[0001] Данное изобретение относится к микробиологическому ферментативному способу производства целевых моносахаридов в свободной форме из сахаров, активированных нуклеотидами.
[0002] Углеводы играют важнейшую роль во всех формах жизни, участвуя в запасании энергии, структурной функции, передаче сигналов, хранении информации и т.д. Для этого в природе существует несколько основных моносахаридов, таких как N-ацетил-глюкозамин, манноза, N-ацетил-маннозамин, фруктоза, фукоза, рибоза, сиаловая кислота, ксилоза и др. и несколько минорных с более узкой специализацией, например, таких как D-аллоза.
[0003] L-фукоза (6-дезокси-L-галактоза) и фукозилированные олиго- и полисахариды представляют большой интерес для химической, косметической и фармацевтической промышленности, поскольку они обладают высоким потенциалом в алиментарных и биомедицинских приложениях (Hauber, H.-P., Schulz, M., Pforte, A., Mack, P., Zabel, P. & Schumacher, U. (2008) Inhalation with fucose and galactose for treatment of Pseudomonas aeroginosa in cyctric fibrosis patients. Int. J. Med. Sci. 5, 371-376.; Isnard, N., Bourles-Dagonet F., Robert, L. & Renard, G. (2005) Studies on corneal wound healing: Effects if fucose on iodine vapor-burnt rabbit corneas. Ophthalmologica 219, 324-333; Robert, L., Fodil-Bourahla, I., Bizbiz, L. & Robert, A.M. (2004) Effect of L-fucose and fucose-rich polysaccharides on elastin biosynthesis, in vivo and in vitro. Biomed. Pharmacother. 58, 123-128; Wild, M. K., Lühn, K., Marquardt, T. & Vestweber, D. (2002) Leukocyte adhesion deficiency II: therapy and genetic defect. Cells Tissues Organs 172, 161-173; Adam, E.C., Mitchell, B.S., Schumacher, D.U., Grant, G. & Schumacher, U. (1997) Pseudomonas aeruginosa II lectin stops human ciliary beating: therapeutic implications of fucose. Am. J. Respir. Care Med. 155, 2102-2104). Известно, что они обладают противовоспалительными, антивирусными и противоопухолевыми свойствами, а также действуют как пребиотики. Благодаря антивозрастному эффекту L-фукоза также представляет интерес в косметологии (Isnard, N., Fodil-Bourathla, I., Robert A.M. & Robert, L. (2004) Pharmacology of skin aging. Stimulation of glycosaminoglycan biosynthesis by L-fucose and fucose rich polysaccharides, effect of in vitro aging of fibroblasts. Biomed. Pharmacother. 58, 202-204). Кроме того, фукозилированные производные известны своими антиаллергенными и эмульгирующими свойствами.
[0004] Тогда как некоторые моносахариды можно получать из природных источников в большом количестве и при разумных затратах (например, глюкозу, N-ацетилглюкозамин и фруктозу), большинство моносахаридов встречаются достаточно редко и могут быть обнаружены в природе только в малых количествах, например, как L-фукоза (6-дезокси-L-галактоза).
[0005] Для коммерческого производства моносахаридов в качестве источников почти исключительно используют олигосахариды естественного происхождения. Осуществляют кислотный гидролиз указанных олигосахаридов и из высвобожденных моносахаридов выделяют отдельные сахара. Из-за большого химического сходства моносахаридов (в основном отличающихся друг от друга только ориентацией отдельных гидроксильных групп) разделение отдельных моносахаридов в чистой форме является достаточно трудоемким и дорогостоящим.
[0006] L-фукоза представляет собой достаточно редкий сахар, который в настоящее время получают путем гидролиза сложных олигосахаридов, происходящих либо из водорослей, либо из бактерий. Для выделения отдельных моносахаридов из комплексных гидролизатов часто используют токсичные химические реагенты, например, такие как ацетат свинца, и избыточное количество органических растворителей (Schweiger, R.G. (1966) Preparation of α-L-fucosides and L-fucose from fucoidan. патент США 3,240,775). Таким образом, выделение отдельных моносахаридов из комплексного гидролизата олигосахаридов является трудным (из-за высокой степени химического сходства высвобождающихся отдельных моносахаридов) и оказывает вредное воздействие на окружающую среду (из-за избыточного применения токсических химических реагентов, таких как карбонат свинца). Кроме того, доступность олигосахаридов, богатых определенными сахарами, в природе может быть достаточно ограниченной и значительно варьировать вследствие сезонных изменений. L-фукоза представляет собой такой редкий моносахарид, который традиционно получают путем кислотного гидролиза полисахаридов, содержащих фукозу. Фукозу в основном получают из полисахарида фукоидан, моносульфата фукана, присутствующего во всех обыкновенных коричневых водорослях, включая семейства Fucaceae и Laminariaceae (Black, W.A.P (1954): The seasonal variation in the combined L-fucose content of the common british Laminariaceae and Fucaceae. J. Sci. Food Agric. 5, 445-448). На сегодняшний день L-фукозу получают в больших количествах в основном путем сбора коричневых водорослей, принадлежащих семейству Fucaceae, которые встречаются во всем мире, но в большом количестве у европейских берегов атлантического океана. Крупномасштабный сбор коричневых водорослей у берегов океана вызывает озабоченность в плане воздействия на окружающую среду и ограничен законами об охране окружающей среды.
[0007] Например, JP 2000351790 описывает способ экстракции фукоидана, а также получения из экстрагированного фукоидана олигосахаридов, содержащих фукозу, и их разделения.
[0008] В недавно опубликованной заявке на патент было продемонстрировано, что помимо гидролиза фукоидана коричневых водорослей L-фукозу также можно получать путем гидролиза бактериальных полисахаридов естественного происхождения, содержащих L-фукозу. WO 2012/034996 A1 описывает штамм, принадлежащий к семейству Enterobacteriaceae, который способен продуцировать внеклеточные полисахариды, содержащие L-фукозу. Для производства L-фукозы выделяют полисахариды, произведенные штаммом, и подвергают гидролизу, например, путем обработки серной кислотой или трифторуксусной кислотой.
[0009] Процесс производства L-фукозы с применением рекомбинантного микроорганизма, обладающего гликозилтрансферазой и гликозидазой, благодаря совместному действию которых синтезируется L-фукоза в свободной форме, впервые был изложен в WO 2014067696 A1. Для данного процесса необходимы два фермента и молекула-акцептор. Гликозилтрансфераза катализирует перенос фукозы от ГДФ-L-фукозы на акцептор, например, лактулозу, для синтеза фукозиллактулозы. Затем гликозидаза осуществляет гидролиз фукозилированного акцептора (например, фукозиллактулозы) до молекулы-акцептора и L-фукозы. После этого акцептор вновь становится доступным для фукозилирования под действием фукозилтрансферазы. L-фукоза затем высвобождается из клетки и поступает в среду, и может быть выделена из надосадочной жидкости. Таким образом, ингибирование пути ГДФ-фукозы по принципу обратной связи можно легко преодолеть и получить существенное количество (несколько г/л) свободной L-фукозы посредством микробиологической ферментации.
[0010] Помимо экстракции L-фукозы из гидролизатов поли- или олигосахаридов разработали несколько путей синтеза L-фукозы из других моносахаридов, таких как L-арабиноза, D-галактоза, L-рамноза, D-манноза и D-глюкоза. Наиболее эффективный путь синтеза, разработанный Defraye et al (1984), начинается от редкого моносахарида L-рамнозы (Defaye, J., Gadelle, A. & Angyal, S. (1984) An efficient synthesis of L-fucose and L-(4-2H)fucose. Carbohydr. Res. 126, 165-169).Как правило, выход такого химического синтеза зачастую оказывается достаточно низким и включает несколько химических стадий. Помимо того, что химический синтез L-фукозы включает несколько стадий синтеза, он требует применения серьезной защиты химических групп. В целом, крупномасштабный химический синтез моносахаридов оказывается экономически невыгодным по сравнению с экстракцией L-фукозы из полисахаридов, взятых из естественных источников.
[0011] Таким образом, в настоящее время получение любых моносахаридов в чистом виде требует значительных усилий по очистке целевого моносахарида от других моносахаридов, зачастую с применением больших объемов органических растворителей и других токсических реагентов. Поэтому избирательное накопление единственного целевого моносахарида, например, L-фукозы, будет очень востребованным. Однако, у большинства микроорганизмов виды моносахаридов, которые они способны утилизировать, ограничены. Кроме того, они зачастую демонстрируют выраженные предпочтения в отношении некоторых моносахаридов, в случае, когда в качестве источника углерода одновременно доступно несколько моносахаридов.
[0012] С учетом указанного выше, задачей данного изобретения является разработка нового способа производства единственного целевого моносахарида в свободной форме, с помощью которого моносахарид можно получать быстро и эффективно, т.е. в большом количестве и с малыми затратами и без отрицательного воздействия на окружающую среду.
[0013] Эта и другие задачи решаются благодаря способу крупномасштабного производства целевого моносахарида в свободной форме с применением микроорганизма, способ включает стадии:
[0014] а) обеспечение микроорганизма для синтеза моносахарида, содержащего фермент, способный катализировать гидролиз моносахарида, активированного нуклеотидом, для высвобождения целевого моносахарида из моносахарида, активированного нуклеотидом, и
[0015] б) культивирование микроорганизма в среде, подходящей для роста микроорганизма, где микроорганизм не способен метаболизировать моносахарид в значительной степени, так что целевой моносахарид продуцируется и накапливается во время стадии культивирования в свободной форме.
[0016] В изобретении предложено следующее решение данной проблемы.
[0017] Способ, предложенный авторами изобретения, не был описан ранее и использует микроорганизм, который содержит и экспрессирует фермент, гидролизующий моносахарид, активированный нуклеотидом, и накапливает свободный моносахарид. Фермент, обладающий гидролизующей активностью в отношении активированных нуклеотидами сахаров, может представлять собой, например, фукозилтрансферазу, предпочтительно, вариант альфа-1,2-фукозилтрансферазы, кодируемой геном wbgL E. coli:O126 (код доступа ADN43847) или 1,2-фукозилтрансферазы futC Helicobacter pylori (код доступа AAD29868). Несмотря на то, что в данном изобретении можно применять немодифицированные микроорганизмы, обладающие описанными выше ферментативными характеристиками, согласно одному аспекту изобретения, микроорганизм представляет собой рекомбинантный микроорганизм, при этом рекомбинантный микроорганизм был трансформирован, чтобы содержать и экспрессировать по меньшей мере одну последовательность нуклеиновой кислоты, которая не встречается у микроорганизма естественным образом и кодирует фермент, способный катализировать гидролиз моносахарида, активированного нуклеотидом.
[0018] В отличие от известных на уровне техники способов, для высвобождения моносахарида используют единственный фермент, способный катализировать гидролиз, т.е. гидролизовать активированные нуклеотидами сахара, т.е. моносахариды, тогда как известные из уровня техники способы применяют по меньшей мере один фермент, переносящий моносахарид от субстрата-донора на субстрат-акцептор, а на следующей стадии происходит высвобождение моносахарида из акцептора при помощи гликозидазы. Указанная стадия не является обязательной в данном изобретении, таким образом, способ производства целевого моносахарида облегчается. Следовательно, в способе, предложенном в данном изобретении, используют фермент, который способен катализировать гидролиз моносахаридов, активированных нуклеотидами, и высвобождать моносахариды в свободной форме «в отсутствие акцептора». Кроме того, согласно изобретению, способ, предложенный в данном изобретении, осуществляют без целенаправленного применения гликозидазы, способной высвобождать моносахариды из субстрата-акцептора.
[0019] Благодаря предложенному новому способу и предложенному новому микроорганизму, рекомбинантному или нет, возможно производство целевого моносахарида в свободной форме и в больших количествах, без необходимости в химических реагентах или сложных стадиях производства. Способ, предложенный в изобретении, представляет способ микробиологической ферментации, подходящий для применения в производстве в промышленных масштабах редких или других моносахаридов, которые могут быть легко выделены из среды, в которой культивируют микроорганизм.
