Изобретение относится к способу получения продукта реакции, катализируемой нитрилазой, включающему стадии (i) обеспечения микроорганизма, содержащего указанную нитрилазу, расположенную на его поверхности, и/или мембранного препарата указанного микроорганизма, и (ii) приведения микроорганизма и/или его мембранного препарата в контакт с одним или более субстратами нитрилазы в условиях, совместимых с активностью нитрилазы. В частности, настоящее изобретение относится к способу получения карбоновой кислоты, включающему стадии (i) обеспечения микроорганизма, содержащего нитрилазу на его поверхности, и/или его мембранного препарата, (ii) приведения микроорганизма и/или его мембранного препарата в контакт с нитрилом таким образом, чтобы нитрилаза могла катализировать конверсию нитрила в карбоновую кислоту. Кроме того, настоящее изобретение относится к способу получения энантиомерно чистой (R)-миндальной кислоты с использованием цельноклеточного биокатализатора с экспонированной на поверхности нитрилазой или его мембранного препарата для процессинга рацемического нитрила миндальной кислоты.
Нитрилы являются важными молекулами-предшественниками для получения множества продуктов, например карбоновых кислот или амидов (Banerjee et al., 2002). Однако вследствие высокой стабильности нитрилов их необходимо подвергать воздействию сильно основных или кислых условий при высоких температурах для химической конверсии нитрилов (Banerjee et al., 2002; Nagasawa et al., 1990). Кроме того, очистка этих продуктов из соответствующей реакционной смеси является очень трудоемкой процедурой.
Нитрилазы (например, EC 3.5.5.1) представляют собой ферменты, способные преобразовывать нитрилы в соответствующую карбоновую кислоту и аммиак за одну стадию (Thimann and Mahadevan, 1964). Применение ферментов нитрилаз вызвало значительный интерес в области промышленности, поскольку, с одной стороны, карбоновые продукты часто используют в качестве промежуточных продуктов в самых разнообразных химических производственных процессах, а с другой стороны, нитрилазы можно использовать для детоксикации отходов (Banerjee et al., 2002; Thuku et al., 2009). Кроме того, нитрилазы высокоизбирательны и обеспечивают получение энантиомерно чистых карбоновых кислот в мягких условиях, без необходимости использования дорогостоящих стадий блокирования и деблокирования, а также катализаторов, необходимых в обычных химических методах (Banerjee et al., 2002). Указанная высокая степень избирательности делает возможным синтез продукта, который невозможно или можно с большим трудом получать общепринятыми методами.
Наиболее важным промышленным применением нитрилаз с точки зрения количества является конверсия рацемического нитрила миндальной кислоты в энантиомерно чистую (R)-миндальную кислоту (Rey et al., 2004). (R)-миндальную кислоту применяют для разделения энантиомеров в рацемате с использованием диастереомерных солей. Кроме того, она является важным хиральным промежуточным продуктом в химическом синтезе новых активных веществ.
Из литературы хорошо известно, что нитрилаза из A. faecalis способна преобразовывать нитрил миндальной кислоты в миндальную кислоту с высокой энантиоселективностью. Хиральные карбоновые кислоты, такие как миндальная кислота, представляют собой соединения, востребованные в органической химии, поскольку они служат в качестве исходных продуктов для множества фармацевтических препаратов или средств защиты растений. Таким образом, например, (R)- или (S)-миндальную кислоту используют для рацемического разделения рацемических аминов. Кроме того, (R)-миндальную кислоту используют в качестве промежуточного продукта для синтеза.
Однако общеизвестно, что нитрилазы являются нестабильными ферментами (Buchholz, Kasche and Bornscheuer, 2005) и, следовательно, редко используются в промышленности в виде очищенных ферментов либо используются в виде цельноклеточных препаратов. Активный центр нитрилаз содержит каталитическую последовательность из трех аминокислотных остатков (каталитическую триаду), содержащую консервативный остаток цистеина активного центра (Kobayashi et al., 1992), подверженный аутоокислению (Kobayashi et al., 1992) и реакционно-способный по отношению ко многим химическим веществам, таким как тиол-содержащие реагенты.
До тех пор пока они находятся внутри клетки, нитрилазы защищены окислительно-восстановительными условиями бактериального цитозоля, которые являются восстановительными (Choi and Lee, 2004), что предотвращает окисление тиоловых групп. Однако показано, что нитрилазы теряют активность под воздействием кислорода, и это наблюдение было связано с восприимчивостью критически важной триады чувствительных к окислению и восстановлению остатков цистеина в активном центре (Mateo et al., 2006).
Были использованы различные стратегии для преодоления недостатков нитрилаз с точки зрения недостаточной активности и/или стабильности.
При одном подходе используют целые экспрессирующие нитрилазу клетки, а не очищенные препараты фермента, при этом имеют дело с внутриклеточными нитрилазами либо в специфичной для штамма, либо в рекомбинантной форме (Kaul et al., 2007). Например, исследователи использовали экспрессирующие нитрилазу клетки Alcaligenes faecalis (Kaul, A. Banerjee and U. C. Banerjee, 2006) в качестве цельноклеточных катализаторов, содержащих нитрилазу, находящуюся внутри клетки. Был достигнут энантиомерный избыток в 97%. Логическое обоснование такого подхода заключается в том, что ферменты остаются в их естественном окружении, бактериальном цитозоле, где эндогенные механизмы сворачивания, а также непрерывный метаболизм и биосинтез белков поддерживают постоянный уровень стабильного и активного белка. Однако цельноклеточные катализаторы следует получать и прикреплять к соответствующей матричной системе (альгинатной) с помощью длительных и трудоемких процедур. Более того, субстраты, подлежащие конверсии, должны пересекать избирательную мембрану бактериальной клетки и, попав в цитозоль, встречаться с целым рядом ферментов с альтернативными активностями и конкурирующими субстратами. И наконец, цельноклеточные катализаторы загрязняют препараты желаемого продукта, и, вследствие этого, исключено использование таких продуктов в чувствительных к этому вариантах применения, например для создания медицинских, косметических и гигиенических продуктов, а также напитков и продуктов питания.
Другой подход к получению высоких уровней функциональных нитрилаз для промышленного применения включает использование шаперонов. Шапероны семейства GroEL являются важными белками, находящимися в цитозоле, которые необходимы для того, чтобы клетки производили, в результате множественных циклов АТФ-зависимого связывания, инкапсуляции и высвобождения субстратных белков, правильно свернутые белки (Hartl, F.U. and Hayer-Hartl, M., 2002). Делеция GroEL в конечном итоге приводит не только к снижению уровней функциональных субстратных белков, но и к гибели бактериальных клеток вследствие накопления больших количеств несвернутого белка. Шапероны, в частности компоненты системы GroEL, были совместно экспрессированы с рекомбинантными нитрилазами в цитозоле E. coli (патент США № 5629190, фигура 9). Этот подход привел к получению более высоких уровней растворимого белка по сравнению с клетками, имеющими только эндогенный уровень шаперонов. Однако необходимым условием такого подхода является использование целых клеток, обеспечивающих механизмы такой укладки, либо добавление отдельно очищенных шаперонов и их кофакторов, а также дополнительных субстратов к препаратам нитрилаз.
В патенте США № 5629190 описано применение клеток E. coli, рекомбинантно экспрессирующих нитрилазы. Однако из-за определенного внутриклеточного набора ферментов данных клеток получаемые реакции являются сложными, а их выход непредсказуемым, даже если белковый агент, такой как GroEL, способствующий сворачиванию полипептидов, которые микроорганизм синтезирует, например рекомбинантно экспрессируемой нитрилазы, экспрессирован рекомбинантно. Как раскрыто в патенте США № 5629190, катализируемая нитрилазой конверсия нитрила в карбоновую кислоту конкурирует с катализируемой нитрилгидратазой реакцией (приводящей к образованию амида кислоты), результатом чего является уменьшение выхода реакции и образование нежелательных побочных продуктов, которые может быть сложно удалять. Другие ферменты, присутствующие в клетке, могут модифицировать нитрил по нитрильной группе или любому другому фрагменту.
Другой подход включает получение и использование перекрестно сшитых ферментных агрегатов, то есть получение растворимого рекомбинантного фермента и последующее сшивание при помощи реакционно-способных бифункциональных химических веществ, таких как глутаральдегид или поли(этиленимин). Однако Kaul et al. (2007) показали, что такая иммобилизация приводит к значительному снижению скоростей реакций и уменьшению константы специфичности. Кроме того, химическое сшивание приводит к изменению субстратной специфичности, наблюдение, связанное с процессом сшивания, который вполне может приводить к образованию белковых молекул, «запертых» в определенной конформации. Кроме того, партии иммобилизованных ферментов, а также продукты их распада и детрит невозможно с легкостью отделять от нужных продуктов реакции.
Yamamoto et al. (1991) тестировали различные изоляты (штаммы дикого типа) в отношении их способности преобразовывать нитрил миндальной кислоты в (R)-(-)-миндальную кислоту. Установлено, что A. faecalis штамм ATCC 8750 обладает самым высоким уровнем активности и энантиоселективности для получения (R)-(-)-миндальной кислоты из рацемического нитрила миндальной кислоты (Yamamoto et al., 1991). В частности, (R)-(-)-миндальная кислота, полученная из нитрила миндальной кислоты с помощью покоящихся клеток, присутствовала в 100% энантиомерном избытке. A. faecalis ATCC 8750 содержит (R)-энантиоселективную нитрилазу и амидазу, используемые для процессинга нитрила миндальной кислоты и амида миндальной кислоты, соответственно. Поскольку (R)-(-)-миндальная кислота производилась из рацемического нитрила миндальной кислоты с выходом 91%, ввиду того что совсем не оставалось (S)-нитрила миндальной кислоты, остающийся в реакционной смеси (S)-(-)-нитрил миндальной кислоты спонтанно рацемизировался из-за химического равновесия (R)- и (S)-изомера нитрила миндальной кислоты с одной стороны и бензальдегида/HCN с другой стороны. Вследствие этого, почти весь нитрил миндальной кислоты потреблялся и преобразовывался в (R)-(-)-миндальную кислоту. Следует отметить, что согласно Yamamoto et al. (1991) для клеток A. faecalis, используемых для производства (R)-миндальной кислоты, необходимы специально оптимизированные условия культивирования.
Между тем, различные нитрилазы бактериального происхождения были клонированы и рекомбинантно экспрессированы в E. coli (Kiziak et al., 2005; Luo et al., 2008; Rustler et al., 2008; патент США № 6180359; Ress-Löschke et al., 1998). Нитрилаза охарактеризована биохимически (Kiziak et al., 2005). Удалось также рекомбинантно экспрессировать ген нитрилазы из A. faecalis ATCC 8750 в E. coli, продукт которого затем использовали в экспериментах по иммобилизации в качестве «перекрестно сшитых ферментных агрегатов» после очистки (Rey et al., 2004). В этом способе нитрилазу нужно выделять из организма-хозяина, то есть E. coli, и очищать. Такой подход является довольно трудоемким. Кроме того, нитрилаза может терять активность во время процедуры.
Нитрилаза из Alcaligenes faecalis подвид faecalis ATCC 8750 описана как гомоолигомерный фермент, состоящий из приблизительно 14 субъединиц с молекулярной массой субъединицы 32 кДа, так что нативный белок имеет общую суммарную молекулярную массу приблизительно около 460 кДа (Yamamoto et al., 1992). Как правило, нитрилазы существуют в виде неактивных димеров и образуют активные олигомеры в результате самоассоциации. Однако количество субъединиц, необходимых для активности, различается от нитрилазы к нитрилазе (Thuku et al., 2009).
Таким образом, нитрилаза имеет ряд недостатков, которые препятствуют использованию нитрилазы в производстве органических соединений:
- нитрилазы чувствительны к окислению, что приводит к снижению активности нитрилаз с течением времени;
- при использовании в виде цельноклеточного биокатализатора нежелательные побочные реакции, катализируемые другими присутствующими в клетке ферментами (такими, как нитрилгидратаза), могут нарушать реакцию, катализируемую нитрилазой;
- нитрилазу невозможно извлекать из реакционной среды.
Аутодисплей представляет собой элегантный инструмент для экспонирования на поверхности бактерий рекомбинантных белков. Эта экспрессионная система основана на механизме секреции семейства белков-аутотранспортеров, относящихся к секреторной системе типа V.
В грамотрицательных бактериях аутотранспортерный путь развивался как для перемещения белков к клеточной поверхности, так и для секреции белков во внеклеточную среду (Jose and Meyer, 2007). Белки-аутотранспортеры синтезируются в виде белков-предшественников, содержащих все необходимые структурные элементы для перемещения к клеточной поверхности (Jose, 2006). Они синтезируются с N-концевым сигнальным пептидом, типичным для пути Sec, который позволяет пересекать внутреннюю мембрану. При нахождении в периплазме после отделения сигнального пептида C-концевая часть предшественника укладывается в наружную мембрану в виде пориноподобной структуры, так называемого β-цилиндра. Через эту пору присоединенный на N-конце домен-«пассажир» переносится на поверхность (Jose, 2002). Там он может быть отщеплен - либо в результате аутопротеолиза, либо дополнительными протеазами - или остается заякоренным на оболочке клетки через транспортерный домен. Замена природного «пассажира» рекомбинантным белком приводит к его правильному переносу на поверхность. Для этой цели методами генетической инженерии можно конструировать искусственный предшественник, состоящий из сигнального пептида, рекомбинантного «пассажира», β-цилиндра и находящейся между ними связующей области, что необходимо для полного доступа к клеточной поверхности. Таким способом аутотранспортер AIDA-I успешно использовали для эффективного экспонирования на поверхности различных доменов-«пассажиров» (Henderson et al., 2004).
В системе аутодисплея наблюдалась самоассоциация субъединиц в активный фермент, например, в случае димерного фермента сорбитолдегидрогеназы (Jose, 2002; Jose and von Schwichow, 2004).
В частности, технология аутодисплея представляет собой метод экспрессии заданных белков на поверхности наружной мембраны E. coli и других грамотрицательных бактерий, в которых система аутодисплея основана на природном механизме секреции белков-аутотранспортеров (Jose and Meyer, 2007). В этом методе рекомбинантный белок-«пассажир» можно с легкостью переносить путем введения его кодирующей последовательности в рамку считывания между сигнальным пептидом и перемещающим доменом аутодисплейного вектора, используя стандартные методы генетической инженерии. Сигнальный пептид можно получать из субъединицы холерного токсина (CTB) и можно объединять с искусственным промотором. В результате этого, белок-«пассажир», который необходимо перенести через наружную мембрану, экспрессируется в виде рекомбинантного белка, слитого с другим белком, называемым аутотранспортером, на наружной мембране E. coli (AIDA-I) (Jose, 2006). C-концевая часть белков-аутотранспортеров образует пориноподобную структуру (β-цилиндр) внутри наружной мембраны E. coli. Эта пориноподобная структура облегчает перемещение рекомбинантного белка-«пассажира» на поверхность наружной мембраны E. coli (Jose, 1995, 2006, 2007).
Задачей настоящего изобретения является предложение способа, позволяющего продуцировать и получать нитрилазы в активном состоянии, которые легко доступны для молекул субстрата. Кроме того, следует избегать присутствия других ферментов, чтобы предотвращать нежелательные побочные реакции. Другой задачей настоящего изобретения является предложение способа производства продуктов реакций, катализируемых нитрилазой (например, карбоновых кислот), с использованием рекомбинантных нитрилаз, в котором проблема снижения активности с течением времени решена, по меньшей мере, частично. Другой задачей настоящего изобретения является предложение способа, в котором продукт реакции, катализируемой нитрилазой (например, карбоновая кислота), можно легко извлекать.
