Область изобретения
Объектом настоящего изобретения является клетка, которая была генетически модифицирована таким образом, что стала способна продуцировать большее количество 2-гидроксиизомасляной кислоты или большее количество полигидроксиалканоатов, содержащих мономерные структурные единицы 2-гидроксиизомасляной кислоты, по сравнению с ее диким типом, отличающаяся тем, что образование 2-гидроксиизомасляной кислоты или полигидроксиалканоатов, содержащих мономерные структурные единицы 2-гидроксиизомасляной кислоты, происходит через ацетоацетил-Коэнзим А в качестве промежуточного продукта и 3-гидроксибутирил-Коэнзим А в качестве прекурсора.
Уровень техники
Метакриловая кислота, ее сложные эфиры и полимеры находят широкое применение при производстве акриловых оконных стекол, продуктов, изготовляемых с применением пресс-формы для литья под давлением, покрытий и многих других продуктов.
Описано множество способов получения метакриловой кислоты. Однако, в основном, во всем мире коммерческое производство базируется на химическом способе, основанном на гидролизе сульфатов метакриламида, которые получают из соответствующих 2-гидроксинитрилов, причем около 1,6 кг серной кислоты необходимо для получения 1 кг метакриловой кислоты.
Химическое превращение 2-гидроксиизомасляной кислоты (2-HIB) в метакриловую кислоту с выходом до 96% описывается в US 3,666,805 и US 5,225,594.
Альтернативный способ получения метакриловой кислоты основывается на гидролизе 2-гидроксинитрилов до 2-гидроксиизомасляной кислоты, с применением нитрилгидролизующих ферментов. При этом речь идет о нитрилазе или комбинации из нитрил гидратазы и амидазы (A.Banerjee, R.Sharrna, U.С.Banerjee, 2002, "The nitrite-degrading enzymes: current status and future prospects", Appl. Microbiol. Biotechnol., 60: 33-44 и US 6,582,943). Серьезным недостатком этого способа является нестабильность нитрилов в нейтральной области рН, необходимой для эффективной нитрилгидролизующей ферментативной активности. Разложение нитрилов в реакционной смеси приводит к накоплению кетонов и цианидов, которые ингибируют нитрилгидролизующую ферментативную активность.
Общий недостаток обоих способов, т.е. доминирующего в настоящее время способа на основе сульфатов метакриламида и ферментативного нитрилгидролизующего способа, состоит в необходимости 2-гидроксинитрилов. Они могут быть получены только из вредных для окружающей среды реагентов, а именно кетонов и цианида.
Альтернативный способ получения 2-гидроксиизомасляной кислоты по ферментативному метаболическому пути, при котором разлагается трет-бутиловый спирт, раскрывается в СА 2,510,657.
Следующий ферментативный способ получения 2-гидроксиизмасляной кислоты раскрывается из РСТ/ЕР2007/052830. В этом способе прекурсор 3-гидроксибутирил-Коэнзим А (3-НВСоА) превращается в 2-гидроксиизомасляную кислоту с помощью мутазы. При практическом применении указанный способ обладает следующими недостатками: указанный способ является периодическим, реагент 3-гидроксиизомасляная кислота (3-НВ) добавляется экзогенно, и условия способа требуют присутствия инертного газа. Степень превращения составляет около 20%.
Поэтому существует необходимость в способе получения 2-гидроксиизомасляной кислоты или полигидроксиалканоатов, содержащих мономерные структурные единицы 2-гидроксиизомасляной кислоты, которому не присущи описанные недостатки.
Таким образом, целью настоящего изобретения является обеспечение способа получения 2-гидроксиизомасляной кислоты, который удовлетворяет потребность в прекурсорах 2-гидроксиизомасляной кислоты или полигидроксиалканоатов, содержащих мономерные структурные единицы 2-гидроксиизомасляной кислоты, которые затем могут быть превращены в метакриловую кислоту, ее сложные эфиры и полимеры.
Описание изобретения
Неожиданно было обнаружено, что образование 2-гидроксиизомасляной кислоты или полигидроксиалканоатов, содержащих мономерные структурные единицы 2-гидроксиизомасляной кислоты через ацетоацетил-Коэнзим А в качестве промежуточного продукта и 3-гидроксибутирил-Коэнзим А в качестве прекурсора, вносит ценный вклад в решение задачи, поставленной в настоящем изобретении.
Термин "прекурсор", как он применяется здесь, означает химическое соединение, которое может быть превращено в целевой продукт по ферментативному пути посредством применения только одного фермента, в то время как термин "промежуточный продукт" означает химическое соединение, которое может быть превращено в целевой продукт по ферментативному пути посредством применения, по меньшей мере, двух ферментов, причем соединения с функциональной группой коэнзима А или без нее должны пониматься как эквивалентные "химические соединения", и, таким образом, формирующие сложный тиоэфир ферменты или расщепляющие сложный тиоэфир ферменты не рассматриваются.
Поэтому объектом настоящего изобретения является клетка, которая была генетически модифицирована таким образом, что стала способна продуцировать большее количество 2-гидроксиизомасляной кислоты или большее количество полигидроксиалканоатов, содержащих мономерные структурные единицы 2-гидроксиизомасляной кислоты, по сравнению с ее диким типом, отличающаяся тем, что образование 2-гидроксиизомасляной кислоты или полигидроксиалканоатов, содержащих мономерные структурные единицы 2-гидроксиизомасляной кислоты, происходит через ацето-ацетил-Коэнзим А в качестве промежуточного продукта и 3-гидроксибутирил-Коэнзим А в качестве прекурсора. Следующими объектами настоящего изобретения являются способ получения клетки по изобретению и способ получения 2-гидроксиизомаслянной кислоты с помощью клетки по изобретению, а также способ получения метакриловой кислоты.
Преимущество настоящего изобретения состоит в том, что 2-гидроксиизомасляная кислота или метакриловая кислота, соответственно, может быть получена как из возобновляемых видов сырья, например из углеводов и/или из глицерина, так и из сырья, получаемого из ископаемых видов топлива, такого как, например, метанол, и, таким образом, можно избежать проблем, связанных с непостоянной доступностью ископаемого сырья.
Следующее преимущество настоящего изобретения состоит в том, что согласно способу по изобретению можно получить метакриловую кислоту при меньшей термической нагрузке и, в общем, с меньшим числом стадий способа по изобретению.
Следующее преимущество настоящего изобретения состоит в возможности избежать многих токсичных или агрессивных веществ, которые накапливаются при обычном химическом способе получения 2-гидроксиизомасляной кислоты.
Далее изобретение описывается с помощью примерных вариантов выполнения изобретения, которые не предназначены для его ограничения.
Все указанные проценты (%), если иного не оговаривается, представляют собой массовые проценты.
Термин "2-гидроксиизомасляная кислота", как он применяется в настоящем документе, всегда обозначает соответствующую С4-карбоновую кислоту в той форме, в которой она существует в зависимости от значения рН после продуцирования соответствующими микроорганизмами. Таким образом, данный термин всегда охватывает чистую кислотную форму (2-гидроксиизомасляная кислота), чистую основную форму (2-гидроксиизобутират), а также смеси из протонированной и депротонированной формы кислоты.
Термин "3-гидроксибутирил-Коэнзим А" охватывает как (R)-, так и (S)-стереоизомеры, причем (R)-стереоизомер особенно предпочтителен.
Фраза "таким образом, что стала способна продуцировать большее количество 2-гидроксиизомасляной кислоты или большее количество полигидроксиалканоатов, содержащих мономерные структурные единицы 2-гидроксиизомасляной кислоты, по сравнению с ее диким типом" относится также к случаю, когда дикий тип генетически модифицированной клетки вообще не способен продуцировать 2-гидроксиизомасляную кислоту или полигидроксиалканоаты, содержащие мономерные структурные единицы 2-гидроксиизомасляной кислоты или, по меньшей мере, никакие обнаруживаемые количества этих соединений, и только после генетической модификации могут продуцироваться обнаруживаемые количества этих компонентов.
Под "диким типом" клетки преимущественно понимается клетка, геном которой находится в том состоянии, в котором он возник естественным образом в процессе эволюции. Это понятие относится как ко всей клетке, так и к отдельным генам. Под понятие "дикий тип" поэтому не попадают, в частности, такие клетки и такие гены, генные последовательности которых были модифицированы человеком, по меньшей мере, частично, посредством рекомбинантных способов.
Из 2-гидроксиизомасляной кислоты можно затем получить метакриловую кислоту посредством щадящей реакции дегидратации. В случае полигидроксиалканоатов, содержащих мономерные структурные единицы 2-гидроксимасляной кислоты, могут быть выделены содержащиеся в клетках граны, наполненные этими полигидроксиалканоатами, и затем полимеры расщепляются с получением 2-гидроксиизомасляной кислоты, которая затем может быть дегидратирована с получением метакриловой кислоты.
При этом согласно настоящему изобретению является предпочтительным, чтобы генетически модифицированная клетка была генетически модифицирована таким образом, что в течение определенного временного интервала, предпочтительно в течение 2 часов, еще более предпочтительно в течение 8 часов, и наиболее предпочтительно в течение 24 часов, она продуцирует, по меньшей мере, в два раза больше, особенно предпочтительно, по меньшей мере, в 10 раз больше, еще более предпочтительно, по меньшей мере, в 100 раз больше, еще более предпочтительно, по меньшей мере, в 1000 раз больше и наиболее предпочтительно, по меньшей мере, в 10000 раз больше 2-гидроксиизомасляной кислоты или полигидроксиалканоатов, содержащих 2-гидроксиизомасляную кислоту, по сравнению с ее диким типом. При этом увеличение в продуцировании продукта можно определить, например, культивируя клетку по изобретению и клетку соответствующего дикого типа по отдельности, при одинаковых условиях (одинаковая клеточная плотность, одинаковая питательная среда, одинаковые условия культивирования), в течение определенного временного интервала, в подходящей питательной среде, и затем определяя количество целевого продукта (2-гидроксиизомасляной кислоты или полигидроксиалканоатов, содержащих 2-гидроксиизомасляную кислоту) либо в клеточном супернатанте, в случае 2-гидроксиизомасляной кислоты, или в клетках, в случае полигидроксиалканоатов, содержащих 2-гидроксиизомасляную кислоту.
Клетки по изобретению могут быть прокариотическими или эукариотическими. При этом речь может идти о клетках млекопитающих (как например, клетки человека), растительных клетках или о микроорганизмах, таких как дрожжи, грибы или бактерии, причем особенно предпочтительны микроорганизмы, и наиболее предпочтительны бактерии и дрожжи.
Среди бактерий, дрожжей или грибов особенно предпочтительными для применения являются бактерии, дрожжи или грибы, хранящиеся в немецком банке микроорганизмов и клеточных культур (DSMZ, Deutsche Sammlung von Mikroorganismn und Zeilkulturen GmbH), Брауншвейг, Германия, в виде штаммов бактерий, дрожжей или грибов. Подходящие согласно настоящему изобретению бактерии принадлежат видам, приведенным по адресу http://www.dsmz.de/species/bacteria.htm,
подходящие согласно настоящему изобретению дрожжи принадлежат видам, приведенным по адресу
http://www.dsmz.de/species/yeasts.htm,
и подходящие согласно настоящему изобретению грибы принадлежат видам, приведенным по адресу
http://www.dsmz.de/species/fungi.htm.
