Изобретение относится к области биотехнологии, конкретно к иммунологии, и направлено на получение лентивирусной конструкции кодирующей Т-клеточный рецептор (TCR), распознающего антигенный эпитоп белка MAGE-A3, и способная индуцировать антиген-специфическое уничтожение клетки-мишени с экспрессией MAGE-A3.
В структуре смертности населения России злокачественные новообразования занимают второе место (15,4%) после болезней системы кровообращения. Ведущими локализациями злокачественных опухолей у обоих полов являются кожа (12,3%, с меланомой - 14,0%), молочная железа (11,4%), трахея, бронхи, легкое (10,5%), желудок (7,0%). При этом, ведущей онкологической патологией у женского населения является рак молочной железы (20,9%), уровень смертности от которого занимает первое место (17,0%) [Под ред. А.Д. Каприна, В.В. Старинского, Г.В. Петровой Злокачественные новообразования в России в 2013 году (заболеваемость и смертность) М.: ФГБУ «МНИОИ им. П.А. Герцена» Минздрава России. - 2015. илл. 250 с. ISBN 978-5-85502-205-6].
Общепринятыми методами лечения рака на сегодняшний день являются хирургия, лучевая терапия и химиотерапия. Использование данных методов позволяет эффективно снижать опухолевую нагрузку за короткий период времени, но не дает возможности элиминировать все опухолевые клетки, диссеминированные в организме пациента. Минимальная опухолевая нагрузка, сохраняющаяся после удаления основной части новообразования, является причиной возникновения рецидивов опухоли и развития метастазов, что в свою очередь приводит к повышению уровня смертности и инвалидизации пациентов с онкологическими заболеваниями.
Таким образом, традиционные подходы к лечению рака обнаруживают недостатки в вопросе устранения минимальной опухолевой нагрузки, и становится очевидной необходимость появления новых технологий, направленных на улучшение процессов распознавания и уничтожения единичных опухолевых клеток, оставшихся в организме пациента после проведения лучевой терапии, химиотерапии или хирургического удаления основной массы опухоли.
Фундаментальные и клинические исследования последних лет подтверждают, что использование потенциала иммунной системы может быть весьма перспективным для устранения опухолевых клеток [Palucka K., Ueno Н., Banchereau J. Recent developments in cancer vaccines. // JImmunol. - 2011. - Vol. 186. - №3. - P. 1325-31; Qian X., Wang X., Jin H. Cell transfer therapy for cancer: past, present, and future. // JImmunolRes. - 2014. doi: 10.1155/2014/525913]. Иммунотерапевтические подходы позволяют активировать клеточное звено иммунитета, нацеливая специфический иммунный ответ против опухолевых клеток, несущих на своей поверхности опухолевые антигены, без повреждения здоровых клеток. В виду вышесказанного целесообразна разработка подходов, позволяющих эффективно генерировать специфический противоопухолевый иммунный ответ в организме пациента.
На сегодняшний день разработано несколько различных иммунотерапевтических подходов, направленных против меланоме, рак яичников и немелкоклеточного рака легкого (НМРЛ), экспрессирующих MAGE-A3 (Zhang B, Ren Z, Zhao J, Zhu Y, Huang B, Xiao C, Zhang Y, Deng J, Mao L, Tang L, Lan D, Gao L, Zhang H, Chen G, Luo OJ. Global analysis of HLA-A2 restricted MAGE-A3 tumor antigen epitopes and corresponding TCRs in non-small cell lung cancer. Theranostics. 2023 Aug 6;13(13):4449-4468. doi: 10.7150/thno.84710). Иммунотерапия против антигена MAGE-A3 направлена на активации эффекторных клеток. С этой целью, в ряде работ использовали вакцинацию MAGE-A3 в сочетании с иммуностимуляторами (AS02B или AS15) (NCT00086866, NCT00849875). С другой стороны, повышение эффективности презентации антигена эффекторным клеткам. Создавали ДНК-конструкции, включающих семейство антигенов MAGE-A, которые трансфицировали дендритные клетки (средство естественной доставки и презентации антигенов эффекторным клеткам) (Lin, L.; Wei, J.; Chen, Y.; Huang, A.; Li, K.K.-W.; Zhang, W. Induction of Antigen-Specific Immune Responses by Dendritic Cells Transduced with a Recombinant Lentiviral Vector Encoding MAGE-A3 Gene. J. Cancer Res. Clin. Oncol. 2014, 140, 281-289.). Этот подход продемонстрировал возможность индуцировать сильный иммунный ответ, который проявлялся в снижении роста опухоли (Duperret, E.K.; Liu, S.; Paik, M.; Trautz, A.; Stoltz, R.; Liu, X.; Ze, K.; Perales-Puchalt, A.; Reed, C.; Yan, J.; et al. A Designer Cross-Reactive DNA Immunotherapeutic Vaccine That Targets Multiple MAGE-A Family Members Simultaneously for Cancer Therapy. Clin. Cancer Res. 2018, 24, 6015-6027.).
Другой иммунотерапевтический подход заключается в разработке адаптивной Т-клеточной терапии, направленной на MAGE-A3, в частности, создание рецепторов Т-клеток (TCR), обладающих высокой аффинностью и специфически связывающий эпитоп MAGE-A3 KVAELVHFL (112-120) на клетке-мишени. Также использование способов и фармацевтических композиций, содержащие модифицированные Т-клетки, для адоптивной терапии (Kageyama, S.; Ikeda, H.; Miyahara, Y.; Imai, N.; Ishihara, M.; Saito, K.; Sugino, S.; Ueda, S.; Ishikawa, T.; Kokura, S.; et al. Adoptive Transfer of MAGE-A4 T-Cell Receptor Gene-Transduced Lymphocytes in Patients with Recurrent Esophageal Cancer. Clin. cancer Res. 2015, 21, 2268-2277; Shah, N.N.; Maatman, T.; Hari, P.; Johnson, B. Multi Targeted CAR-T Cell Therapies for B-Cell Malignancies. Front. Oncol. 2019, 9, 146.; NCT01273181
Основную роль в развитии специфического противоопухолевого иммунного ответа играют цитотоксические Т-лимфоциты. Наличие противоопухолевых цитотоксических Т-лимфоцитов (ЦТЛ), как в отношении их количества, так и нормального функционирования, является необходимым условием для уничтожения опухолевых клеток иммунной системой [Aerts J., Hegmans J. Tumor-specific cytotoxic T cells are crucial for efficacy of immunomodulatory antibodies in patients with lung cancer. // Cancer Res. - 2013. - Vol. 73. - P. 2381-2388]. Иммунотерапевтические подходы, направленные на борьбу с конкретными опухоль-ассоциированными антигенами, также используют активированные антигенспецифические цитотоксические Т-лимфоциты в качестве главного противоопухолевого агента.
