ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Настоящая заявка притязает на приоритет по предварительной заявке за номером 61/030544, поданной 21 февраля 2008 года.
Настоящее изобретение относится к области культивирования клеток млекопитающих и описывает способы и композиции для иммобилизации клеток и культивирования клеток на поверхности твердой подложки, а также открепления клеток от нее, причем данная поверхность подложки содержит от по меньшей мере 0,5% N с суммарным содержанием O и N выше или равно 17,2% и с углом смачивания по меньшей мере приблизительно 13,9 градуса, и не имеет слоя питающих клеток и адгезивного слоя. В одном из осуществлений настоящего изобретения культивируемые клетки обрабатывают соединением, способным ингибировать активность Rho-киназы. В другом осуществлении клетки обрабатывают соединением, способным ингибировать активность Rho.
ПРЕДПОСЫЛКИ СОЗДАНИЯ ИЗОБРЕТЕНИЯ
Культивирование клеток млекопитающих является одним из многих процессов в медико-биологических науках. Для культивирования и анализа опорнозависимых клеток млекопитающих часто используют сосуды из стекла или пластика, например, полистирола, что нередко требует дополнительной обработки поверхности, обеспечивающей закрепление клеток на поверхности сосуда. Подобная обработка может включать нанесение на поверхности адгезивного слоя, например, путем адсорбции, плазмостимулированной полимеризации или привитой сополимеризации. Альтернативно, обработка поверхности может заключаться в химической модификации самой поверхности сосуда, что может быть достигнуто обработкой с использованием коронного разряда в атмосфере, радиочастотной плазменной обработки в вакууме, тлеющего разряда постоянного тока и микроволновой плазменной обработки. Такая обработка приводит к изменению элементного и химического состава поверхности. Протекающие при этом определенные химические процессы зависят от выбранного способа обработки поверхности, энергии и продолжительности обработки, а также газового состава среды, в которой проводится обработка.
Например, в заявке на патент US5449383 описывается подложка из массивного полимерного материала; и тонкого полимерного слоя, способного поддерживать рост клеток, представляющего собой реориентационно-устойчивый полимер, который состоит из полимеризованных в плазме амидных мономеров, предоставляющих свои амидные группы для иммобилизации клеток. При этом указанный амидный мономер выбирают из группы, включающей диметилформамид и амиды с формулой R1-CO-N(R2)R3, где R1 - алифатическая, алициклическая или ароматическая группа, каждая из которых может дополнительно нести в качестве заместителей атомы галогена или гидроксильные группы, а R2 и R3 независимо выбирают из атома водорода или алкильной группы, и указанный тонкий полимерный слой способствует закреплению и пролиферации указанных клеток.
В другом примере, в заявке на патент EP0348969A1 описан способ эндотелиализации полимерной поверхности, который заключается в приведении поверхности полимера в контакт с плазмой, образующейся из азотсодержащего газообразного материала, в результате чего указанная полимерная поверхность модифицируется с появлением поверхностных аминогрупп; затем на указанную модифицированную поверхность наносятся эндотелиальные клетки в количестве, достаточном для образования конфлюэнтного слоя клеток на указанной поверхности, содержащей аминогруппы, причем пролиферация клеток необязательна.
В другом примере, в заявке на патент EP0092302A2 описан способ влияния на рост клеточной культуры в ростовой среде на подложке, которая характеризуется тем, что химическая структура поверхности подложки модифицируется путем обработки поверхности подложки плазмой, образующейся из углерода, водорода, кислорода, азота, серы, фосфора, одного из галогенов, либо соединения любых из перечисленных элементов.
В другом примере, в заявке на патент US 6617152B2 описано устройство для обработки поверхности полимерной подложки, состоящей из следующих компонентов: (а) система подачи газа, микроволновой источник энергии и смесительная камера для подготовки плазменной смеси, причем смесительная камера имеет газовый контакт как с системой подачи газа, так и с микроволновым источником энергии; (b) двухкамерная рабочая область для проведения обработки поверхности, состоящая из внутренней рабочей камеры, находящейся внутри внешней рабочей камеры, причем в указанной внутренней рабочей камере имеется отверстие, обеспечивающее газовый контакт с указанной внешней камерой; (c) указанная смесительная камера для подготовки плазменной смеси через апертуру приводится в газовый контакт с указанной внешней рабочей камерой; (d) система подключения к вакуумной линии, соединенная с указанной внешней камерой; и (e) где указанное отверстие в указанной внутренней рабочей камере ориентировано соосно с указанной апертурой, причем указанное отверстие находится от указанной апертуры на заранее определенном расстоянии.
В одном примере, в заявке на патент US2003/0180903A1 описана полимерная подложка с рабочей поверхностью для культивирования клеток, причем, по данным электронной микроскопии, содержание поверхностного кислорода составляет по меньшей мере 25% для химического анализа на глубине приблизительно 50 Ангстрем.
В одном примере, в заявке на патент WO2006114098 описан микроструктурированный биосовместимый материал для хирургических имплантов и поверхностей тканевых культур, направляющих клетки. Микроструктуру поверхности биоматериала выбирают для стимулирования роста недифференцированных эмбриональных стволовых клеток; стимулирования нейрональной дифференцировки эмбриональных стволовых клеток; или стимулирования дифференцировки эмбриональных стволовых клеток.
В другом примере, в работе Bigdeli et al. (J. Biotechnol. 133:146-153, 2008), описан способ адаптации и/или отбора эмбриональных стволовых клеток человека для культивирования без дифференцировки в условиях отсутствия питающих клеток и без предварительной обработки поверхности твердой подложки белком внеклеточного матрикса. Описанный способ включает следующие этапы: (i) замена культуральной среды, кондиционируемой диплоидными эмбриональными фибробластами из легкого человека, на среду, кондиционируемую неонатальными хондроцитами; (ii) затем ферментативное пассирование клеток с питательного слоя мышиных эмбриональных клеток на обработанные препаратом MatrigelTM носители, затем на носители CostarTM, и, наконец, на носители PrimariaTM; и (iii) снова возврат к первоначально использовавшейся среде. Лишь очень немногие из обработанных таким образом эмбриональных стволовых клеток человека породили устойчивые клеточные линии, что указывает на то, что описанный способ включает отбор эмбриональных стволовых клеток человека по условиям культивирования.
Способы обработки поверхности, которые изменяют элементный и химический состав самой поверхности, успешно применялись для подготовки твердых полимерных подложек для культивирования многих типов клеток млекопитающих. Однако имеются существенные ограничения в отношении неудовлетворительной иммобилизации и/или культивирования при использовании определенных типов клеток млекопитающих, например, плюрипотентных стволовых клеток или клеток мезонефроса человека HEK293.
В работе Graham et al., (J. Gen. Virol. 36:59-72, 1977) описывается происхождение линии клеток HEK293.
Эффективность закрепления клеток линии HEK293 может быть повышена путем нанесения на поверхность твердой подложки адгезивного слоя с помощью, например, белков внеклеточного матрикса, полилизина, полиорнитина или полиэтиленимина перед внесением клеток HEK293 в культуральный сосуд. Однако подготовка адгезивного слоя требует значительного времени и, как правило, приводит к получению нестерильной твердой подложки с меньшим сроком хранения по сравнению с необработанной исходной твердой подложкой. Таким образом, существует значительная потребность в разработке способов и материалов для повышения эффективности закрепления клеток HEK293 на твердой подложке без нанесения адгезивного слоя.
Существующие на сегодня способы культивирования плюрипотентных стволовых клеток, в частности, эмбриональных стволовых клеток (ЭСК), требуют сложных условий культивирования, таких как, например, культивирование эмбриональных стволовых клеток на поверхности твердой подложки с нанесенным слоем питающих клеток или на поверхности твердой подложки с нанесенным адгезивным слоем белка внеклеточного матрикса. В использующих эти способы системах для культивирования нередко применяются питающие клетки или белки внеклеточного матрикса, полученные из организма другого вида по отношению к организму, чьи стволовые клетки культивируются (ксеногенный материал). Для культивирования эмбриональных стволовых клеток могут применяться среды, полученные при введении питающих клеток, то есть, среды, кондиционируемые клетками, отличными от недифференцированных эмбриональных стволовых клеток, причем в эти среды может дополнительно вводиться сыворотка животных.
Например, в работах Reubinoff et al. (Nature Biotechnol. 18:399-404, 2000) и Thompson et al. (Science 282:1145-1147, 1998) описывается способ культивирования линий эмбриональных стволовых клеток из бластоцистов человека с использованием питающего клеточного слоя из мышиных эмбриональных фибробластов.
В другом примере, в работе Xu et al. (Nature Biotechnology 19:971-974, 2001) описывается использование препарата MatrigelTM и ламинина для обработки поверхности твердой подложки перед культивированием эмбриональных стволовых клеток человека без дифференцировки, причем культивирование ведется без использования питающих клеток.
В другом примере, в работе Vallier et al. (J. Cell Sci. 118:4495-4509, 2005) описывается использование эмбриональной бычьей сыворотки для обработки поверхности твердой подложки перед культивированием эмбриональных стволовых клеток человека без дифференцировки, причем культивирование ведется без использования питающих клеток.
В другом примере, в заявке на патент WO2005014799, описывается кондиционированная среда для поддержания, пролиферации и дифференцировки клеток млекопитающих. Заявка на патент US20050244962 гласит: “Приготовленная в соответствии с настоящим изобретением культуральная среда кондиционируется за счет секреторной активности мышиных клеток, в частности, дифференцированных и иммортализованных трансгенных гепатоцитов под названием MMH (Met Murine Hepatocyte).”
В другом примере, в работе Wanatabe et al. (Nature Biotechnol. 35:681-686, 2007), отмечается, что “ингибитор Rho-ассоциированной киназы (ингибитор ROCK) позволяет добиться выживания диссоциированных эмбриональных стволовых клеток человека”, и демонстрируется снижение апоптоза, вызванного диссоциацией, повышение эффективности клонирования (от приблизительно 1% до приблизительно 27%) и облегчение субклонирования после переноса генов при использовании мышиных эмбриональных фибробластов в качестве питающих клеток, коллагена и MatrigelTM в качестве белка внеклеточного матрикса, и Y-27632 или Fasudil в качестве ингибитора ROCK. Более того, диссоциированные эмбриональные стволовые клетки человека, обработанные Y-27632, оказались защищены от апоптоза в несодержащей сыворотки суспензионной культуре.
В другом примере, в работе Peerani et al. (EMBO Journal 26:4744-4755, 2007), отмечается: “Сложность пространственной организации культур эмбриональных стволовых клеток человека (ЭСК) создает гетерогенные микроокружения (ниши), которые определяют дальнейшую судьбу ЭСК. Настоящее исследование демонстрирует, что скорость и траекторию дифференцировки ЭСК можно контролировать путем создания ниш стволовых клеток с заданными свойствами. Размер и состав ниши регулируют соотношение между индуцирующими и ингибирующими дифференцировку факторами. С точки зрения механизма такой регулировки, в результате антагонистических взаимодействий между ЭСК и производной от ЭСК внеэмбриональной энтодермы (ExE) создается пространственный градиент сигнального каскада Smad1, зависящий от размеров ниши. Эти взаимодействия опосредуются локализованной секрецией костного морфогенетического белка BMP2 из ExE и его антагониста, фактора дифференцировки роста GDF3 из ЭСК. Микроструктурирование ЭСК, обработанных малыми интерферирующими РНК против GDF3, BMP2 и Smad1, а также обработка ингибитором Rho-ассоциированной киназы (ROCK) демонстрируют, что независимый контроль активации Smad1 может спасти зависящую от размера колонии дифференцировку ЭСК. Наши результаты впервые показывают роль Smad1 в интеграции пространственной информации и в контроле самообновления и дифференцировки ЭСК в зависимости от размера ниши”.
В другом примере, в работе Koyanagi, M et al (J Neurosci Res. 2007 Sep 7 [Epub ahead of print]), отмечается: “Участие Rho-ГТФазы предполагалось в апоптозе многих типов клеток, включая нейроны, но механизм ее действия до сих пор полностью не выяснен. В настоящей работе мы исследуем роли Rho и ROCK в апоптозе в процессе трансплантации нейронных клеток-предшественников, происходящих от эмбриональных стволовых клеток. Мы обнаружили, что диссоциация предшественников нейронов приводит к активации Rho и индуцирует апоптоз. Обработка ингибитором Rho, внеклеточным ферментом C3, и/или ингибитором ROCK Y-27632 снижает степень апоптоза (аноикоза), вызванного диссоциацией, на 20-30%. Ингибирование ROCK также снижает пузырение мембраны - ранний морфологический признак апоптоза; расщепление каспазы-3 и высвобождение цитохорма C из митохондрий. Полученные результаты дают основание полагать, что диссоциация нейронных клеток-предшественников активирует внутренний каскад смерти клетки, который по меньшей мере частично опосредуется через каскад Rho/ROCK. Более того, в экспериментальной модели трансплантации на животном ингибирование Rho и/или ROCK приводит к подавлению острого апоптоза клеток-трансплантов. После трансплантации вокруг транспланта наблюдается ярко выраженное действие фактора некроза опухоли альфа и профактора роста нервов. Ингибирование ROCK также подавляет апоптоз, которому способствуют эти воспалительные цитокины. В совокупности полученные результаты указывают, что ингибирование каскада Rho/ROCK может повысить выживаемость клеток-трансплантов в заместительной клеточной терапии”.
В другом примере, в работе Yoneda et al (J. Cell Biol. 170: 443-453, August 3, 2005), отмечается: “Гомологичные Rho-киназы млекопитающих (ROCK I и II) считаются функционально избыточными, исходя в основном из сверхэкспрессии киназных конструкций. В качестве нижележащих эффекторов Rho-ГТФаз их основными субстратами являются легкая цепь миозина и миозинфосфатаза. Предполагается участие обеих киназ в сборке пучков микрофиламент и сократительной способности гладких мышц. В настоящей работе анализ адгезии фибробластов к фибронектину показал, что, хотя ROCK II присутствовала в большем количестве, ее активность всегда была ниже, чем активность ROCK I. Специфическое восстановление ROCK I малой интерферирующей РНК приводило к потере тонофибрилл и фокальных адгезий, несмотря на сохранение ROCK II и связанного с гуанинтрифосфатом RhoA. Напротив, при отрицательной модуляции ROCK II наблюдалось усиление микрофиламентного цитоскелета. Поглощение фагоцитов покрытыми бионектином гранулами значительно подавлялось в клетках с удаленной ROCK II, но не в клетках с удаленной ROCK I. Эти эффекты частично обусловлены специфичными липид-связывающими предпочтениями гомологичных доменов с плекстрином ROCK. Киназа ROCK II связывала фосфатидилинозитол 3,4,5P3 и оказалась чувствительной к его уровню, что не наблюдалось в случае киназы ROCK I. Таким образом, эндогенные Rho-ассоциированные киназы специфично регулируются и, в свою очередь, ассоциированы с различными миозиновыми структурами”.
В другом примере, в работе Harb et al (PloS ONE 3(8): e3001. oi:10.1371/journal.pone.0003001, August 2008) описана существенная роль сигнальной оси Rho-Rock-Миозин в регуляции базовых каналов межклеточной коммуникации для эмбриональных стволовых клеток как мыши, так и человека, которая может использоваться для дальнейшего развития [так!] технологии пригодных для медицинских целей свободных от ксенобиотиков сред для плюрипотентных стволовых клеток человека.
Использование ксеногенных материалов может оказаться недопустимым в ряде применений, связанных с использованием плюрипотентных стволовых клеток. В таких случаях можно использовать альтернативные материалы. Например, в работе Stojkovic et al. (Stem Cells 23:895-902, 2005) описано применение сыворотки человека для обработки поверхности твердой подложки перед культивированием эмбриональных стволовых клеток человека без дифференцировки, причем культивирование ведется без использования питающих клеток.
В альтернативной культуральной системе используется не содержащая сыворотки среда с добавлением факторов роста, способных стимулировать пролиферацию эмбриональных стволовых клеток.
Например, в работе Cheon et al. (BioReprod DOI:10.1095/biolreprod.105.046870; 19 Oct 2005) описана не содержащая ни питающих клеток, ни сыворотки культуральная система, в которой эмбриональные стволовые клетки поддерживаются в некондиционированной замещающей сыворотку среде с добавкой различных факторов роста, способных инициировать самообновление эмбриональных стволовых клеток.
В другом примере, в работе Levenstein et al. (Stem Cells 24:568-574, 2006), описаны способы долговременного культивирования эмбриональных стволовых клеток человека в отсутствие фибробластов или кондиционированной среды с использованием среды с добавкой базового фактора роста фибробластов FGF.
В другом примере, в заявке на патент US20050148070, описан способ культивирования эмбриональных стволовых клеток человека в среде с определенной композицией, не содержащей сыворотки и не содержащей питающих клеток - фибробластов. Данный способ заключается в культивировании стволовых клеток в культуральной среде, содержащей альбумин, аминокислоты, витамины, минеральные вещества по меньшей мере один трансферрин или заместитель трансферрина по меньшей мере один инсулин или заместитель инсулина, причем указанная культуральная среда существенно свободна от эмбриональной сыворотки млекопитающих и содержит по меньшей мере приблизительно 100 нг/мл FGF, способного активировать FGF-сигнальный рецептор, где указанный фактор роста поступает из источника, отличного от просто питающего слоя фибробластов, данная среда поддерживает пролиферацию стволовых клеток в недифференцированном состоянии в отсутствие питающих клеток или кондиционированной среды.
В другом примере, в заявке на патент US20050233446, описывается среда с определенной композицией, которая может быть использована при культивировании стволовых клеток, включая недифференцированные зародышевые стволовые клетки приматов. В растворе среда является существенно изотонической относительно культивируемых стволовых клеток. Для каждой культуры конкретно используемая среда состоит из основной среды с добавкой базового FGF, инсулина и аскорбиновой кислоты, необходимых для поддержания существенно недифференцированного роста зародышевых стволовых клеток.
В другом примере, в заявке на патент US6800480, отмечается: “В одном варианте осуществления предлагается культуральная среда для выращивания зародышевых стволовых клеток приматов в существенно недифференцированном состоянии, включающая основную среду с низким содержанием эндотоксина и низким осмотическим давлением, которая эффективно поддерживает рост зародышевых стволовых клеток приматов. Основная среда объединяется с питательной сывороткой, способной эффективно поддерживать рост зародышевых стволовых клеток приматов, и с субстратом, выбранным из группы, включающей питающие клетки и компонент внеклеточного матрикса, полученный из питающих клеток. Указанная среда также включает заменимые аминокислоты, антиоксидант и первый фактор роста, выбранный из группы, включающей нуклеозиды и соль-пируват”.
В другом примере, в заявке на патент US20050244962, отмечается: “В одном аспекте в изобретении предлагается способ культивирования эмбриональных стволовых клеток приматов. Стволовые клетки культивируются в культуре, существенно свободной от эмбриональной сыворотки млекопитающих (предпочтительно также существенно свободной от сыворотки любых животных) и в присутствии фактора роста фибробластов, полученного из источника, иного чем просто питающие клетки-фибробласты. В предпочтительной форме слой питающих фибробластов, ранее необходимый для поддержания культуры стволовых клеток, становится необязательным вследствие добавления достаточного количества фактора роста фибробластов”.
В другом примере, в заявке на патент WO2005065354, описана существенно свободная от питающих клеток и сыворотки изотоническая культуральная среда с определенное композицией, которая включает в себя: а. основную среду; b. базовый фактор роста фибробластов в количестве, достаточном для поддержания роста существенно недифференцированных стволовых клеток млекопитающих; c. инсулин в количестве, достаточном для поддержания роста существенно недифференцированных стволовых клеток млекопитающих; и d. аскорбиновую кислоту в количестве, достаточном для поддержания роста существенно недифференцированных стволовых клеток млекопитающих.
В другом примере, в заявке на патент WO2005086845, описан способ поддержания недифференцированных стволовых клеток, включающий воздействие на стволовые клетки одним из членов семейства белков трансформирующего ростового фактора-бета (TGFβ), одним из членов семейства белков фактора роста фибробластов (FGF) или никотинамидом (NIC) в количестве, достаточном для поддержания клеток в недифференцированном состоянии в течение периода времени, достаточного для получения желаемого результата.
Плюрипотентные стволовые клетки обладают большим потенциалом для научно-исследовательской работы и скрининга лекарственных препаратов. В настоящее время крупномасштабное культивирование линий эмбриональных стволовых клеток человека представляет большие проблемы и сталкивается с существенными трудностями. Возможные пути преодоления этих трудностей заключаются в пассировании и культивировании эмбриональных стволовых клеток человека в виде одиночных клеток. Одиночные клетки более удобны для применения стандартных методик культивирования тканей, таких как, например, подсчет клеток, трансфицирование и т.д.
Например, в работе Nicolas et al. описан способ получения и наработки линий эмбриональных стволовых клеток человека из одиночных клеток, изолированных с помощью флюоресцентной сортировки после генетической модификации с использованием лентивирусных векторов (Stem Cells Dev. 16:109-118, 2007).
В другом примере, в заявке на патент США US2005158852, описан способ “улучшения роста и выживаемости одиночных эмбриональных стволовых клеток человека. Способ включает этапы получения одиночной недифференцированной эмбриональной стволовой клетки человека; смешивания указанной одиночной недифференцированной клетки с внеклеточным матриксом для обволакивания клетки и посева полученной смеси на питающие клетки с питательной средой в ростовом окружении”.
В другом примере, в работе Sidhu et al. (Stem Cells Dev. 15:61-69, 2006), впервые сообщается о трех клонах эмбриональных стволовых клеток человека, hES 3.1, 3.2 и 3.3, полученных из родительской линии hES3 путем сортировки одноклеточных препаратов на проточном цитометре.
Однако пассирование и культивирование эмбриональных стволовых клеток человека в виде одиночных клеток приводит к генетическим аберрациям и к потере плюрипотентности. Для поддержания плюрипотентности и генетической устойчивости большое значение имеют условия культивирования. Как правило, пассирование линий эмбриональных стволовых клеток человека выполняется вручную или с использованием ферментативных агентов, таких как коллагеназа, либераза или диспаза.
Например, в работе Draper et al. отмечается наличие “кариотипических изменений, включающих амплификацию 17q плеча в трех независимых линиях эмбриональных стволовых клеток человека в пяти независимых случаях”. (Nature Biotechnol. 22:53-54, 2004).
В другом примере, в работе Buzzard et al. отмечается: “Мы зафиксировали лишь одно изменение кариотипа... что может быть обусловлено используемыми нами способами культивирования, поскольку наши способы разительно отличаются от способов, используемых большинством других групп. Как правило, мы пассируем эмбриональные стволовые клетки человека через семь дней, сначала подсекая колонию краем разломленной пипетки... При таком способе пассирования не используются никакие ферментативные или химические способы диссоциации клеток. Мы считаем, что именно с этим может быть связана относительная генетическая устойчивость эмбриональных стволовых клеток человека в наших руках”. (Nature Biotechnol. 22:381-382, 2004).
В другом примере, в работе Mitalipova et al., отмечается: “Способы группового пассирования... могут зафиксировать популяции анэуплоидных клеток после многократного пассирования в культуре, но могут применяться на более коротких промежутках времени (по меньшей мере до 15 пассажа) без угрозы изменения кариотипа... возможно удастся поддерживать нормальный кариотип эмбриональных стволовых клеток человека в условиях продолжительного ручного ведения с последующим ограниченным числом групповых пассажей в экспериментах, требующих больших количеств эмбриональных стволовых клеток человека, чем могут обеспечить только способы ручного пассирования”. (Nature Biotechnol. 23:19-20, 2005).
В другом примере, в работе Heng et al., отмечается: “Полученные результаты показали, что второй протокол (трипсинизация с умеренным пипетированием) наносят гораздо меньший ущерб жизнеспособности клеток по сравнению с первым протоколом (обработка коллагеназой и соскребание). Это, в свою очередь, выливается в большую вероятность выживания клеток в циклах заморозки/разморозки”. (Biotechnology and Applied Biochemistry 47:33-37, 2007).
В другом примере, в работе Hasegawa et al., отмечается: “Мы вывели сублинии эмбриональных стволовых клеток человека, переносящих полную диссоциацию. Эти клетки имеют высокую эффективность пересева и высокую эффективность клонирования, при этом они сохраняют свою способность дифференцировки в клетки всех трех зародышевых листков”. (Stem Cells 24:2649-2660, 2006).
