Изобретение относится к области биотехнологии, а именно к получению рекомбинантного белка, который представляет собой рекомбинантную атоксичную форму пневмолизина Streptococcus pneumoniae (S. pneumoniae) для медицинских целей, а также к рекомбинантному штамму Escherichia coli (Е. coli) M15/pSp-raPLY, который нами зарегистрирован в Государственной коллекции патогенных микроорганизмов и клеточных культур «ГКПМ-Оболенск», регистрационный номер 122 и плазмиде для его получения.
Активная иммунизация может быть одним из эффективных средств защиты против условно патогенных микроорганизмов и, в частности, S. pneumoniae. Для профилактики пневмококковой инфекции разработаны полисахаридная и конъюгированные полисахаридные вакцины: Pneumo 23, Prevenar 13, Synflorix (Feldman С., Anderson R. Review: Current and new generation pneumococcal vaccines. Journal of Infection, 2014, 69, 309-325). Эти вакцины успешно себя зарекомендовали, так как их внедрение в практику позволило снизить число пневмококковых заболеваний за счет развития серотипспецифического иммунного ответа на полисахаридные антигены (Lu J., Hou Н., Wang D. et al. Systemic and mucosal immune responses elicited by intranasal immunization with a pneumococcal bacterium-like particle-based vaccine displaying pneumolysin mutant Plym2. Immunology Letters, 2017, 187, 41-46; Wijmenga-Monsuur A.J., van Westen E., Knol M.J. et al. Direct comparison of immunogenicity induced by 10- or 13-valent pneumococcal conjugate vaccine around the 11-month booster in dutch infants. PLoS One, 2015, 10(12): e0144739).
Однако, несмотря на эффективность существующих вакцин, есть ряд нерешенных вопросов, которые остаются актуальными на сегодняшний день. В частности, явление замещения серотипов возбудителя и рост антибиотико-устойчивых штаммов пневмококка (Wijmenga-Monsuur A.J., van Westen Е., Knol M.J. et al. Direct comparison of immunogenicity induced by 10- or 13-valent pneumococcal conjugate vaccine around the 11-month booster in dutch infants. PLoS One, 2015, 10(12): e0144739). По разным причинам в настоящее время для использования в клинической практике не зарегистрировано ни одной серотипнезависимой вакцины на основе белковых антигенов пневмококка.
Streptococcus pneumoniae является одним из основных возбудителей внебольничной пневмонии (Wijmenga-Monsuur A.J., van Westen Е., Knol M.J. et al. Direct comparison of immunogenicity induced by 10- or 13-valent pneumococcal conjugate vaccine around the 11-month booster in dutch infants. PLoS One, 2015, 10(12): e0144739; Steel H.C., Cockeran R., Anderson R., Feldman C. Overview of community-acquired pneumonia and the role of inflammatory mechanisms in the immunopathogenesis of severe pneumococcal disease. Mediators of Inflammation, 2013, Volume 2013, 1-18). Широкое распространение пневмококковая инфекция получила благодаря колонизации микроба на слизистых оболочках верхних дыхательных путей человека, а также возрастающей резистентности невакцинных серотипов пневмококка к химиотерапевтическим средствам и антибиотикам (Brooks L.R.K., Mias G.I. Streptococcus pneumoniae's virulence and host immunity: aging, diagnostics, and prevention. Front. Immunol., 2018, 9: 1366; Brooks L.R.K., Mias G.I. Streptococcus pneumoniae's virulence and host immunity: aging, diagnostics, and prevention. Front. Immunol., 2018, 9: 1366; Weiser J.N., Ferreira D.M., Paton J.C.. Streptococcus pneumoniae: transmission, colonization and invasion. Nat Rev Microbiol., 2018, 16(6): 355-367. doi:10.1038/s41579-018-0001-8). В связи с тем, что белки пневмококка обладают высокой внутривидовой гомологией, они являются перспективными антигенами для разработки вакцин с серотипнезависимой активностью. Существует более 100 пневмококковых белков, однако исследователи сосредоточили свое внимание примерно на 20-30 белках. Наибольший интерес представляют такие белки S. pneumoniae как: пневмолизин, пневмококковый поверхностный белок А, пневмококковый поверхностный белок С, пневмококковый поверхностный антиген А, белки полигистидиновой триады, белки ворсинок пневмококка, белки-ферменты гиалуронидаза, нейраминидаза, стрептокиназа, цинк-зависимая протеаза, белки теплового шока пневмококка (Feldman С, Anderson R. Review: Current and new generation pneumococcal vaccines. Journal of Infection, 2014, 69, 309-325; Feldman C., Anderson R. Review: Current and new generation pneumococcal vaccines. Journal of Infection, 2014, 69, 309-325; Morais V., Texeira E., Suarez N. Next-generation whole-cell pneumococcal vaccine. Vaccines, 2019, 7, 151).
При создании современных вакцинных препаратов все большое внимание уделяется использованию генно-инженерных методов, суть которых заключается во встраивании генов, отвечающих за синтез отдельных протективных антигенов возбудителя, в геном непатогенного микроорганизма (например, Escherichia coli). Во время культивирования полученных продуцентов в большом количестве синтезируются целевые продукты (рекомбинантные белки), которые подвергаются простой очистке и могут быть использованы для приготовления вакцины. Такая стратегия позволяет определить наиболее иммуногенные белковые антигены бактерии. При этом процесс производства вакцин исключает контакт с культурами патогенных микроорганизмов, что является существенным технологическим преимуществом, обеспечивающим безопасность персонала. В нашей стране разработан экспериментальный препарат на основе гибридного (слитного) рекомбинантного белка, включающего аминокислотные последовательности: pspA, spr1895, psaA, защищенный Патентом РФ на изобретение №2012126328/15. Вакцина против пневмонии, вызываемой Streptococcus pneumoniae, на основе гибридного белка. // Суворов А.Н., Духовлинов И.В., Орлов А.И., Байгузин Е.Я. - Заявл. 22.06.2012; опубл. 27.03.2014 // Бюл. №9. - 2012.
