Настоящее изобретение относится к способу скрининга на модулятор экспрессии эндотелиальной NO-синтазы (eNOS), способу диагностирования сердечно-сосудистого заболевания у индивида, применению HEBP1 для идентификации лекарственного средства для профилактики и/или лечения заболевания, связанного с дисфункцией eNOS, в частности сердечно-сосудистого заболевания, применению HEBP1 для обнаружения компонента передачи сигнала при экспрессии eNOS и применению HEBP1 для регуляции активности промотора eNOS.
Синтазы оксида азота (EC 1.14.13.39; NOS) были открыты в 1998, и в то время физиологическое значение их продукта, оксида азота (NO), который является самой маленькой биоактивной молекулой млекопитающих, было неизвестно. Между тем было установлено, что NO является важным мессенджером в регуляции сосудистого тонуса сердечно-сосудистой системы, служит вторичным мессенджером центральной нервной системы и обладает защитным механизмом против бактериальных и опухолевых клеток. В 1989 Нобелевская премия по медицине была присуждена Robert Furchgott, Ferid Murad и Luis J. Ignaro, которые идентифицировали NO в качестве мессенджеров клеток млекопитающих.
NOS являются членами семейства родственных ферментов, кодируемых отдельными генами. NOS является одним из самых регулируемых ферментов биологии. Существует три известных изоформы, две из них являются конститутивными (cNOS), а третья - индуцибельной (iNOS). Клонирование ферментов NOS показало, что гены cNOS включают гены, конститутивные как в головном мозге (NOSl, nNOS или нейронной NOS), так и в эндотелии (NOS3, eNOS, cNOS или эндотелиальной NOS), ген третьей изоформы является индуцибельным геном (NOS2, iNOS или индуцибельной NOS).
В головном мозге nNOS является растворимым ферментом с молекулярной массой 161 кДа и представляет собой самую большую изоформу NOS. Эта изоформа конститутивно экспрессируется в основном в нейронах и в головном мозге и синтезирует лишь малые количества NO. Регуляция ферментативной активности опосредуется Ca2+ через кальмодулин. Однако на ферментативную активность также оказывает влияние фосфорилирование посредством зависимой от Ca2+-кальмодулина протеинкиназы 2, а также протеинкиназы A, C и G. В головном мозге NO важен для регуляции передачи сигнала в синапсе. Периферические кровеносные сосуды и гладкомышечные клетки часто иннервируются нервами, которые вырабатывают NO и действуют антагонистически по отношению к симпатической нервной системе. Кроме того, большие количества nNOS были обнаружены в скелетной мышце, в которой NO контролирует мышечную сократимость и локальный кровоток.
Индуцибельная NOS экспрессируется в макрофагах, но также в других клетках после индукции бактериальным LPS или цитокинами. Как и nNOS, iNOS является по большей части растворимым белком, имеющим молекулярную массу 131 кДа, который вырабатывает огромные количества NO. iNOS регулируется на транскрипционном уровне различными стимулами. После индукции ее экспрессии выработка NO проходит по цитотоксическому принципу макрофагов при разрушении микроорганизмов и паразитов, а также опухолевых клеток. Однако NO может разрушать здоровые клетки организма и вызывать повреждения окружающей ткани. Большая часть воспалительных и аутоиммунных заболеваний характеризуется присутствием огромных количеств активированных макрофагов и нейтрофильных гранулоцитов. Кроме того, iNOS также важна в патологии септического шока, который характеризуется резким расширением артериальных сосудов, гипотонией и повреждениями микрососудов.
Наконец, эндотелиальная NOS ответственна за большую часть синтезируемого в сосудах NO, который защищает от артериосклероза и тромбоза. Интактная eNOS является главным переключателем при физиологическом поддержании сосудистого гомеостаза и играет ключевую роль в патофизиологии сердечно-сосудистой системы. Выработка NO ферментом eNOS в сосудистом эндотелии опосредуется в основных условиях стимуляцией ряда агонистов рецепторов, таких как брадикинин, ацетилхолин и гистамин, а также гемодинамическим давлением протекающей крови. NO приводит к расширению всех типов кровеносных сосудов в результате стимуляции растворимой гуанилилциклазы и увеличения концентрации cGMP в гладкомышечных клетках. Соответственно, NO эндотелиальных клеток является важной эндогенной вызывающей расширение сосудов ответной частью на вазоконстрикцию под действием симпатической нервной системы или ренин-ангиотензиновой системы.
Помимо своего сосудорасширяющего свойства эндотелиальный NO имеет ряд сосудозащитных и антисклеротических свойств. Высвобождаемый в просвет сосудов NO является сильным ингибитором агрегации тромбоцитов и их адгезии к сосудистой стенке. Помимо защиты от тромбоза ингибируется выброс факторов роста, которые могли бы стимулировать пролиферацию гладкомышечных клеток. Было установлено, что у кроликов и мышей генетическое или фармакологическое ингибирование eNOS приводит к прогрессирующему артериосклерозу. Кроме того, эндотелиальный NO может модулировать экспрессию генов, вовлеченных в артериогенез. Это особенно отчетливо видно в случае хемотаксического белка из моноцитов (MCP1), молекул клеточной поверхности, таких как CD11/CD18, P-селектина, адгезивной молекулы 1 сосудистых клеток - VCAM-1 и внутриклеточной адгезивной молекулы - ICAM 1. Соответственно, предотвращается адгезия и инфильтрация липоцитов в сосудистую стенку, предохраняя, тем самым, от ранней фазы артериогенеза. Кроме того, NO ингибирует синтез ДНК, митогенез и пролиферацию сосудистых гладкомышечных клеток. Предполагается, что антипролиферативные эффекты опосредуются cGMP. Кроме того, NO может оказывать прямые эффекты посредством S-нитросилации белков, такие как антиапоптозный эффект в эндотелиальных клетках в результате нитросилации каспазы 3.
Ряд сердечно-сосудистых заболеваний был связан с недостатком биодоступного NO вследствие снижения синтеза и/или увеличения деструкции NO. Другие классические симптомы и заболевания включают гипохолестеринемию, сахарный диабет, гипертонию и неблагоприятные эффекты, опосредованные курением. Как подробно описано выше, патология различных сердечно-сосудистых заболеваний обычно основана на недостатке NO в результате эндотелиальной дисфункции. Недостаток биоактивности NO может быть обусловлен снижением экспрессии и/или активности eNOS, разрывом связей с eNOS, увеличением деструкции NO или ослаблением ответной реакции эффекторных систем для NO.
Консервативная терапия с использованием органических нитратов имеет большое число недостатков из-за выработки огромных количеств NO. В частности, при длительном лечении наблюдается значительное уменьшение эффекта нитрата, которое называют «толерантностью к нитратам». Составляющий 6-8 часов период без нитратов необходим для получения полного эффекта нитрата. Кроме того, обычными неблагоприятными эффектами нитрата являются головная боль, покрасневшая кожа («прилив крови») и риск сильного снижения кровяного давлении с рефлекторной тахикардией. Соответственно, идентификация новых лекарственных средств для долговременной терапии, которые могут индуцировать экспрессию функциональной eNOS и которые могли бы постоянно увеличивать количество биодоступного NO в отличие от нитрата, является интересной и чрезвычайно важной целью исследования. Вследствие своей важной физиологической и патофизиологической функции в организме животного (в том числе человека) важно выявить новые способы модуляции активности eNOS.
Соответственно, целью настоящего изобретения было выявление альтернативных механизмов модулирования активности eNOS.
Неожиданно было обнаружено, что гемсвязывающий белок 1 (HEBP1) вовлечен в активацию экспрессии eNOS. В частности, было установлено, что активность промотора eNOS значительно уменьшена при выключении HEBP1, используя РНК-технологии.
Соответственно, средства и способы взаимодействия с HEBP1 могут быть использованы для регуляции или изменения активности промотора eNOS и/или экспрессии eNOS.
Поэтому в первом аспекте настоящее изобретение относится к способу скрининга на модулятор экспрессии эндотелиальной NO-синтазы (eNOS), который предусматривает
- наличие тест-системы, содержащей гемсвязывающий белок 1 (HEBP1) или его функционально активный вариант,
- приведение тест-системы в контакт с агентом и
- выявление эффекта агента на тест-систему, идентифицируя, тем самым, агент как модулятор экспрессии eNOS.
Как подробно описано выше, eNOS, также известная как синтаза 3 оксида азота (NOS3), синтезирует NO в кровеносных сосудах и вовлечена в регуляцию функционирования сосудов. Этот фермент, в частности, экспрессируется в различных типах эндотелиальных клеток артерий и вен. Однако также было устанолено, что eNOS экспрессируется в плаценте человека, в эпителиальных клетках почечных канальцев и клетках ободочной кишки кролика, и иммунная реакция с участием eNOS была также обнаружена в нейронах гиппокампа и в других областях головного мозга кролика. eNOS конститутивно экспрессируется и вырабатывает малые количества NO по сравнению с iNOS.
Фермент eNOS присутствует в виде гомодимера, причем каждый мономер составлен из нескольких субъединиц (схематическая иллюстрация iNOS представлена в Forstermann and Monzen, 2006, Circulation 113: 1708-1714). C-концевой редуктазный домен связывает никотинамидадениндинуклеотидфосфат (NADPH), флавинмононуклеотид (FMN) и флавинадениндинуклеотид (FAD) и связан с кальмодулинсвязывающим доменом с оксигеназным доменом. Оксигеназный домен имеет простатическую гемовую группу и связывает 6(R)-5,6,7,8-тетрагидробиоптерин (BH4), молекулярный кислород и L-аргинин. Редуктазный домен мономера связан с N-концевым оксигеназным доменом второго мономера. Все изоферменты NOS катализируют флавин-опосредованный электронный перенос со связанного с C-концом NADPH на гем в N-концевом домене. Электронный перенос в редуктазном домене с NADPH на флавин, а также с редуктазного домена на гем оксигеназного домена увеличивается при кальций-индуцированном связывании кальмодулина с NOS. В гемовой группе электроны используются для восстановления и активации молекулярного кислорода. Окисление L-аденина в L-цитролин происходит благодаря двум последовательным реакциям моноокисления, продуцирующим Nω-гидрокси-L-аденин (NOHLA) в качестве промежуточного продукта, синтезируя, тем самым, NO.
Как подробно описано выше, дисфункция эндотелиальной NO-синтазы (eNOS) связана с рядом заболеваний. Можно было подтвердить, что экспрессия eNOS уменьшена при различных заболеваниях, включающих сердечно-сосудистые заболевания, такие как сердечная недостаточность и инфаркт миокарда. Как ни удивительно, было установлено, что гемсвязывающий белок 1 (HEBP1) является фактором модуляции экспрессии eNOS, в частности, посредством взаимодействия с промотором eNOS. Этот факт можно использовать для выявления модулятора экспрессии eNOS, который составляет возможное терапевтическое средство для лечения заболевания, характеризующегося измененной экспрессией eNOS.
Заявленный способ скрининга на модулятор включает обеспечение тест-системы, включающей HEBP1. Как показано в контексте настоящего изобретения, HEBP1 является белком, взаимодействующим с промотором eNOS и, тем самым, изменяющим экспрессию eNOS. HEBP1 также называют гемсвязывающим белком 1, HBP, HEBP или p22HBP. Предполагается, что HEBP1 может связывать свободные порфиногены, которые могут присутствовать в клетке, и, таким образом, способствовать удалению этих потенциально токсичных соединений. Он связывается с высоким сродством с одной молекулой гема или порфиринов, при этом он обладает схожим сродством к металлопорфиринам, свободным порфиринам и N-метилпротопорфирину.
Аминокислотная последовательность белка человека состоит из 189 аминокислот и доступна в PubMed под входящим № NP 057071. Однако белок HEBP1 может также происходить из любого другого вида, и уже была опубликована последовательность белков HEBP1 других видов. Примеры включают Mus musculus (входящий № NP 038574, упоминаемый как Hebpl), Pan Troglodytes (входящий № XP_528742, упоминаемый как LOC473371), Gallus gallus (входящий № NP 001025925, упоминаемый как RCJMB04_2k3), Canis familiaris (входящий № XP_534884, упоминаемый как NOC477690) и Rattus norwegicus (входящий № XP_342776, упоминаемый как HEBP1_предсказанный).
Помимо любого природного варианта HEBP1, такого как видовой вариант или вариант сплайсинга, также могут использоваться модифицированные белки HEBP1. Следует отметить, что модифицированный белок HEBP1 или вариант HEBP1 является функционально активным вариантом в том отношении, что у варианта сохраняется его биологическая функция, заключающаяся во взаимодействии с промотором eNOS и модулировании экспрессии eNOS. Предпочтительно, определяют сохранение биологической функции, например регуляции экспрессии eNOS, составляющее по крайней мере 50%, предпочтительно по крайней мере 60%, более предпочтительно по крайней мере 70%, 80% или 90%, еще предпочтительнее 95% модуляторной активности природного HEBP1. Биологическую активность можно определить как описано в примерах, в частности в примерах 1, 2, 3 или 4 (например, используя линию клеток с геном репортера, находящимся под контролем промотора eNOS, такую как EA.crs03, или меченные биотином энхансеры транскрипции, такие как A012 или A013, или ОТ-ПЦР для определения уровней мРНК, такой как мРНК для eNOS, или гашение триптофановой флуоресценции, или поляризации флуоресценции, или модели на животных).
Вариантом может быть молекула с доменом, состоящим из природного белка HEBP1, и по крайней мере один дополнительный компонент. Например, белок может быть соединен с маркером, таким как метка, используемая для очистки (например, метка в виде 6 His (или гексаHis), Strep-метка, HA-метка, метка в виде c-myc или метка в виде глутатион-S-трансферазы (GST)). В случае необходимости, например, в высокой степени очищенного белка или варианта HEBP1 могут использоваться два или множество маркеров (например, комбинация вышеуказанных маркеров или меток). В этом случае белки очищают с использованием двух или более отдельных хроматографических стадий, в каждом случае используя сродство первой, а затем второй метки. Примерами таких двойных или сдвоенных меток являются GST-His-метка (глутатион-S-трансфераза, слитая с полигистидиновой меткой), 6xHis-Strep-метка (6 остатков гистидина, слитых с Strep-меткой), 6xHis-маркер100-метка (6 остатков гистидина, слитых с 12-аминокислотным пептидом MAP-киназы 2 млекопитающего), 8xHis-HA-метка (8 остатков гистидина, слитых с меткой в виде эпитопа гемагглютинина), His-MBP (His-метка, слитая со связывающим мальтозу белком), FLAG-HA-метка (FLAG-метка, слитая с меткой в виде эпитопа гемагглютинина) и FLAG-Strep-метка. Маркер мог бы использоваться для обнаружения белка с меткой, при этом могли бы использоваться специфические антитела. Подходящие антитела включают антитела против HA (такие как 12CA5 или 3F10), против 6 His, против c-myc и против GST. Кроме того, белок HEBP1 можно было бы соединить с маркером отличного класса, таким как флуоресцентный маркер или радиоактивный маркер, который позволяет обнаружить HEBP1. В другом варианте осуществления HEBP1 мог бы быть частью гибридного белка, в котором вторая часть, такая как белковый компонент, обладающий ферментативной активностью, могла бы использоваться для обнаружения.
В другом варианте осуществления настоящего изобретения вариантом HEBP1 мог бы быть фрагмент HEBP, который сохраняет способность взаимодействовать с промотором eNOS и регулировать экспрессию eNOS. Он может включать белки HEBP1 с короткими C- и/или N-концевыми делециями (например, делециями самое большее 20, 19, 18, 17, 16, 15, 14, 13, 12, 11, 10, 9, 8, 7, 6 5, 4, 3, 2 или 1 аминокислоты). Кроме того, фрагмент HEBP1 может быть, кроме того, модифицированным, как подробно описано выше в отношении белка HEBP1.
Альтернативно или дополнительно, белок HEBP1 или его вариант, описываемый выше, может содержать одну или несколько аминокислотных замен, в частности, в областях, не участвующих во взаимодействии с промотором eNOS и регулировании экспрессии eNOS. Однако предпочтительными являются консервативные аминокислотные замены, при которых аминокислоту замещают химически подобной аминокислотой. Характерные консервативные замены находятся в группе алифатических аминокислот, в группе аминокислот, имеющих боковые цепи с алифатическими гидроксильными группами, в группе аминокислот, имеющих кислотные остатки, в группе амидных производных, в группе аминокислот с основными остатками или в группе аминокислот, имеющих ароматические остатки. Белок или фрагмент HEBP1 или его вариант с заменой может быть модифицирован, как подробно описано выше в отношении белка, или фрагмента, или варианта HEBP1. Ниже в описании настоящего изобретения все детали, приведенные в отношении белка HEBP1, также относятся к его функционально активным вариантам, кроме случаев, оговоренных особо.
Однако наиболее предпочтительно, если белком HEBP1 является природный белок HEBP1, еще предпочтительнее природный белок HEBP1 человека.
Как подробно описано выше, тест-система содержит белок HEBP1 или его функционально активные варианты. Тест-система может также включать дополнительные элементы, такие как средства для выявления эффекта модулятора на тест-систему для идентификации агента в качестве модулятора экспрессии eNOS. Подходящие средства для выявления эффекта модулятора подробно описываются по всему описанию настоящего изобретения. Тест-система может находиться в клеточной системе или бесклеточной системе в зависимости от преобладающих условий.
В случае способа настоящего изобретения тест-систему, содержащую HEBP1 или его функционально активный вариант, приводят в контакт с агентом. Агентом, исследуемым с помощью способа настоящего изобретения, может быть любое исследуемое вещество или исследуемое соединение любой химической природы. Он может быть уже известен в качестве лекарственного средства для заболевания. Альтернативно, он может быть известным химическим соединением, но для которого неизвестно, обладает ли оно терапевтическим эффектом, в другом варианте осуществления, и соединением может быть новое или пока неизвестное химическое соединение. Агентом может быть также смесь исследуемых веществ или исследуемых соединений.
