Изобретение относится к биотехнологии, а именно к получению гибридного рекомбинантного белка F-I наружной мембраны Pseudomonas aeruginosa (далее OprF-OprI) для медицинских целей, а также к рекомбинантным штаммам Escherichia coli (далее Е. coli) и плазмидам для его получения.
Pseudomonas aeruginosa (далее Р. aeruginosa) является одним из основных возбудителей гнойно-воспалительных заболеваний человека, в частности вызывает госпитальные инфекции. Широкое распространение синегнойная инфекция получила благодаря неприхотливости в отношении питательных веществ и условий внешней среды, а также высокой резистентности к химиотерапевтическим средствам и антибиотикам.
Активная иммунизация может быть одним из эффективных средств защиты против условно патогенных микроорганизмов и, в частности, Р. aeruginosa. Для индукции противосинегнойного иммунитета в нашей стране разрабатывались препараты на основе инактивированных бактерий, их компонентов и экзопродуктов:
поливалентная синегнойная корпускулярная вакцина [1-3], пиоиммуноген [4-5], псевдомонас-вакцина [6-9]. По разным причинам в настоящее время для использования в клинической практике не зарегистрировано ни одного препарата, включающего поверхностные антигены синегнойной палочки.
Для иммунопрофилактики и иммунотерапии синегнойных инфекций в НИИ вакцин и сывороток им.Мечникова г.Уфы разработан препарат на основе экзотоксина А Р. aeruginosa - анатоксин синегнойной палочки [10-11]. Но выпуск данного препарата приостановлен в связи с коммерческими трудностями.
Трудности создания эффективных иммунопрепаратов против синегнойной палочки заключаются в том, что патогенез развивающихся с ее участием инфекций обусловлен воздействием целого ряда факторов патогенности, среди которых выделить наиболее значимый для разработки вакцинных препаратов представляется сложной задачей. Во время инфицирования, когда происходит проникновение возбудителя в ткани макроорганизма, первичные иммунные реакции формируются в основном на поверхностные антигены бактериальной клетки (компоненты мембраны и клеточной стенки). При исследовании поверхностных белков выявлено, что наибольшей иммуногенностью обладают некоторые белки наружной мембраны (outer protein - Opr) Р. aeruginosa, которые отвечают за межклеточный транспорт и сохранение структуры бактериальной мембраны. Наиболее иммунологически исследованы белки F и I наружной мембраны (OprF и OprI). Первый белок с молекулярной массой 37,6 кДа формирует поры в плазматической мембране, а второй с молекулярной массой 8,8 кДа, являющийся липопротеином, несет структурную функцию. Было показано, что эти белки обладают наиболее высокой иммуногенностью. Эти данные обусловили исследования по разработке вакцин на основе очищенных мембранных белков [12]. Однако получение подобных препаратов является проблематичным, поскольку содержание белков наружной мембраны незначительно относительно суммарных белков бактериальной клетки.
В последнее время при создании вакцинных препаратов все большое внимание уделяется использованию генно-инженерных методов, суть которых заключается во встраивании генов, отвечающих за синтез отдельных протективных антигенов в возбудителях, в геном непатогенного микроорганизма (например, Е. coli или Saccharomyces cerevisiae). Во время культивирования полученных продуцентов в большом количестве синтезируются целевые продукты (рекомбинантные белки), которые подвергаются простой очистке и могут использоваться для приготовления вакцины. Такая стратегия позволяет вычленить и использовать наиболее иммуногенные компоненты бактерии. При этом процесс производства вакцин исключает контакт с культурами патогенных микроорганизмов, что является существенным технологическим преимуществом, обеспечивающим безопасность персонала.
Технической задачей изобретения является получение рекомбинантной плазмидной ДНК pPA-OPRFI, кодирующей белки OprF и OprI P. aeruginosa и штамма-продуцента гибридного рекомбинантного белка F-I P. aeruginosa, с использованием штамма Е. coli К-12.
Указанная задача решается созданием рекомбинантной плазмидной ДНК pPA-OPRFI и штамма Е. coli PA-OPRFI.