Согласно настоящему изобретению предпочтительными клетками являются клетки, происходящие из рода Aspergillus, Corynebacterium, Brevibacterium, Bacillus, Acinetobacter, Alcaligenes, Lactobacillus, Paracoccus, Lactococcus, Candida, Pichia, Hansenula, Kluyveromyces, Saccharomyces, Escherichia, Zymomonas, Yarrowia, Methylobacterium, Ralstonia, Pseudomonas, Rhodospirillum, Rhodobacter, Burkholdena, Clostridium и Cupriavidus, причем клетки, происходящие из рода Aspergillus nidulans, Aspergillus niger, Alcaligenes latus, Bacillus megaterium, Bacillus subtilis, Brevibacteiium flavum, Brevibacterium lactofermentum, Escherichia coli, Saccharomyces cerevisiae, Kluveromyces lactis, Candida blankii, Candida rugosa, Corynebacterium glutamicum, Corynebacterium efficiens, Zymomonas mobilis, Yarrowia lipolytica, Hansenula polymorpha, Methylobacterium extorquens, Ralstonia eutropha, особенно Ralstonia eutropha H16, Rhodospirillum rubrum, Rhodobacter sphaeroi-des, Paracoccus versutus, Pseudomonas aeruginosa, Pseudomonas putida, Acinetobacter calcoaceticus и Pichia pastoris, являются особенно предпочтительными.
Клетка по настоящему изобретению, которая способна продуцировать, по сравнению с ее диким типом, большее количество 2-гидроксиизомасляной кислоты или большее количество полигидроксиалканоатов, содержащих мономерные структурные единицы 2-гидроксиизомасляной кислоты, через ацетоацетил-Коэнзим А в качестве промежуточного продукта и 3-гидроксибутирил-Коэнзим А в качестве прекурсора, обнаруживает активность фермента Е1, который катализирует превращение ацетоацетил-коэнзима А в 3-гидроксибутирил-коэнзим А.
В случае фермента Е1 речь преимущественно идет о ферменте, выбранном из группы, состоящей из:
3-гидроксиацил-СоА дегидрогеназы (ЕС 1.1.1.35),
ацетоацетил-Коэнзим А редуктазы (ЕС 1.1.1.36),
длинноцепочечной 3-гидроксиацил-СоА дегидрогеназы ((ЕС 1.1.1.211) и 3-гидроксибутирил-Коэнзим А-дегидрогеназы (ЕС 1.1.1.157).
Этот фермент предпочтительно закодирован генами, выбранными из группы, состоящей из phaB, phbB, fabG, phbN1 или phbB2, причем phaB и phbB особенно предпочтительны. Нуклеотидные последовательности этих генов могут быть получены, например, из "Kyoto Encyclopedia of Genes and Genomes" (KEGG-база данных), базы данных национального центра биотехнологической информации (NCBI) национальной медицинской библиотеки (Bethesda, MD, США) или из базы данных нуклеотидных последовательностей европейской лаборатории молекулярной биологии (EMBL, Гейдельберг, Германия и Кембридж, UK).
Преимущество клетки по настоящему изобретению может состоять в том, что клетка по настоящему изобретению, по сравнению с ее диким типом, обнаруживает повышенную активность фермента Е1, который катализирует превращение ацетоацетил-коэнзима А в 3-гидроксибутирил-коэнзим А.
Под фразой "повышенная активность фермента", применяемой выше в отношении фермента Е1 и далее в отношении ферментов Е2 и т.д., предпочтительно нужно понимать повышенную внутриклеточную активность.
Следующие комментарии относительно повышения ферментативной активности в клетках относятся как к повышению активности фермента Е1, так и к повышению активности всех в последующем упоминаемых ферментов, активность которых, при необходимости, может быть повышена.
По существу, повышение ферментативной активности может достигаться посредством того, что повышается число копий генной последовательности или генных последовательностей, которые кодируют фермент, посредством применения сильного промотора, посредством изменения частоты использования кодона, посредством увеличения различными путями периода полураспада мРНК или фермента, посредством модификации регуляции экспрессии гена или посредством применения гена или аллели, кодирующих соответствующий фермент с повышенной активностью, и, при необходимости, посредством комбинирования всех этих действий.
Генетически модифицированные клетки по настоящему изобретению получают, например, трансформацией, трансдукцией, конъюгацией или комбинацией этих способов, применяя вектор, который содержит желаемый ген, аллель этого гена или его части, и промотор, который способствует экспрессии этого гена. Гетерологическая экспрессия достигается, в частности, интеграцией гена или аллелей в хромосому клетки или в экстрахромосомно реплицирующийся вектор.
Обзор возможностей повышения активности ферментов в клетках, на примере пируват-карбоксилазы, рассматривается в источнике DE-A-10031999, который включен в настоящий документ посредством ссылки и объем раскрытия которого относительно возможностей повышения активности ферментов в клетках является частью настоящего изобретения.
Экспрессия вышеназванных ферментов или генов и всех упоминаемых далее ферментов или генов может быть обнаружена с помощью одно- и двумерного фракционирования белков в геле и последующей оптической идентификации концентрации белка в геле с помощью соответствующего программного обеспечения для оценки. Если повышение ферментативной активности базируется исключительно на повышении экспрессии соответствующего гена, то количественное определение указанного повышения ферментативной активности может проводиться простым сравнением одно- или двумерного фракционирования белков клетки дикого типа и генетически модифицированной клетки. Обычный способ получения белковых гелей для коринеформных бактерий и идентификации указанных белков описывается у Hermann et al. (Electrophoresis, 22: 1712.23 (2001). Концентрацию белков можно проанализировать подобным образом с помощью Вестерн-Блот гибридизации со специфическими для анализируемого белка антителами (Sambrook et al., Molecular Cloning: a laboratory manual, 2nd Ed. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, Нью-Йорк, США, 1989) и последующей оптической оценки, с применением соответствующего программного обеспечения для определения концентрации (Lohaus und Meyer (1989) Biospektrum, 5: 32-39; Lottspeich (1999), Ange-wandte Chemie 111: 2630-2647). Активность ДНК-связывающих белков может также быть определена с помощью анализа на замедление подвижности ДНК в геле (также упоминается как торможение в геле) (Wilson et al. (2001) Journal of Bacteriology, 183: 2151-2155). Действие, оказываемое ДНК-связывающими белками на экспрессию других генов, может быть обнаружено различными хорошо описанными способами анализа репортерных генов (Sambrook et al., Molecular Cloning: a laboratory manual, 2nd Ed. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, Нью-Йорк, США, 1989). Внутриклеточные ферментативные активности могут быть определены различными описанными способами (Donahue et al. (2000) Journal of Bacteriology 182 (19): 5624-5627; Ray et al. (2000) Journal of Bacteriology 182 (8): 2277-2284; Freedberg et al. (1973) Journal of Bacteriology 115 (3): 816-823). Если в приведенном далее описании изобретения никакие конкретные способы определения активности определенного фермента не указываются, то определение повышения ферментативной активности, а также определение понижения ферментативной активности происходит преимущественно посредством способов, описанных у Hermann et al., Electophoresis, 22: 1712-23 (2001), Lohaus et al., Biospektrum 5 32-39 (1998), Lottspeich, Angewandte Chemie 111: 2630-2647 (1999) и Wilson et al., Journal of Bacteriology 183: 2151-2155 (2001).
Если повышению ферментативной активности сопутствует мутация эндогенного гена, то такие мутации могут быть достигнуты либо ненаправленно с помощью классических способов, таких как, например, основанных на ультрафиолетовом облучении или вызывающих мутацию мутагенных веществ, либо целенаправленно, с помощью генно-инженерных способов, таких как делеция(ии), инсерция(ии) и/или нуклеотидное замещение(ия). Посредством таких мутаций получают модифицированные клетки. Особенно предпочтительные мутантные ферменты представляют собой, в частности, также такие ферменты, которые больше не ингибируются по типу обратной связи или ингибируются в меньшей степени по сравнению с ферментом дикого типа.
Если повышению ферментативной активности сопутствует повышение синтеза фермента, то к этому случаю относится, например, увеличение числа копий соответствующих генов или мутирование промоторной или регуляторной областей или сайта связывания рибосом, расположенного против хода транскрипции структурного гена. Тем же самым образом действуют экспрессионные кассеты, которые встраиваются против хода транскрипции структурного гена. Кроме того, посредством индуцируемых промоторов можно повышать экспрессию в любой произвольный момент времени. Более того, так называемые "Энхансеры" могут быть включены в ген, кодирующий фермент, в качестве регуляторных последовательностей, которые благодаря повышенному взаимодействию между РНК-полимеразой и ДНК также вызывают повышение экспрессии гена. Экспрессия также улучшается с помощью мер по увеличению продолжительности жизни мРНК. Кроме того, ферментативная активность также усиливается посредством предотвращения расщепления ферментного белка. В этом случае, гены или генные конструкции либо находятся в плазмидах с разным числом копий, либо интегрируются в хромосому и амплифицируются там. Альтернативно, сверхэкспрессия рассматриваемых генов может, кроме того, достигаться путем изменения состава среды и методики культивации. Специалисты в данной области техники находят руководства к действию, среди прочего, у Martin et al. (Bio/Technology 5, 137-146 (1987)), у Guerrero et al. (Gene 138, 35-41 (1994)), Tsuchiya und Morinaga (Bionechnology 6, 428-430 (1988)), у Eikmanns et al. (Gene 102, 93-98 (1991)), в ЕР-А-0472869, в US 4,601,893, у Schwarzer und Punier (Bio/Technology 9, 84-87 (1991), у Reinscheid et al. (Applied and Environmental Microbiology 60, 126-132 (1994)), у LaBarre et al. (Journal of Bacteriology 175, 1001-1007 (1993)), в WO-A-96/15246, у Malumbres et al. (Gene 134, 15-24 (1993), в JP-A-10-229891, у Jensen und Hammer (Biotechnology and Bioengineering 58, 191-195 (1998)) и в известных учебниках по генетике и молекулярной биологии. Вышеописанные действия, так же как и мутации, приводят к генетически модифицированным клеткам.
Для повышения экспрессии соответствующих генов применяются, например, эписомальные плазмиды. Подходящие плазмиды и вектора представляют собой, в принципе, все варианты конструкций, доступные для этой цели специалисту в данной области техники. Такие плазмиды и вектора поставляются, например, фирмами Novagen, Promega, New England Biolabs, Clontech или Gibco BRL. Другие предпочтительные плазмиды и вектора могут быть обнаружены, например, у: Glover, D.М. (1985), DNA cloning: a practical approach, Vol.I-III, IRL Press Ltd., Oxford; Rodriguez, R.L. und Denhardt, D.Т (eds) (1988), Vectors: a survey of molecular cloning vectors and their uses, 179-204, Butterworth, Stoneham; Goeddel, D.V. (1990), Systems for heterologous gene expression, Methods Enzymol. 185, 3-7; Sam-brook, J.; Fritsch, E.F. und Maniatis, T. (1989), Molecular cloning: a laboratory manual, 2nd ed., Cold Spring Harbor Laboratory Press, New York.
Затем плазмидный вектор, содержащий ген, который должен быть амплифицирован, переносится путем конъюгации или трансформации в желаемый штамм. Способ конъюгации описывается, например, у Schäfer et al., Applied and Environmental Microbiology 60: 756-759 (1994). Способы трансформации описываются, например, у Thierbach et al., Applied Microbiology and Biotechnology 29: 356-362 (1988), Dunican und Shivnan, Bio/Technology 7: 1067-1070 (1989) и Tauch et al., FEMS Microbiology Let-ters 123: 343-347 (1994). После гомологической рекомбинации посредством "кроссоверного" события, полученный штамм содержит уже, по меньшей мере, две копии соответствующего гена.