[0020] Выражение «моносахарид», который используют в данном документе и как его обычно понимают в области техники, относится к основной структурной единице углеводов. Моносахариды являются простейшей формой сахаров и обычно представляют собой бесцветные растворимые в воде твердые кристаллические соединения. Примеры моносахаридов включают глюкозу, фруктозу, галактозу, ксилозу, маннозу, фукозу, рамнозу и рибозу. Моносахариды являются строительными блоками дисахаридов, таких как сахароза, и полисахаридов, таких как целлюлоза и крахмал. Термин «олигосахарид», который используют в данном документе и как его обычно понимают в области техники, относится к полимерным сахаридам, содержащим два моносахарида или более.
[0021] Термин «последовательность нуклеиновой кислоты, кодирующая» обычно относится к любому полирибонуклеотиду или полидезоксирибонуклеотиду, который может быть немодифицированной РНК или ДНК или модифицированной РНК или ДНК и обычно представляет ген, который кодирует определенный полипептид или белок. Термин охватывает, без ограничений, одно- и двуцепочечные ДНК, ДНК, представляющие собой смесь одно- и двуцепочечных участков или одно-, двух- и трехцепочечных участков, одно- и двуцепочечные РНК, а также РНК, которые являются смесью одно- и двуцепочечных участков, гибридные молекулы, содержащие ДНК и РНК, которые могут быть одноцепочечными или, чаще, двуцепочечными или трехцепочечными участками, или смесью одно- и двуцепочечных участков. Термин также охватывает полинуклеотиды, которые включают единственный непрерывный участок или несколько дискретных участков, кодирующих полипептид (например, прерывающиеся интегрированным фагом или встроенной последовательностью или редактированием) наряду с дополнительными участками, которые могут также содержать кодирующие и/или некодирующие последовательности.
[0022] В данном контексте термин «полипептид(ы)» относится к любому пептиду или белку, содержащему две или более аминокислот, соединенных друг с другом пептидными связями или модифицированными пептидными связями. «Полипептид(ы)» относятся как к коротким цепям, обычно обозначаемым пептидами, олигопептидами и олигомерами, так и к более длинным цепям, обычно обозначаемым белками. Полипептиды могут содержать аминокислоты, которые не относятся к 20 аминокислотам, кодируемым генами. «Полипептид(ы)» включают и те, которые модифицированы естественным путем, таким как процессинг и другие пост-трансляционные модификации, и путем химической модификации. Следует принимать во внимание, что один и тот же тип модификации одинаковой или разной степени выраженности может встречаться в нескольких сайтах заданного белка, без существенного изменения активности полипептида. Также, заданный полипептид может содержать несколько типов модификаций. Модификации могут возникать в любом участке полипептида, включая полипептидный остов, боковые цепи аминокислот, а также амино- или карбокси-концы.
[0023] В данном изобретении термин «моносахарид, активированный нуклеотидом», используемый в сочетании с «ферментом, способным катализировать гидролиз моносахарида, активированного нуклеотидом» или «ферментом, способным гидролизовать моносахарид, активированный нуклеотидом», описывает фермент, обладающий каталитической активностью в отношении моносахаридов, активированных нуклеотидами; гидролиз приводит к высвобождению целевого моносахарида. В этой связи, термин «гликозилтрансфераза» обозначает и охватывает ферменты, которые катализируют перенос группировок моносахарида от активированного нуклеотидом моносахарида («гликозильного донора») на молекулу гликозильного акцептора. В данном изобретении гликозилтрансфераза, которую применяют в способе и в микроорганизме согласно изобретению, катализирует гидролиз моносахарида, активированного нуклеотидом, в отсутствие молекулы-акцептора. Согласно одному аспекту изобретения, наиболее предпочтительно, если гидролаза активированных нуклеотидами сахаров представляет собой бактериальную фукозилтрансферазу, и предпочтительно, вариант альфа-1,2-фукозилтрансферазы, кодируемой геном wbgL E. coli:O126 (код доступа ADN43847) или другие фукозилтрансферазы, катализирующие гидролиз ГДФ-L-фукозы в отсутствие молекулы-акцептора.
[0024] Соответственно, термин гидролаза сахаров, активированных нуклеотидами, или нуклеиновая кислота/полинуклеотид, кодирующие гидролазу сахаров, активированных нуклеотидами, относится к ферменту, который катализирует гидролитическое расщепление сахаров, активированных нуклеотидами, таких как ГДФ-фукоза, УДФ-галактоза, ГДФ-манноза, ГДФ-рамноза, и других нуклеотид-сахаров естественного происхождения. Предпочтительно, указанная гидролаза сахаров, активированных нуклеотидами, представляет собой гликозилтрансферазу, которая не переносит или преимущественно не переносит моносахарид на молекулу-акцептор. В случае ГДФ-L-фукозы фермент, способный гидролизовать моносахарид, активированный нуклеотидом, представляет собой фукозилтрансферазу, например, альфа-1,2-фукозилтрансферазу, но не ограничивается ей.
[0025] Более конкретно, предпочтительно, если используют альфа-1,2-фукозилтрансферазу WbgL E. coli :126, имеющую замены аминокислотных остатков аспарагина 69 на серин, гистидина 124 на аланин, глутамата 215 на глицин и изолейцина 268 на пролин, или 1,2-фукозилтрансферазу FutC Helicobacter pylori или другие фукозилтрансферазы, проявляющие гидролитическую активность в отношении ГДФ-L-фукозы в отсутствие молекулы-акцептора.
[0026] В рамках данного изобретения указанные термины также охватывают полиморфные варианты нуклеиновых кислот/полинуклеотидов и полипептидов, аллели, мутанты и межвидовые гомологи, имеющие аминокислотную последовательность, которая идентична более чем на 60%, на 65%, 70%, 75%, 80%, 85%, 90%, предпочтительно на 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% или более, предпочтительно на участке протяженностью по меньшей мере 25, 50, 100, 200 или 300 или более аминокислот, с аминокислотными последовательностями альфа-1,2-фукозилтрансфераз, кодируемых геном wbgL E. coli:O126 (номер доступа ADN43847) или futC H. pylori (номер доступа AAD29868).
[0027] Кроме того, полипептид гидролизующего фермента может быть изменен путем вставок или делеций пептидных последовательностей для модификации его активности. Например, полипептидные последовательности могут быть слиты с полипептидом фермента для придания дополнительной ферментативной активности.
[0028] Кроме того, гены, кодирующие фермент, способный гидролизовать моносахарид, активированный нуклеотидом, могут быть изменены таким образом, чтобы генный продукт включал белки или полипептиды, которые представляют функционально эквивалентные генные продукты. Такой эквивалентный генный продукт гидролазы может содержать делеции, вставки или замены аминокислотных остатков в составе аминокислотной последовательности, кодируемой последовательностью гена гидролазы, описанной выше, которые при этом приводят к молчащей замене, таким образом, с образованием функционально-эквивалентного генного продукта, кодирующего фермент, способный гидролизовать моносахарид, активированный нуклеотидом. Аминокислотные замены могут производиться на основании сходства в полярности, заряде, растворимости, гидрофобности, гидрофильности и/или амфипатических свойств задействованых остатков. Например, неполярные (гидрофобные) аминокислоты включают аланин, лейцин, изолейцин, валин, пролин, фенилаланин, триптофан и метионин; полярные незаряженные аминокислоты включают глицин, серин, треонин, цистеин, тирозин, аспарагин и глутамин; положительно заряженные (основные) аминокислоты включают аргинин, лизин и гистидин; и отрицательно заряженные (кислые) аминокислоты включают аспарагиновую кислоту и глутаминовую кислоту.
[0029] В контексте данного изобретения «функционально эквивалентный» относится к полипептиду, способному проявлять in vivo гидролазную активность в отношении сахаров, активированных нуклеотидами, по существу схожую с эндогенной активностью продукта гена гидролазы, кодируемым последовательностью гена гидролазы, описанной выше, если судить по ряду критериев, включая, без ограничений, антигенность, т.е. способность связываться с антителом, направленным против гидролазы сахаров, активированных нуклеотидами, иммуногенность, т.е. способность вырабатывать антитело, которое способно связываться с белком или полипептидом гидролазы моносахаридов, активированных нуклеотидами, а также ферментативную активность. Соответственно, данное изобретение также охватывает ферменты, которые являются функционально эквивалентными непосредственно изложенным в данном документе.
[0030] На основании данного изобретения специалист в области техники также легко может понять, что любые модификации изложенных здесь ферментов можно использовать в способе и микроорганизме, предложенным в данном изобретении, указанные модификации приводят к увеличению гидролизующей активности ферментов, описанных в данном документе. Таким образом, такие модифицированные ферменты, демонстрирующие повышенную гидролизующую активность по сравнению с немодифицированными формами, также входят в объем изобретения.
[0031] В объем изобретения также входят белки, полипептиды и производные (включая фрагменты) гидролазы сахаров, активированных нуклеотидами, которые избирательно модифицированы во время или после трансляции. Кроме того, в качестве замен или вставок в полипептидную последовательность фермента могут быть встроены не являющиеся классическими аминокислоты или химические аналоги аминокислот.
[0032] Согласно предпочтительному воплощению способа, предложенного в изобретении, фермент является вариантом 2-фукозилтрансферазы, кодируемой геном wbgL, или вариантом 1,2-фукозилтрансферазы, кодируемой геном futC Helicobacter pylori, вариант имеет по меньшей мере одну, предпочтительно, по меньшей мере две и, более предпочтительно, более двух модификаций по сравнению с 2-фукозилтрансферазой дикого типа, кодируемой геном wbgL, или с 1,2-фукозилтрансферазой дикого типа, кодируемой геном futC, соответственно, модификация приводит к увеличению гидролизующей активности фермента.
[0033] Фермент, способный гидролизовать сахара, активированные нуклеотидами, можно производить с использованием технологии рекомбинантной ДНК, известной в области техники. Для конструирования экспрессирующих векторов, содержащих последовательности, кодирующие ферменты и соответствующие сигналы, контролирующие транскрипцию, позволяющие синтезировать фермент, катализирующий гидролиз сахаров, активированных нуклеотидами, можно использовать способы, хорошо известные в области техники. Данные способы включают, например, in vitro технологии рекомбинантной ДНК, методы синтеза и генетическую рекомбинацию in vivo. См., например, методы, описанные Sambrook, J. and Russell D.W. (1989) Molecular Cloning: A Laboratory Manual, 2nd Ed., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y.
[0034] Согласно воплощению способа, предложенного в изобретении, по меньшей мере одна модификация представляет собой по меньшей мере одну аминокислотную замену. Согласно другому воплощению модификация представляет собой или содержит по одну, две или более двух, в частности, три, четыре, пять, шесть, семь, восемь, девять или десять аминокислотных замен, где модифицированный фермент, способный гидролизовать моносахарид, активированный нуклеотидом, обладает повышенной гидролизующей активностью в отношении моносахарида, активированного нуклеотидом, по сравнению с немодифицированным ферментом дикого типа.
[0035] Согласно одному воплощению, предпочтительно, если используют альфа-1,2-фукозилтрансферазу WbgL E. coli :126, имеющую замены аминокислотных остатков аспарагина 69 на серин, гистидина 124 на аланин, глутамата 215 на глицин и изолейцина 268 на пролин, или 1,2-фукозилтрансферазу FutC Helicobacter pylori или другие фукозилтрансферазы, проявляющие гидролитическую активность в отношении ГДФ-L-фукозы в отсутствие молекулы-акцептора.
[0036] На основании описания изобретения специалист в области техники поймет, что изобретение охватывает не только конкретные замены, указанные выше, но также и другие модификации этих конкретных описанных ферментов, в частности, другие замены, при условии, что соответственным образом модифицированный фермент обладает повышенной гидролизующей активностью в отношении моносахарида, активированного нуклеотидом, по сравнению с немодифицированным ферментом дикого типа. Альтернативные замены, которые могут оказаться подходящими, также обсуждаются выше в более общем виде, но также могут применяться.
[0037] Здесь и далее в изобретении «рекомбинантная» означает генетически модифицированную ДНК, полученную путем переноса или сплайсинга генов от одних биологических видов в клетки микроорганизма-хозяина, относящегося к другому биологическому виду. Такая ДНК становится частью генетического аппарата хозяина и реплицируется.