Таким образом, существует значительная потребность в способах и композициях для культивирования клеток млекопитающих, включая культивирование плюрипотентных стволовых клеток в отсутствие питающего слоя и адгезивного слоя при сохранении плюрипотентности культивируемых клеток.
КРАТКОЕ ОПИСАНИЕ
В одном осуществлении настоящее изобретение предлагает способы и композиции для иммобилизации клеток и культивирования клеток на поверхности твердой подложки и открепления клеток от нее, причем указанная поверхность подложки содержит от по меньшей мере приблизительно 0,5% N, с суммарным содержанием O и N выше или равно 17,2% и с углом смачивания по меньшей мере приблизительно 13,9 градуса; не имеет слоя питающих клеток и не имеет адгезивного слоя.
В одном осуществлении настоящее изобретение предлагает способ повышения иммобилизации клеток на поверхности, которая содержит от по меньшей мере приблизительно 0,5% N, с суммарным содержанием O и N выше или равно 17,2% и с углом смачивания по меньшей мере приблизительно 13,9 градуса, не имеет слоя питающих клеток и адгезивного слоя, включающий стадии:
а. получения суспензии клеток,
b. обработки суспензии клеток по меньшей мере одним соединением, выбранным из группы, состоящей из: соединения, способного ингибировать активность Rho-киназы, и соединения, способного ингибировать активность Rho, и
c. нанесения обработанной суспензии клеток на поверхность, где они закрепляются.
В одном осуществлении после закрепления на поверхности клетки продолжают поддерживаться в той же культуральной среде. В альтернативном осуществлении по меньшей мере одно из указанных соединений удаляется из культуральной среды.
В одном осуществлении закрепленные клетки снимаются с поверхности путем удаления по меньшей мере одного из указанных соединений из культуральной среды.
В одном осуществлении указанная суспензия клеток представляет собой суспензию кластеров клеток. В альтернативном осуществлении указанная суспензия клеток представляет собой суспензию одиночных клеток.
В одном осуществлении клетки представляют собой плюрипотентные стволовые клетки. В альтернативном осуществлении клетки представляют собой стволовые клетки.
В одном осуществлении настоящее изобретение предлагает способ повышения иммобилизации клеток на поверхности, которая содержит от по меньшей мере приблизительно 0,9% N, с суммарным содержанием O и N выше или равно 22,3% и с углом смачивания по меньшей мере приблизительно 13,9 градуса, не имеет слоя питающих клеток и не имеет адгезивного слоя, включающий стадии:
a. получения суспензии клеток, и
b. нанесения суспензии клеток на поверхность, где они закрепляются.
КРАТКОЕ ОПИСАНИЕ ФИГУР
На фиг.1 приведены фазоконтрастные микрофотографии (4×) клеток линии эмбриональных стволовых клеток человека H1, дважды пассированных как кластеры с использованием ферментативной смеси LIBERASE на поверхностно-модифицированных носителях 2, 3 и 4. На фигуре также приведены изображения клеток линии эмбриональных стволовых клеток человека H1, культивируемых на носителе с поверхностью, обработанной раствором препарата MatrigelTM в соотношении 1:30, и носителе, обработанном по технологии Nunclon DeltaTM.
На фиг.2 показано влияние добавления 10 мкМ Y-27632 на эффективность закрепления эмбриональных стволовых клеток человека на поверхностно-модифицированных носителях. На фигуре приведены фазоконтрастные микрофотографии (4×) клеток линии эмбриональных стволовых клеток человека H1, дважды пассированных как кластеры на поверхностно-модифицированных носителях 3 и 4. Затем клетки пассировали на поверхностно-модифицированные носители 2, 3 и 4 в среде, кондиционированной мышиными эмбриональными фибробластами (MEF) и содержащей 10 мкМ Y-27632. Перед фотографированием клетки культивировали в течение четырех дней. В качестве контроля использовались клетки, культивируемые в отсутствие Y-27632.
На фиг.3 показана динамика обработки различными соединениями эмбриональных стволовых клеток человека, культивируемых на поверхностно-модифицированных носителях, составляющих предмет настоящего изобретения. Клетки линии эмбриональных стволовых клеток человека H1 четырежды пассировали как кластеры с использованием ферментативной смеси LIBERASE на поверхностно-модифицированных носителях 3 и 4 и культивировали в кондиционированной MEF среде. В течение первых двух дней после пассирования клетки обрабатывали либо 10 мкМ ингибитора Rho-киназы Y-27632, либо 0,5 нг/мл ингибитора Rho - проникающей в клетку формы внеклеточного фермента C3 трансферазы. Клетки, которые обрабатывали ингибитором Rho-киназы Y-27632 и затем в течение первых двух дней после каждого пассажа обрабатывали Y-27632 на поверхностно-модифицированном носителе 3, обозначены как “7s”. Клетки, которые обрабатывали ингибитором Rho-киназы Y-27632 и затем в течение первых двух дней после каждого пассажа обрабатывали Y-27632 на поверхностно-модифицированном носителе 4, обозначены как “3s”. Клетки, которые в течение двух дней обрабатывали ингибитором Rho, затем в течение первых двух дней после каждого пассажа обрабатывали ингибитором Rho-киназы Y-27632 и затем в течение первых двух дней после пассажа обрабатывали Y-27632 на поверхностно-модифицированном носителе 3, обозначены как “5s”. Клетки, которые в течение двух дней обрабатывали ингибитором Rho, затем в течение первых двух дней после каждого пассажа обрабатывали ингибитором Rho-киназы Y-27632 и затем в течение первых двух дней после пассажа обрабатывали Y-27632 на поверхностно-модифицированном носителе 4, обозначены как “1s”.
На фиг.4 показаны определенные способом qRT-PCR уровни экспрессии маркеров, ассоциированных с плюрипотентностью и дифференцировкой в эмбриональных стволовых клетках человека, обработанных в соответствии с указанным на фиг.6 протоколом.
На фиг.5 показаны определенные способом проточной цитометрии уровни экспрессии маркеров плюрипотентности в клетках линии эмбриональных стволовых клеток человека H1 на пассаже 4 (p4), пассаже 9 (p9) и затем на пассаже 10, 11 или 12 (p10, p11 или p12).
На фиг.6 приведены иммуно-флюоресцентные фотографии клеток линии эмбриональных стволовых клеток человека H1, которые последовательно пассировали как кластеры с использованием ферментативной смеси LIBERASE на поверхностно-модифицированном носителе 4 и культивировали в среде, кондиционированной MEF. Уровни экспрессии белков, ассоциированных с маркерами плюрипотентности, определялись для клеток, культивированных в течение 11 пассажей на поверхностно-модифицированном носителе 4. Клетки обрабатывали 10 мкМ Y-27632 в течение двух дней после каждого пассажа.
На фиг.7 показана способность эмбриональных стволовых клеток человека образовывать дефинитивную эндодерму после культивирования на поверхностно-модифицированных носителях. Клетки линии эмбриональных стволовых клеток человека H1 пассировали 11 раз как кластеры с использованием ферментативной смеси LIBERASE на поверхностно-модифицированных носителях 3 и 4 и культивировали в среде, кондиционированной MEF. На пассаже 8 (p8) и затем снова на пассаже 10 или 11 (p10-11) клетки в течение двух дней обрабатывали средой DMEM:F12, содержащей 0,5% FBS, 100 нг/мл активина A и 20 нг/мл Wnt3a, и затем обрабатывали средой DMEM:F12, содержащей 2% FBS и 100 нг/мл активина A, в течение еще трех дней. Вертикальная ось на фигуре показывает процент CXCR4-положительных клеток по данным проточной цитометрии. Смотрите также таблицу 5.
На фиг.8 показана способность эмбриональных стволовых клеток человека образовывать панкреатическую эндодерму после культивирования на поверхностно-модифицированных носителях. Клетки линии эмбриональных стволовых клеток человека H1 пассировали восемь раз как кластеры с использованием ферментативной смеси LIBERASE на поверхностно-модифицированных носителях 3 и 4 и культивировали в среде, кондиционированной MEF. На пассаже 8 (p8) клетки дифференцировали в клетки дефинитивной эндодермы путем обработки в течение двух дней средой DMEM:F12, содержащей 0,5% FBS, 100 нг/мл активина A и 20 нг/мл Wnt3a, и затем обработки средой DMEM:F12, содержащей 2% FBS и 100 нг/мл активина A, в течение еще трех дней. Затем клетки дифференцировали далее в клетки зародышевой передней кишки путем обработки в течение четырех дней средой DMEM:F12, содержащей 2% FBS, 100 нг/мл FGF-10 и 1 мкМ циклопамин-KAAD. Затем клетки дифференцировали в клетки панкреатической эндодермы путем обработки в течение четырех дней средой DMEM:F12, содержащей 1% B-27, 100 нг/мл FGF-10, 1 мкМ циклопамин-KAAD и 2 мкМ ретиноевой кислоты. Было проведено иммунофлюоресцентное окрашивание клеток по PDX-1 (зеленый) и E-кадгерину (красный); полное число клеток определяли с помощью красителя Хехст (синий).
На фиг.9 показана способность эмбриональных стволовых клеток человека образовывать эмбриоидные тельца после культивирования на поверхностно-модифицированных носителях.
На фиг.10 показан кариотип эмбриональных стволовых клеток человека, культивированных на поверхностно-модифицированном носителе 4.
На фиг.11 показано влияние обработки ингибиторами Rho-киназы (Y-27632 от компании EMD biosciences, Y-27632 от компании Sigma, Fasudil и Hydroxyfasudil) на эффективность закрепления эмбриональных стволовых клеток человека на поверхностно-модифицированных носителях. Клетки в течение трех дней культивировали в содержащих указанные соединения средах при указанных концентрациях. Затем клетки окрашивали кристаллическим фиолетовым и фотографировали.
На фиг.12 показана зависимость эффективности закрепления эмбриональных стволовых клеток человека на поверхностно-модифицированных носителях от концентрации вводимого в среду Y-27632. На первый день в культуральные среды вводился ингибитор Rho-киназы Y-27632 в указанных концентрациях (0, 1, 2, 4 или 10 мкМ Y-27632). Начиная со второго дня, в течение пяти дней клетки поддерживали в среде, содержащей 10 мкМ Y-27632, ежедневно заменяя среду. На пятый день с носителей удаляли культуральную среду, клетки окрашивали 0,5% кристаллическим фиолетовым и фотографировали.
На фиг.13 показано образование колоний эмбриональных стволовых клеток человека через четыре дня после пассирования на поверхностно-модифицированные носители 2, 3 или 4 с добавлением или без добавления 10 мкМ Y-27632.
На фиг.14 показано образование колоний эмбриональных стволовых клеток человека через четыре дня после пассирования на обработанные препаратом MatrigelTM носители с добавлением или без добавления 10 мкМ Y-27632.
На фиг.15 показано различие между влиянием непрерывной и прерывистой обработки эмбриональных стволовых клеток человека Y-27632 на эффективность закрепления клеток на поверхностно-модифицированном носителе.
На фиг.16 показаны фотографии клеток линии эмбриональных стволовых клеток человека H9, которые пассировали как одиночные клетки, высевая на поверхностно-модифицированный носитель 3 в среде, кондиционированной MEF и содержащей (B) или не содержащей (A) 10 мкМ Y-27632. Фотографии сделаны через 24 часа после посева.
На фиг.17 показаны уровни экспрессии ассоциированных с плюрипотентностью маркеров в клетках линии эмбриональных стволовых клеток человека H9, которые в течение 5 пассажей пассировали как одиночные клетки, используя диссоциирующий фермент TrypLE™ Express, и высевали на поверхностно-модифицированных носителях 3 и 4 с добавлением или без добавления 10 мкМ Y-27632 (Y). По оси Х отложены маркеры плюрипотентности, по оси Y - процент положительных по данному маркеру клеток.
На фиг.18 показано полное число клеток линии эмбриональных стволовых клеток человека H9, которые пассировали как одиночные клетки, высевая на поверхностно-модифицированных носителях 3 и 4. Влияние добавления 10 мкМ Y-27632 (Y) на численность клеток изучали на клетках, пассируемых на обработанные препаратом MatrigelTM носителях (“наивные” клетки, N), и на клетках, 10 раз пассируемых на поверхностно-модифицированных носителях (“акклиматизированные” клетки, A). По оси X отложены различные условия культивирования клеток, по оси Y - число клеток, деленное на 104.
На фиг.19 показана скорость роста клеток линии эмбриональных стволовых клеток человека H9, которые перед данным исследованием пассировали как одиночные клетки на носителях, обработанных препаратом MatrigelTM. Клетки высевали с плотностью 104/см2 и культивировали в среде, кондиционированной MEF, с добавлением или без добавления 10 мкМ Y-27632 на поверхностно-модифицированных носителях 3 и 4. По оси Y отложено количество клеток, собранных на 2, 3 и 4 день после посева (деленное на 104).
На фиг.20 показана скорость роста клеток линии эмбриональных стволовых клеток человека H9, которые перед данным исследованием 10 раз пассировали как одиночные клетки на поверхностно-модифицированных носителях. Клетки высевали с плотностью 104/см2 и культивировали в среде, кондиционированной MEF, с добавлением или без добавления 10 мкМ Y-27632 на поверхностно-модифицированных носителях 3 и 4. По оси Y отложено количество клеток, собранных на 2, 3 и 4 день после посева (деленное на 104).
На фиг.21 приведены фотографии клеток линии эмбриональных стволовых клеток человека H9, которые пассировали как одиночные клетки, высевая на поверхностно-модифицированных носителях 2-4 и 13 в формате 96-луночных планшетов. Кондиционированная MEF культуральная среда содержала 10 мкМ Y-27632. Фотографии сделаны через 48 часов после посева.
На фиг.22 показана способность клеток линии эмбриональных стволовых клеток человека H9, которые пассировали как одиночные клетки, высевая на поверхностно-модифицированных носителях 3 и 4, дифференцироваться в клетки дефинитивной эндодермы. Степень образования дефинитивной эндодермы определяли по измерению уровня экспрессии CXCR на проточном цитометре. Было исследовано влияние добавления 10 мкМ Y-27632 на образование дефинитивной эндодермы. В процессе размножения клетки обрабатывали Y-27632. В качестве контроля использовали клетки, размножавшиеся и дифференцировавшиеся на носителях, обработанных препаратом MatrigelTM. По оси Y отложен процент CXCR4-положительных клеток по данным проточной цитометрии.
На фиг.23 показана способность клеток линии эмбриональных стволовых клеток человека H9, которые пассировали как одиночные клетки, высевая на поверхностно-модифицированных носителях 3 и 4, дифференцироваться в клетки панкреатической эндодермы. Клетки высевали на поверхностно-модифицированные носители и культивировали в среде, кондиционированной MEF, с добавлением 10 мкМ Y-27632, пассируя их 8 раз на поверхностно-модифицированных носителях перед дифференцировкой. По оси Y отложена определенная способом q-PCR кратность возрастания уровня экспрессии маркера панкреатической дифференцировки (Ngn3, Pdx1, инсулин) на стадии поздней передней кишки (PF) и стадии экспрессирующей гормон эндокринной клетки (EN).
На фиг.24 показано закрепление эмбриональных стволовых клеток человека на поверхностно-модифицированных носителях. Клетки линии эмбриональных стволовых клеток человека H9 50-го пассажа высевали в разбавлении 1:2 на следующие носители: поверхностно-модифицированные носители 3 и 4, носители с поверхностью типа CellBINDTM и PrimariaTM. Через 24 часа после посева с носителей удаляли культуральную среду, окрашивали клетки 0,5% кристаллическим фиолетовым и фотографировали. Стрелками на фигуре указаны колонии.
На фиг.25 показано закрепление эмбриональных стволовых клеток человека на поверхностно-модифицированных носителях. Клетки линии эмбриональных стволовых клеток человека H9 50-го пассажа высевали в разбавлении 1:2 в присутствии варьируемых концентраций Y-27632 (0, 1, 2, 4, 10 и 20 микромоль/литр) на следующие носители: поверхностно-модифицированные носители 3 и 4, носители с поверхностью типа CellBINDTM и PrimariaTM. Через 24 часа после посева с носителей удаляли культуральную среду, окрашивали клетки 0,5% кристаллическим фиолетовым и фотографировали. Колонии выглядят как темные точки в лунках. Стрелками на фигуре указаны колонии в необработанных лунках.
На фиг.26 показано закрепление эмбриональных стволовых клеток человека на поверхностно-модифицированных носителях. Клетки линии эмбриональных стволовых клеток человека H9 53-го пассажа высевали в разбавлении 1:3 в отсутствие или в присутствии Y-27632 (0 или 20 микромоль/литр) на следующие носители: поверхностно-модифицированные носители 2-4 и 13, носители с поверхностью типа CellBINDTM и PrimariaTM. Через 48 часов после посева с носителей удаляли культуральную среду, окрашивали клетки 0,5% кристаллическим фиолетовым и фотографировали. Колонии выглядят как темные точки в лунках. Стрелками на фигуре указаны колонии в необработанных лунках.
На фиг.27 показаны первая (октябрь) и вторая (декабрь) попытки иммобилизации клеток линии эмбриональных стволовых клеток человека H9 на поверхностно-модифицированных носителях 14 и 15, а также попытка иммобилизации клеток линии эмбриональных стволовых клеток человека H1 на поверхностно-модифицированных носителях 14 и 15. Клетки линии эмбриональных стволовых клеток человека H9 42-го и 53-го пассажа и клетки линии эмбриональных стволовых клеток человека H1 57-го пассажа высевали в разбавлении 1:2 или 1:3 в присутствии 20 микромоль/литр на поверхностно-модифицированные носители. Через 24-48 часов после посева с носителей удаляли культуральную среду, окрашивали клетки 0,5% кристаллическим фиолетовым и фотографировали. Колонии выглядят как темные точки в лунках. Стрелками на фигуре указаны колонии на носителях.
На фиг.28 показано закрепление эмбриональных стволовых клеток человека на поверхностно-модифицированном носителе 4 в среде определенной композиции mTeSRTM. Клетки линии эмбриональных стволовых клеток человека H9 50-го пассажа высевали в разбавлении 1:2 в отсутствие или в присутствии Y-27632 (0 или 20 микромоль/литр) в необработанные лунки и в лунки, обработанные следующими белками: 0,1% желатина, 2% БСА, 0,34 мг/мл крысиного коллагена, разбавленный 1:1000 MatrigelTM и разбавленный 1:5000 MatrigelTM. Через 48 часов после посева с носителей удаляли культуральную среду, окрашивали клетки 0,5% кристаллическим фиолетовым и фотографировали. Колонии выглядят как темные точки в лунках.
На фиг.29 показаны углы смачивания водой для поверхностно-модифицированных носителей, измерявшиеся в течение 11 недель статическим способом неподвижной капли. Первое измерение выполняли через неделю после обработки поверхности и стерилизации. Каждая точка на графике соответствует среднему углу смачивания (по одному измерению для каждой из семи капель). Углы смачивания для носителей, обработанных по технологии Nuclon DeltaTM и CellBINDTM, измеряли в тех же экспериментальных условиях, что и для поверхностно-модифицированных носителей 1-4 и 13, но обработка поверхности и стерилизация для них проводилась более чем за 12 недель до первого измерения (носитель с поверхностью Nuclon DeltaTM * стерилизовали за неделю до выполнения первого измерения).
На фиг.30 показана плотность отрицательных зарядов на поверхностно-модифицированных носителях, измеренная по реакционной способности поверхностей в отношении положительно заряженных молекул кристаллического фиолетового. Испытывали по три образца каждой поверхности, измеряя оптическое поглощение десорбированного с поверхности каждого образца кристаллического фиолетового в трех повторностях. На фигуре приведены средние значения и их стандартные отклонения по результатам девяти измерений.
На фиг.31 показано влияние поверхностей твердых подложек и Y-27632 на закрепление и рост клеток HEK293 в не содержащей сыворотки среде химически определенной композиции Pro293a-CDMTM (A) и среде EMEM с добавкой 10% бычьей эмбриональной сыворотки (B). Клетки HEK293 высевали на 96-луночные планшеты с поверхностью типа CellBINDTM, поверхностью типа Nunclon DeltaTM или поверхностью 4 по настоящему изобретению. На фигуре показано количество клеток HEK293, закрепившихся на указанных поверхностях, в зависимости от условий культивирования и концентрации Y-27632. Клетки культивировали либо: (i) 96 часов постоянно в среде с присутствующим Y-27632 (Y-27632 96h on); либо (ii) 48 часов постоянно в среде с присутствующим Y-27632 с последующей заменой среды и культивированием в течение 48 часов в среде без Y-27632 (Y-27632 48 часов с/48 часов без). Клетки HEK293 в среде без Y-27632 (No Y-27632) культивировали точно так же, как и клетки в среде с присутствующим Y-27632, то есть, либо выдерживали 96 часов без смены среды, либо заменяли среду через 48 часов. Наличие Y-27632 в культуральной среде повысило эффективность закрепления клеток HEK293 на поверхности 4 и поверхности типа CellBINDTM при использовании концентраций ингибитора 2,0 и 5,0 мкМ. Удаление Y-27632 после 48 часов инкубации привело к откреплению значительной части клеток от поверхности 4 и поверхности типа CellBINDTM. На фигуре приведены средние значения и их стандартные отклонения по результатам трех измерений.
На фиг.32 показано влияние поверхностей твердых подложек и ингибиторов Rho-киназы Y-27632 и H-1152 Y-27632 на рост клеток HEK293 в среде EMEM с добавлением 10% бычьей эмбриональной сыворотки. Клетки HEK293 высевали на 24-луночные планшеты типа Multidish, имеющие либо поверхность типа 4 (A), либо необработанную (но облученную дозой гамма-излучения 25 кГр) полистирольную поверхность (B).
На фиг.33 показано влияние H-1152 и поверхности типа 4 на эффективность закрепления и морфологию клеток HEK293. Клетки HEK293 высевали на 12-луночные планшеты типа Multidish в среде EMEM с добавкой 10% бычьей эмбриональной сыворотки и инкубировали в течение 67 часов в автоматизированном микроскопе-инкубаторе. Кривые роста на фигуре A и микрофотографии на фигуре B демонстрируют общий эффект H-1152 на эффективность закрепления и рост клеток HEK293 на поверхности типа 4, а также влияние замены среды на эффективность закрепления и морфологию клеток HEK293 на поверхности типа 4 в присутствии и в отсутствие H-1152.
На фиг.34 приведены кривые роста клеток HEK293 на поверхности типа 4 и поверхности типа Nunclon DeltaTM в отсутствие или в присутствии 2,5 мкМ Y-27632 в течение трех пассажей. Клетки HEK293 в среде EMEM с добавлением 10% бычьей эмбриональной сыворотки трижды пассировали с использованием трипсинизации.
На фиг.35 показано влияние ингибирования Rho-киназы на эффективность закрепления клеток линии эмбриональных стволовых клеток человека H1 на поверхностно-модифицированных носителях 4, 18 и 19, а также на поверхности типа PrimariaTM. Лунки A&B использовались в качестве контрольных для всех поверхностей. В лунки C&D добавляли 10 мкМ Y-27632. В лунки E&F добавляли 3 мкМ H1152-глицила. В лунки G&H добавляли 10 мкМ H1152-глицила.
На фиг.36 показано влияние ингибирования Rho-киназы на эффективность закрепления клеток линии эмбриональных стволовых клеток человека H1 на поверхностно-модифицированном носителе 30. (--) = без обработки. (RI) = 3 мкМ H1152-глицила. (MG) = адгезивный слой из MATRIGEL в разбавлении 1:30. (MG+RI) = адгезивный слой из MATRIGEL в разбавлении 1:30 + 3 мкМ H1152-глицила.
На фиг.37 показано влияние ингибирования Rho-киназы на эффективность закрепления клеток линии эмбриональных стволовых клеток человека H1 на поверхностно-модифицированном носителе 31. (--) = без обработки. (RI) = 3 мкМ H1152-глицила. (MG) = адгезивный слой из MATRIGEL в разбавлении 1:30. (MG+RI) = адгезивный слой из MATRIGEL в разбавлении 1:30 + 3 мкМ H1152-глицила.
На фиг.38 показано влияние ингибирования Rho-киназы на эффективность закрепления клеток линии эмбриональных стволовых клеток человека H1 на поверхностно-модифицированном носителе 32. (--) = без обработки. (RI) = 3 мкМ H1152-глицила. (MG) = адгезивный слой из MATRIGEL в разбавлении 1:30. (MG+RI) = адгезивный слой из MATRIGEL в разбавлении 1:30 + 3 мкМ H1152-глицила.
