Настоящая заявка истребует приоритет по заявке за номером 61/116452, поданной 20 ноября 2008 г.
ОБЛАСТЬ ПРИМЕНЕНИЯ ИЗОБРЕТЕНИЯ
Настоящее изобретение направлено на создание способов выращивания, размножения и дифференцирования плюрипотентных стволовых клеток на плоских носителях, не имеющих адсорбированного слоя и слоя питающих клеток.
ПРЕДПОСЫЛКИ СОЗДАНИЯ ИЗОБРЕТЕНИЯ
Культивирование клеток млекопитающих является одним из многих процессов в медико-биологических науках. Для культивирования и анализа опорнозависимых клеток млекопитающих часто используют сосуды из стекла или пластика, например полистирола, что нередко требует дополнительной обработки поверхности, обеспечивающей закрепление клеток на поверхности сосуда. Подобная обработка может включать в себя нанесение на поверхности адсорбированного слоя, например, путем адсорбции, плазмостимулированной полимеризации или привитой сополимеризации. Альтернативно, обработка поверхности может включать химическую модификацию самой поверхности сосуда, что может быть достигнуто обработкой с использованием коронного разряда в атмосфере, радиочастотной плазменной обработки в вакууме, тлеющего разряда постоянного тока и микроволновой плазменной обработки.
Существующие на сегодня способы культивирования плюрипотентных стволовых клеток, в частности, эмбриональных стволовых клеток (ЭСК), требуют сложных условий культивирования, таких как, например, культивирование эмбриональных стволовых клеток на поверхности твердого носителя с нанесенным слоем питающих клеток или на поверхности твердого носителя с нанесенным адсорбированным слоем белка внеклеточного матрикса. В использующих эти способы системах для культивирования нередко применяются питающие клетки или белки внеклеточного матрикса, полученные из организма другого вида по отношению к организму, чьи стволовые клетки культивируются (ксеногенный материал). Для культивирования эмбриональных стволовых клеток могут применяться среды, полученные при введении питающих клеток, то есть среды, кондиционируемые клетками, отличными от недифференцированных эмбриональных стволовых клеток, причем в эти среды может дополнительно вводиться сыворотка животных.
Например, в работах Reubinoff et al. (Nature Biotechnol. 18:399-404, 2000) и Thompson et al. (Science 282:1145-1147, 1998) описывается способ культивирования линий эмбриональных стволовых клеток из бластоцистов человека с использованием питающего клеточного слоя из мышиных эмбриональных фибробластов.
В другом примере, работе Xu et al. (Nature Biotechnology 19: 971-974, 2001), описывается использование препарата MATRIGEL® и ламинина для обработки поверхностей твердого носителя перед культивированием эмбриональных стволовых клеток человека без дифференцирования, причем культивирование ведется без использования питающих клеток. В другом примере, в работе Vallier et al. (J. Cell Sci. 118:4495-4509, 2005), описывается использование эмбриональной бычьей сыворотки для обработки поверхностей твердого носителя перед культивированием эмбриональных стволовых клеток человека без дифференцирования, причем культивирование ведется без использования питающих клеток.
В другом примере, в заявке на патент WO2005014799, описывается кондиционированная среда для поддержания, пролиферации и дифференцирования клеток млекопитающих. В заявке на патент US20050244962 говорится: «Приготовленная в соответствии с настоящим изобретением культуральная среда кондиционируется благодаря секреторной активности мышиных клеток, в частности дифференцированных и иммортализованных трангенных гепатоцитов под названием MMH (Met Murine Hepatocyte).»
В другом примере, в работе Wanatabe et al. (Nature Biotechnol. 35:681-686, 2007), говорится, что «ингибитор Rho-ассоциированной киназы (ингибитор ROCK) позволяет добиться выживания диссоциированных эмбриональных стволовых клеток человека», и демонстрируется снижение индуцированного диссоциацией апоптоза, повышение эффективности клонирования (с приблизительно 1% до приблизительно 27%) и облегчение субклонирования после переноса генов при использовании мышиных эмбриональных фибробластов в качестве питающих клеток, коллагена и MATRIGEL® в качестве белка внеклеточного матрикса, и Y-27632 или Fasudil в качестве ингибитора ROCK. Более того диссоциированные эмбриональные стволовые клетки человека, обработанные Y-27632, оказались защищены от апоптоза в не содержащей сыворотки суспензионной культуре.
В другом примере, работе Peerani et al. (EMBO Journal 26:4744-4755, 2007), говорится: «Сложность пространственной организации культур эмбриональных стволовых клеток человека (ЭСК) создает гетерогенные микроокружения (ниши), которые определяют дальнейшую судьбу ЭСК. Настоящее исследование демонстрирует, что скорость и траекторию дифференцирования ЭСК можно контролировать путем создания ниш стволовых клеток с заданными свойствами. Размер и состав ниши регулируют соотношение между индуцирующими и ингибирующими дифференцирование факторами. С точки зрения механизма такой регулировки, в результате антагонистических взаимодействий между ЭСК и производных от ЭСК клеток внеэмбриональной эндодермы (ExE) создается пространственный градиент сигнального каскада Smad1, зависящий от размеров ниши. Эти взаимодействия опосредуются локализованной секрецией костного морфогенетического белка BMP2 из ExE и его антагониста, фактора дифференцирования роста GDF3 из ЭСК. Микроструктурирование ЭСК, обработанных малыми интерферирующими РНК против GDF3, BMP2 и Smad1, а также обработка ингибитором Rho-ассоциированной киназы (ROCK), демонстрируют, что независимый контроль активации Smad1 может спасти зависящее от размера колонии дифференцирование ЭСК. Результаты авторов патента впервые показывают роль Smad1 в интеграции пространственной информации и в контроле самообновления и дифференцирования ЭСК в зависимости от размера ниши».
В другом примере, работе Koyanagi, M et al.(J Neurosci Res. 2008 Feb 1; 86(2): 270-80), говорится: «Участие Rho-ГТФазы предполагалось в апоптозе многих типов клеток, включая нейроны, но механизм ее действия до сих пор полностью не выяснен. В настоящей работе авторы исследуют роль Rho и ROCK в апоптозе в процессе трансплантации нейронных клеток-предшественников, происходящих от эмбриональных стволовых клеток. Авторы обнаружили, что диссоциация предшественников нейронов приводит к активации Rho и индуцирует апоптоз. Обработка ингибитором Rho, внеклеточным ферментом C3, и/или ингибитором ROCK Y-27632 снижает степень индуцированного диссоциацией апоптоза (аноикоза) на 20-30%. Ингибирование ROCK также снижает пузырение мембраны - ранний морфологический признак апоптоза; расщепление каспазы-3; и высвобождение цитохорма c из митохондрий. Полученные результаты дают основание полагать, что диссоциация нейронных клеток-предшественников активирует внутренний каскад смерти клетки, который по меньшей мере частично опосредуется через каскад Rho/ROCK. Более того, в экспериментальной модели трансплантации на животном ингибирование Rho и/или ROCK приводит к подавлению острого апоптоза трансплантированных клеток. После трансплантации вокруг трансплантата наблюдается ярко выраженная экспрессия фактора некроза опухоли альфа и профактора роста нервов. Ингибирование ROCK также подавляет апоптоз, которому способствуют эти воспалительные цитокины. Взятые вместе, полученные результаты указывают на то, что ингибирование каскада Rho/ROCK может повысить выживаемость трансплантированных клеток в заместительной клеточной терапии.»
Использование ксеногенных материалов может оказаться недопустимым в ряде применений, связанных с использованием плюрипотентных стволовых клеток. В таких случаях можно использовать альтернативные материалы. Например, в работе Stojkovic et al. (Stem Cells 23:895-902, 2005) описано применение сыворотки человека для обработки поверхности твердого носителя перед культивированием эмбриональных стволовых клеток человека без дифференцирования, причем культивирование ведется без использования питающих клеток.
В альтернативной культуральной системе используется не содержащая сыворотки среда, обогащенная факторами роста, способными стимулировать пролиферацию эмбриональных стволовых клеток.
Например, в работе Cheon et al. (BioReprod DOI:10.1095/biolreprod.105.046870; 19 Oct 2005) описана не содержащая ни питающих клеток, ни сыворотки культуральная система, в которой эмбриональные стволовые клетки поддерживаются в некондиционированной замещающей сыворотку среде с добавлением различных факторов роста, способных инициировать самообновление эмбриональных стволовых клеток.
В другом примере, в работе Levenstein et al. (Stem Cells 24:568-574, 2006), описаны способы долговременного культивирования эмбриональных стволовых клеток человека в отсутствие фибробластов или кондиционированной среды с использованием среды с добавлением базового фактора роста фибробластов FGF.
В другом примере, заявке на патент US20050148070, описывается способ культивирования эмбриональных стволовых клеток человека в среде с определенным составом, не содержащей сыворотки и не содержащей питающих клеток-фибробластов, включающий: культивирование стволовых клеток в культуральной среде, содержащей альбумин, аминокислоты, витамины, минеральные вещества, по меньшей мере один трансферин или заместитель трансферина, по меньшей мере один инсулин или заместитель инсулина, причем культуральная среда по существу не содержит эмбриональной сыворотки млекопитающих и содержит по меньшей мере приблизительно 100 нг/мл FGF, способного активировать FGF-сигнальный рецептор, где фактор роста поступает из источника, отличного от просто слоя питающих клеток фибробластов, среду, поддерживающую пролиферацию стволовых клеток в недифференцированном состоянии без слоя питающих клеток или кондиционированной среды.
В другом примере, в заявке на патент US20050233446, описывается среда с определенным составом, которая может быть использована при культивировании стволовых клеток, включая недифференцированные зародышевые стволовые клетки приматов. В растворе среда является по существу изотонической относительно культивируемых стволовых клеток. Для каждой культуры конкретно используемая среда состоит из основной среды с добавлением базового FGF, инсулина и аскорбиновой кислоты, необходимых для поддержания по существу недифференцированного роста зародышевых стволовых клеток.
В другом примере, заявке на патент US6800480, говорится: «В одном варианте осуществления предлагается культуральная среда для выращивания зародышевых стволовых клеток приматов в по существу недифференцированном состоянии, включающая основную среду с низким содержанием эндотоксина и низким осмотическим давлением, которая эффективно поддерживает рост зародышевых стволовых клеток приматов. Основная среда объединяется с питательной сывороткой, способной поддерживать рост зародышевых стволовых клеток приматов, и субстратом, выбираемым из группы, состоящей из питающих клеток и внеклеточного матрикса, полученного из питающих клеток. Среда также включает в себя заменимые аминокислоты, антиоксидант и первый фактор роста, выбираемый из группы, состоящей из нуклеозидов и соль-пируватов.»
В другом примере, в заявке на патент US20050244962, говорится: «В одном аспекте изобретения предлагается способ культивирования эмбриональных стволовых клеток приматов. Стволовые клетки культивируются в культуре, по существу свободной от эмбриональной сыворотки млекопитающих (предпочтительно также по существу свободной от сыворотки любых животных) и в присутствии фактора роста фибробластов, полученного из источника, отличного от просто слоя питающих фибробластов. В предпочтительной форме слой питающих фибробластов, ранее необходимый для поддержания культуры стволовых клеток, становится необязательным вследствие добавления достаточного количества фактора роста фибробластов.»
В другом примере, заявке на патент WO2005065354, описывается по существу свободная от питающих клеток и сыворотки изотоническая культуральная среда с определенным составом, содержащая: а. базальную среду; b. базовый фактор роста фибробластов в количестве, достаточном для поддержания роста по существу недифференцированных стволовых клеток млекопитающих; c. инсулин в количестве, достаточном для поддержания роста по существу недифференцированных стволовых клеток млекопитающих; и d. аскорбиновую кислоту в количестве, достаточном для поддержания роста по существу недифференцированных стволовых клеток млекопитающих.
В другом примере, заявке на патент WO2005086845, описан способ поддержания недифференцированных стволовых клеток, причем упомянутый способ включает в себя воздействие на стволовые клетки одним из членов семейства белков трансформирующего ростовой фактор-бета (TGFβ), одним из членов семейства белков фактора роста фибробластов (FGF) или никотинамидом (NIC) в количестве, достаточном для поддержания клеток в недифференцированном состоянии в течение периода времени, достаточного для получения желаемого результата.
Плюрипотентные стволовые клетки обладают большим потенциалом для научно-исследовательской работы и скрининга лекарственных препаратов. В настоящее время крупномасштабное культивирование линий эмбриональных стволовых клеток человека представляет большие проблемы и сталкивается с существенными трудностями. Возможные пути преодоления этих трудностей заключаются в пассировании и культивировании эмбриональных стволовых клеток человека в виде одиночных клеток. Одиночные клетки более удобны для применения стандартных методик культивирования тканей, таких как, например, подсчет клеток, трансфицирование и т.д.
Например, в работе Nicolas et al. описывается способ получения и наработки линий эмбриональных стволовых клеток человека из одиночных клеток, изолированных с помощью флюоресцентной сортировки после генетической модификации с использованием лентивирусных векторов (Stem Cells Dev. 16: 109-118, 2007).
В другом примере, заявке на патент США US2005158852, описан способ «улучшения роста и выживаемости одиночных эмбриональных стволовых клеток человека. Способ включает в себя стадии получения одиночной недифференцированной эмбриональной стволовой клетки человека; смешивания одиночной недифференцированной клетки с внеклеточным матриксом для обволакивания клетки; и посева полученной смеси на питающие клетки с питательной средой в ростовом окружении».
