Настоящее изобретение относится к способу получения фукозилированных олигосахаридов с использованием генетически модифицированной прокариотической клетки хозяина, а также к клетке хозяина, используемой в этом способе, и ее применению для получения фукозилированных олигосахаридов в высоких концентрациях.
ПРЕДШЕСТВУЮЩИЙ УРОВЕНЬ ТЕХНИКИ
Грудное молоко человека представляет собой сложную смесь углеводов, жиров, белков, витаминов, минеральных веществ и микроэлементов. Наиболее преобладающая фракция представлена углеводами, которые в свою очередь могут подразделяться на лактозу и более сложные олигосахариды (олигосахариды грудного молока, ОГМ). В то время как лактоза используется в качестве источника энергии, сложные олигосахариды не метаболизируются в организме младенца. Фракция сложных олигосахаридов составляет до 20% включительно от всей углеводной фракции и состоит из более чем 200 различных олигосахаридов. Наличие и концентрация этих сложных олигосахаридов специфичны для людей, и поэтому они не могут быть обнаружены в больших количествах в молоке других млекопитающих.
К настоящему времени идентифицировано приблизительно 200 различающихся по структуре ОГМ и описаны их многочисленные полезные свойства. ОГМ не перевариваются в организме младенцев, находящихся на грудном вскармливании, но представляют собой ценный источник углерода и энергии для полезных бактерий рода Bifidobacteria, Lactobacillus и Bacteroides в кишечнике, что приводит к превалированию этих бактерий в кишечнике и способствует подавлению ими роста патогенов, предотвращая таким образом инфекции кишечного эпителия. Однако, помимо этого ОГМ напрямую связываются с патогенными бактериями, простейшими и вирусами, блокируя взаимодействия патогена и хозяина путем имитации гликановых цепей рецепторов клеточной поверхности и тем самым защищая ребенка, находящегося на грудном вскармливании, от инфекционных заболеваний.
Наиболее часто встречающимся олигосахаридом является 2'-фукозиллактоза. Другими часто встречающимися ОГМ, входящими в состав грудного молока, являются 3-фукозиллактоза, лакто-N-тетраоза, лакто-N-неотетраоза и лакто-N-фукопентаозы. Помимо этих нейтральных олигосахаридов в грудном молоке могут быть обнаружены кислые ОГМ, такие как, например, 3'-сиалиллактоза, 6'-сиалиллактоза и сиалиллакто-N-тетраоза a, b и с или сиалиллакто-N-фукопентаоза II и так далее. Эти структуры находятся в близком родстве с эпитопами гликоконъюгатов на поверхности эпителиальных клеток, с антигенами гистогрупп крови системы Льюиса, и структурная гомология ОГМ с эпитопами эпителиальных клеток объясняет защитные свойства по отношению к бактериальным патогенам.
Благодаря своим полезным свойствам, ОГМ одобрены к включению в качестве ингредиентов в детские смеси и другие продукты питания, что делает необходимым получение ОГМ в больших количествах вплоть до масштаба многотоннажного производства.
В связи с ограниченным количеством сырья и трудностями в получении очищенных фракций отдельных олигосахаридов грудного молока были разработаны химические методы получения некоторых из этих сложных молекул. Однако, химический и биокаталитический подходы оказались коммерчески несостоятельными, и, кроме того, отдельные химические пути синтеза олигосахаридов грудного молока включают в себя некоторые вредные химические реагенты, обуславливающие риск загрязнения конечного продукта.
В связи с трудностями, возникающими при химическом синтезе олигосахаридов грудного молока, было разработано несколько способов с использованием ферментов и ферментационных подходов. На сегодняшний момент для некоторых из ОГМ, таких как 2'-фукозиллактоза, 3-фукозиллактоза, лакто-N-тетраоза, лакто-N-неотетраоза, лакто-N-фукопентаоза I, лакто-N-дифукогексаоза II, 3'-сиалиллактоза и 6'-сиалиллактоза, разработаны ферментативные подходы с применением по большей части генетически модифицированных бактериальных штаммов, таких как рекомбинантный штамм Escherichia coli.
Однако с использованием даже самых эффективных доступных в настоящее время способов, базирующихся на бактериальной ферментации, не удается достичь или с трудом достигают концентраций ОГМ в культуральной жидкости, составляющих более 20 г/л. Обычно, в случае применения способов в промышленных масштабах должны быть превышены концентрации 50 г/л, хотя более желательной концентрацией является 100 г/л.
Таким образом, задача настоящего изобретения заключается в разработке улучшенного способа ферментации, посредством которого возможно осуществление биосинтеза фукозилированных олигосахаридов, в частности, 2'-фукозиллактозы, в концентрации, превышающей 100 г/л.
КРАТКОЕ ОПИСАНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
Эта и другие задачи решаются путем разработки способа получения фукозилированных олигосахаридов с использованием генетически модифицированной прокариотической клетки хозяина, включающего стадии:
- предоставления клетки хозяина, генетически модифицированной таким образом, что по меньшей мере (1) активность фермента, преобразующего фруктозо-6-фосфат, которая в немодифицированной клетке хозяина находится на обычном уровне, понижена или подавлена; (2) по меньшей мере один ген, кодирующий фермент, необходимый для синтеза ГДФ-фукозы (ГДФ означает гуанозиндифосфат) de novo, сверхэкспрессирован в клетке хозяина; (3) экзогенный ген, кодирующий фукозилтрансферазу, предпочтительно альфа-1,2-фукозилтрансферазу и/или альфа-1,3-фукозилтрансферазу, экспрессирован, предпочтительно сверхэкспрессирован, в клетке хозяина;
- культивирования указанной генетически модифицированной клетки хозяина в культуральной среде, содержащей источник углерода и энергии, выбранный по меньшей мере из одного из следующих источников: глюкозы, сахарозы, глицерина, сукцината, цитрата, пирувата, малата, лактата или этанола; и
- обеспечения подачи лактозы в культуральную среду с лактозой.
На последующей стадии образованный таким образом фукозилированный олигосахарид может быть извлечен или получен из среды, в которой культивируют клетку хозяина.
Стадия выращивания и культивирования генетически модифицированной клетки хозяина и стадия добавления лактозы в культуральную среду могут быть проведены таким образом, что сначала генетически модифицированную клетку хозяина культивируют в течение определенного периода времени и на последующей стадии по окончании периода такого первоначального культивирования осуществляют подачу лактозы, добавляя ее в среду, в которой культивируют клетку хозяина; альтернативно, лактоза может быть добавлена в определенном количестве в начале периода культивирования генетически модифицированной клетки хозяина и может постоянно добавляться в определенном количестве. В альтернативном варианте лактоза может вырабатываться внутри самой среды.
Данная задача также решается посредством генетически модифицированной прокариотической клетки хозяина и посредством ее применения для получения фукозилированного олигосахарида, при этом клетка-хозяин генетически модифицирована таким образом, что по меньшей мере (1) активность фермента, преобразующего фруктозо-6-фосфат, которая в немодифицированной клетке хозяина находится на обычном уровне, понижена или подавлена и/или имеется повышенная активность фермента, катализирующего образование фруктозо-6-фосфата в клетке хозяина; (2) по меньшей мере один ген, кодирующий фермент, необходимый для синтеза ГДФ-фукозы de novo, сверхэкспрессирован; (3) экзогенный ген, кодирующий фукозилтрансферазу, предпочтительно альфа-1,2-фукозилтрансферазу и/или альфа-1,3-фукозилтрансферазу, экспрессирован, предпочтительно сверхэкспрессирован, в клетке хозяина.
Возможно, что клетка-хозяин была подвергнута дальнейшей генетической модификации (4) с целью экспрессирования гена, который кодирует белок, обеспечивающий транспорт или способствующий транспорту желаемого фукозилированного олигосахарида в среду, в которой культивируют клетку хозяина; и/или (5) с целью экспрессирования экзогенного гена, кодирующего бифункциональную L-фукокиназу/L-фукозо-1-фосфат-гуанилилтрансферазу; и/или (6) с целью инактивирования или делетирования генов, кодирующих L-фукозоизомеразу и L-фукулозокиназу; и/или (7) с целью инактивирования или разрушения генов, кодирующих ферменты синтеза колановой кислоты; и/или (8) с целью экспрессирования пермеазы лактозы; и/или (9) с целью инактивирования или делетирования эндогенных генов бета-галактозидазы; и/или (10) с целью экспрессирования гена экзогенно регулируемой бета-галактозидазы; и/или (11) с целью сверхэкспрессирования экзогенных генов, ответственных за метаболизирование галактозы, и/или (12) с целью экспрессирования экзогенного гена, кодирующего фермент, который проявляет фосфатазную активность в отношении фруктозо-1,6-бифосфата.
Кроме того, согласно данному изобретению предложен способ получения фукозилированных олигосахаридов с использованием генетически модифицированной прокариотической клетки хозяина, включающий стадии:
- предоставления прокариотической клетки хозяина, генетически модифицированной таким образом, что по меньшей мере (1) в указанной генетически модифицированной клетке хозяина количество фруктозо-6-фосфата возросло в результате снижения или подавления активности фермента, преобразующего фруктозо-6-фосфат, которая в немодифицированной клетке хозяина находится на обычном уровне или в результате повышения активности фермента, катализирующего образование фруктозо-6-фосфата; что (2) по меньшей мере один ген, кодирующий фермент, необходимый для синтеза ГДФ-фукозы de novo, сверхэкспрессирован в клетке хозяина; (3) экзогенный ген, кодирующий альфа-1,2-фукозилтрансферазу и/или альфа-1,3-фукозилтрансферазу, экспрессирован в клетке хозяина;
- культивирования указанной генетически модифицированной клетки хозяина в культуральной среде, содержащей источник углерода и энергии, выбранный по меньшей мере из одного из следующих источников: глюкозы, сахарозы, глицерина, сукцината, цитрата, пирувата, малата, лактата или этанола; и
- обеспечения подачи лактозы в культуральную среду с лактозой.
С использованием этого подхода полностью решаются задачи, лежащие в основе изобретения.
С использованием способа по изобретению, а также с использованием генетически модифицированной клетки хозяина, применяемой в данном способе, можно получать фукозилированные олигосахариды в концентрации, превышающей 50 г/л, и даже 100 г/л, и даже выше 150 г/л, таким образом предоставляя эффективный инструмент для крупномасштабного и, следовательно, промышленного ферментативного получения фукозилированных олигосахаридов.
В настоящей заявке и согласно общему пониманию существующего уровня техники термин "фукозилированный олигосахарид" означает фукозилированный олигосахарид в той форме, в которой он обнаружен в грудном молоке, т.е. олигосахарид, содержащий остаток фукозы. Предпочтительно, фукозилированный олигосахарид представляет собой олигосахарид, выбранный из 2'-фукозиллактозы, 3-фукозиллактозы или дифукозиллактозы.
Кроме того, в настоящей заявке термин "генетически модифицированная прокариотическая клетка-хозяин" означает прокариотическую клетку, чей генетический материал был изменен с использованием генно-инженерных методов. Например, клетка-хозяин была генетически модифицирована таким образом, что или эндогенные нуклеиновокислотные последовательности природного происхождения, имеющиеся в указанной клетке хозяина, претерпели делетирование, разрыв или подверглись иному воздействию, приводящему к изменению их экспрессии, т.е. подавлению, снижению, супрессии, усилению или им подобному, и/или в клетку хозяина были введены экзогенные нуклеиновые кислоты, т.е. чужеродные для указанной клетки хозяина нуклеиновые кислоты, для осуществления экспрессии в клетке хозяина, например, под контролем управляемого промотора. В этой связи такие генетически модифицированные клетки хозяина также называют "рекомбинантными клетками хозяина". Например, рассматриваемая прокариотическая клетка-хозяин является генетически модифицированной прокариотической клеткой хозяина вследствие введения в подходящую прокариотическую клетку хозяина гетерологичной нуклеиновой кислоты, например, экзогенной нуклеиновой кислоты, являющейся чужеродной для прокариотической клетки хозяина, или рекомбинантной нуклеиновой кислоты, которая в норме не содержится в прокариотической клетке хозяина.
Соответственно, термин "рекомбинантный", использованный в данном описании со ссылкой на клетку бактерии-хозяина указывает на то, что в бактериальной клетке реплицируется гетерологичная нуклеиновая кислота либо экспрессируется пептид или белок, кодируемый гетерологичной нуклеиновой кислотой (т.е. последовательностью, "чужеродной для указанной клетки"). Рекомбинантные клетки могут содержать гены, которые не обнаруживаются в нативной (нерекомбинантной) форме клетки. Рекомбинантные клетки также могут содержать гены, обнаруживаемые в нативной форме клетки, при этом гены модифицируют и возвращают в клетку искусственным способом. Термин также охватывает клетки, которые содержат эндогенную для данной клетки нуклеиновую кислоту, модифицированную без удаления нуклеиновой кислоты из клетки; такие модификации включают модификации, полученные с использованием замены гена, сайт-специфической мутации и схожих методов. Соответственно, "рекомбинантный полипептид" представляет собой полипептид, продуцируемый рекомбинантной клеткой. "Гетерологичная последовательность" или "гетерологичная нуклеиновая кислота", как использовано в данном описании, происходит из источника, чужеродного для данной конкретной клетки хозяина (например, другого вида) или, если происходит из того же источника, то является модифицированной по сравнению со своей исходной формой. Таким образом, гетерологичная нуклеиновая кислота, функционально связанная с промотором, происходит из источника, отличающегося от источника, из которого происходит промотор, или, если происходит из того же источника, то является модифицированной по сравнению со своей исходной формой. Гетерологичная последовательность может быть стабильно введена в геном клетки микроорганизма-хозяина посредством, например, трансфекции, трансформации, конъюгирования или трансдукции, при этом могут быть применены методы, которые будут зависеть от клетки хозяина и подлежащей введению последовательности. Специалисту в данной области известны различные методы, которые описаны, например, в Sambrook et al., Molecular Cloning: A Laboratory Manual, 2nd Ed., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y. (1989).
Соответственно, в настоящей заявке под "генетически модифицированной прокариотической клеткой хозяина" понимают прокариотическую клетку, трансформированную или трансфицированную либо восприимчивую к трансформации или трансфекции экзогенной полинуклеотидной последовательностью.
Нуклеиновокислотные последовательности, использованные в настоящем изобретении, могут, например, входить в состав вектора, которым стабильно трансформируют/трансфицируют клетки микроорганизма-хозяина, или могут быть введены в клетки микроорганизма-хозяина иным образом.
В данном изобретении для экспрессии генов может быть использовано большое разнообразие экспрессирующих систем. Такие векторы включают, среди прочих, векторы на основе хромосом, эписом и вирусов, например, векторы, происходящие из бактериальных плазмид, из бактериофагов, из транспозонов, из эписом дрожжей, из инсерционных элементов, из хромосомных элементов дрожжей, из вирусов, и векторы, происходящие из их комбинаций, как например, векторы, полученные из генетических элементов плазмид и бактериофагов, таких как космиды и фагмиды. Конструкции экспрессирующих систем могут содержать контрольные участки, которые регулируют, а также вызывают экспрессию. Как правило, в этом отношении для экспрессии могут быть использованы любая система или вектор, подходящие для сохранения, увеличения количества или экспрессии полинуклеотидов и для синтеза полипептида в клетке хозяина. Соответствующая последовательность ДНК может быть встроена в экспрессирующую систему любым из множества хорошо известных и рутинных методов, таких как, например, приведенные выше в Sambrook и др.
Данная область техники переполнена патентными и литературными публикациями, имеющими отношение к методологиям "рекомбинантной ДНК" для выделения, синтеза, очистки и амплификации генетического материала, используемого для трансформации выбранных организмов-хозяев. Таким образом, процесс трансформации микроорганизма-хозяина "гибридной" вирусной или кольцевой плазмидной ДНК, включающей в себя выбранные экзогенные (т.е. чужеродные или "гетерологичные") последовательности ДНК, относится к общеизвестным знаниям. Специалист в данной области знаком с множеством способов получения "гибридных" векторов, используемых для трансформации выбранного микроорганизма-хозяина.
Термин "нуклеиновокислотная последовательность, кодирующая…" обычно относится к любому полирибонуклеотиду или полидезоксирибонуклеотиду, который может представлять собой немодифицированную РНК или ДНК либо модифицированную РНК или ДНК и, как правило, представляет собой ген, кодирующий определенный полипептид или белок. Термин включает в себя, без ограничения, одно- и двухцепочечную ДНК, ДНК, представляющую собой смесь из одно- и двухцепочечных участков или одно-, двух- и трехцепочечных участков, одно- и двухцепочечную РНК и РНК, представляющую собой смесь из одно- и двухцепочечных участков, гибридные молекулы, содержащие ДНК и РНК, которые могут представлять собой одноцепочечные, или, в более типичном случае, двухцепочечные, или трехцепочечные участки, либо смесь одно- и двухцепочечных участков. Данный термин также охватывает полинуклеотиды, включающие единый непрерывный участок или прерывающиеся участки, кодирующие полипептид (например, прерванные интегрированным фагом или инсерционной последовательностью, или редактированием), вместе с дополнительными участками, которые также могут содержать кодирующие и/или некодирующие последовательности.
Использованный в данном описании термин "культивирование" означает выращивание и/или инкубирование бактериальной клетки в среде и в условиях, допускающих получение желаемого(ых) олигосахарида(ов) и подходящих для его(их) получения. Специалисту в данной области техники будут легко доступны несколько подходящих клеток бактерии-хозяина, а также сред и условий для их культивирования после прочтения описания данного изобретения с учетом технической и экспертной квалификации этого специалиста.
Следует понимать, что при применении изобретения, описанного в данной заявке, получение одного или более олигосахаридов, определенных в данном описании, возможно при условии, если соответствующие нуклеиновые кислоты, кодирующие релевантные белки/ферменты, описанные в данной заявке, содержатся в клетке(ах).