[0038] «Микроорганизм» здесь обозначает и охватывает любой микроскопический организм, который содержит либо единичную клетку, кластер клеток или многоклеточные, достаточно сложные организмы, которые подходят для использования в способе, предложенном в данном изобретении, и в частности, включают бактерии и дрожжи. Микроорганизм, который применяют согласно изобретению, можно культивировать в жидкой среде, и, как правило, для роста и репликации в среде ему необходим источник углеводов.
[0039] Следовательно, «рекомбинантный микроорганизм-хозяин» означает любой микроорганизм, содержащий последовательности нуклеиновой кислоты, кодирующие гликозилтрансферазу или гидролазу сахаров, активированных нуклеотидами, или кодирующие фукозилтрансферазу или гидролазу ГДФ-L-фукозы, где последовательности нуклеиновой кислоты, кодирующие указанные ферменты, представляют собой последовательности нуклеиновой кислоты, чужеродные/не встречающиеся естественным образом в рекомбинантной клетке (хозяине), где чужеродная/не встречающаяся естественным образом в указанном микроорганизме последовательность интегрирована в геном клетки микроорганизма-хозяина. Таким образом, «не встречающийся естественным образом» означает, что последовательности нуклеиновой кислоты являются чужеродными для клетки микроорганизма-хозяина, т.е. последовательности нуклеиновой кислоты являются гетерологичными по отношению к клетке микроорганизма-хозяина. Гетерологичная последовательность может быть стабильно встроена, например, путем трансфекции, трансформации или трансдукции в геном клетки микроорганизма-хозяина, где можно применять методики, в зависимости от клетки-хозяина, в которую будет встроена последовательность. Специалистам в области техники известны различные методики, они описаны, например, Sambrook et al., 1989, см. выше. Таким образом, клетка-хозяин, в которую была встроена гетерологичная последовательность, будет продуцировать гетерологичные белки, кодируемые последовательностями нуклеиновой кислоты согласно изобретению.
[0040] Для рекомбинантной продукции клетки-хозяева должны быть генетически модифицированы для того, чтобы включать системы экспрессии или их части и последовательности нуклеиновых кислот согласно изобретению. Встраивание последовательности нуклеиновой кислоты в клетку микроорганизма-хозяина можно осуществлять способами, описанными во многих стандартных лабораторных руководствах, таких как Davis et al., Basic Methods in Molecular Biology, (1986) и Sambrook et al., 1989, см. выше.
[0041] Так, последовательности нуклеиновых кислот по изобретению могут, например, входить в состав вектора, которым могут быть стабильно трансформированы/трансфецированы клетки микроорганизма-хозяина.
[0042] Для производства полипептидов по изобретению можно использовать многочисленные разнообразные системы экспрессии. Такие векторы включают, помимо прочих, хромосомные векторы, эписомные векторы и векторы на основе вирусов, например, векторы на основе бактериальных плазмид, бактериофагов, транспозонов, эписом дрожжей, перемещающихся встроенных элементов, элементов хромосом дрожжей, вирусов, а также векторы на основе их комбинаций, такие как на основе плазмид и генетических элементов бактериофагов, такие как космиды и фагмиды. Конструкции систем экспрессии могут содержать контролирующие участки, которые регулируют, а также запускают экспрессию. Как правило, для экспрессии можно применять любую систему или вектор, подходящие для того, чтобы поддерживать, репродуцировать или экспрессировать полинуклеотиды и синтезировать полипептид в клетке-хозяине. Соответствующую последовательность ДНК можно встроить в систему экспрессии посредством любого из многочисленных известных и рутинных способов, например, таких, которые описаны Sambrook et al., см. выше.
[0043] В данном документе термин «выделение» обозначает изолирование, сбор, очистку или иное отделение от культуры микроорганизма моносахарида, произведенного микроорганизмом согласно изобретению.
[0044] Согласно предпочтительному воплощению способа, предложенного в изобретении, микроорганизм дополнительно модифицируют для инактивации или существенного ослабления или отсутствия катаболических путей, ведущих к деградации произведенного моносахарида.
[0045] Согласно другому воплощению, микроорганизм дополнительно модифицируют для инактивации или существенного ослабления или отсутствия генов, вовлеченных в катаболизм L-фукозы.
[0046] Согласно другому воплощению, микроорганизм дополнительно модифицируют для сверхэкспрессии по меньшей мере одного гена, вовлеченного в биосинтез моносахарида, активированного нуклеотидом, для улучшения снабжения моносахарида моносахаридом, активированным нуклеотидом. В этой связи предпочтительно, чтобы по меньшей мере один ген был гетерологичным или гомологичным.
[0047] Согласно другому воплощению способа, предложенного в данном изобретении, по меньшей мере один ген, вовлеченный в биосинтез ГДФ-фукозы, ГДФ-маннозы или ГДФ-рамнозы, сверхэкспрессируется для улучшения снабжения ГДФ-фукозой, ГДФ-маннозой или ГДФ-рамнозой, соответственно. В этой связи предпочтительно, чтобы по меньшей мере один ген был гетерологичным или гомологичным.
[0048] Согласно другому воплощению способа, предложенного в изобретении, микроорганизм дополнительно модифицируют для инактивации или ослабления путей, конкурирующих за моносахарид, активированный нуклеотидом.
[0049] Согласно другому воплощению способа, предложенного в изобретении, микроорганизм дополнительно модифицируют для экспрессии фосфатазы, в случаях, когда моносахарид высвобождается ферментом в фосфорилированной форме.
[0050] В изобретении особенно предпочтительно, если продуцируемый свободный моносахарид выбран из L-фукозы, L-рамнозы или L-маннозы.
[0051] В предпочтительном воплощении микроорганизм культивируют в среде, содержащей источник углерода, который выбран из глицерина, сахарозы, ацетата, глюкозы, фруктозы, мелассы, лактозы, ксилозы, целлюлозы, синтетического газа, двуокиси углерода или окиси углерода. В этом контексте следует понимать, что в качестве источника углерода при культивировании можно использовать любой другой, предпочтительно дешевый субстрат, и специалист в области техники легко сможет использовать источник углерода, подходящий в данном изобретении, для выращивания микроорганизма для крупномасштабного производства целевого моносахарида.
[0052] Согласно одному аспекту изобретения на стадии культивирования микроорганизма, например, рекомбинантного микроорганизма, в среду непрерывно добавляют источник углерода.
[0053] Благодаря непрерывному добавлению источника углерода на стадии культивирования достигается непрерывное и эффективное производство моносахарида.
[0054] Согласно другому аспекту изобретения моносахарид выделяют из надосадочной жидкости культивируемого рекомбинантного микроорганизма-хозяина, которую получают путем центрифугирования культивируемого микроорганизма-хозяина для получения надосадочной жидкости и осадка микроорганизма-хозяина.
[0055] Благодаря предложенному новому способу возможно выделять произведенный моносахарид из среды, в которой культивируют микроорганизм-хозяин, поскольку моносахарид, который продуцируется в клетке микроорганизма, транспортируется в среду, таким образом, делая возможным удобное выделение моносахарида из надосадочной жидкости, после отделения клеток микроорганизма от культуральной среды.
[0056] Другие моно- или олигосахариды, которые могут быть произведены в микроорганизме в ходе синтеза целевого моносахарида, которые ухудшают/препятствуют стадии выделения/очистки целевого моносахарида, могут метаболизироваться микроорганизмом, благодаря чему стадия выделения целевого моносахарида дополнительно улучшается и облегчается. Таким образом, фермент(ы), метаболизирующий(е) сахарид, могут быть добавлены/внесены в среду извне и в конце способа, предложенного в изобретении. В этом случае нежелательные сахара не могут накапливаться и не интерферируют с выделением целевого моносахарида. Гены, кодирующие метаболические пути или ферменты, могут экспрессироваться в микроорганизме для метаболизации нежелательных моносахаридов, которые в противном случае будут мешать, и специалист в области техники, ознакомившись с описанием изобретения, легко выявит другие подходящие пути или ферменты, которые следует дерегулировать/активировать или снабдить, в зависимости от продуцируемого моносахарида.
[0057] Согласно другому аспекту изобретения способ, предложенный в изобретении, включает следующие стадии:
а) обеспечение рекомбинантного микроорганизма-хозяина в среде, подходящей для выращивания микроорганизма, который был трансформирован, чтобы содержать последовательность нуклеиновой кислоты, кодирующую фермент, катализирующий гидролиз моносахарида, активированного нуклеотидом, не существующего в микроорганизме естественным образом, причем микроорганизм не способен метаболизировать продуцируемый моносахарид в существенном количестве,
б) культивирование рекомбинантного микроорганизма-хозяина в указанной среде, в которую моносахарид продуцируется в свободной форме,
в) выделение свободного моносахарида из среды.
[0058] Таким образом, способ, описанный в параграфах выше, включает дополнительную стадию выделения свободного моносахарида из среды.
[0059] Согласно другому аспекту изобретения описан и заявлен микроорганизм.
[0060] Определения, использованные и приведенные выше для конкретных терминов, относящиеся к способу, также относятся и к рекомбинантному микроорганизму, представленному в данном описании.
[0061] Согласно предпочтительному воплощению, микроорганизм, использованный в способе согласно изобретению и заявленный в данном документе, выбран из штамма бактерий или дрожжей, способного синтезировать сахара, активированные нуклеотидами, из которых можно получить целевой моносахарид путем гидролитического расщепления моносахарида, активированного нуклеотидом. Преимущество бактерий Escherichia coli, Corynebacterium glutamicum и дрожжи Saccharomyces sp. состоит в том, что указанные микроорганизмы можно легко и без особых затрат культивировать в лабораторных условиях, и бактерии и дрожжи интенсивно изучали на протяжении многих лет.
[0062] Соответственно, в предпочтительном воплощении микроорганизм-хозяин, используемый в способе согласно изобретению и заявленный в данном документе, выбран из группы, состоящей из бактерий и дрожжей, и предпочтительно представляет собой штамм Escherichia coli.
[0063] В воплощении изобретения также предпочтительно, если рекомбинантный микроорганизм-хозяин дополнительно модифицирован, чтобы не иметь генов, кодирующих ферменты, вовлеченные в метаболизм целевого моносахарида, в случае, когда целевым моносахаридом является L-фукоза, генов, кодирующих L-фукулозозокиназу, L-фукозоизомеразу, фукулозо-1-фосфат-альдолазу и УДФ-глюкозо:ундекапренил-фосфат-глюкозофосфотрансферазу. Кроме того, согласно предпочтительному воплощению, осуществляют делецию генов гликозилтрансфераз, использующих моносахарид, активированный нуклеотидом, в качестве субстрата для синтеза полисахаридов (например, фукозилтрансфераз или ферментов, вовлеченных в синтез фукозилированных олигосахаридов, таких как колановая кислота).
[0064] Кроме того, предпочтительна сверхэкспрессия генов, улучшающих синтез сахаров, активированных нуклеотидами. В случае ГДФ-L-фукозы в соответствующем микроорганизме сверхэкспрессируют гены, кодирующие фосфоманномутазу (manB), маннозо-1-фосфатгуанозилтрансферазу (manC), ГДФ-маннозо-4,6-дегидратазу (gmd) и ГДФ-L-фукозосинтазу (wcaG) E. coli или надлежащие гены других организмов.
[0065] Преимуществом данного воплощения является то, что внутриклеточная деградация полученного моносахарида L-фукозы и продукция колановой кислоты предотвращается и улучшается синтез ГДФ-L-фукозы.
[0066] В другом предпочтительном воплощении рекомбинантный микроорганизм-хозяин дополнительно трансформирован, чтобы содержать гены, позволяющие рекомбинантному микроорганизму-хозяину расти на сахарозе или глицерине в качестве единственного источника углерода, и особенно предпочтительно, если кластер генов csc Escherichia coli W (номер доступа CP0021851) интегрирован в геном микроорганизма-хозяина, где кластер генов содержит гены сахарозопермеазы, фруктокиназы, сахарозогидролазы и репрессора транскрипции (гены cscB, cscK, cscA и cscR, соответственно), что позволяет трансформированному микроорганизму расти на сахарозе как единственном источнике углерода.
[0067] В этой связи следует отметить, что все воплощения, указанные в качестве предпочтительных для способа, предложенного в изобретении, используют заявленный микроорганизм, где это применимо.
