Область изобретения
Настоящее изобретение относится к области биомедицины. В настоящей заявке предложен антигенный полипептид, который может быть использован для продуцирования антител, способных связываться с молекулами, происходящими из PCSK9. Настоящее изобретение также относится к антителу или его функциональному фрагменту, специфически связывающемуся с PCSK9 с высокой аффинностью. В настоящем изобретении также предложена молекула нуклеиновой кислоты, кодирующая антитело по настоящему изобретению или его функциональный фрагмент, вектор экспрессии и клетка-хозяин для экспрессии антитела или его функционального фрагмента по настоящему изобретению, и способ продуцирования антитела по настоящему изобретению или его функционального фрагмента. В изобретении также предложен иммунноконъюгат и фармацевтическая композиция, содержащая антитело по настоящему изобретению или его функциональный фрагмент, и способ лечения различных заболеваний, включающих дислипидемию и родственные заболевания сердечно-сосудистой системы, путем использования антитела в соответствии по настоящему изобретению или его функционального фрагмента.
Предшествующий уровень техники
Наследственная гиперхолестеринемия (FH) представляет собой аутосомальное моногенное доминантное генетическое заболевание, которое является наследственным и имеет различные клинические проявления. FH представляет собой наиболее тяжелое заболевание среди моногенных заболеваний липидного метаболизма и также названо заболеванием рецептора LDL (липопротеинов низкой плотности) или гиперлипидемией типа IIa, которая представляет собой наиболее распространенную наследственную гиперлипидемию у детей, и представляет собой важный фактор риска ишемической болезни сердца. Существенное увеличение холестерина, липопротеина низкой плотности (LDL-C), представляет собой важный симптом заболевания, и таким образом подавление деградации гепатоцитарного LDLR (рецептора LDL) и увеличение захвата LDL посредством LDLR может уменьшать развитие атеросклероза, и риск заболевания может быть уменьшен. Мутации в гене PCSK9 (S127R,F216L) обнаружены у пациентов ADH, у которых были исключены мутации гена LDLR и apoB. Действие PCSK9 на липидный метаболизм было продемонстрировано впервые (Abifadel, Varret et al. 2003). В настоящее время PCSK9 используется в качестве новой терапевтической мишени для дислипидемии и имеет хорошую перспективу для применения.
Пропротеиновая конвертаза субтилизин/кексин 9(PCK9), также называемая как нейтральная регулируемая апоптозом конвертаза 1 (NARC-1), относится к девятому члену субтилизинового семейства кексиноподобной пропротеинконвертазы, и состоит из 692 аминокислотных остатков, находится преимущественно в печени, почках и тонком кишечнике и т.п. Она экспрессируется клетками паренхимы печени, клетками мезенхимы и эпидермальными клетками толстого кишечника и т.п. и находится в крови в виде секреторного белка (Seidah, Benjannet et al. 2003).
Pcsk9 кодируется геном PCSK9, который расположен на человеческой аутосоме LP33-p34.3 и синтезируется в основном в эндоплазматическом ретикулуме. Сначала он образует неактивную структуру предшественника массой 72 кДа и затем расщепляется в положениях 152 и 153 путем автокатализа с образованием домена предшественника массой 14 кДа и зрелого фрагмента массой 57 кДа, содержащего каталитический домен и C-концевой домен (Benjannet, Rhaids et al. 2004). Домен предшественника может связываться с каталитическим доменом не ковалентным образом, что необходимо для правильного свертывания зрелого фрагмента и транспорта из эндоплазматического ретикулума.
Мутации гена PCSK9 можно разделить на тип приобретения функции (D347Y, S127R, F216L, L82X, и Y142X и т.п.) и тип нарушения функции (R46L, Y142X, и C679) (Abifadel, Varret et al. 2003), где вследствие синергии с различными белками мутанты типа приобретения функции могут изменять аффинность PCSK9 или улучшать восприимчивость протеазы к саморасщеплению PCSK9, и улучшать деградацию LDLR со стороны PCSK9, приводя в результате к увеличенному уровню LDL-C в крови, вызывающему ADH или атеросклеротическое заболевание с ранним началом.
Исследования демонстрируют, что PCSK9 не только оказывает важное влияние на дифференцировку клеток печеночной паренхимы и нервных клеток (Seidah, Benjannet et al. 2003), но также корректирует экспрессию рецептора липопротеина низкой плотности (LDLR), за счет этого участвуя в синтезе и метаболизме холестерина. Исследования демонстрируют, что добавление очищенного белка PCSK9 в культуральную среду для клеток HepG2 уменьшало уровень LDLR на клеточной поверхности в зависимости от дозы (Lagace, Curtis et al.2006). Однако уровень LDLR существенно уменьшался у мышей, имеющих клетки печени сверхэкспресссирующие PCSK9, тогда как уровень экспрессии мРНК LDLR не уменьшался (Lambert, Charlton et al. 2009). Таким образом, PCSK9 корректирует уровень LDLR путем посттранскрипционного механизма.
Молекула LDLR состоит из пяти основных доменов: связывающая лиганд обогащенная цистеином область; гомологичный домен предшественника эпидермального фактора роста (EGF), включающий три подобных EGF (эпидермальный фактор роста) последовательности (EGF-A, EGF-B и EGF-C) и структуру бета-спирали, сахаридный домен, трансмембранный домен и цитоплазматическая концевая область, содержащая последовательность, необходимую для интернализации рецептора.
В 2007 году существенный прогресс был достигнут в отношении исследования молекулярного механизма опосредованной PCSK9 деградации LDLR. В этом исследовании продемонстрировали, что PCSK9 секретируется в плазму крови, может связываться с внеклеточным доменом LDLR и вызывать интернализацию, облегчая его деградацию в лизосоме (Zhang, Lagace et al. 2007). Дальнейшее исследование продемонстрировало, что при нейтральной среде pH на клеточной поверхности PCSK9 может связываться с EGF-A на внеклеточном сегменте LDLR, приводя к эндоцитозу в клетке. При клеточной среде с низким pH C-концевой домен PCSK9 может связываться с доменом лиганда в LDLR, таким образом, что LDLR деградирует в лизосоме и не способен вернуться на клеточную мембрану (Fisher, Surdo et al. 2007).
В заключение, в настоящее время PCSK9 становится важной мишенью при дислипидемии и родственных заболеваниях сердечно-сосудистой системы. Моноклональное антитело против него имеет широкие перспективы для применения.
Краткое изложение сущности изобретения
Проблема, решаемая в изобретении
Одна из задач настоящего изобретения заключается в предложении антитела против связанной с дислипидемией мишенью и его функционального фрагмента, способа лечения дислипидемии путем использования указанного антитела или его функционального фрагмента и т.п.
Способ решения технической проблемы
Для достижения вышеупомянутых задач авторы изобретения провели тщательное исследование и завершили настоящее изобретение на основе обнаружения того, что антитело по настоящему изобретению может специфически связываться с PCSK9 и ингибировать связывание PCSK9 с LDLR, за счет этого увеличивая захват LDL-c. Настоящее изобретение включает следующие аспекты:
В одном из аспектов изобретения предложены антитела против PCSK9, способные связываться с пропротеиновой конвертазой субтилизин/кексин 9 (PCK9), и их функциональные фрагменты, содержащие CDR тяжелой цепи, имеющие аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 4, 5, 6, 10, 11, 12, 16, 17, 18, 22, 23, 24, 28, 29 и 30, или из любого из вариантов этой последовательности, и/или CDR легкой цепи, имеющие аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 1, 2, 3, 7, 8, 9, 13, 14, 15, 19, 20, 21, 25, 26 и 27, или из любого из вариантов этой последовательности.
В еще одном аспекте изобретения дополнительно предложены антитела, способные связываться с PCSK9, или их функциональные фрагменты, где аминокислотные последовательности CDR1, CDR2 и CDR3 тяжелой цепи выбраны из одной из групп, состоящих из следующих аминокислотных последовательностей или их вариантов:
и/или аминокислотные последовательности CDR1, CDR2 и CDR3 легкой цепи выбраны из группы, состоящей из следующих аминокислотных последовательностей или их вариантов.
В еще одном аспекте настоящего изобретения предложено антитело, способное связываться с PCSK9, и его функциональный фрагмент, где аминокислотные последовательности CDR1, CDR2 и CDR3 тяжелой цепи и CDR1, CDR2 и CDR3 легкой цепи выбраны из группы, состоящей из следующих аминокислотных последовательностей или их вариантов:
В еще одном аспекте изобретения предложены антитела, способные связываться с PCSK9, или их функциональные фрагменты, которые включают вариабельную область тяжелой цепи, выбранную из группы, состоящей из аминокислотных последовательностей SEQ ID NO: 32, 34, 36, 38 и 40, или из вариантов любой из указанных последовательностей, и/или вариабельную область легкой цепи, выбранную из группы, состоящей из аминокислотных последовательностей SEQ ID NO: 31, 33, 35, 37 и 39, или из вариантов указанных последовательностей.
В еще одном аспекте изобретения предложены антитела, способные связываться с PCSK9, или их функциональные фрагменты, где указанная вариабельная область тяжелой цепи представляет собой SEQ ID NO: 32 или ее вариант и указанная вариабельная область легкой цепи представляет собой SEQ ID NO: 31 или ее вариант, или
вариабельная область тяжелой цепи представляет собой SEQ ID NO: 34 или ее вариант и вариабельная область легкой цепи представляет собой SEQ ID NO: 33 или ее вариант, или
вариабельная область тяжелой цепи представляет собой SEQ ID NO:36 или ее вариант и вариабельная область легкой цепи представляет собой SEQ ID NO:35 или ее вариант, или
вариабельная область тяжелой цепи представляет собой SEQ ID N0:38 или ее вариант и вариабельная область легкой цепи представляет собой SEQ ID NO: 37 или ее вариант, или
вариабельная область тяжелой цепи представляет собой SEQ ID NO: 40 или ее вариант и вариабельная область легкой цепи представляет собой SEQ ID NO: 39 или ее вариант.
В еще одном аспекте изобретения предложены антитела, способные связываться с PCSK9, или их функциональные фрагменты, которые представляют собой химерное антитело, гуманизированное антитело или полностью человеческое антитело.
В еще одном аспекте изобретения предложена выделенная молекула нуклеиновой кислоты, кодирующая антитело, способное связываться с PCSK9, или его функциональный фрагмент, и вектор экспрессии или клетка-хозяин, содержащие указанную молекулу нуклеиновой кислоты.
В еще одном аспекте изобретения предложена фармацевтическая композиция, содержащая антитело, способное связываться с PCSK9, или его функциональный фрагмент, молекулу нуклеиновой кислоты, кодирующую антитело по настоящему изобретению, способное связываться с PCSK9, или его функциональный фрагмент, и вектор экспрессии или клетку-хозяин, или любую их комбинацию, и фармацевтически приемлемый носитель.
В еще одном аспекте настоящего изобретения предложено применение антитела по настоящему изобретению, способного связываться с PCSK9, или его функционального фрагмента, или молекулы нуклеиновой кислоты, кодирующей его, или вектора экспрессии или клетки-хозяина, в изготовлении лекарственного средства для лечения заболеваний, таких как дислипидемия и родственные заболевания сердечно-сосудистой системы, включающие гиперлипидемию или гиперхолестеринемию.
В еще одном аспекте настоящего изобретения предложено применение антитела по настоящему изобретению, способного связываться с PCSK9, или его функционального фрагмента, или молекулы нуклеиновой кислоты, кодирующей его, или вектора экспрессии или клетки-хозяина, в изготовлении лекарственного средства для облегчения клеточного захвата LDL. (Из описания предшествующего уровня техники понятно, что PCSK9 может связываться с LDLR и уменьшать LDLR, за счет чего клеточный захват LDL уменьшается, приводя к увеличению уровня липидов в крови. Антитело против PCSK9 может нейтрализовать PCSK9, за счет этого обращая указанный процесс, улучшая клеточный захват LDL и уменьшая уровень липидов в крови.)
В еще одном аспекте настоящего изобретения предложен имммуноконъюгат, который содержит антитело по настоящему изобретению, способное связываться с PCSK9, или его функциональный фрагмент, конъюгированное(ый) с терапевтическим агентом, где указанный терапевтический агент предпочтительно представляет собой токсин, радиоактивный изотоп, лекарственное средство или цитотоксический агент.
Краткое описание графических материалов
Фиг. 1: Диаграмма электрофореза SDS-PAGE (электрофорез в полиакриламидном геле с додецилсульфатом натрия) человеческого PCSK9;
Фиг. 2: Анализ ELISA (иммуноферментный анализ) связывания человеческого PCSK9 с LDLR;
Фиг. 3: Обнаружение при помощи FACS (сортировка клеток с активацией флуоресценции) связывания человеческого PCSK9 с LDLR;
Фиг. 4: Подтверждение при помощи FACS связывания человеческого PCSK9 с
LDLR;
Фиг. 5: Обнаружение при помощи FACS связывания клеток-кандидатов гибридомы с человеческим PCSK9;
Фиг. 6: Анализ связывания мышиного антитела и PCSK9 D347Y (покрытие антигеном);
Фиг. 7: Анализ связывания мышиного антитела и PCSK9 D347Y (покрытие антителом);
Фиг. 8: Анализ конкурентного связывания мышиного антитела с антигенным эпитопом PCSK D347Y и референсного антитела;
Фиг. 9: Влияние мышиного антитела на захват LDL клетками HepG2;
Фиг. 10: Связывание химерного антитела с человеческим PCSK9;
Фиг. 11: Влияние химерного антитела на захват LDL клетками HepG2;
Фиг. 12: Связывание гуманизированного антитела с человеческим PCSK9;
Фиг. 13: Влияние гуманизированного антитела на захват LDL клетками HepG2;
Фиг. 14: Гуманизированное антитело может эффективно уменьшать уровень LDL-c in vivo у яванского макака (Macaca fascicularis).
Конкретный способ осуществления изобретения
Если не определено иное, то все используемые здесь научные и технологические термины имеют то же самое значение, которое понимает специалист в данной области техники. Для определения и терминологии в области техники специалист в данной области техники может обратиться к Current Protocols In Molecular Biology (Ausubel). Сокращения аминокислотных остатков представляют собой стандартный 3-буквенный и/или 1-буквенный код, принятый в области техники, относящийся к одному из 20 обычных L-аминокислотных остатков.
В изобретении предложено антитело против PCSK9 или его функциональный фрагмент, способное(ый) связываться с пропротеиновой конвертазой субтилизин/кексин 9 (PCK9). Антитело по настоящему изобретению или его функциональный фрагмент обладает по меньшей мере одной из способностей, включающих блокирование взаимодействия между PCSK9 и LDLR с высокой аффинностью; связывание с PCSK9 с высокой специфичностью.
В изобретении дополнительно предложено гуманизированное антитело против PCSK9 и его функциональный фрагмент. Это гуманизированное антитело получают путем осуществления конструирования при помощи компьютерного симулирования мышиного антитела, полученного путем иммунизации мышей, комбинированного с технологией фагового дисплея. Связывающие эпитопы также соответствующим образом идентифицируют в соответствии с характеристиками связывания с белками PCSK9 различных видов. Гуманизированное антитело против PCSK9 по настоящему изобретению и его функциональный фрагмент не только обладают вышеупомянутыми благоприятными характеристиками, но также связываются с человеческим белком PCSK9 или белком PCSK9 яванского макака с высокой аффинностью и взаимодействуют с мышиным белком PCSK9.