На фиг.39 показано влияние ингибирования Rho-киназы на эффективность закрепления клеток линии эмбриональных стволовых клеток человека H1 на поверхностно-модифицированном носителе 33. (--) = без обработки. (RI) = 3 мкМ H1152-глицила. (MG) = адгезивный слой из MATRIGEL в разбавлении 1:30. (MG+RI) = адгезивный слой из MATRIGEL в разбавлении 1:30 + 3 мкМ H1152-глицила.
На фиг.40 показано влияние ингибирования Rho-киназы на эффективность закрепления клеток линии эмбриональных стволовых клеток человека H1 на поверхностно-модифицированном носителе 34. (--) = без обработки. (RI) = 3 мкМ H1152-глицила. (MG) = адгезивный слой из MATRIGEL в разбавлении 1:30. (MG+RI) = адгезивный слой из MATRIGEL в разбавлении 1:30 + 3 мкМ H1152-глицила.
На фиг.41 показаны углы смачивания водой для поверхностно-модифицированных носителей, измерявшиеся в течение 40 недель статическим способом неподвижной капли.
На фиг.42 показаны измеренные статическим способом неподвижной капли углы смачивания водой для поверхностно-модифицированных носителей.
На фиг.43 показана плотность отрицательных зарядов на поверхностно-модифицированных носителях, измеренная по реакционной способности поверхностей в отношении положительно заряженных молекул кристаллического фиолетового.
На фиг.44 показана плотность отрицательных зарядов на поверхностно-модифицированных носителях 4, 22-24 и 29, измеренная по реакционной способности поверхностей в отношении положительно заряженных молекул кристаллического фиолетового. Испытывали по три образца каждой поверхности, измеряя оптическое поглощение десорбированного с поверхности каждого образца кристаллического фиолетового в трех повторностях. Плотность отрицательных зарядов на поверхностях типов 4, 22-24 и 29 нормировали по плотности отрицательных зарядов на поверхности типа Nunclon DeltaTM. На фигуре приведены средние значения и их стандартные отклонения по результатам девяти измерений.
ПОДРОБНОЕ ОПИСАНИЕ
Для ясности описания, а не в ограничение изобретения, подробное описание изобретения разделено на следующие подразделы, которые описывают или иллюстрируют определенные особенности, варианты осуществления или области применения настоящего изобретения.
Определения
Используемый в настоящей заявке термин “адгезивный слой” относится к слою на поверхности твердой подложки, формируемому путем прикрепления молекул к поверхности либо ковалентными (так называемая прививка), либо нековалентными (так называемая адсорбция) связями. Используемые для получения адгезивного слоя молекулы могут быть, например, белковыми и родственными им молекулами, которые могут включать в себя, например, белки внеклеточного матрикса, аминокислоты и т.д., и молекулами небиологической природы, такими как, например, полиэтиленимин.
Используемый в настоящей заявке термин “β-клеточная линия дифференцировки” относится к клеткам, положительным по экспрессии гена транскрипционного фактора PDX-1 и по меньшей мере одного из следующих транскрипционных факторов: NGN-3, Nkx2.2, Nkx6.1, NeuroD, Isl-1, HNF-3 beta, MAFA, Pax4 и Pax6. Клетки с экспрессией маркеров, характерных для β-клеточной линии дифференцировки, включают β-клетки.
Используемый в настоящей заявке термин “клетки с экспрессией маркеров, характерных для линии дифференцировки в клетки дефинитивной эндодермы” относится к клеткам с экспрессией по меньшей мере одного из следующих маркеров: SOX-17, GATA-4, HNF-3 beta, GSC, Cer1, Nodal, FGF-8, Brachyury, смесеобразный белок гомеобокса, FGF-4 CD48, эомезодермин (EOMES), DKK4, FGF-17, GATA-6, CXCR4, C-Kit, CD99 и OTX2. Клетки с экспрессией маркеров, характерных для линии дифференцировки в клетки дефинитивной эндодермы, включают клетки-предшественники клеток первичной полоски, клетки первичной полоски, клетки мезоэндодермы и клетки дефинитивной эндодермы.
Используемый в настоящей заявке термин “клетки с экспрессией маркеров, характерных для линии дифференцировки в клетки панкреатической эндодермы” относится к клеткам с экспрессией по меньшей мере одного из следующих маркеров: PDX-1, HNF-1beta, PTF-1 alpha, HNF-6 и HB9. Клетки с экспрессией маркеров, характерных для линии дифференцировки в клетки панкреатической эндодермы, включают клетки панкреатической эндодермы.
Используемый в настоящей заявке термин “клетки с экспрессией маркеров, характерных для линии дифференцировки в панкреатические эндокринные клетки” относится к клеткам с экспрессией по меньшей мере одного из следующих маркеров: NGN-3, NeuroD, Islet-1, PDX-1, NKX6.1, Pax-4, Ngn-3 и PTF-1 alpha. Клетки с экспрессией маркеров, характерных для линии дифференцировки в панкреатические эндокринные клетки, включают панкреатические эндокринные клетки, панкреатические клетки, экспрессирующие гормоны, и панкреатические клетки, секретирующие гормоны, а также клетки β-клеточной линии дифференцировки.
Используемый в настоящей заявке термин “дефинитивная эндодерма” относится к клеткам, обладающим характерными особенностями клеток, возникающих из эпибласта в ходе гаструляции, и формирующим желудочно-кишечный тракт и его производные. Клетки дефинитивной эндодермы экспрессируют следующие маркеры: CXCR4, HNF-3 beta, GATA-4, SOX-17, Cerberus, OTX2, goosecoid, c-Kit, CD99 и Mixl1.
Используемый в настоящей заявке термин “белки внеклеточного матрикса” относится к белковым и родственным им молекулам, обычно находящимся между клетками в теле или в плаценте. Белки внеклеточного матрикса могут быть получены из ткани, биологических жидкостей, например, крови, или из среды, кондиционированной нерекомбинантными клетками, рекомбинантными клетками или бактериями.
Используемый в настоящей заявке термин “внеэмбриональная эндодерма” относится к популяции клеток, экспрессирующих по меньшей мере один из следующих маркеров: SOX-7, AFP и SPARC.
Используемый в настоящей заявке термин “клетки HEK293” относится к клеточной линии, полученной трансформацией культуры нормальных клеток мезонефроса человека как описано в работе Graham et al. (J. Gen. Virol. 36:59-72, 1977), а также к любым клеткам, полученным из данной родительской линии.
Используемый в настоящей заявке термин “маркеры” означает молекулы нуклеиновых кислот или полипептидов с дифференциальной экспрессией в интересующих клетках. В данном контексте под дифференциальной экспрессией подразумевается повышение уровня экспрессии для положительного маркера и понижение уровня экспрессии для отрицательного маркера. Определяемый уровень маркерной аминокислоты или полипептида в интересующих клетках оказывается значительно выше или ниже по сравнению с другими клетками, что позволяет идентифицировать интересующую клетку и отличить ее от других клеток с помощью любого из множества известных в данной области способов.
Используемый в настоящей заявке термин “матрица” означает носитель с трехмерной структурой, на котором могут закрепиться клетки.
Используемый в настоящей заявке термин “клетка мезэндодермы” относится к клеткам, экспрессирующим по меньшей мере один из следующих маркеров: CD48, эомезодермин (EOMES), SOX-17, DKK4, HNF-3 beta, GSC, FGF-17, GATA-6.
Используемый в настоящей заявке термин “панкреатическая эндокринная клетка” или “панкреатическая клетка, экспрессирующая гормоны” относится к клеткам, способным к экспрессии по меньшей мере одного из следующих гормонов: инсулин, глюкагон, соматостатин и панкреатический полипептид.
Используемый в настоящей заявке термин “панкреатическая клетка, секретирующая гормоны” относится к клеткам, способным к секреции по меньшей мере одного из следующих гормонов: инсулин, глюкагон, соматостатин и панкреатический полипептид.
Используемый в настоящей заявке термин “клетка-предшественник клетки первичной полоски” относится к клеткам, экспрессирующим по меньшей мере один из следующих маркеров: Nodal и FGF-8.
Используемый в настоящей заявке термин “клетка первичной полоски” относится к клеткам, экспрессирующим по меньшей мере один из следующих маркеров: Brachyury, смесеобразный белок гомеобокса и FGF-4.
Используемый в настоящей заявке термин “поверхность” относится к внешнему слою молекул твердой подложки - сосуда или матрицы, предназначенных для культивирования или анализа клеток. Элементный состав, степень гладкости и смачиваемость поверхности могут быть проанализированы способами рентгеновской фотоэлектронной спектроскопии (XPS), атомно-силовой микроскопии (AFM) и измерения угла смачивания, соответственно.
Используемый в настоящей заявке термин “поверхностно-модифицированный носитель” относится к сосуду, содержащему поверхность одного из типов 1-34 описанных в примерах 16, 17 и 26, или содержащему поверхности, поступающие в продажу под торговыми марками Nunclon DeltaTM, CostarTM, FalconTM, CellBINDTM и PrimariaTM. Указанные сосуды могут, например, быть изготовлены из полимера, такого как полистирол (PS), сополимера циклоолефина (COC), поликарбоната (PC), полиметилметакрилата (PMMA) или стирол-акрилонитрильного сополимера (SAN).
Стволовые клетки представляют собой недифференцированные клетки, определяющиеся по их способности на уровне единичной клетки как самообновляться, так и дифференцироваться с образованием клеток-потомков, таких как самообновляющиеся предшественники, необновляющиеся предшественники и окончательно дифференцированные клетки. Стволовые клетки также характеризуются способностью дифференцироваться in vitro в функциональные клетки различных линий, происходящих из нескольких зародышевых листков (эндодермы, мезодермы и эктодермы), а также после трансплантации давать начало тканям, происходящим от нескольких зародышевых листков, и вносить существенный вклад в формирование большинства, если не всех, тканей после инъекции в бластоцисты.
По потенциалу развития стволовые клетки классифицируются следующим образом: (i) тотипотентные, т.е. способные давать начало всем эмбриональным и внеэмбриональным типам клеток; (ii) плюрипотентные, т.е. способные давать начало всем эмбриональным типам клеток; (iii) мультипотентные, т.е. способные давать начало группе клеточных линий в пределах конкретной ткани, органа или физиологической системы (например, гематопоэтические стволовые клетки (ГСК) могут давать таких потомков, как ГСК (самообновление), олигопотентные предшественники, ограниченные клетками крови, и все типы клеток и клеточных элементов (таких как тромбоциты), являющиеся нормальными компонентами крови); (iv) олигопотентные, т.е. способные давать начало более ограниченному набору клеточных линий, чем мультипотентные стволовые клетки; и (v) унипотентные, т.е. способные давать начало единственной клеточной линии (например, сперматогенные стволовые клетки).
Дифференцировка представляет собой процесс, при помощи которого неспециализированная (“некоммитированная”) или менее специализированная клетка приобретает свойства специализированной клетки, например, нервной или мышечной клетки. Дифференцированная клетка или клетка с индуцированной дифференцировкой представляет собой клетку, занявшую более специализированное (“коммитированное”) положение в линии клеточной дифференцировки. Термин “коммитированная” применительно к процессу дифференцировки относится к клетке, дошедшей в ходе процесса дифференцировки до стадии, от которой при нормальных условиях она продолжит дифференцироваться до определенного типа клеток или набора типов клеток и не сможет при нормальных условиях дифференцироваться в иной тип клеток или вернуться к менее дифференцированному типу. Дедифференцировкой называется процесс, в ходе которого клетка возвращается к менее специализированному (или коммитированному) положению в линии дифференцировки. Используемый в настоящей заявке термин “линия дифференцировки” определяет наследственность клетки, то есть определяет, из какой клетки произошла данная клетка и каким клеткам она может дать начало. В линии дифференцировки клетка помещается в наследственную схему развития и дифференцировки. Маркером, специфичным для линии дифференцировки, называется характерная особенность, специфически ассоциированная с фенотипом клеток конкретной линии дифференцировки, которая может быть использована для оценки дифференцировки некоммитированных клеток в клетки данной линии дифференцировки.
Для описания клеток в процессе культивирования используются различные термины. Термин “поддержание” относится в целом к клеткам, помещенным в ростовую среду в условиях, которые способствуют росту и/или делению клеток, в результате чего популяция клеток может увеличиваться или не увеличиваться. Термин “пассирование” означает процесс изъятия клеток из одного культурального сосуда и переноса их в другой культуральный сосуд в условия, которые способствуют росту и/или делению клеток.
Конкретная популяция клеток или клеточная линия иногда описывается или характеризуется числом выполненных с ней пассирований. Например, пассированная 10 раз культивируемая популяция клеток может описываться как культура десятого пассажа, или культура P10. Первичная культура, т.е. первая культура после выделения клеток из ткани, получает обозначение P0. После первого пересева клетки описываются как вторичная культура (P1, или культура первого пассажа). После второго пересева клетки превращаются в третичную культуру (P2, или культура второго пассажа) и т.д. Специалист в данной области определит, что за промежуток времени между последовательными пассированиями популяция клеток может многократно удваиваться; поэтому число удвоений популяций в культуре превышает номер пассажа. Степень размножения клеток (то есть число удваиваний популяции) за промежуток времени между последовательными пассированиями зависит от многих факторов, включая, помимо прочего, плотность посева, тип подложки, тип среды, условия роста и продолжительность времени между пересевами.
В одном осуществлении настоящее изобретение предлагает способ повышения эффективности закрепления клеток на поверхности, которая содержит от по меньшей мере приблизительно 0,9% N, с суммарным содержанием O и N выше или равно 22,3% и с углом смачивания по меньшей мере приблизительно 13,9 градуса, не имеет слоя питающих клеток и адгезивного слоя, включающий стадии:
а. получения суспензии клеток, и
b. нанесения суспензии клеток на поверхность, где они закрепляются.
В одном осуществлении настоящее изобретение предлагает способ повышения эффективности закрепления клеток на поверхности, которая содержит от по меньшей мере приблизительно 0,5% N, с суммарным содержанием O и N выше или равно 17,2% и с углом смачивания по меньшей мере приблизительно 13,9 градуса, не имеет слоя питающих клеток и адгезивного слоя, включающий стадии:
а. получения суспензии клеток,
b. обработки суспензии клеток по меньшей мере одним соединением, выбранным из группы, состоящей из: соединения, способного ингибировать активность Rho-киназы, и соединения, способного ингибировать активность Rho, и
c. нанесение обработанной суспензии клеток на поверхность, где они закрепляются.
В одном осуществлении указанная суспензия клеток представляет собой суспензию кластеров клеток. В альтернативном осуществлении указанная суспензия клеток представляет собой суспензию одиночных клеток.
В одном осуществлении клетки представляют собой плюрипотентные стволовые клетки. В альтернативном осуществлении клетки представляют собой стволовые клетки.
В одном осуществлении указанная поверхность имеет адгезивный слой. В одном осуществлении указанный адгезивный слой состоит из компонента внеклеточного матрикса, например, компонента, полученного из базальной мембраны, или компонента, который может участвовать в лиганд-рецепторном взаимодействии с участием молекулы адгезивного слоя. В одном осуществлении адгезивный слой состоит из препарата MATRIGEL (Becton Dickenson). MATRIGEL® представляет собой растворимый препарат из клеток опухоли Энгельбрета-Холма-Суорма, который при комнатной температуре превращается в гель и образует восстановленную базальную мембрану. Белковый адгезивный слой также может быть образован из ламинина, фибронектина, протеогликана, энтактина, гепарансульфата и т.д., как по отдельности, так и в различных сочетаниях.
В одном осуществлении после закрепления на поверхности клетки продолжают поддерживаться в той же культуральной среде. В альтернативном осуществлении по меньшей мере одно указанное соединение удаляется из культуральной среды после закрепления клеток на поверхности. В одном осуществлении закрепленные клетки снимаются с поверхности путем удаления по меньшей мере одного указанного соединения из культуральной среды.
В одном осуществлении суспензия культивируемых клеток обрабатывается по меньшей мере одним соединением, способным ингибировать активность Rho-киназы. В альтернативном осуществлении суспензия культивируемых клеток обрабатывается по меньшей мере одним соединением, способным ингибировать активность Rho. В альтернативном осуществлении суспензия культивируемых клеток обрабатывается по меньшей мере одним соединением, способным ингибировать активность Rho-киназы, и по меньшей мере одним соединением, способным ингибировать активность Rho.
По меньшей мере одно указанное соединение, способное ингибировать активность Rho-киназы, выбирают из группы, состоящей из следующих соединений: Y-27632, Fasudil и Hydroxyfasudil.
В одном осуществлении по меньшей мере одно указанное соединение, способное ингибировать активность Rho-киназы, представляет собой Y-27632.
По меньшей мере одно указанное соединение, способное ингибировать активность Rho-киназы, может использоваться в концентрации от приблизительно 0,1 мкМ до приблизительно 100 мкМ. В одном осуществлении по меньшей мере одно указанное соединение, способное ингибировать активность Rho-киназы, используется в концентрации приблизительно 10 мкМ.
В одном осуществлении по меньшей мере одно указанное соединение, способное ингибировать активность Rho, представляет собой ингибитор Rho-ГТФазы.
В одном осуществлении по меньшей мере одно указанное соединение, способное ингибировать активность Rho, представляет собой внеклеточный фермент C3 трансферазу.
По меньшей мере одно указанное соединение, способное ингибировать активность Rho, может использоваться в концентрации от приблизительно 0,01 мкг/мл до приблизительно 5 мкг/мл. В одном осуществлении по меньшей мере одно указанное соединение, способное ингибировать активность Rho, используется в концентрации приблизительно 0,5 мкг/мл.
Поверхностно-модифицированные носители
Поверхностно-модифицированные носители в соответствии с настоящим изобретением могут представлять собой сосуды, поверхность которых была модифицирована так, чтобы она содержала от по меньшей мере приблизительно 0,5% N, имела суммарное содержание O и N выше или равно 17,2% и имела угол смачивания по меньшей мере приблизительно 13,9 градуса. Как вариант, указанная поверхность может представлять собой трехмерную матрицу, такую как, например, пористый каркас, на которой могут закрепляться клетки.
В одном осуществлении указанный поверхностно-модифицированный носитель представляет собой носитель, поверхность которого содержит от по меньшей мере приблизительно 0,5% N, с суммарным содержанием O и N выше или равно 17,2% и с углом смачивания по меньшей мере приблизительно 13,9 градуса. В альтернативном осуществлении указанный поверхностно-модифицированный носитель представляет собой носитель, поверхность которого содержит от по меньшей мере приблизительно 0,5% N, с суммарным содержанием O и N выше или равно 19,5% и с углом смачивания по меньшей мере приблизительно 13,9 градуса.
В одном осуществлении указанный поверхностно-модифицированный носитель представляет собой носитель, поверхность которого содержит от по меньшей мере приблизительно 1,3% N, с суммарным содержанием O и N по меньшей мере приблизительно 24,9% и с углом смачивания по меньшей мере приблизительно 20,7 градуса, и который в настоящей заявке именуется “поверхностно-модифицированный носитель 1”.
В одном осуществлении указанный поверхностно-модифицированный носитель представляет собой носитель, поверхность которого содержит от по меньшей мере приблизительно 1,7% N, с суммарным содержанием O и N по меньшей мере приблизительно 29,6% и с углом смачивания по меньшей мере приблизительно 14,3 градуса, и который в настоящей заявке именуется “поверхностно-модифицированный носитель 2”.
В одном осуществлении указанный поверхностно-модифицированный носитель представляет собой носитель, поверхность которого содержит от по меньшей мере приблизительно 2,0% N, с суммарным содержанием O и N по меньшей мере приблизительно 30,7% и с углом смачивания по меньшей мере приблизительно 18,4 градуса, и который в настоящей заявке именуется “поверхностно-модифицированный носитель 3”.
В одном осуществлении указанный поверхностно-модифицированный носитель представляет собой носитель, поверхность которого содержит от по меньшей мере приблизительно 2,1% N, с суммарным содержанием O и N по меньшей мере приблизительно 30,2% и с углом смачивания по меньшей мере приблизительно 17,4 градуса, и который в настоящей заявке именуется “поверхностно-модифицированный носитель 4”.
В одном осуществлении указанный поверхностно-модифицированный носитель представляет собой носитель, поверхность которого содержит от по меньшей мере приблизительно 1,8% N, с суммарным содержанием O и N по меньшей мере приблизительно 28,2% и с углом смачивания по меньшей мере приблизительно 18,8 градуса, и который в настоящей заявке именуется “поверхностно-модифицированный носитель 13”.
В одном осуществлении указанный поверхностно-модифицированный носитель представляет собой носитель, поверхность которого содержит от по меньшей мере приблизительно 1,0% N, с суммарным содержанием O и N по меньшей мере приблизительно 27,8% и с углом смачивания по меньшей мере приблизительно 44,3 градуса, и который поступает в продажу под торговой маркой CELLBIND.
В одном осуществлении указанный поверхностно-модифицированный носитель представляет собой носитель, поверхность которого содержит от по меньшей мере приблизительно 10,2% N, с суммарным содержанием O и N по меньшей мере приблизительно 23,0% и с углом смачивания по меньшей мере приблизительно 39,5 градуса, и который поступает в продажу под торговой маркой PRIMARIA.
Определение характеристик поверхностно-модифицированных носителей
В одном осуществлении элементный состав поверхностно-модифицированных носителей может быть проанализирован способом рентгеновской фотоэлектронной спектроскопии (XPS). Способ XPS, также известный как электронная спектроскопия для химического анализа (ESCA), используется для определения элементов или атомов, присутствующих в поверхности твердой подложки (возможно определение всех элементов в концентрациях менее 0,1 атомного процента, за исключением водорода и гелия), а также для определения химического окружения подобных элементов или атомов. Например, XPS анализ твердого образца полистирола (который содержит только углерод и водород) обычно дает следующие результаты: более 97% углерода, менее 3% кислорода и 0% азота (водород не определяется; определяемые уровни кислорода могут различаться из-за окисления цепей полистирола на поверхности, например, вследствие радиационной стерилизации) (Brevig et al., Biomaterials 26:3039-3053, 2005; Shen and Horbett, J. Biomed. Mater. Res. 57:336-345, 2001).
В одном осуществлении степень гладкости поверхности поверхностно-модифицированных носителей может быть проанализирована способом атомно-силовой микроскопии (AFM). Способ AFM позволяет получать топографию атомов или молекул на поверхности с поперечным разрешением до 1Å и вертикальным разрешением до 0,1Å.
В одном осуществлении смачиваемость поверхности поверхностно-модифицированных носителей может быть проанализирована способом измерения угла смачивания. Например, измерение угла смачивания статическим способом неподвижной капли дает информацию о взаимодействии между поверхностью твердой подложки и жидкостью. Угол смачивания описывает форму капли жидкости, которая покоится на поверхности твердой подложки, и представляет собой контактный угол жидкости на поверхности твердого носителя, измеренный внутри жидкости вдоль линии контакта, где встречаются жидкость, твердая подложка и газ. Поверхность с углом смачивания водой более 90° называется гидрофобной, а поверхность с углом смачивания водой менее 90° - гидрофильной. На очень гидрофильных поверхностях, то есть поверхностях с большим сродством к воде, капля воды полностью растекается по поверхности (эффективный угол смачивания равен 0°).
В одном осуществлении плотность отрицательных зарядов на поверхности поверхностно-модифицированных носителей может быть проанализирована способом измерения реакционной способности поверхности в отношении кристаллического фиолетового. Молекулы кристаллического фиолетового несут положительный заряд, что позволяет им связываться с отрицательно заряженными молекулами и фрагментами молекул, например, отрицательно заряженными функциональными группами, присутствующими на поверхности полимера. Поверхность с высокой реакционной способностью в отношении кристаллического фиолетового имеет большую плотность отрицательных зарядов по сравнению с поверхностью, имеющей низкую реакционную способность в отношении кристаллического фиолетового, при том, что степень гладкости поверхности и, тем самым, ее удельная площадь у них одинакова.
