Настоящее изобретение относится к антителам, к человеческому моноцитарному протеину хемотаксиса (МСР)-1 и к применению указанных антител для лечения заболеваний и нарушений, связанных с миграцией и активацией моноцитов и Т-клеток, например воспалительных заболеваний.
В опубликованной японской заявке на патент JP 05276986 (на имя Sumitomo Electric Co.) описано получение моноклональных антител грызунов к человеческому МСР-1, которые применяют для выявления МСР-1, лечения и диагностики заболеваний, связанных с инфильтрацией макрофагов. В опубликованной японской заявке на патент JP 09067399 (на имя Mitsui Toatsu) описано получение человеческих моноклональных антител к человеческому МСР-1 из человеческих периферических кровяных клеток (гемоцитов), трансформированных EBV (вирус Эпштейна-Барра), которые применяют для лечения воспаления. В опубликованной японской заявке на патент JP 11060502 (на имя Teijin) описано применение ингибитора МСР-1, прежде всего человеческого антитела к МСР-1, для лечения инфаркта головного мозга.
В настоящем описании представлены улучшенные антитела к человеческому МСР-1, предназначенные для лечения заболеваний и нарушений, связанных с миграцией и активацией моноцитов и Т-клеток.
Таким образом, изобретение относится к МСР-1-связывающей молекуле, которая содержит антигенсвязывающий центр антитела, включающий по меньшей мере одну вариабельную область тяжелой цепи (VH) иммуноглобулина, которая состоит из расположенных в указанной последовательности гипервариабельных участков CDR1, CDR2 и CDR3, где CDR1 имеет аминокислотную последовательность His-Tyr-Trp-Met-Ser, CDR2 имеет аминокислотную последовательность Asn-Ile-Glu-Gln-Asp-Gly-Ser-Glu-Lys-Tyr-Tyr-Val-Asp-Ser-Val-Lys-Gly и CDR3 имеет аминокислотную последовательность Asp-Leu-Glu-Gly-Leu-His-Gly-Asp-Gly-Tyr-Phe-Asp-Leu; и ее непосредственным эквивалентам.
Изобретение относится также к МСР-1-связывающей молекуле, которая содержит антигенсвязывающий центр антитела, включающий по меньшей мере одну вариабельную область легкой цепи (vl) иммуноглобулина, которая состоит из расположенных в указанной последовательности гипервариабельных участков CDR1', CDR2' и CDR3', где CDR1' имеет аминокислотную последовательность Arg-Ala-Ser-Gln-Gly-Val-Ser-Ser-Ala-Leu-Ala, CDR2' имеет аминокислотную последовательность Asp-Ala-Ser-Ser-Leu-Glu-Ser и CDR3' имеет аминокислотную последовательность Gln-Gln-Phe-Asn-Ser-Tyr-Pro; и ее непосредственным эквивалентам.
Первым объектом изобретения является один домен МСР-1-связывающей молекулы, содержащий выделенную тяжелую цепь иммуноглобулина, которая включает указанную выше вариабельную область тяжелой цепи (VH).
Вторым объектом изобретения является также МСР-1-связывающая молекула, которая содержит вариабельные области и тяжелой (VH), и легкой цепи (vl), при этом указанная МСР-1-связывающая молекула содержит по меньшей мере один антигенсвязывающий центр антитела, включающий:
а) вариабельную область тяжелой цепи (VH) иммуноглобулина, которая стоит из расположенных в указанной последовательности гипервариабельных участков CDR1, CDR2 и CDR3, где CDR1 имеет аминокислотную последовательность His-Туг-Trp-Met-Ser, CDR2 имеет аминокислотную последовательность Asn-Ile-Glu-Gln-Asp-Gly-Ser-Glu-Lys-Tyr-Tyr-Val-Asp-Ser-Val-Lys-Gly и CDR3 имеет аминокислотную последовательность Asp-Leu-Glu-Gly-Leu-His-Gly-Asp-Gly-Tyr-Phe-Asp-Leu, и
б) вариабельную область легкой цепи (vl) иммуноглобулина, которая состоит из расположенных в указанной последовательности гипервариабельных участков CDR1', CDR2' и CDR3', где CDR1' имеет аминокислотную последовательность Arg-Ala-Ser-Gln-Gly-Val-Ser-Ser-Ala-Leu-Ala, CDR2' имеет аминокислотную последовательность Asp-Ala-Ser-Ser-Leu-Glu-Ser и CDR3' имеет аминокислотную последовательность GIn-Gln-Phe-Asn-Ser-Tyr-Pro; и ее непосредственные эквиваленты.
Если не указано иное, любая представленная в описании полипептидная цепь имеет аминокислотную последовательность, которая начинается на N-конце и заканчивается на С-конце.
Если антигенсвязывающий центр антитела содержит и vh-, и VL-области, то они могут быть расположены на одной и той же полипептидной молекуле или предпочтительно каждая область может располагаться на разных цепях, VH-область может быть частью тяжелой цепи иммуноглобулина или ее фрагмента, а VL-область - частью легкой цепи иммуноглобулина или ее фрагмента.
Понятие «МСР-1-связывающая молекула» относится к любой молекуле, которая обладает способностью связываться с антигеном МСР-1 либо индивидуально, либо в сочетании с другими молекулами. Реакцию связывания можно идентифицировать с помощью стандартных методов (качественные анализы), которые включают, например, биологический анализ ингибирования связывания МСР-1 с его рецептором, т.е. с рецептором хемокина (CCR)-2, например CCR2B, или любой тип анализов связывания, в сравнении с отрицательным контролем, в котором используют антитело неродственной специфичности, но предпочтительно относящееся к тому же изотипу.
Связывание МСР-1-связывающих молекул по изобретению с МСР-1 можно легко продемонстрировать, например, с использованием BIAcore-анализа.
Примеры антигенсвязывающих молекул включают антитела, продуцируемые В-клетками или гибридомами, и химерные антитела, антитела со встроенными путем трансплантации гипервариабельными участками (CDR) или человеческие антитела, или любые их фрагменты, например, F(ab')2- и Fab- фрагменты, а также одну цепь или один домен антитела.
Одноцепочечное антитело состоит из вариабельных областей тяжелых и легких цепей антитела, ковалентно связанных с пептидным линкером, который, как правило, состоит из 10-30 аминокислот, предпочтительно из 15-25 аминокислот. Таким образом, эта структура не включает константную область тяжелых и легких цепей и можно предположить, что небольшой пептидный спейсер должен обладать менее выраженными антигенными свойствами, чем вся константная область. Понятие «химерное антитело» относится к антителу, в котором константные области тяжелых и легких цепей или и тех и других цепей, получают из антитела человека, а вариабельные области как тяжелых, так и легких цепей получают из антитела организма кроме человека (например, мыши) или из антитела организма человека, но которое является другим человеческим антителом. Понятие «антитело со встроенным путем трансплантации гипервариабельным участком» обозначает антитело, в котором гипервариабельные участки (CDR) происходят из антитела-донора, такого как антитело организма кроме человека (например, мыши) или другое человеческое антитело, в то время как все или практически все другие части иммуноглобулина, например, константные области и высококонсервативные участки вариабельных областей, т.е. каркасные участки, происходят из антитела-акцептора, например человеческого антитела. Однако антитело с встроенным путем трансплантации гипервариабельным участком может включать небольшое количество аминокислот последовательности-донора в каркасных участках, например в частях каркасных участков, примыкающим к гипервариабельным участкам. Понятие «человеческое антитело» обозначает антитело, в котором все константные и вариабельные области как тяжелых, так и легких цепей, происходят из антитела человека или практически идентичны последовательностям антитела человека, но необязательно получены из этого же антитела, и включают полученные в мыши антитела, в которых мышиные гены вариабельных и константных областей иммуноглобулина заменены их человеческими копиями, например, как в целом описано в ЕР 0546073 В1, патентах US 5545806. 5569825, 5625126, 5633425,5661016, 5770429, ЕР 0438474 В1 и ЕР 0463151 В1.
Наиболее предпочтительными МСР-1-связывающими молекулами по изобретению являются человеческие антитела, прежде всего антитела AAV293, AAV294 и ABN912, описанные ниже в примерах. (Антитело AAV293 представляет собой человеческое антитело IgG3/κ, a ABN912 представляет собой человеческое антитело IgG4/κ, но они практически идентичны в других аспектах. Антитело AAV294 представляет собой человеческое антитело IgGl/κ, вариабельные области которого идентичны вариабельным областям AAV293 за исключением замен одной аминокислоты в FR1, CDR2 и FR3 vh и CDR1' и FR3' vl, что описано ниже в примерах.)
Таким образом, в предпочтительных химерных антителах вариабельные области как тяжелых, так и легких цепей, получают из антитела человека, например из антитела ABN912, последовательность которого представлена в Seq. Id. No. 1 и Seq. Id. No. 2. Участки константных областей также содержат приемлемые участки человеческих константных областей, например, как это описано в «Sequences of Proteins of Immunological Interest», Kabat E.A. и др., US Department of Health и Human Services, Public Health Service, National Institute of Health.
Гипервариабельные участки могут быть связаны с любым типом каркасных участков, хотя предпочтительно их получают из антител человека. Приемлемые каркасные участки описаны у Kabat E.A. и др., там же. Предпочтительный каркасный участок тяжелой цепи представляет собой каркасный участок человеческой тяжелой цепи, например антитела ABN912, последовательность которого представлена в Seq. Id. No. 1. Она состоит из расположенных в указанной последовательности FR1-, FR2-, FR3- и FR4-областей. Аналогично этому последовательность, приведенная в Seq. Id. No. 2, является предпочтительной в качестве каркасного участка легкой цепи ABN912 и состоит из расположенных в указанной последовательности FR1'-, FR2'-, FR3'- и FR4'-участков. Помимо каркасных участков, приведенных в Seq. Id. No. 1 и Seq. Id. No. 2, можно использовать другие каркасные участки, предпочтительно человеческие каркасные участки, например, описанные у Kabat и др., там же. Несколько аминокислотных остатков каркасных участков, в частности, в тех частях каркасного участка, которые примыкают к гипервариабельным участкам, могут отличаться от аминокислотных остатков соответствующей области каркасного участка, например, для того, чтобы оказывать воздействие на способности к связыванию.
Таким образом, изобретение относится также к МСР-1-связывающей молекуле, которая содержит по меньшей мере один антигенсвязывающий центр антитела, включающий либо первый домен, аминокислотная последовательность которого практически идентична последовательности, представленной в Seq. Id. No. 1, которая начинается с аминокислоты в положении 1 и заканчивается аминокислотой в положении 122, либо описанный выше первый домен и второй домен, аминокислотная последовательность которого практически идентична последовательности, представленной в Seq. Id. No. 2, которая начинается с аминокислоты в положении 1 и заканчивается аминокислотой в положении 109.
Моноклональные антитела, которые вырабатываются против протеина, который встречается у всех людей, как правило, получают в организме кроме человека, например, мышах. Непосредственным результатом этого является то, что полученное с помощью гибридомы ксеногенное антитело при введении людям вызывает нежелательный иммунный ответ, который главным образом опосредуется константной областью ксеногенного иммуноглобулина. Это заметно ограничивает применение таких антител, поскольку их нельзя вводить в течение длительных периодов времени. Вследствие этого особенно предпочтительно применять одноцепочечные, несущие один домен, химерные антитела, антитела со встроенным путем трансплантации гипервариабельным участком или, прежде всего, человеческие антитела, которые при введении людям не должны вызывать выраженную аллогенную реакцию.