Специалист в данной области техники может заменить, добавить и/или удалить одну или более чем одну (например, 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 или более 10) аминокислот в последовательностях, описанных в настоящем изобретении, при условии, что по существу не будет оказываться влияние на активность антитела, за счет этого получая варианты последовательности антитела или его функционального фрагмента, которые считаются включенными в объем настоящего изобретения. Например, аминокислоты в вариабельной области могут быть заменены на аминокислоты, обладающие похожими свойствами. Варианты по настоящему изобретению могут иметь последовательность, имеющую по меньшей мере 95%, 96%, 97%, 98% или 99% идентичность с последовательностью, из которой получен вариант. Идентичность последовательности может быть измерена путем использования программного обеспечения для анализа последовательности, например компьютерной программы BLAST с определенными параметрами, в частности BLASTP или TBLASTN.
Антитело по настоящему изобретению может быть полноразмерным (например, антитело IgG1 или IgG4) или может включать только антигенсвязывающую группировку (например, Fab, F (ab')2 или фрагмент scFv) или может быть модифицировано для изменения функции. Изобретение включает антитело против PCSK9 с модифицированным профилем гликозилирования. В некоторых применениях может быть полезна модификация, удаляющая нежелательные сайты гликозилирования, или приводящая к отсутствию фукозной группировки на олигосахаридной цепи, таким образом, усиливая антитело-зависимую цитотоксичность (ADCC). В других применениях антитело может быть модифицировано галактозильной группировкой, модифицирующей комплемент-зависимую клеточную токсичность (CDC).
Используемый в настоящей заявке термин "функциональный фрагмент" относится к фрагменту антитела, такому как fv, scFv (sc обозначает одиночную цепь), Fab, F(ab')2, Fab', фрагмент scFv-Fc или диатело), или любому фрагменту, который может увеличивать период полувыведения за счет химической модификации или включения в липосому. Химическая модификация представляет собой, например, добавление поли (алкилен) гликоля, такого как полиэтиленгликоль ("ПЭГилирование"), (ПЭГилированные фрагменты, которые называются fv-PEG, scFv-PEG, FAB-PEG, F(ab')2-PEG или Fab'- PEG) (где "ПЭГ" представляет собой полиэтиленгликоль), и фрагмент обладает активностью связывания с EGFR (рецептором эпидермального фактора роста). Предпочтительно, функциональные фрагменты состоят из или содержат частичную последовательность тяжелой или легкой вариабельных цепей антитела-источника, где частичная последовательность достаточна для того, чтобы сохранять ту же самую связывающую специфичность и достаточную аффинность, как его антитело-источник, к PCSK9, аффинность предпочтительно равна по меньшей мере 1/100 аффинности его антитела- источника, более предпочтительно, равна по меньшей мере 1/10. Такие функциональные сегменты содержат по меньшей мере пять аминокислот, предпочтительно 10, 15, 25, 50 и 100 последовательных аминокислот последовательности антитела-источника.
Специалист в данной области техники может клонировать молекулы ДНК, кодирующие антитело против PCSK9 по настоящему изобретению, в вектор, и затем трансформировать вектор в клетку-хозяин. Таким образом, в изобретении дополнительно предложен рекомбинантный ДНК вектор, содержащий молекулу ДНК, кодирующую антитело против PCSK9, раскрытое в изобретении.
Предпочтительно, рекомбинантный ДНК вектор представляет собой вектор экспрессии. Специалист в данной области техники может клонировать молекулу ДНК антитела в вектор экспрессии и трансформировать этот вектор в клетку-хозяина с получением антитела путем индуцируемой экспрессии. Вектор экспрессии по настоящему изобретению содержит последовательность ДНК, кодирующую вариабельную область тяжелой цепи, вариабельную область легкой цепи и/или константную область антитела против PCSK9. Однако два вектора экспрессии могут быть сконструированы по отдельности, где первый содержит вариабельную область тяжелой цепи и константную область, а другой содержит вариабельную область легкой цепи и константную область, для одновременной трансфекции клеток млекопитающих. В предпочтительном воплощении вектор экспрессии дополнительно содержит промотор и последовательность ДНК, кодирующую секреторный сигнальный пептид и по меньшей мере один ген устойчивости к лекарственному средству для скрининга.
Клетка-хозяин по настоящему изобретению может представлять собой прокариотическую клетку-хозяина, эукариотическую клетку-хозяина или фаг. Прокариотические клетки-хозяева могут представлять собой Escherichia coli, Bacillus subtilis, Streptomycete или Proteus mirabilis и т.п. Эукариотическая клетка-хозяин может представлять собой гриб, такой как Pichia pastoris, Saccharomyces cerevisiae и schizochytrium, клетки насекомых, таких как совка луговая и т.п., клетки растений, таких как табак, клетки млекопитающих, такие как клетки BHK, клетки CHO, клетки COS, клетки миеломы и т.п. В некоторых воплощениях клетки-хозяева по настоящему изобретению предпочтительно представляют собой клетки млекопитающих, более предпочтительно клетки BHK, клетки CHO, клетки NSO или клетки COS.
Используемый в настоящей заявке термин "фармацевтическая композиция" относится к комбинации по меньшей мере одного лекарственного средства и возможно фармацевтически приемлемых носителей или вспомогательных агентов, которые комбинируются вместе для достижения некоторой конкретной цели. В некоторых воплощениях фармацевтическая композиция содержит комбинацию, разделенную по времени и/или в пространстве, а также они могут действовать вместе для достижения назначения настоящего изобретения. Например, компоненты в фармацевтической композиции (таких как молекулы антител и нуклеиновых кислот, комбинации молекул нуклеиновых кислот и/или конъюгаты по настоящему изобретению) могут быть введены субъекту в совокупности или по отдельности. Когда компоненты, содержащиеся в фармацевтической композиции, вводят субъекту по отдельности, тогда указанные компоненты могут быть введены одновременно или один за другим. Предпочтительно, фармацевтический носитель представляет собой воду, буферный водный раствор, изотонический солевой раствор, такой как PBS (физиологический раствор, забуференный фосфатом), глюкозу, маннит, декстрозу, лактозу, крахмал, стеарат магния, целлюлозу, карбонат магния, 0,3% глицерин, гиалуроновую кислоту, этанол или полиалкиленгликоль, такой как полипропиленгликоль, триглицериды и т.п. Тип фармацевтически приемлемого носителя главным образом зависит от того, приготовлена ли композиция по изобретению для перорального, назального, подкожного, внутримышечного или внутривенного введения. Композиция по настоящему изобретению может содержать увлажнитель, эмульгатор или буферное вещество в качестве добавок.
Фармацевтическая композиция по настоящему изобретению может быть введена любым подходящим способом, таким как пероральное, назальное, внутрикожное, подкожное, внутримышечное или внутривенное введение.
В одном из связанных аспектов в изобретении предложена фармацевтическая композиция для комбинирования антитела против PCSK9 и второго терапевтического агента. В одном из воплощений второй терапевтический агент представляет собой любой реагент, который благоприятным образом может быть скомбинирован с антителом против PCSK9. Примеры реагентов, которые могут быть благоприятным образом комбинированы с антителами против PCSK9, включают другие реагенты, которые подавляют активность PCSK9 (включая другие антитела или их антигенсвязывающие фрагменты, пептидный ингибитор, низкомолекулярный антагонист и т.п.) и/или реактив для взаимодействия с вышележащими или нижележащими сигнальными путями для PCSK9, но не ограничиваются ими.
Используемый в настоящей заявке термин "устранение, подавление или уменьшение активности PCSK9 для предупреждения или лечения заболеваний или состояний" подразумевает заболевания или состояния, вызываемые экспрессией PCSK9 или имеющие симптомы/особенности вследствие экспрессии PCSK9. В некоторых воплощениях заболевание или состояние выбрано из гиперлипидемии или гиперхолестеринемии.
Используемый в настоящей заявке термин "терапевтически эффективное количество" относится к дозе, достаточной для того, чтобы демонстрировать преимущество для субъекта, которой ее вводят. Реальное количество, а также скорость и время введения будут зависеть от состояния самого пациента и тяжести состояния. Наконец, предписание лечения (такое как определение дозы и т.п.) находится на ответственности практикующего врача и других докторов и зависит от его решения, как правило принимаемого с учетом заболевания, которое лечат, состояния конкретного пациента, места доставки лекарственного средства, способов введения и других факторов, известных докторам.
Используемый в настоящей заявке термин "субъект" относится к млекопитающему, такому как человек, но также может представлять собой других животных, таких как дикие животные (такие как цапля, аист, журавль и т.п.), домашний скот (такие как утки, гуси и т.п.) или экспериментальные животные (такие как орангутанг, обезьяна, крыса, мышь, кролик, морская свинка и т.п.).
С одной стороны, антитело по настоящему изобретению или его функциональный фрагмент содержит CDR тяжелой цепи, имеющие аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 4, 5, 6, 10, 11, 12, 16, 17, 18, 22, 23, 24, 28, 29 и 30, или из любого из вариантов этой последовательности, и/или CDR легкой цепи, имеющие аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 1, 2, 3, 7, 8, 9, 13, 14, 15, 19, 20, 21, 25, 26 и 27, или из любого из вариантов этой последовательности.
В некоторых предпочтительных воплощениях аминокислотные последовательности CDR1, CDR2 и CDR3 CDR тяжелой цепи выбраны из одной из групп, состоящих из следующих аминокислотных последовательностей, или их вариантов:
и/или аминокислотные последовательности CDR1, CDR2 и CDR3 CDR легкой цепи выбраны из группы, состоящей из следующих аминокислотных последовательностей, или их вариантов
В некоторых предпочтительных воплощениях аминокислотные последовательности CDR1, CDR2 и CDR3 тяжелой цепи и CDR1, CDR2 и CDR3 легкой цепи выбраны из группы, состоящей из следующих аминокислотных последовательностей, или их вариантов:
В некоторых воплощениях антитела или их функциональные фрагменты по настоящему изобретению содержат вариабельную область тяжелой цепи, выбранную изгруппы, состоящей из аминокислотных последовательностей SEQ ID NO: 32, 34, 36, 38 и 40, или из вариантов любой из указанных последовательностей, и/или вариабельную область легкой цепи, выбранную из группы, состоящей из аминокислотных последовательностей SEQ ID NO: 31, 33, 35, 37 и 39, или из вариантов указанных последовательностей.
В предпочтительном воплощении вариабельная область тяжелой цепи представляет собой SEQ ID NO: 32 или его вариант; вариабельная область легкой цепи представляет собой SEQ ID NO: 31 или его вариант.
В еще одном предпочтительном воплощении вариабельная область тяжелой представляет собой SEQ ID NO: 34 или его вариант; вариабельная область легкой представляет собой SEQ ID NO: 33 или его вариант.
В еще одном предпочтительном воплощении вариабельная область тяжелой цепи представляет собой SEQ ID NO: 36 или его вариант; вариабельная область легкой цепи представляет собой SEQ ID NO: 35 или его вариант.
В еще одном предпочтительном воплощении вариабельная область тяжелой цепи представляет собой SEQ ID NO: 38 или его вариант; вариабельная область легкой цепи представляет собой SEQ ID NO: 37 или его вариант.
В еще одном предпочтительном воплощении вариабельная область тяжелой цепи представляет собой SEQ ID NO: 40 или его вариант; вариабельная область легкой цепи представляет собой SEQ ID NO: 39 или его вариант.
Антитело или его функциональный фрагмент может представлять собой химерное антитело, гуманизированное антитело или полностью человеческое антитело.
Антитело по настоящему изобретению или его функциональный фрагмент может быть гуманизирован(о). Получение гуманизированного антитела хорошо известно специалистам в данной области техники. Например, последовательности CDR по настоящему изобретению могут быть перенесены в вариабельную область человеческого антитела для получения гуманизированного антитела против PCSK9 по настоящему изобретению. Гуманизированное антитело не вызывает реакцию против антитела (AAR) или реакцию в виде человеческого антитела против мышиного антитела (HAMA), не подвергается быстрому удалению путем нейтрализации направленным против него антителом, и приводит к иммунной эффекторной функции.
В некоторых предпочтительных воплощениях гуманизированное антитело противPCSK9 или его функционального фрагмента содержит вариабельную область тяжелой цепи, выбранную из группы, состоящей из аминокислотных последовательностей SEQ ID NO: 32, 34, 36, 38 и 40, или вариантов любой из указанных последовательностей, и/или вариабельная область легкой цепи, выбранную из группы, состоящей из аминокислотных последовательностей SEQ ID NO: 31, 33, 35, 37 и 39, или вариантов указанных последовательностей.
В одном из предпочтительных воплощений гуманизированного антитела по настоящему изобретению или его функционального фрагмента вариабельная область тяжелой цепи представляет собой SEQ ID NO: 32 или его вариант и указанная вариабельная область легкой цепи представляет собой SEQ ID NO: 31 или его вариант.
В еще одном предпочтительном воплощении гуманизированное антитело по настоящему изобретению или его функциональным фрагментом, вариабельная область тяжелой цепи представляет собой SEQ ID NO: 34 или его вариант и вариабельная область легкой цепи представляет собой SEQ ID NO: 33 или его вариант.
В еще одном предпочтительном воплощении гуманизированного антитела по настоящему изобретению или его функциональным фрагментом вариабельная область тяжелой цепи представляет собой SEQ ID NO: 36 или его вариант и вариабельная область легкой цепи представляет собой SEQ ID NO: 35 или его вариант.
В еще одном предпочтительном воплощении гуманизированного антитела по настоящему изобретению или его функциональным фрагментом, вариабельная область тяжелой цепи представляет собой SEQ ID NO:38 или его вариант и вариабельная область легкой цепи представляет собой SEQ ID NO: 37 или его вариант.
В еще одном предпочтительном воплощении гуманизированного антитела по настоящему изобретению или его функциональным фрагментом, вариабельная область тяжелой цепи представляет собой SEQ ID NO: 40 или его вариант и вариабельная область легкой цепи представляет собой SEQ ID NO: 39 или его вариант.
В еще одном предпочтительном воплощении гуманизированного антитела по настоящему изобретению или его функциональным фрагментом, вариабельная область тяжелой цепи представляет собой SEQ ID NO: 47 или его вариант и вариабельная область легкой цепи представляет собой SEQ ID NO: 45 или его вариант.
В еще одном предпочтительном воплощении гуманизированного антитела по настоящему изобретению или его функциональным фрагментом, вариабельная область тяжелой цепи представляет собой SEQ ID NO: 49 или его вариант и вариабельная область легкой цепи представляет собой SEQ ID NO: 45 или его вариант.
В изобретении также предложена выделенная молекула нуклеиновой кислоты, кодирующая антитело по настоящему изобретению или его функциональный фрагмент. В предпочтительном воплощении молекула нуклеиновой кислоты содержит нуклеотидную последовательность SEQ ID NO: 48, 50 и/или 46; или их комбинацию.
В изобретении также предложен вектор экспрессии, содержащий молекулу нуклеиновой кислоты и клетка-хозяин, содержащая вектор экспрессии.
В изобретении предложен способ получения антитела против PCSK9 или его функционального фрагмента, включающий стадии: культивирования клетки-хозяина по изобретению в условиях, дающих возможности для образования антитела или его функционального фрагмента, и выделения образовавшегося антитела или его функционального фрагмента.
Еще один аспект настоящего изобретения относится к иммуноконъюгату, содержащему антитело или его функциональный фрагмент, конъюгированное(ый) с терапевтическим агентом. Терапевтический агент предпочтительно представляет собой токсин, радиоактивный изотоп, лекарственное средство или цитотоксический агент.
Настоящее изобретение также относится к фармацевтической композиции, содержащей антитело по настоящему изобретению или его функциональный фрагмент и фармацевтически приемлемый носитель.
