Перекрестная ссылка на родственные заявки
В настоящей заявке заявлен приоритет, соответствующий дате приоритета по предварительным заявкам США с серийными № 61/152962, поданной 16 февраля 2009, 61/152966, поданной 16 февраля 2009, и 61/152960, поданной 16 февраля 2009, полное содержание которых включено в настоящее описание в виде ссылки.
Область изобретения
Настоящее изобретение связано с пептидами, содержащими их композициями и способами лечения или предупреждение дислипидемии, сердечно-сосудистого заболевания, эндотелиальной дисфункции, макрососудистого расстройства или микрососудистого расстройства.
Предшествующий уровень техники
Холестерин, циркулирующий в организме человека, переносится липопротеинами плазмы, которые являются частицами комплексной липидной и белковой композиции, которые осуществляют транспорт липидов в крови. Двумя типами липопротеинов плазмы, которые переносят холестерин, являются липопротеины низкой плотности ("ЛПНП") и липопротеины высокой плотности ("ЛПВП"). Считается, что ЛПНП ответственны за доставку холестерина из печени (где он синтезируется или куда доставляется из пищевых источников) к тканям организма, находящимся вне пределов печени. С другой стороны, считается, что ЛПВП способствуют транспорту холестерина из внепеченочных тканей в печень, где холестерин катаболизируется и подвергается элиминации. Такой транспорт холестерина из внепеченочных тканей в печень называется "обратным транспортом холестерина".
Путь обратного транспорта холестерина ("RCT") имеет три основных стадии: (i) выход холестерина, то есть первоначальное удаление холестерина из различных пулов периферических клеток; (ii) этерификацию холестерина под действием лецитин-холестерин-ацилтрансферазы ("LCAT"), таким образом предотвращающую обратный вход вышедшего холестерина в клетку; и (iii) поглощение сложного эфира холестерина липопротеином высокой плотности и доставку комплекса сложного эфира холестерина с ЛПВП к клеткам печени.
Путь RCT опосредован частицами ЛПВП. Каждая частица ЛПВП имеет липидный компонент и белковый компонент. Липидный компонент ЛПВП может быть фосфолипидом, холестерином (или сложным эфиром холестерина) или триглицеридом. Белковый компонент ЛПВП первоначально состоит из ApoA-I. ApoA-I синтезируется печенью и тонким кишечником в виде пре-про-аполипопротеина, который секретируется в виде про-протеина, который быстро расщепляется с получением зрелого полипептида, имеющего 243 аминокислотных остатка. ApoA-I первоначально состоит из 6-8 различных повторяющихся элементов, состоящих из 22 аминокислотных остатков, пространственно разделенных линкерным фрагментом, которым часто служит пролин, а в некоторых случаях он образован несколькими остатками. ApoA-I образует три типа стабильных комплексов с липидами: малые, обедненные липидами комплексы, так называемые ЛПВП пре-β-1; уплощенные дискообразные частицы, содержащие полярные липиды (фосфолипид и холестерин), так называемые ЛПВП пре-β-2; и сферические частицы, содержащие как полярные, так и неполярные липиды, так называемые сферические или зрелые ЛПВП (ЛПВП3 и ЛПВП2).
Были сделаны попытки рекомбинантного продуцирования и введения ApoA-I пациентам с целью защиты от атеросклеротического заболевания. Однако с продуцированием и применением ApoA-I связано множество трудностей, что делает его менее чем идеальным в качестве лекарственного средства; например, ApoA-I является крупным белком, который трудно и дорого производить, и надо будет обойти значительные трудности, связанные с проблемами производства и воспроизведения, в том, что касается стабильности в процессе хранения, доставки активного продукта и времени полужизни in vivo.
С учетом указанных недостатков, были сделаны попытки получения пептидов, которые имитируют активность ApoA-I in vivo. В данной области есть потребность в разработке дополнительных пептидов, которые могли бы имитировать активность ApoA-I in vivo, продукция которых была бы простой и оправданной в отношении расходов.
Краткое описание изобретения
В одном из воплощений настоящее изобретение связано с пептидами, содержащими от 22 до 29 остатков, имеющими формулу I:
R1-Y1-X1-X2-X3-X4-X5-X6-X7-X8-X9-X10-X11-X12-X13-X14-X15-X16-X17-X18-X19-X20-X21-X22-X23-Y2-R2
Формула I
и их фармацевтически приемлемыми солями, в которых
X1 отсутствует или является ахиральным, D- или L-основным аминокислотным остатком;
X2 является ахиральным, D- или L-алифатическим аминокислотным остатком;
X3 является ахиральным, D- или L-алифатическим аминокислотным остатком;
X4 является ахиральным, D- или L-основным аминокислотным остатком;
X5 представляет собой Gln, Asn, D-Gln, D-Asn или основный ахиральный аминокислотный остаток, D- или L-основный аминокислотный остаток;
X6 является ахиральным, D- или L-основным аминокислотным остатком;
X7 является ахиральным, D- или L-гидрофобным аминокислотным остатком;
X8 является ахиральным, D- или L-гидрофобным аминокислотным остатком;
X9 является ахиральным, D- или L-гидрофобным аминокислотным остатком;
X10 представляет собой Leu, Trp, Gly, Nal, D-Leu, D-Trp или D-Nal;
X11 представляет собой Gly или ахиральный, D- или L-алифатический аминокислотный остаток;
X12 является ахиральным, D- или L-гидрофильным аминокислотным остатком;
X13 является ахиральным, D- или L-гидрофильным аминокислотным остатком;
X14 представляет собой Leu, Trp, Gly, D-Leu или D-Trp;
X15 представляет собой Leu, Gly или D-Leu;
X16 является ахиральным, D- или L-кислым аминокислотным остатком;
X17 является ахиральным, D- или L-гидрофильным аминокислотным остатком;
X18 представляет собой Leu, Phe, D-Leu или D-Phe;
X19 представляет собой Leu, Phe, D-Leu или D-Phe;
X20 является ахиральным, D- или L-кислым аминокислотным остатком;
X21 представляет собой Leu, Phe, D-Leu или D-Phe;
X22 является ахиральным, D- или L-алифатическим аминокислотным остатком; и
X23 представляет собой Inp, Nip, azPro, Pip, azPip, D-Nip или D-Pip;
Y1 отсутствует или является аминокислотной последовательностью, содержащей от 1 до 7 остатков;
Y2 отсутствует или является аминокислотной последовательностью, содержащей от 1 до 7 остатков;
R1 представляет собой H или аминозащитную группу;
R2 представляет собой OH или карбоксильную защитную группу; при этом
a) каждый хиральный аминокислотный остаток является L-аминокислотным остатком;
b) каждый хиральный аминокислотный остаток является D- аминокислотным остатком;
c) каждый хиральный аминокислотный остаток является L-аминокислотным остатком, за исключением того, что один или более из каждых хиральных концевых аминокислотных остатков и каждых хиральных аминокислотных остатков, непосредственно к ним примыкающих, является D-аминокислотным остатком; или
d) каждый хиральный аминокислотный остаток является D- аминокислотным остатком, за исключением того, что один или более из каждых хиральных концевых аминокислотных остатков и каждых хиральных аминокислотных остатков, непосредственно к ним примыкающих, является L-аминокислотным остатком.
В другом воплощении настоящее изобретение связано с пептидами, содержащими от 15 до 22 остатков, имеющими следующую формулу II:
R1-Y1-X1-X2-X3-X4-X5-X6-X7-X8-X9-X10-X11-X12-X13-X14-X15-X16-X17-X18-Y2-R2
Формула II
и их фармацевтически приемлемыми солями, в которых
X1 является ахиральным, D- или L-основным аминокислотным остатком;
X2 представляет собой Leu или D-Leu;
X3 является ахиральным, D- или L-основным аминокислотным остатком;
X4 представляет собой Gln, Asn, D-Gln или D-Asn;
X5 представляет собой Leu, D-Leu или ахиральный, D- или L-основный аминокислотный остаток;
X6 представляет собой Leu, Trp, Phe, D-Leu, D-Trp или D-Phe;
X7 является ахиральным, D- или L-кислым аминокислотным остатком;
X8 представляет собой Asn, D-Asn или является ахиральным, D-или L-кислым аминокислотным остатком;
X9 представляет собой Leu, Trp, D-Leu или D-Trp;
X10 представляет собой Leu, Trp, D-Leu или D-Trp;
X11 представляет собой ахиральный, D- или L-кислый аминокислотный остаток;
X12 является ахиральным, D- или L-основным аминокислотным остатком;
X13 представляет собой Leu, Phe, D-Leu или D-Phe;
X14 представляет собой Leu, Phe, D-Leu или D-Phe;
X15 является ахиральным, D- или L-кислым аминокислотным остатком;
X16 представляет собой Leu или D-Leu;
X17 является ахиральным, D- или L-алифатическим аминокислотным остатком;
X18 представляет собой Inp, Nip, azPro, Pip, azPip, D-Nip или D-Pip;
Y1 отсутствует или является аминокислотной последовательностью, содержащей от 1 до 4 остатков;
Y2 отсутствует;
R1 представляет собой H или аминозащитную группу;
R2 представляет собой OH или карбоксильную защитную группу;
причем от нуля до трех остатков от X1 до X17 отсутствуют; и при этом
a) каждый хиральный аминокислотный остаток является L-аминокислотным остатком;
b) каждый хиральный аминокислотный остаток является D- аминокислотным остатком;
c) каждый хиральный аминокислотный остаток является L-аминокислотным остатком, за исключением того, что один или более из каждых хиральных концевых аминокислотных остатков и каждых хиральных аминокислотных остатков, непосредственно к ним примыкающих, является D-аминокислотным остатком; или
d) каждый хиральный аминокислотный остаток является D-аминокислотным остатком, за исключением того, что один или более из каждых хиральных концевых аминокислотных остатков и каждых хиральных аминокислотных остатков, непосредственно к ним примыкающих, является L-аминокислотным остатком.
В другом воплощении настоящее изобретение связано с пептидами, содержащими от 22 до 29 остатков, имеющими формулу III:
R1-Y1-X1-X2-X3-X4-X5-X6-X7-X8-X9-X10-X11-X12-X13-X14-X15-X16-X17-X18-X19-X20-X21-X22-X23-Y2-R2
Формула III
и их фармацевтически приемлемыми солями, в которых
X1 отсутствует или является ахиральным, D- или L-основным аминокислотным остатком;
X2 является ахиральным, D- или L-основным аминокислотным остатком;
X3 представляет собой Leu или D-Leu;
X4 является ахиральным, D- или L-основным аминокислотным остатком;
X5 является ахиральным, D- или L-основным аминокислотным остатком;
X6 представляет собой Gln, Asn, D-Gln или D-Asn;
X7 представляет собой Leu или D-Leu;
X8 представляет собой Ala или D-Ala;
X9 представляет собой Asp или D-Asp;
X10 представляет собой Leu, Phe, Gly, D-Leu или D-Phe;
X11 представляет собой Gly, Leu или D-Leu;
X12 представляет собой Arg или D-Arg;
X13 является ахиральным, D- или L-кислым аминокислотным остатком;
X14 представляет собой Leu, Trp, Gly, D-Leu или D-Trp;
X15 представляет собой Leu или D-Leu;
X16 представляет собой Gln или D-Gln;
X17 представляет собой Glu, Leu, D-Glu или D-Leu;
X18 представляет собой Leu, Phe, D-Leu или D-Phe;
X19 является ахиральным, D- или L-алифатическим аминокислотным остатком;
X20 представляет собой Glu или D-Glu;
X21 представляет собой Leu, Phe, D-Leu или D-Phe;
X22 является ахиральным, D- или L-алифатическим аминокислотным остатком;
X23 представляет собой Inp, Nip, azPro, Pip, azPip, D-Nip или D-Pip;
Y1 отсутствует или является аминокислотной последовательностью, содержащей от 1 до 7 остатков;
Y2 отсутствует или является аминокислотной последовательностью, содержащей от 1 до 7 остатков;
R1 представляет собой H или аминозащитную группу;
R2 представляет собой OH или карбоксильную защитную группу; при этом
a) каждый хиральный аминокислотный остаток является L-аминокислотным остатком;
b) каждый хиральный аминокислотный остаток является D-аминокислотным остатком;
c) каждый хиральный аминокислотный остаток является L-аминокислотным остатком, за исключением того, что один или более из каждых хиральных концевых аминокислотных остатков и каждых хиральных аминокислотных остатков, непосредственно к ним примыкающих, является D-аминокислотным остатком; или
d) каждый хиральный аминокислотный остаток является D-аминокислотным остатком, за исключением того, что один или более из каждых хиральных концевых аминокислотных остатков и каждых хиральных аминокислотных остатков, непосредственно к ним примыкающих, является L-аминокислотным остатком.
Пептид формулы I, II или III, или его фармацевтически приемлемая соль ("миметик ApoA-I") применимы для лечения или предупреждения дислипидемии, сердечно-сосудистого заболевания, эндотелиальной дисфункции, макрососудистого расстройства или микрососудистого расстройства (каждое из которых является "Состоянием").
В другом воплощении настоящее изобретение связано с композициями, содержащими эффективное количество миметика ApoA-I и фармацевтически приемлемый носитель или наполнитель.
В другом воплощении настоящее изобретение связано со способами лечения или предупреждения состояния, предусматривающими введение в случае необходимости эффективного количества миметика ApoA-I млекопитающему.
Краткое описание графического материала
Фиг.1A представляет собой диаграмму Шиффера и Эдмундсона «helical wheel» (диаграмма «Спиральные колеса») идеализированной амфипатической α-спирали, в которой полыми кружочками обозначены гидрофильные аминокислотные остатки, а заполненными кружочками обозначены гидрофобные аминокислотные остатки.
Фиг.1В представляет собой схему-диаграмму спиральной сетки («helical net») идеализированной амфипатической спирали, изображенной на фиг.1A.
Фиг.1С представляет собой цилиндрическую диаграмму амфипатической спирали, изображенной на фиг.1A.
Фиг.2 представляет собой диаграмму Шиффера и Эдмундсона «helical wheel» 22-мерного консенсусного пептида Сегреста (SEQ ID NO:1).
На фиг.3A проиллюстрирована разветвленная сеть миметиков ApoA-I третьего порядка.
На фиг.3B проиллюстрирована разветвленная сеть миметиков ApoA-I четвертого порядка.
На фиг.3C проиллюстрирована разветвленная сеть миметиков ApoA-I смешанного порядка.
На фиг.3D проиллюстрированы примеры разветвленных сетей "Lys-tree" миметиков ApoA-I.
Фиг.4A представляет собой динамическое изображение различных состояний агрегации и пептид-липидных комплексов, которые могут быть получены с помощью миметиков ApoA-I согласно изобретению. Слева: процесс мультимеризации пептидов, происходящий в результате взаимодействия нескольких пептидных спиралей и ведущий к образованию олигомеров в условиях с определенной концентрацией пептида, значениями pH и ионной силы. В центре: взаимодействие миметиков ApoA-I (в любом из указанных состояний агрегации) с липидными компонентами (например, малыми униламеллярными везикулами ("SUV", small unilamellar vesicles)) приводит к реорганизации липидов. Справа: путем изменения липид-пептидного молярного отношения можно получить различные типы пептид-липидных комплексов, от липид-пептидных комицелл при низких липид-пептидных соотношениях до дискообразных частиц и, наконец, до крупных мультиламеллярных комплексов при все более возрастающих липид-пептидных соотношениях.
На фиг.4B проиллюстрирована общепринятая модель дискообразных комплексов миметик ApoA-I-липид, образуемых в определенном интервале соотношений миметик ApoA-I-липид. Каждый миметик ApoA-I, окружающий каемку диска, находится в тесном контакте с двумя своими ближайшими соседями.
Фиг.5 представляет собой репрезентативную хроматограмму проникновения в гель пептид 16/липидного комплекса (липиды представляют собой сфингомиелин, DPPC (дипальмитоилфосфатидилхолин) и DPPG (дипальмитоилфосфатидилглицерин)).
Фиг.6 представляет собой кривую повышения, над исходным уровнем, фракции ЛПВП общего холестерина после введения пептид 16/липидного комплекса (липиды представляют собой сфингомиелин, DPPC и DPPG, и указанные компоненты присутствуют в весовом соотношении пептид 16:сфингомиелин:DPPC:DPPG как 1:1,35:1,35:0,30) кроликам.
Фиг.7 представляет собой кривую повышения фракции ЛПВП свободного холестерина после введения пептид 16/липидного комплекса (указанные липиды представляют собой сфингомиелин, DPPC и DPPG в весовом соотношении 1:1,2125:1,2125:0,075, и весовое соотношение пептид:липид составляет 1:2,5) кроликам.
Фиг.8 представляет собой профиль элюции гельпроникающей хроматографии на исходном уровне (темная линия) и через 20 мин после введения 2,5 мг/кг пептид 16/липидного комплекса (указанные липиды представляют собой сфингомиелин, DPPC и DPPG в весовом соотношении 1:1,2125:1,2125:0,075, и весовое соотношение пептид:липид составляет 1:2,5) кроликам.
Фиг.9 представляет собой кривую повышения уровня фосфолипида в плазме после введения пептид 16/липидного комплекса (указанные липиды представляют собой сфингомиелин, DPPC и DPPG в весовом соотношении 1:1,2125:1,2125:0,075, и весовое соотношение пептид:липид составляет 1:2,5) кроликам на голодный желудок в дозах 0 (квадратики), 10 (треугольники), 20 (кружочки) или 30 (ромбики) мг/кг. Через различные промежутки времени после введения дозы измеряли уровни фосфолипида в плазме. Исходные значения (варьирующие от 0,96 до 1,18 г/л для указанных четырех групп) вычитали для того, чтобы определить степень повышения уровней фосфолипида в плазме. Брали по 3 животных на группу. К 30-34-часовому сроку после введения дозы указанные уровни возвращались к значениям, равным или меньшим, чем исходный уровень.
Фиг.10A представляет собой кривую повышения уровня общего холестерина в плазме после инфузии пептид 16/липидного комплекса (указанные липиды представляют собой сфингомиелин, DPPC и DPPG в весовом соотношении 1:1,2125:1,2125:0,075, и весовое соотношение пептид:липид составляет 1:2,5) кроликам на голодный желудок в дозах 0 (квадратики), 10 (треугольники), 20 (кружочки) или 30 (ромбики) мг/кг. Через различные промежутки времени после введения дозы измеряли уровни общего холестерина в плазме. Исходные значения вычитали для того, чтобы определить степень повышения уровней холестерина в плазме. Исходные значения варьировали от 0,59 до 0,77 г/л. Брали по 3 животных на группу. К 30-34-часовому сроку после введения дозы указанные уровни возвращались к значениям, меньшим, чем исходный уровень, или равным ему.
Фиг.10B представляет собой кривую повышения уровня свободного холестерина в плазме после инфузии пептид 16/липидного комплекса (указанные липиды представляют собой сфингомиелин, DPPC и DPPG в весовом соотношении 1:1,2125:1,2125:0,075, и весовое соотношение пептид:липид составляет 1:2,5) кроликам на голодный желудок в дозах 0 (квадратики), 10 (треугольники), 20 (кружочки) или 30 (ромбики) мг/кг. Через различные промежутки времени после введения дозы измеряли уровни свободного холестерина в плазме. Исходные значения вычитали для того, чтобы определить степень повышения уровней холестерина в плазме. Исходные значения варьировали от 0,21 до 0,27 г/л. Брали по 3 животных на группу. К 30-34-часовому сроку после введения дозы указанные уровни возвращались к значениям, меньшим, чем исходный уровень, или равным ему.
Фиг.10C представляет собой кривую повышения уровня этерифицированного холестерина в плазме после инфузии пептид 16/липидного комплекса (указанные липиды представляют собой сфингомиелин, DPPC и DPPG в весовом соотношении 1:1,2125:1,2125:0,075, и весовое соотношение пептид:липид составляет 1:2,5) кроликам на голодный желудок в дозах 0 (квадратики), 10 (треугольники), 20 (кружочки) или 30 (ромбики) мг/кг. Через различные промежутки времени после введения дозы измеряли уровни этерифицированного холестерина в плазме. Исходные значения вычитали для того, чтобы определить степень повышения уровней холестерина. Исходные значения варьировали от 0,39 до 0,52 г/л. Брали по 3 животных на группу. К 30-34-часовому сроку после введения дозы указанные уровни возвращались к значениям, меньшим, чем исходный уровень, или равным ему.
Фиг.11A представляет собой кривую повышения уровня общего холестерина ЛПВП в плазме после инфузии пептид 16/липидного комплекса (указанные липиды представляют собой сфингомиелин, DPPC и DPPG в весовом соотношении 1:1,2125:1,2125:0,075, и весовое соотношение пептид:липид составляет 1:2,5) кроликам на голодный желудок в дозах 0 (квадратики), 10 (треугольники), 20 (кружочки) или 30 (ромбики) мг/кг. Через различные промежутки времени после введения дозы измеряли уровни общего холестерина ЛПВП. Исходные значения вычитали для того, чтобы определить степень повышения уровней холестерина. Исходные значения общего холестерина ЛПВП варьировали от 0,33 до 0,38 г/л. Брали по 3 животных на группу. К 30-34-часовому сроку после введения дозы указанные уровни возвращались к значениям, меньшим, чем исходный уровень, или равным ему.
Фиг.11B представляет собой кривую повышения уровня свободного холестерина ЛПВП в плазме после инфузии пептид 16/липидного комплекса (указанные липиды представляют собой сфингомиелин, DPPC и DPPG в весовом соотношении 1:1,2125:1,2125:0,075, и весовое соотношение пептид:липид составляет 1:2,5) кроликам на голодный желудок в дозах 0 (квадратики), 10 (треугольники), 20 (кружочки) или 30 (ромбики) мг/кг. Через различные промежутки времени после введения дозы измеряли уровни свободного холестерина ЛПВП. Исходные значения вычитали для того, чтобы определить степень повышения уровней холестерина. Исходные значения свободного холестерина ЛПВП варьировали от 0,11 до 0,13 г/л. Брали по 3 животных на группу. К 30-34-часовому сроку после введения дозы указанные уровни возвращались к значениям, меньшим, чем исходный уровень, или равным ему.
Фиг.11C представляет собой кривую повышения уровня общего холестерина ЛПВП в плазме после инфузии пептид 16/липидного комплекса (указанные липиды представляют собой сфингомиелин, DPPC и DPPG в весовом соотношении 1:1,2125:1,2125:0,075, и весовое соотношение пептид:липид составляет 1:2,5) кроликам на голодный желудок в дозах 0 (квадратики), 10 (треугольники), 20 (кружочки) или 30 (ромбики) мг/кг. Через различные промежутки времени после введения дозы измеряли уровни общего холестерина ЛПВП. Исходные значения вычитали для того, чтобы определить степень повышения уровней холестерина. Исходные значения общего холестерина ЛПВП варьировали от 0,17 до 0,33 г/л. Брали по 3 животных на группу. К 30-34-часовому сроку после введения дозы указанные уровни возвращались к значениям, меньшим, чем исходный уровень, или равным ему.
Фиг.11D представляет собой кривую повышения уровня свободного холестерина ЛПВП после инфузии пептид 16/липидного комплекса (указанные липиды представляют собой сфингомиелин, DPPC и DPPG в весовом соотношении 1:1,2125:1,2125:0,075, и весовое соотношение пептид:липид составляет 1:2,5) кроликам на голодный желудок в дозах 0 (квадратики), 10 (треугольники), 20 (кружочки) или 30 (ромбики) мг/кг. Через различные промежутки времени после введения дозы измеряли уровни свободного холестерина ЛПВП. Исходные значения вычитали для того, чтобы определить степень повышения уровней холестерина. Исходные значения свободного холестерина ЛПВП варьировали от 0,06 до 0,11 г/л. Брали по 3 животных на группу. К 30-34-часовому сроку после введения дозы указанные уровни возвращались к значениям, меньшим, чем исходный уровень, или равным ему.
Фиг.11E представляет собой кривую повышения уровня общего холестерина ЛПВП после инфузии пептид 16/липидного комплекса (указанные липиды представляют собой сфингомиелин, DPPC и DPPG в весовом соотношении 1:1,2125:1,2125:0,075, и весовое соотношение пептид:липид составляет 1:2,5) кроликам на голодный желудок в дозах 0 (квадратики), 10 (треугольники), 20 (кружочки) или 30 (ромбики) мг/кг. Через различные промежутки времени после введения дозы измеряли уровни общего холестерина ЛПВП. Исходные значения вычитали для того, чтобы определить степень повышения уровней холестерина. Исходные значения свободного холестерина ЛПВП варьировали от 0,04 до 0,11 г/л. Брали по 3 животных на группу. К 30-34-часовому сроку после введения дозы указанные уровни возвращались к значениям, меньшим, чем исходный уровень, или равным ему.
Фиг.11F представляет собой кривую повышения уровня свободного холестерина ЛПВП после инфузии пептид 16/липидного комплекса (указанные липиды представляют собой сфингомиелин, DPPC и DPPG в весовом соотношении 1:1,2125:1,2125:0,075, и весовое соотношение пептид:липид составляет 1:2,5) кроликам на голодный желудок в дозах 0 (квадратики), 10 (треугольники), 20 (кружочки) или 30 (ромбики) мг/кг. Через различные промежутки времени после введения дозы измеряли уровни свободного холестерина ЛПВП. Исходные значения вычитали для того, чтобы определить степень повышения уровней холестерина. Исходные значения свободного холестерина ЛПВП варьировали от 0,02 до 0,04 г/л. Брали по 3 животных на группу. К 30-34-часовому сроку после введения дозы указанные уровни возвращались к значениям, меньшим, чем исходный уровень, или равным ему.
Фиг.12 представляет собой кривую повышения уровня триглицеридов в плазме после инфузии пептид 16/липидного комплекса (указанные липиды представляют собой сфингомиелин, DPPC и DPPG в весовом соотношении 1:1,2125:1,2125:0,075, и весовое соотношение пептид:липид составляет 1:2,5) кроликам на голодный желудок в дозах 0 (квадратики), 10 (треугольники), 20 (кружочки) или 30 (ромбики) мг/кг. Через различные промежутки времени после введения дозы измеряли уровни триглицеридов в плазме. Исходные значения (варьировавшие от 0,40 до 0,80 г/л для указанных четырех групп) вычитали для того, чтобы определить степень повышения уровней триглицеридов в плазме. Брали по 3 животных на группу.
Фиг.13 представляет собой кривую повышения уровней свободного холестерина ЛПВП в плазме после инфузии пептид 16/липидного комплекса (указанные липиды представляют собой сфингомиелин, DPPC и DPPG в весовом соотношении 1:1,2125:1,2125:0,075, и весовое соотношение пептид:липид составляет 1:2,5) кроликам на голодный желудок в дозах 0 (квадратики), 2,5 (треугольники), 5 (кружочки) или 10 (ромбики) мг/кг. На исходном уровне и через 5, 20, 40, 60, 90 и 120 минут после инициации инфузии измеряли уровни свободного холестерина ЛПВП в плазме. Исходные значения вычитали для того, чтобы определить степень повышения уровней свободного холестерина ЛПВП в плазме. Брали по 4 животных на группу.
Фиг.14A представляет собой профиль элюции гельпроникающей ВЭЖХ-хроматографии на исходном уровне (темная линия) и через 20 мин после инфузии 2,5 мг/кг пептид 16/липидного комплекса (указанные липиды представляют собой сфингомиелин, DPPC и DPPG в весовом соотношении 1:1,2125:1,2125:0,075, и весовое соотношение пептид:липид составляет 1:2,5). На оси Y отложено поглощение, полученное в результате непрерывного компьютерного анализа свободного холестерина липопротеиновых фракций, элюирующихся с колонки гельпроникающей ВЭЖХ-хроматографии. Пики слева направо соответствуют фракциям ЛПОНП, ЛПНП и ЛПВП.
Фиг.14B представляет собой профиль элюции гельпроникающей ВЭЖХ-хроматографии на исходном уровне (темная линия) и через 20 мин после инфузии 5,0 мг/кг пептид 16/липидного комплекса (указанные липиды представляют собой сфингомиелин, DPPC и DPPG в весовом соотношении 1:1,2125:1,2125:0,075, и весовое соотношение пептид:липид составляет 1:2,5). На оси Y отложено поглощение, полученное в результате непрерывного компьютерного анализа свободного холестерина липопротеиновых фракций, элюирующихся с колонки гельпроникающей ВЭЖХ-хроматографии. Пики слева направо соответствуют фракциям ЛПОНП, ЛПНП и ЛПВП.
Фиг.15 представляет собой кривую повышения уровней свободного холестерина ЛПВП в плазме после инфузии пептид 16/липидного комплекса (указанные липиды представляют собой сфингомиелин, DPPC и DPPG в весовом соотношении 1:1,2125:1,2125:0,075, и весовое соотношение пептид:липид составляет 1:2,5) кроликам на голодный желудок в дозе 20 мг/кг со скоростью 1 мл/мин (треугольники) или 0,2 мл/мин (ромбики). Через различные промежутки времени после введения дозы измеряли уровни свободного холестерина ЛПВП в плазме. Исходные значения вычитали для того, чтобы определить степень повышения уровней свободного холестерина ЛПВП в плазме. Брали по 4 животных на группу, обрабатываемую пептид 16/липидным комплексом, и по 2 животных на группу, обрабатываемую наполнителем.
На фиг.16 проиллюстрированы кривые кинетических профилей пептида 16 (верхние панели), свободного холестерина (средние панели) и фосфолипида (нижние панели) у самцов и самок крыс после введения первой дозы пептид 16/липидного комплекса (указанные липиды представляют собой сфингомиелин, DPPC и DPPG в весовом соотношении 1:1,2125:1,2125:0,075, и весовое соотношение пептид:липид составляет 1:2,5) на день 0. Снижение уровней пептида 16 и фосфолипида в плазме со временем указывает на клиренс пептид 16/липидного комплекса. Представлены кинетики свободного холестерина. Данные в каждой точке представляют собой среднее ± SD (N=3 крысам/группу).
На фиг.17 проиллюстрированы кривые кинетических профилей пептида 16 (верхние панели), свободного холестерина (средние панели) и фосфолипида (нижние панели) у самцов и самок крыс после введения множественной дозы пептид 16/липидного комплекса (указанные липиды представляют собой сфингомиелин, DPPC и DPPG в весовом соотношении 1:1,2125:1,2125:0,075, и весовое соотношение пептид:липид составляет 1:2,5) на день 26. Указанные животные получали пептид 16/липидный комплекс через день в течение 4 недель. Снижение уровней пептида 16 и фосфолипида в плазме со временем указывает на клиренс пептид 16/липидного комплекса. Представлены кинетики свободного холестерина. Данные в каждой точке представляют собой среднее ± SD (N=3 крысам/группу).
На фиг.18 проиллюстрированы кривые кинетических профилей пептида 16 (верхние панели), свободного холестерина (средние панели) и фосфолипида (нижние панели) у самцов и самок макак-крабоедов после введения первой дозы пептид 16/липидного комплекса (указанные липиды представляют собой сфингомиелин, DPPC и DPPG в весовом соотношении 1:1,2125:1,2125:0,075, и весовое соотношение пептид:липид составляет 1:2,5) на день 0. Снижение уровней пептида 16 и фосфолипида в плазме со временем указывает на клиренс пептид 16/липидного комплекса. Представлены кинетики свободного холестерина. Данные в каждой точке представляют собой среднее ± SD (N=3 макакам/группу).
На фиг.19 проиллюстрированы кривые кинетических профилей пептида 16 (верхние панели), свободного холестерина (средние панели) и фосфолипида (нижние панели) у самцов и самок макак-крабоедов после введения множественной дозы пептид 16/липидного комплекса (указанные липиды представляют собой сфингомиелин, DPPC и DPPG в весовом соотношении 1:1,2125:1,2125:0,075, и весовое соотношение пептид:липид составляет 1:2,5) на день 26. Указанные животные получали пептид 16/липидный комплекс через день в течение 4 недель. Снижение уровней пептида 16 и фосфолипида в плазме со временем указывает на клиренс пептид 16/липидного комплекса. Представлены кинетики свободного холестерина. Данные в каждой точке представляют собой среднее ± SD (N=3 макакам/группу).
Фиг.20A представляет собой кривую, отражающую % возрастания, по сравнению с уровнем, наблюдавшимся перед введением дозы, суммарного холестерина плазмы у мышей C57B1/6J после обработки композицией A, B или C. В разных временных точках у животных последовательно отбирали пробы, по 6 животных/на группу.
Фиг.20B представляет собой кривую повышения уровней суммарного холестерина в плазме у мышей C57B1/6J после обработки композицией A, B или C. В разных временных точках у животных последовательно отбирали пробы, по 6 животных/на группу.
Фиг.21A представляет собой кривую, отражающую % возрастания, по сравнению с уровнем, наблюдавшимся перед введением дозы, этерифицированного холестерина у мышей C57B1/6J после обработки композицией A, B или C. В разных временных точках у животных последовательно отбирали пробы, по 6 животных/на группу.
Фиг.21B представляет собой кривую повышения уровней этерифицированного холестерина у мышей C57B1/6J после обработки композицией A, B или C. В разных временных точках у животных последовательно отбирали пробы, по 6 животных/на группу.
Подробное описание изобретения
I. Определения
Термин "приблизительно", который непосредственно предваряет числовое или количественное значение, означает, что указанное числовое или количественное значение имеет разброс величин, составляющий плюс или минус 10%. Например, "приблизительно 1:1" в диапазоне от 0,9:1 до 1,1:1.
Используемый здесь термин "алкил", если не оговорено иначе, относится к необязательно замещенному насыщенному углеводородному радикалу с разветвленной, прямой цепью или циклическому углеводородному радикалу. Типичными алкильными группами являются (C1-C6)-алкильные группы, которые включают в себя, но не ограничены перечисленным, метил, этил, пропил, изопропил, бутил, изобутил, трет-бутил, пентил, изопентил, гексил и т.п. В определенных воплощениях алкильные группы представляют собой (C1-C4)-алкил. Если не оговорено иначе, алкил является незамещенным.
Используемый здесь термин "алкенил", если не оговорено иначе, относится к ненасыщенному углеводородному радикалу с разветвленной, прямой цепью или циклическому неароматическому углеводородному радикалу, имеющему одну или более углерод-углеродных двойных связей. Указанные одна или несколько двойных связей могут быть либо в цис-, либо в транс-конформации. Обычно алкенильные группы включают в себя, но не ограничены перечисленным, этенил, пропенил, изопропенил, бутенил, изобутенил, трет-бутенил, пентенил, гексенил и т.п. В определенных воплощениях алкенильная группа представляет собой (C2-C6)-алкенил.
Используемый здесь термин "алкинил", если не оговорено иначе, относится к ненасыщенному углеводородному радикалу с разветвленной или прямой цепью, имеющему по меньшей мере одну углерод-углеродную тройную связь. Обычно алкинильные группы включают в себя, но не ограничены перечисленным, этинил, пропинил, бутинил, изобутинил, пентинил, гексинил и т.п. В определенных воплощениях алкинильная группа представляет собой (C2-C6)-алкинил.
Используемый здесь термин "арил", если не оговорено иначе, относится к необязательно замещенной системе ароматических колец, в которых каждый атом внутри кольца представляет собой C, O, N или S, таким образом обслуживая гетероциклические ароматические кольца. Обычно арильные группы включают в себя, но не ограничены перечисленным, бензил, фенил, нафтил, антрацил, фуран, имидазол, индазол, индол, изохинолин, изотиазол, изоксазол, пиран, пиразин, пиразол, пиридазин, пиридин, пиримидин, пиррол, пирролизин, хиназолин, хинолин, хинолизин, хиноксалин, тиазол и тиофен. В определенных воплощениях арильная группа представляет собой (C5-C26-арил). В определенных воплощениях гетероарильная группа представляет собой 5-20-членный гетероарил. Если не оговорено иначе, арил является незамещенным.
Используемый здесь термин "аралкил", если не оговорено иначе, относится к алкильной группе, замещенной арильной группой.
Используемый здесь термин "замещенный алкил или арил", если не оговорено иначе, относится к алкильной или арильной группе, в которой один или несколько ее атомов водорода заменены другим заместителем. Типичные заместители включают в себя ORa, -SRa, -NRaRa, -NO2, -CN, галоген, -SO2Ra, -C(O)Ra, -C(O)ORa и -C(O)NRaRa, где каждый Ra независимо является водородом, алкилом или арилом.
Используемый здесь термин "гидрофильная оболочка" если не оговорено иначе, относится к наружной поверхности спирали, имеющей в целом гидрофильный характер.
Используемый здесь термин "гидрофобная оболочка" если не оговорено иначе, относится к наружной поверхности пептида, имеющего в целом гидрофобный характер.
Здесь, когда речь идет о миметике ApoA-I, имеется в виду, что число концевых -NH2-групп равно нулю, когда R1 представляет собой аминозащитную группу, и равно 1, когда R1 является H.
Здесь, когда речь идет о миметике ApoA-I, имеется в виду, что число концевых -COOH-групп равно нулю, когда R1 представляет собой карбоксильную защитную группу, и равно 1, когда R1 является OH.
Используемый здесь термин "млекопитающее", если не оговорено иначе, относится к человеку, мыши, крысе, морской свинке, собаке, кошке, лошади, корове, свинье или не принадлежащему человеческому роду примату, такому как обезьяна, шимпанзе или павиан. В одном из воплощений указанное млекопитающее является человеком.
Когда термин "эффективное количество" используется в связи с миметиком ApoA-I, он означает количество, которое эффективно для лечения или предупреждения какого-либо Состояния.
Используемый здесь термин "свободный холестерин ЛПВП" означает количество такого холестерина, который имеет свободную гидроксильную группу ("свободный холестерин"), которая содержится внутри частиц ЛПВП в сыворотке. Частицы ЛПВП могут образовываться из миметик ApoA-I/липидного комплекса.
Используемый здесь термин "суммарный холестерин ЛПВП" означает количество холестерина, который имеет гидроксильную группу, которая этерифицирована ("этерифицированный холестерин"), которая содержится внутри частиц ЛПВП в сыворотке. Частицы ЛПВП могут образовываться из миметик ApoA-I/липидного комплекса.
Используемый здесь термин "аминокислотный остаток", если не оговорено иначе, включает в себя генетически кодируемые аминокислотные остатки и генетически не кодируемые аминокислотные остатки.
Аббревиатуры, существующие для используемых здесь генетически кодируемых аминокислотных остатков, приведены ниже в таблице 1.
Генетически не кодируемые аминокислотные остатки включают в себя, но не ограничены перечисленным, β-аланин (β-Ala); 2,3-диаминопропионовую кислоту (Dpr); нипекотиновую кислоту (Nip); пипеколиновую кислоту (Pip); орнитин (Orn); цитруллин (Cit); трет-бутилаланин (t-BuA); 2-трет-бутилглицин (t-BuG); N-метилизолейцин (MeIle); фенилглицин (PhG); циклогексилаланин (ChA); норлейцин (Nle); нафтилаланин (Nal); 4-хлорфенилаланин (Phe(4-Cl)); 2-фторфенилаланин (Phe(2-F)); 3-фторфенилаланин (Phe(3-F)); 4-фторфенилаланин (Phe(4-F)); пеницилламин (Pen); l,2,3,4-тетрагидроизохинолин-3-карбоновую кислоту (Tic); β-2-тиенилаланин (Thi); метионинсульфоксид (MSO); гомоаргинин (hArg); N-ацетиллизин (AcLys); 2,4-диаминомасляную кислоту (Dbu); 2,3-диаминомасляную кислоту (Dab); пара-аминофенилаланин (Phe(pNH2)); N-метилвалин (MeVal); гомоцмстемн (hCys), гомофенилаланин (hPhe); гомосерин (hSer); гидроксипролин (Hyp); гомопролин (hPro); и соответствующий D-энантиомер каждого из указанных выше соединений, например, D-β-Ala, D-Dpr, D-Nip, D-Orn, D-Cit, D-t-BuA, D-t-BuG, D-MeIle, D-PhG, D-ChA, D-Nle, D-Nal, D-Phe(4-Cl), D-Phe(2-F), D-Phe(3-F), D-Phe(4-F), D-Pen, D-Tic, D-Thi, D-MSO, D-hArg, D-AcLys, D-Dbu, D-Dab, D-Phe(pNH2), D-MeVal, D-hCys, D-hPhe, D-hSer, D-Hyp и D-hPro. Другие генетически не кодируемые аминокислотные остатки включают в себя 3-аминопропионовую кислоту; 4-аминомасляную кислоту; изонипекотиновую кислоту (Inp); аза-пипеколиновую кислоту (azPip); аза-пролин (azPro); α-аминоизомасляную кислоту (Aib); ε-аминогексановую кислоту (Aha); δ-аминовалериановую кислоту (Ava); N-метилглицин (MeGly).
Термин "хиральный", используемый здесь в отношении аминокислотного остатка, означает аминокислотный остаток, имеющий по меньшей мере один хиральный центр. В одном из воплощений указанный хиральный аминокислотный остаток является L-аминокислотным остатком. Примеры L-аминокислотных остатков включают в себя, не ограничиваясь перечисленным, Ala, Arg, Asn, Asp, Cys, Gln, Glu, His, Ile, Leu, Lys, Met, Phe, Pro, Ser, Thr, Trp, Tyr, Val, β-Ala, Dpr, Nip, Orn, Cit, t-BuA, t-BuG, MeIle, PhG, ChA, NIe, Nal, Phe(4-Cl), Phe(2-F), Phe(3-F), Phe(4-F), Pen, Tic, Thi, MSO, hArg, AcLys, Dbu, Dab, Phe(pNH2), MeVal, hCys, hPhe, hSer, Hyp и hPro. В одном из воплощений указанный хиральный аминокислотный остаток является D-аминокислотным остатком. Примеры D-аминокислотных остатков включают в себя, не ограничиваясь перечисленным, D-Ala, D-Arg, D-Asn, D-Asp, D-Cys, D-Gln, D-Glu, D-His, D-Ile, D-Leu, D-Lys, D-Met, D-Phe, D-Pro, D-Ser, D-Thr, D-Trp, D-Tyr, D-Val, D-β-Ala, D-Dpr, D-Nip, D-Pip, D-Orn, D-Cit, D-t-BuA, D-t-BuG, D-MeIle, D-PhG, D-ChA, D-Nle, D-Nal, D-Phe(4-Cl), D-Phe(2-F), D-Phe(3-F), D-Phe(4-F), D-Pen, D-Tic, D-Thi, D-MSO, D-hArg, D-AcLys, D-Dbu, D-Dab, D-Phe(pNH2), D-MeVal, D-hCys, D-hPhe, D-hSer, D-Hyp и D-hPro.
Термин "ахиральный", используемый здесь в отношении аминокислотного остатка, означает аминокислотный остаток, который не имеет хирального центра. Примеры ахиральных аминокислотных остатков включают в себя, не ограничиваясь перечисленным, Gly, Inp, Aib, Aha, Ava, MeGly, azPip и azPro.
Используемый здесь термин "алифатический аминокислотный остаток", если не оговорено иначе, относится к аминокислотному остатку, имеющему алифатическую углеводородную цепь. Алифатический аминокислотный остаток включает в себя, не ограничиваясь перечисленным, Ala (A), Val (V), Leu (L), Ile (I), Pro (P), azPro, Pip, azPip, β-Ala, Aib, t-BuA, t-BuG, MeIle, ChA, Nle, MeVal, Inp, Nip, hPro, D-Ala, D-Val, D-Leu, D-Ile, D-Pro, D-β-Ala, D-t-BuA, D-t-BuG, D-MeIle, D-Nle, D-MeVal, D-Nip, D-Pip, D-ChA и D-hPro. В одном из воплощений указанный алифатический аминокислотный остаток является L-аминокислотным остатком. В другом воплощении указанный алифатический аминокислотный остаток является D-аминокислотным остатком. В другом воплощении указанный алифатический аминокислотный остаток является ахиральным аминокислотным остатком.
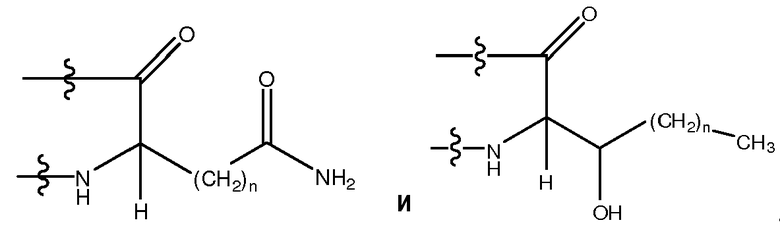
Используемый здесь термин "гидрофильный аминокислотный остаток", если не оговорено иначе, относится к аминокислотному остатку, проявляющему менее чем нулевую гидрофобность, согласно общепринятой нормированной шкале гидрофобности Айзенберга, Eisenberg et al., 1984, J. Mol. Biol. 179:125-142. Гидрофильный аминокислотный остаток включает в себя, не ограничиваясь перечисленным, Pro (P), Gly (G), Thr (T), Ser (S), His (H), Glu (E), Asn (N), Gln (Q), Asp (D), Lys (K), Arg (R), Dpr, Orn, Cit, Pen, MSO, hArg, AcLys, Dbu, Dab, Phe(p-NH2), hCys, hSer, Hyp, D-Pro, D-Thr, D-Ser, D-His, D-Glu, D-Asn, D-Gln, D-Asp, D-Lys, D-Arg, D-Dpr, D-Orn, D-Cit, D-Pen, D-MSO, D-hArg, D-AcLys, D-Dbu, D-Dab, D-Phe(p-NH2), D-hCys, D-hSer и D-Hyp. Другие гидрофильные аминокислотные остатки включают в себя, не ограничиваясь перечисленным, аналоги боковых цепей C1-4, имеющие следующие формулы:
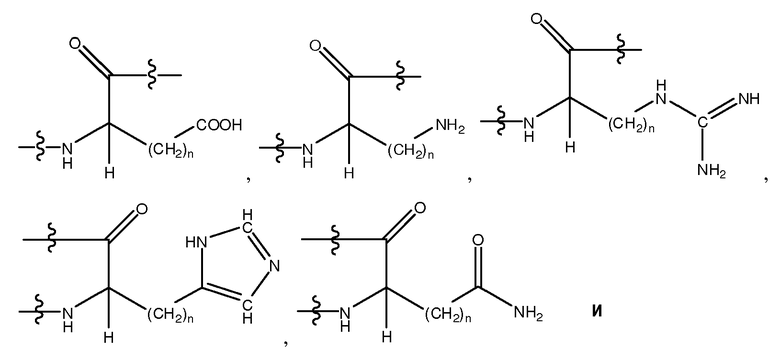
 , где n является целым числом от 1 до 4. В одном из воплощений указанный гидрофильный аминокислотный остаток является L-аминокислотным остатком. В другом воплощении указанный гидрофильный аминокислотный остаток является D-аминокислотным остатком. В другом воплощении указанный гидрофильный аминокислотный остаток является ахиральным аминокислотным остатком. В другом воплощении указанный гидрофильный аминокислотный остаток является кислым L-аминокислотным остатком, кислым D-аминокислотным остатком или кислым ахиральным аминокислотным остатком. В другом воплощении указанный гидрофильный аминокислотный остаток является основным L-аминокислотным остатком, основным D-аминокислотным остатком или основным ахиральным аминокислотным остатком.
, где n является целым числом от 1 до 4. В одном из воплощений указанный гидрофильный аминокислотный остаток является L-аминокислотным остатком. В другом воплощении указанный гидрофильный аминокислотный остаток является D-аминокислотным остатком. В другом воплощении указанный гидрофильный аминокислотный остаток является ахиральным аминокислотным остатком. В другом воплощении указанный гидрофильный аминокислотный остаток является кислым L-аминокислотным остатком, кислым D-аминокислотным остатком или кислым ахиральным аминокислотным остатком. В другом воплощении указанный гидрофильный аминокислотный остаток является основным L-аминокислотным остатком, основным D-аминокислотным остатком или основным ахиральным аминокислотным остатком.
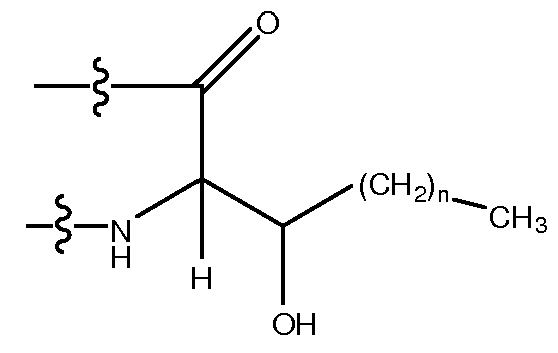
Используемый здесь термин "гидрофобный аминокислотный остаток", если не оговорено иначе, относится к аминокислотному остатку, проявляющему более чем нулевую гидрофобность, согласно общепринятой нормированной шкале гидрофобности Айзенберга, Eisenberg, 1984, J. Mol. Biol. 179:125-142. Гидрофобный аминокислотный остаток включает в себя, не ограничиваясь перечисленным, Ile (I), Phe (F), Val (V), Leu (L), Trp (W), Met (M), Ala (A), Gly (G), Tyr (Y), β-Ala, Nip, t-BuA, t-BuG, MeIle, PhG, ChA, Nle, Nal, Phe(4-Cl), Phe(2-F), Phe(3-F), Phe(4-F), Tic, Thi, MeVal, hPhe, hPro, 3-аминопропионовую кислоту, 4-аминомасляную кислоту, Inp, Aib, Aha, Ava, MeGly, D-Pro, D-Ile, D-Phe, D-Val, D-Leu, D-Trp, D-Met, D-Ala, D-Tyr, D-β-Ala, D-Nip, D-t-BuA, D-t-BuG, D-MeIle, D-PhG, D-ChA, D-Nle, D-Nal, D-Phe(4-Cl), D-Phe(2-F), D-Phe(3-F), D-Phe(4-F), D-Tic, D-Thi, D-MeVal, D-hPhe и D-hPro. Другие гидрофобные аминокислоты включают в себя, не ограничиваясь перечисленным, аналоги боковых цепей C1-4, имеющие следующие формулы:
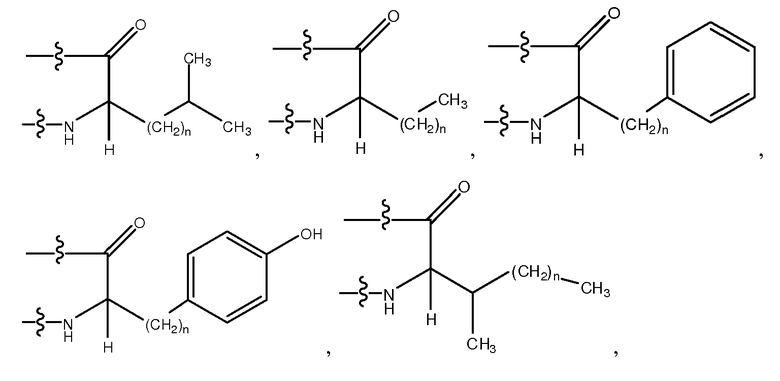
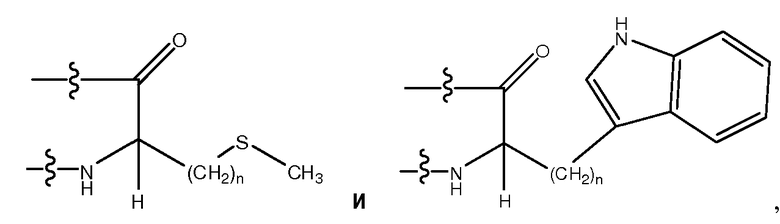
где n является целым числом от 1 до 4. В одном из воплощений указанный гидрофобный аминокислотный остаток является L-аминокислотным остатком. В другом воплощении указанный гидрофобный аминокислотный остаток является D-аминокислотным остатком. В другом воплощении указанный гидрофобный аминокислотный остаток является ахиральным аминокислотным остатком.
Используемый здесь термин "полярный аминокислотный остаток", если не оговорено иначе, относится к гидрофильному аминокислотному остатку, имеющему боковую цепь, которая не заряжена при физиологических значениях pH, но которая содержит по меньшей мере одну связь, в которой пара электронов, обобществленная двумя атомами, удерживается в большей близости к одному из атомов. Полярные аминокислотные остатки включают в себя, не ограничиваясь перечисленным, Asn (N), Gln (Q), Ser (S), Thr (T), Cit, Pen, MSO, AcLys, hCys, hSer, Hyp, D-Asn, D-Gln, D-Ser, D-Thr, D-Cit, D-Pen, D-MSO, D-AcLys, D-hCys, D-hSer и D-Hyp. Другие полярные аминокислоты включают в себя, не ограничиваясь перечисленным, аналоги боковых цепей C1-4, имеющие следующие формулы:
где n является целым числом от 1 до 4. В одном из воплощений указанный полярный аминокислотный остаток является L-аминокислотным остатком. В другом воплощении указанный полярный аминокислотный остаток является D-аминокислотным остатком. В другом воплощении указанный полярный аминокислотный остаток является ахиральным аминокислотным остатком.
Используемый здесь термин "кислый аминокислотный остаток", если не оговорено иначе, относится к гидрофильному аминокислотному остатку, имеющему значение pK боковой цепи, которое составляет менее чем 7. Кислые аминокислотные остатки обычно при физиологических значениях pH имеют отрицательно заряженные боковые цепи в силу утраты иона водорода. Кислые аминокислотные остатки включают в себя, не ограничиваясь перечисленным, Glu (E), Asp (D), D-Glu и D-Asp. Другие кислые аминокислотные остатки включают в себя, не ограничиваясь перечисленным, аналоги боковых цепей C1-4, имеющие следующую формулу:
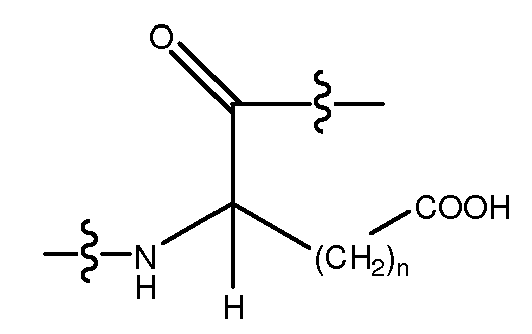 , где n является целым числом от 1 до 4. В одном из воплощений указанный кислый аминокислотный остаток является L-аминокислотным остатком. В другом воплощении указанный кислый аминокислотный остаток является D-аминокислотным остатком. В другом воплощении указанный полярный аминокислотный остаток является ахиральным аминокислотным остатком.
, где n является целым числом от 1 до 4. В одном из воплощений указанный кислый аминокислотный остаток является L-аминокислотным остатком. В другом воплощении указанный кислый аминокислотный остаток является D-аминокислотным остатком. В другом воплощении указанный полярный аминокислотный остаток является ахиральным аминокислотным остатком.
Используемый здесь термин "основный аминокислотный остаток", если не оговорено иначе, относится к гидрофильному аминокислотному остатку, имеющему значение pK боковой цепи, которое составляет более чем 7. Основные аминокислотные остатки обычно при физиологических значениях pH имеют положительно заряженные боковые цепи в силу ассоциации с ионом гидрония. Основные аминокислотные остатки включают в себя, не ограничиваясь перечисленным, His (H), Arg (R), Lys (K), Dpr, Orn, hArg, Dbu, Dab, Phe(p-NH2), D-His, D-Arg, D-Lys, D-Dpr, D-Orn, D-hArg, D-Dbu, D-Dab и D-Phe(p-NH2). Другие основные аминокислотные остатки включают в себя, не ограничиваясь перечисленным, аналоги боковых цепей C1-4, имеющие следующие формулы:
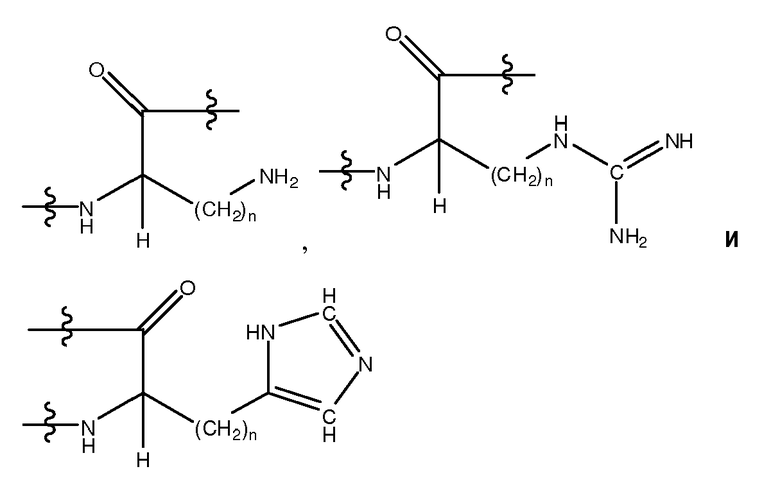 , где n является целым числом от 1 до 4. В одном из воплощений указанный основный аминокислотный остаток является L-аминокислотным остатком. В другом воплощении указанный основный аминокислотный остаток является D-аминокислотным остатком. В другом воплощении указанный основный аминокислотный остаток является ахиральным аминокислотным остатком.
, где n является целым числом от 1 до 4. В одном из воплощений указанный основный аминокислотный остаток является L-аминокислотным остатком. В другом воплощении указанный основный аминокислотный остаток является D-аминокислотным остатком. В другом воплощении указанный основный аминокислотный остаток является ахиральным аминокислотным остатком.
Используемый здесь термин "неполярный аминокислотный остаток", если не оговорено иначе, относится к гидрофобному аминокислотному остатку, имеющему боковую цепь, которая не заряжена при физиологических значениях pH и которая содержит связи, в которых пара электронов, обобществленная двумя атомами, удерживается по существу в одинаковой степени обоими этими атомами (то есть указанная боковая цепь является неполярной). Неполярные аминокислотные остатки включают в себя, не ограничиваясь перечисленным, Leu (L), Val (V), Ile (I), Met (M), Gly (G), Ala (A), Pro (P), azPro, Pip, azPip, β-Ala, Nip, t-BuG, MeIle, ChA, Nle, MeVal, hPro, 3-аминопропионовую кислоту, 4-аминомасляную кислоту, Inp, Aib, Aha, Ava, MeGly, D-Leu, D-Val, D-Ile, D-Met, D-Ala, D-Pro, D-β-Ala, D-Inp, D-t-BuG, D-MeIle, D-ChA, D-Nle, D-MeVal, D-Nip, D-Pip и D-hPro. Другие неполярные аминокислотные остатки включают в себя, не ограничиваясь перечисленным, аналоги боковых цепей C1-4, имеющие следующие формулы:
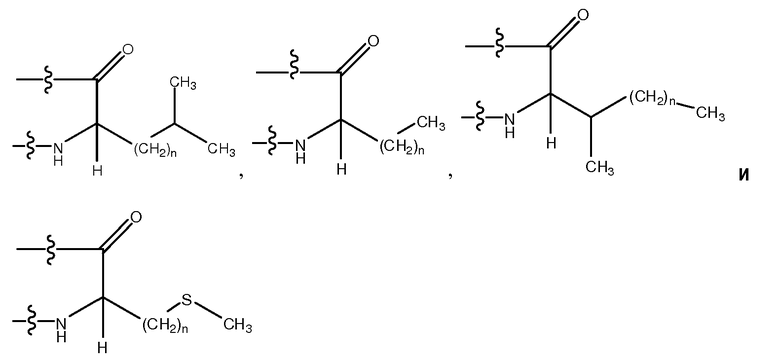
где n является целым числом от 1 до 4. В одном из воплощений указанный неполярный аминокислотный остаток является L-аминокислотным остатком. В другом воплощении указанный неполярный аминокислотный остаток является D-аминокислотным остатком. В другом воплощении указанный неполярный аминокислотный остаток является ахиральным аминокислотным остатком.
Используемый здесь термин "ароматический аминокислотный остаток", если не оговорено иначе, относится к гидрофобному аминокислотному остатку с боковой цепью, имеющей по меньшей мере одно ароматическое или гетероароматическое кольцо. Указанное ароматическое или гетероароматическое кольцо может содержать один или несколько заместителей, таких как -OH, -SH, -CN, -F, -Cl, -Br, -I, -NO2, -NO, -NH2, -NHR, -NRR, -C(O)R, -C(O)OH, -C(O)OR, -C(O)NH2, -C(O)NHR, -C(O)NRR, где каждый R независимо является (C1-C6)-алкилом, замещенным (C1-C6)-алкилом, 5-26-членным арилом и замещенным 5-26-членным арилом. Ароматические аминокислотные остатки включают в себя, не ограничиваясь перечисленным, Phe (F), Tyr (Y), Trp (W), PhG, Nal, Phe(4-Cl), Phe(2-F), Phe(3-F), Phe(4-F), Tic, Thi, hPhe, D-Phe, D-Tyr и D-Trp, D-PhG, D-Nal, D-Phe(4-Cl), D-Phe(2-F), D-Phe(3-F), D-Phe(4-F), D-Tic, D-Thi и D-hPhe. Другие ароматические аминокислотные остатки включают в себя, не ограничиваясь перечисленным, аналоги боковых цепей C1-4, имеющие следующие формулы:
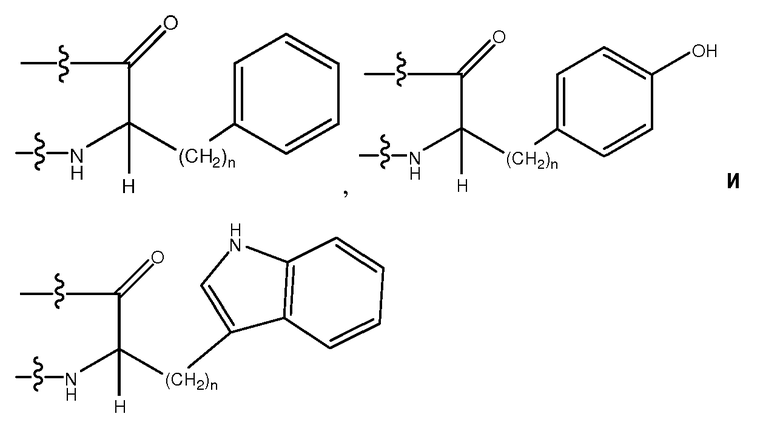 ,
,
где n является целым числом от 1 до 4. В одном из воплощений указанный ароматический аминокислотный остаток является L-аминокислотным остатком. В другом воплощении указанный ароматический аминокислотный остаток является D-аминокислотным остатком. В другом воплощении указанный ароматический аминокислотный остаток является ахиральным аминокислотным остатком.
Классификации генетически кодируемых и генетически не кодируемых аминокислотных остатков в соответствии с указанными выше категориями суммированы ниже в таблице. Следует понимать, что приведенная ниже таблица представлена здесь лишь в иллюстративных целях и не претендует на исчерпывающий список аминокислотных остатков, которые могут быть использованы в описанных здесь миметиках ApoA-I. Другие аминокислотные остатки, специально здесь не описанные, можно с легкостью отнести к той или иной категории на основании наблюдаемых у них физических и химических свойств, с учетом приведенных выше определений. Некоторые классификации аминокислотных остатков включены в таблицу 2 ниже.
Классификации некоторых аминокислотных остатков
Как должно быть очевидно специалистам в данной области, определяемые выше категории не являются взаимоисключающими. Таким образом, аминокислотные остатки, имеющие боковые цепи, проявляющие два или более физико-химических свойств, могут быть включены во множественные категории. Например, аминокислотные боковые цепи, имеющие ароматические фрагменты, которые дополнительно замещены полярными заместителями, такими как Tyr (Y) или его соответствующим D-энантиомером, могут проявлять как гидрофобные ароматические свойства, так и полярные или гидрофильные свойства, и, следовательно, могут быть включены в обе категории, ароматическую и полярную. Отнесение любого аминокислотного остатка к той или иной категории должно быть очевидно специалисту в данной области, в частности, в свете представленного здесь описания.
