Ссылка
Согласно настоящей заявке испрашивается приоритет по международной патентной заявке № PCT/US 2016/053598, поданной 24 сентября 2016 года, которая включена в данный документ посредством ссылки во всей своей полноте.
Предшествующий уровень техники настоящего изобретения
Многие типы клеток можно идентифицировать и распределить по категориям на основании распространенности конкретных наборов белков, эндогенно экспрессирующихся и расположенных на их плазматических мембранах. Это явление обеспечивает возможность изучения клеток с применением процесса, известного как иммунофенотипирование, при котором клетки инкубируют с несущими флуоресцентную метку антителами, которые являются специфическими к известным поверхностным белкам клеток, и клетки связываются с ними. Проточную цитометрию обычно применяют для измерения уровней связанных с поверхностью антител для каждой клетки. Тем не менее, подходы на основе проточной цитометрии ограничиваются количеством флуорофоров, которые можно применять одновременно в одном эксперименте. Более того, количество флуорофоров, которые можно применять одновременно в одном эксперименте при применении подходов на основе проточной цитометрии, ограничивается спектральным перекрытием. Кроме того, проточная цитометрия не пригодна для многих биологически значимых анализов и последующего секвенирования ДНК.
Сущность изобретения
Следовательно, существует потребность в способах характеристики, например, иммунофенотипирования, отдельных клеток без этих ограничений. В отличие от подходов на основе проточной цитометрии в способах, описанных в данном документе, для анализа белков в отдельных клетках применяется считывание данных на основе секвенирования, и они не ограничиваются количеством флуорофоров, которые можно применять в одновременно в одном эксперименте, или их спектральным перекрытием. Кроме того, способы, описанные в данном документе, являются пригодными для многих биологически значимых анализов и последующего секвенирования ДНК. В способах, описанных в данном документе, используются конъюгаты аффинная молекула-олигонуклеотид (например, конъюгаты антитело-олигонуклеотид или конъюгаты тетрамер-олигонуклеотид). Олигонуклеотид в составе конъюгата содержит идентификационную последовательность антигена (AID), которая выступает в роли штрихкода для поверхностного антигена, с которым специфически связывается аффинная часть конъюгата аффинная молекула-олигонуклеотид. Следовательно, при применении способов, описанных в данном документе, антиген (например, поверхностный белок) отдельной клетки можно анализировать без потребности в флуорофорах. Например, поверхностный белок отдельной клетки, который проявляется, можно идентифицировать по идентификационной последовательности антигена. В качестве еще одного примера, Т-клеточный рецептор (TCR) или В-клеточный рецептор (BCR) отдельной клетки, который проявляется, можно идентифицировать по идентификационной последовательности антигена. В соответствии с некоторыми аспектами аффинность связывания TCR или BCR, проявляющегося на поверхности клетки, можно определить с применением конъюгированных аффинной молекулы и олигонуклеотида, описанных в данном документе. Один или несколько из поверхностных белков отдельной клетки можно применять для идентификации, определения характеристик или релевантности отдельной клетки.
Конъюгат аффинная молекула-олигонуклеотид в описанных в данном документе способах, которые можно применять для того, чтобы преодолеть проблемы медленного сортинга клеток, уменьшенного выхода целевых клеток, связанного с сортингом клеток, ограниченного количества выходных потоков и выбранных групп, которые не соответствуют количественно определяемому свойству конъюгата аффинная молекула-олигонуклеотид, такому как аффинность. Изображенный иллюстративный конъюгат аффинная молекула-олигонуклеотид может заменить или улучшить сортинг с использованием измерений связывания тетрамеров на отдельных клетках в резервуарах. Изображенный иллюстративный конъюгат аффинная молекула-олигонуклеотид можно применять в способах, описанных в данном документе, чтобы одновременно обнаруживать последовательности пары TCR, распространенность клона и относительные аффинности тетрамеров. Например, конъюгат аффинная молекула-олигонуклеотид, который представляет собой конъюгат тетрамер-олигонуклеотид, можно применять в способах, описанных в данном документе, чтобы одновременно обнаруживать последовательностей пары TCR, которые связываются с пептидом в составе тетрамера, определять аффинность связывания TCR с комплексом главный комплекс гистосовместимости (МНС)-пептид в составе конъюгата тетрамер-олигонуклеотид и проводить различия между TCR с низкой аффинностью связывания по сравнению с TCR с высокой аффинностью связывания в отношении комплекса МНС-пептид в составе конъюгата тетрамер-олигонуклеотид. В качестве еще одного примера, конъюгат аффинная молекула-олигонуклеотид, который представляет собой конъюгат тетрамер-олигонуклеотид, можно применять в способах, описанных в данном документе, чтобы одновременно обнаруживать последовательности пары BCR, которые связываются с антигеном В-клеточного рецептора в составе тетрамера, определять аффинность связывания BCR с антигеном В-клеточного рецептора в составе конъюгата тетрамер-олигонуклеотид и проводить различия между BCR с низкой аффинностью связывания по сравнению с BCR с высокой аффинностью связывания в отношении антигена В-клеточного рецептора в составе конъюгата тетрамер-олигонуклеотид.
В данном документе описан способ характеристики, например, иммунофенотипирования, клеток в резервуарах (например, в эмульсии) с использованием конъюгатов аффинная молекула-олигонуклеотид. В соответствии с некоторыми вариантами осуществления способ применяют для идентификации клеточных субпопуляций способом, совместимым с анализом отдельных клеток в эмульсионной среде. В соответствии с некоторыми вариантами осуществления способ применяют для идентификации иммунных клеток, специфических в отношении антигена, способом, совместимым с анализом отдельных клеток в эмульсионной среде. В соответствии с некоторыми вариантами осуществления перед анализом клеток антитела, специфические к поверхностному белку, конъюгируют с олигонуклеотидами. В соответствии с некоторыми вариантами осуществления олигонуклеотиды сконструированы таким образом, чтобы они содержали мотив последовательности, который является уникальным для мишенеспецифичности конъюгированного антитела. В соответствии с некоторыми вариантами осуществления олигонуклеотиды сконструированы таким образом, чтобы они содержали мотив последовательности, который является уникальным для мишенеспецифичности конъюгированного тетрамера. Олигонуклеотид может быть конъюгирован с аффинной частью конъюгата аффинная молекула-олигонуклеотид (например, антителом или тетрамером) ковалентно или нековалентно (например, биотин-олигонуклеотид со стрептавидин-антителом).
Способ может включать инкубирование клеток в смеси или растворе с одним или несколькими конъюгатами аффинная молекула-олигонуклеотид. Клетки можно подвергнуть отмывке для удаления несвязанных конъюгатов аффинная молекула-олигонуклеотид. Клетки затем инкапсулируют в резервуарах, например, в эмульсии. Клетки могут присутствовать в резервуарах с плотностью одна клетка на резервуар. Таким образом, конъюгаты аффинная молекула-олигонуклеотид в резервуаре, например, в капле, связываются с клеточной поверхностью, например, посредством специфического взаимодействия антитело-поверхностный белок. Способ может включать прикрепление специфической для резервуара последовательности ДНК (например, уникального штрихкода резервуара) к конъюгированным с аффинными молекулами олигонуклеотидам. Дополнительный анализ клеточной ДНК или мРНК, фенотипические измерения, функциональное исследование, клеточный сортинг или другие реакции можно осуществлять до, одновременно или после мечения штрихкодом конъюгированного с аффинной молекулой олигонуклеотида (например, ДНК-штрихкодом).
Способ может включать инкубирование клеток в смеси или растворе с одним или несколькими конъюгатами тетрамер-олигонуклеотид. Клетки можно подвергнуть отмывке для удаления несвязанных конъюгатов тетрамер-олигонуклеотид. Клетки затем инкапсулируют в резервуарах, например, в эмульсии. Клетки могут присутствовать в резервуарах с плотностью одна клетка на резервуар. Таким образом, конъюгаты тетрамер-олигонуклеотид в резервуаре, например, в капле, связываются с клеточной поверхностью, например, посредством специфического взаимодействия TCR-комплекс МНС-пептид или посредством специфического взаимодействия BCR-антиген В-клеточного рецептора в составе тетрамера. Способ может предполагать прикрепление специфической для резервуара последовательности ДНК (например, уникального штрихкода резервуара) к конъюгированным с тетрамером олигонуклеотидам. Дополнительный анализ клеточной ДНК или мРНК, фенотипические измерения, функциональное исследование, клеточный сортинг или другие реакции можно осуществлять до, одновременно или после мечения штрихкодом конъюгированного с тетрамером олигонуклеотида (например, ДНК-штрихкодом).
Способ может включать экстрагирование нуклеиновых кислот из эмульсии, например, после проведения экспериментов в эмульсии. Экстрагированные нуклеиновые кислоты можно подготовить для секвенирования и секвенировать (например, с применением технологии секвенирования нового поколения). Способ может включать секвенирование молекул полинуклеотида из резервуаров, которые содержат как идентификационную последовательность антигена, так и последовательность специфического для капли штрихкода. Идентификационная последовательность антигена может определять специфический белок клеточной поверхности, с которым связывается конъюгированное с олигонуклеотидом антитело. Идентификационная последовательность антигена может определять специфическое антитело конъюгированного с олигонуклеотидом антитела, которое связывается с конкретным белком клеточной поверхности. Идентификационная последовательность антигена может определять специфический комплекс МНС-пептид в составе конъюгированного с олигонуклеотидом тетрамера, который связывается с конкретным клеточным TCR. Идентификационная последовательность антигена может определять специфический антиген В-клеточного рецептора в составе конъюгированного с олигонуклеотидом тетрамера, который связывается с конкретным клеточным BCR. Таким образом, идентификационная последовательность антигена может указывать на то, какой поверхностный белок экспрессировала анализируемая клетка. В резервуаре, содержащем отдельную клетку, все последовательности, содержащие общую последовательность специфического для капли штрихкода, связаны с одной клеткой. Благодаря этому, отдельную клетку можно анализировать как проявляющую набор поверхностных белков, которые можно применять для ее идентификации, определения характеристик или релевантности. Например, отдельную клетку можно анализировать как проявляющую TCR с определенной аффинностью к комплексу МНС-пептид, в котором также можно идентифицировать последовательность этого TCR, или отдельную клетку можно анализировать как проявляющую BCR с определенной аффинностью к антигену В-клеточного рецептора в составе тетрамера, в котором также можно идентифицировать последовательность этого BCR.
В соответствии с одним аспектом предполагается способ, включающий осуществление реакции во множестве резервуаров, при этом реакция включает прикрепление меченого штрихкодом резервуара полинуклеотида, содержащего последовательность штрихкода резервуара, к олигонуклеотиду в составе конъюгата аффинная молекула-олигонуклеотид, связанного с целевым антигеном отдельной клетки, выделенной в резервуаре из множества резервуаров. В соответствии с одним аспектом предполагается способ, включающий осуществление реакции во множестве резервуаров, при этом реакция включает прикрепление меченого штрихкодом резервуара полинуклеотида, содержащего последовательность штрихкода резервуара, к олигонуклеотиду в составе конъюгата тетрамер-олигонуклеотид, связанного с TCR отдельной клетки, выделенной в резервуаре из множества резервуаров. В соответствии с одним аспектом предполагается способ, включающий осуществление реакции во множестве резервуаров, при этом реакция включает прикрепление меченого штрихкодом резервуара полинуклеотида, содержащего последовательность штрихкода резервуара, к олигонуклеотиду в составе конъюгата тетрамер-олигонуклеотид, связанного с BCR отдельной клетки, выделенной в резервуаре из множества резервуаров.
В соответствии с одним аспектом в данном документе представлен способ, включающий осуществления реакции в резервуаре из множества резервуаров, причем реакция включает прикрепление меченого штрихкодом резервуара полинуклеотида, который содержит последовательность штрихкода резервуара, к олигонуклеотидной части конъюгата аффинная молекула-олигонуклеотид, при этом конъюгат аффинная молекула-олигонуклеотид связывается с целевым антигеном, экспрессирующимся клеткой в резервуаре из множества резервуаров. В соответствии с одним аспектом в данном документе представлен способ, включающий осуществление реакции в резервуаре из множества резервуаров, причем реакция включает прикрепление меченого штрихкодом резервуара полинуклеотида, который содержит последовательность штрихкода резервуара, к олигонуклеотидной части конъюгата тетрамер-олигонуклеотид, при этом конъюгат тетрамер-олигонуклеотид связывается с TCR, экспрессирующимся клеткой в резервуаре из множества резервуаров. В соответствии с одним аспектом в данном документе представлен способ, включающий осуществление реакции в резервуаре из множества резервуаров, причем реакция включает прикрепление меченого штрихкодом резервуара полинуклеотида, который содержит последовательность штрихкода резервуара, к олигонуклеотидной части конъюгата тетрамер-олигонуклеотид, при этом конъюгат тетрамер-олигонуклеотид связывается с BCR, экспрессирующимся клеткой в резервуаре из множества резервуаров.
В соответствии с некоторыми вариантами осуществления клетка представляет собой одну клетку, содержащуюся внутри резервуара. В соответствии с некоторыми вариантами осуществления резервуар содержит два или более резервуара из множества резервуаров. В соответствии с некоторыми вариантами осуществления резервуар содержит каждый резервуар из множества резервуаров. В соответствии с некоторыми вариантами осуществления реакция происходит в двух или более резервуарах из множества резервуаров. В соответствии с некоторыми вариантами осуществления клетка в каждом резервуаре происходит из одного и того же образца. В соответствии с некоторыми вариантами осуществления клетка в резервуаре из первого множества резервуаров в составе двух или более множеств резервуаров происходит из того же образца, что и клетка в резервуаре во втором множестве резервуаров в составе двух или более множеств резервуаров. В соответствии с некоторыми вариантами осуществления олигонуклеотидная часть содержит идентификационную последовательность антигена (AID). В соответствии с некоторыми вариантами осуществления AID выступает в качестве штрихкода для целевого антигена или аффинной части в составе конъюгата аффинная молекула-олигонуклеотид. В соответствии с некоторыми вариантами осуществления AID выступает в качестве штрихкода для TCR с известной специфичностью или комплекса МНС-пептид в составе конъюгата тетрамер-олигонуклеотид. В соответствии с некоторыми вариантами осуществления AID выступает в качестве штрихкода для BCR с известной специфичностью или антигена В-клеточного рецептора в составе конъюгата тетрамер-олигонуклеотид.
В соответствии с некоторыми вариантами осуществления олигонуклеотид дополнительно содержит идентификационную последовательность антигена (AID), выступающую в качестве штрихкода для целевого антигена или аффинной части в составе конъюгата аффинная молекула-олигонуклеотид. В соответствии с некоторыми вариантами осуществления олигонуклеотид дополнительно содержит идентификационную последовательность антигена (AID), выступающую в качестве штрихкода для TCR с известной специфичностью или комплекса МНС-пептид в составе конъюгата тетрамер-олигонуклеотид. В соответствии с некоторыми вариантами осуществления идентификационная последовательность антигена (AID) представляет собой известную последовательность.
В соответствии с некоторыми вариантами осуществления меченый штрихкодом резервуара полинуклеотид происходит из матричного меченого штрихкодом резервуара полинуклеотида в резервуаре.
В соответствии с некоторыми вариантами осуществления способ дополнительно включает секвенирование олигонуклеотида или его ампликона с получением информации о последовательности.
В соответствии с некоторыми вариантами осуществления способ дополнительно включает определение характеристики отдельной клетки на основании информации о последовательности. В соответствии с некоторыми вариантами осуществления информация о последовательности содержит идентификационную последовательность антигена (AID). В соответствии с некоторыми вариантами осуществления способ дополнительно включает определение характеристики отдельной клетки на основании информации о последовательности. В соответствии с некоторыми вариантами осуществления характеристика представляет собой фенотип. В соответствии с некоторыми вариантами осуществления фенотип представляет собой иммунофенотип. В соответствии с некоторыми вариантами осуществления характеристика представляет собой аффинность. В соответствии с некоторыми вариантами осуществления аффинность относится к комплексу МНС-пептид. В соответствии с некоторыми вариантами осуществления аффинность представляет собой относительную аффинность комплекса МНС-пептид (например, TCR с более высокой аффинностью связывания по сравнению с TCR с более низкой аффинностью связывания). В соответствии с некоторыми вариантами осуществления аффинность относится к BCR. В соответствии с некоторыми вариантами осуществления аффинность представляет собой относительную аффинность BCR (например, BCR с более высокой аффинностью связывания по сравнению с BCR с более низкой аффинностью связывания).
В соответствии с некоторыми вариантами осуществления способ дополнительно включает приведение конъюгата аффинная молекула-олигонуклеотид в контакт со множеством клеток, содержащим отдельную клетку. В соответствии с некоторыми вариантами осуществления способ дополнительно включает приведение конъюгата тетрамер-олигонуклеотид в контакт со множеством клеток, содержащим отдельную клетку. В соответствии с некоторыми вариантами осуществления контакт обеспечивают до выделения одной клетки в резервуаре. В соответствии с некоторыми вариантами осуществления способ дополнительно включает отмывку множества клеток после контакта.
В соответствии с некоторыми вариантами осуществления резервуар не содержит конъюгат аффинная молекула-олигонуклеотид, который не связан с целевым антигеном. В соответствии с некоторыми вариантами осуществления резервуар не содержит конъюгат тетрамер-олигонуклеотид, который не связан с TCR. В соответствии с некоторыми вариантами осуществления резервуар не содержит конъюгат тетрамер-олигонуклеотид, который не связан с BCR.
В соответствии с некоторыми вариантами осуществления способ дополнительно включает выделение отдельной клетки в резервуаре. В соответствии с некоторыми вариантами осуществления отдельная клетка связывается с конъюгатом аффинная молекула-олигонуклеотид до выделения.
В соответствии с некоторыми вариантами осуществления способ дополнительно включает лизис отдельной клетки. В соответствии с некоторыми вариантами осуществления лизис осуществляют после выделения отдельной клетки в резервуаре.
В соответствии с некоторыми вариантами осуществления множество клеток представляет собой множество несортированных клеток. В соответствии с некоторыми вариантами осуществления множество клеток представляет собой множество сортированных клеток. В соответствии с некоторыми вариантами осуществления множество клеток представляет собой множество обогащенных клеток.
В соответствии с некоторыми вариантами осуществления последовательность штрихкода резервуара в составе меченого штрихкодом резервуара полинуклеотида или его ампликона в первом резервуаре из множества резервуаров отличается от последовательности штрихкода резервуара в составе меченого штрихкодом резервуара полинуклеотида или его ампликона во втором резервуаре из множества резервуаров. В соответствии с некоторыми вариантами осуществления последовательность штрихкода резервуара в составе каждого меченого штрихкодом резервуара полинуклеотида или его ампликона в отдельном резервуаре из множества резервуаров содержит одинаковую последовательность штрихкода резервуара. В соответствии с некоторыми вариантами осуществления последовательность штрихкода резервуара в составе каждого меченого штрихкодом резервуара полинуклеотида и его ампликона в любом отдельном резервуаре из множества резервуаров является уникальной для последовательности штрихкода резервуара в составе каждого меченого штрихкодом резервуара полинуклеотида и его ампликона в любом другом отдельном резервуаре из множества резервуаров.
В соответствии с некоторыми вариантами осуществления способ дополнительно включает прикрепление меченого штрихкодом резервуара полинуклеотида к клеточному полинуклеотиду из отдельной клетки. В соответствии с некоторыми вариантами осуществления прикрепление меченого штрихкодом резервуара полинуклеотида к олигонуклеотиду в составе конъюгата аффинная молекула-олигонуклеотид и прикрепление меченого штрихкодом резервуара полинуклеотида к клеточному полинуклеотиду из отдельной клетки осуществляют одновременно.
В соответствии с некоторыми вариантами осуществления способ дополнительно включает амплификацию олигонуклеотида или его комплементарной последовательности. В соответствии с некоторыми вариантами осуществления способ дополнительно включает амплификацию клеточного полинуклеотида или его комплементарной последовательности. В соответствии с некоторыми вариантами осуществления амплификацию олигонуклеотида или его комплементарной последовательности и амплификацию клеточного полинуклеотида или его комплементарной последовательности осуществляют одновременно.
В соответствии с некоторыми вариантами осуществления последовательность штрихкода резервуара в составе клеточного полинуклеотида и последовательность штрихкода резервуара в составе олигонуклеотида является одинаковой.
В соответствии с некоторыми вариантами осуществления способ дополнительно включает объединение олигонуклеотидов или их ампликонов из двух или более резервуаров из множества резервуаров. В соответствии с некоторыми вариантами осуществления способ дополнительно включает объединение олигонуклеотидов или их ампликонов и клеточных полинуклеотидов или их ампликонов из двух или более резервуаров из множества резервуаров. В соответствии с некоторыми вариантами осуществления объединение осуществляют до секвенирования.
В соответствии с некоторыми вариантами осуществления конъюгат аффинная молекула-олигонуклеотид содержит множество различных конъюгатов аффинная молекула-олигонуклеотид. В соответствии с некоторыми вариантами осуществления каждый конъюгат аффинная молекула-олигонуклеотид из множества конъюгатов аффинная молекула-олигонуклеотид содержит уникальную идентификационную последовательность антигена (AID). В соответствии с некоторыми вариантами осуществления олигонуклеотид содержит последовательность молекулярного штрихкода аффинной молекулы (АМВ), которая выступает в качестве штрихкода для отдельной молекулы конъюгата аффинная молекула-олигонуклеотид из множества молекул конъюгата аффинная молекула-олигонуклеотид. В соответствии с некоторыми вариантами осуществления каждая молекула конъюгата аффинная молекула-олигонуклеотид из множества молекул конъюгата аффинная молекула-олигонуклеотид содержит уникальную последовательность молекулярного штрихкода аффинной молекулы (АМВ).
В соответствии с некоторыми вариантами осуществления способы и конъюгаты, описанные в данном документе, включает множество различных конъюгатов тетрамер-олигонуклеотид. В соответствии с некоторыми вариантами осуществления каждый конъюгат тетрамер-олигонуклеотид из множества конъюгатов аффинная молекула-олигонуклеотид содержит уникальную идентификационную последовательность антигена (AID). В соответствии с некоторыми вариантами осуществления олигонуклеотид содержит последовательность молекулярного штрихкода аффинной молекулы (АМВ), которая выступает в качестве штрихкода для отдельной молекулы конъюгата тетрамер-олигонуклеотид из множества молекул конъюгата тетрамер-олигонуклеотид. В соответствии с некоторыми вариантами осуществления каждая молекула конъюгата тетрамер-олигонуклеотид из множества молекул конъюгата тетрамер-олигонуклеотид содержит уникальную последовательность молекулярного штрихкода аффинной молекулы (АМВ).
В соответствии с некоторыми вариантами осуществления олигонуклеотид содержит последовательность для слияния, и прикрепление включает прикрепление меченого штрихкодом резервуара полинуклеотида к последовательности для слияния. В соответствии с некоторыми вариантами осуществления олигонуклеотид содержит последовательность для связывания праймера. В соответствии с некоторыми вариантами осуществления олигонуклеотид содержит константную последовательность.
В соответствии с некоторыми вариантами осуществления способ дополнительно включает секвенирование олигонуклеотид а, его комплементарных последовательностей, его амплифицированных продуктов или их комбинации, таким образом получая риды последовательности олигонуклеотида. В соответствии с некоторыми вариантами осуществления способ дополнительно включает сравнение одного или нескольких ридов первой последовательности олигонуклеотида с одним или несколькими ридами второй последовательности олигонуклеотида. В соответствии с некоторыми вариантами осуществления способ дополнительно включает анализ ридов последовательности олигонуклеотида. В соответствии с некоторыми вариантами осуществления способ дополнительно включает анализ последовательностей штрихкода резервуара в ридах последовательности олигонуклеотида. В соответствии с некоторыми вариантами осуществления способ дополнительно включает анализ идентификационных последовательностей антигена (AID) в ридах последовательности олигонуклеотида. В соответствии с некоторыми вариантами осуществления способ дополнительно включает анализ последовательностей молекулярного штрихкода аффинной молекулы (АМВ) в ридах последовательности олигонуклеотида. В соответствии с некоторыми вариантами осуществления анализ включает определение частоты одной или нескольких последовательностей штрихкода резервуара, одной или нескольких последовательностей AID, одной или нескольких последовательностей молекулярного штрихкода аффинной молекулы (АМВ) или их комбинации. В соответствии с некоторыми вариантами осуществления анализ включает сравнение. В соответствии с некоторыми вариантами осуществления способ дополнительно включает сравнение идентификационных последовательностей антигена (AID) в ридах последовательности олигонуклеотида с последовательностями молекулярного штрихкода аффинной молекулы (АМВ) в ридах последовательности олигонуклеотида.
В соответствии с некоторыми вариантами осуществления способ дополнительно включает секвенирование клеточного полинуклеотида, его комплементарных последовательностей, его амплифицированных продуктов или их комбинации, таким образом получая риды последовательности клеточного полинуклеотида. В соответствии с некоторыми вариантами осуществления способ дополнительно включает сравнение ридов последовательности олигонуклеотида с ридами последовательности клеточного полинуклеотида. В соответствии с некоторыми вариантами осуществления способ дополнительно включает сравнение последовательностей штрихкода резервуара в ридах последовательности олигонуклеотида с последовательностями штрихкода резервуара в ридах последовательностей клеточного полинуклеотида. В соответствии с некоторыми вариантами осуществления способ дополнительно включает сравнение ридов последовательности клеточного полинуклеотида. В соответствии с некоторыми вариантами осуществления способ дополнительно включает анализ последовательностей штрихкода резервуара в ридах последовательности клеточного полинуклеотида. В соответствии с некоторыми вариантами осуществления способ дополнительно включает анализ последовательностей молекулярного штрихкода в ридах последовательности клеточного полинуклеотида.
В соответствии с некоторыми вариантами осуществления способ дополнительно включает определение характеристики клетки на основании анализа или сравнения. В соответствии с некоторыми вариантами осуществления способ дополнительно включает выбор антитела, BCR или TCR на основании ридов последовательности олигонуклеотида. В соответствии с некоторыми вариантами осуществления способ включает выбор антитела, BCR или TCR на основании ридов последовательности клеточного полинуклеотида.
В соответствии с некоторыми вариантами осуществления способ дополнительно включает определение аффинности тетрамера в отношении TCR или BCR и/или аффинности TCR или BCR в отношении тетрамера на основании анализа или сравнения. В соответствии с некоторыми вариантами осуществления количество ридов от секвенирования является показателем аффинности тетрамера в отношении TCR или BCR. В соответствии с некоторыми вариантами осуществления большее число ридов последовательности для отдельной клетки указывает на более высокую аффинность связывания у конъюгата тетрамер-олигонуклеотид и/или TCR или BCR по сравнению с отдельной клеткой с меньшим количеством ридов от секвенирования. Аффинность связывания у тетрамера может быть показателем связывания комплекса МНС-пептид в составе тетрамера или антигена В-клеточного рецептора в составе тетрамера.
В соответствии с некоторыми вариантами осуществления способ определения аффинности TCR или BCR в отношении конъюгата тетрамер-олигонуклеотид включает инкубирование одной или множества Т-клеток в присутствии одного или множества конъюгатов тетрамер-олигонуклеотид, при этом каждый из множества конъюгатов тетрамер-олигонуклеотид может содержать уникальные идентификационные штрихкоды. В соответствии с некоторыми вариантами осуществления способ дополнительно включает выделение каждого комплекса клетка-тетрамер и секвенирование по меньшей мере части олигонуклеотида или ее комплементарной последовательности, конъюгированной с тетрамерным комплексом, который описан в данном документе. В соответствии с некоторыми вариантами осуществления способ дополнительно включает подсчет количества ридов от секвенирования, полученных во время секвенирования, для каждой уникальной последовательности штрихкода. В соответствии с некоторыми вариантами осуществления способ дополнительно включает анализ или сравнение информации от секвенирования со стандартной кривой. В соответствии с некоторыми вариантами осуществления способ дополнительно включает определение аффинности TCR или BCR в отношении тетрамера или тетрамера в отношении TCR или BCR на основании секвенирования, или анализа, или сравнения информации от секвенирования. В соответствии с некоторыми вариантами осуществления меченый штрихкодом резервуара полинуклеотид, прикрепленный к олигонуклеотиду, и меченый штрихкодом резервуара полинуклеотид, прикрепленный к клеточному полинуклеотиду, происходят из одного и того же матричного меченого штрихкодом резервуара полинуклеотида в резервуаре. В соответствии с некоторыми вариантами осуществления меченый штрихкодом резервуара полинуклеотид, прикрепленный к олигонуклеотиду, представляет собой продукт амплификации матричного меченого штрихкодом резервуара полинуклеотида.
В соответствии с некоторыми вариантами осуществления меченый штрихкодом резервуара полинуклеотид, прикрепленный к клеточному полинуклеотиду, представляет собой продукт амплификации матричного меченого штрихкодом резервуара полинуклеотида.
В соответствии с некоторыми вариантами осуществления резервуар содержит твердую подложку. В соответствии с некоторыми вариантами осуществления резервуар не содержит твердую подложку. В соответствии с некоторыми вариантами осуществления каждый резервуар из множества резервуаров содержит отдельную клетку. В соответствии с некоторыми вариантами осуществления резервуар представляет собой лунку, эмульсию или каплю. В соответствии с некоторыми вариантами осуществления матричный меченый штрихкодом резервуара полинуклеотид не связан с твердой подложкой. В соответствии с некоторыми вариантами осуществления матричный меченый штрихкодом резервуара полинуклеотид связан с твердой подложкой.
В соответствии с некоторыми вариантами осуществления способ дополнительно включает прикрепление последовательности молекулярного штрихкода в меченом молекулярный штрихкод полинуклеотиде из множества меченых молекулярным штрихкодом полинуклеотидов к клеточному полинуклеотиду, при этом последовательность молекулярного штрихкода выступает в качестве штрихкода для молекулы полинуклеотида отдельной клетки и ее ампликонов.
В соответствии с некоторыми вариантами осуществления прикрепление включает лигирование полинуклеотида в резервуаре с олигонуклеотидом. В соответствии с некоторыми вариантами осуществления прикрепление включает прикрепление полинуклеотида в резервуаре к олигонуклеотиду с использованием фермента. В соответствии с некоторыми вариантами осуществления прикрепление включает гибридизацию полинуклеотида в резервуаре с олигонуклеотидом. В соответствии с некоторыми вариантами осуществления прикрепление дополнительно включает достройку олигонуклеотида. В соответствии с некоторыми вариантами осуществления прикрепление включает амплификацию матричного меченого штрихкодом резервуара полинуклеотида.
В соответствии с некоторыми вариантами осуществления олигонуклеотид является двухцепочечным. В соответствии с некоторыми вариантами осуществления олигонуклеотид является одноцепочечным. В соответствии с некоторыми вариантами осуществления олигонуклеотид представляет собой ДНК. В соответствии с некоторыми вариантами осуществления олигонуклеотид представляет собой РНК.
В соответствии с некоторыми вариантами осуществления клеточный полинуклеотид содержит последовательность вариабельного участка. В соответствии с некоторыми вариантами осуществления способ дополнительно включает спаривание нативных последовательностей цепей, содержащих последовательность вариабельного участка. В соответствии с некоторыми вариантами осуществления клеточный полинуклеотид представляет собой ДНК. В соответствии с некоторыми вариантами осуществления клеточный полинуклеотид представляет собой РНК. В соответствии с некоторыми вариантами осуществления РНК представляет собой мРНК.
В соответствии с некоторыми вариантами осуществления отдельная клетка представляет собой В-клетку. В соответствии с некоторыми вариантами осуществления отдельная клетка представляет собой Т-клетку.
В соответствии с некоторыми вариантами осуществления аффинная часть конъюгата аффинная молекула-олигонуклеотид связывается с внеклеточным антигеном отдельной клетки. В соответствии с некоторыми вариантами осуществления внеклеточный антиген отдельной клетки представляет собой антиген, специфический для иммунной клетки. В соответствии с некоторыми вариантами осуществления внеклеточный антиген отдельной клетки представляет собой антиген, специфический для Т-клетки. В соответствии с некоторыми вариантами осуществления внеклеточный антиген представляет собой CD4. В соответствии с некоторыми вариантами осуществления внеклеточный антиген представляет собой CD8. В соответствии с некоторыми вариантами осуществления внеклеточный антиген отдельной клетки представляет собой антиген, специфический для В-клетки. В соответствии с некоторыми вариантами осуществления внеклеточный антиген представляет собой иммуноглобулин.
В соответствии с некоторыми вариантами осуществления аффинная часть конъюгата аффинная молекула-олигонуклеотид представляет собой антитело или его фрагмент. В соответствии с некоторыми вариантами осуществления аффинная часть конъюгата аффинная молекула-олигонуклеотид представляет собой пептид. В соответствии с некоторыми вариантами осуществления аффинная часть конъюгата аффинная молекула-олигонуклеотид представляет собой белок. В соответствии с некоторыми вариантами осуществления аффинная часть конъюгата аффинная молекула-олигонуклеотид представляет собой аптамер. В соответствии с некоторыми вариантами осуществления аффинная часть конъюгата аффинная молекула-олигонуклеотид представляет собой малую молекулу. В соответствии с некоторыми вариантами осуществления аффинная часть конъюгата аффинная молекула-олигонуклеотид представляет собой лекарственное средство. В соответствии с некоторыми вариантами осуществления аффинная часть конъюгата аффинная молекула-олигонуклеотид представляет собой клетку. В соответствии с некоторыми вариантами осуществления клетка представляет собой антигенпрезентирующую клетку (АРС). В соответствии с некоторыми вариантами осуществления аффинная часть конъюгата аффинная молекула-олигонуклеотид содержит комплекс МНС-пептид. В соответствии с некоторыми вариантами осуществления аффинная часть конъюгата аффинная молекула-олигонуклеотид содержит главный комплекс гистосовместимости (МНС). В соответствии с некоторыми вариантами осуществления МНС присутствует в растворимой и/или мультимерной (например, тетрамерной) форме. В соответствии с некоторыми вариантами осуществления МНС является связанным с пептидом. В соответствии с некоторыми вариантами осуществления пептид представляет собой синтетический пептид. В соответствии с некоторыми вариантами осуществления МНС связывается с Т-клеточным рецептором (TCR) и/или TCR-подобной связывающей молекулой, такой как TCR-подобное антитело, или иммуноглобулин, или химерный антигенный рецептор, например, отдельной клетки.
В соответствии с некоторыми вариантами осуществления аффинная часть конъюгата тетрамер-олигонуклеотид содержит комплекс МНС-пептид. В соответствии с некоторыми вариантами осуществления аффинная часть конъюгата тетрамер-олигонуклеотид содержит главный комплекс гистосовместимости (МНС) в растворимой и/или мультимерной (например, тетрамерной) форме. В соответствии с некоторыми вариантами осуществления МНС связан с белком стрептавидином, который образует комплекс с центральным белком биотином, обеспечивая возможность объединения в мультимерный комплекс до четырех комплексов МНС-стрептавидин (например, как изображено на фиг. 10А). В соответствии с некоторыми вариантами осуществления МНС является связанным с пептидом. В соответствии с некоторыми вариантами осуществления МНС является связанным с пептидом посредством ковалентной или нековалентной связи. В соответствии с некоторыми вариантами осуществления пептид представляет собой синтетический пептид. В соответствии с некоторыми вариантами осуществления пептид представляет собой синтетический пептид. В соответствии с некоторыми вариантами осуществления МНС связывается с TCR и/или TCR-подобной связывающей молекулой, такой как TCR-подобное антитело, или иммуноглобулин, или химерный антигенный рецептор, например, отдельной клетки. В соответствии с некоторыми вариантами осуществления аффинная часть конъюгата тетрамер-олигонуклеотид содержит антиген В-клеточного рецептора в растворимой и/или мультимерной (например, тетрамерной) форме. В соответствии с некоторыми вариантами осуществления антиген В-клеточного рецептора ранее вводили в клетки или субъекту. В соответствии с некоторыми вариантами осуществления антиген В-клеточного рецептора связывается с BCR и/или BCR-подобной связывающей молекулой, такой как химерный антигенный рецептор, например, отдельной клетки.
В соответствии с некоторыми вариантами осуществления аффинная часть специфически связывается с распознающей антиген молекулой и/или иммунорецептором, таким как антитело, или иммуноглобулин, или их часть или гибридный белок, сконструированным иммунорецептором, химерным антигенным рецептором (CAR) или TCR. В соответствии с некоторыми такими вариантами осуществления аффинная часть содержит антиген или эпитоп или их часть, распознаваемые антителом или рецептором, таким как CAR. В соответствии с некоторыми вариантами осуществления аффинная часть содержит антитело или его антигенсвязывающий фрагмент, которые специфически связываются с иммунорецептором. В соответствии с некоторыми аспектами антитело или его антигенсвязывающий фрагмент специфически связываются с вариабельной и/или антигенсвязывающей частью рецептора, такой как идиотоп. В соответствии с некоторыми аспектами аффинная молекула представляет собой антиидиотипическое антитело или его фрагмент.
В соответствии с некоторыми вариантами осуществления аффинная часть конъюгата аффинная молекула-олигонуклеотид содержит главный комплекс гистосовместимости (МНС) или его функциональную или связывающую часть. В соответствии с некоторыми вариантами осуществления аффинная часть содержит мультимер МНС, необязательно, тетрамер МНС. В соответствии с некоторыми вариантами осуществления МНС присутствует в растворимой форме. В соответствии с некоторыми вариантами осуществления МНС связан с пептидом и/или содержит пептид в борозде МНС. В соответствии с некоторыми вариантами осуществления пептид представляет собой синтетический пептид. В соответствии с некоторыми вариантами осуществления МНС связывается с Т-клеточным рецептором (TCR) отдельной клетки. В соответствии с некоторыми вариантами осуществления аффинная часть содержит пептид, который связывается с антителом или химерным антигенным рецептором (CAR), и/или при этом мишень представляет собой антитело или CAR. В соответствии с некоторыми вариантами осуществления аффинная часть представляет собой или содержит антиген или эпитоп, специфически распознаваемый антителом или химерным антигенным рецептором, и/или содержит антитело, которое специфически о связывается с ним, необязательно, антиидиотипическое антитело, которое специфически связывается с его антигенсвязывающей частью.
В соответствии с одним аспектом предполагается композиция, содержащая множество резервуаров, каждый из которых содержит отдельную клетку из образца, содержащего множество клеток, конъюгат аффинная молекула-олигонуклеотид, связанный с целевым антигеном в отдельной клетке, и меченый штрихкодом резервуара полинуклеотид, содержащий последовательность штрихкода резервуара. В соответствии с некоторыми вариантами осуществления меченый штрихкодом резервуара полинуклеотид или его комплементарная последовательность являются прикрепленными к олигонуклеотиду в составе конъюгата аффинная молекула-олигонуклеотид.
В соответствии с одним аспектом предполагается композиция, содержащая множество резервуаров, каждый из которых содержит отдельную клетку из образца, содержащего множество клеток, конъюгат тетрамер-олигонуклеотид, связанный с TCR или BCR отдельной клетки, и меченый штрихкодом резервуара полинуклеотид, содержащий последовательность штрихкода резервуара. В соответствии с некоторыми вариантами осуществления меченый штрихкодом резервуара полинуклеотид или его комплементарная последовательность являются прикрепленными к олигонуклеотиду в составе конъюгата тетрамер-олигонуклеотид. В соответствии с одним аспектом предполагается композиция, содержащая множество резервуаров, каждый из которых содержит отдельную лизированную клетку из образца, содержащего множество клеток, и конъюгат аффинная молекула-олигонуклеотид, связанный с целевым антигеном отдельной лизированной клетки; причем олигонуклеотид в составе конъюгата аффинная молекула-олигонуклеотид содержит последовательность штрихкода резервуара, и при этом клеточный полинуклеотид из отдельной лизированной клетки содержит одну и ту же последовательность штрихкода резервуара.
В соответствии с одним аспектом предполагается композиция, содержащая множество резервуаров, каждый из которых содержит отдельную лизированную клетку из образца, содержащего множество клеток, и конъюгат тетрамер-олигонуклеотид, связанный с TCR или BCR отдельной лизированной клетки; причем олигонуклеотид в составе конъюгата тетрамер-олигонуклеотид содержит последовательность штрихкода резервуара, и при этом клеточный полинуклеотид из отдельной лизированной клетки содержит одну и ту же последовательность штрихкода резервуара. В соответствии с одним аспектом в данном документе представлена композиция, содержащая множество резервуаров, при этом резервуар из множества резервуаров содержит отдельную клетку из образца, содержащего множество клеток, и меченый штрихкодом резервуара полинуклеотид, содержащий последовательность штрихкода резервуара, при этом резервуар дополнительно содержит конъюгат аффинная молекула-олигонуклеотид, который связан с целевым антигеном отдельной клетки, или олигонуклеотидную часть из него.
В соответствии с одним аспектом в данном документе представлена композиция, содержащая множество резервуаров, при этом резервуар из множества резервуаров содержит отдельную клетку из образца, содержащего множество клеток, и меченый штрихкодом резервуара полинуклеотид, содержащий последовательность штрихкода резервуара, при этом резервуар дополнительно содержит конъюгат тетрамер-олигонуклеотид, который связывается с TCR или BCR отдельной клетки, или олигонуклеотидную часть из него.
В соответствии с некоторыми вариантами осуществления реакция происходит в двух или более резервуарах из множества резервуаров. В соответствии с некоторыми вариантами осуществления резервуар содержит каждый резервуар из множества резервуаров. В соответствии с некоторыми вариантами осуществления множество резервуаров содержит два или более множества резервуаров. В соответствии с некоторыми вариантами осуществления клетка в каждом резервуаре происходит из одного и того же образца. В соответствии с некоторыми вариантами осуществления клетка в резервуаре из первого множества резервуаров в составе двух или более множеств резервуаров происходит из того же образца, что и клетка в резервуаре во втором множестве резервуаров в составе двух или более множеств резервуаров. В соответствии с некоторыми вариантами осуществления меченый штрихкодом резервуара полинуклеотид или его комплементарная последовательность являются прикрепленными к олигонуклеотиду в составе конъюгата аффинная молекула-олигонуклеотид. В соответствии с некоторыми вариантами осуществления меченый штрихкодом резервуара полинуклеотид или его комплементарная последовательность являются прикрепленными к олигонуклеотиду в составе конъюгата тетрамер-олигонуклеотид. В соответствии с некоторыми вариантами осуществления отдельную клетку подвергают лизису.
В соответствии с одним аспектом в данном документе представлена композиция, содержащая множество резервуаров, причем резервуар из множества резервуаров содержит отдельную лизированную клетку из образца, содержащего множество клеток, и конъюгат аффинная молекула-олигонуклеотид, содержащий аффинную часть, которая связывается с целевым антигеном отдельной лизированной клетки или олигонуклеотидной частью конъюгата аффинная молекула-олигонуклеотид; при этом олигонуклеотидная часть конъюгата аффинная молекула-олигонуклеотид содержит последовательность штрихкода резервуара, и при этом клеточный полинуклеотид из отдельной лизированной клетки содержит одинаковую последовательность штрихкода резервуара.
В соответствии с одним аспектом в данном документе представлена композиция, содержащая множество резервуаров, причем резервуар из множества резервуаров содержит отдельную лизированную клетку из образца, содержащего множество клеток, и конъюгат тетрамер-олигонуклеотид, содержащий аффинную часть, которая связывается с TCR или BCR отдельной лизированной клетки или олигонуклеотидной частью конъюгата тетрамер-олигонуклеотид; при этом олигонуклеотидная часть конъюгата тетрамер-олигонуклеотид содержит последовательность штрихкода резервуара, и при этом клеточный полинуклеотид из отдельной лизированной клетки содержит одинаковую последовательность штрихкода резервуара.
В соответствии с одним аспектом предполагается набор, содержащий: первый контейнер, содержащий первый олигонуклеотид, содержащий первую идентификационную последовательность антигена (AID), при этом первая последовательность AID представляет собой известную последовательность; второй контейнер, содержащий второй олигонуклеотид, содержащий вторую идентификационную последовательность антигена (AID), при этом вторая последовательность AID представляет собой известную последовательность и является отличной от первой последовательности AID; один или несколько третьих контейнеров, содержащие реактивы, способные к конъюгированию первого олигонуклеотид а с первой аффинной молекулой, и реактивы, способные к конъюгированию второго олигонуклеотида со второй аффинной молекулой; набор инструкций, описывающих, как конъюгировать первый олигонуклеотид с первой аффинной молекулой и второй олигонуклеотид со второй аффинной молекулой.
В соответствии с одним аспектом предполагается набор, содержащий: первый контейнер, содержащий первый олигонуклеотид, содержащий первую идентификационную последовательность антигена (AID), при этом первая последовательность AID представляет собой известную последовательность; второй контейнер, содержащий второй олигонуклеотид, содержащий вторую идентификационную последовательность антигена (AID), при этом вторая последовательность AID представляет собой известную последовательность и является отличной от первой последовательности AID; один или несколько третьих контейнеров, содержащие реактивы, способные к конъюгированию первого олигонуклеотида с первым тетрамером, и реактивы, способные к конъюгированию второго олигонуклеотида со вторым тетрамером; набор инструкций, описывающих, как конъюгировать первый олигонуклеотид с первым тетрамером и второй олигонуклеотид со вторым тетрамером.
В соответствии с одним аспектом предполагается набор, содержащий: первый контейнер, содержащий олигонуклеотид, содержащий идентификационную последовательность антигена (AID), при этом последовательность AID представляет собой известную последовательность; второй контейнер, содержащий реактивы, способные к конъюгированию олигонуклеотида с аффинной молекулой; третий контейнер, содержащий множество меченых штрихкодом резервуара полинуклеотидов; и набор инструкций, описывающий, как прикрепить меченый штрихкодом резервуара полинуклеотид из множества меченых штрихкодом резервуара полинуклеотидов к олигонуклеотиду, когда его конъюгируют с аффинной молекулой.
В соответствии с одним аспектом предполагается набор, содержащий: первый контейнер, содержащий олигонуклеотид, содержащий идентификационную последовательность антигена (AID), при этом последовательность AID представляет собой известную последовательность; второй контейнер, содержащий реактивы, способные к конъюгированию олигонуклеотида с тетрамером; третий контейнер, содержащий множество меченых штрихкодом резервуара полинуклеотидов; и набор инструкций, описывающий, как прикрепить меченый штрихкодом резервуара полинуклеотид из множества меченых штрихкодом резервуара полинуклеотидов к олигонуклеотиду, когда его конъюгируют с тетрамером.
Способы и композиции, раскрытые в данном документе, можно применять для анализа профиля опухолей. Например, способы могут включать связывание клеточных фенотипов с иммунным репертуаром в образцах от пациента, чтобы идентифицировать реагирующие на опухоль TCR. Способы и композиции, раскрытые в данном документе, можно применять для адоптивной клеточной терапии. Например, способы могут включать генетический анализ Т-клеток без сортинга. Например, способы могут включать объединение информации о клонах Т-клеток (с применением TCR) с паттернами экспрессии генов во время производства продукта и лечения. В соответствии с некоторыми вариантами осуществления способы, раскрытые в данном документе, можно применять для отслеживания, характеристики, мониторинга и/или оценки подвергнутых адоптивному переносу клеток, полученных от пациента до, во время прохождения курса адоптивной клеточной терапии или после нее. Способы и композиции, раскрытые в данном документе, можно применять для идентификации TCR к известным мишеням. Например, способы могут включать идентификацию клонов с высокой аффинностью, которые могут сильно реагировать на антиген, но слабо пролиферируют. Способы и композиции, раскрытые в данном документе, можно применять для мультиплексирования образца клеток. Например, эмульсию, содержащую объединенные образцы клеток, приведенные в контакт с одним или несколькими конъюгатами аффинная молекула-олигонуклеотид, можно применять для идентификации исходных образцов клеток при одновременной обработке многочисленных образцов.
Способы и композиции, раскрытые в данном документе, можно применять для мультиплексного измерения аффинности TCR или BCR посредством секвенирования. Способы и композиции, раскрытые в данном документе, можно применять для количественного анализа аффинности связывания TCR или BCR отдельной клетки в отношении тетрамера в составе конъюгата тетрамер-олигонуклеотид. Более того, этот количественный анализ может быть чувствительным к концентрации конъюгата тетрамер-олигонуклеотид, при которой сигнал от аффинности связывания может зависеть от концентрации конъюгата тетрамер-олигонуклеотид (например, снижение концентрации конъюгата тетрамер-олигонуклеотид приводит в результате к соответствующему снижению сигнала от аффинности связывания). Кроме того, способ и композиции, раскрытые в данном документе, можно применять для того, чтобы отличать антигенспецифические клетки от неспецифических клеток. Например, установление различий между клетками с TCR или BCR, которые связываются с тетрамером в составе конъюгата тетрамер-олигонуклеотид, и клетками с TCR или BCR, которые не связываются с тетрамером в составе конъюгата тетрамер-олигонуклеотид. Более того, одним преимуществом способов и композиций, раскрытых в данном документе, является способность отличать популяцию антигенспецифических клеток (т.е. тетрамер + клетки) от неспецифических клеток (т.е. тетрамер - клетки) без применения гейтирования в ходе проточной цитометрии для разделения этих двух популяций. Часто гейтирование в ходе проточной цитометрии может быть «наиболее вероятным предположением» разделения популяций на основании отделения тетрамер + популяции от тетрамер - популяции на графике результатов проточной цитометрии, которое может быть неточным и, вследствие этого, может приводить в результате к потере клеток из каждой популяции или неверной характеристике клетки. Напротив, способы и композиции, описанные в данном документе, не опираются на методики гейтирования для установления различий между популяциями клеток, а вместо этого могут представлять собой способ с высокой пропускной способностью, в котором индивидуально количественно определяют аффинность связывания у каждой клетки в образце. Более того, способы и композиции, раскрытые в данном документе, можно применять для установления различий между клеткой с TCR, которые характеризуются высокой аффинностью связывания в отношении МНС-пептида (т.е. сильная связывающая молекула), и клеткой с TCR, которые характеризуются низкой аффинностью связывания в отношении МНС-пептида (т.е. слабая связывающая молекула), а также для установления различий между клетками с высокой аффинностью связывания и клетками с низкой аффинностью связывания в пределах образца с применением способов с высокой пропускной способностью, описанных в данном документе.
Включение посредством ссылки
Все публикации патенты и патентные заявки, упоминаемые в данном описании, включены в данный документ посредством ссылки во всей своей полноте и с любой целью в той же мере, как если бы было указано, что каждая отдельная публикация, патент или патентная заявка была специально и отдельно включена посредством ссылки.
Краткое описание чертежей
Новые признаки, описанные в данном документе, изложены с характерными деталями в пунктах прилагаемой формулы изобретения. Лучшее понимание признаков и преимуществ признаков, описанных в данном документе, будет достигаться с учетом следующего подробного описания, в котором изложены иллюстративные примеры, в которых используются принципы из признаков, описанных в данном документе, и прилагаемых чертежей, в которых:
На фиг. 1 представлена иллюстративная схема резервуара согласно способам, описанным в данном документе.
На фиг. 2 представлено иллюстративное строение олигонуклеотидной метки, конъюгированной с антителом. Каждый выделенный цветом блок представляет часть полной последовательности олигонуклеотида. Последовательность для слияния применяют для ферментативного прикрепления специфического для капли ДНК-штрихкода внутри эмульсионной реакционной смеси. Показан только один возможный порядок расположения последовательностей, хотя другие порядки расположения являются совместимыми с описанным способом.
На фиг. 3А представлен иллюстративный совместный захват последовательностей иммунного рецептора с дополнительными мРНК- и белками-мишенями. Мишени поверхностные белки количественно определяют с помощью предварительного инкубирования клеток с мечеными ДНК окрашивающими антителами перед секвенированием в эмульсии.
На фиг. 3В представлены иллюстративные результаты измерений мРНК и белка CD4 и CD8 на 3682 парах VαVβ TCR со штрихкодами капель, полученных из Т-клеток здорового человека.
На фиг. 3С представлено иллюстративное соответствие между измерениями мРНК и белка (каждая точка представляет собой штрихкод капли, связанный с парой VαVβ TCR).
На фиг. 3D приведена иллюстративная таблица результатов одновременного выявления мРНК и белка CD4 и CD8 из не подвергнутых сортингу Т-клеток в эмульсии. Из 30000 вводимых Т-клеток было выделено 3682 пары TCR. Частоты пар TCR, называемых CD4+ или CD8+, полученные посредством сравнения мРНК с белком (на основании подсчета молекул, принцип большинства), показаны в виде матрицы.
На фиг. 3Е представлены иллюстративные результаты для 45870 пар TCR из отдельных клеток, полученные с применением конъюгата аффинная молекула-олигонуклеотид, нацеленного на CD4, и конъюгата аффинная молекула-олигонуклеотид, нацеленного на CD8.
На фиг. 4 изображена схема иллюстративного способа применения конъюгата аффинная молекула-олигонуклеотид, нацеленного на CD4, и конъюгата аффинная молекула-олигонуклеотид, нацеленного на CD8.
На фиг. 5 демонстрируются результаты способа мечения штрихкодом отдельной иммунной клетки в эмульсии.
На фиг. 5А представлено иллюстративное изображение двух потоков водной среды, содержащих клетки и лизирующую/реакционную (LR) смесь, пропускаемых в масло, что дает монодисперсную эмульсию с продуктивностью более 8 миллионов капель в час.
На фиг. 5В представлено иллюстративное изображение, показывающее, что клетки в резервуарах подвергают лизису и подвергают прикреплению молекулярного и специфического для капли штрихкода в одной реакции.
На фиг. 5С представлено иллюстративное изображение, показывающее, что целевая мРНК подвергается обратной транскрипции и мечению универсальной адаптерной последовательностью посредством переключения матрицы. Затем происходит ПЦР амплификация матрицы штрихкода капли, первоначально разведенной до количества ~1 молекула на каплю. Амплифицированные штрихкоды прикрепляют к полученным посредством переключения матрицы кДНК посредством комплементарной достройки перекрывающихся последовательностей. Продукты выделяют из эмульсии и очищают с использованием биотина на праймере для RT перед дополнительными стадиями обработки библиотеки и секвенированием с высокой пропускной способностью.
На фиг. 5D представлено иллюстративное изображение, показывающее, что двойное мечение штрихкодами позволяет кластеризацию ридов от секвенирования по молекулам и каплям, из которых они происходят, реконструируя нативные пары цепей рецепторов, при этом сводя к минимуму ошибки при секвенировании и отклонения при амплификации.
На фиг. 6 проиллюстрированы результаты способа выделения BCR из выделенных здоровых В-клеток.
На фиг. 6А представлено иллюстративное изображение капель, в которых 3 миллиона В-клеток переводят в эмульсию в количестве 0,2 клетки/капля, что приводит в результате к тому, что ~90% занятых ячеек содержат отдельные клетки.
На фиг. 6В представлено иллюстративное изображение точности спаривания VHVL. После мечения штрихкодами в эмульсии и секвенирования данные обогащали данными от капель с отдельными клетками и точность спаривания VHVL оценивали с использованием соответствия пар между размноженными клонами.
На фиг. 6С представлен иллюстративный график зависимости процентного содержания штрихкода капли от изотипа Ig. Показаны изотип тяжелой цепи (наиболее распространенный изотип в каждой капле) и использование локуса легкой цепи для 259368 отфильтрованных пар VHVL.
На фиг. 6D представлен иллюстративный график распределенной по рангам распространенности 100 наиболее часто встречающихся клонов тяжелой цепи в каждой из шести независимых фракций эмульсии. Помечена общая частота 0,05%.
На фиг. 6Е представлен иллюстративный график сравнения экспрессии VH и VL в клетках, оцениваемой по количеству захваченных мРНК в каждом штрихкоде капли. 5000 точек показаны для каждого изотипа.
На фиг. 6F представлен иллюстративный график корреляции мутаций в VH относительно VL для пар BCR и распределения по плотности в пределах каждого изотипа.
На фиг. 7 иллюстрируются результаты способа изучения нейтрализующего антитела широкого спектра действия (bNAb) против ВИЧ.
На фиг. 7А представлен иллюстративный график распределения изотипа тяжелых цепей у 38620 выделенных пар VHVL из В-клеток от элитного контролера ВИЧ, введенных в эмульсию. Малая доля цепей IgG хорошо выравнивается с ранее известными bNAb («PGT-подобные»).
На фиг. 7В представлено иллюстративное изображение филогенетических деревьев из полных аминокислотных VDJ последовательностей известных bNAb (черный), а также для вновь выделенных (красный, меченых штрихкодом капли), при этом тяжелые (слева) и легкие цепи (справа) изображены на графике раздельно. Потенциально несоответствующие антитела PGT122 и PGT123 показаны голубым.
На фиг. 7С представлено иллюстративное изображение нейтрализующей активности (IC50, мкг/мл) у 8 вновь выявленных PGT-подобных вариантов в отношении десяти штаммов ВИЧ по сравнению с контрольной партией PGT121.
На фиг. 8 иллюстрируются результаты способа характеристики TIL из опухоли яичника.
На фиг. 8А представлено иллюстративное изображение капель, в которых 400000 несортированных диссоциированных клеток из опухоли яичника были введены в эмульсию и пары BCR и TCR одновременно выделяли посредством мечения штрихкодами в эмульсии.
На фиг. 8В представлен иллюстративный график зависимости штрихкодов капель от комбинаций цепей рецепторов, показывающий количества всех комбинаций VH/VL и Vα/Vβ, наблюдаемых в штрихкодах капель после фильтрования.
На фиг. 8С представлен иллюстративный график зависимости процентного содержания штрихкода капли от распределения изотипов тяжелой цепи в выделенных парах BCR.
На фиг. 8D представлен иллюстративный график корреляции мутаций в VH по сравнению с VL для пар BCR и распределения по плотности в пределах каждого изотипа.
На фиг. 8Е изображены иллюстративные графики количеств захваченных мРНК для пар TCR и пар BCR в целом (вверху) и для различных изотипов (внизу).
На фиг. 8F изображены иллюстративные графики результатов клонального анализа, показывающего упорядоченную по рангам распространенность 30 наиболее часто встречающихся клонов тяжелой цепи BCR (сверху) и 30 наиболее часто встречающихся клонов бета-цепи TCR в каждой из шести независимых фракций эмульсии. Показаны уровни общей частоты 1% и 10%.
На фиг. 9 проиллюстрирован способ иммунофенотипирования с применением конъюгатов антитело-олигонуклеотид.
На фиг. 9А представлено иллюстративное схематическое изображение, показывающее 2 резервуара, каждый из которых содержит отдельную клетку, связанную с конъюгатом антитело-олигонуклеотид. (DB1 - штрихкод 1 капли; DB2 - штрихкод 2 капли; МВ1 - молекулярный штрихкод 1; МВ2 - молекулярный штрихкод 2; AID - идентификационный штрихкод антигена; АМВ1 молекулярный штрихкод 1 антитела; АМВ2 - молекулярный штрихкод 2 антитела).
На фиг. 9В представлено иллюстративное схематическое изображение, показывающее 2 резервуара, каждый из которых содержит молекулы РНК из лизированной клетки из резервуара с фиг. 9А. Молекулы РНК подвергают обратной транскрипции и нематричные нуклеотиды добавляют к концу молекулы кДНК, образованной при обратной транскрипции. Молекулярные штрихкоды гибридизируются с нематричными нуклеотидами, добавленными к концу молекулы кДНК, образованной при обратной транскрипции.
На фиг. 9С представлено иллюстративное схематическое изображение, показывающее 2 резервуара, каждый из которых содержит полинуклеотид, меченый штрихкодом матрицы, который амплифицируется и прикрепляется к кДНК из резервуара с фиг. 9В посредством гибридизации, и кДНК достраивается (вверху). Достроенная кДНК затем амплифицируется (внизу).
На фиг. 9D представлено иллюстративное схематическое изображение, показывающее, что частицы RNA-MB-DB с одинаковым молекулярным штрихкодом (MB), прикрепленным одинаковым идентичным последовательностям РНК, вероятно, являются результатом ПЦР дупликации. Частицы RNA-MB-DB с двумя отличающимися MB, которые прикреплены к одинаковым идентичным последовательностям РНК (RNA1-MB1-DB и RNA1-MB2-DB), представляют собой две независимые молекулы РНК, происходящие из источника, а не являющиеся результатом ПЦР дупликации.
На фиг. 9Е изображено иллюстративное схематическое изображение, на котором показано, что частица DB-AMB-AID с одинаковым молекулярным штрихкодом антитела (АМВ), прикрепленным к последовательности с одинаковым штрихкодом капли (DB) и идентификационным штрихкодом антигена (AID), вероятно, является результатом ПЦР дупликации. Частицы DB1-AMB1-AID1 и DB1-AMB2-AID1 с двумя отличающимися АМВ, прикрепленными к последовательностям с одинаковым штрихкодом капли (DB) и идентификационным штрихкодом антигена (AID), представляют собой две независимые молекулы олигонуклеотида из двух независимых молекул конъюгата антитело-олигонуклеотид, каждая с антителом, которое специфически связывается с одним и тем же целевым антигеном, прикрепленным к одной и той же отдельной клетке в резервуаре, а не являющиеся результатом ПЦР дупликации. Частицы DB1-AMBn-AID1 и частицы DB1-AMBn-AID2 с двумя отличающимися AID, прикрепленными к последовательности с одинаковым штрихкодом капли (DB) и одинаковыми или отличающимися молекулярными штрихкодами антитела (АМВ) представляют собой две независимые молекулы олигонуклеотида из двух независимых молекул конъюгата антитело-олигонуклеотид, прикрепленных к одной и той же отдельной клетке в резервуаре, при этом одна из молекул конъюгата антитело-олигонуклеотид содержит антитело, которое специфически связывается с первым целевым антигеном, а другая молекула конъюгата антитело-олигонуклеотид содержит антитело, которое специфически связывается со вторым целевым антигеном.
На фиг. 10А представлено схематическое изображение иллюстративного конъюгата аффинная молекула-олигонуклеотид согласно способам, описанный в данном документе. Как изображено, иллюстративный конъюгат аффинная молекула-олигонуклеотид может содержать аффинную часть в виде МНС-пептида. В соответствии с некоторыми аспектами такие конъюгаты могут быть названы конъюгатами тетрамер-олигонуклеотид.
На фиг. 10В представлено схематическое изображение иллюстративного конъюгата аффинная молекула-олигонуклеотид согласно способам, описанным в данном документе. В соответствии с некоторыми аспектами такие конъюгаты могут быть названы ко нъюгатами тетрамер-олигонуклеотид.
На фиг. 11А представлен иллюстративный график сигнала от связывания для двух иллюстративных конъюгатов аффинная молекула-олигонуклеотид согласно способам, описанным в данном документе, которые содержат аффинную часть, которая связывается с TCR.
На фиг. 12А представлено иллюстративное схематическое изображение Т-клетки, связанной с иллюстративным конъюгатом аффинная молекула-олигонуклеотид согласно способам, описанным в данном документе.
На фиг. 12В представлено иллюстративное схематическое изображение Т-клетки в капле, связанной с иллюстративным конъюгатом аффинная молекула-олигонуклеотид согласно способам, описанным в данном документе. Нуклеиновые кислоты в капле промаркированы идентификационными последовательностями капли и включены в библиотеку для секвенирования нового поколения.
На фиг. 13 представлено иллюстративное схематическое изображение олигонуклеотидной метки, конъюгированной с иллюстративным конъюгатом аффинная молекула-олигонуклеотид тетрамер. Идентификационная последовательность тетрамера представляет собой короткую константную последовательность ДНК, которая соответствует партии тетрамера и обеспечивает возможность мультиплексного анализа различных мишеней, таких как мишени пептид-МНС, в одном эксперименте. Молекулярный штрихкод представляет собой вырожденную последовательность, которая обеспечивает возможность подсчета молекул для количественного определения связанных тетрамеров.
На фиг. 14 представлены схематические изображения иллюстративного конъюгата аффинная молекула-олигонуклеотид, образованного из реактивов в виде меченого ДНК тетрамера МНС (конъюгат тетрамер-олигонуклеотид). В соответствии с одним вариантом осуществления, связанный с Су5 олигонуклеотид ДНК синтезируют и конъюгируют со стрептавидином или нейтравидином. В соответствии с одним вариантом осуществления нефлуоресцентный олигонуклеотид ДНК конъюгируют с АРС-стрептавидином. В соответствии с одним вариантом осуществления смесь нефлуоресцентного олигонуклеотида ДНК и стрептавидина или нейтравидина конъюгируют с активированным АРС.
На фиг. 15 изображен иллюстративный способ конъюгирования олигонуклеотида с аффинной частью конъюгата аффинная молекула-олигонуклеотид с применением подхода клик-химии.
На фиг. 16 представлено схематическое изображение иллюстративной последовательности операций для получения и характеристики иллюстративных конъюгатов аффинная молекула-олигонуклеотид.
На фиг. 17 изображены результаты иллюстративного способа, описанного в данном документе, в котором применяются 6 различных конъюгатов аффинная молекула-олигонуклеотид, нацеленные на CD3, CD 19, CD4, CD8, HLA-DR и CTLA-4. Каждая точка соответствует штрихкоду капли/отдельной клетке. Идентификацию клеток проводили на основании типа выделенной пары рецепторов (TCR = Т-клетка; Ig = В-клетка).
На фиг. 18 изображен дополнительный вид иллюстративного конъюгата тетрамер-олигонуклеотид. В соответствии с некоторыми вариантами осуществления тетрамер-олигонуклеотид содержит флуорофор (флуоресцентную молекулу). В соответствии с некоторыми вариантами осуществления тетрамер-олигонуклеотид не содержит флуорофор.
На фиг. 19 изображены иллюстративные кривые связывания для двух иллюстративных конъюгатов тетрамер-олигонуклеотид согласно способам, описанным в данном документе, которые содержат аффинную часть в виде МНС-пептида, которая связывается с TCR.
На фиг. 20 изображены графики результатов проточной цитометрии, при которой мишенеспецифические первичные CD8+ Т-клетки или не специфические к мишени первичные CD8+ Т-клетки инкубировали либо с реактивом в виде стандартного тетрамера рМНС, либо с меченым ДНК тетрамером рМНС (конъюгат тетрамер-олигонуклеотид). Меченый ДНК тетрамер рМНС использовали в убывающих концентрациях, которые показаны в виде треугольника над тремя крайними сверху справа секциями.
На фиг. 21 изображено сравнение значения подсчитанного количества конъюгата тетрамер-олигонуклеотид на клетку в клетках, проявляющих не участвующие в связывании TCR (например, TCR, которые не распознаются специфически реактивом в виде конъюгата тетрамер-олигонуклеотид) и участвующие в связывании TCR (например, TCR, которые специфически распознаются реактивом с конъюгатом тетрамер-олигонуклеотид). Использовали возрастающие концентрации конъюгата тетрамер-олигонуклеотид, которые указаны на оси X.
Подробное раскрытие настоящего изобретения
Несколько аспектов описаны ниже со ссылкой на представленные в качестве примера применения для иллюстрации. Следует понимать, что многочисленные конкретные детали, взаимосвязи и способы изложены для обеспечения полного понимания признаков, описанных в данном документе. Квалифицированный специалист в соответствующей области техники, однако, легко поймет, что признаки, описанные в данном документе, могут быть практически реализованы без одной или нескольких из конкретных деталей или с использованием других способов. Признаки, описанные в данном документе, не ограничиваются проиллюстрированным порядком действий или событий, поскольку некоторые действия могут происходить в разном порядке и/или одновременно с другими действиями или событиями. Более того, не все проиллюстрированные действия или события требуются для осуществления метода в соответствии с признаками, описанными в данном документе.
Терминология, используемая в данном документе, предназначена для описания только конкретных случаев и не предполагается как ограничивающая. В контексте данного документа предполагается, что формы единственного числа включают в себя также формы множественного числа, если контекст явно не указывает на иное. Более того, в случае, если термины «в том числе», «включает в себя», «имеющий», «имеет», «с» или их варианты используются и в подробном описании, и/или пунктах формулы изобретения, предполагается, что такие термины являются включительными аналогично термину «содержащий».
Термины «приблизительно» или «примерно» означают пребывание в пределах приемлемого диапазона погрешности для конкретного значения, определяемого средним специалистом в данной области техники, который отчасти будет зависеть от того, как измерено или определено значение, т.е. ограничений измерительной системы. Например, «приблизительно» может означать пребывание в пределах 1 или более чем 1 стандартного отклонения в соответствии с практикой, используемой в данной области техники. В качестве альтернативы, «приблизительно» может означать диапазон до 20%, до 10%, до 5% или до 1% от заданного значения. В качестве альтернативы, в особенности, применительно к биологическим системам или процессам, термин может означать пребывание в пределах порядка величины значения, в пределах величины, 5-кратно и, более предпочтительно, 2-кратно превышающей измеренное значение. При этом конкретные значения описаны в заявке и формуле изобретения, если не указано иное, следует предполагать, что термин «приблизительно» означает пребывание в пределах приемлемого диапазона для конкретного значения.
Целью настоящего изобретения является обеспечение способов и композиций для фенотипирования отдельных клеток (например, иммунных клеток) с применением конъюгатов аффинная молекула-олигонуклеотид (например, конъюгатов антитело-олигонуклеотид) (например, в эмульсиях).
Определения
Термин «антитело» в данном документе, следовательно, используют в наиболее широком смысле, и он включает в себя поликлональные и моноклональные антитела, в том числе интактные антитела и их функциональные (антигенсвязывающие) фрагменты антител, в том числе антигенсвязывающие (Fab) фрагменты, F(ab')2 фрагменты, Fab' фрагменты, Fv фрагменты, фрагменты рекомбинантного IgG (rIgG), одноцепочечные фрагменты антител, в том числе одноцепочечные вариабельные фрагменты (scFv) и фрагменты однодоменных антител (например, sdAb, sdFv, нанотело). Термином охватываются полученные с помощью методов генной инженерии и/или иным образом модифицированные формы иммуноглобулинов, такие как интраантитела, пептидные антитела, химерные антитела, полностью человеческие антитела, гуманизированные антитела и гетероконъюгаты антител, мультиспецифические, например, биспецифические, антитела, диатела, триатела и тетратела, тандемный ди-scFv, тандемный три-scFv. Если не указано иное, термин «антитело» следует понимать как охватывающий функциональные фрагменты этих антител. Термином также охватываются интактные или полноразмерные антитела, в том числе антитела любого класса или подкласса, в том числе IgG и его подклассы, IgM, IgE, IgA, и IgD.
Термины «определяющий комплементарность участок» и «CDR», синонимичные терминам «гипервариабельный участок» или «HVR», являются известными в уровне техники применительно к несмежным последовательностям аминокислот в пределах вариабельных участков антитела, которые придают специфичность в отношении антигена и/или обеспечивают аффинность связывания. В целом, три CDR присутствуют в каждом вариабельном участке тяжелой цепи (CDR-H1, CDR-H2, CDR-H3), и три CDR присутствуют в каждом вариабельном участке легкой цепи (CDR-L1, CDR-L2, CDR-L3). «Каркасные участки» и «FR» являются известными в уровне техники применительно к отличным от CDR частям вариабельных участков тяжелых и легких цепей. В целом, четыре FR присутствуют в каждом полноразмерном вариабельном участке тяжелой цепи (FR-H1, FR-H2, FR-H3 и FR-H4), и четыре FR присутствуют в каждом полноразмерном вариабельном участке легкой цепи (FR-L1, FR-L2, FR-L3 и FR-L4).