В свете вышеизложенного более предпочтительную МСР-1-связывающую молекулу по изобретению выбирают их группы, включающей человеческое антитело к МСР-1, которое содержит по меньшей мере
а) тяжелую цепь иммуноглобулина или ее фрагмент, который включает (I) вариабельную область, состоящую из расположенных в указанной последовательности гипервариабельных участков CDR1, CDR2 и CDR3, и (II) константную область человеческой тяжелой цепи или ее фрагмент, где CDR1 имеет аминокислотную последовательность His-Tyr-Trp-Met-Ser, CDR2 имеет аминокислотную последовательность Asn-Ile-Glu-Gln-Asp-Gly-Ser-Glu-Lys-Tyr-Tyr-Val-Asp-Ser-Val-Lys-Gly и CDR3 имеет аминокислотнаую последовательность Asp-Leu-Glu-Gly-Leu-His-Gly-Asp-Gly-Tyr-Phe-Asp-Leu, и
б) легкую цепь иммуноглобулина или ее фрагмент, который включает (I) вариабельную область, содержащую гипервариабельный участок CDR3' и необязательно также гипервариабельные участки CDR1', CDR2', и (II) константную область человеческой легкой цепи или ее фрагмент, где CDR1' имеет аминокислотную последовательность Arg-Ala-Ser-Gln-Gly-Val-Ser-Ser-Ala-Leu-Ala, CDR2' имеет аминокислотную последовательность Asp-Ala-Ser-Ser-Leu-Glu-Ser и CDR3' имеет аминокислотную последовательность GIn-Gln-Phe-Asn-Ser-Tyr-Pro; и его непосредственные эквиваленты.
В другом варианте МСР-1-связывающую молекулу по изобретению можно выбирать из группы, включающей одноцепочечную связывающую молекулу, которая содержит антигенсвязывающий центр антитела, включающий
а) первый домен, который состоит из расположенных в указанной последовательности гипервариабельных участков CDR1, CDR2 и CDR3, где гипервариабельные участки имеют аминокислотные последовательности, представленные в Seq. Id. No. 1,
б) второй домен, содержащий гипервариабельные участки CDR1', CDR2' и CDR3', где гипервариабельные участки имеют аминокислотные последовательности, представленные в Seq. Id. No. 2, и
в) пептидный линкер, который связан либо с N-концом первого домена и с С-концом второго домена или с С-концом первого домена и с N-концом второго домена; и ее непосредственные эквиваленты.
Как хорошо известно, минорные замены в аминокислотной последовательности, такие как делеция, добавление или замещение одной, небольшого количества или даже нескольких аминокислот, могут приводить к получению аллельной формы исходного протеина, которая обладает практически идентичными свойствами.
Таким образом, понятие «непосредственные эквиваленты» обозначает либо любой один домен МСР-1-связывающей молекулы (молекула X),
(I) в котором гипервариабельные участки CDR1, CDR2 и CDR3, взятые в целом, гомологичны по меньшей мере на 80%, предпочтительно гомологичны по меньшей мере на 90%, более предпочтительно гомологичны по меньшей мере на 95% гипервариабельным участкам, последовательность которых представлена в Seq. Id. No. 1, и
(II) который обладает способностью ингибировать связывание МСР-1 с его рецептором практически в такой же степени, что и молекула, с которой проводится сравнение, несущая каркасные участки, идентичные каркасным участкам молекулы X, но имеющая гипервариабельные участки CDR1, CDR2 и CDR3, идентичные указанным участкам, последовательность которых представлена в Seq. Id. No. I, либо любую МСР-1-связывающую молекулу, имеющую по меньшей мере два домена в сайте связывания (молекула X')
(I) в которой гипервариабельные участки CDR1, CDR2, CDR3, CDR1', CDR2' и CDR3', взятые в целом, гомологичные по меньшей мере на 80%, предпочтительно гомологичны по меньшей мере на 90%, более предпочтительно гомологичны по меньшей мере на 95% гипервариабельным участкам, последовательность которых представлена в Seq. Id. No. 1 и 2, и
(II) которая обладает способностью ингибировать связывание МСР-1 с его рецептором практически в такой же степени, что и молекула, с которой проводится сравнение, несущая каркасные участки и константные области, идентичные каркасным участкам и константным областям молекулы X', но имеющая гипервариабельные участки CDR1, CDR2, CDR3, CDR1', CDR2' и CDR3', идентичные указанным участкам, последовательность которых представлена в Seq. Id. No. 1 и 2.
В контексте настоящего описания подразумевается, что аминокислотные последовательности по меньшей мере на 80% гомологичны друг другу, если они имеют по меньшей мере 80% идентичных аминокислотных остатков в аналогичном положении, по данным оптимального сравнительного анализа, при котором бреши или вставки в аминокислотных последовательностях рассматриваются как неидентичные остатки.
Ингибирование связывания МСР-1 с его рецептором удобно тестировать с помощью различных анализов, включая анализы, которые описаны ниже. Понятие «в такой же степени» обозначает, что молекула, с которой проводится сравнение, и эквивалентная молекула дают практически одинаковые с точки зрения статистики кривые ингибирования связывания МСР-1 при исследовании с помощью одного из указанных выше анализов. Например, для МСР-1-связывающих молекул по изобретению, как правило, получают значения IC50 для ингибирования связывания МСР-1 с его рецептором (CCR2B), которое находится в пределах +/- х 5 (т.е. предпочтительно является практически одинаковым) относительно IC50 соответствующей молекулы, с которой проводится сравнение, при анализе с помощью описанных выше методов.
Например, применяемый анализ может представлять собой анализ конкурентного ингибирования связывания МСР-1 мембранным связывающим МСР-1 рецептором (CCR2B) и МСР-1-связывающими молекулами по изобретению, например, с использованием сцинтилляционного анализа сходства (SPA), описанного ниже в примерах.
Наиболее предпочтительно человеческое антитело к МСР-1 содержит по меньшей мере
а) одну тяжелую цепь, которая несет вариабельную область, аминокислотная последовательность которой практически идентична последовательности, представленной в Seq. Id. No. 1, которая начинается с аминокислоты в положении 1 и заканчивается аминокислотой в положении 122, и константную область человеческой тяжелой цепи; и
б) одну легкую цепь, которая несет вариабельную область, аминокислотная последовательность которой практически идентична последовательности, представленной в Seq. Id. No. 2, которая начинается с аминокислоты в положении 1 и заканчивается аминокислотой в положении 109, и константную область человеческой легкой цепи.
Константная область человеческой тяжелой цепи может быть γ1-, γ2-, γ3-, γ4-, γ-, α1-, α2-, δ- или ε-типа, предпочтительно γ-типа, более предпочтительно γ4-типа, а константная область человеческой легкой цепи может быть κ- или λ-типа (включая подтипы λ1, λ2 и λ3), но предпочтительно κ-типа. Аминокислотные последовательности всех указанных константных областей приведены у Kabat и др., там же.
МСР-1-связывающую молекулу по изобретению можно получать с помощью методов рекомбинантной ДНК. При этом надо сконструировать одну или несколько молекул ДНК, кодирующих связывающую молекулу, поместить под контроль соответствующих последовательностей и трансформировать организм-хозяин, в котором может происходить экспрессия.
Таким образом, в наиболее широком смысле изобретение относится к
(I) молекулам ДНК, кодирующим один домен МСР-1-связывающей молекулы по изобретению, одноцепочечную МСР-1-связывающую молекулу по изобретению, тяжелую или легкую цепь МСР-1-связывающей молекулы по изобретению или их фрагменты и
(II) применению молекул ДНК по изобретению для получения рекомбинантным путем МСР-1-связывающей молекулы по изобретению.
Существующий уровень техники является таким, что позволяет специалистам в данной области синтезировать молекулы ДНК по изобретению на основе представленной в настоящем описании информации, т.е. аминокислотных последовательностей гипервариабельных участков и кодирующих их последовательностей ДНК. Метод конструирования гена вариабельной области описан, например, в EPA 239400 и в целом состоит в следующем: клонируют ген, кодирующий вариабельную область МАт, независимо от его специфичности. Определяют сегменты ДНК, кодирующие каркасный участок и гипервариабельные участки, и сегменты ДНК, кодирующие гипервариабельные участки удаляют таким образом, чтобы сегменты ДНК, кодирующие каркасные участки, сливать в стыках с приемлемыми сайтами рестрикции. Сайты рестрикции можно создавать в требуемых положениях путем мутагенеза молекулы ДНК с использованием стандартных процедур. Кассеты, несущие двухцепочечные синтетические CDR, получают путем синтеза ДНК на основе данных о последовательностях, представленных в Seq. Id. No. 1 или 2. Эти кассеты конструируют так, чтобы они имели «липкие» концы для встраивания путем лигирования в стыки каркаса.
Кроме того, для получения конструкции ДНК, кодирующей МСР-1-связывающие молекулы по изобретению, не требуется использовать мРНК из продуцирующей гибридому линии клеток. Так, в заявке РСТ WO 90/07861 представлены полные инструкции по получению антитела с помощью методов рекомбинантной ДНК на основе только письменной информации, такой как нуклеотидная последовательность гена. Метод заключается в синтезе нескольких олигонуклеотидов, их амплификации с помощью ПЦР и их сплайсинга с получением требуемой последовательности ДНК.
Научной общественности известны экспрессионные векторы, содержащие приемлемые промоторы или гены, кодирующие константные области тяжелой и легкой цепи. Так, после получения молекулы ДНК по изобретению ее можно принятым образом переносить в соответствующий экспрессионный вектор. Молекулы ДНК, кодирующие одноцепочечные антитела, можно также получать стандартными методами, например, описанными в WO 88/1649.
В свете вышеизложенного для удовлетворения критериям полноты описания не требуется использовать депонированные гибридомы или линии клеток.
Конкретным вариантом осуществления изобретения являются описанные ниже первая и вторая конструкции ДНК, предназначенные для получения МСР-1-связывающей молекулы.
Первая конструкция ДНК кодирует тяжелую цепь или ее фрагмент и включает а) первый фрагмент, который кодирует вариабельную область, содержащую альтернативные каркасный и гипервариабельные участки, где гипервариабельные участки состоят из последовательно расположенных CDR1, CDR2 и CDR3, аминокислотные последовательности которых представлены в Seq. Id. No. 1; этот первый фрагмент начинается с кодона, который кодирует первую аминокислоту вариабельной области, и заканчивается кодоном, который кодирует последнюю аминокислоту вариабельной области, и б) второй фрагмент, кодирующий константную область тяжелой цепи или ее фрагмент, который начинается с кодона, который кодирует первую аминокислоту константной области тяжелой цепи, и заканчивается кодоном, который кодирует последнюю аминокислоту константной области или ее фрагмента, после которого расположен стоп-кодон.
Предпочтительно указанный первый фрагмент кодирует вариабельную область, аминокислотная последовательность которой практически идентична аминокислотной последовательности, представленной в SEQ ID No. 1, которая начинается с аминокислоты в положении 1 и заканчивается аминокислотой в положении 122. Более предпочтительно первый фрагмент имеет нуклеотидную последовательность, представленную в SEQ ID No. 1, которая начинается с нуклеотида в положении 1 и заканчивается нуклеотидом в положении 366. Предпочтительно также, чтобы второй фрагмент кодировал константную область человеческой тяжелой цепи, более предпочтительно константную область человеческой цепи γ4-типа. Этот второй фрагмент представляет собой фрагмент ДНК геномного происхождения (который содержит интроны) или фрагмент кДНК (без интронов).
Вторая конструкция ДНК кодирует легкую цепь или ее фрагмент и содержит
а) первый фрагмент, который кодирует вариабельную область, содержащую альтернативные каркасный участок и гипервариабельные участки; где гипервариабельные участки представляют собой CDR1', CDR2' и CDR3', аминокислотные последовательности которых представлены в Seq. Id. No. 2;
этот первый фрагмент начинается с кодона, который кодирует первую аминокислоту вариабельной области и заканчивается кодоном, который кодирует последнюю аминокислоту вариабельной области, и
б) второй фрагмент, кодирующий константную область легкой цепи или ее фрагмент, который начинается с кодона, кодирующего первую аминокислоту константной области легкой цепи, и заканчивающуюся кодоном, который кодирует последнюю аминокислоту константной области или ее фрагмента, после которого расположен стоп-кодон.