Кроме того, аминокислотный остаток Cys (C) или его соответствующий D-энантиомер может образовывать дисульфидные мостики с другими остатками Cys (C) или их соответствующими D-энантиомерами или с другими сульфанил-содержащими аминокислотами. Способность остатков Cys (C) (и других аминокислот, содержащих SH-боковые цепи) существовать в пептиде либо в редуцированной форме со свободными SH-группами, либо в виде окисленной образующей дисульфидные мостики форме, влияет на то, будут ли остатки Cys (C) или их соответствующие D-энантиомеры в результате придавать пептиду гидрофобный или гидрофильный характер. Поскольку Cys (C) или его соответствующий D-энантиомер проявляет гидрофобность, соответствующую 0,29, согласно общепринятой нормированной шкале Айзенберга (Eisenberg, 1984, выше), следует понимать, что в целях осуществления настоящего изобретения Cys (C) и его соответствующий D-энантиомер подпадают под категорию полярных гидрофильных аминокислот, несмотря на общую классификацию, определяемую выше.
II. Миметики ApoA-I
A. Пептиды формулы I
В одном из воплощений настоящее изобретение связано с пептидами, содержащими от 15 до 29 остатков, имеющими формулу I:
R1-Y1-X1-X2-X3-X4-X5-X6-X7-X8-X9-X10-X11-X12-X13-X14-X15-X16-X17-X18-X19-X20-X21-X22-X23-Y2-R2
Формула I
и их фармацевтически приемлемыми солями, в которых
X1 отсутствует или является ахиральным, D- или L-основным аминокислотным остатком;
X2 является ахиральным, D- или L-основным аминокислотным остатком;
X3 является ахиральным, D- или L-алифатическим аминокислотным остатком;
X4 является ахиральным, D- или L-основным аминокислотным остатком;
X5 представляет собой Gln, Asn, D-Gln, D-Asn или ахиральный, D- или L-основный аминокислотный остаток;
X6 представляет собой Gln, Asn, D-Gln, D-Asn или ахиральный, D- или L-основный аминокислотный остаток;
X7 является ахиральным, D- или L-гидрофобным аминокислотным остатком;
X8 является ахиральным, D- или L-гидрофобным аминокислотным остатком;
X9 является ахиральным, D- или L-гидрофильным аминокислотным остатком;
X10 представляет собой Leu, Trp, Gly, Nal, D-Leu, D-Trp или D-Nal;
X11 представляет собой Gly или ахиральный, D- или L-алифатический аминокислотный остаток;
X12 является ахиральным, D- или L-гидрофильным аминокислотным остатком;
X13 является ахиральным, D- или L-гидрофильным аминокислотным остатком;
X14 представляет собой Leu, Trp, Gly, D-Leu или D-Trp;
X15 представляет собой Leu, Gly или D-Leu;
X16 является ахиральным, D- или L-кислым аминокислотным остатком, или ахиральным, D- или L-основным аминокислотным остатком;
X17 является ахиральным, D- или L-гидрофильным аминокислотным остатком;
X18 представляет собой Leu, Phe, D-Leu или D-Phe;
X19 представляет собой Leu, Phe, D-Leu или D-Phe;
X20 является ахиральным, D- или L-кислым аминокислотным остатком;
X21 представляет собой Leu, Phe, D-Leu или D-Phe;
X22 является ахиральным, D- или L-алифатическим аминокислотным остатком; и
X23 представляет собой Inp, Nip, azPro, Pip, azPip, D-Nip или D-Pip;
Y1 отсутствует или является аминокислотной последовательностью, содержащей от 1 до 7 остатков;
Y2 отсутствует или является аминокислотной последовательностью, содержащей от 1 до 7 остатков;
R1 представляет собой H или аминозащитную группу;
R2 представляет собой OH или карбоксильную защитную группу;
причем от нуля до восьми остатков X2-X22 отсутствуют; и
при этом
a) каждый хиральный аминокислотный остаток является L-аминокислотным остатком;
b) каждый хиральный аминокислотный остаток является D-аминокислотным остатком;
c) каждый хиральный аминокислотный остаток является L-аминокислотным остатком, за исключением того, что один или более из каждых хиральных концевых аминокислотных остатков и каждых хиральных аминокислотных остатков, непосредственно к ним примыкающих, является D-аминокислотным остатком; или
d) каждый хиральный аминокислотный остаток является D-аминокислотным остатком, за исключением того, что один или более из каждых хиральных концевых аминокислотных остатков и каждых хиральных аминокислотных остатков, непосредственно к ним примыкающих, является L-аминокислотным остатком.
В другом воплощении настоящее изобретение связано с пептидами, содержащими от 22 до 29 остатков, имеющими формулу I:
R1-Y1-X1-X2-X3-X4-X5-X6-X7-X8-X9-X10-X11-X12-X13-X14-X15-X16-X17-X18-X19-X20-X21-X22-X23-Y2-R2
Формула I
и их фармацевтически приемлемыми солями, в которых
X1 отсутствует или является ахиральным, D- или L-основным аминокислотным остатком;
X2 является ахиральным, D- или L-основным аминокислотным остатком;
X3 является ахиральным, D- или L-алифатическим аминокислотным остатком;
X4 является ахиральным, D- или L-основным аминокислотным остатком;
X5 представляет собой Gln, Asn, D-Gln, D-Asn или ахиральный, D- или L-основный аминокислотный остаток;
X6 является ахиральным, D- или L-основным аминокислотным остатком;
X7 является ахиральным, D- или L-гидрофобным аминокислотным остатком;
X8 является ахиральным, D- или L-гидрофобным аминокислотным остатком;
X9 является ахиральным, D- или L-гидрофильным аминокислотным остатком;
X10 представляет собой Leu, Trp, Gly, Nal, D-Leu, D-Trp или D-Nal;
X11 представляет собой Gly или ахиральный, D- или L-алифатический аминокислотный остаток;
X12 является ахиральным, D- или L-гидрофильным аминокислотным остатком;
X13 является ахиральным, D- или L-гидрофильным аминокислотным остатком;
X14 представляет собой Leu, Trp, Gly, D-Leu или D-Trp;
X15 представляет собой Leu, Gly или D-Leu;
X16 является ахиральным, D- или L-кислым аминокислотным остатком;
X17 является ахиральным, D- или L-гидрофильным аминокислотным остатком;
X18 представляет собой Leu, Phe, D-Leu или D-Phe;
X19 представляет собой Leu, Phe, D-Leu или D-Phe;
X20 является ахиральным, D- или L-кислым аминокислотным остатком;
X21 представляет собой Leu, Phe, D-Leu или D-Phe;
X22 является ахиральным, D- или L-алифатическим аминокислотным остатком; и
X23 представляет собой Inp, Nip, azPro, Pip, azPip, D-Nip или D-Pip;
Y1 отсутствует или является аминокислотной последовательностью, содержащей от 1 до 7 остатков;
Y2 отсутствует или является аминокислотной последовательностью, содержащей от 1 до 7 остатков;
R1 представляет собой H или аминозащитную группу;
R2 представляет собой OH или карбоксильную защитную группу; при этом
a) каждый хиральный аминокислотный остаток является L-аминокислотным остатком;
b) каждый хиральный аминокислотный остаток является D-аминокислотным остатком;
c) каждый хиральный аминокислотный остаток является L-аминокислотным остатком, за исключением того, что один или более из каждых хиральных концевых аминокислотных остатков и каждых хиральных аминокислотных остатков, непосредственно к ним примыкающих, является D-аминокислотным остатком; или
d) каждый хиральный аминокислотный остаток является D-аминокислотным остатком, за исключением того, что один или более из каждых хиральных концевых аминокислотных остатков и каждых хиральных аминокислотных остатков, непосредственно к ним примыкающих, является L-аминокислотным остатком.
В другом воплощении настоящее изобретение связано с пептидами, содержащими от 15 до 21 остатка, имеющими формулу I:
R1-Y1-X1-X2-X3-X4-X5-X6-X7-X8-X9-X10-X11-X12-X13-X14-X15-X16-X17-X18-X19-X20-X21-X22-X23-Y2-R2
Формула I
и их фармацевтически приемлемыми солями, в которых
X1 отсутствует или является ахиральным, D- или L-основным аминокислотным остатком;
X2 является ахиральным, D- или L-основным аминокислотным остатком;
X3 является ахиральным, D- или L-алифатическим аминокислотным остатком;
X4 является ахиральным, D- или L-основным аминокислотным остатком;
X5 представляет собой Gln, Asn, D-Gln, D-Asn или ахиральный, D- или L-основный аминокислотный остаток;
X6 является ахиральным, D- или L-основным аминокислотным остатком;
X7 является ахиральным, D- или L-гидрофобным аминокислотным остатком;
X8 является ахиральным, D- или L-гидрофобным аминокислотным остатком;
X9 является ахиральным, D- или L-гидрофильным аминокислотным остатком;
X10 представляет собой Leu, Trp, Gly, Nal, D-Leu, D-Trp или D-Nal;
X11 представляет собой Gly или ахиральный, D- или L-алифатический аминокислотный остаток;
X12 является ахиральным, D- или L-гидрофильным аминокислотным остатком;
X13 является ахиральным, D- или L-гидрофильным аминокислотным остатком;
X14 представляет собой Leu, Trp, Gly, D-Leu или D-Trp;
X15 представляет собой Leu, Gly или D-Leu;
X16 является ахиральным, D- или L-кислым аминокислотным остатком;
X17 является ахиральным, D- или L-гидрофильным аминокислотным остатком;
X18 представляет собой Leu, Phe, D-Leu или D-Phe;
X19 представляет собой Leu, Phe, D-Leu или D-Phe;
X20 является ахиральным, D- или L-кислым аминокислотным остатком;
X21 представляет собой Leu, Phe, D-Leu или D-Phe;
X22 является ахиральным, D- или L-алифатическим аминокислотным остатком; и
X23 представляет собой Inp, Nip, azPro, Pip, azPip, D-Nip или D-Pip;
Y1 отсутствует или является аминокислотной последовательностью, содержащей от 1 до 7 остатков;
Y2 отсутствует или является аминокислотной последовательностью, содержащей от 1 до 7 остатков;
R1 представляет собой H или аминозащитную группу;
R2 представляет собой OH или карбоксильную защитную группу;
причем от одного до восьми остатков X2-X22 отсутствуют; и при этом
a) каждый хиральный аминокислотный остаток является L-аминокислотным остатком;
b) каждый хиральный аминокислотный остаток является D-аминокислотным остатком;
c) каждый хиральный аминокислотный остаток является L-аминокислотным остатком, за исключением того, что один или более из каждых хиральных концевых аминокислотных остатков и каждых хиральных аминокислотных остатков, непосредственно к ним примыкающих, является D-аминокислотным остатком; или
d) каждый хиральный аминокислотный остаток является D-аминокислотным остатком, за исключением того, что один или более из каждых хиральных концевых аминокислотных остатков и каждых хиральных аминокислотных остатков, непосредственно к ним примыкающих, является L-аминокислотным остатком.
В другом воплощении пептид формулы I или его фармацевтически приемлемая соль имеет 22 аминокислотного остатка в длину, и X1 отсутствует.
Если не оговорено иначе, с имитаторами ApoA-I формулы I связаны следующие воплощения.
В одном из воплощений X2 и X4 оба являются Lys, Orn, D-Lys или D-Orn. В другом воплощении X5 представляет собой GIn, Lys, D-Gln или D-Lys. В другом воплощении X9 представляет собой кислый аминокислотный остаток. В другом воплощении X12 представляет собой Glu, Asn, Gln, Arg, D-Glu, D-Asn, D-Gln или D-Arg. В другом воплощении X13 представляет собой Glu, Asn, Gln, Arg, D-Glu, D-Asn, D-Gln или D-Arg. В другом воплощении X16 представляет собой кислый аминокислотный остаток. В другом воплощении X17 представляет собой Arg, Lys, Orn, D-Arg, D-Lys или D-Orn. В другом воплощении X21 представляет собой Leu или D-Leu. В другом воплощении X22 представляет собой Ala, Val, Leu, D-Ala, D-Val или D-Leu.
В другом воплощении X1 отсутствует; X13 представляет собой кислый аминокислотный остаток, Arg или D-Arg; X14 является основным аминокислотным остатком, Asn, Glu, D-Asn или D-Glu; и X2-X12 и X15-X23 являются такими, как описаны выше в формуле I.
В другом воплощении X1 отсутствует; X2 является Lys, Orn, D-Lys или D-Orn; X3 является Leu или D-Leu; X4 является Lys, Orn, D-Lys или D-Orn; X5 является Lys, Orn, Gln, Asn, D-Lys, D-Orn, D-Gln или D-Asn; X6 представляет собой Lys, Orn, Gln, Asn, D-Lys, D-Orn, D-Gln или D-Asn; X7 представляет собой Leu, Gly, Nal, D-Leu или D-Nal; X8 представляет собой Ala, Trp, Gly, Leu, Phe, Nal, D-Ala, D-Trp, D-Leu, D-Phe или D-Nal; X9 представляет собой Asp, Glu, Gln, Lys, D-Asp, D-Glu, D-Gln или D-Lys; X11 представляет собой Leu, Gly, D-Leu или Aib; X12 представляет собой Asp, Glu, Asn, D-Asp, D-Glu или D-Asn; X13 представляет собой Asn, Gln, Glu, Arg, D-Asn, D-Gln, D-Glu или D-Arg; X16 представляет собой Asp, Arg, Glu, D-Asp, D-Arg или D-Glu; X17 представляет собой Lys, Arg, Orn, Asn, Glu, D-Lys, D-Arg, D-Orn, D-Asn или D-Glu; X20 представляет собой Asp, Glu, D-Asp или D-Glu; и/или X22 представляет собой Ala, Val, Leu, D-Ala, D-Val или D-Leu; и X10, X14, X15, X18, X19, X21 и X23 являются такими, как определено выше в формуле I.
В другом воплощении X1 отсутствует; X9 представляет собой Glu или D-Glu; X12 представляет собой Glu или D-Glu; X13 представляет собой Asn, Glu, D-Asn или D-Glu; X14 представляет собой Leu или D-Leu; X15 представляет собой Leu или D-Leu; X16 представляет собой Glu или D-Glu; X17 представляет собой Arg, Lys, D-Arg или D-Lys; X18 представляет собой Phe или D-Phe; X19 представляет собой Leu или D-Leu; X21 представляет собой Leu или D-Leu; и/или X22 представляет собой Val или D-Val; и X2-X8, X10, X11, X20 и X23 являются такими, как определено выше в формуле I.
В другом воплощении X1 отсутствует; X2 представляет собой Lys, Orn, D-Lys или D-Orn; X3 представляет собой Leu или D-Leu; X4 представляет собой Lys, Orn, D-Lys или D-Orn; X5 представляет собой Lys, Orn, Gln, Asn, D-Lys, D-Orn, D-Gln или D-Asn; X6 представляет собой Lys, Orn, Gln, Asn, D-Lys, D-Orn, D-Gln или D-Asn; X7 представляет собой Leu, Gly, Nal, D-Leu или D-Nal; X8 представляет собой Ala, Trp, Gly, Leu, Phe, Nal, D-Ala, D-Trp, D-Leu, D-Phe или D-Nal; X9 представляет собой Glu или D-Glu; X11 представляет собой Leu, D-Leu, Gly или Aib; X12 представляет собой Glu или D-Glu; X13 представляет собой Asn, Glu, D-Asn или D-Glu; X14 представляет собой Leu или D-Leu; X15 представляет собой Leu или D-Leu; X16 представляет собой Glu или D-Glu; X17 представляет собой Arg, Lys, D-Arg или D-Lys; X18 представляет собой Phe или D-Phe; X19 представляет собой Leu или D-Leu; X20 представляет собой Asp, Glu, D-Asp или D-Glu; X21 представляет собой Leu или D-Leu; и/или X22 представляет собой Val или D-Val; и X10 и X23 являются такими, как определено выше в формуле I.
В другом воплощении X1 отсутствует, только один из X5 и X6 является основным аминокислотным остатком, а другой из X5 и X6 является Gln, Asn, D-Gln или D-Asn.
В другом воплощении Y1 или Y2 отсутствует или является последовательностью, содержащей от одного до семи аминокислотных остатков. В другом воплощении один или более из аминокислотных остатков указанной аминокислотной последовательности являются кислыми аминокислотными остатками. В другом воплощении один или более из аминокислотных остатков указанной аминокислотной последовательности являются основными аминокислотными остатками.
В другом воплощении один из X5 и X6 является Lys, Orn, D-Lys или D-Orn, а другой из X5 и X6 является Gln, Asn, D-Gln или D-Asn.
В другом воплощении каждый из хиральных аминокислотных остатков является L-аминокислотным остатком.
В другом воплощении каждый из хиральных аминокислотных остатков является D-аминокислотным остатком.
В другом воплощении X1 отсутствует; один из X7, X8, X10, X11, X14 и X15 является Gly; и X1-X6, X9, X12, X13, а также X16-X23 являются отличными от Gly.
В другом воплощении X1 отсутствует; X11 является Gly; и каждый из X7, X8, X10, X14 и X15 является отличным от Gly.
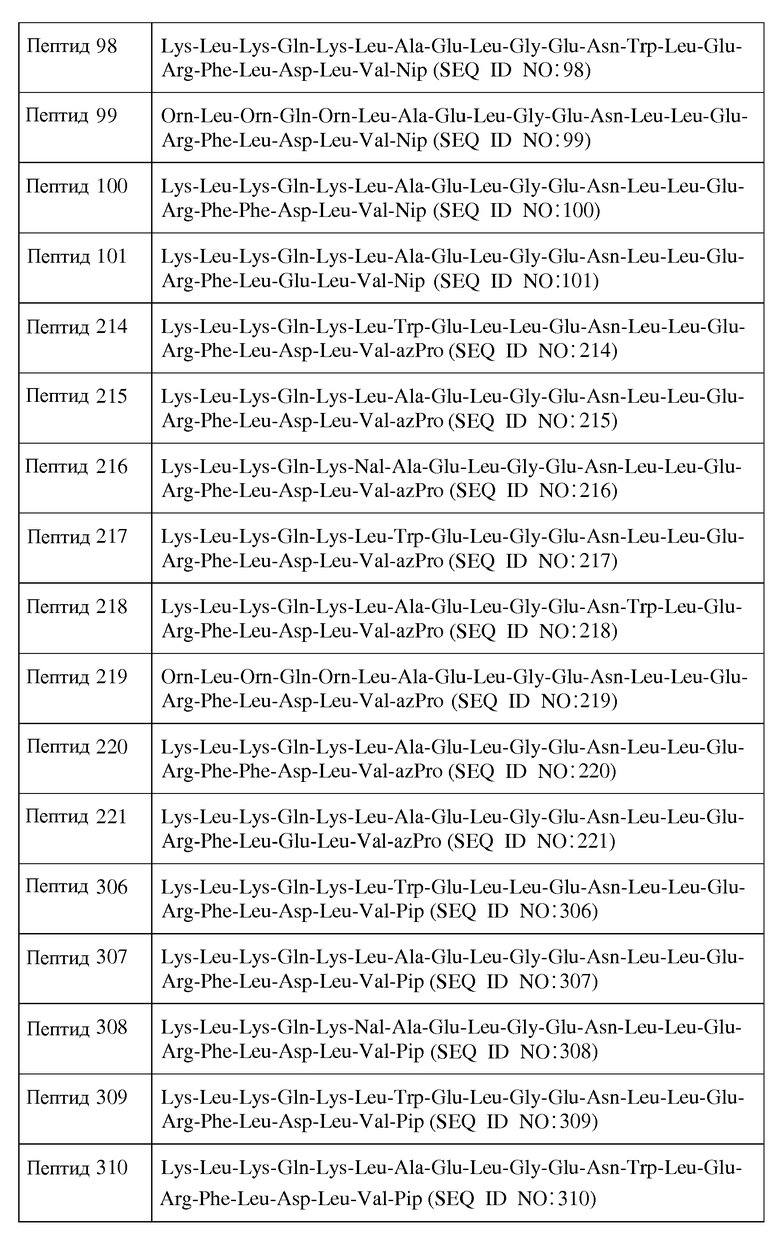
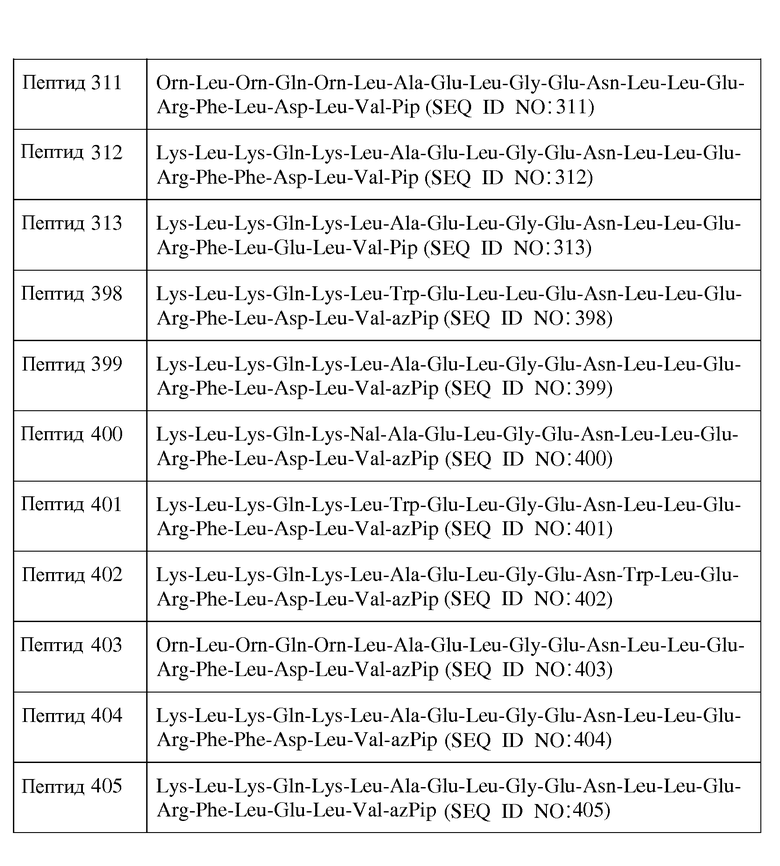
В другом воплощении X1 отсутствует; X2 представляет собой Lys, Orn, D-Lys или D-Orn; X3 представляет собой Leu или D-Leu; X4 является Lys, Orn, D-Lys или D-Orn; X5 является Gln или D-Gln; X6 представляет собой Lys, Orn, D-Lys или D-Orn; X7 представляет собой Leu, Nal, D-Leu или D-Nal; X8 является Ala, Trp, D-Ala или D-Trp; X9 является Glu или D-Glu; X10 представляет собой Leu или D-Leu; X11 является Gly; X12 представляет собой Glu или D-Glu; X13 представляет собой Asn или D-Asn; X14 представляет собой Leu, Trp, D-Leu или D-Trp; X15 является Leu или D-Leu; X16 представляет собой Glu или D-Glu; X17 является Arg или D-Arg; X18 представляет собой Phe или D-Phe; X19 представляет собой Leu, Phe, D-Leu или D-Phe; X20 представляет собой Asp, Glu, D-Asp или D-Glu; X21 является Leu или D-Leu; X22 представляет собой Val или D-Val; и X23 является Inp. В одном из воплощений пептид формулы I является пептидом, представленным в таблице 3 ниже:

или их фармацевтически приемлемой солью.
В другом воплощении X1 отсутствует; X15 является Gly; и каждый из X7, X8, X10, X11 и X14 является отличным от Gly. В одном из воплощений пептид формулы I представляет собой следующее:
или их фармацевтически приемлемую соль.
В другом воплощении X1 отсутствует; X14 является Gly; и каждый из X7, X8, X10, X11 и X15 является отличным от Gly. В одном из воплощений пептид формулы I представляет собой следующее:
или их фармацевтически приемлемую соль.
В другом воплощении X1 отсутствует; X10 является Gly; и каждый из X7, X8, X11, X14 и X15 является отличным от Gly. В одном из воплощений пептид формулы I представляет собой следующее:
или их фармацевтически приемлемую соль.
В другом воплощении X1 отсутствует; X8 является Gly; и каждый из X7, X10, X11, X14 и X15 является отличным от Gly. В одном из воплощений пептид формулы I представляет собой следующее:
или их фармацевтически приемлемую соль.
В другом воплощении X1 отсутствует; X7 является Gly; и каждый из X8, X10, X11, X14 и X15 является отличным от Gly. В одном из воплощений пептид формулы I представляет собой следующее:
или их фармацевтически приемлемую соль.
В другом воплощении X1 отсутствует; и каждый из X7, X8, X10, X11, X14 и X15 является отличным от Gly.
В другом воплощении X1 отсутствует; X2 представляет собой Lys, Orn, D-Lys или D-Orn; X3 представляет собой Leu или D-Leu; X4 является Lys, Orn, D-Lys или D-Orn; один из X5 или X6 является Gln или D-Gln, а другой из X5 или X6 представляет собой Lys, Orn, D-Lys или D-Orn; X7 представляет собой Leu, Nal, D-Leu или D-Nal; X8 является Ala, Leu, Trp, Nal, D-Ala, D-Leu, D-Trp или D-Nal; X9 является Glu или D-Glu; X10 представляет собой Leu, Trp, Nal, D-Leu, D-Trp или D-Nal; X11 является Leu, D-Leu или Aib; X12 представляет собой Glu или D-Glu; X13 представляет собой Asn, Gln, D-Asn или D-Gln; X14 представляет собой Leu, Trp, D-Leu или D-Trp; X15 является Leu или D-Leu; X16 представляет собой Glu или D-Glu; X17 является Arg, Lys, D-Arg или D-Lys; X18 представляет собой Leu, Phe, D-Leu или D-Phe; X19 представляет собой Leu, Phe, D-Leu или D-Phe; X20 представляет собой Asp, Glu, D-Asp или D-Glu; X21 является Leu или D-Leu; X22 представляет собой Val, Leu, D-Val или D-Leu; и X23 является Inp.
В другом воплощении X1 отсутствует; X2 представляет собой Lys или D-Lys; X3 представляет собой Leu или D-Leu; X4 является Lys или D-Lys; X5 представляет собой Gln или D-Gln; X6 является Lys или D-Lys; X7 представляет собой Leu или D-Leu; X8 является Ala или D-Ala; X9 является Glu или D-Glu; X10 представляет собой Leu или D-Leu; X11 является Leu или D-Leu; X12 представляет собой Glu или D-Glu; X13 представляет собой Asn или D-Asn; X14 представляет собой Leu или D-Leu; X15 является Leu или D-Leu; X16 представляет собой Glu или D-Glu; X17 является Arg или D-Arg; X18 представляет собой Phe или D-Phe; X19 представляет собой Leu или D-Leu; X20 представляет собой Asp или D-Asp; X21 является Leu или D-Leu; X22 представляет собой Val или D-Val; и X23 представляет собой Inp.
В другом воплощении X3 является отличным от Lys или D-Lys; X9 является отличным от Trp или D-Trp; X11 является отличным от Glu, Trp, D-Glu или D-Trp; X12 является отличным от Trp, Leu, D-Trp или D-Leu; X13 является отличным от Trp или D-Trp; X15 является отличным от Lys, Trp, D-Lys или D-Trp; X17 является отличным от Trp, Leu, D-Trp или D-Leu; X18 является отличным от Trp или D-Trp; X19 является отличным от Lys, Glu, Trp, Nal, D-Lys, D-Glu, D-Trp или D-Nal; и/или X22 является отличным от Val или D-Val.
В другом воплощении пептид формулы I является одним из пептидов, представленных в таблице 4 ниже:
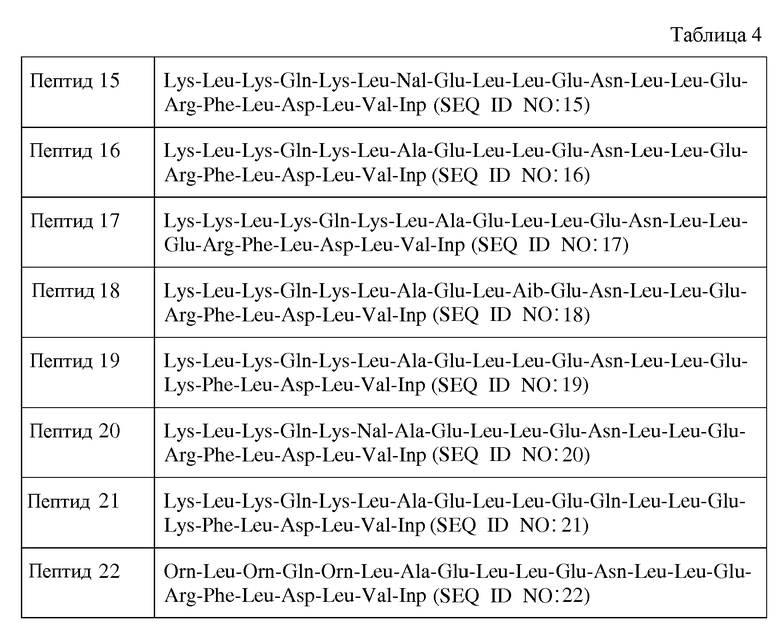
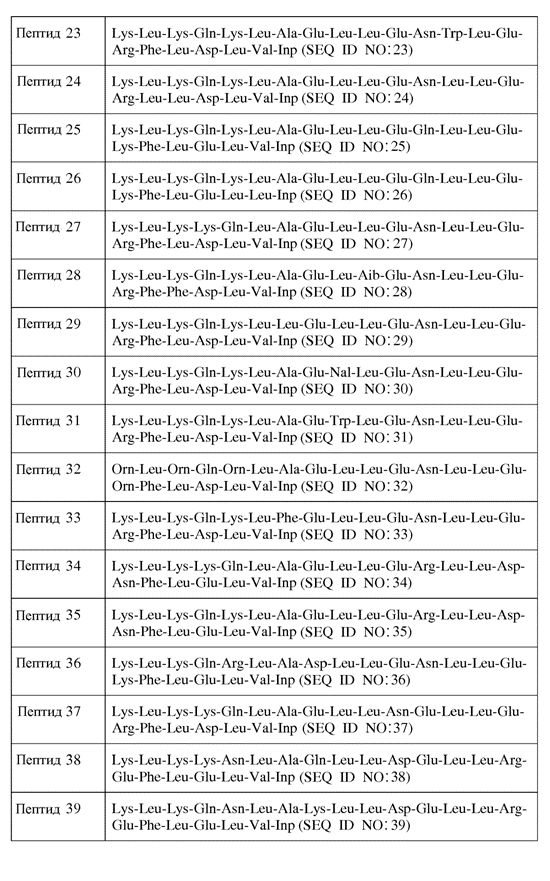
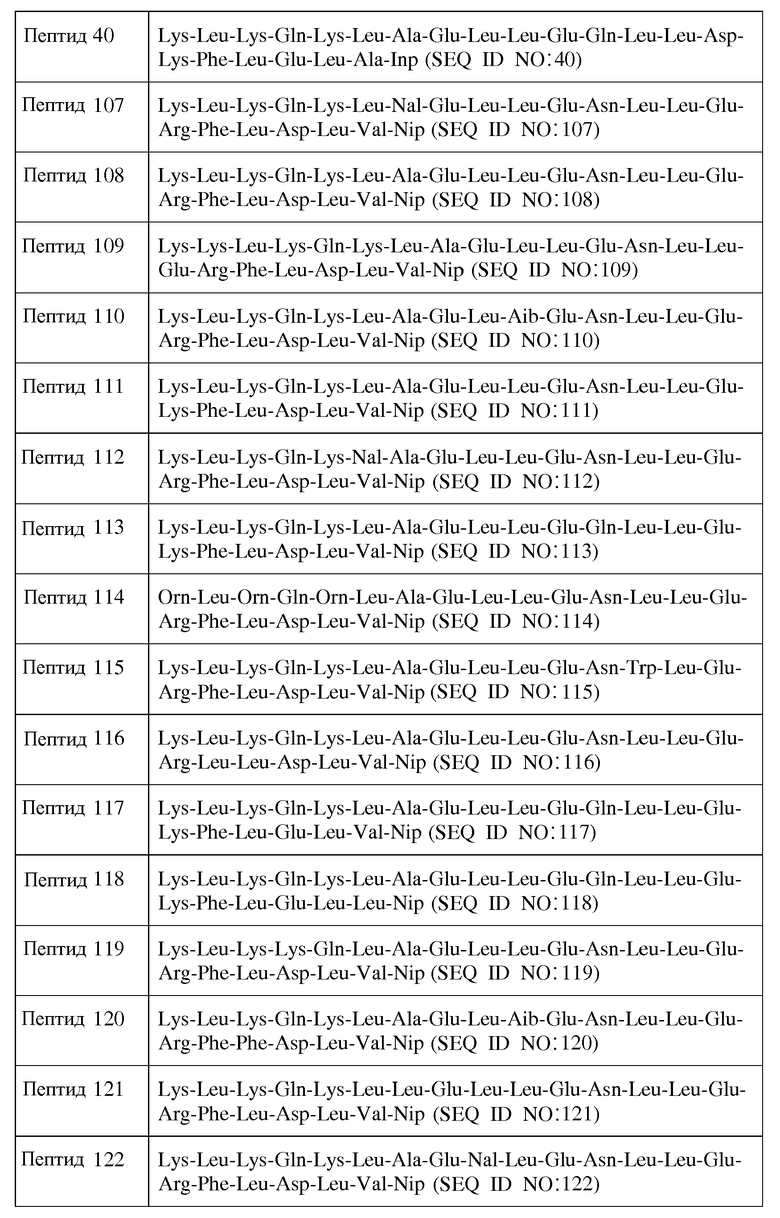
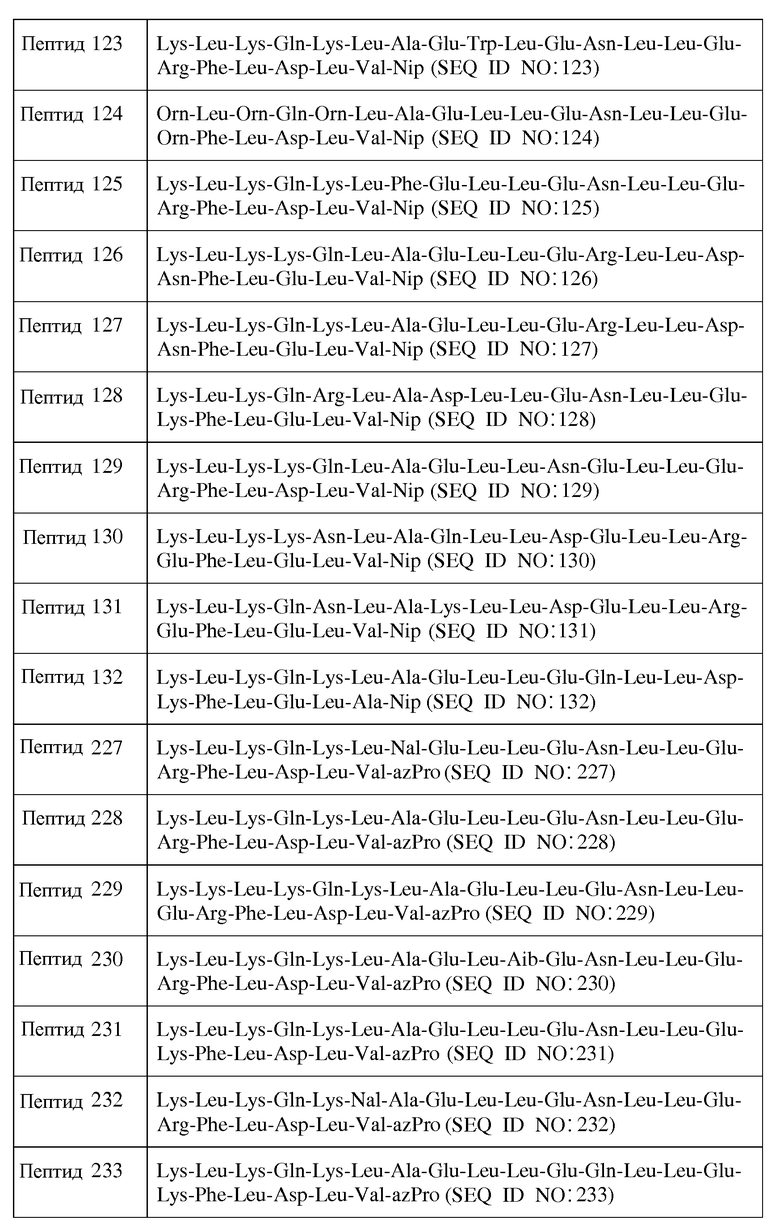
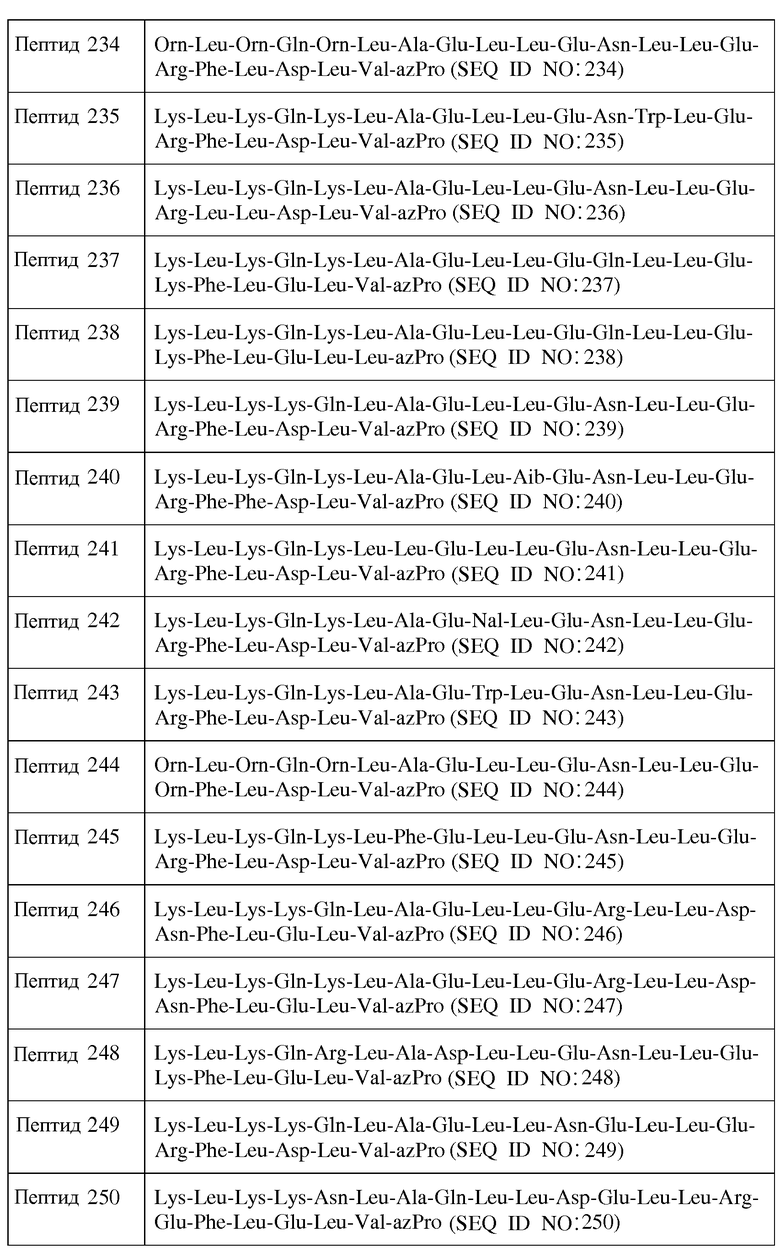

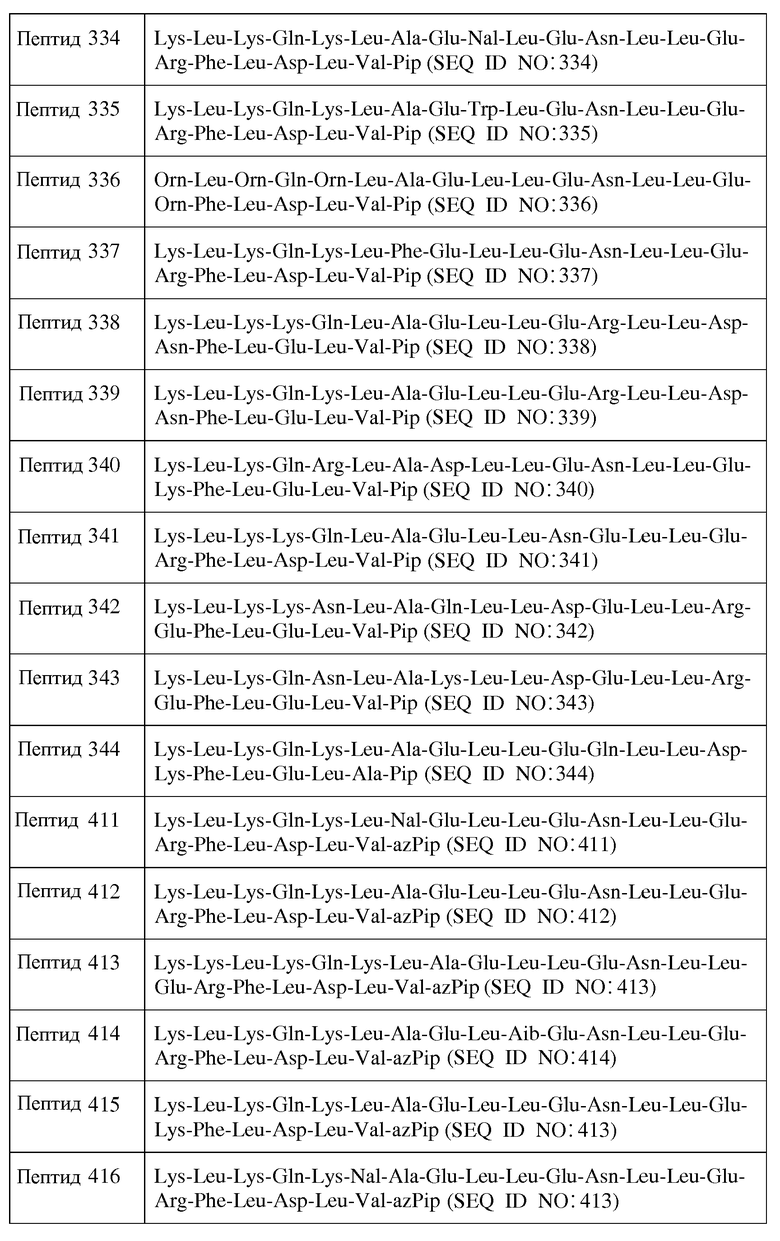
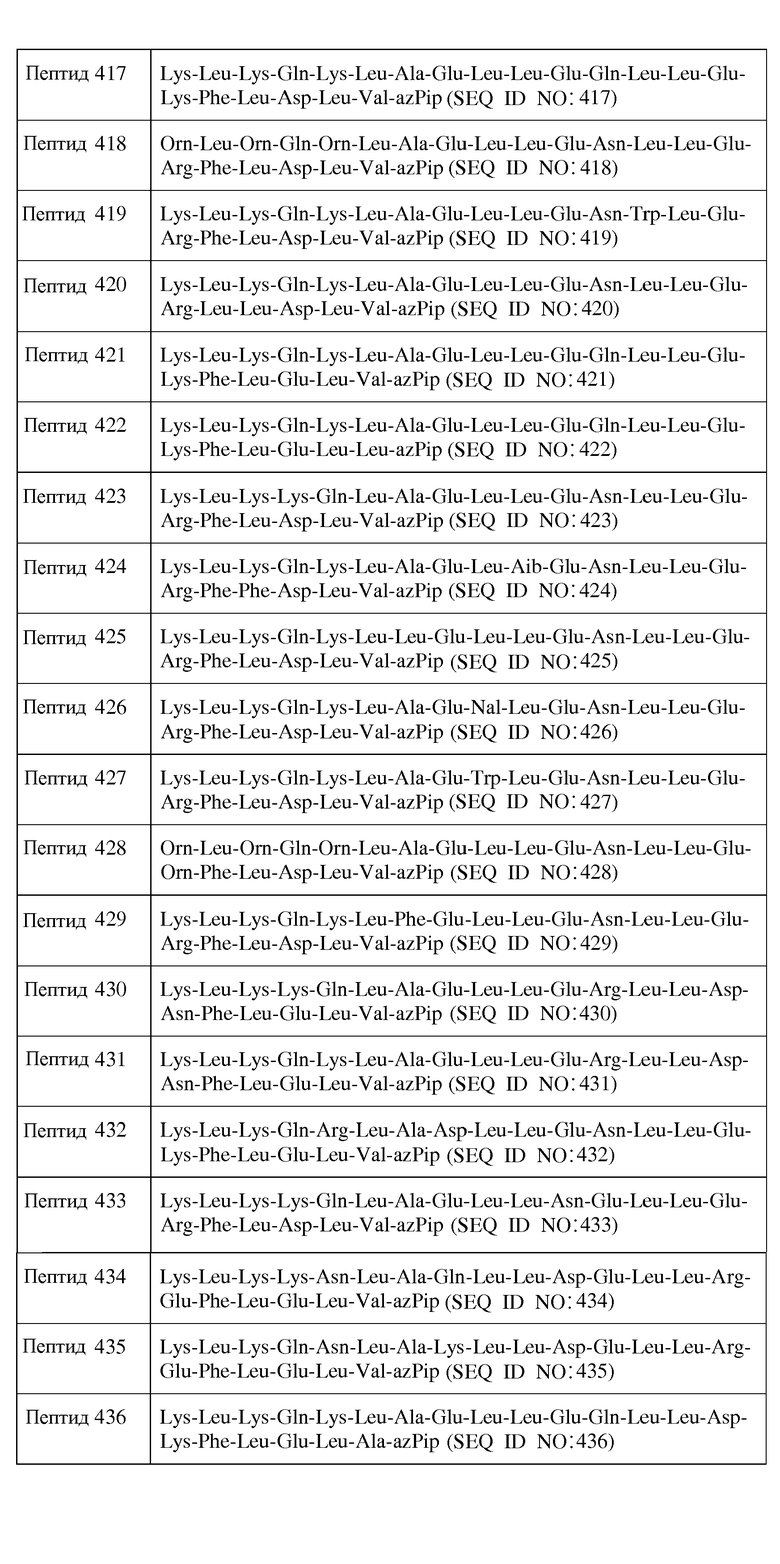 или их фармацевтически приемлемой солью.
или их фармацевтически приемлемой солью.
B. Пептиды формулы II
В одном из воплощений настоящее изобретение связано с пептидами, содержащими от 14 до 22 остатков, имеющими следующую формулу II:
R1-Y1-X1-X2-X3-X4-X5-X6-X7-X8-X9-X10-X11-X12-X13-X14-X15-X16-X17-X18-Y2-R2
Формула II
и их фармацевтически приемлемыми солями, в которых
X1 является ахиральным, D- или L-основным аминокислотным остатком;
X2 представляет собой Leu или D-Leu;
X3 является ахиральным, D- или L-основным аминокислотным остатком;
X4 представляет собой Gln, Asn, D-Gln или D-Asn;
X5 представляет собой Leu, D-Leu или ахиральный, D- или L-основный аминокислотный остаток;
X6 представляет собой Leu, Trp, Phe, D-Leu, D-Trp или D-Phe;
X7 является ахиральным, D- или L-кислым аминокислотным остатком;
X8 представляет собой Asn, D-Asn или является ахиральным, D-или L-кислым аминокислотным остатком;
X9 представляет собой Leu, Trp, D-Leu или D-Trp;
X10 представляет собой Leu, Trp, D-Leu или D-Trp;
X11 представляет собой ахиральный, D- или L-кислый аминокислотный остаток;
X12 является ахиральным, D- или L-основным аминокислотным остатком;
X13 представляет собой Leu, Phe, D-Leu или D-Phe;
X14 представляет собой Leu, Phe, D-Leu или D-Phe;
X15 является ахиральным, D- или L-кислым аминокислотным остатком;
X16 представляет собой Leu или D-Leu;
X17 является ахиральным, D- или L-алифатическим аминокислотным остатком;
X18 представляет собой Inp, Nip, azPro, Pip, azPip, D-Nip или D-Pip;
Y1 отсутствует или является аминокислотной последовательностью, содержащей от 1 до 4 остатков;
Y2 отсутствует или является аминокислотной последовательностью, содержащей от 1 до 4 остатков;
R1 представляет собой H или аминозащитную группу;
R2 представляет собой OH или карбоксильную защитную группу;
причем от нуля до восьми остатков от X1 до X17 отсутствуют; и при этом
a) каждый хиральный аминокислотный остаток является L-аминокислотным остатком;
b) каждый хиральный аминокислотный остаток является D-аминокислотным остатком;
c) каждый хиральный аминокислотный остаток является L-аминокислотным остатком, за исключением того, что один или более из каждых хиральных концевых аминокислотных остатков и каждых хиральных аминокислотных остатков, непосредственно к ним примыкающих, является D-аминокислотным остатком; или
d) каждый хиральный аминокислотный остаток является D-аминокислотным остатком, за исключением того, что один или более из каждых хиральных концевых аминокислотных остатков и каждых хиральных аминокислотных остатков, непосредственно к ним примыкающих, является L-аминокислотным остатком.
В другом воплощении настоящее изобретение связано с пептидами, содержащими от 15 до 22 остатков, имеющими следующую формулу II:
R1-Y1-X1-X2-X3-X4-X5-X6-X7-X8-X9-X10-X11-X12-X13-X14-X15-X16-X17-X18-Y2-R2
Формула II
и их фармацевтически приемлемыми солями, в которых
X1 является ахиральным, D- или L-основным аминокислотным остатком;
X2 представляет собой Leu или D-Leu;
X3 является ахиральным, D- или L-основным аминокислотным остатком;
X4 представляет собой Gln, Asn, D-Gln или D-Asn;
X5 представляет собой Leu, D-Leu или является ахиральным, D-или L-основным аминокислотным остатком;
X6 представляет собой Leu, Trp, Phe, D-Leu, D-Trp или D-Phe;
X7 является ахиральным, D- или L-кислым аминокислотным остатком;
X8 представляет собой Asn, D-Asn или является ахиральным, D-или L-кислым аминокислотным остатком;
X9 представляет собой Leu, Trp, D-Leu или D-Trp;
X10 представляет собой Leu, Trp, D-Leu или D-Trp;
X11 представляет собой ахиральный, D- или L-кислый аминокислотный остаток;
X12 является ахиральным, D- или L-основным аминокислотным остатком;
X13 представляет собой Leu, Phe, D-Leu или D-Phe;
X14 представляет собой Leu, Phe, D-Leu или D-Phe;
X15 является ахиральным, D- или L-кислым аминокислотным остатком;
X16 представляет собой Leu или D-Leu;
X17 является ахиральным, D- или L-алифатическим аминокислотным остатком;
X18 представляет собой Inp, Nip, azPro, Pip, azPip, D-Nip или D-Pip;
Y1 отсутствует или является аминокислотной последовательностью, содержащей от 1 до 4 остатков;
Y2 отсутствует;
R1 представляет собой H или аминозащитную группу;
R2 представляет собой OH или карбоксильную защитную группу;
причем от нуля до трех остатков от X1 до X17 отсутствуют; и при этом
a) каждый хиральный аминокислотный остаток является L-аминокислотным остатком;
b) каждый хиральный аминокислотный остаток является D-аминокислотным остатком;
c) каждый хиральный аминокислотный остаток является L-аминокислотным остатком, за исключением того, что один или более из каждых хиральных концевых аминокислотных остатков и каждых хиральных аминокислотных остатков, непосредственно к ним примыкающих, является D-аминокислотным остатком; или
d) каждый хиральный аминокислотный остаток является D-аминокислотным остатком, за исключением того, что один или более из каждых хиральных концевых аминокислотных остатков и каждых хиральных аминокислотных остатков, непосредственно к ним примыкающих, является L-аминокислотным остатком.
В другом воплощении настоящее изобретение связано с пептидами, содержащими 14 остатков, имеющими следующую формулу II:
R1-Y1-X1-X2-X3-X4-X5-X6-X7-X8-X9-X10-X11-X12-X13-X14-X15-X16-X17-X18-Y2-R2
Формула II
и их фармацевтически приемлемыми солями, в которых
X1 является ахиральным, D- или L-основным аминокислотным остатком;
X2 представляет собой Leu или D-Leu;
X3 является ахиральным, D- или L-основным аминокислотным остатком;
X4 представляет собой Gln, Asn, D-Gln или D-Asn;
X5 представляет собой Leu, D-Leu или является ахиральным, D-или L-основным аминокислотным остатком;
X6 представляет собой Leu, Trp, Phe, D-Leu, D-Trp или D-Phe;
X7 является ахиральным, D- или L-кислым аминокислотным остатком;
X8 представляет собой Asn, D-Asn или является ахиральным, D-или L-кислым аминокислотным остатком;
X9 представляет собой Leu, Trp, D-Leu или D-Trp;
X10 представляет собой Leu, Trp, D-Leu или D-Trp;
X11 представляет собой ахиральный, D- или L-кислый аминокислотный остаток;
X12 является ахиральным, D- или L-основным аминокислотным остатком;
X13 представляет собой Leu, Phe, D-Leu или D-Phe;
X14 представляет собой Leu, Phe, D-Leu или D-Phe;
X15 является ахиральным, D- или L-кислым аминокислотным остатком;
X16 представляет собой Leu или D-Leu;
X17 является ахиральным, D- или L-алифатическим аминокислотным остатком;
X18 представляет собой Inp, Nip, azPro, Pip, azPip, D-Nip или D-Pip;
Y1 отсутствует или является аминокислотной последовательностью, содержащей от 1 до 4 остатков;
Y2 отсутствует;
R1 представляет собой H или аминозащитную группу;
R2 представляет собой OH или карбоксильную защитную группу;
причем от четырех до восьми остатков от X1 до X17 отсутствуют; и при этом
a) каждый хиральный аминокислотный остаток является L-аминокислотным остатком;
b) каждый хиральный аминокислотный остаток является D-аминокислотным остатком;
c) каждый хиральный аминокислотный остаток является L-аминокислотным остатком, за исключением того, что один или более из каждых хиральных концевых аминокислотных остатков и каждых хиральных аминокислотных остатков, непосредственно к ним примыкающих, является D-аминокислотным остатком; или
d) каждый хиральный аминокислотный остаток является D-аминокислотным остатком, за исключением того, что один или более из каждых хиральных концевых аминокислотных остатков и каждых хиральных аминокислотных остатков, непосредственно к ним примыкающих, является L-аминокислотным остатком.
В одном из воплощений пептид формулы II является пептидом, содержащим 18 остатков.
В одном из воплощений пептид формулы II является пептидом, представленным ниже в таблице 5.

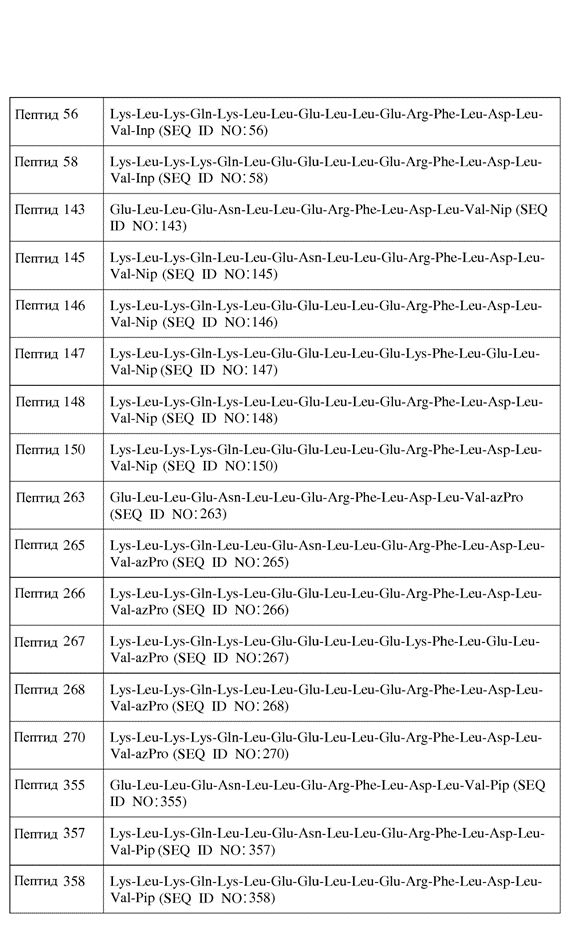
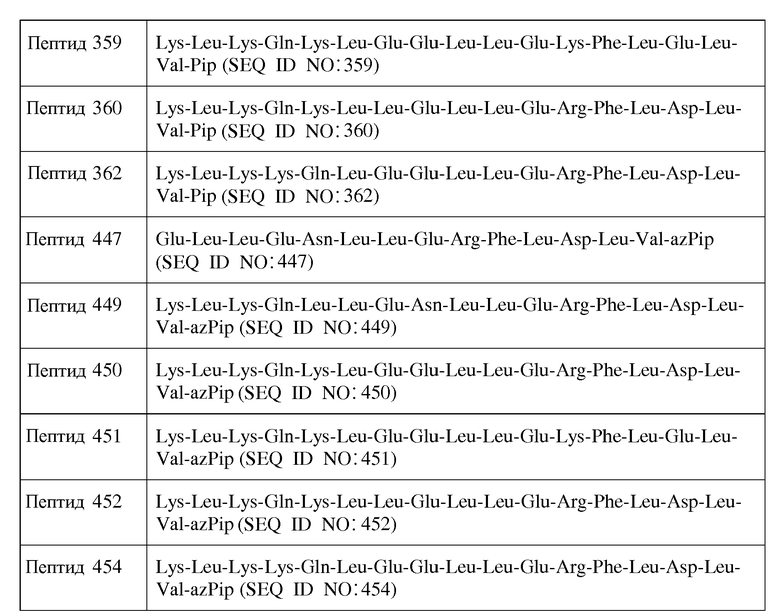
или их фармацевтически приемлемой солью.
C. Пептиды формулы III
В одном из воплощений настоящее изобретение связано с пептидами, содержащими от 15 до 29 остатков, имеющими формулу III:
R1-Y1-X1-X2-X3-X4-X5-X6-X7-X8-X9-X10-X11-X12-X13-X14-X15-X16-X17-X18-X19-X20-X21-X22-X23-Y2-R2
Формула III
и их фармацевтически приемлемыми солями, в которых
X1 отсутствует или является ахиральным, D- или L-основным аминокислотным остатком;
X2 является ахиральным, D- или L-основным аминокислотным остатком;
X3 является ахиральным, D- или L-алифатическим аминокислотным остатком;
X4 является ахиральным, D- или L-основным аминокислотным остатком;
X5 представляет собой Gln, Asn, D-Gln, D-Asn или является ахиральным, D- или L-основным аминокислотным остатком;
X6 представляет собой Gln, Asn, D-Gln или D-Asn, или является ахиральным, D- или L-основным аминокислотным остатком;
X7 является ахиральным, D- или L-гидрофобным аминокислотным остатком;
X8 является ахиральным, D- или L-гидрофобным аминокислотным остатком;
X9 является ахиральным, D- или L-гидрофильным аминокислотным остатком;
X10 представляет собой Leu, Trp, Gly, Nal, D-Leu, D-Trp или D-Nal;
X11 представляет собой Gly или является ахиральным, D- или L-алифатическим аминокислотным остатком;
X12 является ахиральным, D- или L-гидрофильным аминокислотным остатком;
X13 является ахиральным, D- или L-гидрофильным аминокислотным остатком;
X14 представляет собой Leu, Trp, Gly, D-Leu или D-Trp;
X15 представляет собой Leu, Gly или D-Leu;
X16 является ахиральным, D- или L-кислым аминокислотным остатком или ахиральным, D- или L-основным аминокислотным остатком;
X17 является ахиральным, D- или L-гидрофильным аминокислотным остатком;
X18 представляет собой Leu, Phe, D-Leu или D-Phe;
X19 представляет собой Leu, Phe, D-Leu или D-Phe;
X20 является ахиральным, D- или L-кислым аминокислотным остатком;
X21 представляет собой Leu, Phe, D-Leu или D-Phe;
X22 является ахиральным, D- или L-алифатическим аминокислотным остатком; и
X23 представляет собой Inp, Nip, azPro, Pip, azPip, D-Nip или D-Pip;
Y1 отсутствует или является аминокислотной последовательностью, содержащей от 1 до 7 остатков;
Y2 отсутствует или является аминокислотной последовательностью, содержащей от 1 до 7 остатков;
R1 представляет собой H или аминозащитную группу;
R2 представляет собой OH или карбоксильную защитную группу; причем от нуля до восьми остатков X2-X22 отсутствуют; и при этом
a) каждый хиральный аминокислотный остаток является L-аминокислотным остатком;
b) каждый хиральный аминокислотный остаток является D-аминокислотным остатком;
c) каждый хиральный аминокислотный остаток является L-аминокислотным остатком, за исключением того, что один или более из каждых хиральных концевых аминокислотных остатков и каждых хиральных аминокислотных остатков, непосредственно к ним примыкающих, является D-аминокислотным остатком; или
d) каждый хиральный аминокислотный остаток является D-аминокислотным остатком, за исключением того, что один или более из каждых хиральных концевых аминокислотных остатков и каждых хиральных аминокислотных остатков, непосредственно к ним примыкающих, является L-аминокислотным остатком.
В другом воплощении настоящее изобретение связано с пептидами, содержащими от 22 до 29 остатков, имеющими формулу III:
R1-Y1-X1-X2-X3-X4-X5-X6-X7-X8-X9-X10-X11-X12-X13-X14-X15-X16-X17-X18-X19-X20-X21-X22-X23-Y2-R2
Формула III
и их фармацевтически приемлемыми солями, в которых
X1 отсутствует или является ахиральным, D- или L-основным аминокислотным остатком;
X2 является ахиральным, D- или L-основным аминокислотным остатком;
X3 является ахиральным, D- или L-алифатическим аминокислотным остатком;
X4 является ахиральным, D- или L-основным аминокислотным остатком;
X5 представляет собой Gln, Asn, D-Gln, D-Asn или является ахиральным, D- или L-основным аминокислотным остатком;
X6 является ахиральным, D- или L-основным аминокислотным остатком;
X7 является ахиральным, D- или L-гидрофобным аминокислотным остатком;
X8 является ахиральным, D- или L-гидрофобным аминокислотным остатком;
X9 является ахиральным, D- или L-гидрофильным аминокислотным остатком;
X10 представляет собой Leu, Trp, Gly, Nal, D-Leu, D-Trp или D-Nal;
X11 представляет собой Gly или является ахиральным, D- или L-алифатическим аминокислотным остатком;
X12 является ахиральным, D- или L-гидрофильным аминокислотным остатком;
X13 является ахиральным, D- или L-гидрофильным аминокислотным остатком;
X14 представляет собой Leu, Trp, Gly, D-Leu или D-Trp;
X15 представляет собой Leu, Gly или D-Leu;
X16 является ахиральным, D- или L-кислым аминокислотным остатком;
X17 является ахиральным, D- или L-гидрофильным аминокислотным остатком;
X18 представляет собой Leu, Phe, D-Leu или D-Phe;
X19 представляет собой Leu, Phe, D-Leu или D-Phe;
X20 является ахиральным, D- или L-кислым аминокислотным остатком;
X21 представляет собой Leu, Phe, D-Leu или D-Phe;
X22 является ахиральным, D- или L-алифатическим аминокислотным остатком; и
X23 представляет собой Inp, Nip, azPro, Pip, azPip, D-Nip или D-Pip;
Y1 отсутствует или является аминокислотной последовательностью, содержащей от 1 до 7 остатков;
Y2 отсутствует или является аминокислотной последовательностью, содержащей от 1 до 7 остатков;
R1 представляет собой H или аминозащитную группу;
R2 представляет собой OH или карбоксильную защитную группу; при этом
a) каждый хиральный аминокислотный остаток является L-аминокислотным остатком;
b) каждый хиральный аминокислотный остаток является D-аминокислотным остатком;
c) каждый хиральный аминокислотный остаток является L-аминокислотным остатком, за исключением того, что один или более из каждых хиральных концевых аминокислотных остатков и каждых хиральных аминокислотных остатков, непосредственно к ним примыкающих, является D-аминокислотным остатком; или
d) каждый хиральный аминокислотный остаток является D-аминокислотным остатком, за исключением того, что один или более из каждых хиральных концевых аминокислотных остатков и каждых хиральных аминокислотных остатков, непосредственно к ним примыкающих, является L-аминокислотным остатком.