Под применяемой выше и при дальнейшем описании изобретения фразой "повышенная по сравнению с ее диким типом активность фермента Ех" всегда предпочтительно понимается активность соответствующего фермента Ех, которая повысилась в, по меньшей мере, 2 раза, предпочтительно в, по меньшей мере, 10 раз, более предпочтительно в, по меньшей мере, 100 раз, особенно предпочтительно в, по меньшей мере, 1000 раз и наиболее предпочтительно в, по меньшей мере, 10000. Кроме того, клетка по изобретению, обнаруживающая "повышенную активность фермента Ех по сравнению с ее диким типом", в более частном случае также охватывает клетку, дикий тип которой не обнаруживает активность указанного фермента Ех или, по меньшей мере, подтверждаемую активность указанного фермента Ех, и подтверждаемая активность указанного фермента Ех обнаруживается только после повышения указанной ферментативной активности, например, посредством сверхэкспрессии. В этой связи термин "Сверхэкспрессия" или фраза "повышение экспрессии", применяемые далее, охватывают как случай, когда немодифицированная клетка, например клетка дикого типа, не обнаруживает экспрессию, или, по меньшей мере, подтверждаемую экспрессию, и подтверждаемый синтез фермента Ех индуцируется лишь посредством рекомбинантных способов.
Под применяемой далее фразой "пониженная активность фермента Ех" предпочтительно понимается активность фермента, которая была понижена до значения, составляющего, по меньшей мере, 0.5 от первоначального, более предпочтительно, по меньшей мере, 0.1, еще более предпочтительно, по меньшей мере, 0.01, особенно предпочтительно, по меньшей мере, 0.001 и наиболее предпочтительно, по меньшей мере, 0.0001. Фраза "пониженная активность" также охватывает случай отсутствия подтверждаемой активности ("нулевая активность"). Понижение активности определенного фермента может достигаться, например, путем целенаправленной мутации, путем добавления конкурентных или неконкурентных ингибиторов, или другими известными специалисту в данной области техники способами понижения активности конкретного фермента.
Предпочтительно, чтобы для клетки по настоящему изобретению, которая, по сравнению с ее диким типом, способна продуцировать большее количество 2-гидроксиизомасляной кислоты или большее количество полигидроксиалканоатов, содержащих мономерные структурные единицы 2-гидроксиизомасляной кислоты, через ацетоацетил-Коэнзим А в качестве промежуточного продукта и 3-гидроксибутирил-Коэнзим А в качестве прекурсора, в качестве источника углерода могли служить углеводы, глицерин, масла и жиры, диоксид углерода, карбоновые кислоты или метанол.
Кроме того, предпочтительно, чтобы клетка по настоящему изобретению, которая способна продуцировать 2-гидроксиизомасляную кислоту или полигидроксиалканоаты, содержащие мономерные структурные единицы 2-гидроксиизомасляной кислоты, через ацетоацетил-Коэнзим А в качестве промежуточного продукта и 3-гидроксибутирил-Коэнзим А в качестве прекурсора, обнаруживала, в дополнение к активности фермента E1, активность фермента Е2, причем предпочтительно повышенную по сравнению с ее диким типом активность фермента Е2, который катализирует превращение молекул ацетил-Коэнзима А в ацетоацетил-Коэнзим А.
В случае фермента Е2 речь предпочтительно идет о ацетил-СоА С-ацетилтрансферазе (EC-Number 2.3.1.9). Этот фермент предпочтительно кодируется генами, выбранными из группы, состоящей из acat1, acat2, loc484063, loc489421, mgc69098, mgc81403, mgc81256, mgc83664, kat-1, erg10, ygeF, atoB, fadAx, phbA-1, phbA-2, atoB-2, pcaF, pcaF-2, phb-A, bktB, phaA, tioL, thIA, fadA, paaJ, phbAf, pimB, mmgA, yhfS, thi, vraB, th1, mvaC, thiL, paaJ, fadA3, fadA4, fadA5, fadA6, cgl12392, catF, sc8f4.03, thiL1, thiL2, acaB1, acaB2, асаВЗ или асаВ4, причем предпочтительными являются cat1, acat2, atoB, thIA, thIB, phaA и phbA, особенно предпочтительными являются phaA и phbA.
Нуклеотидные последовательности этих генов могут быть получены, например, из "Kyoto Encyclopedia of Genes and Genomes" (KEGG-база данных), базы данных национального центра биотехнологической информации (NCBI) национальной медицинской библиотеки (Bethesda, MD, США) или из базы данных нуклеотидных последовательностей европейской лаборатории молекулярной биологии (EMBL, Гейдельберг, Германия или Кембридж, UK).
Клетка по настоящему изобретению предпочтительно обнаруживает, по меньшей мере, одну активность фермента Е3, предпочтительно повышенную по сравнению с ее диким типом активность фермента Е3, который катализирует превращение 3-гидроксибутирил-Коэнзима А в 2-гидроксиизобутирил-Коэнзим А. В случае фермента Е3 речь преимущественно идет о гидроксил-изобутирил-СоА мутазе, о изобутирил-СоА мутазе (ЕС 5.4.99.13) или о метилмалонил-СоА мутазе (ЕС 5.4.99.2), в каждом случае предпочтительно о коэнзим В12-зависимой мутазе.
Фермент Е3 может быть выделен предпочтительно из микроорганизмов Aquincola tertiaricarbonis L108, DSM18028, DSM18512, Metilibium petroleiphilum PM1, Metilibium sp.R8, Xanthobacter autotrophicus Py2, Rhodobacter sphaeroides (ATCC 17029), Nocardioides sp.JS614, Marinobacter algicola DG893, Sinorhizobium medicae WSM419, Roseovarius sp.217, Pyrococcus furiosus DSM 3638, и, в частности, представляет собой коэнзим В12-зависимую мутазу, описанную в РСТ/ЕР2007/052830, а также один из ферментов, по меньшей мере, одна часть последовательностей которых на аминокислотном уровне идентична аминокислотной последовательности малой или большей субъединицы мутазы, описанной в РСТ/ЕР2007/052830 (номер доступа DQ436457.1 и DQ436456.1), на, по меньшей мере, 60%, предпочтительно на, по меньшей мере, 80%, более предпочтительно на, по меньшей мере, 95%, и особенно предпочтительно на, по меньшей мере, 99%, как было определено с помощью алгоритма blastp, где ограничение на вероятность случайного совпадения равно 10, длина слова равна 3, применяется матрица blosum62, штраф за открытие вставки равен 11, и штраф за продолжение вставки равен 1, и применяется условная установка формы структурного расчета.
В предпочтительном варианте выполнения клетки по изобретению, способной продуцировать 2-гидроксиизомасляную кислоту или полигидроксиалканоаты, содержащие мономерные структурные единицы 2-гидроксиизомасляной кислоты, через ацетоацетил-Коэнзим А в качестве промежуточного продукта и 3-гидроксибутирил-Коэнзим А в качестве прекурсора, клетка по настоящему изобретению в форме ее дикого типа обнаруживает активность фермента Е4, предпочтительно пониженную по сравнению с ее диким типом активность, по меньшей мере, одного фермента Е4, который катализирует превращение 3-гидроксибутирил-Коэнзима А в полигидроксибутират.
В случае фермента Е4 речь преимущественно идет о полигидроксиалканоат синтазе, особенно предпочтительно о полигидроксибутират синтазе. Этот фермент кодируется преимущественно генами phbC и phaC, причем ген phaC особенно предпочтителен.
В следующем предпочтительном варианте выполнения клетки по изобретению, способной продуцировать 2-гидроксиизомасляную кислоту или полигидроксиалканоаты, содержащие мономерные структурные единицы 2-гидроксиизомасляной кислоты, через ацетоацетил-Коэнзим А в качестве промежуточного продукта и 3-гидроксибутирил-Коэнзим А в качестве прекурсора, клетка по настоящему изобретению в форме ее дикого типа обнаруживает активность фермента Es, предпочтительно пониженную по сравнению с ее диким типом активность фермента Ев, который катализирует превращение 3-гидроксибутирил-Коэнзима А в кротонил-Коэнзим А.
В случае фермента Е5 речь идет преимущественно о кротоназе (ЕС-номер 4.2.1.55) или о (3R)-3-Гидроксибутаноил-СоА дегидратазе (ЕС-номер 4.2.1.17). Этот фермент кодируется преимущественно генами, выбранными из группы, состоящей из crt, crt1, crt2, fadB, paaF, причем crt, а также соответствующий ген из Clostridien, являются предпочтительными, причем ген crt из Clostridium acetobutilicum особенно предпочтителен.
Еще в одном предпочтительном варианте клетки по изобретению, способные продуцировать 2-гидроксиизомасляную кислоту или полигидроксиалканоаты, содержащие мономерные структурные единицы 2-гидроксиизомасляной кислоты, через ацетоацетил-Коэнзим А в качестве промежуточного продукта и 3-гидроксибутирил-Коэнзим А в качестве прекурсора, клетка по настоящему изобретению в форме ее дикого типа обнаруживает активность фермента Е6, предпочтительно пониженную по сравнению с ее диким типом активность фермента Е6, который катализирует превращение R-3-гидроксибутирил-Коэнзима А в S-3-гидроксибутирил-Коэнзим А.
Под ферментом Е6 преимущественно понимается 3-гидроксибутирил-СоА эпимераза (ЕС 5.1.2.3). Этот фермент кодируется преимущественно генами, выбранными из группы, состоящей из fadB, fadB1, fadB2, fadJ, fabJ-1, faoA, yfcX, причем fadB, fadJ и yfcX предпочтительны, и fadB и fadJ особенно предпочтительны.
Кроме того, клетка по изобретению, способная продуцировать 2-гидроксиизомасляную кислоту или полигидроксиалканоаты, содержащие мономерные структурные единицы 2-гидроксиизомасляной кислоты, через ацетоацетил-Коэнзим А в качестве промежуточного продукта и 3-гидроксибутирил-Коэнзим А в качестве прекурсора, предпочтительно обнаруживает пониженную по сравнению с ее диким типом активность, по меньшей мере, одного фермента Е7, для которого субстратом служит 3-гидроксибутирил-Коэнзим А.
Вклад в решение поставленных выше задач вносит способ получения 2-гидроксиизомасляной кислоты или полигидроксиалканоатов, содержащих мономерные структурные единицы 2-гидроксиизомасляной кислоты, содержащий следующие стадии:
a) контакт клетки по изобретению с питательной средой, содержащей источник углерода, в условиях, при которых из источника углерода образуется 2-гидроксиизомасляная кислота или полигидроксиалканоаты, содержащие мономерные структурные единицы 2-гидроксиизомасляной кислоты, а также, при необходимости
b) очистка 2-гидроксимасляной кислоты от питательной среды или полигидроксиалканоатов, содержащих мономерные структурные единицы 2-гидроксиизомасляной кислоты, от клеток.