Предпочтительно, указанный первый фрагмент кодирует вариабельную область, аминокислотная последовательность которой практически идентична аминокислотной последовательности, представленной в SEQ ID No. 2, которая начинается с аминокислоты в положении 1 и заканчивается аминокислотой в положении 109. Более предпочтительно первый фрагмент имеет нуклеотидную последовательность, представленную в SEQ ID No. 2, которая начинается с нуклеотида в положении 1 и заканчивается нуклеотидом в положении 327. Предпочтительно также, чтобы второй фрагмент кодировал константную область человеческой легкой цепи, более предпочтительно константную область человеческой цепи κ-типа.
Изобретение относится также к МСР-1-связывающим молекулам, в которых один или более, как правило, лишь небольшое количество остатков CDR1, CDR2, CDR3, CDR1, CDR2' или CDR3' заменены по сравнению с остатками последовательностей, представленных в SEQ ID No. 1 и SEQ ID No. 2; например, в результате мутации, например, сайт направленного мутагенеза соответствующих последовательностей ДНК. Изобретение относится к последовательностям ДНК, кодирующим такие измененные МСР-1-связывающие молекулы. Изобретение относится также к связывающим молекулам, в которых один или более, как правило, лишь небольшое количество остатков каркасных участков заменены по сравнению с остатками последовательностей, представленных в SEQ ID No. 1 и SEQ ID No. 2.
В обеих конструкциях ДНК первый и второй фрагменты могут быть разделены интроном, а также целесообразно, когда в интроне между первым и вторым фрагментом находится энхансер. Наличие энхансера, который является транскрибируемым, но не транслируемым, может повышать эффективность транскрипции. Согласно конкретным вариантам осуществления обе конструкции ДНК содержат энхансер гена тяжелой цепи, предпочтительно человеческого гена.
Каждую конструкцию ДНК помещают под контроль приемлемых контролирующих экспрессию последовательностей, прежде всего, под контроль приемлемого промотора. Можно использовать любой тип промотора при условии, что он адаптирован к организму-хозяину, в который конструкции ДНК должны быть перенесены для экспрессии. Однако, если экспрессия происходит в клетке млекопитающего, то особенно предпочтительно применять промотор гена иммуноглобулина.
Требуемое антитело можно получать в клеточной культуре или в трансгенном животном. Соответствующее трансгенное животное можно получать с помощью стандартных методов, которые включают микроинъекцию в яйцеклетки первой и второй конструкций ДНК, помещенных под контроль приемлемых контролирующих последовательностей, перенос полученных таким образом яйцеклеток в соответствующих псевдобеременных самок и отбор потомства, экспрессирующего требуемое антитело.
Когда цепи антитела получают в клеточной культуре, конструкции ДНК должны быть сначала встроены либо в один экспрессирующий вектор, либо в два различных, но совместимых экспрессирующих вектора, причем последний вариант является предпочтительным.
Таким образом, изобретение относится также к экспрессионному вектору, который может реплицироваться в линии прокариотических или эукариотических клеток и содержит по меньшей мере одну из описанных выше конструкций ДНК.
Каждый экспрессионный вектор, содержащий конструкцию ДНК, затем переносят в приемлемый организм-хозяин. Когда конструкции ДНК встроены индивидуально в два экспрессионных вектора, их можно переносить по отдельности, т.е. по одному типу вектора на клетку, либо посредством котрансформации, указанный последний вариант является предпочтительным. Приемлемый организм-хозяин может представлять собой линию клеток бактерий, дрожжей или млекопитающих, указанный последний вариант является предпочтительным. Более предпочтительно линию клеток млекопитающего получают из лимфомы, например миеломы, гибридомы или здоровых иммортализованных В- клеток, которые предпочтительно не экспрессируют тяжелую или легкую цепи никаких эндогенных антител, например, линию клеток SP 2/0.
Для экспрессии в клетках млекопитающих кодирующие последовательности МСР-1-связывающих молекул предпочтительно интегрируют в ДНК клетки-хозяина в локус, в котором может быть обеспечен высокий уровень экспрессии МСР-1-связывающей молекулы или который является предпочтительным для обеспечения высокого уровня экспрессии МСР-1-связывающей молекулы. Клетки, в которых существуют такие локусы, предпочтительные для интеграции кодирующей последовательности МСР-1-связывающей молекулы, можно выявлять и отбирать на основе данных о уровнях экспрессии МСР-1-связывающей молекулы. Для получения клеток-хозяев, содержащих кодирующую последовательность МСР-1-связывающей молекулы, можно применять любой селектируемый маркер; например, можно использовать ген dhfr/метотрексат, или эквивалентную систему отбора. Системы для экспрессии МСР-1-связывающих молекул по изобретению включают системы амплификации/селекции, основанные на применении GS, например, описанные в ЕР 0256055 В, ЕР 0323997 В и ЕР-А 89303964.4.
Еще одним объектом изобретения является способ получения МСР-1-связывающей молекулы, который предусматривает (I) культивирование организма, трансформированного описанным выше экспрессионным вектором, и (II) выделение МСР-1-связывающей молекулы из культуры.
Наиболее предпочтительно МСР-1-связывающая молекула по изобретению представляет собой человеческое антитело, например, антитела AAV293, AAV294 или ABN912, и ее можно получать культивированием соответствующей линии клеток гибридомы или предпочтительно из рекомбинантной линии клеток, которая содержит ДНК, кодирующую человеческое антитело, включая ДНК, измененную с целью кодирования измененного изотипа антитела или другой функции или свойства антитела.
При создании настоящего изобретения установлено, что AAV294 и более предпочтительно антитела AAV293 и ABN912 дают перекрестную реакцию с рекомбинантным человеческим эотаксином-1. Поскольку эти антитела эффективно взаимодействуют с эотаксином-1, а также с МСР-1, их можно использовать для ингибирования связывания эотаксина-1 с его рецептором наряду с ингибированием связывания МСР-1 с его рецептором. Секреция эотаксина происходит при аллергических заболеваниях и нарушениях, включая аллергические и воспалительные заболевания дыхательных путей, такие как астма. Под объем настоящего изобретения подпадают антитела, прежде всего химерные антитела и антитела со встроенным путем трансплантации гипервариабельным участком, и предпочтительно человеческие антитела, обладающие способностью специфично связываться как с МСР-1, так и с эотаксином, например, с человеческим МСР-1 и человеческим эотаксином, а также применение таких антител для лечения заболеваний, опосредуемых МСР-1 или эотаксином.
Таким образом, еще одним объектом изобретения является антитело к МСР-1, которое дает перекрестную реакцию с эотаксином.
Предпочтительные согласно этому объекту изобретения антитела представляют собой антитела, которые обладают способностью ингибировать связывание МСР-1 с его рецептором и обладают способностью ингибировать связывание эотаксина с его рецептором.
И еще одним объектом изобретение является
I) применение антитела к МСР-1, дающего перекрестную реакцию с эотаксином-1, которое обладает способностью ингибировать связывание МСР-1 и эотаксина-1 с их рецепторам, и для лечения опосредуемых МСР-1 или эотаксином-1 болезни или нарушения;
II) способ лечения опосредуемых МСР-1 или эотаксином болезни или нарушения у пациента, который предусматривает введение пациенту эффективного количества антитела к МСР-1, которое дает перекрестную реакцию с эотаксином и обладает способностью ингибировать связывание МСР-1 и эотаксина с их рецепторами;
III) фармацевтическая композиция, содержащая антитело к МСР-1, которое дает перекрестную реакцию с эотаксином и обладает способностью ингибировать связывание МСР-1 и эотаксина с их рецепторами, в сочетании с фармацевтически приемлемыми эксципиентом, разбавителем или носителем; и
IV) применение антитела к МСР-1, которое дает перекрестную реакцию с эотаксином и обладает способностью ингибировать связывание МСР-1 и эотаксина с их рецепторами, для приготовления лекарственного средства, предназначенного для лечения опосредуемых МСР-1 или эотаксином болезни или нарушения.
В указанных дополнительных объектах антитело к МСР-1 предпочтительно дает перекрестную реакцию с эотаксином-1 и опосредуемые эотаксином болезни предпочтительно представляют собой болезни, опосредуемые эотаксином-1.
Для целей настоящего описания понятие «антитело, обладающее способностью ингибировать связывание МСР-1 и эотаксина с их рецепторами» подразумевает, что антитело обладает способностью ингибировать связывание МСР-1 и эотаксина с их рецепторами практически в такой же степени, что и антитела AAV294, AAV293 или ABN912, где понятие «в такой же степени» имеет указанное выше значение.
В настоящем описании понятие «опосредуемая МСР-1 болезнь» и «опосредуемая эотаксином болезнь» подразумевает все болезни и медицинские состояния, которые либо прямо, либо косвенно связаны с МСР-1 или эотаксином, прежде всего с эотаксином-1, включая этиологию, развитие, прогресс, персистентность или патологию болезни или состояния.
В настоящем описания понятия «лечение» или «лечить» относятся как к профилактическому или превентивному лечению, так и к исцеляющему или изменяющему течение болезни лечению, включая лечения пациента, который имеет риск заражения болезнью или для которого предусматривается возможность заражения болезнью, а также пациентов, которые уже больны или у которых диагностировано наличие болезни или медицинского состояния, и включает подавление клинического рецидива.
Аффинность связывания антител AAV293 и ABN912 с МСР-1 превосходит аффинности связывания ранее описанных антител к МСР-1, например, антител к человеческому МСР-1. Так, для ABN912 значение константы равновесия диссоциации kd, характеризующей связывание с МСР-1, составляет менее примерно 50 пМ, например, примерно 43 пМ. Указанная высокая аффинность связывания обусловливает очень высокую терапевтическую ценность ABN912.
Таким образом, еще одним объектом изобретения является антитело к МСР-1, для которого значение kd, характеризующее связывание с МСР-1, составляет примерно 50пМ или менее. Этот объект изобретения включает также применение таких высокоаффинных антител и содержащих их композиций, аналогичное применению, указанному выше для антител к МСР-1, которые дают перекрестную реакцию с эотаксином.
Кроме того, при создании изобретения было установлено, что антитело ABN912 связывается с антигенным эпитопом МСР-1, который включает остаток аргинина в положении 24 МСР-1. Отсюда ясно, что антитело ABN912 обладает способностью оказывать непосредственное воздействие на связывание МСР-1 с его рецептором (CCR2B); Arg24 является важным остатком МСР-1, который принимает участие в связывании МСР-1 с CCR2B. Помимо этого сайт связывания ABN912 включает остатки Argl8 и Lys49 МСР-1.
Таким образом, еще одним объектом изобретения является антитело к МСР-1, которое связывается с антигенным эпитопом МСР-1, который содержит остаток аргинина в положении 24 МСР-1. Предпочтительно антигенный эпитоп содержит также остаток аргинина в положении 18 и остаток лизина в положении 49 МСР-1. Этот объект изобретения включает также пути применения, способы и композиции, описанные выше для антител к МСР-1, которые дают перекрестную реакцию с эотаксином.
МСР-1-связывающие молекулы, как они определены выше, прежде всего МСР-1 -связывающие молекулы, описанные в первом и втором объектах изобретения; антитела к МСР-1, которые дают перекрестную реакцию с эотаксином, прежде всего антитела, которые обладают способностью ингибировать связывание МСР-1 и эотаксина с их рецепторами; антитела к МСР-1, для которых значение kd, характеризующее связывание МСР-1, составляет примерно 50 пМ или менее; и антитела к МСР-1, которые связываются с антигенным эпитопом МСР-1, который содержит остаток аргинина в положении 24 МСР-1, далее в контексте настоящего описания обозначены как «антитела по изобретению».
Предпочтительно антитела по изобретению представляют собой МСР-1-связывающие молекулы, описанные в первом и втором объектах изобретения. Предпочтительно антитела по изобретение представляют собой человеческие антитела, более предпочтительно антитело ABN912 или его непосредственный эквивалент.
Антитела по изобретению ингибируют воздействие МСР-1 на его клетки-мишени и вследствие этого их можно применять для лечения опосредуемых МСР-1 болезней и нарушений. Эти и другие виды фармакологической активности антител по изобретению можно продемонстрировать с помощью стандартных тестов, например, описанных ниже:
1. Ингибирование связывания МСР-1 с клетками, экспрессирующими CCR2B. Предпосылкой для осуществления сигнальной и эффекторной функций МСР-1 является его взаимодействие с рецептором CCR2B. Для демонстрации того, что антитела по изобретению ингибируют связывание МСР-1 с клеточными мембранами, экспрессирующими указанный рецептор, применяют сцинтилляционный анализ сходства (SPA).