В другом примере, работе Sidhu et al. (Stem Cells Dev. 15:61-69, 2006), впервые сообщается о трех клонах эмбриональных стволовых клеток человека, hES 3.1, 3.2 и 3.3, полученных из родительской линии hES3 путем сортировки одноклеточных препаратов на проточном цитометре.
Однако пассирование и культивирование эмбриональных стволовых клеток человека в виде одиночных клеток приводит к генетическим аберрациям и к потере плюрипотентности. Для поддержания плюрипотентности и генетической устойчивости большое значение имеют условия культивирования. Как правило, пассирование линий эмбриональных стволовых клеток человека выполняется вручную или с использованием ферментативных веществ, таких как коллагеназа, либераза или диспаза.
Например, в работе Draper et al. отмечается наличие «кариотипических изменений, включающих в себя амплификацию 17q плеча в трех независимых линиях эмбриональных стволовых клеток человека в пяти независимых случаях.» (Nature Biotechnol. 22: 53-54, 2004).
В другом пример, работе Buzzard et al., говорится: «мы зафиксировали лишь одно изменение кариотипа…, что может быть обусловлено используемыми нами способами культивирования, поскольку наши способы существенно отличаются от способов, используемых большинством других групп. Как правило, мы пассируем эмбриональные стволовые клетки человека через семь дней, сначала подсекая колонию краем разломленной пипетки… При таком способе пассирования не используются никакие ферментативные или химические способы диссоциации клеток. Мы считаем, что именно с этим может быть связана относительная генетическая устойчивость эмбриональных стволовых клеток человека в наших руках.» (Nature Biotechnol. 22: 381-382, 2004).
В другом примере, работе Mitalipova et al., говорится: «способы группового пассирования... могут зафиксировать популяции анэуплоидных клеток после многократного пассирования в культуре, но могут применяться на более коротких промежутках времени (по меньшей мере до 15 пассажей) без угрозы изменения кариотипа…, возможно удастся поддерживать нормальный кариотип эмбриональных стволовых клеток человека в условиях продолжительного ручного ведения с последующим ограниченным числом групповых пассирований в экспериментах, требующих больших количеств эмбриональных стволовых клеток человека, чем могут обеспечить только способы ручного пассирования». (Nature Biotechnol. 23: 19-20, 2005).
В другом примере, работе Heng et al., говорится: «полученные результаты показали, что второй протокол (трипсинизация с умеренным пипетированием) наносит гораздо меньший ущерб жизнеспособности клеток по сравнению с первым протоколом (обработка коллагеназой и соскребание). Это, в свою очередь, выливается в большую вероятность выживания клеток в циклах заморозки/разморозки.» (Biotechnology and Applied Biochemistry 47:33-37, 2007).
В другом примере, работе Hasegawa et al., говорится: «мы вывели сублинии эмбриональных стволовых клеток человека, переносящих полную диссоциацию. Эти клетки имеют высокую эффективность пересева и высокую эффективность клонирования, при этом они сохраняют свою способность дифференцирования в клетки всех трех зародышевых листков.» (Stem Cells 24:2649-2660, 2006).
В другом примере, заявке на патент США 61/030544, описаны способы и композиции для закрепления клеток на, культивирования клеток на и снятия клеток с поверхности твердого носителя, содержащей от по меньшей мере приблизительно 0,9% азота до по меньшей мере приблизительно 11% азота и от по меньшей мере приблизительно 12% кислорода до по меньшей мере приблизительно 30% кислорода, не имеющей слоя питающих клеток и адсорбированного слоя. В одном из вариантов осуществления настоящего изобретения клетки обрабатывают соединением, способным ингибировать активность Rho-киназы.
Имеется значительная потребность в способах и композициях для культивирования клеток, включая культивирование плюрипотентных стволовых клеток в отсутствие слоя питающих клеток и адсорбированного слоя при сохранении плюрипотентности культивируемых клеток. В настоящем изобретении предложены способы выращивания, размножения и дифференцирования плюрипотентных стволовых клеток на плоских носителях, не имеющих адсорбированного слоя и слоя питающих клеток, причем клетки не требуют обработки соединением, способным ингибировать активность Rho-киназы, для закрепления на плоском носителе.
КРАТКОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
В одном варианте осуществления настоящего изобретения описаны способы закрепления клеток на, культивирования клеток на и снятия клеток с плоского носителя, содержащего до приблизительно 12% N, от по меньшей мере приблизительно 12% O до по меньшей мере приблизительно 55% O, с углом смачивания от приблизительно 18 градусов до приблизительно 32 градусов, не имеющего адсорбированного слоя и слоя питающих клеток.
КРАТКОЕ ОПИСАНИЕ ФИГУР
На фиг.1 показано влияние ингибитора Rho-киназы H-1152 на закрепление эмбриональных стволовых клеток человека линии H1 на плоских носителях. На фиг.1А показано закрепление клеток на мембранах из смешанных эфиров целлюлозы (мембрана № 2 в Таблице 1). На фиг.1В показано закрепление клеток на мембранах из нейлона (мембрана № 4 в Таблице 1). На фиг.1С показано закрепление клеток на мембранах из ацетата целлюлозы (мембрана № 5 в Таблице 1). На фиг.1D показано закрепление клеток на мембранах из поликарбоната (мембрана № 7 в Таблице 1). На фиг.1E показано закрепление клеток на мембранах из полиэтилентерефталата (мембрана № 12 в Таблице 1).
На фиг.2 показано влияние ингибитора Rho-киназы Y-26732 на закрепление эмбриональных стволовых клеток человека линии H9 на мембране из смешанных эфиров целлюлозы (мембрана № 1 в Таблице 1). На фиг.2A показано закрепление клеток в контрольной лунке. На фиг.2B показано закрепление клеток, обработанных 10 мкМ Y-26732. На фиг.2C показано закрепление клеток, обработанных 20 мкМ Y-26732.
На фиг.3 показаны кривые пролиферации эмбриональных стволовых клеток человека линии H1 на покрытой препаратом MATRIGEL® поверхности (сплошная линия) и на мембранах из смешанных эфиров целлюлозы (мембрана № 1 в Таблице 1) (пунктирная линия).
На фиг.4 показаны окрашенные с использованием красителя Гимза хромосомы представительных клеток из эмбриональных стволовых клеток человека линии H1. На фиг.4A показаны хромосомы клеток, культивированных в течение 10 пассажей на покрытых препаратом MATRIGEL® поверхностях. На фиг.4B показаны хромосомы клеток, культивированных в течение 10 пассажей на мембранах из смешанных эфиров целлюлозы (мембрана № 1 в Таблице 1).
На фиг.5 показано влияние ингибитора Rho-киназы Y26732 на закрепление эмбриональных стволовых клеток человека линии H9 на поликарбонатных мембранах (мембрана № 7 в Таблице 1). На фиг.5A показано закрепление клеток в контрольной лунке. На фиг.5B показано закрепление клеток, обработанных 10 мкМ Y-26732. На фиг.5C показано закрепление клеток, обработанных 20 мкМ Y-26732.
На фиг.6 показано влияние ингибитора Rho-киназы H-1152 на закрепление эмбриональных стволовых клеток человека линии H1 на поликарбонатных мембранах (мембрана № 7 в Таблице 1). На фиг.6A показано закрепление клеток в контрольной лунке. На фиг.6B показано закрепление клеток при добавлении в культуральную среду 0,03 мкМ H-1152. На фиг.6C показано закрепление клеток при добавлении в культуральную среду 0,1 мкМ H-1152. На фиг.6D показано закрепление клеток при добавлении в культуральную среду 0,3 мкМ H-1152. На фиг.6E показано закрепление клеток при добавлении в культуральную среду 1 мкМ H-1152. На фиг.6F показано закрепление клеток при добавлении в культуральную среду 3 мкМ H-1152.
На фиг.7 показано открепление эмбриональных стволовых клеток человека линии H1 от поликарбонатных мембран (мембрана № 9 в Таблице 1) после удаления из культуральной среды ингибитора Rho-киназы H-1152. На фиг.7A показано открепление клеток в случае, когда в культуральной среде поддерживалась концентрация H-11523 мкМ. На фиг.7B показано открепление клеток в случае, когда H-1152 был удален из культуральной среды.
На фиг.8 показано влияние размера пор в мембране и обработки ингибитором Rho-киназы на закрепление эмбриональных стволовых клеток человека линии H1 на плоских носителях, представляющих собой: поликарбонатную мембрану № 10 в Таблице 1 для фиг.8A и 8C; и поликарбонатную мембрану № 11 в Таблице 1 для фиг.8B и 8D. На фиг.8A и 8B показано открепление клеток в случае, когда в культуральной среде поддерживалась концентрация H-1152 3 мкМ. На фиг.8C и 8D показано открепление клеток в случае, когда H-1152 был удален из культуральной среды.
На фиг.9 показано сохранение экспрессии ассоциируемых с плюрипотентностью маркеров для эмбриональных стволовых клеток человека линии H1, культивировавшихся в течение трех пассажей на поликарбонатных мембранах (мембрана № 8 в Таблице 1). Уровень экспрессии указанных на фигуре генов определяли с использованием ПЦР в режиме реального времени. Сплошными столбиками показаны данные, полученные для недифференцированных эмбриональных стволовых клеток человека линии H1. Заштрихованными столбиками показаны данные, полученные для клеток, культивировавшихся на поликарбонатных мембранах.
На фиг.10 показана способность эмбриональных стволовых клеток человека линии H1 образовывать эмбриоидные тельца после культивирования в течение 12 пассажей на поликарбонатных мембранах (мембрана № 8 в Таблице 1). На фигуре приведены представительные данные из одного эксперимента.
На фиг.11 показаны полученные на сканирующем электронном микроскопе микрофотографии плоских носителей, составляющих предмет настоящего изобретения.
На фиг.12 показаны полученные на сканирующем электронном микроскопе микрофотографии плоского носителя ULTRAWEBTM.
На фиг.13 показано влияние среды определенного состава mTESRTM на связывание эмбриональных стволовых клеток человека линии H1 с различными плоскими носителями.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Для ясности описания, а не в ограничение изобретения, подробное описание изобретения разделено на следующие подразделы, описывающие или иллюстрирующие определенные особенности, варианты осуществления или области применения настоящего изобретения.
Определения
Используемый в настоящей заявке термин «адсорбированный слой» относится к слою на поверхности твердого носителя, формируемому путем прикрепления молекул к поверхности либо ковалентными (так называемая пересадка), либо нековалентными (так называемая адсорбция) связями. Используемые для получения адсорбированного слоя молекулы могут быть, например, белковыми и родственными им молекулами, которые могут включать в себя, например, белки внеклеточного матрикса, аминокислоты и т.д., или молекулами небиологической природы, такими как, например, полиэтиленимин.
Используемый в настоящей заявке термин «β-клеточная линия дифференцирования» относится к клеткам, положительным по экспрессии гена транскрипционного фактора PDX-1 и по меньшей мере одного из следующих транскрипционных факторов: NGN3, NKX2.2, NKX6.1, NEUROD, ISL1, HNF-3 бета, MAFA, PAX4 или PAX6. Клетки с экспрессией маркеров, характерных для β-клеточной линии дифференцирования, включают в себя β-клетки.
Используемый в настоящей заявке термин «клетки с экспрессией маркеров, характерных для линии дифференцирования в клетки сформированной эндодермы» относится к клеткам, экспрессирующим по меньшей мере один из следующих маркеров: SOX17, GATA4, HNF3 бета, GSC, CER1, Nodal, FGF8, Brachyury, Mix-подобный гомеобоксный белок, FGF4 CD48, эомезодермин (EOMES), DKK4, FGF17, GATA6, CXCR4, C-Kit, CD99 или OTX2. Клетки с экспрессией маркеров, характерных для линии дифференцирования в клетки сформированной эндодермы, включают в себя клетки-предшественники первичной полоски, клетки первичной полоски, клетки мезэндодермы и клетки сформированной эндодермы.
Термин «клетки, экспрессирующие маркеры, характерные для линии панкреатической эндодермы» в настоящем документе обозначает клетки, экспрессирующие по меньшей мере один из следующих маркеров: PDX1, HNF1 бета, PTF1 альфа, HNF6, NKX6.1 или HB9. Клетки с экспрессией маркеров, характерных для линии дифференцирования в клетки панкреатической эндодермы, включают в себя клетки панкреатической эндодермы, клетки первичной кишечной трубки и клетки поздней передней кишки.
Используемый в настоящей заявке термин «сформированная эндодерма» относится к клеткам, обладающим характерными особенностями клеток, происходящих в ходе гаструляции от эпибласта, и формирующим желудочно-кишечный тракт и его производные. Клетки сформированной эндодермы экспрессируют следующие маркеры: HNF3 бета, GATA4, SOX17, церберус, OTX2, гузекоид, C-Kit, CD99 или MIXL1.
Используемый в настоящей заявке термин «панкреатическая эндокринная клетка» или «клетка, экспрессирующая панкреатические гормоны» относится к клеткам, способным к экспрессии по меньшей мере одного из следующих гормонов: инсулина, глюкагона, соматостатина и панкреатического полипептида.
Используемый в настоящей заявке термин «внеэмбриональная эндодерма» относится к популяции клеток, экспрессирующих по меньшей мере один из следующих маркеров: SOX7, AFP или SPARC.
Используемый в настоящей заявке термин «белки внеклеточного матрикса» относится к белковым и родственным им молекулам, обычно находящимся между клетками в теле или в плаценте. Белки внеклеточного матрикса могут быть получены из ткани, биологических жидкостей, например крови, или из среды, кондиционированной нерекомбинантными клетками, рекомбинантными клетками или бактериями.