В еще одном аспекте настоящего изобретения предложен способ предупреждения или лечения заболеваний или состояний путем устранения, ингибирования или уменьшения активности PCSK9, включающий введение нуждающемуся субъекту терапевтически эффективного количества антитела или его функционального фрагмента, нуклеиновой кислоты, вектора экспрессии, клетки хозяина, иммуноконъюгата или фармацевтической композиции по настоящему изобретению.
В настоящем изобретении также предложено применение антитела или его функционального фрагмента, нуклеиновой кислоты, вектора экспрессии, клетки хозяина, иммуноконъюгата или фармацевтической композиции по настоящему изобретению в изготовлении лекарственного средства для лечения заболеваний или состояний.
Следующие примеры приведены для того, чтобы продемонстрировать и дополнительно объяснить некоторые предпочтительные воплощения и аспекты по настоящему изобретению, которые не должны быть интерпретированы как ограничивающие его объем.
Примеры
Пример 1: Клонирование внеклеточной области человеческого PCSK9 и внеклеточной области LDLR в эукариотической системе экспрессии
Внеклеточный фрагмент человеческого PCSK9 амплифицировали при помощи ПЦР (полимеразной цепной реакции) путем использования кДНК гена человеческого PCSK9 (от Beijing Sino Biological Inc.) с известной последовательностью в качестве матрицы (матричная последовательность является следующей), с прямым праймером 5'-GTACACTAGTCACCATGGGCACCGTCAGCTC-3' и обратным праймером 5'-GATCCTCGAGCCTGGAGCTCCTGGGAGG-3'. Амплифицированный продукт подвергали расщеплению двумя ферментами speI и XhoI и клонировали в самостоятельно сконструированную эукариотическую плазмидную систему экспрессии (pSec CAGA2 ECD). Клетку 293E переносили в плазмиду посредством PEI (полиэтиленимин). После 6 суток супернатант культуральной среды собирали, и внеклеточную область белка человеческого PCSK9 очищали при помощи аффинной хроматографии.
Аналогично, внеклеточный фрагмент LDLR амплифицировали путем ПЦР путем использования прямого праймера 5'-GTACGCTAGCCACCATGGGGCCCTGGGGCTG-3' и обратного праймера 5'-GATCCTCGAGCCCTCACGCTACTGG-3'. Амплифицированный продукт подвергали расщеплению двумя ферментами NheI и XhoI и клонировали в самостоятельно сконструированную эукариотическую плазмидную систему экспрессии. Клетку 293E трансфицировали в плазмиду посредством PEI. После 6 суток супернатант культуральной среды собирали, и внеклеточную область LDLR очищали.
Фиг. 1 представляет собой диаграмму электрофореза SDS-PAGE белка внеклеточной области человеческого PCSK9.
Нуклеотидная последовательность внеклеточной области человеческого PCSK9: SEQ ID NO:41
ATGGGCACCG TCAGCTCCAG GCGGTCCTGG TGGCCGCTGC CACTGCTGCT GCTGCTGCTG 61 CTGCTCCTGG GTCCCGCGGG CGCCCGTGCG CAGGAGGACG AGGACGGCGA CTACGAGGAG 121 CTGGTGCTAG CCTTGCGTTC CGAGGAGGAC GGCCTGGCCG AAGCACCCGA GCACGGAACC 181 ACAGCCACCT TCCACCGCTG CGCCAAGGAT CCGTGGAGGT TGCCTGGCAC CTACGTGGTG 241 GTGCTGAAGG AGGAGACCCA CCTCTCGCAG TCAGAGCGCA CTGCCCGCCG CCTGCAGGCC 301 CAGGCTGCCC GCCGGGGATA CCTCACCAAG ATCCTGCATG TCTTCCATGG CCTTCTTCCT 361
GGCTTCCTGG TGAAGATGAG TGGCGACCTG CTGGAGCTGG CCTTGAAGTT GCCCCATGTC 421 GACTACATCG AGGAGGACTC CTCTGTCTTT GCCCAGAGCA TCCCGTGGAA CCTGGAGCGG 481 ATTACCCCTC CACGGTACCG GGCGGATGAA TACCAGCCCC CCGACGGAGG CAGCCTGGTG 541 GAGGTGTATC TCCTAGACAC CAGCATACAG AGTGACCACC GGGAAATCGA GGGCAGGGTC 601 ATGGTCACCG ACTTCGAGAA TGTGCCCGAG GAGGACGGGA CCCGCTTCCA CAGACAGGCC 661 AGCAAGTGTG ACAGTCATGG CACCCACCTG GCAGGGGTGG TCAGCGGCCG GGATGCCGGC 721 GTGGCCAAGG GTGCCAGCAT GCGCAGCCTG CGCGTGCTCA ACTGCCAAGG GAAGGGCACG 781 GTTAGCGGCA CCCTCATAGG CCTGGAGTTT ATTCGGAAAA GCCAGCTGGT CCAGCCTGTG 841 GGGCCACTGG TGGTGCTGCT GCCCCTGGCG GGTGGGTACA GCCGCGTCCT CAACGCCGCC 901 TGCCAGCGCC TGGCGAGGGC TGGGGTCGTG CTGGTCACCG CTGCCGGCAA CTTCCGGGAC 961 GATGCCTGCC TCTACTCCCC AGCCTCAGCT CCCGAGGTCA TCACAGTTGG GGCCACCAAT 1021 GCCCAGGACC AGCCGGTGAC CCTGGGGACT TTGGGGACCA ACTTTGGCCG CTGTGTGGAC 1081 CTCTTTGCCC CAGGGGAGGA CATCATTGGT GCCTCCAGCG ACTGCAGCAC CTGCTTTGTG 1141 TCACAGAGTG GGACATCACA GGCTGCTGCC CACGTGGCTG GCATTGCAGC CATGATGCTG 1201 TCTGCCGAGC CGGAGCTCAC CCTGGCCGAG TTGAGGCAGA GACTGATCCA CTTCTCTGCC 1261 AAAGATGTCA TCAATGAGGC CTGGTTCCCT GAGGACCAGC GGGTACTGAC CCCCAACCTG 1321 GTGGCCGCCC TGCCCCCCAG CACCCATGGG GCAGGTTGGC AGCTGTTTTG CAGGACTGTG 1381 TGGTCAGCAC ACTCGGGGCC TACACGGATG GCCACAGCCA TCGCCCGCTG CGCCCCAGAT 1441 GAGGAGCTGC TGAGCTGCTC CAGTTTCTCC AGGAGTGGGA AGCGGCGGGG CGAGCGCATG 1501 GAGGCCCAAG GGGGCAAGCT GGTCTGCCGG GCCCACAACG CTTTTGGGGG TGAGGGTGTC 1561 TACGCCATTG CCAGGTGCTG CCTGCTACCC CAGGCCAACT GCAGCGTCCA CACAGCTCCA 1621 CCAGCTGAGG CCAGCATGGG GACCCGTGTC CACTGCCACC AACAGGGCCA CGTCCTCACA 1681 GGCTGCAGCT CCCACTGGGA GGTGGAGGAC CTTGGCACCC ACAAGCCGCC TGTGCTGAGG 1741 CCACGAGGTC AGCCCAACCA GTGCGTGGGC CACAGGGAGG CCAGCATCCA CGCTTCCTGC 1801 TGCCATGCCC CAGGTCTGGA ATGCAAAGTC AAGGAGCATG GAATCCCGGC CCCTCAGGAG 1861 CAGGTGACCG TGGCCTGCGA GGAGGGCTGG ACCCTGACTG GCTGCAGTGC CCTCCCTGGG 1921 ACCTCCCACG TCCTGGGGGC CTACGCCGTA GACAACACGT GTGTAGTCAG GAGCCGGGAC 1981 GTCAGCACTA CAGGCAGCAC CAGCGAAGAG GCCGTGACAG CCGTTGCCAT CTGCTGCCGG 2041 AGCCGGCACC TGGCGCAGGC CTCCCAGGAG CTCCAGTGA
Нуклеотидная последовательность внеклеточной области LDLR: SEQ ID NO: 42
ATGGGGCCCT GGGGCTGGAA ATTGCGCTGG ACCGTCGCCT TGCTCCTCGC CGCGGCGGGG ACTGCAGTGG GCGACAGATG CGAAAGAAAC GAGTTCCAGT GCCAAGACGG GAAATGCATC 121 TCCTACAAGT GGGTCTGCGA TGGCAGCGCT GAGTGCCAGG ATGGCTCTGA TGAGTCCCAG 181 GAGACGTGCT TGTCTGTCAC CTGCAAATCC GGGGACTTCA GCTGTGGGGG CCGTGTCAAC 241 CGCTGCATTC CTCAGTTCTG GAGGTGCGAT GGCCAAGTGG ACTGCGACAA CGGCTCAGAC 301 GAGCAAGGCT GTCCCCCCAA GACGTGCTCC CAGGACGAGT TTCGCTGCCA CGATGGGAAG 361 TGCATCTCTC GGCAGTTCGT CTGTGACTCA GACCGGGACT GCTTGGACGG CTCAGACGAG 421 GCCTCCTGCC CGGTGCTCAC CTGTGGTCCC GCCAGCTTCC AGTGCAACAG CTCCACCTGC 481 ATCCCCCAGC TGTGGGCCTG CGACAACGAC CCCGACTGCG AAGATGGCTC GGATGAGTGG 541 CCGCAGCGCT GTAGGGGTCT TTACGTGTTC CAAGGGGACA GTAGCCCCTG CTCGGCCTTC 601 GAGTTCCACT GCCTAAGTGG CGAGTGCATC CACTCCAGCT GGCGCTGTGA TGGTGGCCCC 661 GACTGCAAGG ACAAATCTGA CGAGGAAAAC TGCGCTGTGG CCACCTGTCG CCCTGACGAA 721 TTCCAGTGCT CTGATGGAAA CTGCATCCAT GGCAGCCGGC AGTGTGACCG GGAATATGAC 781 TGCAAGGACA TGAGCGATGA AGTTGGCTGC GTTAATGTGA CACTCTGCGA GGGACCCAAC 841 AAGTTCAAGT GTCACAGCGG CGAATGCATC ACCCTGGACA AAGTCTGCAA CATGGCTAGA 901 GACTGCCGGG ACTGGTCAGA TGAACCCATC AAAGAGTGCG GGACCAACGA ATGCTTGGAC 961 AACAACGGCG GCTGTTCCCA CGTCTGCAAT GACCTTAAGA TCGGCTACGA GTGCCTGTGC 1021 CCCGACGGCT TCCAGCTGGT GGCCCAGCGA AGATGCGAAG ATATCGATGA GTGTCAGGAT 1081 CCCGACACCT GCAGCCAGCT CTGCGTGAAC CTGGAGGGTG GCTACAAGTG CCAGTGTGAG 1141 GAAGGCTTCC AGCTGGACCC CCACACGAAG GCCTGCAAGG CTGTGGGCTC CATCGCCTAC 1201 CTCTTCTTCA CCAACCGGCA CGAGGTCAGG AAGATGACGC TGGACCGGAG CGAGTACACC 1261 AGCCTCATCC CCAACCTGAG GAACGTGGTC GCTCTGGACA CGGAGGTGGC CAGCAATAGA 1321 ATCTACTGGT CTGACCTGTC CCAGAGAATG ATCTGCAGCA CCCAGCTTGA CAGAGCCCAC 1381 GGCGTCTCTT CCTATGACAC CGTCATCAGC AGGGACATCC AGGCCCCCGA CGGGCTGGCT 1441 GTGGACTGGA TCCACAGCAA CATCTACTGG ACCGACTCTG TCCTGGGCAC TGTCTCTGTT 1501 GCGGATACCA AGGGCGTGAA GAGGAAAACG TTATTCAGGG AGAACGGCTC CAAGCCAAGG 1561 GCCATCGTGG TGGATCCTGT TCATGGCTTC ATGTACTGGA CTGACTGGGG AACTCCTGCC 1621 AAGATCAAGA AAGGGGGCCT GAATGGTGTG GACATCTACT CGCTGGTGAC TGAAAACATT 1681 CAGTGGCCCA ATGGCATCAC CCTAGATCTC CTCAGTGGCC GCCTCTACTG GGTTGACTCC 1741 AAACTTCACT CCATCTCAAG CATCGATGTC AACGGGGGCA ACCGGAAGAC CATCTTGGAG 1801 GATGAAAAGA GGCTGGCCCA CCCCTTCTCC TTGGCCGTCT TTGAGGACAA AGTATTTTGG 1861 ACAGATATCA TCAACGAAGC CATTTTCAGT GCCAACCGCC TCACAGGTTC CGATGTCAAC 1921 TTGTTGGCTG AAAACCTACT GTCCCCAGAG GATATGGTTC TCTTCCACAA CCTCACCCAG 1981 CCAAGAGGAG TGAACTGGTG TGAGAGGACC ACCCTGAGCA ATGGCGGCTG CCAGTATCTG 2041 TGCCTCCCTG CCCCGCAGAT CAACCCCCAC TCGCCCAAGT TTACCTGCGC CTGCCCGGAC 2101 GGCATGCTGC TGGCCAGGGA CATGAGGAGC TGCCTCACAG AGGCTGAGGC TGCAGTGGCC 2161 ACCCAGGAGA CATCCACCGT CAGGCTAAAG GTCAGCTCCA CAGCCGTAAG GACACAGCAC 2221 ACAACCACCC GACCTGTTCC CGACACCTCC CGGCTGCCTG GGGCCACCCC TGGGCTCACC 2281 ACGGTGGAGA TAGTGACAAT GTCTCACCAA GCTCTGGGCG ACGTTGCTGG CAGAGGAAAT 2341 GAGAAGAAGC CCAGTAGCGT GAGGG
Пример 2: Анализ ELISA связывания человеческого рекомбинантного белка PCSK9 с LDLR
2.1 Мечение человеческого рекомбинантного белка PCSK9 биотином
Человеческий рекомбинантный белок PCSK9 и биотин-xx-NHS, растворенный в DMSO (диметилсульфоксид), смешивали в отношении 1:10, выдерживали при комнатной температуре в течение 2 часов, и затем реакционную смесь пропускали через колонку для ультрафильтрации 10 kD для разделения меченого биотином человеческого PCSK9 и свободного биотина.
2.2 Обнаружение при помощи ELISA связывания человеческого PCSK9 и LDLR
Для определения связывающей способности человеческого PCSK9 с LDLR 1 мкг/мл LDLR вносили в 96-луночный планшет для ELISA в покрывающем буфере, хранили при 4°C в течение ночи. На следующие сутки раствор из лунок отбирали, и лунки трижды промывали промывающим буфером. Затем раствор PBS, содержащий 2% молока, добавляли для блокирования в течение 60 минут. После трехкратного промывания промывающим буфером в каждую лунку добавляли по 100 мкл меченого биотином PCSK9 в различных концентрациях, инкубировали в течение 1 часа при комнатной температуре, трижды промывали промывающим буфером. Меченые HRP (пероксидаза хрена) козьи антитела против мышиного антитела разбавляли промывающим буфером в отношении 1:10000, инкубировали в течение 1 часа при комнатной температуре, трижды промывали промывающим буфером и добавляли 50 мкл раствора субстрата TMB (тетраметилбензидин) для окрашивания. После проведения реакции в течение 8 минут при комнатной температуре реакцию останавливали при помощи 100 мкл 12M раствора HCl, и поглощение регистрировали при 450 нм.
Как можно видеть на Фиг. 2, человеческий рекомбинантный белок PCSK9 может специфически связываться с LDLR в зависимости от дозы.