Мембраны экспрессирующих CCR2B клеток СНО инкубируют с различными концентрациям антитела-мишени (например, от 10 до 10 М) и остаточное связывание (125-1)-МСР-1 оценивают с помощью SPA, используя гранулы, покрытые агглютинином из проростков пшеницы. При анализе этим методом значение IC50 антител по изобретению, как правило, составляет от примерно 0,1 до примерно 10 нМ, предпочтительно примерно 0,5 нМ (например, 461±206 пМ).
2. Ингибирование опосредуемой МСР-1 сигнализации. Потенциальную способность антител по изобретению ингибировать вызванные МСР-1 физиологические действия оценивают по индуцированной МСР-1 мобилизации внутриклеточного Са2+ в присутствии антитела или без него.
Оценку воздействия на кальций осуществляют с использованием стабильно трансфектированных, экспрессирующих CCR2B клеток линии СНО и линии ТНР-1 с помощью флуоресцентных красителей и анализа с использованием клеточного сортера с возбуждением флуоресценции (FACS), которые описаны ниже в примерах. При анализе этом методом значение IC50 антител по изобретению, как правило, составляет от примерно 0,05 до примерно 10 нМ, предпочтительно примерно 0,5 нМ (например, 390±20 пМ).
Антитела по изобретению предпочтительно дают перекрестную реакцию с эотаксином, прежде всего с эотаксином-1, и вследствие этого предпочтительно могут ингибировать воздействие эотаксина на его клетки-мишени, поэтому их также можно применять для лечения опосредуемых эотаксином болезней и нарушений. Перекрестную реактивность антител по изобретению с эотаксином можно определять с помощью метода, основанного на применении оптического биосенсора, такого как BIAcore® (Karlsson и др., J. Immunol. Meth. 145:229-240 (1991)).
Как указано выше, антитела по изобретению эффективно блокируют воздействия МСР-1 и предпочтительно дают перекрестную реакцию с эотаксином. Таким образом, антитела по изобретению могут найти следующее применение в фармацевтике.
Антитела по изобретению можно применять для профилактики и лечения опосредуемых МСР-1 или эотаксином болезней или медицинских состояний. МСР-1 играет важную роль в передвижении лейкоцитов, прежде всего в миграции моноцитов к местам воспаления, и вследствие этого антитела по изобретению можно применять для ингибирования миграции моноцитов, например, для лечения воспалительных состояний, аллергий и аллергических состояний, аутоиммунных заболеваний, реакции отторжения трансплантата, разных видов рака, включающих инфильтрацию лейкоцитов, стеноза или рестеноза, атеросклероза, ревматоидного артрита и остеоартрита.
Болезни или состояния, которые можно лечить с помощью антител по изобретению включают: воспалительные или аллергические состояния, в том числе респираторные аллергические заболевания, такие как астма, аллергический ринит, ХОЗЛ (хроническое обструктивное заболевание легких), заболевания, связанные с гиперчувствительноостью легкого, гиперчувствительный пневмонит, коллагеноз легкого (КЛ) (например, идиопатический фиброз легкого или КЛ, связанный саутоиммунными заболеваниями, такими как ревматоидный артрит (РА), системная красная волчанка (СКВ) и т.д..); анафилаксия или гиперчувствительные реакции, аллергии на лекарственные средства (например, на пенициллин или цефалоспорины), и аллергии на укусы насекомых; воспалительные заболевания кишечника, такие как болезнь Крона и неспецифический язвенный колит; спондилоартропатии, склеродема; псориаз и воспалительные дерматозы, такие как дерматит, экзема, атонический дерматит, аллергический контактный дерматит, крапивница; васкулит; аутоиммунные заболевания, в частности аутоиммунные заболевания, этиология которых включает воспалительный компонент, такие как артрит (например, ревматоидный артрит, хронический прогрессирующий артрит, псориатический артрит и деформирующий артрит), и ревматоидные заболевания, включая воспалительные состояния и ревматоидные болезни, связанные с потерей костной ткани, связанную с воспалением боль, гиперчувствительность (включая как гиперчувствительность дыхательный путей, так и кожную гиперчувствительность) и аллергии. Специфические аутоиммунные заболевания, для лечения которых можно применять антитела по изобретению, включают аутоиммунные гематологические болезни (такие, например, как гемолитическая анемия, апластическая анемия, истинная анемия эритроцитов и идиопатическая тромбоцитопения), системную красную волчанку, полихондрит, склеродему, грануломатоз Вегенера, дерматомиозит, хронический активный гепатит, тяжелую псевдопаралитическую миастению, псориаз, синдром Стивенса-Джонсона, идоапатическую спру, аутоиммунные воспалительные заболевания кишечника (такие, например, как неспецифический язвенный колит, болезнь Крона и синдром радраженной толстой кишки), аутоиммунный тиреоидит, болезнь Бехчета, эндокринную офтальмопатию, болезнь Грейвса, саркоидоз, рассеянный склероз, первичный билиарный цирроз печени, юношеский диабет (сахарный диабет типа I), увеит (передний и задний), сухой кератоконъюнктивит и весенний кератоконъюнктивит, коллагеноз легкого и гломерулонефрит (без нефротического синдрома или с нефротическим синдромом, например, включая идиопатический нефротический синдром или нефропатию с минимальным изменением); отторжение трансплантата (например, при трансплантации с использованием трансплантатов сердца, легкого, системы сердце-легкое, печени, почки, поджелудочной железы, кожи или роговицы), включая отторжение аллотрансплантата или отторжение ксенотрансплантата или реакцию «трансплантат против хозяина», и связанное с артериосклерозом отторжение трансплантата органа; атеросклероз; рак кожи или органов, связанный с инфильтрацией лейкоцитов; стеноз или рестеноз сосудистой сети, прежде всего артерий, например, коронарной артерии, включая стеноз или рестеноз, возникающие в результате операции на сосудах, а также гиперплазию новой оболочки; и другие заболевания и состояния, такие как воспалительные реакции, включая повреждение, связанное с реперфузией, злокачественные болезни крови, индуцированную цитокином токсичность (например, септический шок или эндотоксический шок), полимиозит, дерматомиозит и грануломатозные заболевания, в том числе саркоидоз.
Антитела по изобретению особенно предпочтительно применять для лечения связанных с метаболизмом заболеваний костной и хрящевой ткани, включая остеоартрит, остеопороз и другие воспалительные повреждения кожи ревматического происхождения, например ревматоидный артрит, и потерю костной ткани в целом, включая связанную с возрастом потерю костной ткани, и, прежде всего, заболевание периодонта.
Соответствующая доза, которую можно применять для лечения указанных выше показаний, естественно, должна варьироваться в зависимости, например, от конкретного применяемого антитела по изобретению, хозяина, пути введения и природы и серьезности состояния, подлежащего лечению. При этом для профилактики удовлетворительные результаты, как правило, получают при использовании доз от примерно 0,05 до примерно 10 мг на кг веса тела, более предпочтительно от примерно 0,1 до примерно 5 мг кг веса тела. Частота введения доз при применении в профилактических целях, как правило, составляет от примерно один раз в неделю до примерно один раз каждые 3 месяца, более предпочтительно от примерно один раз каждые 2 недели до примерно 1 раз каждые 10 недель, например, один раз каждые 4 или 8 недель. Антитело по изобретению целесообразно вводить парентерально, внутривенно, например, в находящуюся впереди от локтевого сустава или другую периферическую вену, внутримышечно или подкожно. Например, профилактическое лечение включает, как правило, введение антитела по изобретению от одного раза в месяц до одного раза каждые 2-3 месяца или еще реже.
Фармацевтические композиции по изобретению можно приготавливать обычным образом. Композиция по изобретению предпочтительно находится в лиофилизированной форме. Для непосредственного введения лиофилизат растворяют в пригодном водном носителе, например, в стерильной воде для инъекций или в стерильном забуференном физиологическом растворе. Если требуется больший объем раствора для введения путем инфузии, например, для быстрого внутривенного введения больших объемов жидкости (болюсная инъекция), целесообразно в физиологический раствор во время приготовления состава добавлять бычий сывороточный альбумин или гепаринизированную собственную кровь пациента. Присутствие избытка такого физиологически инертного протеина препятствует потере антитела в результате адсорбции стенками контейнера и пробирки, которые используются вместе с раствором для инфузии. Если применяют альбумин, то приемлемые концентрации составляют от 0,5 до 4,5% в пересчете на массу физиологического раствора.
Изобретение дополнительно описано с помощью приведенных ниже примеров, представленных только с целью иллюстрации, в которых сделаны ссылки на прилагаемые чертежи: фиг.1:
А - график, иллюстрирующий активность эозинофилпероксидазы (ЕРО) в различных полученных с помощью пункционной биопсии образцах обработанных и необработанных макак-резус;
Б - аналогичный график, иллюстрирующий активности миелопероксидазы (МРО) в полученных с помощью биопсии образцах;
фиг.2 - фотографии, на которых представлено окрашивание эозинофилов в гистологических образцах, взятых у 4 макак-резус до и после лечения с помощью ABN912 и CGP44290 (контрольное антитело);
фиг. 3 - график, иллюстрирующий миграцию Th-клеток (в %) в трансплантатах человеческой кожи при различных режимах обработки, и
фиг. 4 - график, иллюстрирующий относительные аффинности связывания мутантов ABN912 с МСР-1.
Примеры
Для получения антител к человеческому МСР-1 используют трансгенных мышей, созданных с целью экспрессии человеческого спектра антител IgG/κ вместо мышиного спектра иммуноглобулинов (Fishwild и др., Nat. BiotechnoL, 14, 845-851,1996). В-клетки этих мышей иммортализуют с помощью стандартных методов на основе гибридом и получают клетки мышиной гибридомы, которые секретируют человеческое антитело AAV293 подкласса IgG3/κ.
Пример 1: Иммунизация мышей и получение линии клеток гибридомы
Иммунизация
Четырех мышей линии Medarex (мыши No. 16194-16197, фирма Medarex Inc. Аннадал, шт. Нью Джерси, США) иммунизируют рекомбинантным человеческим МСР-1 (фирма R&D Systems, Миннеаполис, шт. Миннесота, США) из расчета 100 мкг протеина на мышь в полном адъюванте Фрейнда в дни 0 и 14 внутрибрюшинно (i.p.) и в день 26 внутривенно (i.v.). Через 41 день ни у одной из мышей не выявлено сывороточное антитело. Мышей дополнительно иммунизируют rhMCP-1 из расчета 100 мкг протеина на мышь подкожно (s.c.) в неполном адъюванте Фрейнда в дни 49 и 65. При анализе через 106 дней в сыворотке одной из мышей (мышь No. 16194) обнаружен выраженный титр антитела к МСР-1. Эту мышь перед осуществлением слияния клеток подвергают 7 дополнительным бустерным инъекциям: 100 мкг протеина на мышь в физиологическом растворе в дни 106 (i.p.), 119 (s.c.) и 135 (i.p.) и 25 мкг протеина на мышь в физиологическом растворе в дни -4 (х2 - i.v. и i.p.) и -3 и -2 (в обоих случаях i.p.).