Используемый в настоящей заявке термин «маркеры» обозначает молекулы нуклеиновых кислот или полипептидов с дифференциальной экспрессией в интересующих клетках. В данном контексте под дифференциальной экспрессией подразумевается повышение уровня экспрессии для положительного маркера и понижение уровня экспрессии для отрицательного маркера. Определяемый уровень маркерной аминокислоты или полипептида в интересующих клетках оказывается значительно выше или ниже по сравнению с другими клетками, что позволяет идентифицировать интересующую клетку и отличить ее от других клеток с помощью любого из множества известных в данной области способов.
Используемый в настоящей заявке термин «клетка мезэндодермы» относится к клеткам, экспрессирующим по меньшей мере один из следующих маркеров: CD48, эомезодермин (EOMES), SOX-17, DKK4, HNF3 бета, GSC, FGF17 или GATA6.
Используемый в настоящей заявке термин «панкреатическая секретирующая гормоны клетка» относится к клеткам, способным к секреции по меньшей мере одного из следующих гормонов: инсулина, глюкагона, соматостатина и панкреатического полипептида.
Используемый в настоящей заявке термин «клетка-предшественник клетки первичной полоски» относится к клеткам, экспрессирующим по меньшей мере один из следующих маркеров: Nodal или FGF8.
Используемый в настоящей заявке термин «клетка первичной полоски» относится к клеткам, экспрессирующим по меньшей мере один из следующих маркеров: Brachyury, Mix-подобный гомеобоксный белок или FGF4.
Стволовые клетки представляют собой недифференцированные клетки, определяемые по их способности на уровне единичной клетки как самообновляться, так и дифференцироваться с образованием клеток-потомков, таких как самообновляющиеся клетки-предшественники, необновляющиеся клетки-предшественники и окончательно дифференцированные клетки. Стволовые клетки также характеризуются способностью дифференцироваться in vitro в функциональные клетки различных клеточных линий дифференцирования из нескольких зародышевых листков (эндодермы, мезодермы и эктодермы), а также после трансплантации давать начало тканям, происходящим от нескольких зародышевых листков, и вносить существенный вклад в формирование большинства, если не всех, тканей после инъекции в бластоцисты.
По потенциалу развития стволовые клетки классифицируют следующим образом: (1) тотипотентные, т.е. способные давать начало всем эмбриональным и внеэмбриональным типам клеток; (2) плюрипотентные, т.е. способные давать начало всем эмбриональным типам клеток; (3) мультипотентные, т.е. способные давать начало группе клеточных линий в пределах конкретной ткани, органа или физиологической системы (например, гематопоэтические стволовые клетки (ГСК) могут давать таких потомков, как ГСК (самообновление), олигопотентные предшественники, ограниченные клетками крови, и все типы клеток и клеточных элементов (таких как тромбоциты), являющиеся нормальными компонентами крови); (4) олигопотентные, т.е. способные давать начало более ограниченному набору клеточных линий, чем мультипотентные стволовые клетки; и (5) унипотентные, т.е. способные давать начало единственной клеточной линии (например, сперматогенные стволовые клетки).
Дифференцирование представляет собой процесс, при помощи которого неспециализированная («некоммитированная») или менее специализированная клетка приобретает свойства специализированной клетки, например нервной или мышечной клетки. Дифференцированная клетка или клетка с индуцированным дифференцированием представляет собой клетку, занявшую более специализированное («коммитированное») положение в линии дифференцирования клетки. Термин «коммитированная» применительно к процессу дифференцирования обозначает клетку, дошедшую в ходе процесса дифференцирования до стадии, от которой в нормальных условиях она продолжит дифференцироваться до определенного типа клеток или набора типов клеток и не сможет в нормальных условиях дифференцироваться в иной тип клеток или вернуться обратно к менее дифференцированному типу. Дедифференцированием называется процесс, в ходе которого клетка возвращается к менее специализированному (или коммитированному) положению в линии дифференцирования. Используемый в настоящей заявке термин «линия дифференцирования» определяет наследственность клетки, то есть определяет, из какой клетки произошла данная клетка и каким клеткам она может дать начало. В линии дифференцирования клетка помещается в наследственную схему развития и дифференцирования. Маркером, специфичным для линии дифференцирования, называется характерная особенность, специфически ассоциированная с фенотипом клеток конкретной линии дифференцирования, которая может использоваться для оценки дифференцирования некоммитированных клеток в клетки данной линии дифференцирования.
Используемый в настоящей заявке термин «поверхность» относится к внешнему слою молекул твердого носителя - сосуда или матрицы, предназначенных для культивирования или анализа клеток. Элементный состав, степень гладкости и смачиваемость поверхности могут быть проанализированы способами рентгеновской фотоэлектронной спектроскопии (РФЭС), атомно-силовой микроскопии (АСМ) и измерения угла смачивания, соответственно.
Для описания клеток в процессе культивирования используются различные термины. Термин «поддержание» относится в целом к клеткам, помещенным в ростовую среду в условиях, которые способствуют росту и/или делению клеток, в результате чего популяция клеток может увеличиваться или не увеличиваться. Термин «пассирование» означает процесс изъятия клеток из одного культурального сосуда и переноса их в другой культуральный сосуд в условиях, которые способствуют росту и/или делению клеток.
Конкретная популяция клеток или клеточная линия иногда описывается или характеризуется числом выполненных с ней пассирований. Например, пассированная 10 раз культивируемая популяция клеток может описываться как культура десятого пассажа, или культура P10. Первичная культура, т.е. первая культура после выделения клеток из ткани, получает обозначение P0. После первого пересева клетки описываются как вторичная культура (P1, или культура первого пассажа). После второго пересева клетки превращаются в третичную культуру (P2, или культура второго пассажа) и т.д. Специалист в данной области определит, что за промежуток времени между последовательными пассированиями популяция клеток может удваиваться многократно; поэтому число удвоений популяций в культуре превышает номер пассажа. Степень размножения клеток (то есть число удваиваний популяции) за промежуток времени между последовательными пассированиями зависит от многих факторов, включая без ограничений плотность посева, тип носителя, тип среды, условия роста и продолжительность времени между пересевами.
Плоские носители настоящего изобретения
Соответствующие целям настоящего изобретения плоские носители могут быть изготовлены из любого материала, способного образовать основу, на которой смогут закрепиться плюрипотентные клетки. Например, плоский носитель может быть изготовлен из поликарбоната. Альтернативно, плоский носитель может быть изготовлен из полиэтилентерефталата (ПЭТФ). Альтернативно, плоский носитель может быть изготовлен из нейлона. Альтернативно, плоский носитель может быть изготовлен из ацетата целлюлозы. Альтернативно, плоский носитель может быть изготовлен из смешанных эфиров целлюлозы. Примеры плоских носителей, соответствующих целям настоящего изобретения, приведены в Таблице 1.
В одном варианте осуществления настоящего изобретения описаны способы закрепления, культивирования и дифференцирования плюрипотентных стволовых клеток на плоском носителе, содержащем до приблизительно 12% N, от по меньшей мере приблизительно 12% O до по меньшей мере приблизительно 55% O, с углом смачивания от приблизительно 18 градусов до приблизительно 32 градусов, не имеющем адсорбированного слоя и слоя питающих клеток. Плоский носитель, содержащий от по меньшей мере приблизительно 8% N до по меньшей мере приблизительно 12% N и от по меньшей мере приблизительно 12% O до по меньшей мере приблизительно 55% O, может представлять собой груботекстурированную волоконную поверхность или, альтернативно, гладкую поверхность.
В одном варианте осуществления настоящего изобретения описан способ закрепления плюрипотентных стволовых клеток на плоском носителе, содержащем до приблизительно 12% N, от по меньшей мере приблизительно 12% O до по меньшей мере приблизительно 55% O, с углом смачивания от приблизительно 18 градусов до приблизительно 32 градусов, не имеющем адсорбированного слоя и слоя питающих клеток, включающий следующие стадии:
а. получение суспензии плюрипотентных стволовых клеток, и
b. добавление суспензии клеток к плоскому носителю с последующим закреплением клеток на носителе.
В одном варианте осуществления после закрепления на поверхности плюрипотентные клетки продолжают поддерживаться в культуральной среде. В одном варианте осуществления после закрепления на поверхности плюрипотентные клетки дифференцируются на плоском носителе.
В одном варианте осуществления эффективность закрепления плюрипотентных стволовых клеток на плоском носителе, содержащем до приблизительно 12% N, от по меньшей мере приблизительно 12% O до по меньшей мере приблизительно 55% O, с углом смачивания от приблизительно 18 градусов до приблизительно 32 градусов, не имеющем адсорбированного слоя и слоя питающих клеток, повышается путем обработки клеток соединением, способным ингибировать активность Rho-киназы. Соединение, способное ингибировать активность Rho-киназы, после закрепления клеток может быть удалено из культуральной среды.
Соединение, способное ингибировать активность Rho-киназы, выбирают из группы, состоящей из: Y-27632, Fasudil, H-1152 и Hydroxyfasudil.
В одном варианте осуществления соединение, способное ингибировать активность Rho-киназы, может использоваться в концентрации от приблизительно 0,1 мкМ до приблизительно 100 мкМ. В одном варианте осуществления по меньшей мере одно соединение, способное ингибировать активность Rho-киназы, используется в концентрации приблизительно 10 мкМ.
Характеризация плоских носителей настоящего изобретения
В одном варианте осуществления элементный состав поверхности описываемых плоских носителей может быть проанализирован способом рентгеновской фотоэлектронной спектроскопии (РФЭС). Способ РФЭС, также известный под названием «электронная спектроскопия для химического анализа» (ЭСХА), используется для определения элементов или атомов, присутствующих в поверхностном слое твердого носителя (возможно определение всех элементов в концентрациях менее 0,1% атом., за исключением водорода и гелия), а также для определения химического окружения подобных элементов или атомов.
В одном варианте осуществления степень гладкости поверхности описываемых плоских носителей может быть проанализирована способом атомно-силовой микроскопии (АСМ). Способ АСМ позволяет получать топографию атомов или молекул на поверхности с поперечным разрешением до 0,1 нм (1Å) и вертикальным разрешением до 0,01 нм (0,1Å).
В одном варианте осуществления смачиваемость поверхности описываемых плоских носителей может быть проанализирована способом измерения угла смачивания. Например, измерение угла смачивания статическим способом неподвижной капли дает информацию о взаимодействии между поверхностью твердой подложки и жидкостью. Угол смачивания описывает форму капли жидкости, которая покоится на поверхности твердой подложки, и представляет собой контактный угол жидкости на поверхности твердого носителя, измеренный внутри жидкости вдоль линии контакта, где встречаются жидкость, твердый носитель и газ. Поверхность с углом смачивания более 90° называется гидрофобной, а поверхность с углом смачивания менее 90° - гидрофильной. На очень гидрофильных поверхностях, то есть поверхностях с большим сродством к воде, капля воды полностью растекается по поверхности (эффективный угол смачивания равен 0°).
В одном варианте осуществления плотность отрицательных зарядов на поверхности описываемых плоских носителей может быть проанализирована способом измерения реакционной способности поверхности в отношении кристаллического фиолетового. Молекулы кристаллического фиолетового несут положительный заряд, что позволяет им связываться с отрицательно заряженными молекулами и фрагментами молекул, например, отрицательно заряженными функциональными группами, присутствующими на поверхности полимера. Поверхность с высокой реакционной способностью в отношении кристаллического фиолетового имеет большую плотность отрицательных зарядов по сравнению с поверхностью, имеющей низкую реакционную способность в отношении кристаллического фиолетового, при той же степени гладкости поверхности и тем самым ее удельной площади.
Плюрипотентные стволовые клетки
Характеристика плюрипотентных стволовых клеток
Плюрипотентные стволовые клетки могут экспрессировать один или более стадийно-специфичных эмбриональных антигенов (SSEA) 3 и 4, а также маркеры, определяемые антителами, обозначенными как Tra-1-60 и Tra-1-81 (Thomson et al., Science 282:1145, 1998). Дифференцирование плюрипотентных стволовых клеток in vitro приводит к потере экспрессии SSEA-4, Tra-1-60 и Tra-1-81 и повышению экспрессии SSEA-1. Недифференцированные полипотентные стволовые клетки, как правило, обладают щелочно-фосфатазной активностью, которую можно определить путем фиксирования клеток 4% раствором параформальдегида с последующим проявлением с использованием красителя Vector Red в качестве субстрата, следуя рекомендациям производителя (компания Vector Laboratories, Бурлингем, Калифорния, США). Недифференцированные плюрипотентные стволовые клетки также, как правило, экспрессируют Oct-4 и TERT, обнаруживаемые способом ОТ-ПЦР.
Другим желательным фенотипическим свойством выращенных плюрипотентных клеток является потенциал дифференцирования в клетки всех трех зародышевых листков: в клетки эндодермальных, мезодермальных и эктодермальных тканей. Плюрипотентность стволовых клеток может быть подтверждена, например, путем инъекции клеток мышам с тяжелым комбинированным иммунодефицитом (SCID), фиксирования образующихся тератом с помощью 4% параформальдегида и их гистологического исследования на предмет наличия клеточных типов, происходящих от трех зародышевых листков. Альтернативно, плюрипотентность можно определить по созданию эмбриоидных телец и анализа их на предмет присутствия маркеров, ассоциирующихся с тремя зародышевыми листками.
Выращенные линии плюрипотентных стволовых клеток могут быть кариотипированы с применением стандартного способа окрашивания с использованием красителя Гимза и сравнения с опубликованными кариотипами соответствующих видов приматов. Желательно получить клетки, имеющие «нормальный кариотип», т.е. эуплоидные клетки, в которых все человеческие хромосомы присутствуют и не имеют видимых изменений.