Данный препарат является вакциной против пневмонии, вызываемой S. pneumoniae, на основе гибридного (слитного) белка, состоящего из комплекса трех белков: pspA, spr1895, psaA. Получаемый гибридный (слитный) белок обладает высокой молекулярной массой и способен вызывать достаточный иммунный ответ, однако белки, входящие в этот комплекс, ответственны только за колонизацию пневмококком верхних дыхательных путей (слизистая носа, носоглотка) и не принимают участия в патогенезе инвазивной пневмококковой инфекции, характерной для нижних дыхательных путей (трахея, бронхи и бронхиолы). Таким образом, защитный иммунный ответ на введение этой вакцины в основном направлен против первичной колонизации пневмококком слизистых верхних дыхательных путей, но не препятствует в дальнейшем развитию более серьезных осложнений, таких как инвазивная пневмония, пневмококковый менингит и сепсис, вызываемых S. pneumoniae.
Известно, что большое значение в развитии пневмококковой пневмонии играет белок микроба пневмолизин, который является токсином (Feldman С., Anderson R. Review: Current and new generation pneumococcal vaccines. Journal of Infection, 2014, 69, 309-32; Nishimoto A.T., Rosch J.W., Tuomanen E.I. Pneumolysin: pathogenesis and therapeutic target. Front. Microbiol., 2020, 11: 1543). Поэтому разрабатываются экспериментальные вакцины на основе детоксицированного пневмолизина (Lu J., Hou Н., Wang D. et al. Systemic and mucosal immune responses elicited by intranasal immunization with a pneumococcal bacterium-like particle-based vaccine displaying pneumolysin mutant Plym2. Immunology Letters, 2017, 187, 41-46). К настоящему времени имеются успешные данные об использовании экспериментальных вакцинных препаратов в разных фазах клинических испытаний.
С. Goulart et al. в 2013 году изучали в доклинических испытаниях действие разных вакцинных препаратов на основе белков пневмококка. Исследователи показали, что смесь белков, состоящая из поверхностного белка А и различных детоксицированых вариантов пневмолизина, работает не хуже, чем слитный белок pspA-dPly, однако отдельно детоксицированный вариант пневмолизина (dPly) работал слабо или полностью не работал (Lu J., Hou Н., Wang D. et al. Systemic and mucosal immune responses elicited by intranasal immunization with a pneumococcal bacterium-like particle-based vaccine displaying pneumolysin mutant Plym2. Immunology Letters, 2017, 187, 41-46. G).
Leroux-Roels et al. в 2014 году участвовали в проведении I и II фазы клинических испытаний с разными вариантами белковых препаратов пневмококка. Исследователи изучали действие dPly и белка полигистидиновой триады D (PhtD) по отдельности и в комбинации. Несмотря на достигнутые положительные результаты в этой работе, следует отметить, что III фаза клинических испытаний так и не была проведена (Lu J., Hou Н., Wang D. et al. Systemic and mucosal immune responses elicited by intranasal immunization with a pneumococcal bacteriumlike particle-based vaccine displaying pneumolysin mutant Plym2. Immunology Letters, 2017, 187, 41-46).
T. Kamtchoua et al. в 2013 году была проведена I фаза клинических испытаний с пневмококковой вакциной, разработанной на основе монопрепарата dPly, который был получен с помощью направленного мутагенеза с заменой 3-х аминокислотных остатков в позициях Т65С, G293C и С428А. Исследователи изучали иммуногенность, используя три дозы белка: 10 мкг, 25 мкг и 50 мкг. Несмотря на успешное выполнение работы, открытые сведения о проведении II и III фазах клинических испытаний в доступной нам литературе отсутствуют (Kamtchoua Т., Bologa М., Hopfer R. et al. Safety and immunogenicity of the pneumococcal pneumolysin derivative PlyD1 in a single-antigen protein vaccine candidate in adults. Vaccine, 2013, 31, 327-333). Вероятно, при проведении II фазы клинических испытаний не была доказана иммуногенность данного препарата, что может быть связано с большим количеством замен аминокислотных остатков в диком штамме пневмолизина.
Таким образом, технический результат нашей заявки на изобретение заключается в получении рекомбинантной плазмидной ДНК pSp-raPLY, кодирующей синтез рекомбинантного атоксичного белка пневмолизина S. pneumoniae и штамма-продуцента рекомбинантной атоксичной формы пневмолизина S. pneumoniae, с использованием штамма Е. coli M15.
Сущность предлагаемой заявки на изобретение:
1. Рекомбинантная плазмидная ДНК pSp-raPLY, кодирующая синтез рекомбинантного атоксичного белка пневмолизина S. pneumoniae, размером 4861 пар оснований, расщепляющаяся в результате гидролиза специфическими нуклеазами рестрикции: XbaI-EcoRI фрагмент ДНК размером 87 пар оснований, содержащего последовательность модифицированного промотора бактериофага Т5 и двух лактозных оперонов; EcoRI-BamHI фрагмент ДНК размером 57 пар оснований, содержащего сайт посадки рибосомы, стартовый ATG-кодон и последовательность, кодирующую шесть гистидинов (Arg-Gly-Ser-His-His-His-His-His-His-Gly-Ser); BamHI-SmaI фрагмент ДНК размером 1413 пар оснований, содержащего последовательность мутантного гена ply S. pneumoniae, полученного с помощью полимеразной цепной реакции (ПЦР); EcoRI-SmaI фрагмент ДНК размером 1470 пар оснований, содержащего сайт посадки рибосомы и последовательность, кодирующую атоксичный рекомбинантный белок пневмолизин S. pneumoniae, (состоит из гистидиновой последовательности и атоксичной формы пневмолизина); HindIII-NhEI фрагмент ДНК размером 120 пар оснований, содержащего лямбда t0 регион остановки транскрипции. Плазмида pSp-raPLY также включает в своем составе регион остановки транскрипции rrnBT1, сайт инициации репликации плазмиды ColE1 и ген, кодирующий устойчивость к ампициллину. В результате экспрессии рекомбинантной плазмиды pSp-raPLY происходит синтез рекомбинантного продукта с молекулярной массой около 53 кДа и его димера с молекулярной массой ~ 106 кДа.
