ПРЕРЕКРЕСТНАЯ ССЫЛКА НА РОДСТВЕННЫЕ ЗАЯВКИ
[0001] Данная заявка заявляет преимущество предварительной заявки на патент США №61/220,344, поданной 25 июня 2009 г., и тем самым включенной ссылкой во всей ее полноте.
ОБЛАСТЬ ТЕХНИКИ
[0002] Описан способ измерения адаптивного иммунитета пациента путем анализа разнообразия генов рецепторов Т-клеток или генов антител с использованием крупномасштабного секвенирования нуклеиновой кислоты, экстрагированной из клеток адаптивной иммунной системы.
ПРЕДПОСЫЛКИ
[0003] Иммунокомпетентность является способностью организма продуцировать нормальный иммунный ответ (т.е. продуцирование антител и/или опосредованный клетками иммунитет) после воздействия патогена, который может быть живым организмом (таким как бактерия или гриб), вируса или специфических антигенных компонентов, выделенных из патогена и введенных в вакцину. Иммунокомпетентность является противоположностью иммунодефициту, или иммунной некомпетентности, или ослабленному иммунитегу. Несколькими примерами могли бы быть новорожденный, у которого пока нет полностью функционирующей иммунной системы, но у которого могут быть переданное матерью антитело (иммуннодефицный); пациент па поздней стадии СПИДа с разрушенной или разрушающейся иммунной системой (иммунная некомпетентность); реципиент с трансплантатом, принимающий лекарство для того, чтобы его организм не отторгал донорский орган (ослабленный иммунитет); связанное с возрастом ослабление функции Т-клеток у лиц пожилого возраста; или индивидуумы, подвергшиеся радиации или химиотерапевтическим лекарственным средствам. Могут быть случаи совмещения, но эти выражения представляют все показатели дисфункциональной иммунной системы. В отношении лимфоцитов иммунокомпетентность означает, что В-клетка или Т-клетка является зрелой и может распознавать антигены и позволять человеку повышать иммунный ответ.
[0004] Иммунокомпетентность зависит от способности адаптивной иммунной системы повышать иммунный ответ, специфический для любых потенциальных чужеродных антигенов, используя высоко полиморфные рецепторы, закодированные В-клетками (иммуноглобулины, Ig) и Т-клетками (Т-клеточные рецепторы, TCR).
[0005] Ig, экспрессированные В-клетками, представляют собой белки, включающие четыре полипсптидные цепи, две тяжелые цепи (Н цепи) и две легкие цепи (L цепи), формирующих HL структуру. Каждая пара Н и L цепей содержит гипервариабельный домен, включающий VL и VH участок, и константный домен. Н цепи Ig бывают нескольких типов: µ, δ, γ, α и β. Разнообразие Ig у индивидуума в основном определено гипервариабельным доменом. V домен Н цепей создается комбинаторным соединением трех типов сегментов гена «germline», сегментами VH, DH и JH. Разнообразие последовательности гипервариабельного домена, кроме того, увеличивается независимым добавлением и делецией нуклеотидов в соединениях VH-DH, DH-JH и VH-JH во время процесса перестройки гена Ig. В этом отношении иммунокомпетентность отражается в разнообразии Ig.
[0006] TCR, экспрессированные αβ Т-клетками, являются белками, включающими две трансмембранные полипептидные цепи (α и β), экспрсссированные от TCRA и TCRB генов, соответственно. Подобные TCR белки экспрессированы в гамма-дельта Т-клетках, из TCRD и TCRG локусов. Каждый TCR пептид содержит вариабельные определяющие комплементарность участки (CDR), а также каркасные участки (FR) и константный участок. Разнообразие последовательностей αβ Т-клеток в значительной степени определено аминокислотной последовательностью петель третьего определяющего комплементарность участка (CDR3) вариабельных доменов α и β цепей, чье разнообразие является результатом рекомбинации между вариабельным (Vβ), разнообразным (Dβ) и соединяющим (Jβ) генными сегментами в локусе β цепи и между аналогичными Vα и Jα генными сегментами в локусе α цепи, соответственно. Существование множества таких генных сегментов в локусах α и β цепей TCR предоставляет большое число отдельных CDR3 последовательностей, подлежащих кодированию. Разнообразие CDR3 последовательностей, кроме того, увеличивается в зависимости от добавления и делении нуклеотидов на Vβ-Dβ, Dβ-Jβ и Vα-Jα соединениях во время процесса перестройки TCR гена. В этом отношении, иммунокомпетентность отражается в разнообразии TCR.
[0007] Существует назревшая потребность в способах оценки или измерения адаптивной иммунной системы пациентов в различных условиях, будь то иммунокомпетентность при ослабленном иммунитете или разрегулированный адаптивный иммунитет при аутоиммунном заболевании. Существует потребность в способах диагностики состояния заболевания или эффекты старения путем оценки иммунокомпетентности пациента. Таким же образом должны быть проконтролированы результаты лечений, которые модифицируют иммунную систему, путем оценки иммунокомпетентности пациента, в процессе прохождения курса лечения. С другой стороны, существует потребность в способах контроля адаптивной иммунной системы в контексте вспышек аутоиммунного заболевании и ремиссий в целях контроля ответа на лечение или потребность начать профилактическое лечение предсимптоматически.
КРАТКОЕ ОПИСАНИЕ
[0008] Одним аспектом данного изобретения является композиция, включающая:
- многообразие праймеров V сегмента, где каждый праймер включает последовательность, которая комплементарна отдельному функциональному V сегменту или небольшому семейству V сегментов; и
- многообразие праймеров J-сегмента, где каждый праймер включает последовательность, которая комплементарна J сегменту;
где праймеры V сегмента и J-сегмента разрешают амплификацию CDR3 участка TCR путем многократной полимеразной цепной реакции (PCR) с продуцированным многообразием амплифицированных молекул ДНК, достаточных для определения разнообразия TCR генов. Одним вариантом осуществления данного изобретения является композиция, где каждый праймер V сегмента включает последовательность, которая комплементарна отдельному Vβ сегменту, а каждый праймер J сегмента включает последовательность, которая комплементарна Jβ сегменту, и где праймеры V сегмента и J-сегмента разрешают амплификацию CDR3 участка TCRβ. Другим вариантом осуществления является композиция, где каждый праймер V-сегмента включает последовательность, которая комплементарна отдельному функциональному Vα сегменту, а каждый праймер J сегмента включает последовательность, которая комплементарна Jα сегменту, и где праймеры V сегмента и J-сегмента разрешают амплификацию CDR3 участка TCRα.
[0009] Другим вариантом осуществления данного изобретения является композиция, где праймеры V сегмента гибридизируются с консервативным сегментом и имеют подобную прочность отжига. В другом варианте осуществления праймер V сегмента заякорен в положении -43 в Vβ сегменте относительно сигнальной последовательности рекомбинации (RSS). В другом варианте осуществления многообразие праймеров V сегмента включает по меньшей мере из 45 праймеров, специфических к 45 различным Vβ генам. В другом варианте осуществления праймеры V сегмента имеют последовательности, которые выбраны из группы, включающей SEQ ID NOS:1-45. В другом варианте осуществления праймеры V сегмента имеют последовательности, которые выбраны из группы, включающей SEQ ID NOS:58-102. Другой вариант осуществления, где праймер V сегмента для каждого Vβ сегмента.
[0010] Другим вариантом осуществления данного изобретения является композиция, где праймеры J сегмента гибридизируются с элементом консервативного каркасного участка Jβ сегмента и имеют подобную прочность отжига. Композиция по пункту 2, где многообразие праймеров J сегмента состоит по меньшей мере из тринадцати праймеров, специфических к тринадцати различным Jβ генам. Другим вариантом осуществления является композиция по п.2, где праймеры J сегмента имеют последовательности, выбранные из группы, включающей SEQ ID NOS:46-57. В другом варианте осуществления праймеры J сегмента имеют последовательности, выбранные из группы, включающей SEQ ID NOS:102-113. Другим вариантом осуществления является праймер J сегмента для каждого Jβ сегмента. В другом варианте осуществления все праймеры J сегмента отжигаются с одинаковым консервативным мотивом.
[0011] Другим вариантом осуществления данного изобретения является композиция, где амплифицированная молекула ДНК начинается с указанного консервативного мотива и амплифицирует соответствующую последовательность для диагностической идентификации J сегмента, и включает CDR3 соединение, и распространяется в V сегмент. В другом варианте осуществления каждый из амплифицированных сегментов Jβ гена имеет уникальный свободный конец из четырех оснований в положениях от +11 до +14 ниже RSS сайта.
[0012] Другим аспектом данного изобретения является композиция, дополнительно включающая совокупность се квенирующих олигонуклеотидов, где секвепирующие олигопуклеотиды гибридизируются с участками в амплифицированных молекулах ДНК. Вариантом осуществления являются гибридизация секвенирующих олигоиуклеотидов рядом со свободным концом из четырех оснований в амплифицированных сегментах Jβ генов в положениях от +11 до +14 ниже RSS сайта. В другом варианте осуществления секвенирующие олигонуклеотиды выбраны из группы, включающей SEG ID NOS:58-70. В другом варианте осуществления V сегмент или J-сегмент выбран так, чтобы включать коррекция ошибок последовательности путем слияния близко родственных последовательностей. Другим вариантом осуществления является композиция, дополнительно включающая универсальный праймер С сегмента для образования кДНК из мРНК.
[0013] Другим аспектом данного изобретения является композиция, включающая:
- многообразие праймеров V сегмента, где каждый праймер V сегмента включает последовательность, которая комплементарна отдельному функциональному V сегменту или небольшому семейству V сегментов; и
- многообразие праймеров J сегмента, где каждый ираймер J сегмента включает последовательность, которая комплементарна J сегменту;
где праймеры V сегмента и J-сегмента разрешают амплификацию CDR3 участка TCRG путем многократной полимеразной цепной реакции (PCR) с получением многообразия амплифицированных молекул ДНК, достаточных для определения разнообразия генов тяжелых цепей антител.
[0014] Другим аспектом данного изобретения является композиция, включающая:
- многообразие праймеров V сегмента, где каждый праймер V сегмента включает последовательность, которая комплементарна отдельному функциональному V сегменту или небольшому семейству V сегментов; и
- многообразие праймеров J сегмента, где каждый праймер J сегмента включает последовательность, которая комплементарна J сегменту;
где праймеры V сегмента и J сегмента разрешают амплификацию CDR3 участка тяжелой цепи (IGII) антитела путем многократной полимеразной цепной реакции (PCR) с продуцированным многообразием амплифицированных молекул ДНК, достаточных для определения разнообразия генов тяжелой цепи антитела.
[0015] Другим аспектом данного изобретения является композиция, включающая:
- многообразие праймеров V сегмента, где каждый праймер V сегмента включает последовательность, которая комплементарна отдельному функциональному V сегменту или небольшому семейству V сегментов; и
- многообразие праймеров J сегмента, где каждый праймер J сегмента включает последовательность, которая комплементарна J сегменту;
где праймеры V сегмента и J сегмента разрешают амплификацию VL участка легкой цепи (IGL) антитела путем многократной полимеразной цепной реакции (PCR) с продуцированным многообразием амплифицированных молекул ДНК, достаточных для определения разнообразия генов легкой цепи антитела.