В одном из вариантов осуществления способа скрининга по настоящему изобретению исследуемое вещество находится в виде библиотеки химических соединений. Библиотеки химических соединений включают множество химических соединений и были составлены из любого из множества источников, в том числе химически синтезированных молекул или природных продуктов, или были созданы с помощью способов комбинаторной химии. Они особенно подходят для высокомасштабного скрининга и могут быть составлены из химических соединений конкретной структуры или соединений конкретного организма, такого как растение. В контексте настоящего изобретения библиотекой химических соединений, предпочтительно, является библиотека, включающая белки и полипептиды или небольшие органические молекулы. Предпочтительно, размер небольшой органической молекулы, в частности, растворимого, неолигомерного, органического соединения составляет менее 500 Д.
В контексте настоящего изобретения тест-систему приводят в контакт с агентом в период времени и в условиях, подходящих для модулирования экспрессии eNOS и его выявления. Подходящие условия включают подходящую температуру и раствор во избежание, например, денатурации включенных белков или для поддержания жизнеспособности клеток, в случае их присутствия. Подходящие условия будут зависеть от конкретной выбранной тест-системы, и квалифицированный специалист способен выбрать такие условия, основываясь на свои общие знания.
После приведения тест-системы в контакт с агентом выявляют эффект агента на тест-систему. В последующем ряд различных систем выявления будет описан подробнее. Однако следует понимать, что они являются примерами, и могут быть также подходящими другие тест-системы.
Если агент оказывает специфический и значительный эффект на тест-систему, агент идентифицируют как модулятор экспрессии eNOS. Модулятор экспрессии eNOS в контексте настоящего изобретения означает агент, изменяющий, либо увеличивающий, либо уменьшающий, экспрессию eNOS. Предпочтительно, экспрессия eNOS увеличивается. В контексте настоящего изобретения экспрессия eNOS является модифицированной, т.е. уменьшенной или предпочтительно увеличенной, по сравнению с контролем, если экспрессия eNOS в соответствующей клетке, приведенной в контакт с модулятором, значительно ниже или выше, соответственно, экспрессии eNOS в контроле (т.е. той же самой клетке, не приведенной в контакт с модулятором). Квалифицированному в данной области специалисту известны статистические способы оценки того, являются две величины статистически отличными друг от друга, такие как проверка по критерию Стьюдента или проверка по критерию хи-квадрат.
В предпочтительном варианте осуществления экспрессия eNOS составляет по крайней мере 110%, предпочтительно по крайней мере 125%, более предпочтительно по крайней мере 150%, 160%, 170%, 180% или 190%, еще предпочтительнее по крайней мере 200% и наиболее предпочтительно по крайней мере 300% от контроля.
Однако для способа по настоящему изобретению не требуется, чтобы эффект модулятора на экспрессию eNOS определялся в этом способе. Следует отметить, что способ скрининга может включать или может не включать стадию, на которой измеряется экспрессия eNOS. Альтернативно измерению экспрессии eNOS, могут использоваться способы выявления, свидетельствующие о модуляции экспрессии eNOS. Особенно в случае высокомасштабного скрининга предпочтительным могло бы быть использование очень легкой и надежной системы выявления, которая включает минимальное количество компонентов. В одном из вариантов осуществления настоящего изобретения тест-система может лишь включать белок HEBP1 (или его функционально активный вариант) и средство для выявления связывания агента/модулятора с белком в отсутствие дополнительных компонентов передачи сигнала, в которую вовлечены HEBP1 и eNOS. Такой системой может быть, например, система, в которой либо исследуемый агент, либо белок HEBP1 или его функционально активный вариант иммобилизован на носителе. Связывание агента с белком HEBP1 или его функционально активным вариантом можно выявить, в силу чего неиммобилизованный партнер по связыванию метят поддающимся обнаружению маркером. Подвергаемый иммобилизации компонент может быть иммобилизован на одном материале или на множестве различных материалов, которые способны к связыванию биомолекулы или ряда биомолекул, исходя из их физических характеристик. Такие материалы включают, но ими не ограничиваются, материалы для анионообменной хроматографии, материалы для катионообменной хроматографии, металлохелаты, пептиды, антитела, полимеры (синтетические или природные), бумагу и т.д.
Иммобилизованный компонент можно привести в контакт с мобильным (т.е. неиммобилизованным) возможным партнером по связыванию, при этом несвязанный мобильный партнер по связыванию удаляют по истечении периода времени, достаточного для создания возможности для связывания. Связывание мобильного и иммобилизованного компонентов можно обнаружить по присутствию маркера мобильного партнера по связыванию в месте иммобилизации иммобилизованного партнера. Например, ряд различных агентов, таких как белки, можно было бы иммобилизовать в многолуночном планшете и можно было бы проинкубировать с меченым белком HEBP1. В тех лунках, в которых обнаруживается маркер, произошло связывание между агентом и белком HEBP1. Соответствующий агент можно идентифицировать как потенциальный модулятор экспрессии eNOS.
Компонент, в частности белок, можно пометить рядом способов, для того чтобы создать возможность для адекватного выявления или очистки. Обычные способы мечения могут использоваться для мечения одной или более функциональных групп на поверхности компонента. В случае белка ими могли бы быть, например, первичные аминогруппы, присутствующие на N-конце каждой полипептидной цепи и в боковой цепи остатков лизина; сульфгидрильные группы, присутствующие на остатках цистеина, сделанные доступными посредством обработки дисульфидных связей восстанавливающим агентом или сделанные имеющимися в наличии посредством модификации остатков лизина с помощью такого реагента, как SATA; или углеводные группы, обычно присутствующие в Fc-области антител, которые могут быть подвергнуты окислению с созданием активных альдегидов для соединения. Компонент или белок можно пометить с использованием ряда различных агентов, таких как биотин (в случае реакции авидина с биотином), ферменты, активированные флуоресцентные красители для мечения аминов, сульфгидрильных или других функциональных групп, например, FITC, флуоресцеином, родамином, синими красителями или Alexa Fluos. Также может использоваться радиоактивная метка, такая как 3H, 32P, 35S, 125I или 14C, а также обычные ферментативные метки, включающие пенициллиназу, пероксидазу хрена и щелочную фосфатазу.
В случае способа настоящего изобретения может использоваться любой подходящий способ выявления. Подходящие способы можно выбрать в зависимости от характеристик тест-системы и исследуемых агентов. Как подробно описано выше, HEBP1 вовлечен в передачу сигнала при экспрессии eNOS. Соответственно, можно определить взаимодействие агентов с HEBP1 (или его вариантом) или компонентом, предшествующим HEBP1 при передаче сигнала. Взаимодействие можно определить непосредственно или опосредованно. «Непосредственно» означает, что определяют связывание агента с HEBP1 (например, используя меченый маркер для выявления комплексов HEBP1/агент). «Опосредовано» означает, что определяют эффект передачи сигнала от HEBP1 (например, активность промотора eNOS, уровень мРНК для eNOS или количество белка eNOS). Подходящие способы подробно описаны, например, в примерах.
В первом случае определяют взаимодействия агента с белком. В данной области известен ряд анализов, в которых может использоваться тест-система и к которым может быть приспособлена тест-система. Это могут быть гетерогенные или гомогенные анализы. Как используется в настоящем описании, гетерогенным анализом является анализ, который включает одну или более стадий промывки, тогда как в гомогенном анализе такие стадии промывки не нужны. Реагенты и соединения только смешивают и измеряют.
В варианте осуществления анализом является ELISA (иммуноферментный твердофазный анализ), DELFIA (усиленный диссоциацией лантанидный флуоресцентный иммуноанализ), SPA (сцинтилляционный анализ сближения), анализ с использованием системы FlashPlate, анализ с использованием FRET (резонансного переноса энергии флуоресценции), анализ с использованием TR-FRET (резонансного переноса энергии флуоресценции с временным разрешением), анализ на основе FP (поляризации флуоресценции), ALPHA (усиленный люминесцентный гомогенный анализ сближения), анализ на основе EFC (комплементации фрагментов фермента), двугибридный скрининг или анализ коиммунопреципитации.
Анализы на основе ELISA (иммуноферментного твердофазного анализа) предлагаются различными компаниями. Он представляет собой биохимический способ, используемый для обнаружения присутствия антитела или антигена в образце. Выполнение ELISA включает использование по крайней мере одного антитела со специфичностью в отношении конкретного антигена (например, сегмента первого или второго белка). Обычно на поверхности иммобилизуют неизвестное количество антигена в образце. Затем всю поверхность заливают конкретным антителом. Это антитело связано с ферментом, который создает поддающийся обнаружению сигнал, такой как изменение цвета или флуоресценция. Например, образец с неизвестным количеством антигена подвергают иммобилизации на твердой подложке (обычно титрационном микропланшете) либо неспецифически (с помощью адсорбции к поверхности), либо специфически (с помощью улавливания другим антителом, специфичным в отношении этого же антигена, в «сэндвич»-ELISA). После иммобилизации антигена добавляют предназначенное для обнаружения антитело, образующее комплекс с антигеном. Предназначенное для обнаружения антитело может быть ковалентно связано с ферментом или само может выявляться с помощью второго антитела, которое связано с ферментом благодаря биоконъюгации. В промежутках между всеми стадиями планшет обычно промывают слабым раствором детергента для удаления любых белков или антител, которые связались неспецифически. После конечной стадии промывки планшет проявляют посредством добавления субстрата для фермента с созданием видимого сигнала, который указывает на количество антигена в образце. В более ранних ELISA используются хромогенные субстраты, хотя в более новых анализах применяются флуорогенные субстраты с намного большей чувствительностью.
Анализы на основе DELFIA (усиленного диссоциацией лантанидного флуоресцентного иммуноанализа) являются твердофазными анализами. Антитело обычно метят европием или другим лантанидом и флуоресценцию европия выявляют после вымывания несвязанных меченных европием антител.
В SPA (сцинтилляционном анализе сближения) и в анализе с использованием системы FlashPlate обычно используются взаимодействия биотин/авидин для улавливания меченных радиоактивным изотопом субстратов. Обычно реакционная смесь включает киназу, биотинилированный пептидный субстрат и γ-[33P]ATP. После реакции биотинилированные пептиды улавливаются стрептавидином. При выявлении с использованием SPA стрептавидин связан с содержащими сцинтиллятор бусинками, тогда как при выявлении с использованием FlashPlate стрептавидин связан с внутренней частью лунки содержащих сцинтиллятор микропланшетов. После иммобилизации меченный радиоактивным изотопом субстрат находится достаточно близко к сцинтиллятору для стимуляции излучения света.
Резонансный перенос энергии флуоресценции (FRET) характеризует перенос энергии между двумя хромофорами без излучения. Хромофор-донор в своем возбужденном состоянии может переносить энергию с помощью механизма безызлучательного дальнодействующего диполь-дипольного взаимодействия на флуорофор-акцептор в непосредственной близости (обычно <10 нм). Поскольку обе молекулы являются флуоресцентными, перенос энергии часто называют «резонансным переносом энергии флуоресценции», хотя на самом деле энергия не переносится с помощью флуоресценции. FRET является способом, подходящим для выявления и количественного анализа взаимодействий белок-агент, взаимодействий белок-белок, взаимодействий белок-ДНК и белок-конформационные изменения. Для анализа связывания белка с агентом, одного белка с другим белком или белка с ДНК одну из молекул метят донором, а другую - акцептором и эти меченные флуорофорами молекулы смешивают. Если они присутствуют в несвязанном состоянии, эмиссия донора выявляется после его возбуждения. После связывания молекул донор и акцептор сближаются, и преимущественно отмечается эмиссия акцептора вследствие межмолекулярного FRET от донора к акцептору. Подходящие соседние элементы для FRET известны в данной области, и квалифицированный специалист сможет выбрать подходящую комбинацию меток для обоих антител. Как используется в настоящем описании в отношении донора и соответствующего акцептора, «соответствующий» относится к акцепторной флуоресцентной составляющей, имеющей спектр эмиссии, который перекрывается со спектром возбуждения донора. Однако оба сигнала должны быть разделимыми друг от друга. Соответственно, максимум для длины волны в спектре эмиссии акцептора должен предпочтительно превышать на по крайней мере 30 нм, более предпочтительно по крайней мере 50 нм, например, по крайней мере 80 нм, по крайней мере 100 нм или по крайней мере 150 нм максимум для длины волны в спектре возбуждения донора (смотри также пример 3.1).
Характерные донорные флуоресцентные составляющие, которые могут использоваться с различными акцепторными флуоресцентными составляющими в технологии FRET, включают флуоресцеин, люцеферовый желтый, B-фикоэритрин, 9-акридинизотиоцианат, люцеферовый желтый VS, производные 4-ацетамидо-4'-изотиоцианатостильбен-2,2'-дисульфокислоты, 7-диэтиламино-3-(4'-изотиоцианатофенил)-4-метилкумарин, сукцинимдил-1-пиренбутират и производные 4-ацетамидо-4'-изотиоцианатостильбен-2,2'-дисульфокислоты. Характерные акцепторные флуоресцентные составляющие, зависящие от используемых донорных флуоресцентных составляющих, включают LC-Red 610, LC-Red 640, LC-Red 670, LC-Red 705, Cy5, Cy5.5, сульфонилхлорид лиссамин-родамина B, тетраметилродамин изотиоцианат, родамин x изотиоцианат, эритрозин изотиоцианат, флуоресцеин, диэтилентриаминпентаацетат или другие хелаторы ионов лантанидов (например, европия или тербия). Донорные и акцепторные флуоресцентные составляющие можно получить, например, от Molecular Probes (Junction City, OR) или Sigma Chemical Co. (St. Louis, MO).
Альтернативно, резонансный перенос энергии флуоресценции с временным разрешением (TR-FRET) может использоваться для тест-системы по настоящему изобретению. В TR-FRET объединены TRF (флуоресценция с временным разрешением) и принцип FRET. Это сочетание объединяет преимущества TRF, заключающиеся в низком уровне фона, и гомогенный формат анализа FRET. Тогда как FRET уже был описан выше, TRF использует преимущества уникальных свойств лантанидов или другого донора с длительной продолжительностью эмиссии. Подходящие для TR-FRET доноры включают, среди прочих, хелаторы лантанидов (криптаты) и некоторые другие металлолигандные комплексы, которые могут иметь продолжительность флуоресценции во временном диапазоне от микро- до миллисекунд и которые поэтому также создают возможность для того, что перенос энергии происходит в измерениях от микро- до миллисекунд. Испускающие флуоресценцию хелаторы лантанидов использовались в качестве доноров энергии в конце семидесятых. Часто используемые лантаниды включают самарий (Sm), европий (Eu), тербий (Tb) и диспрозий (Dy). По причине их специфических фотофизических и спектральных свойств комплексы лантанидов представляют большой интерес для применения флуоресценции в биологии. В частности, они имеют большой стоксов сдвиг и очень длительные продолжительности эмиссии (от микросекунд до миллисекунд) по сравнению с более традиционными флуорофорами.
Обычно органические хромофоры используются в качестве акцепторов. Они включают аллофикоцианин (APC). Соответствующие детали в отношении TR-FRET, а также акцепторов приведены в WO 98/15830.
Анализы на основе поляризации флуоресценции (FP) представляют собой анализы, в которых используется поляризованный свет для возбуждения флуоресцентных пептидов-субстратов в растворе. Эти флуоресцентные пептиды свободны в растворе и поворачиваются, что вызывает превращение излучаемого света в деполяризованный. Если пептид-субстрат связывается с более большой молекулой, то его скорость поворачивания значительно уменьшается, и излучаемый свет остается в высокой степени поляризованным (смотри также пример 4.3).
В другом варианте осуществления настоящего изобретения тест-систему приспосабливают к усиленному люминесцентному гомогенному анализу сближения (ALPHA). ALPHA представляет собой анализ в растворе, который был первоначально разработан Packard BioScience. ALPHA является анализом сближения на основе люминесценции, в котором один партнер по взаимодействию присоединен к донорным бусинкам, тогда как другой партнер соединен с акцепторными бусинками, при этом диаметр обоих видов бусинок составляет лишь приблизительно 250 нм. Соединение фотосенсибилизатор внедрено в донорную бусинку. С помощью этого соединения после освещения лазерным светом при длине волны, составляющей приблизительно 680 нм, кислород окружающей среды превращается в богатый энергией, короткоживущий атомарный кислород. Если вблизи нет акцепторной бусинки, атомарный кислород затухает без создания сигнала. Если донорная и акцепторная бусинки сближены (приблизительно 250 нм) посредством биологического взаимодействия присоединенных биомолекул, атомарный кислород, высвобождаемый донорной бусинкой, вызывает каскад люминесценции/флуоресценции в соседней акцепторной бусинке, приводя к в высокой степени усиленному сигналу в 520-620 нм диапазоне. Люминесцентный сигнал выявляют в подходящем считывающем устройстве. Ради большего количества деталей, в отношении методов ALPHA смотри Ullman et al, 1994, Proc. Natl. Acad. Sci. USA 91, 5426-5430.
Анализы на основе EFC (комплементации фрагментов фермента) или эквивалентные анализы могут использоваться, в частности, для высокомасштабного скрининга соединений. Анализ на основе EFC основан на сконструированном ферменте β-галактозидазе, который состоит из двух фрагментов: ферментного акцептора (EA) и ферментного донора (ED). Когда два фрагмента разделены, β-галактозидазной активности нет, но когда фрагменты находятся вместе, они соединяются (служат дополнением друг друга) с образованием активного фермента. В анализе на основе EFC используется конъюгат ED-аналит, в котором аналит может быть распознан специфическим связывающим белком, таким как антитело или рецептор. В отсутствие специфического связывающего белка конъюгат ED-аналит способен к комплементации EA с образованием активной β-галактозидазы, создавая положительный люминесцентный сигнал. Если конъюгат ED-аналит связывается специфическим связывающим белком, комплементация с помощью EA не допускается, и сигнала нет. В случае предоставления (в образце) свободного аналита он будет конкурировать с конъюгатом ED-аналит за связывание со специфическим связывающим белком. Свободный аналит будет разрешать конъюгату ED-аналита дополнять EA, создавая сигнал, в зависимости от количества свободного аналита, присутствующего в образце.
Двугибридный скрининг представляет собой метод молекулярной биологии, используемый для обнаружения белок-белковых взаимодействий посредством анализа физических взаимодействий (таких как связывание) между двумя белками. Предпосылкой анализа является активация 3' гена(ов) репортера при связывании транскрипционного фактора с 5' активирующей последовательностью. Для целей двугибридного скрининга транскрипционный фактор разделен на два отдельных фрагмента, называемых связывающим доменом (BD) и активирующим доменом (AD). BD является доменом, ответственным за связывание с UAS, а AD является доменом, ответственным за активацию транскрипции.