Использованный в данном описании термин "извлечение" означает выделение, сбор, очистку, накопление или осуществляемое иным образом отделение олигосахарида, продуцируемого микроорганизмом-хозяином по изобретению, от среды культивирования микроорганизма-хозяина.
Согласно одному из воплощений способа и применения по изобретению продуцируемый фукозилированный олигосахарид выбран из меньшей мере из одного из следующего: 2'-фукозиллактозы, 3-фукозиллактозы или дифукозиллактозы.
Согласно одному из воплощений способа по изобретению прокариотическая клетка-хозяин выбрана из группы, состоящей из клеток бактерии-хозяина, предпочтительно выбранных из штамма Escherichia coli, видов Lactobacillus или штамма Corynebacterium glutamicum.
Предпочтительно, чтобы клетка-хозяин представляла собой клетку бактерии-хозяина, выбранную из клеток Escherichia coli, Corynebacterium glutamicum, Bacillus subtilis, Bacillus megaterium, Lactobacillus casei, Lactobacillus acidophilus, Lactobacillus helveticus, Lactobacillus delbrueckii, Lactococcus lactis. Специалист в данной области техники будет осведомлен о дополнительных бактериальных штаммах при прочтении описания настоящего изобретения.
В настоящей заявке и согласно общему пониманию под термином "фукозилтрансфераза" понимают как фермент, который осуществляющий перенос сахара L-фукозы с субстрата-донора ГДФ-фукозы (гуанозин-дифосфат-фукозы) на субстрат-акцептор с образованием фукозилированного олигосахарида. В настоящем изобретении субстратом-акцептором является олигосахарид. Кроме того, фукозилтрансферазы не только катализируют фукозилирование в присутствии гликановых акцепторы, но также могут обуславливать гидролиз ГДФ-L-фукозы в отсутствие субстрата-акцептора.
Соответственно, термины "альфа-1,2-фукозилтрансфераза" или "фукозилтрансфераза" либо нуклеиновая кислота/полинуклеотид, кодирующая(ий) "альфа-1,2-фукозилтрансферазу" или "фукозилтрансферазу", относятся к гликозилтрансферазе, которая катализирует перенос группировки фукозы от субстрата-донора, например, ГДФ-фукозы, на акцепторную молекулу с образованием альфа-1,2-связи. Термины "альфа-1,3-фукозилтрансфераза" или "фукозилтрансфераза" либо нуклеиновая кислота/полинуклеотид, кодирующая(ий) "альфа-1,3-фукозилтрансферазу" или "фукозилтрансферазу", относятся к гликозилтрансферазе, которая катализирует перенос группировки фукозы от субстрата-донора, например, ГДФ-фукозы, на акцепторную молекулу с образованием альфа-1,3-связи. Акцепторной молекулой может быть, например, лактоза, 2'-фукозиллактоза, 3-фукозиллактоза, 3'-сиалиллактоза, 6'-сиалиллактоза, лакто-N-тетраоза, лакто-N-неотетраоза или их производное.
Согласно изобретению экзогенный ген, кодирующий фукозилтрансферазу, выбран из гена, экспрессирующего белок, обладающий альфа-1,2-фукозилтрансферазной активностью, гена, экспрессирующего белок, обладающий альфа-1,3-фукозилтрансферазной активностью, или гена, экспрессирующего белок, обладающий альфа-1,2-фукозилтрансферазной, а также альфа-1,3-фукозилтрансферазной активностью.
Согласно предпочтительным воплощениям, для синтеза 2'-фукозиллактозы экспрессируют подходящую альфа-1,2-фукозилтрансферазу, для синтеза 3-фукозиллактозы экспрессируют подходящую альфа-1,3-фукозилтрансферазу, для синтеза 2',3-дифукозиллактозы экспрессируют как подходящую альфа-1,2-фукозилтрансферазу, так и альфа-1,3-фукозилтрансферазу, или по меньшей мере один ген, кодирующий белок, обладающий альфа-1,2-, а также альфа-1,3-фукозилтрансферазной активностью.
Неограничивающими примерами фукозилтрансфераз, которые могут быть использованы согласно изобретению и которые будут представлять собой часть изобретения, являются, например, бактериальные фукозилтрансферазы и предпочтительно альфа-1,2-фукозилтрансфераза, и более предпочтительно альфа-1,2-фукозилтрансфераза, кодируемая геном wbgL из Е. coli: O126, или альфа-1,2-фукозилтрансфераза, кодируемая геном fucT из Helicobacter pylori, или альфа-1,3-фукозилтрансфераза и более предпочтительно альфа-1,3-фукозилтрансфераза из Akkermansia miciniphila, Bacteroides fragilis, H. pylori или H. hepaticus. Предпочтительно используют гликозилтрансферазу или ее варианты, которые описаны в ЕР 2479263 А1, или в ЕР 2439264, или в WO 2010/142305, содержание которых при этом непосредственно относится к предмету данного изобретения и составляет его.
"Вариант" как термин, использованный в данном описании, относится к полинуклеотиду или полипептиду, который отличается от референсного полинуклеотида или полипептида, в частности, относится к ферменту, упомянутому и использованному в данном описании, соответственно, но сохраняет необходимые (ферментативные) свойства референсного полинуклеотида или полипептида. Типичный вариант полинуклеотида отличается по нуклеотидной последовательности от другого, референсного полинуклеотида. Изменения в нуклеотидной последовательности варианта могут менять или не менять аминокислотную последовательность полипептида, кодируемую референсным полинуклеотидом. Изменения нуклеотидной последовательности могут приводить к аминокислотным заменам, добавлениям, делециям, слияниям и укорочениям в полипептиде, кодируемом референсной последовательностью, как будет рассмотрено ниже. Типичный вариант полипептида отличается по аминокислотной последовательности от другого, референсного полипептида. Как правило, набор различий ограничен, так что последовательности референсного полипептида и варианта в целом очень схожи и во многих участках идентичны. Аминокислотные последовательности варианта и референсного полипептида могут различаться одной или несколькими заменами, одним или несколькими добавлениями, делециями в любой комбинации. Замененный или встроенный аминокислотный остаток может представлять собой или не представлять собой остаток, кодируемый генетическим кодом. Вариант полинуклеотида или полипептида может иметь природное происхождение, как например, аллельный вариант, или может представлять собой вариант, неизвестный в природе. Неприродные варианты полинуклеотидов и полипептидов могут быть созданы методами мутагенеза, методами прямого синтеза и другими методами рекомбинантной технологии, известными специалистам в данной области техники.
В объем настоящего изобретения под этими терминами также включены полиморфные варианты, аллели, мутанты и межвидовые гомологи нуклеиновых кислот/полинуклеотидов и полипептидов, имеющие аминокислотную последовательность/нуклеиновокислотную последовательность, обладающую идентичностью по аминокислотной последовательности более чем примерно на 60%, 65%, 70%, 75%, 80%, 85%, 90%, предпочтительно 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% либо более высокой идентичностью по аминокислотной последовательности, предпочтительно в пределах участка по меньшей мере примерно из 25, 50, 100, 200, 500, 1000 или более аминокислот, с упомянутым в данном описании полипептидом, например, с ферментом, преобразующим фруктозо-6-фосфат, в частности, фосфофруктокиназой А, глюкозо-6-фосфат-изомеразой, фруктозо-6-фосфат-альдолазой, транскетолазой, например tktA, tktB, или трансальдолазой, например, talA, talB, фруктозо-1,6-бифосфат-фосфатазой, использованной в данном описании, с фосфоманномутазой предпочтительно manB, маннозо-1-фосфат-гуанозилтрансферазой, предпочтительно manC, ГДФ-маннозо-4,6-дегидратазой, предпочтительно gmd, и ГДФ-L-фукозо-синтазой, предпочтительно wcaG, с фукозилтрансферазой, использованной в данном описании, с фруктозо-1,6-бифосфат-фосфатазой, предпочтительно функционально активным вариантом фруктозо-1,6-бифосфат-фосфатазы (fbpase) из Pisum sativum, с эффлюксным переносчиком сахара, например, yberc0001_9420 или SetA, и с пермеазой лактозы, например, LacY, используемой в данном описании.
Соответственно, подразумевается, что "функциональный фрагмент" любого из генов/белков, описанных в данной заявке, обозначает варианты последовательности генов/белков, все еще сохраняющих ту же или несколько меньшую активность гена или белка, из которого происходит соответствующий фрагмент.
Использованный в данном описании термин "эндогенный", как определено в данной заявке и как принято в данной области техники, означает, что нуклеиновая кислота, кодирующая представляющий интерес фермент, происходит из бактериальной клетки хозяина, а не введена в указанную клетку хозяина, тогда как "экзогенная" или "рекомбинантная" нуклеиновая кислота введена в указанную клетку хозяина, а не происходит из указанной клетки хозяина.
Согласно другому воплощению нуклеиновая кислота/ген бывают гомологичными или гетерологичными. В настоящей заявке и согласно общему пониманию в релевантной области техники термин "гомологичный" относится к нуклеиновокислотной последовательности/гену, которая(ый) кодирует конкретный(ые) продукт или продукты и происходит из того же вида, в который встроена указанная нуклеиновокислотная последовательность. Соответственно, термин "гетерологичный" относится к нуклеиновокислотной последовательности/гену, кодирующей(ему) конкретный(ые) продукт или продукты и происходящей(ему) из другого вида, чем тот, в который встроены указанная(ый) нуклеиновокислотная последовательность/ген.
Согласно другому воплощению клетка-хозяин по изобретению дополнительно содержит контрольные последовательности, позволяющие осуществлять регулируемую сверхэкспрессию эндогенных или экзогенных/рекомбинантных нуклеиновокислотных последовательностей/генов. Определенный выше термин "контрольная последовательность", который в данном описании используется как синоним выражения "контрольная последовательность экспрессии нуклеиновой кислоты/гена", относится к последовательности, содержащей промоторные последовательности, сигнальную последовательность или ряд сайтов связывания транскрипционных факторов, чьи последовательности влияют на транскрипцию и/или трансляцию нуклеиновокислотной последовательности или гена, функционально связанной(ого) с контрольными последовательностями.
В настоящей заявке термин "функционально связанный", использованный в данном описании, будет означать функциональную связь между контрольной последовательностью экспрессии нуклеиновой кислоты/гена (такой как промоторная, сигнальная последовательность или ряд сайтов связывания транскрипционных факторов) и второй нуклеиновокислотной последовательностью или геном, причем контрольная последовательность экспрессии влияет на транскрипцию и/или трансляцию нуклеиновой кислоты, соответствующей второй последовательности. Соответственно, термин "промотор" означает последовательности ДНК, которые обычно "предшествуют" гену" в полимерной молекуле ДНК и предоставляют сайт инициации транскрипции в мРНК. "Регуляторные" последовательности ДНК, обычно также расположенные "вверх по течению от" гена (т.е. предшествующие гену) в указанной полимерной молекуле ДНК, связывают белки, которые определяют частоту (или степень) инициации транскрипции. Совместно именуемые как "промоторная/регуляторная" или "контрольная" последовательность ДНК, эти последовательности, "предшествующие" выбранному гену (или серии генов) в функциональной полимерной молекуле ДНК, действуют совместно в отношении определения того, будет ли происходить транскрипция (и возможно экспрессия) гена. Последовательности ДНК, которые "следуют" за геном в полимерной молекуле ДНК и обеспечивают наличие сигнала терминации транскрипции в мРНК, именуются как последовательности "терминации" транскрипции.
Как уже было указано выше, нуклеиновокислотная последовательность/ген, используемые согласно изобретению, могут, например, содержаться в векторе, которым стабильно трансформируют/трансфицируют бактериальные клетки хозяина. Определения и подробное описание указанного выше получения рекомбинантов применимы к этому абзацу.
В некоторых воплощениях нуклеиновокислотную последовательность/ген помещают под контроль индуцибельного промотора, представляющего собой промотор, который направляет экспрессию гена, при этом уровень экспрессии может изменяться под действием факторов окружающей среды или связанных с развитием факторов, таких как, например, температура, рН, анаэробные или аэробные условия, свет, транскрипционные факторы и химические реагенты. Такие промоторы обозначаются в данном описании как "индуцибельные" промоторы, которые позволяют регулировать временной режим экспрессии белков, используемых в настоящем изобретении. Специалистам в данной области техники известны индуцибельные промоторы для Е. coli и других бактериальных клеток хозяина.
По всему описанию изобретения подразумевается, что термин "ген" означает линейную последовательность нуклеотидов (или нуклеиновокислотную последовательность; см. выше) располагающуюся в сегменте ДНК, предоставляющую кодируемые "инструкции" для синтеза РНК, трансляция которой в белок приводит к экспрессии белка/пептида. Белок/пептид может, как в настоящем изобретении, иметь определенные ферментативные функции. "Нуклеиновая кислота" относится к состоящему из дезоксирибонуклеотидов или рибонуклеотидов полимеру либо в одно-, либо в двухцепочечной форме и, при отсутствии иных ограничений, охватывает известные аналоги природных нуклеотидов, которые гибридизуются с нуклеиновыми кислотами аналогично природным нуклеотидам. Если не указано иное, взятая в отдельности нуклеиновокислотная последовательность содержит свою комплементарную последовательность. Если не указано иное, конкретная нуклеиновокислотная последовательность подразумевает и комплементарную ей последовательность.
Обычно термин "нуклеиновокислотная последовательность, кодирующая…" или "ген(ы), кодирующий/кодирующие…" относится к любому полирибонуклеотиду или полидезоксирибонуклеотиду, который может представлять собой немодифицированную РНК или ДНК либо модифицированную РНК или ДНК и, как правило, представляет собой ген, который кодирует определенный полипептид или белок. Термин включает в себя, без ограничения, одно- и двухцепочечную ДНК, ДНК, представляющую собой смесь из одно- и двухцепочечных участков или одно-, двух- и трехцепочечных участков, одно- и двухцепочечную РНК и РНК, представляющую собой смесь из одно- и двухцепочечных участков, гибридные молекулы, содержащие ДНК и РНК, которые могут представлять собой одноцепочечные, или, в более типичном случае, двухцепочечные, или трехцепочечные участки, либо смесь одно- и двухцепочечных участков. Данный термин также охватывает полинуклеотиды, включающие в себя единый непрерывный участок или прерывающиеся участки, кодирующие полипептид (например, прерванные интегрированным фагом или инсерционной последовательностью, или редактированием), вместе с дополнительными участками, которые также могут содержать кодирующие и/или некодирующие последовательности.
Соответственно, в настоящем изобретении термины "ген" и "нуклеиновокислотная последовательность" используются взаимозаменяемо.
Кроме того, подразумевается, что использованный в данном описании термин "активность" при ссылке на фермент включает в себя любую молекулу, проявляющую ферментативную активность и действующую как катализатор, чтобы вызвать специфическую биохимическую реакцию, оставаясь при этом неизменной в процессе реакции, в частности, белковую молекулу. В частности, подразумевается, что этим термином должны охватываться белки с ферментативной активностью, способные участвовать в превращении субстрата в продукт.
Молекулы, называемые субстратами в начале ферментативных реакций, превращаются в другие молекулы, называемые продуктами. Почти все химические реакции в биологической клетке нуждаются в ферментах для того, чтобы их протекание осуществлялось со скоростями, достаточными для поддержания жизни. Поскольку ферменты селективны в отношении своих субстратов и ускоряют только некоторые реакции среди множества возможных, набор ферментов, произведенных в клетке, определяет те метаболические пути, которые функционируют в этой клетке.
Соответственно, когда, согласно изобретению, активность фермента "подавлена" или "понижена", фермент не обладает той активностью, которую он имеет, если фермент или его экспрессия не изменены, т.е. в таком случае активность подавлена или понижена по сравнению с неизмененным(ой) ферментом/экспрессией фермента.
Авторы настоящего изобретения смогли предложить способ и генетически модифицированную клетку хозяина, посредством которых при получении фукозилированных олигосахаридов возможно достижение концентрации продукта, превышающей 100 г/л.
Согласно одному из воплощений изобретения, относящихся к способу или клетке хозяина по изобретению, активность фермента, преобразующего фруктозо-6-фосфат, которая в немодифицированной клетке хозяина, там, где это уместно, находится на обычном уровне, понижена или подавлена. В случае использования Е. coli в качестве клетки хозяина предпочтительно, чтобы для способа и клетки хозяина по изобретению фермент, преобразующий фруктозо-6-фосфат, был выбран из группы, состоящей из фосфофруктокиназы, предпочтительно фосфофруктокиназы A (PfKA), глюкозо-6-фосфат-изомеразы, фруктозо-6-фосфат-альдолазы, транскетолазы, предпочтительно tktA, tktB, или трансальдолазы, предпочтительно talA, talB, при этом фермент, преобразующий фруктозо-6-фосфат, будучи иным образом представленным и активным в немодифицированной клетке хозяина Е. coli, был модифицирован таким образом, чтобы его активность была понижена или подавлена.
PfkA эффективно фосфорилирует фруктозо-6-фосфат до фруктозо-1,6-бифосфата. Фруктозо-6-фосфат представляет собой точку ветвления в гликолитическом и глюконеогенном путях и в синтезе ГДФ-L-фукозы, который начинается с ManA-катализируемой изомеризации фруктозо-6-фосфата до маннозо-6-фосфата. В случае выращивания Е. coli на глюконеогенном субстрате, таком как глицерин, фосфорилирование фруктозо-6-фосфата под действием PfkA представляет собой типичную реакцию с большим потреблением АТФ, и в дополнение к этому наблюдается конкуренция с ManA за субстрат.
Согласно одному из воплощений способа и клетки хозяина по изобретению, ферментом, участвующим в образовании фруктозо-6-фосфата, является фруктозо-1,6-бифосфат-фосфатаза, активность которой может быть повышена с целью увеличения количества фруктозо-6-фосфата.
