Перекрестные ссылки на связанные заявки
Настоящая заявка испрашивает приоритет временных патентных заявок США №№ 60/933307, 60/933331 и 60/933312, поданных 6 июня 2007, включенных, таким образом, в качестве ссылки в полном объеме.
Область изобретения
Настоящее изобретение относится к способам конструирования белков с целью оптимизации их эффективности при определенных интересующих окружающих условиях. В некоторых вариантах осуществления настоящее изобретение относится к способам конструирования ферментов с целью оптимизации их каталитической активности при конкретных окружающих условиях. В некоторых предпочтительных вариантах осуществления, настоящее изобретение относится к способам конструирования ферментов с целью оптимизации их каталитической активности и/или стабильности при неблагоприятных окружающих условиях. В некоторых предпочтительных вариантах осуществления настоящее изобретение относится к способам конструирования ферментов с целью оптимизации их стабильности при хранении, конкретно при неблагоприятных окружающих условиях. В некоторых предпочтительных вариантах осуществления настоящее изобретение относится к способам изменения суммарного поверхностного заряда и/или распределения поверхностных зарядов ферментов (например, металлопротеаз) с целью получения вариантов фермента, которые проявляют улучшенную эффективность и/или стабильность в моющих составах по сравнению с начальным или родительским ферментом.
Предпосылки изобретения
Свойства белков, функционирующих вне их естественного окружения, часто являются субоптимальными. Например, ферменты (например, протеазы, липазы, амилазы, целлюлазы и т.д.) часто используются для очистки материи от краски в детергентах для стирки, которые типично включают сложное сочетание активных ингредиентов. Фактически, большинство продуктов для очистки содержат систему поверхностно-активных веществ, отбеливатели, моющие компоненты, гасители мыльной пены, грязесуспендирующие компоненты, грязеотталкивающие компоненты, оптические отбеливатели, смягчители, дисперсанты, соединения, подавляющие перенос красителя, абразивы, бактерициды и отдушки, а также ферменты для очистки. Таким образом, несмотря на сложность современных детергентов, существует множество красок, удалить которые полностью сложно, отчасти из-за субоптимальной эффективности фермента. Несмотря на многие исследования, направленные на разработку ферментов, в данной области сохраняется потребность в способах создания белков для конкретных применений и условий. Фактически, в данной области сохраняется потребность в способах для быстрой и систематической адаптации электростатических свойств других с целью оптимизации их эффективности в коммерческих применениях. В частности, в данной области сохраняется потребность в способах создания промышленно пригодных ферментов, включая в качестве неограничивающих примеров липазы, амилазы, кутиназы, маннаназы, оксидоредуктазы, целлюлазы, пектиназы, протеазы и другие ферменты, для того, чтобы обеспечить улучшенную активность, стабильность и растворимость в очищающих растворах.
Сущность изобретения
Настоящее изобретение относится к способам конструирования белков с целью оптимизации их эффективности при конкретных интересующих окружающих условиях. В некоторых вариантах осуществления настоящее изобретение относится к способам конструирования ферментов с целью оптимизации их каталитической активности при конкретных окружающих условиях. В некоторых предпочтительных вариантах осуществления, настоящее изобретение относится к способам конструирования ферментов с целью оптимизации их каталитической активности и/или стабильности при неблагоприятных окружающих условиях. В некоторых предпочтительных вариантах осуществления, настоящее изобретение относится к способам конструирования ферментов с целью оптимизации их стабильности при хранении, конкретно при неблагоприятных окружающих условиях. В некоторых предпочтительных вариантах осуществления, настоящее изобретение относится к способам изменения суммарного поверхностного заряда и/или распределения поверхностных зарядов ферментов (например, металлопротеаз) с целью получения вариантов фермента, которые проявляют улучшенную эффективность и/или стабильность в моющих составах по сравнению с начальным или родительским ферментом.
Настоящее изобретение относится к способам производства улучшенных вариантов белков, содержащим: тестирование первого свойства множества вариантов белков с единичными заменами в первом тесте и второго свойства во втором тесте, где свойство родительского белка представляет собой заданное значение, равное 1,0, в каждом тесте, благоприятное первое или второе свойство обладает значением, превышающим 1,0, и чрезмерно неблагоприятное первое или второе свойство обладает значением приблизительно менее чем 0,80 или в некоторых предпочтительных вариантах осуществления, приблизительно менее чем 0,60; идентификацию замены, по меньшей мере, в одном из вариантов белков с единичными заменами, которая связана с благоприятным первым свойством и которая не связана с чрезмерно неблагоприятным вторым свойством; идентификацию замены, по меньшей мере, в одном из вариантов белков с единичными заменами, которая связаны с благоприятным вторым свойством и которая не связана с чрезмерно неблагоприятным первым свойством; введение замены из предыдущей стадии в белок для получения варианта белка с множественными заменами. В некоторых вариантах осуществления способы дополнительно содержат тестирование варианта белка с множественными заменами в первом тесте и втором тесте, где улучшенный вариант белка достигает значения, превышающего 1,0, как в первом, так и во втором указанных тестах, или значения, превышающего 1,0, в первом тесте, и значения от 0,80 до 1,0 во втором тесте. В некоторых дополнительных вариантах осуществления, способы дополнительно содержат производство улучшенного варианта белка(ов). В некоторых вариантах осуществления первое и второе свойства обладают отрицательной корреляцией. В некоторых дополнительных вариантах осуществления, благоприятное первое или второе свойство обладает значением, превышающим приблизительно 1,2. В некоторых дополнительных вариантах осуществления, чрезмерно неблагоприятное первое или второе свойство обладает значением приблизительно менее чем 0,40. В некоторых предпочтительных вариантах осуществления, первое свойство представляет собой стабильность, и второе свойство представляет собой эффективность стирки. В некоторых особенно предпочтительных вариантах осуществления, стабильность включает в себя стабильность в детергенте и эффективность стирки включает в себя эффективность отстирывания крови-молока-чернил (КМЧ) в детергенте. В некоторых дополнительных предпочтительных вариантах осуществления, белок представляет собой нейтральную металлопротеазу. В некоторых дополнительных вариантах осуществления, родительский белок представляет собой зрелую форму нейтральной металлопротеазы дикого типа, тогда как в других вариантах осуществления вариант получен из нейтральной металлопротеазы из семейства Bacillaceae. В некоторых особенно предпочтительных вариантах осуществления, вариант получен из нейтральной металлопротеазы рода Bacillus. Еще в дополнительных вариантах осуществления эффективность стирки тестируют в порошке или жидком моющем составе, обладающем pH между 5 и 12,0. В некоторых дополнительных вариантах осуществления эффективность стирки тестируют в холодной воде с жидким детергентом, обладающим основным pH. Еще в дополнительных вариантах осуществления, по меньшей мере, одна из замен содержит изменение суммарного заряда, равное 0, -1 или -2 по отношению к родительской нейтральной металлопротеазе, тогда как в некоторых альтернативных вариантах осуществления, по меньшей мере, одна из замен содержит изменение суммарного заряда, равное +1 или +2 по отношению к родительской нейтральной металлопротеазе. Не планируется, что стадии ограничиваются точным перечисленным выше порядком, так как любой подходящий порядок находит применение в настоящем изобретении. В некоторых предпочтительных вариантах осуществления, улучшенный вариант протеазы обладает изменением суммарного заряда, равным +1 или +2 по отношению к родительской нейтральной металлопротеазе. Еще в дополнительных вариантах осуществления, замены находятся в позициях в родительской нейтральной металлопротеазе, обладающей доступной для растворителя поверхностью (ДРП) приблизительно больше чем 50%. В других дополнительных вариантах осуществления, одна или несколько позиций в родительской нейтральной металлопротеазе представляют собой позиции, обладающие доступной для растворителя поверхностью (ДРП) приблизительно больше чем 65%.
Настоящее изобретение относится к способам производства улучшенных вариантов протеазы, содержащим: тестирование первого свойства множества вариантов протеазы с единичными заменами в первом тесте и второго свойства во втором тесте, где свойство родительской протеазы представляет собой заданное значение, равное 1,0, в каждом тесте, благоприятное первое или второе свойство обладает значением, превышающим 1,0, и чрезмерно неблагоприятное первое или второе свойство обладает значением приблизительно менее чем 0,80 или в некоторых предпочтительных вариантах осуществления приблизительно менее чем 0,60; идентификацию замены, по меньшей мере, в одном из вариантов протеазы с единичными заменами, которые связаны с благоприятным первым свойством и которые не связаны с чрезмерно неблагоприятным вторым свойством; идентификацию замены, по меньшей мере, в одном из вариантов протеазы с единичными заменами, которые связаны с благоприятным вторым свойством и которые не связаны с чрезмерно неблагоприятным первым свойством; введение замены из предыдущей стадии в протеазу, чтобы получить вариант протеазы с множественными заменами. В некоторых вариантах осуществления способы дополнительно содержат тестирование варианта протеазы с множественными заменами в первом тесте и втором тесте, где улучшенный вариант протеазы достигает значения, превышающего 1,0, как в первом, так и во втором тестах, или значения, превышающего 1,0, в первом тесте, и значения от 0,80 до 1,0 во втором тесте. В некоторых дополнительных вариантах осуществления, способы дополнительно содержат производство улучшенного варианта протеазы(з). В некоторых вариантах осуществления первое и второе свойства обладают отрицательной корреляцией. В некоторых дополнительных вариантах осуществления, благоприятное первое или второе свойство обладает значением приблизительно больше чем 1,2. В некоторых дополнительных вариантах осуществления, чрезмерно неблагоприятное первое или второе свойство обладает значением приблизительно менее чем 0,40. В некоторых предпочтительных вариантах осуществления, первое свойство представляет собой стабильность, и второе свойство представляет собой эффективность стирки. В некоторых особенно предпочтительных вариантах осуществления, стабильность содержит стабильность в детергенте и эффективность стирки содержит эффективность отстирывания крови-молока-чернил (КМЧ) в детергенте. В некоторых дополнительных предпочтительных вариантах осуществления, протеаза представляет собой нейтральную металлопротеазу. В некоторых дополнительных вариантах осуществления, родительская протеаза представляет собой зрелую форму нейтральной металлопротеазы дикого типа, тогда как в других вариантах осуществления вариант получен из нейтральной металлопротеазы семейства Bacillaceae. В некоторых особенно предпочтительных вариантах осуществления, вариант получен из нейтральной металлопротеазы рода Bacillus. Еще в дополнительных вариантах осуществления эффективность стирки тестируют в порошке или жидком моющем составе, обладающем pH между 5 и 12,0. В некоторых дополнительных вариантах осуществления, эффективность стирки тестируют в холодной воде с жидким детергентом, обладающим основным pH. Еще в дополнительных вариантах осуществления, по меньшей мере, одна из замен содержит изменение суммарного заряда, равное 0, -1 или -2 по отношению к родительской нейтральной металлопротеазе, тогда как в некоторых альтернативных вариантах осуществления, по меньшей мере, одна из замен содержит изменение суммарного заряда, равное +1 или +2 по отношению к родительской нейтральной металлопротеазе. Не планируется, что стадии ограничиваются точным перечисленным выше порядком, так как любой подходящий порядок находит применение в настоящем изобретении. В некоторых предпочтительных вариантах осуществления, улучшенный вариант протеазы обладает изменением суммарного заряда, равного +1 или +2 по отношению к родительской нейтральной металлопротеазе. Еще в дополнительных вариантах осуществления, замены находятся в позициях в родительской нейтральной металлопротеазе, обладающих доступной для растворителя поверхностью (ДРП) приблизительно больше чем 50%. В других дополнительных вариантах осуществления, одна или несколько позиций в родительской нейтральной металлопротеазе представляют собой позиции, обладающие доступной для растворителя поверхностью (ДРП) приблизительно больше чем 65%. Настоящее изобретение также относится к белкам с множественными заменами, произведенным с помощью способов, изложенных в настоящем документе. В некоторых предпочтительных вариантах осуществления, настоящее изобретение относится к вариантам нейтральной металлопротеазы, произведенных способом, изложенным в настоящем документе. В некоторых особенно предпочтительных вариантах осуществления, настоящее изобретение относится к вариантам протеазы, содержащим замену в позиции остатка, соответствующей позиции остатка 83 нейтральной металлопротеазы Bacillus, приведенной как SEQ ID NO:3. В некоторых дополнительных предпочтительных вариантах осуществления, замена содержит замену L83K. Также предоставлены варианты NprE, содержащие сочетания замен, выбранных из группы, состоящей из: i) 4K-45K-50R-54K-59K-90K-129I-138L-179P-190L-199E-214Q-220E-244S-265P-269H-285R-296E; 45K-50R-59K-90K-1291-138L-179P-190L-199E-214Q-220E-244S-265P-285R; 45K-59K-90K-129I-138L-179P-190L-199E-214Q-220E-265P-285R; и 59K-90K-129I-179P-190L-199E-214Q-220E-265P-285R.
Настоящее изобретение также относится к выделенным полинуклеотидам, кодирующим варианты протеазы, изложенные в настоящем документе. Кроме того, настоящее изобретение относится к экспрессирующим векторам, содержащим полинуклеотид, изложенный в настоящем документе, в функциональном сочетании с промотором. В настоящем изобретении также предоставлены клетки-хозяева, трансформированные экспрессирующим вектором(ами), предоставленным в настоящем документе. Настоящее изобретение также относится к чистящим композициям, содержащим варианты протеазы, произведенные с помощью способов по настоящему изобретению.
Настоящее изобретение относится к способам производства улучшенных вариантов белков, содержащим: a) тестирование множества вариантов белков с единичными заменами в первом тесте первого свойства и второго свойства во втором тесте, где свойство родительского белка представляет собой заданное значение, равное 1,0, в каждом тесте, благоприятное первое или второе свойство обладает значением, превышающим 1,0, и чрезмерно неблагоприятное первое или второе свойство обладает значением приблизительно менее чем 0,80; b) идентификацию замены, по меньшей мере, в одном из вариантов белков с единичными заменами, которая связана с благоприятным первым свойством и которая не связана с чрезмерно неблагоприятным вторым свойством; c) идентификацию замены, по меньшей мере, в одном из вариантов белков с единичными заменами, которая связана с благоприятным вторым свойствам и которая не связана с чрезмерно неблагоприятным первым свойством; и d) введение замены из стадии b) и замены из стадии c) в белок, чтобы получить вариант белка с множественными заменами. В некоторых вариантах осуществления способы дополнительно содержат стадию e) тестирования варианта белка с множественными заменами в первом тесте и втором тесте, где улучшенный вариант белка достигает значения, превышающего 1,0, как в первом, так и во втором тестах, или значения, превышающего 1,0 в первом тесте, и значения от 0,80 до 1,0 во втором тесте.
В некоторых вариантах осуществления белок представляет собой фермент, выбранный из группы, состоящей из протеазы, амилазы, целлюлазы, полиэстеразы, эстеразы, липазы, кутиназы, пектиназы, оксидазы, трансферазы, каталазы и алкалазы. В некоторых предпочтительных вариантах осуществления, фермент представляет собой протеазу или амилазу. В некоторых вариантах осуществления первое и второе интересующие свойства содержат два или более свойства, выбранные из группы, состоящей из связывания субстрата, ингибирования фермента, экспрессии, стабильности в детергенте, тепловой стабильности; скорости реакции; степени завершенности реакции; тепловой активности; разжижения крахмала; разложения биомассы, осахаривания, гидролиза эфира, ферментативного отбеливания, эффективности стирки и модификации текстиля. В некоторых особенно предпочтительных вариантах осуществления, способы дополнительно содержат производство улучшенного варианта белка. В некоторых вариантах осуществления первое и второе свойства обладают отрицательной корреляцией. В некоторых вариантах осуществления настоящего изобретения благоприятные первое или второе свойство обладает значением приблизительно больше чем 1,2, и/или чрезмерно неблагоприятное первое или второе свойство обладает значением менее чем приблизительно 0,60. В некоторых предпочтительных вариантах осуществления, первое свойство представляет собой стабильность, и второе свойство представляет собой эффективность стирки. В поднаборе этих вариантов осуществления, стабильность содержит стабильность в детергенте, и эффективность стирки содержит эффективность отстирывания крови-молока-чернил (КМЧ) в детергенте. В некоторых вариантах осуществления первое свойство представляет собой экспрессию белка, и второе свойство представляет собой ферментативную активность. В поднаборе этих вариантов осуществления, ферментативная активность содержит эффективность отстирывания рисового крахмала в детергенте. В некоторых предпочтительных вариантах осуществления, протеаза выбрана из группы, состоящей из нейтральной металлопротеазы, сериновой протеазы и субтилизина. В поднаборе этих вариантов осуществления, нейтральная металлопротеаза представляет собой нейтральную металлопротеазу семейства Bacillaceae. В некоторых образцовых вариантах осуществления, нейтральная металлопротеаза из рода Bacillus (например, NprE B. subtilis). В дополнительных вариантах осуществления амилаза представляет собой α-амилазу семейства Bacillaceae. В образцовых вариантах осуществления, α-амилаза из рода Bacillus (например, AmyS B. stearothermophilus). В некоторых предпочтительных вариантах осуществления, эффективность стирки тестируют в порошке или жидком моющем составе, обладающем pH между 5 и 12,0, и/или холодной воде с жидким детергентом, обладающим основным pH. В некоторых вариантах осуществления, по меньшей мере, одна из замен содержит изменение суммарного заряда, равное 0, -1 или -2 по отношению к родительскому ферменту. В некоторых вариантах осуществления, по меньшей мере, одна из замен содержит изменение суммарного заряда, равного +1 или +2 по отношению к родительскому ферменту. В некоторых предпочтительных вариантах осуществления, по меньшей мере, одна из замен содержит, по меньшей мере, две замены, первую с изменением суммарного заряда, равным 0, -1 или -2 по отношению к родительскому ферменту; и вторую с изменением суммарного заряда, равным +1 или +2 по отношению к родительскому ферменту. В некоторых вариантах осуществления, по меньшей мере, одна замена содержит от одной до 20 (например, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19 или 20) замен. В некоторых вариантах осуществления улучшенный вариант фермента обладает изменением суммарного заряда, равного -2, -1, 0, +1 или +2 по отношению к родительскому ферменту. В некоторых вариантах осуществления замены находятся в позициях в родительском ферменте, обладающих доступной для растворителя поверхностью (ДРП) приблизительно больше чем 25%, приблизительно больше чем 50% или приблизительно больше чем 65%. Также настоящим изобретением предоставлены выделенные полинуклеотиды, кодирующие варианты фермента, изложенные в настоящем документе. В дополнительных вариантах осуществления настоящее изобретение относится к экспрессирующему вектору, содержащему полинуклеотид в функциональном сочетании с промотором. В некоторых вариантах осуществления предоставлены клетки-хозяева, содержащие экспрессирующий вектор. В дополнительных вариантах осуществления предоставлены чистящие композиции, которые содержат варианты фермента, произведенные с использованием способов по настоящему изобретению.
Настоящее изобретение относится к способам производства улучшенных вариантов белков, содержащим в функциональном порядке: a) тестирование первого свойства множества вариантов белков с единичными заменами в первом тесте и второго свойства во втором тесте, где свойство родительского белка представляет собой заданное значение, равное 1,0, в каждом тесте, благоприятное первое или второе свойство обладает значением, превышающим 1,0, и чрезмерно неблагоприятное первое или второе свойство обладает значением приблизительно менее чем 0,80; b) идентификацию замены, по меньшей мере, в одном из вариантов белков с единичными заменами, которая связана с благоприятным первым свойством и которая не связана с чрезмерно неблагоприятным вторым свойством; c) идентификацию замены, по меньшей мере, в одном из вариантов белков с единичными заменами, которая связана с благоприятным вторым свойством и которая не связана с чрезмерно неблагоприятным первым свойством; и d) введение замены из стадии b) и замены из стадии c) в белок, чтобы получить вариант белка с множественными заменами, где вариант белка с множественными заменами является улучшенным вариантом белка. В некоторых предпочтительных вариантах осуществления, способы дополнительно содержат: стадию e) тестирования варианта белка с множественными заменами в первом тесте и втором тесте, где улучшенный вариант белка достигает значения, превышающего 1,0, как в первом, так и во втором тестах, или значения, превышающего 1,0 в первом тесте, и значения от 0,80 до 1,0 во втором тесте. В некоторых дополнительных вариантах осуществления, родительский белок представляет собой фермент, где улучшенные варианты белков являются ферментами. В некоторых дополнительных вариантах осуществления, фермент выбран из протеаз, амилаз, целлюлаз, полиэстераз, эстераз, липаз, кутиназ, пектиназ, оксидаз, трансфераз и каталаз. В некоторых особенно предпочтительных вариантах осуществления, фермент представляет собой протеазу или амилазу. В некоторых дополнительных вариантах осуществления, первое и второе интересующие свойства содержат два или более из группы, состоящей из связывания субстрата, ингибирования фермента, экспрессии, стабильности в детергенте, тепловой стабильности, скорости реакции, степени завершенности реакции, тепловой активности, разжижения крахмала, разложения биомассы, осахаривания, гидролиза эфира, ферментативного отбеливания, эффективности стирки и модификации текстиля. В некоторых других дополнительных вариантах осуществления, способы дополнительно содержат стадию производства улучшенного варианта белка. В некоторых дополнительных вариантах осуществления, первое и второе свойства обладают отрицательной корреляцией. В некоторых особенно предпочтительных вариантах осуществления, благоприятное первое или второе свойство обладают значением приблизительно больше чем 1,2. В некоторых дополнительных предпочтительных вариантах осуществления, чрезмерно неблагоприятное первое или второе свойство обладает значением приблизительно менее чем 0,60. В некоторых других дополнительных вариантах осуществления, чрезмерно неблагоприятное первое или второе свойство обладает значением приблизительно менее чем 0,40. В некоторых предпочтительных вариантах осуществления, первое свойство представляет собой стабильность, и второе свойство представляет собой эффективность стирки. В некоторых особенно предпочтительных вариантах осуществления, стабильность содержит стабильность в моющих составах и эффективность стирки содержит эффективность отстирывания крови-молока-чернил (КМЧ). В некоторых других дополнительных вариантах осуществления, эффективность стирки тестируют в порошке или жидком моющем составе, обладающем pH между приблизительно 5 и приблизительно 12. В некоторых дополнительных предпочтительных вариантах осуществления, эффективность стирки тестируют в холодной воде с жидким детергентом, обладающим основным pH. В некоторых дополнительных вариантах осуществления, первое свойство представляет собой экспрессию белка, и второе свойство представляет собой ферментативную активность. В некоторых дополнительных вариантах осуществления, протеаза выбрана из нейтральных металлопротеаз и сериновых протеаз. В некоторых особенно предпочтительных вариантах осуществления, сериновая протеаза представляет собой субтилизин. В некоторых дополнительных вариантах осуществления, нейтральная металлопротеаза представляет собой нейтральную металлопротеазу, полученную из члена семейства Bacillaceae. В некоторых альтернативных вариантах осуществления, амилаза представляет собой α-амилазу, полученную из члена семейства Bacillaceae. В некоторых дополнительных вариантах осуществления, по меньшей мере, одна из замен содержит изменение суммарного заряда, равное 0, -1, или -2 по отношению к родительскому ферменту. В некоторых альтернативных вариантах осуществления, по меньшей мере, одна из замен содержит изменение суммарного заряда, равное +1 или +2 по отношению к родительскому ферменту. В некоторых других дополнительных вариантах осуществления, по меньшей мере, одна из замен содержит изменение суммарного заряда, равное 0, -1 или -2 по отношению к родительскому ферменту. В некоторых дополнительных альтернативных вариантах осуществления, по меньшей мере, одна из замен содержит изменение суммарного заряда, равное +1 или +2 по отношению к родительскому ферменту. В некоторых других дополнительных вариантах осуществления, улучшенный вариант фермента обладает изменением суммарного заряда, равным +1 или +2 по отношению к родительскому ферменту. В некоторых альтернативных вариантах осуществления, замены находятся в позициях в родительском ферменте, обладающих доступной для растворителя поверхностью (ДРП) приблизительно больше чем 25%. В некоторых предпочтительных вариантах осуществления, замены находятся в позициях в родительском ферменте, обладающих доступной для растворителя поверхностью (ДРП) приблизительно больше чем 50%, приблизительно больше чем 65%. В некоторых особенно предпочтительных вариантах осуществления, родительский фермент представляет собой фермент дикого типа.
Настоящее изобретение также относится к чистящим композициям, содержащим улучшенный вариант белка, произведенный в соответствии со способами, изложенными в настоящем документе.