Слияние и отбор гибридомы
В день слияния мышь No. 16194 умерщвляют путем ингаляции СО2 и общие клетки селезенки (4,8 × 10 ) с помощью общепринятого метода на основе ПЭГ 4000 подвергают слиянию с клетками линии PAI-0 (5х10), т.е. линиями клеток мышиной миеломы. Слитые клетки высевают в 720 лунок (1 мл/лунку), содержащих питающий подслой мышиных перитонеальных клеток (мыши линии Balb/c, в ГАТ-среде (гипоксантин-аминоптерин-тимидин), дополненной средой RPMI 1640, 10% инактивированной тепловой обработкой фетальной телячьей сыворотки, 5 × 10-3 М β-меркаптоэтанолом, 50 мкг/мл гентамицина. Питательную среду заменяют на каждый 4-й день и через 14 дней ГАТ-среду заменяют ГТ-средой (гипоксантин-тимидин), т.е. средой без аминоптерина. Из 720 первоначально засеянных лунок 461 лунка (64%) является позитивной в отношении роста гибридомы. Супернатанты собирают и подвергают скринингу в отношении МСР-1-реактивных моноклональных антител с помощью ELISA. Выявляют 7 моноклональных антител IgG-подкласса. Осуществляют клонирование с использованием четырех 96-луночных титрационных микропланшетов, засевая каждую лунку из расчета 0,5 клеток/100 мкл. Через 8 дней клоны анализируют с помощью микроскопа, добавляют 100 мкл среды для роста и на следующий день супернатант анализируют с помощью ELISA. Для дальнейшего клонирования и характер из ации отбирают наиболее реактивную гибридому, клон 149. Субклон гибридомы 149-12 отбирают на основе его ингибирующей активности, используя анализ зависящей от rhMCP-1 мобилизации кальция в присутствии продукта моноклонального антитела IgG3/κ, т.е. AAV293.
Чистота и частичные аминокислотные последовательности тяжелой и легкой цепи
Секвенирование аминокислотных последовательностей
Легкую и тяжелую цепи очищенного антитела AAV293 разделяют с помощью ДСН-ПААГ и N-концевые аминокислоты определяют с помощью расщепления по Эдману. Последовательности кДНК, кодирующие вариабельные области тяжелой и легкой цепи, получают с помощью ПЦР-амплификации кДНК, полученной с использованием мРНК из клонированных клеток гибридомы, и полностью секвенируют. N-концевые аминокислотные последовательности вариабельных областей тяжелой и легкой цепи и соответствующие последовательности ДНК представлены ниже в Seq. Id No. 1 и Seq Id No. 2, где CDR выделены жирным шрифтом.
Еще одно моноклональное антитело к МСР-1, в виде продукта моноклонального антитела IgGl/к AAV294, получают также с помощью описанного выше метода. Это антитело связывается с МСР-1 с примерно в 3 раза более низкой аффинностью, чем антитело AAV293, при этом установлено, что оно имеет аминокислотные последовательности vh и vl, которые идентичны аминокислотным последовательностям антитела AAV293, за исключением того, что в vh антитела AAV294 в положении 24 Ala заменен на Val 24, в положении 60 Туг заменен на Phe и в положении 74 Asn заменен на Ser, и в vl антитела AAV294 в положении 30 Ser заменен на Туг и в положении 69 Pro заменен на Thr.
Конструирование экспрессионных векторов для тяжелой и легкой цепи. Клонированные кодирующие последовательности vl и vh амплифицируют с помощью ПЦР и встраивают в соответствующие сайты рестрикции в векторы, несущие кассеты, которые содержат промотор иммуноглобулина, лидерные последовательности антитела RFT2 (Heinrich и др., J. Immunol. 143, 3589-3597 (1989)), фрагмент J-сегментов и сайт донора сплайсированного фрагмента. Кассету для легкой цепи, которая содержит полную vl -область, промотор и лидерную последовательность для секреции, переносят в экспрессионный вектор, содержащий человеческий ген Ck, энхансер тяжелой цепи иммуноглобулина и модифицированную кДНК мышиного гена dhfr для отбора по признаку устойчивости к метотрексату (МТХ).
Кассету для тяжелой цепи переносят соответственно в экспрессионный вектор, несущий человеческий ген IgG4, энхансер тяжелой цепи иммуноглобулина и ген для отбора по признаку устойчивости к неомицину.
Конфигурация и тяжелой, и легкой цепей в экспрессионных векторах соответствует геномной конфигурации перестроенных генов иммуноглобулина, что, по-видимому, имеет решающее значение для достижения высокого уровня экспрессии.
Для получения антитела описанными выше векторами совместно трасфектируют соответствующую линию клеток-хозяев, например клеточную линию SP2/0. Клетки, содержащие векторные последовательности, отбирают с помощь селекции метотрексатом и отобранные клеточные линии культивируют с целью экспрессии антитела ABN912 (человеческое антитело подкласса IgG4/κ к человеческому МСР-1).
Экспрессионные векторы, несущие гены тяжелой и легкой цепей NVP-ABN912, соответственно, линеаризуют и путем электропорации трасфектируют ими клетки Sp2/0. Трансфектированные клетки выращивают в течение 20 ч в неселективной среде RPMI, дополненной фетальной телячьей сывороткой (ФТС), и качестве селектирующего агента добавляют G418 в концентрации 1,4 мг/мл и инкубируют в течение 48-72 ч. Трансфектированные пулы адаптируют к среде RPMI без ФТС, которая содержит общепринятые добавки (пируват, глутамин, человеческий сывороточный альбумин, трансферрин, инсулин). После двух стадий амплификации с использованием метотрексата в концентрациях 200 нМ и 1 мкМ выделяют высокопродуктивные клоны. Стабильность продуктивности клонов и субклонов оценивают в Т175-культурах в течение 4-5 месяцев непрерывного культивирования, осуществляя дополнительные эксперименты с использованием центрифуги. Для высокопродуктивных клонов осуществляют переход от получения лабораторных культур до крупномасштабного культивирования в биореакторах. Получают несколько высокопродуктивных клеточных линий, которые можно применять для производства ABN912, Максимальное количество продукта, накапливаемого в культурах в течение ночи, составляет 504 мг/л.
Пример 2: Результаты биохимических и биологических исследований
Человеческое моноклональное антитело ABN912 связывается с человеческим МСР-1 и нейтрализует его функцию in vitro. Способность моноклонального антитела связываться с рекомбинантным человеческим МСР-1 можно охарактеризовать также с помощью двух независимых биохимических методов, таких как Biacore-анализ и сцинтилляционный анализ сходства (SPA). Специфичность антитела ABN912 в отношении других СС-хемокинов или МСР-1, выделенных из организмов кроме человека, оценивают с помощью BIAcore и с помощью SPA оценивают ингибирование антителом ABN912 связывания МСР-1 с клетками, экспрессирующими CCR2B. Биологическую активность ABN912 в отношении рекомбинантного и продуцируемого в естественных условиях МСР-1 демонстрируют с помощью анализа мобилизации Са2+ в клетках, экспрессирующих CCR2B. Активность ABN912 в отношении природных клеток-мишеней, т.е. моноцитов периферической крови человека (hPBMC), демонстрируют путем оценки вызываемого МСР-1 хемотаксиса.
2.1. BIAcore-анализ
Константы скорости ассоциации и диссоциации для связывания рекомбинантного человеческого МСР-1 с иммобилизованным ABN912 оценивают с помощью BIAcore-анализа и рассчитывают значения kd. ABN912 иммобилизуют на поверхности чипа биосенсора и связывание с рекомбинантным МСР-1 оценивают с помощью поверхностного плазменного резонанса (BIACORE 2000 Instrument Handbook, March 1999 (Version AC); http:www.biacore.com). Полученные результаты приведены ниже в таблице.
ABN912 связывается с рекомбинантным человеческим МСР-1 с очень высокой аффинностью.
2.2. Профили селективности и видоспецифичности в отношении хемокинов
С целью определения специфичности связывания МСР-1 с ABN912 с помощью BIAcore оценивают потенциальную способность связываться с ABN912 нескольких МСР-1, выделенных из организмов кроме человека, и СС-хемокинов.
2.2.1. Взаимодействия с МСР-1, выделенных из организмов кроме приматов
Оценивают связывание с антителом ABN912 МСР-1 из различных обычно применяемых видов лабораторных животных. Опыты осуществляют путем введения 5 нМ раствора хемокина в проточную ячейку устройства BIAcore, предварительно загруженную ABN912. Через 8 мин для каждого из оцениваемых хемокинов определяют количество связанного хемокина. Полученные результаты представлены ниже в таблице в виде процента связывания каждого хемокина по сравнению с рекомбинантным человеческим МСР-1 (100%).
ABN912 обладает специфичностью по отношению к человеческому МСР-1 и не дает перекрестную реакцию с МСР-1 других изученных видов.
2.2.2. Взаимодействия с рекомбинантными хемокинами
Для оценки профиля перекрестной реактивности антитела ABN912 с другими СС-хемокинами с помощью описанного выше метода оценивают их способность связываться с антителом. Ниже в таблице приведены проценты связывания каждого хемокина по сравнению с рекомбинантным человеческим МСР-1.
ABN912 представляет собой специфическое для человеческого МСР-1 антитело и не дает перекрестную реакцию с МСР-2, МСР-3, МСР-4, LEC, RANTES, MIP-1α и MIP-1β, но дает выраженную перекрестную реакцию с человеческим эотаксином, т.е. человеческим эотаксином-1. Установлено, что AAV294 дает также выраженную перекрестную реакцию с эотаксином.
2.3. Ингибирование связывания МСР-1 с клетками, экспрессирующими CCR2B
SPA-технологию (сцинтилляционный анализ сходства) используют для того, чтобы доказать, что антитело ABN912 препятствует связыванию МСР-1 с клеточными мембранами, экспрессирующими рецептор CCR2B. Мембраны СНО-клеток, экспрессирующих CCR2B (СНО №84 - см. ниже), инкубируют с различными концентрациями ABN912 (от 10-14 до 10-8 М) и с помощью SPA измеряют остаточное связывание радиоактивно меченного (125-1)-МСР-1 с использованием гранул, покрытых агглютинином из проростков пшеницы. Среднее значение IC50 [М], полученное по трем независимым экспериментам, составляет (461 ± 206) × 10-12. Таким образом, установлено, что ABN912 препятствует связыванию МСР-1 с клеточными мембранами, которые экспрессируют рецептор CCR2B.
2.4. Ингибирование опосредуемой МСР-1 сигнализации
Ранним признаком индуцируемой МСР-1 сигнализации является мобилизация внутриклеточного Са, которую можно оценивать с помощью флуоресцентных красителей.
Оценку изменения мобилизации кальция осуществляют на линии клеток СНО №84, т.е. линии СНО-клеток, стабильно трансфектированной с целью экспрессии хемокинного рецептора CCR2B. Клетки линии СНО №84 культивируют в не содержащей рибонуклеазу и дезоксирибонуклеазу среде МЕМ-альфа с глутамакс-1 (glutamax-1), дополненной 10% диализированной ФТС, 200 ед./мл пенициллина/стрептомицина и 80 нМ метотрексатом в качестве селективного агента. Когда клеточная культура становится плотной, но до стадии конфлюэнтности, клетки промывают ЗФР и проводят обнаружение, используя кратковременную инкубацию с трипсином-ЭДТК (максимум в течение 1 мин).
Клетки СНО № 84 однократно промывают RPMI путем центрифугирования при 250 × g в течение 7 мин и ресуспендируют 3,5×10 клеток/мл в Hepes-буфере-0,5% БСА, содержащем 0,04% плуроновой кислоты, 1,0 мкМ «fura red» и 0,3 мкМ «fluo-3». Клетки инкубируют с указанными флуоресцентными кальциевыми зондами в течение 1 ч при комнатной температуре в темноте при осторожном периодическом (6-8 раз) перемешивании. Затем клетки собирают дважды в Hepes-буфере-0,5% БАС путем центрифугирования и дебрис ресуспендируют из расчета 1,5-2×106 клеток/мл в Hepes-буфере-0,5% БСА. После такой обработки клетки готовы к стимуляции и оценке кальция и их хранят при комнатной температуре в темноте до использования.
Готовят 20-кратные растворы и антител, и хемокина (МСР-1, фирма R&D Systems, Миннеаполис, шт. Миннесота, США) и перед добавлением в клетки их смешивают при комнатной температуре в течение 5-8 мин. Оценивают зависимость интенсивности как красной, так и зеленой флуоресценции от времени с использованием проточного цитометра (FACS, фирма Becton Dickinson). С целью получения базовых уровней для каждого образца клеток предварительно в течение 15 с оценивают флуоресцентность клеток, в которые введены флуоресцентные Са-зонды. Сбор данных прекращают на короткое время для того, чтобы стимулировать клетки добавлением небольшого объема субстанции, представляющей собой стимул (10-кратная концентрация хемокина с антителом или без него), и продолжают оценку флуоресценции. Общее время оценки флуоресценции составляет 51 с.