Наиболее близким к заявляемому является способ получения человеческого Т-клеточного рецептора, специфичного к эпитопу KVAELVHFL (112-120), полученного из белка MAGE-A3, включающий выделение мононуклеарных клеток (МНК) периферической крови HLA-A02+ доноров, затем МНК культивируют в присутствии рекомбинантного человеческого гранулоцитарно-макрофагального колониестимулирующего фактора (GM-CSF) и интерлейкина-4 (IL-4) в течение 4-х дней; на 5-й день добавляют в качестве фактора праймирования эпитоп KVAELVHFL (112-120) в культуру дендритных клеток с последующей их инкубацией при температуре 37°C; на 6-й день добавляют фактор созревания для завершения созревания дендритных клеток; затем полученные дендритные клетки собирают, промывают питательной средой, подсчитывают их количество, оценивают их жизнеспособность и сокультивируют с аутологичными CD8+ T-лимфоцитами в определенном соотношении, а через 7 дней сокультивирования выделяют антиген-специфические CD8+ T-клетки, затем производят идентификацию последовательности генов TCR с помощью секвенирования полной длины TCR, включая CDR3, выделенной из клонов Т-клеток, при этом цепи TCRα и TCRβ соединяют пептидным линкером 2A (TCRb-P2A-TCRa), а полную конструкцию TCR клонируют в скелет вирусной плазмиды для получения
вирусного вектора несущего ген TCR. Для получения человеческого Т-клеточного рецептора, специфичного к эпитопу KVAELVHFL (112-120) использовали аналог антигенпрезентирующих клеток (клеточная линия Т2) и набор эпитопов MAGE-A3 (Mp1, Mp2, Mp3, Mp4, Mp5, Mp6, Mp7, Mp8, Mp9 и Mp10) и последующий выбор из них тех, которые показали наибольшую иммуногенность в тестах. Противоопухолевой активности полученных TCR-T клеток, специфичных к эпитопу KVAELVHFL (112-120), оценивается по экспрессии маркеров активации (CD69 и CD137) и цитотоксическая активность против клеточной линии опухоли (РС9 и А375), экспрессирующих рецептор MAGE-A3. (Zhang B, Ren Z, Zhao J, Zhu Y, Huang B, Xiao C, Zhang Y, Deng J, Mao L, Tang L, Lan D, Gao L, Zhang H, Chen G, Luo OJ. Global analysis of HLA-A2 restricted MAGE-A3 tumor antigen epitopes and corresponding TCRs in non-small cell lung cancer. Theranostics. 2023 Aug 6;13(13):4449-4468. doi: 10.7150/thno.84710).
Недостатком прототипа является использование аналога антигенпрезентирующих клеток (клеточная линия Т2) и набора эпитопов MAGE-A3 (Mp1, Mp2, Mp3, Mp4, Mp5, Mp6, Mp7, Mp8, Mp9 и Mp10) и последующий выбор из них тех, которые показали наибольшую иммуногенность в тестах. Клеточная линия Т2 не может в полной мере отражать функциональность естественных антигенпрезентирующих клеток (дендритные клетки) и из-за этого и полученные результаты варьируют у разных доноров/пациентов в зависимости от тяжести болезни и стадии противоопухолевой терапии; оценка аффинности полученных TCR с использованием инструмента, который использует только последовательность CDR3β, последовательность эпитопа и информацию об аллеле HLA, чего недостаточно для оценки аффинности; противоопухолевая активность TCR-T клеток оценивается по экспрессии маркеров активации (CD69 и CD137) и цитотоксическая активность против клеточной линии опухоли (РС9 и А375). Данные недостатки не позволяют получить истинную информацию о строении и аффинитете человеческого Т-клеточного рецептора к эпитопу MAGE-A3.
Задачей изобретения является получения в короткий срок точной информации о человеческом Т-клеточном рецепторе, специфичного к эпитопу MAGE-A3 KVAELVHFL (112-120).
Поставленная задача решается тем, что в способе получения человеческого Т-клеточного рецептора, специфичного к эпитопу KVAELVHFL (112-120), полученного из белка MAGE-A3, включающем выделение мононуклеарных клеток (МНК) периферической крови HLA-A02+ доноров, затем МНК культивируют в присутствии рекомбинантного человеческого гранулоцитарно-макрофагального колониестимулирующего фактора (GM-CSF) и интерлейкина-4 (IL-4) в течение 4-х дней; на 5-й день в культуру дендритных клеток добавляют в качестве фактора праймирования эпитоп KVAELVHFL (112-120), с последующей их инкубацией при температуре 37°C; на 6-й день добавляют фактор созревания для завершения созревания дендритных клеток; затем полученные дендритные клетки собирают, промывают питательной средой, подсчитывают их количество, оценивают их жизнеспособность и сокультивируют с аутологичными CD8+ Т-лимфоцитами в определенном соотношении, а через 7 дней сокультивирования выделяют антиген-специфические CD8+ T-клетки, затем производят идентификацию последовательности генов TCR и с помощью секвенирования полной длины TCR, включая CDR3, выделенной из клонов Т-клеток, при этом цепи TCRα и TCRβ соединяют пептидным линкером 2A (TCRb-P2A-TCRa), а полную конструкцию TCR клонируют в скелет вирусной плазмиды для получения вирусного вектора, несущего ген TCR, при этом донорами являются условно-здоровые индивидуумы, из крови которых моноциты выделяли на градиенте фиколла, на 5-й день добавляют в качестве фактора праймирования эпитоп KVAELVHFL (112-120), в дозе 100 мкг/мл и последующей инкубацией в течение 24-х часов, на 6-й день в качестве фактора созревания добавляют TNF-a в концентрации 25 нг/мл в течение 24 часов, а сокультивирование дендритных клеток осуществляется с CD8+ Т-лимфоцитами в присутствии 0,5 мкг/мл антител к CD3 (анти-CD3), 1 мкг/мл антител к CD28 (анти CD28) и IL-2, IL-7, IL-15 (по 10 нг/мл каждый) в соотношении 1:10, через 7 дней сокультивирования выделяют антиген-специфические CD8+ T-клетки с помощью реагентов Flex-T, затем производят идентификацию последовательности генов TCR с помощью секвенирования РНК единичных клеток (Single-cell RNA sequencing (scRNA-seq)) и набор BD Rhapsody TCR/BCR amplification kit, выбор клонов TCR специфичных к эпитопу (112-120) белка MAGE-A3, на основе их аффинитета, фенотипа и специфичности, с помощью нейросетей ERGO-II и NetMHCpan-4.1 и программы TCRscape, причем помимо соединения цепи TCRα и TCRβ пептидным линкером 2A (TCRb-P2A-TCRa) используют 5' и 3' нетранслируемую область бета-глобулина на 5' и 3' концах вставки, в качестве вирусной плазмиды используют лентивирус на основе ВИЧ-1 (pLenti_hPGK), получают один доминирующий TCR с альфа- и бета-цепью, происходящими из одной и той же клетки.
