Настоящее изобретение относится к областям техники генной инженерии, в частности к генной инженерии клеток Bacillus для продукции фукозилированных олигосахаридов и к микробной продукции фукозилированных олигосахаридов с использованием указанных генетически сконструированных клеток Bacillus.
Предшествующий уровень техники
Человеческое грудное молоко обладает оптимальными питательными свойствами для грудных детей. Сахариды, которые присутствуют в человеческом грудном молоке, представляют собой основной компонент человеческого молока, превосходя жиры и белки. Помимо лактозы, которая служит источником энергии, человеческое грудное молоко содержит от примерно 5 до 25 граммов на литр молекул сложных сахаров, а именно, олигосахаридов. Данные олигосахариды обнаружены в значимых концентрациях только в человеческом молоке, и они суммарно известны как олигосахариды грудного молока (ОГМ).
На сегодняшний день идентифицировано приблизительно 200 структурно отличающихся ОГМ. Указанные ОГМ основаны на дисахариде - лактозе (состоящей из группировки глюкозы (Glc) и группировки галактозы (Gal)) и несут дополнительные остатки моносахаридов, которые основаны на N-ацетил-глюкозамине (GlcNAc), фукозе (Fuc), салициловой кислоте (NeuNAc) и галактозе (Gal). Концентрация и состав ОГМ в человеческом грудном молоке варьирует среди индивидуумов и на протяжении периода лактации от вплоть до 20 г/л в молозиве до 5-10 г/л в зрелом молоке.
Молоко женщин, принадлежащее к так называемому «фенотипу секретора», содержит высокое содержание α-1,2-фукозилированных ОГМ. У данных женщин экспрессируется ген FUT2, кодирующий так называемую «фукозилтрансферазу 2». Наиболее распространенными ОГМ в их молоке являются 2'-фукозиллактоза (2'-FL - от англ. 2'-fucosyllactose; Fuc(α1-2)Gal(β1-4)Glc) и лакто-N-фукопентаоза-I (LNPF-I - от англ.lacto-N-fucopentaose-I; Fuc(α1-2)Gal(β1-3)GlcNAc(β1-3)Gal(β1-4)Glc).
Олигосахариды грудного молока не перевариваются во время их прохождения через кишечник грудных детей. За счет их устойчивости в кишечнике грудного ребенка они демонстрируют полезные действия. Более конкретно, ОГМ, как было показано, являются пребиотическими, поскольку они служат источником углерода для комменсальных микроорганизмов родов Bifidobacterium, Bacteroides и Lactobacillus. Таким образом, ОГМ поддерживают пролиферацию данных микроорганизмов в кишечниках грудных детей.
Олигосахариды грудного молока также непосредственно уменьшают колонизацию кишечника грудного ребенка патогенами за счет того, что они предотвращают прилипание указанных патогенов к структурам гликана на поверхности слизистой кишечника. ОГМ функционируют как ловушка, благодаря их структурному сходству с гликанами поверхности эпителия, и ингибируют инвазию патогенов, снижая, вследствие этого, риск инфекций.
Показано, что альфа-1,2-фукозилированные ОГМ защищают от инфицирований Campylobacter jejuni, возбудителем наиболее распространенных бактериальных диарей. α-1,2-Фукозилированные ОГМ также ассоциированы с защитой от диареи, обусловленной термостойким токсином Escherichia coli. Кроме того, риск инфицирований калицивирусами, опосредующими диарею, снижается за счет высокого содержания α-1,2-фукозилированных ОГМ в грудном молоке. ОГМ, главным образом фукозилированный ОГМ лакто-N-фукопентаоза V (LNFP-V; Gal(β1-3)GlcNAc (β1-3)Gal(β1-4)[Fucα1-3]Glc), связывается(ются) с участком связывания токсина А из Clostridium difficile с углеводом, наиболее распространенной причины внутрибольничной диареи. Таким образом, ОГМ, по-видимому, препятствуют взаимодействию токсина toxin А из С. difficile с клеточными рецепторами.
Кроме того, прилипание Pseudomonas aeruginosa к эпителиальным клеткам значимо ингибировалось 2-FL и 3-фукозиллактозой (3-FL - от англ. 3-fucosyllactose; Gal(β1-4)[Fucα1-3]Glc). Связывание норовирусов (вирусов Норуолк, NLV - от англ. Norwalk-like virus), основной причины острого гастроэнтерита, с антигенами группы гисто-крови, предотвращается α-1,2-фукозилированными ОГМ, а также α-1,3-фукозилированными ОГМ. Это указывает на потенциал данных ОГМ в отношении ингибирования связывания капсида норовируса с гликанами рецептора хозяина.
Благодаря известной пользе от ОГМ и главным образом фукозилированных ОГМ экономически целесообразный способ их синтеза является желательным таким образом, чтобы данные олигосахариды или по меньшей мере некоторые из данных олигосахаридов стали доступными в качестве добавки к детской питательной смеси.
Из-за ограниченной поставки и сложностей получения чистых фракций отдельных олигосахаридов грудного молока, были разработаны химические пути синтеза некоторых данных сложных молекул. Однако и химический синтез, и биокаталитические подходы оказались коммерчески нерациональными. Кроме того, в частности химический синтез олигосахаридов грудного молока включает применение нескольких вредных химических веществ, которые накрадывают риск загрязнения конечного продукта.
Из-за проблем, возникающих при химическом синтезе олигосахаридов грудного молока, были разработаны ферментативные подходы к получению ОГМ. На сегодняшний день, раскрыта микробная продукция нескольких ОГМ, таких как 2'-фукозиллактоза, 3-фукозиллактоза, лакто-N-тетраоза, лакто-N-неотетраоза, лакто-N-фукопентаоза I, лакто-N-дифукогексаоза II, 3'-сиалиллактоза и 6'-сиалиллактоза, с использованием главным образом генетически сконструированных штаммов Escherichia coli.
На сегодняшний день рекомбинантные клетки Е. coli используют для микробной продукции ОГМ в промышленном масштабе. Однако род Escherichia coli содержит патогенные члены, а также непатогенные члены. Несмотря на то, что непатогенные штаммы Е. coli используют для микробной продукции ОГМ, такие непатогенные Е. coli не считают безопасными для изготовления продуктов, которые предназначены для потребления человеком во множестве областей. Таким образом, микробные клетки родов, которые расценивались бы как безопасные в таких областях, необходимы для изготовления соединений, которые предназначены для потребления человеком, для получения официального одобрения указанных продуктов для их коммерциализации в указанных областях.
Проблема решается посредством использования бактериальных клеток рода Bacillus, которые расцениваются как безопасные для потребления человеком, и которые способны продуцировать фукозилированный олигосахарид.
Бактерии рода Bacillus являются грамположительными, палочковидными микробными клетками, образующими эндоспоры, или аэробных, или факультативно анаэробных видов. Род Bacillus принадлежит к типу Фирмикуты. Геном членов рода Bacillus имеет тенденцию к парам оснований А-Т в своей частоте использования кодонов. Виды Bacillus почти повсеместно распространены в природе. Например, они могут быть найдены в почве (В. subtilis), а также встречаются в экстремальных условиях окружающей среды, таких как высокий рН (В. alcalophilus), высокая температура (В. thermophilus) или высокое содержание солей (В. halodurans).
Род Bacillus включает 266 видов, имеющих название, которые включают свободноживущие виды, а также паразитарные патогенные виды. Два вида Bacillus считаются с медицинской точки зрения значимыми: В. anthracis, который вызывает сибирскую язву, и В. cereus, который вызывает пищевое отравление. Третий вид, В. thuringiensis, является важным патогеном насекомых, продуцирующим токсин, который может уничтожать насекомых. Таким образом, он используется в качестве инсектицида для контроля над насекомыми-вредителями.
Из-за их статуса GRAS (от англ. generally recognized as safe - общепризнан безопасным), несколько видов Bacillus, например, В. amyloliquefaciens, В. licheniformis и В. subtilis, используются в биотехнологической продукции разных белков и соединений, используемых в пищевой и фармацевтической промышленности.
В. amyloliquefaciens является источником рестриктазы BamHI, и также синтезирует природный белок - антибиотик барназу. Кроме того, В. amyloliquefaciens продуцирует плантазолицин, антибиотик с селективной активностью в отношении B. anthracis. Альфа-амилаза из В. amyloliquefaciens часто используется в гидролизе крахмала. В. amyloliquefaciens также является источником субтилизина, который катализирует распад белков.
В. amyloliquefaciens представляет собой бактерию-колонизатора корней, которая используется для того, чтобы бороться с некоторыми патогенами корней растений в сельском хозяйстве, водной культуре и гидропонике, поскольку она оказывает действие против бактериальных и грибковых патогенов, и может предотвращать инфицирование в результате конкурентного исключения или вытеснения в конкурентной борьбе нежелательного патогена.
Ее высокая способность секретировать щелочную сериновую протеазу сделала В. licheniformis одной из наиболее важных бактерий в промышленном производстве фермента. Субтилизин Carlsberg, секретируемый В. licheniformis, используют в качестве протеазы для моющих средств, и он продается под торговым названием Alcalase®.
В. subtilis представляет собой бактерию, позитивную в отношении каталазы, которая обнаружена в почве и желудочно-кишечном тракте жвачных животных и человека. В. subtilis и вещества, происходящие из данной бактерии, не содержащей эндотоксин, оценивались разными авторитетными органами в отношении их безопасности и пользы в применении в пищевой продукции. В Соединенных Штатах ферменты карбогидраза и протеаза из В. subtilis общепризнаны безопасными (GRAS) Управлением по санитарному надзору за качеством пищевых продуктов и медикаментов США (FDA - от англ. Food and Drug Administration). Bacillus subtilis также присвоен статус «квалифицированной презумпции безопасности» Европейским агентством по безопасности продуктов питания.
Кроме того, нетоксигенные и непатогенные штаммы В. subtilis обычно используются в пищевой промышленности. Например, ферментированные соевые бобы в виде натто, которые обычно потребляют в Японии, содержат вплоть до 10 жизнеспособных клеток В. subtilis на грамм. Ферментированные бобы признают за их вклад в здоровую флору кишечника и поглощение витамина K2. Продукт натто и В. subtilis var. natto в качестве его основного компонента представляют собой FOSHU (от англ. Foods for Specified Health Use - пищевые продукты, специально используемые для поддержания здоровья), одобренные министерством здравоохранения, труда и социальной защиты Японии в качестве эффективных для сохранения здоровья.
С В. subtilis легко работать, она быстро растет, и условия культивирования являются простыми. Рекомбинантные штаммы В. subtilis используют в продукции полигидроксиалканолатов, гиалуроновой кислоты и разных ферментов, таких как амилаза и протеазы.
С природными изолятами В. subtilis дикого типа сложно работать, по сравнению с лабораторными штаммами, которые подвергались процессам одомашнивания на основе мутагенеза и селекции. Данные одомашненные штаммы часто обладают улучшенными способностями к развитию природной компетентности (поглощению и интеграции ДНК окружающей среды), росту и потере способностей, необходимых «в дикой природе». В В. subtilis линейная ДНК, а также мультимерные формы плазмидной ДНК активно поглощаются природными компетентными клетками.
В определенных физиологических условиях маленькая субпопуляция клеток В. subtilis становится компетентной. В В. subtilis природная компетентность регулируется сложной регуляторной сетью. Ключевые регуляторы в данной сети представляют собой, среди прочих, мастер регулятор компетентности ComK и транскрипционный мастер регулятор споруляции Spo0A. Эффективность трансформации клеток В. subtilis и возможно эффективность интеграции ДНК в их геном можно улучшать посредством генной инженерии. Это может быть достигнуто посредством эктопической интеграции экспрессионной кассеты, содержащей регулируемый промотор (например, промотор, индуцируемый маннитом PmtlA) и гены сотK и comS, в геном В. subtilis. Дополнительно, данная стратегия обеспечивает трансформацию В. subtilis за счет природной компетентности с использованием сложной среды (например, LB).
Для продукции фукозилированных сахаридов В. subtilis можно генетически модифицировать разными способами.
Интеграцию генов и/или (одновременную) инактивацию генов посредством нарушения или удаления можно достигать посредством гомологичной рекомбинации. Для эффективной гомологичной рекомбинации по меньшей мере 400-500 п.н. гомологичных плечей необходимы в В. subtilis.
Еще один способ для направленного конструирования генома представляет собой современную систему CRISPR-Cas9. Данный быстрый и безмаркерный инструмент редактирования генома можно использовать для крупномасштабных геномных делеций, маленьких и больших вставок ДНК, сайленсинга генов посредством введения стоп-кодона, а также введения точечных мутаций. Никаких предварительных геномных модификаций не требуется в случае лишенного изъянов редактирования генома посредством CRISPR-Cas9.
Случайная хромосомная интеграция генов и инсерционный мутагенез могут быть выполнены, используя модифицированный транспозон mariner. Данная система не связана с горячими точками в В. subtilis, одновременно демонстрируя высокую эффективность в случайной эктопической интеграции.
Несмотря на то, что виды Bacillus используют для промышленной продукции ферментов, на сегодняшний день не представлены клетки Bacillus для промышленной продукции олигосахаридов, в частности, фукозилированных олигосахаридов.
В китайской патентной заявке CN 108 410 787 А раскрыты рекомбинантные клетки Bacillus subtilis, которые синтезируют лактил-N-неотетраозу. Указанные рекомбинантные клетки В. subtilis имеют ген лактопермеазы, который интегрируют в геном клетки. Кроме того, указанная клетка Bacillus, которая несет плазмиду, содержащую ген бета-1,3-N-глюкозаминтрансферазы и ген β-1,4-галактозилотансферазы. Клетки В. subtilis можно культивировать в присутствии экзогенной лактозы и они синтезируют лактил-N-неотетраозу при титрах вплоть до примерно 1 г/л, что слишком мало для экономически целесообразного промышленного производства.
Несмотря на то, что во множестве патентных заявок упоминается Bacillus в качестве рода, который подходит для продукции олигосахаридов, таких как лакто-N-неотетраоза, никакого коммерческого применения Bacillus для продукций фукозилированных олигосахаридов, в частности фукозилированных олигосахаридов грудного молока, еще не было реализовано, предположительно из-за значительных усилий в модификации метаболизма, которые требуются для реализации необходимых путей биосинтеза для продукции ОГМ в Bacillus. Тогда как указанная выше В. subtilis для продукции LNnT зависит от субстратов-доноров, которые встречаются в природе в клетках В. subtilis, продукция фукозилированного олигосахарида в Bacillus требует реализации гетерологичного метаболического пути в клетке для обеспечения требуемого субстрата-донора - ГДФ (гуанозиндифосфат)-фукозы.
В китайской патентной заявке CN 109 735 479 А раскрыты рекомбинантные клетки Bacillus subtilis для осуществления синтеза 2'-фукозиллактозы, где уровень экспрессии фермента-транспортера лактозы повышен, и где клетка экспрессирует фукозокиназу, фосфатгуанинтрансферазу и фукозилтрансферазу. Сообщалось, что выход 2'-фукозиллактозы в ферментационной среде составлял от 0,424 г/л до 1,042 г/л.
Тем не менее цель настоящего изобретения заключалась в предложении бактериальной клетки рода Bacillus для продукции фукозилированных олигосахаридов.
Цель достигнута посредством предложения клетки Bacillus, в которой имеется лактозопермеаза для импортирования экзогенной лактозы в клетку, путь биосинтеза ГДФ-фукозы de novo для предоставления ГДФ-фукозы и фукозилтрансфераза для переноса группировки фукозы от ГДФ-фукозы на лактозу. Указанную клетку Bacillus можно культивировать в присутствии экзогенной лактозы с получением фукозилированного олигосахарида.
КРАТКОЕ ИЗЛОЖЕНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
Согласно первому аспекту предложена неспорообразующая бактериальная клетка рода Bacillus для получения фукозилированного олигосахарида, где в клетке Bacillus имеется лактозопермеаза, путь биосинтеза ГДФ-фукозы и фукозилтрансфераза.
Согласно второму аспекту предложено применение неспорообразующей бактериальной клетки рода Bacillus согласно первому аспекту для получения фукозилированного олигосахарида.
Согласно третьему аспекту предложен способ получения фукозилированного олигосахарида, включающий:
- предоставление неспорообразующей бактериальной клетки рода Bacillus, где в указанной клетке Bacillus имеется лактозопермеаза, путь биосинтеза ГДФ-фукозы и фукозилтрансфераза, акцептирующая лактозу;
- культивирование клетки Bacillus в культуральной среде, содержащей лактозу, и в условиях, которые являются пермиссивными в отношении клетки Bacillus, с продукцией фукозилированного олигосахарида, и
- возможно, извлечение фукозилированного олигосахарида из культуральной среды и/или клетки Bacillus.
Согласно четвертому аспекту предложены фукозилированные олигосахариды, которые были продуцированы клеткой Bacillus, в которой имеется лактозопермеаза, путь биосинтеза ГДФ-фукозы и фукозилтрансфераза.
Согласно пятому аспекту предложено применение фукозилированного олигосахарида, который был продуцирован клеткой Bacillus, в которой имеется лактозопермеаза, путь биосинтеза ГДФ-фукозы и фукозилтрансфераза, для изготовления питательной композиции.
Согласно шестому аспекту предложены питательные композиции, содержащие по меньшей мере один фукозилированный олигосахарид, который продуцирован клеткой Bacillus, в которой имеется лактозопермеаза, путь биосинтеза ГДФ-фукозы и фукозилтрансфераза.
Подробное описание
Настоящее изобретение относится к клеткам Bacillus для продукции фукозилированных олигосахаридов, их применению и способам получения фукозилированных олигосахаридов посредством культивирования указанных клеток Bacillus в культуральной среде, содержащей лактозу, и в условиях, которые являются пермиссивными в отношении продукции указанного фукозилированного олигосахарида указанной клеткой Bacillus.
Для того, чтобы иметь способность продуцировать фукозилированный олигосахарид, клетка Bacillus должна предоставлять фукозилтрансферазе субстрат-донор, содержащий группировку фукозы, и субстрат-акцептор, представляющий собой дисахарид или олигосахарид, таким образом, чтобы фукозилтрансфераза могла переносить группировку фукозы от субстрата-донора на указанный субстрат-акцептор, с образованием, вследствие этого, фукозилированного олигосахарида.
Следует понимать, что фукозилированный олигосахарид, который предназначена продуцировать указанная клетка Bacillus, представляет собой желательный фукозилированный олигосахарид, тогда как другие фукозилированные олигосахариды, которые могут быть образованы за счет разнородности фукозилтрансферазы во время продукции желательного фукозилированного олигосахарида, считаются нежелательными фукозилированными олигосахаридами или побочными продуктами. Подходящий субстрат-донор представляет собой ГДФ-L-фукозу, и подходящий субстрат-акцептор для образования фукозиллактозы представляет собой дисахарид - лактозу. Полученный желательный фукозилированный олигосахарид представляет собой 2'-фукозиллактозу, 3-фукозиллактозу или 2',3-дифукозиллактозу.
Клетки Bacillus дикого типа как не синтезируют лактозу внутриклеточно, так и не поглощают и не метаболизируют экзогенную лактозу. Однако лактоза представляет собой субстрат-акцептор для группировки фукозы под действием фукозилтрансферазы, акцептирующей лактозу, в образовании некоторых фукозилированных олигосахаридов. Следовательно, с возможностью продуцировать фукозилированный олигосахарид, такой как 2'-фукозиллактоза, 3-фукозиллактоза или 2',3-дифукозиллактоза, клетка Bacillus должна обладать способностью предоставления лактозы фукозилтрансферазе, акцептирующей лактозу, или посредством образования лактозы внутриклеточно и/или посредством поглощения лактозы извне.
В одном воплощении клетка Bacillus для продуцирования фукозилированного олигосахарида может поглощать экзогенную лактозу, имея лактозопермеазу. Термин «экзогенный» в отношении лактозы, в том виде, в котором он используется в данном документе, относится к лактозе, которая не происходит из клетки Bacillus, а именно, являясь внутриклеточно синтезированной клеткой Bacillus, а которая происходит снаружи по отношению к клетке Bacillus, и которую добавляют к культуральной среде, в которой выращивают клетку Bacillus для получения фукозилированного олигосахарида.
В дополнительном и/или альтернативном воплощении клетка Bacillus генетически сконструирована с возможностью поглощать экзогенную лактозу, а именно для того, чтобы иметь лактозопермеазу. Таким образом, в клетке Bacillus для продукции фукозилированного олигосахарида имеется гетерологичная лактозопермеаза. Подходящая лактозопермеаза представляет собой LacY Е. coli или ее функциональный вариант.
Термин «гетерологичный», в том виде, в котором он используется в данном документе, в отношении белков, полипептидов, ферментов и транспортеров, а также в отношении молекул нуклеиновой кислоты и/или нуклеотидных последовательностей, относится к молекуле, которая не является нативной в отношении вида клетки, которая содержит указанную молекулу. Термин «ненативный» указывает на то, что указанная молекула отсутствует в клетке-предшественнике, встречающейся в природе или дикого типа, а именно, клетке того же вида, который наиболее часто встречается в природе, клетки Bacillus.
Термин «функциональный вариант», в том виде, в котором он используется в данном документе, в отношении ферментов и/или транспортных молекул, относится к белкам или полипептидам, обладающим такой же активностью (ферментативной, каталитической или транслоцирующей), как и фермент или транспортер, на который ссылаются, но который имеет аминокислотную последовательность, отличную от аминокислотной последовательности молекулы фермента или транспортера, на которую ссылаются. Таким образом, типичный вариант белка/полипептида отличается по аминокислотной последовательности от белка/полипептида, на который ссылаются. Вариант и белок/полипептид, на который ссылаются, может отличаться по аминокислотной последовательности одной или более заменами, присоединениями и/или делециями в любой комбинации. Следовательно, термин «функциональный вариант» включает усеченные версии белка/полипептида, на который ссылаются, которые обладают такой же активностью, как и белок/полипептид, на который ссылаются. Замещенный или вставленный аминокислотный остаток может представлять собой или может не представлять собой аминокислотный остаток, кодируемый генетическим кодом. Вариант белка/полипептида может представлять собой встречающийся в природе, как например, аллельный вариант, или он может представлять собой вариант, который не известно, чтобы встречался в природе. Варианты белков/полипептидов, не встречающиеся в природе, могут быть созданы посредством методик мутагенеза, посредством прямого синтеза или посредством других методов генной инженерии, известных специалистам в данной области. В пределах объема настоящего раскрытия в термин «вариант» также включены белки и межвидовые гомологи, которые имеют аминокислотную последовательность/последовательность нуклеиновой кислоты, которая обладает более чем примерно 60%-ной идентичностью аминокислотных последовательностей, 65%-ной, 70%-ной, 75%-ной, 80%-ной, 85%-ной, 90%-ной, предпочтительно 91%-ной, 92%-ной, 93%-ной, 94%-ной, 95%-ной, 96%-ной, 97%-ной, 98%-ной или 99%-ной или большей идентичностью аминокислотных последовательностей, предпочтительно, на протяжении области из по меньшей мере примерно 25, 50, 100, 200, 500, 1000 или более аминокислот, с референсным полипептидом.
Термин «такая же активность», в том виде, в котором он используется в данном документе, относится к ферментативной, каталитической активности или активности транспортировки белка/полипептида лишь с точки зрения качества. Следовательно, «функциональный вариант» также включает варианты, которые обладают повышенной или пониженной активностью, по сравнению с активностью белка/полипептида, на который ссылаются.
В дополнительном и/или альтернативном воплощении клетка Bacillus генетически сконструирована для того, чтобы содержать или экспрессировать нуклеотидную последовательность, которая кодирует лактозопермеазу, предпочтительно нуклеотидную последовательность, которая кодирует лактозопермеазу LacY Е. coli или ее функциональный вариант.