В качестве источника углерода могут применяться углеводы [как например, моносахариды (например, глюкоза, фруктоза, галактоза, арабиноза, ксилоза), олигосахариды (например, мальтоза, сахароза, лактоза), и полисахариды (например, крахмал, гидролизованный крахмал, целлюлоза, гидролизованная целлюлоза, гемицеллюлоза, гидролизованная гемицеллюлоза)], также как и продукты их реакции, такие как, например, сахарные спирты и полигидроксикислоты;
диоксид углерода;
органические моно-, ди- и трикарбоновые кислоты, необязательно содержащие одну или более, например 1, 2, 3 или 4, гидроксильные группы, например уксусная кислота, винная кислота, итаконовая кислота, янтарная кислота, пропионовая кислота, молочная кислота, 3-гидроксипропионовая кислота, фумаровая кислота, малеиновая кислота, 2,5-фурандикарбоновая кислота, глутаровая кислота, левулиновая кислота, глюконовая кислота, аконитовая кислота, янтарная кислота и диаминопимелиновая кислота, лимонная кислота;
липиды;
масла или жиры, такие как, например, рапсовое масло, соевое масло, пальмовое масло, подсолнечное масло, арахисовое масло и кокосовое масло;
насыщенные и ненасыщенные жирные кислоты с предпочтительно 10-22 атомами углерода, например γ-линоленовая кислота, дигомо-γ-линоленовая кислота, арахидоновая кислота, пальмитиновая кислота, стеариновая кислота, линолевая кислота, эйкозапентаеновая кислота и докозагексаеновая кислота;
углеводороды, как например метан;
спирты, например, с 1-22 атомами углерода, например бутанол, метанол, этанол;
диолы с предпочтительно 3-8 атомами углерода, например пропандиол и бутандиол;
многоатомные спирты (также упоминаемые как высшие спирты) с тремя или более, например 3, 4, 5 или 6, ОН-группами, например глицерин, сорбитол, маннитол, ксилитол и арабинитол;
кетоны с, предпочтительно, 1-10 атомами углерода, и при необходимости 1 или более гидроксильными группами, например ацетон и ацетоин;
лактоны, например γ-бутиролактон, циклодекстрины, биополимеры, например полигидроксиацетат, сложные полиэфиры, например полилактид, полисахариды, полиизопреноиды, полиамиды;
ароматические соединения, например ароматические амины, ванилин и индиго;
белки, например ферменты, такие как амилазы, пектиназы, кислые, гибридные или нейтральные целлюлазы, эстеразы, такие как липазы, панкреазы, протеазы, ксиланазы, и оксидоредуктазы, такие как лакказа, каталаза и пероксидаза, глюканазы, фитазы; каротиноиды, например ликопин, β-каротин, астаксантин, зеаксантин и кантаксантин;
протеиногенные и непротеиногенные аминокислоты, например лизин, глютамат, метионин, фенилаланин, аспарагиновая кислота, триптофан и треонин;
пуриновые и пиримидиновые основания;
нуклеозиды и нуклеотиды, например никотинамидадениндинуклеотид (NAD) и аденозин-5'-монофосфат (AMP);
а также прекурсоры и производные вышеупомянутых соединений, например соли упомянутых кислот.
Эти вещества могут применяться как по отдельности, так и в виде смеси. Особенно предпочтительным является применение углеводов, в частности моносахаридов, олигосахаридов или полисахаридов, как это описано в US 6136576, например C5-сахаров или глицерина.
Метанол является предпочтительно применяемым спиртом, так как его можно получить из множества различных источников, таких как, например, биогаз, биомасса, природный газ или уголь.
Источники углерода могут применяться в различных формах (чистые или в виде раствора/суспензии) и в виде различных композиций (очищенные или в виде сырья) после различных стадий обработки (например, сок сахарного тростника, сироп, патока, тростниковый сахар, белый сахарный песок; кукурузное зерно, мука, крахмал, декстрин, глюкоза), перед или после обработки (обработка паром, предварительная обработка кислотой, предварительная ферментативная обработка).
Предпочтительно, в альтернативном варианте выполнения настоящего изобретения, в качестве источника углерода применяется СО2 или СО, в частности, синтез-газ. Применяемыми в этом случае клетками по изобретению являются клетки ацетогенов, как например, виды рода Acetobacterium, такие как A. woodii и Clostridium aceticum. Более конкретно, применяемыми клетками являются клетки ацетогенов, выбранных из группы, состоящей из Thermoanaerobacter kivui, Acetobacterium woodii, Acetoanaerobium notera, Clostridium aceticum, Butyribacterium methylotrophicum, Clostridium acetobutylicum, Moorella thermoacetica, Eubacterium limosum, Peptostreptococcus productus, Clostridium ljungdahlii и Clostridium carboxidivorans. Особенно подходящими в этом отношении являются Clostridium carboxidivorans, особенно штаммы "Р7" и "Р11". Такие клетки описываются, например, в US 2007/0275447 и US 2008/0057554. Другие особенно подходящие в этом отношении клетки представляют собой Clostridium ljungdahlii, особенно штаммы, выбранные из группы, состоящей из Clostridium ljungdahlii РЕТС, Clostridium ljungdahlii ERI2, Clostridium ljungdahlii C01 и Clostridium ljungdahlii O-52, и описанные в WO 98/00558 и WO 00/68407.
Генетически модифицированные клетки по настоящему изобретению могут вступать в контакт с питательной средой и, таким образом, культивироваться непрерывно при непрерывном способе или периодически при периодическом способе (культивирование партиями) или культивирование периодическим способом с подпиткой (воздушно-приточный способ), или культивирование повторяющимся периодическим способом с подпиткой (повторяющийся воздушно-приточный способ), с целью получения 2-гидроксиизомасляной кислоты или полигидроксиалканоатов, содержащих мономерные структурные единицы 2-гидроксиизомасляной кислоты. Также возможен полунепрерывный процесс, как описывается, например, в GB-A-1009370. Описание подходящих способов культивирования приводится в учебнике Chmiel ("Bioprozesstechnik 1. Einführung in die Bioverfahrenstechnik" (Gustav Fischer Verlag, Штутгарт, 1991)) или в учебнике Storhas ("Bioreaktoren und periphere Einrichtungen", Vieweg Verlag, Брауншвейг/Висбаден, 1994).
Применяемая культуральная среда должна подходящим образом удовлетворять требованиям соответствующих штаммов. Описания культуральных сред для различных микроорганизмов приводится в руководстве "Manual of Methods for General Bacteriology" американского биотехнологического общества (Washington D.С., США, 1981).
В качестве источника азота могут применяться органические азотосодержащие соединения, такие как пептон, дрожжевой экстракт, мясной экстракт, солодовый экстракт, жидкий кукурузный экстракт, соевая мука и мочевина, или неорганические соединения, такие как сульфат аммония, хлорид аммония, фосфат аммония, карбонат аммония и нитрат аммония. Источники азота могут применяться по отдельности или в виде смеси.
В качестве источника фосфора могут применяться фосфорная кислота, дигидрофосфат калия или гидрофосфат дикалия или соответствующие натриевые соли. Культуральная среда должна, кроме того, содержать соль металла, например, сульфат магния или сульфат железа, которые необходимы для роста. Наконец, в дополнение к вышеупомянутым веществам могут применяться важные для роста вещества, такие как аминокислоты и витамины. Кроме того, культуральная среда может содержать подходящие прекурсоры. Указанные вещества при применении могут однократно добавляться в культуральную среду или подходящим образом вноситься в процессе культивирования.
Для контроля рН культуры подходящим образом вводятся основные соединения, такие как гидроксид натрия, гидроксид калия, аммиак или нашатырный спирт, или кислотные соединения, такие как фосфорная кислота или серная кислота. Для контроля ценообразования могут применяться антивспенивающие агенты, такие как, например, сложные полигликолевые эфиры жирных кислот. Для поддержания стабильности плазмиды в среду вносятся подходящие селективно действующие агенты, такие как, например, антибиотики. Для поддержания аэробных условий в культуру вносят кислород или кислородсодержащие газовые смеси, такие как, например, воздух. Температура культивации лежит обычно в интервале от 20°С до 45°С и предпочтительно в интервале от 25°С до 40°С. Предпочтение отдается применению клеток, таких как клетки, описанные в US 6803218, особенно если субстратом для указанных клеток служит глицерин. В этом случае культивирование клеток может происходить при температуре в интервале от 40 до 100°С.
Очистка 2-гидроксиизомасляной кислоты от питательной среды предпочтительно осуществляется непрерывно, причем в этой связи также предпочтительно проводить непрерывное получение 2-гидроксиизомасляной кислоты с помощью ферментации, так чтобы весь процесс получение 2-гидроксиизомасляной кислоты, вплоть до ее очистки от ферментационного бульона, проводился непрерывно. Для непрерывной очистки получаемой 2-гидроксиизомасляной кислоты от ферментационного бульона ее необходимо непрерывно пропускать через устройство для удаления применяемых при ферментации микроорганизмов, предпочтительно через фильтр, способный отделять вещества с размером в интервале от 20 до 200 кДа, где имеет место твердое/жидкое разделение. Также возможно применение центрифуги, подходящего устройства для седиментации или комбинации этих устройств, причем особенно предпочтительным является сначала отделить, по меньшей мере, часть микроорганизмов путем седиментации, а затем подвергнуть ферментационный бульон, частично освобожденный от микроорганизмов, ультрафильтрации или центрифугированию.
После удаления микроорганизмов, продукт ферментации, обогащенный 2-гидроксиизомасляной кислотой, подается в разделительную установку, предпочтительно многоступенчатую разделительную установку. В этой разделительной установке предусмотрены несколько последовательно соединенных ступеней разделения, из которых, в каждом случае, выходят рециркуляционные линии, идущие к ферментационному чану. Далее из соответствующих ступеней разделения выходят отводящие линии. Отдельные ступени разделения могут работать по принципу электродиализа, обратного осмоса, ультрафильтрации или нанофильтрации. Как правило, отдельные ступени разделения содержат мембранные разделительные устройства. Выбор отдельных ступеней разделения зависит от природы и объема побочных продуктов ферментации и остаточных субстратов.
Наряду с выделением 2-гидроксиизомасляной кислоты посредством электродиализа, обратного осмоса, ультрафильтрации или нанофильтрации, в ходе которого в качестве конечного продукта получают водный раствор 2-гидроксиизомасляной кислоты, 2-гидроксиизомасляную кислоту можно также выделить с помощью экстракционных способов из свободного от микроорганизмов ферментационного раствора, причем в этом случае в конце можно получить чистую 2-гидроксиизомасляную кислоту. Для выделения 2-гидроксиизомасляной кислоты экстракцией к ферментационному раствору могут добавляться, например, соединения аммония или амины, с получением аммонийной соли 2-гидроксиизомасляной кислоты. Эта аммонийная соль затем может быть выделена из ферментационного раствора путем добавления органического экстракционного агента и последующего нагревания полученной смеси, вследствие чего аммонийная соль накапливается в органической фазе. 2-гидроксиизомасляная кислота может затем быть выделена из этой фазы, например, на следующих стадиях экстракции, с получением чистой 2-гидроксиизомасляной кислоты. Детали данного способа разделения раскрываются в WO-A-02/090312, объем раскрытия которого, в отношении выделения гидроксикарбоновых кислот из ферментационных растворов, включен в настоящий документ в виде ссылки и образует часть настоящего изобретения.
В зависимости от способа выделения 2-гидроксиизомасляной кислоты из ферментационного раствора получают либо водный раствор 2-гидроксиизомасляной кислоты, содержащий от 2 до 90 мас.%, предпочтительно от 7,5 до 50 мас.% и особенно предпочтительно от 10 до 25 мас.% 2-гидроксиизомасляной кислоты, либо чистую 2-гидроксиизомасляную кислоту.
При увеличении концентраций 2-гидроксиизомасляная кислота имеет склонность к образованию ее циклического димера (Тетраметилгликолид, TMG). На стадии дегидратации способа по настоящему изобретению с этим димером обращаются таким же образом, как и с 2-гидроксиизомасляной кислотой, и, таким образом, в случае данной стадии способа по изобретению, термин "2-гидроксиизомасляная кислота" охватывает этот димер.
Далее полученную с помощью способа по изобретению 2-гидроксиизомасляную кислоту можно нейтрализовывать перед, во время или после очистки, причем для этого могут применяться, например, основания, такие как, например, гидроксиды щелочных или щелочноземельных металлов, как например гидроксид кальция или гидроксид натрия, или, например, NH3 или NH4OH.