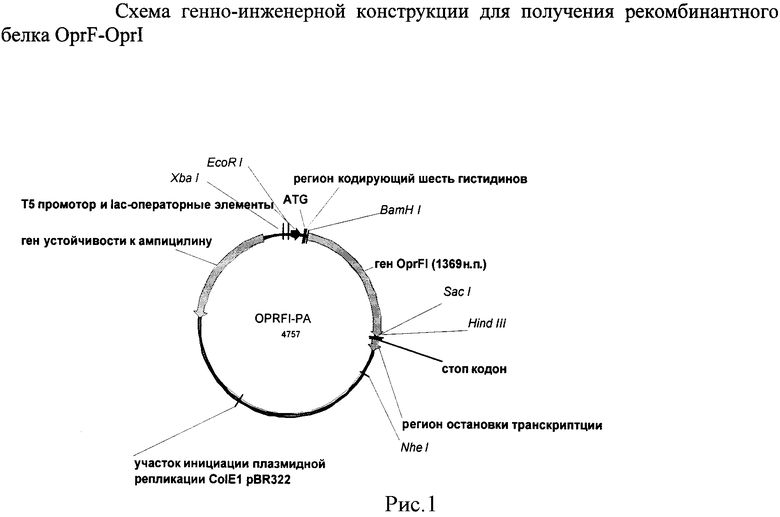
Плазмида pPA-OPRFI имеет 4757 пар оснований, характеризуется наличием следующих фрагментов:
Xba I - EcoR I фрагмента ДНК размером 87 пар оснований, содержащего последовательность модифицированного промотора бактериофага Т5 и двух лактозных оперонов.
EcoR I - BamH I фрагмента ДНК размером 57 пар оснований, содержащего сайт посадки рибосомы, стартовый ATG-кодон и последовательность, кодирующую шесть гистидинов (Arg-Gly-Ser-His-His-His-His-His-His-Gly-Ser).
BamH I - Sad фрагмента ДНК размером 1312 пар оснований, содержащего слитые полные последовательности генов oprF и oprI P. aeruginosa.
EcoR I - Sac I фрагмента ДНК размером 1369 пар оснований, содержащего сайт посадки рибосомы и последовательность, кодирующую OprF-OprI (состоит из гистидиновой последовательности, белка OprF и белка OprI).
Hind III - NhE I фрагмента ДНК размером 1116 пар оснований, содержащего лямбда t0 регион остановки транскрипции.
Плазмида pPA-OPRFI также включает в своем составе регион остановки транскрипции rrnBT1, сайт инициации репликации плазмиды ColEl и ген, кодирующий устойчивость к ампициллину.
На рисунке 1 представлена схема плазмиды pPA-OPRFI.
На рисунке 2 приведена аминокислотная последовательность OprF-OprI, закодированная в плазмиде pPA-OPRFI.
Штамм Е. coli PA-OPRFI получен трансформацией клеток родительского штамма Е. coli К-12, содержащих плазмиду pREP4, с использованием традиционной генно-инженерной технологии.
Штамм Е. coli PA-OPRFI характеризуется следующими признаками:
Культурально-морфологические признаки.
Клетки мелкие, прямые, утолщенной палочковидной формы, грамотрицательные, неспороносные.
Клетки хорошо растут на простых питательных средах. При росте на агару "Дифко" колонии круглые, гладкие, выпуклые, мутные, блестящие, серые, края ровные. При росте в жидких средах (на минимальной среде с глюкозой или LB-бульоне) образуют интенсивную ровную муть.
Физико-биологические признаки.
Аэроб. Температурный диапазон роста 4-42°С при оптимуме pH 6,5-7,5.
В качестве источника азота используют как минеральные соли в аммонийной и нитратной формах, так и органические соединения в виде аминокислот, пептона, триптона, дрожжевого экстракта и т.д.
В качестве источника углерода используют аминокислоты, глицерин, углеводы.
Устойчивость к антибиотикам. Клетки проявляют устойчивость к ампицилину (до 200 мкг/мл) и к канамицину (до 100 мкг/мл).
Штамм Е. coli PA-OPRFI - продуцент OprF-OprI.
Способ, условия и состав среды для хранения штамма:
L-бульон с 10%-ным глицерином и антибиотиками при -70°С в ампулах и в лиофилизированном состоянии в ампулах при 4°С.