Пример 3: Клеточный анализ связывания PCSK9 с LDLR
3.1 Конструирование стабильно трансфицируемого клеточного штамма 293F-LDLR
Сконструированную эукариотическую экспрессирующуюся плазмиду с полноразмерной последовательностью LDLR с пуромициновой системой скрининга трансфицировали в адгезивные клетки 293F с использованием PEI (полиэтиленимин). Через 24 часа после трансфекции осуществляли скрининг с использованием пуромицина (2 мкг/мл) до формирования стабильно трансфицируемого клеточного пула 293F-LDLR. В то же самое время, 96-луночный планшет засеивали 0,8 клетками/лунку в предельных разведениях. Через 15 суток моноклональное антитело 293F-LDLR отделяли и клетки пересевали с образованием стабильно трансфицируемого клеточного штамма 293F-LDLR.
3.2 Связывание меченного биотином человеческого PCSK9 D347Y со стабильно трансфицируемым клеточным штаммом 293F-LDLR
Различные концентрации меченого биотином человеческого рекомбинантного белка PCSK9 D347Y смешивали со стабильно трансфицируемой клеточной линией 293F-LDLR, инкубировали в течение 15 минут при температуре 4°C. После трехкратного промывания буфером FACS (20 мМ Tris, 100 мМ NaCl, 2 мМ Ca2+, 1% FBS (эмбриональная бычья сыворотка), pH 7,4) добавляли стрептавидин-аллофикоцианин (SA-APC, 2 мкг/мл), и инкубировали в течение 20 мин при 4°C. После трехкратного промывания буфером FACS обнаружение осуществляли путем проточной цитофлуориметрии.
Как показано на Фиг. 3, сконструированный человеческий рекомбинантный белок PCSK9 D347Y может специфически связываться с LDLR на клетках 293T-LDLR, демонстрируя зависимое от дозы действие.
3.3 Связывание меченного биотином человеческого PCSK9 с LDLR на клетках HepG2
Для дополнительного подтверждения связывающей способности hPCSK9 с LDLR различные концентрации меченого биотином человеческого рекомбинантного белка PCSK9 D347Y смешивали с клетками HepG2, инкубировали в течение 15 минут при температуре 4°C. После трехкратного промывания PBS добавляли стрептавидин- аллофикоцианин (SA-APC, 2 мкг/мл) и инкубировали в течение 20 мин при 4°C. После трехкратного промывания буфером FACS обнаружение осуществляли путем проточной цитофлуориметрии.
Как показано на Фиг. 4 результат согласуется с вышеизложенным. Человеческий рекомбинантный белок PCSK9 может специфически связываться с LDLR на клетках HepG2.
Пример 4: Получение мышиного антитела против PCSK9
4.1 Иммунизация животных:
Человеческий рекомбинантный белок PCSK9 в качестве антигена смешивали с равным количеством иммунного адъюванта (адъювант Фрейнда), и иммунизировали 5 шестинедельных самок мышей FVB. После первой иммунизации повторную иммунизацию осуществляли раз в неделю, при этом всего осуществляли 4 иммунизации.
4.2 Слияние клеток
После последней повторной иммунизации лимфатические узлы в основании бедра мыши собирали и измельчали в нормальном физиологическом растворе, и обогащенную лимфоцитами суспензию собирали, сливали с клетками SP20 в соответствии с традиционным способом электротрансфекции. Слитые клетки распределяли по 96 лункам с использованием полной культуральной среды RPMI-1640, содержащей HAT, и выращивали при 37°C с 5% CO2.
Пример 5: Анализ блокирования лиганда и рецептора
1220 клонов, которые могут секретировать антитела, связывающиеся с человеческим белком PCSK9, подвергали скринингу из 20000 различных моноклональных клеток гибридомы путем реакции ELISA. 15 из этих 1220 антител могут ингибировать связывание меченного биотином человеческого PCSK9 с LDLR на HepG2. Авторы изобретения сфокусировались на последующих экспериментах с 5 антителами, обладающими самой большой ингибирующей способностью.
Супернатант вышеуказанных пяти клеток гибридомы смешивали с меченным биотином человеческим PCSK9 (400 нг/мл) и инкубировали при комнатной температуре в течение 20 мин. Затем эту смесь и стабильно трансфицируемый клеточный штамм 293T-LDLR инкубировали при 37°C в течение 15 минут. После трехкратного промывания PBS добавляли 0,2 мкг/мл SA-APC и инкубировали при 4°C в течение 15 минут. После трехкратного промывания PBS проточный цитофлуориметр использовали для подтверждения того, может ли антитело, секретируемое клетками гибридомы, подавлять связывание человеческого PCSK9 с LDLR на поверхности клеток 293F-LDLR. Аналогично, с использованием референсного моноклонального антитела против PCSK9 (Merck) в качестве контроля осуществляли вышеприведенный анализ с концентрацией 4 мкг/мл.
Как показано на Фиг. 5 результат проточной цитофлуориметрии демонстрирует, что по сравнению с референсным антителом все 5 моноклональных антител-кандидатов 3G12, 1A5, 1B5, 1B11 и 2B12 могут быть эффективно связаны с человеческим PCSK9 так,
чтобы подавлять деградацию LDLR.
Пример 6: Перекрестная реакция между PCSK у различных видов Перекрестную реакцию между моноклональными мышиными антителами- кандидатами 3G12, 1A5 и 1B5, 1B11 и 2B12; человеческим PCSK9; мышиным PCSK9 и PCSK9 яванского макака определяли при помощи ELISA. Результаты эксперимента показывают, что мышиное антитело против PCSK9, полученное авторами изобретения, может перекрестно реагировать с антителами против PCSK9, о которых сообщали другие компании, что будет облегчать эксперименты на животных и экономить время. Результаты эксперимента представлены в таблице 1:
Таблица 1
ч: полученные от человека; м: полученные от мыши; ям: полученные от яванского макака.
Пример 7: Определение константы связывания с человеческим рекомбинантным белком PCSK9
Как показано в таблице 2, определяли константу комбинирования между различными мышиными антителами и HPCSK9 при помощи прибора ForteBio, что свидетельствует о том, что полученное мышиное антитело может специфически связываться с человеческим PCSK9.
Таблица 2
Пример 8: Связывание мышиного антитела-кандидата с человеческим PCSK9 96-луночный планшет для ELISA засевали, покрывали 4 мкг/мл стрептавидина, инкубировали при 37°C в течение 90 мин. Затем раствор из лунок отбирали, и лунки трижды промывали промывающим буфером, и раствор PBS, содержащий 2% молока, добавляли для блокирования в течение 60 минут. После трехкратного промывания промывающим буфером в каждую лунку добавляли 2 мкг/мл меченого биотином человеческого PCSK9, инкубировали в течение 1 часа при 37°C и трижды промывали промывающим буфером. Затем добавляли химерные антитела, которые разбавляли в различных отношениях, инкубировали при 37°C в течение 1 часа. Затем добавляли химерное антитело в различных разведениях и инкубировали в течение 1 часа при 37°C. После трехкратного промывания промывающим буфером меченые HRP мышиные антитела против человеческого IgG (H+L) разбавляли промывающим буфером в отношении 1:5000, инкубировали в течение 1 часа при комнатной температуре. После трехкратного промывания промывающим буфером добавляли 100 мкл раствора субстрата TMB для окрашивания, осуществляя реакцию в течение 10 минут при комнатной температуре. Реакцию останавливали при помощи 100 мкл 12M раствора HCl, и поглощение регистрировали при 450 нм.
Как показано на Фиг. 6, мышиное антитело может специфически связываться с человеческим PCSK9. По сравнению с референсным антителом 2B12 и 3G12 обладают лучшей связывающей способностью.
Для подтверждения специфического связывания антитела с человеческим PCSK9 по аналогии с вышеприведенным экспериментом антитело (1 мкг/мл) высевали в 96- луночный планшет, инкубировали в течение 60 минут при 37°C. Затем раствор из лунок отбирали и лунки трижды промывали промывающим буфером. Затем раствор PBS, содержащий 2% молоко, добавляли для блокирования в течение 60 минут. После трехкратного промывания промывающим буфером добавляли 100 мкл меченого биотином человеческого PCSK9 в различных концентрациях, инкубировали в течение 1 часа при 37°C. После трехкратного промывания промывающим буфером HRP-стрептавидин разбавляли промывающим буфером в отношении 1:5000, инкубировали в течение 1 часа при комнатной температуре. После трехкратного промывания промывающим буфером добавляли 100 мкл раствора субстрата TMB для окрашивания и реакцию осуществляли в течение 10 минут при комнатной температуре. Реакцию останавливали при помощи 100 мкл 12M раствора HCl, и поглощение регистрировали при 450 нм. Как показано на Фиг. 7, профиль связывания мышиного антитела с человеческим PCSK9 согласуется с профилем в предыдущем эксперименте.
На основании этого дополнительно сравнивали сходства и различия между связыванием различных сконструированных мышиных антител с антигенными эпитопами человеческого PCSK9. Покрывали 4 мкг/мл стрептавидина и инкубировали при постоянной температуре 37°C в течение 90 минут. Затем раствор из лунок отбирали, и лунки трижды промывали промывающим буфером. Затем раствор PBS, содержащий 2% молоко, добавляли для блокирования в течение 60 минут. После трехкратного промывания промывающим буфером в каждую лунку добавляли 100 мкл меченого биотином человеческого PCSK9 в концентрации 2 мкг/мл, инкубировали в течение 1 часа при 37°C и трижды промывали промывающим буфером. Затем добавляли химерные антитела, которые разбавляли в различных отношениях с 200 нг/мл референсного антитела, инкубировали при 37°C в течение 90 минут. После трехкратного промывания промывающим буфером меченые HRP козьи антитела против человеческого IgG-Fc разбавляли промывающим буфером в отношении 1:5000, инкубировали в течение 1 часа при комнатной температуре. После трехкратного промывания промывающим буфером добавляли 100 мкл раствора субстрата TMB для окрашивания, и реакцию осуществляли в течение 10 минут при комнатной температуре. Реакцию останавливали при помощи 100 мкл 12M раствора HCl, и поглощение регистрировали при 450 нм.
Как показано на Фиг. 8, 1B11 и 2B12 не могут значимо конкурировать с референсным антителом за связывание с антигенным эпитопом человеческого PCSK9; тогда как 1A5 и 1B5, 3G12 могут значимо конкурентно связываться с антигенными эпитопами на человеческом PCSK, которые в ином случае могут связываться с референсным моноклональным антителом. Предполагается, что конкурирующая связывающая способность 1A5, 1B5 и 3G12 значимо сильнее, чем у референсного антитела.
Пример 9: Действие захвата LDL на клетки HepG2
Антитела в различных разведениях инкубировали с меченым биотином человеческим PCSK9-D347Y (2,5 мкг/мл) в течение 1 часа при комнатной температуре и затем смешивали с FL-LDL (5 мкг/мл), инкубировали в течение 3 часов при 37°C и трижды промывали PBS, и затем флуоресценцию обнаруживали с использованием Tecan Safire 2 (возбуждение при 514 нм/испускание при 570 нм).
Как можно видеть, исходя из результатов на Фиг. 9 и в таблице 3, различные мышиные моноклональные антитела могут связываться с человеческим PSCK9 для подавления его действия в отношении захвата LDL с IC50 (средняя ингибирующая концентрация) приблизительно 1мкг/мл.
Таблица 3
Пример 10: Получение последовательностей-кандидатов вариабельной области
Кандидаты-клетки гибридомы выращивали, центрифугировали при 1000 об./мин и тотальную РНК экстрагировали при помощи тризола. После синтеза первую цепь кДНК использовали в качестве последующей матрицы для амплификации соответствующей последовательности ДНК вариабельной области клеток гибридомы (Jones and Bendig 1991). В 50 мкл реакционной системы добавляли 1 мкл кДНК, 5 мкл 10X буфера для ПЦР, по 1 мкл прямых и обратных праймеров, соответственно (25 пмоль), 1 мкл dNTP (дезоксирибонуклеозидтрифосфатов), 1 мкл 25 ммоль PL MgCl2, 39 мкл H2O, предварительная денатурация при 95°C в течение 10 мин, и 1 мкл фермента Taq. Осуществляли температурные циклы и амплификацию путем ПЦР Условия реакции представляли собой: денатурация при 94°C в течение 1 мин, отжиг при 58°C в течение 1 мин, элонгация при 72°C в течение 15 с, всего 32 цикла, и затем поддерживали при 72°C в течение 10 мин.
После секвенирования амплифицированных продуктов последовательности вариабельной области тяжелой цепи и легкой цепи в гибридоме-кандидате являются следующими: 1A5 LC SEQ ID NO:31
DIQMTQSPSSLSASLGDKVTITCKASQDINKYIDWYQHKPGKGPRLLIHYTSTLQP GIP SRF SGSGS GRDYSF SISNLEPEDIATYYCVQYDDLWTF GGGTKLEIK SEQ ID NO:1 LCDR1: ASQDINKYID SEQ ID NO:2 LCDR2: YTSTLQP SEQ ID NO:3 LCDR3: VQYDDLWT 1A5 HC SEQ ID NO:32
QVQLKQSGPSLVQPSQSLSITCTVSGFSLTSYGVHWVRQSPGKGLEWLGVIWRG GSTDYNAAFMSRLSITKDNSKSQVFFKMNSLQADDTAIYYCANHRDWGQGTLVTVSA SEQ ID NO:4 HCDR1: SYGVH
SEQ ID NO:5 HCDR2: VIWRGGSTDYNAAFMS SEQ ID NO:6 HCDR3: HRD 3G12 LC SEQ ID NO:33
DIQMTQSPSSLSASLGGKVTITCKASQDINKYIDWYQHKPGKSPRLLIHYASTLQP GIPSRFSGSGSGRDYSFSISNLEPEDIATYYCLQYDDLWTFGGGTKLEIK SEQ ID NO:7 LCDR1: ASQDINKYID SEQ ID NO:8 LCDR2: YASTLQP SEQ ID NO:9 LCDR3: LQYDDLWT 3G12 HC SEQ ID NO:34
QVQLKQSGPSLLQPSQSLSITCTVSGFSLTSYGIHWVRQSPGKGLEWLGVIWRGGI TDYNAPFMSRLNITKDNSKNQVFFKMNSLQVDDTAIYYCANHRDWGQGTLVTVSA SEQ ID NO:10 HCDR1: SYGIH SEQ ID NO:11 HCDR2: VIWRGGITDYNAPFMS SEQ ID NO:12 HCDR3: HRD 1B5 LC SEQ ID NO:35
DIVLTQSPASLVVSLGQRATISCRASESVDNYGISFMNWFQQKPGQPPKLLIYTTS NQGSGVRARLSGSGCGTDFSLNIHPMEEDDSAMYFCQQSKEVPYTFGGGTKLEIK SEQ ID NO:13 LCDR1: RASESVDNYGISFMN SEQ ID NO:14 LCDR2: TTSNQGS SEQ ID NO:15 LCDR3: QQSKEVPYT 1B5 HC SEQ ID NO:36
DVQLQESGPGLVKPSQSLSLTCTVTGYSITSDYAWNWIRQFPGNKVEWMGYISY SGSSSYNPSLKGRISITRDTSKNQFFLQLNSVTTEDTATYYCARFYYRFDAYFDSWGQGT TLTVSS
SEQ ID NO:16 HCDR1: SDYAWN SEQ ID NO:17 HCDR2: YISYSGSSSYNPSLKG SEQ ID NO:18 HCDR3: FYYRFDAYFDS 2B12 LC
SEQ ID NO:37
DIVLTQSPASLAVSLGQRATISCRASESVEYYGTSLMHWYQHKPGQTPKLLIYSG SNVESGVPARF SGSGS GTDF SLNIHPVEEDDIAMYFCQQSREVPSTFGGGTKLEIK SEQ ID NO:19 LCDR1: RASESVEYYGTSLMH SEQ ID NO:20 LCDR2: SGSNVES SEQ ID NO:21 LCDR3: QQSREVPST 2B12 HC SEQ ID NO:38
DVQLQESGPGLVKPSQSLSLTCSVTGFSITSDYAWNWIRQFPGNKLEWMGYISYS GTTNYNPSLKSRISITRDTSKNQFFLHLNSVITEDTATYYCARREGHYSWFPYWGQGTLV TVSA
SEQ ID NO:22 HCDR1: SDYAWN
SEQ ID NO:23 HCDR2: YISYSGTTNYNPSLKS
SEQ ID NO:24 HCDR3: REGHYSWFPY
1B11 LC
SEQ ID NO:39
DIQMTQSPASLSASVGETVTITCRASENIYSYLAWYQQKQENSPQLLVYNAYTLA DGVPSRFSGSGSGTQFSLKIISLQPEDFGNYYCQHHYRTPPTFGGGTKLEIK SEQ ID NO:25 LCDR1: RASENIYSYLA SEQ ID NO:26 LCDR2: NAYTLAD SEQ ID NO:27 LCDR3: QHHYRTPPT 1B11 HC SEQ ID NO:40
QVQLQQSGAVLVRPGTSIKVSCKASGYAFTNYLIEWIKKRPGQGLEWIGMINPGS GDTNFNEKFKAKATLTADKSSTTAYMQLNSLTFDDSAVYFCARSSQLGLPYWGQGTLV TVSA
SEQ ID NO:28 HCDR1: NYLIE
SEQ ID NO:29 HCDR2: MINPGSGDTNFNEKFKA
SEQ ID NO:30 HCDR3: SSQLGLPY
Пример 11: Конструирование вектора экспрессии для химерного антитела
Fc фрагмент константной области тяжелой цепи и k/£ константную область легкой цепи клонировали из клеток человеческой крови (Beijing Blood Institute) и затем лигировали в плазмиду pCDNA3.1 для конструирования. Вышеприведенные фрагменты последовательности тяжелой цепи и легкой цепи синтезировали в Genscript, причем тяжелую цепь, расщепленную при помощи BspqI, и легкую цепь, расщепленную при помощи BspqI, соответственно, лигировали в сконструированные плазмиды pCDNA3.1, и секвенировали для получения правильных клонов. Последующие экспериментальные материалы получали путем трансфекции этих серий плазмид в клетки и очистки. По аналогии с вышеприведенными экспериментами тяжелую цепь и легкую цепь клонировали в сконструированную плазмиду pCDNA3.1, содержащую Fc фрагмент мышиной константной области тяжелой цепи и k/£ константную область легкой цепи.