[0016] Другим аспектом данного изобретения является способ, включающий:
- отбор многообразия праймеров V сегмента, где каждый праймер V сегмента включает последовательность, которая комплементарна отдельному функциональному V сегменту или небольшому семейству V сегментов; и
- отбор многообразия праймеров J сегмента, где каждый праймер J сегмента включает последовательность, которая комплементарна J сегменту;
- комбинирование праймеров V сегмента и J сегмента с образцом геномной ДНК для разрешения амплификации CDR3 участка путем многократной полимеразной цепной реакции (PCR) с получением многообразия амплифицированных молекул ДНК, достаточных для определения разнообразия TCR генов.
[0017] Одним вариантом осуществления данного изобретения является способ, где каждый праймер V сегмента включает последовательность, которая комплементарна отдельному функциональному Vβ сегменту, а каждый праймер J сегмента включает последовательность, которая комплементарна Jβ сегменту; и где комбинирование праймеров V сегмента и J сегмента с образцом геномной ДНК разрешает амплификацию CDR3 участка TCR путем многократной полимеразной цепной реакции (PCR) и продуцирует многообразие амплифицированных молекул ДНК. В другом варианте осуществления каждый праймер V сегмента включает последовательность, которая комплементарна отдельному функциональному Vα сегменту, а каждый праймер J сегмента включает последовательность, которая комплементарна Jα сегменту; и где комбинирование праймеров V сегмента и J сегмента с образцом геномной ДНК разрешает амплификацию CDR3 участка TCR путем многократной полимеразной цепной реакции (PCR) и продуцирует многообразие амплифицированных молекул ДНК.
[0018] Другим вариантом осуществления данного изобретения является способ, дополнительно включающий этап секвенирования амплифицированных молекул ДНК. В другом варианте осуществления на этапе секвенирования используется совокупность секвенирующих олигонуклеотидов, которые гибридизируются с участками в амплифицированных молекулах ДНК. Другим вариантом осуществления является способ, дополнительно включающий этап расчета общего разнообразия CDR3 последовательностей TCRβ среди амплифицированных молекул ДНК. В другом варианте осуществления способ показывает, что общее разнообразие нормального человека составляет более 1*106 последовательностей, более 2*106 последовательностей или более 3*106 последовательностей.
[0019] Другим аспектом данного изобретения является способ диагностики иммунодефицита у пациента-человека, включающий измерение разнообразия CDR3 последовательностей TCR пациента, и сравнение разнообразия субъекта с разнообразием, полученным от нормального субъекта. Вариантом осуществления данного изобретения является способ, где измерение разнообразия последовательностей TCR включает этапы:
- отбора многообразия праймеров V сегмента, где каждый праймер V сегмента включает последовательность, которая комплементарна отдельному функциональному V сегменту или небольшому семейству V сегментов; и
- отбор многообразия праймеров J сегмента, где каждый праймер J сегмента включает последовательность, которая комплементарна J сегменту;
- комбинирование праймеров V сегмента и J сегмента с образцом геномной ДНК для разрешения амплификации CDR3 участка TCR путем многократной полимеразной цепной реакции (PCR) с продуцированным многообразием амплифицированных молекул ДНК;
- секвенирование амплифицированных молекул ДНК;
- расчет общего разнообразия CDR3 последовательностей TCR среди амплифицированных молекул ДНК.
[0020] Вариантом осуществления данного изобретения является способ, где сравнение разнообразия определяется расчетом с использованием следующего уравнения:
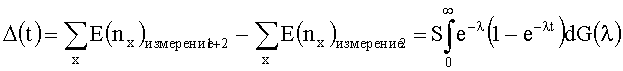
где G(λ) является эмпирической функцией распределения параметров λ1, …, λS, nx является числом клонотипов, секвенированных строго x раз, и
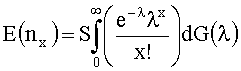
[0021] Другим вариантом осуществления данного изобретения является способ, где сравнивается разнообразие по меньшей мере двух образцов геномной ДНК. В другом варианте осуществления один образец геномной ДНК является образцом от пациента, а другой образец - от нормального субъекта. В другом варианте осуществления один образец геномной ДНК является образцом от пациента перед терапевтическим курсом лечения, а другой образец - от пациента после курса лечения. В другом варианте осуществления два образца геномной ДНК являются образцами от одного и того же пациента в различные точки времени во время курса лечения. В другом варианте осуществления заболевание диагностируется на основе сравнения разнообразия среди образцов геномной ДНК. В другом варианте осуществления иммунокомпетентность пациента-человека оценивается путем сравнения.
ПОДРОБНОЕ ОПИСАНИЕ ИЛЛЮСТРАТИВНЫХ ВАРИАНТОВ ОСУЩЕСТВЛЕНИЯ
[0022] TCR и гены Ig могут образовывать миллионы отличных белков путем соматической мутации. Вследствие этого создающего разнообразие механизма гипервариабельные определяющие комплементарность участки этих генов могут кодировать последовательности, которые могут взаимодействовать с миллионами лигандов, и эти участки связаны с константным участком, который может передавать сигнал клетке, показывающей связывание родственного лиганда белка.
[0023] Адаптивная иммунная система применяет несколько стратегий для создания спектра антигенных рецепторов Т- и В-клеток с достаточным разнообразием для распознавания группы потенциальных патогенов. В αβ и γβ Т-клетках, которые, прежде всего, распознают пеитидпые антигены, представленные молекулами МНС, большая часть из разнообразия рецепторов содержится в третьем определяющем комплементарность участке (CDR3) α и β цепей (или γ и δ цепей) Т-клеточного рецептора (TCR). Хотя было приблизительно оценено, что адаптивная иммунная система может образовывать до 1018 отличных TCR αβ пар, прямая экспериментальная оценка CDR3 разнообразия TCR не удалась.
[0024] В данном документе описывается новый способ измерения CDR3 разнообразия TCR, который основан на секвенировании отдельной молекулы ДНК, и применение этого подхода к последовательности CDR3 участков в миллионах перестроенных TCRβ генов, выделенных из Т-клеток периферической крови двух здоровых взрослых людей.
[0025] Способность адаптивной иммунной системы повышать иммунный ответ, специфический для любого из большого количества потенциальных чужеродных антигенов, которым индивидуум может быть подвергнут, зависит от высоко полиморфных рецепторов, закодированных В-клетками (иммуноглобулинами) и Т-клетками (Т-клеточными рецепторами; TCR). TCR, экспрессированные αβ Т-клетками, которые, прежде всего, распознают пептидные антигены, представленные молекулами главного комплекса гистосовместимости (МНС) класса I и II, являются гетеродимерными белками, включающими две трансмембранные полипептидные цепи (α и β), причем каждая содержит один вариабельный и один константный домен. Пептидная специфичность αβ Т-клеток в значительной части определяется аминокислотной последовательностью, закодированной в петлях третьего определяющего комплементарность участка (CDR3) вариабельных доменов α и β цепи. CDR3 участки β и α цепей сформированы рекомбинацией между неперекрывающимися вариабельным (Vβ), разнообразным (Dβ) и соединяющим (Jβ) сегментами гена в локусе β цепи и между аналогичными Vα и Jα сегментами гена в локусе α цепи, соответственно. Существование множества таких генных сегментов в локусах α и β цепи TCR разрешает большое число отличных CDR3 последовательностей, подлежащих кодированию. Разнообразие CDR3 последовательностей, кроме того, увеличивается независимым от матрицы добавлением и делецией нуклеотидов на Vβ-Dβ, Dβ-Jβ и Vα-Jα соединениях во время процесса генной перестройки TCR.
[0026] Предыдущие попытки оценки разнообразия рецепторов в спектре αβ Т-клеток взрослого человека зависели от исследования перестроенных генов α и β цепи TCR, экспрессированных в небольших хорошо определенных подсовокупностей спектра с последующей экстраполяцией разнообразия, присутствующего в этих подсовокупностях, на полный спектр, с оценкой приблизительно 106 уникальных TCRβ цепи CDR3 последовательностей на индивидуум, с 10-20% этих уникальных TCRβ CDR3 последовательностей, экспрессированными клетками в «обученной» антигеном CD45RO+ категории. Точность и четкость этой оценки весьма ограничены необходимостью экстраполировать разнообразие, наблюдаемое в сотнях последовательностей, на полный спектр, и вполне возможно, что реальное число уникальных TCRβ цепи CDR3 последовательностей в αβ Т-клеточном спектре значительно больше, чем 1×106.
[0027] Недавние достижения в высокопроизводительной технологии ДНК секвенирования сделали возможным значительно более глубокое секвенирование, чем основанные на капиллярах технологии. Комплексная библиотека матричных молекул, несущих универсальные PCR адаптерные последовательности на каждом конце, гибридизирована с «газоном» комплементарных олигонуклеотидов, иммобилизированных на твердой поверхности. Твердофазная PCR используется для амплификации гибридизированной библиотеки, что дает в результате миллионы матричных кластеров на поверхности, причем каждый включает множество (~1000) идентичных копий отдельной молекулы ДНК из оригинальной библиотеки. Интервал 30-54 пар оснований в молекулах в каждом кластере секвенирован с использованием химической технологии обратимой терминации с использованием красителей, для разрешения одновременного секвенирования из геномной ДНК перестроенных CDR3 участков TCRβ цепи, выполняемого в миллионах Т-клеток. Такой подход дает возможность прямого секвенирования значительной доли уникально перестроенных CDR3 участков TCRβ в популяциях αβ Т-клеток, что тем самым разрешает оценить относительную частоту каждой CDR3 последовательности в популяции.
[0028] Точная оценка разнообразия CDR3 последовательностей TCRβ в полном αβ Т-клеточном спектре из разнообразия, измеренного в конечной выборке Т-клеток, требует оценки числа CDR3 последовательностей, присутствующих в спектре, которые не наблюдались в образце. CDR3 разнообразие TCRβ цепи в полном αβ Т-клеточном спектре было оценено с использованием прямых измерений числа уникальных CDR3 последовательностей TCRR, наблюдаемых в образцах крови, содержащих миллионы αβ Т-клеток. Результаты в данном документе идентифицируют нижнюю границу для CDR3 разнообразия TCRβ в CD4+ и CD8+ Т-клеточных категориях, которая в несколько раз выше, чем предыдущие оценки. Кроме того, результаты в данном документе демонстрируют, что имеется по меньшей мере 1,5×106 уникальных CDR3 последовательностей TCRβ в CD45RO+ категории «обученных» антигеном Т-клеток, большая доля которых присутствует при низкой относительной частоте. Существование такой разнообразной популяции CDR3 последовательностей TCRβ в «обученных» антигеном клетках ранее не было продемонстрировано.
[0029] Разнообразный пул TCRβ цепей в каждом здоровом индивидуум является образцом из оцененного теоретического предела более 1011 возможных последовательностей. Однако реализованная совокупность перестроенных TCR не равномерно отобрана из этого теоретического предела. Различные Vβ и Jβ обнаружены с более чем тысячекратной разностью частот. Кроме того, степени инсерции нуклеотидов являются сильно смещенными. Этот уменьшенный предел полученных TCRβ последовательностей ведет к вероятности обладания людьми общими β цепями. С данными о последовательностях, сгенерированными способами, описанными в данном документе, могут быть вычислены in vivo применение J, применение V, моно- и динуклеотидные смещения и зависимое от положения применение аминокислот. Эти смещения значительно уже размера предела последовательностей, из которого выбраны TCRβ, что указывает на то, что различные индивидуумы имеют общие TCRβ цепи с идентичными аминокислотными последовательностями. Результаты в данном документе показывают, что многие тысячи таких идентичных последовательностей являются попарно общими в геномах отдельных людей.