Штамм Е. coli PA-OPRFI идентифицирован по Определителю Берги (1974) как штамм вида Е. coli.
Особенностью заявляемого способа является разработка технологии, позволяющей получать высокоочищенный белок OprF-OprI, состоящий из полноразмерных высокоиммуногенных аминокислотных последовательностей мембранных белков OprF и OprI P. aeruginosa.
Способ включает следующие этапы:
Культивирование в питательной среде штамма Е. coli PA-OPRFI;
Осаждение биомассы центрифугированием;
Разрушение клеток микроорганизма при растворении их в денатурирующем буфере, содержащем 8 М мочевину.
Очистка с помощью аффинной хроматографии на никель-активированных сорбентах;
Ренатурация очищенного OprF-OprI с помощью диализа.
Оценка протективных свойств очищенного OprF-OprI.
Оптимальными условиями проведения отдельных стадий получения рекомбинантного OprF-OprI являются следующие:
Разрушение клеток осуществляют в результате растворения биомассы в 8 М буферном растворе мочевины;
Удаление нерастворимых компонентов клетки осуществляют центрифугированием;
Ренатурацию проводят при температуре 4°С путем ступенчатого диализа против 50 мМ буферного раствора Tris-HCl (pH 9,0).
Выход рекомбинантного OprF-OprI в результате применения способа в оптимальном режиме составляет не менее 40 мг очищенного OprF с 1 л культуральной среды, уровень экспрессии OprF-OprI - не менее 100 мг с 1 л культуральной среды.
Сущность и преимущества заявляемой группы изобретений иллюстрируются следующими примерами.
Пример 1. Получение рекомбинантной плазмиды pPA-OPRFI.
Технология получения плазмиды pPA-OPRFI включает следующие этапы:
Выделение геномной ДНК,
Амплификация генов OprF и oprIP. aeruginosa с помощью ПЦР,
Встраивание амплифицированных последовательностей в плазмидный вектор.
Для выделения геномной ДНК можно использовать любой штамм синегнойной палочки.
Культуру клеток Р. aeruginosa хранили в лиофилизированном состоянии. Для получения живой культуры ампулу вскрывали и добавляли 1 мл бульона Мюллер-Хинтона (кислотный гидролизат казеина - 17,5 г/л, вытяжка из говядины - 300,0 г/л, растворимый крахмал - 1,5 г/л). Суспензию переносили в пробирку объемом 10 мл и инкубировали 4 часа при 37°С. Затем с использованием микробиологической петли инокулировали штрихом пробирки со скошенной агаризованной (1,5% бактоагара) средой (бульон Мюллер-Хинтона). Пробирки с культурой инкубировали в течение 16 часов при 37°С.
Полученную биомассу аккуратно снимали микробиологической петлей с поверхности агаризованной среды и переносили в микроцентрифужную пробирку. Бактериальную биомассу растворяли в 10 мМ буферного раствора Tris-HCl (pH 8,0). Затем добавляли раствор (10%) саркозила до конечной концентрации 0,5% и раствор (20 мг в мл) протеиназы К до конечной концентрации 20 мкг в мл. Смесь инкубировали 1 час при температуре +50°С с помешиванием.
Для осаждения ДНК из раствора добавляли 2,5 объема этанола и 1/10 объема 3 М раствора ацетата натрия. Пробирку инкубировали при температуре -70°С 15 минут, а затем центрифугировали 10 минут при 16000 g и температуре +4°С. Супернатант удаляли и тщательно промывали осадок 70%-ным спиртом. После промывки осадок высушивали на воздухе.
После высыхания на воздухе осадок растворял в ТЕ-буфере (10 мМ Tris-HCl, 1 мМ EDTA, pH 8,0).
При ПЦР каждого гена использовали пару праймеров, один (прямой) из которых соответствовал началу, а второй (обратный) был комплиментарен концу гена.
Прямой праймер для oprF имел следующую структуру (5'-3'): AAG GAT CCA TGA ААС TGA AGA АСА ССТ TAG, а обратный: ААА GAT CTT TTG GCT ТСА GCT ТСТ АС. Прямой праймер содержал на 5'-конце сайт рестрикции ВатН I, а обратный сайт рестрикции Bgl II.