Источники плюрипотентных стволовых клеток
К типам плюрипотентных стволовых клеток, которые можно использовать, относятся устойчивые линии плюрипотентных клеток, получаемые из формируемой после вынашивания плода ткани, в том числе из преэмбриональной ткани (такой как бластоциста), эмбриональной ткани или ткани плода, взятой в любой момент в ходе вынашивания, как правило, но не обязательно, до срока приблизительно 10-12 недель беременности. Не ограничивающими настоящее изобретение примерами являются устойчивые линии человеческих эмбриональных стволовых клеток или человеческих эмбриональных зародышевых клеток, например, клеточные линии человеческих эмбриональных стволовых клеток H1, H7 и H9 (WiCell). Также возможно использование описываемых в настоящей заявке составов в ходе первоначального установления или стабилизации таких клеток, в этом случае исходными клетками являются первичные плюрипотентные клетки, взятые напрямую из тканей-источников. Также соответствуют целям настоящего изобретения клетки, взятые из популяции плюрипотентных стволовых клеток, уже культивированных в отсутствие питающих клеток. Также соответствуют целям настоящего изобретения клетки мутантных линий эмбриональных стволовых клеток человека, таких как, например, BG01v (компания BresaGen, Атенс, Джорджия, США). Также соответствуют целям настоящего изобретения плюрипотентные стволовые клетки, полученные из неплюрипотентных клеток, таких как, например, соматических клеток взрослого.
В одном варианте осуществления эмбриональные стволовые клетки человека готовят, как описано в следующих публикациях: Thomson et al. (патент США № 5843780; Science 282:1145, 1998; Curr. Top. Dev. Biol. 38:133 ff., 1998; Proc. Natl. Acad. Sci. U.S.A. 92:7844, 1995).
Культивирование плюрипотентных стволовых клеток
В одном осуществлении перед культивированием в соответствии со способами, составляющими предмет настоящего изобретения, плюрипотентные стволовые клетки культивируют на слое питающих клеток или белка внеклеточного матрикса, которые различным образом поддерживают плюрипотентные стволовые клетки. Например, плюрипотентные стволовые клетки культивируют на слое питающих клеток, который поддерживает пролиферацию плюрипотентных стволовых клеток без их существенного дифференцирования. Рост плюрипотентных стволовых клеток на слое питающих клеток без дифференцирования поддерживают путем использования (i) культурального сосуда с имеющимся слоем питающих клеток; и (ii) среды, кондиционированной путем предварительного культивирования клеток иного типа, или некондиционированной среды, например, не содержащей сыворотки среды или даже среды с химически определенным составом.
В другом примере плюрипотентные стволовые клетки культивируют в культуральной системе, по существу свободной от питающих клеток, но тем не менее способной поддерживать пролиферацию плюрипотентных стволовых клеток без существенного дифференцирования. Рост плюрипотентных стволовых клеток в свободной от питающих клеток культуральной системе без дифференцирования поддерживают путем использования (i) адгезивного слоя на твердом носителе, содержащего один или более белков внеклеточного матрикса; и (ii) среды, кондиционированной путем предварительного культивирования клеток иного типа, или некондиционированной среды, например не содержащей сыворотки среды или даже среды с химически определенным составом.
В альтернативном варианте осуществления плюрипотентные стволовые клетки культивируют на состоящем из смешанных эфиров целлюлозы плоском носителе в среде, кондиционированной путем предварительного культивирования клеток иного типа, или некондиционированной среде, например, не содержащей сыворотки среде или даже среде с химически определенным составом.
Культуральная среда. Пример соответствующей целям настоящего изобретения клеточной культуральной среды приведен в заявке на патент США US20020072117. Другой пример соответствующей целям настоящего изобретения клеточной культуральной среды приведен в заявке на патент США US6642048. Другой пример соответствующей целям настоящего изобретения клеточной культуральной среды приведен в заявке на патент WO2005014799. Другой пример соответствующей целям настоящего изобретения клеточной культуральной среды приведен в работе Xu et al. (Stem Cells 22: 972-980, 2004). Другой пример соответствующей целям настоящего изобретения клеточной культуральной среды приведен в заявке на патент США US20070010011. Другой пример соответствующей целям настоящего изобретения клеточной культуральной среды приведен в работе Cheon et al. (BioReprod DOI:10.1095/biolreprod.105.046870; 19 Oct. 2005). Другой пример соответствующей целям настоящего изобретения клеточной культуральной среды приведен в работе Levenstein et al. (Stem Cells 24: 568-574, 2006). Другой пример соответствующей целям настоящего изобретения клеточной культуральной среды приведен в заявке на патент США US20050148070. Другой пример соответствующей целям настоящего изобретения клеточной культуральной среды приведен в заявке на патент США US20050233446. Другой пример соответствующей целям настоящего изобретения клеточной культуральной среды приведен в заявке на патент США US6800480. Другой пример соответствующей целям настоящего изобретения клеточной культуральной среды приведен в заявке на патент США US20050244962. Другой пример соответствующей целям настоящего изобретения клеточной культуральной среды приведен в заявке на патент WO2005065354. Другой пример соответствующей целям настоящего изобретения клеточной культуральной среды приведен в заявке на патент WO2005086845.
Культуральная среда, соответствующая целям настоящего изобретения, может также быть приготовлена из указанных ниже компонентов, таких как, например, модифицированная по способу Дульбекко среда Игла (DMEM), Gibco № 11965-092; нокаут-среда Игла, модифицированная по способу Дульбекко (KO DMEM), Gibco № 10829-018; основная среда Хэма F12/50% DMEM; 200 мM L-глутамина, Gibco № 15039-027; раствор заменимых аминокислот, Gibco 11140-050; β-меркаптоэтанол, Sigma № M7522; человеческий рекомбинантный основной фактор роста фибробластов (bFGF), Gibco № 13256-029.
Дифференцирование плюрипотентных стволовых клеток
В одном осуществлении настоящего изобретения плюрипотентные стволовые клетки размножают в культуре с сохранением их плюрипотентности. Изменения в плюрипотентности культивируемых клеток во времени могут быть определены по изменению уровней экспрессии ассоциированных с плюрипотентностью маркеров. Альтернативно, за изменениями плюрипотентности можно следить по изменениям уровней экспрессии маркеров, ассоциированных с дифференцированием, или маркеров, ассоциированных с клетками иных типов.
В альтернативном осуществлении плюрипотентные стволовые клетки размножают в культуре и затем обрабатывают таким образом, чтобы индуцировать их дифференцирование в клетки иного типа. К таким клеткам иного типа могут относиться клетки с экспрессией маркеров, характерных для линии дифференцирования сформированной эндодермы. Альтернативно, к таким клеткам иного типа могут относиться клетки с экспрессией маркеров, характерных для линии дифференцирования панкреатической эндодермы. Альтернативно, к таким клеткам иного типа могут относиться клетки с экспрессией маркеров, характерных для линии дифференцирования панкреатических эндокринных клеток. Альтернативно, к таким клеткам иного типа могут относиться клетки с экспрессией маркеров, характерных для линии дифференцирования β-клеток.
Плюрипотентные стволовые клетки, обработанные в соответствии со способами, составляющими предмет настоящего изобретения, могут быть дифференцированы в различные клетки иных типов с использованием любых соответствующих способов, известных специалистам в данной области.
Например, плюрипотентные стволовые клетки, обработанные в соответствии со способами, составляющими предмет настоящего изобретения, могут быть дифференцированы в нервные клетки, сердечные клетки, гепатоциты и т.д.
Например, плюрипотентные стволовые клетки, обработанные в соответствии со способами, составляющими предмет настоящего изобретения, могут быть дифференцированы в нервные клетки-предшественники и кардиомиоциты в соответствии со способами, описанными в заявке на патент WO2007030870.
В другом примере плюрипотентные стволовые клетки, обработанные в соответствии со способами, составляющими предмет настоящего изобретения, могут быть дифференцированы в гепатоциты в соответствии со способами, описанными в заявке на патент США № 6458589.
Например, плюрипотентные стволовые клетки могут быть дифференцированы в клетки с экспрессией маркеров, характерных для линии дифференцирования сформированной эндодермы, в соответствии со способами, описанными в работе D'Amour et al., Nature Biotechnol. 23:1534-1541, 2005.
Например, плюрипотентные стволовые клетки могут быть дифференцированы в клетки с экспрессией маркеров, характерных для линии дифференцирования сформированной эндодермы, в соответствии со способами, описанными в работе Shinozaki et al., Development 131:1651-1662, 2004.
Например, плюрипотентные стволовые клетки могут быть дифференцированы в клетки с экспрессией маркеров, характерных для линии дифференцирования сформированной эндодермы, в соответствии со способами, описанными в работе McLean et al., Stem Cells 25:29-38, 2007.
Например, плюрипотентные стволовые клетки могут быть дифференцированы в клетки с экспрессией маркеров, характерных для линии дифференцирования сформированной эндодермы, в соответствии со способами, описанными в работе D'Amour et al., Nature Biotechnol. 24:1392-1401, 2006.
Маркеры, характерные для линии дифференцирования сформированной эндодермы, выбирают из группы, состоящей из следующих маркеров: SOX17, GATA4, HNF3 бета, GSC, CER1, NODAL, FGF8, Brachyury, Mix-подобный гомеобоксный белок, FGF4 CD48, эомезодермин (EOMES), DKK4, FGF17, GATA6, CXCR4, C-Kit, CD99 и OTX2. Целям настоящего изобретения соответствуют клетки с экспрессией по меньшей мере одного из маркеров, характерных для линии дифференцирования сформированной эндодермы. В одном аспекте настоящего изобретения клетка с экспрессией маркеров, характерных для линии дифференцирования сформированной эндодермы, представляет собой клетку-предшественницу первичной полоски. В другом аспекте настоящего изобретения клетка с экспрессией маркеров, характерных для линии дифференцирования сформированной эндодермы, представляет собой мезэндодермальную клетку. В другом аспекте настоящего изобретения клетка с экспрессией маркеров, характерных для линии дифференцирования сформированной эндодермы, представляет собой клетку сформированной эндодермы.
Например, плюрипотентные стволовые клетки могут быть дифференцированы в клетки с экспрессией маркеров, характерных для линии дифференцирования панкреатической эндодермы, в соответствии со способами, описанными в работе D'Amour et al., Nature Biotechnol. 24:1392-1401, 2006.
Маркеры, характерные для линии дифференцирования панкреатической эндодермы, выбирают из группы, состоящей из следующих маркеров: PDX1, HNF1 бета, PTF1 альфа, HNF6, HB9 и PROX1. Целям настоящего изобретения соответствуют клетки с экспрессией по меньшей мере одного из маркеров, характерных для линии дифференцирования панкреатической эндодермы. В одном аспекте настоящего изобретения клетка с экспрессией маркеров, характерных для линии дифференцирования панкреатической эндодермы, представляет собой клетку панкреатической эндодермы.
Плюрипотентные стволовые клетки могут быть дифференцированы в клетки с экспрессией маркеров, характерных для линии дифференцирования панкреатических эндокринных клеток, с использованием любых соответствующих способов, известных специалистам в данной области.
Например, плюрипотентные стволовые клетки могут быть дифференцированы в клетки с экспрессией маркеров, характерных для линии дифференцирования панкреатических эндокринных клеток, в соответствии со способами, описанными в работе D'Amour et al., Nature Biotechnol. 24:1392-1401, 2006.
Например, плюрипотентные стволовые клетки могут быть дифференцированы в клетки с экспрессией маркеров, характерных для линии дифференцирования панкреатических эндокринных клеток, в соответствии со способами, описанными в работе D'Amour et al., Nature Biotechnol. 24: 1392-1401, 2006.
Маркеры, характерные для линии дифференцирования панкреатических эндокринных клеток, выбирают из группы, состоящей из следующих маркеров: NGN3, NEUROD, ISL1, PDX1, NKX6.1, PAX4, NGN3 и PTF-1 альфа. В одном осуществлении панкреатическая эндокринная клетка способна экспрессировать по меньшей мере один из следующих гормонов: инсулин, глюкагон, соматостатин и панкреатический полипептид. Целям настоящего изобретения соответствуют клетки с экспрессией по меньшей мере одного из маркеров, характерных для линии дифференцирования панкреатических эндокринных клеток. В одном аспекте настоящего изобретения клетка с экспрессией маркеров, характерных для линии дифференцирования панкреатических эндокринных клеток, представляет собой панкреатическую эндокринную клетку. Панкреатическая эндокринная клетка может представлять собой панкреатическую клетку, экспрессирующую гормоны. Альтернативно, панкреатическая эндокринная клетка может представлять собой панкреатическую клетку, секретирующую гормоны.
В одном аспекте настоящего изобретения панкреатическая эндокринная клетка представляет собой клетку с экспрессией маркеров, характерных для линии дифференцирования β-клеток. Клетка с экспрессией маркеров, характерных для линии дифференцирования β-клеток, экспрессирует Pdx1 и по меньшей мере один из следующих транскрипционных факторов: NGN3, NKX2.2, NKX6.1, NEUROD, ISL1, HNF3 бета, MAFA, PAX4 и PAX6. В одном аспекте настоящего изобретения клетка с экспрессией маркеров, характерных для линии дифференцирования β-клеток, представляет собой β-клетку.
Настоящее изобретение иллюстрируют, но не ограничивают, следующие примеры.
ПРИМЕРЫ
Пример 1: Закрепление эмбриональных стволовых клеток человека на плоских носителях настоящего изобретения.
