Настоящее изобретение относится к способам получения фукозилированных соединений и связанных с ними клеток.
Женское молоко, состоящее из сложной смеси углеводов, белков, липидов, гормонов и микроэлементов, обеспечивает всеми необходимыми питательными веществами развитие младенцев. Кроме того, женское молоко содержит некоторые защитные вещества. Помимо иммуноглобулинов женское молоко содержит набор сложных олигосахаридов, обладающих защитными свойствами. Фракция олигосахаридов женского молока (НМО) включает, помимо основного углеводного компонента лактозы, более 130 различных сложных олигосахаридов. Это структурное разнообразие сложных олигосахаридов и присутствие их в большом количестве характерно только для человека. Напротив, в коровьем молоке обнаруживают только следовые количества значительно более сложных олигосахаридов, и вследствие этого в обычно применяемой молочной смеси не хватает указанных олигосахаридов.
Данные клинических исследований показывают, что у вскармливаемых грудью младенцев реже наблюдаются случаи диареи, респираторных заболеваний и отита среднего уха по сравнению с младенцами, вскормленными молочной смесью. Длительное время указанные защитные эффекты женского молока приписывали присутствию секретируемых иммуноглобулинов, однако, в настоящее время установлено, что НМО могут представлять собой главную линию защиты от патогенов для вскармливаемых грудью младенцев.
Многие из сложных НМО демонстрируют гомологию с гликоконъюгатами клеточной поверхности, такими как антиген тканей и групп крови системы Льюис × (Lex), Gal(β1-4)[Fuc-(α1-3)]GlcNAc(β1) (Newburg, 2001), который часто служит в качестве рецепторов патогенов. Таким образом, экскретируя растворимые ловушки, имитирующие структуры гликоконъюгатов клеточной поверхности, природа создала здесь эффективный механизм предупреждения инфекций. Например, было показано, что НМО могут радикально снижать вирулентность патогенных Escherichia coli (Cravioto et al., 1991), Vibrio cholerae (Coppa et al., 2006), Streptococcus pneumoniae (Andersson et al., 1986) или Campylobacter jejuni (Ruiz-Palacios et al., 2003), а также способны нейтрализовать токсины, подобные термостабильному энтеротоксину E.coli (Crane et al., 1994). Помимо упомянутых местных эффектов в кишечнике, НМО, попадая в системный кровоток, также способны вызывать системные эффекты у младенцев (Gnoth et al., 2001).
Влияние НМО на белково-углеводные взаимодействия, например на связывание селектин-лейкоцит, может модулировать иммунные ответы и снижать воспалительные ответы (Bode, 2006, Kunz & Rudloff, 2006).
Сложные олигосахариды представляют собой третий по величине компонент женского молока, после лактозы и жиров. Практически все они в совокупности имеют лактозу на восстанавливающем конце и несут фукозу и/или сиаловую кислоту на невосстанавливающем конце. Они построены из моносахаридов от 3 вплоть до 32 единиц, и большинство из них содержит фукозу от 1 до 15 единиц фукозы. Таким образом, фукозилированные олигосахариды демонстрируют высокий потенциал в качестве биоактивных компонентов продуктов питания с противоинфекционными и пребиотическими свойствами.
Фукозилтрансферазы (FucT), катализирующие перенос остатков фукозы от донора, гуанозин-дифосфат-активированной L-фукозы (GDP-L-фукоза), к некоторым молекулам-акцепторам, экспрессируются в животных, растениях, грибах и бактериях (Ма et al., 2006). Их характеризуют по месту присоединения фукозы, следовательно, различают α1,2, α1,3/4 и α1,6 FucT. Помимо FucT человека, которая исходно ответственна за биосинтез НМО и антигенов группы крови, описано несколько бактериальных FucT. Активность FucT наилучшим образом исследована для желудочного патогена человека Helicobacter pylori, у которого к его липополисахариду (LPS) присоединены фукозосодержащие антигены Льюиса (Wang et al,, 2000). Точная роль указанных структур антигенов Льюиса в процессе инфекции Н. pylori не ясна, но обсуждают роль молекулярной мимикрии в осуществлении обмана иммунной системы хозяина, адгезии и заселении (Bergman et al., 2006).
Благодаря мощному потенциалу НМО, выступающих в качестве способствующих укреплению здоровья биологически активных добавок, существует серьезная заинтересованность в их рентабельном широкомасштабном производстве. Получение с помощью катализа в процессах бактериальной ферментации представляет собой наиболее выгодный способ по сравнению с экстракцией НМО из женского молока и химическим синтезом, который представляет собой трудоемкий процесс и требует многочисленных стадий защиты и снятия защиты (Kretzschmar & Stahl, 1998). За последние десять лет были сообщения о нескольких попытках синтеза НМО с применением как ферментации с рекомбинантной E.coli, так и ферментативного превращения in vitro (Albermann et al., 2001, Dumon et al., 2006, Dumon et al., 2001, Dumon et al., 2004, Koizumi et al., 2000). Однако доступность донора, сахаронуклеотида GDP-фукозы, представляет собой узкое место в получении фукозилированных олигосахаридов. Эту высокоэнергетическую молекулу нельзя в настоящее время эффективно или рентабельно получить с помощью химического или энзиматического синтеза. Большинство публикаций сообщают о системах производства фукозилированных соединений на основе пула эндогенной GDP-фукозы E.coli, который, однако, чрезвычайно ограничен и применим только для индуцируемого синтеза содержащего фукозу экзополисахарида, колановой кислоты (Grant etal., 1970).
Например, Albermann et al. (2001) применили в энзиматическом синтезе рекомбинантные ферменты. GDP-β-L-фукозу получают путем превращения GDP-D-маннозы в ООР-4-кето-6-дезокси-D-маннозу. Полученное соединение под действием ОВР-4-кето-6-дезокси-D-манноза-3,5-эпимеразы-4-редуктазы превращают в GDP-β-L-фукозу, которую очищают с помощью препаративной HPLC.
Другой подход, предложенный Koizumi и соавторами для синтеза LeX из N-ацетиллактозамина (LacNAc), включает следующую комбинацию: образования GTP из добавленной GMP с помощью Corynebacterium ammoniagenes, синтез GDP-фукозы через GDP-маннозу и фукозилирование LacNAc путем оверэкспрессии α1,3-FucT H. pylori в разных штаммах E.coli (Koizumi et а1., 2000). Поскольку для этого подхода, связанного с бактериями, необходимо применять пермеабилизацию и, следовательно, уничтожение клеток, то не существует возможности с помощью указанного выбранного подхода осуществить продолжительный и крупномасштабный ферментационный процесс.
Все еще существует потребность в способах получения фукозилированных соединений, которые помогли бы преодолеть, по меньшей мере, некоторые из недостатков существующего уровня техники.
Одно из воплощений изобретения представляет собой способ получения генетически модифицированной клетки, обладающей способностью продуцировать фукозилированные соединения, включающий следующие стадии:
- трансформирование клетки для экспрессии фукозокиназы;
- трансформирование клетки для экспрессии фукозо-1-фосфатгуанилилтрансферазы;
- трансформирование клетки для экспрессии фукозилтрансферазы.
В соответствии со способом изобретения получают генетически модифицированную клетку. Ее трансформируют для экспрессии фукозокиназы, фукозо-1-фосфатгуанилилтрансферазы и фукозилтрансферазы.
Способы введения генов в клетку известны специалистам.
В предпочтительном воплощении, генетически модифицированная клетка представляет собой микроорганизм, которые выбирают из группы, состоящей из родов Escherichia, Klebsiella, Helicobacter, Bacillus, Lactobacillus, Streptococcus, Lactococcus, Pichia, Saccharomyces и Kluyveromyces.
В предпочтительном воплощении изобретения, активность фукозакиназы и фукозо-1-фосфатгуанилилтрансферазы объединяют в бифункциональном ферменте. Подходящие гены для трансформации, кодирующие фукозокиназу, фукозо-1-фосфатгуанилилтрансферазу, и/или бифункциональные гены, кодирующие фукозокиназу/фукозо-1-фосфатгуанилилтрансферазу, могут быть получены из родов Bacteroides, Lentisphaera, Ruminococcus, Solibacter, Arabidopsis, Oryza, Physcomitrella, Vitis, Danio, Bos, Equus, Macaco, Pan, Homo, Rattus, Mus и Xenopus.
Подходящие гены фукозилтрансферазы могут быть получены из организмов, которые выбирают из группы родов Helicobacter, Escherichia, Yersinia, Enterococcus, Shigella, Klebsiella, Salmonella, Bacteroides, Dictyostelium, Arabidopsis, Drosophila, Homo, Bos, Mus, Rattus, Gallus, Canis и Sus.
В зависимости от источника гена и клетки, примененной для экспрессии, для усиления экспрессии может быть полезна оптимизация кодонов.
Некоторые клетки имеют катаболический путь для фукозы. В этом случае рекомендуется инактивировать этот катаболический путь. Подходящие способы включают инактивацию одного или нескольких генов, которые выбирают из группы, состоящей из гена фукозо-1-фосфат-альдолазы, гена фукозоизомеразы и гена фуколозокиназы.
Подходящие соединения, производные фукозы, которые могут быть получены с помощью генетически модифицированных клеток настоящего изобретения, представляют собой фукозиллактозы, предпочтительно 2'-фукозиллактозу, 3-фукозиллактозу или лактодифукотетраозу.