Коиммунопреципитация может использоваться для идентификации белковых комплексов посредством преципитации одного белка, который, как полагают, находится в комплексе, добавочные члены комплекса также улавливаются и могут быть идентифицированы. Белковые комплексы после связывания со специфическим антителом удаляют из основной массы раствора посредством захвата связывающимся с антителом белком, присоединенным к твердой подложке, такой как агарозный шарик. Эти связывающиеся с антителами белки (белок A, белок G, белок L) были исходно выделены из бактерий и распознают широкий ряд антител. После первоначального захвата белка или белкового комплекса твердую подложку промывают несколько раз для удаления любых белков, связавшихся благодаря антителу неспецифически и непрочно. После промывки подвергнутый преципитации белок(и) элюируют и анализируют, используя гель-электрофорез, масс-спектрометрию, Вестерн-блоттинг или любое количество других способов для идентификации составных частей комплекса. Таким образом, коиммунопреципитация является стандартным способом для оценки белок-белкового взаимодействия. Подходящая тест-система, в которой вызывается коиммунопреципитация, описана в примере 1.
В другом предпочтительном варианте осуществления настоящего изобретения средство для выявления взаимодействия между первым и вторым белком может быть приспособлено к определению одного или нескольких компонентов, следующих за HEBP1 в каскаде передачи сигнала. Определение может включать определение концентрации мРНК для eNOS или белка eNOS, или белка-репортера или мРНК для него. Концентрация может определяться в ответ на возможный модулятор, описываемый выше. Средства и способы для определения концентраций одной(го) или нескольких нуклеиновых кислот или белков хорошо известны квалифицированному специалисту и включают те, которые включают ОТ-ПЦР, масс-спектрометрии или FRET (смотри также примеры).
Приводимые в качестве примеров тест-системы и их применение описаны в примерах 1-4.
Предпочтительно, способ адаптируют к высокомасштабному скринингу. В этом способе большое количество соединений скринируют относительно агентов в либо бесклеточных анализах, либо анализах на основе цельных клеток. Как правило, эти скрининги выполняют в 96-луночных планшетах, используя технологии, базируемые в автоматизированных, роботизированных местах, или в форматах ранжированных рядов («чипов») с высокой плотностью размещения.
В конкретном варианте осуществления настоящего изобретения тест-система настоящего изобретения включает
- промотор eNOS и/или
- один или несколько транскрипционных факторов для промотора eNOS.
Промотор eNOS представляет собой регуляторный район гена eNOS, который делает возможной транскрипцию гена. Как и в случае многих других конститутивно экспрессируемых генов, в промоторе eNOS отсутствует классический TATA-бокс. Однако может быть идентифицирован ряд консервативных цис-элементов для Sp1, Ets, GATA, NF-1, AP-1, гемодинамического давления и стерола. Известными стимулами транскрипции eNOS являются, например, гемодинамическое давление протекающей крови, гипоксия и такие агенты как эстроген и лизофосфатилхолин. Транскрипция eNOS может уменьшаться из-за оксигенированного липопротеина низкой плотности (oxLDL) и фактора α некроза опухолей.
Схематическая иллюстрация сайтов связывания транскрипционных факторов в генах eNOS человека представлена Searles (Searles, 2006, Am. J. Physiol. Cell Physiol. 291: C803-C860). С помощью детального анализа проксимального канонического промотора гена eNOS человека были идентифицированы два положительных регуляторных домена (PRDI и II) в положениях -104/-95 и -144/- 115, соответственно, относительно начала транскрипции. Члены семейства Ets, Sp1, варианты Sp3, MAZ и YY1 были идентифицированы в качестве регулирующих транскрипцию факторов в пределах этого района. В PRDI и II можно было выявить положительные в отношении транскрипции eNOS белок-ДНК и белок-белковые взаимодействия, что означает, что транскрипция eNOS точно регулируется сложным взаимодействием транскрипционных факторов. Помимо того в положении -230/-227 находится связывающий сайт GATA, который важен для базальной транскрипции eNOS. Кроме этих цис-элементов была идентифицирована последовательность энхансера 269NT, находящаяся на расстоянии 4,9 т.п.о. в направлении 3'-5' от начала транскрипции, функционирование которой регулируется AP2- MAZ-, Sp1- и Ets-родственными факторами в нуклеопротеиновых комплексах. Однако описанные выше транскрипционные факторы являются в основном повсеместно экспрессируемыми белками, не подходящими для избирательной регуляции или контроля eNOS.
На посттранскрипционном уровне eNOS может регулироваться действующими в цис-положении РНК-элементами, кальмодулином и внутриклеточным Ca2+, действующим в качестве активатора синтеза NO. Кроме того, eNOS может также активироваться при аллостерическом связывании белка 90 теплового шока, индуцируемого, например, под действием гистамина, VEGF или гемодинамического давления. Активность eNOS может быть также модулирована фосфорилированием остатков серина или треонина. Вовлеченными в это протеинкиназами являются, например, протеинкиназа A, протеинкиназа C, активируемая аденозинмонофосфатом протеинкиназа, Ca2+/CaM-зависимая протеинкиназа и серин/треонинкиназа AKT. Фосфорилирование Ser1177 может индуцировать гемодинамическое давление, VEGF и эстрадиол, и оно увеличивает активность eNOS. Однако фосфорилирование Thr497 производит уменьшение активности. Процессы дефосфорилирования опосредуются фосфатазами PPA2 и PP1.
Кроме того, было установлено, что на активность eNOS может отрицательно влиять белок-белковое взаимодействие, например, благодаря связи с C-концевым доменом связанных с G-белками рецепторов (например, рецептора брадикинина B2). В анализах с использованием дрожжевой двугибридной системы был идентифицирован белок с мол. м. 34 кДа, который был назван NOSIP (взаимодействующим с eNOS белком). NOSIP связывается с оксигеназным доменом eNOS и активирует транслокацию фермента из кавеул во внутриклеточные области, что приводит к снижению синтеза NO. Оксигеназный домен eNOS использовался в качестве «приманки» (насадки) для идентификации взаимодействующего с eNOS белка NOSTRIN (индуктора перемещения eNOS). Сверхэкспрессия NOSTRIN приводит к транслокации eNOS из плазматической мембраны в сосудистую структуру и к снижению выброса NO. Можно было установить, что NOSTRIN образует трехкомпонентный комплекс с eNOS и кавеолином-1. Кроме того, он ответственен за привлечение белков-медиаторов, таких как динамин-2.
Промоторы eNOS эндотелиальных клеток человека, быка, мыши и свиньи были клонированы и продемонстрировали высокую степень гомологии последовательностей. Ген eNOS состоит из 26 экзонов и охватывает приблизительно 21 т.п.о. геномной ДНК на хромосоме 7Q35-36. мРНК для eNOS 4052NT конститутивно экспрессируется в эндотелиальных клетках и является очень стабильной.
Кроме HEBP1 может присутствовать один или несколько транскрипционных факторов для промотора eNOS и/или один или несколько вышеотмеченных факторов, необходимых для осуществления транскрипции.
Тест-система по настоящему изобретению может включать клетку, в частности клетку млекопитающего, особенно клетку человека. Примеры подходящих клеток включают эндотелиальные клетки. Эти клетки могут быть, например, «первичными» клетками, такими как эндотелиальная клетка пупочной вены человека (HUVEC) (смотри пример 1), или такой линией клеток, как клетки EA.hy926 (смотри пример 1). Однако можно использовать любую другую клетку или линию клеток, необязательно генетически модифицированную для включения белка HEBP1 или компонентов, необходимых для выявления эффекта.
В предпочтительном способе настоящего изобретения эффект определяют с помощью флуоресценции. Подходящие способы подробно описаны выше и могут включать флуоресцентный маркер, FRET, поляризацию флуоресценции, подробно описанные в настоящем описании.
В другом предпочтительном варианте осуществления настоящего изобретения способ используется для отбора лекарственного средства для профилактики и/или лечения заболевания, связанного с дисфункцией eNOS, в частности сердечно-сосудистого заболевания, такого как инфаркт миокарда и/или сердечная недостаточность.
Приводимые в качестве примеров заболевания, связанные с дисфункцией eNOS, подробно описаны выше. Однако предпочтительными являются сердечно-сосудистые заболевания, особенно инфаркт миокарда и/или сердечная недостаточность. В соответствии с настоящим изобретением термин «профилактика заболевания» относится к снижению риска развития преобладающего заболевания, тогда как термин «лечение заболевания» относится к уменьшению интенсивности симптомов преобладающего заболевания, замедлению течения заболевания и т.д. Профилактика или предупредительная мера является способом предупреждения повреждения, болезни или заболевания в первую очередь. Лечение используется после того, как проблема со здоровьем уже возникла. Лечение используется в отношении проблемы со здоровьем и может привести к ее устранению, но чаще лечение лишь уменьшает проблему до тех пор, пока лечение продолжится. Лечение с полным излечением является подмножеством лечения, при котором полностью обращают в течение заболевания или навсегда прекращают проблемы со здоровьем.
Другой объект настоящего изобретения относится к способу диагностирования сердечно-сосудистого заболевания у индивида, включающему
- определение уровня мРНК для HEBP1 или белка HEBP1 в образце, полученном от индивида,
причем повышенный или пониженный уровень мРНК для HEBP1 или белка HEBP1 относительно контроля служит признаком сердечно-сосудистого заболевания.
Как показано в примерах, измененный уровень белка HEBP1 связан с сердечно-сосудистым заболеванием, в частности сердечной недостаточностью и/или инфарктом миокарда. Соответственно, уровень мРНК для HEBP1 или белка HEBP1 в образце, полученном от индивида, можно определить для выявления уровня, отличающегося от контрольного уровня (например, у здорового индивида или уровня, определенного в группе здоровых индивидов), который служит признаком вышеуказанного заболевания.
Термины «образец от индивида» и «исследуемый образец» относятся ко всем биологическим жидкостям и экскрециям, выделенным от любого конкретного индивида, в частности человека. В контексте настоящего изобретения такие образцы включают, но ими не ограничиваются, кровь, сыворотку крови, плазму крови, аспират из соска молочной железы, мочу, сперму, семенную жидкость, семенную плазму, простатическую жидкость, экскрет, слезы, слюну, пот, биопсию, асцитическую жидкость, спинномозговую жидкость, молоко, лимфу, образцы жидкости после промывки бронхов и другие образцы промывной жидкости или образцы тканевых экстрактов. Обычно кровь или образцы тканей сердечно-сосудистой системы являются предпочтительными исследуемыми образцами для применения в связи с настоящим изобретением.
Уровень мРНК для HEBP1 или белка HEBP1 можно определить с помощью ряда способов, в том числе способами, описанными в настоящем документе, в частности, также в связи со способом скрининга настоящего изобретения и в примерах.
В альтернативном варианте может использоваться масс-спектрометрия. Термин «масс-спектрометрия» относится к использованию источника ионизации для образования ионов в газовой фазе на основе образца на поверхности и выявлению ионов в газовой фазе с помощью масс-спектрометрии. Термин «масс-спектрометрия с использованием лазерной десорбции» относится к использованию лазера в качестве источника ионизации для образования ионов в газовой фазе на основе образца на поверхности и выявлению ионов в газовой фазе с помощью масс-спектрометрии. Предпочтительным способом масс-спектрометрии для биомолекул, таких как HEBP1, является масс-спектрометрия с использованием лазерной десорбции/ионизации из матрицы или MALDI. При использовании MALDI аналит обычно смешивают с матричным материалом, который при высыхании кристаллизуется вместе с аналитом. Матричный материал поглощает энергию от источника энергии, которая в противном случае разрушила бы неустойчивые биомолекулы или аналиты. Другим предпочтительным способом является масс-спектрометрия с использованием усиленной поверхности лазерной десорбции/ионизации или SELDI. При использовании SELDI поверхность, на которую наносят аналит, играет активную роль в захвате аналита и/или его десорбции. В контексте настоящего изобретения образец включает биологический образец, который, возможно, был подвергнут хроматографической или другой химической обработке, и подходящий матричный субстрат.
В масс-спектрометрии «кажущаяся молекулярная масса» относится к значению отношения молекулярной массы (в дальтонах) к заряду, m/z, выявленных ионов. То, каким образом получают величину кажущейся молекулярной массы, зависит от типа используемого масс-спектрометра. При использовании времяпролетного масс-спектрометра кажущаяся молекулярная масса является зависимостью времени от ионизации до выявления. Термин «сигнал» относится к любому ответу, порожденному исследуемой биомолекулой. Например, термин «сигнал» относится к ответу, порожденному биомолекулой при ударе о детектор масс-спектрометра. Интенсивность сигнала коррелирует с количеством или концентрацией биомолекулы. Сигнал определяется двумя величинами: величиной кажущейся молекулярной массы и величиной интенсивности, порожденной, как описано. Величина массы является атомарной характеристикой биомолекулы, тогда как величина интенсивности соответствует определенному количеству или концентрации биомолекулы с соответствующей величиной кажущейся молекулярной массы. Таким образом, «сигнал» всегда относится к свойствам биомолекулы.
Альтернативно, присутствие и количество HEBP1 в исследуемом образце можно получить и количественно определить, используя обычные методы, известные квалифицированным в данной области специалистам. Квалифицированному специалисту в данной области хорошо известны, например, способы количественного определения антигенов или антител в исследуемых образцах. Например, присутствие и количество HEPB1 в исследуемом образце можно определить, используя иммуноанализ. Иммуноанализ обычно включает: (a) обеспечение антитела (или антигена), которое специфически связывается с биомаркером; (b) приведение исследуемого образца в контакт с антителом или антигеном и (c) выявление присутствия комплекса антитела, связанного с антигеном, в исследуемом образце или комплекса антигена, связанного с антителом, в исследуемом образце. Приводимое в качестве примера антитело описано в примере. Однако может быть продуцировано альтернативное антитело в соответствии со способами, известными квалифицированному специалисту.
После получения антитела HEBP1 можно выявить и/или количественно определить, используя любой из ряда общепризнанных иммунологических анализов связывания. Анализы, которые могут использоваться в настоящем изобретении, включают, например, иммуноферментный твердофазный анализ (ELISA), который также известен как «сэндвич-анализ», иммуноферментный анализ (EIA), радиоиммуноанализ (RIA), иммунофлуоресцентный анализ (FIA), иммунохемилюминесцентный анализ (CLIA), импульсный иммуноанализ (CIA), иммуноферментный анализ на фильтрационном материале (MEIA), твердофазный иммуноанализ с флуоресцентным усилением (FLISA), реакции агглютинации и мультиплексные иммунофлуоресцентные анализы (такие как LuminexTM LabMAP) и т.д. Ради обзора обычных иммуноанализов смотри также Methods in Cell Biology: Antibodies in Cell Biology, volume 37 (Asai, ed. 1993); Basic and Clinical Immunology (Stites & Terr, eds., 7th ed. 1991).
Обычно исследуемый образец, полученный от индивида, можно привести в контакт с антителом, которое специфически связывает антиген. Перед приведением антитела в контакт с исследуемым образцом антитело можно необязательно закрепить на твердой подложке для облегчения промывки и последующего выделения комплекса. Примеры твердых подложек включают стекло или пластмассу в форме, например, титрационного микропланшета, предметного стекла или покровного стекла, палочки, бусинки или микробусинки.
После инкубации образца с антителами смесь отмывают и выявляют образованный комплекс антитело-антиген. Это можно осуществить посредством инкубации отмытой смеси с реагентом для детектирования. Этот реагент для детекции может быть, например, вторым антителом, которое мечено поддающейся обнаружению меткой. Что касается обнаруживаемой метки, то любая обнаруживаемая метка, известная в данной области, может быть использована. Например, обнаруживаемая метка может быть радиоактивной меткой (такая как, например, 3H, 125I, 35S, 14C, 32P и 35P), ферментативной меткой (такая как, например, пероксидаза хрена, щелочная фосфатаза, глюкозо-6-фосфатдегидрогеназа и т.п.), хемилюминесцентной меткой (такая как, например, эфиры акридиния, тиоэфиры акридиния, сульфонамиды акридиния, эфиры фенантридиния, люминал, изолюминол и т.п.), флуоресцентной меткой (такая как, например, флуоресцеин (например, 5-флуоресцеин, 6-карбоксифлуоресцеин, 3'6-карбоксифлуоресцеин, 5(6)-карбоксифлуоресцеин, 6-гексахлорфлуоресцеин, 6-тетрахлорфлуоресцеин, флуоресцеин изотиоцианат и т.п.)), родамином, фикобилипротеинами, R-фикоэритрином, квантовыми примесями (например, покрытый сульфидом цинка селенид кадмия), термометрической меткой или иммунологической меткой для полимеразной цепной реакции.
При проведении анализов могут потребоваться стадии инкубации и/или промывки после каждого объединения реагентов. Стадии инкубации могут применяться в зависимости от приблизительно 5 секунд до нескольких часов, предпочтительно от приблизительно 5 минут до приблизительно 24 часов. Однако время инкубации будет зависеть от формата анализа, биомаркера (антигена), объема раствора, концентраций и т.п. Обычно анализы проводят при температуре окружающей среды, хотя их можно проводить во всем диапазоне температур, например 10-40°С.
Предпочтительно, если уровень мРНК для HEBP1 или белка HEBP1 в полученном от индивида образце является пониженным по сравнению с контролем.
Образцом может быть любой образец, подходящий для выявления измененного уровня мРНК для HEBP1 или белка HEBP1. Однако предпочтительно уровень определяют в образце сердечно-сосудистой системы, таком как эндотелиальная клетка, особенно сердечно-сосудистой системы, или клетка сердца.
Как подробно описано выше, дисфункция eNOS и изменения в HEBP1 имеют отношение, в частности, к заболеванию сердечно-сосудистой системы. Предпочтительно сердечно-сосудистым заболеванием является сердечная недостаточность и/или инфаркт миокарда.
