Область техники
Изобретение относится к области генной инженерии и предназначено для лечения широкого спектра злокачественных новообразований
В генной терапии действующим компонентом является генетическая конструкция, которая обеспечивает экспрессию одного или нескольких биологически активных веществ (БАВ), индуцирующих гибель опухолевых клеток. Генетические конструкции, содержащие так называемые «гены-убийцы» (от англ. suicide genes), используются для лечения опухолей в сочетании с пролекарствами. Данный подход получил название ген-направленной энзиматической пролекарственной терапии или ГНЭПТ (от англ. Gene-Directed Enzyme-Prodrug Therapy), (Connors ТА. Gene Ther. 1995. 2(10):702-9). Механизм действия ГНЭПТ заключается во введении в опухолевые клетки «гена-убийцы», который кодирует целевой фермент, катализирующий превращение малотоксичного пролекарства в токсичный для опухолевых клеток агент. При этом, агент действует не только на те клетки, в которых произошло превращение, но также проникает по градиенту концентрации в соседние клетки. Этот феномен носит название «эффекта свидетеля» (от англ. bystander effect), что значительно увеличивает эффективность ГНЭПТ (Kuriyama S., et al. Hepatology. 1995. 22(6): 1838-46). Этот феномен может объяснить то, что трансдукция генетической конструкцией менее 10% клеток может оказаться достаточной для уничтожения всей популяции опухолевых клеток (Huber BE., et al. Proc Natl Acad Sci USA. 1994. 91(17): 8302-6). Следует добавить, что высокий уровень локальной экспрессии «гена-убийцы» в опухолевых клетках позволяет существенно повысить специфичность терапевтического воздействия и снизить токсичность БАВ в нормальных тканях.
Уровень техники
Система ГНЭПТ на основе генетической конструкции, кодирующей цитозиндезаминазу (CD), и пролекарства 5-фторцитозин (5-FC) является перспективной метаболической парой целевой фермент / пролекарство. Цитозиндезаминаза катализирует превращение 5-FC в 5-фторурацил (5-FU), цитостатический эффект которого достигается тремя разными путями: 1) одно из производных 5-FU, 5-фтордезоксиуридин-5-монофосфат, необратимо ингибирует тимидилатсинтазу, что приводит к торможению синтеза ДНК;
2) другие производные 5-FU, 5-фторуридин-5-трифосфат (5-FUTO) и 5-фтордезоксиуридин-5-трифосфат, встраиваются в нуклеотидную последовательность при репликации ДНК, что останавливает синтез ДНК; 3) 5-фторуридин-трифосфат предотвращает процессинг РНК и индуцирует апоптоз (Twitty CG., et al. Hum Gene Ther Methods. 2016. 27(1): 17-31). Таким образом, система ГНЭПТ на основе CD/5-FC действует не только на делящиеся, но и на непролиферирующие клетки.
Доклинические и клинические исследования генотерапевтических препаратов на основе гена цитозиндеаминазы и генов иммунного ответа
Препарат MVA-FCU1 представляет собой модифицированный вирус осповакцины Анкара (MVA), несущий химерный ген FCU1, с системным введением 5-фторцитозина, который хорошо переносит и обладает противоопухолевым действием: у восьми из 16 пациентов наблюдалась стабилизация опухоли первичных или вторичных опухолей печени (Erbs P., et al. Cancer Gene Ther, 2008. 15(1): 18-28; Husseini F., et al. Ann Oncol. 2017. 28(1): 169-174). Препарат Toca 511 на основе гаммаретровируса, кодирующий ген CD, стабильно реплицируется в ксенотрансплантатах опухолей человека и после введения 5-фторцитозина вызывает полную регрессию таких ксенотрансплантатов (Perez O.D., et al. Mol Ther. 2012. 20(9): 1689-98). В КИ NCT01470794 фазы III наблюдали многолетние стойкие ответы у пациентов с глиобластомой на фоне 5-фторцитозина (Cloughesy T.F., et al. Neuro Oncol. 2018. 20(10): 1383-1392). Лечение плазмидной конструкцией на основе аденовирусного вектора, кодирующей ген CD, в присутствии пролекарства приводит к полной регрессии плевральной эффузии у пациентов (Zarogoulidis P., et al. Cancer Gene Ther. 2012. 19(9): 593-600).
ГНЭПТ системы на основе невирусных векторов можно дополнить генами-активаторами противоопухолевого иммунного ответа. Так, экспрессия генов CD и IL-12 в присутствии 5-фторцитозина позволяет существенно увеличить торможение роста опухолей у мышей (Hwang K.S., et al. ВМС Cancer. 2005. 5: 51). Воздействие на этап иммунного цикла можно осуществлять посредством «сигналов опасности», которые представляют собой молекулы / макромолекулекулы, ассоциированные с повреждениями (от англ. damage-associated molecular patterns, DAMPs). Они выделяются поврежденными клетками в ответ на воздействие патогенов, токсинов, механических повреждений и т.д. и активируют иммунный ответ через стимуляцию антиген-представляющих клеток. Активацию «сигналов опасности» можно запустить также посредством генной терапии, что было продемонстрировано в работах Koyama, Wang и др. (Koyata Y., et al. Biotechnol Lett. 2016. 38(11): 1857-1866; Wang W., et al. Oncotarget. 2014. 5(1): 150-160; Li, J.L., et al. Gene Ther. 2009. 16(3): 376-382; Liu Z., et al. Hum Gene Ther. 2017. 28(8): 667-680).
В качестве «сигнала опасности» используют структурный компонент жгутиков бактерий - флагеллин - и белок внеклеточного матрикса - декорин. Флагеллин является мощным активатором системы врожденного иммунитета благодаря специфического рецептора TLR5, который находится на поверхности клеток иммунной системы. Этот рецептор относится к системе Толл-подобных рецепторов (Toll-Like Receptor, TLR) (Hayashi F., et al. Nature. 2001. 410(6832): 1099-1103). Geng и соавторы (Geng D et al., Cancer Res. 2015; 75(10), 1959-1971) получили модифицированные Т-клетки, секретирующие флагеллин. Введение таких клеток мышам с меланомой обеспечивало высокий противоопухолевый эффект по сравнению с контролем. Gudkov AV и соавторы (Yang Н., Oncotarget. 2016. 7(3), 2936-50) разработали рекомбинантную форму флагеллина Salmonella enterica (Entholimod), свойства которой изучались в клиническом исследовании с привлечением 40 участников (https://clinicaltrials.gov/ct2/show/NCT02715882; Burdelya L.G., et al. Science. 2008. 320(5873), 226-30).
Техническая проблема
Множество факторов влияют на эффективность и безопасность генной терапии на основе ГНЭПТ как инновационного способа лечения онкологических заболеваний. К ним можно отнести способы введения генотерапевтического препарата, продолжительность курса лечения, тип вектора и способ его адресной доставки, комбинацию терапевтических генов в составе плазмидной конструкции, тип клеток или тканей-мишеней. В двухкомпонентной системе фермент/пролекарство фермент должен иметь более высокое сродство к пролекарству, чем к его натуральному субстрату, иначе для достижения требуемого терапевтического эффекта может потребоваться гораздо более высокая концентрация пролекарства, что увеличит риск связанных с приемом пролекарства нежелательных явлений. С другой стороны, важными эндогенными факторами, влияющими на безопасность генной терапии, являются стабильность экспрессируемого in vivo «гена-убийцы» и иммуногенность. В физиологических условиях пролекарство должно также быть стабильным и отличаться низкой токсичностью до активации ферментом, но проявлять высокую токсичность для раковых клеток после активации в качестве лекарства. Более того, такое лекарство должно обладать высоким локальным и генерализованным «эффектом свидетеля», чтобы преодолеть недостатки, ассоциированные с векторами с низким уровнем трансфекции клеток. В этом контексте выбор типа вектора также является критичным для разработки эффективной ГНЭПТ плазмидной конструкции. Как правило, вирусные векторы более эффективны в трансфекции клеток, однако их использование сопряжено с вероятностью неспецифической интеграции в геном, возникновения генотоксичности и вторичной канцерогенности. В клинических испытаниях чаще всего используются аденовирусные векторы, а также ретровирусные и лентивирусные векторы, терапевтическая эффективность которых ограничена из-за их неспособности распространяться и трансфицировать опухолевые клетки, непосредственно активируя иммунный ответ. Поэтому выбор невирусных и не интегрирующихся в геном векторов является более целесообразным для дизайна плазмидной конструкции с точки зрения снижения вышеуказанных рисков, присущих вирусным векторам. Другой проблемой хорошо трансфицирующих клетки как вирусных, так и невирусных векторов является их побочная экспрессия в не-опухолевых клетках, приводящая не только к созданию избыточной метаболической перегрузки трансфицированных клеток, но и к возрастанию рисков нежелательных явлений. Поэтому ее решение реализуется путем включения в состав вектора ткане- или опухоль-специфичного промотора или рационализации способа введения генотерапевтического препарата и пролекарства (например, внутриопухолевое введение препарата и системное введение в организм малотоксичного пролекарства). Поскольку существуют объективные ограничения эффективности экзогенного вектора в клеточной системе вследствие его уязвимости к клеточным ДНКзам, целесообразно «упаковывать» плазмидную конструкцию в полимерные наноносители, например, самособирающиеся наночастицы, имеющие высокий тропизм к опухоль-специфичным поверхностным рецепторам. Наконец, высокая гетерогенность популяции как опухолевых клеток внутри одной опухоли индивидуума, так и клеток того же типа опухоли у разных индивидуумов также детерминируют адаптацию композиции генотерапевтического препарата и его нацеливание на клеточную субпопуляцию, которая в максимальной степени может быть ответственна за прогрессию опухоли, резистентность к химиотерапии/радиотерапии, метастазирование и рецидив рака. Например, устойчивые к определенному типу воздействий стволовые опухолевые клетки, контролирующие процессы самообновления и дифференцировки опухолевой массы, гиперэкспрессируют на своей поверхности специфические маркеры (CD24, CD44, CD133) и альдегиддегидрогеназу (ALDH) (Ysar SB & Jimeno A. Mol Cancer Ther. 2010. 9(9), 2450-2457). С другой стороны, возникновение резистентных клеточных субпопуляций опухоли к цитотоксическому агенту, происходящему из системно вводимого пролекарства, а также данные об умеренной эффективности ГНЭПТ плазмидных конструкций (Cloughesy et al., Neuro Oncol. 2018. 20(10), 1383-1392; Freytag et al., Int J Radiat Oncol Biol Phys. 2014. 89(2), 268-276; Husseini F., et al., Ann Oncol. 2017. 28(1), 169-174) указывают на необходимость создания лекарств двойного действия путем введения дополнительных «генов-убийц» в состав вектора (Hu W & Liu W. Vitro Cell Dev Biol Anim. 2010. 46(6), 497-501), либо введения генов, ассоциированных с апоптозом, либо генов-активаторов иммунного ответа, не требующих использования пролекарств, что было бы хорошим вариантом решения этой проблемы и повышения терапевтической эффективности ГНЭПТ, а также расширения спектра ЗНО для генной терапии.
Сведения о близких аналогах FCU1-BsFm/PP
Изобретение, раскрытое в патенте WO2015080631 (RU2014000893), представляет собой бицистронную плазмидную конструкцию на основе аденовирусного вектора, содержащую ген TLR5 (toll-подобный рецептор 5) и ген, кодирующий агонист TLR5-негликозилированную форму флагеллина. Изобретение призвано обеспечить специфическое уничтожение раковых клеток путем NF-κВ-опосредованного апоптоза. К важным отличиям аналога от FCU1-BsFm/PP относятся отсутствие в плазмидной ДНК элементов системы ГНЭПТ, наличие последовательности промоторной области гена, кодирующего убиквитин С, а также существенная разница нуклеотидных последовательностей гена флагеллина. Недостатками данного изобретения являются повышенная вероятность интеграции аденовирусного вектора в геном и отсутствие способности оказывать прямое цитотоксическое действие на опухолевые клетки, вследствие чего противоопухолевый эффект in vivo может быть недостаточным для достижения существенной регрессии опухоли.
В патенте RU2539764C2 (от 27.01.2015) (с.8, строки 41-48, фигура 5, пример 7) раскрывается экспрессионная конструкция PhTSurv269-FCU1-mGM-CSF-pGL3, включающая: PhTERT - фрагмент промоторного региона гена обратной транскриптазы теломеразы человека (hTERT); PhSurv269 - фрагмент промоторного региона гена сурвивина человека; FCU1 - гибридный ген цитозиндезаминазы/урацилфосфорибозилтрансферазы Saccharomyces cerevisiae; IRES - участок посадки рибосомы вируса энцефаломиокардита человека; mGMCSF - ген гранулоцитарно-макрофагального колониестимулирующего фактора мыши; SV40 late poly(A) - сигнал полиаденилирования вируса SV40; Amp(r) - ген р-лактамазы. Отличиями объекта техники от аналога являются: наличие в нем гена, кодирующего секретируемый мутантный флагеллин Bacillus subtitilis; использование в качестве носителя блок-сополимера на основе ПЭГ-ПЭИ-ТАТ, нековалентно связывающего терапевтическую плазмидную ДНК.
В публикации (Безбородова О.А. и др. Противоопухолевая эффективность комбинированной генной и лучевой терапии у животных. Доклады Академии Наук, 2016, т. 470, №4, с. 471-474) описаны результаты исследования противоопухолевого действия комбинации генной терапии с использованием конструкции CMV-FCU1-IRESmGM-CSF-pGL3 и 5-фторцитозина (5-FC) в комплексе с блок-сополимером ПЭГ-ПЭИ-ТАТ (FCU1-mGM/5-FC) и лучевой терапии (однократное облучение опухоли в дозе 5 Гр) на мышиной модели аденокарциномы толстой кишки СТ26.
В патенте RU2476596C1 (от 27.02.2013) (п.2-12, 15-20) раскрывается формула генотерапевтического лекарственного средства, включающего плазмидную конструкцию на основе pGL3-pCMV-FCU1, несущую ген рекомбинантной цитозиндезаминазы под контролем сильного неспецифичного промотора цитомегаловируса, или конструкцию pG13-pBIRC5-1.5-FCU1, содержащую ген цитозиндезаминазы под контролем опухоль-специфичного промотора гена сурвивина человека, которые вводят в сочетании с 5-фторцитозином, что приводит к гибели раковых клеток (пример 12, фигура 8). Плазмидные конструкции вводятся внутриопухолево в составе полиплексов с использованием блок-сополимера ПЭИ-ПЭГ-ТАТ (см. пример 14, пп. 5-12 Формулы изобретения). Отличиями объекта техники от аналога являются наличие в нем гена, кодирующего секретируемый мутантный флагеллин Bacillus subtitilis.
В формуле аналогичного изобретения, защищенного патентом RU2580220C2, есть три признака сходства с FCU1-BsFm/PP. Так, по п. 1 аналог представляет собой «рекомбинантную плазмиду pCpG-CytDA/upp размером 4981 н.п., обеспечивающую экспрессию гибридного белка цитозиндезаминаза Е. coli - урацилфосфорибозил трансферазу…»; по п. 2 - «рекомбинантную плазмиду pCpG-CytDA/upp/VP22 размером 5876 н.п., обеспечивающую экспрессию гибридного белка цитозиндезаминазы Е. coli -урацилфосфорибозил трансфераза-белка VP22 герпесвируса человека…»; по п. 3 - «способ осуществления терапии группы онкологических заболеваний, представляющих собой солидные опухоли, путем введения композиции… в организм пациента интратуморально, после чего также введения 5-фторцитозина в терапевтически эффективных количествах». Тем не менее, различие заключается в источнике происхождения нуклеотидных последовательностей химерного гена FCU1, включенного в плазмидную конструкцию: в аналоге (патент RU2580220C2) это Е. coli, тогда как FCU1 в FCU1-BsFm/PP - это ген дрожжей. Еще одно различие - использование в сравниваемых плазмидных конструкциях разных векторов. Недостатком аналога является отсутствие данных о терапевтической активности (помимо мышей с перевитой меланомой), а также данных о продолжительности экспрессии целевых генов плазмидной конструкции in vivo. Кроме того, можно констатировать очень слабое доказательство противоопухолевой эффективности pCpG-CytDA/upp/VP22 на животной модели меланомы В16, которая является низкочувствительной к системе FCUl/5-фторцитозин, если считать, что терапевтический эффект является значимым при ТРО≥75%, и УПЖ≥35%. Исследование противоопухолевой активности FCU1-BsFm/PP показало, что даже добавление в плазмидную конструкцию «сигнала опасности» (гена мутантного флагеллина или декорина) не приводит к значимому усилению активности несмотря на то, что меланома является иммуногенной опухолью.
В статье Akbulut и соавторов (Akbulut Н., et al. Hum Gene Ther. 2019. 30(8): 999-1007) описана бицистронная плазмидная конструкция на основе аденовирусного вектора, кодирующая гены CD и GM-CSF под контролем CMV промотора (Ad-CD-GMCSF). Показана эффективность конструкции как in vitro на опухолевых клеточных линиях (HEK293, трансформированные клетки почки человека, CRL-2638, клетки карциномы толстой кишки мыши, CCL-51 клетки рака молочной железы мыши), так и in vivo - на мышиной модели с привитой опухолью рака толстой кишки. Плазмидная конструкция экспрессировала в клетках как цитозиндезаминазу, так и иммуногенный белок GM-CSF, значительно увеличивая количество опухолеспецифичных Т-клеток и Т-клеточную цитотоксичность. У мышей, получивших Ad-CD-GMCSF и 5-фторцитозин, значимо тормозился рост опухоли и возрастала продолжительность жизни по сравнению с мышами, получившими конструкцию с геном CD (Ad-CD) и нативным контролем. Препарат FCU1-BsFm/PP отличается от описанного продукта тем, что имеет в своем составе более совершенную конструкцию химерного гена FCU1 (вместо гена CD), что имеет большее преимущество в продукции цитотоксических агентов в опухолевых клетках и потенциал усиления «эффекта свидетеля».