Настоящее изобретение представляет собой синтез в клетке, который начинается с фукозы, вместо препаративного синтеза с рекомбинантными ферментами, начинающегося с GDP-D-маннозы, как описано у Albermann et al. (2001).
Дополнительное воплощение изобретения представляет собой генетически модифицированные клетка, получаемая по способу изобретения. Для получения фукозилированных соединений генетически модифицированную клетку изобретения культивируют в подходящих условиях культивирования в среде, включающей фукозу и субстрат-акцептор.
Подходящие субстраты-акцепторы представляют собой, например, моно-, ди- или олигосахарид или пептид, например, лактозу, 2'-фукозиллактозу или 3-фукозиллактозу.
Предпочтительные фукозилированные соединения, полученные с помощью способа получения, представляют собой фукозиллактозы, предпочтительно 2'-фукозиллактозу или 3-фукозиллактозу или лактодифукотетраозу.
В этом документе впервые сообщают об эффективном синтезе GDP-фукозы в E.coli из добавленной извне L-фукозы и, таким образом, о создании фукозного «реутилизационного» пути в E.coli. Однако этот подход также применен к другим легко культивируемым организмам, представляющим интерес для пищевой или фармацевтической промышленности (например, к Lactobacillus spp.). Применение указанного недавно открытого пути открывает совершенно новые перспективы получения олигосахаридов, помимо 2'-фукозиллактозу и 3-фукозиллактозу, в которых нет необходимости зависеть от дорогостоящего и трудоемкого получения GDP-фукозы (in vitro) или эндогенных, сложно регулируемых, биосинтетических путей образования GDP-фукозы (in vivo).
В так называемом «пути реутилизации фукозы» фукозу исходно фосфорилируют до фукозо-1-фосфата под действием фермента фукозокиназы. Фукозо-1-фосфат превращают в GDP-фукозу под действием фермента фукозо-1-Р-гуанилилтрансферазы. Недавно был описан первый бактериальный фермент, Fkp, обладающий обеими активностями, фукозокиназы и L-фукозо-1-Р-гуанилилтрансферазы (Coyne et al., 2005). Кишечная бактерия Bacteroides fragilis использует этот фермент для продукции GDP-фукозу, которая служит для навешивания остатков фукозы на полисахариды и гликопротеины капсулы.
Краткое описание чертежей
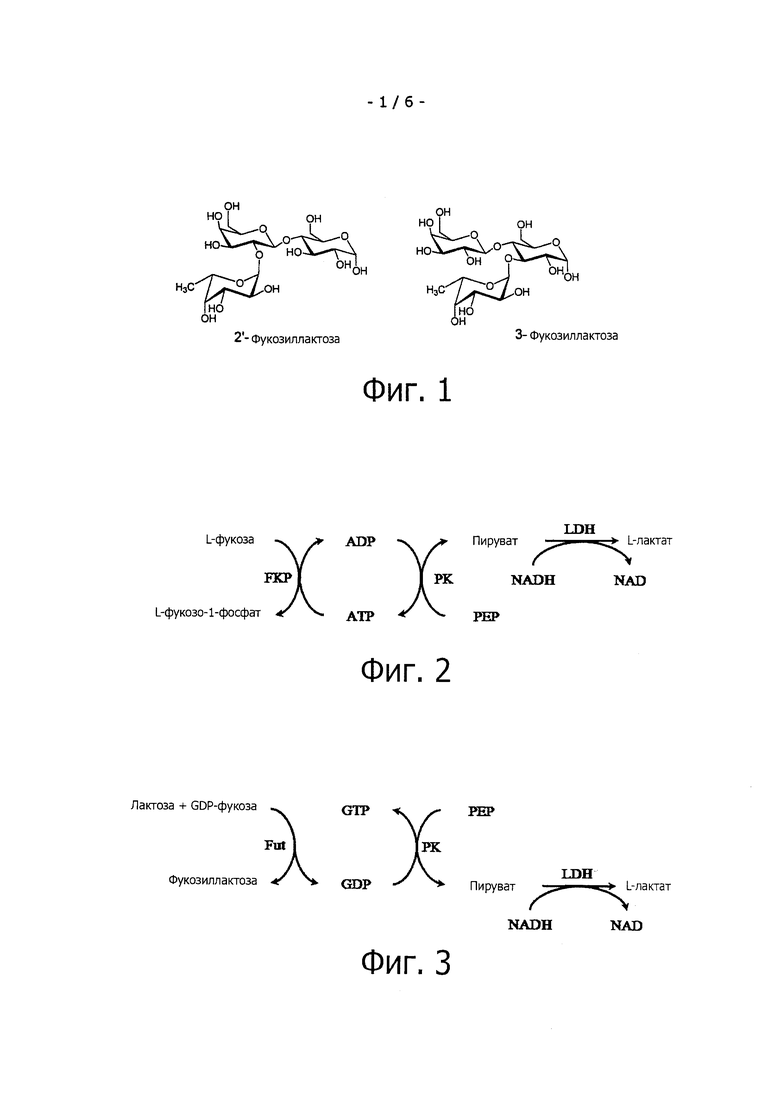
Фиг.1 раскрывает структуры исключительно сложных олигосахаридов женского молока (НМО) 2'-фукозиллактозы и 3-фукозиллактозы.
На фиг.2 приведена схема фотометрического анализа для определения активности Fkp в ферментативных реакциях, сопряженных с окислением NADH; Fkp = бифункциональная фукозокиназа / фукозо-1-фосфатгуанилилтрансфераза, РК=пируваткиназа, LDH = L-лактатдегидрогеназа, PEP=фосфоенолпируват.
На фиг.3 приведена схема фотометрического анализа для определения активности FucT в ферментативных реакциях, сопряженных с окислением NADH; NADH; FucT = фукозилтрансфераза, РК = пируваткиназа, LDH = L-лактатдегидрогеназа, PEP = фосфоенолпируват.
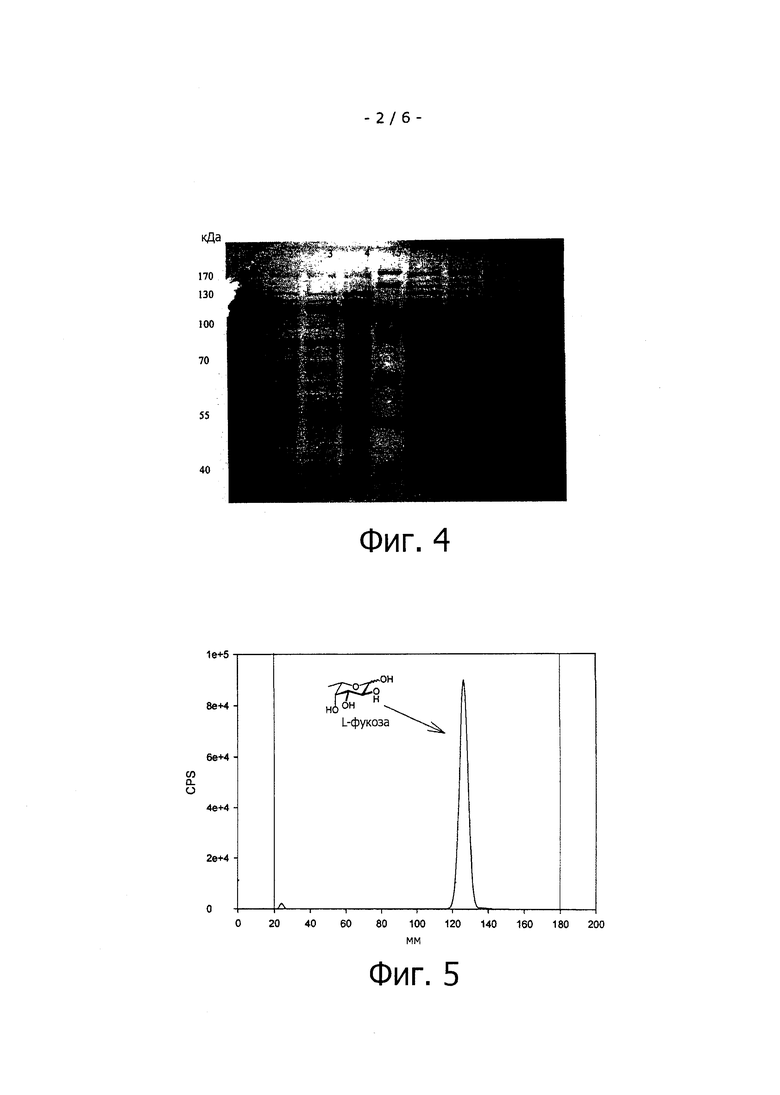
На фиг.4 показано образование белка после индукции. Дорожки 1-4: экспрессия растворимого Fkp (105,7 kDa) и/или FutAco (49,3 kDa) или FucT2 (35,9 kDa), в неочищенных экстрактах E.coli BW25113 bfucA (DE3) pCOLA-fkp-fucP (дорожка 1), Е.coli BW25113 bfucA (DE3) pET-futAco (дорожка 2), E. coli BW 25113 bfucA (DE3) pCOLA-fkp-fucP + pETfutAco (дорожка 3) и E. coli BW 25113 ΔfucA (DE3) pCOLA-fkp-fucP + pCAW55 (дорожка 4); дорожка 5: предварительно окрашенный белковый маркер «PageRuler™» («Fermentas», Германия); дорожки 6-9: экспрессия нерастворимого Fkp и/или FutAco или FucT2, в клеточном дебрисе, ресуспендированном в 6 М мочевине, из E. coli BW25113 ΔfucA (ВЕ3) pCOLA-fkp-fucP (дорожка 6), E. coli BW 25113 ΔfucA (DE3) pET-futAco (дорожка 7), E. coli BW 25113 ΔfucA (DE3) pCOLA-fkp-fucP + pETfotAco (дорожка 8) и E. coli BW 25113 ΔfucA (DE3) pCOLA-fkp-fucP + pCAW55 (дорожка 9).