Решить поставленные выше задачи позволяет, в частности, также способ получения метакриловой кислоты или сложных эфиров метакриловой кислоты, содержащий следующие стадии:
IA) получение 2-гидроксиизомасляной кислоты согласно способу, описанному выше, и, при необходимости, очистка и/или нейтрализация 2-гидроксиизомасляной кислоты,
IB) дегидратация 2-гидроксиизомасляной кислоты с получением метакриловой кислоты, а также, при необходимости, этерификация метакрилата или метакриловой кислоты.
Согласно стадии способа IB) 2-гидроксиизомасляная кислота дегидратируется с получением метакриловой кислоты, для этой реакции возможно применять либо чистую 2-гидроксиизомасляную кислоту, выделенную из ферментационного раствора, либо водный раствор 2-гидроксиизомасляной кислоты, который был выделен в процессе обработки ферментационного раствора, и при необходимости, также концентрирован, например, посредством дистилляции, при необходимости, в присутствии подходящих азеотропообразователей, еще до стадии дегидратации.
Стадия дегидратации, в принципе, может проводиться в жидкой фазе или в газовой фазе. Согласно изобретению является предпочтительным, чтобы стадия дегидратации проводилась в присутствии катализатора, причем тип применяемого катализатора зависит от того, проводится ли газофазная дегидратация или дегидратация в жидкой фазе.
В качестве подходящих катализаторов дегидратации рассматриваются как кислотные, так и щелочные катализаторы. Кислотные катализаторы предпочтительны, в частности, благодаря их незначительной склонности к образованию олигомеров. В качестве катализатора дегидратации может применяться как гомогенный, так и гетерогенный катализатор. Если в качестве катализатора дегидратации применяется гетерогенный катализатор, то предпочтительно, чтобы катализатор дегидратации находился в контакте с носителем х. В качестве соответствующего носителя х специалисты в данной области техники могут рассматривать все подходящие твердые материалы. В этой связи предпочтительно, чтобы указанные твердые материалы обладали подходящим объемом пор, необходимым для хорошего связывания с катализатором дегидратации и насыщения им. Кроме того, общий объем пор, согласно стандарту DIN 66133, предпочтительно лежит в интервале от 0,01 до 3 мл/г, и особенно предпочтительно общий объем пор лежит в интервале от 0,1 до 1,5 мл/г. Кроме того, твердые материалы, подходящие в качестве носителя х, предпочтительно имеют площадь поверхности в интервале от 0,001 до 1000 м2/г, предпочтительно в интервале от 0,005 до 450 м2/г и особенно предпочтительно в интервале от 0,01 до 300 м2/г, как определено с помощью теста BET согласно стандарту DIN 66131. В качестве носителя для катализатора дегидратации может применяться сыпучий материал, со средним диаметром частиц в интервале от 0,1 до 40 мм, предпочтительно в интервале от 1 до 10 мм и наиболее предпочтительно в интервале от 1,5 до 5 мм. Кроме того, в качестве носителя для катализатора дегидратации могут служить стены реактора, в котором проходит дегидратация.
Кроме того, носитель может быть кислотным или основным, либо кислотный или основный катализатор дегидратации может доставляться на инертном носителе. В качестве способов нанесения упоминаются, в частности, погружение или пропитывание, или включение в матрицу носителя.
Подходящие носители х, которые также могут обладать свойствами катализатора дегидратации, представляют собой, в частности, природные или синтетические силикатные материалы, такие как, в частности, морденит, монтмориллонит, кислые цеолиты; носители, покрытые одноосновными, двухосновными или полиосновными неорганическими кислотами, в частности фосфорной кислотой, или кислыми солями неорганических кислот, такими как вещества оксидного или силикатного типа, как например, Al2O3, TiO2; оксиды и смешанные оксиды, такие как, например, γ-Al2O3 и ZnO-Al2O3 смешанные оксиды гетерополикислот.
Согласно одному варианту выполнения настоящего изобретения, носитель х, по меньшей мере, частично состоит из соединения оксидного типа. Такие соединения оксидного типа должны содержать, по меньшей мере, один из элементов, выбранных из Si, Ti, Zr, Al, P или комбинацию, по меньшей мере, двух этих элементов. Такие носители благодаря их кислотным или основным свойствам могут сами действовать в качестве катализатора дегидратации. Предпочтительный класс соединений, действующих как носитель х и как катализатор дегидратации содержит кремний/алюминий/ фосфор оксиды. Предпочтительные основные вещества, которые действуют как в качестве катализатора дегидратации, так и в качестве носителя х содержат щелочные металлы, щелочноземельные металлы, лантан, лантаноиды или комбинации, по меньшей мере, двух из перечисленных веществ в форме их оксидов. Такие кислотные или основные катализаторы дегидратации коммерчески доступны от Evonik Degussa GmbH и от Sudchemie AG. Ионообменные вещества представляют собой следующий класс. Они также могут быть основными или кислотными.
Подходящими гомогенными катализаторами дегидратации являются, в частности, неорганические кислоты, предпочтительно фосфорсодержащие кислоты, более того, предпочтительно фосфорная кислота. Эти неорганические кислоты могут наноситься на носитель х путем погружения или пропитки.
В частности, применение гетерогенных катализаторов является особенно предпочтительным при газофазной дегидратации. Однако при дегидратации в жидкой фазе применяются как гомогенные, так и гетерогенные катализаторы дегидратации.
Более того, предпочтительно, чтобы при выполнении способа по изобретению применялся катализатор дегидратации со значением НО в интервале от +1 до -10, предпочтительно в интервале от +2 до -8,2 и, более предпочтительно, в случае дегидратации в жидкой фазе, в интервале от +2 до -3, и, в случае газофазной дегидратации, в интервале от -3 до -8,2. Значение НО соответствует функции кислотности Гаммета и определяется с помощью так называемого аминного титрования и применения индикаторов или по поглощению газообразного основания (смотрите "Studies in Surface Science and Catalytics", Vol.51, 1989: "New solid Acids and Bases, their catalytic Properties", K.Tannabe et al.).
Согласно особенно предпочтительному варианту выполнения способа по настоящему изобретению, в качестве кислотного твердого катализатора применяется носитель с пористой структурой, находящийся в контакте с неорганической кислотой, предпочтительно с фосфорной кислотой или с суперкислотами, такими как, например, сульфатированный или фосфатированный оксид циркония, и который основан, по меньшей мере, на 90 мас.%, предпочтительно, по меньшей мере, на 95 мас.% и особенно предпочтительно, по меньшей мере, на 99 мас.%, на оксиде кремния, предпочтительно SiO2. Контакт носителя с пористой структурой с неорганической кислотой осуществляется предпочтительно путем пропитывания указанного носителя указанной кислотой, причем кислота составляет предпочтительно от 10 до 70 мас.%, особенно предпочтительно от 20 до 60 мас.% и наиболее предпочтительно от 30 до 50 мас.% от массы носителя, после установления контакта проводят сушку. После сушки неорганическую кислоту фиксируют путем нагревания носителя предпочтительно до температуры в интервале от 300 до 600°С, более предпочтительно в интервале от 400 до 500°С.
Согласно особенно предпочтительному варианту выполнения способа по настоящему изобретению дегидратация проводится в газовой фазе. При этом могут применяться обычные устройства, известные специалисту в области газофазных реакций, например трубчатый реактор. Особенно предпочтительным является применение кожухотрубных теплообменников, а также реакторов, которые содержат в качестве теплообменника тепловые панели.
Согласно одному варианту выполнения настоящего изобретения, при проведении газофазной дегидратации чистая 2-гидроксиизомасляная кислота поступает в реактор, содержащий неподвижный слой одного из вышеупомянутых катализаторов. Согласно другому варианту выполнения настоящего изобретения, 2-гидроксиизомасляная кислота поступает в реактор в виде водного раствора 2-гидроксиизомасляной кислоты, содержащего от 2 до 80 мас.%, особенно предпочтительно от 5 до 50 мас.% и наиболее предпочтительно от 10 до 25 мас.% 2-гидроксиизомасляной кислоты относительно общей массы водного раствора. Условия давления и условия температуры внутри реактора выбираются таким образом, чтобы 2-гидроксиизомасляная кислота или ее водный раствор на входе в реактор находились в газообразной форме. Дегидратация в газовой фазе предпочтительно осуществляется при температуре в интервале от 200 до 400°С, особенно предпочтительно от 250 до 350°С. Давление внутри реактора при газофазной дегидратации предпочтительно лежит в интервале от 0,1 до 50 бар, особенно предпочтительно в интервале от 0,2 до 10 бар и наиболее предпочтительно в интервале от 0,5 до 5 бар.
В случае газофазной дегидратации количество 2-гидроксиизомасляной кислоты, поданной в реактор, предпочтительно лежит в интервале от 10 до 100 об.%, особенно предпочтительно в интервале от 20 до 100 об.% и наиболее предпочтительно в интервале от 30 до 100 об.%.
Согласно другому предпочтительному варианту выполнения способа по изобретению, дегидратация проводится в жидкой фазе. Дегидратация в жидкой фазе также может проводиться в любом устройстве, известном специалисту в данной области техники, в котором жидкость может быть нагрета до желаемой температуры реакции, причем возможно поддерживать в устройстве давление, достаточное, чтобы компоненты реакции оставались жидкими при желательных температурных условиях.
Согласно конкретному варианту выполнения способа по изобретению, способ дегидратации в жидкой фазе включает первую стадию, на которой в реактор подается чистая 2-гидроксиизомасляная кислота или водный раствор, содержащий от 5 до 100 мас.%, особенно предпочтительно от 20 до 100 мас.% и наиболее предпочтительно от 50 до 100 мас.% 2-гидроксиизомасляной кислоты относительно общей массы водного раствора. Условия давления и условия температуры внутри реактора выбираются таким образом, чтобы на входе в реактор указанная 2-гидроксиизомасляная кислота или указанный водный раствор находились в жидкой форме.
Согласно особенно предпочтительному варианту выполнения способа по изобретению, при котором дегидратация проводится в жидкой фазе, 2-гидроксиизомасляную кислоту или ее водный раствор пропускают над неподвижным слоем катализатора внутри реактора дегидратации таким образом, чтобы жидкая фаза протекала над поверхностью частиц катализатора. Такой способ может осуществляться, например, в реакторе с орошаемым слоем.
Дегидратация в жидкой фазе выполняется при температуре предпочтительно в интервале от 200 до 350°С, особенно предпочтительно в интервале от 250 до 300°С. Давление внутри реактора в случае дегидратации в жидкой фазе предпочтительно лежит в интервале от 1 до 50 бар, особенно предпочтительно в интервале от 2 до 25 бар и наиболее предпочтительно в интервале от 3 до 10 бар.
Катализ дегидратации, как при газофазной дегидратации, так и при дегидратации в жидкой фазе, может происходить гомогенно или гетерогенно.
При гомогенном катализе, катализатор, который в данном случае имеет форму неорганической кислоты, такой как, например, фосфорная кислота или серная кислота, сначала вступает в контакт с чистой 2-гидроксиизомасляной кислотой или с водным раствором, содержащим 2-гидроксиизомасляную кислоту. Затем полученная композиция поступает в реактор и превращается в метакриловую кислоту при требуемых условиях давления и температуры. По возможности, в реактор нужно также подводить неорганическую кислоту независимо от 2-гидроксиизомасляной кислоты или ее водного раствора. В этом случае, реактор содержит, по меньшей мере, две подводящие линии, одна для 2-гидроксиизомасляной кислоты или водного раствора, содержащего 2-гидроксиизомасляную кислоту, и вторая для катализатора. Если реакция дегидратации проводится в жидкой фазе в реакторе с орошаемым слоем, то это предпочтительно, чтобы катализатор вместе с 2-гидроксиизомасляной кислотой или водным раствором, содержащим 2-гидроксиизомасляную кислоту, поступал в верхнюю часть реактора.