В соответствии с настоящим изобретением HEBP1 может использоваться для идентификации лекарственного средства для профилактики и/или лечения заболевания, связанного с дисфункцией eNOS, в частности сердечно-сосудистого заболевания. Следовательно, другим объектом настоящего изобретения является применение HEBP1 для идентификации лекарственного средства для профилактики и/или лечения заболевания, связанного с дисфункцией eNOS, в частности сердечно-сосудистого заболевания.
Однако эндотелиальная дисфункция вследствие снижения оксида азота в эндотелиальных клетках является одним из основных механизмов развития сосудистых заболеваний и часто приводит к атеросклерозу. Она является очень часто встречающейся у пациентов с сахарным диабетом, гипертензией или другими хроническими патофизиологическими состояниями. Соответственно, модулятор может использоваться при профилактике или лечении побочных эффектов хронических патофизиологических состояний, связанных с эндотелиальной дисфункцией, таких как сахарный диабет или гипертензия.
Предпочтительно, лекарственное средство изменяет, предпочтительно увеличивает, экспрессию eNOS.
Также в соответствии с настоящим изобретением HEBP1 может использоваться для выявления компонента передачи сигнала при экспрессии eNOS. Следовательно, в еще одном варианте осуществления настоящее изобретение относится к применению HEBP1 для выявления компонента передачи сигнала при экспрессии eNOS. Выявление дополнительных компонентов может осуществляться с использованием описанных в примерах способов, таких как использование HEBP1 в качестве «приманки», ранжированных микрорядов белков, киРНК (коротких интерферирующих РНК), репортерных систем, масс-спектрометрии, аффинной очистки, электрофореза в SDS-ПААГ и т.д. (смотри, в частности, примеры 1 и 2, в которых HEBP1 должен использоваться в качестве «приманки»). В этих способах HEBP1 является предпочтительно HEBP1 человека. HEBP1 может использоваться для выявления партнеров по связыванию и/или все еще неизвестных компонентов, предшествующих HEBP1 или следующих за ним при передаче сигнала. В зависимости от компонента способы, описанные выше в связи со способом скрининга настоящего изобретения, могут использоваться для количественного анализа или выявления эффектов выявляемых компонентов.
Кроме того, другой объект настоящего изобретения относится к применению HEBP1 для регулирования активности промотора eNOS. В соответствии с настоящим изобретением HEBP1 может использоваться для регулирования активности промотора eNOS. Регулирование включает увеличение, активирование, уменьшение, ингибирование и блокирование активности промотора eNOS. Регулирование активности промотора eNOS может использоваться для регуляции экспрессии eNOS в клетке, ткани или органе. Необязательно в регуляцию могут быть вовлечены дополнительные компоненты, такие как компоненты, специфически регулирующие HEBP1 или промотор eNOS, такие как AVE3085, AVE9488 или вещество 9257 (смотри примеры). Однако промотор eNOS может также быть функционально связанным (например, в результате генетического конструирования) с отличным геном (например, геном репортера или любым другим геном) для регуляции экспрессии этого гена промотором eNOS и HEBP1. Эта конструкция может использоваться для моделирования экспрессии eNOS в клетках сердечно-сосудистой системы, в частности эндотелиальных клетках, с исследовательской целью, лечебной целью или любой другой целью. Например, эта конструкция может использоваться в модели сердечно-сосудистого заболевания для исследования экспрессии eNOS или активности промотора eNOS и его регуляции. Промотор eNOS и HEBP1 могут использоваться в качестве индуцибельного или тканеспецифического промотора с базальной активностью, как это известно квалифицированному специалисту.
Настоящее изобретение не ограничивается конкретной методикой, протоколами и реагентами, представленными в настоящем описании, поскольку они могут меняться. Кроме того, используемая в настоящем описании терминология предназначена лишь для описания конкретных вариантов осуществления и, как предполагается, не ограничивает объем настоящего изобретения. Используемые в настоящем описании и прилагаемой формуле изобретения формы единственного числа включают ссылку на множественное число, если только из контекста ясно не следует иное. Также слова «включать», «содержать» и «охватывать» должны интерпретироваться как включительно, а не исключительно.
Кроме особо оговоренных случаев, все технические и научные термины и все акронимы, используемые в настоящем описании, имеют значение, одинаковое со значением, в котором они обычно понимаются специалистом со средним уровнем компетентности в области техники настоящего изобретения. Хотя любые способы и материалы, схожие с описанными в настоящем описании способами и материалами или эквивалентные им, могут использоваться при осуществлении на практике настоящего изобретения, здесь описываются предпочтительные способы и материалы.
Ниже настоящее изобретение проиллюстрировано следующими фигурами и примерами, хотя будет понятно, что эти фигуры и примеры включены только с целью иллюстрации и, как предполагается, не ограничивают объем настоящего изобретения, кроме особо оговоренных случаев.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
На фиг.1 демонстрируется валидация различных HEBP1-киРНК на уровне мРНК. Относительную экспрессию HEBP1, приведенную к таковой GAPDH, определяли через 24 ч после трансфекции клеток EA.csr03 (96-луночный формат, n=4) с использованием специфических TaqMan®-зондов с помощью количественной ОТ-ПЦР (*p<0,05 в сравнение с siLV2-контролем). киРНК 25 для ингибирования HEBP1 способна к ингибированию транскрипции гена HEBP1.
На фиг.2 демонстрируется выключение HEBP1, используя киРНК, и последующее измерение активации промотора под действием AVE3085. Этот эксперимент выполняли в 96-луночном формате (n=4). Измеренную хемилюминесценцию приводили к содержанию общего клеточного белка. киРНК 25 для ингибирования HEBP1 способна к ингибированию активации промотора eNOS.
На фиг.3 демонстрируются исследования связывания гемина и PPIX с гексаHis-HEBP1 человека с помощью гашения триптофановой флуоресценции. К раствору 0,5 мкМ HEBP1 добавляли гемин и PPIX в концентрациях от 30 нМ до 10 мкМ. После инкубации в течение 10 мин измеряли триптофановую флуоресценцию (λвозбуждения=295 нм/λэмиссии=340 нм). Представлены средние значения ± среднеквадратическое отклонение 6 экспериментов, *p<0,05 в сравнение с 30 нМ гемином/PPIX. Гемин и PPIX способны гасить триптофановую флуоресценцию гексаHis-HEBP1 человека дозозависимым образом.
На фиг.4 демонстрируется влияние энхансера транскрипции eNOS - 9257 на связывание гемина с 6xHis-huHEBP1 на основе измерения гашения триптофановой флуоресценции. Раствор 0,5 мкМ HEBP1 предварительно инкубировали с 9257 (10 мкМ) на льду в течение 10 мин. После добавления гемина в различных концентрациях измеряли триптофановую флуоресценцию (λвозбуждения=295 нм/λэмиссии=340 нм). Представлены средние значения ± среднеквадратическое отклонение 6 экспериментов, *p<0,05 в сравнение с DMSO-контролем. 9257 оказывает влияние на связывание гемина с гексаHis-HEBP1.
На фиг.5 демонстрируется исследование специфического связывания A300 с HEBP1 с помощью измерения поляризации флуоресценции. Вещество A300 (30 нМ) инкубировали с различными концентрациями гексаHis-HEBP1 или BSA при комнатной температуре в течение 10 мин. Впоследствии измеряли поляризацию флуоресценции (λвозбуждения=530 нм/λэмиссии=585 нм). Представлены средние значения ± среднеквадратическое отклонение 5 экспериментов, *p<0,05 в сравнение с DMSO-контролем без белка. Являющееся энхансером транскрипции eNOS вещество A300 связывается специфически с гексаHis-HEBP1.
На фиг.6 демонстрируется экспрессия HEBP1_предсказанного в ткани сердца крыс с хронической сердечной недостаточностью после инфаркта миокарда. Крысы были разделены на три группы. Одна группа была ложно оперированной (группа 1), тогда как в группах 2 и 3 хроническая сердечная недостаточность была вызвана инфарктом миокарда. Только последнюю из названных групп подвергали лечению AVE3085 (10 мг/(кг день)) в течение 9 недель. Впоследствии в ткани сердца определяли экспрессию мРНК (HEBP1 по сравнению с GAPDH). Представлены средние значения ± среднеквадратическое отклонение 7 экспериментов, *p<0,05 в сравнение с ложно оперированными. Этот эксперимент наводит на мысль о том, что HEBP1 вовлечен в патогенез хронической сердечной недостаточности.
ПРИМЕРЫ
ПРИМЕР 1
Очистка потенциальных мишеней энхансеров транскрипции eNOS, используя аффинную хроматографию
Принцип аффинной очистки являющихся потенциальными мишенями белков действует так, что фармакофор, белок-мишень которого должен быть очищен, иммобилизуют на матрице посредством химического связывания (так называемого линкера). Если к материалу со специфическим сродством добавляются белковые лизаты клеточных культур, фармакофор действует в качестве разновидности «приманки» и извлекает белки со сродством к фармакофору. Этот способ был использован в настоящей работе для очистки/амплификации молекул-мишеней. Были использованы специфические материалы для колонки в активной и неактивной конформации фармакофора (смотри ниже). 3×107 клеток EA.csr03 (смотри ниже) использовались для аффинной хроматографии и хранили в виде клеточных осадков при -70°C. Для лизиса осадки ресуспендировали в буфере DPBS с 0,5% Tween 20 (1 мл/100 мг клеточного осадка) и разрушали с использованием ультразвука (3×30 с, импульс 5, интенсивность 20%). Контролирование лизиса клеток осуществляли с помощью микроскопии. Клеточный дебрис отделяли с помощью двух стадий центрифугирования (первой - в течение 10 мин при 1000×g/4°C, второй - в течение 30 мин при 70000×g/4°C). Концентрацию белка в супернатанте определяли с помощью способа с использованием BCA. Приблизительно 5 мг белка было получено из 3×107 клеток.
Выполняли последующую препаративную аффинную хроматографию с использованием лизатов клеток EA.csr03 на специфических связанных с сефарозой материалах для колонки и идентификацию связанных белков с помощью масс-спектрометрии. После нескольких стадий промывки связанные белки элюировали с помощью добавления SDS и разделяли, используя электрофорез в полиакриламидном геле. После окрашивания геля серебром всю дорожку всех элюированных белков разрезали на приблизительно 60 кусочков геля. Их по отдельности подвергали расщеплению трипсином до фрагментов белков, разделяли на C18-колонке и идентифицировали, используя MS-MS/MS.
Электрофорез в полиакриламидном геле в денатурирующих условиях (электрофорез в SDS-ПААГ) представляет собой способ гель-электрофореза, при котором различные белки разделяются в соответствии со своей молекулярной массой. Эта возможность является следствием добавления анионного поверхностно-активного вещества додецилсульфата натрия (SDS) к белковой смеси, подвергаемой разделению. SDS присоединяется к белкам и маскирует собственный заряд белков. Образуются отрицательно заряженные комплексы SDS-белок с постоянным отношением заряда к массе, составляющим приблизительно 1,4 г SDS/г белка (~1 молекула SDS/3 аминокислоты). Благодаря дополнительному нагреванию образцов вторичные и третичные структуры белков разрушаются. Кроме того, в образцы добавляют восстанавливающие агенты, например β-меркаптоэтанол и DTT, для расщепления дисульфидных мостиков. Средой для разделения, используемой при электрофорезе в SDS-ПААГ, является гелевая матрица из полиакриламида, который является результатом сшивания акриламида с метиленбисакриламидом. При применении электрического поля комплексы SDS-белок перемещаются по матрице для разделения и разделяются в соответствии со своим размером благодаря так называемому «эффекту молекулярного сита».
Для получения образцов различных клеток для гель-электрофореза клетки тщательно дважды промывали DPBS (37°C), а затем лизировали с использованием 1× буфера для образцов с SDS (50 мМ Trizma Base (pH 6,8), 1,6% (в отношении веса к объему - вес./об.) SDS; 4% (вес./об.) глицерина, 0,01% (вес./об.) бромфенолового синего, 5% (об./об.) β-меркаптоэтанола, 325Е Benzonase®, ингибиторы протеаз и вода до 10 мл). Затем добавляли фермент Benzonase® и образцы встряхивали в течение 15 мин при 37°C. Benzonase® является генетически сконструированной эндонуклеазой, деструктурирующей РНК и ДНК в клеточном лизате, значительно уменьшая, тем самым, вязкость образцов и обеспечивая лучшее разделение смеси белков при электрофорезе. Затем белковые образцы подвергают денатурации посредством нагревания при 70°C в течение 20 мин. Образцы либо использовали непосредственно для электрофореза, либо хранили при -20°C до использования. В экспериментах по состоянию фосфорилирования MAP-киназами, кроме того, ингибиторы фосфатазы (смеси 1 и 2) в соотношении 1:100 добавлялись в DPBS и буфер для образцов. Для гель-элетрофореза была использована система Novex® Midi Gel System от Invitrogen (Karlsruhe, Германия). При использовании этой готовой гелевой системы возможно внесение в каждый гель вплоть до 26 образцов. Кроме того, в объединенной камере XCell4 SureLockTM Midi-Cell электрофорезу может одновременно подвергаться вплоть до четырех гелей. Использовались 4-12% Bis-Tris полиакриламидные гели с 26 карманами для образцов. Они являются так называемыми «градиентными гелями», т.е. концентрация полиакриламида возрастает с увеличением хода разделения и поэтому позволяет одновременно разделять небольшие и большие белки. Во всех экспериментах в гель вносился также стандарт подходящих белков для оценки молекулярных масс исследуемых белков. В зависимости от требуемого диапазона разделения делался выбор между подвижным буфером MES-SIDES и подвижным буфером MOPS-SIDES. Кроме того, в подвижный буфер верхней камеры добавляли 435 мкл антиоксиданта NuPAGE® на гель. Гель-электрофорез проводили при постоянном напряжении, составляющем 200 В, в течение 40 мин при использовании подвижного буфера MES-SIDES и в течение 55 мин при использовании подвижного буфера MOPS-SIDES.
В случае окрашивания гелей с белками серебром окрашивание белков растворами серебра классически основано на том принципе, что ионы Ag+ образуют комплексы с остатками глютаминовой кислоты, аспарагиновой кислоты и цистеина. Восстановление ионов Ag+ дает атомарное серебро, вызывая коричневатое окрашивание белковых полос. Преимущество окрашивания серебром над другими способами, например окрашиванием Кумасси, заключается в высокой чувствительности способа. Поэтому можно визуализировать даже количества белка, начиная с 5 нг белка/0,5 см полосы, что является большим преимуществом, особенно в случае характеристических исследований. В случае характеристического исследования белков в полиакриламидных гелях окрашивание серебром осуществляли, используя наборы II для окрашивания SilverSNAP® от компании Pierce (Rockford, США), следуя стандартным инструкциям производителя. Принцип этой процедуры основан на фиксации белков в геле посредством использования раствора этанола и уксусной кислоты (30% (об./об.) этанола, 10% (об./об.) уксусной кислоты). За этим следует инкубация с использованием раствора соли серебра и восстановление ионов серебра до атомарного серебра, которое окрашивает белковые полосы.
«Вестерн-блоттинг» означает перенос белков после разделения с помощью гель-электрофореза на полимерный опорный слой. Таким образом белки становятся более легко доступными для антител при последующей иммунодетекции. В качестве опорных материалов пригодны, по существу, различные полимеры, например нейлон, PVDF и нитроцеллюлоза. Электрическое напряжение, применяемое перпендикулярно к гелю и мембране, вызывает миграцию белков из геля на мембрану. Рисунок полос предшествующего разделения сохраняется в этом процессе. В способе блоттинга, который был использован, с использованием нитроцеллюлозы в качестве опорного материала связывание белков с мембраной основано на гидрофобных взаимодействиях. Для приготовления геля для блоттинга после гель-электрофореза его уравновешивали в течение 20 мин в 2× буфере для переноса (50 мл 20× буфера для переноса NuPAGE®, 50 мл метанола, 500 мкл антиоксиданта NuPAGE® и воды до 500 мл). Нитроцеллюлозную мембрану промывали недолго водой и инкубировали вместе с шестью листами фильтровальной бумаги на каждый гель в 2× буфере для переноса. Сам перенос осуществляли в «Semi-Dry-Blotter» (Biostep, Jahnsdorf, Германия). Гель и мембрану помещали между пропитанными листами фильтровальной бумаги над ними и под ними по «принципу сэндвича» и применяли постоянное напряжение, составляющее 20 В, в течение 60 мин.
В предшествующих исследованиях два низкомолекулярных соединения с родственными структурами усиливали активность промотора eNOS зависящим от концентрации образом:
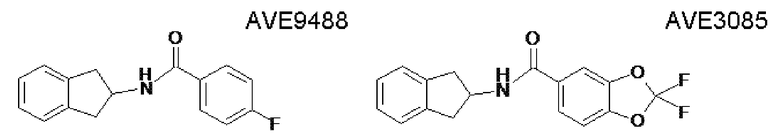
AVE9488: 4-фтор-N-индан-2-илбензамид; CAS №: 291756-32-6
AVE3085: индан-2-иламид 2,2-дифторбензо[1,3]диоксол-5-карбоновой кислоты; CAS №: 450348-85-3
AVE3085 использовали в качестве «приманки» для идентификации потенциальных мишеней. Были синтезированы четыре различных материала для колонок для обогащения потенциальных мишеней, используя аффинную хроматографию (таблица 1). Для синтеза фармакофор подвергали реакции с активированной с помощью N-гидроксисукцинимидного эфира (NHS) сефарозой и связывали ковалентно благодаря образованию эфирного мостика. Сайты связывания NHS-сефарозы, которые не были заполнены, заполняли посредством реакции с этаноламином.
контрольный
активный
Активный
неактивный
Материал A095 представляет собой материал для колонки без фармакофора в качестве «приманки» на линкере. Назначением этого контроля служит идентификация и исключение белков, которые связываются неспецифически с материалом колонки, в перечне являющихся потенциальными мишенями молекул.
В других материалах для колонок фармакофор ковалентно присоединен через линкер к частицам сефарозы. Таким образом, материал A093 является материалом, в котором фармакофор соединен с матрицей через линкер с помощью трех полиамидных связей. В случае материалов A092 и A094 этот линкер является немного длиннее, поскольку он содержит дополнительную эфирную связь. Кроме того, фармакофор находился в активной конформации в материале A092 и в неактивной конформации в A094.