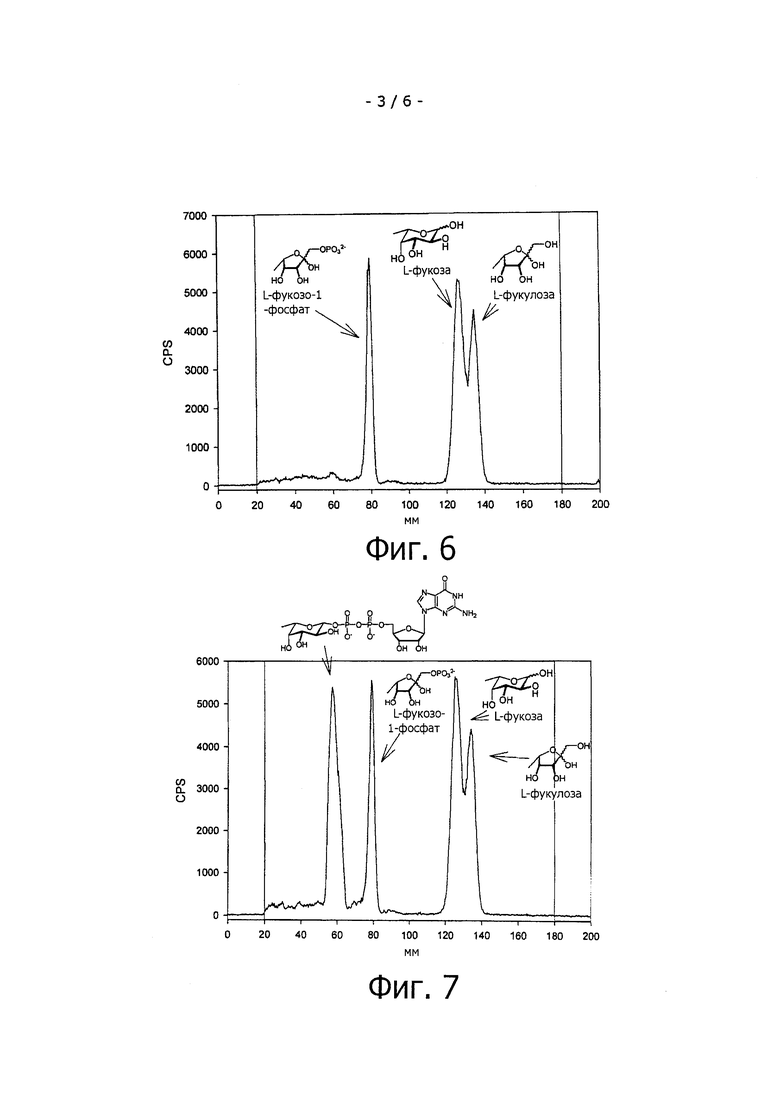
На фиг.5 показана тонкослойная хроматография с радиометрическим детектированием (радио-TLC) 3Н-фукозы, проведенная в системе бутанол:ацетон:уксусная кислота:вода (35:35:7:23) и проанализированная с помощью радио-TLC детектора.
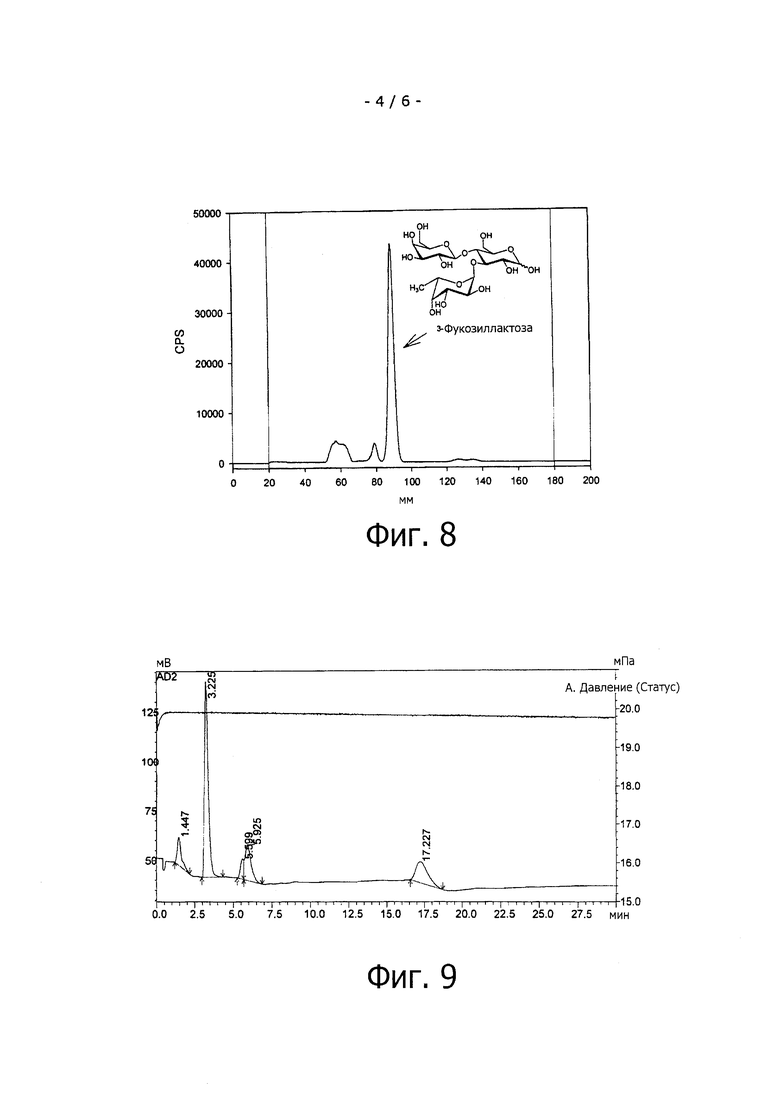
На фиг.6 показана радио-ТLС клеточного экстракта из E. coli BW 25113 ΔfucA (DE3) pCOLADuet-1 pETDuet-1, демонстрирующая фукозу и фукулезу и фукулозо-1-фосфат, однако, распад фукулозо-1-фосфаа ингибируется из-за генного нокаута (гена фукулозо- 1-фосфат-альдолазы (fucA}.
На фиг.7 показана радио-ТLС клеточного экстракта из E. coli BW 25113 ΔfucA (DE3) pCOLA-fkp-fucP, демонстрирующая накопление GDP-фукозы, продуцируемой бифункциональной фукозокиназой / фукозо-1-фосфатгуанилилтрансферазой, Fkp, из Bacteroides fragilis, а также фукозы и продуктов деградации фукулозы и фукулозо-1-фосфата.
На фиг.8 показана радио-TLC клеточного экстракта из E. coli BW 25113 ΔfucA (DE3) pCOLA-fkp-fucP pET-futAco, демонстрирующая накопление 3-фукозиллактозы, продуцируемой фукозилтрансферазой Helicobacter pylori с оптимизированным кодоном через GDP-фукозу, обеспечиваемую бифункциональной фукозокиназой / фукозо-1-фосфатгуанилилтрансферазой (Fkp). Фукоза и продукты деградации фукулозы и фукулозо-1-фосфата присутствуют только в минимальном количестве; количество GDP-фукозы существенно снижено вследствие продукции 3-фукозиллактозы.
На фиг.9 показан HPAED-анализ клеточного лизата из негативного контрольного штамма E. coli BW 25113 ΔfucA (DE3) pCOLADuet-1 pETDuet-1, демонстрирующий наличие внутриклеточной L-фукозы, лактозы, глицерина и L-рамнозы, но отсутствие фукозиллактозы.