2. Штамм бактерии Е. coli M15/pSp-raPLY - продуцент рекомбинантной атоксичной формы пневмолизина S. pneumoniae, полученный путем трансформации родительского штамма бактерий Е. coli M15 рекомбинантной плазмидной ДНК pSp-raPLY по п. 1.
3. Получение рекомбинантного атоксичного белка пневмолизина S. pneumoniae с использованием штамма Е. coli M15/pSp-raPLY по п. 2, включающий его культивирование в питательной среде, разрушение клеток микроорганизма и очистку полученного продукта с использованием аффинной хроматографии на никель-активированных сорбентах и растворением с ренатурацией очищенного препарата в нативных условиях при помощи диализа.
Нами была создана рекомбинантная плазмидная ДНК pSp-raPLY и штамм Е. coli M15/pSp-raPLY.
Плазмида pSp-raPLY имеет 4861 пар оснований и характеризуется наличием следующих фрагментов:
XbaI - EcoRI фрагмент ДНК размером 87 пар оснований, содержащего последовательность модифицированного промотора бактериофага Т5 и двух лактозных оперонов.
EcoRI - BamHI фрагмент ДНК размером 57 пар оснований, содержащего сайт посадки рибосомы, стартовый ATG-кодон и последовательность, кодирующую шесть гистидинов (Arg-Gly-Ser-His-His-His-His-His-His-Gly-Ser).
BamHI - SmaI фрагмент ДНК размером 1413 пар оснований, последовательность мутантного гена ply S. pneumoniae, полученного с помощью полимеразной цепной реакции (ПНР).
EcoRI - SmaI фрагмент ДНК размером 1470 пар оснований, содержащего сайт посадки рибосомы и последовательность, кодирующую атоксичный рекомбинантный белок пневмолизин S. pneumoniae, (состоит из гистидиновой последовательности и атоксичной формы пневмолизина).
HindIII - NhEI фрагмент ДНК размером 120 пар оснований, содержащего лямбда t0 регион остановки транскрипции.
На фиг. 1 представлена схема плазмиды pSp-raPLY для получения атоксичного рекомбинантного белка пневмолизина.
Заявленный рекомбинантный белок, представляющий собой атоксичную форму пневмолизина S. pneumoniae имеет аминокислотную последовательность, представленную на фиг. 2.
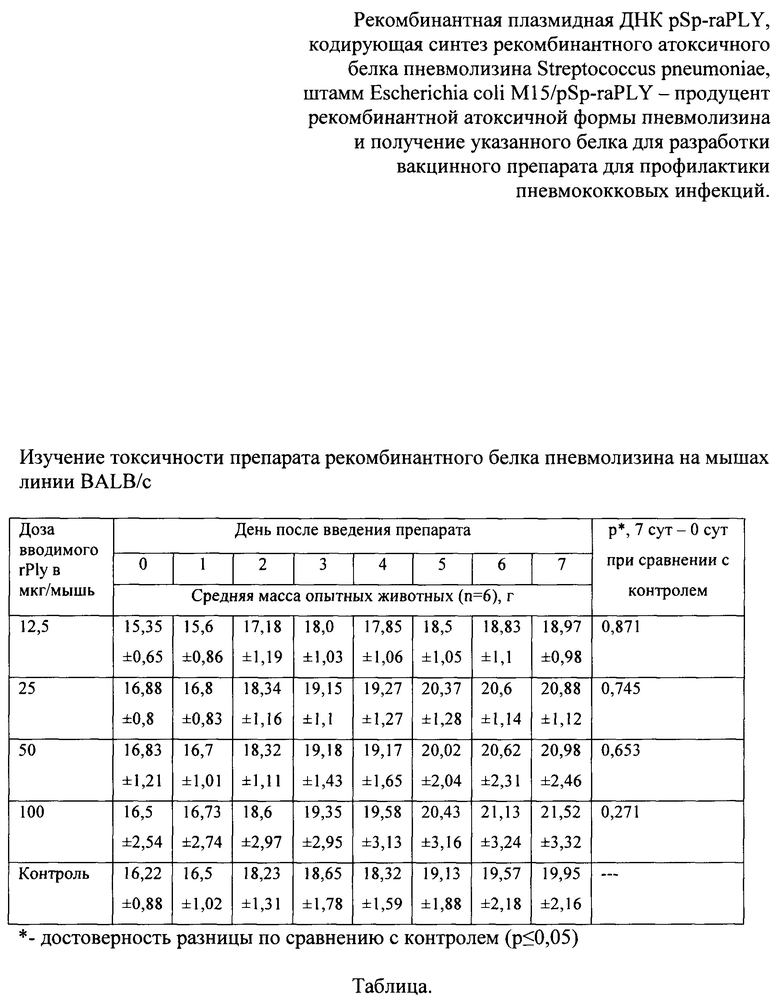
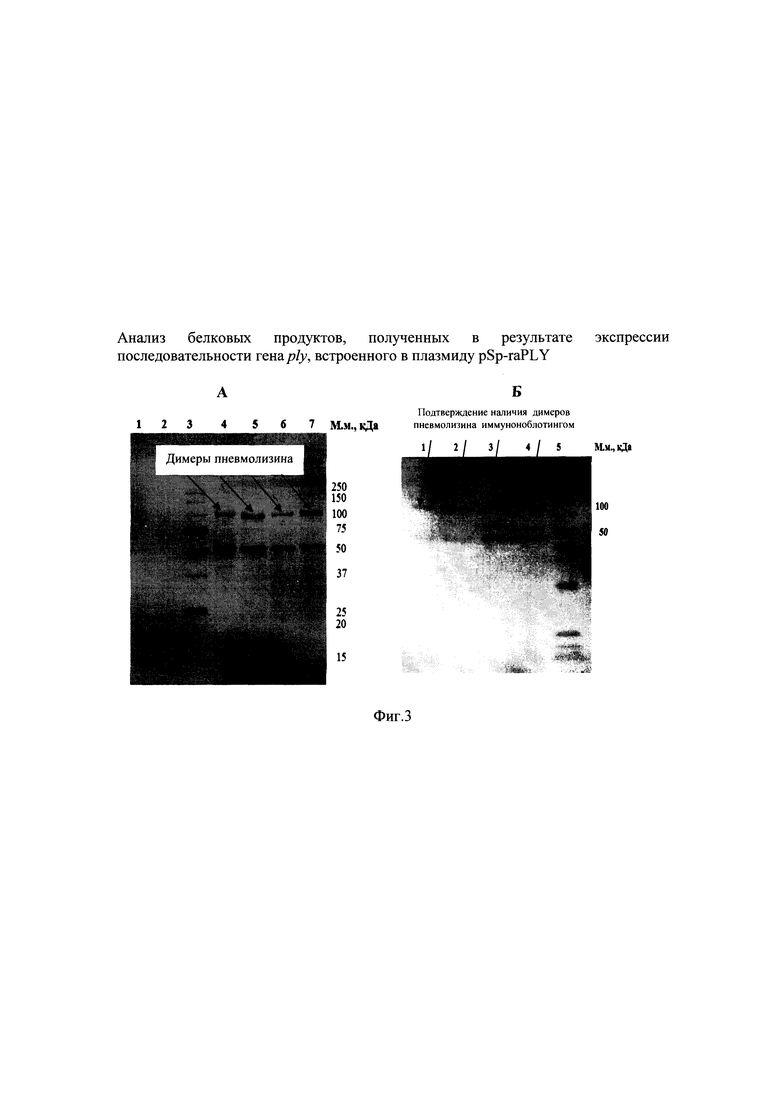
На фиг. 3 представлен анализ белковых продуктов, полученных в результате экспрессии в клетках Е. coli последовательности рекомбинантного гена ply, встроенного в плазмиду pSp-raPLY.