Аффинные очистки являющихся потенциальными мишенями белков из лизатов цельных клеток EA.csr03 были выполнены с использованием всех четырех материалов для колонок, и были идентифицированы обогащенные таким образом белки, после трипсинолизиса, посредством масс-спектрометрии. Однако, кроме того, была также определена экспрессия в клетках EA.hy926 и HUVEC.
Клетки EA.csr03 являются клетками стабильной линии клеток с геном люциферазы под контролем промотора eNOS. Она была получена посредством трансфекции клеток EA.hy926 конструкцией репортера - люциферазы светлячка с добавлением фрагмента размером 3,5 т.п.о. промотора eNOS. Культивирование этих клеток выполняли с использованием IMDM с GlutaMAXTM I, дополненной 100 Е/мл пенициллина, 100 мкг/мл стрептомицина, 10% FCS и 0,4 мг/мл генетицина в качестве антибиотика для отбора. Клетки EA.hy926 являются стабильной линий клеток человека, которая была получена посредством слияния HUVEC с линией клеток гибридомы A549. Эта линия клеток характеризуется экспрессией специфических маркеров эндотелиальных клеток, например, «фактора Виллебранда». Используемыми клетками является исходная культура Cora Jean Edgell пассажа 32. Клетки культивировали в IMDM с GlutaMAXTM I, дополненной 100 Е/мл пенициллина, 100 мкг/мл стрептомицина, добавкой к среде HAT Hybri-MaxTM (гипоксантина, аминоптерина и тимидина) и протективной среды Biotect.
Перечень являющихся потенциальными мишенями молекул включает белки
- которые были идентифицированы не везде с использованием по крайней мере четырех фрагментов белков;
- которые не связывались с материалом колонки без какого-либо фармакофора, предлагаемого в качестве «приманки»;
- которые связывались предпочтительно с материалами колонок с фармакофором в активной конформации.
В таблице 2 представлен сводный перечень потенциальных мишеней, полученных таким образом. В этой таблице представлены, для каждого идентифицированного белка, уникальный номер доступа в банки данных UniProtKB/Swiss-Prot, теоретическая молекулярная масса и число фрагментов белков, с помощью которых он был идентифицирован.
Кроме гемсвязывающего белка 1 (HEBPl), перечисленные белки являются белками, которые связывались исключительно с активной конформацией материала колонки. HEBP1 также связывался, в виде единственного белка, с неактивной конформацией материала колонки. Однако в этом эксперименте его можно было идентифицировать с использованием лишь двух фрагментов белков в сравнении с 18 в случае активной конформации.
ПРИМЕР 2
Идентификация являющихся потенциальными мишенями белков, используя ранжированные микроряды белков
Эксперименты с использованием ранжированных микрорядов белков были выполнены, используя ProtoArrays® версии 3.0 (Invitrogen, Karlsruhe, Германия). Они представляли собой покрытые нитроцеллюлозой стеклянные пластинки, на которых было иммобилизовано приблизительно 5000 рекомбинантных белков человека, в двух повторах.
Для исследования сродства соединений, являющихся энхансерами транскрипции eNOS, с использованием указанных ранжированных микрорядов использовали меченные биотином энхансеры транскрипции eNOS A012 (в активной конформации) и A012 (в неактивной конформации) и связывание этих веществ с белками выявляли и подвергали количественному анализу посредством измерения флуоресценции, используя меченный IRDye®680 стрептавидин.
На первой стадии ранжированные микроряды инкубировали в течение одного часа при 4°C с буфером для блокирования MOPS с 0,1% (об./об.) игепала и 1% (вес./об.) BSA. Затем ранжированные ряды инкубировали с меченными биотином энхансерами транскрипции eNOS A012 и A013, иногда в присутствии немеченого вещества 9257, при 4°C в течение 90 мин без встряхивания, в различных условиях испытания в буфере для образцов MOPS с 1% (об./об.) игепала и 1% (вес./об.) BSA.
Вещество 9257
После промывки три раза, в каждом случае в течение 1 мин, чистым буфером для образцов MOPS на льду их инкубировали в течение 30 мин с меченным IRDye®)680 стрептавидином, разведенным 1/1000, в буфере для образцов MOPS вдали от света, также на льду. Меченные RDye®680 и IRDye® вторые антитела, которые распознают константную Fc-часть первых антител, использовались исключительно для иммунодетекции определенных белков. Флуоресцентная метка, используемая в настоящем описании, обладает свойством испускания ближней части инфракрасного диапазона (длин волн λ=700-800 нм) после возбуждения лазерным светом. Преимущество этого диапазона длин волн состоит в отличном отношении сигнала к шуму благодаря низкому уровню собственной флуоресценции. Для выявления используется ODYSSEYTM от LI-COR Biosciences (Bad Homburg, Германия). Используя этот прибор, можно выявить обе длины волны эмиссии на одной мембране одновременно. Благодаря одновременному выявлению белка-мишени и контрольного белка нормализация отдельных образцов возможна всего лишь в одну стадию. Выявление с использованием ODYSSEYTM (LI-COR Biosciences, Bad Homburg, Германия) следовало за тремя дополнительными стадиями промывки (каждая в течение 1 мин) с использованием 0,1% (об./об.) игепала в PBS.
После переноса мембрану блокировали в течение одного часа в буфере для блокирования ODYSSEY, разведенном 1:1 PBS. Назначением этой стадии является заполнение любых остающихся свободными сайтов связывания белков на мембране, а значит предотвращение неспецифического связывания антител. Мембрану инкубировали в течение ночи при 4°C с 1-2 первыми антителами в буфере для связывания антител (буфере для блокирования ODYSSEY (разведенном 1:1 PBS), 0,25% (об./об.) Tween 20, 0,02% (вес./об.) натрия азида). Инкубация со вторыми антителами следовала за промывкой мембраны (4×5 мин) с использованием PBST (PBS с 0,1% Tween 20) для удаления не связавшегося антитела. После инкубации в течение одного часа со вторыми антителами в буфере для связывания антител (с защитой от света, при комнатной температуре) мембрану снова промывали 4×5 мин с использованием PBST с последующим выявлением.
Для того чтобы иметь возможность исследовать сродство соединений, являющихся энхансерами транскрипции eNOS, используя эти ранжированные микроряды, эти соединения должны быть либо непосредственно помечены, либо должна была существовать возможность их мечения опосредованно. В экспериментах авторов настоящего изобретения меченные биотином энхансеры транскрипции eNOS использовались в активной (A012) и неактивной конформации (A013) (таблица 3). Их можно выявить на основе флуоресценции благодаря связыванию с высоким сродством меченного IRDye®680 стрептавидина с остатками биотина на веществах.
активный
неактивный
Для того чтобы иметь возможность использовать меченные биотином вещества, являющиеся энхансерами транскрипции eNOS, - A012 и A013 в экспериментах с использованием ранжированных микрорядов, было необходимо принять меры для того, чтобы остаток биотина не мешал действию фармакофора и чтобы существовала достаточная активность. С этой целью осуществляли контроль веществ в клетках при анализе транскрипции eNOS. При сравнении A012 с контрольным веществом AVE9488 является очевидным, что несмотря на введение линкера с биотиновой меткой, A012 (EC50=1,2 мкМ) является даже немного активнее в клетках, чем контрольный AVE9488 (EC50=3,0 мкМ). Меченное биотином вещество A013 (в неактивной конформации) теперь имело какую-либо едва значительную клеточную активность (EC50 в клетках >10 мкМ). Таким образом, два вещества A012 и A013 подходили для применения в экспериментах с использованием мечения и ранжированных микрорядов.
В эксперименте с использованием ранжированных микрорядов белков один ранжированный ряд в каждом случае инкубировали в трех различных условиях: A: активное меченное биотином вещество, являющееся энхансером транскрипции eNOS, - A012 (100 мкМ), B: A012 (100 мкМ) в присутствии активного немеченого вещества 9257 (25 мкМ) и C: неактивное меченное биотином вещество A013 (100 мкМ).
Мечение трех различных белковых дубликатов (A: числа 1-3) активным меченным биотином веществом A012 было легко различимым. Анализ релевантного конкурентного эксперимента с использованием вещества 9257 (B) показывает, что связывание A012 было заметно слабее, поскольку он вытеснялся 9257 из сайта связывания. Когда ранжированный микроряд инкубировали с неактивным меченным биотином веществом A013, не было связывания вещества с белками (C), которые ранее метились активным биотин-веществом.
Эти анализы мечения (A) и конкурентные анализы (B) оценивали посредством количественного определения флуоресцентных сигналов. Белки, в случае которых могла быть продемонстрирована конкуренция, были включены в перечень потенциальных мишеней (таблица 4).
Поскольку исследованные белки были продуцированы исключительно с помощью рекомбинантных методов, было невозможно подтвердить их функцию. Поскольку эти белки были идентифицированы исключительно на основе аффинностей связывания, требовалось подтверждение их функции в качестве белков - возможных мишеней.
ПРИМЕР 3
Валидация потенциальных мишеней из экспериментов с использованием аффинной хроматографии и ранжированных микрорядов белков
1. Профиль экспрессии в эндотелиальных клетках потенциальных мишеней из аффинной хроматографии.
Широкий профиль экспрессии всех потенциальных мишеней был создан на уровне мРНК в различных типах эндотелиальных клеток, чтобы сделать возможной лучшую оценку релевантности белков, обогащенных при аффинной хроматографии. В основу этого исследования было положено представление, что обогащение слабо экспрессируемых белков на материале колонки могло бы указывать на более высокую специфичность связывания по сравнению с белками, которые присутствуют в большем количестве в клетках. С этой целью определяли экспрессию всех являющихся потенциальными мишенями белков, используя количественную ОТ-ПЦР и специфические TaqMan®-зонды.
В количественной ПЦР с использованием обратной транскриптазы в режиме реального времени (количественной ОТ-ПЦР) обратную транскрипцию и ПЦР в режиме реального времени выполняют последовательно при постановке реакции. Ферментную смесь двух обратных транскриптаз (Omniscript/Sensiscript обратную транскриптазу) и HOTStarTaq ДНК-полимеразу добавляли к реакции. В начале реакции ДНК-полимераза все еще находится в неактивной форме, и происходит только обратная транскрипция мРНК в кДНК при 50°C. Затем обратные транскриптазы инактивируют, а ДНК-полимераза активируется при нагревании при 95°C с последующими стадиями амплификации кДНК.
Амплифицированную в режиме реального времени кДНК подвергают количественному анализу посредством пары специфических в отношении последовательностей праймеров и флуоресцентно меченого олигонуклеотидного зонда, так называемого TaqMan®-зонда. Он присоединяется к ДНК во время амплификации между двумя праймерами. TaqMan®-зонд помечен на одном конце флуоресцентной меткой, а на другом конце - гасителем. Флуоресценцию нельзя выявить на основе резонансного переноса энергии флуоресценции (FRET) в начале ПЦР. Однако если во время амплификации Taq-полимераза приблизится к зонду, проявится ее 5'→3' экзонуклеазная активность на двойной цепи. Зонд расщепляется, в результате чего флуорофор отделяется от гасителя. Этот процесс приводит к поддающейся обнаружению флуоресценции, которая увеличивается пропорционально количеству образованного продукта ПЦР.
Обычная постановка реакции в объеме 20 мкл в 96-луночном формате обычно состояла из 10 мкл 2× смеси Quantitect RT Mastermix, 1 мкл 20× смеси для анализа (праймера и TaqMan-зонда от Applied Biosystems), 0,2 мкл ОТ-смеси, 5 мкл РНК и 3,8 мкл воды, лишенной нуклеаз. Реакции проводили в системе детекции ПЦР iCycler® от BioRad-Laboratories (Munich, Германия) в соответствии со следующим протоколом амплификации.
Количественный анализ мРНК мишени проводили посредством приведения к количеству мРНК нерегулируемого гена GAPDH (гена «домашнего хозяйства») и расчета с использованием метода ΔΔct. Этот метод основывается на расчете порогового значения флуоресценции (ct) относительно фоновой флуоресценции в начале, когда еще продукт ПЦР не присутствует. Относительную экспрессию определенного гена мишени по сравнению с геном, не подвергнутым стимуляции контролем, приведенную к GAPDH, рассчитывают, следовательно, следующим образом:
ΔΔct = ctгена мишени - ctGAPDH
ΔΔct = ctобразца - ctконтроля
2-ΔΔct = ×-кратная экспрессия
Используемыми образцами клеток были сначала препараты РНК из не подвергнутых обработке клеток EA.csr03, лизаты которых также использовались при аффинной хроматографии. Профиль экспрессии сравнивали с таковым EA.hy926 и первичных HUVEC. Первичные эндотелиальные клетки пупочной вены человека (HUVEC) готовились заново для каждого эксперимента с помощью стандартных способов и использовались лишь вплоть до пассажа 3. Среда для роста этих клеток состояла из IMDM с GlutaMAXTM I, которая была дополнена 100 Е/мл пенициллина, 100 мкг/мл стрептомицина и 20% FCS. Культивирование выполняли на материалах для культивирования клеток с покрытием коллагеном I при 37°C и 5% CO2.
Установленные для потенциальных мишеней экспрессии были разделены на три класса (таблица 5).
Что касается линии клеток EA.csr03, использованной при хроматографии, высокая экспрессия была установлена для исследованных генов IDH3A, PPP2CA, GLO1, HTATIP2 и HEBP1. Средние уровни экспрессии были также отмечены в случае PIR, PDXK, LGALS3, MAPK3 и ADK, и очень низкая экспрессия была установлено только для PTCD1 и PCBP3.
При сравнении уровней экспрессии отдельных генов между линией клеток с репортером EA.csr03, линией родительских для них клеток EA.hy926 и первичными HUVEC различия обычно не обнаруживаются. Только в случае PIR и MAPK3, более низкий уровень экспрессии обнаруживается в линиях клеток по сравнению с первичными HUVEC.
2. Профиль экспрессии в эндотелиальных клетках являющихся потенциальными мишенями белков из экспериментов с использованием ранжированных микрорядов белков.
Белки в ранжированных микрорядах белков (смотри выше) были исключительно рекомбинантными белками. Поэтому было важно проверить, какие из них экспрессируются в эндотелиальных клетках. С этой целью определяли с помощью количественной ОТ-ПЦР относительную экспрессию потенциальных мишеней относительно GAPDH в препаратах РНК из не подвергнутых обработке клеток EA.csr03, клеток EA.hy926 и первичных HUVEC. Результаты представлены в таблице 6.
Экспрессия большей части потенциальных мишеней была низкой в исследованных типах эндотелиальных клеток (TBPL1, TFEB, MAPK7, PSAT1). Средняя экспрессия наблюдалась лишь для PRKAA1, а высокая - для CMAS.
В этом эксперименте, кроме одного исключения (PSAT1), не было различий в уровнях экспрессии отдельных мишеней между линиями клеток и первичными HUVEC.
3. Валидация потенциальных мишеней, используя технологию киРНК (коротких интерферирующих РНК).
На основе экспериментов с использованием аффинной хроматографии и ранжированных микрорядов белков был представлен сводный перечень всего из 18 являющихся потенциальными мишенями белков. Идентификация этих белков основывалась на сродстве к различным энхансерам транскрипции eNOS в бесклеточных системах. По этой причине все кандидаты были подвергнуты дальнейшей валидации в клетках.
РНК-интерференция означает специфическую в отношении последовательностей супрессию на посттранскрипционном уровне некоторых генов у животных и растений, инициируемую двухцепочечной РНК (дцРНК), которая гомологична гену-мишени. В Caenorhabditis elegans и Drosophila melanogaster смогли установить разрушение длинной, специфической в отношении последовательности дцРНК цитоплазматическим, RNase III-подобным белком «дайсером» в качестве механизма, на котором основана РНК-интерференция. Происходит образование коротких фрагментов длиной 19-25 нуклеотидов, так называемых «коротких интерферирующих РНК» (киРНК). Результирующие киРНК затем включаются в РНК-белковые комплексы, названные «индуцированным РНК комплексом сайленсинга» (RISC), и служат в качестве медиаторов последующей деструкции РНК. Разделение двух цепей киРНК вызывает активацию RISC. Активированный комплекс теперь может связываться с гомологичной мРНК и разрезать ее в месте, находящемся на расстоянии приблизительно 12 нуклеотидов от 3'-конца киРНК.
В настоящей работе технология РНК-интерференции представляла собой ключевой метод валидации влияния являющихся потенциальными мишенями энхансеров транскрипции eNOS молекул на промотор eNOS. Для переноса молекул киРНК использовались препараты катионных липосом, Lipofectin® в случае HUVEC и LipofectamineTM 2000 в случае EA.hy926 или EA.csr03. Они обладают свойством образования комплекса отрицательно заряженных нуклеиновых кислот на своей положительно заряженной поверхности. Благодаря липидоподобной природе липосом они способны к взаимодействию с клеточной мембраной и переносу нуклеиновых кислот в комплексе внутрь клетки.
Для трансфекции с использованием липосом киРНК в эндотелиальные клетки клетки EA.hy926 или EA.csr03 засевали с плотностью, составляющей 50000 клеток в каждую лунку, в объеме 100 мкл среды для роста в 96-луночные планшеты. После инкубации в течение 24 ч в термостате клетки промывали стерильным DPBS и покрывали в каждом случае 100 мкл/лунку специальной среды Opti-MEM® I без сыворотки и антибиотиков. Затем используемые киРНК (смотри ниже) предварительно инкубировали с липофектамином 2000 в соответствии с инструкциями производителя и, наконец, к клетками добавляли 50 мкл/лунку смеси для трансфекции. Кроме особо оговоренных случаев, киРНК использовали в конечных концентрациях, составляющих 100 нМ. На каждую лунку использовали, после предшествующей оптимизации, 0,8 мкл липофектамина 2000 для трансфекции клеток EA.hy926 и 0,5 мкл для клеток EA.csr03. После проведения инкубации клеток в термостате в течение 6 часов среду для трансфекции замещали стандартной средой для роста и следующие экспериментальные стадии выполняли в заданном интервале времени, составляющем 24-72 часа после трансфекции. Первичные HUVEC засевали с плотностью, составляющей 100000 клеток в каждой лунке, в 6-луночные планшеты с покрытием коллагеном I и культивировали в стандартной среде для роста вплоть до 60-70% конфлюэнтности. Перед трансфекцией клетки промывали стерильным DPBS и покрывали в каждом случае 800 мкл/лунку специальной среды Opti-MEM® I без сыворотки и антибиотиков. На лунку использовали 6 мкл липофектина и киРНК в конечной концентрации, составляющей 100 нМ. Предварительную инкубацию киРНК и липофектина проводили в соответствии с инструкциями производителя в отношении HUVEC. После добавления к клеткам 200 мкл/лунку препарата для трансфекции инкубацию проводили в термостате в течение 6 часов. Затем среду для трансфекции замещали стандартной средой для роста. Последующие экспериментальные стадии выполняли между 24-72 часами после трансфекции.