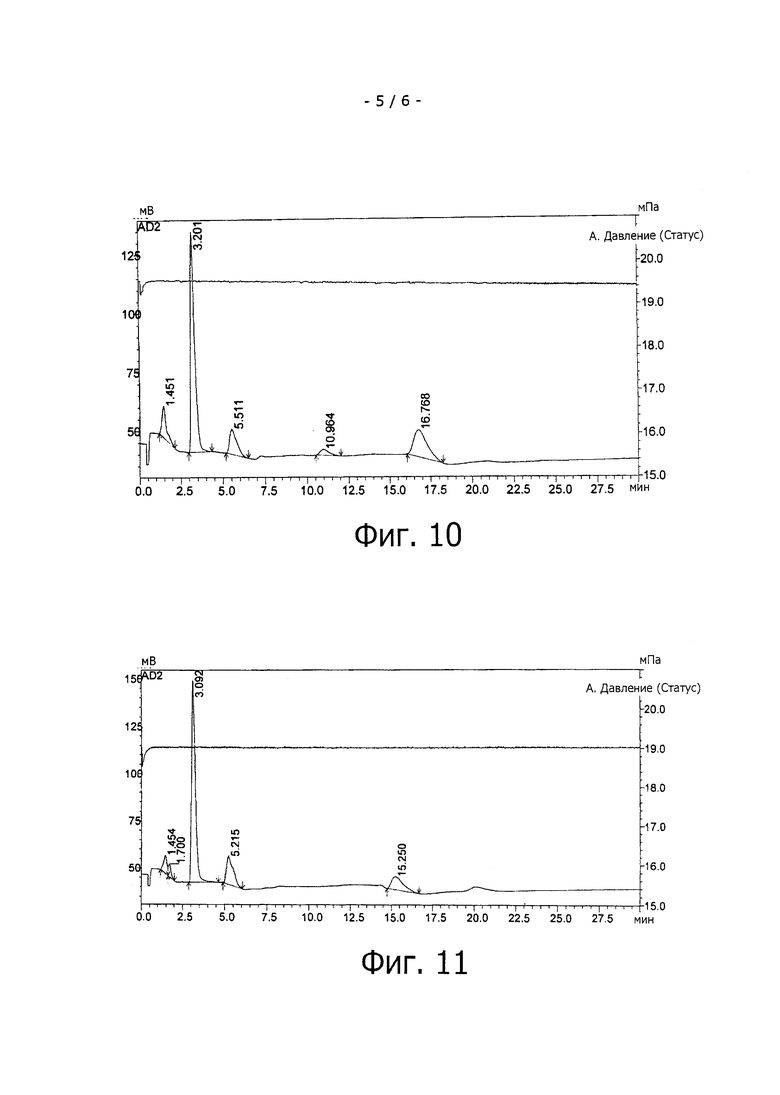
На фиг.10 показан клеточный лизат штамма E. coli BW 25113 ΔfucA (DE3) pCOLA-fkp-fucP pET-futAco, продуцирующий 3-фукозиллактозу (время удержания равно примерно 11 мин); кроме того, можно видеть пики L-фукозы, лактозы, глицерина и L-рамнозы.
На фиг.11 показан HPAED-анализ клеточного лизата из штамма E.coli BW 25113 ΔfucA (DE3) pCOLA-fkp-fucP pCAW55, демонстрирующий продукцию 2'-фукозиллактозы (время удержания равно примерно 22 мин). Кроме того, можно видеть L-фукозу, лактозу, глицерин и L-рамнозу.
На фиг.12а и b показан HPLC-анализ с электрохимической регистрацией экспрессии GDP-фукозы в E.coli JM109 (ВЕ3) ΔfucA (фиг.12а) и E.coli JM109 (DE3) ΔfucA pCOLA-fkp-fucP (фиг.12b).
Примеры
Это изобретение дополнительно объяснено следующими, нелимитирующими примерами:
Пример 1
Конструирование экспрессирующих плазмид и получение продуцирующих штаммов
Для успешного предотвращения деградации добавленной извне фукозы ген fucA, кодирующий ключевой фермент катаболизма, фукулозо-1-фосфат-альдолазу, должен быть удален из генома штамма BW 25113 E.coli. Для создания делеции fucA применяют методику, предложенную в работе (Datsenko & Wanner, 2000). Для экспрессии гетерологичного гена, с помощью Т7-промотора, индуцибельную Т7 RNA-полимеразу включают в несущий делецию штамм E.coli BW 25113 ΔfucA с помощью λDE3 набора для лизогенизации («Novagen»). Полученный штамм после этого называют E.coli BW 25113 ΔfucA (ВЕ3). Плазмиды pCOLA-fkp-fucP и pET-futAco конструируют с помощью pCOLADuet-1 и pETDuet-1 векторов экспрессии («Novagen»). Все праймеры, примененные для конструирования, перечислены в таблице 2. Ген fkp (код доступа в Банке генов AY 849806) амплифицируют с помощью PCR с праймерами flcp-NcoI-прямой и fkp-NotI-обратный, применяя геномную DNA Bacteroides fragilis ATCC 25285D. Ген fucP (код доступа в Банке генов СР 000948) Escherichia coli К12 амплифицируют из геномной DNA E.coli ТОР10 («Invitrogen», США), применяя праймеры FucP-NdeI-прямой и FucP-XhoI-обратный. Оба. fkp nfucP вставляют в первый и второй сайт множественного клонирования (MCS) pCOLADuet-1, соответственно, применяя указанные сайты рестрикциии. Полученную плазмиду обозначают как pCOLA-fkp-fucP. Ген futA (код доступа в Банке генов АЕ 000511) штамма 26695 Н. pylori был кодон-оптимизирован для экспрессии в E.coli и получен синтетическим способом в компании «GenScript Corporation» («Piscataway», NJ, США). Ген амплифицируют, применяя праймеры FutAco-Ncol-прямой и FutAco-BamHI-обратный, и вставляют в первый MCS pETDuet-1, получая pET-fritAco. Правильность вставки клонированных генов проверяют с помощь ретрикционного анализа и секвенирования с помощью рекомендованных праймеров pACYCDuetUPl, pET-Upstream, DuetDOWN-1, DuetUP2 и T7-Terminator, перечисленных в руководстве по применению векторов Duet «Duet Vectors Manual» («Novagen»). Плазмида pCAW55, содержащая ген fucT2, который кодирует α1,2-фукозилтрансферазу из Helicobacter pylori NCTC364 была предоставлена С. Albermann (Институт микробиологии, Университет Штутгарта) и основана на векторе pJOE2702 (Stumpp et al., 2000). Ген fucT2 был вставлен через сайты рестрикции NdeI/PstI и находился под контролем L-рамнозо-индуцибельного промотор rhaРBАD. Клетки E.coli BW 25113 ΔfucA (DE3) трансформируют вектором экспрессии путем электропорации (Dower et al., 1988). Все бактериальные штаммы, примененные в этом эксперименте, перечислены в таблице 1.
pET-futAco
GGAATCCCTTATCAGATAACG
Пример 2
Условия культивирования и получение клеточных экстрактов Штаммы E.coli инокулируют в 10 мл 2xYT бульона (Sambrook & Russell, 2001), содержащего 100 мкг мл-1 ампициллина и/или 50 мкг мл-1 канамицина, и инкубируют в течение ночи в ротационном шейкере при 37°С. На следующий день, в 30 мл свежего 2xYT бульона, дополненного соответствующими антибиотиками, инокулируют 1/100 из выросшей в течение ночи культуры, и инкубируют при 37°С в ротационном шейкере с хорошей аэрацией. Когда оптическая плотность (ОD600 нм) культуры достигает значения, равного приблизительно 0,5, добавляют индукторы: изопропил-1-тио-β-D-галактопиранозид (IPTG) и/или L-рамнозу, в концентрации, равной 0,1 мМ и 0,1% соответственно. Культуры дополнительно инкубируют при 28°С в течение ночи (приблизительно 15 ч) при постоянном перемешивании. Для измерения активности фотометрическим способом отбирают аликвоту культуры клеток, клетки осаждают и ресуспендируют в пятикратном (масса/объем) 50 мМ Tris-HCl pH 7,5. Добавляют стеклянные шарики, по массе в четыре раза превышающие осадок клеток, и полученную суспензию перемешивают на вортексе два раза, каждый раз в течение пяти минут, и в промежутках хранят на льду в течение дополнительных пяти минут. Дебрис клеток удаляют центрифугированием (13200 rpm, 5 мин, 4°С) и полученный неочищенный экстракт хранят 4°С.
Для продуцирования фукозиллактозы in vivo, клетки промывают одним объемом, по отношению к объему культуры, солевого раствора с фосфатным буфером рН 7,4 (PBS) (Sambrook & Russell, 2001) и ресуспендируют в 30 мл модифицированной минеральной среды М9; по стандартной для М9 прописи (Sambrook & Russell, 2001), добавляют следующие соединения: 20 мМ L-фукозу, 20 мМ лактозу, 0,5% глицерин, 0,5 мМ гуанозин и 1x GIBCO MEM раствор витаминов (100Х) («Invitrogen», США). Индукторы, L-рамнозу (0,1%) и IPTG (0,1 мМ), также добавляют ко всем культурам не зависимо от того, какой штамм культивируют, для того, чтобы избежать различий в условиях культивирования. Снова, культуры инкубируют при 28°С в течение ночи (приблизительно 15 ч) при постоянном перемешивании. Культуры центрифугируют и супернатанты декантируют и хранят при -20°С. Клетки последовательно промывают с PBS, ресуспендируют в дистиллированной воде, и пермеабилизуют с помощью автоклавирования (100°С, 5 мин). Для удаления клеточного дебриса, образцы центрифугируют (8500 rpm, 30 мин) и прозрачный клеточный лизат хранят при -20°С.
Пример 3
SDS-PAGE (SDS-электрофорез в полиакриламидном геле)
Экспрессию гетерологичных белков контролируют с помощью SDS-PAGE (Sambrook & Russell, 2001). Экстракты белков готовили в 1x SDS-буфере для нанесения на гель, и полиакриламидные гели прокрашивали Кумаси бриллиантовым синим.