Пример 12: Связывание химерного антитела с человеческим PCSK9
96-луночный планшет для ELISA засевали, покрывали 4 мкг/мл стрептавидина и инкубировали при 37°C в течение 90 мин. Затем раствор из лунок отбирали и лунки трижды промывали промывающим буфером. Раствор PBS, содержащий 2% молока, добавляли для блокирования в течение 60 минут. После трехкратного промывания промывающим буфером в каждую лунку добавляли по 100 мкл меченного биотином PCSK9 в концентрации 2 мкг/мл, инкубировали при 37°C в течение 1 часа и трижды промывали промывающим буфером. Затем добавляли химерные антитела, которые разбавляли в различных отношениях, инкубировали при 37°C в течение 1 часа. После трехкратного промывания промывающим буфером меченые HRP мышиные антитела против человеческого IgG (H+L) разбавляли промывающим буфером в отношении 1:5000, инкубировали в течение 1 часа при комнатной температуре. После трехкратного промывания промывающим буфером добавляли 100 мкл раствора субстрата TMB для окрашивания и реакцию осуществляли в течение 10 минут при комнатной температуре. Реакцию останавливали при помощи 100 мкл 12M раствора HCl, и поглощение регистрировали при 450 нм.
Как показано на Фиг. 10, как химерное антитело 3G12, так и мышиное антитело могут специфически связываться с человеческим PCSK9.
Пример 13: Влияние химерного антитела на захват LDL клетками HepG2
Сконструированное химерное антитело (10 мкг/мл) и меченный биотином человеческий PCSK9 (400 нг/мл) смешивали и инкубировали при комнатной температуре в течение 60 мин. Смесь добавляли к клеткам HepG2 и затем добавляли FL-LDL (5
мкг/мл), инкубировали при 37°C в течение 3 часов. После 3 промываний с использованием PBS образцы загружали и детектировали при помощи проточной цитофлуориметрии.
Как показано на Фиг. 11, сконструированное химерное антитело 3G12 может подавлять действие человеческого PCSK9 на захват LDL.
Пример 14: Гуманизация антитела
Гуманизацию осуществляли в соответствии с последовательностями вариабельной области антител, секретируемых полученными выше клетками гибридомы. Вкратце, процесс гуманизации включал следующие стадии: A, выравнивание последовательностей генов антител, секретируемых каждой из клеток гибридомы, с последовательностю гена человеческой эмбриональной линии антитела, и выявление последовательности с высокой гомологией; Б, анализ аффинности HLA-DR для отбора каркасной последовательности человеческой эмбриональной линии с низкой аффинностью; В, использование технологии компьютерного модулирования для получения предполагаемого пространственного-3D профиля связывания путем анализа вариабельной области и соседней каркасной аминокислотной последовательности при помощи молекулярного докинга. Путем расчета факторов, таких как электростатическая сила, сила Ван-дер-Ваальса, гидрофильность, гидрофобность и энтропия, анализа отдельных критических аминокислот, которые могут взаимодействовать с PD-1 и поддерживать пространственную конструкцию в каждой последовательности гена антитела, секретируемого клетками гибридомы, их сшивают с геном выбранной каркасной цепи человеческой эмбриональной линии, и на основе этого метят аминокислотные сайты каркасной цепи, которые необходимо сохранить. Синтезируют случайные праймеры и готовят фаговую библиотеку для скрининга библиотеки гуманизированного антитела (Pini, A.et al.(1998).
Design and Use of a Phage Display Library: HUMAN ANTIBODIES WITH 10 SUBNANOMOLAR AFFINITY AGAINST A MARKER OF ANGIOGENESIS ELUTED FROM A TWO-DIMENSIONAL GEL., Journal of Biological Chemistry, 273(34): 2176921776). На основе вышеприведенного авторы изобретения получили следующие различные гуманизированные антитела, включая следующие клоны: 32 и 77, которые имеют одну и ту же легкую цепь.
32/77-LC
(SEQ ID NO:45, вариабельная область легкой цепи)
DIQMTQSPSSLSASVGDRVTITCQASQDINKYIDWYQHKPGKAPKLLIHYASTLO PGVPSRFSGSGSGRDYTFTISSLQPEDIATYYCLQYDDLWTFGQGTKVEIK
(SEQ ID NO:46, нуклеотидная последовательность вариабельной области легкой
цепи)
GACATCCAGATGACCCAGAGCCCCAGCAGCCTGAGCGCCAGCGTGGGCGAC AGAGTGACCATCACCTGCCAGGCCAGCCAGGACATCAACAAGTACATCGACTGGTA CCAGCACAAGCCCGGCAAGGCCCCCAAGCTGCTGATCCACTACGCCAGCACCCTGC AGCCCGGCGTGCCCAGCAGATTCAGCGGCAGCGGCAGCGGCAGAGACTACACCTTC ACCATCAGCAGCCTGCAGCCCGAGGACATCGCCACCTACTACTGCCTGCAGTACGA CGACCTGTGGACCTTCGGCCAGGGCACCAAGGTGGAGATCAAG 32-HC(SEQ ID NO:47, вариабельная область тяжелой цепи)
QVQLQESGPGLVKPSQTLSLTCTVSGFSISSYGIHWIRQSPGKGLEWIGVIWRGGI TDYNAPFMSRVTISKDNSKNQVSFKLSSVTAADTAVYYCANHRDWGQGTLVTVSS
(SEQ ID NO:48, нуклеотидная последовательность вариабельной области тяжелой
цепи)
CAGGTGCAGCTGCAGGAAAGCGGCCCGGGCCTGGTGAAACCGAGCCAGACC CTGAGCCTGACCTGCACCGTGAGCGGCTTTAGCATTAGCAGCTATGGCATTCATTGG ATTCGCCAGAGCCCGGGCAAAGGCCTGGAATGGATTGGCGTGATTTGGCGCGGCGG CATTACCGATTATAACGCGCCGTTTATGAGCCGCGTGACCATTAGCAAAGATAACAG CAAAAACCAGGTGAGCTTTAAACTGAGCAGCGTGACCGCGGCGGATACCGCGGTGT ATTATTGCGCGAACCATCGCGATTGGGGCCAGGGCACCCTGGTGACCGTGAGCAGC 77-HC
(SEQ ID NO:49, вариабельная область тяжелой цепи)
QVQLQESGPGLVKPSQTLSLTCTVSGFSISSYGVHWIRQSPGKGLEWIGVIWRGG STDYNAAFMSRVTISKDNSKNQVSFKLSSVTAADTAVYYCANHRDWGQGTLVTVS S
(SEQ ID NO:50, нуклеотидная последовательность вариабельной области тяжелой
цепи)
CAGGTGCAGCTGCAGGAAAGCGGCCCGGGCCTGGTGAAACCGAGCCAGACCCT GAGCCTGACCTGCACCGTGAGCGGCTTTAGCATTAGCAGCTATGGCGTGCATTGGAT TCGCCAGAGCCCGGGCAAAGGCCTGGAATGGATTGGCGTGATTTGGCGCGGCGGCA GCACCGATTATAACGCGGCGTTTATGAGCCGCGTGACCATTAGCAAAGATAACAGC AAAAACCAGGTGAGCTTTAAACTGAGCAGCGTGACCGCGGCGGATACCGCGGTGTA
TTATTGCGCGAACCATCGCGATTGGGGCCAGGGCACCCTGGTGACCGTGAGCAGC
Пример 15: Связывание гуманизированного антитела с человеческим PCSK9
96-луночный планшет для ELISA засевали, покрывали 0,1 мкг/мл стрептавидина и инкубировали при 37°C в течение 60 мин. Затем раствор из лунок отбирали и лунки трижды промывали промывающим буфером. Раствор PBS, содержащий 2% BSA, добавляли для блокирования в течение 60 минут. После трехкратного промывания промывающим буфером в каждую лунку добавляли по 100 мкл меченого биотином PCSK9 в концентрации 0,2 мкг/мл, инкубировали при 37°C в течение 1 часа и трижды промывали промывающим буфером. Затем добавляли гуманизированные антитела, которые разбавляли в различных отношениях, инкубировали при 37°C в течение 1 часа. После трехкратного промывания промывающим буфером меченые HRP мышиные антитела против человеческого IgG (H+L) разбавляли промывающим буфером в отношении 1:10000, инкубировали в течение 1 часа при комнатной температуре. После трехкратного промывания промывающим буфером добавляли 100 мкл раствора субстрата TMB для окрашивания и реакцию осуществляли в течение 10 минут при комнатной температуре. Реакцию останавливали при помощи 100 мкл 12M раствора HCl, и поглощение регистрировали при 450 нм.
Как показано на Фиг. 12, после гуманизации связывающая способность с человеческим PSK9 не изменяется с величиной EC50 (средняя эффективная концентрация), составляющей приблизительно 5 нг/мл.
Пример 16: Влияние гуманизированного антитела на захват LDL клетками HepG2
Гуманизированное антитело и референсное антитело (градиент разбавления с различными концентрациями с начальной концентрацией 10 мкг/мл и 2-кратным градиентом концентрации) и HuPCSK9-D347Y (2,5 мкг/мл) инкубировали при комнатной температуре в течение 30 мин и добавляли к клеткам HepG2, инкубировали при 37°C с 7% CO2 в течение 1 ч. Затем добавляли Dil-LDL (5 мкг/мл), инкубировали при 37°C с 7% CO2 в течение 5 часов. После четырехкратного промывания при помощи PBS Tecan M1000 Pro использовали для обнаружения при помощи флуорисценции (возбуждение 554 нм/испускание 571 нм).
Как показано на Фиг. 14, гуманизированное антитело может эффективно подавлять связывание PCSK9 с LDLR для улучшения захвата LDL клетками HepG2 с величиной IC50
приблизительно 1 мкг/мл.
Пример 17: Гуманизированное антитело может эффективно уменьшать in vivo уровень LDL-c у яванского макака (Macaca fascicularis)
Яванского макака использовали в качестве животной модели с подкожным введением одиночной дозы. В различные моменты времени изменения уровня LDL-c в сыворотке крови и концентрации антитела были обнаружены при различных дозах (3, 10, 30 мг/кг).
Как показано на Фиг. 14, при детектировании можно видеть, что при различных вводимых дозах количество LDL-c в сыворотке крови существенно уменьшается. На D7 (седьмые сутки) после введения обнаружено 70% уменьшение уровня LDL-c. В то же самое время, период полувыведения гуманизированного антитела по сравнению с химерным антителом значительно увеличивался.
Ссылки
Abifadel, M., et al. (2003). "Mutations in PCSK9 cause autosomal dominant hypercholesterolemia." Nat Genet 34(2): 154-156.
Abifadel, M., et al. (2003). "Mutations in PCSK9 cause autosomal dominant hypercholesterolemia." Nat Genet 34(2): 154-156.
Benjannet, S., et al. (2004). "NARC-1/PCSK9 and its natural mutants zymogen cleavage and effects on the low density lipoprotein (LDL) receptor and LDL cholesterol." Journal of Biological Chemistry 279(47): 48865-48875.
Fisher, T. S., et al. (2007). "Effects of pH and Low Density Lipoprotein (LDL) on PCSK9-dependent LDL Receptor Regulation." Journal of Biological Chemistry 282(28): 2050220512.
Jones, S. T. and M. M. Bendig (1991). "Rapid PCR-cloning of full-length mouse immunoglobulin variable regions." Bio/Technology 9(1): 88-89.
Lagace, T. A., et al. (2006). "Secreted PCSK9 decreases the number of LDL receptors in hepatocytes and inlivers of parabiotic mice." Journal of Clinical Investigation 116(11): 29953005.
Lambert, G., et al. (2009). "Molecular basis of PCSK9 function." Atherosclerosis 203(1):
1-7.
Seidah, N. G., et al. (2003). "The secretory proprotein convertase neural apoptosis- regulated convertase 1 (NARC-1): liver regeneration and neuronal differentiation." Proceedings
of the National Academy of Sciences 100(3): 928-933.
Zhang, D.-W., et al. (2007). "Binding of Proprotein Convertase Subtilisin/Kexin Type 9 to Epidermal Growth Factor-like Repeat A of Low Density Lipoprotein Receptor Decreases Receptor Recycling and Increases Degradation." Journal of Biological Chemistry 282(25): 18602-18612
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ.