Удивительно, но авторы настоящего изобретения обнаружили, что нитрилазу можно экспрессировать на поверхности клетки-хозяина, такого как микроорганизм, при этом нитрилаза является полностью функциональной, то есть способной катализировать конверсию нитрилов, приводящую к образованию карбоновой кислоты.
Первым аспектом настоящего изобретения является способ получения продукта реакции, катализируемой нитрилазой, включающий стадии:
(i) обеспечения микроорганизма, содержащего указанную нитрилазу, расположенную на его поверхности, и/или мембранного препарата указанного микроорганизма,
(ii) приведения микроорганизма и/или его мембранного препарата в контакт с одним или более субстратами нитрилазы в условиях, совместимых с активностью нитрилазы.
В способе по настоящему изобретению можно использовать любой известный субстрат нитрилаз. Продукт может представлять собой любое соединение, являющееся продуктом реакции, катализируемой нитрилазой. Используемый в данном документе термин «условия, совместимые с активностью нитрилазы» включает условия, в которых нитрилаза является активной, то есть в которых нитрилаза способна преобразовывать один или более субстратов в продукты. Можно использовать любые известные условия, в которых нитрилаза является активной.
Предпочтительно, если один или, по меньшей мере, один субстрат нитрилазы, используемый в способе по настоящему изобретению, представляет собой нитрил. Также предпочтительно, если продукт, получаемый способом по настоящему изобретению, представляет собой карбоновую кислоту. «Условия, совместимые с активностью нитрилазы» могут быть условиями, в которых нитрилаза катализирует конверсию нитрила, приводящую к образованию карбоновой кислоты. Предпочтительный вариант осуществления настоящего изобретения представляет собой способ получения карбоновой кислоты, включающий стадии:
(i) обеспечения микроорганизма, содержащего нитрилазу, расположенную на его поверхности, и/или его мембранного препарата,
(ii) приведения микроорганизма и/или его мембранного препарата в контакт с нитрилом таким образом, что нитрилаза катализирует конверсию нитрила, приводящую к образованию карбоновой кислоты.
Способ по настоящему изобретению необязательно может дополнительно включать стадию
(iii) извлечения микроорганизма, используемого на стадии (ii).
При использовании в данном документе ссылка на «стадию (i)», «стадию (ii)» или «стадию (iii)» означает ссылку на соответствующую стадию способа для получения продукта реакции, катализируемой нитрилазой, и предпочтительного способа получения карбоновой кислоты, описанного в данном документе.
Настоящее изобретение основано на том удивительном открытии, что бактериальные нитрилазы, будучи рекомбинантным методом слиты с подходящей аутотранспортерной системой, переносятся на поверхность бактериальной клетки и интегрируются в конформации, соответствующей активному состоянию, в бактериальную мембрану таким образом, что они остаются каталитически компетентными в течение нескольких дней и могут быть использованы для катализа реакций в промышленных масштабах. Это особенно удивительно, поскольку наружная мембрана бактериальных клеток лишена механизма сворачивания белков, подходящего для цитозольных белков. Кроме того, что самое удивительное, авторы настоящего изобретения обнаружили, что фермент не теряет активность после или во время переноса чувствительного к окислению и восстановлению фермента из восстановительных условий бактериального цитозоля на наружную мембрану, где кислород присутствует в повышенных количествах, создавая сильно окислительную среду.
Не желая быть связанными какой-либо теорией, авторы настоящего изобретения предположили, что нитрилазы, будучи иммобилизованы на поверхности бактериальных клеток путем аутодисплея, образуют агрегаты или трехмерные структуры, которые защищают реакционно-способные каталитические аминокислотные остатки от дезактивации, даже при воздействии повышенных уровней кислорода и сильных механических сил, связанных с энергичным встряхиванием в рамках рутинных методов, используемых для выращивания большого количества бактериальных клеток.
Цельноклеточный биокатализатор, описанный в данном документе, впервые позволяет получать активную нитрилазу на поверхности клетки грамотрицательной бактерии, такой как E. coli, при этом клетку можно использовать для конверсии нитрила и для получения соответствующей карбоновой кислоты. В частности, цельноклеточный биокатализатор с экспонированной нитрилазой, описанный в данном документе, можно применять для биотехнологической конверсии нитрилов.
Важно то, что указанную реакцию проводят в мягких условиях, с хорошей производительностью и без образования токсичных побочных продуктов. В частности, нитрилаза, рекомбинантно экспрессированная на поверхности микроорганизма, способна катализировать конверсию нитрилов в мягких условиях, например, при pH приблизительно 7,0.
Используемый в данном документе термин «цельноклеточный биокатализатор» означает целую клетку, в частности микроорганизм, содержащую природный катализатор, такой как ферментный белок, в частности нитрилазу, для осуществления химической трансформации органических соединений, в частности нитрилов, причем природный катализатор предпочтительно расположен на поверхности клетки. Согласно изобретению, катализатор может быть рекомбинантно экспрессирован.
Бактериальная клетка содержит ряд отсеков, отделенных друг от друга гидрофобными мембранами. Грамположительная бактериальная клетка содержит плазматическую мембрану, ограничивающую цитозоль, внутреннюю часть клетки. Плазматическая мембрана окружена слоем пептидогликанов. Напротив, у грамотрицательных бактерий в дополнение к плазматической мембране имеется другая мембрана, называемая наружной мембраной. Используемый в данном документе термин «поверхность» предпочтительно означает слой микроорганизма, контактирующий с окружающей средой, например жидкой культуральной средой. В предпочтительном варианте осуществления настоящего изобретения поверхность представляет собой сторону грамотрицательной бактериальной клетки, контактирующую с жидкой культуральной средой. Используемые в данном документе выражения «экспонированная на поверхности» и «экспрессированная на поверхности» используются взаимозаменяемо и означают локализацию нитрилазы на поверхности клетки.
Используемый в данном документе термин «микроорганизм» означает клетку-хозяина. Клетка-хозяин по изобретению может представлять собой прокариотический или эукариотический микроорганизм, предпочтительно прокариотический микроорганизм, более предпочтительно бактериальную клетку, даже более предпочтительно грамотрицательную бактериальную клетку. Наиболее предпочтительным микроорганизмом является клетка E. coli. Термины «микроорганизм» и «клетка-хозяин» используются в данном документе взаимозаменяемо.
Используемый в данном документе термин «нитрилаза» означает фермент нитрилазу, его каталитически активную часть, производное или аналог, которые катализируют конверсию нитрилов в карбоновые кислоты и аммиак. Следует отметить, что используемые в данном документе термины «производное» или «аналог» молекулы означают часть или модифицированную версию молекулы. В настоящем изобретении нитрилаза может представлять собой любую нитрилазу, такую как нитрилаза, способная катализировать конверсию нитрила, приводящую к образованию карбоновой кислоты. Предпочтительная нитрилаза является ферментом согласно EC 3.5.5.1 (другое название «нитриламиногидролаза»).
Нитрилаза может проявлять только нитрилазную активность. Нитрилаза может также быть многофункциональным ферментом, который в дополнение к другим активностям проявляет нитрилазную активность.
Квалифицированному специалисту известен целый ряд видов, являющихся источником нитрилаз, которые включают прокариот и эукариот. Например, нитрилазу, используемую в настоящем изобретении, можно получать из Bordetella, Klebsiella, Aspergillus, Alcaligenes, Saccharomyces, Burkholderia, Neurospora, Lachancea, Debaryomyces, Yarrowia, Candida, Kluyveromyces, Rhodococcus, Nocardia и/или Rhizobium. Примерами видов, являющихся источником нитрилазы, которую можно использовать по настоящему изобретению, являются Bordetella bronchiseptica, Klebsiella pneumoniae, Aspergillus niger, Alcaligenes faecalis, Saccharomyces cerevisiae, Burkholderia multivorans, Aspergillus fumigatus, Neurospora crassa, Lachancea thermotolerans, Debaryomyces hansenii, Yarrowia lipolytica, Candida glabrata, Kluyveromyces lactis, Rhodococcus rhodochrous, Nocardia sp., Rhizobium leguminosarum и/или Nocardia farcinica.
Нитрилазу по настоящему изобретению предпочтительно получают из Alcaligenes, Klebsiella и/или Saccharomyces. Более предпочтительную нитрилазу выбирают из нитрилаз, полученных из Alcaligenes faecalis, Klebsiella pneumoniae и Saccharomyces cerevisiae. Alcaligenes faecalis может представлять собой Alcaligenes faecalis подвид faecalis. Klebsiella pneumoniae может представлять собой Klebsiella pneumoniae подвид ozaenae. Еще более предпочтительной нитрилазой является нитрилаза Alcaligenes faecalis.
Нитрилазу по настоящему изобретению можно выбирать из группы, включающей нитрилазы P_887662 [GI: 33600102, 01-май-2009, Bordetella bronchiseptica RB50], AAA25057 [GI: 149175, 26-апрель-1993, Klebsiella pneumoniae], NP_943299 [GI: 38639530, 30-апрель-2009, Klebsiella pneumoniae], P10045 [GI: 115192, 20-январь-2009, Klebsiella pneumoniae подвид ozaenae], XP_001389617 [GI: 145230706, 28-февраль-2008, Aspergillus niger], ACS13754 [GI: 239738518, 15-июнь-2009, Alcaligenes sp. ECU0401], BAA02684 [GI: 216203, 16-февраль-2008, Alcaligenes faecalis], CAK46957 [GI: 134083480, 24-март-2007, Aspergillus niger], EDV09642 [GI: 190406375, 16-июнь-2008, Saccharomyces cerevisiae RM11-1a], YP_001945058 [GI: 189349430, 07-май-2009, Burkholderia multivorans ATCC 17616], NP_012102 [GI: 6322027, 16-июнь-2008, Saccharomyces cerevisiae RM11-1a], 1F89B [GI: 16975400, 19-ноябрь-2001, Saccharomyces cerevisiae], NP_012409 [GI: 6322335, 05-ноябрь-2009, Saccharomyces cerevisiae], EDN63257 [GI: 151945002, 13-июль-2007, Saccharomyces cerevisiae YJM789], EDN59257 [GI: 151940875, 13-июль-2007, Saccharomyces cerevisiae YJM789], CAY80342 [GI: 259147089, 23-сентябрь-2009, Saccharomyces cerevisiae EC1118], EEU05439 [GI: 256270219, 20-август-2009, Saccharomyces cerevisiae JAY291], P40447 [GI: 731891, 03-ноябрь-2009, Saccharomyces cerevisiae], XP_751200 [GI: 70992703, 27-февраль-2008, Aspergillus fumigatus Af293], CAD71250 [GI: 28950282, 14-ноябрь-2006, Neurospora crassa], CAK48039 [GI: 134075478, 24-март-2007, Aspergillus niger), CAR23067 [GI: 238934886, 08-октябрь-2009, Lachancea thermotolerans], CAG86637 [GI: 199431326, 10-сентябрь-2008, Debaryomyces hansenii], XP_500602 [GI: 50546150, 29-октябрь-2008, Yarrowia lipolytica], CAG78819 [GI: 49651877, 23-октябрь-2008, Yarrowia lipolytica], CAG59341 [GI: 49525722, 16-декабрь-2008, Candida glabrata], XP_454637 [GI: 50309261, 18-апрель-2008, Kluyveromyces lactis], P20960 [GI: 417386, 05-май-2009, Alcaligenes faecalis], ABO46008 [GI: 134034945, 05-ноябрь-2009, Rhodococcus rhodochrous], AAX18182 [GI: 60280369, 02-март-2005, Nocardia sp. C-14-1], BAA02127 [GI: 216932, 16-февраль-2008, Rhodococcus rhodochrous], Q02068 [GI: 417382, 20-январь-2009, Rhodococcus rhodochrous], CAK02877 [GI: 115259785, 13-май-2009, Rhizobium leguminosarum bv. viciae 3841], CAF05970 [GI: 40882143, 10-октябрь-2009, Neurospora crassa], CAK08726 [GI: 115257629, 13-май-2009, Rhizobium leguminosarum bv. viciae 3841], BAD58116 [GI: 54016746, 10-май-2008, Nocardia farcinica IFM 10152] и Q5Z1U0 [GI: 81603033, 03-ноябрь-2009, Nocardia farcinica]. В этом списке использован формат «регистрационный номер Genbank [идентификатор Genbank, дата записи в базу данных, вид/штамм]». Последовательности нитрилаз этой группы можно определять по регистрационному номеру Genbank, идентификатору Genbank (GI), дате записи в базу данных Genbank, а также виду и/или штамму, из которого последовательность получена.
Используемый в данном документе термин «нитрилаза» включает любую нитрилазу, имеющую идентичность по меньшей мере 70%, по меньшей мере 80%, по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 98% или по меньшей мере 99% с нитрилазой, раскрытой в данном документе. Квалифицированному специалисту известны подходящие методы определения степени идентичности на уровне аминокислотной последовательности, например, при помощи BLAST или PBLAST.
Как указано выше, активная нитрилаза может представлять собой гомоолигомерный фермент, состоящий из приблизительно 14 субъединиц. Как правило, нитрилазы существуют в виде неактивных димеров. В настоящем изобретении нитрилаза, экспонированная на поверхности клетки-хозяина, может быть гомомультимером, включающим три или более идентичные субъединицы. В частности, гомомультимер может содержать от 7 до 16 идентичных субъединиц, например 7, 8, 9, 10, 11, 12, 13, 14, 15 или 16 идентичных субъединиц. Предпочтительным является число, равное приблизительно 14 субъединиц, то есть 11, 12, 13, 14, 15 или 16 субъединиц. Гомомультимеры (также называемые в данном документе «гомоолигомеры») могут быть образованы в результате спонтанной ассоциации нескольких идентичных полипептидных субъединиц, экспонированных на мембране клетки-хозяина.
Используемый в данном документе термин «нитрил» означает любое соединение, содержащее по меньшей мере одну -CN функциональную группу (нитрильную группу). Нитрил может также содержать одну, две, три, четыре или даже больше нитрильных групп.
Нитрильная группа может быть связана с ароматическим фрагментом, например, арильной группой, содержащей от 6 до 14 атомов углерода, предпочтительно от 6 до 10 атомов углерода. Арильная группа может содержать одно, два или три конденсированных ароматических кольца. Более предпочтительно, арильная группа может быть фенильной группой. Арильная группа может быть замещена одним или более заместителями, например одним, двумя, тремя, четырьмя, пятью или даже более заместителями.
Нитрильная группа может быть связана с алифатическим фрагментом, например алкильной группой, содержащей от 1 до 8 атомов углерода, от 1 до 6 атомов углерода, от 1 до 4 атомов углерода или от 1 до 3 атомов углерода. Алкильная группа может быть с прямой или разветвленной цепью. Алкильная группа может содержать циклический фрагмент. Алкильная группа может быть замещена одним или более заместителями, например одним, двумя, тремя, четырьмя, пятью или даже более заместителями.
Нитрил может содержать алкиларильную группу, при этом алкил и арил имеют значения, описанные в данном документе. По меньшей мере одна нитрильная группа может быть связана с алкильной группой и/или арильной группой. Алкильный и/или арильный фрагмент может быть замещен одним или более заместителями, например одним, двумя, тремя, четырьмя, пятью или даже более заместителями.