В другом воплощении настоящее изобретение связано с пептидами, содержащими от 15 до 21 остатка, имеющими формулу III:
R1-Y1-X1-X2-X3-X4-X5-X6-X7-X8-X9-X10-X11-X12-X13-X14-X15-X16-X17-X18-X19-X20-X21-X22-X23-Y2-R2
Формула III
и их фармацевтически приемлемыми солями, в которых
X1 отсутствует или является ахиральным, D- или L-основным аминокислотным остатком;
X2 является ахиральным, D- или L-основным аминокислотным остатком;
X3 является ахиральным, D- или L-алифатическим аминокислотным остатком;
X4 является ахиральным, D- или L-основным аминокислотным остатком;
X5 представляет собой Gln, Asn, D-Gln, D-Asn или является ахиральным, D- или L-основным аминокислотным остатком;
X6 является ахиральным, D- или L-основным аминокислотным остатком;
X7 является ахиральным, D- или L-гидрофобным аминокислотным остатком;
X8 является ахиральным, D- или L-гидрофобным аминокислотным остатком;
X9 является ахиральным, D- или L-гидрофильным аминокислотным остатком;
X10 представляет собой Leu, Trp, Gly, Nal, D-Leu, D-Trp или D-Nal;
X11 представляет собой Gly или является ахиральным, D- или L-алифатическим аминокислотным остатком;
X12 является ахиральным, D- или L-гидрофильным аминокислотным остатком;
X13 является ахиральным, D- или L-гидрофильным аминокислотным остатком;
X14 представляет собой Leu, Trp, Gly, D-Leu или D-Trp;
X15 представляет собой Leu, Gly или D-Leu;
X16 является ахиральным, D- или L-кислым аминокислотным остатком;
X17 является ахиральным, D- или L-гидрофильным аминокислотным остатком;
X18 представляет собой Leu, Phe, D-Leu или D-Phe;
X19 представляет собой Leu, Phe, D-Leu или D-Phe;
X20 является ахиральным, D- или L-кислым аминокислотным остатком;
X21 представляет собой Leu, Phe, D-Leu или D-Phe;
X22 является ахиральным, D- или L-алифатическим аминокислотным остатком; и
X23 представляет собой Inp, Nip, azPro, Pip, azPip, D-Nip или D-Pip;
Y1 отсутствует или является аминокислотной последовательностью, содержащей от 1 до 7 остатков;
Y2 отсутствует или является аминокислотной последовательностью, содержащей от 1 до 7 остатков;
R1 представляет собой H или аминозащитную группу;
R2 представляет собой OH или карбоксильную защитную группу; причем от одного до восьми остатков X2-X22 отсутствуют; и при этом
a) каждый хиральный аминокислотный остаток является L-аминокислотным остатком;
b) каждый хиральный аминокислотный остаток является D-аминокислотным остатком;
c) каждый хиральный аминокислотный остаток является L-аминокислотным остатком, за исключением того, что один или более из каждых хиральных концевых аминокислотных остатков и каждых хиральных аминокислотных остатков, непосредственно к ним примыкающих, является D-аминокислотным остатком; или
d) каждый хиральный аминокислотный остаток является D-аминокислотным остатком, за исключением того, что один или более из каждых хиральных концевых аминокислотных остатков и каждых хиральных аминокислотных остатков, непосредственно к ним примыкающих, является L-аминокислотным остатком.
В другом воплощении пептид формулы III содержит 22 аминокислотных остатка в длину, и X1 отсутствует.
В одном из воплощений пептид формулы III является пептидом, представленным ниже в таблице 6.

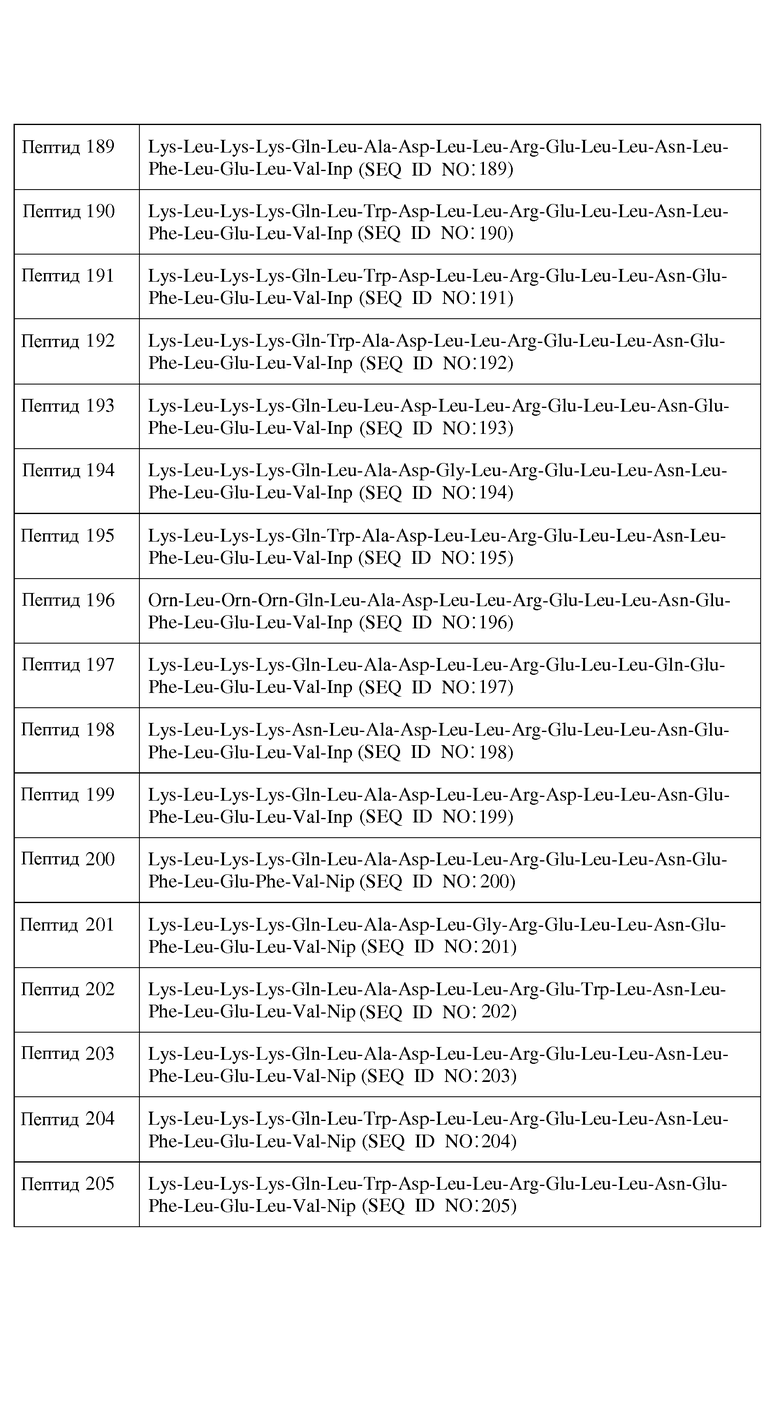
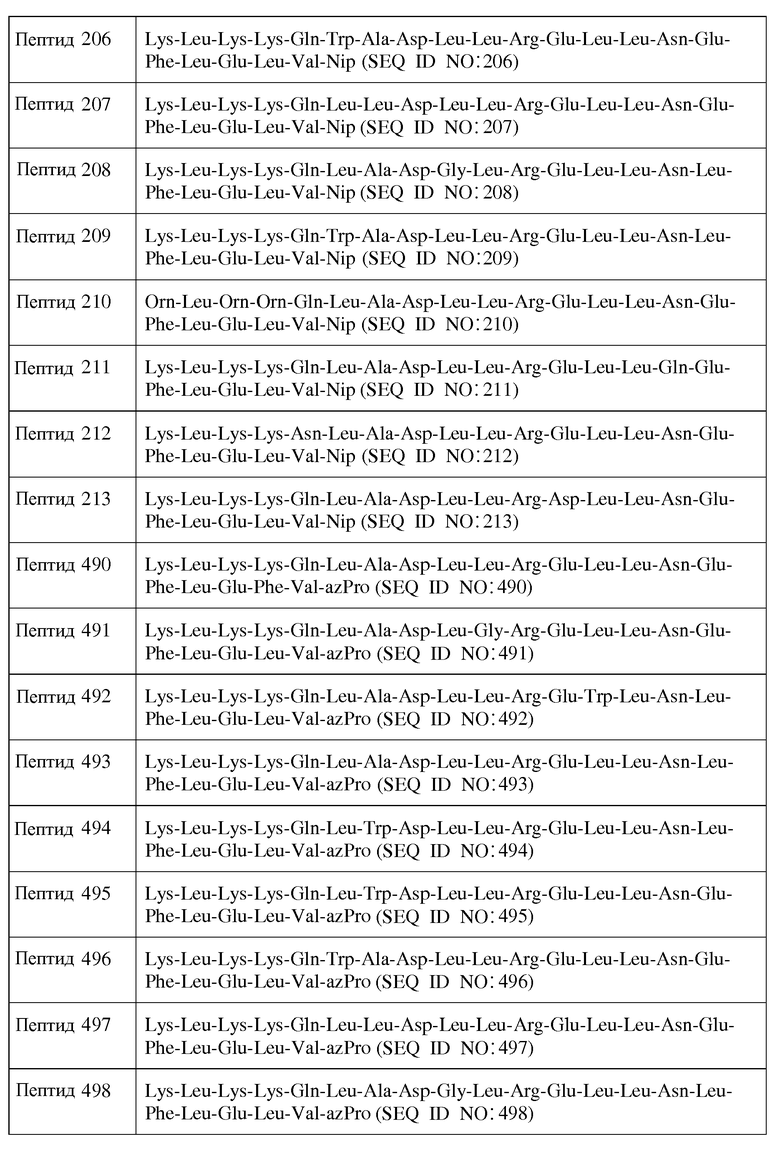
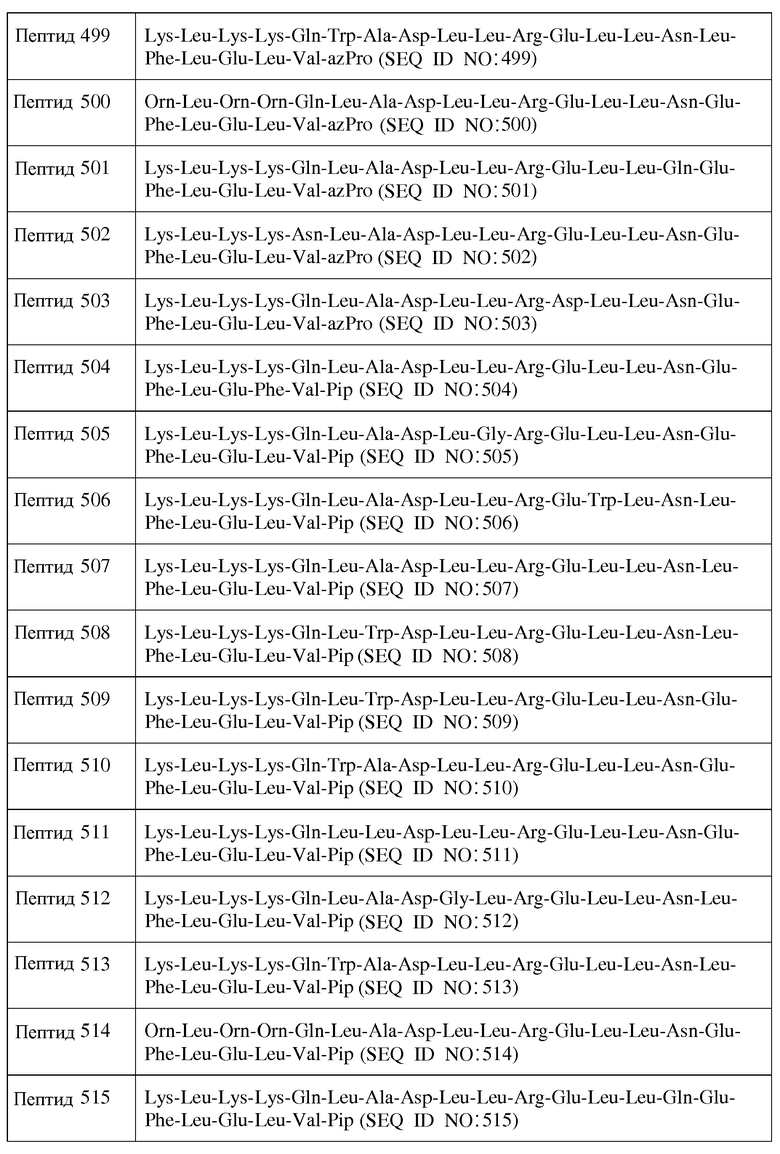
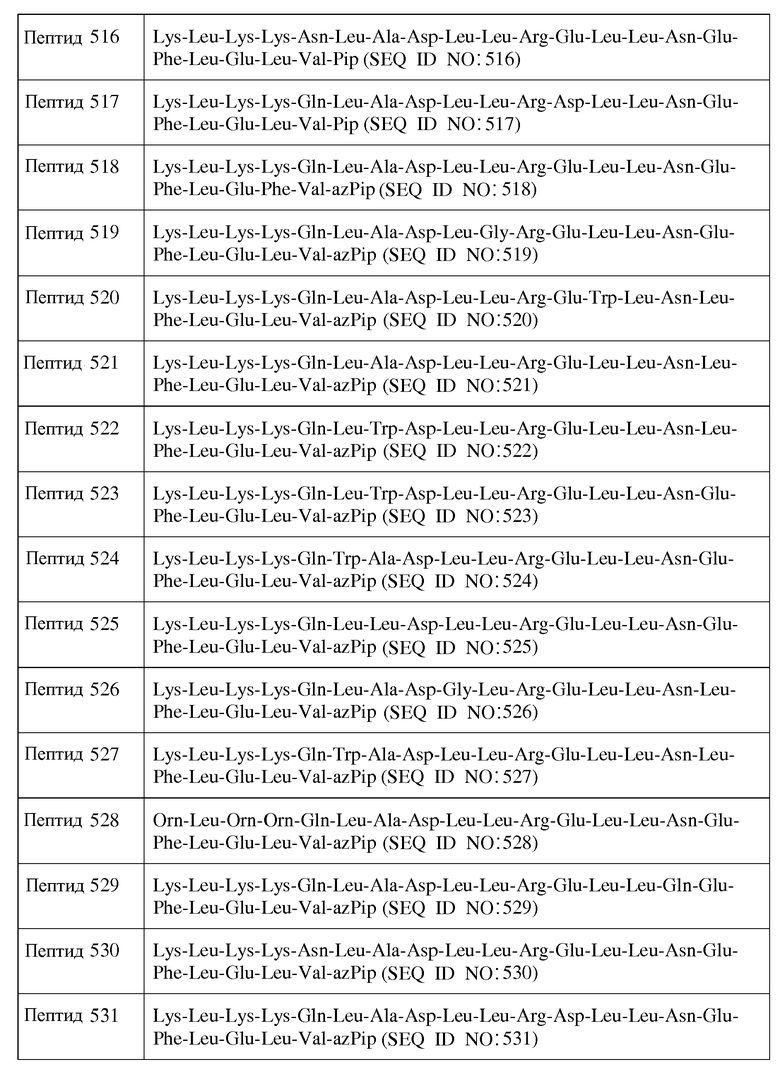
или их фармацевтически приемлемой солью.
В других воплощениях настоящее изобретение включает в себя миметики ApoA-I, в которых одна или более из их амидных связей необязательно заменена связью, отличной от амидной, включая, но не ограничиваясь замещенным амидом или изостерой амида. Таким образом, хотя различные остатки X1-X23, Y1 и Y2 в формулах I, II и III описаны в терминах аминокислот, в определенных воплощениях настоящего изобретения, вместо одной или нескольких амидных связей присутствует неамидная связь.
В другом воплощении атом азота одной или нескольких из амидных связей миметиков ApoA-I является замещенным, так, что замещенная амидная связь имеет формулу -C(O)NR'-, где R' представляет собой (C1-C6)-алкил, (C2-C6)-алкенил, (C2-C6)-алкинил, (C5-C20)-арил, (C6-C26)-аралкил, 5-20-членный гетероарил или 6-26-членный алкгетероарил. В другом воплощении R' замещен -OR, -SR, -NRR, -NO2, -CN, галогеном, -SO2R, -C(O)R, -C(O)OR и -C(O)NRR, где каждый из R независимо является водородом, алкилом или арилом.
В другом воплощении неамидная связь заменяет одну или несколько амидных связей миметиков ApoA-I и включает в себя, но не ограничена перечисленным, -CH2NH-, -CH2S-, -CH2CH2-, -CH=CH- (цис- и транс-), -C(O)CH2-, -CH(OH)CH2- и -CH2SO-. Соединения, имеющие такие неамидные связи, а также способы получения таких соединений, хорошо известны специалистам в данной области (см., например, Spatola, March 1983, Vega Data Vol.1, Issue 3; Spatola, 1983, "Peptide Backbone Modifications" In: Chemistry and Biochemistry of Amino Acids Peptides and Proteins, Weinstein, ed., Marcel Dekker, New York, p.267 (обзор общего характера); Morley, 1980, Trends Pharm. Sci. 1: 463-468; Hudson et al., 1979, Int. J. Prot. Res. 14: 177-185 (-CH2NH-, -CH2CH2-); Spatola et al., 1986, Life Sci. 38: 1243-1249 (-CH2-S); Hann, 1982, J. Chem. Soc. Perkin Trans. I. 1: 307-314 (-CH=CH-, цис- и транс-); Almquist et al., 1980, J. Med. Chem. 23: 1392-1398 (-COCH2-); Jennings-White et al., Tetrahedron. Lett. 23: 2533 (-COCH2-); European Patent Application EP 45665 (1982) CA 97: 39405 (-CH(OH)CH2-); Holladay et al., 1983, Tetrahedron Lett. 24: 4401-4404 (-C(OH)CH2-); а также Hruby, 1982, Life Sci. 31: 189-199 (-CH2-S-).
Кроме того, одна или несколько амидных связей миметиков ApoA-I могут быть заменены одним или несколькими пептидомиметиками или имитаторами амидных фрагментов, которые не вызывают значительных нарушений структуры или активности указанных пептидов. Подходящие имитаторы амидных фрагментов описаны, например, у Olson et al., 1993, J. Med. Chem. 36: 3039-3049.
В определенных воплощениях миметик ApoA-I представлен в виде фармацевтически приемлемой соли. Соль может быть образована на C-конце или на N-конце, или же на боковой цепи кислотного или основного аминокислотного остатка.
В определенных воплощениях фармацевтически приемлемая соль является солью металла, солью органического амина или солью присоединения кислот.
Соли металла могут образовываться при добавлении неорганического основания к пептиду формул I, II или III. Неорганическое основание состоит из катиона металла, спаренного с противоионом основания, таким, например, как гидроксид, карбонат, бикарбонат или фосфат. Металл может быть щелочным металлом, щелочноземельным металлом, переходным металлом или металлом основной группы. В определенных воплощениях металлом является литий, натрий, калий, церий, магний, марганец, железо, кальций, алюминий или цинк.
Органические соли амина могут образовываться при добавлении органического амина к пептиду формул I, II или III. В определенных воплощениях органическим амином является триэтиламин, этаноламин, диэтаноламин, триэтаноламин, морфолин, пиперидин, N-метилпиперидин, N-этилпиперидин, дибензиламин, пиперазин, пиридин, пиразин или пипиразин.
Соли присоединения кислот могут образовываться при добавлении кислоты к пептиду формул I, II или III. В определенных воплощениях кислота является органической. В определенных воплощениях кислота является неорганической. В других воплощениях кислота является хлористоводородной кислотой, бромистоводородной кислотой, йодистоводородной кислотой, азотной кислотой, серной кислотой, сернистой кислотой, фосфорной кислотой, изоникотиновой кислотой, молочной кислотой, салициловой кислотой, винной кислотой, аскорбиновой кислотой, гентизиновой кислотой, глюконовой кислотой, глюкуроновой кислотой, сахарной кислотой, муравьиной кислотой, бензойной кислотой, глутаминовой кислотой, пантотеновой кислотой, уксусной кислотой, фумаровой кислотой, янтарной кислотой, метансульфоновой кислотой, этансульфоновой кислотой, бензолсульфоновой кислотой, пара-толуолсульфоновой кислотой, лимонной кислотой или малеиновой кислотой. В определенных других воплощениях солью присоединения кислот является гидрохлорид, гидробромид, гидройодид, нитрат, сульфат, сульфит, бисульфат, фосфат, кислый фосфат, изоникотинат, лактат, салицилат, тартрат, битартрат, аскорбат, гентизинат, глюконат, глюкаронат, сахарат, формат, бензоат, глутамат, пантотенат, ацетат, фумарат, сукцинат, метансульфонат, этансульфонат, бензолсульфонат, пара-толуилсульфонат, цитрат или малеат.
В определенных воплощениях R1 представляет собой аминозащитную группу. В определенных воплощениях указанной аминозащитной группой являются (C1-C6)-алкил, (C2-C6)-алкенил, (C2-C6)-алкинил, (C5-C26)-арил, (C6-C26-аралкил), 5-20-членный гетероарил или 6-26-членный алкгетероарил; -C(O)R; -C(O)OR; -SO2R; или -SR, где R является H или (C1-C6)-алкилом, (C2-C6)-алкенилом, (C2-C6)-алкинилом, (C5-C26)-арилом, (C6-C26)-аралкилом 5-20-членным гетероарилом или 6-26-членным алкгетероарилом. В других воплощениях (C1-C6)-алкил, (C2-C6)-алкенил, (C2-C6)-алкинил, (C5-C26)-арил, (C6-C26-аралкил), 5-20-членный гетероарил или 6-26-членный алкгетероарил замещены одним или несколькими ORa, -SRa, -NRaRa, -NO2, -CN, галогеном, -SO2Ra, -C(О)Ra, -C(O)ORa и -C(O)NRaRa, где каждый Ra независимо является водородом, алкилом или арилом. Когда R1 является H, число аминозащитных групп в миметике ApoA-I равно нулю; а когда R1 является аминозащитной группой, число аминозащитных групп в миметике ApoA-I равно 1.
В других воплощениях аминозащитной группой являются дансил; метоксикарбонил; этоксикарбонил; 9-фторенилметоксикарбонил; 2-хлорэтоксикарбонил; 2,2,2-трихлорэтоксикарбонил; 2-фенилэтоксикарбонил; трет-бутоксикарбонил; бензилоксикарбонил; пара-метоксибензилоксикарбонил; пара-нитробензилоксикарбонил; o- нитробензилоксикарбонил; пара-бромбензилоксикарбонил; пара-хлорбензилоксикарбонил; пара-йодбензилоксикарбонил; 2,4-дихлорбензилоксикарбонил; дифенилметоксикарбонил; 3,5-диметоксибензилоксикарбонил; феноксикарбонил; 2,4,6-три-трет-бутилфеноксикарбонил; 2,4,6-триметилбензилоксикарбонил; формил; ацетил; хлорацетил; трихлорацетил; трифторацетил; фенилацетил; пиколиноил; бензоил; пара-фенилбензоил; фталоил; метил; трет-бутил; аллил; [2-(триметилсилил)этокси]метил; 2,4-диметоксибензил; 2,4-динитрофенил; бензил; 4-метоксибензил; дифенилметил; трифенилметил; бензолсульфенил; o-нитробензолсульфенил; 2,4-динитробензолсульфенил; пара-толуолсульфонил; бензолсульфонил; 2,3,6-триметил-4-метоксибензолсульфонил; 2,4,6-триметоксибензолсульфонил; 2,6-диметил-4-метоксибензолсульфонил; пентаметилбензолсульфонил; 4-метоксибензолсульфонил; 2,4,6-триметилбензолсульфонил; или бензилсульфонил. В других воплощениях аминозащитной группой является ацетил, формил или дансил.
В определенных воплощениях R2 представляет собой H или карбоксильную защитную группу. В определенных воплощениях карбоксильной защитной группой являются О-(C1-C6)-алкил, О-(C2-C6)-алкенил, О-(C2-C6)-алкинил, О-(C5-C26)-арил, О-(C5-C26-аралкил), O-(5-20-членный гетероарил) или О-(6-26-членный алкгетероарил); или -NRR, где R представляет собой H или (C1-C6)-алкил, (C2-C6)-алкенил, (C2-C6)-алкинил, (C5-C26)-арил, (C6-C26-аралкил), 5-20-членный гетероарил или 6-26-членный алкгетероарил. В других воплощениях (C1-C6)-алкил, (C2-C6)-алкенил, (C2-C6)-алкинил, (C5-C26)-арил, (C6-C26)-аралкил, 5-20-членный гетероарил или 6-26-членный алкгетероарил замещены одним или несколькими из ORa, -SRa, -NRaRa, -NO2, -CN, галогена, -SO2Ra, -C(О)Ra, -C(O)ORa и -C(O)NRaRa, где каждый Ra независимо является водородом, алкилом или арилом. Когда R1 является H, число карбоксильных защитных групп в миметике ApoA-I равно нулю; а когда R1 является карбоксильной защитной группой, число карбоксильных защитных групп в миметике ApoA-I равно 1.
В других воплощениях карбоксильной защитной группой являются метокси; этокси; 9-фторенилметокси; метоксиметокси; метилтиометокси; тетрагидропиранокси; тетрагидрофуранокси; метоксиэтоксиметокси; бензилоксиметокси; фенацилокси; пара-бромфенацилокси; α-метилфенацилокси; пара-метоксифенацилокси; дезилокси; 2-хлорэтокси; 2,2,2-трихлорэтокси, 2-метилтиоэтолкси; 2-(пара-толуолсульфонил)метокси; трет-бутокси; циклопентокси; циклогексокси; аллилокси; металлилокси; циннамокси; α-метилциннамокси; фенокси; 2,6-диметилфенокси; 2,6-диизопропилфенокси; бензилокси; трифенилметокси; дифенилметокси; 2,4,6-триметилбензилокси; пара-бромбензилокси; o-нитробензилокси; N,N-диметиламидо; пирролидинил; или пиперидинил.
В объем настоящего изобретения входят также защищенные формы миметиков ApoA-I, то есть формы миметиков ApoA-I, в которых одна или несколько из их -NH2- или COOH-групп защищены защитной группой. В одном из воплощений одна или несколько -NH2-групп защищены аминозащитной группой, как описано выше. В другом воплощении одна или несколько COOH-групп защищены, как описано выше, карбоксильной защитной группой.
В одном из воплощений миметики ApoA-I обладают способностью образовывать амфипатическую α-спираль в присутствии одного или нескольких липидов. Под термином "амфипатическая, или амфифильная" подразумевается, что такая α-спираль имеет противоположные гидрофильную и гидрофобную поверхности, ориентированные вдоль ее длинной оси, то есть на одной стороне такой спирали спроецированы в основном гидрофильные боковые цепи, тогда как на противоположной стороне спирали спроецированы в основном гидрофобные боковые цепи. На фиг.1A и 1B представлены два наглядных изображения противоположных гидрофильной и гидрофобной сторон типовой идеализированной амфипатической α-спирали. На фиг.1A представлена диаграмма Шиффера-Эдмундсона «helical wheel» («Спиральные колеса») (Schiffer and Edmundson, 1967, Biophys. J. 7: 121-135). В таком «колесе» длинная ось спирали перпендикулярна странице. Начиная от N-конца, следующие друг за другом аминокислотные остатки (представленные кружочками) радиально расположены по периметру кольца с интервалами в 100°. Таким образом, аминокислотный остаток n+1 расположен под углом поворота в 100° относительно остатка n, остаток n+2 расположен под углом поворота в 100° относительно остатка n+1, и т.д. Расположение под углом поворота в 100° дает в результате 3,6 аминокислотного остатка на виток, что обычно и наблюдается в идеализированной α-спирали. На фиг.1A четко видны противоположные гидрофильная и гидрофобная поверхности спирали; гидрофильные аминокислотные остатки обозначены полыми кружочками, а заполненными кружочками обозначены гидрофобные аминокислотные остатки.
На фиг.1В представлена схема-диаграмма спиральной сетки идеализированной амфипатической спирали, изображенной на фиг.1A. (Lim, 1978, FEBS Lett. 89: 10-14). В типичной схеме-диаграмме спиральной сетки α-спираль представлена в виде цилиндра, который разрезан вдоль центра его гидрофильной поверхности и выправлен. Таким образом, центр гидрофобной поверхности, определяемый гидрофобным моментом спирали (Eisenberg et al., 1982, Nature 299: 371-374), лежит в центре этой фигуры и сориентирован таким образом, что выходит из плоскости страницы. Иллюстрация спирального цилиндра до разрезания и его выправление показаны на фиг.1C. Путем разрезания цилиндра в разных плоскостях дает возможность наблюдать разные картины одной и той же алифатической спирали, и таким путем можно получить различную информацию о свойствах спирали.
Хоть авторы изобретения и не желают связывать себя какой бы то ни было конкретной теорией, тем не менее, они полагают, что определенные структурные и/или физические свойства амфипатической спирали, образуемой миметиками ApoA-I, могут быть важны для их активности. Такие свойства включают в себя степень амфифильности, суммарную гидрофобность, среднюю гидрофобность, гидрофобные и гидрофильные углы, гидрофобный момент, средний гидрофобный момент и суммарный заряд α-спирали.
Степень амфифильности (степень асимметрии гидрофобности) амфипатической спирали, образуемой миметиками ApoA-I, удобно рассчитывать путем вычисления гидрофобного момента (µH) спирали. Способы расчета µH для конкретной пептидной последовательности хорошо известны в данной области и описаны, например, у Eisenberg, 1984, Ann. Rev. Biochem. 53: 595-623. Фактическое значение µH, полученное для конкретного пептида, будет зависеть от общего числа аминокислотных остатков, составляющих данный пептид. Таким образом, обычно непосредственное сравнение значений µH для пептидов разной длины не является информативным.
Амфифильность пептидов различной длины можно напрямую сравнивать на уровне среднего гидрофобного момента (<µH>). Средний гидрофобный момент может быть получен путем деления µH на число остатков в спирали (то есть <µH> = µH/N). Обычно миметики ApoA-I, проявляющие <µH> в области от 0,45 до 0,65, определяемое с помощью общепринятой нормированной шкалы гидрофобности Айзенберга (Eisenberg, 1984, J. Mol. Biol. 179: 125-142), считаются входящими в объем настоящего изобретения. В одном из воплощений <µH> находится в области от 0,50 до 0,60.
Суммарную, или тотальную, гидрофобность (Ho) пептида удобно рассчитать, взяв алгебраическую сумму гидрофобностей каждого аминокислотного остатка в пептиде (то есть 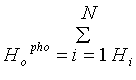
Тотальная гидрофобность гидрофобной поверхности (Ho pho) амфипатической спирали можно получить, взяв сумму гидрофобностей гидрофобных аминокислотных остатков, которые попадают в область гидрофобного угла, как описано ниже (то есть 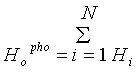
Гидрофобный угол (pho-угол) обычно определяется как угол или дуга, покрываемые наиболее длинным непрерывным отрезком гидрофобных аминокислотных остатков, когда пептид рассматривают согласно представлениям Шиффера-Эдмундсона «helical wheel» («Спиральные колеса») (то есть число смежных гидрофобных остатков на «колесе» умножают на 20°). Гидрофильный угол (phi-угол) представляет собой разницу между 360° pho-углом (то есть 360° pho-угол). Специалистам в данной области должно быть понятно, что pho- и phi-углы могут отчасти зависеть от числа аминокислотных остатков в пептиде. Например, со ссылкой на фиг.2, можно увидеть, что только 18 аминокислотных остатков подходят для одного поворота спирального «колеса» Шиффера-Эдмундсона для 22-членного консенсус-пептида (Segrest's consensus 22-mer), Pro-Val-Leu-Asp-Glu-Phe-Arg-Glu-Lys-Leu-Asn-Glu-Glu-Leu-Glu-Ala-Leu-Lys-Gln-Lys-Leu-Lys (SEQ ID NO:1). Меньшее количество аминокислотных остатков оставляют брешь в колесе; большее количество аминокислотных остатков являются причиной того, что определенные положения в таком колесе должны быть заняты более чем одним аминокислотным остатком.
В случае пептидов, насчитывающих 15 или более аминокислотных остатков, таких как миметик ApoA-I, содержащий от 15 до 29 остатков, "непрерывный" отрезок из гидрофобных аминокислотных остатков означает, что по меньшей мере один аминокислотный остаток в положениях вдоль такого колеса, содержащих два или более аминокислотных остатков, является гидрофобным аминокислотным остатком. Таким образом, со ссылкой на фиг.2, pho-угол является дугой, покрываемой остатками 16, 2, 6, 17, 10, 3 и 14, несмотря на наличие гидрофильного остатка в положении 20, поскольку остаток в положении 2, который занимает то же самое положение на колесе, является гидрофобным остатком. Обычно миметики ApoA-I, имеющие pho-угол в области от 160° до 220°, считаются входящими в объем настоящего изобретения. В определенных воплощениях pho-угол заключен в области от 180° до 200°.
В пептиде 16 (Lys-Leu-Lys-Gln-Lys-Leu-Ala-Glu-Leu-Leu-Glu-Asn-Leu-Leu-Glu-Arg-Phe-Leu-Asp-Leu-Val-Inp (SEQ ID NO:16)) или его фармацевтически приемлемой соли, наглядном миметике ApoA-I, положительно заряженные аминокислотные остатки кластеризованы у последнего N-концевого поворота спирали. Хоть авторы изобретения и не желают связывать себя какой бы то ни было конкретной теорией, тем не менее они полагают, что кластер из основных остатков на N-конце стабилизирует спираль в результате электростатических взаимодействий диполя заряд (NH3 +)-спираль. Подразумевается также, что стабилизация происходит в результате гидрофобных взаимодействий между боковыми цепями лизина и центральной частью (кором) спирали (см., Groebke et al., 1996, Proc. Natl. Acad. Sci. U.S.A. 93: 4025-4029; Esposito et al., 1997, Biopolymers 41: 27-35).
За исключением положительно заряженного N-концевого кластера, отрицательные заряды в пептиде 16 или в его фармацевтически приемлемой соли расположены в оставшейся части гидрофильной поверхности по меньшей мере с одним отрицательно заряженным (кислым) аминокислотным остатком на виток, давая в результате непрерывный отрезок отрицательных зарядов вдоль гидрофильной поверхности спирали. Один положительный заряд локализован на остатке 16, который потенциально вносит вклад в стабильность спирали путем образования солевого мостика с кислым остатком на соседнем витке спирали.
Считается, что с помощью ЯМР-исследования пептида 16 или его фармацевтически приемлемой соли можно было бы продемонстрировать, что аминокислотные остатки 13, 14, 17 и 20 указанного пептида образуют гидрофобный кластер вблизи C-конца спирали. Phe-17 располагается в центре этого кластера, и, как считается, играет важную роль в стабилизации данного гидрофобного кластера.
Хоть авторы изобретения и не желают связывать себя какой бы то ни было конкретной теорией, тем не менее они полагают, что гидрофобный кластер, образованный остатками 13, 14, 17 и 20 пептида 16 или его фармацевтически приемлемой соли, является существенным в реализации связывания липида и активации LCAT. Считается, что амфифильные пептиды будут связываться с фосфолипидами, ориентируя свои гидрофобные поверхности в направлении алкильных цепей липидных фрагментов. Таким образом, считается, что указанный высокогидрофобный кластер вносит свой вклад в обеспечение высокого сродства к липидам, наблюдаемое в отношении миметиков ApoA-I согласно изобретению. Поскольку связывание липидов является необходимым предварительным условием для активации лецитин-холестерин-ацилтрансферазы (LCAT), считается, что указанный гидрофобный кластер также важен для активации LCAT.
Ароматические остатки могут быть важны в заякоривании пептидов и белков к липидам (De Kruijff, 1990, Biosci. Rep. 10: 127-130; O'Neil and De Grado, 1990, Science 250: 645-651; Blondelle et al., 1993, Biochim. Biophys. Acta 1202: 331-336). Таким образом, дополнительно считается, что Phe-17, который расположен в центре гидрофобного кластера, также может играть важную роль в заякоривании пептида 16 или его фармацевтически приемлемой соли к липидам.
Длинная ось α-спирали, образуемой миметиками ApoA-I, в целом имеет обычно искривленную конфигурацию. В типичных амфифильных спиралях обнаружено, что длины водородных связей гидрофильных и гидрофобных поверхностей варьируют таким образом, что гидрофобная сторона спирали является вогнутой (Barlow and Thornton, 1988, J. Mol. Biol. 201: 601-619; Zhou et al., 1992, J. Am. Chem. Soc. 33: 11174-11183; Gesell et al., 1997, J. Biomol. NMR 9: 127-135). Хоть авторы изобретения и не желают связывать себя какой бы то ни было конкретной теорией, тем не менее они полагают, что общая кривизна гидрофобной поверхности спирали может быть важна для связывания дискообразных комплексов искривленная спираль позволяет пептиду лучше "соответствовать" каемкам вокруг дискообразных частиц, тем самым увеличивая стабильность комплекса пептида с диском.
В общепринятой структурной модели ApoA-I амфифильные α-спирали сгруппированы вокруг каемки дискообразных ЛПВП (см. фиг.4B). В данной модели предполагается, что спирали выстроены вдоль их гидрофобных поверхностей и направлены на липидные ацильные цепи (Brasseur et al., 1990, Biochim. Biophys. Acta 1043: 245-252). Спирали расположены в антипараллельном порядке, и предполагается, что кооперативный эффект между спиралями вносит свой вклад в стабильность дискообразного комплекса ЛПВП (Brasseur et al., выше). Было высказано предположение, что одним из факторов, который вносит свой вклад в стабильность дискообразного комплекса ЛПВП, является существование ионных взаимодействий между кислыми и основными остатками, приводящих к образованию межмолекулярных солевых мостиков или водородных связей между остатками на соседних антипараллельных спиралях. В данной модели пептиды рассматриваются не как отдельный объект, а во взаимодействии по меньшей мере с двумя другими соседними пептидными молекулами (фиг.4B).
Общепринятым является также и то, что внутримолекулярная водородная связь или солевой мостик, образованный между кислым и основным остатками, соответственно, в положениях i и i+3 спирали, стабилизирует спиральную структуру (Marqusee et al:, 1985, Proc. Natl. Acad. Sci. USA 84(24): 8898-8902).
Таким образом, миметики ApoA-I обладают способностью образовывать межмолекулярные водородные связи друг с другом, когда они выстроены в антипараллельном порядке и их гидрофобные поверхности повернуты в одном и том же направлении, так же, как и в случае, когда пептиды связываются с липидами. Миметики ApoA-I обладают также способностью образовывать внутримолекулярные водородные связи или солевые мостики вблизи N- и C-концов спирали.
Более того, при расположении в указанном антипараллельном порядке спирали оказываются плотно упакованными; нет никаких стерических ограничений, препятствующих тесному контакту между спиралями. Миметики ApoA-I обладают способностью к плотной упаковке и ионному взаимодействию, с образованием внутри- и/или межмолекулярных солевых мостиков и/или водородных связей при связывании с липидами в антипараллельном порядке.
Миметики ApoA-I способны к самоассоциации. Феномен самоассоциации зависит от условий pH, концентрации пептида и ионной силы и может приводить в результате к нескольким состояниям ассоциации, от мономерной до нескольких вариантов мультимерных форм (фиг.4A). Гидрофобное ядро пептидных агрегатов благоприятствует гидрофобным взаимодействиям с липидами. Способность пептидов агрегировать даже при очень низких концентрациях может способствовать их связыванию с липидами. Предполагается, что в центре пептидных агрегатов возникают также пептид-пептидные взаимодействия, и они могут конкурировать с липид-пептидными взаимодействиями.
Гидрофобное ядро агрегатов миметиков ApoA-I способствует взаимодействиям с липидами. Способность миметиков ApoA-I к агрегации даже при очень низких концентрациях может способствовать их связыванию с липидами. Взаимодействия между миметиками ApoA-I и липидами приводит к образованию пептид-липидных комплексов. Как проиллюстрировано на фиг.4A, полученный тип комплекса (комицеллы, диски, везикулы или мультислои) может зависеть от липид:пептидного молярного отношения, при этом комицеллы обычно образуются при низких липид:пептидных молярных отношениях, а с увеличением молярных отношений образуются дискообразные и везикулярные или многослойные комплексы. Мицеллы обычно образуются приблизительно при соотношениях 2 моль липидов приблизительно к 1 моль ApoA-I или приблизительно 2 моль липидов приблизительно к 6-10 моль миметика ApoA-I. Дискообразные комплексы обычно образуются при соотношениях, приблизительно составляющих 50-100 моль липида: приблизительно к 1 моль ApoA-I или приблизительно от 6 примерно до 10 моль миметика ApoA-I. Везикулярные комплексы обычно образуются при соотношениях, приблизительно составляющих от 200 примерно до 300 моль липида: приблизительно к 1 моль ApoA-I или приблизительно от 6 примерно до 10 моль миметика ApoA-I. Указанное свойство было описано для амфифильных пептидов (Epand, The Amphipathic Helix, 1993) и для ApoA-I (Jones, 1992, Structure and Function of Apolipoproteins, Chapter 8, pp.217-250). Липид-пептидное молярное отношение определяет также размер и состав таких комплексов.
D. Измененные формы пептидов формулы I, II и III и их фармацевтически приемлемых солей
В других воплощениях миметики ApoA-I содержат 22 или менее аминокислотных остатков. Безусловно, усеченные или внутренне укороченные формы формулы I, II или III, содержащие 21, 20, 19, 18, 17, 16 или даже 15 аминокислотных остатков, которые по существу сохраняют суммарные характеристики и свойства амфифильных спиралей, образованных миметиками ApoA-I, считаются входящими в объем, охватываемый настоящим изобретением.
В одном из воплощений настоящего изобретения усеченные формы миметиков ApoA-I получают путем делеции одного или нескольких аминокислотных остатков с N- и/или C-конца. Укороченные за счет внутренних областей формы миметиков ApoA-I получают путем делеции одного или нескольких аминокислотных остатков, занимающих положения внутри миметиков ApoA-I. Подвергнутые делеции внутренние аминокислотные остатки могут быть последовательными остатками или не следующими друг за другом остатками.
Специалистам в данной области должно быть понятно, что делеция внутреннего аминокислотного остатка в миметике ApoA-I может вызвать на месте делеции поворот плоскости раздела гидрофильно-гидрофобной поверхности спирали на 100°. Поскольку такие ротации могут значительно изменить амфифильные свойства образовавшейся в результате этого спирали, в одном из воплощений настоящего изобретения один или нескольких аминокислотных остатков подвергают делеции, так, чтобы по существу сохранилась ориентация плоскости гидрофильно-гидрофобной поверхности вдоль все оси спирали.
Это удобно осуществлять путем делеции достаточного количества последовательных или не следующих друг за другом аминокислотных остатков, с тем, чтобы делеции был подвергнут один полный виток спирали. Идеализированная α-спираль имеет 3,6 остатков на виток. Таким образом, в одном из воплощений делеции подвергают группы из 3-4 последовательных или не следующих друг за другом аминокислотных остатков. Вопрос о том, следует ли подвергнуть делеции 3 аминокислотных остатка или 4 аминокислотных остатка, может зависеть от положения внутри спирали первого подвергаемого делеции остатка. Определение приблизительного количества последовательных или не следующих друг за другом аминокислотных остатков, которые составляют один полный поворот спирали от любой конкретной начальной точки внутри амфипатической спирали, полностью находится в пределах компетенции специалистов в данной области.
Миметики ApoA-I могут быть также удлинены с одного или с обоих концов, или же за счет внутренних областей путем добавления аминокислотных остатков, которые не вызывают существенных помех, а в определенных воплощениях даже усиливают структурные и/или функциональные свойства пептидов. Безусловно, удлиненные миметиков ApoA-I, содержащие 23, 24, 25, 26, 27, 28 или 29 аминокислотных остатков, считаются входящими в объем, охватываемый настоящим изобретением. Такие удлинения миметиков ApoA-I могут по существу сохранять суммарную амфифильность и другие свойства миметиков ApoA-I. Специалистам в данной области буде, безусловно, понятно, что добавление аминокислотных остатков во внутренние области может вызвать ротацию плоскости гидрофильно-гидрофобной поверхности на месте включения добавочного остатка, наподобие того, что было описано выше для случая внутренних делеций. Таким образом, соображения, обсуждаемые выше в связи с внутренними делециями, применимы также и к добавлениям аминокислотных остатков во внутренние области.
В одном из воплощений миметики ApoA-I удлиняют на их N-и/или C-конце, добавляя аминокислотную последовательность, содержащую от 1 до 7 остатков.
В одном из воплощений миметики ApoA-I удлиняют на их N-и/или C-конце по меньшей мере на один виток спирали. Такие удлинения стабилизируют вторичную структуру спирали в присутствии липидов, так же как и описанные ранее концевые аминокислотные остатки и сегменты.
В другом воплощении миметики ApoA-I удлиняют на N-конце на один основный аминокислотный остаток, такой как Lys (K). В одном из воплощений X1 представляет собой Lys, X2 представляет собой Lys, X3 представляет собой Leu, X4 представляет собой Lys, X5 представляет собой Gln, X6 представляет собой Lys, X7 представляет собой Leu, X8 представляет собой Ala, X9 представляет собой Glu, X10 представляет собой Leu, X11 представляет собой Leu, X12 представляет собой Glu, X13 представляет собой Asn, X14 представляет собой Leu, X15 представляет собой Leu, X16 представляет собой Glu, X17 представляет собой Arg, X18 представляет собой Phe, X19 представляет собой Leu, X20 представляет собой Asp, X21 представляет собой Leu, X22 представляет собой Val и X23 представляет собой Inp.
В объем настоящего изобретения входят также "защищенные" формы миметиков ApoA-I, то есть формы миметиков ApoA-I, в которых R1 представляет собой аминозащитную группу и/или R2 представляет собой карбоксильную защитную группу. Считается, что удаление N- и/или C-концевых зарядов миметиков ApoA-I, содержащих 18 или менее аминокислотных остатков (путем синтезирования N-ацилированных пептидных амидов/сложного эфира/гидразидов/спиртов и их заместителей) может привести к образованию миметиков, которые приближаются, а в определенных воплощениях даже превосходят, активность незащищенной формы такого миметика. В определенных воплощениях, когда такие миметики содержат 22 или более аминокислотных остатков, считается, что блокирование N- или C-конца может привести к образованию миметиков ApoA-I, которые проявляют более низкую активность, чем неблокированные формы. Однако защита обоих, N- и C-концов, миметиков ApoA-I, содержащих 22 или более аминокислотных остатков, может восстановить активность. Таким образом, в одном из воплощений настоящего изобретения защищены или N-, и/или C-конец (в другом воплощении оба конца) миметиков ApoA-I, содержащих 18 или менее аминокислотных остатков, тогда как N- и C-концы пептидов, содержащих 22 или более аминокислотных остатков, являются либо оба защищенными, либо оба незащищенными. Обычно N-концевые блокирующие группы включают в себя RC(O)-, где R представляет собой -H, (C1-C6)-алкил, (C2-C6)-алкенил, (C2-C6)-алкинил, (C5-C20)-арил, (C6-C26)-аралкил, 5-20-членный гетероарил или 6-26-членный алкгетероарил. Конкретные N-концевые блокирующие группы включают в себя ацетил, формил и дансил.
Типичные C-концевые блокирующие группы включают в себя -C(O)NRR и -C(O)OR, где каждый R независимо является таким, как определено выше. Конкретные С-концевые блокирующие группы включают в себя такие, в которых каждый R независимо является метилом. Хоть и не желая связывать себя какой бы то ни было конкретной теорией, авторы изобретения тем не менее полагают, что такие концевые блокирующие группы стабилизируют α-спираль в присутствии липидов (см., например, Venkatachelapathi et al., 1993, PROTEINS: Structure, Function and Genetics 15: 349-359).
E. Димеры, тримеры, тетрамеры и мультимеры пептидов формулы I, II или III и их фармацевтически приемлемые соли
Структура нативного ApoA-I содержит семь спиральных элементов, которые, как считается, сообща участвуют в связывании липидов (Nakagawa et al., 1985, J. Am. Chem. Soc. 107: 7087-7092; Anantharamaiah et al., 1985, J. Biol. Chem. 260: 10248-10262; Vanloo et al., 1991, J. Lipid Res. 32: 1253-1264; Mendez et al., 1994, J. Clin. Invest. 94: 1698-1705; Palgunari et al., 1996, Arterioscler. Thromb. Vase. Biol. 16: 328-338; Demoor et al., 1996, Eur. J. Biochem. 239:74-84). Таким образом, в настоящее изобретение включены также димеры, тримеры, тетрамеры и даже полимеры более высокого порядка («мультимеры») миметиков ApoA-I. Такие мультимеры могут быть представлены в виде тандемных повторов, разветвленных сетей или их сочетаний. Миметики ApoA-I могут быть напрямую присоединены друг к другу или же разделены одним или несколькими линкерами.
Миметики ApoA-I, которые содержат мультимеры, могут быть пептидами формулы I, II или III, аналогами формулы I, II или III, измененными формами формулы I, II или III, усеченными формами или формами с внутренними делециями формулы I, II или III, удлиненными формами формулы I, II или III и/или их сочетаниями. Миметики ApoA-I могут быть соединены по принципу голова-к-хвосту (то есть N-концом к C-концу), по принципу голова-к-голове, (то есть N-концом к N-концу), по принципу хвост-к-хвосту (то есть С-концом к C-концу), или в различных их комбинациях и в виде их фармацевтически приемлемых солей.
В одном из воплощений настоящего изобретения мультимеры являются тандемными повторами двух, трех, четырех и приблизительно до десяти миметиков ApoA-I. В одном из воплощений мультимеры являются тандемными повторами от 2 до 8 пептидов. Таким образом, в одном из воплощений настоящее изобретение связано с мультимерами, имеющими следующую структурную формулу:
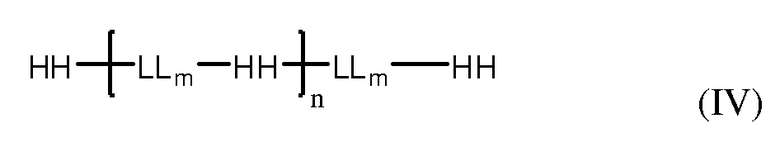
в которой
каждое m независимо является целым числом от 0 до 1, и в одном из воплощений m равно 1; n является целым числом от 0 до 10, и в одном из воплощений n является целым числом от 0 до 8; каждый "HH" независимо является радикалом, полученным из миметика ApoA-I; и каждый "LL" независимо представляет собой линкер.
В структуре (IV) линкер LL может быть любой бифункциональной молекулой, способной к ковалентному связыванию двух пептидов друг с другом. Таким образом, подходящими линкерами являются бифункциональные молекулы, в которых функциональные группы способны ковалентно присоединиться к N-и/или C-концу пептида. Функциональные группы, подходящие для присоединения к N- или C-концу пептидов, хорошо известны специалистам в данной области и имеют удобный химизм для обеспечения образования такой ковалентной связи.
Линкер может быть гибким, жестким или полужестким, в зависимости от требуемых свойств мультимера. Подходящие линкеры включают в себя, например, аминокислотные остатки, такие как Pro, azPro, Pip, azPip или Gly, или же пептидные сегменты, содержащие приблизительно от 2 примерно до 5, 10, 15 или 20, или даже более аминокислотных остатков, бифункциональные органические соединения, такие как H2N(CH2)nCOOH, HO(CH2)nCOOH и HO(CH2CH2O)nCH2CH2COOH, где n является целым числом от 1 до 12, и т.п. Примеры таких линкеров, также как и способы получения таких линкеров и соединений, включающих такие линкеры, хорошо известны в данной области (см., например, Hunig et al., 1974, Chem. Ber. 100: 3039-3044; Basak et al., 1994, Bioconjug. Chem. 5(4): 301-305).
В одном из воплощений настоящего изобретения тандемные повторы внутренне прерываются единственным пролиновым остатком. В тех случаях, когда миметики ApoA-I не содержат N- или C-концевой пролиновый остаток, LL может быть Pro, D-Pro, azPro, Pip, D-Pip или azPip, и m равно 1.
В определенных воплощениях настоящего изобретения может быть необходимо использование расщепляемых линкеров, которые обеспечивают высвобождение одного или нескольких спиральных сегментов (HH) в определенных условиях. Подходящие расщепляемые линкеры включают в себя пептиды, имеющие последовательности аминокислотных остатков, которые распознаются протеиназами, олигонуклеотиды, которые могут быть расщеплены эндонуклеазами, и органические соединения, которые могут расщепляться химическим путем, например, в кислотных, щелочных или иных условиях. Обычно условия расщепления бывают относительно мягкими, с тем, чтобы не вызвать денатурацию или чтобы как-нибудь иначе не вызвать деградацию спиральных сегментов и/или нерасщепленных линкеров, образующих мультимеры.
Пептидные и олигонуклеотидные линкеры, которые могут селективно расщепляться, также как и способы расщепления указанных линкеров, хорошо известны в данной области и должны быть очевидны для специалистов. Подходящие линкеры в виде органических соединений, которые могут селективно расщепляться, хорошо известны специалистам в данной области и включают таковые, описанные, например, в заявке WO 94/08051, а также в цитируемых в ней публикациях.
В одном из воплощений используемыми линкерами являются пептиды, которые представляют собой субстраты для эндогенных циркулирующих ферментов, обеспечивающие таким образом возможность мультимерам селективно расщепляться in vivo. Эндогенным ферментом, подходящим для расщепления линкеров, является, например, про-аполипопротеин A-I-пропептидаза. Соответствующие ферменты, также как и пептидные сегменты, которые выступают в качестве субстратов для таких ферментов, хорошо известны в данной области (см., например, Edelstein et al., 1983, J. Biol. Chem. 258: 11430-11433; Zanis, 1983, Proc. Natl. Acad. Sci. USA 80: 2574-2578).
В одном из воплощений используют линкеры удовлетворительной длины и гибкости, с тем, чтобы обеспечить антипараллельную ориентацию спиральных сегментов (HH) структуры (II) и образование межмолекулярных водородных связей или солевых мостиков в присутствии липидов. Линкеры удовлетворительной длины и гибкости включают в себя, но не ограничены перечисленным, остаток или радикал Pro, D-Pro, azPro, Pip, D-Pip, azPip, Gly, Cys-Cys, H2N(CH2)nCOOH, HO(CH2)nCOOH или HO(CH2CH2O)nCH2CH2COOH, где n принимает значения от 1 до 12 или от 4 до 6; H2N-арил-COOH и углеводороды.
Альтернативно, поскольку нативные аполипопротеины обеспечивают возможность кооперативного связывания между антипараллельными спиральными сегментами, пептидные линкеры, которые по первичной последовательности соответствуют пептидным сегментам, соединяющим соседние спирали нативных аполипопротеинов, включая, например, ApoA-I, ApoA-II, ApoA-IV, ApoC-I, ApoC-II, ApoC-III, ApoD, ApoE и ApoJ, могут быть удобным образом использованы для связывания миметиков ApoA-I формулы I. Такие последовательности хорошо известны в данной области (см., например, Rosseneu et al., "Analysis of the Primary and of the Secondary Structure of the Apolipoproteins," In: Structure and Function of Lipoproteins, Ch.6, 159-183, CRC Press, Inc., 1992).
Другие линкеры, которые обеспечивают образование межмолекулярных водородных связей или солевых мостиков между тандемными повторами антипараллельных спиральных сегментов, включают в себя пептиды с разворотами в обратную сторону, такие как β-изгибы и γ-изгибы, а также органические молекулы, которые имитируют структуры пептидных β-изгибов и/или γ-изгибов. Обычно развороты в обратную сторону являются сегментами пептида, которые меняют направление полипептидной цепи на противоположное, так, чтобы создать возможность отдельной полипептидной цепи объединиться с областями антипараллельной β-складчатой структуры или антипараллельной α-спиральной структуры. β-изгибы обычно состоят из четырех аминокислотных остатков, а γ-изгибы обычно состоят из трех аминокислотных остатков.
Конформации и последовательности многих пептидных β-изгибов подробно описаны в данной области и включают в себя, лишь для примера, а не в порядке ограничения, тип-I, тип-I', тип-II, тип-II', тип-III, тип-III', тип-IV, тип-V, тип-V', тип-VIa, тип-VIb, тип-VII и тип-VIII (см. Richardson, 1981, Adv. Protein Chem. 34: 167-339; Rose et al., 1985, Adv. Protein Chem. 37: 1-109; Wilmot et al., 1988, J. Mol. Biol. 203: 221-232; Sibanda et al., 1989, J. Mol. Biol. 206: 759-777; Tramontano et al., 1989, Proteins: Struct. Funct. Genet. 6: 382-394).
Специфические конформации коротких пептидных изгибов, таких как β-изгибы, зависят прежде всего от положений определенных аминокислотных остатков в изгибе (обычно Gly, Asn или Pro). Как правило, β-изгиб типа-I совместим с любым аминокислотным остатком в положениях от 1 до 4 такого изгиба, с тем лишь исключением, что Pro не может находиться в положении 3. Gly преобладает в положении 4, а Pro преобладает в положении 2 изгибов обоих типов, типа-I и типа-II. Остатки Asp, Asn, Ser и Cys зачастую встречаются в положении 1, где их боковые цепи часто образуют водородную связь с NH остатка 3.
В изгибах типа-II остатки Gly и Asn встречаются наиболее часто в положении 3, поскольку они наиболее легко принимают углы остова. В идеальном случае в изгибах типа-I' Gly находится в положениях 2 и 3, а в изгибах типа-II' Gly находится в положении 2. Изгибы типа-III обычно могут иметь наибольшее количество аминокислотных остатков, но изгибы типа-III', как правило, требуют наличия Gly в положениях 2 и 3. Изгибы типа-VIa и VIb обычно содержат цис-пептидную связь и Pro в качестве внутреннего остатка. В качестве обзора разных типов и последовательностей β-изгибов в белках и пептидах читатель может обратиться к публикации Wilmot et al., 1988, J. Mol. Biol. 203: 221-232.
Конформации и последовательности многих пепидных γ-изгибов также подробно описаны в данной области (см., например, Rose et al., 1985, Adv. Protein Chem. 37: 1-109; Wilmer-White et al., 1987, Trends Biochem. Sci. 12: 189-192; Wilmot et al., 1988, J. Mol. Biol. 203: 221-232; Sibanda et al., 1989, J. Mol. Biol. 206: 759-777; Tramontano et al., 1989, Proteins: Struct. Funct. Genet. 6: 382-394). Все из указанных типов структур β-изгибов и γ-изгибов и соответствующие им последовательности, также как и открытые позже структуры и последовательности пептидных β-изгибов и γ-изгибов, специально включены в настоящее описание.
Альтернативно, линкер (LL) может содержать органическую молекулу или фрагмент, который имитирует структуру пептидного β-изгиба или γ-изгиба. Такие имитирующие β-изгиб и/или γ-изгиб фрагменты, также как способы синтезирования пептидов, содержащих такие фрагменты, хорошо известны в данной области и включают в себя, помимо прочего, таковые, описанные у Giannis and Kolter, 1993 Angew. Chem. Intl. Ed. Eng. 32: 1244-1267; Kahn et al., 1988, J. Molecular Recognition 1: 75-79; и Kahn et al., 1987, Tetrahedron Lett. 28: 1623-1626.
Еще в одном из воплощений настоящего изобретения мультимеры представлены в виде разветвленных сетей (см., например, фиг.3). Такие сети удобно получать путем использования мультифункциональных связующих фрагментов, которые обеспечивают присоединение к простому связующему фрагменту более чем двух спиральных элементов. Таким образом, в разветвленных сетях используются молекулы, имеющие три, четыре или даже больше функциональных групп, которые способны ковалентно связываться с N- и/или C-концом пептида. Подходящие связующие фрагменты включают в себя, например, остатки аминокислот, имеющих боковые цепи, несущие функциональные группы гидроксила, сульфанила, амино, карбоксила, амида и/или сложного эфира, таких, например, как Ser (S), Thr (T), Cys (C), Tyr (Y), Asn (N), Gln (Q), Lys (K), Arg (R), Orn, Asp (D) и Glu (E); а также соответствующий D-энантиомер каждого из указанных выше; или остатки других органических молекул, содержащие такие функциональные группы.
Спиральные сегменты, присоединенные к отдельному связующему фрагменту, необязательно должны быть присоединены одинаковыми концами. На самом деле в определенных воплощениях спиральные сегменты присоединяют к отдельному связующему фрагменту таким образом, чтобы они были организованы в антипараллельную структуру, то есть чтобы некоторые из спиралей были прикреплены через свои N-концы, а остальные через свои C-концы.
Спиральные сегменты могут быть присоединены непосредственно к связующему фрагменту или, как было описано ранее, могут быть пространственно отделены от связующего фрагмента одним или несколькими бифункциональными линкерами (LL).
Если обратиться к фиг.3A и 3B, можно увидеть, что разветвленная сеть может быть описана в терминах числа «узлов», образующих сеть, где каждый мультифункциональный связующий фрагмент образует узел. На фиг.3A и 3B спиральные сегменты (то есть миметики ApoA-I) проиллюстрированы в виде цилиндров, а мультифункциональные связующие фрагменты (или узлы) - в виде кружочков (●), где число линий, выходящих из кружочка, указывает на "порядок" (или число функциональных групп) мультифункционального связующего фрагмента.
Число узлов в сети будет обычно зависеть от суммарно требуемого числа спиральных сегментов и, как правило, будет составлять приблизительно от 1 до 2. Конечно, было бы желательно, чтобы для данного числа требуемых спиральных сегментов, сети, имеющие связующие фрагменты более высокого порядка, имели бы меньше узлов. Например, если обратиться к фиг.3A и 3B, сеть третьего порядка (то есть сеть, имеющая трифункциональные связующие фрагменты) из семи спиральных элементов имеет три узла (фиг.3A), тогда как сеть четвертого порядка (то есть сеть, имеющая тетрафункциональные связующие фрагменты) из семи спиральных элементов имеет только два узла (фиг.3B).
Сети могут быть униформного (одинакового) порядка, то есть они могут быть сетями, в которых все узлы являются, например, трифункциональными или тетрафункциональными связующими фрагментами, или могут быть сетями смешанного порядка, например, сетями, в которых узлы являются смешанными узлами, являясь, например, трифункциональными или тетрафункциональными связующими фрагментами. Конечно, следует понимать, что даже в случае сетей униформного порядка нет необходимости в том, чтобы связующие фрагменты были идентичными. В сетях третьего порядка можно использовать, например, два, три, четыре или даже больше различных трифункциональных связующих фрагментов.