В патенте WO2013115673A1 описан наиболее близкий аналог FCU1-BsFm/PP, который совпадает с ним по признакам наличия элементов системы ГНЭПТ: химерный ген FCU1, элементы системы доставки на основе ПЭИ/ПЭГ, конъюгированные с ТАТ-пептидом, а также в области техники (лечение широкого спектра ЗНО). Экспрессионная плазмидная конструкция способна превращать внутри раковой клетки малотоксичное пролекарство 5-фторцитозин во внутриклеточный токсин. Первое отличие препарата FCU1-BsFm/PP заключается в том, что промоторный регион плазмидной конструкции представлен последовательностью сильного конститутивного, но неспецифичного промотора цитомегаловируса человека, тогда как аналог имеет в своем составе «многопрофильный промотор с тандемной комбинацией BIRC5 и TERT промоторов или производное тандемной комбинации BIRC5 и TERT промоторов, образованное в результате делеции, замены, инсерции или иной мутации промотора, не влияющих на активность промотора, но при этом меняющих его структуру». Другим положительным отличием препарата FCU1-BsFm/PP является наличие в составе плазмидной конструкции гена, кодирующего мутантный ген секретируемого флагеллина Bacillus subtilis. Экспрессия флагеллина в опухолевых клетках может приводить к активации локального и генерализованного противоопухолевого иммунного ответа. Таким образом, кардинальные отличия от аналога реализованы в иной комбинации генов в составе плазмидной ДНК и разной композиции нуклеотидных последовательностей промоторных областей для инициации транскрипции терапевтических генов.
Список иллюстраций:
Фигура 1 - Схема плазмидной конструкции FCU1-BsFm под контролем сильного конститутивного неспецифического промотора CMV. Обозначения: FCU1 - химерный ген цитозиндезаминазы и урацилфосфорибозилтрансферазы дрожжей, BsFm - ген секретируемого мутантного флагеллина Bacillus subtilis, Р2А - последовательность 2А-пептида, CMV - промотор цитомегаловируса человека, Amp(r) - ген устойчивости к ампициллину (селективный маркер), SV40 late poly(A) - сигнал полиаденилирования из генома вируса SV40. Обозначены сайты узнавания эндонуклеаз рестрикции SmaI и FseI, фланкирующие последовательность целевого гена.
Фигура 2 - Определение трансфекционной активности образца блок-сополимера на клетках линии СТ26. Верхняя панель - результаты анализа нетрансфицированных клеток, установка гейтов. Нижняя панель - трансфекция клеток плазмидной ДНК pEGFP-N1. Ось X - интенсивность флуоресценции зеленого флуоресцентного белка. Ef=(NLR+NUR)/NLL*100, где Ef - эффективность трансфекции в %; NLR, NUR, NLL -количество зарегистрированных клеток в нижнем правом, верхнем правом и нижнем левом квадрантах соответственно. Процент трансфицированных клеток в примере равен 26,77%.
Фигура 3 - Определение содержания плДНК в клетках С26, трансфицированных FCU1-BsFm/PP, в зависимости от времени после трансфекции. А - калибровочная кривая для определения содержания гДНК в препаратах тотальной ДНК, выделенных из трансфицированных клеток С26 мыши. Представлены средние значения, полученные для трех независимых серий разведений калибровочных образцов со стандартным отклонением. Б - калибровочная кривая для определения содержания плДНК в препаратах тотальной ДНК, выделенных из трансфицированных клеток С26 мыши. Представлены средние значения, полученные для трех независимых серий разведений калибровочных образцов со стандартным отклонением. В - содержание плДНК FCU1-BsFm в трансфицированных клетках С26 в зависимости от времени после трансфекции. Для каждой временной точки результат представлен в повторе. К - контрольный образец ДНК, выделенный из интактных клеток С26. Представлены средние значения в трипликатах со стандартным отклонением.
Фигура 4 - Определение содержания плДНК в опухолях мышей в зависимости от времени после однократного внутриопухолевого введения препарата FCU1-BsFm/PP. А - калибровочная кривая для определения содержания гДНК в препаратах тотальной ДНК, выделенной из опухолей мышей. Представлены средние значения, полученные для трех независимых серий разведений калибровочных образцов со стандартным отклонением. Б - калибровочная кривая для определения содержания плДНК в препаратах тотальной ДНК, выделенных из опухолей мышей. Представлены средние значения, полученные для трех независимых серий разведений калибровочных образцов со стандартным отклонением. В - содержание плДНК FCU1-BsFm в опухолях мышей в зависимости от времени после введения препарата FCU1-BsFm/PP. Для каждой временной точки результат представлен в повторе. К - контрольный образец ДНК, выделенный из опухоли мыши, не подвергшейся воздействию ЛС.Представлены средние значения в трипликатах со стандартным отклонением.
Фигура 5 - Относительный уровень транскрипции кассеты FCU1-BsFm в клетках линии С26. Высота столбцов отражает значение Q±s.e.m, где Q - относительный уровень транскрипции. Для конечных результатов по каждой временной точке все трипликаты по дням рассчитали как отдельные значения, количество повторов для одного образца от 5 до 15. К - контрольные нетрансфицированные клетки.
Фигура 6 - Относительный уровень транскрипции кассеты FCU1-BsFm в опухолях мышей.
Фигура 7 - Вестерн-блот анализ содержания белка FCU1 в лизатах клеток линии С26, трансфицированных FCU1-BsFm/PP и собранных через 24 часа, 48 часов, 3 дня, 5 дней и 7 дней после трансфекции. К - контроль, лизат нетрансфицированных клеток.
Фигура 8 - Результаты количественной денситометрической обработки результатов вестерн-блот анализа содержания белка FCU1 в лизатах клеток линии С26, трансфицированных препаратом FCU1-BsFm/PP и собранных через 24 часа, 48 часов, 3 дня, 5 дней и 7 дней после трансфекции. Для каждой временной точки анализировался трипликат в двух биологический повторностях А и Б (N=6). К - контроль, лизат нетрансфицированных клеток; ND - не детектируется.
Фигура 9 - Депарафинизированные окрашенные срезы тканей опухоли, фиксированных формалином. Г&Э - окрашивание гематоксилином и эозином. ПкАт - иммунопероксидазное окрашивание с поликлональными антителами, докрашивание гематоксилином. Ig - иммунопероксидазное окрашивание с Ig кролика (контроль), докрашивание гематоксилином.
Фигура 10 - Схема вариантов исследования эффективности FCU1-BsFm/PP.
Фигура 11 - Динамика роста саркомы S37 у мышей F1 (самок), леченных препаратом в сочетании с 5-FC с разной кратностью введения препарата (в опытных группах n=9, в контрольной группе n=11). Лечение начинали на 7-е сутки роста опухоли (объем опухоли ≈ 50 мм3).
FCU1-BsFm вводили внутриопухолево однократно, двукратно, трехкратно и четырехкратно в разовой дозе 0,04 мкг ДНК/мм3 (курсовые - 0,04 ДНК/мм3, 0,08 мкг ДНК/мм3, 0,12 мкг ДНК/мм3 и 0,16 ДНК/мм3). 5-FC в составе препарата Анкотил вводили внутрибрюшинно в разовой дозе 250 мг/кг дважды в сутки (суточная доза - 500 мг/кг, курсовые дозы - 3500 мг/кг, 5500 мг/кг, 7500 мг/кг и 9500 мг/кг) (интервал - 8-10 часов).
Фигура 12 - Динамика роста саркомы S37 у мышей F1 (самок), получавших лечение системой FCU1-BsFm/PP+5-FC в разном режиме (в опытных группах n=9, в контрольной группе n=11). Лечение начинали на 7-е сутки роста опухоли (объем опухоли ≈ 50 мм3).
Фигура 13 - Объемы лимфоузлов с метастатическим поражением у мышей F1 (самок) с опухолью S37, получавших лечение системой FCU1-BsFm/PP+5-FC в разной дозе и разном режиме (в опытных группах n=9, в контрольной группе n=11).
Фигура 14 - Выживаемость мышей (самок) с привитой саркомой S37, получавших FCU1-BsFm/PP в разных режимах введения (в опытных группах n=9, в контрольной группе -n=11). Лечение начинали на 7-е сутки роста опухоли (объем опухоли ≈ 50 мм3).
Фигура 15 - Дизайн исследования по оценке эффективности выбранного режима введения FCU1-BsFm/PP на мышах с привитой карциномой С26.
Фигура 16 - Схема оценки противоопухолевого иммунного ответа на лечение генотерапевтическим ЛС посредством повторного привития опухоли излеченным животным (tumor rechallenge).
Фигура 17 - Динамика роста саркомы S37 после повторного привития излеченным животным.
Фигура 18 - Динамика роста карциномы СТ26 после повторного привития излеченным животным.
Фигура 19 - Динамика роста карциномы легкого СМТ 167 при воздействии ЛС FCU1-BsFm/PP и пролекарства 5-FC. FCU1-BsFm/PP вводили трехкратно интратуморально с интервалом 5 дней в разовой дозе 0,04 мкг ДНК/мм3. 5-FC в составе препарата Анкотил вводили внутрибрюшинно в разовой дозе 250 мг/кг, дважды в день с интервалом 8-10 часов (суточная доза от 500 мг/кг) в течение 15 дней по схеме (курсовая доза 7500 мг/кг). Животным контрольной группы вводили интратуморально и внутрибрюшинно фосфатный буфер (PBS) в режиме и объеме, соответствующим режиму и объему введения препарата в опытной группе.
Фигура 20 - Динамика роста карциномы шейки матки РШМ5 при воздействии FCU1-BsFm/PP и пролекарства 5-FC. FCU1-BsFm/PP вводили трехкратно интратуморально с интервалом 5 дней в разовой дозе 0,04 мкг ДНК/мм3. 5-FC в составе препарата Анкотил вводили внутрибрюшинно в разовой дозе 250 мг/кг, дважды в день с интервалом 8-10 часов (суточная доза от 500 мг/кг) в течение 15 дней по схеме (курсовая доза 7500 мг/кг). Животным контрольной группы вводили интратуморально и внутрибрюшинно фосфатный буфер (PBS) в режиме и объеме, соответствующим режиму и объему введения препарата в опытной группе.
Фигура 21 - Динамика роста карциномы молочной железы 4Т1 у мышей при воздействии препарата FCU1-BsFm/PP и пролекарства 5-FC. FCU1-BsFm/PP вводили трехкратно интратуморально с интервалом 5 дней в разовой дозе 0,04 мкг ДНК/мм3. 5-FC в составе препарата Анкотил вводили внутрибрюшинно в разовой дозе 250 мг/кг, дважды в день с интервалом 8-10 часов (суточная доза от 500 мг/кг) в течение 15 дней по схеме (курсовая доза 7500 мг/кг). Животным контрольной группы вводили интратуморально и внутрибрюшинно фосфатный буфер (PBS) в режиме и объеме, соответствующим режиму и объему введения препарата в опытной группе.
Фигура 22 - Динамика роста аденокарциномы яичников человека SC-OV-3 при воздействии ЛС FCU1-BsFm/PP и пролекарства 5-FC. FCU1-BsFm/PP вводили трехкратно интратуморально с интервалом 5 дней в разовой дозе 0,04 мкг ДНК/мм3. 5-FC вводили внутрибрюшинно в разовой дозе 250 мг/кг, дважды в день с интервалом 8-10 часов (суточная доза от 500 мг/кг) в течение 15 дней по схеме (курсовая доза 7500 мг/кг). Животным контрольной группы вводили интратуморально и внутрибрюшинно фосфатный буфер (PBS) в режиме и объеме, соответствующим режиму и объему введения препарата в опытной группе.
Фигура 23 - Динамика роста колоректальной аденокарциномы человека НСТ116 при воздействии препарата FCU1-BsFm/PP и 5-FC. ЛС FCU1-BsFm/PP вводили трехкратно интратуморально с интервалом 5 дней в разовой дозе 0,04 мкг ДНК/мм3. 5-FC в составе препарата Анкотил вводили внутрибрюшинно в разовой дозе 250 мг/кг, дважды в день с интервалом 8-10 часов (суточная доза от 500 мг/кг) в течение 15 дней по схеме (курсовая доза 7500 мг/кг). Животным контрольной группы вводили интратуморально и внутрибрюшинно PBS в режиме и объеме, соответствующим режиму и объему введения препаратов в опытных группах.
Фигура 24 - Динамика роста плоскоклеточного рака кожи А431 при воздействии FCU1-BsFm/PP и пролекарства 5-фторцитозин. FCU1-BsFm/PP вводили трехкратно интратуморально с интервалом 5 дней в разовой дозе 0,04 мкг ДНК/мм3. 5-FC вводили внутрибрюшинно в разовой дозе 250 мг/кг, дважды в день с интервалом 8-10 часов (суточная доза от 500 мг/кг) в течение 15 дней по схеме (курсовая доза 7500 мг/кг). Животным контрольной группы вводили интратуморально и внутрибрюшинно фосфатный буфер (PBS) в режиме и объеме, соответствующим режиму и объему введения препарата в опытной группе.
Раскрытие изобретения
Общие сведения
Изобретение относится к генотерапевтическим препаратам для лечения широкого спектра солидных злокачественных новообразований, при котором препарат вводится внутриопухолево в первичный или вторичный опухолевый очаг (метастаз), а малотоксичное пролекарство вводится посредством инфузий в системный кровоток.
Задача состояла в создании плазмидной конструкции для адресной доставки в опухолевый очаг, экспрессия целевых генов которой должна обеспечивать достижение противоопухолевого эффекта, опосредованного через цитотоксическое и иммуностимулирующее воздействие, а также в тестировании подобного эффекта в доклинических исследованиях на животных.
Техническим результатом было создание препарата FCU1-BsFm/PP на основе плазмидной конструкции невирусной природы, кодирующей целевые терапевтические гены FCU1 и BsFm, упакованной в специальный блок-сополимер (РР). Блок-сополимер обеспечивает наиболее эффективный способ доставки плазмидной ДНК внутрь клеток. Экспрессированный химерный белок FCU1 в присутствии экзогенного малотоксичного пролекарства 5-фторцитозин (МНН) вызывает гибель опухолевых клеток за счет превращения пролекарства в цитотоксический агент 5-фторурацил. Экспрессированный мутантный флагеллин опосредует активацию локального и генерализованного противоопухолевого иммунного ответа.
Препарат представляет собой генотерапевтическую плазмидную конструкцию, упакованную в блок-сополимер. Принцип действия препарата основан на трансфекции опухолевых клеток, где происходит запуск экспрессии нескольких терапевтических генов, которые закодированы в плазмидной ДНК, что приводит к ограниченному по времени накоплению белковых продуктов, которые в норме отсутствуют в клетках. При определенных условиях препарат оказывает двойное фармакологическое действие. Во-первых, функционирование одного из белковых продуктов в присутствии специального малотоксичного пролекарства способствует накоплению внутри клеток ряда цитотоксических метаболитов, которые ингибируют процесс репликации активно делящихся клеток. Цитотоксические метаболиты обладают способностью высвобождаться из клеток-продуцентов, которые были трансфицированы плазмидной ДНК, и диффундировать в соседние опухолевые клетки, оказывая так называемый «эффект свидетеля». Во-вторых, другой белковый продукт является регулятором локального и генерализованного противоопухолевого иммунного ответа.
Более конкретно, действующим началом разрабатываемого препарата является модифицированный ген цитозиндезаминазы дрожжей (FCY1), который при введении в раковые клетки превращает пролекарство 5-фторцитозин в антиметаболит 5-фторурацил, который затем конвертиртируется клеточными ферментами в цитотоксичные метаболиты. Для увеличения эффективности конверсии пролекарства в цитотоксичные метаболиты использован химерный ген FCY1/FUR1, где ген FUR1 кодирует урацилфосфорибозилтрансферазу дрожжей, что позволяет добиться более выраженного цитотоксического действия на опухолевые клетки, а также более сильного локального «эффекта свидетеля».
Использование флагеллина, который является агонистом Толл-подобных рецепторов (TLR) и представителем так называемых «сигналов опасности», ставит своей целью активацию иммунной системы организма. Флагеллин связывается с TLR-рецепторами на поверхности антиген-представляющих клеток (АПК). В результате взаимодействия происходит привлечение в опухоль и активация АПК, которые захватывают опухолевые антигены, процессируют и представляют их Т-клеткам. Рецепторы, способные распознать флагеллин, расположены на поверхности клеток, поэтому генетически кодируемый флагеллин должен секретироваться из продуцирующих его клеток во внеклеточную среду.
Описание генотерапевтической плазмидной конструкции FCUl-BsFm
FCU1-BsFm представляет собой бицистронную плазмидную конструкцию на основе невирусного вектора pGL3. В единую экспрессионную кассету под контролем сильного неспецифического промотора цитомегаловируса человека (CMV) входит химерный ген FCU1 цитозиндезаминазы (FCY1) и урацилфосфорибозилтрансферазы (FUR1) Saccharomyces cerevisiae arg4 и ген секретируемого мутантного флагеллина (BsFm) Bacillus subtilis. Кодирующие последовательности терапевтических генов FCU1 и BsFm в плазмидной конструкции соединены последовательностью, кодирующей 2А-пептид тешовируса свиней (Р2А). Схема плазмидной конструкции FCUl-BsFm показана на фигуре 1. Нуклеотидная последовательность плазмидной конструкции соответствует SEQ ID №1.
Ген FCU1, кодирующий химерный белок FCY1/FUR1, был сконструирован путем объединения кодирующих последовательностей двух генов - FCY1 (471 пар оснований, chrXVI:677,165-677,635) и FUR1 (642 пар оснований, chrVIII:362,124-362,765), - которые были получены с помощью амплификации на матрице геномной ДНК штамма дрожжей Saccharomyces cerevisiae arg4. Кодоны инициации и терминации транскрипции были удалены олигонуклеотидным сайт-направленным мутагенезом. Фрагменты генов были разделены кодоном, соответствующим аминокислоте аланину, в результате чего образующийся химерный белок содержал аланиновый линкер между белками FCY1 и FURL Кроме того, позднее в структуру химерного гена была введена консенсусная последовательность Козака CGCC для оптимизации экспрессии в клетках млекопитающих. Нуклеотидная последовательность гена FCU1 соответствует SEQ ID №2.