Пример 4
Энзиматические фотометрические анализы
Пример 4а
Для определения активности Fkp, фукозокиназную активность фермента измеряют по количеству ADP, возникающего из АТР, используемого в качестве субстрата пируваткиназой (РК) в процессе дефосфорилирования фосфоенолпирувата (PEP), при этом полученный пируват затем превращается в L-лактат под действием L-лактатдегидрогеназы (LDH), что сопровождается поглощением NADH. Соответствующие реакции суммированы на фиг.2. Реакцию проводили в 1000 мкл 65 мМ MOPS-буфера (рН 7,5) содержащего 10 мМ L-фукозу, 15 мМ PEP, 5 мМ MgSO4, по 0,2 мМ каждого АТР и NADH, и 5 U каждого РК и LDH. После добавления 25 мкл неочищенного экстракта, окисление NADH до NAD регистрируют по убыли поглощения при 340 нм, с помощью спектрофотометра V-630 Bio (JASCO GmbH, Германия).
Пример 4b
Аналогично, активность FucT (как показано на фиг.3) измеряют по появлению GDP (из донора GDP-L-фукозы), который фосфорилируется до GTP под действием РК при превращении PEP в пируват. LDH катализирует конечную реакцию восстановления пируват до L-лактата, сопровождающуюся поглощением NADH. Клеточные экстракты (25 мкл) тестируют в 1000 мкл реакционной смеси, содержащей 10 мМ лактозы, 100 мкМ GDP-L-фукозу, 5 мМ MgSO4, по 0,2 мМ каждого АТР и NADH, и по U каждого РК и LDH в 50 мМ Tris-HCl буфере (рН 7,5). Убыль NADH регистрируют при 340 нм.
Пример 5
Анализ олигосахаридов
Образцы анализируют с помощью высокоэффективной анионообменной хроматографии (HPAED), применяя импульсный амперометрический детектор «Decade II» от компании «Antec Leyden» (Нидерланды) и колонку «CarboPac PA20» («Dionex», Германия), соединенную системой HPLC («Shimadzu», Германия). Чувствительность детектора устанавливают на 50 мкА с приложенным импульсным потенциалом, равным 0,05 V. Моно-, ди- и олигосахариды элюируют 10 мМ гидроксидом натрия со скоростью элюции, равной 0,4 мл мин-1. После 30 мин изократического элюирования 10 мМ NaOH колонку промывают в течение 20 мин 200 мМ NaOH для получения постоянных времен удерживания и после этого регенерируют 10 мМ NaOH в течение 20 мин.
Пример 6
Введение Н-фукозы
Клетки E.coli BW 25113 ΔfucA (DE3) трансформируют векторами pCOLADuet-1, pETDuet-1, pCOLA-fkp-fucP и pET-futAco для создания следующих штаммов:
E.coli BW 25113 ΔfucA (DE3) pCOLADuet-1 pETDuet-1;
E.coli BW 25113 ΔfucA (DE3) pCOLA-fkp-fucP;
E.coli BW 25113 ΔfucA (DE3) pCOLA-fkp-fucP pET-futAco.
В экспериментах по введению, штамм E.coli BW 25113 ΔfucA (DE3) pCOLADuet-1 pETDuet-1 служит в качестве пустого вектора-контроля. Все три штамма затем применяют в экспериментах по введению меченой тритием фукозы. Для экспериментов по введению, клетки культивируют в 3 мл среды 2xYT, содержащей 20 мкл L-5,6-3Н-фукозы (40-60 Ci/ммоль и 1 MCi/мл), 50 мМ лактозу и 1 мМ IPTG. В соответствии с примененными векторами экспрессии среду 2xYT дополняют ампициллином (100 мкг мл-1) и/или канамицином (50 мкг мл-1). По 3 мл культуры E.coli инкубируют при комнатной температуре в течение ночи. Затем клетки собирают центрифугированием и отделяют от культуральной среды, полученные осадки клеток ресуспендируют в 200 мкл ddH2O и кипятят 5 мин. После охлаждения на льду в течение 10 мин клеточный дебрис собирают центрифугированием при 13000 rpm в течение 10 мин. Из полученных таким способом клеточных супернатантов E.coli по 20 мкл каждой культуры наносят на силикагелевые пластины для TLC («Silica gel 60»). Для проведения TLC на пластине, применяют смеси растворителей, состоящие из бутанола: ацетона: уксусной кислоты: воды (35:35:7:23). Затем проводят радио-TLC анализ с помощью радио-TLC детектора («Raytest»). Для определения Rf-величин нерадиоактивного контрольного материала платины для TLC опрыскивают раствором анисового альдегида (5 мл конц. H2SO4, 100 мл этанола, 1,5 мл уксусной кислоты, 2 мл анисового альдегида) и нагревают.
Пример 7
Создание эффективного пути «реутилизации» L-фукозы в E.coli. Поскольку в качестве субстрата-акцептора для фукозилтрансферазы применяют лактозу, дефицитный по β-галактозидазе (lacZ-) штамм E.coli BW 25113 был выбран для преодоления проблемы быстрой деградации лактозы (Datsenko & Wanner, 2000). L-Фукоза также может быть эффективно превращена диким типом E.coli путем изомеризации в фукулозу, путем фосфорилирования в фукулозо-1-фосфат и путем последующего ретроальдольного расщепления фукулозо-1-фосфата в глицерин-3-фосфат и L-лактальдегид. Для предупреждения деградации добавленной фукозы ген fucA, кодирующий ключевой катаболический фермент пути деградации фукозы фукулозо-1-фосфат-альдолазу (FucA), удаляют из геномного штамма E.coli BW 25113. Полученный штамм E.coli BW 25113 ΔfucA не способен расти как на фукозе, так и на лактозе в качестве единственного источника углерода на чашках с минимальной М9. Лизогенизация с рекомбинантным фагом λDE3 приводит к образованию штамма E.coli BW 25113 ΔfucA (DE3), для которого могут быть применены вектора экспрессии, стимулируемые Т7-промотором. Способность нуклеотидов активировать фукозу до GDP-фукозы очень ограничена в природе и также в течение длительного времени известна только у некоторых млекопитающих (человека, свиньи, мыши). Активация нуклеотидами фукозы опосредуется в этом случае двумя последовательными энзиматическими стадиями, сначала фосфорилирование фукозы до фукозо-1-фосфата, катализируемое фукозокиназой, и затем превращение фукозо-1-фосфата в GDP-фукозу, катализируемое гуанилилтрансферазой, соответственно. Хотя у млекопитающих путь реутилизации фукозы включает две отдельные ферментативные реакции, недавно обнаружены бактериальные и растительные белки, которые включают обе энзиматические активности. Гетерологичная экспрессия фукозокиназы человека в E.coli приводит только к едва детектируемой активности (Hinderlich et al., 2002). Биохимические исследования показали, что фукокиниза млекопитающих представляет собой высоко регулируемый фермент (Park et al., 1998). Для того чтобы изучить, не представляет ли собой Fkp, недавно открытый в В. Fragilis, более подходящий фермент для активации фукозы и для эффективного обеспечения GDP-фукозы в синтезе фукозилированных олигосахаридов в E.coli, мы амплифицировали ген из геномной DNA В. fragilis и клонировали его в бактериальный вектор экспрессии для гетерологичной экспрессии.
Для синтеза 2'- и 3-фукозиллактозы для совместной экспрессии выбирают следующие фукозилтрансферазы: ген futA из Н. pylori 26695 (Appelmelk et al., 1999), кодирующий α1,3-фукозилтрансферазу, и ген α1,2'-фукозилтрансферазы fucT2 из Н. pylori NCTC364 (Albermann et al., 2001). Перед началом процесса клонирования, частоту использования кодона futA оптимизируют для экспрессии в E.coli, и затем синтезируют ген в компании «GenScript corporation» (США). Полученный ген futAco вставляют в вектор экспрессии pETDuet-1, и экспрессию тестируют с коэкспрессией Fkp и FucP и без нее. Применяя стандартные условия для индукции, проводят совместную экспрессию Fkp, FucP и FutAco или FucT2. Образование белка проверяют после индукции с IPTG и/или L-рамнозой с помощью SDS-PAGE (смотри фиг.4), регистрируя явно выраженную продукцию растворимого белка Fkp, тогда как индукция локализованного в мембране белка фукозопермеазы (FucP), как ожидали, не была обнаружена в цитоплазме клетки, согласно SDS-PAGE. Однако было подтверждено, что генные продукты futAco и fucT2 прежде всего локализованы в тельцах включения, регистрируемая растворимая фракция была незначительной.
Пример 8
Фотометрическая регистрация энзиматтеской активности
Неочищенные экстракты, полученные из индуцированных культур, тестируют на фукозокиназную и фукозилтрансферазную активность, с помощью вспомогательных ферментов в сопряженном энзиматическом анализе, как описано выше. Очевидно, существует значительная базовая как NADH-оксидазная, так и/или фосфатазная активность в E.coli BW 25113 ΔfucA (DE3), которые ответственны за невоспроизводимые результаты и низкую определяемую фукозокиназную и фукозилтрансферазную активность различных штаммов. Вследствие этого, было решено определять энзиматическую активность, регистрируя образование внутриклеточного продукта (GDP-фукозы и фукозиллактозы).