На фиг. 3А - ПААГ окрашенный Кумасси R-250, Фиг. 3Б - нитроцеллюлозная мембрана после иммуноблоттинга с моноклональными антителами к пневмолизину.
На фиг. 3А треки: 1 и 2 - продукты неиндуцированного продуцента рекомбинантного белка (слабая полоса); 3 - маркер молекулярной массы (#26614; тяжелые фрагменты 250, 150 и 100 кДа); 4, 5, 6 и 7 - продукты индуцированного продуцента рекомбинантного белка;
На фиг. 3Б треки: 1, 2, 3 и 4 - продукты индуцированного продуцента рекомбинантного белка; 5 - маркер молекулярной массы.
В таблице представлены данные по анализу токсичности полученной рекомбинантной формы пневмолизина S. pneumoniae.
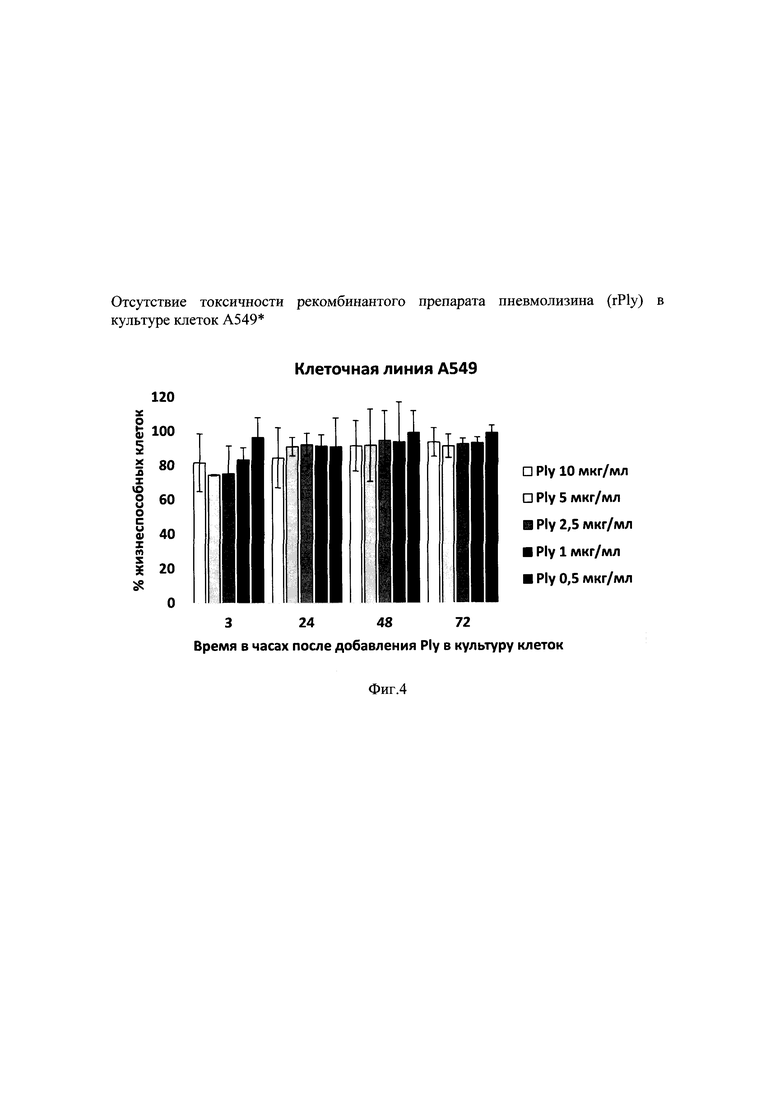
На фиг. 4 приведены результаты опыта по анализу токсичности полученной рекомбинантной формы пневмолизина S. pneumoniae в культуре клеток А-549 in vitro.
Штамм Е. coli M15/pSp-raPLY получен трансформацией клеток родительского штамма Е. coli M15, с использованием традиционной генно-инженерной технологии.
Штамм Е. coli M15/pSp-raPLY характеризуется следующими признаками.
Культурально-морфологические признаки: клетки мелкие, прямые, утолщенной палочковидной формы, грамотрицательные, неспороносные. Микроб хорошо растет на простых питательных средах. При росте на агаре «Дифко» колонии круглые, гладкие, выпуклые, мутные, блестящие, светло-бежевые, края ровные. При росте в жидких средах (на минимальной среде с глюкозой или LB-бульоне) образуют интенсивную ровную муть.
Физико-биологические признаки: факультативный анаэроб, температурный диапазон роста 35-37°С при оптимуме рН в пределах 6,5-7,5.