Флагеллин бактерий не содержит в своей структуре сигналов секреции из клеток млекопитающих, поэтому на N-конец этого белка был добавлен сигнальный пептид из незрелого белка IL-2 человека (MYRMQLLSCIALSLALVTNS) (Zhang et al, J Gene Med. 2005; 7(3), 354-65), который способен инициировать секрецию различных белков из клеток млекопитающих. Таким образом, белок флагеллин Bacillus subtilis дикого типа был модифицирован в химерный белок (BsF), включивший в себя последовательность сигнального пептида. В процессе секреции сигнальный пептид обычно отщепляется от целевого белка, что нивелирует его влияние на биологическую активность флагеллина во внеклеточной среде. Была сконструирована мутантная форма BsF, в четырех предсказанных сайта N-гликозилирования были введены замены кодонов N/Q. Кроме того, кодирующая последовательность BsF была оптимизирована по кодонному составу для увеличения продукции целевого белка в клетках млекопитающих. Белок BsF с измененной последовательностью был назван BsFm. Нуклеотидная последовательность мутантного гена BsFm соответствует SEQ ID №3.
Описание способа получения генотерапевтической плазмидной ДНК
В качестве плазмидного вектора использовали плазмиду pGL3-Basic (Promega, США), в которую был клонирован промотор цитомегаловируса (CMV) человека (pGL3-CMV). Из плазмиды pGL3-CMV вырезали ген люциферазы (Luc) по уникальным сайтам рестрикции HindIII и FseI. Плазмиду pGL3-CMV (2,5 мкг) обрабатывали рестриктазой HindIII (15 единиц) при 37°С в течение ночи в объеме реакционной смеси 50 мкл. Расщепленную плазмиду очищали с помощью набора Cleanup Mini (Евроген, Россия). Линейную плазмиду обрабатывали рестриктазой FseI (15 единиц) при 37°С в течение ночи в объеме реакционной смеси 50 мкл. Расщепленную плазмиду очищали с помощью набора Cleanup Mini (Евроген, Россия). Образец линейной плазмиды наносили на агарозный гель, проводили электрофорез и вырезали из геля полосу, соответствующую размеру линейной плазмиды (фрагмент, равный 3700 п. н.). При этом удаляли ген люциферазы (фрагмент, равный 1700 п. н.).
Получали плазмидную конструкцию на основе плазмиды pGL3, содержащую экспрессионную кассету гена FCU1 и гена декарина мыши (mDCN) под контролем промотора CMV (FCU1-mDCN). Амплификацию гена mDCN проводили с помощью специфичных праймеров (DCN_XmaI_for - AT-CCCGGG-CCT-ATGAAGGCAACTCTCATCTTCTTCCT и DCN_FseI_rev - TA-GGCCGGCC-TAA-CTTGTAGTTTCCAAGTTGAATGGCAGA), используя в качестве матрицы ДНК плазмиды pCMV6-Entry (Origen, США). Праймеры содержали сайты XmaI и FseI для направленного клонирования. Праймер направления Rev вводил стоп-кодон ТАА, который отсутствует на конце гена mDCN в плазмиде pCMV6-Entry, поскольку плазмида кодирует слитой белок декарина с тэгом Мус-DDK. Праймер направления For вводил три последние нуклеотида ССТ от Р2А-пептида, которые удалялись при рестрикции вектора по сайту XmaI. Для препаративной наработки фрагмента ДНК проводили 8 циклов амплификации и после завершения циклов инкубировали реакционную смесь в объеме 50 мкл с 300 нг матрицы в течение 10 мин при 72°С. Фрагмент гена mDCN очищали в агарозном геле и вырезали фрагмент, равный 1065 п. н. ДНК выделяли из геля с помощью набора Cleanup Mini (Евроген, Россия), при этом элюцию проводили 20 мкл буфера для элюции. Фрагмент обрабатывали рестриктазой XmaI (10 единиц) при 37°С в течение ночи в объеме реакционной смеси 30 мкл. Смесь после рестрикции очищали с помощью набора Cleanup Mini. Далее фрагмент ДНК обрабатывали второй рестриктазой FseI (10 единиц) при 37°С в течение ночи. Смесь после рестрикции очищали с помощью набора Cleanup Mini. Для создания целевой конструкции использовали плазмидную конструкцию, содержащую экспрессионную кассету гена FCU1 и гена hGMCSF под контролем промотора CMV (FCU1-hGMCSF), которая была создана ранее. Из плазмидной конструкции удаляли ген hGMCSF и кодон от Р2А-пептида по уникальным сайтам рестрикции XmaI и FseI. Плазмиду FCU1-hGMCSF (2,5 мкг) обрабатывали рестриктазой XmaI (15 единиц) при 37°С в течение ночи в объеме реакционной смеси 50 мкл. Расщепленную плазмиду очищали при помощи набора Cleanup Mini. Линейную плазмиду наносили на агарозный гель и проводили электрофорез. Затем вырезали фрагмент геля, соответствующий по размеру линейной плазмиде (5200 п. н). При этом удаляли ген hGMCSF и кодон от Р2А (фрагмент 450 п. н.). Линейный вектор выделяли из геля с помощью набора Cleanup Mini. Для лигирования вектора со вставкой брали 50 нг вектора и 25-40 нг вставки, что в молярном соотношении вектор:вставка составляет 1:2,5. Реакцию лигирования проводили в течение ночи при 14°С с использованием 1 мкл (1 Weiss unit) Т4 ДНК лигазы. Полученной лигазной смесью трансформировали компетентные клетки Е. coli штамм DH5α. Пробирку с клетками (200 мкл) извлекали из морозильной камеры и инкубировали на льду в течение 15 мин. К клеткам добавляли 1/3 объема лигазной смеси и инкубировали на льду в течение 30 мин. Далее прогревали клетки в течение 30 с при 42°С. Образцы помещали в лед и сразу добавляли среду LB до конечного объема 1 мл и инкубировали при 37°С в течение 1-1,5 ч. Аликвоту объемом 100 мкл рассевали на чашки Петри с твердым LB-агаром, содержащим ампициллин (100 мкг/мл). Далее проводили скрининг колоний на наличие целевой вставки (ген mDCN). Колонии, содержащие вставку, выращивали в жидкой среде LB с добавлением ампициллина (100 мкг/мл) при инкубации на шейкере при скорости 150 об/мин и 37°С в течение 20 ч. Пробирки с выросшими клетками центрифугировали при 3000 об/мин. Плазмиду из полученного осадка очищали с помощью набора Plasmid Miniprep (Евроген, Россия) и хранили при температуре - 20°С. Точность нуклеотидной последовательности полученной конструкции подтверждали секвенированием с использованием праймеров: SV40polyA-pGL3 (CGATTTTACCACATTTGTAGAGGTTT) и FCU1F100 (ACCATGCCGCCTTCCCAGA).
Последовательность гена BsFm в составе коммерческого вектора pUC57 была получена путем синтеза гена в компании Proteogenix (Франция). Далее целевая последовательность была вырезана по фланкирующим сайтам рестрикции AgeI и XbaI, введенным в ходе синтеза этого гена, и клонирована в плазмиду CMV-41-BBL-pGL3, разрезанную по сайтам AgeI и XbaI (при этом была удалена кассета 41-BBL). Наличие и ориентация вставки были подтверждены с помощью рестрикционного анализа, а последовательность вставки - посредством секвенирования по Сэнгеру. Амплификацию гена BsFm проводили с помощью специфических праймеров BSFm_for (ATCCCGGGCCTATGAGAATGCAGCTGCTGCTG) и BSFm_rev (ATGGCCGGCCTCACCTCAGCAGCTGCAGCAC), используя в качестве матрицы ДНК ранее полученную плазмиду BsFm-pGL3 под контролем промотора CMV. Праймеры содержат сайты SmaI и FseI для направленного клонирования. Праймер направления BSFm_for вводит три последние нуклеотида кодирующей последовательности 2А пептида (ССТ). Полученный продукт амплификации обрабатывали рестриктазами SmaI/ FseI и лигировали в полученную ранее плазмиду FCU1-mDCN-pGL3 под контролем промотора CMV, гидролизованную по сайтам SmaI и FseI (при этом был удален ген mDCN). Наличие и ориентация вставки были подтверждены с помощью рестрикционного анализа, корректность нуклеотидной последовательности вставки была подтверждена при помощи секвенирования по Сэнгеру.
Для создания рекомбинантного штамма-продуцента целевой плазмидной ДНК был использован E.coli DH5 fhuA2 А2 Δ(argF-lacZ)U169 phoA glnV44 Ф80 Δ(lacZ)M15 gyrA96 recA1 relA1 endA1 thi-1 hsdR17. Для этого 1 нг плазмидной конструкции смешивали с 200 мкл заранее приготовленных компетентных клеток E.coli штамма DH5a, инкубировали в ледяной бане в течение 30 минут и подвергали тепловому шоку в течение 30 сек при температуре 42°С. К смеси добавляли 1 мл среды LB без антибиотика и инкубировали в течение 1 часа при 37°С. 100 мкл смеси высевали на чашку Петри с агаризованной средой LB с ампициллином (100 мкг/мл) и инкубировали в течение 12-16 часов при температуре 37°С до появления колоний. Отдельные колонии пересевали на сектора на чашки Петри и проверяли на наличие требуемой плазмидной конструкции. Колонии, содержащие плазмидную конструкцию, выращивали в LB среде, содержащей ампициллин, до достижения оптической плотности 1 при длине волны 660 нм, добавляли глицерин до 15% и замораживали в криопробирках (Nunc, США) при -70°С.
Наращивание биомассы Е. coli, содержащей плазмидную ДНК, проводилось в ферментере объемом 10 литров. Для этого материал для посева из замороженной культуры рекомбинантного штамма Е. coli высевали на чашку Петри с агаризованной средой LB с ампициллином (100 мкг/мл) и проводили инкубацию в течение 12-16 часов при температуре 37°С до образования одиночных колоний. Засевали 10 мл среды LB, содержащей ампициллин (100 мкг/мл), одиночной колонией с чашки Петри. Выращивали начальную культуру 8 часов при 37°С с интенсивным встряхиванием (200-250 об/мин). Пересевали 10 мл начальной культуры в 500 мл среды LB с ампициллином и выращивали промежуточную культуру в тех же условиях. 500 мл промежуточной культуры пересевали в ферментер, содержащий 9 л LB среды с ампициллином, и выращивали в течение 10 часов при температуре 37°С с интенсивной аэрацией и перемешиванием. Биомассу собирали на проточной центрифуге на 6000 g и 40°С.
Выделение и очистка проходили в три стадии:
1 стадия - гель-фильтрация осветленного и сконцентрированного бактериального лизата - выполнялась на смоле Separose 6 Fast Flow. При этом происходило отделение плазмидной ДНК от массы бактериальной РНК;
2 стадия - адсорбционная хроматография на тиофильной ароматической смоле PlasmidSelect Xtra для отделения сверхспирализованной плазмидной ДНК от кольцевой и линейной форм;
3 стадия - ионообменная хроматография на смоле SOURCE 30Q для завершающей очистки плазмидной ДНК (final polishing).
Смену хроматографического буфера и концентрирование плазмидной ДНК производили на половолоконных картриджах (Hollow fibers) компании GE Healthcare. На первом этапе выделения плазмидной ДНК проводили щелочной лизис бактериальной массы согласно стандартной методике. Осажденную бактериальную массу ресуспендировали в 100 мл буфера (25 мМ трис-HCl, 10 мМ ЭДТА, 50 мМ глюкозы, рН 7,5, 100 мкг/мл РНКаза А), далее добавляли 100 мл лизирующего буфера (200 мМ NaOH, 1% SDS (додецилсульфат натрия)). Через 4 минуты после добавления лизирующего буфера проводили нейтрализацию с помощью добавления охлажденного ацетатного буфера (3 М CH3CO2K). Полученный раствор фильтровали и добавляли изопропанол в соотношении 1:0,7, далее смесь центрифугировали при 14 тыс.об/мин в течение 30 мин при 4°С. Супернатант сливали, осадок промывали 70% этанолом, центрифугировали 15 мин при тех же условиях. Полученный осадок растворяли в 35 мл трис-HCl буфера (50 мМ трис-HCl, 10 мМ ЭДТА, рН 7,5). Далее проводилась очистка плазмидной ДНК на колонках «PlasmidSelect Xtra Starter Kit» (GE Healthcare Life Sciences, США).
Описание свойств генотерапевтической плазмидной ДНК
1) Плазмидная конструкция накапливается в количестве 2.5 × 108 копий плазмидной ДНК/мкг гДНК через 24 часа после трансфекции культуры опухолевых клеток с последующим плавным снижением в течение семидневного периода наблюдения.
2) Максимальная относительная транскрипция экспрессионной кассеты FCU1-BsFm происходит через 24 часа после трансфекции культуры опухолевых клеток. Уровень транскрипции плавно снижается (временные точки 48 часов и 72 часа) в два раза каждые сутки, затем резко пададает до полного исчезновения транскриптов в течение семидневного периода наблюдения.
3) Относительный уровень транскрипции экспрессионной кассеты FCU1-BsFm в опухолевой ткани низкий, с тенденцией к увеличению количества транскриптов к третьим суткам эксперимента и дальнейшему плавному падению в течение семидневного периода наблюдения.
4) Максимальное накопление белка FCU1 в культуре опухолевых клеток имеет место через 24 часа после трансфекции, а затем оно снижается почти вдвое через 48 часов и не детектируется на 5 день после трансфекции.
5) По данным иммуногистохимического анализа срезов опухолевого материала, белок FCU1 окрашивается максимально интенсивно через 48 часов после введения плазмидной конструкции с постепенным снижением к 5-7 суткам наблюдения.
6) В трансфицированных плазмидной конструкцией опухолевых клетках уровень продукции цитокинового белка IL-8 значимо превышает контроль, что косвенно свидетельствовует о факте экспрессии мутантного гена флагеллина в составе плазмиды.
7) Наиболее выраженный ингибирующий эффект вызывает использование препарата FCU1-BsFm/PP с конечной концентрацией блок-сополимера 12,5 мкМ при введении его с интервалом 5 дней (торможение роста опухоли после окончания лечения составило 71%).
8) На моделях перевиваемых опухолей саркомы S37 при трехкратном внутриопухолевом введении FCU1-BsFm/PP в разовой дозе 0,04 мкг ДНК/мм3 и пролекарства 5-фторцитозин (внутрибрюшинно) через 14 дней после окончания лечения (в сравнению с контролем) торможение роста опухоли составило 72-80%, задержка роста опухоли - 10,4-10,6 дней, индекс торможения метастазирования 60,5-63,4%, показатель погибших клеток - 0,7. Медиана выживаемости в группе с введением FCU1-BsFm/PP составила 50,0-53,0 суток против 32,0 суток в контрольной группе.
9) На моделях перевиваемых опухолей колоректальной карциномы СТ26 при трехкратном внутриопухолевом введении FCU1-BsFm/PP в разовой дозе 0,04 мкг ДНК/мм3 и пролекарства 5-фторцитозин (внутрибрюшинно) через 14 дней после окончания лечения (в сравнению с контролем) торможение роста опухоли составило 74%, задержка роста опухоли - 16,7 суток, показатель погибших клеток - 1,1 (эффект «++» по рейтингу противоопухолевой активности). Медиана выживаемости в группе с введением FCU1-BsFm/PP составила 44,1 суток против 33,0 суток в контрольной группе.
10) После повторной перевивки опухоли излеченным FCU1-BsFm/PP животным (tumor rechallenge) опухоль не развилась в течение 60 дней, что свидетельствует о стойком приобретенном противоопухолевом иммунитете.
11) Препарат FCU1-BsFm/PP вводится путем инъекции внутрь опухолевого очага, с целью его равномерного распределения по площади поражения. Доза препарата FCU1-BsFm/PP рассчитывается в зависимости от объема опухоли.
Описание способа приготовления ГЛФ препарата FCU1-BsFm/PP
Двухкомпонентная жидкая готовая лекарственная форма (ГЛФ), состоит из двух флаконов. Флакон А представляет раствор плазмидной ДНК в HEPES-глюкозном буфере объемом 1 мл. Флакон Б представляет собой раствор блок-сополимера РР в боратном буфере. Размораживают при комнатной температуре в течение 10-15 минут флакон А, тщательно перемешивают. Размораживают при комнатной температуре в течение 10-15 минут флакон Б, тщательно перемешивают. Непосредственно перед применением содержимое флакона А добавляли к содержимому флакона Б, выдерживали при комнатной температуре в течение 5-10 минут и тщательно перемешивали. Полученный раствор выдерживали 40 минут при комнатной температуре. Таким образом, готовый к использованию генотерапевтический препарат содержит 2 мл раствора (80 мкг/мл сверхскрученной плазмидной ДНК, 12,5 мкМ блок-сополимера, 5 мМ HEPES рН 7,4, 5% глюкозы, 12,5 мМ боратного буфера рН 7,5).
Описание способа доставки плазмидной конструкции
Блок-сополимер РР включает в себя линейные поликатионы полиэтиленимин (ПЭИ) и полиэтиленгликоль (ПЭГ), конъюгированные с пептидом GRKKKRRQRC (ТАТ-пептид), который увеличивает способность препарата проникать в клетки. Для двухстадийного синтеза конъюгатов использовали линейный ПЭИ с молекулярной массой 25000 г/моль, бифункциональный ПЭГ MAL-dPEG™24-NHS ester (Quanta BioDesign, Ltd., США) с молекулярной массой 1394,55 г/моль и синтетический ТАТ-пептид. На первой стадии бифункциональный ПЭГ взаимодействовал при рН 5,5 с аминогруппой ПЭИ. На второй стадии пептид за сульфгидрильную группу N-концевого цистеина присоединяли к второму концу ПЭГ, содержащему реакционноспособный малеимид, при рН 7,0. Ожидаемая пришивка ПЭГ/ПЭИ в блок-сополимерах - 1,1-1,5. Ожидаемая пришивка пептида ТАТ/ПЭИ - 0,8-1,4. Блок-сополимер имеет мольное соотношение ПЭГ/ПЭИ от 1,1 до 1,5 (определяемое спектрофотометрически), ТАТ/ПЭИ - 0,5-0,9 (определяется флуориметрически), гидродинамический диаметр 3-6 нм (99,99% частиц) (определяется методом динамического светорассеивания), специфическая активность составляет не менее 20% трансфекции в культуре клеток СТ26 in vitro.