Пример 9
Изучение утилизации введенной извне 3H-L-фукозы при образовании GDP-фукозы и 3-фукозиллактозы в рекомбинантной E.coli
Цель этого эксперимента заключается в подтверждении образования 3-фукозиллактозы из фукозы и лактозы через образование GDP-фукозы под действием бифункционального фермента Fkp пути реутилизации фукозы из Bacteroides fragilis. Негативные контрольные штаммы E.coli BW 25113 ΔfucA (DE3) pCOLADuet-1 pETDue-1, а также штаммы E.coli BW 25113 ΔfucA (DE3) pCOLA-fkp-fucP, экспрессирующие Fkp и фукозопермеазу, и штаммы E.coli BW 25113 ΔfucA (DE3) pCOLA-fkp-fucP pET-futAco, экспрессирующие Fkp, фукозопермеазу и α1,3-фукозилтрансферазу, обрабатывают, как описано выше. Клеточные экстракты, полученные из указанных штаммов, наносят на пластинку для TLC, которую проводят, как описано выше, и анализируют с помощью радио-TLC детектора. Кроме того, стандартную 3H-меченую L-фукозу наносят на пластинку для TLC и хроматографируют (см. фиг.5). Нерадиоактивные стандарты для L-фукозы и L-фукулозо-1-фосфата, GDP-L-фукозы, а также для 3-фукозиллактозы анализируют аналогичным способом с помощью TLC и последующего окрашивания раствором анисового альдегида (данные не приведены).
Результаты эксперимента с негативным контролем (смотри фиг.6) демонстрируют продукты первой и второй катаболических стадий метаболизма фукозы, т.е. L-фукулозу (полученную из фукозы под действием фукозоизомеразы) и L-фукулозо-1-фосфат (полученную из фукулозы под действием фуколозокиназы). Дальнейшую деградацию фукозы эффективно ингибируют нокдауном гена fucA, который кодирует фермент фукулозо-1-фосфат-альдолазу, катализирующую реакцию ретроальдольного расщепления фукулозо-1-фосфата до L-лактальдегида и дигидроксиацетонфосфата.
E.coli клетки, коэкспрессирующие бифункциональные фукозокиназу / фукозо-1-фосфатгуанилилтрансферазу, Fkp из Bacteroides fragilis демонстрируют образование GDP-фукозы (см. фиг.7), которая несомненно накапливается в клетках и может только минимально отклоняться в сторону других метаболических путей, продукты которых в противном случае могли бы появиться на радио-TLC.
Клеточные экстракты из штамма E.coli BW 25113 ΔfucA (DE3) pCOLA-fkp-fucP pET-futAco демонстрируют образование 3-фукозиллактозы, и только небольшое количество GDP-фукозы (см. фиг.8). Этот результат соответствует исходной цели эксперимента, а именно, продемонстрировать образование 3-фукозиллактозы через GDP-фукозу, которую поставляет бифункциональный фермент, Fkp, пути реутилизации из Bacteroides fragilis. Количество продуктов деградации фукозы, фукулозы и фукулозо-1-фосфата, также значительно снижается, благодаря потреблению GDP-фукозы при образовании фукозиллактозы и возникшему сдвигу равновесия реакции от фукулозо-1-фосфата и фукулозы к фукозе, которая постоянно выводится из реакции в результате образования GDP-фукозы.
Пример 10
Изучение образования 2'-фукозиллактозы и 3-фукозиллактозы под действием рекомбинантной E.coli
Штамм E.coli BW 25113 ΔfucA (БЕЗ), несущий pCOLA-fkp-fucP, а также ген futAco или fuсТ2 в отдельном векторе экспрессии, а также E.coli BW 25113 ΔfucA (DE3), несущий пустые векторы pCOLADuet-1 и pETDuet-1 (негативный контроль) растят в 2xYT бульоне, и экспрессию белка индуцируют с помощью IPTG и/или L-рамнозы течение 15 часов при 28°С. Клетки последовательно промывают PBS и ресуспендируют в модифицированной среде М9, дополненной L-фукозой, лактозой и гуанозином, IPTG и L-рамнозой. После фазы ферментации (28°С, 15 часов) клетки собирают, супернатанты объединяют, и клеточные лизаты получают, как описано выше.
Анализ с помощью HPAED показал, что времена удержания на примененной HPLC-колонке, равные приблизительно 3 мин для стандарта L-фукозы, приблизительно 17 мин для стандарта лактозы, приблизительно 11 мин для стандарта 3-фукозиллактозы и приблизительно 22 мин для примененного стандарта 2'-фукозиллактозы (данные не приведены). Глицерин, который в качестве источника углерода представляет собой часть культуральной среды, регистрировали со временем удержания равным приблизительно 1,5 мин, и индуктор, L-рамнозу, со временем удержания равным 5,5 мин. Оба вещества регистрировали внутриклеточно в ходе анализа клеточных лизатов.
Клеточные лизаты из E.coli BW 25113 ΔfucA (ВЕЗ) pCOLADuet-1 pETDuet-1 негативного контрольного штамма показали наличие внутриклеточной L-фукозы и лактозы, но, как ожидали, отсутствие фукозиллактозы (см. фиг.9). Помимо, вышеуказанных молекул при анализе обнаружили также внесенный в среду источник углерода глицерин и индуктор транскрипции L-рамнозу.
HPAED-анализ клеточного лизата из штамма E.coli BW 25113 ΔfucA (DE3) pCOLA-fkp-fucP pET-futAco, коэкспрессирующего ген fkp B.fragilis и ген фукозопермеазы E.coli в комбинации с геном α1,3-фукозилтрансферазы из Helicobacter pylori с оптимизированным кодоном, показал внутриклеточное образование 3-фукозиллактозы (пик приблизительно на 11 мин, см. фиг.10). L-фукоза и лактоза, как глицерин и L-рамноза, также представляют собой компоненты клеточного лизата.
Клеточный лизат из штамма E.coli BW 25113 ΔfucA (DE3) pCOLA-fkp-fucP pCAW55 продемонстрировал внутриклеточное образование 2'-фукозиллактозы (см. фиг.11), благодаря коэкспрессии α1,2-фукозилтрансферазы FucT2. Кроме того, L-фукозу, лактозу, глицерин и L-рамнозу можно видеть в клеточном лизате, точно также как в клеточном лизате из негативного контроля и из продуцирующего 3-фукозиллактозу штамма E.coli BW 25113 ΔfucA (DE3) pCOLA-fkp-fucP pET-futAco.
Указанные результаты ясно демонстрируют образование 3- и 2'-фукозиллактозы в рекомбинантных клетках E.coli из добавленных извне L-фукозы и лактозы. С помощью гетерологичной экспрессии белка Fkp из В. fragilis, катализирующего двухстадийную реакцию фосфорилирования фукозы и переноса гуанилила на фукозо-1-фосфата, получают эффективное образование GDP-фукозы. α1,3-фукозилтрансфераза FutAco с оптимизированным кодоном, исходно полученная из Helicobacter pylori, или α1,2-фукозилтрансфераза FucT2 из Helicobacter pylori, соответственно, могут превращать поставляемую таким путем GDP-фукозу в 2'- и 3-фукозиллактозу.
Пример 11
Экспрессия GDP-фукозы в клетках E.coli JM109
Увеличение внутриклеточного содержания GDP-фукозы в результате экспрессии Fkp было показано путем параллельной культивации штамма E.coli, экспрессирующего Fkp из плазмиды, и штамма E.coli, не содержащего копии Fkp. Штамм E.coli JM109 (DE3) ΔfucA в этом случае применяют в качестве контрольного штамма без Fkp. Штамм, экспрессирующий Fkp, представлял собой тот же штамм E.coli JM109 (DE3) ΔfucA, на этот раз содержащий плазмиду pCOLA-fkp-fucP, и, следовательно, несущий гены, кодирующие фукозокиназу / фукозо-1-фосфатгуанилилтрансферазу, Fkp, и фукозопермеазу, FucP. Поскольку гены были клонированы в сайты множественного клонирования (MCS) 1 и 2 вектора pCOLADuet-1 («Novagen», UK), экспрессия обоих генов может быть индуцирована добавлением IPTG, поскольку оба MCS фланкированы Т7-промотором/оператором на 5'-стороне.
Оба штамма культивируют в двойном повторе в 30 мл 2YT среды, дополненной канамицином для штамма с pCOLA-fkp-fucP для сохранения плазмиды, при 37°С и 220 rpm. Индукцию экспрессии Fkp начинают при OD600=0,5 добавлением 1 мМ IPTG и к обоим штаммам добавляют 20 мМ фукозу и затем культивируют дополнительные 3 часа при 37°С и 220 rpm. Клетки осаждают центрифугированием, и осадки ресуспендируют в дистиллированной воде (5 масс/объем). Указанные клеточные суспензии инкубируют при 95°С течение 10 минут для лизирования клеток. Клеточный дебрис удаляют центрифугированием и супернатанты анализируют с помощью HPLC.
HPLC-анализ проводят путем электрохимической регистрации с помощью импульсного амперометрического детектора «Decade II» (Antec Leyden, Нидерланды). 20 мМ гидрохлорида натрия + 825 мМ ацетата натрия применяют в качестве элюента на колонке «CarboPac PA20» («Dionex», США). GDP-фукозу элюируют со временем удержания, равным 16,0 минут.