Прямой праймер для opr1 имел следующую структуру (5'-3'): АА GGA ТСС ААС ААС GTT CTG ААА ТТС ТС, а обратный TTG AGC ТСТ ТАС TTG CGG CTG GCT ТТТ ТСС. Прямой праймер содержал на 5'-конце сайт рестрикции BamH I, a обратный сайт рестрикции Sac I.
ПЦР проводили при следующих реакционных условиях: 50 mM KCl, 10 тМ Tris-HCl, 0,1% Triton X-100, 1,5 mM MgCl, 0,120 mM dNTP, (pH 9,0). На 50 мкл реакционной смеси использовали 3 ед. TAQ-полимеразы (Fermentas) и 0,1 мкг геномной ДНК.
Реакцию проводили в амплификаторе "Терцик". Процесс амплификации гена oprF состоял из следующих стадий: прогревания при 94°С в течении 3 мин и 30-ти циклов ПЦР (94°С - 1 мин, 30 сек 65°С - 1 мин, 72°С - 1 мин) и инкубации при 72°С в течение 2 мин. Процесс амплификации гена oprI состоял из следующих стадий: прогревания при 94°С в течение 3 мин и 30-ти циклов ПЦР (94°С - 1 мин, 30 сек 65°С - 1 мин, 72°С - 20 сек) и инкубации при 72°С в течение 40 сек.
В первом случае полученный фрагмент ДНК имел размер около 1 kb, а во втором около 0,25 kb. Первый амплификат обработали рестриктазами BamH I и Bgl II, а второй BamH I и Sac I. Параллельно рестриктазами BamH I и Sac I был обработан вектор pQE-30 (QIAGEN).
Рестрикты очистили в агарозном геле и объединили в одну конструкцию при реакции лигирования. При отборе рекомбинантных клонов, который проводился с использованием рестриктного анализа и секвенирования, отбирали образцы с конструкциями, представляющие собой ген oprF, слитый с геном oprI, расположенный в прямой ориентации в векторной плазмиде. В результате была получена рекомбинантная плазмида pPA-OPRFI размером 4757 пар оснований, в которой ДНК слитого гена фланкирована сайтами рестрикции BamH I и Sac I (рис.1).
Пример 2. Получение штамма E.coli PA-OPRFI - продуцента OprF-OprI.
Штамм-продуцент E.coli PA-OPRFI получали путем трансформации клеток родительского штамма E.coli K12, содержащего плазмиду pREP4, рекомбинантной плазмидой pPA-OPRFI с последующим отбором рекомбинантных клонов на среде с ампициллином и канамицином при 37°С.
Первичный отбор рекомбинантных плазмид проводили с использованием рестриктного анализа и секвенирования.
Далее отбор продуцентов проводился по способности контролируемого синтеза рекомбинантного белка в клетках. Для этого инокулировали бактериальной колонией 3 мл среды LB, содержащей ампициллин и канамицин в концентрациях 100 и 50 мкг в мл соответственно. Выращивали в термостатированном шейкере со скоростью орбитального вращения 250 об/мин в течение 12-16 часов и температуре 37°С. Затем в 5 мл свежей среды вносили 100 мкл выросшей культуры и инкубировали в прежних условиях в течение 2-3 часов до достижения культурой активной фазы роста (0,6 оптических единиц при длине волны 600 нм). Добавляли 5 мкл 1 М водного раствора химического индуктора изопропил-β-d-тиогалактопиранозид (ИПТГ). Инкубацию продолжали в прежних условиях 4 часа, после чего осаждали полученную биомассу.