Описание способа доставки препарата FCUl-BsFm/PP в опухолевые очаги
Препарат может вводится шприцом в пальпируемые опухолевые очаги, расположенные подкожно и внутрикожно, при этом выбор типа иглы и шприца зависит от размера и локализации опухоли. В опухоли небольшого размера используется «веерная» техника введения препарата, которая предполагает использование одной точки входа в опухолевый очаг с последующим введением по нескольким путям внутри очага для достижения наиболее полного и равномерного охвата. Препарат распределяют посредством вытягивания иглы на себя (не выходя из очага) и последующего перенаправления иглы столько раз, сколько необходимо для распределения всей дозы препарата. При наличии опухолевого очага большого размера, когда «веерная» техника введения препарата не совсем подходит, используется техника равномерного введения препарата в несколько разных точек входа в опухолевый очаг.
Примеры осуществления изобретения
Пример 1. Анализ эффективности трансфекции эукариотических клеток препаратом блок-сополимера РР
Оценку специфической активности блок-сополимера осуществляли методом транзиентной трансфекции клеточной линии СТ26. На фигуре 2 приведены результаты тестирования полученного блок-сополимера. Измерение эффективности трансфекции проводили с использованием плазмиды pEGFP-N1, содержащей ген зеленого флуоресцентного белка под контролем промотора ранних генов цитомегаловируса. Для проведения анализа использовали проточный цитофлуориметр FACS Scan Analyzer (Бектон Дикинсон, США) с длиной света возбуждающего лазера 488 нм и фильтров принимающих детекторов 530 нм (канал FL1, детекция FITC, GFP), 585 нм (канал FL2, детекция РЕ/PI, RFP) и >670 nm (канал FL3< детекция PerCP). После проведения трансфекции клетки, получившие плазмиду pEGFP-N1, продуцировали зеленый флуоресцирующий белок, наличие которого в клетке детектировалось по увеличению сигнала над базовым уровнем по каналу FL1. При этом не происходило увеличения интенсивности сигнала в красной области спектра (канал FL2) (см. фигуру 2, правый график распределения клеток по интенсивности свечения - зафиксирован сдвиг части клеточной популяции вправо по сравнению с исходной).
Пример 2. Оценка длительности присутствия плазмидной конструкции в клетках С26 и в опухоли С26 после воздействия FCU1-BsFm/PP
Клетки линии С26 трансфицировали препаратом FCU1-BsFm/PP. Культивированные клетки собирали через 24 часа, 48 часов, 3 дня, 5 дней и 7 дней после трансфекции. Для in vitro оценки длительности присутствия плазмидной ДНК (плДНК) FCU1-BsFm в трансфицированных клетках использовали метод количественной ПЦР-амплификации в режиме реального времени с применением специфических праймеров: комплементарные плДНК и гДНК мыши: pGLplfor (5'-CTCATAGCTCACGCTGTAGG-3')/pGLp1rev (5'-ATACCTCGCTCTGCTAATCC-3') для амплификации фрагмента плДНК размером 194 п. н. и Ginfg_v2_for (5'-CTCAAGTGGCATAGATGTGG-3')/Ginfg_v2_rev (5'-AGAAACAACAGGGAAACTGG-3') для амплификации фрагмента уникального гена γ-интерферона мыши размером 107 п.н. Для определения количества гДНК и плДНК использовали калибровочные кривые. Для построения калибровочной кривой для определения количества гДНК готовили три независимые серии четырехкратных разведений гДНК, выделенной из нетрансфицированных клеток С26. Было использовано по четыре разведения на каждую серию, начиная с 12.5 нг гДНК на реакцию. Калибровочные образцы амплифицировали с праймерами Ginfg_v2_for/Ginfg_v2_rev, комплементарными гену γ-интерферона. Для построения калибровочной кривой для определения количества плДНК готовили три независимые серии четырехкратных разведений плДНК, начиная с 0,2 нг плДНК на реакцию. При этом в каждый калибровочный образец добавляли по 25 нг гДНК, выделенной из нетрансфицированных клеток С26, чтобы учесть возможное влияние гДНК на эффективность амплификации плДНК. Калибровочные образцы амплифицировали с праймерами pGLp1for/pGLp1rev, комплементарными фрагменту терапевтической плДНК. Калибровочные кривые, а также гистограмма, демонстрирующая зависимость содержания плДНК в трансфицированных клетках С26 от времени после трансфекции, приведены на фигуре 3. Представленные данные свидетельствуют о высоком содержании плДНК в трансфицированных клетках С26 в начале эксперимента, через 24 часа после трансфекции полученное значение составило 2.5 × 108 копий плазмиды/мкг гДНК. Далее наблюдали плавное снижение значений до 1.5 × 108 копий плазмиды/мкг гДНК на 3 день эксперимента и резкое снижение на 5 день эксперимента. К концу эксперимента (7 день) количество плДНК снижалось в 50 раз по сравнению с 1 днем эксперимента.
Выделение и амплификацию плазмидной ДНК FCU1-BsFm из опухолевой массы осуществляли так же, как и в случае трансфицированных клеток. Для построения калибровочной кривой для определения количества плДНК готовили три независимые серии четырехкратных разведений плДНК начиная с 0,002 нг плДНК на реакцию. При этом в каждый калибровочный образец добавляли по 25 нг гДНК, выделенной из нетрансфицированных клеток С26, чтобы учесть возможное влияние гДНК на эффективность амплификации плДНК. Калибровочные образцы амплифицировали с праймерами pGLp1for/pGLp1rev, комплементарными фрагменту терапевтической плДНК, также строили график зависимости количества амплифицированной матрицы от флуоресценции красителя SYBR Green, выраженной в условных единицах.
Калибровочные кривые, а также гистограмма, демонстрирующая зависимость содержания плДНК в опухолях мышей С26 от времени после введения ЛС, приведены на фигуре 4. Полученные результаты свидетельствуют о высоком содержании плДНК в опухолях мышей после введения ЛС на протяжении всего эксперимента. В целом наблюдали снижение количества плДНК в опухолях с течением времени. Так, на седьмой день эксперимента содержание плазмиды в опухолях было в 1,7 и более раз ниже, чем через 24 часа после введения препарата FCU1-BsFm/PP.
Пример 3. Оценка уровня экспрессии плазмидной конструкции FCU1-BsFm на уровне РНК методом ОТ-ПЦР
Для определения уровня и продолжительности транскрипции генов FCU1 и BsFm в составе плазмидной конструкции FCU1-BsFm in vitro и in vivo использовали метод ОТ-ПЦР в режиме реального времени. Для амплификации целевых фрагментов терапевтических генов FCU1 и BsFm были подобраны пары праймеров: FCU1-Forward (5'-TATTGGTGAGGCTTCGGAAC-3')/FCU1-Reverse (5'-CGATTCACCAGCTCTGACAA-3') для амплификации целевого фрагмента гена FCU1 размером 274 п.н. и BsFm-Forward (5'-GAGATGGCCAGCAAGAACTC-3')/BsFm-Reverse (5'-ACCTTGTAGGTGCCATCCAG-3') для амплификации целевого фрагмента гена BsFm размером 274 п. о. Уровень транскрипции гена FCU1 в образцах определяли в относительных единицах по сравнению с уровнем транскрипции референсного гена рибосомальной 18S РНК. Для амплификации целевого фрагмента рибосомальной 18S РНК мыши была подобрана пара праймеров m18S-Forward (5'-TGCAATTATTCCCCATGAACG-3')/m18S-Reverse (5'-GCCTCACTAAACCATCCAATC-3'), размер ампликона 123 п.н. Таким образом, было показано, что максимальная относительная транскрипция экспрессионной кассеты FCU1-BsFm наблюдается через 24 часа после трансфекции. Далее в течение двух суток уровень транскрипции плавно снижался (временные точки - 48 часов и 72 часа) в два раза каждые сутки, а затем резко падал до полного исчезновения транскриптов через 7 дней после трансфекции (фигура 5). Также можно отметить, что относительный уровень транскрипции кассеты FCU1-BsFm в опухолях был низким, с тенденцией к увеличению количества транскриптов к третьим суткам эксперимента и дальнейшему плавному падению (фигура 6).
Пример 4. Оценка уровня и продолжительности экспрессии плазмидной конструкции FCU1-BsFm на уровне белка
Для проведения вестерн-блоттинга образцы трансфицированных клеток и образцы опухолей С26 лизировали в RIPA-буфере с добавлением ингибитора протеаз. Клеточные лизаты подвергали электрофоретическому разделению в полиакриламидном геле по методу Лэммли в присутствии додецилсульфата натрия (SDS) в трис-глициновой буферной системе. Белки переносили на поливинилиденфторид-содержащую мембрану и проводили иммунодетекцию антигена, используя в качестве первичных антител овечьи IgG (ab35251, Abcam, Англия), связывающиеся с N-концевой областью белка FCU1, и вторичные ослиные антитела к овечьим IgG (sc-2473, SantaCruz, США), конъюгированные с пероксидазой хрена. Дополнительно проводили иммунодетекцию белка «домашнего хозяйства» бета-актина мыши. Для этого мембрану отмывали и гибридизовали с антителами повторно, используя в качестве первичных кроличьи моноклональные антитела к актину (А2103, Merck, Германия), а в качестве вторичных - конъюгат козлиных антител (к кроличьим IgG) с пероксидазой хрена (7074S, Cell Signalling Technology, Нидерланды). Белки на мембране визуализировали с помощью набора Immun-Star HRP Chemiluminescent (BIO-RAD, США) и регистрировали на приборе Bio-Rad VersaDoc МР4000, используя компьютерную программу Quantity One. Трансфицированные клетки С26 были получены в двух биологических повторностях (повторы А и Б) путем постановки двух независимых трансфекций для каждой из временных точек. В качестве отрицательного контроля использовали нетрансфицированные клетки С26. Полученные результаты представлены на фигуре 7.
Несмотря на то, что на каждую дорожку было нанесено 120 тысяч клеток каждого образца, количество детектируемого актина в разных образцах не одинаково. Такие различия могут возникать из-за погрешностей при сборе клеточного материала, пробоподготовке и при нанесении образцов на гель. Обычно метод вестерн-блот анализа применяется для качественной оценки присутствия белка, однако в случае денситометрической обработки полученных изображений вестерн-блот может рассматриваться как количественный метод. При денситометрической обработке полученных результатов значения FCU1 нормировались на значения актина для каждого из анализируемых образцов, таким образом наблюдаемые различия по актину были учтены при обработке результатов (фигура 8). Количественная денситометрическая обработка проводилась по трипликатам для каждого из образцов в двух биологических повторах А и Б (N=6) с помощью программы ImageJ (National Institutes of Health (NIH), США). Методом вестерн-блоттинга было показано, что в образцах клеток линии карциномы кишки мыши С26, собранных через различные промежутки времени после трансфекции, максимальное количество белка FCU1 накапливается через 24-72 часа после трансфекции, а затем постепенно снижается и перестает детектироваться на 5 день после трансфекции. Транскрипция генов FCU1 и BsFm происходит в составе одной матричной РНК, при трансляции которой образуются дискретные целевые белки в одинаковом соотношении. Поэтому, если каким-либо методом показана экспрессия одного белка кассеты, можно считать, что второй белок также синтезируется, поскольку гены находятся в одной рамке считывания.
Дополнительно был выявлен целевой белок FCU1 с помощью иммуногистохимического окрашивания на серийных срезах тканей опухоли, фиксированных 10% нейтральным формалином и заключенных в парафин. Срезы получали через 48, 120 и 168 часов после внутриопухолевого введения ЛС. После гидратации срезов эндогенную пероксидазу инактивировали в 3% растворе Н2О2, приготовленном на забуференном физиологическом растворе (ЗФР), рН 7,4. Неспецифические взаимодействия блокировали 3% раствором бычьего сывороточного альбумина в ЗФР. Выявление целевого белка проводили методом непрямого иммунопероксидазного окрашивания с использованием кроличьих поликлональных антител к цитозиндезаминазе (Cytosine deaminase Polyclonal Antibody, Bioss Antibodies Inc), антивидовых антител, меченных биотином (Santa Cruz Biotechnology), стрептавидина, конъюгированного с пероксидазой хрена (BD Bioscience), с последующим хромогенным окрашиванием (Liquid DAB+Chromogen substrate System; Dako Cytomation). В качестве контроля использовали неспецифические иммуноглобулины кролика. Срезы докрашивали гематоксилином, обезвоживали и заключали в канадский бальзам. Анализ препаратов проводили под микроскопом Olympus ВХ51. При анализе срезов ткани опухоли было выявлено специфическое окрашивание. Было показано, что максимальное иммунопероксидазное окрашивание наблюдается в опухоли через 48 часов после введения препарата FCU1-BsFm/PP, интенсивность окрашивания снижалась к 5-7 суткам наблюдения (фигура 9).
Пример 5. Оценка продукции флагеллина трансфицированными клетками
Флагеллин способен связываться с TLR5 рецепторами на поверхности клеток, что приводит к активации различных сигнальных путей и продукции ряда цитокинов во внеклеточную среду. Так, стимулированные флагеллином дендритные клетки усиленно продуцируют TNF-α, IL-8, IL-1β, МСР-1, MIP-1α, MIP-1β, RANTES. Взаимодействие с рецептором TLR-5 высокоспецифично и происходит при низких концентрациях флагеллина - до 8,5×10-10 М. Была использована способность клеток линии А-549 секретировать интерлейкин-8 (IL-8) при добавлении в культуральную среду. Для этого предварительно проводили транзиентную трансфекцию клеток линии СТ26 плазмидной конструкцией FCU1-BsFm, а также контрольных конструкций CMV-BsFm (положительный контроль), CMV-FCU1 (отрицательный контроль). Через 48 часов после трансфекции собирали кондиционированные среды. Среды очищали от клеток с помощью центрифугирования, готовили разведения в 10, 100 и 500 раз в смеси со свежей средой DMEM/F12 с 10% сывороткой, после чего добавляли разведенные кондиционированные среды к клеткам А-549. Клетки инкубировали 24 часа при 37°С в атмосфере 5% CO2, после чего отбирали среды, кондиционированные клетками А-549, и определяли в них концентрацию IL-8 с помощью твердофазного иммуноферментного анализа (ELISA), при этом использовали коммерческий набор ELISA для человеческого IL8/CXC (HUMAN IL-8 / CXCL8 ELISA KIT, Sigma-Aldrich, США). В лунки 96-луночного ИФА-планшета с иммобилизованными первичными антителами к IL-8 вносили образцы кондиционированной среды, разбавленной в 10, 100 или 500 раз, а также концентрационные стандарты. Затем добавляли детектирующие биотинилированные антитела против IL-8 и далее раствор меченного пероксидазой хрена стрептавидина. После отмывки добавляли проявляющий субстрат, после развития окраски на спектрофотометре Benchmark™ Plus Microplate Reader (Bio-Rad, США) определяли оптическое поглощение при 450 нм. С целью подбора оптимальных условий содержание IL-8 в образцах определяли в два этапа.
На основании калибровочного графика (зависимость оптической плотности при λ=450 нм раствора окрашенного продукта реакции от концентрации белка, полученного для концентрационных стандартов) определяли уровень продукции IL-8 клетками А-549, инкубированными с различными разведениями кондиционированных сред клеток СТ26, трансфицированных целевой и контрольными плазмидными конструкциями. В таблице 1 показано, что среды клеток СТ26, трансфицированных FCU1-BsFm и CMV-BsFm, содержащим ген флагеллина, индуцировали высокий уровень продукции уровень продукции белка IL-8 клетками А-549 при разведениях от 10 до 500 раз, что свидетельствует о высоком уровне продукции флагеллина клетками СТ26. При этом, по-видимому, бицистронная конструкция FCU1-BsFm обеспечивала немного меньший уровень продукции флагеллина, т.к. при разведении среды уже в 100 раз клетки А549 отвечали более низким уровнем продукции IL-8, чего не наблюдалось на положительном контроле (CMV-BsFm). Интересно, что при небольшом разведении (в 10 раз) кондиционированная среда, полученная от клеток, трансфицированных плазмидной конструкцией CMV-FCU1, также индуцировала небольшую продукцию IL-8, однако при большем разведении этот эффект пропадал. Это, вероятно, связано с накоплением какого-либо другого фактора в среде трансфицированных клеток СТ26, также способного запускать ответ в клетках А549. Таким образом, бицистронная плазмидная конструкция FCU1-BsFm в клетках СТ26 обеспечивает высокий уровень биологической активности флагеллина в кондиционированной среде, хотя и несколько меньший по сравнению с моноцистронной конструкцией pCMV-BsFm. Это согласуется с выводами работы о том, что экспрессия второго цистрона в бицистронной конструкции на основе Р2А может быть снижена по сравнению с моноцистронным вектором. Таким образом, созданная бицистронная плазмидная конструкция FCU1-BsFm обладает требуемой биологической активностью и может быть использована в экспериментах на животных.
Пример 6. Терапевтическая эффективность препарата FCU1-BsFm/PP в зависимости от концентрации блок-сополимера
Сравнительную оценку терапевтической эффективности плазмидной конструкции проводили на мышах линии BALB/c (самки, возраст 6-8 недель, масса 18-20 г. ) с привитой карциномой толстой кишки СТ26. Животные были получены из питомника «Пущино» ФГБУН ФИБХ РАН. Оценку эффективности противоопухолевого лечения животных в работе проводили на основе мультипараметрического анализа показателей: объема опухоли (V, мм3), торможения роста опухоли (ТРО, %), степени торможения опухоли (Т/С, %), задержки роста опухоли (Т-С, сутки), количества погибших клеток (lgn), медианы выживаемости по кривой, построенной по методу Каплан-Мейера, и увеличения продолжительности жизни животных (УПЖ, %). Минимальные значения показателей, характеризующие лечение как эффективное: ТРО≥70%, Т/С≤30%, УПЖ≥50%, lgn≥0,7 (Руководство по экспериментальному (доклиническому) изучению новых фармакологических веществ. Часть 1. М.: Гриф и К, 2012; Cytotoxic Anticancer Drugs: Models and Concepts for Drug Discovery and Development. Proceedings of the Twenty-Second Annual Cancer Symposium Detroit, Michigan, USA - April 26-28, 1990).