Плюрипотентные стволовые клетки
Определение характеристики плюрипотентных стволовых клеток
Плюрипотентные стволовые клетки могут экспрессировать один или несколько стадиеспецифических эмбриональных антигенов (SSEA) 3 и 4, а также маркеры, определяемые антителами, обозначаемыми как Tra-1-60 и Tra-1-81 (Thomson et al., Science 282:1145 1998). Дифференцировка плюрипотентных стволовых клеток in vitro приводит к потере экспрессии SSEA-4, Tra- 1-60 и Tra-1-81 (при ее наличии) и повышению экспрессии SSEA-1. Недифференцированные плюрипотентные стволовые клетки, как правило, обладают щелочно-фосфатазной активностью, которую можно определить путем фиксирования клеток 4% раствором параформальдегида с последующим проявлением с использованием красителя Vector Red в качестве субстрата, следуя рекомендациям производителя (компания Vector Laboratories, Берлингейм, Калифорния, США). Недифференцированные плюрипотентные стволовые клетки также, как правило, экспрессируют Oct-4 и TERT, обнаруживаемые способом RT-PCR.
Другим желательным фенотипическим свойством выращенных плюрипотентных клеток является потенциал дифференцировки в клетки всех трех зародышевых листков: в эндодермальные, мезодермальные и эктодермальные ткани. Плюрипотентность стволовых клеток может быть подтверждена, например, путем инъекции клеток мышам с тяжелым комбинированным иммунодефицитом (SCID), фиксирования образующихся тератом с помощью 4% параформальдегида и их гистологического исследования на предмет наличия клеточных типов, происходящих от трех зародышевых листков. Как вариант, плюрипотентность можно определить путем создания эмбриоидных телец и анализа их на предмет присутствия маркеров, ассоциируемых с тремя зародышевыми листками.
Выращенные линии плюрипотентных стволовых клеток могут быть кариотипированы с применением стандартного способа окрашивания красителем Гимза (G-banding) и сравнения с опубликованными кариотипами соответствующих видов приматов. Желательно получать клетки, имеющие “нормальный кариотип”, т.е. эуплоидные клетки, в которых присутствуют все человеческие хромосомы и не имеют видимых изменений.
Источники плюрипотентных стволовых клеток
К типам плюрипотентных стволовых клеток, которые можно использовать, относятся стабильные линии плюрипотентных клеток, получаемых из ткани, формирующейся после наступления беременности, в том числе из преэмбриональной ткани (например, бластоциста), эмбриональной ткани или ткани плода, взятой в любой момент в ходе беременности, как правило, но не обязательно, до срока приблизительно 10-12 недель беременности. Не ограничивающими настоящее изобретение примерами являются стабильные линии человеческих эмбриональных стволовых клеток или человеческих эмбриональных зародышевых клеток, например, клеточные линии человеческих эмбриональных стволовых клеток H1, H7 и H9 (WiCell). Также возможно использование описываемых в настоящей заявке композиций в ходе первоначального установления или стабилизации таких клеток; в этом случае исходными клетками являются первичные плюрипотентные клетки, взятые непосредственно из тканей-источников. Также пригодны для целей настоящего изобретения клетки, взятые из популяции плюрипотентных стволовых клеток, уже культивируемых в отсутствие питающих клеток, или из популяции плюрипотентных стволовых клеток, уже культивируемых в присутствии питающих клеток. Также пригодны для целей настоящего изобретения клетки мутантных линий эмбриональных стволовых клеток человека, таких как, например, BG01v (компания BresaGen, Атенс, Джорджия, США). Также пригодны для целей настоящего изобретения клетки, полученные из соматических клеток взрослого человека, такие как, например, клетки, описанные в работе Takahashi et al, Cell 131: 1-12 (2007).
В одном осуществлении эмбриональные стволовые клетки человека готовят, как описано в следующих публикациях: Thomson et al. ( Патент США № 5,843,780; Science 282:1145, 1998; Curr. Top. Dev. Biol. 38:133 ff., 1998; Proc. Natl. Acad. Sci. U.S.A. 92:7844, 1995).
Культивирование плюрипотентных стволовых клеток
В одном осуществлении перед культивированием в соответствии со способами, составляющими предмет настоящего изобретения, плюрипотентные стволовые клетки культивируют на слое питающих клеток или белка внеклеточного матрикса, которые различным образом поддерживают плюрипотентные стволовые клетки. Например, плюрипотентные стволовые клетки культивируют на слое питающих клеток, который поддерживает пролиферацию плюрипотентных стволовых клеток без их существенной дифференцировки. Рост плюрипотентных стволовых клеток на слое питающих клеток без дифференцировки поддерживается с помощью (i) культурального сосуда с имеющимся слоем питающих клеток и (ii) среды, кондиционированной путем предварительного культивирования клеток иного типа, или некондиционированной среды, например, не содержащей сыворотки или даже среды с химически определенной композицией.
В другом примере плюрипотентные стволовые клетки культивируют в культуральной системе, существенно свободной от питающих клеток, но, тем не менее, способной поддерживать пролиферацию плюрипотентных стволовых клеток без существенной дифференцировки. Рост плюрипотентных стволовых клеток в свободной от питающих клеток культуральной системе без дифференцировки поддерживается за счет (i) адгезивного слоя на поверхности твердой подложки, содержащего один или более белков внеклеточного матрикса; и (ii) среды, кондиционированной путем предварительного культивирования клеток иного типа, или некондиционированной среды, например, не содержащей сыворотки, или даже среды с химически определенной композицией.
В альтернативном осуществлении плюрипотентные стволовые клетки культивируют на поверхностно-модифицированном носителе, содержащем от по меньшей мере приблизительно 0,5% N, с суммарным содержанием O и N выше или равно 17,2% и с углом смачивания по меньшей мере приблизительно 13,9 градуса, в среде, кондиционированной путем предварительного культивирования клеток иного типа, или некондиционированной среде, например, не содержащей сыворотки или даже среде с химически определенной композицией.
Культуральная среда: Пример клеточной культуральной среды, соответствующей целям настоящего изобретения, приведен в заявке на патент США US20020072117. Другой пример клеточной культуральной среды, соответствующей целям настоящего изобретения, приведен в заявке на патент США US6642048. Другой пример клеточной культуральной среды, соответствующей целям настоящего изобретения, приведен в заявке на патент WO2005014799. Другой пример клеточной культуральной среды, соответствующей целям настоящего изобретения, приведен в работе Xu et al (Stem Cells 22: 972-980, 2004). Другой пример клеточной культуральной среды, соответствующей целям настоящего изобретения, приведен в заявке на патент США US20070010011. Другой пример клеточной культуральной среды, соответствующей целям настоящего изобретения, приведен в работе Cheon et al. (BioReprod DOI:10.1095/biolreprod.105.046870; 19 Oct 2005). Другой пример клеточной культуральной среды, соответствующей целям настоящего изобретения, приведен в работе Levenstein et al. (Stem Cells 24: 568-574, 2006). Другой пример клеточной культуральной среды, соответствующей целям настоящего изобретения, приведен в заявке на патент США US20050148070. Другой пример клеточной культуральной среды, соответствующей целям настоящего изобретения, приведен в заявке на патент США US20050233446. Другой пример клеточной культуральной среды, соответствующей целям настоящего изобретения, приведен в заявке на патент США US6800480. Другой пример клеточной культуральной среды, соответствующей целям настоящего изобретения, приведен в заявке на патент США US20050244962. Другой пример клеточной культуральной среды, соответствующей целям настоящего изобретения, приведен в заявке на патент WO2005065354. Другой пример клеточной культуральной среды, соответствующей целям настоящего изобретения, приведен в заявке на патент WO2005086845.
Культуральная среда, соответствующая целям настоящего изобретения, может также быть приготовлена из указанных ниже компонентов, таких как, например, модифицированная по способу Дульбекко среда Игла (DMEM), Gibco # 11965-092; нокаут-среда Игла, модифицированная по способу Дульбекко (KO DMEM), Gibco #10829-018; основная среда Хэма F12/50% DMEM; 200 мМ L-глутамина, Gibco # 15039-027; раствор заменимых аминокислот, Gibco 11140-050; β-меркаптоэтанол, Sigma # M7522; человеческий рекомбинантный основной фактор роста фибробластов (bFGF), Gibco # 13256-029.
Дифференцировка плюрипотентных стволовых клеток
В одном осуществлении настоящего изобретения плюрипотентные стволовые клетки размножают в культуре с сохранением их плюрипотентности. Изменения в плюрипотентности культивируемых клеток во времени могут быть определены по изменению уровней экспрессии ассоциированных с плюрипотентностью маркеров. Альтернативным образом, за изменениями плюрипотентности можно следить по изменениям уровней экспрессии маркеров, ассоциированных с дифференцировкой, или маркеров, ассоциированных с клетками иных типов.
В альтернативном осуществлении плюрипотентные стволовые клетки размножают в культуре и затем обрабатывают таким образом, чтобы индуцировать их дифференцировку в клетки иного типа. К таким клеткам иного типа могут относиться клетки с экспрессией маркеров, характерных для линии дифференцировки дефинитивной эндодермы. Альтернативным образом, к таким клеткам иного типа могут относиться клетки с экспрессией маркеров, характерных для линии дифференцировки панкреатической эндодермы. Альтернативным образом, к таким клеткам иного типа могут относиться клетки с экспрессией маркеров, характерных для линии дифференцировки панкреатической эндокринных клеток. Альтернативным образом, к таким клеткам иного типа могут относиться клетки с экспрессией маркеров, характерных для линии дифференцировки β-клеток.
Плюрипотентные стволовые клетки, обработанные в соответствии со способами, составляющими предмет настоящего изобретения, могут быть дифференцированы в различные клетки иных типов с использованием любых соответствующих способов, известных специалистам в данной области.
Например, плюрипотентные стволовые клетки, обработанные в соответствии со способами, составляющими предмет настоящего изобретения, могут быть дифференцированы в нервные клетки, сердечные клетки, гепатоциты и т.д.
Например, плюрипотентные стволовые клетки, обработанные в соответствии со способами, составляющими предмет настоящего изобретения, могут быть дифференцированы в нервные клетки-предшественники и кардиомиоциты в соответствии со способами, описанными в заявке на патент WO2007030870.
В другом примере, плюрипотентные стволовые клетки, обработанные в соответствии со способами, составляющими предмет настоящего изобретения, могут быть дифференцированы в гепатоциты в соответствии со способами, описанными в заявке на патент США 6458589.
Например, плюрипотентные стволовые клетки могут быть дифференцированы в клетки с экспрессией маркеров, характерных для линии дифференцировки дефинитивной эндодермы, в соответствии со способами, описанными в работе D'Amour et al., Nature Biotechnol. 23:1534-1541, 2005.
Например, плюрипотентные стволовые клетки могут быть дифференцированы в клетки с экспрессией маркеров, характерных для линии дифференцировки дефинитивной эндодермы, в соответствии со способами, описанными в работе Shinozaki et al., Development 131:1651-1662, 2004.
Например, плюрипотентные стволовые клетки могут быть дифференцированы в клетки с экспрессией маркеров, характерных для линии дифференцировки дефинитивной эндодермы, в соответствии со способами, описанными в работе McLean et al., Stem Cells 25:29-38, 2007.
Например, плюрипотентные стволовые клетки могут быть дифференцированы в клетки с экспрессией маркеров, характерных для линии дифференцировки дефинитивной эндодермы, в соответствии со способами, описанными в работе D'Amour et al., Nature Biotechnol. 24:1392-1401, 2006.
Маркеры, характерные для линии дифференцировки дефинитивной эндодермы, выбирают из группы, включающей SOX17, GATA4, Hnf-3beta, GSC, Cer1, Nodal, FGF-8, Brachyury, смесеобразный белок гомеобокса, FGF-4 CD48, эомезодермин (EOMES), DKK4, FGF-17, GATA6, CXCR4, C-Kit, CD99 и OTX2. Для целей настоящего изобретения пригодны клетки с экспрессией по меньшей мере одного из маркеров, характерных для линии дифференцировки дефинитивной эндодермы. В одном аспекте настоящего изобретения клетка с экспрессией маркеров, характерных для линии дифференцировки дефинитивной эндодермы, представляет собой клетку-предшественницу первичной полоски. В другом аспекте настоящего изобретения клетка с экспрессией маркеров, характерных для линии дифференцировки дефинитивной эндодермы, представляет собой мезэндодермальную клетку. В другом аспекте настоящего изобретения клетка с экспрессией маркеров, характерных для линии дифференцировки дефинитивной эндодермы, представляет собой клетку дефинитивной эндодермы.
Например, плюрипотентные стволовые клетки могут быть дифференцированы в клетки с экспрессией маркеров, характерных для линии дифференцировки панкреатической эндодермы, в соответствии со способами, описанными в работе D'Amour et al., Nature Biotechnol. 24:1392-1401, 2006.
Маркеры, характерные для линии дифференцировки панкреатической эндодермы, выбирают из группы, включающей Pdx1, HNF-1beta, PTF1a, HNF-6, HB9 и PROX1. Для целей настоящего изобретения пригодны клетки с экспрессией по меньшей мере одного из маркеров, характерных для линии дифференцировки панкреатической эндодермы. В одном аспекте настоящего изобретения клетка с экспрессией маркеров, характерных для линии дифференцировки панкреатической эндодермы, представляет собой клетку панкреатической эндодермы.
Плюрипотентные стволовые клетки могут быть дифференцированы в клетки с экспрессией маркеров, характерных для линии дифференцировки панкреатических эндокринных клеток, с использованием любых соответствующих способов, известных специалистам в данной области.
Например, плюрипотентные стволовые клетки могут быть дифференцированы в клетки с экспрессией маркеров, характерных для линии дифференцировки панкреатических эндокринных клеток, в соответствии со способами, описанными в работе D'Amour et al., Nature Biotechnol. 24:1392-1401, 2006.
Например, плюрипотентные стволовые клетки могут быть дифференцированы в клетки с экспрессией маркеров, характерных для линии дифференцировки панкреатических эндокринных клеток, в соответствии со способами, описанными в работе D'Amour et al., Nature Biotechnol. 24:1392-1401, 2006.
Маркеры, характерные для линии дифференцировки панкреатических эндокринных клеток, выбирают из группы, включающей NGN-3, NeuroD, Islet-1, Pdx-1, NKX6.1, Pax-4, Ngn-3 и PTF-1 alpha. В одном осуществлении настоящего изобретения панкреатическая эндокринная клетка способна к экспрессии по меньшей мере одного из следующих гормонов: инсулин, глюкагон, соматостатин и панкреатический полипептид. Для целей настоящего изобретения пригодны клетки с экспрессией по меньшей мере одного из маркеров, характерных для линии дифференцировки панкреатических эндокринных клеток. В одном аспекте настоящего изобретения клетка с экспрессией маркеров, характерных для линии дифференцировки панкреатических эндокринных клеток, представляет собой панкреатическую эндокринную клетку. Указанная панкреатическая эндокринная клетка может представлять собой панкреатическую клетку, экспрессирующую гормоны. Альтернативно, указанная панкреатическая эндокринная клетка может представлять собой панкреатическую клетку, секретирующую гормоны.
В одном аспекте настоящего изобретения панкреатическая эндокринная клетка представляет собой клетку с экспрессией маркеров, характерных для линии дифференцировки β-клеток. Клетка с экспрессией маркеров, характерных для линии дифференцировки β-клеток, экспрессирует Pdx1 и по меньшей мере один из следующих транскрипционных факторов: NGN-3, Nkx2.2, Nkx6.1, NeuroD, Isl-1, HNF-3 beta, MAFA, Pax4 и Pax6. В одном аспекте настоящего изобретения клетка с экспрессией маркеров, характерных для линии дифференцировки β-клеток, представляет собой β-клетку.
Настоящее изобретение иллюстрируют, но не ограничивают, следующие примеры.
ПРИМЕРЫ
Пример 1
Пассирование и поддержание эмбриональных стволовых клеток человека в виде кластеров клеток
Клетки линий эмбриональных стволовых клеток человека H1 и H9 изначально поддерживали на инактивированных митомицином C первичных мышиных эмбриональных фибробластах (MEF). Эмбриональные стволовые клетки человека несколькими пассажами перевели с питающих клеток MEF на носитель с поверхностью, обработанной препаратом MatrigelTM (компания Becton-Dickinson, Бедфорд, Массачусетс, США).
Обработка поверхности препаратом MatrigelTM: Препарат MatrigelTM с пониженным содержанием факторов роста (Growth Factor Reduced Matrigel) размораживали при температуре 4°C и затем разводили в холодной среде DMEM/F12 (компания Invitrogen, Карлсбад, Калифорния, США) в соотношении 1:30. На каждую чашку размером 6 см или на каждую лунку 6-луночного планшета наносили достаточный для полного покрытия поверхности объем приготовленной жидкости (соответственно 2 мл и 1 мл) и инкубировали носители в течение одного часа при комнатной температуре. Обработанные таким образом носители использовали в течение нескольких часов или хранили при температуре 4°C до двух недель.
Культивирование эмбриональных стволовых клеток человека: Колонии недифференцированных эмбриональных стволовых клеток человека (линий H9 или H1) собирали с питающего слоя путем инкубации в растворе 1 мг/мл коллагеназы IV (компания Sigma-Aldrich, Сент-Луис, Миссури, США) в среде DMEM/F12 в течение 10 минут с последующим соскребанием сколом пипетки. Скопления клеток осаждали центрифугированием при 600 g в течение четырех минут, полученные сгустки клеток осторожно диспергировали 2-мл пипеткой для разделения колоний на небольшие кластеры клеток. Полученные таким образом кластеры клеток высевали на обработанные препаратом MatrigelTM чашки в кондиционированной мышиными эмбриональными фибробластами среде (MEF-CM) с добавлением bFGF (8 нг/мл; компания R&D Systems, Миннеаполис, Миннесота, США) с плотностью 50-150 колоний на чашку размером 6 см в 5 мл ростовой среды. Среду заменяли ежедневно. Колонии, культивируемые на поверхности MatrigelTM в среде MEF-CM, пересевали по мере укрупнения, когда они занимали 70-80% площади поверхности, приблизительно каждые 3-4 дня. Эмбриональные стволовые клетки человека в колониях имели высокое соотношение объема ядра и объема цитоплазмы и выраженные нуклеоли, подобно эмбриональным стволовым клеткам человека, поддерживаемым на питающем слое (смотрите фиг.1). Дифференцированные клетки составляли менее 5% от общего числа клеток в культуре.
Для пассирования клеток в установленном порядке в среде MEF-CM на поверхности MatrigelTM клетки инкубировали в растворе 1 мг/мл коллагеназы IV в среде DMEM/F12 до 60 минут и снимали их с поверхности чашек с помощью мощной струи среды DMEM/F12 и соскабливания. Собранные клетки центрифугировали, диспергировали и высевали в соотношении 1:3 или 1:4.
Пример 2
Пассирование эмбриональных стволовых клеток человека в виде одиночных клеток
Эмбриональные стволовые клетки человека линии H9 растили как одиночные клетки в соответствии со способами, описанными в заявке на патент США LFS5163USPSP, права на который принадлежат компании LifeScan Inc. Клетки пассировали путем обработки ферментом TrypLE™ Express в течение пяти минут при температуре 37°C и высевали с плотностью 10 000 клеток/см2 на поверхность подложки.
Пример 3
Иммобилизация, культивирование и поддержание плюрипотентности эмбриональных стволовых клеток человека на поверхностно-модифицированных носителях, не имеющих на своей поверхности белков/компонентов внеклеточного матрикса и питающих клеток
Перед началом исследования эмбриональные стволовые клетки человека линии H1 на 49-ом пассаже поддерживали в среде, кондиционированной MEF, на носителях с поверхностью Nunclon DeltaTM, обработанных препаратом MatrigelTM с пониженным содержанием факторов роста в разбавлении 1:30. Для пассирования клетки снимали с поверхности путем инкубации в растворе 1 мг/мл коллагеназы или путем соскабливания вручную.
Затем приготовленные таким образом клетки высевали в две необработанные лунки поверхностно-модифицированных носителей в формате 6-луночных планшетов. Кроме того, одну из лунок каждого планшета обрабатывали свободным от ксеногенного материала 0,1% желатином человека в качестве контроля. Клетки также высевали непосредственно на необработанные и обработанные желатином лунки 6-луночных планшетов с поверхностями типа CostarTM (кат. № 3516; компания Corning, Корнинг, Нью-Йорк, США), FalconTM (кат. № 351146; компания Becton Dickinson, Франклин Лэйкс, Нью-Джерси, США) и Nunclon DeltaTM (кат. № 140675; компания Thermo Fisher Scientific, Роскиль, Дания), используемые в качестве отрицательных контролей, и лунки тех же планшетов, обработанные препаратом MatrigelTM с пониженным содержанием факторов роста в разбавлении 1:30, используемые в качестве положительных контролей. При всех обработках клетки поддерживали в среде, кондиционированной MEF.
Мы обнаружили, что после двух пассажей поверхностно-модифицированные носители 2, 3 и 4 имели на своей поверхности закрепленные колонии эмбриональных стволовых клеток, которые после ферментативной диссоциации снова закрепились на поверхности носителя и продолжали рост. Никаких ярко выраженных различий в эффективности закрепления или скорости роста клеток на обработанных желатином или необработанных лунках поверхностно-модифицированных носителей 2, 3 и 4 не наблюдалось.
Клетки, механически снятые с носителей, обработанных препаратом MatrigelTM с пониженным содержанием факторов роста в разбавлении 1:30, плохо закреплялись на поверхностно-модифицированных носителях 2, 3 и 4, тогда как клетки, диссоциированные ферментативно раствором 1 мг/мл коллагеназы, хорошо закреплялись на обработанных желатином или необработанных лунках поверхностно-модифицированных носителей 2, 3 и 4.
Эмбриональные стволовые клетки линии H1 49 пассажа (H1p49), внесенные на поверхностно-модифицированные носители 1 и 5-12, а также на необработанные и обработанные желатином носители с поверхностями типов Nunclon DeltaTM, FalconTM и CostarTM, на перечисленных поверхностях не закреплялись. Однако те же самые клетки успешно закреплялись на носителях, обработанных препаратом MatrigelTM с пониженным содержанием факторов роста в разбавлении 1:30, что указывает на способность клеток закрепляться на поверхности подложки.
Как правило, промежуток времени между последовательными пересевами эмбриональных стволовых клеток линии H1, культивируемых на носителях, обработанных препаратом MatrigelTM с пониженным содержанием факторов роста в разбавлении 1:30, составлял 3-4 дня, однако клетки, культивируемые на поверхностно-модифицированных носителях 2, 3 и 4, созревали для следующего пересева лишь на седьмой день. Это, вероятно, связано с пониженной эффективностью закрепления клеток на обработанных поверхностях, поскольку непосредственно после пересева на обработанных препаратом MatrigelTM поверхностях наблюдалось больше исходных колоний, чем на поверхностях носителей 2, 3 и 4.
Клетки 50-го пассажа разделяли в соотношении 1 к 2, половину образца собирали для выделения РНК и проверяли на уровень экспрессии маркеров плюрипотентности (смотрите таблицу 1). Вторую половину каждого образца высевали на поверхностно-модифицированные носители. Образовавшимся на этом пассаже (p51) колониям также потребовалось 7 дней культивирования для созревания к следующему пересеву, и маленькие колонии, которые развивались лишь после 4 дней культивирования, показаны на фиг.1. Эти колонии сохраняли классическую морфологию колоний эмбриональных стволовых клеток.
Культивирование на поверхностно-модифицированных носителях 2, 3 и 4 остановили после четырех пассажей; образцы проанализировали способом qRT-PCR на уровень экспрессии маркеров плюрипотентности (смотрите таблицу 2) и дифференцировали в линию дефинитивной эндодермы (DE). На четвертом пассаже клетки сохраняли экспрессию классических маркеров плюрипотентности: Oct4, Nanog, Sox2 и TERT. Более того, клетки были способны дифференцироваться в клетки линии дефинитивной эндодермы при обработке средой, содержащей DMEM/F12, 100 нг/мл активина A, 20 нг/мл Wnt3a и 0,5-2,0% FBS (смотрите таблицу 3), что указывает на сохранение плюрипотентности клеток вплоть до четвертого пассажа.
Пример 4
Иммобилизация, культивирование и поддержание плюрипотентности эмбриональных стволовых клеток человека на поверхностно-модифицированных носителях, не имеющих на своей поверхности белков/компонентов внеклеточного матрикса и питающих клеток: Эффект от ингибирования Rho и ингибирования Rho-киназы
Перед началом исследования эмбриональные стволовые клетки человека линии H1 на 49-ом пассаже поддерживали в среде, кондиционированной MEF, на носителях с поверхностью Nunclon DeltaTM, обработанных препаратом MatrigelTM с пониженным содержанием факторов роста в разбавлении 1:30. Для пассирования клетки снимали с поверхности путем инкубации в растворе 1 мг/мл коллагеназы.