Наподобие линейных мультимеров, спиральные элементы, содержащие разветвленную сеть, могут быть, но необязательно, идентичными.
Пример такой разветвленной сети смешанного порядка проиллюстрирован на фиг.3C. На фиг.3C спиральные сегменты (то есть миметики ApoA-I) проиллюстрированы в виде цилиндров, а мультифункциональные связующие фрагменты - в виде кружочков (●), где число линий, выходящих из кружочка, указывает на "порядок" (или число функциональных групп) мультифункционального связующего фрагмента. Линии, соединяющие спиральные сегменты, представляют собой, как было описано выше, бифункциональные линкеры LL. Спиральные сегменты, которые содержатся в разветвленных сетях, могут, как было описано выше, быть тандемными повторами миметиков ApoA-I.
В одном из иллюстративных воплощений разветвленные сети согласно изобретению описываются следующей формулой:
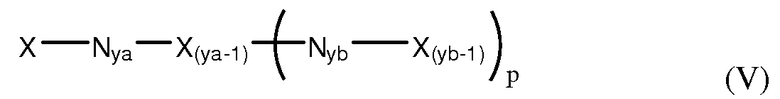
в которой
каждый X независимо является радикалом, полученным из мультимера следующей формулы:
в которой
каждый HH независимо является радикалом, полученным из миметика ApoA-I;
каждый LL является бифункциональным линкером;
каждое m независимо является целым числом от 0 до 1;
каждое n независимо является целым числом от 0 до 8;
каждый из Nya и Nyb независимо является мультифункциональным связующим фрагментом, где where ya и yb представляют собой число функциональных групп, соответственно, на Nya и Nyb; каждое ya и yb независимо является целым числом от 3 до 8; и p является целым числом от 0 до 7.
В одном из воплощений разветвленная сеть содержит "Lys tree" («лизиновое дерево»), то есть сеть, в которой мультифункциональный связующий фрагмент представляет собой один или более остатков Lys (K) (см., например, фиг.3D).
В одном из иллюстративных воплощений разветвленные сети указанного «лизинового дерева» ("Lys tree") согласно изобретению описываются следующими формулами:
в которой
каждый X независимо является радикалом, полученным из мультимера следующей формулы:
в которой
каждый HH независимо является радикалом, полученным из миметика ApoA-I формулы I;
каждый LL является бифункциональным линкером;
каждое n независимо является целым числом от 0 до 8;
каждое m независимо является целым числом от 0 до 1;
R1 представляет собой -OR или -NRR; и
каждый R независимо представляет собой -H, (C1-C6)-алкил, (C2-C6)-алкенил, (C2-C6)-алкинил или (C5-C20)-арил.
Некоторые дополнительные иллюстративные миметики ApoA-I представлены ниже в таблице 7:
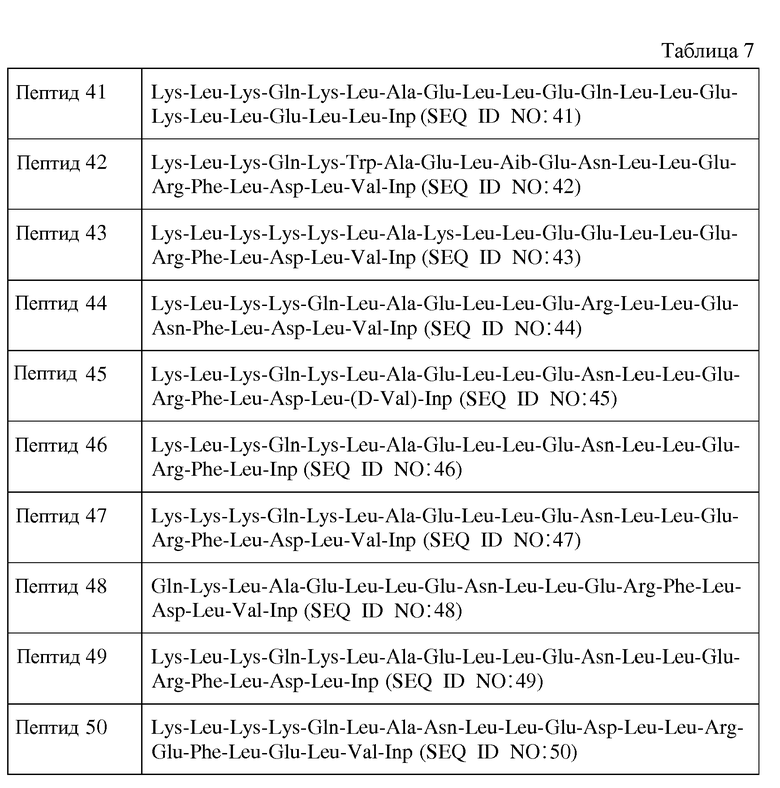

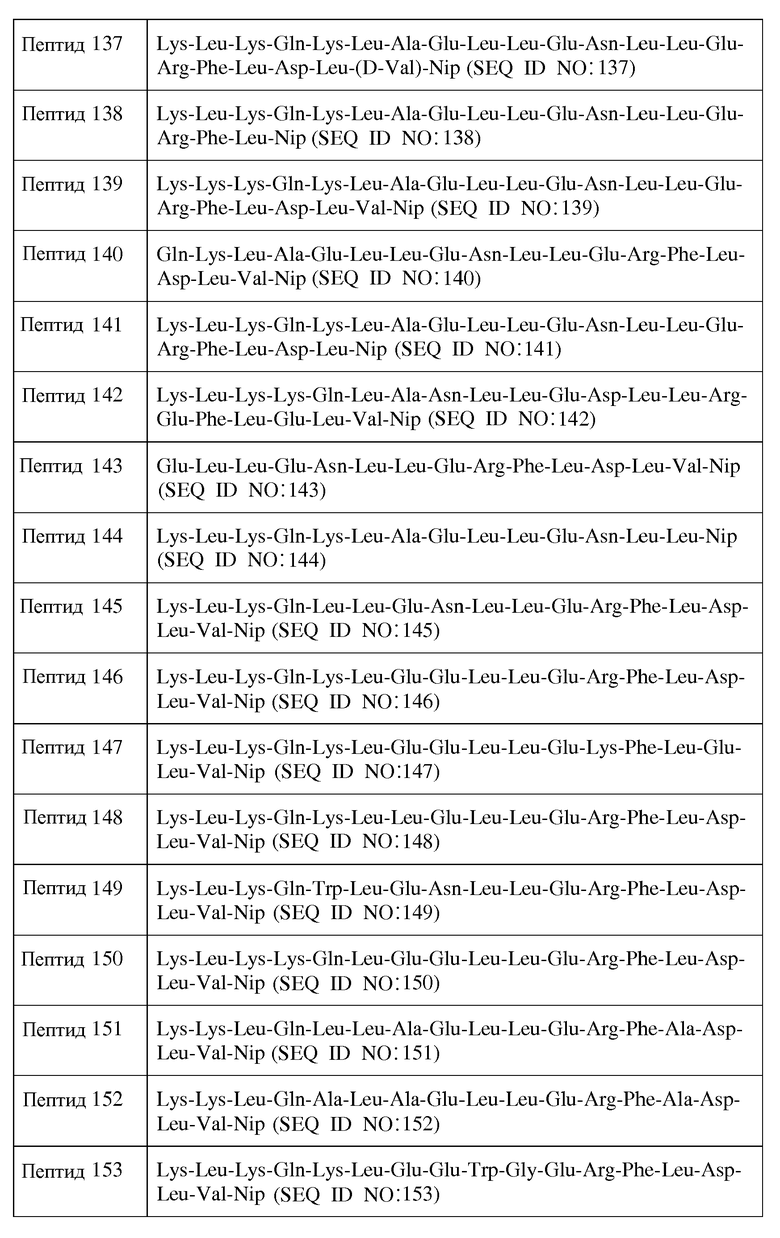
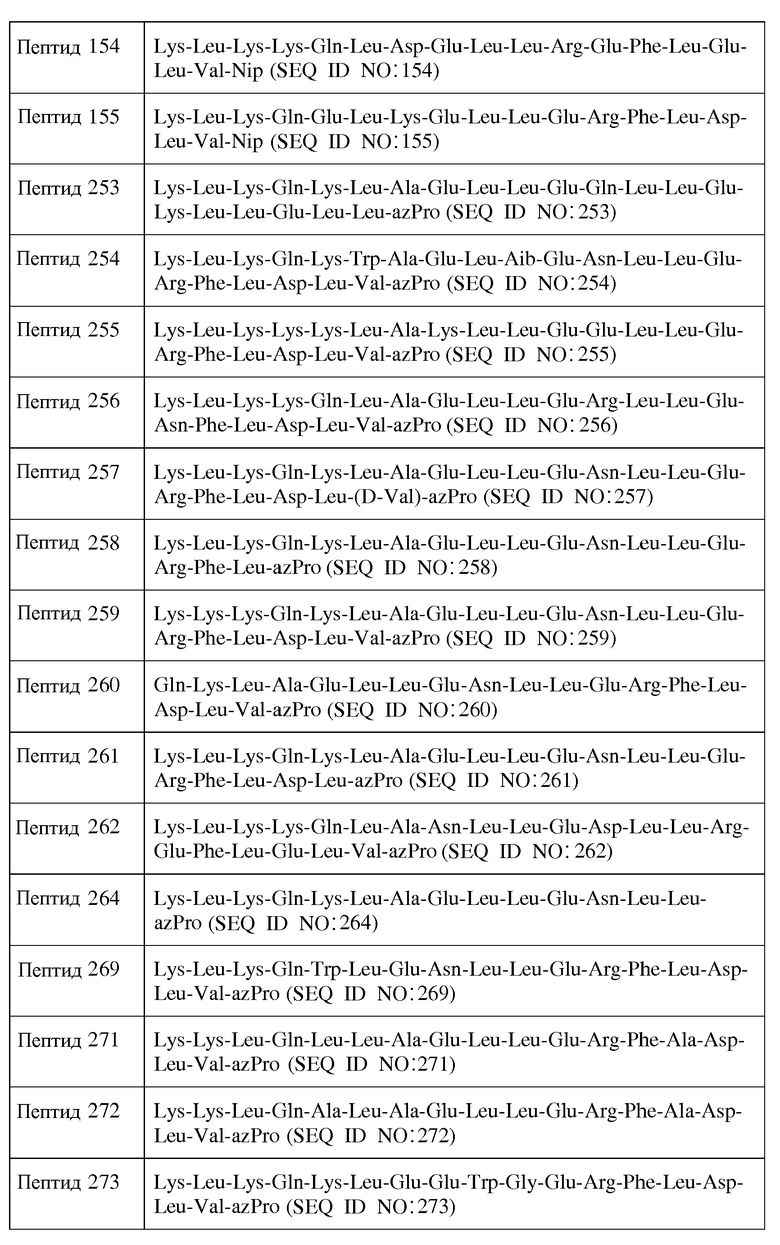
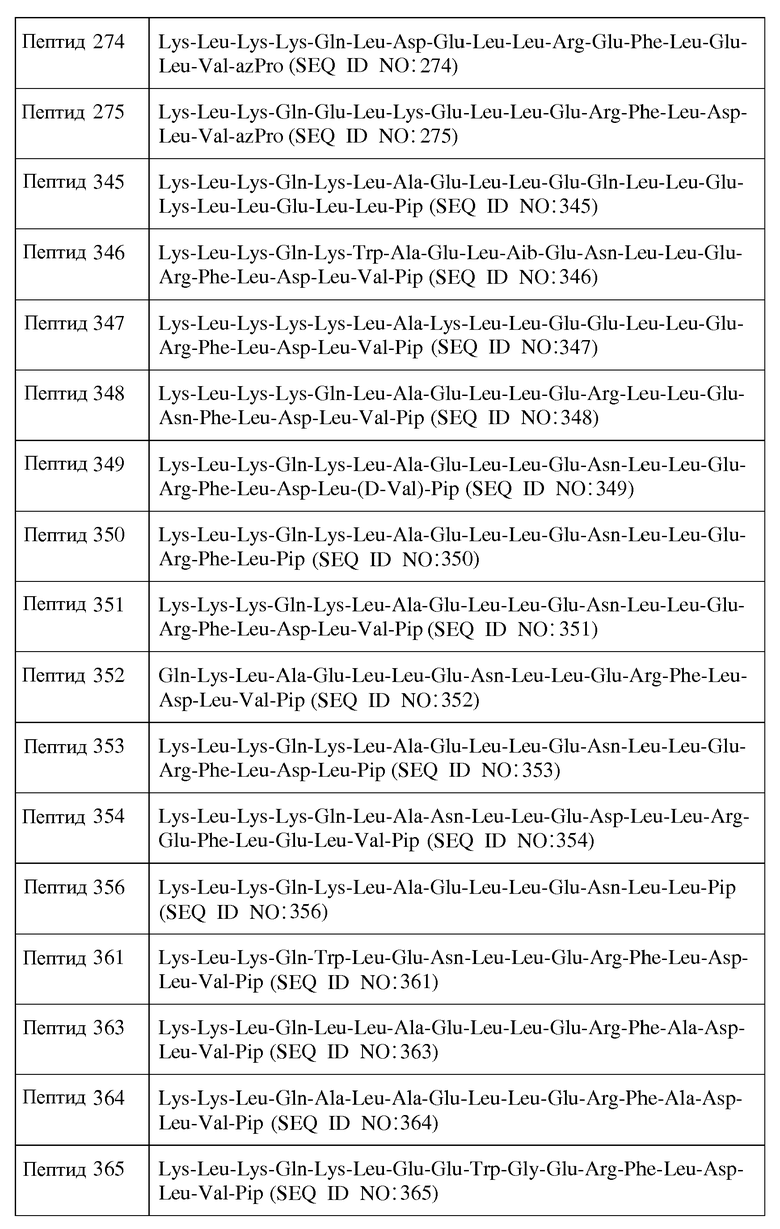
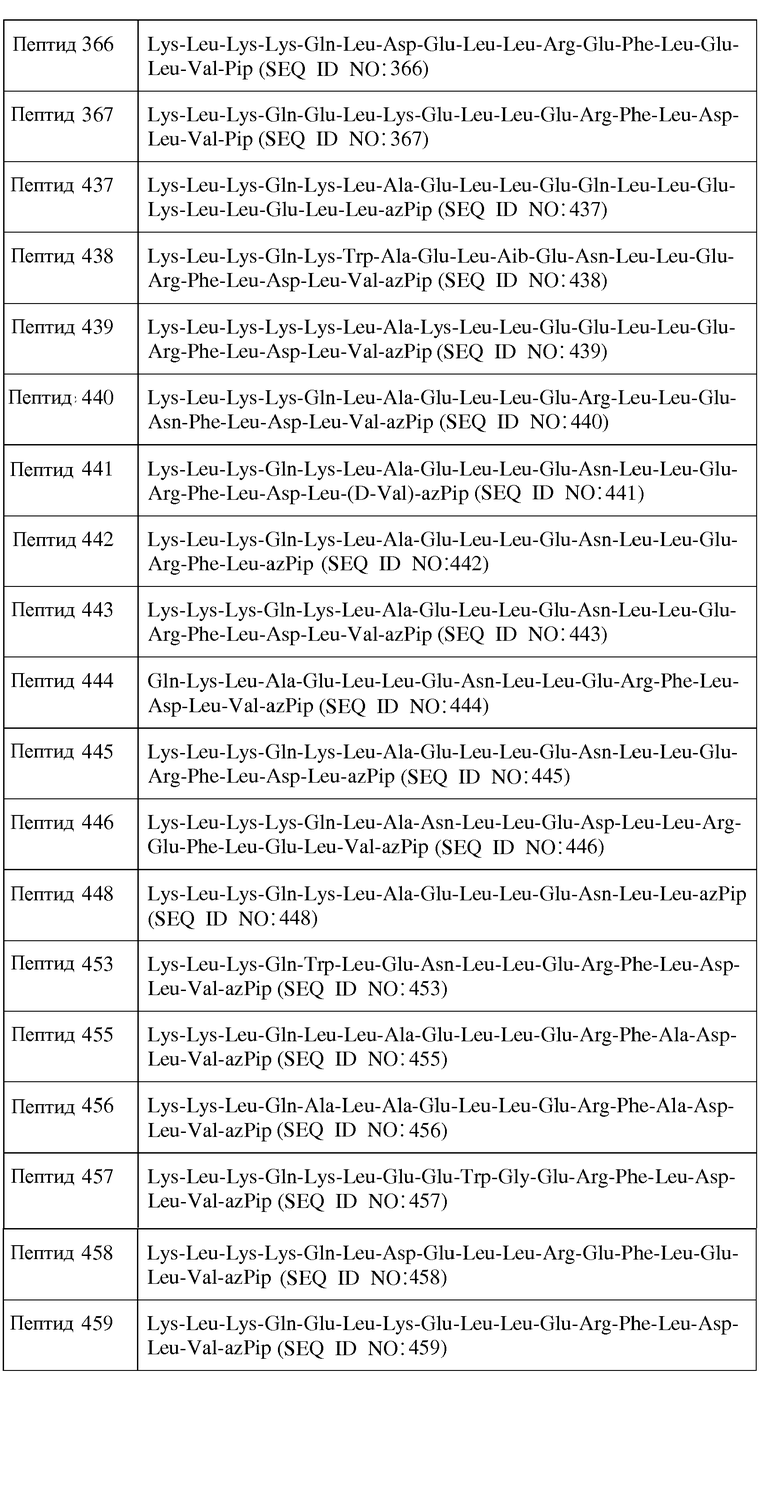
или их фармацевтически приемлемая соль.
Некоторые иллюстративные миметики ApoA-I, имеющие ацетилированный N-конец и амидированный C-конец, представлены ниже в таблицах 8 и 9:
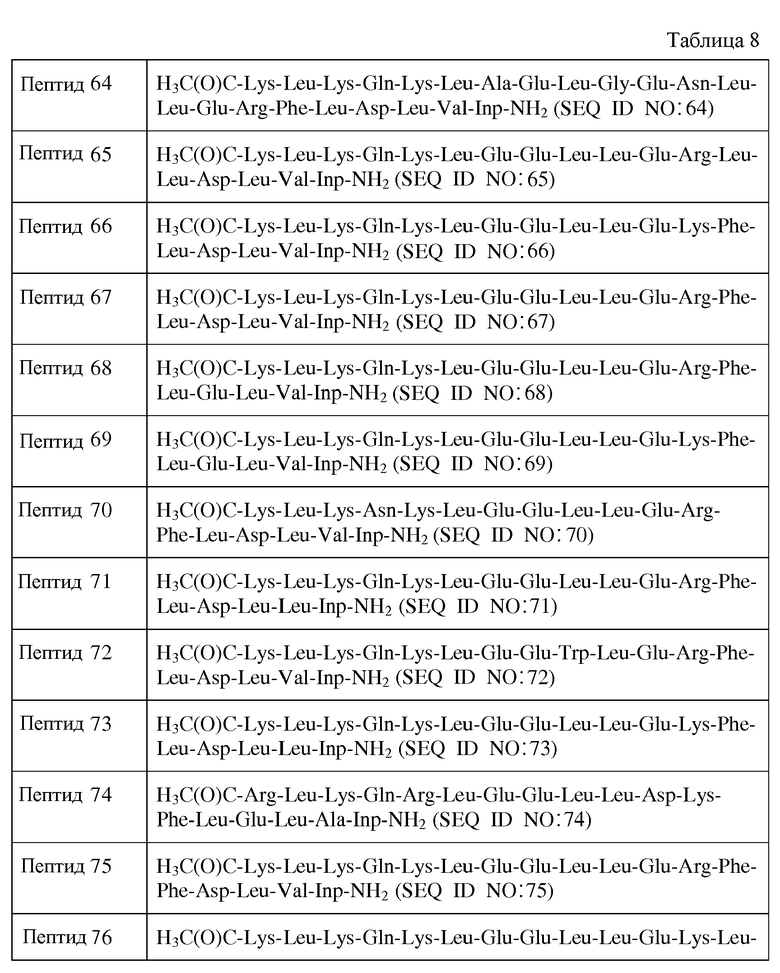
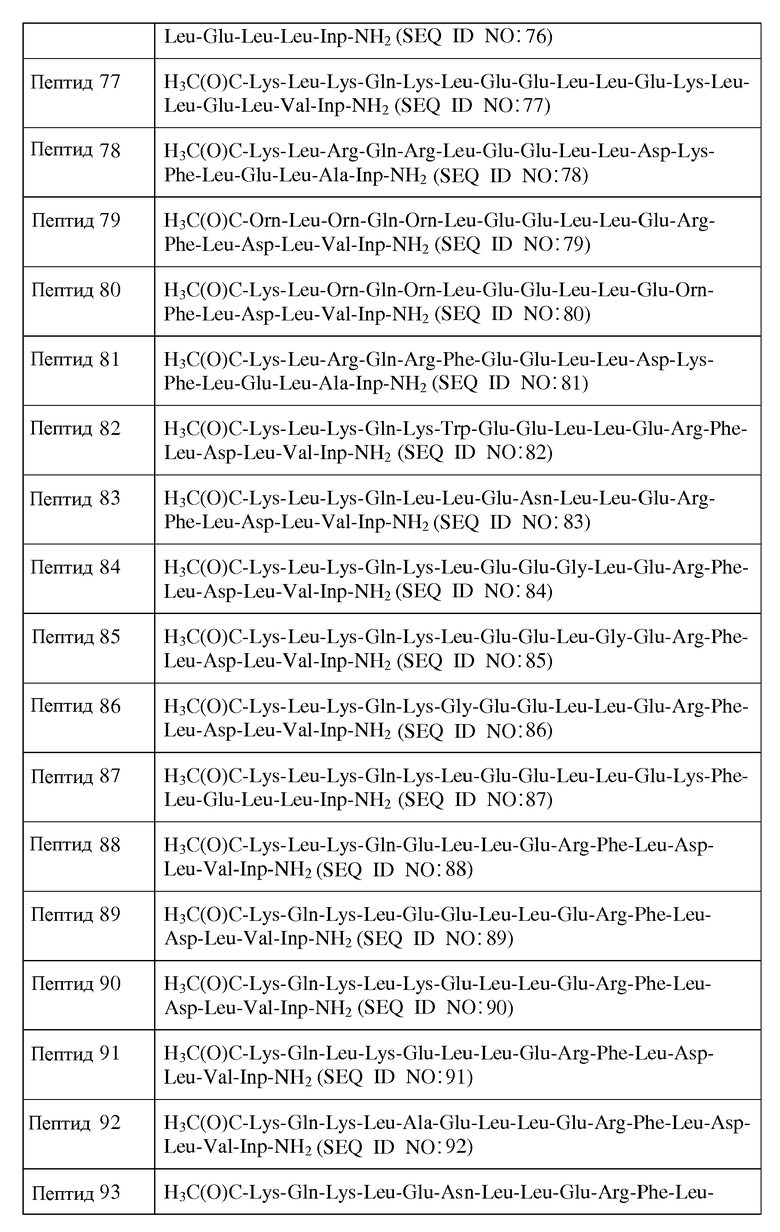
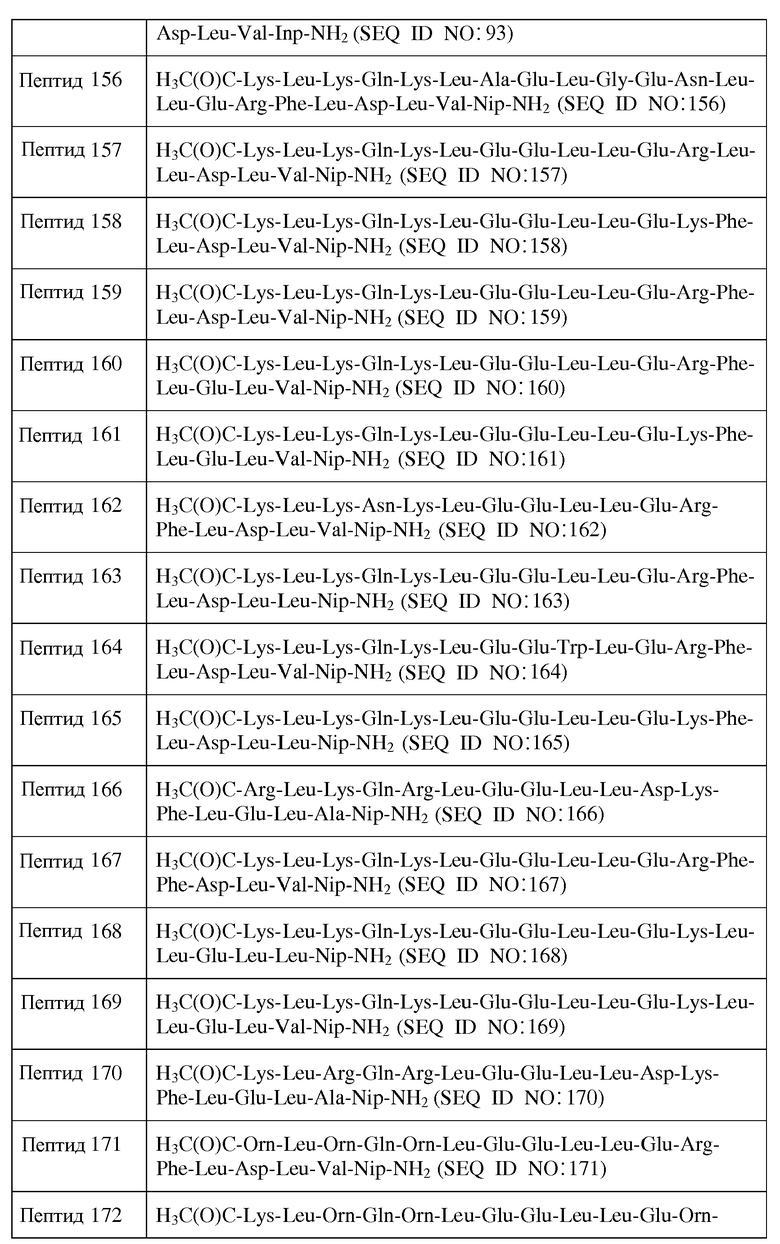
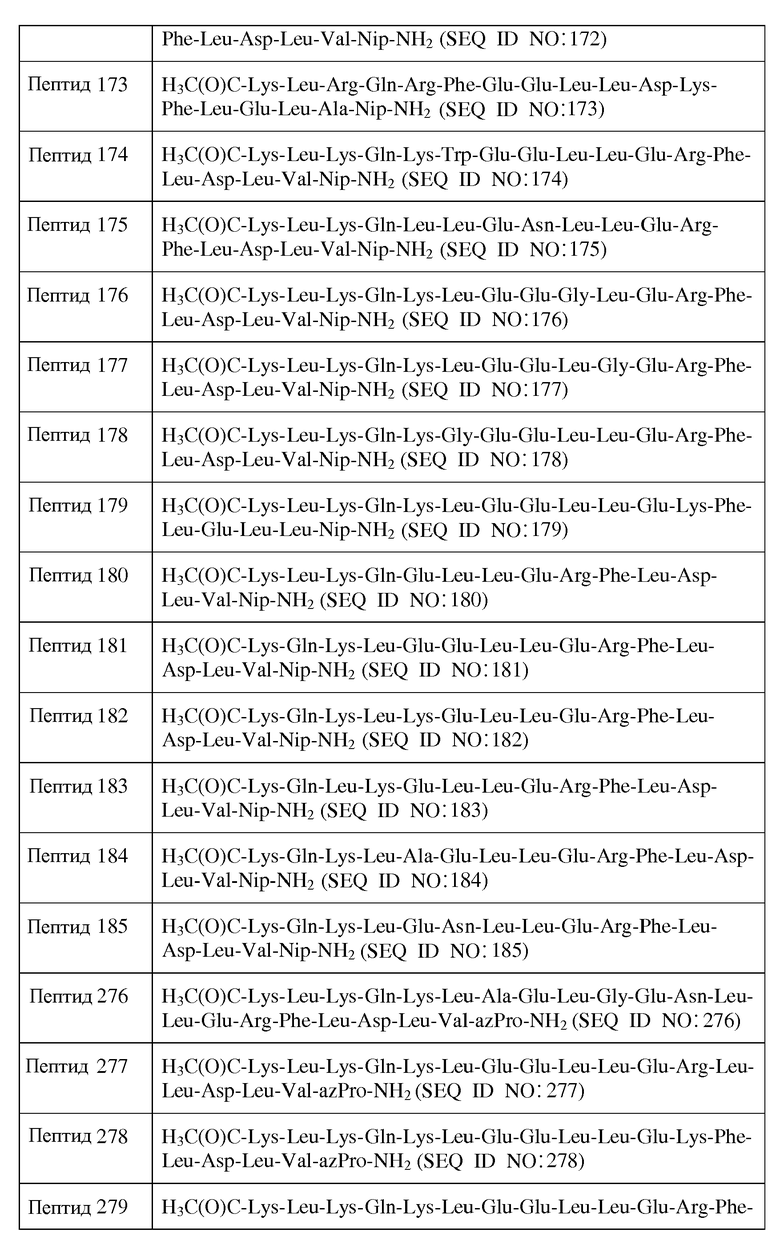
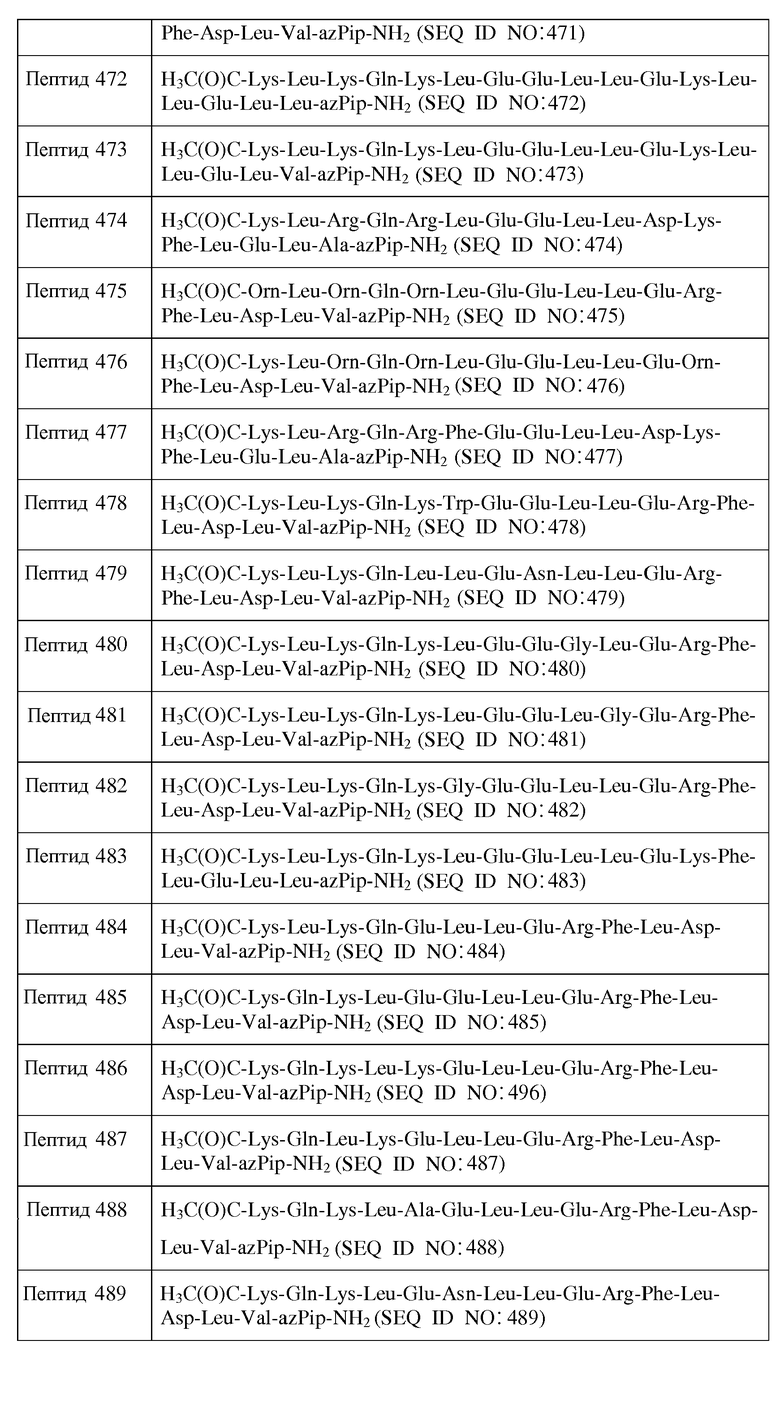
или их фармацевтически приемлемая соль.
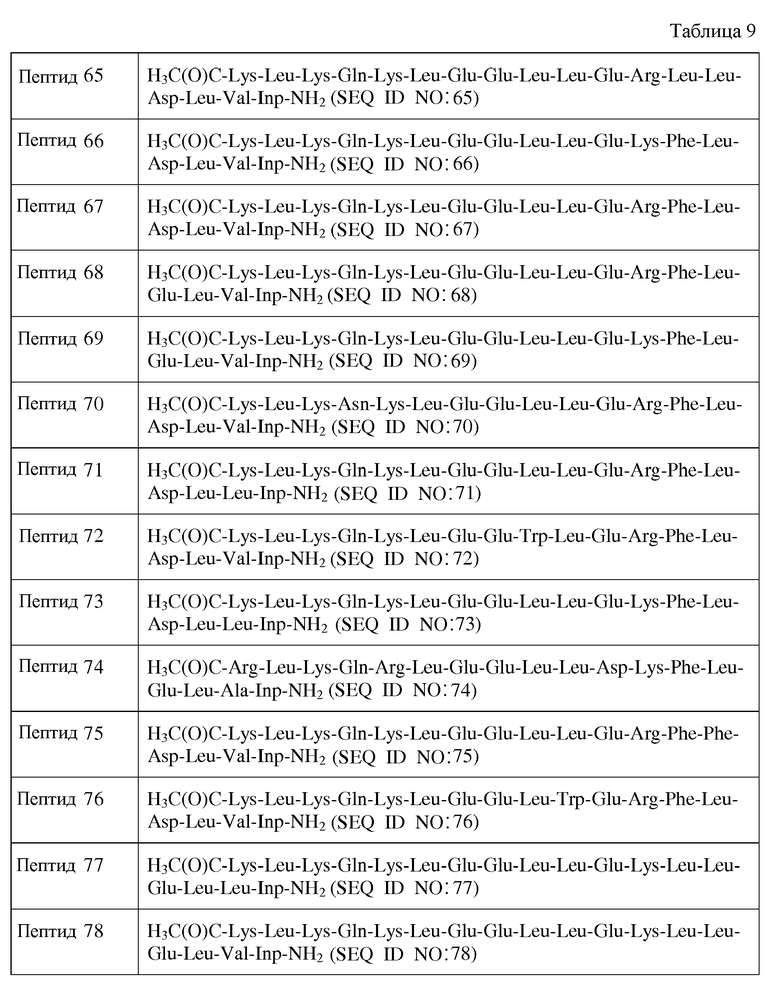
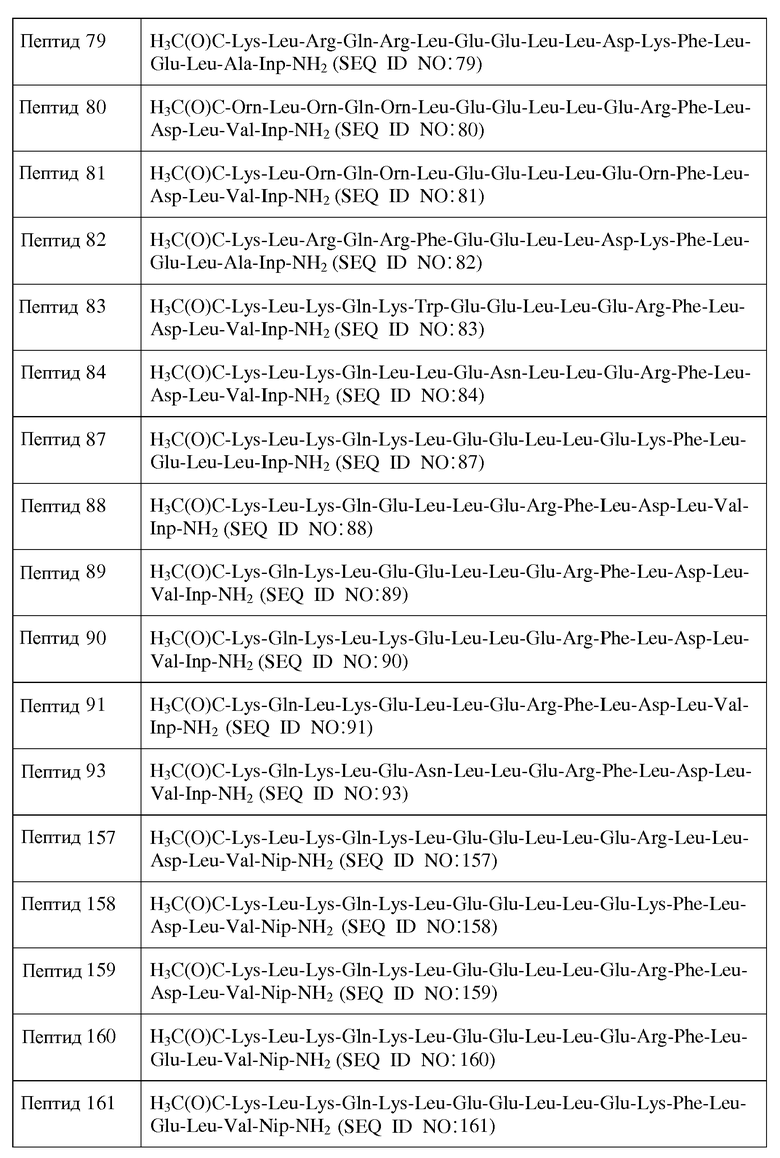
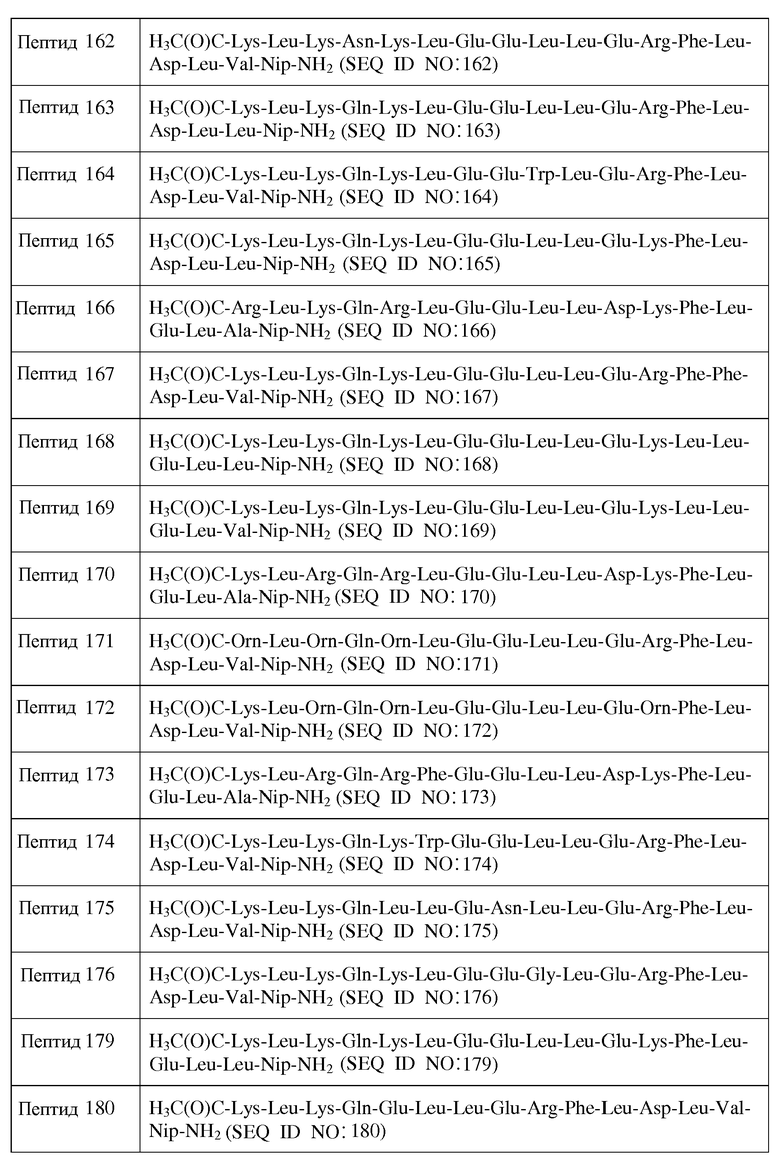
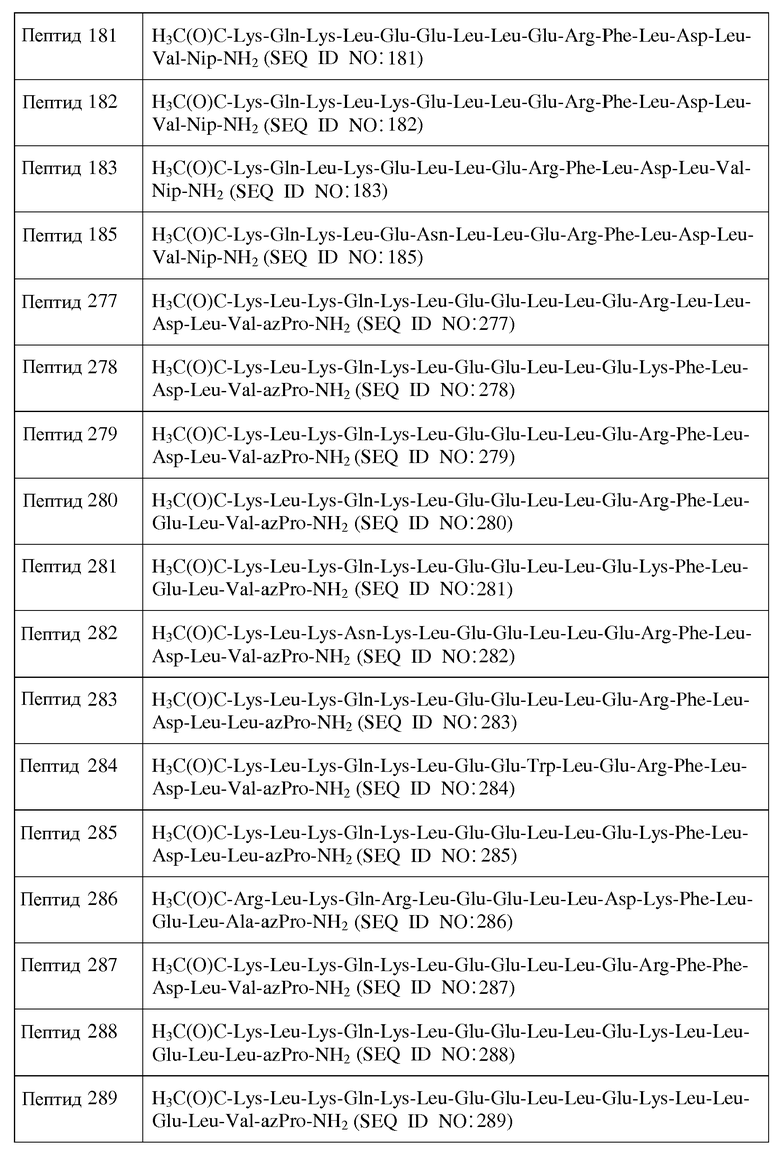
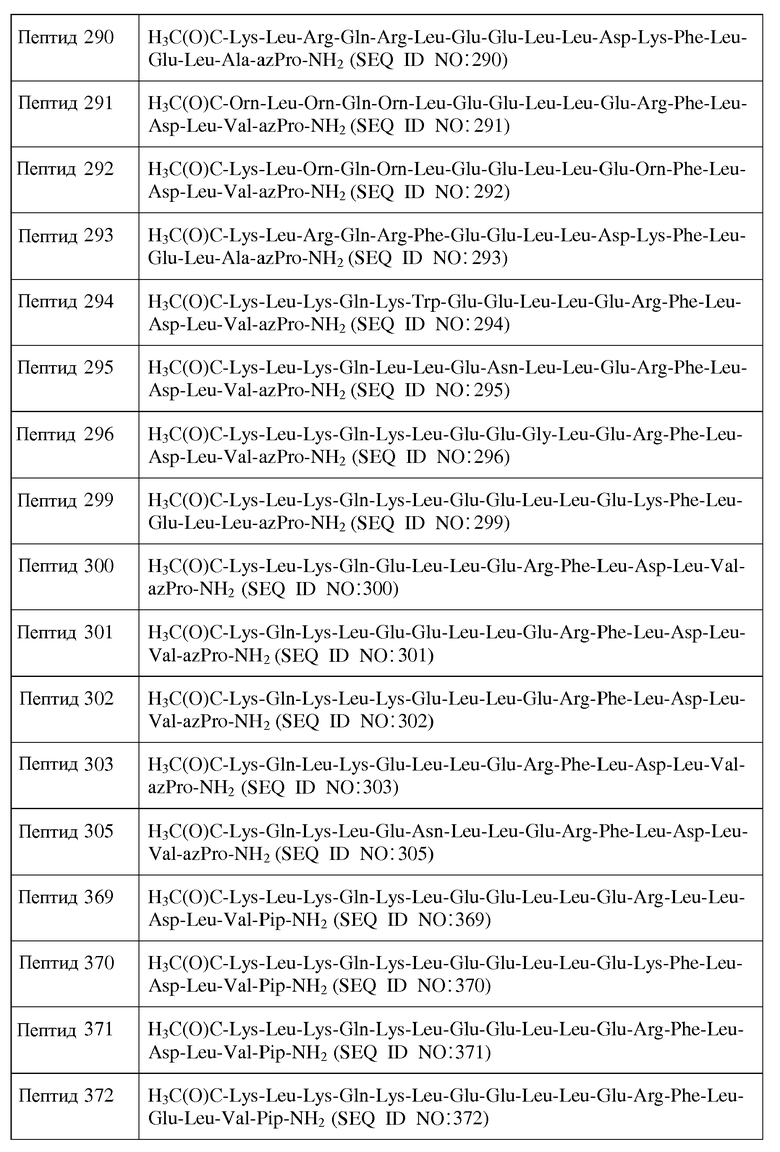
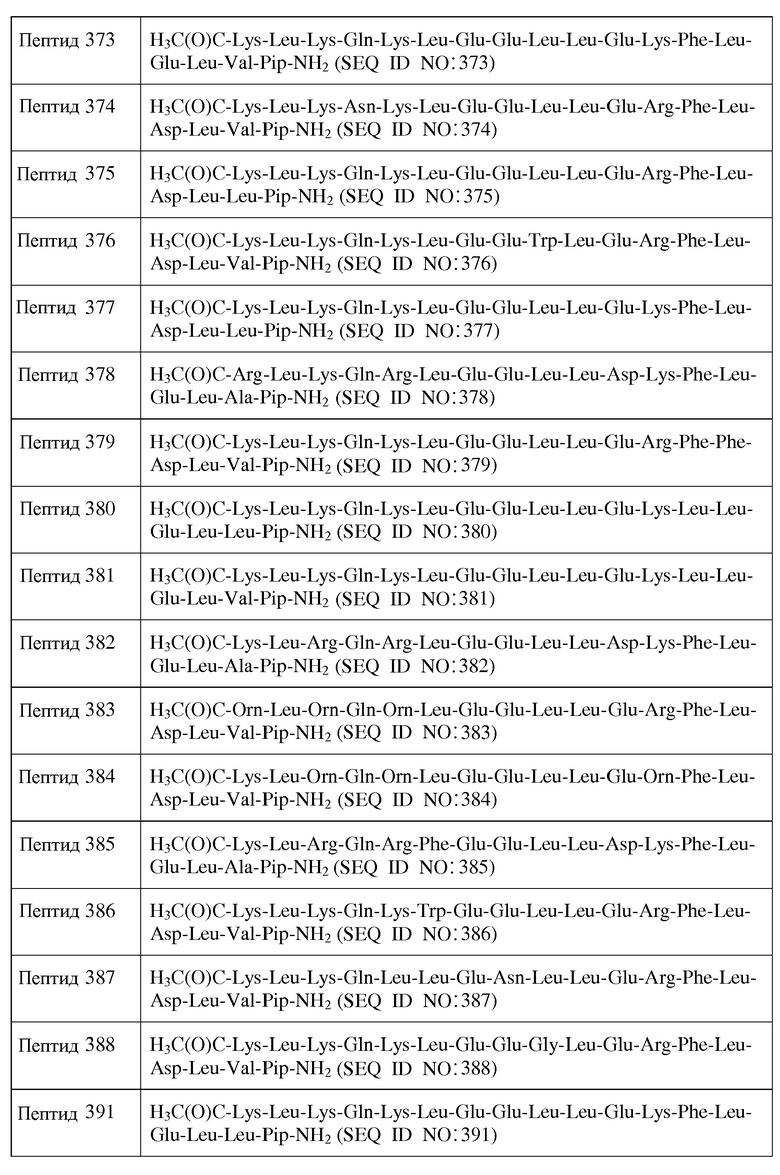
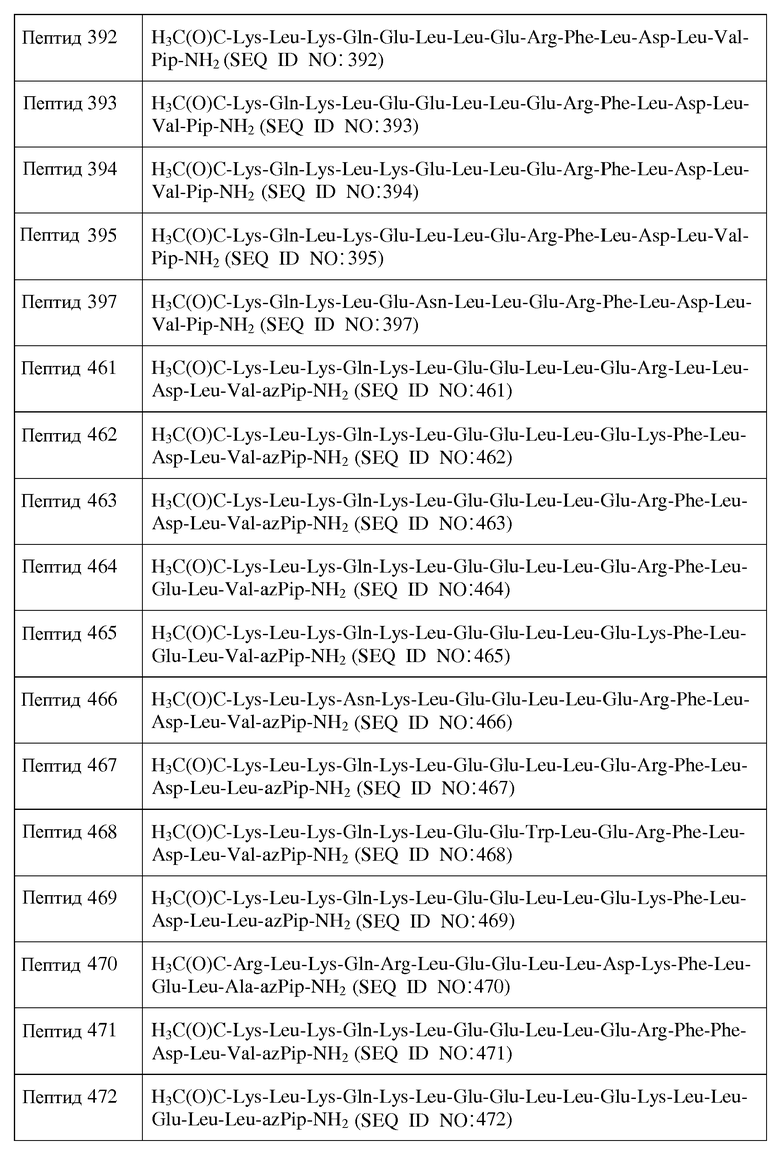
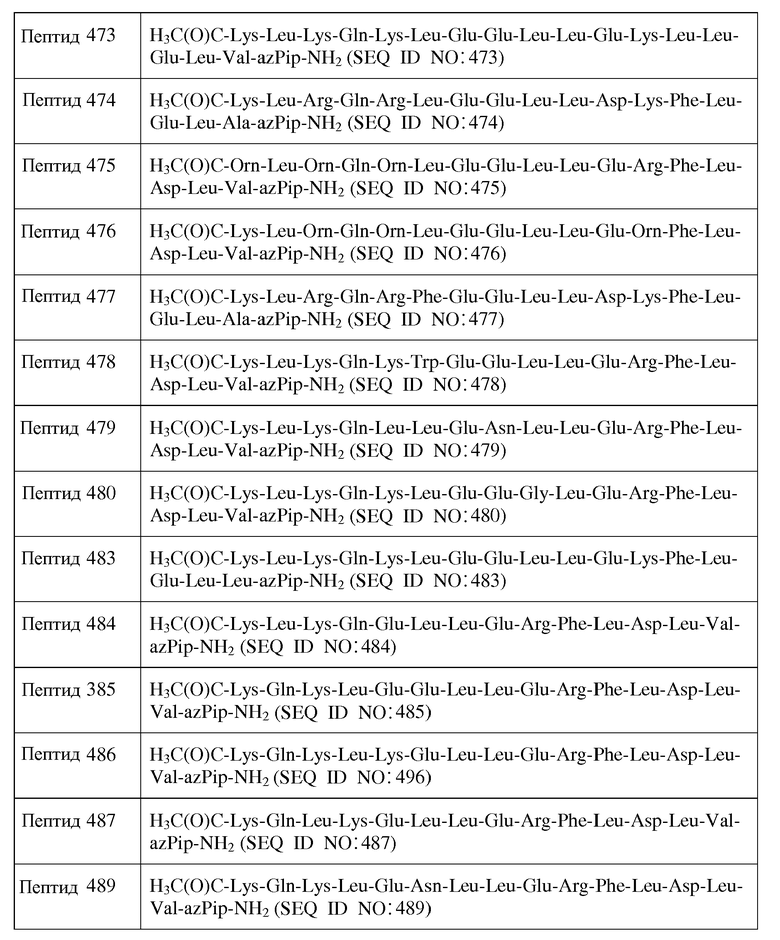
или их фармацевтически приемлемая соль.
III. Синтез миметиков ApoA-I
Миметики ApoA-I могут фактически быть получены с использованием любой известной в данной области технологии получения пептидов. Например, миметики ApoA-I могут быть получены стандартными способами ступенчатого пептидного синтеза в растворе или твердофазного пептидного синтеза, или же с использованием технологий рекомбинантной ДНК.
A. Химический синтез
Миметики ApoA-I могут быть получены с использованием стандартных способов ступенчатого пептидного синтеза в жидкой фазе или твердофазного пептидного синтеза (см., например, Chemical Approaches to the Synthesis of Peptides and Proteins, Williams et al., Eds., 1997, CRC Press, Boca Raton Fla., и приведенные там ссылки; Solid Phase Peptide Synthesis: A Practical Approach, Atherton & Sheppard, Eds., 1989, IRL Press, Oxford, England, и приведенные там ссылки).
Альтернативно, миметики ApoA-I могут быть получены путем сегментной конденсации пептидов, как описано, например, у Liu et al., 1996, Tetrahedron Lett. 37(7): 933-936; Baca, et al., 1995, J. Am. Chem. Soc. 117: 1881-1887; Tarn et al., 1995, Int. J. Peptide Protein Res. 45: 209-216; Schnolzer and Kent, 1992, Science 256: 221-225; Liu and Tarn, 1994, J. Am. Chem. Soc. 116(10): 4149-4153; Liu and Tarn, 1994, Proc. Natl. Acad. Sci. USA 91: 6584-6588; Yamashiro and Li, 1988, Int. J. Peptide Protein Res. 31: 322-334). Это уместно, в частности, в случае пептидов, имеющих глициновый остаток. Другие способы, используемые для синтеза миметиков ApoA-I, описаны у Nakagawa et al., 1985, J. Am. Chem. Soc. 107: 7087-7092.
Миметики ApoA-I, имеющие N- и/или C-концевые блокирующие группы, могут быть получены с использованием стандартных технологий органической химии. Например, способы ацилирования N-конца пептида или амидирования, или этерификации C-конца пептида хорошо известны в данной области. Способы осуществления других модификаций на N- и/или C-конце знакомы специалистам в данной области, также как и способы защиты любых функциональных групп боковых цепей, которые могут быть необходимы для присоединения концевых блокирующих групп.
Фармацевтически приемлемые соли (противоионы) могут быть получены стандартными способами путем ионообменной хроматографии или другими хорошо известными в данной области способами.
Миметики ApoA-I, которые представлены в виде тандемных мультимеров, могут быть синтезированы стандартными способами путем добавления линкера (линкеров) к пептидной цепи на соответствующей стадии синтеза. Альтернативно, можно синтезировать спиральные сегменты, и каждый сегмент вводить в реакцию с указанным линкером. Безусловно, конкретный способ синтеза будет зависеть от состава линкера. Подходящие системы защиты и соответствующий химизм хорошо известны и будут очевидны специалистам в данной области.
Миметики ApoA-I, которые представлены в виде разветвленных сетей, могут быть синтезированы стандартными способами с использованием тримерных и тетрамерных смол и химических технологий, описываемых Tarn, 1988, Proc. Natl. Acad. Sci. USA 85: 5409-5413 и Demoor et al., 1996, Eur. J. Biochem. 239: 74-84. Модифицированные синтетические смолы и стратегии синтеза разветвленных сетей более высокого или более низкого порядков, или же таких, которые содержат комбинации различных спиральных сегментов миметиков ApoA-I, находятся в пределах компетенции специалистов в области пептидной химии и/или органической химии.
Образование, в случае необходимости, дисульфидных связей может быть осуществлено в присутствии умеренно окисляющих средств. Могут быть использованы химические окисляющие средства, или же миметики ApoA-I могут быть просто экспонированы в атмосферный воздух с целью образования таких связей. В данной области известны разные способы, включая и способы, описанные, например, Tarn et al., 1979, Synthesis 955-957; Stewart et al., 1984, Solid Phase Peptide Synthesis, 2d Ed., Pierce Chemical Company Rockford, 111; Ahmed et al., 1975, J. Biol. Chem. 250: 8477-8482; а также Pennington et al., 1991 Peptides 1990, 164-166, Giralt and Andreu, Eds., ESCOM Leiden, The Netherlands. Дополнительная альтернатива описана Kamber et al., 1980, Helv. Chim. Acta 63: 899-915. A method conducted on solid supports is described by Albericio, 1985, Int. J. Peptide Protein Res. 26: 92-97. Любой из указанных способов может быть использован для образования дисульфидных мостиков в пептидах согласно изобретению.
Миметики ApoA-I, содержащие один или несколько внутренних остатков глицина, можно синтезировать с относительно высоким выходом путем сегментной конденсации пептидов, обеспечивая таким образом преимущественные возможности для широкомасштабной продукции. Сегментная конденсация, то есть объединение малых цепей составляющих пептидов с образованием более крупной пептидной цепи используется для получения многих биологически активных пептидов, включая миметики ApoA-I, насчитывающие 44 аминокислотных остатка (см., например, Nakagawa et al., 1985, J. Am Chem. Soc. 107: 7087-7083; Nokihara et al., 1989, Peptides 1988: 166-168; Kneib-Cordonnier et al., 1990, Int. J. Pept. Protein Res. 35: 527-538).
Преимущества синтеза через сегментную конденсацию включают в себя возможность конденсировать предварительно образованные сегменты в жидкой фазе и легкость очистки конечного продукта. Недостатки указанного способа включают в себя низкие уровни эффективности взаимодействия и выхода на стадии конденсации, а также низкую растворимость определенных пептидных последовательностей. Эффективность взаимодействия на стадии конденсации можно повысить путем увеличения времени взаимодействия. Обычно увеличение времени взаимодействия приводит к повышенной рацемизации продукта (Sieber et al., 1970, Helv. Chim. Acta 53: 2135-2150). Однако поскольку глицин лишен хирального центра, он не подвергается рацемизации (пролиновые остатки, в силу стерических препятствий, также мало подвержены или же совсем не подвержены рацемизации при длительном времени взаимодействия). Таким образом, при вариантах реализации, которые связаны с наличием внутренних глициновых остатков, синтез через сегментную конденсацию может осуществляться в больших количествах и с большим выходом путем синтезирования избирательных сегментов, что дает преимущество в связи с тем, что глициновые остатки не подвергаются рацемизации. Таким образом, миметики ApoA-I, содержащие один или более внутренних глициновых остатков, обеспечивают преимущество при синтезе в силу возможности получения крупномасштабных объемов препарата.
B. Рекомбинантный синтез
Если миметик ApoA-I полностью состоит из генетически кодируемых аминокислотных остатков или если часть его состоит из них, тогда такой миметик ApoA-I или соответствующая его часть может также быть синтезирована с помощью стандартной рекомбинантной технологии генной инженерии.
Для получения рекомбинантной продукции полинуклеотидную последовательность, кодирующую соответствующий пептид, встраивали в подходящий экспрессирующий вектор, то есть в вектор, который содержит элементы, необходимые для транскрипции и трансляции встроенной кодирующей последовательности, или, в случае вектора РНК-вируса, элементы, необходимые для репликации и трансляции. Затем экспрессирующий носитель трансфицируют в подходящую хозяйскую клетку, которая будет экспрессировать указанный пептид. Затем, в зависимости от того, какую экспрессирующую систему используют, экспрессируемый пептид выделяют тем или иным из хорошо известных в данной области способов. Способы продуцирования рекомбинантных белков и пептидов хорошо известны в данной области (см., например, публикации Sambrook et al., 1989, Molecular Cloning A Laboratory Manual, Cold Spring Harbor Laboratory, N. Y.; а также Ausubel et al., 1989, Current Protocols in Molecular Biology, Greene Publishing Associates and Wiley Interscience, N. Y., каждая из которых в полном объеме включена в настоящее описание в виде ссылки).
Для повышения эффективности продукции полинуклеотид может быть сконструирован таким образом, чтобы он кодировал множество элементов пептида, разделенных участками ферментативного расщепления либо гомополимеров (повторяющиеся пептидные элементы), либо гетерополимеров (различные пептиды, присоединенные друг к другу). Полученный в результате полипептид может быть расщеплен (например, путем обработки соответствующим ферментом) с тем, чтобы восстановить пептидные элементы. Это может повысить выход пептидов, находящихся под контролем одного промотора. В одном из воплощений полицистронный полинуклеотид может быть сконструирован таким образом, чтобы транскрибировалась единственная мРНК, которая кодирует множество пептидов (то есть гомополимеры или гетерополимеры), при этом чтобы каждая кодирующая область была оперативно связана с кэп-независимой последовательностью, осуществляющей контроль трансляции; например, с участком внутренней посадки рибосомы (IRES). При использовании в подходящих вирусных экспрессирующих системах трансляцию каждого пептида, кодируемого мРНК, направляют внутренне в транскрипт, например, с помощью IRES. Таким образом, полицистронная конструкция направляет транскрипцию единственной крупной полицистронной мРНК, которая, в свою очередь, направляет трансляцию множественных индивидуальных пептидов. Такой подход исключает продуцирование и ферментативный процессинг полипротеинов и может значительно повысить выход пептида, находящихся под контролем одного промотора.