В качестве источника азота пневмококк использует минеральные соли в аммонийной и нитратной формах и органические соединения в виде аминокислот, пептона, триптона, дрожжевого экстракта и т.д.
В качестве источника углерода микроб использует аминокислоты, глицерин, углеводы.
Клетки проявляют устойчивость к ампициллину (до 200 мкг/мл) и канамицину (до 70 мкг/мл).
Штамм Е. coli M15/pSp-raPLY - продуцент атоксичной формы пневмолизина S. pneumoniae.
Способ, условия и состав среды для хранения штамма:
LB-бульон с 10% глицерином и антибиотиками при минус 70°С в ампулах или в лиофилизированном состоянии в ампулах при 4°С.
Штамм Е. coli M15/pSp-raPLY идентифицирован по «определителю Берги» (1974) как штамм вида Е. coli.
Особенностью заявляемого штамма Е. coli M15/pSp-raPLY является получение высокоочищенного рекомбинантного белка пневмолизина S. pneumoniae сравнительно более простым способом и в более короткие сроки, по сравнению с традиционными методами получения бактериальных белков. При этом в получаемом белковом препарате отсутствует токсичность, что позволяет его характеризовать как перспективный вакцинный кандидат.
Получение рекомбинантного белка включает следующие этапы:
культивирование в питательной среде штамма Е. coli M15/pSp-raPLY;
осаждение биомассы центрифугированием;
разрушение клеток микроорганизма при растворении их в денатурирующем буфере, содержащем 8 М мочевину;
очистка с помощью аффинной хроматографии на никель-активированных сорбентах;
ренатурация очищенного рекомбинантного белка (атоксичной формы пневмолизина S. pneumoniae) выполнена с помощью диализа.
Оптимальными условиями проведения отдельных стадий получения рекомбинантного белка (атоксичной формы пневмолизина S. pneumoniae):
разрушение клеток осуществляют в результате растворения биомассы в 8 М буферном растворе мочевины;
удаление нерастворимых компонентов клетки осуществляют центрифугированием;
ренатурацию проводят при температуре 4°С путем диализа против 50 мМ буферного раствора Tris-HCl (рН=9,0).
Выход рекомбинантного белка (атоксичной формы пневмолизина S. pneumoniae) в результате применения способа в оптимальном режиме составляет не менее 120 мг рекомбинантного белка с 16 л культуральной среды, уровень синтеза рекомбинантного белка - не менее 7,5 мг с 1 л культуральной среды.
Сущность и преимущества заявляемой группы изобретений иллюстрируется следующими примерами.
Пример 1. Получение рекомбинантной плазмиды pSp-raPLY.
Технология получения плазмиды pSp-raPLY включает следующие этапы:
выделение геномной ДНК S. pneumoniae;
амплификация гена ply S. pneumoniae с помощью ПЦР;
встраивание амплифицированной последовательности в плазмидный вектор.
Для выделения геномной ДНК использовали штамм, полученный из коллекции центра коллективного пользования НИИВС им. И.И. Мечникова S. pneumoniae серотипа 19F, депонированный в Государственной коллекции патогенных микроорганизмов III-IV групп патогенности ФГБУ «Научный центр экспертизы средств медицинского применения» Минздрава России под регистрационным номером 298.
Культуру клеток S. pneumoniae хранили в лиофилизированном состоянии. Для восстановления жизнеспособности культуры ампулу с лиофилизированным штаммом S. pneumoniae вскрывали стерильно над лотком в боксе биологической безопасности. После вскрытия ампулы, лиофилизат растворяли путем добавления 0,3 мл сердечно-мозгового бульона. Содержимое ампулы переносили пастеровской пипеткой в пробирку с 2 мл сердечно-мозгового бульона и инкубировали при 37°С в 5,5% CO2 среде в течение 2 ч. После этапа подращивания, культуру высевали на питательный агар с 5% содержанием лошадиной крови, из расчета 100 мкл на одну чашку и инкубировали при 37°С в 5,5% CO2 среде в течение 18-24 ч. В работе использовали культуру S. pneumoniae после двух восстановительных пассажей. Чистоту выросшей культуры определяли визуально и микроскопически в мазках, окрашенных по Граму.
Полученную биомассу аккуратно снимали микробиологической петлей с поверхности агаризованной среды и переносили в микроцентрифужную пробирку. Осажденную бактериальную биомассу S. pneumoniae растворяли в 200 мкл ТЕ-буфера, рН=8,0. После этого в пробирку добавляли 10 мкл протеиназы К (10 мг/мл) и 200 мкл лизирующего раствора. Смесь инкубировали при температуре 65°С в течение 15 мин, периодически перемешивая содержимое пробирки. Затем прибавляли 200 мкл осаждающего раствора и переносили весь объем пробирки на колонку «К-сорб-100» (Синтол) и проводили выделение геномной ДНК согласно протоколу производителя. Полученные растворы ДНК хранили при минус 20°С.
При ПЦР использовали пару праймеров, один из которых соответствовал началу (прямой), а второй (обратный) был комплементарен концу гена ply S. pneumoniae. Прямой праймер содержал на 5'-конце сайт рестрикции BamHI, а обратный сайт рестрикции SmaI.
Прямой праймер имел следующую структуру (5'-3'): GGATCACTTAGTCCAACCAC, а обратный: GCAAACATTCTTCTCTCCTA.
ПЦР проводили при следующих реакционных условиях: буфер х5 для Phu-полимеразы, содержащий 1,5 mM MgCl2, 0,2 mM dNTP, праймеры Forward и Reverse по 0,01 mM каждый. На 50 мкл реакционной смеси использовали 1 ед. Phu-полимеразы (Fermentas) и 1 мкг геномной ДНК.
Реакцию проводили в амплификаторе "Biometra". Процесс амплификации состоял из следующих стадий: прогревания при 98°С в течении 1 мин, и 30-ти циклов ПЦР (98°С - 15 сек., 55-65°С - 20 сек, 72°С - 50 сек, финальная элонгация - 5 мин) и инкубации при 72°С в течении 2 мин.