Затем приготовленные таким образом клетки высевали в необработанные лунки поверхностно-модифицированных носителей в формате 6-луночных планшетов. Клетки также высевали непосредственно на необработанные и обработанные желатином лунки 6-луночных планшетов с поверхностями типа CostarTM, FalconTM и Nunclon DeltaTM, используемые в качестве отрицательных контролей, и лунки тех же планшетов, обработанные препаратом MatrigelTM с пониженным содержанием факторов роста в разбавлении 1:30, используемые в качестве положительных контролей. При всех обработках клетки поддерживали в среде, кондиционированной MEF.
Эмбриональные стволовые клетки линии H1 49 пассажа, нанесенные на поверхностно-модифицированные носители 1 и 5-12, а также на необработанные и обработанные желатином носители с поверхностями типов Nunclon DeltaTM и CostarTM, на перечисленных поверхностях не закреплялись, однако они закреплялись на поверхностно-модифицированных носителях 2, 3 и 4. Те же самые клетки успешно закреплялись на носителях, обработанных препаратом MatrigelTM с пониженным содержанием факторов роста в разбавлении 1:30, что указывает на способность клеток закрепляться на поверхности подложки.
Как правило, промежуток времени между последовательными пересевами эмбриональных стволовых клеток линии H1, культивируемыми на носителях, обработанных препаратом MatrigelTM с пониженным содержанием факторов роста в разбавлении 1:30, составлял 3-4 дня, однако клетки, культивируемые на поверхностно-модифицированных носителях 2, 3 и 4, созревали для следующего пересева лишь на седьмой день. Это, вероятно, связано с пониженной эффективностью закрепления клеток на поверхностно-модифицированных носителях, поскольку непосредственно после пересева на обработанных препаратом MatrigelTM поверхностях наблюдалось больше исходных колоний, чем на поверхностно-модифицированных носителях 2, 3 и 4.
Клетки 50-го пассажа разделяли в соотношении 1 к 2, половину образца собирали для выделения РНК и проверяли на уровень экспрессии маркеров плюрипотентности (смотрите таблицу 1). Вторую половину каждого образца высевали на поверхностно-модифицированные носители. Образовавшимся на этом пассаже (p51) колониям также потребовалось 7 дней культивирования для созревания к следующему пересеву, и маленькие колонии, которые развивались лишь после 4 дней культивирования, показаны на фиг.1. Эти колонии сохраняли классическую морфологию колоний эмбриональных стволовых клеток.
Из-за задержки в пассировании клетки разделяли в соотношении 1 к 2, и половину образца 4-го пассажа высевали в среде, кондиционированной MEF, или в среде, кондиционированной MEF с добавлением ингибитора Rho-киназы (ROCK), Y-27632, в концентрации 10 мкМ в попытке улучшить кинетику роста клеток. После пассажа клетки в течение 48 часов выдерживали в исходной среде, после чего среду заменяли на свежую среду, кондиционированную MEF, без добавки ингибитора.
Добавление Y-27632 в концентрации 10 мкМ существенно повысило эффективность посева клеток 52-го пассажа (p52), и через четыре дня после пересева улучшение роста колоний было явно выражено (смотрите фиг.2). Альтернативным образом, перед коллагеназной диссоциацией эмбриональные стволовые клетки человека линии H1 дополнительно обрабатывали проникающей в клетку формой ингибитора Rho, C3 экзотрансферазы, в концентрации 0,5 нг/мл, что также повысило эффективность посева клеток.
Если клетки, высеянные в среде с добавкой 10 мкМ Y-27632, были готовы к следующему пассированию через 4 дня после посева, клетки, высеянные в среде без ингибитора ROCK не были готовы к разделению через 4 дня после посева. Клетки, обработанные ингибитором Rho, C3 экзотрансферазой, также не были готовы к пассированию через четыре дня после посева; кроме того, эти клетки отличались повышенной степенью дифференцировки в фибробластообразную морфологию. По этой причине клетки, обработанные ингибитором Rho на пассаже 4, на всех последующих пассажах обрабатывали Y-27632 (смотрите фиг.3).
Клетки пассировали далее по меньшей мере до пассажа 10 на поверхностно-модифицированных носителях 3 и 4 и проверили на наличие маркеров, ассоциированных с плюрипотентностью: генов - способом qRT-PCR; экспрессию маркеров клеточной поверхности - способом проточной цитометрии; а также иммунофлюоресценцию поверхности клетки я ядерных белков (смотрите фиг.4-6). Плюрипотентность клеток также подтвердили, проверив способность клеток к дифференцировке в клетки линий дефинитивной эндодермы, панкреатической эндодермы, а также к образованию эмбриоидных телец, состоящих из всех трех зародышевых листков (смотрите фиг.7-9). Клетки также проверили на кариотипическую устойчивость, и мы обнаружили, что клетки были способны сохранять нормальный кариотип (смотрите фиг.10).
Пример 5
Закрепление и открепление эмбриональных стволовых клеток человека путем ингибирования Rho-киназы
Перед началом исследования эмбриональные стволовые клетки человека линии H9 на 40-ом пассаже поддерживали в среде, кондиционированной MEF, на носителях с поверхностью Nunclon DeltaTM, обработанных препаратом MatrigelTM, с пониженным содержанием факторов роста в разбавлении 1:30. Для пассирования клетки снимали с поверхности путем инкубации в растворе 1 мг/мл коллагеназы или путем соскабливания вручную.
Затем приготовленные таким образом клетки высевали на поверхностно-модифицированные носители 2, 3, 4 и 13 в формате 12-луночных планшетов в присутствии возрастающих концентраций одного из следующих ингибиторов Rho-киназы:Y-27632 (компании Sigma, Сент-Луис, Миссури, США или компании EMD, Сан-Диего, Калифорния, США), Fasudil (Sigma), или Hydroxyfasudil (Sigma), и поддерживали в течение трех дней, ежедневно заменяя среду и ингибитор. К концу третьего дня культуральную среду удаляли и носители окрашивали кристаллическим фиолетовым (0,5% раствор в воде) для визуализации колоний.
Мы обнаружили, что на третий день поверхностно-модифицированные носители 2, 3, 4 и 13 в присутствии возрастающих концентраций ингибитора Rho-киназы имели на своей поверхности закрепленные колонии эмбриональных стволовых клеток. Наилучшие результаты были получены при использовании Y-27632 (10 мкМ), хотя некоторые колонии закреплялись и росли и в присутствии других ингибиторов Rho-киназы, Fasudil и Hydroxyfasudil (смотрите фиг.11).
Мы попытались определить оптимальную дозировку Y-27632 для стимулирования закрепления клеток на поверхности путем обработки клеток варьируемой в некотором диапазоне концентрацией Y-27632 в культуральной среде в течение первого дня после пересева. После первого дня культивирования клетки в последующие дни обрабатывали Y-27632 в концентрации 10 мкМ. По нашим наблюдениям, максимальная концентрация для стимулирования закрепления и роста эмбриональных стволовых клеток составила 10 мкМ (смотрите фиг.12), что имело место на поверхностно-модифицированных носителях 2, 3, 4, 13, а также носителях с поверхностью типа CellBINDTM (компания Corning, Корнинг, Нью-Йорк, США).
Также проверили влияние непрерывной обработки одиночной дозой Y-27632 на закрепление и рост клеток. Клетки получали 0, 1, 4 или 10 мкМ Y-27632 в течение 4 дней. Хотя частичное закрепление клеток наблюдалось и на поверхностно-модифицированных носителях без обработки (0 мкМ), оптимальная концентрация для стимулирования закрепления и роста эмбриональных стволовых клеток составила 10 мкМ Y-27632 (смотрите таблицу 4) на поверхностно-модифицированных носителях 2, 3, 4, 13.
Поскольку введение ингибитора ROCK существенно повысило эффективность посева и ускорило рост клеток на поверхностно-модифицированных носителях 2, 3 и 4 по сравнению с необработанными клетками (смотрите фиг.13), мы решили выяснить, является ли это следствием должного закрепления клеток на поверхности или следствием повышенной пролиферации клеток. Мы обнаружили, что ингибирование Rho-киназы не повышает пролиферацию клеток, поскольку обработанные Y-27632 клетки растут практически с той же плотностью, что и необработанные клетки (смотрите фиг.14). Вместо этого обработка Y-27632 способствует закреплению клеток на поверхности и позволяет им расти с нормальной кинетикой пролиферации (смотрите фиг.15). Удаление ингибитора Rho-киназы из ростовой среды эмбриональных стволовых клеток, пассированных в присутствии ингибитора Rho-киназы, приводит к откреплению клеток с поверхности. При культивировании эмбриональных стволовых клеток в суспензии происходит формирование эмбриоидных телец с дифференцировкой во все три зародышевые линии. По этой причине, хотя повторное введение ингибитора Rho-киназы и восстанавливало закрепление клеток на поверхности (смотрите фиг.15), как и ожидалось, в тех образцах, где в ходе культивирования ингибитор Rho-киназы удаляли на 24 часа и позволяли клеткам открепиться от поверхности, расти в суспензии в течение 24 часов, а затем опять вводили ингибитор Rho-киназы, наблюдалась значительная дифференцировка культуры эмбриональных стволовых клеток.
Пример 6
Эмбриональные стволовые клетки человека линии H9, пассируемые с использованием фермента TrypLE™ Express на поверхностно-модифицированных носителях, демонстрируют повышенную адгезию в присутствии Y-27632
Исходное пассирование эмбриональных стволовых клеток человека линии H9 на поверхностно-модифицированные носители. Адгезия клеток к поверхности повышается при постоянном наличии 10 мкМ Y-27632. Это справедливо для всех четырех испытанных поверхностно-модифицированных носителей: 2, 3, 4 и 13. Фотографии клеток линии H9 через 24 часа после посева на поверхностно-модифицированном носителе 3 приведены на фиг.16.
Пример 7
Одиночные эмбриональные стволовые клетки человека линии H9, пассируемые с использованием фермента TrypLE™ Express на поверхностно-модифицированных носителях, сохраняют плюрипотентность
Эмбриональные стволовые клетки человека плюрипотентны и могут дифференцироваться в клетки всех линий. Данное плюрипотентное состояние клеток должна поддерживать поверхность, на которой они растут. Чтобы определить, способны ли поверхностно-модифицированные носители поддерживать плюрипотентность эмбриональных стволовых клеток человека, последние пассировали 38 раз с использованием коллагеназы и 38 раз с использованием фермента TrypLE™ Express с последующими 5 пассажами на поверхностно-модифицированном носителе 3 (поверхность 3), поверхностно-модифицированном носителе 4 (поверхность 4) или носителе с поверхностью, обработанной препаратом MatrigelTM в разбавлении 1:30. В культуральную среду для указанных образцов добавляли 10 мкМ Y-27632. Уровень экспрессии маркеров плюрипотентности Tra-1-60, Tra-1-81, SSEA-3 и SSEA-4 оценивали способом проточной цитометрии. Полученные результаты приведены на фиг.17. По оси Y отложен процент положительных по маркерам клеток. Одиночные эмбриональные стволовые клетки человека на поверхностно-модифицированных носителях могут сохранять свою плюрипотентность.
Пример 8
Ингибирование Rho-киназы стимулирует адгезию и рост клеток линии эмбриональных стволовых клеток человека H9, выращиваемых как одиночные клетки на поверхностно-модифицированных носителях при переносе с поверхности типа Matrigel TM
Роль Y-27632 в адгезии и росте эмбриональных стволовых клеток человека была изучена в связи с использованием поверхностно-модифицированных носителей. Эмбриональные стволовые клетки человека линии H9 пассировали 38 раз с использованием коллагеназы и 50 раз с использованием фермента TrypLE™ Express с последующим высеванием на поверхностно-модифицированные носители 3 и 4 (“наивные” клетки). Альтернативным образом, эмбриональные стволовые клетки человека линии H9 пассировали 38 раз с использованием коллагеназы и 38 раз с использованием фермента TrypLE™ Express с последующими 9 пассажами на поверхностно-модифицированном носителе 3 (“акклиматизированные” клетки, Поверхность 3) и поверхностно-модифицированном носителе 4 (“акклиматизированные” клетки, Поверхность 4). Клетки высевали с плотностью 104/см2 в среде, кондиционированной MEF, и выращивали два дня в присутствии 10 мкМ Y-27632 или в отсутствие ингибитора. Полученные результаты приведены на фиг.18. Добавление Y-27632 улучшает закрепление “наивных” клеток на поверхностно-модифицированных носителях 3 и 4. Добавление Y-27632 не улучшило закрепление “акклиматизированных” клеток на поверхностно-модифицированных носителях 3 и 4. Поверхностно-модифицированный носитель 3 имел лучшие показатели закрепления и/или роста “наивных” клеток. Поверхностно-модифицированный носитель 4 имел лучшие показатели закрепления и/или роста “акклиматизированных” эмбриональных стволовых клеток человека. За клетками наблюдали в течение 4 дней (смотрите фиг.19 и 20). “Наивные” клетки имели повышенную скорость роста при культивировании в присутствии 10 мкМ Y-27632, причем некоторое преимущество имел поверхностно-модифицированный носитель 3 (фиг.19). “Акклиматизированные” клетки имели повышенную скорость роста при культивировании в отсутствие Y-27632 (смотрите фиг.20).
Пример 9
Поверхностно-модифицированные носители могут использоваться для скрининга соединений
Поверхностно-модифицированные носители в формате 96-луночного планшета и в стандартном формате Общества скрининга биомолекул (Society for Biomolecular Screening, SBS) можно использовать для выращивания одиночных эмбриональных стволовых клеток человека в присутствии 10 мкМ Y-27632. Фотографии одиночных клеток линии H9 на 96-луночном планшете приведены на фиг.21. Благодаря этому скрининг соединений можно проводить непосредственно на 96-луночном планшете без нанесения дополнительных адгезивных слоев или питающих клеток, таких как MatrigelTM и мышиные эмбриональные фибробласты, соответственно.
Пример 10
Культивируемые на поверхностно-модифицированных носителях одиночные эмбриональные стволовые клетки способны дифференцироваться в клетки дефинитивной эндодермы
Одна из целей заключается в дифференцировке эмбриональных стволовых клеток человека в клетки различных линий. Чтобы определить, способны ли поверхностно-модифицированные носители поддерживать дифференцировку клеток, эмбриональные стволовые клетки человека пассировали 38 раз с использованием коллагеназы и 38 раз с использованием фермента TrypLE™ Express с последующими 9 пассажами на поверхностно-модифицированном носителе 3 (поверхность 3) и поверхностно-модифицированном носителе 4 (поверхность 4). В качестве положительного контроля эмбриональные стволовые клетки человека растили на носителе, обработанном препаратом MatrigelTM в разбавлении 1:30. В процессе роста указанных образцов клеток в питающую среду вводили 10 мкМ Y-27632. После выращивания проверяли способность культивированных клеток к образованию дефинитивной эндодермы. Коротко говоря, 70% конфлюэнтных культур в течение двух дней обрабатывали смесью, содержащей 100 нг/мл активина A, 10 нг/мл Wnt3a и 0,5% FBS в среде DMEM-F12. Затем клетки в течение трех дней обрабатывали смесью, содержащей 100 нг/мл активина A и 2% FBS в среде DMEM/F12. Дифференцированные в дефинитивную эндодерму клетки определяли по уровню экспрессии белка CXCR4 на проточном цитометре (смотрите фиг.22). Процент положительных по данному маркеру клеток отложен по оси Y. Эмбриональные стволовые клетки человека, культивируемые как одиночные клетки, способны дифференцироваться в клетки дефинитивной эндодермы в присутствии или в отсутствие Y-27632 на поверхностно-модифицированных носителях 3 и 4.
Пример 11
Культивируемые на поверхностно-модифицированных носителях одиночные эмбриональные стволовые клетки способны дифференцироваться в клетки панкреатической эндодермы
После выполнения протокола дифференцировки в клетки дефинитивной эндодермы клетки в течение трех дней инкубировали в смеси, содержащей FGF-7 (50 нг/мл; R&D Systems), ингибитор белка “ежик Соник” (sHh), KAAD циклопамина (2,5 мкМ; Sigma-Aldrich) и 2% FBS в среде DMEM-F12. На этой стадии клетки, не получавшие Y-27632 в процессе роста, откреплялись с поверхности поверхностно-модифицированных носителей 3 и 4. Клетки, которые в процессе роста обрабатывали Y-27632, далее в течение четырех дней инкубировали в смеси, содержащей FGF-7 (50 нг/мл), KAAD циклопамин (2,5 мкМ), ретиноевую кислоту (1 мкМ; Sigma-Aldrich) и 1% B27 (Invitrogen) в среде DMEM-F12 (стадия поздней передней кишки, PF). Затем в течение четырех дней клетки инкубировали в смеси, содержащей Exendin 4 (50 нг/мл; Sigma-Aldrich), DAPT (1 мкМ; Calbiochem) и 1% B27 в среде DMEM-F12. Дифференцировка при этом перешла в стадию панкреатической эндодермы (EN). Затем следовала трехдневная обработка в среде CMRL (Invitrogen), содержащей 50 нг/мл HGF, IGF (R&D Systems), Exendin 4 (50 нг/mL) и 1% B27. На стадиях PF и EN из одной из лунок поверхностно-модифицированных носителей 3 и 4 отбирали образцы РНК и анализировали отобранные на этих стадиях образцы способом PCR в режиме реального времени на уровни следующих панкреатических маркеров: Pdx1, Nkx6.1, Nkx2.2, Pax4, NeuroD, HNF3b, Ptf1a, инсулин и AFP. Анализ на уровни перечисленных панкреатических маркеров повторили на данной стадии. Параллельно с обработанными образцами также проводили PCR в режиме реального времени на РНК образцов необработанных эмбриональных стволовых клеток той же линии. Обработанные образцы нормировали по наборам необработанных контролей до кратности изменения равной единице. Велось отслеживание и сравнение уровней экспрессии Pdx1 и инсулина для разных поверхностно-модифицированных носителей.
В результате было обнаружено индуцирование маркеров панкреатической эндодермы в клетках, обработанных на поверхностно-модифицированных носителях 3 и 4, хотя уровень экспрессии был несколько выше у клеток, обработанных на поверхностно-модифицированном носителе 3 (смотрите фиг.23). Оба поверхностно-модифицированных носителя в присутствии Y-27632 в процессе роста способны поддерживать дифференцировку одиночных эмбриональных стволовых клеток человека в клетки поздней передней кишки и панкреатической эндодермы. Одиночные клетки, не обрабатывавшиеся Y-27632 в процессе роста, откреплялись от поверхности до начала дифференцировки в клетки поздней передней кишки.
Пример 12
Эмбриональные стволовые клетки человека линий H1 и H9 закрепляются на поверхностно-модифицированных носителях, причем адгезия улучшается при обработке клеток Y-27632
Эмбриональные стволовые клетки человека линии H9 49-го пассажа, предварительно находившиеся на пластиковом носителе с поверхностью, обработанной препаратом MatrigelTM в разбавлении 1:30, и культивируемые в среде, кондиционированной MEF с добавлением 8 нг/мл bFGF, обработали ферментативной смесью LIBERASE и перенесли на поверхностно-модифицированные носители в среду, кондиционированную MEF с добавлением 8 нг/мл bFGF, без какой-либо дополнительной обработки или введения дополнительных реагентов при возрастающих концентрациях Y-27632. Мы обнаружили, что через 24 и 48 часов после посева эмбриональных стволовых клеток человека линии H9 на поверхностно-модифицированные носители после окраски кристаллическим фиолетовым можно было наблюдать небольшие колонии на поверхности следующих носителей: поверхности 2-4 и 13, CellBINDTM и PrimariaTM (кат. № 353846, компания Becton Dickinson, Франкоин Лэйкс, Нью-Джерси, США) (смотрите фиг.24-26). Более того, степень адгезии колоний эмбриональных стволовых клеток человека линии H9 повышалась при введении в среду Y-27632, и эффективность ингибитора зависела от его концентрации (смотрите фиг.25). При низких концентрациях Y-27632 (от 1 до 2 микромоль на литр) наблюдалось минимальное повышение эффективности закрепления эмбриональных стволовых клеток на поверхности носителя по сравнению с необработанными клетками (смотрите рисунок 25), тогда как более высокие концентрации Y-27632 (от 4 до 20 микромоль на литр) стимулировали закрепление клеток на поверхностно-модифицированных носителях, как показывает окраска кристаллическим фиолетовым (смотрите фиг.25 и 26).
В дополнение к динамической регуляции эффективности закрепления эмбриональных стволовых клеток человека при добавлении Y-27632 в культуральную среду, мы также обнаружили разные степени адгезии клеток к различным поверхностно-модифицированным носителям в присутствии Y-27632. Например, клетки показали меньшую степень адгезии к носителям с поверхностью типа CellBINDTM и большую вероятность открепления от поверхности типа CellBINDTM с течением времени даже при наличии постоянно присутствующего Y-27632, тогда как клетки более эффективно закреплялись на поверхностно-модифицированных носителях 3, 4 или 13, а также поверхности типа PrimariaTM с меньшей вероятностью открепления при обработке ингибитором Rho-киназы Y-27632 (смотрите фиг.25 и 26).
Пример 13
Эмбриональные стволовые клетки человека линий H1 и H9 закрепляются и образуют колонии с разными скоростями на поверхностно-модифицированных носителях в присутствии Y-27632
Эмбриональные стволовые клетки человека линии H1 и H9, предварительно находившиеся на пластиковом носителе с поверхностью, обработанной препаратом MatrigelTM в разбавлении 1:30, и культивируемые в среде, кондиционированной MEF с добавлением 8 нг/мл bFGF, обработали ферментативной смесью LIBERASE и перенесли на поверхностно-модифицированные носители в среду, кондиционированную MEF с добавлением 8 нг/мл bFGF, без какой-либо дополнительной обработки или с введением 20 мкМ Y-27632. Через сорок восемь часов после посева эмбриональных стволовых клеток человека линии H9 на поверхностно-модифицированные носители 14 и 15 можно было наблюдать небольшие колонии в образцах со средой, в которую добавили 20 мкМ Y-27632 (степень закрепления и эффективность образования колоний изменялись от эксперимента к эксперименту) (смотрите фиг.27). Эмбриональные стволовые клетки человека линии H1 также закреплялись и образовывали колонии на обоих поверхностно-модифицированных носителях 14 и 15 при введении в среду 20 мкМ Y-27632, причем это было выражено в большей степени по сравнению со связыванием у клеток линии H9. Полученные данные указывают, что степень закрепления и эффективность образования колоний эмбриональных стволовых клеток человека на поверхности твердых подложек меняются от линии к линии.
Пример 14
Закрепление эмбриональных стволовых клеток человека на поверхностно-модифицированных носителях с помощью среды определенной композиции
Эмбриональные стволовые клетки человека линии Н9 49-го пассажа дважды пассировали в среду определенной композиции, mTeSRTM, на пластиковом носителе, обработанном MatrigelTM. Затем клетки обрабатывали ферментативной смесью LIBERASE и высевали на поверхностно-модифицированный носитель Nunc4 в среде mTeSRTM. Клетки высевали в среду с добавлением или без добавления 20 мкМ Y-27632. Лунки перед посевом клеток также обрабатывали различными белками в течение 30 минут (без обработки, 0,1% желатина, 2% БСА, 0,34 мг/мл крысиного коллагена I, 1:1000 MatrigelTM или 1:5000 MatrigelTM), чтобы определить, могут ли эти белки способствовать адгезии эмбриональных стволовых клеток человека в среде определенной композиции с Y-27632 или без него (фиг.28). По нашим наблюдениям, в отсутствие Y-27632 эмбриональные стволовые клетки человека, посеянные на поверхностно-модифицированном носителе в среде определенной композиции, не закреплялись даже в присутствии белков внеклеточной матрицы, таких как коллаген I или 1:1000 MatrigelTM. Тем не менее, при добавлении в среду определенной композиции mTeSRTM 20 мкМ Y-27632 эмбриональные стволовые клетки человека закреплялись на поверхности Nunc4. Кроме того, их закрепление было одинаковым и в необработанных лунках, и в лунках, обработанных 0,1% желатина, 2% БСА и 0,34 мг/мл крысиного коллагена I. Незначительное повышение прикрепляемости эмбриональных стволовых клеток человека было отмечено в лунках с низкой концентрацией MatrigelTM (в разбавлении 1:1000 и 1:5000), однако эти концентрации MatrigelTM были недостаточны для улучшения адгезии в присутствии Y-27632. Эти результаты показывают, что в присутствии ингибитора ROCK, Y-27632 эмбриональные стволовые клетки человека можно культивировать и на модифицированных пластиковых подложках в среде определенной композиции, и что низкие концентрации MatrigelTM, в соотношении примерно 1:1000 или 1:5000, могут улучшать адгезию.