(г. Дармштадт)
(г. Дармштадт)
(г. Дармштадт)
(г. Дармштадт)
(г. Дармштадт)
(г. Дармштадт)
(г. Дармштадт)
(г. Дармштадт)
(г. Дармштадт)
(г. Дармштадт)
(г. Дармштадт)
(г. Дармштадт)
(г. Дармштадт)
(г. Дармштадт)
(г. Дармштадт)
(г. Дармштадт)
(г. Дармштадт)
(г. Дармштадт)
(г. Дармштадт)
(г. Дармштадт)
(г. Дармштадт)
(г. Дармштадт)
(г. Дармштадт)
(г. Дармштадт)
(г. Дармштадт)
(г. Дармштадт)
(г. Дармштадт)
(г. Дармштадт)
(г. Дармштадт)
(г. Дармштадт)
(г. Дармштадт)
(г. Дармштадт)
(г. Дармштадт)
(г. Дармштадт)
(г. Дармштадт)
(г. Дармштадт)
(г. Дармштадт)
(г. Дармштадт)
(г. Дармштадт)
(г. Дармштадт)
(г. Дармштадт)
(г. Дармштадт)
(г. Дармштадт)
(г. Дармштадт)
(г. Дармштадт)
(г. Дармштадт)
(г. Дармштадт)
(г. Дармштадт)
(г. Дармштадт)
(г. Дармштадт)
(г. Дармштадт)
(г. Дармштадт)
(г. Дармштадт)
(г. Дармштадт)
(г. Дармштадт)
(г. Дармштадт)
Функциональную релевантность всех являющихся потенциальными мишенями белков проверяли, используя технологию киРНК в линии клеток с геном репортера под контролем промотора eNOS EA.csr03. Для успешного использования этой технологии при анализе транскрипции eNOS использовали 3-4 имеющихся в продаже киРНК с различными целевыми последовательностями, которые отличались очень заметно по своей эффективности в исключении являющихся их мишенями белков, для каждой мишени-кандидата.
Для идентификации лучших киРНК их трансфецировали в клетки EA.hy926 (в конечной концентрации, составляющей 100 нМ, кроме особо оговоренных случаев) и спустя 24 часа выделяли и очищали РНК. Остающееся количество РНК соответствующей мишени определяли количественно в ОТ-ПЦР по сравнению с GAPDH. В случае наличия функционального антитела против белка-кандидата экспрессию белка по сравнению с GAPDH анализировали в отдельных постановках реакций через 48 часов после трансфекции киРНК посредством лизиса клеток буфером для образцов с SDS и последующего Вестерн-блоттинга.
Момент времени для получения образцов выбирали на основе предварительно проведенных кинетических исследований различных киРНК (не представленные данные), в которых выключение достигало своего максимума на уровне мРНК через 24 ч после трансфекции и на уровне белка на основании последующей трансляции через 48 ч.
После идентификации самой эффективной киРНК для каждой потенциальной мишени ее затем использовали в клетках EA.csr03 при анализе транскрипции eNOS. Таким образом, стало возможным, благодаря почти полному исключению этих белков, изучение их роли в индуцируемом веществом усилении транскрипции eNOS. Если бы исследуемый белок имел ключевую функцию в индуцируемом веществом усилении транскрипции eNOS, AVE3085 не должен бы дольше быть в состоянии активировать промотор eNOS.
4. Общие сведения обо всех подвергнутых валидации киРНК и проверка самых эффективных из них в анализе транскрипции eNOS
Исследование эффективностей различных киРНК в отношении белков-мишеней было опробовано. Соответствующую остаточную мРНК определяли с помощью количественной ОТ-ПЦР через 24 часа после трансфекции эндотелиальных клеток человека (EA.hy926/EA.csr03). Затем самые эффективные киРНК были отобраны и использованы в анализе транскрипции eNOS. Через 48 часов после трансфекции клетки EA.csr03 обрабатывали AVE3085 (5 мкМ) в течение дополнительных 18 ч. После лизиса клеток определяли люциферазную активность и содержание общего клеточного белка.
Поскольку для проверки всех киРНК было необходимо использовать несколько 96-луночных планшетов для культивирования клеток, AVE3085-индуцируемую активность гена репортера определяли относительно люциферазной активности в DMSO-контроле. Благодаря выключению HEBP1 промотор eNOS мог быть теперь значимо активирован лишь в 1,6 по сравнению с активацией в 2,4 раза в siLV2-контроле. Следовательно, белок HEBP1, по-видимому, играет роль в AVE3085-опосредованном усилении транскрипции eNOS.
Все другие киРНК были исследованы или оценены в анализе транскрипции eNOS. Наблюдаемые эффекты на состояние базальной активации промотора и на AVE3085-индуцируемую активацию промотора суммированы в таблице 7.
(siLV2=100%)
(siLV2=100%)
(siLV2=100%)
Только в случае HEBP1 продемонстрировано, после его выключения с помощью киРНК, значительное снижение активации промотора eNOS с помощью AVE3085, составляющее 64% в сравнении с siLV2-контролем. Поэтому он является единственным белком-кандидатом, который мог бы играть роль мишени в случае AVE3085-опосредованного усиления транскрипции.
5. Отбор эффективных киРНК и использование в анализе транскрипции eNOS с использованием примера гемсвязывающего белка 1 (HEBP1)
Как уже отмечалось, HEBP1 был единственным белком, в случае которого, после выключения благодаря киРНК, промотор eNOS не мог дольше быть активирован до степени, одинаковой с таковой в случае siLV2-контроля. Поэтому HEBP1 должен быть подтвержден еще раз в качестве примера кандидата на усиление транскрипции eNOS с помощью AVE3085.
Для анализа транскрипции в случае активации промотора eNOS засевали 20000 клеток линии клеток с геном репортера EA.csr03 в каждую лунку в прозрачном 96-луночном планшете для культивирования клеток. Спустя 24 часа готовили различные разведения вещества в среде для культивирования клеток, начиная с базовых растворов в DMSO, и их предварительно инкубировали в течение 10 мин при 37°C в водяной бане. Затем эти инкубационные среды (100 мкл/лунку) добавляли к клеткам. После инкубации в течение 18 часов клетки дважды промывали DPBS (100 мкл/лунку) и добавляли буфер для лизиса с высвобождением люциферазы (50 мкл/лунку). Спустя 10 мин к клеточным лизатам добавляли растворенный субстрат для люциферазы люциферин (100 мкл/лунку). Во время катализируемого люциферазой окисления люциферина до оксилюциферина происходит испускание хемилюминесценции. Испускаемый свет является показателем люциферазной активности, а следовательно, опосредованно, показателем активации промотора eNOS. Испускаемые световые единицы определяли в считывающем устройстве для микропланшетов Genios от Tecan Deutschland (Crailsheim, Германия).
В экспериментах, в которых клетки EA.csr03 обрабатывали различными киРНК до измерения люцеферазной активности, иногда отмечались незначительные изменения числа клеток вследствие токсических эффектов. Поэтому для стандартизации люциферазной активности было необходимо определить содержание общего белка в клеточных лизатах. Для этого после обработки клетки дважды промывали с использованием 100 мкл/лунку DPBS и растворяли в 60 мкл/лунку буфера для лизиса с CHAPS, состоящего из HEPES (20 мМ, pH 7,4), натрия хлорида (150 мМ) и 1,1% CHAPS, и встряхивали в течение 20 мин при комнатной температуре. Из каждого образца 15 мкл этого лизата отбирали, переносили в планшет для определения белка и разводили 135 мкл воды в соотношении 1:10. Содержание общего белка определяли с помощью микротеста для определения белка с использованием BCA в соответствии с инструкциями производителя. В каждом случае к остающимся 45 мкл клеточного лизата/лунку добавляли 45 мкл 2× реагента для лизиса клеток с высвобождением люциферазы и люциферазную активность определяли, как описано ранее.
Статистический анализ экспериментальных результатов выполняли, используя программное обеспечение GraphPad Prism 4.03. Все результаты представляли как среднее значение ± среднеквадратическое отклонение. Значимость результатов оценивали с помощью «проверки по критерию Стьюдента». При p-значении, составляющем <0,05, данные считались значимыми и отмечены на графиках с помощью «*» или «#».
Как и при ранее представленных валидациях различных киРНК, самая эффективная HEBP1-киРНК была идентифицирована на уровне мРНК, используя количественную ОТ-ПЦР (фиг.1). В случае клеток, обработанных siHEBPl 12 и siHEBPl 20, 69% и 40% мРНК для HEBP1, соответственно, все еще можно было выявить через 24 часа после трансфекции. Напротив, после обработки клеток siHEBP1 25 остаточное количество мРНК для HEBP1, на уровне 10% в сравнение с siLV2-котролями, было очень небольшим. Таким образом, siHEBP1 25 обеспечивает очень эффективную киРНК для выключения HEBP1 в клеточной среде и очень подходит для дальнейшего применения в анализе транскрипции eNOS. Для этой потенциальной мишени валидация на уровне белка была невозможной, так как в наличии не было функционального антитела.
Затем используемые условия испытания применяли в анализе транскрипции eNOS (фиг.2). В обработанных siLV2 контрольных клетках возможно активирование промотора eNOS под действием AVE3085 в 2,4 раза в сравнение с DMSO-контролем. Однако в случае выключения HEBP1, используя siHEBP1 25, промотор значимо активируется в сравнение с siLV2 лишь в 1,6 раз. Основываясь на этом результате, HEBP1 является, вероятно, белком, который играет основную роль в AVE3085-опосредованном усилении транскрипции eNOS.
ПРИМЕР 4
Биохимическая валидация гемсвязывающего белка 1 (HEBP1) в качестве потенциальной мишени в случае усиления транскрипции eNOS
Гемсвязывающий белок 1 (HEBP1) был единственным белком-кандидатом, в случае которого, после исключения белка с помощью технологии киРНК, активация промотора eNOS под действием AVE3085 была уменьшена.
Исходная ситуация для дальнейшей биохимической проверочной работы в отношении HEBP1 была непростой. Единственное антитело, имеющееся в продаже в тот момент времени (Abnova GmbH, Heidelberg, Германия, каталожный № H00050865-A01), продемонстрировало значительную неспецифичность в Вестерн-блоттинге (не представленные данные). В продаже также не было рекомбинантного белка человека. Поэтому исходным переключением на дальнейшую проверочную работу была рекомбинантная экспрессия и очистка HEBP1 человека.
1. Экспрессия и очистка гемсвязывающего белка 1 человека
Для улучшения очистки HEBP1 был обеспечен с аффинной меткой гексаHis на N-конце и экспрессирован в E. coli с помощью рекомбинантных методов, используя ранее клонированную экспрессионную плазмиду. Клеточные осадки от всего 4 литров культуры E. coli использовались для каждой серии очистки. ГексаHis-HEBP1 очищали, как описано далее, используя материал с аффинностью связывания с гексаHis (металлоаффинную колонку TALONTM Superflow).
Поскольку гемсвязывающего белка 1 (HEBP1) человека нет в продаже, но он требовался для его дальнейшей валидации в качестве потенциальной мишени, его экспрессировали в E. coli с помощью рекомбинантных методов, а затем выделили. Для клонирования ГексаHis-HEBP1 человека в вектор для экспрессии в E. coli ген HEBP1 человека амплифицировали с помощью ПЦР посредством использования имеющегося в наличии вектора для экспрессии в клетках млекопитающих «huHEBPl-pcDNA3.1», используя ДНК-полимеразу PhusionTM Hot Start High-Fidelity. В гене HEBP1 человека обеспечивался сайт расщепления EcoRI на 5'-конце благодаря разработке праймера. Используемыми праймерами были
прямой-EcoRI: 5'-CGTGAATTCGATCAAGAACTCGCTGTTCG-S' SEQ ID NO: 24
обратный: 5'-TAAACGGGCCCTCTAGACTC-S' SEQ ID NO: 25
Полученный продукт ПЦР выделяли с использованием системы для очистки продуктов ПЦР QIAquick и встраивали через расщепление в сайтах для EcoRI и XhoI в «сайт множественного клонирования» (MCS) исходного вектора ChampionTM pET302/NT-His с помощью направленного лигирования. Используемый исходный вектор уже несет на 5'-конце MCS генетический код для аффинной метки ГексаHis и для присоединения желаемого белка к N-концу.
Полученным вектором трансформировали компетентные клетки JM 109 E. coli, там его амплифицировали и плазмидную ДНК очистили с использованием системы для выделения плазмид Pure YieldTM. Наконец, концентрацию и чистоту ДНК определяли в объеме 2 мкл, используя спектрофотометр NanoDrop® ND-1000 (NanoDrop Technologies, Wilmington, США).
Экспрессию белка ГексаHis-HEBP1 человека осуществляли, используя описанный экспрессионный вектор для ГексаHis-HEBP1. Чтобы достигнуть наивысшего из возможных уровней экспрессии белка в E. coli, был выбран химически компетентный штамм BL21 StarTM (DE3). Этот штамм характеризуется мутацией в гене РНКазы, которая в основном ответственна за деструкцию транскриптов мРНК. Благодаря результирующему дефициту РНКазы накапливаются большие количества мРНК, приводя, тем самым, к более высокому выходу белка. Для экспрессии белка 50 мкл-аликвоту компетентных клеток E. coli для каждой серии трансформации оттаивали на льду и добавляли 10 нг плазмидной ДНК в объеме 5 мкл. За инкубацией на льду в течение 30 мин следовал тепловой шок в течение 30 с при 42°C в водяной бане. Затем клетки немедленно помещали на лед, добавляли 250 мкл среды для трансформации S.O.C. и смесь встряхивали в течение одного часа при 37°C. Затем в каждом случае 100 мкл высевали на чашку с ампициллином-агаром для отбора трансформированных клеток благодаря гену устойчивости к ампициллину, присутствующему в экспрессионной плазмиде. Чашку с агаром инкубировали в течение ночи при 37°C, затем четыре различных клона засевали в 5 мл среды Луриа Бертани (10 г триптона; 5 г дрожжевого экстракта; 10 г натрия хлорида и воды до 1000 мл) с карбенициллином (50 мкг/мл) и снова культивировали в течение ночи. В каждом случае добавляли глицерин к 1 мл этих культур и смесь хранили в качестве штоковых культур при -70°C. Остающиеся 4 мл предварительной культуры культивировали в 100 мл среды LB с карбенициллином (50 мкг/мл) вплоть до оптической плотности, составляющей 0,6 при длине волны λ=600 нм (ОП600), и добавляли до 1 мМ изопропил-β-D-тиогалактопиранозида (IPTG), индуктор lac-оперона в E. coli. IPTG взаимодействует с репрессором LacR, а значит предотвращает его связывание с оператором. Достигается транскрипция релевантного гена. После индукции экспрессии белка с помощью IPTG клетки культивировали в течение дополнительных 4 ч. Для сбора клеток суспензию клеток центрифугировали несколькими частями в течение 15 мин при 3000×g/4°C, среду отбрасывали, а осадки хранили при -20°C до дальнейшего использования.
Предварительно экспрессированный в E. coli гексаHis-HEBP1 очищали хроматографически, используя «AKTApurifier» (GE Healthcare Europe, Freiburg, Германия). Используемым для аффинной очистки материалом для колонки была металлоаффинная колонка TALONTM Superflow (Clontech Laboratories, Mountain View, США). Он представляет собой аффинный материал с положительно заряженными ионами кобальта, закрепленными на гранулах сефарозы с помощью хелатообразующих агентов, на своей поверхности. Благодаря высокоспецифическому взаимодействию остатков гистидина с ионами кобальта рекомбинантные белки, предоставляемые с меткой ГексаHis, обратимо связываются с материалом колонки, а следовательно, обогащаются. Этот принцип называется «аффинной хроматографией с использованием иммобилизованных металлов» (IMAC). Нативные белки подвергают элюированию с помощью имидазола, который конкурирует с остатками гистидина за связывание с ионами кобальта.
В каждом случае 20 мл буфера TALON®xTractor на грамм клеточного осадка использовали для лизиса клеток E. coli. Клетки тщательно ресуспендировали и центрифугировали в течение 20 мин при 12000×g/4°C для осветления лизата. Супернатант удаляли и пропускали через 0,2 мкм стерильный фильтр для полного удаления всех частиц.
Для очистки гексаHis-HEBP1 клеточный лизат вносили в колонку TALON, предварительно промытую буфером для уравновешивания (pH 7,0), состоящим из натрия фосфата (50 мМ) и натрия хлорида (300 мМ), со скоростью потока 0,5 мл/мин. Колонку снова промывали с использованием 10-20 объемов колонки буфера для уравновешивания и связанный ГексаHis-меченный HEBP1 смывали с колонки содержащим имидазол буфером для элюирования (pH 7,0), состоящим из натрия фосфата (50 мМ), натрия хлорида (300 мМ) и имидазола (150 мМ). Элюат собирали фракциями объемом 2 мл в коллекторе фракций. С помощью УФ-детектора с регулируемыми длинами волн, соединенного с колонкой, было возможно слежение как за введением клеточного лизата, так и за элюированием ГексаHis-HEBP1.
Проточный материал и полученные фракции, которые объединяли попарно в каждом случае, концентрировали посредством центрифугирования при 4000×g в колонках Amicon с размером отсечки 10 кДа. Посредством промывки в каждом случае 8 мл буфера для диализа pH 7,4, состоящего из HEPES (20 мМ) и натрия хлорида (100 мМ), и дополнительных стадий центрифугирования выполняли, кроме того, буферный обмен в образцах, в заключение концентрируя до конечного объема, составляющего приблизительно 0,5 мл. Отбирали небольшие количества концентратов и готовили образцы с SDS для анализа с помощью электрофореза в SDS-ПААГ и окрашивания серебром или Вестерн-блоттинга. Остаток либо использовали немедленно в последующих экспериментах, либо быстро замораживали в жидком азоте и хранили при -20°C до дальнейшего использования.