Используемые в данном документе один или более заместители могут быть независимо выбраны из -OH, йода, брома, хлора, фтора, арила, алкила, алкокси и тому подобного, причем алкил и арил имеют значения, описанные в данном документе, и алкокси содержит от 1 до 8 атомов углерода, от 1 до 6 атомов углерода, от 1 до 4 атомов углерода или от 1 до 3 атомов углерода. Предпочтительной алкокси-группой является метокси. Алкокси-группа может быть с прямой или разветвленной цепью. Алкокси-группа может содержать циклический фрагмент.
Алкильная группа, ароматическая группа и/или алкокси-группа могут независимо содержать по меньшей мере один гетероатом, например один, два, три, четыре или даже более гетероатомов, так что по меньшей мере один гетероатом замещает атом углерода. Гетероатомы можно независимо выбирать из N, O и S.
Типичные нитрилы для использования в качестве субстратов нитрилазы по изобретению представляют собой ароматические нитрилы, такие как нитрил миндальной кислоты, бензонитрил, фенилпропионитрил, фенилглицинонитрил, бромоксинил, йоксинил, хлороксинил, анизонитрил, 3-бром-4-гидроксибензонитрил, 3-фтор-4-гидроксибензонитрил, 4-гидрокси-3,5-диметобензонитрил и пруназин; а также алифатические нитрилы, такие как н-бутиронитрил, н-валеронитрил, изобутиронитрил и сукцинонитрил. Особенно предпочтительным субстратом по изобретению является нитрил миндальной кислоты.
В соответствии с настоящим изобретением, предпочтительными субстратами для нитрилазы из Alcaligenes faecalis являются нитрил миндальной кислоты и пруназин. Предпочтительными субстратами для нитрилазы из Klebsiella pneumoniae являются бромоксинил, йоксинил, хлороксинил, анизонитрил, 3-бром-4-гидроксибензонитрил, 3-фтор-4-гидроксибензонитрил и 4-гидрокси-3,5-диметобензонитрил. Предпочтительными субстратами для нитрилазы из Saccharomyces cerevisiae являются бензонитрил, фенилпропионитрил, нитрил миндальной кислоты, фенилглицинонитрил, н-бутиронитрил, н-валеронитрил, изобутиронитрил и сукцинонитрил.
В одном аспекте изобретения ген, кодирующий нитрилазу, амплифицировали из целой ДНК Alcaligenes faecalis методом ПЦР и клонировали. Ген может быть слит надлежащим образом с рамкой считывания аутотранспортера при помощи подходящих сайтов рестрикции, присоединенных к праймеру для ПЦР. Экспрессию слитого белка можно продемонстрировать стандартными лабораторными методами. Фермент, экспрессированный на поверхности и иммобилизованный на ней, способен внеклеточно преобразовывать добавленный субстрат (в данном случае рацемический нитрил миндальной кислоты).
При соблюдении применяемых в настоящее время условий культивирования и экспрессии можно добиться коэффициента трансформации приблизительно 50%, предпочтительно приблизительно 60%, более предпочтительно приблизительно 70% и даже более предпочтительно приблизительно 80% через 5 дней. Соединения, имеющие асимметричный атом С, можно получать способом по настоящему изобретению в энантиомерном избытке. Энантиомерный избыток (%ee) может составлять, по меньшей мере, приблизительно 50%, 60%, 70%, 80%, 90%, 95%, 96%, 97%, 98% или 99%, предпочтительно, по меньшей мере, приблизительно 99% или выше для продукта, в частности, для карбоновой кислоты. В частности, для (R)-миндальной кислоты был достигнут коэффициент трансформации приблизительно 80% через 5 дней и энантиомерный избыток (%ee) выше чем 99%, как показано методом хиральной ВЭЖХ.
При использовании в данном документе, энантиомерный избыток (ee) вещества представляет собой меру оптической чистоты вещества. Энантиомерный избыток можно определять в растворе соединения, которое будет проанализировано. Соответствующие растворители (такие как гидрофобные и гидрофильные растворители) и их смеси известны.
Энантиомерный избыток можно рассчитывать по следующей формуле:
ee%=100*([R]-[S])/([R]+[S]),
где [R], [S] указывают на концентрацию продуктов и где R и S означают (R)- и (S)-изомеры, соответственно. Таким образом, в рацемической смеси, то есть [R]=[S], ee%=100*0/([R]+[S])=0%, то есть нет избытка R над S или наоборот. Таким образом, энантиомерный избыток рацемической смеси равен 0%, тогда как энантиомерный избыток энантиомерно чистого соединения равен 100%. Концентрацию (R)- и (S)-изомеров можно определять любым методом, известным в данной области для определения соединений, имеющих ассиметричные атомы C, таким как, но не ограничиваясь ими, определение удельного вращения смеси, хиральная колоночная хроматография (например, хиральная ВЭЖХ) и ЯМР-спектроскопия.
Изобретение, описанное в данном документе, имеет то преимущество, что мягкие условия реакции во время гидролиза нитрила сочетаются с легким методом очистки продукта, поскольку для этой цели клетки нужно только удалить центрифугированием из порции реакционной смеси.
Нитрилаза может быть рекомбинантно экспрессирована в микроорганизме и перенесена к клеточной поверхности. Микроорганизм размножается самостоятельно и производит фермент сам, так что больше нет необходимости в сложной очистке биокатализатора. Примером такого микроорганизма является штамм E. coli, описанный в примере 1 в данном документе.
Используемый в данном документе термин «рекомбинантная» означает последовательность нуклеиновой кислоты, составленную из по меньшей мере двух последовательностей нуклеиновой кислоты, функционально связанных друг с другом, которые естественным образом не связаны друг с другом. По меньшей мере две последовательности можно получать из одного и того же организма. Например, рекомбинантная конструкция может содержать сигнальную последовательность из вида Alcaligenes и кодирующую нитрилазу последовательность из того же вида Alcaligenes, которая обычно не связана с указанной сигнальной последовательностью. По меньшей мере две последовательности можно получать из по меньшей мере двух различных организмов. Например, сигнальный пептид из E. coli, ген нитрилазы из Alcaligenes, трансмембранный линкер из E. coli и аутотранспортерный домен из E. coli можно функционально связывать. Другим примером является рекомбинантный полипептид, в котором сигнальный пептид получен из β-субъединицы холерного токсина (CTB), аутотранспортерный домен и трансмембранный линкер происходят из E. coli, и нитрилаза происходит из Alcaligenes.
В предпочтительном варианте осуществления настоящего изобретения используемый в данном документе термин «экспрессируемая рекомбинантно», также используемый как синоним термина «экспрессированная рекомбинантным образом» или тому подобного, означает экспрессию рекомбинантной последовательности нуклеиновой кислоты. Например, экспрессия в E. coli, да и вообще в любом организме, последовательности нуклеиновой кислоты, содержащей сигнальный пептид E. coli и последовательность нитрилазы Alcaligenes, представляет собой рекомбинантную экспрессию.
Предпочтительно, если нитрилаза слита с транспортерным доменом аутотранспортера. Транспортерный домен аутотранспортера по изобретению может быть любым транспортерным доменом аутотранспортера и предпочтительно способен образовывать структуру β-цилиндра. Подробное описание структуры β-цилиндра и предпочтительные примеры аутотранспортеров с β-цилиндром описаны в WO97/35022, включенном в данный документ посредством ссылки. В статье Henderson et al. (2004) описаны аутотранспортерные белки, содержащие подходящие аутотранспортерные домены (обобщенная информация представлена в таблице 1 статьи Henderson et al., 2004). Информация из статьи Henderson et al. (2004) включена в данный документ посредством ссылки. Например, транспортерный домен аутотранспортера можно выбирать из Ssp (P09489, S. marcescens), Ssp-h1 (BAA33455, S. marcescens), Ssp-h2 (BAA11383, S. marcescens), PspA (BAA36466, P. fluorescens), PspB (BAA36467, P. fluorescens), Ssa1 (AAA80490, P. haemolytica), SphB1 (CAC44081, B. pertussis), AspA/NaIP (AAN71715, N. meningitidis), VacA (Q48247, H. pylori), AIDA-I (Q03155, E. coli), IcsA (AAA26547, S. flexneri), MisL (AAD16954, S. enterica), TibA (AAD41751, E. coli), Ag43 (P39180, E. coli), ShdA (AAD25110, S. enterica), AutA (CAB89117, N. meningitidis), Tsh (I54632, E. coli), SepA (CAC05786, S. flexneri), EspC (AAC44731, E. coli), EspP (CAA66144, E. coli), Pet (AAC26634, E. coli), Pic (AAD23953, E. coli), SigA (AAF67320, S. flexneri), Sat (AAG30168, E. coli), Vat (AAO21903, E. coli), EpeA (AAL18821, E. coli), EatA (AAO17297, E. coli), EspI (CAC39286, E. coli), EaaA (AAF63237, E. coli), EaaC (AAF63038, E. coli), пертактина (P14283, B. pertussis), BrkA (AAA51646, B. pertussis), Tef (AAQ82668, B. pertussis), Vag8 (AAC31247, B. pertussis), PmpD (O84818, C. trachomatis), Pmp20 (Q9Z812, C. pneumoniae), Pmp21 (Q9Z6U5, C. pneumoniae), протеазы IgA1 (NP_283693, N. meningitidis), App (CAC14670, N. meningitidis), протеазы IgA1 (P45386, H. influenzae), Hap (P45387, H. influenzae), rOmpA (P15921, R rickettsii), rOmpB (Q53047, R rickettsii), ApeE (AAC38796, S. enterica), EstA (AAB61674, P. aeruginosa), Lip-1 (P40601, X. luminescens), McaP (AAP97134, M. catarrhalis), BabA (AAC38081, H. pylori), SabA (AAD06240, H. pylori), AlpA (CAB05386, H. pylori), Aae (AAP21063, A. actinomycetemcomitans), NanB (AAG35309, R haemolytica), а также вариантов этих аутотранспортеров. Для каждого из типичных аутотранспортерных белков в скобках приведены примеры соответствующих регистрационных номеров Genbank и видов, из которых можно получать аутотранспортер. Предпочтительно транспортерный домен аутотранспортера представляет собой белок AIDA-I E. coli или его вариант, такой как, например, описанный в статье Niewert U., Frey A., Voss T., Le Bouguen C., Baljer G., Franke S., Schmidt MA. The AIDA Autotransporter System is Associated with F18 and Stx2e in Escherichia coli Isolates from Pigs Diagnosed with Edema Disease and Postweaning Diarrhea. Clin. Diagn. Lab. Immunol. 2001 Jan, 8(1):143-149;9.
Варианты вышеуказанных последовательностей аутотранспортеров можно, например, получать, изменяя аминокислотную последовательность в петлевых структурах β-цилиндра, которые не являются частью трансмембранных фрагментов. Необязательно, части нуклеиновой кислоты, кодирующие поверхностные петли, можно полностью удалять. Более того, в амфипатическом β-листе могут иметь место консервативные аминокислотные замены, то есть замены гидрофильной аминокислоты на другую гидрофильную аминокислоту и/или замены гидрофобной аминокислоты на другую гидрофобную аминокислоту. Предпочтительно, вариант имеет идентичность последовательности, равную по меньшей мере 70%, по меньшей мере 80%, по меньшей мере 90%, по меньшей мере 95% или по меньшей мере 98% на аминокислотном уровне с соответствующей природной последовательностью аутотранспортерного домена, особенно в области β-листов.
Предпочтительно, если стадия (i) способа по изобретению включает стадии:
(a) обеспечения микроорганизма, содержащего последовательность нуклеиновой кислоты, функционально связанную с последовательностью контроля экспрессии, причем указанная последовательность нуклеиновой кислоты содержит:
(1) часть, кодирующую сигнальный пептид,
(2) часть, кодирующую рекомбинантную нитрилазу, которая будет экспонирована,
(3) необязательно, часть, кодирующую сайт узнавания протеазы,
(4) часть, кодирующую трансмембранный линкер, и
(5) часть, кодирующую транспортерный домен аутотранспортера,
(b) культивирования микроорганизма в условиях, при которых последовательность нуклеиновой кислоты со стадии (a) экспрессируется, и продукт экспрессии последовательности нуклеиновой кислоты экспонируется на поверхности микроорганизма.
Специалист в данной области способен подобрать условия, при которых последовательность нуклеиновой кислоты экспрессируется, и для этого необходимы лишь рутинные эксперименты, например тестирование различных температур, сред, плотностей клеток и/или концентраций химических веществ, индуцирующих экспрессию.
Микроорганизм, получаемый на стадии (a), можно получать трансформацией последовательностью нуклеиновой кислоты, функционально связанной с последовательностью контроля экспрессии, при этом указанная нуклеиновая кислота содержит:
(1) часть, кодирующую сигнальный пептид,
(2) часть, кодирующую рекомбинантную нитрилазу, которая будет экспонирована,
(3) необязательно, часть, кодирующую сайт узнавания протеазы,
(4) часть, кодирующую трансмембранный линкер, и
(5) часть, кодирующую транспортерный домен аутотранспортера,
где компоненты (1)-(5) представляют собой последовательности нуклеиновой кислоты, описанные ниже в данном документе.
Можно использовать любой известный метод трансформации. Используемый в данном документе термин «трансформация» предпочтительно означает введение гетерологичной конструкции в клетку, предпочтительно бактериальную клетку, которая после этого называется трансформированной. Квалифицированному специалисту в данной области известны протоколы и методы трансформации, например электротрансформация или трансформация химически компетентных клеток.
В другом варианте осуществления патентоспособный способ дополнительно включает стадию (c) получения мембранного препарата из микроорганизма со стадии (b).
Мембранный препарат, используемый по настоящему изобретению, содержит каталитически активную нитрилазу. Используемый в данном документе термин «мембранный препарат» предпочтительно означает продукт, обогащенный мембранными компонентами. Специалист в данной области знаком с протоколами и методами, которые можно использовать для получения мембранных препаратов. Например, бактериальные клетки можно собирать из культуры и подвергать лизису, например, путем периодического замораживания-оттаивания, разрушения ультразвуком, ресуспендирования в лизирующих буферах или тому подобного, с последующим дифференциальным центрифугированием для выделения мембранных фракций клеток. В предпочтительном варианте осуществления настоящего изобретения мембранный препарат представляет собой препарат наружной мембраны, то есть препарат, обогащенный компонентами наружной мембраны по сравнению с компонентами других мембран и компартментов, таких как цитозоль, внутренняя мембрана и периплазма. Специалист в данной области знаком с протоколами и методами, которые можно использовать для выделения наружной мембраны, например обработкой лизоцимом бактериальных клеток и последующими стадиями центрифугирования. В предпочтительном варианте осуществления настоящего изобретения мембранный препарат представляет собой препарат наружной мембраны, содержащий каталитически активную нитрилазу, при этом мембранный препарат получают из клеток, которые синтезируют указанную нитрилазу в цитозоле, а затем переносят ее на наружную мембрану. В другом варианте осуществления указанный мембранный препарат можно соответствующим образом обрабатывать, например, встраивая мембранные белки, такие как слитый белок нитрилазы, в искусственные везикулы. Специалист в данной области владеет методами функционального воссоздания мембранных белков в везикулах, например, с использованием подходящих детергентов.
Стадия (a) способа по настоящему изобретению относится к получению микроорганизма. Микроорганизм может быть любым микроорганизмом, описанным в данном документе. На стадии (a) получают клетку-хозяина, в частности хозяина-бактерии, которая содержит последовательность нуклеиновой кислоты, функционально связанную с последовательностью контроля экспрессии, то есть промотором и, необязательно, дополнительными последовательностями, необходимыми для экспрессии гена в соответствующей клетке-хозяине. Используемая в данном документе фраза «последовательность нуклеиновой кислоты, функционально связанная с последовательностью контроля экспрессии» означает функциональное взаимодействие последовательности нуклеиновой кислоты с последовательностью контроля экспрессии. Квалифицированному специалисту известны соответствующие промоторы и последовательности контроля экспрессии. Промотор и/или последовательность контроля экспрессии могут быть гомологичными или гетерологичными для клетки-хозяина.