Изобретение на наш взгляд, является новым. Существенным в данном способе является получение в короткий срок более точной информации о строении и аффинитете человеческого Т-клеточного рецептора, специфичного к эпитопу MAGE-A3 KVAELVHFL (112-120), обладающего высоким аффинитетом к целевому пептиду и минимальным аффинитетом к нецелевым пептидам, что определяет меньший риск развития побочных реакций.
Это достигается, в частности, использованием точного метода анализа транскриптома клеток (секвенирование РНК единичных клеток), позволяющего получить полную информацию о полной длине конкретного Т-клеточного рецептора (α-цепи и β-цепи) на каждой клетке и использующего для этого значительно меньшее количество целевых клеток, что позволяет получать необходимую информацию используя биологический материал доноров. Данный метод позволяет не проводить дополнительные процедуры (вакцинация доноров клетками, нагруженные целевым антигеном, и длительное культивирование исследуемых клеток для получения необходимого количества) и сократить время получение Т-клеточного рецептора. Использование нейросетей ERGO-II, NetMHCpan-4.1 и программы TCRscape (https://github.com/Perik-Zavodskii/TCRscape) позволяет выбрать клоны TCR к антигену MAGE-A3, обладающего высоким аффинитетом к целевому пептиду и минимальным аффинитетом к нецелевым пептидам. Использование Flex-T и одновременно двух флуорохромов для выделения позитивных клонов является более специфичным методом обнаружения антигенспецифичных клеток.
Способ осуществляется следующим образом.
1. Получение антиген-специфических Т-клеток.
Мононуклеарные клетки (МНК) получают из периферической крови условно-здоровых доноров с генотипом HLA-A02. МНК разделяют на прилипающую и неприлипающую фракции посредством адгезии на пластике в течение 30 минут при +37° в условиях СО2-инкубатора. Получение зрелых дендритных клеток (ДК) из прилипающей фракции проводили с использованием GM-CSF (100 нг/мл) и IL-4 (50 нг/мл). Для нагрузки антигеном использовали пептид KVAELVHFL (112-120), полученный из белка MAGE-A3, в дозе 100 мкг/мл на 5 сутки культивирования, с последующим созреванием ДК с помощью TNF-alpha (25 нг/мл) на 6 сутки культивирования. Клетки неприлипшей фракции МНК были культивированы в течении 6 суток. На 7 день проводилось объединение культур ДК и CD8+ клеток из неприлипшей фракции (соотношение 1:10) с добавлением 0,5 мкг/мл антител к CD3 (анти-CD3), 1 мкг/мл антител к CD28 (анти-CD28) и IL-2, IL-7, IL-15 (по 10 нг/мл каждый) для поддержания жизнеспособности и пролиферации CD8+ цитотоксических Т-лимфоцитов.
Через 7-8 дней совместного культивирования проводили выделение антиген-специфических Т-клеток из совместной культуры ДК и CD8+ клеток с помощью реагентов Flex-T и соответствующих пептидов с одновременным окрашиванием позитивных специфических клеток двумя флюорохромами и антителами против CD8+ для идентификации методом проточной сортировки.
После полного цикла протокола получения АГ-специфических Т-клеток было получено 2 образца специфичных для эпитопа KVAELVHFL (112-120), антигена MAGE-A3. Жизнеспособность полученных клеток оценивали с помощью красителей 7AAD и кальцеина. Содержание 7AAD-негативных/кальцеин-позитивных клеток в пробе составляло не менее 80%. Полученные клетки были помечены антителами для мультиплексирования биологических образцов SampleTag для использования на платформе для исследования мультиома единичных клеток BD Rhapsody.
Помеченные антителами для мультиплексирования Т-клетки в количестве 27 000 были загружены в станцию BD Rhapsody Express, в которой было произведено добавление улавливающих поли-А-несущие молекулы (мРНК, включая альфа- и бета-цепи Т-клеточного рецептора (TCR), и SampleTag) металлических бус с молекулярными штрихкодами, лизис клеток и гибридизация поли-А-несущие молекул лизированных клеток с металлическими бусами. Поли-А-несущие молекулы затем были обратно транскрибированы с получением кДНК. К кДНК добавили Template Switch Oligo (TSO) и провели ещё один раунд обратной транскрипции, что позволяет производить прочтения с 5’-конца кДНК. кДНК была амплифицирована в двух раундах полугнездовой ПЦР. Затем, для TCR библиотеки был произведен случайный отжиг праймеров и элонгация. С целью получения финальных библиотек иммунного транскриптома (379 генов связанные с иммунным ответом), TCR и SampleTag, продукты второй полугнездовой ПЦР библиотек SampleTag и иммунного транскриптома и продукты случайного отжига и элонгации библиотеки TCR были амплифицированны в гнездовой ПЦР с праймерами, содержащими адаптеры для секвенаторов Illumina. Полученные библиотеки были пулированы из расчета 27000 загруженных клеток и 10000 прочтений на клетку для библиотеки иммунного транскриптома, 15000 прочтений на клетку для библиотеки TCR, 1200 прочтений на клетку для библиотеки SampleTag и секвенированы на приборе NovaSeq 6000 на ячейке S2 c 150 парными прочтениями.
Полученные FASTQ файлы были обработаны при помощи пайплайна BD версии 1.11.1L, который проводит контроль качества прочтений, их выравнивание на референсный транскриптом, в том числе и Т-клеточных рецепторов, строит библиотеки клеточных индексов, устраняет ПЦР эффекты при помощи UMI RSEC (unique molecular identifier recursive substitution error correction) и UMI DBEC (unique molecular identifier distribution-based error correction), строит библиотеки клеток, устраняет технический шум при помощи анализа второй производной и выдает финальные матрицы экспрессии генов и Adaptive Immune Receptor Repertoire (AIRR) - матрицы последовательностей альфа- и бета-цепей Т-клеточного рецептора.
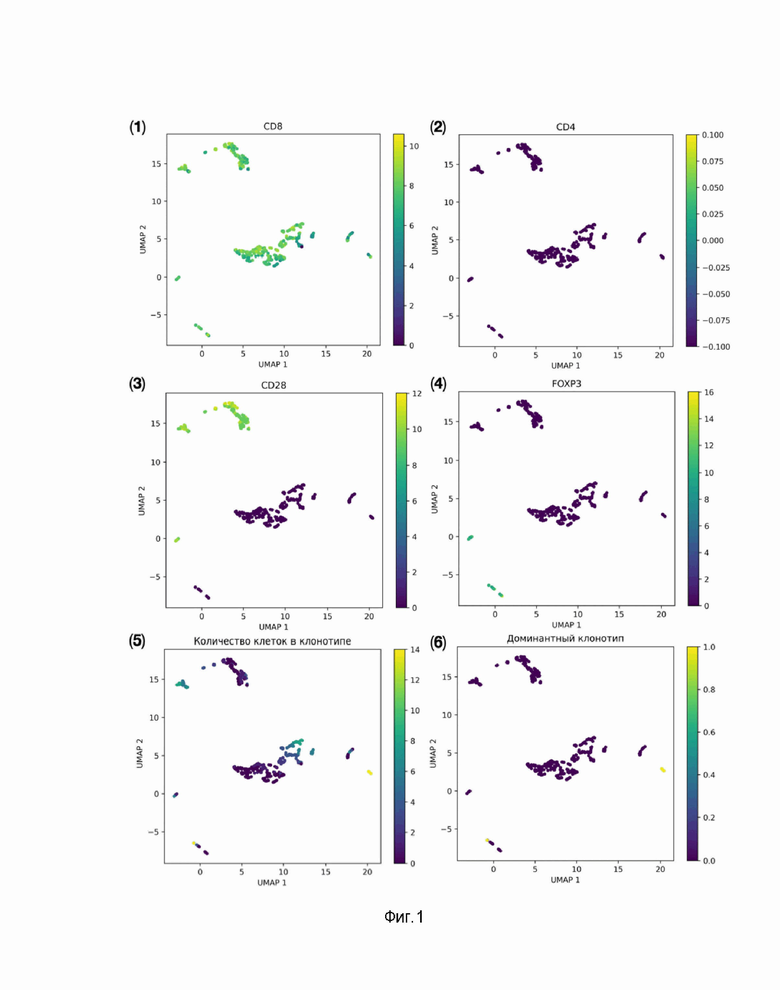
Нами было получено 7700 единичных клеток. Биоинформатический анализ данных секвенирования производился с помощью программного инструмента TCRscape (https://github.com/Perik-Zavodskii/TCRscape). TCRscape - это инструмент для определения доминантных (по количеству единичных клеток) по CDR3-петле или по полным аминокислотным последовательностям клонотипов Т-клеточных рецепторов и оценка фенотипа антиген-специфических TCR T-клеток. TCRscape использует матрицы экспрессии генов единичных клеток и матрицу репертуара рецепторов адаптивного иммунитета (AIRR), сгенерированные инструментом для обработки сырых данных секвенирования РНК единичных клеток BD Rhapsody. Для исследуемой матрицы экспрессии генов выполняется log2(Counts Per Million) нормализация, а с использованием матрицы AIRR обнаруживаются и подсчитываются клонотипы Т-клеточных рецепторов (TCR). Избранные гены нормализованной матрицы экспрессии генов, связанные со специфичностью и аффинностью TCR, объединяются с матрицей клонотипов наряду с результатами оценки связывания нейронной сети ERGO-II, далее для объединенной матрицы выполняется анализ главных компонентов (PCA) с целью оценки размерности данных, выполняется уменьшение размерности Uniform Manifold Approximation and Projection (UMAP) с учетом размерности данных, а затем ищутся кластеры с использованием метода HDBSCAN. Для эпитопа MAGE-A3 обнаружено более 3000 клонотипов, среди них 191 с 2 или более клетками на клонотип.
Для прогнозирования специфичности связывания полученных TCR была использована нейронная сеть ERGO-II. После загрузки репозитория нейросети, выполнялся запуск инструмента из терминала с выбором входного файла и базы данных (McPAS), на основе которой была обучена модель. McPAS-TCR - вручную курируемая и основанная на опубликованной литературе база данных, содержащая информацию примерно о сорока тысячах последовательностях Т-клеточных рецепторов, антигенов, с которыми они связываются, типе Т-клеток (CD4+/CD8+) и типе MHC (MHC-I/MHC-II класса). В McPAS-TCR включается информация о Т-лимфоцитах, экспансирующихся при различных патологических состояниях человека или мыши (в т.ч. вирусные инфекции, онкологические заболевания и аутоиммунные реакции). Входной CSV-файл для запуска ERGO-II содержит информацию о последовательности TCR CDR3α и CDR3β, последовательности пептида, типе MHC (MHC-I/MHC-II класса), генах V и J, а также типе Т-клеток (CD4+/CD8+). Выходной файл содержит значение прогнозирования вероятности связывания Т-клеточного рецептора с комплексом пептид/MHC (нормализованная оценка аффинитета - score), которое варьируется от 0 до 1, где 0 - минимальный аффинитет, а 1 - максимальный аффинитет. Используя ERGO-II, мы можем предсказать аффинитет полученных TCR к пептиду, что облегчает выбор между кандидатами из пула TCR в зависимости от желаемого применения.
Известно, что многие CAR-Т и TCR-T клетки имеют нетаргетный эффект, так как могут взаимодействовать с родственными белками. Чтобы снизить данный эффект, был произведён компьютерный анализ взаимодействия полученных TCR c пептидами родственных белков. Так с помощью нейросети ERGO-II in silico была предиктирована вероятность связывания TCR комплексом пептид/MHC (пептиды из белков подсемейства MAGE, белка Titin).
Нетаргетные пептиды выбирали в результате анализа литературы и анализировали при помощи нейросети NetMHCpan-4.1 cо следующими параметрами: длина пептида - 8-11 аминокислот, аллель - HLA-A*02:01, взвешенный критерий достоверности предиктированного связывания (rank) - меньше 2%.
На основании анализа доминантности клонотипов и фенотипического анализа антигенспецифических Т-клеток с использованием TCRscape, аффинитета клонотипов к целевому пептиду и минимального аффинитета к нецелевым пептидам при помощи нейросетей ERGO-II и NetMHCpan-4.1 был выбран доминантный клон с наибольшим числом клеток. На фиг.1 представлен анализ клонотипов специфичных к MAGE-A3 CD8 Т-клеток с помощью TCRscape, а именно UMAP-визуализация клонотипов MAGE-A3-специфичных CD8 Т-клеток с использованием TCRscape; 1), 2), 3) и 4) представляют экспрессию маркерных генов в каждом кластере; 5) представляют количество клеток для каждого клонотипа; 6) представляют доминирующий клонотип.
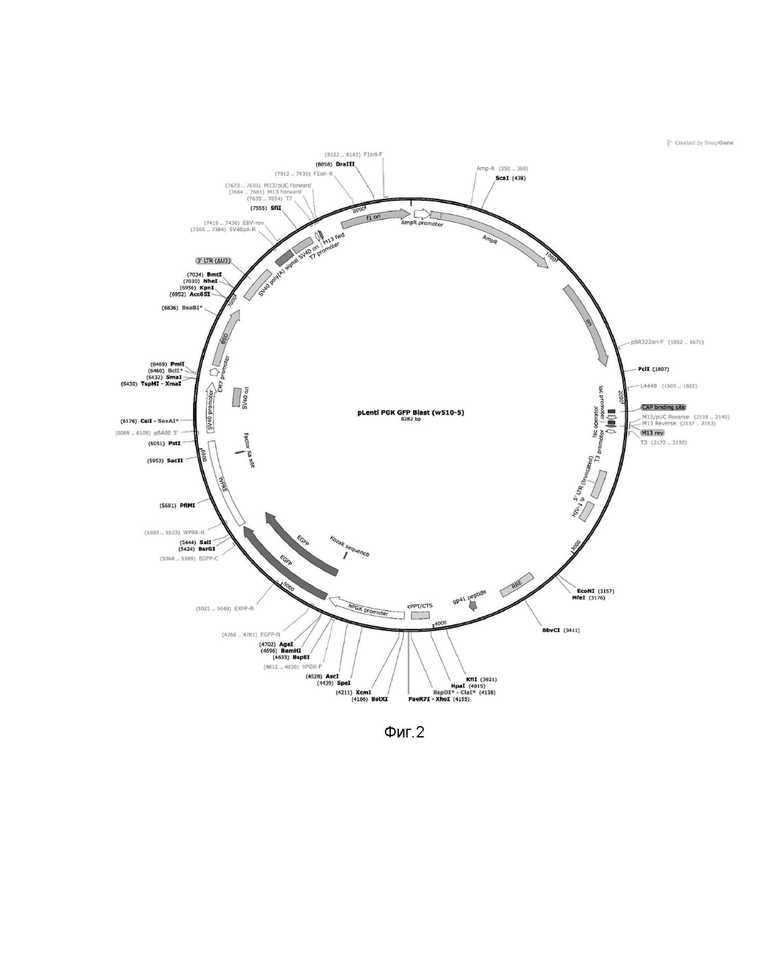
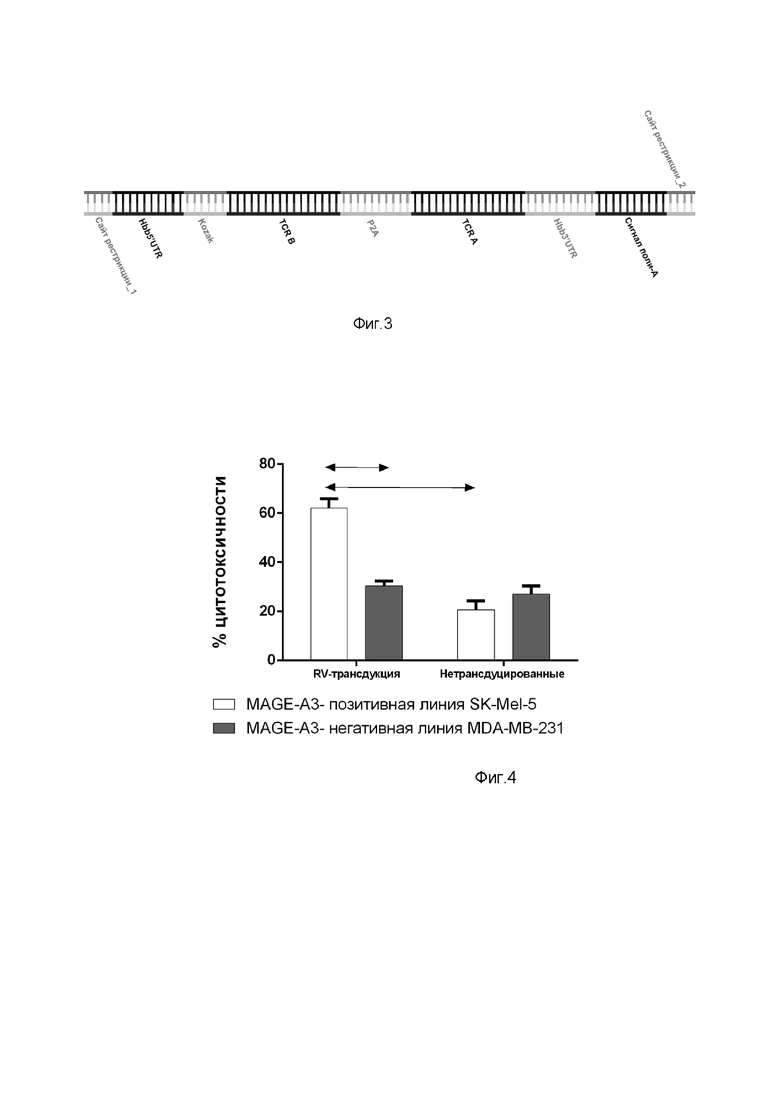
Далее был сконструирован лентивирусный вектор на основе ВИЧ-1 (pLenti_hPGK_gfp) с вставкой вместо гена EGFP, схема плазмиды pLenti_hPGK, показана на фиг.2. Последовательность вставка представлена на диске CD-RW. Сессия записи диска закрыта для последующей записи на него информации. конструкция была использована для получения вирусного вектора, несущего ген TCR, который специфичен к эпитопу опухоль-ассоциированных антигенов MAGE-A3. В состав трансферной плазмиды лентивирусного вектора включены последовательности TCRa и TCRb в единой рамке считывания, разделенные при помощи сигнала для сброса полипептида с полисом P2A и 5' и 3' нетранслируемую область бета-глобулина на 5' и 3' концах вставки. Cхема конструирования вставки для плазмиды показана на фиг. 3.
Для синтеза и клонирования специфического гена, кодирующего TCR, специфичный к эпитопу опухолеассоциированного антигена MAGE-A3, был использован следующий протокол:
1. Синтез гена методом ПЦР (полимеразная цепная реакция).
Ген, кодирующий TCR, был синтезирован путем амплификации с помощью ПЦР. Сначала, были синтезированы праймеры для получения полной последовательности нуклеотидов вставки в плазмиду. Перечень последовательности прилагается.
ПЦР выполнена с использованием набора ThermoFisherScientific (#F549), который включает ДНК-полимеразу Phusion Hot Start II, систему 5-кратных буферов HF и GC, 50 мМ раствор MgCl2, ДМСО, была проведена ПЦР на амплификаторе Bio Rad Thermal cycler C-1000. Фланкирующие праймеры For (GGATCCACATTTGCTTCTGA) и Rev (TGTACATTTATTGCAATGAAAAT) использовали с концентрацией 10 мкМ. Остальные праймеры использовали с концентрацией 2 мкМ. Реакционную смесь для первой ПЦР готовили следующим образом (конечный объем - 50 мл): 10 мкл 5X буфера GC, 10 мкл эквимолярного раствора праймеров, 0,5 мкл смесь дНТФ (25 мМ каждый), 1 мкл полимеразы Phusion HS II, 28,5 мкл mQ. Режим: начальная денатурация (1 цикл): 98°C - 2 мин; амплификация (15 циклов): 98°C - 10 сек, 60°C - 15 сек, 72°C - 2.5 мин; досинтез (1 цикл): 72°C - 5 мин; хранение при + 4°C.
После завершения первой ПЦР была проведена вторая ПЦР, направленная на получение ампликонов целевой длины (2029 п. н.). Реакционную смесь для второй ПЦР готовили следующим образом (конечный объем - 50 мл): 10 мкл 5X буфера GC, 2 мкл рабочего раствора фланкирующих праймеров (10 мкМ каждый), 6 мкл реакционной смеси, полученной после ПЦР 1 этапа, 0,5 мкл смесь дНТФ (25 мМ каждый), 1 мкл полимеразы Phusion HS II, 30,5 мкл mQ. Режим: начальная денатурация (1 цикл): 98°C- 2 мин; амплификация (25 циклов): 98°C - 15 сек, 50°C - 15 сек, 72°C - 2,5 сек; досинтез (1 цикл): 72°C - 5 мин; хранение при + 4°C.
После второй ПЦР проводили горизонтальный гель-электрофорез в 1% агарозном геле при напряженности 6 В/см в течение не менее 30 минут. В геле были видны фрагменты ПЦР длиной 2029 п. н. Фрагмент вырезали из геля и очищали с помощью коммерческого набора LumiPure для выделения ДНК из агарозного геля.
2. Клонирование в промежуточный вектор pUC57:
Вектор pUC57, линеаризованный по сайту EcoRV, был использован в качестве вектора клонирования. Реакции лигирования проводили с использованием ДНК-лигазы Т4 (5 единиц/мкл) Thermo Scientific™ (#EL0011). Соотношение вектор: очищенный ПЦР-продукт 1:3. Инкубация 16-18 ч при температуре +4°C.
Лигазная смесь была трансформирована в клетки E. coli XL1-blue для химической трансформации Eurogene CC001 по методике производителя. Затем эти трансформированные клетки высевали на чашки Петри с LB-агаром, содержащим X-gal, IPTG и ампициллин в дозе 100 мкг/мл. Инкубация чашек в течение 16-18ч при +37°С. Верификацию белых клонов проводили методом ПЦР-скрининга с использованием специфических праймеров для вставки (M13 for (5-GTAAAACGACGGCCAGT-3), M13_rev (5-AGCGGATAACAATTTCACACAGGA-3).). Для скрининга использовали реакционную смесь для ПЦР Basic, 2x Lumiprobe # 5024. Ожидаемый размер продукта ПЦР составил 2175 п. н. Клоны, давшие положительные результаты ПЦР, культивировали в жидкую LB-среду (5 мл, ампициллин 100 мкг/мл). Выделение плазмид проводили на следующий день (через 16 часов) с помощью набора LumiPure для выделения плазмидной ДНК (Spin Miniprep), Lumiprobe #1583. После чего подходящие клоны отбирали путем секвенирования.
3. Переклонирование искомого гена в конечный лентивирусный вектор pLenti_hPGK_gfp:
Вставку подготавливали для переклонирования путем вырезания плазмиды pUC57_M с последовательной реакцией рестрикции, сначала по сайту BstAUI (Сибэнзим) (1 ч при 37°С в буфере Fast Digest от ThermoScientific, #B72), потом по BamHI (Сибэнзим) (также в буфере Fast Digest от ThermoScientific, #B72). Затем проводили горизонтальный гель-электрофорез в 1% агарозном геле реакционной смеси при напряжении 5-6 В/см не менее 45 мин, и вырезание из геля срезы с желаемым фрагментом размером (2023 п. н.). Фрагмент был очищен с помощью набора LumiPuree для выделения ДНК из агарозного геля, Lumiprobe, №5793.
pLenti_hPGK_gfp вектор подготавливали для переклонирование путем удаления гена, кодирующего gfp, из вектора с одновременной рестрикцией по сайтам BstAUI (Сибэнзим) и BamHI (сибэнзим) Fast Digest от ThermoScientific, #B72, 1 ч при 37°С. Затем проводили горизонтальный гель-электрофорез в 1% агарозном геле реакционной смеси при напряжении 5-6 В/см не менее 45 мин, и вырезание из геля срезы с желаемым фрагментом размером 7594 п. н., соответствующему вектору без вставки. Фрагмент был очищен с помощью набора LumiPuree для выделения ДНК из агарозного геля, Lumiprobe, №5793.
4. Клонирование гена в вектор pLenti_hPGK_gfp сайтами BstAUI, BamHI:
Постановка реакции лигирования проводилась с помощью T4 DNA Ligase (5 U/μL) Thermo Scientific™ (#EL0011). Соотношение вектор: фрагмент 1:5. Инкубация в течение 16-18ч при +4°С.
Лигазная смесь была трансформирована в клетки E. coli XL1-blue для химической трансформации Eurogene CC001 по методике производителя. Затем эти трансформированные клетки высевали на чашки Петри с LB-агаром, содержащим ампициллин в дозе 100 мкг/мл. Инкубация чашек ночь при +37°С. Верификацию клонов проводили методом ПЦР-скрининга с использованием специфических праймеров hPGK-F (5- GTGTTCCGCATTCTGCAAG-3), WPRE-R (5- CATAGCGTAAAAGGAGCAACA-3), для скрининга использовалась реакционная смесь для ПЦР Basic, 2x Lumiprobe # 5024. Ожидаемый размер продукта ПЦР составил 2147 п. н. Клоны, давшие положительные результаты ПЦР, культивировали в жидкую LB-среду (5 мл, ампициллин 100 мкг/мл, ночная культура). Выделение плазмид проводили на следующий день (через 16 часов) с помощью набора LumiPure для выделения плазмидной ДНК (Spin Miniprep), Lumiprobe #1583. После чего подходящие клоны отбирали путем секвенирования.
Трансферная плазмида, плазмида, кодирующая VSV-G, и упаковочные плазмиды лентивируса третьего поколения (Gag-pol и Rev) были наработаны в E. coli (штамм NEB Stable), полученные плазмиды были проверены с помощью рестрикционных ферментов и гель-электрофореза. Вышеперечисленные плазмиды были доставлены при помощи липофектамина 2000 в упаковочные клетки HEK-293T (Human embryonic kidney 293T).
Наработанные лентивирусы были концентрированы при помощи коммерческого набора TransLv™ Lentivirus Precipitation Solution (5×) и титрованы при помощи количественной (со стандартами в разведении) PCR на провирусную ДНК (TransLvTM Lentivirus qPCR Titration Kit) в HEK-293T клетках. Для целевых клеток была подобрана необходимая множественность инфекции (Multiplicity Of Infection, MOI) для трансдукции наибольшего количества клеток с наибольшей витальностью и с наименьшим числом маркеров истощения. Затем Т-клетки-мишени трансдуцировали полученными лентивирусными частицами в соответствии с оптимальным MOI, после чего отсортировали CD8 клетки. Для оценки цитотоксической активности трансдуцированных TCR-Т клеток их сокультивировали с клетками опухолевых линий (соотношение 1:5), отличающихся по экспрессии MAGE-A3 (SK-MEL-5 - MAGE-A3-позитивная, HCT 116 - MAGE-A3-позитивная и MDA-MB-231 - MAGE-A3-негативная). Контролем служили нетрансдуцированные Т-клетки.
На фиг.4 показан процент гибели опухолевых клеток MAGE-A3-позитивной линии (SK-MEL-5 и HCT 116) и MAGE-A3-негативной линии (MDA-MB-231) при сокультивировании их с CD8+Т-клетками, трансдуцированные лентивирусного вектора, несущего ген, кодирующий T клеточный рецептор, специфичный к эпитопу MAGE-A3, и с нетрансдуцированными CD8+Т-клетками (контроль), n=6. Стрелками обозначены статистически значимые различия между экспериментальными и контрольной группами (р≤0,05).
Таким образом, предложенный способ получения человеческого Т-клеточного рецептора, специфичного для эпитопа KVAELVHFL (112-120) белка MAGE-A3 позволяет создать в короткий срок, без дополнительных длительных процедур (получение дендритноклеточных вакцин, вакцинация доноров), и имеющее точное описание строения T-клеточного рецептора, специфичного для эпитопа KVAELVHFL (112-120) MAGE-A3. Проведенные исследования in vitro CD8+ клеток, трансдуцированных лентивирусного вектора, несущего ген, кодирующий разработанный Т-клеточный рецептор, специфичный к эпитопу KVAELVHFL (112-120) MAGE-A3, с использованием прямого цитотоксического теста против опухолевых клеток, экспрессирующих антиген MAGE-A3 продемонстрировали высокий показатель цитотоксичности против опухолевых клеток, экспрессирующих антиген MAGE-A3, в отличие от соответствующего (антиген-негативного) контроля.