Для наблюдения за процессом элюирования измеряли оптическую плотность при длинах волн λ=280 нм и λ=405 нм. Оптическая плотность при λ=280 нм является обычным измерением белка. Гемсвязывающий белок демонстрирует дополнительное поглощение света при λ=405 нм.
Элюирование ГексаHis-HEBP1 после добавления имидазола происходило только после определенного времени удержания. Начиная с фракции 10 наблюдалось резкое увеличение белка и содержание гема во фракциях. Элюирование основной части белка можно было регистрировать вплоть до фракции 16, после которой наблюдалось непрерывное уменьшение содержания белка. Начиная с этой фракции на хроматограмме выявлялась разновидность «плеча» при λ=280 нм, высота которого продолжала уменьшаться и к фракции 30 почти возвращалась к базальному уровню.
Все фракции объединяли попарно, а затем осуществляли обмен буфера с использованием 20 мМ HEPES (pH 7,4) и 100 мМ натрия хлорида для удаления любого имидазола, все еще присутствующего. Образцы концентрировали и отбирали небольшие количества для получения образцов с SDS, при этом остаток использовали в последующих экспериментах.
Образцы с SDS, созданные на основе фракций, подвергали гель-электрофорезу. С целью контроля также использовали образец проточного материала колонки - не связавшихся белков. Затем параллельно выполняли окрашивание серебром белка в геле или иммунодетекцию ГексаHis-метки HEBP1 в Вестерн-блоттинге.
При окрашивании серебром довольно большие количества белка с мол. м. приблизительно 23 кДа различимы во фракциях 12-28, и, начиная с фракции 16, он находится в почти полностью чистой форме. Наблюдаемая молекулярная масса хорошо согласуется с теоретическим значением 21 кДа для HEBP1. Вторая полоса белка размером, составляющим приблизительно 40 кДа, различима как незначительная «примесь» в этих фракциях и могла бы являться димером HEBP1.
Результат окрашивания серебром был подтвержден при иммунологическом детектировании аффинной метки ГексаHis в Вестерн-блоттинге. Очень большие количества ГексаHis-меченного HEBP1 с молекулярной массой, составляющей ~23 кДа, были обнаружены во фракциях 12-28. И опять также незначительная «примесь» с мол. м. приблизительно 40 кДа была обнаружена с помощью антитела против ГексаHis-метки, что могло бы быть дополнительным свидетельством присутствия димера. В проточном материале антитело не распознало ГексаHis-HEBP1, т.е. связывание белка с материалом колонки происходило в количественном соотношении.
Основываясь на результатах окрашивания серебром и Вестерн-блоттинга, фракции 16-28 с чистотой HEBP1, составляющей >95%, использовали в последующих анализах. Содержание белка в отдельных фракциях определяли, используя способ с использованием BCA. Обычно получали всего 8 мг очищенного ГексаHis-HEBP1 из одной очистки.
2. Исследования связывания на 6xHis-huHEBP1 посредством гашения триптофановой флуоресценции
Для проведения исследований связывания с использованием HEBP1 человека можно использовать собственную флуоресценцию остатков триптофана при λ=340 нм, используя длину волны возбуждения λ=295 нм. Результирующую флуоресценцию можно «погасить» посредством связывания порфиринов вблизи, в пространственном отношении, остатков триптофана. Это приводит к уменьшению собственной флуоресценции, которое, следовательно, обеспечивает меру связывания лиганда. Для оптимизации экспериментальных условий для этих измерений гашения сначала было необходимо оценить спектроскопические свойства ГексаHis-HEBP1.
2.1. Спектроскопическая характеристика ГексаHis-HEBP1
Спектр поглощения HEBP1 (35 мкМ) с использованием фотометра Nanodrop® показал основное сильное поглощение при λ=290 нм (не представленные данные). Основываясь на этом результате, затем забуференный HEPES раствор HEBP1 (0,5 мкМ) подвергали возбуждению при длине волны λ=290 нм и определяли результирующую эмиссию на различных длинах волн.
Определение спектра эмиссии показало максимум для длины волны λ=340 нм. Для создания спектра возбуждения возбуждение осуществляли при различных длинах волн и эмиссию измеряли на постоянной длине волны λ=340 нм. Этот спектр показал максимальное возбуждение при λ=295 нм.
Спектроскопическая характеристика ГексаHis-HEBP1 человека хорошо согласуется с последующими определениями триптофановой флуоресценции при λвозбуждения=295 нм и λэмиссии=340 нм.
2.2 Связывание гемина и протопорфирина IX с ГексаHis-HEBP1
Затем были проведены исследования связывания гемина и протопорфирина IX (PPIX) с ГексаHis-HEBP1 человека в присутствии энхансеров транскрипции eNOS посредством гашения триптофановой флуоресценции. Исследования связывания с использованием HEBP1 человека проводили, используя собственную флуоресценцию остатков триптофана, которые в нем содержатся. Флуоресценцию, возникающую на λ=340 нм после возбуждения длиной волны λ=295 нм, можно «погасить» благодаря связыванию порфиринов вблизи остатков триптофана. Измеряемая флуоресценция уменьшается в зависимости от концентрации лигандов. Используя нелинейную регрессию можно рассчитать параметры связывания.
Гемин и PPIX в концентрациях от 30 нМ до 10 мкМ добавляли в буферный раствор HEBP1 (500 нМ) и после инкубации в течение 10 мин измеряли триптофановую флуоресценцию (фиг.3). Было возможно использовать более высокие концентрации порфиринов вследствие низкой растворимости в водосодержащих системах.
Зависимое от концентрации уменьшение флуоресценции наблюдали как в случае гемина, так и в случае PPIX. Использование нелинейной регрессии значений дало значение IC50=3 мкМ для гемина и = 13 мкМ для PPIX. Следовательно, выяснено, что гемин является в 10 раз более сильным, чем PPIX, и он использовался в последующих экспериментах.
2.3. Влияние энхансеров транскрипции eNOS на связывание гемина с гексаHis-HEBP1 человека.
Затем исследовали влияние энхансеров транскрипции eNOS на гашение триптофановой флуоресценции гемином. В экспериментах рекомбинантный HEBP1 человека в концентрации, составляющей 500 нМ, предварительно инкубировали на льду в течение 30 мин с веществом, являющимся энхансером транскрипции eNOS, - 9257 в сравнение с DMSO-контролем в буфере для связывания (pH 7,4), состоящем из HEPES (20 мМ), натрия хлорида (100 мМ) и DTT (1 мМ). Черные 96-луночные планшеты использовали в качестве резервуара для реакций. Затем добавляли гемин или протопорфирин IX в различных концентрациях благодаря добавлению 200-кратного концентрированного исходного раствора в DMSO и инкубировали в течение 10 мин при комнатной температуре. В заключение измеряли триптофановую флуоресценцию после предварительной оптимизации при λвозбуждения=295 нм и λэмиссии=340 нм в считывающем устройстве для микропланшетов Safire2 (Tecan Deutschland, Crailsheim, Германия).
Используемым прототипом было вещество 9257, поскольку относительно AVE9488 и AVE3085 оно обладает большей эффективностью и растворимостью в воде, а следовательно, приводит к меньшим помехам в этом анализе.
Сначала авторы настоящего изобретения исследовали, может ли вещество 9257 гасить триптофановую флуоресценцию HEBP1. Уменьшения флуоресценции нельзя было определить (не представленные данные). Поэтому связывание 9257 исследовали опосредованно через конкуренцию с гемином. Для этого HEBP1 (500 нМ) предварительно инкубировали с 9257 (10 мкМ) на льду в сравнении с DMSO-контролем. Через 30 мин добавляли гемин в концентрациях, меняющихся от 0,1 до 10 мкМ, и осуществляли инкубацию при комнатной температуре в течение дополнительных 10 мин до измерения триптофановой флуоресценции (фиг.4).
На фиг.4 демонстрируется сдвиг кривой связывания гемина вправо во время предварительной инкубации с 9257. Расчет значения IC50 для обеих кривых с использованием линейной регрессии дал 3,4 мкМ для 9257 в сравнении с 1,1 мкМ для DMSO-контроля. Значение IC50 было увеличенным в 3,1 раза благодаря присутствию 9257. Энхансер транскрипции eNOS - 9257 вытесняет гемин из кармана для связывания без гашения им самим триптофановой флуоресценции.
Анализ триптофановой флуоресценции оказался не очень подходящим для более детальных и широких исследований. Поэтому авторы настоящего изобретения затем создали альтернативный способ выполнения экспериментов по связыванию энхансеров транскрипции eNOS с HEBP1.
3. Эксперименты по связыванию энхансеров транскрипции eNOS с гексаHis-HEBP1 человека с использованием измерения поляризации флуоресценции.
Дальнейшие исследования связывания энхансеров транскрипции eNOS с HEBP1 человека проводили, используя способ поляризации флуоресценции. Для этой цели вещество A300 было использовано и снабжено ковалентно родаминовой меткой через линкер.
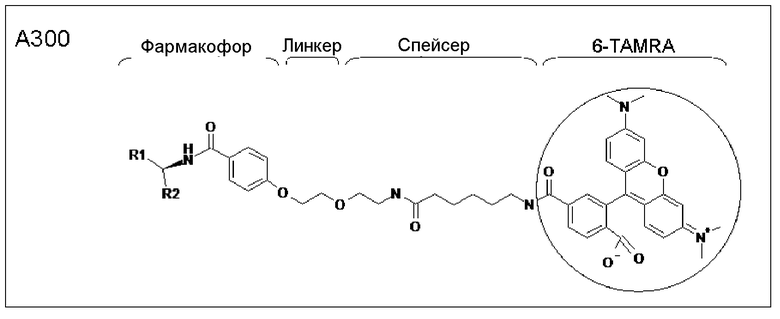
Флуорофоры обычно испускают линейно-поляризованный свет в случае их возбуждения также линейно-поляризованным светом. Однако изменение этой поляризации флуоресценции зависит от степени, в которой флуорофор может поворачиваться между моментом времени возбуждения и таковым эмиссии. Если флуорофор может поворачиваться в пространстве и не прикреплен к партеру по связыванию, он испускает «изотропно» линейно-поляризованный свет благодаря собственному повороту на различные углы. Однако если флуорофор прикреплен и больше не способен к повороту, он испускает линейно-поляризованный под меньшим углом свет («анизотропия»). Это явление можно использовать для измерения связывания флуоресцентно меченой молекулы с белком. При использовании этого принципа измерения до возбуждения в приборе для измерения падающий свет линейно поляризуется поляризатором. Интенсивность флуоресценции I испускаемого света измеряют во втором поляризаторе, так называемом «анализаторе», в двух положениях относительно первого поляризатора (параллельном и перпендикулярном). Поляризацию P определяют с использованием следующего уравнения:
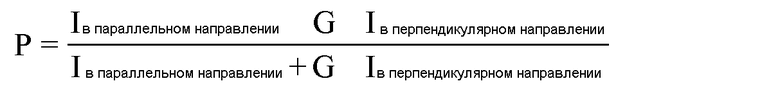
P: поляризация
Iв параллельном направлении: интенсивность флуоресценции, измеренная, когда поляризатор и анализатор расположены параллельно друг к другу.
Iв перпендикулярном направлении: интенсивность флуоресценции, измеренная, когда поляризатор и анализатор расположены перпендикулярно друг к другу.
G: весовой множитель (множитель, специфичный для прибора)
Полученная поляризация P является безмерной величиной, которую часто указывают в мП (в единицах миллиполяризации).
В настоящей работе метод поляризации флуоресценции использовали для проведения экспериментов по связыванию меченного родамином вещества, являющегося энхансером транскрипции eNOS, - A300 с рекомбинатным гексаHis-HEBP1 человека. Для приготовления образцов различные концентрации вещества A300 вместе с гексаHis-HEBP1 или BSA в буфере для связывания (pH 7,4), состоящем из HEPES (20 мМ), натрия хлорида (100 мМ) и DTT (1 мМ), помещали на лед. После инкубации при комнатной температуре в течение 10 мин поляризацию измеряли при длине волны возбуждения λ=530 нм и длине волны эмиссии λ=585 нм в считывающем устройстве для микропланшетов Safire2 (Tecan Deutschland, Crailsheim, Германия). Для проводимых в настоящем описании экспериментов использовали исключительно черные 96-луночные титрационные микропланшеты со стеклянным дном, поскольку они характеризуются особенно низкой фоновой флуоресценцией.
3.1. Активность меченного родамином вещества A300 в анализе транскрипции eNOS
Перед фактическими биохимическими экспериментами вещество A300 проверяли на активность в клетках в анализе транскрипции eNOS в сравнение с AVE3085 в качестве контроля посредством определения зависимостей эффекта от концентрации исследуемого вещества. Используя нелинейную регрессию, были рассчитаны значения EC50, составляющие 160 нМ для A300 и 400 нМ для AVE3085. Следовательно, меченное родамином вещество, являющееся энхансером транскрипции, - A300 имело в 2,5 раза более высокую активность в клетках, чем контрольное вещество и очень подходило для применения в экспериментах по связыванию с использованием способов поляризации флуоресценции.
3.2 Спектроскопическая характеристика меченного родамином вещества, являющегося энхансером транскрипции eNOS, - A300
Сначала регистрировали спектр поглощения для вещества A300 (100 нМ) в буферном растворе, в котором максимум поглощения был обнаружен для длины волны, составляющей 555 нм.
Техническим недостатком считывающего устройства для микропланшетов Safire2 является то, что имеются лишь четыре определенных длины волны возбуждения для измерения поляризации флуоресценции. Однако эмиссия возможна при любой желаемой длине волны.
Длиной волны возбуждения, ближайшей к максимуму возбуждения A300 и имеющейся в Safire2, была λ=530 нм. По этой причине для валидации и контроля в качестве последующих измерений снова регистрировали спектр эмиссии с использованием возбуждения при λ=530 нм.
Из зарегистрированного спектра было легко понять, что даже при использовании этой не полностью оптимальной длины волны возбуждения можно было с высокой чувствительностью измерить эмиссию на 585 нм. Следовательно, эту систему можно было использовать для проведения анализов связывания веществ, являющихся энхансерами транскрипции eNOS, с гексаHis-HEBP1 человека, используя способ поляризации флуоресценции.
3.3. Исследования связывания меченного родамином вещества, являющегося энхансером транскрипции eNOS, - A300 с HEBP1 человека методом поляризации флуоресценции
В исследованиях связывания использовали A300 в качестве флуоресцентного образца в постоянной концентрации, составляющей 30 нМ, к которому добавляли увеличивающиеся концентрации HEBP1. Для определения доли неспецифического связывания при связывании с обычным белком проводились параллельные эксперименты с использованием равных концентраций бычьего сывороточного альбумина (BSA). Долю специфического связывания можно подсчитать на основе различия между связыванием A300 с HEBP1 и неспецифическим связыванием с BSA (фиг.5).
Кривая связывания A300 с HEBP1 представляет собой типичную кривую насыщения, достигающую плато при увеличении концентраций белков. Напротив, в случае связывания A300 с BSA получена линейная кривая, указывающая на связывание, которое не становится предельным. В результате обнаружения различия в значениях поляризации в случаях связывании A300 с HEBPl или BSA, кривую можно подвергнуть оценке, при которой можно подсчитать долю специфического связывания. Используя нелинейную регрессию, значение KD, составляющее 11,7 мкМ, можно рассчитать для специфического связывания A300 с гексаHis-HEBP1 человека.
4. Предварительные исследования экспрессии HEBP1 в модели патологического состояния на животных
В другом контексте было проведено исследование на животных, в котором у крыс развивалась хроническая сердечная недостаточность после вызова инфаркта миокарда. На основе имеющихся образцов РНК из ткани сердца можно было определить экспрессию HEBP1_предсказанного на уровне мРНК. Последовательность гена HEBP1 еще не была выверена у крысы, и она была идентифицирована посредством совмещения последовательностей от других видов, поэтому все еще используется обозначение HEBP1_предсказанный (NM_001108651).
Выделение РНК из используемых типов клеток выполняли, используя способ минивыделения с использованием RNeasy®. На первой стадии к клеткам добавляли буфер, содержащий гуанидин тиоцианат и β-меркаптоэтанол (буфер RLT), который очень эффективно денатурирует белки и, в частности, инактивирует РНКазы. Образцы клеток в 6-луночном формате разлагали в каждом случае в 600 мкл буфера для лизиса, а вязкость уменьшали посредством центрифугирования в течение 2 мин при 14000×g в колонках QIAshredder. При добавлении 70% (об./об.) этанола любая присутствующая РНК осаждается и связывается с мембраной из диоксида кремния миниколонки RNeasy®. За несколькими стадиями промывки различными буферами следует элюирование очищенной тотальной РНК с использованием 50 мкл воды без нуклеаз. Образцы клеток в 96-луночном формате разлагали в каждом случае с использованием 140 мкл буфера для лизиса, и РНК затем выделяли методом минивыделения с использованием RNeasy® в BIOROBOT 8000 от Qiagen (Hilden, Германия) в соответствии с инструкциями производителя. Концентрацию и чистоту приготовленной РНК определяли, используя спектрофотометр NanoDrop® ND-1000 (NanoDrop Technologies Inc., Wilmington, США). При этом на основе общего объема, составляющего 2 мкл, оптическую плотность образцов измеряли при длине волны λ=260 нм (A260) для нуклеиновых кислот. Превращение измеренной оптической плотности в концентрацию раствора РНК осуществляли с использованием коэффициента экстинкции, составляющего 40 нг×см-1×мкл-1, по закону Ламберта-Бера. Вывод о чистоте РНК делали при дополнительном измерении оптической плотности при длине волны λ=280 нм (A280), при которой поглощают белки, фенолы или другие примеси. Отношение A260/A280 должно быть как можно ближе к значению 2 для «чистой» РНК. Полученную РНК затем разводили в соотношении 1:5 и либо использовали немедленно в количественной ОТ-ПЦР, либо хранили при -80°C для дальнейшего использования.