[0068] Соответственно, данное изобретение также относится к применению микроорганизма, обладающего ферментом, катализирующим гидролиз моносахарида, активированного нуклеотидами, где микроорганизм не способен метаболизировать моносахарид, и изобретение дополнительно относится к применению рекомбинантного микроорганизма согласно изобретению для производства моносахарида, в частности, L-фукозы.
[0069] Следует отметить, что определения, приведенные выше для описания некоторых терминов, относящихся к способу, предложенному в изобретении, относятся к микроорганизму, рекомбинантному или немодифицированному, как заявлено и описано в данном документе.
[0070] В альтернативном варианте, способ производства моносахаридов можно применять в бесклеточных системах, где фермент согласно изобретению и подходящие субстраты смешивают в водной реакционной среде. Фермент можно применять в свободной форме в растворе или он может быть связан или иммобилизован на носителе, таком как полимер, а субстрат можно добавлять к носителю. Носителем можно, например, заполнять колонку.
[0071] В частности, данное изобретение относится к способу, в котором в качестве рекомбинантного микроорганизма-хозяина используют рекомбинантный штамм Escherichia coli, где в рекомбинантном штамме Escherichia coli осуществили делецию гена L-фукозоизомеразы, гена L-фукулозокиназы и УДФ-глюкозо:ундекапренил-фосфат-глюкозофосфотрансферазы, и где рекомбинантный штамм Escherichia coli был трансформирован, чтобы содержать a) гены, позволяющие штамму E. coli расти на сахарозе или глицерине в качестве единственного источника углерода, гены, кодирующие, соответственно, сахарозопермеазу, фруктокиназу, сахарозогидролазу и репрессор транскрипции, б) гены, кодирующие фосфоманномутазу, маннозо-1-фосфатгуанозилтрансферазу, ГДФ-маннозо-4,6-дегидратазу и ГДФ-L-фукозосинтазу E. coli или других организмов, в) ген, кодирующий фермент, катализирующий гидролиз моносахаридов, активированных нуклеотидами, например, фукозилтрансферазу, которая гидролизует ГДФ-L-фукозу в отсутствие молекулы-акцептора. Описание воплощений и приведенные графические материалы подтверждают следующие преимущества.
[0072] Не требует доказательств, что характеристики, упомянутые выше, а также характеристики, которые объясняются ниже, могут применяться не только в соответствующих приведенных комбинациях, но также и в других комбинациях или сами по себе, при этом оставаясь в рамках данного изобретения. Также следует отметить, что конкретные признаки, представленные в зависимых пунктах, могут быть скомбинированы друг с другом в рамках изобретения иным способом, таким образом, изобретение также охватывает другие воплощения, имеющие любую возможную комбинацию признаков, представленных в зависимых пунктах.
[0073] Несколько воплощений изобретения проиллюстрированы графическими материалами и более подробно объясняются в описании, приведенном ниже. Графические материалы.
[0074] Фиг. 1. Определение свободной L-фукозы в культуральной надосадочной жидкости штамма-продуцента E. coli BL21(DE3), содержащего библиотеку pDEST:wbgL после случайного мутагенеза wbgL в результате ПЦР пониженной точности. Надосадочную жидкость культур штамма-продуцента E. coli BL21(DE3), несущего клоны pDEST:wbgL (после ПЦР пониженной точности), использовали в колориметрическом L-фукозодегидрогеназном тесте (подробное описание анализа приведено в параграфе [0089] ниже). Лунки, обозначенные «WT», содержали надосадочную жидкость штамма-продуцента E. coli BL21(DE3) культуры pDEST:wbgL, экспрессирующие немодифицированный ген wbgL. Лунки, обозначенные «D1» и «H2», содержали культуральную надосадочную жидкость клонов, синтезирующих варианты WbgL (H124A E215G) (клон D1) и WbgL (N69S I268P) (клон H2);
[0075] Фиг. 2. Анализ культуральной надосадочной жидкости штамма-продуцента E. coli BL21(DE3), несущего pDEST:wbgL(H124A E215G) (клон D1) и pDEST:wbgL(N69S I268P) (клон H2) с использованием жидкостной хроматографии с тандемной масс-спектрометрией (LC-MS/MS). Клон D1 и клон H2 продуцировали 4,3 г/л и 2,5 г/л L-фукозы, соответственно.
[0076] Фиг. 3. Определение L-фукозы, высвобожденной из ГДФ-L-фукозы вследствие ГДФ-L-фукозогидролазной активности. Клеточные лизаты E. coli BL21(DE3) pDEST:futC и E. coli BL21(DE3) pDEST:wbgL (H124A E215G) использовали in vitro в ГДФ-L-фукозогидролазных тестах при содержании ГДФ-L-фукозы5 мМ. Свободную L-фукозу определяли с помощью колориметрического L-фукозодегидрогеназного теста (описание анализа приведено в параграфе [0089] ниже). В лунке 1 содержалось 0,27 единиц FutC, в лунку 2 вносили 0,32 единицы WbgL (H124A E215G). Для подтверждения стабильности ГДФ-L-фукозы проводили анализ с бычьим сывороточным альбумином (лунка 3).
[0077] Фиг. 4. Последовательность <Ptet-manCB-PT5-gmd> (SEQ ID No. 1), wcaG- dhfr (A), <cscB-cscK-cscA-cscR> (SEQ ID No. 2) (B), <wbgL> (SEQ ID No. 3) (C) и <futC> (SEQ ID No. 4)
[0078] ОПИСАНИЕ ПРИМЕРОВ ОСУЩЕСТВЛЕНИЯ ИЗОБРЕТЕНИЯ
[0079] Конструирование штамма E. coli, продуцирующего фукозу
[0080] Для конструирования штамма, продуцирующего фукозу, методом генной инженерии использовали Escherichia coli BL21 (DE3) (Novagen, Дармштадт, Германия). Поскольку фукоза образуется в результате гидролиза ГДФ-L-фукозы, синтез ГДФ-L-фукозы усиливается при интеграции в геном и сверхэкспрессии генов, кодирующих фосфоманномутазу (manB), маннозо-1-фосфатгуанозилтрансферазу (manC), ГДФ-маннозо-4,6-дегидратазу (gmd) и ГДФ-L-фукозосинтазу (wcaG) E. coli K12 DH5α. Оперон manC-manB находился под транскрипционным контролем промотора Ptet и оперон gmd-wcaG экспрессировался под контролем промотора PT5. Кластер генов <Ptet-manCB-PT5-gmd, wcaG- dhfr > (SEQ ID No. 1; Фиг. 4A) также содержал ген dhfr, кодирующий дигидрофолатредуктазу, придающую устойчивость к триметоприму клеткам, интегрировавшим ген. Для интеграции кластера в геном E. coli BL21(DE3) посредством транспозиции кластер фланкирован инвертированными концевыми повторами, специфически распознающимися мобильным генетическим элементом (транспозоном) Himar1 семейства mariner. Кроме того, в геном организма-хозяина встраивали кластер генов csc E. coli W. Кластер генов содержит гены сахарозопермеазы (cscB), фруктокиназы (cscK), сахарозогидролазы (cscA) и репрессора транскрипции (cscR). Интеграция кластера <cscB-cscK-cscA-cscR> (SEQ ID No. 2; Фиг. 4B), фланкированного инвертированными концевыми повторами, специфически распознающимися Himar1, осуществлялась посредством транспозиции Himar1 и придавала организму-хозяину способность расти, используя сахарозу в качестве единственного источника углерода (Choi, K.-H. and Kim, K.-J. (2009) Applications of transposon-based gene delivery system in bacteria. J. Microbiol. Biotechnol. 19, 217-228).
[0081] Для предотвращения истощения ГДФ-L-фукозы при образовании колановой кислоты осуществляли делецию гена wcaJ, который, как предполагают, кодирует УДФ-глюкозо:ундекапренил-фосфат-глюкозо-1-фосфаттрансферазу в геноме E. coli BL21(DE3) согласно способу, предложенному Datsenko and Warner (Datsenko, K.A. and Warner B.L. (2000) One-step inactivation of chromosomal genes in Escherichia coli K-12 using PCR products. Proc. Natl. Acad. Sci. USA 97, 6640-6645). УДФ-глюкозо:ундекапренил-фосфат-глюкозо-1-фосфаттрансфераза кодирует первую стадию синтеза колановой кислоты (Stevenson, G., Andrianopoulos, K., Hobbs, M. and Reeves, P. R. (1996) Organization of the Escherichia coli K-12 gene cluster responsible for production of the extracellular polysaccharide colonic acid. J. Bacteriol. 178, 4885-4893). Кроме того, для ингибирования деградации L-фукозы инактивировали гены пути катаболизма фукозы fucI и fucK, кодирующие фукозоизомеразу и фукулозокиназу, соответственно, путем геномного нокаута.
[0082] Клонирование генов 2-фукозилтрансфераз wbgL и futC и мутагенез wbgL
[0083] Ген 2-фукозилтрансферазы wbgL (SEQ ID No. 3; Фиг. 4C) E. coli:O126 (номер доступа AND43847) был кодон-оптимизирован и получен синтетическим способом в компании GenScript (Пискатауэй, США). Ген futC (SEQ ID No. 4, Фиг. 4D), кодирующий 1,2-фукозилтрансферазу Helicobacter pylori (номер доступа AAD29868) был также синтезирован и кодон-оптимизирован для экспрессии в E. coli. Для клонирования в вектор pDEST14 гены амплифицировали с использованием праймеров 6128 (SEQ ID No. 5) и 6129 (SEQ ID No. 6) для wbgL и праймеров 6195 (SEQ ID No. 7) и 6196 (SEQ ID No. 8) для futC, соответственно; последовательности праймеров приведены в Таблице 1 ниже:
[0084] Таблица 1. Перечень олигонуклеотидов, использованных в полимеразной цепной реакции
[0085] Мутации были внедрены в ген wbgL в результате нескольких раундов ПЦР пониженной точности с использованием набора Diversify® PCR Random Mutagenesis (Clonetech, Маунтин-Вью, США) согласно инструкциям производителя и праймеров 6128 и 6129. Очищенные продукты ПЦР пониженной точности клонировали в вектор pDEST14 с получением pDEST:wbgL (после ПЦР пониженной точности). Клонирование в вектор pDEST14 обычно осуществляли с использованием технологии Gateway (руководство по технологии Gateway® (Life Technologies, Карлсбад, США)). Плазмиды секвенировали в компании LGC Genomics (Берлин, Германия). Рекомбинантными плазмидами трансформировали подходящие хозяева E. coli путем электропорации.
[0086] Питательная среда и культивирование клеток
[0087] Штамм-продуцент E. coli BL21(DE3), несущий плазмидную библиотеку pDEST:wbgL (после ПЦР пониженной точности) выращивали в среде с минеральными солями с добавлением 1% (об/об) глицерина и 1% (об/об) сахарозы в качестве источников углерода. Среда содержала 2 г/л NH4H2PO4, 7 г/л K2HPO4, 2 г/л KOH, 0,3 г/л лимонной кислоты, 0,98 г/л MgSO4 × 7 H2O и 0,02 г/л CaCl2 × 6 H2O. В нее добавляли один миллилитр раствора микроэлементов (54,4 г/л железа аммонийного цитрат, 9,8 г/л MnCl2 × 4 H2O, 1,6 г/л CoCl2 × 6 H2O, 1 г/л CuCl2 × 2 H2O, 1,9 г/л H3BO3, 9 г/л ZnSO4 × 7 H2O, 1,1 г/л Na2MoO4 × 2 H2O, 1,5 г/л Na2SeO3, 1,5 г/л NiSO4 × 6 H2O). Для селекции добавляли 10 мкг/мл триметоприма и 100 мкг/мл ампициллина. Клетки культивировали в 96-луночных планшетах для микротитрования в течение 24 ч при 30°C со встряхиванием. 50 мкл начальной культуры переносили в 96-луночные планшеты с 400 мкл свежей среды, забуференной HEPES (100 мМ), содержащей 0,3 мМ IPTG для индукции экспрессии генов wbgL в pDEST. Индуцированные культуры выращивали в течение двух суток при 30°C при встряхивании. Клетки осаждали центрифугированием и использовали надосадочную жидкость для определения свободной L-фукозы.