<110> JUNMENG BIOSCIENCE CO. LTD
<120> АНТИТЕЛО ПРОТИВ PCSK9 И ЕГО ПРИМЕНЕНИЕ
<130> 20150829
<160> 50
<170> PatentIn version 3.5
<210> 1
<211> 10
<212> PRT
<213> Mus musculus
<400> 1
Ala Ser Gln Asp Ile Asn Lys Tyr Ile Asp
1 5 10
<210> 2
<211> 7
<212> PRT
<213> Mus musculus
<400> 2
Tyr Thr Ser Thr Leu Gln Pro
1 5
<210> 3
<211> 8
<212> PRT
<213> Mus musculus
<400> 3
Val Gln Tyr Asp Asp Leu Trp Thr
1 5
<210> 4
<211> 5
<212> PRT
<213> Mus musculus
<400> 4
Ser Tyr Gly Val His
1 5
<210> 5
<211> 16
<212> PRT
<213> Mus musculus
<400> 5
Val Ile Trp Arg Gly Gly Ser Thr Asp Tyr Asn Ala Ala Phe Met Ser
1 5 10 15
<210> 6
<211> 6
<212> PRT
<213> Mus musculus
<400> 6
His Arg Asp
1
<210> 7
<211> 10
<212> PRT
<213> Mus musculus
<400> 7
Ala Ser Gln Asp Ile Asn Lys Tyr Ile Asp
1 5 10
<210> 8
<211> 7
<212> PRT
<213> Mus musculus
<400> 8
Tyr Ala Ser Thr Leu Gln Pro
1 5
<210> 9
<211> 8
<212> PRT
<213> Mus musculus
<400> 9
Leu Gln Tyr Asp Asp Leu Trp Thr
1 5
<210> 10
<211> 5
<212> PRT
<213> Mus musculus
<400> 10
Ser Tyr Gly Ile His
1 5
<210> 11
<211> 16
<212> PRT
<213> Mus musculus
<400> 11
Val Ile Trp Arg Gly Gly Ile Thr Asp Tyr Asn Ala Pro Phe Met Ser
1 5 10 15
<210> 12
<211> 3
<212> PRT
<213> Mus musculus
<400> 12
His Arg Asp
1
<210> 13
<211> 15
<212> PRT
<213> Mus musculus
<400> 13
Arg Ala Ser Glu Ser Val Asp Asn Tyr Gly Ile Ser Phe Met Asn
1 5 10 15
<210> 14
<211> 7
<212> PRT
<213> Mus musculus
<400> 14
Thr Thr Ser Asn Gln Gly Ser
1 5
<210> 15
<211> 9
<212> PRT
<213> Mus musculus
<400> 15
Gln Gln Ser Lys Glu Val Pro Tyr Thr
1 5
<210> 16
<211> 6
<212> PRT
<213> Mus musculus
<400> 16
Ser Asp Tyr Ala Trp Asn
1 5
<210> 17
<211> 16
<212> PRT
<213> Mus musculus
<400> 17
Tyr Ile Ser Tyr Ser Gly Ser Ser Ser Tyr Asn Pro Ser Leu Lys Gly
1 5 10 15
<210> 18
<211> 11
<212> PRT
<213> Mus musculus
<400> 18
Phe Tyr Tyr Arg Phe Asp Ala Tyr Phe Asp Ser
1 5 10
<210> 19
<211> 15
<212> PRT
<213> Mus musculus
<400> 19
Arg Ala Ser Glu Ser Val Glu Tyr Tyr Gly Thr Ser Leu Met His
1 5 10 15
<210> 20
<211> 7
<212> PRT
<213> Mus musculus
<400> 20
Ser Gly Ser Asn Val Glu Ser
1 5
<210> 21
<211> 9
<212> PRT
<213> Mus musculus
<400> 21
Gln Gln Ser Arg Glu Val Pro Ser Thr
1 5
<210> 22
<211> 6
<212> PRT
<213> Mus musculus
<400> 22
Ser Asp Tyr Ala Trp Asn
1 5
<210> 23
<211> 6
<212> PRT
<213> Mus musculus
<400> 23
Ser Asp Tyr Ala Trp Asn
1 5
<210> 24
<211> 10
<212> PRT
<213> Mus musculus
<400> 24
Arg Glu Gly His Tyr Ser Trp Phe Pro Tyr
1 5 10
<210> 25
<211> 11
<212> PRT
<213> Mus musculus
<400> 25
Arg Ala Ser Glu Asn Ile Tyr Ser Tyr Leu Ala
1 5 10
<210> 26
<211> 7
<212> PRT
<213> Mus musculus
<400> 26
Asn Ala Tyr Thr Leu Ala Asp
1 5
<210> 27
<211> 9
<212> PRT
<213> Mus musculus
<400> 27
Gln His His Tyr Arg Thr Pro Pro Thr
1 5
<210> 28
<211> 5
<212> PRT
<213> Mus musculus
<400> 28
Asn Tyr Leu Ile Glu
1 5
<210> 29
<211> 17
<212> PRT
<213> Mus musculus
<400> 29
Met Ile Asn Pro Gly Ser Gly Asp Thr Asn Phe Asn Glu Lys Phe Lys
1 5 10 15
Ala
<210> 30
<211> 8
<212> PRT
<213> Mus musculus
<400> 30
Ser Ser Gln Leu Gly Leu Pro Tyr
1 5
<210> 31
<211> 106
<212> PRT
<213> Mus musculus
<400> 31
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Leu Gly
1 5 10 15
Asp Lys Val Thr Ile Thr Cys Lys Ala Ser Gln Asp Ile Asn Lys Tyr
20 25 30
Ile Asp Trp Tyr Gln His Lys Pro Gly Lys Gly Pro Arg Leu Leu Ile
35 40 45
His Tyr Thr Ser Thr Leu Gln Pro Gly Ile Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Arg Asp Tyr Ser Phe Ser Ile Ser Asn Leu Glu Pro
65 70 75 80
Glu Asp Ile Ala Thr Tyr Tyr Cys Val Gln Tyr Asp Asp Leu Trp Thr
85 90 95
Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 32
<211> 111
<212> PRT
<213> Mus musculus
<400> 32
Gln Val Gln Leu Lys Gln Ser Gly Pro Ser Leu Val Gln Pro Ser Gln
1 5 10 15
Ser Leu Ser Ile Thr Cys Thr Val Ser Gly Phe Ser Leu Thr Ser Tyr
20 25 30
Gly Val His Trp Val Arg Gln Ser Pro Gly Lys Gly Leu Glu Trp Leu
35 40 45
Gly Val Ile Trp Arg Gly Gly Ser Thr Asp Tyr Asn Ala Ala Phe Met
50 55 60
Ser Arg Leu Ser Ile Thr Lys Asp Asn Ser Lys Ser Gln Val Phe Phe
65 70 75 80
Lys Met Asn Ser Leu Gln Ala Asp Asp Thr Ala Ile Tyr Tyr Cys Ala
85 90 95
Asn His Arg Asp Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ala
100 105 110
<210> 33
<211> 106
<212> PRT
<213> Mus musculus
<400> 33
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Leu Gly
1 5 10 15
Gly Lys Val Thr Ile Thr Cys Lys Ala Ser Gln Asp Ile Asn Lys Tyr
20 25 30
Ile Asp Trp Tyr Gln His Lys Pro Gly Lys Ser Pro Arg Leu Leu Ile
35 40 45
His Tyr Ala Ser Thr Leu Gln Pro Gly Ile Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Arg Asp Tyr Ser Phe Ser Ile Ser Asn Leu Glu Pro
65 70 75 80
Glu Asp Ile Ala Thr Tyr Tyr Cys Leu Gln Tyr Asp Asp Leu Trp Thr
85 90 95
Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 34
<211> 111
<212> PRT
<213> Mus musculus
<400> 34
Gln Val Gln Leu Lys Gln Ser Gly Pro Ser Leu Leu Gln Pro Ser Gln
1 5 10 15
Ser Leu Ser Ile Thr Cys Thr Val Ser Gly Phe Ser Leu Thr Ser Tyr
20 25 30
Gly Ile His Trp Val Arg Gln Ser Pro Gly Lys Gly Leu Glu Trp Leu
35 40 45
Gly Val Ile Trp Arg Gly Gly Ile Thr Asp Tyr Asn Ala Pro Phe Met
50 55 60
Ser Arg Leu Asn Ile Thr Lys Asp Asn Ser Lys Asn Gln Val Phe Phe
65 70 75 80
Lys Met Asn Ser Leu Gln Val Asp Asp Thr Ala Ile Tyr Tyr Cys Ala
85 90 95
Asn His Arg Asp Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ala
100 105 110
<210> 35
<211> 111
<212> PRT
<213> Mus musculus
<400> 35
Asp Ile Val Leu Thr Gln Ser Pro Ala Ser Leu Val Val Ser Leu Gly
1 5 10 15
Gln Arg Ala Thr Ile Ser Cys Arg Ala Ser Glu Ser Val Asp Asn Tyr
20 25 30
Gly Ile Ser Phe Met Asn Trp Phe Gln Gln Lys Pro Gly Gln Pro Pro
35 40 45
Lys Leu Leu Ile Tyr Thr Thr Ser Asn Gln Gly Ser Gly Val Arg Ala
50 55 60
Arg Leu Ser Gly Ser Gly Cys Gly Thr Asp Phe Ser Leu Asn Ile His
65 70 75 80
Pro Met Glu Glu Asp Asp Ser Ala Met Tyr Phe Cys Gln Gln Ser Lys
85 90 95
Glu Val Pro Tyr Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 36
<211> 120
<212> PRT
<213> Mus musculus
<400> 36
Asp Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1 5 10 15
Ser Leu Ser Leu Thr Cys Thr Val Thr Gly Tyr Ser Ile Thr Ser Asp
20 25 30
Tyr Ala Trp Asn Trp Ile Arg Gln Phe Pro Gly Asn Lys Val Glu Trp
35 40 45
Met Gly Tyr Ile Ser Tyr Ser Gly Ser Ser Ser Tyr Asn Pro Ser Leu
50 55 60
Lys Gly Arg Ile Ser Ile Thr Arg Asp Thr Ser Lys Asn Gln Phe Phe
65 70 75 80
Leu Gln Leu Asn Ser Val Thr Thr Glu Asp Thr Ala Thr Tyr Tyr Cys
85 90 95
Ala Arg Phe Tyr Tyr Arg Phe Asp Ala Tyr Phe Asp Ser Trp Gly Gln
100 105 110
Gly Thr Thr Leu Thr Val Ser Ser
115 120
<210> 37
<211> 111
<212> PRT
<213> Mus musculus
<400> 37
Asp Ile Val Leu Thr Gln Ser Pro Ala Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Gln Arg Ala Thr Ile Ser Cys Arg Ala Ser Glu Ser Val Glu Tyr Tyr
20 25 30
Gly Thr Ser Leu Met His Trp Tyr Gln His Lys Pro Gly Gln Thr Pro
35 40 45
Lys Leu Leu Ile Tyr Ser Gly Ser Asn Val Glu Ser Gly Val Pro Ala
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Ser Leu Asn Ile His
65 70 75 80
Pro Val Glu Glu Asp Asp Ile Ala Met Tyr Phe Cys Gln Gln Ser Arg
85 90 95
Glu Val Pro Ser Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 38
<211> 119
<212> PRT
<213> Mus musculus
<400> 38
Asp Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1 5 10 15
Ser Leu Ser Leu Thr Cys Ser Val Thr Gly Phe Ser Ile Thr Ser Asp
20 25 30
Tyr Ala Trp Asn Trp Ile Arg Gln Phe Pro Gly Asn Lys Leu Glu Trp
35 40 45
Met Gly Tyr Ile Ser Tyr Ser Gly Thr Thr Asn Tyr Asn Pro Ser Leu
50 55 60
Lys Ser Arg Ile Ser Ile Thr Arg Asp Thr Ser Lys Asn Gln Phe Phe
65 70 75 80
Leu His Leu Asn Ser Val Ile Thr Glu Asp Thr Ala Thr Tyr Tyr Cys
85 90 95
Ala Arg Arg Glu Gly His Tyr Ser Trp Phe Pro Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ala
115
<210> 39
<211> 107
<212> PRT
<213> Mus musculus
<400> 39
Asp Ile Gln Met Thr Gln Ser Pro Ala Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Glu Thr Val Thr Ile Thr Cys Arg Ala Ser Glu Asn Ile Tyr Ser Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Gln Glu Asn Ser Pro Gln Leu Leu Val