Предпочтительно, последовательность нуклеиновой кислоты находится в рекомбинантном векторе, например плазмидном векторе.
Последовательность нуклеиновой кислоты на стадии (a) содержит (1) часть, кодирующую сигнальный пептид, предпочтительно часть, кодирующую сигнальный пептид грамотрицательной бактерии, делающий возможным транспорт в периплазму через внутреннюю мембрану клетки. Сигнальный пептид может быть сигнальным пептидом, гомологичным для клетки-хозяина. Сигнальный пептид может также быть сигнальным пептидом, гетерологичным для клетки-хозяина.
Далее, последовательность нуклеиновой кислоты содержит (2) часть, кодирующую нитрилазу, которая будет экспонирована (= «полипептид-пассажир» или «пассажир»). Можно использовать нуклеиновую кислоту, кодирующую любую нитрилазу, описанную в данном документе.
Последовательность нуклеиновой кислоты необязательно содержит (3) часть, кодирующую сайт узнавания протеазы. Используемый в данном документе термин «сайт узнавания протеазы» означает специфическую аминокислотную последовательность, узнаваемую специфической протеазой, которая впоследствии расщепляет полипептид путем гидролиза амидной связи, характерной для сайта узнавания протеазы. Сайт узнавания протеазы может быть сайтом узнавания для собственной протеазы, то есть протеазы, естественным образом существующей в клетке-хозяине, либо для добавленной извне протеазы. Например, добавленная извне протеаза может быть протеазой IgA (см. также EP-A-0 254 090), тромбином или фактором X. Собственная протеаза, например, может быть выбрана из OmpT, OmpK или протеазы X. Сайт узнавания протеазы может быть гомологичным для клетки-хозяина. Сайт узнавания протеазы может быть также гетерологичным для клетки-хозяина.
Кроме того, последовательность нуклеиновой кислоты содержит (4) часть, кодирующую трансмембранный линкер, который необходим для презентирования полипептида-пассажира, то есть рекомбинантной нитрилазы (2) на внешней поверхности наружной мембраны клетки-хозяина. Можно использовать домен трансмембранного линкера, который гомологичен по отношению к аутотранспортеру, то есть домен трансмембранного линкера кодируется фрагментом нуклеиновой кислоты, непосредственно слитым с 5'-концом последовательности нуклеиновой кислоты, кодирующей аутотранспортерный домен. Также можно использовать домен трансмембранного линкера, который гетерологичен по отношению к аутотранспортеру. Длина трансмембранного линкера предпочтительно составляет 30-160 аминокислот.
Кроме того, последовательность нуклеиновой кислоты содержит (5) часть, кодирующую транспортерный домен аутотранспортера. В контексте настоящего изобретения аутодисплей может представлять собой рекомбинантный дисплей белков или полипептидов на поверхности при помощи аутотранспортера в любой грамотрицательной бактерии. Транспортерный домен может быть любым транспортерным доменом, описанным в данном документе.
Компоненты (1)-(5) в последовательности нуклеиновой кислоты по настоящему изобретению предпочтительно ориентированы в направлении от 5'-конца к 3'-концу. В продукте экспрессии, полученном на стадии (b), аминокислотные последовательности, кодируемые последовательностями нуклеиновой кислоты (1)-(5), предпочтительно расположены от N-конца к C-концу.
Стадия (b) в способе по настоящему изобретению относится к культивированию микроорганизма в условиях, при которых последовательность нуклеиновой кислоты стадии (a) экспрессируется и продукт экспрессии, содержащий рекомбинантную нитрилазу, экспонируется на поверхности микроорганизма. Специалисту в данной области известны подходящие условия культивирования. Способ по изобретению делает возможной эффективную экспрессию белков-пассажиров на поверхности микроорганизмов, в частности E. coli или других грамотрицательных бактериальных клеток, в количестве вплоть до 100 000 или более молекул на клетку при использовании жидкой среды следующего состава: от 5 г/л до 20 г/л, предпочтительно приблизительно 10 г/л, триптона, от 2 г/л до 10 г/л, предпочтительно приблизительно 5 г/л, дрожжевого экстракта, от 5 г/л до 20 г/л, в частности приблизительно 10 г/л, NaCl и оставшаяся часть - вода. По возможности, среда должна содержать минимально возможное количество двухвалентных катионов, так что предпочтительно используют Aqua bidest или высокоочищенную воду, например воду, очищенную Millipore. Жидкая среда предпочтительно может дополнительно содержать ЭДТА в концентрации от 2 мкМ до 20 мкМ, в частности 10 мкМ. Кроме того, она предпочтительно может содержать восстанавливающие реагенты, такие как 2-меркаптоэтанол или дитиотреитол или дитиоэритритол в предпочтительной концентрации от 2 мМ до 20 мМ. Восстанавливающие реагенты способствуют поддержанию несвернутой структуры полипептида в процессе транспортировки. Кроме того, жидкая среда может дополнительно содержать источники C, предпочтительно глюкозу, например, в количестве вплоть до 10 г/л, для того, чтобы способствовать секреции, то есть переносу «пассажира» в окружающую среду. Для поверхностного дисплея предпочтительно не добавлять дополнительный источник C. Предпочтительные условия культивирования для клеток грамотрицательных бактерий, таких как E. coli, описаны в примерах.
Стадию (ii) можно проводить в аэробных и/или окислительных условиях. В предпочтительном варианте осуществления термин «аэробные условия» означает условия, характеризующиеся наличием кислорода. В другом предпочтительном варианте осуществления термин «аэробные условия» означает наличие повышенных уровней кислорода по сравнению с уровнями, нормальными для данной температуры и жидкости. В другом предпочтительном варианте осуществления термин «аэробные условия» означает условия, характеризующиеся повышенным газообменом между жидкостью, например культуральной средой, и окружающей газовой средой, содержащей кислород, например воздухом. Специалист в данной области способен измерять уровни кислорода, например, путем использования подходящих чувствительных к кислороду электродов. Кроме того, специалист в данной области способен создавать аэробные условия, например, продувая буферы или среды воздухом или кислородом, добавляя высвобождающие кислород химические вещества или просто энергично встряхивая такие буферы или среды в присутствии кислорода или воздуха. С другой стороны, специалист в данной области способен создавать анаэробные условия, например, интенсивно продувая буферы или среды аргоном или азотом либо герметизируя культуральные сосуды, чтобы свести к минимуму газообмен.
В предпочтительном варианте осуществления используемый в данном документе термин «окислительные условия» означает добавление в среды или буферы либо наличие окислителей, то есть химических веществ, вызывающих высвобождение электронов другими веществами, при этом ферменты имеют тенденцию окисляться. Например, восстановленные окислительно-восстановительные ферменты, как правило, окисляются в присутствии кислорода. С другой стороны, используемый в данном документе термин «восстановительные» или «восстановительные условия» означает добавление в среды или буферы либо наличие восстановителей, то есть химических веществ, вызывающих поглощение электронов другими веществами. Специалист в данной области может создавать желаемые окислительно-восстановительные условия, например, путем добавления окислителей, таких как кислород, и добавления восстановителей, например тиоловых реагентов, таких как меркаптоэтанол или ДТТ. Кроме того, специалист в данной области способен измерять окислительно-восстановительный потенциал растворов, например, с помощью окислительно-восстановительных электродов.
В грамотрицательной бактериальной клетке-хозяине, такой как E. coli, после перемещения рекомбинантного «пассажира» «пассажир» остается прикрепленным к поверхности наружной мембраны за счет β-цилиндра, который служит якорем в наружной мембране. Благодаря контролируемой интеграции β-цилиндра в наружной мембране C-концевая часть β-цилиндра направлена к внутренней стороне наружной мембраны, в то время как N-концевая часть линкера, с которой рекомбинантный белок-«пассажир» ковалентно связан, направлена к внешней поверхности наружной мембраны, то есть в окружающую среду.
Необязательная стадия (c) относится к получению мембранного препарата микроорганизма со стадии (b). Подходящие методы получения мембранных препаратов хорошо известны специалистам в данной области.
Используемый по данному документу мембранный препарат предпочтительно является мембранным препаратом наружной мембраны.
Стадия (ii) способа по настоящему изобретению относится к созданию контакта микроорганизма и/или его мембранного препарата с одним или более субстратами нитрилазы в условиях, совместимых с активностью нитрилазы. Как указано выше, в предпочтительном варианте осуществления стадия (ii) относится к созданию контакта микроорганизма и/или его мембранного препарата с нитрилом в условиях, при которых нитрилаза катализирует конверсию нитрила, приводящую к образованию карбоновой кислоты. Можно использовать любые подходящие условия, известные в данной области. В частности, реакцию проводят в жидкой среде. В жидкой среде микроорганизм может образовывать суспензию. Микроорганизм также может быть иммобилизованным. При использовании мембранного препарата мембранный препарат может образовывать частицы, которые могут быть диспергированы в жидкой среде. Реакцию можно проводить при рН в диапазоне от приблизительно 5,0 до приблизительно 9,0, предпочтительно от приблизительно 6,0 до приблизительно 8,0 и более предпочтительно от приблизительно 6,5 до приблизительно 7,5.
Стадия (iii) способа по настоящему изобретению относится к извлечению микроорганизма, используемого на стадии (ii). Как показано на примерах настоящего изобретения, после реакционного цикла, например в течение 24 часов, бактериальные клетки по-прежнему проявляют значительную активность. Таким образом, клетки можно использовать в следующем цикле реакций. Извлечение можно проводить путем центрифугирования реакционной смеси. При необходимости клетки можно промывать.
Другой аспект изобретения относится к применению микроорганизма, экспрессирующего нитрилазу на поверхности, и/или его мембранного препарата для получения продукта реакции, катализируемой нитрилазой, в частности для получения карбоновой кислоты. Предпочтительно, если получаемая карбоновая кислота представляет собой миндальную кислоту, в частности (R)-миндальную кислоту.
Другой аспект изобретения относится к микроорганизму, экспонирующему рекомбинантную нитрилазу на поверхности, причем микроорганизм представляет собой микроорганизм, описанный в данном документе, и нитрилаза представляет собой нитрилазу, описанную в данном документе. В частности, микроорганизм экспонирует слитый полипептид, содержащий сигнальный пептид, рекомбинантную нитрилазу, необязательно, сайт узнавания протеазы, трансмембранный линкер и транспортерный домен аутотранспортера, на своей поверхности, при этом компоненты слитых белков являются такими, как описано в данном документе.
Другой аспект изобретения относится к мембранному препарату, содержащему нитрилазу. Мембранный препарат можно получать из микроорганизма, описанного в данном документе, например, на стадии (c) способа по настоящему изобретению. Нитрилаза может представлять собой любую нитрилазу, описанную в данном документе.
Еще один аспект изобретения относится к способу получения микроорганизма, экспонирующего рекомбинантную нитрилазу на своей поверхности, включающему введение последовательности нуклеиновой кислоты в указанный микроорганизм, при этом указанная последовательность нуклеиновой кислоты содержит:
(1) часть, кодирующую сигнальный пептид,
(2) часть, кодирующую рекомбинантную нитрилазу, которая будет экспонирована,
(3) необязательно, часть, кодирующую сайт узнавания протеазы,
(4) часть, кодирующую трансмембранный линкер, и
(5) часть, кодирующую транспортерный домен аутотранспортера,
при этом нуклеиновая кислота функционально связана с последовательностью контроля экспрессии. Компоненты (1)-(5) являются последовательностями нуклеиновой кислоты, описанными выше в данном документе. Микроорганизм может быть любым микроорганизмом, описанным в данном документе, таким как бактерия, более конкретно, грамотрицательная бактерия, даже более конкретно E. coli. Нитрилаза может быть любой нитрилазой, описанной в данном документе. «Введение нуклеиновой кислоты в указанный микроорганизм» включает трансформацию, описанную в данном документе.
Далее изобретение иллюстрируется следующими фигурами и примерами, которые не должны рассматриваться как ограничивающие объем изобретения.
Фигура 1: Схема аутотранспортерного слитого белка, кодируемого pAT-NitAf. Окружение сайтов слияния приведено в виде последовательностей. Две аминокислоты на N-конце, которые были добавлены в результате процедуры клонирования, выделены курсивом и жирным шрифтом. Указан сайт расщепления сигнальной пептидазой. SP, сигнальная пептидаза.
Фигура 2: SDS-ПААГ фракции наружной мембраны E. coli BL21 (DE3) pAT-NitAf, содержащей избыточно экспрессированный аутотранспортерный слитый белок AFP (1), продукта расщепления целых клеток, содержащих избыточно экспрессированный аутотранспортер, протеиназой K (2) и набором маркеров (M).
Фигура 3: Проявление активности нитрилазы
Образование (R)-миндальной кислоты из (R,S)-нитрила миндальной кислоты. Реакционная смесь содержала в общем объеме 1 мл: Na-фосфатный буфер (50 мМ, рН 7,5), (R,S)-нитрил миндальной кислоты (10 мМ) и клетки E. coli BL21 (DE3) pAT-NitAf, соответствующие OD578=10. Точки данных представляют собой средние значения из троекратно повторенных экспериментов; стандартные ошибки, отмеченные чертами, слишком малы, чтобы быть видимыми. Черные символы представляют индукцию при OD578=1, и белые символы представляют индукцию при OD578=0,5. Трис-HCl буфер pH 7 (▼/∇), фосфатный буфер рН 7,5 (■/□).
Фигура 4: Определение методом хиральной ВЭЖХ (R)-миндальной кислоты. Чистая (R)-миндальная кислота (10 мМ) в метаноле (впереди), экстракция миндальной кислоты после конверсии рацемического нитрила миндальной кислоты E. coli BL21 pAT-NitAf (сзади).
Фигура 5: рН-зависимость коэффициента конверсии (R,S)-нитрила миндальной кислоты в R-миндальную кислоту. Реакцию проводили либо в 50 мМ ацетатном буфере (белые столбики), либо в 50 мМ Трис-HCl буфере (черные столбики). Время реакции составляло 48 час при 37°C.
Фигура 6: Стабильность цельноклеточного биокатализатора в процессе хранения при -18°C. Клетки замораживали в среде, содержащей 20% глицерина, и после оттаивания начинали конверсию (получение миндальной кислоты) при помощи E. coli BL21 (DE3) pAT-NitAf в 50 мМ фосфатном буфере рН 7,5, 10 мМ нитриле миндальной кислоты на протяжении 120 час при 37°C.
Фигура 7: Возможность повторного использования E. coli BL21 (DE3) pAT-NitAf в различных буферах и температурных условиях. Каждый цикл соответствует периоду конверсии в 24 часа. Затем биокатализатор удаляли центрифугированием (60 с при 13000 об/мин), ресуспендировали в свежем буфере, объединяли со свежим субстратом (конечная концентрация 10 мМ) и инкубировали в течение дополнительных 24 часов. Черные символы представляют Трис-HCl буфер (50 мМ, pH 7), белые символы представляют натрий-фосфатный буфер (50 мМ, pH 7,5). 45°C (●/○), 42°C (□/■), 37°C (▲/∆), 30°C (▼/∇).
Фигура 8: Рестрикционное расщепление плазмидной ДНК с использованием KpnI и XhoI. Дорожка 1) 1-kb маркер, 2) - 12) плазмидная ДНК из трансформантов, расщепленная с использованием KpnI/XhoI, 13) 1-kb маркер.
