Настоящее изобретение относится к клетке, стабильно культивируемой в Среде, где клетка адаптирована для продукции олигосахаридов, клетка трансформирована таким образом, что она содержит по меньшей мере одну последовательность нуклеиновых кислот, кодирующую фермент, задействованный в синтезе олигосахарида.
Такие клетки известны, например, из Dumon и др., 2001.
Давно известно, что грудное молоко у людей содержит, кроме лактозы, комплексную смесь олигосахаридов, называемую олигосахариды грудного молока (Human Milk Oligosaccharides (HMO)). Эта фракция олигосахаридов материнского молока является уникальной по своему составу и качеству. В отличие от других млекопитающих, грудное молоко у людей содержит олигосахариды в концентрации от 7 до 12 г/л, что в 10-100 раз выше, чем у большинства других млекопитающих (Boehm & Stahl, 2007, Kunz и др., 2000, Newburg & Neubauer, 1995).
В настоящее время, структурно было охарактеризовано более 80 соединений, относящихся к НМО. В целом, НМО характеризуются, в отличие от других олигосахаридов, обнаруженных в организме человека, лактозным компонентом на восстанавливающем конце, и фукозой и/или сиаловой кислотой на невосстанавливающем конце.
Различают два основных типа: Олигосахариды со структурой I типа имеют фукозу α1,4-связанную с GlcNAc, тогда как олигосахариды со структурой II типа проявляют α1,3-фукозилирование GlcNAc или глюкозы; любой тип может содержать α1,2-связанную фукозу с галактозой. Наиболее важными олигосахаридами являются 2'-фукозиллактоза и 3-фукозиллактоза.
Эти структуры являются близкородственными эпитопам гликоконъюгатов поверхности эпителиальных клеток, гисто-антигенов групп крови Льюиса, таких как Льюис x (Lex(Gal(β1-4)[Fuc-(α1-3)]GlcNAc(β1)) (Newburg, 2001). Структурная гомология НМО с эпителиальными этипопами обуславливает защитные свойства по отношению к бактериальным патогенам.
Например, вирулентность патогенных Escherichia coli (Cravioto и др., 1991), Vibrio cholerae (Coppa и др., 2006), Streptococcus pneumoniae (Andersson и др., 1986) или Campylobacterjejuni (Ruiz-Palacios и др., 2003) может быть значительно уменьшена путем связывания патогенов с НМО вместо гликанов поверхности слизистой оболочки человека, и также связывания токсинов, подобных термостабильному энтеротоксину Е. coli (Crane и др., 1994).
Кроме указанных местных эффектов в желудочно-кишечном тракте, НМО также способны вызывать системные эффекты у младенцев при поступлении в большой круг кровообращения (Gnoth и др., 2001). Влияние НМО на белок-углеводные взаимодействия, например, связывание селектин-лейкоцит может модулировать иммунные ответные реакции и уменьшать воспалительные ответные реакции (Bode, 2006, Kunz & Rudloff, 2006).
Для уменьшения частоты инфекций, обнаруживаемых у новорожденных и, следовательно, также детской смертности в течение первого года жизни, несомненно, является чрезвычайно важным в целом обеспечить новорожденных питанием, включающим НМО. Это может быть легко осуществимо в сообществах, где грудное вскармливание является общепринятым и широко распространено. Тем не менее, это не является обычной практикой.
Существует много медицинских причин, таких как возможная передача инфекционных заболеваний от матери к ребенку, которые при определенных условиях выступают против грудного вскармливания. Во многих странах Африки, например, грудное вскармливание может быть основной причиной ВИЧ-инфицирования в грудном возрасте.
Также культурные особенности могут приводить к отмене грудного вскармливания, как в случаях в больших промышленно-развитых странах, таких как, например, США.
Вследствие того факта, что НМО могут быть обнаружены в естественных источниках, таких как в молоке других млекопитающих, только в низких концентрациях, экстрагирование олигосахаридов из природных источников неприемлемо для удовлетворения потребности в НМО.
Химический синтез олигосахаридов является трудоемким и требует многих стадий защиты и снятия защиты (Kretzschmar & Stahl, 1998), следовательно, в целом является относительно дорогостоящим и характеризуется низкой воспроизводимостью.
Таким образом, ферментативное получение олигосахаридов с использованием биотехнологически сконструированных организмов является перспективным альтернативным источником для синтеза НМО в промышленном масштабе.
За последнее десятилетие, было опубликовано несколько успешных попыток синтеза НМО, используя либо ферментацию с рекомбинантными Е. coli или ферментативное превращение in vitro. Эти попытки главным образом были сконцентрированы на синтезе фукозилированных соединений, относящихся к или очень сходных с НМО.
Например, в нескольких публикациях описан синтез структур Льюиса лакто-N-неофукопентаозы, лакто-N-неодифукогексаозы и лакто-N-неодифукооктаозы, а также 2′- и 3-фукозиллактозы (Albermann и др., 2001, Dumon и др., 2006, Dumon и др., 2001, Dumon и др., 2004, Koizumi и др., 2000). В этих случаях, ферментативное фукозилирование эдуктов, таких как, например, лактоза, осуществляют с помощью фукозилтрансферазы (FucTs).
Большинство публикаций, описывающих получение фукозилированных соединений, в этой связи описывает применение FucTs, имеющих происхождение из Helicobacter pylori. В целом, FucTs человека также можно использовать для этой цели. Тем не менее, при сверхэкспрессии в бактериальных клетках, FucTs из бактериальных источников менее подвержены таким проблемам, как неправильная укладка и нерастворимость.
Кроме того, большинство опубликованных систем для синтеза фукозилированных соединений основываются на эндогенном пуле GDP-фукозы Е. coli, который обычно используется для синтеза фукозо-содержащей экзополисахаридной колониевой кислоты (Grant и др., 1970). В этих случаях, доступность GDP-фукозы является, очевидно, критическим элементом, ограничивающим эффективность синтеза.
Недавно, изобретатели настоящего изобретения описали новый цельноклеточный процесс получения, используя фермент Fkp (Parkot и др., 2008), полное содержание этой патентной заявки включено в данную заявку путем ссылки.
Fkp (Coyne и др., 2005), который имеет происхождение из Bacteroidesfragilis, представляет собой бифункциональный фермент, который обладает как фукокиназной, так и L-фукоза-1-Р-гуанилилтрансферазной активностью. В данном случае, экзогенно поставляемая фукоза сначала фосфорилируется и затем нуклеотид-активируется с образованием важной молекулы предшественника GDP-фукоза. «Реутилизационный» путь L-фукозы на основе Fkp успешно используется для синтеза фукозилированных олигосахаридов (Parkot и др., 2008).
Даже при том, что на уровне биохимического синтеза как такового, были разработаны важные стратегии, известные методы для синтеза in vivo олигосахаридов все несут определенные преимущества, ингибируя, в большинстве случаев, массовую продукцию олигосахаридов.
Реальная сложность при продукции, в клетках, олигосахаридов с высокой производительностью, с одной стороны, состоит в массивном внутриклеточном обогащении продуцируемых олигосахаридов и нуклеотидов побочными продуктами, и, с другой стороны, экстрагирование продуцированных олигосахаридов.