Согласно другому воплощению изобретения, по меньшей мере один ген, кодирующий фермент, необходимый для синтеза ГДФ-фукозы de novo, сверхэкспрессирован в клетке хозяина.
ГДФ-фукоза, которая упомянута выше, служит в качестве донора L-фукозы для реакции опосредованного фукозилтрансферазой переноса L-фукозы на субстрат-акцептор с образованием фукозилированного олигосахарида.
В случае прокариотической клетки хозяина, генетически модифицированной для собственного продуцирования ГДФ-фукозы, не требуется добавления извне L-фукозы, которая может быть превращена в ГДФ-фукозу посредством "реутилизационного" пути (salvage pathway), поскольку клетка-хозяин эффективно продуцирует ГДФ-фукозу, необходимую для процесса фукозилирования желаемого олигосахарида.
По меньшей мере один ген, кодирующий фермент, необходимый для синтеза ГДФ-фукозы de novo и сверхэкспрессируемый в клетке хозяина, может представлять собой эндогенный ген или экзогенный ген, который может быть интегрирован в геном клетки хозяина.
В одном из воплощений настоящего изобретения экзогенными генами, кодирующими ферменты, необходимые для синтеза ГДФ-фукозы de novo, являются: ген, кодирующий фосфоманномутазу, предпочтительно manB, ген, кодирующий маннозо-1-фосфат-гуанозилтрансферазу, предпочтительно manC, ген, кодирующий ГДФ-маннозо-4,6-дегидратазу, предпочтительно gmd, и ген, кодирующий ГДФ-L-фукозосинтазу, предпочтительно wcaG.
Согласно одному из воплощений изобретения и также как упомянуто выше, по меньшей мере один экзогенный ген, кодирующий фукозилтрансферазу, выбран из генов, экспрессия которых приводит к получению белка, проявляющего альфа-1,2-фукозилтрансферазную активность и/или альфа-1,3-фукозилтрансферазную активность. В этой связи особенно предпочтителен случай, если альфа-1,2-фукозилтрансфераза выбрана из группы, состоящей из wbgL из Е. coli: O126 или альфа-1,2-фукозилтрансферазы, кодируемой геном fucT 2 из Helicobacter pylori, и если ген, кодирующий альфа-1,3-фукозилтрансферазу, выбран из группы, состоящей из гена, кодирующего альфа-1,3-фукозилтрансферазу из видов Akkermansia muciniphila и Bacteroides fragilis, Helicobacter pylori или Helicobacter hepaticus.
Согласно другому воплощению изобретения предпочтительно, если клетку хозяина подвергают дальнейшей генетической модификации: (1) с целью экспрессирования гена, предпочтительно экзогенного гена, который кодирует белок, обеспечивающий экспорт или способствующий экспорту желаемого фукозилированного олигосахарида в культуральную среду; и/или (2) с целью экспрессирования экзогенного гена, кодирующего бифункциональную L-фукокиназу/L-фукозо-1-фосфат-гуанилилтрансферазу; и/или (3) с целью проявления генов fucI и fucK, имеющих мутации или делеции, в результате чего наблюдают понижение или подавление активностей L-фукозоизомеразы (FucI) и L-фукулозокиназы (FucK), и/или (4) с целью инактивирования или разрушения генов, кодирующих ферменты синтеза колановой кислоты; и/или (5) с целью экспрессирования, предпочтительно сверхэкспрессирования, эндогенной и/или экзогенной пермеазы для импорта лактозы; и/или (6) с целью инактивирования или делетирования эндогенных генов бета-галактозидазы; и/или (7) с целью экспрессирования гена, кодирующего бета-галактозидазу, предпочтительно экзогенно регулируемую бета-галактозидазу; и/или (8) с целью сверхэкспрессирования эндогенного и/или экзогенного гена, кодирующего фруктозо-1,6-бифосфат-фосфатазу.
С использованием дополнительной генетической модификации, как указано выше, можно еще более усовершенствовать способ получения фукозилированного олигосахарида.
С использованием делетирования или инактивирования генов, кодирующих L-фукозоизомеразу (например, FucI) и L-фукулозокиназу (например, FucK), можно избежать катаболизма внутриклеточной фукозы.
При разрушении, делетировании или инактивировании генов, кодирующих ферменты биосинтеза колановой кислоты (например, гена wcaJ, катализирующего первую стадию синтеза колановой кислоты, при использовании Е. coli в качестве клетки хозяина), предотвращается внутриклеточное продуцирование колановой кислоты, которое в противном случае может конкурировать в реакции с участием фукозилтрансферазы за субстрат ГДФ-L-фукозу.
Согласно предпочтительному воплощению, белком, который обеспечивает экспорт или способствует экспорту желаемого фукозилированного олигосахарида в культуральную среду, является эффлюксный переносчик сахара, предпочтительно выбранный из yberc0001_9420 и SetA из Е. coli.
Согласно предпочтительному воплощению, геном, кодирующим бифункциональную L-фукокиназу/L-фукозо-1-фосфат-гуанилилтрансферазу, является ген fkp из Bacteroides fragilis.
Согласно предпочтительному воплощению, фруктозо-1,6-бифосфат-фосфатаза кодируется геном, представляющим собой функционально активный вариант гена фруктозо-1,6-бифосфат-фосфатазы (fbpase) из Pisum sativum.
Согласно предпочтительному воплощению пермеазой лактозы является LacY из Е. coli.
В случае экспрессии гена, кодирующего бифункциональную L-фукокиназу/L-фукозо-1-фосфат-гуанилилтрансферазу, которая катализирует синтез ГДФ-L-фукозы, например, гена fkp из Bacteroides fragilis, предотвращается образование свободной L-фукозы, которая может накапливаться как побочный продукт вследствие гидролиза ГДФ-L-фукозы, тем самым сохраняя свободную L-фукозу для синтеза желаемого фукозилированного олигосахарида.
Согласно предпочтительному воплощению, экзогенными генами, ответственными за метаболизирование галактозы, являются гены, содержащие оперон galETKM и/или galP из Е. coli.
Согласно одному из воплощений изобретения, гены, которые модифицированы в клетке хозяина или которыми модифицируют клетку хозяина, являются эндогенными или экзогенными генами.
По всему описанию изобретения и применительно к каждому(ой) гену/нуклеиновой кислоте, которые были экзогенно введены в клетку хозяина, предпочтительно, согласно одному из воплощений способа и клетки хозяина по изобретению, чтобы по меньшей мере один из экзогенных генов, предпочтительно интегрированный в геном клетки хозяина, был сверхэкспрессирован, предпочтительно в результате эндогенного или экзогенного индуцирования либо конститутивным образом.
Соответственно, предпочтительно, если по меньшей мере один ген или несколько из следующих далее генов сверхэкспрессирован(ы): (1) экзогенные гены, кодирующие ферменты, необходимые для синтеза ГДФ-фукозы de novo; (2) экзогенный ген, кодирующий фукозилтрансферазу; (3) экзогенный ген, кодирующий эффлюксный переносчик сахара; (4) экзогенный ген, кодирующий бифункциональную L-фукокиназу/L-фукозо-1-фосфат-гуанилилтрансферазу; (5) ген, кодирующий пермеазу лактозы; (6) экзогенно регулируемый ген, кодирующий бета-галактозидазу; и/или (7) экзогенные гены, ответственные за метаболизирование галактозы; (8) экзогенный ген, кодирующий фруктозо-1,6-бисфосфат-фосфатазу; при этом сверхэкспрессия может быть вызвана, например, посредством регулируемого промотора, который инициирует транскрипцию данного(ых) гена(ов), либо в определенный момент времени, либо в течение промежутка времени в ходе культивирования, либо в течение всего времени культивирования.
Кроме того, согласно предпочтительному воплощению, экзогенные гены, которые подлежат введению в клетку хозяина, используемую в способе по изобретению, интегрируют в геном клетки хозяина.
Кроме того, согласно одному из аспектов изобретения и если не указано иное, гены, которые модифицированы в клетке хозяина/которыми модифицируют клетку хозяина, также могут, согласно изобретению, быть эндогенными генами, и их экспрессия может быть усилена, или повышена, или может наблюдаться сверхэкспрессия, либо в противном случае, может быть подавлена или снижена.
В случае инактивирования или делетирования эндогенного(ых) гена(ов) бета-галактозидазы расщепление добавляемой извне лактозы предотвращается; однако, поскольку желательно иметь расщепленную лактозу, которая не метаболизируется, и которая, в противном случае, затрудняет очистку желаемого фукозилированного олигосахарида, также предпочтительно, если экзогенно регулируемый ген, кодирующий бета-галактозидазу или мутированную форму бета-галактозидазы, экспрессируется в клетке хозяина. Например, может быть экспрессирован фрагмент lacZΩ гена lacZ, экспрессия которого, например, может регулироваться посредством репрессора, например, посредством чувствительного к температуре транскрипционного репрессора, например cI857. B этом случае синтез Ω-фрагмента бета-галактозидазы может быть инициирован путем повышения температуры до 42°С.
Репрессор, в настоящей заявке и согласно общему пониманию в релевантной области техники, представляет собой ДНК- или РНК-связывающий белок, который ингибирует экспрессию одного или более генов, связываясь с оператором или соответствующими сайленсерами. ДНК-связывающий репрессор блокирует присоединение РНК-полимеразы к промотору, предотвращая таким образом транскрипцию генов с образованием матричной РНК.
В качестве промотора для альфа-фрагмента экзогенной бета-галактозидазы можно использовать, например, промотор PgbA из Е. coli BL21 (DE3). α- и Ω-фрагменты бета-галактозидазы объединяют для получения активной бета-галактозидазы в клетке.
В предпочтительном воплощении лактозу предоставляют путем добавления лактозы от начала культивирования в концентрации по меньшей мере 5 мМ, более предпочтительно в концентрации, превышающей 10, 20, 30, 40, 50, 60, 70, 80, 90 или 100 мМ, либо еще более предпочтительно в концентрации 300 мМ или выше, чем 300 или 400 мМ.
Согласно еще одному воплощению лактозу предоставляют путем добавления лактозы в культуральную среду в такой концентрации, что в течение фазы образования продукта при культивировании достигается концентрация лактозы, составляющая по меньшей мере 5 мМ, более предпочтительно по меньшей мере 10, 20 или 30 мМ.
Альтернативно, лактоза может продуцироваться клеткой хозяина внутриклеточно, как описано в патенте ЕР 1927316 А1, содержание которого при этом однозначно упоминается и включено посредством ссылки.
В способе по изобретению предпочтительно, если клетки хозяина культивируют в течение по меньшей мере примерно 60, 80, 100 или примерно 120 часов либо в непрерывном режиме.
Так, согласно одному из аспектов изобретения, т.е. в случае способа непрерывного культивирования, источник углерода постоянно добавляют в среду при прохождении стадии культивирования клетки хозяина. В результате постоянного добавления источника углерода при прохождении стадии культивирования осуществляется непрерывное и эффективное продуцирование олигосахарида.
Согласно другому аспекту способ по изобретению представляет собой или включает метод периодического культивирования с подпиткой, при этом используют либо подпитываемую культуру постоянного объема, когда подпитку субстратом осуществляют без разбавления культуры, либо подпитываемую культуру вариабельного объема, когда ферментируемый объем изменяется в зависимости от продолжительности ферментирования вследствие подпитки субстратом.
Как упомянуто выше, настоящее изобретение также относится к генетически модифицированной прокариотической клетке хозяина, при этом клетка-хозяин генетически модифицирована таким образом, что (1) активность фермента, преобразующего фруктозо-6-фосфат, которая в немодифицированной клетке хозяина находится на обычном уровне, понижена или подавлена; (2) по меньшей мере один ген, кодирующий фермент, необходимый для синтеза ГДФ-фукозы de novo, сверхэкспрессирован; (3) экзогенный ген, кодирующий фукозилтрансферазу, предпочтительно ген, кодирующий альфа-1,2-фукозилтрансферазу и/или альфа-1,3-фукозилтрансферазу, экспрессирован в данной клетке.
Как упомянуто для приведенного выше способа, клетка-хозяин предпочтительно выбрана из штамма Escherichia coli, штамма Lactobacillus или штамма Corynebacterium.
Согласно одному из воплощений клетки хозяина по изобретению, внутриклеточное количество фруктозо-6-фосфата повышают посредством (1) понижения или подавления активности фермента, преобразующего фруктозо-6-фосфат, выбранного из группы фосфофруктокиназы, глюкозо-6-фосфат-изомеразы, фруктозо-6-фосфат-альдолазы, транскетолазы, например, tktA, tktB, или трансальдолазы, например, talA, talB, или (2) повышения активности фруктозо-1,6-бифосфат-фосфатазы.
В предпочтительном воплощении гены, кодирующие ферменты, необходимые для синтеза ГДФ-фукозы de novo, сверхэкспрессированы.
Еще в одном другом предпочтительном воплощении экзогенными генами, кодирующими по меньшей мере одну фукозилтрансферазу, являются гены, кодирующие альфа-1,2-фукозилтрансферазы и/или альфа-1,3-фукозилтрансферазы, и они выбраны из гена wbgL из Е. coli O126 или гена fucT2 из Helicobacter pylori, когда речь идет об альфа-1,2-фукозилтрансферазах, и из генов видов Akkermansia muciniphila, Bacteroides fragilis, Helicobacter pylori или Helicobacter hepaticus, когда речь идет об альфа-1,3-фукозилтрансферазах.
Согласно одному из воплощений клетка-хозяин, описанная выше, возможно подвергнута дальнейшей генетической модификации (4) с целью экспрессирования экзогенного гена, кодирующего эффлюксный переносчик сахара; и/или (5) с целью экспрессирования экзогенного гена, кодирующего бифункциональную L-фукокиназу/L-фукозо-1-фосфат-гуанилилтрансферазу; и/или (6) с целью инактивирования или делетирования генов, кодирующих L-фукозоизомеразу и L-фукулозокиназу; и/или (7) с целью инактивирования или разрушения генов, кодирующих УДФ(уридин-дифосфат)-глюкоза:ундекапренил-фосфат-глюкозо-1-фосфат-трансферазу; и/или (8) с целью экспрессирования экзогенной пермеазы лактозы; и/или (9) с целью инактивирования или делетирования эндогенных генов бета-галактозидазы; и/или (10) с целью экспрессирования экзогенно регулируемого гена, кодирующего бета-галактозидазу; и/или (11) с целью экспрессирования экзогенных генов, ответственных за метаболизирование галактозы; и/или с целью экспрессирования экзогенного гена, кодирующего фруктозо-1,6-бисфосфат-фосфатазу.
Настоящее изобретение также относится к применению генетически модифицированной прокариотической клетки хозяина по изобретению для получения фукозилированного олигосахарида.
Другие преимущества следуют из описания воплощений и приложенных графических материалов.
Очевидно, что вышеупомянутые признаки и признаки, которые еще подлежат разъяснению ниже, можно использовать не только в соответственно конкретизированных сочетаниях, но также и в других сочетаниях или самостоятельно, без отклонения от объема настоящего изобретения.
КРАТКОЕ ОПИСАНИЕ ФИГУР
Некоторые воплощения изобретения проиллюстрированы на фигурах и разъяснены более подробно в следующей далее части описания. На фигурах показаны:
Фиг. 1. Схематичная типичная иллюстрация генетически модифицированной клетки хозяина, подлежащей применению в способе по изобретению;
Фиг. 2. Результаты анализов супернатантов из глицерин-содержащих культур, в которых выращивали 2'-фукозиллактоза-продуцирующие штаммы Е. coli, с использованием высокоэффективной жидкостной хроматографии (HPLC); и
Фиг. 3. SEQ ID No. 1-7.
ПОДРОБНОЕ ОПИСАНИЕ ФИГУР И ВОПЛОЩЕНИЙ
На Фиг. 1 показана типичная иллюстрация клетки хозяина по изобретению, подлежащей применению в способе по изобретению, при этом изображены типичные пути ферментирования типичных фукозилированных олигосахаридов - 2'-фукозиллактозы. На Фиг. 1 показана типичная бактериальная клетка-хозяин, генетически модифицированная согласно изобретению в отношении продуцирования 2'-фукозиллактозы.
Как можно видеть на Фиг. 1, обычно в качестве источника углерода используют глицерин, в то время как лактозу добавляют извне. Лактоза транспортируется в клетку хозяина посредством пермеазы (например, LacY). Глицерин переносится в клетку хозяина облегченной диффузией посредством GlpF. Внутри прокариотической клетки хозяина глицерин превращается в глицеральдегид-3-фосфат, который превращается в фруктозо-6-фосфат (1) предпочтительно в результате сверхэкспрессии экзогенного гена, кодирующего Fbpase, и (2) благодаря ингибированию обратной реакции посредством инактивации фосфофруктокиназы A (PfkA). Благодаря сверхэкспрессии экзогенных ферментов, необходимых для синтеза ГДФ-фукозы de novo, т.е. фосфоманномутазы, ManB, маннозо-1-фосфат-гуанозилтрансферазы, ManC, ГДФ-маннозо-4,6-дегидратазы, Gmd, и ГДФ-L-фукозосинтазы, WcaG, продуцируется ГДФ-L-фукоза.
На следующей стадии ГДФ-L-фукоза под действием альфа-1,2-фукозилтрансферазы, например, WbgL, взаимодействует с интернализированной лактозой с получением 2-фукозиллактозы, которая экспортируется посредством эффлюксного переносчика, например TPYb, в среду, в которой культивируют клетку хозяина.
На Фиг. 2 показаны результаты анализов супернатантов из глицерин-содержащих культур, в которых выращивали 2'-фукозиллактоза-продуцирующие штаммы Е. coli, с использованием HPLC.