Применяемые индикаторы Са при связывании с кальцием сдвигают интенсивность флуоресценции в противоположные стороны; флуоресценция красителя fura red понижается, в то время как флуоресценция красителя fluo-3 повышается. В каждом эксперименте оценивают красную и зеленую флуоресценцию, рассчитывают соотношение между красной и зеленой флуоресценцией и строят график в зависимости от времени. «Базовое соотношение» и «соотношение после стимуляции» соответственно определяют как среднее значение соотношений, полученных непосредственно перед стимуляцией, и среднее значение максимальных величин, полученных после стимуляции. Интенсивность реакции оценивают количественно с помощью коэффициента стимуляции (S.I.), представляющего собой «соотношение после стимуляции» деленное на «базовое соотношение». Коэффициенты стимуляции, полученные в присутствии антитела, выражают в виде процента S.I., полученного в присутствии только растворителя.
Ингибирование (%) антителом А1, растворенном в растворителе S1, выражают формулой:
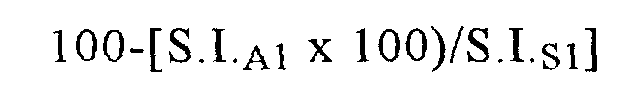
Ингибирование антителом ABN912 сравнивают с ингибированием антителом-предшественником, т.е. AAV293, и неродственным имеющимся в продаже мышиным антителом к МСР-1 (фирма R&D Systems). Кроме того, поскольку применяемый для получения AAV293 иммуноген представляет собой негликозилированный рекомбинантный человеческий МСР-1 из Е. coli, для стимуляции клеток СНО №84 также применяют супернатант МСР-1 из стимулированной TNFα линии клеток HUVEC (эндотелиальные клетки пупочной вены человека) и согласно описанному выше методу определяют антагонистическое действие ABN912 в отношении мобилизации Са2+. Полученные результаты приведены ниже в таблице.
AAV293
(IC50 в нМ)
(фирма R&D
Systems)
(IC50 в нМ)
0,39 ± 0,02 1,53 ± 0,47
0,66 ± 0,16
2,12 ± 0,06
ABN912 специфично ингибирует индуцируемую МСР-1 мобилизацию Са в клетках СНО №84, но при этом обладает одинаковой эффективностью в отношении МСР-1, полученного как из Е. coli, так и из HUVEC.
2.5. Ингибирование индуцируемого МСР-1 хемотаксиса
Способность ABN912 ингибировать хемотактическое действие МСР-1 в отношении hPBMC (моноциты периферическое крови человека) оценивают в опытах по оценке хемотаксиса с использованием камеры Бойдена (Boyden). HPBMC получают с помощью устройства типа Lymphoprep и в качестве исходной популяции используют 2×106 моноцитов на мл. В нижний и верхний отсек камеры добавляют ABN912 или неродственное антитело (отрицательный контроль) в указанных молярных соотношениях. Хемотаксис индуцируют, добавляя в нижний отсек 5,7 нМ раствор рекомбинантного человеческого МСР-1 или 1 нМ fMIFL (отрицательный контроль). Клеткам дают возможность мигрировать из верхнего отсека в нижний отсек в течение 90 мин при комнатной температуре. Количество прошедших в нижний отсек клеток оценивают путем окрашивания и последующего подсчета. Установлено, что ABN912 ингибирует индуцируемый МСР-1 хемотаксис hPBMC в зависимости от дозы. Воздействие ABN912 на индуцируемый МСР хемотаксис является специфическим; неродственное антитело не оказывает воздействия на индуцируемый МСР хемотаксис, а ABN912 не оказывает воздействия на индуцируемый fMIFL хемотаксис.
2.6. Ингибирование эмиграции лейкоцитов у макак-резус
Указанную механистическую модель используют для решения вопроса о том, можно ли ингибировать индуцируемую МСР-1 эмиграцию лейкоцитов в кожу макак-резус с помощью NVP-ABN912 при его быстром внутривенном введении (в виде болюса).
Самцам макак-резус вводят 30 мкг/кг IL-3 в течение 13 дней (дважды в день, i.v.) для того, чтобы повысить количество лейкоцитов и примировать эндотелиальные клетки для оптимального рекрумента. На 13-й день обезьян анестезируют и МСР-1 (10 мкг) вводят в трех повторностях интрадермально в правую часть грудного и брюшного отдела. Через 4 ч обезьян вновь анестезируют и с помощью пункционной биопсии получают образцы из мест, в которые инъецировали МСР-1. Затем осуществляют внутривенное введение в виде болюса либо NVP-ABN912, либо гуманизированного применяемого в качестве отрицательного контроля антитела к человеческому карциноэмбриональному антигену совместимого изотипа (CGP44290), в каждом случае до достижения дозы 5 мг/кг. И, наконец, вновь инъецируют МСР-1 (10 мкг), в трех повторностях, интрадермально в левую часть грудного и брюшного отдела. Эти места инъекций располагают симметрично относительно мест проведенных ранее инъекций. Через 4 ч обезьян вновь анестезируют и с помощью пункционной биопсии получают образцы из мест, в которые инъецировали МСР-1. Все полученные с помощью пункционной биопсии образцы затем разрезают пополам и одну половину замораживают в жидком азоте для ферментного анализа. Вторую половину хранят в формалине для последующего гистологического анализа. В эозинофилах и нейтрофилах оценивают активность эозинофилпероксидазы (ЕРО) и миелопероксидазы (МРО) соответственно. Параллельно в качестве контроля у каждой обезьяны берут образцы из мест, в которые инъецировали ЗФР, а также из мест, в которые не осуществляли инъекцию для стимуляции инфильтрации. Во всех контрольных вариантах не обнаружено выраженных опосредуемых антителом воздействий.
Ферментативная активность в местах инъекций МСР-1 до и после обработки NVP-ABN912 и CGP44290 (IgG4, применяемый в качестве отрицательного контроля совместимый изотип), соответственно, представлена в виде среднего значения ± СКО. Для каждого варианта каждой обезьяне делают инъекции в несколько различных мест (в 3 разных места). Объединенные данные, полученные в двух независимых экспериментах, представлены на фиг.1, где на фиг.1А представлены данные об эмиграции эозинофилов, а на фиг.1Б - об эмиграции нейтрофилов.
Стандартизованные данные об активности ЕРО (А) или МРО (Б) в образцах пункционной биопсии до (МСР-1, без обработки, 100%) и после обработки антителами представлены в виде среднего значения ± СКО.
В том случае, когда эмиграцию лейкоцитов в кожу макак-резус индуцируют с помощью 10 мкг МСР-1, в присутствии NVP-ABN912 наблюдают 85,5%-ное ингибирование эмиграции эозинофилов и 81,4%-ное ингибирование эмиграции нейтрофилов по сравнению с эмиграцией лейкоцитов перед введением антитела. При применении CGP44290 наблюдают только 20%-ное ингибирование обоих типов клеток. Учитывая, что инъекция только ЗФР приводит к рекрументу 13,0% эозинофиллов и 22,7% нейтрофилов относительно уровня реакции, обнаруженной при использовании 10 мкг МСР-1, введение NVP-ABN912 снижает уровень эмиграции лейкоцитов до базового уровня, который наблюдается при стимуляции ЗФР. Различия между результатами, полученными при использовании NVP-ABN912 и контрольного антитела, являются статистически высоко достоверными (р<0,01), и, принимая во внимание уровень действия, они, вероятно, имеют определенное биологическое значение. Репрезентативные гистологические данные об эмиграции эозинофилов представлены на фиг.2.
Данные гистологического анализа образцов пункционной биопсии, полученных в эксперименте по индуцируемой МСР-1 эмиграции эозинофилов, представлены для четырех обезьян (2279, 2280, 2281 и 2282).
(A), (Б), (Д), (Е) - окрашивание эозинофилов до введения антител обезьянам 2279, 2280, 2281 и 2282 соответственно.
(B), (Г), (Ж), (3) - окрашивание эозинофилов после введения антител обезьянам 2279, 2280, 2281 и 2282 соответственно.
Обезьян 2279 и 2280 обрабатывают применяемым в качестве отрицательного контроля антителом совместимого изотипа CGP44290 (5 мг/кг), а обезьян 2281 и 2282 обрабатывают NVP-ABN912 (5 мг/кг). Эозинофилы окрашиваются в красный цвет (они видны как темные пятна на черно-белой фотографии), и стрелками на панели (Ж) и (3) выделено несколько эозинофилов, присутствующих у двух обработанных NVP-ABN912 обезьян.
При использовании CGP 44290 не обнаружено различий между образцами, полученными до и после обработки, в то время как у обезьян, обработанных NVP-ABN912, обнаружено практически полное ингибирование эмиграции эозинофилов.
2.7 Ингибирование Т-клеточной инфильтрации у мышей линии SCID-hu
Указанную механистическую модель применяют для решения вопроса о том, можно ли ингибировать индуцируемую МСР-1 инфильтрацию Th-клеток в кожу мышей линии SCID-hu Skin с помощью внутрибрюшной инъекции NVP-ABN912.
Мышам линии SCID трансплантируют два небольших кусочка кожи взрослого человека (получая линию SCID-hu Skin). Качество трансплантатов оценивают в течение 5-6 недель после трансплантации и затем отбирают мышей, трансплантация которых считается успешной (в целом >85%) для экспериментов по миграции in vivo. Для оценки опосредуемой хемокином миграции РВМС выделяют стандартным методом разделения в градиенте плотности из образцов лейкоцитной пленки и соответственно вводят (i.p. 1×108 клеток/мышь, объем 500 мкл) мышам линии SCID-hu Skin, которым предварительно трансплантируют (5-8 недель) два кусочка кожи человека (в правую и левую стороны верхней области спины). Перенос клеток осуществляют в день 0. МСР-1 (фирма R&D Systems, Миннеаполис, шт. Минессота) и ЗФР вводят внутрикожно в человеческие трансплантаты в экспериментальные дни 1, 2, 4 и 6. Контрольным мышам (обработанным CGP44290) вводят 500 нг МСР-1 в правый кожный трансплантат и такой же объем (30 мкл) ЗФР в левый трансплантат. Мышам, обработанным NVP-ABN912, инъецируют 500 нг МСР-1 и в правый, и в левый кожный трансплантат. NVP-ABN912 и контрольный изотип CGP44290 вводят i.p. (100 мкг/мышь, 4 мг/кг, объем 500 мкл) в день 0 (за 5 ч до переноса клеток) и в экспериментальные дни 2 и 5. На 8-й день всех мышей умерщвляют и человеческие кожные трансплантаты выделяют для дальнейшего анализа. Суспензии отдельных клеток каждого человеческого кожного трансплантата получают с помощью устройства DAKO Medimachine согласно инструкции производителя. Эти клеточные суспензии окрашивают с помощью МАт к человеческим CD3 и CD4, конъюгированного с ФИТЦ и ПЭ, и анализируют с использованием проточного цитометра типа FACSCalibur (фирма Becton Dickinson, Сан Хосе, Калифорния). Результаты представлены на фиг.3. Установлено, что 0,73±0,15% введенных путем инфузии человеческих Th-клеток проникает в кожный трансплантат группы, обработанной ABN912 (5 мышей, n=10), по сравнению с 3,73±1,03% в соответствующей группе, обработанной CGP44290 (3 мыши, n=3). У мышей, у которых миграцию клеток индуцируют ЗФР, в человеческий кожный трансплантат (3 мыши, n=3) проникает 0,34±0,17% Th-клеток.
Статистический анализ осуществляют на основе однонаправленного вариационного анализа с использованием post hoc-критерия Дуннетта для множественного сравнения. Снижение Th-клеточной инфильтрации у животных, обработанных ABN912, в сравнении с животными, обработанными CGP44290, соответственно оказалось высоко достоверным (р<0,01, **).