Настоящее изобретение также относится к выделенным вариантам нейтральной металлопротеазы, обладающим аминокислотной последовательностью, содержащей, по меньшей мере, одну замену аминокислоты, выполненную в позиции, эквивалентной позиции в нейтральной металлопротеазе, содержащей аминокислотную последовательность, изложенную в SEQ ID NO:3. В некоторых предпочтительных вариантах осуществления, по меньшей мере, одна замена выполнена в позиции, эквивалентной позиции 83 аминокислотной последовательности, изложенной в SEQ ID NO:3. В некоторых особенно предпочтительных вариантах осуществления, замена представляет собой L83K.
Краткое описание чертежей
На фиг.1A показана относительная специфическая гидролитическая активность для BODIPY-крахмала по отношению к экспрессии комбинаторной зарядовой библиотеки (КЗБ) AmyS-S242Q во встряхиваемых пробирках.
На фиг.1B показана относительная активность очистки на микрообразцах с рисовым крахмалом в TIDE 2× по отношению к экспрессии КЗБ AmyS-S242Q во встряхиваемых пробирках.
На фиг.2A показана относительная экспрессия во встряхиваемых пробирках по отношению к относительному изменению суммарного заряда для КЗБ AmyS-S242Q.
На фиг.2B показана относительная гидролитическая активность для BODIPY-крахмала по отношению к относительному изменению суммарного заряда для КЗБ AmyS-S242Q.
На фиг.3A показана относительная экспрессия во встряхиваемых пробирках по отношению к относительному изменению суммарного заряда для КЗБ AmyS-S242Q.
На фиг.3B показана активность очистки микрообразцов с рисовым крахмалом по отношению к относительному изменению суммарного заряда для КЗБ AmyS-S242Q.
Общее описание изобретения
Настоящее изобретение относится к способам конструирования белков с целью оптимизации их эффективности при конкретных интересующих окружающих условиях. В некоторых вариантах осуществления настоящее изобретение относится к способам конструирования ферментов с целью оптимизации их каталитической активности при конкретных окружающих условиях. В некоторых предпочтительных вариантах осуществления, настоящее изобретение относится к способам конструирования ферментов с целью оптимизации их каталитической активности и/или стабильности при неблагоприятных окружающих условиях. В некоторых предпочтительных вариантах осуществления, настоящее изобретение относится к способам конструирования ферментов с целью оптимизации их стабильности при хранении, конкретно при неблагоприятных окружающих условиях. В некоторых предпочтительных вариантах осуществления, настоящее изобретение относится к способам изменения суммарного поверхностного заряда и/или распределения поверхностных зарядов ферментов (например, металлопротеаз) с целью получения вариантов фермента, которые проявляют улучшенную эффективность и/или стабильность в моющих составах по сравнению с начальным или родительским ферментом.
Протеаза субтилизин является основным ферментом, используемым в детергентах для стирки и возможно наиболее широко используемым ферментом в мире. Отмечалось, что поверхностные электростатические эффекты могут модулировать каталитическую активность субтилизина (см., например, Russell and Fersht, Nature 328:496-500 [1987]). Совсем недавно были открыты мутации, которые участвуют в изменении суммарного заряда субтилизина и имеют сильное влияние на эффективность стирки в детергентах (см., например, EP патент № 0479870 B1). Полагалось, что это положительное воздействие является результатом сдвига pI (изоэлектрической точки) субтилизина в сторону pH моющего раствора. Однако последняя работа показала, что это заключение не всегда является применимым (см., например, патент США № 6673590 B1). Как указано в этом патенте, влияние зарядовых мутаций в субтилизине сильно зависит от концентрации детергента, причем мутации, снижающие pI родительского субтилизина, предоставляют фермент, который более эффективен при низкой концентрации детергента, и мутации, повышающие pI, предоставляют фермент, который более эффективен при высокой концентрации детергента. Это обладает значительной выгодой, так как концентрация детергента в моющих растворах значительно меняется в мировом масштабе. Таким образом, специалистам в данной области становится понятно, что существует оптимальная pI субтилизина для эффективной стирки, которая зависит от pH и концентрации детергента в моющем растворе. Описывались дальнейшие попытки увеличения активности субтилизина в детергентах для стирки (см. патентную заявку США № 2005/0221461). К удивлению было обнаружено, что варианты субтилизина, обладающие таким же суммарным электростатическим зарядом, как у родительского субтилизина, обладают повышенной эффективностью стирки в условиях мытья как с высокой, так и с низкой концентрацией детергента.
Если не указано иначе, осуществление настоящего изобретения на практике затрагивает общепринятые способы, которые являются общеупотребительными в конструировании белков, молекулярной биологии, микробиологии и рекомбинантной ДНК, которые известны специалистам в данной области. Такие способы известны специалистам в данной области и описаны во множестве текстов и справочных работ, хорошо известных профессионалам в данной области. Все патенты, патентные заявки, статьи и публикации, отмеченные в настоящем документе как выше, так и ниже, настоящим явно включены в настоящий документ в качестве ссылки по причине того, что они содержат уместные сведения, относящиеся к способам и композициям, предоставленным настоящим изобретением.
Пока в настоящем документе не установлено обратное, все технические и научные термины, использованные в настоящем документе, обладают таким же значением, в каком обычно их понимает специалист в области, к которой принадлежит это изобретение. Хотя любые способы и материалы, схожие или эквивалентные способам и материалам, описываемым в настоящем документе, находят применение в осуществлении настоящего изобретения на практике, некоторые предпочтительные способы и материалы описаны в настоящем документе. Таким образом, термины, которые определены непосредственно ниже, описаны более полно по отношению к описанию в целом.
Так же, как используется в настоящем документе, единственное число включает множественное число до тех пор, пока в контексте не будет ясно указано обратное. Числовые диапазоны включают в себя числа, определяющие этот диапазон. Если не указано иначе, нуклеиновые кислоты записаны слева направо в направлении от 5' к 3'; аминокислотные последовательности записаны слева направо в направлении от амино- к карбокси-концу соответственно. Следует понимать, что это изобретение не ограничено конкретными описанными способами, протоколами и реактивами, так как они могут меняться в зависимости от контекста, в котором их используют специалисты в данной области.
Предполагается, что каждое максимальное числовое ограничение, заданное на всем протяжении этого описания, включает каждое нижнее числовое ограничение, как если бы такие нижние числовые ограничения были явно прописаны в настоящем документе. Каждое минимальное числовое ограничение, заданное на всем протяжении этого описания, будет включать каждое верхнее числовое ограничение, как если бы такие верхние числовые ограничения были явно прописаны в настоящем документе. Каждый числовой диапазон, заданный на всем протяжении этого описания, будет включать каждый более узкий числовой диапазон, который попадает в пределы такого более широкого числового диапазона, как если бы все такие более узкие числовые диапазоны были явно прописаны в настоящем документе.
Кроме того, заголовки, предоставленные в настоящем документе, не являются ограничениями для различных аспектов или вариантов осуществления изобретения, которые могут иметь место по отношению к описанию в целом. Таким образом, термины, определенные непосредственно ниже, более полно определены по отношению к описанию в целом. Тем не менее, для того чтобы облегчить понимание изобретения, определенное количество терминов определено ниже.
Определения
Как используется в настоящем документе, термины «протеаза» и «протеолитическая активность» относятся к белку или пептиду, проявляющему способность к гидролизу пептидов или субстратов, обладающих пептидными связями. Для измерения протеолитической активности существует множество хорошо известных процедур (см., например, Kalisz, «Microbial Proteases», In: Fiechter (ed.), Advances in Biochemical Engineering/Biotechnology [1988]). Например, протеолитическую активность можно установить с помощью сравнительных анализов, в которых анализируют способность соответствующей протеазы гидролизовать коммерческий субстрат. Образцовые субстраты, пригодные для такого анализа протеазы или протеолитической активности, включают в качестве неограничивающих примеров диметилказеин (Sigma C-9801), бычий коллаген (Sigma C-9879), бычий эластин (Sigma E-1625) и бычий кератин (ICN Biomedical 902111). Колориметрические тесты, использующие эти субстраты, хорошо известны в данной области (см., например, WO 99/3401 1; и патент США № 6376450). The pNA assay (см., например, Del Mar et al, Anal. Biochem, 99:316-320 [1979]) также находят применение при определении активной концентрации фермента для фракций, собранных в процессе градиентного элюирования. В этом анализе измеряют скорость, с которой p-нитроанилин высвобождается при гидролизе ферментом растворимого синтетического субстрата, сукцинил-аланин-аланин-пролин-фенилаланин-p-нитроанилида (sAAPF-pNA). Скорость развития желтой окраски в результате реакции гидролиза измеряют при 410 нм на спектрофотометре и она пропорциональна активной концентрации фермента. Кроме того, измерение оптической плотности при 280 нм можно использовать для определения общей концентрации белка. Отношение активного фермента к общему белку дает чистоту фермента.
Как используется в настоящем документе, термины «протеаза ASP», «протеаза ASP» и «ASP», относятся к сериновым протеазам, описываемым в настоящем документе и описанным в патенте США, серийный № заявки 10/576331). В некоторых предпочтительных вариантах осуществления, протеаза ASP представляет собой протеазу, обозначенную в настоящем документе как протеаза 69B4, полученная из штамма 69B4 Cellulomonas. Таким образом, в предпочтительных вариантах осуществления термин «протеаза 69B4» относится к встречающейся в природе зрелой протеазе, полученной из штамма 69B4 Cellulomonas (DSM 16035). В альтернативных вариантах осуществления, настоящее изобретение относится к фрагментам протеазы ASP.
Термин «гомологи протеазы Cellulomonas» относится к встречающимся в природе протеазам, обладающим аминокислотными последовательностями, в значительной степени идентичными зрелой протеазе, полученной из штамма 69B4 Cellulomonas, или полинуклеотидной последовательности, которая кодирует такие встречающиеся в природе протеазы, и эти протеазы сохраняют функциональные характеристики сериновой протеазы, кодируемой такими нуклеиновыми кислотами. В некоторых вариантах осуществления эти гомологи протеазы обозначают как «целлюломонадины».
Как используются в настоящем документе, термины «вариант ASP», «вариант протеазы ASP» и «вариант протеазы 69B» используют по отношению к протеазам, которые схожи с ASP дикого типа, в частности их функцией, но обладают мутациями в своей аминокислотной последовательности, которые отличают их последовательность от протеазы дикого типа.
Как используется в настоящем документе, «Cellulomonas ssp.» относится ко всем видам рода «Cellulomonas», которые являются грамположительными бактериями, систематизированными в качестве членов семейства Cellulomonadaceae, подотряда Micrococcineae, отряда Actinomycetales, класса Actinobacteria. Установлено, что род Cellulomonas продолжает подвергаться таксономической реорганизации. Таким образом, предполагается, что род включает виды, которые были пересистематизированы.
Как используется в настоящем документе, «Streptomyces ssp.» относится ко всем видам рода «Streptomyces», которые являются грамположительными бактериями, систематизированным в качестве членов семейства Streptomycetaceae, подотряда Streptomycineae, отряда Actinomycetales, класса Actinobacteria. Установлено, что род Streptomyces продолжает подвергаться таксономической реорганизации. Таким образом, предполагается, что род включает в себя виды, которые были пересистематизированы.
Как используется в настоящем документе, «род Bacillus» включает все виды рода «Bacillus», известные специалистам в данной области, включая в качестве неограничивающих примеров B. subtilis, B. licheniformis, B. lentus, B. brevis, B. stearothermophilus, B. alkalophilus, B. amyloliquefaciens, B. clausii, B. halodurans, B. megaterium, B. coagulans, B. circulans, B. lautus и B. thuringiensis. Установлено, что род Bacillus продолжает подвергаться таксономической реорганизации. Таким образом, предполагается, что род включает в себя виды, которые были пересистематизированы, включая в качестве неограничивающих примеров такие организмы, как B. stearothermophilus, который теперь называется «Geobacillus stearothermophilus». Производство устойчивых эндоспор в присутствии кислорода сочли определяющим признаком рода Bacillus, хотя эта характеристика также применима к недавно названным Alley clobacillus, Amphibacillus, Aneurinibacillus, Anoxybacillus, Brevibacillus, Filobacillus, Gracilibacillus, Halobacillus, Paenibacillus, Salibacillus, Thermobacillus, Ureibacillus и Virgibacillus.
Термины «полинуклеотид» и «нуклеиновая кислота», используемые в настоящем документе взаимозаменяемо, относятся к полимерной форме нуклеотидов любой длины, или рибонуклеотидов, или дезоксирибонуклеотидов. Эти термины включают в качестве неограничивающих примеров одно-, двух- или трехцепочечные ДНК, геномную ДНК, кДНК, РНК, ДНК-РНК гибриды или полимер, содержащий пуриновые и пиримидиновые основания, или другие природные, химически, биохимически модифицированные, ненатуральные нуклеотидные основания или их производные. Приведены неограничивающие примеры полинуклеотидов: гены, фрагменты генов, фрагменты хромосом, EST, экзоны, интроны, мРНК, тРНК, рРНК, рибозимы, кДНК, рекомбинантные полинуклеотиды, разветвленные полинуклеотиды, плазмиды, векторы, выделенные ДНК с любой последовательностью, выделенные РНК с любой последовательностью, зонды из нуклеиновых кислот и праймеры. В некоторых вариантах осуществления полинуклеотиды содержат модифицированные нуклеотиды, такие как метилированные нуклеотиды и нуклеотидные аналоги, урацил, другие сахара и связывающие группы, такие как фторрибоза и тиоат, и ветвящиеся нуклеотиды. В альтернативных вариантах осуществления, последовательность нуклеотидов прерывается ненуклеотидными компонентами.
Как используется в настоящем документе, термины «ДНК конструкция» и «трансформирующая ДНК» используют взаимозаменяемо по отношению к ДНК, используемой для введения последовательностей в клетку-хозяина или организм. ДНК может быть создана in vitro с помощью ПЦР или любого другого пригодного способа(ов), известного для этих целей в данной области. В особенно предпочтительных вариантах осуществления ДНК конструкция содержит интересующую последовательность (например, в виде вводимой последовательности). В некоторых вариантах осуществления последовательность функционально связана с дополнительными элементами, такими как управляющие элементы (например, промоторы и т.д.). ДНК конструкция может дополнительно содержать селективный маркер. Она может дополнительно содержать вводимую последовательность, фланкированную гомологичными фрагментами. В дополнительном варианте осуществления трансформирующая ДНК содержит другие негомологичные последовательности, добавленные на концах (например, спейсерные последовательности или фланкирующие последовательности). В некоторых вариантах осуществления концы вводимой последовательности замкнуты так, чтобы трансформирующая ДНК образовывала замкнутое кольцо. Трансформирующие последовательности могут быть дикого типа, мутантными или модифицированными. В некоторых вариантах осуществления ДНК конструкция содержит последовательности, гомологичные хромосоме клетки-хозяина. В других вариантах осуществления ДНК конструкция содержит негомологичные последовательности. Когда ДНК конструкция собрана in vitro, ее можно использовать для: 1) вставки гетерологичных последовательностей в требуемую последовательность-мишень клетки-хозяина; и/или 2) мутирования области хромосомы клетки-хозяина (т.е. замещения эндогенной последовательности гетерологичной последовательностью) и/или 3) удаления намеченных генов; и/или введения реплицирующейся плазмиды в хозяйский организм.
Как используется в настоящем документе, термины «экспрессирующая кассета» и «экспрессирующий вектор» относятся к конструкции из нуклеиновой кислоты, созданной рекомбинантно или синтетически, с рядом определенных элементов из нуклеиновых кислот, которые позволяют транскрипцию конкретной нуклеиновой кислоты в клетке-мишени. Рекомбинантная экспрессирующая кассета может быть встроена в плазмиду, хромосому, митохондриальную ДНК, пластидную ДНК, вирус или фрагмент нуклеиновой кислоты. Типично, часть экспрессирующего вектора с рекомбинантной экспрессирующей кассетой содержит, среди прочих последовательностей, последовательность нуклеиновой кислоты, подлежащую транскрипции, и промоторы. В предпочтительных вариантах осуществления экспрессирующие векторы обладают способностью встраивать и экспрессировать гетерологичные ДНК фрагменты в клетке-хозяине. Многие прокариотические и эукариотические экспрессирующие векторы коммерчески доступны. Выбор подходящих экспрессирующих векторов находится в компетенции специалистов в данной области. Термин «экспрессирующая кассета» используется взаимозаменяемо в настоящем документе с «ДНК конструкцией» и ее грамматическими эквивалентами. Выбор подходящих экспрессирующих векторов находится в компетенции специалистов в данной области.
Как используется в настоящем документе, термин «вектор» относится к полинуклеотидной конструкции, спроектированной для введения нуклеиновых кислот в один или несколько типов клеток. Векторы включают клонирующие векторы, экспрессирующие векторы, челночные векторы, плазмиды, кассеты и т.п. В некоторых вариантах осуществления полинуклеотидная конструкция содержит последовательность ДНК, кодирующую протеазу (например, предшественник или зрелую протеазу), которая функционально связана с подходящей пропоследовательностью (например, секреторной и т.д.), которая допускает осуществление экспрессии ДНК в пригодном хозяйском организме.
Как используется в настоящем документе, термин «плазмида» относится к кольцевой двухцепочечной (дц) ДНК конструкции, используемой в качестве клонирующего вектора, который образует внехромосомный самореплицирующийся генетический элемент в некоторых эукариотах и прокариотах, или интегрируется в хромосому хозяина.
Как используется в настоящем документе в контексте относительно введения последовательности нуклеиновой кислоты в клетку, термин «введенный» относится к любому способу, пригодному для переноса последовательности нуклеиновой кислоты в клетку. Такие способы для введения включают в качестве неограничивающих примеров слияние протопластов, трансфекцию, трансформацию, конъюгирование и трансдукцию (см., например, Ferrari et al., «Genetics», in Hardwood et al., (eds.), Bacillus. Plenum Publishing Corp., стр.57-72 [1989]).
Как используется в настоящем документе, термины «трансформированный» и «стабильно трансформированный» относятся к клетке, которая имеет негативную (гетерологичную) полинуклеотидную последовательность, интегрированную в ее геном или в виде эписомной плазмиды, которая сохраняется, по меньшей мере, в течение двух поколений.
Как используется в настоящем документе, термин «нуклеотидная последовательность, кодирующая селективный маркер» относится к нуклеотидной последовательности, которая делает возможной экспрессию в клетках-хозяевах и где экспрессия селективного маркера дает клеткам, содержащим экспрессированный ген, способность к росту в присутствии соответствующего селективного средства или при недостатке необходимого питательного вещества.
Как используется в настоящем документе, термины «селективный маркер» и «постоянный селективный маркер» относятся к нуклеиновой кислоте (например, гену), которая допускает экспрессию в клетке-хозяине, которая позволяет просто отбирать те хозяйские организмы, которые содержат вектор. Примеры таких селективных маркеров включают в качестве неограничивающих примеров противомикробные вещества. Таким образом, термин «селективный маркер» относится к генам, которые обеспечивают индикацию того, что клетка-хозяин захватила входящую интересующую ДНК, или наступление какой-нибудь другой реакции. Типично, селективные маркеры представляют собой гены, которые дают устойчивость к противомикробным средствам или метаболическое преимущество клетке-хозяину, чтобы позволить отличить клетки, содержащие экзогенную ДНК, от клеток, которые не приняли никакую экзогенную последовательность в процессе трансформации. «Постоянный селективный маркер» представляет собой маркер, который расположен на хромосоме микроорганизма, подлежащего трансформации. Постоянный селективный маркер кодирует ген, который отличается от селективного маркера в трансформирующей ДНК конструкции. Селективные маркеры хорошо известны специалистам в данной области. Как отмечено выше, предпочтительно, маркер представляет собой маркер устойчивости к противомикробному средству (например, ampR; phleoR; specR; kanR; eryR; tetR; cmpR и neoR (см., например, Guerot-Fleury, Gene, 167:335-337 [1995]; Palmeros et al, Gene 247:255-264 [2000]; и Trieu-Cuot et al., Gene, 23:331-341 [1983])). Другие пригодные в соответствии с изобретением маркеры включают в качестве неограничивающих примеров ауксотрофные маркеры, такие как триптофан; и маркеры обнаружения, такие как β-галактозидаза.
Как используется в настоящем документе, термин «промотор» относится к последовательности нуклеиновой кислоты, которая направляет транскрипцию гена, расположенного в направлении 5'-3'. В предпочтительных вариантах осуществления промотор соответствует клетке-хозяину, в которой намеченный ген должен быть экспрессирован. Промотор, вместе с другими транскрипционными и трансляционными регуляторными последовательностями нуклеиновой кислоты (также называемыми «управляющими последовательностями»), необходим для экспрессии заданного гена. В основном, транскрипционные и трансляционные регуляторные последовательности включают в качестве неограничивающих примеров промоторные последовательности, участки связывания рибосомы, транскрипционные стартовые и останавливающие последовательности, трансляционные стартовые и останавливающие последовательности и энхансерные или активаторные последовательности.
Нуклеиновая кислота «функционально связана», когда она расположена в функциональной взаимосвязи с другой последовательностью нуклеиновой кислоты. Например, ДНК, кодирующая секреторную лидерную последовательность (т.е. сигнальный пептид), функционально связана с ДНК для полипептида, если она экспрессирована в виде белка-предшественника, который участвует в секреции полипептида; промотор или энхансер функционально связан с кодирующей последовательностью, если он влияет на транскрипцию последовательности; или сайт связывания рибосомы функционально связан с кодирующей последовательностью, если он расположен так, чтобы облегчать трансляцию. Как правило, «функционально связан» обозначает, что последовательности ДНК соединены смежно, и, в случае секреторной лидерной последовательности, смежны и находятся в одной фазе считывания. Однако энхансеры не должны быть смежными. Связывание выполняется посредством лигирования в пригодных участках рестрикции. Если такие сайты отсутствуют, синтетические олигонуклеотидные адаптеры или линкеры используют в соответствии с состоянием техники.
Как используется в настоящем документе, термин «ген» относится к полинуклеотиду (например, сегменту ДНК), который кодирует полипептид и содержит области, которые идут до и после кодирующих областей, а также промежуточные последовательности (интроны) между отдельными кодирующими сегментами (экзонами).
Как используется в настоящем документе, «гомологичные гены» относятся к паре генов из различных, но обычно родственных видов, которые соответствуют друг другу и которые идентичны или очень схожи друг с другом. Термин охватывает гены, которые разделились посредством видообразования (т.е. развития новых видов) (например, ортологичные гены), а также гены, которые разделились генетическим удвоением (например, паралогичные гены).
Как используется в настоящем документе, «ортолог» и «ортологичные гены» относятся к генам в различных видах, которые развились из общего предкового гена (т.е. гомологичный ген) посредством видообразования. Типично, ортологи сохраняют одну функцию в процессе эволюции. Идентификация ортологов находит применение при надежном предсказании функции гена во вновь секвенированных геномах.
Как используется в настоящем документе, «паралог» и «паралогичные гены» относятся к генам, которые связаны через дупликацию в пределах генома. Тогда как ортологи сохраняют одну функцию в процессе эволюции, паралоги развивают новые функции, даже если некоторые функции часто связаны с исходной функцией. Примеры паралогичных генов включают в качестве неограничивающих примеров гены, кодирующие трипсин, химотрипсин, эластазу и тромбин, все они являются сериновыми протеазами и встречаются вместе в одних и тех же видах.
Как используется в настоящем документе, «гомология» относится к сходству или идентичности последовательности, причем идентичность является предпочтительной. Эту гомологию устанавливают с помощью известных в данной области способов (см., например, Smith and Waterman, Adv. Appl. Math., 2:482 [1981]; Needleman and Wunsch, J. Mol. Biol., 48:443 [1970]; Pearson and Lipman, Proc. Natl. Acad.Sci. USA, 85:2444 [1988]; программы, такие как GAP, BESTFIT, FASTA и TFASTA в пакете программного обеспечения Wisconsin Genetics, Genetics Computer Group, Madison, WI; и Devereux et ah, Nucl. Acid Res., 12:387-395 [1984]).
Как используется в настоящем документе, «аналогичная последовательность» представляет собой последовательность, в которой функция гена в основном является такой же, так как ген основан на родительском гене (например, протеаза штамма 69B4 Cellulomonas). Дополнительно, аналогичные гены включают, по меньшей мере, приблизительно 45%, приблизительно 50%, приблизительно 55%, приблизительно 60%, приблизительно 65%, приблизительно 70%, приблизительно 75%, приблизительно 80%, приблизительно 85%, приблизительно 90%, приблизительно 95%, приблизительно 97%, приблизительно 98%, приблизительно 99% или приблизительно 100% идентичность последовательности с последовательностью родительского гена. Альтернативно, аналогичные последовательности обладают выравниванием с 70-100% генов, найденных в области родительского гена (например, протеаза штамма 69B4 Cellulomonas) и/или имеют, по меньшей мере, 5-10 генов, найденных в области, выровненной с генами в хромосоме, содержащей родительский ген (например, хромосома штамма 69B4 Cellulomonas). В дополнительных вариантах осуществления более чем одно из вышеуказанных свойств относится к последовательности. Аналогичные последовательности устанавливают известными способами для выравнивания последовательностей. Общеупотребительным способом выравнивания является BLAST, хотя, как указано выше и ниже, существуют другие способы, которые также находят применение при выравнивании последовательностей.