[0030] Технология оценивания использует два пула праймеров для обеспечения высоко мультиплексной реакции PCR. «Прямой» пул имеет праймер, специфический к каждому V сегменту в гене (несколько праймеров, направленных на высоко консервативный участок, используются, чтобы одновременно захватить множество V сегментов). Праймеры «обратного» пула отжигаются с консервативной последовательностью в соединяющем ("J") сегменте. Пул амплифицированного сегмента включает соответствующую последовательность для идентификации каждого J сегмента, а также для разрешения отжига J-сегмент-специфического праймера для ресеквенирования. Это дает возможность непосредственного наблюдения большой фракции соматических перестроек, присутствующих у индивидуума. Это, в свою очередь, позволяет быстро сравнить TCR спектр у индивидуумов с аутоиммунным нарушением (или показанием другого целевого заболевания) с TCR спектром контролей.
[0031] Адаптивная иммунная система теоретически может генерировать огромное разнообразие CDR3 последовательностей Т-клеточных рецепторов - гораздо больше, чем вероятно будет экспрессировано у какого-либо индивидуума в любой момент времени. Предыдущие попытки измерить, какая фракция этого теоретического разнообразия фактически используется в αβ Т-клеточном спектре у взрослого, однако, не предоставили четкую оценку разнообразия. Как описывалось в данном документе, разработка нового подхода к этому вопросу, основанная на секвенировании отдельной молекулы ДНК, и аналитический вычислительный подход к оцениванию разнообразия спектра с использованием измерений разнообразия в конечных выборках. Анализ продемонстрировал, что ряд уникальных CDR3 TCRβ последовательностей в спектре взрослого значительно превышает предыдущие оценки, основанные на исчерпывающем капиллярном секвенировании небольших сегментов спектра. Разнообразие TCRβ цепи в CD45RO- популяции (обогащенной наивными Т-клетками), наблюдаемое с использованием способов, описанных в данном документе, в пять раз больше, чем сообщалось ранее. Главное открытие заключается в числе уникальных CDR3 TCRβ последовательностей, экспрессированных в «обученных» антигеном CD45RO+ Т-клетках, результаты в данном документе показывают, что это число в 10-20 раз больше, чем ожидалось на основе предыдущих результатов других. Распределение частот CDR3 последовательностей в CD45RO+ клетках позволяет предположить, что Т-клеточный спектр содержит большое число клонов с небольшим размером клонов.
[0032] Результаты в данном документе показывают, что полученная совокупность TCRβ цепей отбирается неравномерно из огромного потенциального предела последовательностей. В частности, последовательности Р цепи, более близкие к зародышевой линии (несколько инсерций и делеций на границах V-D и D-J), по-видимому, созданы с относительно высокой частотой. TCR последовательности, близкие к зародышевой линии, являются общими у разных людей, потому что последовательность зародышевой линии для V, D и J является общей, по модулю небольшое число полиморфизмов, среди человеческой популяции.
[0033] Т-клеточные рецепторы, экспрессированные зрелыми αβ Т-клетками, являются гетеродимерами, у которых две составные цепи сгенерированы событиями независимой перестройки вариабельных локусов α и β цепей TCR. α цепь имеет меньшее разнообразие, чем β цепь, так, что более высокая фракция α является общей у индивидуумов, и сотни точных αβ рецепторов TCR являются общими у любой пары индивидуумов.
Клетки
[0034] В-клетки и Т-клетки могут быть полученными из ряда образцов ткани, включая костный мозг, тимус, лимфатические железы, периферические ткани и кровь, но периферическая кровь наиболее легкодоступна. Образцы периферической крови получаются путем флеботомии у субъектов. Мононуклеарные клетки периферической крови (РВМС) выделяются способами, известными специалисту в данной области, например, Ficoll-Hypaque® разделением по градиенту плотности. Предпочтительно, для анализа используются цельные РВМС. В- и/или Т-лимфоциты, вместо, могут быть проточно отсортированы на множество категорий для каждого субъекта: например, CD8+CD45RO+/- и CD4+CD45RO+/- с использованием флуоресцентно меченных античеловеческих антител, например, CD4 FITC (клон М-Т466, Miltenyi Biotec), CD8 РЕ (клон RPA-T8, BD Biosciences), CD45RO ECD (клон UCHL-1, Beckman Coulter) и CD45RO АРС (клон UCIIL-1, BD Biosciences). Окрашивание общих РВМС может быть выполнено с подходящей комбинацией антител с последующим промыванием клеток перед анализом. Лимфоцитные подсовокупности могут быть выделены FACS сортингом, например, с помощью системы сортировки клеток BD FACSAria™ (BD Biosciences), и анализом результатов с программным обеспечением FlowJo (Treestar Inc.), а также концептуально подобными способами, включающими специфические антитела, иммобилизированные на поверхностях или гранулах.
Выделение нуклеиновой кислоты
[0035] Общая геномная ДНК экстрагируется из клеток, например, с использованием QIAamp® DNA blood Mini Kit (QIAGEN®). Приблизительная масса отдельного гаплоидпого генома составляет 3 пг.Предпочтительно, по меньшей мере 100000-200000 клеток используются для анализа разнообразия, т.е. от около 0,6 до 1,2 мкг ДНК из диплоидных Т-клеток. С использованием РВМС в качестве источника, число Т-клеток может быть оценено в около 30% всех клеток.
[0036] Альтернативно, из клеток может быть выделена общая нуклеиновая кислота, включая и геномную ДНК, и мРНК. Если разнообразие оценивается по мРНК в экстракте нуклеиновой кислоты, мРНК должна быть превращена в кДНК до измерения. Это легко может быть выполнено способами, известными специалисту в данной области.
Амплификация ДНК
[0037] Система мультиплексной PCR используется для амплификации перестороенных TCR локусов из геномной ДНК, предпочтительно из CDR3 участка, более предпочтительно из CDR3 участка TCRα, TCRγ или TCRδ, наиболее предпочтительно из CDR3 участка TCRβ.
[0038] В целом, система мультиплексной PCR может использовать по меньшей мере 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24 или 25, предпочтительно 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38 или 39, наиболее предпочтительно 40, 41, 42, 43, 44 или 45 «прямых» праймеров, из которых каждый «прямой» праймер является специфическим к последовательности, соответствующей одному или нескольким сегментам TRB V участка, показанным в SEQ ID NOS:114-248; и по меньшей мере 3, 4, 5, 6, или 7, предпочтительно 8, 9, 10, 11, 12 или 13 «обратных» праймеров, из которых каждый «обратный» праймер является специфическим к последовательности, соответствующей одному или нескольким сегментам TRB J участка, показанным в SEQ ID N08:249-261. Наиболее предпочтительно существует праймер J сегмента для каждого J сегмента.
[0039] Предпочтительно праймеры сконструированы так, чтобы не пересекать границу интрона/экзона. «Прямые» праймеры предпочтительно должны отжигаться с V сегментами на участке относительно сильного сохранения последовательности между V сегментами так, чтобы максимизировать сохранение последовательности среди этих праймеров. Следовательно, это минимизирует потенциал для свойств дифференциального отжига каждого праймера, и, таким образом, амплифицированный участок между V и J праймерами содержит достаточно информации TCR V последовательности для идентификации используемого специфического сегмента V гена.
[0040] Предпочтительно, праймеры J сегментов гибридизируются с консервативным элементом J сегмента и имеют подобную прочность отжига. Наиболее предпочтительно, все праймеры J сегментов отжигаются с одинаковым мотивом консервативного каркасного участка. И «прямые», и «обратные» праймеры предпочтительно модифицированы на 5' конце с последовательностью универсального «прямого» праймера, совместимой с ДНК секвенатором.
[0041] Например, система мультиплексной PCR может использовать 45 «прямых» праймеров (Таблица 1), каждый специфический к функциональному TCR Vβ сегменту, и тринадцать «обратных» праймеров (Таблица 2), каждый специфический к TCR Jβ сегменту. Xn и Yn соответствуют полинуклеотидам с длинами пит, соответственно, которые были бы специфическими для технологии секвенирования отдельной молекулы, используемой для считывания в оценивании.
[0042] 45 «прямых» PCR праймеров Таблицы 1 являются комплементарными к каждому из 48 функциональных вариабельных сегментов, а тринадцать «обратных» PCR праймеров из Таблицы 2 комплементарны к каждому из функциональных соединяющих (J) генных сегментов из TRB локуса (TRBJ). Сегменты TRB V участка идентифицированы в Перечне последовательностей в SEQ ID NOS:114-248, а сегменты TRB J участка - в SEQ ID NOS:249-261. Праймеры сконструированы так, что соответствующая информация представлена в амплифицированной последовательности для однозначной идентификации и V, и J генов (>40 пар оснований последовательности вверх по сигнальной последовательности рекомбинации (RSS) V гена, и >30 пар оснований вниз по RSS J гена). Альтернативные праймеры могут быть выбраны специалистом из генов V и J участков каждой субъединицы TCR.
[0043] «Прямые» праймеры модифицированы на 5' конце с последовательностью универсального «прямого» праймера, совместимой с ДНК секвенатором (Xn Таблицы 1). Подобным образом, все из обратных праймеров модифицированы последовательностью универсального «обратного» праймера (Ym Таблицы 2). Один пример таких универсальных праймеров показан в Таблицах 3 и 4 для секвенирующей системы с одноконцевым считыванием Illumina GAII. 45 TCR Vβ «прямых» праймеров отжигаются с Vβ сегментами на участке относительно сильного сохранения последовательности между Vβ сегментами так, чтобы максимизировать сохранение последовательности среди этих праймеров.
[0044] Общий продукт PCR для перестроенного CDR3 участка TCRR с использованием этой системы, как ожидали, имеет длину приблизительно 200 пар оснований. Геномные матрицы амплифицированы PCR с использованием пула из 45 TCR Vβ F праймеров ("VF пул") и пула из двенадцати TCR Jβ R праймеров ("JR пул"). Например, 50 мкл реакционных смесей PCR могут быть использованы с 1,0 мкМ VF пула (22 пМ для каждого уникального TCR Vβ F праймера), 1,0 мкМ JR пула (77 пМ для каждого уникального TCRBJR праймера), 1X смеси QIAGEN Multiplex PCR master (QIAGEN инвентарный номер 206145), 10% 0-раствора (QIAGEN) и 16 нг/мкл гДНК.
[0045] Совокупность праймеров IGH была сконструирована, чтобы попытаться вместить потенциал для соматической гипермутации в перестроенных IGH генах, как наблюдали после начальной стимуляции наивных В-клеток. Следовательно, все праймеры были сконструированы, чтобы быть немного длиннее нормального, и чтобы заякорить 3' концы каждого праймера в высоко консервативных последовательностях трех или более нуклеотидов, которые должны быть устойчивы и к функциональным, и к нефункциональным соматическим мутациям.
[0046] «Обратные» праймеры IGHJ были сконструированы, чтобы заякорить 3' конец каждого PCR праймера в мотиве высоко консервативной GGGG последовательности в IGHJ сегментах. Эти последовательности показаны в Таблице 5. Подчеркнутые последовательности имеют десять пар оснований из RSS, которые могут быть удалены. Они были исключены из штрихкодовой схемы. Выделенная жирным шрифтом последовательность является обратно-комплементарной последовательностью IGH J «обратных» PCR праймеров. Выделенная курсивом последовательность является штрихкодом для J идентичности (восемь штрихкодов показывают шесть генов и два аллеля в генах). Следующая последовательность в подчеркнутом сегменте может показывать дополнительные аллельные идентичности.