Пример 15
Поверхностно-модифицированные носители в форме колбы могут способствовать закреплению эмбриональных стволовых клеток человека и дифференцировке в дефинитивную эндодерму и панкреатическую эндодерму
Эмбриональные стволовые клетки человека линий H1 и H9, ранее высеянные на пластиковом носителе, обработанном 1:30 MatrigelTM и выращенные в среде, кондиционированной MEF с добавлением 8 нг/мл bFGF были обработаны ферментативной смесью LIBERASE и посеяны в колбы T25, T75, T150 и T175 с плотностью 1:2 или 1:3; при этом колбы были различного размера и имели модифицированную поверхность. Клетки посеяли в среде, кондиционированной MEF с добавлением 8 нг/мл bFGF и 20 мкМ Y-27632. Колонии эмбриональных стволовых клеток человека затем выращивали с ежедневной заменой среды, кондиционированной MEF с добавлением 8 нг/мл bFGF и 20 мкМ Y-27632, пока носители не заполнились примерно на 50%. На данном этапе среду заменили на среду DMEM/F12, содержащую 2% БСА, 100 нг/мл активина A, 20 нг/мл Wnt3a и 20 мкМ Y-27632 и поддерживали клетки в этой среде в течение 2 дней с ежедневной заменой среды. На третий и четвертый день среду меняли на среду DMEM/F12, содержащую 2% БСА, 100 нг/мл активина-A и 20 мкМ Y-27632. Затем клетки снимали с поверхности с помощью TrypLE и проводили анализ способом проточной цитометрии на экспрессию маркера поверхности дефинитивной эндодермы (DE), CXCR4. По нашим наблюдениям, при этих условиях эмбриональные стволовые клетки человека дифференцировались в популяцию, высоко положительную по CXCR4, достигавшем почти 90% CXCR4+; это говорит о том, что клетки в основном дифференцировались в дефинитивную эндодерму (таблица 5). Кроме того, закрепление клеток на поверхности культуры в процессе роста или в процессе дифференцировки зависело от поддержания ингибирования ROCK, поскольку удаление Y-27632 из культуральной среды приводило к откреплению клеток от пластика.
Мы хотели определить, может ли панкреатическая эндодерма формироваться из дефинитивной эндодермы, получаемой на поверхностно-модифицированных носителях в форме колбы. Для этого мы инкубировали клетки в течение дополнительных четырех дней с Y-27632 (20 мкМ), FGF-7 (50 нг/мл), циклопамином KAAD (2,5 мкМ), и 1% B27 (Invitrogen) в среде DMEM-F12, а затем еще четыре дня в этой среде, дополненной ретиноевой кислотой (1 мкМ; Sigma-Aldrich) для дифференцировки клеток до стадии панкреатической эндодермы. Затем были взяты образцы РНК и выполнен их анализ способом PCR в режиме реального времени на уровень панкреатический маркера Pdx1. Обработанные образцы нормировали по набору необработанных контролей до кратности изменения равной единице. По нашим наблюдениям, в образцах повысился уровень PDX1 по сравнению с недифференцированными эмбриональными стволовыми клетками человека, при этом уровень мРНК был по меньшей мере в 256 раз выше в дифференцированных клетках, чем в недифференцированных эмбриональных стволовых клетках человека.
Пример 16
Обработка поверхности и поверхностно-модифицированные носители
Поверхностно-модифицированные носители приготавливали способом плазменной обработки коронным разрядом или микроволновой плазменной обработки изделий, полученных литьем под давлением (таблица 6). Для изготовления изделий способом литься под давлением применяли такие полимеры, как полистирол, поликарбонат, смесь поликарбоната и полистирола и сополимер циклоолефина. Поверхностно-модифицированные носители упаковывали в отдельные полиэтиленовые пакеты, затем стерилизовали гамма-излучением (25 кГр) и, наконец, хранили при комнатной температуре до использования при культивировании клеток или в экспериментах по определению характеристик поверхности. Поверхностно-модифицированные носители 18, 30 и 31-32 были изготовлены способом литья из тех же полимеров, что и поверхностно-модифицированные носители 19, 33 и 34, соответственно, но не проходили обработку плазмой. Поверхности 14 и 31 не облучались гамма-излучением.
Плазменная обработка коронным разрядом проводилась в металлической вакуумной камере, внутри которой находился только один электрод, электрически изолированный от внутреннего объема камеры (C-Lab Plasma; Vetaphone A/S, Дания). Металлические стенки служили вторым электродом (заземлением). Самонастраивающийся генератор коронного разряда создавал электрическое поле, дававшее достаточно энергии для возникновения плазмы во всем объеме камеры. Обрабатываемое изделие помещали на дно камеры. Камеру закрывали и вакуумировали до давления 1 Па (10-2 мбар). При этом давлении клапан вакуумного насоса закрывали и включали генератор коронного разряда. Генератор был установлен в режим выходной мощности 2000 Вт. Через плазму пропускали ток в течение 5-60 секунд. Затем открывали клапан впуска газа (воздуха), и давление в камере возвращалось к атмосферному значению.
Обработку микроволновой плазмой выполняли в кварцевых вакуумных камерах (модель 300-E для поверхностно-модифицированных носителей 5-12 и модель 440 для поверхностно-модифицированных носителей 14 и 15; обе - производства Technics Plasma GmbH, Германия). Энергия для генерирования плазмы поступала от 2,43 ГГц микроволнового генератора, расположенного вне камеры. Обрабатываемое изделие помещали на стеклянный поддон внутри камеры. Камеру закрывали и вакуумировали до давления от 30 до 50 Па (от 0,3 до 0,5 мбар). Клапан вакуумного насоса оставался открытым, и давление поддерживали на нужном уровне, регулируя расход газа (воздуха или кислорода) клапаном впуска газа. Затем включали микроволновой генератор. Генератор был установлен в режим мощности от 500 до 600 Вт. Затем клапан насоса закрывали и открывали клапан впуска воздуха, чтобы давление в камере вернулось к атмосферному значению.
В таблице 6 показаны мощность, время, давление и газы, используемые при обработке поверхностно-модифицированных носителей плазмой коронного разряда и микроволновой плазмой.
Пример 17
Определение характеристик поверхностно-модифицированных носителей, составляющих предмет настоящего изобретения
Углы смачивания водой
Поверхностно-модифицированные носители 1-4 и 13 упаковывали в отдельные полиэтиленовые пакеты, стерилизовали и хранили при комнатной температуре на протяжении всего периода проведения анализов. Углы смачивания сначала измеряли через неделю после обработки поверхности и стерилизации, а затем повторно в моменты времени, указанные на фиг.29. Все измерения углов смачивания выполняли статическим способом неподвижной капли и с помощью измерительной головки PG-X производства компании FIBRO Systems AB, Швеция [гониометр, состоящий из видеокамеры и программного обеспечения (v. 3.1)]. Углы смачивания определяли по тангенсу угла наклона. Капли воды MilliQ в количестве 4,0 мкл наносили автоматической пипеткой в статическом режиме согласно инструкциям производителя. Угол смачивания каждой капли измеряли один раз (для каждого образца на каждый момент времени наносилось 7 капель). Для каждого момента времени использовали новый образец, чтобы не допустить влияния предыдущих измерений. Измерения на поверхностях Nunclon DeltaTM и CellBINDTM проводили при тех же экспериментальных условиях, что и измерения на Поверхностях 1-4 и 13, но обработку поверхности и стерилизацию производили более чем за 12 недель до первого измерения (Nunclon DeltaTM* стерилизовали за одну неделю до первого измерения). На фиг.29 показано, что поверхностно-модифицированные носители 1-4 и 13 обладали схожей гидрофильностью и были более гидрофильными (имели меньшие углы смачивания водой), чем поверхности Nunclon DeltaTM и CellBINDTM. Гидрофильность поверхностно-модифицированных носителей 1-4 и 13 была стабильной по меньшей мере 12 недель после обработки и стерилизации поверхности.
CellBINDTM ранее описывалась как имеющая угол смачивания 13,4 градуса (стандартное отклонение 4 градуса) [Технический отчет Corning (2005), Поверхность Corning® CellBIND® Усовершенствованная поверхность для улучшения закрепления клеток (CLS-AN-057 REV1) на http://catalog2.corning.com/Lifesciences/media/pdf/t_CellBIND_Improved_Surface_CLS_AN_057.pdf].
Плотность отрицательных зарядов
Плотность отрицательных зарядов определяли на поверхностно-модифицированных носителях 1-4 и 13, поверхности Nunclon DeltaTM, поверхности CellBINDTM, поверхности PrimariaTM, поверхности FalconTM и необработанной (но стерилизованной) полистирольной поверхности (все в формате 3-см чашки). 3 мл водного раствора кристаллического фиолетового (0,015% в/о) добавляли на каждую чашку и инкубировали чашки при мягком потряхивании (50 об./мин) в течение 60 минут при комнатной температуре. Для удаления кристаллического фиолетового, не связанного с поверхностями, чашки трижды промыли 3 мл воды MilliQ и сушили в течение ночи при 60°C. Кристаллический фиолетовый, связанный с поверхностью, десорбировали добавлением 1,5 мл раствора 0,1 M HCl в EtOH (99%) и инкубировали чашки при плавном потряхивании (50 об./мин.) в течение 2 минут при комнатной температуре. Поглощение раствора HCl:EtOH с десорбированным кристаллическим фиолетовым измеряли при 590 нм с помощью считывателя микропланшетов EnVision 2100 (Perkin Elmer; Уолтэм, Массачусетс, США). Были внесены поправки в значения поглощения на фоновое поглощение раствора HCl:EtOH. Плотность отрицательных зарядов измеряли на трех чашках для каждой поверхности, проводя по три измерения поглощения для каждой чашки.
Плотность отрицательного заряда для поверхностно-модифицированных носителей показана на фиг.30. Плотность отрицательных зарядов поверхностей 1-4 и 13 была схожей, но более длительная обработка поверхности в интервале 5-60 секунд могла приводить к более низким значениям плотности отрицательных зарядов поверхности. Поверхности 1-4 и 13 имели значительно более низкую плотность отрицательных зарядов, чем поверхность CellBINDTM и поверхность Nunclon DeltaTM обработанные в 2007 году. Поверхности 1-4 и 13 имели плотности отрицательных зарядов того же уровня, что и поверхность Nunclon DeltaTM, обработанная в 2005 году, и значительно более высокую плотность отрицательных зарядов, чем поверхность PrimariaTM, поверхность FalconTM и необработанная (но стерилизованная) полистирольная поверхность. Более низкие значения плотности отрицательных зарядов поверхности Nunclon DeltaTM, обработанной в 2005 году, чем у поверхности Nunclon DeltaTM, обработанной в 2007 году, указывают на то, что полистирол с обработанной поверхностью становится со временем немного менее отрицательно заряженным. Высокая плотность отрицательных зарядов поверхности CellBINDTM не связана с большей грубостью поверхности и ее большей площадью (смотрите анализ AFM в данном примере).
Рентгеновская фотоэлектронная спектроскопия (XPS)
Был проведен анализ поверхностно-модифицированных носителей 1-4 и 13-15 и носителей с поверхностями Nunclon DeltaTM, CostarTM, FalconTM, CellBINDTM и PrimariaTM способом XPS. Образцы в виде вырезанных из носителя секций подвергали рентгеновскому излучению; при этом секции фиксировали пружинными зажимами на держателе образца из нержавеющей стали. Образцы облучали Al kα излучением (1486 эВ). Угол между образцом и анализатором при проведении анализа составлял 45°. Спектры изучали способом подгонки с помощью программного обеспечения, предоставленного производителем прибора - компанией Physical Electronics. Для обработки данных использовали программное обеспечение с приложением MatlabTM. Для анализа использовали рентгеновский фотоэлектронный спектрометр Physical Electronics, модель 5400 Х. Анализировали от двух до пяти самых внешних нанометров в глубину в области диаметром примерно один миллиметр на обработанной поверхности носителя для каждого из двух планшетов с данной поверхностью.
Элементный состав поверхности в атомных процентах показан в таблице 7. Все поверхностно-модифицированные носители содержат углерод, кислород и азот (при XPS водород не определяется) на поверхности. Поверхности 1-4, поверхность 13 и поверхность CellBINDTM содержали больше кислорода, чем другие проанализированные поверхности. Поверхности 1-4 и поверхности 13-15 содержали меньше азота, чем поверхность PrimariaTM, о больше азота, чем поверхности носителей Nunclon DeltaTM, CostarTM, FalconTM, и CellBINDTM. Уровни кислорода и азота положительно коррелировали с более длительным временем обработки поверхности (поверхности 1-4 и 13), а самые высокие уровни обоих этих элементов достигались при 30 или 60-секундной обработке плазмой коронного разряда (поверхность 3 и поверхность 4, соответственно). Поверхности 3 и 4 имели схожий элементный состав. Поверхности 2 и 13 имели схожий элементный состав и по своему элементному составу были ближе к поверхностям 3 и 4, чем к поверхности 1.
Пики спектров C1s исследовали способом подгонки (способом наилучших хи-квадратов) для определения и количественной оценки сред связывания углерода в поверхностях по ширине пиков и энергетическим положениям элементов согласно литературным данным (таблица 8). Концентрации указаны в атомных процентах, которые получали перемножением площади в процентах и атомной концентрации. Поверхности 2-4 и 13 были схожи с точки зрения углерод-связывающих сред. Доля углерода в связывающей среде C*-C-O-C-C* была ниже в поверхностях 2-4 и 13, чем в других проанализированных поверхностях. Доля углерода в связывающей среде O-[C=O]-O была выше в поверхностях 2-4 и 13, чем в других проанализированных поверхностях. Также была обнаружена идентичность поверхностей 2-4 и 13 и поверхности 1, поверхности CellBINDTM, и/или поверхности PrimariaTM. Доля углерода в связывающей среде C-O-C или в связывающей среде C-NH3+ (та же точка энергия в спектре) была выше в поверхностях 1-4 и 13, чем в других проанализированных поверхностях. Доля углерода в связывающей среде C-O-C*=O была выше в поверхностях 2-4, поверхности 13, и поверхности PrimariaTM, чем в других проанализированных поверхностях. Доля углерода в связывающей среде CO3 - была выше в поверхностях 2-4, поверхности 13 и поверхности CellBINDTM, чем в других проанализированных поверхностях. Доля углерода в связывающей среде C=O была выше в поверхностях 1-4, поверхности 13 и поверхности CellBINDTM, чем в других проанализированных поверхностях. Доля углерода в связывающей среде C-[O]-C была выше в поверхностях 1-4, поверхности 13, поверхности CellBINDTM и поверхности PrimariaTM, чем в других проанализированных поверхностях. Пик потери энергии связан с ароматическим переходом Π→Π* и является индикатором ароматичности поверхности.
Пики спектров O1s были почти гауссовой формы, и их подгонка была невозможна. Пики спектров N1s были исследованы способом подгонки (способом наилучших хи-квадратов) для определения и количественной оценки сред связывания азота в поверхностях по ширине пиков и энергетическим положениям элементов согласно литературным данным (таблица 9). Концентрации указаны в атомных процентах, которые получали перемножением площади в процентах и атомной концентрации. Сигналы N1s поверхностей Nunclon DeltaTM, CellBINDTM, CostarTM, и FalconTM были слабыми, поэтому было невозможно определить среды связывания азота для этих поверхностей. Спектры N1s для поверхностно-модифицированных носителей 1-4 и 13 были неразличимыми, поэтому показаны данные подгонки двух репрезентативных спектров. Доля азота в связывающей среде -NH3 + была выше в поверхностях 1-4 и 13, чем в поверхностях 14 и 15 и поверхности PrimariaTM. Азот в связывающей среде -NH2 был обнаружен только в поверхностях 14 и 15 и поверхности PrimariaTM. Азот в связывающей среде -NO2 был обнаружен только в поверхностях 1-4 и 13 и в одном образце поверхности 15. Азот в связывающей среде -NO3 был обнаружен только в поверхности 15 и поверхности PrimariaTM.
CellBINDTM ранее была описана как поверхность, имеющая в элементном составе 70,4% углерода, 29,0% кислорода, 0,6% азота и <0,01% других элементов и относительно высокую концентрацию групп C-[O]-C, C=O и COOH/R по данным анализа ESCA [технический отчет Corning (2005), Поверхность Corning® CellBIND®: Усовершенствованная поверхность для улучшенного закрепления клеток (Improved Surface for Enhanced Cell Attachment) (CLS-AN-057 REV1) на http://catalog2.corning.com/Lifesciences/media/pdf/t_CellBIND_Improved_Surface_CLS_AN_057.pdf].
PrimariaTM ранее была описана как поверхность, имеющая в элементном составе 74,6% углерода, 14,1% кислорода, 11,1% азота и 0,2% других элементов, преимущественно с нитрильными (C≡N) и мочевинными [HN(C=O)NH] средами связывания углерода и азота по данным анализа ESCA.
Атомно-силовая микроскопия (АСМ)
Был проведен анализ поверхностно-модифицированных носителей 1-4 и 13 и носителей с поверхностями Nunclon DeltaTM and CellBINDTM способом AFM. Образцы анализировали с помощью многорежимного атомно-силового микроскопа Digital Instruments в полуконтактном режиме. Использовался наконечник для полуконтактного режима типа TESP7. Образцы крепили к дискам для образцов двойной клейкой лентой. Анализировались зоны 10 мкМ × 10 мкМ и 500 нм × 500 нм на обработанной части поверхности носителей. Средняя шероховатость поверхности (Ra) и максимальная высота (Rmax) в нанометрах показаны в таблице 10. Как и носители с поверхностями Nunclon DeltaTM и CellBINDTM, поверхностно-модифицированные носители 1-4 и 13 были относительно гладкими, а Ra и Rmax не коррелировали со временем обработки поверхности ни в одной из двух сканограмм. Анализ поверхностей из необработанного полистирола и окисленного полистирола, предназначенных для культивирования клеток, и поверхности PrimariaTM были описаны в работе Shen and Horbett (J. Biomed. Mater. Res. 57:336-345, 2001): шероховатость поверхности примерно 4 нм для всех трех поверхностей.
Пример 18
Элементный состав поверхности и угол смачивания с точки зрения закрепления эмбриональных стволовых клеток человека и образования колоний
Сводные результаты XPS анализа элементного состава поверхности, измерения угла смачивания поверхности и экспериментов по закреплению эмбриональных стволовых клеток человека и образованию колоний показана в таблице 11.
Закрепление эмбриональных стволовых клеток человека и образование колоний (не менее 15 колоний на 10 см2 поверхности) на поверхности твердой подложки в отсутствие соединения, способного ингибировать Rho или Rho-киназу, наблюдались только на поверхностно-модифицированных носителях 2-4 и 13, носителях CellBINDTM и носителях PrimariaTM (клетки наносили на поверхность в виде суспензии кластеров клеток). Поверхностно-модифицированные носители 2-4 и 13 поддерживали закрепление клеток, образование колоний и пассирование. После примерно трех пассирований скорость роста эмбриональных стволовых клеток человека на поверхностно-модифицированных носителях 2-4 и 13 внезапно упала (только в отсутствие ингибирования Rho и ингибирования Rho-киназы), хотя морфология клеток указывала на то, что клетки не дифференцировались. Кроме того, экспрессия маркера плюрипотентности сохранялась в клетках, которые пассировали четыре раза на поверхности 3. Носители CellBINDTM поддерживали закрепление эмбриональных стволовых клеток человека и образование колоний, но перед первым пассированием наблюдалась дифференциация клеток. Наблюдения за морфологией клеток показали, что носители PrimariaTM поддерживают закрепление эмбриональных стволовых клеток человека и образование колоний без признаков дифференциации (пассирование не тестировалось). Содержание как кислорода (например, поверхность 2 по сравнению с поверхностью 14), так и азота (например, поверхность PrimariaTM по сравнению с поверхностью CostarTM; и поверхности 2 и 13 по сравнению с поверхностью CellBINDTM) в поверхностях влияло на способность поверхностей поддерживать закрепление эмбриональных стволовых клеток человека и образование колоний в отсутствие ингибирования Rho и ингибирования Rho-киназы. Поверхности с содержанием азота не менее чем примерно 0,9%, суммарным содержанием азота и кислорода не менее чем примерно 22,3% и углом смачивания водой не менее чем примерно 13,9 градусов поддерживали закрепление ЭСК человека и образование колоний в отсутствие ингибирования Rho и ингибирования Rho-киназы.
Закрепление эмбриональных стволовых клеток человека и образование колоний (не менее 15 колоний на 10 см2 поверхности) на поверхности твердой подложки в присутствии соединения, способного ингибировать Rho или Rho-киназу, наблюдались на поверхностно-модифицированных носителях 1-15, поверхностно-модифицированном носителе 19, поверхностно-модифицированном носителе 33, поверхностно-модифицированном носителе 34, носителях CellBINDTM и PrimariaTM (клетки наносили на поверхности в виде суспензии кластеров клеток). Мы заметили, что поверхности 2-4, 13 и PrimariaTM лучше содействовали закреплению эмбриональных стволовых клеток человека и образованию колоний, чем поверхности 1, 19, 33, 34 и CellBINDTM, которые, в свою очередь, были лучше поверхностей 5-12, 14 и 15. На поверхностно-модифицированных носителях 3 и 4 и в присутствии ингибитора Rho-киназы эмбриональные стволовые клетки человека прикреплялись и образовывали колонии, которые разрастались и пассирование которых можно было произвести не менее 10 раз, давая начало плюрипотентным клеткам с нормальным кариотипом (кариотип проверялся только у клеток, выращенных на поверхности 4). Содержание как кислорода (например, CellBINDTM по сравнению с Nunclon DeltaTM), так и азота (например, PrimariaTM по сравнению с CostarTM; и поверхности 2 и 13 по сравнению с CellBINDTM) в поверхностях влияло на способность поверхностей поддерживать закрепление эмбриональных стволовых клеток человека и образование колоний в присутствии ингибитора Rho-киназы. Поверхности с содержанием азота по меньшей мере примерно 0,5%, суммарным содержанием азота и кислорода по меньшей мере примерно 17,2% и углом смачивания водой по меньшей мере примерно 13,9 градусов поддерживали прикрепление эмбриональных стволовых клеток человека и образование колоний в присутствии ингибитора Rho-киназы. Поверхности с содержанием азота по меньшей мере примерно 0,5%, суммарным содержанием азота и кислорода по меньшей мере примерно 17,3%, но менее 19,9%, и углом смачивания с водой по меньшей мере примерно 9,4 градусов в некоторых случаях (поверхность 14) поддерживали прикрепление эмбриональных стволовых клеток человека и образование колоний в присутствии Rho-киназы, а в некоторых случаях (поверхности 22-24) - не поддерживали.
Мы заметили, что удаление ингибитора Rho-киназы из культур эмбриональных стволовых клеток человека, выращенных на поверхностно-модифицированном носителе 4, привело к откреплению эмбриональных стволовых клеток человека от поверхности твердой подложки. После этого клетки можно было снова закрепить на поверхности повторной обработкой ингибитором Rho-киназы. Поскольку ферментативное пассирование эмбриональных стволовых клеток человека является потенциальным стрессором и может приводить к кариотипической нестабильности, благодаря временному удалению ингибитора Rho-киназы для пассирования эмбриональных стволовых клеток человека можно устранить стрессы ферментативного пассирования.
Закрепление эмбриональных стволовых клеток человека и образование колоний также были продемонстрированы на поверхностно-модифицированном носителе в среде без животных компонентов, с ингибированием Rho-киназы. Предварительная обработка поверхностно-модифицированного носителя 4 белками внеклеточного матрикса привела к росту числа колоний, но только в присутствии ингибитора Rho-киназы.