Для экспрессии миметиков ApoA-I могут быть использованы различные векторные экспрессирующие системы хозяйских клеток. Таким системы включают в себя, но не ограничены перечисленным, микроорганизмы, такие как бактерии, трансформированные рекомбинантной ДНК бактериофага или плазмидной ДНК, содержащими соответствующую кодирующую последовательность; дрожжи или нитчатые грибы, трансформированные рекомбинантным дрожжевым или грибковым экспрессирующими векторами, содержащими соответствующую кодирующую последовательность; системы клеток насекомых, инфицированных векторами, экспрессирующими рекомбинантный вирус (например, бакуловирус), содержащий соответствующую кодирующую последовательность; системы клеток растений, инфицированных векторами, экспрессирующими рекомбинантный вирус (например, вирус мозаики цветной капусты), или трансформированных векторами, экспрессирующими рекомбинантную плазмиду (например, Ti-плазмиду), содержащую соответствующую кодирующую последовательность; или системы клеток животных.
Элементы экспрессии экспрессирующих систем могут варьировать по эффективности и специфичностям. В зависимости от используемой системы хозяина/вектора, любой из числа подходящих элементов транскрипции и трансляции, включая конститутивные и индуцибельные промоторы, может быть использован в экспрессирующем векторе. Например, при клонировании в бактериальные системы могут быть использованы индуцибельные промоторы, такие как pL бактериофага λ, plac, ptrp, ptac (гибридный промотор ptrp-lac) и т.п.; при клонировании в системы клеток насекомых могут быть использованы такие промоторы как бакуловирусный полиэдронный промотор; при клонировании в системы клеток растений могут быть использованы промоторы, полученные из генома клеток растений (например, промоторы [белков] теплового шока; промотор малой субъединицы RUBISCO; промотор a/b-связывающего белка хлорофилла) или из вирусов растений (например, промотор 35S-РНК вируса CaMV; промотор белка оболочки ВТМ); при клонировании в системы клеток млекопитающих могут быть использованы промоторы, полученные из генома клеток млекопитающих (например, промотор металлотионеина) или из вирусов млекопитающих (например, поздний промотор аденовируса; промотор вируса коровьей оспы в 7,5 т.н.); при создании клеточных линий, которые содержат множественные копии экспрессируемого продукта, могут быть использованы векторы на основе SV40, BPV и EBV с соответствующим селективным маркером.
В случаях, когда используются экспрессирующие векторы растений, экспрессия последовательностей, кодирующих миметики ApoA-I, может быть под управлением любого из целого ряда промоторов. Например, могут быть использованы вирусные промоторы, такие как промоторы 35S-РНК и 19S-РНК вируса CaMV (Brisson et al., 1984, Nature 310: 511-514), или промотор белка оболочки ВТМ (Takamatsu et al., 1987, EMBO J. 6: 307-311); альтернативно, могут быть использованы промоторы растений, такие как промотор малой субъединицы RUBISCO (Coruzzi et al., 1984, EMBO J. 3: 1671-1680; Broglie et al., 1984, Science 224: 838-843) или промоторы [белков] теплового шока, например, hspl7.5-E или hspl7.3-B соевых бобов (Gurley et al., 1986, Mol. Cell. Biol. 6: 559-565). Указанные конструкции могут быть встроены в клетки растений с помощью Ti-плазмид, Ri-плазмид, векторов вирусов растений, путем прямой ДНК-трансформации, микроинъекции, электропорации и т.д. В качестве обзора таких технологий см., например, Weissbach & Weissbach, 1988, Methods for Plant Molecular Biology, Academic Press, N. Y., Section VIII, pp.421-463; а также Grierson & Corey, 1988, Plant Molecular Biology, 2d Ed., Blackie, London, Ch.7-9.
В одной из экспрессирующих систем насекомых, которые могут быть использованы для продуцирования миметиков ApoA-I, Autographa californica, вирус ядерного полиэдроза (AcNPV) используется в качестве вектора для экспрессии чужеродных генов. Указанный вирус растет в клетках Spodoptera frugiperda. Кодирующая последовательность может быть клонирована в области, не являющиеся жизненно важными (например, ген полиэдрона) областями вируса и помещены под контроль промотора AcNPV (например, полиэдронного промотора). Успешное встраивание кодирующей последовательности приведет к инактивации гена полиэдрона и продукции неокклюдированного рекомбинантного вируса (то есть вируса, лишенного белковой оболочки, кодируемой полиэдронным геном). Затем такие рекомбинантные вирусы используются для инфицирования клеток Spodoptera frugiperda, в которых встроенный ген экспрессируется (см., например, Smith et al., 1983, J. Virol. 46: 584; Smith, патент США № 4215051). Дополнительные примеры такой системы экспрессии можно найти в Current Protocols in Molecular Biology, Vol.2, Ausubel et al., eds., Greene Publish. Assoc. & Wiley Interscience.
В хозяйских клетках млекопитающих может быть использован целый ряд экспрессирующих систем на основе вируса. В случаях, когда в качестве экспрессирующего вектора используется, кодирующая последовательность может быть лигирована с комплексом, осуществляющим контроль транскрипции/трансляции аденовируса, например, с последовательностью позднего промотора и трехкомпонентной лидерной последовательностью. Затем такой химерный ген может быть встроен в аденовирусный геном путем рекомбинации in vitro или in vivo. В результате встраивания в область, не являющуюся жизненно важной областью вирусного генома (например, в область El или E3), будет получен рекомбинантный вирус, который будет жизнеспособным и будет способен к экспрессированию пептида в инцированных хозяйских клетках (см., например, Logan & Shenk, 1984, Proc. Natl. Acad. Sci. (USA) 81: 3655-3659). Альтернативно, может быть использован промотор вакцины 7,5 К (см., например, Mackett et al., 1982, Proc. Natl. Acad. Sci. (USA) 79: 7415-7419; Mackett et al., 1984, J. Virol. 49: 857-864; Panicali et al., 1982, Proc. Natl. Acad. Sci. 79: 4927-4931).
Другие экспрессирующие системы для продуцирования миметиков ApoA-I будут очевидны для специалистов в данной области.
C. Очистка
Миметики ApoA-I могут быть очищены с использованием известных в данной области технологий, таких как хроматография с обращенной фазой, высокоэффективная жидкостная хроматография, ионообменная хроматография, гель-электрофорез, аффинная хроматография и т.п. Конкретные условия, используемые для очистки конкретного миметика ApoA-I, могут отчасти зависеть от стратегии синтеза и таких факторов как суммарный заряд, гидрофобность, гидрофильность и т.д., и также будут очевидны для специалистов в данной области. Мультимерные разветвленные пептиды могут быть очищены, например, путем ионообменной хроматографии или эксклюзионной хроматографии.
Для очистки посредством аффинной хроматографии может быть использовано любое антитело, которое специфически связывается с миметиком ApoA-I. Для продуцирования антител можно иммунизировать различных животных-хозяев, включая, но ими не ограничиваясь, кроликов, мышей, крыс и т.д., путем инъекции пептида. Пептид может быть привязан к соответствующему носителю, такому как БСА, через функциональную группу боковой цепи или посредством линкеров, прикрепленных к функциональной группе боковой цепи. Для усиления иммунологического ответа могут быть использованы различные адъюванты, в зависимости от вида хозяина, включая, но ими не ограничиваясь, адъювант Фрейнда (полный и неполный), минеральные гели, такие как гидроокись алюминия, поверхностно-активные вещества, такие как лизолецитин, поверхностно-активные полиолы, полианионы, пептиды, масляные эмульсии, гемоцианин лимфы улитки, динитрофенол, а также потенциально полезные человеческие адъюванты, такие как БЦЖ (бацилла Кальметта-Герена) и Corynebacterium parvum.
Моноклональные антитела против миметика ApoA-I могут быть получены с использованием любой технологии, которая обеспечивает продуцирование молекул антитела стабильными клеточными линиями в культуре. Такие технологии включают в себя, но ими не ограничиваются, гибридомную технику, описанную Kohler и Milstein, 1975, Nature 256: 495-497, или Kaprowski, патент США № 4376110, который включен в настоящее описание в виде ссылки; технологию человеческих B-клеточных гибридом) Kosbor et al., 1983, Immunology Today 4: 72; Cote et al., 1983, Proc. Natl. Acad. Sci. U.S.A. 80: 2026-2030); и технологию EBV-гибридом (Cole et al., 1985, Monoclonal Antibodies and Cancer Therapy, Alan R. Liss, Inc., pp.77-96 (1985)). Кроме того, могут быть получены использованы технологии, разработанные для продуцирования "химерных антител" (Morrison et al., 1984, Proc. Natl. Acad. Sci. U.S.A. 81: 6851-6855; Neuberger et al., 1984, Nature 312: 604-608; Takeda et al., 1985, Nature 314: 452-454, Boss, патент США № 4816397; Cabilly, патент США № 4816567, которые включены в настоящее описание в виде ссылки) путем сплайсинга генов молекулы мышиного антитела с подходящей антигенной специфичностью с генами молекулы человеческого антитела с подходящей биологической активностью. Или же могут быть получены "гуманизированные" антитела (см., например, Queen, патент США № 5585089, который включен в настоящее описание в виде ссылки). Альтернативно, технологии, описанные для продуцирования одноцепочечных антител (патент США № 4946778), могут быть адаптированы для продуцирования пептид-специфических одноцепочечных антител.
Фрагменты антител, которые содержат делеции специфических связывающих участков, могут быть получены с использованием известных технологий. Например, такие фрагменты включают в себя, но не ограничены F(ab')2-фрагментами, которые можно получить путем обработки пепсином молекул антитела и Fab-фрагментов, которые могут быть получены путем восстановления дисульфидных мостиков F(ab')2-фрагментов. Альтернативно, могут быть сконструированы экспрессирующие Fab библиотеки (Huse et al., 1989, Science 246: 1275-1281), позволяющие быстро и с легкостью идентифицировать моноклональные Fab-фрагменты с требуемой специфичностью представляющего интерес пептида.
Антитело или фрагмент антитела, специфичный в отношении требуемого миметика ApoA-I, может быть присоединен, например, к агарозе, и комплекс антитело-агароза использован в иммунохроматографии для очистки пептидов согласно изобретению. См. Scopes, 1984, Protein Purification: Principles and Practice, Springer-Verlag New York, Inc., N. Y., Livingstone, 1974, Methods In Enzymology: Immunoaffinity Chromatography of Proteins 34: 723-731.
IV. Композиции
В одном из воплощений настоящее изобретение связано с композициями, содержащими эффективное количество миметика ApoA-I и фармацевтически приемлемого носителя или наполнителя.
Композиции могут быть составлены для введения млекопитающему путем инъекции. Инъецируемые препараты включают в себя стерильные суспензии, растворы или эмульсии активного ингредиента в водном или масляном носителях. Композиции могут также включать в себя рецептурные агенты, такие как суспендирующий, стабилизирующий и/или диспергирующий агенты. Композиции для инъекции могут быть представлены в единичной дозированной форме, например, в ампулах, или в контейнерах для многократных доз, и могут содержать добавочные консерванты. Альтернативно, композиция для инъекции может быть представлена в виде порошка, предусматривающая перед применением восстановление подходящим носителем, включая, но не ограничиваясь стерильной апирогенной водой, буфером, раствором декстрозы и т.д. Для таких целей миметик ApoA-I может быть лиофилизирован, или же может быть получен со-лиофилизированный пептид-липидный комплекс. Предназначенные для хранения препараты могут поставляться в единичных дозированных формах и восстанавливаться перед применением in vivo.
Для пролонгированной доставки указанная композиция может быть составлена в виде препарата пролонгированного действия для введения путем имплантации; например, путем подкожной, внутрикожной или внутримышечной инъекции. Таким образом, например, миметик ApoA-I может быть составлен с соответствующими полимерными или гидрофобными веществами (например, в виде эмульсии в приемлемом масле) или ионообменными смолами, или же в виде медленно растворимых производных; например, в виде медленно растворимой солевой формы миметика ApoA-I.
В другом воплощении, композиции вводят внутривенно. Альтернативно, могут быть использованы системы трансдермальной доставки для чрескожной абсорбции, производимые в виде адгезивного диска или пластыря, из которых медленно высвобождается активный ингредиент. С этой целью могут быть использованы усилители проницаемости для облегчения трансдермального проникновения миметика ApoA-I. Благоприятное воздействие может быть достигнуто, в частности, путем включения миметика ApoA-I в нитроглицериновый пластырь для применения в отношении млекопитающего, имеющего такое Состояние, как ишемическая болезнь сердца или гиперхолестеринемия.
Для перорального введения композиции могут быть представлены, например, в виде таблеток или капсул, полученных стандартным способом в составе с фармацевтически приемлемыми эксципиентами, такими как связующие средства (например, прежелатинизированный кукурузный крахмал, поливинилпирролидон или гидроксипропилметилцеллюлоза); наполнители (например, лактоза, микрокристаллическая целлюлоза или дикальций-фосфат); смазывающие вещества (например, стеарат магния, тальк или двуокись кремния); разрыхлители (например, картофельный крахмал или натрия гликолят крахмал); или увлажнители (например, лаурилсульфат натрия). Таблетки могут быть покрыты способами, хорошо известными в данной области. Жидкие препараты для перорального введения могут быть представлены, например, в виде растворов, сиропов или суспензий, или же они могут быть представлены в виде сухого продукта для его составления с водой или другим подходящим носителем перед применением. Такие жидкие препараты могут быть получены стандартными способами с фармацевтически приемлемыми добавками, такими как суспендирующие средства (например, сироп сорбита, производные целлюлозы или гидрогенизированные пищевые жиры); эмульгирующие средства (например, лецитин или акация); неводные носители (например, миндальное масло, жирные сложные эфиры, этиловый спирт или фракционированные овощные масла); и консерванты (например, метил или пропил-пара-гидроксибензоаты или сорбиновая кислота). Такие препараты могут, если это желательно, содержать также буферные соли, ароматизаторы, красители и подсластители. Препараты для перорального введения могут быть составлены таким образом, чтобы было обеспечено контролируемое высвобождение миметика ApoA-I.
Для буккального введения композиции могут быть представлены в виде таблеток или леденцов, составленных стандартным способом. Для ректального и вагинального путей введения активный ингредиент может быть составлен в виде растворов (для удерживающих клизм), суппозиториев или мазей.
Для введения путем ингаляции миметик ApoA-I удобно доставлять в виде аэрозольного спрея из герметичных упаковок или небулайзера, с применением подходящего пропеллента, например, дихлордифторметана, трихлорфторметана, дихлортетрафторэтана, двуокиси углерода или другого подходящего газа. В случае находящегося под давлением аэрозоля дозированную единицу можно определить, если использовать клапан для доставки отмеренного количества. Капсулы и картриджи, например, желатиновые, для использования в ингаляторе или инсуффляторе могут быть составлены таким образом, чтобы они содержали порошковую смесь миметика ApoA-I и подходящей порошковой основы, такой как лактоза или крахмал.
Композиции, если это желательно, могут быть представлены в упаковке или в распылительном устройстве, которое может содержать одну или более единиц дозированных форм, содержащих миметик ApoA-I. Такая упаковка может содержать, например, металлическую или пластиковую пленку, такую как в блистерной упаковке. Указанные упаковка или распылительное устройство могут быть снабжены инструкциями по применению.
В определенных воплощениях миметик ApoA-I можно составить или ввести в виде комплекса с липидом. Соответственно, настоящее изобретение включает в себя миметик ApoA-I/липидные комплексы, содержащие их композиции и способы их введения. Указанные комплексы могут иметь несколько преимуществ, поскольку они могут иметь увеличенное время полужизни в циркуляторном русле, в частности, тогда, когда такой комплекс имеет плотность и размер, сходные с ЛПВП, и в особенности с популяциями пре-β-1 или пре-β-2 ЛПВП. Такие комплексы удобно получать любым из целого ряда описанных ниже способов. Стабильные препараты, имеющие относительно длительный срок хранения, могут быть получены путем лиофилизации, при этом описанный ниже способ солиофилизации приведен здесь лишь в качестве одного из воплощений. Лиофилизированные комплексы могут быть использованы для получения основной массы материала для составления фармацевтических дозированных форм или для получения индивидуальных аликвот или дозированных форм, которые могут быть восстановлены путем регидратации стерильной водой или соответствующим буферным раствором перед введением субъекту.
Для получения указанных комплексов могут быть использованы различные способы, хорошо знакомые специалистам в данной области. Например, может быть использован целый ряд доступных технологий получения липосом или протеолипосом. Например, для образования комплекса миметик ApoA-I может быть подвергнут действию ультразвука (с использованием ванночки или ультразвукового аппарата) совместно с соответствующими липидами. Альтернативно, миметик ApoA-I может быть скомбинирован с заранее составленными липидными везикулами с получением спонтанно составленных пептид-липидных комплексов. Еще в одном альтернативном воплощении указанные комплексы могут быть образованы путем детергентного диализа; например, смесь миметика ApoA-I, липида и детергента подвергают диализу для удаления детергента и восстановления или образования комплексов (см., например, Jonas et al., 1986, Methods in Enzymol. 128: 553-582).
Альтернативно, указанные комплексы могут быть получены способами, описанными в патенте США № 6004925 ("патент '925"), полное описание которого включено в настоящее описание в виде ссылки. В способах, описанных в патенте '925, миметик ApoA-I и липид скомбинированы в системе растворителя, в которой совместно растворяется каждый ингредиент и которая может быть полностью удалена путем лиофилизации. С этой целью выбирают пару растворителей для обеспечения совместной растворимости как миметика ApoA-I, так и липида. В одном из воплощений миметик ApoA-I такого комплекса может быть растворен в водном или органическом растворителе или смеси растворителей (растворитель 1). Липидный, такой как фосфолипидный, компонент растворяют в водном или органическом растворителе или в смеси растворителей (растворитель 2), которые смешивается с растворителем 1, и два указанных раствора смешивают друг с другом. Альтернативно, миметик ApoA-I и липид могут быть встроены в систему совместных растворителей; то есть в смесь смешивающихся растворителей. Альтернативно, миметик ApoA-I и липид могут быть суспендированы в растворителе или в смеси растворителей. В одном из воплощений указанная смесь растворителей является смесью органического растворителя и воды. Примеры органических растворителей включают в себя, не ограничиваясь перечисленным, уксусную кислоту, ксилен, циклогексан и метанол. Примеры смесей растворителей включают в себя, не ограничиваясь перечисленным, уксусную кислоту и ксилен, уксусную кислоту и циклогексан, а также метанол и ксилен. Подходящее соотношение миметика ApoA-I и липидов может быть прежде всего определено эмпирически, так, чтобы полученные в результате комплексы обладали подходящими физическими и химическими свойствами; то есть чтобы обычно (но необязательно) они были сходными по размеру с ЛПВП. Полученную в результате смесь замораживают и лиофилизируют досуха. Иногда для облегчения лиофилизации к смеси добавляют дополнительный растворитель. Такой лиофилизированный продукт можно хранить в течение длительных периодов, и обычно он остается при этом стабильным.
Альтернативно, указанные комплексы могут быть получены путем совместной лиофилизации миметика ApoA-I с пептидом в растворах или суспензиях. Выбранный гомогенный раствор пептида и фосфолипидов в органическом растворителе или в смеси органических растворителей может быть лиофилизирован, и пептид/фосфолипидные комплексы могут быть образованы спонтанно в результате гидратации лиофилизированного порошка водным буфером.
Лиофилизированный продукт может быть восстановлен с целью получения раствора или суспензии комплекса. Для этого лиофилизированный порошок подвергают повторной гидратации водным раствором до нужного объема (часто 5-20 мг пептида/мл, что является обычным для внутривенной инъекции). В одном из воплощений лиофилизированный порошок подвергают повторной гидратации забуференным фосфатом физиологическим раствором, двууглекислым физиологическим раствором или же физиологическим раствором. Показатель pH указанной смеси может быть доведен до 7,5-8,5. Смесь можно встряхивать или перемешивать на вортексе для облегчения регидратации, и в большинстве случаев стадию восстановления можно проводить при температуре, равной или превышающей температуру фазового перехода липидного компонента комплекса. В течение минут получают препарат восстановленных липид-белковых комплексов.
Можно определить свойства аликвоты полученного в результате восстановленного препарата для подтверждения того, что комплексы в составе препарата имеют требуемую классификацию по размерам; например, классификацию по размерам ЛПВП. Определение свойств восстановленного препарата может быть предпринято с использованием любого способа, известного в данной области, включая, но не ограничиваясь фильтрационной эксклюзионной по размеру хроматографией, гель-фильтрационной хроматографией, фильтрационной колоночной хроматографией, гель-проникающей хроматографией и нативным ПААГ-электрофорезом. В одном из воплощений свойства восстановленного препарата определяют путем гель-фильтрационной хроматографии. Размер полученных в результате комплексов может быть определен по их эффективности. В описанных ниже примерах используется система гель-фильтрационной хроматографии Pharmacia Superose 6 FPLC. Буфер, который используется, содержит 150 мМ NaCl в 50 мМ фосфатном буфере, pH приблизительно от 7,0 примерно до 9, в одном из воплощений, 7,5-8,5, в другом воплощении - 7,4. Типичный объем образца составляет от 20 до 200 микролитров комплексов, содержащих 5 мг пептида/мл. Скорость протекания колонки составляет 0,5 мл/мин. Серия белков с известной молекулярной массой и диаметром Стокса, а также человеческие ЛПВП используются в качестве стандартов для калибровки колонки комплексов белков и липопротеинов в отношении поглощения или рассеяния света с длиной волны 254 или 280 нм.
Восстановленный препарат также может быть охарактеризован в отношении определения концентрации, конечного значения pH и осмоляльности полученного раствора, а также концентрации и целостности пептида и индивидуальных липидов. Концентрация миметика ApoA-I и липида комплексов может быть измерена любым из способов, известных в данной области, включая, но не ограничиваясь белковым и фосфолипидным анализами, а также хроматографическими методами, такими как высокоэффективная жидкостная хроматография ("ВЭЖХ"), гель-фильтрационная хроматография, газовая хроматография ("ГХ"). Хроматографы могут быть соединены с различными детекторами, включая, но не ограничиваясь детекторами масс-спектрометрии, УФ или диодной матрицы, флуоресценции и эластическими детекторами светового рассеяния. Целостность миметика ApoA-I и липида в указанных комплексах может быть определена описанной выше хроматографической технологии, а также путем аминокислотного анализа, с помощью тонкослойной хроматографии и стандартных анализов по определению окисления липидов.
Липид миметик ApoA-I/липидного комплекса может представлять собой один или несколько различных липидов включая, но не ограничиваясь, насыщенными, ненасыщенными, природными и синтетическими липидами и фосфолипидами, а также их фармацевтически приемлемыми солями. Типичные соли включают в себя, но не ограничиваются натриевой, кальциевой, магниевой и калиевой солями.
Подходящие липиды комплексов миметик ApoA-I/липид включают в себя, но не ограничены перечисленным, фосфолипиды (C1-C6)-алкильной цепи, фосфатидилхолин (ФХ), яичный фосфатидилхолин, фосфатидилхолин соевых бобов, дипальмитоилфосфатидилхолин, димиристоилфосфатидилхолин, дистеароилфосфатидилхолин-1-миристоил-2-пальмитоилфосфатидилхолин, 1-пальмитоил-2-миристоилфосфатидилхолин, 1-пальмитоил-2-стеароилфосфатидилхолин, 1-стеароил-2-пальмитоилфосфатидилхолин, 1-пальмитоил-2-олеоилфосфатидилхолин, 1-олеоил-2-пальмитилфосфатидилхолин, диолеоилфосфатидилхолин, диолеоилфосфатидилэтаноламин, дилауроилфосфатидилглицеринфосфатидилхолин, фосфатидилсерин, фосфатидилэтаноламин, фосфатидилинозит, сфингомиелин, сфинголипиды, фосфатидилглицерин, дифосфатидилглицерин, димиристоилфосфатидилглицерин, дипальмитоилфосфатидилглицерин, дистеароилфосфатидилглицерин, диолеоилфосфатидилглицерин, димиристоилфосфатидную кислоту, дипальмитоилфосфатидную кислоту, димиристоилфосфатидилэтаноламин, дипальмитоилфосфатидилэтаноламин, димиристоилфосфатидилсерин, дипальмитоилфосфатидилсерин, фосфатидилсерин мозга, сфингомиелин, сфингомиелин мозга, дипальмитоилсфингомиелин, дистеароилсфингомиелин, фосфатидную кислоту, галактоцереброзид, ганглиозиды, цереброзиды, дилауроилфосфатидилхолин, (1,3)-D-маннозил-(1,3)диглицерид, аминофенилгликозид, 3-холестерил-6'-(гликозилтио)гексилэфир-гликолипиды и холестерин, а также их производные.
В одном из воплощений липид миметик ApoA-I/липидного комплекса является нейтральным фосфолипидом. Нейтральный фосфолипид может быть любым фосфолипидом, который имеет суммарный заряд, приблизительно равный нулю при физиологическом pH. В определенных воплощениях нейтральный фосфолипид является цвиттерионом, который имеет суммарный заряд, приблизительно равный нулю при физиологическом pH.
В другом воплощении нейтральный фосфолипид представляет собой лецитин (известный также как фосфатидилхолин). В определенных воплощениях нейтральный фосфолипид представляет собой смесь нейтральных фосфолипидов, которая содержит приблизительно от 5 примерно до 100 вес.% лецитина. В других воплощениях смесь нейтральных фосфолипидов содержит приблизительно 100 вес.% лецитина. В определенных воплощениях нейтральный фосфолипид представляет собой смесь нейтральных фосфолипидов, которая содержит приблизительно от 5 примерно до 100 моль.% лецитина. В других воплощениях смесь нейтральных фосфолипидов содержит приблизительно 100 моль.% лецитина.
В другом воплощении нейтральный фосфолипид представляет собой сфингомиелин. В определенных воплощениях нейтральный фосфолипид представляет собой смесь нейтральных фосфолипидов, которая содержит приблизительно от 5 примерно до 100 вес.% сфингомиелина. В другом воплощении нейтральный фосфолипид представляет собой смесь нейтральных фосфолипидов, которая содержит приблизительно 100 вес.% сфингомиелина. В определенных воплощениях нейтральный фосфолипид представляет собой смесь нейтральных фосфолипидов, которая содержит приблизительно от 5 примерно до 100 моль.% сфингомиелина. В других воплощениях нейтральный фосфолипид представляет собой смесь нейтральных фосфолипидов, которая содержит приблизительно 100 моль.% сфингомиелина.
В другом воплощении нейтральный фосфолипид миметик ApoA-I/липидного комплекса представляет собой смесь нейтральных фосфолипидов, которая содержит лецитин и сфингомиелин. Молярное отношение лецитина к сфингомиелину может варьировать, но, как правило, изменяется в пределах приблизительно от 20: примерно к 1 и до приблизительно 1: примерно к 20. В определенных воплощениях молярное отношение лецитина к сфингомиелину изменяется в пределах приблизительно от 10: примерно к 3 и до приблизительно 10: примерно к 6. В других воплощениях молярное отношение лецитина к сфингомиелину изменяется в пределах приблизительно от 1: примерно к 20 и до приблизительно 3: примерно к 10.
В другом воплощении нейтральный фосфолипид миметик ApoA-I/липидного комплекса представляет собой смесь нейтральных фосфолипидов, которая содержит лецитин, сфингомиелин и один или несколько дополнительных нейтральных фосфолипидов. Обычно дополнительный нейтральный фосфолипид составляет приблизительно от 5 примерно до 100 вес.% смеси.
В другом воплощении липид миметик ApoA-I/липидного комплекса представляет собой заряженный фосфолипид. Подходящие заряженные фосфолипиды включают в себя, не ограничиваясь перечисленным, фосфатидилинозит, фосфатидилсерин, фосфатидилглицерин и фосфатидную кислоту.
В одном из воплощений липид миметик ApoA-I/липидного комплекса представляет собой смесь по меньшей мере одного нейтрального фосфолипида и по меньшей мере одного заряженного фосфолипида. Суммарное количество заряженного фосфолипида (фосфолипидов) в липидной смеси может варьировать, но, как правило, изменяется в пределах приблизительно от 0,2 примерно до 10 вес.% липидной смеси. В определенных воплощениях суммарное количество заряженного фосфолипида (фосфолипидов) в липидной смеси составляет приблизительно от 0,2 примерно до 2 вес.%, приблизительно от 0,2 примерно до 3 вес.%, приблизительно от 0,2 примерно до 4 вес.%, приблизительно от 0,2 примерно до 5 вес.%, приблизительно от 0,2 примерно до 6 вес.%, приблизительно от 0,2 примерно до 7 вес.%, приблизительно от 0,2 примерно до 8 вес.% или приблизительно от 0,2 примерно до 9 вес.% липидной смеси. В определенных воплощениях суммарное количество заряженного фосфолипида (фосфолипидов) в липидной смеси составляет приблизительно 0,2, приблизительно 0,3, приблизительно 0,4, приблизительно 0,5, приблизительно 0,6, приблизительно 0,7, приблизительно 0,8, приблизительно 0,9, приблизительно 1,0, приблизительно 1,1, приблизительно 1,2, приблизительно 1,3, приблизительно 1,4, приблизительно 1,5, приблизительно 1,6, приблизительно 1,7, приблизительно 1,8, приблизительно 1,9, приблизительно 2,0, приблизительно 2,1, приблизительно 2,2, приблизительно 2,3, приблизительно 2,4, приблизительно 2,5, приблизительно 2,6, приблизительно 2,7, приблизительно 2,8, приблизительно 2,9 или приблизительно 3,0, приблизительно 4, приблизительно 5, приблизительно 6, приблизительно 7, приблизительно 8, приблизительно 9 или приблизительно 10 вес.% липидной смеси. Суммарное количество нейтрального фосфолипида (фосфолипидов) в липидной смеси также может варьировать и может зависеть от количества заряженного фосфолипида (фосфолипидов) и любых других липидов, включенных в ее состав. В одном из воплощений суммарное количество нейтрального фосфолипида (фосфолипидов) в липидной смеси приблизительно составляет приблизительно от 90 примерно до 99,8 вес.% липидной смеси. В одном из воплощений липид миметик ApoA-I/липидного комплекса представляет собой смесь сфингомиелина и заряженного фосфолипида. В другом воплощении липид миметик ApoA-I/липидного комплекса представляет собой смесь сфингомиелина, дипальмитоилфосфатидилхолина ("DPPC") и заряженного фосфолипида.
В одном из воплощений липид миметик ApoA-I/липидного комплекса представляет собой сфингомиелин. В другом воплощении сфингомиелин получен из молока, яиц или мозга, или же получен синтетическим путем. В другом воплощении липид миметик ApoA-I/липидного комплекса представляет собой аналог или производное сфингомиелина. Подходящие аналоги или производные сфингомиелина включают в себя, но не ограничены перечисленным, альмитоилсфингомиелин, стеароилсфингомиелин, D-эритроза-сфингомиелин и D-эритроза-дигидросфингомиелин.
В другом воплощении сфингомиелин является искусственно обогащенным в одной конкретной насыщенной или ненасыщенной ацильной цепи. Например, сфингомиелин молока (Avanti Phospholipid, Alabaster, Ala.) имеет длинные насыщенные ацильные цепи. Сфингомиелин молока содержит приблизительно 20% C16:0 (углерод 16, насыщенный) ацильной цепи по сравнению с яичным сфингомиелином, который содержит 80% C16:0. С использованием экстракции растворителя можно обогатить сфингомиелин молока в конкретной ацильной цепи с получением композиции, имеющей концентрацию ацильной цепи, сравнимую, например, с яичным сфингомиелином. Ацильные цепи, которые могут быть использованы в настоящем изобретении, включают в себя, не ограничиваясь перечисленным, насыщенные ацильные цепи (такие как дипальмитоильные, дистеароильные, диарахидонильные и дибензоильные ацильные цепи), ненасыщенные цепи (такие как диолкоильные цепи), смешанные цепи насыщенных и ненасыщенных ацильных цепей (такие как пальмитоильные или олеоильные цепи), насыщенные и/или ненасыщенные цепи смешанных длин, а также эфирные аналоги насыщенных и ненасыщенных ацильных цепей.
Сфингомиелин может быть полусинтетическим, так, чтобы он имел конкретную ацильную цепь. Например, сфингомиелин молока может прежде всего быть очищен из молока, затем одна конкретная ацильная цепь, например, ацильная цепь C16:0, может быть расщеплена и заменена другой ацильной цепью (такой как пальмитиновая кислота или олеиновая кислота).
Сфингомиелин может быть также полностью синтезирован, например, путем крупномасштабного синтеза. См., например, Dong et al., патент США № 5220043; Weis, 1999, Chem. Phys. Lipids 102(1-2): 3-12. В одном из воплощений предопределенный уровень насыщения и состав жирных кислот выбирают из синтетического сфингомиелина.
В другом воплощении липид миметик ApoA-I/липидного комплекса представляет собой смесь сфингомиелина и другого липида. В этом воплощении сфингомиелин, как правило, составляет приблизительно от 25 примерно до 75 вес.% указанной смеси.
В другом воплощении, липид миметик ApoA-I/липидного комплекса представляет собой смесь сфингомиелина и DPPC. В другом воплощении липид миметик ApoA-I/липидного комплекса представляет собой смесь сфингомиелина, DPPC и дипальмитоилфосфатидилглицерина ("DPPG"). В одном из воплощений DPPG присутствует в количестве приблизительно от 0 примерно до 10 моль.% или вес.% смеси. В другом воплощении DPPG присутствует в количестве приблизительно от 2 примерно до 4 моль или вес.% смеси. В другом воплощении сфингомиелин и DPPG присутствуют в смеси в весовом или молярном отношении приблизительно 1: примерно к 1. В другом воплощении сфингомиелин, DPPC и DPPG присутствуют в смеси в весовом или молярном отношении, соответственно, приблизительно 1: примерно к 1: приблизительно к 0,06. В другом воплощении сфингомиелин, DPPC и DPPG присутствуют в смеси в молярном отношении, соответственно, 1,04:1:0,061. В другом воплощении сфингомиелин, DPPC и DPPG присутствуют в смеси в весовом отношении, соответственно, 1:1:0,062. В другом воплощении смесь содержит приблизительно 48,5 моль или вес.% сфингомиелина, приблизительно 48,5 моль или вес.% DPPC и приблизительно 3 молярных или весовых % DPPG.
В другом воплощении комплекс миметик ApoA-I/липидный комплекс содержит один или несколько дополнительных пептидов. В одном из воплощений дополнительным пептидом является ApoA-I.
В одном из воплощений весовое отношение суммарного количества пептида к липиду в каждом миметик АроА-I/липидном комплексе составляет от приблизительно 1: примерно к 0,5 до приблизительно 1: примерно к 5. В другом воплощении весовое отношение суммарного количества пептида к липиду в каждом миметик ApoA-I/липидном комплексе составляет от приблизительно 1: примерно к 1 до приблизительно 1: примерно к 5. В другом воплощении весовое отношение суммарного количества пептида к липиду в каждом миметик АроА-I/липидном комплексе составляет от приблизительно 1: примерно к 2 до приблизительно 1: примерно к 5. В другом воплощении весовое отношение суммарного количества пептида к липиду в каждом миметик АроА-I/липидном комплексе составляет приблизительно 1: примерно к 2,5. В другом воплощении весовое отношение суммарного количества пептида к липиду в каждом миметик АроА-I/липидном комплексе составляет от приблизительно 1: примерно к 3 до 1: приблизительно к 5. В другом воплощении молярное отношение суммарного количества пептида к липиду в каждом миметик АроА-I/липидном комплексе составляет от приблизительно 1: примерно к 2,5 до приблизительно 1: примерно к 20. В другом воплощении молярное отношение суммарного количества пептида к липиду в каждом миметик АроА-I/липидном комплексе составляет приблизительно 1: примерно к 9,2.
Когда липид миметик ApoA-I/липидного комплекса является смесью сфингомиелина, DPPC и DPPG, весовое отношение пептид: сфингомиелин: DPPC: DPPG обычно составляет, соответственно, приблизительно 1: приблизительно 1: приблизительно 1: приблизительно 0,08. В одном из воплощений весовое отношение пептид:сфингомиелин:DPPC:DPPG составляет, соответственно, 1:1,2125:1,2125:0,075. Молярное отношение пептид: сфингомиелин:DPPC:DPPG обычно составляет, соответственно, приблизительно 1: приблизительно 4: приблизительно 4: приблизительно 0,03. В одном из воплощений молярное отношение пептид: сфингомиелин:DPPC:DPPG составляет, соответственно, 1:4,55:4,36:0,27.
В другом воплощении, миметик ApoA-I/липидный комплекс содержит приблизительно от 40 примерно до 85 вес.% липида и приблизительно от 15 примерно до 60 вес.% пептида.
В другом воплощении каждый миметик ApoA-I/липидный комплекс составляет приблизительно от 2 примерно до 12 нм в диаметре.
V. Способы лечения или предупреждения состояния
Хоть авторы изобретения и не желают связывать себя какой бы то ни было конкретной теорией, тем не менее они полагают, что спираль, образуемая миметиками ApoA-I согласно изобретению, точно имитирует структурные и функциональные свойства амфипатических спиральных областей нативного ApoA-I, которые важны для осуществления активации связывания липида, оттока холестерина и/или активации LCAT, таким образом приводя к образованию пептидов, которые проявляют высокую ApoA-I-подобную активность. В одном из воплощений миметики ApoA-I функционируют путем образования амфипатических спиралей (в присутствии липидов), связывания липидов, образования пре-β-подобных или ЛПВП-подобных комплексов, активации LCAT, повышения концентрации ЛПВП в сыворотке и активации оттока холестерина.
В одном из воплощений миметики ApoA-I активируют LCAT. В другом воплощении миметики ApoA-I не активируют LCAT. В другом воплощении миметики ApoA-I активируют LCAT, но только в такой степени, которая не приводит к ускорению этерификации холестерина. В другом воплощении миметики ApoA-I активируют LCAT и, таким образом, ускоряют этерификацию холестерина, но при этом ускорение этерификации холестерина только за счет активации LCAT является недостаточным для лечения или предупреждения состояния.
В одном из воплощений настоящее изобретение связано со способами лечения или предотвращения состояния, предусматривающими введение млекопитающему, в случае необходимости, эффективного количества миметика ApoA-I.
Примеры дислипидемии включают в себя любое расстройство, при котором повышение концентрации ЛПВП в сыворотке, активации LCAT и активация оттока холестерина и RCT являются благотворными. Такие расстройства включают в себя, но не ограничены перечисленным, гиперпротеинемию (такую как гиперхиломикронемия), высокую концентрацию ЛПНП в сыворотке, высокую концентрацию ЛПОНП в сыворотке, гиперлипидемию (такую как гиперхолестеринемия или гиперглицеридемия (такую как гипертриглицеридемия)), низкую концентрацию ЛПВП в сыворотке, гипохолестеринемию, A-бета-липопротеинемию, дефицит ApoA-I и болезнь Танжера.
Примеры сердечно-сосудистых заболеваний включают в себя, но не ограничены перечисленным, метаболический синдром, ишемическую болезнь сердца, атеросклероз, рестеноз (например, профилактика или лечение атеросклеротических бляшек, которые развиваются в результате медицинских процедур, таких как баллонная ангиопластика), эндотоксемию (которая часто является результатом септического шока), застойную сердечную недостаточность (такую как хроническая или острая сердечная недостаточность), циркуляторный шок, кардиомиопатию, пересадку сердца, инфаркт миокарда, сердечную аритмию (такую как фибрилляция предсердия), суправентрикулярную тахикардию, трепетание предсердий, пароксизмальную предсердную тахикардию, аневризму, стенокардию, острое нарушение мозгового кровообращения (инсульт), болезнь периферических сосудов, цереброваскулярную болезнь, болезнь почек, атерогенез, атеросклероз, острый панкреатит и болезнь коронарных артерий.
Эндотелиальная дисфункция представляет собой любой дисбаланс между сосудорасширяющими и сосудосуживающими факторами и ингибирующими рост и стимулирующими рост факторами, продуцируемыми эндотелием. Эндотелиальная дисфункция обычно ослабляет способность кровеносных сосудов к расслаблению.
Примеры макрососудистых расстройств включают в себя любое расстройство крупного кровеносного сосуда. Такие расстройства включают в себя, но не ограничены перечисленным, преходящее нарушение мозгового кровообращения, инсульт, стенокардию, инфаркт миокарда, сердечную недостаточность и болезнь периферических сосудов.
Примеры микрососудистых расстройств включают в себя любое расстройство мелкого кровеносного сосуда. Такие расстройства включают в себя, но не ограничены перечисленным, диабетическую ретинопатию (непролиферативную, пролиферативную, макулярную эдему), микроальбуминурию, макроальбуминурию, конечную стадию почечного заболевания, эректильную дисфункцию, автономную невропатию, периферическую невропатию, остеомиелит и ишемию нижней конечности.
Миметики ApoA-I можно вводить отдельно или в сочетании с одним или несколькими другими средствами, которые применимы для лечения состояния. Такая терапия включает в себя, но не ограничена перечисленным, одновременное или последовательное введение рассматриваемых лекарственных средств.
В одном из воплощений способы лечения или предупреждения состояния могут дополнительно предусматривать введение одного или нескольких лекарственных средств из одного или нескольких из следующих классов: ингибиторов ACE (ангиотензинпревращающий фермент), бета-блокаторов, нитратов, блокаторов кальциевых каналов, диуретиков, тромболитических средств и средств, снижающих уровень холестерина в крови. В другом воплощении способы лечения или предупреждения состояния дополнительно предусматривают введение одного или нескольких из следующих лекарственных средств: холестирамина, колестипола, колесевелама, гемфиброзила, ципрофибрата, клофибрата, фенофибрата, безафибрата, эзетимиба, рамиприла, верапамила, никардипина, дилтиазема, карведилола, надопола, изосорбида мононитрата, пропранолола, изосорбида динитрата, дигоксина, фуросемида, метопролола тартрата, трандолаприла, нитроглицерина, амлодипина безилата, оксикодона, клопидогрела, нифедипина, атенолола, лизиноприла, ампирина и ланоксина.
Еще в одном из воплощений, способы лечения или предупреждения состояния могут дополнительно включать в себя введение одного или нескольких понижающих уровень холестерина лекарственных средств, известных специалистам в данной области; например, секвестранты желчных кислот, ниацин, и/или статины, такие как аторвастатин, симвастатин, правастатин, флувастатин и питавастатин. Такая схема лечения может вызвать особенно благотворные терапевтические эффекты, поскольку каждое лекарственное средство действует на разные мишени в системах синтеза и транспорта холестерина; то есть секвестранты желчных кислот действуют на рециркуляцию холестерина, на популяцию хиломикронов и ЛПНП; ниацин прежде всего действует на популяцию ЛПОНП и ЛПНП; статины ингибируют синтез холестерина, снижая уровень популяции ЛПНП (и, возможно, повышая экспрессию рецептора ЛПНП); тогда как миметики ApoA-I действуют на RCT, повышают уровень ЛПВП, повышают активность LCAT и активность оттока холестерина.
В другом воплощении способы лечения или предупреждения состояния могут дополнительно включать в себя введение фибрата, такого как клинофибрат, симбифибрат, фенофибрат и бензафибрат.
Еще в одном из воплощений, способы лечения или предупреждения состояния могут дополнительно включать в себя введение противомикробного средства и/или противовоспалительного средства, например, такого, которое используют для лечения септического шока, вызванного эндотоксином.
Миметики ApoA-I можно вводить любым из подходящих путей, которые обеспечивают биодоступность в системе кровообращения. Этого можно достичь парентеральными путями введения, включая внутривенную (в/в), внутримышечную (в/м), внутрикожную, подкожную (п/к) и внутрибрюшинную (в/б) инъекции. Однако могут быть использованы также и другие пути введения. Например, поглощение через желудочно-кишечный тракт может быть осуществлено через пероральный путь введения (включая, без ограничения, глотательный, буккальный и сублингвальный пути), при условии, что используются соответствующие композиции (например, с энтерическими покрытиями), чтобы избежать или минимизировать деградацию пептидов, например, в жестких условиях слизистой рта, желудка и/или тонкого кишечника. Альтернативно, может быть использовано введение через ткани слизистой, такие как вагинальный и ректальный пути введения, чтобы избежать или минимизировать деградацию в желудочно-кишечном тракте. В качестве еще одной альтернативы, композиции согласно изобретению могут быть введены чрескожно (например, трансдермально), глазным путем или путем ингаляции. Специалистам должно быть понятно, что выбранный путь введения может варьировать, в зависимости от состояния, возраста и податливости реципиента.
Фактическая применяемая доза миметика ApoA-I может варьировать в зависимости от пути введения и может быть скорректирована с целью достижения концентраций миметика ApoA-I в плазме крови, составляющих от 100 мг/л до 2 г/л. В одном из воплощений дозу миметика ApoA-I корректируют с целью достижения уровня свободного миметика ApoA-I или входящего в состав комплекса миметика ApoA-I в сыворотке по меньшей мере в течение 24 часов после введения, составляющего приблизительно на 10 мг/дл - 300 мг/дл выше, чем базовый (исходный) уровень до введения.
Миметики ApoA-I можно вводить по самым разным терапевтическими схемам. В одном из воплощений миметик ApoA-I вводят путем инъекции в дозе от 0,5 мг/кг до 100 мг/кг один раз в неделю. В другом воплощении необходимые уровни в сыворотке могут поддерживаться путем непрерывной инфузии или путем перемежающейся инфузии, обеспечивающей приблизительно от 0,5 мг/кг/час до 100 мг/кг/час миметика ApoA-I. В одном из воплощений миметик ApoA-I вводят в дозе приблизительно 20 мг/кг.
В другом воплощении миметик ApoA-I вводят путем внутривенной инъекции один или более раз в день. В другом воплощении миметик ApoA-I вводят путем инъекции по одному разу каждые 3-15 дней, по одному разу каждые 5-10 дней или по одному разу каждые 10 дней. В другом воплощении миметик ApoA-I вводят в виде серии поддерживающих инъекций, при этом указанные серии поддерживающих инъекций вводят по одному разу каждые 6 месяцев до одного года. Серии поддерживающих инъекций можно вводить, например, в течение целого дня (перфузия для поддержания определенно установленного уровня комплексов в плазме), нескольких дней (например, четыре инъекции в течение восьмидневного периода) или нескольких недель (например, четыре инъекции в течение четырехнедельного периода).
Еще в одном из воплощений можно вводить повышающиеся дозы миметика ApoA-I, начиная приблизительно с 1 до 5 доз в количестве, составляющем приблизительно от 50 мг приблизительно до 200 мг на введение, с последующими повторяющимися дозами приблизительно от 200 мг приблизительно до 1 г на введение. В зависимости от нужд пациента, введение может быть произведено путем медленной инфузии продолжительностью более одного часа, путем быстрой инфузии продолжительностью в один час или менее, или же путем однократной болюсной инъекции.
Токсичность и терапевтическая эффективность миметиков ApoA-I может быть определена с помощью стандартных фармацевтических процедур в клеточной культуре или на экспериментальных животных, с определением показателей LD50 (доза, летальная для 50% популяции) и ED50 (доза, терапевтически эффективная для 50% популяции). Отношение доз, оказывающих токсический и терапевтический эффекты, представляет собой терапевтический индекс, и он может быть выражен в виде отношения LD50/ED50. В одном из воплощений миметики ApoA-I имеют широкие терапевтические индексы.
VI. Способы анализа
Миметики ApoA-I могут быть исследованы в отношении их способности образовывать α-спирали в присутствии липидов, связываться с липидами, образовывать комплексы с липидами, активировать LCAT, активировать отток холестерина и пр.
Способы анализов для исследования структуры и/или функции миметиков ApoA-I хорошо известны в данной области. Несколько таких способов приведено ниже. Например, приведенные далее в примерах методы циркулярного, или кругового, дихроизма (CD) и ядерного магнитного резонанса (ЯМР) могут быть использованы для анализа структуры миметиков ApoA-I и, в частности, степени спиральности в присутствии липидов. Способность связываться с липидами может быть определена с помощью флуоресцентного спектроскопического анализа, описанного ниже в примерах. Способность пептидов и/или пептидных аналогов активировать LCAT может быть с легкостью определена с помощью способа активации LCAT, описанного ниже в примерах. Анализы in vitro и in vivo, описанные ниже в примерах, могут быть использованы для оценки времени полужизни, распределения, оттока холестерина и эффектов в отношении RCT.
В целом миметики ApoA-I согласно изобретению, которые проявляют свойства, перечисленные ниже в таблице 10, считаются особенно эффективными.
Способность миметика ApoA-I образовывать α-спираль в присутствии липидов может быть продемонстрирована с помощью описанного ниже метода CD. Те пептиды, которые спирализованы по меньшей мере на 40% (неблокированные пептиды, имеющие 18 или менее аминокислотных остатков) или спирализованы по меньшей мере на 60% (блокированные пептиды, имеющие 18 или менее аминокислотных остатков; неблокированные пептиды, имеющие 22 или более аминокислотных остатка) и которые связываются с липидами (в концентрации, приблизительно составляющей 5 мкМ и с липид:пептидным молярным отношением, приблизительно составляющим 30), в частности, те миметики ApoA-I, которые содержат флуоресцентный остаток Trp (W) или Nal, могут быть идентифицированы с помощью описанного далее флуоресцентного анализа. Однако для миметиков ApoA-I, которые не содержат флуоресцентных остатков, связывание с липидами наблюдается тогда, когда степень спиральности в присутствии липидов повышается.
В одном из воплощений настоящего изобретения миметики ApoA-I, в частности, те, которые проявляют свойство связывания с липидами в присутствии SUV (пептиды в 0,5-10 мкМ; с липид:пептидным молярным отношением в области от 1 до 50), подвергают скринингу на предмет определения их способности активировать LCAT, поскольку пептиды, способные активировать LCAT, являются особенно эффективными в описанных здесь способах. В одном из воплощений у миметиков ApoA-I проявляется по меньшей мере приблизительно 38% способность активировать LCAT по сравнению с нативным человеческим ApoA-I (что определяется с помощью описанного здесь анализа активации LCAT). В другом воплощении у миметиков ApoA-I проявляется по меньшей мере приблизительно 50%, 60%, 70%, 80% или даже 90% способность активировать LCAT по сравнению с нативным человеческим ApoA-I.
VII. Прочие применения
Миметики ApoA-I используются в анализах in vitro для определения сывороточных ЛПВП, например, в диагностических целях. Поскольку обычно миметики ApoA-I ассоциированы с ЛПВП-компонентом сыворотки, такие миметики могут быть использованы в качестве "маркеров" для популяции ЛПВП. Соответственно, настоящее изобретение связано также со способами измерения сывороточной концентрации ЛПВП, предусматривающими приведение сывороточных ЛПВП в контакт с количеством миметика ApoA-I, которое ассоциировано с сывороточными ЛПВП, и расчет количества ApoA-I-ассоциированного ЛПВП. Более того, миметики ApoA-I могут быть использованы в качестве "маркеров" для субпопуляции ЛПВП, которая эффективна в обратном транспорте холестерина ("RCT"). С этой целью миметик ApoA-I может быть добавлен или смешан с образцом сыворотки млекопитающего, содержащим ЛПВП; после соответствующего времени инкубации компонент ЛПВП может быть исследован путем детектирования встроенного миметика ApoA-I. Это может быть осуществлено с помощью меченого миметика ApoA-I (например, радиоактивными метками, флуоресцентными метками, ферментативными метками, красителями и т.д.) или же с помощью иммуноанализов с использованием антител (или фрагментов антител), специфичных в отношении миметика ApoA-I.
Альтернативно, меченые миметики ApoA-I используются в процедурах визуализации (например, CAT-сканы, MRI-сканы) для визуализации системы циркуляции или для мониторинга RCT, или с целью визуализации накопления ЛПВП в жировых полосках, в очагах атеросклеротических повреждений и т.д. (там, где ЛПВП должны быть активны в оттоке холестерина).
Далее настоящее изобретение включает в себя следующие неограничивающие иллюстративные примеры.
Примеры
Пример 1
Синтез миметиков ApoA-I
Миметики ApoA-I получают путем твердофазного пептидного синтеза (SPPS) с использованием химии Fmoc(9-фторенилметилоксикарбонила). C-концевой остаток ковалентно связывают с 4-метилбензгидриламиновой (MBHA) смолой. Затем присоединяют остальные аминокислотные остатки путем циклически повторяющегося удаления защитной группы Fmoc и присоединения защищенной аминогруппы. После твердофазной сборки пептида этот пептид отделяют от смолы трифторуксусной кислотой (ТФУ). Неочищенный пептид получали путем преципитации и сушки. Идентичность неочищенного пептида подтверждали масс-спектроскопически и путем аминокислотного анализа.
Пример 2
Очистка миметиков ApoA-I
Очистку миметиков ApoA-I предпринимали в соответствии с примером 1 путем препаративной обращеннофазной ВЭЖХ со стационарной фазой C18 (привитой диоксид кремния, размер частиц 15 мкм, размер пор 120 Å) с использованием градиента вода/ацетонитрил (0,1% противоион ТФУ). Элюирующиеся фракции детектировали по УФ-поглощению при 220 нм. В результате каждого прогона производится приблизительно 15 г неочищенного пептида, очищенные фракции собираются и концентрируются на роторном испарителе. Затем раствор пептида очищают на колонке C18 ВЭЖХ, используемой на первой стадии очистки. После этого раствор пептида концентрируют на ротороном испарителе для удаления ацетонитрила и лиофилизируют.
Далее, лиофилизированный пептидный порошок вновь солюбилизируют в смеси 90% воды/10% ацетонитрила, и в процессе ионообменной хроматографии противоион обменивается на ацетат (смола Dowex, элюирующая среда 90% воды/10% ацетонитрила). Очищенный пептид с ацетатным противоионом фильтруют через стерильную мембрану с диаметром пор 0,22 микрометра и лиофилизируют.
Пример 3
Синтез пептида 16
Пептид 16 был синтезирован на твердофазной подложке с использованием химии Fmoc(9-фторенилметилоксикарбонила). C-концевой изонипекотинильный остаток был ковалентно связан со смолой линкером типа линкера Ванга (4-метилбензгидриламиновой (MBHA) смолой). Используемыми защитными группами аминокислот были следующие: трет-бутильная группа в случае Glu и Asp, Boc-группа в случае Lys, Pbf-группа в случае Arg, Trt-группа в случае Asn и Gln.
Твердофазную сборку пептида осуществляли вручную в реакторе 601, снабженном пористым диском, механической мешалкой и устройством для барботирования азотом. Смолу, пара-метилбензгидриламиновую смолу (полистирол-l%-дивинилбензол), размачивали и промывали дихлорметан (DCM)/диметилформамидом (ДМФ) (95/5). Встраивание C-концевого остатка достигалось путем связывания C-концевой изонипекотиновой кислоты, этерифицированной на линкере MPPA (линкер типа линкера Ванга). Реакцию связывания проводили с 1,35 экв. Fmoc-Inp-MPPA-линкера, 1,35 экв. N-гидроксибензотриазола (HOBt), бензотриазол-1-ил-окситрипирролидинофосфония гексафторфосфата (PyBOP) и 4,05 экв. диизопропилэтиламина (DIEA) в ДМФ/ДХМ(50/50). После связывания смолу 3 раза промывали ДМФ.
Пептидную цепочку собирали на смоле путем циклически повторяющегося удаления защитной группы Fmoc и присоединения защищенной аминогруппы. Все аминокислотные остатки присоединяли, следуя одному и тому же циклу: сначала защитную группу Fmoc удаляли в пиперидине (35% в ДМФ) в трех повторных циклах. (Реакция снятия защиты Fmoc занимала приблизительно 16 мин). После удаления защитной группы Fmoc смолу в девяти повторных циклах промывали ДМФ. Затем Fmoc-защищенные аминокислотные остатки (2 экв.) соединяли с 2 экв. N,N'- диизопропилкарбодиимида (DIC)/HOBt в смеси ДМФ и ДХМ (50/50). (Стадия связывания занимала приблизительно от одного часа до целого дня). Для того, чтобы определить, окончена ли реакция связывания, использовали нингидриновый тест. Если нингидриновый тест указывает на то, что реакция связывания не завершена, стадию связывания повторяли с меньшим избытком (0,5-1 экв.) аминокислоты, PYBOP, HOBt в ДМФ/ДХМ и DIEA. После стадии связывания смолу промывали ДМФ в трех повторных циклах.
Затем пептид отделяли от смолы и убирали защитные группы. Отделение от смолы и снятие защиты осуществляли партиями в смеси ТФУ/вода/анизол (90/5/5 об./об./об.) при концентрации 5 мл/г пептида по отношению к смоле в течение 2,5 часов при комнатной температуре. В процессе постепенного добавления смолы к смеси реагентов температуру регулировали таким образом, чтобы она оставалась ниже 25°C. Пептид был растворим в ТФУ и экстрагировался от смолы путем фильтрации через пористый диск. После концентрирования на роторном испарителе пептид подвергали преципитации в холодном метил-трет-бутиловом эфире (MTBE) (0±5°C) и фильтровали. Неочищенный пептид промывали MTBE и сушили при пониженном давлении в печи при 30°C.
После удаления последней защитной группы Fmoc пептид обрабатывали ТФУ/H2O для отщепления и удаления защитных групп боковых цепей. Затем неочищенный пептид осаждали из холодного эфира и собирали путем фильтрации.
Пример 4
Очистка пептида 16
Пептид 16, полученный в соответствии с примером 3, очищали путем препаративной обращеннофазной ВЭЖХ (высокоэффективная жидкостная хроматография) со стационарной фазой C18 с использованием градиента вода/ацетонитрил (противоион ТФУ). Колонку Prochrom LC110 набивали новой или специализированной стационарной фазой C18 (привитой диоксид кремния, размер частиц 15 мкм, размер пор 120 ангстрем). Набивку колонки контролировали с помощью SST для определенного числа слоев и коэффициента асимметрии пика.
На колонке Prochrom LC110 количество пептида, инъецируемого на каждый пробег, приблизительно составляло 15 г неочищенного пептида, растворенного в воде/ацетонитриле (80/20) при концентрации, приблизительно составляющей 75 г/л. Через колонку пропускали градиент буфера B в буфере A (со скоростью тока приблизительно в 450 мл/мин и УФ-детектированием при 220 нм): буфер A=0,1% ТФУ в воде; буфер B = ацетонитрил/0,1% ТФУ в воде (80/20), при следующих условиях:
при скорости тока 1 мл/мин
Элюируемые фракции анализировали путем аналитической ВЭЖХ и собирали в составе четырех категорий: обедненной, передней примесной, чистой и задней примесной, в соответствии с заранее установленными стандартами. Установленные стандарты для чистоты в процессе выполнения ВЭЖХ для классификации фракций по категориям являются следующими:
Для обеспечения большего выхода продукта примесные фракции, примыкающие к чистым (переднюю примесную и заднюю примесную) подвергали повторному пропусканию через ту же самую колонку. "Чистые пулы" концентрировали на роторном испарителе с целью удаления ацетонитрила.
Пример 5
Обмен противоионами и сушка пептида 16
Чистые пулы из примера 4 смешивали и ставили на мешалку для гомогенизации. Концентрирование чистого пептида 16 предпринимали с помощью обращеннофазной ВЭЖХ на препаративной колонке, которая служила для очистки. На колонке Prochrom LC110 объем чистого пептида, инъецированного на каждый пробег, приблизительно составлял 20 л при концентрации, приблизительно составляющей 5 г/л. Через колонку пропускали градиент буфера B в буфере A (со скоростью тока приблизительно в 450 мл/мин и УФ-детектированием при 220 нм): буфер A=0,1% ТФУ в воде; буфер B = ацетонитрил/0,1% ТФУ в воде (80/20). Через колонку пропускали ступенчатый градиент буфера B в буфере A (со скоростью тока приблизительно в 450 мл/мин и УФ-детектированием при 220 нм): буфер A=0,1% ТФУ в воде; буфер B = ацетонитрил/0,1% ТФУ в воде (80/20).
Объем растворителя для смыва полного пика собирали, концентрировали на роторном испарителе для удаления ацетонитрила и лиофилизировали на бутылочном лиофилизаторе. Полученные в результате лиофилизированные пулы очищенного пептида смешивали в воде/ацетонитриле (90/10) в концентрации 80 г/л и перемешивали до полного растворения перед ионообменной хроматографией на ацетате Dowex, строго основном, со смолой в 50-100 меш. (Ацетат Dowex был получен путем обработки смолы Dowex C1 1н NaOH, затем промывки очищенной водой, обработки AcOH/H2O (25/75) и промывки очищенной водой.) Образец элюировали водой/ ацетонитрилом (90/10). Объем растворителя для смыва полного пика собирали и концентрировали на роторном испарителе в случае, если элюционный объем был слишком большим. Очищенный раствор пептида фильтровали через стерильную фильтрационную капсулу (0,22 микрометра) и лиофилизировали на shelf-лиофилизаторе.
Пример 6
Анализ чистоты пептида 16
Чистоту пептида 16 определяли с помощью анализа путем аналитической ВЭЖХ с обращенной фазой. Чистоту устанавливали путем интегрирования площадей всех пиков (нормирование площади). Анализ предпринимали с помощью системы Waters Alliance HPLC с модулем 2695, состоящим из насоса с двойным клапаном с возвратно-поступательным движением, дегазатора, автоматической инъекционной системы, с регулируемым колоночным термостатом Пельтье; с УФ-детекторным модулем 2487; и программным обеспечением Empower Pro Version 5.00. Используемой колонкой являлась колонка Symmetry C18 (5 мкм) или эквивалентная ей колонка, 250×4,6 мм. Температура колонки составляла 60°C. Инъекции элюировали при градиентном профиле со скоростью тока, составляющей 1 мл/мин. Элюентом A являлась 0,1% ТФУ (например, Acros 13972) в воде milli-Q, тогда как элюентом B является 0,1% ТФУ в ацетонитриле в градиенте категории ВЭЖХ (например, SDS 00637G). Градиентный профиль показан ниже:
Пептид 16 детектировали по УФ-поглощению при 210 нм. Время протекания составляло 45 мин, со временем удерживания в 22 мин между моментом инъекции и вымыванием с колонки. Пептид 16 перевешивали во флаконе ВЭЖХ и растворяли в очищенной воде с получением концентрации, приблизительно составляющей 1,2 мг/мл. Инъецировали по 20 мкл растворов пептида.
Пример 7
Определение свойств миметиков ApoA-I посредством LC-MS (жидкостной хроматограф-масс-спектрометр)
Стандартный коммерчески доступный трехстадийный квадрупольный масс-спектрометр (модель TSQ 700; Finnigan MAT, San Jose Calif., USA) использовали для определения массы. Интерфейс с пневматически оснащенным электрораспылением (ESI) использовали для внесения образца в источник ионизации с атмосферным давлением масс-спектрометра. Интерфейсный [межфазный] распылитель работал при положительном потенциале в 4,5 кВ. Температуру стального капилляра поддерживали на уровне 200°C, тогда как температура распределителя составляла 70°C. Положительные ионы, вырабатываемые в результате процесса, происходящего в таком ионном испарителе, поступают в анализатор масс-спектрометра. Умножитель установлен на 1000 В. Анализаторный компартмент масс-спектрометра установлен на 4E-6. Все данные получали при разрешении <1 мкм.
Миметики ApoA-I анализировали путем прямой инфузии очищенных миметиков ApoA-I с использованием системы микроколонок ABI (Applied Biosystems), состоящей из шприцевого насоса (модель 140B), УФ-детектора (модель 785A) и духового шкафа/инжектора (модель 112A). Система растворителей состоит из воды (растворитель A) и ацетонитрила (растворитель B), каждый из которых содержит 0,1% ТФУ. Миметики ApoA-I инфузируют либо с использованием градиента, либо в изократических условиях и элюируют с колонки Aquapore C18. Скорость тока обычно составляет 300 мкл/мин. Концентрация каждого миметика ApoA-I приблизительно составляет 0,03 мг/мл, 20 мкл из которых инъецируют (например, 30 пмоль).