2.8. Характеристика и механизм действия ABN912-связывающего эпитопа на человеческом МСР-1
Трехмерную структуру комплекса МСР-1 с Fab-фрагментом NVP-ABN912 определяют с помощью кристаллографии в рентгеновских лучах.
Fab-фрагмент NVP-ABN912 получают протеолитическим расщеплением из полного антитела и очищают с использованием протеин А-хроматографии с последующей гель-фильтрацией. Комплекс, включающий Fab и рекомбинантный человеческий МСР-1, очищают с помощью протеин G-хроматографии и гель-фильтрации и концентрируют с помощью ультрафильтрации до 26 мг/л в 50 мМ Трис-HCl, рН 8,0, 0,1М NaCl. Кристаллы выращивают при комнатной температуре с использованием метода диффузии паров в висящих каплях в буфере, содержащем 20% (мас./об.) ПЭГ 4000, 10 об.% изопропанола, 0,1М Na Hepes pH 7,5. Они относятся к пространственной группе P212121 с размерами элементарной ячейки а=63,90Е, b=86,08Е, с=321,64Е и содержат по три комплекса на асимметричную ячейку. Перед получением данных один кристалл погружают примерно на 3 мин в буфер, содержащий 17% (мас./об.) ПЭГ 4000, 8,5 об.% изопропанола, 15% глицерина, 85 мМ Na Hepes, рН 7,5. Затем кристалл закрепляют с помощью найлоновой петли CryoLoop и сразу же замораживают в потоке азота при 120К. Диффракционную картину оценивают с помощью системы плоского изображения MAR 345 с использованием дающего SNBL-пучок устройства типа European Synchrotron Radiation Facility (λ=0,90Е). В целом проводят 242617 наблюдений, что соответствует 68948 отдельным отражениям (Rsym=0,050), при разрешении 20,0 и 2,33Е (полнота 89,3%). Структуру определяют путем молекулярной замены, используя в качестве исходных моделей рентгенограммы Fab-фрагмента моноклонального антитела 3D6 (RCSB-номер 1DFB; Не и др., 1992) и человеческого МСР-1 (RCSB-номер IDOL; Lubkowski и др., 1997). Структуру уточняют с учетом динамики угла поворота и минимизации энергии, используя программу CNX, до достижения конечного R-фактора, составляющего 0,224 (Rfree=0,261) для всех отражений между 20 и 2,33А. Конечная модель включает L1- L214 и HI-Н222-локусы антитела ABN912 и остатки 10-71 человеческого антитела МСР-1. Модель имеет правильное геометрическое строение, характеризующееся среднеквадратичным отклонением (СКО), составляющим 0,006А для длин связей и 1,36° для углов между связями.
Для комплекса Fab-фрагмент NVP-ABN912/человеческий МСР-1 характерна большая внутренняя поверхность связывания (площадь объединенной внутренней поверхности - 1,590Е2), включающая множество гидрофобных и полярных связей, а также несколько имеющих решающее значение электростатических взаимодействий. Основные контакты с антигеном опосредуются длинной H-CDR3-петлей антитела NVP-ABN912, которая охватывает центр объединенного с антигеном сайта и в результате этого в значительной степени погружается в комплекс. Связывающий эпитоп человеческого МСР-1 содержит аминокислотные остатки Asn 14, Thr 16, Asn 17, Arg 18, Lys 19, Ile 20, Ser 21, Gln 23, Arg 24, Lys 49, Glu 50, Ile 51 и Cys 52. Остатки Arg 18, Arg 24 и Lys 49 принимают участие в электростатических взаимодействиях с остатками антитела Glu L55, Asp H99 и Glu HI 01 и в связанных с Н взаимодействиях с Туг L94, Trp H33, Asn H50, Gln H53, Asp H99 и Tyr Н108. Кроме того, гуанидиниевый фрагмент Arg 18 и Arg 24 МСР-1 является π-ориентированным относительно ароматических колец Trp H33 и Tyr Н32 соответственно. Дополнительные взаимодействия между NVP-ABN912 и Туг 13, Gin 17, Ser 21, Lys 19, Arg 24 и Glu 50 антигена опосредованы 8 молекулами воды, погруженными во внутреннюю поверхность протеина. Поскольку Tyr 13 и Arg 24 МСР-1 принимают участие в связывании с хемокинным рецептором CCR2B (Hemmerich и др., Biochemistry, 18; 13013-13025, 1999), вероятно, что NVP-ABN912 непосредственно конкурирует с рецептором за связывание с антагонистом.
2.9. Картирование эпитопа ABN912 на МСР-1 с помощью сайтнаправленного мутагенеза
С целью выявления остатков МСР-1, которые являются функционально важными для распознавания NVP-ABN912, осуществляют анализ с использованием аланинсканирующего мутагенеза (Cunningham В. С. и Wells J. A. Science 244, 1081-1085 (1989)). Первоначально визуально оценивают трехмерную структуру МСР-1 (Handel Т.М. и Domaille, Р. J. Biochemistry 35, 6569-6584 (1996); Lubkowski J., Bujacz G., Boque L., Domaille P. J., Handel Т. М. и Wlodawer, A., Nature Struct. Biol. 4, 64-69 (1997)) с целью идентификации выступающих над поверхностью остатков с использованием пакета программ WebLab Viewer. В целом 39 остатков, которые потенциально могут взаимодействовать с ABN912, индивидуально подвергают мутации с заменой на аланин или лизин (D3K, А48К) с использованием набора для сайтнаправленного мутагеназа QuikChange Site-Directed Mutagenesis (Papworth С., Bauer J. С., Braman J. и Wright D. A. Strategies 9 (3), 3-4 (1996)). Полученные в результате мутантные гены экспрессируют в клетках линии HEK.EBNA сначала путем трансфекции клеток 2 мкг экспрессионных плазмид, несущих мутантные последовательности МСР-1, с реагентом для трансфекции Geneporter (фирма Gene Therapy Systems). После трансфекции клетки инкубируют в течение 3 дней при 37°С, затем супернатанты собирают, центрифугируют в течение 5 мин и очищают с помощью аффинной хроматографии. Концентрации мутантного МСР-1 определяют с помощью анализа протеинов (фирма Biorad), используя очищенный МСР-1 в качестве стандарта. Как правило, из 3 мл культуры получают по 40 мкг очищенного человеческого МСР-1 или мутантных МСР-1.
Аффинность МСР-1 и мутантного МСР-1 к NVP-ABN912 оценивают с помощью поверхностного плазменного резонанса с использованием устройства BIAcore (фирма BIAcore). Первоначально поверхность сенсорного элемента активируют и инъецируют 30 мкг/мл раствора антитела к Fcγ для ковалентного связывания с элементом. Антитело NVP-ABN912 накапливают на модифицированной с помощью антитела к Fcγ поверхностью, инъецируя 5 мкг/мл раствора. Готовят разбавления растворов МСР-1 или мутантных МСР-1, получая конечные концентрации от 0,75 до 4 нМ и инъецируют для связывания с иммобилизованным NVP-ABN912 на поверхности сенсорного элемента. Определяют ассоциацию и диссоциацию в течение 5 мин, регистрируя при этом сигнал поверхностного плазменного резонанса. Затем анализируют титрованные серии с помощью пакета программ BIAevaluation 3.0 (Software part of the instrument installation; Handbook Cat.No. BR-1002-29; версия 7-97). Результаты представлены на фиг.4, где продемонстрировано связывание МСР-1 и мутантных МСР-1 CNVP-ABN912.