Вследствие внутриклеточного обогащения, продукты реакций синтеза могут постепенно развивать продукт-ингибирующее действие на синтетические ферменты. В определенной точке, синтез таким образом становится неэффективно медленным. Кроме того, продукты могут достигать цитотоксических концентраций, вызывая лизис клеток или по меньшей мере метаболическую останову. В любом случае, не представляется возможной непрерывная внутриклеточная продукция олигосахаридов.
Кроме того, можно ожидать, что накопление избыточных количеств олигосахаридов в конечном итоге будет приводить к лизису клеток и клеточной гибели. Этот клеточный лизис или позже осуществленный клеточный лизис, используемый для экстрагирования синтезированных олигосахаридов из клетки, будет приводить к комплексной смеси целевых олигосахаридов и клеточных компонентов (метаболиты, дебрис). Очистка целевого олигосахарида из этой комплексной смеси является дорогостоящей и, таким образом, для большинства олигосахаридов, экономически не выгодной.
Учитывая эти обстоятельства, биотехнологическая продукция олигосахаридов является чрезвычайно неэффективной и тяжело поддается контролю, в особенности, поскольку известную биотехнологическую продукцию олигосахаридов осуществляют, используя периодическое культивирование, такое периодическое культивирование является, с экономической точки зрений, чрезвычайно неудовлетворительным.
С учетом изложенного выше, задачей изобретения является улучшение способов биотехнологической продукции олигосахаридов таким образом, чтобы облегчить продукцию и сделать ее более контролируемой, и в такой способ в целом увеличить выход олигосахаридов.
В соответствии с изобретением, эта и другие задачи решаются путем обеспечения типа клетки, указанной в начале, где клетка дополнительно трансформирована таким образом, что она содержит по меньшей мере одну последовательность нуклеиновых кислот, кодирующую белок семейства эффлюксного переносчика сахара, ее функциональный гомолог или производное.
Таким образом задача изобретения решается полностью.
В соответствии с изобретением, под олигосахаридами поминают короткие полимеры моносахаридов, содержащие по меньшей мере 2 субъединицы сахара. Олигосахариды могут быть либо разветвленными или образовывать линейную цепь из субъединиц. Кроме того, сахарные субъединицы олигосахаридов могут характеризоваться количеством химических модификаций. Таким образом, олигосахариды в соответствии с настоящим изобретением могут содержать один или больше несахарных компонентов.
Последовательность нуклеиновых кислот, в соответствии с изобретением, описывает генетический код, представленный полимером нуклеиновых кислот, таким как, например, полимер дезоксирибонуклеиновых кислот или полимер рибонуклеиновых кислот. Генетический код, таким образом, может включать кодирующие последовательности, включая информацию для формирования белка, или некодирующие участки, содержащие, например, промоторные участки, участки для прикрепления регуляторных или вспомогательных соединений, спейсеры и структурные последовательности, оказывающие влияния на вторичную или третичную структуру самого полимера нуклеиновых кислот и/или участия в его процессинге.
″Трансформированы таким образом, что содержат″, в соответствии с настоящим изобретением, относится к любому методу инсерции по меньшей мере одной дополнительной последовательности нуклеиновых кислот в клетки, последовательность нуклеиновых кислот после этого присутствует внутри клетки либо в виде плазмиды или интегрирована в хромосому/хромосомы клеток. Известные методы трансформации включают, например, химическую трансформацию или электропорацию. Стабильный трансгенез, даже при отсутствии селективных агентов, может в целом достигаться путем хромосомной интеграции по меньшей мере одной дополнительной последовательности нуклеиновых кислот. Для этого, клетка может инфицироваться вирусом или фагом. Альтернативно, можно применять другие способы гомологичной и негомологичной рекомбинации, например, системы на основе вирусов или транспозонов.
Для биотехнологических применений, экспорт более крупных олигосахаридов из клетки является комплексной проблемой. Это является одной из основных проблем, поскольку было идентифицировано только несколько клеточных механизмов, относящихся к такому транспорту. Причиной редких случаев экспорта олигосахаридов из клетки является тот факт, что синтез олигосахарид потребляет большое количество клеточных ресурсов. Следовательно, потеря этих компонентов в целом неблагоприятна для клетки.
Кроме того, большинство известных механизмов для экспорта олигосахаридов задействует химические модификации олигосахаридов, например, путем их связывания с липидными компонентами (Alaimo и др. 2006). Вследствие этого, олигосахариды являются не только мембранно-связанными, ингибируя их высвобождение в среду, а также химически связанными с липидными компонентами, уменьшая их растворимость в водной среде. Следовательно, такие механизмы плохо подходят для применения для получения олигосахаридов в промышленном масштабе.
Семейство эффлюксного переносчика сахара (SET), впервые описанное Liu с соавторами (Liu и др., 1999а) для Е. coli, содержит белки SetA, SetB и SetC. Гомологи (аминокислотная идентичность >50%) белков переносчика обнаружена изначально в Enterobacteriaceae (Liu и др. 1999а).
Кроме глюкозы и лактозы, SET экспортерные белки проявляют субстратную специфичность к определенным моно- и ди-сахаридам, а также, например, к индукторной молекуле изопропил-β-D-тиогалактозид (IPTG) и токсическому аналогу сахара о-нитрофенил-β-D-тиогалактозид (ONPG) (Liu и др., 1999b). Тем не менее, биохимические исследования показали, что например, SetA проявляет очень низкую до нуля транспортную активность для больших или великообъемных молекул, таких как гептозы или трисахариды.
Учитывая вышеизложенные причины, не можно было предполагать, что экспортерные белки семейства SET будут пригодны вообще для транспорта олигосахаридов.
Тем не менее, изобретателями было обнаружено, неожиданно, что, сверхэкспрессия SET экспортерных белков приводит к чрезвычайно эффективному экспорту олигосахаридов.
Кроме того, было показано, что SET белки экспортируют лактозу, один из эдуктов реакций синтеза. Следовательно, можно предположить, что синтез олигосахаридов, полученных в модифицированных клетках, будет осуществляться чрезвычайно неэффективно и медленно вследствие постоянного отвода эдукта из клеток.
В отличие от этого, изобретатели смогли показать впервые, что, несмотря на сверхэкспрессию SET экспортерных белков, синтез олигосахаридов, полученных в модифицированных клетках, является чрезвычайно продуктивным.
В целом, является предпочтительным, если клетку выбирают из группы, включающей клетки бактерий, грибов, животных и растений. Следовательно, особенно предпочтительно, когда клетка представляет собой клетку Escherichia coli.
Преимуществом в этом случае является то, что клетки Е. coli обладают высокой метаболической активностью и высоким коэффициент воспроизводства. Дополнительно, Е. coli является одним из наиболее хорошо охарактеризованных организмов для молекулярно-биологических и биотехнологических целей. Многие техники для трансформации и культивирования бактерий, известные в данной области, были специально адаптированы для Е. coli. Дополнительно, коммерчески доступны штаммы Е. coli с различным генетическим фоном.