Ранее было показано, что ингибитор Rho-киназы Y26732 повышает эффективность закрепления эмбриональных стволовых клеток человека на поверхностно-модифицированных носителях (см. заявку на патент США № 61/030544). Цель исследований, составляющих предмет настоящего изобретения, состояла в определении способности эмбриональных стволовых клеток человека к закреплению на других плоских поверхностях. Испытанные в рамках настоящего изобретения плоские поверхности перечислены в Таблице 1.
Перед тестированием клетки линии эмбриональных стволовых клеток человека H1 нарабатывали на культуральных планшетах, покрытых препаратом MATRIGEL® со сниженным содержание факторов роста в разбавлении 1:30. Клетки высевали на культуральные чашки размером 100 мм в 10 мл кондиционированной MEF среде с добавлением 20 нг/мл bFGF (MEF-CM/bFGF). Клетки культивировали при температуре 37°C в увлажненной атмосфере, содержащей 5% CO2. Культуральную среду ежедневно заменяли на свежую порцию MEF-CM/bFGF. При достижении степени слияния приблизительно 80% клетки пассировали путем обработки раствором 1 мг/мл препарата LIBERASE в течение 5 минут при температуре 37°C. Расщепление останавливали путем удаления фермента из чашки и промывания клеток средой MEF-CM/bFGF. Клетки собирали соскабливанием вручную в 10 мл среды MEF-CM/bFGF и переносили в коническую пробирку объемом 50 мл. Затем клетки центрифугировали при 200× g (1000 об/мин) на настольной центрифуге с получением дебриса. После удаления супернатанта клетки ресуспендировали в 40 мл среды MEF-CM/bFGF и равномерно распределяли по четырем культуральным чашкам размером 100 мм, покрытых препаратом MATRIGEL® со сниженным содержание факторов роста в разбавлении 1:30.
Клетки линии эмбриональных стволовых клеток человека H1 высевали на различные плоские носители, перечисленные в Таблице 1, с плотностью 100000 клеток/см2. Используемые плоские носители не имели адсорбированного слоя и слоя питающих клеток. Клетки культивировали в среде MEF-CM/bFGF, как описано выше. Определяли влияние ингибитора Rho-киназы H-1152 на закрепление клеток на плоских носителях. В используемую для посева клеток среду добавляли 3 мкМ H-1152. Затем клетки выдерживали в течение 24 часов для закрепления. По истечении указанного периода клетки фиксировали 4% раствором параформальдегида в течение 5 минут при комнатной температуре. Затем клетки окрашивали 1% раствором гематоксилина и определяли численность клеток оптической микроскопией. В качестве контрольного образца использовали лунки с чистой средой.
На следующих носителях клетки линии эмбриональных стволовых клеток человека H1 закреплялись независимо от присутствия в среде ингибитора Rho-киназы: мембраны из смешанных эфиров целлюлозы (мембрана № 2, фиг.1A); мембраны из нейлона (мембрана № 4, фиг.1B) и мембраны из ацетата целлюлозы (мембрана № 5, фиг.1C). Эффективность закрепления клеток на перечисленных мембранах повышалась при добавлении в среду 3 мкМ H-1152 (см. фиг.1A-1C).
На следующих плоских носителях клетки линии эмбриональных стволовых клеток человека H1 закреплялись только в присутствии 3 мкМ H-1152: мембраны из поликарбоната (мембрана № 7, фиг.1D) и мембрана из полиэтилентерефталата (мембрана № 12, фиг.1E). Удаление H-1152 из культуральной среды приводило к откреплению клеток линии H1 от мембран обоих указанных типов. В отсутствие H-1152 клетки на указанных мембранах не закреплялись.
Пример 2: Воздействие обработки ингибитором Rho-киназы на закрепление эмбриональных стволовых клеток человека на плоских носителях, состоящих из смешанных эфиров целлюлозы (мембрана № 1).
Перед экспериментом клетки линии эмбриональных стволовых клеток человека H9 культивировали на покрытых препаратом MATRIGEL® чашках. Клетки высевали на мембраны из смешанных ацетатов целлюлозы (мембрана № 1) с плотностью 150000 клеток/см2 в кондиционированной MEF среде. Используемый плоский носитель не имел адсорбированного слоя и слоя питающих клеток. Изучили воздействие обработки ингибитором Rho-киназы на закрепление клеток на плоском носителе. Клетки обрабатывали 0, 10 или 20 мкМ Y26732. Через 24 часа клетки фиксировали 4% раствором параформальдегида, промывали фосфатным буфером, сушили на воздухе и окрашивали кристаллическим фиолетовым. Численность клеток определяли оптической микроскопией. В качестве контрольного образца использовали лунки с чистой средой.
Клетки закреплялись на плоском носителе в отсутствие Y26732 (фиг.2A). Добавление Y26732 в концентрации 10 и 20 мкМ повысило эффективность закрепления клеток на плоском носителе (фиг.2B-2C). Удаление Y26732 из культуральной среды на 24 часа не привело к откреплению клеток от плоского носителя.
Пример 3: Воздействие культивирования на мембране плоского носителя № 1 на скорость пролиферации эмбриональных стволовых клеток человека.
Сравнивали скорости пролиферации клеток линии эмбриональных стволовых клеток человека, культивируемых на покрытых препаратом MATRIGEL® чашках, и клеток, культивируемых на мембране № 1. Клетки высевали с равной плотностью на оба носителя. Чтобы получить одноклеточную суспензию для определения численности клеток, носители с клетками обрабатывали отделяющим препаратом TrypLE. Образцы с клетками отбирали через промежутки времени, указанные на фиг.3. Пролиферация клеток происходила со сравнимыми скоростями. Период удвоения составил приблизительно 1,151 дня и 1,138 дня для клеток на препарате MATRIGEL® и на мембране № 1, соответственно.
Пример 4: Эмбриональные стволовые клетки человека сохраняют свою плюрипотентность в течение трех пассажей на плоских носителях, состоящих из смешанных эфиров целлюлозы (мембрана № 1).
Клетки линии эмбриональных стволовых клеток человека H1 высевали на плоских носителях, состоящих из смешанных эфиров целлюлозы (мембрана № 1), с плотностью 75000 клеток/см2 в среде MEF-CM с добавлением 20 нг/мл bFGF. Перед пассированием клетки культивировали в течение 5 или 6 дней до достижения степени слияния приблизительно 75-90%, следуя описанным выше способам. Культуральную среду заменяли ежедневно. После культивирования в течение 3 пассажей клетки собирали и определяли уровень экспрессии ассоциированных с плюрипотентностью маркеров с помощью проточной цитометрии. Как показано в Таблице 2, более 95% клеток сохранило экспрессию ассоциированных с плюрипотентностью маркеров клеточной поверхности, включая Tra1-60, Tra1-81, SSEA-3 и SSEA-4, что указывает на сохранение плюрипотентности клеток.
Пример 5: Эмбриональные стволовые клетки человека сохраняют устойчивый кариотип в течение десяти пассажей на плоских носителях, состоящих из смешанных эфиров целлюлозы (мембрана № 1).
Клетки линии эмбриональных стволовых клеток человека линии H1 в течение 10 пассажей культивировали на покрытых препаратом MATRIGEL® культуральных планшетах и на мембранах из смешанных эфиров целлюлозы. Клетки культивировали, следуя описанным выше способам. Искомый кариотип определяли с помощью цитогенетического анализа, анализируя двадцать метафазных клеток, окрашенных с использованием красителя Гимза. Как показано на фиг.4, окрашенные хромосомы представительной клетки из культуры, выращенной на покрытых препаратом MATRIGEL® культуральных планшетах (фиг.4A), и другой клетки из культуры, выращенной на мембране из смешанных эфиров целлюлозы (фиг.4B), демонстрируют нормальный мужской кариотип.
Кариотип также определяли путем анализа двухсот интерфазных ядер способом флуоресцентной гибридизации in situ (FISH), используя зонды на хромосому 12p и 17q для идентификации очень малых популяций клеток с изменениями в числе копий хромосом 12 и 17, которые не могут быть обнаружены стандартными способами цитогенетики. В клетках, культивировавшихся на препарате MATRIGEL® и на мембранах из смешанных эфиров целлюлозы, не было обнаружено аномальных клеток с трисомией по 12 и (или) 17 хромосоме.
Пример 6: Эмбриональные стволовые клетки человека способны к дифференцированию в продуцирующие инсулин клетки на плоских носителях, состоящих из смешанных эфиров целлюлозы (мембрана № 1).
Клетки линии эмбриональных стволовых клеток человека H1 высевали на плоских носителях, состоящих из смешанных эфиров целлюлозы (мембрана № 1), с плотностью 150000 клеток/см2 в кондиционированной MEF среде с добавлением 20 нг/мл bFGF. Клетки дифференцировали в продуцирующие инсулин клетки, обрабатывая клетки в соответствии с изложенным в Таблице 3 протоколом дифференцирования. Клетки культивировали в кондиционированной MEF среде с добавлением 20 нг/мл bFGF в течение 3-4 дней до достижения степени слияния приблизительно 75-90%. Клетки обрабатывали в среде DMEM-F12 с добавлением 2% не содержащего жирных кислот бычьего сывороточного альбумина (FAF-BSA), 100 нг/мл активина A и 20 нг/мл Wnt3A в течение двух дней с последующей обработкой в среде DMEM-F12 с добавлением 2% не содержащего жирных кислот бычьего сывороточного альбумина (FAF-BSA) и 100 нг/мл активина в течение еще двух дней. Затем клетки обрабатывали в среде DMEM-F12 с добавлением 2% BSA, 20 нг/мл FGF7 и 250 нМ циклопамин-KAAD в течение трех дней с последующей обработкой в среде DMEM-F12 с добавлением 1% препарата B27, 20 нг/мл FGF7, 250 нМ циклопамина-KAAD, 2 мкМ M ретиноевой кислоты (RA) и 100 нг/мл ногина (Noggin) в течение 4 дней. Затем клетки обрабатывали в среде DMEM-F12 с добавлением 1% препарата B27, 1 мкМ ингибитора ALK5 № 2 (номер по каталогу Axxora: ALX-270-445-M001), 100 нг/мл ногина (Noggin), 100 нг/мл нетрина-4, 50 нг/мл экстендина-4 и 1 мкМ DAPT в течение 3 дней. Клетки культивировали в среде DMEM-F12 с добавлением 1% препарата B27 и 1 мкМ ингибитора ALK5 № 2 в течение 7 дней и затем в среде DMEM-F12 с добавлением 1% препарата B27 в течение еще 7 дней.
По завершении протокола дифференцирования отобрали образцы РНК для определения уровней экспрессии маркеров, характерных для линии дифференцирования панкреатических эндокринных клеток. Полученная КТ плотность для инсулина составляла приблизительно 17. Соответствующая величина КТ плотности для GAPDH составила приблизительно 19; полученные данные свидетельствуют о том, что клетки после обработки имеют высокие уровни экспрессии инсулина.
Пример 7: Эмбриональные стволовые клетки человека закрепляются на плоских носителях, состоящих из поликарбонатных мембран, зависимым от присутствия ингибитора Rho-киназы образом.
Клетки линии эмбриональных стволовых клеток человека H9 высевали на плоских поликарбонатных носителях (мембрана № 7), с плотностью 150000 клеток/см2 в кондиционированной MEF среде. Изучили влияние обработки ингибитором Rho-киназы на закрепление клеток: в культуральную среду добавляли ингибитор Rho-киназы Y26732 в концентрации 0, 10 и 20 мкМ. Через 24 часа клетки фиксировали 4% раствором параформальдегида, промывали фосфатным буфером, сушили на воздухе и окрашивали кристаллическим фиолетовым. Численность клеток определяли оптической микроскопией. В качестве контрольного образца использовали лунки с чистой средой.
Клетки не закреплялись на мембране в контрольных чашках (фиг.5A). Добавление Y26732 привело к закреплению клеток на мембранах (фиг.5B-5C).
В отдельном эксперименте определили влияние ингибитора Rho-киназы H-1152 на закрепление клеток линии эмбриональных стволовых клеток человека H1 на мембране № 7. Клетки высевали на плоских носителях, состоящих из поликарбонатных мембран (мембрана № 7), с плотностью 150000 клеток/см2 в кондиционированной MEF среде с добавлением 20 нг/мл bFGF. В культуральную среду добавляли ингибитор Rho-киназы H-1152 в концентрациях 0; 0,03; 0,1; 0,3; 1 и 3 мкМ. Через 24 часа клетки фиксировали 4% раствором параформальдегида, промывали фосфатным буфером, сушили на воздухе и окрашивали кристаллическим фиолетовым. Численность клеток определяли оптической микроскопией. В качестве контрольного образца использовали лунки с чистой средой.
Клетки не закреплялись на мембране в контрольной чашке (фиг.6A) и в чашках с добавлением 0,03 и 0,1 мкМ H-1152 (фиг.6B-6C). Однако закрепление наблюдалось в культурах, обработанных 0,3; 1 и 3 мкМ H-1152 (фиг.6D-6F).
Пример 8: Удаление ингибитора Rho-киназы из культуральной среды приводит к откреплению эмбриональных стволовых клеток человека от плоских носителей, состоящих из поликарбонатных мембран.
Клетки линии эмбриональных стволовых клеток человека H1 высевали на плоских поликарбонатных носителях (мембрана № 9), с плотностью 100000 клеток/см2, в кондиционированной MEF среде с добавлением 20 нг/мл bFGF и 3 мкМ ингибитора Rho-киназы H-1152. Клетки культивировали в течение 24 часов и затем заменяли культуральную среду на кондиционированную MEF среду с добавлением 20 нг/мл bFGF, но без H-1152. Через 24 часа клетки на мембране фиксировали 4% раствором параформальдегида, промывали фосфатным буфером, сушили на воздухе и окрашивали кристаллическим фиолетовым. Численность клеток определяли оптической микроскопией. В качестве контрольного образца использовали лунки с добавлением H-1152. Удаление H-1152 из культуральной среды привело к откреплению клеток от плоского носителя (фиг.7).