На Фиг. 2 изображен HPLC-профиль образца бульона после ферментации 2'-фукозиллактоза-продуцирующего штамма, несущего ген yberc0001_9420, кодирующий гетерологичный переносчик (черный цвет), и HPLC-профиль образца бульона после ферментации того же штамма, но с делецией гена yberc0001_9420, кодирующего гетерологичный переносчик (серый цвет). Ферментацию обоих штаммов проводили в течение 111 ч при 28°С, используя глицерин в качестве источника углерода и энергии.
Пример 1
Конструирование штамма Е. coli BL21(DE3) для получения 2'-фукозиллактозы
Используя Е. coli BL21(DE3) в качестве исходного штамма-хозяина, конструировали штамм для получения 2'-фукозиллактозы методом биосинтеза с применением цельных клеток. Геномное конструирование такого штамма включало события разрушения и делетирования генов и интеграцию гетерологичных генов.
Поскольку 2'-фукозиллактоза синтезируется из лактозы, которую применяют при культивировании бактерий, и из ГДФ-L-фукозы, которая продуцируется живыми клетками, сначала копию гена lacZ дикого типа, кодирующего эндогенную β-галактозидазу, инактивировали посредством мутагенеза с использованием ошибочно спаривающихся олигонуклеотидов (см. Ellis et al., "High efficiency mutagenesis, repair, and engineering of chromosomal DNA using single-stranded oligonucleotides", Proc. Natl. Acad. Sci. USA, 98: 6742-6746 (2001)). С использованием этого же способа осуществляли разрушение гена, кодирующего арабиноза-изомеразу, araA.
Ген фрагмента lacZΩ вводили под контролем чувствительного к температуре транскрипционного репрессора cI857. Ген фрагмента lacZα экспрессируется под контролем промотора PgbA из Е. coli BL21 (DE3) в этом штамме, с получением в результате этого штамма LacZ+.
Геномные делетирования осуществляли с использованием опосредуемой фагом λ Red-зависимой рекомбинации в соответствии со способом Datsenko и Warner (см. "One-step inactivation of chromosomal genes in Escherichia coli K-12 using PCR products", Proc. Natl. Acad. Sci. USA, 97: 6640-6645 (2000)). Гены fucI и fucK, кодирующие L-фукозоизомеразу и L-фукулозокиназу, соответственно, были делетированы для предотвращения деградации L-фукозы. Также были делетированы гены wzxC-wcaJ. По всей вероятности ген wcaJ кодирует УДФ-глюкоза:ундекапренил-фосфат-глюкозо-1-фосфат-трансферазу, катализирующую первую стадию синтеза колановой кислоты (см. Stevenson et al., "Organization of the Escherichia coli K-12 gene cluster responsible for production of the extracellular polysaccharide colonic acid", J. Bacteriol., 178: 4885-4893, (1996)); в случае ГДФ-фукозы получение колановой кислоты будет осуществлено с применением реакции с участием фукозилтрансферазной активности.
Интегрирование в геном гетерологичных генов выполняли посредством транспозиции. Осуществляли интегрирование в геном больших генных кластеров, опосредованное гиперактивной С9-мутантной формой транспозазы mariner Himar1 (см. Lampe et al., "Hyperactive transposase mutants of the Himar1 mariner transposon", Proc. Natl. Acad. Sci. USA, 96: 11428-11433 (1999)), ген которой был встроен в плазмиду pEcomar под транскрипционным контролем промотора Para. Чтобы усилить синтез ГДФ-фукозы de novo, гены, кодирующие фосфоманномутазу (manB), маннозо-1-фосфат-гуанозилтрансферазу (manC), ГДФ-маннозо-4,6-дегидратазу (gmd) и ГДФ-L-фукозосинтазу (wcaG) из Е. coli K12 DH5α сверхэкспрессировали в штамме Е. coli BL21(DE3); оперон manCB помещали под контроль конститутивного промотора Ptet, оперон gmd, wcaG транскрибируется, начиная с конститутивного промотора РТ5. Транспозонную кассету <Ptet-manCB-РТ5-gmd, wcaG-FRT-dhfr-FRT> (SEQ ID No. 1), включающую в себя ген дигидрофолатредуктазы (dhfr) для приобретения устойчивости к триметоприму, фланкированную инвертированными концевыми повторами, специфично распознаваемыми mariner-подобным элементом транспозазы Himar1, встраивали в геном Е. coli от pEcomar C9-manCB-gmd, wcaG-dhfr.
Для хромосомной интеграции одиночных генов использовали транспозазу EZ-Tn5™ (Epicentre, USA). Чтобы получить EZ-Tn5 транспосомы, представляющий интерес ген вместе с FRT-сайтом, фланкированным кассетой устойчивости к антибиотикам, амплифицировали вместе с праймерами, которые несли на обоих сайтах состоящие из 19 пар оснований (п.о.) сайты распознавания - мозаичные концы (5'-CTGTCTCTTATACACATCT (SEQ ID No. 8)) для транспозазы EZ-Tn5. Используя транспозазу EZ-Tn5™, интегрировали ген, кодирующий импортер лактозы LacY из Е. coli K12 TG1 (номер доступа ABN72583), ген 2-фукозилтрансферазы, wbgL, из Е. coli: O126 (номер доступа ADN43847) и ген yberc0001_9420, кодирующий эффлюксный переносчик сахара из главного транспортерного суперсемейства из Yersinia bercovieri, АТСС (Американская коллекция типовых структур) 43970 (номер доступа EEQ08298) с применением соответствующих интеграционных кассет: <Ptet-lacY-FRT-aadA-FRT> (SEQ ID No. 2), <Ptet-wbgLcoFRT-neoFRT> (SEQ ID No. 3) и Ptet-yberc0001_9420coFRT-cat-FRT> (SEQ ID No. 4), получая штамм. Гены wbgL и yberc0001_9420 были получены путем синтеза и оптимизированы по кодонам (одновременно) компанией GenScript Cooperation (USA). После успешного интегрирования гена lacY ген устойчивости удаляли из стрептомицин-устойчивых клонов под действием рекомбиназы FLP, кодируемой плазмидой рСР20 (Datsenko and Warner, "One-step inactivation of chromosomal genes in Escherichia coli K-12 using PCR products", Proc. Natl. Acad. Sci. USA, 97: 6640-6645 (2000)).
Поскольку в E. coli BL21(DE3) отсутствует функциональный gal-оперон, в штамм В интегрировали естественным образом регулируемую копию оперона galETKM из Е. coli K посредством EZ-транспозиции с использованием интеграционной кассеты <Pgal-galE-galT-galK-galM> (SEQ ID No. 5). Прошедшие интегрирование образцы отбирали из агара Мак-Конки, содержащего 1% галактозы, в виде колоний красного цвета. Полученный штамм способен метаболизировать моносахариды глюкозу и галактозу, образующиеся при гидролизе лактозы.
Пример 2
Проверка усиленного экспорта 2'-фукозиллактозы под действием эффлюксного переносчика сахара из Yersinia bercovieri, АТСС 43970
Нокаут гена yberc0001_9420
Чтобы продемонстрировать функциональность гетерологичного переносчика сахара из Yersinia bercovieri, АТСС 43970, ген yberc0001_9420 делетировали из штамма Е. coli BL21(DE3) lacZ-, araA-, fucI-, fucK-, wcaJ-, который содержал хромосомные интеграции генов manB, manC, gmd, wcaG, lacY, wbgL; и yberc0001_9420 делетировали посредством гетерологичной рекомбинации согласно Datsenko и Wanner (2000; см. выше) с использованием кассеты устойчивости к гентамицину аасС1 из плазмиды pBBR-MCS5 (Kovach, Elzer et al. 1995, "Four new derivatives of the broad-host-range cloning vector pBBR1MCS, carrying different antibiotic-resistance cassettes", Gene, 166, 175-176), которую встраивали в ген yberc0001_9420, получая штамм Δyberc0001_9420.
Условия культивирования для получения 2'-фукозиллактозы
Штамм Е. coli BL21 (DE3), несущий ген yberc0001_9420, кодирующий гетерологичный переносчик, и штамм Δyberc0001_9420 культивировали при 28°С в ферментерах емкостью 3 л (New Brunswick, Edison, USA), исходя из 800 мл среды с минеральными солями, содержащей NH4H2PO4 - 7 г/л, K2HPO4 - 7 г/л, KOH - 2 г/л, лимонную кислоту - 0,3 г/л, MgSO4×7H2O - 2 г/л и CaCl2×6H2O - 0,015 г/л, дополненной раствором микроэлементов из расчета 1 мл/л (содержащим цитрат аммония-железа(III) - 54,4 г/л, MnCl2×4H2O - 9,8 г/л, CoCl2×6H2O - 1,6 г/л, CuCl2×2H2O - 1 г/л, H3BO3 - 1,9 г/л, ZnSO4×7H2O - 9 г/л, Na2MoO4×2H2O -1,1 г/л, Na2SeO3 - 1,5 г/л, NiSO4×6H2O - 1,5 г/л), содержащей 1,5% глицерина в качестве источника углерода и антибиотики триметоприм (10 мкг/мл) и канамицин (15 мкг/мл). Культивирование начинали, используя 2,5% (об./об.) инокулята из прекультуры, растущей в той же глицерин-содержащей среде. Лактозу в качестве акцептора в реакции с участием фукозилтрансферазы добавляли в течение семи часов, получая концентрацию 30 мМ в культуральной смеси, начиная при значении оптической плотности при 660 нм (OD660 нм), составляющем примерно 10. Затем вручную корректировали содержание лактозы для поддержания избытка акцепторной молекулы; глицерин добавляли в непрерывном режиме.
Анализ культурального супернатанта и обнаружение 2'-фукозиллактозы посредством HPLC
Анализ посредством высокоэффективной жидкостной хроматографии (HPLC) проводили, используя рефрактометрический детектор (RID-10A) (Shimadzu, Germany) и колонку ReproSil Carbohydrate, 5 мкм (250 мм × 4,6 мм) (Dr. Maisch GmbH, Germany), присоединенные к HPLC-системе (Shimadzu, Germany). Элюирование выполняли в изократическом режиме с использованием смеси ацетонитрил : H2O (68/32 (об./об.)) в качестве элюента при 35°С и скорости потока 1,4 мл/мин. На колонку наносили по 20 мкл образца. Концентрацию 2'-фукозиллактозы рассчитывали из стандартной кривой. С этой целью в HPLC-образцы добавляли 10% (об./об.) 100 мМ раствора сахарозы в качестве внутреннего стандарта, после чего фильтровали (размер пор 0,22 мкм) и очищали посредством твердофазной экстракции на ионообменной матрице (Strata ABW, Phenomenex).
Обнаружение 2'-фукозиллактозы в супернатантах культур Е. coli BL21(DE3)
Через 111 ч ферментирования при 28°С в среде с минеральными солями и глицерином в качестве источника углерода посредством HPLC обнаруживали 2'-фукозиллактозу в концентрации 73 мМ (35,6 г/л) и 25 мМ (12,2 г/л) в культуральном супернатанте штаммов, у которых имелся и отсутствовал ген yberc0001_9420, кодирующий переносчик, см. Фиг 2: на Фиг. 2 изображен HPLC-профиль для бульона после ферментации 2'-фукозиллактоза-продуцирующего штамма, несущего ген yberc0001_9420, кодирующий гетерологичный переносчик (черный цвет), и HPLC-профиль для бульона после ферментации того же штамма, но с делецией гена yberc0001_9420, кодирующего гетерологичный переносчик (серый цвет). Делетирование в штамме гена yberc0001_9420, кодирующего гетерологичный переносчик сахара, уменьшает обнаруживаемое количество 2'-фукозиллактозы в супернатанте. Это свидетельствует о том, что белок-переносчик действительно усиливает получение 2'-фукозиллактозы посредством более быстрого транспорта данного трисахарида за пределы клетки, поскольку генетическое окружение за исключением гена yberc0001_9420, идентично в обоих штаммах. Кроме того, более низкой плотности клеток достигали в случае клеток, не имеющих переносчика 2'-фукозиллактозы, вероятно из-за осмотического стресса, вызванного существенным накоплением сахара внутри клеток. Как показано на Фиг. 2, количество 2',3-дифукозиллактозы, обнаруживаемое в культуре штамма Δyberc0001_9420, примерно в два раза больше, чем в бульоне с исходным штаммом. Повышенное продуцирование 2',3-дифукозиллактозы, когда L-фукоза переносится на 2'-фукозиллактозу в катализируемой фукозилтрансферазой реакции, также свидетельствует о более высоких внутриклеточных концентрациях акцепторной молекулы 2'-фукозиллактозы в штамме с нокаутом гена yberc0001_9420 по сравнению со штаммом, сверхэкспрессирующим yberc0001_9420.
На Фиг. 2 более светлыми линиями, т.е. серыми линиями, изображен супернатант штамма Δyberc0001_9420 Е. coli BL21(DE3) Δyberc0001_9420, черными линиями, изображен супернатант из культуры содержащего ген yberc0001_9420 штамма Е. coli BL21(DE3). Образцы отбирали через 111 ч ферментирования при 28°С в среде с минеральными солями с использованием глицерина в качестве источника углерода.
Пример 3
Получение 2'-фукозиллактозы Ферментативным способом
Процессы ферментации проводили в ферментерах емкостью 3 л при 30°С и при рН 7,0; значение рН регулировали титрованием 25%-ным раствором аммиака. Штамм, описанный в примере 2, культивировали в среде с минеральными солями, описанной в примере 2, используя глицерин в качестве источника углерода и энергии. Содержимое ферментера с начальным объемом 1 л инокулировали прекультурой, культивированной в этой же среде. После расходования 2% глицерина, содержащегося в данной порции, глицерин (60%, об./об.) подавали в непрерывном режиме. Лактозу в концентрации 0,66 М добавляли тремя порциями (с интервалом в один час) по 10 мл каждая, когда достигали значение OD600 нм, равное 6. После этого в непрерывном режиме подавали лактозу для поддержания концентрации лактозы в ферментере по меньшей мере 10 мМ. Через 86 ч культивирования достигали конечной концентрации 2'-фукозиллактозы, составляющей 91,3 мМ (44,6 г/л). В результате подъема температуры до 42°С экспрессируется ген β-галактозидазы, и лактоза и продукты ее расщепления глюкоза и галактоза метаболизируются штаммом, продуцирующим 2'-фукозиллактозу.
Пример 4
HPLC-анализ культурального супернатанта
Анализ посредством HPLC проводили, используя рефрактометрический детектор (RID-10A) (Shimadzu, Germany) и колонку с амидным сорбентом XBridge, 3,5 мкм (250×4,6 мм) от Waters (Eschborn, Germany), соединенные с системой HPLC (Shimadzu, Germany). Элюирование выполняли в изократическом режиме, используя в качестве элюента смесь 30% А: 50% (об./об.) ACN (ацетонитрил) в деионизованной воде (ddH2O), с 0,1% (об./об.) NH4OH, и 70% В: 80% (об./об.) ACN в ddH2O, с 0,1% (об./об.) NH4OH, при 35°С и со скоростью потока 1,4 мл/мин. На колонку наносили 10 мкл образца и концентрацию 2'-фукозиллактозы рассчитывали по стандартной кривой. С этой целью в HPLC-образцы добавляли 10% (об./об.) 100 мМ раствора сахарозы в качестве внутреннего стандарта перед фильтрованием (размер пор 0,22 мкм) и очисткой посредством твердофазной экстракции на ионообменной матрице (Strata ABW, Phenomenex). С использованием тех же условий анализа также обнаруживали побочные продукты, такие как L-фукоза, 3-фукозиллактоза, 2',3-дифукозиллактоза и фукозилгалактоза.
Пример 5
Улучшение 2'-фукозиллактоза-продуцирующего штамма с использованием метаболического конструирования
Дальнейшего улучшения, касающегося синтеза 2'-фукозиллактозы с использованием штамма Е. coli, достигали посредством делетирования гена pfkA, кодирующего фосфофруктокиназу А. При культивировании Е. coli на глюконеогенном субстрате, таком как глицерин, фосфорилирование фруктозо-6-фосфата под действием PfkA представляет собой типичную реакцию с большим потреблением АТФ, и в дополнение к этому наблюдается конкуренция с ManA за субстрат. Ген pfkA делетировали с использованием гетерологичной рекомбинации согласно Datsenko и Wanner (2000, см. выше), используя кассету устойчивости к гентамицину (аасС1), фланкированную сайтами lox71/66 (см. Lambert, Bongers et al., 2007, "Cre-lox-based system for multiple gene deletions and selectable-marker removal in Lactobacillus plantarum", Appl. Environ. Microbial., 73, 1126-113). После успешного делетирования гена pfkA, из генома Е. coli удаляли ген устойчивости к антибиотику, используя рекомбиназу Cre (см. Abremski, Hoess et al., 1983, "Studies on the properties of P1 site-specific recombination: evidence for topological-L unlinked products following recombination", Cell, 32, 1301-1311), которая была клонирована под контролем промотора Para в каркасе плазмиды pKD46 (см. Datsenko и Wanner, 2000).
Для различных фукозилтрансфераз помимо трансферазной активности демонстрировали ГДФ-L-фукозогидролазную активность. Кроме того, такая гидролитическая активность была показана для wbgL, альфа-1,2-фукозилтрансферазы, используемой для синтеза 2'-фукозиллактозы (см. ЕР 3050973 А1). Чтобы сохранить свободную L-фукозу для получения 2'-фукозиллактозы и устранить загрязняющую L-фукозу из культурального бульона, ген fkp, кодирующий бифункциональную L-фукокиназу/L-фукозо-1-фосфат-гуанилилтрансферазу из Bacteroides fragilis, под транскрипционным контролем промотора Ptet, вместе с геном аасС1, фланкированным lox71/66, интегрировали в хромосому штамма, описанного в примере 1, посредством транспозиции с использованием транспозазы EZ-Tn5™; <Ptet-fkp-lox-aacC1-lox> (SEQ ID 6). После успешного интегрирования ген устойчивости к гентамицину удаляли из генома так, как описано выше.
Пример 6
Оптимизированный способ ферментации для получения 2'-фукозиллактозы
Используя оптимизированную среду с минеральными солями, которая содержит KH2PO4 - 3 г/л, K2HPO4 - 12 г/л, (NH4)2SO4 - 5 г/л, лимонную кислоту - 0,3 г/л, MgSO4×7H2O - 2 г/л, NaCl - 0,1 г/л и CaCl2×6H2O - 0,015 г/л, вместе с раствором микроэлементов из расчета 1 мл/л (содержащим цитрат аммония-железа(III) - 54,4 г/л, MnCl2×4H2O - 9,8 г/л, CoCl2×6H2O - 1,6 г/л, CuCl2×2H2O - 1 г/л, Н3ВО3 - 1,9 г/л, ZnSO4×7H2O - 9 г/л, Na2MoO4×2H2O - 1,1 г/л, Na2SeO3 - 1,5 г/л, NiSO4×6H2O - 1,5 г/л), и 2% глицерина в качестве источника углерода, штамм Е. coli, описанный в примере 5, культивировали в ферментере емкостью 3 л при 33°С. Значение рН поддерживали на уровне 7,0 путем титрования 25%-ным раствором аммиака. Содержимое ферментера инокулировали прекультурой с OD600нм 0,1, растущей в той же среде. Лактозу добавляли, когда для культуры наблюдали значение OD600нм, равное 5, с получением концентрации 30 мМ. На протяжении всего процесса ферментации поддерживали концентрацию лактозы 20-30 мМ, что контролировали с использованием HPLC-анализов. Подпитку глицерином (60%, об./об.) начинали после расходования глицерина в содержимом ферментера со скоростями потока 4,5 мл/л/ч в течение 20 часов, после чего подпитку осуществляли в течение 33 часов при 5,7 мл/л/ч и в течение 18 часов при 7,1 мл/л/ч в течение периода, составляющего 18 часов (скорости подпитки относятся к начальному объему). В целом через 93 ч достигали концентрации 2'-фукозиллактозы 106,5 г/л (217 мМ).
Пример 7
Конструирование улучшенного продуцирующего 2'-фукозиллактозу штамма посредством стимулирования метаболической активности
Чтобы усилить поток метаболизируемого источника углерода - глицерина через глюконеогенный путь от триозофосфатов к фруктозо-6-фосфату для подпитки биосинтеза ГДФ-L-фукозы, гены, кодирующие фруктозо-1,6-бифосфат-альдолазу (fbaB) и гетерологичный ген фруктозо-1,6-бифосфат-фосфатазы (fbpase) из Pisum sativum сверхэкспрессировали в штамме, описанном в примере 5. Выполняли слияние гена fbaB из Е. coli BL21 (DE3) с промотором Ptet. Активность FBPase из хлоропластов P. sativum аллостерически регулируется посредством дисульфид-дитиольного обмена, обусловленного восстановлением с участием тиоредоксинов. Замена остатка цистеина 153 на серии приводит к получению конститутивно активного фермента. Ген, кодирующий FBPase из хлоропластов P. sativum (номер доступа AAD10213), оптимизированный по кодонам для экспрессии в Е. coli, с гексагистидиновой меткой на N-конце и модифицированный для кодирования C153S-варианта фермента, приобретали у Genescript. Ген fbpase транскрибируется с промотора Т7. Кассету <Ptet-fbaB-PT7-His6-fbpase-lox-aacC1-lox> (SEQ ID No. 7) использовали для опосредуемого транспозазой EZ-Tn5™ интегрирования в штамм хозяина. После удаления гена устойчивости к гентамицину из генома Е. coli этот штамм использовали для получения 2'-фукозиллактозы.
Пример 8
Получение 2'-фукозиллактозы в концентрации 150 г/л ферментативным способом
Штамм, продуцирующий 2'-фукозиллактозу, генетически модифицированный так, как описано в примере 7, культивировали в той же среде при 33°С, как описано в примере 5. Кроме того, первоначально в среду для ферментации добавляли порцию 60 мМ лактозы в 2%-ом глицерине. Непрерывную подпитку лактозой с концентрацией по лактозе 0,66 М начинали при значении OD600нм, составляющем примерно 10. Кроме того, добавление лактозы осуществляли, используя 1 М концентрированный раствор. Концентрацию лактозы поддерживали при значении приблизительно 30 мМ. После окончания фазы порционной подпитки, на что указывало повышение уровня растворенного кислорода, начинали подпитку глицерином (60%, об./об.) со скоростью потока 6,9 мл/л/ч в течение 37 часов (относительно начального объема). После этого подпитку снижали до 9,4 мл/л/ч в течение 19 часов и затем снова повышали до 7,3 мл/л/ч в течение 19 часов. Через 93 часа после внесения затравки в ферментер достигали концентрации 2'-фукозиллактозы, составляющей 150,2 г/л.
Пример 9
Получение 3-фукозиллактозы из глицерина
С использованием Е. coli BL21 (DE3) с lacZ ΔwcaJ ΔfucIK с хромосомной интеграцией генов, кодирующих ферменты для синтеза ГДФ-фукозы de novo (ManB, ManC, Gmd, WcaG), конструировали 3-фукозиллактоза-продуцирующий штамм.
Ген, кодирующий альфа-1,3-фукозилтранферазу из Bacteroides fragilis (ЕР 2439264 А1), вместе с геном, кодирующим эффлюксный переносчик сахара SetA из Е. coli (US 2014/0120611 А1), и геном, придающим устойчивость к гентамицину, интегрировали в геном Е. coli. Ферментацию штамма с целью продуцирования 3 фукозиллактозы проводили в условиях, описанных в примере 6. Подпитку глицерином начинали после окончания фазы порционной подпитки со скоростью подпитки 7,4 мл/л/ч (относительно начального объема). В культуральную смесь добавляли лактозу до концентрации 33 мМ, когда достигали значения OD600нм, составляющего 30. На протяжении всего процесса добавляли лактозу, чтобы поддерживать ее концентрацию в супернатанте по меньшей мере 10 мМ. Через 88 ч процесс останавливали при концентрации 3-фукозиллактозы в супернатанте 30 г/л.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> ЙЕННЕВАЙН БИОТЕХНОЛОГИ ГМБХ
<120> Улучшенный способ получения фукозилированных
олигосахаридов
<130> 2827P112EP
<140> EP 16 196 486.1
<141> 2016-10-29
<160> 8
<170> PatentIn version 3.5
<210> 1
<211> 6783
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Транспозонная кассета
<400> 1
gccagatgat taattcctaa tttttgttga cactctatca ttgatagagt tattttacca 60
ctccctatca gtgatagaga aaagtgaaat gaatagttcg acaaaaatct agaaataatt 120
ttgtttaact ttaagaagga gatatacaat ttcgtcgaca cacaggaaac atattaaaaa 180
ttaaaacctg caggagtttg aaggagatag aaccatggcg cagtcgaaac tctatccagt 240
tgtgatggca ggtggctccg gtagccgctt atggccgctt tcccgcgtac tttatcccaa 300
gcagttttta tgcctgaaag gcgatctcac catgctgcaa accaccatct gccgcctgaa 360
cggcgtggag tgcgaaagcc cggtggtgat ttgcaatgag cagcaccgct ttattgtcgc 420
ggaacagctg cgtcaactga acaaacttac cgagaacatt attctcgaac cggcagggcg 480
aaacacggca cctgccattg cgctggcggc gctggcggca aaacgtcata gcccggagag 540
cgacccgtta atgctggtat tggcggcgga tcatgtgatt gccgatgaag acgcgttccg 600
tgccgccgtg cgtaatgcca tgccatatgc cgaagcgggc aagctggtga ccttcggcat 660
tgtgccggat ctaccagaaa ccggttatgg ctatattcgt cgcggtgaag tgtctgcggg 720
tgagcaggat atggtggcct ttgaagtggc gcagtttgtc gaaaaaccga atctggaaac 780
cgctcaggcc tatgtggcaa gcggcgaata ttactggaac agcggtatgt tcctgttccg 840
cgccggacgc tatctcgaag aactgaaaaa atatcgcccg gatatcctcg atgcctgtga 900
aaaagcgatg agcgccgtcg atccggatct caattttatt cgcgtggatg aagaagcgtt 960
tctcgcctgc ccggaagagt cggtggatta cgcggtcatg gaacgtacgg cagatgctgt 1020
tgtggtgccg atggatgcgg gctggagcga tgttggctcc tggtcttcat tatgggagat 1080
cagcgcccac accgccgagg gcaacgtttg ccacggcgat gtgattaatc acaaaactga 1140
aaacagctat gtgtatgctg aatctggcct ggtcaccacc gtcggggtga aagatctggt 1200
agtggtgcag accaaagatg cggtgctgat tgccgaccgt aacgcggtac aggatgtgaa 1260
aaaagtggtc gagcagatca aagccgatgg tcgccatgag catcgggtgc atcgcgaagt 1320
gtatcgtccg tggggcaaat atgactctat cgacgcgggc gaccgctacc aggtgaaacg 1380
catcaccgtg aaaccgggcg agggcttgtc ggtacagatg caccatcacc gcgcggaaca 1440
ctgggtggtt gtcgcgggaa cggcaaaagt caccattgat ggtgatatca aactgcttgg 1500
tgaaaacgag tccatttata ttccgctggg ggcgacgcat tgcctggaaa acccggggaa 1560
aattccgctc gatttaattg aagtgcgctc cggctcttat ctcgaagagg atgatgtggt 1620
gcgtttcgcg gatcgctacg gacgggtgta aacgtcgcat caggcaatga atgcgaaacc 1680
gcggtgtaaa taacgacaaa aataaaattg gccgcttcgg tcagggccaa ctattgcctg 1740
aaaaagggta acgatatgaa aaaattaacc tgctttaaag cctatgatat tcgcgggaaa 1800
ttaggcgaag aactgaatga agatatcgcc tggcgcattg gtcgcgccta tggcgaattt 1860
ctcaaaccga aaaccattgt gttaggcggt gatgtccgcc tcaccagcga aaccttaaaa 1920
ctggcgctgg cgaaaggttt acaggatgcg ggcgttgacg tgctggatat tggtatgtcc 1980
ggcaccgaag agatctattt cgccacgttc catctcggcg tggatggcgg cattgaagtt 2040
accgccagcc ataatccgat ggattataac ggcatgaagc tggttcgcga gggggctcgc 2100
ccgatcagcg gagataccgg actgcgcgac gtccagcgtc tggctgaagc caacgacttt 2160
cctcccgtcg atgaaaccaa acgcggtcgc tatcagcaaa tcaacctgcg tgacgcttac 2220
gttgatcacc tgttcggtta tatcaatgtc aaaaacctca cgccgctcaa gctggtgatc 2280
aactccggga acggcgcagc gggtccggtg gtggacgcca ttgaagcccg ctttaaagcc 2340
ctcggcgcgc ccgtggaatt aatcaaagtg cacaacacgc cggacggcaa tttccccaac 2400
ggtattccta acccactact gccggaatgc cgcgacgaca cccgcaatgc ggtcatcaaa 2460
cacggcgcgg atatgggcat tgcttttgat ggcgattttg accgctgttt cctgtttgac 2520
gaaaaagggc agtttattga gggctactac attgtcggcc tgttggcaga agcattcctc 2580
gaaaaaaatc ccggcgcgaa gatcatccac gatccacgtc tctcctggaa caccgttgat 2640
gtggtgactg ccgcaggtgg cacgccggta atgtcgaaaa ccggacacgc ctttattaaa 2700
gaacgtatgc gcaaggaaga cgccatctat ggtggcgaaa tgagcgccca ccattacttc 2760
cgtgatttcg cttactgcga cagcggcatg atcccgtggc tgctggtcgc cgaactggtg 2820
tgcctgaaag ataaaacgct gggcgaactg gtacgcgacc ggatggcggc gtttccggca 2880
agcggtgaga tcaacagcaa actggcgcaa cccgttgagg cgattaaccg cgtggaacag 2940
cattttagcc gtgaggcgct ggcggtggat cgcaccgatg gcatcagcat gacctttgcc 3000
gactggcgct ttaacctgcg cacctccaat accgaaccgg tggtgcgcct gaatgtggaa 3060
tcgcgcggtg atgtgccgct gatggaagcg cgaacgcgaa ctctgctgac gttgctgaac 3120
gagtaaaaac gcggccgcga tatcgttgta aaacgacggc cagtgcaaga atcataaaaa 3180
atttatttgc tttcaggaaa atttttctgt ataatagatt cataaatttg agagaggagt 3240
ttttgtgagc ggataacaat tccccatctt agtatattag ttaagtataa atacaccgcg 3300
gaggacgaag gagatagaac catgtcaaaa gtcgctctca tcaccggtgt aaccggacaa 3360
gacggttctt acctggcaga gtttctgctg gaaaaaggtt acgaggtgca tggtattaag 3420
cgtcgcgcat cgtcattcaa caccgagcgc gtggatcaca tttatcagga tccgcacacc 3480
tgcaacccga aattccatct gcattatggc gacctgagtg atacctctaa cctgacgcgc 3540
attttgcgtg aagtacagcc ggatgaagtg tacaacctgg gcgcaatgag ccacgttgcg 3600
gtctcttttg agtcaccaga atataccgct gacgtcgacg cgatgggtac gctgcgcctg 3660
ctggaggcga tccgcttcct cggtctggaa aagaaaactc gtttctatca ggcttccacc 3720
tctgaactgt atggtctggt gcaggaaatt ccgcagaaag agaccacgcc gttctacccg 3780
cgatctccgt atgcggtcgc caaactgtac gcctactgga tcaccgttaa ctaccgtgaa 3840
tcctacggca tgtacgcctg taacggaatt ctcttcaacc atgaatcccc gcgccgcggc 3900
gaaaccttcg ttacccgcaa aatcacccgc gcaatcgcca acatcgccca ggggctggag 3960
tcgtgcctgt acctcggcaa tatggattcc ctgcgtgact ggggccacgc caaagactac 4020
gtaaaaatgc agtggatgat gctgcagcag gaacagccgg aagatttcgt tatcgcgacc 4080
ggcgttcagt actccgtgcg tcagttcgtg gaaatggcgg cagcacagct gggcatcaaa 4140
ctgcgctttg aaggcacggg cgttgaagag aagggcattg tggtttccgt caccgggcat 4200
gacgcgccgg gcgttaaacc gggtgatgtg attatcgctg ttgacccgcg ttacttccgt 4260
ccggctgaag ttgaaacgct gctcggcgac ccgaccaaag cgcacgaaaa actgggctgg 4320
aaaccggaaa tcaccctcag agagatggtg tctgaaatgg tggctaatga cctcgaagcg 4380
gcgaaaaaac actctctgct gaaatctcac ggctacgacg tggcgatcgc gctggagtca 4440
taagcatgag taaacaacga gtttttattg ctggtcatcg cgggatggtc ggttccgcca 4500
tcaggcggca gctcgaacag cgcggtgatg tggaactggt attacgcacc cgcgacgagc 4560
tgaacctgct ggacagccgc gccgtgcatg atttctttgc cagcgaacgt attgaccagg 4620
tctatctggc ggcggcgaaa gtgggcggca ttgttgccaa caacacctat ccggcggatt 4680
tcatctacca gaacatgatg attgagagca acatcattca cgccgcgcat cagaacgacg 4740
tgaacaaact gctgtttctc ggatcgtcct gcatctaccc gaaactggca aaacagccga 4800
tggcagaaag cgagttgttg cagggcacgc tggagccgac taacgagcct tatgctattg 4860
ccaaaatcgc cgggatcaaa ctgtgcgaat catacaaccg ccagtacgga cgcgattacc 4920
gctcagtcat gccgaccaac ctgtacgggc cacacgacaa cttccacccg agtaattcgc 4980
atgtgatccc agcattgctg cgtcgcttcc acgaggcgac ggcacagaat gcgccggacg 5040
tggtggtatg gggcagcggt acaccgatgc gcgaatttct gcacgtcgat gatatggcgg 5100
cggcgagcat tcatgtcatg gagctggcgc atgaagtctg gctggagaac acccagccga 5160
tgttgtcgca cattaacgtc ggcacgggcg ttgactgcac tatccgcgag ctggcgcaaa 5220
ccatcgccaa agtggtgggt tacaaaggcc gggtggtttt tgatgccagc aaaccggatg 5280
gcacgccgcg caaactgctg gatgtgacgc gcctgcatca gcttggctgg tatcacgaaa 5340
tctcactgga agcggggctt gccagcactt accagtggtt ccttgagaat caagaccgct 5400
ttcggggggg gagctaacgc gccatttaaa tcaacctcag cggtcatagc tgtttcctgt 5460
gactgagcaa taactagcat aaccccttgg ggcctctaaa cgggtcttga ggggtttttt 5520
gctgaaacca atttgcctgg cggcagtagc gcggtggtcc cacctgaccc catgccgaac 5580
tcagaagtga aacgccgtag cgccgatggt agtgtggggt ctccccatgc gagagtaggg 5640
aactgccagg catcaaataa aacgaaaggc tcagtcgaaa gactgggcct ttcgggatcc 5700
aggccggcct gttaacgaat taatcttccg cggcggtatc gataagcttg atatcgaatt 5760
ccgaagttcc tattctctag aaagtatagg aacttcaggt ctgaagagga gtttacgtcc 5820
agccaagcta gcttggctgc aggtcgtcga aattctaccg ggtaggggag gcgcttttcc 5880
caaggcagtc tggagcatgc gctttagcag ccccgctggg cacttggcgc tacacaagtg 5940
gcctctggcc tcgcacacat tccacatcca ccggtaggcg ccaaccggct ccgttctttg 6000
gtggcccctt cgcgccacct tctactcctc ccctagtcag gaagttcccc cccgccccgc 6060
agctcgcgtc gtgcaggacg tgacaaatgg aagtagcacg tctcactagt ctcgtgcaga 6120
tggacagcac cgctgagcaa tggaagcggg taggcctttg gggcagcggc caatagcagc 6180
tttgctcctt cgctttctgg gctcagaggc tgggaagggg tgggtccggg ggcgggctca 6240
ggggcgggct caggggcggg gcgggcgccc gaaggtcctc cggaggcccg gcattctgca 6300
cgcttcaaaa gcgcacgtct gccgcgctgt tctcctcttc ctcatctccg ggcctttcga 6360
cctgcagcct gttgacaatt aatcatcggc atagtatatc ggcatagtat aatacgacaa 6420
ggtgaggaac taaaccatgg gtcaaagtag cgatgaagcc aacgctcccg ttgcagggca 6480
gtttgcgctt cccctgagtg ccacctttgg cttaggggat cgcgtacgca agaaatctgg 6540
tgccgcttgg cagggtcaag tcgtcggttg gtattgcaca aaactcactc ctgaaggcta 6600
tgcggtcgag tccgaatccc acccaggctc agtgcaaatt tatcctgtgg ctgcacttga 6660
acgtgtggcc taatgagggg atcaattctc tagagctcgc tgatcagaag ttcctattct 6720
ctagaaagta taggaacttc gatggcgcct catccctgaa gccaataggg ataacagggt 6780
aat 6783
<210> 2
<211> 2851
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Интеграционная кассета
<400> 2
tggccagatg attaattcct aatttttgtt gacactctat cattgataga gttattttac 60
cactccctat cagtgataga gaaaagtgaa atgaatagtt cgacaaaaat ctagaaataa 120
ttttgtttaa ctttaagaag gagatataca aatgtactat ttaaaaaaca caaacttttg 180
gatgttcggt ttattctttt tcttttactt ttttatcatg ggagcctact tcccgttttt 240
cccgatttgg ctacatgaca tcaaccatat cagcaaaagt gatacgggta ttatttttgc 300
cgctatttct ctgttctcgc tattattcca accgctgttt ggtctgcttt ctgacaaact 360
cgggctgcgc aaatacctgc tgtggattat taccggcatg ttagtgatgt ttgcgccgtt 420
ctttattttt atcttcgggc cactgttaca atacaacatt ttagtaggat cgattgttgg 480
tggtatttat ctaggctttt gttttaacgc cggtgcgcca gcagtagagg catttattga 540
gaaagtcagc cgtcgcagta atttcgaatt tggtcgcgcg cggatgtttg gctgtgttgg 600
ctgggcgctg tgtgcctcga ttgtcggcat catgttcacc atcaataatc agtttgtttt 660
ctggctgggc tctggctgtg cactcatcct cgccgtttta ctctttttcg ccaaaacgga 720
tgcgccctct tctgccacgg ttgccaatgc ggtaggtgcc aaccattcgg catttagcct 780
taagctggca ctggaactgt tcagacagcc aaaactgtgg tttttgtcac tgtatgttat 840
tggcgtttcc tgcacctacg atgtttttga ccaacagttt gctaatttct ttacttcgtt 900
ctttgctacc ggtgaacagg gtacgcgggt atttggctac gtaacgacaa tgggcgaatt 960
acttaacgcc tcgattatgt tctttgcgcc actgatcatt aatcgcatcg gtgggaaaaa 1020
cgccctgctg ctggctggca ctattatgtc tgtacgtatt attggctcat cgttcgccac 1080
ctcagcgctg gaagtggtta ttctgaaaac gctgcatatg tttgaagtac cgttcctgct 1140
ggtgggctgc tttaaatata ttaccagcca gtttgaagtg cgtttttcag cgacgattta 1200
tctggtctgt ttctgcttct ttaagcaact ggcgatgatt tttatgtctg tactggcggg 1260
caatatgtat gaaagcatcg gtttccaggg cgcttatctg gtgctgggtc tggtggcgct 1320
gggcttcacc ttaatttccg tgttcacgct tagcggcccc ggcccgcttt ccctgctgcg 1380
tcgtcaggtg aatgaagtcg ctgggagcta agcggccgcg tcgacacgca aaaaggccat 1440
ccgtcaggat ggccttctgc ttaatttgat gcctggcagt ttatggcggg cgtcctgccc 1500
gccaccctcc gggccgttgc ttcgcaacgt tcaaatccgc tcccggcgga tttgtcctac 1560
tcaggagagc gttcaccgac aaacaacaga taaaacgaaa ggcccagtct ttcgactgag 1620
cctttcgttt tatttgatgc ctggcagttc cctactctcg catggggaga ccccacacta 1680
ccatcatgta tgaatatcct ccttagttcc tattccgaag ttcctattct ctagaaagta 1740
taggaacttc ggcgcgtcct acctgtgaca cgcgtgccgc agtctcacgc ccggagcgta 1800
gcgaccgagt gagctagcta tttgtttatt tttctaaata cattcaaata tgtatccgct 1860
catgagacaa taaccctgat aaatgcttca ataatattga aaaaggaaga gtatgaggga 1920
agcggtgatc gccgaagtat cgactcaact atcagaggta gttggcgtca tcgagcgcca 1980
tctcgaaccg acgttgctgg ccgtacattt gtacggctcc gcagtggatg gcggcctgaa 2040
gccacacagt gatattgatt tgctggttac ggtgaccgta aggcttgatg aaacaacgcg 2100
gcgagctttg atcaacgacc ttttggaaac ttcggcttcc cctggagaga gcgagattct 2160
ccgcgctgta gaagtcacca ttgttgtgca cgacgacatc attccgtggc gttatccagc 2220
taagcgcgaa ctgcaatttg gagaatggca gcgcaatgac attcttgcag gtatcttcga 2280
gccagccacg atcgacattg atctggctat cttgctgaca aaagcaagag aacatagcgt 2340
tgccttggta ggtccagcgg cggaggaact ctttgatccg gttcctgaac aggatctatt 2400
tgaggcgcta aatgaaacct taacgctatg gaactcgccg cccgactggg ctggcgatga 2460
gcgaaatgta gtgcttacgt tgtcccgcat ttggtacagc gcagtaaccg gcaaaatcgc 2520
gccgaaggat gtcgctgccg actgggcaat ggagcgcctg ccggcccagt atcagcccgt 2580
catacttgaa gctagacagg cttatcttgg acaagaagaa gatcgcttgg cctcgcgcgc 2640
agatcagttg gaagaatttg tccactacgt gaaaggcgag atcaccaagg tagtcggcaa 2700
ataatgtcta acaattcgtt caagccgagg ggccgcaaga tccggccacg atgacccggt 2760
cgtcgggtac cggcagggcg gggcgtaagg cgcgccattt aaatgaagtt cctattccga 2820
agttcctatt ctctagaaag tataggaact t 2851
<210> 3
<211> 2858
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Интеграционная кассета
<400> 3
ggccagatga ttaattccta atttttgttg acactctatc attgatagag ttattttacc 60
actccctatc agtgatagag aaaagtgaaa tgaatagttc gacaaaaatc tagaaataat 120
tttgtttaac tttaagaagg agatatacaa atgggcagca ttattcgtct gcagggtggt 180
ctgggtaatc agctgtttca gtttagcttt ggttatgccc tgagcaaaat taatggtaca 240
ccgctgtatt tcgacattag ccattatgcc gaaaacgatg atcatggtgg ttatcgtctg 300
aataatctgc agattccgga agaatatctg cagtattata ccccgaaaat taataatatt 360
tataaactgc tggtgcgtgg cagccgtctg tatccggata tttttctgtt tctgggcttt 420
tgcaacgaat ttcatgccta tggctacgat tttgaatata ttgcccagaa atggaaaagc 480
aaaaaataca ttggctactg gcagagcgaa cacttttttc ataaacatat tctggacctg 540
aaagaatttt ttattccgaa aaatgtgagc gaacaggcaa atctgctggc agcaaaaatt 600
ctggaaagcc agagcagcct gagcattcat attcgtcgtg gcgattatat taaaaacaaa 660
accgcaaccc tgacacatgg tgtttgtagc ctggaatatt ataaaaaagc cctgaacaaa 720
atccgcgatc tggcaatgat tcgtgatgtg tttatcttta gcgacgatat cttctggtgc 780
aaagaaaata ttgaaaccct gctgagcaaa aaatataata tttattatag cgaagatctg 840
agccaagaag aggatctgtg gctgatgagc ctggcaaatc atcatattat tgccaatagc 900
agctttagtt ggtggggtgc atatctgggt agcagcgcaa gccagattgt tatttatccg 960
accccgtggt atgatattac cccgaaaaac acctatatcc cgattgtgaa ccattggatc 1020
aacgttgata aacatagcag ctgctaagcg gccgcgtcga cacgcaaaaa ggccatccgt 1080
caggatggcc ttctgcttaa tttgatgcct ggcagtttat ggcgggcgtc ctgcccgcca 1140
ccctccgggc cgttgcttcg caacgttcaa atccgctccc ggcggatttg tcctactcag 1200
gagagcgttc accgacaaac aacagataaa acgaaaggcc cagtctttcg actgagcctt 1260
tcgttttatt tgatgcctgg cagttcccta ctctcgcatg gggagacccc acactaccat 1320
catgtatgaa tatcctcctt agttcctatt ccgaagttcc tattctctag aaagtatagg 1380
aacttcggcg cgtcctacct gtgacacgcg tcaagatccc ctcacgctgc cgcaagcact 1440
cagggcgcaa gggctgctaa aggaagcgga acacgtagaa agccagtccg cagaaacggt 1500
gctgaccccg gatgaatgtc agctactggg ctatctggac aagggaaaac gcaagcgcaa 1560
agagaaagca ggtagcttgc agtgggctta catggcgata gctagactgg gcggttttat 1620
ggacagcaag cgaaccggaa ttgccagctg gggcgccctc tggtaaggtt gggaagccct 1680
gcaaagtaaa ctggatggct ttcttgccgc caaggatctg atggcgcagg ggatcaagat 1740
ctgatcaaga gacaggatga ggatcgtttc gcatgattga acaagatgga ttgcacgcag 1800
gttctccggc cgcttgggtg gagaggctat tcggctatga ctgggcacaa cagacaatcg 1860
gctgctctga tgccgccgtg ttccggctgt cagcgcaggg gcgcccggtt ctttttgtca 1920
agaccgacct gtccggtgcc ctgaatgaac tgcaggacga ggcagcgcgg ctatcgtggc 1980
tggccacgac gggcgttcct tgcgcagctg tgctcgacgt tgtcactgaa gcgggaaggg 2040
actggctgct attgggcgaa gtgccggggc aggatctcct gtcatctcac cttgctcctg 2100
ccgagaaagt atccatcatg gctgatgcaa tgcggcggct gcatacgctt gatccggcta 2160
cctgcccatt cgaccaccaa gcgaaacatc gcatcgagcg agcacgtact cggatggaag 2220
ccggtcttgt cgatcaggat gatctggacg aagagcatca ggggctcgcg ccagccgaac 2280
tgttcgccag gctcaaggcg cgcatgcccg acggcgagga tctcgtcgtg acccatggcg 2340
atgcctgctt gccgaatatc atggtggaaa atggccgctt ttctggattc atcgactgtg 2400
gccggctggg tgtggcggac cgctatcagg acatagcgtt ggctacccgt gatattgctg 2460
aagagcttgg cggcgaatgg gctgaccgct tcctcgtgct ttacggtatc gccgctcccg 2520
attcgcagcg catcgccttc tatcgccttc ttgacgagtt cttctgagcg ggactctggg 2580
gttcgaaatg accgaccaag cgacgcccaa cctgccatca cgagatttcg attccaccgc 2640
cgccttctat gaaaggttgg gcttcggaat cgttttccgg gacgccggct ggatgatcct 2700
ccagcgcggg gatctcatgc tggagttctt cgcccacccc agcttcaaaa gcgctctcgg 2760
taccggcagg gcggggcgta aggcgcgcca tttaaatgaa gttcctattc cgaagttcct 2820
attctctaga aagtatagga acttcgaagc agctccag 2858
<210> 4
<211> 2631
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Интеграционная кассета
<400> 4
ggccagatga ttaattccta atttttgttg acactctatc attgatagag ttattttacc 60
actccctatc agtgatagag aaaagtgaaa tgaatagttc gacaaaaatc tagaaataat 120
tttgtttaac tttaagaagg agatatacaa atgaagtcgg cactgacctt ttcccgtcgc 180
atcaatccgg tgtttctggc gttctttgtc gttgcttttc tgagcggtat cgcaggcgca 240
ctgcaggctc cgaccctgag tctgtttctg tccacggaag tgaaagttcg tccgctgtgg 300
gttggtctgt tctataccgt caacgcaatc gctggcatta cggttagctt tatcctggcg 360
aaacgttcag attcgcgcgg tgaccgtcgc aagctgatta tggtgtgcta tctgatggcg 420
gttggcaact gtctgctgtt tgccttcaat cgtgattacc tgaccctgat cacggcaggt 480
gtgctgctgg cgagcgttgc caacaccgca atgccgcaga ttttcgcgct ggcccgtgaa 540
tatgccgaca gctctgcacg cgaagtggtt atgtttagtt ccatcatgcg cgctcaactg 600
agtctggcat gggtgattgg tccgccgctg tcctttatgc tggcgctgaa ttacggtttt 660
accctgatgt tctcaatcgc ggccggcatt ttcgttctgt cggccctggt cgtgtggttt 720
atcctgccga gtgtcccgcg tgcagaaccg gttgtcgatg caccggtggt tgtccagggt 780
tcactgttcg cagacaaaaa cgttctgctg ctgtttatcg cgtcgatgct gatgtggacc 840
tgcaatacga tgtatattat cgatatgccg ctgtacatta ccgcaagcct gggtctgccg 900
gaacgtctgg ctggtctgct gatgggtacc gcagctggcc tggaaattcc gatcatgctg 960
ctggcgggtt attctgtgcg ttactttggc aaacgcaaga ttatgctgtt cgctgttctg 1020
gcgggtgtcc tgttttatac cggcctggtt ctgtttaaat tcaagacggc cctgatgctg 1080
ctgcagatct ttaacgcaat tttcatcggt attgtggctg gcattggtat gctgtacttc 1140
caagatctga tgccgggtcg tgcaggtgca gcaaccacgc tgtttaccaa tagcatctct 1200
acgggtgtca ttctggcagg cgtgctgcaa ggcggtctga ccgaaacgtg gggccatgac 1260
agcgtctatg tgatggcgat ggtcctgtct attctggccc tgattatctg tgcacgtgtg 1320
cgcgaagctt aaatcgatac tagcataacc ccttggggcc tctaaacgcg tcgacacgca 1380
aaaaggccat ccgtcaggat ggccttctgc ttaatttgat gcctggcagt ttatggcggg 1440
cgtcctgccc gccaccctcc gggccgttgc ttcgcaacgt tcaaatccgc tcccggcgga 1500
tttgtcctac tcaggagagc gttcaccgac aaacaacaga taaaacgaaa ggcccagtct 1560
ttcgactgag cctttcgttt tatttgatgc ctggcagttc cctactctcg catggggaga 1620
ccccacacta ccatcatgta tgaatatcct ccttagttcc tattccgaag ttcctattct 1680
ctagaaagta taggaacttc ggcgcgtcct acctgtgacg gaagatcact tcgcagaata 1740
aataaatcct ggtgtccctg ttgataccgg gaagccctgg gccaactttt ggcgaaaatg 1800