Кроме того, является предпочтительным, если фермент выбирают из группы, включающей гликозилтрансферазу, гликозилтрансферазу Leioir типа, гликозилтрансферазу не-Leloir типа, сиалилтрансферазу, галактозилтрансферазу, фукозилтрансферазу, маннозилтрансферазу, N-ацетилглюкозаминилтрансферазу, N-ацетилгалактозаминилтрансферазу.
В соответствии с одним вариантом осуществления, фермент представляет собой фукозилтрансферазу.
Кроме того, является предпочтительным, если эффлюксный переносчик сахара представляет собой SetA или его производное.
Преимуществом в этом случае является то, что SetA, из биохимически охарактеризованных представителей семейства SET экспортерных белков, характеризуется наибольшей субстратной специфичностью. В этом случае, наибольшее разнообразие олигосахаридов может, по крайней мере потенциально, экспортироваться и таким образом продуцироваться, используя SetA.
Также является предпочтительным, если клетка дополнительно транформирована таким образом, что она содержит по меньшей мере одну последовательность нуклеиновых кислот, кодирующую белок, облегчающий или промотирующий импорт эдуктов, необходимых для синтеза олигосахарида.
Следовательно, является благоприятным, что концентрация эдуктов внутри клетки повышается. В нормальных условиях, импорт эдуктов, таких как фукоза или лактоза, ограничивается доступностью соответствующих импортерных белков. Однако, в том случае, когда клетка модифицируется для синтеза олигосахаридов в промышленном масштабе, импорт эдуктов, рассчитанный на эндогенных уровнях импортерных белков, может являться недостаточным для постоянного пополнений реакций синтеза эдуктами. Эти проблему можно решить путем сверхэкспрессии соответствующих импортерных белков.
Значимые импортерные белки в данном случае представляют собой главным образом импортеры для моно- или дисахаридов, такие как импортеры лактозы, например, Е. coli β-галактозид пермеаза (LacY), или импортеры фукозы, например Е. coli фукоза пермеаза (FucP), а также могут содержать импортеры для нуклеотидов и других эдуктов.
Является особенно предпочтительным, если клетка трансформирована таким образом, что она содержит по меньшей мере одну последовательность нуклеиновых кислот, кодирующую белок, выбранный из группы, включающей переносчик лактозы, переносчик фукозы, переносчик сиаловой кислоты, переносчик галактозы, переносчик маннозы, N-ацетилглюкозамин переносчик, N-ацетилгалактозамин переносчик, АВС-переносчик, переносчик для нуклеотид-активированного сахара и переносчик для нуклеинового основания, нуклеозида или нуклеотида.
В этой связи, нуклеотид-активированный сахар может представлять собой, но не ограничиваясь только ими, GDP-фукозу, СМР-сиаловую кислоту, UDP-галактозу, UDP-глюкозу, GDP-маннозу, UDP-глюкозамин или UDP-галактозамин.
Кроме того, термин нуклеиновое основание обозначает гуанин, цитозин, аденин, тимин и урацил. Нуклеозид обозначает гуанозин, цитидин, аденозин, тимидин и уридин, тогда как нуклеотид может представлять собой моно-, ди- или трифосфат гуанозина, цитидина, аденозина, тимидина или уридина.
Дополнительно, является предпочтительным, если клетка дополнительно трансформирована таким образом, что она содержит по меньшей мере одну последовательность нуклеиновых кислот, кодирующую белок, выбранный из группы, включающей нуклеотидилтрансферазу, гуанилилтрансферазу, уридилилтрансферазу, Fkp, L-фукозокиназу, фукоза-1-фосфат гуанилилтрансферазу, синтетазу СМР-сиаловой кислоты, галактозокиназу, галактоза-1-фосфат уридилилтрансферазу, глюкозокиназу, глюкозу-1-фосфат уридилилтрансферазу, маннозокиназу, манноза-1-фосфат гуанилилтрансферазу, GDP-4-кето-6-дезокси-D-манноза редуктазу, глюкозамин киназу, глюкозамин-фосфат ацетилтрансферазу, N-ацетил-глюкозамин-фосфат уридилилтрансферазу, UDP-N-ацетилглюкозамин 4-эпимеразу, UDP-N-ацетилглюкозамин 2-эпимеразу.
В этой связи, термин нуклеотидил трансфераза в целом относится к ферментам, которые могут переносить нуклеотиды на фосфосахара, которые представляют собой встречающиеся в природа сахара или искусственные сахара.
Преимущество здесь состоит в том, как описано также в Parkot и др. 2008, включенную в настоящую заявку путем ссылки, что внутриклеточные пулы нуклеотид-активированных Сахаров, таких как GDP-фукоза, могут быть пополнены. В данном случае, синтез олигосахаридов становится более эффективным.
Кроме того, в целом является предпочтительным, катаболический путь клетки для выбранных моно-, ди- или олигосахаридов, задействованных в и/или необходимых для синтеза олигосахаридов, по меньшей мере частично инактивирован.
В данном случае, преимущество заключается в том, что суммарная эффективность синтеза может быть повышена. Это является важным обстоятельством, поскольку меньше эдуктов, предназначенных для синтеза олигосахаридов, расходуется при эндогенном метаболизме клетки.
В варианте осуществления изобретения, описанном ниже, например, применяются клетки, которые являются дефицитными по разложению лактозы. Это может осуществляться путем фермента β-галактозидазы, который кодируется геном lacZ. Эта генетическая модификация предотвращает внутриклеточное расщепление лактозы на быстро метаболизируемые моносахариды глюкозу и галактозу. Таким образом, лактоза присутствует в более высокой концентрации в качестве акцепторной молекулы для последующих реакций гликозилирования/фукозилирования.
В альтернативном варианте осуществления, клетки, используемые для синтеза олигосахаридов, являются, самостоятельно или дополнительно к дефициту lacZ, описанному выше, дефицитны по распаду L-фукозы. Это можно осуществлять путем инактивации гена fucA, кодирующего ключевой катаболический фермент фукулоза-1-фосфат альдолаза (FucA) пути распада фукозы.
Несомненно, также доступны другие техники, от ферментативных ингибиторов до стабильно трансфектированных конструктов и РНК, которые могут применяться по меньшей мере для частичной инактивации катаболических путей.
В объеме настоящего изобретения, в целом является предпочтительным, если олигосахариды содержат по меньшей мере три субъединицы и/или характеризуются молекулярным весом по меньшей мере приблизительно 480 г/моль.
Все важные НМО имеют молекулярный вес выше указанного предела.
Настоящее изобретение также охватывает способ продукции олигосахаридов, который включает стадии:
a) обеспечение клетки в соответствии с изобретением,
b) культивирование клетки в среде в условиях, предоставляющих возможность продукции указанных олигосахаридов,
c) экстрагирование указанных олигосахаридов из культуральной среды.
В объеме настоящего изобретения, под «условиями, предоставляющими возможность для» понимают условия, относящиеся к физическим или химическим параметрам, включая, но не ограничиваясь только ими, температуру, рН, давление, осмотическое давление и концентрацию продукта/эдукта.
В предпочтительном варианте осуществления изобретения, «условия, предоставляющие возможность для» могут включать температуру в диапазоне 30+/-20°С, значение рН в диапазоне 7+/-3.