В дополнение к пассированию эмбриональных стволовых клеток человека ферментативными способами, сохраняющими характерные для колонии условия культивирования за счет пассирования клеток как кластеров, эмбриональные стволовые клетки человека можно было также пассировать как одиночные клетки с помощью таких ферментов, как TrypLE™ или Accutase™. Как в присутствии, так и в отсутствие ингибитора Rho-киназы колонии эмбриональных стволовых клеток человека диссоциировали в суспензию отдельных клеток с помощью TrypLE™, прикрепленного к поверхностно-модифицированным носителям 3 и 4, и формировали колонии, которые можно было пассировать не менее пяти раз и получать клетки с маркерами плюрипотентности.
Удаление ингибитора Rho-киназы из культур эмбриональных стволовых клеток человека, полученных пассированием клеток как суспензии одиночных клеток, не приводило к откреплению эмбриональных стволовых клеток человека от поверхности твердой подложки, а приводило к образованию колоний, которые росли быстрее, чем когда ингибитор Rho-киназы не удалялся.
Пример 19
Обработка Y-27632 для улучшения закрепления клеток HEK293 на поверхностно-модифицированных носителях
Клетки мезонефроса человека 293 (HEK293, ECACC № 85120602) поддерживали в минимальной эссенциальной среде Игла (EMEM; Lonza, Вервьер, Бельгия), с добавлением 10% эмбриональной бычьей сыворотки (FBS; Lonza). Клетки адаптировали к среде Pro293a-CDM (Lonza) - среде с определенным составом без сыворотки, оптимизированной для культивирования адгезивных HEK293 посредством постепенного, в несколько пассажей, применения среды ЕМЕМ с добавлением сыворотки и среды Pro293a-CDM в последовательности соотношений 3:1, 1:1, 1:3, 1:7 и, наконец, 0:1. Для поддержания и адаптации клетки HEK293 высевали в количестве 2,0×104 клеток/см2 в колбе объемом 75 см2 с поверхностью Nunclon DeltaTM (Thermo Fisher Scientific, Роскильд, Дания) и пассировали при уровне конфлюэнтности 70-80% с помощью трипсина/ЭДТК для диссоциации.
Среду Pro293a-CDM (100 мкл) с добавлением Y-27632 (Sigma Chemical Co., Сент-Луис, Миннесота) в концентрации 1,0, 4,0 или 10 мкМ помещали в 96-луночные планшеты с плоским дном с поверхностью 4, поверхностью Nunclon DeltaTM или поверхностью CellBINDTM. В лунки добавляли еще 100 мкл среды Pro293a-CDM с клетками HEK293 (4,0×104 клеток/см2). Затем культуры инкубировали при температуре 37°C в увлажненной атмосфере 5% CO2 в воздухе в течение: (i) 96 часов; или (ii) 48 часов с последующим однократным промыванием культур 200 мкл физиологического раствора, забуференного фосфатом Дульбекко (DPBS; Lonza), а затем добавляли 200 мкл среды Pro293a-CDM без Y-27632 и, наконец, инкубировали культуры еще 48 часов.
Затем определяли количество жизнеспособных клеток в лунках с помощью набора для определения активности лактатдегидрогеназы (ЛДГ) производства компании Roche, Швейцария. Коротко говоря, лунки промывали средой Pro293a-CDM, и адгезивные клетки лизировали в 100 мкл DPBS с 2% (об./об.) Triton X-100 (Sigma Chemical Co.) в течение 30-минутной инкубации при температуре 37°C. Лизат и 100-мкл смеси катализатора и реактива смешивали и инкубировали в темноте при температуре 25°C в течение 30 минут. Реакцию прекращали добавлением 50 мкл 1,0 M HCl и измеряли поглощение на 490 нм в считывателе микропланшетов (Genios Pro; Tecan, Австрия). Число клеток подсчитывали с помощью значений A490 для этих образцов и стандартов, содержащих ЛДГ по известному количеству клеток.
Влияние поверхностей твердых подложек и Y-27632 на закрепление и рост клеток HEK293 в среде Pro293a-CDM показано на фиг.31a, где 96-часовое непрерывное воздействие Y-27632 отмечено как “Y-27632 96 ч on”, а 48-часовое непрерывное воздействие Y-27632 с последующей заменой среды и 48-часовым инкубированием в отсутствие Y-27632 отмечено как “Y-27632 48 ч on/48 ч off”. В отсутствие Y-27632 клетки HEK293 закрепляются на всех трех поверхностях. Замена среды через 48 часов инкубирования привела к тому, что в культурах стало значительно меньше клеток по данным измерения через 96 часов инкубирования. Улучшение закрепления клеток HEK293 на поверхности 4 и поверхности CellBINDTM было отмечено при применении Y-27632 в концентрации 2,0 и 5,0 мкМ. Удаление Y-27632 через 48 часов инкубирования привело к откреплению значительного количества клеток от всех трех поверхностей.
Аналогичный эксперимент был проведен с использованием неадаптированных клеток НЕК293 в количестве 2,0×104 на см2 и EMEM с добавлением 10% эмбриональной бычьей сыворотки. Влияние поверхностей твердых подложек и Y-27632 на закрепление и рост клеток HEK293 в EMEM с добавлением 10% эмбриональной бычьей сыворотки показано на фиг.31b, где 96-часовое непрерывное воздействие Y-27632 отмечено как “Y-27632 96 ч on”, а 48-часовое непрерывное воздействие Y-27632 с последующей заменой среды и 48-часовым инкубированием в отсутствие Y-27632 отмечено как “Y-27632 48 ч on/48 ч off". В отсутствие Y-27632, клетки HEK293 закреплялись на всех трех поверхностях. Замена среды после 48 часов инкубирования приводила к тому, что в культурах стало значительно меньше клеток по данным измерения через 96 часов инкубирования. Улучшение закрепления клеток HEK293 на поверхности 4 и поверхности CellBINDTM было отмечено при применении Y-27632 в концентрации 2,0 и 5,0 мкМ. Удаление Y-27632 через 48 часов инкубирования привело к откреплению значительного количества клеток от поверхности 4 и CellBINDTM.
Пример 20
Обработка Y-27632 и H-1152 улучшает рост клеток HEK293 на поверхностно-модифицированных носителях
Клетки HEK293 поддерживали в EMEM (Lonza) с добавлением 10% эмбриональной бычьей сыворотки (Lonza). Клетки пассировали при уровне конфлюэнтности 70-80% с помощью трипсина/ЭДТК для диссоциации и высевали в количестве 2,0×104 клеток/см2 в колбе объемом 75 см2 с поверхностью Nunclon DeltaTM (Thermo Fisher Scientific, Роскильде, Дания).
EMEM (500 мкл) с добавлением 10% эмбриональной бычьей сыворотки с 1,0, 5,0, 10, 15 или 20 мкМ Y-27632 (Sigma Chemical Co.) или 0,4, 1,2, 1,6, 2,4 или 2,8 мкМ H-1152 (Calbiochem, EMD Chemicals Inc., Дармштадт, Германия) наносили на 24-луночные планшеты Multidish с поверхностью 4 или необработанной (но прошедшей гамма-облучение; 25 кГр) полистирольной поверхностью. В лунки добавляли еще 500 мкл EMEM с добавлением 10% эмбриональной бычьей сыворотки и с клетками HEK293 (2,0×104 клеток/см2). Культуры помещали в аппарат IncuCyteTM Plus (Essen Instruments, Мичиган, США) и инкубировали при температуре 37°C в увлажненной атмосфере 5% CO2 в воздухе. IncuCyteTM Plus представляет собой автоматизированную платформу для получения изображений, конфигурация которой допускает установку внутри CO2 инкубатора и обеспечивает кинетическое неинвазивное получение изображений живых клеток способом снятия фазоконтрастных изображений клеток; при этом пользователь определяет моменты времени и точки съемки в культурах. Первичная измеряемая величина прибора - конфлюэнтность культуры, то есть, доля поверхности, покрытой клетками. Клетки HEK293 инкубировали в течение 72 часов без манипуляций и получали изображения каждые два часа в 9 позициях в трех экземплярах культур. Конфлюэнтность культуры определяли с помощью программного обеспечения IncuCyteTM Plus software (v. 3.4.1.25966).
Повышение концентрации Y-27632 и H-1152 улучшает закрепление и рост клеток HEK293 на поверхности 4 (фиг.32a). Влияние необработанной поверхности культуры клеток и Y-27632 или H-1152 на закрепление и рост клеток HEK293 показано на фиг.32b. Рост и закрепление клеток HEK293 немного улучшались в присутствии 10 мкМ Y-27632 и 0,6-1,2 мкМ H-1152. Тем не менее, улучшение роста и прикрепления клеток HEK293 на необработанной поверхности культуры клеток незначительно по сравнению с поверхностью 4.
Пример 21
Обработка H-1152 улучшает рост и прикрепление клеток HEK293 к поверхностно-модифицированным носителям
Клетки HEK293 поддерживали в EMEM (Lonza) с добавлением 10% эмбриональной бычьей сыворотки (Lonza). Клетки пассировали при уровне конфлюэнтности 70-80% с помощью трипсина/ЭДТК для диссоциации и высевали в количестве 2,0×104 клеток/см2 в колбе объемом 75 см2 с поверхностью Nunclon DeltaTM (Thermo Fisher Scientific, Роскильде, Дания).
EMEM (1,0 мл) с добавлением 10% эмбриональной бычьей сыворотки с 0,4, 0,8, 1,2, 1,6, 2,0, 2,4 или 2,8 мкМ H-1152 наносили на 12-луночные планшеты Multidish с поверхностью 4. В лунки добавляли еще 1,0 мл EMEM с добавлением 10% эмбриональной бычьей сыворотки и с клетками HEK293 (4,0×104 клеток/см2). Культуры помещали в аппарат IncuCyteTM Plus и инкубировали при температуре 37°C в увлажненной атмосфере 5% CO2 в воздухе в течение 42 часов (изображения получали каждые 6 часов). Затем удаляли 1 мл культурной среды пипеткой и добавляли 1,0 мл EMEM с добавлением 10% эмбриональной бычьей сыворотки с 0,2, 0,4, 0,6, 0,8, 1,0, 1,2 и 1,4 мкМ H-1152. Культуры снова помещали в аппарат IncuCyteTM Plus и получали изображения каждый час в течение следующих 25 часов. Изображения получали в 9 положениях в трех экземплярах культур и определяли конфлюэнтность культур с помощью программного обеспечения IncuCyteTM Plus. Изображения IncuCyteTM Plus, полученные в определенных положениях в культурах клеток HEK293, выращенных в отсутствие или в присутствии H-1152 (0,6 мкМ), считывали и представляли в виде фазоконтрастных микрофотографий для сравнения с морфологией культуры HEK293 в следующие моменты времени: в начале инкубации (0 часов), непосредственно перед заменой среды (42 часа), через 1 час после замены среды (43 часа) и, наконец, после 52 часов инкубации.
В отсутствие H-1152 и в присутствии 0,2 мкМ или 0,4 мкМ H-1152 замена 50% среды через 42 часа инкубации приводила к значительному снижению конфлюэнтности культуры (фиг.33a). В присутствии 0,6 мкМ, 0,8 мкМ или 1,4 мкМ H-1152 влияние замены среды было минимальным. Клетки HEK293, выращиваемые на поверхности 4 в присутствии H-1152, покрывали поверхности твердых подложек более равномерно, чем клетки HEK293, выращиваемые на поверхности 4 в отсутствие H-1152 (фиг.33b). В отсутствие H-1152 клетки HEK293 образовывали крупные кластеры, тогда как клетки HEK293 в присутствии H-1152 образовывали кластеры меньшего размера с более низкой плотностью клеток.
Пример 22
Обработка Y-27632 улучшает рост клеток HEK293 в процессе трех пассирований на поверхностно-модифицированных носителях
EMEM (500 мкл) с добавлением 10% эмбриональной бычьей сыворотки с 5,0 мкМ Y-27632 помещали в лунки 24-луночных планшетов Multidish с поверхностью 4 или поверхностью Nunclon DeltaTM. В лунки добавляли еще 500 мкл EMEM с добавлением 10% эмбриональной бычьей сыворотки с клетками HEK293 (2,0×104 клеток/см2) и инкубировали культуры при 37°C в увлажненной атмосфере 5% CO2 в воздухе в течение трех 3 дней. Клетки пассировали обработкой трипсином/ЭДТК (Lonza, Вервье, Бельгия) в течение двух минут при температуре 37°C и определяли общее число клеток с помощью цитометра NucleoCount (Chemometec A/S, Аллерод, Дания). Для последовательных пассирований клетки HEK293 высевали в количестве 2,0×104 клеток/см2. Рост клеток HEK293 на поверхности 4 и поверхности Nunclon DeltaTM улучшался в присутствии 2,5 мкМ Y-27632 (фиг.34).
Пример 23
Иммобилизация, культивирование и поддержание эмбриональных стволовых клеток человека с использованием поверхностно-модифицированных носителей 4, 18 и 19 без белков/компонентов внеклеточного матрикса и питающих клеток
Эмбриональные стволовые клетки человека линии Н1 42-го пассажа, поддерживаемые в пластиковой посуде с покрытием 1:30 MATRIGEL в среде, кондиционированной MEF с добавлением 8 нг/мл bFGF, снимали обработкой ферментативной смесью LIBERASE™ и помещали на поверхностно-модифицированные носители в формате 96-луночных планшетов, в разбавлении 1 к 2 в среде, кондиционированной MEF, с добавлением 8 нг/мл bFGF. Клетки помещали на модифицированные поверхности 4, 18, 19 или Primaria™. Для определения влияния ингибирования Rho-киназы на связывание с модифицированной поверхностью мы обрабатывали клетки либо 10 мкМ ингибитора Rho-киназы Y-27632, либо 3 или 10 мкМ ингибитора Rho-киназы H-1152 глицила. Необработанные клетки использовали в качестве контролей. После 24 часов в культуре из лунок удаляли жидкость, клетки высушивали, а в лунки добавляли кристаллический фиолетовый.
По нашим наблюдениям, после 24 часов в культуре колонии эмбриональных стволовых клеток закреплялись и распространялись в случае обработки ингибиторами Rho-киназы на поверхностно-модифицированных носителях 4 и 19 и носителе Primaria™, однако такого эффекта не наблюдалось на поверхностно-модифицированном носителе 18 (фиг.35).
Пример 24
Иммобилизация, культивирование и поддержание эмбриональных стволовых клеток человека с использованием поверхностно-модифицированных носителей 30, 31, 32, 33 и 34 без белков/компонентов внеклеточного матрикса и питающих клеток
Эмбриональные стволовые клетки человека линии H1 47-го пассажа, поддерживаемые в пластиковой посуде с покрытием 1:30 MATRIGEL в среде, кондиционированной MEF с добавлением 8 нг/мл bFGF, снимали обработкой ферментативной смесью TrypLE™ и помещали на поверхностно-модифицированные носители в формате 96-луночных планшетов, в разбавлении 1 к 3 в среде, кондиционированной MEF с добавлением 8 нг/мл bFGF. Клетки помещали на модифицированные поверхности 30, 31, 32, 33 или 34. Для определения влияния ингибирования Rho-киназы на связывание с модифицированной поверхностью мы обрабатывали клетки 3 мкМ ингибитора Rho-киназы H-1152 глицила. Необработанные клетки использовали в качестве контролей. Кроме того, клетки высевали в лунки на поверхностно-модифицированных носителях, предварительно обработанных MatrigeTM. Через 24 часа после высевания среду меняли на свежую MEF-кондиционированную среду с добавлением 8 нг/мл bFGF, а в случае клеток, посеянных в присутствии ингибитора Rho-киназы, в среду добавляли 3 мкМ H-1152 глицила. После 48 часов в культуре из лунок удаляли жидкость, клетки высушивали, а в лунки добавляли кристаллический фиолетовый.
По нашим наблюдениям, после 48 часов в культуре колонии эмбриональных стволовых клеток закреплялись и распространялись в случае обработки ингибиторами Rho-киназы на поверхностно-модифицированных носителях 33 и 34 (фиг.39 и 40, соответственно), однако такого эффекта не наблюдалось на поверхностно-модифицированных носителях 30, 31 или 32 (фиг.36-40, соответственно).
Пример 25
Иммобилизация, культивирование и поддержание эмбриональных стволовых клеток человека с использованием поверхностно-модифицированных носителей 22, 23, 24 или 29 без белков/компонентов внеклеточного матрикса и питающих клеток
Эмбриональные стволовые клетки человека линии Н1 46-го пассажа, поддерживаемые в пластиковой посуде с покрытием 1:30 MATRIGEL в среде, кондиционированной MEF с добавлением 8 нг/мл bFGF, снимали обработкой ферментативной смесью Liberase™ и помещали на 60 мм чашке с модифицированной поверхностью в разбавлении 1 к 3 в среде, кондиционированной MEF с добавлением 8 нг/мл bFGF. Клетки помещали на поверхностно-модифицированные носители 3, 4, 22, 23, 24 и 29. Для определения влияния ингибирования Rho-киназы на связывание с модифицированной поверхностью мы обрабатывали клетки 3 мкМ ингибитора Rho-киназы H-1152 глицила для высевания клеток. Среду меняли на свежую MEF-кондиционированную среду с 8 нг/мл bFGF и 1 мкМ ингибитора Rho-киназы H-1152 глицила через 24 часа после высевания клеток. Клетки, посеянные на модифицированную поверхность 3, 4 или пластиковую поверхность, покрытую MatrigelTM, использовали в качестве контролей. Носители исследовали способом фазоконтрастной микроскопии через 24 и 48 часов после высевания. По нашим наблюдениям, после 48 часов в культуре колонии эмбриональных стволовых клеток не закреплялись на поверхностно-модифицированным носителях 22, 23, 24 и 29 с ингибитором Rho-киназы или без него, тогда как клетки, помещенные на поверхностно-модифицированный носитель 3 или 4 в присутствии ингибитора Rho-киназы закреплялись и распространялись.
Пример 26
Дальнейшее определение характеристик поверхностно-модифицированных носителей, составляющих предмет настоящего изобретения
Углы смачивания водой
Поверхностно-модифицированные носители 1-4 и 13 упаковывали в отдельные полиэтиленовые пакеты, стерилизовали и хранили при комнатной температуре в течение 40-недельного периода проведения анализа. Углы смачивания сначала измеряли через одну неделю после обработки поверхности и стерилизации, а затем повторно в моменты времени, указанные на фиг.41. Все измерения углов смачивания выполнялись, как описано в примере 17. Измерения на поверхностях Nunclon DeltaTM и CellBINDTM проводились в тех же экспериментальных условиях, что и измерения на поверхностях 1-4 и 13, только обработка поверхности и стерилизация проводились более чем за 12 недель до первого измерения (Nuclon DeltaTM* стерилизовали за одну неделю до первого измерения). На фиг.41 показано, что поверхностно-модифицированные носители 1-4 и 13 имели схожую гидрофильность и были более гидрофильными (имели меньшие углы смачивания водой), чем поверхности Nunclon DeltaTM и CellBINDTM. Гидрофильные свойства поверхностно-модифицированных носителей 1-4 и 13 были стабильными не менее чем 41 неделю после обработки поверхности и стерилизации.
Углы смачивания измеряли также на поверхностно-модифицированных носителях 5-12, 22-24, 29, 30 и 33, которые были упакованы в полиэтиленовые пакеты, простерилизованы как описано в примере 16 и хранились при комнатной температуре 9 недель (кроме поверхностно-модифицированного носителя 29, который хранился 28 недель). Поверхностно-модифицированные носители 18, 19, 32 и 34 были в формате одиночной микролунки и поэтому не могли использоваться для измерения углов смачивания. Поверхностно-модифицированные носители 30 и 33 имели формат микролуночного планшета, и измерение углов смачивания производилось на обратной стороне планшета, а не внутри лунок. Измерения углов смачивания проводились, как описано в примере 17 (в случае поверхностно-модифицированного носителя 29, имеющего высокую гидрофильность, наносили небольшую каплю 2,5 мкл воды MilliQ), но анализировали по три образца с нанесением 7 капель на каждый образец. Измерения на носителях с поверхностями CostarTM, FalconTM, PrimariaTM и Nunclon DeltaTM проводились при тех же экспериментальных условиях, но обработка поверхности и стерилизация проводились более чем за 12 недель до первого измерения. На фиг.42 показано, что поверхностно-модифицированные носители 5-12 были более гидрофильными (имели меньшие углы смачивания водой), чем поверхности Nunclon DeltaTM, CostarTM и FalconTM. Гидрофильность поверхностей 5-12 была сравнима с гидрофильностью поверхности PrimariaTM и выше, чем гидрофильность поверхностей 1-4 и 13 (показано на фиг.41). Гидрофильность поверхностно-модифицированных носителей 22-24 и 33 была сравнима с гидрофильностью поверхностей 1-4 и 13 (показано на фиг.41), тогда как гидрофильность поверхности 30 была сравнима с гидрофильностью поверхностей Nunclon DeltaTM, CostarTM и FalconTM. Поверхностно-модифицированный носитель 29 был значительно более гидрофильным, чем другие анализировавшиеся поверхности.
Плотность отрицательных зарядов
Плотность отрицательных зарядов определяли на поверхностно-модифицированных носителях 5-12 (все в формате 5-см чашки), 18, 19, 30, 32, 33 и 34 (все в формате микролунок), поверхностно-модифицированных носителях 22-24 и 29 (все в формате 6-см чашки) и поверхности CellBINDTM (в формате 3-см чашки), поверхности PrimariaTM (в формате Multidish-6) и поверхности Nunclon DeltaTM (в формате 3-см чашки). Водный раствор кристаллического фиолетового (0,015% в/о) в избытке добавляли в каждый формат (0,34 мл/см2 для формата чашек и 0,13 мл/см2 для формата микролунок) и инкубировали при плавном потряхивании (50 об./мин.) в течение 60 минут при комнатной температуре. Для удаления кристаллического фиолетового, не связанного с поверхностями, тарелки трижды промывали 3 мл воды MilliQ для формата чашек и трижды 350 мкл воды MilliQ для формата микролунок, а затем высушивали в течение ночи при температуре 60°C. Кристаллический фиолетовый, связанный с поверхностью, десорбировали добавлением 0,17 мл/см2 раствора 0,1 M HCl в EtOH (99%) и инкубировали чашки при плавном потряхивании (50 об./мин.) в течение 2 минут при комнатной температуре). Измеряли поглощение раствора HCl:EtOH с десорбированным кристаллическим фиолетовым на длине волны 590 нм с помощью считывателя микропланшетов EnVision 2100 (Perkin Elmer; Уолтэм, Массачусетс, США). В значения поглощения вносились поправки на фоновое поглощение раствора HCl:EtOH. Плотность отрицательных зарядов измеряли на трех чашках с поверхностно-модифицированными носителями 5-12, 22-24, 29, CellBINDTM, PrimariaTM и Nunclon DeltaTM с проведением трех измерений поглощения для каждой тарелки. Для поверхностно-модифицированных носителей 18, 19, 30, 32, 33 и 34 проводилось три измерения на одном образце.
Показатели плотности отрицательных зарядов поверхностно-модифицированных носителей 5-12 были схожи, и эти поверхности имели значительно более низкие плотности отрицательных зарядов, чем поверхность CellBINDTM и Nunclon DeltaTM, но значительно более высокие показатели плотности отрицательных зарядов, чем поверхность PrimariaTM (фиг.43). Показатели плотности отрицательных зарядов поверхностно-модифицированных носителей 19, 33 и 34 были значительно выше, чем показатели плотности отрицательных зарядов поверхностно-модифицированных носителей 18, 30 и 32, которые являлись соответствующими необработанными поверхностями из того же полимерного материала. Показатели плотности отрицательных зарядов поверхностно-модифицированных носителей 22-24 и 29 нормировали относительно плотности отрицательных зарядов поверхности Nunclon DeltaTM, и, как видно из фиг.44, поверхностно-модифицированные носители 2-24 имели большую плотность отрицательных зарядов, чем поверхность Nunclon DeltaTM, тогда как плотность отрицательных зарядов поверхностно-модифицированного носителя 29 была значительно ниже, чем плотность отрицательных зарядов поверхности Nunclon DeltaTM (и поверхности 4).