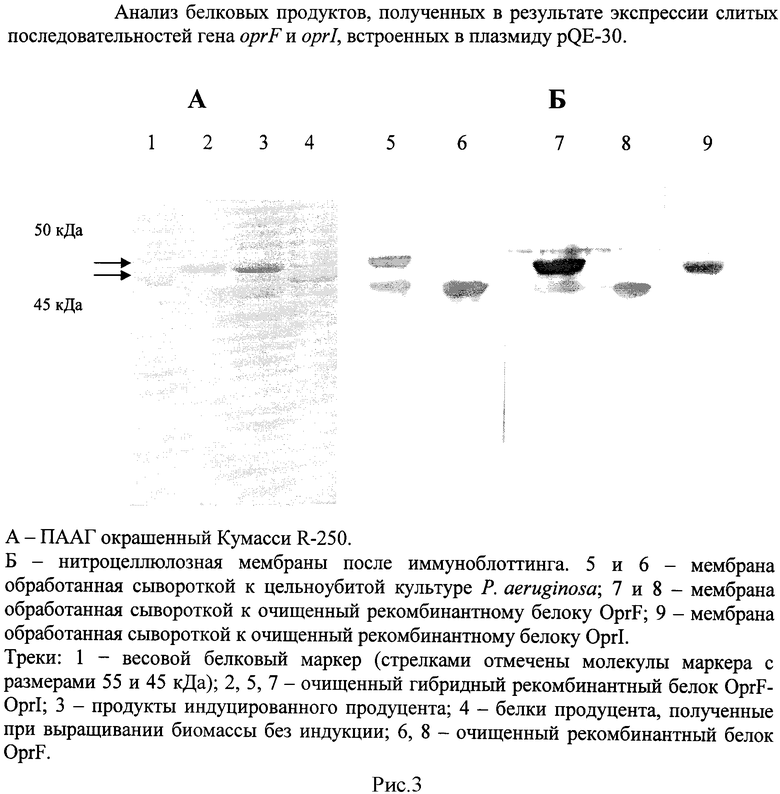
В качестве контроля использовали бактериальную культуру продуцента, инкубированную при тех же условиях, но без добавления ИПТГ. Белковые продукты анализировали электрофорезом в полиакриламидном геле (ПААГ) по методу Лэммли. (рис.3).
При анализе биомассы, полученной после индукции экспрессии рекомбинантного гена, выявлено наличие специфичного продукта, который отсутствовал в клетках продуцента, выращенного без добавления ИПТГ. Молекулярная масса синтезированного рекомбинантного продукта составила между 45 и 55 кДа. Расчетная масса для рекомбинантного OprF-OprI соответствовала 47,9 кДа. По результатам электрофореза установлено, что количество рекомбинантного белка составляло не менее одной трети относительно всех клеточных белков.
Рекомбинантные белки проявили высокую специфичность при взаимодействии с поликлональной гипериммунной кроличьей сывороткой к Р. aeruginosa и с поликлональными гипериммунным кроличьими сыворотками к очищенным рекомбинантным белкам OprF и OprI (рис.3).
Пример 3. Получение OprF-OprI из штамма Е. coli PA-OPRFI.
Получение OprF-OprI проводили в 4 этапа:
1 этап. Культивирование штамма E.coli PA-OPRFI с индукцией экспрессии рекомбинантного гена.
2 этап. Получения клеточного лизата штамма E.coli PA-OPRFI после экспрессии рекомбинантного гена.
3 этап. Хроматографическая очистка OprF-OprI.
4 этап. Растворение и ренатурация OprF-OprI.
Культивирование штамма E.coli PA-OPRFI с индукцией экспрессии рекомбинантного гена.
Выращивали посевной материал штамма E.coli PA-OPRFI в 500 мл среды LB, содержащей ампициллин и канамицин в концентрациях 100 и 50 мкг в мл соответственно. Культивирование проводили течение 12 ч при температуре 37°С в термостатированном шейкере со скоростью орбитального вращения 250 об/мин.
Полученную культуру асептически вносили в лабораторный ферментер с 10 л стерильной среды LB, содержащей ампициллин и канамицин в концентрациях 100 и 50 мкг в мл соответственно. Выращивание в ферментере проводили при температуре 37°С, концентрации растворенного кислорода 30%, оборотов мешалки 300 об/мин.
При достижении плотности культуры 0,6 оптических единиц при длине волны 600 нм в ферментер вносили 10 мл 1 М водного раствора ИПТГ. Инкубацию продолжали в прежних условиях 4 часа.
Получения клеточного лизата штамма E.coli PA-OPRFI после экспрессии рекомбинантного гена.