Лактозопермеаза LacY Е. coli кодируется областью, кодирующей белок (а именно, открытая рамка считывания) гена lacY Е. coli (номер доступа Gen Bank: NP_414877.1). Следовательно, в одном воплощении клетка Bacillus генетически сконструирована для того, чтобы содержать и экспрессировать нуклеотидную последовательность, которая кодирует лактозопермеазу LacY Е. coli или ее функциональный вариант.
В одном дополнительном и/или альтернативном воплощении частота использования кодона нуклеотидной последовательности, кодирующей гетерологичную лактозопермеазу, приспособлена к частоте использования кодона Bacillus. Частота использования кодона В. subtilis, например, необычна тем, что суммарное содержание GC ниже примерно 45%, содержание GC первой буквы данных кодонов выше примерно 51%, содержание CG второй буквы данных кодонов ниже примерно 36,1% и содержание CG третьей буквы данных кодонов ниже примерно 46%.
Для экспрессии лактозопермеазы клетка Bacillus содержит рекомбинантный ген лактозопермеазы, где нуклеотидная последовательность, которая кодирует лактозопермеазу, функционально связана с последовательностями контроля экспрессии, которые опосредуют экспрессию открытой рамки считывания лактозопермеазы.
Термин «функционально связанный», в том виде, в котором он используется в данном документе, будет означать функциональную связь между последовательностью контроля экспрессии нуклеотидной кислоты/гена (такой как промотор, сигнальная последовательность или ряд сайтов связывания транскрипционных факторов) и второй последовательностью нуклеиновой кислоты (обычно называемой «областью, кодирующей белок», «открытой рамкой считывания» и иногда даже «геном»), где последовательность контроля экспрессии влияет на транскрипцию и/или трансляцию нуклеотидной последовательности, соответствующей второй последовательности. Соответственно, термин «промотор» обозначает последовательности ДНК, которые обычно «предшествуют» открытой рамке считывания в ДНК-полимере и обеспечивают сайт инициации транскрипции в мРНК. «Регуляторные» последовательности ДНК, также обычно расположенные «выше» (то есть, предшествуя) открытой рамки считывания в данном ДНК-полимере, связывают белки, которые определяют частоту (или скорость) инициации транскрипции. В совокупности называемые «промоторной/регуляторной» или «контрольной» ДНК-последовательностью, данные последовательности, которые предшествуют выбранной открытой рамке считывания (или серии открытых рамок считывания) в функциональном ДНК-полимере способствуют определению того, будет ли происходить транскрипция (и в конечном итоге экспрессия) открытой рамки считывания. Последовательности ДНК, которые «следуют за» геном в ДНК-полимере и обеспечивают сигнал для терминации транскрипции в мРНК, называются последовательностями, «терминирующими» транскрипцию.
Рекомбинантный ген лактозопермеазы может быть интегрирован в хромосому Bacillus или присутствовать в виде эписомальной версии на плазмиде в клетке Bacillus.
Экспрессия гетерологичного гена лактозопермеазы в клетке Bacillus позволяет полученной клетке Bacillus поглощать извне поставляемую лактозу из культуральной среды. Поглощенная лактоза может затем служить субстратом-акцептором для фукозилтрансферазы (см. ниже в данном документе).
Клетка Bacillus для продукции фукозилированного олигосахарида должна иметь способность предоставлять субстрат-донор для переноса группировки фукозы на субстрат-акцептор. Субстрат-донор для группировки фукозы представляет собой ГДФ-фукозу. Следовательно, клетка Bacillus должна иметь способность внутриклеточно продуцировать ГДФ-фукозу. Для внутриклеточного биосинтеза ГДФ-фукозы, в клетке Bacillus имеется путь биосинтеза ГДФ-фукозы. Bacillus генетически сконструирована для того, чтобы иметь путь биосинтеза ГДФ-фукозы. Путь биосинтеза ГДФ-фукозы может представлять собой или путь биосинтеза ГДФ-фукозы de novo и/или реутилизационный путь ГДФ-фукозы.
В одном воплощении в клетке Bacillus имеется путь биосинтеза ГДФ-фукозы de novo для внутриклеточного биосинтеза ГДФ-фукозы. Клетка Bacillus генетически сконструирована для того, чтобы иметь путь биосинтеза ГДФ-фукозы de novo.
Путь биосинтеза ГДФ-фукозы de novo включает активности следующих ферментов:
1. фруктозо-6-фосфатизомераза;
2. фосфоманномутаза;
3. ГДФ:маннозо-1-фосфатгуанилилтрансфераза;
4. ГДФ-маннозо-4,6-дегидратаза; и
5. ГДФ-фукозосинтаза.
Следовательно, в клетке Bacillus для получения фукозилированного олигосахарида имеется фруктозо-6-фосфатизомераза (обычно также обозначаемая маннозо-6-фосфатизомеразой), фосфоманномутаза, ГДФ:маннозо-1-фосфатгуанилилтрансфераза, ГДФ-маннозо-4,6-дегидратаза и ГДФ-фукозосинтаза.
В дополнительном и/или альтернативном воплощении клетка Bacillus генетически сконструирована для того, чтобы иметь фруктозо-6-фосфатизомеразу, фосфоманномутазу, ГДФ:маннозо-1-фосфатгуанилилтрансферазу, ГДФ-маннозо-4,6-дегидратазу и ГДФ-фукозосинтазу.
Путь биосинтеза ГДФ-фукозы de novo начинается с изомеризации фруктозо-6-фосфата до маннозо-6-фосфата, реакции, которая катализируется фруктозо-6-фосфатизомеразой. В дополнительном и/или альтернативном воплощении клетка Bacillus генетически сконструирована для того, чтобы содержать и экспрессировать нуклеотидную последовательность, которая кодирует фруктозо-6-фосфатизомеразу. Подходящий фруктозо-6-фосфат представляет собой ManA Е. coli или его функциональный вариант. Типичная нуклеотидная последовательность представляет собой нуклеотидную последовательность, которая кодирует ManA Е. coli.
Фруктозо-6-фосфатизомераза ManA Е. coli кодируется областью, кодирующей белок, гена тапА Е. coli (номер доступа GenBank: NP_416130.3). Таким образом, в дополнительном воплощении клетка Bacillus генетически сконструирована для того, чтобы содержать и экспрессировать нуклеотидную последовательность, которая кодирует ManA Е. coli или его функциональный вариант.
Фруктозо-6-фосфатизомераза ManA Е. coli кодируется открытой рамкой считывания гена тапА Е. coli (номер доступа GenBank: NP_416130.3). Таким образом, в дополнительном и/или альтернативном воплощении клетка Bacillus содержит ген тапА Е. coli или его функциональный вариант.
В дополнительном и/или альтернативном воплощении частота использования кодона нуклеотидной последовательности, кодирующей фруктозо-6-фосфатизомеразу, приспособлена к частоте использования кодона Bacillus.
Другие подходящие фруктозо-6-фосфатизомеразы представляют собой ManA В. subtilis и белки-паралоги GmuF В. subtilis (YdhS) и Pmi В. subtilis (Yvyl) или их функциональные варианты. Иллюстративные нуклеотидные последовательности представляют собой нуклеотидные последовательности, которые кодируют ManA В. subtilis, GmuF В. subtilis или Pmi В. subtilis. Фруктозо-6-фосфатизомеразу ManA В. subtilis кодирует открытая рамка считывания гена тапА В. subtilis (номер доступа Gen Bank: NP_389084.1).
Фруктозо-6-фосфатизомераза ManA В. subtilis кодируется областью, кодирующей белок, гена тапА В. subtilis (номе доступа GenBank: (NP_389084.1)). Таким образом, в дополнительном воплощении клетка Bacillus генетически сконструирована для того, чтобы содержать и экспрессировать нуклеотидную последовательность, которая кодирует ManA В. subtilis или его функциональный вариант. Таким образом, в дополнительном и/или альтернативном воплощении клетка Bacillus содержит ген тапА В. subtilis или его функциональный вариант.В еще одном дополнительном и/или альтернативном воплощении клетка Bacillus генетически сконструирована для того, чтобы содержать и экспрессировать нуклеотидную последовательность, которая кодировала ген gmuF В. subtilis или его функциональный вариант, и/или нуклеотидную последовательность, которая кодирует ген pmi В. subtilis или его функциональный вариант.
В дополнительном и/или альтернативном воплощении экспрессия нативного гена тапА, нативного гена gmuF и/или нативного гена pmi может быть повышена. Экспрессия нативного гена тапА, нативного гена gmuF и/или нативного гена pmi может быть повышена по той причине, что эндогенный промотор по меньшей мере одного из нативного гена тапА, нативного гена gmuF и нативного гена pmi заменен более сильным промотором, а именно промотором, опосредующим повышенный уровень экспрессии, по сравнению с нативным промотором гена тапА, гена gmuF и гена pmi, соответственно. В еще одном дополнительном или альтернативном воплощении экспрессия нативного гена тапА, нативного гена gmuF и/или нативного гена pmi может быть усилена по той причине, что дополнительные копии нативного гена тапА, нативного гена gmuF и/или нативного гена pmi размножаются в клетке Bacillus.
В еще одном дополнительном и/или альтернативном воплощении экспрессия нативного гена тапА может усиливаться в результате делеции или функциональной инактивации гена тапР, кодирующего транспортер маннозы, приводящей к конститутивной экспрессии нативного гена тапА.
В еще одном дополнительном или альтернативном воплощении экспрессия нативного гена gmuF может быть усилена в результате делеции или функциональной инактивации гена gmuR, кодирующего соответствующий репрессор транскрипции.
Для экспрессии фруктозо-6-фосфатизомеразы клетка Bacillus содержит рекомбинантный ген фруктозо-6-фосфатизомеразы, в котором нуклеотидная последовательность, которая кодирует фруктозо-6-фосфатизомеразу, функционально связана с последовательностями контроля экспрессии.
Рекомбинантный ген фруктозо-6-фосфатизомеразы может быть интегрирован в хромосому Bacillus или присутствовать в виде эписомальной версии на плазмиде в клетке Bacillus.
На второй стадии биосинтеза de novo ГДФ-фукозы маннозо-6-фосфат превращается в маннозо-1-фосфат под действием ферментативной активности фосфоманномутазы. В дополнительном и/или альтернативном воплощении клетка Bacillus генетически сконструирована для того, чтобы содержать и экспрессировать нуклеотидную последовательность, которая кодирует фосфоманномутазу.
Подходящая фосфоманномутаза представляет собой фосфоманномутазу ManB Е. coii. Таким образом, в дополнительном воплощении клетка Bacillus генетически сконструирована для того, чтобы содержать и экспрессировать нуклеотидную последовательность, которая кодирует ManB Е. coli или ее функциональный вариант.
Фосфоманномутаза ManB Е. coli кодируется нуклеотидной последовательностью области, кодирующей белок, ген тапВ Е. coli (номер доступа Gen Bank: NP_416552.1). Таким образом, в дополнительном и/или альтернативном воплощении клетка Bacillus содержит ген тапВ Е. coli или его функциональный вариант.
В дополнительном и/или альтернативном воплощении частота использования кодона нуклеотидной последовательности, кодирующей фосфоманномутазу, приспособлена к частоте использования кодона Bacillus.
Для экспрессии фосфоманномутазы клетка Bacillus содержит рекомбинантный ген фосфоманномутазы, в котором нуклеотидная последовательность, которая кодирует фосфоманномутазу, функционально связана с последовательностями контроля экспрессии.
Рекомбинантный ген фосфоманномутазы может быть интегрирован в хромосому Bacillus или присутствовать в клетке Bacillus в виде эписомальной версии на плазмиде.
Следующая стадия в пути биосинтеза ГДФ-фукозы cfe novo представляет собой образование ГДФ-маннозы посредством следующей реакции:
α-D-маннозо-1-фосфат+ГТФ+Н+=>дифосфат+ГДФ-α-D-манноза
Превращение маннозо-1-фосфата в ГДФ-маннозу опосредовано ГДФ:маннозо-1-фосфатгуанилилтрансферазой. Таким образом, в клетке Bacillus для продукции фукозилированного олигосахарида имеется гетерологичная ГДФ:маннозо-1-фосфатгуанилилтрансфераза. Подходящая ГДФ:маннозо-1-фосфатгуанилилтрансфераза представляет собой ManC Е. coli или ее функциональный вариант.
В дополнительном и/или альтернативном воплощении клетка Bacillus генетически сконструирована, чтобы содержать и экспрессировать нуклеотидную последовательность, которая кодирует ГДФ:маннозо-1-фосфатгуанилилтрансферазу. Иллюстративная нуклеотидная последовательность представляет собой нуклеотидную последовательность, которая кодирует ManC Е. coli. Таким образом, в дополнительном воплощении клетка Bacillus генетически сконструирована для того, чтобы содержать и экспрессировать нуклеотидную последовательность, которая кодирует ManC Е. coli или ее функциональный вариант.
ГДФ:маннозо-1-фосфатгуанилилтрансфераза ManC Е. coli кодируется открытой рамкой считывания гена тапС Е. coli (номер доступа Gen Bank: NP_416553.1). Таким образом, в дополнительном и/или альтернативном воплощении клетка Bacillus содержит ген тапС Е. coli или его функциональный вариант.
В дополнительном и/или альтернативном воплощении частота использования кодона нуклеотидной последовательности, кодирующей ГДФ:маннозо-1-фосфатгуанилилтрансферазу, приспособлена к частоте использования кодона Bacillus.
В случае экспрессии ГДФ:маннозо-1-фосфатгуанилилтрансферазы в клетке Bacillus содержится рекомбинантный ген ГДФ:маннозо-1-фосфатгуанилилтрансеразы, в котором нуклеотидная последовательность, которая кодирует ГДФ:маннозо-1-фосфатгуанилилтрансферазу, функционально связана с последовательностями контроля экспрессии.
Рекомбинантный ген ГДФ:маннозо-1-фосфатгуанилилтрансферазы может быть интегрирован в хромосому Bacillus или находиться в виде эписомальной версии на плазмиде.
На последующей стадии пути ГДФ-фукозы de novo, катализируемой ГДФ-маннозо-4,6-дегидратазой, ГДФ-D-манноза превращается в ГДФ-4-дегидро-6-дезокси-D-маннозу.
Таким образом, в клетке Bacillus для получения фукозилированного олигосахарида имеется ГДФ-маннозо-4,6-дегидратаза. Подходящей ГДФ-маннозо-4,6-дегидратазой является Gmd Е. coll.
В дополнительном и/или альтернативном воплощении клетка Bacillus генетически сконструирована для того, чтобы содержать и экспрессировать нуклеотидную последовательность, которая кодирует ГДФ-маннозо-4,6-дегидратазу. Иллюстративная нуклеотидная последовательность представляет собой нуклеотидную последовательность, которая кодирует Gmd Е. coll. Таким образом, в дополнительном воплощении клетка Bacillus генетически сконструирована для того, чтобы содержать и экспрессировать нуклеотидную последовательность, которая кодирует Gmd Е. coli или ее функциональный вариант.
ГДФ-маннозо-4,6-дегидратаза Gmd Е. coli кодируется открытой рамкой считывания гена gmd Е. coli (номер доступа Gen Bank: NP_416557.1). Таким образом, в дополнительном и/или альтернативном воплощении в клетке Bacillus содержится ген gmd Е. coli или его функциональный вариант.
В дополнительном и/или альтернативном воплощении частота использования кодона нуклеотидной последовательности, кодирующей ГДФ-маннозо-4,6-дегидратазу, приспособлена к частоте использования кодона Bacillus.
В случае экспрессии ГДФ-маннозо-4,6-дегидратазы, в клетке Bacillus содержится рекомбинантный ген ГДФ-маннозо-4,6-дегидратазы, в котором нуклеотидная последовательность, которая кодирует ГДФ-маннозо-4,6-дегидратазу, функционально связана с последовательностями контроля экспрессии.
Рекомбинантный ген ГДФ-маннозо-4,6-дегидратазы может быть интегрирован в хромосому Bacillus или находиться в виде эписомальной версии на плазмиде.
В конечной реакции пути биосинтеза de novo ГДФ-фукозы ГДФ-4-дегидро-6-дезокси-D-манноза превращается в ГДФ-фукозу в результате двухстадийной NADPH (от англ. reduced nicotinamide adenine dinucleotide phosphate восстановленный никотинамидадениндинуклеотидфосфат)-зависимой реакции. Данное превращение опосредовано ГДФ-фукозосинтазой и включает эпимеразную реакцию и редуктазную реакцию. Указанная эпимеразная реакция превращает ГДФ-4-кето-6-дезоксиманнозу в ГДФ-4-кето-6-дезоксигалактозу, которая затем восстанавливается до ГДФ-фукозы.
Таким образом, в клетке Bacillus для продукции фукозилированного олигосахарида имеется ГДФ-фукозосинтаза. Подходящей ГДФ-фукозосинтазой является WcaG Е. coli или ее функциональный вариант.
В дополнительном и/или альтернативном воплощении клетка Bacillus генетически сконструирована для того, чтобы содержать и экспрессировать нуклеотидную последовательность, которая кодирует ГДФ-фукозосинтазу. Иллюстративная нуклеотидная последовательность представляет собой нуклеотидную последовательность, которая кодирует WcaG Е. coli. Таким образом, в дополнительном воплощении клетка Bacillus генетически сконструирована для того, чтобы содержать и экспрессировать нуклеотидную последовательность, которая кодирует WcaG Е. coli или ее функциональный вариант.
ГДФ-фукозосинтаза WcaG Е. coli кодируется открытой рамкой считывания гена wcaG Е. coli (номер доступа Gen Bank: NP_416556.1). Таким образом, в дополнительном и/или альтернативном воплощении в клетке Bacillus содержится ген wcaG Е. coli или его функциональный вариант.
В дополнительном и/или альтернативном воплощении частота использования кодона нуклеотидной последовательности, кодирующей ГДФ-фукозосинтазу, приспособлена к частоте использования кодона Bacillus.
В случае экспрессии ГДФ-фукозосинтазы, в клетке Bacillus содержится рекомбинантный ген ГДФ-фукозосинтазы, в котором нуклеотидная последовательность, которая кодирует ГДФ-фукозосинтазу, функционально связана с последовательностями контроля экспрессии.
Рекомбинантный ген ГДФ-фукозосинтазы может быть интегрирован в хромосому Bacillus или находиться в виде эписомальной версии на плазмиде.
В дополнительном и/или альтернативном воплощении в клетке Bacillus имеется реутилизационный путь ГДФ-фукозы для внутриклеточного биосинтеза ГДФ-фукозы. В пути реутилизации ГДФ-L-фукозы свободная цитоплазматическая фукоза фосфорилирована под действием L-фукокиназы с образованием L-фукозо-L-фосфата, который затем дополнительно превращается в ГДФ-L-фукозу.
Реутилизационный путь для биосинтеза ГДФ-фукозы включает следующие ферменты:
I. фукозокиназа; и
II. L-фукозо-1-фосфатгуанилилтрансфераза.
Следовательно, в клетке Bacillus для продукции фукозилированного олигосахарида, в которой имеется реутилизационный путь ГДФ-фукозы, имеется фукозокиназа и L-фукозо-1-фосфатгуанилилтрансфераза.
В дополнительном и/или альтернативном воплощении клетка Bacillus генетически сконструирована для того, чтобы иметь фукозокиназу и L-фукозо-1-фосфатгуанилилтрансферазу.
Фукозокиназа, также называемая фукокиназой, АТФ:6-дезокси-L-галактозо-1-фосфотрансфераза, АТФ:β-L-фукозо-1-фосфотрансфераза или L-фукокиназная активность, L-фукозокиназная активность катализирует реакцию L-фукоза+АТФ → β-L-фукозо-1-фосфат+АДФ+2 Н+.
Затем L-фукозо-1-фосфатгуанилилтрансфераза или ГДФ-L-фукозопирофосфорилаза превращает указанный β-L-фукозс-1-фосфат в ГДФ-L-фукозу.
В дополнительном и/или альтернативном воплощении клетка Bacillus генетически сконструирована для того, чтобы содержать и экспрессировать нуклеотидную последовательность, которая кодирует фукозокиназу или ее функциональный вариант, и L-фукозо-1-фосфатгуанилилтрансферазу или ее функциональный вариант.
В дополнительном и/или альтернативном воплощении фукозокиназа и L-фукозо-1-фосфатгуанилилтрансфераза объединены в одном единственном полипептиде. Подходящие гены для трансформации, кодирующие фукозокиназу, фукозо-1-фосфатгуанилилтрансферазу и/или бифункциональную L-фукозо-1-фосфатгуанилилтрансферазу, могут быть получены из родов Bacteroides, Lentisphaera, Ruminococcus, Solibacter, Arabidopsis, Oryza, Physcomitrella, Vitis, Danio, Bos, Equus, Macaca, Pan, Homo, Rattus, Mus и Xenopus. Пример бифункциональной фукозокиназы/L-фукозо-1-фосфатгуанилилтрансферазы найден в Bactericides fragilis.
В В. fragilis бифункциональная фукозокиназа/L-фукозо-1-фосфатгуанилилтрансфераза Fkp кодируется геном fkp В. fragilis (номер доступа GenBank AY849806).
В дополнительном и/или альтернативном воплощении клетка Bacillus генетически сконструирована для того, чтобы содержать и экспрессировать нуклеотидную последовательность, которая кодирует Fkp В. fragilis или ее функциональный вариант.
В дополнительном и/или альтернативном воплощении частота использования кодона нуклеотидных последовательностей, кодирующих фукозокиназу, фукозо-1-фосфатгуанилилтрансферазу и/или бифункциональную фукозокиназу/L-фукозо-1-фосфатгуанилилтрансферазу, приспособлена к частоте использования кодона Bacillus.
В случае экспрессии фукозокиназы и фукозо-1-фосфатгуанилилтрансферазы, в клетке Bacillus содержится по меньшей мере один рекомбинантный ген, в котором область, кодирующая белок, кодирующая фукозокиназу и L-фукозо-1-фосфатгуанилилтрансферазу и/или бифункциональную фукозокиназу/L-фукозо-1-фосфатгуанилилтрансферазу фукозокиназу, функционально связана с последовательностями контроля экспрессии.
Рекомбинантный ген фукозокиназы, ген фукозо-1-фосфатгуанилилтрансферазы и/или ген бифункциональной фукозокиназы/L-фукозо-1-фосфатгуанилилтрансферазы, может быть интегрирован в хромосому Bacillus или находиться в виде эписомальной версии на плазмиде.
В случае внутриклеточного образования фукозиллактозы в клетке Bacillus имеется фукозилтрансфераза. Ферментативная активность фукозилтрансферазы переносит группировку фукозы с субстрата-донора на субстрат-акцептор. Для биосинтеза фукозиллактозы указанный субстрат-акцептор представляет собой лактозу. Следовательно, фукозилтрансфераза представляет собой фукозилтрансферазу, акцептирующую лактозу.
Фукозилтрансфераза выбрана из группы, состоящей из α-1,2-фукозилтрансфераз для биосинтеза 2'-фукозиллактозы и α-1,3-фукозилтрансфераз для биосинтеза 3-фукозиллактозы.
В дополнительном и/или альтернативном воплощении клетка Bacillus генетически сконструирована для того, чтобы содержать и экспрессировать по меньшей мере одну нуклеотидную последовательность, которая кодирует фукозилтрансферазу.
Для получения 2'-фукозиллактозы (2'-FL) α-1,2-фукозилтрансферазы WbgL из O126 Е. coli и FucT2 из Helicobacter pylori (ЕР 2 479 263 В1), α-1,2-фукозилтрансферазы WblA из Vibrio cholera O22, FutD из Н. bilis АТСС 437879, FutE из Н cinaede CCUG 18818, FutN из Bacteroides vulgatus АТСС 8482, FutO из Bacteroides ovatus АТСС 8483, WbgN из E. coli O55:H7, Bft1 и Bft3 из Bacteroides fragilis NCTC 9343 (WO 2014/018596 A2), и α-1,2-фукозилтрансферазы FucT2 из H. pylori для синтеза сахаридов Lewis Y и Lewis В (US 6670160 B2) были описаны и являются подходящими α-1,2-фукозилтрансферазами для биосинтеза 2'-FL в клетках Bacillus.