Способ, описанный выше, обеспечивает преимущество, состоящее в том, что олигосахариды могут быть экстрагировали непосредственно из культуральной среды, тогда как для других способов необходим лизис клеток и последующая экстракция олигосахаридов из полученного лизата.
В этой связи, является предпочтительным, если стадию b) осуществляют, используя биореактор с непрерывным потоком.
Преимущество состоит в том, что при использовании биореактора с непрерывным потоком, количество продуцируемых олигосахаридов легко может быть повышено. Это обусловлено тем, что синтез происходит непрерывно на относительно высоком уровне.
Также является предпочтительным, если среда на стадии b) содержит одно или несколько веществ, выбранных из группы, включающей основные добавочные компоненты для поддержания роста и размножения клеток, селективные агенты, эффекторы активности генов и эдукты, необходимые для синтеза олигосахаридов.
Типические добавочные компоненты для поддержания роста и размножения клеток широко известны из уровня техники и описаны, например, в Sambrook и Russel, 2001. Такие основные добавочные компоненты отвечают за общую потребность культивируемых клеток в питательных веществах, и включают, например, белки, углеводы, липиды и минералы.
Также из уровня техники известны селективные агенты, такие как антибиотики, например, из Sambrook и Russel, 2001. Такие агенты можно использовать для защиты культур генетически модифицированных клеток от остаточного загрязнения конкурирующими микроорганизмами, такими как грибы или бактерии. Дополнительно, например, в бактериальных популяциях, селективные агенты могут использоваться для стабилизации генетической информации, содержащейся в плазмиде.
Эффекторы активности генов можно использовать для селективной индукции или репрессии активности определенных генов или установки генов в клетке. Такие эффекторы включают от простых химических соединений, таких как изопропил-1-тио-β-D-галактопиранозид (IPTG) до более комплексных соединений, таких как гормоны. Такие эффекторы активности генов хорошо известны из уровня техники, например, из Sambrook и Russel, 2001.
Кроме того, является предпочтительным, если эдукты выбирают из группы, включающей арабинозу, треозу, эритрозу, рибозу, рибулозу, ксилозу, глюкозу, D-2-дезокси-2-амино-глюкозу, N-ацетилглюкозамин, глюкозамин, фруктозу, маннозу, галактозу, N-ацетилгалактозамин, галактозамин, сорбозу, фукозу, N-ацетилнейраминовую кислоту, гликозид, искусственный сахар, нуклеиновое основание, нуклеозид, нуклеотид и любой возможный их ди- или полимер.
В целом является предпочтительным, если олигосахариды содержат по меньшей мере три субъединицы и/или характеризуются молекулярным весом по меньшей мере приблизительно 480 г/моль.
В соответствии с вариантом осуществления настоящего изобретения, олигосахарид, продуцируемый с помощью способа, описанного выше, представляет собой фукозиллактозу, преимущество в этом случае заключается в том, что фукозиллактоза является одним из наиболее перспективных соединений, присутствующих в НМО.
Дальнейшие преимущества становятся понятными из описания вариантов осуществления изобретения и приложенных фигур.
Совершенно очевидно, что вышеуказанные характерные особенности и характерные особенности, представленные ниже, можно использовать не только в соответствующих указанных комбинациях, а также в других комбинациях или самостоятельно, без отклонения от сущности заявляемого изобретения.
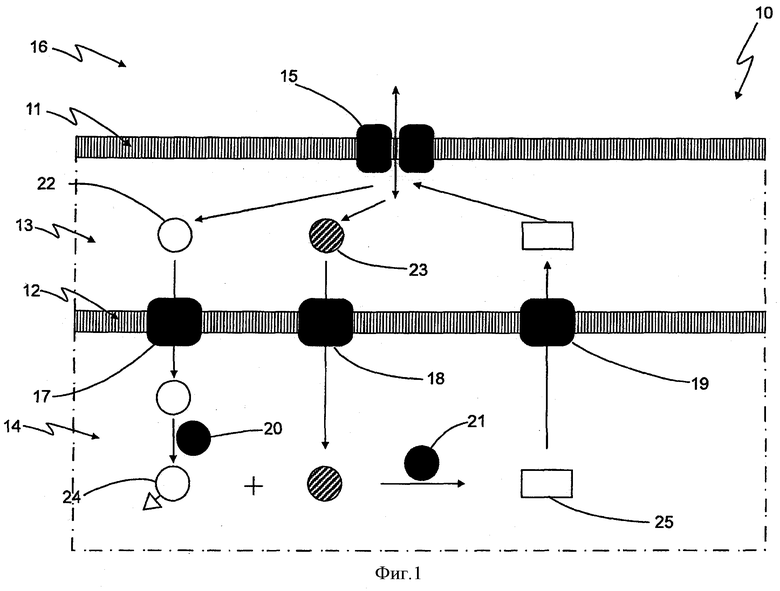
Определенные варианты осуществления изобретения иллюстрируются на фигурах и поясняются более подробно в последующем описании. На фигурах:
Фиг.1 показан схематический обзор синтеза олигосахаридов и транспорта внутри грамотрицательной бактериальной клетки, модифицированной в соответствии с изобретением;
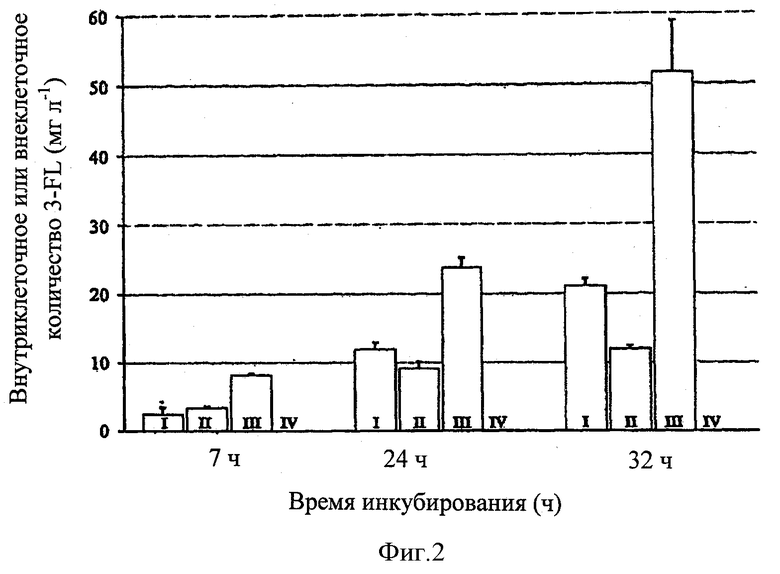
Фиг.2 представлены результаты измерений 3-фукозиллактозы в клеточной влажной массе и супернатантах бактериальных культур, сравнивая различные генотипы; и
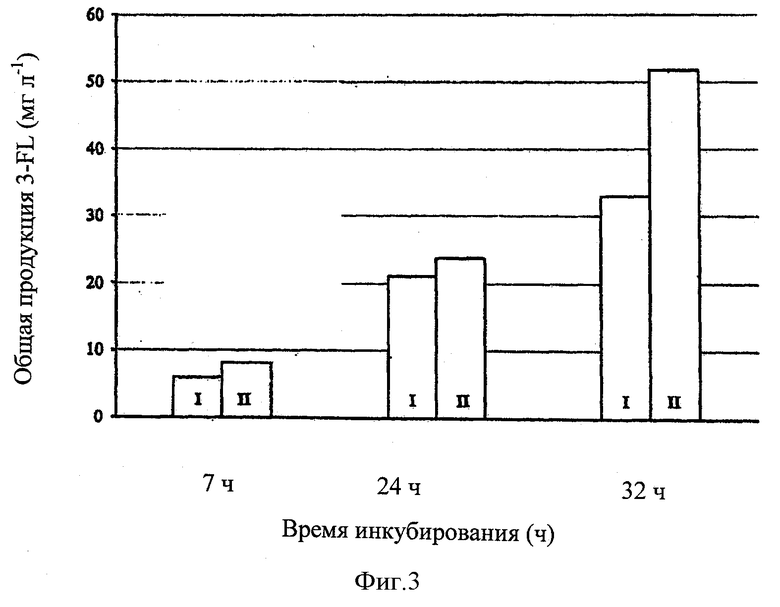
Фиг.3 представлено сравнение между количествами 3-фукозиллактозы, синтезированной в бактериальных культурах с различными генотипами.