Одним из подходящих примеров является алгоритм PILEUP. PILEUP создает множественное выравнивание последовательностей из группы родственных последовательностей с использованием прогрессивных, парных выравниваний. Также он может построить дерево, на котором показаны родственные связи, использованные для создания выравнивания. PILEUP использует упрощенный способ прогрессивного выравнивания авторов Feng и Doolittle (Feng and Doolittle, J. Mol. Evol., 35:351-360 [1987]). Способ схож со способом, описанным авторами Higgins и Sharp (Higgins and Sharp, CABIOS 5:151-153 [1989]). Полезные параметры PILEUP включают в себя default gap weight, равный 3,00, default gap length weight, равный 0,10, и weighted end gaps.
Другим примером пригодного алгоритма является алгоритм BLAST, описанный авторами Altschul et al. (Altschul et al., J. Mol. Biol., 215:403-410 [1990]; и Karlin et al., Proc. Natl. Acad. Sci., USA, 90:5873-5787 [1993]). В частности, полезной программной реализацией BLAST является программа WU-BLAST-2 (см., Altschul et al., Meth. Enzymol., 266:460-480 [1996]). WU-BLAST-2 использует несколько параметров поиска, большая часть которых установлена на значения по умолчанию. Устанавливали следующие значения регулируемых параметров: overlap span = 1, overlap fraction = 0,125, word threshold (T)=11. Параметры HSP S и HSP S2 являются динамическими величинами и устанавливаются самой программой в зависимости от состава конкретной последовательности и состава конкретной базы данных, против которой проводится поиск интересующей последовательности. Однако величины могут быть скорректированы для увеличения чувствительности. Величину % идентичности аминокислотных последовательностей устанавливают делением количества совпадающих идентичных остатков на общее количество остатков в «более длинной» последовательности в выровненной области. «Более длинная» последовательность представляет собой последовательность, которая содержит большее количество истинных остатков в выровненной области (пропуски, введенные программой WU-Blast-2 для максимизации оценки выравнивания, игнорируются).
Таким образом, «процент идентичности (%) последовательности нуклеиновой кислоты» определяется как процентное содержание нуклеотидных остатков в испытываемой последовательности, которые идентичны нуклеотидным остаткам стартовой последовательности (т.е. интересующей последовательности). В предпочтительном способе используется модуль BLASTN программы WU-BLAST-2, в котором параметры установлены по умолчанию, причем overlap span и overlap fraction установлены на 1 и 0,125 соответственно.
Как используется в настоящем документе, термин «гибридизация» относится к процессу, с помощью которого цепочка нуклеиновой кислоты соединяется с комплементарной цепочкой посредством спаривания оснований, которое известно в данной области.
Последовательность нуклеиновой кислоты считается «поддающейся избирательной гибридизации» с эталонной последовательностью нуклеиновой кислоты, если две последовательности специфически гибридизуются друг с другом при условиях гибридизации и отмывки от средних до крайне строгих. Условия гибридизации основаны на температуре плавления (Tm) связывающего комплекса или зонда из нуклеиновой кислоты. Например, «максимальная строгость» типично наступает приблизительно при Tm-5°C (на 5°C ниже Tm зонда); «крайняя строгость» приблизительно при 5-10°C ниже Tm; «средняя строгость» приблизительно при 10-20°C ниже Tm зонда; и «низкая строгость» приблизительно при 20-25°C ниже Tm. Функционально, максимально строгие условия можно использовать для идентификации последовательности, обладающей строгой идентичностью или почти строгой идентичностью с зондом для гибридизации; тогда как гибридизацию при средней или низкой строгости условий можно использовать для обнаружения или установления гомологов полинуклеотидной последовательности.
Умеренные и крайне строгие условия гибридизации хорошо известны в данной области. Пример условий с высокой строгостью включает гибридизацию приблизительно при 42°C в 50% формамиде, 5× SSC, 5× растворе Денхардта, 0,5% SDS и 100 мкг/мл денатурированной ДНК-носителя с последующей двукратной отмывкой в 2× SSC и 0,5% SDS при комнатной температуре и двумя дополнительными разами в 0,1× SSC и 0,5% SDS при 42°C. Пример умеренно строгих условий включает инкубирование в течение ночи при 37°C в растворе, содержащем 20% формамида, 5× SSC (150 мМ NaCl, 15 мМ трицитрат натрия), 50 мМ фосфат натрия (pH 7,6), 5× раствор Денхардта, 10% сульфат декстрана и 20 мг/мл денатурированной порезанной ДНК спермы лосося, с последующей отмывкой фильтров в 1× SSC приблизительно при 37-50°C. Специалисты в данной области знают, как подбирать температуру, ионную силу и т.д. по мере необходимости, чтобы приспособиться к таким факторам, как длина зонда и т.п.
Как используется в настоящем документе, «рекомбинантный» включает ссылку на клетку или вектор, который был модифицирован введением гетерологичной последовательности нуклеиновой кислоты, или клетку, полученную из клетки, модифицированной таким образом. Таким образом, например, рекомбинантные клетки экспрессируют гены, которые не найдены в идентичной форме в природной (нерекомбинантной) форме клетки, или экспрессируют нативные гены, которые в других отношениях экспрессируются ненормально, недостаточно экспрессируются или не экспрессируются вовсе в результате преднамеренного вмешательства человека. «Рекомбинация», «рекомбинирование» и создание «рекомбинированной» нуклеиновой кислоты, как правило, представляет собой сборку из двух или более фрагментов нуклеиновых кислот, где сборка дает начало химерному гену.
В предпочтительном варианте осуществления мутантные последовательности ДНК создают сайт-насыщающим мутагенезом, по меньшей мере, в одном кодоне. В другом предпочтительном варианте осуществления сайт-насыщающий мутагенез выполняют для двух или более кодонов. В дополнительном варианте осуществления мутантные последовательности ДНК обладают более чем 50%, более чем 55%, более чем 60%, более чем 65%, более чем 70%, более чем 75%, более чем 80%, более чем 85%, более чем 90%, более чем 95% или более чем 98% гомологией с последовательностью дикого типа. В альтернативных вариантах осуществления, мутантная ДНК создана in vivo с помощью любой известной процедуры мутагенеза, например, такой как облучение, нитрозогуанидин и т.п. Затем требуемую последовательность ДНК выделяют и используют в способах, предоставленных в настоящем документе.
Как используется в настоящем документе, термин «последовательность-мишень» относится к последовательности ДНК в клетке-хозяине, которая кодирует последовательность, где в геном клетки-хозяина требуется встроить вводимую последовательность. В некоторых вариантах осуществления последовательность-мишень кодирует функциональный ген дикого типа или оперон, тогда как в других вариантах осуществления последовательность-мишень кодирует функционально мутантный ген или оперон, или нефункциональный ген или оперон.
Как используется в настоящем документе, «фланкирующая последовательность» относится к любой последовательности, которая расположена или в 3'-5' направлении или в 5'-3' направлении относительно рассматриваемой последовательности (например, для генов A-B-C, ген B фланкирован генными последовательностями A и C). В предпочтительном варианте осуществления вводимая последовательность фланкирована гомологичными фрагментами с каждой стороны. В другом варианте осуществления вводимая последовательность и гомологичные фрагменты содержат блок, который фланкирован спейсерными последовательностями с каждой стороны. В некоторых вариантах осуществления фланкирующая последовательность присутствует только с одной стороны (или 3' или 5'), но в предпочтительных вариантах осуществления она находится с каждой стороны от фланкируемой последовательности. В некоторых вариантах осуществления фланкирующая последовательность присутствует только с одной стороны (или 3' или 5'), тогда как в предпочтительных вариантах осуществления она присутствует с каждой стороны от фланкируемой последовательности.
Как используется в настоящем документе, термин «спейсерная последовательность» относится к любой дополнительной ДНК, которая фланкирует гомологичные фрагменты (типично векторные последовательности). Однако термин охватывает любую негомологичную последовательность ДНК. Не ограничиваясь какой-либо теорией, спейсерная последовательность предоставляет некритическую мишень для клетки для инициации поглощения ДНК.
Как используется в настоящем документе, термины «амплификация» и «амплификация гена» относятся к процессу, с помощью которого специфическая последовательность ДНК непропорционально реплицируется из условия, чтобы амплифицированный ген стал присутствовать в повышенном числе копий, чем изначально присутствовало в геноме. В некоторых вариантах осуществления отбор клеток посредством роста в присутствии лекарственного средства (например, ингибитора для поддающегося ингибированию фермента) приводит к амплификации или эндогенного гена, кодирующего продукт гена, необходимый для роста в присутствии лекарственного средства, или к амплификации экзогенной (т.е. вводимой) последовательности, кодирующей этот продукт гена, или как к тому, так и к другому.
«Амплификация» является особым случаем репликации нуклеиновой кислоты, использующим специфичность к матрице. Она отличается от неспецифической репликации матрицы (т.е. репликации, которая зависит от матрицы, но не зависит от специфической матрицы). Здесь специфичность матрицы различается по точности репликации (т.е. синтезу правильной полинуклеотидной последовательности) и нуклеотидной (рибо- или дезоксирибо-) специфичности. Специфичность матрицы часто описывают в терминах специфичности «мишени». Последовательности-мишени представляют собой «мишени» в том смысле, что происходит поиск и отсортировывание «мишени» от других нуклеиновых кислот. Изначально способы амплификации были созданы для такого отсортировывания.
Как используется в настоящем документе, термин «соамплификация» относится к введению в одну клетку амплифицируемого маркера в сочетании с другими генными последовательностями (т.е. содержащими один или несколько не подлежащий отбору генов, таких как гены, содержащиеся в экспрессирующем векторе) и применению соответствующего давления отбора из условия, чтобы клетки амплифицировали как амплифицируемый маркер, так и другие, не подлежащие отбору генные последовательности. Амплифицируемый маркер может быть физически связан с другими генными последовательностями или альтернативно в одну клетку могут быть введены два раздельных фрагмента ДНК, один содержит амплифицируемый маркер и другой содержит не подлежащий отбору маркер.
Как используется в настоящем документе, термины «амплифицируемый маркер», «амплифицируемый ген» и «амплифицируемый вектор» относятся к гену или вектору, кодирующему ген, который допускает амплификацию этого гена при соответствующих условиях роста.
В большинстве способов амплификации «специфичность матрицы» достигается посредством выбора фермента. Ферменты амплификации представляют собой ферменты, которые в условиях их использования будут обрабатывать только специфические последовательности нуклеиновых кислот в гетерологичной смеси нуклеиновых кислот. Например, в случае репликазы Qβ, РНК MDV-1 представляет собой специфическую матрицу для репликазы (см., например, Kacian et al., Proc. Natl. Acad. Sci. USA 69:3038 [1972]), и другие нуклеиновые кислоты не реплицируются этим ферментом амплификации. Подобным образом, в случае РНК полимеразы T7, этот фермент амплификации обладает строгой специфичностью к своим собственным промоторам (см., Chamberlin et al., Nature 228:227 [1970]). В случае ДНК лигазы T4, фермент не будет лигировать два олигонуклеотида или полинуклеотида, если имеет место несовпадение между олигонуклеотидным или полинуклеотидным субстратом и матрицей в точке лигирования (см., Wu and Wallace, Genomics 4:560 [1989]). В конечном счете, было обнаружено, что Taq и Pfu полимеразы, благодаря их способности работать при высокой температуре, проявляют высокую специфичность к последовательностям, связанным и, таким образом, обозначенным с помощью праймеров; высокая температура приводит к термодинамическим условиям, которые способствуют гибридизации праймеров с последовательностями-мишенями, но не гибридизации не с последовательностями-мишенями.
Как используется в настоящем документе, термин «амплифицируемая нуклеиновая кислота» относится к нуклеиновым кислотам, которые могут быть амплифицированы любым способом амплификации. Предполагается, что «амплифицируемая нуклеиновая кислота» обычно будет содержать «матрицу образца».
Как используется в настоящем документе, термин «матрица образца» относится к нуклеиновой кислоте, которая происходит из образца, который анализируют на присутствие «мишени» (определена ниже). В отличие от этого, «фоновая матрица» используется в отношении нуклеиновой кислоты, отличной от матрицы образца, которая может присутствовать или отсутствовать в образце. Фоновая матрица чаще всего является произвольной. Это может быть результатом переноса или это может быть из-за присутствия загрязнителей из нуклеиновых кислот, которые должны быть найдены и удалены из образца. Например, нуклеиновые кислоты из организмов, отличные от тех, что должны быть обнаружены, могут присутствовать в качестве фона в тестируемом образце.
Как используется в настоящем документе, термин «праймер» относится к олигонуклеотиду, который или встречается в природе в виде очищенного продукта расщепления рестриктазами, или создан синтетически, который способен действовать в качестве точки инициации синтеза, когда он помещен в условия, в которых индуцирован синтез продукта праймерного удлиняющегося сегмента, который комплементарен цепочке нуклеиновой кислоты (т.е. в присутствии нуклеотидов и индуцирующего агента, такого как ДНК полимераза и при подходящих температуре и pH). Для максимальной эффективности амплификации праймер предпочтительно является одноцепочечным, но альтернативно может быть двухцепочечным. Если праймер является двухцепочечным, то сначала его обрабатывают для разделения его цепочек перед использованием для изготовления продуктов элонгации. Предпочтительно, праймер представляет собой олигодезоксирибонуклеотид. Праймер должен быть достаточно длинным для того, чтобы направлять синтез продуктов элонгации в присутствии индуцирующего агента. Точные длины праймеров будут зависеть от многих факторов, включая температуру, источник праймера, и применяемого способа.
Как используется в настоящем документе, термин «зонд» относится к олигонуклеотиду (т.е. последовательности нуклеотидов), который, или встречается в природе в виде очищенного продукта расщепления рестриктазами, или создан синтетически, рекомбинантно, или с помощью ПЦР амплификации, который поддается гибридизации с другим интересующим олигонуклеотидом. Зонд может быть одноцепочечным или двухцепочечным. Зонды пригодны для обнаружения, идентификации и выделения определенных генетических последовательностей. Предполагается, что любой зонд, использованный в настоящем изобретении, будет мечен любой «репортерной молекулой» для того, чтобы его можно было обнаружить в любой системе обнаружения, включая в качестве неограничивающих примеров ферментативные (например, ELISA, а также гистохимические анализы, основанные на ферменте), флуоресцентные, радиоактивные и люминесцентные системы. Не планируется, что настоящее изобретение будет ограничиваться любой конкретной системой обнаружения или меткой.
Как используется в настоящем документе, термин «мишень» при использовании в отношении полимеразной цепной реакции, относится к области нуклеиновой кислоты, связанной праймерами, используемыми в полимеразной цепной реакции. Таким образом, происходит поиск и отсортировывание «мишени» от других последовательностей нуклеиновых кислот. «Сегмент» определен как область нуклеиновой кислоты внутри последовательности-мишени.
Как используется в настоящем документе, термин «полимеразная цепная реакция» («ПЦР») относится к способам из патентов США №№ 4683195, 4683202 и 4965188, которые включают способы для повышения концентрации сегмента последовательности-мишени в смеси геномной ДНК без клонирования или очистки, которые известны специалистам в данной области. Так как требуемые амплифицированные сегменты последовательности-мишени становятся преобладающими последовательностями (в пересчете на концентрацию) в смеси, о них говорят, что они «ПЦР амплифицированы».
Как используется в настоящем документе, термин «реактивы для амплификации» относится к тем реактивам (дезоксирибонуклеотид трифосфаты, буфер и т.д.), которые необходимы для амплификации, за исключением праймеров, матрицы из нуклеиновой кислоты и фермента амплификации. Типично, реактивы для амплификации, наряду с другими компонентами реакции, помещают и содержат в реакционном сосуде (пробирке, микролунке и т.д.).
С помощью ПЦР можно амплифицировать одну копию специфической последовательности-мишени в геномной ДНК до уровня, который можно определить с помощью нескольких различных способов (например, гибридизации с меченым зондом; встраивание биотинилированных праймеров с последующим обнаружением фермент-авидинового конъюгата; встраивание дезоксинуклеотид трифосфатов, меченных 32P, таких как dCTP или dATP, в амплифицированный сегмент). В дополнение к геномной ДНК, любая олигонуклеотидная или полинуклеотидная последовательность может быть амплифицирована с помощью соответствующего набора праймерных молекул. В частности, амплифицированные сегменты, созданные самим процессом ПЦР, сами являются эффективными матрицами для последующих ПЦР амплификаций.
Как используется в настоящем документе, термины «ПЦР продукт», «ПЦР фрагмент» и «продукт амплификации» относятся к полученной смеси соединений после завершения двух или более циклов ПЦР из стадий денатурации, отжига и элонгации. Эти термины охватывают случай, в котором происходила амплификация одного или нескольких сегментов одной или нескольких последовательностей-мишеней.
Как используется в настоящем документе, термин «RT-ПЦР» относится к репликации и амплификации РНК последовательностей. В этом способе, в котором обратная транскрипция объединена с ПЦР, наиболее часто используется процедура с одним ферментом, в которой применяется термостабильная полимераза, как описано в патенте США № 5322770. В RT-ПЦР, РНК матрица превращается в кДНК благодаря обратной транскриптазной активности полимеразы и затем амплифицируется с использованием полимеразной активности полимеразы (т.е. как в других способах ПЦР).
Как используется в настоящем документе, термины «рестрикционные эндонуклеазы» и «рестрикционные ферменты» относятся к бактериальным ферментам, каждый из которых режет двухцепочечную ДНК непосредственно в или рядом с конкретной нуклеотидной последовательностью.
«Участок рестрикции» относится к нуклеотидной последовательности, распознаваемой и разрезаемой с помощью заданной рестрикционной эндонуклеазы и часто представляет собой сайт для встраивания фрагментов ДНК. В определенных вариантах осуществления участки рестрикции создают внутри селективного маркера и в 5'- и 3'-концах ДНК конструкции.
Как используется в настоящем документе, термин «хромосомная интеграция» относится к процессу, при помощи которого вводимую последовательность вводят в хромосому клетки-хозяина. Гомологичные области трансформирующей ДНК выравниваются с гомологичными областями хромосомы. Впоследствии последовательность между гомологичными фрагментами замещается вводимой последовательностью через двойной кроссинговер (т.е. гомологичную рекомбинацию). В некоторых вариантах осуществления настоящего изобретения, гомологичные участки инактивирующего хромосомного сегмента ДНК конструкции выравниваются с фланкирующими гомологичными областями естественной хромосомной области хромосомы Bacillus. Впоследствии естественная хромосомная область в двойном кроссинговере (т.е. гомологичной рекомбинации) удаляется с помощью ДНК конструкции.
«Гомологичная рекомбинация» обозначает обмен фрагментами ДНК между двумя молекулами ДНК или спаренными хромосомами в сайте с идентичными или почти идентичными нуклеотидными последовательностями. В предпочтительном варианте осуществления хромосомная интеграция представляет собой гомологичную рекомбинацию. «Гомологичные последовательности», как используется в настоящем документе, обозначают последовательность нуклеиновой кислоты или полипептида, обладающую 100%, 99%, 98%, 97%, 96%, 95%, 94%, 93%, 92%, 91%, 90%, 88%, 85%, 80%, 75% или 70% идентичностью последовательности с другой последовательностью нуклеиновой кислоты или полипептида при оптимальном выравнивании для сравнения. В некоторых вариантах осуществления гомологичные последовательности обладают от 85% до 100% идентичностью последовательности, тогда как в других вариантах осуществления имеет место от 90% до 100% идентичность последовательности, и в более предпочтительных вариантах осуществления имеет место 95% и 100% идентичность последовательности.
Как используется в настоящем документе, «аминокислота» относится к пептидным или белковым последовательностям или их фрагментам. Термины «белок», «пептид» и «полипептид» используют взаимозаменяемо.
Как используется в настоящем документе, «интересующий белок» и «интересующий полипептид» относятся к требуемому и/или оцениваемому белку/полипептиду. В некоторых вариантах осуществления «интересующий белок» представляет собой «родительский белок» (т.е. стартовый белок). В некоторых вариантах осуществления родительский белок представляет собой фермент дикого типа, который используется в качестве стартовой точки в конструировании/проектировании белка. В некоторых вариантах осуществления интересующий белок экспрессирован внутриклеточно, тогда как в других вариантах осуществления он представляет собой секретируемый полипептид. В особенно предпочтительных вариантах осуществления эти ферменты включают сериновые протеазы и металлопротеазы, описываемые в настоящем документе. В некоторых вариантах осуществления интересующий белок представляет собой секретируемый полипептид, слитый с сигнальным пептидом (т.е. амино-концевым удлиняющим сегментом на белке, подлежащем секреции). Почти все секретируемые белки используют амино-концевой белковый удлиняющий сегмент, который играет ключевую роль при направлении и перемещении белков-предшественников через мембрану. Этот удлиняющий сегмент протеолитически удаляется с помощью сигнальной пептидазы во время или сразу после прохождения через мембрану.
Как используется в настоящем документе, термин «гетерологичный белок» относится к белку или полипептиду, который в природе не встречается в клетке-хозяине. Примеры гетерологичных белков включают ферменты, такие как гидролазы, включая протеазы. В некоторых вариантах осуществления гены, кодирующие белки, представляют собой встречающиеся в природе гены, тогда как в других вариантах осуществления используются мутировавшие и/или синтетические гены.
Как используется в настоящем документе, «гомологичный белок» относится к белку или полипептиду, который является нативным или встречается в клетке в природе. В предпочтительных вариантах осуществления клетка представляет собой грамположительную клетку, тогда как в особенно предпочтительных вариантах осуществления клетка представляет собой клетку-хозяина Bacillus. В альтернативных вариантах осуществления, гомологичный белок представляет собой нативный белок, синтезируемый другими организмами, включая в качестве неограничивающих примеров E. coli, Cellulomonas, Bacillus, Streptomyces, Trichoderma и Aspergillus. Данное изобретение относится к клеткам-хозяевам, продуцирующим гомологичный белок посредством технологии рекомбинантных ДНК.
Как используется в настоящем документе, «область оперона» содержит группу смежных генов, которые транскрибируются в виде одной транскрипционной единицы из общего промотора и таким образом подвергаются корегуляции. В некоторых вариантах осуществления оперон содержит ген-регулятор. В наиболее предпочтительных вариантах осуществления, используются опероны, которые обладают повышенной экспрессией, судя по измерениям уровней РНК, но обладают неизвестной или излишней функцией.
Как используется в настоящем документе, «противомикробная область» представляет собой область, содержащую, по меньшей мере, один ген, который кодирует противомикробный белок.
Говорят, что полинуклеотид «кодирует» РНК или полипептид, если в своем нативном состоянии или при манипуляциях посредством способов, известных специалистам в данной области, он может быть транскрибирован и/или транслирован для синтеза РНК, полипептида или его фрагмента. Об антисмысловой цепи такой нуклеиновой кислоты также говорят, что она кодирует последовательности.
Как известно в данной области, ДНК может быть транслирована с помощью РНК полимеразы для образования РНК, но РНК может быть обратно транскрибирована с помощью обратной транскриптазы для образования ДНК. Таким образом, ДНК может кодировать РНК и наоборот.
Термины «регуляторный сегмент», или «регуляторная последовательность», или «последовательность, контролирующая экспрессию» относятся к полинуклеотидной последовательности ДНК, которая функционально связана с полинуклеотидной последовательностью ДНК, которая кодирует аминокислотную последовательность полипептидной цепи, чтобы влиять на экспрессию кодируемой аминокислотной последовательности. Регуляторная последовательность может ингибировать, подавлять или способствовать экспрессии функционально связанной полинуклеотидной последовательности, кодирующей аминокислоту.
«Хозяйский штамм» или «клетка-хозяин» относится к хозяйскому организму, пригодному для экспрессии вектора, содержащего ДНК по настоящему изобретению.
Фермент «сверхэкспрессирован» в клетке-хозяине, если фермент экспрессирован в клетке при более высоком уровне, чем уровень, при котором он экспрессирован в соответствующей клетке дикого типа.
В настоящем документе термины «белок» и «полипептид» используются взаимозаменяемо. На протяжении этого раскрытия используется трехбуквенный код для аминокислот, определенный в соответствии с IUPAC-IUB Объединенной комиссией по биохимической номенклатуре (JCBN). Также следует понимать, что полипептид может кодироваться более чем одной нуклеотидной последовательностью по причине вырожденности генетического кода.
«Пропоследовательность» представляет собой аминокислотную последовательность между сигнальной последовательностью и зрелой протеазой, которая необходима для секреции протеазы. Отщепление пропоследовательности приведет к образованию зрелой активной протеазы.