Основные детерминанты МСР-1 для NVP-ABN912, выявленные с помощью описанного выше метода, представляют собой остатки Т16, R18, R24 и К49. Замена Т16, R18 и R24 на аланин приводит к полному отсутствию связывания с NVP-ABN912, в то время как замена К49 на аланин приводит к 133-кратному снижению аффинности.
| название | год | авторы | номер документа |
|---|---|---|---|
| АНТИТЕЛА К ЧЕЛОВЕЧЕСКОМУ IL-1БЕТА | 2001 |
|
RU2286351C2 |
| РЕКОМБИНАНТНОЕ АНТИТЕЛО К ЧЕЛОВЕЧЕСКОМУ ИНТЕРЛЕЙКИНУ 1β | 2001 |
|
RU2264413C2 |
| АНТАГОНИСТИЧЕСКИЕ АНТИТЕЛА К IL-17 | 2005 |
|
RU2426741C2 |
| НОВОЕ ПРИМЕНЕНИЕ СОЕДИНЕНИЙ ИЛ-1β | 2012 |
|
RU2571563C2 |
| ТЕРАПЕВТИЧЕСКИЕ СВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ | 2002 |
|
RU2328506C2 |
| МОНОКЛОНАЛЬНЫЕ АНТИТЕЛА К TIM-3 ЧЕЛОВЕКА | 2019 |
|
RU2777336C1 |
| СПОСОБ ПОЛУЧЕНИЯ БИСПЕЦИФИЧЕСКИХ АНТИТЕЛ, БИСПЕЦИФИЧЕСКИЕ АНТИТЕЛА И ТЕРАПЕВТИЧЕСКОЕ ПРИМЕНЕНИЕ ТАКИХ АНТИТЕЛ | 2018 |
|
RU2795240C2 |
| АНТИТЕЛО К SIRPα И ЕГО ПРИМЕНЕНИЕ | 2021 |
|
RU2822496C1 |
| НОВОЕ ПРИМЕНЕНИЕ СОЕДИНЕНИЙ ИЛ-1β | 2006 |
|
RU2468817C2 |
| АНТИ-C5 АНТИТЕЛА И ИХ ПРИМЕНЕНИЕ | 2018 |
|
RU2774716C2 |
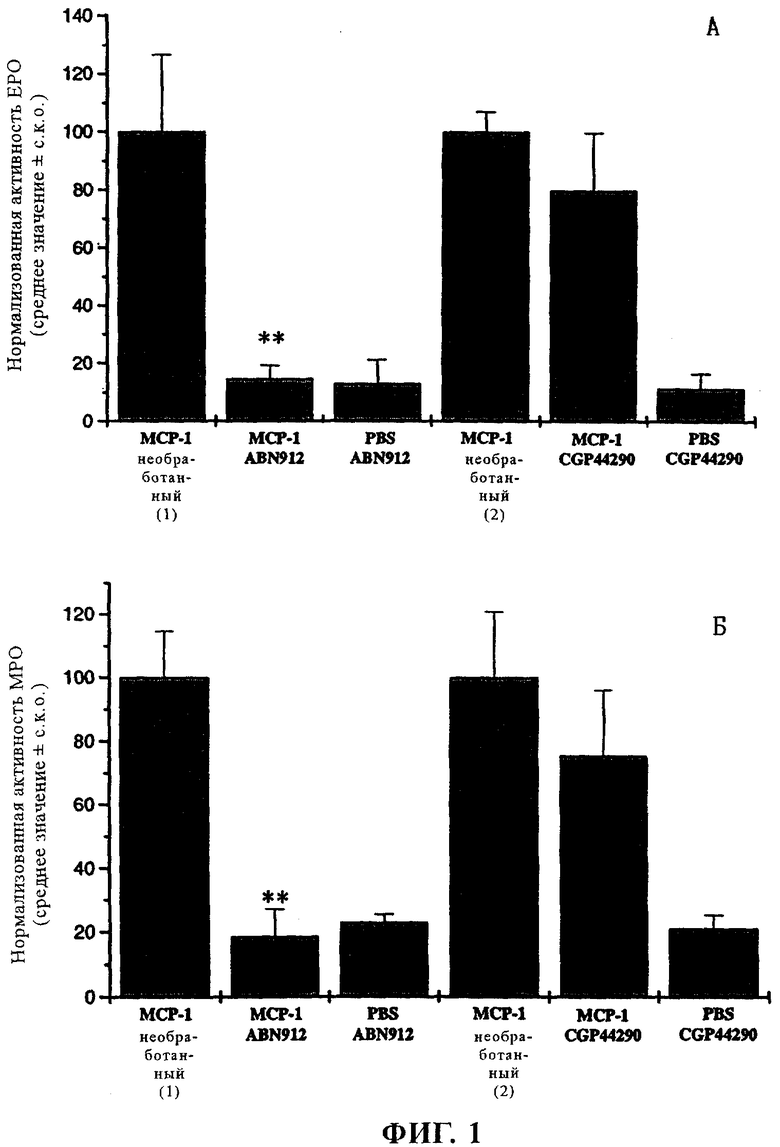
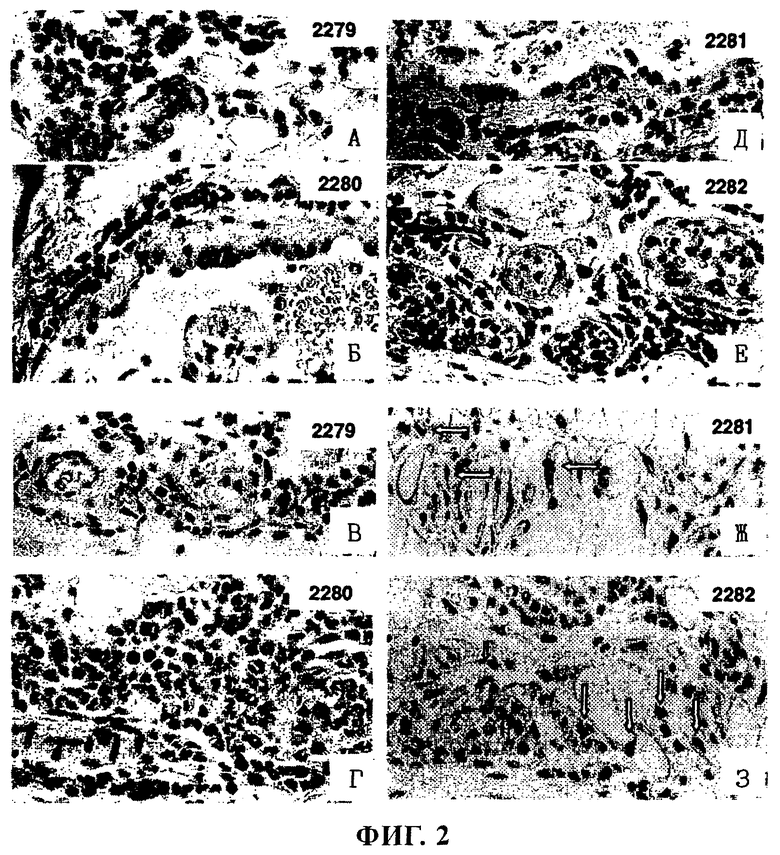
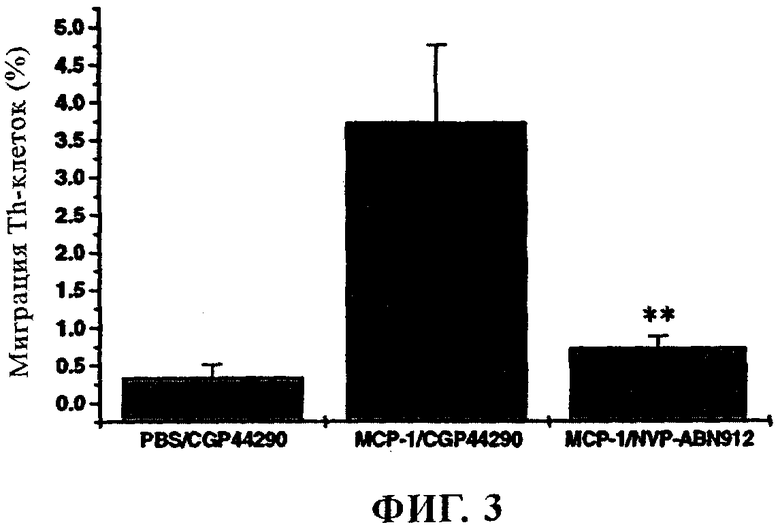
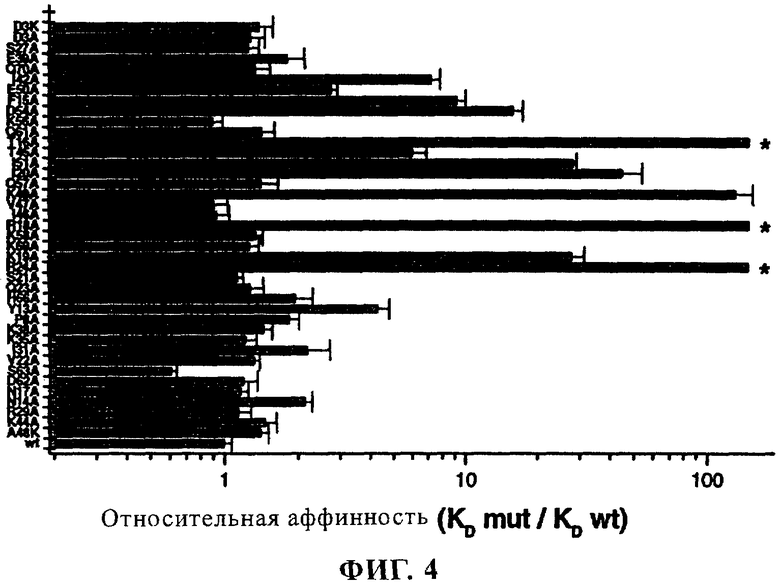






Настоящее изобретение относится к иммунологии и биотехнологии. Описаны варианты МСР-1-связывающих молекул. Одна из МСР-1-связывающих молекул содержит, по меньшей мере, одну вариабельную область тяжелой цепи (VН) иммуноглобулина, которая состоит из гипервариабельных участков CDR1, CDR2 и CDR3, тогда как другие содержат и легкую и тяжелую цепи. Предложены ДНК конструкции, кодирующие указанные МСР-1 связывающие молекулы и экспрессирующий вектор, несущий по меньшей мере одну из конструкций ДНК. Описан способ получения МСР-1 связывающей молекулы. Раскрыт также способ лечения опосредуемых МСР-1 или эотаксином-1 заболевания или нарушения на основе антитела к МСР-1, которое перекрестно связывает эотаксин-1. Описана фармацевтическая композиция на основе антитела к МСР-1, которое перекрестно связывает эотаксин-1, для лечения опосредуемых МСР-1 или эотаксином-1 болезни или нарушения у пациента. МСР-1-связывающие молекулы по изобретению ингибируют связывание МСР-1 с его рецептором. Иммобилизованное полное антитело по изобретению является высокоспецифичным, поскольку связывает рекомбинантный МСР-1 человека с KD(43±2,9)×10-12 и может найти применение далее в медицине. 9 н. и 4 з.п. ф-лы, 4 ил., 4 табл.
(а) CDR1 - His-Tyr-Trp-Met-Ser, CDR2 -Asn-Ile-Glu-Gln-Asp-Gly-Ser-Glu-Lys-Tyr-Tyr-Val-Asp-Ser-Val-Lys-Gly и CDR3 - Asp-Leu-Glu-Gly-Leu-His-Gly-Asp-Gly-Tyr-Phe-Asp-Leu; или
(б) гипервариабельные участки CDR1, CDR2 и CDR3, взятые в целом, гомологичные по меньшей мере на 80% гипервариабельным участкам CDR1, CDR2 и CDR3, представленным в Seq.ID No.l.
(а) вариабельную область тяжелой цепи (VH) иммуноглобулина, которая включает последовательности гипервариабельных участков CDR1, CDR2 и CDR3, где CDR1 имеет аминокислотную последовательность His-Tyr-Trp-Met-Ser, CDR2 имеет аминокислотную последовательность Asn-Ile-Glu-Gln-Asp-Gly-Ser-Glu-Lys-Tyr-Tyr-Val-Asp-Ser-Val-Lys-Gly и CDR3 имеет аминокислотную последовательность Asp-Leu-Glu-Gly-Leu-His-Gly-Asp-Gly-Tyr-Phe-Asp-Leu; или
(б) вариабельную область легкой цепи (VL) иммуноглобулина, которая включает последовательности гипервариабельных участков CDR1', CDR2' и CDR3', где CDR1' имеет аминокислотную последовательность Arg-Ala-Ser-Gln-Gly-Val-Ser-Ser-Ala-Leu-Ala, CDR2' имеет аминокислотную последовательность Asp-Ala-Ser-Ser-Leu-Glu-Ser и CDR3' имеет аминокислотную последовательность GIn-Gln-Phe-Asn-Ser-Tyr-Pro;
(в) гипервариабельные участки CDR1, CDR2, CDR3, CDR1', CDR2' и CDR3', взятые в целом, гомологичны по меньшей мере на 80% гипервариабельным участкам, представленным в Seq.ID No. 1 и 2.
а) первый фрагмент, который кодирует вариабельную область, содержащую альтернативные каркасный и гипервариабельные участки, где гипервариабельные участки состоят из последовательных участков CDR1, CDR2 и CDR3, аминокислотные последовательности которых представлены в SEQ ID No. 1; этот первый фрагмент начинается с кодона, который кодирует первую аминокислоту вариабельной области, и заканчивается кодоном, который кодирует последнюю аминокислоту вариабельной области, и
б) второй фрагмент, кодирующий константную область тяжелой цепи или ее фрагмент, который начинается с кодона, кодирующего первую аминокислоту константной области тяжелой цепи, и заканчивается кодоном, который кодирует последнюю аминокислоту константной области или его фрагмента, после которого расположен стоп-кодон.
а) первый фрагмент, который кодирует вариабельную область, содержащую альтернативные каркасный и гипервариабельные участки, где гипервариабельные участки состоят из последовательных участков CDR1, CDR2 и CDR3, аминокислотные последовательности которых представлены в SEQ ID No. 1; этот первый фрагмент начинается с кодона, который кодирует первую аминокислоту вариабельной области, и заканчивается кодоном, который кодирует последнюю аминокислоту вариабельной области, за которым следует стоп-кодон,
б) второй фрагмент, который кодирует вариабельную область, содержащую альтернативные каркасный участок и гипервариабельные участки, где гипервариабельные участки представляют собой CDR1', CDR2' и CDR3', аминокислотные последовательности которых представлены в SEQ ID No. 2; этот первый фрагмент начинается с кодона, который кодирует первую аминокислоту вариабельной области и заканчивается кодоном, который кодирует последнюю аминокислоту вариабельной области, за которым следует стоп-кодон.
| Разборный с внутренней печью кипятильник | 1922 |
|
SU9A1 |
| BLESS N.M | |||
| et al | |||
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| J | |||
| Immunol | |||
| ЩИТОВОЙ ДЛЯ ВОДОЕМОВ ЗАТВОР | 1922 |
|
SU2000A1 |
| ХИМЕРНОЕ МОНОКЛОНАЛЬНОЕ АНТИТЕЛО, ВЗАИМОДЕЙСТВУЮЩЕЕ С ЧЕЛОВЕЧЕСКОЙ ПЛАЦЕНТАРНОЙ ЩЕЛОЧНОЙ ФОСФАТАЗОЙ, ВАРИАБЕЛЬНАЯ ОБЛАСТЬ ТЯЖЕЛОЙ ГАММА-ЦЕПИ ХИМЕРНОГО МОНОКЛОНАЛЬНОГО АНТИТЕЛА, ВАРИАБЕЛЬНАЯ ОБЛАСТЬ ЛЕГКОЙ K-ЦЕПИ ХИМЕРНОГО МОНОКЛОНАЛЬНОГО АНТИТЕЛА, ФРАГМЕНТ КДНК, КОДИРУЮЩИЙ ВАРИАБЕЛЬНУЮ ОБЛАСТЬ ТЯЖЕЛОЙ ГАММА-ЦЕПИ ХИМЕРНОГО МОНОКЛОНАЛЬНОГО АНТИТЕЛА И ФРАГМЕНТ КДНК, КОДИРУЮЩИЙ ВАРИАБЕЛЬНУЮ ОБЛАСТЬ ЛЕГКОЙ K-ЦЕПИ ХИМЕРНОГО МОНОКЛОНАЛЬНОГО АНТИТЕЛА | 1990 |
|
RU2102479C1 |