Пример 1: Синтез олигосахаридов и транспорт внутри грамотрицательной бактериальной клетки
На фигуре 1 показа фрагмент грамотрицательной бактериальной клетки 10. Грамотрицательная бактериальная клетка 10 в соответствии с изобретением содержит внешнюю мембрану 11, плазматическую мембрану 12, периплазматическое пространство 13, расположенное между указанной внешней мембраной 11 и указанной плазматической мембраной 12, и цитозоль 14, заключенный в указанной плазматической мембране 12. Внешняя мембрана содержит порины, через которые водорастворимые соединения могут проходить из среды 16 в периплазматическое пространство 13 и наоборот.
В соответствии с одним вариантом осуществления настоящего изобретения, плазматическая мембрана содержит FucP, служащий в качестве первого эдукта-переносчик а17, LacY, служащий в качестве второго эдукта переносчика 18, и SetA, служащий в качестве экспортера продукта.
Кроме того, Fkp, служащий в качестве нуклеотидилтрансферазы 20, и FutAco, служащий в качестве гликозилтрансферазы 21, содержатся в цитозоле 14.
Когда, в соответствии с этим вариантом осуществления, фукозу, в качестве первого эдукта 22, и лактозу, в качестве второго эдукта 23, подают в среду 16, то они входят в периплазматическое пространство 13 через порин 15. Затем, первый эдукт 22 транспортируется с помощью переносчика первого эдукта 17 в цитозоль 14. В цитозоле, первый эдукт 22 модифицируется с помощью нуклеотидилтрансферазы 20, что приводит к получению нуклеотидилированного первого эдукта 24, GDP-фукозы.
Второй эдукт 23, импортируется в цитозоль 14 с помощью импортера второго эдукта 15.
Затем гликозилтрансфераза 21 катализирует реакцию между нуклеотидилированным первым эдуктом, GDP-фукоза, и вторым эдуктом, лактозой, что приводит к получению олигосахарида 25, 3-фукозиллактозы, и GDP (не показано).
После этого, олигосахарид 25 экспортируется из цитозоля 14 с помощью экспортера продукта 19 (SetA) в периплазматическое пространство 13 и может покидать периплазматическое пространство 13 через порины 15, поступая в среду 16.
Пример 2: Материалы и методы
2.1 Конструирование экспрессионных плазмид и разработка штаммов Е. coli
Е. coli JM109(DE3) (Promega; www.promega.com) использовали в качестве исходного хозяйского штамма для разработки продуцирующего штамма Е. coli. Все олигонуклеотидные праймеры, используемые для процедур клонирования, перечислены в Таблице 1. Плазмиды pACYC-lacY и pACYC-lacY-setA конструировали следующим образом: Гены lacY (в соответствии с номером доступа GenBank АСВ02461) (GenBank; www.ncbi.nlm.nih.gov) и setA (в соответствии с номером доступа GenBank YP_025293) (GenBank) амплифицировали из геномной ДНК Е. coli TOP 10 (Invitrogen; www.invitrogen.com), используя праймеры lacY NcoI прямой/lacY EcoRI обратный и setA NdeI прямой/setA XhoI обратный. ПЦР продукты подвергали рестрикционному расцеплению с помощью указанных ферментов, и лигировали с соответствующе расщепленным экспрессионным вектором pACYCDuet-1 (Novagen; www.merckbiosciences.co.uk).
Полученные плазмиды проверяли с помощью рестриктазных фрагментов, электрофореза в агарозном геле, а также секвенирования с праймерами pACYCduetUP1, DuetDOWN-1-Праймер, DuetUP2-Праймер и Т7-Терминатор-Праймер для корректной инсерции генов (данные не представлены). Используемые плазмиды pCOLA-fkp-fucP и рЕТ-futAco были сконструированы ранее (Parkot и др., 2008). Для получения штаммов JM00, JM01 и JM02, различные комбинации плазмид вводили в Е. coli JM109(DE3) путем электропорации (Dower и др., 1988). Все плазмиды и бактериальные штаммы перечислены в Таблице 2.
2.2 Инактиваыия катаболизма фукозы в Е. coli:
Для предотвращения разложения вводимой из внешних источников фукозы, ген fucA, кодирующий катаболический фермент L-фукулоза-1-фосфат альдолаза, делетировали из хромосомы Е. coli JM109 (DE3). Все олигонуклеотидные праймеры, используемые для процедур мутагенеза, перечислены в Таблице 1. Для конструирования делетированного мутанта fucA, применяли методологию Datsenko и Wanner (Datsenko и Wanner, 2000), используя праймеры fucA-knock-f и fucA-knock-r. Корректную делению fucA подтверждали с помощью ПЦР, используя праймеры fucA-контроль-f и fucA-контроль-r, фланкирующие хромосомный инсерционный сайт, и фукозо-отрицательный фенотип верифицировали путем помещения бактерии в М9 минимальный агар (Sambrook и Russell, 2001) с дополненной фукозой в качестве единственного источника углерода (данные не представлены).
ApR резидентный к ампициллину; KmR, резидентный к канамицину; CmR, резистентный к хлорамфениколу.
2.3 Условия культивирования и приготовление клеточных экстрактов
Е. coli штаммы инокулировали 1:100 из выращиваемых в течение ночи культур в 100 мл минеральной среды (Samain и др., 1999), содержащей 7,0 г л-1 NH4H2PO4, 7,0 г л-1 K2HPO4, 1,0 г л-1 MgSO4×7H2O, 0,5 г л-1 лимонной кислоты, 2,0 г л-1 КОН, 0,0045 г л-1 триамин·HCl и 7,5 мл л-1 раствора микроэлементов. Маточный раствор микроэлементов содержал 70 мМ нитрилотриацетата (рН 6,5), 7,5 г л-1 лимоннокислого железа, 1,3 г л-1 MnCl2×4H2O, 0,21 г л-1 CoCl2×6H2O, 0,13 г л-1 CuCl2×2H2O, 0,25 г л-1 Н3ВО3, 1,2 г л-1 ZnSO4×7H2O и 0,15 г л-1 Na2MoO4×2H2O. Среду дополняли 0,1% глюкозой и 1% глицерином в качестве источника углерода, и с 100 мкг мл-1 ампициллина, 50 мкг мл-1 канамицина и/или 20 мкг мл-1 хлорамфеникола перед инкубированием на роторном шейкере при 37°С, обеспечивая хорошую аэрацию.