В качестве модели солидной опухоли использовали карциному толстой кишки СТ26 (АТСС® CRL-2638™). СТ26 культивировали в пластиковых флаконах с поверхностью роста клеток 75 см2 (Costar, США) в стандартных условиях (37°С, 5% СО2) в среде RPMI1640 с добавлением 12,5% FCS и антибиотиков - 60 мкг/мл пеницилина, 100 мкг/мл стрептомицина и 0,25 мкг/мл амфотерицина. Для прививки животным использовали клетки 2-4 пассажа. Клеточную суспензию вводили мышам подкожно по 0,1×106 клеток/мышь на боковую поверхность тела в объеме 0,1 мл. После инокуляции опухолевых клеток животные находились под ежедневным мониторингом с оценкой их общего состояния и состояния опухоли в месте прививки. Воздействие начинали на 13-й день после инокуляции опухолевых клеток, когда средний объем опухоли в группе составлял 70±15 мм3.
Препарат FCU1-BsFm/PP вводили внутриопухолево 3 раза за время эксперимента с интервалом 3 и 5 дней. Внутрибрюшинное введение пролекарства 5-фторцитозин (5-FC) в составе препарата Анкотил (MEDA PHARMACEUTICALS SWITZERLAND, GmbH, Швейцария) начинали через 24 часа после введения препарата FCU1-BsFm/PP в разовой дозе 250 мг/кг, 2 раза в сутки с интервалом 8-10 часов в течение 15 дней (курсовая доза -7500 мг/кг). Выбор дозы пролекарства осуществляли на основании данных литературы, где указано, что в качестве оптимальной дозы пролекарства 5-FC является 500 мг/кг (Kim et al., Transl Oncol, 2018. 11(1), 74-85; Mitchell et al., Neuro Oncol, 2017. 19(7), 930-939). Поскольку 5-FC при парентеральном введении имеет короткий период полувыведения (до 4-6 часов), для поддержания его концентрации в крови 5-FC вводили дважды в сутки с интервалом 8-10 часов. 5-фторцитозин вводили внутрибрюшинно, так как в соответствии с российскими и международными рекомендациями данный вид введения является альтернативным для парентерального введения. Результаты исследования представлены в таблице 2.
Проведенные исследования показали, что наиболее выраженный ингибирующий эффект наблюдается в случае использования препарата FCU1-BsFm/PP с конечной концентрацией блок-сополимера РР 12,5 мкМ при введении с интервалом 5 дней: ТРО после окончания лечения составило 71%. Уменьшение интервала введения препарата с концентрацией РР 12.5 мкМ до 3 дней приводило к снижению противоопухолевого действия: после окончания лечения (28-й день роста опухоли) ТРО составляло 58%, через 7 дней после окончания лечения (35-й день роста опухоли) ТРО составляло 47%. При сохранении режима трехкратного введения с интервалом 5 дней, но уменьшении конечной концентрации РР до 7,5 мкМ было отмечено отсутствие лечебного действия: динамика роста опухоли не отличалась от таковой в контрольной группе (показатель ТРО=15% свидетельствует об отсутствии противоопухолевого эффекта). Таким образом, в терапевтической системе было показано, что конечная концентрация РР 12,5 мкМ в FCU1-BsFm/PP является наиболее эффективной. Все дальнейшие исследования по подбору эффективной дозы и схемы введения разрабатываемого препарата FCU1-BsFm/PP проводились с использованием концентрации блок-сополимера, равной 12,5 мкМ.
Пример 7. Терапевтическая эффективность с варьированием кратности введения FCU1-BsFm/PP и интервалов между введениями
Интервал внутриопухолевого введения препарата FCU1-BsFm/PP варьировали от 5 до 7 дней, пролекарство 5-фторцитозин во всех опытных группах мышей вводили внутрибрюшинно дважды в день (интервал 8-10 часов) в суточной дозе 500 мг/кг или 400 мкг/кг.Длительность введения 5-фторцитозина зависела от кратности введения FCU1-BsFm/PP: каждая группа получала внутрибрюшинные инъекции 5-фторцитозина в течение 7 дней после последнего введения FCU1-BsFm/PP. Таким образом, при двукратном применении FCU1-BsFm/PP 5-фторцитозин вводили в течение 11 дней, при трехкратном - 15, при четырехкратном - 19. Всего было сформировано 6 экспериментальных групп. День 1 -день начала лечения мышей. Дизайн исследования представлен на фигуре 10.
Терапевтический эффект возрастал при переходе от двукратного введения (интервал 5 дней) к трехкратному и четырехкратному введению (см. фигуру 11). При двукратном введении мышам FCU1-BsFm/PP в разовой дозе 0,04 мкг ДНК/мм3 с интервалом 5 дней (группа FCU1-BsFm (0,04×2)/5-FC (500) ТРО составило 61% после окончания лечения (21-й день роста опухоли), конечная величина Т/С составила 67% (Т/С для контрольных животных составило 100%), задержка роста опухоли составила 4,3 дня.
Увеличение кратности введения FCU1-BsFm/PP в разовой дозе 0,04 мкг ДНК/мм3 до трех раз с интервалом 5 дней (группа FCU1-BsFm (0.04×3)/5-FC (500) привело к повышению противоопухолевого эффекта: после окончания лечения (25-е сутки наблюдения) ТРО составило 80%, конечный показатель Т/С уменьшился до 28% («++»), а задержка роста опухоли увеличилась до 10,6 дней. Этот режим оказался эффективным по показателю логарифма числа погибших опухолевых клеток (lgn=0,7) и индексу торможения метастазирования, который составлял 72,8%. При сохранении режима трехкратного введения, но при увеличении интервала введения FCU1-BsFm/PP до 7 дней (группа FCU1-BsFm (0,04×3/7)/5-FC (500) была отмечена слабо выраженная тенденция к снижению эффективности лечения мышей с саркомой S37: Т/С составило 42% («+»); Т-С - 6,3 дней (см. фигуру 12). Увеличение кратности введения в разовой дозе 0,04 мкг ДНК/мм3 с трех до четырех раз с интервалом 5 дней (группа FCU1-BsFm (0,04×4)/5-FC (500) в сочетании с 5-FC (суточная доза 500 мг/кг, курсовая - 9500 мг/кг) значимо не привело к увеличению противоопухолевой эффективности: после окончания лечения (30-е сутки наблюдения) ТРО составило 74%, конечный показатель Т/С - 26%, lgn - 0,7, ТМ - 60,5% (фигура 13).
Сравнение медианы выживаемости в группах выявило слабо выраженную тенденцию к ее увеличению (фигура 14): 53,0 суток против 50,0 суток в группе животных, которым вводили 5-FC в курсовой дозе 7600 мг/кг. Эти данные могут свидетельствовать о некотором снижении токсичности лечения. Торможение метастазирования составляло 63,9%. Сравнение результатов лечения мышей с саркомой S37 при трехкратном и четырехкратном введении препарата в разовой дозе 0,04 мкг ДНК/мм3 выявило сходную высокую эффективность лечения в этих группах животных. Так, через 14 дней после окончания лечения ТРО составило 72-80%, конечный показатель Т/С - 20-28%, задержка роста опухоли - 10,4-10,6 дней соответственно, lgn - 0,7, ТМ - 60,5-63,4% во всех группах животных. При этом характер кривой зависимости «доза-эффект» свидетельствовал, что достигнут максимально возможный эффект (по lgn), а потому дальнейшее повышение дозы препарата является нецелесообразным. Полученный противоопухолевый эффект был устойчивым: медиана выживаемости составила 50,0-53,0 суток, против 32,0 суток в контрольной группе животных.
Противоопухолевая эффективность двух схем введения ЛС (двукратное введение и трехкратное введение с интервалом 5 дней) на модели СТ26 была проанализирована для того, чтобы оценить воспроизводимость результатов, полученных на модели саркомы S37. Сравнение эффективности двух схем лечения ЛС проводили на половозрелых мышах BALB/c (возраст 6-8 недель, масса 18-20 г.) с привитой карциномой толстой кишки СТ26 (АТСС® CRL-2638™). Животные были получены из питомника «Пущино» ФГБУН ФИБХ РАН. Лечение начинали, когда средний объем опухоли в группе достигал 50±10 мм3. Дизайн исследования представлен на фигуре 15.
Оценку эффективности противоопухолевого лечения проводили по показателям ТРО, Т/С, Т-С, lgn, медианы выживаемости и увеличению продолжительности жизни животных. Результаты исследования представлены в таблицах 3 и 4. Проведенные исследования показали, что трехкратное внутриопухолевое введение мышам с привитой СТ26 ЛС в дозе 0,04 мкг ДНК/мм3 опухоли с интервалом 5 дней на фоне введения пролекарства 5-FC в суточной дозе 500 мг/кг в течение 15 дней (курсовая доза 7500 мг/кг) эффективно (рисунки 22 и 23): ТРО возрастало по мере увеличения срока наблюдения и составило 74%, задержка роста опухоли - 16,7 суток, показатель lgn достигал максимально возможного значения 1,1 («эффект++» по рейтингу противоопухолевой активности). Медиана выживаемости животных в опытной и контрольной группах различались на 11,1 суток - 33,0 и 44,1 суток соответственно. Двукратное введение препарата FCU1-BsFm/PP (группа сравнения) мышам в разовой дозе 0,04 мкг ДНК/мм3 с интервалом 5 дней было менее эффективным: ТРО составило 50%, показатель Т/С - 50%, задержка роста опухоли - 11,1 дня, а интегральный показатель lgn=0,7, что также характеризует наличие противоопухолевого эффекта.
Таким образом, в результате проведенных исследований на двух опухолевых моделях различного генеза было показано, что наиболее оптимальной является трехкратное за курс (с интервалом 5 дней) внутриопухолевое введение лекарственного средства в разовой дозе 0,04 мкг ДНК/мм3.
Клеточные линии меланомы в условиях in vivo обладают низкой чувствительностью к цитотоксическому агенту 5-фторурацилу, который образуется в опухоли в результате экспрессии цитозиндезаминазы дрожжей в составе плазмидной конструкции в присутствии 5-фторцитозина. Как следствие опухолевые модели меланомы мыши, в частности модель B16F0, являются низкочувствительными к воздействию системы FCU1/5-FC. Был проведен эксперимент по оценке эффективности на модели B16F0 с расчетом на то, что добавление к FCU1/5-FC гена флагелина может увеличить противоопухолевый эффект системы. В качестве групп сравнения использовали мышей с B16F0, которым вводили моногенные плазмидные конструкции, содержащие гены FCU1 и BsFm. Пролекарство 5-FC вводили внутрибрюшинно через 48 часов после введения генных конструкций в разовой дозе 500 мг/кг, 1 раз в сутки в течение 15 дней с интервалами между введениями генной конструкции (курсовая доза 7500 мг/кг). Данные по динамике роста опухолей, ТРО, СПЖ и УПЖ представлены в таблице 5. Эффективность лечения меланомы B16F0 при применении плазмидных конструкций на фоне введения пролекарства у животных была ниже, чем на моделях опухолей S37 и СТ26.
При трехкратном внутриопухолевом введении моногенной конструкции с вставкой гена BsFm (х3) с последующим ежедневным внутрибрюшинным введением PBS в течение 15 дней динамика роста меланомы B16F0 незначимо отличалась от таковой в контрольной группе. Трехкратное введение моногенной конструкции с вставкой гена FCU1 (х3) на фоне введения пролекарства не приводило к значимому увеличению противоопухолевой эффективности: ТРО составило 48% после окончания лечения, при этом было отмечено биологически незначимое увеличение продолжительности жизни до 23%. Применение бицистронной плазмидной конструкции FCU1-BsFm (х3) в комбинации с пролекарством было (так же, как и применение конструкции FCU1) малоэффективно: отмечено краткосрочное торможение первичного очага (ТРО=50%) на 16 сутки наблюдения, которое было сравнимо с таковым в группе сравнения, в которой мыши получали конструкцию с геном FCU1. Таким образом, проведенные исследования на модели меланомы B16F0 с использованием плазмидной конструкции FCU1-BsFm не выявили значимого противоопухолевого эффекта в режиме их трехкратного введения в дозе 0,08 мкг ДНК/мм3 (курсовая доза - 0,04 × (v1+v2+v3) мкг ДНК/мм3, где v1, v2 и v3 - объемы опухоли, мм3, в момент 1-го, 2-го и 3-го введений) на фоне 5-FC, который вводили через 48 часов после введения генных конструкций в разовой дозе 500 мг/кг 1 раз в сутки в течение 15 дней с интервалами между введениями плазмидной конструкции (курсовая доза 7500 мг/кг). Эти данные подтверждают предыдущие результаты о неэффективности воздействия на меланому B16F0 посредством системы FCU1/5-FC как в монорежиме, так и в комбинации с системами, активирующими противоопухолевый иммунный ответ несмотря на то, что меланома является иммуногенной опухолью.
Пример 8. Оценка долгосрочных противоопухолевых эффектов введения FCU-BsFm/PP
Для оценки развития стойкого противоопухолевого иммунитета в ответ на лечение противораковыми препаратами используют метод повторной перевивки опухоли ранее леченным животным (tumor rechallenge). В большинстве исследований в эксперимент берут животных, которые находятся в состоянии полной ремиссии 60 дней (исчезновение опухолевого узла) (Serebrovskaya et al., Cytokine, 2016. 84, 10-16; Wu et al., J Pharmacol Sci, 2016. 132(1), 24-30; Luo et al., Nat Nanotechnol, 2017. 12(7), 648-654). Животным на противоположный от регрессировавшей опухоли бок повторно прививают опухоль и наблюдают за ее развитием. Ингибирование роста привитой опухоли считается признаком развития противоопухолевого иммунитета. Оценку стойкости приобретенного противоопухолевого иммунитета проводили на мышах линии BALB/c с карциномой кишечника СТ26 и на мышах F1 с саркомой S37, излеченных после трехкратного внутриопухолевого введения препарата FCU1-BsFm в разовой дозе 0,04 мкг ДНК/мм3 опухоли в сочетании с 5-фторцитозином, который вводили внутрибрюшинно в разовой дозе 250 мг/кг дважды в сутки (суточная доза - 500 мг/кг, курсовая - 7500 мг/кг).
Формировали две группы мышей: опытная группа - излеченные мыши, группа сравнения - нативные мыши (нелеченые). Раковые клетки СТ26 прививали подкожно по 0,1×106 клеток/мышь; штамм саркомы S37 прививали подкожно по 1×106 клеток/мышь F1. После инокуляции опухолевых клеток животные не подвергались никакому воздействию и находились под ежедневным мониторингом с оценкой их общего состояния и состояния опухоли в месте прививки. Наблюдение продолжали в течение 60 дней. Схема эксперимента представлена на фигуре 16, повторное привитие опухоли мышам, излечившимся от карциномы СТ26, проводили через 60 дней.
У всех мышей в группе сравнения наблюдали образование опухоли - саркомы S37 (фигура 17). У мышей после излечения (полная ремиссия) в результате воздействия FCU1-BsFm/PP наблюдали образование опухолевых очагов, однако с задержкой, которая составляла от 4 до 9 дней. Неуспех данного эксперимента может быть обусловлен увеличением времени (125 дней вместо 60 дней) между констатацией полной ремиссии у животных до проведения процедуры повторного привития опухоли (125 дней), поскольку, согласно стандартным международным протоколам, данный интервал должен составлять 60 дней.
В случае повторной прививки опухоли СТ26 излечившимся мышам развитие опухолевых образований наблюдали только в контрольных группах (первичное привитие опухоли нативным животным) (фигура 18), тогда как у мышей после излечения в результате введения препарата FCU1-BsFm/PP (полная ремиссия) опухоль не развилась в течение 60 дней после повторной прививки, что может свидетельствовать о стойком приобретенном противоопухолевом иммунитете. Таким образом, было показано, что лечение FCU1-BsFm/PP в сочетании с 5-фторцитозином может обеспечивать защиту излеченных животных от рецидивирования и возврата к клиническим проявлениям заболевания.
Пример 9. Сравнительная оценка противоопухолевой эффективности генотерапевтического препарата и пролекарства 5-фторцитозин на опухолях разного гистогенеза
На следующий день после того, как средний объем опухолей СМТ167 у мышей С57 В1/6 составил 50±20 мм3, было инициировано лечение животных препаратом FCU1-BsFm/PP в дозе 0,04 мкг ДНК/мм3 опухоли в сочетании с пролекарством 5-фторцитозин. Результаты исследований представлены на фигуре 19. Было установлено, что после окончания лечения (24-31 дни) в группах мышей, которым вводили FCU1-BsFm/PP на фоне пролекарства, наблюдалось торможение роста опухоли, максимальное значение которого достигало 52%. На 35 день наблюдения (через 10 дней после окончания курса терапии) показатель ТРО снижался до 42%. Средняя продолжительность жизни в группе мышей, получавших лечение, была сравнима с контрольной группой: 47±6 суток vs 40±5 дней. Таким образом, выраженный противоопухолевый эффект в отношении карциномы легкого СМТ 167 при применении FCU1-BsFm/PP был слабо выраженным.
В день, когда средний объем опухолей РШМ5 у мышей F1 (C57B1/6j × СВА) составил 60±10 мм3 было инициировано лечение животных препаратом FCU1-BsFm/PP в дозе 0,04 мкг ДНК/мм3 опухоли в сочетании с пролекарством 5-фторцитозин. Результаты исследований представлены на фигуре 20. Было установлено, что эффективность лечения рака шейки матки РШМ5 при применении у животных FCU1-BsFm/PP в комбинации с пролекарством 5-FC проявлялась в торможении роста опухоли в диапазоне 48-68%, достигая максимального значения 68% на 21 сутки и снижаясь до 48% к 32 суткам наблюдения. Т/С на момент окончания лечения достигала 33%. Однако отмеченное торможение роста первичного очага не приводило к существенному увеличению продолжительности жизни животных, которое составило 13%.