Фигура 9: Выделенная наружная мембрана после расщепления трипсином клонов E. coli UT 5600 (DE3) pAT-NitAf и pAT-NitSc. Дорожка 1) pAT-NitSc, расщепленная трипсином, 2) pAT-NitSc, нерасщепленная, 3) pAT-NitAf, расщепленная протеиназой K, 4) pAT-NitAf, нерасщепленная, 5) маркеры молекулярной массы (кДа).
Фигура 10: Анализ методом ВЭЖХ следующих конверсий нитрила миндальной кислоты при помощи E. coli BL21 (DE3) pAT-NitAf.
A) Отрицательный контроль. E. coli BL21 (DE3), OD578=10, время инкубации: 120 ч/30°C.
1) Фронт растворителя ВЭЖХ, 2) нитрил миндальной кислоты, 3) бензальдегид
B) Конверсия при помощи E. coli BL21 (DE3) pAT-NitAf, OD578=10, время инкубации: 120 ч/30°C. 1) Фронт растворителя ВЭЖХ, 2) нитрил миндальной кислоты, 3) бензальдегид.
Фигура 11: Миндальная кислота, полученная с использованием рекомбинантного цельноклеточного биокатализатора E. coli BL21 (DE3) pAT-NitAf.
Фигура 12: Производство миндальной кислоты при помощи E. coli BL21 (DE3) pAT-NitAf при 30°C и 37°C, соответственно, в фосфатном буфере, pH 7,5. При OD578=1 клетки индуцировали с помощью 1 мМ ИПТГ в течение 1 часа при 30°C. Концентрация субстрата в момент времени t=0 составляла 10 мМ нитрила миндальной кислоты.
Фигура 13: Производство миндальной кислоты при помощи E. coli BL21 (DE3) pAT-NitAf при 37°C в фосфатном буфере, pH 7,5 и с использованием различных концентраций субстрата. При OD578=1 клетки индуцировали с помощью 1 мМ ИПТГ в течение 1 часа при 30°C. Последующая конверсия занимала 120 часов.
Фигура 14: Производство миндальной кислоты при помощи E. coli BL21 (DE3) pAT-NitAf при 37°C (исключения указаны на фигуре) в различных буферных системах. При OD578=1 клетки индуцировали с помощью 1 мМ ИПТГ в течение 1 часа при 30°C. Концентрация субстрата в момент времени t=0 составляла 10 мМ нитрила миндальной кислоты и конверсия занимала 120 часов.
Фигура 15: Определение производства и идентичности (R)-миндальной кислоты, полученной с помощью E. coli BL21 (DE3) pAT-NitAf за период времени 5 дней. Хиральная-OM колонка (компания CS-Chromatographie Service) со смесью гексан:2-пропанол:ТФУ (90:10:0,1), поток: 0,5 мл/мин. Детекция при 210 нм. A) Анализ клеточного супернатанта E. coli BL21 (DE3) pAT-NitAf, экстрагированного этилацетатом. B) Рацемическая миндальная кислота, объединенная с продуктом конверсии из A).
Фигура 16: Производство миндальной кислоты после различного времени хранения при 6°C (фигура 16). Конверсия при помощи E. coli BL21 (DE3) pAT-NitAf в 50 мМ фосфатном буфере рН 7,5, 10 мМ нитрила миндальной кислоты, 120 часов при 37°C.
Фигура 17: Производство миндальной кислоты при помощи E. coli BL21 (DE3) pAT-NitAf в различных буферах и при различных температурах, в каждом случае в течение 24 часов, с использованием 10 мМ нитрила миндальной кислоты при 1000 об/мин в термомиксере.
Фигура 18: Определение производства 3,5-дибромо-4-гидроксибензойной кислоты при помощи E. coli JK321 pAT-NitKp. Время инкубации 120 часов, OD578=10, субстрат 2,5 мМ бромоксинил.
Примеры
Следующие примеры демонстрируют, что авторы настоящего изобретения смогли идентифицировать три нитрилазных гена из различных организмов и клонировать указанные гены в систему аутодисплея.
Пример 1: Нитрилаза из Alcaligenes faecalis
Цельноклеточные биокатализаторы являются привлекательными технологическими инструментами для производства большого количества редких химических веществ, особенно при необходимости регио- или энантиоселективного синтеза. В настоящем исследовании был создан цельноклеточный биокатализатор для синтеза (R)-миндальной кислоты с использованием нитрила миндальной кислоты в качестве субстрата. С этой целью нитрилазу из Alcaligenes faecalis подвид faecalis ATCC 8750 экспрессировали на поверхности Escherichia coli (E. coli) методом аутодисплея. Система аутодисплея представляет собой эффективную систему поверхностного дисплея для грамотрицательных бактерий и основана на аутотранспортерном пути. С помощью этой технологии стало возможным экспрессировать мультигомомерную нитрилазу в активной форме на поверхности E. coli и получать энантиомерно чистую (R)-миндальную кислоту с энантиомерным избытком >99%. Внедрение слитого белка нитрилаза-аутотранспортер в наружную мембрану контролировали методом SDS-ПААГ, а экспонирование фермента на поверхности можно было проверять с помощью теста на доступность для протеазы. При оптимальных условиях можно было производить 2,6 мМ (R)-миндальной кислоты за 24 часа, используя бактериальную суспензию с OD578=10 в биотрансформации небольшого масштаба (1 мл).
В качестве модели мультигомомерного фермента использовали нитрилазу из Alcaligenes faecalis подвид faecalis ATCC 8750.
Нитрилаза была доступна для расщепления протеазой при расщеплении целых клеток и демонстрировала такое же субстратное ингибирование в отношении бензальдегида, что и свободный фермент.
Результаты
Конструкция слитого белка
Ген нитрилазы амплифицировали полимеразной цепной реакцией (ПЦР) из геномной ДНК Alcaligenes faecalis подвид faecalis ATCC 8750 с помощью олигонуклеотидов, выведенных из опубликованных данных о последовательностях (см. экспериментальную часть). ПЦР-праймеры добавляли сайт XhoI на 5'-конце, а сайт KpnI на 3'-конце кодирующей нитрилазу области, что необходимо для слияния нитрилазы в рамке считывания с аутотранспортерными доменами, нужными для аутодисплея. В качестве векторной основы, кодирующей каркас аутотранспортера, использовали pET-Adx, как описано ранее (Jose, 2001). Полученный продукт ПЦР лигировали в расщепленный XhoI/KpnI pET-Adx, заменяя прежний «пассажир» Adx, чтобы получить pAT-NitAf. Полученный аутотранспортерный слитый белок обладает типичной структурой, содержащей сигнальный пептид, нитрилазу в качестве «пассажира», трансмембранный линкер и β-цилиндр (фигура 1). Сигнальный пептид, происходящий из β-субъединицы холерного токсина (CTB), использовали для переноса через внутреннюю мембрану. β-цилиндр (транспортерный домен аутотранспортера) для перенесения «пассажира» через наружную мембрану и связующая область (трансмембранный линкер) для обеспечения полного доступа к поверхности происходили из AIDA-I (Jose, 2001).
Полученный зрелый слитый белок, кодируемый pAT-NitAf, имеет предсказанную молекулярную массу приблизительно 90 кДа после процессинга сигнальной пептидазой. pAT-NitAf переносили трансформацией в E. coli BL21 (DE3), отрицательный по OmpT мутант, что предотвращало расщепление экспонированной нитрилазы. Этот штамм получил название E. coli BL21 (DE3) pAT-NitAf.
Экспрессия слитого белка нитрилаза-аутотранспортер
Белковый биосинтез искусственного слитого белка начинали путем добавления 1 мМ ИПТГ, как описано в экспериментальной части. Для исследования того, экспонирована ли нитрилаза полностью на поверхности или в сторону периплазмы, проводили цельноклеточное расщепление интактных клеток E. coli BL21 (DE3) pAT-NitAf. Поскольку наружная мембрана E. coli непроницаема для протеаз, подобных протеиназе K или трипсину, разрушение белка добавленной извне протеазой должно происходить вследствие его экспонирования на поверхности. Экспрессию и цельноклеточное расщепление контролировали электрофорезом в полиакриламидном геле с додецилсульфатом натрия (SDS-ПААГ). Последующее окрашивание Кумасси бриллиантовым синим показало, что добавление протеиназы K приводило к сильному уменьшению полосы слитого белка (фигура 2).
Чувствительным маркером целостности анализируемых клеток является интенсивность белковых полос, соответствующих Omp F/C и OmpA. Не должно быть никаких изменений в соотношении двух полос Omp в расщепляемом и нерасщепляемом образце. Если полоса OmpA исчезает, целостность наружной мембраны нарушена и периплазматический фрагмент OmpA разрушен протеиназой K. В экспериментах авторов изобретения размер и количество OmpA были одинаковы и в обработанных протеиназой и в не обработанных клетках.
В итоге авторы изобретения пришли к выводу, что почти весь фермент-«пассажир» экспрессирован на поверхности бактерии.
Ферментативная активность экспонированной на поверхности нитрилазы
Для анализа клетки E. coli BL21 (DE3), несущие плазмиду pAT-NitAf, доводили до оптической плотности при 578 нм (OD578), равной 10, в 50 мМ натрий-фосфатном буфере pH 7,5 в общем объеме 1 мл. Поскольку фермент описан для демонстрации субстратной специфичности в отношении гидроксинитрилов, рацемический нитрил миндальной кислоты использовали в качестве субстрата в конечной концентрации 10 мМ. Биоконверсию анализировали методом ВЭЖХ и подсчитывали количество полученной миндальной кислоты. Первые биоконверсии проводили с клетками, индуцированными ИПТГ при OD578=0,5. Это приводило к продукции 2,1 мМ миндальной кислоты после биоконверсии в течение 72 часов. Дальнейшая инкубация вплоть до 120 часов не приводила к дальнейшему увеличению продукции миндальной кислоты. Биоконверсию можно было значительно улучшить в два раза путем индукции ИПТГ при OD578=1. С такой модификацией биоконверсия не стабилизировалась через 72 часа, но, напротив, увеличивалась на протяжении периода наблюдения, составляющего 120 часов, до общей величины 6,6 мМ миндальной кислоты (фигура 3). Количество добавленного субстрата было ограничено в связи с появлением ингибирования фермента, вызванного свободным бензальдегидом, который образовывался спонтанно в результате распада нитрила миндальной кислоты (Yamamoto et al., 1992).
Конверсия нитрила миндальной кислоты в миндальную кислоту
Был разработан метод ВЭЖХ для отделения субстрата (нитрила миндальной кислоты) от продукта (миндальной кислоты) и для ее обнаружения. При использовании E. coli BL21 (DE3) pAT-NitAf была подтверждена конверсия нитрила миндальной кислоты в миндальную кислоту (фигура 10). Анализ методом ВЭЖХ также выявил, что в водной системе нитрил миндальной кислоты присутствует в равновесии с бензальдегидом (+HCN), как и ожидалось (фигура 10). Изменение некоторых параметров реакции привело к получению концентрации продукта 2,4 мМ, что соответствует массе 0,36 мг в 1 мл образца, через 120 часов, начиная с концентрации субстрата 5 мМ при использовании клеточной суспензии с OD578=10 (фигура 11).
Энантиомерный избыток
Следующим шагом была проверка энантиомерного избытка полученной миндальной кислоты. С этой целью миндальную кислоту экстрагировали из водного буфера растворителем с этилацетатом. Экстрагированную миндальную кислоту растворяли в метаноле и непосредственно использовали для анализа методом хиральной ВЭЖХ. Определенный энантиомерный избыток (ee) составлял ee > 99% для биоконверсии при 37°C (фигуры 4 и 15A). Это показало, что экспонированная на поверхности нитрилаза из A. faecalis способна продуцировать (R)-миндальную кислоту с высокой энантиомерной чистотой. Из фигуры 15A очевидно, что был обнаружен почти исключительно пик, характерный для времени удержания (R)-энантиомера миндальной кислоты. Рассчитанная энантиомерная чистота составляла более чем 99%. На фигуре 15B показана идентичность (R)-миндальной кислоты, определенная по пикам чистых (R)- и (S)-энантиомеров с конверсией из фигуры 15A.
Оптимальный рН
Хотя ранее было опубликовано, что оптимальная величина pH для нитрилазы из Alcaligenes faecalis составляет pH 7,5, авторы изобретения изучили, не происходит ли смещение оптимального значения pH в результате экспонирования на поверхности. Клетки ресуспендировали в ацетатном буфере или в Трис-HCl буфере (оба 50 мМ) до конечного значения OD578=10. Через 120 часов инкубации при 37°C супернатант анализировали методом ВЭЖХ и рассчитывали количество конверсионной миндальной кислоты. Авторы изобретения определили оптимальное значение pH, равное 7, при конверсии 5,5 мМ миндальной кислоты. Была обнаружена сильная потеря активности в диапазоне от pH 7 до pH 8. Использование дополнительно исследованного фосфатного буфера pH 7,5 привело к конверсии только 4,4 мМ миндальной кислоты при прочих одинаковых условиях (данные не представлены) (фигура 5).
При оптимизации температуры реакции и величины рН стало возможно получать вплоть до 2,6 мМ миндальной кислоты в течение 24 часов. Амид миндальной кислоты никогда не обнаруживали.
Кинетика
В серии экспериментов анализировали несколько параметров с точки зрения их влияния на производство миндальной кислоты в лабораторных условиях. На фигуре 12 представлена кинетика производства миндальной кислоты при различных температурах. При 37°C было обнаружено возрастание концентрации продукта приблизительно на 20% по сравнению с конверсией при 30°C. Прогрессирование конверсии при обоих значениях температуры, 30°C и 37°C, также показало, что концентрация полученной миндальной кислоты лишь медленно возрастает после конверсии в течение приблизительно 144 часов.
Концентрация субстрата
Далее анализировали влияние концентрации субстрата на производство миндальной кислоты. В водном растворе субстрат нитрил миндальной кислоты находится в равновесии с бензальдегидом и HCN. Однако при избыточных концентрациях бензальдегид оказывает токсическое воздействие на клетки, тем самым ограничивая увеличение концентрации субстрата, как показано на фигуре 13. Дальнейшее снижение концентрации нитрила до менее 10 мМ не приводило к дальнейшему увеличению производительности, из чего следует, что 10 мМ нитрила миндальной кислоты является оптимальной концентрацией субстрата.
Различные буферные системы
Сравнительное изучение производства миндальной кислоты при использовании цельноклеточного катализатора E. coli BL21 (DE3) pAT-NitAf также проводили, используя различные буферные системы. На фигуре 14 показано, что наилучшие результаты были достигнуты при использовании раствора в Трис-буфере с pH 6,5 по сравнению с любыми другими используемыми буферами.
Стабильность при хранении
Для определения стабильности при хранении сконструированного цельноклеточного биокатализатора, индуцированные ИПТГ клетки E. coli BL21 (DE3) pAT-NitAf доводили до OD578=50 в 50 мМ натрий-фосфатном буфере pH 7,5, содержащем 20% (v/v) глицерина, и хранили при -18°C. Глицерин добавляли в образцы при -18°C в качестве антифриза, чтобы предотвратить лизис клеток. В разные промежутки времени образцы промывали один раз в фосфатном буфере и доводили до OD578=10. Альтернативно, после индукции ИПТГ аликвоты клеток хранили при 6°C в холодильнике и подвергали тесту на нитрилазу после различных сроков хранения. Реакцию начинали добавлением 10 мМ нитрила миндальной кислоты и образцы инкубировали в течение 120 часов при 37°C. После хранения 35 дней при -18°C клетки все еще сохраняли 72% их исходной активности (фигура 6). Через одну неделю активность цельноклеточного катализатора при +6°C составляла только приблизительно 50% от исходной активности (фигура 16). В данных экспериментах это также коррелировало с наблюдаемым лизисом клеток. Здесь, опять же, очевидно, что часть клеток проявляет относительно стабильную остаточную активность на протяжении измеренного периода времени.