Когда культуры достигали оптической плотности (ОП600 нм) приблизительно 1,0, индуктор изопропил-1-тио-β-D-галактопиранозид (IPTG) добавляли в концентрации 0,5 мМ, и культуры инкубировали в течение ночи при 28°С в условиях постоянного встряхивания. Приблизительно через 16 часов, добавляли 40 мМ L-фукозы и 20 мМ лактозы. После этого культуры непрерывно инкубировали при 28°С при постоянном встряхивании.
В определенные периоды времени, 20 мл образцов культур отбирали, и клетки собирали путем центрифугирования. Супернатанты культур отделяли и сразу анализировали с помощью высокоэффективной анионообменной хроматографии (НРАЕС) или хранили при -20°С. Осадки клеток после центрифугирования промывали с помощью PBS (Sambrook и Russell, 2001), ресуспедировали в дистиллированной воде в 5-ти кратном количестве по весу осадков после центрифугирования, и лизировали путем кипячения в течение 10 минут. Для получения внутриклеточных фракций, клеточный дебрис отделяли путем центрифугирования и прозрачный лизат клеток хранили при -20°С или сразу анализировали с помощью НРАЕС.
2.4 SDS-PAGE (электрофорез в полиакриламидном геле в присутствии додецилсульфата натрия)
Экспрессию гетерологичных белков проверяли с помощью SDS-PAGE (Sambrook и Russell, 2001) (данные не представлены). Белковые экстракты приготавливали в 1×SDS гель-загрузочном буфере, и полиакриламидные гели окрашивали с помощью кумасси бриллиантового голубоого.
2.5 Определение олигосахаридов с помощью высокоэффективной анионообменной хроматографии с импульсным амперометрическим детектированием (HPAEC-PAD)
Образцы анализировали с помощью высокоэффективной анионообменной хроматографии (НРАЕС), используя Decade II импульсный амперометрический детектор (PAD) (Antec Leyden; www.antec-leyden.nl) и колонку CarboPac PA20 (Dionex; www.dionex.com), присоединенную к ВЭЖХ системе (Shimadzu; www.shimadzu.eu). Чувствительность детектора устанавливала на 50 мкА с применяемым 0,05 В импульсным напряжением.
Моно-, ди-, и олигосахариды элюировали с помощью 10 мМ гидроксида натрия при скорости потока 0,4 мл мин-1. После изократического элюирования в течение 30 минут с 10 мМ NaOH колонку промывали в течение 20 минут с помощью 200 мМ NaOH для получения постоянного времени удерживания и после этого регенерировали с помощью 10 мМ NaOH в течение 20 минут. Для всех анализируемых образцов, 20 мкл 1:2 dH2O-разведенных растворов использовали для НРАЕС анализа. Анализы с помощью HPAEC-PAD показали время удерживания на используемой ВЭЖХ колонке приблизительно 3,5 минуты для стандарта L-фукоза, прибл. 15 минут для стандарта лактоза, и прибл. 11-12 минут для 3-фукозиллактозного стандарта (данные не представлены). Стандарты для веществ глицерин и глюкоза, которые добавляли в культуральную среду в качестве источника углерода, записывали со временем удерживания прибл. 1,5 минуты и 7-8 минут, соответственно.
Пример 3: Продукция 3-фукозиллактозы и SetA-зависимая секреция в культуральную среду с помощью рекомбинантных Е. coli
Задачей этого исследования был анализ SetA-опосредованного экспорта внутриклеточной 3-фукозиллактозы. Для ферментационных экспериментов использовали штаммы Е. coli JM01 и JM02 (см. Таблицу 2). Штаммы JM01 и JM02, которые оба экспрессируют ферменты Fkp и FutAco (α1,3-фукозилтрансфераза), а также белки-переносчики FucP и LacY, отличались только по экспрессии SetA переносчик. JM01 не сверхпродуцирует SetA, a JM02 сверхпродуцирует SetA.
На фигуре 2 показаны количества 3-фукозиллактозы во влажной клеточной массе и супернатанте Е. coli культур JM01 и JM02, определенные с помощью HPAEC-PAD анализа.
Для определения влияния сверхэкспрессии SetA, измерения осуществляли через 7, 24 и 32 часов после индукции экспрессии fkp, fucP, lacY, futAco и setA.
На фигуре, внеклеточные фракции 3-фукозиллактозы, измеренные в Е. coli JM01 (SetA не сверхэкспрессируется) представлены в колонках I, внутриклеточные фракции представлены в колонках II. В случае Е. coli JM02 (SetA-сверхэкспрессируется), внеклеточные фракции 3-фукозиллактозы представлены в колонках III и внутриклеточные фракции представлены в колонках IV. Все значения представляют собой средние значения из двойных экспериментов, планки погрешностей увязывают соответствующие стандартные отклонения.
В этих измерениях было показано, что штамм JM01, который не сверхэкспрессирует SetA, накапливает 3-фукозиллактозу в цитозольной фракции.
В отличие от этого, внутриклеточная концентрация 3-фукозиллактозы в случае штамма JM002, сверхэкспрессирующего setA, находится ниже предела обнаружения.
Кроме того, супернатанты из JM01- и JM02- культур проявляют определенное содержание 3-фукозиллактозы. Посредством этого, однако, содержание 3-фукозиллактозы, обнаруженное в супернатанте JM02 культуры, значительно повышено по сравнению с содержанием 3-фукозиллактозы, обнаруженном в супернатанте JM01 культуры. Тогда как через 32 ч, концентрация 3-фукозиллактозы в супернатанте JM01 составляет приблизительно 21 мг л-1, то концентрация 3-фукозиллактозы в случае JM02 составляет выше 51 мг л-1.
Сравнение общих количеств 3-фукозиллактозы в культурах Е. coli JM01 и JM02 представлено на фигуре 3.
В данном случае, общее количество 3-фукозиллактозы в Е. coli JM01 культурах представлено в колонках I и общее количество 3-фукозиллактозы в Е. coli JM02 культурах представлено в колонках II. Аналогичным образом, представлены средние значения для двойных экспериментов.
После инкубирования в течение 32 часов, штамм JM02 продуцирует 51,68 мг л-1, приблизительно на 57% больше 3-фукозиллактозы в целом, чем штамм JM01 (32,99 мг л-1).
Пример 4: Обсуждение
Экспериментальные результаты для HPAEC-PAD анализа показали, что сильные различия в синтезе и транспорте 3-фукозиллактозы существуют между штаммами JM01, без сверхэкспрессии SetA, и JM02, сверхэкспрессирующего SetA.
Начиная с того, что 3-фукозиллактоза не определяется в клеточной влажной массе JM02 культуры, тогда как более высокая концентрация 3-фукозиллактозы может быть измерена в супернатанте.
Это очевидно свидетельствует о том, что сверхэкспрессия SetA в Е. coli приводит к чрезвычайно эффективному экспорту 3-фукозиллактозы из клетки.