Для получения 3-фукозиллактозы описаны α-1,3-фукозилтрансфераза Amuc из Akkermansia muciniphila и FucT6 и FucT7 из Bacteroides fragilis (ЕР 2 439 264 А1), α-1,3-фукозилтрансфераза FutA из Н. pylori (US 2014/ 0120611 А1), и они являются подходящими α-1,3-фукозилтрансферазами для биосинтеза 3-FL в клетках Bacillus. Кроме того, в WO 2016/040531 А1 раскрыта α-1,3-фукозилтрансфераза CafC из В. nordii CL02T12C05 для синтеза 3-фукозиллактозы и лактодифукотетраозы, и CafD из Н. hepaticus АТСС51449 для продукции LNnFP-III.
Дополнительные фукозилтрансферазы, которые могут быть экспрессированы в клетках Bacillus, для продукции фукозилированных сахаридов раскрыты в WO 2019/0088133 А1, который включен в данный документ посредством ссылки.
В дополнительном и/или альтернативном воплощении частота использования кодона нуклеотидной последовательности, кодирующей фукозилтрансферазу, приспособлена к частоте использования кодона Bacillus.
В случае экспрессии фукозилтрансферазы клетка Bacillus содержит по меньшей мере один рекомбинантный ген, в котором область, кодирующая белок, кодирующая фукозилтрансферазу, функционально связана с последовательностями контроля экспрессии.
Рекомбинантный ген фукозилтрансферазы может быть интегрирован в хромосому Bacillus или находиться в виде эписомальной версии на плазмиде.
В дополнительном и/или альтернативном воплощении клетка Bacillus для продукции фукозилированного олигосахарида может поглощать L-фукозу. Способность поглощать L-фукозу является преимуществом клеток Bacillus, в которых имеется реутилизационный путь ГДФ-фукозы.
В дополнительном и/или альтернативном воплощении клетка Bacillus генетически сконструирована для того, чтобы сделать возможным или улучшить поглощение L-фукозы. Таким образом, в клетке Bacillus имеется гетерологичная L-фукозопермеаза. Подходящая L-фукозопермеаза представляет собой FucP Е. coli или ее функциональный вариант.
В дополнительном и/или альтернативном воплощении клетка Bacillus генетически сконструирована для того, чтобы содержать и экспрессировать нуклеотидную последовательность, которая кодирует L-фукозопермеазу, предпочтительно нуклеотидную последовательность, которая кодирует FucP Е. coli или ее функциональный вариант.
L-фукозопермеаза Е. coli кодируется областью, кодирующей белок, гена fucP Е. coli (номер доступа Gen Bank: NP_417281.1). Следовательно, клетка Bacillus генетически сконструирована для того, чтобы содержать и экспрессировать нуклеотидную последовательность, которая кодирует FucP Е. coli или ее функциональный вариант.
В дополнительном и/или альтернативном воплощении частота использования кодона нуклеотидной последовательности, кодирующей L-фукозопермеазу, приспособлена к частоте использования кодона Bacillus.
Для экспрессии L-фукозопермеазы в клетке Bacillus содержится рекомбинантный ген L-фукозопермеазы, в котором нуклеотидная последовательность, которая кодирует L-фукозопермеазу, функционально связана с последовательностями контроля экспрессии.
Рекомбинантный ген L-фукозопермеазы может быть интегрирован в хромосому Bacillus или находиться в виде эписомальной версии на плазмиде.
В дополнительном и/или альтернативном воплощении клетка Bacillus для продукции фукозилированного олигосахарида не обладает никакой β-галактозидазной активностью или обладает сниженной β-галактозидазной активностью, по сравнению с клеткой дикого типа того же вида.
Внутриклеточный биосинтез фукозилированных олигосахаридов требует импортирования лактозы в качестве субстрата-акцептора для фукозилтрансферазы, акцептирующей лактозу. Любая внутриклеточная ферментативная активность, которая гидролизует поглощенную лактозу, будет воздействовать на эффективность образования фукозиллактозы, поскольку пул внутриклеточной лактозы будет уменьшен. Таким образом, будет преимущественным, если клетка Bacillus для продукции фукозилированного олигосахарида не будет обладать или будет обладать по меньшей мере, по сравнению с клеткой Bacillus дикого типа, сниженной активностью бета-галактозидазы.
В дополнительном и/или альтернативном воплощении клетка Bacillus генетически сконструирована для устранения или по меньшей мере уменьшения активности β-галактозидазы клетки.
В дополнительном и/или альтернативном воплощении клетка Bacillus генетически сконструирована посредством делеции или функциональной инактивации гена дапА. В еще одном воплощении клетка Bacillus генетически сконструирована для снижения уровня экспрессии гена дапА, по сравнению с клеткой Bacillus дикого типа.
Ген ganA Bacillus также называется yvfH или 1асА. Он представляет собой ген регулона GanR, который содержит гены, кодирующие ферменты, участвующие в утилизации галактана. Ген ganA кодирует бета-галактозидазу, которая участвует в утилизации галактана Bacillus.
Делеция или функциональная инактивация гена ganA устраняет GanA-опосредуемую β-галактозидазную активность в клетке Bacillus, в то время как уменьшение уровня экспрессии ganA снижает количество GanA в клетке Bacillus и, следовательно, β-галактозидазную активность, которая могла бы препятствовать биосинтезу фукозилированного олигосахарида.
В дополнительном и/или альтернативном воплощении клетка Bacillus генетически сконструирована посредством делеции или функциональной инактивации гена yesZ. Ген yesZ Bacillus кодирует бета-галактозидазу YesZ, которая играет роль в деградации рамногалактуронана, происходящего из клеточных стенок растений. Экспрессия гена yesZ Bacillus индуцируется рамногалактуронаном I. В еще одном воплощении клетка Bacillus генетически сконструирована для того, чтобы уменьшать уровень экспрессии гена yesZ, по сравнению с клеткой Bacillus дикого типа.
Делеция или функциональная инактивация гена yesZ устраняет YesZ-опосредуемую β-галактозидазную активность в клетке Bacillus, тогда как уменьшение уровня экспрессии yesZ снижает количество YesZ в клетке Bacillus и, следовательно, β-галактозидазную активность, которая могла бы препятствовать биосинтезу фукозилированного олигосахарида.
Когда В. subtilis поступает в постэкспоненциальную фазу роста, они (начинают) продуцируют большие количества внеклеточных протеаз. Чужеродные белки часто являются чувствительными к протеазам. Таким образом, штамм, не содержащий экзопротеазу, является желательным для повышения стабильности гетерологичных белков и для обеспечения аккумуляции высоких уровней чужеродных белков. Геном Bacillus кодирует по меньшей мере восемь внеклеточных протеаз, а именно, nprE, aprE, epr, bpr, mpr, nprB, vpr и wprA. Таким образом, в дополнительном и/или альтернативном воплощении клетка генетически сконструирована в том смысле, что по меньшей мере один ген, кодирующий внеклеточную протеазу, был удален или функционально инактивирован, предпочтительно, по меньшей мере один из генов, выбранных из группы, состоящей из nprE, aprE, epr, bpr, mpr, nprB, vpr и wprA. Предпочтительно, два, три, четыре, пять, шест, семь или восемь генов, выбранных из группы, состоящей из nprE, aprE, epr, bpr, mpr, nprB, vpr и wprA, удалены или функционально инактивированы.
В. subtilis синтезирует пульхерриминовую кислоту при росте в средах, содержащих углевод, такой как глюкоза или лактоза. Выделяемая пульхерриминовая кислота образует красный пигмент пульхерримин, соль пульхерриминовой кислоты (хелат железа (III)), где железо находится в среде для выращивания. Образование данного нежелательного побочного продукта во время процессов ферментации можно предотвратить/аннулировать посредством делеции или нарушения генов yvmC и/или сурХ. Ген yvmC кодирует цикподипептидсинтазу, и ген сурХ кодирует цитохром Р450 цикло-l-лейцил-l-лейцилдипептидоксидазу.
Таким образом, в дополнительном и/или альтернативном воплощении клетка Bacillus генетически сконструирована так, чтобы по меньшей мере один из генов yvmC и сурХ удален или функционально инактивирован.
В ризобактерии В. subtilis имеются гены для синтеза более чем 20 антибиотиков. Среди них имеются пептидные антибиотики, подобно лантибиотикам Bacillus subtilis и лантибиотик-подобным пептидам (субтилин, эрицин S, мерсацидин, субланцин 168, субтилозин А), и нерибосомно синтезируемые (пептидные) антибиотики (сурфактин, итурин, бацилломицин, микосубтилин, коринебактин/ бациллибактин, фенгицин, плипастатин, микобациллин, TL-119, бацилизин, бацилизоцин, амикумацин, 3,3'-неотрегалосадиамин, диффицидин, ризоктицин).
Для продукции фукозилированного олигосахарида предпочтительно использовать клетку Bacillus, которая не продуцирует антибиотик. Таким образом, в дополнительном и/или альтернативном воплощении клетка Bacillus не синтезирует один или более антибиотиков, выбранных из группы, состоящей из лантибиотиков и лантибиотик-подобных пептидов, таких как субтилин, эрицин S, мерсацидин, субланцин 168, субтилозин А; нерибосомно синтезируемых (пептидных) антибиотиков, таких как сурфактин, итурин, бацилломицин, микосубтилин, коринебактин/бациллибактин, фенгицин, плипастатин, микобациллин, TL-119, бацилизин, бацилизоцин, амикумацин, 3,3'-неотрегалозадиамин, диффицидин и ризоктицин. Клетка Bacillus может быть генетически сконструирована с получением клетки Bacillus, которая не синтезирует один или более из указанных антибиотиков.
Клетки Bacillus дикого типа могут образовывать споры. Споруляцию, а именно процесс образования спор, у бактерий рассматривают как реакцию бактериальной клетки, которая инициирует программу развития, приводящую к образованию дочерних клеток отличной морфологии и направления развития. Споруляцию Bacillus исследовали как основную модель дифференцировки клеток. Во время споруляции палочковидная клетка Bacillus делится асимметрично, что приводит вследствие этого к получению двух генетически идентичных клеток с разной морфологией и направлениями развития.
Однако при промышленном получении нежелательно, если бактериальный штамм - продуцент образует споры во время ферментации. Таким образом, предпочтительно использовать клетки Bacillus для продукции фукозилированных олигосахаридов, которые не способны образовывать споры. Такие клетки Bacillus называются «неспорообразующими».
Предпочтительно, неспорообразующая клетка Bacillus, способная продуцировать фукозилированный олигосахарид, происходила из одного из штаммов В. Subtilis, перечисленных в Таблице 1.

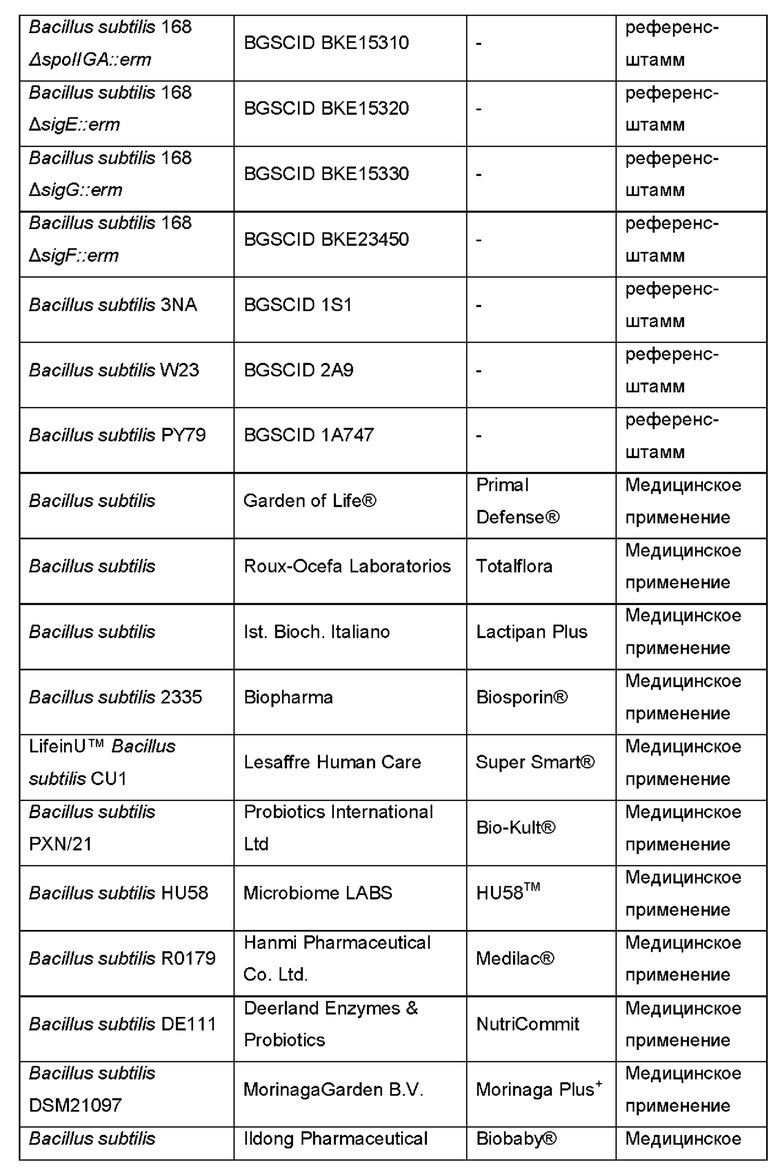
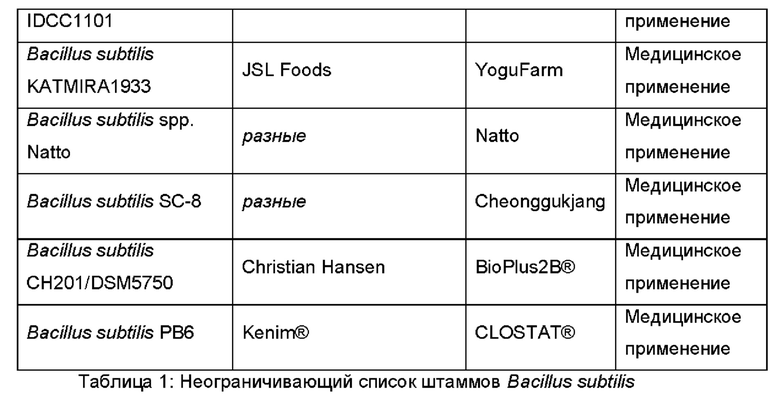
В дополнительном и/или альтернативном воплощении клетка Bacillus генетически сконструирована посредством делеции или функциональной инактивации Spo0A. Подходящая функциональная инактивация Spo0A включает делецию участка фосфорилирования, где фосфотрансферазы Spo0F и Spo0B фосфорилируют Spo0A.
В дополнительном и/или альтернативном воплощении клетка Bacillus генетически сконструирована посредством делеции или функциональной инактивации генов, кодирующих фактор сигма SigE (sigE) и/или фактор сигма SigF (sigF).
Клетка Bacillus по изобретению способна продуцировать фукозилированный олигосахарид, при культивации в присутствии лактозы в среде и в условиях, которые являются пермиссивными в отношении клетки Bacillus для продукции фукозилированного олигосахарида.
Согласно второму аспекту предложено применение клетки Bacillus, как ранее описано в данном документе, для продукции фукозилированного олигосахарида, предпочтительно, фукозилированного олигосахарида грудного молока, такого как 2'-FL, 3-FL или 2',3-DiFL.
Поскольку Bacillus обычно расценивают как безопасную, продукция фукозилированных олигосахаридов для потребления человеком с использованием организма-продуцента, который в общем считается безопасным, также будет считаться безопасной, при условии, что изменения в клетках Bacillus для продукции фукозилированного олигосахарида не влияют на безопасность штамма-продуцента в отношении здоровья человека и окружающей среды. Следовательно, ожидают, что будет меньше проблем, влияющих на официальное одобрение фукозилированного олигосахарида для потребления человеком, в частности, фукозилированных олигосахаридов грудного молока для потребления человеком, более конкретно, для детской питательной смеси, которая состоит из или содержит фукозилированный олигосахарид, который был продуцирован посредством использования клетки Bacillus, как описано в данном документе. Таким образом, одобрение продуцируемых микробами фукозилированных олигосахаридов в детской питательной смеси и питательных композициях органами регулирования, а также среди потребителей, должно быть лучше.
Согласно третьему аспекту предложен способ получения фукозилированного олигосахарида, который включает:
- предоставление неспорообразующей клетки Bacillus, которая генетически сконструирована для того, чтобы иметь лактозопермеазу, путь биосинтеза ГДФ-фукозы и фукозилтрансферазу;
- культивирование клетки Bacillus в культуральной среде, содержащей лактозу, и в условиях, которые являются пермиссивными в отношении продукции фукозилированного олигосахарида, и
возможно извлечение фукозилированного олигосахарида из культуральной среды и/или из клетки Bacillus.
Неспорообразующая клетка Bacillus, которая предложена, представляет собой клетку Bacillus, как описано в данном документе.
В дополнительном и/или альтернативном воплощении фукозилированный олигосахарид выбран из группы, состоящей из 2'-фукозиллактозы (2'-FL), 3-фукозиллактозы (3-FL), 2',3-фукозиллактозы (DiFL - от англ. 2',3-difucosyllactose), лакто-N-фукопентаозы I (LNFP I - от англ. lacto-N-fucopentaose I), лакто-N-неофукопентаозы I (LNnFP I - от англ. lacto-N-neofucopentaose I), лакто-N-фукопентаозы II (LNFP II - от англ. lacto-N-fucopentaose II), лакто-N-фукопентаозы III (LNFP III - от англ. lacto-N-fucopentaose III), лакто-N-фукопентаозы V (LNFP V - от англ. lacto-N-fucopentaose V), лакто-N-неофукопентаозы V (LNnFP V - от англ. lacto-N-neofucopentaose V), лакто-N-дифукогексаозы I (LNDH I - от англ. lacto-N-difucohexaose I) и лакто-N-дифукогексаозы II (LND - от англ. lacto-N-difucohexaose II).
В дополнительном и/или альтернативном воплощении культуральная среда содержит L-фукозу, в частности, для культивирования клетки Bacillus для продукции фукозилированного олигосахарида, в которой имеется реутилизационный путь ГДФ-фукозы для обеспечения ГДФ-фукозы в качестве субстрата-донора для фукозилтрансферазы.
Изобретение также относится к фукозилированным олигосахаридам, которые были продуцированы клеткой Bacillus, и/или способу, как описано в данном документе, к применению указанных фукозилированных олигосахаридов для изготовления питательной композиции, предпочтительно, детской питательной смеси, добавки к рациону или продукта лечебного питания. Кроме того, настоящее изобретение также относится к питательным композициям, содержащим фукозилированный олигосахарид, который был продуцирован клеткой Bacillus, и/или способу, как описано в данном документе.
Настоящее изобретение будет описано относительно конкретных воплощений и со ссылкой на графические материалы, но данное изобретение не ограничивается ими, а только формулой изобретения. Кроме того, термины первый, второй и тому подобное в описании и в формуле изобретения используются для проведения различия между похожими элементами и не обязательно для описания последовательности, во времени, в пространстве, по рангу или любым другим образом. Следует понимать, что термины, используемые таким образом, являются взаимозаменяемыми в соответствующих обстоятельствах, и что воплощения изобретения, описанные в данном документе, способны работать в последовательностях, отличных от описанных или проиллюстрированных в данном документе.
Следует отметить, что термин «содержащий», используемый в формуле изобретения, не следует считать ограничивающимся средствами, перечисленными в дальнейшем; он не исключает других элементов или стадий. Таким образом, его следует считать определяющим наличие заявленных признаков, целых чисел, стадий или компонентов, на которые ссылаются, но он не исключает наличие или добавление одного или более других признаков, целых чисел, стадий или компонентов или их групп. Таким образом, объем выражения «устройство, содержащее средства А и В» не следует ограничивать устройствами, состоящими только из компонентов А и В. Оно означает, что в отношении настоящего изобретения, единственными релевантными компонентами устройства являются А и В.
Ссылка на всем протяжении данного описания изобретения на «одно воплощение» или «воплощение» означает, что конкретный признак, структура или характеристика, описанные в связи с данным воплощением, включены в по меньшей мере одно воплощение настоящего изобретения. Таким образом, появления фраз «в одном воплощении» или «в воплощении» в разных местах по всему объему данного описания изобретения не обязательно все относятся к одному и тому же воплощению, но могут. Кроме того, конкретные признаки, структуры или характеристики могут быть объединены любым подходящим образом, как будет очевидно среднему специалисту в данной области из данного раскрытия, в одном или более воплощениях.
Аналогично, следует понимать, что в описании иллюстративных воплощений изобретения разные признаки изобретения иногда сгруппированы вместе в одном единственном воплощении, фигуре или его описании в целях упрощения раскрытия и помощи в понимании одного или более из разных аспектов изобретения. Данный способ раскрытия, однако, не нужно считать отражающим мысль, что заявленное изобретение требует больше признаков, чем явным образом перечислены в каждом пункте. Скорее, как отражено в приведенной ниже формуле изобретения, аспекты изобретения заключаются меньше чем во всех признаках одного вышеизложенного раскрытого воплощения. Таким образом, формула изобретения после подробного описания явным образом включена тем самым в данное подробное описание, причем каждый пункт отдельно стоит в виде отдельного воплощения данного изобретения.
Кроме того, в то время как некоторые воплощения, описанные в данном документе, включают некоторые, но не все признаки, включенные в другие воплощения, подразумевается, что комбинации признаков разных воплощений находятся в объеме изобретения и образуют разные воплощения, как будет понятно специалистам в данной области. Например, в приведенной ниже формуле изобретения любое из заявленных воплощений можно использовать в любой комбинации.
Кроме того, некоторые из воплощений описаны в данном документе как способ или комбинация элементов способа, которые могут быть реализованы посредством процессора компьютерной системы или с помощью других средств выполнения функции. Таким образом, процессор с необходимыми инструкциями для осуществления такого способа или элемента способа образует средство осуществления способа или элемента способа. Кроме того, описанный в данном документе элемент воплощения аппарата представляет собой пример средства осуществления функции, выполняемой элементом, с целью осуществления изобретения.
В описании и графических материалах, предоставленных в данном документе, изложены многочисленные конкретные подробности. Однако понятно, что воплощения изобретения можно осуществлять на практике без данных конкретных подробностей. В других примерах хорошо известные способы, структуры и методики не были показаны подробно для того, чтобы не затруднять понимание данного описания. Теперь изобретение будет описано с помощью подробного описания нескольких воплощений изобретения. Ясно, что другие воплощения изобретения могут быть скомпонованы в соответствии со знаниями специалистов в данной области, не отклоняясь от истинной сущности или технической идеи изобретения, причем изобретение ограничено только условиями прилагаемой формулы изобретения.
ПРИМЕРЫ
Пример 1: Трансформация Bacillus subtilis
Bacillus subtilis можно подвергать генетическим манипуляциям посредством разных методик. Для трансформации В, subtilis компетентные клетки готовили посредством модифицированного протокола двухстадийного способа (Anagnostopoulos, С.and Spizizen, J. (1961) J Bacteriol 81 (5): 741-746). Ночную культуру инокулировали в среде MG1 и встряхивали при 37°С. Среда MG1 представляет собой минимальную среду Спицайзена, которая дополнена 0,5% глюкозой, 5 мМ MgSO4 и 0,02% казаминовыми кислотами (возможно, дополнительно дополнена биотином и/или L-триптофаном). На следующее утро данную культуру разводили 1:20 в свежей среде MG1 и инкубировали при 37°С в течение приблизительно 6 ч. 1 мл культуры разводили в 8 мл среды MG2, которая отличается от среды MG1 по концентрации казаминовых кислот (0,01% вместо 0,02%). В укороченном протоколе ночную культуру непосредственно разводят в среде MG2. После инкубации в течение еще 90 мин порцию культуры, 1 мл, смешивали с 1-3 мкг мультимерной плазмидной ДНК или линейной ДНК и инкубировали при 37°С в течение 30-60 мин при встряхивании. Мультимерную плазмидную ДНК получали или посредством использования штамма Е. coli NM538 для размножения плазмидной ДНК или посредством линеаризации плазмиды в результате расщепления рестриктазой с одним разрезом, которая осуществляет расщепление в пределах каркаса с последующим повторным лигированием Т4 ДНК лигазой.