В день, когда средний объем опухоли 4Т1 у мышей BALB/c составил 50±10 мм3, было инициировано лечение животных препаратом FCU1-BsFm/PP в дозе 0,04 мкг ДНК/мм3 опухоли в сочетании с пролекарством 5-фторцитозин. 4Т1 - крайне агрессивная метастазирующая опухоль, имитирующая трижды негативный рак молочной железы 4 стадии человека. Результаты исследований представлены на фигуре 21. Противоопухолевая эффективность препарата FCU1-BsFm/PP в сочетании с 5-FC на модели рака молочной железы 4Т1 проявлялась в статистически значимом (р=0,008) торможении роста опухоли в течение 8 дней (65-73% на 17-24 сутки наблюдения). После окончания лечения конечный показатель Т/С составил 28%, задержка роста опухоли - 14 дней (время, необходимое для достижения объема опухоли 100 мм3). Показатель средней продолжительности жизни животных в опытной группе, получавших лечение препаратом FCU1-BsFm/PP и пролекарством 5-FC, был значительно выше, чем в контрольной группе: УПЖ составило 42%.
На следующий день после того, как средний объем опухоли SC-OV-3 у мышей Nu/j составил 50±10 мм3, было инициировано лечение животных препаратом FCU1-BsFm/PP в дозе 0,04 мкг ДНК/мм3 опухоли в сочетании с пролекарством 5-фторцитозин. Результаты исследования представлены на фигуре 22. Представленные данные демонстрируют значимый лечебный эффект генотерапевтического препарата и пролекарства в отношении опухоли человека SC-OV-3: ТРО составило более 70% в течение 27 дней после окончания лечения, причем максимальное ТРО (85%) отмечалось на 52 день. Также наблюдали задержку роста опухоли, которая составляла 29 дней (время, необходимое для достижения объема опухоли 300 мм3). Показатель Т/С на этот момент составил 15%, что классифицируется как высокая противоопухолевая активность (++++). Медиана выживаемости животных в опытной группе превысила таковую в контрольной группе на 33 дня (96 дней и 63 дня соответственно). Таким образом, в отношении ксенографта SC-OV-3 был получен значимый лечебный эффект.
В день, когда средний объем опухоли НСТ 116 у мышей Nu/j составил 50±10 мм3, было инициировано лечение животных препаратом FCU1-BsFm/PP в дозе 0,04 мкг ДНК/мм3 опухоли в сочетании с пролекарством 5-фторцитозин. Результаты исследования представлены на фигуре 23. Было установлено, что введение препарата FCUl-BsFm/PP и пролекарства 5-FC мышам с НСТ116 (колоректальная аденокарцинома) имело высокую эффективность. ТРО возрастало по мере увеличения срока наблюдения и составило 70-78% на 5-20 сутки после окончания лечения. Задержка роста опухоли составила 31 день (время, необходимое для достижения объема опухоли 300 мм3). Показатель Т/С на момент окончания лечения составил 20%. Медианы средней продолжительности жизни животных в опытной и контрольной группах различались на 35 дней - 148 дней и 83 дня соответственно. Таким образом, в отношении ксенографта НСТ116 был получен значимый лечебный эффект.
Иммунодефицитные мыши линии Nu/j, самки с привитыми клетками А431 (плоскоклеточный рак кожи) были распределены в 2 группы: опытную и контрольную. В каждой группе было по 10 мышей. На следующий день после того, как средний объем опухолей у животных в группах составил 80±10 мм3, мышам опытной группы начинали внутриопухолевое введение генотерапевтического препарата FCU1-BsFm/PP в дозе 0,04 мкг ДНК/мм3 опухоли. Результаты исследования представлены на фигуре 24. ТРО составило 62-75% в течение 18 суток после окончания лечения. Также наблюдали задержку роста опухоли, которая составляла 23 дня (время, необходимое для достижения объема опухоли 100 мм3). Показатель Т/С на этот момент составил 25%, излеченность на этой модели ксенографта достигла 20%.
Основные выводы на основе примеров 7-9:
1. Наиболее эффективный режим введения генотерапевтического препарата FCU1-BsFm/PP - 3-х кратное за курс (с интервалом 5 дней) внутриопухолевое введение FCU1-BsFm/PP в разовой дозе 0,04 мкг ДНК/мм3 (курсовая доза - 0,04 × (v1+v2+v3) мкг ДНК/мм3, где v1, v2 и v3 - объемы опухоли, мм3, в момент 1-го, 2-го и 3-го введений FCU1-BsFm/PP) на фоне инфузий 5-фторцитозина в диапазоне суточной дозы 500 мг/кг по схеме: введение FCU1-BsFm/PP в Дни 1, 6, 11; введение 5-фторцитозина в Дни 2-5, 7-10, 12-18.
2. На моделях перевиваемых опухолей различного гистогенеза у иммунокомпетентных мышей был определен спектр противоопухолевой активности препарата FCU1-BsFm/PP при системном введении 5-фторцитозина. Высокий по критериям эффективности противоопухолевый эффект лечения наблюдали на моделях саркомы S37, опухолях толстой кишки С26 и СТ26, карциномы молочной железы 4Т1, умеренный - на модели рака шейки матки РШМ5, низкую эффективность лечения наблюдали на модели СМТ167. В отношении меланомы М3 лечение оказалось неэффективным. Таким образом, противоопухолевая эффективность комбинации препарата FCU1-BsFm/PP и пролекарства 5-фторцитозин зависит от гистологической формы опухоли и снижается в ряду S37>С26 ~ СТ26>4Т1>РМШ5>СМТ 167>>М3. На моделях ксенографтов опухолей человека различного гистогенеза у иммунодефицитных мышей была выявлена противоопухолевая активность препарата FCU1-BsFm/PP при системном введении 5-фторцитозина. Высокий по критериям эффективности противоопухолевый эффект лечения наблюдали на модели колоректальной карциномы человека НСТ116, менее выраженный, но биологически значимый эффект - на модели плоскоклеточного рака кожи человека А431.

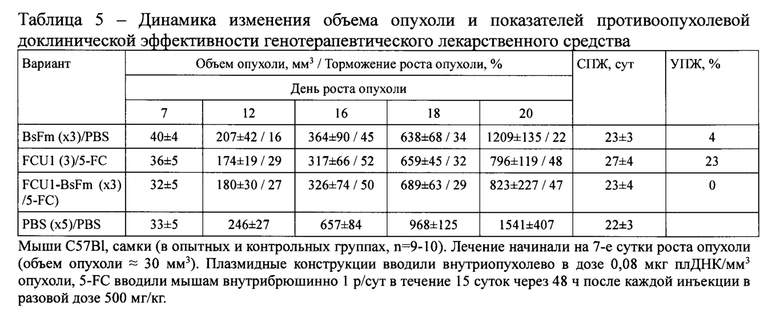
--->
<?xml version="1.0" encoding="UTF-8"?>
<!DOCTYPE ST26SequenceListing PUBLIC "-//WIPO//DTD Sequence Listing
1.3//EN" "ST26SequenceListing_V1_3.dtd">
<ST26SequenceListing originalFreeTextLanguageCode="ru"
nonEnglishFreeTextLanguageCode="ru" dtdVersion="V1_3" fileName="PP
для лечения солидных опухолей.xml" softwareName="WIPO Sequence"
softwareVersion="2.2.0" productionDate="2023-02-28">
<ApplicantFileReference>ABC1</ApplicantFileReference>
<ApplicantName languageCode="ru">Федеральное государственное
бюджетное учреждение "Центр стратегического планировния и
управления медико-биологическими рисками здоровью" Федерального
медико-биологического агентства</ApplicantName>
<ApplicantNameLatin>Federal State Budgetary Institution "Center
for Strategic Planning and Management of Biomedical Health
Risks" of the Federal Medical Biological
Agency</ApplicantNameLatin>
<InventionTitle languageCode="ru">Генотерапевтический препарат
FCU1-BsFm/PP для лечения солидных опухолей</InventionTitle>
<SequenceTotalQuantity>3</SequenceTotalQuantity>
<SequenceData sequenceIDNumber="1">
<INSDSeq>
<INSDSeq_length>6224</INSDSeq_length>
<INSDSeq_moltype>DNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..6224</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other DNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q2">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>Saccharomyces cerevisiae, Bacillus
subtilis</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>ggtaccgagctcttacgcgtgctagccctaatagtaatcaattacgggg
tcattagttcatagcccatatatggagttccgcgttacataacttacggtaaatggcccgcctggctgac
cgcccaacgacccccgcccattgacgtcaataatgacgtatgttcccatagtaacgccaatagggacttt
ccattgacgtcaatgggtggagtatttacggtaaactgcccacttggcagtacatcaagtgtatcatatg
ccaagtacgccccctattgacgtcaatgacggtaaatggcccgcctggcattatgcccagtacatgacct
tatgggactttcctacttggcagtacatctacgtattagtcatcgctattaccatggtgatgcggttttg
gcagtacatcaatgggcgtggatagcggtttgactcacggggatttccaagtctccaccccattgacgtc
aatgggagtttgttttggcaccaaaatcaacgggactttccaaaatgtcgtaacaactccgccccattga
cgcaaatgggcggtaggcgtgtacggtgggaggtctatataagcagagctggtttagtgaaccgtcagat
ccgctagcgctaccggactcagatcgggctcgagctcaagctttattgcggtagtttatcacagttaaat
tgctaacgcagtcagtgcttctgacacaacagtctcgaacttaagctgcagaagttggtcgtgaggcact
gggcaggtaagtatcaaggttacaagacaggtttaaggagaccaatagaaactgggcttgtcgagacaga
gaagactcttgcgtttctgataggcacctattggtcttactgacatccactttgcctttctctccacagg
tgtccactcccagttcaattacagctcttaaggctagagtacttaatacgactcactataggctagcctc