MS-эксперименты полномасштабного сканирования получены путем сканирования квадруполя 1 из m/z 500-1500 в 4 s. Данные получали с помощью системы Alpha DEC и обрабатывали с использованием программного обеспечения, поставляемого Finnigan MAT (BIOWORKS).
Пример 8
Определение свойств пептида 16 посредством LC-MS
Масс-спектральный анализ осуществляли с помощью инструментария Thermo-Finnigan LCQ Advantage. Источником служила электрораспылительная ионизация (ESI-MS). Параметры MS: газовый поток азота = 30 условных единиц, напряжение при электростатическом напылении = 5,2 В, температура в капилляре = 270°C, напряжение в капилляре = 38 В, на объективе в колене электронно-лучевой трубки (Tube Lens Offset)=55 В. Тестируемый раствор в 100 мкг/мл раствора пептида 16 анализировали в растворе метанол/вода/муравьиная кислота 47/47/6 об./об./об. (прямая инфузия в MS при скорости инъекции 5 мкл/мин посредством 500 мкл шприца). Результат, полученный после обратной фильтрации, совпадал с теоретическим значением.
Пример 9
Аминокислотный анализ миметиков ApoA-I
Аминокислотный анализ производили с помощью аминокислотного анализатора ABI 420 (Applied Biosystems). Указанная система состоит из трех модулей: гидролизующего и дериватизирующего инструмента, обращеннофазовой ВЭЖХ и системы данных. Образцы пептидов наносили (3 раза в трех повторах) на пористые предметные стекла и последовательно подвергали гидролизу в условиях газовой фазы (155°C, 90 мин). После удаления HCl, полученные в результате аминокислоты превращали в PTC-AA (фенилтиокарбамоил-аминокислоты) с использованием PITC (фенилизотиоцианат). После переноса образца в ячейку ВЭХ полученные в результате смеси фракционировали на колонке Aquapore C18 в градиентном режиме (растворитель A: 50 ммоль ацетата аммония (NH4Ac), pH 5,4, в воде; растворитель B: 32 ммоль ацетата натрия (NaOAc) в водном ацетонитриле) в условиях контролируемой температуры. Данные ВЭЖХ обрабатывали с использованием пакета программ, обеспечиваемых компанией Applied Biosystems. Количественную оценку осуществляли относительно стандартных пептидов, поставляемых компанией Applied Biosystems.
Пример 10
Аминокислотный анализ пептида 16
Пептид 16 (приблизительно 700 мкг) гидролизовали 100 микролитрами 6н HCl (например, Pierce 24308) при 110°C в течение 20 часов на составные аминокислоты, которые после дериватизации разделяли и количественно оценивали против стандартной смеси аминокислот (аминокислотный стандарт H, например, Pierce 20088). Аминокислоты дериватизировали с помощью o-фталальдегида (OPA-реагент, например, Fluka 5061-3335) и 9-фторенилметилхлорфомиата (Fmoc-реагент, например, Fluka 5061-3337), затем инъецировали на колонку C-18 ВЭЖХ. Для анализа использовали Agilent 1100 ВЭЖХ с УФ-детектором и программное обеспечение Chemstation. Используемой колонкой была колонка Hypersil ODS 200×2,1 мм, 5 мкм. Используемый градиент представлял собой 0-60% B за 17 мин до 100% B за 7 мин при скорости тока 0,45 мл/мин. Буфер A=2,3 г ацетата натрия в 1000 мл H2O + 180 мкл триэтиламина, pH доводили до 7,2 с помощью 2% раствора уксусной кислоты + 3,3 мл тетрагидрофурана. Буфер В=2,3 г ацетата натрия в 200 мл H2O, pH доводили до 7,2 с помощью 2% раствора уксусной кислоты + 400 мл ацетонитрила + 400 мл метанола. Оценку аминокислот проводили в трех повторах, при этом аминокислоты детектировали по УФ-поглощению при 368 и 262 нм. Стандартный раствор Pierce инъецировали как перед, так и после инъецирования пептидного образца в трех параллельных пробах.
Пример 11
Получение пептид/липидных комплексов путем со-лиофилизации
50 мг миметика ApoA-I растворяли в 1 мл ледяной уксусной кислоты в 1 миллилитровом прозрачном стеклянном пузырьке с крышечкой. Растворению пептида содействовали периодическим перемешиванием на вортексе в течение 10-минутных периодов при комнатной температуре. 50 мг дипальмитоилфосфатидилхолина (DPPC; липиды Avanti Polar, 99% чистоты, продукт #850355) и 50 мг яичного сфингомиелина (NOF) растворяли в 1 мл ледяной уксусной кислоты. DPPG растворяли в смеси 90% ледяной уксусной кислоты и 10% воды (об./об.) в концентрации 10 мг/мл. Растворению DPPG способствовала инкубация при 37°C. Миметик ApoA-I, сфингомиелин, растворы DPPC и DPPG смешивали с получением весового отношения миметик ApoA-I:сфингомиелин:DPPC:DPPG, составляющего, соответственно, 1:1,35:1,35:0,30. Полученный в результате раствор замораживали при -20°C и лиофилизировали в течение более чем 12 часов.
Лиофилизированный порошок подвергали гидратации в двууглекислом солевом буфере (20 мМ бикарбонат натрия, 130 мМ NaCl, pH 8,2) с получением конечной концентрации миметика ApoA-I, составляющей 10 мг/мл. Смесь перемешивали для облегчения регидратации. После гидратации pH доводили до pH 7,4, используя раствор 1н. NaOH. Для содействия комплексообразованию гидратированный порошок инкубировали в водяной бане при 50°C в течение 15 минут, с последующим выдерживанием при комнатной температуре в течение 15 минут. Нагревание и охлаждение повторяли до тех пор, пока не добивались получения очищенного прозрачного раствора миметик ApoA-I/фосфолипидные комплексы в буфере.
Пример 12
Получение пептид 16/липидного комплекса путем гомогенизации
Получали натрий-фосфатный буфер (12 мМ, pH 8,2) и нагревали его до 50°C.
Дисперсию DPPG получали путем диспергирования DPPG в буфере в концентрации 45 мг/мл. Раствор пептида получали путем растворения пептида 16 в буфере в концентрации 30 мг/мл. pH пептидного раствора доводили приблизительно до 8,2 путем добавления NaOH. Затем пептидный раствор объединяли с дисперсией DPPG и инкубировали при 50°C до тех пор, пока не добивались получения прозрачного раствора пептид/DPPG.
Дисперсию сфингомиелин/DPPC получали путем диспергирования сфингомиелина и DPPC в буфере в концентрации 38,3 мг/мл каждого из компонентов, сфингомиелина и DPPC. Затем дисперсию сфингомиелин/DPPC перемешивали путем интенсивного горизонтального встряхивания перемешивания.
Растворы пептид/DPPG и сфингомиелин/DPPC объединяли и гомогенизировали, используя гомогенизатор высокого давления (Avestin C3), до тех пор, пока раствор не становился прозрачным и не начинали образовываться комплексы. После гомогенизации добавляли изотонический агент (130 мМ NaCl). Затем раствор стерилизовали путем пропускания через стерильный фильтр и заполняли им стеклянные флаконы. Конечная концентрация пептида 16 в растворе составляла 15 мг/мл.
Пример 13
Анализ пептид 16/липидного комплекса
a. Распределение комплексов по размеру
Подтверждали идентичность пептид 16/липидного комплекса, полученного в соответствии с примером 12, и распределение комплексов по размеру определяли с помощью гель-проникающей хроматографии (GPC). Для разделения использовали Tosoh TSK-GEL G3000SWXL (ID 7,8 мм, длина 30 см). Инъекции элюировали 6 мМ фосфатным буфером, содержащим 150 мМ NaCl (pH 7,4), и при изократической скорости тока, составляющей 1 мл/мин. Образцы получали путем 20x разведения мобильной фазой, при этом используемый объем инъекции составлял 100 мкл. Пропускную способность колонки проверяли перед каждым нанесением путем инъецирования четырех стандартов с известными молекулярными весами. Комплекс детектировали посредством УФ при длине волны 220 нм. Идентичность подтверждали путем сравнения времени удерживания комплекса со ссылочным стандартом. Распределение комплексов по размеру выражали как процент от суммарного пика в хроматограмме. GPC-хроматограмма для пептид 16/липидного комплекса, полученного в соответствии с примером 12, представлена на фиг.5.
b. Идентичность, чистота и содержание пептида 16 комплекса
Идентичность, чистоту и содержание пептида 16 комплекса определяли с помощью сверхэффективной жидкостной хроматографии ("UPLC") с УФ-детектированием при длине волны 215 нм. Для данного разделения использовали 100-миллиметровую колонку Acquity BEH C18 с I.D. в 2,1 мм и размером частиц в 1,7 мкм. Инъекции элюировали с помощью мобильной фазы бинарного градиента 0,1% (об./об.) ТФУ при соотношении метанол/ацетонитрил/вода как 52,5/22,5/22 об./об./об. и 0,1% (об./об.) ТФУ при соотношении метанол/ацетонитрил/вода как 56/24/20 об./об./об. Образцы получали путем 20x разведения и инъецировали, используя объем инъекции в 7,5 мкл. Комбинация органических растворителей мобильной фазы способствовала растворению комплексов и отделению пептида 16 от липида комплекса. Идентичность пептида 16 подтверждали путем сравнения его времени удерживания со ссылочным стандартом. Чистоту пептида 16 выражали как процент от суммарного пика в хроматограмме. Содержание пептида 16 вычисляли с помощью калибровочной кривой, построенной на основании ссылочных стандартов разбавленных растворов пептида 16.
c. Определение липидного содержания в комплексе
Липидное содержание пептид 16/липидного комплекса, полученного в соответствии с примером 12, определяли с помощью ферментативного анализа с использованием метода DAOS. Набор для анализа производится компанией Wako Pure Chemical Industries, Ltd (Phospholipids C kit). Образцы растворяли в 75x, используя фосфатный буфер. Ферменты в указанном наборе для анализа гидролизовали сфингомиелин и DPPC с высвобождением холина, который затем вступал в реакцию с несколькими другими ферментами, активируя голубой пигмент. Голубой пигмент определяли спектрофотометрически. Проводили количественное определение образцов из калибровочной кривой, полученной на основании разведений холата натрия и голубого пигмента. Гидролизованный сфингомиелин и DPPC содержали холин и, таким образом, их рассчитывали указанным методом.
Пример 14
Гель-фильтрационная хроматография человеческих ЛПВП на суперозе 6
Человеческий ЛПВП2 (HDL2) получен следующим образом: 300 мл замороженной человеческой плазмы (Mannheim Blutspendzentrale #1185190) подвергали оттаиванию, ее плотность доводили до 1,25 с помощью твердого бромида калия и центрифугировали в течение 45 часов при 40,000 об/мин на роторе Ti45 (Beckman) при 20°C. Полученный в результате несвязанный верхний слой собирали, диализовали против дистиллированной воды, плотность доводили до 1,07 с помощью твердого бромида калия и центрифугировали, как описано выше, в течение 70 часов. Нижний слой (на уровне 1 см от дна пробирки) собирали, добавляли 0,01% азид натрия, и этот слой хранили при 4°C в течение 4 дней. 20 мкл ЛПВП2 наносили на суперозу 6, Pharmacia, в системе гель-фильтрационной хроматографии, с использованием 0,9% NaCl в качестве колоночного элюата. Скорость протекания через колонку составляла 0,5 мл/мин. Проводили мониторинг колоночного элюата по поглощению или рассеянию света при длине волны 254 нм. Серию белков с известными молекулярными весами и диаметрами Стокса использовали в качестве стандартов для калибровки колонки для расчета диаметров Стокса частиц (Pharmacia Gel Filtration Calibration Kit Instruction Manual, Pharmacia Laboratory Separation, Piscataway, N.J., revised April 1985).
Пример 15
Определение пептида 16 в плазме у крысы и обезьяны с использованием преципитации путем детектирования с помощью жидкостной хроматографии и тандемной масс-спектрометрии (LC-MS/MS)
Концентрации пептида 16 определяли в плазме крыс или обезьян в интервале концентраций от 1 до 500 мкг/мл с использованием холостой пробы матрикса. Меченный изотопом пептид 16 использовали в качестве раствора для внутреннего стандарта и добавляли к оттаявшим образцам плазмы. Затем в указанных образцах осаждали белок, используя воду: ацетонитрил: ТФУ (70:20:10 об./об./об.), с последующим смешиванием и центрифугированием. Супернатант переносили в чистый 96-луночный планшет, и к каждой лунке добавляли воду: ацетонитрил: ТФУ (70:30:0,1 об./об./об.), с последующим смешиванием и центрифугированием перед анализом LC-MS/MS. Условия ЖХ (LC) были следующими: Acquity UPLC и турбо-ион-распылительная масс-спектросукопия (положительный ион) (MS/MS), использование колонки BEH Shield RP18 с пропускаемым градиентом вода:ацетонитрил: ТФУ 0,1%.
Концентрации пептида 16 в калибровочных стандартах и образцах QC были определены с помощью линейной регрессии, построенной по принципу наименьших квадратов, с величиной, обратной концентрации (1/x), в качестве весового коэффициента.
Пример 16
Измерение фармакокинетики пептид 16/липидного комплекса у крыс
Фармакокинетику пептид 16/липидного комплекса (где липиды представлены сфингомиелином, DPPC и DPPG в весовом отношении 1:1,2125:1,2125:0,075, а пептид: липидное весовое отношение составляет 1:2,5) оценивали у крыс Windstar.
По девять животных каждого пола на группу были взяты для оценки фармакокинетики. Животные в контрольной группе носителя получали внутривенно по 20 мл/кг 130-миллимолярного хлорида натрия в 12 мМ фосфатном буфере, pH 8,2. Животным в группах, обрабатываемых пептид 16/липидным комплексом, вводили его по 15, 30 или 60 мг/кг через день путем внутривенной инфузии. Приблизительно 0,5 мл крови отбирали из ретроорбитального синуса под изофлорановой анестезией и собирали в пробирки, содержащие Na3ЭДТА в качестве антикоагулянта, из когорт по 3 животных на когорту в начальной точке и через 0,0833, 0,5, 1, 2, 6, 12, 24 и 48 часов после введения дозы на день 0 и день 26. Таким образом, у каждой когорты животных кровь отбирали в трех разных временных точках. Плазму отделяли в результате последующего центрифугирования и замораживали при -20°C на период до проведения анализа. Уровни пептида анализировали методом LC-MS/MS, как описано в примере 8. Фармакокинетические параметры из индивидуальных концентраций в плазме определяли путем фармакокинетического анализа без компартментализации с использованием кинетики 4.4.1. После введения дозы уровни пептида 16 в плазме быстро повышались, затем в течение 6 часов после окончания введения инфузии оставались поддающимися количественному определению у животных, которым вводили пептид 16/липидный комплекс в дозах 15 и 30 мг/кг. Детектируемые уровни пептида 16 наблюдались вплоть до 12 часов у животных обоего пола, обработанных 60 мг/кг. Как и ожидалось, в случае внутривенного введения Tmax достигался сразу же после введения. Было установлено, что время полужизни циркулирующих уровней пептида 16 составляет от 0,5 до 5 часов у крыс обоего пола, и оно повышается с повышением вводимой дозы. Клиренс и объем распределения снижались с повышением вводимой дозы. На основе объема распределения можно было сделать заключение, что пептид 16/липидный комплекс обычно распределялся по всему объему плазмы.
Пример 17
Измерение фармакокинетики пептид 16/липидного комплекса у обезьян
Фармакокинетику пептид 16/липидного комплекса (где липиды представлены сфингомиелином, DPPC и DPPG в весовом отношении 1:1,2125:1,2125:0,075, а пептид: липидное весовое отношение составляет 1:2,5) оценивали у макак-крабоедов.
Животным в контрольной группе носителя внутривенно вводили по 10 мл/кг 130-миллимолярного хлорида натрия в 12 мМ фосфатном буфере, pH 8,2. Животным в группах, обрабатываемых пептид 16/липидным комплексом, вводили его по 15, 30 или 60 мг/кг через день путем внутривенной инфузии. Кровь отбирали в пробирки, содержащие Na3ЭДТА в качестве антикоагулянта, в начальной точке, в конце инфузирования, а затем через 1, 2, 6, 12, 24 и 48 часов после введения дозы. В каждой временной точке приблизительно 1 мл крови отбирали из бедренного сосуда, при этом животных фиксировали, и обходились без анестезирования. Плазму отделяли в результате последующего центрифугирования и замораживали при -20°C на период до проведения анализа. Уровни пептида 16 анализировали методом LC-MS/MS, как описано в примере 8. Фармакокинетические параметры из индивидуальных концентраций плазмы определяли путем фармакокинетического анализа без компартментализации с использованием кинетики 4.4.1. Пептид 16 определяли в плазме через 12 часов после окончания инфузии у животных обоего пола, которым вводили пептид 16/липидный комплекс в дозе 15 мг/кг. Детектируемые уровни пептида 16 наблюдались вплоть до 24 часов у животных, обработанных 30 и 60 мг/кг. Уровни фосфолипидов также возрастали после введения дозы, а затем возвращались к исходным уровням через промежутки времени, сравнимые с таковыми для пептида 16. Как и ожидалось, в случае внутривенного введения Tmax достигался сразу же после введения дозы. Было установлено, что время полужизни циркулирующих уровней пептида 16 составляет от 2 до 7 часов у обезьян обоего пола, и оно повышается с повышением вводимой дозы. Клиренс и объем распределения снижались с повышением вводимой дозы. На основе объема распределения можно было сделать заключение, что пептид 16/липидный комплекс обычно распределялся прежде всего в плазменном компартменте.
Пример 18
Мобилизация холестерина у кроликов
a. Получение пептид 16/липидного комплекса
Пептид 16 был синтезирован посредством F-moc-синтеза в соответствии с примером 3 и очищен путем обращеннофазовой хроматографии, согласно описанному в примере 4.
Затем пептид 16 образовывал комплекс в смеси со сфингомиелином, DPPG и DPPC в результате совместной лиофилизации растворов пептида 16, сфингомиелина, DPPG и DPPC в смеси ледяная уксусная кислота: вода. Полученный в результате лиофилизированный порошок восстанавливали буфером (натрий-фосфатный буфер, 12 мМ, pH 8,2) с образованием суспензии пептид 16/липидного комплекса, имеющего весовое отношение пептид 16:сфингомиелин:DPPC:DPPG, составляющее 1:1,35:1,35:0,30.
b. Введение пептид 16/липидного комплекса кроликам
Новозеландских кроликов-самцов, весящих от 3 до 4 кг, использовали для того, чтобы продемонстрировать мобилизацию холестерина пептид 16/липидным комплексом.
Условия в помещении, в котором содержали животных, были следующими: температура, 22±2°C; относительная влажность, 55+15%; а также 12-часовой цикл света/12-часовой цикл темноты.
Животные проходили акклиматизацию по меньшей мере в течение 7 дней перед началом исследований. Животные находились на повседневной неограниченной контролируемой брикетированной диете. Неограниченный доступ к воде животные имели на протяжение всего исследования.
Перед введением пептид 16/липидного комплекса животные в течение ночи были лишены доступа к пище. Животных взвешивали непосредственно перед введением пептид 16/липидного комплекса. Пептид 16/липидный комплекс вводили внутривенно в дозе 20 мг/кг. Вводимый объем устанавливали в соответствии с весом. Питание возобновлялось приблизительно через 6 часов после введения пептид 16/липидного комплекса.
c. Анализ образцов крови
Перед сбором образцов крови животных в течение ночи лишали доступа к пище. Кровь отбирали на исходном уровне, затем через 5 мин, 15 мин, 30 мин, 1 час, 2 часа, 4 часа, 6 часов, 8 часов, 30 часов и 34 часа после начала инфузии. Образцы крови отбирали из яремной вены или из ушной маргинальной вены. Кровь отбирали из яремной вены шприцем, снабженным иглой с ЭДТА (приблизительно 1 мл крови за время отбора образца). Непосредственно после отбора крови образцы крови хранили приблизительно при 4°C, чтобы избежать изменений в образцах крови. Образцы крови центрифугировали (3500 g в течение 10 минут приблизительно при 5°C). Образцы плазмы отделяли, брали из них аликвоты (по 3 аликвоты по меньшей мере объемом в 200 мкл (аликвоты A, B, C)) и хранили приблизительно при -80°C. Оставшийся сгусток крови отбрасывали.
Сывороточный фосфолипид (Фосфолипид B, Набор # 990-54009, Wako Chemicals GmbH, Neuss, Germany), триглицериды (Триглицериды, Набор # 1488872, Boehringer Mannheim Corporation, Indianapolis, Ind.), суммарный холестерин и неэтерифицированный холестерин определяли, используя коммерчески доступные наборы для автоматического анализатора Hitachi 912 (Roche Diagnostics Corporation, Indianapolis, Ind.).
Профили липопротеина анализировали с помощью гель-фильтрационной хроматографии на колонке 1×30 см сефарозы 6HR, снабженной детектором суммарного или свободного холестерина в режиме on-line, как описано Kieft et al. (J Lipid Res 1991; 32: 859-866, 1991). Область под пиками, соответствующую липопротеинам размером с ЛПОНП, ЛПНП и ЛПВП, интегрировали. Фракцию свободного или суммарного холестерина каждого пика умножали на суммарный холестерин в плазме или свободный холестерин, определяемый посредством автоматического анализатора для определения ЛПОНП, ЛПНП и ЛПВП свободного и суммарного холестерина. Этерифицированный холестерин в сыворотке и в липопротеиновых фракциях ЛПОНП, ЛПНП и ЛПВП вычисляли путем вычитания показателей свободного холестерина из уровня суммарного холестерина.
Повышение фракции ЛПВП суммарного холестерина после инфузии комплексов наносили как функцию от времени, и это проиллюстрировано на фиг.6. Суммарный ЛПВП холестерин повышался при введении пептид 16/липидного комплекса, что указывает на тканевую мобилизацию холестерина и смещение в сторону ЛПВП.
Пример 19
Мобилизация холестерина у кроликов
a. Получение пептид 16/липидного комплекса
Пептид 16/липидный комплекс получали в соответствии с примером 12. Пептид 16/липидный комплекс имел весовое отношение пептид 16:сфингомиелин:DPPC:DPPG, составляющее 1:1,2125:1,2125:0,075 и весовое отношение пептид:липид, составляющее 1:2,5.
b. Введение пептид 16/липидного комплекса кроликам
Новозеландских кроликов-самцов, весящих от 3 до 4 кг, использовали для того, чтобы продемонстрировать повышение уровней ЛПВП у кроликов под действием пептид 16/липидного комплекса.
Условия в помещении, в котором содержали животных, были следующими: температура, 22±2°C; относительная влажность, 55+15%; а также 12-часовой цикл света/12-часовой цикл темноты.
Животные проходили акклиматизацию по меньшей мере в течение 7 дней перед началом исследований. Животные находились на неограниченной повседневной контролируемой брикетированной диете. Неограниченный доступ к воде животные имели на протяжении всего исследования.
Перед введением пептид 16/липидного комплекса животные в течение ночи были лишены доступа к пище. Животных взвешивали непосредственно перед введением пептид 16/липидного комплекса. Для изучения минимальной дозы, при которой может быть зарегистрирована мобилизация холестерина, животным вводили 2,5, 5 и 10 мг/кг пептид 16/липидного комплекса или забуференного фосфатом физиологического раствора в контроле. Для исследования каждой дозы брали по четыре животных на группу. Питание возобновлялось приблизительно через 6 часов после введения пептид 16/липидного комплекса или забуференного фосфатом физиологического раствора в контроле.
c. Анализ образцов крови
Перед сбором образцов крови животных в течение ночи лишали доступа к пище. Кровь отбирали на исходном уровне, затем через 5 мин, 15 мин, 30 мин, 1 час, 2 часа, 4 часа, 6 часов, 8 часов, 30 часов и 34 часа после начала инфузии. Образцы крови отбирали из яремной вены или из ушной маргинальной вены. Кровь отбирали из яремной вены шприцем, снабженным иглой с ЭДТА (приблизительно 1 мл крови за время отбора образца). Непосредственно после отбора крови образцы крови хранили приблизительно при 4°C, чтобы избежать изменений в образцах крови. Образцы крови центрифугировали (3500 g в течение 10 минут приблизительно при 5°C). Образцы плазмы отделяли, брали из них аликвоты (по 3 аликвоты объемом по меньшей мере в 200 мкл (аликвоты A, B, C)) и хранили приблизительно при -80°C. Оставшийся сгусток крови отбрасывали.
Сывороточный фосфолипид (Фосфолипид B, Набор # 990-54009, Wako Chemicals GmbH, Neuss, Germany), триглицериды (Триглицериды, Набор # 1488872, Boehringer Mannheim Corporation, Indianapolis, Ind.), суммарный холестерин и неэтерифицированный холестерин определяли, используя коммерчески доступные наборы для автоматического анализатора Hitachi 912 (Roche Diagnostics Corporation, Indianapolis, Ind.).
Профили липопротеина анализировали с помощью гель-фильтрационной хроматографии на колонке 1×30 см сефарозы 6HR, снабженной детектором суммарного или свободного холестерина в режиме on-line, как описано Kieft et al. (J Lipid Res 1991; 32: 859-866, 1991). Область под пиками, соответствующая липопротеинам размером с ЛПОНП, ЛПНП и ЛПВП, интегрировали. Фракцию свободного или суммарного холестерина каждого пика умножали на суммарный холестерин в плазме или свободный холестерин, определяемый посредством автоматического анализатора для определения ЛПОНП, ЛПНП и ЛПВП свободного и суммарного холестерина. Этерифицированный холестерин в сыворотке и в липопротеиновых фракциях ЛПОНП, ЛПНП и ЛПВП вычисляли путем вычитания показателей свободного холестерина из уровня суммарного холестерина.
Повышение фракции ЛПВП свободного холестерина после инфузии комплексов наносили в виде функции от времени, и это проиллюстрировано на фиг.7. Четкое повышение фракции ЛПВП свободного холестерина на фоне исходного уровня наблюдалось при введении дозы в 2,5 мг/кг, что указывает на высокий потенциал пептид 16/липидного комплекса. Через пять и 20 минут после начала инфузии уровень холестерина повышался на 30% над исходным уровнем. Такое увеличение было статистически достоверным (p<0,05, согласно парному двупараметрическому T-критерию Стьюдента). В отличие от этого, никаких изменений от базового уровня в группе плацебо обнаружено не было.
Фармакологический эффект пептид 16/липидного комплекса при дозе в 2,5 мг/кг был дополнительно подтвержден путем сравнения исходных сканов липопротеиновых фракций, элюирующихся с колонки эксклюзионной по размеру ВЭЖХ, что проиллюстрировано на фиг.8. На хроматограммах ВЭЖХ после инъекции наблюдается четкое повышение фракции ЛПВП свободного холестерина относительно базового уровня.
Пример 20
Доза-ответ пептид 16/липидного комплекса
Доза-ответ пептид 16/липидного комплекса (где липиды представлены сфингомиелином, DPPC и DPPG в весовом отношении 1:1,2125:1,2125:0,075, а пептид: липидное весовое отношение составляет 1:2,5) оценивали у белых новозеландских кроликов.
Некормленным белым новозеландским кроликам, используемым в качестве модели мобилизации холестерина, пептид 16/липидный комплекс в концентрациях 5, 10 или 15 мг/мл (на основе концентрации пептида) или забуференный фосфатом физиологический раствор в качестве контроля носителя, вводили внутривенно со скоростью 1 мл/мин некормленным животным при объеме инфузии, составляющем 2 мл/кг. В каждой группе использовали по три животных на дозу. Конечные дозы составляли 0, 10, 20 или 30 мг/кг. Кровь отбирали на исходном уровне, затем через 5 мин, 15 мин, 30 мин, 1 час, 2 часа, 4 часа, 6 часов, 8 часов, 30 часов и 34 часа после начала инфузии. Затем определяли уровни липида и липопротеина в плазме. Уровни липопротеина определяли с помощью эксклюзионного по размеру фракционирования ВЭЖХ, со встроенным детектором свободного и суммарного холестерина, в соответствии со способом, описанным Usui, S., Hara, Y., Hosaki, S., and Okazaki, M., A new on-line dual enzymatic method for simultaneous quantification of cholesterol and triglycerides in lipoproteins by HPLC. J. Lipid Res. 43, 805-814 (2002). Область под основными пиками, соответствующая липопротеинам размером с ЛПОНП, ЛПНП и ЛПВП, интегрировали. Фракцию свободного или суммарного холестерина в каждом пике умножали на уровень суммарного холестерина в плазме или свободного холестерина, чтобы определить уровень холестерина в каждой фракции. Уровни сложного эфира холестерина в каждой фракции определяли путем вычитания показателей свободного холестерина из уровня суммарного холестерина в каждой фракции. В данной модели повышение уровней ЛПВП холестерина в плазме является показателем тканевой мобилизации холестерина и смещения в сторону ЛПВП.
На фиг.9 показано дозо-зависимое повышение фосфолипидов в плазме после инфузии кроликам пептид 16/липидного комплекса. Такое повышение отражает уровни циркуляции пептид 16/липидного комплекса, поскольку фосфолипид является компонентом пептид 16/липидного комплекса. Уровни пептида 16 образовывали пик в течение первых 30 минут, а затем снижались до исходных уровней. Наблюдалось также дозо-зависимое увеличение в мобилизации холестерина. Это следовало из того, что повышались как уровни суммарного холестерина в плазме (фиг.10A), так и уровни суммарного холестерина ЛПВП (фиг.11A). Основная часть повышенного холестерина была представлена в виде свободного холестерина (фиг.10 и 11).
Повышение суммарного и свободного холестерина во фракции ЛПНП (фиг.11C и 11D) наблюдалось при двух самых высоких дозах. Повышение уровня свободного холестерина было приблизительно равно таковому суммарного холестерина, что указывает на малое увеличение сложного эфира холестерина в указанной фракции. Повышение свободного холестерина во фракции ЛПНП, в отсутствие повышения сложного эфира холестерина, указывает на то, что такое повышение не может отражать увеличение уровня типичных ЛПНП, богатых сложным эфиром холестерина. Комплексы, появляющиеся в этой фракции липопротеина, скорее всего являются продуктом инфузированного пептид 16/липидного комплекса, который вызвал повышение холестерина в результате процесса мобилизации холестерина. Наблюдаемое повышение холестерина ЛПОНП было распределено между этерифицированной и неэтерифицированной фракциями холестерина. Уровни триглицеридов транзитно повышались в течение первых четырех-шести часов после введения всех доз пептид 16/липидного комплекса (фиг.12). Не обнаружено очевидной взаимосвязи между введенной дозой и повышением уровня триглицеридов.
Пример 21
Минимальная эффективная доза пептид 16/липидного комплекса
Минимальная эффективная доза пептид 16/липидного комплекса (где липиды представлены сфингомиелином, DPPC и DPPG в весовом отношении 1:1,2125:1,2125:0,075, а пептид: липидное весовое отношение составляет 1:2,5) оценивали у белых новозеландских кроликов.
Изучали минимальную дозу, при которой можно регистрировать мобилизацию холестерина. Животным вводили 0, 2,5, 5 и 10 мг/кг пептид 16/липидного комплекса. Для исследования каждой дозы брали по четыре животных на группу. Фармакологический эффект был наиболее очевиден при повышении уровня свободного холестерина во фракции ЛПВП по сравнению с исходными уровнями (фиг.13). Этого и следовало ожидать, поскольку основная часть повышенного холестерина сначала, после инфузии пептид 16/липидного комплекса, представлена свободным холестерином во фракции ЛПВП. Кроме того, свободным холестерином представлена приблизительно одна треть суммарного ЛПВП холестерина, что облегчает детектирование повышения уровня этой фракции. Наблюдалось четкое повышение свободного ЛПВП холестерина над базовым уровнем при дозе в 2,5 мг/кг. Через пять и 20 минут после начала инфузии уровень холестерина повышался на 30% над базовым уровнем. Такое повышение было статистически достоверным (p<0,05, согласно парному двупараметрическому T-критерию Стьюдента). В отличие от этого, никаких изменений от базового уровня в контрольной группе обнаружено не было.
Фармакологический эффект пептид 16/липидного комплекса при дозе в 2,5 мг/кг или 5 мг/кг был дополнительно подтвержден путем сравнения исходных сканов липопротеиновых фракций, элюирующихся с колонки эксклюзионной по размеру ВЭЖХ. Как можно видеть в этих двух примерах, на фиг.14, на хроматограммах ВЭЖХ имеет место четкое повышение свободного холестерина относительно базового уровня во фракции ЛПВП. Это показано по увеличению площади под пиком ЛПВП для образца, взятого через 20 минут после начала инфузии пептид 16/липидного комплекса (светлая линия на фиг.14), по сравнению с площадью под пиком ЛПВП на базовом уровне (темная линия на фиг.14).
Пример 22
Влияние скорости инфузии на эффективность пептид 16/липидного комплекса
Влияние скорости инфузии на эффективность пептид 16/липидного комплекса (где липиды представлены сфингомиелином, DPPC и DPPG в весовом отношении 1:1,2125:1,2125:0,075, а пептид:липидное весовое отношение составляет 1:2,5) оценивали у белых новозеландских кроликов.
Изучали влияние скорости инфузии пептид 16/липидного комплекса на мобилизацию холестерина. Пептид 16/липидный комплекс в концентрации 10 мг/мл (на основе концентрации пептида) или забуференный фосфатом физиологический раствор в качестве контроля носителя вводили в объемной дозе 2 мл/кг со скоростью либо 1 мл/мин, либо 0,2 мл/мин. Конечная доза пептид 16/липидного комплекса составляла 20 мг/кг. По четыре животных изучали в группах пептид 16/липидного комплекса и по два животных в контрольных группах носителя. Вес кроликов варьировал от 2,2 до 2,8 кг.
Скорость увеличения уровня фосфолипида в плазме в результате инфузии пептид 16/липидного комплекса и скорость увеличения уровня холестерина в плазме в результате последующей мобилизации холестерина была более медленной у животных, которым пептид 16/липидный комплекс вводили с меньшей скоростью. Однако максимальные уровни мобилизации фосфолипида и холестерина были сходными. На фиг.15 показано, что увеличение свободного ЛПВП холестерина после инфузии пептид 16/липидного комплекса со скоростью 1 мл/мин или 0,2 мл/мин было сходным. Таким образом, в настоящей модели скорость инфузии пептид 16/липидного комплекса, превышающая тестируемые скорости, мало отражается или же вовсе не отражается на мобилизации холестерина.
Пример 23
Фармакокинетические исследования пептид 16/липидного комплекса на крысах и обезьянах
Фармакокинетика пептид 16/липидного комплекса (где липиды представлены сфингомиелином, DPPC и DPPG в весовом отношении 1:1,2125:1,2125:0,075, а пептид: липидное весовое отношение составляет 1:2,5) оценивали у крыс и обезьян.
a. Методология исследования
Концентрации пептид 16/липидного комплекса в плазме крыс и обезьян определяли с помощью жидкостной хроматографии с использованием оснащения техникой тандемной масс-спектрометрии (LC-MS/MS). Пептид 16, компонент пептид 16/липидного комплекса, экстрагировали из плазмы, содержащей ЭДТА, после преципитации белковой фракции ацетонитрилом. Указанный способ позволяет измерить экстрагированный пептид 16 и внутренний стандарт меченного изотопом пептида 16. Экстракты ресуспендировали, и пептид изучали методом сверхэффективной жидкостной хроматографии в сочетании с тандемным масс-спектрометром (MS/MS). Калибровочный интервал указанного метода составляет 1-500 мкг/мл для образца объемом в 25 мкл. Способы экстракции и LC-MS/MS были основаны на общих рекомендациях по осуществлению биоаналитического метода установления достоверности и в соответствии с инструкциями GLP (Good Laboratory Practice). Проверка достоверности данных показала, что указанные методы являются достаточно чувствительными, специфичными, надежными и точными для определения пептид 16/липидного комплекса в плазме крыс и обезьян.
b. Фармакокинетические исследования у крыс
По 9 крыс каждого пола на группу включали для оценки токсикокинетики пептид 16/липидного комплекса после введения дозировки на день 0 (первая доза) и день 26. Животные в контрольной группе носителя получали внутривенно по 20 мл/кг 130-миллимолярного хлорида натрия в 12 мМ фосфатном буфере, pH 8,2. Животным в группах, обрабатываемых пептид 16/липидным комплексом, вводили его по 15, 30 или 60 мг/кг через день путем внутривенной инфузии. Приблизительно 0,5 мл крови отбирали из ретроорбитального синуса под изофлорановой анестезией и собирали в пробирки, содержащие Na3ЭДТА в качестве антикоагулянта, из когорт по 3 животных на группу в начальной точке и через 0,0833, 0,5, 1, 2, 6, 12, 24 и 48 часов после введения дозы на день 0 и день 26. Таким образом, у каждой когорты животных кровь отбирали в трех разных временных точках. Плазму отделяли в результате последующего центрифугирования и замораживали при -20°C на период вплоть до проведения анализа [на установке] Covance (UK). Уровни пептида анализировали методом LC-MS/MS. Токсикокинетические параметры определяли из индивидуальных концентраций в плазме посредством фармакокинетического анализа без компартментализации с использованием кинетики 4.4.1.
Как показано на фиг.16 и 17, уровни пептида 16 в плазме после введения дозировки быстро повышались, затем в течение 6 часов после окончания введения инфузии оставались поддающимися количественному определению у животных, которым вводили пептид 16/липидный комплекс в дозах 15 и 30 мг/кг. Детектируемые уровни пептида наблюдались вплоть до 12 часов у животных обоего пола, обработанных дозой в 60 мг/кг. Уровни фосфолипидов возрастали после введения дозы, а затем возвращались к исходным уровням через промежутки времени, сходные с таковыми для указанного пептида. Уровень свободного холестерина (неэтерифицированного) повышался после инфузии дозо-зависимым образом, что является показателем мобилизации холестерина. После этого происходило снижение уровня холестерина, что свидетельствует о том, что частицы пептид 16/липидного комплекса эффективно удаляют холестерин из циркуляции. Сходную картину наблюдали на день 0 и день 26.
Средние значения токсикокинетических параметров для пептид 16/липидного комплекса на день 0 (первая доза) и день 26 (последняя доза) представлены ниже в таблице 11:
Токсикокинетические параметры пептид 16/липидного комплекса у крысы†
Tmax достигался сразу же после введения дозы. Установленное время полужизни циркулирующих уровней пептида 16 составляло от 0,835 до 2,56 часов у крыс обоего пола, и оно повышалось с повышением вводимой дозы. Клиренс и объем распределения снижались с повышением вводимой дозы. На основе объема распределения можно было сделать заключение, что пептид 16/липидный комплекс обычно распределялся прежде всего в плазменном компартменте (ссылочный объем плазмы у крыс = 30 мл/кг). См. Davies, B and Morris, T. Physiological parameters in laboratory animals and human, Pharmaceutical Research, 10, 1093-1095, 1993.
Повышение показателей AUC(0-12 ч) и Cmax с увеличением дозы (на основе дозы в 15 мг/кг) представлено в таблице 12. Значения Cmax были пропорциональны дозе у обоих полов, тогда как значения AUC(0-12 ч) возрастали более чем пропорционально дозе, что свидетельствует о больших временах удерживания пептид 16/липидного комплекса в циркуляции с повышением дозы.
Повышение AUC и Cmax с увеличением дозы пептид 16/ липидного комплекса
Не наблюдалось существенных связанных с полом различий в фармакокинетических профилях, значениях AUC или Cmax после введения одиночной дозы и множественных доз. На основании показателей Cmax и AUC, в течение 4-недельного периода введения не удалось обнаружить накопления пептида 16 или пептид 16/липидного комплекса.
c. Фармакокинетические исследования у обезьян
Токсикокинетику пептид 16/липидного комплекса оценивали после введения дозировки на день 0 (первая доза) и день 26. Животные в контрольной группе носителя получали внутривенно по 10 мл/кг 130-миллимолярного хлорида натрия в 12 мМ фосфатном буфере, pH 8,2. Животным в группах, обрабатываемых пептид 16/липидным комплексом, вводили его по 15, 30 или 60 мг/кг через день путем внутривенной инфузии. Кровь отбирали в пробирки, содержащие Na3ЭДТА в качестве антикоагулянта, в начальной точке, в конце инфузирования, а затем через 1, 2, 6, 12, 24 и 48 часов после введения дозы на день 0 и день 26. В каждой временной точке приблизительно 1 мл крови отбирали из бедренного сосуда, при этом животных фиксировали, и обходились без анестезирования. Плазму отделяли после центрифугирования и замораживали при -20°C на период вплоть до проведения анализа [на установке] Covance (UK). Уровни пептида анализировали методом LC-MS/MS. Токсикокинетические параметры определяли из индивидуальных концентраций в плазме посредством фармакокинетического анализа без компартментализации с использованием кинетики 4.4.1.
Как показано на фиг.18 и 19, уровни пептида 16 в плазме могли детектироваться вплоть до 12 часов после последней инфузии у животных обоего пола, обработанных дозой пептид 16/липидного комплекса в 15 мг/кг. Детектируемые уровни пептида наблюдались вплоть до 24 часов у животных, обработанных 30 и 60 мг/кг. Уровни фосфолипидов также возрастали после введения дозы, а затем возвращались к исходным уровням через промежутки времени, сходные с таковыми для указанного пептида. Уровень свободного холестерина (неэтерифицированного) повышался после инфузии дозо-зависимым образом, что является показателем мобилизации холестерина. После этого происходило снижение уровня холестерина, что свидетельствует о том, что частицы пептид 16/липидного комплекса эффективно удаляют холестерин из циркуляции. Сходную картину наблюдали на день 0 и день 26.
Средние значения токсикокинетических параметров для пептид 16/липидного комплекса на день 0 (первая доза) и день 26 (последняя доза) представлены ниже в таблице 13:
Токсикокинетические параметры пептид 16/липидного комплекса у обезьян†
Т=0 соответствует окончанию инфузии.
Tmax достигался сразу же после введения дозы. Установленное время полужизни циркулирующих уровней пептида 16 составляло от 2,11 до 4,58 часов у крыс обоего пола, и оно повышалось дозо-зависимым образом. Клиренс и объем распределения снижались с повышением вводимой дозы. На основе объема распределения можно было сделать заключение, что пептид 16/ липидный комплекс обычно распределялся прежде всего в плазменном компартменте (ссылочный объем плазмы у крыс = 45 мл/кг). См. Davies, B and Morris, T. Physiological parameters in laboratory animals and human, Pharmaceutical Research, 10, 1093-1095, 1993.
Повышение показателей AUC(0-24 ч) и Cmax с увеличением дозы (на основе дозы в 15 мг/кг) представлено в таблице 14. Значения Cmax были пропорциональны дозе у обоих полов, тогда как значения AUC(0-24 ч) возрастали более чем пропорционально дозе, что свидетельствует о больших временах удерживания пептид 16/липидного комплекса в циркуляции с повышением дозы.
Повышение AUC и Cmax с увеличением дозы пептид 16/липидного комплекса
Не наблюдалось существенных связанных с полом различий в фармакокинетических профилях, значениях AUC или Cmax после введения одиночной дозы и множественных доз.
На основании показателей Cmax и AUC, в течение 4-недельного периода введения не удалось обнаружить накопления пептида 16 или пептид 16/липидного комплекса.
Пример 24
Фармакокинетические исследования пептида 16 и пептид 16/липидных комплексов у мышей
После инъекции одной из трех пептидных композиций в плазме измеряли суммарный холестерин, неэтерифицированный холестерин и сложный эфир холестерина (как разницу между показателями суммарного и неэтерифицированного холестеринов).
Пептидные композиции: (A) пептид 16; (B) комплекс пептид 16/DPPC (весовое отношение 1:2); (C) комплекс пептид 16/DPPC (весовое отношение 1:2,5). Каждая из композиций A, B и C была получена в виде растворов в концентрации 15 мг/мл.
20 мышам C57B1/6J ограничивали питание по меньшей мере в течение двух недель, в соответствии с установленной диетой для грызунов, содержащей 60% ккал% жира (Reseach diets - D12492). В питьевую воду добавляли 5% глюкозы. После 3 часов голодания им вводили пептидные композиции в дозе 30 мг/кг (внутривенные инъекции по 50 мкл), после чего образцы крови отбирали через 15, 30, 60, 120 и 240 минут. Один образец крови брали перед инъекцией.
Образцы плазмы анализировали на предмет определения суммарного холестерина (наборы производства Biolabo-CEROOX-SOP002, CER00X-SOP003). Сложный эфир холестерина рассчитывали в виде разницы между суммарным холестерином и неэтерифицированным холестерином.
Полученные результаты представлены на фиг.21 и 22.
В данной заявке приведен целый ряд ссылок, каждая из которых в полном объеме включена в настоящее описание в качестве ссылки.
Далее следуют некоторые иллюстративные воплощения настоящего изобретения:
1. Пептид, содержащий от 22 до 29 остатков, имеющий следующую формулу I:
R1-Y1-X1-X2-X3-X4-X5-X6-X7-X8-X9-X10-X11-X12-X13-X14-X15-X16-X17-X18-X19-X20-X21-X22-X23-Y2-R2
Формула I
или его фармацевтически приемлемая соль, где
X1 отсутствует или является ахиральным аминокислотным остатком, D-основным аминокислотным остатком или L-основным аминокислотным остатком;
X2 является ахиральным аминокислотным остатком, D-основным аминокислотным остатком или L-основным аминокислотным остатком;
X3 является алифатическим ахиральным аминокислотным остатком, алифатическим D-аминокислотным остатком или алифатическим L-аминокислотным остатком;
X4 является ахиральным основным аминокислотным остатком, D-основным аминокислотным остатком или L-основным аминокислотным остатком;
X5 представляет собой Gln, Asn, D-Gln, D-Asn или ахиральный основный аминокислотный остаток, D-основный аминокислотный остаток или L-основный аминокислотный остаток;
X6 является ахиральным основным аминокислотным остатком, D-основным аминокислотным остатком или L-основным аминокислотным остатком;
X7 является ахиральным гидрофобным аминокислотным остатком, D-гидрофобным аминокислотным остатком или L-гидрофобным аминокислотным остатком;
X8 является ахиральным гидрофобным аминокислотным остатком, D-гидрофобным аминокислотным остатком или L-гидрофобным аминокислотным остатком;
X9 является ахиральным гидрофильным аминокислотным остатком, D-гидрофильным аминокислотным остатком или L-гидрофильным аминокислотным остатком;
X10 представляет собой Leu, Trp, Gly, Nal, D-Leu, D-Trp или D-Nal;
X11 представляет собой Gly или ахиральный алифатический аминокислотный остаток, D-алифатический аминокислотный остаток или L-алифатический аминокислотный остаток;
X12 является ахиральным гидрофильным аминокислотным остатком, D-гидрофильным аминокислотным остатком или L-гидрофильным аминокислотным остатком;
X13 является ахиральным гидрофильным аминокислотным остатком, D-гидрофильным аминокислотным остатком или L-гидрофильным аминокислотным остатком;
X14 представляет собой Leu, Trp, Gly, D-Leu или D-Trp;
X15 представляет собой Leu, Gly или D-Leu;
X16 является ахиральным кислым аминокислотным остатком, D-кислым аминокислотным остатком или L-кислым аминокислотным остатком;
X17 является ахиральным гидрофильным аминокислотным остатком, D-гидрофильным аминокислотным остатком или L-гидрофильным аминокислотным остатком;
X18 представляет собой Leu, Phe, D-Leu или D-Phe;
X19 представляет собой Leu, Phe, D-Leu или D-Phe;
X20 является ахиральным кислым аминокислотным остатком, D-кислым аминокислотным остатком или L-кислым аминокислотным остатком;
X21 представляет собой Leu, Phe, D-Leu или D-Phe;
X22 является ахиральным алифатическим аминокислотным остатком, D-алифатическим аминокислотным остатком или L-алифатическим аминокислотным остатком; и
X23 представляет собой Inp, Nip, azPro, Pip, azPip, D-Nip или D-Pip;
Y1 отсутствует или является аминокислотной последовательностью, содержащей от 1 до 7 остатков, где каждый остаток последовательности независимо является ахиральным, D- или L-аминокислотным остатком;
Y2 отсутствует или является последовательностью, содержащей от 1 до 7 аминокислотных остатков, где каждый остаток последовательности независимо является ахиральным, D- или L-аминокислотным остатком;
R1 представляет собой H или аминозащитную группу; и
R2 представляет собой OH или карбоксильную защитную группу; и при этом
a) все аминокислотные остатки, не являющиеся концевыми аминокислотными остатками и остатками, непосредственно примыкающими к концевым аминокислотным остаткам, являются ахиральными или L-аминокислотными остатками; или
b) все аминокислотные остатки, не являющиеся концевыми аминокислотными остатками и остатками, непосредственно примыкающими к концевым аминокислотным остаткам, являются ахиральными или D-аминокислотными остатками.
2. Пептид или фармацевтически приемлемая соль указанного пептида, согласно воплощению 1, где
X3 представляет собой Leu или D-Leu;
X7 представляет собой Leu, Gly, Nal, D-Leu или D-Nal;
X8 представляет собой Ala, Nal, Trp, Gly, Leu, Phe, D-Ala, D-Nal, D-Trp, D-Leu или D-Phe;
X11 представляет собой Leu, Gly, Aib или D-Leu; и
X22 представляет собой Ala, Leu, Val, D-Ala, D-Leu или D-Val.
3. Пептид или фармацевтически приемлемая соль пептида, согласно воплощению 1, где
X1 отсутствует или представляет собой Lys или D-Lys;
X2 представляет собой Lys, Orn, D-Lys или D-Orn;
X4 представляет собой Lys, Orn, D-Lys или D-Orn;
X5 представляет собой Gln, Asn, Lys, Orn, D-Gln, D-Asn, D-Lys или D-Orn;
X6 представляет собой Gln, Asn, Lys, Orn, D-Gln, D-Asn, D-Lys или D-Orn;
X9 представляет собой Asp, Glu, D-Asp или D-Glu;
X12 представляет собой Glu, Asp, D-Asp или D-Glu;
X13 представляет собой Asn, Gln, D-Asn или D-Gln;
X16 представляет собой Asp, Glu, D-Asp или D-Glu;
X17 представляет собой Lys, Arg, Orn, D-Lys, D-Arg или D-Orn; и
X20 представляет собой Asp, Glu, D-Asp или D-Glu.
4. Пептид или фармацевтически приемлемая соль указанного пептида, согласно воплощению 3, где X18 представляет собой Phe или D-Phe.
5. Пептид или фармацевтически приемлемая соль указанного пептида, согласно воплощению 1, где указанный пептид является пептидом, содержащим 22 или 23 пептидных остатка.
6. Пептид или фармацевтически приемлемая соль указанного пептида, согласно воплощению 5, где R1 представляет собой H и R2 представляет собой OH.
7. Пептид или фармацевтически приемлемая соль указанного пептида, согласно воплощению 5, где
X1 отсутствует, представляет собой Lys или D-Lys;
X2 представляет собой Lys, Orn, D-Lys или D-Orn;
X3 представляет собой Leu или D-Leu;
X4 представляет собой Lys, Orn, D-Lys или D-Orn;
X5 представляет собой Gln, Asn, Lys, Orn, D-Gln, D-Asn, D-Lys или D-Orn;
X6 представляет собой Lys, Orn, D-Lys или D-Orn;
X7 представляет собой Gly, Leu, Nal, D-Leu или D-Nal;
X8 представляет собой Ala, Nal, Trp, Leu, Phe, Gly, D-Ala, D-Nal, D-Trp, D-Leu или D-Phe;
X9 представляет собой Asp, Glu, D-Asp или D-Glu;
X11 представляет собой Gly, Leu, Aib или D-Leu;
X12 представляет собой Glu, Asp, D-Glu или D-Asp;
X13 представляет собой Asn, Gln, D-Asn или D-Gln;
X16 представляет собой Asp, Glu, D-Asp, или D-Glu;
X17 представляет собой Lys, Arg, Orn, D-Lys, D-Arg или D-Orn;
X20 представляет собой Asp, Glu, D-Asp или D-Glu; и
X22 представляет собой Ala, Val, Leu, D-Ala, D-Val или D-Leu.
8. Пептид или фармацевтически приемлемая соль указанного пептида, согласно воплощению 7, где X5 представляет собой Gln, Asn, D-Gln или D-Asn, а X6 представляет собой Lys, Orn, D-Lys или D-Orn; или X5 представляет собой Lys, Orn, D-Lys или D-Orn, а X6 представляет собой Gln, Asn, D-Gln или D-Asn.
9. Пептид или фармацевтически приемлемая соль указанного пептида, согласно воплощению 7, где X1 отсутствует, а пептид является пептидом, содержащим 22 пептидных остатка. D
10. Пептид или фармацевтически приемлемая соль указанного пептида, согласно воплощению 9, где каждый из X7, X8, X10, X11, X14 и X15 является отличным от Gly.
11. Пептид или фармацевтически приемлемая соль указанного пептида, согласно воплощению 9, где только один из X7, X8, X10, X11, X14 и X15 является Gly.
12. Пептид или фармацевтически приемлемая соль указанного пептида, согласно воплощению 9, где
X2 и X4 представляют собой Lys, Orn, D-Lys или D-Orn;
X5 представляет собой Gln, Lys, D-Gln или D-Lys;
X9 является ахиральным кислым аминокислотным остатком, D-кислым аминокислотным остатком или L-кислым аминокислотным остатком;
X12 представляет собой Glu, Asn, GIn, Arg, D-Glu, D-Asn, D-Gln или D-Arg;
X13 представляет собой Glu, Asn, Gln, Arg, D-Glu, D-Asn, D-Gln или D-Arg;
X16 является ахиральным кислым аминокислотным остатком, D-кислым аминокислотным остатком или L-кислым аминокислотным остатком;
X17 представляет собой Arg, Lys, Orn, D-Arg, D-Lys или D-Orn;
X21 представляет собой Leu или D-Leu; и
X22 представляет собой Ala, Val, Leu, D-Ala, D-Val или D-Leu.
13. Пептид или фармацевтически приемлемая соль указанного пептида, согласно воплощению 1, где пептид представляет собой
Lys-Leu-Lys-Gln-Lys-Leu-Trp-Glu-Leu-Leu-Glu-Asn-Leu-Leu-Glu-Arg-Phe-Leu-Asp-Leu-Val-Inp (SEQ ID NO:2);
Lys-Leu-Lys-Gln-Lys-Leu-Ala-Glu-Leu-Gly-Glu-Asn-Leu-Leu-Glu-Arg-Phe-Leu-Asp-Leu-Val-Inp (SEQ ID NO:3);
Lys-Leu-Lys-Gln-Lys-Nal-Ala-Glu-Leu-Gly-Glu-Asn-Leu-Leu-Glu-Arg-Phe-Leu-Asp-Leu-Val-Inp (SEQ ID NO:4);
Lys-Leu-Lys-Gln-Lys-Leu-Trp-Glu-Leu-Gly-Glu-Asn-Leu-Leu-Glu-Arg-Phe-Leu-Asp-Leu-Val-Inp (SEQ ID NO:5);
Lys-Leu-Lys-Gln-Lys-Leu-Ala-Glu-Leu-Gly-Glu-Asn-Trp-Leu-Glu-Arg-Phe-Leu-Asp-Leu-Val-Inp (SEQ ID NO:6);
Orn-Leu-Orn-Gln-Orn-Leu-Ala-Glu-Leu-Gly-Glu-Asn-Leu-Leu-Glu-Arg-Phe-Leu-Asp-Leu-Val-Inp (SEQ ID NO:7);
Lys-Leu-Lys-Gln-Lys-Leu-Ala-Glu-Leu-Gly-Glu-Asn-Leu-Leu-Glu-Arg-Phe-Phe-Asp-Leu-Val-Inp (SEQ ID NO:8);
Lys-Leu-Lys-Gln-Lys-Leu-Ala-Glu-Leu-Gly-Glu-Asn-Leu-Leu-Glu-Arg-Phe-Leu-Glu-Leu-Val-Inp (SEQ ID NO:9);
Lys-Leu-Lys-Gln-Lys-Leu-Ala-Glu-Leu-Leu-Glu-Asn-Leu-Gly-Glu-Arg-Phe-Leu-Asp-Leu-Val-Inp (SEQ ID NO:10);
Lys-Leu-Lys-Gln-Lys-Leu-Ala-Glu-Leu-Leu-Glu-Asn-Gly-Leu-Glu-Arg-Phe-Leu-Asp-Leu-Val-Inp (SEQ ID NO:11);
Lys-Leu-Lys-Gln-Lys-Leu-Ala-Glu-Gly-Leu-Glu-Asn-Leu-Leu-Glu-Arg-Phe-Leu-Asp-Leu-Val-Inp (SEQ ID NO:12);
Lys-Leu-Lys-Gln-Lys-Leu-Gly-Glu-Leu-Leu-Glu-Asn-Leu-Leu-Glu-Arg-Phe-Leu-Asp-Leu-Val-Inp (SEQ ID NO:13);
Lys-Leu-Lys-Gln-Lys-Gly-Ala-Glu-Leu-Leu-Glu-Asn-Leu-Leu-Glu-Arg-Phe-Leu-Asp-Leu-Val-Inp (SEQ ID NO:14);
Lys-Leu-Lys-Gln-Lys-Leu-Nal-Glu-Leu-Leu-Glu-Asn-Leu-Leu-Glu-Arg-Phe-Leu-Asp-Leu-Val-Inp (SEQ ID NO:15);
Lys-Leu-Lys-Gln-Lys-Leu-Ala-Glu-Leu-Leu-Glu-Asn-Leu-Leu-Glu-Arg-Phe-Leu-Asp-Leu-Val-Inp (SEQ ID NO:16);
Lys-Leu-Lys-Gln-Lys-Leu-Ala-Glu-Leu-Aib-Glu-Asn-Leu-Leu-Glu-Arg-Phe-Leu-Asp-Leu-Val-Inp (SEQ ID NO:18);
Lys-Leu-Lys-Gln-Lys-Leu-Ala-Glu-Leu-Leu-Glu-Asn-Leu-Leu-Glu-Lys-Phe-Leu-Asp-Leu-Val-Inp (SEQ ID NO:19);
Lys-Leu-Lys-Gln-Lys-Nal-Ala-Glu-Leu-Leu-Glu-Asn-Leu-Leu-Glu-Arg-Phe-Leu-Asp-Leu-Val-Inp (SEQ ID NO:20);
Lys-Leu-Lys-Gln-Lys-Leu-Ala-Glu-Leu-Leu-Glu-Gln-Leu-Leu-Glu-Lys-Phe-Leu-Asp-Leu-Val-Inp (SEQ ID NO:21);
Orn-Leu-Orn-Gln-Orn-Leu-Ala-Glu-Leu-Leu-Glu-Asn-Leu-Leu-Glu-Arg-Phe-Leu-Asp-Leu-Val-Inp (SEQ ID NO:22);
Lys-Leu-Lys-Gln-Lys-Leu-Ala-Glu-Leu-Leu-Glu-Asn-Trp-Leu-Glu-Arg-Phe-Leu-Asp-Leu-Val-Inp (SEQ ID NO:23);
Lys-Leu-Lys-Gln-Lys-Leu-Ala-Glu-Leu-Leu-Glu-Asn-Leu-Leu-Glu-Arg-Leu-Leu-Asp-Leu-Val-Inp (SEQ ID NO:24);
Lys-Leu-Lys-Gln-Lys-Leu-Ala-Glu-Leu-Leu-Glu-Gln-Leu-Leu-Glu-Lys-Phe-Leu-Glu-Leu-Val-Inp (SEQ ID NO:25);
Lys-Leu-Lys-Gln-Lys-Leu-Ala-Glu-Leu-Leu-Glu-Gln-Leu-Leu-Glu-Lys-Phe-Leu-Glu-Leu-Leu-Inp (SEQ ID NO:26);
Lys-Leu-Lys-Gln-Lys-Leu-Ala-Glu-Leu-Aib-Glu-Asn-Leu-Leu-Glu-Arg-Phe-Phe-Asp-Leu-Val-Inp (SEQ ID NO:28);
Lys-Leu-Lys-Gln-Lys-Leu-Leu-Glu-Leu-Leu-Glu-Asn-Leu-Leu-Glu-Arg-Phe-Leu-Asp-Leu-Val-Inp (SEQ ID NO:29);
Lys-Leu-Lys-Gln-Lys-Leu-Ala-Glu-Nal-Leu-Glu-Asn-Leu-Leu-Glu-Arg-Phe-Leu-Asp-Leu-Val-Inp (SEQ ID NO:30);
Lys-Leu-Lys-Gln-Lys-Leu-Ala-Glu-Trp-Leu-Glu-Asn-Leu-Leu-Glu-Arg-Phe-Leu-Asp-Leu-Val-Inp (SEQ ID NO:31);
Orn-Leu-Orn-Gln-Orn-Leu-Ala-Glu-Leu-Leu-Glu-Asn-Leu-Leu-Glu-Orn-Phe-Leu-Asp-Leu-Val-Inp (SEQ ID NO:32);
Lys-Leu-Lys-Gln-Lys-Leu-Phe-Glu-Leu-Leu-Glu-Asn-Leu-Leu-Glu-Arg-Phe-Leu-Asp-Leu-Val-Inp (SEQ ID NO:33);
Lys-Leu-Lys-Gln-Arg-Leu-Ala-Asp-Leu-Leu-Glu-Asn-Leu-Leu-Glu-Lys-Phe-Leu-Glu-Leu-Val-Inp (SEQ ID NO:36);
Lys-Leu-Lys-Gln-Lys-Leu-Ala-Glu-Leu-Leu-Glu-Gln-Leu-Leu-Asp-Lys-Phe-Leu-Glu-Leu-Ala-Inp (SEQ ID NO:40);
Lys-Leu-Lys-Gln-Lys-Leu-Trp-Glu-Leu-Leu-Glu-Asn-Leu-Leu-Glu-Arg-Phe-Leu-Asp-Leu-Val-Nip (SEQ ID NO:94);
Lys-Leu-Lys-Gln-Lys-Leu-Ala-Glu-Leu-Gly-Glu-Asn-Leu-Leu-Glu-Arg-Phe-Leu-Asp-Leu-Val-Nip (SEQ ID NO:95);
Lys-Leu-Lys-Gln-Lys-Nal-Ala-Glu-Leu-Gly-Glu-Asn-Leu-Leu-Glu-Arg-Phe-Leu-Asp-Leu-Val-Nip (SEQ ID NO:96);
Lys-Leu-Lys-Gln-Lys-Leu-Trp-Glu-Leu-Gly-Glu-Asn-Leu-Leu-Glu-Arg-Phe-Leu-Asp-Leu-Val-Nip (SEQ ID NO:97);
Lys-Leu-Lys-Gln-Lys-Leu-Ala-Glu-Leu-Gly-Glu-Asn-Trp-Leu-Glu-Arg-Phe-Leu-Asp-Leu-Val-Nip (SEQ ID NO:98);
Orn-Leu-Orn-Gln-Orn-Leu-Ala-Glu-Leu-Gly-Glu-Asn-Leu-Leu-Glu-Arg-Phe-Leu-Asp-Leu-Val-Nip (SEQ ID NO:99);
Lys-Leu-Lys-Gln-Lys-Leu-Ala-Glu-Leu-Gly-Glu-Asn-Leu-Leu-Glu-Arg-Phe-Phe-Asp-Leu-Val-Nip (SEQ ID NO:100);
Lys-Leu-Lys-Gln-Lys-Leu-Ala-Glu-Leu-Gly-Glu-Asn-Leu-Leu-Glu-Arg-Phe-Leu-Glu-Leu-Val-Nip (SEQ ID NO:101);
Lys-Leu-Lys-Gln-Lys-Leu-Ala-Glu-Leu-Leu-Glu-Asn-Leu-Gly-Glu-Arg-Phe-Leu-Asp-Leu-Val-Nip (SEQ ID NO:102);
Lys-Leu-Lys-Gln-Lys-Leu-Ala-Glu-Leu-Leu-Glu-Asn-Gly-Leu-Glu-Arg-Phe-Leu-Asp-Leu-Val-Nip (SEQ ID NO:103);
Lys-Leu-Lys-Gln-Lys-Leu-Ala-Glu-Gly-Leu-Glu-Asn-Leu-Leu-Glu-Arg-Phe-Leu-Asp-Leu-Val-Nip (SEQ ID NO:104);
Lys-Leu-Lys-Gln-Lys-Leu-Gly-Glu-Leu-Leu-Glu-Asn-Leu-Leu-Glu-Arg-Phe-Leu-Asp-Leu-Val-Nip (SEQ ID NO:105);
Lys-Leu-Lys-Gln-Lys-Gly-Ala-Glu-Leu-Leu-Glu-Asn-Leu-Leu-Glu-Arg-Phe-Leu-Asp-Leu-Val-Nip (SEQ ID NO:106);
Lys-Leu-Lys-Gln-Lys-Leu-Nal-Glu-Leu-Leu-Glu-Asn-Leu-Leu-Glu-Arg-Phe-Leu-Asp-Leu-Val-Nip (SEQ ID NO:107);
Lys-Leu-Lys-Gln-Lys-Leu-Ala-Glu-Leu-Leu-Glu-Asn-Leu-Leu-Glu-Arg-Phe-Leu-Asp-Leu-Val-Nip (SEQ ID NO:108);
Lys-Leu-Lys-Gln-Lys-Leu-Ala-Glu-Leu-Aib-Glu-Asn-Leu-Leu-Glu-Arg-Phe-Leu-Asp-Leu-Val-Nip (SEQ ID NO:110);
Lys-Leu-Lys-Gln-Lys-Leu-Ala-Glu-Leu-Leu-Glu-Asn-Leu-Leu-Glu-Lys-Phe-Leu-Asp-Leu-Val-Nip (SEQ ID NO:111);
Lys-Leu-Lys-Gln-Lys-Nal-Ala-Glu-Leu-Leu-Glu-Asn-Leu-Leu-Glu-Arg-Phe-Leu-Asp-Leu-Val-Nip (SEQ ID NO:112);
Lys-Leu-Lys-Gln-Lys-Leu-Ala-Glu-Leu-Leu-Glu-Gln-Leu-Leu-Glu-Lys-Phe-Leu-Asp-Leu-Val-Nip (SEQ ID NO:113);
Orn-Leu-Orn-Gln-Orn-Leu-Ala-Glu-Leu-Leu-Glu-Asn-Leu-Leu-Glu-Arg-Phe-Leu-Asp-Leu-Val-Nip (SEQ ID NO:114);
Lys-Leu-Lys-Gln-Lys-Leu-Ala-Glu-Leu-Leu-Glu-Asn-Trp-Leu-Glu-Arg-Phe-Leu-Asp-Leu-Val-Nip (SEQ ID NO:115);
Lys-Leu-Lys-Gln-Lys-Leu-Ala-Glu-Leu-Leu-Glu-Asn-Leu-Leu-Glu-Arg-Leu-Leu-Asp-Leu-Val-Nip (SEQ ID NO:116);
Lys-Leu-Lys-Gln-Lys-Leu-Ala-Glu-Leu-Leu-Glu-Gln-Leu-Leu-Glu-Lys-Phe-Leu-Glu-Leu-Val-Nip (SEQ ID NO:117);
Lys-Leu-Lys-Gln-Lys-Leu-Ala-Glu-Leu-Leu-Glu-Gln-Leu-Leu-Glu-Lys-Phe-Leu-Glu-Leu-Leu-Nip (SEQ ID NO:118);
Lys-Leu-Lys-Gln-Lys-Leu-Ala-Glu-Leu-Aib-Glu-Asn-Leu-Leu-Glu-Arg-Phe-Phe-Asp-Leu-Val-Nip (SEQ ID NO:120);
Lys-Leu-Lys-Gln-Lys-Leu-Leu-Glu-Leu-Leu-Glu-Asn-Leu-Leu-Glu-Arg-Phe-Leu-Asp-Leu-Val-Nip (SEQ ID NO:121);
Lys-Leu-Lys-Gln-Lys-Leu-Ala-Glu-Nal-Leu-Glu-Asn-Leu-Leu-Glu-Arg-Phe-Leu-Asp-Leu-Val-Nip (SEQ ID NO:122);
Lys-Leu-Lys-Gln-Lys-Leu-Ala-Glu-Trp-Leu-Glu-Asn-Leu-Leu-Glu-Arg-Phe-Leu-Asp-Leu-Val-Nip (SEQ ID NO:123);
Orn-Leu-Orn-Gln-Orn-Leu-Ala-Glu-Leu-Leu-Glu-Asn-Leu-Leu-Glu-Orn-Phe-Leu-Asp-Leu-Val-Nip (SEQ ID NO:124);
Lys-Leu-Lys-Gln-Lys-Leu-Phe-Glu-Leu-Leu-Glu-Asn-Leu-Leu-Glu-Arg-Phe-Leu-Asp-Leu-Val-Nip (SEQ ID NO:125);
Lys-Leu-Lys-Gln-Arg-Leu-Ala-Asp-Leu-Leu-Glu-Asn-Leu-Leu-Glu-Lys-Phe-Leu-Glu-Leu-Val-Nip (SEQ ID NO:128); или
Lys-Leu-Lys-Gln-Lys-Leu-Ala-Glu-Leu-Leu-Glu-Gln-Leu-Leu-Asp-Lys-Phe-Leu-Glu-Leu-Ala-Nip (SEQ ID NO:132), или
фармацевтически приемлемую соль одного из вышеперечисленных [соединений].