[0088] E. coli BL21(DE3), содержащие pDEST: wbgL (H124A E215G) и pDEST:futC, соответственно, выращивали в среде 2YT (Sambrook, J. and Russell D. W. (1989) Molecular Cloning: A Laboratory Manual, 2nd Ed., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y.) при 30°C до OD600 нм 0,3 в присутствии 100 мкг/мл ампициллина. Транскрипцию wbgL (H124A E215G) и futC индуцировали добавлением 0,3 мМ IPTG. Клетки собирали центрифугированием через 20 ч после индукции. Клетки использовали для анализа активности ГДФ-L-фукозогидролазы in vitro.
[0089] Ферментативный анализ для определения свободной L-фукозы и активности ГДФ-L-фукозогидролазы in vitro
[0090] Свободную L-фукозу определяли колориметрическим способом, используя L-фукозодегидрогеназу (FuDH) Pseudomonas sp. No.1143 (номер доступа D32042), которая катализирует НАДФ-зависимую трансформацию L-фукозы в L-фуконо-1,5-лактон. При колориметрическом определении нитросиний тетразолий в присутствии феназина метосульфата восстанавливается в сине-фиолетовый формазан под действием НАДФ-Н. Образование формазана определяют при 571 нм. (Mayer, K. M. and Arnold, F. H. (2002) A Colorimetric Assay to Quantify Dehydrogenase Activity in Crude Cell Lysates. J. Biomol. Screen. 7, 135-140).
[0091] Ген fuDH Pseudomonas sp. No.1143 сверхэкспрессировали в E. coli BL21(DE3). Рекомбинантный белок FuDH, имеющий N-концевой His6-таг, обогащали из неочищенного экстракта при помощи аффинной хроматографии с использованием иммобилизованных металлов на колонках Ni Sepharose™ 6 Fast Flow (GE Healthcare, Поллардс Вуд, Великобритания).
[0092] Для детекции свободной L-фукозы в культурах штамма-продуцента фукозы E. coli BL21(DE3), несущего pDEST:wbgL (после ПЦР пониженной точности) каждые 200 мкл реакционной смеси для L-фукозодегидрогеназного теста содержали 50 мкл надосадочной жидкости культуры клеток и 150 мкл раствора реагента, содержащего 0,8 мМ НАДФ-Н, 0,3 мМ нитросиний тетразолий, 0,03 мМ феназина метосульфат и 4,7 единиц His6-FuDH в 50 мМ Tris (pH 8,0) с добавлением 0,13 % (мас/об) желатина (все химические реагенты приобретали в компании Sigma Aldrich, Сент Луис, США). Образование сине-фиолетового формазана определяли через 10 мин инкубации при комнатной температуре при 571 нм.
[0093] Для определения активности ГДФ-L-фукозогидролазы в клеточных лизатах клетки E. coli BL21(DE3) pDEST:wbgL (H124A E215G) и E. coli BL21(DE3) pDEST:futC ресуспендировали в 50 мМ буфере HEPES (pH 7,5) с добавлением 5 мМ MnCl2 и разрушали с использованием стеклянных шариков и гомогенизатора Mini-Beatbeater (BioSpec Producs, Бартлсвиль, США). В ГДФ-L-фукозогидролазном тесте L-фукоза отщеплялась от ГДФ-L-фукозы. 200 мкл реакционной смеси для определения гидролазы содержали 12,5 мкл 100 мМ ГДФ-L-фукозы (Sigma Aldrich, Сент-Луис, США) в 50 мМ буфера HEPES (pH 7,5), 5 мМ MnCl2 и 50 мкл бесклеточного экстракта.
[0094] Для проверки стабильности ГДФ-L-фукозы гидролазный тест также проводили, взяв вместо неочищенного экстракта 50 мкл бычьего сывороточного альбумина (30 мг/мл). Концентрации белка оценивали по Бредфорду, используя доступный для приобретения раствор красителя (Roti-Quant®, Carl Roth, Карлсруэ, Германия). После инкубации в течение 1 часа при 30°C ферменты в тестах с ГДФ-L-фукозой инактивировали путем нагревания до 95°C в течение 10 мин. Для определения свободной L-фукозы с помощью L-фукозодегидрогеназного теста использовали 50 мкл смеси после ГДФ-L-фукозогидролазной реакции. Анализ проводили, как описано выше, и инкубировали в течение 24 ч при комнатной температуре.
[0095] Анализ методом жидкостной хроматографии с тандемной масс-спектрометрией (LC-MS/MS)
[0096] Масс-спектрометрический анализ выполняли с использованием мониторинга множественных реакций (MRM, от англ. multiple reaction monitoring) и тройной квадрупольной масс-спектрометрической системы детекции (Shimadzu LC-MS 8050) (Shimadzu Corporation, Киото, Япония). Ионы-предшественники отбирают и анализируют в квадруполе 1, фрагментацию осуществляют в ячейке соударений, используя для диссоциации, индуцированной соударениями, газ аргон, отбор фрагментированных ионов осуществляют в квадруполе 3.
[0097] Хроматографическое разделение фукозы и мальтотриозы после разведения культуральной надосадочной жидкости и реакционной смеси из анализа in vitro, соответственно, 1:100 водным раствором мальтотриозы (категории LC/MS (для жидкостной хроматографии/масс-спектрометрии)) с концентрацией 10 мкг/мл, выполняли на колонках для ВЭЖХ XBridge Amide HPLC (3,5 мкм, 2,1 × 50 мм (Waters, США) с картриджем XBridge Amide guard (3,5 мкм, 2,1 × 10 мм) (Waters, США). Система ВЭЖХ состоит из автоматического пробоотборника Shimadzu Nexera X2 SIL-30ACMP, у которого устанавливали температуру 8°C, насоса Shimadzu LC-20AD и колоночного термостата Shimadzu CTO-20AC, у которого устанавливали температуру 30°C (Shimadzu Corporation, Киото, Япония). Подвижная фаза состояла из ацетонитрила:H2O (62:38 % (об/об)) с 10 мМ ацетата аммония. В прибор инжектировали 1 мкл образца; прогон осуществляли в течение 3 мин со скоростью тока 300 мкл/мин. L-фукозу и мальтотриозу (добавляемую в качестве внутреннего стандарта для нормализации) анализировали, с помощью мониторинга множественных реакций при ионизации электрораспылением в режиме регистрации отрицательных ионов. Масс-спектрометр эксплуатировали в режиме низкого разрешения. Фукоза образует ион с m/z 163,2 [M-H], а мальтотриоза - ион с m/z 503,2 [M-H]. Ион-предшественник L-фукозы далее фрагментировался в ячейке соударений на фрагменты ионов с m/z 88,9, m/z 70,8 и m/z 58,9 Молекулярный ион мальтотриозы (m/z 503,2) образовывал фрагменты с m/z 341,1, m/z 161,05 и m/z 100,9. Энергию соударений, Q1 и Q3 Pre Bias оптимизировали для каждого аналита в отдельности.
[0098] Результаты
[0099] Синтез свободной L-фукозы штаммом-продуцентом E. coli BL21(DE3), экспрессирующим ген фукозилтрансферазы
[00100] Для усиления синтеза ГДФ-L-фукозы в геном E.coli BL21(DE3) были интегрированы и сверхэкспрессированы гетерологичные гены, кодирующие фосфоманномутазу, маннозо-1-фосфат-гуанозилтрансферазу, ГДФ-маннозо-4,6-дегидратазу и ГДФ-L-фукозосинтазу. Кроме того, для придания штамму BL21(DE3) способности расти на сахарозе в геном интегрировали кластер генов csc, кодирующих сахарозопермеазу, фруктокиназу, сахарозогидролазу и репрессор транскрипции E. coli W.
[00101] 2-фукозилтрансфераза WbgL катализирует перенос L-фукозы от молекулы-донора ГДФ-L-фукозы на олигосахарид-акцептор. Однако, при выращивании штамма-продуцента E. coli BL21(DE3), несущего pDEST:wbgL, в соответствующей среде в отсутствие молекулы-акцептора в надосадочной жидкости бактериальных культур может определяться свободная L-фукоза. Свободная L-фукоза высвобождается из ГДФ-фукозы благодаря ГДФ-L-фукозогидролазной активности WbgL.
[00102] Для дальнейшего улучшения ГДФ-L-фукозогидролазной активности WbgL осуществляли случайный мутагенез гена wbgL с помощью ПЦР пониженной точности. Библиотеку почти из 5000 штаммов-продуцентов E. coli BL21(DE3), несущих клоны pDEST:wbgL(после ПЦР пониженной точности), анализировали на предмет увеличения ГДФ-L-фукозогидролазной активности. Как показал анализ культуральной надосадочной жидкости с помощью L-фукозодегидрогеназного теста, два клона (обозначенные клонами D1 и H2) демонстрировали повышенную продукцию свободной L-фукозы (Фиг. 1). У каждого из двух клонов были обнаружены аминокислотные замены последовательности WbgL. Вариант WbgL клона D1 содержал аминокислотные замены гистидина 124 на аланин и глутамата 215 на глицин, у варианта WbgL клона H2 остаток аспарагина 69 был заменен на серин, а изолейцин 268 - на пролин.
[00103] Кроме того, надосадочную жидкость штамма-продуцента E. coli BL21(DE3), несущего плазмиды pDEST:wbgL, pDEST:wbgL(H124A E215G) D1 и pDEST:wbgL(N69S I268P) H2, анализировали с использованием LC-MS/MS. В каждом образце обнаруживали L-фукозу с помощью MRM-анализа. Количество свободной L-фукозы определяли, используя мальтотриозу в качестве внутреннего стандарта для нормализации. У штамма, экспрессирующего ген wbgL дикого типа, в культуральной надосадочной жидкости определялось 0,12 г/л L-фукозы. Очевидно, что гидролитическая активность вариантов WbgL (H124A, E215G) и (N69S, I268P) была повышена. Клоны D1 и H2 продуцировали 0,43 г/л и 0,25 г/л L-фукозы, соответственно (Фиг. 2).
[00104] Определение активности ГДФ-L-фукозогидролазы в клеточных лизатах микроорганизмов, экспрессирующих гены фукозилтрансферазы
[00105] Активность ГДФ-L-фукозогидролазы анализировали в бесклеточных экстрактах E. coli BL21(DE3) pDEST:wbgL (H124A E215G) и E. coli BL21(DE3) pDEST:futC, культивируемых в присутствии индуктора транскрипции IPTG. Описано, что 2-фукозилтрансфераза FutC гидролизует ГДФ-L-фукозу в отсутствие олигосахаридного субстрата (Stein, D.B., Lin Y.-N., Lin, C.-H. (2008) Characterization of Helicobacter pylori α1,2-fucosyltransferase for enzymatic synthesis of tumor-associated antigens. Adv. Synth. Catal. 350, 2313-2321). ГДФ-L-фукоза расщеплялась в гидролазном тесте где реакционная смесь содержала 5 мМ ГДФ-L-фукозы и клеточные лизаты соответствующих штаммов. Свободную L-фукозу определяли с помощью L-фукозодегидрогеназного теста. Когда в тесте вместо неочищенного экстракта использовали бычий сывороточный альбумин, свободная L-фукоза не определялась, что свидетельствует о стабильности ГДФ-L-фукозы (Фиг. 3).
[00106] Для количественного определения L-фукозы, высвобожденной в результате гидролиза ГДФ-L-фукозы in vitro, после гидролазной реакции смеси просветляли при помощи твердофазной экстракции с использованием ионообменных картриджей (Strata ABW, Phenomenex, Ашаффенбург, Германия) и анализировали при помощи LC-MS/MS. В анализах, где использовали клеточные лизаты штаммов, экспрессирующих futC и wbgL(H124A E215G), через 1 час инкубации определялось 0,38 г/л и 0,32 г/л L-фукозы, соответственно, что соответствовало специфической активности ГДФ-L-фукозогидролазы 0,18 Ед/мг для FutC и 0,21 Ед/мг для WbgL (H124A E215G).