В данной заявке, изобретателями было показано, что SetA, в отличие от того, что можно было предполагать на основе известного уровня техники, может эффективно экспортировать большие олигосахариды, характеризирующийся, в данном случае, тремя субъединицами и молекулярным весом 488 г/моль.
В отличие от этого, во влажной клеточной массе JM01, обнаруживается значительное количество 3-фукозиллактозы, тогда как только сравнительно небольшое количество 3-фукозиллактозы присутствует в супернатанте.
Эти результаты свидетельствуют о том, что при отсутствии сверхэкспрессии SetA, 3-фукозиллактоза значительно накапливается в цитозоле бактериальных клеток. Тот факт, что также в этих условиях 3-фукозиллактозу обнаруживается в супернатанте, может, в соответствии с современным уровнем знаний, объясняться повышенным лизисом бактериальных клеток, возникающих вследствие высоких внутриклеточных концентраций 3-фукозиллактозы.
При сравнении суммарных количеств 3-фукозиллактозы в JM01- и JM02-культурах, становится очевидным, что сверхэкспрессия SetA не только приводит к повышенной концентрации 3-фукозиллактозы в супернатанте, но также повышает общее количество синтезированной 3-фукозиллактозы (см. Фигуру 3).
Такое повышение общей эффективности синтеза может быть обусловлено более высокой жизнеспособностью клеток, которые сверхэкспрессируют SetA, жизнеспособность которых будет являться результатом избегания цитотоксичности продукта. Альтернативно или дополнительно, повышение общей эффективности синтеза также может быть обусловлено избеганием ингибирующих эффектов продукта на синтетические ферменты вследствие экспорта продукта, обусловленного сверхэкспрессией SetA.
В данной заявке, изобретателями было показано, что сверхэкспрессия SetA переносчика является эффективным путем для усиления выхода синтеза олигосахаридов, выполняемого с помощью биотехнологических методов, использующих культивируемые клетки. Кроме того, можно практически полностью избежать ингибирования продуктом релевантных синтетических ферментов и цитотоксичности продукта синтеза, продукция облегчается и становится более контролируемой.
Перечень ссылок
Alaimo, С., Catrein, I., Morf, L., Marolda, C.L., Callewaert, N., Valvano, M.A., Feldman, M.F. & M. Aebi, (2006) Two distinct but interchangeable mechanisms for flipping oflipid-linked oligosaccharides. EMBO J 25(5):967-976.
Albermann, C., W. Piepersberg & U.F. Wehmeier, (2001) Synthesis of the milk oligosaccharide 2'-fucosyllactose using recombinant bacterial enzymes. Carbohydr Res 334: 97-103.
Andersson, В., O. Porras, L.A. Hanson, T. Lagergard & C. Svanborg-Eden, (1986) Inhibition of attachment of Streptococcus pneumoniae and Haemophilus influenzae by human milk and receptor oligosaccharides. J Infect Dis 153: 232-237.
Bode, L., (2006) Recent advances on structure, metabolism, and function of human milk oligosaccharides. JNutr 136:2127-2130.
Boehm, G. & B. Stahl, (2007) Oligosaccharides from milk. J Nutr 137: 847S-849S.
Coppa, G.V., L. Zampini, T. Galeazzi, B. Facinelli, L. Ferrante, R. Capretti & G. Orazio, (2006) Human milk oligosaccharides inhibit the adhesion to Caco-2 cells of diarrea pathogens: Escherichia coli, Vibrio cholerae, and Salmonella fyris. Pediatr Res 59: 377-382.
Coyne, M.J., B. Reinap, M.M. Lee & L.E. Comstock, (2005) Human symbionts use a host-like pathway for surface fucosylation. Science 307: 1778-1781.
Crane, J.K., S.S. Azar, A. Stam & D.S. Newburg, (1994) Oligosaccharides from human milk block binding and activity of the Escherichia coli heat-stable enterotoxin (STa) in T84 intestinal cells. J Nutr 124: 2358-2364.
Cravioto, A., A.Tello, H. Villafan, J. Ruiz, S. del Vedovo & J.R. Neeser, (1991) Inhibition of localized adhesion of enteropathogenic Escherichia coli to HEp-2 cells by immunoglobulin and oligosaccharide fractions of human colostrum and breast milk. J Infect Dis 163: 1247-1255.
Datsenko, К.A., and B.L. Wanner. 2000. One-step inactivation of chromosomal genes in Escherichia coli K-12 using PCR products. Proc Nati Acad Sci USA 97:6640-5.
Dower, W.J., J.F. Miller, and C.W. Ragsdale. 1988. High efficiency transformation of E. coli by high voltage electroporation. Nucleic Acids Res 16:6127-45.
Dumon, C, C. Bosso, J.P. Utille, A. Heyraud & E.Samain, (2006) Production of Lewis x tetrasaccharides by metabolically engineered Escherichia coli. Chembiochem 7:359-365.
Dumon, C, B. Priem, S.L. Martin, A. Heyraud, C. Bosso & E. Samain, (2001) In vivo fucosylation of lacto-N-neotetraose and lacto-N-neohexaose by heterologous expression of Helicobacter pylori alpha-1,3 fucosyltransferase in engineered Escherichia coli. Glycoconj J 18:465-474.
Dumon, C, E.Samain & B. Priem, (2004) Assessment of the two Helicobacter pylori alpha-1,3-fucosyltransferase ortholog genes for the large-scale synthesis of LewisX human milk oligosaccharides by metabolically engineered Escherichia coli. Biotechnol Prog 20:412-419.
Gnoth, M.J., S. Rudloff, C. Kunz & R.K. Kinne, (2001) Investigations of the in vitro transport of human milk oligosaccharides by a Caco-2 monolayer using a novel high performance liquid chromatography-mass spectrometry technique. J Biol Chem 276:34363-34370.
Grant, W.D., I.W. Sutherland & J.F. Wilkimson, (1970) Control ofcolanic acid synthesis. J Bacterial 103:89-96.
Koizumi, S., Т. Endo, K. Tabata, H. Nagano, J. Ohnishi & A. Ozaki, (2000) Large-scale production of GDP-fucose and Lewis X by bacterial coupling. J Ind Microbiol Biotechnol 25:213-217.
Kretzschmar, G. & W. Stahl, (1998) Large scale synthesis of linker-modified sialyl-Lewis(X), Lewis(X) and N-acetyllactosamine. Tetrahedron 54:6341-6358.
Kunz, С. & S. Rudloff, (2006) Health promoting aspects of milk oligosaccharides. Int Dairy 716:1341-1346.
Kunz, C., S. Rudloff, W. Baier, N. Klein & S. Strobel, (2000) Oligosaccharides in human milk: structural, functional, and metabolic aspects. Annu Rev Nutr 20: 699-722.
Liu, J.Y., P.F. Miller, M. Gosink & E.R. Olson, (1999a) The identification of a new family of sugar efflux pumps in Escherichia coli. Mol Microbiol 31:1845-1851.
Liu, J.Y., P.F. Miller, J. Willard & E.R. Olson, (1999b) Functional and biochemical characterization of Escherichia coli sugar efflux transporters. Journal of Biological Chemistry 214:22977-22984.