Температурная зависимость цельноклеточного катализатора
Известно, что оптимальная температура для фермента, выделенного из A. faecalis, составляет 40-45°C. Эти выводы подтверждаются данными фигуры 17, на которой представлены температурные тесты в диапазоне от 30°C до 45°C. Использование фосфатного буфера или Трис-буфера в реакционной смеси, судя по всему, не имело особого значения.
Возможность повторного использования цельноклеточного катализатора (фигура 7)
Для изучения возможности повторного использования и стабильности цельноклеточного биокатализатора проводили 5 циклов реакций конверсии по 24 часа с использованием различных буферов и температурных условий. «Цикл» представляет собой 24-часовую реакцию конверсии с последующим центрифугированием клеток E. coli и добавлением свежего субстрата для следующего цикла. Наибольший выход в течение 24 часов был достигнут в случае E. coli BL21 (DE3) pAT-NitAf в натрий-фосфатном буфере (pH 7,5, 45°C) с 2,62 мМ миндальной кислоты, однако активность быстро падала до минимума всего в 10% от исходной активности через 5 циклов. При использовании Трис-HCl буфера (pH 7, 45°C) можно было достичь конверсии, составляющей 2,53 мМ миндальной кислоты в течение 24 часов, через 5 циклов можно было получить более чем 33% от исходного коэффициента конверсии. Активность клеток в Трис-HCl (37°C) падала от 2,24 мМ в первом цикле до 1,25 мМ через 5 циклов, это составляет более чем 55% остаточной активности. Во время первого цикла имели место лишь незначительные различия в производительности между клетками, ресуспендированными в натрий-фосфатном буфере или в Трис-HCl буфере, однако общая производительность, напротив, была всегда лучше в тестируемых образцах с Трис-HCl буфером.
Экспериментальная часть
Бактериальные штаммы и плазмиды
Для целей субклонирования использовали E. coli TOP10 [F- mcrA Δ(mrr-hsd RMS-mcrBC) Ф80lacZΔM15 ΔlacX74 recA1 araD139 Δ(ara-leu)7697 galU galK rpsL (StrR) endA1 nupG] от Invitrogen (Carlsbad, CA, USA). Escherichia coli BL21 (DE3) [B, F-, dcm, ompT, Ion, hsdS (rB- mB-), gal, λ (DE3)] использовали для экспрессии аутотранспортерных слитых белков.
Штаммы E. coli обычно выращивали при 37°C в среде Лурия-Бертани (LB), содержащей ампициллин (100 мкг/мл) при энергичном встряхивании (200 об/мин). Твердую среду готовили путем добавления агара (1,5%, w/v) к среде.
Плазмиду pCR®4-TOPO® от Invitrogen (Carlsbad, CA, USA) использовали для субклонирования гена нитрилазы. Конструкция плазмиды pET-Adx04, кодирующей каркас аутотранспортера AIDA-I, описана ранее (Rustler et al., 2008; патент США № 6180359). На фигуре 8 представлены плазмиды, выделенные из 11 трансформантов, в которых правильный фрагмент ДНК размером приблизительно 1 kb, соответствующий ПЦР-фрагменту, был обнаружен внутри правильной аутотранспортерной плазмиды (приблизительно 7 kb) в 7 клонах после расщепления при помощи XhoI и KpnI.
Методы рекомбинантной ДНК
Ген, кодирующий нитрилазу, амплифицировали из геномной ДНК Alcaligenes faecalis подвид faecalis ATCC 8750 (LGC Standards GmbH, Wesel, Germany).
Праймеры для амплификации гена нитрилазы были выведены на основании информации об уже описанной последовательности гена нитрилазы A. faecalis ATCC 8750 (Kaul et al., 2007). Прямой праймер Nit-XhoI (5'-CCGCTCGAGCAGACAAGAAAAATCGTCC) и обратный праймер Nit-KpnI (3'-GGGGTACCGGACGGTTCTTGCACC) использовали для амплификации фрагмента 1073 bp, содержащего ген нитрилазы. Для облегчения клонирования продуктов ПЦР добавляли сайты рестрикции XhoI и KpnI (подчеркнутые участки в последовательности праймеров представляют собой сайты рестрикции для соответствующих ферментов). Реакционная смесь для ПЦР содержала 10 нг геномной ДНК, соответствующие количества смеси Eppendorf PCR Mastermix от Eppendorf (Hamburg, Germany), 1 мкл (10 мкМ маточного раствора) прямого и обратного праймеров (Sigma-Aldrich Chemical Company, Milwaukee, USA) в объеме 10 мкл. Реакцию ПЦР проводили в амплификаторе Eppendorf Mastercycler gradient от Eppendorf (Hamburg, Germany). Протокол ПЦР состоял из первоначальной стадии денатурации в течение 5 мин при 95°C, последующих 30 циклов по 30 сек при 94°C, 1 мин при 64°C, 1 мин при 72°C и, в заключение, последней стадии элонгации в течение 6 мин при 72°C.
Продукт ПЦР непосредственно использовали для субклонирования в вектор pCR®4-TOPO® в соответствии с инструкциями производителя. Получение плазмидной ДНК проводили, как описано в другом месте. Полученную плазмиду расщепляли XhoI/KpnI и очищали электрофорезом в агарозном геле при помощи набора Qiagen Gel Extraction Kit (Qiagen, Hilden, Germany).
Полученный фрагмент встраивали в расщепленный XhoI/KpnI вектор pET-Adx04, где он заменял ген, кодирующий Adx04, и получали pAT-NitAf.
Препарат наружной мембраны и SDS-ПААГ
Клетки E. coli выращивали в течение ночи и соответствующее количество культуры использовали для инокуляции в среду LB, содержащую 2-меркаптоэтанол (10 мМ) и ампициллин (100 мкг/мл). Клетки культивировали при 37°C (200 об/мин) до достижения оптической плотности (OD578), равной 0,5 или 1. Для индуцируемой экспрессии добавляли ИПТГ (конечная концентрация 1 мМ) от компании Roth (Karlsruhe, Germany). Через 60 мин при 30°C клетки собирали и промывали Трис-HCl (0,2 М, pH 8,2). Для теста на доступность для протеазы клетки ресуспендировали в 2 мл Трис-HCl (0,2 М, pH 6,8), содержащем 25 мкл раствора протеиназы K (2%), и инкубировали в течение 60 мин в водяной бане при 37°C. Дифференциальное фракционирование клеток проводили по модифицированному методу Hantke (Hantke, 1981).
Для SDS-ПААГ изоляты наружной мембраны разводили 1:2 буфером для образцов, состоящим из Трис-HCl (100 мМ, pH 6,8), додецилсульфата натрия (4%), бромфенолового синего (0,2%), 2-меркаптоэтанола (2%), дитиотреитола (0,2 М) и глицерина (20%), кипятили в течение 5 мин и анализировали в 10% полиакриламидных гелях. Белки окрашивали Кумасси бриллиантовым синим и для определения размеров использовали набор неокрашенных белков Pageruler от компании Fermentas (St. Leon-Rot, Germany).
Проведение стандартной конверсии
Стандартный эксперимент проводили следующим образом: предварительную культуру рекомбинантной E. coli BL21 (DE3) штамма pAT-NitAf использовали для инокуляции основной культуры в соотношении 1:100 и культивировали при 37°C и 200 об/мин в инкубаторе-шейкере. По достижении OD578=1 добавляли для индукции ИПТГ в конечной концентрации 1 мМ. Затем происходила индукция в течение одного часа при 30°C и 200 об/мин. После этого клетки промывали один раз 50 мМ фосфатным буфером, pH 7,5, и в конечном итоге доводили до OD578=10, используя тот же буфер. Затем 1-мл аликвоты указанной клеточной суспензии объединяли с 20 мкл нитрила миндальной кислоты (0,5 М) и инкубировали при различных температурах (в частности, 37°C) в термомиксере при 1000 об/мин или 200 об/мин. Образцы отбирали в различные интервалы времени, центрифугировали (6000×g, 5 мин) и стерильно фильтровали, используя 0,2-мкм мембранный фильтр из полиэфирсульфона от компании VWR (Darmstadt, Germany). Супернатант непосредственно анализировали методом ВЭЖХ. После стерильной фильтрации никакой дальнейшей биоконверсии не наблюдали.
Аналитические методы
Конверсию нитрила анализировали на приборе для ВЭЖХ (Millenium Chromatography Manager 3.2), снабженном диодно-матричным детектором 996 и 2695 сепарационным модулем, от компании Waters (MA, USA). Для ахирального анализа использовали колонку с обращенной фазой [100×4,6 мм (внутренний диаметр), заполненную Hypersil ODS с диаметром частиц 5 мкм] от компании Hewlett Packard (Böblingen, Germany) для определения продуктов конверсии, которые обнаруживали спектрофотометрически при 210 нм. Метанол (40% v/v) и фосфорную кислоту (0,1%) в воде использовали в качестве подвижной фазы для разделения ВЭЖХ. Для разделения энантиомеров миндальной кислоты использовали колонку с хиральной фазой [150×4,6 мм (внутренний диаметр), заполненную Reprosil Chiral-OM с диаметром частиц 5 мкм] от компании CS-Chromatographie Service (Langerwehe, Germany).
Подвижная фаза состояла из гексана (90% v/v), изопропилового спирта (10% v/v) и трифторуксусной кислоты (0,1% v/v). Миндальную кислоту экстрагировали из образцов после биоконверсии, как описано в другом месте, с той незначительной модификацией, что этилацетат использовали вместо дихлорметана (Yamamoto, 1991).
Пример 2: Нитрилаза из Klebsiella pneumoniae
Ген нитрилазы из K. pneumoniae амплифицировали методом ПЦР и встраивали в соответствующий вектор для аутодисплея после промежуточного клонирования в стандартный вектор E. coli и анализа последовательности, как описано в примере 1. Конструкция для аутодисплея, кодирующая слитый белок, содержащий нитрилазу K. pneumoniae, аутотранспортерный домен, трансмембранный линкер и сигнальный пептид, как описано для pAT-NitAf в примере 1, получила название pAT-NitKp. Соответствующий штамм E. coli, экспонирующий слитый полипептид на поверхности, назван E. coli BL21 (DE3) pAT-NitKp.
Обнаружение экспрессии и поверхностной локализации нитрилазы Klebsiella pneumoniae проводили, как в случае с нитрилазой Alcaligenes faecalis (см. также пример 1).
Из литературы хорошо известно, что указанная нитрилаза проявляет очень высокую субстратную специфичность в отношении бромоксинила. Указанное токсичное соединение широко используется в качестве гербицида в сельском хозяйстве и каждый год применяется в Германии в промышленных масштабах. Соответствующая карбоновая кислота в природе проявляет низкую токсичность и стабильность.
Производство 3,5-дибромо-4-гидроксибензойной кислоты из бромоксинила при помощи E. coli JK321 pAT-NitKp показано на фигуре 18. На этой фигуре продемонстрировано, что нитрилаза K. pneumonia, экспрессированная на поверхности клетки E. coli, способна преобразовывать нитрил в соответствующую ему карбоновую кислоту.
Пример 3: Нитрилаза из Saccharomyces cerevisiae
Ген нитрилазы из S. cerevisiae амплифицировали методом ПЦР и клонировали, как описано в примере 1. Анализ последовательности выявил 100% идентичность с последовательностью нитрилазы S. cerevisiae, полученной из базы данных. Нитрилазу клонировали в E. coli в виде слитого белка, содержащего аутотранспортерный домен, трансмембранный линкер и сигнальный пептид, как описано в примере 1. И вновь анализ последовательности выявил 100% идентичность с последовательностью из базы данных. Определение экспрессии проводили аналогично примеру 1.
Конструкция для аутодисплея, кодирующая слитый белок, содержащий нитрилазу S. cerevisiae, аутотранспортерный домен, трансмембранный линкер и сигнальный пептид, как описано для pAT-NitAf в примере 1, получила название pAT-NitSc. Соответствующий штамм E. coli, экспонирующий слитый полипептид на поверхности, назван E. coli BL21 (DE3) pAT-NitSc.
Установлено, что после индукции ИПТГ слитый белок, содержащий нитрилазу S. cerevisiae, был локализован на поверхности. Проводили расщепление трипсином целых клеток, как описано в примере 1. Результаты представлены на фигуре 9.