Точные границы аминокислотных последовательностей заданного CDR или FR можно легко определить с применением любой из ряда хорошо известных схем, в том числе описанных в Kabat et al. (1991), "Sequences of Proteins of Immunological Interest," 5th Ed. Public Health Service, National Institutes of Health, Bethesda, MD (схема нумерации «Kabat»), Al-Lazikani et al., (1997) JMB 273,927-948 (схема нумерации «Chothia»), MacCallum et al., J. Mol. Biol. 262:732-745 (1996), "Antibody-antigen interactions: Contact analysis and binding site topography," J. Mol. Biol. 262, 732-745." (схема нумерации на основе «контактных остатков»), Lefranc MP et al., "IMGT unique numbering for immunoglobulin and T cell receptor variable domains and Ig superfamily V-like domains," Dev Comp Immunol, 2003 Jan; 27(1):55-77 (схема нумерации «IMGT»), и Honegger A and Pltickthun A, "Yet another numbering scheme for immunoglobulin variable domains: an automatic modeling and analysis tool," J Mol Biol, 2001 Jun 8; 309(3):657-70, (схема нумерации «Aho»).
Границы заданного CDR или FR могут варьировать в зависимости от схемы, используемой для идентификации. Например, схема по Kabat основывается на выравниваниях структур, тогда как схема по Chothia основывается на информации о структуре. Схемы нумерации как по Kabat, так и по Chothia основываются на длинах наиболее распространенных последовательностей участков антител со вставками, которым предоставляются буквенные обозначения вставок, например, «30а», и делециями, возникающими в некоторых антителах. В двух схемах определенные вставки и делеции («инделы») находятся в различных положениях, что приводит в результате к отличающейся нумерации. Схема на основе контактных остатков основывается на анализе кристаллических структур комплексов и является подобной во многих отношениях схеме нумерации по Chothia.
В таблице А ниже приведены иллюстративные границы положения CDR-L1, CDR-L2, CDR-L3 и CDR-H1, CDR-H2, CDR-H3, идентифицированные с помощью схемы по Kabat, Chothia и схемы на основе контактных остатков, соответственно. Для CDR-H1 приведена нумерация остатков согласно схемам нумерации как по Kabat, так и по Chothia. FR располагаются между CDR, например, FR-L1 располагается между CDR-L1 и CDR-L2 и т.д. Следует отметить, что поскольку показанная схема нумерации по Kabat помещает вставки в Н35А и Н35 В, конец петли CDR-H1 по Chothia при нумерации с использованием показанной системы нумерации по Kabat варьирует между Н32 и Н34 в зависимости от длины петли.
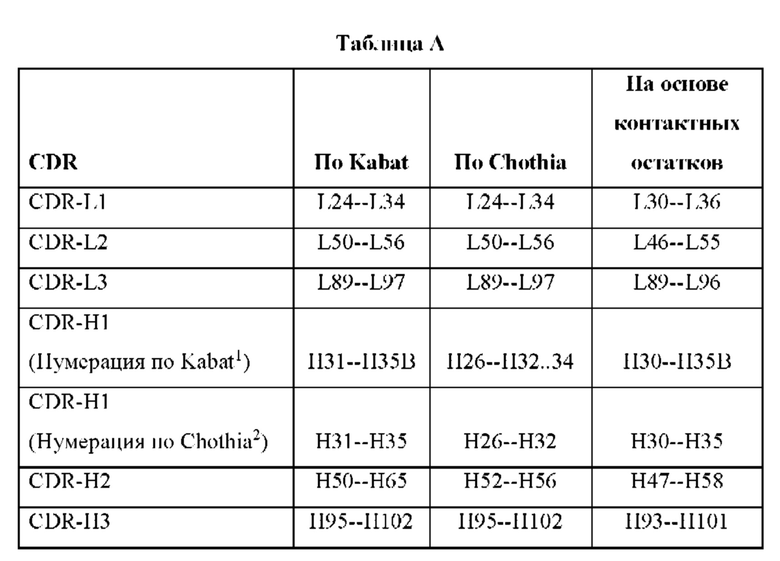
Таким образом, если не определено иное, «CDR» или «определяющий комплементарность участок» или отдельные определенные CDR (например, CDR-H1, CDR-Н2) заданного антитела или его участка, такого как его вариабельный участок, следует понимать как охватывающие (или специфически определяющие) определяющий комплементарность участок, который определяется с помощью любой из вышеупомянутых схем. Например, в случае, когда указывается, что конкретный CDR (например, CDR-H3) содержит аминокислотную последовательность соответствующего CDR в заданной аминокислотной последовательности VH или VL, понятно, что такой CDR имеет последовательность соответствующего CDR (например, CDR-H3) в пределах вариабельного участка, который определяется с помощью любой из вышеупомянутых схем. В соответствии с некоторыми вариантами осуществления определенные последовательности CDR являются определенными.
Подобным образом, если не определено иное, FR или отдельный определенный FR (отдельные определенные FR) (например, FR-H1, FR-H2) заданного антитела или его участка, такого как его вариабельный участок, следует понимать как охватывающий (или специфически определяющий) каркасный участок, который определяется с помощью любой из известных схем. В некоторых случаях определяется схема для идентификации конкретного CDR, FR или нескольких FR или CDR, как например, для CDR, который определен по Kabat, Chothia или с помощью способа на основе контактных остатков. В других случаях задана конкретная аминокислотная последовательность CDR или FR.
Термин «вариабельный участок» или «вариабельный домен» относится к домену тяжелой или легкой цепи антитела, который задействован в связывании антитела с антигеном. Вариабельные домены тяжелой цепи и легкой цепи (VH и VL, соответственно) нативного антитела обычно имеют подобные структуры, при этом каждый домен содержит четыре консервативных каркасных участка (FR) и три CDR. (См., например, Kindt et al. Kuby Immunology, 6th ed., W.H. Freeman and Co., page 91 (2007). Одного VH или VL домена может быть достаточно для придания специфичности связывания с антигеном. Более того, антитела, которые связываются с конкретным антигеном, могут быть выделены с применением VH или VL домена из антитела, которое связывается с антигеном, для скрининга библиотеки соответствующих VL или VH доменов, соответственно. См., например, Portolano et al., J. Immunol. 150:880-887 (1993); Clarkson et al., Nature 352:624-628 (1991).
В числе обеспеченных антител находятся фрагменты антител. «Фрагмент антитела» относится к молекуле, отличной от интактного антитела, которая содержит часть интактного антитела, которая связывается с антигеном, с которым связывается интактное антитело. Примеры фрагментов антител включают в себя, без ограничения, Fv, Fab, Fab', Fab'-SH, F(ab')2; диатела; линейные антитела; одноцепочечные молекулы антител (например, scFv) и мультиспецифические антитела, образованные из фрагментов антител. В соответствии с конкретными вариантами осуществления антитела представляют собой одноцепочечные фрагменты антитела, содержащие вариабельный участок тяжелой цепи и/или вариабельный участок легкой цепи, такие как scFv.
Если не указано иное, термин «TCR» следует понимать как охватывающий полные TCR, а также их антигенсвязывающие части или антигенсвязывающие фрагменты (также называемые фрагментами, связывающими МНС-пептид). В соответствии с некоторыми вариантами осуществления TCR представляет собой интактный или полноразмерный TCR. В соответствии с некоторыми вариантами осуществления TCR представляет собой антигенсвязывающую часть, которая является меньшей, чем полноразмерный TCR, но которая связывается со специфическим антигенным пептидом, связанным с (т.е. находящимся в контексте) молекулой МНС, т.е. с комплексом МНС-пептид. В некоторых случаях антигенсвязывающая часть или фрагмент TCR могут содержать только часть структурных доменов полноразмерного или интактного TCR, но при этом все еще способны связываться с эпитопом (например, комплексом МНС-пептид), с которым связывается полный TCR. В некоторых случаях антигенсвязывающая часть или фрагмент TCR содержит вариабельные домены TCR, такие как вариабельный домен α-цепи и вариабельный домен β-цепи TCR, достаточные для образования сайта связывания для связывания специфического комплекса МНС-пептид, такого как обычно, в котором каждая цепь содержит три определяющих комплементарность участка. Включены полипептиды или белки, имеющие связывающий домен, который представляет собой антигенсвязывающий домен или является гомологичным антигенсвязывающему домену. Антитела с привитым определяющим комплементарность участком (CDR) и TCR, а также другие гуманизированные антитела и TCR (в том числе модификации CDR и модификации каркасных участков) также предполагаются этими терминами. Следует отметить, что несмотря на то, что упоминаться могут только цепи иммуноглобулинов {например, тяжелые цепи и легкие цепи), раскрытое изобретение можно применять ко многим другим отличающимся типам парных последовательностей, например, парам цепей Т-клеточного рецептора (цепей TCRα и TCRβ и цепей TCRγ и TCRδ), и оно не ограничивается иммуноглобулинами.
Способность Т-клеток распознавать антигены, ассоциированные с различными злокачественными опухолями или организмами-возбудителями инфекций, обеспечивается им TCR, который состоит из обеих из альфа(α)-цепи и бета(β)-цепи или обеих из гамма(γ)- и дельта(δ)-цепи. Белки, из которых состоят эти цепи, кодируются ДНК, которая использует уникальный механизм для создания огромного разнообразия TCR. Этот мультисубъединичный распознающий иммунорецептор связывается с комплексом CD3 и связывает пептиды, презентируемые белками МНС класса I и II на поверхности антигенпрезентирующих клеток (АРС). Связывание TCR с антигенным пептидом на АРС является главным событием в активации Т-клеток, которая происходит в иммунологическом синапсе в точке контакта между Т-клеткой и АРС.
Каждый TCR содержит определяющие комплементарность участки (CDR) вариабельного домена, а также каркасные участки (FR). Аминокислотная последовательность петель третьего определяющего комплементарность участка (CDR3) вариабельных доменов α- и β-цепи в значительной мере определяет разнообразие последовательности αβ Т-клеток, возникающих в результате рекомбинации между вариабельными (Vβ), обеспечивающими разнообразие (Dβ) и соединительными (Jβ) генными сегментами в локусе β-цепи и между аналогичными Vα и Jα генными сегментами в локусе α-цепи, соответственно. Существование нескольких таких генных сегментов в локусах α- и β-цепей TCR обеспечивает возможность того, чтобы они кодировали большое количество отличающихся последовательностей CDR3. Независимое добавление и делеция нуклеотидов в соединениях Vβ-Dβ, Dβ-Jβ и Vα-Jα во время процесса перестройки гена TCR дополнительно повышает разнообразие последовательности CDR3. В этом отношении иммунологическая компетентность отражается в разнообразии TCR.
Иммуноглобулины (Ig), экспрессируемые В-клетками, в некоторых аспектах представляют собой белки, состоящие из четырех полипептидных цепей, двух тяжелых цепей (IgHs) и двух легких цепей (IgL), образующих структуру H2L2. Каждая пара IgH- и IgL-цепей содержит гипервариабельный домен, состоящий из VL и VH участка, и константный домен. Цепи IgH у Ig относятся к нескольким типам: μ, δ, γ, α и β. Разнообразие Ig у индивида преимущественно определяется гипервариабельным доменом. Подобно TCR V-домен в цепях IgH образован посредством комбинаторного соединения VH, DH и JH генных сегментов. Независимое добавление и делеция нуклеотидов в соединениях VH-DH, DH-JH и VH-JH в ходе процесса перестройки гена Ig дополнительно повышает разнообразие последовательности гипервариабельного домена. В данном случае иммунологическая компетентность отражается в разнообразии Ig.
Термин «вариабельный участок» или «вариабельный домен» относится к домену тяжелой или легкой цепи антитела, который задействован в связывании антитела с антигеном. Вариабельные домены тяжелой цепи и легкой цепи (VH и VL, соответственно) нативного антитела обычно имеют подобные структуры, при этом каждый домен содержит четыре консервативных каркасных участка (FR) и три CDR. (См., например, Kindt et al. Kuby Immunology, 6th ed., W.H. Freeman and Co., page 91 (2007). Одного VH или VL домена может быть достаточно для придания специфичности связывания с антигеном. Более того, антитела, которые связываются с конкретным антигеном, могут быть выделены с применением VH или VL домена из антитела, которое связывается с антигеном, для скрининга библиотеки соответствующих VL или VH доменов, соответственно. См., например, Portolano et al., J. Immunol. 150:880-887 (1993); Clarkson et al., Nature 352:624-628 (1991).
«Аффинная часть» относится к части конъюгата аффинная молекула-олигонуклеотид, которая взаимодействует с целевым антигеном. Иллюстративные аффинные части включают в себя антитела, пептиды, белки, аптамеры, малые молекулы, лекарственные средства, клетки, МНС и другие.
«Гипервариабельный участок» относится к аминокислотным остаткам в антителе или TCR, которые являются ответственными за связывание антигена. Гипервариабельный участок содержит аминокислотные остатки из определяющего комплементарность участка или CDR. Каркасные или FR остатки представляют собой остатки в вариабельном домене, отличные от остатков в гипервариабельном участке, которые определены в данном документе.
В число обеспеченных антител входят фрагменты антител. «Фрагмент антитела» относится к молекуле, отличной от интактного антитела, которая содержит часть интактного антитела, которая связывается с антигеном, с которым связывается интактное антитело. Примеры фрагментов антител включают в себя, без ограничения, Fv, Fab, Fab', Fab'-SH, F(ab')2; диатела; линейные антитела; одноцепочечные молекулы антител (например, scFv) и мультиспецифические антитела, образованные из фрагментов антител. В соответствии с конкретными вариантами осуществления антитела представляют собой одноцепочечные фрагменты антител, содержащие вариабельный участок тяжелой цепи и/или вариабельный участок легкой цепи, такие как scFv.
Однодоменные антитела представляют собой фрагменты антител, содержащие весь вариабельный домен тяжелой цепи антитела или его часть или весь вариабельный домен легкой цепи антитела или его часть. В соответствии с определенными вариантами осуществления однодоменное антитело представляет собой человеческое однодоменное антитело.
Фрагменты антител могут быть получены с помощью разнообразных методик, в том числе, без ограничения, с помощью протеолитического расщепления интактного антитела, а также с помощью продуцирования рекомбинантными клетками-хозяевами. В соответствии с некоторыми вариантами осуществления антитела представляют собой фрагменты, полученные с помощью методик на основе рекомбинации, такие как фрагменты, содержащие расположение в порядке, который не встречается в естественных условиях, как например, фрагменты, у которых два или более участков или цепей антитела соединены синтетическими линкерами, например, пептидными линкерами, и/или фрагменты, которые не могут быть получены с помощью расщепления ферментом интактного антитела, встречающегося в естественных условиях. В соответствии с некоторыми аспектами фрагменты антитела представляют собой scFv.
Также предполагаются фрагменты TCR, в том числе антигенсвязывающие фрагменты. В соответствии с некоторыми вариантами осуществления TCR представляет собой его антигенсвязывающую часть, такую как вариант полноразмерного TCR, не содержащий его трансмембранного и/или цитоплазматического участка(участков), который можно назвать полным растворимым TCR. В соответствии с некоторыми вариантами осуществления TCR представляет собой димерный TCR (dTCR). В соответствии с некоторыми вариантами осуществления TCR представляет собой одноцепочечный TCR (scTCR), такой как scTCR, имеющий структуру, которая описана в РСТ публикациях международных патентных заявок WO 03/020763, WO 04/033685 или WO 2011/044186. В соответствии с определенными вариантами осуществления TCR представляет собой фрагмент одноцепочечного TCR, содержащий вариабельный участок альфа-цепи, связанный с вариабельным участком бета-цепи, такой как scTv. В соответствии с некоторыми вариантами осуществления scTv также называется scFv.
Одноцепочечный Fv или scFv в некоторых аспектах относится к фрагментам антител или TCR, которые содержат вариабельный домен тяжелой цепи (VH) и вариабельный домен легкой цепи (VL) антитела или вариабельный домен альфа- или гамма-цепи (Vα или Vγ) и вариабельный домен бета- или дельта-цепи (Vβ или Vδ) TCR, при этом эти домены присутствуют в одной полипептидной цепи. Обычно полипептид Fv дополнительно содержит полипептидный линкер между VH и VL доменами, или Vα и Vβ доменами, или Vγ и Vδ доменами, который позволяет sFv образовывать желательную структуру для связывания антигена.
Диатело в некоторых аспектах относится к небольшим фрагментам антител и/или TCR с двумя сайтами связывания антигена, при этом фрагменты содержат VH, соединенный с VL в той же полипептидной цепи (VH-VL), или Vα, соединенный с Vβ в той же полипептидной цепи (Vα-Vβ), или Vγ, соединенный с Vδ в той же полипептидной цепи (Vγ-Vδ). Посредством применения линкера, который является слишком коротким, чтобы обеспечить возможность спаривания между двумя доменами на одной и той же цепи, домены заставляют спариваться с комплементарными доменами другой цепи и создавать два сайта связывания антигена. Иллюстративные диатела описаны более полно, например, в патентных документах ЕР 404097 и WO 93111161.
Биспецифическое антитело или биспецифический TCR в некоторых аспектах относится к антителу или TCR, которые проявляют специфичность в отношении двух различных типов антигенов. Термины, которые используют в данном документе, в частности, включают в себя, без ограничения, антитела и TCR, которые проявляют специфичность связывания в отношении антигена-мишени и в отношении другой мишени, что облегчает доставку в конкретную ткань. Подобным образом, мультиспецифические антитела и TCR характеризуются двумя или более специфичностями связывания.
Линейное антитело или «линейный TCR» в некоторых аспектах относится к паре тандемно расположенных Fd сегментов (например, VH-CH1-VH-CH1 или Vα-Cα1-Vα-Cα1), которые образуют пару антигенсвязывающих участков. Линейные антитела и TCR могут быть биспецифическими или моноспецифическими, например, как описано в Zapata et al., Protein Eng. 8(10): 1057-1062 (1995).
Антигенсвязывающий домен в некоторых аспектах относится к одному или нескольким фрагментам антитела или TCR, которые сохраняют способность к специфическому связыванию с антигеном. Неограничивающие примеры фрагментов антител, включенные в такие термины, включают в себя, без ограничения, (i) Fab фрагмент, моновалентный фрагмент, состоящий из VL, VH, CL и CH1 доменов; (ii) F(ab')2 фрагмент, бивалентный фрагмент, содержащий два Fab фрагмента, связанные дисульфидным мостиком в шарнирном участке; (iii) Fd фрагмент, состоящий из VH и CH1 доменов; (iv) Fv фрагмент, содержащий VL и VH домены одного плеча антитела, в том числе scFv, (v) dAb фрагмент (Ward et al., (1989) Nature 341:544 546), который содержит VH домен; и (vi) выделенный CDR. В это определение дополнительно включены антитела, содержащие одну тяжелую цепь и одну легкую цепь, или TCR с одной альфа-цепью или одной бета-цепью.
«F(ab')2» и «Fab'» фрагменты можно получить посредством обработки Ig протеазой, такой как пепсин и папаин, и включают в себя фрагменты антител, полученные посредством расщепления иммуноглобулина вблизи дисульфидных связей, существующих между шарнирным участком в каждой из двух тяжелых цепей. Например, папаин расщепляет IgG выше дисульфидных связей, существующих между шарнирными участками в каждой из двух тяжелых цепей, с получением двух гомологичных фрагментов антитела, в которых легкая цепь, состоящая из VL и CL, и фрагмент тяжелой цепи, состоящий из VH и CHγ1 (участок γ1 в константном участке тяжелой цепи), соединены в своих С-концевых участках посредством дисульфидной связи. Каждый из этих двух гомологичных фрагментов антител называют 'Fab'. Пепсин также расщепляет IgG ниже дисульфидных связей, существующих между шарнирными участками в каждой из двух тяжелых цепей, чтобы получить фрагмент антитела, немного больший, чем фрагмент, в котором два вышеупомянутых 'Fab' соединены в шарнирном участке. Этот фрагмент антитела называется F('ab')2. Fab фрагмент также содержит константный домен легкой цепи и первый константный домен (CH1) тяжелой цепи. 'Fab' фрагменты отличаются от Fab фрагментов добавлением нескольких остатков на карбоксильном конце CH1 домена тяжелой цепи, в том числе одного или нескольких цистеинов из шарнирного участка антитела. Fab'-SH представляет собой обозначение в данном документе для Fab', в котором цистеиновый остаток(цистеиновые остатки) в константных доменах несут свободную тиольную группу. F(ab')2 фрагменты антител первоначально получают в виде пар Fab' фрагментов, которые имеют между собой цистеины из шарнирного участка.
Fv в некоторых аспектах относится к антителу или фрагменту TCR, который содержит полный сайт распознавания антигена и сайт связывания антигена. Этот участок состоит из димера вариабельного домена одной тяжелой цепи и одной легкой цепи, или одной TCRα-цепи и одной TCRβ-цепи, или одной TCRγ-цепи и одной TCRδ-цепи в жесткой нековалентной ассоциации. Для этой конфигурации характерно, что три CDR в каждом вариабельном домене взаимодействуют, определяя сайт связывания антигена на поверхности димера VH-VL, или димера Vα-Vβ, или димера Vγ-Vδ. В совокупности, комбинация одного или нескольких из CDR из каждой из VH и VL цепей, или Vα-Vβ цепей, или Vγ-Vδ цепей придает антителу или TCR специфичность связывания антигена. Например, будет понятно, что CDRH3 и CDRL3, например, могут быть достаточными для придания антителу или TCR специфичности связывания антигена при переносе VH и VL цепей, или Vα и Vβ цепей, или Vγ-Vδ цепей реципиентного выбранного антитела, TCR или их антигенсвязывающего фрагмента, и эту комбинацию CDR можно исследовать в отношении связывания, аффинности и т.д. Даже отдельный вариабельный домен (или половина Fv, содержащая только три CDR, специфических в отношении антигена) характеризуется способностью к распознаванию и связыванию антигена, хотя и, возможно, с более низкой аффинностью, чем при объединении со вторым вариабельным доменом. Более того, несмотря на то что два домена в Fv фрагменте (VL и VH, или Vα и Vβ, или Vγ и Vδ) кодируются отдельными генами, их можно соединить с применением способов на основе рекомбинации посредством синтетического линкера, который обеспечивает возможность их получения в виде единой белковой цепи, в которой VL и VH, или Vα и Vβ, или Vγ и Vδ участки цепи спариваются с образованием моновалентных молекул (известных как одноцепочечный Fv (scFv); Bird et al. (1988) Science 242:423-426; Huston et al. (1988) Proc. Nail. Acad. Sci. USA 85:5879-5883; и Osbourn et al. (1998) Nat. Biotechnol. 16:778). Предполагается, что такие scFv также охватываются термином «антигенсвязывающая часть» антитела. Последовательности любых VH и VL в конкретном scFv можно связать с кДНК или геномными последовательностями Fc участка с целью создания экспрессионных векторов, кодирующих полные молекулы Ig (например, IgG) или другие изотипы. VH и VL также можно применять в создании Fab, Fv или других фрагментов Ig с применением либо технологии химии белков, либо технологии на основе рекомбинантной ДНК.
Антигенсвязывающие полипептиды также включают в себя димеры тяжелых цепей, такие как, например, антитела верблюдовых и акул. Антитела верблюдовых и акул содержат гомодимерную пару из двух цепей V-подобных и C-подобных доменов (ни та, ни другая не имеет легкой цепи). Поскольку VH участок у IgG из димера тяжелых цепей у верблюдовых не должен осуществлять гидрофобные взаимодействия с легкой цепью, участок в тяжелой цепи, который в нормальных условиях контактирует с легкой цепью, изменен у верблюдовых на гидрофильные аминокислотные остатки. VH домены у IgG из димера тяжелых цепей называют VHH доменами. Ig-NAR у акул содержат гомодимер одного вариабельного домена (называемый V-NAR доменом) и пять С-образных константных доменов (C-NAR домены). У верблюдовых разнообразие репертуара антител определяется CDR 1, 2 и 3 в VH или VHH участках. CDR3 в верблюжьем VHH участке характеризуется сравнительно большой длиной, составляющей в среднем 16 аминокислот (Muyldermans et al., 1994, Protein Engineering 7(9): 1129).
«Гуманизированное» антитело представляет собой антитело, в котором все или практически все аминокислотные остатки в CDR получены из CDR, отличных от человеческих, и все или практически все аминокислотные остатки в FR получены из человеческих FR. Гуманизированное антитело необязательно может включать в себя по меньшей мере часть константного участка антитела, полученного из человеческого антитела. «Гуманизированная форма» антитела, отличного от человеческого, относится к варианту антитела, отличного от человеческого, которое подверглось гуманизации, как правило, с целью снижения иммуногенности у людей, при этом сохраняя специфичность и аффинность исходного антитела, отличного от человеческого. В соответствии с некоторыми вариантами осуществления некоторые остатки в FR у гуманизированного антитела заменены на соответствующие остатки из антитела, отличного от человеческого (например, антитела, из которого получают остатки CDR), например, для восстановления или улучшения специфичности или аффинности антитела.
В числе обеспеченных антител находятся человеческие антитела. «Человеческое антитело» представляет собой антитело с аминокислотной последовательностью, соответствующей таковой у антитела, продуцируемого человеком или человеческой клеткой, или из источника, отличного от человека, который использует человеческие репертуары антител или другие последовательности, кодирующие человеческие антитела, в том числе библиотеки человеческих антител. Термин исключает гуманизированные формы антител, отличных от человеческих, которые содержат антигенсвязывающие участки, отличные от человеческих, такие как антигенсвязывающие участки, в которых все или практически все CDR являются отличными от человеческих.
Человеческие антитела можно получить посредством введения иммуногена трансгенному животному, которое было модифицировано с целью продуцирования интактных человеческих антител или интактных антител с человеческими вариабельными участками в ответ на антигенный стимул. Такие животные, как правило, содержат все локусы человеческого иммуноглобулина или их часть, которые заменяют эндогенные локусы иммуноглобулина, или которые присутствуют внехромосомно или произвольно интегрированы в хромосомы животного. У таких трансгенных животных эндогенные локусы иммуноглобулина обычно были инактивированы. Человеческие антитела также можно получить из библиотек человеческих антител, в том числе библиотек для фагового дисплея и бесклеточных библиотек, содержащих кодирующие антитело последовательности, полученные из репертуара человеческих антител.
В число обеспеченных антител входят моноклональные антитела, в том числе фрагменты моноклональных антител. Термин «моноклональное антитело» в контексте данного документа относится к антителу, полученному из или находящемуся в составе популяции практически однородных антител, т.е. отдельные антитела, содержащиеся в популяции, являются идентичными за исключением возможных вариантов, содержащих встречающиеся в естественных условиях мутации, или возникающих во время получения препарата моноклональных антител, такие варианты обычно присутствуют в незначительных количествах. В отличие от препаратов поликлональных антител, которые, как правило, включают в себя различные антитела, направленные на различные эпитопы, каждое моноклональное антитело в препарате моноклональных антител направлено на один эпитоп на антигене. Термин не следует толковать как требующий получения антитела с помощью какого-либо конкретного способа. Моноклональное антитело может быть получено с помощью ряда методик, в том числе, без ограничения, получение из гибридомы, способы на основе рекомбинантной ДНК, фаговый дисплей и другие способы дисплея антител.
Термины «полипептид» и «белок» используются взаимозаменяемо, они относятся к полимеру из аминокислотных остатков и не ограничиваются минимальной длиной. Полипептиды, в том числе представленные антитела и цепи антител, а также другие пептиды, например, линкеры и связывающие пептиды, могут включать в себя аминокислотные остатки, в том числе природные аминокислотные остатки и/или аминокислотные остатки, отличные от природных. Термины также включают в себя модификации полипептида после экспрессии, например, гликозилирование, сиалилирование, ацетилирование, фосфор илирование и т.п. В соответствии с некоторыми аспектами полипептиды могут содержать модификации относительно нативной или природной последовательности при условии, что белок сохраняет желаемую активность. Эти модификации могут быть преднамеренными, как в случае с айт-направленного мутагенеза, или они могут быть случайными, как например, вследствие мутаций у хозяев, которые продуцируют белки, или ошибок вследствие ПЦР амплификации.
«Последовательность зародышевого типа» относится к генной последовательности из зародышевой линии (гаплоидные гаметы и те диплоидные клетки, из которых они образовываются). ДНК зародышевого типа содержит многочисленные генные сегменты, которые кодируют отдельную тяжелую или легкую цепь Ig, или отдельную цепь TCRα или TCRβ, или отдельную цепь TCRγ или TCRδ. Эти генные сегменты хранятся в зародышевых клетках, но не могут быть транскрибированы и транслированы до тех пор, пока они не подвергнуться перестройке в функциональные гены. Во время дифференцировки В-клеток и Т-клеток в костном мозге эти генные сегменты перетасовываются в произвольном порядке за счет динамической генетической системы, способной создавать более чем 108 специфичностей. Большинство из этих генных сегментов опубликованы и собраны в базу данных зародышевых последовательностей.
Аффинность относится к константе равновесия для обратимого связывания двух средств и выражена в виде KD. Аффинность связывающего белка в отношении лиганда, как например, аффинность антитела в отношении эпитопа или, например, аффинность TCR в отношении комплекса МСН-пептид, может составлять, например, от приблизительно 100 наномоль (нМ) до приблизительно 0,1 нМ, от приблизительно 100 нМ до приблизительно 1 пикомоль (пМ) или от приблизительно 100 нМ до приблизительно 1 фемтомоль (фМ). Термин «авидность» относится к устойчивости комплекса из двух или более средств к диссоциации после разведения.
Эпитоп в соответствии с некоторыми аспектами относится к части антигена или другой макромолекулы, способной к осуществлению связывающего взаимодействия со связывающим карманом в вариабельном участке антитела или TCR. Такие связывающие взаимодействия могут проявляться в виде межмолекулярного контакта с одним или несколькими аминокислотными остатками в одном или нескольких CDR. В связывании антигена могут быть задействованы, например, CDR3, пара CDR3 или в некоторых случаях взаимодействия вплоть до всех шести CDR VH и VL цепей. Эпитоп может представлять собой линейную пептидную последовательность (т.е. «непрерывный») или может состоять из несмежных аминокислотных последовательностей (т.е. «конформационный» или «прерывающийся»). Антитело или TCR могут распознавать одну или несколько аминокислотных последовательностей; таким образом, эпитоп может определять более чем одну отдельную аминокислотную последовательность. В соответствии с некоторыми аспектами TCR может распознавать одну или несколько аминокислотных последовательностей или эпитопов в контексте МНС. Эпитопы, распознаваемые антителами и TCR, можно определить с помощью методик составления пептидных карт и анализа последовательности, хорошо известных специалисту в данной области техники. Связывающие взаимодействия проявляются в виде межмолекулярных контактов с одним или несколькими аминокислотными остатками в CDR.
В соответствии с некоторыми вариантами осуществления упоминание антитела или TCR со специфическим связыванием относится к ситуации, в которой антитело или TCR не будет проявлять какое-либо существенное связывание с молекулами, отличными от антигена, содержащего эпитоп, распознаваемый антителом или TCR. Термин также является применимым в случаях, когда, например, антигенсвязывающий домен является специфическим в отношении конкретного эпитопа, который несут ряд антигенов, в этом случае выбранное антитело, TCR или их антигенсвязывающий фрагмент, несущий антигенсвязывающий домен, будет способен к связыванию с различными антигенами, несущими эпитоп. Термины «преимущественно связывается» или «специфически связывается» означают, что антитела, TCR или их фрагменты связываются с эпитопом с большей аффинностью, чем они связываются с неродственными аминокислотными последовательностями, и, если они являются перекрестно реактивными с другими полипептидами, содержащими эпитоп, они не являются токсичными при уровнях, в которых их составляют для введения с целью применения у человека. В соответствии с одним аспектом такая аффинность является по меньшей мере 1-кратно большей, по меньшей мере 2-кратно большей, по меньшей мере 3-кратно большей, по меньшей мере 4-кратно большей, по меньшей мере 5-кратно большей, по меньшей мере 6-кратно большей, по меньшей мере 7-кратно большей, по меньшей мере 8-кратно большей, по меньшей мере 9-кратно большей, 10-кратно большей, по меньшей мере 20-кратно большей, по меньшей мере 30-кратно большей, по меньшей мере 40-кратно большей, по меньшей мере 50-кратно большей, по меньшей мере 60-кратно большей, по меньшей мере 70-кратно большей, по меньшей мере 80-кратно большей, по меньшей мере 90-кратно большей, по меньшей мере 100-кратно большей или по меньшей мере 1000-кратно большей, чем аффинность у антитела, TCR или их фрагмента в отношении неродственных аминокислотных последовательностей. Термин «связывание» относится к непосредственной ассоциации между двумя молекулами, обусловленной, например, ковалентными, электростатическими, гидрофобными и ионными взаимодействиями и/или взаимодействиями при образовании водородных связей в физиологических условиях, и оно включает в себя такие взаимодействия, как солевые мостики и водные мостики, а также любые другие общепринятые значения связывания.
Термин «связывание» относится к непосредственной ассоциации между двумя молекулами, обусловленной, например, ковалентными, электростатическими, гидрофобными и ионными взаимодействиями и/или взаимодействиями при образовании водородных связей в физиологических условиях, и оно включает в себя такие взаимодействия, как солевые мостики и водные мостики, а также любые другие общепринятые значения связывания.
В соответствии с некоторыми аспектами термин «тетрамер» может относиться к комплексу, содержащему четыре субъединицы, связанные с одной молекулой стрептавидина, который может связываться с популяцией клеток и, следовательно, идентифицировать ее. Субъединица может представлять собой комплекс МНС-пептид. Субъединица может представлять собой МНС без ассоциированного пептида. Субъединица может представлять собой антиген В-клеточного рецептора. Популяция клеток, идентифицируемых с помощью тетрамера, может представлять собой популяцию, которая экспрессирует рецептор, такой как TCR или BCR, который связывается с субъединицей тетрамера. Популяция клеток может представлять собой антигенспецифические Т-клетки. Популяция клеток может представлять собой антигенспецифические В-клетки. Тетрамер может нести флуоресцентную метку. В контексте данного документа тетрамер МНС-пептид можно использовать взаимозаменяемо с рМНС.
«Фармацевтически приемлемый» относится к молекулярным частицам и композициям, которые являются физиологически переносимыми и, как правило, не дают аллергическую или подобную нежелательную реакцию, такую как расстройство желудка, головокружение и т.п., при введении человеку.
«Предупреждение» относится к профилактике, предупреждению появления симптомов, предупреждению развития заболевания или нарушения, ассоциированного с избыточными уровнями белка или коррелирующего с активностью белка.
«Ингибирование», «лечение» и «осуществление лечения» используются взаимозаменяемо и относятся, например, к остановке развития симптомов, продлению жизни, частичному или полному облегчению симптомов и частичной или полной ликвидации состояния, заболевания или нарушения, ассоциированного с избыточными уровнями белка или коррелирующего с активностью белка. Например, лечение рака включает в себя, без ограничения, остановку развития, частичное или полное устранение роста злокачественных клеток или опухоли. Лечение или частичное устранение включает в себя, например, кратное снижение роста опухоли или ее размера и/или объема, как например, приблизительно 2-кратное, приблизительно 3-кратное, приблизительно 4-кратное, приблизительно 5-кратное, приблизительно 10-кратное, приблизительно 20-кратное, приблизительно 50-кратное или любое кратное снижение в диапазоне между приведенными значениями. Подобным образом, лечение или частичное устранение может включать в себя процентное снижение роста опухоли или ее размера и/или объема, составляющее приблизительно 1%, 2%, 3%, 4%, 5%, 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 95%, или любое процентное снижение в диапазоне между приведенными значениями.
Нейтрализующее антитело или нейтрализующий TCR в соответствии с некоторыми аспектами относится к любому антителу или TCR, которые ингибируют размножение патогена, такого как вирус или бактерия, вне зависимости от механизма, посредством которого достигается нейтрализация.
Репертуар антител или репертуар TCR относится к набору антител, TCR или их фрагментов. Репертуар антител можно применять, например, для выбора конкретного антитела или скрининга в отношении конкретного свойства, такого как способность к связыванию, специфичность связывания, способность транспорту через желудочно-кишечный тракт, стабильность, аффинность и т.п. Термин, в частности, включает в себя библиотеки антител и TCR, в том числе все формы комбинаторных библиотек, такие как, например, библиотеки для фагового дисплея антител, в том числе, без ограничения, библиотеки для фагового дисплея одноцепочечных Fv (scFv) и Fab антител из любого источника, в том числе библиотеки не подвергнутых какому-либо воздействию молекул, синтетические и полусинтетические библиотеки.
«Целевая молекула нуклеиновой кислоты», «целевой полинуклеотид», «целевая молекула полинуклеотида» относятся к любой нуклеиновой кислоте, представляющей интерес.
Полимеразная цепная реакция (ПЦР) относится к реакции амплификации последовательностей полинуклеотидов in vitro посредством одновременной достройки праймеров комплементарных цепей двухцепочечного полинуклеотида. Реакции ПЦР продуцируют копии матричного полинуклеотида, фланкированные сайтами для связывания праймеров. При использовании двух праймеров результатом является экспоненциальное увеличение количества копий матричного полинуклеотида обеих цепей с каждым циклом, поскольку с каждым циклом реплицируются обе цепи. Дуплекс полинуклеотида имеет концы, соответствующие концам применяемых праймеров. ПЦР может включать один или несколько повторов денатурирования матричного полинуклеотида, гибридизации праймеров с сайтами связывания для праймеров и достройки праймеров ДНК- или РНК-полимеразой в присутствии нуклеотидов. Конкретные температуры, продолжительности на каждой стадии и скорости смены стадий зависят от многих факторов, хорошо известных квалифицированному специалисту в данной области техники. (McPherson et al., IRL Press, Oxford (1991 and 1995)). Например, в традиционной ПЦР с применением ДНК-полимеразы Taq двухцепочечный матричный полинуклеотид можно денатурировать при температуре >90°С, праймеры могут гибридизироваться при температуре в диапазоне 50-75°С, и праймеры могут достраиваться при температуре в диапазоне 72-78°С. В соответствии с некоторыми вариантами осуществления ПЦР включает ПЦР с обратной транскрипцией (RT-PCR), ПЦР в реальном времени, гнездовую ПЦР, количественную ПЦР, мультиплексную ПЦР или подобное. В соответствии с некоторыми вариантами осуществления ПЦР не предусматривает RT-PCR. (патенты США №№5168038, 5210015, 6174670, 6569627 и 5925517; Mackay et al., Nucleic Acids Research, 30: 1292-1305 (2002)). RT-PCR включает, что реакции ПЦР предшествует реакция обратной транскрипции, и амплификации подвергается полученная в результате кДНК, гнездовая ПЦР включает двухстадийную ПЦР, при этом ампликон первой реакции ПЦР с применением первого набора праймеров становится образцом для второй реакции ПЦР с применением второго набора праймеров, по меньшей мере один из которых связывается с имеющим внутреннее расположение участком в ампликоне первой реакции ПЦР. Мультиплексная ПЦР включает реакцию ПЦР, при которой множество последовательностей полинуклеотидов подвергаются ПЦР в одной и той же реакционной смеси одновременно. Объемы реакционной смеси для ПЦР могут составлять от 0,2 пл до 1000 мкл. Количественная ПЦР включает реакцию ПЦР, предназначенную для измерения абсолютного или относительного количества, распространенности или концентрации одной или нескольких последовательностей в образце. Количественные измерения могут включать в себя сравнение одной или нескольких эталонных последовательностей или стандартов с последовательностью полинуклеотида, представляющей интерес.(Freeman et al., Biotechniques, 26: 112-126 (1999); Becker-Andre et al., Nucleic Acids Research, 17: 9437-9447 (1989); Zimmerman et al., Biotechniques, 21: 268-279 (1996); Diviacco et al., Gene, 122: 3013-3020 (1992); Becker-Andre et al., Nucleic Acids Research, 17: 9437-9446 (1989)).
В контексте данного документа «нуклеотид», «нуклеозид», «нуклеотидный остаток» и «нуклеозидный остаток» могут означать дезоксирибонуклеотидный или рибонуклеотидный остаток или другой подобный нуклеозидный аналог, способный служить в качестве компонента праймера, подходящего для применения в реакции амплификации (например, реакции ПЦР). Такие нуклеозиды и их производные можно применять в качестве строительных блоков для праймеров, описанных в данном документе, за исключением тех случаев, когда указано иное. Ничто в данной заявке не говорит о невозможности использования производных нуклеозидов или оснований, которые были химически модифицированы с целью повышения их стабильности или применимости в реакции амплификации, при условии, что химическая модификация не препятствует их распознаванию полимеразой как дезоксигуанина, дезоксицитозина, дезокситимидина или дезоксиаденина в соответствующих случаях. В соответствии с некоторыми вариантами осуществления нуклеотидные аналоги могут стабилизировать образование гибрида. В соответствии с некоторыми вариантами осуществления нуклеотидные аналоги могут дестабилизировать образование гибрида. В соответствии с некоторыми вариантами осуществления нуклеотидные аналоги могут повышать специфичность гибридизации. В соответствии с некоторыми вариантами осуществления нуклеотидные аналоги могут снижать специфичность гибридизации.
«Нуклеиновая кислота» или грамматические эквиваленты данного термина относятся либо к отдельному нуклеотиду, либо по меньшей мере к двум нуклеотидам, ко валентно связанным вместе.
«Полинуклеотид» или грамматические эквиваленты данного термина относятся по меньшей мере к двум нуклеотидам, ковалентно связанным вместе. Полинуклеотид содержит молекулу, содержащую два или более нуклеотидов. Полинуклеотид содержит полимерную форму нуклеотидов любой длины либо из рибонуклеотидов, либо из дезоксирибонуклеотидов, либо из пептидонуклеиновых кислот (PNA), которые содержат пуриновые и пиримидиновые основания или другие натуральные, химически или биохимически модифицированные, ненатуральные нуклеотидные основания или производные нуклеотидных оснований. Остов полинуклеотида может содержать сахара и фосфатные группы или модифицированные или замещенные сахарные или фосфатные группы. Полинуклеотид может содержать модифицированные нуклеотиды, такие как метилированные нуклеотиды и нуклеотидные аналоги. Последовательность нуклеотидов может прерываться ненуклеотидными компонентами. Полинуклеотид может включать в себя другие молекулы, такие как другой гибридизированный Полинуклеотид. Полинуклеотиды включают в себя последовательности дезоксирибонуклеиновой кислоты (ДНК), рибонуклеиновой кислоты (РНК) или обеих из них. Неограничивающие примеры полинуклеотидов включают в себя ген, фрагмент гена, экзон, интрон, межгенную ДНК (в том числе, без ограничения, гетерохроматиновую ДНК), матричную РНК (мРНК), транспортную РНК, рибосомальную РНК, рибозимы, малые интерферирующие РНК (siRNA), кДНК, рекомбинантные полинуклеотиды, разветвленные полинуклеотиды, плазмиды, векторы, выделенную ДНК из последовательности, выделенную РНК из последовательности, зонды на основе нуклеиновой кислоты и праймеры. Полинуклеотиды могут быть выделенными из природных источников, рекомбинантными или искусственно синтезированными.
Полинуклеотиды могут включать в себя нестандартные нуклеотиды, такие как нуклеотидные аналоги или модифицированные нуклеотиды. В соответствии с некоторыми вариантами осуществления нестандартные нуклеотиды могут стабилизировать образование гибрида. В соответствии с некоторыми вариантами осуществления нестандартные нуклеотиды могут дестабилизировать образование гибрида. В соответствии с некоторыми вариантами осуществления нестандартные нуклеотиды могут повышать специфичность гибридизации. В соответствии с некоторыми вариантами осуществления нестандартные нуклеотиды могут снижать специфичность гибридизации. Примеры нестандартных модификаций нуклеотидов включают в себя 2'-O-Ме, 2'-O-аллил, 2'-O-пропаргил, 2'-O-алкил, 2'-фтор, 2'-арабино, 2'-ксило, 2'-фторарабино, фосфоротиоат, фосфородитиоат, фосфороамидаты, 2'-амино, 5-алкил-замещенный пиримидин, 3'-дезоксигуанозин, 5-галоген-замещенный пиримидин, алкил-замещенный пурин, галоген-замещенный пурин, бициклические нуклеотиды, 2'МОЕ, молекулы PNA, молекулы LNA, LNA-подобные молекулы, диаминопурин, S2T, 5-фторурацил, 5-бромурацил, 5-хлорурацил, 5-йодурацил, гипоксантин, ксантин, 4-ацетилцитозин, 5-(карбоксигидроксилметил)урацил, 5-карбоксиметиламинометил-2-тиоуридин, 5-карбоксиметиламинометилурацил, дигидроурацил, бета-D-галактозилквеуозин, инозин, N6-изопентениладенин, 1-метилгуанин, 1-метилинозин, 2,2-диметилгуанин, 2-метиладенин, 2-метилгуанин, 3-метилцитозин, 5-метилцитозин, N6-аденин, 7-метилгуанин, 5-метиламинометилурацил, 5-метоксиаминометил-2-тиоурацил, бета-D-маннозилквеуозин, 5'-метоксикарбоксиметилурацил, 5-метоксиурацил, 2-метилтио-D46-изопентениладенин, урацил-5-оксиуксусную кислоту (v), вибутоксозин, псевдоурацил, квеуозин, 2-тиоцитозин, 5-метил-2-тиоурацил, 2-тиоурацил, 4-тиоурацил, 5-метилурацил, метиловый эфир урацил-5-оксиуксусной кислоты, урацил-5-оксиуксусную кислоту (v), 5-метил-2-тиоурацил, 3-(3-амино-3-N-2-карбоксипропил)урацил, (acp3)w, 2,6-диаминопурин и их производные.
«Субъект», «индивид», «хозяин» или «пациент» относится к живым организмам, таким как млекопитающие. Примеры субъектов и хозяев включают в себя, без ограничения, лошадей, коров, верблюдов, овец, свиней, коз, собак, кошек, кроликов, морских свинок, крыс, мышей (например, гуманизированных мышей), песчанок, отличных от человека приматов (например, макаков), людей и т.п., животных, отличных от млекопитающих, в том числе, например, позвоночных, отличных от млекопитающих, таких как птицы (например, курицы или утки), рыбы (например, акулы) или лягушки (например, Xenopus), и беспозвоночных, отличных от млекопитающих, а также их трансгенные разновидности. В соответствии с определенными аспектами субъект относится к отдельному организму (например, человеку). В соответствии с определенными аспектами предполагается группа индивидов, состоящая из небольшой когорты, имеющей или общий иммунный фактор для изучения, и/или заболевание, и/или когорты индивидов без заболевания (например, отрицательный/нормальный контроль). Субъект, от которого получают образцы, может быть поражен любым заболеванием и/или нарушением (например, одной или несколькими аллергиями, инфекциями, раком или аутоиммунными нарушениями или т.п.), и его можно сравнить с субъектом отрицательного контроля, который не поражен заболеванием.
«Набор» относится к системе доставки для доставки материалов или реактивов для осуществления способа, раскрытого в данном документе. В соответствии с некоторыми вариантами осуществления наборы включают в себя системы, которые обеспечивают возможность хранения, транспортировки или доставки реактивов для реакции (например, зондов, ферментов и т.д. в соответствующих контейнерах) и/или вспомогательных материалов (например, буферов, письменных инструкций по осуществлению анализа и т.д.) из одного места в другое. Например, наборы включают в себя одну или несколько емкостей (например, коробок), содержащих соответствующие реактивы для реакции и/или вспомогательные материалы. Такое содержимое может доставляться целевому получателю вместе или раздельно. Например, первый контейнер может содержать фермент для применения в анализе, в то время как второй контейнер содержит множество праймеров.
Полипептид в соответствии с некоторыми аспектами относится к молекуле, содержащей по меньшей мере две аминокислоты. В соответствии с некоторыми вариантами осуществления полипептид состоит из одного пептида. В соответствии с некоторыми вариантами осуществления полипептид содержит два или более пептидов. Например, полипептид может содержать по меньшей мере приблизительно 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 30, 40, 50, 60, 70, 80, 90, 100, 200, 300, 400, 500, 600, 700, 800, 900 или 1000 пептидов или аминокислот. Примеры полипептидов включают в себя, без ограничения, цепи из аминокислот, белки, пептиды, гормоны, полипептиды с сахаридами, липидами, гликолипидами, фосфолипидами, антитела, ферменты, киназы, рецепторы, транскрипционные факторы и лиганды.
Образец в соответствии с некоторыми аспектами относится к биологическому, экологическому, медицинскому образцу, образцу от субъекта или пациента или образцу, содержащему полинуклеотид, такой как целевой полинуклеотид.
Конъюгаты аффинная молекула-олигонуклеотид
Конъюгат аффинная молекула-олигонуклеотид содержит часть в виде аффинной молекулы (например, антитело или комплекс МНС-пептид) и олигонуклеотидную часть. Идентификационную последовательность антигена в олигонуклеотиде в составе конъюгата аффинная молекула-олигонуклеотид можно применять для идентификации одного или нескольких антигенов, с которыми специфически взаимодействует конъюгат аффинная молекула-олигонуклеотид. В соответствии с некоторыми вариантами осуществления олигонуклеотид ковалентно прикреплен к аффинной части конъюгата. В соответствии с некоторыми вариантами осуществления олигонуклеотид нековалентно прикреплен к аффинной части конъюгата. Конъюгат аффинная молекула-олигонуклеотид может представлять собой конъюгат тетрамер-олигонуклеотид. Конъюгат тетрамер-олигонуклеотид может содержать тетрамер (например, тетрамер антигена B-клеточного рецептора или тетрамер комплекса МНС-пептид) и олигонуклеотидную часть. Идентификационную последовательность антигена в олигонуклеотиде в составе конъюгата тетрамер-олигонуклеотид можно применять для идентификации одного или нескольких антигенов, с которым специфически взаимодействует конъюгат тетрамер-олигонуклеотид. В соответствии с некоторыми вариантами осуществления олигонуклеотид ковалентно прикреплен к аффинной части или тетрамеру в составе конъюгата. В соответствии с некоторыми вариантами осуществления олигонуклеотид нековалентно прикреплен к аффинной части или тетрамеру в составе конъюгата.
В соответствии с некоторыми вариантами осуществления конъюгат аффинная молекула-олигонуклеотид содержит одну аффинную часть. В соответствии с некоторыми вариантами осуществления конъюгаты аффинная молекула-олигонуклеотид представляют собой мультивалентные конъюгаты аффинная молекула-олигонуклеотид. Например, мультивалентные конъюгаты аффинная молекула-олигонуклеотид могут содержать антигенсвязывающие домены по меньшей мере двух аффинных молекул, конъюгированные с одним или несколькими олигонуклеотидами. Например, мультивалентные конъюгаты аффинная молекула-олигонуклеотид могут содержать антигенсвязывающие домены по меньшей мере приблизительно 2, 3, 4, 5, 6, 7, 8, 9, 10, 20, 50, 100, 200, 500 или 1000 аффинных молекул, конъюгированные с одним или несколькими олигонуклеотидами. В соответствии с некоторыми вариантами осуществления конъюгат аффинная молекула-олигонуклеотид содержит один олигонуклеотид. В соответствии с некоторыми вариантами осуществления конъюгат аффинная молекула-олигонуклеотид содержит 2 или более олигонуклеотидов. Например, конъюгат аффинная молекула-олигонуклеотид может содержать по меньшей мере приблизительно 2, 3, 4, 5, 6, 7, 8, 9, 10, 20, 50, 100, 200, 500 или 1000 олигонуклеотидов, конъюгированных с одной или несколькими аффинными молекулами (например, антителом или комплексом МНС-пептид). В соответствии с некоторыми вариантами осуществления конъюгат аффинная молекула-олигонуклеотид содержит 2 или более олигонуклеотидов, содержащих одинаковую идентификационную последовательность антигена (AID). Например, конъюгат аффинная молекула-олигонуклеотид может содержать по меньшей мере приблизительно 2, 3, 4, 5, 6, 7, 8, 9, 10, 20, 50, 100, 200, 500 или 1000 олигонуклеотидов, содержащих одинаковую последовательность AID.
Аффинная часть конъюгатов аффинная молекула-олигонуклеотид
Аффинная часть (или домен) конъюгата аффинная молекула-олигонуклеотид содержит участок, молекулу, домен, часть, фрагмент или долю конъюгата аффинная молекула-олигонуклеотид, которые связываются с целевым антигеном. Таким образом, аффинная часть придает способность к связыванию или специфическому связыванию с заданным целевым антигеном, таким как внеклеточный домен белка клеточной поверхности. В соответствии с некоторыми вариантами осуществления аффинная часть не взаимодействует значительным образом с антигеном в составе другого конъюгата аффинная молекула-олигонуклеотид, содержащего отличающуюся идентификационную последовательность антигена. В соответствии с некоторыми вариантами осуществления аффинная часть представляет собой молекулу, которая может содержать нуклеиновую кислоту, или к которой может быть прикреплен олигонуклеотид без значительного ухудшения связывания аффинной части с целевым антигеном.
Аффинная часть конъюгата аффинная молекула-олигонуклеотид может представлять собой молекулу нуклеиновой кислоты, или она может являться белковой. Аффинные части включают в себя, без ограничения, РНК, ДНК, гибриды РНК-ДНК, малые молекулы (например, лекарственные средства), аптамеры, полипептиды, белки, антитела и их фрагменты, TCR и их фрагменты, вирусы, вирусные частицы, клетки, их фрагменты и их комбинации. (См., например, Fredriksson et al., (2002) Nat Biotech 20:473-77; Gullberg et al., (2004) PNAS, 101:8420-24). Например, аффинная часть может представлять собой одноцепочечную РНК, двухцепочечную РНК, одноцепочечную ДНК, двухцепочечную ДНК, ДНК или РНК, содержащую один или несколько двухцепочечных участков и один или несколько одноцепочечных участков, гибрид РНК-ДНК, малую молекулу, аптамер, полипептид, белок, антитело, фрагмент антитела, TCR, фрагмент TCR, MHC, комплекс МНС-пептид, вирусную частицу, клетку или любую их комбинацию.
В соответствии с некоторыми вариантами осуществления аффинная часть конъюгата аффинная молекула-олигонуклеотид нацелена на клетку. Например, аффинная часть конъюгата аффинная молекула-олигонуклеотид может быть нацелена на Т-клетку или В-клетку. В соответствии с некоторыми вариантами осуществления аффинная часть конъюгата аффинная молекула-олигонуклеотид нацелена на конкретный тип клеток или субпопуляцию клеток. Например, аффинная часть конъюгата аффинная молекула-олигонуклеотид может быть нацелена на CD4+ Т-клетку или CD8+ Т-клетку. Например, аффинная часть конъюгата аффинная молекула-олигонуклеотид может быть нацелена на Т-клетку, содержащую TCR, который специфически распознает конкретный антиген. Например, аффинная часть конъюгата аффинная молекула-олигонуклеотид может быть нацелена на Т-клетку, содержащую TCR, который специфически распознает конкретный комплекс МНС-пептид.
В соответствии с некоторыми вариантами осуществления аффинная часть конъюгата аффинная молекула-олигонуклеотид нацелена на внеклеточный домен мишени на клетке. Например, аффинная часть конъюгата аффинная молекула-олигонуклеотид может быть нацелена на внеклеточный внеклеточный домен рецептора клетки, например, Т-клеточного рецептора (TCR). Например, аффинная часть конъюгата аффинная молекула-олигонуклеотид может быть нацелена на гликозилированный участок внеклеточного домена рецептора на клетке. Например, аффинная часть конъюгата аффинная молекула-олигонуклеотид может быть нацелена на лиганд-связывающий участок внеклеточного домена рецептора на клетке. Например, аффинная часть конъюгата аффинная молекула-олигонуклеотид может быть нацелена на участок внеклеточного домена рецептора на клетке, которая не связывается с лигандом.
В соответствии с некоторыми вариантами осуществления аффинная часть конъюгата тетрамер-олигонуклеотид содержит комплекс МНС-пептид. В соответствии с некоторыми вариантами осуществления комплекс МНС-пептид нацелен на TCR. В соответствии с некоторыми вариантами осуществления аффинная часть конъюгата тетрамер-олигонуклеотид содержит антиген В-клеточного рецептора. В соответствии с некоторыми вариантами осуществления антиген В-клеточного рецептора нацелен на BCR.
Белки
В соответствии с некоторыми вариантами осуществления аффинная часть представляет собой полипептид, белок или любой его фрагмент. В соответствии с некоторыми вариантами осуществления аффинная часть конъюгата аффинная молекула-олигонуклеотид представляет собой белок. В соответствии с некоторыми вариантами осуществления аффинная часть конъюгата аффинная молекула-олигонуклеотид представляет собой пептид. Например, аффинная часть конъюгата аффинная молекула-олигонуклеотид может представлять собой антитело, такое как связывающий домен антитела. Например, аффинная часть конъюгата аффинная молекула-олигонуклеотид может представлять собой комплекс МНС-пептид. Например, аффинная часть может представлять собой очищенный полипептид, выделенный полипептид, меченый посредством слияния последовательности полипептид, полипептид, прикрепленный к мембране клетки, или вирусу, или вириону или охватывающий их, цитоплазматический белок, внутриклеточный белок, внеклеточный белок, киназу, фосфатазу, ароматазу, хеликазу, протеазу, оксидоредуктазу, редуктазу, трансферазу, гидролазу, лиазу, изомеразу, гликозилазу, белок внеклеточного матрикса, лигазу, переносчик ионов, канал, пору, апоптотический белок, белок клеточной адгезии, патогенный белок, аберрантно экспрессируемый белок; транскрипционный фактор, транскрипционный регулятор, белок, участвующий в трансляции, шаперон, секретируемый белок, лиганд, гормон, цитокин, хемокин, ядерный белок, рецептор, трансмембранный рецептор, белок, участвующий в передаче сигнала, антитело, мембранный белок, интегральный мембранный белок, периферический мембранный белок, белок клеточной стенки, глобулярный белок, волокнистый белок, гликопротеин, липопротеин, хромосомный белок, любой их фрагмент или любую их комбинацию. В соответствии с некоторыми вариантами осуществления аффинная часть представляет собой гетерологичный полипептид. В соответствии с некоторыми вариантами осуществления аффинная часть представляет собой белок, сверхэкспрессированный в клетке с применением методик молекулярной биологии, таких как трансфекция. В соответствии с некоторыми вариантами осуществления аффинная часть представляет собой рекомбинантный полипептид. Например, аффинная часть может содержать образцы, продуцируемые в бактериальных (например, Е. coli), дрожжевых клетках, клетках млекопитающих или насекомых (например, белки, сверхэкспрессируемые организмами). В соответствии с некоторыми вариантами осуществления аффинная часть представляет собой полипептид, содержащий мутацию, вставку, делецию или полиморфизм. В соответствии с некоторыми вариантами осуществления аффинная часть представляет собой антиген, такой как полипептид, применяемый для иммунизации организма или для генерирования иммунной реакции у организма, как например, для выработки антител.
В соответствии с некоторыми вариантами осуществления аффинная часть конъюгата тетрамер-олигонуклеотид представляет собой тетрамер. В соответствии с некоторыми вариантами осуществления субъединица тетрамера представляет собой белок. В соответствии с некоторыми вариантами осуществления субъединица тетрамера представляет собой пептид. В соответствии с некоторыми вариантами осуществления субъединица тетрамера представляет собой комплекс белков и/или пептидов. В соответствии с некоторыми вариантами осуществления комплекс белков и/или пептидов может быть ковалентно или нековалентно связан и/или ассоциирован. Например, аффинная часть конъюгата тетрамер-олигонуклеотид может представлять собой комплекс МНС-пептид. Некоторые неограничивающие примеры пептида в комплексе МНС-пептид могут представлять собой пептид из очищенного полипептида, выделенный полипептид, меченый посредством слияния последовательности полипептид, полипептид, прикрепленный к мембране клетки, или вирусу, или вириону или охватывающий их, цитоплазматический белок, внутриклеточный белок, внеклеточный белок, киназу, фосфатазу, ароматазу, хеликазу, протеазу, оксидоредуктазу, редуктазу, трансферазу, гидролазу, лиазу, изомеразу, гликозилазу, белок внеклеточного матрикса, лигазу, переносчик ионов, канал, пору, апоптотический белок, белок клеточной адгезии, патогенный белок, аберрантно экспрессируемый белок, транскрипционный фактор, транскрипционный регулятор, белок, участвующий в трансляции, шаперон, секретируемый белок, лиганд, гормон, цитокин, хемокин, ядерный белок, рецептор, трансмембранный рецептор, белок, участвующий в передаче сигнала, антитело, мембранный белок, интегральный мембранный белок, периферический мембранный белок, белок клеточной стенки, глобулярный белок, волокнистый белок, гликопротеин, липопротеин, хромосомный белок, любой их фрагмент или любую их комбинацию. В соответствии с некоторыми вариантами осуществления пептид получен из ассоциированного с заболеванием антигена. Например, в соответствии с некоторыми аспектами пептид представляет собой опухолеассоциированный антиген, в том числе универсальный опухолевый антиген, ассоциированный с вирусом антиген, ассоциированный с аутоиммунным заболеванием антиген, ассоциированный с воспалительным заболеванием антиген, ассоциированный с бактерией антиген или любую их комбинацию, или может быть получен из них. В соответствии с некоторыми вариантами осуществления аффинная часть представляет собой белок, сверхэкспрессированный в клетке с применением методик молекулярной биологии, таких как трансфекция. В соответствии с некоторыми вариантами осуществления аффинная часть представляет собой рекомбинантный полипептид. Например, аффинная часть может содержать образцы, продуцируемые в бактериальных (например, Е. coli), дрожжевых клетках, клетках млекопитающих или насекомых (например, белки, сверхэкспрессируемые организмами). В соответствии с некоторыми вариантами осуществления аффинная часть представляет собой полипептид, содержащий мутацию, вставку, делецию или полиморфизм. В соответствии с некоторыми вариантами осуществления аффинная часть представляет собой антиген, такой как полипептид, применяемый для иммунизации организма или для генерирования иммунной реакции у организма, как например, для выработки антител. В качестве другого примера, аффинная часть конъюгата тетрамер-олигонуклеотид может представлять собой антиген В-клеточного рецептора. Антиген В-клеточного рецептора может представлять собой субъединицу тетрамера.
Антитела
В соответствии с некоторыми вариантами осуществления аффинная часть представляет собой антитело, например, связывающий фрагмент антитела. Антитело может специфически связываться с другой молекулой в конкретной пространственной и полярной организации. Например, антитело может представлять собой очищенное антитело, выделенное антитело, фрагмент антитела или антитело, меченное посредством слияния последовательности. В соответствии с некоторыми вариантами осуществления антитело сверхэкспрессируют в клетке с применением методик молекулярной биологии, таких как трансфекция. В соответствии с некоторыми вариантами осуществления антитело представляет собой рекомбинантное антитело. Антитело может специфически связываться с другой молекулой в конкретной пространственной и полярной организации, такой как молекула клеточной поверхности. Антитело может представлять собой моноклональное, поликлональное или рекомбинантное антитело и может быть получено с помощью методик, которые являются хорошо известными в уровне техники, таких как иммунизация хозяина и сбор сыворотки (поликлональные), или посредством получения непрерывных гибридных клеточных линий и сбора секретированного белка (моноклональные), или посредством клонирования и экспрессии нуклеотидных последовательностей или их подвергнутых мутагенезу вариантов, кодирующих по меньшей мере аминокислотные последовательности, требующиеся для специфического связывания природных антител. Кроме того, агрегаты, полимеры и конъюгаты иммуноглобулинов или их фрагменты можно применять в случае, когда это уместно, при условии, что сохраняется аффинность связывания в отношении конкретной молекулы. Примеры фрагментов антител включают в себя Fab фрагмент, моновалентный фрагмент, состоящий из VL, VH, CL и CH1 доменов; F(ab)2 фрагмент, бивалентный фрагмент, содержащий два Fab фрагмента, связанные дисульфидным мостиком в шарнирном участке; Fd фрагмент, состоящий из VH и CH1 доменов; Fv фрагмент, состоящий из VL и VH доменов одного плеча антитела; фрагмент однодоменного антитела (dAb) (Ward et at., (1989) Nature 341:544-46), которое состоит из VH домена; и выделенный CDR и одноцепочечный фрагмент (scFv), в котором пара VL и VH участков образует моновалентные молекулы (известные как одноцепочечный Fv (scFv); см., например. Bird et al., (1988) Science 242:423-26; и Huston et al., (1988) PNAS 85:5879-83). Таким образом, фрагменты антител включают в себя Fab, F(ab)2, scFv, Fv, dAb и т.п. Несмотря на то что два домена VL и VH кодируются отдельными генами, их можно соединить с применением способов на основе рекомбинации с помощью искусственного пептидного линкера, который позволяет получать их в виде единой белковой цепи. Такие одноцепочечные антитела включают в себя один или несколько антигенсвязывающих фрагментов. Эти фрагменты антител можно получить с применением общепринятых методик, известных специалисту в данной области техники, и фрагменты могут быть подвергнуты скринингу в отношении полезности таким же образом, как и интактные антитела. Антитела могут быть человеческими, гуманизированными, химерными, выделенными, собачьими, кошачьими, ослиными, овечьими, могут быть получены из любого растения, животного или млекопитающего.
MHC
Распознавание антигенных структур клеточным звеном иммунной системы в некоторых случаях опосредуется экспрессируемыми на поверхности молекулами главного комплекса гистосовместимости (МНС). Клетки, такие как антигенпрезентирующие клетки (АРС), в соответствии с некоторыми аспектами перерабатывают белки, такие как антигены, в короткие пептиды, которые могут презентироваться в специальной пептид-связывающей складке в молекуле МНС и в соответствии с некоторыми аспектами в результате этого могут распознаваться Т-клетками. Специфическое распознавание эпитопа (фрагмента пептида) Т-клеточным рецептором (TCR) обычно требует одновременного взаимодействия с молекулой МНС. Стабильный мультимерный комплекс может быть получен с субъединицами белка МНС, содержащими связанный пептид. Комплекс МНС-антиген может образовывать стабильную структуру с Т-клетками, распознающими комплекс через их антигенный рецептор, таким образом обеспечивая связывание с Т-клетками, которые специфически распознают антиген. Аффинная часть конъюгата аффинная молекула-олигонуклеотид или конъюгата тетрамер-олигонуклеотид может быть нацелена на Т-клетку. Аффинная часть конъюгата аффинная молекула-олигонуклеотид может быть специфически нацелена на Т-клетку. Аффинная часть конъюгата аффинная молекула-олигонуклеотид или конъюгата тетрамер-олигонуклеотид может быть нацелена на Т-клеточный рецептор или TCR-подобную молекулу, такую как TCR-подобный CAR. Аффинная часть конъюгата аффинная молекула-олигонуклеотид может быть специфически нацелена на Т-клеточный рецептор. Например, аффинная часть конъюгата аффинная молекула-олигонуклеотид или конъюгата тетрамер-олигонуклеотид может содержать молекулу МНС. Например, аффинная часть конъюгата аффинная молекула-олигонуклеотид или конъюгата тетрамер-олигонуклеотид может содержать комплекс МНС-пептид (МНС-р). Аффинная часть конъюгата аффинная молекула-олигонуклеотид или конъюгата тетрамер-олигонуклеотид может иметь формулу (А-В-Р)n, в которой А представляет собой α-цепь белка МНС I класса или МНС II класса, В представляет собой β-цепь белка МНС II класса или β2 микроглобулина для белка МНС I класса, и Р представляет собой пептид. В соответствии с некоторыми вариантами осуществления n равно 1. В соответствии с некоторыми вариантами осуществления n является большим или равным 2. Субъединицы белка МНС могут присутствовать в растворимой форме. Например, растворимые субъединицы белка МНС могут быть получены из нативных субъединиц белка МНС посредством делеции трансмембранного домена или его части. В соответствии с некоторыми вариантами осуществления субъединицы белка МНС не содержат цитоплазматический домен. В соответствии с некоторыми вариантами осуществления субъединицы белка МНС не содержат трансмембранный домен.
Пептид (Р) может иметь длину от приблизительно 6 до 12 аминокислот в случае комплексов с белками МНС I класса, например, приблизительно от 8 до 10 аминокислот. Пептид может иметь длину от приблизительно 6 до 20 аминокислот в случае комплексов с белками МНС II класса, например, приблизительно от 10 до 18 аминокислот. Пептиды могут иметь последовательность, полученную из широкого спектра белков. Пептиды могут представлять собой Т-клеточные эпитопы. Последовательности эпитопов из ряда антигенов являются известными в уровне техники. В качестве альтернативы, последовательность эпитопа может быть определена эмпирически посредством выделения и секвенирования пептидов, связанных с нативными белками МНС, посредством синтеза ряда пептидов из целевой последовательности с последующим анализом реактивности Т-клеток в отношении различных пептидов или посредством получения ряда связывающих комплексов с различными пептидами и количественного определения связывания с Т-клетками. Получение фрагментов, идентификация последовательностей и идентификация минимальной последовательности описана в патенте США №5019384 и в источниках, цитируемых в нем. Пептиды можно получать с помощью ряда способов. Их можно подходящим образом синтезировать с помощью общепринятых методик с использованием автоматических синтезаторов, или их можно синтезировать вручную. В качестве альтернативы, могут быть получены последовательности ДНК, которые кодируют конкретный пептид, и их можно клонировать и экспрессировать с обеспечением желаемого пептида. В этом случае первой аминокислотой может быть метионин. Кроме того, пептиды можно получать с помощью способов на основе рекомбинации в виде гибридных последовательностей с белками, которые представляют один из специфических связывающих элементов в составе пары, обеспечивая возможность очистки гибридного белка с помощью аффинных реактивов, с последующим протеолитическим расщеплением, обычно по сконструированному сайту, и с получением на выходе желаемого пептида (см., например, Driscoll et al. (1993) J. Mol. Bio. 232:342-350). Пептид также можно выделить из природных источников и очищать с помощью известных методик, в том числе, например, хроматографии на ионообменных материалах, разделения по размеру, иммуноаффинной хроматографии и электрофореза.
В соответствии с некоторыми вариантами осуществления α- и β-субъединицы получают отдельно и обеспечивают возможность их ассоциации in vitro с образованием стабильного комплекса в виде гетеродуплекса (см., например, Altman et al. (1993), или Garboczi et al. (1992)). В соответствии с некоторыми вариантами осуществления α- и β-субъединицы экспрессируют вместе в одной клетке. В соответствии с некоторыми вариантами осуществления применяют единую молекулу, имеющую α- и β-субъединицы. Например, одноцепочечный гетеродимер можно создать посредством слияния двух субъединиц с применением короткого пептидного линкера, например, пептида или линкера из 15-25 аминокислот (см., например, Bedzyk et al. (1990) J. Biol. Chem. 265:18615). Растворимые гетеродимеры также могут быть получены посредством выделения нативного гетеродимера и расщепления протеазой, например, папаином, с получением растворимого продукта.
Растворимые субъединицы могут независимо экспрессироваться с ДНК-конструкции, кодирующей усеченный белок. С целью экспрессии последовательности ДНК можно вставить в соответствующий вектор экспрессии, где может использоваться нативный участок инициации транскрипции или экзогенный участок инициации транскрипции, например, промотор, отличающийся от промотора, который ассоциирован с геном в хромосоме в нормальных условиях. Промотор можно вводить с помощью способов на основе рекомбинации in vitro или в результате гомологичной интеграции последовательности в хромосому. Участки инициации транскрипции известны для широкого спектра экспрессионных хозяев. Экспрессионные хозяева могут включать прокариотов или эукариотов, в частности, Е. coli, В. subtilis,, клетки млекопитающего, такого как клетки СНО, клетки COS, клетки почки обезьяны, лимфоидные клетки, человеческие клеточные линии и т.д.