Последовательности IGH J обратных PCR праймеров показаны в Таблице 6.
[0047] V праймеры были сконструированы в консервативном участке FR2 между двумя консервативными триптофановыми (W) кодонами.
[0048] Последовательности праймеров заякорены на 3' конце в триптофановом кодоне для всех IGHV семейств, которые сохраняют этот кодон. Он позволяет последним трем нуклеотидам (TGG триптофана) заякориться на последовательности, которая, как ожидали, будет устойчива к соматической гипермутации, обеспечивая 3' якорь пяти из шести нуклеотидов для каждого праймера. Последовательность протягивается вверх дальше, чем нормальная, и включает вырожденные нуклеотиды, чтобы разрешить ошибочные спаривания, индуцированные гипермутацией (или между близко родственными IGH V семействами) без значительного изменения характеристик отжига праймера, как показано в Таблице 7. Последовательностями V генных сегментов являются SEQ ID NOS:262-420.
[0049] Условия тепловых циклов могут вытекать из способов, известных специалисту в данной области. Например, с использованием термоциклера PCR Express (Hybaid, Ashford, UK), могут быть использованы следующие условия циклов: 1 цикл при 95°С в течение 15 минут, 25-40 циклов при 94°С в течение 30 секунд, 59°С в течение 30 секунд и 72°С в течение 1 минуты с последующим одним циклом при 72°С в течение 10 минут.
Секвенирование
[0050] Секвенирование выполняется с использованием совокупности секвенирующих олигонуклеотидов, которые гибридизируются с определенным участком в амплифицированных молекулах ДНК.
[0051] Предпочтительно, каждый амплифицированный J генный сегмент имеет уникальный свободный конец из четырех оснований в положениях от +11 до +14 ниже RSS сайта. Следовательно, секвенирующие олигопуклеогиды гибридизируются рядом со свободным концом из четырех оснований в амплифицированных Jβ генных сегментах в положениях от +11 до +14 ниже RSS сайта.
[0052] Например, секвенируемые олигонуклеотиды для TCRB могут быть сконструированы для отжига с консенсусным нуклеотидным мотивом, наблюдаемого только ниже этого "свободного конца", так, что первые четыре основания считанной последовательности будут однозначно идентифицировать J сегмент (Таблица 8).
[0053] Информация, используемая для определения J и V сегмента считанной последовательности, полностью вмещена в амплифицированной последовательности и не зависит от идентичности PCR праймеров. Эти секвенируемые олигонуклеотиды были выбраны так, что беспорядочное примирование реакции секвенирования для одного J сегмента олигонуклеотидом, специфическим к другому J сегменту, будет генерировать данные последовательности, начиная точно с такого же нуклеотида, что и данные последовательности от корректного секвенируемого олигонуклеотида. Таким образом, беспорядочный отжиг секвенируемых олигонуклеотидов не повлиял на качество генерируемых данных последовательности.
[0054] Средняя длина CDR3 участка, определенного как нуклеотиды между вторым консервативным цистеином V сегмента и консервативным фенилаланином J сегмента, составляет 35+/-3, поэтому последовательности, начинающиеся от свободного конца Jβ сегмента, почти всегда будут захватывать полное V-D-J соединение в считанных 50 парах оснований.
[0055] βj генные сегменты TCR имеют длину приблизительно 50 пар оснований. PCR праймеры, которые отжигаются и продлеваются с ошибочно спаренными последовательностями, называются беспорядочные праймеры. Jβ «обратные» PCR праймеры TCR были сконструированы для минимизации перекрывания с секвенирующими олигонуклеотидами, чтобы минимизировать беспорядочное примирование в контексте мультиплексной PCR. 13 Jβ «обратных» праймеров TCR заякорены на 3' конце в мотиве консенсусного сплайс-сайта с минимальным перекрыванием секвенируемых праймеров. Jβ праймеры TCR обеспечивают постоянную температуру отжига с использованием программы для секвенатора с параметрами по умолчанию.
[0056] Для реакции секвенирования IGHJ секвенируемые праймеры удлиняются на три нуклеотида по консервативным CAG последовательностям, как показано в Таблице 9.
Последовательность обработки данных
[0057] Для быстрого анализа результатов секвенирования специалистом в данной области может быть разработан алгоритм. Предпочтительным способом является следующее.
[0058] Применение этапа PCR для амплификации CDR3 участков TCRβ перед секвенированием потенциально может ввести систематическое смещение в предполагаемом относительном изобилии последовательностей из-за различий в эффективности PCR амплификации CDR3 участков с использованием разных сегментов Vβ и Jβ генов. Каждый цикл PCR амплификации потенциально вводит смещение средней величины 1,51/15=1,027. Таким образом, 25 циклов PCR вводят общее смещение средней величины 1,02725=1,95 в предполагаемое относительное изобилие отличных последовательностей CDR3 участка.
[0059] Секвенированные считывания были отфильтрованы по тем, которые включают CDR3 последовательности. Обработка данных секвенатора включает серии этапов для устранения ошибок в первичной последовательности каждого считывания и для сжатия данных. Фильтр сложности удаляет приблизительно 20% последовательностей, которые прочитаны неправильно, из секвенатора. Затем последовательности должны были иметь совпадение минимум шести оснований и для одного из тринадцати J-участков TCRB, и для одного из 54 V-участков. При применении фильтра для контрольной линии, содержащей фаговую последовательность, в среднем только одна последовательность из 7-8 миллионов прошла эти этапы. Наконец, был применен алгоритм ближайшего соседа, чтобы разрушить данные уникальных последовательностей путем слияния близкородственных последовательностей, для устранения и ошибки PCR, и ошибки секвенирования.
[0060] При анализе данных соотношение последовательностей в продукте PCR должно быть получено в обратном направлении от данных последовательности перед оцениванием истинного распределения клонотипов в крови. Для каждой последовательности, наблюдаемой данное число раз в данных в данном документе, оценивается вероятность того, что эта последовательность была отобрана из пула PCR конкретного размера. Поскольку секвенированные CDR3 участки отбираются случайным образом из огромного пула продуктов PCR, число наблюдений для каждой последовательности получены из распределений Пуассона. Параметры Пуассона квантованы в соответствии с числом Т-клеточных геномов, которые предоставили матрицу для PCR. Простая смешанная модель Пуассона обеих оценок этих параметров и места попарной вероятности для каждой последовательности являются полученными из каждого распределения. Это метод максимизации ожидаемого результата, который воспроизводит изобилие каждой последовательности, которая была получена из крови.
[0061] Для оценки разнообразия применяется формула "ненаблюдаемых видов". Для употребления этой формулы клонотипы уникальных адаптивных иммунных рецепторов (например, TCRB) занимают место видов. Математическое решение предоставляет, что для общего числа "видов" или клонотипов TCRβ, S, при эксперименте секвенирования наблюдается xs копий последовательности s. Для всех из ненаблюдаемых клонотипов xs равняется 0, и каждый TCR клонотип "захвачен" в крови, полученной согласно пуассоновскому способу с параметром λS. Число Т-клеточных геномов секвенировали в первом измерении 7 и во втором измерении. Поскольку имеется большое число уникальных последовательностей, интеграл будет означать сумму. Если G(X) является эмпирической функцией распределения параметров λ1, …, λS, а nx является числом клонотипов секвенированных строго х раз, тогда общее число клонотипов, т.е. измерение разнообразия Е, выводится из следующей формулы:
[0062] Для данного эксперимента, где Т-клетки отбираются из некого произвольного источника (например, полученная кровь), формула используется для оценки (estimate) общего разнообразия видов во всем источнике. Идея заключается в том, что отобранное число клонотипов при каждом размере вмещает достаточную информацию для оценки исходного распределения клонотипов во всем источнике. Для вывода формулы было оценено число ожидаемых новых видов, если точное измерение повторялось. Лимит формулы такой, как если бы повторяли измерения бесконечное число раз. Результатом является ожидаемое число видов в общей популяции исходного источника. Значение Д(1), число новых клонотипов, наблюдаемых во втором измерении, должно быть определено предпочтительно с использованием следующего уравнения:
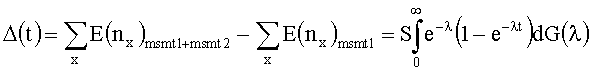
в котором msmt1 и msmt2 являются числом клонотипов из измерения 1 и 2, соответственно. Разложение Тейлора 1-е-λt дает A(t)=E(x1)t-E(x2)t2++E(x3)t3-…, которое может быть округленным, путем замещения ожидаемых результатов E(nx) наблюдаемыми числами в первом измерении. С использованием в числах, наблюдаемых в первом измерении, эта формула предсказывает, что 1,6*105 новых уникальных последовательностей должны наблюдаться во втором измерении. Фактическое значение второго измерения составило 1,8*105 новых последовательностей TCRβ, откуда следует, что прогноз предоставил действительную нижнюю границу общего разнообразия. Было использовано преобразование Эйлера для упорядочивания Δ(t) для продуцирования нижней границы Δ(∞).
Использование измерения разнообразия для диагностики заболевания
[0063] Измерение разнообразия может быть использовано для диагностики заболевания или эффекта курса лечения следующим образом. Спектры Т-клеточных и/или В-клеточных рецепторов могут быть измерены в различные точки времени, например, после курса лечения лейкемии трансплантацией гематопоэтических стволовых клеток (HSCT). И изменение разнообразия, и полное разнообразие спектра TCRB может быть использовано для измерения иммунокомпетентности. Может быть использован стандарт для ожидаемой степени иммунной реконструкция после трансплантации. Степень изменения разнообразия между любыми двумя точками времени может быть использована для интенсивной модификации курса лечения. Общее разнообразие в фиксированной точке времени также является важным показателем, поскольку этот стандарт может быть использован для сравнения различных пациентов. В частности, общее разнообразие является показателем, который должен коррелироваться с клиническим определением иммунной реконструкции. Эта информация может быть использована для модификации режимов приема профилактических лекарственных средств - антибиотиков, противовирусных препаратов и противогрибковых препаратов, например, после HSCT.
[0064] Оценка иммунной реконструкции после аллогенной трансплантации гематопоэтических клеток может быть определена путем измерения изменений разнообразия. Эти способы также расширят анализ того, как разнообразие лимфоцитов снижается с возрастом, если судить по анализу ответов Т-клеток на вакцинацию. Кроме того, способы данного изобретения обеспечивают средство оценки исследуемых терапевтических средств (например, интерлейкин-7 (IL-7)), которые оказывают прямое воздействие па воспроизведение, рост и развитие αβ Т-клеток. Более того, применение этих способов для изучения популяций тимусных Т-клеток обеспечит понимание процессов как генной перестройки Т-клеточного рецептора, так и положительной и отрицательной селекции тимоцитов.
[0065] Новорожденный, у которого еще нет полностью функционирующей иммунной системы, но у которого может быть переданное матерью антитело, является иммунодефицитным. Новорожденный восприимчив к ряду заболеваний, пока его иммунная система автономно развивается, и измерения адаптивной иммунной системы по данному изобретению, вероятно, могут оказаться полезными для новорожденных пациентов.
[0066] Разнообразие лимфоцитов может быть оценено при других состояниях врожденного или приобретенного иммунодефицита. Больные СПИДом пациенты с нарушенной или недостаточной иммунной системой могут быть проконтролированы для определения стадии заболевания и измерения ответа пациента на лечения, направленные на восстановление иммунокомпетентности.