Биомассу осаждали в центрифуге Beckman J2-21 (ротор JA-14) со скоростью вращения 3000 об/мин в течение 10 мин. Осадок собирали и растворяли в одном литре следующего раствора: 8 М мочевина; 0,1 М NaH2PO4; 0,01 М Tris-HCl; pH - 8,0. Растворение биомассы проводили в орбитальном шейкере при вращении 250 об/мин в течение 12 ч при комнатной температуре.
Затем лизат освобождали от нерастворенных фрагментов клетки в центрифуге Beckman J2-21 (ротор JA-14) со скоростью вращения 15000 об/мин при комнатной температуре в течение часа. Супернатант переносили в чистую стерильную колбу.
Хроматографическая очистка OprF-OprI.
К осветленному лизату добавляли к нему 20 мл суспензии Ni-агарозы (QIAGEN) и инкубировали 1 час на комнатной температуре в орбитальном шейкере при легком покачивании (40 об/мин).
Полученную суспензию пятью этапами пропускали через колонку для хроматографии (Bio-Rad) объемом 250 мл и диаметром 5 см. В результате из колонки удалялась жидкая фаза лизата и оставался сорбент со связанным рекомбинантным белком.
Далее через колонку пропускали два литра промывочного раствора следующего состава: 8 М мочевина; 0,1 М NaH2PO4; 0,01 М Tris-HCl; pH - 6,3.
Для эллюции рекомбинантного белка через колонку пропускали десять раз по 15 мл первого эллюционного раствора и десять раз по 15 мл второго эллюционного раствора, собирая каждую фракцию в отдельную пробирку. Растворы для эллюции имели тот же состав, что и раствор для промывки, только первый эллюционный раствор имел pH - 5,9, а второй - 4,5.
С аликвотами от собранных фракций проводили электрофорез в полиакриламидном геле. После анализа результатов электрофореза отбирали фракции, содержащие очищенный белок.
Растворение и ренатурация OprF-OprI.
Очищенный рекомбинантный белок растворяли в 50 мМ Tris-HCl (pH 9,0) с помощью ступенчатого диализа при температуре 4°С и перемешивании буфера на магнитной мешалке. Для этого фракции, содержащие очищенный белок, объединяли и переносили в диализный мешок.
Завязанный диализный мешок помещали в емкость, содержащую 1 литр следующего раствора: 6 М мочевина; 50 мМ Tris-HCl; pH - 9,0. Через два часа поочередно меняли содержимое на буферные растворы с содержанием 4, 2 и 1 М мочевины.
Затем в емкость для диализа наливали чистый буферный раствор (50 мМ Tris-HCl, pH 9,0). Его трижды меняли каждые два часа.
Содержание белков в препарате определяли спектрофотометрически при длине волны 280 нм. При расчете концентрации рекомбинантных белков использовали коэффициент экстинкции 0,59, рассчитанный в программе OMIGA.
Препараты очищенного OprF-OprI хранили в аликвотах по 5 мл в стерильных пробирках при -70°С.
Пример 4. Протективные свойства OprF-OprI.
Протективные свойства очищенного OprF-OprI оценивали на мышиной модели.
Для проведения иммунизации к препарату очищенного рекомбинантного белка добавляли гель гидроокиси алюминия (ГУП "Иммунопрепарат") из расчета: 1 мг белка - 3 мг гидроокиси алюминия. Концентрацию белка в препарате доводили до требуемого значения фосфатно-солевым буфером. Сорбцию проводили в течение суток при 4-7°С.
В качестве модельных животных использовали белых беспородных мышей массой 18-22 г. Иммунизационная доза составила 50 мкг белка на одно животное. Объем вводимого препарата составлял 0,5 мл. Препараты животным вводили внутрибрюшинно. Использована схема двукратной иммунизации с двухнедельным интервалом. В качестве контроля культуры были использованы неиммунизированные мыши той же партии. Кроме того, в качестве препарата сравнения использовался рекомбинантный белок OprF P. aeruginosa.