--->
SEQUENCE LISTING
<110> Jennewein Biotechnologies GmbH
<120> Fermentation process for producing monosaccharides in free form
from nucleotide-activated sugars
<130> 2827P110EP
<140> EP15153383.3
<141> 2015-01-30
<160> 8
<170> PatentIn version 3.5
<210> 1
<211> 6741
<212> DNA
<213> Artificial Sequence
<220>
<223> <Ptet-manCB-PT5-gmd, wcaG-dhfr>
<400> 1
ggccagatga ttaattccta atttttgttg acactctatc attgatagag ttattttacc 60
actccctatc agtgatagag aaaagtgaaa tgaatagttc gacaaaaatc tagaaataat 120
tttgtttaac tttaagaagg agatatacaa tttcgtcgac acacaggaaa catattaaaa 180
attaaaacct gcaggagttt gaaggagata gaaccatggc gcagtcgaaa ctctatccag 240
ttgtgatggc aggtggctcc ggtagccgct tatggccgct ttcccgcgta ctttatccca 300
agcagttttt atgcctgaaa ggcgatctca ccatgctgca aaccaccatc tgccgcctga 360
acggcgtgga gtgcgaaagc ccggtggtga tttgcaatga gcagcaccgc tttattgtcg 420
cggaacagct gcgtcaactg aacaaactta ccgagaacat tattctcgaa ccggcagggc 480
gaaacacggc acctgccatt gcgctggcgg cgctggcggc aaaacgtcat agcccggaga 540
gcgacccgtt aatgctggta ttggcggcgg atcatgtgat tgccgatgaa gacgcgttcc 600
gtgccgccgt gcgtaatgcc atgccatatg ccgaagcggg caagctggtg accttcggca 660
ttgtgccgga tctaccagaa accggttatg gctatattcg tcgcggtgaa gtgtctgcgg 720
gtgagcagga tatggtggcc tttgaagtgg cgcagtttgt cgaaaaaccg aatctggaaa 780
ccgctcaggc ctatgtggca agcggcgaat attactggaa cagcggtatg ttcctgttcc 840
gcgccggacg ctatctcgaa gaactgaaaa aatatcgccc ggatatcctc gatgcctgtg 900
aaaaagcgat gagcgccgtc gatccggatc tcaattttat tcgcgtggat gaagaagcgt 960
ttctcgcctg cccggaagag tcggtggatt acgcggtcat ggaacgtacg gcagatgctg 1020
ttgtggtgcc gatggatgcg ggctggagcg atgttggctc ctggtcttca ttatgggaga 1080
tcagcgccca caccgccgag ggcaacgttt gccacggcga tgtgattaat cacaaaactg 1140
aaaacagcta tgtgtatgct gaatctggcc tggtcaccac cgtcggggtg aaagatctgg 1200
tagtggtgca gaccaaagat gcggtgctga ttgccgaccg taacgcggta caggatgtga 1260
aaaaagtggt cgagcagatc aaagccgatg gtcgccatga gcatcgggtg catcgcgaag 1320
tgtatcgtcc gtggggcaaa tatgactcta tcgacgcggg cgaccgctac caggtgaaac 1380
gcatcaccgt gaaaccgggc gagggcttgt cggtacagat gcaccatcac cgcgcggaac 1440
actgggtggt tgtcgcggga acggcaaaag tcaccattga tggtgatatc aaactgcttg 1500
gtgaaaacga gtccatttat attccgctgg gggcgacgca ttgcctggaa aacccgggga 1560
aaattccgct cgatttaatt gaagtgcgct ccggctctta tctcgaagag gatgatgtgg 1620
tgcgtttcgc ggatcgctac ggacgggtgt aaacgtcgca tcaggcaatg aatgcgaaac 1680
cgcggtgtaa ataacgacaa aaataaaatt ggccgcttcg gtcagggcca actattgcct 1740
gaaaaagggt aacgatatga aaaaattaac ctgctttaaa gcctatgata ttcgcgggaa 1800
attaggcgaa gaactgaatg aagatatcgc ctggcgcatt ggtcgcgcct atggcgaatt 1860
tctcaaaccg aaaaccattg tgttaggcgg tgatgtccgc ctcaccagcg aaaccttaaa 1920
actggcgctg gcgaaaggtt tacaggatgc gggcgttgac gtgctggata ttggtatgtc 1980
cggcaccgaa gagatctatt tcgccacgtt ccatctcggc gtggatggcg gcattgaagt 2040
taccgccagc cataatccga tggattataa cggcatgaag ctggttcgcg agggggctcg 2100
cccgatcagc ggagataccg gactgcgcga cgtccagcgt ctggctgaag ccaacgactt 2160
tcctcccgtc gatgaaacca aacgcggtcg ctatcagcaa atcaacctgc gtgacgctta 2220
cgttgatcac ctgttcggtt atatcaatgt caaaaacctc acgccgctca agctggtgat 2280
caactccggg aacggcgcag cgggtccggt ggtggacgcc attgaagccc gctttaaagc 2340
cctcggcgcg cccgtggaat taatcaaagt gcacaacacg ccggacggca atttccccaa 2400
cggtattcct aacccactac tgccggaatg ccgcgacgac acccgcaatg cggtcatcaa 2460
acacggcgcg gatatgggca ttgcttttga tggcgatttt gaccgctgtt tcctgtttga 2520
cgaaaaaggg cagtttattg agggctacta cattgtcggc ctgttggcag aagcattcct 2580
cgaaaaaaat cccggcgcga agatcatcca cgatccacgt ctctcctgga acaccgttga 2640
tgtggtgact gccgcaggtg gcacgccggt aatgtcgaaa accggacacg cctttattaa 2700
agaacgtatg cgcaaggaag acgccatcta tggtggcgaa atgagcgccc accattactt 2760
ccgtgatttc gcttactgcg acagcggcat gatcccgtgg ctgctggtcg ccgaactggt 2820
gtgcctgaaa gataaaacgc tgggcgaact ggtacgcgac cggatggcgg cgtttccggc 2880
aagcggtgag atcaacagca aactggcgca acccgttgag gcgattaacc gcgtggaaca 2940
gcattttagc cgtgaggcgc tggcggtgga tcgcaccgat ggcatcagca tgacctttgc 3000
cgactggcgc tttaacctgc gcacctccaa taccgaaccg gtggtgcgcc tgaatgtgga 3060
atcgcgcggt gatgtgccgc tgatggaagc gcgaacgcga actctgctga cgttgctgaa 3120
cgagtaaaaa cgcggccgcg atatcgttgt aaaacgacgg ccagtgcaag aatcataaaa 3180
aatttatttg ctttcaggaa aatttttctg tataatagat tcataaattt gagagaggag 3240
tttttgtgag cggataacaa ttccccatct tagtatatta gttaagtata aatacaccgc 3300
ggaggacgaa ggagatagaa ccatgtcaaa agtcgctctc atcaccggtg taaccggaca 3360
agacggttct tacctggcag agtttctgct ggaaaaaggt tacgaggtgc atggtattaa 3420
gcgtcgcgca tcgtcattca acaccgagcg cgtggatcac atttatcagg atccgcacac 3480
ctgcaacccg aaattccatc tgcattatgg cgacctgagt gatacctcta acctgacgcg 3540
cattttgcgt gaagtacagc cggatgaagt gtacaacctg ggcgcaatga gccacgttgc 3600
ggtctctttt gagtcaccag aatataccgc tgacgtcgac gcgatgggta cgctgcgcct 3660
gctggaggcg atccgcttcc tcggtctgga aaagaaaact cgtttctatc aggcttccac 3720
ctctgaactg tatggtctgg tgcaggaaat tccgcagaaa gagaccacgc cgttctaccc 3780
gcgatctccg tatgcggtcg ccaaactgta cgcctactgg atcaccgtta actaccgtga 3840
atcctacggc atgtacgcct gtaacggaat tctcttcaac catgaatccc cgcgccgcgg 3900
cgaaaccttc gttacccgca aaatcacccg cgcaatcgcc aacatcgccc aggggctgga 3960
gtcgtgcctg tacctcggca atatggattc cctgcgtgac tggggccacg ccaaagacta 4020
cgtaaaaatg cagtggatga tgctgcagca ggaacagccg gaagatttcg ttatcgcgac 4080
cggcgttcag tactccgtgc gtcagttcgt ggaaatggcg gcagcacagc tgggcatcaa 4140
actgcgcttt gaaggcacgg gcgttgaaga gaagggcatt gtggtttccg tcaccgggca 4200
tgacgcgccg ggcgttaaac cgggtgatgt gattatcgct gttgacccgc gttacttccg 4260
tccggctgaa gttgaaacgc tgctcggcga cccgaccaaa gcgcacgaaa aactgggctg 4320
gaaaccggaa atcaccctca gagagatggt gtctgaaatg gtggctaatg acctcgaagc 4380
ggcgaaaaaa cactctctgc tgaaatctca cggctacgac gtggcgatcg cgctggagtc 4440
ataagcatga gtaaacaacg agtttttatt gctggtcatc gcgggatggt cggttccgcc 4500
atcaggcggc agctcgaaca gcgcggtgat gtggaactgg tattacgcac ccgcgacgag 4560
ctgaacctgc tggacagccg cgccgtgcat gatttctttg ccagcgaacg tattgaccag 4620
gtctatctgg cggcggcgaa agtgggcggc attgttgcca acaacaccta tccggcggat 4680
ttcatctacc agaacatgat gattgagagc aacatcattc acgccgcgca tcagaacgac 4740
gtgaacaaac tgctgtttct cggatcgtcc tgcatctacc cgaaactggc aaaacagccg 4800
atggcagaaa gcgagttgtt gcagggcacg ctggagccga ctaacgagcc ttatgctatt 4860
gccaaaatcg ccgggatcaa actgtgcgaa tcatacaacc gccagtacgg acgcgattac 4920
cgctcagtca tgccgaccaa cctgtacggg ccacacgaca acttccaccc gagtaattcg 4980
catgtgatcc cagcattgct gcgtcgcttc cacgaggcga cggcacagaa tgcgccggac 5040
gtggtggtat ggggcagcgg tacaccgatg cgcgaatttc tgcacgtcga tgatatggcg 5100
gcggcgagca ttcatgtcat ggagctggcg catgaagtct ggctggagaa cacccagccg 5160
atgttgtcgc acattaacgt cggcacgggc gttgactgca ctatccgcga gctggcgcaa 5220
accatcgcca aagtggtggg ttacaaaggc cgggtggttt ttgatgccag caaaccggat 5280
ggcacgccgc gcaaactgct ggatgtgacg cgcctgcatc agcttggctg gtatcacgaa 5340
atctcactgg aagcggggct tgccagcact taccagtggt tccttgagaa tcaagaccgc 5400
tttcgggggg ggagctaacg cgccatttaa atcaacctca gcggtcatag ctgtttcctg 5460
tgactgagca ataactagca taaccccttg gggcctctaa acgggtcttg aggggttttt 5520
tgctgaaacc aatttgcctg gcggcagtag cgcggtggtc ccacctgacc ccatgccgaa 5580
ctcagaagtg aaacgccgta gcgccgatgg tagtgtgggg tctccccatg cgagagtagg 5640
gaactgccag gcatcaaata aaacgaaagg ctcagtcgaa agactgggcc tttcgggatc 5700
caggccggcc tgttaacgaa ttaatcttcc gcggcggtat cgataagctt gatatcgaat 5760
tccgaagttc ctattctcta gaaagtatag gaacttcagg tctgaagagg agtttacgtc 5820
cagccaagct agcttggctg caggtcgtcg aaattctacc gggtagggga ggcgcttttc 5880
ccaaggcagt ctggagcatg cgctttagca gccccgctgg gcacttggcg ctacacaagt 5940
ggcctctggc ctcgcacaca ttccacatcc accggtaggc gccaaccggc tccgttcttt 6000
ggtggcccct tcgcgccacc ttctactcct cccctagtca ggaagttccc ccccgccccg 6060
cagctcgcgt cgtgcaggac gtgacaaatg gaagtagcac gtctcactag tctcgtgcag 6120
atggacagca ccgctgagca atggaagcgg gtaggccttt ggggcagcgg ccaatagcag 6180