[0067] Другое применение способов данного изобретения заключается в обеспечении диагностических показателей для реципиентов трансплантата паренхиматозного органа, принимающих лекарство так, что их организмы не будут отторгать орган донора. Обычно эти пациенты находятся под иммуносупрессивным лечением. Контролирование иммунокомпетентности хозяина поможет перед и после трансплантации.
[0068] Индивидуумы, подвергшиеся воздействию радиации или химиотерапевтических лекарственных средств, подлежат трансплантации костного мозга или, в противном случае, требуют пополнения популяций Т-клеток, наряду с соответствующей иммунокомпетентностью. Способы данного изобретения обеспечивают средство для качественной и количественной оценки трансплантата костного мозга или восстановления лимфоцитов в ходе этих курсов лечения.
[0069] Одним методом определения разнообразия является сравнение по меньшей мере двух образцов геномной ДНК, из которых предпочтительно один образец геномной ДНК является образцом от пациента, а другой образец взят у нормального субъекта, или, альтернативно, из которых один образец геномной ДНК является образцом от пациента перед терапевтическим курсом лечения, а другой образец - от пациента после курса лечения, или из которых два образца геномной ДНК являются образцами одного и того же пациента в различные точки времени во время курса лечения. Другой метод диагностики может быть основан на сравнении разнообразия среди образцов геномной DNA, например, в которых иммунокомпетентность пациента-человека оценивается путем сравнения.
Биомаркеры
[0070] Общие последовательности TCR у индивидуумов представляют новый класс потенциальных биомаркеров для ряда заболеваний, включая различные типы рака, аутоиммунные заболевания и инфекционные заболевания. Имеются "общие" Т-клетки, которые были описаны для множества заболеваний человека. TCR могут использоваться в качестве биомаркеров, поскольку Т-клетки являются результатом клепального расселения, с помощью которого иммунная система амплифицирует эти биомаркеры посредством быстрого деления клеток. После амплификацияи TCR легко диагностируются, даже если мишень небольшая (например, опухоль ранней стадии). TCR также могут использоваться в качестве биомаркеров, поскольку во многих случаях Т-клетки, кроме того, могут способствовать заболеванию и поэтому могут создавать мишень для лекарственного средства. Т-клеточные самовзаимодействия, как полагают, играют важную роль в некоторых заболеваниях, связанных с аутоиммунитетом, например рассеянный склероз, диабет I типа и ревматоидный артрит.
ПРИМЕРЫ
[0071] Пример 1: Получение образца, выделение РВМС, сортировка FACS и экстракция геномной ДНК
[0072] Образцы периферической крови двух здоровых доноров мужского пола возрастом 35 и 37 лет были получены с письменным информированным согласием с использованием форм, одобренных Экспертным советом организации Центра онкологических исследований Фреда Хатчинсона (FIICRC). Мононуклеарные клетки периферической крови (РВМС) были выделены разделением по градиенту плотности Ficoll-Hypaque®. Т-лимфоциты были проточно отсортированы на четыре категории для каждого субъекта: CD8+CD45RO+/- и CD4+CD45RO+/-. Для характеристики лимфоцитов были использованы следующие конъюгированные античеловеческие антитела: CD4 FITC (клон М-Т466, Miltenyi Biotec), CD8 РЕ (клон RPA-T8, BD Biosciences), CD45RO ECD (клон UCHL-1, Beckman Coulter) и CD45RO АРС (клон UCIIL-1, BD Biosciences). Окрашивание общих РВМС было выполнено с подходящей комбинацией антител в течение 20 минут при 4°С, и окрашенные клетки были один раз промыты перед анализом. Подсовокупности лимфоцитов были выделены FACS сортировкой в системе сортировки клеток BD FACSAria™ (BD Biosciences). Данные были проанализированы на программном обеспечении FlowJo (Treestar Inc.).
[0073] Общая геномная ДНК была экстрагирована из отсортированных клеток с использованием набора QIAamp® DNA blood Mini Kit (QIAGEN®). Приблизительная масса отдельного гаплоидного генома составляет 3 пг. Для того чтобы миллионы образцов перестроенных TCRB в каждой категории Т-клеток, было получено 6-27 микрограмм матричной ДНК из каждой категории (смотри Таблицу 10).
[0074] Пример 2: Виртуальное спектральное типирование β цепи Т-клеточного рецептора
[0075] Виртуальное спектральное типирование β цепи TCR было выполнено следующим образом. Комплементарная ДНК была синтезирована из РНК, экстрагированной из отсортированных Т-клеточных популяций, и использовалась в качестве матрицы для мультиплексной PCR амплификации перестроенного CDR3 участка β цепи TCR. Каждая мультиплексная реакция содержала 6-РАМ-меченый антисмысловой праймер, специфический для константного участка β цепи TCR, и от двух до пяти вариабельных ген-специфических смысловых праймеров β цепи TCR (TRBV). Были исследованы все 23 функциональных Vβ семейства. PCR реакции были выполнены на термоциклере Hybaid PCR Express (Hybaid, Ashford, UK) при следующих условиях цикла: 1 цикл при 95°С в течение 6 минут, 40 циклов при 94°С в течение 30 секунд, 58°С в течение 30 секунд и 72°С в течение 40 секунд, с последующим 1 циклом при 72°С в течение 10 минут. Каждая реакционная среда содержала матрицу кДНК, 500 мкМ dNTP, 2 мМ MgCl2 и 1 единицу ДНК-полимеразы AmpliTaq Gold (Perkin Elmer) в буфере AmpliTaq Gold, в окончательном объеме 20 мкл. После завершения аликвота PCR продукта была разбавлена 1:50 и проанализирована с использованием анализатора ДНК. Выход анализатора ДНК был превращен в распределение интенсивности флуоресценции против длины путем сравнения со следом интенсивности флуоресценции эталонного образца, содержащего стандарты известного размера.
[0076] Пример 3: Мультиплексная PCR амплификация CDR3 участков TCRβ
[0077] CDR3 участок соединения был оперативно определен следующим образом. Соединение начинается со второго консервативного цистенпа V-участка и заканчивается на консервативном фенилаланине J-участка. С получением обратно-комплементарных наблюдаемых последовательностей и транслированием фланкирующих участков были идентифицированы аминокислоты, определяющие границы соединения. Число нуклеотидов между этими границами определяет длину и, поэтому, рамку CDR3 участка. Для того, чтобы образовать матричную библиотеку для секвенирования система мультиплексной PCR была выбрана для амплификации перестроенных TCRβ локусов из геномной ДНК. Система мультиплексной PCR использует 45 «прямых» праймеров (Таблица 3), каждый специфичен к функциональному Vβ сегменту TCR, и тринадцать «обратных» праймеров (Таблица 4), каждый специфичен к Jβ сегменту TCR. Праймеры были выбраны для предоставления того, что адекватная информация представлена в амплифицированной последовательности для однозначного идентифицирования, и V, и J генов (>40 пар оснований последовательности вверх по сигнальной последовательности рекомбинации (RSS) V гена и >30 пар оснований вниз но RSS J гена).
[0078[«Прямые» праймеры модифицированы на 5' конце с последовательностью универсального «прямого» праймера, совместимой с твердофазовой PCR на кластерной станции Illumina GA2. Сходным образом, все из «обратных» праймеров модифицированы с последовательностью универсального «обратного» праймера GA2. 3' конец каждого «прямого» праймера заякорен в положении -43 в Vβ сегменте, относительно сигнальной последовательности рекомбинации (RSS), тем самым обеспечивая уникальную Vβ последовательность свободного конца в амплифицированном участке. Тринадцать «обратных» праймеров, специфических к каждому Jβ сегменту, заякорены на 3' интроне, причем 3' конец каждого праймера пересекает соединение интрон/экзон. Были сконструированы тринадцать секвенируемых праймеров, комплементарных Jβ сегментам, которые комплементарны амплифицированной части Jβ сегмента так, что несколько первых оснований сгенерированной последовательности будут захватывать уникальную Jβ последовательность свободного конца.
[0079] В среднем J делециями были 4 пар оснований +/-2,5 пары оснований, что подразумевает, что J делеции более 10 нуклеотидов встречаются в менее 1% последовательностей. Каждый из тринадцати различных Jβ генных сегментов TCR имел уникальный свободный конец из четырех оснований в положениях от +11 до +14 вниз по RSS сайту. Таким образом, были сконструированы секвенируемые олигонуклеотиды для отжига с консенсусным нуклеотидным мотивом, наблюдаемым только вниз по этому "свободному концу", так, что первые четыре основания считанной последовательности будут однозначно идентифицировать J сегмент (Таблица 5).
[0080] Использованная информация для определения J и V сегмента считанной последовательности целиком содержится в пределах амплифицированной последовательности и не зависит от идентичности PCR праймеров. Эти секвенируемые олигонуклеотиды были выбраны так, что беспорядочное примирование реакции секвенирования для одного J сегмента олигонуклеотидом, специфическим к другому J сегменту, будет генерировать данные последовательности, начиная с точно такого же нуклеотида, что и данные последовательности от корректного секвенируемого олигонуклеотида. Таким образом, беспорядочный отжиг секвенируемых олигонуклеотидов не влияет на качество генерированных данных о последовательностях.
[0081] Средняя длина CDR3 участка, определенного, следуя правилу, как нуклеотиды между вторым консервативным цистеином V сегмента и консервативным фенилаланином J сегмента, составляет 35+/-3, поэтому последовательности, начинающиеся с Jβ сегментного свободного конца, будут почти всегда захватывать полное VNDNJ соединение в считанных 50 парах оснований.
[0082] βJ генные сегменты TCR имеют длину приблизительно 50 пар оснований. PCR праймеры, которые отжигаются и удлиняются с ошибочно спаренными последовательностями, называются беспорядочными праймерами. Вследствие риска беспорядочного примирования в контексте мультиплексной PCR, особенно в контексте генного семейства, были сконструированы «обратные» PCR праймеры TCR Jβ для минимизации перекрывания с секвенируемыми олигонуклеотидами. Таким образом, 13 TCR Jβ «обратных» праймеров заякорены на 3' конце на мотиве консенсусного сплайс-сайта с минимальным перекрыванием секвенируемых праймеров. Были сконструированы TCR Jβ праймеры для постоянной температуры отжига (58 градусов в 50 мМ соли) с использованием программы OligoCalc с параметрами по умолчанию (http://www.basic. northwestern.edu/biotools/onnrocalc.html).
[0083] Были сконструированы 45 TCR Vβ «прямых» праймеров для отжига с Vβ сегментами на участке относительно сильного сохранения последовательности между Vβ сегментами для двух точных целей. Во-первых, увеличение до максимума сохранения последовательности среди этих праймеров минимизирует потенциал для свойств дифференциального отжига каждого праймера. Во-вторых, праймеры были выбраны так, что амплифицированный участок между V и J праймерами будет включать достаточную информацию последовательности TCR Vβ для идентификации используемого специфического Vβ генного сегмента. Это устраняет риск ошибочного распределения TCR Vβ генного сегмента в событии беспорядочного примирования TCR Vβ праймерами. TCR Vβ «прямые» праймеры были сконструированы для всех известных непсевдогенов в TCRβ локусе.