Изобретение относится к области биотехнологии и иммунологии. Предложен способ получения человеческого Т-клеточного рецептора, специфичного к эпитопу KVAELVHFL (112-120), полученному из белка MAGE-A3. Выделяют мононуклеарные клетки периферической крови условно-здоровых HLA-A02+ доноров, разделяют их на прилипающую и неприлипающую фракции. Прилипающую фракцию культивируют с рекомбинантным человеческим GM-CSF и IL-4. Получают культуру дендритных клеток, инкубируют ее с добавлением фактора праймирования – эпитопа KVAELVHFL. Добавляют TNF-α для завершения созревания дендритных клеток, которые затем сокультивируют с аутологичными CD8+ T-клетками, анти-CD3 и анти-CD28 антителами, IL-2, IL-7 и IL-15. Выделяют антиген-специфические CD8+ T-клетки, отбирают клоны TCR на основе их аффинитета, фенотипа и специфичности. Идентифицируют последовательность генов TCR с помощью секвенирования полной длины TCR, включая CDR3. Конструируют лентивирусный вектор на основе ВИЧ-1, несущий ген TCR, для получения одного доминирующего TCR с альфа- и бета-цепью из одной и той же клетки. Изобретение обеспечивает получение в короткий срок более точной информации о строении и аффинитете человеческого TCR, обладающего высоким аффинитетом и специфичностью к целевому пептиду, что уменьшает риск развития побочных реакций. 4 ил.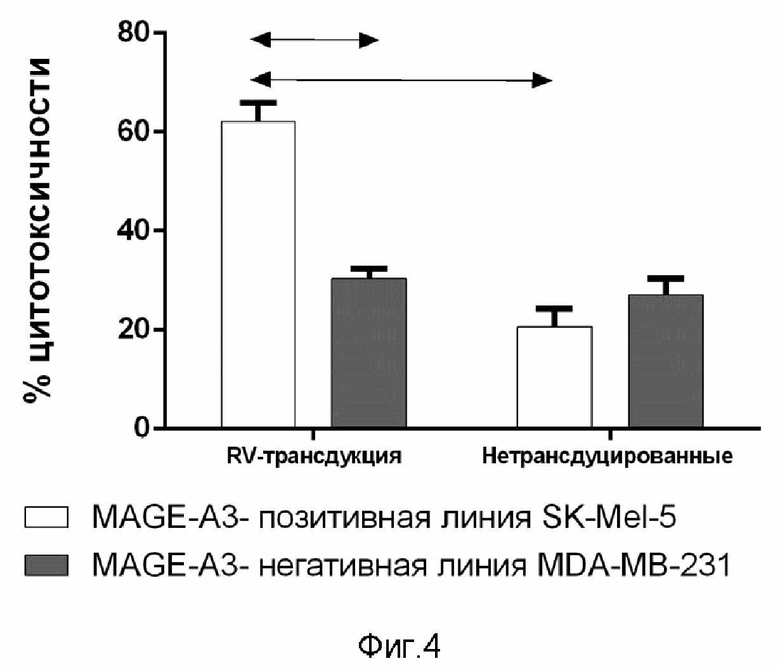
Способ получения человеческого Т-клеточного рецептора, специфичного к эпитопу KVAELVHFL (112-120), полученному из белка MAGE-A3, включающий выделение мононуклеарных клеток (МНК) периферической крови HLA-A02+ доноров, их последующее разделение на прилипающую и неприлипающую фракции посредством адгезии на пластике в течение 30 мин при +37°С в условиях СО2-инкубатора, затем прилипающую фракцию МНК культивируют в присутствии рекомбинантного человеческого гранулоцитарно-макрофагального колониестимулирующего фактора (GM-CSF) и интерлейкина-4 (IL-4) в течение 4 дней; на 5-й день добавляют в качестве фактора праймирования эпитоп KVAELVHFL (112-120) в культуру дендритных клеток с последующей их инкубацией при температуре 37°C; на 6-й день добавляют фактор созревания для завершения созревания дендритных клеток; затем полученные дендритные клетки собирают, промывают питательной средой, подсчитывают их количество, оценивают их жизнеспособность и сокультивируют с аутологичными CD8+ T-лимфоцитами в определенном соотношении, а через 7 дней сокультивирования выделяют антиген-специфические CD8+ T-клетки, затем производят идентификацию последовательности генов TCR с помощью секвенирования полной длины TCR, включая CDR3, выделенной из клонов Т-клеток, при этом цепи TCRα и TCRβ соединяют пептидным линкером 2A (TCRb-P2A-TCRa), а полную конструкцию TCR клонируют в скелет вирусной плазмиды для получения вирусного вектора, несущего ген TCR, отличающийся тем, что донорами являются условно-здоровые индивидуумы, из крови которых моноциты выделяли на градиенте фиколла, на 5-й день добавляют в качестве фактора праймирования эпитоп KVAELVHFL (112-120) в дозе 100 мкг/мл и с последующей инкубацией в течение 24 ч, на 6-й день в качестве фактора созревания добавляют TNF-α в концентрации 25 нг/мл в течение 24 ч, а сокультивирование дендритных клеток осуществляют с CD8+ T-лимфоцитами в присутствии 0,5 мкг/мл антител к CD3 (анти-CD3), 1 мкг/мл антител к CD28 (анти-CD28) и IL-2, IL-7, IL-15 (по 10 нг/мл каждый) в соотношении 1:10, через 7 дней сокультивирования выделяют антиген-специфические CD8+ T-клетки с помощью реагентов Flex-T, затем производят идентификацию последовательности генов TCR с помощью секвенирования РНК единичных клеток (Single-cell RNA sequencing (scRNA-seq)) и набора BD Rapsody TCR/BCR amplification kit, выбор клонов TCR специфичного к эпитопу KVAELVHFL (112-120) белка MAGE-A3, на основе их аффинитета, фенотипа и специфичности, с помощью нейросетей ERGO-II и NetMHCpan-4.1 и программы TCRscape, причем помимо соединения цепи TCRα и TCRβ пептидным линкером 2A (TCRb-P2A-TCRa) используют 5' и 3' нетранслируемую область бета-глобулина на 5' и 3' концах вставки, в качестве вирусной плазмиды используют лентивирус на основе ВИЧ-1 (pLenti_hPGK), получают один доминирующий TCR с альфа- и бета-цепью, происходящими из одной и той же клетки.
| ZHANG B | |||
| et al | |||
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |
| Насос | 1917 |
|
SU13A1 |
| MARIJE A.J | |||
| DE ROOIJ et al | |||
| A library of cancer testis specific T cell receptors for T cell receptor gene therapy, Molecular Therapy - Oncolytics, Volume 28, | |||