ctttgctcct tcgctttctg ggctcagagg ctgggaaggg gtgggtccgg gggcgggctc 6240
aggggcgggc tcaggggcgg ggcgggcgcc cgaaggtcct ccggaggccc ggcattctgc 6300
acgcttcaaa agcgcacgtc tgccgcgctg ttctcctctt cctcatctcc gggcctttcg 6360
acctgcagcc tgttgacaat taatcatcgg catagtatat cggcatagta taatacgaca 6420
aggtgaggaa ctaaaccatg ggtcaaagta gcgatgaagc caacgctccc gttgcagggc 6480
agtttgcgct tcccctgagt gccacctttg gcttagggga tcgcgtacgc aagaaatctg 6540
gtgccgcttg gcagggtcaa gtcgtcggtt ggtattgcac aaaactcact cctgaaggct 6600
atgcggtcga gtccgaatcc cacccaggct cagtgcaaat ttatcctgtg gctgcacttg 6660
aacgtgtggc ctaatgaggg gatcaattct ctagagctcg ctgatcagaa gttcctattc 6720
tctagaaagt ataggaactt c 6741
<210> 2
<211> 5160
<212> DNA
<213> Artificial Sequence
<220>
<223> <cscB-cscK-cscA-cscR>
<400> 2
gcgactgtac cagaacatga atgaggcgtt tggattaggc gattattagc agggctaagc 60
attttactat tattattttc cggttgaggg atatagagct atcgacaaca accggaaaaa 120
gtttacgtct atattgctga aggtacaggc gtttccataa ctatttgctc gcgtttttta 180
ctcaagaaga aaatgccaaa tagcaacatc aggcagacaa tacccgaaat tgcgaagaaa 240
actgtctggt agcctgcgtg gtcaaagagt atcccagtcg gcgttgaaag cagcacaatc 300
ccaagcgaac tggcaatttg aaaaccaatc agaaagatcg tcgacgacag gcgcttatca 360
aagtttgcca cgctgtattt gaagacggat atgacacaaa gtggaacctc aatggcatgt 420
aacaacttca ctaatgaaat aatccagggg ttaacgaaca gcgcgcagga aaggatacgc 480
aacgccataa tcacaactcc gataagtaat gcattttttg gccctacccg attcacaaag 540
aaaggaataa tcgccatgca cagcgcttcg agtaccacct ggaatgagtt gagataacca 600
tacaggcgcg ttcctacatc gtgtgattcg aataaacctg aataaaagac aggaaaaagt 660
tgttgatcaa aaatgttata gaaagaccac gtccccacaa taaatatgac gaaaacccag 720
aagtttcgat ccttgaaaac tgcgataaaa tcctcttttt ttacccctcc cgcatctgcc 780
gctacgcact ggtgatcctt atctttaaaa cgcatgttga tcatcataaa tacagcgcca 840
aatagcgaga ccaaccagaa gttgatatgg ggactgatac taaaaaatat gccggcaaag 900
aacgcgccaa tagcatagcc aaaagatccc caggcgcgcg ctgttccata ttcgaaatga 960
aaatttcgcg ccattttttc ggtgaagcta tcaagcaaac cgcatcccgc cagatacccc 1020
aagccaaaaa atagcgcccc cagaattaga cctacagaaa aattgctttg cagtaacggt 1080
tcataaacgt aaatcataaa cggtccggtc aagaccagga tgaaactcat acaccagatg 1140
agcggtttct tcagaccgag tttatcctga acgatgccgt agaacatcat aaatagaatg 1200
ctggtaaact ggttgaccga ataaagtgta cctaattccg tccctgtcaa ccctagatgt 1260
cctttcagcc aaatagcgta taacgaccac cacagcgacc aggaaataaa aaagagaaat 1320
gagtaactgg atgcaaaacg atagtacgca tttctgaatg gaatattcag tgccataatt 1380
acctgcctgt cgttaaaaaa ttcacgtcct atttagagat aagagcgact tcgccgttta 1440
cttctcacta ttccagttct tgtcgacatg gcagcgctgt cattgcccct ttcgccgtta 1500
ctgcaagcgc tccgcaacgt tgagcgagat cgataattcg tcgcatttct ctctcatctg 1560
tagataatcc cgtagaggac agacctgtga gtaacccggc aacgaacgca tctcccgccc 1620
ccgtgctatc gacacaattc acagacattc cagcaaaatg gtgaacttgt cctcgataac 1680
agaccaccac cccttctgca cctttagtca ccaacagcat ggcgatctca tactcttttg 1740
ccagggcgca tatatcctga tcgttctgtg tttttccact gataagtcgc cattcttctt 1800
ccgagagctt gacgacatcc gccagttgta gcgcctgccg caaacacaag cggagcaaat 1860
gctcgtcttg ccatagatct tcacgaatat taggatcgaa gctgacaaaa cctccggcat 1920
gccggatcgc cgtcatcgca gtaaatgcgc tggtacgcga aggctcggca gacaacgcaa 1980
ttgaacagag atgtaaccat tcgccatgtc gccagcaggg caagtctgtc gtctctaaaa 2040
aaagatcggc actggggcgg accataaacg taaatgaacg ttccccttga tcgttcagat 2100
cgacaagcac cgtggatgtc cggtgccatt catcttgctt cagatacgtg atatcgactc 2160
cctcagttag cagcgttctt tgcattaacg caccaaaagg atcatccccc acccgaccta 2220
taaacccact tgttccgcct aatctggcga ttcccaccgc aacgttagct ggcgcgccgc 2280
caggacaagg cagtaggcgc ccgtctgatt ctggcaagag atctacgacc gcatccccta 2340
aaacccatac tttggctgac atttttttcc cttaaattca tctgagttac gcatagtgat 2400
aaacctcttt ttcgcaaaat cgtcatggat ttactaaaac atgcatattc gatcacaaaa 2460
cgtcatagtt aacgttaaca tttgtgatat tcatcgcatt tatgaaagta agggacttta 2520
tttttataaa agttaacgtt aacaattcac caaatttgct taaccaggat gattaaaatg 2580
acgcaatctc gattgcatgc ggcgcaaaac gccctagcaa aacttcatga gcaccggggt 2640
aacactttct atccccattt tcacctcgcg cctcctgccg ggtggatgaa cgatccaaac 2700
ggcctgatct ggtttaacga tcgttatcac gcgttttatc aacatcatcc gatgagcgaa 2760
cactgggggc caatgcactg gggacatgcc accagcgacg atatgatcca ctggcagcat 2820
gagcctattg cgctagcgcc aggagacgat aatgacaaag acgggtgttt ttcaggtagt 2880
gctgtcgatg acaatggtgt cctctcactt atctacaccg gacacgtctg gctcgatggt 2940
gcaggtaatg acgatgcaat tcgcgaagta caatgtctgg ctaccagtcg ggatggtatt 3000
catttcgaga aacagggtgt gatcctcact ccaccagaag gaatcatgca cttccgcgat 3060
cctaaagtgt ggcgtgaagc cgacacatgg tggatggtag tcggggcgaa agatccaggc 3120
aacacggggc agatcctgct ttatcgcggc agttcgttgc gtgaatggac cttcgatcgc 3180
gtactggccc acgctgatgc gggtgaaagc tatatgtggg aatgtccgga ctttttcagc 3240
cttggcgatc agcattatct gatgttttcc ccgcagggaa tgaatgccga gggatacagt 3300
taccgaaatc gctttcaaag tggcgtaata cccggaatgt ggtcgccagg acgacttttt 3360
gcacaatccg ggcattttac tgaacttgat aacgggcatg acttttatgc accacaaagc 3420
tttttagcga aggatggtcg gcgtattgtt atcggctgga tggatatgtg ggaatcgcca 3480
atgccctcaa aacgtgaagg atgggcaggc tgcatgacgc tggcgcgcga gctatcagag 3540
agcaatggca aacttctaca acgcccggta cacgaagctg agtcgttacg ccagcagcat 3600
caatctgtct ctccccgcac aatcagcaat aaatatgttt tgcaggaaaa cgcgcaagca 3660
gttgagattc agttgcagtg ggcgctgaag aacagtgatg ccgaacatta cggattacag 3720
ctcggcactg gaatgcggct gtatattgat aaccaatctg agcgacttgt tttgtggcgg 3780
tattacccac acgagaattt agacggctac cgtagtattc ccctcccgca gcgtgacacg 3840
ctcgccctaa ggatatttat cgatacatca tccgtggaag tatttattaa cgacggggaa 3900
gcggtgatga gtagtcgaat ctatccgcag ccagaagaac gggaactgtc gctttatgcc 3960
tcccacggag tggctgtgct gcaacatgga gcactctggc tactgggtta acataatatc 4020
aggtggaaca acggatcaac agcgggcaag ggatccgcgt cactcttccc ccttcacgac 4080
cttcaataat atgcaatgca gcttcccgcc cgataatgtc atgtggaagc tgaattgtgg 4140
tcagcggcgg taaaaacaga tgcccgacgc caaccagatt atcaaagccc attacggcga 4200
catcctgcgg gattcgtacc cccttcgcca gaagaacctg ataagccaca aaggctgcgc 4260
gatcgttacc acatatcaga acatcaaaat ctggtttgcc cggtttgaag tgggcattga 4320
gtaaacttgc gagatcggtg tagtgatcat cacctgttgc catgtgaaat tgtttcacct 4380
cagccagatc tcgtccagca tcacgccagg cctgctcaaa tccctgccga cgataccctg 4440
ttgccaacgc actttccggt agccagaagc ataacggttg acgatagccc gccgcgagca 4500
aatgctgtgt tgattcatat tgtgcagtgt aatcatcagg gatataactg ggtaacgctg 4560
ggtcatccgc cacacagttc gccaatacaa tattttcacc atacagagac tcaggcagcg 4620
tgatatgtcg cagccccatt gtagtataga taatgccatc cggacggtgg gcaagcagct 4680
gacgtgccgc gcgggcagcg tcatcttcag aaaaaatatt gattaaaaaa ctattccagc 4740
cgaactcgct ggcggtttgc tcaatggcaa gcagaatatc aacagagaaa ggagtggtag 4800
ccgtgtcctg cgccagcacg gcgagagtcg acggcttacg tccttgagcg cgcatcttac 4860
gggcggaaag atcaggaaca taattcaggg tctggattgc ctgcaatacg cggtcacgcg 4920
ttgcaggacg cacagattct gcattatgca tcacccggga gactgtcatc atcgacactc 4980
ccgccaggcg tgcgacatcc tttaatgaag ccatacccaa gccgtttgcc gtaaaacggg 5040
cactgtagca gaaacagacg tcactggcga gatccaacgc cctatcacct gacacagcaa 5100
tacaataaaa aataacaata attcccggac aattgtcccc aattccgcct ctgttctcgc 5160
<210> 3
<211> 897
<212> DNA
<213> Artificial Sequence
<220>
<223> wbgL gene codon optimized
<400> 3
atgggcagca ttattcgtct gcagggtggt ctgggtaatc agctgtttca gtttagcttt 60
ggttatgccc tgagcaaaat taatggtaca ccgctgtatt tcgacattag ccattatgcc 120
gaaaacgatg atcatggtgg ttatcgtctg aataatctgc agattccgga agaatatctg 180
cagtattata ccccgaaaat taataatatt tataaactgc tggtgcgtgg cagccgtctg 240
tatccggata tttttctgtt tctgggcttt tgcaacgaat ttcatgccta tggctacgat 300
tttgaatata ttgcccagaa atggaaaagc aaaaaataca ttggctactg gcagagcgaa 360
cacttttttc ataaacatat tctggacctg aaagaatttt ttattccgaa aaatgtgagc 420
gaacaggcaa atctgctggc agcaaaaatt ctggaaagcc agagcagcct gagcattcat 480
attcgtcgtg gcgattatat taaaaacaaa accgcaaccc tgacacatgg tgtttgtagc 540
ctggaatatt ataaaaaagc cctgaacaaa atccgcgatc tggcaatgat tcgtgatgtg 600
tttatcttta gcgacgatat cttctggtgc aaagaaaata ttgaaaccct gctgagcaaa 660