Затем, клетки распределяли по чашкам с 2xYT и агаром, содержащим соответствующий антибиотик. Антибиотики добавляли в соответствующих концентрациях: 5 мкг⋅мл-1 эритромицина, 5 мкг⋅мл-1 хлорамфеникола, 10 мкг⋅мл-1 канамицина,100 мкг⋅мл-1 пектиномицина.
В качестве альтернативы, для трансформации протопластов (Romero, D. et al. (2006) Journal of Microbiological Methods 66:556-559) клетки выращивали в 20 мл бульона Penassay (РАВ - от англ. Penassay broth) при 37°С до начала стационарной фазы роста (OD600 (от англ. optical density - оптическая фаза) равна 1,7-2). Затем клетки осаждали центрифугированием и ресуспендировали в 10 мл среды SMPP (0,3%-ный бычий сывороточный альбумин, 5%-ная 2 М сахароза, 25% 4×РАВ, 50% 2×SMM), состоящей из 2×SMM, представляющего собой 1 М сахарозу, 0,04 М гидрат двунатриевой соли малеиновой кислоты и 0,04 М MgCl2 (рН 6,5). После добавления лизоцима (10 мг мл-1) и мутанолизина (75 Ед мл-1) смесь инкубировали при 37°С при встряхивании с образованием протопластов. Образование протопластов проверяли посредством микроскопа. Затем протопласты аккуратно собирали посредством центрифугирования при 5200×g и 4°С в течение 5 мин, два раза промывали охлажденным льдом промывочным буфером для электротрансформации (1×SMM) и, в конечном итоге, суспендировали в данном растворе. Плазмидную ДНК (1-3 мкг) добавляли к 120 мкл суспензии протопластов, и смесь держали на льду в течение по меньшей мере 5 мин. Смесь для трансформации переносили в кювету, объемом 0,2 см, и один импульс для электропорации прикладывали на уровне 25 мкФ, 400 Ω и 0,7 кВ. Сразу после разряда электропорации 1 мл восстанавливающей среды (равные объемы 4×РАВ и 2×SMM, свежеприготовленные перед применением) добавляли в кювету. Затем реакционную смесь для трансформации переносили в 2 мл пробирку и инкубировали при 37°С при встряхивании в течение 12 ч. Для регенерации клеточную суспензию распределяли по чашкам с DM3 и агаром (Chang, S. and Cohen, S. (1979) MGG 168(1):111-115) и инкубировали при 37°C в течение 48 ч. Регенерирующая смесь DM3 содержала следующие стерильные растворы на литр: 200 мл 4%-ного агара, 100 мл 5%-ных казаминовых кислот, 50 мл 10%-ого дрожжевого экстракта, 100 мл 3,5%-ного K2HPO4 и 1,5%-ного KH2PO4, 25 мл 20%-ной глюкозы, 20 мл 1 М MgCl2, 500 мл 0,5 М сорбита и 5 мл простерилизованного посредством фильтрации 2%-ного бычьего сывороточного альбумина (добавляемые к смеси, когда температура ниже 55°С), и ее дополняли соответствующим антибиотиком.
Электропорацию В. subtilis проводили в соответствии с модифицированным протоколом от Zhang et al. (2011), предоставленным MoBiTec GmbH (Zhang, G., Bao, P., Zhang. Y., Deng, A. Chen. N. и Wen, T. (2011) Anal. Biochem., 409:130-137). Ночную культуру 2×YT разводили в 100 раз свежей средой 2×YT, и культуру выращивали до OD600 0,2 при 37°С на ротационной качалке. Затем культуру дополняли 1%-ным DL-треонином, 2%-ным глицином, 0,1%-ным триптофаном и 0,03% Tween 80. После культивирования в течение еще 60 мин, клеточную суспензию охлаждали на льду в течение 20 мин, центрифугировали при 5000×g в течение 10 мин при 4°С и два раза промывали буфером для электропорации (0,5 М трегалоза, 0,5 М сорбит, 0,5 М маннит, 0,5 мМ MgCl2, 0,5 мМ K2HPO4, 0,5 мМ KH2PO4, рН 7,4, стерилизовали посредством фильтрации и хранили в замороженном виде). Наконец, клетки ресуспендировали в буфере для электропорации в соотношении 1/100 исходного объема культуры, и 100 мкл клеточной суспензии смешивали с ДНК. Смесь для трансформации переносили в 0,1-см кювету, и электропорацию проводили при 1,8 кВ с помощью одного импульса, доставляемого посредством прибора MicroPulser™ (Bio-Rad). Сразу после переноса импульса, 1 мл бульона 2×YT, содержащего 0,5 М сорбит и 0,38 М маннит, добавляли в кювету. Суспензию для трансформации переносили в пробирку, объемом 2 мл, и инкубировали при 37°С в течение 3 ч на ротационной качалке. Клетки распределяли по чашке с 2×YT и агаром и инкубировали при 37°С в течение ночи.
Используя альтернативный протокол электропорации (Xue, G. P., J. S. Johnson, и В. P. Dalrymple: 1999; Journal of Microbiological Methods 34:183-191), 5 мл LB, содержащей 0,5 M глюцит, инокулировали В. subtilis и инкубировали в течение ночи при 37°С. Далее, ночную культуру разводили (1:16) 75 мл LB, содержащей 0,5 М глюцит, и инкубировали до получения OD600 0,85 - 0,95. Затем клетки осаждали посредством центрифугирования в течение 10 мин при 4°С при 5,000×g и четыре раза промывали буфером для электропорации, охлажденным посредством льда (10% глицерин, 0,5 М глюцит, 0,5 М маннит). Наконец, клетки ресуспендировали в 1-2 мл буфера для электропорации. Электропорацию проводили, используя 60 мкл компетентных клеток с ДНК в охлажденной кювете для электропорации (расстояние между электродами 1 мм). Смесь клетки-ДНК подвергали воздействию одного электрического импульса на уровне 25 мкФ, 200 Ω и 21 кВ/см. Наконец, 1 мл бульона для восстановления (LB, содержащая 0,5 М глюцит и 0,38 М маннит) добавляли к электропермеабилизированным клеткам, и бактериальную культуру инкубировали в течение 3 ч при 37°С с последующим посевом на LB с агаром с добавлением антибиотика.
Использовали две разные обогащенные среды, а именно бульон Luria (LB) и 2×YT. Среда бульон Luria (LB) состояла из 1%-ного триптона, 0,5%-ого дрожжевого экстракта и 0,5% NaCl (рН 7,2).
Среда 2×YT состояла из 1,6%-ного триптона, 1%-ого дрожжевого экстракта и 0,5% NaCl (рН 7,5).
Для получения чашек с обогащенной средой с агаром добавляли 15 г л-1 агара.
Для экспериментов с встряхиваемыми колбами использовали минимальную среду Спицайзена (Spizizen, J. 1958 Proc. Natl. Acad. Sci. U. S. A. 44(10):1072-1078).
Минимальная среда Спицайзена содержит следующие соли: 2 г/л (NH4)2SO4, 14 г/л K2HPO4, 6 г/л KH2PO4, 1 г/л цитрата Na3 × 2⋅H2O и 0,2 г/л MgSO4 × 7⋅H2O.
Предкультуральная среда, состоящая из минимальных солей Спицайзена, дополненная 2% D-глюкозой, 0,05% казаминовыми кислотами и MgSO4, до конечной концентрации 2 мМ (возможно дополнительно дополненная биотином и/или L-триптофаном).
Основная культуральная среда состояла из минимальных солей Спицайзена, дополненная 2%-ной D-глюкозой, 0,05%-ыми казаминовыми кислотами, MgSO4 до конечной концентрации 2 мМ и 0,5 мл⋅л-1 1000×раствора микроэлементов (возможно дополнительно дополненная биотином и/или L-триптофаном).
Раствор микроэлементов (1000х) состоял из 100,6 г⋅л-1 C6H9NO6, 56,4 г⋅л-1 цитрата железа (III)-аммония, 9,8 гл-1 MnCl2 × 4⋅H2O, 1,6 г⋅л-1 CoCl2 × 6 H2O, 1 г⋅л-1 CuCl2 × 2⋅H2O, 1,9 г⋅л-1 Н3 ВО3, 9 г⋅л-1 ZnSO4 × 7⋅H2O, 1,1 г⋅л-1 Na2MoO4 × 2⋅H2O, 1,5 г⋅л-1 Na2SeO3, 1,5 г⋅л-1 NiSO4 × 6⋅H2O.
При необходимости, соответствующий(ие) антибиотик(и) добавляли к среде для того, чтобы сделать ее селективной.
Штаммы В. subtilis исходно выращивали на чашках с обогащенными средами с агаром с получением одиночных колоний. Данные чашки выращивали в течение 1 суток при 30-37°С. В экспериментах со встряхиваемыми колбами 20 мл предварительной культуры инокулировали одиночной колонией и выращивали в течение ночи при 30-37°С на ротационной качалке. Следующие 20 мл основных культур инокулировали данной предварительной культурой до начальной OD600 примерно 0,1 и инкубировали при 30-37°С на ротационной качалке. Если требовалась индукция, 40-60 мл основной культуры разбивали на порции по 20 мл в момент времени индукции. Объем культуры не превышал 20% емкости встряхиваемой колбы.
Пример 2: Конструирование штамма-продуцента Bacillus subtilis для 2'-фукозиллактозы
Модификации метаболизма штамма Bacillus subtilis с дефектом спорообразования (таблица 1) достигали посредством интеграции гетерологичных генов тапС Е. coli, тапВ Е. coli и тапА Е. coli и одновременного удаления эндогенного гена lacA посредством гомологичной рекомбинации. Ген дапА (yvfN, lacA) В. subtilis, который расположен в пределах галактозного оперона, кодирует бета-галактозидазу.
Для синтеза ГДФ-маннозы открытые рамки считывания тапС, тапВ и тапА функционально связаны с конститутивным промотором Р43 В, subtilis (часть репозитория iGem: ID последовательности: ВВа_K143013) в виде оперона. Ген тапС (номер доступа Gen Bank: NP_416553.1) кодирует ГДФ:маннозо-1-фосфатгуанилилтрансферазу из Е. coli. Ген тапВ (номер доступа Gen Bank: NP_416552.1) кодирует фосфоманномутазу Е. coli и ген тапА (номер доступа Gen Bank: NP_389084.1) кодирует фруктозо-6-фосфатизомеразу В. subtilis. Каждый из генов, упомянутых выше, сливали in silico с последовательностью RBS В. subtilis. Описанную в данном документе экспрессионную кассету <Р43-тапСВА> подвергали оптимизации кодонов для экспрессии в В. subtilis и получали синтетически посредством GenScript Corp. Далее, кассету полной интеграции собирали и клонировали в pBR322 (New England Biolabs GmbH, Франкфурт, Германия) с созданием суицидной плазмиды <pBR322 flank дапА up-lox71-erm-lox66-P43-manCBA-flank ganA down> (SEQ ID NO: 1). Затем, В. subtilis трансформировали данной плазмидой за счет ее природной компетентности. Клетки распределяли по чашкам с 2х YT и агаром, содержащим соответствующий антибиотик (5 мкг мл-1 эритромицина). Интеграцию экспрессионной кассеты <Р43-тапСВА> в локус ganA генома В. subtilis, с получением штамма А, проверяли посредством ПЦР (полимеразная цепная реакция) на колониях. Экспрессию гена подтверждали посредством направленной протеомики и/или посредством ПЦР в реальном времени.
Для продукции 2'-фукозиллактозы из лактозы и ГДФ-маннозы в полученном штамме для интеграции <Р43-тапСВА> конструировали экспрессионную плазмиду (SEQ ID NO: 2), которая содержит все необходимые гены под контролем индуцибельного промотора Pgrac100. Таким образом, экспрессионный вектор рНТ253 В. subtilis (MoBiTec GmbH, Геттинген, Германия) использовали в качестве каркаса. Открытую рамку считывания гена lacY Е. coli (номер доступа Gen Bank: NP_414877.1) амплифицировали посредством ПЦР из хромосомной ДНК. Открытую рамку считывания гена wbgL, кодирующего α-1,2-фукозилтрансферазы O126 Е. coli, подвергали оптимизации кодонов для экспрессии в В. subtilis и получали синтетически посредством GenScript Corp. Кроме того, открытые рамки считывания генов gmd Е. coli (номер доступа Gen Bank: NP_416557.1; кодирующий ГДФ-маннозо-4,6-дегидратазу) и wcaG (номер доступа Gen Bank: NP_416556.1; кодирующий ГДФ-фукозосинтазу) подвергали оптимизации кодонов для экспрессии в В. subtilis и получали синтетически посредством GenScript Corp. Каждый ген, заключенный в индуцибельной экспрессионной кассете, был связан с последовательностью RBS В, subtilis. Кроме того, подходящую терминирующую последовательность В. subtilis из части репозитория iGem (ID последовательности: ВВа_В0015) вводили в конечную экспрессионную плазмиду <рНТ253 Pgrac100-wbgL-gmd-wcaG-lacY-terminator> (SEQ ID NO: 2). Штамм интеграции <P43-manCBA> трансформировали данной экспрессионной плазмидой (SEQ ID NO: 2) за счет его природной компетентности. Экспрессию генов подтверждали посредством направленной протеомики и/или посредством ПЦР в реальном времени. Трансформанты культивировали в условиях, которые являются пермиссивными в отношении В. Subtilis, для продукции 2'-фукозиллактозы в присутствии экзогенной лактозы.
Пример 3: Конструирование штамма-продуцента Bacillus subtilis для 2'-фукозиллактозы
Штамм А, который описан в примере 2, использовали в качестве родительского штамма. Для продукции 2'-фукозиллактозы из лактозы и ГДФ-маннозы, дополнительно, гены lacY, gmd, wcaG и wbgL Е. coli интегрировали в эндогенный локус атуЕ (атуА) штамма А (кодирующий альфа-амилазу).
Открытую рамку считывания гена wbgL (кодирующего α-1,2-фукозилтрансферазу) подвергали оптимизации кодонов для экспрессии в В. subtilis и получали синтетически посредством GenScript Corp. Кроме того, открытые рамки считывания генов gmd (номер доступа Gen Bank: NP_416557.1; кодирующего ГДФ-маннозо-4,6-дегидратазу) и wcaG (номер доступа Gen Bank: NP_416556.1; кодирующего ГДФ-фукозосинтазу) подвергали оптимизации кодонов для экспрессии в В. subtilis (проводилось GenScript Corp), помещали под контроль сильного конститутивного промотора PlepA В. subtilis в виде оперона из двух генов и получали синтетически с помощью GenScript Corp. Открытую рамку считывания гена lacY Е. coli (номер доступа Gen Bank: NP_414877.1), кодирующего лактозопермеазу, амплифицировали посредством ПЦР из хромосомной ДНК.
Все необходимые гены для продукции 2'-фукозиллактозы из лактозы и ГДФ-маннозы собирали в конститутивную экспрессионную кассету <P43-wbgL-lacY-PlepA-gmd-wcaG>, в которой каждый ген был связан с последовательностью RBS В. subtilis. Дополнительно, подходящую терминирующую последовательность В. subtilis из части репозитория iGem (ID последовательности: ВВа_В0015) помещали ниже данной экспрессионной кассеты. Описанную выше конструкцию клонировали в pBR322 (New England Biolabs GmbH, Франкфурт, Германия) с получением <pBR322 flank amyE up-lox71-aacd9-lox66-P43-wbgL-lacY-PlepA-gmd wcaG-ferminator-flank amyE down> (SEQ ID NO: 3).
Полученный вектор для интеграции amyE (SEQ ID NO: 3) использовали для трансформации В. subtilis за счет ее природной компетентности. Проводили селекцию трансформантов посредством спектиномицина (100 мкг мл-1). Интеграцию экспрессионной кассеты <P43-wbgL-lacY-PlepA-gmd wcaG-terminator> в локус атуЕ генома В. subtilis проверяли посредством ПЦР на колониях. Экспрессию генов подтверждали посредством направленной протеомики и/или посредством ПЦР в реальном времени.
Пример 4: Конструирование штамма-продуцента Bacillus subtilis для 2'-фукозиллактозы
Для получения 2'-фукозиллактозы из ГДФ-фукозы и экзогенной лактозы, исходно, ген lacY Е. coli интегрировали в эндогенный локус атуЕ (атуА) (кодирующий альфа-амилазу). Открытую рамку считывания гена lacY Е. coli (номер доступа Gen Bank: NP_414877.1), кодирующего лактозопермеазу, амплифицировали посредством ПЦР из хромосомной ДНК. Кассету для интеграции <flank атуЕ up-lox71-aad9-lox66-P43-lacY-flank атуЕ down> (SEQ ID NO: 4) конструировали и клонировали в pBR322 (New England Biolabs GmbH, Франкфурт, Германия). В. subtilis трансформировали полученной плазмидой за счет ее природной компетентности. Интеграцию экспрессионной кассеты <Р43-lacY> в локус атуЕ генома В. Subtilis, приводящую к получению штамма В, проверяли посредством ПЦР на колониях. Экспрессию генов подтверждали посредством направленной протеомики и/или посредством ПЦР в реальном времени.
Для синтеза ГДФ-фукозы из экзогенной L-фукозы конструировали экспрессионную плазмиду В. subtilis, содержащую все необходимые гены. Плазмиду pDW1 использовали в качестве каркаса. Данную плазмиду получали синтетически посредством GenScript Corp, и она содержит гены bla и erm для селекции в Е. coli и В. subtilis, соответственно. Дополнительно, она содержит репликон рМВ1 для размножения в Е. coli и репликон pUB110 для размножения в В. subtilis.
Ген fkp В. fragilis (номер доступа GenBank AY849806) кодирует бифункциональную фукозокиназу/L-фукозо-1-фосфатгуанилилтрансферазу, ген wbgL Е. coli кодирует α-1,2-фукозилтрансферазы и ген fucP Е. coli (номер доступа Gen Bank: NP_417281.1) кодирует L-фукозопермеазу. Каждую из открытых рамок считывания данных генов подвергали оптимизации кодонов для экспрессии в В. subtilis и получали синтетически посредством GenScript Corp. Конструировали экспрессионную кассету, которая содержит гены fkp, wbgL и fucP под контролем сильного конститутивного промотора Р436. subtilis.
Каждый ген, заключенный в экспрессионную кассету, был связан с последовательностью RBS В. subtilis. Кроме того, подходящую терминирующую последовательность В. subtilis из части репозитория iGem (ID последовательности: ВВа_В0015) вводили в конечную экспрессионную плазмиду <pDW1-P43-fkp-wbgL-fucP-terminator> (SEQ ID NO: 5). Штамм В трансформировали данной экспрессионной плазмидой (SEQ ID NO: 5) за счет его природной компетентности. Экспрессию генов подтверждали посредством направленной протеомики и/или посредством ПЦР в реальном времени. Трансформанты культивировали в условиях, которые являются пермиссивными в отношении В. Subtilis, с получением 2'-фукозиллактозы в присутствии экзогенной лактозы и L-фукозы.
Пример 5: Получение 2'-фукозиллактозы с использованием штаммов Bacillus subtilis с модифицированным метаболизмом
Предварительную культуру инокулировали штаммом Bacillus subtilis с модифицированным метаболизмом, подходящим для биосинтеза 2'-фукозиллактозы (как описано в примере 2, 3 и 4, соответственно).