Субъединицы можно сверхэкспрессировать в подходящей клетке-хозяине, и, если необходимо, солюбилизировать. Две субъединицы можно объединить с пептидом и обеспечить возможность фолдинга in vitro с образованием стабильного комплекса в виде гетеродимера с доменами, связанными дисульфидными связями между цепями. Пептид может быть включен в первоначальную реакцию фолдинга, или его можно присоединять к пустому гетеродимеру на более поздней стадии. МНС-связывающий сайт может не содержать пептидов до присоединения пептида. Исключением будут являться те случаи, когда желательно пометить Т-клетки с природным комплексом пептид-МНС, таким как тот, который может присутствовать на поверхности клеток, являющихся мишенью для аутоиммунной атаки и т.д. Гетеродимер МНС будет связываться с пептидом в борозде, образованной двумя удаленными от мембраны доменами, либо α2 и α1 в случае I класса, либо α1 и β1 в случае II класса. Условия, которые позволяют фолдинг и ассоциацию субъединиц и пептида являются известными в уровне техники (см., например, Altman et al. (1993), и Garboczi et al. (1992)). В качестве одного примера пермиссивных условий примерно эквимолярные количества солюбилизированных α- и β-субъединиц смешивают в растворе мочевины. Рефолдинг инициируют посредством разведения или диализа в буферный раствор без мочевины. Пептиды загружают в пустые гетеродимеры II класса при рН приблизительно 5-5,5 в течение приблизительно 1-3 суток с последующей нейтрализацией, концентрированном и заменой буфера. Тем не менее, специалисту в данной области техники будет хорошо понятно, что конкретные условия фолдинга не являются критически важными для практического осуществления настоящего изобретения.
В соответствии с некоторыми вариантами осуществления мономерный комплекс (α-β-Р) может быть мультимеризованным. Например, мультимер может быть образован посредством связывания мономеров с мультивалентной частицей через специфические сайты прикрепления на α- или β-субъединице. В соответствии с некоторыми вариантами осуществления мультимер образуется при химическом сшивании мономеров. В уровне техники известен ряд реактивов, способных к сшиванию белков, в том числе, без ограничения, азидобензоилгидразид, N-[4-(пара-азидосалициламино)бутил]-3'-[2'-пиридилдитио]пропионамид), бис-сульфосукцинимидил-суберат, диметиладипимидат, дисукцинимидилтартрат, N-γ-малеимидобутирилоксисукцинимидовый сложный эфир, N-гидрокси-сульфосукцинимидил-4-азидобензоат, N-сукцинимидил[4-азидофенил]-1,3'-дитиопропионат, N-сукцинимидил[4-йодацетил]аминобензоат, глутаровый альдегид, формальдегид и сукцинимидил-4-[N-малеимидометил]циклогексан-1-карбоксилат. Сайт прикрепления для связывания с мультивалентной частицей может быть встречающимся в естественных условиях, или его можно ввести с помощью генной инженерии. Сайт может представлять собой специфический связывающий элемент в составе пары или сайт, который модифицирован с целью обеспечения специфического связывающегося элемента в составе пары, где комплементарная пара имеет множество сайтов для специфического связывания. Связывание с комплементарным связывающим элементом может представлять собой химическую реакцию, связывание эпитоп-рецептор или связывание гаптен-рецептор, где гаптен связан с цепью в субъединице. В соответствии с предпочтительным вариантом осуществления одна из субъединиц является слитой с аминокислотной последовательностью, обеспечивающей сайт распознавания для модифицирующего фермента. Последовательность для распознавания обычно будет включена посредством слияния вблизи карбокси-конца последовательности субъединицы во избежание возможного создания препятствия в сайте связывания антигенного пептида. Экспрессионная кассета будет подходящим образом включать в себя последовательность, кодирующую сайт распознавания.
Представляющие интерес модифицирующие ферменты включают в себя BirA, различные гликозилазы, фарнезил-протеинтрансферазу, протеинкиназы и т.п. Можно обеспечить возможность реакции субъединицы с модифицирующим ферментом в любое удобное время, обычно после образования мономера. Группа, вводимая с помощью модифицирующего фермента, например, биотин, сахар, фосфат, фарнезил и т.д., обеспечивает комплементарный связывающий элемент в составе пары или уникальный сайт для дополнительной модификации, такой как химическое сшивание, биотинилирование и т.д., который будет обеспечивать комплементарный связывающий элемент в составе пары. Альтернативной стратегией является введение неспаренного цистеинового остатка в субъединицу, таким образом вводя уникальный и химически реакционноспособный сайт для связывания. Сайт прикрепления также может представлять собой встречающийся в естественных условиях или введенный эпитоп, где мультивалентный партнер по связыванию будет представлять собой антитело, например, IgG, IgM и т.д. Любая модификация будет находиться в сайте, например, близком к С-концу, который не будет препятствовать связыванию. Примером образования мультимера является введение распознаваемой последовательности для фермента BirA, который катализирует биотинилирование белкового субстрата. Мономер с биотинилированной субъединицей затем связывают с мультимерным партнером по связыванию, например, стрептавидином или авидином, с которыми биотин связывается с крайне высокой аффинностью. Стрептавидин имеет валентность 4, обеспечивая мультимер (α-β-Р)4. Мультивалентный партнер по связыванию может присутствовать в свободной форме в растворе или может быть прикреплен к нерастворимой подложке. Примеры подходящих нерастворимых подложек включают в себя гранулы, например, магнитные гранулы, мембраны и микротитровальные планшеты. Они, как правило, выполнены из стекла, пластика (например, полистирола), полисахаридов, нейлона или нитроцеллюлозы. Прикрепление к нерастворимой подложке является полезным в случае, когда связывающий комплекс нужно применять с целью разделения Т-клеток.
Антиген В-клеточного рецептора
В-клетки, такие как небольшие популяции В-клеток или субпопуляции В-клеток, могут распознавать антигены с помощью В-клеточных рецепторов (BCR). Тетрамеры с субъединицами из антигенов В-клеточного рецептора можно применять для идентификации и изучения этих В-клеток или субпопуляций В-клеток. Антигены В-клеточного рецептора включают в себя любой антиген с эпитопом, способным к индукции иммунной реакции посредством BCR. Например, антиген В-клеточного рецептора может представлять собой антиген, который был применен для иммунизации субъекта. Антиген В-клеточного рецептора может быть превращен в тетрамер посредством применения небольшой линейной пептидной части антигена В-клеточного рецептора, в котором пептидная часть содержит эпитоп BCR. Небольшую линеаризованную пептидную часть можно затем экспрессировать с сайтом биотинилирования в экспрессионном векторе и очистить в виде мономера, подлежащего применению в тетрамере. Мономер с биотинилированной субъединицей затем можно связать с мультимерным партнером по связыванию, например, стрептавидином или авидином, с которыми биотин связывается с крайне высокой аффинностью. Стрептавидин имеет валентность 4, обеспечивая тетрамер антигена В-клеточного рецептора. Получаемый в результате тетрамер можно очистить с применением колоночной гель-хроматографии.
Клетки
В соответствии с некоторыми вариантами осуществления аффинная часть конъюгата аффинная молекула-олигонуклеотид представляет собой клетку. Например, аффинная часть может представлять собой интактную клетку, клетку, обработанную соединением (например, лекарственным средством), зафиксированную клетку, лизированную клетку или любую их комбинацию. В соответствии с некоторыми вариантами осуществления аффинная часть представляет собой отдельную клетку. Например, аффинная часть конъюгата аффинная молекула-олигонуклеотид может представлять собой Т-клетку или В-клетку. В соответствии с некоторыми вариантами осуществления аффинная часть представляет собой множество клеток. В соответствии с некоторыми вариантами осуществления аффинная часть представляет собой Т-клетку. В соответствии с некоторыми вариантами осуществления аффинная часть представляет собой В-клетку. В соответствии с некоторыми вариантами осуществления аффинная часть представляет собой антигенпрезентирующую клетку (АРС). В соответствии с некоторыми вариантами осуществления аффинная часть конъюгата аффинная молекула-олигонуклеотид представляет собой конкретный тип клеток или субпопуляцию клеток. Например, аффинная часть конъюгата аффинная молекула-олигонуклеотид может представлять собой CD4+ Т-клетку или CD8+ Т-клетку. Например, аффинная часть конъюгата аффинная молекула-олигонуклеотид может представлять собой Т-клетку, содержащую TCR, который специфически распознает конкретный антиген. Например, аффинная часть конъюгата аффинная молекула-олигонуклеотид может представлять собой Т-клетку, содержащую TCR, который специфически распознает конкретный комплекс МНС-пептид. В соответствии с некоторыми вариантами осуществления аффинная часть представляет собой клетку.
Малые молекулы
В соответствии с некоторыми вариантами осуществления аффинная часть конъюгата аффинная молекула-олигонуклеотид представляет собой малую молекулу, такую как лекарственное средство. Например, малая молекула может представлять собой макроциклическую молекулу, ингибитор, лекарственное средство или химическое соединение. В соответствии с некоторыми вариантами осуществления малая молекула содержит не более чем пять доноров водородной связи. В соответствии с некоторыми вариантами осуществления малая молекула содержит не более чем десять акцепторов водородной связи. В соответствии с некоторыми вариантами осуществления малая молекула характеризуется молекулярной массой, составляющей 500 дальтон или менее. В соответствии с некоторыми вариантами осуществления малая молекула характеризуется молекулярной массой, составляющей от приблизительно 180 до 500 дальтон. В соответствии с некоторыми вариантами осуществления малая молекула характеризуется коэффициентом разделения октанол-вода lop P, составляющим не более чем пять. В соответствии с некоторыми вариантами осуществления малая молекула характеризуется коэффициентом разделения log P, составляющим от -0,4 до 5,6. В соответствии с некоторыми вариантами осуществления малая молекула характеризуется молярной преломляющей способностью, составляющей от приблизительно 40 до 130. В соответствии с некоторыми вариантами осуществления малая молекула содержит от приблизительно 20 до приблизительно 70 атомов. В соответствии с некоторыми вариантами осуществления малая молекула имеет площадь полярной поверхности, составляющую 140 ангстрем2 или менее.
Нуклеиновые кислоты
В соответствии с некоторыми вариантами осуществления аффинная часть представляет собой полимерную форму рибонуклеотидов и/или дезоксирибонуклеотидов (аденина, гуанина, тимина или цитозина), такую как ДНК или РНК (например, мРНК). ДНК включает в себя двухцепочечную ДНК, находящуюся в линейных молекулах ДНК (например, рестрикционных фрагментах), вирусах, плазмидах и хромосомах. В соответствии с некоторыми вариантами осуществления аффинная часть полинуклеотида представляет собой одноцепочечную, двухцепочечную, малую интерферирующую РНК (siRNA), информационную РНК (мРНК), транспортную РНК (тРНК), хромосому, ген, некодирующую геномную последовательность, геномную ДНК (например, фрагментированную геномную ДНК), очищенный полинуклеотид, выделенный полинуклеотид, гибридизированный полинуклеотид, сайт связывания для транскрипционного фактора, митохондриальную ДНК, рибосомную РНК, эукариотический полинуклеотид, прокариотический полинуклеотид, синтезированный полинуклеотид, лигированный полинуклеотид, рекомбинантный полинуклеотид, полинуклеотид, содержащий аналог нуклеиновой кислоты, метилированный полинуклеотид, деметилированный полинуклеотид, любой их фрагмент или любую их комбинацию. В соответствии с некоторыми вариантами осуществления аффинная часть представляет собой полинуклеотид, содержащий двухцепочечный участок и конец, который не является двухцепочечным (например, участок 5' или 3' «липкого» конца). В соответствии с некоторыми вариантами осуществления аффинная часть представляет собой рекомбинантный полинуклеотид. В соответствии с некоторыми вариантами осуществления аффинная часть представляет собой гетерологичный полинуклеотид. Например, аффинная часть может содержать полинуклеотиды, продуцируемые в бактериальных (например, Е. coli), дрожжевых клетках, клетках млекопитающих или насекомых (например, полинуклеотиды, гетерологические по отношению к организмам). В соответствии с некоторыми вариантами осуществления аффинная часть представляет собой полинуклеотид, содержащий мутацию, вставку, делецию или полиморфизм.
В соответствии с некоторыми вариантами осуществления аффинная часть представляет собой аптамер. Аптамер представляет собой выделенную молекулу нуклеиновой кислоты, которая связывается с целевым анализируемым веществом, таким как белок, с высокой специфичностью и аффинностью. Аптамер представляет собой трехмерную структуру, удерживаемую в определенной(определенных) конформации(конформациях), которая(которые) обеспечивает(обеспечивают) химические контакты для специфического связывания с ее заданной мишенью. Несмотря на то что аптамеры представляют собой молекулы, имеющие в основе нуклеиновую кислоту, существует фундаментальное различие между аптамерами и другими молекулами нуклеиновой кислоты, такими как гены и мРНК. В последнем случае структура нуклеиновой кислоты кодирует информацию в своей линейной последовательности оснований, и, следовательно, эта последовательность является значимой для функции хранения информации. Как полная противоположность, функция аптамера, которая основывается на специфическом связывании с целевой молекулой, не зависит полностью от консервативной линейной последовательности оснований (некодирующая последовательность), а, скорее, зависит от конкретной вторичной/третичной/четвертичной структуры. Любой кодирующий потенциал, которым может обладать аптамер является полностью случайным и не играет какой бы то ни было роли в связывании аптамера с его когнатной мишенью. Аптамеры также нужно отличать от встречающихся в естественных условиях последовательностей нуклеиновой кислоты, которые связываются с определенными белками. Эти последние последовательности представляют собой встречающиеся в естественных условиях последовательности, заключенные в геноме организма, которые связываются со специализированной подгруппой белков, которые вовлечены в транскрипцию, трансляцию и транспортировку встречающихся в естественных условиях нуклеиновых кислот (например, белки, связывающиеся с нуклеиновыми кислотами). С другой стороны, аптамеры представляют собой короткие, выделенные, не встречающиеся в естественных условиях молекулы нуклеиновой кислоты. Несмотря на то что можно идентифицировать аптамеры, которые связываются с белками, связывающимися с нуклеиновыми кислотами, в большинстве случаев такие аптамеры характеризуются незначительной или отсутствующей идентичностью последовательности с последовательностями, которые распознаются белками, связывающимися с нуклеиновыми кислотами, в естественных условиях. Что более важно, аптамеры могут связываться практически с любым белком (а нет только с белками, связывающимися с нуклеиновыми кислотами), а также с почти любой мишенью, представляющей интерес, в том числе с малыми молекулами, углеводами, пептидами и т.д. Для большинства мишеней, даже для белков, встречающейся в естественных условиях последовательности нуклеиновой кислоты, с которой она связывается, не существует. Для тех мишеней, которые имеют такую последовательность, например, белки, связывающиеся с нуклеиновой кислотой, такие последовательности будут отличаться от аптамеров в результате относительно низкой аффинности связывания, которая используется в естественных условиях, по сравнению с крепко связывающимися аптамерами. Аптамеры способны к специфическому связыванию с выбранными мишенями и модулированию активности или связывающих взаимодействий у мишеней, например, посредством связывания аптамеры могут блокировать у своей мишени способность к функционированию. Функциональное свойство специфического связывания с мишенью является характерным свойством аптамера. Типичный аптамер имеет размер 6-35 кДа (20-100 нуклеотидов), связывается со своей мишенью с аффинностью, имеющей величину порядка микромолей или даже менее наномолей, и может различать близкородственные мишени (например, аптамеры могут селективно связываться с родственными белками из одного и того же семейства генов). Аптамеры способны использовать обычно наблюдаемые межмолекулярные взаимодействия, такие как образование водородных связей, взаимодополняющие электростатические взаимодействия, гидрофобные контакты и стерическое исключение, для связывания с конкретной мишенью. Аптамеры имеют ряд желательных характеристик для применения в качестве терапевтических и диагностических средств, в том числе высокую специфичность и аффинность, низкую иммуногенность, биологическую эффективность и отличные фармакокинетические свойства. Аптамер может содержать молекулярную структуру типа стебель и петля, образованную в результате гибридизации комплементарных полинуклеотидов, которые связаны ковалентно (например, структура петли «шпилька»). Стебель содержит гибридизированные полинуклеотиды, и петля представляет собой участок, который ковалентно связывает два комплементарных полинуклеотида.
В соответствии с некоторыми вариантами осуществления аффинная часть представляет собой множество аффинных частей, такое как смесь или библиотека аффинных частей. В соответствии с некоторыми вариантами осуществления аффинная часть представляет собой множество различных аффинных частей. Например, аффинная часть может содержать множество по меньшей мере приблизительно из 2, 3, 4, 5, 6, 7, 8, 9, 10, 15, 20, 30, 40, 50, 60, 70, 80, 90, 100, 200, 500, 1000, 2000, 3000, 4000, 5000, 6000, 7000, 8000, 9000, 10000, 11000, 12000, 13000, 14000, 15000, 16000, 17000, 18000, 19000, 20000, 25000 или 30000 аффинных частей.
Олигонуклеотидная часть конъюгатов аффинная молекула-олигонуклеотид
Олигонуклеотидная часть конъюгата аффинная молекула-олигонуклеотид представляет собой нуклеиновую кислоту, которая связана с аффинной частью конъюгата аффинная молекула-олигонуклеотид. В соответствии с некоторыми вариантами осуществления Олигонуклеотидная часть конъюгата тетрамер-олигонуклеотид представляет собой нуклеиновую кислоту, которая связана с аффинной частью конъюгата тетрамер-олигонуклеотид. В соответствии с некоторыми вариантами осуществления Олигонуклеотидная часть конъюгата тетрамер-олигонуклеотид представляет собой нуклеиновую кислоту, которая связана с одним или несколькими элементами кора тетрамерного комплекса. Например, нуклеиновая кислота может быть конъюгирована с мономерным или тетрамерным стрептавидиновым кором. Иллюстративные схематические изображения конъюгата тетрамер-олигонуклеотид представлены на фиг. 10А, 10В, 13, 14 и 18. В соответствии с некоторыми вариантами осуществления олигонуклеотид непосредственно связан с аффинной частью. В соответствии с некоторыми вариантами осуществления олигонуклеотид опосредованно связан с аффинной частью. В соответствии с некоторыми вариантами осуществления олигонуклеотид связан с аффинной частью нековалентно. В соответствии с некоторыми вариантами осуществления олигонуклеотид связан с аффинной частью ковалентно. В соответствии с некоторыми вариантами осуществления олигонуклеотид представляет собой синтезированный олигонуклеотид. В соответствии с предпочтительным вариантом осуществления олигонуклеотид не осуществляет значимого взаимодействия напрямую с целевым анализируемым веществом в составе аффинной части.
Олигонуклеотид, связанный с аффинной частью конъюгата аффинная молекула-олигонуклеотид или конъюгата тетрамер-олигонуклеотид, может содержать одну или несколько последовательностей штрихкода. Например, олигонуклеотид, связанный с аффинной частью конъюгата аффинная молекула-олигонуклеотид или конъюгата тетрамер-олигонуклеотид, может содержать идентификационную последовательность антигена (AID) и последовательность молекулярного штрихкода антигена (АМВ). Олигонуклеотид может содержать идентификационную последовательность антигена (AID), последовательность для слияния, сайт для праймера, последовательность молекулярного штрихкода, константную последовательность или любую их комбинацию.
Олигонуклеотид может содержать химическую модификацию для обеспечения возможности конъюгирования с аффинной частью конъюгата аффинная молекула-олигонуклеотид (например, амин, тиол или биотин) или конъюгата тетрамер-олигонуклеотид.
Олигонуклеотид может содержать множество олигонуклеотидов. Множество олигонуклеотидов может содержаться во множестве конъюгатов аффинная молекула-олигонуклеотид или во множестве конъюгатов тетрамер-олигонуклеотид. Например, олигонуклеотид может содержать множество по меньшей мере приблизительно из 2, 3, 4, 5, 6, 7, 8, 9, 10, 15, 20, 30, 40, 50, 60, 70, 80, 90, 100, 200, 500, 1000, 2000, 3000, 4000, 5000, 6000, 7000, 8000, 9000, 10000, 11000, 12000, 13000, 14000, 15000, 16000, 17000, 18000, 19000, 20000, 25000 или 30000 олигонуклеотидов. Например, множество по меньшей мере приблизительно из 2, 3, 4, 5, 6, 7, 8, 9, 10, 15, 20, 30, 40, 50, 60, 70, 80, 90, 100, 200, 500, 1000, 2000, 3000, 4000, 5000, 6000, 7000, 8000, 9000, 10000, 11000, 12000, 13000, 14000, 15000, 16000, 17000, 18000, 19000, 20000, 25000 или 30000 олигонуклеотидов может содержаться во множестве по меньшей мере приблизительно из 2, 3, 4, 5, 6, 7, 8, 9, 10, 15, 20, 30, 40, 50, 60, 70, 80, 90, 100, 200, 500, 1000, 2000, 3000, 4000, 5000, 6000, 7000, 8000, 9000, 10000, 11000, 12000, 13000, 14000, 15000, 16000, 17000, 18000, 19000, 20000, 25000 или 30000 конъюгатов аффинная молекула-олигонуклеотид или конъюгатов тетрамер-олигонуклеотид.
Олигонуклеотид может содержать последовательность штрихкода в олигонуклеотиде, последовательность для слияния в олигонуклеотиде, последовательность для связывания праймера в олигонуклеотиде, константную последовательность в олигонуклеотиде или любую их комбинацию.
Идентификационная последовательность антигена (AID) в олигонуклеотиде
Олигонуклеотид может содержать последовательность штрихкода антигена в олигонуклеотиде или ее комплементарную последовательность. Штрихкод антигена в олигонуклеотиде может обеспечивать возможность идентификации комплекса аффинная молекула-олигонуклеотид или комплекса тетрамер-олигонуклеотид, содержащего штрихкод антигена в олигонуклеотиде. Штрихкод антигена в олигонуклеотиде может обеспечивать возможность идентификации аффинной части, к которой прикреплен штрихкод антигена в олигонуклеотиде. Штрихкод антигена в олигонуклеотиде можно применять для идентификации аффинной части из множества различных аффинных частей, которые связываются с различными целевыми анализируемыми веществами. Штрихкод антигена в олигонуклеотиде может служить в качестве штрихкода исключительно для комплекса аффинная молекула-олигонуклеотид или исключительно для комплекса тетрамер-олигонуклеотид. Штрихкод антигена в олигонуклеотиде может служить в качестве штрихкода исключительно для аффинной части. Таким образом, последовательность штрихкода антигена в олигонуклеотиде может служить в качестве штрихкода для конкретной аффинной части.
Штрихкод антигена в олигонуклеотиде может представлять собой уникальную последовательность штрихкода. Например, любой штрихкод антигена в олигонуклеотиде из множества штрихкодов антигена в олигонуклеотиде может представлять собой уникальную последовательность штрихкода. Количество теоретически возможных различных последовательностей штрихкода антигена может напрямую зависеть от длины последовательности штрихкода. Например, если можно применять ДНК-штрихкод из произвольно собранных аденинового, тимидинового, гуанозинового и цитидинового нуклеотидов, теоретическое максимальное количество возможных последовательностей штрихкода может составлять 1048576 в случае длины в десять нуклеотидов и может составлять 1073741824 в случае длины в пятнадцать нуклеотидов. Последовательность штрихкода антигена в олигонуклеотиде может содержать последовательность по меньшей мере из 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 25, 30, 40, 45 или 50 или большего количества последовательных нуклеотидов. Олигонуклеотид может содержать две или более последовательностей штрихкода антигена в олигонуклеотиде или их комплементарные последовательности. Последовательность штрихкода антигена в олигонуклеотиде может содержать собранную в произвольном порядке последовательность нуклеотидов. Последовательность штрихкода антигена в олигонуклеотиде может представлять собой вырожденную последовательность. Последовательность штрихкода антигена в олигонуклеотиде может представлять собой известную последовательность. Последовательность штрихкода антигена в олигонуклеотиде может представлять собой предварительно определенную последовательность. В соответствии с предпочтительным вариантом осуществления последовательность штрихкода антигена в олигонуклеотиде представляет собой известную уникальную последовательность, которая выступает в качестве штрихкода для аффинной части, с которой она связана, в результате чего сигнал, содержащий штрихкод антигена в олигонуклеотиде (например, рид последовательности) или его комплементарную последовательность, можно применять для идентификации аффинной части из множества различных аффинных частей, которые взаимодействуют с различными целевыми анализируемыми веществами.
Например, олигонуклеотид, связанный с аффинной частью конъюгата аффинная молекула-олигонуклеотид, может содержать штрихкод, который представляет собой идентификационную последовательность антигена (AID). Последовательность AID может служить в качестве штрихкода для аффинной части конъюгата аффинная молекула-олигонуклеотид. Последовательность AID может служить в качестве штрихкода для антигена, на который нацелена аффинная часть. Последовательность AID можно применять для идентификации аффинной части конъюгата аффинная молекула-олигонуклеотид и/или антигена, на который нацелена аффинная часть. Например, последовательность AID может служить в качестве штрихкода для антитела в составе конъюгата антитело-олигонуклеотид. Например, последовательность AID может служить в качестве штрихкода для антигена, на который нацелено антитело в составе конъюгата антитело-олигонуклеотид. Например, последовательность AID можно применять для иммунофенотипирования клеток. Например, последовательность AID может служить в качестве штрихкода для пептида в составе комплекса МНС-пептид.
Последовательность AID может быть уникальной для каждого антигена, на который нацелена аффинная часть конъюгатов аффинная молекула-олигонуклеотид. Последовательность AID может быть уникальной для аффинной части конъюгатов аффинная молекула-олигонуклеотид. Например, последовательность AID может быть уникальной для каждого антитела, которое специфически связывается с отличающимся целевым антигеном клетки. В соответствии с некоторыми вариантами осуществления последовательность AID представляет собой определенную последовательность. В соответствии с некоторыми вариантами осуществления последовательность AID представляет собой известную последовательность. Последовательность AID для каждого олигонуклеотида может быть определена посредством секвенирования олигонуклеотида или продуктов амплификации олигонуклеотида, например, с помощью секвенирования нового поколения.
Последовательность молекулярного штрихкода в олигонуклеотиде
Олигонуклеотид может содержать последовательность молекулярного штрихкода в олигонуклеотиде или ее комплементарную последовательность. Штрихкод олигонуклеотида может обеспечивать возможность идентификации молекулы комплекса аффинная молекула-Олигонуклеотид, содержащего штрихкод олигонуклеотида. Молекулярный штрихкод в олигонуклеотиде может служить в качестве штрихкода исключительно для молекулы комплекса аффинная молекула-олигонуклеотид. Молекулярный штрихкод в олигонуклеотиде может служить в качестве штрихкода исключительно для молекулы аффинной части. Таким образом, последовательность молекулярного штрихкода в олигонуклеотиде может служить в качестве штрихкода для конкретной молекулы в составе аффинной части.
Молекулярный штрихкод в олигонуклеотиде может представлять собой уникальную последовательность штрихкода. Например, любой молекулярный штрихкод в олигонуклеотиде из множества молекулярных штрихкодов в олигонуклеотиде может представлять собой уникальную последовательность штрихкода. Количество теоретически возможных различных последовательностей молекулярного штрихкода может напрямую зависеть от длины последовательности штрихкода. Например, если можно применять ДНК-штрихкод из произвольно собранных аденинового, тимидинового, гуанозинового и цитидинового нуклеотидов, теоретическое максимальное количество возможных последовательностей штрихкода может составлять 1048576 в случае длины в десять нуклеотидов и может составлять 1073741824 в случае длины в пятнадцать нуклеотидов. Последовательность молекулярного штрихкода в олигонуклеотиде может содержать последовательность по меньшей мере из 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 25, 30, 40, 45 или 50 или большего количества последовательных нуклеотидов. Олигонуклеотид может содержать две или более последовательности молекулярного штрихкода в олигонуклеотиде или их комплементарные последовательности. Последовательность молекулярного штрихкода в олигонуклеотиде может содержать собранную в произвольном порядке последовательность нуклеотидов. Последовательность молекулярного штрихкода в олигонуклеотиде может представлять собой вырожденную последовательность. Последовательность молекулярного штрихкода в олигонуклеотиде может представлять собой известную последовательность. Последовательность молекулярного штрихкода в олигонуклеотиде может представлять собой предварительно определенную последовательность. В соответствии с предпочтительным вариантом осуществления последовательность молекулярного штрихкода в олигонуклеотиде представляет собой уникальную последовательность, которую можно применять для идентификации молекулы в составе аффинной части из множества молекул в составе аффинной части, которые взаимодействуют с целевым анализируемым веществом.
Например, олигонуклеотид, связанный с аффинной частью конъюгата аффинная молекула-олигонуклеотид, может содержать штрихкод, который представляет собой последовательность молекулярного штрихкода антигена (АМВ). Последовательность молекулярного штрихкода антигена (АМВ) может быть уникальной для каждой молекулы олигонуклеотида в составе конъюгата аффинная молекула-олигонуклеотид. Последовательность АМВ может обеспечивать возможность подсчета количества молекул олигонуклеотида в составе конъюгата аффинная молекула-олигонуклеотид, которые связались с антигеном, таким как антиген отдельной клетки в резервуаре, например, капле эмульсии. Последовательность АМВ для каждого олигонуклеотида может быть определена посредством секвенирования олигонуклеотида или продуктов амплификации олигонуклеотида, например, с помощью секвенирования нового поколения.
Последовательность для слияния в олигонуклеотиде
Олигонуклеотид, связанный с аффинной частью конъюгата аффинная молекула-олигонуклеотид или конъюгата тетрамер-олигонуклеотид, может содержать последовательность для слияния. Последовательность для слияния может обеспечивать возможность достройки при ПЦР последовательности специфического для капли штрихкода на олигонуклеотиде в составе конъюгата аффинная молекула-олигонуклеотид, например, связанного с клеточной поверхностью конъюгата аффинная молекула-олигонуклеотид, или конъюгата тетрамер-олигонуклеотид, например, связанного с TCR или BCR конъюгата тетрамер-олигонуклеотид. Последовательность для слияния в каждом олигонуклеотиде из множества олигонуклеотидов может быть идентичной. Последовательность для слияния может содержать последовательность, которая является комплементарной последовательности штрихкода капли. В соответствии с некоторыми вариантами осуществления последовательность для слияния располагается на конце олигонуклеотида. В соответствии с некоторыми вариантами осуществления последовательность для слияния на конце олигонуклеотида не конъюгирована напрямую с аффинной частью конъюгата антитело-олигонуклеотид или конъюгата тетрамер-олигонуклеотид. В соответствии с некоторыми вариантами осуществления последовательность для слияния на конце олигонуклеотида содержит свободный конец.
Последовательность для слияния может содержать участок, комплементарный участку 3'-метящего полинуклеотида, такого как полинуклеотид, содержащий штрихкод резервуара. Последовательность для слияния может содержать участок, комплементарный комплементарной последовательности участка полинуклеотида, такого как полинуклеотид, содержащий штрихкод резервуара. Например, последовательность для слияния может содержать 3' участок, такой как 3'-концевой участок, который является комплементарным 3'-метящему полинуклеотиду или его комплементарной последовательности, содержащим штрихкод, такой штрихкод резервуара.
3'-Метящий полинуклеотид может представлять собой полинуклеотид, применяемый для добавления нуклеиновых кислот к 3' концу целевого полинуклеотида, такого как олигонуклеотид в составе конъюгата аффинная молекула-олигонуклеотид или конъюгата тетрамер-олигонуклеотид. 3'-Метящий полинуклеотид может представлять собой полинуклеотид, применяемый в качестве матрицы для добавления нуклеиновых кислот к 3' концу целевого полинуклеотида, такого как олигонуклеотид в составе конъюгата аффинная молекула-олигонуклеотид или конъюгата тетрамер-олигонуклеотид. 3'-Метящий полинуклеотид может представлять собой полинуклеотид, который гибридизируется с 3' концом целевого полинуклеотида, такого как олигонуклеотид в составе конъюгата аффинная молекула-олигонуклеотид или конъюгата тетрамер-олигонуклеотид. 3'-Метящий полинуклеотид может представлять собой полинуклеотид, содержащий 3' участок, такой как 3'-концевой участок, который гибридизируется с 3' концом целевого полинуклеотида, такого как олигонуклеотид в составе конъюгата аффинная молекула-олигонуклеотид или конъюгата тетрамер-олигонуклеотид.
В соответствии с некоторыми вариантами осуществления 3'-метящий полинуклеотид представляет собой меченый штрихкодом резервуара полинуклеотид. Штрихкод резервуара может быть добавлен к олигонуклеотиду в составе конъюгата аффинная молекула-олигонуклеотид или конъюгата тетрамер-олигонуклеотид. Например, штрихкод резервуара может гибридизироваться с олигонуклеотидом в составе конъюгата аффинная молекула-олигонуклеотид или конъюгата тетрамер-олигонуклеотид. Меченый штрихкодом резервуара полинуклеотид может содержать 3' участок, такой как 3'-концевой участок, который гибридизируется с 3' концом олигонуклеотида в составе конъюгата аффинная молекула-олигонуклеотид или конъюгата тетрамер-олигонуклеотид.
В соответствии с некоторыми вариантами осуществления 3'-метящий полинуклеотид представляет собой амплифицированный продукт. В соответствии с некоторыми вариантами осуществления 3'-метящий полинуклеотид представляет собой амплифицированный продукт, происходящий из одной молекулы. В соответствии с некоторыми вариантами осуществления 3'-метящий полинуклеотид представляет собой амплифицированный продукт меченого штрихкодом резервуара полинуклеотида. В соответствии с некоторыми вариантами осуществления 3'-метящий полинуклеотид представляет собой амплифицированный продукт, происходящий из одного меченого штрихкодом резервуара полинуклеотида. Участок, расположенный в направлении 5' относительно 3'-участка, который гибридизируется с 3' концом олигонуклеотида в составе конъюгата аффинная молекула-олигонуклеотид, может содержать участок, комплементарный праймеру или его комплементарной последовательности. Участок, расположенный в направлении 5' относительно 3'-участка, который гибридизируется с 3' концом олигонуклеотида в составе конъюгата аффинная молекула-олигонуклеотид, может содержать участок, комплементарный праймеру, который может применяться для амплификации олигонуклеотида в составе конъюгата аффинная молекула-олигонуклеотид или конъюгата тетрамер-олигонуклеотид. Например, набор праймеров, содержащий первый праймер, который является комплементарным участку, расположенному в направлении 5' относительно 3'-участка, который гибридизируется с 3' концом олигонуклеотида в составе конъюгата аффинная молекула-олигонуклеотид, конъюгата тетрамер-олигонуклеотид или его комплементарной последовательностью, и второй праймер, который является комплементарным сайту для праймера в олигонуклеотиде в составе конъюгата аффинная молекула-олигонуклеотид или конъюгата тетрамер-олигонуклеотид, можно применять для амплификации олигонуклеотида в составе конъюгата аффинная молекула-олигонуклеотид или конъюгата тетрамер-олигонуклеотид.
Участок, расположенный в направлении 5' относительно 3'-участка, который гибридизируется с 3' концом олигонуклеотида в составе конъюгата аффинная молекула-олигонуклеотид, может содержать участок, комплементарный праймеру или его комплементарной последовательности, которые применяли для амплификации меченого штрихкодом резервуара полинуклеотида.
Последовательность для слияния в олигонуклеотиде может составлять по меньшей мере приблизительно 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 25, 30, 35, 40, 45, 50, 55, 60, 65, 70, 75, 80, 85, 90, 95, 100 или больше последовательных нуклеотидов. Последовательность для слияния в олигонуклеотиде может представлять собой последовательность известной длины. Последовательность для слияния в олигонуклеотиде может представлять собой известную последовательность. Последовательность для слияния в олигонуклеотиде может представлять собой предварительно определенную последовательность. Последовательность для слияния в олигонуклеотиде может представлять собой неизвестную последовательность известной длины. Последовательность для слияния в олигонуклеотиде может представлять собой известную последовательность известной длины.
Константная последовательность в олигонуклеотиде
Олигонуклеотид, связанный с аффинной частью конъюгата аффинная молекула-олигонуклеотид или конъюгата тетрамер-олигонуклеотид, может содержать константную последовательность. Константная последовательность является необязательной. Константная последовательность в каждом олигонуклеотиде из множества конъюгатов аффинная молекула-олигонуклеотид или конъюгатов тетрамер-олигонуклеотид может быть идентичной.
Константную последовательность в олигонуклеотиде можно применять для увеличения длины олигонуклеотида или для отделения одного или нескольких из штрихкода в олигонуклеотиде, последовательности для слияния в олигонуклеотиде и сайта для связывания праймера в олигонуклеотиде друг от друга. В соответствии с некоторыми вариантами осуществления олигонуклеотид не содержит константную последовательность в олигонуклеотиде. Например, олигонуклеотид может быть связан с аффинной частью на конце олигонуклеотида, содержащем сайт для связывания праймера в олигонуклеотиде.
В соответствии с некоторыми вариантами осуществления константная последовательность в олигонуклеотиде прикреплена к аффинной части комплекса аффинная молекула-олигонуклеотид или комплекса тетрамер-олигонуклеотид. В соответствии с некоторыми вариантами осуществления константная последовательность в олигонуклеотиде располагается выше последовательности для связывания праймера в олигонуклеотиде. Например, константная последовательность в олигонуклеотиде может располагаться в направлении 5' относительно последовательности для связывания праймера в олигонуклеотиде. В соответствии с некоторыми вариантами осуществления константная последовательность в олигонуклеотиде располагается ниже последовательности для связывания праймера в олигонуклеотиде. Например, константная последовательность в олигонуклеотиде может располагаться в направлении 3' относительно последовательности для связывания праймера в олигонуклеотиде. В соответствии с некоторыми вариантами осуществления константная последовательность в олигонуклеотиде располагается выше штрихкода в олигонуклеотиде. Например, константная последовательность в олигонуклеотиде может располагаться в направлении 5' относительно штрихкода в олигонуклеотиде. В соответствии с некоторыми вариантами осуществления константная последовательность в олигонуклеотиде располагается ниже штрихкода в олигонуклеотиде. Например, константная последовательность в олигонуклеотиде может располагаться в направлении 3' относительно штрихкода в олигонуклеотиде. В соответствии с некоторыми вариантами осуществления константная последовательность в олигонуклеотиде располагается выше последовательности для слияния в олигонуклеотиде. Например, константная последовательность в олигонуклеотиде может располагаться в направлении 5' относительно последовательности для слияния в олигонуклеотиде.
В соответствии с некоторыми вариантами осуществления константная последовательность в олигонуклеотиде располагается между последовательностью для связывания праймера в олигонуклеотиде и аффинной частью комплекса аффинная молекула-олигонуклеотид или комплекса тетрамер-олигонуклеотид. Например, константная последовательность в олигонуклеотиде может располагаться в направлении 5' относительно последовательности для связывания праймера в олигонуклеотиде и прикреплена к аффинной части олигонуклеотида. В соответствии с некоторыми вариантами осуществления константная последовательность в олигонуклеотиде располагается между последовательностью для связывания праймера в олигонуклеотиде и штрихкодом в олигонуклеотиде. Например, константная последовательность в олигонуклеотиде может располагаться в направлении 3' относительно последовательности для связывания праймера в олигонуклеотиде и в направлении 5' относительно штрихкода в олигонуклеотиде. В соответствии с некоторыми вариантами осуществления константная последовательность в олигонуклеотиде располагается между последовательностью для слияния в олигонуклеотиде и штрихкодом в олигонуклеотиде. Например, константная последовательность в олигонуклеотиде может располагаться в направлении 3' относительно штрихкода в олигонуклеотиде и в направлении 5' относительно последовательности для слияния в олигонуклеотиде.
Константная последовательность в олигонуклеотиде может составлять по меньшей мере 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 25, 30, 35, 40, 45, 50, 55, 60, 65, 70, 75, 80, 85, 90, 95, 100, 200, 250, 300, 400, 500 или большее количество последовательных нуклеотидов. Константная последовательность в олигонуклеотиде может содержать неслучайную последовательность нуклеотидов. Константная последовательность в олигонуклеотиде может представлять собой последовательность известной длины. Константная последовательность в олигонуклеотиде может представлять собой известную последовательность. Константная последовательность в олигонуклеотиде может представлять собой предварительно определенную последовательность. Константная последовательность в олигонуклеотиде может представлять собой неизвестную последовательность известной длины. Константная последовательность в олигонуклеотиде может представлять собой известную последовательность известной длины.
Сайт для связывания праймера в олигонуклеотиде
Олигонуклеотид, связанный с аффинной частью конъюгата аффинная молекула-олигонуклеотид или конъюгата тетрамер-олигонуклеотид, может содержать сайт для праймера. Сайт для праймера может содержать последовательность, которая является комплементарной праймеру, такому как праймер для амплификации. Последовательность для связывания праймера в олигонуклеотиде можно применять в качестве сайта для связывания праймера для реакции, такой как амплификация или секвенирование. Последовательность для связывания праймера в олигонуклеотиде может представлять собой первую последовательность для связывания праймера для пары праймеров, применяемых для реакции, такой как амплификация или секвенирование. Например, последовательность для связывания праймера в олигонуклеотиде может представлять собой сайт для связывания прямого праймера. Например, сайт для связывания праймера в олигонуклеотиде может представлять собой сайт для связывания обратного праймера. Например, сайт для связывания праймера в олигонуклеотиде может представлять собой сайт для связывания прямого праймера, и последовательность для связывания праймера в меченом штрихкодом резервуара полинуклеотиде, прикрепленном к олигонуклеотиду, может представлять собой последовательность для связывания обратного праймера. В соответствии с некоторыми вариантами осуществления последовательность для связывания праймера в олигонуклеотиде представляет собой последовательность для связывания универсального праймера.
Последовательность для связывания праймера в олигонуклеотиде и последовательность для связывания праймера в полинуклеотиде, прикрепленном к олигонуклеотиду, (например, в меченом штрихкодом резервуара полинуклеотиде) могут характеризоваться температурами плавления, которые отличаются не более чем на 6, 5, 4, 3, 2 или 1 градус Цельсия. Нуклеотидная последовательность последовательности для связывания праймера в олигонуклеотиде и последовательности для связывания праймера в полинуклеотиде, прикрепленном к олигонуклеотиду, может отличаться, в результате чего полинуклеотид, который гибридизируется с последовательностью для связывания праймера в олигонуклеотиде; не гибридизируется с последовательностью для связывания праймера в полинуклеотиде, прикрепленном к олигонуклеотиду. Нуклеотидная последовательность последовательности для связывания праймера в олигонуклеотиде и последовательности для связывания праймера в полинуклеотиде, прикрепленном к олигонуклеотиду, может отличаться, в результате чего полинуклеотид, который гибридизируется с последовательностью для связывания праймера в полинуклеотиде, прикрепленном к олигонуклеотиду, не гибридизируется с последовательностью для связывания праймера в олигонуклеотиде.
Расположение элементов в олигонуклеотиде
Элементы олигонуклеотида могут быть расположены в таком порядке, чтобы последовательность для слияния в олигонуклеотиде располагалась на одном конце олигонуклеотида. Элементы олигонуклеотида могут быть расположены в таком порядке, чтобы он содержал штрихкод в олигонуклеотиде выше последовательности для слияния в олигонуклеотиде. Элементы олигонуклеотида могут быть расположены в таком порядке, чтобы он содержал последовательность для связывания праймера в олигонуклеотиде выше штрихкода в олигонуклеотиде. Элементы олигонуклеотида могут быть расположены в таком порядке, чтобы константная последовательность в олигонуклеотиде располагалась выше или ниже последовательности для связывания праймера в олигонуклеотиде. Элементы олигонуклеотида могут быть расположены в таком порядке, чтобы константная последовательность в олигонуклеотиде располагалась выше последовательности штрихкода олигонуклеотида. Элементы олигонуклеотида могут быть расположены в таком порядке, чтобы константная последовательность в олигонуклеотиде располагалась на одном конце олигонуклеотида, например, на конце олигонуклеотида, который не содержит последовательность для слияния в олигонуклеотиде. Например, элементы олигонуклеотида могут располагаться в следующем порядке: последовательность для слияния в олигонуклеотиде, последовательность штрихкода в олигонуклеотиде, последовательность для связывания праймера в олигонуклеотиде и константная последовательность в олигонуклеотиде. Например, элементы олигонуклеотида могут располагаться в следующем порядке: последовательность для слияния в олигонуклеотиде, последовательность штрихкода в олигонуклеотиде, последовательность для связывания праймера в олигонуклеотиде и константная последовательность в олигонуклеотиде, продолжающаяся по направлению к аффинной части. Например, элементы олигонуклеотида могут располагаться в следующем порядке: последовательность для слияния в олигонуклеотиде, последовательность штрихкода в олигонуклеотиде, последовательность для связывания праймера в олигонуклеотиде и константная последовательность в олигонуклеотиде от 5' конца к 3' концу или от 3' конца к 5' концу. Например, олигонуклеотид может содержать 5'-концевую последовательность для слияния в олигонуклеотиде, уникальную последовательность штрихкода в олигонуклеотиде, последовательность для связывания обратного праймера в олигонуклеотиде и 3' константную последовательность в олигонуклеотиде, прикрепленную к аффинной части (например, через группу первичного амина, прикрепленную к 3' концу) в таком порядке. Например, элементы олигонуклеотида, прикрепленного к аффинной части, могут располагаться, продолжаясь в направлении аффинной части, в следующем порядке: последовательность для слияния в олигонуклеотиде, последовательность штрихкода в олигонуклеотиде, константная последовательность в олигонуклеотиде и последовательность сайта для связывания праймера в олигонуклеотиде.
Элементы олигонуклеотида могут располагаться в таком порядке, чтобы последовательность для слияния располагалась на одном конце олигонуклеотида. Элементы олигонуклеотида могут располагаться в таком порядке, чтобы последовательность для слияния располагалась ниже последовательности AID. Элементы олигонуклеотида могут располагаться в таком порядке, чтобы последовательность для слияния располагалась ниже последовательности АМВ. Элементы олигонуклеотида могут располагаться в таком порядке, чтобы последовательность для слияния располагалась ниже константной последовательности. Элементы олигонуклеотида могут располагаться в таком порядке, чтобы последовательность для слияния располагалась ниже сайта для праймера.
Элементы олигонуклеотида могут располагаться в таком порядке, чтобы сайт для праймера располагался на одном конце олигонуклеотида. Элементы олигонуклеотида могут располагаться в таком порядке, чтобы сайт для праймера располагался выше последовательности AID. Элементы олигонуклеотида могут располагаться в таком порядке, чтобы сайт для праймера располагался выше последовательности АМВ. Элементы олигонуклеотида могут располагаться в таком порядке, чтобы сайт для праймера располагался выше константной последовательности. Элементы олигонуклеотида могут располагаться в таком порядке, чтобы сайт для праймера располагался выше последовательности для слияния.
Элементы олигонуклеотида могут располагаться в таком порядке, чтобы он содержал последовательность AID выше последовательности для слияния. Элементы олигонуклеотида могут располагаться в таком порядке, чтобы он содержал последовательность AID ниже сайта для праймера. Последовательность AID может быть расположена выше или ниже последовательности АМВ. Последовательность AID может быть расположена выше или ниже константной последовательности. Элементы олигонуклеотида могут располагаться в таком порядке, чтобы он содержал последовательность AID между последовательностью для слияния и сайтом для праймера.
Элементы олигонуклеотида могут располагаться в таком порядке, чтобы он содержал последовательность АМВ выше последовательности для слияния. Элементы олигонуклеотида могут располагаться в таком порядке, чтобы он содержал последовательность АМВ ниже сайта для праймера. Последовательность АМВ может быть расположена выше или ниже последовательности АМВ. Последовательность АМВ может быть расположена выше или ниже константной последовательности. Элементы олигонуклеотида могут располагаться в таком порядке, чтобы он содержал последовательность АМВ между последовательностью для слияния и сайтом для праймера.
Элементы олигонуклеотида могут располагаться в таком порядке, чтобы он содержал константную последовательность выше последовательности для слияния. Элементы олигонуклеотида могут располагаться в таком порядке, чтобы он содержал константную последовательность ниже сайта для праймера. Константная последовательность может быть расположена выше или ниже последовательности AID. Константная последовательность может быть расположена выше или ниже последовательности АМВ. Элементы олигонуклеотида могут располагаться в таком порядке, чтобы он содержал константную последовательность между последовательностью для слияния и сайтом для праймера.
Элементы олигонуклеотида могут располагаться в таком порядке, чтобы последовательность АМВ и/или последовательность AID не располагалась на одном конце олигонуклеотида, например, на конце олигонуклеотида, который содержит последовательность для слияния или сайт для праймера. Например, элементы олигонуклеотида могут располагаться в следующем порядке: последовательность для слияния, последовательность AID, последовательность АМВ и сайт для праймера. Например, элементы олигонуклеотида могут располагаться в следующем порядке: последовательность для слияния, последовательность АМВ, последовательность AID и сайт для праймера.
Например, элементы олигонуклеотида могут располагаться в следующем порядке: последовательность для слияния, последовательность AID, последовательность АМВ, константная последовательность и сайт для праймера. Например, элементы олигонуклеотида могут располагаться в следующем порядке: последовательность для слияния, последовательность АМВ, последовательность AID, константная последовательность и сайт для праймера. Например, элементы олигонуклеотида могут располагаться в следующем порядке: последовательность для слияния, константная последовательность, последовательность AID, последовательность АМВ и сайт для праймера. Например, элементы олигонуклеотида могут располагаться в следующем порядке: последовательность для слияния, константная последовательность, последовательность АМВ, последовательность AID и сайт для праймера. Например, элементы олигонуклеотида могут располагаться в следующем порядке: последовательность для слияния, последовательность AID, константная последовательность, последовательность АМВ и сайт для праймера. Например, элементы олигонуклеотида могут располагаться в следующем порядке: последовательность для слияния, последовательность АМВ, константная последовательность, последовательность AID и сайт для праймера.
Например, элементы олигонуклеотида могут располагаться в следующем порядке: последовательность для слияния, последовательность AID, последовательность АМВ, константная последовательность и сайт для праймера, продолжаясь по направлению к аффинной части конъюгата аффинная молекула-олигонуклеотид. Например, элементы олигонуклеотида могут располагаться в следующем порядке: последовательность для слияния, последовательность AID, последовательность АМВ, константная последовательность и сайт для праймера, продолжаясь по направлению к аффинной части конъюгата аффинная молекула-олигонуклеотид от 5' конца к 3' концу олигонуклеотида. Например, олигонуклеотид может содержать 5'-концевую последовательность для слияния, последовательность AID, последовательность АМВ, константную последовательность и 3' сайт для праймера, прикрепленный к аффинной части конъюгата аффинная молекула-олигонуклеотид {например, через группу первичного амина в антителе, прикрепленном к 3' концу олигонуклеотида), в таком порядке.
Получение конъюгата аффинная молекула-олигонуклеотид
Конъюгаты аффинная молекула-олигонуклеотид или конъюгаты тетрамер-олигонуклеотид, используемые в способах и композициях, описанных в данном документе, можно получить с применением любого удобного способа. Аффинная часть может быть прямо или непрямо (например, через линкер) связана с олигонуклеотидом. Аффинная часть может быть связана ковалентно (например, с помощью химического сшивания) или нековалентно (например, посредством взаимодействия стрептавидин-биотин) с олигонуклеотидом. Конструирование и получение конъюгатов аффинная молекула-олигонуклеотид широко описано в уровне техники, в том числе различные отличающиеся аффинные части, которые можно применять, конструирование олигонуклеотидов для анализов методом близкого лигирования и связывание таких олигонуклеотидов с аффинными частями с образованием конъюгатов аффинная молекула-олигонуклеотид. Детали и принципы, описанные в уровне техники, можно применять к конструированию конъюгатов аффинная молекула-олигонуклеотид для применения в способах согласно настоящему изобретению (см., например, международную заявку WO 2007107743, и патенты США №7306904 и №6878515).
Можно использовать реакцию прямого связывания между олигонуклеотидом и аффинной частью, например, в которой каждый из них обладает функциональной группой (например, заместителем или группой для химической обработки), способной реагировать с функциональной группой на другой молекуле. Функциональные группы могут присутствовать на олигонуклеотиде или аффинной части или могут быть введены в эти компоненты (например, посредством реакций окисления, реакций восстановления, реакций расщепления и т.п.). Были описаны способы получения конъюгатов нуклеиновая кислота/полипептид (см., например, патент США №5733523).
Функциональные группы в антителе или полипептиде, которые можно применять для связывания с олигонуклеотидом, включают в себя, без ограничения, углеводы, тиольные группы (HS-) аминокислот, аминогруппы аминокислот (H2N-) и карбокси-группы аминокислот. Например, углеводные структуры могут окисляться до альдегидов и реагировать с соединением, содержащим H2NNH-группу, с образованием функциональной группы -C=NH-NH-. Например, можно обеспечивать реакцию тиольных групп с группой, реагирующей с тиолом, с образованием тиоэфира или дисульфида. Например, свободные тиольные группы белков можно вводить в белки посредством тиолирования или расщепления дисульфидов в нативных цистеиновых остатках. Например, можно обеспечить реакцию аминогруппы (например, на амино-конце или омега-аминогруппы в лизиновом остатке) с электрофильной группой (например, активированной карбокси-группой) с образованием амидной группы. Например, карбокси-группу (например, карбокси-конец или карбокси-группу двухосновной альфа-аминокислоты) можно активировать и привести в контакт с аминогруппой с образованием амидной группы. Другие иллюстративные функциональные группы включают в себя, например, SPDP, карбодиимид, глутаровый альдегид и т.п.
В соответствии с иллюстративным вариантом осуществления олигонуклеотид ковалентно связывают с аффинной частью с применением коммерческого набора (универсальный набор для конъюгации антител и нуклеотидов "All-in-One Antibody-Oligonucleotide Conjugation Kit"; Solulink, Inc.). Например, во-первых, 3'-амино-олигонуклеотид можно дериватизировать сульфо-S-4FB. Во-вторых, аффинную часть можно модифицировать S-HyNic группой. В-третьих, можно обеспечить реакцию HyNic-модифицированной аффинной части с 4FB-модифицированным олигонуклеотидом с получением на выходе конъюгата аффинная молекула-олигонуклеотид с бис-арилгидразоном в качестве промежуточной части. Избыток 4FB-модифицированного олигонуклеотида можно в дальнейшем удалить с помощью магнитной аффинной матрицы. В целом, выделенная аффинная часть может быть по меньшей мере приблизительно на 95%, 96%, 97%, 98%, 99% или 100% свободной от HyNic-модифицированной аффинной части и 4FB-модифицированного олигонуклеотида. Бис-арилгидразоновая связь может быть устойчивой как к температуре {например, 94°С), так и к рН {например, 3-10).
В случае, когда используют связывающие группы, такие линкеры могут быть выбраны, чтобы обеспечить ковалентное прикрепление или нековалентное прикрепление аффинной части и олигонуклеотида через связывающую группу. Ряд подходящих линкеров являются известными в уровне техники. В соответствии с некоторыми вариантами осуществления линкер имеет молекулярную массу по меньшей мере приблизительно 50 или 100 дальтон. В соответствии с некоторыми вариантами осуществления линкер имеет молекулярную массу не более чем приблизительно 300; 500; 1000; 10000 или 100000 дальтон. Линкер может содержать функциональную группу на любом из концов, причем реакционноспособная функциональная группа способна связываться с олигонуклеотидом. Линкер может содержать функциональную группу на любом из концов, причем реакционноспособная функциональная группа способна связываться с аффинной частью. Функциональные группы могут присутствовать на олигонуклеотиде, аффинной части и/или линкере или могут быть введены в эти компоненты {например, посредством реакций окисления, реакций восстановления, реакций расщепления и т.п.).
Иллюстративные линкеры включают в себя полимеры, цепи алифатических углеводородов, ненасыщенные углеводородные цепи, полипептиды, полинуклеотиды, циклические линкеры, ациклические линкеры, углеводороды, эфиры, полиамины и другие известные в уровне техники. Иллюстративные функциональные группы линкеров включают в себя нуклеофильные функциональные группы {например, амины, аминогруппы, гидрокси-группы, сульфгидрильные группы, аминогруппы, спирты, тиолы и гидразиды), электрофильные функциональные группы {например, альдегиды, сложные эфиры, винилкетоны, эпоксиды, изоцианаты и малеимиды) и функциональные группы, способные к реакциям циклоприсоединения, образующие дисульфидные связи или связывающиеся с металлами. Например, функциональные группы линкеров могут представлять собой первичные амины, вторичные амины, гидроксамовые кислоты, N-гидроксисукцинимидиловые сложные эфиры, N-гидроксисукцинимидил-карбонаты, оксикарбонилимидазолы, нитрофенильные сложные эфиры, трифторэтиловые сложные эфиры, глицидиловые эфиры, винилсульфоны, малеимиды, азидобензоилгидразид, N-[4-(пара-азидосалициламино)бутил]-3'-[2'-пиридилдитио]пропионамид), бис-сульфосукцинимидилсуберат, диметиладипимидат, дисукцинимидилтартрат, N-малеимидобутирилоксисукцинимидный сложный эфир, N-гидроксисульфосукцинимидил-4-азидобензоат, N-сукцинимидил-[4-азидофенил]-1,3'-дитиопропионат, N-сукцинимидил-[4-йодацетил]аминобензоат, глутаровый альдегид и сукцинимидил-4-[N-малеимидометил]циклогексан-1-карбоксилат, N-гидроксисукцинимидный сложный эфир 3-(2-пиридилдитио)пропионовой кислоты (SPDP), N-гидроксисукцинимидный сложный эфир 4-(N-малеимидометил)-циклогексан-1-карбоновой кислоты (SMCC) и т.п.
В соответствии с другими вариантами осуществления конъюгаты аффинная молекула-олигонуклеотид или конъюгаты тетрамер-олигонуклеотид можно получать с применением in vitro протоколов, которые дают на выходе конъюгаты аффинная молекула-олигонуклеотид или конъюгаты тетрамер-олигонуклеотид, таких как получение аффинной части in vitro из векторов, которые кодируют аффинную часть. Примеры таких представляющих интерес in vitro протоколов включают в себя: протоколы на основе RepA (см., например, Fitzgerald et at, Drug Discov Today (2000) 5:253-58 и WO 9837186), протоколы на основе рибосомного дисплея (см., например, Hanes et at, PNAS (1997) 94:4937-42; Roberts et at, Curr Opin Chem Biol (1999) Jun; 3: 268-73; Schaffitzel et at, J Immunol Methods (1999) Dec 10; 231:119-35; и международная заявка WO 9854312) и т.д.
Методики для конъюгирования молекул нуклеиновой кислоты с антителами являются хорошо известными в уровне техники (см., например, Anion et al, "Monoclonal Antibodies For Immunotargeting Of Drugs In Cancer Therapy," в Monoclonal Antibodies And Cancer Therapy (Reisfeld et al. eds., Alan R. Liss, Inc., 1985); Hellstrom et al, "Antibodies For Drug Delivery," в Controlled Drug Delivery (Robinson et al. eds., Marcel Deiker, Inc., 2nd ed. 1987); Thorpe, "Antibody Carriers Of Cytotoxic Agents hi Cancer Therapy: A Review," в Monoclonal Antibodies '84: Biological And Clinical Applications (Pinchera et al. eds., 1985); "Analysis, Results, and Future Prospective of the Therapeutic Use of Radiolabeled Antibody hi Cancer Therapy," в Monoclonal Antibodies For Cancer Detection And Therapy (Baldwin et al. eds., Academic Press, 1985); и Thorpe et al, 1982, Immunol. Rev. 62: 119-58. См. также, например, PCT публикацию международной заявки WO 89/12624.) Например, молекула нуклеиновой кислоты может быть ковалентно прикреплена к лизинам или цистеинам на антителе, как например, через функциональную группу N-гидроксисукцинимидного сложного эфира или малеимида, соответственно.
Целевые антигены
Целевым антигеном для аффинной части может быть молекула нуклеиновой кислоты или им может быть белковая молекула, такая как целевой белок или пептид. Целевой антиген может представлять собой соединение или композицию, которые присутствуют на клетке в образце. В соответствии с некоторыми вариантами осуществления целевой антиген может представлять собой соединение или композицию, способные вызывать клеточно-опосредованную иммунную реакцию (то есть адаптивную иммунную реакцию), особенно, у млекопитающего, такого как человек. В соответствии с некоторыми вариантами осуществления целевой антиген может распознаваться Т-клеткой в контексте молекулы МНС. Целевые антигены включают в себя, без ограничения, клетки, тканевые экстракты, тканевые или клеточные лизаты, белки, отдельно или в виде смеси, множество белков, пептидов, смесей пептидов, липиды, углеводы, сахара и т.п. Целевой антиген может быть характерным для заболевания, такого как инфекционное заболевание, аутоиммунное заболевание или рак. Целевой антиген может представлять собой, например, вирусный антиген, бактериальный антиген, раковый антиген и т.д.
В соответствии с некоторыми вариантами осуществления целевой антиген представляет собой вирусный антиген. Вирусные антигены включают в себя, например, белок оболочки вируса, антиген вируса гриппа, антиген ВИЧ, антиген вируса гепатита В или антиген вируса гепатита С.
В соответствии с некоторыми вариантами осуществления целевой антиген представляет собой раковый антиген (например, белок, пептид, липид, углевод и т.д.), который исключительно или преимущественно экспрессируется или сверхэкспрессируется опухолевой клеткой или раковой клеткой, в результате чего антиген ассоциируется с опухолью или раком. Раковый антиген может представлять собой раковый антиген только из одного типа рака или опухоли, в результате чего раковый антиген ассоциируется только с одним типом рака или опухоли или является характерным для них. В качестве альтернативы, раковый антиген может представлять собой раковый антиген из более чем одного типа рака или опухоли (например, может быть характерным для них). Например, раковый антиген может экспрессироваться как клетками злокачественной опухоли молочной железы, так и клетками злокачественной опухоли предстательной железы и совсем не экспрессируется нормальными, не опухолевыми или не раковыми клетками или экспрессируется лишь минимально. Раковый антиген может представлять собой раковый антиген меланомы или антиген рака молочной железы. Иллюстративные раковые антигены включают в себя антигены из группы, состоящей из gp100. MART-1, NY-ESO-1, члена семейства белков MAGE, например, MAGE-A1, мезотелина, тирозиназы, TRP-1, TRP-2, PMSA, Her-2 и р53.
Целевой антиген может быть получен естественным, искусственным образом, посредством синтеза или методов на основе рекомбинации. Таким образом, целевой антиген может представлять собой синтетический, рекомбинантный, выделенный и/или очищенный белок, полипептид или пептид. Способы создания или получения таких антигенов являются известными в уровне техники. Например, подходящие способы синтеза de novo полипептидов и белков (например, антигенных полипептидов и белков) описаны в Chan et al., Fmoc Solid Phase Peptide Synthesis, Oxford University Press. Oxford, United Kingdom, 2005; Peptide and Protein Drug Analysis, ed. Reid, R., Marcel Dekker, Inc., 2000; Epitope Mapping, ed. Westwood et al., Oxford University Press. Oxford. United Kingdom, 2000; и в патенте США №5449752. Также, полипептиды и белки (например, антигенные полипептиды и белки) можно получать с помощью способов на основе рекомбинации с применением нуклеиновых кислот, которые кодируют полипептид или белок, с применением стандартных способов на основе рекомбинации. См., например, Sambrook et al., Molecular Cloning: A Laboratory Manual, 3rd ed. Cold Spring Harbor Press, Cold Spring Harbor, N.Y. 2001; и Ausubel et al., Current Protocols in Molecular Biology. Greene Publishing Associates and John Wiley & Sons, NY, 1994. Нуклеотидные последовательности многих антигенов являются известными в уровне техники, и они доступны из базы данных GenBank на веб-сайте Национального центра биотехнологической информации (NCBI). Кроме того, антиген можно выделить из и/или очистить от источника, такого как растение, бактерия, насекомое, млекопитающее, например, крыса, человек и т.д. Способы выделения и очистки являются хорошо известными в уровне техники.
Антиген может представлять собой свободный антиген, например, несвязанный антигенный пептид (например, свободный пептид), или может представлять собой связанный антиген, например, тетрамер МНС-пептид или антигенный пептид, презентируемый клеткой-носителем, которая была активирована пептидом.
В соответствии с некоторыми вариантами осуществления целевое анализируемое вещество представляет собой мембраносвязанный белок. В соответствии с одним вариантом осуществления мембраносвязанный белок представляет собой CD4, классический мембранный белок I типа с одним трансмембранным (ТМ) доменом. (Carr et at, (1989) J. Biol. Chem. 264:21286-95). В соответствии с другим вариантом осуществления мембраносвязанный белок представляет собой GPR77, несколько раз пересекающий мембрану, сопряженный с G-белком рецепторный (GPCR) мембранный белок. (Cain & Monk, (2002) J. Biol. Chem. 277:7165-69).
Дополнительные иллюстративные мембраносвязанные белки включают в себя, без ограничения, GPCR (например, адренергические рецепторы, ангиотензиновые рецепторы, холе цистокинино вые рецепторы, мускариновые ацетилхолиновые рецепторы, нейротензиновые рецепторы, галаниновые рецепторы, дофаминовые рецепторы, опиоидные рецепторы, серотониновые рецепторы, соматостатиновые рецепторы и т.д.), ионные каналы (например, никотиновые ацетилхолиновые рецепторы, натриевые каналы, калиевые каналы и т.д.), рецепторные тирозин-киназы, рецепторные серин/треонин-киназы, рецепторные гуанилатциклазы, рецепторы факторов роста и гормонов (например, рецептор эпидермального фактора роста (EGF)) и другие. Также можно применять мутантные или модифицированные варианты мембраносвязанных белков. Например, некоторые единичные или множественные точковые мутации GPCR сохраняют функцию и вовлечены в заболевание (см., например, Stadel et al., (1997) Trends in Pharmacological Review 18:430-37).
Характеристика отдельных клеток, мечение клеток полинуклеотидом со штрихкодом и спаривание цепей
Способы, описанные в данном документе, могут включать характеристику клеток с использованием конъюгатов аффинная молекула-олигонуклеотид или конъюгатов тетрамер-олигонуклеотид. Множество клеток могут быть приведены в контакт с одним или несколькими конъюгатами аффинная молекула-олигонуклеотид или одним или несколькими конъюгатами тетрамер-олигонуклеотид. Клетки можно отмывать для удаления несвязанных конъюгатов. Клетки можно выделять в резервуарах в виде отдельных клеток. Конъюгаты аффинная молекула-олигонуклеотид, связанные с выделенными клетками, можно модифицировать таким образом, чтобы они содержали последовательность штрихкода резервуара, как например, посредством прикрепления меченого штрихкодом резервуара полинуклеотида к олигонуклеотиду в составе конъюгатов. Конъюгаты тетрамер-олигонуклеотид, связанные с выделенными клетками, можно модифицировать таким образом, чтобы они содержали последовательность штрихкода резервуара, как например, посредством прикрепления меченого штрихкодом резервуара полинуклеотида к олигонуклеотиду в составе конъюгатов.
Полинуклеотид, несущий штрихкод резервуара, также можно вводить во время образования резервуаров. Эти меченые штрихкодом резервуара полинуклеотиды могут нести вырожденные штрихкоды, в результате чего каждый олигонуклеотид, содержащий штрихкод резервуара, содержит уникальный идентификационный код, соответствующий резервуару, в котором они находятся.
Олигонуклеотиды можно амплифицировать и амплифицированные продукты реакции можно выделить из резервуаров. Амплифицированные продукты могут быть обогащены с помощью ПЦР с целью добавления меток для секвенирования нового поколения (NGS). Библиотеку можно секвенировать с применением платформы для секвенирования с высокой пропускной способностью с последующим анализом последовательностей штрихкода резервуара, и/или последовательностей AID, и/или последовательностей АМВ. Поскольку каждая отдельная клетка является выделенной в своем соответствующем резервуаре, каждый штрихкода резервуара наблюдается дважды, когда секвенированные амплифицированные продукты олигонуклеотида происходят из одного и того же резервуара и, следовательно, из уникальной отдельной клетки. Поскольку каждая AID в олигонуклеотиде выступает в качестве штрихкода для аффинной части конъюгата аффинная молекула-олигонуклеотид или конъюгатов тетрамер-олигонуклеотид, к которым она прикреплена, и каждая отдельная клетка является выделенной в своем соответствующем резервуаре, каждая AID наблюдается в случае последовательностей, содержащих одинаковый штрихкод резервуара, когда секвенированные амплифицированные продукты олигонуклеотида происходят из конкретного конъюгата аффинная молекула-олигонуклеотид, связанного с отдельной клеткой, в одном и том же резервуаре. Каждый отличающийся АМВ отдельно наблюдают в наборе последовательностей, все из которых содержат одинаковый штрихкод резервуара, когда амплифицированные продукты олигонуклеотида, имеющего АМВ в наборе секвенированных, происходят из отличающейся (по сравнению с другими отдельными АМВ) одной олигонуклеотидной части одной молекулы конъюгата аффинная молекула-олигонуклеотид, связанной с клеткой в одном и том же резервуаре, например, в случаях, в которых применяют резервуары с отдельными клетками, связанной с отдельной клеткой в резервуаре. Каждый отдельный АМВ наблюдают, когда все амплифицированные продукты олигонуклеотида с последовательностями, содержащими одинаковый штрихкод резервуара, происходят из олигонуклеотидной части одной (одной и той же) молекулы конъюгата аффинная молекула-олигонуклеотид или молекулы конъюгата тетрамер-олигонуклеотид (например, представляют собой дубликаты или ампликоны в результате ПЦР). Таким образом, каждый олигонуклеотид, наблюдаемый с заданной комбинацией конкретного АМВ и конкретного штрихкода резервуара, указывает на отдельную молекулу конъюгата аффинная молекула-олигонуклеотид, связавшегося с отдельной клеткой, или конъюгата тетрамер-олигонуклеотид, связавшегося с отдельной клеткой; следовательно, выявление заданного количества (например, 2, 3, 4 или более) из многочисленных секвенированных олигонуклеотидов с такой комбинацией АМВ/штрихкод резервуара является показателем количества (например, 2, 3, 4 или более) конъюгатов аффинная молекула-олигонуклеотид, связавшихся с отдельной клеткой, или конъюгатов тетрамер-олигонуклеотид, связавшихся с отдельной клеткой, например, в анализируемой клеточной популяции или образце. Таким образом, такое количество может быть показателем количества молекул на клетке, для связывания с которой предназначена данная аффинная часть конъюгата аффинная молекула-олигонуклеотид, например, количества копий, экспрессирующихся на или в отдельной клетке.