Таблица 3: Содержание внутриклеточной GDP-фукозы в E.coli JM109 (DE3) ΔfucA с экспрессией и без экспрессии фукозокиназы / фукозо-1-фосфатгуанилилтрансферазы, Fkp, из pCOLA-fkp-fucP.
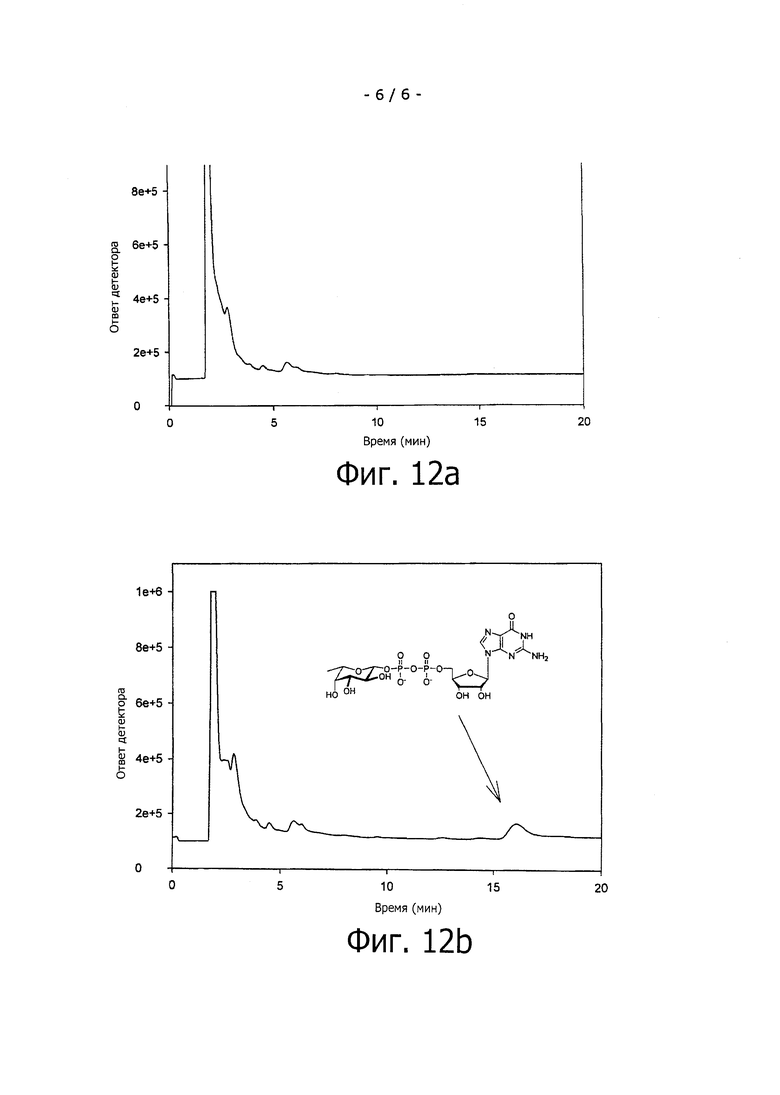
На фиг.12а показан HPLC-анализ клетки E.coli JM109 (DE3) ΔfucA для экспрессии GDP-фукозы без экспрессии белка FKP.
Фиг.12b представляет собой анализ клеток Е, coli JM109 (DE3) ΔfucA pCOLA-fkp-fucP, коэкспрессирующих белок Fkp вместе с импортером фукозы FucP. Пик на 16,0 мин соответствует GDP-фукозе, как подтверждено с помощью достоверного стандарта.
Ссылки
Albermann, С., W.Piepersberg & U.F.Wehmeier, (2001) Synthesis of the milk oligosaccharide 2'-fucosyllactose using recombinant bacterial enzymes. Carbohydr Res 334: 97-103.
Andersson, В., O.Porras, L.A.Hanson, T.Lagergard & C.Svanborg-Eden, (1986) Inhibition of attachment of Streptococcus pneumoniae and Haemophilus influenzae by human milk and receptor oligosaccharides. JInfect Dis 153: 232-237.
Appelmelk, В.J., S.L.Martin, M.A.Monteiro, C.A.Clayton, A.A.McColm, P.Zheng, T.Verboom, J.J.Maaskant, D.H. van den Eijnden, C.H.Hokke, M.B.Perry, C.M.Vandenbroucke-Grauls & J.G.Kusters, (1999) Phase variation in Helicobacter pylori lipopolysaccharide due to changes in the lengths of poly(C) tracts in alpha3-rucosyltransferase genes. Infect Immun 67: 5361-5366.
Appelmelk, В.J., M.C.Martino, E.Veenhof, M.A.Monteiro, J.J.Maaskant, R.Negrini, F.Lindh, M.Perry, G.Del Giudice & C.M.Vandenbroucke-Grauls, (2000) Phase variation in H type I and Lewis a epitopes of Helicobacter pylori lipopolysaccharide. Infect Immun 68: 5928-5932.
Bergman, M., G. Del Prete, Y. van Kooyk & B.Appelmelk, (2006) Helicobacter pylori phase variation, immune modulation and gastric autoimmunity. Nat Rev Microbiol 4:151-159.
Bode, L., (2006) Recent advances on structure, metabolism, and function of human milk oligosaccharides. JNutr 136: 2127-2130.
Coppa, G.V., L.Zampini, T.Galeazzi, B.Facinelli, L.Fen-ante, R.Capretti & G.Orazio (2006) Human milk oligosaccharides inhibit the adhesion to Caco-2 cells ofdian-heal pathogens: Escherichia coli. Vibrio cholerae, and Salmonella fyris. Pediatr Res 59: 377-382.
Coyne, M.J., B.Reinap, M.M.Lee & L.E.Comstock, (2005) Human symbionts use a host-like pathway for surface fucosylation. Science 307: 1778-1781.
Crane, J.K., S.S.Azar, A.Stam & D.S.Newburg, (1994) Oligosaccharides from human milk block binding and activity of the Escherichia coli heat-stable enterotoxin (STa) in T84 intestinal cells. JNutr 124: 2358-2364.
Cravioto, A., A.Tello, H.Villafan, J.Ruiz, S. del Vedovo & J.R.Neeser, (1991) Inhibition of localized adhesion of enteropathogenic Escherichia coli to HEp-2 cells by immunoglobulin and oligosaccharide fractions of human colostrum and breast milk. J Infect Dis 163: 1247-1255.
Datsenko, К.А. & B.L.Wanner, (2000) One-step inactivation of chromosomal genes in Escherichia coli K-12 using PCR products. Proc NatlAcadSci USA 97: 6640-6645.
Dower, W.J., J.F.Miller & C.W.Ragsdale, (1988) High efficiency transformation of E.coli by high voltage electroporation. Nucleic Acids Res 16: 6127-6145.
Dumon, С., С.Bosso, J.P.Utille, A.Heyraud & E.Samain, (2006) Production of Lewis x tetrasaccharides by metabolically engineered Escherichia coli. Chembiochem 7: 359-365.
Dumon, С., В.Priem, S.L.Martin, A.Heyraud, C.Bosso & E.Samain, (2001) In vivo fucosylation of lacto-N-neotetraose and lacto-N-neohexaose by heterologous expression of Helicobacter pylori alpha-1,3 fucosyltransferase in engineered Escherichia coli. Glycoconj.718:465-474.
Dumon, С., E.Samain & В.Priem, (2004) Assessment of the two Helicobacter pylori alpha-1,3-fucosyltransferase ortholog genes for the large-scale synthesis of LewisX human milk oligosaccharides by metabolically engineered Escherichia coli. Biotechnol Prog 20: 412-419.
Ge, Z., N.W.Chan, M.M.Palcic & D.E.Taylor, (1997) Cloning and heterologous expression of an alphal,3-fucosyltransferase gene from the gastric pathogen Helicobacter pylori. JBiol Chem 272: 21357-21363.
Gnoth, M.J., S.Rudloff, C.Kunz & R.K.Kinne, (2001) Investigations of the in vitro transport of human milk oligosaccharides by a Caco-2 monolayer using a novel high performance liquid chromatography-mass spectrometry technique. J Biol Chem 276: 34363-34370.
Grant, W.D., I.W.Sutherland & J.F.Wilkimson, (1970) Control of colanic acid synthesis. J Bacterial 103: 89-96.
Hinderlich S, Berger, M., Blume, A., Chen, H., Ghaderi, D. & Bauer, C. (2002) Identification of human L-fucose kinase amino acid sequence. Biochem. Biophys. Res. Commun. 294, 650-654.
Koizumi, S., T.Endo, K.Tabata, H.Nagano, J.Ohnishi & A.Ozaki, (2000) Large-scale production of GDP-fucose and Lewis X by bacterial coupling. J Ind Microbiol Biotechnol 25:213-217.
Kretzschmar, G. & W.Stahl, (1998) Large scale synthesis of linker-modified sialyl-Lewis(X), Lewis(X) and N-acetyllactosamine. Tetrahedron 54: 6341-6358.
Kunz, C. & S.Rudloff, (2006) Health promoting aspects of milk oligosaccharides. Int Dairy J 16: 1341-1346.
Li, M., X.W.Liu, J.Shao, J.Shen, Q.Jia, W.Yi, J.K.Song, R.Woodward, C.S.Chow & P.G.Wang, (2008) Characterization of a novel alpha 1,2-fucosyltransferase of Escherichia coli O128:b12 and functional investigation of its common motif. Biochemistry 47: 378-387.