При гетерогенном катализе, катализатор находится в форме твердой подложки, расположенной в пространстве реактора, например, в форме неподвижного слоя, в форме покрытых катализатором панелей, предпочтительно тепловых панелей, которые расположены внутри реактора, или же в виде покрытых катализатором стен реактора. Возможные реакторы описываются, например, в DE-A-19848208, DE-A-10019381 и ЕР-A-I234612. В случае гетерогенного катализа, предпочтительными катализаторами являются носители, находящиеся в контакте с неорганическими кислотами, предпочтительно пропитанные пористые носители. Затем 2-гидроксиизомасляную кислоту или водный раствор, содержащий 2-гидроксиизомасляную кислоту, в жидкой или парообразной форме, приводят в контакт с поверхностью твердого каталитического материала.
Согласно особенно предпочтительному варианту выполнения способа по изобретению, дегидратация 2-гидроксиизомасляной кислоты проводится в жидкой фазе, при давлении в интервале от 200 до 500 мбар, при температуре в интервале от 160 до 300°С, предпочтительно от 200 до 240°С, и в присутствии ионов щелочного металла в качестве катализатора.
При данных условиях реакции полученная метакриловая кислота может быть подвергнута процессу перегонки в газообразной форме вместе с водой и затем конденсирована в примыкающий сосуд в виде водного раствора, с получением, по возможности, водного раствора метакриловой кислоты, не содержащего какие-либо компоненты катализатора.
Согласно предпочтительному варианту выполнения способа по изобретению, раствор метакриловой кислоты, полученный таким образом, при необходимости, может быть этерифицирован без последующей обработки. В этом случае раствор указанной метакриловой кислоты при нагревании вводится в контакт с соответствующими спиртами и подходящими катализаторами этерификации, известными специалисту в данной области техники, такими как, например, концентрированные кислоты, с превращением, таким образом, метакриловой кислоты в соответствующие сложные эфиры.
Предпочтительные спирты, среди прочего, представляют собой спирты с, по меньшей мере, одним атомом углерода, предпочтительно с 2-12 атомами углерода, и особенно предпочтительно с 4-9 атомами углерода. Указанные спирты могут иметь линейную, разветвленную или циклическую структуру. Кроме того, спирты могут содержать ароматические группы или заместители, такие как, например, атомы галогенов. К предпочтительным спиртам в частности относятся: метанол, этанол, н-пропанол, изо-пропанол, н-бутанол, 1-метилпропанол, 2-метилпропанол, трет-бутанол, н-пентанол, 1-метилбутанол, 2-метилбутанол, 3-метилбутанол, 2,2-диметилпропанол, н-гексанол, 1-метилпентанол, 2-метилпентанол, 3-метилпентанол, 4-метилпентанол, 1,1-диметил-бутанол, 2,2-диметилбутанол, 3,3-диметилбутанол, 1,2-диметилбутанол, н-гептанол, 1-метилгексанол, 2-метилгексанол, 3-метилгексанол, 4-метилгексанол, 1,2-диметилпентанол, 1,3-диметилпентанол, 1,1-диметилпентанол, 1,1,2,2-тетраметилпропанол, бензиловый спирт, н-октанол, 2-этилгексанол, н-нонанол, 1-метилоктанол, 2-метилоктанол, н-деканол, н-ундеканол, 1-метилдеканол, 2-метилдеканол, н-додеканол, 2,4-диэтилоктанол, циклопентанол, циклогексанол, 4-трет-бутилциклогексанол, циклогептанол, циклододеканол, 2-(диметиламино)этанол, 3-(диметиламино)пропанол, 4-(диметиламино)бутанол, 5-(диметиламино)пентанол, 6-(диметиламино)гексанол, 8-(диметиламино)октанол, 10-(диметиламино)деканол, 12-(диметиламино)додеканол, 2-(диэтиламино)этанол, 3-(диэтиламино)пропанол, 4-(диэтиламино)бутанол, 5-(диэтиламино)пентанол, 6-(диэтиламино)гексанол, 8-(диэтиламино)октанол, 10-(диэтиламино)деканол, 12-(диэтиламино)-додеканол, 2-(ди-(изо-пропил)амино)этанол, 3-(ди-(изо-пропил)-амино)пропанол, 4-(ди-(изо-пропил)амино)бутанол, 5-(ди-(изо-пропил) амино)пентанол, 6-(ди-(изо-пропил)амино)гексанол, 8-(ди-(изо-пропил)амино)октанол, 10-(ди-(изо-пропил)амино)деканол, 12-(ди-(изо-пропил)амино)додеканол, 2-(дибутиламино)этанол, 3-(дибутиламино)пропанол, 4-(дибутиламино)бутанол, 5-(дибутиламино)пентанол, 6-(дибутиламино)гексанол, 8-(дибутиламино)октанол, 10-(дибутиламино)деканол, 12-(дибутиламино)додеканол, 2-(дигексиламино)этанол, 3-(дигексиламино)пропанол, 4-(дигексиламино)бутанол, 5-(дигексиламино)пентанол, 6-(дигексиламино)гексанол, 8-(дигексиламино)октанол, 10-(дигексиламино)деканол, 12-(дигексиламино)додеканол, 2-(метилэтиламино)этил-, 2-(метил-пропиламино)этанол, 2-(метил-изо-пропиламино)этанол, 2-(метил-бутиламино)этанол, 2-(метилгексиламино)этанол, 2-(метилоктил-амино)этанол, 2-(этилпропиламино)этанол, 2-(этил-изо-пропиламино)этанол, 2-(этилбутиламино)этанол, 2-(этилгексиламино)-этанол, 2-(этилоктиламино)этанол, 3-(метилэтиламино)пропанол, 3-(метилпропиламино)пропанол, 3-(метил-изо-пропиламино)пропанол, 3-(метилбутиламино)пропанол, 3-(метилгексиламино)пропанол, 3-(метилоктиламино)пропанол, 3-(этилпропиламино)пропанол, 3-(этил-изо-пропиламино)пропанол, 3-(этилбутиламино)пропанол, 3-(этилгексиламино)пропанол, 3-(этилоктиламино)пропанол, 4-(метилэтиламино)бутанол, 4-(метилпропиламино)бутанол, 4-(метил-изо-пропиламино)бутанол, 4-(метилбутиламино)бутанол, 4-(метилгексиламино)-бутанол, 4-(метилоктиламино)бутанол, 4-(этилпропиламино)бутанол, 4-(этил-изо-пропиламино)бутанол, 4-(этилбутиламино)бутанол, 4-(этилгексиламино)бутанол, 4-(этилоктиламино)бутанол, 2-(N-пиперидинил)этанол, 3-(N-пиперидинил)пропанол, 4-(N-пиперидинил)-бутанол, 5-(N-пиперидинил)пентанол, 6-(N-пиперидинил)гексанол, 8-(N-пиперидинил)октанол, 10-(N-пиперидинил)деканол, 12-(N-пиперидинил)додеканол, 2-(N-пирролидинил)этанол, 3-(N-пирролидинил)пропанол, 4-(N-пирролидинил)бутанол, 5-(N-пирролидинил)пентил-, 6-(N-пирролидинил)гексанол, 8-(N-пирролидинил)октанол, 10-(N-пирролидинил)деканол, 12-(N-пирролидинил)додеканол, 2-(N-морфолино)этанол, 3-(N-морфолино)-пропанол, 4-(N-морфолино)бутанол, 5-(N-морфолино)пентанол, 6-(N-морфолино)гексанол, 8-(N-морфолино)октанол, 10-(N-морфолино)-деканол, 12-(N-морфолино)додеканол, 2-(N'-этил-N-пиперазинил) этанол, 3-(N'-метил-N-пиперазинил)пропанол, 4-(N'-метил-N-пиперазинил)бутанол, 5-(N'-метил-N-пиперазинил)пентанол, 6-(N'-метил-N-пиперазинил)гексанол, 8-(N'-метил-N-пиперазинил)октанол, 10-(N'-метил-N-пиперазинил)деканол, 12-(N'-метил-N-пиперазинил)додеканол, 2-(N'-этил-N-пиперазинил)этанол, 3-(N'-этил-N-пиперазинил)пропанол, 4-(N'-этил-N-пиперазинил)бутанол, 5-(N'-этил-N-пиперазинил)пентанол, 6-(N'-этил-N-пиперазинил)гексанол, 8-(N'-этил-N-пиперазинил)октанол, 10-(N'-этил-N-пиперазинил)деканол, 12-(N'-этил-N-пиперазинил)додеканол, 2-(N'-изо-пропил-N-пиперазинил)этанол, 3-(N'-изо-пропил-N-пиперазинил)пропанол, 4-(N'-изо-пропил-N-пиперазинил)бутанол, 5-(N'-изо-пропил-N-пиперазинил)пентанол, 6-(N'-изо-пропил-N-пиперазинил)гексанол, 8-(N'-изо-пропил-N-пиперазинил)октанол, 10-(N'-изо-пропил-N-пиперазинил)деканол, 12-(N'-изо-пропил-N-пиперазинил)додеканол, 3-оксабутанол, 3-оксапентанол, 2,2-диметил-4-оксапентанол, 3,6-диоксагептанол, 3,6-диоксаоктанол, 3,6,9-триоксадеканол, 3,6,9-триоксаундеканол, 4-оксапентанол, 4-оксагексанол, 4-оксагептанол, 4,8-диоксанонанол, 4,8-диоксадеканол, 4,8-диоксаундеканол, 5-оксагексанол или 5,10-диоксаундеканол.
Кроме того, возможно применять в качестве растворителей этоксилированные и/или пропоксилированные спирты, а также смешанные этоксилированные/пропоксилированные спирты, в частности Ra-(O-CH2-CH2)x-OH или Ra-(O-CH(CH3)-CH2)x-OH, или Ra(O-CH2-CH(CH3))x-OH, где Ra представляет собой C1-С20-алкил, и х представляет собой целое число от 10 до 20, или этоксилированные и/или пропоксилированные аминоспирты, например Rb 2N(-CH2-CH2-O)y-H или Rb 2N(-CH(CH3)-CH2-O)y-H, или Rb 2N(-CH2CH(CH3)-O)y-H, где у представляет собой целое число от 1 до 4. Rb представляет собой алкильную группу с 1-6 атомами углерода, причем атом азота вместе с заместителями Rb может образовывать пяти- или семичленное кольцо. При необходимости кольцо может содержать в качестве заместителей одну или более алкильную группу с короткими цепями, такую как, например, метил, этил или пропил.
Тем не менее, очистка метакриловой кислоты перед этерификацией может быть предпочтительна, причем, в принципе, возможно применять любой известный специалисту в данной области техники способ очистки, который обычно применяется для очистки загрязненной метакриловой кислоты, полученной путем каталитического газофазного окисления пропилена.
Если реакция дегидратации проводилась в газовой фазе, то предпочтительно сначала конденсировать метакриловую кислоту, с получением водного раствора метакриловой кислоты. При этом может применяться любой известный специалисту в данной области техники способ конденсации, например фракционная конденсация, как раскрывается в WO-A-2004/035 514, WO-A-03/014 172 или ЕР-А-ЕР А 1163201, или тотальная конденсация, как раскрывается в ЕР-А-0695736. По возможности, в ходе конденсации необходимо добавлять дополнительные растворители, в частности воду, чтобы поглощение метакриловой кислоты было по возможности более полным.