14. Пептид или фармацевтически приемлемая соль пептида, согласно воплощению 5, где указанный пептид является пептидом, содержащим 23 пептидных остатка.
15. Пептид или фармацевтически приемлемая соль пептида, согласно воплощению 14, где указанный пептид представляет собой
Lys-Lys-Leu-Lys-Gln-Lys-Leu-Ala-Glu-Leu-Leu-Glu-Asn-Leu-Leu-Glu-Arg-Phe-Leu-Asp-Leu-Val-Inp (SEQ ID NO:17); или
Lys-Lys-Leu-Lys-Gln-Lys-Leu-Ala-Glu-Leu-Leu-Glu-Asn-Leu-Leu-Glu-Arg-Phe-Leu-Asp-Leu-Val-Nip (SEQ ID NO:109), или
фармацевтически приемлемую соль одного из вышеперечисленных [соединений].
16. Пептид или фармацевтически приемлемая соль указанного пептида, согласно воплощению 5, где X1 отсутствует, а пептид является пептидом, содержащим 22 пептидных остатка.
17. Пептид или фармацевтически приемлемая соль указанного пептида, согласно воплощению 16, где пептид представляет собой
Lys-Leu-Lys-Lys-Gln-Leu-Ala-Glu-Leu-Leu-Glu-Arg-Leu-Leu-Asp-Asn-Phe-Leu-Glu-Leu-Val-Inp (SEQ ID NO:34);
Lys-Leu-Lys-Gln-Lys-Leu-Ala-Glu-Leu-Leu-Glu-Arg-Leu-Leu-Asp-Asn-Phe-Leu-Glu-Leu-Val-Inp (SEQ ID NO:35);
Lys-Leu-Lys-Lys-Gln-Leu-Ala-Glu-Leu-Leu-Glu-Arg-Leu-Leu-Asp-Asn-Phe-Leu-Glu-Leu-Val-Nip (SEQ ID NO:126); или
Lys-Leu-Lys-Gln-Lys-Leu-Ala-Glu-Leu-Leu-Glu-Arg-Leu-Leu-Asp-Asn-Phe-Leu-Glu-Leu-Val-Nip (SEQ ID NO:127) или
фармацевтически приемлемую соль одного из вышеперечисленных [соединений].
18. Пептид или фармацевтически приемлемая соль указанного пептида, согласно воплощению 16, где
X9 представляет собой Gln, Lys, D-Gln, D-Lys, ахиральный кислый аминокислотный остаток, D-кислый аминокислотный остаток или L-кислый аминокислотный остаток;
X12 представляет собой Asn, D-Asn, ахиральный кислый аминокислотный остаток, D-кислый аминокислотный остаток или L-кислый аминокислотный остаток; и
X17 представляет собой Asn, Glu, D-Asn, D-Glu, ахиральный основный аминокислотный остаток, D-основный аминокислотный остаток или L-основный аминокислотный остаток.
19. Пептид или фармацевтически приемлемая соль указанного пептида, согласно воплощению 16, где
X9 представляет собой Gln, Lys, D-Gln, D-Lys, ахиральный кислый аминокислотный остаток, D-кислый аминокислотный остаток или L-кислый аминокислотный остаток;
X12 представляет собой Asn, D-Asn, ахиральный кислый аминокислотный остаток, D-кислый аминокислотный остаток или L-кислый аминокислотный остаток; и
X17 представляет собой Asn, Glu, D-Asn, D-Glu, ахиральный основный аминокислотный остаток, D-основный аминокислотный остаток или L-основный аминокислотный остаток.
20. Пептид или фармацевтически приемлемая соль указанного пептида, согласно воплощению 16, где
X2 представляет собой Lys, Orn, D-Lys или D-Orn;
X3 представляет собой Leu или D-Leu;
X4 представляет собой Lys, Orn, D-Lys или D-Orn;
X5 представляет собой Lys, Orn, GIn, Asn, D-Lys, D-Orn, D-Gln или D-Asn;
X6 представляет собой Lys, Orn, D-Lys или D-Orn;
X7 представляет собой Leu, Gly, Nal, D-Leu или D-Nal;
X8 представляет собой Ala, Trp, Gly, Leu, Phe, Nal, D-Ala, D-Trp, D-Leu, D-Phe или D-Nal;
X9 представляет собой Asp, Glu, Gln, Lys, D-Asp, D-Glu, D-Gln или D-Lys;
X11 представляет собой Leu, Gly, Aib или D-Leu;
X12 представляет собой Asp, Glu, Asn, D-Asp, D-Glu или D-Asn;
X13 представляет собой Asn, Gln, Glu, Arg, D-Asn, D-Gln, D-Glu или D-Arg;
X16 представляет собой Asp, Glu, D-Asp или D-Glu;
X17 представляет собой Lys, Arg, Orn, Asn, Glu, D-Lys, D-Arg, D-Orn, D-Asn или D-Glu;
X20 представляет собой Asp, Glu, D-Asp или D-Glu; и
X22 представляет собой Ala, Val, Leu, D-AIa, D-Val или D-Leu.
21. Пептид или фармацевтически приемлемая соль указанного пептида, согласно воплощению 16, где
X9 представляет собой Glu или D-Glu;
X12 представляет собой Glu или D-Glu;
X13 представляет собой Asn, Glu, D-Asn или D-Glu;
X14 представляет собой Leu или D-Leu;
X15 представляет собой Leu или D-Leu;
X16 представляет собой Glu или D-Glu;
X17 представляет собой Arg, Lys, D-Arg или D-Lys;
X18 представляет собой Phe или D-Phe;
X19 представляет собой Leu или D-Leu;
X21 представляет собой Leu или D-Leu; и
X22 представляет собой Val или D-Val.
22. Пептид или фармацевтически приемлемая соль указанного пептида, согласно воплощению 16, где X11 представляет собой Gly, а каждый из X7, X8, X10, X14 и X15 является отличным от Gly.
23. Пептид или фармацевтически приемлемая соль указанного пептида, согласно воплощению 16, где
X2 представляет собой Lys, Orn, D-Lys или D-Orn;
X3 представляет собой Leu или D-Leu;
X4 является Lys, Orn, D-Lys или D-Orn;
X5 представляет собой Gln или D-Gln;
X6 представляет собой Lys, Orn, D-Lys или D-Orn;
X7 представляет собой Leu, Nal, D-Leu или D-Nal;
X8 представляет собой Ala, Trp, D-Ala или D-Trp;
X9 представляет собой Glu или D-Glu;
X10 представляет собой Leu или D-Leu;
X11 является Gly;
X12 представляет собой Glu или D-Glu;
X13 представляет собой Asn или D-Asn;
X14 представляет собой Leu, Trp, D-Leu или D-Trp;
X15 представляет собой Leu или D-Leu;
X16 представляет собой Glu или D-Glu;
X17 представляет собой Arg или D-Arg;
X18 представляет собой Phe или D-Phe;
X19 представляет собой Leu, Phe, D-Leu или D-Phe;
X20 представляет собой Asp, Glu, D-Asp или D-Glu;
X21 представляет собой Leu или D-Leu; и
X22 представляет собой Val или D-Val.
24. Пептид или фармацевтически приемлемая соль указанного пептида, согласно воплощению 20, где указанный пептид представляет собой
Lys-Leu-Lys-Gln-Lys-Leu-Trp-Glu-Leu-Leu-Glu-Asn-Leu-Leu-Glu-Arg-Phe-Leu-Asp-Leu-Val-Inp (SEQ ID NO:2);
Lys-Leu-Lys-Gln-Lys-Leu-Ala-Glu-Leu-Gly-Glu-Asn-Leu-Leu-Glu-Arg-Phe-Leu-Asp-Leu-Val-Inp (SEQ ID NO:3);
Lys-Leu-Lys-Gln-Lys-Nal-Ala-Glu-Leu-Gly-Glu-Asn-Leu-Leu-Glu-Arg-Phe-Leu-Asp-Leu-Val-Inp (SEQ ID NO:4);
Lys-Leu-Lys-Gln-Lys-Leu-Trp-Glu-Leu-Gly-Glu-Asn-Leu-Leu-Glu-Arg-Phe-Leu-Asp-Leu-Val-Inp (SEQ ID NO:5);
Lys-Leu-Lys-Gln-Lys-Leu-Ala-Glu-Leu-Gly-Glu-Asn-Trp-Leu-Glu-Arg-Phe-Leu-Asp-Leu-Val-Inp (SEQ ID NO:6);
Orn-Leu-Orn-Gln-Orn-Leu-Ala-Glu-Leu-Gly-Glu-Asn-Leu-Leu-Glu-Arg-Phe-Leu-Asp-Leu-Val-Inp (SEQ ID NO:7);
Lys-Leu-Lys-Gln-Lys-Leu-Ala-Glu-Leu-Gly-Glu-Asn-Leu-Leu-Glu-Arg-Phe Phe-Asp-Leu-Val-Inp (SEQ ID NO:8);
Lys-Leu-Lys-Gln-Lys-Leu-Ala-Glu-Leu-Gly-Glu-Asn-Leu-Leu-Glu-Arg-Phe-Leu-Glu-Leu-Val-Inp (SEQ ID NO:9);
Lys-Leu-Lys-Gln-Lys-Leu-Trp-Glu-Leu-Leu-Glu-Asn-Leu-Leu-Glu-Arg-Phe-Leu-Asp-Leu-Val-Nip (SEQ ID NO:94);
Lys-Leu-Lys-Gln-Lys-Leu-Ala-Glu-Leu-Gly-Glu-Asn-Leu-Leu-Glu-Arg-Phe-Leu-Asp-Leu-Val-Nip (SEQ ID NO:95);
Lys-Leu-Lys-Gln-Lys-Nal-Ala-Glu-Leu-Gly-Glu-Asn-Leu-Leu-Glu-Arg-Phe-Leu-Asp-Leu-Val-Nip (SEQ ID NO:96);
Lys-Leu-Lys-Gln-Lys-Leu-Trp-Glu-Leu-Gly-Glu-Asn-Leu-Leu-Glu-Arg-Phe-Leu-Asp-Leu-Val-Nip (SEQ ID NO:97);
Lys-Leu-Lys-Gln-Lys-Leu-Ala-Glu-Leu-Gly-Glu-Asn-Trp-Leu-Glu-Arg-Phe-Leu-Asp-Leu-Val-Nip (SEQ ID NO:98);
Orn-Leu-Orn-Gln-Orn-Leu-Ala-Glu-Leu-Gly-Glu-Asn-Leu-Leu-Glu-Arg-Phe-Leu-Asp-Leu-Val-Nip (SEQ ID NO:99);
Lys-Leu-Lys-Gln-Lys-Leu-Ala-Glu-Leu-Gly-Glu-Asn-Leu-Leu-Glu-Arg-Phe-Phe-Asp-Leu-Val-Nip (SEQ ID NO:100); или
Lys-Leu-Lys-Gln-Lys-Leu-Ala-Glu-Leu-Gly-Glu-Asn-Leu-Leu-Glu-Arg-Phe-Leu-Glu-Leu-Val-Nip (SEQ ID NO:101), или
фармацевтически приемлемую соль одного из вышеперечисленных [соединений].
25. Пептид или фармацевтически приемлемая соль указанного пептида, согласно воплощению 16, где X15 является Gly, а каждый из X7, X8, X10, X11 и X14 является отличным от Gly.
26. Пептид или фармацевтически приемлемая соль указанного пептида, согласно воплощению 25, где указанный пептид представляет собой
Lys-Leu-Lys-Gln-Lys-Leu-Ala-Glu-Leu-Leu-Glu-Asn-Leu-Gly-Glu-Arg-Phe-Leu-Asp-Leu-Val-Inp (SEQ ID NO:10); или
Lys-Leu-Lys-Gln-Lys-Leu-Ala-Glu-Leu-Leu-Glu-Asn-Leu-Gly-Glu-Arg-Phe-Leu-Asp-Leu-Val-Nip (SEQ ID NO:102), или
фармацевтически приемлемую соль одного из вышеперечисленных [соединений].
27. Пептид или фармацевтически приемлемая соль указанного пептида, согласно воплощению 16, где X14 является Gly, а каждый из X7, X8, X10, X11 и X15 является отличным от Gly.
28. Пептид или фармацевтически приемлемая соль указанного пептида, согласно воплощению 27, где указанный пептид представляет собой
Lys-Leu-Lys-Gln-Lys-Leu-Ala-Glu-Leu-Leu-Glu-Asn-Gly-Leu-Glu-Arg-Phe-Leu-Asp-Leu-Val-Inp (SEQ ID NO:11); или
Lys-Leu-Lys-Gln-Lys-Leu-Ala-Glu-Leu-Leu-Glu-Asn-Gly-Leu-Glu-Arg-Phe-Leu-Asp-Leu-Val-Nip (SEQ ID NO:103), или
фармацевтически приемлемую соль одного из вышеперечисленных [соединений].
29. Пептид или фармацевтически приемлемая соль указанного пептида, согласно воплощению 16, где X10 является Gly, а каждый из X7, X8, X11, X14 и X15 является отличным от Gly.
30. Пептид или фармацевтически приемлемая соль указанного пептида, согласно воплощению 29, где указанный пептид представляет собой
Lys-Leu-Lys-Gln-Lys-Leu-Ala-Glu-Gly-Leu-Glu-Asn-Leu-Leu-Glu-Arg-Phe-Leu-Asp-Leu-Val-Inp (SEQ ID NO:12); или
Lys-Leu-Lys-Gln-Lys-Leu-Ala-Glu-Gly-Leu-Glu-Asn-Leu-Leu-Glu-Arg-Phe-Leu-Asp-Leu-Val-Nip (SEQ ID NO:104), или
фармацевтически приемлемую соль одного из вышеперечисленных [соединений].
31. Пептид или фармацевтически приемлемая соль указанного пептида, согласно воплощению 16, где X8 является Gly, а каждый из X7, X10, X11, X14 и X15 является отличным от Gly.
32. Пептид или фармацевтически приемлемая соль указанного пептида, согласно воплощению 31, где указанный пептид представляет собой
Lys-Leu-Lys-Gln-Lys-Leu-Gly-Glu-Leu-Leu-Glu-Asn-Leu-Leu-Glu-Arg-Phe-Leu-Asp-Leu-Val-Inp (SEQ ID NO:13); или
Lys-Leu-Lys-Gln-Lys-Leu-Gly-Glu-Leu-Leu-Glu-Asn-Leu-Leu-Glu-Arg-Phe-Leu-Asp-Leu-Val-Nip (SEQ ID NO:105), или
фармацевтически приемлемую соль одного из вышеперечисленных [соединений].
33. Пептид или фармацевтически приемлемая соль указанного пептида, согласно воплощению 16, где X7 является Gly, а каждый из X8, X10, X11, X14 и X15 является отличным от Gly.
34. Пептид или фармацевтически приемлемая соль указанного пептида, согласно воплощению 33, где указанный пептид представляет собой
Lys-Leu-Lys-Gln-Lys-Gly-Ala-Glu-Leu-Leu-Glu-Asn-Leu-Leu-Glu-Arg-Phe-Leu-Asp-Leu-Val-Inp (SEQ ID NO:14); или
Lys-Leu-Lys-Gln-Lys-Gly-Ala-Glu-Leu-Leu-Glu-Asn-Leu-Leu-Glu-Arg-Phe-Leu-Asp-Leu-Val-Nip (SEQ ID NO:106), или
фармацевтически приемлемую соль одного из вышеперечисленных [соединений].
35. Пептид или фармацевтически приемлемая соль указанного пептида, согласно воплощению 1, где указанный пептид представляет собой
Lys-Leu-Lys-Gln-Lys-Leu-Ala-Glu-Leu-Leu-Glu-Asn-Leu-Leu-Glu-Arg-Phe-Leu-Asp-Leu-Val-Inp (SEQ ID NO:16); или
Lys-Leu-Lys-Gln-Lys-Leu-Ala-Glu-Leu-Leu-Glu-Asn-Leu-Leu-Glu-Arg-Phe-Leu-Asp-Leu-Val-Nip (SEQ ID NO:108), или
фармацевтически приемлемую соль одного из вышеперечисленных [соединений].
36. Пептид, содержащий от 15 до 22 остатков, имеющий следующую формулу II:
R1-Y1-X1-X2-X3-X4-X5-X6-X7-X8-X9-X10-X11-X12-X13-X14-X15-X16-X17-X18-Y2-R2
Формула II
или его фармацевтически приемлемая соль, где
X1 является ахиральным, D- или L-основным аминокислотным остатком;
X2 представляет собой Leu или D-Leu;
X3 является ахиральным, D- или L-основным аминокислотным остатком;
X4 представляет собой Gln, Asn, D-Gln или D-Asn;
X5 представляет собой Leu, D-Leu или является ахиральным, D- или L-основным аминокислотным остатком;
X6 представляет собой Leu, Trp, Phe, D-Leu, D-Trp или D-Phe;
X7 является ахиральным, D- или L-кислым аминокислотным остатком;
X8 представляет собой Asn, D-Asn или является ахиральным, D- или L-кислым аминокислотным остатком;
X9 представляет собой Leu, Trp, D-Leu или D-Trp;
X10 представляет собой Leu, Trp, D-Leu или D-Trp;
X11 представляет собой ахиральный, D- или L-кислый аминокислотный остаток;
X12 является ахиральным, D- или L-основным аминокислотным остатком;
X13 представляет собой Leu, Phe, D-Leu или D-Phe;
X14 представляет собой Leu, Phe, D-Leu или D-Phe;
X15 является ахиральным, D- или L-кислым аминокислотным остатком;
X16 представляет собой Leu или D-Leu;
X17 является ахиральным, D- или L-алифатическим аминокислотным остатком;
X18 представляет собой Inp, Nip, azPro, Pip, azPip, D-Nip или D-Pip;
Y1 отсутствует или является аминокислотной последовательностью, содержащей от 1 до 4 остатков;
Y2 отсутствует;
R1 представляет собой H или аминозащитную группу;
R2 представляет собой OH или карбоксильную защитную группу;
причем от нуля до трех остатков от X1 до X17 отсутствуют; и при этом
a) каждый хиральный аминокислотный остаток является L-аминокислотным остатком;
b) каждый хиральный аминокислотный остаток является D-аминокислотным остатком;
c) каждый хиральный аминокислотный остаток является L-аминокислотным остатком, за исключением того, что один или более из каждых хиральных концевых аминокислотных остатков и каждых хиральных аминокислотных остатков, непосредственно к ним примыкающих, является D-аминокислотным остатком; или
d) каждый хиральный аминокислотный остаток является D-аминокислотным остатком, за исключением того, что один или более из каждых хиральных концевых аминокислотных остатков и каждых хиральных аминокислотных остатков, непосредственно к ним примыкающих, является L-аминокислотным остатком.
37. Пептид или фармацевтически приемлемая соль указанного пептида, согласно воплощению 36, где
X17 представляет собой Ala, Leu, Val, D-Ala, D-Leu или D-Val.
38. Пептид или фармацевтически приемлемая соль указанного пептида, согласно воплощению 36, где
X1 представляет собой His, Lys, Arg, D-His, D-Lys или D-Arg;
X3 представляет собой Lys, Arg, Orn, D-Lys, D-Arg или D-Orn;
X5 представляет собой Lys, Arg, Orn, D-Lys, D-Arg или D-Orn;
X7 представляет собой Glu или D-Glu;
X8 представляет собой Asn, Glu, D-Asn или D-Glu;
X11 представляет собой Asp, Glu, D-Asp или D-Glu;
X12 представляет собой Arg, Lys, Orn, D-Arg, D-Lys или D-Orn; и
X15 представляет собой Asp, Glu, D-Asp или D-Glu.
39. Пептид или фармацевтически приемлемая соль указанного пептида, согласно воплощению 38, где
X13 представляет собой Phe или D-Phe.
40. Пептид или фармацевтически приемлемая соль указанного пептида, согласно воплощению 36, где указанный пептид является пептидом, содержащим 18 остатков.
41. Пептид или фармацевтически приемлемая соль указанного пептида, согласно воплощению 40, где R1 представляет собой H, и R2 представляет собой OH.
42. Пептид или фармацевтически приемлемая соль указанного пептида, согласно воплощению 41, где
X1 представляет собой Arg, Lys, Orn, D-Arg, D-Lys или D-Orn;
X3 представляет собой Arg, Lys, Orn, D-Arg, D-Lys или D-Orn;
X5 представляет собой Arg, Lys, Orn, D-Arg, D-Lys или D-Orn;
X7 представляет собой Glu или D-Glu;
X8 представляет собой Glu, Asn, D-Glu или D-Asn;
X11 представляет собой Glu, Asp, D-Glu или D-Asp;
X12 представляет собой Arg, Lys, Orn, D-Arg, D-Lys или D-Orn;
X15 представляет собой Asp, Glu, D-Asp или D-Glu; и
X17 представляет собой Ala, VaI, Leu, D-AIa, D-Val или D-Leu.
43. Пептид или фармацевтически приемлемая соль указанного пептида, согласно воплощению 36, где указанный пептид представляет собой
Lys-Leu-Lys-Gln-Leu-Leu-Glu-Asn-Leu-Leu-Glu-Arg-Phe-Leu-Asp-Leu-Val-Inp (SEQ ID NO:53);
Lys-Leu-Lys-Gln-Lys-Leu-Glu-Glu-Leu-Leu-Glu-Arg-Phe-Leu-Asp-Leu-Val-Inp (SEQ ID NO:54);
Lys-Leu-Lys-Gln-Leu-Leu-Glu-Asn-Leu-Leu-Glu-Arg-Phe-Leu-Asp-Leu-Val-Nip (SEQ ID NO:145); или
Lys-Leu-Lys-Gln-Lys-Leu-Glu-Glu-Leu-Leu-Glu-Arg-Phe-Leu-Asp-Leu-Val-Nip (SEQ ID NO:146), или
фармацевтически приемлемую соль одного из вышеперечисленных [соединений].
44. Пептид или фармацевтически приемлемая соль указанного пептида, согласно воплощению 36, где указанный пептид представляет собой
H3C(O)C-Lys-Leu-Lys-Gln-Lys-Leu-Glu-Glu-Leu-Leu-Glu-Arg-Leu-Leu-Asp-Leu-Val-Inp-NH2 (SEQ ID NO:65);
H3C(O)C-Lys-Leu-Lys-Gln-Lys-Leu-Glu-Glu-Leu-Leu-Glu-Lys-Phe-Leu-Asp-Leu-Val-Inp-NH2 (SEQ ID NO:66);
H3C(O)C-Lys-Leu-Lys-Gln-Lys-Leu-Glu-Glu-Leu-Leu-Glu-Arg-Phe-Leu-Asp-Leu-Val-Inp-NH2 (SEQ ID NO:67);
H3C(O)C-Lys-Leu-Lys-Gln-Lys-Leu-Glu-Glu-Leu-Leu-Glu-Arg-Phe-Leu-Glu-Leu-Val-Inp-NH2 (SEQ ID NO:68);
H3C(O)C-Lys-LeU-Lys-Gln-Lys-LeU-Glu-Glu-Leu-Leu-Glu-Lys-Phe-Leu-Glu-Leu-Val-Inp-NH2 (SEQ ID NO:69);
H3C(O)C-Lys-Leu-Lys-Asn-Lys-Leu-Glu-Glu-Leu-Leu-Glu-Arg-Phe-Leu-Asp-Leu-Val-Inp-NH2 (SEQ ID NO:70);
H3C(O)C-Lys-Leu-Lys-Gln-Lys-Leu-Glu-Glu-Leu-Leu-Glu-Arg-Phe-Leu-Asp-Leu-Leu-Inp-NH2 (SEQ ID NO:71);
H3C(O)C-Lys-Leu-Lys-Gln-Lys-Leu-Glu-Glu-Trp-Leu-Glu-Arg-Phe-Leu-Asp-Leu-Val-Inp-NH2 (SEQ ID NO:72);
H3C(O)C-Lys-Leu-Lys-Gln-Lys-Leu-Glu-Glu-Leu-Leu-Glu-Lys-Phe-Leu-Asp-Leu-Leu-Inp-NH2 (SEQ ID NO:73);
H3C(O)C-Arg-Leu-Lys-Gln-Arg-Leu-Glu-Glu-Leu-Leu-Asp-Lys-Phe-Leu-Glu-Leu-Ala-Inp-NH2 (SEQ ID NO:74);
H3C(O)C-Lys-Leu-Lys-Gln-Lys-Leu-Glu-Glu-Leu-Leu-Glu-Arg-Phe-Phe-Asp-Leu-Val-Inp-NH2 (SEQ ID NO:75);
H3C(O)C-Lys-Leu-Lys-Gln-Lys-Leu-Glu-Glu-Leu-Trp-Glu-Arg-Phe-Leu-Asp-Leu-Val-Inp-NH2 (SEQ ID NO:76);
H3C(O)C-Lys-Leu-Lys-Gln-Lys-Leu-Glu-Glu-Leu-Leu-Glu-Lys-Leu-Leu-Glu-Leu-Leu-Inp-NH2 (SEQ ID NO:77);
H3C(O)C-Lys-Leu-Lys-Gln-Lys-Leu-Glu-Glu-Leu-Leu-Glu-Lys-Leu-Leu-Glu-Leu-Val-Inp-NH2 (SEQ ID NO:78);
H3C(O)C-Lys-Leu-Arg-Gln-Arg-Leu-Glu-Glu-Leu-Leu-Asp-Lys-Phe-Leu-Glu-Leu-Ala-Inp-NH2 (SEQ ID NO:79);
H3C(O)C-Orn-Leu-Orn-Gln-Orn-Leu-Glu-Glu-Leu-Leu-Glu-Arg-Phe-Leu-Asp-Leu-Val-Inp-NH2 (SEQ ID NO:80);
H3C(O)C-Lys-Leu-Orn-Gln-Orn-Leu-Glu-Glu-Leu-Leu-Glu-Orn-Phe-Leu-Asp-Leu-Val-Inp-NH2 (SEQ ID NO:81);
H3C(O)C-Lys-Leu-Arg-Gln-Arg-Phe-Glu-Glu-Leu-Leu-Asp-Lys-Phe-Leu-Glu-Leu-Ala-Inp-NH2 (SEQ ID NO:82);
H3C(O)C-Lys-Leu-Lys-Gln-Lys-Trp-Glu-Glu-Leu-Leu-Glu-Arg-Phe-Leu-Asp-Leu-Val-Inp-NH2 (SEQ ID NO:83);
H3C(O)C-Lys-Leu-Lys-Gln-Leu-Leu-Glu-Asn-Leu-Leu-Glu-Arg-Phe-Leu-Asp-Leu-Val-Inp-NH2 (SEQ ID NO:84);
H3C(O)C-Lys-Leu-Lys-Gln-Lys-Leu-Glu-Glu-Leu-Leu-Glu-Lys-Phe-Leu-Glu-Leu-Leu-Inp-NH2 (SEQ ID NO:87);
H3C(O)C-Lys-Leu-Lys-Gln-Glu-Leu-Leu-Glu-Arg-Phe-Leu-Asp-Leu-Val-Inp-NH2 (SEQ ID NO:88);
H3C(O)C-Lys-Gln-Lys-Leu-Glu-Glu-Leu-Leu-Glu-Arg-Phe-Leu-Asp-Leu-Val-Inp-NH2 (SEQ ID NO:89);
H3C(O)C-Lys-Leu-Lys-Gln-Lys-Leu-Glu-Glu-Leu-Leu-Glu-AIg-Leu-Leu-Asp-Leu-Val-Nip-NH2 (SEQ ID NO:157);
H3C(O)C-Lys-Leu-Lys-Gln-Lys-Leu-Glu-Glu-Leu-Leu-Glu-Lys-Phe-Leu-Asp-Leu-Val-Nip-NH2 (SEQ ID NO:158);
H3C(O)C-Lys-Leu-Lys-Gln-Lys-Leu-Glu-Glu-Leu-Leu-Glu-Arg-Phe-Leu-Asp-Leu-Val-Nip-NH2 (SEQ ID NO:159);
H3C(O)C-Lys-Leu-Lys-Gln-Lys-Leu-Glu-Glu-Leu-Leu-Glu-Arg-Phe-Leu-Glu-Leu-Val-NiP-NH2 (SEQ ID NO:160);
H3C(O)C-Lys-Leu-Lys-Gln-Lys-Leu-Glu-Glu-Leu-Leu-Glu-Lys-Phe-Leu-Glu-Leu-Val-NiP-NH2 (SEQ ID NO:161);
H3C(O)C-Lys-Leu-Lys-Asn-Lys-Leu-Glu-Glu-Leu-Leu-Glu-Arg-Phe-Leu-Asp-Leu-Val-Nip-NH2 (SEQ ID NO:162);
H3C(O)C-Lys-Leu-Lys-Gln-Lys-Leu-Glu-Glu-Leu-Leu-Glu-Arg-Phe-Leu-Asp-Leu-Leu-Nip-NH2 (SEQ ID NO:163);
H3C(O)C-Lys-Leu-Lys-Gln-Lys-Leu-Glu-Glu-Trp-Leu-Glu-Arg-Phe-Leu-Asp-Leu-Val-Nip-NH2 (SEQ ID NO:164);
H3C(O)C-Lys-Leu-Lys-Gln-Lys-Leu-Glu-Glu-Leu-Leu-Glu-Lys-Phe-Leu-Asp-Leu-Leu-Nip-NH2 (SEQ ID NO:165);
H3C(O)C-Arg-Leu-Lys-Gln-Arg-Leu-Glu-Glu-Leu-Leu-Asp-Lys-Phe-Leu-Glu-Leu-Ala-Nip-NH2 (SEQ ID NO:166);
H3C(O)C-Lys-Leu-Lys-Gln-Lys-Leu-Glu-Glu-Leu-Leu-Glu-Arg-Phe-Phe-Asp-Leu-Val-Nip-NH2 (SEQ ID NO:167);
H3C(O)C-Lys-LeU-Lys-Gln-Lys-Leu-Glu-Glu-Leu-Leu-Glu-Lys-Leu-Leu-Glu-Leu-Leu-Nip-NH2 (SEQ ID NO:168);
H3C(O)C-Lys-Leu-Lys-Gln-Lys-Leu-Glu-Glu-Leu-Leu-Glu-LyS-LeU-Leu-Glu-Leu-Val-Nip-NH2 (SEQ ID NO:169);
H3C(O)C-Lys-Leu-Arg-Gln-Arg-Leu-Glu-Glu-Leu-Leu-Asp-Lys-Phe-Leu-Glu-Leu-Ala-Nip-NH2 (SEQ ID NO:170);
H3C(O)C-Orn-Leu-Orn-Gln-Orn-Leu-Glu-Glu-Leu-Leu-Glu-Arg-Phe-Leu-Asp-Leu-Val-Nip-NH2 (SEQ ID NO:171);
H3C(O)C-Lys-Leu-Orn-Gln-Orn-Leu-Glu-Glu-Leu-Leu-Glu-Orn-Phe-Leu-Asp-Leu-Val-Nip-NH2 (SEQ ID NO:172);
H3C(O)C-Lys-Leu-Arg-Gln-Arg-Phe-Glu-Glu-Leu-Leu-Asp-Lys-Phe-Leu-Glu-Leu-Ala-Nip-NH2 (SEQ ID NO:173);
H3C(O)C-Lys-Leu-Lys-Gln-Lys-Trp-Glu-Glu-Leu-Leu-Glu-Arg-Phe-Leu-Asp-Leu-Val-Nip-NH2 (SEQ ID NO:174);
H3C(O)C-Lys-Leu-Lys-Gln-Leu-Leu-Glu-Asn-Leu-Leu-Glu-Arg-Phe-Leu-Asp-Leu-Val-Nip-NH2 (SEQ ID NO:175);
H3C(O)C-Lys-Leu-Lys-Gln-Lys-Leu-Glu-Glu-Gly-Leu-Glu-Arg-Phe-Leu-Asp-Leu-Val-Nip-NH2 (SEQ ID NO:176);
H3C(O)C-Lys-Leu-Lys-Gln-Lys-Leu-Glu-Glu-Leu-Leu-Glu-Lys-Phe-Leu-Glu-Leu-Leu-Nip-NH2 (SEQ ID NO:179);
H3C(O)C-Lys-Leu-Lys-Gln-Glu-Leu-Leu-Glu-Arg-Phe-Leu-Asp-Leu-Val-Nip-NH2 (SEQ ID NO:180); или
H3C(O)C-Lys-Gln-Lys-Leu-Glu-Glu-Leu-Leu-Glu-Arg-Phe-Leu-Asp-Leu-Val-Nip-NH2 (SEQ ID NO:181), или
фармацевтически приемлемую соль одного из вышеперечисленных [соединений].
45. Пептид, содержащий от 22 до 29 остатков, имеющий следующую формулу III:
R1-Y1-X1-X2-X3-X4-X5-X6-X7-X8-X9-X10-X11-X12-X13-X14-X15-X16-X17-X18-X19-X20-X21-X22-X23-Y2-R2
Формула III
или его фармацевтически приемлемая соль, где
X1 отсутствует или является ахиральным, D- или L-основным аминокислотным остатком;
X2 является ахиральным, D- или L-основным аминокислотным остатком;
X3 представляет собой Leu или D-Leu;
X4 является ахиральным, D- или L-основным аминокислотным остатком;
X5 является ахиральным, D- или L-основным аминокислотным остатком;
X6 представляет собой Gln, Asn, D-Gln или D-Asn;
X7 представляет собой Leu или D-Leu;
X8 представляет собой Ala или D-AIa;
X9 представляет собой Asp или D-Asp;
X10 представляет собой Leu, Phe, Gly, D-Leu или D-Phe;
X11 представляет собой Gly, Leu или D-Leu;
X12 представляет собой Arg или D-Arg;
X13 является ахиральным, D- или L-кислым аминокислотным остатком;
X14 представляет собой Leu, Trp, Gly, D-Leu или D-Trp;
X15 представляет собой Leu или D-Leu;
X16 представляет собой Gln или D-Gln;
X17 представляет собой Glu, Leu, D-Glu или D-Leu;
X18 представляет собой Leu, Phe, D-Leu или D-Phe;
X19 является ахиральным, D- или L-алифатическим аминокислотным остатком;
X20 представляет собой Glu или D-Glu;
X21 представляет собой Leu, Phe, D-Leu или D-Phe;
X22 является ахиральным, D- или L-алифатическим аминокислотным остатком;
X23 представляет собой Inp, Nip, azPro, Pip, azPip, D-Nip или D-Pip;
Y1 отсутствует или является аминокислотной последовательностью, содержащей от 1 до 7 остатков;
Y2 отсутствует или является аминокислотной последовательностью, содержащей от 1 до 7 остатков;
R1 представляет собой H или аминозащитную группу;
R2 представляет собой OH или карбоксильную защитную группу;
при этом
a) каждый хиральный аминокислотный остаток является L-аминокислотным остатком;
b) каждый хиральный аминокислотный остаток является D-аминокислотным остатком;
c) каждый хиральный аминокислотный остаток является L-аминокислотным остатком, за исключением того, что один или более из каждых хиральных концевых аминокислотных остатков и каждых хиральных аминокислотных остатков, непосредственно к ним примыкающих, является D-аминокислотным остатком; или
d) каждый хиральный аминокислотный остаток является D-аминокислотным остатком, за исключением того, что один или более из каждых хиральных концевых аминокислотных остатков и каждых хиральных аминокислотных остатков, непосредственно к ним примыкающих, является L-аминокислотным остатком.
46. Пептид или фармацевтически приемлемая соль указанного пептида, согласно воплощению 45, где указанный пептид является пептидом, содержащим 22 или 23 остатка.
47. Пептид или фармацевтически приемлемая соль указанного пептида, согласно воплощению 46, где указанный пептид является пептидом, содержащим 22 остатка.
48. Пептид или фармацевтически приемлемая соль указанного пептида, согласно воплощению 47, где X22 представляет собой Val, Leu, D-Val или D-Leu.
49. Пептид или фармацевтически приемлемая соль указанного пептида, согласно воплощению 47, где
X2 представляет собой Lys или D-Lys;
X4 представляет собой Lys или D-Lys;
X5 представляет собой Lys или D-Lys;
X13 представляет собой Glu или D-Glu;
X18 представляет собой Phe или D-Phe;
X19 представляет собой Leu или D-Leu; и
X22 представляет собой Ala, Leu, Val, D-Ala, D-Leu или D-Val.
50. Пептид или фармацевтически приемлемая соль указанного пептида, согласно воплощению 47, где
X2 представляет собой Lys или D-Lys;
X4 представляет собой Lys или D-Lys;
X5 представляет собой Lys или D-Lys;
X13 представляет собой Glu или D-Glu;
X18 представляет собой Phe или D-Phe;
X19 представляет собой Leu или D-Leu; и
X22 представляет собой Ala, Leu, Val, D-Ala, D-Leu или D-Val.
51. Пептид или фармацевтически приемлемая соль указанного пептида, согласно воплощению 47, где X13 и X17 представляют собой Glu или D-Glu.
52. Пептид или фармацевтически приемлемая соль указанного пептида, согласно воплощению 46, где
X1 отсутствует;
X2 представляет собой Lys или D-Lys;
X4 представляет собой Lys или D-Lys;
X5 представляет собой Lys или D-Lys;
X18 представляет собой Phe или D-Phe;
X19 представляет собой Leu или D-Leu; и
X22 представляет собой Val или D-Val.
53. Пептид или фармацевтически приемлемая соль указанного пептида, согласно воплощению 47, где X13 или X17 представляют собой Glu или D-Glu.
54. Пептид или фармацевтически приемлемая соль указанного пептида, согласно воплощению 47, где X22 представляет собой Val или D-Val и X6 представляет собой Gln или D-Gln.
55. Пептид или фармацевтически приемлемая соль указанного пептида, согласно воплощению 47, где X22 представляет собой Val или D-Val или X6 представляет собой Gln или D-Gln.
56. Пептид или фармацевтически приемлемая соль указанного пептида, согласно воплощению 47, где только один из X10, X11 и X14 представляет собой Gly.
57. Пептид или фармацевтически приемлемая соль указанного пептида, согласно воплощению 45, где указанный пептид представляет собой
Lys-Leu-Lys-Lys-Gln-Leu-Ala-Asp-Leu-Leu-Arg-Glu-Leu-Leu-Gln-Glu-Phe-Leu-Glu-Leu-Val-Inp (SEQ ID NO:197); или
Lys-Leu-Lys-Lys-Gln-Leu-Ala-Asp-Leu-Leu-Arg-Glu-Leu-Leu-Gln-Glu-Phe-Leu-Glu-Leu-Val-Nip (SEQ ID NO:211), или
фармацевтически приемлемую соль одного из вышеперечисленных [соединений].
58. Пептид или фармацевтически приемлемая соль указанного пептида, согласно воплощению 47, где X10 представляет собой Gly, а X17 представляет собой Glu или D-Glu.
59. Пептид или фармацевтически приемлемая соль указанного пептида, согласно воплощению 47, где каждый из X10, X11 и X14 является отличным от Gly.
60. Пептид или фармацевтически приемлемая соль указанного пептида, согласно воплощению 47, где X17 представляет собой Leu или D-Leu.
61. Пептид или фармацевтически приемлемая соль указанного пептида, согласно воплощению 60, где X14 представляет собой Trp или D-Trp и X10 представляет собой Leu, Phe, D-Leu или D-Phe.
62. Пептид или фармацевтически приемлемая соль указанного пептида, согласно воплощению 60, где X14 представляет собой Trp или D-Trp или X10 представляет собой Leu, Phe, D-Leu или D-Phe.
63. Пептид или фармацевтически приемлемая соль указанного пептида, согласно воплощению 45, где R1 представляет собой H и R2 представляет собой OH.
64. Пептид согласно любому из воплощений 1-63, где пептид представлен в виде фармацевтически приемлемой соли.
65. Пептид согласно воплощению 64, где указанная соль является солью металла или органической солью амина.
66. Пептид согласно воплощению 65, где указанный металл является щелочным металлом или щелочноземельным металлом.
67. Пептид согласно воплощению 65, где указанный металл является литием, натрием, калием, магнием, кальцием, алюминием или цинком.
68. Пептид согласно воплощению 65, где указанный амин представляет собой триэтиламин, этаноламин, диэтаноламин, триэтаноламин, морфолин, N-метилпиперидин, N-этилпиперидин или дибензиламин.
69. Пептид согласно воплощению 64, где указанная соль представляет собой соль присоединения кислоты.
70. Пептид согласно воплощению 69, где указанной солью присоединения кислоты является гидрохлорид, гидробромид, гидройодид, нитрат, сульфат, сульфит, бисульфат, фосфат, кислый фосфат, изоникотинат, лактат, салицилат, тартрат, битартрат, аскорбат, гентизинат, глюконат, глюкаронат, сахарат, формат, бензоат, глутамат, пантотенат, ацетат, фумарат, сукцинат, метансульфонат, этансульфонат, бензолсульфонат, пара-толуилсульфонат, цитрат или малеат.
71. Пептид или фармацевтически приемлемая соль указанного пептида, согласно любому из воплощений 1-63, где R1 представляет собой аминозащитную группу.
72. Пептид или фармацевтически приемлемая соль указанного пептида, согласно воплощению 71, где указанной аминозащитной группой является дансил; метоксикарбонил; этоксикарбонил; 9-фторенилметоксикарбонил; 2-хлорэтоксикарбонил; 2,2,2-трихлорэтоксикарбонил; 2-фенилэтоксикарбонил; трет-бутоксикарбонил; бензилоксикарбонил; пара-метоксибензилоксикарбонил; пара-нитробензилоксикарбонил; o- нитробензилоксикарбонил; пара-бромбензилоксикарбонил; пара-хлорбензилоксикарбонил; пара-йодбензилоксикарбонил; 2,4-дихлорбензилоксикарбонил; дифенилметоксикарбонил; 3,5- диметоксибензилоксикарбонил; феноксикарбонил; 2,4,6-три-трет-бутилфеноксикарбонил; 2,4,6-триметилбензилоксикарбонил; формил; ацетил; хлорацетил; трихлорацетил; трифторацетил; фенилацетил; пиколиноил; бензоил; пара-фенилбензоил; фталоил; метил; трет-бутил; аллил; [2-(триметилсилил)этокси]метил; 2,4-диметоксибензил; 2,4-динитрофенил; бензил; 4-метоксибензил; дифенилметил; трифенилметил; бензолсульфенил; o-нитробензолсульфенил; 2,4-динитробензолсульфенил; пара-толуолсульфонил; бензолсульфонил; 2,3,6-триметил-4-метоксибензолсульфонил; 2,4,6-триметоксибензолсульфонил; 2,6-диметил-4-метоксибензолсульфонил; пентаметилбензолсульфонил; 4-метоксибензолсульфонил; 2,4,6-триметилбензолсульфонил; или бензилсульфонил.
73. Пептид или фармацевтически приемлемая соль указанного пептида, согласно любому из воплощений 1-63, где R2 представляет собой карбоксильную защитную группу.
74. Пептид или фармацевтически приемлемая соль указанного пептида, согласно воплощению 73, где указанной карбоксильной защитной группой является метокси; этокси; 9-фторенилметокси; метоксиметокси; метилтиометокси; тетрагидропиранокси; тетрагидрофуранокси; метоксиэтоксиметокси; бензилоксиметокси; фенацилокси; пара-бромфенацилокси; α-метилфенацилокси; пара-метоксифенацилокси; дезилокси; 2-хлорэтокси; 2,2,2-трихлорэтокси, 2-метилтиоэтолкси; 2-(пара-толуолсульфонил)метокси; трет-бутокси; циклопентокси; циклогексокси; аллилокси; металлилокси; циннамокси; α-метилциннамокси; фенокси; 2,6-диметилфенокси; 2,6-диизопропилфенокси; бензилокси; трифенилметокси; дифенилметокси; 2,4,6-триметилбензилокси; пара-бромбензилокси; o-нитробензилокси; N,N-диметиламидо; пирролидинил; или пиперидинил.
75. Пептид или фармацевтически приемлемая соль указанного пептида, согласно любому из воплощений 1-63, где одна или более из -NH2- или COOH-групп указанного пептида являются защищенными защитной группой.
76. Композиция, содержащая эффективное количество пептида или фармацевтически приемлемой соли указанного пептида, согласно любому из воплощений 1-75, и фармацевтически приемлемый носитель или наполнитель.
77. Способ лечения или предупреждения дислипидемии, предусматривающий введение млекопитающему, в случае необходимости, эффективного количества пептида или фармацевтически приемлемой соли указанного пептида, согласно любому из воплощений 1-75.
78. Способ согласно воплощению 77, где указанная дислипидемия представляет собой гиперпротеинемию, высокую концентрацию в сыворотке липопротеина низкой плотности, высокую концентрацию в сыворотке липопротеина очень низкой плотности, гиперлипидемию, низкую концентрацию липопротеина высокой плотности в сыворотке, гипохолестеринемию, A-бета-липопротеинемию, дефицит ApoA-I и болезнь Танжера.
79. Способ согласно воплощению 77, где указанная дислипидемия представляет собой гиперлипидемию, гиперхолестеринемию, дефицит ApoA-I или гипертриглицеридемию.
80. Способ согласно воплощению 77, где указанное лечение предусматривает повышение концентрации липопротеина высокой плотности в сыворотке.
81. Способ лечения или предупреждения сердечно-сосудистого заболевания, предусматривающий введение млекопитающему, в случае необходимости, эффективного количества пептида или фармацевтически приемлемой соли указанного пептида, согласно любому из воплощений 1-75.
82. Способ согласно воплощению 81, где сердечно-сосудистое заболевание представляет собой метаболический синдром, ишемическую болезнь сердца, атеросклероз, рестеноз, эндотоксемию, застойную сердечную недостаточность, циркуляторный шок, кардиомиопатию, пересадку сердца, инфаркт миокарда, сердечную аритмию, суправентрикулярную тахикардию, трепетание предсердий, пароксизмальную предсердную тахикардию, аневризму, стенокардию, острое нарушение мозгового кровообращения, болезнь периферических сосудов, цереброваскулярную болезнь, болезнь почек, атерогенез, атеросклероз, острый панкреатит и болезнь коронарных артерий.
83. Способ согласно воплощению 81, где сердечно-сосудистое заболевание представляет собой атеросклероз, рестеноз или метаболический синдром.
84. Способ лечения или предупреждения эндотелиальной дисфункции, предусматривающий введение млекопитающему, в случае необходимости, эффективного количества пептида или фармацевтически приемлемой соли указанного пептида, согласно любому из воплощений 1-75.
85. Способ лечения или предупреждения макрососудистого расстройства, предусматривающий введение млекопитающему, в случае необходимости, эффективного количества пептида или фармацевтически приемлемой соли указанного пептида, согласно любому из воплощений 1-75.
86. Способ согласно воплощению 85, где указанное макрососудистое расстройство представляет собой преходящее нарушение мозгового кровообращения, инсульт, стенокардию, инфаркт миокарда, сердечную недостаточность или болезнь периферических сосудов.
87. Способ лечения или предупреждения микрососудистого расстройства, предусматривающий введение млекопитающему, в случае необходимости, эффективного количества пептида или фармацевтически приемлемой соли указанного пептида, согласно любому из воплощений 1-75.
88. Способ согласно воплощению 87, где указанное микрососудистое расстройство представляет собой диабетическую ретинопатию, микроальбуминурию, макроальбуминурию, конечную стадию почечного заболевания, эректильную дисфункцию, автономную невропатию, периферическую невропатию, остеомиелит или ишемию нижней конечности.
89. Способ согласно любому из воплощений 77-88, согласно которому млекопитающее является человеком.
90. Способ согласно любому из воплощений 77-89, согласно которому введение осуществляют перорально, внутривенно, внутримышечно, интратекально, подкожно, сублингвально, назально, накожно, трансдермально, глазным путем или путем ингаляции.