Ma, В., J.L.Simala-Grant & D.E.Taylor, (2006) Fucosylation in prokaryotes and eukaryotes. Glycobiology 16: 158R-184R.
Martin, S.L., M.R.Edbrooke, T.C.Hodgman, D.H. van den Eijnden & M.I.Bird, (1997) Lewis X biosynthesis in Helicobacter pylori. Molecular cloning of an alpha(1,3)-fucosyltransferase gene. JBiol Chem 272: 21349-21356.
Newburg, D.S., (2001) Bioactive components of human milk: evolution, efficiency, and protection. Adv Exp MedBiol 501: 3-10.
Newburg, D.S. & S.H.Neubauer, (1995) Carbohydrates in milk. In: Handbook of Milk Composition. R.G.Jensen (ed). San Diego, CA: Academic Press, pp.273-349.
Rasko, D.A., G.Wang, M.M.Palcic & D.E.Taylor, (2000) Cloning and characterization of the alpha(1,3/4) fucosyltransferase of Helicobacter pylori. J Biol Chem 275:4988-4994.
Ruiz-Palacios, G.M., L.E.Cervantes, P.Ramos, B.Chavez-Munguia & D.S.Newburg, (2003) Campylobacter jejuni binds intestinal H(0) antigen (Fuc alpha 1, 2Gal beta 1, 4GlcNAc), and fucosyloligosaccharides of human milk inhibit its binding and infection. J Biol Chem 278:14112-14120.
Sambrook, J. & D.W.Russell, (2001) Molecular cloning: a laboratory manual. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY.
Stumpp, Т., В.Wilms & J.Altenbuchner, (2000) Ein neues L-Rhamnose-induzierbares Expressionssystem fur Escherichia coli. BIOspektrum 6: 33-36.
Wang, G., Z.Ge, D.A.Rasko & D.E.Taylor, (2000) Lewis antigens in Helicobacter pylori: biosynthesis and phase variation. Mol Microbiol 36: 1187-1196.
Wang, G., D.A.Rasko, R.Sherbume & D.E.Taylor, (1999) Molecular genetic basis for the variable expression of Lewis Y antigen in Helicobacter pylori: analysis of the alpha (1,2) fucosyltransferase gene. Mol Microbiol 31: 1265-1274.
Park, S.H., I.Pastuszak, R.Drake & A.D.Elbein, (1998). Purification to apparent homogenicity and properties of pig kidney L-fucose kinase. JBiol Chem 273: 5685-5691.
| название | год | авторы | номер документа |
|---|---|---|---|
| СИНТЕЗ НМО | 2009 |
|
RU2517602C2 |
| НОВЫЕ ФУКОЗИЛТРАНСФЕРАЗЫ И ИХ ПРИМЕНЕНИЕ | 2011 |
|
RU2642307C2 |
| ПРОДУКЦИЯ ФУКОЗИЛИРОВАННЫХ ОЛИГОСАХАРИДОВ В BACILLUS | 2020 |
|
RU2810729C2 |
| НОВЫЕ ФУКОЗИЛТРАСФЕРАЗЫ И ИХ ПРИМЕНЕНИЯ | 2011 |
|
RU2628307C2 |
| ФУКОЗИЛТРАНСФЕРАЗЫ И ИХ ПРИМЕНЕНИЕ ДЛЯ ПОЛУЧЕНИЯ ФУКОЗИЛИРОВАННЫХ ОЛИГОСАХАРИДОВ | 2018 |
|
RU2818835C2 |
| УЛУЧШЕННЫЙ СПОСОБ ПОЛУЧЕНИЯ ФУКОЗИЛИРОВАННЫХ ОЛИГОСАХАРИДОВ | 2017 |
|
RU2790445C2 |
| ФЕРМЕНТАТИВНЫЙ СПОСОБ ПРОИЗВОДСТВА МОНОСАХАРИДОВ В СВОБОДНОЙ ФОРМЕ ИЗ САХАРОВ, АКТИВИРОВАННЫХ НУКЛЕОТИДАМИ | 2016 |
|
RU2746739C1 |
| Ферментативная продукция углеводов микробными клетками с использованием смешанного сырья | 2020 |
|
RU2809122C2 |
| ПРИМЕНЕНИЕ ГЛИКОЗИДАЗ В ПОЛУЧЕНИИ ОЛИГОСАХАРИДОВ | 2019 |
|
RU2810730C2 |
| Ферментативное получение олигосахаридов посредством общей ферментации с использованием смешанного сырья | 2019 |
|
RU2801231C2 |
Группа изобретений относится к генетически модифицированной клетке для продуцирования фукозилированных олигосахаридов, способу ее получения и способу получения фукозилированных олигосахаридов. Генетически модифицированную клетку получают трансформированием клетки геном, кодирующим фукозокиназу, геном, кодирующим фукозо-1-фосфатгуанилилтрансферазу, геном, кодирующим фукозилтрансферазу. Катаболический путь для фукозы инактивирован в указанной клетке путем инактивации одного или нескольких генов, которые выбирают из группы, состоящей из гена фукозо-1-фосфат-альдолазы, гена фукозоизомеразы и гена фуколозокиназы. При этом клетка представляет собой микроорганизм, который выбирают из группы, состоящей из родов Escherichia, Klebsiella, Helicobacter, Bacillus, Lactobacillus, Streptococcus, Lactococcus, Pichia, Saccharomyces и Kluyveromyces. С использованием указанной генетически модифицированной клетки получают фукозилированные олигосахариды путем ее культивирования в подходящих условиях в среде, включающей фукозу и субстрат-акцептор, в котором субстрат-акцептор представляет собой моно-, ди- или олигосахарид или пептид. 3 н. и 6 з.п. ф-лы, 12 ил., 3 табл., 11 пр.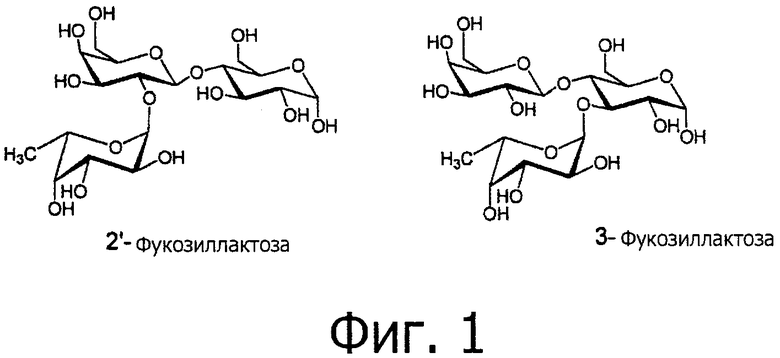
1. Способ получения генетически модифицированной клетки, обладающей способностью продуцировать фукозилированные олигосахариды, включающий следующие стадии:
- трансформирование клетки геном, кодирующим фукозокиназу;
- трансформирование клетки геном, кодирующим фукозо-1-фосфатгуанилилтрансферазу;
- трансформирование клетки геном, кодирующим фукозилтрансферазу,
в котором генетически модифицированная клетка представляет собой микроорганизм, который выбирают из группы, состоящей из родов Escherichia, Klebsiella, Helicobacter, Bacillus, Lactobacillus, Streptococcus, Lactococcus, Pichia, Saccharomyces и Kluyveromyces, и
в котором катаболический путь для фукозы инактивирован в указанной клетке путем инактивации одного или нескольких генов, которые выбирают из группы, состоящей из гена фукозо-1-фосфат-альдолазы, гена фукозоизомеразы и гена фуколозокиназы.
2. Способ по п.1, в котором ген, кодирующий фукозилтрансферазу получают из организма, который выбирают из группы, состоящей из родов Helicobacter, Escherichia, Yersinia, Enterococcus, Shigella, Klebsiella, Salmonella, Bacteroides, Dictyostelium, Arabidopsis, Drosophila, Homo, Bos, Mus, Rattus, Gallus, Canis и Sus.
3. Способ по п.1, в котором фукозилированный олигосахарид представляет собой фукозиллактозу, предпочтительно 2′-фукозиллактозу, 3-фукозиллактозу или лактодифукотетраозу.
4. Способ по п.1, в котором фукозокиназу и фукозо-1-фосфатгуанилилтрансферазу объединяют в бифункциональный фермент, полученный из Bacteroides.
5. Способ по п. 1, в котором фукозилированный олигосахарид представляет собой человеческий молочный олигосахарид.
6. Генетически модифицированная клетка для продуцирования фукозилированных олигосахаридов, полученная способом по любому из пп. 1-5.
7. Способ получения фукозилированных олигосахаридов, включающий стадии культивирования клетки по п.6 в подходящих условиях культивирования в среде, включающей фукозу и субстрат-акцептор, в котором субстрат-акцептор представляет собой моно-, ди- или олигосахарид или пептид.
8. Способ по п.7, в котором субстрат-акцептор представляет собой лактозу, 2′-фукозиллактозу или 3-фукозиллактозу.
9. Способ по п.7, в котором фукозилированный олигосахарид представляет собой фукозиллактозу, предпочтительно 2′-фукозиллактозу или 3-фукозиллактозу или лактодифукотетраозу.