35 40 45
Tyr Asn Ala Tyr Thr Leu Ala Asp Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Gln Phe Ser Leu Lys Ile Ile Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Gly Asn Tyr Tyr Cys Gln His His Tyr Arg Thr Pro Pro
85 90 95
Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 40
<211> 117
<212> PRT
<213> Mus musculus
<400> 40
Gln Val Gln Leu Gln Gln Ser Gly Ala Val Leu Val Arg Pro Gly Thr
1 5 10 15
Ser Ile Lys Val Ser Cys Lys Ala Ser Gly Tyr Ala Phe Thr Asn Tyr
20 25 30
Leu Ile Glu Trp Ile Lys Lys Arg Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Met Ile Asn Pro Gly Ser Gly Asp Thr Asn Phe Asn Glu Lys Phe
50 55 60
Lys Ala Lys Ala Thr Leu Thr Ala Asp Lys Ser Ser Thr Thr Ala Tyr
65 70 75 80
Met Gln Leu Asn Ser Leu Thr Phe Asp Asp Ser Ala Val Tyr Phe Cys
85 90 95
Ala Arg Ser Ser Gln Leu Gly Leu Pro Tyr Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ala
115
<210> 41
<211> 2079
<212> DNA
<213> Homo sapiens
<400> 41
atgggcaccg tcagctccag gcggtcctgg tggccgctgc cactgctgct gctgctgctg 60
ctgctcctgg gtcccgcggg cgcccgtgcg caggaggacg aggacggcga ctacgaggag 120
ctggtgctag ccttgcgttc cgaggaggac ggcctggccg aagcacccga gcacggaacc 180
acagccacct tccaccgctg cgccaaggat ccgtggaggt tgcctggcac ctacgtggtg 240
gtgctgaagg aggagaccca cctctcgcag tcagagcgca ctgcccgccg cctgcaggcc 300
caggctgccc gccggggata cctcaccaag atcctgcatg tcttccatgg ccttcttcct 360
ggcttcctgg tgaagatgag tggcgacctg ctggagctgg ccttgaagtt gccccatgtc 420
gactacatcg aggaggactc ctctgtcttt gcccagagca tcccgtggaa cctggagcgg 480
attacccctc cacggtaccg ggcggatgaa taccagcccc ccgacggagg cagcctggtg 540
gaggtgtatc tcctagacac cagcatacag agtgaccacc gggaaatcga gggcagggtc 600
atggtcaccg acttcgagaa tgtgcccgag gaggacggga cccgcttcca cagacaggcc 660
agcaagtgtg acagtcatgg cacccacctg gcaggggtgg tcagcggccg ggatgccggc 720
gtggccaagg gtgccagcat gcgcagcctg cgcgtgctca actgccaagg gaagggcacg 780
gttagcggca ccctcatagg cctggagttt attcggaaaa gccagctggt ccagcctgtg 840
gggccactgg tggtgctgct gcccctggcg ggtgggtaca gccgcgtcct caacgccgcc 900
tgccagcgcc tggcgagggc tggggtcgtg ctggtcaccg ctgccggcaa cttccgggac 960
gatgcctgcc tctactcccc agcctcagct cccgaggtca tcacagttgg ggccaccaat 1020
gcccaggacc agccggtgac cctggggact ttggggacca actttggccg ctgtgtggac 1080
ctctttgccc caggggagga catcattggt gcctccagcg actgcagcac ctgctttgtg 1140
tcacagagtg ggacatcaca ggctgctgcc cacgtggctg gcattgcagc catgatgctg 1200
tctgccgagc cggagctcac cctggccgag ttgaggcaga gactgatcca cttctctgcc 1260
aaagatgtca tcaatgaggc ctggttccct gaggaccagc gggtactgac ccccaacctg 1320
gtggccgccc tgccccccag cacccatggg gcaggttggc agctgttttg caggactgtg 1380
tggtcagcac actcggggcc tacacggatg gccacagcca tcgcccgctg cgccccagat 1440
gaggagctgc tgagctgctc cagtttctcc aggagtggga agcggcgggg cgagcgcatg 1500
gaggcccaag ggggcaagct ggtctgccgg gcccacaacg cttttggggg tgagggtgtc 1560
tacgccattg ccaggtgctg cctgctaccc caggccaact gcagcgtcca cacagctcca 1620
ccagctgagg ccagcatggg gacccgtgtc cactgccacc aacagggcca cgtcctcaca 1680
ggctgcagct cccactggga ggtggaggac cttggcaccc acaagccgcc tgtgctgagg 1740
ccacgaggtc agcccaacca gtgcgtgggc cacagggagg ccagcatcca cgcttcctgc 1800
tgccatgccc caggtctgga atgcaaagtc aaggagcatg gaatcccggc ccctcaggag 1860
caggtgaccg tggcctgcga ggagggctgg accctgactg gctgcagtgc cctccctggg 1920
acctcccacg tcctgggggc ctacgccgta gacaacacgt gtgtagtcag gagccgggac 1980
gtcagcacta caggcagcac cagcgaagag gccgtgacag ccgttgccat ctgctgccgg 2040
agccggcacc tggcgcaggc ctcccaggag ctccagtga 2079
<210> 42
<211> 2365
<212> DNA
<213> Homo sapiens
<400> 42
atggggccct ggggctggaa attgcgctgg accgtcgcct tgctcctcgc cgcggcgggg 60
actgcagtgg gcgacagatg cgaaagaaac gagttccagt gccaagacgg gaaatgcatc 120
tcctacaagt gggtctgcga tggcagcgct gagtgccagg atggctctga tgagtcccag 180
gagacgtgct tgtctgtcac ctgcaaatcc ggggacttca gctgtggggg ccgtgtcaac 240
cgctgcattc ctcagttctg gaggtgcgat ggccaagtgg actgcgacaa cggctcagac 300
gagcaaggct gtccccccaa gacgtgctcc caggacgagt ttcgctgcca cgatgggaag 360
tgcatctctc ggcagttcgt ctgtgactca gaccgggact gcttggacgg ctcagacgag 420
gcctcctgcc cggtgctcac ctgtggtccc gccagcttcc agtgcaacag ctccacctgc 480
atcccccagc tgtgggcctg cgacaacgac cccgactgcg aagatggctc ggatgagtgg 540
ccgcagcgct gtaggggtct ttacgtgttc caaggggaca gtagcccctg ctcggccttc 600
gagttccact gcctaagtgg cgagtgcatc cactccagct ggcgctgtga tggtggcccc 660
gactgcaagg acaaatctga cgaggaaaac tgcgctgtgg ccacctgtcg ccctgacgaa 720
ttccagtgct ctgatggaaa ctgcatccat ggcagccggc agtgtgaccg ggaatatgac 780
tgcaaggaca tgagcgatga agttggctgc gttaatgtga cactctgcga gggacccaac 840
aagttcaagt gtcacagcgg cgaatgcatc accctggaca aagtctgcaa catggctaga 900
gactgccggg actggtcaga tgaacccatc aaagagtgcg ggaccaacga atgcttggac 960
aacaacggcg gctgttccca cgtctgcaat gaccttaaga tcggctacga gtgcctgtgc 1020
cccgacggct tccagctggt ggcccagcga agatgcgaag atatcgatga gtgtcaggat 1080
cccgacacct gcagccagct ctgcgtgaac ctggagggtg gctacaagtg ccagtgtgag 1140
gaaggcttcc agctggaccc ccacacgaag gcctgcaagg ctgtgggctc catcgcctac 1200
ctcttcttca ccaaccggca cgaggtcagg aagatgacgc tggaccggag cgagtacacc 1260
agcctcatcc ccaacctgag gaacgtggtc gctctggaca cggaggtggc cagcaataga 1320
atctactggt ctgacctgtc ccagagaatg atctgcagca cccagcttga cagagcccac 1380
ggcgtctctt cctatgacac cgtcatcagc agggacatcc aggcccccga cgggctggct 1440
gtggactgga tccacagcaa catctactgg accgactctg tcctgggcac tgtctctgtt 1500
gcggatacca agggcgtgaa gaggaaaacg ttattcaggg agaacggctc caagccaagg 1560
gccatcgtgg tggatcctgt tcatggcttc atgtactgga ctgactgggg aactcctgcc 1620
aagatcaaga aagggggcct gaatggtgtg gacatctact cgctggtgac tgaaaacatt 1680
cagtggccca atggcatcac cctagatctc ctcagtggcc gcctctactg ggttgactcc 1740
aaacttcact ccatctcaag catcgatgtc aacgggggca accggaagac catcttggag 1800
gatgaaaaga ggctggccca ccccttctcc ttggccgtct ttgaggacaa agtattttgg 1860
acagatatca tcaacgaagc cattttcagt gccaaccgcc tcacaggttc cgatgtcaac 1920
ttgttggctg aaaacctact gtccccagag gatatggttc tcttccacaa cctcacccag 1980
ccaagaggag tgaactggtg tgagaggacc accctgagca atggcggctg ccagtatctg 2040
tgcctccctg ccccgcagat caacccccac tcgcccaagt ttacctgcgc ctgcccggac 2100
ggcatgctgc tggccaggga catgaggagc tgcctcacag aggctgaggc tgcagtggcc 2160
acccaggaga catccaccgt caggctaaag gtcagctcca cagccgtaag gacacagcac 2220
acaaccaccc gacctgttcc cgacacctcc cggctgcctg gggccacccc tgggctcacc 2280
acggtggaga tagtgacaat gtctcaccaa gctctgggcg acgttgctgg cagaggaaat 2340
gagaagaagc ccagtagcgt gaggg 2365
<210> 43
<211> 31
<212> DNA
<213> Искусственная последовательность
<220>
<223> Праймер
<400> 43
gtacgctagc caccatgggg ccctggggct g 31
<210> 44
<211> 25
<212> DNA
<213> Искусственная последовательность
<220>
<223> Праймер
<400> 44
gatcctcgag ccctcacgct actgg 25
<210> 45
<211> 106
<212> PRT
<213> Искусственная последовательность
<220>
<223> Вариабельная область легкой цепи
<400> 45
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Gln Ala Ser Gln Asp Ile Asn Lys Tyr
20 25 30
Ile Asp Trp Tyr Gln His Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
His Tyr Ala Ser Thr Leu Gln Pro Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Arg Asp Tyr Thr Phe Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Ile Ala Thr Tyr Tyr Cys Leu Gln Tyr Asp Asp Leu Trp Thr
85 90 95
Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 46
<211> 318
<212> DNA
<213> Искусственная последовательность
<220>
<223> Нуклеотидная последовательность вариабельной области легкой цепи
<400> 46
gacatccaga tgacccagag ccccagcagc ctgagcgcca gcgtgggcga cagagtgacc 60
atcacctgcc aggccagcca ggacatcaac aagtacatcg actggtacca gcacaagccc 120
ggcaaggccc ccaagctgct gatccactac gccagcaccc tgcagcccgg cgtgcccagc 180
agattcagcg gcagcggcag cggcagagac tacaccttca ccatcagcag cctgcagccc 240
gaggacatcg ccacctacta ctgcctgcag tacgacgacc tgtggacctt cggccagggc 300
accaaggtgg agatcaag 318
<210> 47
<211> 111
<212> PRT
<213> Искусственная последовательность
<220>
<223> Вариабельная область тяжелой цепи
<400> 47
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Phe Ser Ile Ser Ser Tyr
20 25 30