Пример 9: Пористость плоского носителя влияет на эффективность закрепления эмбриональных стволовых клеток человека.
Клетки линии эмбриональных стволовых клеток человека H1 42-го пассажа высевали на следующие плоские носители: мембрану № 10 (размер пор 0,4 мкм); и мембрану № 11 (размер пор 3 мкм). Клетки высевали с плотностью 100000 клеток/см2 в кондиционированной MEF среде с добавлением 20 нг/мл bFGF. Также изучили воздействие ингибирования Rho-киназы на закрепление клеток на плоских носителях. В культуральную среду добавили 3 мкМ H-1152. Через 24 часа культуральную среду заменяли на кондиционированную MEF среду с добавлением 20 нг/мл bFGF, но без H-1152. Еще через 24 часа культивирования клетки на мембране фиксировали 4% раствором параформальдегида, промывали фосфатным буфером, сушили на воздухе и окрашивали кристаллическим фиолетовым. В качестве контрольного образца использовали лунки с добавлением 1 мкМ Η-1152. Численность клеток определяли оптической микроскопией. В качестве контрольного образца использовали лунки с чистой средой.
Обнаружили, что на мембране № 10 (фиг.8A) закрепилось большее количество клеток, чем на мембране № 11 (фиг.8B). Присутствие 1 мкМ H-1152 в культуральной среде необходимо для поддержания закрепления клеток линии H1 на мембранах (фиг.8A-8B). Удаление H-1152 из культуральной среды привело к откреплению клеток от мембраны № 10 и мембраны № 11 (фиг.8C-8D).
Пример 10: Эмбриональные стволовые клетки человека сохраняют свою плюрипотентность в течение многих пассажей на плоских носителях, состоящих из поликарбонатных мембран.
Клетки линии эмбриональных стволовых клеток человека H1 высевали на плоских носителях, состоящих из поликарбонатных мембран (мембрана № 8). Клетке культивировали в кондиционированной MEF среде с добавлением 20 нг/мл bFGF и 3 мкМ H-1152. Культуральную среду заменяли ежедневно. Клетки пассировали путем удаления H-1152 из среды, используя для открепления клеток от носителя осторожное помешивание. Клетки культивировали в течение 3 пассажей и затем собирали для проведения проточной цитометрии и количественного ОТ-ПЦР анализа. Как показано в Таблице 4, по результатом анализа на проточном цитометре более 95% клеток сохранило экспрессию ассоциированных с плюрипотентностью маркеров клеточной поверхности, включая Tra1-60, Tra1-81, SSEA-3 и SSEA-4. На фиг.9 приведены результаты количественного анализа ОТ-ПЦР, показывающие, что уровни экспрессии ряда генов в клетках линии H1, культивировавшихся на поликарбонатных мембранах в течение 3 пассажей, сравнимы с уровнями экспрессии для недифференцированных клеток линии H1.
В отдельном исследовании клетки линии эмбриональных стволовых клеток человека H1 высевали на плоском носителе - поликарбонатной мембране (мембрана № 8). Клетки культивировали в кондиционированной MEF среде с добавлением 20 нг/мл bFGF и 1 мкМ H-1152. Культуральную среду заменяли ежедневно. Клетки пассировали путем удаления H-1152 из среды, используя для открепления клеток от носителя осторожное помешивание. Клетки культивировали в течение 9 пассажей и затем собирали для проведения проточной цитометрии. Как показано в Таблице 5, более 95% клеток сохранило экспрессию ассоциированных с плюрипотентностью маркеров клеточной поверхности, включая Tra1-60, Tra1-81, SSEA-3 и SSEA-4.
Альтернативный способ оценки плюрипотентности связан со способностью клеток образовывать эмбриоидные тельца. Клетки линии эмбриональных стволовых клеток человека H1 высевали на плоских поликарбонатных носителях (мембрана № 8). Клетки культивировали в кондиционированной MEF среде с добавлением 20 нг/мл bFGF и 3 мкМ H-1152. Культуральную среду заменяли ежедневно. Клетки пассировали путем удаления H-1152 из среды, используя для открепления клеток от носителя осторожное помешивание. Клетки культивировали в течение 12 пассажей.
Для формирования эмбриоидных телец использовали следующий протокол. Клетки линии H1 собирали и культивировали в среде DMEM/F12 с добавлением 20% эмбриональной бычьей сыворотки на планшетах с ультранизким закреплением (номер по каталогу Corning: 3471). Для питания клеток культуральную среду через день на 50% заменяли на свежую. Эмбриоидные тельца сформировались через 14 дней (фиг.10).
Пример 11: Эмбриональные стволовые клетки человека способны к образованию сформированной эндодермы после культивирования на плоских носителях, состоящих из поликарбонатных мембран.
Клетки линии эмбриональных стволовых клеток человека H1 высевали на плоский носитель - поликарбонатную мембрану (мембрана № 8). Сначала клетки культивировали в кондиционированной MEF среде с добавлением 20 нг/мл bFGF и 3 мкМ H-1152. Затем до начала эксперимента клетки в течение 10 пассажей культивировали в кондиционированной MEF среде с добавлением 20 нг/мл bFGF и 1 мкМ H-1152.
Затем клетки высевали на культуральные планшеты размером 100 мм, покрытые препаратом MATRIGEL® в разбавлении 1:30. Клетки культивировали в кондиционированной MEF среде с добавлением 20 нг/мл bFGF в течение 3 дней. Затем клетки обрабатывали в среде DMEM/F12 с добавлением 2% не содержащего жирных кислот бычьего сывороточного альбумина, 100 нг/мл активина A и 20 нг/мл Wnt3A в течение двух дней с последующей обработкой в среде DMEM/F12 с добавлением 2% не содержащего жирных кислот бычьего сывороточного альбумина и 100 нг/мл активина в течение еще двух дней. После этого клетки сняли с носителя с помощью обработки препаратом TRYPLE и получили одноклеточную суспензию. Уровни экспрессии маркеров, характерных для линии дифференцирования сформированной эндодермы, определяли с помощью проточной цитометрии.
Как показано в Таблице 6, более 90% клеток оказались дважды положительны по CD99 и CXCR4 (CD184), и 12% клеток оказались положительны по CD9 и отрицательны по CXCR4. Полученные данные свидетельствуют о сохранении способности клеток к дифференцированию по линии сформированной эндодермы.
Пример 12: Физические характеристики плоских носителей настоящего изобретения.
Определили химическую структуру поверхности плоских носителей, составляющих предмет настоящего изобретения. В Таблицах 7-10 приведены результаты анализа способом рентгеновской фотоэлектронной спектроскопии (РФЭС) и результаты измерения контактного угла. При проведении РФЭС использовали глубину анализа приблизительно 5-10 нм (50-100 Å). Как правило, 95% наблюдаемого сигнала формируется в пределах слоя указанной толщины.
В мембранах 1-3 обнаружено близкое содержание кислорода, углерода (в основном в виде C-O и C-(C,H), возможно O-C-O) и азота (в виде NO3, NO2 и возможно C-N и R4-N+). В мембране 3 также содержались следовые количества Na+ и SOx и повышенные концентрации C-(C,H). Мембрана 4 содержала C-(C,H), C-(O,N) и (O,N)-C=O, а также возможно следовые количества натрия. Мембрана 5 содержала в основном C-O, C-(C,H), O-C-O и/или O-C=O. Также были обнаружены следовые количества Na+ и SOx. Мембраны 6-11 содержали C-(C,H), C-O, O-C=O, C-N, CO3, p-p*, а также следовые количества R4-N+, SOx и либо Na+, либо Cr3+. Поверхность мембраны 6 также может содержать следовые количества хлора. Следовые количества хрома были обнаружены только на мембранах 10 и 11, на мембранах 6-9 был обнаружен Na+. Поверхность мембраны 12 содержала C-(C,H), C-O, O-C=O и pi-pi* в согласии со структурой ПЭТФ. Также были обнаружены следовые количества азота и натрия.
На фиг.11 показаны полученные на сканирующем электронном микроскопе микрофотографии плоских носителей, составляющих предмет настоящего изобретения. Наблюдали два типа морфологии поверхности. Первый тип характеризовался наличием открытой сети волокон. Первый тип характеризовался наличием гладкой листовой поверхности с разбросанными по ней круглыми отверстиями.
В Таблице 10 приведены результаты измерения контактного угла для поверхностей, составляющих предмет настоящего изобретения. Поверхности 1-5 имели контактный угол в диапазоне от приблизительно 18° до приблизительно 32°. Для закрепления на поверхностях 1-5 плюрипотентные стволовые клетки не требовали присутствия ингибитора активности Rho-киназы.
Поверхности 6-12 имели контактный угол более 32°. Для закрепления на этих поверхностях плюрипотентные стволовые клетки требовали присутствия ингибитора активности Rho-киназы.
Пример 13: Закрепление плюрипотентных стволовых клеток на плоском носителе, состоящем из полиамина.
Состоящий из полиамина плоский носитель был изготовлен в соответствии со способами, изложенными в патенте США US6743273 и работе Schindler M et al, Biomaterials 26(28): 5624-5631; 2005. Указанный плоский носитель доступен в продаже под торговой маркой ULTRAWEBTM. Синтетические поверхности носителя ULTRAWEB™ состоят из случайным образом ориентированных нановолокон полиамида со средним диаметров волокна 280 нм, нанесенных способом электропрядения. Распределение по размерам нановолокон находится в диапазоне от 200 до 400 нм. Первая тестированная поверхность ULTRAWEB™ была несколько гидрофильной (номер по каталогу 3870XX1), вторая поверхность (номер по каталогу 3871XX1) была несколько гидрофильной и имела покрытие из полиаминового материала, который служил для нановолокон источником свободных аминовых групп и положительного заряда. Обе поверхности имеют высокую эффективность адсорбции белков, обусловленной гидрофобными взаимодействиями. Полученные на сканирующем электронном микроскопе микрофотографии носителей с разрешением 5 микрон при увеличении 10000× показаны на фиг.12. Однако клетки линии эмбриональных стволовых клеток человека H1 оказались неспособными закрепиться на обеих тестированных поверхностях UTRAWEBTM.
Пример 14: Влияние использования среды с химически определенным составом на закрепление плюрипотентных клеток на плоских носителях настоящего изобретения.
Клетки линии эмбриональных стволовых клеток человека H1 высевали на следующие плоские носители: мембрану 1 (смешанные эфиры целлюлозы), мембрану 4 (нейлон), мембрану 5 (ацетат целлюлозы) и нитроцеллюлозу. Клетки высевали в разбавлении 1:3 в среде определенного состава mTESRTM и культивировали в течение 24 часов. В качестве контрольного образца использовали параллельное культивирование в кондиционируемой MEF среде. Культивирование клеток в среде mTESRTM не сказалось на способности клеток к закреплению на плоских носителях. Клетки были способны к закреплению на мембранах 1, 4 и 5 и на нитроцеллюлозе. Наибольшую эффективность связывания клеток при использовании среды mTESRTM показала мембрана 4, затем шла мембрана 5 с такой же эффективностью связывания, как у нитроцеллюлозы, и затем мембрана 1.