Полученный фрагмент ДНК размером 1413 пар оснований обработали рестриктазами BamHI и SmaI. Параллельно этими же рестриктазами был обработан вектор pQE-30 (QIAGEN). Оба рестрикта очищали в агарозном геле и объединили в одну конструкцию при реакции лигирования. В результате была получена рекомбинантная плазмида pSp-raPLY размером 4861 пар оснований, в которой ДНК гена ply была фланкирована сайтами рестрикции BamHI и SmaI (фиг. 1). По результатам секвенирования расшифрована аминокислотная последовательность полученного рекомбинантного белка пневмолизина (фиг. 2).
Пример 2. Получение штамма Е. coli M15/pSp-raPLY - продуцента атоксичной формы пневмолизина S. pneumoniae.
Штамм-продуцент Е. coli M15/pSp-raPLY получали путем трансформации клеток родительского штамма Е. coli M15, содержащего плазмиду pREP4, рекомбинантной плазмидой pSp-raPLY с последующим отбором рекомбинантных клонов, выращенных при температуре 37°С на среде LB, содержащей ампициллин и канамицин.
Первичный отбор рекомбинантных плазмид проводили с использованием рестриктного анализа и секвенирования.
Далее отбор продуцентов проводился по способности контролируемого синтеза рекомбинантного белка в клетках штамма-продуцента. Для этого добавляли бактериальной петлей одну колонию в 1 мл среды LB, содержащей ампициллин и канамицин в концентрациях 200 и 70 мкг в мл соответственно. Выращивали в термостатированном шейкере со скоростью орбитального вращения 200 об/мин в течение 12-16 ч при температуре 37°С. Затем в 5 мл свежей среды вносили 100 мкл выросшей культуры и инкубировали в прежних условиях в течение 2-3 ч до достижения культурой активной фазы роста (0,6 оптических единиц при длине волны 600 нм). Добавляли 5 мкл 1 М водного раствора химического индуктора изопропил-β-d-тиогалактопиранозид (ИПТГ). Инкубацию продолжали в прежних условиях 4 ч, отбирая пробы для электрофореза в течение каждого часа, после завершения культивирования осаждали полученную биомассу.
В качестве контроля использовали бактериальную культуру продуцента, инкубированную при тех же условиях, но без добавления ИПТГ. Белковые продукты анализировали электрофорезом в полиакриламидном геле (ПААГ) по методу Лэммли (фиг. 3А).
При анализе биомассы, полученной после индукции экспрессии рекомбинантного гена, выявлено наличие специфичного продукта, который отсутствовал или очень слабо экспрессировался в клетках продуцента, выращенного без добавления ИПТГ. Молекулярная масса синтезированного рекомбинантного продукта составила примерно 53 кДа и 106 кДа. Расчетная масса для рекомбинантной атоксичной формы пневмолизина S. pneumoniae составила 53 кДа, а для его димера - 106 кДа. По результатам электрофореза установлено, что количество рекомбинантного белка составляло не менее одной пятой относительно всех клеточных белков.
Рекомбинантные белки проявили высокую специфичность при взаимодействии с коммерческими моноклональными антителами к пневмолизину (фиг. 3Б).
Пример 3. Получение рекомбинантного атоксичного белка пневмолизина S. pneumoniae из штамма Е. coli M15/pSp-raPLY.
Получение рекомбинантного атоксичного белка пневмолизина S. pneumoniae проводили в 4 этапа.
1 этап: культивирование штамма Е. coli M15/pSp-raPLY с индукцией экспрессии рекомбинантного гена ply.
2 этап: получение клеточного лизата штамма Е. coli M15/pSp-raPLY после экспрессии рекомбинантного гена ply.
3 этап: хроматографическая очистка рекомбинантного атоксичного белка пневмолизина S. pneumoniae.
4 этап: растворение и ренатурация рекомбинантного атоксичного белка пневмолизина S. pneumoniae.
Культивирование штамма Е. coli M15/pSp-raPLY с индукцией экспрессии рекомбинантного гена ply.
Выращивали посевной материал штамма Е. coli M15/pSp-raPLY в 200 мл среды LB содержащей ампициллин и канамицин в концентрациях 200 и 70 мкг в мл соответственно. Культивирование проводили в течение 12 ч при температуре 37°С в термостатированном шейкере со скоростью орбитального вращения 200 об/мин.
Полученную культуру асептически вносили в колбы суммарным объемом 4 л стерильной среды LB, содержащей ампициллин и канамицин в концентрациях 200 и 70 мкг в мл соответственно. Проводили культивирование в тех же условиях 1,5-2 ч.
При достижении плотности культуры 0,6 оптических единиц при длине волны 600 нм в термостатированном шейкере вносили 4 мл 1 М водного раствора ИПТГ. Инкубацию продолжали в прежних условиях 4 ч.
Получение клеточного лизата штамма Е. coli M15/pSp-raPLY после экспрессии рекомбинантного гена ply.
Биомассу осаждали в центрифуге Beckman J2-21 (ротор JA-14) со скоростью вращения 3000 об/мин в течение 30 мин, замораживали и хранили при минус 20°С. Осадок, собранный с 16 л. культуры растворяли в 500 мл следующего раствора: 8 М мочевина; 0,1 М NaH2PO4; 0,01 М Tris-HCl; рН=8,0. Растворение биомассы проводили в орбитальном шейкере при вращении 100 об/мин в течение 12 ч при комнатной температуре.
Затем лизат освобождали от нерастворенных фрагментов клетки в центрифуге Eppendorf 5804 R со скоростью вращения 10000 об/мин при комнатной температуре в течение 1 ч. Супернатант переносили в чистую стерильную колбу.
Хроматографическая очистка рекомбинантного атоксичного белка пневмолизина S. pneumoniae.
К осветленному лизату добавляли 25 мл суспензии Ni-агарозы (QIAGEN), и инкубировали 1 ч при комнатной температуре в орбитальном шейкере при легком покачивании (60 об/мин).
Полученную суспензию пропускали через колонку для хроматографии (Bio-Rad) объемом 250 мл и диаметром 5 см. В результате из колонки удалялась жидкая фаза лизата и оставался сорбент со связанным рекомбинантным белком.