Из водного раствора метакриловой кислоты, полученного после конденсации, или же из водного раствора метакриловой кислоты, полученного после дегидратации в жидкой фазе, может затем быть удалена вода и другие примеси на следующих стадиях очистки. При этом возможно сначала удалять воду с помощью азеотропной дистилляции в присутствии азеотропообразователей, как это описывается, например, в DE-A-19853064. Применение высококипящих органических растворителей для поглощения метакриловой кислоты также возможно, как это описывается, например, в ЕР-А-0974574. Наряду с этими способами дистилляции, также могут применяться мембраны для обезвоживания, как это предлагается, например, в DE-A-4401405. Кроме того, очистку водного раствора метакриловой кислоты, полученного с помощью дегидратации в жидкой фазе или конденсации, можно проводить способом процесса кристаллизации.
Полученная после удаления воды метакриловая кислота может быть затем очищена на следующих стадиях способа. Таким образом могут быть удалены содержащиеся высококипящие примеси, которые все еще присутствуют на следующих стадиях дистилляции. Тем не менее, особенно предпочтительно, если последующая очистка метакриловой кислоты, полученной после удаления воды, проводится с помощью способов кристаллизации, как описывается, например, в DE-A-10149353.
Полученная таким образом очищенная метакриловая кислота затем, при необходимости, может быть подвергнута этерификации.
Вклад в решение поставленных выше задач, кроме того, вносит способ получения метакриловой кислоты или сложных эфиров метакриловой кислоты, содержащий следующие стадии:
IIA) получение полигидроксиалканоатов, содержащих мономерные структурные единицы 2-гидроксиизомасляной кислоты, согласно вышеописанному способу,
IIB) расщепление полигидроксиалканоатов, содержащих мономерные структурные единицы 2-гидроксиизомасляной кислоты, с получением 2-гидроксиизомасляной кислоты, а также, при необходимости, нейтрализация 2-гидроксиизомасляной кислоты и/или очистка 2-гидроксиизомасляной кислоты,
IIC) дегидратация 2-гидроксиизомасляной кислоты с получением метакриловой кислоты, а также, при необходимости, этерификация метакрилата или метакриловой кислоты.
Вклад в решение вышеупомянутых задач также вносит способ получения полиметакриловой кислоты или сложных эфиров полиметакриловой кислоты, содержащий следующие стадии:
IIIA) получение метакриловой кислоты согласно вышеописанному способу,
IIIB) радикальная полимеризация метакриловой кислоты,
причем, при необходимости, карбоксильные группы метакриловой кислоты, до или после радикальной полимеризации, могут быть, по меньшей мере, частично этерифицированы.
Приведенные далее примеры служат для иллюстрации настоящего изобретения и возможных вариантов выполнения настоящего изобретения, и никоим образом не предназначены для ограничения сущности и объема настоящего изобретения до упомянутых в них вариантов выполнения изобретения, где сущность и объем настоящего изобретения определяются полным описанием и формулой изобретения.
Следующие фигуры относятся к примерам:
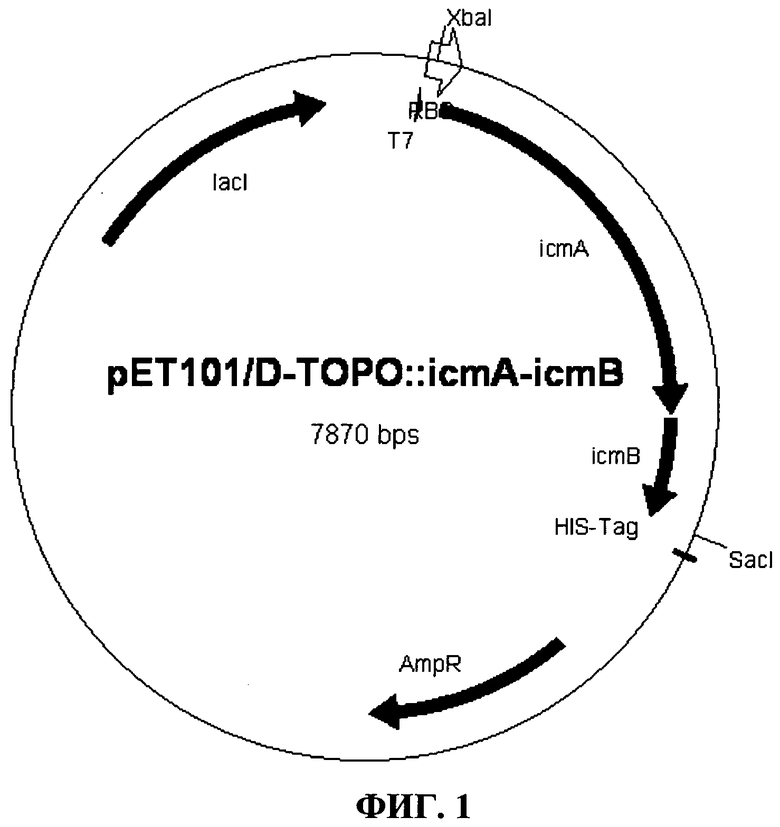
Фиг.1: гибридная плазмида pET101/D-TOPO::icmA-icmB
Фиг.2: гибридная плазмида pBBR1MCS-2::icmA-icmB
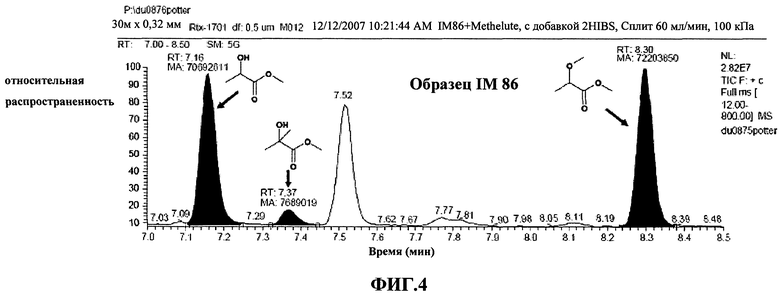
Фиг.3: количественное определение 2-гидроксиизомасляной кислоты в образце IM-86 было проведено после добавления к образцу 2-гидроксиизомасляной кислоты. Пик метиллактата (время удерживания (Rt.) равно 7,16 минут) применялся в качестве внутреннего стандарта. Приведены участки ГХ-МС-хроматограмм для первоначального образца и образца с добавкой.
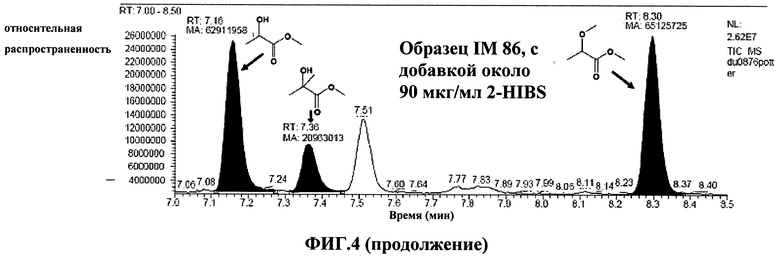
Фиг.4: добавление 2-гидроксиизомасляной кислоты к образцу IM-89. Приводятся участки ЯМР-спектров для первоначального образца (А) и образца с добавкой(В).
Примеры
1. Выделение геномной ДНК и амплификация фрагментов icmA и icmB
Геномную ДНК выделили из штамма Aquincola tertiaricarbonis (A. tertiaricarbonis DSMZ 18512) с помощью набора Kit DNeasy Blood & Tissue (Qiagen GmbH, Хильден, Германия), согласно указаниям производителя, и применяли в качестве матрицы для ПЦР при амплификации фрагментов icmA (1,7 т.п.н.; DQ436456) и icmB (0,4 т.п.н.; DQ436457). Эти фрагменты кодируют фермент Е3. Олигонуклеотиды 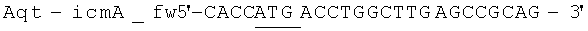
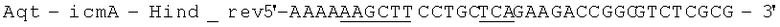
(обратный праймер; стоп-кодон и сайт рестрикции Hind III подчеркнуты) применяются при амплификации icmA, и Олигонуклеотиды 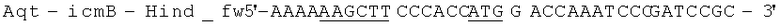
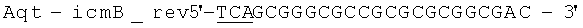
Полимеразную цепную реакцию (ПЦР, согласно SAIKI et al., 1985, Enzymatic amplification of β-globin genomic sequences and restriction site analysis for diagnosis of sickle cell anemia. Science 230: 1350-1354.) проводили с помощью Pfu-полимеразы (Promega, Мэдисон, США). Проводилось 35 циклов, где в течение 60 секунд поддерживалась температура 95°С, в течение 30 секунд поддерживалась температура 65°С, и в течение 4 минут поддерживалась температура 72°С. ПЦР проводили в термоциклере (Primus 96 advanced; PEQLAB Biotechnologie GMBH, Эрланген, Германия).
Фрагменты очистили с помощью набора QIAquick PCR Purification Kit (Qiagen GmbH, Хильден), согласно указаниям производителя, и затем подвергли рестрикции с помощью Hindlll. Обе смеси были лигированы по сайту рестрикции HindIII.
Продукт лигирования icmA-icmB (2,1 т.п.н.) применяли в качестве матрицы для Pfu-ПЦР, с применением олигонуклеотидов 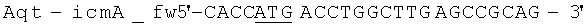

2. Получение экспрессионного вектора Ralstonia eutropha
Очищенный ПЦР-фрагмент icmA-icmB (2,1 т.п.н.) лигировали в вектор pET101/D-TOPO (Invitrogen GmbH, Карлсруэ, Германия) согласно указаниям производителя. Полученную гибридную плазмиду pET101/D-TOPO::icmA-icmB (Фиг.1, Seq. ID No.1) перенесли в компетентные DH5α клетки Е. coli (New England Biolabs, Франкфурт) и контролировали посредством рестрикции и секвенирования.
Для достижение экспрессии в штаммах R. eutropha, дикий тип которых проявляют активности ферментов E1, E2 и Е4, конструкция должна быть клонирована в подходящий вектор с широким кругом хозяев. Вектор, применяемый в pBBR1MCS-2, описанный у KOVACH et al. (1995). Four new derivatives of the broad-host-range cloning vector pBBRIMCS carrying different antibiotic-resistance cassettes. Gene, 166: 175-176.
С этой целью плазмиды pET101/D-TOPO::icmA-icmB и pBBR1MCS-2 подвергли рестрикции ферментами XbaI и SacI, и фрагмент icmA-icmBQ был лигирован в целевой вектор pBBR1MCS-2, и компетентные DH5α клетки Е. coli (New England Biolabs, Франкфурт) были трансформированы с помощью полученной гибридной плазмиды pBBR1MCS-2::icmA-icmB (Фиг.2, Seq. ID No.2).
Плазмиду проверили с помощью рестрикции и секвенирования, и перенесли в компетентные S17-1 клетки Е. coli, штамм, который делает возможным конъюгационный перенос плазмид в, среди прочего, штаммы Ralstonia eutropha. Для этого проводится конъюгация с точечным скрещиванием (как описывается у FRIEDRICH et al.,1981, Naturally occurring genetic transfer of hydrogen-oxidizing ability between strains of Alcaligenes eutrophus. J Bacteriol 147: 198-205), с применением E. coli S17-1 pBBR1MCS-2::icmA-icmB в качестве донора, и как R. eutropha H16 (переклассифицировали как Cupnavidus necator, DSMZ 428), так и R. eutropha PHB-4 (переклассифицировали как Cupnavidus necator, DSMZ 541) в качестве реципиента.
Таким образом могут быть получены трансконъюганты, которые несут плазмиду pBBR1MCS-2::icmA-icmB.