Термины «сигнальная последовательность» или «сигнальный пептид» относятся к любой последовательности нуклеотидов и/или аминокислот, которые участвуют в секреции зрелой или прекурсорной формы белка. Это определение сигнальной последовательности является функциональным, что говорит о том, что включены все те аминокислотные последовательности, кодируемые N-концевой частью белкового гена, которые участвуют в осуществлении секреции белка. Они часто, но не всегда, связаны с N-концевой частью белка или с N-концевой частью белка-предшественника. Сигнальная последовательность может быть эндогенной или экзогенной. Сигнальная последовательность может быть естественным образом связана с белком (например, протеазой) или может быть из другого гена, кодирующего другой секретируемый белок. Одна образцовая экзогенная сигнальная последовательность содержит первые семь аминокислотных остатков сигнальной последовательности из субтилизина B. subtilis, слитого с остатком от сигнальной последовательности субтилизина из B. lentus (ATCC 21536).
Термин «гибридная сигнальная последовательность» относится к сигнальным последовательностям, в которых часть последовательности, полученная из экспрессирующего хозяйского организма, слита с сигнальной последовательностью гена, подлежащего экспрессии. В некоторых вариантах осуществления применяются синтетические последовательности.
Термин «в значительной степени одинаковая сигнальная активность» относится к сигнальной активности, на которую указывает значительной степени одинаковая секреция протеазы в сбраживаемую среду, например уровень протеазы в сбраживаемой среде составляет, по меньшей мере, 50%, по меньшей мере, 60%, по меньшей мере, 70%, по меньшей мере, 80%, по меньшей мере, 90%, по меньшей мере, 95%, по меньшей мере, 98% от уровней секретируемой протеазы в сбраживаемой среде, которая обеспечена сигнальной последовательностью.
Термин «зрелая» форма белка или пептида относится к конечной функциональной форме белка или пептида. В качестве примера, зрелая форма протеазы NprE по настоящему изобретению, по меньшей мере, содержит аминокислотную последовательность из SEQ ID NO:3.
Термин «прекурсорная» форма белка или пептида относится к зрелой форме белка, которая имеет пропоследовательность, функционально связанную с амино- или карбокси-концом белка. Прекурсор также может обладать «сигнальной» последовательностью, функционально связанной с амино-концом пропоследовательности. Прекурсор также может содержать дополнительные полинуклеотиды, которые участвуют в посттрансляционной активности (например, полинуклеотиды отщепляются оттуда, чтобы покинуть зрелую форму белка или пептида).
«Встречающийся в природе фермент» и «встречающийся в природе белок» указывают на фермент или белок, обладающий немодифицированной аминокислотной последовательностью, идентичной последовательности, найденной в природе. Встречающиеся в природе ферменты включают нативные ферменты, такие ферменты естественным образом экспрессированы или найдены в конкретном микроорганизме.
Термины «выделенный из» и «полученный из» относятся не только к ферменту (например, протеазе), синтезированному или синтезируемому штаммом рассматриваемого организма, но также к ферменту, кодируемому последовательностью ДНК, выделенной из такого штамма, и продуцируемому в хозяйском организме, содержащем такую последовательность ДНК. Дополнительно, термин относится к ферменту, который кодируется последовательностью ДНК синтетического и/или кДНК происхождения и который обладает идентификационными характеристиками рассматриваемого фермента.
«Производное» в рамках этого определения, как правило, сохраняет характеристическую протеолитическую активность, наблюдаемую у дикого типа, нативной или родительской формы в случае, если производное пригодно для целей, сходных с целями применения дикого типа, нативной или родительской формы. Функциональные производные фермента охватывают встречающиеся в природе, синтетически или рекомбинантно созданные пептиды или фрагменты пептидов, обладающие основными характеристиками родительского фермента.
Термин «функциональное производное» относится к производному нуклеиновой кислоты, имеющему функциональные характеристики нуклеиновой кислоты, кодирующей фермент. Функциональные производные нуклеиновой кислоты, которые кодируют ферменты, предоставленные в настоящем документе, охватывают встречающиеся в природе, синтетически или рекомбинантно созданные нуклеиновые кислоты или фрагменты. Нуклеиновая кислота дикого типа, кодирующая ферменты по настоящему изобретению, включает встречающиеся в природе аллели и гомологи, основанные на вырожденности генетического кода, известной в данной области.
Термин «идентичный» в контексте двух последовательностей нуклеиновых кислот или полипептидов относится к остаткам в двух последовательностях, которые являются одинаковыми при выравнивании с максимальным совпадением, которое измеряется с использованием одного из следующих алгоритмов сравнения последовательностей или анализа.
Термин «оптимальное выравнивание» относится к выравниванию, дающему наивысшее значение процента идентичности.
«Процент идентичности последовательностей», «процент идентичности аминокислотных последовательностей», «процент идентичности генных последовательностей» и/или «процент идентичности последовательности нуклеиновых кислот/полинуклеотидов» по отношению к двум аминокислотным, полинуклеотидным и/или генным последовательностям (в соответствующих случаях) относятся к процентному содержанию остатков, которые идентичны в двух последовательностях, когда эти последовательности оптимально выровнены. Таким образом, 80% идентичность аминокислотных последовательностей обозначает, что 80% аминокислот в двух оптимально выровненных полипептидных последовательностях идентичны.
Таким образом, фраза «в значительной степени идентичны» в контексте двух нуклеиновых кислот или полипептидов относится к полинуклеотиду или полипептиду, который обладает, по меньшей мере, 70% идентичностью последовательности, предпочтительно по меньшей мере, 75%, предпочтительно по меньшей мере, 80%, предпочтительно по меньшей мере, 85%, предпочтительно по меньшей мере, 90%, предпочтительно по меньшей мере, 95%, предпочтительно по меньшей мере, 97%, предпочтительно по меньшей мере, 98% и предпочтительно по меньшей мере, 99% идентичностью последовательности по сравнению с эталонной последовательностью при использовании программ или алгоритмов (например, BLAST, ALIGN, CLUSTAL) со стандартными параметрами. Одним признаком того, что два полипептида в значительной степени идентичны, является то, что первый полипептид обладает иммунологической перекрестной реактивностью со вторым полипептидом. Типично, полипептиды, которые отличаются консервативными аминокислотными заменами, иммунологически кросс-реактивны. Таким образом, полипептид в значительной степени идентичен второму полипептиду, например, когда два пептида отличаются только консервативной заменой. Другой признак того, что две последовательности нуклеиновой кислоты в значительной степени идентичны, заключается в том, что две молекулы гибридизуются друг с другом при строгих условиях (например, в диапазоне от средне до крайне строгих).
Термин «выделенный» или «очищенный» относится к материалу, который удален из своего исходного окружения (например, природного окружения, если он встречается в природе). Например, о материале говорят, что он «очищен», когда в конкретной композиции он присутствует в большей или меньшей концентрации, чем встречается в существующем в природе организме, или в организме дикого типа, или в сочетании с компонентами, которые обычно не присутствуют при экспрессии во встречающемся в природе организме или в организме дикого типа. Например, встречающийся в природе полинуклеотид или полипептид, присутствующий в живом организме, не является выделенным, но тот же полинуклеотид или полипептид, отделенный от некоторых или всех материалов, сосуществующих с ним в естественной системе, является выделенным. В некоторых вариантах осуществления такие полинуклеотиды являются частью вектора и/или такие полинуклеотиды или полипептиды являются частью композиции, и все же выделены в таком векторе или композиции, которые не являются частью их естественной среды. В некоторых предпочтительных вариантах осуществления, говорят, что нуклеиновая кислота или белок очищены, например, если они являются источником в основном одной полосы на электрофоретическом геле или в блоттинге.
Термин «выделенный», при использовании в отношении последовательности ДНК, относится к последовательности ДНК, которая была удалена из ее естественного генетического окружения и таким образом свободна от других посторонних или нежелательных кодирующих последовательностей, и существует в форме, пригодной для использования в генетически сконструированных системах продуцирования белка. Такие выделенные молекулы представляют собой молекулы, которые отделены от их естественного окружения и включают кДНК и геномные клоны. Выделенные молекулы ДНК по настоящему изобретению свободны от других генов, с которыми они, как правило, связаны, но могут содержать встречающиеся в природе 5'- и 3'-нетранслируемые области, такие как промоторы и терминаторы. Идентификация связанных областей будет очевидна специалистам в данной области (см., например, Dynan and Tijan, Nature 316:774-78, 1985). Термин «выделенная последовательность ДНК» альтернативно обозначается как «клонированная последовательность ДНК».
Термин «выделенный», при использовании в отношении белка, относится к белку, который найден в состоянии, отличающемся от естественной среды. В предпочтительной форме, выделенный белок в значительной степени свободен от других белков, конкретно от других гомологичных белков. Выделенный белок обладает более чем 10% чистотой, предпочтительно более чем 20% чистотой и даже более предпочтительно более чем 30% чистотой, которую определяют с помощью SDS-PAGE. Дополнительные аспекты изобретения охватывают белок в форме с высокой степенью очистки (т.е. более чем 40% чистоты, более чем 60% чистоты, более чем 80% чистоты, более чем 90% чистоты, более чем 95% чистоты, более чем 97% чистоты и даже более чем 99% чистоты), которую определяют с помощью SDS-PAGE.
Как используется в настоящем документе, термин «комбинаторный мутагенез» относится к способам, в которых создают библиотеки вариантов стартовых последовательностей. В этих библиотеках варианты содержат одну или несколько мутаций, выбранных из предопределенного набора мутаций. Кроме того, способы предоставляют средства для введения случайных мутаций, которые не являются членами предопределенного набора мутаций. В некоторых вариантах осуществления способы содержат способы, изложенные в заявке США № 09/699250, поданной 26 октября 2000 г. В альтернативных вариантах осуществления, способы комбинаторного мутагенеза охватывают коммерчески доступные наборы (например, QUIKCHANGE® Multisite, Stratagene, La Jolla, CA).
Как используется в настоящем документе, термин «вариант» относится к белку, который был получен из белка-предшественника (например, «родительского» белка) посредством добавления, замены или делеции одной или нескольких аминокислот. В некоторых вариантах осуществления вариант содержит, по меньшей мере, одну модификацию, которая содержит изменения заряда по сравнению с белком-предшественником. В некоторых предпочтительных вариантах осуществления, белок-предшественник представляет собой родительский белок, который представляет собой белок дикого типа.
Как используется в настоящем документе, термин «библиотека мутантов» относится к популяции клеток, которые идентичны в большей части своих геномов, но включают различные гомологи одного или нескольких генов. Такие библиотеки можно использовать, например, для идентификации генов или оперонов с улучшенными характеристиками.
Как используется в настоящем документе, термин «стартовый ген» относится к интересующему гену, который кодирует интересующий белок, который должен быть улучшен и/или изменен с использованием настоящего изобретения.
Как используется в настоящем документе, термины «множественное выравнивание последовательностей» и «МВП» относятся к последовательностям множества гомологов стартового гена, которые выровнены с использованием алгоритма (например, Clustal W).
Как используется в настоящем документе, термины «консенсусная последовательность» и «каноническая последовательность» относятся к первоначальной аминокислотной последовательности, с которой сравнивают все варианты конкретного белка или интересующей последовательности. Термины также относятся к последовательности, в которой приведены нуклеотиды, которые наиболее часто присутствуют в интересующей последовательности ДНК. Для каждой позиции в гене консенсусная последовательность дает аминокислоту, которая наиболее часто встречается в этой позиции в МВП.
Как используется в настоящем документе, термин «консенсусная мутация» относится к различиям в последовательности стартового гена и консенсусной последовательности. Консенсусные мутации устанавливают сравнением последовательности стартового гена и консенсусной последовательности, полученной из МВП. В некоторых вариантах осуществления консенсусные мутации введены в стартовый ген из условия, чтобы он стал больше похож на консенсусную последовательность. Консенсусные мутации также включают аминокислотные замены, которые меняют аминокислоту в стартовом гене на аминокислоту, которая более часто встречается в МВП в этой позиции по отношению к частоте этой аминокислоты в стартовом гене. Таким образом, термин консенсусная мутация включает все единичные аминокислотные замены, которые заменяют аминокислоту стартового гена на аминокислоту, которая встречается более часто, чем аминокислота в МВП.
Как используется в настоящем документе, термин «исходный вариант с благоприятными свойствами» относится к варианту, который был идентифицирован посредством скрининга библиотеки комбинаторного консенсусного мутагенеза. В предпочтительных вариантах осуществления исходные варианты с благоприятными свойствами обладают улучшенными эксплуатационными характеристиками по сравнению со стартовым геном.
Как используется в настоящем документе, термин «улучшенный вариант с благоприятными свойствами» относится к варианту, который был установлен посредством скрининга расширенной библиотеки комбинаторного консенсусного мутагенеза.
Как используется в настоящем документе, термины «улучшающая мутация» и «мутация, повышающая эффективность» относятся к мутации, которая ведет к улучшенной эффективности, когда ее вводят в стартовый ген. В некоторых предпочтительных вариантах осуществления, эти мутации идентифицируют посредством секвенирования вариантов с благоприятными свойствами, установленных в ходе стадии скрининга данного способа. В большинстве вариантов осуществления вероятно, что мутации, которые чаще находят в вариантах с благоприятными свойствами, являются улучшающими мутациями по сравнению с нескринированной библиотекой комбинаторного консенсусного мутагенеза.
Как используется в настоящем документе, термин «обогащенная библиотека комбинаторного консенсусного мутагенеза» относится к библиотеке ККМ, которая спроектирована и сконструирована на основе результатов скрининга и/или секвенирования из раннего цикла ККМ мутагенеза и скрининга. В некоторых вариантах осуществления обогащенная библиотека ККМ основана на последовательности исходного варианта с благоприятными свойствами, полученного из раннего цикла ККМ. В дополнительных вариантах осуществления, обогащенная ККМ спроектирована из условия, чтобы мутациям, которые часто наблюдаются в исходных вариантах с благоприятными свойствами из ранних циклов мутагенеза и скрининга, отдавалось предпочтение. В некоторых предпочтительных вариантах осуществления это осуществляется посредством пропуска праймеров, которые кодируют мутации, снижающие эффективность, или посредством увеличения концентрации праймеров, которые кодируют мутации, повышающие эффективность, по отношению к другим праймерам, которые использовались в ранних библиотеках ККМ.
Как используется в настоящем документе, термин «мутации, снижающие эффективность» относится к мутациям в библиотеке комбинаторного консенсусного мутагенеза, которые реже находят в вариантах с благоприятными свойствами в результате скрининга по сравнению с нескринированной библиотекой комбинаторного консенсусного мутагенеза. В предпочтительных вариантах осуществления процесс скрининга удаляет и/или снижает относительное содержание вариантов, которые содержат «мутации, снижающие эффективность».
Как используется в настоящем документе, термин «функциональный анализ» относится к анализу, который обеспечивает измерение активности белка. В особенно предпочтительных вариантах осуществления термин относится к аналитическим системам, в которых белок анализируют на предмет его способности функционировать при его обычной нагрузке. Например, в случае ферментов, функциональный анализ касается определения эффективности фермента в катализе реакции.
Как используется в настоящем документе, термин «намеченное свойство» относится к свойству стартового гена, которое должно быть изменено. Не планируется, что настоящее изобретение ограничивается любым конкретным намеченным свойством. Однако, в некоторых предпочтительных вариантах осуществления, намеченное свойство представляет собой стабильность продукта гена (например, устойчивость к денатурации, протеолизу или другим разрушающим факторам), тогда как в других вариантах осуществления изменен уровень продукции в продуцирующем хозяйском организме. Фактически, предполагается, что любое свойство стартового гена найдет применение в настоящем изобретении.
Термин «свойство» или его грамматические эквиваленты в контексте нуклеиновой кислоты, как используется в настоящем документе, относится к любой характеристике или свойству нуклеиновой кислоты, которое можно выбрать или обнаружить. Эти свойства включают в качестве неограничивающих примеров свойство воздействовать на связывание с полипептидом, свойство, придаваемое клетке, содержащей конкретную нуклеиновую кислоту, свойство воздействовать на транскрипцию гена (например, сила промотора, распознание промотора, регулирование промотора, энхансерная функция), свойство воздействовать на процессинг РНК (например, сплайсинг РНК, стабильность РНК, конформацию РНК и посттранскрипционную модификацию), свойство воздействовать на трансляцию (например, уровень, регулирование, связывание мРНК с рибосомальными белками, посттрансляционная модификация). Например, в нуклеиновой кислоте сайт связывания фактора транскрипции, полимеразы, регуляторного фактора и т.д. может быть изменен для создания требуемых характеристик или для идентификации нежелательных характеристик.
Термин «свойство» или его грамматические эквиваленты в контексте полипептида, как используется в настоящем документе, относится к любой характеристике или свойству полипептида, которое можно выбрать или обнаружить. Эти свойства включают в качестве неограничивающих примеров окислительную стабильность, субстратную специфичность, каталитическую активность, тепловую стабильность, стабильность в щелочах, pH профиль активности, устойчивость к протеолитической деградации, KM, kcat, отношение kcat/KM, укладку белка, стимулирование иммунного ответа, способность связываться с лигандом, способность связываться с рецептором, способность секретироваться, способность экспонироваться на поверхности клетки, способность к олигомеризации, способность передавать сигналы, способность стимулировать клеточную пролиферацию, способность ингибировать клеточную пролиферацию, способность индуцировать апоптоз, способность модифицироваться фосфорилированием или гликозилированием, способность лечить заболевание.
Как используется в настоящем документе, термин «скрининг» имеет свое обычное значение в данной области и, в основном, представляет собой многостадийный процесс. На первой стадии предоставляют мутантную нуклеиновую кислоту или вариант ее полипептида. На второй стадии определяют свойство мутантной нуклеиновой кислоты или варианта полипептида. На третьей стадии свойство, которое определили, сравнивают со свойством соответствующей родительской нуклеиновой кислоты, со свойством соответствующего встречающегося в природе полипептида или со свойством стартового материала (например, начальной последовательности) для создания мутантной нуклеиновой кислоты.
Профессионалу в данной области будет ясно, что процедура скрининга для получения нуклеиновой кислоты или белка с измененным свойством зависит от свойства стартового материала, модификация которого предназначена для облегчения создания мутантной нуклеиновой кислоты. Поэтому профессионал оценит то, что изобретение не ограничено любым определенным свойством, подлежащим скринингу, и что следующее описание свойств перечисляет только пояснительные примеры. Способы скрининга конкретного свойства, как правило, описаны в данной области. Например, можно измерить связывание, pH, специфичность и т.д. до и после мутации, где перемена указывает на изменение. Предпочтительно, скрининг осуществляется высокопроизводительным способом, одновременно включая множество образцов, подлежащих скринингу, включая в качестве неограничивающих примеров анализы с использованием чипов, фаговый дисплей и множество субстратов и/или индикаторов.
Как используется в настоящем документе, в некоторых вариантах осуществления скрининг охватывает стадии отбора, на которых интересующие варианты обогащают из совокупности вариантов. Примеры этих вариантов осуществления включают отбор вариантов, которые дают хозяйскому организму преимущество в росте, а также фаговый дисплей или любой другой способ выявления, где варианты могут быть захвачены из совокупности вариантов на основании их связывающих или каталитических свойств. В предпочтительном варианте осуществления библиотека вариантов подвергается внешнему воздействию (нагревание, протеаза, денатурация) и впоследствии варианты, которые остаются интактными, идентифицируют в скрининге или обогащают посредством отбора. Предполагается, что термин охватывает любые пригодные средства для отбора. Фактически, не планируется, что настоящее изобретение будет ограничено любым конкретным способом скрининга.
Как используется в настоящем документе, термин «нацеленный случайный отбор» относится к процессу, в котором создается множество последовательностей, в которых одна или несколько позиций были рандомизированы. В некоторых вариантах осуществления рандомизация является полной (т.е. все четыре нуклеотида, A, T, G и C, могут встречаться в рандомизированной позиции). В альтернативных вариантах осуществления, рандомизация нуклеотида ограничена поднабором четырех нуклеотидов. Нацеленный случайный отбор может быть применен к одному или нескольким кодонам последовательности, кодирующим один или несколько интересующих белков. После экспрессии из полученных библиотек создают белковые популяции, в которых одна или несколько аминокислотных позиций могут содержать смесь из всех 20 аминокислот или поднабора аминокислот, который определяется схемой рандомизации рандомизированного кодона. В некоторых вариантах осуществления отдельные члены популяции, полученной в результате нацеленного случайного отбора, отличаются по количеству аминокислот, из-за нацеленной или случайной вставки или делеции кодонов. В дополнительных вариантах осуществления синтетические аминокислоты включены в созданные белковые популяции. В некоторых предпочтительных вариантах осуществления, большая часть членов популяции, полученной в результате нацеленного случайного отбора, проявляет более высокую гомологию последовательности по отношению к консенсусной последовательности, чем стартовый ген. В некоторых вариантах осуществления последовательность кодирует один или несколько интересующих белков. В альтернативных вариантах осуществления, белки имеют различные биологические функции. В некоторых дополнительных предпочтительных вариантах осуществления, вводимая последовательность содержит, по меньшей мере, один селективный маркер. Эта последовательность может кодировать один или несколько интересующих белков. Они могут обладать другой биологической функцией(ями). Во многих случаях вводимая последовательность будет содержать селективный маркер, такой как ген, который обеспечивает устойчивость к антибиотику.
Термины «модифицированная последовательность» и «модифицированные гены», используемые в настоящем документе взаимозаменяемо, относятся к последовательности, которая содержит делецию, вставку или нарушение встречающейся в природе последовательности нуклеиновой кислоты. В некоторых предпочтительных вариантах осуществления, продукт экспрессии модифицированной последовательности представляет собой усеченный белок (например, если модификация представляет собой делецию или нарушение последовательности). В некоторых особенно предпочтительных вариантах осуществления, усеченный белок сохраняет биологическую активность. В альтернативных вариантах осуществления, продукт экспрессии модифицированной последовательности представляет собой удлиненный белок (например, модификации, содержащие вставку в последовательность нуклеиновой кислоты). В некоторых вариантах осуществления вставка приводит к усеченному белку (например, когда вставка приводит к образованию стоп-кодона). Таким образом, вставка может приводить или к усеченному белку, или к удлиненному белку в качестве продукта экспрессии.
Как используется в настоящем документе, термины «мутантная последовательность» и «мутантный ген» используют взаимозаменяемо и они относятся к последовательности, которая обладает изменением, по меньшей мере, в одном кодоне, встречающемся в последовательности клетки-хозяина дикого типа. Продукт экспрессии мутантной последовательности представляет собой белок с измененной аминокислотной последовательностью по отношению к дикому типу. Продукт экспрессии может обладать измененными функциональными возможностями (например, улучшенной ферментативной активностью).
Термины «мутагенный праймер» или «мутагенный олигонуклеотид» (используются в настоящем документе взаимозаменяемо) предназначены для обозначения олигонуклеотидных структур, которые соответствуют части матричной последовательности и которые поддаются гибридизации с ней. Что касается мутагенных праймеров, праймер не будет точно совпадать с матричной нуклеиновой кислотой, несовпадение или несовпадения в праймере используются для введения требуемой мутации в библиотеку нуклеиновых кислот. Как используется в настоящем документе, «немутагенный праймер» или «немутагенный олигонуклеотид» относится к олигонуклеотидным структурам, которые точно совпадают с матричной нуклеиновой кислотой. В одном из вариантов осуществления изобретения используются только мутагенные праймеры. В другом предпочтительном варианте осуществления изобретения, праймеры спроектированы для того, чтобы, по меньшей мере, в одной области, в которую мутагенный праймер был включен, также имел место немутагенный праймер, включенный в олигонуклеотидную смесь. Добавляя смесь мутагенных праймеров и немутагенных праймеров, соответствующих, по меньшей мере, одному из мутагенных праймеров, в результате можно создать библиотеку нуклеиновых кислот, в которой представлено множество комбинаторных мутационных шаблонов. Например, если требуется, чтобы некоторые члены библиотеки мутантных нуклеиновых кислот сохранили свою родительскую последовательность в определенных позициях, тогда как другие члены являлись мутантами в таких сайтах, немутагенные праймеры предоставляют способность получить конкретный уровень немутантных членов в библиотеке нуклеиновых кислот для заданного остатка. Способы по изобретению используют мутагенные и немутагенные олигонуклеотиды, которые, как правило, обладают длиной между 10-50 основаниями, более предпочтительно длиной приблизительно 15-45 оснований. Однако может быть необходимо использовать праймеры, которые или короче чем 10 оснований, или длиннее чем 50 оснований, для получения требуемого результата мутагенеза. В отношении соответствующих мутагенных и немутагенных праймеров не обязательно, чтобы соответствующие олигонуклеотиды были одинаковой длины, но только чтобы происходило перекрытие в области, соответствующей вводимой мутации.