Далее через колонку пропускали 1 л первого промывочного раствора следующего состава: 8 М мочевина; 0,1 М NaH2PO4; 0,01 М Tris-HCl; рН=6,3. Затем через колонку пропускали 0,5 л второго промывочного раствора следующего состава: 8 М мочевина; 0,1 М NaH2PO4; 0,01 М Tris-HCl; рН=5,9.
Для элюции рекомбинантного белка использовали раствор следующего состава: следующего состава: 8 М мочевина; 0,1 М NaH2PO4; 0,01 М Tris-HCl; рН=4,5. С полученным элюатом проводили оценочный электрофорез в ПААГ.
Растворение и ренатурация рекомбинантного атоксичного белка пневмолизина S. pneumoniae.
Очищенный рекомбинантный белок растворяли в 50 мМ Tris-HCl (рН=9,0) с помощью диализа при температуре 4°С и перемешивании буфера на магнитной мешалке. Для этого препарат очищенного рекомбинантного белка переносили в диализный мешок.
Завязанный диализный мешок помещали в емкость, содержащую 1 л следующего раствора: 50 мМ Tris-HCl; рН=9,0. Через 4 ч проводили смену буферного раствора. Процедуру повторяли пять раз.
Содержание белка в препарате определяли спектрофотометрически при длине волны 280 нм. При расчете концентрации рекомбинантных белков использовали коэффициент экстинкции 0,7, рассчитанный в программе Vector NTI Advance 11.0.
Препараты очищенного рекомбинантного белка (атоксичная форма пневмолизина S. pneumoniae) хранили в аликвотах по 10 мл в стерильных пробирках при минус 70°С.
Пример 4. Оценка токсичности рекомбинантного белка пневмолизина S. pneumoniae.
Препараты очищенного рекомбинантного белка пневмолизина вводили однократно внутрибрюшинно животным в дозах 12,5 мкг/мышь, 25 мкг/мышь, 50 мкг/мышь и 100 мкг/мышь. Число животных в каждой группе было десять (n=10). Наблюдение за мышами проводили в течение 7 дней. Учитывали среднюю прибавку веса мышей в каждой группе на протяжении всего эксперимента по сравнению с контрольной группой мышей, которым однократно вводили стерильный физ. раствор. Значимой разницы (р<0,05) в изменении прибавки веса животных по сравнению с контролем не было выявлено (таблица).
В следующем опыте изучали токсичность очищенного рекомбинантного белка пневмолизина в культуре клеток А-549 in vitro. Рекомбинантный белок добавляли в концентрациях от 0,5 мкг/мл до 10 мкг/мл. Ни одна из взятых концентраций пневмолизина не оказывала токсичного эффекта на жизнеспособность клеток карциномы легкого человека (А-549) по сравнению с контролем, который принимали условно за 100% жизнеспособных клеток (фиг. 4).
| название | год | авторы | номер документа |
|---|---|---|---|
| ИММУНОБИОЛОГИЧЕСКОЕ СРЕДСТВО НА ОСНОВЕ РЕКОМБИНАНТНОГО БЕЛКА ДЛЯ ПРОФИЛАКТИКИ ПНЕВМОКОККОВОЙ ИНФЕКЦИИ | 2024 |
|
RU2823752C1 |
| ВАКЦИНА ПРОТИВ ПНЕВМОНИИ, ВЫЗЫВАЕМОЙ Streptococcus pneumoniae, НА ОСНОВЕ ГИБРИДНОГО БЕЛКА | 2012 |
|
RU2510281C2 |
| СПОСОБ ПОЛУЧЕНИЯ ИММУНОГЕННОЙ КОМПОЗИЦИИ НА ОСНОВЕ ГИБРИДНОГО БЕЛКА pESAT6-DBD И ДЕКСТРАНА, РЕКОМБИНАНТНАЯ ПЛАЗМИДА pESAT6-DBD, ШТАММ Escherichia coli, ХИМЕРНЫЙ БЕЛОК ESAT6-DBD И ИХ ПРИМЕНЕНИЕ | 2013 |
|
RU2520737C1 |
| СПОСОБ ПОЛУЧЕНИЯ ИММУНОГЕННОЙ КОМПОЗИЦИИ НА ОСНОВЕ ГИБРИДНОГО БЕЛКА Ag85A-DBD И ДЕКСТРАНА, РЕКОМБИНАНТНАЯ ПЛАЗМИДА pAg85A-DBD, ШТАММ Escherichia coli [pREP4, pAg85A-DBD], ХИМЕРНЫЙ БЕЛОК Ag85A-DBD | 2013 |
|
RU2520078C1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДА pESAT6-CFP10-DBD, РЕКОМБИНАНТНЫЙ ШТАММ Escherichia coli M15 [pREP4, pESAT6-CFP10-DBD], СПОСОБ ПОЛУЧЕНИЯ, ИММОБИЛИЗАЦИИ, КОНЦЕНТРИРОВАНИЯ И ОЧИСТКИ РЕКОМБИНАНТНОГО БЕЛКА ESAT6-CFP10-DBD НА ДЕКСТРАНЕ, РЕКОМБИНАНТНЫЙ БЕЛОК ESAT6-CFP10-DBD И ИММУНОГЕННАЯ КОМПОЗИЦИЯ, СОДЕРЖАЩАЯ БЕЛОК ESAT6-CFP10-DBD | 2013 |
|
RU2539026C1 |
| Рекомбинантная ДНК, обеспечивающая получение рекомбинантного белка Cov1, обладающего иммуногенными свойствами в отношении вируса SARS-CoV-2 | 2021 |
|
RU2776484C1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК, КОДИРУЮЩАЯ СИНТЕЗ РЕКОМБИНАНТНОГО БЕЛКА TUL4spCBD, ШТАММ ESCHERICHIA COLI M15 [pREP4, pTUL4spCBD] - ПРОДУЦЕНТ РЕКОМБИНАНТНОГО БЕЛКА TUL4spCBD, РЕКОМБИНАНТНЫЙ БЕЛОК TUL4spCBD И СПОСОБ ЕГО ПОЛУЧЕНИЯ, СПОСОБ ПОЛУЧЕНИЯ СПЕЦИФИЧЕСКИХ АНТИТЕЛ К БЕЛКУ TUL4spCBD | 2004 |
|
RU2270249C1 |
| СПОСОБ ПОЛУЧЕНИЯ ИММУНОГЕННОЙ КОМПОЗИЦИИ НА ОСНОВЕ ГИБРИДНОГО БЕЛКА CFP10-DBD И ДЕКСТРАНА, РЕКОМБИНАНТНАЯ ПЛАЗМИДА pCFP10-DBD, ШТАММ Escherichia coli [pREP4, pCFP10-DBD], ХИМЕРНЫЙ БЕЛОК CFP10-DBD И ИХ ПРИМЕНЕНИЕ | 2013 |
|
RU2546875C1 |