aaatataata tttattatag cgaagatctg agccaagaag aggatctgtg gctgatgagc 720
ctggcaaatc atcatattat tgccaatagc agctttagtt ggtggggtgc atatctgggt 780
agcagcgcaa gccagattgt tatttatccg accccgtggt atgatattac cccgaaaaac 840
acctatatcc cgattgtgaa ccattggatc aacgttgata aacatagcag ctgctaa 897
<210> 4
<211> 903
<212> DNA
<213> Artificial Sequence
<220>
<223> futC codon optimized
<400> 4
atggccttta aagtggttca gatctgcggc ggtctgggca atcagatgtt tcaatatgcg 60
ttcgccaaat cactgcaaaa acattcgaac accccggttc tgctggatat cacgagtttt 120
gattggtccg accgtaaaat gcagctggaa ctgttcccga ttgatctgcc gtacgcaagc 180
gctaaagaaa tcgcgattgc caaaatgcag cacctgccga aactggtccg tgatgcactg 240
aaatgtatgg gctttgaccg cgtttcacaa gaaattgtct tcgaatatga accgaaactg 300
ctgaaaccgt cgcgtctgac ctatttcttt ggttactttc aggatccgcg ctacttcgac 360
gctatctctc cgctgattaa acaaaccttt acgctgccgc cgccgccgga aaacaacaaa 420
aacaacaaca aaaaagaaga agaatatcag tgcaaactga gtctgatcct ggcggccaaa 480
aattccgtct ttgtgcatat tcgtcgcggc gattacgtgg gcatcggttg tcagctgggt 540
attgactatc agaaaaaagc actggaatac atggctaaac gtgtgccgaa tatggaactg 600
tttgttttct gcgaagatct ggaatttacc cagaacctgg acctgggcta tccgttcatg 660
gatatgacca cgcgcgacaa agaagaagaa gcgtactggg atatgctgct gatgcagtca 720
tgtcaacatg gtattatcgc caatagcacg tattcttggt gggcagctta cctgattgaa 780
aacccggaaa aaattatcat tggcccgaaa cattggctgt ttggtcacga aaatatcctg 840
tgcaaagaat gggtcaaaat tgaaagccac ttcgaagtga aatctcagaa atataacgcg 900
taa 903
<210> 5
<211> 74
<212> DNA
<213> Artificial Sequence
<220>
<223> Primer 6128
<400> 5
ggggacaagt ttgtacaaaa aagcaggctt cgaaggagat acaacatggg cagcattatt 60
cgtctgcagg gtgg 74
<210> 6
<211> 60
<212> DNA
<213> Artificial Sequence
<220>
<223> Primer 6129
<400> 6
ggggaccact ttgtacaaga aagctgggtt tagcagctgc tatgtttatc aacgttgatc 60
<210> 7
<211> 75
<212> DNA
<213> Artificial Sequence
<220>
<223> Primer 6195
<400> 7
ggggacaagt ttgtacaaaa aagcaggctt cgaaggaggt agaacatggc ctttaaagtg 60
gttcagatct gcggc 75
<210> 8
<211> 64
<212> DNA
<213> Artificial Sequence
<220>
<223> Primer 6196
<400> 8
ggggaccact ttgtacaaga aagctgggtc ctattacgcg ttatatttct gagatttcac 60
ttcg 64
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| УЛУЧШЕННЫЙ СПОСОБ ПОЛУЧЕНИЯ ФУКОЗИЛИРОВАННЫХ ОЛИГОСАХАРИДОВ | 2017 |
|
RU2790445C2 |
| ФУКОЗИЛТРАНСФЕРАЗЫ И ИХ ПРИМЕНЕНИЕ ДЛЯ ПОЛУЧЕНИЯ ФУКОЗИЛИРОВАННЫХ ОЛИГОСАХАРИДОВ | 2018 |
|
RU2818835C2 |
| Ферментативная продукция углеводов микробными клетками с использованием смешанного сырья | 2020 |
|
RU2809122C2 |
| Ферментативное получение олигосахаридов посредством общей ферментации с использованием смешанного сырья | 2019 |
|
RU2801231C2 |
| ПРОДУКЦИЯ ФУКОЗИЛИРОВАННЫХ ОЛИГОСАХАРИДОВ В BACILLUS | 2020 |
|
RU2810729C2 |
| СИНТЕЗ ФУКОЗИЛИРОВАННЫХ СОЕДИНЕНИЙ | 2009 |
|
RU2584599C2 |
| ПРИМЕНЕНИЕ ГЛИКОЗИДАЗ В ПОЛУЧЕНИИ ОЛИГОСАХАРИДОВ | 2019 |
|
RU2810730C2 |
| СИНТЕЗ НМО | 2009 |
|
RU2517602C2 |
| НОВЫЕ ФУКОЗИЛТРАСФЕРАЗЫ И ИХ ПРИМЕНЕНИЯ | 2011 |
|
RU2628307C2 |
| ПРОДУКЦИЯ ГЛИКОПРОТЕИНОВ С МОДИФИЦИРОВАННЫМ ФУКОЗИЛИРОВАНИЕМ | 2008 |
|
RU2479629C2 |

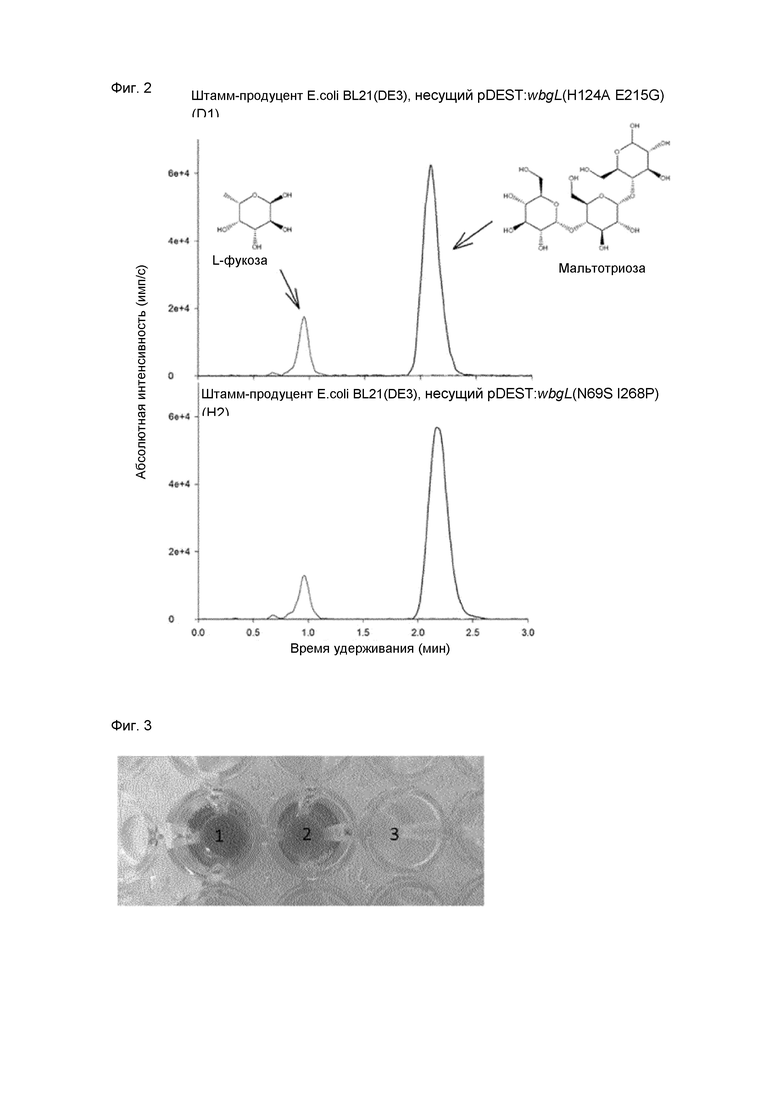







Группа изобретений относится к способу производства L-фукозы в свободной форме с использованием рекомбинантного микроорганизма и рекомбинантному микроорганизму для производства L-фукозы в свободной форме. Способ предусматривает культивирование рекомбинантного микроорганизма в среде, подходящей для роста микроорганизма, где этот микроорганизм не способен метаболизировать L-фукозу, так что L-фукоза накапливается во время стадии культивирования, и выделение свободной L-фукозы из среды. При этом рекомбинантный микроорганизм для синтеза L-фукозы содержит гетерологичный фермент, катализирующий гидролиз ГДФ-L-фукозы для высвобождения L-фукозы из ГДФ-L-фукозы, где указанный фермент представляет собой вариант альфа-1,2-фукозилтрансферазы, кодируемой геном wbgL из E. coli:O126, содержащий две аминокислотные замены, выбранные из группы, состоящей из (i) замен гистидина 124 на аланин и глутамата 215 на глицин, и (ii) замен аспарагина 69 на серин и изолейцина 268 на пролин. Группа изобретений обеспечивает эффективное получение целевого моносахарида. 2 н. и 7 з.п. ф-лы, 4 ил., 1 табл.
1. Способ производства L-фукозы в свободной форме с использованием рекомбинантного микроорганизма, при котором:
а) берут рекомбинантный микроорганизм для синтеза L-фукозы, содержащий гетерологичный фермент, катализирующий гидролиз ГДФ-L-фукозы для высвобождения L-фукозы из ГДФ-L-фукозы, где указанный фермент представляет собой вариант альфа-1,2-фукозилтрансферазы, кодируемой геном wbgL из E. coli:O126, где указанный вариант содержит две аминокислотные замены, где указанные две аминокислотные замены выбраны из группы, состоящей из (i) замен гистидина 124 на аланин и глутамата 215 на глицин, и (ii) замен аспарагина 69 на серин и изолейцина 268 на пролин, и
б) культивируют этот рекомбинантный микроорганизм в среде, подходящей для роста микроорганизма, где этот микроорганизм не способен метаболизировать L-фукозу, так что L-фукоза накапливается во время стадии культивирования, и
в) выделяют свободную L-фукозу из среды.
2. Способ согласно п. 1, отличающийся тем, что по меньшей мере один ген, вовлеченный в биосинтез ГДФ-L-фукозы, сверхэкспрессируется для улучшения снабжения ГДФ-L-фукозой.
3. Способ согласно п. 2, отличающийся тем, что по меньшей мере один ген является гетерологичным или гомологичным.
4. Способ согласно любому из предшествующих пунктов, отличающийся тем, что микроорганизм культивируют в среде, содержащей источник углерода, который выбран из группы, состоящей из глицерина, сахарозы, глюкозы, фруктозы, мелассы, ксилозы, целлюлозы, синтетического газа, кукурузного сиропа и лактозы.
5. Способ согласно любому из пп. 1-4, отличающийся тем, что микроорганизм-хозяин представляет собой штамм Escherichia coli, Corynebacterium spp., Clostridium spp., Bacillus spp., Pseudomonas spp., Lactobacillus spp. или Saccharomyces sp.
6. Рекомбинантный микроорганизм для производства L-фукозы, содержащий гетерологичный фермент, катализирующий гидролиз ГДФ-L-фукозы и высвобождать L-фукозу из ГДФ-L-фукозы, где указанный фермент представляет собой вариант альфа-1,2-фукозилтрансферазы, кодируемой геном wbgL из E. coli:O126, где указанный вариант содержит две аминокислотные замены, где указанные две аминокислотные замены выбраны из группы, состоящей из (i) замен гистидина 124 на аланин и глутамата 215 на глицин, и (ii) замен аспарагина 69 на серин и изолейцина 268 на пролин, и где этот микроорганизм не способен метаболизировать L-фукозу.
7. Рекомбинантный микроорганизм согласно п. 6, отличающийся тем, что последовательность нуклеиновой кислоты, кодирующей указанный гетерологичный фермент, интегрирована в геном указанного микроорганизма.
8. Рекомбинантный микроорганизм согласно п. 6 или 7, отличающийся тем, что он дополнительно модифицирован, чтобы содержать гетерологичные гены, кодирующие фосфоманномутазу, маннозо-1-фосфат-гуанозилтрансферазу, ГДФ-маннозо-4,6-дегидратазу и ГДФ-L-фукозосинтазу.
9. Рекомбинантный микроорганизм согласно любому из пп. 6-8, отличающийся тем, что микроорганизм представляет собой штамм Escherichia coli, Corynebacterium spp., Clostridium spp., Bacillus spp., Pseudomonas spp., Lactobacillus spp. или Saccharomyces sp.
| WO 2014067696 A1, 08.05.2014 | |||
| STEIN D | |||
| ET AL | |||
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| Adv | |||
| Synth | |||
| Catal | |||
| Станок для изготовления деревянных ниточных катушек из цилиндрических, снабженных осевым отверстием, заготовок | 1923 |
|
SU2008A1 |
| Опускной разборный снаряд для подводного наращивания свай | 1924 |
|
SU2313A1 |
| PETTIT N | |||
| ET AL | |||
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |