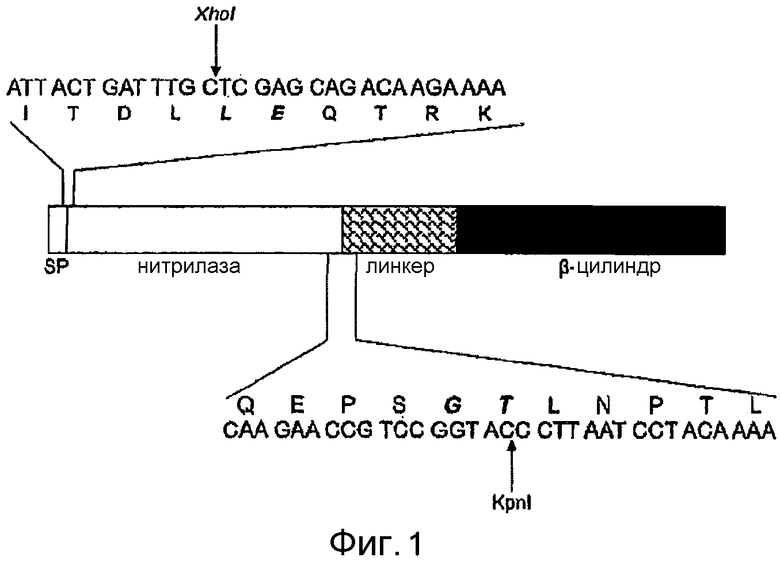
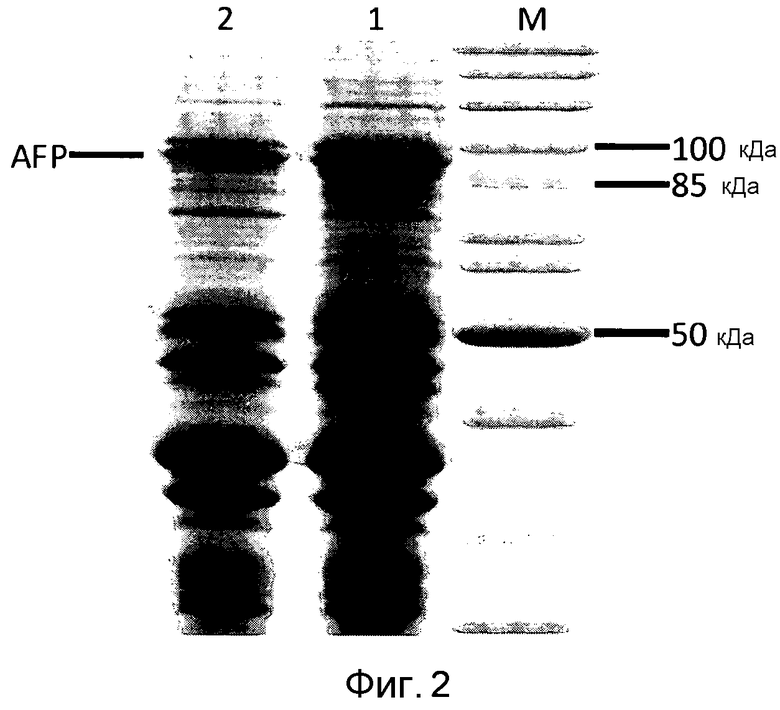
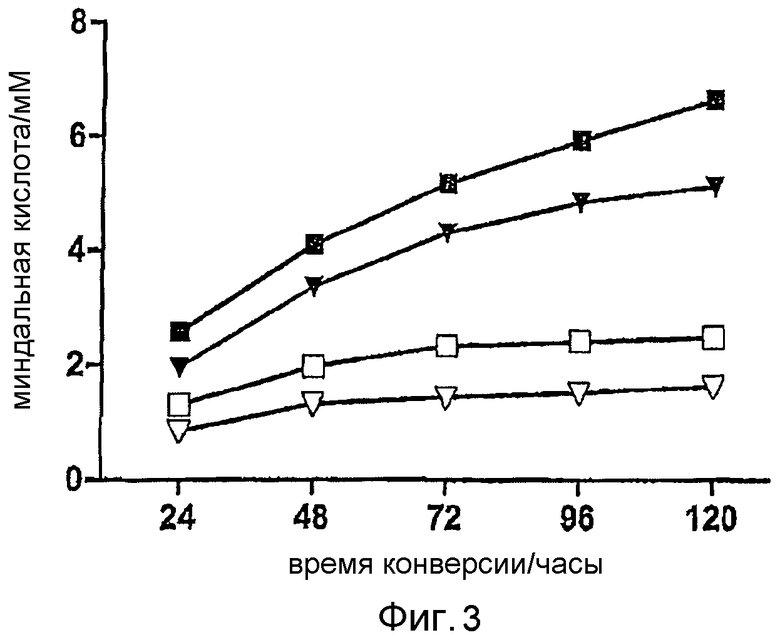
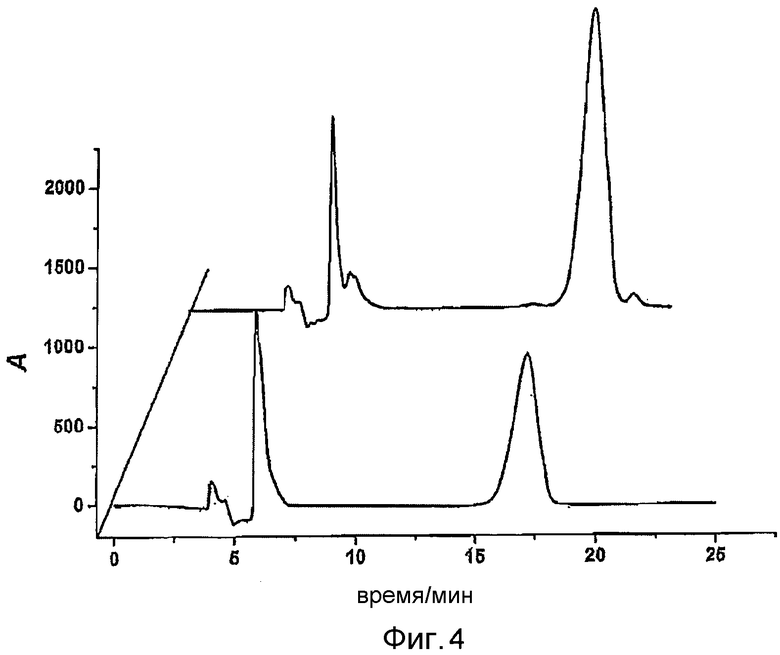
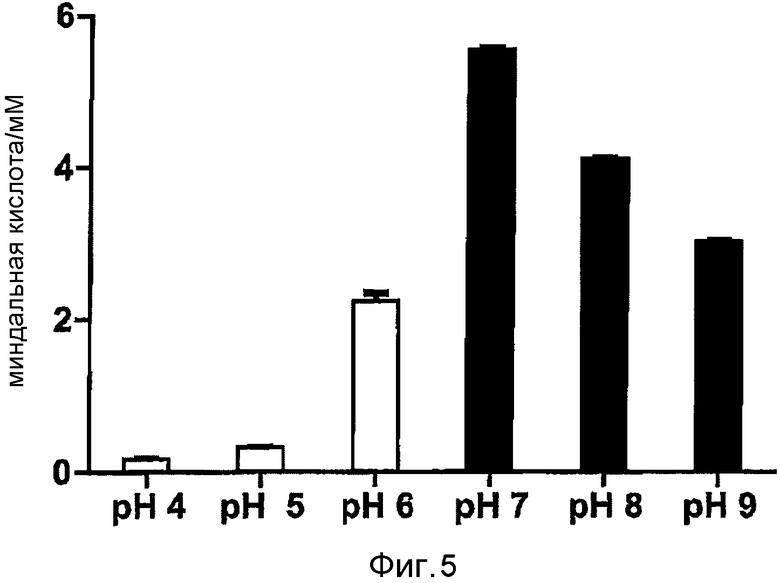
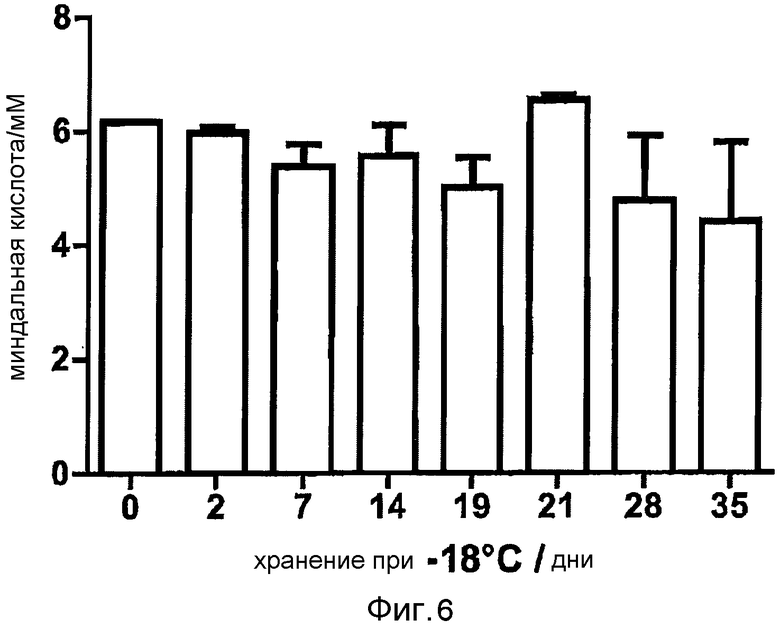
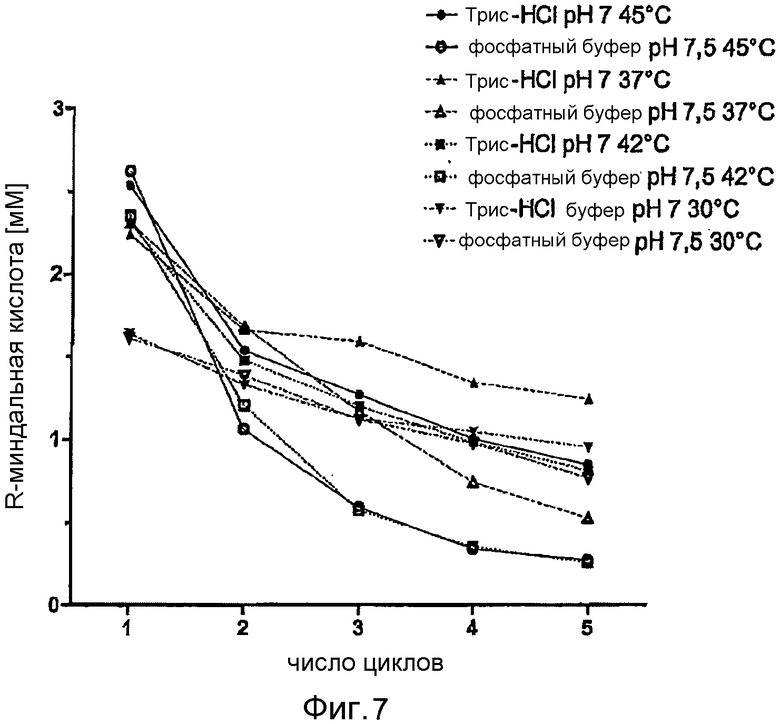
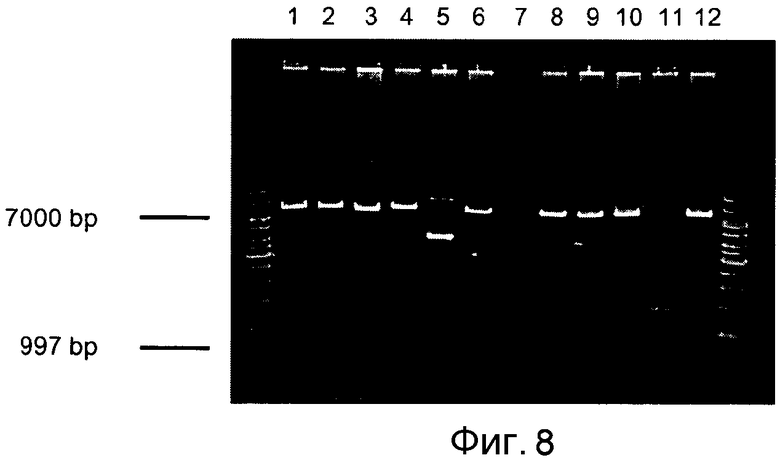
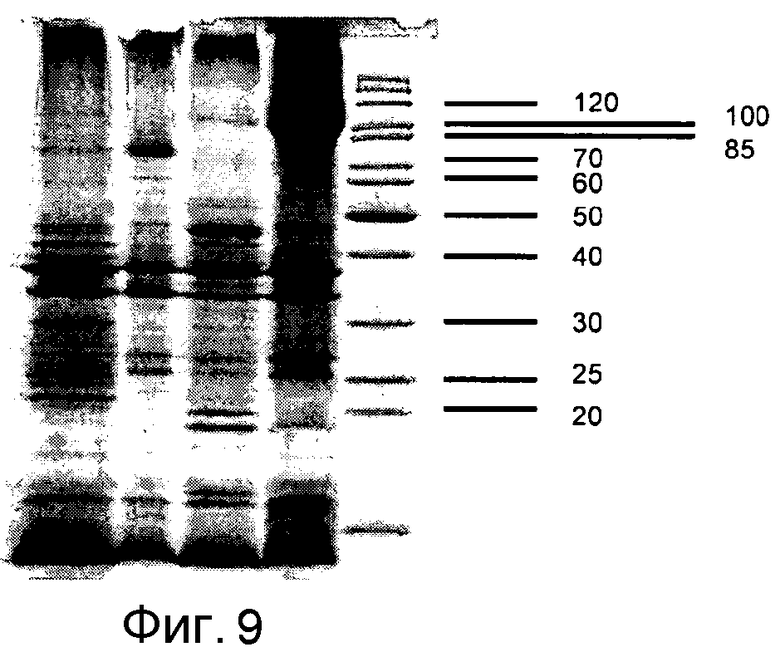
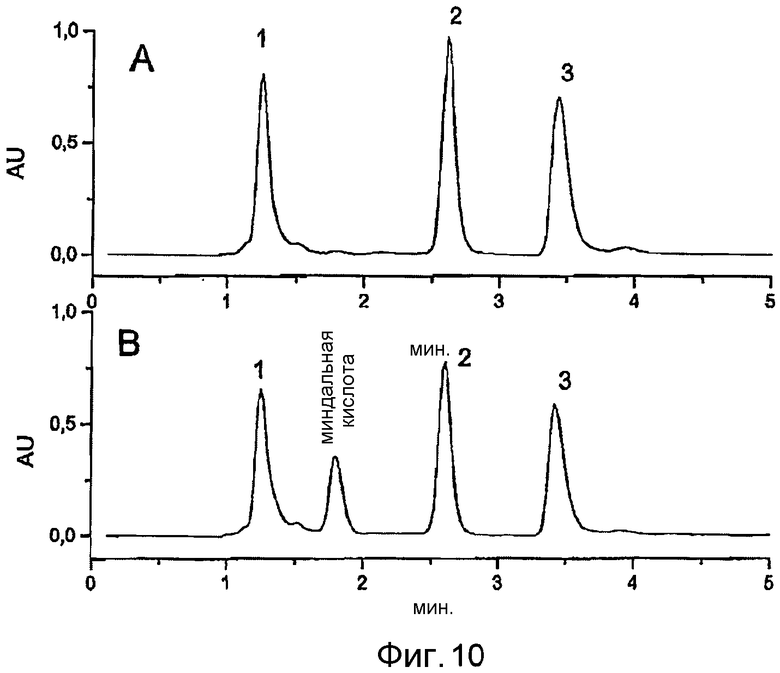
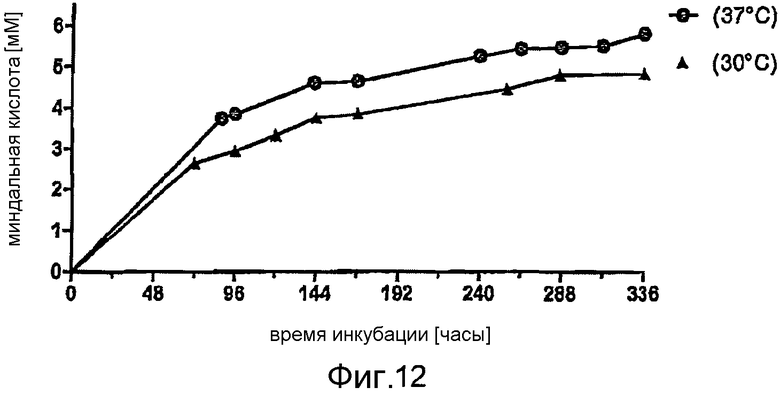
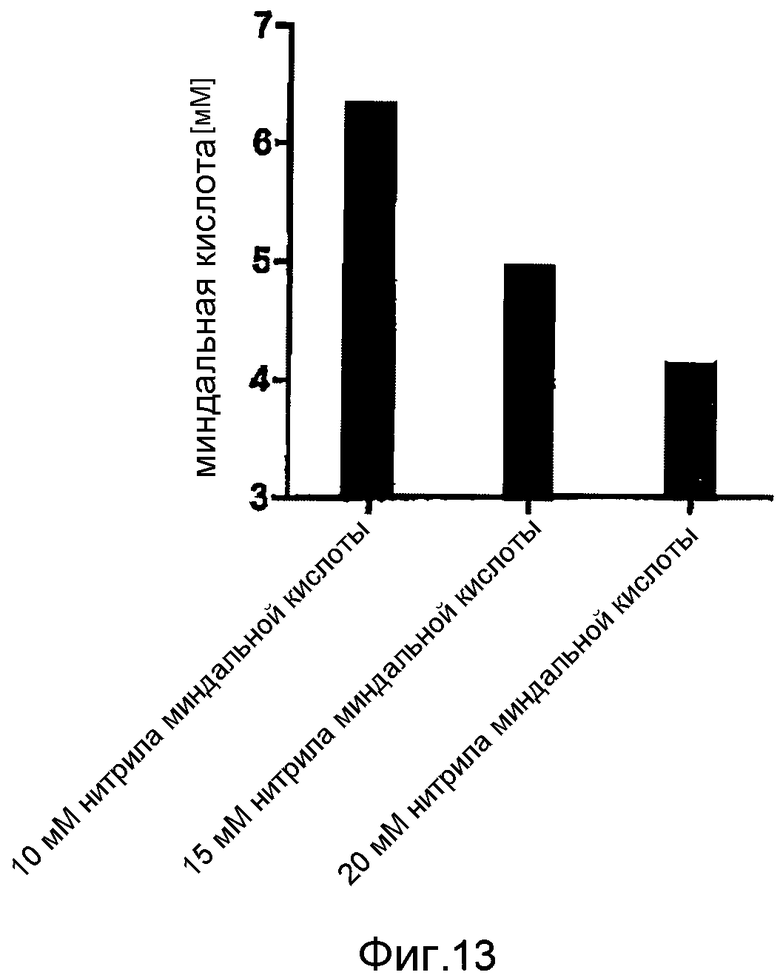
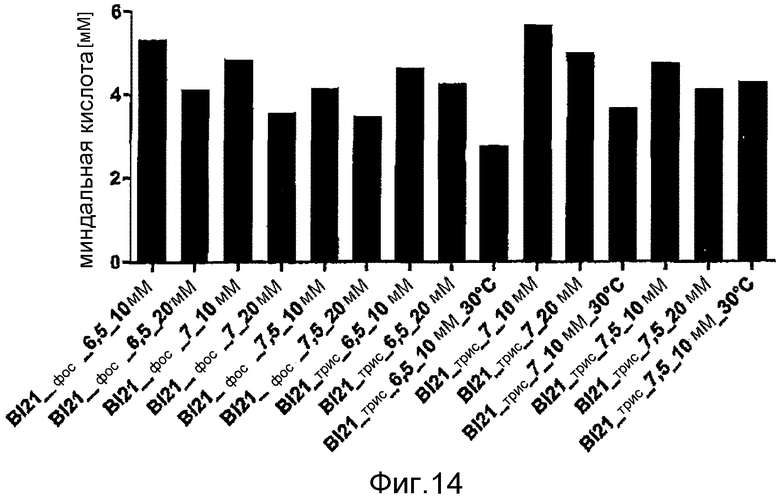
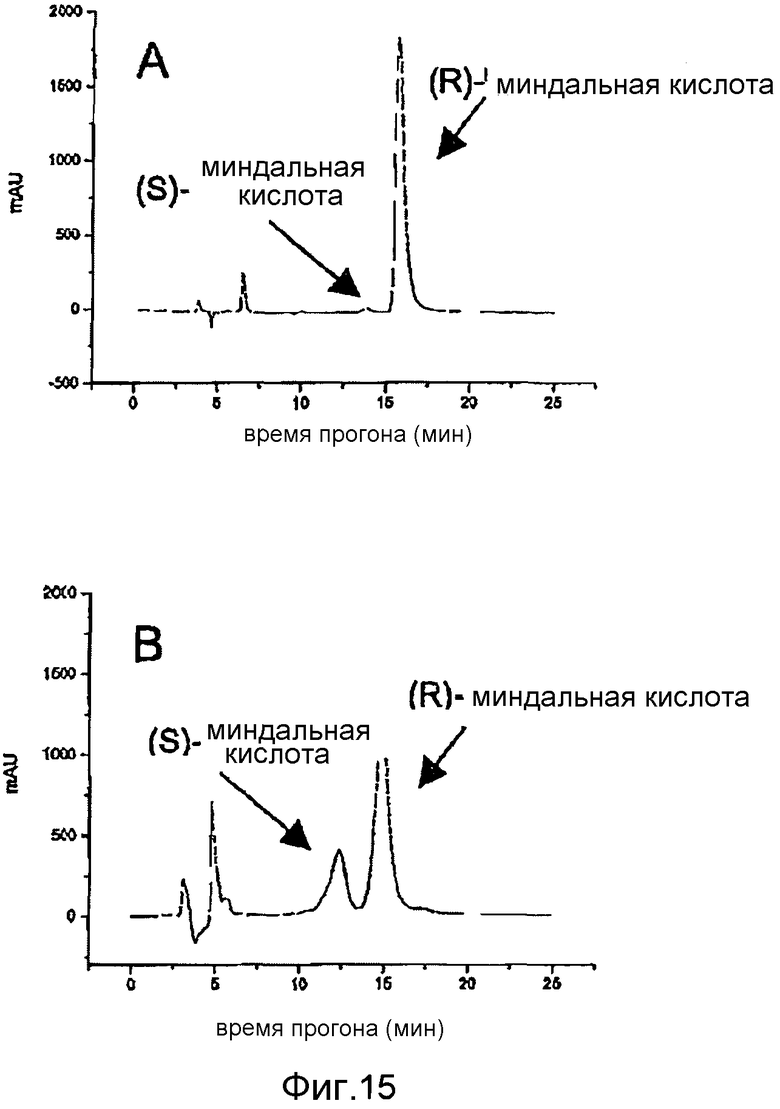
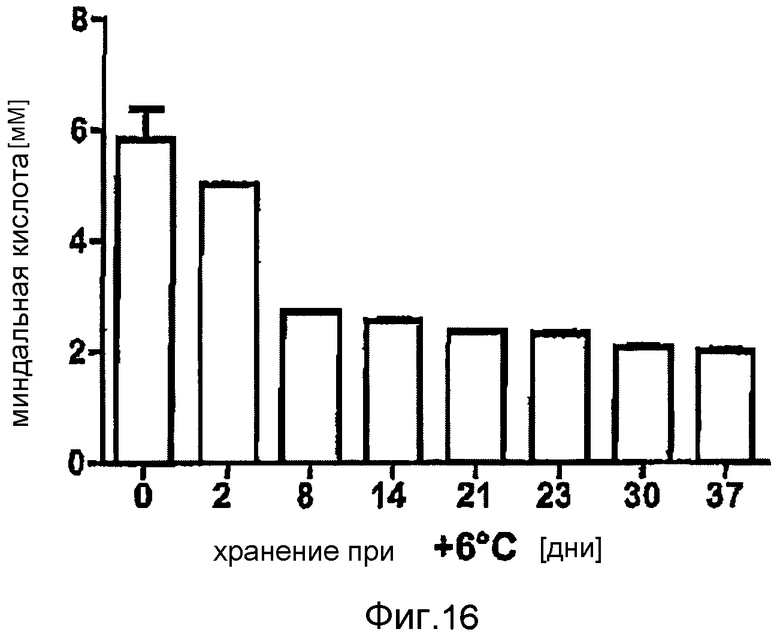
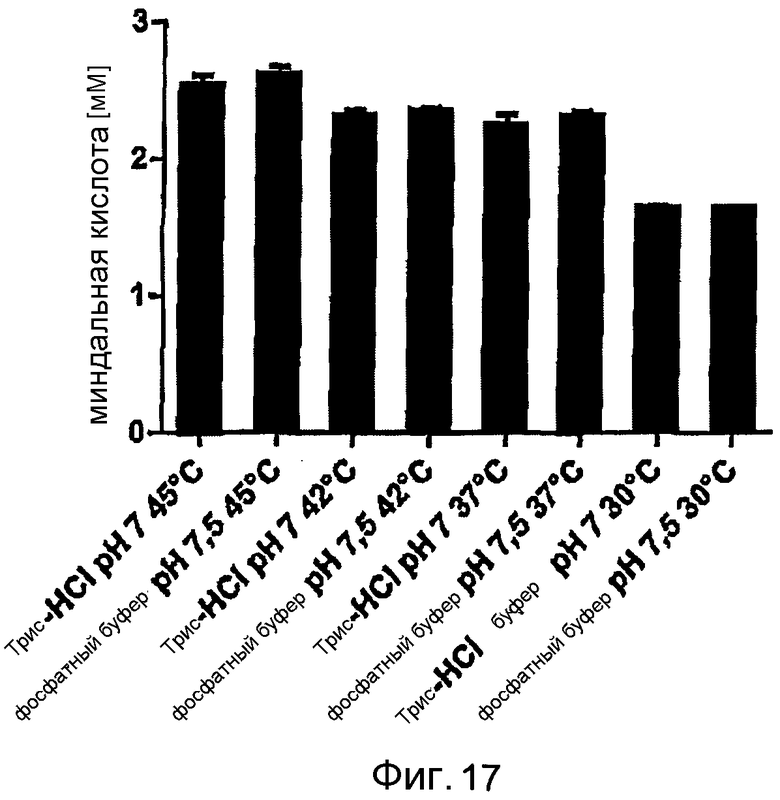
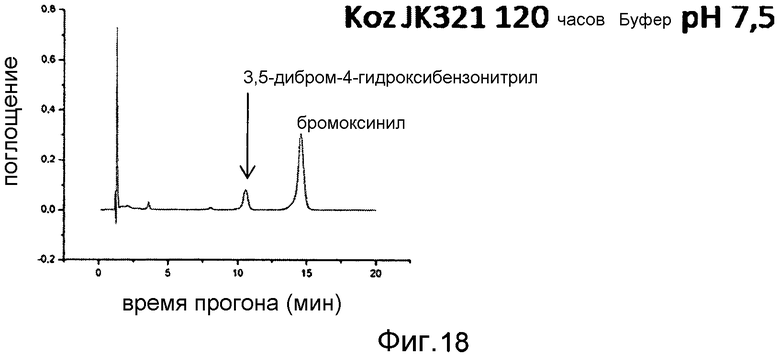
Изобретение относится к биохимии, в частности к способу получения продукта реакции, катализируемой нитрилазой, слитой с транспортным доменом аутотранспортера, где способ включает стадии (i) обеспечения грамотрицательной бактерии, содержащей указанную нитрилазу, расположенную на его поверхности, и/или мембранного препарата указанной грамотрицательной бактерии, и (ii) приведения грамотрицательной бактерии и/или ее мембранного препарата в контакт с одним или более субстратами нитрилазы в условиях, совместимых с активностью нитрилазы. Изобретение позволяет повысить стабильность нитрилазы и увеличить выход продукта реакции. 5 н. и 12 з.п. ф-лы, 18 ил., 3 пр.
1. Способ получения продукта реакции, катализируемой нитрилазой, включающий стадии:
(i) обеспечения грамотрицательной бактерии, содержащей указанную нитрилазу, расположенную на ее поверхности, и/или мембранного препарата указанной грамотрицательной бактерии,
(ii) приведения указанных грамотрицательной бактерии и/или ее мембранного препарата в контакт с одним или более субстратами нитрилазы в условиях, совместимых с активностью нитрилазы, причем нитрилаза слита с транспортерным доменом аутотранспортера.
2. Способ по п.1, в котором продукт реакции, катализируемой нитрилазой, представляет собой карбоновую кислоту и, по меньшей мере, один субстрат нитрилазы представляет собой нитрил.
3. Способ по п.1 или 2, в котором нитрилаза рекомбинантно экспрессирована в указанной грамотрицательной бактерии и перенесена на ее поверхность.
4. Способ по п.1 или 2, в котором стадия (i) включает
(а) обеспечение грамотрицательной бактерии, содержащей последовательность нуклеиновой кислоты, функционально связанную с последовательностью контроля экспрессии, причем указанная последовательность нуклеиновой кислоты содержит:
(1) часть, кодирующую сигнальный пептид,
(2) часть, кодирующую рекомбинантную нитрилазу, которая будет экспонирована,
(3) необязательно, часть, кодирующую сайт узнавания протеазы,
(4) часть, кодирующую трансмембранный линкер, и
(5) часть, кодирующую транспортерный домен аутотранспортера,
(b) культивирование указанной грамотрицательной бактерии в условиях, при которых последовательность нуклеиновой кислоты со стадии (а) экспрессируется и продукт экспрессии последовательности нуклеиновой кислоты экспонируется на поверхности указанной грамотрицательной бактерии.
5. Способ по п.4, дополнительно включающий стадию
(c) получения мембранного препарата из грамотрицательной бактерии со стадии (b).