В некоторых вариантах осуществления праймеры добавляют в предопределенном соотношении. Например, если требуется, чтобы полученная библиотека имела значительный уровень определенной специфической мутации и меньшее количество отличающейся мутации в том же или другом сайте, посредством подбора количества добавляемого праймера можно создать требуемую смещенную библиотеку. Альтернативно, добавляя большие или меньшие количества немутагенных праймеров, можно регулировать частоту, с которой соответствующая мутация(и) образуется в библиотеке мутантных нуклеиновых кислот.
Как используется в настоящем документе, фраза «смежные мутации» относится к мутациям, которые присутствуют в одном олигонуклеотидном праймере. Например, смежные мутации могут быть прилегающими или близкими друг к другу, однако они будут введены в конечные мутантные матричные нуклеиновые кислоты с помощью одного праймера.
Как используется в настоящем документе, фраза «несмежные мутации» относится к мутациям, которые представлены отдельными олигонуклеотидными праймерами. Например, несмежные мутации будут введены в конечные мутантные матичные нуклеиновые кислоты с помощью раздельно изготовленных олигонуклеотидных праймеров.
Термины «последовательность дикого типа», «последовательность нуклеиновой кислоты дикого типа» и «ген дикого типа», используемые в настоящем документе взаимозаменяемо, относятся к последовательности, которая является нативной или встречающейся в природе в клетке-хозяине. В некоторых вариантах осуществления последовательность дикого типа относится к интересующей последовательности, которая представляет собой начальную точку проекта по конструированию белка. Последовательность дикого типа может кодировать или гомологичный, или гетерологичный белок. Гомологичный белок представляет собой белок, который клетка-хозяин будет продуцировать без вмешательства. Гетерологичный белок представляет собой белок, который клетка-хозяин не будет продуцировать без вмешательства.
Термин «окислительно устойчивый» относится к протеазам по настоящему изобретению, которые сохраняют определенное количество ферментативной активности в течение заданного периода времени в условиях, преобладающих в процессе протеолиза, гидролиза, очистки или другого процесса по изобретению, например когда они подвергаются или приводятся в контакт с отбеливателями или окисляющими средствами. В некоторых вариантах осуществления протеазы сохраняют, по меньшей мере, 50%, 60%, 70%, 75%, 80%, 85%, 90%, 92%, 95%, 96%, 97%, 98% или 99% протеолитической активности после контакта с отбеливающим или окисляющим средством в течение заданного периода времени, например, по меньшей мере, 1 минута, 3 минуты, 5 минут, 8 минут, 12 минут, 16 минут, 20 минут и т.д.
Термин «хелатор-стабильный» относится к протеазам по настоящему изобретению, которые сохраняют определенное количество ферментативной активности в течение заданного периода времени в условиях, преобладающих в ходе протеолиза, гидролиза, очистки или другого процесса по изобретению, например когда они подвергаются или приводятся в контакт с хелирующими агентами. В некоторых вариантах осуществления протеазы сохраняют, по меньшей мере, 50%, 60%, 70%, 75%, 80%, 85%, 90%, 92%, 95%, 96%, 97%, 98% или 99% протеолитической активности после контакта с хелатирующим агентом в течение заданного периода времени, например, по меньшей мере, 10 минут, 20 минут, 40 минут, 60 минут, 100 минут и т.д.
Термины «термически стабильный» и «термостабильный» относятся к ферментам по настоящему изобретению, которые сохраняют определенное количество ферментативной активности после воздействия установленных температур в течение заданного периода времени в условиях, преобладающих в процессе ферментативной обработки, гидролиза, очистки или другого процесса по изобретению, например когда их подвергают воздействию измененных температур. Измененные температуры включают повышенные или пониженные температуры. В некоторых вариантах осуществления протеазы сохраняют, по меньшей мере, 50%, 60%, 70%, 75%, 80%, 85%, 90%, 92%, 95%, 96%, 97%, 98% или 99% протеолитической активности после воздействия измененных температур в течение заданного периода времени, например, по меньшей мере, 60 минут, 120 минут, 180 минут, 240 минут, 300 минут и т.д.
Термины «увеличенная стабильность» или «уменьшенная стабильность» в контексте окисления, хелатирования, термически и/или pH-стабильного фермента относятся к повышенной сохраненной ферментативной активности со временем по сравнению с другими сериновыми протеазами (например, субтилизиновыми протеазами) и/или ферментами дикого типа.
Термины «увеличенная стабильность» или «уменьшенная стабильность» в контексте окисления, хелатирования, термически и/или pH-стабильного фермента относятся к меньшей сохраненной ферментативной активности со временем по сравнению с другими сериновыми протеазами (например, субтилизиновыми протеазами) и/или ферментами дикого типа.
Как используется в настоящем документе, термин «чистящий состав» включает, если не указано иначе, гранулированные или порошковые моющие средства общего назначения или для жестких условий, в частности чистящие детергенты; жидкие, гелеобразные или пастообразные моющие средства общего назначения, в частности, так называемые жидкости для жестких условий; жидкие детергенты для деликатной материи; средства для ручного мытья посуды или облегченные средства для мытья посуды, в частности средства с обильным пенообразованием; средства для автоматического мытья посуды, включая различные таблетированные, гранулированные, жидкие средства и средства для ополаскивания посуды для домашнего и институционального применения; жидкие чистящие и дезинфицирующие средства, включая антибактериальные средства для ручного мытья, чистящие плитки, полоскания для рта, очищающие средства для зубов и полости рта, шампуни для автомобилей и ковров, чистящие средства для ванных комнат; шампуни и ополаскиватели для волос; гели для душа и пены для ванны и средства для очистки металла; а также вспомогательные чистящие средства, такие как отбеливающие добавки и средства для «stain-stick» или предварительной обработки.
Если не указано иначе, концентрации всех компонентов или композиции находятся в соответствии с активной концентрацией того компонента или композиции, за исключением примесей, например, остаточных растворителей или побочных продуктов, которые могут присутствовать в коммерчески доступных источниках.
Массы ферментативных компонентов основаны на общем активном белке. Все процентные содержания и отношения вычисляются по массе, если не указано иначе. Все процентные содержания и отношения вычисляются на основе целой композиции, если не указано иначе.
Следует понимать, что каждое максимальное числовое ограничение, даваемое на всем протяжении этого описания, включает каждое нижнее числовое ограничение, как если бы такие нижние числовые ограничения были явно указаны в настоящем документе. Каждое минимальное числовое ограничение, даваемое на всем протяжении этого описания, будет включать каждое верхнее числовое ограничение, как если бы такие верхние числовые ограничения были явно указаны в настоящем документе. Каждый числовой диапазон, указанный на всем протяжении этого описания, будет включать каждый числовой диапазон, который входит в пределы такого широкого числового диапазона, как если бы более узкие числовые диапазоны были явно указаны в настоящем документе.
Термин «активность очистки» относится к эффективности очистки, достигаемой ферментом в условиях, преобладающих в ходе ферментативной обработки, гидролиза, очистки или другого процесса по изобретению. В некоторых вариантах осуществления эффективность очистки определяют посредством применения различных чистящих тестов, относящихся к чувствительным к ферменту краскам, например траве, крови, молоку или яичному белку, которая определяется различными хроматографическими, спектрофотометрическими или другими количественными способами после того, как краски подвергают стандартным условиям мытья. Образцовые анализы включают в качестве неограничивающих примеров анализы, описанные в WO 99/34011 и патенте США № 6605458, а также те способы, которые включены в примеры.
Термин «эффективное для очистки количество» протеазы относится к количеству протеазы, описываемой в настоящем документе, перед которым достигается требуемый уровень ферментативной активности в конкретном чистящем составе. Такие эффективные количества легко определяются специалистом в данной области и основаны на многих факторах, таких как конкретная используемая протеаза, чистящее применение, конкретная композиция чистящего состава и требуется ли жидкая или сухая (например, гранулированная, кусковая) композиция и т.д.
Термин «вспомогательные очищающие вещества», как используется в настоящем документе, обозначает любое жидкое, твердое или газообразное вещество, выбранное для конкретного требуемого типа чистящего состава и формы продукта (например, жидкость, гранулы, порошок, кусок, паста, спрей, таблетка, гель; или пенная композиция), эти вещества также предпочтительно совместимы с протеолитическим ферментом, используемым в композиции. В некоторых вариантах осуществления гранулированные композиции находятся в «компактной» форме, тогда как в других вариантах осуществления жидкие композиции находятся в «концентрированной» форме.
Термин «увеличенная эффективность» в контексте активности очистки относится к увеличенной или повышенной активности очистки определенных чувствительных к ферменту красок, таких как яйца, молоко, трава или кровь, которую определяют посредством обычной оценки после стандартного цикла стирки и/или множества циклов отмывки.
Термин «уменьшенная эффективность» в контексте активности очистки относится к сниженной или меньшей активности очистки конкретных чувствительных к ферменту красок, таких как яйца, молоко, трава или кровь, которую определяют посредством обычной оценки после стандартного цикла стирки.
Термин «сравнительная эффективность» в контексте активности очистки относится, по меньшей мере, к 60%, по меньшей мере, 70%, по меньшей мере, 80%, по меньшей мере, 90%, или, по меньшей мере, 95% активности очистки сравниваемой протеазы (например, коммерчески доступных протеаз). Эффективность очистки может быть определена посредством сравнения протеаз по настоящему изобретению с другими протеазами в различных чистящих тестах, касающихся чувствительных к ферменту красок, таких как кровь, молоко и/или чернила (КМЧ), которую определяют с помощью обыкновенных спектрофотометрических или аналитических способов после цикла стирки при стандартных условиях.
Как используется в настоящем документе, система «с низкой концентрацией детергента» содержит детергенты, в которых приблизительно менее чем 800 м.д. моющих компонентов присутствуют в воде для мытья. Японские детергенты типично рассматриваются как системы с низкой концентрацией детергента, так как обычно они содержат приблизительно 667 м.д. моющих компонентов, присутствующих в воде для стирки.
Как используется в настоящем документе, системы «со средней концентрацией детергента» содержат детергенты, где между приблизительно 800 м.д. и приблизительно 2000 м.д. моющих компонентов присутствуют в воде для стирки. Как правило, североамериканские детергенты рассматриваются как системы со средней концентрацией детергента, так как они обычно содержат приблизительно 975 м.д. моющих компонентов, присутствующих в воде для стирки. Бразильские детергенты типично содержат приблизительно 1500 м.д. моющих компонентов, присутствующих в воде для стирки.
Как используется в настоящем документе, системы «с высокой концентрацией детергента» содержат детергенты, где приблизительно больше чем 2000 м.д. моющих компонентов присутствует в воде для стирки. Европейские детергенты, как правило, рассматриваются как системы с высокой концентрацией детергента, так как они содержат приблизительно 3000-8000 м.д. моющих компонентов в воде для стирки.
Как используется в настоящем документе, «чистящие композиции для материи» включают моющие составы для ручной и машинной стирки, включая добавочные композиции для стирки и композиции, пригодные для использования при замачивании и/или предварительной обработке окрашенной материи (например, одежды, полотен и других текстильных материалов).
Как используется в настоящем документе, «чистящие композиции не для материи» включают чистящие композиции для нетекстильных (т.е. не материи) поверхностей, включая в качестве неограничивающих примеров моющие составы для мытья посуды, чистящие композиции для полости рта, чистящие композиции для зубов и персональные чистящие композиции.
«Компактная» форма чистящих композиций в настоящем документе лучше всего отражена в плотности и, с точки зрения композиции, в количестве наполнителей из неорганических солей. Наполнители из неорганических солей представляют собой традиционные ингредиенты моющих составов в порошковой форме. В традиционных моющих составах, солевые наполнители присутствуют в существенных количествах, типично 17-35% по массе от общей композиции. В отличие от этого, в компактных композициях, солевые наполнители присутствуют в количествах, не превышающих 15% от общей композиции. В некоторых вариантах осуществления солевые наполнители присутствуют в количествах, которые не превышают 10% или более предпочтительно 5% по массе композиции. В некоторых вариантах осуществления наполнители из неорганических солей выбраны из сульфатов и хлоридов щелочных и щелочноземельных металлов. Предпочтительный солевой наполнитель представляет собой сульфат натрия.
Подробное описание изобретения
Настоящее изобретение относится к способам конструирования белков с целью оптимизации их эффективности при конкретных интересующих окружающих условиях. В некоторых вариантах осуществления настоящее изобретение относится к способам конструирования ферментов с целью оптимизации их каталитической активности при конкретных окружающих условиях. В некоторых предпочтительных вариантах осуществления, настоящее изобретение относится к способам конструирования ферментов с целью оптимизации их каталитической активности и/или стабильности при неблагоприятных окружающих условиях. В некоторых предпочтительных вариантах осуществления, настоящее изобретение относится к способам конструирования ферментов с целью оптимизации их стабильности при хранении, конкретно при неблагоприятных окружающих условиях. В некоторых предпочтительных вариантах осуществления настоящее изобретение относится к способам изменения суммарного поверхностного заряда и/или распределения поверхностных зарядов ферментов (например, металлопротеаз) с целью получения вариантов фермента, которые проявляют улучшенную эффективность и/или стабильность в моющих составах по сравнению с начальным или родительским ферментом.
В некоторых вариантах осуществления настоящее изобретение относится к способам конструирования фермента для одновременной оптимизации его каталитической активности и стабильности при неблагоприятных окружающих условиях, даже когда эти два свойства обладают отрицательной корреляцией при анализе влияния отдельных мутаций. В частности, настоящее изобретение относится к способам изменения суммарного поверхностного заряда и/или распределения поверхностных зарядов металлопротеазы с целью получения вариантов фермента, проявляющих улучшенную эффективность в моющих составах.
Настоящее изобретение относится к способам и композициям, содержащим, по меньшей мере, один вариант нейтральной металлопротеазы, который обладает улучшенной эффективностью стирки и/или стабильностью в моющем составе(ах). В некоторых особенно предпочтительных вариантах осуществления настоящее изобретение относится к вариантам нейтральной металлопротеазы Bacillus amyloliquefaciens. Настоящее изобретение находит конкретное использование в применениях, включая в качестве неограничивающих примеров очистку, отбеливание и дезинфицирование. Дополнительно, настоящее изобретение относится к способам конструирования фермента для оптимизации его каталитической активности при неблагоприятных окружающих условиях. В частности, настоящее изобретение относится к способам изменения суммарного поверхностного заряда и/или распределения поверхностных зарядов металлопротеазы с целью получения вариантов фермента, проявляющих улучшенную эффективность и/или стабильность в моющих составах.
Многие белки и ферменты крайне восприимчивы к денатурации и подлежат необратимой денатурации при хранении в детергентах для стирки. Известно, что детергенты для стирки содержат анионные, катионные и неионные поверхностно-активные средства, где поверхностно-активное вещество классифицируется на основании его ионных (электрический заряд) свойств в воде. Эти ингредиенты взаимодействуют с поверхностным зарядом белковой молекулы, что приводит к денатурации белка (например, к потере структуры и функции). Было показано, что NprE (нейтральная металлопротеаза) нестабильна при хранении в моющих составах, содержащих поверхностно-активное вещество, такое как LAS. LAS представляет собой анионное поверхностно-активное вещество, в котором общий отрицательный заряд усиливает взаимодействие с положительно заряженными боковыми цепями аминокислот, расположенных на поверхности белка. Такие электростатические взаимодействия воздействуют на важную стабильность белка, ослабляя или нарушая стабилизирующие электростатические взаимодействия. Затем нарушается укладка дестабилизированного белка и он становится неактивным. В процессе разработки настоящего изобретения было обнаружено, что поверхностный заряд фермента серьезно влияет на эффективность стирки и/или стабильность в детергенте. Дополнительно было обнаружено, что распределение заряженных остатков на поверхности протеазы сильно влияет на эффективность стирки и/или стабильность. Способы конструирования белков по настоящему изобретению эффективно оптимизируют протеазы для увеличения эффективности одного или нескольких свойства в моющих составах посредством оптимизации суммарного поверхностного заряда и/или распределения поверхностных зарядов.
В кратком изложении, в некоторых вариантах осуществления настоящего изобретения способы затрагивают создание библиотек для оценки сайтов с участием множества аминокислотных остатков интересующего фермента и тестирование вариантов фермента на предмет интересующих свойств. Это делает возможной идентификацию полезных, нейтральных и вредоносных мутаций, а также оптимального изменения заряда (по отношению к родительскому ферменту) для интересующего свойства(в). В некоторых альтернативных вариантах осуществления заряд сканирует все остатки для создания вариантов с мутациями, которые изменяют заряд в каждом сайте (например, мутирует нейтральные остатки в положительные и/или отрицательные заряды, и мутирует заряженные остатки в противоположно заряженные и/или нейтральные остатки). В некоторых дополнительных предпочтительных вариантах осуществления, способы касаются создания комбинаторных «сбалансированных по зарядам» библиотек вариантов, которые содержат полезные мутации, которые меняют заряд фермента в требуемом направлении и полезные или нейтральные мутации, которые меняют заряд в противоположном направлении, и затем тестирования сбалансированной по зарядам библиотек на предмет интересующего свойства(в). Таким образом, поверхностный заряд фермента и распределение поверхностного заряда оптимизированы одновременно и можно идентифицировать варианты фермента, обладающие улучшениями множества свойств.
Способы по настоящему изобретению находят применение при улучшении эффективности различных классов ферментов, а также протеаз (например, амилаз, целлюлаз, оксидаз, кутиназ, маннаназ, пектиназ, амилаз, липаз и т.п.). Фактически, не планируется, что настоящее изобретение должно ограничиваться ни любым конкретным ферментом, ни классом ферментов. Кроме того, настоящее изобретение находит применение при оптимизации неферментативных свойств белка, которые требуют конкретный поверхностный заряд и распределение зарядов (например, экспрессия, связывание с клеточной поверхностью, податливость для образования состава и т.д.).
I. Производство вариантов протеазы с улучшенными свойствами
Большое количество библиотек для оценки сайтов сконструировали для NprE, в которой каждая аминокислота зрелого белка была замещена большинством других аминокислот (см., заявку на патент США, серия № 10/576331 и WO 2005/052146). Проводили скрининг этих библиотек на предмет стабильности в детергенте и эффективности очистки от КМЧ. Затем данные скрининга анализировали в отношении эффекта изменения заряда, внесенного мутациями. При конструировании вариантов NprE желательны как увеличенная стабильность, так и хорошая эффективность очистки от КМЧ (кровь-молоко-чернила), которые с самого начала казались взаимоисключающими свойствами. Настоящее изобретение относится к средствам для производства более стабильного варианта, который проявляет хорошую эффективность очистки от КМЧ.
Таким образом, настоящее изобретение относится к способам для установления мутаций, которые повышают стабильность или эффективность очистки от КМЧ без чрезмерного уменьшения другого параметра. Как используется в настоящем документе, фраза «чрезмерно неблагоприятный» относится к свойствам белка, обладающим менее чем требуемыми величинами. Этот термин охватывает некоторые белки с низкой эффективностью, обладающие нейтральными мутациями (менее чем 80% величины эффективности родительского белка или белка дикого типа); белки с низкой эффективностью, обладающие невредоносными мутациями (менее чем 50%); и в основном неактивные белки, обладающие вредоносным мутациями (менее чем 5%). В некоторых вариантах осуществления величины относительной эффективность выражены в виде показателя эффективности (ПЭ), который представляет собой отношение эффективности варианта белка к эффективности родительского белка. Затем зарядовые мутации уравновешивают для того, чтобы конечный вариант был от +1 до +3 по отношению к ферменту дикого типа. Кроме того, настоящее изобретение относится к средствам для выбора аминокислотных остатков, которые, по-видимому, не взаимодействуют в трехмерной структуре фермента, таким образом минимизируя несуммируемость между множеством мутаций.
Как описано в настоящем документе, четыре варианта NprE сконструировали с помощью способов по настоящему изобретению. Эти варианты содержали от десяти до восемнадцати мутаций. Как описано в мелких подробностях в примерах, эти варианты проявляли повышенную стабильность и эффективность очистки от КМЧ, похожие на фермент дикого типа.
II. Основные способы производства полезных вариантов фермента
Как описано в настоящем документе, определяли взаимосвязь между эффективностью стирки в анализе с микрообразцами КМЧ и общим зарядом на поверхности фермента. Способы по настоящему изобретению находят применение при повышении эффективности различных ферментов и белков (например, амилаз, целлюлаз, оксидаз, кутиназ, маннаназ, пектиназ, липаз, протеаз и других ферментов). В кратком изложении, идентифицировали аминокислотные остатки, расположенные на поверхности фермента дикого типа, которые приблизительно больше, чем на 25% доступны для растворителя, приблизительно больше, чем на 50% доступны для растворителя или приблизительно больше, чем на 65% доступны для растворителя, и создавали библиотеки для оценки сайтов, в которых каждый остаток дикого типа замещали множеством других встречающихся в природе аминокислот. В некоторых вариантах осуществления конструирование белков на поверхности молекулы касается замещения боковых цепей нейтральных аминокислот кислыми или основными боковыми цепями и/или замещения положительно заряженных боковых цепей нейтральными или отрицательно заряженными боковыми цепями или наоборот. Кроме того, изменение суммарного заряда вариантов фермента, которые проявляли улучшенную эффективность отстирывания от КМЧ, отмечали для того, чтобы определить эту структурно-функциональную взаимосвязь. В дополнительных вариантах осуществления, когда для заданного фермента устанавливали оптимальное значение заряда, проводили скрининг природных изолятов для того, чтобы идентифицировать варианты фермента с оптимальным значением заряда/распределением зарядов.
III. Производство вариантов амилазы с улучшенными свойствами
Сконструировали комбинаторную зарядовую библиотеку для AmyS-S242Q посредством введения комбинаций замен в четыре позиции в зрелом ферменте. Проводили скрининг этой библиотеки на предмет гидролиза BODIPY-крахмала, эффективности очистки микрообразцов с рисовым крахмалом и экспрессии фермента. Затем данные скрининга анализировали в отношении эффекта изменения заряда, внесенного мутациями. При конструировании вариантов AmyS-S242Q желательны как увеличенная экспрессия белка, так и хорошая эффективность фермента, несмотря на то, что было обнаружено, что эти свойства обладают отрицательной корреляцией. Настоящее изобретение относится к средствам производства варианта с более высоким уровнем экспрессии, который проявляет хорошую эффективность очистки от рисового крахмала. Таким образом, настоящее изобретение относится к способам идентификации мутаций, которые повышают экспрессию или ферментативную активность без чрезмерного уменьшения другого параметра.
Экспериментальная часть
Следующие примеры предоставлены для того, чтобы продемонстрировать и дополнительно проиллюстрировать определенные предпочтительные варианты осуществления и аспекты настоящего изобретения и не должны рассматриваться в качестве ограничивающих его область.