[0084] Общий PCR продукт для успешно пререстроенного CDR3 участка TCRβ с использованием этой системы, как ожидается, имеет длину приблизительно 200 пар оснований. Геномные матрицы были PCR амплифицированы с использованием эквимолярного пула 45 TCR Vβ F праймеров ("VF пул") и эквимолярного пула тринадцати TCR Jβ R праймеров ("JR пул"). 50 мкл реакционной смеси PCR было установлено при 1,0 мкМ VF пула (22 нМ для каждого уникального TCR Vβ F праймера), 1,0 мкМ JR пула (77 пМ для каждого уникального TCRBJR праймера), 1X смеси QIAGEN Multiplex PCR master (QIAGEN инвентарный номер 206145), 10% Q-раствора (QIAGEN) и 16 нг/мкл гДНК. Были использованы следующие условия термического цикла на термоциклере PCR Express (Hybaid, Ashford, UK) при следующих условиях цикла: 1 цикл при 95°С в течение 15 минут, 25-40 циклов при 94°С в течение 30 секунд, 59°С в течение 30 секунд и 72°С в течение 1 минуты, с последующим одним циклом при 72°С в течение 10 минут. 12-20 лунок PCR были выполнены для каждой библиотеки для того, чтобы отобрать от сотен тысяч до миллионов перестроенных TCRβ CDR3 локусов.
[0085] Пример 4: Предварительная обработка данных последовательности
[0086] Обработка данных секвенатора включает серии этапов для устранения ошибок в первичной последовательности каждого считывания и сжатия данных. Сначала фильтр сложности удаляет из секвенатора приблизительно 20% последовательностей, которые прочитаны неправильно. Затем последовательности должны были иметь совпадение минимум шести оснований, и для одного из тринадцати J-участков, и для одного из 54 V-участков. При употреблении фильтра для контрольной линии, содержащей фаговую последовательность, в среднем только одна последовательность из 7-8 миллионов прошла эти этапы без ложного распознавания сигнала. Наконец, был применен алгоритм ближайшего соседа, чтобы разрушить данные уникальных последовательностей путем слияния близкородственных последовательностей, для устранения, и ошибки PCR, и ошибки секвенирования (смотри Таблицу 10).
[0087] Пример 5: Оценивание относительного изобилия последовательности CDR3 в PCR пулах и образцах крови
[0088] После разрушения данных, исходное распределение Т-клеточных последовательностей в восстановлении крови было получено из данных последовательности. В способе используются три этапа: 1) проточная сортировка Т-клеток, полученных из периферической крови, 2) PCR амплификация и 3) секвенирование. При анализе данных соотношение последовательностей в PCR продукте должно быть получено в обратном направлении от данных последовательности перед оцениванием истинного распределения клонотипов в крови.
[0089] Для каждой последовательности, наблюдаемой данное число раз в данных в данном документе, оценивается вероятность того, что эта последовательность была отобрана из пула PCR конкретного размера. Поскольку секвенированные CDR3 участки отбираются случайным образом из огромного пула PCR продуктов, число наблюдений для каждой последовательности получены из распределений Пуассона. Параметры Пуассона квантованы в соответствии с числом Т-клеточных геномов, которые предоставили матрицу для PCR. Простая смешанная модель Пуассона, и оценок этих параметров, и места попарной вероятности для каждой последовательности являются полученными из каждого распределения. Это способ максимизации ожидаемого результата, который воспроизводит изобилие каждой последовательности, которая была взята из крови.
[0090] Пример 6: Модель ненаблюдаемых видов для оценивания истинного разнообразия
[0091] Смешанная модель может воссоздавать частоту каждого TCRβ CDR3 вида, полученного из крови, но возникает важный вопрос, сколько уникальных CDR3 видов присутствовало у донора? Это фундаментальный вопрос, который нуждается в ответе, поскольку у каждого донора доступный образец ограничен, и в будущем будет иметь более важное значение, так как эти способы экстраполированы на меньшие объемы крови, которые обоснованно могут быть получены от пациентов, подлежащих курсу лечению.
[0092] Математическое решение предоставляет то, что для общего числа "видов" или клонотипов TCRβ, S, при эксперименте секвенирования наблюдается xs копий последовательности s. Для всех из ненаблюдаемых клонотипов xs равняется 0, и каждый клонотип TCR "захвачен" в крови, полученной согласно пуассоновскому способу с параметром λS. Число Т-клеточных геномов секвенировали в первом измерении 1 и во втором измерении. Поскольку имеется большое число уникальных последовательностей, интеграл будет означать сумму. Если G(λ) является эмпирической функцией распределения параметров λ1, …, λS, а nx является числом клонотипов секвенированных строго х раз, тогда
[0093] Значение A(t) является числом новых клонотипов, наблюдаемых во втором эксперименте секвенирования.
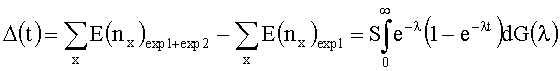
[0094] Разложение Тейлора l-e-λt дает Δ(t)=E(x1)t-E(x2)t2+E(x3)t3-…, которое может быть округленным, путем замещения ожидаемых результатов (Е(nx)) наблюдаемыми числами в первом измерении. С использованием в числах, наблюдаемых в первом измерении, эта формула предсказывает, что 1,6*105 новых уникальных последовательностей должны наблюдаться во втором измерении. Фактическое значение второго измерения составило 1,8* 105 новых последовательностей TCRβ, которые предполагают, что прогноз предоставил действительную нижнюю границу общего разнообразия. Было использовано преобразование Эйлера для упорядочения Δ(t) для продуцирования нижней границы Δ(∞).
[0095] Пример 7: Коррекция ошибки и оценка смещения
[0096] Ошибка последовательности в данных первичной последовательности получается прежде всего из двух источников: (1) ошибка включения нуклеотида, которая происходит во время амплификации TCRβ CDR3 матричных последовательностей с помощью PCR, и (2) ошибки восстановлений последовательности оснований, введенных во время секвенирования PCR-амплифицированпой библиотеки CDR3 последовательности. Большое количество данных разрешает выполнять прямой корректирующий ошибки код для коррекции большинства ошибок в данных первичной последовательности, относимых к этим двум источникам. После коррекции ошибок число уникальных в рамке CDR3 последовательностей и число наблюдений каждой уникальной последовательности были сведены в таблицу для каждой из четырех проточно отсортированных Т-клеточных популяций от двух доноров. Относительная частота распределения CDR3 последовательностей в четырех определенных проточной цитометрией популяциях продемонстрировала, что «обученные» антигеном CD45RO+ популяции содержали значительно больше уникальных CDR3 последовательностей с высокой относительной частотой, чем CD45RO- популяции. Гистограммы частоты CDR3 последовательностей TCRβ, наблюдаемых в четырех различных Т-клеточных подсовокупностях, отличающихся экспрессией CD4, CD8 и CD45RO и присутствующих в крови, показали, что каждая из десяти уникальных последовательностей наблюдались 200 раз в CD4+CD45RO+ («обученном» антигеном) Т-клеточном образце, который наблюдался чаще, в два раза, чем в CD4+CD45RO- популяциях.
[0097] Использование этапа PCR для амплификации CDR3 участков TCRR перед секвенированием могло бы потенциально ввести систематическое смещение в подразумеваемое относительное изобилие последовательностей из-за различий в эффективности PCR амплификации CDR3 участков с использованием различных Vβ и Jβ генных сегментов. Для оценки величины какого-либо такого смещения CDR3 участки TCRR из образца приблизительно 30000 уникальных CD4+CD45RO+ Т-лимфоцитных геномов было амплифицировано с помощью 25 циклов PCR, при которой точка PCR продукта была разделена на две половины. Половина совокупности была отложена, а другая половина PCR продукта была амплифицирована при дополнительных 15 циклах PCR, всего 40 циклов амплификации. Затем PCR продукты, амплифицированные с помощью 25 и 40 циклов, были секвенированы и сравнены. Более 95% последовательностей из 25 циклов также были обнаружены в образце из 40 циклов: линейная корреляция наблюдается при сравнении частоты последовательностей у этих образцов. Для последовательностей, наблюдаемых данное число раз в линии 25 циклов, комбинация смещения PCR и отбор ряд входит в ряд по среднему числу наблюдений при 40 циклах. Консервативно приписывая среднее отклонение по линии (в 1,5 раза) полностью смещению PCR, каждый цикл PCR амплификации потенциально вводит смещение средней величины 1,51/15=1,027. Таким образом, 25 циклов PCR вводят общее смещение средней величины 1,02725=1,95 в полученную относительную представленность отличных последовательностей CDR3 участка.
[0098] Пример 8: Использование Jβ генного сегмента
[0099] CDR3 участок в каждой β цепи TCR включает последовательность, полученную из одного из тринадцати Jβ генных сегментов. Анализ CDR3 последовательностей в четырех различных Т-клеточных популяциях от двух доноров продемонстрировал, что фракция общих последовательностей, которые включали последовательности, полученные из тринадцати различных Jβ генных сегментов, изменилась более чем в 20 раз. Использование Jβ среди четырех различных Т, определенных проточной цитометрией Т-клеток от отдельного донора, было относительно константным у данного донора. Более того, паттерны использования Jβ, наблюдаемые у двух доноров, которые были подрозумеваемы из анализа геномной ДНК из Т-клеток, секвенированных с использованием GA, качественно подобны таковым, наблюдаемым в Т-клетках из пуповинной крови и от здоровых взрослых доноров, оба из которых были подрозумеваемы из анализа кДНК из Т-клеток, секвенированных с использованием исчерпывающих, основанных на капиллярах способов.
[0100] Пример 9: Смещение нуклеотидной инсерции
[0101] Большая часть разнообразия на CDR3 соединениях в α и β цепях TCR создается инсерциями нематричных нуклеотидов с помощью фермента концевой дезоксинуклеотидил-трансферазы (TdT). Однако in vivo выбор играет значительную роль в создании спектра TCR, что приводит к непредсказуемости. Частоты TdT нуклеотидной инсерции, независимые от выбора, были рассчитаны с использованием TCR последовательностей рамки. Эти последовательности являются нефункциональными перестройками, которые выполняются на одном аллеле в Т-клетках, где второй аллель имеет функциональную перестройку. Смещение мононуклеотидной инсерции TdT способствует С и G (Таблица 11).
[0102] Подобные частоты нуклеотидов наблюдаются в последовательностях рамки (Таблица 12).
[0103] N участки из последовательностей TCR вне рамки были использованы для показателя динуклеотидного смещения. Для выделения маргинального вклада динуклеотидного смещения, динуклеотидные частоты были разделены на мононуклеотидные частоты каждого из двух оснований. Показатель представляет собой
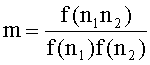
[0104] Матрица для m представлена в Таблице 13.
[0105] Многие из динуклеотидов являются недо- или сверхпоказанными. Например, вероятности обнаружения GG пары являются очень высокими. Поскольку кодоны GGN транслируются в глицин, на CDR3 участках ожидается множество глицинов.
[0106] Пример 10: Распределения аминокислот на CDR3 участках
[0107] Распределение аминокислот на CDR3 участках β цепей TCR является общим для зародышевых последовательностей для V, D и J участков, смещения инсерции TdT и выбора. Распределение аминокислот в этом участке для четырех различных Т-клеточных подкатегорий очень похоже у различных подтипов Т-клеток. Разделение последовательностей в β цепях фиксированной длины, зависимое от положения распределение среди аминокислот, которые сгруппированы по шести химическим признакам: небольшие, специфические и крупные гидрофобные, нейтральные полярные, кислотные и основные. Распределения практически идентичны, за исключением CD8+«обученных» антигеном Т-клеток, которые имеют более высокую долю кислотных оснований, особенно в положении 5.