Newburg, D.S., (2001) Bioactive components of human milk: evolution, efficiency, and protection. Adv Exp Med Biol 501:3-10.
Newburg, D.S. & S.H. Neubauer, (1995) Carbohydrates in milk. In: Handbook of Milk Composition. R.G. Jensen (ред.). San Diego, CA: Academic Press, cc. 273-349.
Parkot, J., E. Hiifner & S. Jennewein, (2008) Synthesis of fucosylated compounds. Европейская патентная публикация ЕР 08172267.
Ruiz-Palacios, G.M., L.E. Cervantes, P. Ramos, B. Chavez-Munguia & D.S. Newburg, (2003) Campylobacterjejuni binds intestinal H(O) antigen (Fuc alpha 1, 2 Gal beta 1, 4 GlcNAc), and fucosyloligosaccharides of human milk inhibit its binding and infection. JBiol Chem 278:14112-14120.
Samain, E., V. Chazalet & R.A. Geremia, (1999) Production of O-acetylated and sulfated chitooligosaccharides by recombinant Escherichia coli strains harboring different combinations of nod genes. J Biotechnol 72:33-47.
Sambrook, J. & D.W. Russell, (2001) Molecular cloning: a laboratory manual. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY.


Изобретение относится к биотехнологии, в частности к бактериальной клетке, стабильно культивируемой в среде, где клетка адаптирована для продукции олигосахаридов. Причем клетка трансформирована таким образом, что она содержит по меньшей мере одну последовательность нуклеиновых кислот, кодирующую фермент, задействованный в синтезе олигосахарида. При этом дополнительно клетка трансформирована таким образом, что она содержит по меньшей мере одну последовательность нуклеиновых кислот, кодирующую белок семейства эффлюксного переносчика сахара или его функциональный гомолог. Кроме того, изобретение относится к способу продукции олигосахаридов, задействующего вышеуказанную клетку. Изобретение позволяет получать олигосахариды с высокой степенью эффективности. 2 н. и 11 з.п. ф-лы, 3 ил., 2 табл., 4 пр.
1. Бактериальная клетка, стабильно культивируемая в среде, где клетка адаптирована для продукции олигосахаридов, клетка трансформирована таким образом, что она содержит по меньшей мере одну последовательность нуклеиновых кислот, кодирующую фермент, задействованный в синтезе олигосахарида, которая характеризуется тем, что бактериальная клетка дополнительно трансформирована таким образом, что она содержит по меньшей мере одну последовательность нуклеиновых кислот, кодирующую белок семейства эффлюксного переносчика сахара или его функциональный гомолог.
2. Бактериальная клетка по п.1, отличающаяся тем, что клетка представляет собой клетку Escherichia coli.
3. Бактериальная клетка по п.1 или 2, отличающаяся тем, что фермент выбирают из группы, включающей гликозилтрансферазу, гликозилтрансферазу Leloir типа, гликозилтрансферазу не-Leloir типа, фукозилтрансферазу, сиалилтрансферазу, галактозилтрансферазу, маннозилтрансферазу, N-Ацетилглюкозаминилтрансферазу, N-Ацетилгалактозаминилтрансферазу.
4. Бактериальная клетка по п.3, отличающаяся тем, что фермент представляет собой фукозилтрансферазу.
5. Бактериальная клетка по любому из пп.1-4, отличающаяся тем, что эффлюксный переносчик сахара представляет собой SetA.
6. Бактериальная клетка по любому из пп.1-5, отличающаяся тем, что она дополнительно трансформирована таким образом, что она содержит по меньшей мере одну последовательность нуклеиновых кислот, кодирующую белок, облегчающий или промотирующий импорт эдуктов, необходимых для синтеза олигосахарида, где белок выбран из группы, включающей переносчик лактозы, переносчик фукозы, переносчик сиаловой кислоты, переносчик галактозы, переносчик маннозы, переносчик N-ацетилглюкозамина, переносчик N-ацетилгалактозамина, АВС-переносчик, переносчик для нуклеотид-активированного сахара и переносчик для нуклеинового основания, нуклеозида или нуклеотида.
7. Бактериальная клетка по любому из пп.1-6, отличающаяся тем, что она дополнительно трансформирована таким образом, что она содержит по меньшей мере одну последовательность нуклеиновых кислот, кодирующую белок, выбранный из группы, включающей нуклеотидилтрансферазу, гуанилилтрансферазу, уридилилтрансферазу, Fkp, L-фукозокиназу, фукоза-1-фосфат гуанилилтрансферазу, синтетазу СМР-сиаловой кислоты, галактозокиназу, галактоза-1-фосфат уридилилтрансферазу, глюкозокиназу, глюкозу-1-фосфат уридилилтрансферазу, маннозокиназу, манноза-1-фосфат гуанилилтрансферазу, GDР-4-кето-6-дезокси-D-манноза редуктазу, глюкозамин киназу, глюкозамин-фосфат ацетилтрансферазу, N-ацетил-глюкозамин-фосфат уридилилтрансферазу, UDP-N-ацеталглюкозамин 4-эпимеразу, UDP-N-ацетил-глюкозамин 2-эпимеразу.
8. Бактериальная клетка по любому из пп.1-7, отличающаяся тем, что олигосахариды содержат по меньшей мере три субъединицы и/или характеризуются молекулярным весом по меньшей мере приблизительно 480 г/моль.
9. Способ продукции олигосахаридов, который включает стадии:
а) обеспечение бактериальной клетки по любому из пп.1-8,
b) культивирование бактериальной клетки в среде в условиях, предоставляющих возможность продукции указанных олигосахаридов,
c) экстрагирование указанных олигосахаридов из культуральной среды.
10. Способ по п.9, отличающийся тем, что стадию b) осуществляют, используя биореактор с непрерывным потоком.
11. Способ по п.9 или 10, отличающийся тем, что среда на стадии b) содержит эдукты, необходимые для синтеза олигосахаридов, где эдукты выбирают из группы, включающей арабинозу, треозу, эритрозу, рибозу, рибулозу, ксилозу, глюкозу, D-2-дезокси-2-амино-глюкозу, N-ацетилглюкозамин, глюкозамин, фруктозу, маннозу, галактозу, N-ацетилгалактозамин, галактозамин, сорбозу, фукозу, лактозу, N-ацетилнейраминовую кислоту, гликозид, искусственный сахар, нуклеиновое основание, нуклеозид, нуклеотид и любой возможный их ди- или полимер.
12. Способ по любому из пп.9-11, отличающийся тем, что олигосахариды содержат по меньшей мере три субъединицы и/или характеризуются молекулярным весом по меньшей мере приблизительно 480 г/моль.
13. Способ по любому из пп.9-12, отличающийся тем, что олигосахарид представляет собой фукозиллактозу.
| Прибор, замыкающий сигнальную цепь при повышении температуры | 1918 |
|
SU99A1 |
| DUMON C et.al | |||
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| Glycoconj J | |||
| Перекатываемый затвор для водоемов | 1922 |
|
SU2001A1 |
| LIU JY et.al | |||
| Functional and biochemical characterization of Escherichia coli sugar efflux | |||