Предварительную культуру инкубировали при 30-37°С в течение ночи и затем разводили до исходной OD600 примерно 0,1 в свежей основной культуральной среде. Когда основная культура достигала OD600 приблизительно 0,5, 2 мМ лактозу добавляли к среде для выращивания. Когда для экспрессии гена использовали индуцибельный промотор Pgrac100, индукцию осуществляли посредством лактозы (2 мМ) или как лактозы (2 мМ), так и IPTG (от англ. isopropylthiogalactoside - изопропилтиогалактозид) (1 мМ). Для утилизирующего биосинтеза ГДФ-фукозы, дополнительно, добавляли 2 мМ L-фукозу. Культивирование прекращали после 24 ч/48 ч, и внутриклеточную и внеклеточную 2'-фукозиллактозу анализировали посредством тонкослойной хроматографии и/или ВЭЖХ (высокоэффективной жидкостной хроматографии) и/или масс-спектрометрии (как описано в WO 2017/042382 А или WO 2019/008133 А). Выявляли биосинтез существенных количеств 2'-фукозиллактозы.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Jennewein Biotechnologie GmbH
<120> ПРОДУКЦИЯ ФУКОЗИЛИРОВАННЫХ ОЛИГОСАХАРИДОВ В BACILLUS
<130> P 1903 WO
<160> 5
<170> PatentIn version 3.5
<210> 1
<211> 11270
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Суицидная плазмида
<400> 1
ttctcatgtt tgacagctta tcatcgataa gctttaatgc ggtagtttat cacagttaaa
60
ttgctaacgc agtcaggcac cgtgtatgaa atctaacaat gcgctcatcg tcatcctcgg
120
caccgtcacc ctggatgctg taggcatagg cttggttatg ccggtactgc cgggcctctt
180
gcgggatatc gtccattccg acagcatcgc cagtcactat ggcgtgctgc tagcgctata
240
tgcgttgatg caatttctat gcgcacccgt tctcggagca ctgtccgacc gctttggccg
300
ccgcccagtc ctgctcgctt cgctacttgg agccactatc gactacgcga tcatggcgac
360
cacacccgtc ctgtggatcc tctacgccgg acgcatcgtg gccggcatca ccggcgccac
420
aggtgcggtt gctggcgcct atatcgccga catcaccgat ggggaagatc gggctcgcca
480
cttcgggctc atgagcgctt gtttcggcgt gggtatggtg gcaggccccg tggccggggg
540
actgttgggc gccatctcct tgcatgcacc attccttgcg gcggcggtgc tcaacggcct
600
caacctacta ctgggctgct tcctaatgca ggagtcgcat aagggagagc gtcgaccgat
660
gcccttgaga gccttcaacc cagtcagctc cttccggtgg gcgcggggca tgactatcgt
720
cgccgcactt atgactgtct tctttatcat gcaactcgta ggacaggtgc cggcagcgct
780
ctgggtcatt ttcggcgagg accgctttcg ctggagcgcg acgatgatcg gcctgtcgct
840
tgcggtattc ggaatcttgc acgccctcgc tcaagccttc gtcactggtc ccgccaccaa
900
acgtttcggc gagaagcagg ccattatcgc cggcatggcg gccgacgcgc tgggctacgt
960
cttgctggcg ttcgcgacgc gaggctggat ggccttcccc attatgattc ttctcgcttc
1020
cggcggcatc gggatgcccg cgttgcaggc catgctgtcc aggcaggtag atgacgacca
1080
tcagggacag cttcaaggat cgctcgcggc tcttaccagc ctaacttcga tcactggacc
1140
gctgatcgtc acggcgattt atgccgcctc ggcgagcaca tggaacgggt tggcatggat
1200
tgtaggcgcc gccctatacc ttgtctgcct ccccgcgttg cgtcgcggtg catggagccg
1260
ggccacctcg acctgaatgg aagccggcgg cacctcgcta acggattcac cactccaaga
1320
attggagcca atcaattctt gcggagaact gtgaatgcgc aaaccaaccc ttggcagaac
1380
atatccatcg cgtccgccat ctccagcagc cgcacgcggc gcatctcggg cagcgttggg
1440
tcctggccac gggtgcgcat gatcgtgctc ctgtcgttga ggacccggct aggctggcgg
1500
ggttgcctta ctggttagca gaatgaatca ccgatacgcg agcgaacgtg aagcgactgc
1560
tgctgcaaaa cgtctgcgac ctgagcaaca acatgaatgg tcttcggttt ccgtgtttcg
1620
taaagtctgg aaacgcggaa gtcagcgccc tgcaccatta tgttccggat ctgcatcgca
1680
ggatgctgct ggctaccctg tggaacacct acatctgtat taacgaagcg ctggcattga
1740
ccctgagtga tttttctctg gtcccgccgc atccataccg ccagttgttt accctcacaa
1800
cgttccagta accgggcatg ttcatcatca gtaacccgta tcgtgagcat cctctctcgt
1860
ttcatcggta tcattacccc catgaacaga aatccccctt acacggaggc atcagtgacc
1920
aaacaggaaa aaaccgccct taacatggcc cgctttatca gaagccagac attaacgctt
1980
ctggagaaac tcaacgagct ggacgcggat gaacaggcag acatctgtga atcgcttcac
2040
gaccacgctg atgagcttta ccgcagctgc ctcgcgcgtt tcggtgatga cggtgaaaac
2100
ctctgacaca tgcagctccc ggagacggtc acagcttgtc tgtaagcgga tgccgggagc
2160
agacaagccc gtcagggcgc gtcagcgggt gttggcgggt gtcggggcgc agccatgacc
2220
cagtcacgta gcgatagcgg agtgtatact ggcttaacta tgcggcatca gagcagattg
2280
tactgagagt gcaccagcgg gcaaggaaag ccttcaatat gtgcagtggt atgtcaactc
2340
tatgaagatc agcctgttta caatggcagg gtctttgctc tgtgtgacgt ttacggccta
2400
tgcgttttcg cgctttcggt ttaaagggag gaaatacgct ttaacgctct ttttattgct
2460
gcagatgatt cctcagtttt cagctttaat tgccttgttt gtgctggcgc aaatcttggg
2520
aatgatcaat agccactggc tgctaatctt gctttatatc ggcggcctga tcccgatgaa
2580
tacgtatttg atgaaagggt acatggattc cattccgatg gatttagacg aaagcgccaa
2640
gattgacgga gccagcagca ccagaatctt cttccagatc attctgccat tatcaaaacc
2700
gatggcggca gtcgtggcca tgaacggctt taccggtccg ctcggagatt ttgtgctgtc
2760
ctcaaccata ttgagaacgc ctgaatcata tacattgccc gtcggtctat tcaatttagt
2820
gaatgatgtc atgggggcca gctatacgac atttgcggcc ggagccctgc ttatcagcat
2880
accggttgcc gtcatcttta ttatgctgca aaagaatttt gtgtccggat taaccgcagg
2940
cggaacgaag ggctaagaga acaaggagga gaatgtgatg tcaaagcttg aaaaaacgca
3000
cgtaacaaaa gcaaaattta tgctccatgg gggagactac aaccccgatc agtggctgga
3060
tcggcccgat attttagctg acgatatcaa actgatgaag ctttctcata cgaatacgtt
3120
ttctgtcggc accagtgaat tcgagctcgg tacctaccgt tcgtataatg tatgctatac
3180
gaagttatta cgcgttaacc cgggcccgcg gatgcatatg atcagatcct ttaactctgg
3240
caaccctcaa aattgaatga gacatgctac acctccggat aataaatata tataaacgta
3300
tatagatttc ataaagtcta acacactaga cttatttact tcgtaattaa gtcgttaaac
3360
cgtgtgctct acgaccaaaa ctataaaacc tttaagaact ttcttttttt acaagaaaaa
3420
agaaattaga taaatctctc atatctttta ttcaataatc gcatccgatt gcagtataaa
3480
tttaacgatc actcatcatg ttcatattta tcagagctcg tgctataatt atactaattt
3540
tataaggagg aaaaaatatg ggcattttta gtatttttgt aatcagcaca gttcattatc
3600
aaccaaacaa aaaataagtg gttataatga atcgttaata agcaaaattc atataaccaa
3660
attaaagagg gttataatga acgagaaaaa tataaaacac agtcaaaact ttattacttc
3720
aaaacataat atagataaaa taatgacaaa tataagatta aatgaacatg ataatatctt
3780
tgaaatcggc tcaggaaaag gccattttac ccttgaatta gtaaagaggt gtaatttcgt
3840
aactgccatt gaaatagacc ataaattatg caaaactaca gaaaataaac ttgttgatca
3900
cgataatttc caagttttaa acaaggatat attgcagttt aaatttccta aaaaccaatc
3960
ctataaaata tatggtaata taccttataa cataagtacg gatataatac gcaaaattgt
4020
ttttgatagt atagctaatg agatttattt aatcgtggaa tacgggtttg ctaaaagatt
4080
attaaataca aaacgctcat tggcattact tttaatggca gaagttgata tttctatatt
4140
aagtatggtt ccaagagaat attttcatcc taaacctaaa gtgaatagct cacttatcag
4200
attaagtaga aaaaaatcaa gaatatcaca caaagataaa caaaagtata attatttcgt
4260
tatgaaatgg gttaacaaag aatacaagaa aatatttaca aaaaatcaat ttaacaattc
4320
cttaaaacat gcaggaattg acgatttaaa caatattagc tttgaacaat tcttatctct
4380
tttcaatagc tataaattat ttaataagta agttaaggga tgcataaact gcatccctta
4440
acttgttttt cgtgtgccta ttttttgtga atcgattata acttcgtata atgtatgcta
4500
tacgaacggt aggatcctct agagtcgacc tgcaggcatt ttacattttt agaaatgggc
4560
gtgaaaaaaa gcgcgcgatt atgtaaaata taaagattaa ctaataagga ggacaaacat
4620
ggcacaatct aaactgtatc cggttgtgat ggcgggcgga tctggatcaa gactgtggcc
4680
gctttcacgc gttctttacc cgaaacaatt tctgtgtctg aaaggagatt taacaatgct
4740
gcagacaaca atttgcagat taaatggcgt ggaatgtgaa tctccggtcg ttatttgcaa
4800
cgaacagcat agatttatcg tcgctgaaca acttcgccag ctgaacaaac tgacagaaaa
4860
catcatcctg gaaccggccg gcagaaatac agcaccggcg attgctcttg cagcgttagc
4920
tgccaaacgc cattcaccgg aaagcgatcc gcttatgtta gtgctggcag cggatcatgt
4980
cattgcggat gaagatgctt ttagagctgc cgttcgcaac gctatgccgt atgccgaagc
5040
aggcaaactt gtgacatttg gaattgtccc ggatttaccg gaaacaggct atggatatat
5100
cagacgcggc gaagtgtcag cgggagaaca agatatggtt gcctttgaag tggcacagtt
5160
tgtcgaaaaa ccgaatctgg aaacagcgca agcgtatgtt gcttcaggcg aatattattg
5220
gaacagcgga atgtttcttt ttagagccgg ccgctatctg gaagaactta aaaaatacag
5280
accggatatt ttagatgcat gtgaaaaagc gatgtcagct gtcgatccgg atttaaactt
5340
tatcagagtt gatgaagaag cctttctggc atgcccggaa gaaagcgttg attatgcagt
5400
gatggaacgc acagccgatg cagtggtcgt tccgatggat gcgggctggt ctgatgttgg
5460
atcatggtca agcctttggg aaattagcgc gcatacagct gaaggcaatg tttgtcatgg
5520
agatgtgatc aaccataaaa cagaaaacag ctatgtttac gcggaatctg gcctggtgac
5580
aacagtcgga gttaaagatt tagtggtcgt tcagacaaaa gatgcggtgt taattgctga
5640
tagaaacgcc gtgcaagatg ttaagaaagt tgtcgaacag atcaaagcag atggaagaca
5700
tgaacataga gtccatcgcg aagtttatcg cccgtgggga aaatatgata gcattgatgc
5760
gggcgatcgc tatcaagtca aacgcatcac agttaaaccg ggcgaaggac tgtctgttca
5820
gatgcatcat catcgcgcag aacattgggt tgtggtcgcc ggcacagcaa aagtgacaat
5880
tgatggcgat attaaactgc ttggagaaaa cgaatctatt tatatcccgt taggagctac
5940
acattgcctg gaaaacccgg gcaaaattcc gcttgattta atcgaagtca gaagcggatc
6000
ttatcttgaa gaagatgatg ttgtgcgctt tgccgataga tatggccgcg tttaagctct
6060
taaggaggat tttagaatga aaaaacttac atgctttaaa gcgtatgata ttcgcggcaa
6120
acttggagaa gaattaaacg aagatattgc ttggagaatc ggccgcgcct atggagaatt
6180
tctgaaaccg aaaacaatcg tgctgggcgg agatgtcaga cttacatcag aaacacttaa
6240
actggctctg gccaaaggcc tgcaagatgc tggagttgat gtgcttgata ttggcatgag
6300
cggaacagaa gaaatctatt ttgccacatt tcatttagga gtcgatggcg gaatcgaagt
6360
tacagcaagc cataatccga tggattataa cggaatgaaa ctggttagag aaggcgcaag
6420
accgatttct ggcgatacag gattaagaga tgtgcagcgc ctggcagaag cgaacgattt
6480
tccgccggtc gatgaaacaa aaagaggacg ctatcaacag attaatttac gcgatgcgta
6540
tgttgatcat ctttttggct acatcaacgt taaaaacctg acaccgctta aactggtgat
6600
caattctggc aacggagcag cgggaccggt tgtggatgct attgaagcca gatttaaagc
6660
acttggcgca ccggttgaat taatcaaagt gcataataca ccggatggca attttccgaa
6720
cggcattccg aacccgctgc ttccggaatg cagagatgat acacgcaatg cggttattaa
6780
acatggcgct gatatgggaa tcgcctttga tggagatttt gatcgctgtt ttctttttga
6840
tgaaaaaggc caatttatcg aaggatacta catcgtgggc ttactggctg aagcatttct
6900
ggagaaaaat ccgggagcga aaattatcca tgatccgaga ctttcttgga atacagtcga
6960
tgtcgttaca gctgccggcg gaacaccggt tatgtcaaaa acaggccatg cttttatcaa
7020
agaaagaatg cgcaaagaag atgcaatcta tggcggagaa atgagcgcgc atcattactt
7080
tagagatttt gcttactgcg attctggcat gatcccgtgg cttttagtcg cggaactggt
7140
ttgtcttaaa gataaaacac tgggagaact ggtcagagat cgcatggcag cgtttccggc
7200
ttcaggcgaa attaatagca aactggcaca accggtggaa gcgatcaaca gagtcgaaca
7260
gcatttttca cgcgaagcac tggcggttga tagaacagat ggaattagca tgacatttgc
7320
cgattggaga tttaatcttc gcacatctaa cacagaaccg gtggtcagac ttaacgttga
7380
atcacgcggc gatgtgccgt taatggaagc aagaacacgc acactgctta cattactgaa
7440
tgaataaaaa ggaggaacta ctatgacaac agaaccgctg tttttcaaac cggtctttaa
7500
agaaagaatt tggggcggaa cagctctggc cgattttggc tatacaatcc cgtctcaacg
7560
cacaggagaa tgctgggctt ttgcagcgca tcaaaatggc cagagcgttg tgcagaacgg
7620
catgtacaaa ggatttacac tgtctgaact gtgggaacat catagacatc tttttggcca
7680
attagaagga gatcgctttc cgctgcttac aaaaatttta gatgctgatc aggatctgtc
7740
agttcaggtt catccgaatg atgaatatgc caatatccat gaaaacggcg aacttggaaa
7800
aacagaatgc tggtatatta tcgattgtca gaaagatgct gaaattatct atggccataa
7860
cgccacaaca aaagaagaat taacaacaat gatcgaaaga ggagaatggg atgaattact
7920
gagacgcgtg aaagtcaaac cgggcgattt cttttatgtc ccgtcaggaa cagttcatgc
7980
aattggcaaa ggaatccttg cgttagaaac acaacagaat agcgatacaa catacagact
8040
gtacgattac gatcgcaaag atgcagaagg caaacttaga gaactgcatc ttaaaaaatc
8100
aatcgaagtt atcgaagtgc cgagcattcc ggaacgccat acagttcatc atgaacaaat
8160
cgaagattta ctgacaacaa cactgatcga atgcgcatat tttagcgtgg gcaaatggaa
8220
cctttcagga agcgcgtctt taaaacaaca gaaaccgttt ctgcttattt ctgtgatcga
8280
aggcgaagga cgcatgattt caggcgaata tgtctatccg tttaagaaag gcgatcacat
8340
gttactgccg tatggcctgg gagaatttaa acttgaagga tatgcggaat gtatcgtttc
8400
tcatctttaa gcgaggcagg atcaggacaa tgattatatt tttgtcatga atttcacgga
8460
agaaaaacag ctggtcacgt ttgatcagag tgtgaaggac ataatgacag gagacatatt
8520
gtcaggcgac ctgacgatgg aaaagtatga agtgagaatt gtcgtaaaca cacattaggc
8580
tgatgctccg ctcgatatgg gcggattctt ttttctatag aatgaaaacg cttgctaagt
8640
cttgggggga tgaaatcatg aaaagcaaag tgaaaatgtt ctttgcggct gccatcgtgt
8700
ggagtgcatg tagttcaaca ggatatgccg ctgccattga gaaggagaag cacgtgtcag
8760
agcttcgggc agaggatctt tttgttaaaa aagtagaggg gatgaacaag gattttatca
8820
aaggggcaga tgtatccagc gttattgctt tggaaaacag cggtgtcacc ttttacaata
8880
caaacggaaa acgccaggat atctttacaa ctttaaaaca ggctggggtc aactatgttc
8940
gcgtccgcat ctggaatcac ccgtatgatt caaatggcaa cgggtatggc gggggaaaca
9000
atgatgttca aaaagccatc gaaatcggaa aaagagcgac agcgaacgga atgaaggtgc
9060
tggccgactt tcactactct gatttctggg ccgatccagc gaaacaaaag gtgcccaaag
9120
cctgggcgaa tctcagcttt gaagcaaaaa aagcaaagct ctatgagtat acgaaacaaa
9180
gcctgcaaaa gatgatcaag gaaggcgtgc ggtgtgaaat accgcacaga tgcgtaagga
9240
gaaaataccg catcaggcgc tcttccgctt cctcgctcac tgactcgctg cgctcggtcg
9300
ttcggctgcg gcgagcggta tcagctcact caaaggcggt aatacggtta tccacagaat
9360
caggggataa cgcaggaaag aacatgtgag caaaaggcca gcaaaaggcc aggaaccgta
9420
aaaaggccgc gttgctggcg tttttccata ggctccgccc ccctgacgag catcacaaaa
9480
atcgacgctc aagtcagagg tggcgaaacc cgacaggact ataaagatac caggcgtttc
9540
cccctggaag ctccctcgtg cgctctcctg ttccgaccct gccgcttacc ggatacctgt
9600
ccgcctttct cccttcggga agcgtggcgc tttctcatag ctcacgctgt aggtatctca
9660
gttcggtgta ggtcgttcgc tccaagctgg gctgtgtgca cgaacccccc gttcagcccg
9720
accgctgcgc cttatccggt aactatcgtc ttgagtccaa cccggtaaga cacgacttat
9780
cgccactggc agcagccact ggtaacagga ttagcagagc gaggtatgta ggcggtgcta
9840
cagagttctt gaagtggtgg cctaactacg gctacactag aaggacagta tttggtatct
9900
gcgctctgct gaagccagtt accttcggaa aaagagttgg tagctcttga tccggcaaac
9960
aaaccaccgc tggtagcggt ggtttttttg tttgcaagca gcagattacg cgcagaaaaa
10020
aaggatctca agaagatcct ttgatctttt ctacggggtc tgacgctcag tggaacgaaa
10080
actcacgtta agggattttg gtcatgagat tatcaaaaag gatcttcacc tagatccttt
10140
taaattaaaa atgaagtttt aaatcaatct aaagtatata tgagtaaact tggtctgaca
10200
gttaccaatg cttaatcagt gaggcaccta tctcagcgat ctgtctattt cgttcatcca
10260
tagttgcctg actccccgtc gtgtagataa ctacgatacg ggagggctta ccatctggcc
10320
ccagtgctgc aatgataccg cgagacccac gctcaccggc tccagattta tcagcaataa
10380
accagccagc cggaagggcc gagcgcagaa gtggtcctgc aactttatcc gcctccatcc
10440
agtctattaa ttgttgccgg gaagctagag taagtagttc gccagttaat agtttgcgca
10500
acgttgttgc cattgctgca ggcatcgtgg tgtcacgctc gtcgtttggt atggcttcat
10560
tcagctccgg ttcccaacga tcaaggcgag ttacatgatc ccccatgttg tgcaaaaaag
10620
cggttagctc cttcggtcct ccgatcgttg tcagaagtaa gttggccgca gtgttatcac
10680
tcatggttat ggcagcactg cataattctc ttactgtcat gccatccgta agatgctttt
10740
ctgtgactgg tgagtactca accaagtcat tctgagaata gtgtatgcgg cgaccgagtt
10800
gctcttgccc ggcgtcaaca cgggataata ccgcgccaca tagcagaact ttaaaagtgc
10860
tcatcattgg aaaacgttct tcggggcgaa aactctcaag gatcttaccg ctgttgagat
10920
ccagttcgat gtaacccact cgtgcaccca actgatcttc agcatctttt actttcacca
10980
gcgtttctgg gtgagcaaaa acaggaaggc aaaatgccgc aaaaaaggga ataagggcga
11040
cacggaaatg ttgaatactc atactcttcc tttttcaata ttattgaagc atttatcagg
11100
gttattgtct catgagcgga tacatatttg aatgtattta gaaaaataaa caaatagggg
11160
ttccgcgcac atttccccga aaagtgccac ctgacgtcta agaaaccatt attatcatga
11220
cattaaccta taaaaatagg cgtatcacga ggccctttcg tcttcaagaa
11270
<210> 2
<211> 12427
<212> ДНК
<213> Искусственная последовательность
<220>
<223> экспрессионная плазмида
<400> 2
ttaagttatt ggtatgactg gttttaagcg caaaaaaagt tgctttttcg tacctattaa
60
tgtatcgttt tagaaaaccg actgtaaaaa gtacagtcgg cattatctca tattataaaa
120
gccagtcatt aggcctatct gacaattcct gaatagagtt cataaacaat cctgcatgat
180
aaccatcaca aacagaatga tgtacctgta aagatagcgg taaatatatt gaattacctt
240
tattaatgaa ttttcctgct gtaataatgg gtagaaggta attactatta ttattgatat
300
ttaagttaaa cccagtaaat gaagtccatg gaataataga aagagaaaaa gcattttcag
360
gtataggtgt tttgggaaac aatttccccg aaccattata tttctctaca tcagaaaggt
420
ataaatcata aaactctttg aagtcattct ttacaggagt ccaaatacca gagaatgttt
480
tagatacacc atcaaaaatt gtataaagtg gctctaactt atcccaataa cctaactctc
540
cgtcgctatt gtaaccagtt ctaaaagctg tatttgagtt tatcaccctt gtcactaaga
600
aaataaatgc agggtaaaat ttatatcctt cttgttttat gtttcggtat aaaacactaa
660
tatcaatttc tgtggttata ctaaaagtcg tttgttggtt caaataatga ttaaatatct
720
cttttctctt ccaattgtct aaatcaattt tattaaagtt catttgatat gcctcctaaa
780
tttttatcta aagtgaattt aggaggctta cttgtctgct ttcttcatta gaatcaatcc
840
ttttttaaaa gtcaatatta ctgtaacata aatatatatt ttaaaaatat cccactttat
900
ccaattttcg tttgttgaac taatgggtgc tttagttgaa gaataaagac cacattaaaa
960
aatgtggtct tttgtgtttt tttaaaggat ttgagcgtag cgaaaaatcc ttttctttct
1020
tatcttgata ataagggtaa ctattgccga tcgtccattc cgacagcatc gccagtcact
1080
atggcgtgct gctagcgcca ttcgccattc aggctgcgca actgttggga agggcgatcg
1140
gtgcgggcct cttcgctatt acgccagctg gcgaaagggg gatgtgctgc aaggcgatta
1200
agttgggtaa cgccagggtt ttcccagtca cgacgttgta aaacgacggc cagtgaattc
1260
gagctcaggc cttaactcac attaattgcg ttgcgctcac tgcccgcttt ccagtcggga
1320
aacctgtcgt gccagctgca ttaatgaatc ggccaacgcg cggggagagg cggtttgcgt
1380
attgggcgcc agggtggttt ttcttttcac cagtgagacg ggcaacagct gattgccctt
1440
caccgcctgg ccctgagaga gttgcagcaa gcggtccacg ctggtttgcc ccagcaggcg