В соответствии с некоторыми вариантами осуществления способы, описанные в данном документе, дополнительно включают мечение штрихкодом полинуклеотидов, полученных из клеток, и/или полинуклеотидов в резервуаре, которые являются отличными от конъюгатов аффинная молекула-олигонуклеотид или конъюгатов тетрамер-олигонуклеотид и отличными от их частей или копий. Например, отдельные клетки, инкапсулированные в резервуарах, с которыми связываются или связались конъюгаты аффинная молекула-олигонуклеотид или конъюгаты тетрамер-олигонуклеотид, можно подвергнуть лизису, и дополнительные полинуклеотиды, такие как полинуклеотиды из отдельной клетки или в отдельной клетке, можно пометить штрихкодом. В соответствии с некоторыми вариантами осуществления олигонуклеотидную часть конъюгата аффинная молекула-олигонуклеотид или конъюгата тетрамер-олигонуклеотид в резервуаре, например, такого, который связывается или связался с отдельной клеткой в резервуаре, и/или ее копию или амплифицированный продукт можно пометить штрихкодом с использованием последовательности штрихкода резервуара. В соответствии с некоторыми вариантами осуществления один или несколько клеточных полинуклеотидов из отдельной клетки можно пометить штрихкодом с использованием одинаковой последовательности штрихкода резервуара; такие дополнительные один или несколько клеточных полинуклеотидов в соответствии с некоторыми вариантами осуществления являются дополнительно помеченными с использованием молекулярных штрихкодов.
Как пары цепей Т-клеточного рецептора, так и пары иммуноглобулиновых цепей антитела представляют собой типы иммунных рецепторов. В соответствии с некоторыми вариантами осуществления клеточный полинуклеотид представляет собой полинуклеотид, кодирующий иммуноглобулиновую цепь (или часть) антитела; в соответствии с некоторыми вариантами осуществления, он представляет собой или содержит полинуклеотид, кодирующий TCR (или часть). В соответствии с некоторыми вариантами осуществления антиген, связанный с конъюгатом аффинная молекула-олигонуклеотид, представляет собой TCR или антитело или их цепь или часть. В соответствии с одним аспектом способы, описанные в данном документе, дополнительно включают создание библиотек полинуклеотидов для секвенирования с высокой пропускной способностью и диагностики. В соответствии с одним аспектом способы, описанные в данном документе, дополнительно включают разработку панелей полученных от человека библиотек для обнаружения антитела и/или TCR от пациента или когорт со специфическими общими свойствами. Раскрытое изобретение можно применять к многочисленным отличающимся типам парных вариабельных последовательностей, например, парам цепей Т-клеточного рецептора и парам иммуноглобулиновых цепей антитела, вместе с характеристикой отдельных клеток с применением конъюгатов аффинная молекула-олигонуклеотид. Например, полинуклеотиды, комплементарные клеточным полинуклеотидам, как например, полинуклеотидам тяжелой и/или легкой цепи, например, VH и/или VL цепей антител, и/или альфа, и/или бета, и/или гамма, и/или дельта цепей, например, Vα/Vβ и Vγ/Vδ цепей Т-клеточного рецептора (TCR) (как например, полученные из их каркасных частей), можно вводить во время образования резервуаров (или включать в них). Полинуклеотид, несущий штрихкод резервуара, также можно вводить во время образования резервуара (или включать в него). Эти меченые штрихкодом резервуара полинуклеотиды могут нести вырожденные штрихкоды, в результате чего каждый клеточный полинуклеотид, содержащий штрихкод резервуара, содержит уникальный идентификационный код, соответствующий резервуару, в котором он находится во время реакции(реакций). Таким образом, в соответствии с некоторыми такими вариантами осуществления, множество полинуклеотидов с одинаковым уникальным идентификационным кодом считаются происходящими из одного и того же резервуара и в соответствии с некоторыми аспектами, следовательно, из одной клетки. Множество полинуклеотидов, несущих молекулярный штрихкод, также можно вводить во время образования резервуаров или включать в них. Эти содержащие молекулярный штрихкод полинуклеотиды могут нести вырожденные штрихкоды, в результате чего каждая молекула клеточного полинуклеотида, содержащая молекулярный штрихкод, содержит уникальный идентификационный код, соответствующий молекуле полинуклеотида одной клетки, из которой они произошли. Миллионы отдельных иммунных клеток можно лизировать внутри эмульсии, и клеточные транскрипты, такие как транскрипты VH и VL, и/или Vα/Vβ, и/или Vγ/Vδ цепей, можно подвергать обратной транскрипции или копировать с применением праймеров с последующим мечением штрихкодом резервуара и молекулярным штрихкодом и ПЦР амплификацией меченых штрихкодом полинуклеотидов. Каждую из VH и VL и/или Vα/Vβ и/или Vγ/Vδ цепей, происходящих из одной иммунной клетки (например, В-клетки или Т-клетки), можно виртуально связать друг с другом с использованием идентификации по одинаковому штрихкоду резервуара.
VH и VL и/или Vα/Vβ и/или Vγ/Vδ цепи можно затем выделить из резервуаров и подвергнуть обогащению с помощью ПЦР с целью добавления меток для секвенирования нового поколения (NGS). Библиотеку можно подвергнуть секвенированию с применением платформы для секвенирования с высокой пропускной способностью с последующим анализом разнообразия репертуара, частоты антитела, характеристики CDR3, филогенетического анализа соматических гипермутаций и т.д. Базу данных корректно подобранных пар VH и VL, и/или Vα/Vβ, и/или Vγ/Vδ можно создать посредством деконволюции последовательностей штрихкода резервуара и молекулярного штрихкода. Поскольку каждая отдельная иммунная клетка является выделенной в своем соответствующем резервуаре, каждый штрихкод резервуара наблюдается дважды, когда секвенированные транскрипты происходят из одних и тех же капель эмульсии и, следовательно, из уникальной отдельной клетки. В случае каждого отличающегося наблюдаемого молекулярного штрихкода последовательности содержат одинаковый штрихкод резервуара, когда секвенированные транскрипты происходят из отличающейся молекулы транскрипта из отдельной клетки. В случае каждого одинакового наблюдаемого молекулярного штрихкода последовательности содержат одинаковый штрихкод резервуара, когда секвенированные транскрипты происходят из одной и той же молекулы транскрипта из отдельной клетки (например, дубликаты в результате ПЦР).
Параллельно с секвенированием библиотеку VH и VL и/или Vα/Vβ и/или Vγ/Vδ цепей, выделенных из резервуаров, можно клонировать в экспрессионные векторы для антител и котрансфицировать для скрининга посредством дисплея на клетках дрожжей. Клонирование этого пула идентичных библиотек является предпочтительным способом по сравнению с разделением биологического образца вначале, поскольку лишь некоторые редкие иммунные клетки будут охватываться в одном или другом анализе. Библиотеку полученных от человека VH и VL и/или Vα и Vβ, и/или Vγ и Vδ цепей можно экспрессировать независимо от корректного или некорректного соответствия пар, как в случае классических анализов методом дисплея. Затем можно осуществлять дисплей на клетках дрожжей в отношении одного или нескольких антигенов-мишеней с целью обогащения потенциальными антителами-кандидатами.
Положительные антитела-кандидаты, выявляемые в результате технологий дисплея, таких как дисплей на клетках дрожжей, можно подвергнуть секвенированию и использовать в качестве последовательности запроса относительно базы данных штрихкодов соответствующих пар. Каждая подвергшаяся дисплею на клетках дрожжей VH и/или Vα и/или Vγ цепь может снова образовывать пару со своей соответствующей VL или Vβ или Vδ цепью, соответственно, и каждая подвергшаяся дисплею на клетках дрожжей VL и/или Vβ и/или Vδ цепь моэет снова образовывать пару со своей соответствующей VH или Vα или Vγ цепью, соответственно. Эти правильно спаренные кандидаты могут быть подвергнуты синтезу гена и их можно экспрессировать в линиях клеток млекопитающих и подтвердить их функцию в отношении мишени, представляющей интерес. Эти кандидаты могут представлять собой полностью человеческие антитела и/или TCR.
Образцы
В соответствии с некоторыми вариантами осуществления любой образец, содержащий полинуклеотиды, можно применять в способах, описанных в данном документе. Любой образец, содержащий клетку, обычно можно применять в способах, описанных в данном документе. Например, образец может представлять собой биологический образец от субъекта или из образца, полученного от него, содержащий РНК или ДНК. Полинуклеотиды можно экстрагировать из биологического образца, или образец можно сразу подвергнуть воздействию способов без экстракции или очистки полинуклеотидов. Образец может представлять собой экстрагированную или выделенную ДНК или РНК. Образец также может представлять собой общую РНК или ДНК, экстрагированную из биологического образца, библиотеку кДНК, вирусную или геномную ДНК. В соответствии с одним вариантом осуществления полинуклеотиды являются выделенными из биологического образца, содержащего ряд других соединений, таких как белки, липиды и нематричные нуклеиновые кислоты. Молекулы матрицы нуклеиновой кислоты могут быть получены из любого клеточного материала, полученного от животного, растения, бактерии, гриба или любого другого клеточного организма. В соответствии с определенными вариантами осуществления полинуклеотиды получены из отдельной клетки. Полинуклеотиды можно получить непосредственно из организма или из биологического образца, полученного из организма. Любой образец ткани или жидкости организма можно применять в качестве источника нуклеиновой кислоты для применения в настоящем изобретении. Полинуклеотиды также могут быть выделены из культивируемых клеток, таких как первичная клеточная культура или клеточная линия. Клетки или ткани, из которых получены матричные нуклеиновые кислоты, могут быть инфицированными вирусом или другим внутриклеточным патогеном.
В соответствии с определенными вариантами осуществления иммунные клетки, продуцирующие антитело или TCR, можно выделить из крови или других биологических образцов от субъекта или хозяина, такого как человек или другое животное, такого как человек или другое животное, которые были иммунизированы, или которые страдают от инфекции, рака, аутоиммунного состояния или любых других заболеваний, для идентификации специфического для патогена, опухоли и/или заболевания антитела или TCR, имеющего возможную клиническую значимость. Например, у человека может быть диагностировано заболевание, могут проявляться симптомы заболевания, может не диагностироваться заболевание или могут не проявляться симптомы заболевания. Например, человек может представлять собой человека, который подвергался воздействию возбудителя инфекции (например, вирусов, бактерий, паразитов, прионов и т.д.), антигена или заболевания, и/или который может получить пользу от антител или TCR против возбудителя инфекции (например, вирусов, бактерий, паразитов, прионов и т.д.), антигена или заболевания. Например, животное может представлять собой животное, которое подвергалось воздействию возбудителя инфекции (например, вирусов, бактерий, паразитов, прионов и т.д.), антигена или заболевания, и/или животное, которое может получить пользу от антител или TCR против возбудителя инфекции (например, вирусов, бактерий, паразитов, прионов и т.д.), антигена или заболевания. Определенные иммунные клетки от иммунизированных хозяев вырабатывают антитела или TCR против одного или нескольких рассматриваемых целевых антигенов и/или одного или нескольких неизвестных антигенов. Согласно настоящему изобретению пул лимфоцитов может быть обогащен желательными иммунными клетками с помощью любого подходящего способа, такого как скрининг и сортинг клеток с применением флуоресцентно-активированного клеточного сортинга (FACS), магнитно-активированного клеточного сортинга (MACS), пэннинга или другого способа скрининга для получения множества иммунных клеток из образца, такого как библиотека иммунных клеток, до секвенирования цепей антитела, получения антитела или получения экспрессионной библиотеки. В отличие от способов обогащения из предшествующего уровня техники, которые обеспечивают только небольшое количество субпопуляций иммунных клеток, экспрессирующих различные антитела, и, следовательно, только небольшое количество встречающихся в естественных условиях комбинаций вариабельных доменов, библиотека иммунных клеток согласно настоящему изобретению содержит по меньшей мере 2 субпопуляции иммунных клеток или отдельные иммунные клетки, экспрессирующие различные антитела или TCR. Например, библиотека иммунных клеток согласно настоящему изобретению может содержать по меньшей мере 5, 10, 100, 250, 500, 750, 1000, 2500, 5000, 10000, 25000, 50000, 75000, 10000, 250000, 500000, 750000, 1000000, 2500000, 5000000, 7500000 или 10000000 субпопуляций или отдельных иммунных клеток, экспрессирующих отличающиеся антитела или TCR. Способы согласно настоящему изобретению максимально повышают выделение иммунных клеток и обеспечивают очень высокое разнообразие.
Т-клетки могут быть получены из ряда источников, в том числе мононуклеарных клеток периферической крови, костного мозга, тимуса, биоптата ткани, опухоли, ткани лимфатического узла, ассоциированной с кишечником лимфоидной ткани, ассоциированной со слизистой лимфоидной ткани, ткани селезенки или любой другой лимфоидной ткани и опухолей. Т-клетки могут быть получены из линий Т-клеток и из аутологичных или аллогенных источников. Т-клетки могут быть получены из отдельного индивида или популяции индивидов, например, из популяции индивидов, все из которых страдают от одинакового заболевания, такого как рак или инфекционное заболевание. В соответствии с некоторыми вариантами осуществления клетки из циркулирующей крови у индивида получают с помощью афереза или лейкафереза. Продукт афереза, как правило, содержит лимфоциты, в том числе Т-клетки, моноциты, гранулоциты, В-клетки, другие ядерные лейкоциты, эритроциты и тромбоциты. В соответствии с одним вариантом осуществления клетки, собранные с помощью афереза или лейкафереза, могут подвергать отмывке для удаления фракции плазмы и для помещения клеток в соответствующий буфер или среду для последующих стадий обработки. В соответствии с одним вариантом осуществления настоящего изобретения клетки отмывают фосфатно-буферным солевым раствором (PBS). В соответствии с альтернативным вариантом осуществления в отмывочном растворе отсутствует кальций и может отсутствовать магний или могут отсутствовать многие, если не все двухвалентные катионы. Как будет хорошо понятно квалифицированному специалисту в данной области техники, стадию отмывки можно осуществлять с помощью способов, известных специалистам в данной области техники, как например, с применением полуавтоматической «проточной» центрифуги. После отмывки клетки можно ресуспендировать в ряде биологически совместимых буферах, таких как, например, не содержащий Ca++/Mg++ PBS. В качестве альтернативы, нежелательные компоненты образца от афереза можно удалить, и клетки непосредственно ресуспендировать в среде культивирования. В соответствии с другими вариантами осуществления Т-клетки выделяют из лимфоцитов периферической крови посредством лизиса эритроцитов и посредством центрифугирования в градиенте плотности PERCOLL™. Конкретную субпопуляцию Т-клеток, такую как CD28+, CD4+, CD8+, CD45RA+ и CD45RO+ Т-клетки, можно дополнительно выделить с помощью методик положительной или отрицательной селекции. Например, CD3+, CD28+ Т-клетки могут быть подвергнуты положительной селекции с применением конъюгированных с CD3/CD28 магнитных гранул (например, DYNABEADS® М-450 CD3/CD28 Т Cell Expander). В соответствии с некоторыми вариантами осуществления обогащение Т-клеточной популяции посредством отрицательной селекции можно осуществлять с использованием комбинации антител, направленных на поверхностные маркеры, уникальные для подвергающихся отрицательной селекции клеток. Один такой способ представляет собой клеточный сортинг и/или селекцию посредством методов отрицательной магнитной иммуноадгезии или проточной цитометрии, в которых применяется коктейль моноклональных антител, направленных на маркеры клеточной поверхности, присутствующие на клетках, подвергаемых отрицательной селекции. Например, в случае обогащения CD4+ клетками посредством отрицательной селекции коктейль моноклональных антител, как правило, включает в себя антитела к CD14, CD20, CD11b, CD16, HLA-DR и CD8. Еще один способ получения Т-клеток для стимуляции заключается в замораживании клеток после стадии отмывки, что не требует стадии удаления моноцитов. Не вдаваясь в какую-либо теорию, замораживание и последующая стадия оттаивания обеспечивает более однородный продукт посредством удаления гранулоцитов и, в некоторой мере, моноцитов из клеточной популяции. После стадии отмывки, в ходе которой удаляют плазму и тромбоциты, клетки можно суспендировать в растворе для замораживания. Несмотря на то что многие растворы для замораживания и параметры замораживания являются известными в уровне техники и будут пригодными в этом контексте, один способ включает применение PBS, содержащего 20% DMSO и 8% человеческого сывороточного альбумина (HSA), или другой подходящей среды для замораживания клеток. Ее затем разводят в соотношении 1:1 средой для того, чтобы конечная концентрация DMSO и HSA составляла 10% и 4%, соответственно. Клетки затем замораживают до -80°С со скоростью, составляющей 1°С в минуту, и хранят в паровой фазе в цистерне для хранения с жидким азотом.
В соответствии с некоторыми вариантами осуществления используют иммунные клетки от неиммунизированных доноров-людей или доноров, отличных от человека. Наивный репертуар у животного (репертуар до антигенной стимуляции) обеспечивает животное с антителами или TCR, которые могут связываться с умеренной аффинностью (KA, составляющая приблизительно от 1×10-6 до 1×10-7 М) практически с любой «не-своей» молекулой. Разнообразие последовательности сайтов связывания антитела или TCR не закодировано напрямую в генах зародышевого типа, а собирается на комбинаторно из сегментов V-гена. Иммунизации запускают пролиферацию (клональное размножение) любой иммунной клетки, образующей комбинацию VH-VL, или Vα-Vβ, или Vγ-Vδ, которая связывается с иммуногеном, и секрецию ею соответствующего антитела, как отмечено выше. Тем не менее, применение клеток селезенки и/или иммунных клеток или других лимфоцитов периферической крови (PBL) от неиммунизированного субъекта может обеспечивать лучшее представление о возможном репертуаре антител или TCR, а также позволяет конструировать последующую библиотеку B-клеточных или T-клеточных антител или TCR с использованием животного любого вида.
В некоторых случаях для того, чтобы получить достаточное количество нуклеиновой кислоты для исследования, забирают объем крови, составляющий по меньшей мере 0,001, 0,005, 0,01, 0,05, 0,1, 0,5, 1, 2, 3, 4, 5, 10, 20, 25, 30, 35, 40, 45 или 50 мл.
В некоторых случаях исходный материал представляет собой периферическую кровь. Клетки периферической крови могут быть обогащены конкретным типом клеток (например, мононуклеарными клетками; эритроцитами; CD4+-клетками; CD8+-клетками; иммунными клетками; Т-клетками, NK-клетками или т.п.). Клетки периферической крови также могут быть селективно обеднены конкретным типом клеток (например, мононуклеарными клетками; эритроцитами; CD4+-клетками; CD8+-клетками; иммунными клетками; Т-клетками, NK-клетками или т.п.).
В некоторых случаях, исходный материал может представлять собой образец ткани, содержащий плотную ткань, неограничивающие примеры которых включают в себя ткань головного мозга, печени, легкого, почки, предстательной железы, яичника, селезенки, лимфатического узла (в том числе небной миндалины), щитовидной железы, поджелудочной железы, сердца, скелетной мышцы, кишечника, гортани, пищевода и желудка. В других случаях исходный материал может представлять собой клетки, содержащие нуклеиновые кислоты, иммунные клетки и, в частности, В-клетки или Т-клетки. В некоторых случаях исходный материал может представлять собой образец, содержащий нуклеиновые кислоты из любого организма, из которого можно получить генетический материал. В некоторых случаях образец представляет собой жидкость, например, кровь, слюну, лимфу или мочу.
Образец может быть взят у субъекта с состоянием. В некоторых случаях субъект, у которого берут образец, может являться пациентом, например, пациентом с раком или пациентом с подозрением на рак. Субъект может представлять собой млекопитающее, например, человека, и может являться самцом или самкой. В некоторых случаях самка является беременной. Образец может представлять собой опухолевый биоптат. Биопсию может осуществлять, например, медицинский работник, в том числе врач, помощник врача, медсестра, ветеринар, стоматолог, мануальный терапевт, фельдшер скорой помощи, дерматолог, онколог, гастроэнтеролог или хирург.
В некоторых случаях отличные от нуклеиновой кислоты материалы можно удалять из исходного материала с применением ферментативных обработок (таких как расщепление протеазой).
В некоторых случаях кровь можно собирать в аппарате, содержащем хелатор магния, в том числе, без ограничения, EDTA, и хранить при 4°С. Необязательно, можно добавлять хелатор кальция, в том числе, без ограничения, EGTA. В другом случае в кровь добавляют ингибитор лизиса клеток, в том числе, без ограничения, формальдегид, производные формальдегида, формалин, глутаровый альдегид, производные глутарового альдегида, сшивающее средство для белков, сшивающее средство для нуклеиновых кислот, сшивающее средство для белков и нуклеиновых кислот, сшивающие средства, реагирующие с первичным амином, сшивающие средства, реагирующие с сульфгидрильной группой, средства, присоединяющие сульфгидрильную группу, или средства, восстанавливающие дисульфидную группу, сшивающие средства, реагирующие с углеводами, сшивающие средства, реагирующие с карбоксильными группами, сшивающие средства с фотореакционной активностью или расщепляемые сшивающие средства.
В некоторых случаях, когда экстрагируемый материал содержит одноцепочечную РНК, двухцепочечную РНК или гибрид ДНК-РНК, эти молекулы можно превратить в двухцепочечную ДНК с применением методик, известных в данной области техники. Например, обратную транскриптазу можно использовать для синтеза ДНК из молекул РНК. В некоторых случаях превращение РНК в ДНК может потребовать предшествующей стадии лигирования, чтобы лигировать линкерный фрагмент с РНК, таким образом обеспечивая возможность применения универсальных праймеров для инициации обратной транскрипции. В других случаях поли-А хвост в молекуле мРНК, например, можно применять для инициации обратной транскрипции. После преобразования в ДНК способы, подробно описанные в данном документе, в некоторых случаях можно применять для того, чтобы дополнительно захватить, выбрать, пометить или выделить желательную последовательность.
Молекулы нуклеиновой кислоты включают в себя дезоксирибонуклеиновую кислоту (ДНК) и/или рибонуклеиновую кислоту (RNA). Молекулы нуклеиновой кислоты могут быть синтетическими или могут быть получены из природных источников. В соответствии с одним вариантом осуществления молекулы нуклеиновой кислоты являются выделенными из биологического образца, содержащего ряд других соединений, таких как белки, липиды и нематричные нуклеиновые кислоты. Молекулы матрицы нуклеиновой кислоты могут быть получены из любого клеточного материала, полученного от животного, растения, бактерии, гриба или любого другого клеточного организма. В соответствии с определенными вариантами осуществления молекулы нуклеиновой кислоты получены из отдельной клетки. Биологические образцы для применения в настоящем изобретении включают в себя вирусные частицы или препараты. Молекулы нуклеиновой кислоты могут быть получены непосредственно из организма или из биологического образца, полученного от организма, например, из крови, мочи, спинномозговой жидкости, семенной жидкости, слюны, мокроты, кала и ткани. Любой образец ткани или жидкости организма можно применять в качестве источника нуклеиновой кислоты для применения в настоящем изобретении. Молекулы нуклеиновой кислоты также могут быть выделены из культивируемых клеток, таких как первичная клеточная культура или клеточная линия. Клетки или ткани, из которых получены матричные нуклеиновые кислоты, могут быть инфицированными вирусом или другим внутриклеточным патогеном.
Образец также может представлять собой общую РНК, экстрагированную из биологического образца, библиотеку кДНК, вирусную или геномную ДНК. В соответствии с определенными вариантами осуществления молекулы нуклеиновой кислоты являются связанными с другими целевыми молекулами, такими как белки, ферменты, субстраты, антитела, связывающие средства, гранулы, малые молекулы, пептиды или любая другая молекула. Обычно нуклеиновую кислоту можно экстрагировать из биологического образца с помощью ряда методик, таких как описанные в Sambrook and Russell, Molecular Cloning: A Laboratory Manual, Third Edition, Cold Spring Harbor, N.Y. (2001). Молекулы нуклеиновой кислоты могут быть одноцепочечными, двухцепочечными или двухцепочечными с одноцепочечными участками (например, структурами типа «стебель» и «петля»).
Способы экстрагирования ДНК являются хорошо известными в уровне техники. Классический протокол выделения ДНК основывается на экстрагировании с применением органических растворителей, таких как смесь фенола и хлороформа, с последующим осаждением с использованием этанола (J. Sambrook et at., "Molecular Cloning: A Laboratory Manual," 1989, 2nd Ed., Cold Spring Harbour Laboratory Press: New York, N.Y.). Другие способы включают в себя: экстрагирование ДНК высаливанием (P. Sunnucks et at., Genetics, 1996, 144: 747-756; S.M. Aljanabi et al., Nucl. Acids Res. 1997, 25: 4692-4693), экстрагирование ДНК с использованием солей бромида триметиламмония (S. Gustincich et at., BioTechniques, 1991, 11: 298-302) и экстрагирование ДНК с использованием тиоцианата гуанидиния (J.В.W. Hammond et al., Biochemistry, 1996, 240: 298-300). Ряд наборов коммерчески доступны для экстрагирования ДНК из биологических образцов (например, BD Biosciences Clontech (Пало-Альто, Калифорния): Epicentre Technologies (Мадисон, Висконсин); Gentra Systems, Inc. (Миннеаполис, Миннесота); MicroProbe Corp. (Ботелл, Вашингтон); Organon Teknika (Дарем, Северная Каролина); и Qiagen Inc. (Валенсия, Калифорния)).
Способы экстрагирования РНК также являются хорошо известными в уровне техники (например, J. Sambrook et al., "Molecular Cloning: A Laboratory Manual" 1989, 211d Ed., Cold Spring Harbour Laboratory Press: New York), и наборы для экстрагирования РНК из жидкостей организма являются коммерчески доступными (например, Ambion, Inc. (Остин, Техас); Amersham Biosciences (Пискатауэй, Нью-Джерси); BD Biosciences Clontech (Пало-Альто, Калифорния); BioRad Laboratories (Геркулес, Калифорния); Dynal Biotech Inc. (Лейк Саксесс, Нью-Йорк); Epicentre Technologies (Мадисон, Висконсин); Gentra Systems, Inc. (Миннеаполис, Миннесота); GIBCO BRL (Гейтерсберг, Мэриленд); Invitrogen Life Technologies (Карлсбад, Калифорния); MicroProbe Corp. (Ботелл, Вашингтон); Organon Teknika (Дарем, Северная Каролина); Promega, Inc. (Мадисон, Висконсин) и Qiagen Inc. (Валенсия, Калифорния)).
Один или несколько образцов могут происходить из одного или нескольких источников. Один или несколько из образцов могут происходить из двух или более источников. Один или несколько из образцов могут происходить из одного или нескольких субъектов. Один или несколько из образцов могут происходить из двух или более субъектов. Один или несколько из образцов могут происходить из одного и того же субъекта. Один или несколько субъектов могут происходить из одного и того же вида. Один или несколько субъектов могут происходить из разных видов. Один или несколько субъектов могут быть здоровыми. Один или несколько субъектов могут быть поражены заболеванием, нарушением или состоянием.
В соответствии с некоторыми вариантами осуществления образец представляет собой жидкость, такую как кровь, слюна, лимфа, моча, спинномозговая жидкость, семенная жидкость, мокрота, кал или гомогенаты тканей.
Образец может быть взят у субъекта с состоянием. В соответствии с некоторыми вариантами осуществления субъект, у которого берут образец, может являться пациентом, например, пациентом с раком или пациентом с подозрением на рак. Субъект может представлять собой млекопитающее, например, человека, и может являться самцом или самкой. В соответствии с некоторыми вариантами осуществления самка является беременной. Образец может представлять собой опухолевый биоптат. Биопсию может осуществлять, например, медицинский работник, в том числе врач, помощник врача, медсестра, ветеринар, стоматолог, мануальный терапевт, фельдшер скорой помощи, дерматолог, онколог, гастроэнтеролог или хирург.
В соответствии с некоторыми вариантами осуществления полинуклеотиды связаны с другими целевыми молекулами, такими как белки, ферменты, субстраты, антитела, связывающие средства, гранулы, малые молекулы, пептиды или любая другая молекула. В соответствии с некоторыми вариантами осуществления полинуклеотиды не связаны с твердой подложкой. Нуклеиновые кислоты можно экстрагировать из биологического образца с помощью ряда методик (Sambrook et at., Molecular Cloning: A Laboratory Manual, Third Edition, Cold Spring Harbor, N.Y. (2001)).
В соответствии с некоторыми вариантами осуществления образец представляет собой слюну. В соответствии с некоторыми вариантами осуществления образец представляет собой цельную кровь. В соответствии с некоторыми вариантами осуществления для того чтобы получить достаточное количество полинуклеотидов для исследования, забирают объем крови, составляющий по меньшей мере приблизительно 0,001, 0,005, 0,01, 0,05, 0,1, 0,5, 1, 2, 3, 4, 5, 10, 20, 25, 30, 35, 40, 45 или 50 мл. В соответствии с некоторыми вариантами осуществления кровь можно собирать в аппарате, содержащем хелатор магния, в том числе, без ограничения, EDTA, и хранить при 4°С. Необязательно, можно добавлять хелатор кальция, в том числе, без ограничения, EGTA.
В соответствии с некоторыми вариантами осуществления в кровь добавляют ингибитор лизиса клеток, в том числе, без ограничения, формальдегид, производные формальдегида, формалин, глутаровый альдегид, производные глутарового альдегида, сшивающее средство для белков, сшивающее средство для нуклеиновых кислот, сшивающее средство для белков и нуклеиновых кислот, сшивающие средства, реагирующие с первичным амином, сшивающие средства, реагирующие с сульфгидрильной группой, средства, присоединяющие сульфгидрильную группу, или средства, восстанавливающие дисульфидную группу, сшивающие средства, реагирующие с углеводами, сшивающие средства, реагирующие с карбоксильными группами, сшивающие средства с фотореакционной активностью или расщепляемые сшивающие средства. В соответствии с некоторыми вариантами осуществления отличные от нуклеиновой кислоты материалы можно удалять из исходного материала с применением ферментативных обработок (таких как расщепление протеазой).
Множество образцов может содержать по меньшей мере 2, 3, 4, 5, 10, 20, 30, 40, 50, 60, 70, 80, 90 или 100 или большее количество образцов. Множество образцов может содержать по меньшей мере приблизительно 100, 200, 300, 400, 500, 600, 700, 800, 900 или 1000 или большее количество образцов. Множество образцов может содержать по меньшей мере приблизительно 1000, 2000, 3000, 4000, 5000, 6000, 7000, 8000 образцов, 9000 или 10000 образцов, или 100000 образцов, или 1000000 или большее количество образцов. Множество образцов может содержать по меньшей мере приблизительно 10000 образцов.
Один или несколько полинуклеотидов в первом образце могут отличаться от одного или нескольких полинуклеотидов во втором образце. Один или несколько полинуклеотидов в первом образце могут отличаться от одного или нескольких полинуклеотидов во множестве образцов. Один или несколько полинуклеотидов в образце могут характеризоваться по меньшей мере приблизительно 80%, 85%, 90%, 95%, 96%, 97%, 98%, 99% или 100% идентичностью последовательности. В соответствии с некоторыми вариантами осуществления один или несколько полинуклеотидов в образце могут отличаться менее чем приблизительно 100, 90, 80, 70, 60, 50, 40, 30, 25, 20, 25, 10, 9, 8, 7, 6, 5, 4, 3, 2 или 1 нуклеотидом или парой оснований. Множество полинуклеотидов в одном или нескольких образцах из множества образцов могут содержать две или более идентичных последовательностей. По меньшей мере приблизительно 1%, 2%, 3%, 4%, 5%, 6%, 7%, 8%, 9%, 10%, 15%, 20%, 25%, 30%, 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95%, 97%, 98%, 99% или 100% от общего количества полинуклеотидов в одном или нескольких из множества образцов могут содержать одинаковую последовательность. Множество полинуклеотидов в одном или нескольких образцах из множества образцов может содержать по меньшей мере две отличающиеся последовательности. По меньшей мере приблизительно 5%, 10%, 15%, 20%, 25%, 30%, 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% от общего количества полинуклеотидов в одном или нескольких из множества образцов могут содержать по меньшей мере две отличающиеся последовательности. В соответствии с некоторыми вариантами осуществления один или несколько полинуклеотидов представляют собой варианты друг друга. Например, один или несколько полинуклеотидов могут содержать однонуклеотидные полиморфизмы или другие типы мутаций. В другом примере один или несколько полинуклеотидов представляют собой сплайс-варианты.
Первый образец может содержать одну или несколько клеток, и второй образец может содержать одну или несколько клеток. Одна или несколько клеток в первом образце могут относиться к тому же типу клеток, что и одна или несколько клеток во втором образце. Один или несколько клеток в первом образце может относиться к типу клеток, отличному от одной или нескольких отличающихся клеток из множества образцов.
Множество образцов можно получать параллельно. Множество образцов можно получать в одно и то же время. Множество образцов можно получать последовательно. Множество образцов можно получать в течение периода порядка лет, например, 100 лет, 10 лет, 5 лет, 4 лет, 3 лет, 2 лет или 1 года от получения одного или нескольких отличающихся образцов. Один или несколько образцов можно получать в пределах приблизительно одного года от получения одного или нескольких отличающихся образцов. Один или несколько образцов можно получать в пределах 12 месяцев, 11 месяцев, 10 месяцев, 9 месяцев, 8 месяцев, 7 месяцев, 6 месяцев, 4 месяцев, 3 месяцев, 2 месяцев или 1 месяца от получения одного или нескольких отличающихся образцов. Один или несколько образцов можно получить в пределах 30 суток, 28 суток, 26 суток, 24 суток, 21 суток, 20 суток, 18 суток, 17 суток, 16 суток, 15 суток, 14 суток, 13 суток, 12 суток, 11 суток, 10 суток, 9 суток, 8 суток, 7 суток, 6 суток, 5 суток, 4 суток, 3 суток, 2 суток или 1 суток от получения одного или нескольких отличающихся образцов. Один или несколько образцов можно получать в пределах приблизительно 24 часов, 22 часов, 20 часов, 18 часов, 16 часов, 14 часов, 12 часов, 10 часов, 8 часов, 6 часов, 4 часов, 2 часов или 1 часа от получения одного или нескольких отличающихся образцов. Один или несколько образцов можно получать в пределах приблизительно 60 секунд, 45 секунд, 30 секунд, 20 секунд, 10 секунд, 5 секунд, 2 секунд или 1 секунды от получения одного или нескольких отличающихся образцов. Один или несколько образцов можно получать в пределах менее чем одной секунды от получения одного или нескольких отличающихся образцов.
Отличающиеся полинуклеотиды в образце могут присутствовать в образце в различных концентрациях или количествах (например, в различном количестве молекул). Например, концентрация или количество одного полинуклеотида могут быть большими, чем концентрация или количество другого полинуклеотида в образце. В соответствии с некоторыми вариантами осуществления концентрация или количество по меньшей мере одного полинуклеотида в образце являются по меньшей мере приблизительно в 1,5, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 20, 25, 30, 35, 40, 45, 50, 60, 70, 80, 90, 100, 200, 300, 400, 500, 600, 700, 800, 900, 1000 или более раз большими, чем концентрация или количество по меньшей мере одного другого полинуклеотида в образце. В еще одном примере, концентрация или количество одного полинуклеотида являются меньшими, чем концентрация или количество другого полинуклеотида в образце. Концентрация или количество по меньшей мере одного полинуклеотида в образце могут являться по меньшей мере приблизительно в 1,5, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 20, 25, 30, 35, 40, 45, 50, 60, 70, 80, 90, 100, 200, 300, 400, 500, 600, 700, 800, 900, 1000 или более раз меньшими, чем концентрация или количество по меньшей мере одного другого полинуклеотида в образце.
В соответствии с некоторыми вариантами осуществления два или более образца могут содержать отличающиеся количества или концентрации полинуклеотидов. В соответствии с некоторыми вариантами осуществления концентрация или количество одного полинуклеотида в одном образце могут быть большими, чем концентрация или количество того же полинуклеотида в отличающемся образце. Например, образец крови может содержать большее количество конкретного полинуклеотида, чем образец мочи. В качестве альтернативы один образец можно разделить на две или более части образца. Части образца могут содержать отличающиеся количества или концентрации одного и того же полинуклеотида. Концентрация или количество по меньшей мере одного полинуклеотида в одном образце могут быть по меньшей мере приблизительно в 1,5, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 20, 25, 30, 35, 40, 45, 50, 60, 70, 80, 90, 100, 200, 300, 400, 500, 600, 700, 800, 900, 1000 или более раз большими, чем концентрация или количество того же полинуклеотида в другом образце. В качестве альтернативы, концентрация или количество одного полинуклеотида в одном образце могут быть меньшими, чем концентрация или количество того же полинуклеотида в отличающемся образце. Например, концентрация или количество по меньшей мере одного полинуклеотида в одном образце могут быть по меньшей мере приблизительно в 1,5, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 20, 25, 30, 35, 40, 45, 50, 60, 70, 80, 90, 100, 200, 300, 400, 500, 600, 700, 800, 900, 1000 или более раз меньшими, чем концентрация или количество того же полинуклеотида в другом образце.
Целевые полинуклеотиды
В некоторых случаях представленные в данном документе способы направлены на амплификацию и секвенирование молекулы целевого полинуклеотида, такой как молекула полинуклеотида из клетки, олигонуклеотид в составе конъюгата аффинная молекула-олигонуклеотид или их ампликоны. В некоторых случаях представленные в данном документе способы направлены на амплификацию и секвенирование по меньшей мере одного участка молекулы целевого полинуклеотида. В некоторых случаях представленные в данном документе способы направлены на амплификацию и секвенирование по меньшей мере одной молекулы целевого полинуклеотида. В соответствии с одним аспектом целевые полинуклеотиды представляют собой олигонуклеотиды в составе конъюгатов аффинная молекула-олигонуклеотид. В соответствии с одним аспектом целевые полинуклеотиды представляют собой РНК. В соответствии с некоторыми вариантами осуществления целевые полинуклеотиды РНК представляют собой мРНК. В соответствии с некоторыми вариантами осуществления целевые полинуклеотиды РНК являются полиаденилированными. В соответствии с некоторыми вариантами осуществления полинуклеотиды РНК не являются полиаденилированными. В соответствии с некоторыми вариантами осуществления целевые полинуклеотиды представляют собой полинуклеотиды ДНК. Например, целевые полинуклеотиды включают в себя кДНК. Полинуклеотиды ДНК могут представлять собой геномную ДНК. Полинуклеотиды ДНК могут содержать экзоны, интроны, нетранслируемые участки или любую их комбинацию.
В соответствии с одним аспектом целевые полинуклеотиды представляют собой геномные нуклеиновые кислоты. ДНК, полученная из генетического материала в хромосомах конкретного организма, может представлять собой геномную ДНК. В соответствии с некоторыми вариантами осуществления целевые полинуклеотиды включают в себя последовательности, содержащие вариабельные участки антитела или TCR, продуцируемых иммунной клеткой. В соответствии с некоторыми вариантами осуществления целевые полинуклеотиды включают в себя последовательности, содержащие вариабельный участок тяжелой цепи антитела, продуцируемого иммунной клеткой. В соответствии с некоторыми вариантами осуществления целевые полинуклеотиды включают в себя последовательности, содержащие вариабельный участок легкой цепи антитела, продуцируемого иммунной клеткой. В соответствии с некоторыми вариантами осуществления целевые полинуклеотиды включают в себя последовательности, содержащие вариабельный участок альфа-цепи TCR, продуцируемого иммунной клеткой. В соответствии с некоторыми вариантами осуществления целевые полинуклеотиды включают в себя последовательности, содержащие вариабельный участок бета-цепи TCR, продуцируемого иммунной клеткой. В соответствии с некоторыми вариантами осуществления целевые полинуклеотиды включают в себя последовательности, содержащие вариабельный участок гамма-цепи TCR, продуцируемого иммунной клеткой. В соответствии с некоторыми вариантами осуществления целевые полинуклеотиды включают в себя последовательности, содержащие вариабельный участок дельта-цепи TCR, продуцируемого иммунной клеткой. Например, целевые полинуклеотиды могут включать в себя полинуклеотид-матрицу, применяемую для образования продуктов реакции обратной транскрипции или реакции достройки праймеров, а также включают в себя сами продукты реакции обратной транскрипции или продукты реакции достройки праймера. Например, целевые полинуклеотиды включают в себя представляющие интерес полинуклеотиды, которые могут подвергаться реакции обратной транскрипции или реакции достройки праймера.
В соответствии с некоторыми вариантами осуществления целевые полинуклеотиды включают в себя последовательности, содержащие AID олигонуклеотидов в составе конъюгатов аффинная молекула-олигонуклеотид или конъюгатов тетрамер-олигонуклеотид. В соответствии с некоторыми вариантами осуществления целевые полинуклеотиды включают в себя последовательности, содержащие AMD олигонуклеотидов в составе конъюгатов аффинная молекула-олигонуклеотид или конъюгатов тетрамер-олигонуклеотид. Например, целевые полинуклеотиды включают в себя РНК или ДНК. Например, целевые полинуклеотиды включают в себя синтезированные олигонуклеотиды. Например, целевые полинуклеотиды включают в себя олигонуклеотиды, содержащие AID и/или АМВ.
Целевые полинуклеотиды могут быть получены практически из любого источника и могут быть подготовлены с применением способов, известных в уровне техники. Например, целевые полинуклеотиды можно непосредственно выделять без амплификации с применением способов, известных в уровне техники, в том числе, без ограничения, экстрагирования фрагмента геномной ДНК или мРНК из организма или клетки (например, иммунной клетки) с получением целевых полинуклеотидов. Целевой полинуклеотид также может охватывать кДНК, образуемую из РНК (такой как мРНК) посредством ПЦР с обратной транскрипцией. В некоторых случаях целевой полинуклеотид представляет собой молекулу РНК. В некоторых случаях целевой полинуклеотид представляет собой молекулу мРНК или кДНК, полученную из молекулы мРНК. В некоторых случаях целевой полинуклеотид представляет собой молекулу мРНК или молекулу кДНК, полученную из молекулы мРНК из отдельной иммунной клетки. В некоторых случаях целевые полинуклеотиды представляют собой молекулы мРНК или молекулы кДНК, полученные из молекул мРНК из отдельных иммунных клеток. В некоторых случаях целевые полинуклеотиды представляют собой молекулы мРНК, кодирующие последовательность антитела из отдельной иммунной клетки. В некоторых случаях целевые полинуклеотиды представляют собой молекулы мРНК, кодирующие последовательности тяжелой цепи антитела из отдельных иммунных клеток. В некоторых случаях целевые полинуклеотиды представляют собой молекулы мРНК, кодирующие последовательность тяжелой цепи антитела из отдельной иммунной клетки. В некоторых случаях целевые полинуклеотиды представляют собой молекулы мРНК, кодирующие последовательности легкой цепи антитела из отдельных иммунных клеток. В некоторых случаях целевые полинуклеотиды представляют собой молекулы мРНК, кодирующие последовательность легкой цепи антитела из отдельной иммунной клетки. В некоторых случаях целевые полинуклеотиды представляют собой молекулы мРНК, кодирующие последовательности вариабельного домена антитела из отдельных иммунных клеток. В некоторых случаях целевые полинуклеотиды представляют собой молекулы мРНК, кодирующие последовательность вариабельного домена антитела из отдельной иммунной клетки. В некоторых случаях целевые полинуклеотиды представляют собой молекулы мРНК, кодирующие последовательности вариабельного домена легкой цепи антитела из отдельных иммунных клеток. В некоторых случаях целевые полинуклеотиды представляют собой молекулы мРНК, кодирующие последовательность вариабельного домена легкой цепи антитела из отдельной иммунной клетки. В некоторых случаях целевые полинуклеотиды представляют собой молекулы мРНК, кодирующие последовательности вариабельного домена тяжелой цепи антитела из отдельных иммунных клеток. В некоторых случаях целевые полинуклеотиды представляют собой молекулы мРНК, кодирующие последовательность вариабельного домена тяжелой цепи антитела из отдельной иммунной клетки. В некоторых случаях целевой полинуклеотид может представлять собой бесклеточную нуклеиновую кислоту, например, ДНК или РНК. В некоторых случаях целевые полинуклеотиды представляют собой молекулы мРНК, кодирующие последовательности вариабельного участка альфа-, бета-, гамма- и/или дельта-цепи TCR из отдельных иммунных клеток. В некоторых случаях целевые полинуклеотиды представляют собой молекулы мРНК, кодирующие последовательности VH и/или VL BCR из отдельных иммунных клеток.
Способы, описанные в данном документе, можно применять для создания библиотеки полинуклеотидов из одного или нескольких целевых полинуклеотидов для секвенирования. В соответствии с некоторыми вариантами осуществления библиотеки можно создать из двух или более участков целевого полинуклеотида. В соответствии с некоторыми вариантами осуществления способов библиотеки можно создать из двух или более целевых полинуклеотидов. В соответствии с некоторыми вариантами осуществления целевые полинуклеотиды представляют собой геномные нуклеиновые кислоты или ДНК, полученную из хромосом. В соответствии с некоторыми вариантами осуществления целевые полинуклеотиды включают в себя последовательности, содержащие вариант, такой как полиморфизм или мутация. В соответствии с некоторыми вариантами осуществления целевые полинуклеотиды включают в себя ДНК, а не РНК. В соответствии с некоторыми вариантами осуществления целевые полинуклеотиды включают в себя РНК, а не ДНК. В соответствии с некоторыми вариантами осуществления целевые полинуклеотиды включают в себя ДНК и РНК. В соответствии с некоторыми вариантами осуществления целевой полинуклеотид представляет собой одноцепочечный полинуклеотид. В соответствии с некоторыми вариантами осуществления целевой полинуклеотид представляет собой двухцепочечный полинуклеотид. В соответствии с некоторыми вариантами осуществления целевой полинуклеотид представляет собой одну цепь двухцепочечного полинуклеотида.
Целевые полинуклеотиды можно синтезировать или получать из любого биологического образца и подготовить с применением способов, известных в уровне техники. В соответствии с некоторыми вариантами осуществления целевые полинуклеотиды выделяют непосредственно без амплификации. Способы непосредственного выделения являются известными в уровне техники. Неограничивающие примеры включают в себя экстрагирование геномной ДНК или мРНК из биологического образца, организма или клетки. В соответствии с некоторыми вариантами осуществления один или несколько целевых полинуклеотидов очищают от биологического образца. В соответствии с некоторыми вариантами осуществления целевой полинуклеотид не очищают от биологического образца, в котором он содержится. В соответствии с некоторыми вариантами осуществления целевой полинуклеотид является выделенным из биологического образца. В соответствии с некоторыми вариантами осуществления целевой полинуклеотид не выделяют из биологического образца, в котором он содержится. В соответствии с некоторыми вариантами осуществления целевой полинуклеотид может представлять собой бесклеточную нуклеиновую кислоту. В соответствии с некоторыми вариантами осуществления целевой полинуклеотид может представлять собой фрагментированную нуклеиновую кислоту. В соответствии с некоторыми вариантами осуществления целевой полинуклеотид может представлять собой транскрибированную нуклеиновую кислоту. В соответствии с некоторыми вариантами осуществления целевой полинуклеотид представляет собой модифицированный полинуклеотид. В соответствии с некоторыми вариантами осуществления целевой полинуклеотид представляет собой немодифицированный полинуклеотид.
В соответствии с некоторыми вариантами осуществления целевой полинуклеотид представляет собой олигонуклеотид из конъюгата аффинная молекула-олигонуклеотид или конъюгата тетрамер-олигонуклеотид. В соответствии с некоторыми вариантами осуществления множество целевых полинуклеотидов содержит множество олигонуклеотидов из множества конъюгатов аффинная молекула-олигонуклеотид или множества конъюгатов тетрамер-олигонуклеотид. В соответствии с некоторыми вариантами осуществления множество целевых полинуклеотидов содержит 2, 3, 4, 5, 6, 7, 8, 9, 10, 20, 30, 40, 50, 60, 70, 80, 90, 100, 150, 200, 250, 300, 350, 400, 450, 500, 550, 600, 650, 700, 750, 800, 850, 900 или 1000 или большее количество олигонуклеотидов из множества конъюгатов аффинная молекула-олигонуклеотид или конъюгатов тетрамер-олигонуклеотид. В соответствии с некоторыми вариантами осуществления множество целевых полинуклеотидов содержит множество олигонуклеотидов из 2, 3, 4, 5, 6, 7, 8, 9, 10, 20, 30, 40, 50, 60, 70, 80, 90, 100, 150, 200, 250, 300, 350, 400, 450, 500, 550, 600, 650, 700, 750, 800, 850, 900, или 1000 или большего количества конъюгатов аффинная молекула-олигонуклеотид или конъюгатов тетрамер-олигонуклеотид. В соответствии с некоторыми вариантами осуществления множество целевых полинуклеотидов содержит 2, 3, 4, 5, 6, 7, 8, 9, 10, 20, 30, 40, 50, 60, 70, 80, 90, 100, 150, 200, 250, 300, 350, 400, 450, 500, 550, 600, 650, 700, 750, 800, 850, 900 или 1000 или большее количество олигонуклеотидов из 2, 3, 4, 5, 6, 7, 8, 9, 10, 20, 30, 40, 50, 60, 70, 80, 90, 100, 150, 200, 250, 300, 350, 400, 450, 500, 550, 600, 650, 700, 750, 800, 850, 900, или 1000 или большего количества конъюгатов аффинная молекула-олигонуклеотид или конъюгатов тетрамер-олигонуклеотид.
В соответствии с некоторыми вариантами осуществления целевой полинуклеотид содержит последовательность AID. В соответствии с некоторыми вариантами осуществления множество целевых полинуклеотидов содержит 2, 3, 4, 5, 6, 7, 8, 9, 10, 20, 30, 40, 50, 60, 70, 80, 90, 100, 150, 200, 250, 300, 350, 400, 450, 500, 550, 600, 650, 700, 750, 800, 850, 900 или 1000 или большее количество последовательностей AID. В соответствии с некоторыми вариантами осуществления целевой полинуклеотид содержит последовательность АМВ. В соответствии с некоторыми вариантами осуществления множество целевых полинуклеотидов содержит по меньшей мере 10, 20, 30, 40, 50, 60, 70, 80, 90, 100, 150, 200, 250, 300, 350, 400, 450, 500, 550, 600, 650, 700, 750, 800, 850, 900, 1000, 1500, 2000, 3000, 4000, 5000, 6000, 7000, 8000, 9000, 10000, 11000, 12000, 13000, 14000, 15000, 16000, 17000, 18000, 19000, 20000, 25000, 30000, 35000, 40000, 45000, 50000, 60000, 70000, 80000, 90000, 100000, 200000, 300000, 400000, 500000, 600000, 700000, 800000, 900000, 1×106, 2×106, 3×106, 4×106, 5×106, 6×106, 7×106, 8×106, 9×106, 1×107, 2×107, 3×107, 4×107, 5×107, 6×107, 7×107, 8×107, 9×107, 1×108, 2×108, 3×108, 4×108, 5×108, 6×108, 7×108, 8×108, 9×108, 1×109, 2×109, 3×109, 4×109, 5×109, 6×109, 7×109, 8×109, 9×109, 1×1010, 2×1010, 3×1010, 4×1010, 5×1010, 6×1010, 7×1010, 8×1010, 9×1010, 1×1011, 2×1011, 3×1011, 4×1011, 5×1011, 6×1011, 7×1011, 8×1011, 9×1011, 1×1012, 2×1012, 3×1012, 4×1012, 5×1012, 6×1012, 7×1012, 8×1012 или 9×1012 или большее количество последовательностей АМВ.
В соответствии с некоторыми вариантами осуществления целевой полинуклеотид представляет собой полинуклеотид из отдельной клетки. В соответствии с некоторыми вариантами осуществления целевые полинуклеотиды происходят из отдельных клеток. В соответствии с некоторыми вариантами осуществления целевой полинуклеотид представляет собой полинуклеотид из образца, содержащего множество клеток.
В соответствии с некоторыми вариантами осуществления целевой полинуклеотид кодирует последовательность биологического маркера. В соответствии с некоторыми вариантами осуществления целевой полинуклеотид кодирует две или более последовательностей биологического маркера. В соответствии с некоторыми вариантами осуществления множество целевых полинуклеотидов кодирует последовательность биологического маркера. В соответствии с некоторыми вариантами осуществления множество целевых полинуклеотидов кодирует две или более последовательностей биологического маркера. В соответствии с некоторыми вариантами осуществления множество целевых полинуклеотидов кодирует 3, 4, 5, 6, 7, 8, 9, 10, 20, 30, 40, 50, 60, 70, 80, 90 или 100 или большее количество последовательностей биологического маркера.
В соответствии с некоторыми вариантами осуществления множество целевых полинуклеотидов содержит панель олигонуклеотидных последовательностей. В соответствии с некоторыми вариантами осуществления множество целевых полинуклеотидов содержит панель последовательностей иммуноглобулинов. Например, панель последовательностей иммуноглобулинов может представлять собой последовательности VH и/или VL. В соответствии с некоторыми вариантами осуществления множество целевых полинуклеотидов содержит панель последовательностей TCR. В соответствии с некоторыми вариантами осуществления панель последовательностей иммуноглобулинов или TCR содержит 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 последовательностей иммуноглобулинов или TCR. В соответствии с некоторыми вариантами осуществления множество целевых полинуклеотидов содержит панель последовательностей BCR. Например, панель последовательностей BCR может представлять собой последовательности VH и/или VL. В соответствии с некоторыми вариантами осуществления панель последовательностей иммуноглобулинов, BCR или TCR содержит по меньшей мере приблизительно 10, 20, 30, 40, 50, 60, 70, 80, 90, 100, 150, 200, 250, 300, 350, 400, 450, 500, 550, 600, 650, 700, 750, 800, 850, 900, 1000, 1500, 2000, 3000, 4000, 5000, 6000, 7000, 8000, 9000, 10000, 11000, 12000, 13000, 14000, 15000, 16000, 17000, 18000, 19000, 20000, 25000, 30000, 35000, 40000, 45000, 50000, 60000, 70000, 80000, 90000, 100000, 200000, 300000, 400000, 500000, 600000, 700000, 800000, 900000, 1×106, 2×106, 3×106, 4×106, 5×106, 6×106, 7×106, 8×106, 9×106, 1×107, 2×107, 3×107, 4×107, 5×107, 6×107, 7×107, 8×107, 9×107, 1×108, 2×108, 3×108, 4×108, 5×108, 6×108, 7×108, 8×108, 9×108, 1×109, 2×109, 3×109, 4×109, 5×109, 6×109, 7×109, 8×109, 9×109, 1×1010, 2×1010, 3×1010, 4×1010, 5×1010, 6×1010, 7×1010, 8×1010, 9×1010, 1×1011, 2×1011, 3×1011, 4×1011, 5×1011, 6×1011, 7×1011, 8×1011, 9×1011, 1×1012, 2×1012, 3×1012, 4×1012, 5×1012, 6×1012, 7×1012, 8×1012 или 9×1012 последовательностей иммуноглобулинов, BCR или TCR. В соответствии с некоторыми вариантами осуществления панель последовательностей иммуноглобулинов, BCR или TCR содержит не более чем приблизительно 10, 20, 30, 40, 50, 60, 70, 80, 90, 100, 150, 200, 250, 300, 350, 400, 450, 500, 550, 600, 650, 700, 750, 800, 850, 900, 1000, 1500, 2000, 3000, 4000, 5000, 6000, 7000, 8000, 9000, 10000, 11000, 12000, 13000, 14000, 15000, 16000, 17000, 18000, 19000, 20000, 25000, 30000, 35000, 40000, 45000, 50000, 60000, 70000, 80000, 90000, 100000, 200000, 300000, 400000, 500000, 600000, 700000, 800000, 900000, 1×106, 2×106, 3×106, 4×106, 5×106, б×106, 7×106, 8×106, 9×106, 1×107, 2×107, 3×107, 4×107, 5×107, 6×107, 7×107, 8×107, 9×107, 1×108, 2×108, 3×108, 4×108, 5×108, 6×108, 7×108, 8×108, 9×108, 1×109, 2×109, 3×109, 4×109, 5×109, 6×109, 7×109, 8×109, 9×109, 1×1010, 2×1010, 3×1010, 4×1010, 5×1010, 6×1010, 7×1010, 8×1010, 9×1010, 1×1011, 2×1011, 3×1011, 4×1011, 5×1011, 6×1011, 7×1011, 8×1011, 9×1011, 1×1012, 2×1012, 3×1012, 4×1012, 5×1012, 6×1012, 7×1012, 8×1012 или 9×1012 последовательностей иммуноглобулинов, BCR или TCR. В соответствии с некоторыми вариантами осуществления панель последовательностей иммуноглобулинов, BCR или TCR содержит приблизительно 10-20, 10-30, 10-40, 10-30, 10-40, 10-50, 10-60, 10-70, 10-80, 10-90, 10-100, 50-60, 50-70, 50-80, 50-90, 50-100, 100-200, 100-300, 100-400, 100-300, 100-400, 100-500, 100-600, 100-700, 100-800, 100-900, 100-1000, 500-600, 500-700, 500-800, 500-900, 500-1000, 1000-2000, 1000-3000, 1000-4000, 1000-3000, 1000-4000, 1000-5000, 1000-6000, 1000-7000, 1000-8000, 1000-9000,1000-10000, 5000-6000, 5000-7000, 5000-8000, 5000-9000, 5000-10000, 1-1×105, 1-2×105, 1-3×105, 1-4×105, 1-5×105, 1-6×105, 1-7×105, 1-8×105, 9×105, 1-1×106, 1-2×106, 1-3×106, 1-4×106, 1-5×106, 1-6×106, 1-7×106, 1-8×106, 9×106, 1×107, 1-2×107, 1-3×107, 1-4×107, 1-5×107, 1-6×107, 1-7×107, 1-8×107, 1-9×107, 1-1×108, 1-2×108, 1-3×108, 1-4×108, 1-5×108, 1-6×108, 1-7×108, 1-8×108, 1-9×108, 1-1×109, 1-2×109, 1-3×109, 1-4×109, 1-5×109, 1-6×109, 1-7×109, 1-8×109, 1-9×109, 1-1×1010, 1-2×1010, 1-3×1010, 1-4×1010, 1-5×1010, 1-6×1010, 1-7×1010, 1-8×1010, 1-9×1010, 1-1×1011, 1-2×1011, 1-3×1011, 1-4×1011, 1-5×1011, 1-6×1011, 1-7×1011, 1-8×1011, 1-9×1011, 1-1×1012, 1-2×1012, 1-3×1012, 1-4×1012, 1-5×1012, 1-6×1012, 1-7×1012, 1-8×1012 или 1-9×1012 последовательностей иммуноглобулинов, BCR или TCR.
В соответствии с некоторыми вариантами осуществления целевой полинуклеотид имеет длину приблизительно 10, 20, 30, 40, 50, 60, 70, 80, 90, 100, 150, 200, 250, 300, 350, 400, 450, 500, 550, 600, 650, 700, 750, 800, 850, 900, 1000, 1500, 2000, 3000, 4000, 5000, 6000, 7000, 8000, 9000, 10000, 11000, 12000, 13000, 14000, 15000, 16000, 17000, 18000, 19000 или 20000 оснований или пар оснований. В соответствии с некоторыми вариантами осуществления целевой полинуклеотид имеет длину по меньшей мере приблизительно 10, 20, 30, 40, 50, 60, 70, 80, 90, 100, 150, 200, 250, 300, 350, 400, 450, 500, 550, 600, 650, 700, 750, 800, 850, 900, 1000, 1500, 2000, 3000, 4000, 5000, 6000, 7000, 8000, 9000, 10000, 11000, 12000, 13000, 14000, 15000, 16000, 17000, 18000, 19000 или 20000 оснований или пар оснований. В соответствии с некоторыми вариантами осуществления целевой полинуклеотид имеет длину не более приблизительно 10, 20, 30, 40, 50, 60, 70, 80, 90, 100, 150, 200, 250, 300, 350, 400, 450, 500, 550, 600, 650, 700, 750, 800, 850, 900, 1000, 1500, 2000, 3000, 4000, 5000, 6000, 7000, 8000, 9000, 10000, 11000, 12000, 13000, 14000, 15000, 16000, 17000, 18000, 19000 или 20000 оснований или пар оснований. В соответствии с некоторыми вариантами осуществления целевой полинуклеотид имеет длину приблизительно 10-20, 10-30, 10-40, 10-30, 10-40, 10-50, 10-60, 10-70, 10-80, 10-90, 10-100, 50-60, 50-70, 50-80, 50-90, 50-100, 100-200, 100-300, 100-400, 100-300, 100-400, 100-500, 100-600, 100-700, 100-800, 100-900, 100-1000, 500-600, 500-700, 500-800, 500-900, 500-1000, 1000-2000, 1000-3000, 1000-4000, 1000-3000, 1000-4000, 1000-5000, 1000-6000, 1000-7000, 1000-8000, 1000-9000, 1000-10000, 5000-6000, 5000-7000, 5000-8000, 5000-9000 или 5000-10000 оснований или пар оснований. В соответствии с некоторыми вариантами осуществления средняя длина целевых полинуклеотидов или их фрагментов может составлять менее чем приблизительно 100, 200, 300, 400, 500 или 800 пар оснований, или менее чем приблизительно 5, 10, 20, 30, 40, 50, 60, 70, 80, 90, 100, 110, 120, 130, 140, 150, 160, 170, 180, 190 или 200 нуклеотидов, или менее чем приблизительно 1, 2, 5, 10, 20, 30, 40, 50, 60, 70, 80, 90, 100 килобаз. В соответствии с некоторыми вариантами осуществления целевая последовательность из относительно короткой матрицы, такой как образец, содержащий целевой полинуклеотид, составляет приблизительно 40, 45, 50, 55, 60, 65, 70, 75, 80, 85, 90, 95 или 100 оснований. В соответствии с определенными вариантами осуществления данные от секвенирования выравнивают с известными или ожидаемыми последовательностями с применением базы данных, содержащей последовательности иммуноглобулина или последовательности TCR, ассоциированные с заболеванием или состоянием.
В соответствии с некоторыми вариантами осуществления способ дополнительно включает определение последовательности зародышевого типа у первого клеточного полинуклеотида, второго клеточного полинуклеотида или обоих из них, причем первый клеточный полинуклеотид содержит последовательность IgH или VH, и при этом второй клеточный полинуклеотид содержит последовательность IgL или VL или любую их комбинацию. В соответствии с некоторыми вариантами осуществления способ дополнительно включает определение отличий последовательности IgL, IgH, VH, VL или любой их комбинации от таких последовательностей зародышевого типа. В соответствии с некоторыми вариантами осуществления способ дополнительно включает определение по меньшей мере одной из общего количества уникальных последовательностей IgH; общего количества уникальных последовательностей IgL; общего количества уникальных последовательностей IgH и IgL; общего количества уникальных спаренных последовательностей IgL и IgH; частоты последовательности IgH или последовательности IgL или частоты комбинации последовательности IgH и последовательности IgL относительно одной или нескольких других.
В соответствии с некоторыми вариантами осуществления способ дополнительно включает определение последовательности зародышевого типа у первого клеточного полинуклеотида, второго клеточного полинуклеотида или обоих из них, причем первый клеточный полинуклеотид содержит последовательность TCRα или Vα, и при этом второй клеточный полинуклеотид содержит последовательность TCRβ или Vβ или любую их комбинацию. В соответствии с некоторыми вариантами осуществления способ дополнительно включает определение отличий последовательности TCRα, TCRβ, Vα, Vβ или любой их комбинации от таких последовательностей зародышевого типа.
В соответствии с некоторыми вариантами осуществления способ дополнительно включает определение по меньшей мере одной из общего количества уникальных последовательностей TCRα; общего количества уникальных последовательностей TCRβ; общего количества уникальных последовательностей TCRα и TCRβ; общего количества уникальных спаренных последовательностей TCRβ и TCRα; частоты последовательности TCRα или последовательности TCRβ или частоты комбинации последовательности TCRα и последовательности TCRβ относительно одной или нескольких других. В соответствии с некоторыми вариантами осуществления способ дополнительно включает определение последовательности зародышевого типа у первого клеточного полинуклеотида, второго клеточного полинуклеотида или обоих из них, причем первый клеточный полинуклеотид содержит последовательность TCRγ или Vγ, и при этом второй клеточный полинуклеотид содержит последовательность TCRδ или Vδ или любую их комбинацию. В соответствии с некоторыми вариантами осуществления способ дополнительно включает определение отличий последовательности TCRγ, TCRδ, Vγ, Vδ или любой их комбинации от таких последовательностей зародышевого типа. В соответствии с некоторыми вариантами осуществления способ дополнительно включает определение по меньшей мере одной из общего количества уникальных последовательностей TCRγ; общего количества уникальных последовательностей TCRδ; общего количества уникальных последовательностей TCRγ и TCRδ; общего количества уникальных спаренных последовательностей TCRδ и TCRγ; частоты последовательности TCRγ или последовательности TCRδ или частоты комбинации последовательности TCRγ и последовательности TCRδ относительно одной или нескольких других. В соответствии с некоторыми вариантами осуществления способ дополнительно включает определение по меньшей мере одного из общего количества последовательностей из первого гена; общего количества последовательностей из второго гена; общего количества уникальных последовательностей из первого гена; общего количества уникальных последовательностей из второго гена или частоты последовательности из первого гена или последовательности из второго гена.
В соответствии с некоторыми вариантами осуществления способ дополнительно включает выбор антитела или TCR на основании общего количества одной или нескольких пар индивидуально спаренных последовательностей IgL и IgH, или последовательностей TCRα и TCRβ, или последовательностей TCRγ и TCRδ и отличия от зародышевого типа. В соответствии с некоторыми вариантами осуществления способ дополнительно включает выбор антитела или TCR на основании одной или нескольких последовательностей IgL или IgH, последовательностей TCRα и TCRβ или последовательностей TCRγ и TCRδ и отличия от зародышевого типа. В соответствии с некоторыми вариантами осуществления способ дополнительно включает выбор антитела или TCR на основании одного или нескольких из паттернов последовательностей, анализа отличий, динамических характеристик или частоты. В соответствии с некоторыми вариантами осуществления способ дополнительно включает выбор антитела или TCR на основании частоты.
Клонирование и экспрессия антител, BCR и TCR
«Экспрессионная библиотека антител», или «экспрессионная библиотека BCR», или «экспрессионная библиотека TCR», или «экспрессионная библиотека» в контексте данного документа могут относиться к набору молекул (т.е. двум или более молекулам) либо на уровне нуклеиновых кислот, либо на уровне белков. Следовательно, эти термины могут относиться к набору экспрессионных векторов, которые кодируют множество молекул антитела, BCR или TCR (т.е. на уровне нуклеиновых кислот), или могут относиться к набору молекул антитела, BCR или TCR после того, как они были экспрессированы в соответствующей экспрессионной системе (т.е. на уровне белков). В качестве альтернативы экспрессионные векторы/экспрессионная библиотека могут содержаться в подходящих клетках-хозяевах, в которых они могут экспрессироваться. Молекулы антитела, которые закодированы или экспрессируются в экспрессионных библиотеках согласно настоящему изобретению, могут присутствовать в любом соответствующем формате, например, они могут представлять собой молекулы целого антитела, BCR или TCR или могут представлять собой фрагменты антител, фрагменты BCR или фрагменты TCR, например, одноцепочечные антитела (например, scFv антитела), Fv антитела, Fab' антитела, (Fab')2 фрагменты, диатела и т.д. Термины «кодирующий» и «который кодирует» в случае последовательности нуклеиновой кислоты, «кодирующей»/«которая кодирует» или кодирующей последовательности ДНК конкретного фермента или нуклеотидной последовательности «кодирующей»/«которая кодирует» конкретный фермент, а также другие синонимичные термины относятся к последовательности ДНК, которая транскрибируется и транслируется в фермент при помещении под контроль соответствующих регуляторных последовательностей. «Промоторная последовательность» представляет собой регуляторный участок ДНК, способный к связыванию РНК-полимеразы в клетке и инициации транскрипции нижележащей (в направлении 3') кодирующей последовательности. Промотор является частью последовательности ДНК. Этот участок последовательности имеет старт-кодон на своем 3' конце. Промоторная последовательность включает в себя минимальное количество оснований с элементами, необходимыми для инициации транскрипции на уровнях, которые могут выявляться как превышающие фоновый уровень. Тем не менее, после того как РНК-полимераза связывается с последовательностью, и транскрипция инициируется на старт-кодоне (на 3' конце с промотором), транскрипция продолжается далее в направлении 3'. В пределах промоторной последовательности будет обнаруживаться сайт инициации транскрипции (подходящим образом определяемый посредством картирования с использованием нуклеазы S1), а также белок-связывающие домены (консенсусные последовательности), ответственные за связывание РНК-полимеразы.
Молекулы антител, BCR или TCR, идентифицированные с помощью, полученные из, выбранные из или получаемые из экспрессионных библиотек антител, BCR или TCR согласно настоящему изобретению образуют еще один дополнительный аспект настоящего изобретения. Опять же, эти молекулы антитела, BCR или TCR могут представлять собой белки или нуклеиновые кислоты, кодирующие молекулы антитела, BCR или TCR, причем эти нуклеиновые кислоты, в свою очередь, могут быть включены в соответствующий экспрессионный вектор и/или могут содержаться в подходящей клетке-хозяине.
Пул кДНК можно подвергнуть реакции ПЦР с полинуклеотидами, которые гибридизируются с константным участком генов тяжелой цепи антитела или BCR, и полинуклеотидами, которые гибридизируются с 5' концом генов участка VH-, или Vα-, или Vγ-цепи антитела, BCR или TCR. Пул кДНК можно подвергнуть реакции ПЦР с полинуклеотидами, которые гибридизируются с константным участком генов тяжелой цепи или альфа или гамма цепи антитела, BCR или TCR, и полинуклеотидам, которые гибридизируются с участком, расположенным в направлении 5' относительно 5' конца участка VH-, или Vα-, или Vγ-цепи меченого штрихкодом полинуклеотида, содержащего последовательность антитела, BCR или TCR. Реакция ПЦР также может быть настроена для амплификации пула VL-, или Vβ-, или Vδ-цепей, например, каппа и лямбда классов. Пул кДНК можно подвергнуть реакции ПЦР с полинуклеотидами, которые гибридизируются с константным участком генов легкой цепи антитела, и полинуклеотидами, которые гибридизируются с 5' концом генов участка VL-, или Vβ-, или Vδ-цепи антитела, BCR или TCR. Пул кДНК можно подвергнуть реакции ПЦР с полинуклеотидами, которые гибридизируются с константным участком генов легкой цепи антитела, и полинуклеотидами, которые гибридизируются с участком, находящимся в положении 5' относительно 5' конца участка VL-, или Vβ-, или Vδ-цепи меченого штрих кодом полинуклеотида, содержащего последовательность антитела, BCR или TCR. Такие олигонуклеотиды или праймеры можно сконструировать на основе известной и общедоступной информации из базы данных о последовательности гена иммуноглобулина, BCR или TCR.
В соответствии с некоторыми вариантами осуществления последовательности VH и VL, или Vα и Vβ, или Vγ и Vδ можно подходящим образом получать из библиотеки последовательностей VH и VL, или Vα и Vβ, или Vγ и Vδ, полученных с помощью ПЦР амплификации с применением одного или нескольких праймеров, которые не являются специфическими к генам тяжелой или легкой цепи и, в частности, к одному или обоим концевым участкам полинуклеотидов VH и VL, или Vα и Vβ, или Vγ и Vδ. В соответствии с некоторыми вариантами осуществления последовательности VH и VL можно подходящим образом получить из библиотеки последовательностей VH и VL, или Vα и Vβ, или Vγ и Vδ, полученных с помощью ПЦР амплификации с применением праймеров, специфических к участку меченого штрихкодом резервуара полинуклеотида. В соответствии с некоторыми вариантами осуществления последовательности VH и VL можно подходящим образом получить из библиотеки последовательностей VH и VL, или Vα и Vβ, или Vγ и Vδ, полученной с помощью ПЦР амплификации с применением праймеров, специфических к семейству С-генов, или праймеров, специфических к С-гену. В соответствии с некоторыми вариантами осуществления последовательности VH и VL можно подходящим образом получить из библиотеки последовательностей VH и VL, или Vα и Vβ, или Vγ и Vδ, полученных с помощью ПЦР амплификации с применением набора праймеров с первым праймером, специфическим к участку меченого штрихкодом резервуара полинуклеотида, и вторым праймером или множеством вторых праймеров, которые представляют собой праймеры, специфические к семейству С-генов, или праймеры, специфические к С-гену. В соответствии с некоторыми вариантами осуществления последовательности VH и VL, или Vα и Vβ, или Vγ и Vδ можно подходящим образом получить из библиотеки последовательностей VH и VL, или Vα и Vβ, или Vγ и Vδ, полученных с помощью ПЦР амплификации с применением набора праймеров с первым праймером, специфическим к участку меченого штрихкодом резервуара полинуклеотида, и вторым праймером, специфическим к универсальной последовательности.