В следующем раскрытии экспериментов использованы следующие сокращения: °C (градусы по Цельсию); об/мин (обороты в минуту); H2O (вода); HCl (соляная кислота); а.к. и А.К. (аминокислота); п.о. (пара оснований); т.п.н. (тысяча пар оснований); кДа (килодальтон); г (грамм); мкг (микрограмм); мг (миллиграмм); нг (нанограмм); мкл (микролитр); мл (миллилитр); мм (миллиметр); нм (нанометр); мкм (микрометр); M (молярный); мМ (миллимолярный); мкМ (микромолярный); ЕД (единица); В (вольт); ММ (молекулярная масса); сек (секунда); мин(ы) (минута/минуты); час(ы) (час/часы); MgCl2 (хлорид магния); NaCl (хлорид натрия); OD280 (оптическая плотность при 280 нм ); OD405 (оптическая плотность при 405 нм); OD600 (оптическая плотность при 600 нм); PAGE (электрофорез в полиакриламидном геле); EtOH (этанол); PBS (фосфатно-солевой буфер [150 мМ NaCl, 10 мМ натрий-фосфатный буфер, pH 7,2]); LAS (лаурилсульфонат натрия); SDS (додецилсульфат натрия); Трис (трис(гидроксиметил)аминометан); TAED (N,N,N'N'-тетраацетилэтилендиамин); BES (полиэфирсульфон); MES (2-морфолиноэтансульфоновая кислота, моногидрат; общая масса 195,24; Sigma # M-3671); CaCl2 (хлорид кальция, безводный; общая масса 110,99; Sigma # C-4901); DMF (N,N-диметилформамид, общая масса 73,09, d=0,95); Abz-AGLA-Nba (2-аминобензоил-L-аланилглицил-L-лейцил-L-аланино-4-нитробензиламид, общая масса 583,65; Bachem # H-6675, VWR catalog H 100040-598); SBG 1% («Супер бульон с глюкозой»; 6 г сойтона [Difco], 3 г дрожжевого экстракта, 6 г NaCl, 6 г глюкозы); pH доводили до 7,1 с помощью NaOH перед стерилизацией с помощью известных в данной области способов; масс./об. (масса к объему); об./об. (объем к объему); Npr и npr (нейтральная металлопротеаза); SEQUEST® (программа поиска в базе данных SEQUEST, University of Washington); Npr и npr (ген нейтральной металлопротеазы); nprE и NprE (нейтральная металлопротеаза B. amyloliquefaciens); PMN (очищенная металлопротеаза MULTIFECT®); ПМТ (планшет микротитратора); MS (масс-спектроскопия); ИУК (индекс удаления краски); TIGR (The Institute for Genomic Research, Rockville, MD); AATCC (American Association of Textile and Coloring Chemists); Procter & Gamble (Procter & Gamble, Inc., Cincinnati, OH); Beckman (Beckman Coulter, Inc., Fullerton, CA); Amersham (Amersham Life Science, Inc. Arlington Heights, IL); ICN (ICN Pharmaceuticals, Inc., Costa Mesa, CA); Pierce (Pierce Biotechnology, Rockford, IL); EMPA (Eidgenossische Material Prufungs und Versuch Anstalt, St. Gallen, Switzerland); CFT (Center for Test Materials, Vlaardingen, The Netherlands); Amicon (Amicon, Inc., Beverly, MA); ATCC (American Type Culture Collection, Manassas, VA); Becton Dickinson (Becton Dickinson Labware, Lincoln Park, NJ); Perkin-Elmer (Perkin-Elmer, Wellesley, MA); Rainin (Rainin Instrument, LLC, Woburn, MA); Eppendorf (Eppendorf AG, Hamburg, Germany); Waters (Waters, Inc., Milford, MA); Geneart (Geneart GmbH, Regensburg, Germany); Perseptive Biosystems (Perseptive Biosystems, Ramsey, MN); Molecular Probes (Molecular Probes, Eugene, OR); BioRad (BioRad, Richmond, CA); Clontech (CLONTECH Laboratories, Palo Alto, CA); Difco (Difco Laboratories, Detroit, MI); GIBCO BRL или Gibco BRL (Life Technologies, Inc., Gaithersburg, MD); Epicentre (Epicentre Biotechnologies, Madison, WI); Zymo Research (Zymo Research Corp., Orange, CA); Integrated DNA Technologies (Integrated DNA Technologies, Inc., Coralville, IA): New Brunswick (New Brunswick Scientific Company, Inc., Edison, NJ); Thermoelectron (Thermoelectron Corp., Waltham, MA); BMG (BMG Labtech, GmbH, Offenburg, Germany); Greiner (Greiner Bio-One, Kremsmuenster, Austria); Novex (Novex, San Diego, CA); Finnzymes (Finnzymes OY, Finland) Qiagen (Qiagen, Inc., Valencia, CA); Invitrogen (Invitrogen Corp., Carlsbad, CA); Sigma (Sigma Chemical Co., St. Louis, MO); DuPont Instruments (Asheville, NY); Global Medical Instrumentation или GMI (Global Medical Instrumentation; Ramsey, MN); MJ Research (MJ Research, Waltham, MA); Infors (Infors AG, Bottmingen, Switzerland); Stratagene (Stratagene Cloning Systems, La Jolla, CA); Roche (Hoffmann La Roche, Inc., Nutley, NJ); Ion Beam Analysis Laboratory (Ion Bean Analysis Laboratory, The University of Surrey Ion Beam Centre (Guildford, UK); TOM (Terg-o-Meter); КМЧ (кровь-молоко-чернила); BaChem (BaChem AG, Bubendorf, Switzerland); Molecular Devices (Molecular Devices, Inc., Sunnyvale, CA); MicroCal (Microcal, Inc., Northhampton, MA); Chemical Computing (Chemical Computing Corp., Montreal, Canada); NCBI (National Center for Biotechnology Information); GE Healthcare (GE Healthcare, UK).
Пример 1
Анализы
Следующие анализы использовали в описанных ниже примерах. Любое отклонение от приведенных ниже протоколов отмечено в примерах. В этих экспериментах спектрофотометр использовали для измерения оптической плотности продуктов, образованных после завершения реакций. Рефрактометр использовали для измерения коэффициента отражения образцов.
A. Определение содержания белка
1. Анализ с БХК (бицинхониновая кислота) для определения содержания белка
В этих анализах анализ с БХК (Pierce) использовали для определения концентрации белка в образцах протеазы в масштабе планшета микротитратора (ПМТ). В этой аналитической системе использовали следующие химические растворы и растворы реактивов: реактив для анализа белка с БКХ и буфер для разведения Pierce (50 мМ MES, pH 6,5, 2 мМ CaCl2, 0,005% TWEEN®-80). Использовали оборудование ПМТ-ридер SpectraMAX (type 340). ПМТ были получены из Costar (type 9017).
В тесте по 200 мкл реактива с БХК раскапывали в каждую лунку, а потом добавляли по 20 мкл разбавленного белка. После тщательного перемешивания, ПМТ инкубировали в течение 30 минут при 37°C. Удаляли пузырьки воздуха и считывали оптическую плотность (OD) раствора в лунках при 562 нм. Для определения концентрации белка, фоновые показания прибора вычитали из показаний прибора для образцов. По величинам OD562 для белковых стандартов (очищенная протеаза) построили калибровочную кривую. Концентрации белка в образцах экстраполировали из калибровочной кривой.
2. Анализ Бредфорда для определения содержания белка
В этих анализах анализ с окрашивающим реактивом Бредфорда (Quick Start) использовали для определения концентрации белка в образцах протеазы в масштабе ПМТ.
В этой аналитической системе использовали следующие химические растворы и растворы реактивов: окрашивающий реактив Бредфорда Quick Start (BIO-RAD кат. № 500-0205), буфер для разведения (10 мМ NaCl, 0,1 мМ CaCl2, 0,005% TWEEN®-80). Использовали оборудование Biomek FX Robot (Beckman) и ПМТ-ридер SpectraMAX (type 340; Molecular Devices). ПМТ были из Costar (type 9017).
В тесте по 200 мкл окрашивающего реактива Бредфорда раскапывали в каждую лунку, а потом добавляли по 15 мкл буфера для разведения. В конце добавляли в лунки по 10 мкл фильтрованного культурального бульона.
После тщательного перемешивания ПМТ инкубировали, по меньшей мере, в течение 10 минут при комнатной температуре. Удаляли пузырьки воздуха и OD лунок считывали при 595 нм. Для определения концентрации белка фоновое показание прибора (т.е. от неинокулированных лунок) вычитали из показаний прибора для образцов. Полученные величины OD595 давали относительную меру содержания белка в образцах.
B. Анализ с микрообразцами для тестирования эффективности протеазы
Использованное оборудование включало Eppendorf Thermomixer и ПМТ-ридер SpectraMAX (type 340). ПМТ были получены из Costar (type 9017). Препараты детергента (жидкий детергент для стирки TIDE® 2× Ultra Clean Breeze (Procter & Gamble); условия мытья США).
Воду Milli-Q доводили до жесткости воды 6 gpg (Ca/Mg=3/1) и добавляли 0,78 г/л детергента TIDE® 2× Ultra Clean Breeze. Детергент предварительно подвергали тепловой обработке при 95°C в течение одного часа для инактивирования всех ферментов, присутствующих в составе. Моющий раствор взбалтывали в течение 15 минут. Затем добавляли 5 мМ HEPES (свободная кислота) и pH доводили до 8,2.
Микрообразцы
Круглые микрообразцы 0,25 дюйма в диаметре получали из CFT Vlaardingen. Перед вырезанием образцов материю (EMPA 116) промывали водой. В каждую лунку 96-луночного планшета микротитратора помещали по одному микрообразцу.
Способ тестирования
Требуемый моющий раствор изготавливали, как описано выше. После уравновешивания Thermomixer при 25°C, 190 мкл моющего раствора добавляли в каждую лунку ПМТ, содержащую микрообразец. К этой смеси добавляли по 10 мкл разбавленного раствора фермента для того, чтобы конечная концентрация фермента составляла 1 мкг/мл (установлена в анализе с БХК). ПМТ запечатывали лентой и помещали в инкубатор на 30 минут при встряхивании при 1400 об/мин. После инкубирования при соответствующих условиях по 100 мкл раствора из каждой лунки переносили в свежий ПМТ. Новый ПМТ, содержащий по 100 мкл раствора в лунке, считывали при 405 нм с помощью ПМТ-ридера SpectraMax. Также были включены пустые контроли, а также контроль, содержащий микрообразец и детергент, но не фермент.
Вычисление эффективности КМЧ
Полученное значение оптической плотности корректировали по пустому значению (т.е. полученному после инкубирования микрообразцов в отсутствие фермента). Полученная оптическая плотность давала меру гидролитической активности тестируемого фермента.
C. Анализ стабильности с TIDE®
Стабильность дикого типа и вариантов протеазы измеряли после стадии инкубирования в присутствии 25% термообработанного жидкого детергента для стирки TIDE® 2× Ultra Clean Breeze. Начальную и остаточную активность определяли с использованием описанного ниже AGLA-анализа, флуоресцентного 96-луночного планшетного ридера, инкубатора/шейкера (iEMS; Thermoelectron) и инкубатора/шейкера (Innova; New Brunswick (type 4230). ПМТ были из Costar (type 9017) и из Greiner (черные планшеты, type 655076).
Химикаты и реактивы
В этой аналитической системе использовали следующие химические растворы и растворы реактивов:
TIDE® 2× Ultra Clean Breeze
125 г TIDE® 2× Ultra Clean Breeze (термообработанный при 95°C, как указано выше), растворенные в смеси из 50 г 50 мМ HEPES pH 8,2 и 275 мл воды; концентрация TIDE® составляла 27,7%, после разбавления супернатантом 25% (ниже называемый как «TIDE®»)
Буфер для разведения MES
52,6 мМ MES/NaOH, 2,6 мМ CaCl2, 0,005% TWEEN®-80, pH 6,5
Субстрат AGLA, BaChem, кат. № H-6675 или American Peptide Co., кат. № 81-0-31
Раствор субстрата AGLA
451 мг AGLA растворяли в 16 мл N,N-диметилформамида; этот раствор выливали в 304 мл MES-буфера (52,6 мМ MES/NaOH, 2,6 мМ CaCl2, 0,005% TWEEN®-80, pH 6,5) при встряхивании.
Способы тестирования
Бесстрессовые условия
Сначала 20 мкл фильтрованного культурального бульона разводили в 180 мкл MES буфера для разведения. Затем 20 мкл этого разбавленного бульона разводили в 180 мкл MES буфера для разведения. Затем 10 мкл этого разведения разводили в 190 мкл раствора субстрата AGLA в планшете, предварительно прогретом до 25°C. Удаляли все присутствующие пузырьки и планшет измеряли в соответствии с протоколом протеазного анализа с AGLA.
Стрессовые условия
Сначала 20 мкл фильтрованного культурального бульона разводили в 180 мкл моющего раствора TIDE® и после предварительного перемешивания в шейкере iEMS в течение 5 минут, дополнительно инкубировали в шейкере Innova. Планшет инкубировали всего в течение 60 минут при 32°C, при 200 об/мин. Кроме того, 20 мкл фильтрованного культурального бульона разбавляли в 180 мкл моющего раствора TIDE® и после предварительного перемешивания в шейкере iEMS в течение 5 минут, дополнительно инкубировали в шейкере Innova. Планшет инкубировали всего в течение 40 минут при 20°C, при 200 об/мин. Затем 20 мкл любого из этих растворов разбавляли в 180 мкл буфера для разведения MES и 10 мкл этого разведения разбавляли в 190 мкл раствора субстрата AGLA в планшете, предварительно прогретом до 25°C. Все присутствующие пузырьки воздуха удаляли и планшет измеряли в соответствии с протоколом протеазного анализа с AGLA.
Вычисления
Измерения флуоресценции проводили при возбуждении на 350 нм и испускании на 415 нм. Программное обеспечение спектрофотометра вычисляло реакционные скорости увеличения флуоресценции для каждой лунки для линии линейной регрессии мЕОФ/мин:
Процент остаточной активности: (Наклон в стрессовых условиях)*100/ (Наклон в бесстрессовых условиях)
D. Протеазный анализ с 2-аминобензоил-L-аланилглицил-L-лейцил-L-аланино-4-нитробензиламидом (Abz-AGLA-Nba)
Предоставленный ниже способ обеспечивает определенную степень технических подробностей, которые делают данные воспроизводимого протеазного анализа независящими от времени и места. Хотя анализ является приспосабливаемым к заданным лабораторным условиям, любые данные, полученные с помощью модифицированной процедуры, следует приводить в соответствии с результатами, полученными исходным способом.
Нейтральные металлопротеазы расщепляют пептидную связь между глицином и лейцином в 2-аминобензоил-L-аланилглицил-L-лейцил-L-аланино-4-нитробензиламиде (Abz-AGLA-Nba). Свободный 2-аминобензоил-L-аланилглицин (Abz-AG) в растворе обладает флуоресценцией с максимумом испускания при 415 нм и максимумом возбуждения при 340 нм. Флуоресценцию Abz-AG гасили нитробензиламидом в интактной молекуле Abz-AGLA-Nba.
В этих экспериментах высвобождение Abz-AG с помощью протеолитического расщепления Abz-AGLA-Nba отслеживали посредством флуоресцентной спектроскопии (Возб. 340/Исп. 415). Скорость проявления Abz-AG являлась мерой протеолитической активности. Анализы выполняли без ограничения начальной скорости субстратом.
Перемешиватель для микропланшетов с контролем температуры (например, Eppendorf Thermomixer) требовался для воспроизводимости результатов анализов. Анализируемые растворы инкубировали до требуемой температуры (например, 25°C) в перемешивателе для микропланшетов перед добавлением фермента. Растворы ферментов добавляли в планшет в перемешивателе, энергично перемешивали и быстро переносили в планшет-ридер.
Требовался спектрофотометр с возможностью непрерывной записи данных и линейно-регрессионного анализа, наряду с контролем температуры (например, SpectraMax M5, Gemini EM, Molecular Devices). Ридер всегда поддерживал требуемую температуру (например, 25°C). Ридер настраивали для обнаружения top-read флуоресценции и устанавливали возбуждение на 350 нм и испускание на 415 нм без применения фильтра с ограниченной полосой пропускания. PMT устанавливали на среднюю чувствительность и 5 считываний для каждой лунки. Включали автокалибрование, но только для калибрования перед первым считыванием. Анализ измеряли в течение 3 минут при минимальном интервале считывания в соответствии с количеством лунок, выбранных для отслеживания. Ридер настраивали для вычисления скорости мЕОФ/мин (тысячные доли единицы относительной флуоресценции в минуту). Количество считываний, использованное для вычисления скорости (точек Vmax), устанавливали на количество, эквивалентное 2 минутам, которое определялось с помощью интервала считывания (например, при считывании каждые 10 секунд будут использоваться 12 точек для вычисления скорости). Максимум ЕОФ устанавливали на 50000.
Все пипетирование основных растворов фермента и субстрата выполняли поршневыми пипетками (Rainin Microman). Рабочие растворы буфера, фермента и рабочий раствор для анализа пипетировали одно- или многоканальными пипетками с вытеснением воздухом (Rainin LTS) из пробирок, емкостей с реактивами или микропланшетов с основными растворами. Для минимизации потерь реактивов удобна пипетка с репитером (Eppendorf) при переносе раствора для анализа в лунки микропланшета, когда используют только несколько лунок. Автоматизированные пипетторы, такие как Beckman FX или Cybio Cybi-well, также удобны при переносе растворов ферментов из микропланшета со стоковым раствором в микропланшет для анализа для того, чтобы одновременно инокулировать целый микропланшет.
Реактивы и растворы
52,6 мМ MES/NaOH, 2,6 мМ CaCl2, pH 6,5 - MES буфер
MES кислоту (10,28 г) и 292 мг безводного CaCl2 растворяли приблизительно в 900 мл очищенной воды. Раствор титровали с помощью NaOH до pH 6,5 (при 25°C или с чувствительным к температуре pH зондом). pH-отрегулированный буфер доводили до общего объема 1 л. Конечный раствор фильтровали через 0,22 мкм стерильный фильтр и хранили при комнатной температуре.
48 мМ Abz-AGLA-Nba в DMF - основной раствор Abz-AGLA-Nba
Приблизительно 28 мг Abz-AGLA-Nba помещали в маленькую пробирку. Его растворяли в DMF (объем меняется в зависимости от массы Abz-AGLA-Nba) и встряхивали в течение нескольких минут. Раствор хранили при комнатной температуре в защищенном от света месте.
50 мМ MES, 2,5 мМ CaCl2, 5% DMF, 2,4 мМ Abz-AGLA-Nba pH 6,5 - раствор для анализа
Один мл основного раствора Abz-AGLA-Nba добавляли в 19 мл MES буфера и встряхивали. Раствор хранили при комнатной температуре в защищенном от света месте.
50 мМ MES, 2,5 мМ CaCl2, pH 6,5 - буфер для разведения фермента
Этот буфер изготавливали добавлением 5 мл очищенной вода к 95 мл MES буфера.
50 мМ MES, 2,5 мМ CaCl2, 5% DMF, pH 6,5 - буфер для разведения субстрата
5 мл чистого DMF добавляли к 95 мл MES буфера. Этот буфер использовали для определения кинетических параметров.
Растворы ферментов
Основные растворы фермента разбавляли буфером для разведения фермента до концентрации приблизительно 1 м.д. (1 мкг/мл). Нейтральную протеазу MULTIFECT® (NprE дикого типа) разбавляли до концентрации ниже 6 м.д. (6 мкг/мл). Серийные разведения были предпочтительными. При комнатной температуре растворы были стабильны в течение 1 часа, но для более длительных периодов хранения растворы хранили во льду.
Процедура
Сначала изготавливали все буферные, основные и рабочие растворы. Каждое разведение фермента анализировали трижды, если не указано иначе. При неполном заполнении лунки в стоковом микропланшете заполняли рабочим раствором фермента так, чтобы образовывались полные вертикальные колонки, начиная с левой части планшета (чтобы подстроиться под планшет-ридер). Соответствующий планшет для анализа заполняли сходным образом. Спектрофотометр для микропланшетов настраивали так, как описывалось ранее.
Сначала 200-мкл аликвоту раствора для анализа помещали в лунки 96-луночного микропланшета. Планшет инкубировали в течение 10 мин при 25°C в перемешивателе микропланшетов с контролем температуры, защищенном от света. Анализ начинали с переноса 10 мкл рабочих растворов фермента из микропланшета с основным раствором в микропланшет для анализа в перемешивателе. Оптимально, использовали 96-луночную головку пипеттора, или в некоторых экспериментах, 8-луночную многоканальную пипетку использовали для переноса сначала из крайней левой колонки. Растворы энергично перемешивали в течение 15 секунд (900 об/мин в Eppendorf Thermomixer). Микропланшет для анализа незамедлительно переносили в спектрофотометр для микропланшетов и начинали записывать измерения флуоресценции при возбуждении при 350 нм и испускании при 415 нм. Программное обеспечение спектрофотометра пересчитывало скорости реакций по увеличению флуоресценции для каждой лунки в прямую линейной регрессии в мЕОФ/мин. В некоторых экспериментах для выравнивания температуры в перемешиватель для микропланшетов помещали второй планшет, тогда как начинали считывать первый планшет.
Уровни начальных скоростей линейно зависели от концентрации продукта (т.е. флуоресценции высвобожденного 2-аминобензоила) до 0,3 мМ продукта, что соответствует приблизительно 50000 ЕОФ в растворе при начальных 2,3 мМ Abz-AGLA-Nba при фоновой флуоресценции приблизительно 22000 ЕОФ. Abz-AGLA-Nba растворяли в DMF и использовали в день его изготовления.
ПРИМЕР 2
Производство протеазы NprE в B. subtilis
В этом примере описаны проводившиеся эксперименты по производству протеазы NprE в B. subtilis. В частности, предоставлены способы, которые использовали для трансформации плазмиды pUBnprE в B. subtilis. Трансформацию выполняли, как известно в данной области (см., например, WO 02/14490, и заявку на патент США серия № 11/581,102). Предоставленная ниже последовательность ДНК (лидерная последовательность nprE, пропоследовательность nprE и зрелая последовательность ДНК nprE из B.amyloliquefaciens) кодирует белок-предшественник NprE.


В приведенной выше последовательности жирным обозначена ДНК, которая кодирует зрелую протеазу NprE, стандартный шрифт обозначает лидерную последовательность (лидерная последовательность nprE) и подчеркивание обозначает пропоследовательности (пропоследовательности nprE). Предоставленная ниже аминокислотная последовательность (лидерная последовательность NprE, пропоследовательность NprE и зрелая последовательность ДНК NprE) (SEQ ID NO:2), соответствует полноразмерному белку NprE. В этой последовательности подчеркивание обозначает пропоследовательность и жирный обозначает зрелую протеазу NprE.


Зрелая последовательность NprE приведена в SEQ ID NO:3. Эту последовательность использовали в качестве основы при изготовлении библиотек вариантов, описываемых в настоящем документе.

Экспрессирующий вектор pUBnprE сконструировали посредством амплификации гена nprE из хромосомной ДНК B. amyloliquefaciens с помощью ПЦР с использованием двух специфических праймеров:
Oligo AB 1740: CTGCAGGAATTCAGATCTTAACATTTTTCCCCTATCATTTTTCCCG (SEQ ID NO:4); и
Oligo AB 1741: GGATCCAAGCTTCCCGGGAAAAGACATATATGATCATGGTGAAGCC (SEQ ID NO:5)
ПЦР выполняли в термоциклере с высокоточной ДНК полимеразой Phusion (Finnzymes). ПЦР смесь содержала 10 мкл 5× буфера (Finnzymes Phusion), 1 мкл 10 мМ dNTP, 1,5 мкл ДМСО, по 1 мкл каждого праймера, 1 мкл ДНК полимеразы Phusion Finnzymes, 1 мкл раствора хромосомной ДНК 50нг/мкл, 34,5 мкл воды MilliQ. Использовали следующий протокол ПЦР: 1) 30 сек при 98°C; 2) 10 сек при 98°C; 3) 20 сек при 55°C; 4) 1 мин при 72°C; 5) 25 циклов из стадий со 2 до 4; и 6) 5 мин при 72°C.
В результате получили фрагмент ДНК 1,9 т.п.н., который расщепляли ДНК рестриктазами BglII и BclI. Многокопийный вектор pUB110 Bacillus (см., например, Gryczan, J Bacteriol, 134:318-329 [1978]) расщепляли рестриктазой BamHI, затем ПЦР фрагмент × BglII × BclI лигировали в вектор pUB110 × BamHI для образования экспрессирующего вектора pUBnprE.
pUBnprE трансформировали в штамм B. subtilis (∆aprE, ∆nprE, oppA, ∆spoIIE, degUHy32, ∆amyE::(xylR,pxylA-comK). Трансформацию в B. subtilis выполняли, как описано в WO 02/14490. Избирательный рост трансформантов B. subtilis, несущих вектор pUBnprE, осуществляли во встряхиваемых колбах, содержащих 25 мл среды MBD (среда определенного состава на основе MOPS), с 20 мг/л неомицина. Среду MBD изготавливали, в основном, как известно в данной области (см., Neidhardt et al., J Bacteriol, 119:736-747 [1974]), за исключением того, что NH4Cl2, FeSO4 и CaCl2 исключали из основной среды, использовали 3 мМ K2HPO4 и в основную среду добавляли 60 мМ мочевины, 75 г/л глюкозы и 1% сойтона. Также питательные микроэлементы изготавливали в виде 100× основного раствора, содержащего в одном литре 400 мг FeSO4∙7H2O, 100 мг MnSO4∙H2O, 100 мг ZnSO4∙7H2O, 50 мг CuCl2∙2H2O, 100 мг CoCl2∙6H2O, 100 мг NaMoO4∙2H2O, 100 мг Na2B4O7∙10H2O, 10 мл 1 М CaCl2 и 10 мл 0,5 M цитрата натрия. Культуру инкубировали в течение трех дней при 37°C в инкубаторе/шейкере (Infors). Это культивирование привело к образованию секретированной NprE протеазы с протеолитической активностью, которую продемонстрировали с помощью протеазных анализов. Анализ гелей выполняли с использованием гелей NuPage Novex 10% Bis-Tris (Invitrogen, кат. № NP0301BOX). Чтобы подготовить образцы для анализа, 2 объема супернатанта смешивали с 1 объемом 1 М HCl, 1 объемом 4× LDS буфера для образцов (Invitrogen, кат. № NP0007) и 1% PMSF (20 мг/мл) и впоследствии нагревали в течение 10 минут при 70°C. Затем по 25 мкл каждого образца загружали в гель вместе с 10 мкл предварительно окрашенных белковых стандартов SeeBlue Plus2 (Invitrogen, кат. № LC5925). Результаты ясно показывали, что стратегия клонирования nprE, описанная в этом примере, пригодна для производства активной NprE в B. subtilis.