1500
aaaatcctgt ttgatggtgg ttgacggcgg gatataacat gagctgtctt cggtatcgtc
1560
gtatcccact accgagatat ccgcaccaac gcgcagcccg gactcggtaa tggcgcgcat
1620
tgcgcccagc gccatctgat cgttggcaac cagcatcgca gtgggaacga tgccctcatt
1680
cagcatttgc atggtttgtt gaaaaccgga catggcactc cagtcgcctt cccgttccgc
1740
tatcggctga atttgattgc gagtgagata tttatgccag ccagccagac gcagacgcgc
1800
cgagacagaa cttaatgggc ccgctaacag cgcgatttgc tggtgaccca atgcgaccag
1860
atgctccacg cccagtcgcg taccgtcttc atgggagaaa ataatactgt tgatgggtgt
1920
ctggtcagag acatcaagaa ataacgccgg aacattagtg caggcagctt ccacagcaat
1980
ggcatcctgg tcatccagcg gatagttaat gatcagccca ctgacgcgtt gcgcgagaag
2040
attgtgcacc gccgctttac aggcttcgac gccgcttcgt tctaccatcg acaccaccac
2100
gctggcaccc agttgatcgg cgcgagattt aatcgccgcg acaatttgcg acggcgcgtg
2160
cagggccaga ctggaggtgg caacgccaat cagcaacgac tgtttgcccg ccagttgttg
2220
tgccacgcgg ttgggaatgt aattcagctc cgccatcgcc gcttccactt tttcccgcgt
2280
tttcgcagaa acgtggctgg cctggttcac cacgcgggaa acggtctgat aagagacacc
2340
ggcatactct gcgacatcgt ataacgttac tggtttcatc aaaatcgtct ccctccgttt
2400
gaatatttga ttgatcgtaa ccagatgaag cactctttcc actatcccta cagtgttatg
2460
gcttgaacaa tcacgaaaca ataattggta cgtacgatct ttcagccgac tcaaacatca
2520
aatcttacaa atgtagtctt tgaaagtatt acatatgtaa gatttaaatg caaccgtttt
2580
ttcggaagga aatgatgacc tcgtttccac cggaattagc ttggtaccaa aggaggtaag
2640
gatcactaga aaatttttta aaaaatctct tgacattgga agggagatat gttattataa
2700
gaattgcgga attgtgagcg gataacaatt cccatataga ttaactaata aggaggacaa
2760
acatgggttc tatcatcaga cttcagggtg gtttaggcaa tcaactgttt caatttagct
2820
ttggttatgc tctttcaaaa atcaatggca caccgctgta ctttgatatt tcacattacg
2880
cagaaaacga tgatcatggc ggatatagac tgaataacct tcaaatcccg gaagaatatc
2940
ttcagtacta cacaccgaaa atcaacaaca tctacaaact gcttgtcaga ggcagccgct
3000
tatatccgga tatttttctg tttcttggct tttgcaacga atttcatgcg tatggctatg
3060
attttgaata catcgcgcaa aaatggaaaa gcaaaaaata catcggatac tggcagtctg
3120
aacatttctt tcataaacat attctggatc tgaaagaatt tttcatcccg aaaaacgtta
3180
gcgaacaagc taacttactg gcagcgaaaa tcttagaatc tcagtcaagc ctgtcaattc
3240
atatcagacg cggcgattac atcaaaaaca aaacagccac actgacacat ggagtttgct
3300
ctcttgaata ctacaagaaa gcactgaaca aaatcagaga tttagccatg attcgcgatg
3360
tgtttatctt ttctgatgat attttctggt gtaaagaaaa catcgaaaca cttctgagca
3420
aaaaatacaa catctactac tcagaagatt taagccaaga agaagattta tggctgatgt
3480
ctctggctaa tcatcatatt atcgccaact cttcattttc atggtggggc gcatatcttg
3540
gaagctctgc gagccagatt gttatctatc cgacaccgtg gtatgatatt acaccgaaaa
3600
acacatacat tcctatcgtg aaccattgga tcaacgtgga caaacattcc tcctgctaag
3660
gagaaaagct agcgattaac taataaggag gaactgaaat gtcaaaagtg gcgcttatta
3720
caggcgtcac aggacaagat ggcagctatt tagctgaatt tctgcttgaa aaaggatatg
3780
aagtgcatgg cattaaaaga cgcgcatcat catttaacac agaaagagtc gatcatatct
3840
atcaagatcc gcatacatgc aacccgaaat ttcatctgca ttatggcgat ctgagcgata
3900
catctaacct gacaagaatt ttacgcgaag tccagccgga tgaagtttat aacctgggcg
3960
cgatgtctca tgtcgctgtt tcatttgaaa gcccggaata tacagccgat gtcgatgcaa
4020
tgggaacact tagattactg gaagccattc gctttttagg cctggaaaag aaaacacgct
4080
tttatcaggc atctacatca gaactgtatg gacttgttca agaaatcccg cagaaagaaa
4140
caacaccgtt ttatccgaga agcccgtatg cggtcgctaa actttacgcg tactggatca
4200
cagttaatta tcgcgaatct tatggaatgt atgcttgcaa tggcatctta tttaaccatg
4260
aatcaccgag acgcggagaa acatttgtta caagaaaaat cacacgcgcc attgcaaata
4320
tcgcccaagg actggaaagc tgtctttatt taggcaacat ggattctctt agagattggg
4380
gccatgcaaa agattacgtt aaaatgcaat ggatgatgtt acaacaggaa cagccggaag
4440
attttgtgat cgccacagga gtgcaatatt cagtcagaca gtttgttgaa atggcagcgg
4500
ctcagcttgg cattaaactg cgctttgaag gcacaggagt ggaagaaaaa ggaatcgttg
4560
tgagcgttac aggccatgat gcgccgggag tgaaaccggg cgatgtgatt atcgccgtcg
4620
atccgagata ttttcgcccg gcagaagtcg aaacactttt aggagatccg acaaaagcgc
4680
atgaaaaact gggctggaaa ccggaaatta cactgagaga aatggttagc gaaatggtgg
4740
ctaacgattt agaagccgcg aaaaaacatt ctctgcttaa atcacatggc tatgatgtgg
4800
cgatcgctct ggaatcttaa aaaggaggaa ctactatgag caaacaaaga gtttttatcg
4860
ctggccatcg cggaatggtg ggctctgcca ttagacgcca attagaacag agaggagatg
4920
tggaacttgt cttaagaaca cgcgatgaac tgaatctgct tgattcaaga gctgtccatg
4980
atttctttgc cagcgaacgc atcgatcagg tttatcttgc agcggctaaa gttggcggaa
5040
ttgtggcaaa taacacatat ccggcggatt ttatctacca aaacatgatg atcgaatcta
5100
acattatcca tgccgcacat cagaatgatg ttaacaaact gctgtttctg ggatcaagct
5160
gcatctatcc gaaacttgca aaacaaccga tggcggaatc agaactttta cagggcacac
5220
tggaaccgac aaatgaaccg tatgcaattg cgaaaatcgc gggaatcaaa ctttgtgaat
5280
cttacaacag acaatacggc agagattatc gctcagtgat gccgacaaac ttatatggac
5340
cgcatgataa ttttcatccg tctaactcac atgtcattcc ggcactgctt agacgctttc
5400
atgaagctac agcccagaac gcgccggatg ttgtggtctg gggctcagga acaccgatgc
5460
gcgaatttct gcatgttgat gatatggcgg ctgccagcat ccatgtgatg gaacttgctc
5520
atgaagtctg gttagaaaat acacaaccga tgcttagcca tattaacgtc ggcacaggag
5580
ttgattgcac aatcagagaa ttagctcaga caattgccaa agttgtggga tataaaggcc
5640
gcgtcgtttt tgatgcctct aaaccggatg gcacaccgag aaaattactg gatgtgacac
5700
gcctgcatca acttggatgg tatcatgaaa ttagcttaga agcaggcctg gcgtctacat
5760
accaatggtt tttagaaaac caggatagat ttcgcggcgg atcataaagt gatagcggta
5820
ccattatagg taagagagga atgtacacat gtactattta aaaaacacaa acttttggat
5880
gttcggttta ttctttttct tttacttttt tatcatggga gcctacttcc cgtttttccc
5940
gatttggcta catgacatca accatatcag caaaagtgat acgggtatta tttttgccgc
6000
tatttctctg ttctcgctat tattccaacc gctgtttggt ctgctttctg acaaactcgg
6060
gctgcgcaaa tacctgctgt ggattattac cggcatgtta gtgatgtttg cgccgttctt
6120
tatttttatc ttcgggccac tgttacaata caacatttta gtaggatcga ttgttggtgg
6180
tatttatcta ggcttttgtt ttaacgccgg tgcgccagca gtagaggcat ttattgagaa
6240
agtcagccgt cgcagtaatt tcgaatttgg tcgcgcgcgg atgtttggct gtgttggctg
6300
ggcgctgtgt gcctcgattg tcggcatcat gttcaccatc aataatcagt ttgttttctg
6360
gctgggctct ggctgtgcac tcatcctcgc cgttttactc tttttcgcca aaacggatgc
6420
gccctcttct gccacggttg ccaatgcggt aggtgccaac cattcggcat ttagccttaa
6480
gctggcactg gaactgttca gacagccaaa actgtggttt ttgtcactgt atgttattgg
6540
cgtttcctgc acctacgatg tttttgacca acagtttgct aatttcttta cttcgttctt
6600
tgctaccggt gaacagggta cgcgggtatt tggctacgta acgacaatgg gcgaattact
6660
taacgcctcg attatgttct ttgcgccact gatcattaat cgcatcggtg ggaaaaacgc
6720
cctgctgctg gctggcacta ttatgtctgt acgtattatt ggctcatcgt tcgccacctc
6780
agcgctggaa gtggttattc tgaaaacgct gcatatgttt gaagtaccgt tcctgctggt
6840
gggctgcttt aaatatatta ccagccagtt tgaagtgcgt ttttcagcga cgatttatct
6900
ggtctgtttc tgcttcttta agcaactggc gatgattttt atgtctgtac tggcgggcaa
6960
tatgtatgaa agcatcggtt tccagggcgc ttatctggtg ctgggtctgg tggcgctggg
7020
cttcacctta atttccgtgt tcacgcttag cggccccggc ccgctttccc tgctgcgtcg
7080
tcaggtgaat gaagtcgctt aaggatccat gtctagagtc gacgtccccg gggcagcccg
7140
cctaatgagc gggctttttt cacgtcccag gcatcaaata aaacgaaagg ctcagtcgaa
7200
agactgggcc tttcgtttta tctgttgttt gtcggtgaac gctctctact agagtcacac
7260
tggctcacct tcgggtgggc ctttctgcgt ttatacccgg ggcagcccgc ctaatgagcg
7320
ggcttttttc acgtcacgcg tccatggaga tctttgtctg caactgaaaa gtttatacct
7380
tacctggaac aaatggttga aacatacgag gctaatatcg gcttattagg aatagtccct
7440
gtactaataa aatcaggtgg atcagttgat cagtatattt tggacgaagc tcggaaagaa
7500
tttggagatg acttgcttaa ttccacaatt aaattaaggg aaagaataaa gcgatttgat
7560
gttcaaggaa tcacggaaga agatactcat gataaagaag ctctaaaact attcaataac
7620
cttacaatgg aattgatcga aagggtggaa ggttaatggt acgaaaatta ggggatctac
7680
ctagaaagcc acaaggcgat aggtcaagct taaagaaccc ttacatggat cttacagatt
7740
ctgaaagtaa agaaacaaca gaggttaaac aaacagaacc aaaaagaaaa aaagcattgt
7800
tgaaaacaat gaaagttgat gtttcaatcc ataataagat taaatcgctg cacgaaattc
7860
tggcagcatc cgaagggaat tcatattact tagaggatac tattgagaga gctattgata
7920
agatggttga gacattacct gagagccaaa aaacttttta tgaatatgaa ttaaaaaaaa
7980
gaaccaacaa aggctgagac agactccaaa cgagtctgtt tttttaaaaa aaatattagg
8040
agcattgaat atatattaga gaattaagaa agacatggga ataaaaatat tttaaatcca
8100
gtaaaaatat gataagatta tttcagaata tgaagaactc tgtttgtttt tgatgaaaaa
8160
acaaacaaaa aaaatccacc taacggaatc tcaatttaac taacagcggc caaactgaga
8220
agttaaattt gagaagggga aaaggcggat ttatacttgt atttaactat ctccatttta
8280
acattttatt aaaccccata caagtgaaaa tcctctttta cactgttcct ttaggtgatc
8340
gcggagggac attatgagtg aagtaaacct aaaaggaaat acagatgaat tagtgtatta
8400
tcgacagcaa accactggaa ataaaatcgc caggaagaga atcaaaaaag ggaaagaaga
8460
agtttattat gttgctgaaa cggaagagaa gatatggaca gaagagcaaa taaaaaactt
8520
ttctttagac aaatttggta cgcatatacc ttacatagaa ggtcattata caatcttaaa
8580
taattacttc tttgattttt ggggctattt tttaggtgct gaaggaattg cgctctatgc
8640
tcacctaact cgttatgcat acggcagcaa agacttttgc tttcctagtc tacaaacaat
8700
cgctaaaaaa atggacaaga ctcctgttac agttagaggc tacttgaaac tgcttgaaag
8760
gtacggtttt atttggaagg taaacgtccg taataaaacc aaggataaca cagaggaatc
8820
cccgattttt aagattagac gtaaggttcc tttgctttca gaagaacttt taaatggaaa
8880
ccctaatatt gaaattccag atgacgagga agcacatgta aagaaggctt taaaaaagga
8940
aaaagagggt cttccaaagg ttttgaaaaa agagcacgat gaatttgtta aaaaaatgat
9000
ggatgagtca gaaacaatta atattccaga ggccttacaa tatgacacaa tgtatgaaga
9060
tatactcagt aaaggagaaa ttcgaaaaga aatcaaaaaa caaataccta atcctacaac
9120
atcttttgag agtatatcaa tgacaactga agaggaaaaa gtcgacagta ctttaaaaag
9180
cgaaatgcaa aatcgtgtct ctaagccttc ttttgatacc tggtttaaaa acactaagat
9240
caaaattgaa aataaaaatt gtttattact tgtaccgagt gaatttgcat ttgaatggat
9300
taagaaaaga tatttagaaa caattaaaac agtccttgaa gaagctggat atgttttcga
9360
aaaaatcgaa ctaagaaaag tgcaataaac tgctgaagta tttcagcagt tttttttatt
9420
tagaaatagt gaaaaaaata taatcaggga ggtatcaata tttaatgagt actgatttaa
9480
atttatttag actggaatta ataattaaca cgtagactaa ttaaaattta atgagggata
9540
aagaggatac aaaaatatta atttcaatcc ctattaaatt ttaacaaggg ggggattaaa
9600
atttaattag aggtttatcc acaagaaaag accctaataa aatttttact agggttataa
9660
cactgattaa tttcttaatg ggggagggat taaaatttaa tgacaaagaa aacaatcttt
9720
taagaaaagc ttttaaaaga taataataaa aagagctttg cgattaagca aaactcttta
9780
ctttttcatt gacattatca aattcatcga tttcaaattg ttgttgtatc ataaagttaa
9840
ttctgttttg cacaaccttt tcaggaatat aaaacacatc tgaggcttgt tttataaact
9900
cagggtcgct aaagtcaatg taacgtagca tatgatatgg tatagcttcc acccaagtta
9960
gcctttctgc ttcttctgaa tgtttttcat atacttccat gggtatctct aaatgatttt
10020
cctcatgtag caaggtatga gcaaaaagtt tatggaattg atagttcctc tctttttctt
10080
caactttttt atctaaaaca aacactttaa catctgagtc aatgtaagca taagatgttt
10140
ttccagtcat aatttcaatc ccaaatcttt tagacagaaa ttctggacgt aaatcttttg
10200
gtgaaagaat ttttttatgt agcaatatat ccgatacagc accttctaaa agcgttggtg
10260
aatagggcat tttacctatc tcctctcatt ttgtggaata aaaatagtca tattcgtcca
10320
tctacctatc ctattatcga acagttgaac tttttaatca aggatcagtc ctttttttca
10380
ttattcttaa actgtgctct taactttaac aactcgattt gtttttccag atctcgaggg
10440
taactagcct cgccgatccc gcaagaggcc cggcagtcag gtggcacttt tcggggaaat
10500
gtgcgcggaa cccctatttg tttatttttc taaatacatt caaatatgta tccgctcatg
10560
agacaataac cctgataaat gcttcaataa tattgaaaaa ggaagagtat gagtattcaa
10620
catttccgtg tcgcccttat tccctttttt gcggcatttt gccttcctgt ttttgctcac
10680
ccagaaacgc tggtgaaagt aaaagatgct gaagatcagt tgggtgcacg agtgggttac
10740
atcgaactgg atctcaacag cggtaagatc cttgagagtt ttcgccccga agaacgtttt
10800
ccaatgatga gcacttttaa agttctgcta tgtggcgcgg tattatcccg tattgacgcc
10860
gggcaagagc aactcggtcg ccgcatacac tattctcaga atgacttggt tgagtactca
10920
ccagtcacag aaaagcatct tacggatggc atgacagtaa gagaattatg cagtgctgcc
10980
ataaccatga gtgataacac tgcggccaac ttacttctga caacgatcgg aggaccgaag
11040
gagctaaccg cttttttgca caacatgggg gatcatgtaa ctcgccttga tcgttgggaa
11100
ccggagctga atgaagccat accaaacgac gagcgtgaca ccacgatgcc tgtagcaatg
11160
gcaacaacgt tgcgcaaact attaactggc gaactactta ctctagcttc ccggcaacaa
11220
ttaatagact ggatggaggc ggataaagtt gcaggaccac ttctgcgctc ggcccttccg
11280
gctggctggt ttattgctga taaatctgga gccggtgagc gtgggtctcg cggtatcatt
11340
gcagcactgg ggccagatgg taagccctcc cgtatcgtag ttatctacac gacggggagt
11400
caggcaacta tggatgaacg aaatagacag atcgctgaga taggtgcctc actgattaag
11460
cattggtaac tgtcagacca agtttactca tatatacttt agattgattt aaaacttcat
11520
ttttaattta aaaggatcta ggtgaagatc ctttttgata atctcatgac caaaatccct
11580
taacgtgagt tttcgttcca ctgagcgtca gaccccgtag aaaagatcaa aggatcttct
11640
tgagatcctt tttttctgcg cgtaatctgc tgcttgcaaa caaaaaaacc accgctacca
11700
gcggtggttt gtttgccgga tcaagagcta ccaactcttt ttccgaaggt aactggcttc
11760
agcagagcgc agataccaaa tactgtcctt ctagtgtagc cgtagttagg ccaccacttc
11820
aagaactctg tagcaccgcc tacatacctc gctctgctaa tcctgttacc agtggctgct
11880
gccagtggcg ataagtcgtg tcttaccggg ttggactcaa gacgatagtt accggataag
11940
gcgcagcggt cgggctgaac ggggggttcg tgcacacagc ccagcttgga gcgaacgacc
12000
tacaccgaac tgagatacct acagcgtgag ctatgagaaa gcgccacgct tcccgaaggg
12060
agaaaggcgg acaggtatcc ggtaagcggc agggtcggaa caggagagcg cacgagggag
12120
cttccagggg gaaacgcctg gtatctttat agtcctgtcg ggtttcgcca cctctgactt
12180
gagcgtcgat ttttgtgatg ctcgtcaggg gggcggagcc tatggaaaaa cgccagcaac
12240
gcggcctttt tacggttcct ggccttttgc tggccttttg ctcacatgtt ctttcctgcg
12300
ttatcccctg attctgtgga taaccgtatt accgcctttg agtgagctga taccgctcgc
12360
cgcagccgaa cgaccgagcg cagcgagtca gtgagcgagg aagcggaaga gcgcccaata
12420
cgcatgc
12427
<210> 3
<211> 12330
<212> ДНК
<213> Искусственная последовательность
<220>
<223> интеграционный вектор
<400> 3
ttctcatgtt tgacagctta tcatcgataa gctttaatgc ggtagtttat cacagttaaa
60
ttgctaacgc agtcaggcac cgtgtatgaa atctaacaat gcgctcatcg tcatcctcgg
120
caccgtcacc ctggatgctg taggcatagg cttggttatg ccggtactgc cgggcctctt
180
gcgggatatc gtccattccg acagcatcgc cagtcactat ggcgtgctgc tagcgctata
240
tgcgttgatg caatttctat gcgcacccgt tctcggagca ctgtccgacc gctttggccg
300
ccgcccagtc ctgctcgctt cgctacttgg agccactatc gactacgcga tcatggcgac
360
cacacccgtc ctgtggatcc tctacgccgg acgcatcgtg gccggcatca ccggcgccac
420
aggtgcggtt gctggcgcct atatcgccga catcaccgat ggggaagatc gggctcgcca
480
cttcgggctc atgagcgctt gtttcggcgt gggtatggtg gcaggccccg tggccggggg
540
actgttgggc gccatctcct tgcatgcacc attccttgcg gcggcggtgc tcaacggcct
600
caacctacta ctgggctgct tcctaatgca ggagtcgcat aagggagagc gtcgaccgat
660
gcccttgaga gccttcaacc cagtcagctc cttccggtgg gcgcggggca tgactatcgt
720
cgccgcactt atgactgtct tctttatcat gcaactcgta ggacaggtgc cggcagcgct
780
ctgggtcatt ttcggcgagg accgctttcg ctggagcgcg acgatgatcg gcctgtcgct
840
tgcggtattc ggaatcttgc acgccctcgc tcaagccttc gtcactggtc ccgccaccaa
900
acgtttcggc gagaagcagg ccattatcgc cggcatggcg gccgacgcgc tgggctacgt
960
cttgctggcg ttcgcgacgc gaggctggat ggccttcccc attatgattc ttctcgcttc
1020
cggcggcatc gggatgcccg cgttgcaggc catgctgtcc aggcaggtag atgacgacca
1080
tcagggacag cttcaaggat cgctcgcggc tcttaccagc ctaacttcga tcactggacc
1140
gctgatcgtc acggcgattt atgccgcctc ggcgagcaca tggaacgggt tggcatggat
1200
tgtaggcgcc gccctatacc ttgtctgcct ccccgcgttg cgtcgcggtg catggagccg
1260
ggccacctcg acctgaatgg aagccggcgg cacctcgcta acggattcac cactccaaga
1320
attggagcca atcaattctt gcggagaact gtgaatgcgc aaaccaaccc ttggcagaac
1380
atatccatcg cgtccgccat ctccagcagc cgcacgcggc gcatctcggg cagcgttggg
1440
tcctggccac gggtgcgcat gatcgtgctc ctgtcgttga ggacccggct aggctggcgg
1500
ggttgcctta ctggttagca gaatgaatca ccgatacgcg agcgaacgtg aagcgactgc
1560
tgctgcaaaa cgtctgcgac ctgagcaaca acatgaatgg tcttcggttt ccgtgtttcg
1620
taaagtctgg aaacgcggaa gtcagcgccc tgcaccatta tgttccggat ctgcatcgca
1680
ggatgctgct ggctaccctg tggaacacct acatctgtat taacgaagcg ctggcattga
1740
ccctgagtga tttttctctg gtcccgccgc atccataccg ccagttgttt accctcacaa
1800
cgttccagta accgggcatg ttcatcatca gtaacccgta tcgtgagcat cctctctcgt
1860
ttcatcggta tcattacccc catgaacaga aatccccctt acacggaggc atcagtgacc
1920
aaacaggaaa aaaccgccct taacatggcc cgctttatca gaagccagac attaacgctt
1980
ctggagaaac tcaacgagct ggacgcggat gaacaggcag acatctgtga atcgcttcac
2040
gaccacgctg atgagcttta ccgcagctgc ctcgcgcgtt tcggtgatga cggtgaaaac
2100
ctctgacaca tgcagctccc ggagacggtc acagcttgtc tgtaagcgga tgccgggagc
2160
agacaagccc gtcagggcgc gtcagcgggt gttggcgggt gtcggggcgc agccatgacc
2220
cagtcacgta gcgatagcgg agtgtatact ggcttaacta tgcggcatca gagcagattg
2280
tactgagagt gcaccatatg acccgacatc cggcgttctc atggcggtgc ttgccgccag
2340
cggtattccg tatgtcaagt ggctgcggtt tatggtgccg cttgctctga tttggttctt
2400
gatcgggctt gtctttatcg tgatcggagt catgatcaat tgggggccgt tttaacgatt
2460
gctgcccgcc ggcttgtacg gcgggctttt gagttattca ttgcagaagc gcaggctgtt
2520
attgtaacat gtaagccata agccattcgt aaaagtgcgg gaggaaggtc atgaataatc
2580
tgcgtaatag actttcaggc gtgaatggga aaaataagag agtaaaagaa aaagaacaaa
2640
aaatctggtc ggagattggg atgatagcgg gagcatttgc gctgcttgat gtgatcatcc
2700
gcggcattat gtttgaattt ccgtttaaag aatgggctgc aagccttgtg tttttgttca
2760