| название | год | авторы | номер документа |
|---|---|---|---|
| ПЕПТИДНЫЙ АНАЛОГ АЦИЛИРОВАННОГО ОКСИНТОМОДУЛИНА | 2018 |
|
RU2752787C1 |
| Композиция для лечения гиперлипидемии, содержащая производное оксинтомодулина | 2013 |
|
RU2768853C1 |
| ПЕПТИДНЫЕ ИНГИБИТОРЫ РЕЦЕПТОРА ИНТЕРЛЕЙКИНА-23 ДЛЯ ПЕРОРАЛЬНОГО ПРИЕМА И ИХ ПРИМЕНЕНИЕ ДЛЯ ЛЕЧЕНИЯ ВОСПАЛИТЕЛЬНЫХ ЗАБОЛЕВАНИЙ КИШЕЧНИКА | 2015 |
|
RU2736637C2 |
| АНАЛОГИ ЭФР(А) С ЗАМЕСТИТЕЛЯМИ - ЖИРНЫМИ КИСЛОТАМИ | 2017 |
|
RU2747877C2 |
| СОЕДИНЕНИЯ-АГОНИСТЫ GIP И СПОСОБЫ | 2015 |
|
RU2716985C2 |
| СПОСОБЫ ЛЕЧЕНИЯ ГИПЕРЛИПИДЕМИИ У ПАЦИЕНТОВ С ДИАБЕТОМ ПОСРЕДСТВОМ ВВЕДЕНИЯ ИНГИБИТОРА PCSK9 | 2018 |
|
RU2772712C2 |
| Фармацевтическая композиция, содержащая инсулин и глюкагон | 2019 |
|
RU2823246C2 |
| Слитые молекулы, происходящие от Cholix-токсина, для пероральной доставки биологически активных нагрузок | 2015 |
|
RU2723178C2 |
| ЛЕЧЕНИЕ КОЖНЫХ ЗАБОЛЕВАНИЙ | 2021 |
|
RU2840027C1 |
| ГЛИКАНЗАВИСИМЫЕ ИММУНОТЕРАПЕВТИЧЕСКИЕ МОЛЕКУЛЫ | 2016 |
|
RU2754661C2 |
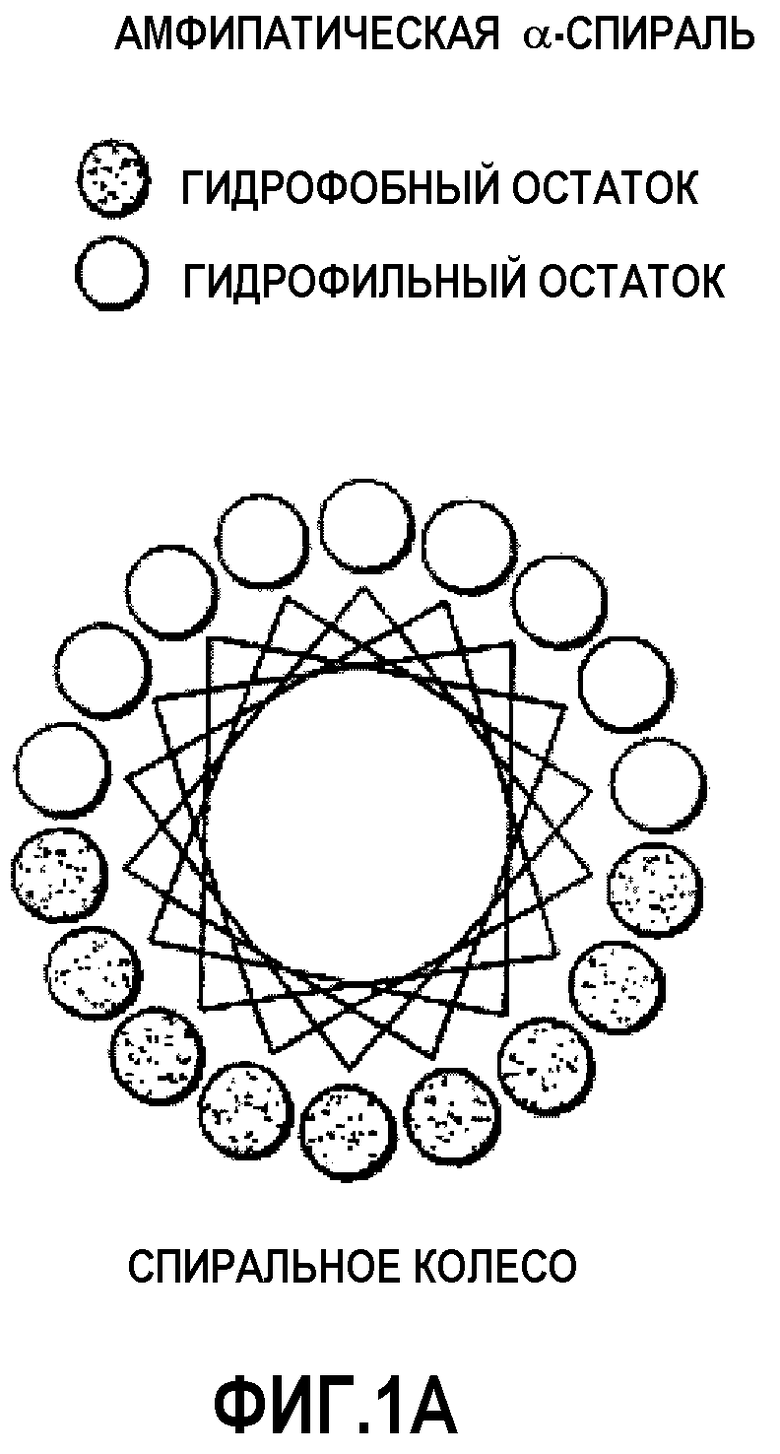
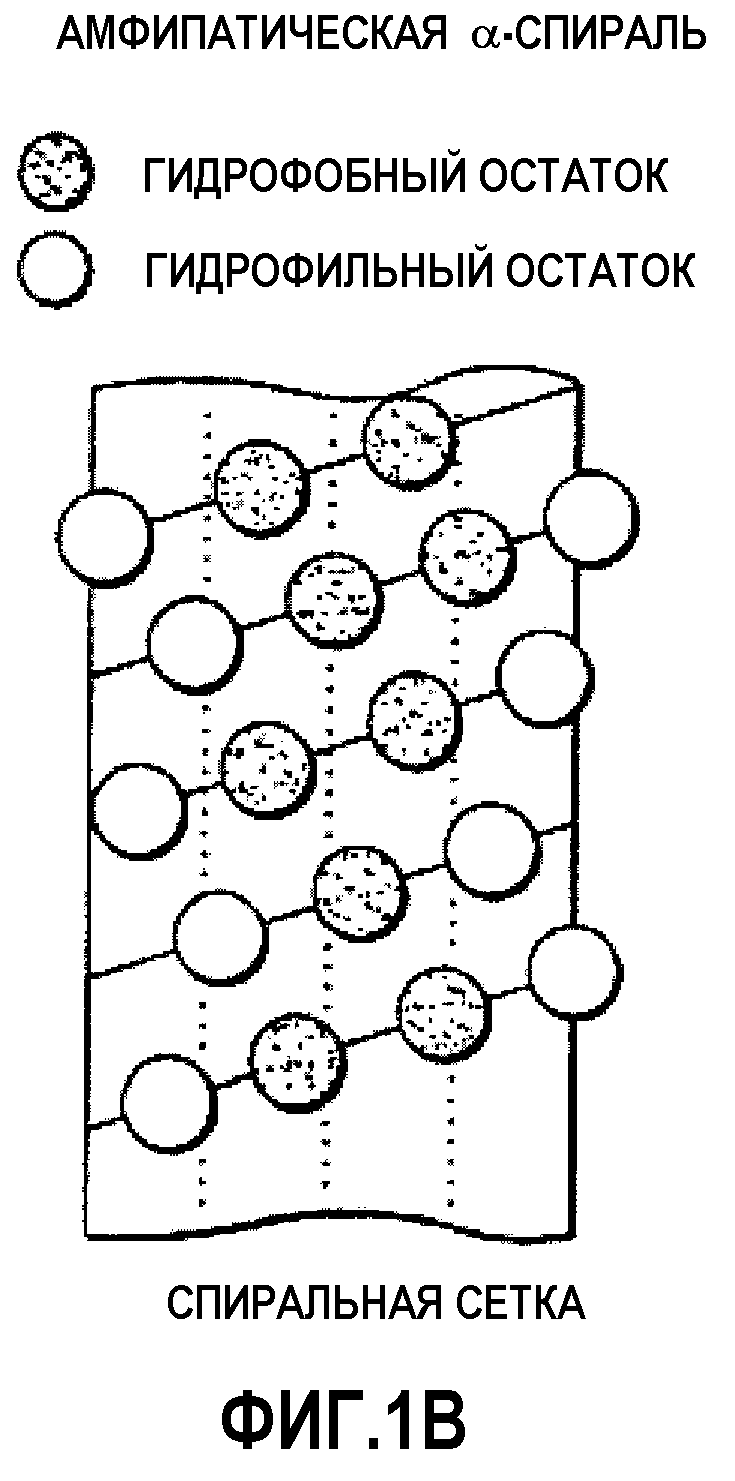
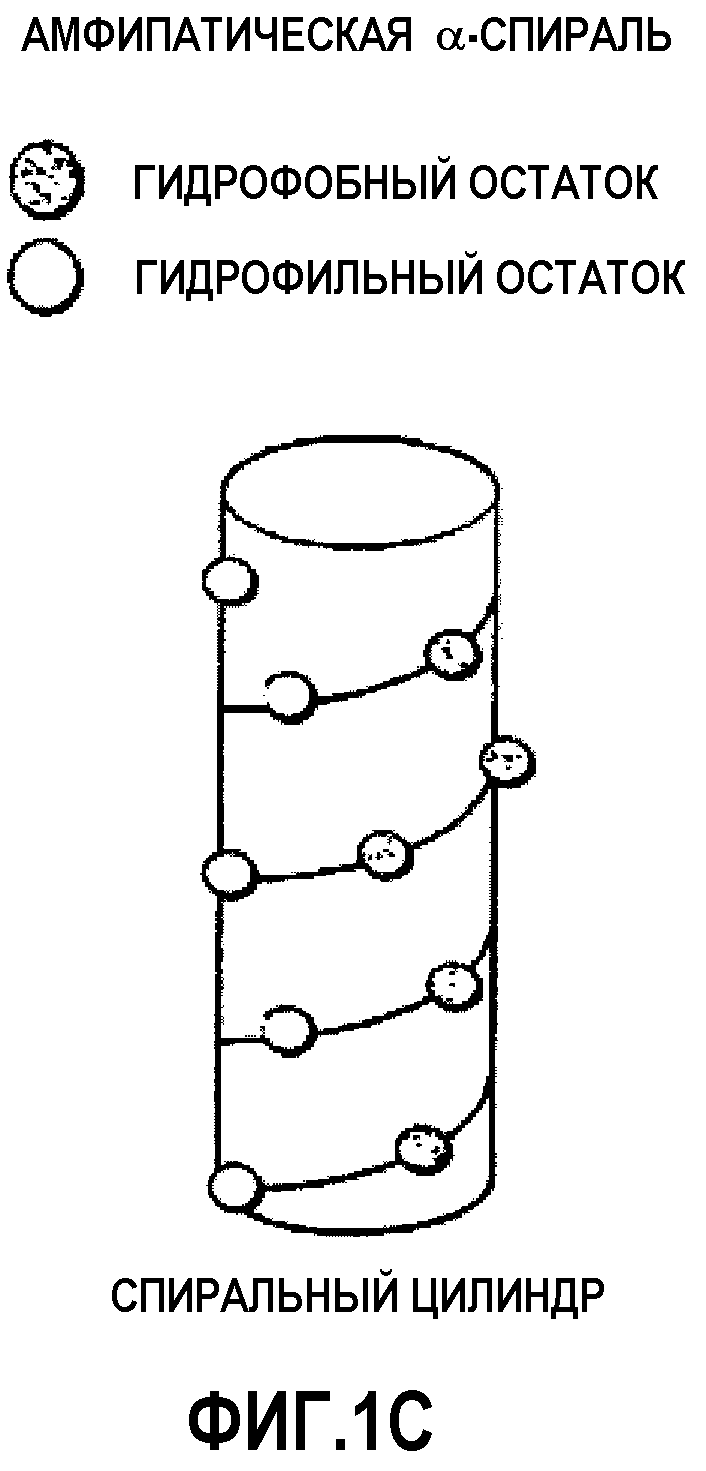
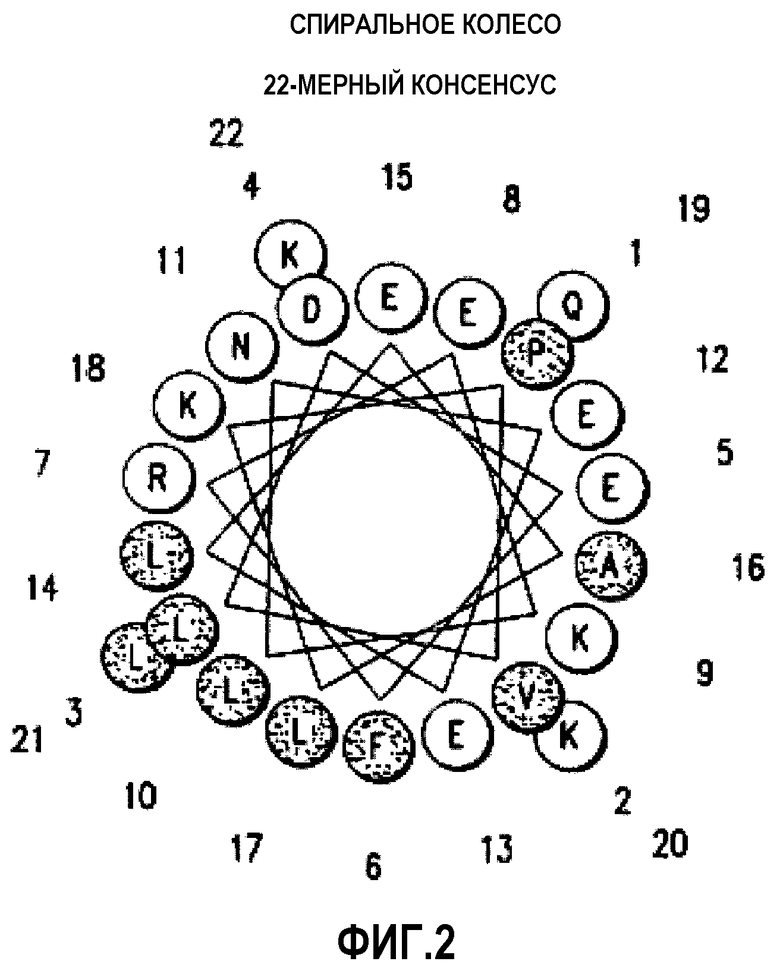
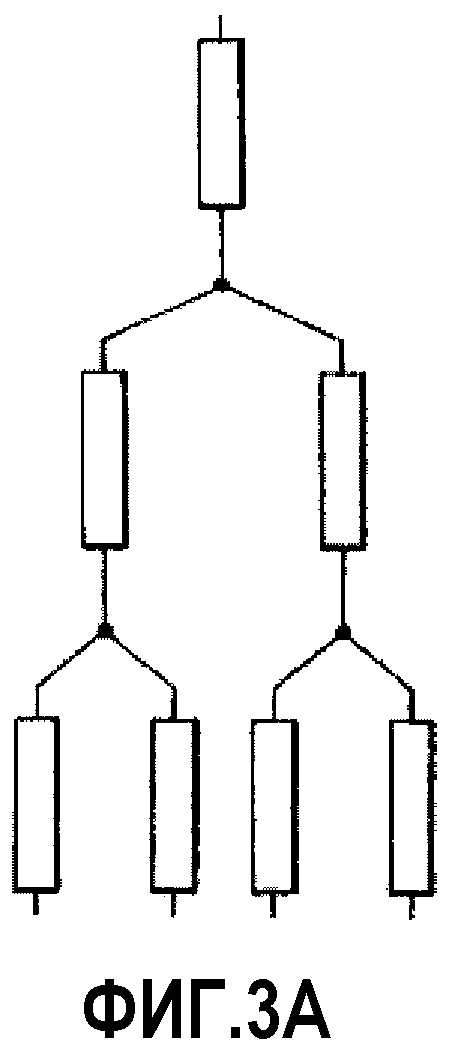
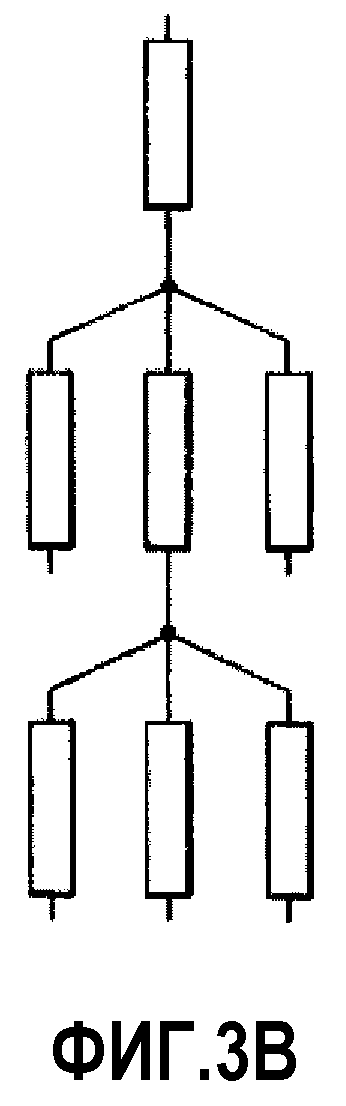
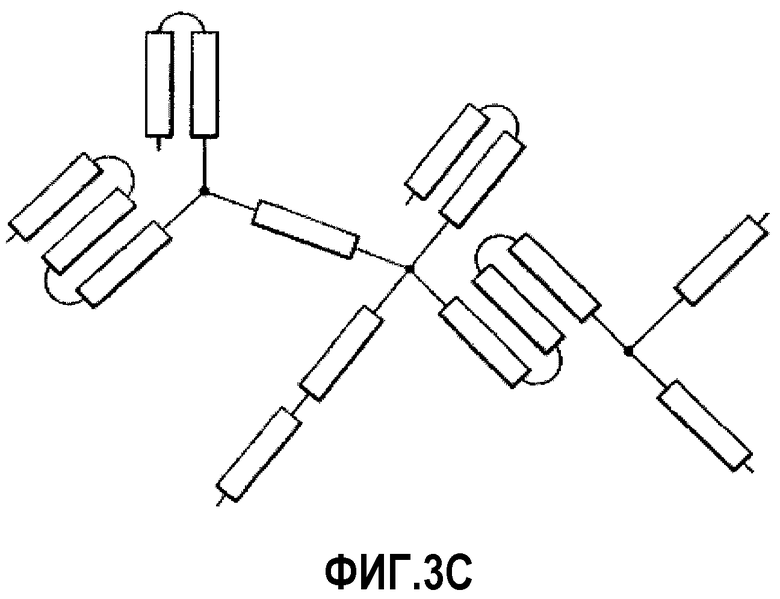
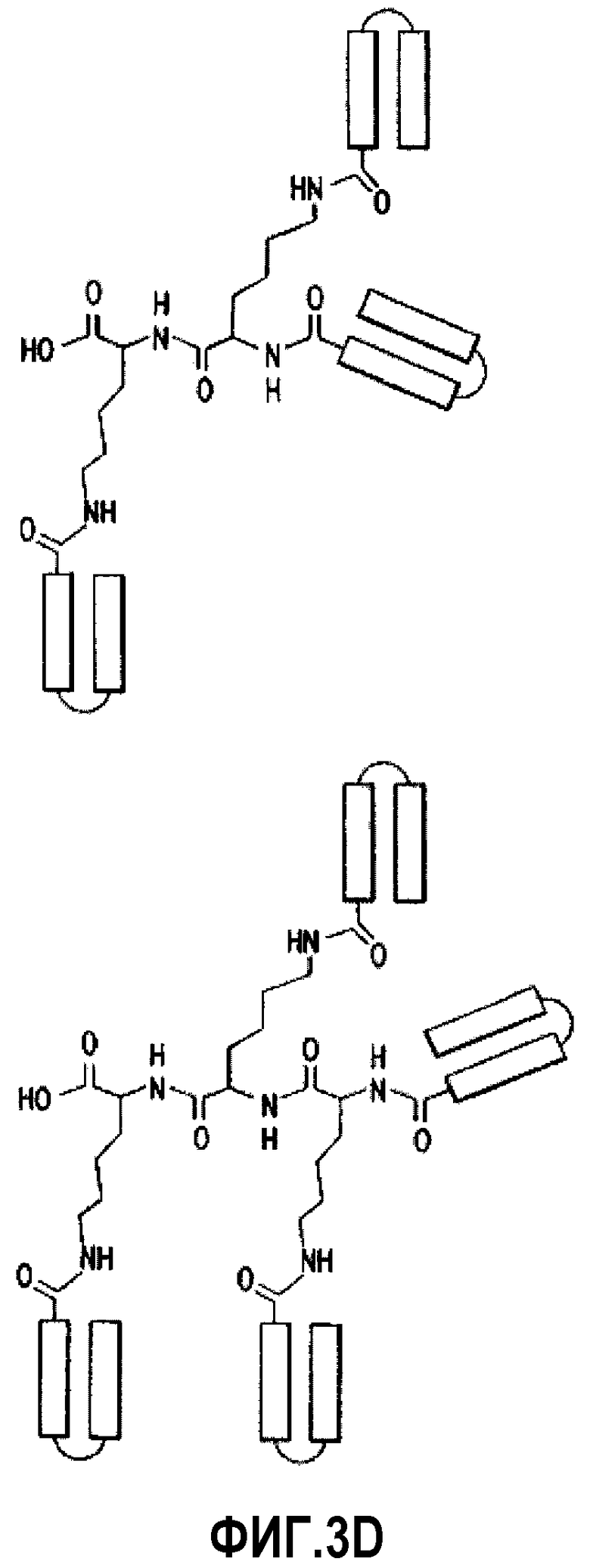
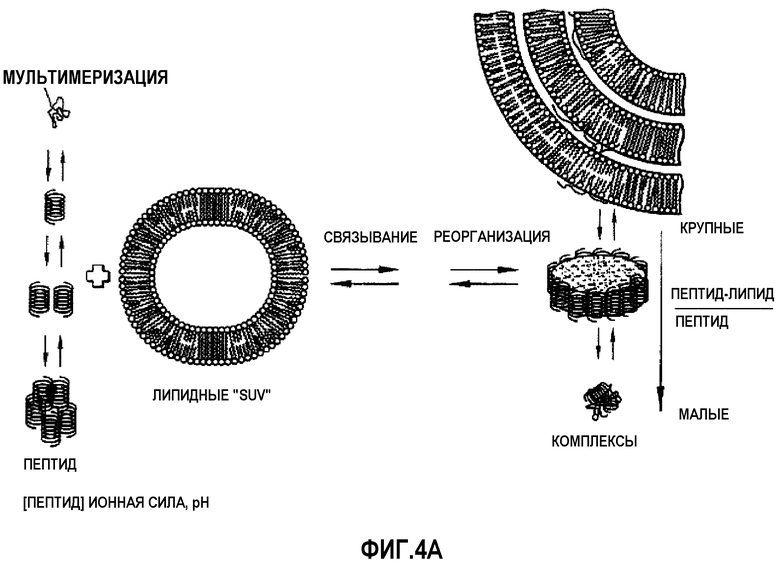
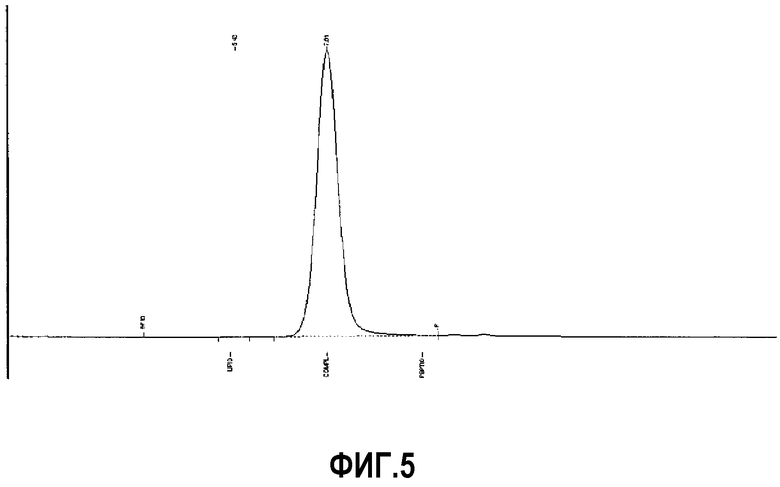
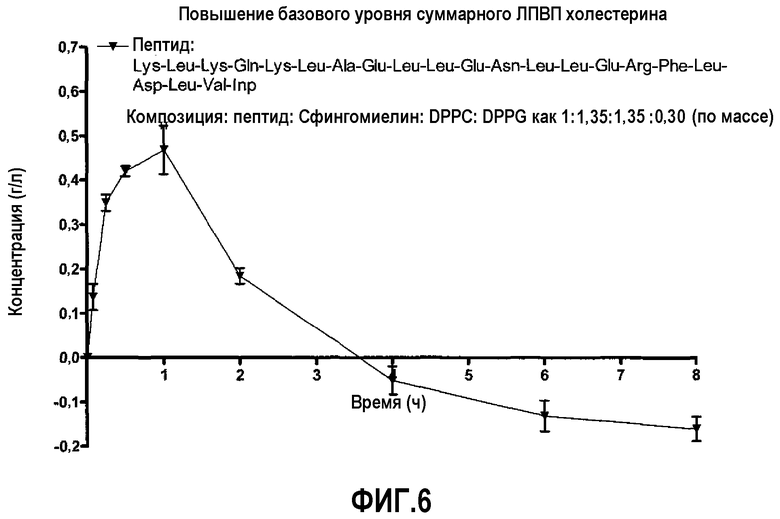
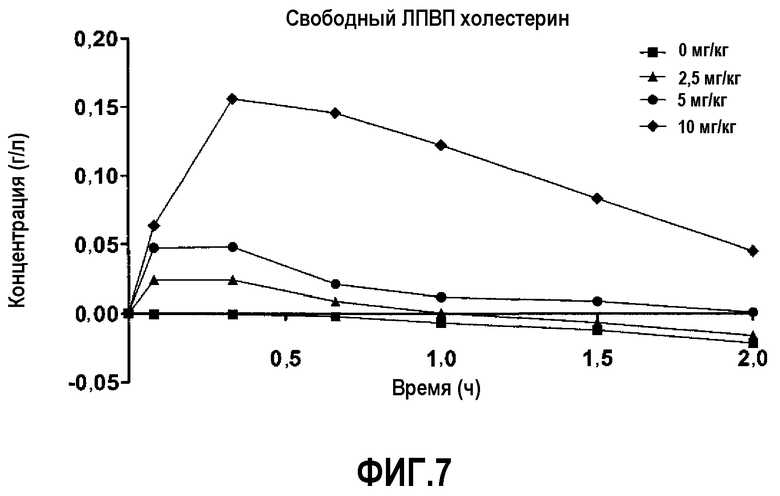
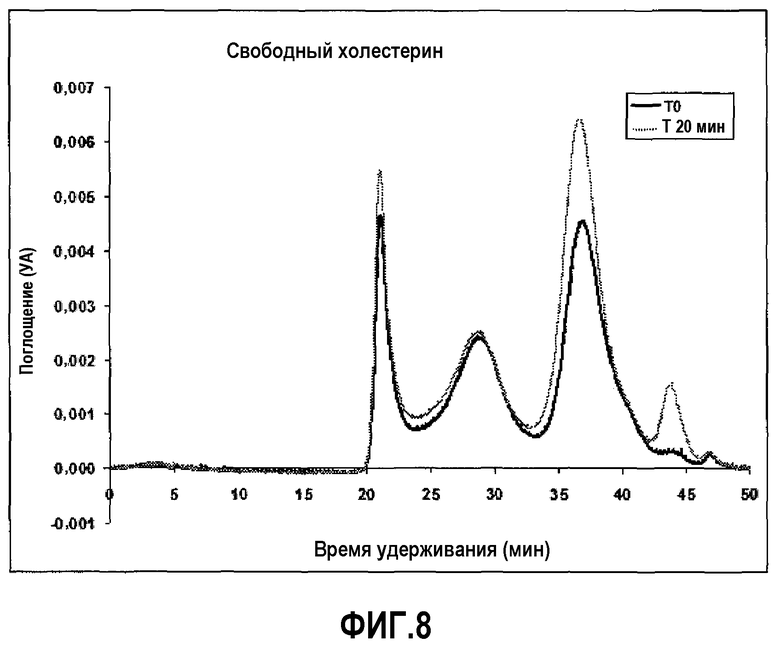
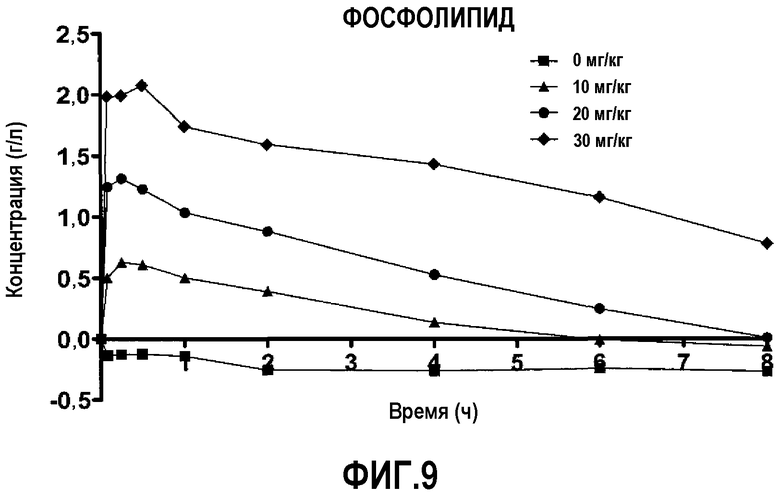
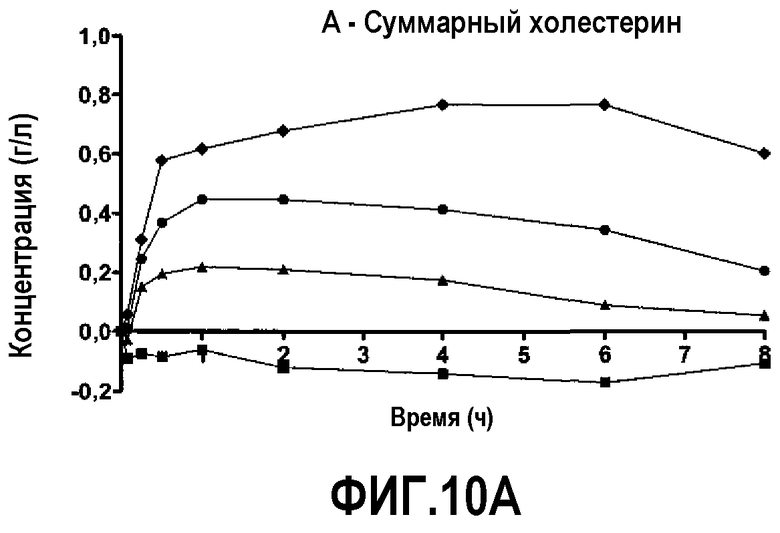
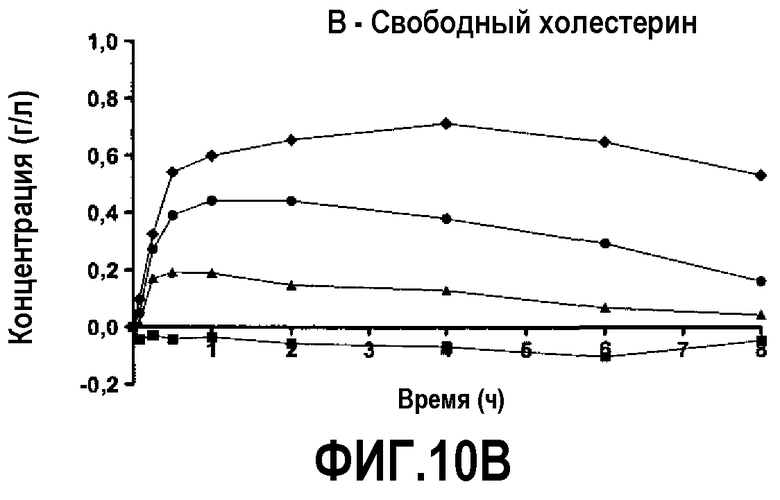
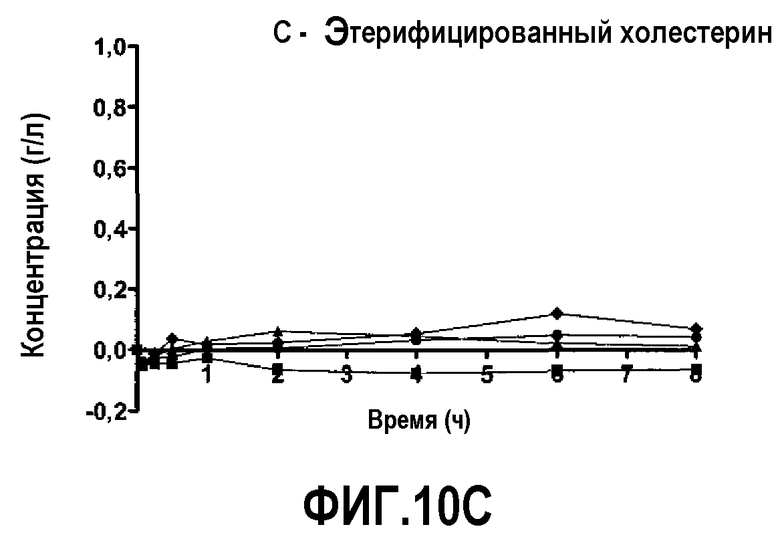
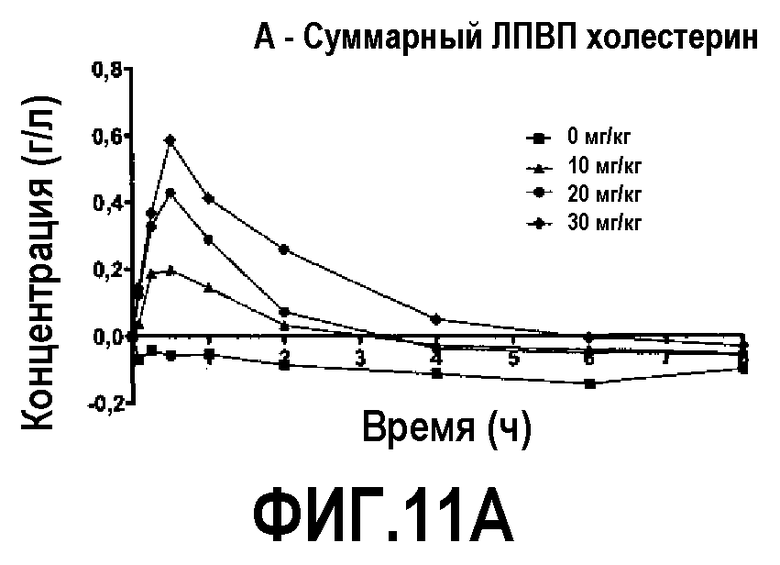
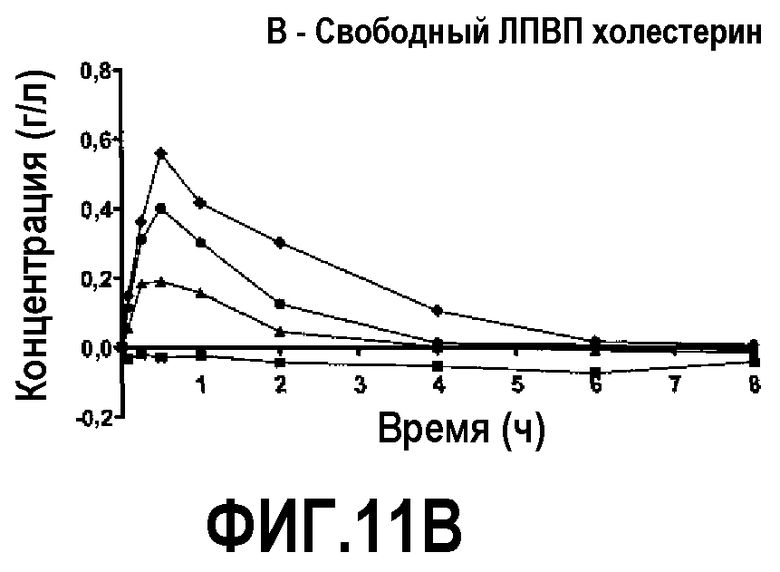
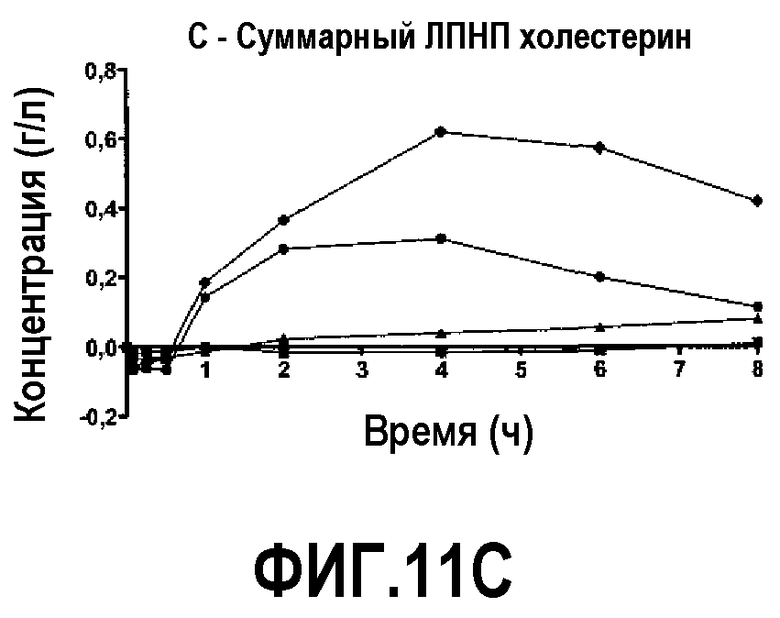
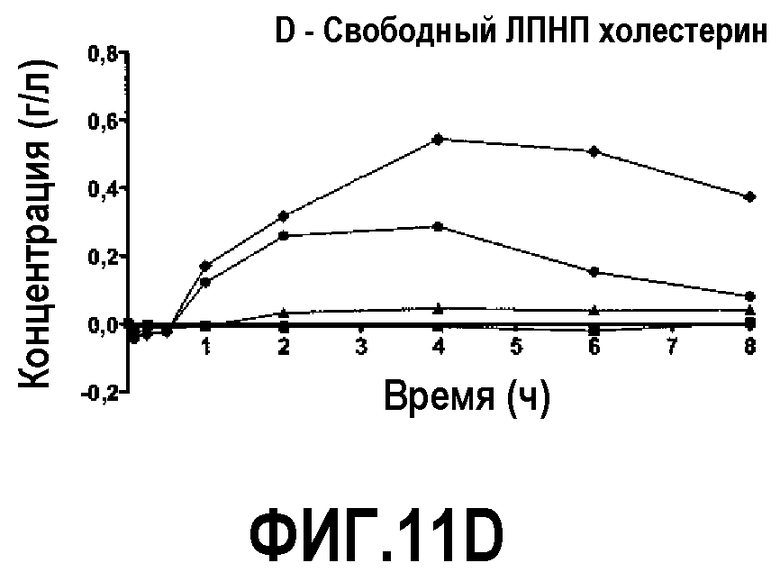
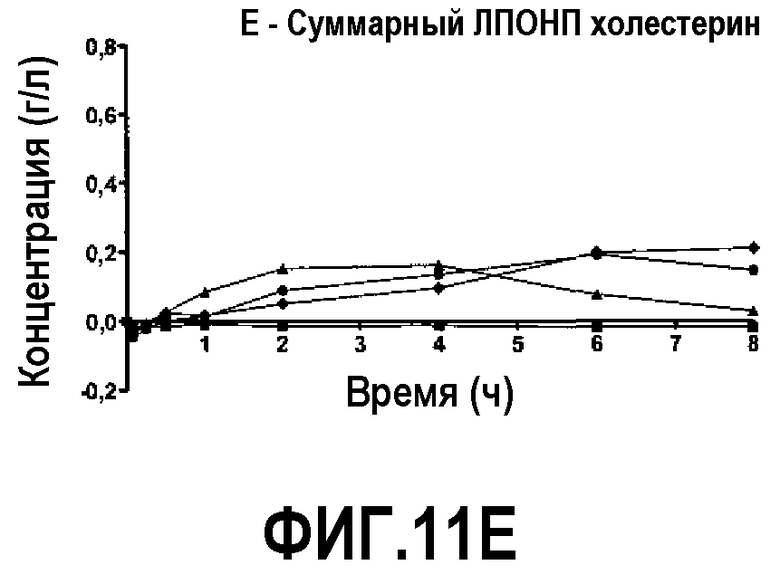
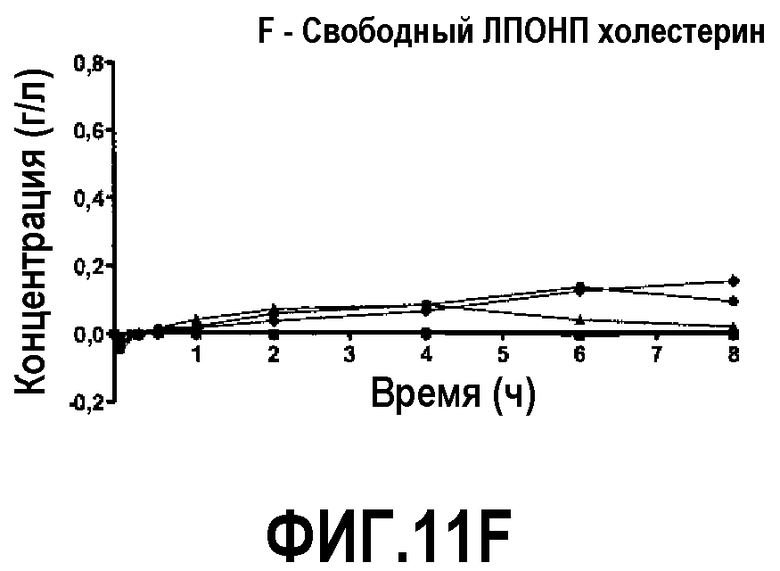
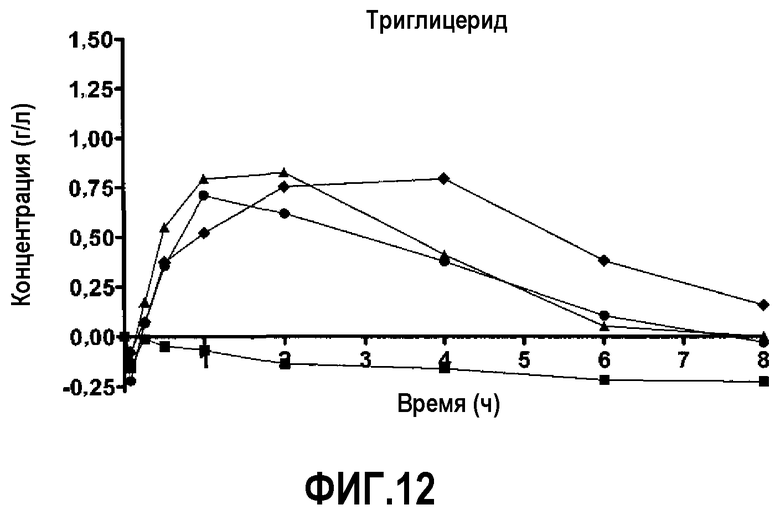
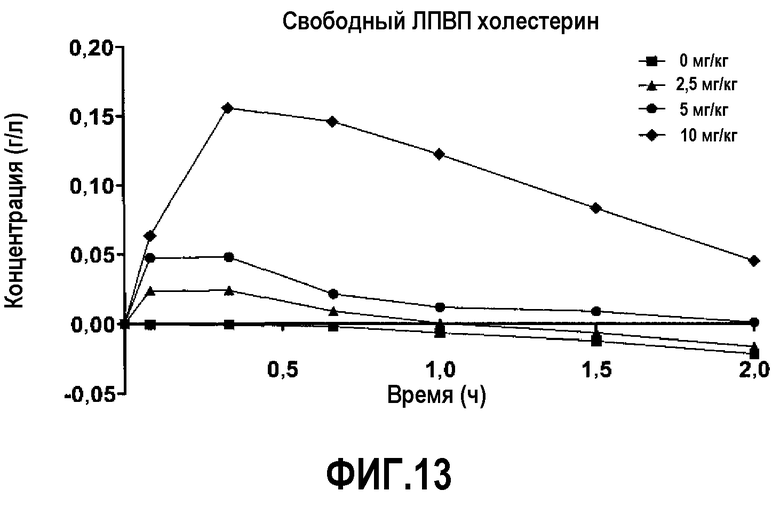
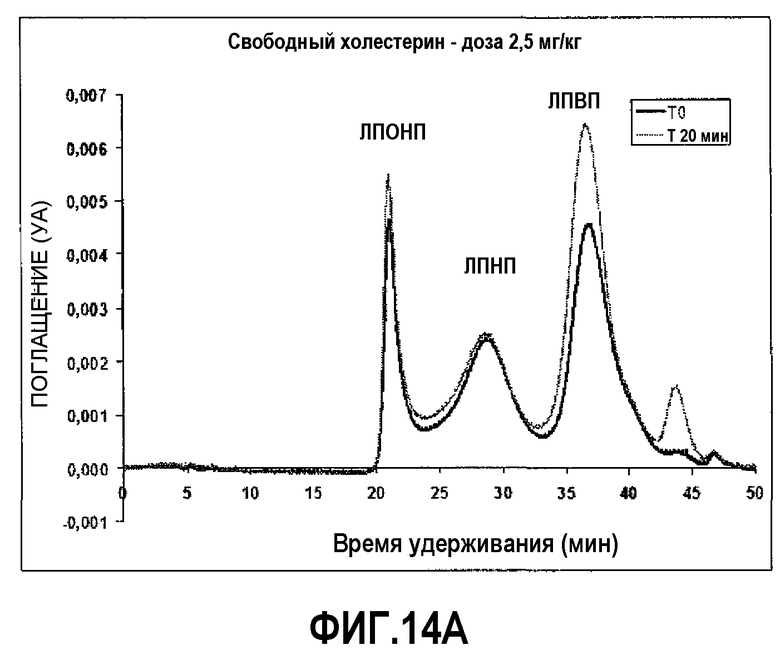
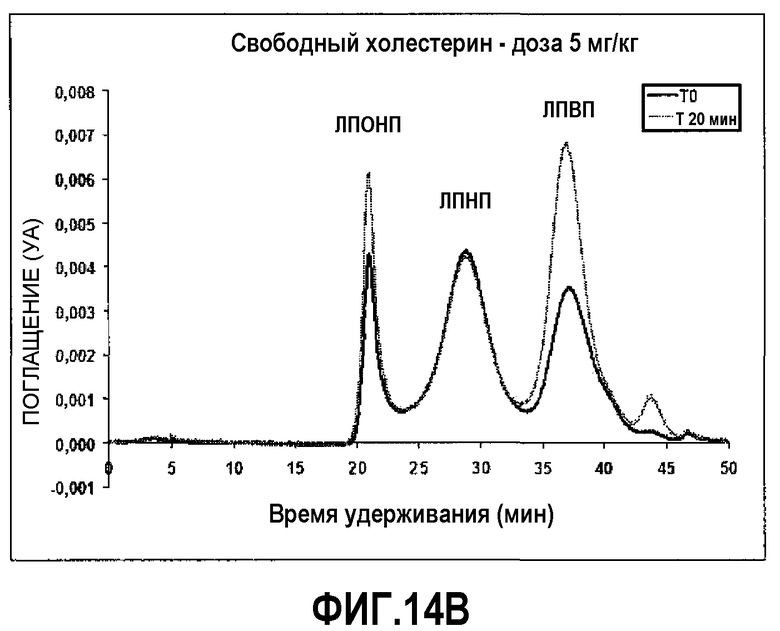
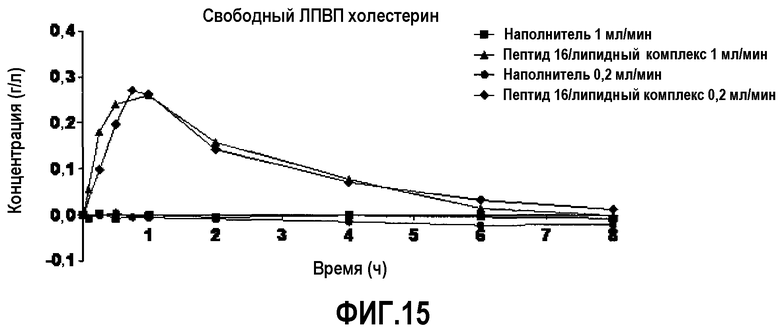
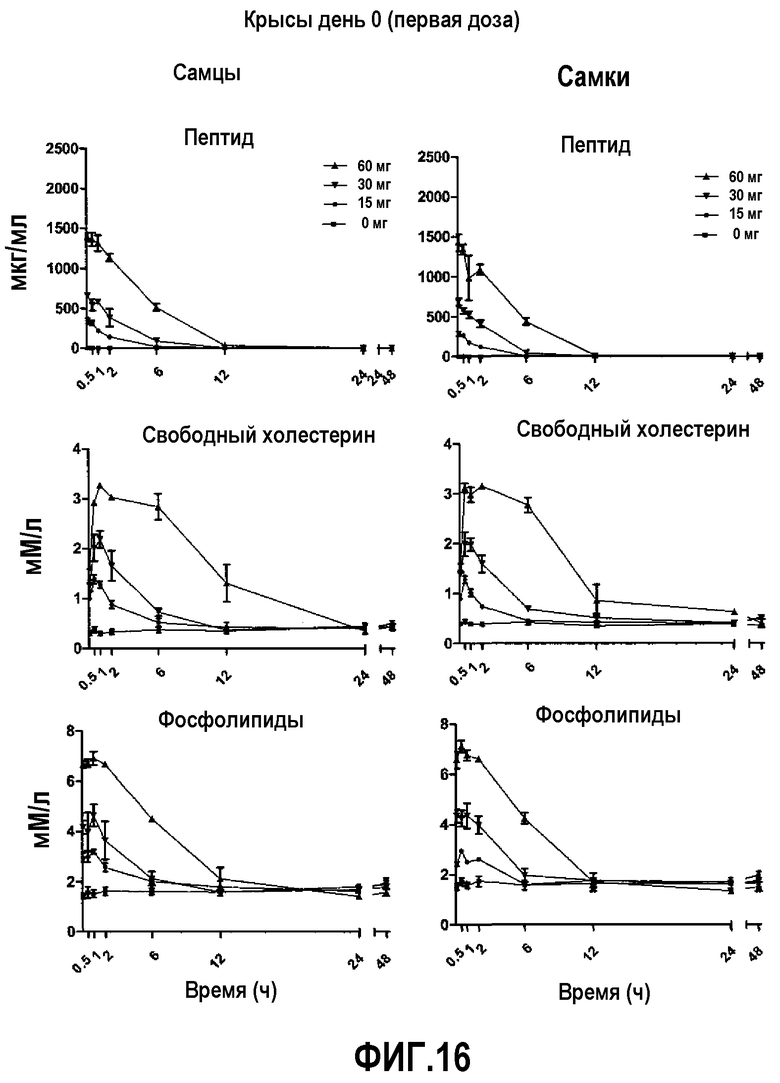
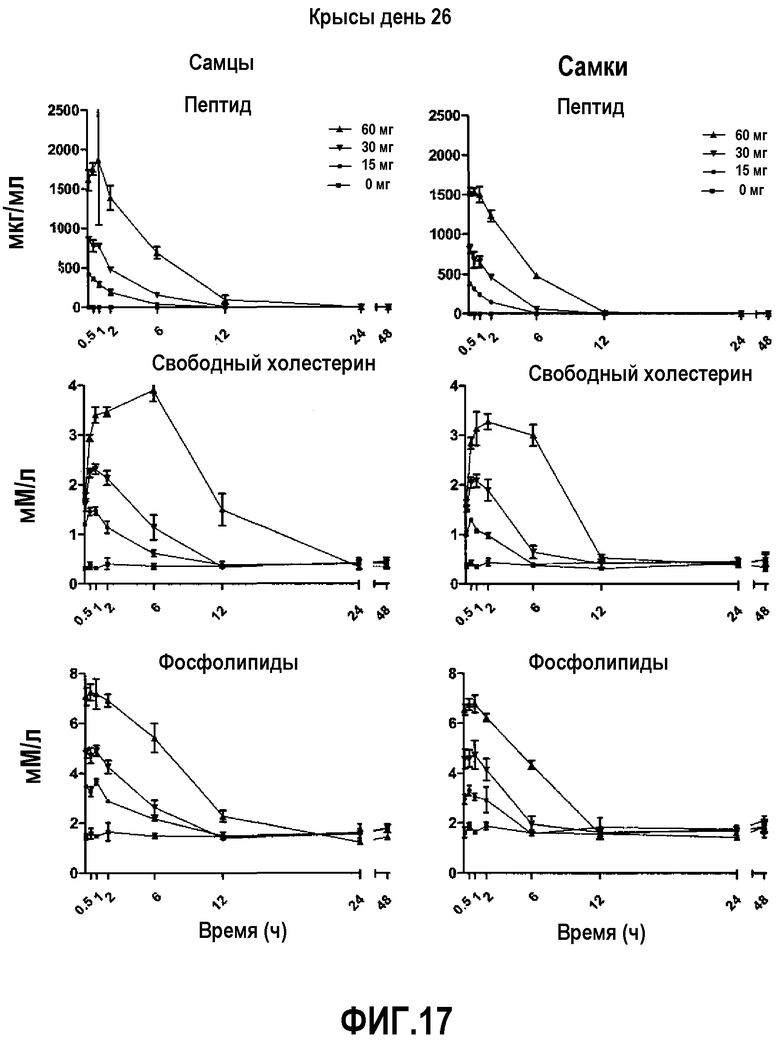
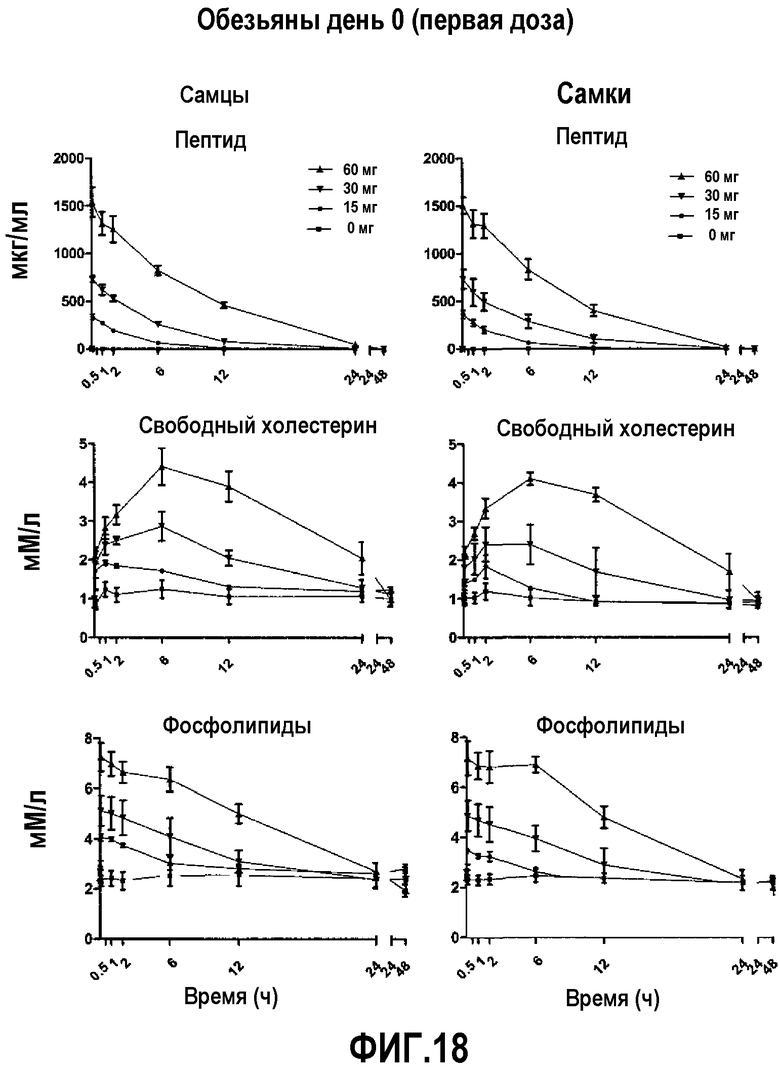
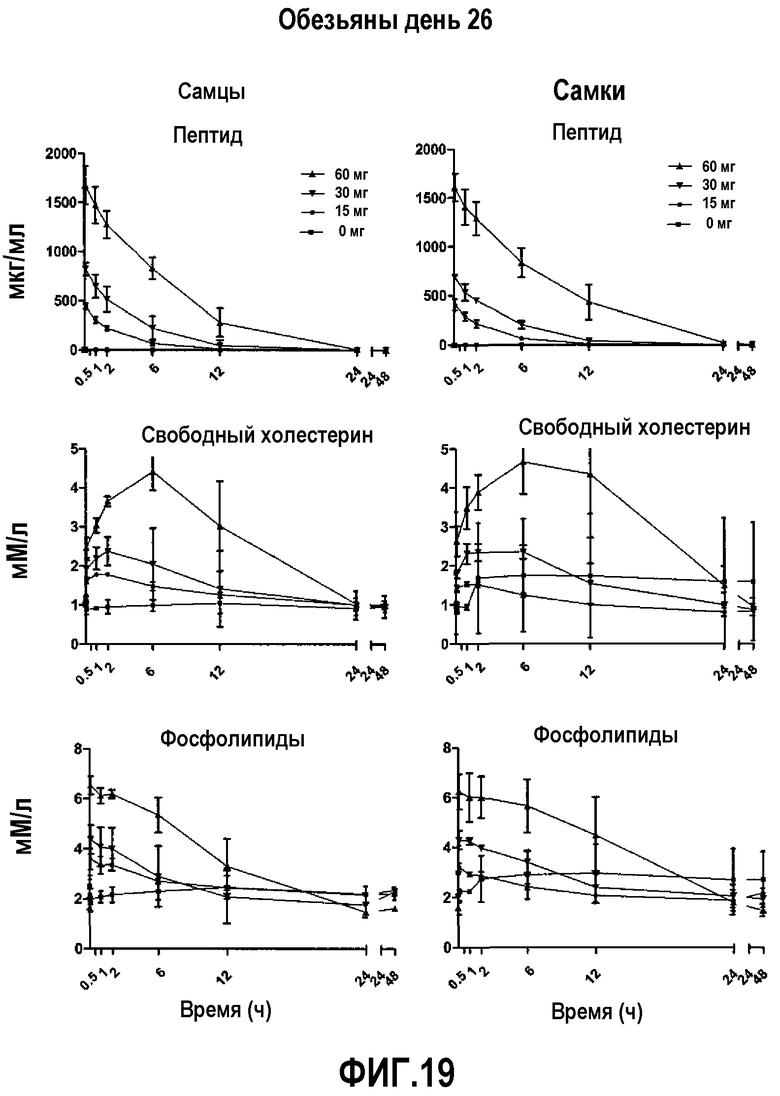
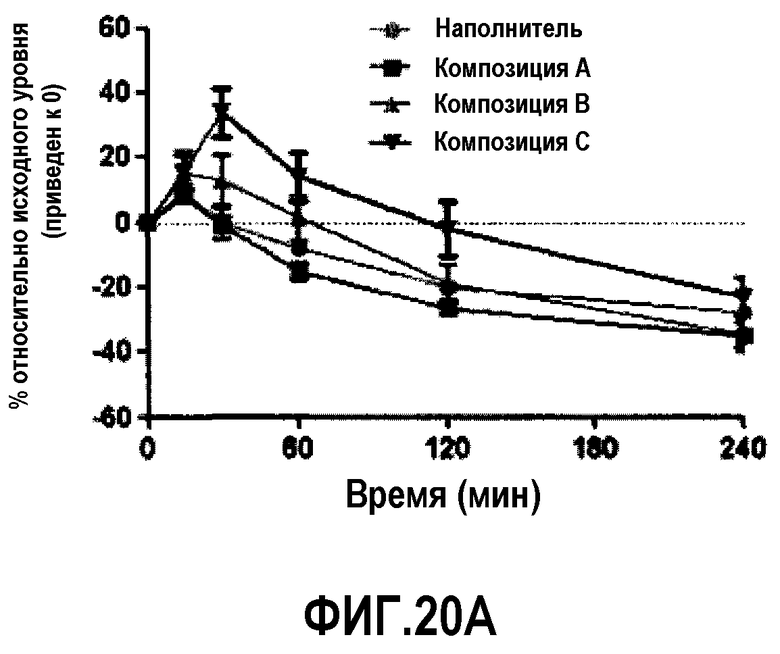
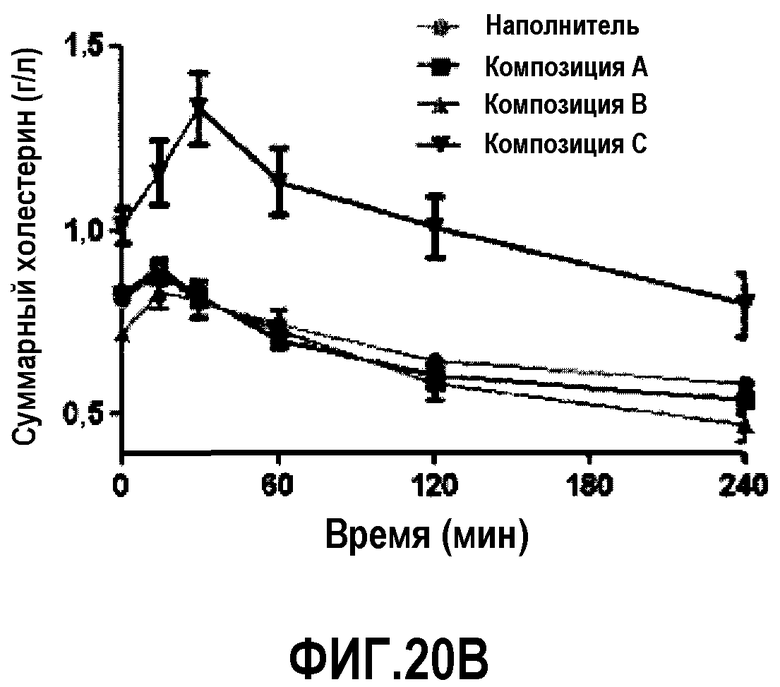
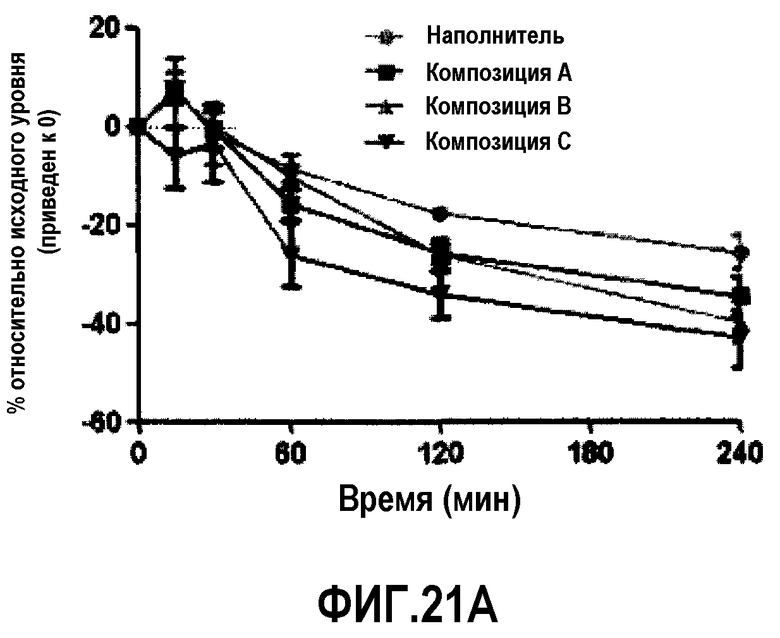
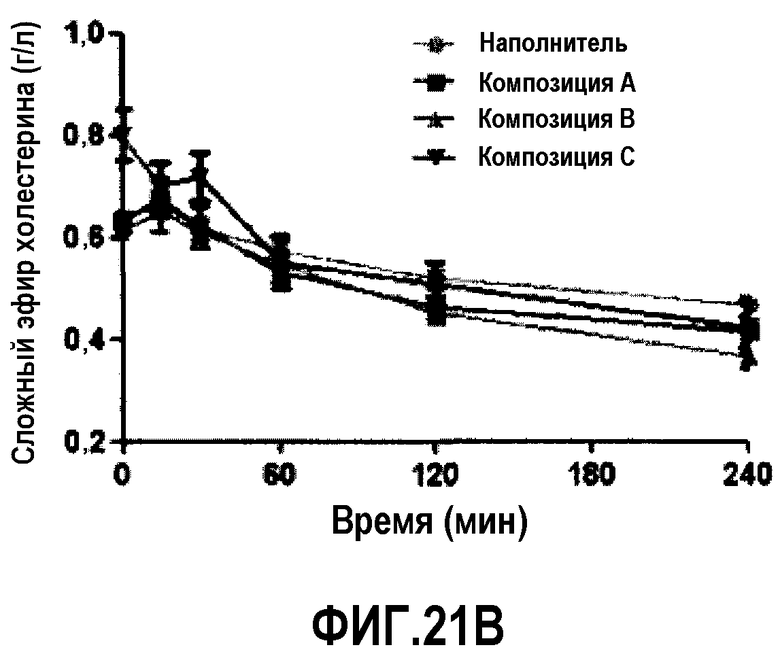
Изобретение относится к области биотехнологии, конкретно к получению пептида, являющегося миметиком ApoA-I, и может быть использовано в медицине. Указанный пептид характеризуется последовательностью Lys-Leu-Lys-Gln-Lys-Leu-Ala-Glu-Leu-Leu-Glu-Asn-Leu-Leu-Glu-Arg-Phe-Leu-Asp-Leu-Val-Inp (SEQ ID NO: 16). Изобретение также относится к пептид/липидному комплексу на основе вышеуказанного пептида, в котором фосфолипид представляет собой один или более из сфингомиелина, DPPC и DPPG, фармацевтической композиции, его включающей, и способам лечения дислипидемии, сердечно-сосудистого заболевания, эндотелиальной дисфункции, макрососудистых расстройств и микрососудистых расстройств с его использованием. Изобретение позволяет получить миметик ApoA-I более стабильный по сравнению с ApoA-I и простой в получении. 8 н. и 6 з.п. ф-лы, 21 ил., 14 табл., 24 пр.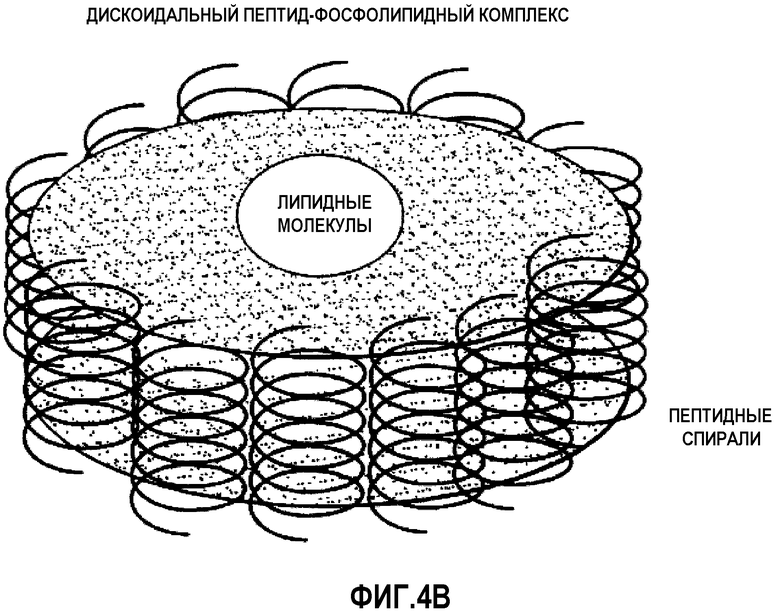
1. Пептид, являющийся миметиком ApoA-I, имеющий следующую аминокислотную последовательность:
Lys-Leu-Lys-Gln-Lys-Leu-Ala-Glu-Leu-Leu-Glu-Asn-Leu-Leu-Glu-Arg-Phe-Leu-Asp-Leu-Val-Inp (SEQ ID NO: 16),
или его фармацевтически приемлемая соль.
2. Пептид/липидный комплекс, предназначенный для лечения дислипидемии, сердечно-сосудистого заболевания, эндотелиальной дисфункции, макрососудистых расстройств и микрососудистых расстройств, содержащий фосфолипид и пептид или фармацевтически приемлемую соль пептида по п.1, где фосфолипид представляет собой один или более из сфингомиелина, DPPC и DPPG и отношение суммарного количества пептида к фосфолипиду составляет от примерно 1 : примерно 0,5 до примерно 1 : примерно 5.
3. Пептид/липидный комплекс по п.2, где фосфолипид представляет собой смесь сфингомиелина и DPPC или DPPG.
4. Пептид/липидный комплекс по п.2, где фосфолипид представляет собой смесь сфингомиелина, DPPC и DPPG.
5. Пептид/липидный комплекс по п.4, где массовое соотношение пептид : сфингомиелин : DPPC : DPPG составляет 1 : 1,2125 : 1,2125 : 0,075.
6. Пептид/липидный комплекс по п.4, где массовое соотношение сфингомиелин : DPPC : DPPG составляет 48,5 : 48,5 : 3.
7. Пептид/липидный комплекс по п.2, где массовое соотношение суммарного количества пептида к фосфолипиду составляет 1 : 2,5.
8. Пептид/липидный комплекс по п.2, где фосфолипид представляет собой смесь сфингомиелина, DPPC и DPPG, массовое соотношение пептид : сфингомиелин : DPPC : DPPG составляет 1 : 1,2125 : 1,2125 : 0,075, и массовое соотношение пептида к липиду составляет 1 : 2,5.
9. Композиция, предназначенная для лечения или предупреждения дислипидемии, сердечно-сосудистого заболевания, эндотелиальной дисфункции, макрососудистого расстройства или микрососудистого расстройства, содержащая пептид/липидный комплекс по любому из пп. 2-8 в количестве, эффективном для лечения или предупреждения дислипидемии, сердечно-сосудистого заболевания, эндотелиальной дисфункции, макрососудистого расстройства или микрососудистого расстройства, и фармацевтически приемлемый носитель или наполнитель.
10. Способ лечения или предупреждения дислипидемии, предусматривающий введение нуждающемуся в этом млекопитающему пептид/липидного комплекса по любому из пп.2-8 в количестве, эффективном для лечения дислипидемии.
11. Способ лечения или предупреждения сердечно-сосудистого заболевания, предусматривающий введение нуждающемуся в этом млекопитающему пептид/липидного комплекса по любому из пп.2-8 в количестве, эффективном для лечения и предупреждения сердечно-сосудистого заболевания.
12. Способ лечения или предупреждения эндотелиальной дисфункции, предусматривающий введение нуждающемуся в этом млекопитающему пептид/липидного комплекса по любому из пп.2-8 в количестве, эффективном для лечения и предупреждения эндотелиальной дисфункции.
13. Способ лечения или предупреждения макрососудистого расстройства, предусматривающий введение нуждающемуся в этом млекопитающему пептид/липидного комплекса по любому из пп.2-8 в количестве, эффективном для лечения или предупреждения макрососудистого расстройства.
14. Способ лечения или предупреждения микрососудиствого расстройства, предусматривающий введение нуждающемуся в этом млекопитающему пептид/липидного комплекса по любому из пп.2-8 в количестве, эффективном для лечения или предупреждения микрососудистого расстройства.
| WO 9916409 A2, 08.04.1999 | |||
| АГОНИСТЫ АПОЛИПОПРОТЕИНА А-I И ИХ ИСПОЛЬЗОВАНИЕ ДЛЯ ЛЕЧЕНИЯ ЗАБОЛЕВАНИЙ, СВЯЗАННЫХ С ДИСЛИПИДЕМИЕЙ | 1998 |
|
RU2215747C2 |
| COPELAND T.D | |||
| et al, Substitution of proline with pipecolic acid at the scissile bond converts a peptide substrate of HIV proteinase into a selective inhibitor, Biochem Biophys Res Commun, 1990, v | |||
| Универсальный двойной гаечный ключ | 1920 |
|
SU169A1 |
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| Приспособление для съемки жилетно-карманным фотографическим аппаратом со штатива | 1921 |
|
SU310A1 |
| HATANO M | |||
| et al., Properties of a toxic phospholipids in the northern | |||