[0108] Особенный интерес представляет сравнение между CD8+ и CD4+ последовательностями TCR, поскольку они связываются с пептидами, представленными HLA молекулами класса I и класса II, соответственно. CD8+ «обученные» антигеном Т-клетки имеют несколько положений с более высокой долей кислотных аминокислот. Это могло бы выполнить связывание с основным остатком, обнаруженным в HLA молекулах класса I, но не класса II.
[0109] Пример 11: β цепи TCR с идентичными аминокислогными последовательностями, обнаруженные у разных людей
[0110] Последовательности β цепи TCR были транслированы в аминокислоты, а затем попарно сравнивались у двух доноров. Наблюдалось несколько тысяч точных совпадений последовательностей. Например, при сравнении CD4+CD45RO- подкатегорий приблизительно 8000 из 250000 уникальных аминокислотных последовательностей от донора 1 точно совпадали с таковыми у донора 2. Многие из этих совпадающих последовательностей на аминокислотном уровне имеют множество нуклеотидных различий в положениях третьего кодона. Из примера, упомянутого выше, 1500/8000 идентичных аминокислотных совпадений имели >5 ошибочных спариваний нуклеотидов. Между любыми двумя Т-клеточными подтипами 4-5% уникальных последовательностей TCRβ, как было обнаружено, имеют идентичные аминокислотные совпадения.
[0111] Были рассмотрены две возможности: что 1) отбор при развитии TCR продуцирует эти общие последовательности, и 2) большое смещение в частоте нуклеотидной инсерции с помощью TdT создает сходные нуклеотидные последовательности. Попарные совпадения в рамке были сравнены с попарными совпадениями вне рамки (смотри Примеры 1-4, выше). Изменяющиеся рамки зафиксировали все из признаков генетического кода, и поэтому должно быть найдено одинаковое число совпадений, если смещение последовательности отвечало за все наблюдение. Однако было обнаружено почти в два раза больше совпадений в рамке, чем совпадений вне рамки, что позволяет предположить, что отбор на уровне белка играет значительную роль.
[0112] Для подтверждения этого обнаружения тысяч идентичных аминокислотных последовательностей β цепи TCR были сравнены два донора в отношении CD8+CD62L+CD45RA+ (подобных наивным) TCR от третьего донора - 44-летней женщины кавказской национальности CMV+. Были обнаружены идентичные попарные совпадения нескольких тысяч последовательностей на аминокислотном уровне между третьим донором и каждым из первоначальных двух доноров. В отличие от этого, 460 последовательностей были общими у всех трех доноров. Большое отклонение в общем числе уникальных последовательностей между донорами является продуктом исходного материала и отклонений загрузки в секвенатор, а не является показательным для отклонения в истинном разнообразии в крови доноров.
[0113] Пример 12: Клонотипы более высокой частоты являются более близкими к зародышевым
[0114] Отклонение в числе копий между различными последовательностями в каждой Т-клеточной подкатегории изменялось с коэффициентом более 10000 раз. Единственным свойством, коррелированным с числом копий, было (число инсерций плюс число делеций), которое обратно зависимо. Результаты анализа показали, что делеции играют меньшую роль, чем инсерции в обратной зависимости с числом копий.
[0115] Последовательности с меньшим количеством инсерций и делеций имеют рецепторные последовательности, более близкие к зародышевой линии. Одной из возможностей увеличения числа последовательностей, более близких к зародышевой линии, является то, что они создаются множество раз во время развития Т-клеток. Так как последовательности зародышевой линии общие у людей, общие TCRβ цепи, вероятно, создаются с помощью TCR с небольшим числом инсерций и делеций.
[0116] Пример 13: Анализ "спектрального типа" CDR3 последовательностей TCRβ с помощью использования V генного сегмента и CDR3 длины
[0117] TCR разнообразие главным образом оценивали с использованием способа спектрального типирования TCR, способ, основанный на PB-PCR, которая не позволяет оценить разнообразие CDR3 TCR на уровне последовательностей, а оценивает разнообразие длины CDR3 TCRα или TCRR, выраженные как мРНК в субпопуляциях αβ Т-клеток, которые используют одинаковый Vα или Vβ генный сегмент. Спектральные типы поликлональных Т-клеточных популяций с разнообразными спектрами CDR3 последовательностей TCR, например, рассматриваемых в пуповинной крови или в периферической крови здоровых молодых людей, обычно содержат CDR3 последовательности 8-10 различных длин, которые являются множествами из трех нуклеотидов, отражая отбор по транскриптам в рамке. Спектральное типирование также обеспечивает приблизительную количественную информацию об относительной частоте CDR3 последовательностей с каждой специфической длиной. Чтобы оценить, сможет ли прямое секвенирование CDR3 участков TCRβ из Т-клеточной геномной ДНК с использованием секвенатора точно захватить все из разнообразия длин CDR3, которые идентифицированы спектральным типированием, "виртуальные" спектральные типы TCRβ (смотри Примеры выше) были сгенерированы из данных последовательности и сравнивались со спектральными типами TCRβ, сгенерированными с использованием традиционных PCR способов. Виртуальные спектральные типы вмещали все из длин CDR3 и информацию об относительной частоте, представленную в традиционных спектральных типах. Прямое секвенирование CDR3 TCRβ захватывает всю информацию разнообразия TCR, представленную в традиционном спектральном типе. Сравнение данных стандартного спектрального типа TCRβ и вычисленных распределений длин CDR3 TCRβ для последовательностей с использованием типичных Vβ генных сегментов TCR и присутствием в CD4+CD45RO+ клетках от донора 1. Сокращение информации, вмещенной в последовательности данных, на гистограмме частоты уникальных последовательностей CDR3 с различными длинами в каждом Vβ семействе легко воспроизводит всю информацию, содержащуюся в данных спектрального типа. Кроме того, виртуальные спектральные типы выявили присутствие в каждом Vβ семействе редкие последовательности CDR3 и с очень короткой, и с очень протяженной длинами CDR3, которые не были обнаружены традиционным спектральным тонированием на основе PCR.
[0118] Пример 14: Оценивание общего разнообразия последовательностей CDR3
[0119] После коррекции ошибок число уникальных последовательностей CDR3, наблюдаемых в каждой линии проточной ячейки секвенатора, обычно превышала 1×105. Учитывая, что PCR продукты, секвенированные в каждой линии, обязательно были получены из небольшой фракции Т-клеточных геномов, присутствующих у каждого из двух доноров, общее число уникальных последовательностей CDR3 TCRβ во всем Т-клеточном спектре каждого индивидуума, вероятно, будет намного выше. Оценивание числа уникальных последовательностей во всем спектре, следовательно, требует оценки числа дополнительных уникальных последовательностей CDR3, которые существуют в крови, но не наблюдались в образце. Оценивание общего разнообразия видов в большой, комплексной популяции с использованием измерений разнообразия видов, присутствующего в выборе конечного объема, исторически назвали "проблемой ненаблюдаемых видов" (смотри Примеры выше). Решение начинается с определения числа новых видов или CDR3 последовательностей TCRβT, которые наблюдаются, если эксперимент повторяется, т.е. если секвенирование повторяется на идентичном образце Т-клеток периферической крови, например, на идентично полученной библиотеке PCR продуктов CDR3 TCRβ в различных линиях проточной ячейки секвенатора, и подсчитывается число новых последовательностей CDR3. Для CD8+CD45RO- клеток от донора 2 предсказано и обнаружено несколько новых последовательностей CDR3 во второй линии в пределах 5% (смотри Примеры выше), что предполагает, что это аналитическое решение может, в самом деле, быть использовано для оценки общего числа уникальных последовательностей CDR3 TCRβ во всем спектре.
[0120] Полученные в результате оценки общего числа уникальных последовательностей CDR3 TCRβ в четырех определенных проточной цитометрией Т-клеточных категориях показаны в Таблице 14.
[0121] Примечательно, что общее разнообразие TCRβ в этих популяциях составляет 3-4 миллиона уникальных последовательностей в периферической крови. Удивительно, но CD45RO+ или «обученная» антигеном категория вмещает приблизительно 1,5 миллиона из этих последовательностей. Это по меньшей мере на порядок больше, чем ожидалось. Это расхождение, скорее всего, связано с большим числом этих последовательностей, наблюдаемых при низкой относительной частоте, что может быть выявлено только путем глубокого секвенирования. Оцененные размеры спектра CDR3 TCRβ каждой категории у двух доноров находятся в пределах 20% друг от друга.
[0122] Результаты в данном документе демонстрируют, что реализованное разнообразие рецептора TCRβ по меньшей мере в пять раз выше, чем предварительные оценки (~4*106 отличных CDR3 последовательностей), и, в частности, предполагают значительно большее разнообразие TCRβ среди CD45RO+ «обученных» антигеном αβ Т-клеток, чем это ранее сообщалось (~1,5*106 отличных CDR3 последовательностей). Однако биоинформатический анализ данных последовательностей TCR показывает сильные смещения в содержании моно- и динуклеотидов, что подразумевает, что использованные последовательности TCR отбираются из распределения гораздо меньшего, чем теоретический размер. При большом разнообразии цепей TCRβ у каждого человека, отобранных из сильно сокращенного предела последовательностей, у каждого человека можно ожидать перекрывание пулов последовательностей TCR. Фактически, результаты показали, что около 5% CD8+ наивных цепей TCRβ с точными аминокислотными совпадениями являются общими у каждой пары трех разных индивидуумов. Поскольку TCRα пул, как ранее было измерено, существенно меньше, чем теоретическое разнообразие TCRβ, эти результаты показывают, что могут быть обнаружены сотни и тысячи достоверных общих αβ TCR.