gagaattcacgcgtggtacctctagagtcgacgaattccaccatggtgacagggggaatggcaagcaagt
gggatcagaagggtatggacattgcctatgaggaggcggccttaggttacaaagagggtggtgttccaat
tggcggatgtcttatcaataacaaagacggaagtgttctcggtcgtggtcacaacatgagatttcaaaag
ggatccgccacactacatggtgagatctccactttggaaaactgtgggagattagagggcaaagtgtaca
aagataccactttgtatacgacgctgtctccatgcgacatgtgtacaggtgccatcatcatgtatggtat
tccacgctgtgttgtcggtgagaacgttaatttcaaaagtaagggcgagaaatatttacaaactagaggt
cacgaggttgttgttgttgacgatgagaggtgtaaaaagatcatgaaacaattcatcgatgaaagacctc
aggattggtttgaagatattggtgaggcttcggaaccatttaagaacgtctacttgctacctcaaacaaa
ccaattgctgggtttgtacaccatcatcagaaataagaatacaactagacctgatttcattttctactcc
gatagaatcatcagattgttggttgaagaaggtttgaaccatctacctgtgcaaaagcaaattgtggaaa
ctgacaccaacgaaaacttcgaaggtgtctcattcatgggtaaaatctgtggtgtttccattgtcagagc
tggtgaatcgatggagcaaggattaagagactgttgtaggtctgtgcgtatcggtaaaattttaattcaa
agggacgaggagactgctttaccaaagttattctacgaaaaattaccagaggatatatctgaaaggtatg
tcttcctattagacccaatgctggccaccggtggtagtgctatcatggctacagaagtcttgattaagag
aggtgttaagccagagagaatttacttcttaaacctaatctgtagtaaggaagggattgaaaaataccat
gccgccttcccagaggtcagaattgttactggtgccctcgacagaggtctagatgaaaacaagtatctag
ttccagggttgggtgactttggtgacagatactactgtgttaccggttccggagccacgaacttctctct
gctgaagcaagcaggagacgtggaggagaatcccgggcctatgagaatgcagctgctgctgctgatcgcc
ctgtccctggccctggtgacaaatagcatgagaatcaatcacaatatcgccgccctgaatacactgaaca
ggctgagctcccagaacagcgccagccagaagaatatggagaagctgagctccggcctgaggatcaatag
agccggcgatgacgccgccggcctggctattagcgagaagatgaggggccagatcagaggcctggagatg
gccagcaagaactcccaggacggcatcagcctgatccagacagccgagggcgccctgacagagacacacg
ccatcctgcagagggtgagggagctggtggtgcaggccggcaatacaggcacccaggataaggccaccga
tctgcagtccatccaggacgagatctccgccctgaccgatgagatcgacggcatctcccagagaaccgag
tttaacggcaagaagctgctggatggcacctacaaggtggacaccgccacccccgccaaccagaagaatc
tggtgtttcagatcggcgcccaggccacacagcagatctccgtgaatatcgaggatatgggcgccgacgc
cctgggcatcaaggaggctgatggcagcatcgccgccctccactccgtgaacgatctggatgtgaccaag
tttgccgacaatgccgccgacaccgccgatatcggctttgacgcccagctgaaggtggtggatgaggcca
tcaaccaggtgagctcccagagggccaagctgggcgccgtgcagaacagactggagcacaccatcaataa
tctgagcgcctccggcgagcagctgaccgctgctgagagcaggatcagagacgtggacatggccaaggag
atgtccgagtttaccaagaataatatcctgagccaggcctcccaggccatgctggcccaggctaaccagc
agccccagaatgtgctgcagctgctgaggtgaggccggccgcttcgagcagacatgataagatacattga
tgagtttggacaaaccacaactagaatgcagtgaaaaaaatgctttatttgtgaaatttgtgatgctatt
gctttatttgtaaccattataagctgcaataaacaagttaacaacaacaattgcattcattttatgtttc
aggttcagggggaggtgtgggaggttttttaaagcaagtaaaacctctacaaatgtggtaaaatcgataa
ggatccgtcgaccgatgcccttgagagccttcaacccagtcagctccttccggtgggcgcggggcatgac
tatcgtcgccgcacttatgactgtcttctttatcatgcaactcgtaggacaggtgccggcagcgctcttc
cgcttcctcgctcactgactcgctgcgctcggtcgttcggctgcggcgagcggtatcagctcactcaaag
gcggtaatacggttatccacagaatcaggggataacgcaggaaagaacatgtgagcaaaaggccagcaaa
aggccaggaaccgtaaaaaggccgcgttgctggcgtttttccataggctccgcccccctgacgagcatca
caaaaatcgacgctcaagtcagaggtggcgaaacccgacaggactataaagataccaggcgtttccccct
ggaagctccctcgtgcgctctcctgttccgaccctgccgcttaccggatacctgtccgcctttctccctt
cgggaagcgtggcgctttctcatagctcacgctgtaggtatctcagttcggtgtaggtcgttcgctccaa
gctgggctgtgtgcacgaaccccccgttcagcccgaccgctgcgccttatccggtaactatcgtcttgag
tccaacccggtaagacacgacttatcgccactggcagcagccactggtaacaggattagcagagcgaggt
atgtaggcggtgctacagagttcttgaagtggtggcctaactacggctacactagaagaacagtatttgg
tatctgcgctctgctgaagccagttaccttcggaaaaagagttggtagctcttgatccggcaaacaaacc
accgctggtagcggtggtttttttgtttgcaagcagcagattacgcgcagaaaaaaaggatctcaagaag
atcctttgatcttttctacggggtctgacgctcagtggaacgaaaactcacgttaagggattttggtcat
gagattatcaaaaaggatcttcacctagatccttttaaattaaaaatgaagttttaaatcaatctaaagt
atatatgagtaaacttggtctgacagttaccaatgcttaatcagtgaggcacctatctcagcgatctgtc
tatttcgttcatccatagttgcctgactccccgtcgtgtagataactacgatacgggagggcttaccatc
tggccccagtgctgcaatgataccgcgagacccacgctcaccggctccagatttatcagcaataaaccag
ccagccggaagggccgagcgcagaagtggtcctgcaactttatccgcctccatccagtctattaattgtt
gccgggaagctagagtaagtagttcgccagttaatagtttgcgcaacgttgttgccattgctacaggcat
cgtggtgtcacgctcgtcgtttggtatggcttcattcagctccggttcccaacgatcaaggcgagttaca
tgatcccccatgttgtgcaaaaaagcggttagctccttcggtcctccgatcgttgtcagaagtaagttgg
ccgcagtgttatcactcatggttatggcagcactgcataattctcttactgtcatgccatccgtaagatg
cttttctgtgactggtgagtactcaaccaagtcattctgagaatagtgtatgcggcgaccgagttgctct
tgcccggcgtcaatacgggataataccgcgccacatagcagaactttaaaagtgctcatcattggaaaac
gttcttcggggcgaaaactctcaaggatcttaccgctgttgagatccagttcgatgtaacccactcgtgc
acccaactgatcttcagcatcttttactttcaccagcgtttctgggtgagcaaaaacaggaaggcaaaat
gccgcaaaaaagggaataagggcgacacggaaatgttgaatactcatactcttcctttttcaatattatt
gaagcatttatcagggttattgtctcatgagcggatacatatttgaatgtatttagaaaaataaacaaat
aggggttccgcgcacatttccccgaaaagtgccacctgacgcgccctgtagcggcgcattaagcgcggcg
ggtgtggtggttacgcgcagcgtgaccgctacacttgccagcgccctagcgcccgctcctttcgctttct
tcccttcctttctcgccacgttcgccggctttccccgtcaagctctaaatcgggggctccctttagggtt
ccgatttagtgctttacggcacctcgaccccaaaaaacttgattagggtgatggttcacgtagtgggcca
tcgccctgatagacggtttttcgccctttgacgttggagtccacgttctttaatagtggactcttgttcc
aaactggaacaacactcaaccctatctcggtctattcttttgatttataagggattttgccgatttcggc
ctattggttaaaaaatgagctgatttaacaaaaatttaacgcgaattttaacaaaatattaacgcttaca
atttgccattcgccattcaggctgcgcaactgttgggaagggcgatcggtgcgggcctcttcgctattac
gccagcccaagctaccatgataagtaagtaatattaaggtacgggaggtacttggagcggccgcaataaa
atatctttattttcattacatctgtgtgttggttttttgtgtgaatcgatagtactaacatacgctctcc
atcaaaacaaaacgaaacaaaacaaactagcaaaataggctgtccccagtgcaagtgcaggtgccagaac
atttctctatcgata</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="2">
<INSDSeq>
<INSDSeq_length>1119</INSDSeq_length>
<INSDSeq_moltype>DNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..1119</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other DNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q4">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>Saccharomyces
cerevisiae</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>atggtgacagggggaatggcaagcaagtgggatcagaagggtatggaca
ttgcctatgaggaggcggccttaggttacaaagagggtggtgttccaattggcggatgtcttatcaataa
caaagacggaagtgttctcggtcgtggtcacaacatgagatttcaaaagggatccgccacactacatggt
gagatctccactttggaaaactgtgggagattagagggcaaagtgtacaaagataccactttgtatacga
cgctgtctccatgcgacatgtgtacaggtgccatcatcatgtatggtattccacgctgtgttgtcggtga
gaacgttaatttcaaaagtaagggcgagaaatatttacaaactagaggtcacgaggttgttgttgttgac
gatgagaggtgtaaaaagatcatgaaacaattcatcgatgaaagacctcaggattggtttgaagatattg
gtgaggcttcggaaccatttaagaacgtctacttgctacctcaaacaaaccaattgctgggtttgtacac
catcatcagaaataagaatacaactagacctgatttcattttctactccgatagaatcatcagattgttg
gttgaagaaggtttgaaccatctacctgtgcaaaagcaaattgtggaaactgacaccaacgaaaacttcg
aaggtgtctcattcatgggtaaaatctgtggtgtttccattgtcagagctggtgaatcgatggagcaagg
attaagagactgttgtaggtctgtgcgtatcggtaaaattttaattcaaagggacgaggagactgcttta
ccaaagttattctacgaaaaattaccagaggatatatctgaaaggtatgtcttcctattagacccaatgc
tggccaccggtggtagtgctatcatggctacagaagtcttgattaagagaggtgttaagccagagagaat
ttacttcttaaacctaatctgtagtaaggaagggattgaaaaataccatgccgccttcccagaggtcaga
attgttactggtgccctcgacagaggtctagatgaaaacaagtatctagttccagggttgggtgactttg
gtgacagatactactgtgtt</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="3">
<INSDSeq>
<INSDSeq_length>972</INSDSeq_length>
<INSDSeq_moltype>DNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..972</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other DNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q6">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>Bacillus subtilis</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>atgagaatgcagctgctgctgctgatcgccctgtccctggccctggtga
caaatagcatgagaatcaatcacaatatcgccgccctgaatacactgaacaggctgagctcccagaacag
cgccagccagaagaatatggagaagctgagctccggcctgaggatcaatagagccggcgatgacgccgcc
ggcctggctattagcgagaagatgaggggccagatcagaggcctggagatggccagcaagaactcccagg
acggcatcagcctgatccagacagccgagggcgccctgacagagacacacgccatcctgcagagggtgag
ggagctggtggtgcaggccggcaatacaggcacccaggataaggccaccgatctgcagtccatccaggac
gagatctccgccctgaccgatgagatcgacggcatctcccagagaaccgagtttaacggcaagaagctgc
tggatggcacctacaaggtggacaccgccacccccgccaaccagaagaatctggtgtttcagatcggcgc