Пример 3
Создание библиотек для оценки сайтов (БОС)
В этом примере описаны способы, использованные при создании БОС nprE.
Вектор pUBnprE, содержащий описанную выше экспрессирующую кассету nprE, служил в качестве ДНК матрицы. Этот вектор содержал уникальный участок рестрикции BglII, который использовали при создании библиотек для оценки сайтов. В кратком изложении, для создания библиотеки для оценки сайтов nprE проводили три ПЦР реакции, включая два ПЦР-мутагенеза для введения интересующего мутировавшего кодона в зрелую ДНК последовательность nprE, и третью ПЦР для объединения двух ПЦР-мутагенезов для того, чтобы сконструировать экспрессирующий вектор pUBnprE, содержащий требуемый мутировавший кодон в зрелой последовательности nprE.
Способ мутагенеза основывался на подходе с кодон-специфичной мутацией, в котором создание всех возможных мутаций одновременно в конкретном ДНК триплете выполняли с использованием прямого и обратного олигонуклеотидных праймеров длиной от 25 до 45 нуклеотидов, включающих специально разработанную триплетную последовательность ДНК NNS (N=A, C, T или G; и S=C или G), которая соответствовала последовательности кодона, подлежащего мутации, и гарантировала случайное встраивание нуклеотидов в конкретный кодон зрелой nprE. Число, указанное в имени праймера, соответствует положению конкретного кодона зрелой nprE. Оцениваемые сайты включали в себя: 4, 12, 13, 14, 23, 24, 33, 45, 46, 47, 49, 50, 54, 58, 59, 60, 65, 66, 87, 90, 96, 97, 100, 186, 196, 211, 214, 228 и 280. Образцовый список последовательностей праймеров описан в заявке на патент США, серия № 11/581,102.
Для создания библиотек для оценки сайтов использовали два дополнительных праймера, содержащих участок рестрикции BglII вместе с частью ДНК последовательности pUBnprE, фланкирующей участок рестрикции BglII. Эти праймеры производили в Invitrogen (масштаб 50 наномоль, обессоленные):
pUB-BglII-FW GTCAGTCAGATCTTCCTTCAGGTTATGACC (SEQ ID NO:6); и
pUB-BglII-RV GTCTCGAAGATCTGATTGCTTAACTGCTTC (SEQ ID NO:7).
Создание каждой БОС начинали с двух первичных ПЦР амплификаций с использованием праймера pUB-BglII-FW и специфического обратного праймера nprE для мутагенеза. Для второй ПЦР использовали праймер pUB-BglII-RV и специфический прямой праймер nprE для мутагенеза (праймеры для прямого и обратного мутагенеза, соответствующие позициям кодона зрелой nprE).
Введение мутаций в зрелую последовательность nprE выполняли с использованием высокоточной ДНК полимеразы Phusion (Finnzymes; кат. № F-530L). Все ПЦР проводили в соответствии с протоколом Finnzymes, приложенным к полимеразе. Условия ПЦР для первичных ПЦР составляли:
для первичной ПЦР 1:
праймер pUB-BglII-FW и специфический обратный праймер NPRE для мутагенеза - оба по 1 мкл (10 мкМ);
для первичной ПЦР 2:
праймер pUB-BglII-RV и специфический прямой праймер NPRE для мутагенеза - оба по 1 мкл (10 мкМ); вместе с
ПЦР программа: 30 секунд при 98°C, 30× (10 секунд при 98°C, 20 секунд при 55°C, 1,5 минуты при 72°C) и 5 мин при 72°C выполняли в термоциклере Peltier PTC-200 (MJ Research). Эксперименты с ПЦР привели к образованию двух фрагментов приблизительно от 2 до 3 т.п.о., которые обладали перекрывающимся участком приблизительно в 30 пар нуклеотидов в области интересующего кодона зрелой NprE. Фрагменты соединяли в третьей ПЦР реакции с использованием этих двух указанных выше фрагментов и прямого и обратного праймеров BglII. Соединяющую ПЦР реакцию проводили в следующем растворе: праймер pUB-BglII-FW и праймер pUB-BglII-RV - оба по 1 мкл (10 мкМ) вместе с
Программа для соединяющей ПЦР была следующей: 30 секунд при 98°C, 30× (10 секунд при 98°C, 20 секунд при 55°C, 2:40 минут при 72°C) и 5 мин при 72°C в термоциклере Peltier PTC-200 (MJ Research).
Амплифицированный линейный фрагмент 6,5 т.п.о. очищали с помощью QIAQUICK® PCR purification kit (Qiagen, кат. № 28106) и расщепляли рестриктазой BglII для образования липких концов на обоих концах соединяющего фрагмента:
- 35 мкл очищенного линейного фрагмента ДНК
- 4 мкл буфера REACT® 3 (Invitrogen)
- 1 мкл BglII, 10 единиц/мл (Invitrogen)
Условия реакции: 1 час, 30°C.
Лигирование фрагмента, расщепленного с помощью BglII и очищенного с использованием QIAQUICK® PCR purification kit (Qiagen, кат. № 28106), привело к образованию кольцевой и мультимерной ДНК, содержащей требуемую мутацию:
- 30 мкл очищенного, расщепленного с помощью BglII фрагмента ДНК
- 8 мкл буфера для T4 ДНК лигазы (Invitrogen кат. № 46300-018)
- 1 мкл T4 ДНК лигазы, 1 единица/мкл (Invitrogen кат. № 15224-017)
Условия реакции: 16-20 часов, при 16°C.
Впоследствии легирующую смесь трансформировали в штамм B. subtilis (∆aprE, ∆nprE, oppA, ∆spoIIE, degUHy32, ∆amyE::(xylR,pxylA-comK). Трансформацию в B. subtilis выполняли, как описано в WO 02/14490. Для каждой библиотеки собирали по 96 отдельных колоний и выращивали их в среде MOPS с неомицином и 1,25 г/л дрожжевого экстракта для анализа последовательности (BaseClear) и целей скрининга. Каждая библиотека включала не более чем 19 сайт-специфичных вариантов nprE.
Варианты создавали посредством выращивания трансформантов из БОС B. subtilis в 96-луночных ПМТ при 37°C в течение 68 часов в среде MBD с 20 мг/л неомицина и 1,25 г/л дрожжевого экстракта.
Пример 4
Создание вариантов протеазы с помощью мутагенеза QUICKHANGE®
В этом примере описаны альтернативные способы для создания БОС nprE, хотя предоставленные в настоящем документе способы пригодны для производства БОС других интересующих ферментов (например, Asp). Как в вышеприведенном примере 3, вектор pUBnprE, содержащий экспрессирующую кассету nprE, служил в качестве источника ДНК матрицы для создания БОС nprE и вариантов NprE. Основное различие между двумя способами состоит в том, что этот способ требует амплификации целого вектора с использованием комплементарных праймеров для сайт-направленного мутагенеза.
Материалы
Штамм Bacillus, содержащий вектор pUBnprE
Qiagen Plasmid Midi Kit (Qiagen кат. № 12143)
Ready-Lyse Lysozyme (Epicentre кат. № R 1802M)
dam Methylase Kit (New England Biolabs кат. № M0222L)
Zymoclean Gel DNA Recovery Kit (Zymo Research кат. № D4001)
Сайт-специфические праймеры nprE для мутагенеза, масштаб 100 нмоль, 5' фосфорилированные, очищенные в PAGE (Integrated DNA Technologies)
QUIKCHANGE® Multi Site-Directed Mutagenesis Kit (Stratagene кат. № 200514)
MJ Research PTC-200 Peltier Thermal Cycler (Bio-Rad Laboratories)
1,2% агарозные E-gels (Invitrogen кат. № G5018-01)
TempliPhi Amplification Kit (GE Healthcare кат. № 25-6400-10)
Компетентные клетки B. subtilis (∆aprE, ∆nprE, oppA, ∆spoIIE, degUHy32, ∆amyE::(xylR,pxylA-comK)
Способы
Для получения плазмид pUBnprE, содержащих одну мутацию (идентифицированную посредством скрининга БОС nprE, как описано выше в примере 4 и заявке на патент США, серия № 11/581,102), отдельную колонию каждого интересующего штамма Bacillus использовали для инокулирования пробирки с 5 мл LB + 10 м.д. неомицин (например, стартовая культура). Культуру выращивали при 37°C, при встряхивании при 225 об/мин в течение 6 часов. Затем 100 мл свежей LB+10 м.д. неомицина инокулировали 1 мл стартовой культуры. Эту культуру выращивали в течение ночи при 37°C, при встряхивании при 225 об/мин. После этого инкубирования, клеточный осадок собирали с помощью достаточного центрифугирования для предоставления клеточного осадка. Клеточный осадок ресуспендировали в 10 мл буфера P1 (Qiagen Plasmid Midi Kit). Затем 10 мкл Ready-Lyse Lysozyme добавляли к ресуспендированному клеточному осадку и инкубировали при 37°C в течение 30 мин. Протокол Qiagen Plasmid Midi Kit продолжали с использованием 10 мл буферов P2 и P3, чтобы учесть увеличенный объем клеточной культуры. После выделения из Bacillus каждой плазмиды pUBnprE, содержащей отдельную мутацию в nprE, определяли концентрацию каждой плазмиды. Затем плазмиды dam метилировали с помощью dam Methylase Kit (New England Biolabs) по инструкциям изготовителя, чтобы метилировать приблизительно по 2 мкг каждой плазмиды pUBnprE на пробирку. Для очистки и концентрирования dam-метилированных плазмид pUBnprE использовали Zymoclean Gel DNA recovery kit. Затем определяли количество dam-метилированных плазмид pUBnprE и разводили их до рабочей концентрации по 50 нг/мкл каждой. Смесь сайт-специфичных праймеров для мутагенеза изготавливали для каждой реакции отдельно. Например, при использовании плазмиды pUBnprE T14R в качестве источника матрицы, пробирка со смешанными сайт-специфичными праймерами для мутагенеза будет содержать 10 мкл nprE-S23R, 10 мкл nprE-G24R, 10 мкл nprE-N46K и 10 мкл nprE-T54R (все праймеры по 10 мкМ каждого). ПЦР реакцию с использованием QuikChange Multi Site-Directed Mutagenesis Kit (Stratagene) выполняли по инструкциям производителя, например, 1 мкл dam-метилированной плазмиды pUBnprE, содержащей одну мутацию (50 нг/мкл), 2 мкл nprE сайт-специфичных праймеров для мутагенеза (10 мкМ), 2,5 мкл 10× буфер QuikChange Multi Reaction, 1 мкл смеси dNTP, 1 мкл смеси ферментов QuikChange Multi (2,5 Ед/мкл), и 17,5 мкл дистиллированной, автоклавированной воды, для предоставления конечного объема реакционной смеси 25 мкл. Библиотеки вариантов NprE амплифицировали с использованием следующих условий: 95°C в течение 1 мин (только 1-й цикл), затем 95°C в течение 1 мин, 55°C в течение 1 мин, 65°C в течение 13,5 мин и 29 повторений цикла. Продукт реакции хранили при 4°C в течение ночи. Затем реакционную смесь подвергали обработке рестриктазой DpnI (поставляемой с QUIKCHANGE® Multi Site-Directed Mutagenesis Kit) для расщепления родительской плазмиды pUB-nprE, используя протокол производителя, т.е. 1,5 мкл рестрикционного фермента DpnI добавляли в каждую пробирку и инкубировали при 37°C в течение 3 часов; затем 2 мкл расщепленного с помощью DpnI продукта ПЦР реакции анализировали на 1,2% E-gel, чтобы гарантировать, что ПЦР реакция прошла и что родительская матрица была разрушена. Затем для создания большого количества ДНК для увеличения размера библиотеки мультивариантов nprE использовали TempliPhi амплификацию по типу катящегося кольца, используя протокол производителя, т.е. 1 мкл обработанной в DpnI QuikChange Multi Site-Directed Mutagenesis ПЦР, 5 мкл буфера для образцов TempliPhi, 5 мкл реакционного буфера TempliPhi и 0,2 мкл смеси ферментов TempliPhi, для образования конечного объема реакции ~11 мкл; инкубировали при 30°C в течение 3 часов; реакцию TempliPhi разводили добавлением 200 мкл дистиллированной, автоклавированной воды и, в кратком изложении, интенсивно перемешивали. Затем 1,5 мкл разбавленного материала TempliPhi трансформировали в компетентные клетки B. subtilis, и мультиварианты nprE отбирали для использования планшетов с LA+10 м.д. неомицина + 1,6% снятого молока. Колонии отбирали и затем секвенировали для идентификации различных сочетаний вариантов библиотеки NprE.
В таблице 4-1 приведены названия праймеров и последовательности, использованные в этих экспериментах. В Integrated DNA Technologies синтезировали все праймеры (масштаб 100 нмоль, 5'-фосфорилированные и очищенные в PAGE). Дополнительные праймеры для мутагенеза описаны в заявке на патент США, сер. № 11/581102). Оцениваемые сайты включали в себя: 4, 12, 13, 23, 45, 49, 50, 54, 59, 60, 65, 82, 90, 110, 119, 128, 129, 130, 135, 136, 137, 138, 139, 140, 151, 152, 155, 179, 190, 197, 198, 199, 204, 205, 214, 216, 217, 218, 219, 220, 221, 222, 224, 243, 244, 260, 261, 263, 265, 269, 273, 282, 285, 286, 289, 293, 296, 297 и 299.

Пример 5
Экспрессия, ферментация, очистка и определение характеристик вариантов протеаз
Этот пример описывает способы, использованные для экспрессии, ферментации и очистки протеаз трансформированного B. subtilis из предыдущих примеров.
Рекомбинантный Bacillus subtilis культивировали с помощью традиционной периодической ферментации в питательной среде. Один глицериновый сосуд культуры B. subtilis, содержащий нейтральную металлопротеазу B. amyloliquefaciens, использовали для инокулирования 600 мл 1% среды SBG, содержащей 200 мг/л хлорамфеникола. Культуры выращивали в течение 36-48 часов при 37°C, после этого времени культуральную жидкость собирали посредством центрифугирования при 12000 об/мин, как известно в данной области. Эту процедуру выполняли дважды. Конечные концентрации фермента, полученные для 48-часовых культур, находились в диапазоне приблизительно между 1,4 и 2 г/л. Через 36 часов инкубирования при 37°C, ферментационный бульон собирали и центрифугировали при 12000 об/мин (центрифуга SORVALL®, модель RC5B). Секретированные нейтральные металлопротеазы выделяли из культуральной жидкости и концентрировали приблизительно 10-кратно с помощью Amicon filter system 8400 с BES (полиэфирсульфон) с порогом 10 кДа.
Концентрированный супернатант диализировали в течение ночи при 4°C против 25 мМ MES буфера, pH 5,4, содержащего 10 мМ NaCl. Затем диализат загружали в катионообменную колонку Poros HS20 (общий объем ~83 мл; связывающая емкость ~4,5 г белка/мл колонки; воды), как описано ниже. Предварительно колонку уравновешивали с использованием 25 мМ MES буфера, pH 5,4, содержащего 10 мМ NaCl. Затем приблизительно 200-300 мл образца загружали в колонку. Связанный белок элюировали с использованием градиента pH от 5,4 до 6,2 через объем MES буфера, равный 10 колонкам. Элюирование белка было между pH 5,8 и 6,0, и оценивалось с использованием протеолитической активности, как описано в настоящем документе, и 10% (масс./об.) NUPAGE® SDS-PAGE (Novex). Затем объединяли фракции, содержащие нейтральную протеазу. Хлориды кальция и цинка в соотношении 3:1 добавляли перед тем, как довести pH до 5,8. Для очистки белка использовали Perceptive Biosystems BIOCAD® Vision (GMI).
Определяли гомогенность очищенного белка, оцененного с использованием 10% (масс./об.) NUPAGE® SDS-PAGE, при чистоте, превышающей 95%. Типично, очищенные препараты показывали ничтожную активность сериновой протеазы при оценке с использованием стандартного протеазного анализа с субстратом N-сукцинил-L-Ala-L-Ala-L-Pro-L-Phe-p-нитроанилидом (Bachem). Этот анализ выполняли в формате планшета микротитратора (ПМТ) (96 лунок) с использованием 100 мМ Трис-HCl буфера, pH 8,5, содержащего 10 мМ CaCl2 и 0,005% TWEEN®-80. Субстрат (p-AAPF NA) изготавливали посредством изготовления основного раствора 160 мМ в ДМСО (диметилсульфоксид) (100 мг/мл) и разведения этого основного раствора в 100 раз Трис-HCl буфером, содержащим CaCl2 и 0,005% TWEEN®-80. Затем 10 мкл разбавленного раствора протеазы (разведения изготавливали с использованием 100 мМ Трис-HCl буфера, pH 8,5, содержащего 10 мМ CaCl2 и 0,005% TWEEN®-80) добавляли к 190 мкл 1 мг/мл раствора p-AAPF NA. Пробу перемешивали в течение 5 минут и при 410 нм считывали кинетические изменения в течение от 2 до 5 минут. Угловой коэффициент отклика измеряли и использовали в качестве индикатора количества активности сериновой протеазы. Изготавливали состав для хранения белка с применением 25 мМ MES буфера, pH 5,8, содержащего 1 мМ хлорида цинка, 4 мМ хлорида кальция и 40% пропиленгликоля.
Пример 6
Компенсация влияния мутаций на активность и стабильность протеаз
Этот пример описывает варианты протеазы с множественными заменами, сконструированные для оптимизации двух противоречащих свойств фермента. В таблице 6-1 показано, как вычисляли изменение заряда:
Вычисление изменения заряда
Как установили в процессе разработки настоящего изобретения, усредненная стабильность снижалась при увеличении положительного заряда. Однако эффективность очистки от КМЧ увеличивалась при увеличении положительного заряда. Оптимальное значение эффективности очистки от КМЧ при условиях тестирования достигали при изменении заряда приблизительно на +1.
Увеличенная стабильность и эффективность очистки от КМЧ являются желательными в сконструированном варианте NprE. Однако эти свойства, по всей видимости, являются противоречащими свойствами. Как установлено в процессе разработки настоящего изобретения, при применении способов по настоящему изобретению возможно создавать более стабильный вариант без существенного компромисса с эффективностью очистки от КМЧ посредством избирательного объединения единичных мутаций. Стратегию, описываемую в настоящем документе, успешно применяли для производства вариантов NprE с множественными заменами, обладающих улучшениями первого свойства (например, стабильности в качестве основного свойства), при одновременном улучшении или не ухудшении второго свойства (например, эффективности очистки от КМЧ в качестве вторичного свойства). В частности, для выбора интересующих замен применяли следующие критерии. Отбирали мутации, которые предоставляют повышенную стабильность в детергенте или эффективность очистки от КМЧ без чрезмерного уменьшения другого параметра. Кроме того, компенсировались зарядовые мутации, так что конечный вариант обладал суммарным зарядом от +1 до +3 по отношению к ферменту дикого типа. Кроме того, аминокислотные остатки, которые казались не участвующими в трехмерной структуре, для минимизации несуммируемости множественных мутаций.
В процессе разработки настоящего изобретения сконструировали четыре варианта, каждый содержал от десяти до восемнадцати замен. Эти варианты приведены в таблице 6-3. Важно, что эти варианты с множественными заменами обладали увеличенной стабильностью в детергенте и сходной эффективностью очистки по сравнению с ферментом дикого типа. Это осуществляли посредством введения отрицательных и нейтральных стабилизирующих заряд мутаций, которые не были слишком вредоносными для эффективности очистки от КМЧ, при компенсации положительными зарядовыми мутациями, влияющими на эффективность, которые не слишком сильно влияли на стабильность. Сконструировали дополнительный набор пар вариантов. Первый из каждой пары обладал стабилизирующей отрицательной зарядовой мутацией, которая снижала эффективность очистки от КМЧ, и второй из каждой пары обладал компенсирующей положительной зарядовой мутацией, которая восстанавливала эффективность очистки от КМЧ и одновременно поддерживала стабильность на уровне вышеуказанного дикого типа. Значения эффективности очистки и стабильности для этих вариантов также приведены в таблице 6-4.
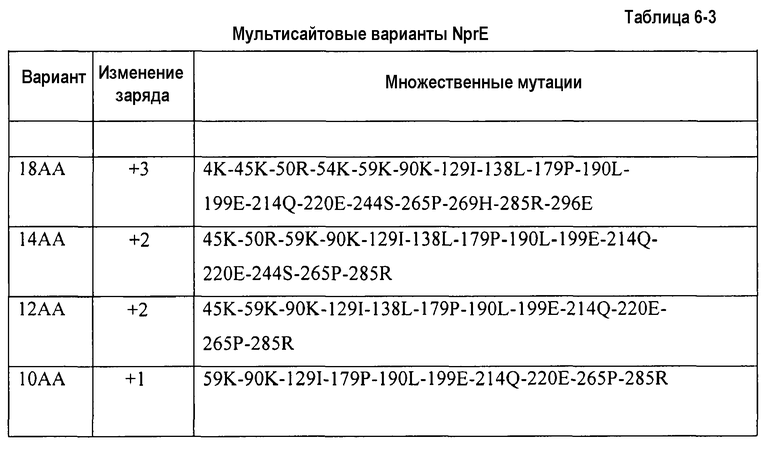
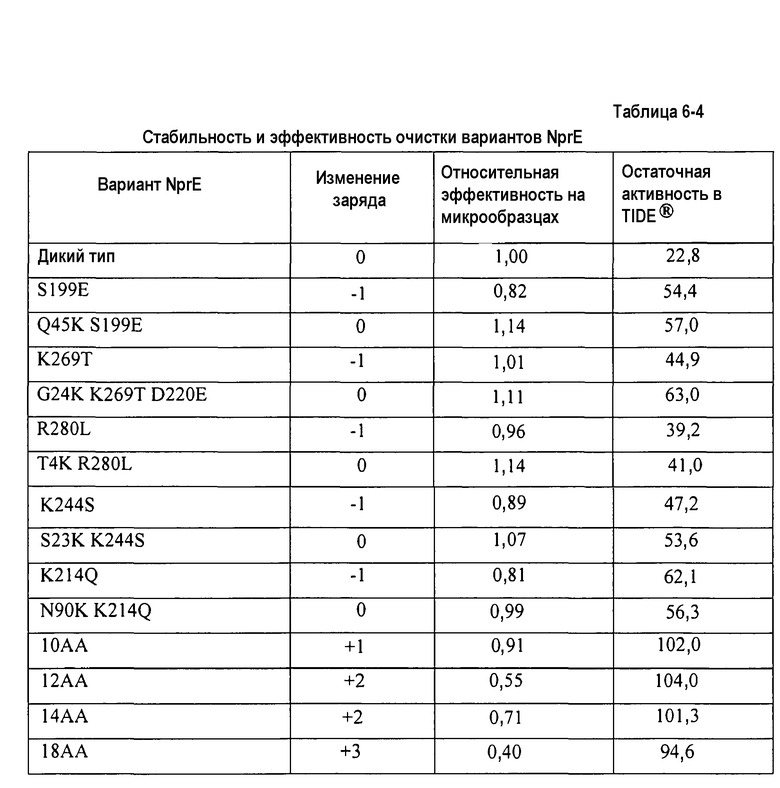
Пример 7
Компенсация влияния мутаций на активность и экспрессию амилазы
В этом примере проиллюстрировано, что два противоречащих свойства фермента могут быть оптимизированы одновременно посредством введения множественных замен аминокислот.
В этом примере описано, как проводили эксперименты по производству α-амилазы Bacillus stearothermophilus (также упоминаемой в настоящем документе как AmyS), мутантной усеченной формы амилазы AmyS (S242Q обладающей делецией 29 аминокислот, также упоминаемой в настоящем документе как S242Q), ее вариантов в B. subtilis. Трансформацию выполняли, как известно в данной области (см., например, WO 02/14490). В кратком изложении, ген, кодирующий родительские амилазы, клонировали в экспрессирующий вектор pHPLT, который содержит последовательность промотора LAT (PLAT), кодирующую сигнальный пептид LAT (preLAT), за которым идут участки рестрикции Pstl и Hpal для клонирования.