Большую часть препаратов РНК можно было использовать в количественной ОТ-ПЦР непосредственно без удаления геномной ДНК, которая все еще присутствует, поскольку имеющиеся в наличии зонды обычно гибридизуются в месте соединения экзона с интроном, а значит может быть распознана лишь результирующая кДНК. В случае зондов, не удовлетворявших этому требованию, выполняли, в соответствии с инструкциями производителя, расщепление ДНКазой с использованием набора RNase-Free® DNAse на миниколонке RNeasy®, в сочетании с выделением РНК.
Были исследованы три различные группы, в каждой из которых было семь животных, на экспрессию HEBP1_предсказанного: одна группа была ложно оперированной, т.е. была открыта лишь брюшная полость, без вызова инфаркта миокарда. У четырнадцати животных (двух групп, в каждой из которых было семь животных) развивалась хроническая сердечная недостаточность после вызова инфаркта миокарда. Одну группу подвергали лечению в течение 9 недель энхансером транскрипции eNOS - AVE3085 в дозе 10 мг/кг/день по сравнению с плацебо.
Тотальную РНК выделяли из сердца и определяли относительную экспрессию HEBP1_предсказанного по сравнению с GAPDH с помощью количественной ОТ-ПЦР (фиг.6). Данные по экспрессии показывают, что при сравнении животных, у которых развилась сердечная недостаточность после инфаркта миокарда, с группой, подвергшейся ложной операции, было увеличение экспрессии HEBP1 _предсказанного в 2 раза. В отличие от этого, лечение AVE3085 вообще не продемонстрировало какого-либо влияния на экспрессию HEBP1_предсказанного. HEBPl_предсказанный, по-видимому, играет роль в патологии этой модели, поскольку его экспрессия не изменялась.
| название | год | авторы | номер документа |
|---|---|---|---|
| НОВЫЕ ЭУКАРИОТИЧЕСКИЕ КЛЕТКИ И СПОСОБЫ ИХ ПОЛУЧЕНИЯ ДЛЯ РЕКОМБИНАНТНОЙ ЭКСПРЕССИИ ПРЕДСТАВЛЯЮЩЕГО ИНТЕРЕС ПРОДУКТА | 2014 |
|
RU2712507C2 |
| ПРИМЕНЕНИЕ ИНГИБИТОРА РАК ДЛЯ ЛЕЧЕНИЯ ЗАБОЛЕВАНИЯ СУСТАВОВ | 2004 |
|
RU2360696C2 |
| АНТИТЕЛА ПРОТИВ КОМПЛЕКСА ANGPTL3/8 И СПОСОБЫ ИХ ИСПОЛЬЗОВАНИЯ | 2019 |
|
RU2799796C2 |
| ЛЕЧЕНИЕ ЗАБОЛЕВАНИЙ, СВЯЗАННЫХ С НЕЙТРОФИЧЕСКИМ ФАКТОРОМ ГОЛОВНОГО МОЗГА (BDNF), ПУТЕМ ИНГИБИРОВАНИЯ ПРИРОДНОГО АНТИСМЫСЛОВОГО ТРАНСКРИПТА BDNF | 2013 |
|
RU2661104C2 |
| ЛЕЧЕНИЕ РАКА С ИСПОЛЬЗОВАНИЕМ ХИМЕРНОГО АНТИГЕНСПЕЦИФИЧЕСКОГО РЕЦЕПТОРА НА ОСНОВЕ ГУМАНИЗИРОВАННОГО АНТИТЕЛА ПРОТИВ EGFRvIII | 2014 |
|
RU2708032C2 |
| ГУМАНИЗИРОВАННЫЕ АНТИТЕЛА К CXCR4 ДЛЯ ЛЕЧЕНИЯ РАКА | 2011 |
|
RU2595394C2 |
| СОЕДИНЕНИЯ ДЛЯ ЛЕЧЕНИЯ БЛОКАДЫ РЕМИЕЛИНИЗАЦИИ ПРИ ЗАБОЛЕВАНИЯХ, СВЯЗАННЫХ С ЭКСПРЕССИЕЙ БЕЛКА ОБОЛОЧКИ HERV-W | 2013 |
|
RU2636355C2 |
| МОНОКЛОНАЛЬНОЕ АНТИТЕЛО К IL-5Rα | 2017 |
|
RU2698048C2 |
| НОВЫЕ ЭУКАРИОТИЧЕСКИЕ КЛЕТКИ И СПОСОБЫ ИХ ПОЛУЧЕНИЯ ДЛЯ РЕКОМБИНАНТНОЙ ЭКСПРЕССИИ ПРЕДСТАВЛЯЮЩЕГО ИНТЕРЕС ПРОДУКТА | 2014 |
|
RU2720525C2 |
| РЕГИСТРАЦИЯ РАХ2 ДЛЯ ДИАГНОСТИКИ РАКА МОЛОЧНОЙ ЖЕЛЕЗЫ | 2009 |
|
RU2522485C2 |





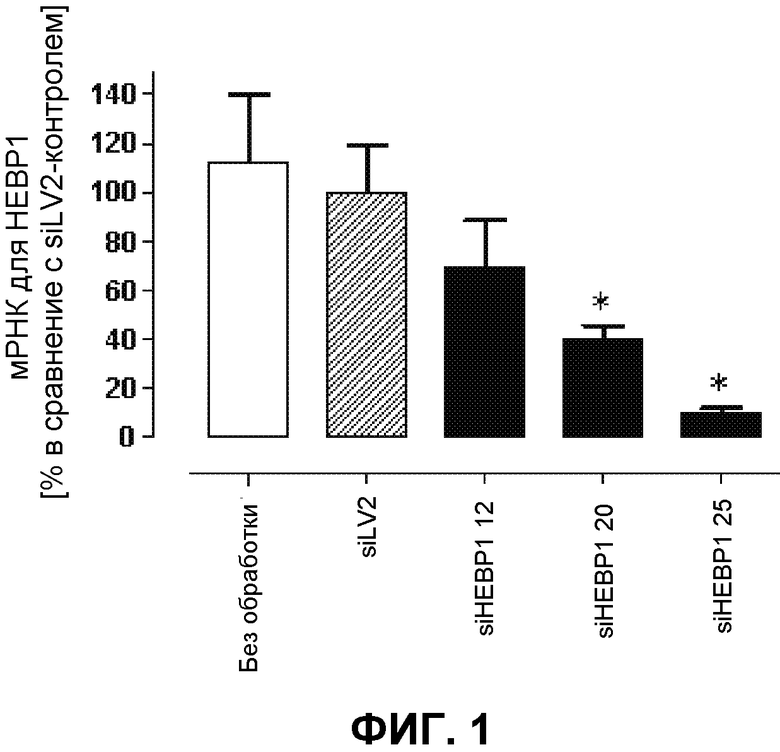
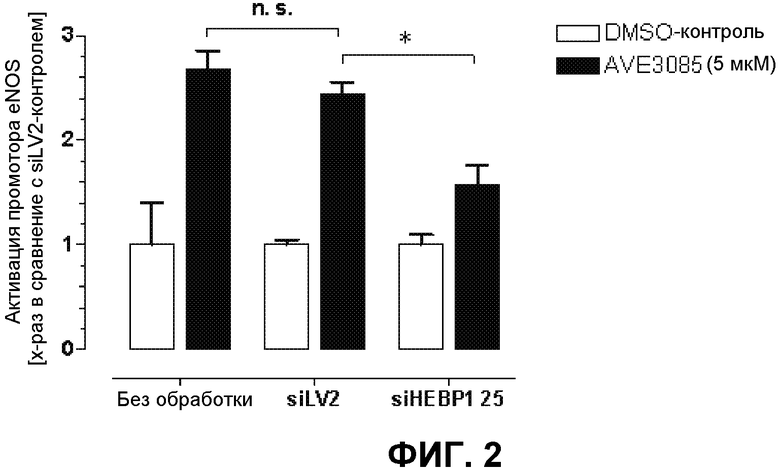
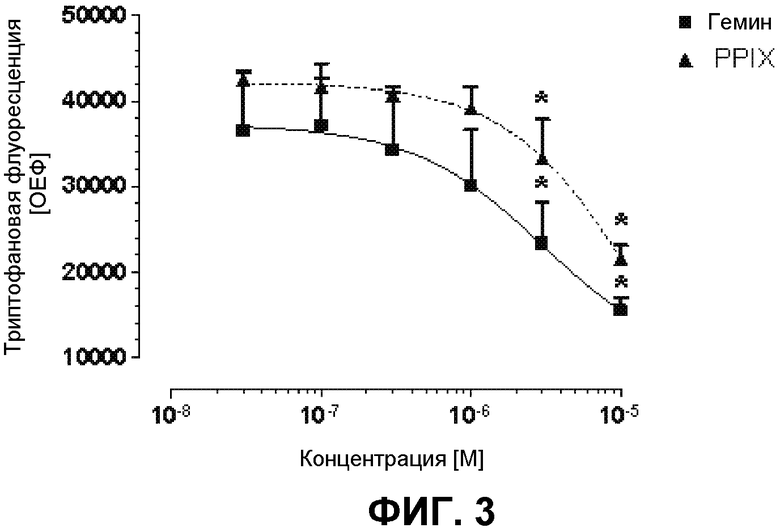
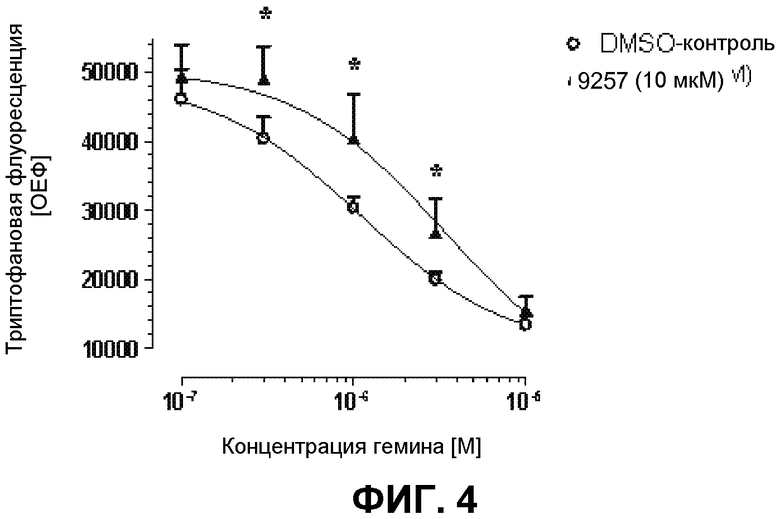
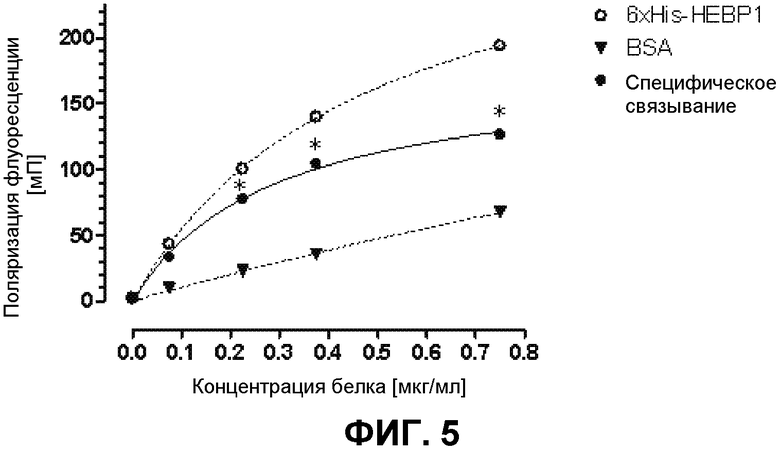
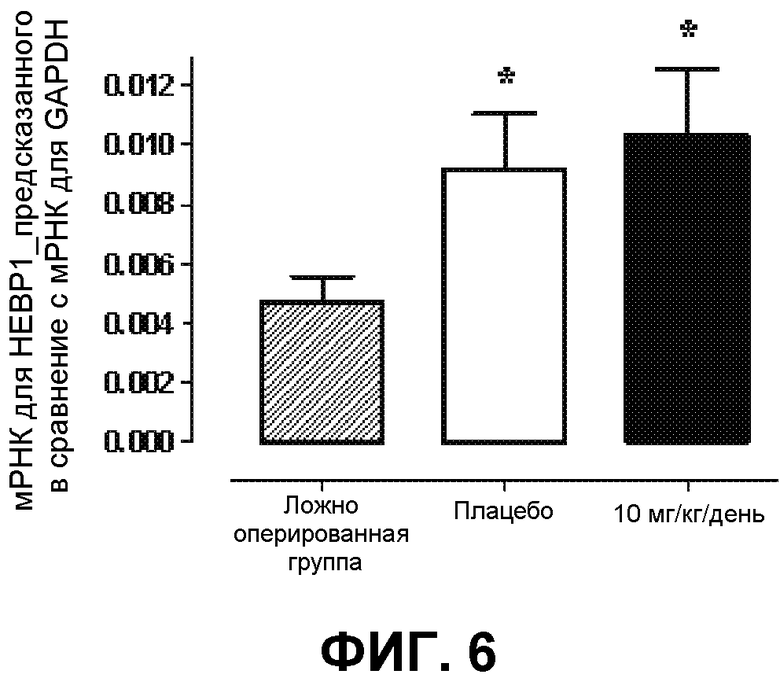
Группа изобретений основана на выявлении механизмов модулирования активности eNOS и относится к способу скрининга на модулятор экспрессии эндотелиальной NO-синтазы (eNOS), способу диагностирования сердечно-сосудистого заболевания у индивида, применению HEBP1 для идентификации лекарственного средства для профилактики и/или лечения заболевания, связанного с дисфункцией eNOS, в частности сердечно-сосудистого заболевания, применению HEBP1 для обнаружения компонента передачи сигнала при экспрессии eNOS и применению HEBP1 для регуляции активности промотора eNOS. Группа изобретений эффективна в лечении и профилактике заболеваний, связанных с дисфункцией eNOS. 5 н. и 20 з.п. ф-лы, 6 ил., 7 табл., 4 пр.
1. Способ скрининга на модулятор экспрессии эндотелиальной NO-синтазы (eNOS), предусматривающий
- наличие тест-системы, включающей гемсвязывающий белок 1 (HEBP1) или его функционально активный вариант,
- приведение тест-системы в контакт с агентом и
- выявление эффекта агента на тест-систему, идентифицируя тем самым агент в качестве модулятора экспрессии eNOS.
2. Способ по п.1, в котором тест-система дополнительно включает
- промотор eNOS и/или
- один или несколько транскрипционных факторов для промотора eNOS.
3. Способ по п.1 или 2, в котором тест-система включает клетку.
4. Способ п.3, где клетка является клеткой млекопитающего.
5. Способ по п.4, где клетка является клеткой человека.
6. Способ по п.1 или 2, где эффект определяется флуоресценцией.
7. Способ по п.1 или 2, где способ используется для скрининга лекарственного средства для профилактики и/или лечения заболевания, связанного с дисфункцией eNOS.
8. Способ по п.3, где способ используется для скрининга лекарственного средства для профилактики и/или лечения заболевания, связанного с дисфункцией eNOS.
9. Способ по п.4, где способ используется для скрининга лекарственного средства для профилактики и/или лечения заболевания, связанного с дисфункцией eNOS.
10. Способ по п.7, где заболевание, связанное с дисфункцией eNOS, представляет собой сердечно-сосудистое заболевание.
11. Способ по п.8, где заболевание, связанное с дисфункцией eNOS, представляет собой сердечно-сосудистое заболевание.
12. Способ по п.9, где заболевание, связанное с дисфункцией eNOS, представляет собой сердечно-сосудистое заболевание.
13. Способ диагностирования сердечно-сосудистого заболевания у индивида, включающий
- определение уровня мРНК для НЕВР1 или белка HEBP1 в образце, полученном от индивида,
причем повышенный или пониженный уровень мРНК для HEBP1 или белка HEBP1 относительно контроля служит признаком сердечно-сосудистого заболевания, где контролем является индивид или группа индивидов, не страдающих сердечно-сосудистым заболеванием.
14. Способ по п.13, в котором уровень понижен относительно контроля.
15. Способ по п.13 или 14, в котором уровень определяют в клетке сердца.
16. Способ по любому из пп.13 или 14, в котором сердечно-сосудистым заболеванием является сердечная недостаточность и/или инфаркт миокарда.
17. Способ по п.15, в котором сердечно-сосудистым заболеванием является сердечная недостаточность и/или инфаркт миокарда.
18. Способ по п.1, 2, 13 или 14, где HEBP1 является HEBP1 человека.
19. Применение НЕВР1 для идентификации лекарственного средства для профилактики и/или лечения заболевания, связанного с дисфункцией eNOS.
20. Применение по п.19, где заболевание, связанное с дисфункцией eNOS, является сердечно-сосудистым заболеванием.
21. Применение по п.19 или 20, где лекарственное средство значительно изменяет экспрессию eNOS по сравнению с контролем, который не контактирует с лекарственным средством.
22. Применение по п.21, где лекарственное средство значимо повышает экспрессию eNOS.
23. Применение HEBP1 для выявления компонента передачи сигнала при экспрессии eNOS.
24. Применение по любому из пп.19-23, где HEBP1 является HEBP1 человека.
25. Применение НЕВР1 для регулирования активности промотора eNOS.
| Способ подсочки хвойных пород | 1988 |
|
SU1607745A1 |
| US2002048581 A1, 2002-04-25 | |||
| СПОСОБ ОПРЕДЕЛЕНИЯ РИСКА РАЗВИТИЯ СЕРДЕЧНО-СОСУДИСТЫХ ОСЛОЖНЕНИЙ У БОЛЬНЫХ ГИПЕРТОНИЧЕСКОЙ БОЛЕЗНЬЮ | 2006 |
|
RU2334466C2 |
| WO2004041170 A2, 2004-05-21 | |||
| ANNALISA DEL PRETE et al., Migration of dendritic cells across blood and lymphatic endothelial Barriers, Vascular Cell Signalling, 2006 Schattauer GmbH, Stuttgart, pp | |||
| Машина для добывания торфа и т.п. | 1922 |
|
SU22A1 |