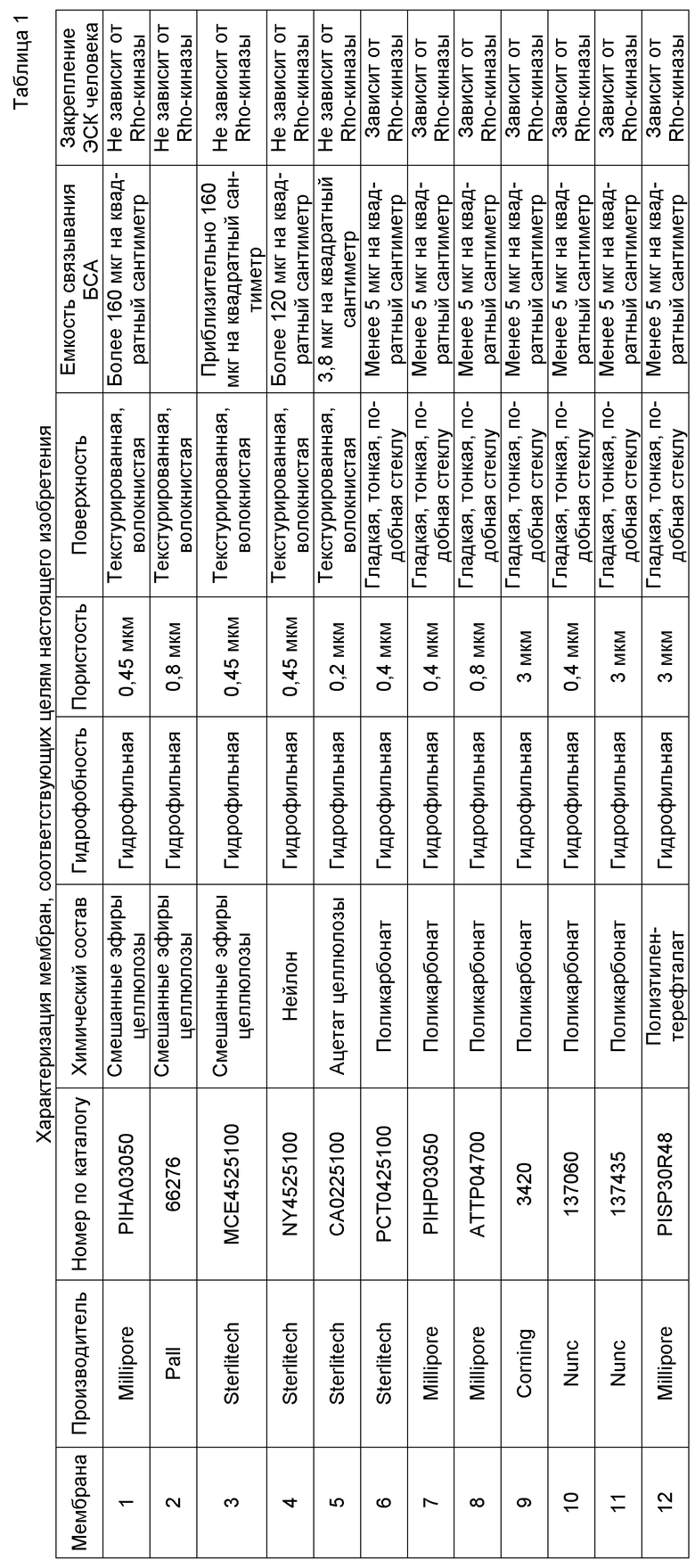
Уровни экспрессии ассоциированных с плюрипотентностью маркеров клеточной поверхности эмбриональных стволовых клеток человека линии H1 после выращивания на мембранах из смешанных эфиров целлюлозы в течение 3 пассажей по результатам проточной цитометрии
Протокол обработки эмбриональных стволовых клеток человека для индуцирования дифференцирования в продуцирующие инсулин клетки
2% не содержащего жирных кислот бычьего сывороточного альбумина (FAF-BSA)
100 нг/мл активина A
20 нг/мл Wnt3A
2% не содержащего жирных кислот бычьего сывороточного альбумина (FAF-BSA)
100 нг/мл активина A
2% БСА
20 нг/мл FGF7
250 нМ циклопамин-KAAD
1% препарата B27
20 нг/мл FGF7
250 нМ циклопамин-KAAD
2 мкМ ретиноевой кислоты (RA)
100 нг/мл ногина (Noggin)
1% препарата B27
1 мкМ ингибитора ALK5 №2
100 нг/мл ногина (Noggin)
100 нг/мл нетрина-4
50 нг/мл экстендина-4
1 мкМ DAPT
1% препарата B27
1 мкМ ингибитора ALK5 № 2
1% препарата B27
Уровни экспрессии ассоциированных с плюрипотентностью маркеров клеточной поверхности эмбриональных стволовых клеток человека линии H1 после выращивания на поликарбонатных мембранах в течение 3 пассажей по результатам проточной цитометрии
Уровни экспрессии ассоциированных с плюрипотентностью маркеров клеточной поверхности эмбриональных стволовых клеток человека линии H1 после выращивания на поликарбонатных мембранах в течение 9 пассажей по результатам проточной цитометрии
Уровни экспрессии маркеров клеточной поверхности эмбриональных стволовых клеток человека линии H1, ассоциированных с дифференцированием по линии сформированной эндодермы. Клетки выращивали на поликарбонатных мембранах в течение 10 пассажей и обрабатывали для индуцирования дифференцирования по линии сформированной эндодермы
Химическая структура поверхности (атомные концентрации в %)
Концентрации (в %) углеродных функциональных групп
Концентрации (в %) азотных функциональных групп
Результаты измерения контактного угла для планшетов настоящего изобретения
Все цитируемые в настоящем документе публикации полностью включены в настоящий документ путем ссылки. Хотя различные аспекты изобретения иллюстрируются выше ссылками на примеры и предпочтительные варианты осуществления, подразумевается, что область изобретения определяется не упомянутым выше описанием, а следующими пунктами формулы изобретения, составленными в соответствии с принципами патентного законодательства.
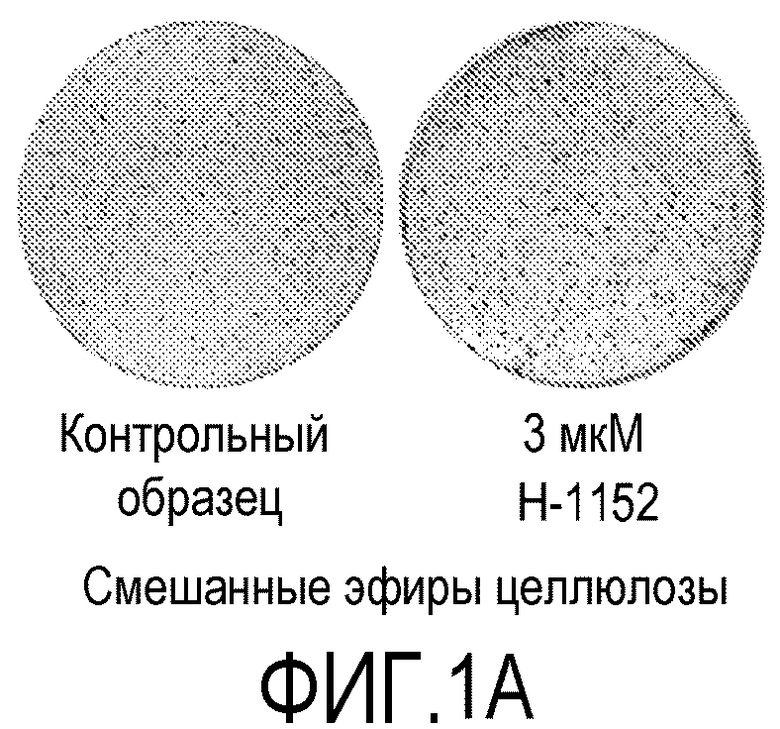
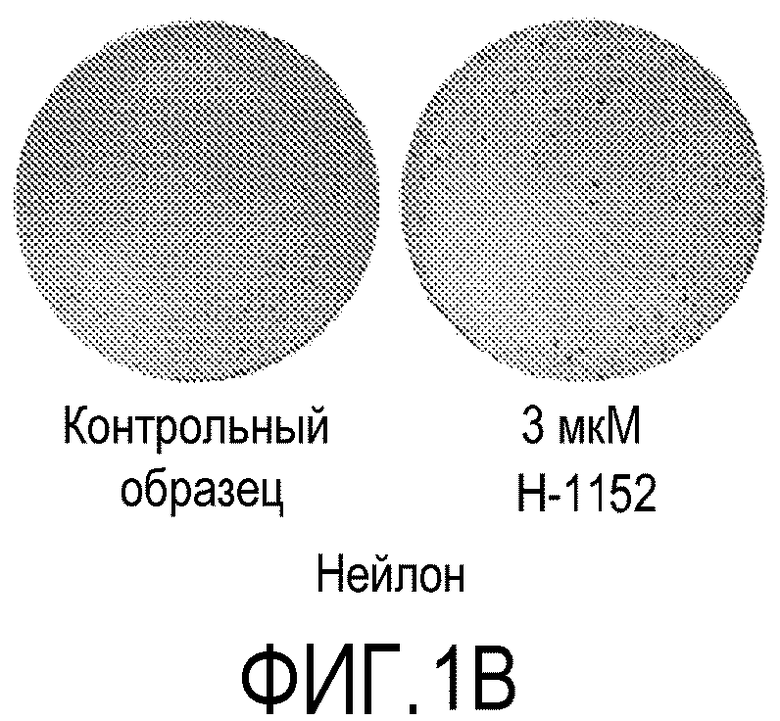
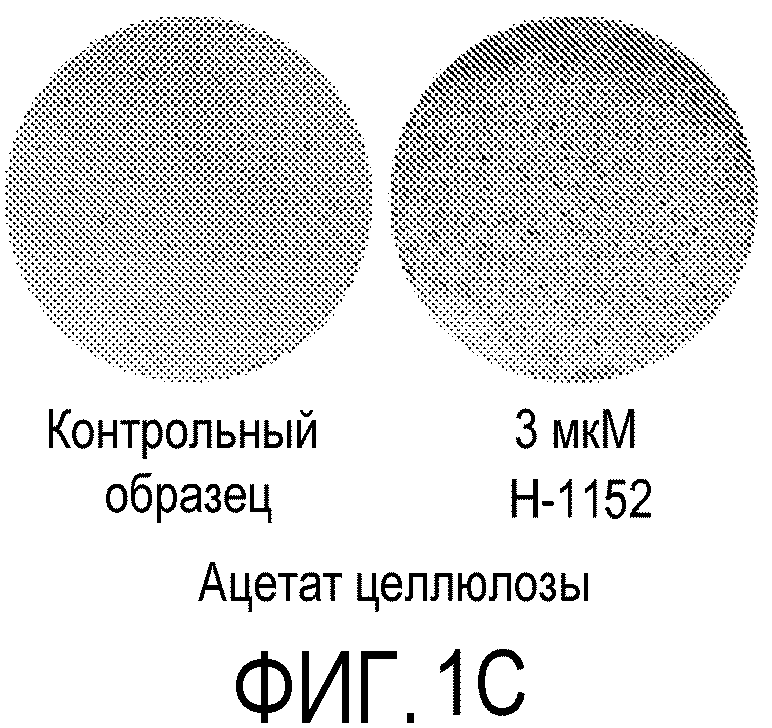
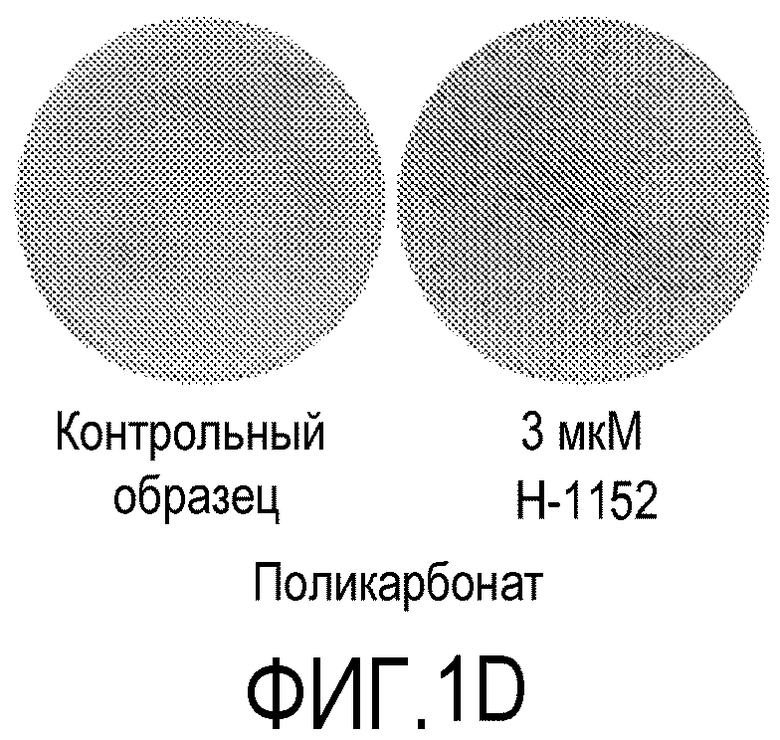




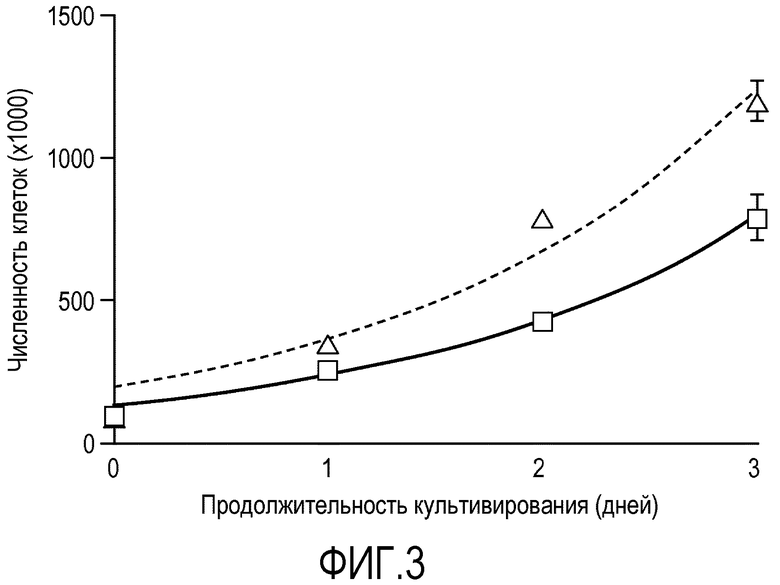
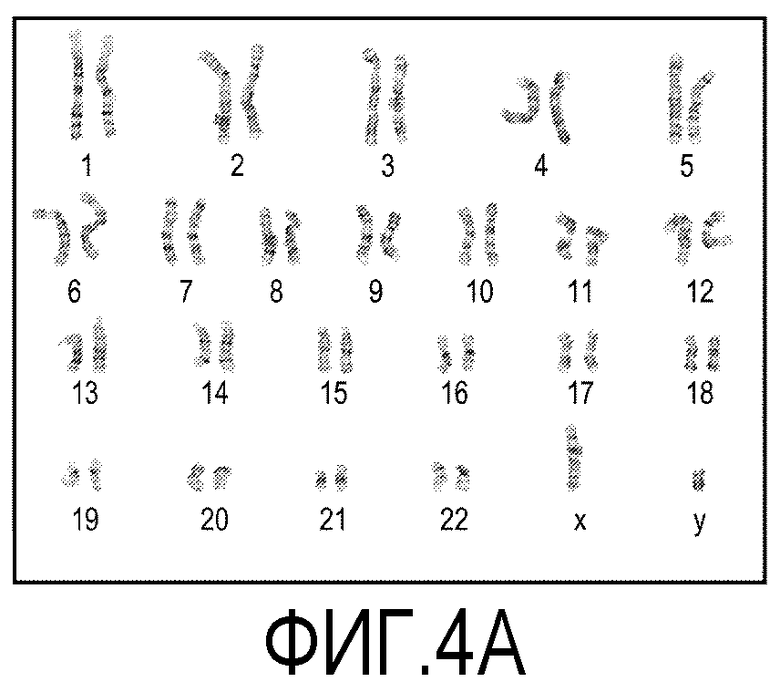
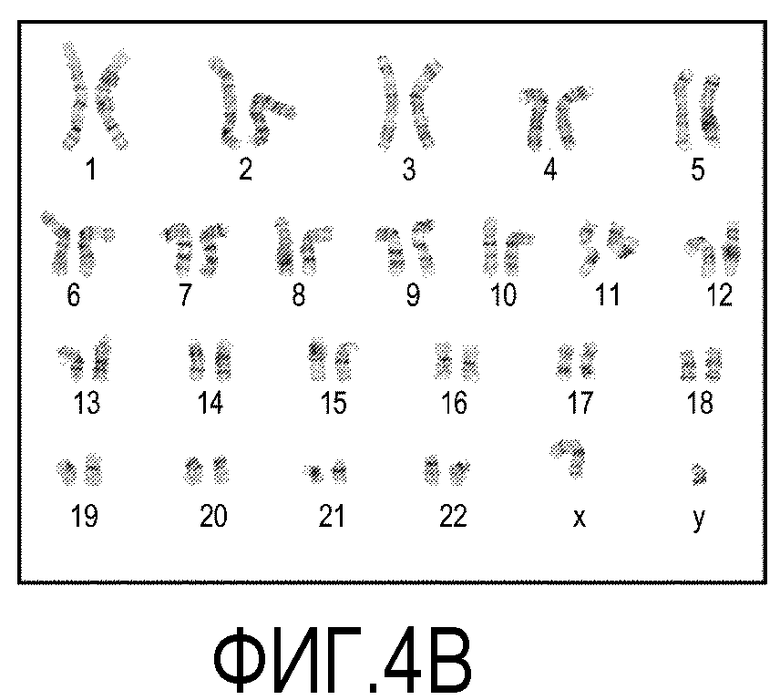












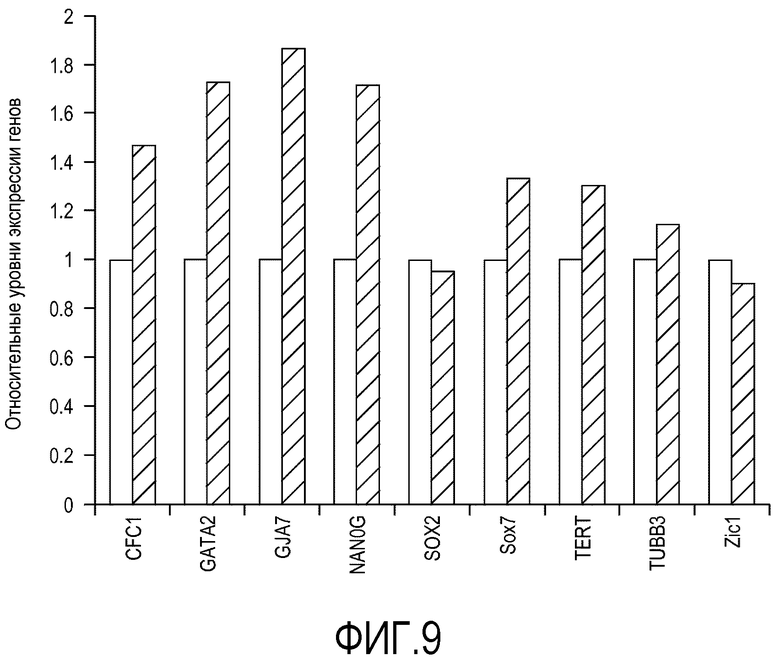
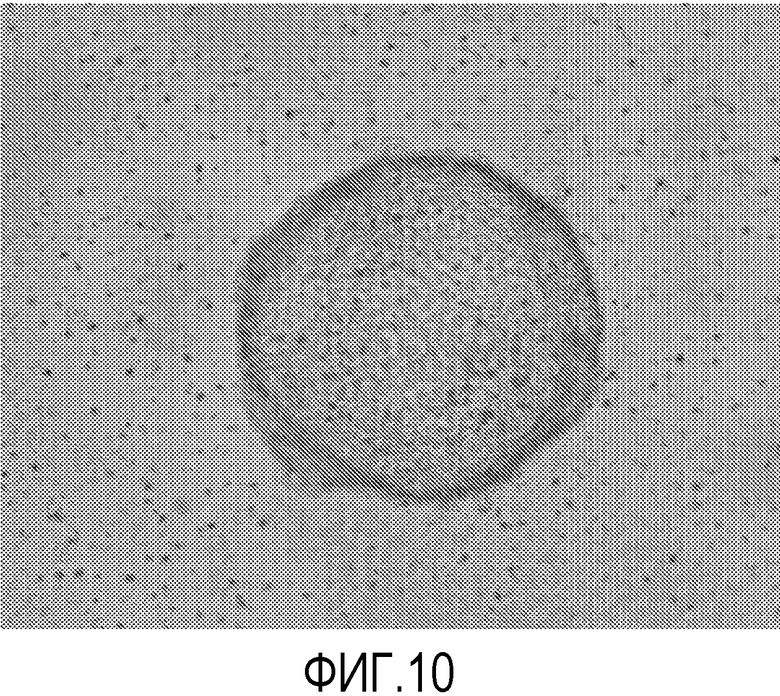
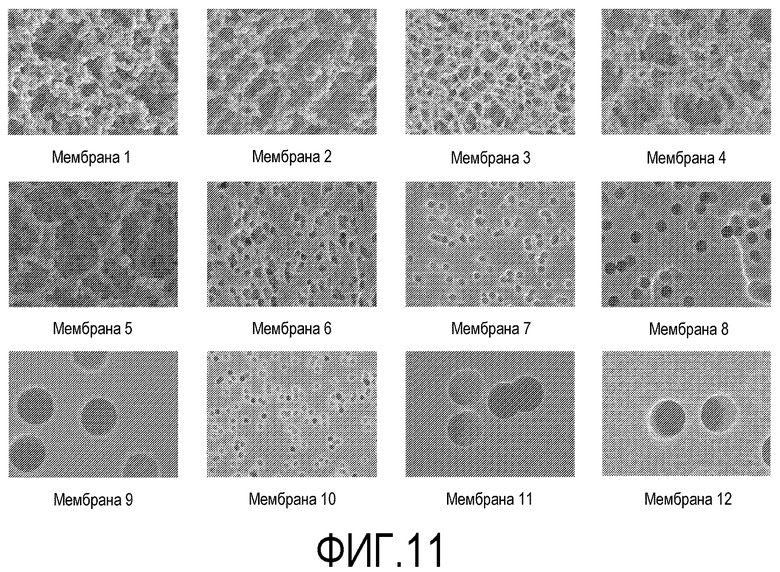


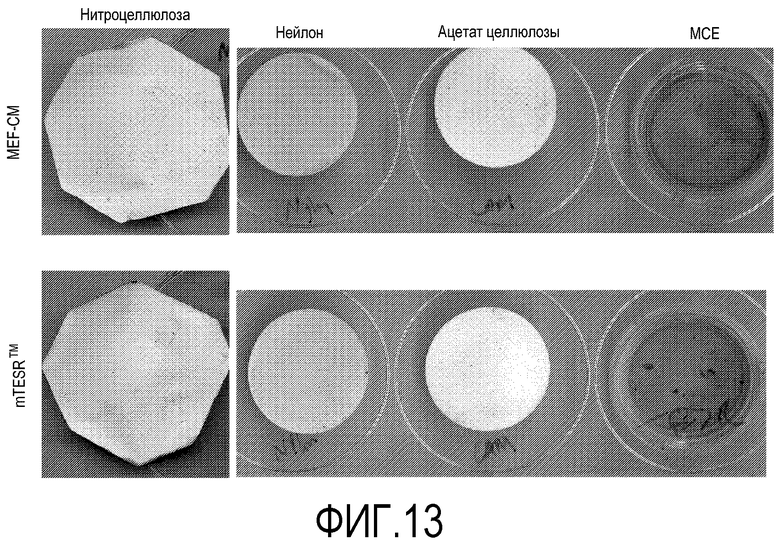
Изобретение относится к области биотехнологии. Описан способ закрепления плюрипотентных стволовых клеток человека на плоском носителе. Способ предусматривает получение суспензии плюрипотентных стволовых клеток и добавление к суспензии соединения, способного ингибировать активность Rho-киназы. Далее проводят добавление суспензии клеток к плоскому носителю и проводят удаление соединения после закрепления клеток. 2 н. и 12 з.п. ф-лы, 32 ил., 10 табл., 14 пр.
1. Способ закрепления плюрипотентных стволовых клеток человека на плоском носителе, содержащем до 12% N, от по меньшей мере 12% O до по меньшей мере 55% O, с углом смачивания от 18 градусов до 32 градусов, и не имеющем адсорбированного слоя и слоя питающих клеток, включающий следующие стадии:
a. получение суспензии плюрипотентных стволовых клеток;
b. добавление к суспензии плюрипотентных стволовых клеток соединения, способного ингибировать активность Rho-киназы;
c. добавление суспензии клеток (b) к плоскому носителю;