tcattatctt atattactgc atcagggctg cggcatccgg aatgctcatg ccgagaatag
2820
acaccaaaga agaactgcaa aaacgggtga agcagcagcg aatagaatca attgcggtcg
2880
cctttgcggt agtggtgctt acgatgtacg acagggggat tccccataca ttcttcgctt
2940
ggctgaaaat gattcttctt tttatcgtct gcggcggcgt tctgtttctg cttcggtatg
3000
tgattgtgaa gctggcttac agaagagcgg taaaagaaga aataaaaaag aaatcatctt
3060
ttttgtttgg aaagcgaggg aagcgttcac agtttcgggc agcttttttt ataggaacat
3120
tgatttgtat tcactctgcc aagttgtttt gatagagtga ttgtgataat tttaaatgta
3180
agcgttaaca aaattctcca gtcttcacat cggtttgaaa ggaggaagcg gaagaatgaa
3240
gtaagaggga tttttgactc cgaagtaagt cttcaaaaaa tcaaataagg agtgtcaaga
3300
ccagtgaatt cgagctcggt acctaccgtt cgtataatgt atgctatacg aagttatgat
3360
aaaaaattta gaagccaatg aaatctataa ataaactaaa ttaagtttat ttaattaaca
3420
actatggata taaaataggt actaatcaaa atagtgagga ggatatattt gaatacatac
3480
gaacaaatta ataaagtgaa aaaaatactt cggaaacatt taaaaaataa ccttattggt
3540
acttacatgt ttggatcagg agttgagagt ggactaaaac caaatagtga tcttgacttt
3600
ttagtcgtcg tatctgaacc attgacagat caaagtaaag aaatacttat acaaaaaatt
3660
agacctattt caaaaaaaat aggagataaa agcaacttac gatatattga attaacaatt
3720
attattcagc aagaaatggt accgtggaat catcctccca aacaagaatt tatttatgga
3780
gaatggttac aagagcttta tgaacaagga tacattcctc agaaggaatt aaattcagat
3840
ttaaccataa tgctttacca agcaaaacga aaaaataaaa gaatatacgg aaattatgac
3900
ttagaggaat tactacctga tattccattt tctgatgtga gaagagccat tatggattcg
3960
tcagaggaat taatagataa ttatcaggat gatgaaacca actctatatt aactttatgc
4020
cgtatgattt taactatgga cacgggtaaa atcataccaa aagatattgc gggaaatgca
4080
gtggctgaat cttctccatt agaacatagg gagagaattt tgttagcagt tcgtagttat
4140
cttggagaga atattgaatg gactaatgaa aatgtaaatt taactataaa ctatttaaat
4200
aacagattaa aaaaattata aaaaaattga aaaaatggtg gaaacacttt tttcaatttt
4260
tttgttttat tatttaatat ttgggaaata ttcattctaa tataacttcg tataatgtat
4320
gctatacgaa cggtaggatc ctctagagtc gacctgcagg cattttacat ttttagaaat
4380
gggcgtgaaa aaaagcgcgc gattatgtaa aatataaaga ttaactaata aggaggacaa
4440
acatgggttc tatcatcaga cttcagggtg gtttaggcaa tcaactgttt caatttagct
4500
ttggttatgc tctttcaaaa atcaatggca caccgctgta ctttgatatt tcacattacg
4560
cagaaaacga tgatcatggc ggatatagac tgaataacct tcaaatcccg gaagaatatc
4620
ttcagtacta cacaccgaaa atcaacaaca tctacaaact gcttgtcaga ggcagccgct
4680
tatatccgga tatttttctg tttcttggct tttgcaacga atttcatgcg tatggctatg
4740
attttgaata catcgcgcaa aaatggaaaa gcaaaaaata catcggatac tggcagtctg
4800
aacatttctt tcataaacat attctggatc tgaaagaatt tttcatcccg aaaaacgtta
4860
gcgaacaagc taacttactg gcagcgaaaa tcttagaatc tcagtcaagc ctgtcaattc
4920
atatcagacg cggcgattac atcaaaaaca aaacagccac actgacacat ggagtttgct
4980
ctcttgaata ctacaagaaa gcactgaaca aaatcagaga tttagccatg attcgcgatg
5040
tgtttatctt ttctgatgat attttctggt gtaaagaaaa catcgaaaca cttctgagca
5100
aaaaatacaa catctactac tcagaagatt taagccaaga agaagattta tggctgatgt
5160
ctctggctaa tcatcatatt atcgccaact cttcattttc atggtggggc gcatatcttg
5220
gaagctctgc gagccagatt gttatctatc cgacaccgtg gtatgatatt acaccgaaaa
5280
acacatacat tcctatcgtg aaccattgga tcaacgtgga caaacattcc tcctgctaaa
5340
gtgatagcgg taccattata ggtaagagag gaatgtacac atgtactatt taaaaaacac
5400
aaacttttgg atgttcggtt tattcttttt cttttacttt tttatcatgg gagcctactt
5460
cccgtttttc ccgatttggc tacatgacat caaccatatc agcaaaagtg atacgggtat
5520
tatttttgcc gctatttctc tgttctcgct attattccaa ccgctgtttg gtctgctttc
5580
tgacaaactc gggctgcgca aatacctgct gtggattatt accggcatgt tagtgatgtt
5640
tgcgccgttc tttattttta tcttcgggcc actgttacaa tacaacattt tagtaggatc
5700
gattgttggt ggtatttatc taggcttttg ttttaacgcc ggtgcgccag cagtagaggc
5760
atttattgag aaagtcagcc gtcgcagtaa tttcgaattt ggtcgcgcgc ggatgtttgg
5820
ctgtgttggc tgggcgctgt gtgcctcgat tgtcggcatc atgttcacca tcaataatca
5880
gtttgttttc tggctgggct ctggctgtgc actcatcctc gccgttttac tctttttcgc
5940
caaaacggat gcgccctctt ctgccacggt tgccaatgcg gtaggtgcca accattcggc
6000
atttagcctt aagctggcac tggaactgtt cagacagcca aaactgtggt ttttgtcact
6060
gtatgttatt ggcgtttcct gcacctacga tgtttttgac caacagtttg ctaatttctt
6120
tacttcgttc tttgctaccg gtgaacaggg tacgcgggta tttggctacg taacgacaat
6180
gggcgaatta cttaacgcct cgattatgtt ctttgcgcca ctgatcatta atcgcatcgg
6240
tgggaaaaac gccctgctgc tggctggcac tattatgtct gtacgtatta ttggctcatc
6300
gttcgccacc tcagcgctgg aagtggttat tctgaaaacg ctgcatatgt ttgaagtacc
6360
gttcctgctg gtgggctgct ttaaatatat taccagccag tttgaagtgc gtttttcagc
6420
gacgatttat ctggtctgtt tctgcttctt taagcaactg gcgatgattt ttatgtctgt
6480
actggcgggc aatatgtatg aaagcatcgg tttccagggc gcttatctgg tgctgggtct
6540
ggtggcgctg ggcttcacct taatttccgt gttcacgctt agcggccccg gcccgctttc
6600
cctgctgcgt cgtcaggtga atgaagtcgc ttaaagtcaa tgtatgaatg gatacgggat
6660
atgaatcaat aagtacgtga aagagaaaag caacccagat atgataggga acttttctct
6720
ttcttgtttt acattgaatc tttacaatcc tattgatata atctaagcta gtgtattttg
6780
cgtttaatag tggagaaaag ctagcgatta actaataagg aggaactgaa atgtcaaaag
6840
tggcgcttat tacaggcgtc acaggacaag atggcagcta tttagctgaa tttctgcttg
6900
aaaaaggata tgaagtgcat ggcattaaaa gacgcgcatc atcatttaac acagaaagag
6960
tcgatcatat ctatcaagat ccgcatacat gcaacccgaa atttcatctg cattatggcg
7020
atctgagcga tacatctaac ctgacaagaa ttttacgcga agtccagccg gatgaagttt
7080
ataacctggg cgcgatgtct catgtcgctg tttcatttga aagcccggaa tatacagccg
7140
atgtcgatgc aatgggaaca cttagattac tggaagccat tcgcttttta ggcctggaaa
7200
agaaaacacg cttttatcag gcatctacat cagaactgta tggacttgtt caagaaatcc
7260
cgcagaaaga aacaacaccg ttttatccga gaagcccgta tgcggtcgct aaactttacg
7320
cgtactggat cacagttaat tatcgcgaat cttatggaat gtatgcttgc aatggcatct
7380
tatttaacca tgaatcaccg agacgcggag aaacatttgt tacaagaaaa atcacacgcg
7440
ccattgcaaa tatcgcccaa ggactggaaa gctgtcttta tttaggcaac atggattctc
7500
ttagagattg gggccatgca aaagattacg ttaaaatgca atggatgatg ttacaacagg
7560
aacagccgga agattttgtg atcgccacag gagtgcaata ttcagtcaga cagtttgttg
7620
aaatggcagc ggctcagctt ggcattaaac tgcgctttga aggcacagga gtggaagaaa
7680
aaggaatcgt tgtgagcgtt acaggccatg atgcgccggg agtgaaaccg ggcgatgtga
7740
ttatcgccgt cgatccgaga tattttcgcc cggcagaagt cgaaacactt ttaggagatc
7800
cgacaaaagc gcatgaaaaa ctgggctgga aaccggaaat tacactgaga gaaatggtta
7860
gcgaaatggt ggctaacgat ttagaagccg cgaaaaaaca ttctctgctt aaatcacatg
7920
gctatgatgt ggcgatcgct ctggaatctt aaaaaggagg aactactatg agcaaacaaa
7980
gagtttttat cgctggccat cgcggaatgg tgggctctgc cattagacgc caattagaac
8040
agagaggaga tgtggaactt gtcttaagaa cacgcgatga actgaatctg cttgattcaa
8100
gagctgtcca tgatttcttt gccagcgaac gcatcgatca ggtttatctt gcagcggcta
8160
aagttggcgg aattgtggca aataacacat atccggcgga ttttatctac caaaacatga
8220
tgatcgaatc taacattatc catgccgcac atcagaatga tgttaacaaa ctgctgtttc
8280
tgggatcaag ctgcatctat ccgaaacttg caaaacaacc gatggcggaa tcagaacttt
8340
tacagggcac actggaaccg acaaatgaac cgtatgcaat tgcgaaaatc gcgggaatca
8400
aactttgtga atcttacaac agacaatacg gcagagatta tcgctcagtg atgccgacaa
8460
acttatatgg accgcatgat aattttcatc cgtctaactc acatgtcatt ccggcactgc
8520
ttagacgctt tcatgaagct acagcccaga acgcgccgga tgttgtggtc tggggctcag
8580
gaacaccgat gcgcgaattt ctgcatgttg atgatatggc ggctgccagc atccatgtga
8640
tggaacttgc tcatgaagtc tggttagaaa atacacaacc gatgcttagc catattaacg
8700
tcggcacagg agttgattgc acaatcagag aattagctca gacaattgcc aaagttgtgg
8760
gatataaagg ccgcgtcgtt tttgatgcct ctaaaccgga tggcacaccg agaaaattac
8820
tggatgtgac acgcctgcat caacttggat ggtatcatga aattagctta gaagcaggcc
8880
tggcgtctac ataccaatgg tttttagaaa accaggatag atttcgcggc ggatcataag
8940
gatccatgtc tagagtcgac gtccccgggg cagcccgcct aatgagcggg cttttttcac
9000
gtcccaggca tcaaataaaa cgaaaggctc agtcgaaaga ctgggccttt cgttttatct
9060
gttgtttgtc ggtgaacgct ctctactaga gtcacactgg ctcaccttcg ggtgggcctt
9120
tctgcgttta tagaattcat attacttaga ggatactatt gagagagcta ttgataagat
9180
ggttgagaca ttacctgaga gccaaaaaac tttttatgaa tatgaattaa aaaaaagaac
9240
caacaaaggc tgagacagac tccgggcaag gctagacggg acttaccgaa agaaaccatc
9300
aatgatggtt tcttttttgt tcataaatca gacaaaactt ttctcttgca aaagtttgtg
9360
aagtgttgca caatataaat gtgaaatact tcacaaacaa aaagacatca aagagaaaca
9420
taccctggaa ggatgattaa tgatgaacaa acatgtaaat aaagtagctt taatcggagc
9480
gggttttgtt ggaagcagtt atgcatttgc gttaattaac caaggaatca cagatgagct
9540
tgtggtcatt gatgtaaata aagaaaaagc aatgggcgat gtgatggatt taaaccacgg
9600
aaaggcgttt gcgccacaac cggtcaaaac atcttacgga acatatgaag actgcaagga
9660
tgctgatatt gtctgcattt gcgccggagc aaaccaaaaa cctggtgaga cacgccttga
9720
attagtagaa aagaacttga agattttcaa aggcatcgtt agtgaagtca tggcgagcgg
9780
atttgacggc attttcttag tcgcgacaaa tccggttgat atcctgactt acgcaacatg
9840
gaaattcagc ggcctgccaa aagagcgggt gattggaagc ggcacaacac ttgattctgc
9900
gagattccgt ttcatgctga gcgaatactt tggcgcagcg cctcaaaacg tacacgcgca
9960
tattatcgga gagcacggcg acacagagct tcctgtttgg agccacgcga atgtcggcgg
10020
tgtgccggtc agtgaactcg ttgagaaaaa cgatgcgtac aaacaagagg agctggacca
10080
aattgtagat gatgtgaaaa acgcagctta ccatatcatt gagaaaaaag gcgcgactta
10140
ttatggggtt gcgatgagtc ttgctcgcat tacaaaagcc attcttcata atgaaaacag
10200
catattaact gtcagcacat atttggacgg gcaatacggt gcagatgacg tgtacatcgg
10260
tgtcatatgc ggtgtgaaat accgcacaga tgcgtaagga gaaaataccg catcaggcgc
10320
tcttccgctt cctcgctcac tgactcgctg cgctcggtcg ttcggctgcg gcgagcggta
10380
tcagctcact caaaggcggt aatacggtta tccacagaat caggggataa cgcaggaaag
10440
aacatgtgag caaaaggcca gcaaaaggcc aggaaccgta aaaaggccgc gttgctggcg
10500
tttttccata ggctccgccc ccctgacgag catcacaaaa atcgacgctc aagtcagagg
10560
tggcgaaacc cgacaggact ataaagatac caggcgtttc cccctggaag ctccctcgtg
10620
cgctctcctg ttccgaccct gccgcttacc ggatacctgt ccgcctttct cccttcggga
10680
agcgtggcgc tttctcatag ctcacgctgt aggtatctca gttcggtgta ggtcgttcgc
10740
tccaagctgg gctgtgtgca cgaacccccc gttcagcccg accgctgcgc cttatccggt
10800
aactatcgtc ttgagtccaa cccggtaaga cacgacttat cgccactggc agcagccact
10860
ggtaacagga ttagcagagc gaggtatgta ggcggtgcta cagagttctt gaagtggtgg
10920
cctaactacg gctacactag aaggacagta tttggtatct gcgctctgct gaagccagtt
10980
accttcggaa aaagagttgg tagctcttga tccggcaaac aaaccaccgc tggtagcggt
11040
ggtttttttg tttgcaagca gcagattacg cgcagaaaaa aaggatctca agaagatcct
11100
ttgatctttt ctacggggtc tgacgctcag tggaacgaaa actcacgtta agggattttg
11160
gtcatgagat tatcaaaaag gatcttcacc tagatccttt taaattaaaa atgaagtttt
11220
aaatcaatct aaagtatata tgagtaaact tggtctgaca gttaccaatg cttaatcagt
11280
gaggcaccta tctcagcgat ctgtctattt cgttcatcca tagttgcctg actccccgtc
11340
gtgtagataa ctacgatacg ggagggctta ccatctggcc ccagtgctgc aatgataccg
11400
cgagacccac gctcaccggc tccagattta tcagcaataa accagccagc cggaagggcc
11460
gagcgcagaa gtggtcctgc aactttatcc gcctccatcc agtctattaa ttgttgccgg
11520
gaagctagag taagtagttc gccagttaat agtttgcgca acgttgttgc cattgctgca
11580
ggcatcgtgg tgtcacgctc gtcgtttggt atggcttcat tcagctccgg ttcccaacga
11640
tcaaggcgag ttacatgatc ccccatgttg tgcaaaaaag cggttagctc cttcggtcct
11700
ccgatcgttg tcagaagtaa gttggccgca gtgttatcac tcatggttat ggcagcactg
11760
cataattctc ttactgtcat gccatccgta agatgctttt ctgtgactgg tgagtactca
11820
accaagtcat tctgagaata gtgtatgcgg cgaccgagtt gctcttgccc ggcgtcaaca
11880
cgggataata ccgcgccaca tagcagaact ttaaaagtgc tcatcattgg aaaacgttct
11940
tcggggcgaa aactctcaag gatcttaccg ctgttgagat ccagttcgat gtaacccact
12000
cgtgcaccca actgatcttc agcatctttt actttcacca gcgtttctgg gtgagcaaaa
12060
acaggaaggc aaaatgccgc aaaaaaggga ataagggcga cacggaaatg ttgaatactc
12120
atactcttcc tttttcaata ttattgaagc atttatcagg gttattgtct catgagcgga
12180
tacatatttg aatgtattta gaaaaataaa caaatagggg ttccgcgcac atttccccga
12240
aaagtgccac ctgacgtcta agaaaccatt attatcatga cattaaccta taaaaatagg
12300
cgtatcacga ggccctttcg tcttcaagaa
12330
<210> 4
<211> 4254
<212> ДНК
<213> Искусственная последовательность
<220>
<223> интеграционная кассета
<400> 4
gcaggctgtt attgtaacat gtaagccata agccattcgt aaaagtgcgg gaggaaggtc
60
atgaataatc tgcgtaatag actttcaggc gtgaatggga aaaataagag agtaaaagaa
120
aaagaacaaa aaatctggtc ggagattggg atgatagcgg gagcatttgc gctgcttgat
180
gtgatcatcc gcggcattat gtttgaattt ccgtttaaag aatgggctgc aagccttgtg
240
tttttgttca tcattatctt atattactgc atcagggctg cggcatccgg aatgctcatg
300
ccgagaatag acaccaaaga agaactgcaa aaacgggtga agcagcagcg aatagaatca
360
attgcggtcg cctttgcggt agtggtgctt acgatgtacg acagggggat tccccataca
420
ttcttcgctt ggctgaaaat gattcttctt tttatcgtct gcggcggcgt tctgtttctg
480
cttcggtatg tgattgtgaa gctggcttac agaagagcgg taaaagaaga aataaaaaag
540
aaatcatctt ttttgtttgg aaagcgaggg aagcgttcac agtttcgggc agcttttttt
600
ataggaacat tgatttgtat tcactctgcc aagttgtttt gatagagtga ttgtgataat
660
tttaaatgta agcgttaaca aaattctcca gtcttcacat cggtttgaaa ggaggaagcg
720
gaagaatgaa gtaagaggga tttttgactc cgaagtaagt cttcaaaaaa tcaaataagg
780
agtgtcaaga atgtttgcaa aacgattcaa aacctcttta ctgccgttat tcgctggatt
840
tttattgctg tttcatttgg ttctggcagg accggcggct gcgagtgctg aaacggcgaa
900
caaatcgaat gagcttacag caccgtcgat caaaagcgga accattcttc atgcatggcc
960
agtgaattcg agctcggtac ctaccgttcg tataatgtat gctatacgaa gttatgataa
1020
aaaatttaga agccaatgaa atctataaat aaactaaatt aagtttattt aattaacaac
1080
tatggatata aaataggtac taatcaaaat agtgaggagg atatatttga atacatacga
1140
acaaattaat aaagtgaaaa aaatacttcg gaaacattta aaaaataacc ttattggtac
1200
ttacatgttt ggatcaggag ttgagagtgg actaaaacca aatagtgatc ttgacttttt
1260
agtcgtcgta tctgaaccat tgacagatca aagtaaagaa atacttatac aaaaaattag
1320
acctatttca aaaaaaatag gagataaaag caacttacga tatattgaat taacaattat
1380
tattcagcaa gaaatggtac cgtggaatca tcctcccaaa caagaattta tttatggaga
1440
atggttacaa gagctttatg aacaaggata cattcctcag aaggaattaa attcagattt
1500
aaccataatg ctttaccaag caaaacgaaa aaataaaaga atatacggaa attatgactt
1560
agaggaatta ctacctgata ttccattttc tgatgtgaga agagccatta tggattcgtc
1620
agaggaatta atagataatt atcaggatga tgaaaccaac tctatattaa ctttatgccg
1680
tatgatttta actatggaca cgggtaaaat cataccaaaa gatattgcgg gaaatgcagt
1740
ggctgaatct tctccattag aacataggga gagaattttg ttagcagttc gtagttatct
1800
tggagagaat attgaatgga ctaatgaaaa tgtaaattta actataaact atttaaataa
1860
cagattaaaa aaattataaa aaaattgaaa aaatggtgga aacacttttt tcaatttttt
1920
tgttttatta tttaatattt gggaaatatt cattctaata taacttcgta taatgtatgc
1980
tatacgaacg gtaggatcct ctagagtcga cctgcaggca ttttacattt ttagaaatgg
2040
gcgtgaaaaa aagcgcgcga ttatgtaaaa tataaagtga tagcggtacc attataggta
2100
agagaggaat gtacacatgt actatttaaa aaacacaaac ttttggatgt tcggtttatt
2160
ctttttcttt tactttttta tcatgggagc ctacttcccg tttttcccga tttggctaca
2220
tgacatcaac catatcagca aaagtgatac gggtattatt tttgccgcta tttctctgtt
2280
ctcgctatta ttccaaccgc tgtttggtct gctttctgac aaactcgggc tgcgcaaata
2340
cctgctgtgg attattaccg gcatgttagt gatgtttgcg ccgttcttta tttttatctt
2400
cgggccactg ttacaataca acattttagt aggatcgatt gttggtggta tttatctagg
2460
cttttgtttt aacgccggtg cgccagcagt agaggcattt attgagaaag tcagccgtcg
2520
cagtaatttc gaatttggtc gcgcgcggat gtttggctgt gttggctggg cgctgtgtgc
2580
ctcgattgtc ggcatcatgt tcaccatcaa taatcagttt gttttctggc tgggctctgg
2640
ctgtgcactc atcctcgccg ttttactctt tttcgccaaa acggatgcgc cctcttctgc
2700
cacggttgcc aatgcggtag gtgccaacca ttcggcattt agccttaagc tggcactgga
2760
actgttcaga cagccaaaac tgtggttttt gtcactgtat gttattggcg tttcctgcac
2820
ctacgatgtt tttgaccaac agtttgctaa tttctttact tcgttctttg ctaccggtga
2880
acagggtacg cgggtatttg gctacgtaac gacaatgggc gaattactta acgcctcgat
2940
tatgttcttt gcgccactga tcattaatcg catcggtggg aaaaacgccc tgctgctggc
3000
tggcactatt atgtctgtac gtattattgg ctcatcgttc gccacctcag cgctggaagt
3060
ggttattctg aaaacgctgc atatgtttga agtaccgttc ctgctggtgg gctgctttaa
3120
atatattacc agccagtttg aagtgcgttt ttcagcgacg atttatctgg tctgtttctg
3180
cttctttaag caactggcga tgatttttat gtctgtactg gcgggcaata tgtatgaaag
3240
catcggtttc cagggcgctt atctggtgct gggtctggtg gcgctgggct tcaccttaat
3300
ttccgtgttc acgcttagcg gccccggccc gctttccctg ctgcgtcgtc aggtgaatga
3360
agtcgcttaa gcaatcaatg tcggatgcca gcctggcttt gattacgtgc taaatggttt
3420
atataatgac tcgggcttaa gcggttctct tccccattga gggcaaggct agacgggact
3480
taccgaaaga aaccatcaat gatggtttct tttttgttca taaatcagac aaaacttttc
3540
tcttgcaaaa gtttgtgaag tgttgcacaa tataaatgtg aaatacttca caaacaaaaa
3600
gacatcaaag agaaacatac cctggaagga tgattaatga tgaacaaaca tgtaaataaa
3660
gtagctttaa tcggagcggg ttttgttgga agcagttatg catttgcgtt aattaaccaa
3720
ggaatcacag atgagcttgt ggtcattgat gtaaataaag aaaaagcaat gggcgatgtg
3780
atggatttaa accacggaaa ggcgtttgcg ccacaaccgg tcaaaacatc ttacggaaca
3840
tatgaagact gcaaggatgc tgatattgtc tgcatttgcg ccggagcaaa ccaaaaacct
3900
ggtgagacac gccttgaatt agtagaaaag aacttgaaga ttttcaaagg catcgttagt
3960
gaagtcatgg cgagcggatt tgacggcatt ttcttagtcg cgacaaatcc ggttgatatc
4020
ctgacttacg caacatggaa attcagcggc ctgccaaaag agcgggtgat tggaagcggc
4080
acaacacttg attctgcgag attccgtttc atgctgagcg aatactttgg cgcagcgcct
4140