ccaggccacacagcagatctccgtgaatatcgaggatatgggcgccgacgccctgggcatcaaggaggct
gatggcagcatcgccgccctccactccgtgaacgatctggatgtgaccaagtttgccgacaatgccgccg
acaccgccgatatcggctttgacgcccagctgaaggtggtggatgaggccatcaaccaggtgagctccca
gagggccaagctgggcgccgtgcagaacagactggagcacaccatcaataatctgagcgcctccggcgag
cagctgaccgctgctgagagcaggatcagagacgtggacatggccaaggagatgtccgagtttaccaaga
ataatatcctgagccaggcctcccaggccatgctggcccaggctaaccagcagccccagaatgtgctgca
gctgctgaggtga</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
</ST26SequenceListing>
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| МНОГОПРОФИЛЬНЫЙ ПРОМОТОР, ЭКСПРЕССИРУЮЩИЙ ВЕКТОР И СПОСОБ ИЗБИРАТЕЛЬНОГО УБИЙСТВА РАКОВЫХ КЛЕТОК С ИХ ИСПОЛЬЗОВАНИЕМ | 2012 |
|
RU2476596C1 |
| УНИВЕРСАЛЬНЫЕ РАКОВОСПЕЦИФИЧНЫЕ ПРОМОТОРЫ И ИХ ИСПОЛЬЗОВАНИЕ В ПРОТИВООПУХОЛЕВОЙ ТЕРАПИИ | 2013 |
|
RU2539764C2 |
| ФАРМАКОЛОГИЧЕСКАЯ КОМБИНАЦИЯ ПОЛИКАТИОННОГО НОСИТЕЛЯ ПЭГ-ПЭИ-ТАТ С ЗАКЛЮЧЕННОЙ В НЕМ ПЛАЗМИДОЙ, НЕСУЩЕЙ ТЕРАПЕВТИЧЕСКИЕ ГЕНЫ HSVtk И GM-CSF ДЛЯ ЦЕЛЕЙ ГЕНОТЕРАПИИ ОПУХОЛЕВЫХ ЗАБОЛЕВАНИЙ | 2013 |
|
RU2575077C2 |
| Лекарственная комбинация для ген-иммунной терапии | 2022 |
|
RU2792683C1 |
| ПОКСВИРУСНЫЕ ОНКОЛИТИЧЕСКИЕ ВЕКТОРЫ | 2008 |
|
RU2503717C2 |
| ПОКСВИРУСНЫЕ ОНКОЛИТИЧЕСКИЕ ВЕКТОРЫ | 2008 |
|
RU2557312C2 |
| ПОКСВИРУСНЫЕ ОНКОЛИТИЧЕСКИЕ ВЕКТОРЫ | 2008 |
|
RU2508401C2 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК pGEM-Puro-DS-Apo, СОДЕРЖАЩАЯ СИНТЕТИЧЕСКИЙ ГЕН АПОПТИНА, ФЛАНКИРОВАННЫЙ ПОСЛЕДОВАТЕЛЬНОСТЯМИ ГЕНОМА ВИРУСА ОСПОВАКЦИНЫ ИЗ РАЙОНА C10L-C12L, И РЕКОМБИНАНТНЫЙ ШТАММ VVdGF-ApoS24/2 ВИРУСА ОСПОВАКЦИНЫ, ПРОДУЦИРУЮЩИЙ АПОПТИН | 2012 |
|
RU2492238C1 |
| УНИВЕРСАЛЬНЫЙ ПРОМОТОР ДЛЯ ЭКСПРЕССИИ ТЕРАПЕВТИЧЕСКИХ ГЕНОВ В КЛЕТКАХ МЛЕКОПИТАЮЩИХ | 2013 |
|
RU2551784C1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДА рСрG-СytDA/upp ДЛЯ ЭКСПРЕССИ ГИБРИДНОГО БЕЛКА ЦИТОЗИНДЕЗАМИНАЗА-УРАЦИЛФОСФОРИБОЗИЛ ТРАНСФЕРАЗА, РЕКОМБИНАНТНАЯ ПЛАЗМИДА рСрG-СytDA/upp/VP22 ДЛЯ ЭКСПРЕССИИ ГИБРИДНОГО БЕЛКА ЦИТОЗИНДЕЗАМИНАЗА-УРАЦИЛФОСФОРИБОЗИЛ ТРАНСФЕРЕЗА-VP22, ЛЕЧЕБНАЯ КОМПОЗИЦИЯ ДЛЯ ТЕРАПИИ ОНКОЛОГИЧЕСКИХ ЗАБОЛЕВАНИЙ И СПОСОБ ЕЕ ПРИМЕНЕНИЯ | 2012 |
|
RU2580220C2 |
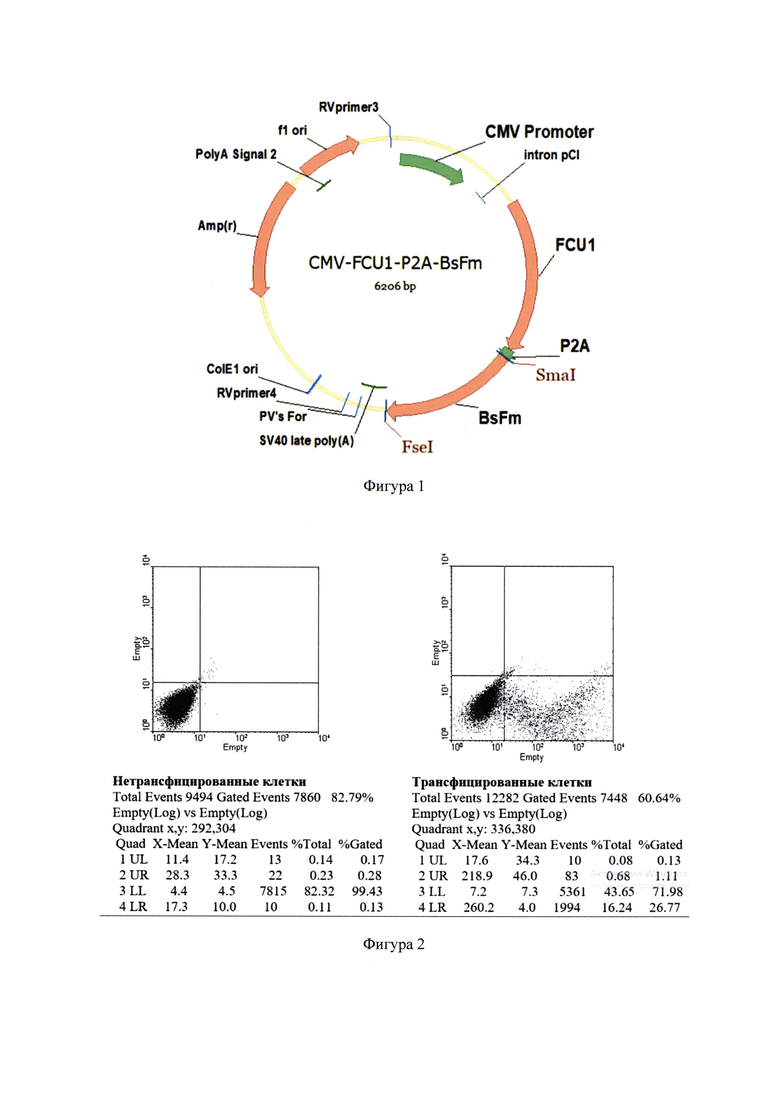

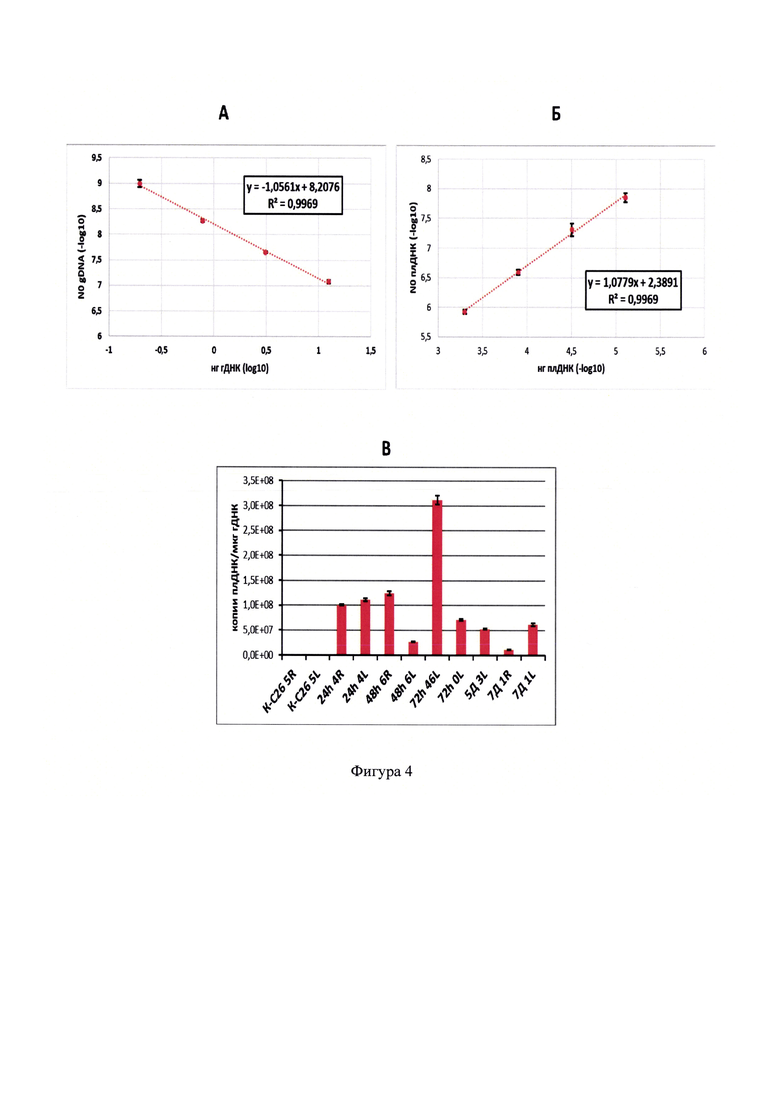
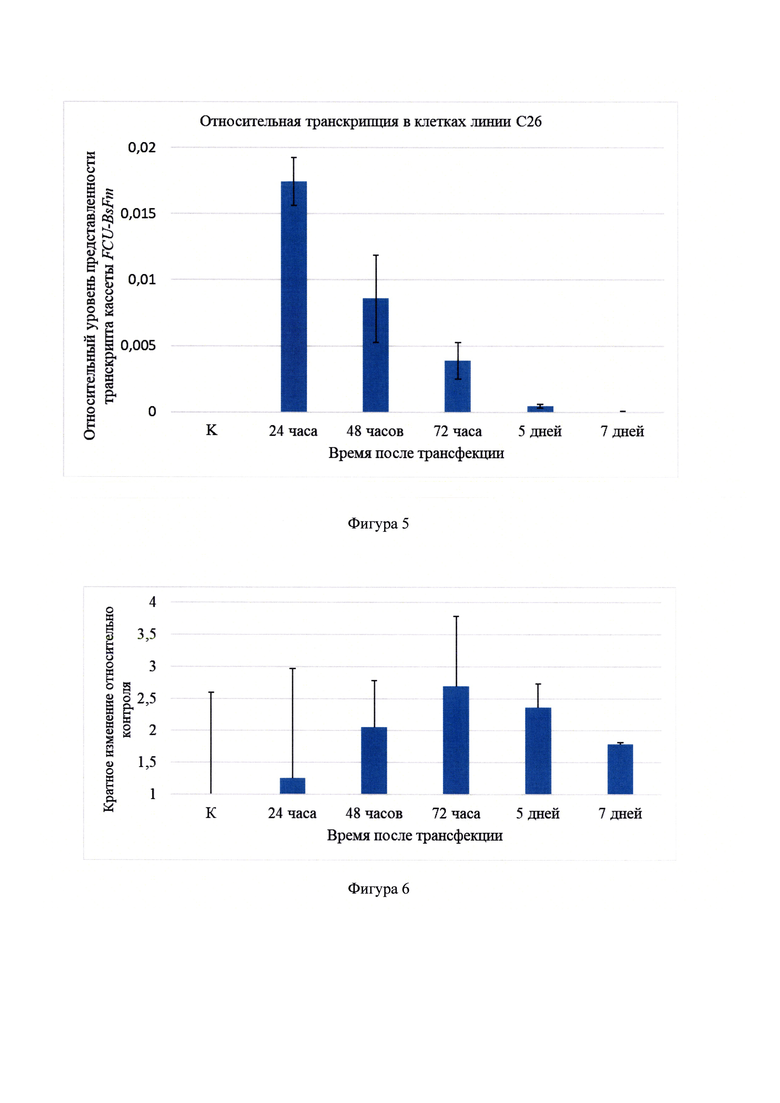
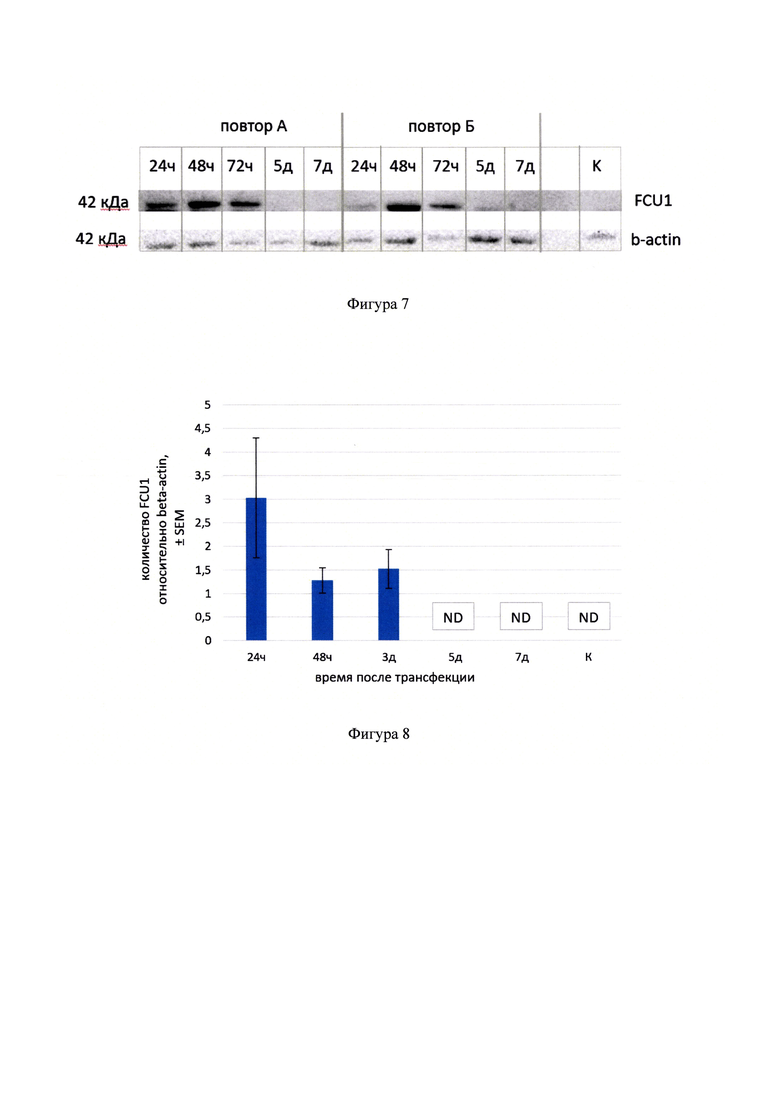
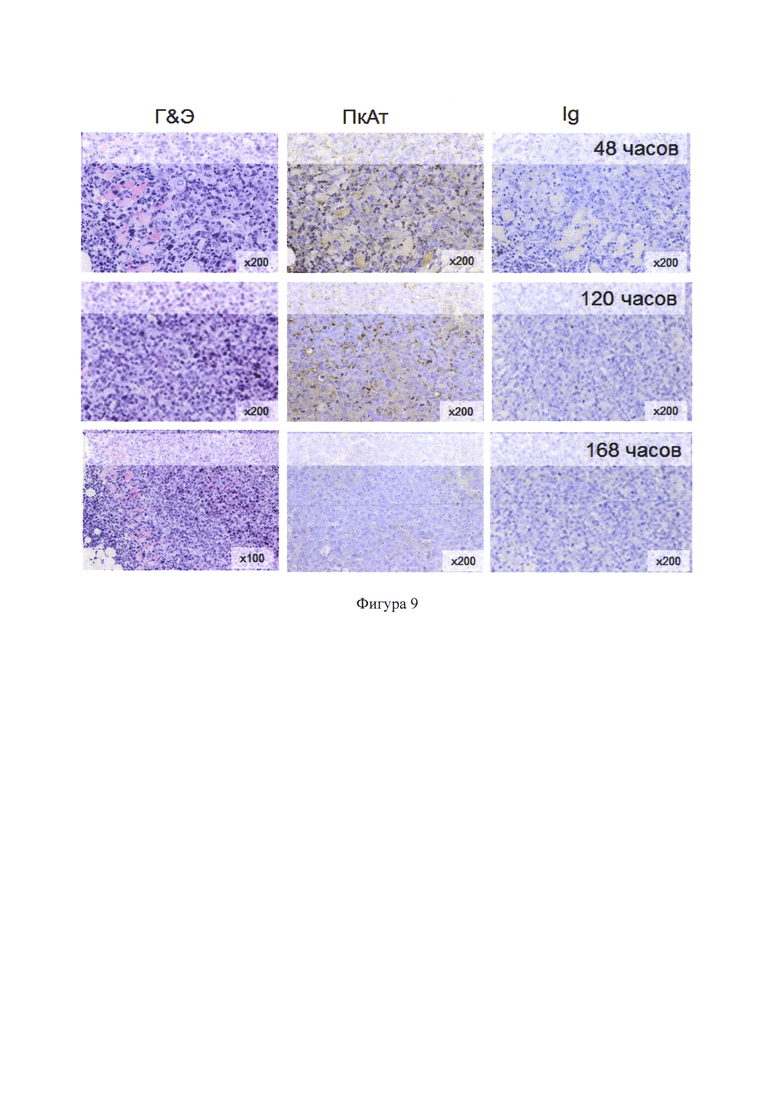

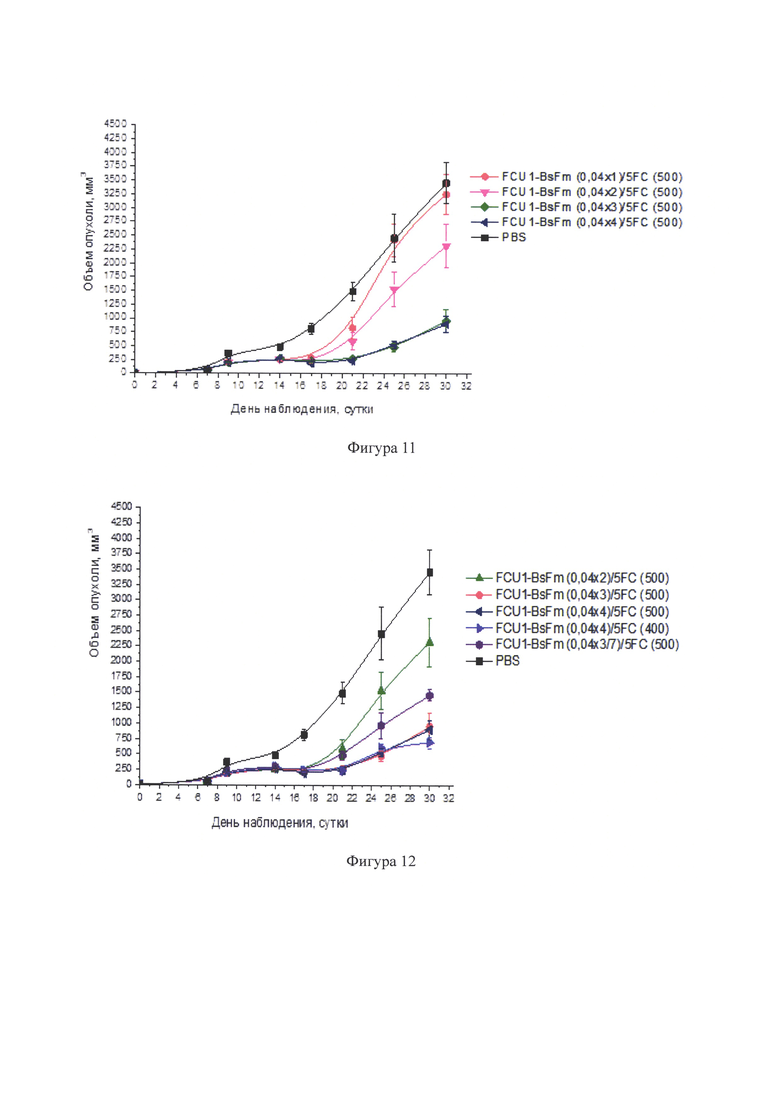




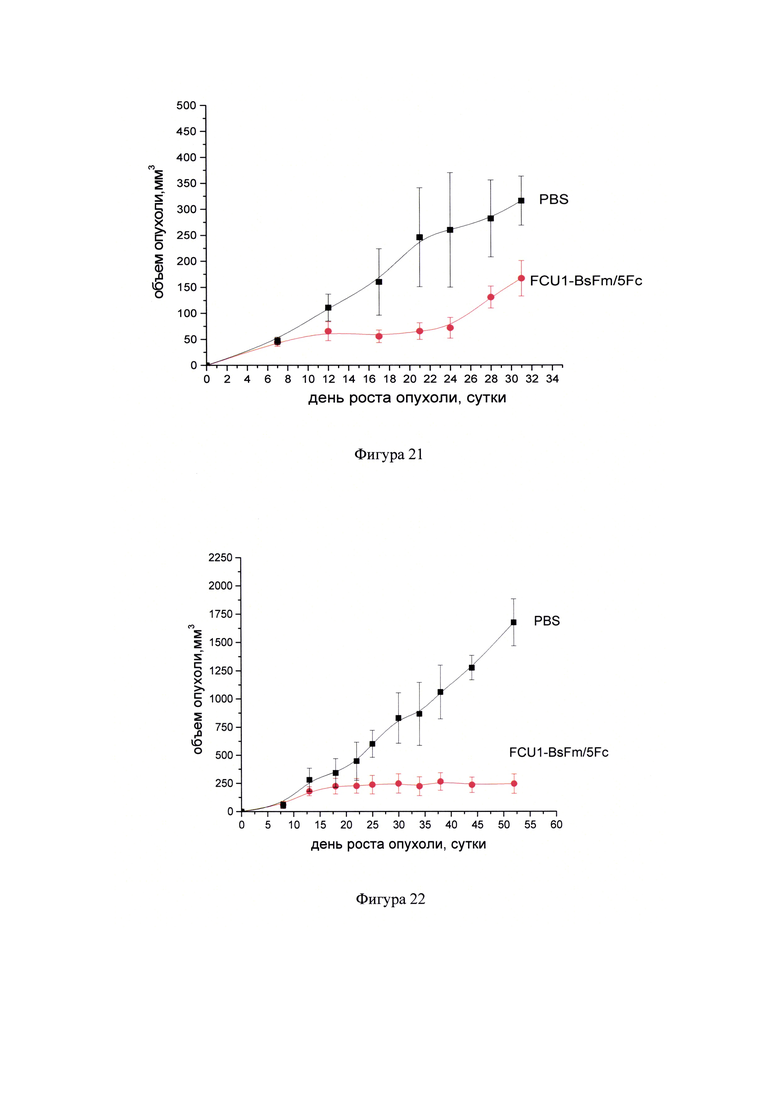
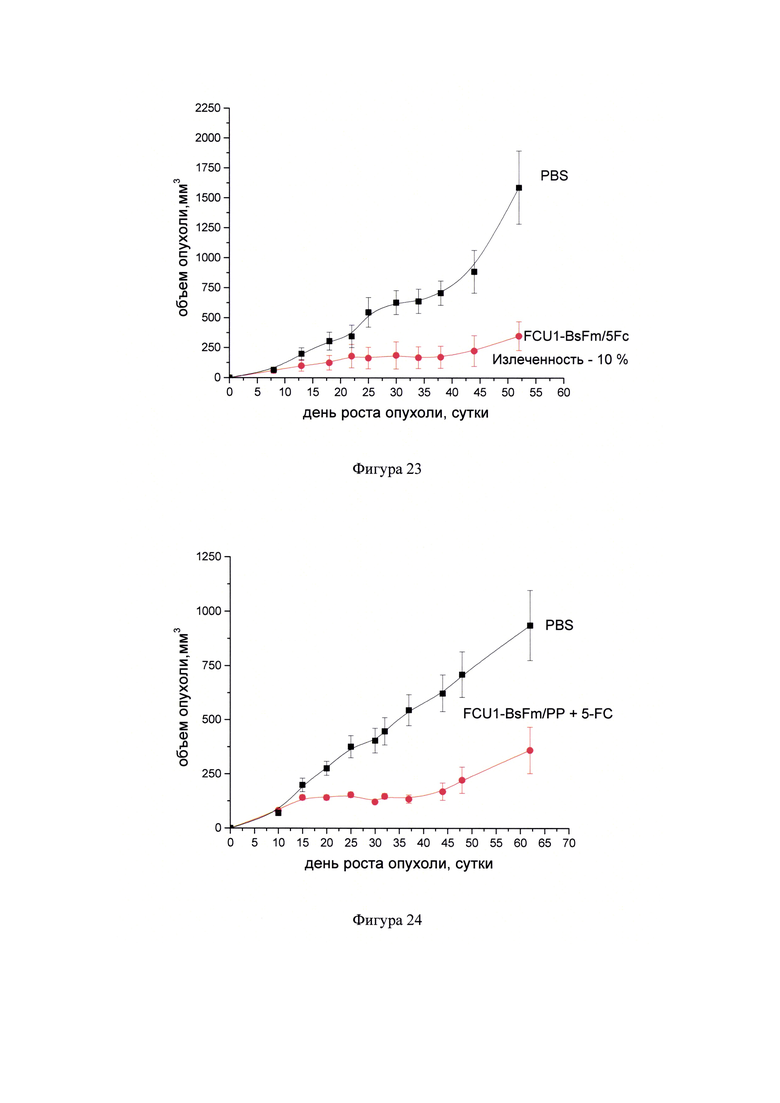
Группа изобретений относится к области генной инженерии. Предложены бицистронная плазмидная конструкция FCU1-BsFm, способ приготовления противоопухолевого препарата FCU1-BsFm/PP, способ его доставки, способы ингибирования репликации опухолевых клеток и лечения злокачественных опухолей. Плазмидная конструкция FCU1-BsFm кодирует химерный ген цитозиндезаминазы и урацилфосфорибозилтрансферазы дрожжей и ген секретируемого мутантного флагеллина Bacillus subtilis. Для получения препарата FCU1-BsFm/PP плазмидную конструкцию «упаковывают» в носитель на основе полиэтиленгликоля, полиэтиленимина и ТАТ-пептида. Способ лечения злокачественных опухолей предусматривает внутриопухолевое введение препарата FCU1-BsFm/PP в сочетании с введением пролекарства 5-фторцитозин. Экспрессия конструкции FCU1-BsFm в опухолевом очаге обеспечивает ингибирование репликации опухолевых клеток. Опухоли представляют собой саркому, опухоль толстой кишки, аденокарциному легкого, рак шейки матки, рак молочной железы, аденокарциному яичника или плоскоклеточный рак кожи. Изобретения обеспечивают эффективный способ доставки плазмидной ДНК в опухолевый очаг и внутрь клеток, обеспечивают достижение противоопухолевого эффекта, опосредованного через цитотоксическое и иммуностимулирующее воздействие. 5 н. и 1 з.п. ф-лы, 24 ил., 5 табл., 9 пр.
1. Бицистронная плазмидная конструкция FCU1-BsFm на основе невирусного вектора pGL3, обладающая противоопухолевой активностью, где опухоль представляет собой саркому, или опухоль толстой кишки, или аденокарциному легкого, или рак шейки матки, или рак молочной железы, или аденокарциному яичника, или плоскоклеточный рак кожи, содержащая экспрессионную кассету с химерным геном FCU1 цитозиндезаминазы (FCY1) и урацилфосфорибозилтрансферазы (FUR1) Saccharomyces cerevisiae arg4 и геном секретируемого мутантного флагеллина BsFm Bacillus subtilis, представленная на SEQ ID No.1.
2. Способ приготовления противоопухолевого препарата FCU1-BsFm/PP, предусматривающий смешивание бицистронной плазмидной конструкции по п.1 с блок-сополимером РР на основе полиэтиленимина (ПЭИ) и полиэтиленгликоля (ПЭГ), при этом ПЭИ-ПЭГ конъюгирован с ТАТ-пептидом с аминокислотной последовательностью GRKKKRRQRC для облегчения проникновения плазмидной ДНК внутрь опухолевых клеток, концентрация блок-сополимера в составе противоопухолевого препарата достигает 12,5 мкМ, с использованием линейных ПЭИ-ПЭГ, причем используют линейные ПЭИ с молекулярной массой 25000 г/моль и бифункциональный ПЭГ MAL-dPEG™24-NHS сложный эфир с молекулярной массой 1394,55 г/моль, при этом мольное соотношение ПЭГ/ПЭИ составляет от 1,1 до 1,5.
3. Способ доставки противоопухолевого препарата FCU1-BsFm/PP, полученного способом по п.2, в опухолевые очаги, где опухоль представляет собой саркому, или опухоль толстой кишки, или аденокарциному легкого, или рак шейки матки, или рак молочной железы, или аденокарциному яичника, или плоскоклеточный рак кожи, предусматривающий внутриопухолевое введение препарата FCU1-BsFm/PP с помощью шприца в пальпируемые опухолевые очаги, при этом препарат распределяют путем вытягивания иглы на себя, но не выводя ее из очага, и перенаправляют иглу в опухоль с разных сторон столько раз, сколько необходимо для введения всей дозы препарата, количество которого зависит от объема опухоли.
4. Способ ингибирования репликации опухолевых клеток посредством экспрессии бицистронной плазмидной конструкции по п.1 в опухолевом очаге, который представлен саркомой, или опухолью толстой кишки, или аденокарциномой легкого, или раком шейки матки, или раком молочной железы, или аденокарциномой яичника, или плоскоклеточным раком кожи, в сочетании с введением пролекарства 5-фторцитозин.
5. Способ лечения злокачественных опухолей, где опухоли представляют собой саркому, или опухоль толстой кишки, или аденокарциному легкого, или рак шейки матки, или рак молочной железы, или аденокарциному яичника, или плоскоклеточный рак кожи, предусматривающий введение способом по п.3 противоопухолевого препарата FCU1-BsFm/PP в виде двухкомпонентной жидкой готовой лекарственной формы (ГЛФ), где первый компонент ГЛФ представлен плазмидной конструкцией FCU1-BsFm в HEPES-глюкозном буфере объемом 1 мл, смешивается перед введением со вторым компонентом ГЛФ, представляющим собой блок-сополимер РР в боратном буфере, в сочетании с введением пролекарства 5-фторцитозин.
6. Способ лечения злокачественных опухолей по п.5, который предусматривает режим введения FCU1-BsFm/PP и пролекарства 5-фторцитозин: трехкратное за курс внутриопухолевое введение FCU1-BsFm/PP в разовой дозе 0,04 мкг плазмидной ДНК/мм3 в дни 1, 6, 11 с интервалом 5 дней и инфузии 5-фторцитозина в суточной дозе 500 мг/кг в дни 2-5, 7-10, 12-18.
| МНОГОПРОФИЛЬНЫЙ ПРОМОТОР, ЭКСПРЕССИРУЮЩИЙ ВЕКТОР И СПОСОБ ИЗБИРАТЕЛЬНОГО УБИЙСТВА РАКОВЫХ КЛЕТОК С ИХ ИСПОЛЬЗОВАНИЕМ | 2012 |
|
RU2476596C1 |
| ALEKSEENKO I | |||
| ET AL | |||
| Step-by-Step Immune Activation for Suicide Gene Therapy Reinforcement | |||
| Int J Mol Sci | |||
| Способ регенерирования сульфо-кислот, употребленных при гидролизе жиров | 1924 |
|
SU2021A1 |
| MIREL D | |||
| B., CHAMBERLIN M | |||
| J | |||
| Видоизменение прибора с двумя приемами для рассматривания проекционные увеличенных и удаленных от зрителя стереограмм | 1919 |
|
SU28A1 |
| Journal of bacteriology | |||
| Механизм для сообщения поршню рабочего цилиндра возвратно-поступательного движения | 1918 |
|
SU1989A1 |