В соответствии с некоторыми вариантами осуществления после обратной транскрипции полученные в результате последовательности кДНК можно амплифицировать с помощью ПЦР с применением одного или нескольких праймеров, специфических к генам иммуноглобулинов или к генам BCR и, в частности, к одному или обоим концевым участкам полинуклеотидов VH и VL, или Vα и Vβ, или Vγ и Vδ. В соответствии с некоторыми вариантами осуществления последовательности VH и VL можно получить из библиотеки последовательностей VH и VL, или Vα и Vβ, или Vγ и Vδ, полученных с помощью ПЦР амплификации с применением праймеров, специфических к семейству V-генов, или праймеров, специфических к V-гену (Nicholls et al., J. Immunol. Meth., 1993, 165:81; международная заявка WO 93/12227), или их можно сконструировать в соответствии со стандартными способами, известными в уровне техники, которые основываются на доступной информации о последовательности. (Последовательности VH и VL, или Vα и Vβ, или Vγ и Vδ можно лигировать обычно с использованием промежуточной спейсерной последовательности (например, кодирующей гибкий пептидный спейсер в составе рамки считывания) с образованием кассеты, кодирующей одноцепочечное антитело). Последовательности V-участка можно подходящим образом клонировать в виде кДНК или продуктов ПЦР амплификации в случае экспрессирующих иммуноглобулины клеток. Участки VH и VL, или Vα и Vβ, или Vγ и Vδ секвенируют, необязательно, в способах, описанных в данном документе и, в особенности, после определенных стадий, которые отмечены (например, после ПЦР на отдельных клетках; после дисплея на поверхности клетки млекопитающего или другой клетки, после скрининга с применением FACS и т.п.). Секвенирование можно применять, помимо прочих причин, для подтверждения того, что разнообразие находится на приемлемом уровне. Секвенирование может включать в себя секвенирование с высокой пропускной способностью, глубокое секвенирование (при котором секвенируют одинаковый ген из множества отдельных образцов для идентификации различий в последовательностях) или комбинации этих двух методов.
В соответствии с некоторыми вариантами осуществления нецелесообразно физически связывать комбинации природных VH и VL, или Vα и Vβ, или Vγ и Vδ с применением способов, описанных в данном документе. В соответствии с некоторыми вариантами осуществления кДНК, полинуклеотиды со штрихкодами или подвергшиеся ПЦР амплификации кДНК со штрихкодами не являются физически связанными. В соответствии с некоторыми вариантами осуществления кДНК, полинуклеотиды со штрихкодами или подвергшиеся ПЦР амплификации кДНК со штрихкодами не являются физически связанными в одной и той же реакционной смеси или резервуаре.
В соответствии с некоторыми вариантами осуществления комбинации природных VH и VL, или Vα и Vβ, или Vγ и Vδ физически связывают, применяя дополнительно к праймерам для кДНК один праймер или множество праймеров для 5' конца гена VH, или Vα, или Vγ и другой праймер или множество праймеров для 5' конца гена VL, или Vβ, или Vδ. Эти праймеры также содержат комплементарные «хвосты» из дополнительной последовательности, чтобы обеспечить возможность самосборки генов VH и VL, или Vα и Vβ, или Vγ и Vδ. После ПЦР амплификации и связывания вероятность получения смешанных продуктов, иными словами, смешанных вариабельных участков является минимальной, поскольку реакции амплификации и связывания осуществлялись в каждой клетке. Риск смешивания можно дополнительно снизить за счет использования реактивов, заполняющих пространство, таких как меченные дигоксигенином нуклеотиды, чтобы дополнительно гарантировать, что пары кДНК V-участков не покинут клеточный компартмент и перемешаются между собой, а останутся внутри клетки для ПЦР амплификации и связывания. Амплифицированные последовательности связываются посредством гибридизации комплементарных концевых последовательностей. После связывания последовательности могут быть выделены из клеток для применения в дополнительных стадиях способа, описанных в данном документе. Например, выделенную ДНК можно подвергнуть ПЦР амплификации с применением концевых праймеров, если необходимо, и клонировать в векторы, которые могут представлять собой плазмиды, фаги, космиды, фагмиды, вирусные векторы или их комбинации, которые подробно описываются ниже. Сайты для удобного рестрикционного фермента можно включить в гибридизированные последовательности для облегчения клонирования. Эти векторы также можно сохранить в виде библиотеки связанных вариабельных участков для последующего применения.
Для обеспечения дополнительных комбинаций VH и VL, или Vα и Vβ, или Vγ и Vδ можно выбрать экспрессионную систему. Например, экспрессионные системы на основе бактериофага обеспечивают возможность случайной рекомбинации последовательностей тяжелой и легкой цепей. Другие подходящие экспрессионные системы являются известными специалисту в данной области техники.
Следует отметить, что в случае последовательностей VH и VL, или Vα и Vβ, или Vγ и Vδ, полученных из организмов, отличных от человека, в соответствии с некоторыми вариантами осуществления может быть предпочтительно химеризовать эти последовательности с полностью человеческим Fc. В контексте данного документа «химеризованный» относится к иммуноглобулину, BCR или TCR, в которых вариабельные участки тяжелой и легкой цепей или участки Vα и Vβ или Vγ и Vδ происходят не из человека, и в которых константные участки тяжелой и легкой цепей или цепи Vα и Vβ или Vγ и Vδ происходят из человека. Ее осуществляют посредством амплификации и клонирования вариабельных доменов в человеческий Fc. Человеческий Fc может являться частью вектора или может содержаться в отдельной молекуле, а также можно применять библиотеку Fc. В соответствии с предпочтительным вариантом осуществления химеризованные молекулы, выращиваемые в клетках млекопитающего, таких как СНО клетки, дважды подвергают скринингу с использованием FACS с целью обогащения клеточной популяции клетками, экспрессирующими антитело, представляющее интерес. Химеризованные антитела, BCR или TCR характеризуют либо посредством секвенирования с последующей характеристикой функциональных свойств, либо с помощью непосредственной характеристики функциональных свойств или определения кинетических характеристик. Выращивание, скрининг и характеристика подробно описаны ниже.
Важно отметить, что вышеописанные реакции ПЦР описаны для клонирования антител в форме IgG. Они являются предпочтительными, поскольку они обычно ассоциированы с более зрелой иммунной реакцией и, в целом, проявляют более высокую аффинность, чем антитела IgM, что, таким образом, делает их более желательными для определенных терапевтических и диагностических применений. Тем не менее, ясно, что можно сконструировать полинуклеотиды, которые будут обеспечивать возможность клонирования одной или нескольких из других форм молекул иммуноглобулинов, например, IgM, IgA, IgE и IgD, если это является желательным или подходящим.
После того как антитело, BCR или TCR были идентифицированы, и соответствующие популяции клеток были выделены в соответствующее время и, необязательно, обогащены, как описано выше, экспрессионные библиотеки для антитела, BCR или TCR не обязательно нужно создавать немедленно, можно обеспечить сохранение генетического материала, содержащегося в клетках, интактным, тем самым обеспечивая возможность получения библиотеки в дальнейшем. Благодаря этому, например, клетки, клеточный лизат или нуклеиновую кислоту, например, РНК или ДНК, полученную из них, можно хранить до дальнейшего использования с помощью подходящих способов, например, с помощью замораживания, а экспрессионные библиотеки получают в дальнейшем, если желательно.
После того как была создана библиотека экспрессионных векторов кодируемые молекулы антител можно затем экспрессировать в соответствующей экспрессионной системе и подвергать скринингу с использованием соответствующих методик, которые являются хорошо известными и документально подтвержденными в уровне техники. Таким образом, определенный выше способ согласно настоящему изобретению может включать дополнительные стадии осуществления экспрессии библиотеки экспрессионных векторов в соответствующей экспрессионной системе и скрининга экспрессированной библиотеки в отношении антител с желательными свойствами.
Как указано в данном документе, полинуклеотиды, полученные с помощью способов согласно настоящему раскрытию, которые содержат полинуклеотид, кодирующий последовательности антитела, BCR или TCR, может включать в себя, без ограничения, полинуклеотиды, кодирующие саму аминокислотную последовательность фрагмента антитела, BCR или TCR, некодирующую последовательность полного антитела, BCR или TCR или ее часть, кодирующую последовательность для фрагмента или части антитела, BCR или TCR, а также дополнительные последовательности, такие как кодирующая последовательность по меньшей мере одной сигнальной лидерной последовательности или слитого пептида, с или без вышеупомянутых дополнительных кодирующих последовательностей, таких как по меньшей мере один интрон, вместе с дополнительными некодирующими последовательностями, в том числе, без ограничения, некодирующими 5' и 3' последовательностями, такими как транскрибируемые нетранслируемые последовательности, которые играют роль в транскрипции, процессинге мРНК, в том числе сигналы сплайсинга и полиаденилирования (например, связывание рибосом и стабильность мРНК); дополнительную кодирующую последовательность, которая кодирует дополнительные аминокислоты, такие как те, которые обеспечивают дополнительные функциональные группы. Следовательно, последовательность, кодирующая антитело, может быть слита с маркерной последовательностью, такой как последовательность, кодирующая пептид, который облегчает очистку слитого антитела или TCR, содержащего фрагмент или часть антитела или TCR.
Первичные ПЦР продукты можно затем необязательно подвергать вторичной реакции ПЦР с новыми наборами полинуклеотидов, которые гибридизируются с 5' и 3' концами вариабельных доменов антитела, BCR или TCR VH, VL каппа и VL лямбда, или Vα и Vβ, или Vγ и Vδ (если это необходимо в зависимости от того, предназначена ли первая реакция ПЦР, с которой применяют новые наборы полинуклеотидов, для амплификации частей генов тяжелой или легкой цепи антитела, или генов Vα или Vβ TCR, или генов Vγ или Vδ TCR). Эти полинуклеотиды преимущественно включают в себя последовательности ДНК, специфические для определенного набора рестрикционных ферментов (т.е. сайты для рестрикционного фермента), для последующего клонирования. Выбранные рестрикционные ферменты нужно выбирать таким образом, чтобы они не производили разрезы в пределах сегментов V-гена человеческого антитела, BCR или TCR. Такие полинуклеотиды можно конструировать на основании известной и общедоступной последовательности гена иммуноглобулина или TCR и информации из базы данных рестрикционных ферментов. Тем не менее, предпочтительные сайты для рестрикционного фермента для включения представляют собой NcoI, Hind III, MluI и NotI. Продукты таких вторичных реакций ПЦР представляют собой репертуары различных фрагментов/доменов V-тяжелой, V-легкой каппа и V-легкой лямбда цепи антитела. Вследствие этого этот тип вторичной реакции ПЦР обычно осуществляют, когда представляющим интерес форматом экспрессионной библиотеки является формат scFv или Fv, при котором присутствуют только VH и VL, или Vα и Vβ, или Vγ и Vδ домены антитела или TCR.
ПЦР продукты также можно подвергать реакции ПЦР с новыми наборами праймеров, которые гибридизируются с 5' и 3' концами меченых штрихкодами полинуклеотидов. Эти полинуклеотиды преимущественно могут включать в себя последовательности ДНК, специфические для определенного набора рестрикционных ферментов (т.е. сайты для рестрикционного фермента), для последующего клонирования. Выбранные рестрикционные ферменты нужно выбирать таким образом, чтобы они не производили разрезы в пределах сегментов V-гена человеческого антитела или TCR. Такие полинуклеотиды можно конструировать на основании известной и общедоступной последовательности гена иммуноглобулина, BCR или TCR и информации из базы данных рестрикционных ферментов. Тем не менее, предпочтительные сайты для рестрикционного фермента для включения представляют собой NcoI, Hind III, MluI и NotI. Продукты таких вторичных реакций ПЦР представляют собой репертуары различных фрагментов/доменов VH, VL каппа и VL лямбда цепи антитела или Vα и Vβ или Vγ и Vδ фрагментов/доменов TCR.
Fv или Fab фрагменты или одноцепочечные антитела тяжелой или легкой цепи, или Vα или Vβ цепи, или Vγ или Vδ цепи, BCR или TCR также можно применять с этой системой. Тяжелую или легкую цепь, или Vα или Vβ цепь, или Vγ или Vδ цепь можно подвергнуть мутагенезу с последующим добавлением комплементарной цепи в раствор. Двум цепям затем позволяют объединиться и образовать функциональный фрагмент антитела. Добавление произвольных неспецифических последовательностей легкой или тяжелой цепи, или Vα или Vβ цепи, или Vγ или Vδ цепи обеспечивает возможность получения комбинаторной последовательности для создания библиотеки из разнообразных членов.
Библиотеки таких репертуаров клонированных фрагментов, содержащих участки вариабельного домена тяжелой цепи, или Vα-цепи или Vγ-цепи или их фрагменты, и/или участки вариабельного домена легкой цепи или Vβ-цепи или Vδ-цепи или их фрагменты, гены антитела, BCR или TCR, полученные из В- или Т-лимфоцитов хозяев, подвергшихся иммунной стимуляции, как определено в данном документе, образуют дополнительные аспекты настоящего изобретения. Эти библиотеки, содержащие клонированные вариабельные участки, необязательно могут быть вставлены в экспрессионные векторы с образованием экспрессионных библиотек.
В соответствии с некоторыми вариантами осуществления реакции ПЦР можно настроить таким образом, чтобы сохранить все или часть константных участков цепей различных антител, BCR или TCR, содержащихся в выделенной популяции иммунных клеток. Желательно, когда форматом экспрессионной библиотеки является формат Fab, при этом компонент тяжелой, или альфа-, или гамма-цепи содержит домены VH, или Vα, или Vγ и CH, или Сα, или Сγ, а компонент легкой цепи, или Vβ цепи, или Vδ цепи содержит домены VL или Vβ или Vδ цепи и CL, или Сβ, или Сδ. Опять же, библиотеки таких клонированных фрагментов, содержащие все или часть константных участков цепей антитела, BCR или TCR, образуют дополнительные аспекты настоящего изобретения.
Эти нуклеиновые кислоты могут подходящим образом содержать последовательности в дополнение к полинуклеотиду согласно настоящему изобретению. Например, множественный сайт клонирования, содержащий один или несколько рестрикционных сайтов для эндонуклеазы, может быть вставлен в нуклеиновую кислоту, чтобы помочь в выделении полинуклеотида. Транслируемые последовательности также могут быть вставлены для помощи в выделении транслированного полинуклеотида согласно настоящему изобретению. Например, гексагистидиновая маркерная последовательность обеспечивает удобное средство для очистки белков согласно настоящему изобретению. Нуклеиновая кислота согласно настоящему изобретению за исключением кодирующей последовательности необязательно представляет собой вектор, адаптер или линкер для клонирования и/или экспрессии полинуклеотида согласно настоящему изобретению.
Дополнительные последовательности можно добавлять к таким последовательностям для клонирования и/или экспрессии с целью оптимизации их функционирования при клонировании и/или экспрессии, чтобы помочь в выделении полинуклеотида или чтобы улучшить введение полинуклеотида в клетку. Применение векторов клонирования, векторов экспрессии, адаптеров и линкеров является хорошо известным в уровне техники. (См., например, Ausubel, выше; или Sambrook, выше).
Библиотеки, раскрытые в данном документе, можно применять в ряде применений. В контексте данного документа библиотека содержит множество молекул. В соответствии с некоторыми вариантами осуществления библиотека содержит множество полинуклеотидов. В соответствии с некоторыми вариантами осуществления библиотека содержит множество праймеров. В соответствии с некоторыми вариантами осуществления библиотека содержит множество ридов последовательностей от одного или нескольких полинуклеотидов, ампликонов или наборов ампликонов. Библиотеку можно хранить и применять много раз для получения образцов для анализа. Некоторые применения включают в себя, например, генотипирование полиморфизмов, изучение процессинга РНК и выбор типичных представителей клонов для выполнения секвенирования в соответствии со способами, представленными в данном документе. Можно получить библиотеки, содержащие множество полинуклеотидов, таких как праймеры, или библиотеки для секвенирования или амплификации, при этом множество полинуклеотидов содержит по меньшей мере приблизительно 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 20, 25, 30, 35, 40, 45, 50, 60, 70, 80, 90, 100, 200, 300, 400, 500, 600, 700, 800, 900, 1000, 1500, 2000, 3000, 4000, 5000, 6000, 7000, 8000, 9000, 10000, 11000, 12000, 13000, 14000, 15000, 16000, 17000, 18000, 19000, 20000, 30000, 40000, 50000, 60000, 70000, 80000, 90000, 100000, 200000, 300000, 400000, 500000, 600000, 700000, 800000, 900000, 1000000, 50000000, 100000000 или большее количество молекулярных штрихкодов или штрихкодов резервуара. В соответствии с некоторыми вариантами осуществления библиотеки полинуклеотидов содержат множество по меньшей мере приблизительно из 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 20, 25, 30, 35, 40, 45, 50, 60, 70, 80, 90, 100, 200, 300, 400, 500, 600, 700, 800, 900, 1000, 1500, 2000, 3000, 4000, 5000, 6000, 7000, 8000, 9000, 10000, 11000, 12000, 13000, 14000, 15000, 16000, 17000, 18000, 19000, 20000, 30000, 40000, 50000, 60000, 70000, 80000, 90000, 100000, 200000, 300000, 400000, 500000, 600000, 700000, 800000, 900000, 1000000, 50000000, 100000000 или большего количества уникальных полинуклеотидов, при этом каждый уникальный полинуклеотид содержит один или несколько молекулярных штрихкодов и штрихкодов резервуара.
Штрихкоды
Молекулярный штрихкод, такой как молекулярный штрихкод антигена, содержит информацию, которая является уникальной для отдельной молекулы, такой как отдельный олигонуклеотид в составе комплекса аффинная молекула-олигонуклеотид или комплекса тетрамер-олигонуклеотид или молекула полинуклеотида из отдельной клетки или из отдельного резервуара. Штрихкод резервуара содержит информацию, которая является уникальной для полинуклеотидов из отдельной клетки или присутствует в одном резервуаре по сравнению с полинуклеотидами из отличающейся отдельной клетки или присутствующими в отличающемся отдельном резервуаре. В соответствии с некоторыми вариантами осуществления уникальная информация содержит уникальную последовательность нуклеотидов. Например, последовательность молекулярного штрихкода или штрихкод резервуара может определяться идентификационными данными и порядком уникальной или произвольной последовательности нуклеотидов, содержащей молекулярный штрихкод или штрихкод резервуара. В соответствии с некоторыми вариантами осуществления уникальную информацию нельзя применять для идентификации последовательности целевого полинуклеотида. Например, молекулярный штрихкод может быть прикреплен к одному целевому полинуклеотиду, но молекулярный штрихкод нельзя применять для определения целевого полинуклеотида, к которому он прикреплен. В соответствии с некоторыми вариантами осуществления, уникальная информация не представляет собой известную последовательность, связанную с идентификационной информацией последовательности целевого полинуклеотида. Например, штрихкод резервуара может быть прикреплен к одному или нескольким целевым полинуклеотидам, но штрихкод резервуара нельзя применять для определения того, к какому из одного или нескольких целевых полинуклеотидов он прикреплен. В соответствии с некоторыми вариантами осуществления уникальная информация содержит произвольную последовательность нуклеотидов. В соответствии с некоторыми вариантами осуществления уникальная информация содержит одну или несколько уникальных последовательностей нуклеотидов на полинуклеотиде. В соответствии с некоторыми вариантами осуществления уникальная информация содержит вырожденную нуклеотидную последовательность или вырожденный штрихкод. Вырожденный штрихкод может характеризоваться переменным составом или последовательностью нуклеотидных оснований. Например, вырожденный штрихкод может представлять собой произвольную последовательность. В соответствии с некоторыми вариантами осуществления комплементарная последовательность молекулярного штрихкода или штрихкода резервуара также является последовательностью молекулярного штрихкода или штрихкода резервуара.
Штрихкод может характеризоваться любой длиной нуклеотидов. Например, любой из штрихкодов, описанных в данном документе, может иметь длину в пределах диапазона от 2 до 36 нуклеотидов, от 4 до 36 нуклеотидов, или от 6 до 30 нуклеотидов, или от 8 до 20 нуклеотидов, от 2 до 20 нуклеотидов, от 4 до 20 нуклеотидов, или от 6 до 20 нуклеотидов. В соответствии с определенными аспектами температуры плавления штрихкодов в составе набора находятся в пределах 10°С относительно друг друга, в пределах 5°С относительно друг друга или в пределах 2°С относительно друг друга. В соответствии с определенными аспектами температуры плавления штрихкодов в составе набора не находятся в пределах 10°С относительно друг друга, в пределах 5°С относительно друг друга или в пределах 2°С относительно друг друга. В соответствии с некоторыми аспектами штрихкоды являются членами набора с минимальной кросс-гибридизацией. Например, нуклеотидная последовательность у каждого члена такого набора может в значительной мере отличаться от таковой у каждого другого члена набора, благодаря чему ни один член не может образовывать стабильный дуплекс с комплементарной цепью любого другого члена при жестких условиях гибридизации. В соответствии с некоторыми вариантами осуществления нуклеотидная последовательность каждого члена минимально кросс-гибридизующегося набора отличается от последовательности каждого другого члена по меньшей мере двумя нуклеотидами. Технологии с применением штрихкодов описаны в Winzeler et al. (1999) Science 285:901; Brenner (2000) Genome Biol. 1:1 Kumar et al. (2001) Nature Rev. 2:302; Giaever et al. (2004) Proc. Natl. Acad. Sci. USA 101:793; Eason et al. (2004) Proc. Natl. Acad. Sci. USA 101:11046; и Brenner (2004) Genome Biol. 5:240.
Например, штрихкод может содержать по меньшей мере приблизительно 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49, 50, 60, 70, 80, 90, 100, 200, 500 или 1000 нуклеотидов. Например, штрихкод может содержать не более чем приблизительно 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49, 50, 60, 70, 80, 90, 100, 200, 500 или 1000 нуклеотидов. В соответствии с некоторыми вариантами осуществления штрихкод имеет конкретную длину, выраженную в нуклеотидах. Например, штрихкод может иметь длину приблизительно 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49, 50, 60, 70, 80, 90, 100, 200, 500 или 1000 нуклеотидов.
В соответствии с некоторыми вариантами осуществления каждый штрихкод во множестве штрихкодов имеет по меньшей мере приблизительно 2 нуклеотида. Например, каждый штрихкод во множестве штрихкодов может иметь длину по меньшей мере приблизительно 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49, 50, 60, 70, 80, 90, 100, 200, 500 или 1000 нуклеотидов. В соответствии с некоторыми вариантами осуществления каждый штрихкод во множестве штрихкодов имеет не более чем приблизительно 1000 нуклеотидов. Например, каждый штрихкод во множестве штрихкодов может иметь длину не более чем приблизительно 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49, 50, 60, 70, 80, 90, 100, 200, 500 или 1000 нуклеотидов.
Количество молекулярных штрихкодов может превышать общее количество молекул, которые нужно пометить, во множестве резервуаров. Количество штрихкодов резервуара может превышать общее количество молекул, которые нужно пометить, во множестве резервуаров. Например, количество молекулярных штрихкодов или штрихкодов резервуара может быть по меньшей мере приблизительно в 2, 3, 4, 5, 6, 7, 8, 9, 10, 15, 20, 30, 40, 50, 60, 70, 80, 90 или 100 раз большим, чем общее количество молекул, которые нужно пометить, во множестве резервуаров.
Количество различных молекулярных штрихкодов может превышать общее количество молекул, которые нужно пометить, во множестве резервуаров. В соответствии с некоторыми вариантами осуществления количество различных молекулярных штрихкодов является по меньшей мере приблизительно в 1, 1,5, 2, 2,5, 3, 3,5, 4, 4,5, 5, 6, 7, 8, 9, 10, 15, 20, 30, 40, 50, 60, 70, 80, 90 или 100 раз большим, чем общее количество молекул, которые нужно пометить, во множестве резервуаров.
Количество различных молекулярных штрихкодов в отдельном резервуаре может превышать количество различных молекул, которые нужно пометить, в отдельном резервуаре. В соответствии с некоторыми вариантами осуществления количество различных молекулярных штрихкодов в отдельном резервуаре является по меньшей мере приблизительно в 1, 1,5, 2, 2,5, 3, 3,5, 4, 4,5, 5, 6, 7, 8, 9, 10, 15, 20, 30, 40, 50, 60, 70, 80, 90 или 100 раз большим, чем количество различных молекул, которые нужно пометить, в отдельном резервуаре.
Количество различных штрихкодов резервуара может быть меньшим, чем общее количество молекул, которые нужно пометить, во множестве резервуаров. В соответствии с некоторыми вариантами осуществления количество различных штрихкодов резервуара является по меньшей мере приблизительно в 1, 1,5, 2, 2,5, 3, 3,5, 4, 4,5, 5, 6, 7, 8, 9, 10, 15, 20, 30, 40, 50, 60, 70, 80, 90 или 100 раз меньшим, чем общее количество молекул, которые нужно пометить, во множестве резервуаров.
Количество молекул амплифицированного продукта из молекулы меченого штрихкодом резервуара полинуклеотида в отдельном резервуаре может превышать количество различных молекул, которые нужно пометить, в отдельном резервуаре. В соответствии с некоторыми вариантами осуществления количество молекул амплифицированного продукта из молекулы меченого штрихкодом резервуара полинуклеотида в отдельном резервуаре является по меньшей мере приблизительно в 1, 1,5, 2, 2,5, 3, 3,5, 4, 4,5, 5, 6, 7, 8, 9, 10, 15, 20, 30, 40, 50, 60, 70, 80, 90 или 100 раз большим, чем количество различных молекул, которые нужно пометить, в отдельном резервуаре.
Количество молекул меченого штрихкодом резервуара полинуклеотида в отдельном резервуаре может быть меньшим, чем количество различных молекул, которые нужно пометить, в отдельном резервуаре. В соответствии с некоторыми вариантами осуществления количество молекул меченого штрихкодом резервуара полинуклеотида в отдельном резервуаре является по меньшей мере приблизительно в 1, 1,5, 2, 2,5, 3, 3,5, 4, 4,5, 5, 6, 7, 8, 9, 10, 15, 20, 30, 40, 50, 60, 70, 80, 90 или 100 раз меньшим, чем количество различных молекул, которые нужно пометить, в отдельном резервуаре.
Количество молекул меченого штрихкодом резервуара полинуклеотида в отдельном резервуаре может составлять одну молекулу. Количество молекул неамплифицированного меченого штрихкодом резервуара полинуклеотида в отдельном резервуаре может составлять одну молекулу.
В соответствии с некоторыми вариантами осуществления по меньшей мере приблизительно 1%, 2%, 3%, 4%, 5%, 6%, 7%, 8%, 9%, 10%, 15%, 20%, 25%, 30%, 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95%, 97% или 100% отличающихся молекулярных штрихкодов имеют одинаковую концентрацию. В соответствии с некоторыми вариантами осуществления по меньшей мере приблизительно 1%, 2%, 3%, 4%, 5%, 6%, 7%, 8%, 9%, 10%, 15%, 20%, 25%, 30%, 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95%, 97% или 100% отличающихся штрихкодов резервуара имеют одинаковую концентрацию.
В соответствии с некоторыми вариантами осуществления по меньшей мере приблизительно 1%, 2%, 3%, 4%, 5%, 6%, 7%, 8%, 9%, 10%, 15%, 20%, 25%, 30%, 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95%, 97% или 100% отличающихся молекулярных штрихкодов имеют отличающуюся концентрацию. В соответствии с некоторыми вариантами осуществления по меньшей мере приблизительно 1%, 2%, 3%, 4%, 5%, 6%, 7%, 8%, 9%, 10%, 15%, 20%, 25%, 30%, 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95%, 97% или 100% отличающихся штрихкодов резервуара имеют отличающуюся концентрацию.
Молекулярные штрихкоды или штрихкоды резервуара в популяции молекулярных штрихкодов или штрихкодов резервуара могут иметь по меньшей мере 10, 15, 20, 25, 30, 35, 40, 45, 50, 60, 70, 80, 90, 100, 200, 300, 400, 500, 600, 700, 800, 900, 1000 или большее количество отличающихся последовательностей. Например, молекулярные штрихкоды или штрихкоды резервуара в популяции могут иметь по меньшей мере 2000, 3000, 4000, 5000, 6000, 7000, 8000, 9000, 10000, 15000, 20000, 25000, 30000, 35000, 40000, 45000, 50000, 60000, 70000, 80000, 90000, 100000, 200000, 300000, 400000, 500000, 600000, 700000, 800000, 900000, 1000000 или большее количество отличающихся последовательностей. Таким образом, множество молекулярных штрихкодов или штрихкодов резервуара можно применять для создания по меньшей мере 10, 15, 20, 25, 30, 35, 40, 45, 50, 60, 70, 80, 90, 100, 200, 300, 400, 500, 600, 700, 800, 900, 1000 или большего количества отличающихся последовательностей из одного или нескольких полинуклеотидов, таких как целевые полинуклеотиды. Например, множество молекулярных штрихкодов или штрихкодов резервуара можно применять для создания по меньшей мере 2000, 3000, 4000, 5000, 6000, 7000, 8000, 9000, 10000, 15000, 20000, 25000, 30000, 35000, 40000, 45000, 50000, 60000, 70000, 80000, 90000, 100000, 200000, 300000, 400000, 500000, 600000, 700000, 800000, 900000, 1×106, 2×106, 3×106, 4×106, 5×106, 6×106, 7×106, 8×106, 9×106, 1×107, 2×107, 3×107, 4×107, 5×107, 6×107, 7×107, 8×107, 9×107, 1×108, 2×108, 3×108, 4×108, 5×108, 6×108, 7×108, 8×108, 9×108, 1×109, 2×109, 3×109, 4×109, 5×109, 6×109, 7×109, 8×109, 9×109, 1×1010, 2×1010, 3×1010, 4×1010, 5×1010, 6×1010, 7×1010, 8×1010, 9×1010, 1×1011, 2×1011, 3×1011, 4×1011, 5×1011, 6×1011, 7×1011, 8×1011, 9×1011, 1×1012, 2×1012, 3×1012, 4×1012, 5×1012, 6×1012, 7×1012, 8×1012, 9×1012 или большего количества отличающихся последовательностей из одного или нескольких полинуклеотидов, таких как целевые полинуклеотиды. Например, множество молекулярных штрихкодов или штрихкодов резервуара можно применять для создания по меньшей мере приблизительно 10, 15, 20, 25, 30, 35, 40, 45, 50, 60, 70, 80, 90, 100, 200, 300, 400, 500, 600, 700, 800, 900, 1000, 2000, 3000, 4000, 5000, 6000, 7000, 8000, 9000, 10000, 15000, 20000, 25000, 30000, 35000, 40000, 45000, 50000, 60000, 70000, 80000, 90000, 100000, 200000, 300000, 400000, 500000, 600000, 700000, 800000, 900000, 1×106, 2×106, 3×106, 4×106, 5×106, 6×106, 7×106, 8×106, 9×106, 1×107, 2×107, 3×107, 4×107, 5×107, 6×107, 7×107, 8×107, 9×107, 1×108, 2×108, 3×108, 4×108, 5×108, 6×108, 7×108, 8×108, 9×108, 1×109, 2×109, 3×109, 4×109, 5×109, 6×109, 7×109, 8×109, 9×109, 1×1010, 2×1010, 3×1010, 4×1010, 5×1010, 6×1010, 7×1010, 8×1010, 9×1010, 1×1011, 2×1011, 3×1011, 4×1011, 5×1011, 6×1011, 7×1011, 8×1011, 9×1011, 1×1012, 2×1012, 3×1012, 4×1012, 5×1012, 6×1012, 7×1012, 8×1012, 9×1012 или большего количества различных последовательностей из по меньшей мере приблизительно 10, 15, 20, 25, 30, 35, 40, 45, 50, 60, 70, 80, 90, 100, 200, 300, 400, 500, 600, 700, 800, 900, 1000, 2000, 3000, 4000, 5000, 6000, 7000, 8000, 9000, 10000, 15000, 20000, 25000, 30000, 35000, 40000, 45000, 50000, 60000, 70000, 80000, 90000, 100000, 200000, 300000, 400000, 500000, 600000, 700000, 800000, 900000, 1×106, 2×106, 3×106, 4×106, 5×106, 6×106, 7×106, 8×106, 9×106, 1×107, 2×107, 3×107, 4×107, 5×107, 6×107, 7×107, 8×107, 9×107, 1×108, 2×108, 3×108, 4×108, 5×108, 6×108, 7×108, 8×108, 9×108, 1×109, 2×109, 3×109, 4×109, 5×109, 6×109, 7×109, 8×109, 9×109, 1×1010, 2×1010, 3×1010, 4×1010, 5×1010, 6×1010, 7×1010, 8×1010, 9×1010, 1×1011, 2×1011, 3×1011, 4×1011, 5×1011, 6×1011, 7×1011, 8×1011, 9×1011, 1×1012, 2×1012, 3×1012, 4×1012, 5×1012, 6×1012, 7×1012, 8×1012, 9×1012 или большего количества целевых полинуклеотидов.
В соответствии с некоторыми вариантами осуществления один или несколько молекулярных штрихкодов применяют, чтобы объединить последовательности в группу или категорию. В соответствии с некоторыми вариантами осуществления один или несколько молекулярных штрихкодов применяют, чтобы объединить последовательности в группу или категорию, при этом последовательности в каждой категории содержат одинаковый молекулярный штрихкод. В соответствии с некоторыми вариантами осуществления один или несколько молекулярных штрихкодов или штрихкодов резервуара применяют, чтобы объединить последовательности в группу или категорию, при этом последовательности в каждой категории содержат набор ампликонов. В соответствии с некоторыми вариантами осуществления один или несколько молекулярных штрихкодов применяют, чтобы объединить последовательности в группу или категорию, при этом последовательности в каждой категории содержат множество последовательностей, при этом полинуклеотиды, из которых было образовано множество последовательностей, были получены из одной и той же молекулы полинуклеотида в реакции амплификации.
В соответствии с некоторыми вариантами осуществления один или несколько штрихкодов резервуара применяют, чтобы объединить последовательности в группу или категорию. В соответствии с некоторыми вариантами осуществления один или несколько штрихкодов резервуара применяют, чтобы объединить последовательности в группу или категорию, при этом последовательности в каждой категории содержат один и тот же штрихкод резервуара. В соответствии с некоторыми вариантами осуществления один или несколько штрихкодов резервуара применяют, чтобы объединить последовательности в группу или категорию, при этом последовательности в каждой категории содержат один или несколько наборов ампликонов. В соответствии с некоторыми вариантами осуществления один или несколько штрихкодов резервуара применяют, чтобы объединить последовательности в группу или категорию, при этом последовательности в каждой категории содержат множество последовательностей, при этом полинуклеотиды, из которых было образовано множество последовательностей, были получены из полинуклеотидов из одного и того же резервуара или отдельной клетки.
В соответствии с некоторыми вариантами осуществления одну или несколько последовательностей AID применяют, чтобы объединить последовательности в группу или категорию. В соответствии с некоторыми вариантами осуществления одну или несколько последовательностей AID применяют, чтобы объединить последовательности в группу или категорию, при этом последовательности в каждой категории содержат одинаковую последовательность AID. В соответствии с некоторыми вариантами осуществления одну или несколько последовательностей AID применяют, чтобы объединить последовательности в группу или категорию, при этом последовательности в каждой категории содержат один или несколько наборов ампликонов. В соответствии с некоторыми вариантами осуществления одну или несколько последовательностей AID применяют, чтобы объединить последовательности в группу или категорию, при этом последовательности в каждой категории содержат множество последовательностей, при этом полинуклеотиды, из которых было образовано множество последовательностей, были получены из полинуклеотидов из одного и того же резервуара или отдельной клетки.
В соответствии с некоторыми вариантами осуществления одну или несколько последовательностей АМВ применяют, чтобы объединить последовательности в группу или категорию. В соответствии с некоторыми вариантами осуществления одну или несколько последовательностей АМВ применяют, чтобы объединить последовательности в группу или категорию, при этом последовательности в каждой категории содержат одинаковую последовательность АМВ. В соответствии с некоторыми вариантами осуществления одну или несколько последовательностей АМВ применяют, чтобы объединить последовательности в группу или категорию, при этом последовательности в каждой категории содержат один или несколько наборов ампликонов. В соответствии с некоторыми вариантами осуществления одну или несколько последовательностей АМВ применяют, чтобы объединить последовательности в группу или категорию, при этом последовательности в каждой категории содержат множество последовательностей, при этом полинуклеотиды, из которых было образовано множество последовательностей, были получены из полинуклеотидов из одного и того же резервуара или отдельной клетки.
В соответствии с некоторыми вариантами осуществления одну или несколько последовательностей AID и последовательностей АМВ применяют, чтобы объединить последовательности в группу или категорию. В соответствии с некоторыми вариантами осуществления одну или несколько последовательностей AID и последовательностей АМВ применяют, чтобы объединить последовательности в группу или категорию, при этом последовательности в каждой категории содержат одинаковую последовательность AID. В соответствии с некоторыми вариантами осуществления одну или несколько последовательностей AID и последовательностей АМВ применяют, чтобы объединить последовательности в группу или категорию, при этом последовательности в каждой категории содержат одинаковую последовательность AID и последовательность АМВ. В соответствии с некоторыми вариантами осуществления одну или несколько последовательностей AID и последовательностей АМВ применяют, чтобы объединить последовательности в группу или категорию, при этом последовательности в каждой категории содержат одинаковую последовательность AID и одинаковую последовательность АМВ. В соответствии с некоторыми вариантами осуществления одну или несколько последовательностей AID и последовательностей АМВ применяют, чтобы объединить последовательности в группу или категорию, при этом последовательности в каждой категории содержат один или несколько наборов ампликонов. В соответствии с некоторыми вариантами осуществления одну или несколько последовательностей AID и последовательностей АМВ применяют, чтобы объединить последовательности в группу или категорию, при этом последовательности в каждой категории содержат множество последовательностей, при этом полинуклеотиды, из которых было образовано множество последовательностей, были получены из одного и того же полинуклеотида в реакции амплификации и из одной и той же отдельной клетки или резервуара.
В соответствии с некоторыми вариантами осуществления один или несколько штрихкодов резервуара и последовательностей АМВ применяют, чтобы объединить последовательности в группу или категорию. В соответствии с некоторыми вариантами осуществления один или несколько штрихкодов резервуара и последовательностей АМВ применяют, чтобы объединить последовательности в группу или категорию, при этом последовательности в каждой категории содержат одинаковый штрихкод резервуара. В соответствии с некоторыми вариантами осуществления один или несколько штрихкодов резервуара и последовательностей АМВ применяют, чтобы объединить последовательности в группу или категорию, при этом последовательности в каждой категории содержат одинаковый штрихкод резервуара и последовательность АМВ. В соответствии с некоторыми вариантами осуществления один или несколько штрихкодов резервуара и последовательностей АМВ применяют, чтобы объединить последовательности в группу или категорию, при этом последовательности в каждой категории содержат одинаковый штрихкод резервуара и одинаковую последовательность АМВ. В соответствии с некоторыми вариантами осуществления один или несколько штрихкодов резервуара и последовательностей АМВ применяют, чтобы объединить последовательности в группу или категорию, при этом последовательности в каждой категории содержат один или несколько наборов ампликонов. В соответствии с некоторыми вариантами осуществления один или несколько штрихкодов резервуара и последовательностей АМВ применяют, чтобы объединить последовательности в группу или категорию, при этом последовательности в каждой категории содержат множество последовательностей, при этом полинуклеотиды, из которых было образовано множество последовательностей, были получены из одного и того же полинуклеотида в реакции амплификации и из одной и той же отдельной клетки или резервуара.
В соответствии с некоторыми вариантами осуществления одну или несколько последовательностей AID и штрихкодов резервуара применяют, чтобы объединить последовательности в группу или категорию. В соответствии с некоторыми вариантами осуществления одну или несколько последовательностей AID и штрихкодов резервуара применяют, чтобы объединить последовательности в группу или категорию, при этом последовательности в каждой категории содержат одинаковую последовательность AID. В соответствии с некоторыми вариантами осуществления одну или несколько последовательностей AID и штрихкодов резервуара применяют, чтобы объединить последовательности в группу или категорию, при этом последовательности в каждой категории содержат одинаковую последовательность AID и одинаковый штрихкод резервуара. В соответствии с некоторыми вариантами осуществления одну или несколько последовательностей AID и штрихкодов резервуара применяют, чтобы объединить последовательности в группу или категорию, при этом последовательности в каждой категории содержат один или несколько наборов ампликонов. В соответствии с некоторыми вариантами осуществления одну или несколько последовательностей AID и штрихкодов резервуара применяют, чтобы объединить последовательности в группу или категорию, при этом последовательности в каждой категории содержат множество последовательностей, при этом полинуклеотиды, из которых было образовано множество последовательностей, были получены из одного и того же полинуклеотида в реакции амплификации и из одной и той же отдельной клетки или резервуара.
В соответствии с некоторыми вариантами осуществления один или несколько молекулярных штрихкодов и штрихкодов резервуара применяют, чтобы объединить последовательности в группу или категорию. В соответствии с некоторыми вариантами осуществления один или несколько молекулярных штрихкодов и штрихкодов резервуара применяют, чтобы объединить последовательности в группу или категорию, при этом последовательности в каждой категории содержат одинаковый молекулярный штрихкод и одинаковый штрихкод резервуара. В соответствии с некоторыми вариантами осуществления один или несколько молекулярных штрихкодов и штрихкодов резервуара применяют, чтобы объединить последовательности в группу или категорию, при этом последовательности в каждой категории содержат один или несколько наборов ампликонов. В соответствии с некоторыми вариантами осуществления один или несколько молекулярных штрихкодов и штрихкодов резервуара применяют, чтобы объединить последовательности в группу или категорию, при этом последовательности в каждой категории содержат множество последовательностей, при этом полинуклеотиды, из которых было образовано множество последовательностей, были получены из одного и того же полинуклеотида в реакции амплификации и из одной и той же отдельной клетки или резервуара. В соответствии с некоторыми вариантами осуществления один или несколько молекулярных штрихкодов и штрихкодов резервуара не применяют для выравнивания последовательностей.
В соответствии с некоторыми вариантами осуществления один или несколько молекулярных штрихкодов не применяют для выравнивания последовательностей. В соответствии с некоторыми вариантами осуществления один или несколько молекулярных штрихкодов применяют для выравнивания последовательностей. В соответствии с некоторыми вариантами осуществления один или несколько молекулярных штрихкодов применяют, чтобы объединить последовательности в группу или категорию, а мишенеспецифический участок применяют для выравнивания последовательностей. В соответствии с некоторыми вариантами осуществления один или несколько штрихкодов резервуара не применяют для выравнивания последовательностей. В соответствии с некоторыми вариантами осуществления один или несколько штрихкодов резервуара применяют для выравнивания последовательностей. В соответствии с некоторыми вариантами осуществления один или несколько штрихкодов резервуара применяют, чтобы объединить последовательности в группу или категорию, а мишенеспецифический участок применяют для выравнивания последовательностей. В соответствии с некоторыми вариантами осуществления один или несколько молекулярных штрихкодов и штрихкодов резервуара применяют для выравнивания последовательностей. В соответствии с некоторыми вариантами осуществления один или несколько молекулярных штрихкодов и штрихкодов резервуара применяют, чтобы объединить последовательности в группу или категорию, а мишенеспецифический участок применяют для выравнивания последовательностей.
В соответствии с некоторыми вариантами осуществления выравниваемые последовательности содержат одинаковую AID. В соответствии с некоторыми вариантами осуществления выравниваемые последовательности содержат одинаковый АМВ. В соответствии с некоторыми вариантами осуществления выравниваемые последовательности содержат одинаковые AID и АМВ. В соответствии с некоторыми вариантами осуществления выравниваемые последовательности содержат одинаковые AID и штрихкод резервуара. В соответствии с некоторыми вариантами осуществления выравниваемые последовательности содержат одинаковые АМВ и штрихкод резервуара. В соответствии с некоторыми вариантами осуществления выравниваемые последовательности содержат одинаковый молекулярный штрихкод. В соответствии с некоторыми вариантами осуществления выравниваемые последовательности содержат одинаковый штрихкод резервуара. В соответствии с некоторыми вариантами осуществления выравниваемые последовательности содержат одинаковые молекулярный штрихкод и штрихкод резервуара. В соответствии с некоторыми вариантами осуществления один или несколько молекулярных штрихкодов или штрихкодов резервуара применяют для выравнивания последовательностей, при этом выравниваемые последовательности содержат две или более последовательностей из набора ампликонов. В соответствии с некоторыми вариантами осуществления один или несколько молекулярных штрихкодов или штрихкодов резервуара применяют для выравнивания последовательностей, причем выравниваемые последовательности содержат множество последовательностей, при этом полинуклеотиды, из которых было образовано множество последовательностей, были получены из одной и той же молекулы полинуклеотида в реакции амплификации. В соответствии с некоторыми вариантами осуществления один или несколько молекулярных штрихкодов или штрихкодов резервуара применяют для выравнивания последовательностей, причем выравниваемые последовательности содержат множество последовательностей, при этом полинуклеотиды, из которых было образовано множество последовательностей, были получены из отдельной клетки или отдельного резервуара.
Получение капель
Разделение образца со множеством клеток на малые реакционные объемы в сочетании с мечением молекулярными штрихкодами и штрихкодами резервуара полинуклеотидов, происходящих из или полученных из отдельной клетки из множества клеток, могут обеспечить секвенирование с высокой пропускной способностью репертуара последовательностей, таких как последовательности биологического маркера.
Разделение образца со множеством клеток на малые реакционные объемы в сочетании с мечением молекулярными штрихкодами и штрихкодами резервуара полинуклеотидов, происходящих из или полученных из отдельной клетки из множества клеток, могут обеспечить секвенирование с высокой пропускной способностью репертуара последовательностей, таких как последовательности, составляющие процент транскриптома организма. Например, репертуар последовательностей может содержать множество последовательностей, составляющих по меньшей мере приблизительно 0,00001%, 0,00005%, 0,00010%, 0,00050%, 0,001%, 0,005%, 0,01%, 0,05%, 0,1%, 0,5%, 1%, 2%, 2,5%, 3%, 3,5%, 4%, 4,5%, 5%, 6%, 7%, 8%, 9%, 10%, 15%, 20%, 30%, 35%, 40%, 45, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95%, 98%, 99% или 100% транскриптома организма.
Разделение образца иммунных клеток на малые реакционные объемы в сочетании с мечением молекулярными штрихкодами и штрихкодами резервуара полинуклеотидов, происходящих из или полученных из отдельной иммунной клетки из множества иммунных клеток, могут обеспечить секвенирование с высокой пропускной способностью репертуара последовательностей тяжелых и легких цепей. Эти способы также могут обеспечивать возможность спаривания тяжелых и легких цепей после секвенирования на основании последовательностей со штрихкодами. Разделение образца на небольшие объемы реакционной смеси, как описано в данном документе, также может обеспечивать возможность применения уменьшенных количеств реактивов, таким образом снижая затраты материалов в анализе.
В некоторых случаях реакцию обратной транскрипции и/или реакцию амплификации (например, ПЦР) осуществляют в каплях, как например, цифровую ПЦР в каплях. В соответствии с определенными аспектами настоящим изобретением предполагаются, что замкнутые объемы жидкости содержат весь целевой материал или его часть. В соответствии с некоторыми вариантами осуществления замкнутый объем представляет собой каплю. Несмотря на то что в данной заявке делается ссылка на «капли», этот термин используется взаимозаменяемо с замкнутым объемом жидкости и ячейкой с жидкостью, если не указано иное. За исключением случаев, когда определено иное, термин «капля» используется из соображения удобства, и можно применять любую ячейку или замкнутый объем жидкости. Капли в контексте данного документа могут включать в себя эмульсионные композиции (или смеси двух или более несмешивающихся жидкостей), такие как описанные в патенте США №7622280. Капли можно получать с помощью устройств, описанных в международной заявке WO/2010/036352. Термин «эмульсия» в контексте данного документа может относиться к смеси несмешивающихся жидкостей (таких как масло и вода). Эмульсии в масляной фазе и/или эмульсии типа вода в масле обеспечивают возможность компартментализации реакционных смесей в водяных каплях. Эмульсии могут содержать водяные капли в непрерывной масляной фазе. Эмульсии, представленные в данном документе, могут представлять собой эмульсии типа масло в воде, при этом капли представляют собой масляные капли в пределах непрерывной водной фазы. Капли, представленные в данном документе, предназначены для предотвращения смешивания между замкнутыми объемами, при этом каждый замкнутый объем защищает свое содержимое от испарения и слияния с содержимым других замкнутых объемов.
Смеси или эмульсии, описанные в данном документе, могут быть стабильными или нестабильными. Эмульсии могут быть относительно стабильными и характеризоваться минимальной коалесценцией. Коалесценция происходит, когда небольшие капли объединяются с образованием все более крупных. В некоторых случаях менее чем 0,00001%, 0,00005%, 0,00010%, 0,00050%, 0,001%, 0,005%, 0,01%, 0,05%, 0,1%, 0,5%, 1%, 2%, 2,5%, 3%, 3,5%, 4%, 4,5%, 5%, 6%, 7%, 8%, 9% или 10% капель, образованных генератором капель, сливаются с другими каплями. Эмульсии также могут характеризоваться ограниченной флокуляцией, процессом, при котором дисперсная фаза переходит из суспензии в хлопья.
Могут быть образованы капли, характеризующиеся средним диаметром, равным приблизительно, меньшим чем приблизительно, или большим чем приблизительно, или составляющим по меньшей мере приблизительно 0,001, 0,01, 0,05, 0,1, 1, 5, 10, 20, 30, 40, 50, 60, 70, 80, 100, 120, 130, 140, 150, 160, 180, 200, 300, 400 или 500 микрон. Капли могут иметь средний диаметр, составляющий от приблизительно 0,001 до приблизительно 500, от приблизительно 0,01 до приблизительно 500, от приблизительно 0,1 до приблизительно 500, от приблизительно 0,1 до приблизительно 100, от приблизительно 0,01 до приблизительно 100 или от приблизительно 1 до приблизительно 100 микрон. Микропотоковые способы получения капель эмульсии с применением микроканального фокусирования в перекрестном потоке или физического перемешивания известны для получения либо монодисперсных, либо полидисперсных эмульсий. Капли могут представлять собой монодисперсные капли. Капли можно образовывать таким образом, чтобы размер капель не изменялся более чем на плюс или минус 5% от среднего размера капель. В некоторых случаях капли можно образовывать таким образом, чтобы размер капель не изменялся более чем на плюс или минус 2% от среднего размера капель. Генератор капель может образовывать популяцию капель из одного образца, при этом ни одна из капель не варьирует по размеру более чем на плюс или минус приблизительно 0,1%, 0,5%, 1%, 1,5%, 2%, 2,5%, 3%, 3,5%, 4%, 4,5%, 5%, 5,5%, 6%, 6,5%, 7%, 7,5%, 8%, 8,5%, 9%, 9,5% или 10% от среднего размера в общей популяции капель.
Более высокая механическая стабильность может быть полезной для микропотоковых манипуляций и обработки жидкостей с использованием более высокого сдвигового усилия (например, в капиллярах микропотокового устройства или посредством поворотов на 90 градусов, таких как клапаны, на пути жидкости). Капли или капсулы, подвергнутые предварительной и последующей термической обработке, могут являться механически устойчивыми к стандартным манипуляциям с помощью пипетки и центрифугированию.
Каплю можно образовывать посредством пропускания потока масляной фазы через водный образец. Водная фаза может содержать буферный раствор и реактивы для проведения реакции амплификации, в том числе клетки, нуклеотиды, нуклеотидные аналоги, полинуклеотиды с молекулярными штрихкодами, праймеры полинуклеотидов со штрихкодами резервуара, нуклеиновые кислоты-матрицы и ферменты, такие как ДНК-полимераза, РНК-полимераза и/или обратная транскриптаза.
Водная фаза может содержать буферный раствор и реактивы для осуществления реакции амплификации с использованием твердой поверхности, такой как гранула, или без нее. Буферный раствор может содержать приблизительно, больше чем приблизительно или меньше чем приблизительно 1, 5, 10, 15, 20, 30, 50, 100 или 200 мМ Tris. В некоторых случаях концентрация хлорида калия может составлять приблизительно, больше чем приблизительно или меньше чем приблизительно 10, 20, 30, 40, 50, 60, 80, 100, 200 мМ. Буферный раствор может содержать приблизительно 15 мМ Tris и 50 мМ KCl. Нуклеотиды могут содержать молекулы дезоксинуклеотидтрифосфатов, в том числе dATP, dCTP, dGTP и dTTP, в концентрациях, составляющих приблизительно, больших чем приблизительно или меньших чем приблизительно 50, 100, 200, 300, 400, 500, 600 или 700 мкМ каждого из них. В некоторых случаях dUTP добавляют в водную фазу до концентрации, составляющей приблизительно, большей чем приблизительно или меньшей чем приблизительно 50, 100, 200, 300, 400, 500, 600 или 700, 800, 900 или 1000 мкМ. В некоторых случаях хлорид магния или ацетат магния (MgCl2) добавляют в водную фазу в концентрации, составляющей приблизительно, большей чем приблизительно или меньшей чем приблизительно 1,0, 2,0, 3,0, 4,0 или 5,0 мМ. Концентрация MgCl2 может составлять приблизительно 3,2 мМ. В некоторых случаях применяют ацетат магния или магний. В некоторых случаях применяют сульфат магния.
Можно применять неспецифическое блокирующее средство, такое как BSA или желатин из бычьей кожи, при этом желатин или BSA присутствует в диапазоне концентраций примерно 0,1-0,9% масса/объем. Другие возможные блокирующие средства могут включать в себя бета-лактоглобулин, казеин, сухое молоко или другие обычные блокирующие средства. В некоторых случаях предпочтительные концентрации BSA и желатин составляют приблизительно 0,1% масса/объем.
Праймеры для амплификации в водной фазе могут иметь концентрацию, составляющую приблизительно, большую чем приблизительно или меньшую чем приблизительно 0,05, 0,1, 0,2, 0,3, 0,4, 0,5, 0,6, 0,7, 0,8, 0,9, 1,0, 1,2, 1,5, 1,7 или 2,0 мкМ. Концентрация праймера в водной фазе может составлять от приблизительно 0,05 до приблизительно 2, от приблизительно 0,1 до приблизительно 1,0, от приблизительно 0,2 до приблизительно 1,0, от приблизительно 0,3 до приблизительно 1,0, от приблизительно 0,4 до приблизительно 1,0 или от приблизительно 0,5 до приблизительно 1,0 мкМ. Концентрация праймеров может составлять приблизительно 0,5 мкМ. Пригодные диапазоны для концентраций целевой нуклеиновой кислоты при ПЦР включают в себя, без ограничения, от приблизительно 1 пг до приблизительно 500 нг.
В некоторых случаях водная фаза также может содержать добавки, в том числе, без ограничения, неспецифические фоновые/блокирующие нуклеиновые кислоты (например, ДНК спермы лосося), биологические консерванты (например, азид натрия), усиливающие добавки для ПЦР (например, бетаин, трегалоза и т.д.) и ингибиторы (например, ингибиторы РНКазы). Другие добавки могут включать в себя, например, диметилсульфоксид (DMSO), глицерин, (моно)гидрат бетаина (N,N,N-триметилглицин = [карбокси-метил] триметиламмоний), трегалозу, 7-деаза-2'-дезоксигуанозинтрифосфат (dC7GTP или 7-деаза-2'-dGTP), BSA (бычий сывороточный альбумин), формамид (метанамид), хлорид тетраметиламмония (ТМАС), другие производные тетраалкиламмония (например, хлорид тетраэтиламмония (ТЕА-Cl) и хлорид тетрапропиламмония (TPrA-Cl), неионный детергент (например, Triton Х-100, Tween 20, Nonidet Р-40 (NP-40)) или PREXCEL-Q. В некоторых случаях водная фаза может содержать 0, 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 различных добавок. В других случаях водная фаза может содержать по меньшей мере 0, 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 различных добавок.
В некоторых случаях неионный блоксополимер этиленоксида/пропиленоксида можно добавлять в водную фазу в концентрации, составляющей приблизительно 0,1%, 0,2%, 0,3%, 0,4%, 0,5%, 0,6%, 0,7%, 0,8%, 0,9% или 1,0%. Общепринятые поверхностно-активные вещества биологического происхождения включают в себя неионные поверхностно-активные вещества, такие как Pluronic F-68, Tetronics и Zonyl FSN. Pluronic F-68 может присутствовать в концентрации, составляющей приблизительно 0,5% масса/объем.
В некоторых случаях сульфат магния может заменять хлорид магния в подобных концентрациях. Широкий спектр распространенных коммерческих буферов для ПЦР от разнообразных поставщиков может заменять буферный раствор.
Эмульсию можно составить таким образом, чтобы получать капли с высокой степенью монодисперсности, имеющие подобную жидкости пленку на границе раздела, которые можно превратить посредством нагревания в микрокапсулы, имеющие подобную твердому веществу пленку на границе раздела; такие микрокапсулы могут вести себя как биореакторы, способные сохранять свое содержимое в течение процесса реакции, такой как ПЦР амплификация. Превращение в форму микрокапсулы может происходить при нагревании. Например, такое превращение может происходить при температуре, большей чем приблизительно 50°С, 60°С, 70°С, 80°С, 90°С или 95°С. В некоторых случаях это нагревание производится с применением термоциклера. Во время процесса нагревания верхний слой из жидкого или минерального масла можно применять для предотвращения испарения. Перед нагреванием избыток масла в непрерывной фазе можно удалить или не удалять. Биологически совместимые капсулы могут быть устойчивыми к коалесценции и/или флокуляции в широком диапазоне условий термической и механической обработки. После превращения капсулы можно хранить при температуре, составляющей приблизительно, большей чем приблизительно или меньшей чем приблизительно 3°С, 4°С, 5°С, 6°С, 7°С, 8°С, 9°С, 10°С, 15°С, 20°С, 25°С, 30°С, 35°С или 40°С. Эти капсулы могут быть полезны в биомедицинских применениях, таких как стабильная, контролируемая цифровым устройством инкапсуляция макромолекул, в особенности, водных биологических жидкостей, содержащих смесь нуклеиновых кислот или белка, или оба из них вместе; доставка лекарственных средств и вакцин; библиотеки биологических молекул; клинические применения для визуализации и другие.
Микрокапсулы могут содержать один или несколько полинуклеотидов и могут противостоять коалесценции, в особенности, при высоких температурах. Соответственно, реакции ПЦР амплификации могут происходить с очень высокой плотностью (например, количество реакций на единицу объема). В некоторых случаях более чем 100000, 500000, 1000000, 1500000, 2000000, 2500000, 5000000 или 10000000 отдельных реакций могут происходить в мл. В некоторых случаях реакции происходят в отдельной лунке, например, лунке микротитровального планшета, без перемешивания между реакционными объемами. Микрокапсулы также могут содержать другие компоненты, необходимые для обеспечения возможности протекания обратной транскрипции, достройки праймеров и/или реакции ПЦР, например, праймеры, зонды, dNTP, ДНК- или РНК-полимеразы и т.д. Эти капсулы проявляют устойчивость к коалесценции и флокуляции в широком диапазоне условий термической и механической обработки.
В некоторых случаях стадию амплификации осуществляют посредством проведения цифровой ПЦР, такой как цифровая ПЦР с использованием микропотокового устройства или цифровая ПЦР в капле.
Капли можно создавать с применением микропотоковых систем или устройств. В контексте данного документа префикс «микро-» (например, «микроканал» или «микропотоковый»), в целом, относится к элементам или изделиям, имеющим ширину или диаметр менее чем приблизительно 1 мм и в некоторых случаях менее чем приблизительно 100 микрон (микрометров). В некоторых случаях элемент или изделие включает в себя канал, по которому может течь жидкость. Кроме того, «микропотоковый» в контексте данного документа относится к устройству, аппарату или системе, которые включают в себя по меньшей мере один микроканал.
Микропотоковые системы и устройства были описаны в ряде контекстов, как правило, в контексте миниатюризированного лабораторного (например, клинического) анализа. Другие применения также были описаны. Например, публикации международных патентных заявок №№ WO 01/89788; WO 2006/040551; WO 2006/040554; WO 2004/002627; WO 2008/063227; WO 2004/091763; WO 2005/021151; WO 2006/096571; WO 2007/089541; WO 2007/081385 и WO 2008/063227.
Капля обычно включает в себя количество первой жидкости с образцом во второй жидкости-носителе. Любую методику, известную в уровне техники для образования капель, можно применять со способами согласно настоящему изобретению. Иллюстративный способ включает обеспечение протекания потока жидкости с образцом, содержащим целевой материал (например, иммунную клетку), таким образом, чтобы он пересекался с двумя противоположными потоками текущей жидкости-носителя. Жидкость-носитель является несмешивающейся с жидкостью с образцом. Пересечение жидкости с образцом с двумя противоположными потоками текущей жидкости-носителя приводит в результате к разделению жидкости с образцом на отдельные капли с образцом, содержащие целевой материал.
Жидкость-носитель может представлять собой любую жидкость, которая является несмешивающейся с жидкостью с образцом. Иллюстративная жидкость-носитель представляет собой масло. В соответствии с определенными вариантами осуществления жидкость-носитель включает в себя поверхностно-активное вещество.
Тот же способ можно применять для создания отдельных капель, которые содержат другие реактивы, такие как реактивы для реакции амплификации, такой как полимеразная цепная реакция (ПЦР), или реакции амплификации, не основывающейся на ПЦР, такой как амплификация со множественным замещением цепей, или других способов, известных квалифицированному специалисту в данной области техники. Подходящие реактивы для проведения реакций амплификации на основе ПЦР являются известными квалифицированным специалистам в данной области техники и включают в себя, без ограничения, ДНК-полимеразы, прямые и обратные праймеры, дезоксинуклеотидтрифосфаты (dNTP) и один или несколько буферов.
В соответствии с определенными вариантами осуществления замкнутые объемы жидкости образуют посредством обеспечения первой ячейки с жидкостью (например, капли), содержащей целевой материал (например, иммунную клетку и/или твердую подложку, такую как гранула), и второй жидкости (например, в виде потока жидкости или в каплях). Первая и вторая жидкости объединяются с образованием капли. Объединение можно осуществлять посредством воздействия электрическим полем на две жидкости. В соответствии с определенными вариантами осуществления вторая жидкость содержит реактивы для проведения реакции амплификации, такой как полимеразная цепная реакция или реакция амплификации.
В соответствии с определенными аспектами настоящее изобретение относится к способу получения библиотеки меченых уникальным штрихкодом последовательностей тяжелой и легкой цепи антитела, и/или последовательностей альфа- и бета-цепи TCR, и/или последовательностей гамма- и дельта-цепи TCR, включающему получение множества конструкций нуклеиновой кислоты, в котором каждая конструкция включает в себя уникальный N-мер и функциональный N-мер. Функциональный N-мер может представлять собой произвольный N-мер, ПЦР праймер, универсальный праймер, антитело, «липкий» конец или любую другую последовательность. Способ может включать в себя получение М наборов из количества N замкнутых объемов жидкости, каждый из которых содержит одну или несколько копий уникальной конструкции. С помощью способа можно создавать библиотеки штрихкодов с более высокой сложностью, добавляя дополнительную конструкцию к каждому замкнутому объему в наборе и повторяя это для каждого набора с получением N×M замкнутых объемов, каждый из которых содержит уникальную пару конструкций. Пары можно гибридизировать или лигировать с получением новых конструкций. В каждой конструкции в библиотеке штрихкодов каждый уникальный N-мер можно адаптировать для идентификации с помощью секвенирования, гибридизации с зондом, других способов или комбинации способов.
Библиотеки капель
В целом, библиотеки капель составляют из ряда элементов библиотеки, которые объединяют вместе в одном наборе. Библиотеки могут варьировать по сложности от одного элемента библиотеки до 1×1015 или большего количества элементов библиотеки. Каждый элемент библиотеки представляет собой один или несколько заданных компонентов с фиксированной концентрацией. Элемент может представлять собой, без ограничения, клетки, гранулы, аминокислоты, белки, полипептиды, нуклеиновые кислоты, полинуклеотиды или химические соединения-малые молекулы. Элемент может содержать идентификатор, такой как молекулярный штрихкод, штрихкод резервуара или оба из них.
Элемент библиотеки клеток может включать в себя, без ограничения, гибридомы, В-клетки, Т-клетки, первичные клетки, культивируемые клеточные линии, раковые клетки, стволовые клетки или любой другой тип клеток. Элементы клеточной библиотеки получают посредством инкапсулирования ряда клеток в количестве от одной до десятков тысяч в отдельных каплях. Количество инкапсулированных клеток обычно определяется пуассоновским статистическим распределением, исходя из численной плотности клеток и объема капли. Тем не менее, в некоторых случаях количество отклоняется от пуассоновского статистического распределения, как описано в Edd et al., Lab Chip, 8(8):1262-1264, 2008. Небольшие размеры клеток обеспечивают возможность получения библиотек в массе со множеством вариантов клеток, таких как иммунные клетки, каждая из которых продуцирует одно антитело или TCR, при этом все они присутствуют в одной исходной среде, а затем эту среду разделяют на капсулы в виде отдельных капель, которые содержат не более чем одну клетку. Клетки внутри капсул в виде отдельных капель затем лизируют, полинуклеотиды тяжелой цепи и легкой цепи, и/или полинуклеотиды альфа- и бета-цепи, и/или полинуклеотиды гамма- и дельта-цепи из лизированных клеток метят молекулярными штрихкодами и штрихкодами резервуара и амплифицируют, а затем объединяют или собирают в пул с образованием библиотеки, состоящей из элементов библиотеки в виде тяжелой и легкой цепи, и/или альфа- и бета-цепи, и/или гамма- и дельта-цепи.
Элемент библиотеки, имеющий в основе гранулу, содержит одну или несколько гранул и также может содержать другие реактивы, такие как антитела, ферменты или другие белки. В случае, когда все элементы библиотеки содержат отличающиеся типы гранул, но одинаковую окружающую среду, все элементы библиотеки могут быть получены из одной исходной жидкости, или они имеют различные исходные жидкости. В случае клеточных библиотек, полученных в массе из набора вариантов, элементы библиотеки будут получены из различных исходных жидкостей. Желательно, чтобы они имели именно одну клетку в капле и при этом лишь небольшое количество капель, содержащих более чем одну клетку, начиная с множества клеток. В некоторых случаях могут быть достигнуты отклонения от пуассоновского статистического распределения, чтобы обеспечить улучшенную загрузку капель, в результате чего присутствует большее количество капель именно с одной клеток в капле и небольшое количество исключений в виде пустых капель или капель, содержащих более чем одну клетку.
В соответствии с некоторыми вариантами осуществления желательно иметь именной один меченый штрихкодом резервуара полинуклеотид на каплю и лишь небольшое количество капель, содержащих более чем один меченый штрихкодом резервуара полинуклеотид, начиная со множества меченых штрихкодом резервуара полинуклеотидов. В некоторых случаях могут быть достигнуты отклонения от пуассоновского статистического распределения, чтобы обеспечить улучшенную загрузку капель, в результате чего присутствует большее количество капель именно с одним меченым штрихкодом резервуара полинуклеотидом на каплю и небольшое количество исключений в виде пустых капель или капель, содержащих более чем один меченый штрихкодом резервуара полинуклеотид.
Примерами библиотек капель являются наборы капель, которые имеют отличающиеся содержимое, варьирующее от гранул, клеток, малых молекул, ДНК, праймеров, антител и меченых штрихкодом полинуклеотидов. Капли варьируют по размеру от примерно 0,5 микрона до 500 микрон в диаметре, что соответствует объему от приблизительно 1 пиколитра до 1 нанолитра. Тем не менее, капли могут иметь размер от 5 микрон и до 500 микрон. Предпочтительно, капли имеют диаметр менее 100 микрон, от приблизительно 1 микрона до приблизительно 100 микрон. Наиболее предпочтительный размер составляет приблизительно 20-40 микрон в диаметре (10-100 пиколитров). Предпочтительные оцениваемые свойства библиотек капель включают в себя баланс осмотического давления, однородный размер и диапазоны размеров.
Капли, содержащиеся в библиотеке капель, обеспечиваемой с помощью настоящего изобретения, предпочтительно являются однородными по размеру. То есть диаметр любой капли в составе библиотеки будет варьировать менее чем на 5%, 4%, 3%, 2%, 1% или 0,5% по сравнению с диаметром других капель в составе той же библиотеки. Однородный размер капель в библиотеке может являться критически важным для поддержания стабильности и целостности капель, а также может быть необходимым для последующего применения капель в составе библиотеки для различных биологических и химических анализов, описанных в данном документе.
Настоящее изобретение относится к библиотеке капель, содержащей множество водяных капель в несмешивающейся жидкости, при этом каждая капля предпочтительно является практически однородной по размеру и содержит отличающийся элемент библиотеки. Настоящее изобретение относится к способу образования библиотеки капель, включающему обеспечение одной водной жидкости, содержащей отличающиеся элементы библиотеки, инкапсулирование каждого элемента библиотеки в водяную каплю в несмешивающейся жидкости.
В соответствии с определенными вариантами осуществления различные типы элементов (например, клетки или гранулы) объединяют в одном источнике, содержащемся в одной и той же среде. После первоначального объединения элементы затем инкапсулируют в капли с получением библиотеки капель, в которой каждая капля с отличающимся типом гранулы или клеткой представляет собой отличающийся элемент библиотеки. Разведение исходного раствора обеспечивает возможность процесса инкапсулирования. В соответствии с некоторыми вариантами осуществления образованные капли либо будут содержать один элемент, либо не будут содержать ничего, т.е. являются пустыми. В соответствии с другими вариантами осуществления образованные капли будут содержать несколько копий элемента библиотеки. Инкапсулируемые элементы обычно представляют собой варианты типа. В одном примере элементы представляют собой иммунные клетки в образце крови, и каждая иммунная клетка является инкапсулированной для амплификации и мечения штрихкодом последовательностей антитела из нуклеотидов в иммунных клетках.
Например, в одном типе эмульсионной библиотеки присутствуют элементы библиотеки, которые содержат различные частицы, т.е. клетки или меченые штрихкодом полинуклеотиды, в отличающейся среде, и которые являются инкапсулированными до объединения в пул. В одном примере определенное количество элементов библиотеки, т.е. количество n различных клеток или меченых штрихкодом полинуклеотидов, содержится в различных средах. Каждый из элементов библиотеки отдельно эмульгируют и собирают в пул, в этот момент каждый из количества n собранных отличающихся элементов библиотеки объединяют и собирают в один пул. Полученный в результате пул содержит множество капель эмульсии типа вода в масле, каждая из которых содержит отличающийся тип частицы.
В соответствии с некоторыми вариантами осуществления образованные капли либо будут содержать один элемент библиотеки, либо не будут содержать ничего, т.е. являются пустыми. В соответствии с другими вариантами осуществления образованные капли будут содержать несколько копий элемента библиотеки. Содержимое гранул следует пуассоновскому распределению, при котором существует дискретное распределение вероятности, которое выражает вероятность ряда событий, происходящих в фиксированный период времени, если эти события происходят с известной средней скоростью и независимо от времени с момента последнего события. Масла и поверхностно-активные вещества, применяемые для создания библиотек, предотвращают обмен содержимым библиотеки между каплями.
Праймеры
Обычно одну или несколько пар праймеров можно применять в реакции амплификации; один праймер в паре праймеров может представлять собой прямой праймер, и один праймер в паре праймеров может представлять собой обратный праймер.
В некоторых случаях первую пару праймеров можно применять в реакции амплификации; один праймер в первой паре может представлять собой прямой праймер, комплементарный последовательности в молекуле первого целевого полинуклеотида, и один праймер в первой паре может представлять собой обратный праймер, который может быть комплементарным второй последовательности в молекуле первого целевого полинуклеотида, и первый целевой локус может находиться между первой последовательностью и второй последовательностью. В соответствии с некоторыми вариантами осуществления первый целевой локус содержит последовательность VH, или Vα, или Vγ. В соответствии с некоторыми вариантами осуществления первый целевой локус содержит последовательность AID и/или последовательность АМВ.
В некоторых случаях вторую пару праймеров можно применять в реакции амплификации; один праймер во второй паре может представлять собой прямой праймер, комплементарный первой последовательности в молекуле второго целевого полинуклеотида, и один праймер во второй паре может представлять собой обратный праймер, комплементарный второй последовательности в молекуле второго целевого полинуклеотида, и второй целевой локус может находиться между первой последовательностью и второй последовательностью. В соответствии с некоторыми вариантами осуществления второй целевой локус содержит последовательность VL, или Vβ, или Vδ.
В некоторых случаях третью пару праймеров можно применять в реакции амплификации; один праймер в третьей паре может представлять собой прямой праймер, комплементарный первой последовательности в молекуле третьего целевого полинуклеотида, и один праймер в третьей паре может представлять собой обратный праймер, комплементарный второй последовательности в молекуле третьего целевого полинуклеотида, и третий целевой локус может находиться между первой последовательностью и второй последовательностью. В соответствии с некоторыми вариантами осуществления третий целевой локус содержит штрихкод, такой как молекулярный штрихкод или штрихкод резервуара.
Длина прямого праймера и обратного праймера может зависеть от последовательности целевого полинуклеотида и целевого локуса. Например, длину и/или TM прямого праймера и обратного праймера можно оптимизировать. В некотором случае праймер может иметь длину, приблизительно равную, приблизительно большую чем или приблизительно меньшую чем 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49, 50, 51, 52, 53, 54, 55, 56, 57, 58, 59 или 60 нуклеотидов. В некоторых случаях праймер имеет длину от приблизительно 15 до приблизительно 20, от приблизительно 15 до приблизительно 25, от приблизительно 15 до приблизительно 30, от приблизительно 15 до приблизительно 40, от приблизительно 15 до приблизительно 45, от приблизительно 15 до приблизительно 50, от приблизительно 15 до приблизительно 55, от приблизительно 15 до приблизительно 60, от приблизительно 20 до приблизительно 25, от приблизительно 20 до приблизительно 30, от приблизительно 20 до приблизительно 35, от приблизительно 20 до приблизительно 40, от приблизительно 20 до приблизительно 45, от приблизительно 20 до приблизительно 50, от приблизительно 20 до приблизительно 55 или от приблизительно 20 до приблизительно 60 нуклеотидов.
Праймер может представлять собой одноцепочечную ДНК до связывания с матричным полинуклеотидом. В некоторых случаях праймер изначально содержит двухцепочечную последовательность. Подходящая длина праймера может зависеть от предполагаемого применения праймера, но может находиться в диапазоне от приблизительно 6 до приблизительно 50 нуклеотидов или от приблизительно 15 до приблизительно 35 нуклеотидов. Молекулам коротких праймеров обычно могут требоваться меньшие температуры для образования достаточно стабильных гибридных комплексов с матрицей. В соответствии с некоторыми вариантами осуществления праймер не обязательно должен отражать точную последовательность нуклеиновой кислоты матрицы, но может являться в достаточной мере комплементарным для гибридизации с матрицей. В некоторых случаях праймер может являться частично двухцепочечным до связывания с матричным полинуклеотидом. Праймер с двухцепочечной последовательностью может иметь петлю «шпилька» с длиной, равной приблизительно, большей чем приблизительно или меньшей чем приблизительно 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19 или 20 оснований. Двухцепочечная часть праймера может иметь длину, равную приблизительно, большую чем приблизительно, меньшую чем приблизительно или составляющую по меньшей мере приблизительно 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49 или 50 пар оснований. Конструирование подходящих праймеров для амплификации заданной целевой последовательности является хорошо известным в уровне техники.
Праймеры могут включать дополнительные элементы, которые обеспечивают возможность выявления или иммобилизации праймера, но не изменяют основного свойства праймера (например, действие в качестве точки инициации синтеза ДНК). Например, праймеры могут содержать дополнительную последовательность нуклеиновой кислоты на 5' конце, которая не гибридизируется с целевой нуклеиновой кислотой, но которая облегчает клонирование, или дополнительную амплификацию, или секвенирование амплифицированного продукта. Например, дополнительная последовательность может содержать сайт для связывания праймера, такой как сайт для связывания универсального праймера. Участок праймера, который является в достаточной мере комплементарным матрице, чтобы гибридизироваться, можно назвать в данном документе участком гибридизации.
В другом случае праймер, используемый в способах и композициях, описанных в данном документе, может содержать один или несколько универсальных нуклеозидов. Неограничивающие примеры универсальных нуклеозидов представляют собой 5-нитроиндол и инозин, которые описаны в публикациях заявок на патент США №2009/0325169 и №2010/0167353.
Праймеры могут быть сконструированы в соответствии с известными параметрами для исключения образования вторичных структур и гибридизации молекулы самой с собой. Различные пары праймеров могут гибридизироваться и плавиться приблизительно при одинаковых температурах, например, в пределах 1°С, 2°С, 3°С, 4°С, 5°С, 6°С, 7°С, 8°С, 9°С или 10°С относительно другой пары праймеров. В некоторых случаях первоначально применяют более чем 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 15, 20, 25, 30, 35, 40, 45, 50, 100, 200, 500, 1000, 5000, 10000 праймеров или более. Такие праймеры могут гибридизироваться с целевыми полинуклеотидами, описанными в данном документе.
Праймеры могут быть получены с помощью ряда способов, в том числе, без ограничения, с помощью клонирования соответствующих последовательностей и прямого химического синтеза с применением способов, хорошо известных в уровне техники (Narang et al., Methods Enzymol. 68:90 (1979); Brown et al., Methods Enzymol. 68:109 (1979)). Праймеры также могут быть получены из коммерческих источников. Праймеры могут характеризоваться идентичной температурой плавления. Праймеры могут характеризоваться неидентичными температурами плавления. Длины праймеров можно нарастить или сократить на 5' конце или 3' конце для получения праймеров с желаемыми температурами плавления. Один из праймеров в паре праймеров может быть длиннее, чем другой праймер. Длины гибридизирующихся 3' участков праймеров в составе пары праймеров могут отличаться. Кроме того, гибридизирующееся положение в каждой паре праймеров можно определять таким образом, чтобы последовательность и длина пар праймеров давала на выходе желаемую температуру плавления. Уравнением для определения температуры плавления у праймеров, меньших чем 25 пар оснований, является правило Уоллеса (TM=2(A+T)+4(G+C)). Для конструирования праймеров также можно применять компьютерные программы. TM (температура плавления или гибридизации) для каждого праймера можно рассчитать с применением компьютерных программ. Температуру гибридизации для праймеров можно пересчитать и повысить после любого цикла амплификации, в том числе, без ограничения, цикла 1, 2, 3, 4, 5, циклов 6-10, циклов 10-15, циклов 15-20, циклов 20-25, циклов 25-30, циклов 30-35 или циклов 35-40. После первоначальных циклов амплификации 5' половина праймеров может включаться в продукты из каждого локуса, представляющего интерес; следовательно, TM можно пересчитать, исходя из последовательностей как 5' половины, так и 3' половины каждого праймера.
Сайт для праймера включает в себя область матрицы, с которой гибридизируется праймер. В соответствии с некоторыми вариантами осуществления праймеры являются способными действовать в качестве точки инициации для управляемого матрицей синтеза нуклеиновой кислоты. Например, праймеры могут инициировать управляемый матрицей синтез нуклеиновой кислоты при наличии четырех отличающихся нуклеотидов и обеспечивающего полимеризацию средства или фермента, такого как ДНК- или РНК-полимераза или обратная транскриптаза. Пара праймеров включает в себя 2 праймера: первый праймер с 5' участком, расположенным против хода транскрипции, который гибридизируется с 5' концом последовательности матрицы, и второй праймер с 3' участком, расположенным по ходу транскрипции, который гибридизируется с комплементарной последовательностью 3' конца последовательности матрицы. Набор праймеров включает в себя два или более праймеров: первый праймер или первое множество праймеров с 5' участком, расположенным против хода транскрипции, который гибридизируется с 5' концом последовательности матрицы или множества последовательностей матриц, и второй праймер или второе множество праймеров с 3' участком, расположенным по ходу транскрипции, который гибридизируется с комплементарной последовательностью 3' конца последовательности матрицы или множества последовательностей матриц. В соответствии с некоторыми вариантами осуществления праймер содержит мишенеспецифическую последовательность. В соответствии с некоторыми вариантами осуществления праймер содержит последовательность штрихкода образца. В соответствии с некоторыми вариантами осуществления праймер содержит универсальную праймерную последовательность. В соответствии с некоторыми вариантами осуществления праймер содержит последовательность ПЦР-праймера. В соответствии с некоторыми вариантами осуществления праймер содержит последовательность ПЦР-праймера, применяемую для инициации амплификации полинуклеотида. (Dieffenbach, PCR Primer: A Laboratory Manual, 2nd Edition (Cold Spring Harbor Press, New York (2003)). Сайт или последовательность для связывания универсального праймера обеспечивает возможность прикрепления универсального праймера к полинуклеотиду и/или ампликону. Универсальные праймеры являются хорошо известными в уровне техники и включают в себя, без ограничения, -47F (M13F), alfaMF, АОХ3', АОХ5', BGHr, CMV-30, CMV-50, CVMf, LACrmt, лямбда gt10F, лямбда gt10R, лямбда gt11F, лямбда gt11R, М13 rev, M13Forward(-20), M13Reverse, male, p10SEQPpQE, pA-120, pet4, pGAP Forward, pGLRVpr3, pGLpr2R, pKLAC14, pQEFS, pQERS, pucU1, pucU2, reversA, seqIREStam, seqIRESzpet, seqori, seqPCR, seqpIRES-, seqpIRES+, seqpSecTag, seqpSecTag+, seqretro+PSI, SP6, T3-prom, T7-prom и T7-termInv. В контексте данного документа прикрепление может относиться обоим или одному из ковалентных взаимодействий и нековалентных взаимодействий. Прикрепление универсального праймера у сайту для связывания универсального праймера можно применять для амплификации, выявления и/или секвенирования полинуклеотида и/или ампликона. Сайт связывания универсального праймера может содержать по меньшей мере приблизительно 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 30, 40, 50, 60, 70, 80, 90, 100, 200, 300, 400, 500, 600, 700, 800, 900 или 1000 нуклеотидов или пар оснований. В еще одном примере сайт связывания универсального праймера содержит по меньшей мере приблизительно 1500, 2000, 2500, 3000, 3500, 4000, 4500, 5000, 5500, 6000, 6500, 7000, 7500, 8000, 8500, 9000, 9500 или 10000 нуклеотидов или пар оснований. В соответствии с некоторыми вариантами осуществления сайт связывания универсального праймера содержит 1-10, 10-20, 10-30 или 10-100 нуклеотидов или пар оснований. В соответствии с некоторыми вариантами осуществления сайт связывания универсального праймера содержит приблизительно 1-90, 1-80, 1-70, 1-60, 1-50, 1-40, 1-30, 1-20, 1-10, 2-90, 2-80, 2-70, 2-60, 2-50, 2-40, 2-30, 2-20, 2-10, 1-900, 1-800, 1-700, 1-600, 1-500, 1-400, 1-300, 1-200, 1-100, 2-900, 2-800, 2-700, 2-600, 2-500, 2-400, 2-300, 2-200, 2-100, 5-90, 5-80, 5-70, 5-60, 5-50, 5-40, 5-30, 5-20, 5-10, 10-90, 10-80, 10-70, 10-60, 10-50, 10-40, 10-30, 10-20, 10-10, 5-900, 5-800, 5-700, 5-600, 5-500, 5-400, 5-300, 5-200, 5-100, 10-900, 10-800, 10-700, 10-600, 10-500, 10-400, 10-300, 10-200, 10-100, 25-900, 25-800, 25-700, 25-600, 25-500, 25-400, 25-300, 25-200, 25-100, 100-1000, 100-900, 100-800, 100-700, 100-600, 100-500, 100-400, 100-300, 100-200, 200-1000, 200-900, 200-800, 200-700, 200-600, 200-500, 200-400, 200-300, 300-1000, 300-900, 300-800, 300-700, 300-600, 300-500, 300-400, 400-1000, 400-900, 400-800, 400-700, 400-600, 400-500, 500-1000, 500-900, 500-800, 500-700, 500-600, 600-1000, 600-900, 600-800, 600-700, 700-1000, 700-900, 700-800, 800-1000, 800-900 или 900-1000 нуклеотидов или пар оснований.
Праймеры могут иметь длину, совместимую с применением в синтезе продуктов достройки праймеров. Праймер может представлять собой полинуклеотид, который имеет длину 8-200 нуклеотидов. Длина праймера может зависеть от последовательности матричного полинуклеотида и матричного локуса. Например, длину и/или температуру плавления (TM) праймера или набора праймеров можно оптимизировать. В некотором случае праймер может иметь длину, приблизительно равную, приблизительно большую чем или приблизительно меньшую чем 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49, 50, 51, 52, 53, 54, 55, 56, 57, 58, 59 или 60 нуклеотидов. В соответствии с некоторыми вариантами осуществления праймеры имеют длину приблизительно 8-100 нуклеотидов, например, 10-75, 15-60, 15-40, 18-30, 20-40, 21-50, 22-45, 25-40, 7-9, 12-15, 15-20, 15-25, 15-30, 15-45, 15-50, 15-55, 15-60, 20-25, 20-30, 20-35, 20-45, 20-50, 20-55 или 20-60 нуклеотидов в длину и любую длину между этими значениями. В соответствии с некоторыми вариантами осуществления праймеры имеют длину не более приблизительно 10, 12, 15, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 35, 40, 45, 50, 55, 60, 65, 70, 75, 80, 85, 90, 95 или 100 нуклеотидов.
Обычно одну или несколько пар праймеров можно применять в экспоненциальной реакции амплификации; один праймер в паре праймеров может представлять собой прямой праймер, и один праймер в паре праймеров может представлять собой обратный праймер. В соответствии с некоторыми вариантами осуществления первую пару праймеров можно применять в реакции экспоненциальной амплификации; один праймер в составе первой пары может представлять собой прямой праймер, комплементарный последовательности молекулы первого матричного полинуклеотида, и один праймер в составе первой пары может представлять собой обратный праймер, комплементарный второй последовательности молекулы первого матричного полинуклеотида, и первый матричный локус может находиться между первой последовательностью и второй последовательностью. В соответствии с некоторыми вариантами осуществления вторую пару праймеров можно применять в реакции амплификации; один праймер во второй паре может представлять собой прямой праймер, комплементарный первой последовательности в молекуле второго целевого полинуклеотида, и один праймер во второй паре может представлять собой обратный праймер, комплементарный второй последовательности в молекуле второго целевого полинуклеотида, и второй целевой локус может находиться между первой последовательностью и второй последовательностью. В соответствии с некоторыми вариантами осуществления второй целевой локус содержит последовательность вариабельного домена легкой цепи антитела. В соответствии с некоторыми вариантами осуществления третью пару праймеров можно применять в реакции амплификации; один праймер в третьей паре может представлять собой прямой праймер, комплементарный первой последовательности в молекуле третьего матричного полинуклеотида, и один праймер в третьей паре может представлять собой обратный праймер, комплементарный второй последовательности в молекуле третьего матричного полинуклеотида, и третий матричный локус может находиться между первой последовательностью и второй последовательностью.
Один или несколько праймеров могут гибридизироваться по меньшей мере с частью из множества матричных полинуклеотидов. Один или несколько праймеров могут гибридизироваться с 3' концом и/или 5' концом из множества матричных полинуклеотидов. Один или несколько праймеров могут гибридизироваться с внутренним участком из множества матричных полинуклеотидов. Внутренний участок может представлять собой по меньшей мере приблизительно 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49, 50, 100, 150, 200, 220, 230, 240, 250, 260, 270, 280, 290, 300, 310, 320, 330, 340, 350, 360, 370, 380, 390, 400, 410, 420, 430, 440, 450, 460, 470, 480, 490, 500, 510, 520, 530, 540, 550, 560, 570, 580, 590, 600, 650, 700, 750, 800, 850, 900 или 1000 нуклеотидов с 3' концов или 5' концов множества матричных полинуклеотидов. Один или несколько праймеров могут содержать фиксированную панель праймеров. Один или несколько праймеров могут содержать по меньшей мере один или несколько специально созданных праймеров. Один или несколько праймеров могут содержать по меньшей мере один или несколько контрольных праймеров. Один или несколько праймеров могут содержать по меньшей мере один или несколько праймеров генов «домашнего хозяйства». Один или несколько праймеров могут содержать универсальный праймер. Универсальный праймер может гибридизироваться с сайтом связывания для универсального праймера. В соответствии с некоторыми вариантами осуществления один или несколько специально созданных праймеров гибридизируются с SBC, мишенеспецифическим участком, их комплементарными последовательностями или любой их комбинацией. Один или несколько праймеров могут содержать универсальный праймер. Один или несколько праймеров могут быть предназначены для амплификации или осуществления достройки праймера, обратной транскрипции, линейной достройки, неэкспоненциальной амплификации, экспоненциальной амплификации, ПЦР или любого другого способа амплификации одного или нескольких целевых или матричных полинуклеотидов.
Мишенеспецифический участок может содержать по меньшей мере приблизительно 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49, 50, 100, 150, 200, 220, 230, 240, 250, 260, 270, 280, 290, 300, 310, 320, 330, 340, 350, 360, 370, 380, 390, 400, 410, 420, 430, 440, 450, 460, 470, 480, 490, 500, 510, 520, 530, 540, 550, 560, 570, 580, 590, 600, 650, 700, 750, 800, 850, 900 или 1000 нуклеотидов или пар оснований. В другом примере мишенеспецифический участок содержит по меньшей мере приблизительно 1500, 2000, 2500, 3000, 3500, 4000, 4500, 5000, 5500, 6000, 6500, 7000, 7500, 8000, 8500, 9000, 9500 или 10000 нуклеотидов или пар оснований. В соответствии с некоторыми вариантами осуществления мишенеспецифический участок содержит приблизительно 5-10, 10-15, 10-20, 10-30, 15-30, 10-75, 15-60, 15-40, 18-30, 20-40, 21-50, 22-45, 25-40, 7-9, 12-15, 15-20, 15-25, 15-30, 15-45, 15-50, 15-55, 15-60, 20-25, 20-30, 20-35, 20-45, 20-50, 20-55, 20-60, 2-900, 2-800, 2-700, 2-600, 2-500, 2-400, 2-300, 2-200, 2-100, 25-900, 25-800, 25-700, 25-600, 25-500, 25-400, 25-300, 25-200, 25-100, 100-1000, 100-900, 100-800, 100-700, 100-600, 100-500, 100-400, 100-300, 100-200, 200-1000, 200-900, 200-800, 200-700, 200-600, 200-500, 200-400, 200-300, 300-1000, 300-900, 300-800, 300-700, 300-600, 300-500, 300-400, 400-1000, 400-900, 400-800, 400-700, 400-600, 400-500, 500-1000, 500-900, 500-800, 500-700, 500-600, 600-1000, 600-900, 600-800, 600-700, 700-1000, 700-900, 700-800, 800-1000, 800-900 или 900-1000 нуклеотидов или пар оснований.
Праймеры могут быть сконструированы в соответствии с известными параметрами для исключения образования вторичных структур и гибридизации молекулы самой с собой. В соответствии с некоторыми вариантами осуществления различные пары праймеров могут гибридизироваться и плавиться приблизительно при одинаковых температурах, например, в пределах 1°С, 2°С, 3°С, 4°С, 5°С, 6°С, 7°С, 8°С, 9°С или 10°С относительно другой пары праймеров. В соответствии с некоторыми вариантами осуществления один или несколько праймеров во множестве праймеров могут гибридизироваться и плавиться приблизительно при одинаковых температурах, например, в пределах 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10°С относительно другого праймера во множестве праймеров. В соответствии с некоторыми вариантами осуществления один или несколько праймеров во множестве могут гибридизироваться и плавиться при температурах, отличных от температур для другого праймера во множестве праймеров.
Множество праймеров для одной или нескольких стадий способов, описанных в данном документе, может содержать множество праймеров, содержащих приблизительно, не более чем приблизительно или по меньшей мере приблизительно 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 30, 40, 50, 60, 70, 80, 90, 100, 200, 300, 400, 500, 600, 700, 800, 900, 1000, 1500, 2000, 3000, 4000, 5000, 6000, 7000, 8000, 9000, 10000, 11000, 12000, 13000, 14000, 15000, 16000, 17000, 18000, 19000, 20000, 30000, 40000, 50000, 60000, 70000, 80000, 90000, 100000, 200000, 300000, 400000, 500000, 600000, 700000, 800000, 900000, 1000000, 50000000, 100000000 различных праймеров. Например, каждый праймер во множестве праймеров может содержать отличающийся мишене- или матрицеспецифический участок или последовательность.
Обратная транскрипция
В некоторых случаях целевые полинуклеотиды получают из РНК посредством обратной транскрипции. В некоторых случаях целевые полинуклеотиды получают из ДНК посредством достройки праймера, как например, с применением полимеразы.
Способы, описанные в данном документе, можно применять в сочетанных обратной транскрипции и ПЦР (обратная транскрипция-ПЦР). Например, обратную транскрипцию и ПЦР можно осуществлять в две отдельных стадии. Первую копию кДНК из образца мРНК можно синтезировать с применении одного из полинуклеотид-dT-праймера, специфического к последовательности праймера, универсального праймера или любого праймера, описанного в данном документе.
Обратную транскрипцию и ПЦР можно осуществлять в реакционной смеси в одном закрытом резервуаре. Например, можно использовать три праймера: один для обратной транскрипции и два для ПЦР. Праймер для обратной транскрипции может связываться с мРНК в положении 3' относительно положения ПЦР ампликона. Хотя это и не является необходимым, праймер для обратной транскрипции может включать в себя остатки РНК или модифицированные аналоги, такие как основания 2'-O-метил-РНК, которые не будут образовывать субстрат для РНКазы Н при гибридизации с мРНК.
Температура для проведения реакции обратной транскрипции зависит от применяемой обратной транскриптазы. В некоторых случаях применяют термостабильную обратную транскриптазу, и реакцию обратной транскрипции осуществляют при температуре от приблизительно 37°С до приблизительно 75°С, от приблизительно 37°С до приблизительно 50°С, от приблизительно 37°С до приблизительно 55°С, от приблизительно 37°С до приблизительно 60°С, от приблизительно 55°С до приблизительно 75°С, от приблизительно 55°С до приблизительно 60°С, при приблизительно 37°С или при приблизительно 60°С. В некоторых случаях применяют обратную транскриптазу, которая переносит 3 или более концевых нуклеотидов, не относящихся к матрице, на конец транскрибированного продукта.
Реакцию обратной транскрипции и реакцию ПЦР, описанную в данном документе, можно осуществлять в различных форматах, известных в уровне техники, как например, в пробирках, микротитровальных планшетах, микропотоковых устройствах или, предпочтительно, в каплях.
Реакцию обратной транскрипции можно осуществлять в объемах, находящихся в диапазоне от 5 мкл до 100 мкл или в объемах реакционной смеси от 10 мкл до 20 мкл. В каплях объемы реакционной смеси могут находиться в диапазоне от 1 пл до 100 нл или от 10 пл до 1 нл. В некоторых случаях реакцию обратной транскрипции осуществляют в капле, имеющей объем, который составляет приблизительно 1 нл или меньше. В некоторых случаях реакция ПЦР проходит в капле, имеющей объем реакционной смеси в диапазоне от 1 пл до 100 нл, предпочтительно, от 10 пл до 1 нл. В некоторых случаях реакцию ПЦР осуществляют в капле, имеющей объем, который составляет приблизительно 1 нл или меньше. В некоторых случаях реакцию обратной транскрипции и реакцию ПЦР осуществляют в одной и той же капле, имеющей объем реакционной смеси в диапазоне от 1 пл до 100 нл или от 10 пл до 1 нл. В некоторых случаях реакцию обратной транскрипции и реакцию ПЦР осуществляют в капле, имеющей объем, который составляет приблизительно 1 нл или меньше, или объем, который составляет приблизительно 1 пл или меньше. В некоторых случаях реакцию обратной транскрипции и реакцию ПЦР осуществляют в разных каплях. В некоторых случаях реакцию обратной транскрипции и реакцию ПЦР осуществляют во множестве капель, каждая из которых имеет объем реакционной смеси в диапазоне от 1 пл до 100 нл или от 10 пл до 1 нл. В некоторых случаях реакцию обратной транскрипции и реакцию ПЦР осуществляют во множестве капель, каждая из которых имеет объем, приблизительно равный или меньший чем 1 нл.
В некоторых случаях первая реакция ПЦР проходит в первой капле, имеющей реакционный объем в диапазоне от 1 пл до 100 нл, предпочтительно, от 10 пл до 1 нл, и вторая реакция ПЦР проходит во второй капле, имеющей реакционный объем в диапазоне от 1 пл до 100 нл, предпочтительно, от 10 пл до 1 нл. В некоторых случаях первая реакция ПЦР проходит в первой капле, имеющей объем, который является приблизительно равным или меньшим чем 1 нл, и вторая реакция ПЦР проходит во второй капле, имеющей объем, который является приблизительно равным или меньшим чем 1 нл.
В некоторых случаях первую реакцию ПЦР и вторую реакцию ПЦР осуществляют во множестве капель, каждая из которых имеет объем реакционной смеси в диапазоне от 1 пл до 100 нл или от 10 пл до 1 нл. В некоторых случаях первую реакцию ПЦР и вторую реакцию ПЦР осуществляют во множестве капель, каждая из которых имеет объем, приблизительно равный или меньший чем 1 нл.
Целевые полинуклеотиды, такие как РНК, могут быть подвергнуты обратной транскрипции в кДНК с применением одного или нескольких праймеров для обратной транскрипции. Один или несколько праймеров для обратной транскрипции могут содержать участок, комплементарный участку РНК, такому как константный участок (например, константный участок тяжелой или легкой цепи или поли-А-хвост мРНК). В соответствии с некоторыми вариантами осуществления праймеры для обратной транскрипции могут содержать первый праймер для обратной транскрипции с участком, комплементарным константному участку первой РНК, и второй праймер для обратной транскрипции с участком, комплементарным константному участку второй РНК. В соответствии с некоторыми вариантами осуществления праймеры для обратной транскрипции могут содержать первый праймер для обратной транскрипции с участком, комплементарным константному участку первой РНК, и один или несколько праймеров для обратной транскрипции с участком, комплементарным константному участку одной или нескольких РНК, соответственно.
В соответствии с некоторыми вариантами осуществления праймеры для обратной транскрипции не содержат штрихкод.
Праймеры для обратной транскрипции могут дополнительно содержать участок, который не является комплементарным участку в РНК. В соответствии с некоторыми вариантами осуществления участок, который не является комплементарным участку РНК, располагается в направлении 5' относительно участка праймеров, который является комплементарным РНК. В соответствии с некоторыми вариантами осуществления участок, который не является комплементарным участку РНК, располагается в направлении 3' относительно участка праймеров, который является комплементарным РНК. В соответствии с некоторыми вариантами осуществления участок, который не является комплементарным участку РНК, представляет собой участок 5' «липкого» конца. В соответствии с некоторыми вариантами осуществления участок, который не является комплементарным участку РНК, содержит инициирующий сайт для амплификации и/или реакции секвенирования. При применении одного или нескольких праймеров, описанных в данном документе, молекулы РНК подвергаются обратной транскрипции с применением подходящих реактивов, известных в уровне техники.
После осуществления реакций обратной транскрипции молекул РНК полученные в результате молекулы кДНК можно пометить молекулярным штрихкодом и штрихкодом резервуара и амплифицировать с помощью одной или нескольких реакций ПЦР, таких как первая и/или вторая реакция ПЦР. В первой и/или второй реакции ПЦР могут использоваться пары праймеров или множество пар праймеров. В первой и/или второй реакции ПЦР может использоваться множество прямых/обратных праймеров и обратный праймер. В первой и/или второй реакции ПЦР может использоваться множество прямых/обратных праймеров и прямой праймер. Первый и/или второй праймер из множества прямых/обратных праймеров может представлять собой прямой/обратный праймер, содержащий участок, комплементарный молекулам кДНК или молекулам меченой штрихкодом кДНК. Первый и/или второй праймер из множества прямых/обратных праймеров может представлять собой прямой/обратный праймер, содержащий участок, комплементарный молекулам меченой штрихкодом кДНК.
В соответствии с некоторыми вариантами осуществления множество прямых/обратных праймеров содержит один или несколько прямых/обратных праймеров, при этом каждый из прямых/обратных праймеров во множестве прямых/обратных праймеров содержит участок, комплементарный одному или нескольким участкам, расположенным выше или ниже относительно V-сегмента кДНК или меченой штрихкодом кДНК. Например, множество прямых/обратных праймеров содержит прямой/обратный праймер, содержащий участок, комплементарный участку, расположенному выше или ниже относительно V-сегмента кДНК или меченой штрихкодом кДНК, и один или несколько других прямых/обратных праймеров, содержащих участок, комплементарный одному или нескольким другим участкам, расположенным выше или ниже относительно V-сегмента кДНК или меченых штрихкодами кДНК. Например, множество прямых/обратных праймеров содержит первый и/или второй прямой/обратный праймер, содержащий участок, комплементарный первому и/или второму участку, расположенному выше или ниже относительно V-сегмента кДНК или меченой штрихкодом кДНК, и второй прямой/обратный праймер, содержащий участок, комплементарный второму участку, расположенному выше или ниже относительно V-сегмента кДНК или меченых штрихкодом кДНК. Например, множество прямых/обратных праймеров содержит первый и/или второй прямой/обратный праймер, содержащий участок, комплементарный первому и/или второму участку, расположенному выше или ниже относительно V-сегмента кДНК или меченых штрихкодом кДНК, второй прямой/обратный праймер, содержащий участок, комплементарный второму участку, расположенному выше или ниже относительно V-сегмента кДНК или меченых штрихкодами кДНК, и третий прямой/обратный праймер, содержащий участок, комплементарный третьему участку, расположенному выше или ниже относительно V-сегмента кДНК или меченых штрихкодом кДНК, и т.д. Праймеры во множестве прямых/обратных праймеров можно применять для гибридизации со всеми возможными участками, расположенными выше или ниже относительно всех V-сегментов, экспрессируемых клетками, такими как иммунные В-клетки или Т-клетки, в образце.
В соответствии с некоторыми вариантами осуществления множество прямых/обратных праймеров содержит один или несколько прямых/обратных праймеров, при этом каждый из прямых/обратных праймеров во множестве прямых/обратных праймеров содержит участок, комплементарный одному или нескольким участкам, расположенным выше или ниже относительно С-сегмента кДНК или меченых штрихкодами кДНК. Например, множество прямых/обратных праймеров содержит прямой/обратный праймер, содержащий участок, комплементарный участку, расположенному выше или ниже относительно С-сегмента кДНК или меченых штрихкодами кДНК, и один или несколько других прямых/обратных праймеров, содержащих участок, комплементарный одному или нескольким другим участкам, расположенным выше или ниже относительно С-сегмента кДНК или меченых штрихкодами кДНК. Например, множество прямых/обратных праймеров содержит первый и/или второй прямой/обратный праймер, содержащий участок, комплементарный первому и/или второму участку, расположенному выше или ниже относительно С-сегмента кДНК или меченых штрихкодами кДНК, и второй прямой/обратный праймер, содержащий участок, комплементарный второму участку, расположенному выше или ниже относительно С-сегмента кДНК или меченых штрихкодами кДНК. Например, множество прямых/обратных праймеров содержит первый и/или второй прямой/обратный праймер, содержащий участок, комплементарный первому и/или второму участку, расположенному выше или ниже относительно С-сегмента кДНК или меченых штрихкодами кДНК, второй прямой/обратный праймер, содержащий участок, комплементарный второму участку, расположенному выше или ниже относительно С-сегмента кДНК или меченых штрихкодами кДНК, и третий прямой/обратный праймер, содержащий участок, комплементарный третьему участку, расположенному выше или ниже относительно С-сегмента кДНК или меченых штрихкодами кДНК, и т.д. Праймеры во множестве прямых/обратных праймеров можно применять для гибридизации со всеми возможными участками, расположенными выше или ниже относительно всех С-сегментов, экспрессируемых клетками, такими как иммунные В-клетки или Т-клетки, в образце.
В соответствии с некоторыми вариантами осуществления множество прямых/обратных праймеров содержит один или несколько прямых/обратных праймеров, при этом каждый из прямых/обратных праймеров во множестве прямых/обратных праймеров содержит участок, комплементарный одному или нескольким участкам, расположенным выше или ниже относительно молекулярного штрихкода в меченых штрихкодами кДНК. Например, множество прямых/обратных праймеров содержит прямой/обратный праймер, содержащий участок, комплементарный участку, расположенному выше или ниже относительно молекулярного штрихкода в меченых штрихкодами кДНК, и один или несколько других прямых/обратных праймеров, содержащих участок, комплементарный одному или нескольким другим участкам, расположенным выше или ниже относительно молекулярного штрихкода в меченых штрихкодами кДНК. Например, множество прямых/обратных праймеров содержит первый и/или второй прямой/обратный праймер, содержащий участок, комплементарный первому и/или второму участку, расположенному выше или ниже относительно молекулярного штрихкода в меченых штрихкодами кДНК, и второй прямой/обратный праймер, содержащий участок, комплементарный второму участку, расположенному выше или ниже относительно молекулярного штрихкода в меченых штрихкодами кДНК. Например, множество прямых/обратных праймеров содержит первый и/или второй прямой/обратный праймер, содержащий участок, комплементарный первому и/или второму участку, расположенному выше или ниже относительно молекулярного штрихкода в меченых штрихкодами кДНК, второй прямой/обратный праймер, содержащий участок, комплементарный второму участку, расположенному выше или ниже относительно молекулярного штрихкода в меченых штрихкодами кДНК, и третий прямой/обратный праймер, содержащий участок, комплементарный третьему участку, расположенному выше или ниже относительно молекулярного штрихкода в меченых штрихкодами кДНК, и т.д. Множество прямых/обратных праймеров можно применять для гибридизации со всеми возможными участками, расположенными выше или ниже относительно всех молекулярных штрихкодов, экспрессируемых клетками, такими как иммунные В-клетки или Т-клетки, в образце.
В соответствии с некоторыми вариантами осуществления множество прямых/обратных праймеров содержит один или несколько прямых/обратных праймеров, при этом каждый из прямых/обратных праймеров во множестве прямых/обратных праймеров содержит участок, комплементарный одному или нескольким участкам, расположенным выше или ниже относительно штрихкода резервуара в меченых штрихкодами кДНК. Например, множество прямых/обратных праймеров содержит прямой/обратный праймер, содержащий участок, комплементарный участку, расположенному выше или ниже относительно штрихкода резервуара в меченых штрихкодами кДНК, и один или несколько других прямых/обратных праймеров, содержащих участок, комплементарный одному или нескольким другим участкам, расположенным выше или ниже относительно штрихкода резервуара в меченых штрихкодами кДНК. Например, множество прямых/обратных праймеров содержит первый и/или второй прямой/обратный праймер, содержащий участок, комплементарный первому и/или второму участку, расположенному выше или ниже относительно штрихкода резервуара в меченых штрихкодами кДНК, и второй прямой/обратный праймер, содержащий участок, комплементарный второму участку, расположенному выше или ниже относительно штрихкода резервуара в меченых штрихкодами кДНК. Например, множество прямых/обратных праймеров содержит первый и/или второй прямой/обратный праймер, содержащий участок, комплементарный первому и/или второму участку, расположенному выше или ниже относительно штрихкода резервуара в меченых штрихкодами кДНК, второй прямой/обратный праймер, содержащий участок, комплементарный второму участку, расположенному выше или ниже относительно штрихкода резервуара в меченых штрихкодами кДНК, и третий прямой/обратный праймер, содержащий участок, комплементарный третьему участку, расположенному выше или ниже относительно штрихкода резервуара в меченых штрихкодами кДНК, и т.д. Праймеры во множество прямых/обратных праймеров можно применять для гибридизации со всеми возможными участками, расположенными выше или ниже относительно всех штрихкодов резервуара, экспрессируемых клетками, такими как иммунные В-клетки или Т-клетки, в образце.
Прямые/обратные праймеры во множестве прямых/обратных праймеров дополнительно содержат участок, которые не является комплементарным участку РНК. В соответствии с некоторыми вариантами осуществления участок, который не является комплементарным участку РНК располагается в направлении 5' относительно участка прямых/обратных праймеров, который является комплементарным РНК (т.е. участкам, расположенным выше или ниже относительно V-сегмента). В соответствии с некоторыми вариантами осуществления участок, который не является комплементарным участку РНК располагается в направлении 3' относительно участка прямых/обратных праймеров, который является комплементарным РНК. В соответствии с некоторыми вариантами осуществления участок, который не является комплементарным участку РНК, представляет собой участок 5' «липкого» конца. В соответствии с некоторыми вариантами осуществления участок, который не является комплементарным участку РНК содержит инициирующий сайт для амплификации и/или второй реакции секвенирования. В соответствии с некоторыми вариантами осуществления участок, который не является комплементарным участку РНК содержит инициирующий сайт для амплификации и/или третьей реакции секвенирования. В соответствии с некоторыми вариантами осуществления участок, который не является комплементарным участку РНК содержит инициирующий сайт для второй и третьей реакции секвенирования. В соответствии с некоторыми вариантами осуществления последовательность инициирующего сайта для второй и третьей реакции секвенирования является одинаковой. При применении одного или нескольких прямых/обратных праймеров и обратного праймера, которые описаны в данном документе, молекулы кДНК амплифицируют с применением подходящих реактивов, известных в уровне техники. В соответствии с некоторыми вариантами осуществления участок является комплементарным участку РНК, такому как константный участок или поли-А-хвост мРНК.
В соответствии с некоторыми вариантами осуществления множество прямых/обратных праймеров содержит один или несколько прямых/обратных праймеров, при этом каждый из прямых/обратных праймеров во множестве прямых/обратных праймеров содержит участок, комплементарный одному или нескольким участкам, расположенным выше или ниже относительно штрихкода резервуара в меченых штрихкодом олигонуклеотидах в составе конъюгатов аффинная молекула-олигонуклеотид или конъюгатов тетрамер-олигонуклеотид. В соответствии с некоторыми вариантами осуществления множество прямых/обратных праймеров содержит один или несколько прямых/обратных праймеров, при этом каждый из прямых/обратных праймеров во множестве прямых/обратных праймеров содержит участок, комплементарный одному или нескольким участкам, расположенным выше или ниже относительно AID в меченых штрихкодом олигонуклеотидах в составе конъюгатов аффинная молекула-олигонуклеотид или конъюгатов тетрамер-олигонуклеотид. В соответствии с некоторыми вариантами осуществления множество прямых/обратных праймеров содержит один или несколько прямых/обратных праймеров, при этом каждый из прямых/обратных праймеров во множестве прямых/обратных праймеров содержит участок, комплементарный одному или нескольким участкам, расположенным выше или ниже относительно АМВ в меченых штрихкодом олигонуклеотидах в составе конъюгатов аффинная молекула-олигонуклеотид или конъюгатов тетрамер-олигонуклеотид. Например, множество прямых/обратных праймеров содержит прямой/обратный праймер, содержащий участок, комплементарный участку, расположенному выше или ниже относительно штрихкода резервуара, AID и/или АМВ в меченых штрихкодом олигонуклеотидах в составе конъюгатов аффинная молекула-олигонуклеотид или конъюгатов тетрамер-олигонуклеотид, и один или несколько других прямых/обратных праймеров, содержащих участок, комплементарный одному или нескольким другим участкам, расположенным выше или ниже относительно штрихкода резервуара, AID и/или АМВ в меченых штрихкодом олигонуклеотидах в составе конъюгатов аффинная молекула-олигонуклеотид или конъюгатов тетрамер-олигонуклеотид. Например, множество прямых/обратных праймеров содержит первый и/или второй прямой/обратный праймер, содержащий участок, комплементарный первому и/или второму участку, расположенному выше или ниже относительно штрихкода резервуара, AID и/или АМВ в меченых штрихкодом олигонуклеотидах в составе конъюгатов аффинная молекула-олигонуклеотид или конъюгатов тетрамер-олигонуклеотид, и второй прямой/обратный праймер, содержащий участок, комплементарный второму участку, расположенному выше или ниже относительно штрихкода резервуара, AID и/или АМВ в меченых штрихкодом олигонуклеотидах в составе конъюгатов аффинная молекула-олигонуклеотид или конъюгатов тетрамер-олигонуклеотид. Например, множество прямых/обратных праймеров содержит первый и/или второй прямой/обратный праймер, содержащий участок, комплементарный первому и/или второму участку, расположенному выше или ниже относительно штрихкода резервуара, AID и/или АМВ в меченых штрихкодом олигонуклеотидах в составе конъюгатов аффинная молекула-олигонуклеотид или конъюгатов тетрамер-олигонуклеотид, второй прямой/обратный праймер, содержащий участок, комплементарный второму участку, расположенному выше или ниже относительно штрихкода резервуара, AID и/или АМВ в меченых штрихкодом олигонуклеотидах в составе конъюгатов аффинная молекула-олигонуклеотид или конъюгатов тетрамер-олигонуклеотид, и третий прямой/обратный праймер, содержащий участок, комплементарный третьему участку, расположенному выше или ниже относительно штрихкода резервуара, AID и/или АМВ в меченых штрихкодом олигонуклеотидах в составе конъюгатов аффинная молекула-олигонуклеотид или конъюгатов тетрамер-олигонуклеотид, и т.д.
Амплификация
После того как штрихкод резервуара был добавлен к целевому полинуклеотиду, целевой полинуклеотид можно амплифицировать. Например, после того как штрихкод резервуара был добавлен к олигонуклеотиду в составе конъюгата аффинная молекула-олигонуклеотид или конъюгата тетрамер-олигонуклеотид, олигонуклеотид можно амплифицировать. Например, после того как штрихкод резервуара был добавлен к клеточному полинуклеотиду, меченый штрихкодом резервуара клеточный полинуклеотид можно амплифицировать.
Реакционная смесь для реакции амплификации может содержать одну или несколько добавок. В некоторых случаях одна или несколько добавок представляют собой диметилсульфоксид (DMSO), глицерин, (моно)гидрат бетаина (N,N,N-триметилглицин = [карбокис-метил] триметиламмоний), трегалозу, 7-деаза-2'-дезоксигуанозинтрифосфат (dC7GTP или 7-деаза-2'-dGTP), BSA (бычий сывороточный альбумин), формамид (метанамид), хлорид тетраметиламмония (ТМАС), другие производные тетраалкиламмония (например, хлорид тетраэтиламмония (ТЕА-Cl) и хлорид тетрапропиламмония (TPrA-Cl), неионный детергент (например, Triton Х-100, Tween 20, Nonidet Р-40 (NP-40)) или PREXCEL-Q. В некоторых случаях реакционная смесь для реакции амплификации содержит 0, 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 различных добавок. В других случаях реакционная смесь для реакции амплификации содержит по меньшей мере 0, 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 различных добавок.
Реакции термоциклирования можно осуществлять на образцах, содержащихся в реакционных объемах (например, каплях). Капли могут быть полидисперсными или предпочтительно монодисперсными, полученными посредством перемешивания, ультразвуковой обработки или с помощью микропотокового устройства с Т-образным соединением каналов или с помощью других средств, известных специалисту, знакомому с данной областью. Плотности могут превышать 20000 капель/40 мкл (капли объемом 1 нл), 200000 капель/40 мкл (капли объемом 100 пл). Капли могут оставаться интактными во время термоциклирования. Капли могут оставаться интактными во время термоциклирования при плотностях, больших чем приблизительно 10000 капель/мкл, 100000 капель/мкл, 200000 капель/мкл, 300000 капель/мкл, 400000 капель/мкл, 500000 капель/мкл, 600000 капель/мкл, 700000 капель/мкл, 800000 капель/мкл, 900000 капель/мкл или 1000000 капель/мкл. В других случаях две или более капель не сливаются во время термоциклирования. В других случаях более чем 100 или более чем 1000 капель не сливаются во время термоциклирования.
Можно применять любую ДНК-полимеразу, которая катализирует достройку праймера, в том числе, без ограничения, ДНК-полимеразу Е. coli, фрагмент Кленова ДНК-полимеразы 1 Е. coli, ДНК-полимеразу Т7, ДНК-полимеразу Т4, Taq полимеразу, ДНК-полимеразу Pfu, ДНК-полимеразу Vent, полимеразу бактериофага 29, REDTaq™, геномную ДНК-полимеразу или секвеназу. В некоторых случаях применяют термостабильную ДНК-полимеразу. Также можно осуществлять ПЦР с «горячим стартом», при которой реакционную смесь нагревают до 95°С в течение двух минут перед добавлением полимеразы, или полимеразу могут сохранять неактивной до первой стадии нагревания в цикле 1. ПЦР с «горячим стартом» можно применять для сведения к минимуму неспецифической амплификации. Любое количество циклов ПЦР можно применять для амплификации ДНК, например, количество, приблизительно равное, большее чем приблизительно или меньшее чем приблизительно 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44 или 45 циклов. Количество циклов амплификации может составлять приблизительно 1-45, 10-45, 20-45, 30-45, 35-45, 10-40, 10-30, 10-25, 10-20, 10-15, 20-35, 25-35, 30-35 или 35-40.
Амплификацию целевых нуклеиновых кислот можно осуществлять с помощью любых средств, известных в уровне техники. Целевые нуклеиновые кислоты можно амплифицировать с помощью полимеразной цепной реакции (PCR) или изотермической амплификации ДНК. Примеры методик ПЦР, которые можно применять, включают в себя, без ограничения, количественную ПЦР, количественную флуоресцентную ПЦР (QF-PCR), мультиплексную флуоресцентную ПЦР (MF-PCR), ПЦР в реальном времени (ПЦР с обратной транскрипции), ПЦР в отдельной клетке, ПЦР полиморфизма длины рестрикционных фрагментов (PCR-RFLP), PCR-RFLP/обратная транскрипция-PCR-RFLP, ПЦР с «горячим стартом», гнездовую ПЦР, ПЦР методом молекулярных колоний in situ, амплификацию по типу катящегося кольца in situ (RCA), цифровую ПЦР (dPCR), цифровую ПЦР в каплях (ddPCR), мостиковую ПЦР, ПЦР в объемах порядка пиколитров и эмульсионную ПЦР. Другие подходящие способы амплификации включают в себя лигазную цепную реакцию (LCR), транскрипционную амплификацию, ПЦР с инвертируемым молекулярным зондом (MIP), самоподдерживающуюся репликацию последовательностей, селективную амплификацию последовательностей целевого полинуклеотида, запускаемую консенсусной последовательностью полимеразную цепную реакция (CP-PCR), запускаемую произвольным праймером полимеразную цепную реакцию (AP-PCR), запускаемую вырожденным полинуклеотидом ПЦР (DOP-PCR) и изотермическую амплификацию последовательности нуклеиновой кислоты (NABSA). Другие способы амплификации, которые можно использовать в данном документе, включают в себя описанные в патентах США №№5242794; 5494810; 4988617 и 6582938, а также включают в себя амплификацию РНК, опосредованную репликазой Q-бета. Амплификация может представлять собой изотермическую амплификацию, например, изотермическую линейную амплификацию.
В соответствии с некоторыми вариантами осуществления амплификация не происходит на твердой подложке. В соответствии с некоторыми вариантами осуществления амплификация не происходит на твердой подложке в капле. В соответствии с некоторыми вариантами осуществления амплификация не происходит на твердой подложке, когда амплификация не происходит в капле.
Секвенирование
После осуществления одного или нескольких из способов или стадий способа, описанных в данном документе, библиотеку образованных полинуклеотидов можно секвенировать.
Секвенирование можно осуществлять с помощью любого способа секвенирования, известного в уровне техники. В соответствии с некоторыми вариантами осуществления секвенирование можно осуществлять с высокой пропускной способностью. Подходящие технологии секвенирования нового поколения включают в себя платформу 454 Life Sciences (Roche, Branford, CT) (Margulies et al., Nature, 437, 376-380 (2005)); геномный анализатор от Illumina, анализ метилирования GoldenGate или анализы метилирования Infinium, т.е. матрица с гранулами для анализа метилирования Infinium HumanMethylation 27K BeadArray или матрица для анализа метилирования VeraCode GoldenGate methylation array (Illumina, Сан-Диего, Калифорния; Bibkova et al., Genome Res. 16, 383-393 (2006); и патенты США №№6306597, 7598035, 7232656), или секвенирование ДНК с помощью лигирования, систему SOLiD (Applied Biosystems/Life Technologies; патенты США №№6797470, 7083917, 7166434, 7320865, 7332285, 7364858 и 7429453); или технологию секвенирования отдельных молекул ДНК Helicos True (Harris et al., Science, 320, 106-109 (2008); и патенты США №№7037687, 7645596, 7169560 и 7769400), технологию секвенирования отдельных молекул в реальном времени (SMRTTm) от Pacific Biosciences и секвенирование (Soni et al., Clin. Chem. 53, 1996-2001 (2007)). Эти системы обеспечивают возможность мультиплексного параллельного секвенирования многих полинуклеотидов, выделенных из образца (Dear, Brief Funct. Genomic Proteomic, 1(4), 397-416 (2003), и McCaughan et al., J. Pathol., 220, 297-306 (2010)). В соответствии с некоторыми вариантами осуществления, полинуклеотиды секвенируют посредством секвенирования с помощью лигирования модифицированных красителем зондов, пиросеквенирования или секвенирования отдельных молекул. Определение последовательности полинуклеотида можно осуществлять с помощью способов секвенирования, таких как секвенирование отдельных молекул Helioscope™, секвенирование ДНК Nanopore, массивно-параллельное секвенирование молекулярных сигнатур (MPSS) от Lynx Therapeutics, пиросеквенирование 454, секвенирование отдельных молекул в реальном времени (RNAP), секвенирование Illumina (Solexa), секвенирование SOLiD, Ion Torrent™, ионное полупроводниковое секвенирование, SMRT™ секвенирование отдельных молекул, секвенирование методом молекулярных колоний, секвенирование ДНК на наносферах и подход VisiGen Biotechnologies. В качестве альтернативы, при определении последовательности полинуклеотидов можно применять платформы для секвенирования, в том числе, без ограничения, геномный анализатор IIx, HiSeq и MiSeq, предлагаемый Illumina, технологию секвенирования отдельных молекул в реальном времени (SMRT™), как например, систему PacBio RS, предлагаемую Pacific Biosciences (Калифорния) и секвенатор Solexa, технологию секвенирования отдельных молекул True Single Molecule Sequencing (tSMS™), как например, секвенатор HeliScope™, предлагаемый Helicos Inc. (Кембридж, Массачусетс). Секвенирование может предусматривать секвенирование MiSeq. Секвенирование может предусматривать секвенирование HiSeq. В соответствии с некоторыми вариантами осуществления определение последовательности полинуклеотида предусматривает секвенирование спаренных концов, нанопоровое секвенирование, секвенирование с высокой пропускной способностью, секвенирование методом «дробовика», секвенирование с использованием красителя-терминатора, секвенирование ДНК со множественными праймерами, праймер-опосредованную загрузку, дидезокси-секвенирование по Сенгеру, секвенирование по Максаму-Гилберту, пиросеквенирование, секвенирование единичных молекул или любую их комбинацию. В качестве альтернативы, последовательность полинуклеотида можно определить с помощью электронной микроскопии или матрицы из химически чувствительных полевых транзисторов (chemFET).
Способ может дополнительно включать секвенирование одного или нескольких полинуклеотидов в библиотеке. Способ может дополнительно включать выравнивание одной или нескольких последовательностей полинуклеотида, ридов последовательностей, последовательностей ампликона или набора последовательностей ампликона в библиотеке друг с другом.
Выравнивание может включать сравнение исследуемой последовательности, такой как рид последовательности, с одной или несколькими другими исследуемыми последовательностями, эталонными последовательностями или их комбинацией. В соответствии с некоторыми вариантами осуществления выравнивание можно применять для определения консенсусной последовательности из множества последовательностей или выравниваемых последовательностей. В соответствии с некоторыми вариантами осуществления выравнивание включает определение консенсусной последовательности из множества последовательностей, каждая из которых имеет идентичный молекулярный штрихкод или штрихкод резервуара. В соответствии с некоторыми вариантами осуществления длина последовательности, выравниваемой с целью сравнения, составляет по меньшей мере 30%, по меньшей мере 40%, по меньшей мере 50%, по меньшей мере 60%, по меньшей мере 70%, по меньшей мере 80%, по меньшей мере 90% или по меньшей мере 95% от длины эталонной последовательности. Фактическое сравнение двух или более последовательностей можно осуществлять с помощью хорошо известных способов, например, с применением математического алгоритма. Неограничивающий пример такого математического алгоритма описан в Karlin, S. and Altschul, S., Proc. Natl. Acad. Sci. USA, 90- 5873-5877 (1993). Такой алгоритм встроен в программы NBLAST и XBLAST (версия 2.0), которые описаны в Altschul, S. et al., Nucleic Acids Res., 25:3389-3402 (1997). При использовании программ BLAST и Gapped BLAST можно использовать любые подходящие параметры соответствующих программ (например, NBLAST). Например, можно установить следующие параметры для сравнения последовательностей: оценка = 100, длина слова = 12, или их можно варьировать (например, W=5 или W=20). Другие примеры включают в себя алгоритм из Myers and Miller, CABIOS (1989), ADVANCE, ADAM, BLAT и FASTA. В соответствии с некоторыми вариантами осуществления процентную идентичность между двумя аминокислотными последовательностями можно установить с применением, например, программы GAP в составе пакета программного обеспечения GCG (Accelrys, Кембридж, Великобритания).
Секвенирование может включать секвенирование по меньшей мере приблизительно 10, 20, 30, 40, 50, 60, 70, 80, 90 100 или большего количества нуклеотидов или пар оснований в полинуклеотидах. В соответствии с некоторыми вариантами осуществления секвенирование включает секвенирование по меньшей мере приблизительно 200, 300, 400, 500, 600, 700, 800, 900, 1000 или большего количества нуклеотидов или пар оснований в полинуклеотидах. В других случаях секвенирование включает секвенирование по меньшей мере приблизительно 1500, 2000, 3000, 4000, 5000, 6000, 7000, 8000, 9000, 10000 или большего количества нуклеотидов или пар оснований в полинуклеотидах.
Секвенирование может включать по меньшей мере приблизительно 200, 300, 400, 500, 600, 700, 800, 900 1000 или большее количество ридов от секвенирования за проход. В контексте данного документа рид последовательности содержит последовательность нуклеотидов, определенную из последовательности или потока данных, получаемого с помощью методики секвенирования. В соответствии с некоторыми вариантами осуществления секвенирование включает секвенирование по меньшей мере приблизительно 1500, 2000, 3000, 4000, 5000, 6000, 7000, 8000, 9000, 10000 или большего количества ридов от секвенирования за проход. Секвенирование может включать количество ридов, большее чем, меньшее чем или равное приблизительно 1000000000 ридов от секвенирования за проход. Секвенирование может предусматривать количество ридов, большее чем, меньшее чем или равное приблизительно 200000000 ридов за проход.
Ферменты
Способы и наборы, раскрытые в данном документе, могут предусматривать один или несколько ферментов. Примеры ферментов включают в себя, без ограничения, лигазы, обратные транскриптазы, полимеразы и рестрикционные нуклеазы.
В соответствии с некоторыми вариантами осуществления прикрепление адаптера к полинуклеотидам предусматривает применение одной или нескольких лигаз. Примеры лигаз включают в себя, без ограничения, ДНК-лигазы, такие как ДНК-лигаза I, ДНК-лигаза III, ДНК-лигаза IV и ДНК-лигаза Т4, и РНК-лигазы, такие как РНК-лигаза I Т4 и РНК-лигаза II Т4.
Способы и наборы, раскрытые в данном документе, могут дополнительно предусматривать применение одной или нескольких обратных транскриптаз. В соответствии с некоторыми вариантами осуществления обратная транскриптаза представляет собой обратную транскриптазу ВИЧ-1, обратную транскриптазу M-MLV, обратную транскриптазу AMV и теломеразную обратную транскриптазу. В соответствии с некоторыми вариантами осуществления обратная транскриптаза представляет собой обратную транскриптазу M-MLV.
В соответствии с некоторыми вариантами осуществления способы и наборы, раскрытые в данном документе, предусматривают применение одной или нескольких протеаз.
В соответствии с некоторыми вариантами осуществления способы и наборы, раскрытые в данном документе, предусматривают применение одной или нескольких полимераз. Примеры полимераз включают в себя, без ограничения, ДНК-полимеразы и РНК-полимеразы. В соответствии с некоторыми вариантами осуществления ДНК-полимераза представляет собой ДНК-полимеразу I, ДНК-полимеразу II, холофермент ДНК-полимеразы III и ДНК-полимеразу IV. Коммерчески доступные ДНК-полимеразы включают в себя, без ограничения, ДНК-полимеразу Bst 2.0, ДНК-полимеразу Bst 2.0 WarmStart™, ДНК-полимеразу Bst, ДНК-полимеразу IV Sulfolobus, ДНК-полимеразу Taq, ДНК-полимеразу 9°N™m, (экзо-) ДНК-полимеразу Deep VentR™, ДНК-полимеразу Deep VentR™, Hemo KlenTaq™, ДНК-полимеразу Taq LongAmp®, ДНК-полимеразу OneTaq®, ДНК-полимеразу Phusion®, ДНК-полимеразу Q5™ High-Fidelity, ДНК-полимеразу γ Therminator™, ДНК-полимеразу Therminator™, ДНК-полимеразу II Therminator™, ДНК-полимеразу III Therminator™, ДНК-полимеразу VentR®, (экзо-) ДНК-полимеразу VentR®, ДНК-полимеразу Bsu, ДНК-полимеразу phi29, ДНК-полимеразу T4, ДНК-полимеразу Т7, концевую трансферазу, Taq полимеразу Titanium®, ДНК-полимеразу Taq KAPA и ДНК-полимеразу Taq KAPA для использования в методиках с «горячим стартом».
В соответствии с некоторыми вариантами осуществления полимераза представляет собой РНК-полимеразы, такие как РНК-полимераза I, РНК-полимераза II, РНК-полимераза III, поли(А)-полимераза Е. coli, РНК-полимераза phi6 (RdRP), поли(U)-полимераза, РНК-полимераза SP6 и РНК-полимераза Т7.
Дополнительные реактивы
Способы и наборы, раскрытые в данном документе, могут предусматривать применение одного или нескольких реактивов. Примеры реактивов включают в себя, без ограничения, реактивы для ПЦР, реактивы для лигирования, реактивы для обратной транскрипции, реактивы-ферменты, реактивы для гибридизации, реактивы для подготовки образца, реактивы для аффинного захвата, твердые подложки, такие как гранулы, и реактивы для очистки и/или выделения нуклеиновых кислот.
В соответствии с другими вариантами осуществления способы, наборы и композиции, раскрытые в данном документе, могут предусматривать подложку. В соответствии с некоторыми вариантами осуществления способы, наборы и композиции, раскрытые в данном документе, не предусматривают подложку. Как правило, твердая подложка содержит один или несколько материалов, содержащих одну или несколько жестких или полужестких поверхностей. В соответствии с некоторыми вариантами осуществления подложка представляет собой подложку, отличную от твердой. Подложка или субстрат могут содержать мембрану, бумагу, пластик, поверхность с покрытием, плоскую поверхность, стекло, микроскопический препарат, чип или любую их комбинацию. В соответствии с некоторыми вариантами осуществления один или несколько поверхностей подложки являются практически плоскими, хотя в соответствии с некоторыми вариантами осуществления может быть желательным физическое разделение участков синтеза для различных соединений, например, с использованием лунок, приподнятых участков, штырьков, вытравленных канавок или подобного. В соответствии с некоторыми вариантами осуществления твердые подложки содержат гранулы, смолы, гели, микросферы или другие геометрические конфигурации. В качестве альтернативы, твердые подложки могут содержать чипы из диоксида кремния, микрочастицы, наночастицы, планшеты и матрицы. Твердая подложка может предусматривать применение гранул, которые самособираются в микролунках. Например, твердая подложка предусматривает технологию BeadArray от Illumina. В качестве альтернативы, твердая подложка предусматривает технологию Bead Array от Abbott Molecular и систему Applied Microarray FlexiPlex™. В других случаях твердая подложка представляет собой планшет. Примеры планшетов включают в себя, без ограничения, мультиматричные планшеты MSD, MSD Multi-Spot® планшеты, микропланшет, микропланшет ProteOn, AlphaPlate, планшет DELFIA, IsoPlate и LumaPlate. В соответствии с некоторыми вариантами осуществления подложка может содержать множество гранул. В соответствии с некоторыми вариантами осуществления подложка может содержать матрицу. В соответствии с некоторыми вариантами осуществления подложка может содержать микроскопический препарат на стекле. Способы, субстраты и методики, применимые к полимерам (патенты США №№5744305, 5143854, 5242974, 5252743, 5324633, 5384261, 5405783, 5424186, 5451683, 5482867, 5491074, 5527681, 5550215, 5571639, 5578832, 5593839, 5599695, 5624711, 5631734, 5795716, 5831070, 5837832, 5856101, 5858659, 5936324, 5968740, 5974164, 5981185, 5981956, 6025601, 6033860, 6040193, 6090555, 6136269, 6269846 и 6428752; публикации заявок на патент США №№20090149340, 20080038559, 20050074787; и в РСТ публикациях международных заявок №№ WO 00/58516, WO 99/36760 и WO 01/58593). Прикрепление полинуклеотидов к подложке может предусматривать амин-тиольное сшивание, малеимидное сшивание, применение N-гидроксисукцинимида или N-гидроксисульфосукцинимида, Zenon или SiteClick. Прикрепление меченых нуклеиновых кислот к подложке может предусматривать прикрепление биотина ко множеству полинуклеотидов и покрытие одной или нескольких гранул стрептавидином. В соответствии с некоторыми вариантами осуществления твердая подложка представляет собой гранулу. Примеры гранул включают в себя, без ограничения, гранулы со стрептавидином, гранулы с агарозой, магнитные гранулы, Dynabeads®, микрогранулы для MACS®, гранулы с конъюгированным антителом (например, микрогранулу с антителом к иммуноглобулину), гранулы с конъюгированным белком А, гранулы с конъюгированным белком G, гранулы с конъюгированным белком A/G, гранулы с конъюгированным белком L, гранулы с конъюгированным полинуклеотид-dT, гранулы диоксида кремния, гранулы из материала, подобные диоксида кремния, микрогранулу с антителом к биотину, микрогранулу с антителом к флуорохрому, и BcMag™ магнитные гранулы с карбоксильными концевыми группами. Диаметр гранул может составлять приблизительно 5 мкм, 10 мкм, 20 мкм, 25 мкм, 30 мкм, 35 мкм, 40 мкм, 45 мкм или 50 мкм. Твердая подложка может представлять собой чип или микрочип. Твердая подложка может содержать дискретные участки. Твердая подложка может представлять собой чип, например, чип с адресацией.
Твердая подложка может содержать практически любой нерастворимый или твердый материал и часто композиция твердой подложки выбрана таким образом, чтобы она была нерастворимой в воде. Например, твердая подложка может содержать или состоять, по существу, из силикагеля, стекла (например, стекла с контролируемым размером пор (CPG)), нейлона, Sephadex®, Sepharose®, целлюлозы, поверхности металла (например, стали, золота, серебра, алюминия, кремния и меди), магнитного материала, пластичного материала (например, полиэтилена, полипропилена, полиамида, сложного полиэфира, поливинилидендифторида (PVDF)) и т.п. Примеры гранул для применения в соответствии с вариантами осуществления могут включать в себя аффинный фрагмент, который позволяет грануле взаимодействовать с молекулой нуклеиновой кислоты. Твердая фаза (например, гранула) может содержать связывающий элемент в составе пары (например, авидин, стрептавидин или его производное). Например, гранула может представлять собой покрытую стрептавидином гранулу, и молекула нуклеиновой кислоты для иммобилизации на грануле может включать в себя биотиновый фрагмент. В некоторых случаях каждая молекула полинуклеотида может включать в себя два аффинных фрагмента, таких как биотин, чтобы дополнительно стабилизировать полинуклеотид. Гранулы могут включать в себя дополнительные элементы для применения в иммобилизации нуклеиновых кислот, или их можно применять в последующих процессах скрининга или селекции. Например, гранула может включать в себя аффинную часть, флуоресцентную метку или гаситель флуоресценции. В некоторых случаях гранула может являться магнитной. В некоторых случаях твердая подложка представляет собой гранулу. Примеры гранул включают в себя, без ограничения, гранулы со стрептавидином, гранулы с агарозой, магнитные гранулы, Dynabeads®, микрогранулы для MACS®, гранулы с конъюгированным антителом (например, микрогранулу с антителом к иммуноглобулину), гранулы с конъюгированным белком А, гранулы с конъюгированным белком G, гранулы с конъюгированным белком A/G, гранулы с конъюгированным белком L, гранулы с конъюгированным полинуклеотид-dT, гранулы диоксида кремния, гранулы из материала, подобные диоксида кремния, микрогранулу с антителом к биотину, микрогранулу с антителом к флуорохрому, и BcMag™ магнитные гранулы с карбоксильными концевыми группами. Гранулы или частицы могут поддаваться разбуханию (например, полимерные гранулы, такие как смола Ванга) или не поддаваться разбуханию (например, CPG). В соответствии с некоторыми вариантами осуществления твердая фаза является, по существу, гидрофильной. В соответствии с некоторыми вариантами осуществления твердая фаза (например, гранула) является, по существу, гидрофобной. В соответствии с некоторыми вариантами осуществления твердая фаза содержит связывающий элемент в составе пары (например, авидин, стрептавидин или их производное) и является, по существу, гидрофобной или, по существу, гидрофильной. В соответствии с некоторыми вариантами осуществления твердая фаза содержит связывающий элемент в составе пары (например, авидин, стрептавидин или их производное) и характеризуется связывающей способностью, большей чем приблизительно 1350 пикомоль несвязанного захватывающего средства (например, несвязанного биотина) на мг твердой подложки. В соответствии с некоторыми вариантами осуществления связывающая способность твердой фазы, содержащей связывающий элемент в составе пары, является большей, чем 800, 900, 1000, 1100, 1200, 1250, 1300, 1350, 1400, 1450, 1500, 1600, 1800, 2000 пикомоль несвязанного захватывающего средства на мг твердой подложки. Другими примерами гранул, которые являются подходящими для настоящего изобретения, являются коллоидные формы золота или гранулы, такие как полистирольные гранулы или гранулы диоксида кремния. Можно применять частицы практически любых радиусов. Примеры гранул могут включать в себя гранулы, имеющие радиус, находящийся в диапазоне от 150 нанометров до 10 микрон. Также можно применять другие размеры.
Способы и наборы, раскрытые в данном документе, могут предусматривать применение одного или нескольких буферов. Примеры буферов включают в себя, без ограничения, отмывочные буферы, буферы для лигирования, буферы для гибридизации, буферы для амплификации и буферы для обратной транскрипции. В соответствии с некоторыми вариантами осуществления буфер для гибридизации представляет собой коммерчески доступный буфер, такой как Hyb раствор рТМАС, гибридизационный раствор SSPE и буфер для гибридизации ECONO™. Буферы, раскрытые в данном документе, могут содержать один или несколько детергентов.
Способы и наборы, раскрытые в данном документе, могут предусматривать применение одного или нескольких носителей. Носители могут усиливать или улучшать эффективность одной или нескольких реакций, раскрытых в данном документе (например, реакции лигирования, обратной транскрипции, амплификации, гибридизации). Носители могут снижать или предотвращать неспецифическую потерю молекул или любых их продуктов (например, полинуклеотида и/или ампликона). Например, носитель может снижать неспецифическую потерю полинуклеотида вследствие абсорбции на поверхностях. Носитель может снижать аффинность полинуклеотида к поверхности или субстрату (например, контейнеру, пробирке Эппендорфа, наконечнику пипетки). В качестве альтернативы, носитель может повышать аффинность полинуклеотида к поверхности или субстрату (например, грануле, матрице, стеклу, микроскопическому препарату, чипу). Носители могут защищать полинуклеотид от расщепления. Например, носители могут защищать молекулу РНК от рибонуклеаз. В качестве альтернативы, носители могут защищать молекулу ДНК от ДНКазы. Примеры носителей включают в себя, без ограничения, полинуклеотиды, такие как ДНК и/или РНК, или полипептиды. Примеры ДНК-носителей включают в себя плазмиды, векторы, полиаденилированную ДНК и полинуклеотиды ДНК. Примеры РНК-носителей включают в себя полиаденилированную РНК, фаговую РНК, РНК фага MS2, РНК E.coli, РНК дрожжей, тРНК дрожжей, РНК млекопитающего, тРНК млекопитающего, короткие полиаденилированные синтетические рибонуклеотиды и полинуклеотиды РНК. РНК-носитель может представлять собой полиаденилированную РНК. В качестве альтернативы, РНК-носитель может представлять собой не-полиаденилированную РНК. В соответствии с некоторыми вариантами осуществления носитель происходит из бактерии, дрожжей или вируса. Например, носитель может представлять собой полинуклеотид или полипептид, полученный из бактерии, дрожжей или вируса. Например, носитель представляет собой белок из Bacillus subtilis. В другом примере носитель представляет собой полинуклеотид из Escherichia coli. В качестве альтернативы, носитель представляет собой полинуклеотид или пептид из млекопитающего (например, человека, мыши, козы, крысы, коровы, овцы, свиньи, собаки или кролика), птицы, амфибии или рептилии.
Способы и наборы, раскрытые в данном документе, могут предусматривать применение одного или нескольких контрольных средств. Контрольные средства могут включать в себя контрольные полинуклеотиды, неактивные ферменты, неспецифические конкуренты. В качестве альтернативы, контрольные средства содержат контроли яркости сигнала от гибридизации, контроли яркости сигнала от зонда, матрицы нуклеиновой кислоты, контроли в виде добавляемых стандартных образцов, контроли для ПЦР амплификации. Контроли для ПЦР амплификации могут представлять собой положительные контроли. В других случаях контроли для ПЦР амплификации представляют собой отрицательные контроли. Контроли матрицы нуклеиновой кислоты могут иметь известные концентрации. Контрольные средства могут содержать одну или несколько меток.
Контроли в виде добавляемых стандартных образцов могут представлять собой матрицы, которые добавляют в реакционную смесь или образец. Например, добавляемую в виде стандартного образца матрицу можно добавлять в реакционную смесь для реакции амплификации. Добавляемую в виде стандартного образца матрицу можно добавлять в реакционную смесь для реакции амплификации в любое время после первого цикла амплификации. В соответствии с некоторыми вариантами осуществления добавляемую в виде стандартного образца матрицу добавляют в реакционную смесь для реакции амплификации после цикла номер 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 20, 25, 30, 35, 40, 45 или 50. Добавляемую в виде стандартного образца матрицу можно добавлять в реакционную смесь для реакции амплификации в любое время до последнего цикла амплификации. Добавляемая в виде стандартного образца матрицу может содержать один или несколько нуклеотидов или пар оснований нуклеиновой кислоты. Добавляемая в виде стандартного образца матрица может содержать ДНК, РНК или любую их комбинацию. Добавляемая в виде стандартного образца матрица может содержать одну или несколько меток.
В данном документе раскрыты молекулы, материалы, композиции и компоненты, которые можно применять для способов и композиций, раскрытых в данном документе, можно применять совместно со способами и композициями, раскрытыми в данном документе, можно применять в подготовке способов и композиций, раскрытых в данном документе, или они являются продуктами способов и композиций, раскрытых в данном документе. Понятно, что в случае, когда раскрыты комбинации, субпопуляции, взаимодействия, группы и т.д. этих материалов, и хотя конкретное упоминание каждой разнообразной отдельной молекулы и соединения и их совокупных комбинаций и преобразование этих молекул и соединений не могут быть полностью раскрыты, каждый из элементов специально предполагается и описывается в данном документе. Например, если раскрыт и обсуждается нуклеотид или нуклеиновая кислота, и обсуждается ряд модификаций, которые можно произвести в ряде молекул, в том числе в нуклеотиде или нуклеиновой кислоте, специально предполагаются каждая и любая комбинация и преобразование нуклеотида или нуклеиновой кислоты и модификации, которые возможны, если специально не указано иное. Эта идея распространяется на все аспекты данной заявки, в том числе, без ограничения, стадии в способах получения и применение раскрытых способов и композиций. Таким образом, если присутствует ряд дополнительных стадий, которые можно осуществлять, понятно, что каждую из этих дополнительных стадий можно осуществлять с любым конкретным вариантом осуществления или комбинацией вариантов осуществления раскрытых способов, и что каждая такая комбинация специально предусматривается и должна считаться раскрытой.
Несмотря на то некоторые варианты осуществления, описанные в данном документе, были показаны и описаны в данном документе, такие варианты осуществления представлены только в качестве примера. Специалистам в данной области техники встретятся многочисленные варианты, изменения и замены, не отступающие от раскрытия, представленного в данном документе. Следует понимать, что различные альтернативы вариантам осуществления, описанным в данном документе, могут быть использованы при практическом осуществлении способа, описанного в данном документе.
Если это не объясняется иным образом, все технические и научные термины, применяемые в данном документе, имеют такое же значение, которое обычно понятно квалифицированному специалисту в данной области техники, к которой относится настоящее раскрытие. Следующие источники содержат варианты осуществления способов и композиций, которые можно применять в данном документе: The Merck Manual of Diagnosis and Therapy, 18th Edition, опубликованный Merck Research Laboratories, 2006 (ISBN 0-9119102); Benjamin Lewin, Genes IX, опубликованный Jones & Bartlett Publishing, 2007 (ISBN-13: 9780763740634); Kendrew et al. (eds.), The Encyclopedia of Mol. Biology, опубликованный Blackwell Science Ltd., 1994 (ISBN 0-632-02182-9); и Robert A. Meyers (ed.), Mol. Biology and Biotechnology: a Comprehensive Desk Reference, опубликованный VCH Publishers, Inc., 1995 (ISBN 1-56081-569-8).
Стандартные процедуры согласно настоящему раскрытию являются описанными, например, в Maniatis et al., Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y., USA (1982); Sambrook et al., Molecular Cloning: A Laboratory Manual (2 ed.), Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y., USA (1989); Davis et al., Basic Methods in Molecular Biology, Elsevier Science Publishing, Inc., New York, USA (1986); или в Methods in Enzymology: Guide to Molecular Cloning Techniques Vol. 152, S.L. Berger and A.R. Kimmerl (eds.), Academic Press Inc., San Diego, USA (1987)). Current Protocols in Molecular Biology (CPMB) (Fred M. Ausubel, et al. ed., John Wiley and Sons, Inc.), Current Protocols in Protein Science (CPPS) (John E. Coligan, et al., ed., John Wiley and Sons, Inc.), Current Protocols in Immunology (CPI) (John E. Coligan, et al., ed. John Wiley and Sons, Inc.), Current Protocols in Cell Biology (CPCB) (Juan S. Bonifacino et al. ed., John Wiley and Sons, Inc.), Culture of Animal Cells: A Manual of Basic Technique by R. Ian Freshney, Publisher: Wiley-Liss; 5th edition (2005), и Animal Cell Culture Methods (Methods in Cell Biology, Vol. 57, Jennie P. Mather and David Barnes editors, Academic Press, 1st edition, 1998).
ПРИМЕРЫ
Пример 1. Иммунотипирование отдельных Т-лимфоцитов в популяции Т-клеток
Способы иммунофенотипирования, описанные в данном документе, подтверждали посредством анализа экспрессии как мРНК CD4 и CD8, так и поверхностных белков в человеческих Т-лимфоцитах (фиг. 1). Канонически, ожидается, что зрелые Т-клетки могут относится или к подтипу CD4, или к подтипу CD8. Подтип CD4 должен экспрессировать мРНК CD4 и белок CD4 вместе, но должен экспрессировать либо мРНК CD8, либо белок CD8. Подтип CD8 должен экспрессировать мРНК CD8 и белок CD8 вместе, но должен экспрессировать либо мРНК CD4, либо белок CD4.
30000 Т-клеток инкубировали с CD4-специфическим конъюгатом антитело-олигонуклеотида, содержащим идентификационную последовательность антигена CD4, и CD8-специфическим конъюгатом антитело-олигонуклеотид, содержащим идентификационную последовательность антигена CD8. Содержание мРНК CD4, CD8 и TCR Т-клеток также анализировали в эксперименте с эмульсией отдельных клеток. мРНК Т-клеточного рецептора альфа, Т-клеточного рецептора бета, CD4 и CD8 подвергали обратной транскрипции в кДНК, и обеспечивали слияние последовательности специфического для капли штрихкода как с кДНК, так и с конъюгированными с антителом олигонуклеотидами посредством полимеразной цепной реакции. Эту ДНК экстрагировали из эмульсии и анализировали с использованием секвенирования нового поколения.
Почти все штрихкоды капель (относящиеся к отдельной клетке) ассоциировали либо с CD4-специфическим идентификатором антигена, либо CD8-специфическим идентификатором антигена. Обнаружили значительное соответствие между результатами анализов для мРНК, и поверхностных белков (фиг. 3).
Пример 2
Секвенирование иммунного репертуара имеет широкий спектр применений в общей иммунологии, при аутоиммунных заболеваниях, инфекционных заболеваниях и онкологических заболеваниях. Несмотря на то что во многих исследованиях изучали разнообразие BCR и TCR в циркулирующей крови, существует растущий интерес к иммунным рецепторам TIL, чьи функции тесно связаны с ростом или регрессией злокачественной опухоли и все еще являются многообразными и часто неохарактеризованными. Критически важным шагом к лучшему пониманию TIL будет выделение и функциональная характеристика их BCR и TCR, поскольку они могут обеспечить возможность идентификации новых опухолеассоциированных антигенов. Опухолевые антигены крайне необходимы для разработки противораковых вакцин, понимания роли ингибиторов иммунологических контрольных точек и совершенствования терапии Т-клетками с химерными антигенными рецепторами (CAR-T) в солидных опухолях. Тем не менее, несмотря на десятилетия технического прогресса в секвенирование иммунных молекул, ни одно исследование не выявило полноразмерных спаренных в нативном формате BCR (тяжелые и легкие цепи) и TCR (альфа- и бета-цепи) из неоднородного образца, такого как опухоль без культивирования in vitro или сортинга клеток, стадий, которые ограничивают и вносят погрешность в наблюдаемом репертуаре. Техническая трудность является особенно большой, поскольку первичные, не подвергающиеся культивированию иммунные клетки могут содержать в 100 раз меньше РНК рецептора, чем в стимулируемых in vitro клетках. Чтобы обеспечить возможность всестороннего анализа спаренных в нативном формате BCR и TCR из сложных неоднородных образцов был разработан способ на основе микропотоковой обработки в эмульсии для параллельного выделения и мечения ДНК-штрихкодами больших количеств отдельных клеток. Вплоть до миллиона клеток в час выделяют в отдельные капли эмульсии объемом ~65 пиколитров. Внутри капель клетки лизируют, целевую мРНК подвергают обратной транскрипции с использованием мишенеспецифических праймеров и в ходе двухстадийного процесса мечения ДНК-штрихкодами прикрепляют к кДНК как специфические для молекулы, так и специфические для капли штрихкоды. После последующего выделения и секвенирования нового поколения стратегия двойного мечения штрихкодами обеспечивает возможность кластеризации ридов последовательностей как по молекулам, так и по клеткам, из которых они происходят. Это обеспечивает возможность существенной коррекции ошибок и отклонений при амплификации, подсчета клонов как на уровне мРНК, так и на уровне клеток, определение изотипа тяжелой цепи и, что важно, выделение полноразмерных спаренных в нативном формате V(D)J последовательностей BCR и TCR одновременно с крайне высокой пропускной способностью.
Опухоль-инфильтрирующие лимфоциты (TIL) являются критически важными для противораковых иммунных реакций, но их изучение представляет сложности вследствие их непредсказуемой распространенности, фенотипов и функции. Способ глубокой характеристики TIL без необходимости в сортинге, стимуляции или культивировании клеток был разработан авторами настоящего изобретения. В способе на основе мечения штрихкодами отдельных клеток в эмульсии захватывают последовательности спаренных в нативном формате В-клеточных и Т-клеточных рецепторов (BCR и TCR) с лимфоцитов среди миллионов вводимых клеток. В отличие от предыдущих подходов, выделенные вариабельные участки являются полноразмерными и могут быть связанными с дополнительными мРНК-мишенями и белками-мишенями. Способ подтверждали с использованием 3 миллионов В-клеток из крови здорового человека и 350000 В-клеток от пациента с ВИЧ перед обработкой 400000 несортированных клеток из аденокарциномы яичника, выделением спаренных BCR и TCR из более чем 11000 TIL. Результаты, полученные авторами настоящего изобретения, представляют наиболее глубокую выборку репертуара любых спаренных BCR или TCR, а также первую демонстрацию одновременного секвенирования РНК и измерения белков из отдельных клеток в эмульсии.
Пример 3. Крупномасштабное выделение пар VHVL В-клеток из образца крови здорового человека
Первоначально разработали технологию, и способность к спариванию и пропускную способность оценивали с использованием 3 миллионов В-клеток, выделенных с помощью отрицательного обогащения на гранулах из периферической крови здорового добровольца. Эмульсию разделяли на шесть раздельных фракций, которые подвергали обработке параллельно и не смешивали повторно перед секвенированием. Клетки загружали в эмульсию в количестве 0,2 клеток на каплю при условии пуассоновской вероятности, что ~90% занятых капель содержат отдельные клетки, что согласовывалось с наблюдениями капель эмульсии (фиг. 5А). После разрушения эмульсии и осуществляли стадии дополнительной обработки библиотек с помощью секвенирования спаренных концов 325+300 п.о. С использованием Illumina MiSeq. Для обработки данных от секвенирования штрихкоды капель и молекулярные штрихкоды использовали совместно для сбора ридов в результате ПЦР репликации от каждой исходной молекулы мРНК и определяли консенсусную последовательность для каждой мРНК, сохраняя только последовательности мРНК, построенные по меньшей мере из двух ридов. Прямые и обратные риды сшивали с получением полноразмерных продуктов, содержащих 5' UTR, полную V(D)J последовательность и константный участок, достаточный для определения изотипа. Подвергшиеся перестройке последовательности тяжелой и легкой цепей иммуноглобулина аннотировали с использованием IMGT High-VQuest и/или IgBLAST.
Полученный в результате набор данных содержал 324988 штрихкодов капель, которые были ассоциированы с мРНК по меньшей мере одной тяжелой цепи (VH) и одной легкой цепи (VL), причем в нем присутствовали 229869 отличающихся клональных линий VH по результатам оценки с помощью кластерного анализа тяжелой цепи. Поскольку этот необработанный набор включает в себя данные от многих клеток, а также капель с отдельными клетками, данные от капель с отдельными клетками обогащали посредством отфильтровывания штрихкодов капель, связанных с не объединенными вместе V(D)J последовательности тяжелой или легкая цепь (фиг. 5В). Эта стадия становится возможным благодаря большому разнообразию типичных иммунных репертуаров, в которых мРНК VH или VL из двух произвольных клеток практически никогда не будут совпадать. Полученный в результате обогащенный набор данных содержал 259368 штрихкодов капель VHVL и содержал 182745 клональных линий VH, удобно представляя наиболее глубокую выборку репертуара спаренных иммунных молекул на данный момент. Точность спаривания VHVL посредством идентификации показателей распространенности для клонального размножения оценивали непосредственно, поскольку клонально родственные клетки должны проявлять соответствия в спаривании их VHVL. Идентифицировали 2604 клонов VH, которые наблюдали более чем в одной фракции эмульсии, что с высокой достоверностью указывает на клетки, образованные в результате клонального размножения. Соответствие последовательностей VL, спаренных с этими VH клонами во фракциях было очень высоким и указывало на точность спаривания 96,1%, обеспечивая высокую достоверности в полном профильтрованном наборе данных от 259368 пар VHVL. Последовательности VH и VL, находящиеся в разных фракциях, были неизменно связаны с разными штрихкодами капли и молекулярными штрихкодами в каждой фракции и, следовательно, не представляли перекрестное загрязнение библиотек. Анализ может недооценивать точность спаривания, поскольку известно, что некоторые В-клетки экспрессируют несколько легких цепей. 75,0% из 259368 профильтрованных штрихкодов капель VHVL или «пар VHVL» содержали IgM и/или IgD (фиг. 5С), которые часто наблюдали совместно, что ожидается при условии типичного IgM+IgD+ фенотипа наивных В-клеток. Также обнаружили меньшие, но значительные фракции пар VHVL для IgA (18,3%) и IgG (6,6%). Все изотипы VH спаривались или с Igκ, или с Igλ в соотношении ~3:2. Из числа 182745 клональных линий VH размножение клонов оценивали двумя способами: по количеству штрихкодов капель, связанных с клоном, и по наблюдению клона во фракциях эмульсии. Клоны, наблюдаемые по многочисленным штрихкодам капель, могут отражать клональное размножение или капли с несколькими штрихкодами, которые ожидаются у ~37% капель при условии исходного пуассоновского рассеивания штрихкодов в капли при λ=1. Тем не менее, любой клон, представленный >8 штрихкодами капли, вероятно, является действительно размноженным (пуассоновская вероятность в одной капле <10-6). В то время как в общей сложности 6,0% клонов наблюдали в более чем одной фракции, в случае клонов, наблюдаемых с более чем 8 штрихкодами капель (в общей сложности 0,7%), 99% из них наблюдали в более чем одной фракции. Все 100 наиболее часто наблюдаемых клонов (30-137 штрихкодов капли для каждого, фиг. 5D) наблюдали по меньшей мере в пяти из шести фракций. Комбинация подсчета штрихкодов и независимого анализа фракций, следовательно, позволяет выявление редких размноженных линий из числа многочисленного фона неразмноженных клонов. Тем не менее, примечательно, что даже наиболее распространенный размноженный клон присутствовал в менее чем одной клетки из тысячи, иллюстрируя большое разнообразие репертуаров периферических органов иммунной системы человека.
Количество захваченных мРНК каждой VH и VL цепи в парах представляет собой оценку уровня экспрессии (фиг. 5Е). Обычно захватывали менее десяти мРНК тяжелой цепи (среднее значение 2,0) и легкой цепи (среднее значение 4,0) на штрихкод капли, наблюдали небольшую популяцию штрихкодов капли с захваченными мРНК тяжелой и легкой цепи в количестве от дюжин до сотен на клетку, происходящую почти исключительно из экспрессирующих IgG и IgA клеток. Интересно, что степень мутации VH и VL в парах сильно коррелировала как в пределах каждого изотипа (например, VH относительно VL для IgG), так и между изотипами (например, IgG относительно IgM) (фиг. 2F). Более того, почти все пары IgG и IgA были в значительной степени подвержены мутациям в обеих из своих VH и VL цепях, тогда как пары IgM и IgD преимущественно показывали небольшую мутацию VH или VL. Эти результаты согласуются с механизмом активации В-клеток, приводящим к переключению класса с IgM и IgD на IgG или IgA, повышенной экспрессии иммуноглобулинов и соматической гипермутации, которая воздействует на локусы как тяжелой, так и легкой цепи в клетке. Помимо этого наблюдения, что сильно мутированные VH цепи проявляют тенденцию к спариванию с сильно мутированными VL цепями, этот способ может создавать большие количества полноразмерных, спаренных в нативном формате BCR из репертуаров покоящихся человеческих В-клеток.
Пример 4. Выделение известных пар VHVL с низкой частотой из элитного контроллера ВИЧ
В качестве дополнительного подтверждения чувствительности спаривания и точности анализа обработке подвергали образец, в котором уже присутствует и общеизвестно образование нескольких редких (<1 клетка на 10000) нативных пар VHVL. Получали периферические В-клетки от пациента-элитного контролера ВИЧ, чьи В-клетки памяти подвергались интенсивному изучению в последние годы в поиске антител, проявляющих нейтрализующую активность в отношении ВИЧ. 350000 В-клеток подвергали обработке с получением в общей сложности 38620 фильтрованных пар VHVL. Интересно, что этот индивид демонстрировал большую долю IgG, чем предшествующий образец в здоровом состоянии (фиг. 3А) или типичные репертуары здоровых периферических В-клеток. Последовательности VH из этого набора данных сравнивали со всеми описанными нейтрализующими антителами широкого спектра действия (bNAb) от этого индивида, в том числе PGT121, и обнаружили восемь близких или идентичных последовательностей VH, указывающих на то, что это семейство bNAb составляет менее чем 0,03% от циркулирующих В-клеток. Важно то, что все легкие цепи, спаренные с этими тяжелыми цепями, относились к ожидаемой и столь же редкой линии bNAb, проявляя одинаковую перестановку Igλ-V3-21/J3 и характерную вставку тройного кодона, как ранее описывалось, подтверждая высокую точность и чувствительность данного способа. Более того, на филогенетическом дереве всех известных и новообразованных PGT121-подобных пар VHVL от этого индивида (фиг. 3В) деревья VH и VL показывают поразительно подобную топологию, в которой спаренные последовательностями VH и VL занимают зеркальные положения, что, вероятно, отражает общую филогенетическую историю. Вариантные пары, исследуемые в данном случае, хорошо согласуются с этим правилом. Интересно, что два опубликованных антитела PGT122 и PGT123 выступают в качестве исключений; подтверждает тот факт, что для этих двух антител пары не обнаруживались, а вместо этого были обнаружены PGT122VH:PGT123VL-подобные и PGT123VH:PGT122VL-подобные пары, указывая на неподтвержденное спаривание в исходном отчете. ДНК, кодирующая полные V(D)J участки, 8 новых PGT-подобных пар VHVL синтезировали, экспрессировали антитела в виде полного IgG и исследовали их способность к нейтрализации нескольких псевдоштаммов ВИЧ (фиг. 6С). Антитела экспрессировались хорошо и все из них проявляли сильную нейтрализующую активность в отношении вируса, демонстрируя пользу данного подходя в быстром получении спаренных в нативном формате функциональных вариантов антител из соответствующего биологического образца.
Пример 5. Пары В-клеточных и Т-клеточных рецепторов из опухоль-инфильтрирующих лимфоцитов
Подтвердив мечение штрихкодами в эмульсии для выделения спаренных рецепторов с высокой пропускной способностью, иммунные рецепторы выделяли напрямую из опухоли. Отбирали диссоциированный с использованием протеазы образец иссеченной аденокарциномы яичника и вводили 400000 несортированных клеток в эмульсию. Окрашивание на CD3/CD19 отдельной аликвоты образца указывало на значительные количества инфильтрирующих В- (~5%) и Т-клеток (~20%) в материале. Распределение отдельных клеток в эмульсии было подобно очищенным клеткам, хотя и с некоторым ограниченным образованием видимых коньков и избыточной вариацией в размере и форме клеток в каплях, что ожидалось вследствие неоднородности типов клеток в образце (фиг. 7А).
Праймеры, нацеленные на константные участки альфа и бета цепей Т-клеточного рецептора вместе, использовали с праймерами для BCR, используемыми ранее, и после секвенирования и жесткого отфильтровывания выделяли тысячи штрихкодов капель, связанных с продуктами в виде BCR или TCR. Для оценки точности на отдельных клетках подсчитывали все возможные комбинации четырех целевых локусов (VH, VL, Vα, Vβ) со штрихкодами капель (фиг. 7В). Подавляющее большинство (97,9%) штрихкодов капель с более чем одной целевой цепью содержали ожидаемые с биологической точки зрения пары VH+VL BCR или Vα+Vβ TCR, при этом лишь 2,1% содержали смешанные комбинации BCR-TCR. Поскольку мечение штрихкодами продуктов является неискаженным по отношению к целевой цепи, этот результат обеспечивает высокую степень достоверности у полученных в результате 6056 пар VHVL BCR и 5217 пар VαVβ TCR. BCR показывали выраженное преобладание IgG (>80%) по сравнению с другими изотипами (фиг. 7С), хотя все они присутствовали (IgE лишь <0,05%). Каппа- и лямбда-легкие цепи присутствовали в соотношениях, подобных наборам данных для периферической крови.
Подобно периферической крови наблюдали корреляцию между изотипом BCR и уровнем мутаций как в VH, так и VL цепях, причем пары IgG и IgA показывают больший уровень мутаций VH и VL, чем IgD и IgM, и общую корреляцию мутаций между VH и VL в каждом изотипе (фиг. 7D). Интересно, что в то время как пары IgD, IgM и IgA показывали очень подобные распределения мутаций между наборами данных для опухоли и периферической крови (фиг. 5F), фракция IgG в опухоли также содержала значительную долю последовательностей, содержащих малое количество мутаций или не содержащих мутаций, чего не наблюдали в периферической крови. Для TCR и для BCR, содержащих IgD, наблюдали подобные количества захваченных мРНК на штрихкод капли с результатами для BCR из периферической крови (фиг. 7Е, обычно <10 на штрихкод капли). Разительно отличаясь от этого, полученные из опухолей пары IgM, IgA и IgG показывали повышенный в 10-100 раз средний уровень экспрессии с сотнями или тысячами целевых мРНК, захваченных по многим из штрихкодов капель. Затем оценивали разнообразие репертуаров захваченных из TIL TCR и BCR (фиг. 7F). Из общего количества 5217 пар TCR наблюдали 2423 отдельных клона TCR бета. Семь клонов присутствовали с частотой >1%, причем наиболее распространенный клон составлял 16,9% от всех штрихкодов капель. Из общего количества 6056 пар BCR наблюдали 1518 отдельных клонов тяжелой цепи, причем 15 клонов встречались с частотой >1%, но ни один не встречался с частотой >5%. Хотя это составляет значительно более ограниченное разнообразие, чем репертуар BCR периферической крови здорового человека (в котором не присутствовало ни одного клона с частотой более 0,06%), присутствие столь большого количества подвергнутых переключению класса, мутированных и экспрессированных на высоком уровне клонов в образце из опухоли демонстрирует необходимость подхода с глубокой и чувствительной подготовкой образца для характеристики TIL. Этот способ обеспечивает возможность быстрого извлечения больших количеств пар иммунорецепторов TIL как из В-, так и Т-клеток одновременно без необходимости предварительной сортировки или экзогенной активации определенных популяций TIL.
Пример 6. Захват дополнительных фенотипических маркеров, представляющих интерес
Спаривание цепей рецепторов посредством мечения штрихкодами капель потенциально обеспечивает возможность захвата дополнительных мишеней помимо иммунных рецепторов. Для изучения этой возможности здоровые Т-клетки разделяли на CD4+ и CD8+ популяции с помощью обогащения на магнитных гранулах и вводили 20000 клеток каждого типа в отдельные серии эмульсий с праймерами, нацеленными на мРНК альфа- и бета-цепей TCR, а также CD4 и CD8. После секвенирования 47,0% из 3861 штрихкодов капель, содержащих Vα и Vβ TCR («пары TCR») из CD4+ выделенных клеток, были связаны с мРНК CD4, в то время как только 0,3% были связаны с мРНК CD8. Напротив, 50,6% из 2235 пар TCR из CD8+ выделенных клеток были ассоциированы с мРНК CD8, в то время как только 0,6% были связаны с мРНК CD4. Это демонстрирует высокую специфичность, но ограниченную чувствительность подхода на основе мРНК для фенотипирования клеток подобно предыдущему сообщению. В отличие от этого, белки, такие как рецепторы клеточной поверхности обычно присутствуют в намного больших количествах (1000-100000 на клетку), чем кодирующие их мРНК, что потенциально делает их выявление более простым, а также потенциально более прямо связано с фенотипом клеток. Для измерения уровней целевого белка на каждой клетке специально созданные метки из олигонуклеотидов ДНК конъюгировали с антителами к человеческим CD4 и CD8 и инкубировали меченные антитела с не подвергнутой разделению смесью CD4+ и CD8+ Т-клеток перед введением 30000 клеток в эмульсию (фиг. 8А). ДНК-метки несут антитело-специфические метки в последовательности, а также молекулярные штрихкоды и последовательность, комплементарную амплифицированным штрихкодам капель, что обеспечивает возможность мечения эмульсии штрихкодами капель и подсчета молекул, аналогичного выполняемому для мРНК. ДНК-метки определяли как мишени одновременно с мРНК TCR, CD4 и CD8. После секвенирования и отфильтровывания 3682 штрихкодов капель идентифицировали с высокой достоверностью как пары VαVβ TCR. В соответствии с предыдущим экспериментом примерно половине (52%) пар TCR можно присвоить статус CD4 или CD8 на основании мРНК (фиг. 8В). Тем не менее, более 95% штрихкодов капли можно отнести к CD4 или CD8 на основании статуса белков, при этом среднее количество подсчитанных молекул на каплю в значительной степени выше для белков CD4/8 (среднее значение 20,5), чем для мРНК CD4/8 (среднее значение 1,0). Соответствие между сигналами от мРНК и белка было высоким (фиг. 8С): 96,0% капель давали согласующиеся результаты как мРНК, так и белка. В некоторых редких случаях выявляли белки как CD4, так и CD8, что, вероятно, было результатом того, что капели содержали две или более клеток. Мечение штрихкодами в эмульсии впервые позволяет непосредственно связывать иммунные рецепторы отдельной клетки с маркерными мРНК и белками, представляющими интерес, и все это с высокой пропускной способностью. Применение этого подхода к TIL с расширенным набором связанных онкологическими заболеваниями иммунных маркеров, таких как антитело к PD-1 и антитело к CTLA-4, обеспечено.
Пример 7. Образцы от человека
Образец крови для подтверждения здорового репертуара собирали с одобрения проекта «Персональный геном». РВМС для эксперимента с bNAb для ВИЧ получали от донора 17, инфицированного ВИЧ-1 донора из когорты G согласно протоколу IAVI. Все образцы от людей с ВИЧ собирали с письменного информированного согласия согласно клиническим протоколам, одобренным Национальным комитетом по этике Республики Руанда, Наблюдательным советом по вопросам биоэтики университета Эмори, Комиссии по этическим вопросам в исследованиях университета Замбии, Комиссией по этическим вопросам в исследованиях больницы Чаринг-Кросс, Комиссией по научным и этическим вопросам UVRI, Комитетом по этическим вопросам в исследованиях университета Нового Южного Уэльса, больницей св. Винсента и территориальной службой здравоохранения Восточного Сиднея, Комиссией по вопросам этики и исследований Национальной больницы Кениата, Кейптаунским комитетом по этическим вопросам в исследованиях, Международным экспертным советом, комиссией по этике университета Махидол, Наблюдательным советом Армейского научно-исследовательского института им. Уолтера Рида (WRAIR) и комитетом Республики Кот-д'Ивуар "National d'Ethique des Sciences de la Vie et de la Sante" (CNESVS). Криоконсервированную диссоциированную иссеченную аденокарциному от отдельного донора получали от Conversant Biologies с письменного информированного согласия согласно одобренному IRB протоколу.
Пример 8. Подготовка клеток
Для исследования 3 миллионов здоровых В-клеток 50 мл крови забирали в пробирки для клеточных препаратов Vacutainer СРТ с гепарином натрия (BD), центрифугировали в течение 20 мин. при 1800×g, отмывали дважды в буфере для для подготовки клеток (1×PBS, дополненный 2% фетальной бычьей сывороткой и 2 мМ EDTA) с использованием центрифугирования при 200×g для удаления тромбоцитов и полученные в результате РВМС криоконсервировали в среде RPMI-1640 (Life Technologies) + 20% фетальной бычьей сыворотки + 10% DMSO при температуры -80°С до востребования. Перед образованием эмульсии РВМС подвергали оттаиванию, отмывали дважды в буфере для подготовки клеток и подсчитывали. В-клетки выделяли с использованием набора для обогащения В-клеток на основе отрицательной селекции (Stem Cell Technologies) в соответствии с инструкциями производителя. Клетки пропускали через 20 мкм сито для клеток и разводили до концентрации 6,2×106 клеток/мл (эксперимент с 3 миллионами В-клеток) или 3,1×106 клеток/мл (эксперименты с PGT от донора и опухолью яичника) в буфере для пролиферации клеток.
Пример 9. Мечение штрихкодом иммунных рецепторов в эмульсии
Платформа для образования эмульсии состояла из трех насосов Mitos P-Pumps (Dolomite Microfluidics), приводимых в действие одним воздушным компрессором, каждый с сенсором расхода Mitos Flow Rate sensor, чтобы обеспечить возможность контролируемого компьютером потока двух водных фаз и одной флуорофильной масляной непрерывной фазы в покрытый флуорофильным материалом кварцевый чип для двух реактивов Dolomite Small 2-Reagent chip. Один вводной канал с водной жидкостью содержал клетки с требуемой плотностью желаемого уровня заполнения клетками капель, в то время как второй канал с водной жидкостью содержал реактивы смесь для лизиса и реакции, состоящую из реакционного буфера AbPair™ и олигонуклеотидов (www.abvitro.com/catalog AV2070-1S и AV2080-1S), 5 единиц/мл обратной транскриптазы на основе MuMLV (Thermo Scientific) и 0,1 единиц/мл полимеразы Herculase II PCR. Шприц Hamilton Microliter на 100 мкл использовали для вертикальной загрузки трубчатой PEEK петли для образца с внутренним диаметром 100 мкм смесью LR в две инъекции, по ~100 мкл каждая. Шприц Hamilton Gastight на 100 мкл использовали для загрузки ~110 мкл клеточной суспензии в трубчатую FEP петлю с внутренним диаметром 0,2 мм и объемом ~100 мкл. Эмульсию образовывали посредством направления струй сфокусированного потока водных фаз с идентичными скоростями потока через чип для 2 реактивов с одновременным потоком масла из двух каналов для масла в чипе. Эмульсия, покидающая выходной канал из чипа, стекала по каплям в пробирки для ПЦР-стрипов объемом 0,2 мл (Eppendorf) на холодном блоке, после чего избыточное масло удаляли посредством пипетирования со дна пробирки, добавляли 40 мкл наслаиваемого раствора (25 мМ Na-EDTA, рН 8,0) и пробирки переносили в стандартный термоциклер для реакции мечения транскрипта. В течение 45 мин. стадии обратной транскрипции (RT) РНК подвергают обратной транскрипции при 42°С с мишенеспецифическими праймерами для RT с основанным на переключении праймера добавлением универсальной адаптерной последовательности, содержащей рандомизированный молекулярный штрихкод. После RT эмульсии подвергали 40 циклам термоциклирования (каждый цикл: 82°С в течение 10 с, 65°С в течение 25 с) для осуществления ПЦР амплификации матриц со штрихкодами капель, который разводили в первоначальной смеси для лизиса и реакции до 30000 копий/мкл, получая концентрацию в конечной смеси 15000 копий/мкл или ~1 на каплю объемом ~65 пл. Один конец штрихкода капли содержит сайт для праймера Illumina read 2 («Р7»), в то время как другой конец совпадает с общей последовательностью универсального адаптерного олигонуклеотида. Таким образом, во время ПЦР подвергнутые переключению матрицы кДНК могут гибридизироваться с амплифицированными цепями штрихкодов капель и становятся сплайсированными посредством достройки перекрывающихся последовательностей с получением полноразмерных продуктов, содержащих мишень, последовательности молекулярного штрихкода и штрихкода капли.
Пример 10. Разрушение эмульсии, очистка, последующие ПЦР, объединение и секвенирование
После термоциклирования наслаиваемый раствор удаляли посредством пипетирования и 40 мкл разрушающего эмульсию раствора (1:1 FC-40 : перфтороктанол) добавляли вместе с 15 мкл раствора для очистки лизата (12,5 мкл протеазы Qiagen, 2,5 мкл 0,5 М Na-EDTA, рН 8,0). После 10-кратного переворачивания для разрушения эмульсии смесь инкубировали в течение 15 минут при 50°С и 3 минут при 95°С для инактивации протеазы. После центрифугирования при 15000×g в течение 1 мин. для выделения водной фазы выделенный материал тщательно очищали для удаления олигонуклеотидов, реактивов и избыточных ПЦР продуктов со штрихкодами капель. Поскольку полноразмерные продукты содержат биотин вследствие 5' биотинилирования праймера для RT, их можно эффективно отделить от избыточных ПЦР продуктов со штрихкодами капель посредством очистки на гранулах со стрептавидином, таким образом сводя к минимуму артефакты от рекомбинации при последующей ПЦР, распространенную проблему при подходах с достройкой при перекрытии. Первые продукты очищали с помощью гранул AMPure ХР (Agencourt) с использованием инструкций производителя в соотношении 1:1 с последующей очисткой с помощью гранул со стрептавидином (New England Biolabs) также с использованием инструкций производителя за которой следует элюирование в деионизированной воде при 95°С с последующей второй очисткой с гранулами AMPure ХР в соотношении 1:1. Продукты затем вносят в ПЦР для обогащения мишени, при которой праймеры, специфические к константным участкам В- или Т-клеточных рецепторов-мишеней использовали вместе с праймером, специфическим к универсальному концу последовательность штрихкода капли. Этот обратный праймер также содержал индекс в виде штрихкода из шести оснований для мультиплексного секвенирования на инструменте MiSeq в соответствии с инструкциями производителя. Таким образом, только полноразмерные меченые штрихкодом целевые последовательности амплифицируются на этой стадии. Все мишени вначале амплифицировали вместе в течение семи циклов: 98°С в течение 10 секунд; 64°С в течение 20 секунд; 72°С в течение 15 секунд с использованием полимеразы Q5 с «горячим стартом» (New England Biolabs) в рекомендуемых производителем условиях, включая стадию активации полимеразы в течение 2 минут при 98°С вначале реакции. За этим следовала очистка AMPure ХР с соотношением гранулы : ПЦР продукты, составляющим 1,5:1. Затем осуществляли второе целенаправленное воздействие из семи циклов в отношении каждой цепи (VH, VL, Vα, Vβ) отдельно с использованием тех же условий термоциклирования, что и до этого, с последующей очисткой с помощью AMPure ХР. Осуществляли окончательную ПЦР с теми же условиями термоциклирования, а затем 5-15 циклов (в зависимости от выхода, оцениваемого с помощью qPCR) для добавления полноразмерных адаптеров для секвенирования Illumina и получения достаточного количества материала для количественного определения TapeStation D1000 (Agilent). Библиотеки затем объединяли и секвенировали на платформе MiSeq (Illumina) V3 2×300 п.о.
Пример 11. Модификации на платформе MiSeq
Реконструкция полного вариабельного V(D)J участка BCR или TCR требуют сшивания ридов Illumina с двумя спаренными концами. Для улучшения этого процесса рид от прямой последовательности в наборе 2×300 п.о. достраивали до 325 п.о. Добавляемый в виде стандартного образца 10% phiX использовали для уменьшения проблем ограниченного разнообразия библиотеки, поскольку библиотеки иммунных рецепторов имеют ограниченное разнообразие в сайтах для праймера в константном участке.
Пример 12. Обзор обработки ридов с помощью методов биоинформатики
Риды Illumina MiSeq подвергали обработке с использованием специально разработанных программных конвейеров, построенных вокруг пакета pRESTO (версия 0.4) для создания полноразмерных консенсусных последовательностей для молекул мРНК из каждой капли, аннотировали с использованием IgBLAST и/или IMGT/HighV-QUEST и дополнительно выравнивали, фильтровали и подвергали обработке специально разработанными скриптами и пакетом Change-O с получением статистических данных.
Пример 13. Обработка и аннотирование ридов, определение изотипа
Обработка сырых ридов, аннтотирование V(D)J и определение клона осуществляли с использованием специально разработанных программных конвейеров, использующих пакеты pRESTO и Change-O. Вкратце, сырые риды от Illumina со спаренными концами 325+300 п.о. подвергали контролю качества, подрезке праймера и специфические к капле (DB) и специфические к молекуле штрихкоды (MB) идентифицировали посредством нечеткого совпадения сайтов для праймера. Вместе, DB и MB уникально определяют происхождение молекулы, и этот уникальный молекулярный идентификатор (UMI) использовали для того, чтобы объединить в группы соответствующие реплицированные в результате ПЦР риды (минимум из двух), происходящие из одной и той же молекулы, для создания консенсусной последовательности для каждой последовательности мРНК. Специфическое для изотипа инициирование подтверждали посредством нечеткого совпадения известных специфических к изотипу константных участков в пределах последовательностей с обрезанными праймерами. Сегменты генов зародышевого типа V(D)J и перестроенную структуру определяли с использованием IgBLAST и подтверждали с использованием IMGT/HighV-QUEST, когда это требовалось, интерпретировали с помощью Change-O и специально разработанных скриптов.
Клоны устанавливали посредством кластеризации методом одиночной связи в пределах групп функциональных V(D)J последовательностей, имеющих совпадающий ген IGHV, ген IGHJ и длину участка соединения, как реализовано в Change-O. Для набора данных от 3 миллионов циркулирующих клеток использовали взвешенное внутриклональное расстояние 4,0 с использованием симметризованной модели транзиции/трансверсии в качестве ограничения между ближайшими соседями у клонов.
Пример 14. Фильтрование включения иммунного рецептора в каплю и расчет точности совпадения при спаривании
Точность выделения В-клеточной последовательности из капель можно оценить двумя способами с использованием этого способа мечения штрихкодами: с использованием соответствия последовательностей мРНК внутри капель и по соответствию спаривания между фракциями. В каждой капле захватывают несколько мРНК на локус; экспрессируемые V(D)J последовательности из одной клетки должны совпадать. Наличие более чем одной продуктивной VDJ и одной продуктивной VJ последовательности на каплю отмечают с помощью методов биоинформатики как предполагаемое включение иммунного рецептора или занятие нескольких клеток при использовании ограничения 2% разнообразия последовательности (средние различия пар нуклеотидов pi55 <0,02) у нескольких выравниваемых V(D)J сегментов для определения соответствия последовательности. Консенсусные последовательности тяжелой и легкой цепи строили для каждой капли с аллельным исключением и их использовали для определения клона и анализа спаривания между фракциями. Для набора из 3 миллионов циркулирующих В-клеток каждая линия VH ассоциирована с одним (в идеальном случае) из >20000 клонов легкой цепи в наборе данных. Среди 259368 капель, исключенных по иммунному локусу, с парами VHVL, 10870 кластеров с перестройкой локуса VDJ тяжелой цепи присутствовали по меньшей мере в двух из шести физически разделенных фракций эмульсии. Эти кластеры представляют собой либо размноженные линии, либо независимую, но подобную перестройку одинаковых экзонов VJ. В случае, когда подвергшийся перестройке VDJ спаривается с соответствующим подвергшимся перестройке VJ в двух параллелях, оба эксперимента независимо давали истинно положительные результаты (33157 из 35922 возможных парных сравнений для 2604 клонов с более редкими перестройками). Следовательно, точность для каждой параллели составляет 96,1% (0,923^0,5).
Пример 15. Изучение последовательности-кандидата bNAb против ВИЧ
Новые спаренные в нативном формате нейтрализующие антитела широкого спектра действия (BNAb) к HIV изучали посредством анализа наших 38620 пар VHVL в отношении сходства с известными последовательностями bNAb HCDR3s, VDJ последовательностями и последовательностями линий от донора 17 отбирали из литературы с использованием tblastx, MUSCLE, и PhyML с последующим изучением в ручном режиме филогенетических деревьев полной аминокислотной последовательности V(D)J для выбора антител-кандидатов, перемежающиеся с известными последовательностями bNAb.
Пример 16. Экспрессия и очистка белка bNAb против ВИЧ
Последовательности антитела синтезировали и клонировали в ранее описанные векторы тяжелой и легкой цепей. Плазмиды тяжелой и легкой цепи подвергали совместной трансфекции (соотношение 1:1) в клетки 293 FreeStyle с использованием 293fectin (Invitrogen) в соответствии с протоколом производителя. Супернатанты антител собирали через четверо суток после трансфекции и очищали с помощью аффинной хроматографии с белком-А. Очищенные антитела подвергали замене буфера на PBS перед использованием в дополнительных анализах.
Пример 17. Получение псевдовируса и анализы нейтрализации
Псевдовирусы получали посредством трансфекции клеток 293Т с использованием экспрессионной плазмиды для Env ВИЧ-1 и дефектной по Env плазмиды в виде геномного остова (pSG3ΔEnv). Псевдовирусы собирали через 72 часа после трансфекции для применения в анализах нейтрализации. Нейтрализующую активность оценивали с использованием одного раунда анализа репликации псевдовируса и клеток-мишеней TZM-В1. Вкратце, клетки TZM-B1 высевали в 96-луночный планшет с плоским дном. В этот планшет добавляли псевдовирус, который предварительно инкубировали с серийными разведениями антитела в течение 1 часа при 37°С. Экспрессию репортерного гена люциферазы количественно определяли спустя 72 часа после инфекции при лизисе и добавлении субстрата люциферазы Bright-Glo (Promega). Для определения значений IC50 кривые доза-ответ подбирали с помощью нелинейной регрессии.
Пример 18. Идентификация целевых цепей в опухоли яичника
После одновременного захвата BCR и TCR из диссоциированной ткани опухоли яичника в эмульсии риды фильтровали с использованием молекулярных штрихкодов и штрихкодов капель, как описано ранее, но затем проводили поиск в отношении присутствия каждого из четырех возможных типов целевой цепи (BCR VH, BCR VL, TCR Vα, TCR Vβ). Целевые цепи сохраняли, если они обосновывались по меньшей мере двумя мРНК, каждая с по меньшей мере двумя ридами от секвенирования. Все штрихкоды капель, содержащие только пары VHVL BCR или VαVβ TCR, анализировали дополнительно. Клоны тяжелой цепи BCR и бета-цепи TCR называли на основании отличающихся аминокислотных последовательностей CDR3.
Пример 19. Выявление белка в эмульсии посредством окрашивания с помощью меченого ДНК антитела
Конструировали одноцепочечные олигонуклеотиды ДНК из 200 п.о., содержащие уникальные идентификационные последовательности антигена из 5 п.о., и их модифицировали 5' аминогруппой (Integrated DNA Technologies). Мышиные моноклональные антитела к человеческому CD4 (BioLegend, #300516) и CD8a (BioLegend, #301018) конъюгировали с олигонуклеотидными метками ДНК с использованием набора Thunder-Link (hinova Biosciences) в соответствии с протоколом производителя. Для мечения клеток перед переносом в эмульсию два миллиона подвергнутых отрицательной селекции Т-клеток из периферической крови разводили в 400 мкл буфера для клеток + 2 мМ EDTA + 0,05% азида натрия. Одноцепочечную ДНК из спермы лосося добавляли к клеткам до конечной концентрации 200 мкг/мл и клетки перемешивали вращательным движением при комнатной температуре в течение пяти минут. Смесь меченых ДНК антител к CD4 и CD8a (каждое в конечной концентрации 5 нМ) добавляли к клеткам и инкубировали при комнатной температуре в течение 30 минут. Клетки промывали три раза буфером для клеток + 2 мМ EDTA + 0,05% азида натрия + 200 мкг/мл одноцепочечной ДНК из спермы лосося. Клетки ресуспендировали в буфере для клеток + 0,05% азида натрия перед введением в анализ в эмульсии. 30000 клеток использовали для секвенирования эмульсии.
Пример 20. Окрашивание с помощью конъюгата тетрамер-олигонуклеотид
Получали конъюгат тетрамер-олигонуклеотид, состоящий из четырех комплексов МНС-пептид, каждый со стрептавидиновым кором, и флуорофора. Каждый конъюгат тетрамер-олигонуклеотид дополнительно содержал молекулярный штрихкод и/или идентификационный код тетрамера для использования в количественном определении, как описано ниже. Иллюстративный меченый флуоресцентной молекулой конъюгат тетрамер-олигонуклеотид показан на фиг. 18.
Иллюстративный способ определения аффинности двух отличающихся TCR описан в данном документе. Две популяции Т-клеток с известной отличающейся аффинностью в отношении МНС-пептида инкубируют с различными концентрациями конъюгата тетрамер-олигонуклеотид, содержащего тетрамер МНС-пептид (рМНС). Несвязавшийся тетрамер-олигонуклеотид смывают, а клетки вводят в анализ в эмульсии и секвенируют. Сигнал от связывания рассчитывают на основании ридов от последовательности для олигонуклеотида в составе конъюгата тетрамер-олигонуклеотид для каждой Т-клетки при каждой концентрации конъюгата тетрамер-олигонуклеотид (фиг. 19). Ожидается, что Т-клетки, инкубируемые при более высокой концентрации конъюгата тетрамер-олигонуклеотид, показывают более высокий сигнал от связывания. Ожидается, что Т-клетки с TCR, которые характеризуются более высокой аффинностью в отношении МНС-пептида в составе конъюгата тетрамер-олигонуклеотид, проявляют более высокий сигнал от связывания по сравнению с Т-клетками с TCR, которые характеризовались более низкой аффинностью в отношении МНС-пептид в составе конъюгата тетрамер-олигонуклеотид при одинаковой концентрации конъюгата тетрамер-олигонуклеотид в пределах подходящего для обработки диапазона результатов анализа. Например, на фиг. 19 между концентрациями log[тетрамер] -8 и -4, сигнал от связывания для TCR1 выше, чем для TCR2, указывая на то, что в этом анализе TCR1, как ожидается, будет характеризоваться более высокой аффинностью в отношении применяемого реактива в виде тетрамера.
Пример 21. Окрашивание с помощью конъюгата тетрамер-олигонуклеотид в сравнении с традиционным окрашиванием с помощью тетерамера МНС-пептид
Мишенеспецифические Т-клетки получали для обеспечения положительного контроля для исследования специфичности связывания у иллюстративных реактивов конъюгата тетрамер-олигонуклеотид, как описано в данном документе. Иллюстративный способ получения мишенеспецифических Т-клеток, в целом, описан в Но et al., J. Immunol. Methods, 310:1-2, 40-52. Вкратце, дендритные клетки получают из адгезивных фракций в образцах мононуклеарных клеток периферической крови (РВМС), полученных от нормальных людей-доноров с HLA-A02:01, посредством культивирования в течение двух суток в присутствии GM-CSF и IL-4 с последующим инкубированием, начиная с Дня 3, в присутствии провоспалительных цитокинов с получением зрелых дендритных клеток. В День 4 полученные в результате зрелые дендритные клетки собирают, отмывают и стимулируют желаемым полученным из мишени пептидом, подлежащим использованию в тетрамере МНС-пептид (рМНС). В День 5 аутологичные CD8+ Т-клетки от нормальных людей-доноров инкубируют с простимулированными пептидом дендритными клетками. В День 8 IFNγ в культурах измеряют в качестве индикатора для культур, содержащих антигенспецифические Т-клетки. Клетки из реагирующих совместных культур отбирают и повторно стимулируют два или три раза простимулированными пептидом дендритными клетками для обогащения антигенспецифическими Т-клетками. В соответствии с некоторыми вариантами осуществления эти антигенспецифические Т-клетки получают из таких обогащенных популяций посредством клеточного сортинга и/или клонирования методом серийных разведений, по существу, как описано в Но et al. 2006. CD8+ Т-клетки, которые не являются мишенеспецифическими, использовали в качестве контрольной Т-клеточной популяции.
Мишенеспецифические CD8+ Т-клетки и контрольную популяцию неспецифических CD8+ Т-клеток инкубировали либо с меченым флуоресцентной молекулой стандартным тетрамером МНС-пептид, либо с различными уменьшающимися концентрациями меченого флуоресцентной молекулой конъюгата тетрамер-олигонуклеотид. После инкубирования присутствие клеток, окрашивающихся положительно стандартным тетрамером МНС-пептид или конъюгатом тетрамер-олигонуклеотид, идентифицировали с помощью проточной цитометрии. Как стандартный тетрамер МНС-пептид, так и конъюгат тетрамер-олигонуклеотид, инкубируемые с неспецифическими CD8+ Т-клетками, проявляли незначительное положительное окрашивание или его отсутствие (фиг. 21, верхний ряд графиков проточной цитометрии). Стандартный тетрамер МНС-пептид, инкубируемый с мишенеспецифическими CD8+ Т-клетками, показывал популяцию хорошо окрашивающихся, положительных по двум маркерам МНС-пептид тетрамер+ CD8+ клеток (фиг. 21, нижний ряд, левый график проточной цитометрии). Конъюгат тетрамер-олигонуклеотид, инкубируемый с мишенеспецифическими CD8+ Т-клетками, показывал популяцию хорошо окрашивающихся, положительных по двум маркерам конъюгат тетрамер-олигонуклеотид+ CD8+ клеток, при этом эта положительная по двум маркерам популяция уменьшалась при пониженной концентрации конъюгата тетрамер-олигонуклеотид (фиг. 21, нижний ряд, три правых графика проточной цитометрии). Это указывает на то, что конъюгаты тетрамер-олигонуклеотид связываются с мишенеспецифическими Т-клетками аналогично стандартным тетрамерам МНС-пептид, и что связывание конъюгатов тетрамер-олигонуклеотид с мишенеспецифическими Т-клетками было зависимым от концентрации конъюгата тетрамер-олигонуклеотид.
Пример 22. Сигнал от связывания конъюгата тетрамер-олигонуклеотид
Мишенеспецифические (участвующие в связывании TCR) или неспецифические Т-клетки (не участвующие в связываниии TCR), которые в общем описаны в примере 21, также использовали для оценки количества связавшихся конъюгатов тетрамер-олигонуклеотид на клетку. Популяцию клеток, не участвующих в связывании TCR, и популяцию клеток, участвующих в связывании TCR, раздельно инкубировали с различными концентрациями конъюгата тетрамер-олигонуклеотид, содержащего тетрамер МНС-пептид, распознаваемый TCR в составе популяции клеток, участвующих в связывании TCR, а также по меньшей мере одну уникальную последовательность олигонуклеотида (например, молекулярный штрихкод, идентификационный код тетрамера и/или идентификационный код антигена). Несвязавшийся тетрамер-олигонуклеотид смывали, а клетки вводили в анализ в эмульсии и секвенировали. Сигнал от связывания рассчитывали на основании количества ридов от последовательности олигонуклеотида для каждого тетрамера при каждой концентрации конъюгата тетрамер-олигонуклеотид, что представлено в графической форме на фиг. 21. Результаты, показанные на фиг. 21, указывают на то, что конъюгаты тетрамер-олигонуклеотид являются способными к специфическому распознаванию и связыванию с TCR на первичных Т-клетках-мишенях дозозависимо.