| название | год | авторы | номер документа |
|---|---|---|---|
| КОМПОЗИЦИИ И СПОСОБЫ ИЗМЕРЕНИЯ И КАЛИБРОВКИ СИСТЕМАТИЧЕСКОЙ ОШИБКИ АМПЛИФИКАЦИИ В МУЛЬТИПЛЕКСНЫХ ПЦР-РЕАКЦИЯХ | 2013 |
|
RU2631797C2 |
| КОНЪЮГАТЫ АФФИННАЯ МОЛЕКУЛА-ОЛИГОНУКЛЕОТИД И ИХ ПРИМЕНЕНИЯ | 2017 |
|
RU2763554C2 |
| Vβ-Dβ-Jβ ОЛИГОНУКЛЕОТИД Т-КЛЕТОЧНОГО РЕЦЕПТОРА, ПАРА ПРАЙМЕРОВ, ОЛИГОНУКЛЕОТИДНЫЙ ЗОНД, СПОСОБ ОБНАРУЖЕНИЯ КЛОНА MBP83-99Vβ13.1 Т-КЛЕТОК, ЭКСПРЕССИРУЮЩИХ МОТИВ LGRAGLTY Т-КЛЕТОЧНОГО РЕЦЕПТОРА, НАБОР, СПОСОБ ЛЕЧЕНИЯ АУТОИММУННОГО ЗАБОЛЕВАНИЯ И СПОСОБ ЕГО МОНИТОРИНГА | 2000 |
|
RU2251552C2 |
| ГЕНЕТИЧЕСКИ МОДИФИЦИРОВАННЫЕ В ОТНОШЕНИИ Т-КЛЕТОЧНОГО РЕЦЕПТОРА МЫШИ | 2012 |
|
RU2661106C2 |
| СПОСОБ ОПРЕДЕЛЕНИЯ МАРКЕРОВ НАЛИЧИЯ ОПУХОЛЕВЫХ Т-ЛИМФОБЛАСТОВ | 2016 |
|
RU2674994C2 |
| Способ получения человеческого Т-клеточного рецептора, специфичного к эпитопу 112-120 (KVAELVHFL) рецептора MAGE-A3 | 2024 |
|
RU2840503C1 |
| КОНЪЮГАТЫ АФФИННАЯ ЧАСТЬ-ОЛИГОНУКЛЕОТИД И ИХ ПРИМЕНЕНИЯ | 2016 |
|
RU2778754C2 |
| СПОСОБ ИЗМЕРЕНИЯ ИЗМЕНЕНИЯ В ИММУННОМ РЕПЕРТУАРЕ ИНДИВИДУУМА | 2016 |
|
RU2714752C2 |
| СЕКВЕНИРОВАНИЕ ПОЛИНУКЛЕОТИДНЫХ БИБЛИОТЕК С ВЫСОКОЙ ПРОПУСКНОЙ СПОСОБНОСТЬЮ И АНАЛИЗ ТРАНСКРИПТОМОВ | 2018 |
|
RU2790291C2 |
| ОПОСРЕДОВАННЫЕ ГУМАНИЗИРОВАННЫМИ Т-КЛЕТКАМИ ИММУННЫЕ ОТВЕТЫ У НЕ ОТНОСЯЩИХСЯ К ЧЕЛОВЕКУ ЖИВОТНЫХ | 2016 |
|
RU2732628C2 |











































































































































































Изобретение относится к области молекулярной биологии, биохимии, иммунологии и генетической инженерии. Предложена композиция для определения разнообразия CDR3 последовательностей TCRB или IGH в образце. Изобретение может быть использовано для оценки адаптивной иммунной системы пациентов в медицине. 35 з.п. ф-лы, 14 табл., 14 пр.
1. Композиция для определения разнообразия CDR3 последовательностей TCRB или IGH в образце, включающая:
(a) множество праймеров V-сегмента и
(b) множество праймеров J-сегмента;
при этом каждый из указанного множества праймеров V-сегмента и указанного множества праймеров J-сегмента состоит из 15-50 нуклеотидов,
где указанное множество праймеров V-сегмента содержит праймеры V-сегмента TCRB или праймеры V-сегмента IGH,
где указанное множество праймеров J-сегмента содержит праймеры J-сегмента TCRB или праймеры J-сегмента IGH,
где каждое из указанного множества праймеров V-сегмента содержит первую последовательность и вторую последовательность, причем указанная первая последовательность комплементарна части первого участка сегмента V-участка TCRB или сегмента V-участка тяжелой цепи иммуноглобулина (IGH), причем указанный первый участок расположен непосредственно на 5′ по отношению ко второму участку указанного сегмента V-участка TCRB или IGH, причем во время перестройки генов TCRB или IGH возникают нематричные делеции, причем указанный второй участок указанного сегмента V-участка TCRB или IGH находится рядом и на 5′ по отношению к сигнальной последовательности V-рекомбинации (V-RSS) указанного сегмента V-участка TCRB или IGH, причем указанная первая последовательность расположена на 3′ по отношению ко второй последовательности на указанном праймере V-сегмента,
где указанная вторая последовательность каждого из указанных праймеров V-сегмента TCRB не является комплементарной указанному первому участку указанного сегмента V-участка TCRB,
где указанная вторая последовательность каждого из указанных праймеров V-сегмента IGH не является комплементарной указанному первому участку указанного сегмента V-участка IGH,
где каждый из указанного множества праймеров J-сегмента имеет первую последовательность и вторую последовательность, причем указанная первая последовательность комплементарна части первого участка сегмента J-участка TCRB или сегмента J-участка IGH, причем указанный первый участок расположен непосредственно на 3′ по отношению ко второму участку указанного сегмента J-участка TCRB или IGH, причем во время перестройки генов TCRB или IGH возникают нематричные делеции, при этом указанный второй участок указанного сегмента J-участка TCRB или IGH находится рядом и на 3′ по отношению к сигнальной последовательности J-рекомбинации (J-RSS) указанного сегмента J-участка TCRB или указанного сегмента J-участка IGH, причем указанная первая последовательность расположена на 3′ по отношению ко второй последовательности на указанном праймере J-сегмента,
где указанная вторая последовательность указанного праймера J-сегмента TCRB не является комплементарной указанному первому участку указанного сегмента J-участка TCRB,
где указанная вторая последовательность указанного праймера J-сегмента IGH не является комплементарной указанному первому участку указанного сегмента J-участка IGH, и
где амплификация перестроенных молекул нуклеиновой кислоты путем одиночной многократной полимеразной цепной реакции (PCR) из образца, содержащего лимфоциты, полученного от субъекта млекопитающего, с использованием указанных праймеров V-сегмента и J-сегмента производит по меньшей мере 106 отличных ампликонов, представляющих разнообразие перестроенных последовательностей CDR3 TCRB или IGH, присутствующих в указанном образце.
2. Композиция по п.1, где множество праймеров V-сегмента имеет сходную прочность отжига.
3. Композиция по п.1, где все праймеры J-сегмента отжигаются с одинаковым мотивом консервативного каркасного участка.
4. Композиция по п.1, где ампликон содержит последовательность, которая начинается с консервативного мотива, включает CDR3 соединение и распространяется в V-сегмент, и где ампликон может быть использован для диагностического определения J-сегмента.
5. Композиция по п.1, дополнительно включающая набор олигонуклеотидов для секвенирования, где указанные олигонуклеотиды для секвенирования комплементарны указанной второй последовательности указанных праймеров V-сегмента и указанной второй последовательности указанных праймеров J-сегментов.
6. Композиция по п.1, где каждый из ампликонов охватывает V-D-J-соединение.
7. Композиция по п.1, где каждый из множества праймеров V-сегмента способен к гибридизации с множеством сегментов V-участка, и каждый из множества праймеров J-сегмента способен к гибридизации с множеством сегментов J-участка.
8. Композиция по п.1, дополнительно включающая универсальный праймер С-сегмента для образования кДНК из мРНК.
9. Композиция по п.1, где 3′ конец каждого из указанных праймеров V-сегмента заякорен в положении -43 в сегменте V-участка TCRB относительно сигнальной последовательности рекомбинации (V-RSS).
10. Композиция по п.1, где множество праймеров V-сегмента TCRB состоит по меньшей мере из 14 праймеров, специфичных к 14 различным генам V TCRB.
11. Композиция по п.1, где указанные первые последовательности указанного множества праймеров V-сегмента TCRB содержат последовательности, выбранные из группы, включающей SEQ ID NOS:1-45.
12. Композиция по п.1, где указанные первые последовательности указанного множества праймеров V-сегмента TCRB содержат последовательности, выбранные из группы, включающей SEQ ID NOS:58-102.
13. Композиция по п.1, где каждый V-сегмент комплементарен по меньшей мере одному праймеру V-сегмента.
14. Композиция по п.1, где указанные праймеры не пересекают границу интрона/экзона.
15. Композиция по п.1, где указанное множество праймеров J-сегмента TCRB гибридизируется с консервативным элементом указанного сегмента J-участка TCRB и характеризуется сходной прочностью отжига.
16. Композиция по п.1, где множество праймеров J-сегмента TCRB включает по меньшей мере пять праймеров, специфичных к пяти различным генам J TCRB.
17. Композиция по п.1, где указанное множество праймеров J-сегмента TCRB содержит последовательности, выбранные из группы, включающей SEQ ID NOS:46-57 и 483.
18. Композиция по п.1, где указанное множество праймеров J-сегмента TCRB содержит последовательности, выбранные из группы, включающей SEQ ID NOS:103-113, 468 и 484.
19. Композиция по п.1, где имеется праймер J-сегмента TCRB для каждого сегмента J-участка TCRB.
20. Композиция по п.1, где каждый амплифицированный сегмент J-участка TCRB имеет уникальную метку из четырех оснований в положениях от +11 до +14 в 5′-3′ направлении сайта J-RSS.
21. Композиция по п.20, дополнительно включающая набор олигонуклеотидов для секвенирования, которые гибридизируются рядом с меткой из четырех оснований в амплифицированных сегментах J-участка TCRB в положениях от +11 до +14 в 5′-3′ направлении сайта J-RSS.
22. Композиция по п.20, где указанный набор олигонуклеотидов для секвенирования выбран из группы, включающей SEQ ID NOS:470-482.
23. Композиция по п.1, где образец содержит геномную ДНК, полученную от субъекта-млекопитающего.
24. Композиция по п.1, где образец содержит кДНК, транскрибированную от мРНК, полученной от субъекта-млекопитающего.
25. Композиция по п.5, где олигонуклеотиды для секвенирования выбраны из группы, включающей SEQ ID NOS:470-482.
26. Композиция по п.1, где указанная вторая последовательность каждого из указанных праймеров V-сегмента содержит последовательность олигонуклеотида для секвенирования.
27. Композиция по п.26, где указанный олигонуклеотид для секвенирования выбран из группы, включающей SEQ ID NOS:470-482.
28. Композиция по п.1, где указанная вторая последовательность каждого из указанных праймеров J-сегмента содержит последовательность олигонуклеотида для секвенирования.
29. Композиция по п.28, где указанная последовательность олигонуклеотида для секвенирования выбрана из группы, включающей SEQ ID NOS:470-482.
30. Композиция по п.1, где указанный первый участок указанного V-сегмента составляет по меньшей мере 40 оснований от консенсусного мотива на 5′-конце V-RSS.
31. Композиция по п.1, где каждый из указанных праймеров J-сегмента имеет 3′-конец, местоположение которого определяется по меньшей мере 14 парами оснований от консенсусного мотива на 3′-конце J-RSS.
32. Композиция по п.1, где длина ампликонов, полученных от указанного множества праймеров V-сегмента и указанного множества праймеров J-сегмента, составляет приблизительно 200 оснований.
33. Композиция по п.1, где указанные первые последовательности указанного множества праймеров V-сегмента TCRB включают SEQ ID NOS:1-45.
34. Композиция по п.1, где указанные первые последовательности указанного множества праймеров V-сегмента TCRB включают SEQ ID NOS:58-102.
35. Композиция по п.1, где указанные первые последовательности указанного множества праймеров V-сегмента IGH содержат последовательности, которые выбраны из группы, включающей SEQ ID NOS:443-451.
36. Композиция по п.1, где указанные первые последовательности указанного множества праймеров J-сегмента IGH содержат последовательности, которые выбраны из группы, включающей SEQ ID NOS:421-431.
| СПОСОБ РАЗВЕРТКИ ЛАЗЕРНОГО ПУЧКА В ОПОРНУЮ СВЕТОВУЮ ПЛОСКОСТЬ И УСТРОЙСТВО ДЛЯ ЕГО ОСУЩЕСТВЛЕНИЯ | 1993 |
|
RU2062982C1 |
| US 20060234234 A1, 19.10.2006 | |||
| US 0006416948 B1, 09.07.2002 | |||
| US 20020110807 A1, 15.08.2002 | |||
| АНТИТЕЛА К ЧЕЛОВЕЧЕСКОМУ МСР-1 | 2001 |
|
RU2314316C2 |