Создание КЗБ AmyS-S242Q B. stearothermophilus
Плазмидную ДНК AmyS-S242Q выделяли из трансформированного штамма B. subtilis (генотип: ∆aprE, ∆nprE, amyE::xylRPxylAcomK-phleo) и посылали в DNA2.0 Inc. в качестве матрицы для конструирования КЗБ. Выполняли запрос DNA2.0 Inc. (Mountain View, CA) на создание позиционной библиотеки в каждом из четырех сайтов амилазы AmyS-S242Q (S242Q). Варианты поставлялись в виде глицериновых основных растворов в 96-луночных планшетах. Комбинаторную зарядовую библиотеку AmyS S242Q проектировали посредством идентификации следующих четырех остатков: Gln-97, Gln-319, Gln-358 и Gln-443. Для четырех сайтов создавали КЗБ из 81 члена, выполняя все сочетания трех возможностей в каждом сайте: дикого типа, аргинин или аспарагиновая кислота.
Аминокислотную последовательность зрелой усеченной амилазы S242Q с замененной аминокислотой, показанную курсивом, использовали в качестве основы при изготовлении библиотек вариантов, описываемых в настоящем документе:

Экспрессия амилазы - масштаб 2 мл
Клоны B. subtilis, содержащие экспрессирующие векторы AmyS, S242Q или AmyTS23t, реплицировали с помощью стального 96-луночного репликатора из глицериновых основных растворов в 96-луночные культуральные планшеты (BD, 353075), содержащие 150 мкл среды LB+10 мкг/мл неомицина, которые выращивали в течение ночи при 37°C, 220 об/мин в увлажненной камере. Аликвоту 100 мкл из ночной культуры использовали для инокулирования 2000 мкл среды определенного состава + 10 мкг/мл неомицина в 5 мл пластиковых культуральных пробирках. Культуральную среду обогащали средой полуопределенного состава, основанной на буфере MOPS, с мочевиной в качестве основного источника азота, глюкозой в качестве основного источника углерода и добавлением 1% сойтона и 5 мМ кальция для активного роста клеток. Культуральные пробирки инкубировали при 37°C, 250 об/мин, в течение 72 часов. После этого инкубирования культуральные бульоны центрифугировали в течение 10 минут при 3000×g. Раствор супернатанта декантировали в 15 мл полипропиленовые конические пробирки и забирали аликвоты по 80 мкл каждого образца в 96-луночные планшеты для количественного определения белка.
Определение концентрации амилазы титрованием антител
Как описано в настоящем документе, концентрацию и специфическую активность α-амилазы определяли титрованием с ингибирующим поликлональным антителом. Было обнаружено, что поликлональные антитела, индуцированные к α-амилазе Bacillus stearothermophilus (AmyS), сильно ингибируют AmyS и α-амилазу из TS23 Bacillus sp. (например, связывание достаточно сильное для того, чтобы давать линейную потерю активности при титровании). Следовательно, это антитело можно использовать для измерения концентрации фермента, которую в свою очередь будут использовать для вычисления специфической активности. В кратком изложении, измеряют степень ингибирования фермента, вызванного несколькими известными концентрациями антитела. Исходя из этой информации, экстраполируют концентрацию антитела, необходимую для полного ингибирования, которая эквивалентна концентрации фермента в образце. Активность и ингибирование α-амилазы измеряли с использованием анализа с флуорогенным BODIPY-крахмалом. Буфер представлял собой 50 мМ MOPS, pH 7,0, содержащий 0,005% Tween-80.
Поликлональное антитело, направленное против очищенной AmyS, индуцировали в кролике и очищали стандартными способами. Эмпирическое значение «видимой концентрации» основного раствора антитела определяли посредством измерения ингибирования образца AmyS с известной специфической активностью. Затем образец антитела использовали для определения концентрации и специфической активности вариантов AmyS и TS23t. Эти величины использовали для создания нормализованных 96-луночных основных планшетов с ферментом, в которых все варианты разводили до общей концентрации.
Анализы с BODIPY-крахмалом для определения активности амилазы
Анализ с BODIPY-крахмалом выполняли с использованием EnzChek® Ultra Amylase Assay Kit (E33651, Invitrogen). Основной раствор крахмального субстрата DQ 1 мг/мл изготавливали растворением содержимого сосуда, содержащего лиофилизированный субстрат в 100 мкл 50 мМ натрий-ацетатного буфера при pH 4,0. Сосуд энергично перемешивали в течение приблизительно 20 секунд и оставляли при комнатной температуре, в темноте, при периодическом перемешивании до растворения. Добавляли 900 мкл буфера для анализа (50 мМ ацетата натрия с 2,6 мМ CaCl2 pH 5,8) и сосуд энергично перемешивали в течение 20 секунд. Раствор субстрата хранили при комнатной температуре, в темноте, до тех пор, пока он не будет готов к использованию, или при 4°C. Для анализа 100 мкг/мл рабочего раствора субстрата DQ изготавливали из 1 мг/мл раствора субстрата в буфере для анализа. 190 мкл 100 мкг/мл раствора субстрата добавляли в каждую лунку в 96-луночном плоскодонном планшете микротитратора. По 10 мкл образцов фермента добавляли в лунки, перемешивали в течение 30 секунд с использованием перемешивателя с термостатом при 800 об/мин. Путой образец, который содержал только буфер и субстрат (без фермента), включали в анализ. Скорость изменения интенсивности флуоресценции измеряли (возбуждение: 485 нм, испускание: 520 нм) во флуоресцентном ридере для планшетов микротитратора при 25°C в течение 5 минут.
Связывание α-амилазы
Варианты амилазы инкубировали с микрообразцами с рисовым крахмалом CS-28 или без них при стандартных условиях мытья в течение 30 мин. Количество свободного фермента измеряли с помощью анализа с BODIPY-крахмалом. Фракцию фермента, связанного с микрообразцами, вычисляли следующим образом:
Связанная фракция = (Активность фермента в отсутствие образца - Активность фермента в присутствии образца)/(Активность фермента в отсутствие образца)
Результаты
Как установлено в процессе разработки настоящего изобретения, среднее значение экспрессии AmyS-242Q снижалось с увеличением положительного заряда. Однако специфический гидролиз BODIPY-крахмала увеличивался с увеличением положительного заряда. Увеличенные экспрессия рекомбинантной амилазы и гидролиз крахмала являются желательными в сконструированном варианте AmyS-242Q, пригодном для разжижения крахмала при производстве топливного этанола или, например, в очистке в моющих применениях. Однако эти свойства, по всей видимости, являются противоречащими свойствами. Как установлено в процессе разработки настоящего изобретения, при использовании способов по настоящему изобретению можно создать вариант амилазы с более высокой экспрессией без существенного компромисса с гидролизом крахмала посредством избирательного объединения отдельных мутаций. Стратегию, описываемую в настоящем документе, успешно применяли для создания и отбора вариантов AmyS-242Q с множественными заменами, обладающих улучшениями первого свойства (например, экспрессии в качестве первичного свойства), при одновременном улучшении или не ухудшении второго свойства (например, гидролиза крахмала в качестве вторичного свойства).
Кроме того, в противоположность среднему значению экспрессии вариантов AmyS-242Q, очистка микрообразцов с рисовым крахмалом увеличивалась при увеличении положительного заряда. Увеличенная экспрессия и эффективность очистки рекомбинантной амилазы являются желательными при конструировании варианта AmyS- 242Q. Однако, по всей видимости, эти свойства также являются противоречащими свойствами. Как установлено в процессе разработки настоящего изобретения, при применении способов по настоящему изобретению можно создать вариант амилазы с более высокой экспрессией без существенного компромисса с эффективностью очистки посредством избирательного объединения отдельных мутаций. Описываемую в настоящем документе стратегию успешно применяли для создания и отбора вариантов AmyS-242Q с множественными заменами, обладающих улучшениями первого свойства (например, экспрессии в качестве первичного свойства), при одновременном улучшении или не ухудшении второго свойства (например, очистка микрообразцов с рисовым крахмалом в качестве вторичного свойства).
В частности, восемнадцать членов комбинаторной зарядовой библиотеки (КЗБ) AmyS-S242Q, содержащей варианты, обладающие сочетаниями из от одной до четырех замен заряженных остатков, тестировали на экспрессию во встряхиваемых пробирках, гидролиз BODIPY-крахмала и активность очистки от рисового крахмала. Победители AmyS-S242Q приведены в таблицах 7-1 и 7-2. Важно, что варианты с множественными заменами из таблицы 7-1 обладают равной или улучшенной экспрессией и равным или улучшенным гидролизом BODIPY-крахмала по сравнению с родительским ферментом. Сходным образом, варианты с множественными заменами из таблицы 7-2 обладают равной или улучшенной экспрессией и равной или улучшенной активностью очистки от рисового крахмала по сравнению с родительским ферментом.
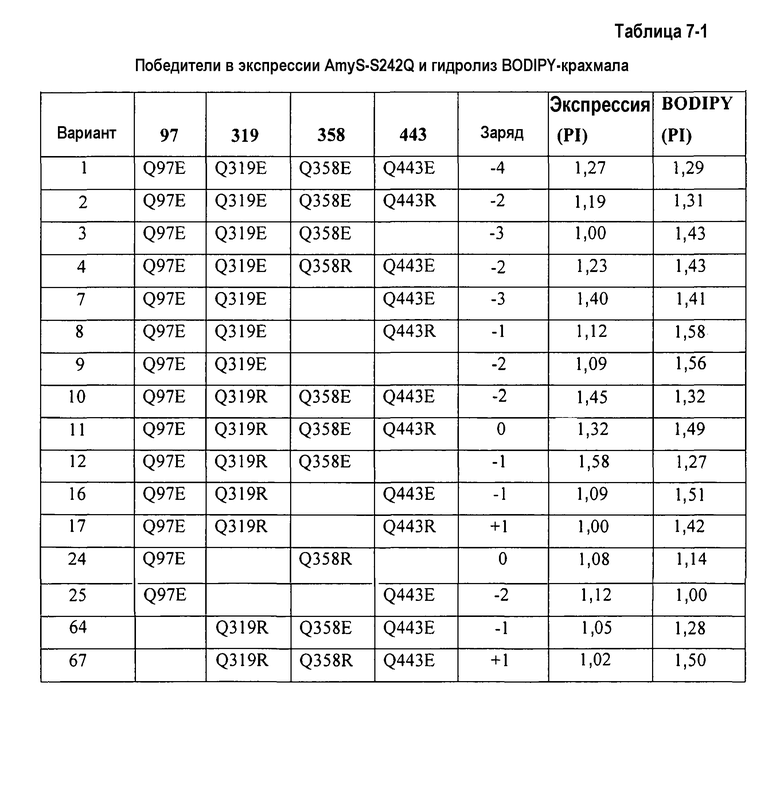
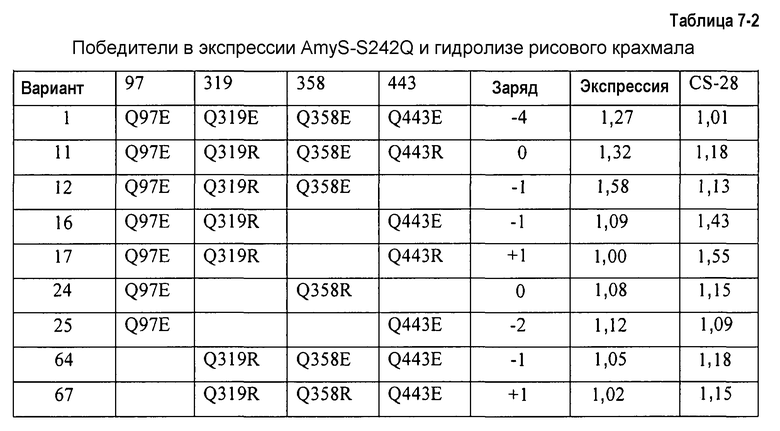
В общем, так как активность фермента и синтез фермента обладают различными зависимостями от заряда (см. фиг.2A, 2B, 3A и 3B), они обладают отрицательной корреляцией (см. фиг.1A и 1B). Однако существует множество вариантов, которые обладают как улучшенной экспрессией, так и улучшенной активностью, и анализ библиотеки, таким образом, делает возможной их идентификацию.
Несмотря на то что способ продемонстрировали на примере амилазы, он применим к другим классам ферментов, таким как протеазы, липазы, целлюлазы, трансферазы и пектиназы. Кроме того, любое сочетание из двух или более свойств может анализироваться одновременно, например экспрессия, активность, связывание, тепловая стабильность, стабильность в присутствии детергента и/или хелатора.
Все патенты и публикации, отмеченные в описании, показывают уровень специалистов в области, к которой принадлежит изобретение. Специалисты в данной области легко оценят то, что настоящее изобретение хорошо приспособлено для выполнения задач и получения оговоренных результатов и преимуществ, а также того, что с ними связано. Композиции и способы, описываемые в настоящем документе, показывают образцовые предпочтительные варианты осуществления и они не предназначены для ограничения объема изобретения. Профессионалам в данной области совершенно очевидно, что варьирование замен и модификаций может быть выполнено в описываемом в настоящем документе изобретении без отступления от области и сущности изобретения.
Иллюстративно описанное в настоящем документе изобретение может быть осуществлено на практике надлежащим образом при отсутствии любого элемента или элементов, ограничения или ограничений, которые специально не описаны в настоящем документе. Термины и выражения, которые были использованы, используются в качестве терминов описания, но не для ограничения, и не имеет места намерение при использовании таких терминов и выражений исключать эквиваленты показанных и описанных свойств или их фрагментов, но установлено, что различные модификации возможны в рамках заявленного объема изобретения. Таким образом, следует понимать, что несмотря на то что настоящее изобретение было раскрыто конкретно с помощью предпочтительных вариантов осуществления и необязательных свойств, специалисты в данной области могут прибегнуть к модификации и изменению идей, раскрытых в настоящем документе, и что такие модификации и вариации считают попадающими в объем данного изобретения, который определен в настоящем документе.
В настоящем документе изобретение было описано широко и в общем. Все более узкие разновидности и менее общая группировка попадает в пределы общего раскрытия, также образуя часть изобретения. Это включает общее описание изобретения с исключением или негативным ограничением, удаляющим любой объект из рода, независимо от того, определенно указан исключенный объект в настоящем документе или нет.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБЫ УЛУЧШЕНИЯ МНОЖЕСТВЕННЫХ СВОЙСТВ БЕЛКА | 2008 |
|
RU2553550C2 |
| СПОСОБЫ УЛУЧШЕНИЯ ЭФФЕКТИВНОСТИ БЕЛКОВ | 2008 |
|
RU2569106C2 |
| ПРОДУЦИРОВАНИЕ ТЕРМОЛИЗИНА И ЕГО ВАРИАНТОВ И ЕГО ИСПОЛЬЗОВАНИЕ В ЖИДКИХ ДЕТЕРГЕНТАХ | 2008 |
|
RU2536255C2 |
| ПРОДУЦИРОВАНИЕ ТЕРМОЛИЗИНА И ЕГО ВАРИАНТОВ И ЕГО ИСПОЛЬЗОВАНИЕ В ЖИДКИХ ДЕТЕРГЕНТАХ | 2014 |
|
RU2733541C2 |
| ВАРИАНТЫ АЛЬФА-АМИЛАЗЫ С ИЗМЕНЕННЫМИ СВОЙСТВАМИ | 2008 |
|
RU2499044C2 |
| КОМПОЗИЦИИ И СПОСОБЫ, ВКЛЮЧАЮЩИЕ В СЕБЯ ПРИМЕНЕНИЕ ВАРИАНТНЫХ МИКРОБНЫХ ПРОТЕАЗ | 2009 |
|
RU2575073C2 |
| ПРИМЕНЕНИЕ И ПОЛУЧЕНИЕ СТАБИЛЬНОЙ ПРИ ХРАНЕНИИ НЕЙТРАЛЬНОЙ МЕТАЛЛОПРОТЕИНАЗЫ | 2006 |
|
RU2433182C2 |
| ВАРИАНТЫ СЕРИНОВОЙ ПРОТЕАЗЫ С МНОЖЕСТВЕННЫМИ МУТАЦИЯМИ | 2007 |
|
RU2558261C2 |
| ПРИМЕНЕНИЕ БЕЛКОВЫХ ГИДРОЛИЗАТОВ ДЛЯ СТАБИЛИЗАЦИИ ДЕТЕРГЕНТНЫХ СОСТАВОВ МЕТАЛЛОПРОТЕАЗЫ | 2008 |
|
RU2484138C2 |
| КОМПОЗИЦИИ И СПОСОБЫ, ВКЛЮЧАЮЩИЕ ВАРИАНТЫ СУБТИЛИЗИНА | 2009 |
|
RU2575602C2 |
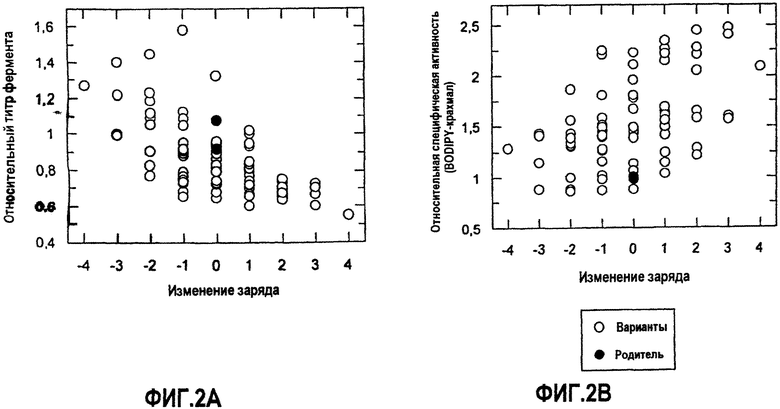
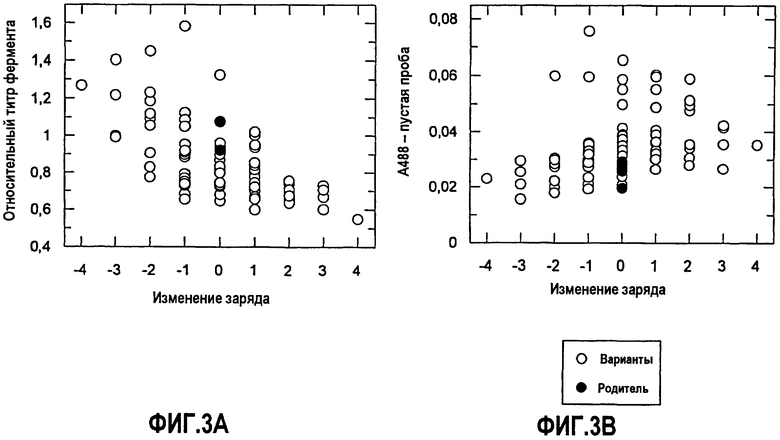
Изобретение относится к способу идентификации улучшенных вариантов белка. Способ включает тестирование множества вариантов белка с единичными заменами в первом тесте первого свойства и во втором тесте второго свойства. Свойству родительского белка присваивается значение 1,0 в каждом тесте. Благоприятное первое или второе свойство обладает значением, превышающим 1,0, и чрезмерно неблагоприятное первое или второе свойство обладает значением менее чем приблизительно 0,80. Указанный родительский белок представляет собой протеазу. Первым свойством является стабильность, вторым свойством является эффективность стирки. Осуществляют идентификацию замены, введение замены в белок для получения варианта белка с множественными заменами. При этом замена не взаимодействует в трехмерной структуре указанного родительского белка, и одна из замен содержит изменение суммарного заряда, равное 0, -1 или -2 по отношению к родительскому ферменту, и, по меньшей мере, одна из замен содержит изменение суммарного заряда, равное +1 или +2 по отношению к родительскому ферменту. Тестируют вариант белка с множественными заменами в первом и втором тесте и идентифицируют указанный вариант белка. Изобретение позволяет получить вариант белка с улучшенной стабильностью и эффективностью стирки. 13 з.п. ф-лы, 6 ил., 6 табл., 7 пр.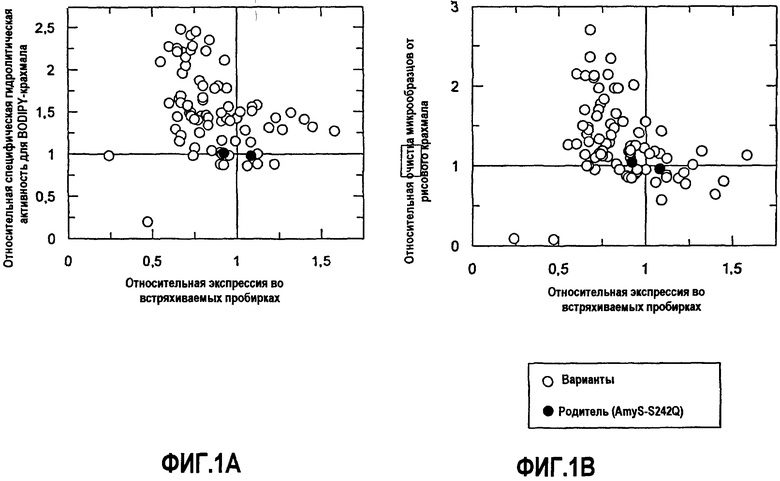
1. Способ идентификации улучшенных вариантов белка, включающий в функциональном порядке:
a) тестирование множества вариантов белка с единичными заменами в первом тесте первого свойства и во втором тесте второго свойства, где свойству родительского белка присваивается значение 1,0 в каждом тесте, благоприятное первое или второе свойство обладает значением, превышающим 1,0, и чрезмерно неблагоприятное первое или второе свойство обладает значением менее чем приблизительно 0,80, где указанный родительский белок представляет собой протеазу, указанное первое свойство представляет собой стабильность и указанное второе свойство представляет собой эффективность стирки;
b) идентификацию замены, по меньшей мере, в одном из вариантов белка с единичными заменами, которая связана с благоприятным первым свойством и которая не связана с чрезмерно неблагоприятным вторым свойством;
c) идентификацию замены, по меньшей мере, в одном из вариантов белка с единичными заменами, которая связана с благоприятным вторым свойством и которая не связана с чрезмерно неблагоприятным первым свойством; и
d) введение замены из стадии b) и замены из стадии c) в белок для получения варианта белка с множественными заменами, где
(i) замена из стадии b) и замена из стадии c) не взаимодействуют в трехмерной структуре указанного родительского белка, и
(ii) по меньшей мере, одна из замен содержит изменение суммарного заряда, равное 0, -1 или -2 по отношению к родительскому ферменту, и, по меньшей мере, одна из замен содержит изменение суммарного заряда, равное +1 или +2 по отношению к родительскому ферменту;
е) тестирование указанного варианта белка с множественными заменами в указанном первом тесте и в указанном втором тесте; и
f)идентификацию указанного варианта белка с множественными заменами как варианта белка с улучшенной стабильностью и эффективностью стирки, если он достигает значения, превышающего 1,0 как в первом, так и во втором указанных тестах, или значения, превышающего 1,0 в первом тесте, и значения от 0,80 до 1,0 во втором тесте.
2. Способ по п.1, где указанное благоприятное первое или второе свойство обладает значением больше чем приблизительно 1,2.
3. Способ по п.1, где указанное чрезмерно неблагоприятное первое или второе свойство обладает значением менее чем приблизительно 0,60.
4. Способ по п.3, где указанное чрезмерно неблагоприятное первое или второе свойство обладает значением менее чем приблизительно 0,40.
5. Способ по п.1, где указанная стабильность включает стабильность в моющих составах и эффективность стирки включает эффективность отстирывания крови-молока-чернил (КМЧ).
6. Способ по п.1, где указанную эффективность стирки тестируют в порошке или жидком моющем составе, обладающем pH между приблизительно 5 и приблизительно 12.
7. Способ по п.1, где указанную эффективность стирки тестируют в холодной воде в жидком детергенте, обладающем основным pH.
8. Способ по п.1, где указанная протеаза выбрана из нейтральной металлопротеазы и сериновой протеазы.
9. Способ по п.8, где указанная сериновая протеаза представляет собой субтилизин.
10. Способ по п.8, где указанная нейтральная металлопротеаза представляет собой нейтральную металлопротеазу, полученную из члена семейства Bacillaceae.
11. Способ по п.1, где замены находятся в родительском ферменте в положениях, обладающих доступной для растворителя поверхностью (ДРП) больше чем приблизительно 25%.
12. Способ по п.1, где замены находятся в родительском ферменте в положениях, обладающих доступной для растворителя поверхностью (ДРП) больше чем приблизительно 50% или больше чем приблизительно 65%.
13. Способ по п.1, где указанный родительский фермент представляет собой фермент дикого типа.
| WO 03076580 A2, 18.09.2003 | |||
| ZHANG W | |||
| ET AL | |||
| Способ восстановления хромовой кислоты, в частности для получения хромовых квасцов | 1921 |
|
SU7A1 |
| Environ | |||
| Microbiol | |||
| Способ обработки целлюлозных материалов, с целью тонкого измельчения или переведения в коллоидальный раствор | 1923 |
|
SU2005A1 |
| HAMAMATSU N | |||
| ET AL | |||
| Directed evolution by accumulating tailored mutations: thermostabilization of lactate oxidase with less trade-off with catalytic | |||