| название | год | авторы | номер документа |
|---|---|---|---|
| КОНЪЮГАТЫ АФФИННАЯ ЧАСТЬ-ОЛИГОНУКЛЕОТИД И ИХ ПРИМЕНЕНИЯ | 2016 |
|
RU2778754C2 |
| СЕКВЕНИРОВАНИЕ ПОЛИНУКЛЕОТИДНЫХ БИБЛИОТЕК С ВЫСОКОЙ ПРОПУСКНОЙ СПОСОБНОСТЬЮ И АНАЛИЗ ТРАНСКРИПТОМОВ | 2018 |
|
RU2790291C2 |
| КОМПОЗИЦИИ И СПОСОБЫ ИЗМЕРЕНИЯ И КАЛИБРОВКИ СИСТЕМАТИЧЕСКОЙ ОШИБКИ АМПЛИФИКАЦИИ В МУЛЬТИПЛЕКСНЫХ ПЦР-РЕАКЦИЯХ | 2013 |
|
RU2631797C2 |
| СПОСОБ И КОМПОЗИЦИЯ ДЛЯ АНАЛИЗА КЛЕТОЧНЫХ КОМПОНЕНТОВ | 2016 |
|
RU2717491C2 |
| ИДЕНТИФИКАЦИЯ АССОЦИИРОВАННЫХ С ОБРАЗЦОМ ПОЛИНУКЛЕОТИДОВ | 2012 |
|
RU2644681C2 |
| ВЫСОКОПРОИЗВОДИТЕЛЬНЫЕ БИБЛИОТЕКИ ОДИНОЧНЫХ ЯДЕР И ОДИНОЧНЫХ КЛЕТОК И СПОСОБЫ ИХ ПОЛУЧЕНИЯ И ИСПОЛЬЗОВАНИЯ | 2020 |
|
RU2838545C2 |
| СПОСОБЫ И СРЕДСТВА ПОЛУЧЕНИЯ БИБЛИОТЕКИ ДЛЯ СЕКВЕНИРОВАНИЯ | 2019 |
|
RU2815513C2 |
| КОМПОЗИЦИИ ДЛЯ Т-КЛЕТОЧНОЙ ТЕРАПИИ С ХИМЕРНЫМ АНТИГЕННЫМ РЕЦЕПТОРОМ И ИХ ПРИМЕНЕНИЯ | 2018 |
|
RU2819805C2 |
| СКОНСТРУИРОВАННЫЕ ВЫСОКОАФФИННЫЕ T-КЛЕТОЧНЫЕ РЕЦЕПТОРЫ ЧЕЛОВЕКА | 2014 |
|
RU2729383C2 |
| СКОНСТРУИРОВАННЫЕ ВЫСОКОАФФИННЫЕ T-КЛЕТОЧНЫЕ РЕЦЕПТОРЫ ЧЕЛОВЕКА | 2014 |
|
RU2740648C2 |
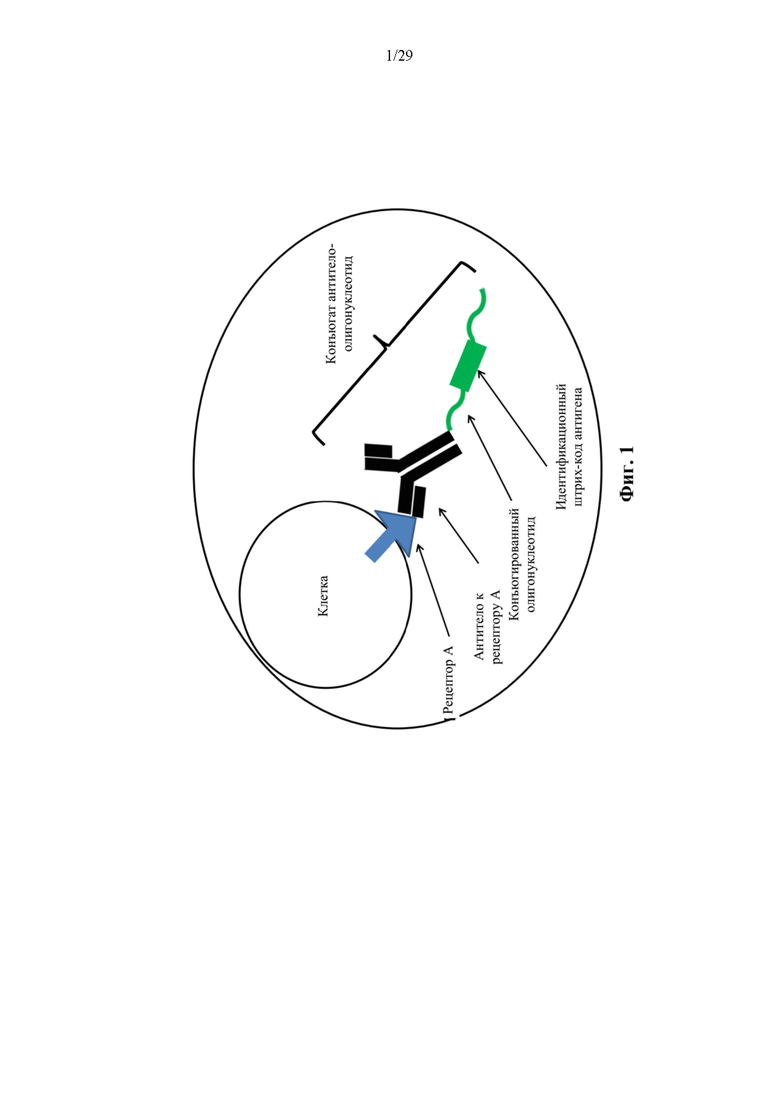
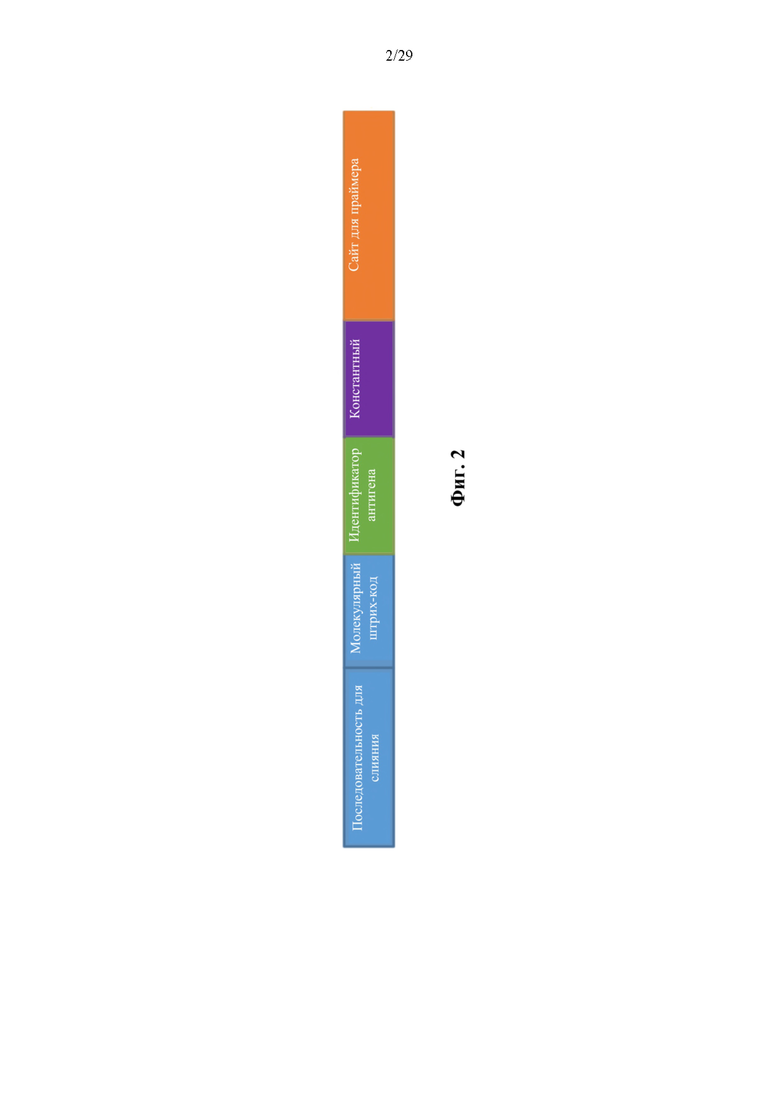
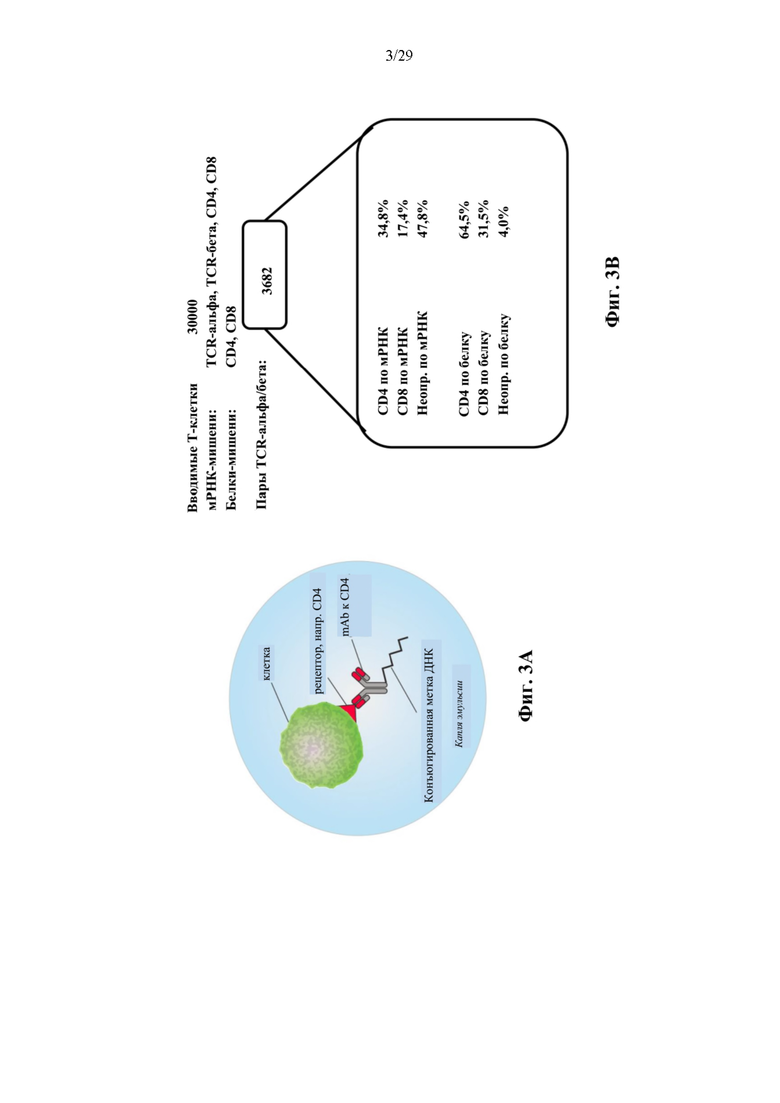
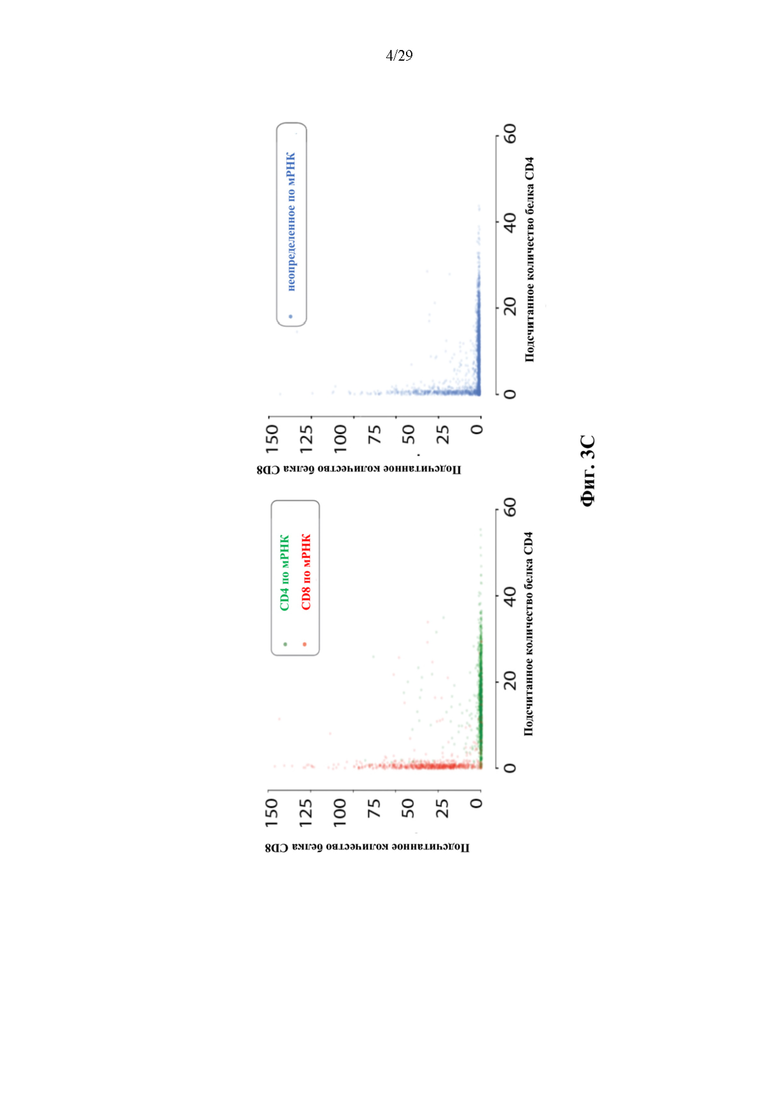
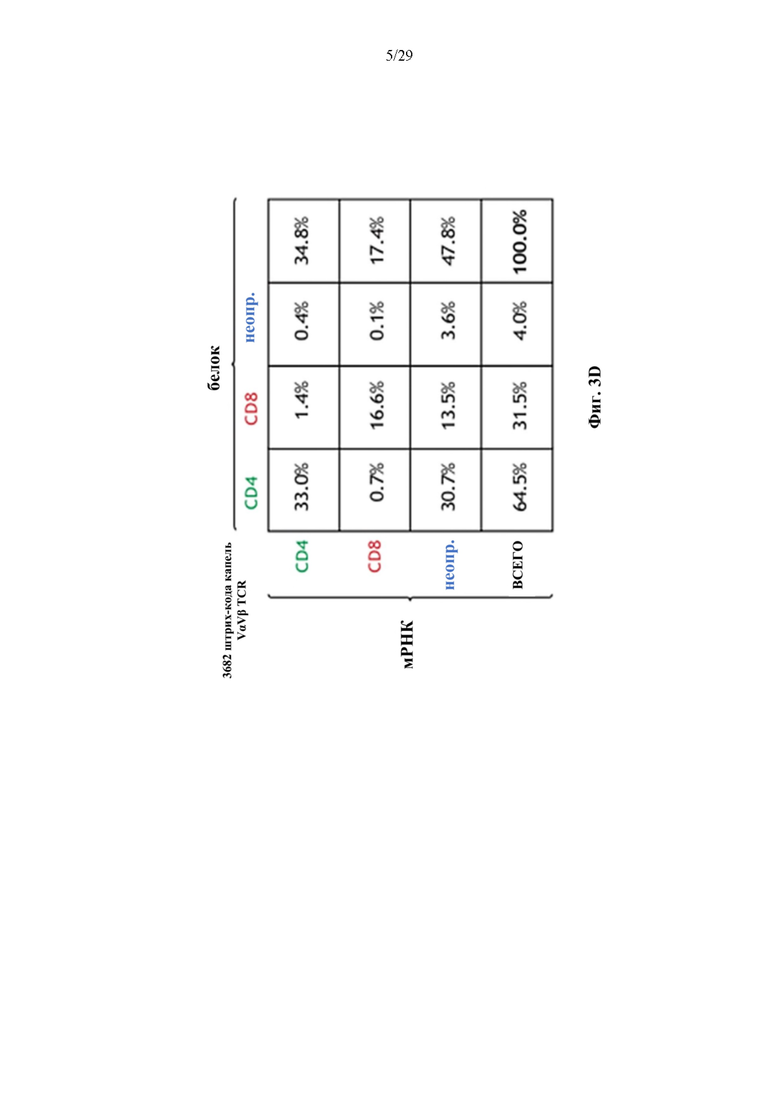
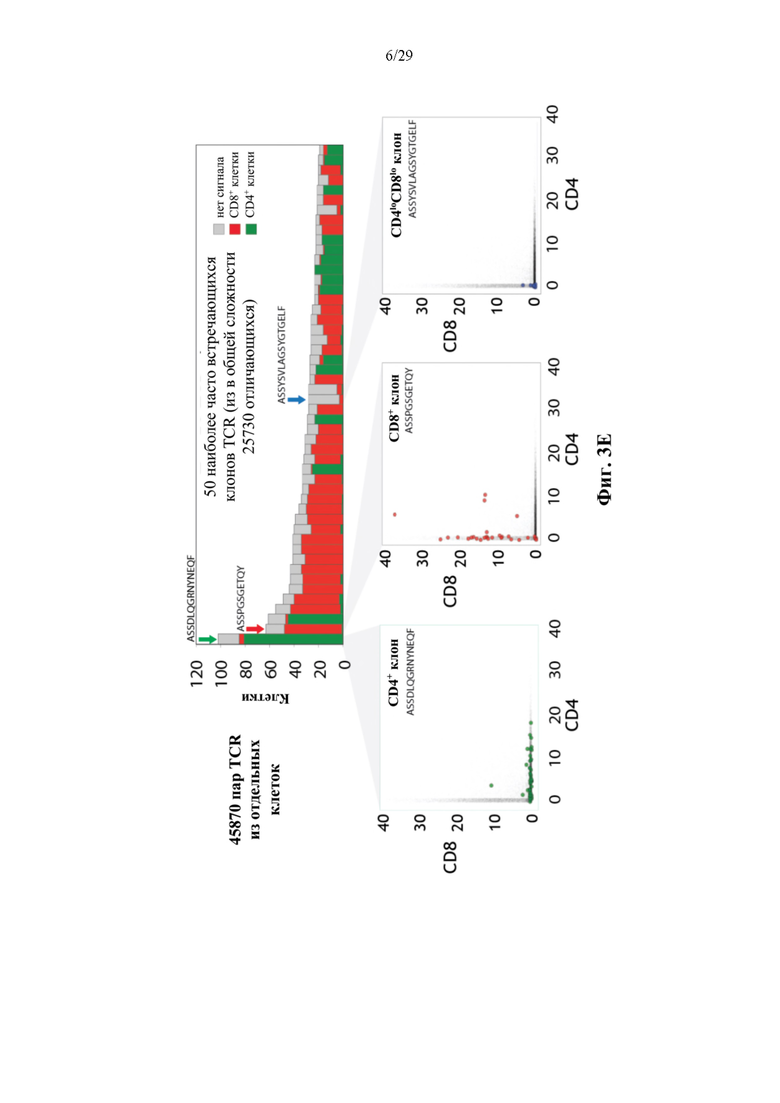
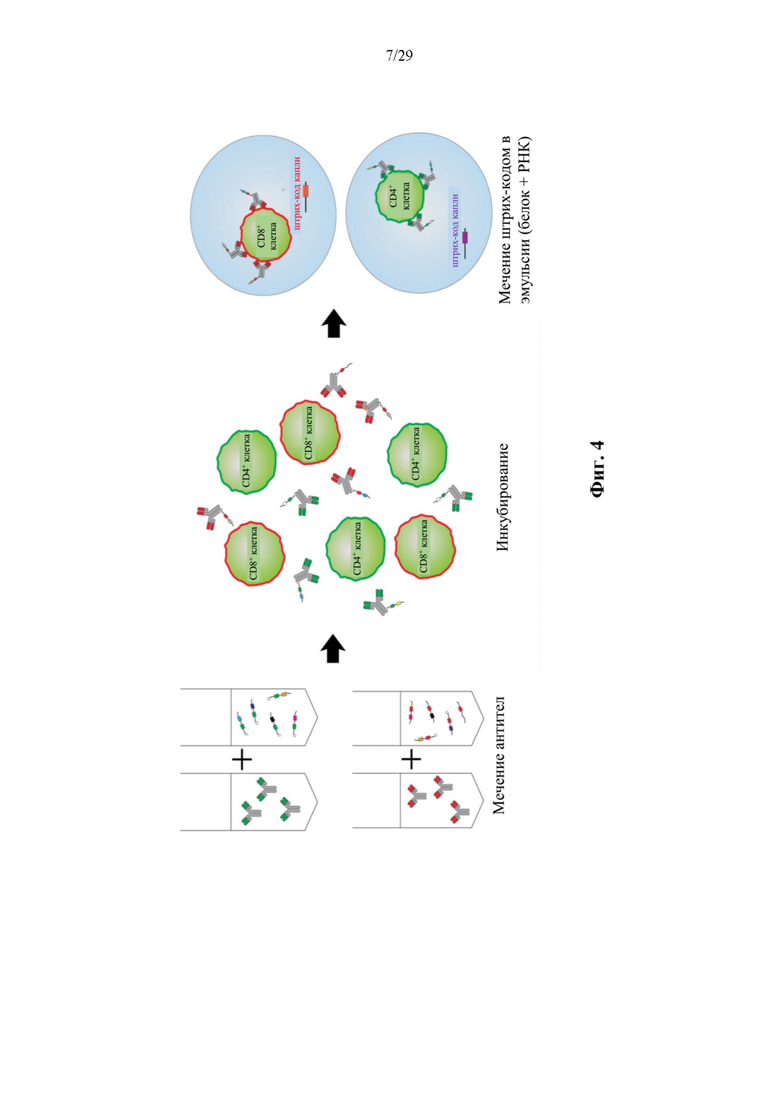

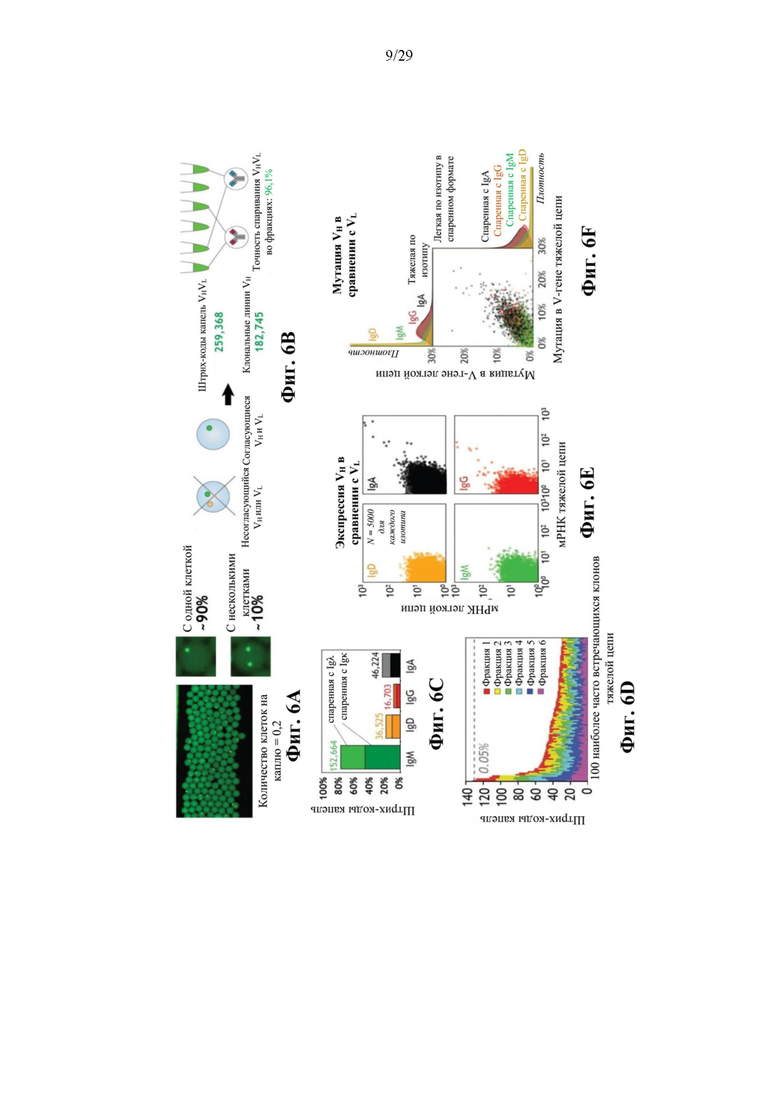
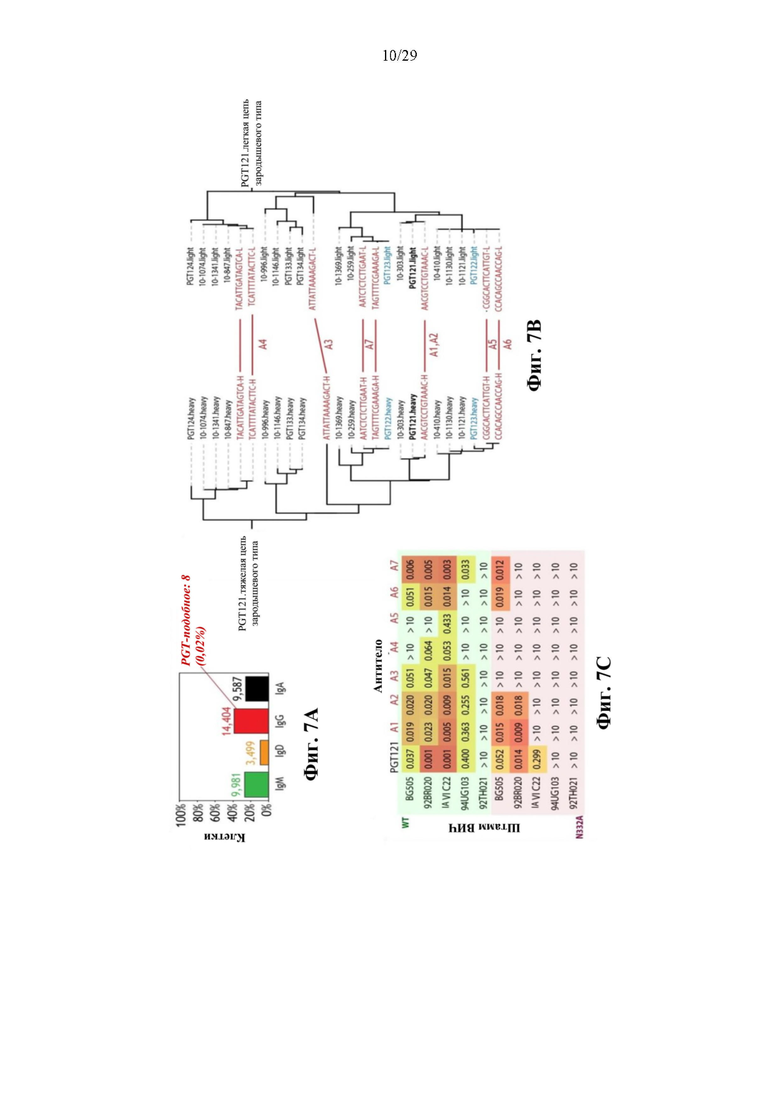
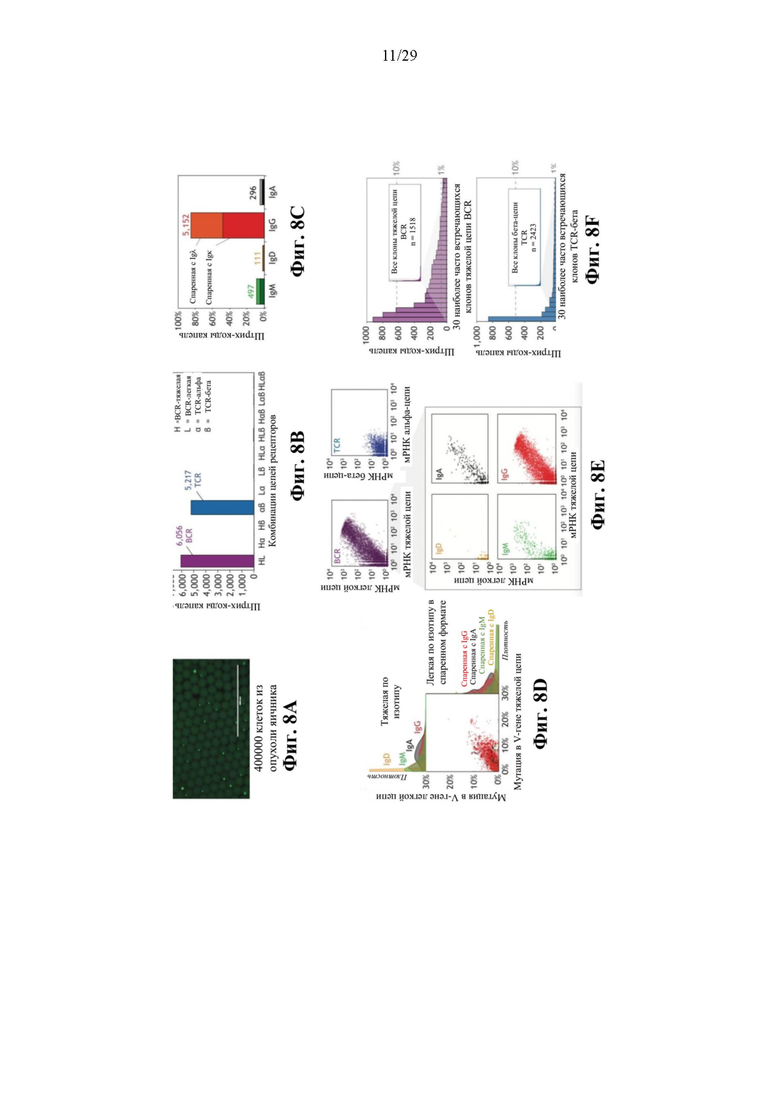
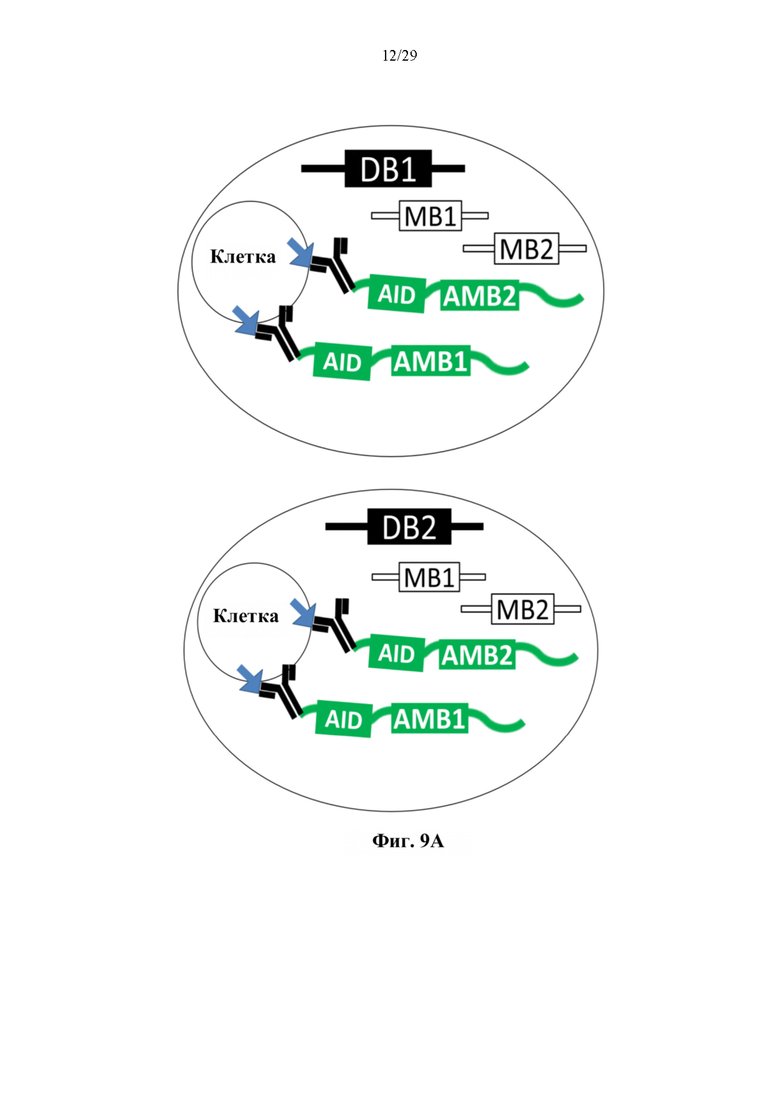


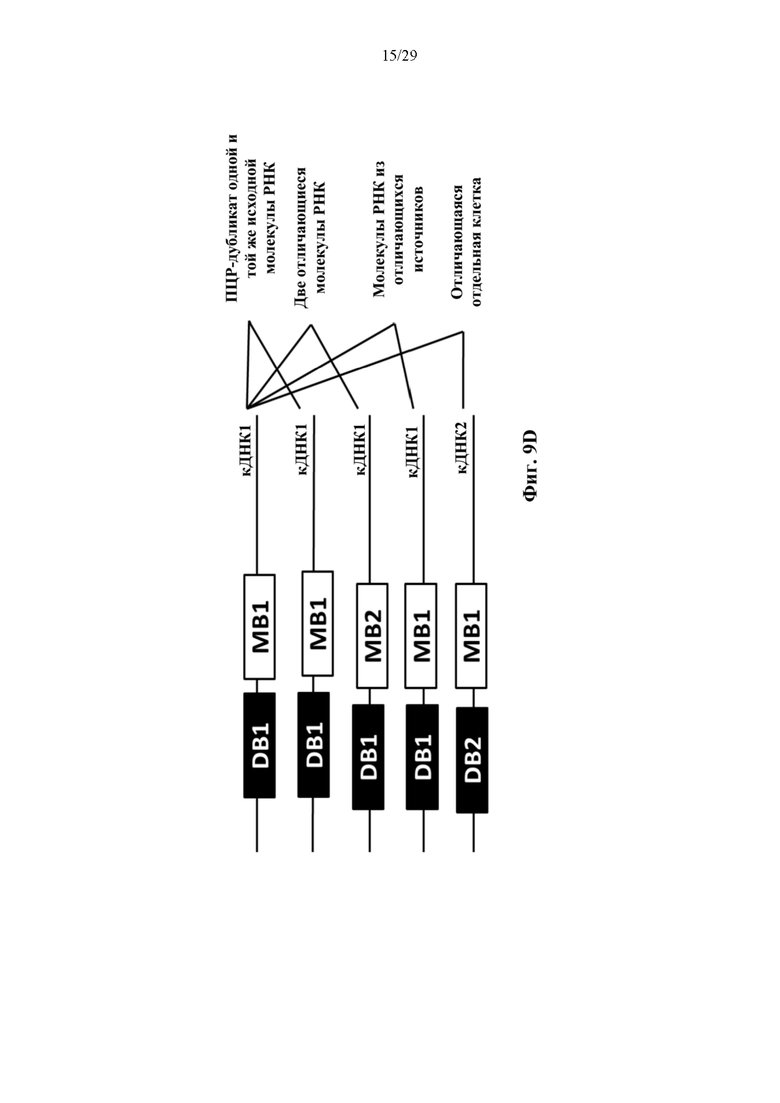
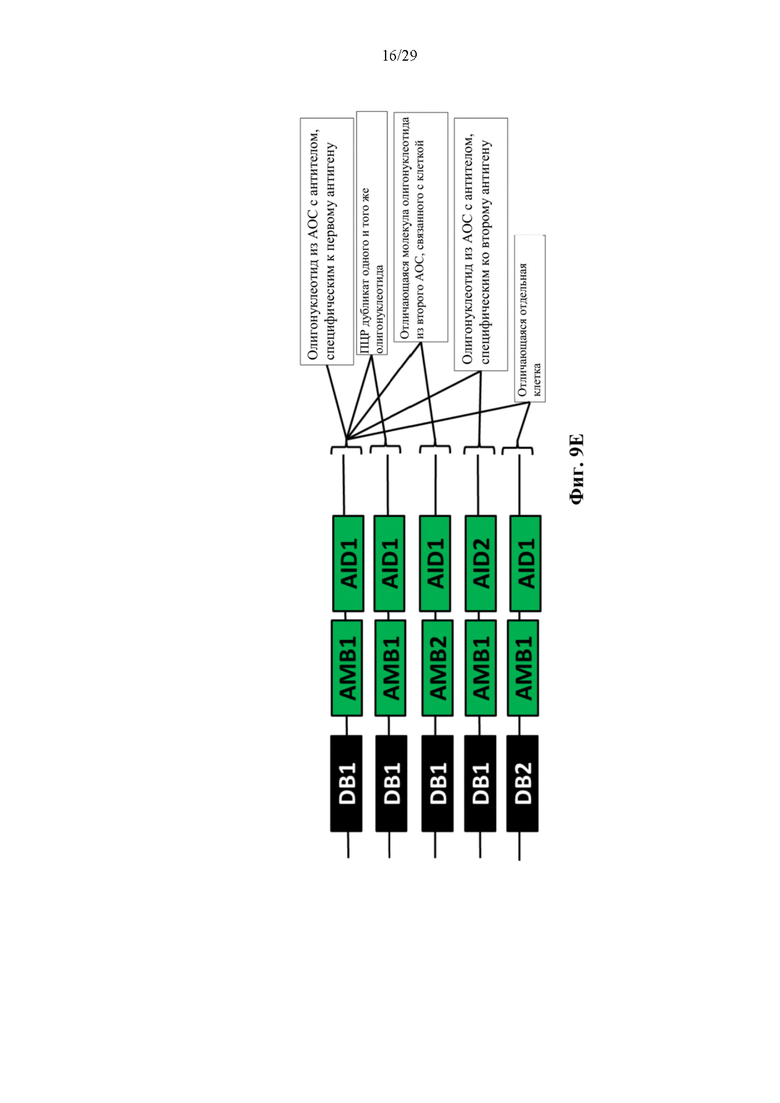
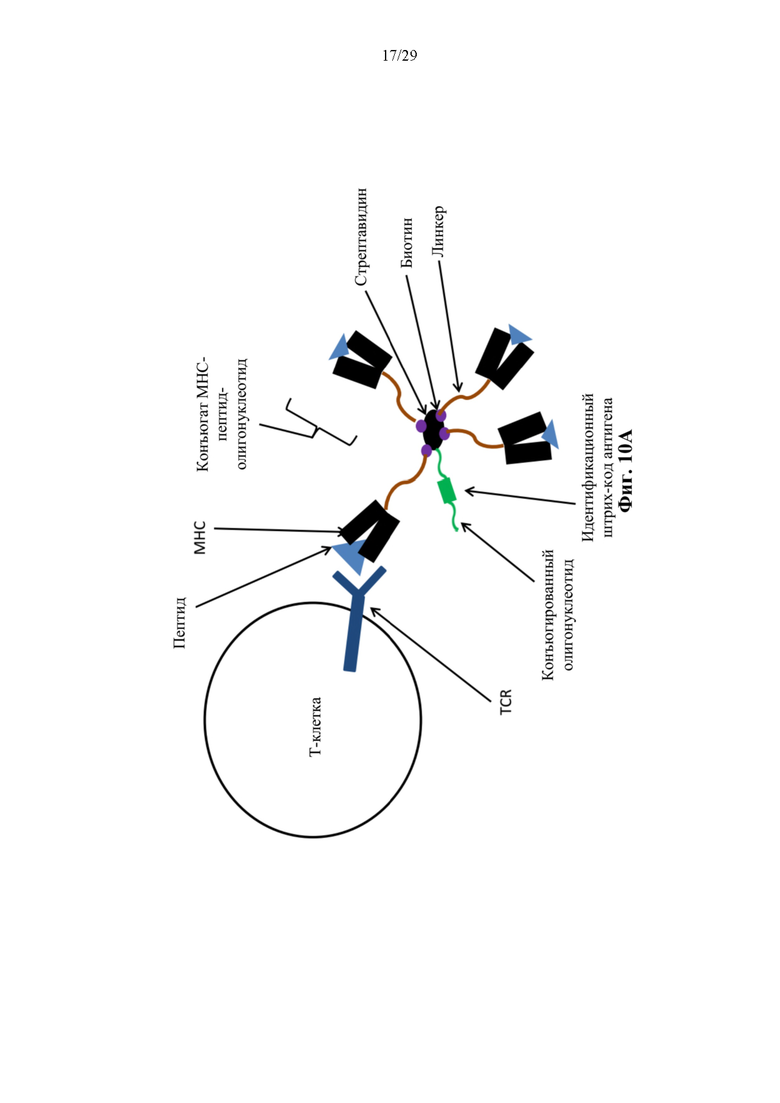
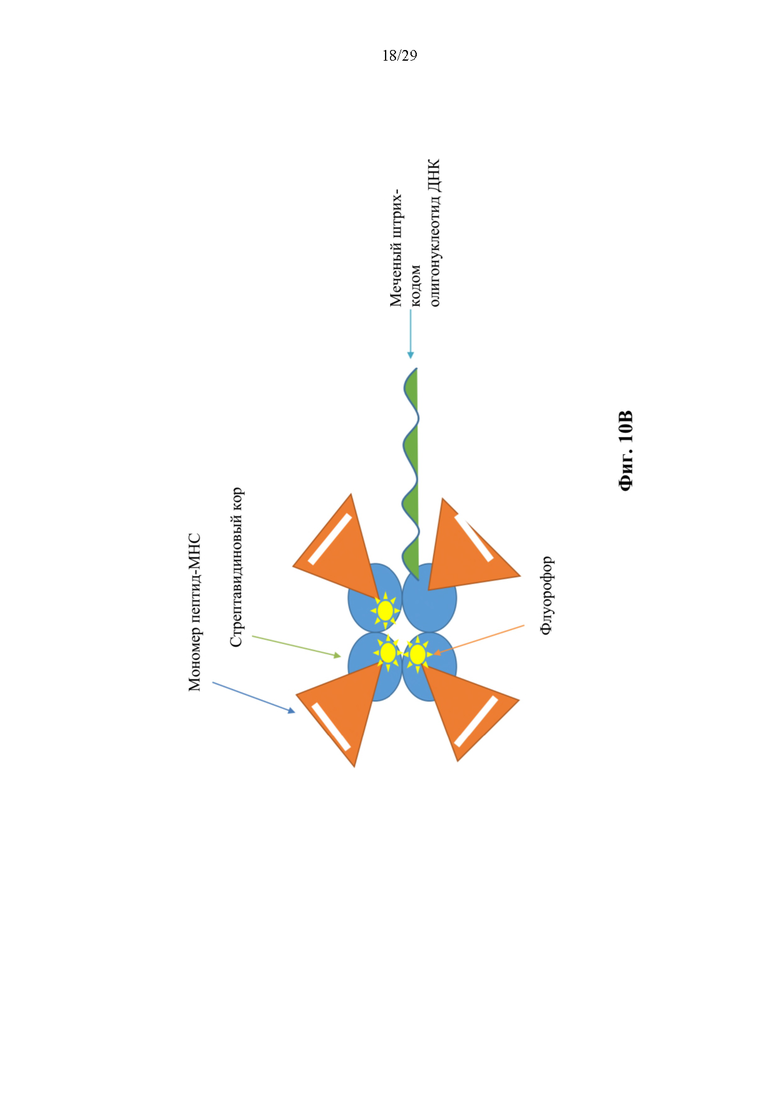
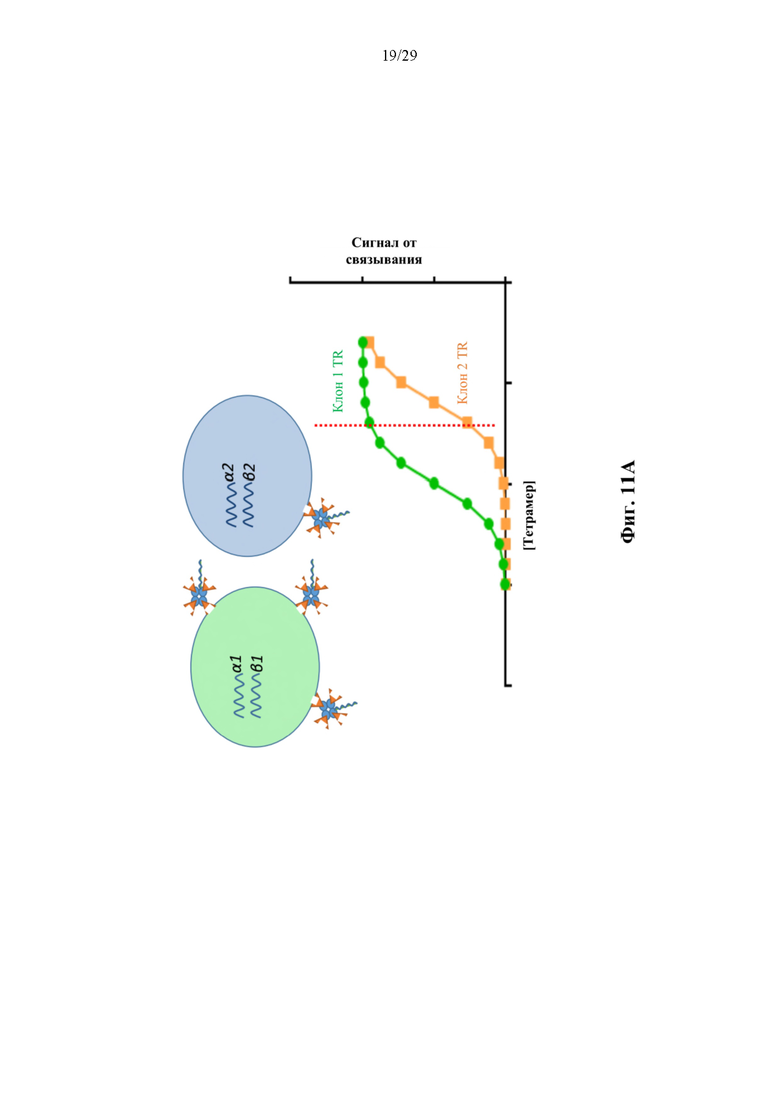

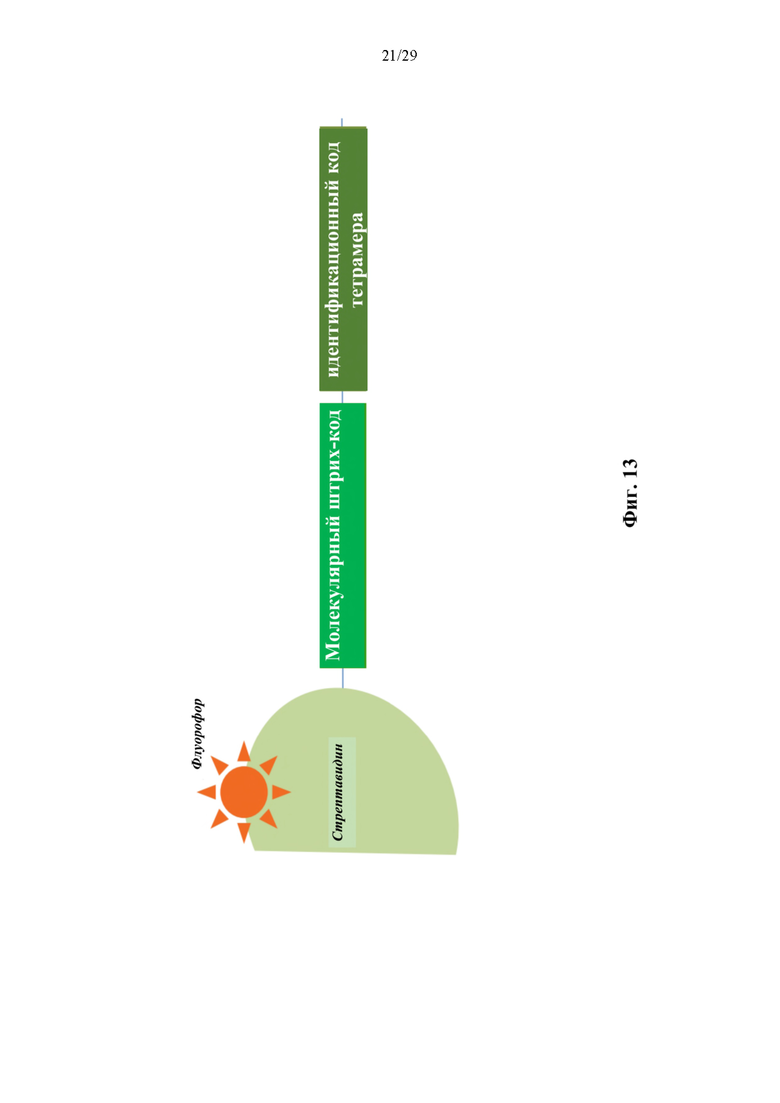

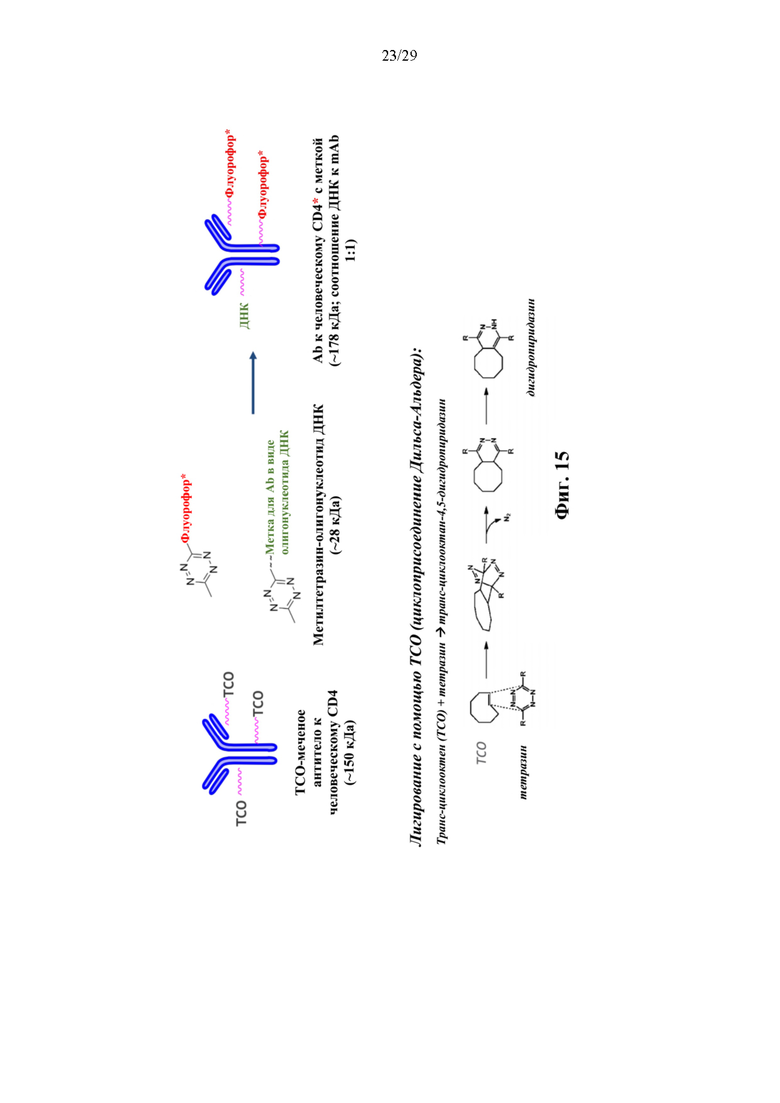
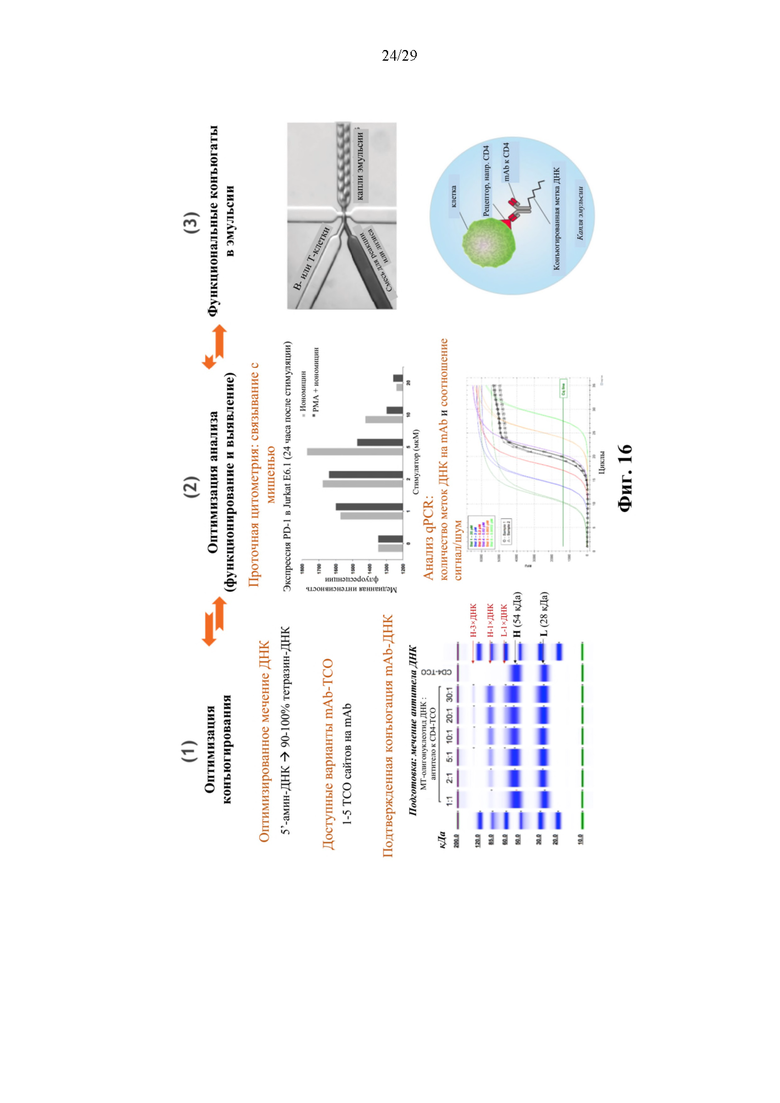
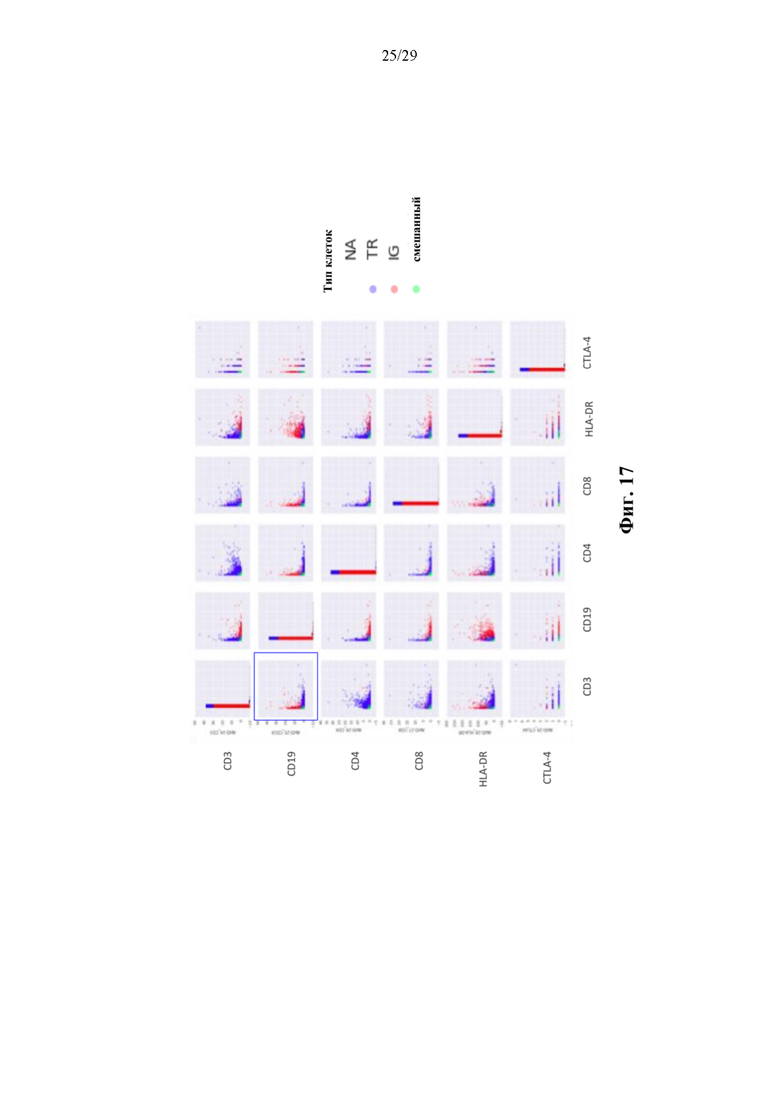
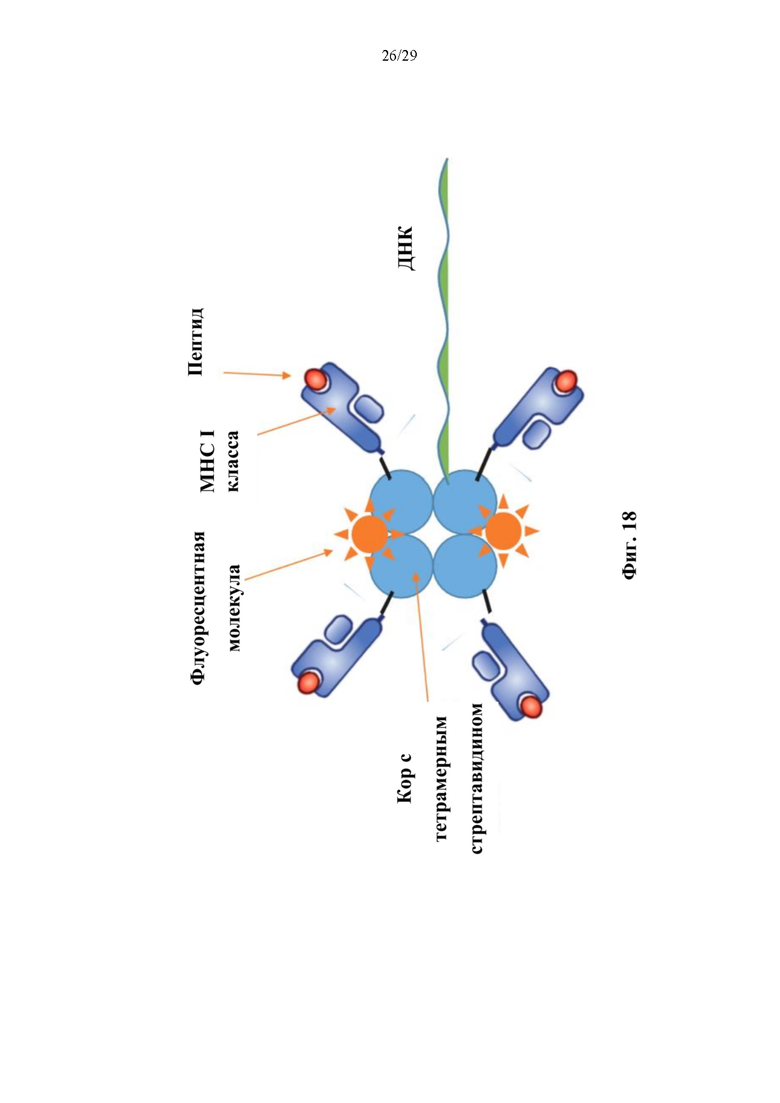
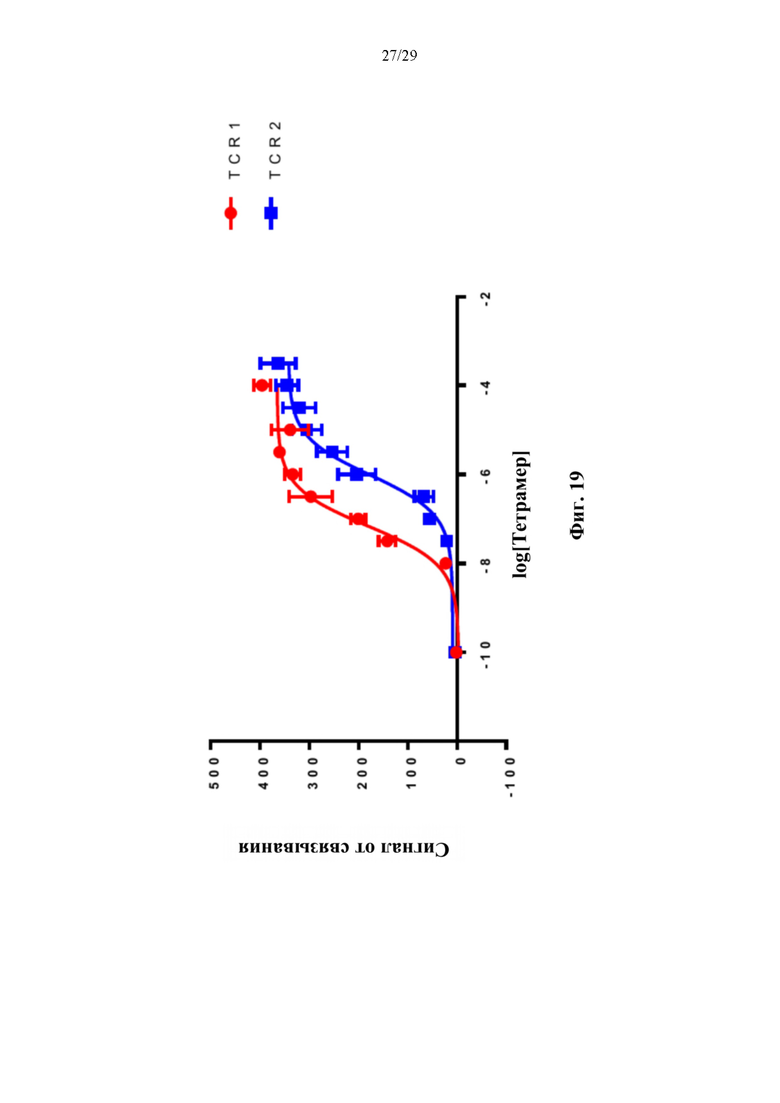
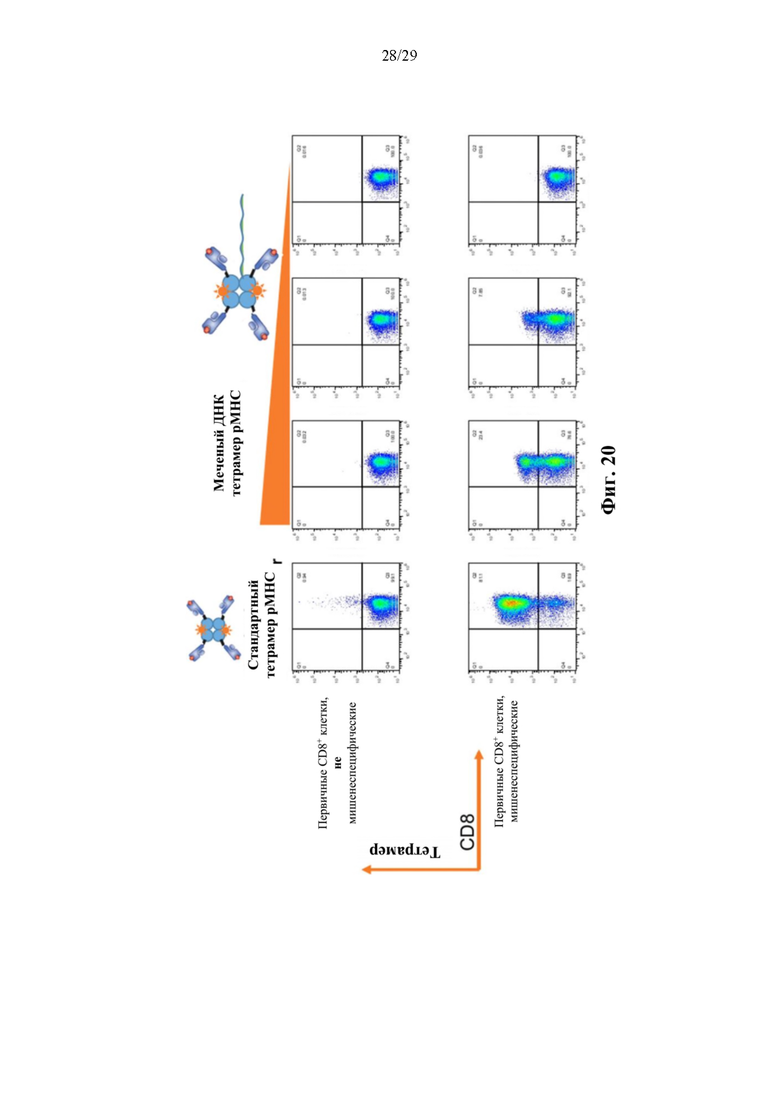
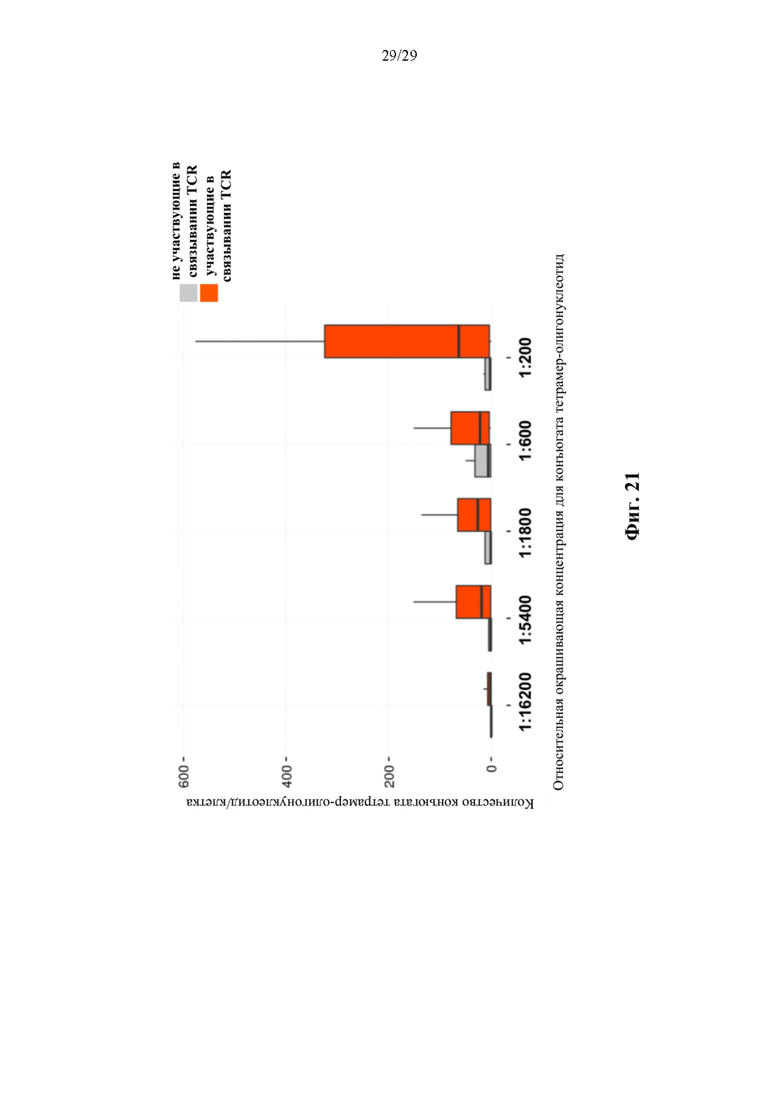
Изобретение относится к области биотехнологии, конкретно к способу выбора T-клеточного рецептора (TCR). Предложенный способ, включающий прикрепление меченого штрихкодом капли полинуклеотида к олигонуклеотидной части молекулы конъюгата pMHC-олигонуклеотид в капле и полинуклеотиду TCR из отдельной T-клетки из множества T-клеток, позволяет идентифицировать отдельные Т-клетки для их дальнейшего секвенирования. 6 н. и 87 з.п. ф-лы, 21 ил., 1 табл., 22 пр.
1. Способ выбора T-клеточного рецептора (TCR), включающий
(а) приведение множества молекул конъюгата pMHC-олигонуклеотид в контакт со множеством T-клеток;
(b) прикрепление меченого штрихкодом капли полинуклеотида или его комплементарной последовательности к
(i) олигонуклеотидной части молекулы конъюгата pMHC-олигонуклеотид в капле из множества капель и
(ii) полинуклеотиду TCR из отдельной T-клетки из множества T-клеток в капле;
(c) секвенирование олигонуклеотидной части или ее комплементарной последовательности и полинуклеотида TCR или его комплементарной последовательности, таким образом получая информацию о последовательности;
и
(d) выбор TCR на основании информации о последовательности.
2. Способ выбора T-клеточного рецептора (TCR), включающий
(a) приведение множества молекул конъюгата pMHC-олигонуклеотид в контакт со множеством T-клеток, при этом молекулы конъюгата pMHC-олигонуклеотид содержат выявляемую часть;
(b) получение обогащенной популяции T-клеток из множества T-клеток из (a) на основании выявления выявляемой части;
(c) выделение отдельной T-клетки, связанной с молекулой конъюгата pMHC-олигонуклеотид, из обогащенной популяции в первую каплю из множества капель;
(d) прикрепление меченого штрихкодом капли полинуклеотида или его комплементарной последовательности к
(i) олигонуклеотидной части молекулы конъюгата pMHC-олигонуклеотид в первой капле и
(ii) полинуклеотиду TCR из отдельной T-клетки в первой капле;
(e) секвенирование олигонуклеотидной части или ее комплементарной последовательности и полинуклеотида TCR или его комплементарной последовательности, таким образом получая информацию о последовательности;
и
(f) выбор TCR на основании информации о последовательности.
3. Способ по п. 2, при этом обогащенная популяция T-клеток содержит соотношение T-клеток, связанных с молекулой конъюгата pMHC-олигонуклеотид, к общему количеству T-клеток, которое является большим, чем соотношение T-клеток, связанных с молекулой конъюгата pMHC-олигонуклеотид, к общему количеству T-клеток во множестве T-клеток после контакта.
4. Способ выбора T-клеточного рецептора (TCR), включающий
(a) приведение первого множества молекул конъюгата pMHC-олигонуклеотид в контакт с первым множеством T-клеток при первом соотношении молекул конъюгата pMHC-олигонуклеотид и клеток;
(b) приведение второго множества молекул конъюгата pMHC-олигонуклеотид в контакт со вторым множеством T-клеток при втором соотношении молекул конъюгата pMHC-олигонуклеотид и клеток, которое является более низким, чем первое соотношение молекул конъюгата pMHC-олигонуклеотид и клеток;
(c) прикрепление первого меченого штрихкодом капли полинуклеотида или его комплементарной последовательности к
(i) олигонуклеотидной части молекулы конъюгата pMHC-олигонуклеотид в первой капле из множества капель и
(ii) первому полинуклеотиду TCR из отдельной T-клетки из первой популяции T-клеток в первой капле;
(d) прикрепление второго меченого штрихкодом капли полинуклеотида или его комплементарной последовательности к
(i) олигонуклеотидной части молекулы конъюгата pMHC-олигонуклеотид во второй капле из множества капель и
(ii) второму полинуклеотиду TCR из отдельной T-клетки из второй популяции T-клеток во второй капле;
(e) секвенирование олигонуклеотидной части молекулы конъюгата pMHC-олигонуклеотид в первой капле или ее комплементарной последовательности и первого полинуклеотида TCR или его комплементарной последовательности, таким образом получая первый набор информации о последовательности;
(f) секвенирование олигонуклеотидной части молекулы конъюгата pMHC-олигонуклеотид во второй капле или ее комплементарной последовательности и второго полинуклеотида TCR или его комплементарной последовательности, таким образом получая второй набор информации о последовательности; и
(g) определение первого сигнала от связывания TCR, кодируемого первым полинуклеотидом TCR, с молекулой конъюгата pMHC-олигонуклеотид на основании первого набора информации о последовательности, и определение второго сигнала от связывания TCR, кодируемого вторым полинуклеотидом TCR, с молекулой конъюгата pMHC-олигонуклеотид на основании второго набора информации о последовательности, и при этом первый полинуклеотид TCR и второй полинуклеотид TCR кодируют один и тот же TCR.
5. Способ по п. 1 или 2, дополнительно включающий определение аффинности связывания TCR, кодируемого полинуклеотидом TCR, с молекулой конъюгата pMHC-олигонуклеотид.
6. Способ по п. 4, дополнительно включающий определение аффинности связывания одного и того же TCR, кодируемого первым полинуклеотидом TCR и вторым полинуклеотидом TCR, на основании первого сигнала от связывания и второго сигнала от связывания.
7. Способ по п. 4, при этом молекулы конъюгата pMHC-олигонуклеотид из первого и второго множества содержат идентичный пептид.
8. Способ по п. 4 или 7, при этом MHC в составе молекул конъюгата pMHC-олигонуклеотид из первого и второго множества относятся к идентичному подтипу.
9. Способ выбора T-клеточного рецептора (TCR), включающий
(a) приведение множества молекул конъюгата pMHC-олигонуклеотид в контакт с множеством T-клеток;
(b) прикрепление
(i) первого меченого штрихкодом капли полинуклеотида или его комплементарной последовательности к олигонуклеотидной части молекулы конъюгата pMHC-олигонуклеотид в первой капле из множества капель, и первому полинуклеотиду TCR из первой отдельной T-клетки из популяции T-клеток в первой капле, и
(ii) второго меченого штрихкодом капли полинуклеотида или его комплементарной последовательности к олигонуклеотидной части молекулы конъюгата pMHC-олигонуклеотид во второй капле из множества капель и полинуклеотиду TCR из второй отдельной T-клетки из популяции T-клеток во второй капле;
(c) секвенирование
(i) олигонуклеотидной части молекулы конъюгата pMHC-олигонуклеотид в первой капле или ее комплементарной последовательности и первого полинуклеотида TCR или его комплементарной последовательности, таким образом получая первый набор информации о последовательности, содержащий риды последовательности штрихкода капли полинуклеотиду TCR и риды последовательности штрихкода капли олигонуклеотида, и
(ii) олигонуклеотидной части молекулы конъюгата pMHC-олигонуклеотид во второй капле или ее комплементарной последовательности и второго полинуклеотида TCR или его комплементарной последовательности, таким образом получая второй набор информации о последовательности, содержащий риды последовательности штрихкода капли полинуклеотида TCR и риды последовательности штрихкода капли олигонуклеотида;
(d) сравнение аффинности связывания TCR, кодируемого первым полинуклеотидом TCR, и аффинности связывания TCR, кодируемого вторым полинуклеотидом TCR, при этом аффинность связывания TCR, кодируемого первым полинуклеотидом TCR, определяют на основании первого набора информации о последовательности, и при этом определение аффинности связывания TCR, кодируемого вторым полинуклеотидом TCR, осуществляют на основании второго набора информации о последовательности; и
(e) выбор TCR, кодируемого первым полинуклеотидом TCR, на основании сравнения, при этом выбранный TCR, кодируемый первым полинуклеотидом TCR, характеризуется более сильной аффинностью связывания, чем TCR, кодируемый вторым полинуклеотидом TCR.
10. Способ по п. 9, при этом первая отдельная T-клетка происходит из пациента с заболеванием или состоянием.
11. Способ по п. 10, при этом вторая отдельная T-клетка происходит из пациента без заболевания или состояния.
12. Способ по любому из пп. 1 или 4-11, при этом молекулы конъюгата pMHC-олигонуклеотид содержат выявляемую часть.
13. Способ по п. 12, дополнительно включающий получение обогащенной популяции T-клеток на основании выявления выявляемой части.
14. Способ по п. 12 или 13, при этом выявляемая часть содержит флуорофор.
15. Способ по любому из пп. 1-14, при этом последовательность штрихкода капли в меченом штрихкодом капли полинуклеотиде или его комплементарной последовательности в первой капле отличается от последовательности штрихкода капли в меченом штрихкодом капли полинуклеотиде или его ампликоне во второй капле из множества капель.
16. Способ по любому из пп. 1-8, при этом информация о последовательности содержит риды последовательности штрихкода капли полинуклеотида TCR и риды последовательности штрихкода капли олигонуклеотида.
17. Способ по любому из пп. 1 или 4-16, при этом способ дополнительно включает после контакта получение обогащенной популяции T-клеток, содержащей более высокий процент T-клеток, связанных с молекулой конъюгата pMHC-олигонуклеотид, чем процент T-клеток, связанных с молекулой конъюгата pMHC-олигонуклеотид, во множестве T-клеток после контакта.
18. Способ по любому из пп. 1-17, при этом способ дополнительно включает определение количества молекул конъюгата pMHC-олигонуклеотид, связанных с отдельной клеткой.
19. Способ по любому из пп. 1-18, при этом олигонуклеотидная часть молекулы конъюгата pMHC-олигонуклеотид содержит последовательность молекулярного штрихкода тетрамера (TMB), которая является уникальной для отдельной молекулы конъюгата pMHC-олигонуклеотид из множества молекул конъюгата pMHC-олигонуклеотид.
20. Способ по п. 19, при этом информация о последовательности содержит риды последовательности молекулярного штрихкода тетрамера.
21. Способ по п. 19 или 20, при этом количество молекул конъюгата pMHC-олигонуклеотид, связанных с отдельной T-клеткой, определяют из информации о последовательности.
22. Способ по любому из пп. 20 или 21, при этом количество молекул конъюгата pMHC-олигонуклеотид, связанных с отдельной T-клеткой, определяют из ридов последовательности молекулярного штрихкода тетрамера.
23. Способ по любому из пп. 19-22, при этом каждая молекула конъюгата pMHC-олигонуклеотид из множества молекул конъюгата pMHC-олигонуклеотид содержит уникальную последовательность TMB.
24. Способ по любому из пп. 1-23, при этом конъюгат pMHC-олигонуклеотид содержит мультимер главного комплекса гистосовместимости (MHC), где, необязательно, мультимер представляет собой тетрамер.
25. Способ по п. 24, при этом MHC присутствует в растворимой форме.
26. Способ по п. 24, при этом pMHC часть молекулы конъюгата pMHC-олигонуклеотид содержит синтетический пептид.
27. Способ по п. 24, при этом pMHC часть молекулы конъюгата pMHC-олигонуклеотид связывается с TCR отдельной T-клетки.
28. Способ по любому из пп. 1-27, при этом олигонуклеотидная часть содержит идентификационную последовательность тетрамера (TID).
29. Способ по п. 28, при этом TID выступает в качестве штрихкода для пептида или pMHC части молекулы конъюгата pMHC-олигонуклеотид.
30. Способ по любому из пп. 1-29, при этом меченый штрихкодом капли полинуклеотид происходит из матричного меченого штрихкодом капли полинуклеотида в капле.
31. Способ по любому из пп. 1-30, при этом способ дополнительно включает определение характеристики отдельной T-клетки в капле на основании информации о последовательности.
32. Способ по п. 31, при этом информация о последовательности содержит идентификационную последовательность тетрамера (TID) или ее комплементарную последовательность и/или содержит количество копий молекул, имеющих последовательность TID.
33. Способ по любому из пп. 1-32, при этом способ дополнительно включает отмывку множества T-клеток после контакта.
34. Способ по любому из пп. 1 или 4-33, при этом способ дополнительно включает выделение отдельной T-клетки в капле.
35. Способ по п. 34, при этом отдельная T-клетка связывается с молекулой конъюгата pMHC-олигонуклеотид до выделения.
36. Способ по любому из пп. 1-35, при этом способ дополнительно включает лизис отдельной T-клетки.
37. Способ по п. 36, при этом лизис осуществляют после выделения отдельной T-клетки в капле.
38. Способ по любому из пп. 1-37, при этом множество T-клеток представляет собой множество несортированных T-клеток.
39. Способ по любому из пп. 1-38, при этом последовательность штрихкода капли в каждом меченом штрихкодом капли полинуклеотиде или его комплементарной последовательности в отдельной капле из множества капель содержит одинаковую последовательность штрихкода капли.
40. Способ по любому из пп. 1-39, при этом последовательность штрихкода капли в каждом меченом штрихкодом капли полинуклеотиде или его комплементарной последовательности в любой отдельной капле из множества капель является уникальной для капли.
41. Способ по любому из пп. 1-40, при этом прикрепление меченого штрихкодом капли полинуклеотида к олигонуклеотидной части и прикрепление меченого штрихкодом капли полинуклеотида к полинуклеотиду TCR из отдельной T-клетки осуществляют одновременно.
42. Способ по любому из пп. 1-41, при этом способ дополнительно включает амплификацию олигонуклеотидной части или ее комплементарной последовательности.
43. Способ по любому из пп. 1-42, при этом способ дополнительно включает амплификацию клеточного полинуклеотида TCR или его комплементарной последовательности.
44. Способ по п. 43, при этом амплификацию олигонуклеотидной части или ее комплементарной последовательности и амплификацию полинуклеотида TCR или его комплементарной последовательности осуществляют одновременно.
45. Способ по любому из пп. 41-44, при этом последовательность штрихкода капли меченого штрихкодом капли полинуклеотида, присоединенного к полинуклеотиду TCR, и последовательность штрихкода капли меченого штрихкодом капли полинуклеотида, присоединенного к олигонуклеотидной части, является одинаковой.
46. Способ по любому из пп. 1-45, при этом способ дополнительно включает объединение олигонуклеотидов, их амплифицированных продуктов или их комплементарных последовательностей из двух или более капель из множества капель.
47. Способ по п. 46, при этом способ дополнительно включает объединение олигонуклеотидов, их амплифицированных продуктов или их комплементарных последовательностей и полинуклеотидов TCR, их амплифицированных продуктов или их комплементарных последовательностей из двух или более капель из множества капель.
48. Способ по п. 46 или 47, при этом объединение осуществляют до секвенирования.
49. Способ по любому из пп. 1-48, при этом молекула конъюгата pMHC-олигонуклеотид содержит множество отличающихся молекул конъюгата pMHC-олигонуклеотид.
50. Способ по п. 49, при этом каждая молекула конъюгата pMHC-олигонуклеотид из множества молекул конъюгата pMHC-олигонуклеотид содержит уникальную идентификационную последовательность тетрамера (TID).
51. Способ по любому из пп. 1-50, при этом олигонуклеотидная часть содержит последовательность для слияния, и прикрепление включает прикрепление меченого штрихкодом капли полинуклеотида к последовательности для слияния.
52. Способ по любому из пп. 1-51, при этом олигонуклеотидная часть содержит последовательность для связывания праймера.
53. Способ по любому из пп. 1-52, при этом олигонуклеотидная часть содержит константную последовательность.
54. Способ по любому из пп. 1-53, при этом способ дополнительно включает анализ последовательностей штрихкода капли в ридах последовательности олигонуклеотида.
55. Способ по любому из пп. 1-54, при этом способ дополнительно включает анализ идентификационных последовательностей тетрамера (TID) в ридах последовательности олигонуклеотида.
56. Способ по любому из пп. 1-55, при этом способ дополнительно включает анализ последовательностей молекулярного штрихкода тетрамера (TMB) в ридах последовательности олигонуклеотида.
57. Способ по п. 56, при этом анализ включает определение частоты одной или более последовательностей штрихкода капли, одной или более последовательностей TID, одной или более последовательностей молекулярного штрихкода тетрамера (TMB) или их комбинации.
58. Способ по п. 56 или 57, при этом анализ включает сравнение.
59. Способ по любому из пп. 54-58, при этом способ дополнительно включает сравнение идентификационных последовательностей тетрамера (TID) в ридах последовательности олигонуклеотида с последовательностями молекулярного штрихкода тетрамера (TMB) в ридах последовательности олигонуклеотида.
60. Способ по любому из пп. 41-59, при этом способ дополнительно включает секвенирование полинуклеотида TCR, его комплементарных последовательностей, его амплифицированных продуктов или их комбинации, таким образом получая риды последовательности полинуклеотида TCR.
61. Способ по п. 60, при этом способ дополнительно включает сравнение ридов последовательности олигонуклеотида с ридами последовательности полинуклеотида TCR.
62. Способ по п. 60 или 61, при этом способ дополнительно включает сравнение последовательностей штрихкода капли в ридах последовательности олигонуклеотида с последовательностями штрихкода капли в ридах последовательности полинуклеотида TCR.
63. Способ по любому из пп. 60-62, при этом способ дополнительно включает сравнение ридов последовательности полинуклеотида TCR.
64. Способ по любому из пп. 60-63, при этом способ дополнительно включает анализ последовательностей штрихкода капли в ридах последовательности полинуклеотида TCR.
65. Способ по любому из пп. 60-64, при этом способ дополнительно включает анализ последовательностей молекулярного штрихкода в ридах последовательности полинуклеотида TCR.
66. Способ по любому из пп. 61-65, при этом способ дополнительно включает определение характеристики клетки на основании анализа или сравнения.
67. Способ по любому из пп. 54-66, при этом способ дополнительно включает выбор TCR на основании ридов последовательности олигонуклеотида.
68. Способ по любому из пп. 60-67, при этом способ дополнительно включает выбор TCR на основании ридов последовательности полинуклеотида TCR.
69. Способ по любому из пп. 1-68, при этом меченый штрихкодом капли полинуклеотид, прикрепленный к олигонуклеотиду, и меченый штрихкодом капли полинуклеотид, прикрепленный к полинуклеотиду TCR, происходят из одного и того же матричного меченого штрихкодом капли полинуклеотида в капле.
70. Способ по любому из пп. 1-69, при этом меченый штрихкодом капли полинуклеотид, прикрепленный к олигонуклеотиду, представляет собой продукт амплификации матричного меченого штрихкодом капли полинуклеотида.
71. Способ по п. 69 или 70, при этом меченый штрихкодом капли полинуклеотид, прикрепленный к полинуклеотиду TCR, представляет собой продукт амплификации матричного меченого штрихкодом капли полинуклеотида.
72. Способ по любому из пп. 1-71, при этом капля содержит твердую подложку.
73. Способ по любому из пп. 1-71, при этом капля не содержит твердую подложку.
74. Способ по любому из пп. 1-73, при этом каждая капля из множества капель содержит отдельную клетку.
75. Способ по любому из пп. 69-71, при этом матричный меченый штрихкодом капли полинуклеотид не связан с твердой подложкой.
76. Способ по любому из пп. 69-71, при этом матричный меченый штрихкодом капли полинуклеотид связан с твердой подложкой.
77. Способ по любому из пп. 1-76, при этом способ дополнительно включает прикрепление последовательности молекулярного штрихкода в меченом молекулярным штрихкодом полинуклеотиде из множества меченых молекулярным штрихкодом полинуклеотидов к полинуклеотиду TCR, при этом последовательность молекулярного штрихкода выступает в качестве штрихкода для отдельной молекулы полинуклеотида TCR и ее амплифицированных продуктов.
78. Способ по любому из пп. 1-77, при этом прикрепление включает лигирование меченого штрихкодом капли полинуклеотида или его комплементарной последовательности с олигонуклеотидной частью.
79. Способ по любому из пп. 1-78, при этом прикрепление включает прикрепление меченого штрихкодом капли полинуклеотида или его комплементарной последовательности к олигонуклеотидной части с помощью фермента.
80. Способ по любому из пп. 1-77, при этом прикрепление включает гибридизацию меченого штрихкодом капли полинуклеотида или его комплементарной последовательности с олигонуклеотидной частью.
81. Способ по п. 80, при этом прикрепление дополнительно включает достройку олигонуклеотида.
82. Способ по любому из пп. 1-81, при этом прикрепление включает амплификацию матричного меченого штрихкодом капли полинуклеотида.
83. Способ по любому из пп. 1-82, при этом олигонуклеотидная часть является двухцепочечной.
84. Способ по любому из пп. 1-82, при этом олигонуклеотидная часть является одноцепочечной.
85. Способ по любому из пп. 1-84, при этом олигонуклеотидная часть представляет собой ДНК.
86. Способ по любому из пп. 1-84, при этом олигонуклеотидная часть представляет собой РНК.
87. Способ по любому из пп. 1-86, при этом полинуклеотид TCR содержит последовательность вариабельного участка.
88. Способ по любому из пп. 1-87, при этом способ дополнительно включает спаривание последовательностей нативных цепей TCR, содержащих последовательность вариабельного участка.
89. Способ по любому из пп. 1-88, при этом полинуклеотид TCR представляет собой ДНК.
90. Способ по любому из пп. 1-88, при этом полинуклеотид TCR представляет собой РНК.
91. Способ по п. 90, при этом РНК представляет собой мРНК.
92. Эмульсия для выбора T-клеточного рецептора (TCR), причем эмульсия содержит множество капель, при этом по меньшей мере одна капля из множества капель содержит
(a) отдельную T-клетку из образца, содержащего множество T-клеток; и
(b) меченый штрихкодом капли полинуклеотид, содержащий последовательность штрихкода капли;
при этом капля дополнительно содержит молекулу конъюгата pMHC-олигонуклеотид, которая связывается с TCR отдельной T-клетки.
93. Эмульсия для выбора T-клеточного рецептора (TCR), причем эмульсия содержит множество капель, при этом по меньшей мере одна капля из множества капель содержит
(a) отдельную лизированную T-клетку из образца, содержащего множество T-клеток; и
(b) молекулу конъюгата pMHC-олигонуклеотид, содержащую pMHC часть, которая связывается с TCR отдельной лизированной T-клетки;
при этом олигонуклеотидная часть конъюгата pMHC-олигонуклеотид содержит последовательность штрихкода капли, и при этом полинуклеотид TCR из отдельной лизированной T-клетки содержит одинаковую последовательность штрихкода капли.
| BENTZEN A | |||
| K | |||
| et al | |||
| Large-scale detection of antigen-specific T cells using peptide-MHC-I multimers labeled with DNA barcodes, Nature biotechnology, 2016, V | |||
| Нивелир для отсчетов без перемещения наблюдателя при нивелировании из средины | 1921 |
|
SU34A1 |
| Печь-кухня, могущая работать, как самостоятельно, так и в комбинации с разного рода нагревательными приборами | 1921 |
|
SU10A1 |
| WO 2015185067 A1, 10.12.2015 | |||
| WO 2014145047 A1, 18.09.2014 | |||
| HAN A | |||
| et al | |||
| Linking T-cell receptor sequence to functional phenotype at the single-cell level, Nature | |||