Через две недели после последней иммунизации для индукции экспериментальной инфекции мышей внутрибрюшинно заражали в объеме 0,5 мл различными дозами (с двукратным шагом) живой вирулентной культуры Р. aeruginosa (штамм РА-170015), выращенной на агаризованной среде Хоттингера. Иммунизированных мышей заражали следующими дозами: 50, 100, 200, 400 и 800 млн микробных клеток (м.к.)/0,5 мл. Заражение контрольных мышей осуществляли в более низких дозах: 25, 50, 100, 200 млн м.к./0,5 мл. Подсчет погибших и выживших животных проводили в течение пяти дней с последующим определением ЛД50, которую вычисляли по формуле Кербера в модификации Ашмарина-Воробьева: ЛД50=lgA-lg2 (B1/C1+…+Bn/Cn)-0,5, где А - максимальная инфекционная доза в опыте, В - количество животных, павших в группе, С - первоначальное количество животных в группе, n - количество групп (табл. 1).
Рекомбинантные белки в результате иммунизации защищали мышей от экспериментальной внутрибрюшинной инфекции. При этом гибридный рекомбинантный белок (OprF-OprI) показал лучшие результаты. Индекс эффективности (ИЭ) защитных свойств рекомбинантного белка OprF-OprI составил 4,9, что оказалось в полтора раза выше аналогичного результата для белка OprF (ИЭ 3,0).
Источники информации
1. Мороз А.Ф., Константинов Б.А., Анциферова Н.Г., Радкевич С.А., Ковырялкина О.В. Применение поливалентной синегнойной вакцины для профилактики послеоперационных гнойно-септических осложнений в кардиохирургии // Современная госпитальная инфекция. Тезисы докладов. Л. - 1980. - С.92.
2. Мороз А.Ф. Применение поливалентной вакцины с защитным эффектом против Pseudomonas aeruginosa // I Всесоюзная конференция по ранам и раневой инфекции. М. - 1977. - С.73-74.
3. Титова Т.И., Сидорова Т.Н., Радкевич С.А. и др. Получение и изучение свойств поливалентной корпускулярной синегнойной вакцины // Журн. микробиол. - 1985. - №8. - С.80-84
4. Станиславский Е.С., Joo I. Вакцины против Pseudomonas aeruginosa инфекции: итоги и перспективы исследований // Бактериальные антигены. Сборник трудов - Москва. - 1982. - С.3-14.
5. Станиславский Е.С., Joo I., Северцева М.К. и др. Иммунологическая эффективность и безвредность в эксперименте пиоиммуногена-вакцины против инфекции Pseudomonas aeruginosa // Журн. микробиол. - 1982. - №5. - С.70-75
6. Бандман О.А., Едвабная Л.С., Булк В.Ф. и др. Бесклеточная вакцина псевдомонас: иммунохимическая характеристика и иммуногенность // Журн. микробиол. - 1987. - №11. - С.44-47.
7. Едвабная Л.С., Зайднер И.Г., Макаренко Т.А. и др. Белковые протективные антигены Pseudomonas aeruginosa // Журн. микробиол. - 1985. - №11. - С.18-23.
8. Макаренко Т.А., Балаян С.С., Сергиенко А.И. и др. Иммунологический ответ у доноров-добровольцев, иммунизированных вакциной псевдомонас // Журн. микробиол. - 1991. - №11. - С.39-41.
9. Макаренко Т.А., Станиславский Е.С. Иммунологическое изучение белков клеточной стенки Pseudomonas aeruginosa // Журн. микробиол. - 1996. - №2. - С.7-9.
10. Пат. 1410527 С СССР, С12Р 1/04. Способ получения экзотоксина А синегнойной палочки // Н.А.Михайлова, Т.Н.Кузнецова, Ф.А.Шаймухаметов, А.Ф.Мороз, И.А.Баснакьян. - Заявл. 10.09.86; Опубл. 09.06.95 // БИПМ. - 1995.
11. Пат. 1481962 С СССР, А61К 35/74. Способ обезвреживания очищенного экзотоксина А синегнойной палочки // Н.А.Михайлова, Ф.А.Шаймухаметов, Т.Н.Кузнецова, А.Ф.Мороз. - Заявл. 15.07.87; Опубл. 09.06.95 // БИПМ. - 1995.