caaaacgtac acgcgcatat tatcggagag cacggcgaca cagagcttcc tgtttggagc
4200
cacgcgaatg tcggcggtgt gccggtcagt gaactcgttg agaaaaacga tgcg
4254
<210> 5
<211> 10340
<212> ДНК
<213> Искусственная последовательность
<220>
<223> экспрессионная плазмида
<400> 5
tcgcgcgttt cggtgatgac ggtgaaaacc tctgacacat gcagctcccg gagacggtca
60
cagcttgtct gtaagcggat gccgggagca gacaagcccg tcagggcgcg tcagcgggtg
120
ttggcgggtg tcggggctgg cttaactatg cggcatcaga gcagattgta ctgagagtgc
180
accatatgcc ttcaatcctt ttaataacaa ttatagcatc taatcttcaa caaactggcc
240
cgtttgttga actactcttt aataaaataa tttttccgtt cccaattcca cattgcaata
300
atagaaaatc catcttcatc ggctttttcg tcatcatctg tatgaatcaa atcgccttct
360
tctgtgtcat caaggtttaa ttttttatgt atttctttta acaaaccacc ataggagatt
420
aaccttttac ggtgtaaacc ttcctccaaa tcagacaaac gtttcaaatt cttttcttca
480
tcatcggtca taaaatccgt atcctttaca ggatattttg cagtttcgtc aattgccgat
540
tgtatatccg atttatattt atttttcggt cgaatcattt gaacttttac atttggatca
600
tagtctaatt tcattgcctt tttccaaaat tgaatccatt gtttttgatt cacgtagttt
660
tctgtattct taaaataagt tggttccaca cataccaata catgcatgtg ctgattataa
720
gaattatctt tattatttat tgtcacttcc gttgcacgca taaaaccaac aagattttta
780
ttaatttttt tatattgcat cattcggcga aatccttgag ccatatctga caaactctta
840
tttaattctt cgccatcata aacattttta actgttaatg tgagaaacaa ccaacgaact
900
gttggctttt gtttaataac ttcagcaaca accttttgtg actgaatgcc atgtttcatt
960
gctctcctcc agttgcacat tggacaaagc ctggatttac aaaaccacac tcgatacaac
1020
tttctttcgc ctgtttcacg attttgttta tactctaata tttcagcaca atcttttact
1080
ctttcagcct ttttaaattc aagaatatgc agaagttcaa agtaatcaac attagcgatt
1140
ttcttttctc tccatggtct cacttttcca ctttttgtct tgtccactaa aacccttgat
1200
ttttcatctg aataaatgct actattagga cacataatat taaaagaaac ccccatctat
1260
ttagttattt gtttagtcac ttataacttt aacagatggg gtttttctgt gcaaccaatt
1320
ttaagggttt tcaatacttt aaaacacata cataccaaca cttcaacgca cctttcagca
1380
actaaaataa aaatgacgtt atttctatat gtatcaagat aagaaagaac aagttcaaaa
1440
ccatcaaaaa aagacacctt ttcaggtgct ttttttattt tataaactca ttccctgatc
1500
tcgacttcgt tcttttttta cctctcggtt atgagttagt tcaaattcgt tctttttagg
1560
ttctaaatcg tgtttttctt ggaattgtgc tgttttatcc tttaccttgt ctacaaaccc
1620
cttaaaaacg tttttaaagg cttttaagcc gtctgtacgt tccttaagat tttacatttt
1680
tagaaatggg cgtgaaaaaa agcgcgcgat tatgtaaaat ataaagatta actaataagg
1740
aggacaaaca tgcaaaaact gctttctctg ccgtcaaatc ttgtccagtc ttttcatgaa
1800
cttgaaagag ttaaccgcac agattggttt tgcacatcag atccggttgg caaaaaactg
1860
ggctcaggcg gaggcacaag ctggttactg gaagaatgtt acaacgaata cagcgatgga
1920
gcgacatttg gcgaatggct ggaaaaagaa aaacgcattc tgttacatgc tggaggccaa
1980
agcagacgcc ttccgggata tgccccgtct ggcaaaatct taacaccggt cccggttttt
2040
agatgggaaa gaggacaaca tttaggccag aacctgctta gcttacaact gccgctttat
2100
gaaaaaatta tgtctctggc tccggataaa ctgcatacac tgattgcctc tggcgatgtg
2160
tatatccgca gcgaaaaacc gcttcagtct atcccggaag ctgatgttgt gtgctatgga
2220
ttatgggtcg atccgtctct ggcaacacat catggcgttt ttgcgtcaga tagaaaacat
2280
ccggaacaac tggattttat gctgcagaaa ccgtcacttg cagaattaga aagcctgtct
2340
aaaacacatt tatttctgat ggatattggc atctggttac tgtcagatag agcggtcgaa
2400
attctgatga aacgcagcca taaagaatca agcgaagaac ttaaatacta cgatctgtac
2460
tcagattttg gacttgcttt aggcacacat ccgcgcattg aagatgaaga agtgaataca
2520
cttagcgtcg caatcctgcc tcttccggga ggcgaatttt accattacgg aacatcaaaa
2580
gaactgattt cttcaacact tagcgttcaa aacaaagtgt atgatcagag acgcatcatg
2640
catagaaaag tgaaaccgaa tccggcaatg tttgtccaaa acgcggtcgt tcgcattccg
2700
ctttgcgctg aaaatgccga tttatggatt gaaaacagcc atatcggacc gaaatggaaa
2760
atcgcatcaa gacatattat cacaggcgtg ccggaaaatg attggtcact tgcagtcccg
2820
gcgggagtgt gtgtcgatgt ggtcccgatg ggagataaag gctttgttgc gagaccgtat
2880
ggattagatg atgtgtttaa aggcgatctg cgcgatagca aaacaacact tacaggcatt
2940
ccgtttggcg aatggatgtc taaaagagga ttatcataca cagatttaaa aggccgcaca
3000
gatgatctgc aggctgtttc agtgtttccg atggtgaaca gcgtcgaaga attaggcctg
3060
gtccttagat ggatgctttc tgaaccggaa ttagaagaag gaaaaaatat ttggctgcgc
3120
tctgaacatt tttcagcaga tgaaatctca gctggcgcca accttaaaag actgtacgcg
3180
caaagagaag aatttcgcaa aggaaattgg aaagcacttg cggttaacca tgaaaaatca
3240
gtgttttatc agctggatct ggcagatgca gcggaagatt ttgttcgcct tggcttagat
3300
atgccggaac ttttaccgga agatgctctg caaatgtcac gcattcataa tcgcatgctt
3360
agagcccgca tcctgaaact tgatggaaaa gattatagac cggaagaaca ggctgccttt
3420
gatctgctta gagatggatt actggatggc attagcaacc gcaaatctac accgaaactg
3480
gatgtctatt ctgatcaaat tgtttggggc agatcaccgg tgcgcatcga tatggcaggc
3540
ggctggacag atacaccgcc gtatagcctt tattctggag gcaatgttgt gaaccttgcg
3600
attgaattaa atggccaacc gccgcttcag gtctacgtta aaccgtgcaa agattttcat
3660
atcgttctga gatcaatcga tatgggagct atggaaatcg tgagcacatt tgatgaactg
3720
caagattaca agaaaattgg ctcaccgttt agcatcccga aagcagcgtt aagcctggcc
3780
ggatttgctc cggccttttc tgctgtcagc tatgcatcac tggaagaaca gcttaaagat
3840
tttggcgcag gaattgaagt tacactttta gctgccatcc cggcgggctc tggattaggc
3900
acaagctcta ttctggcttc aacagtgctt ggagccatca atgatttttg cggcctggca
3960
tgggataaaa acgaaatttg tcaaagaaca cttgtcttag aacagctgct tacaacaggc
4020
ggcggctggc aagatcagta tggcggagtc ctgcaaggag ttaaactgct gcagacagaa
4080
gctggctttg cccagtcacc gttagttcgc tggctgccgg atcatctttt tacacatccg
4140
gaatacaaag attgtcatct tttatactac acaggaatta caagaacagc aaaaggcatt
4200
ttagcggaaa tcgtgtcaag catgtttctg aatagctcac tgcatcttaa cctgcttagc
4260
gaaatgaaag cacatgcgct tgatatgaat gaagctattc aaagaggctc ttttgtcgaa
4320
tttggacgct tagttggcaa aacatgggaa cagaataaag ccctggatag cggaacaaac
4380
ccgccggcag tggaagcgat catcgatctg atcaaagatt acacactggg atacaaactg
4440
cctggagcag gcggcggcgg ctatctgtat atggttgcga aagatccgca agcagctgtg
4500
agaattagaa aaatcctgac agaaaatgct ccgaacccga gagcccgctt tgtggaaatg
4560
acactttcag ataaaggatt tcaggtctca agaagctaag ctcttaagga ggattttaga
4620
atgggttcta tcatcagact tcagggtggt ttaggcaatc aactgtttca atttagcttt
4680
ggttatgctc tttcaaaaat caatggcaca ccgctgtact ttgatatttc acattacgca
4740
gaaaacgatg atcatggcgg atatagactg aataaccttc aaatcccgga agaatatctt
4800
cagtactaca caccgaaaat caacaacatc tacaaactgc ttgtcagagg cagccgctta
4860
tatccggata tttttctgtt tcttggcttt tgcaacgaat ttcatgcgta tggctatgat
4920
tttgaataca tcgcgcaaaa atggaaaagc aaaaaataca tcggatactg gcagtctgaa
4980
catttctttc ataaacatat tctggatctg aaagaatttt tcatcccgaa aaacgttagc
5040
gaacaagcta acttactggc agcgaaaatc ttagaatctc agtcaagcct gtcaattcat
5100
atcagacgcg gcgattacat caaaaacaaa acagccacac tgacacatgg agtttgctct
5160
cttgaatact acaagaaagc actgaacaaa atcagagatt tagccatgat tcgcgatgtg
5220
tttatctttt ctgatgatat tttctggtgt aaagaaaaca tcgaaacact tctgagcaaa
5280
aaatacaaca tctactactc agaagattta agccaagaag aagatttatg gctgatgtct
5340
ctggctaatc atcatattat cgccaactct tcattttcat ggtggggcgc atatcttgga
5400
agctctgcga gccagattgt tatctatccg acaccgtggt atgatattac accgaaaaac
5460
acatacattc ctatcgtgaa ccattggatc aacgtggaca aacattcctc ctgctaaaaa
5520
ggaggaacta ctatgggcaa tacatcaatt caaacacaga gctatagagc agttgataaa
5580
gatgcgggac aaagccgctc ttatattatc ccgtttgcac tgctttgctc tctgtttttc
5640
ctgtgggctg tggccaataa cctgaatgat attttactgc cgcagtttca acaggcgttt
5700
acacttacaa actttcaagc tggattaatt cagagcgcct tttactttgg ctactttatc
5760
atcccgatcc cggctggaat ccttatgaaa aaactgtctt acaaagctgg aattatcaca
5820
ggcttatttc tgtatgcact gggagcagct ctgttttggc ctgctgccga aatcatgaat
5880
tatacactgt ttctggtcgg actgtttatt atcgcagcgg gactgggctg tcttgaaaca
5940
gctgccaacc cgtttgtcac agttcttgga ccggaatcaa gcggccattt tagactgaac
6000
cttgcacaaa catttaactc atttggagca atcatcgcgg ttgtgtttgg ccagagctta
6060
attctgtcta atgtcccgca tcaaagccag gatgttctgg ataaaatgtc tccggaacaa
6120
ctttcagcgt ataaacattc acttgtgtta agcgtccaga caccgtatat gattatcgtt
6180
gcaatcgtgc ttttagtcgc gctgcttatt atgttaacaa aatttccggc actgcaatct
6240
gataaccatt cagatgcgaa acagggctca tttagcgctt ctttatcaag actggccaga
6300
attagacatt ggcgctgggc tgtgttagcc caattttgct atgtcggagc acagacagcg
6360
tgttggtcat atctgatccg ctatgctgtc gaagaaattc cgggaatgac agccggcttt
6420
gcagcgaatt atcttacagg cacaatggtt tgctttttca tcggaagatt tacaggcaca
6480
tggttaattt ctcgctttgc accgcataaa gtgcttgctg cctatgcgtt aatcgcaatg
6540
gcgctgtgcc ttatttcagc ttttgccggc ggacatgtcg gccttattgc attaacactg
6600
tgtagcgcgt ttatgtctat ccaatacccg acaatcttta gcctgggaat taaaaacctt
6660
ggccaggata caaaatacgg atcttcattt atcgttatga caattatcgg cggaggcatt
6720
gttacaccgg tgatgggatt tgtgagcgat gcagcgggca atatcccgac agctgaatta
6780
attccggctc tgtgttttgc cgttatcttt atctttgcta gatttcgctc tcaaacagcc
6840
acaaactaat ctagagtcga cgtccaggca tcaaataaaa cgaaaggctc agtcgaaaga
6900
ctgggccttt cgttttatct gttgtttgtc ggtgaacgct ctctactaga gtcacactgg
6960
ctcaccttcg ggtgggcctt tctgcgttta tatacgcgtt aacccgggcc cgcggatgga
7020
tatgatcaga tcctttaact ctggcaaccc tcaaaattga atgagacatg ctacacctcc
7080
ggataataaa tatatataaa cgtatataga tttcataaag tctaacacac tagacttatt
7140
tacttcgtaa ttaagtcgtt aaaccgtgtg ctctacgacc aaaactataa aacctttaag
7200
aactttcttt ttttacaaga aaaaagaaat tagataaatc tctcatatct tttattcaat
7260
aatcgcatcc gattgcagta taaatttaac gatcactcat catgttcata tttatcagag
7320
ctcgtgctat aattatacta attttataag gaggaaaaaa tatgggcatt tttagtattt
7380
ttgtaatcag cacagttcat tatcaaccaa acaaaaaata agtggttata atgaatcgtt
7440
aataagcaaa attcatataa ccaaattaaa gagggttata atgaacgaga aaaatataaa
7500
acacagtcaa aactttatta cttcaaaaca taatatagat aaaataatga caaatataag
7560
attaaatgaa catgataata tctttgaaat cggctcagga aaaggccatt ttacccttga
7620
attagtaaag aggtgtaatt tcgtaactgc cattgaaata gaccataaat tatgcaaaac
7680
tacagaaaat aaacttgttg atcacgataa tttccaagtt ttaaacaagg atatattgca
7740
gtttaaattt cctaaaaacc aatcctataa aatatatggt aatatacctt ataacataag
7800
tacggatata atacgcaaaa ttgtttttga tagtatagct aatgagattt atttaatcgt
7860
ggaatacggg tttgctaaaa gattattaaa tacaaaacgc tcattggcat tacttttaat
7920
ggcagaagtt gatatttcta tattaagtat ggttccaaga gaatattttc atcctaaacc
7980
taaagtgaat agctcactta tcagattaag tagaaaaaaa tcaagaatat cacacaaaga
8040
taaacaaaag tataattatt tcgttatgaa atgggttaac aaagaataca agaaaatatt
8100
tacaaaaaat caatttaaca attccttaaa acatgcagga attgacgatt taaacaatat
8160
tagctttgaa caattcttat ctcttttcaa tagctataaa ttatttaata agtaagttaa
8220
gggatgcata aactgcatcc cttaacttgt ttttcgtgtg cctatttttt gtgaatcgat
8280
tcagctgcat taatgaatcg gccaacgcgc ggggagaggc ggtttgcgta ttgggcgctc
8340
ttccgcttcc tcgctcactg actcgctgcg ctcggtcgtt cggctgcggc gagcggtatc
8400
agctcactca aaggcggtaa tacggttatc cacagaatca ggggataacg caggaaagaa
8460
catgtgagca aaaggccagc aaaaggccag gaaccgtaaa aaggccgcgt tgctggcgtt
8520
tttccatagg ctccgccccc ctgacgagca tcacaaaaat cgacgctcaa gtcagaggtg
8580
gcgaaacccg acaggactat aaagatacca ggcgtttccc cctggaagct ccctcgtgcg
8640
ctctcctgtt ccgaccctgc cgcttaccgg atacctgtcc gcctttctcc cttcgggaag
8700
cgtggcgctt tctcatagct cacgctgtag gtatctcagt tcggtgtagg tcgttcgctc
8760
caagctgggc tgtgtgcacg aaccccccgt tcagcccgac cgctgcgcct tatccggtaa
8820
ctatcgtctt gagtccaacc cggtaagaca cgacttatcg ccactggcag cagccactgg
8880
taacaggatt agcagagcga ggtatgtagg cggtgctaca gagttcttga agtggtggcc
8940
taactacggc tacactagaa gaacagtatt tggtatctgc gctctgctga agccagttac
9000
cttcggaaaa agagttggta gctcttgatc cggcaaacaa accaccgctg gtagcggtgg
9060
tttttttgtt tgcaagcagc agattacgcg cagaaaaaaa ggatctcaag aagatccttt
9120
gatcttttct acggggtctg acgctcagtg gaacgaaaac tcacgttaag ggattttggt
9180
catgagatta tcaaaaagga tcttcaccta gatcctttta aattaaaaat gaagttttaa
9240
atcaatctaa agtatatatg agtaaacttg gtctgacagt taccaatgct taatcagtga
9300
ggcacctatc tcagcgatct gtctatttcg ttcatccata gttgcctgac tccccgtcgt
9360
gtagataact acgatacggg agggcttacc atctggcccc agtgctgcaa tgataccgcg
9420
agacccacgc tcaccggctc cagatttatc agcaataaac cagccagccg gaagggccga
9480
gcgcagaagt ggtcctgcaa ctttatccgc ctccatccag tctattaatt gttgccggga
9540
agctagagta agtagttcgc cagttaatag tttgcgcaac gttgttgcca ttgctacagg
9600
catcgtggtg tcacgctcgt cgtttggtat ggcttcattc agctccggtt cccaacgatc
9660
aaggcgagtt acatgatccc ccatgttgtg caaaaaagcg gttagctcct tcggtcctcc
9720
gatcgttgtc agaagtaagt tggccgcagt gttatcactc atggttatgg cagcactgca
9780
taattctctt actgtcatgc catccgtaag atgcttttct gtgactggtg agtactcaac
9840
caagtcattc tgagaatagt gtatgcggcg accgagttgc tcttgcccgg cgtcaatacg
9900
ggataatacc gcgccacata gcagaacttt aaaagtgctc atcattggaa aacgttcttc
9960
ggggcgaaaa ctctcaagga tcttaccgct gttgagatcc agttcgatgt aacccactcg
10020
tgcacccaac tgatcttcag catcttttac tttcaccagc gtttctgggt gagcaaaaac
10080
aggaaggcaa aatgccgcaa aaaagggaat aagggcgaca cggaaatgtt gaatactcat
10140
actcttcctt tttcaatatt attgaagcat ttatcagggt tattgtctca tgagcggata
10200
catatttgaa tgtatttaga aaaataaaca aataggggtt ccgcgcacat ttccccgaaa
10260
agtgccacct gacgtctaag aaaccattat tatcatgaca ttaacctata aaaataggcg
10320
tatcacgagg ccctttcgtc
10340
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| Ферментативная продукция углеводов микробными клетками с использованием смешанного сырья | 2020 |
|
RU2809122C2 |
| СИНТЕЗ ФУКОЗИЛИРОВАННЫХ СОЕДИНЕНИЙ | 2009 |
|
RU2584599C2 |
| ФУКОЗИЛТРАНСФЕРАЗЫ И ИХ ПРИМЕНЕНИЕ ДЛЯ ПОЛУЧЕНИЯ ФУКОЗИЛИРОВАННЫХ ОЛИГОСАХАРИДОВ | 2018 |
|
RU2818835C2 |
| УЛУЧШЕННЫЙ СПОСОБ ПОЛУЧЕНИЯ ФУКОЗИЛИРОВАННЫХ ОЛИГОСАХАРИДОВ | 2017 |
|
RU2790445C2 |
| Ферментативное получение олигосахаридов посредством общей ферментации с использованием смешанного сырья | 2019 |
|
RU2801231C2 |
| ПРОДУКЦИЯ СИАЛИРОВАННЫХ ОЛИГОСАХАРИДОВ В КЛЕТКАХ BACILLUS | 2020 |
|
RU2816764C1 |
| ПРИМЕНЕНИЕ ГЛИКОЗИДАЗ В ПОЛУЧЕНИИ ОЛИГОСАХАРИДОВ | 2019 |
|
RU2810730C2 |
| НОВЫЕ ФУКОЗИЛТРАНСФЕРАЗЫ И ИХ ПРИМЕНЕНИЕ | 2011 |
|
RU2642307C2 |
| НОВЫЕ ФУКОЗИЛТРАСФЕРАЗЫ И ИХ ПРИМЕНЕНИЯ | 2011 |
|
RU2628307C2 |
| СИНТЕЗ НМО | 2009 |
|
RU2517602C2 |
Изобретение относится к биотехнологии. Предложена неспорообразующая клетка Bacillus subtilis для получения 2'-фукозиллактозы. При этом в указанной клетке нарушена споруляция в результате делеции или функциональной инактивации одного или более генов, 5 кодирующих Spo0A, сигма E и сигма F. Также указанная клетка содержит лактозопермеазу LacY E. coli и α-1,2-фукозилтрансферазу WbgL E. coli. Также клетка генетически сконструирована для экспрессии генов пути биосинтеза ГДФ-фукозы de novo, представляющих собой гены E. coli manA, E. coli manB, E. coli manC, E. coli gmd и E. coli wcaG; и причем указанная клетка Bacillus не обладает какой-либо β-галактозидазной активностью или обладает пониженной β-галактозидазной активностью по сравнению с клеткой-предшественником Bacillus дикого типа того же вида в результате делеции или функциональной инактивации ganA. Также предложены применение указанной клетки Bacillus для получения 2'-фукозиллактозы и способ получения 2'-фукозиллактозы с использованием указанной клетки Bacillus. Изобретение может быть использовано для эффективной продукции 2'-фукозиллактозы. 3 н. и 1 з.п. ф-лы, 1 табл., 5 пр.
1. Неспорообразующая клетка Bacillus subtilis для получения 2'-фукозиллактозы, где нарушена споруляция в результате делеции или функциональной инактивации одного или более генов, кодирующих Spo0A, сигма E и сигма F, причем указанная клетка генетически сконструирована, чтобы иметь лактозопермеазу, где лактозопермеаза представляет собой LacY E. coli, генетически сконструирована для экспрессии генов пути биосинтеза ГДФ-фукозы de novo, представляющих собой гены E. coli manA, E. coli manB, E. coli manC, E. coli gmd и E. coli wcaG; генетически сконструирована, чтобы иметь α-1,2-фукозилтрансферазу WbgL E. coli, и причем указанная клетка Bacillus не обладает какой-либо β-галактозидазной активностью или обладает пониженной β-галактозидазной активностью по сравнению с клеткой-предшественником Bacillus дикого типа того же вида в результате делеции или функциональной инактивации ganA.
2. Неспорообразующая клетка Bacillus subtilis по п. 1, где клетка Bacillus subtilis дополнительно генетически сконструирована в результате делеции или функциональной инактивации yesZ.
3. Применение клетки Bacillus по п. 1 или 2 для получения 2'-фукозиллактозы.
4. Способ получения 2'-фукозиллактозы, включающий:
- предоставление неспорообразующей клетки Bacillus, охарактеризованной в п. 1 или 2;
- культивирование указанной клетки Bacillus в среде, которая содержит лактозу, и в условиях, которые являются пермиссивными в отношении получения фукозилированного олигосахарида; и возможно
- извлечение 2'-фукозиллактозы из среды и/или клетки Bacillus.
| US 2012208181 A1, 16.08.2012 | |||
| CN 109735479 A, 10.05.2019 | |||
| US 2014031541 A1, 30.01.2014 | |||
| СИНТЕЗ ФУКОЗИЛИРОВАННЫХ СОЕДИНЕНИЙ | 2009 |
|
RU2584599C2 |
| ПИТАТЕЛЬНАЯ КОМПОЗИЦИЯ ДЛЯ МЛАДЕНЦЕВ И/ИЛИ ДЕТЕЙ МЛАДШЕГО ВОЗРАСТА, СОДЕРЖАЩАЯ ОЛИГОСАХАРИДЫ | 2017 |
|
RU2742510C2 |
| US 20170081353 A1, 23.03.2017 | |||
| WO 2003040352 A1, 15.05.2003. | |||