Gly Ile His Trp Ile Arg Gln Ser Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Val Ile Trp Arg Gly Gly Ile Thr Asp Tyr Asn Ala Pro Phe Met
50 55 60
Ser Arg Val Thr Ile Ser Lys Asp Asn Ser Lys Asn Gln Val Ser Phe
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Asn His Arg Asp Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
100 105 110
<210> 48
<211> 333
<212> DNA
<213> Искусственная последовательность
<220>
<223> Нуклеотидная последовательность вариабельной области тяжелой цепи
<400> 48
caggtgcagc tgcaggaaag cggcccgggc ctggtgaaac cgagccagac cctgagcctg 60
acctgcaccg tgagcggctt tagcattagc agctatggca ttcattggat tcgccagagc 120
ccgggcaaag gcctggaatg gattggcgtg atttggcgcg gcggcattac cgattataac 180
gcgccgttta tgagccgcgt gaccattagc aaagataaca gcaaaaacca ggtgagcttt 240
aaactgagca gcgtgaccgc ggcggatacc gcggtgtatt attgcgcgaa ccatcgcgat 300
tggggccagg gcaccctggt gaccgtgagc agc 333
<210> 49
<211> 111
<212> PRT
<213> Искусственная последовательность
<220>
<223> Вариабельная область тяжелой цепи
<400> 49
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Phe Ser Ile Ser Ser Tyr
20 25 30
Gly Val His Trp Ile Arg Gln Ser Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Val Ile Trp Arg Gly Gly Ser Thr Asp Tyr Asn Ala Ala Phe Met
50 55 60
Ser Arg Val Thr Ile Ser Lys Asp Asn Ser Lys Asn Gln Val Ser Phe
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Asn His Arg Asp Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
100 105 110
<210> 50
<211> 333
<212> DNA
<213> Искусственная последовательность
<220>
<223> Нуклеотидная последовательность вариабельной области тяжелой цепи
<400> 50
caggtgcagc tgcaggaaag cggcccgggc ctggtgaaac cgagccagac cctgagcctg 60
acctgcaccg tgagcggctt tagcattagc agctatggcg tgcattggat tcgccagagc 120
ccgggcaaag gcctggaatg gattggcgtg atttggcgcg gcggcagcac cgattataac 180
gcggcgttta tgagccgcgt gaccattagc aaagataaca gcaaaaacca ggtgagcttt 240
aaactgagca gcgtgaccgc ggcggatacc gcggtgtatt attgcgcgaa ccatcgcgat 300
tggggccagg gcaccctggt gaccgtgagc agc 333
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| АНТИ-PCSK9 АНТИТЕЛО, ЕГО АНТИГЕНСВЯЗЫВАЮЩИЙ ФРАГМЕНТ И ИХ МЕДИЦИНСКОЕ ПРИМЕНЕНИЕ | 2016 |
|
RU2739208C2 |
| АНТИТЕЛО ПРОТИВ IL-17A И ЕГО ПРИМЕНЕНИЕ | 2019 |
|
RU2816204C2 |
| АНТИТЕЛА К TIGIT И ИХ ПРИМЕНЕНИЕ | 2020 |
|
RU2827445C2 |
| АНТИТЕЛО К CLDN-18.2 И ЕГО ПРИМЕНЕНИЕ | 2021 |
|
RU2829997C1 |
| IL-5-СВЯЗЫВАЮЩАЯ МОЛЕКУЛА, СПОСОБ ЕЕ ПОЛУЧЕНИЯ И ЕЕ ПРИМЕНЕНИЕ | 2021 |
|
RU2811518C1 |
| АНТИТЕЛО К GIPR И ЕГО СЛИТЫЙ С GLP-1 БЕЛОК, А ТАКЖЕ ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ НА ЕГО ОСНОВЕ И ЕГО ПРИМЕНЕНИЕ | 2019 |
|
RU2800370C2 |
| СПОСОБ ЛЕЧЕНИЯ ЗАБОЛЕВАНИЙ, СВЯЗАННЫХ С ХОЛЕСТЕРИНОМ | 2021 |
|
RU2832333C1 |
| Антитела к TSLP человека и их применение | 2021 |
|
RU2825460C1 |
| ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ, СОДЕРЖАЩАЯ АНТИТЕЛА К PCSK-9, И ЕЕ ПРИМЕНЕНИЕ | 2018 |
|
RU2782792C2 |
| СЛИТЫЙ БЕЛОК НА ОСНОВЕ АНТИТЕЛА К ЕТА И TGF-β TRAP, СОДЕРЖАЩАЯ ЕГО ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ И ИХ ПРИМЕНЕНИЕ | 2020 |
|
RU2832190C2 |

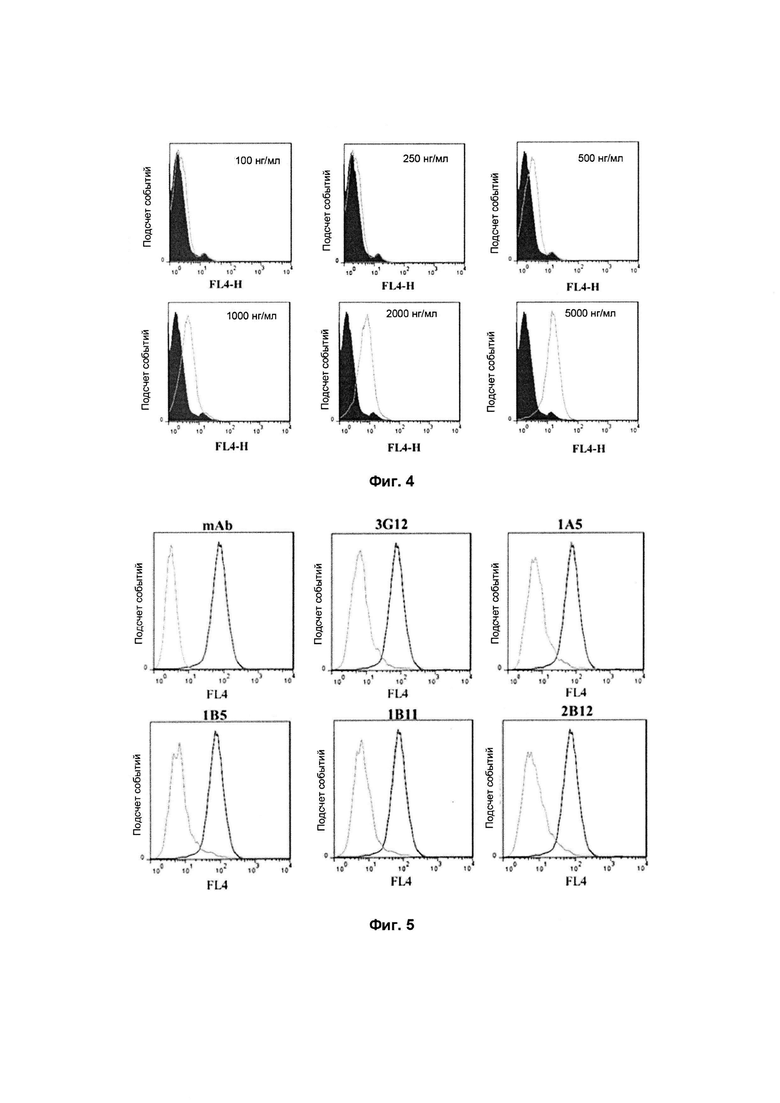
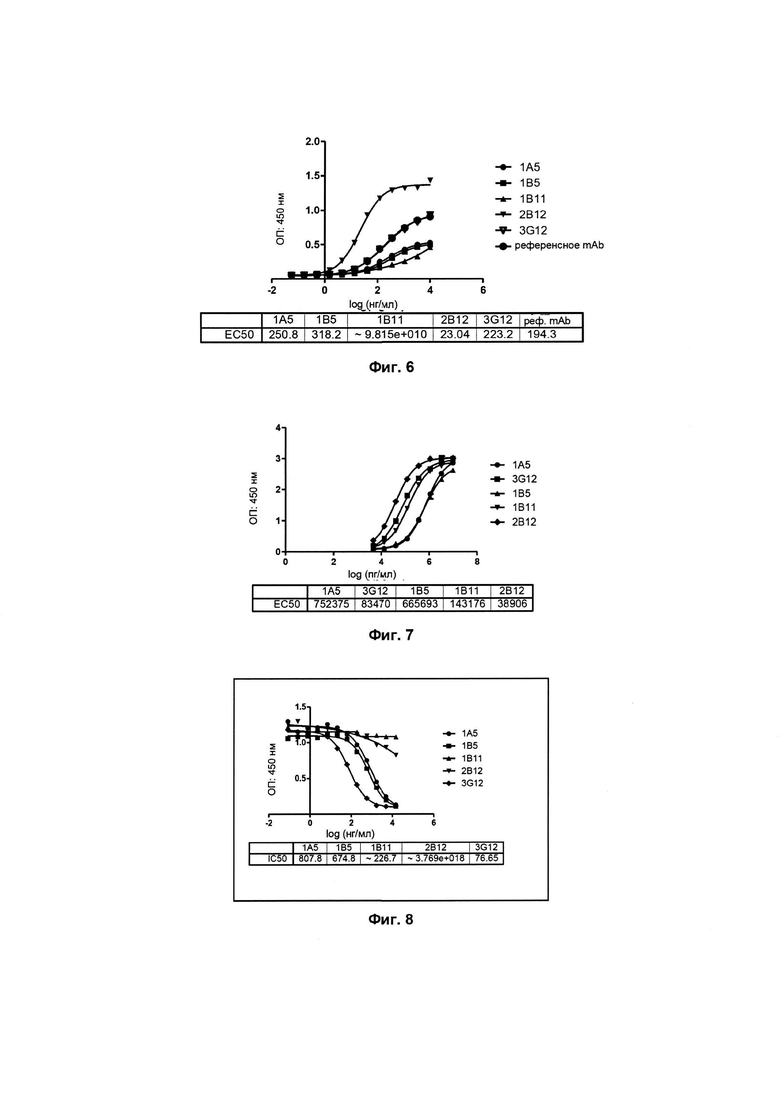
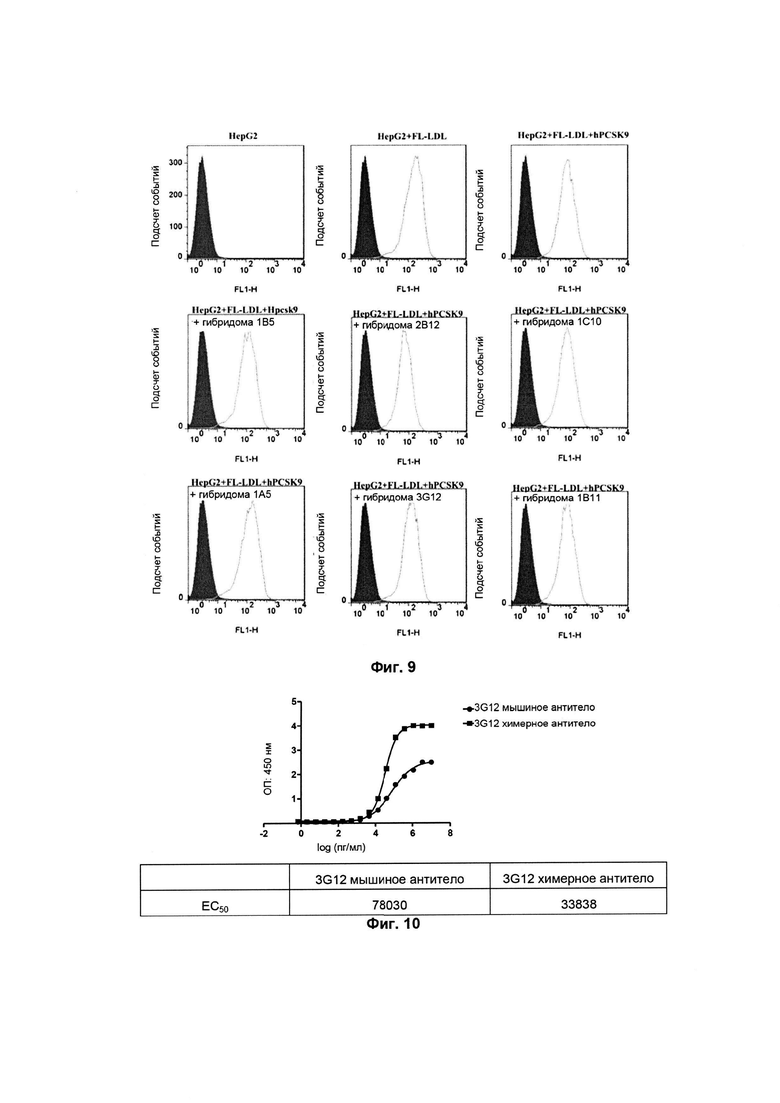
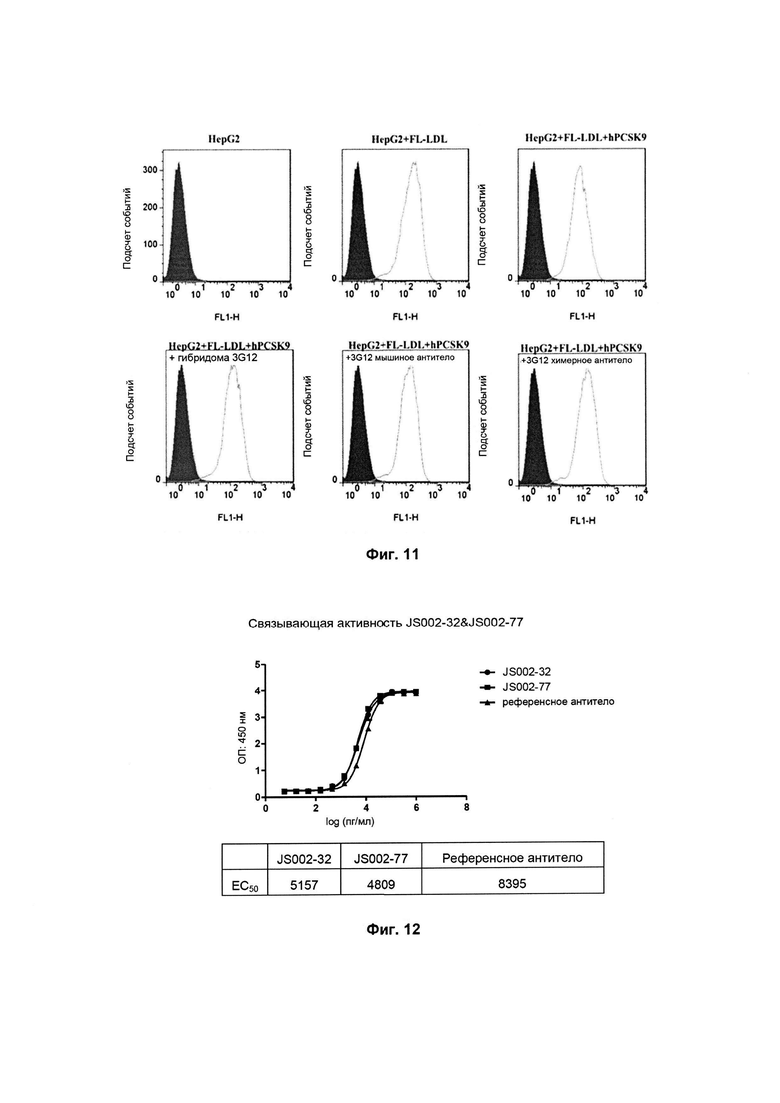
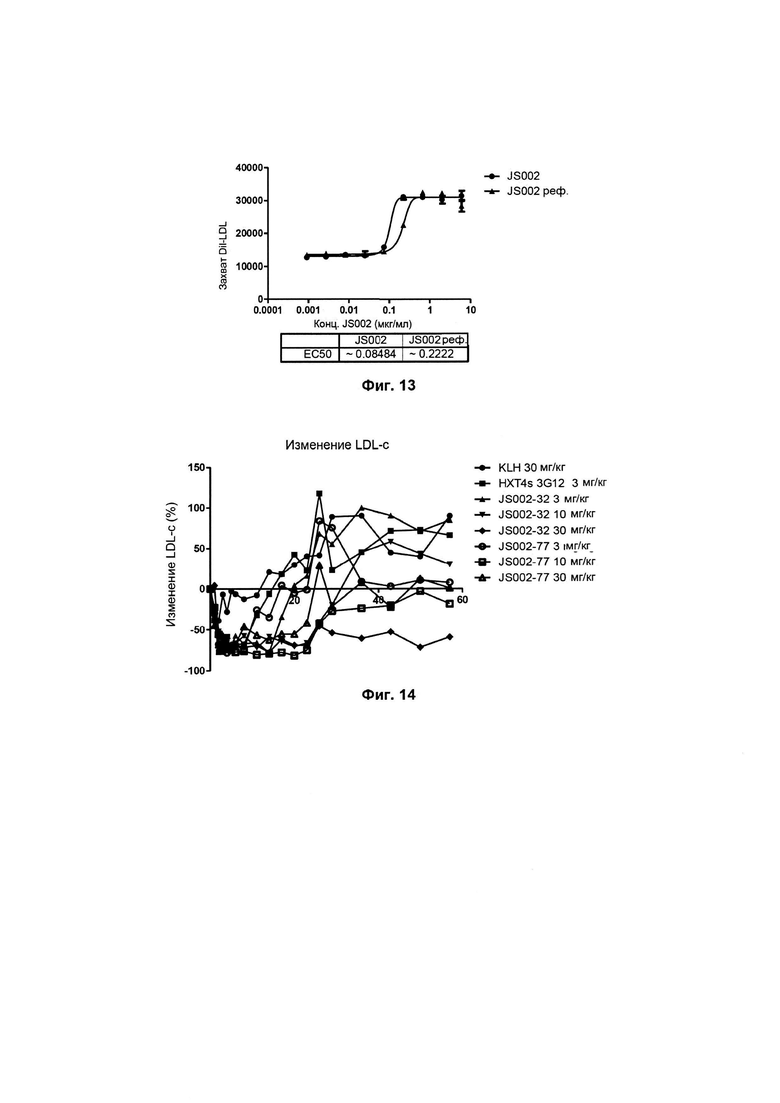
Изобретение относится к области биохимии, в частности к антителу, которое специфично связывается с PCSK9. Также раскрыты фармацевтическая композиция, содержащая указанное антитело; молекула нуклеиновой кислоты, кодирующей указанное антитело; вектор и клетка-хозяин, содержащие указанную молекулу нуклеиновой кислоты; иммуноконъюгат, содержащий указанное антитело. Раскрыто применение указанного антитела в изготовлении лекарственного средства для лечения дислипидемии или средства для облегчения клеточного захвата LDL (липопротеинов низкой плотности). Изобретение обладает способностью эффективно лечить заболевания, ассоциированные с PCSK9. 9 н. и 6 з.п. ф-лы, 14 ил., 3 табл., 17 пр.
1. Антитело для специфического связывания с PCSK9 или его функциональный фрагмент, содержащее(ий) участки CDR1, CDR2 и CDR3 тяжелой цепи и CDR1, CDR2 и CDR3 легкой цепи, где аминокислотные последовательности CDR1, CDR2 и CDR3 тяжелой цепи и CDR1, CDR2 и CDR3 легкой цепи выбраны из одной из групп, состоящих из следующих аминокислотных последовательностей:
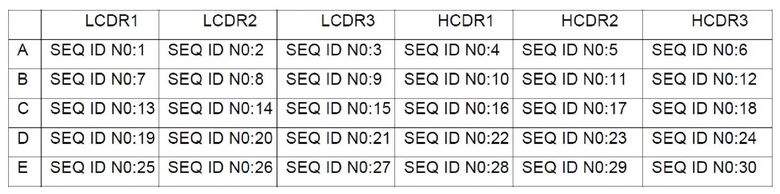
2. Антитело или его функциональный фрагмент по п. 1, которое(ый) содержит вариабельную область тяжелой цепи, выбранную из группы, состоящей из аминокислотных последовательностей SEQ ID NO: 32, 34, 36, 38 и 40, и вариабельную область легкой цепи, выбранную из группы, состоящей из аминокислотных последовательностей SEQ ID NO: 31, 33, 35, 37 и 39.
3. Антитело или его функциональный фрагмент по п. 2, где указанная вариабельная область тяжелой цепи представляет собой SEQ ID NO: 32 и указанная вариабельная область легкой цепи представляет собой SEQ ID NO: 31, или
вариабельная область тяжелой цепи представляет собой SEQ ID NO: 34 и вариабельная область легкой цепи представляет собой SEQ ID NO: 33, или
вариабельная область тяжелой цепи представляет собой SEQ ID NO: 36 и вариабельная область легкой цепи представляет собой SEQ ID NO: 35, или
вариабельная область тяжелой цепи представляет собой SEQ ID NO: 38 и вариабельная область легкой цепи представляет собой SEQ ID NO: 37, или
вариабельная область тяжелой цепи представляет собой SEQ ID NO: 40 и вариабельная область легкой цепи представляет собой SEQ ID NO: 39.
4. Антитело или его функциональный фрагмент по любому из пп. 1-3, представляющее собой химерное антитело, гуманизированное антитело или полностью человеческое антитело.
5. Антитело для специфического связывания с PCSK9 или его функциональный фрагмент, содержащее(ий) вариабельную область тяжелой цепи, выбранную из группы, состоящей из аминокислотных последовательностей SEQ ID NO: 47 и SEQ ID NO: 49, и вариабельную область легкой цепи, выбранную из группы, состоящей из аминокислотной последовательности SEQ ID NO: 45,
или
вариабельную область тяжелой цепи, кодируемую нуклеотидом, кодирующим аминокислотную последовательность SEQ ID NO: 47 или SEQ ID NO: 49, и вариабельную область легкой цепи, кодируемую нуклеотидом, кодирующим аминокислотную последовательность SEQ ID NO: 45.
6. Выделенная молекула нуклеиновой кислоты, кодирующая антитело или его функциональный фрагмент по любому из пп. 1-5.
7. Выделенная молекула нуклеиновой кислоты по п. 6, выбранная из группы, состоящей из SEQ ID NO: 46, 48 и 50.
8. Вектор экспрессии, содержащий молекулу нуклеиновой кислоты по п. 6 или 7.
9. Клетка-хозяин для экспрессии антитела, способного к специфическому связыванию с PCSK9, содержащая молекулу нуклеиновой кислоты по п. 6 или 7.
10. Фармацевтическая композиция для лечения заболеваний, ассоциированных с фактором PSCK9, содержащая терапевтически эффективное количество антитела или его функционального фрагмента по любому из пп.1-5 и фармацевтически приемлемый носитель.
11. Применение антитела или его функционального фрагмента по любому из пп. 1-5 в изготовлении лекарственного средства для лечения дислипидемии или родственных заболеваний сердечно-сосудистой системы.
12. Применение по п. 11, где указанное заболевание представляет собой гиперлипидемию или гиперхолестеринемию.
13. Применение антитела или его функционального фрагмента по любому из пп. 1-5 в изготовлении лекарственного средства для облегчения клеточного захвата LDL (липопротеинов низкой плотности).
14. Иммуноконъюгат для лечения заболеваний, ассоциированных с фактором PSCK9, содержащий антитело или его функциональный фрагмент по любому из пп.1-5, конъюгированный с терапевтическим агентом.
15. Иммуноконъюгат по п. 14, где указанный терапевтический агент представляет собой токсин, радиоизотоп, лекарственное средство или цитотоксический агент.
| RU 2015109166 A, 10.08.2015 | |||
| EA 201270019 A1, 29.06.2012 | |||
| US 7928189 B2, 19.04.2011 | |||
| WO 2014107739 A1, 10.07.2014. |