agacgttgat cggcacgtaa gaggttccaa ctttcaccat aatgaaataa gatcactacc 1860
gggcgtattt tttgagttgt cgagattttc aggagctaag gaagctaaaa tggagaaaaa 1920
aatcactgga tataccaccg ttgatatatc ccaatggcat cgtaaagaac attttgaggc 1980
atttcagtca gttgctcaat gtacctataa ccagaccgtt cagctggata ttacggcctt 2040
tttaaagacc gtaaagaaaa ataagcacaa gttttatccg gcctttattc acattcttgc 2100
ccgcctgatg aatgctcatc cggaattacg tatggcaatg aaagacggtg agctggtgat 2160
atgggatagt gttcaccctt gttacaccgt tttccatgag caaactgaaa cgttttcatc 2220
gctctggagt gaataccacg acgatttccg gcagtttcta cacatatatt cgcaagatgt 2280
ggcgtgttac ggtgaaaacc tggcctattt ccctaaaggg tttattgaga atatgttttt 2340
cgtctcagcc aatccctggg tgagtttcac cagttttgat ttaaacgtgg ccaatatgga 2400
caacttcttc gcccccgttt tcaccatggg caaatattat acgcaaggcg acaaggtgct 2460
gatgccgctg gcgattcagg ttcatcatgc cgtttgtgat ggcttccatg tcggcagatg 2520
cttaatgaat acaacagtac tgcgatgagt ggcagggcgg ggcgtaaggc gcgccattta 2580
aatgaagttc ctattccgaa gttcctattc tctagaaagt ataggaactt c 2631
<210> 5
<211> 4259
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Интеграционная кассета
<400> 5
ttactcagca ataaactgat attccgtcag gctggaatac tcttcgccag gacgcaggaa 60
gcagtccggt tgcggccatt cagggtggtt cgggctgtcc ggtagaaact cgctttccag 120
agccagccct tgccagtcgg cgtaaggttc ggttccccgc gacggtgtgc cgccgaggaa 180
gttgccggag tagaattgca gagccggagc ggtggtgtag accttcagct gcaatttttc 240
atctgctgac cagacatgcg ccgccacttt cttgccatcg cctttggcct gtaacaagaa 300
tgcgtgatcg taacctttca ctttgcgctg atcgtcgtcg gcaagaaact cactggcgat 360
gattttggcg ctgcggaaat caaaagacgt tccggcgaca gatttcaggc cgtcgtgcgg 420
aatgccgcct tcatcaaccg gcagatattc gtccgccaga atctgcaact tgtgattgcg 480
cacgtcagac tgctcgccgt caagattgaa atagacgtga ttagtcatat tcaccgggca 540
aggtttatca actgtggcgc gataagtaat ggagatacgg ttatcgtcgg tcagacgata 600
ttgcaccgtc gcgccgagat tacccgggaa gccctgatca ccatcatctg aactcagggc 660
aaacagcacc tgacgatcgt tctggttcac aatctgccag cgacgtttgt cgaacccttc 720
cggcccgccg tgcagctggt taacgccctg acttggcgaa agcgtcacgg tttcaccgtc 780
aaaggtataa cggctattgg cgatacggtt ggcataacga ccaatagagg cccccagaaa 840
cgcggcctga tcctgatagc attccgggct ggcacagccg agcagcgcct cgcggacgct 900
gccatcggaa agcggaatac gggcggaaag taaagtcgca ccccagtcca tcagcgtgac 960
taccatccct gcgttgttac gcaaagttaa cagtcggtac ggctgaccat cgggtgccag 1020
tgcgggagtt tcgttcagca ctgtcctgct ccttgtgatg gtttacaaac gtaaaaagtc 1080
tctttaatac ctgtttttgc ttcatattgt tcagcgacag cttgctgtac ggcaggcacc 1140
agctcttccg ggatcagcgc gacgatacag ccgccaaatc cgccgccggt catgcgtacg 1200
ccacctttgt cgccaatcac agctttgacg atttctacca gagtgtcaat ttgcggcacg 1260
gtgatttcga aatcatcgcg catagaggca tgagactccg ccatcaactc gcccatacgt 1320
ttcaggtcgc cttgctccag cgcgctggca gcttcaacgg tgcgggcgtt ttcagtcagt 1380
atatgacgca cgcgttttgc cacgatcggg tccagttcat gcgcaacagc gttgaactct 1440
tcaatggtga catcacgcag ggctggctgc tggaagaaac gcgcaccggt ttcgcactgt 1500
tcacgacggg tgttgtattc gctgccaacc agggtacgtt tgaagttact gttgatgatg 1560
acgacagcca cacctttggg catggaaact gctttggtcc ccagtgagcg gcaatcgatc 1620
agcaaggcat gatctttctt gccgagcgcg gaaattagct gatccatgat cccgcagtta 1680
cagcctacaa actggttttc tgcttcctga ccgttaagcg cgatttgtgc gccgtccagc 1740
ggcagatgat aaagctgctg caatacggtt ccgaccgcga cttccagtga agcggaagaa 1800
cttaacccgg caccctgcgg cacattgccg ctgatcacca tgtccacgcc gccgaagctg 1860
ttgttacgca gttgcagatg tttcaccacg ccacgaacgt agttagccca ttgatagttt 1920
tcatgtgcga caatgggcgc atcgagggaa aactcgtcga gctgattttc ataatcggct 1980
gccatcacgc gaactttacg gtcatcgcgt ggtgcacaac tgatcacggt ttgataatca 2040
atcgcgcagg gcagaacgaa accgtcgttg tagtcggtgt gttcaccaat caaattcacg 2100
cggccaggcg cctgaatggt gtgagtggca gggtagccaa atgcgttggc aaacagagat 2160
tgtgtttttt ctttcagact catttcttac actccggatt cgcgaaaatg gatatcgctg 2220
actgcgcgca aacgctctgc tgcctgttct gcggtcaggt ctcgctgggt ctctgccagc 2280
atttcataac caaccataaa tttacgtacg gtggcggagc gcagcagagg cggataaaag 2340
tgcgcgtgca gctgccagtg ttgattctct tcgccattaa atggcgcgcc gtgccagccc 2400
atagagtagg ggaaggagca ctggaagagg ttgtcataac gactggtcag ctttttcaac 2460
gccagcgcca gatcgctgcg ctgggcgtcg gtcaaatcgg tgatccgtaa aacgtgggct 2520
ttgggcagca gtagcgtttc gaacggccag gcagcccagt aaggcacgac ggctaaccag 2580
tgttcggttt cgacaacggt acggctaccg tctgccagct cgcgctgaac ataatccacc 2640
agcattggtg atttctgttc ggcaaaatat tctttttgca ggcggtcttc gcgctcagct 2700
tcgttaggca ggaagctatt tgcccaaatc tgaccgtgcg gatgcgggtt agagcagccc 2760
atcgccgcgc ctttgttttc aaaaacctgc acccatgggt acgttttccc cagttctgcg 2820
gtttgctcct gccaggtttt gacgatttcc gtcaatgctg caacgctgag ctctggcagc 2880
gttttactgt gatccggtga aaagcagatc acccggctgg tgccgcgcgc gctctggcaa 2940
cgcatcagcg gatcgtgact ttctggcgca tctggcgtgt cagacatcaa agccgcaaag 3000
tcattagtga aaacgtaagt cccggtgtaa tcggggtttt tatcgcctgt cacccgcaca 3060
ttacctgcgc agaggaagca atctggatcg tgcgcaggta acacctgttt ggctggcgtt 3120
tcctgcgccc cctgccaggg gcgcttagcg cggtgcggtg aaaccagaat ccattgcccg 3180
gtgagcgggt tgtagcggcg atgtggatga tcaacgggat taaattgcgt catggtcgtt 3240
ccttaatcgg gatatccctg tggatggcgt gactgccagt gccaggtgtc ctgcgccatt 3300
tcatcgagtg tgcgcgttac gcgccagttc agttcacggt cggctttgct ggcgtccgcc 3360
cagtaggccg gaaggtcgcc ctcgcgacgc ggtgcaaaat gataattaac cggtttgccg 3420
caggctttgc tgaaggcatt aaccacgtcc agcacgctgt tgcctacgcc agcgccgagg 3480
ttgtagatgt gtacgcctgg cttgttcgcc agtttttcca tcgccacgac gtgaccgtcc 3540
gccagatcca ttacgtggat gtaatcgcgt acgccagtac catcttcggt cggataatcg 3600
ttaccaaaaa tcgccagcga gtcgcgacgg cctacagcaa cctgggcgat gtatggcatc 3660
aggttattcg gaatgccttg cggatcttcg cccatatcgc ccgacggatg cgcgccaacc 3720
gggttgaagt agcgcagcag ggcaatgctc cagtccggct gggctttttg cagatcggtg 3780
aggatctgtt ccaccatcag cttgcttttg ccgtaagggc tttgcggtgt gccggtcggg 3840
aagctttcaa cgtatggaat tttgggctga tcgccataaa cggtggcgga ggagctaaaa 3900
ataaagtttt tgacgttagc ggcgcgcatg gcgctaatca ggcgcagagt gccgttgaca 3960
ttgttgtcgt aatattccag cggtttttgt accgattcgc ccacggcttt cagcccggcg 4020
aagtggatca cggtgtcgat agcgtgatcg tgcaggatct cggtcatcaa cgcttcgtta 4080
cgaatatcgc cttcaacaaa cgttggatgt ttgccgccta aacgctcgat aacaggcagt 4140
acgctgcgct tactgttaca gaggttatca agaatgatga catcatgacc gttttgcagt 4200
aattgcacac aggtatgact tccaatgtaa ccgctaccac cggtaaccag aactctcat 4259
<210> 6
<211> 4223
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Интеграционная кассета
<400> 6
tggccagatg attaattcct aatttttgtt gacactctat cattgataga gttattttac 60
cactccctat cagtgataga gaaaagtgaa atgaatagtt cgacaaaaat ctagaaataa 120
ttttgtttaa ctttaagaag gagatataca aatgcaaaaa ctactatctt taccgtccaa 180
tctggttcag tcttttcatg aactggagag ggtgaatcgt accgattggt tttgtacttc 240
cgacccggta ggtaagaaac ttggttccgg tggtggaaca tcctggctgc ttgaagaatg 300
ttataatgaa tattcagatg gtgctacttt tggagagtgg cttgaaaaag aaaaaagaat 360
tcttcttcat gcgggtgggc aaagccgtcg tttacccggc tatgcacctt ctggaaagat 420
tctcactccg gttcctgtgt tccggtggga gagagggcaa catctgggac aaaatctgct 480
ttctctgcaa cttcccctat atgaaaaaat catgtctttg gctccggata aactccatac 540
actgattgcg agtggtgatg tctatattcg ttcggagaaa cctttgcaga gtattcccga 600
agcggatgtg gtttgttatg gactgtgggt agatccgtct ctggctaccc atcatggcgt 660
gtttgcttcc gatcgcaaac atcccgaaca actcgacttt atgcttcaga agccttcgtt 720
ggcagaattg gaatctttat cgaagaccca tttgttcctg atggacatcg gtatatggct 780
tttgagtgac cgtgccgtag aaatcttgat gaaacgttct cataaagaaa gctctgaaga 840
actaaagtat tatgatcttt attccgattt tggattagct ttgggaactc atccccgtat 900
tgaagacgaa gaggtcaata cgctatccgt tgctattctg cctttgccgg gaggagagtt 960
ctatcattac gggaccagta aagaactgat ttcttcaact ctttccgtac agaataaggt 1020
ttacgatcag cgtcgtatca tgcaccgtaa agtaaagccc aatccggcta tgtttgtcca 1080
aaatgctgtc gtgcggatac ctctttgtgc cgagaatgct gatttatgga tcgagaacag 1140
tcatatcgga ccaaagtgga agattgcttc acgacatatt attaccgggg ttccggaaaa 1200
tgactggtca ttggctgtgc ctgccggagt gtgtgtagat gtggttccga tgggtgataa 1260
gggctttgtt gcccgtccat acggtctgga cgatgttttc aaaggagatt tgagagattc 1320
caaaacaacc ctgacgggta ttccttttgg tgaatggatg tccaaacgcg gtttgtcata 1380
tacagatttg aaaggacgta cggacgattt acaggcagtt tccgtattcc ctatggttaa 1440
ttctgtagaa gagttgggat tggtgttgag gtggatgttg tccgaacccg aactggagga 1500
aggaaagaat atctggttac gttccgaaca tttttctgcg gacgaaattt cggcaggtgc 1560
caatctgaag cgtttgtatg cacaacgtga agagttcaga aaaggaaact ggaaagcatt 1620
ggccgttaat catgaaaaaa gtgtttttta tcaacttgat ttggccgatg cagctgaaga 1680
ttttgtacgt cttggtttgg atatgcctga attattgcct gaggatgctc tgcagatgtc 1740
acgcatccat aaccggatgt tgcgtgcgcg tattttgaaa ttagacggga aagattatcg 1800
tccggaagaa caggctgctt ttgatttgct tcgtgacggc ttgctggacg ggatcagtaa 1860
tcgtaagagt accccaaaat tggatgtata ttccgatcag attgtttggg gacgtagccc 1920
cgtgcgcatc gatatggcag gtggatggac cgatactcct ccttattcac tttattcggg 1980
aggaaatgtg gtgaatctag ccattgagtt gaacggacaa cctcccttac aggtctatgt 2040
gaagccgtgt aaagacttcc atatcgtcct gcgttctatc gatatgggtg ctatggaaat 2100
agtatctacg tttgatgaat tgcaagatta taagaagatc ggttcacctt tctctattcc 2160
gaaagccgct ctgtcattgg caggctttgc acctgcgttt tctgctgtat cttatgcttc 2220
attagaggaa cagcttaaag atttcggtgc aggtattgaa gtgactttat tggctgctat 2280
tcctgccggt tccggtttgg gcaccagttc cattctggct tctaccgtac ttggtgccat 2340
taacgatttc tgtggtttag cctgggataa aaatgagatt tgtcaacgta ctcttgttct 2400
tgaacaattg ctgactaccg gaggtggatg gcaggatcag tatggaggtg tgttgcaggg 2460
tgtgaagctt cttcagaccg aggccggctt tgctcaaagt ccattggtgc gttggctacc 2520
cgatcattta tttacgcatc ctgaatacaa agactgtcac ttgctttatt ataccggtat 2580
aactcgtacg gcaaaaggga tcttggcaga aatagtcagt tccatgttcc tcaattcatc 2640
gttgcatctc aatttacttt cggaaatgaa ggcgcatgca ttggatatga atgaagctat 2700
acagcgtgga agttttgttg agtttggccg tttggtagga aaaacctggg aacaaaacaa 2760
agcattggat agcggaacaa atcctccggc tgtggaggca attatcgatc tgataaaaga 2820
ttataccttg ggatataaat tgccgggagc cggtggtggc gggtacttat atatggtagc 2880
gaaagatccg caagctgctg ttcgtattcg taagatactg acagaaaacg ctccgaatcc 2940
gcgggcacgt tttgtcgaaa tgacgttatc tgataaggga ttccaagtat cacgatcata 3000
actgaaacca atttgcctgg cggcagtagc gcggtggtcc cacctgaccc catgccgaac 3060
tcagaagtga aacgccgtag cgccgatggt agtgtggggt ctccccatgc gagagtaggg 3120
aactgccagg catcaaataa aacgaaaggc tcagtcgaaa gactgggcct ttcgggatcc 3180
aggccggcct gttaagacgg ccagtgaatt cgagctcggt acctaccgtt cgtataatgt 3240
atgctatacg aagttatcga gctctagaga atgatcccct cattaggcca cacgttcaag 3300
tgcagcgcac accgtggaaa cggatgaagg cacgaaccca gttgacataa gcctgttcgg 3360
ttcgtaaact gtaatgcaag tagcgtatgc gctcacgcaa ctggtccaga accttgaccg 3420
aacgcagcgg tggtaacggc gcagtggcgg ttttcatggc ttgttatgac tgtttttttg 3480
tacagtctat gcctcgggca tccaagcagc aagcgcgtta cgccgtgggt cgatgtttga 3540
tgttatggag cagcaacgat gttacgcagc agcaacgatg ttacgcagca gggcagtcgc 3600
cctaaaacaa agttaggtgg ctcaagtatg ggcatcattc gcacatgtag gctcggccct 3660
gaccaagtca aatccatgcg ggctgctctt gatcttttcg gtcgtgagtt cggagacgta 3720
gccacctact cccaacatca gccggactcc gattacctcg ggaacttgct ccgtagtaag 3780
acattcatcg cgcttgctgc cttcgaccaa gaagcggttg ttggcgctct cgcggcttac 3840
gttctgccca ggtttgagca gccgcgtagt gagatctata tctatgatct cgcagtctcc 3900
ggcgagcacc ggaggcaggg cattgccacc gcgctcatca atctcctcaa gcatgaggcc 3960
aacgcgcttg gtgcttatgt gatctacgtg caagcagatt acggtgacga tcccgcagtg 4020
gctctctata caaagttggg catacgggaa gaagtgatgc actttgatat cgacccaagt 4080
accgccacct aacaattcgt tcaagccgag atcgtagaat ttcgacgacc tgcagccaag 4140
cataacttcg tataatgtat gctatacgaa cggtaggatc ctctagagtc gacctgcagg 4200
catgagatgt gtataagaga cag 4223
<210> 7
<211> 3792
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Интеграционная кассета