3. Получение 2-гидроксиизомасляной кислоты в рекомбинантных клетках R. eutropha
Продуцирование 2-гидоксиизомасляной кислоты было изучено путем выращивания несущих плазмиду штаммов R. eutropha, описанных в примере 2, в 50 мл MSM-среде Фолльбрехта ((NH4)2HPO4, 2.0 г; KH2РО4, 2.1 г; MgSO4×7H2O, 0.2 г; FeCl3×6H2O, 6 мг; CaCl2×2H2О, 10 мг; раствор микроэлементов (Pfennig and Lippert, 1966), 0.1 мл). В среду дополнительно ввели глюконат натрия (15 г/л), канамицин (50 мкг/мл) и Коэнзим В12 (60 мкг/мл). Клетки инкубировали на термостатированном шейкере при 30°С и 160 об/мин. Через 30 часов дополнительно добавили глюконат натрия (1,5%, мас./об.) и Коэнзим В12 [60 мкг/мл]. Урожай культуры собрали после 52 часов культивирования с помощью центрифугирования при 5000 об/мин (4°С). Супернатан культуры хранили при -20°С до проведения анализа.
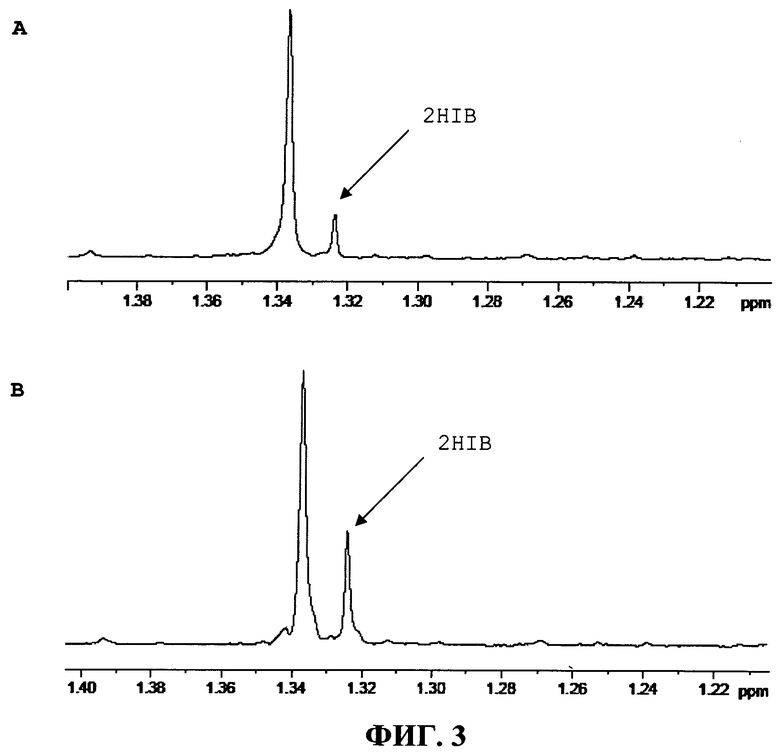
Обнаружение и количественное определение 2-гидроксиизомасляной кислоты провели с помощью количественной 1H-ЯМР спектроскопии. Образцы количественно концентрировали. Для остатка определили 1Н-ЯМР спектры и вычислили содержание относительно TSP (трисилилпропионовой кислоты) в качестве внутреннего стандарта. В спектре 2-гидроксиизомасляной кислоты обнаруживается синглет при около 1,36 частей на миллион (ppm), и чистая 2-гидроксиизомасляная кислота добавляется к остатку для получения доказательств (Фигура. 3).
В проанализированных образцах были обнаружены концентрации 2-гидроксиизомасляной кислоты до 0,72 ммоль/кг. Напротив, в соответствующих контрольных смесях, содержащих пустую плазмиду, содержание 2-гидроксиизомасляной кислоты не было обнаружено. ЯМР измерения были качественно и количественно подтверждены с помощью ГХ-МС и добавления чистой 2-гидроксиизомасляной кислоты (Фиг.4). В этом случае, хроматографическое разделение осуществляется на капиллярной колонке 30 м Rtx-1701 (Fisher Scientific, Питсбург, США). После лиофилизации образцы ресуспендировали с применением реагента дериватизации "Methelute" (Pierce, Рокфорд, США). 0,5 мкп этого раствора непосредственно нанесли с помощью инжектора для ввода проб с делением потока/без деления потока. Пики 2-гидроксиизомасляной кислоты были идентифицированы путем сравнения полученного масс-спектра с базой данных спектров. Содержание 2-гидроксиизомасляной кислоты оценили путем добавления к образцам определенного количества сравнительного вещества 2-гидроксиизомасляной кислоты. В проанализированных образцах были обнаружены концентрации до 44 мкг/мл. В контрольных смесях, содержащих пустую плазмиду, 2-гидроксиизомасляная кислота не была обнаружена. Подобным образом также возможно обнаружить 2-гидроксиизомасляную кислоту, при синтезе которой в качестве источника углерода выступает фруктоза (1,5%, мас./об.).
4. Дегидратация 2-гидроксиизомасляной кислоты с получением метакрилата
5 мл раствора 2-гидроксиизомасляной кислоты (0,2 г/л), полученного согласно примеру 3, смешали при перемешивании с NaOH (0.06). Раствор инкубировали при перемешивании и охлаждении с помощью обратного холодильника при 185-195°С, при пониженном давлении (300 Торр). В течение 5 часов каждый час добавляли еще 0,5 мг 2-гидроксиизомасляной кислоты в 5 мл, причем указанный раствор также содержал 0,4 мас.% п-метоксифенола для предотвращения полимеризации метакрилата. После 24 часов инкубирования реакция завершилась. Превращение 2-гидроксиизомасляной кислоты в метакрилат составило более 90%. Выделение метакриловой кислоты из реакционной смеси осуществляли посредством дистилляции.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ПОЛУЧЕНИЯ МЕТАКРИЛОВОЙ КИСЛОТЫ ИЛИ СЛОЖНЫХ ЭФИРОВ МEТАКРИЛОВОЙ КИСЛОТЫ | 2008 |
|
RU2491346C9 |
| СПОСОБ ФЕРМЕНТАТИВНОГО ПОЛУЧЕНИЯ 2-ГИДРОКСИ-2-МЕТИЛКАРБОНОВЫХ КИСЛОТ | 2007 |
|
RU2459871C2 |
| СПОСОБ ПОЛУЧЕНИЯ АЛКИЛ(МЕТ)АКРИЛАТОВ | 2006 |
|
RU2409552C2 |
| ФЕРМЕНТАТИВНОЕ ПОЛУЧЕНИЕ 1-БУТАНОЛА | 2006 |
|
RU2429295C2 |
| СПОСОБ ПОЛУЧЕНИЯ МЕТИЛМЕТАКРИЛАТА | 2017 |
|
RU2758142C2 |
| СПОСОБЫ ПОЛУЧЕНИЯ РАМНОЛИПИДОВ | 2015 |
|
RU2694392C2 |
| МОДИФИЦИРОВАННЫЕ МИКРООРГАНИЗМЫ И СПОСОБЫ ПОЛУЧЕНИЯ БУТАДИЕНА С ИХ ПРИМЕНЕНИЕМ | 2012 |
|
RU2639564C2 |
| ФЕРМЕНТАТИВНОЕ ПОЛУЧЕНИЕ ЧЕТЫРЕХУГЛЕРОДНЫХ СПИРТОВ | 2006 |
|
RU2394913C2 |
| СПОСОБ ПОЛУЧЕНИЯ АМИДА КАРБОНОВОЙ КИСЛОТЫ ИЗ КАРБОНИЛЬНОГО СОЕДИНЕНИЯ И ЦИАНИСТОВОДОРОДНОЙ КИСЛОТЫ | 2009 |
|
RU2552619C9 |
| Новые способы дисплея циклических пептидов на частицах бактериофага | 2015 |
|
RU2702087C2 |




Изобретение относится к рекомбинантной клетке Ralstonia eutropha, предназначенной для получения 2-гидроксиизомасляной кислоты. Клетка трансформирована плазмидой с последовательностью SEQ ID NO:2. Клетка, несущая указанную плазмиду, продуцирует 2-гидроксиизомасляную кислоту в концентрации до 0,72 ммоль/кг. 4 ил., 4 пр.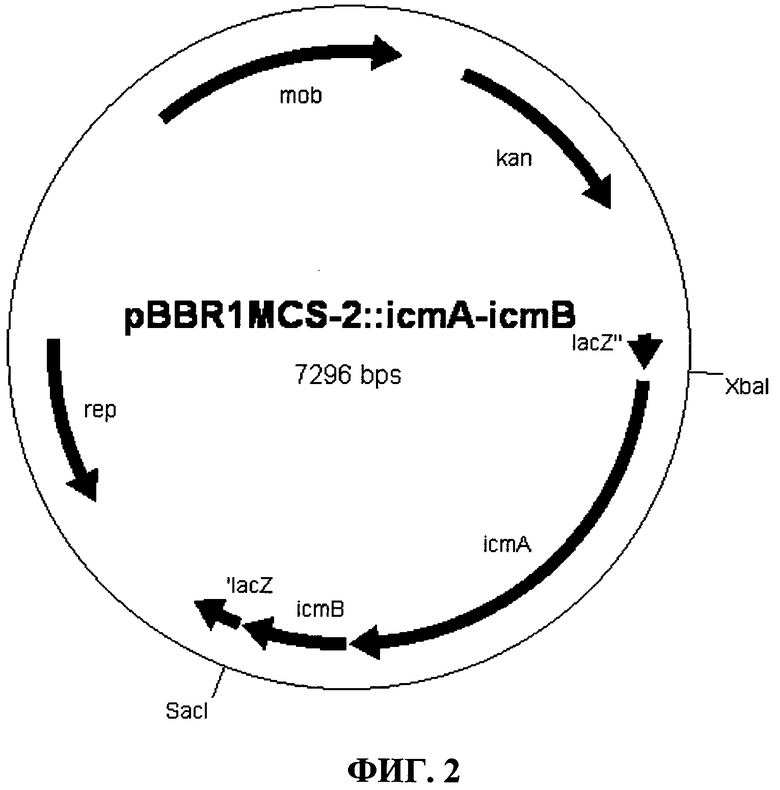
Рекомбинантная клетка Ralstonia eutropha, предназначенная для получения 2-гидроксиизомасляной кислоты, содержащая плазмиду с последовательностью SEQ ID NO:2.
| WO 2007110394 A2, 04.10.2007 | |||
| WO 2007141208 A2, 13.12.2007 | |||
| СПОСОБ ГЕНЕТИЧЕСКОЙ МОДИФИКАЦИИ ИНТЕРЕСУЮЩЕГО ЭНДОГЕННОГО ГЕНА ИЛИ ХРОМОСОМНОГО ЛОКУСА (ВАРИАНТЫ) И ЕГО ИСПОЛЬЗОВАНИЕ | 2001 |
|
RU2290441C2 |
| REN Q | |||
| ET AL | |||
| Переносная печь для варки пищи и отопления в окопах, походных помещениях и т.п. | 1921 |
|
SU3A1 |
| Приспособление для соединения пучка кисти с трубкою или втулкою, служащей для прикрепления ручки | 1915 |
|
SU66A1 |
| Очаг для массовой варки пищи, выпечки хлеба и кипячения воды | 1921 |
|
SU4A1 |
| Устройство для выемки отдельных труб в экономайзерах | 1925 |
|
SU1311A1 |
| FUKUI T | |||
| ET AL | |||
| Engineering of Ralstonia eutropha for production of | |||