12. Lee NG, Jung SB, Ahn BY, Kim YH, Kim JJ, Kim DK, Kim IS, Yoon SM, Nam SW, Kim HS, Park WJ. Immunization of bum-patients with a Pseudomonas aeruginosa outer membrane protein vaccine elicits antibodies with protective efficacy // Vaccine - 2000 - №18 - Р.1952-1961.

Изобретение относится к области биотехнологии и касается рекомбинантной плазмидной ДНК pPA-OPRFI, кодирующей гибридный рекомбинантный белок F-I наружной мембраны Pseudomonas aeruginosa, штамма бактерий E.coli PA-OPRFI, продуцента такого гибридного белка и способа получения указанного рекомбинантного белка. Представленная плазмидная ДНК содержит фрагмент ДНК, содержащий последовательность модифицированного промотора бактериофага Т5 и двух лактозных оперонов; фрагмент ДНК, содержащий сайт посадки рибосомы, стартовый ATG-кодон и последовательность, кодирующую шесть гистидинов; фрагмент ДНК, содержащий полноразмерные последовательности генов oprF и opri P.aeruginosa; фрагмент ДНК, содержащий сайт посадки рибосомы, и фрагмент ДНК, содержащий лямбда t0 регион остановки транскрипции. Представленные изобретения позволяют получать гибридный рекомбинантный белок F-I наружной мембраны P.aeruginosa для его иммунобиологического тестирования при разработке вакцины против синегнойной палочки, а также для получения донорских иммуноглобулинов, предназначенных для терапии активных форм синегнойной инфекции. 3 н.п. ф-лы, 3 ил., 1 табл., 4 пр.
1. Рекомбинантная плазмидная ДНК pPA-OPRFI, кодирующая гибридный рекомбинантный белок F-I наружной мембраны Р.aeruginosa размером 4757 пар оснований, содержащая: Xba I - EcoR I фрагмент ДНК размером 87 пар оснований, содержащий последовательность модифицированного промотора бактериофага Т5 и двух лактозных оперонов; EcoR I - BamH I фрагмент ДНК размером 57 пар оснований, содержащий сайт посадки рибосомы, стартовый ATG-кодон и последовательность, кодирующую шесть гистидинов (Arg-Gly-Ser-His-His-His-His-His-His-Gly-Ser); BamH I - Sac I фрагмент ДНК размером 1312 пар оснований, содержащий полноразмерные последовательности генов oprF и opri P.aeruginosa, полученного с помощью полимеразной цепной реакции (ПЦР); EcoR I - Sac I фрагмент ДНК размером 1369 пар оснований, содержащий сайт посадки рибосомы и последовательность, кодирующую белки OprF и OprI (состоит из гистидиновой последовательности и собственно белков OprF и OprI); Hind III-NhE I фрагмент ДНК размером 1116 пар оснований, содержащий лямбда t0 регион остановки транскрипции.
2. Штамм бактерий Е.coli PA-OPRFI - продуцент гибридного рекомбинантного белка F-I наружной мембраны Р.aeruginosa, полученный путем трансформации родительского штамма бактерий Escherichia coli К12 рекомбинантной плазмидной ДНК pPA-OPRFI по п.1.
3. Способ получения гибридного рекомбинантного белка F-I наружной мембраны Р.aeruginosa с использованием трансформированного методами генетической инженерии штамма бактерий Е.coli PA-OPRFI по п.2, включающий его культивирование в питательной среде, разрушение клеток микроорганизма, очистку полученного продукта с использованием аффинной хроматографии на никель-активированных сорбентах и растворением с ренатурацией очищенного препарата в нативных условиях при помощи ступенчатого диализа.
| US 0006300102 B1, 09.10.2001 | |||
| Полимерная композиция для тепло- и гидроизоляции теплопроводов | 1976 |
|
SU717106A1 |
| Baumann U et al., Recombinant OprF-OprI as a vaccine against Pseudomonas aeruginosa infections, Vaccine, 2004 Feb 17, Vol.22, No.7, p.p.840-7 | |||
| Yong-Mei Han et al., High-level expression, purification, polyclonal antibody preparation against recombinant OprD from Pseudomonas aeruginosa, | |||