| Иммуногенная композиция на основе рекомбинантных псевдоаденовирусных частиц, а также на основе белковых антигенов и способ получения иммуногенной композиции | 2018 |
|
RU2691302C1 |
| Штамм вируса гриппа ИнфлюБакт-H7/PspA для производства комбинированной вакцины против вируса гриппа А и бактериальной пневмонии, вызываемой Streptococcus pneumoniae | 2021 |
|
RU2776196C1 |


Изобретение относится к биотехнологии и представляет собой рекомбинантную плазмидную ДНК pSp-raPLY, содержащую ген для экспрессии рекомбинантного атоксичного пневмолизина S. pneumoniae, экспрессия которого находится под контролем промотора фага Т5, двух лактозных оперонов, лямбда t0 региона остановки транскрипции, терминатора транскрипции rrnBT1. Плазмида pSp-raPLY содержит ген устойчивости к ампициллину, участок инициации репликации плазмиды ColE1. Данной плазмидой трансформируют клетки реципиентного штамма Escherichia coli M15. Реципиентный штамм Е. coli M15 содержит плазмиду pREP4, которая включает ген lac белка, способного связываться с lac-оперонами, и ген устойчивости к канамицину. В своем геноме штамм Е. coli M15 содержит высокоэкспрессируемый ген РНК-полимеразы бактериофага Т5. В результате получен штамм Е. coli M15/pSp-raPLY с продуктивностью не менее 5 мг рекомбинантной атоксичной формы пневмолизина S. pneumoniae на 1 л культуральной среды или 5-10 г влажной биомассы клеток штамма. 3 н.п. ф-лы, 4 ил., 1 табл., 4 пр.
1. Рекомбинантная плазмидная ДНК pSp-raPLY, кодирующая синтез рекомбинантного атоксичного белка пневмолизина S. pneumoniae, размером 4861 пар оснований, расщепляющаяся в результате гидролиза специфическими нуклеазами рестрикции: XbaI-EcoRI фрагмент ДНК размером 87 пар оснований, содержащего последовательность модифицированного промотора бактериофага Т5 и двух лактозных оперонов; EcoRI-ВатHI фрагмент ДНК размером 57 пар оснований, содержащего сайт посадки рибосомы, стартовый ATG-кодон и последовательность, кодирующую шесть гистидинов Arg-Gly-Ser-His-His-His-His-His-His-Gly-Ser, BamHl-Smal фрагмент ДНК размером 1413 пар оснований, содержащего последовательность мутантного гена ply S. pneumoniae, полученного с помощью полимеразной цепной реакции, EcoRI-SmaI фрагмент ДНК размером 1470 пар оснований, содержащего сайт посадки рибосомы и последовательность, кодирующую атоксичную форму пневмолизина S. Pneumonia, состоящий из гистидиновой последовательности и атоксичной формы пневмолизина, HindIII-NhEI фрагмент ДНК размером 120 пар оснований, содержащего лямбда t0 регион остановки транскрипции; плазмида pSp-raPLY также включает в своем составе регион остановки транскрипции rrnBT1, сайт инициации репликации плазмиды ColE1 и ген, кодирующий устойчивость к ампициллину; в результате экспрессии рекомбинантной плазмиды pSp-raPLY происходит синтез рекомбинантного продукта с молекулярной массой 50 кДа и его димера с молекулярной массой 100 кДа.
2. Штамм бактерии Е. coli M15/pSp-raPLY - продуцент рекомбинантной атоксичной формы пневмолизина S. pneumoniae, полученный путем трансформации родительского штамма бактерий Е. coli Ml5 рекомбинантной плазмидной ДНК pSp-raPLY по п. 1.
3. Способ получения рекомбинантного атоксичного белка пневмолизина S. pneumoniae с использованием штамма Е. coli M15/pSp-raPLY по п. 2, являющегося нетоксичным димером с молекулярной массой от 50 до 100 кДа, формирующего иммунный ответ для профилактики пневмококковых инфекций, включающий следующие этапы: культивирование в питательной среде штамма Е. coli M15/pSp-raPLY; осаждение биомассы центрифугированием; разрушение клеток микроорганизма при растворении их в денатурирующем буфере, содержащем 8М мочевину; очистка с помощью аффинной хроматографии на никель-активированных сорбентах; ренатурация очищенной атоксичной формы пневмолизина S. pneumoniae, выполненная с помощью диализа.
| МОДИФИЦИРОВАННЫЙ ПНЕВМОЛИЗИН, РЕКОМБИНАНТНАЯ ПЛАЗМИДА, СПОСОБ ПОЛУЧЕНИЯ МОДИФИЦИРОВАННОГО ПНЕВМОЛИЗИНА, ВАКЦИНА | 1989 |
|
RU2121481C1 |
| Рекомбинантная плазмида, экспрессирующая клонированный ген гемолизина Vibrio cholerae, и штамм Escherichia coli - суперпродуцент прогемолизина Vibrio cholerae | 2017 |
|
RU2671099C1 |
| ВАКЦИНА ПРОТИВ ПНЕВМОНИИ, ВЫЗЫВАЕМОЙ Streptococcus pneumoniae, НА ОСНОВЕ ГИБРИДНОГО БЕЛКА | 2012 |
|
RU2510281C2 |
| LU J.et al | |||
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |
| Immunology Letters, 2017, 187, 41-46. | |||