<400> 7
gggaattgat tctggtacca aatgagtcga ccggccagat gattaattcc taatttttgt 60
tgacactcta tcattgatag agttatttta ccactcccta tcagtgatag agaaaagtga 120
aatgaatagt tcgacaaaaa tctagaaata attttgttta actttaagaa ggagatatac 180
aaatgattac ccgcaaaagg cgggccagga caatccatag ccgatatcca atcggaattt 240
acgggagcat agtaatgaca gatattgcac agttgcttgg caaagacgcc gacaaccttt 300
tacagcaccg ttgtatgact attccttctg accagcttta tctccccgga catgactacg 360
tagaccgcgt gatgattgac aataatcgcc cgccagcggt gttacgtaat atgcagacgt 420
tgtacaacac tgggcgtctg gctggcacag gatatctttc tattctgccg gttgaccagg 480
gcgttgagca ctctgccgga gcttcatttg ctgctaaccc gctctacttt gacccgaaaa 540
acattgttga actggcgatc gaagcgggct gtaactgtgt ggcatcaact tacggcgtgt 600
tggcgtcggt atcgcggcgc tatgcgcatc gcattccatt cctcgtcaaa cttaatcaca 660
acgagacgct aagttacccg aacacctacg atcaaacgct gtatgccagc gtggagcagg 720
ccttcaacat gggcgcggtg gcggttggtg cgactatcta ttttggttcg gaagagtcac 780
gtcgccagat tgaagaaatt tctgcggctt ttgaacgtgc gcacgagctg ggcatggtga 840
cagtgctgtg ggcctatttg cgtaactccg cctttaagaa agatggcgtt gattaccatg 900
tttccgccga cctgaccggt caggcaaacc atctggcggc gaccataggt gcagatatcg 960
tcaaacaaaa aatggcggaa aataacggcg gctataaagc aattaattac ggttataccg 1020
acgatcgcgt gtacagcaag ttaaccagcg aaaacccgat tgatctggtg cgttatcagt 1080
tagctaactg ctatatgggc cgggccgggt tgataaactc cggcggtgct gcaggcggtg 1140
aaactgacct cagcgatgca gtgcgtactg cggttatcaa caaacgcgct ggcggaatgg 1200
ggctgattct tggacgtaag gcgttcaaga aatcgatggc tgacggcgtg aaactgatta 1260
acgccgtgca ggatgtttat ctcgatagca aaattactat cgcctaagag gatcgagatc 1320
tcgatcccgc gaaattaata cgactcacta taggggaatt gtgagcggat aacaattccc 1380
ctctagaaat aattttgttt aactttaaga aggagatata ccatgggcca tcatcatcat 1440
catcatcatc atcatcacag cagcggccat atcgaaggtc gtcatatggc ggtgaaagaa 1500
gcgaccagcg agaccaagaa gcgtagcggt tacgagatca ttaccctgac cagctggctg 1560
ctgcaacaag aacagaaggg tatcattgac gcggaactga ccatcgttct gagcagcatt 1620
agcatggcgt gcaaacagat cgcgagcctg gtgcaacgtg cgaacattag caacctgacc 1680
ggtacccaag gcgcggttaa catccagggt gaagaccaaa agaaactgga tgttattagc 1740
aacgaggtgt tcagcaactg cctgcgtagc agcggtcgta ccggcatcat tgcgagcgag 1800
gaagaggacg tggcggttgc ggtggaagag agctacagcg gtaactatat cgtggttttt 1860
gacccgctgg atggcagcag caacctggat gcggctgtga gcaccggtag catcttcggc 1920
atttacagcc cgaacgacga gagcctgccg gattttggtg acgatagcga cgataacacc 1980
ctgggcaccg aagagcaacg ttgcatcgtt aacgtgtgcc aaccgggtag caacctgctg 2040
gcggcgggct actgcatgta tagcagcagc gttgcgttcg tgctgaccat tggcaagggc 2100
gttttcgtgt ttaccctgga cccgctgtac ggtgaattcg tgctgaccca ggagaacctg 2160
caaatcccga agagcggtga aatttacagc tttaacgagg gcaactataa actgtgggat 2220
gaaaacctga agaaatatat cgacgatctg aaggaaccgg gtccgagcgg taaaccgtac 2280
agcgcgcgtt atatcggtag cctggttggc gacttccacc gtaccctgct gtacggtggc 2340
atttacggtt atccgcgtga taagaaaagc aagaacggca aactgcgtct gctgtatgaa 2400
tgcgcgccga tgagctttat tgttgagcag gcgggtggca aaggtagcga cggccaccag 2460
cgtgtgctgg atatccaacc gaccgaaatt caccagcgtg ttccgctgta cattggtagc 2520
accgaagagg ttgaaaaagt tgaaaagtat ctggcgtaat cgagtctggt aaagaaaccg 2580
ctgctgcgaa atttgaacgc cagcacatgg actcgtctac tagcgcagct taattaacct 2640
aggctgctgc caccgctgag caataactag cataacccct tggggcctct aaacgggtct 2700
tgaggggttt tttgctgaaa ggaggaacta tatccggatt ggcgaatggg acgcgccctg 2760
tagcggcgca ttaagcgcgg cgggtggacg gccagtgaat tcgagctcgg tacctaccgt 2820
tcgtataatg tatgctatac gaagttatcg agctctagag aatgatcccc tcattaggcc 2880
acacgttcaa gtgcagcgca caccgtggaa acggatgaag gcacgaaccc agttgacata 2940
agcctgttcg gttcgtaaac tgtaatgcaa gtagcgtatg cgctcacgca actggtccag 3000
aaccttgacc gaacgcagcg gtggtaacgg cgcagtggcg gttttcatgg cttgttatga 3060
ctgttttttt gtacagtcta tgcctcgggc atccaagcag caagcgcgtt acgccgtggg 3120
tcgatgtttg atgttatgga gcagcaacga tgttacgcag cagcaacgat gttacgcagc 3180
agggcagtcg ccctaaaaca aagttaggtg gctcaagtat gggcatcatt cgcacatgta 3240
ggctcggccc tgaccaagtc aaatccatgc gggctgctct tgatcttttc ggtcgtgagt 3300
tcggagacgt agccacctac tcccaacatc agccggactc cgattacctc gggaacttgc 3360
tccgtagtaa gacattcatc gcgcttgctg ccttcgacca agaagcggtt gttggcgctc 3420
tcgcggctta cgttctgccc aggtttgagc agccgcgtag tgagatctat atctatgatc 3480
tcgcagtctc cggcgagcac cggaggcagg gcattgccac cgcgctcatc aatctcctca 3540
agcatgaggc caacgcgctt ggtgcttatg tgatctacgt gcaagcagat tacggtgacg 3600
atcccgcagt ggctctctat acaaagttgg gcatacggga agaagtgatg cactttgata 3660
tcgacccaag taccgccacc taacaattcg ttcaagccga gatcgtagaa tttcgacgac 3720
ctgcagccaa gcataacttc gtataatgta tgctatacga acggtaggat cctctagagt 3780
cgacctgcag gc 3792
<210> 8
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер
<400> 8
ctgtctctta tacacatct 19
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| ФУКОЗИЛТРАНСФЕРАЗЫ И ИХ ПРИМЕНЕНИЕ ДЛЯ ПОЛУЧЕНИЯ ФУКОЗИЛИРОВАННЫХ ОЛИГОСАХАРИДОВ | 2018 |
|
RU2818835C2 |
| Ферментативное получение олигосахаридов посредством общей ферментации с использованием смешанного сырья | 2019 |
|
RU2801231C2 |
| ПРОДУКЦИЯ ФУКОЗИЛИРОВАННЫХ ОЛИГОСАХАРИДОВ В BACILLUS | 2020 |
|
RU2810729C2 |
| Ферментативная продукция углеводов микробными клетками с использованием смешанного сырья | 2020 |
|
RU2809122C2 |
| НОВЫЕ ФУКОЗИЛТРАСФЕРАЗЫ И ИХ ПРИМЕНЕНИЯ | 2011 |
|
RU2628307C2 |
| НОВЫЕ ФУКОЗИЛТРАНСФЕРАЗЫ И ИХ ПРИМЕНЕНИЕ | 2011 |
|
RU2642307C2 |
| ФЕРМЕНТАТИВНЫЙ СПОСОБ ПРОИЗВОДСТВА МОНОСАХАРИДОВ В СВОБОДНОЙ ФОРМЕ ИЗ САХАРОВ, АКТИВИРОВАННЫХ НУКЛЕОТИДАМИ | 2016 |
|
RU2746739C1 |
| СИНТЕЗ ФУКОЗИЛИРОВАННЫХ СОЕДИНЕНИЙ | 2009 |
|
RU2584599C2 |
| СИНТЕЗ НМО | 2009 |
|
RU2517602C2 |
| ПРИМЕНЕНИЕ ГЛИКОЗИДАЗ В ПОЛУЧЕНИИ ОЛИГОСАХАРИДОВ | 2019 |
|
RU2810730C2 |
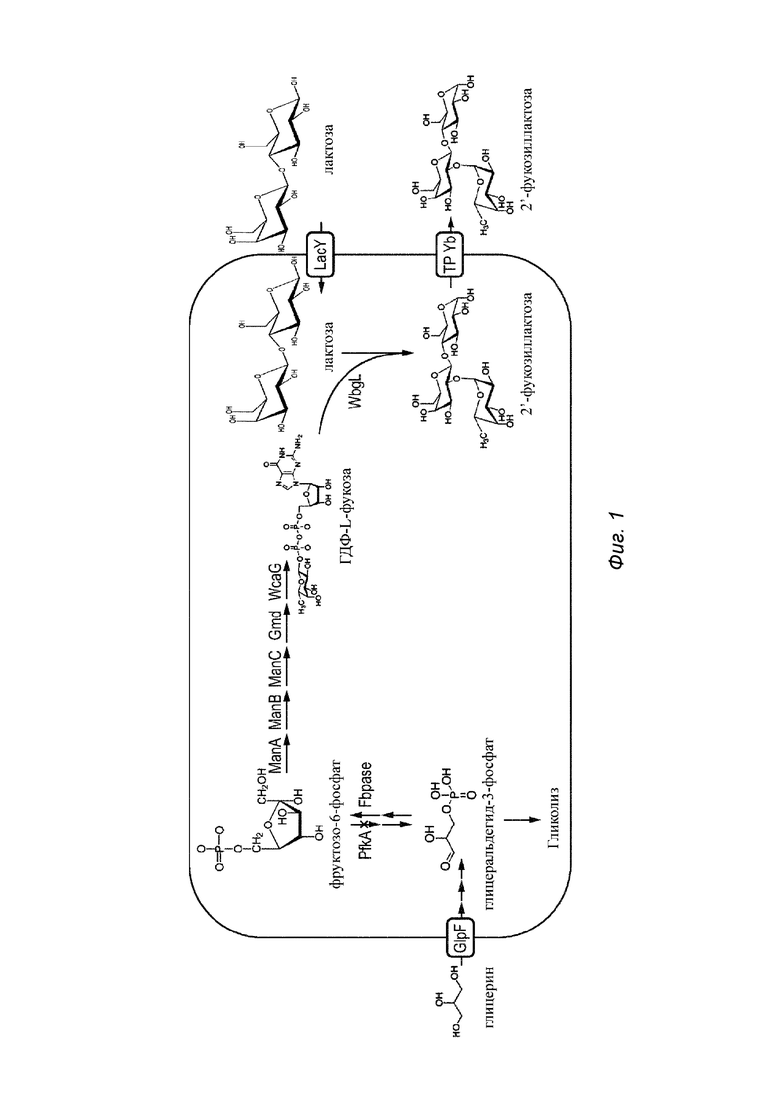
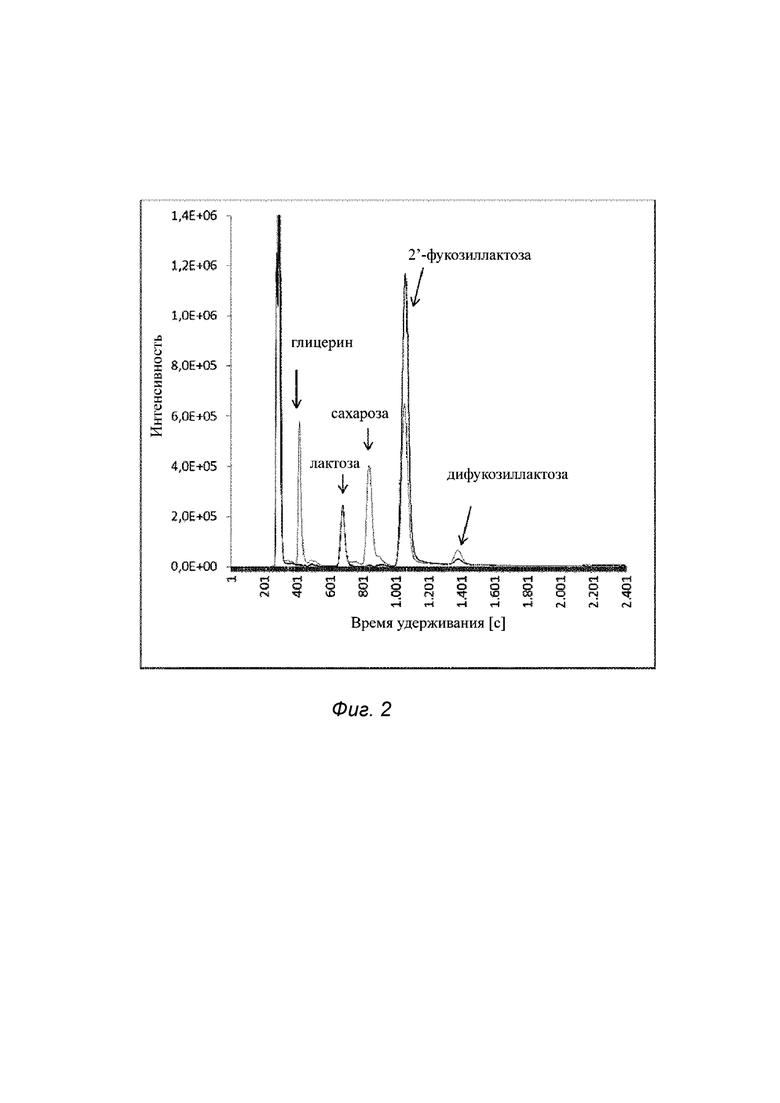












Изобретение относится к области биотехнологии, в частности к получению 2'-фукозиллактозы с использованием генетически модифицированной прокариотической клетки-хозяина. Клетку-хозяина генетически модифицируют таким образом, что по меньшей мере (1) активность фермента, преобразующего фруктозо-6-фосфат, понижена или подавлена и/или путем повышения активности фруктозо-1,6-бифосфат-фосфатазы; (2) по меньшей мере один ген, кодирующий фермент, необходимый для синтеза ГДФ-фукозы de novo, сверхэкспрессирован в клетке-хозяине; (3) экзогенный ген, кодирующий альфа-1,2-фукозилтрансферазу, экспрессирован в клетке-хозяине; и (4) ген, кодирующий фруктозо-1,6-бифосфат-альдолазу, сверхэкспрессирован. Культивируют указанную клетку-хозяина в культуральной среде, содержащей глицерин, и обеспечивают подачу лактозы. Получают фукозилированный олигосахарид, который может быть извлечен из среды. Изобретение позволяет осуществлять биосинтез 2'-фукозиллактозы в концентрации, превышающей 100 г/л. 14 з.п. ф-лы, 3 ил., 9 пр.
1. Способ получения 2'-фукозиллактозы с использованием генетически модифицированной прокариотической клетки-хозяина, включающий стадии:
- предоставления прокариотической клетки-хозяина, генетически модифицированной таким образом, что по меньшей мере (1) активность фермента, преобразующего фруктозо-6-фосфат, которая в немодифицированной клетке-хозяине находится на обычном уровне, понижена или подавлена, где указанный фермент, преобразующий фруктозо-6-фосфат, представляет собой фосфофруктокиназу, и/или путем повышения активности фруктозо-1,6-бифосфат-фосфатазы; (2) по меньшей мере один ген, кодирующий фермент, необходимый для синтеза ГДФ-фукозы de novo, сверхэкспрессирован в клетке-хозяине, где указанный по меньшей мере один ген, кодирующий фермент, необходимый для синтеза ГДФ-фукозы de novo, представляет собой ген, кодирующий фермент ManA, катализирующий изомеризацию фруктозо-6-фосфата до маннозо-6-фосфата, ген, кодирующий фосфоманномутазу, ген, кодирующий маннозо-1-фосфат-гуaнозилтрансферазу, ген, кодирующий ГДФ-маннозо-4,6-дегидратазу, или ген, кодирующий ГДФ-L-фукозосинтазу; (3) экзогенный ген, кодирующий альфа-1,2-фукозилтрансферазу, экспрессирован в клетке-хозяине; и (4) ген, кодирующий фруктозо-1,6-бифосфат-альдолазу, сверхэкспрессирован;
- культивирования и/или выращивания указанной генетически модифицированной клетки-хозяина в культуральной среде, содержащей источник углерода и/или энергии, который представляет собой глицерин; и
- обеспечения подачи лактозы в культуральную среду; с получением тем самым фукозилированного олигосахарида, который может быть извлечен из среды, в которой культивируют клетку-хозяина.
2. Способ по п. 1, где клетка-хозяин выбрана из группы, состоящей из бактериальных клеток-хозяев.
3. Способ по п. 1 или 2, где количество фруктозо-6-фосфата в клетке возрастает в результате возрастания активности фруктозо-1,6-бифосфат-фосфатазы.
4. Способ по любому из пп. 1-3, где ген, кодирующий фосфоманномутазу, представляет собой manB, ген, кодирующий маннозо-1-фосфат-гуaнозилтрансферазу, представляет собой manC, ген, кодирующий ГДФ-маннозо-4,6-дегидратазу, представляет собой gmd, и ген, кодирующий ГДФ-L-фукозосинтазу, представляет собой wcaG.
5. Способ по п. 4, где ген, кодирующий альфа-1,2-фукозилтрансферазу, выбран из группы, состоящей из гена wbgL из E. coli O126 или гена fucT2 из Helicobacter pylori.
6. Способ по любому из пп. 1-5, где клетку-хозяина подвергают дальнейшей генетической модификации с целью экспрессирования гена, кодирующего белок, который обеспечивает экспорт или способствует экспорту желаемого фукозилированного олигосахарида в культуральную среду.
7. Способ по любому из пп. 1-6, где эндогенная или экзогенная пермеаза для импорта лактозы сверхэкспрессирована.
8. Способ по любому из пп. 1-7, где гены, которые модифицированы в клетке-хозяине или которыми модифицируют клетку-хозяина, являются эндогенными или экзогенными генами.
9. Способ по любому из пп. 1-8, где по меньшей мере один из генов, которые модифицированы в клетке-хозяине или которыми модифицируют клетку-хозяина, сверхэкспрессирован в результате эндогенного или экзогенного индуцирования или конститутивным образом.
10. Способ по п. 9, где ген, кодирующий белок, который обеспечивает экспорт или способствует экспорту желаемой 2'-фукозиллактозы, представляет собой ген, кодирующий эффлюксный переносчик сахара.
11. Способ по любому из пп. 3-10, где фруктозо-1,6-бифосфат-фосфатаза кодируется геном, представляющим собой функционально активный вариант гена фруктозо-1,6-бифосфат-фосфатазы (fbpase) из Pisum sativum как присутствует в составе последовательности SEQ ID No. 7.
12. Способ по любому из пп. 7-10, где пермеаза лактозы представляет собой LacY из E. coli.
13. Способ по любому из пп. 1-12, где предоставление лактозы осуществляют путем добавления лактозы от начала культивирования в концентрации по меньшей мере 5 мМ.
14. Способ по любому из пп. 1-13, где предоставление лактозы осуществляют путем добавления лактозы в культуральную среду в такой концентрации, что в течение фазы образования продукта при культивировании достигается концентрация лактозы, составляющая по меньшей мере 5 мМ.
15. Способ по любому из пп. 1-14, где клетки-хозяева культивируют в течение по меньшей мере 60 часов.
| US 2013122553 A1, 16.05.2013 | |||
| CHIN Y.-W | |||
| et al | |||
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |
| Biotechnology and Bioengineering, 23 May 2016, 113(11), p.2443-2452 | |||
| Печь-кухня, могущая работать, как самостоятельно, так и в комбинации с разного рода нагревательными приборами | 1921 |
|
SU10A1 |
| WO 2010142305 A1, 16.12.2010 | |||
| WO 2015150328 A1, 08.10.2015 | |||
| WO 2016040531 | |||