6. Способ по п.1 или 2, в котором транспортерный домен аутотранспортера выбирают из группы, включающей Ssp, Ssp-h1, Ssp-h2, PspA, PspB, Ssa1, SphB1, AspA/NaIP, VacA, AIDA-I, IcsA, MisL, TibA, Ag43, ShdA, AutA, Tsh, SepA, EspC, EspP, Pet, Pic, SigA, Sat, Vat, EpeA, EatA, EspI, EaaA, EaaC, пертактин, BrkA, Tef, Vag8, PmpD, Pmp20, Pmp21, протеазу IgA1, App, Hap, rOmpA, rOmpB, ApeE, EstA, Lip-1, McaP, BabA, SabA, AlpA, Aae, NanB, a также их варианты.
7. Способ по п.1 или 2, в котором нитрилазу выбирают из группы, включающей нитрилазы Alcaligenes, нитрилазы Klebsiella и нитрилазы Saccharomyces.
8. Способ по п.1 или 2, в котором указанная грамотрицательная бактерия представляет собой Escherichia coli.
9. Способ по п.1 или 2, в котором стадию (ii) выполняют при pН в диапазоне от приблизительно 6,5 до приблизительно 7,5.
10. Способ по п.1 или 2, в котором стадию (ii) выполняют в аэробных и/или окислительных условиях.
11. Способ по п.2, в котором нитрил выбирают из группы, включающей нитрил миндальной кислоты, пруназин, бромоксинил, йоксинил, хлороксинил, анизонитрил, 3-бром-4-гидроксибензонитрил, 3-фтор-4-гидроксибензонитрил, 4-гидрокси-3,5-диметобензонитрил, бензонитрил, фенилпропионитрил, фенилглицинонитрил, н-бутиронитрил, н-валеронитрил, изобутиронитрил и сукцинонитрил.
12. Способ по п.1 или 2, в котором продукт реакции, катализируемой нитрилазой, представляет собой карбоновую кислоту и указанная карбоновая кислота производится в энантиомерном избытке, по меньшей мере, приблизительно 70%.
13. Способ по п.1 или 2, дополнительно включающий стадию (iii) извлечения грамотрицательной бактерии, используемой на стадии (ii).
14. Грамотрицательная бактерия, экспонирующая рекомбинантную нитрилазу на своей поверхности, причем нитрилаза слита с транспортерным доменом аутотранспортера.
15. Мембранный препарат, содержащий рекомбинантную нитрилазу, полученный из грамотрицательной бактерии по п.14.
16. Применение грамотрицательной бактерии по п.14 и/или мембранного препарата по п.15 для получения продукта реакции, катализируемой нитрилазой.
17. Способ получения грамотрицательной бактерии, экспонирующей рекомбинантную нитрилазу на своей поверхности, включающий введение последовательности нуклеиновой кислоты в указанную грамотрицательную бактерию, причем последовательность нуклеиновой кислоты содержит:
(1) часть, кодирующую сигнальный пептид,
(2) часть, кодирующую рекомбинантную нитрилазу, которая будет экспонирована,
(3) необязательно, часть, кодирующую сайт узнавания протеазы,
(4) часть, кодирующую трансмембранный линкер, и
(5) часть, кодирующую транспортерный домен аутотранспортера, причем последовательность нуклеиновой кислоты функционально связана с последовательностью контроля экспрессии.
| BANERJEE A et.al | |||
| Enantioselective nitrilase from Pseudomonas putida: cloning, heterologous expression, and bioreactor studies, Mol Biotechnol | |||
| Колосоуборка | 1923 |
|
SU2009A1 |
| Печь-кухня, могущая работать, как самостоятельно, так и в комбинации с разного рода нагревательными приборами | 1921 |
|
SU10A1 |
| Станок для изготовления деревянных ниточных катушек из цилиндрических, снабженных осевым отверстием, заготовок | 1923 |
|
SU2008A1 |
| SINGH R et.al | |||
| Ударно-вращательная врубовая машина | 1922 |
|
SU126A1 |