d. обеспечение возможности закрепления клеток на плоском носителе; и
e. удаление соединения, способного ингибировать активность Rho-киназы, после того как клетки закрепились на поверхности носителя;
где соединение, способное ингибировать активность Rho-киназы, усиливает или требуется для закрепления суспензии клеток на плоском носителе, и
где закрепление включает культивирование клетки и носителя в условиях, которые способствуют росту без дифференцирования,
где указанные плюрипотентные стволовые клетки человека получены из клеточной линии человеческих эмбриональных стволовых клеток H1, H7 или H9 (WiCeIl).
2. Способ по п. 1, в котором клетки поддерживают в культуре после их закрепления на поверхности плоского носителя.
3. Способ по п. 1, в котором клетки далее дифференцируются после их закрепления на поверхности плоского носителя.
4. Способ по п. 1, в котором соединение, способное ингибировать активность Rho-киназы, выбрано из группы, состоящей из Y-27632, fasudil, H-1152 и hydroxyfasudil.
5. Способ по п. 1, в котором ингибитор активности Rho-киназы добавляют в количестве от 0,1 мкМ до 100 мкМ.
6. Способ по п. 1, в котором по меньшей мере 95% клеток поддерживает экспрессию маркеров, связанных с плюрипотентностью.
7. Способ усиления закрепления плюрипотентных стволовых клеток человека на поверхности плоского носителя, содержащего до 12% Ν, от по меньшей мере 12% O до по меньшей мере 55% O, с углом смачивания от 18 градусов до 32 градусов, и не имеющем адсорбированного слоя и слоя питающих клеток, включающий следующие стадии:
a. обработка суспензии плюрипотентных стволовых клеток соединением, способным ингибировать активность Rho-киназы;
b. приведение плоского носителя в контакт с обработанной суспензией плюрипотентных стволовых клеток человека;
c. обеспечение возможности закрепления клеток на поверхности плоского носителя; и
d. удаление соединения, способного ингибировать активность Rho-киназы после того, как клетки закрепились на поверхности плоского носителя;
где соединение, способное ингибировать активность Rho-киназы, усиливает закрепление суспензии клеток на плоском носителе,
где указанные плюрипотентные стволовые клетки человека получены из клеточной линии человеческих эмбриональных стволовых клеток H1, H7 или H9 (WiCeIl).
8. Способ по п. 7, в котором клетки поддерживают в культуре после их закрепления на поверхности плоского носителя.
9. Способ по п. 7, в котором клетки далее дифференцируются после их закрепления на поверхности плоского носителя.
10. Способ по п. 7, в котором соединение, способное ингибировать активность Rho-киназы, удаляют после того, как плюрипотентные стволовые клетки закрепляются на поверхности плоского носителя.
11. Способ по п. 7, в котором соединение, способное ингибировать активность Rho-киназы, выбрано из группы, состоящей из Y-27632, fasudil, H-1152 и hydroxyfasudil.
12. Способ по п. 7, где способ включает от 0,1 мкМ до 100 мкМ соединения, способного ингибировать активность Rho-киназы.
13. Способ по п. 7, в котором удаление соединения, способного ингибировать активность Rho-киназы, не приводит к отделению клеток.
14. Способ по п. 7, в котором по меньшей мере 95% клеток поддерживает экспрессию маркеров, связанных с плюрипотентностью.
| HARB N | |||
| et al., "The Rho-Rock-Myosin signaling axis determines cell-cell integrity of self-renewing pluripotent stem cells", PLoS One | |||
| Станок для изготовления деревянных ниточных катушек из цилиндрических, снабженных осевым отверстием, заготовок | 1923 |
|
SU2008A1 |
| VAN KOOTEN T.G | |||
| et al., "Plasma-treated polystyrene surfaces: model surfaces for studying cell-biomaterial interactions", Biomaterials | |||
| Способ приготовления мыла | 1923 |
|
SU2004A1 |
| WO 2008035110 A1, 27.03.2008 | |||
| СПОСОБ КУЛЬТИВИРОВАНИЯ МЕЗЕНХИМАЛЬНЫХ СТВОЛОВЫХ КЛЕТОК ЧЕЛОВЕКА EX VIVO | 2006 |
|
RU2323252C1 |