Рентгеновская фотоэлектронная спектроскопия (XPS)
Поверхностно-модифицированные носители 5-12, 18, 19, 22-24, 29, 30, 31-34 анализировали способом XPS, как описано в примере 17. Элементный состав поверхности в атомных процентах показан в таблице 12. Все поверхности содержали углерод, кислород и азот (в XPS водород не определяется), кроме поверхностно-модифицированных носителей 31 и 32 (не обработанных плазмой), которые не содержали азота. Поверхностно-модифицированные носители 5-12 содержали меньше кислорода, чем поверхностно-модифицированные носители 1-4 и 13, но значительно больше кислорода, чем поверхности CostarTM, FalconTM и Nunclon DeltaTM (показано в таблице 7). Поверхностно-модифицированные носители 5-12 подготавливали микроволновой обработкой плазмой, в то время как поверхностно-модифицированные носители 1-4 и 13 обрабатывали плазмой коронного разряда. Поверхностно-модифицированные носители 19, 33 и 34, которые подготавливали обработкой плазмой коронного разряда, но изготовляли литьем под давлением из полимерных материалов кроме полистирола (использовавшегося в изготовлении поверхностно-модифицированных носителей 1-4 и 13), содержали кислород в количествах, сравнимых с его количеством в поверхностно-модифицированных носителях 1-4 и 13. Поверхностно-модифицированные носители 22-24 содержали меньше кислорода, чем поверхностно-модифицированные носители 1-4 и 13. Поверхностно-модифицированный носитель 29 содержал кислород в количестве, сравнимом с его количеством в поверхностно-модифицированных носителях 1-4 и 13. Поверхностно-модифицированные носители 5-12, 19, 33 и 34 содержали меньше азота, чем поверхностно-модифицированные носители 1-4 и 13, но больше, чем поверхности CostarTM, FalconTM и Nunclon DeltaTM (показано в таблице 7). Поверхностно-модифицированный носитель 29 содержал значительно больше азота, чем другие анализировавшиеся поверхности, в том числе поверхность PrimariaTM.
Пики спектров C1s исследовали способом подгонки (способом наилучших хи-квадратов) для определения и количественной оценки сред связывания углерода на поверхностно-модифицированных носителях по ширине пиков и энергетическим положениям элементов согласно литературным данным (таблица 13). Концентрации указаны в атомных процентах, которые получали перемножением площади в процентах и атомной концентрации. Все поверхности, обработанные плазмой, кроме поверхностей 10, 22-24 и 29, были схожи с точки зрения сред, связывающих углерод. Доля углерода в связывающей среде C-[O]-C была значительно выше в поверхностях 19, 33 и 34, чем в поверхностях 5-12, 18, 30 и 32 и поверхностях 1-4 и 13 (показано в таблице 8). Доля углерода в связывающей среде O-[C=O]-O была ниже в поверхностях 5-12, 19, 33 и 34, чем в поверхностях 1-4 и 13. Доля углерода в связывающей среде C*-C-O-C-C* была значительно выше в поверхностях 5-9, 11, 12, 19, 33 и 34, чем в поверхностях 1-4 и 13, но сравнима с долей углерода в поверхностях Nunclon DeltaTM и CellBINDTM. Доля углерода в связывающей среде C-O-C или C-NH3 + (в том же энергетическом положении спектра) была ниже в поверхностях 5-12, 19, 33 и 34, чем в поверхностях 1-4 и 13, но выше, чем в поверхностях CostarTM, FalconTM, CellBINDTM и PrimariaTM. Доля углерода в связывающей среде C-O-C*=O была выше в поверхностях 19, 33 и 34, чем в поверхностях 5-12, и сравнима с его количеством в поверхностях 1-4 и 13. Доля углерода в связывающей среде C=O была выше в поверхностях 5-12, чем в поверхностях 19, 33 и 34, но ниже, чем в поверхностях 1-4 и 13. Доля углерода в связывающей среде CO3 - была выше в поверхностях 5-12, чем в поверхностях 19, 33 и 34, и сравнима с его долей в поверхностях 1-4 и 13. Поверхности 22-24 были схожи с точки зрения сред, связывающих углерод. Доля углерода в связывающих средах C-[O]-C, O-[C=O]-O, C-O-C или C-NH3 +, C-O-C*=O и C=O для поверхностей 22-24 были значительно ниже, чем для поверхностей 1-4 и 13. Доля углерода в связывающих средах CO3 - и связывающих средах C*-C-O-C-C* была выше в поверхностях 22-24, чем в поверхностях 1-4 и 13. Связывающая углерод среда поверхности 29 отличалась от связывающей углерод среды всех остальных поверхностей, обработанных плазмой. Доля углерода в связывающей среде C-[O]-C была сравнима с его долей поверхностях 1-4 и 13. Доля углерода в связывающих средах O-[C=O]-O, CO3 - и C*-C-O-C-C* была ниже в поверхности 29, чем в поверхностях 1-4 и 13. Доля углерода в связывающих средах C-O-C или C-NH3 +, C-O-C*=O и в связывающих средах C=O была выше для поверхности 29, чем для поверхностей 1-4 и 13. Пик потери энергии был обусловлен ароматическим переходом P®P* и является индикатором ароматичности поверхности.
Пики спектров O1s имели почти гауссовой формы, и их подгонка была невозможна. Пики спектров N1s были исследованы способом подгонки (способом наилучших хи-квадратов) для определения и количественной оценки сред связывания азота в поверхностях по ширине пиков и энергетическим положениям элементов согласно литературным данным (таблица 14). Концентрации указаны в атомных процентах, получаемых перемножением площади в процентах и атомной концентрации. Доля азота в связывающей среде -NH3 + во всех поверхностях, кроме поверхности 9, была ниже, чем в поверхностях 1-4 и 13. Доля азота в связывающей среде -NH2 в поверхностях 5-12, 19, 33 и 34 варьировалась, но была выше, чем в поверхностях 1-4 и 13. Доля азота в связывающей среде -NO2 в поверхностях 5-12, 19, 33 и 34 варьировалась, но была ниже, чем в поверхностях 1-4 и 13. Доля азота в связывающей среде -NO3 в поверхностях 5-12, 19, 33 и 34 варьировалась, но была выше, чем в поверхностях 1-4 и 13. Связывающие азот среды поверхностей 22-24 и 29 отличались от сред других поверхностей, обработанных плазмой. Доля азота в связывающей среде -NH2 в поверхностях 22-24 и 29 варьировалась, но была значительно выше, чем в поверхностях 1-4 и 13. Доля азота в связывающей среде -NO2 была ниже в поверхностях 22-24 и 29, чем в поверхностях 1-4 и 13. Доля азота в связывающей среде O=C-N-C=O была сравнима в поверхностях 22-24 и 29 и поверхностях 1-4 и 13.
Публикации, цитируемые в данном документе, полностью включаются в настоящий документ в форме ссылок. Хотя различные аспекты изобретения иллюстрируются выше ссылками на примеры и предпочтительные варианты осуществления, подразумевается, что область изобретения ограничивается не указанным выше описанием, а следующими пунктами формулы изобретения, составленными в соответствии с принципами патентного законодательства.
ТАБЛИЦЫ
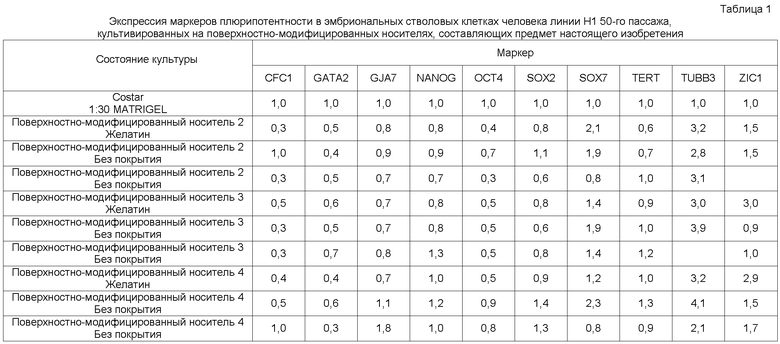
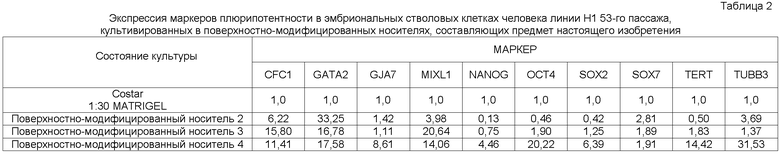

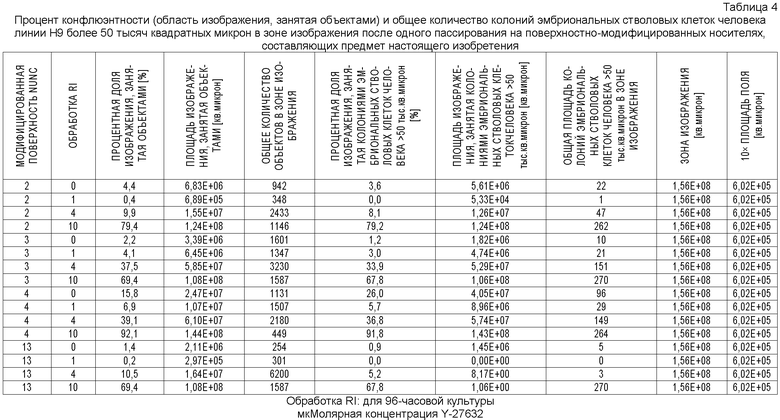


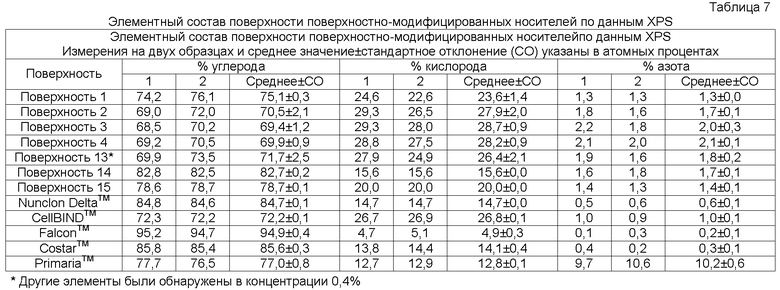
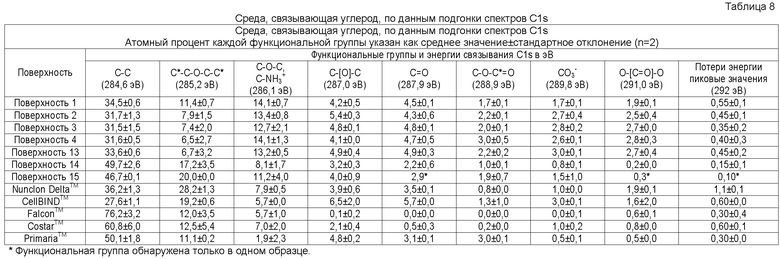

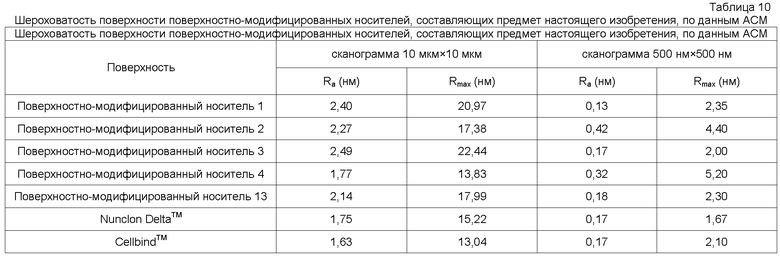

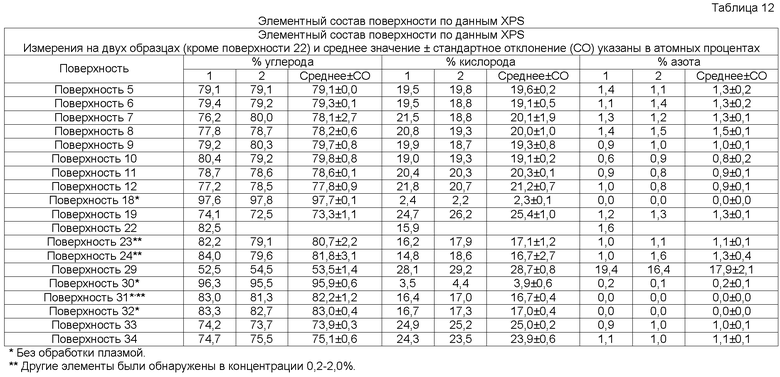
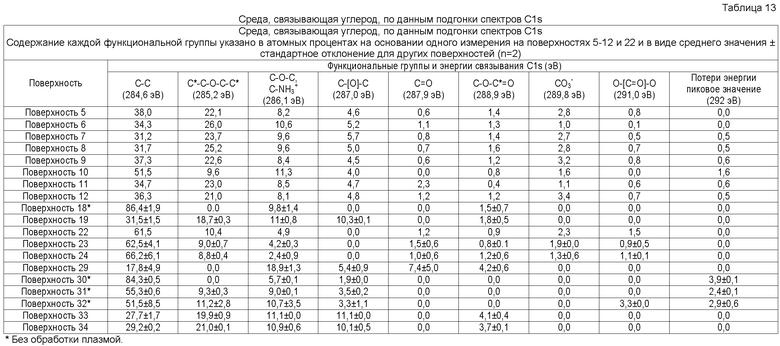
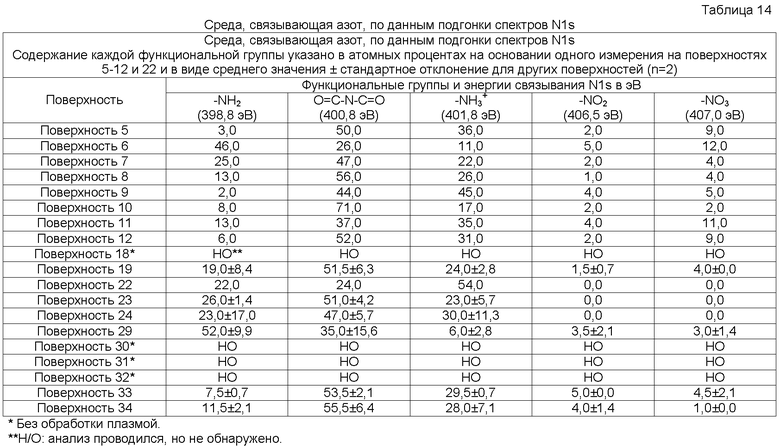

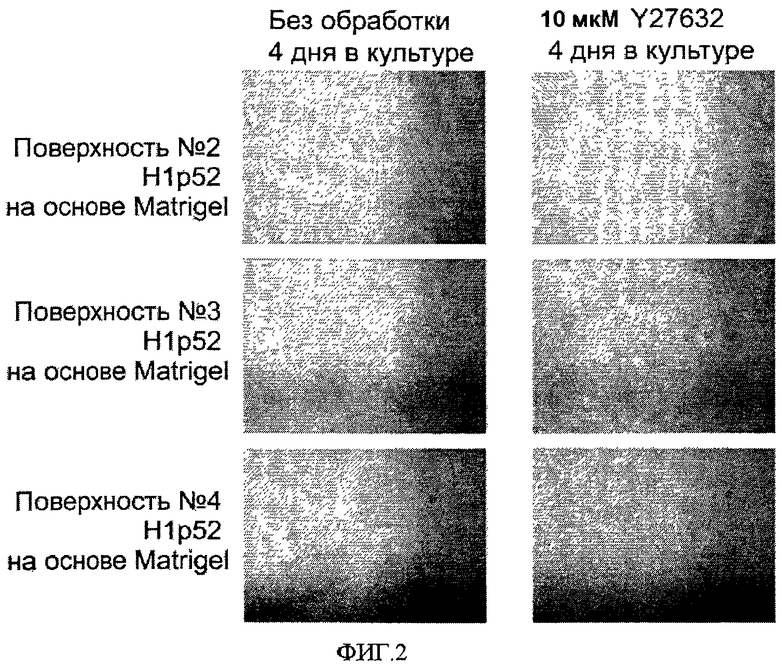
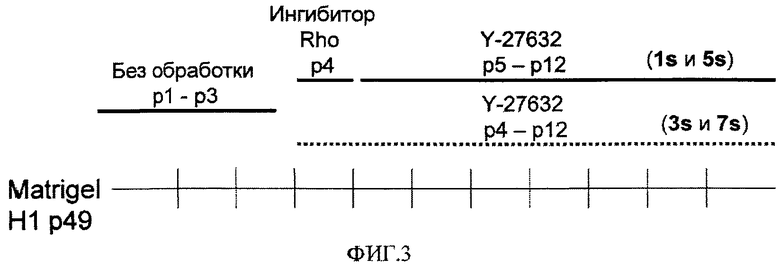
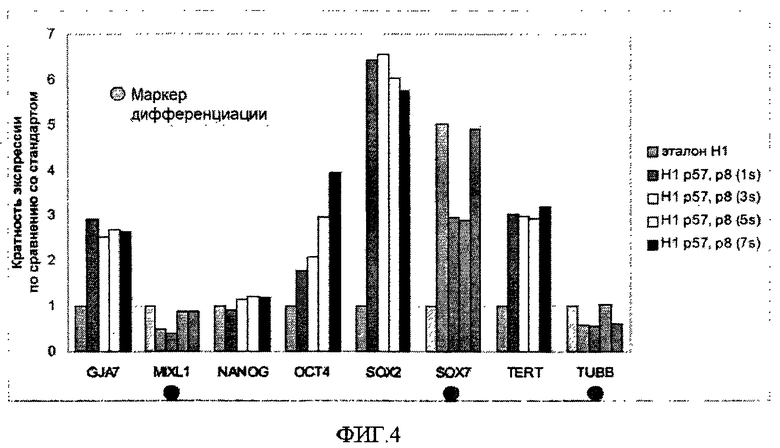

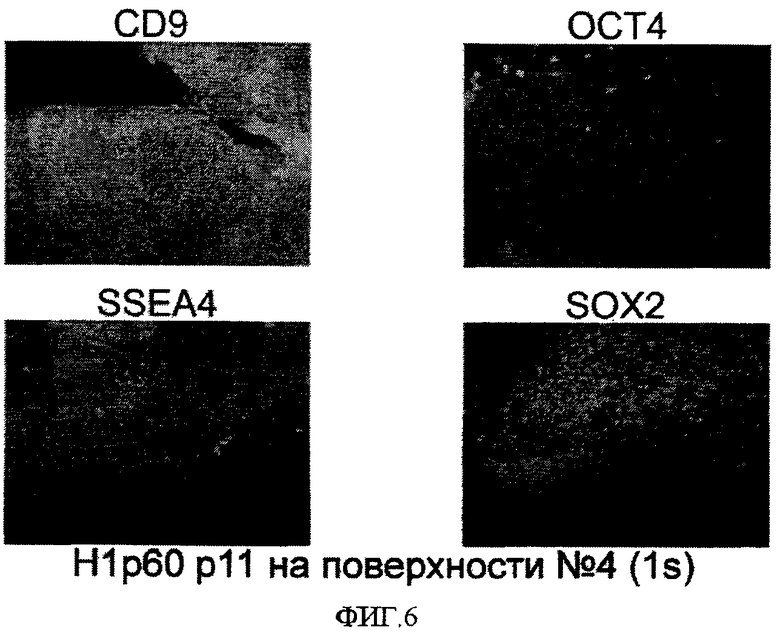
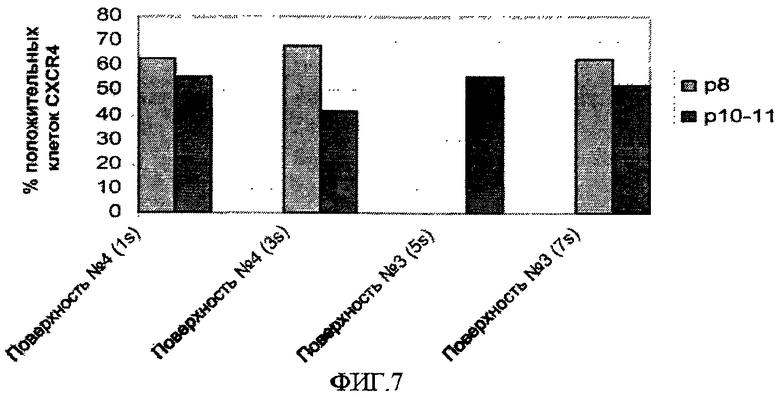
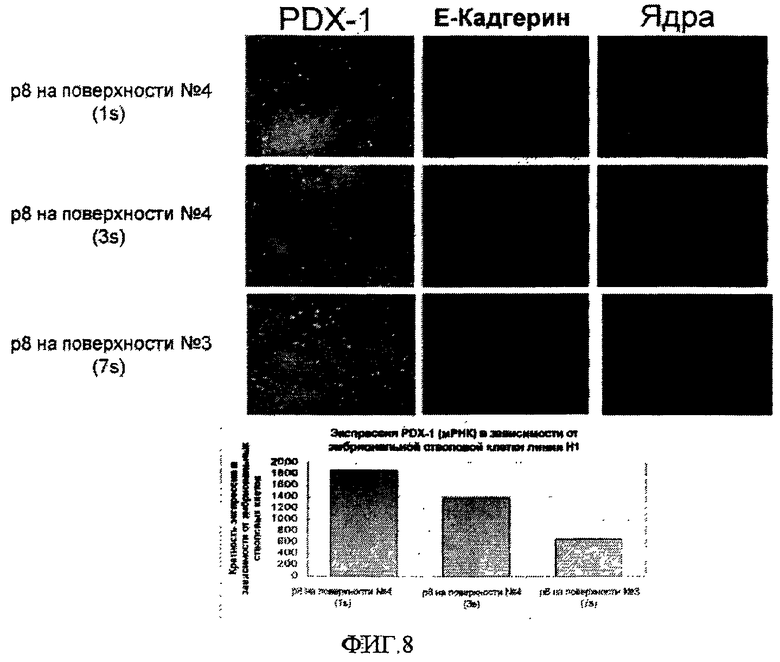
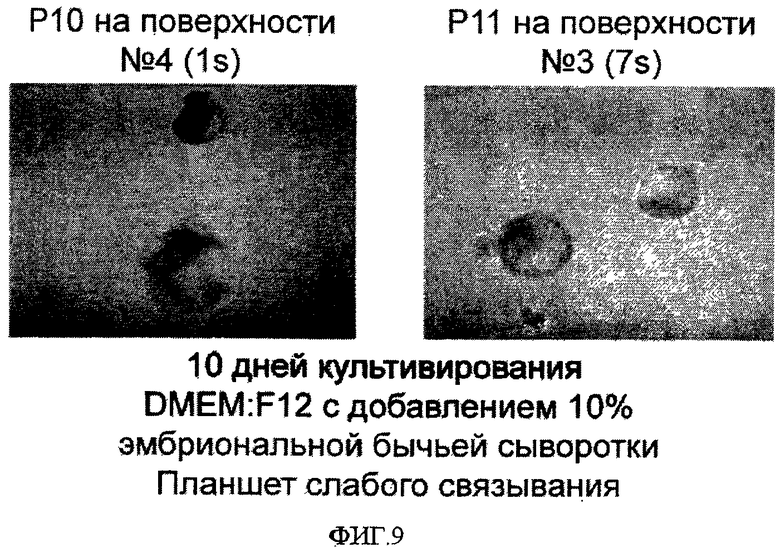
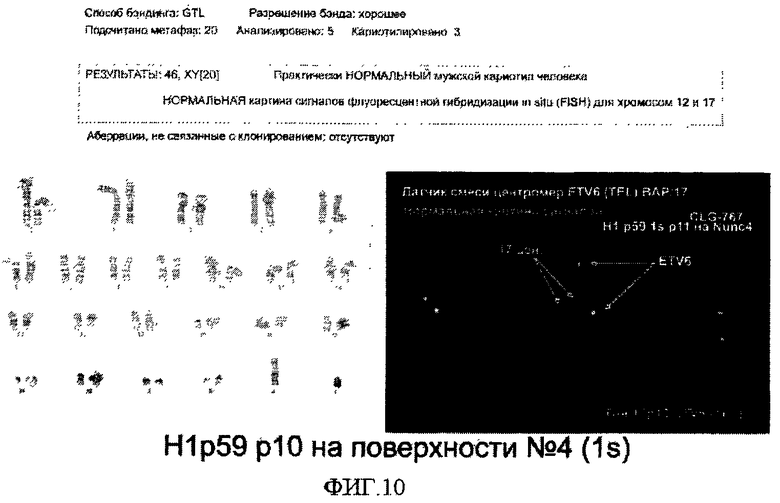
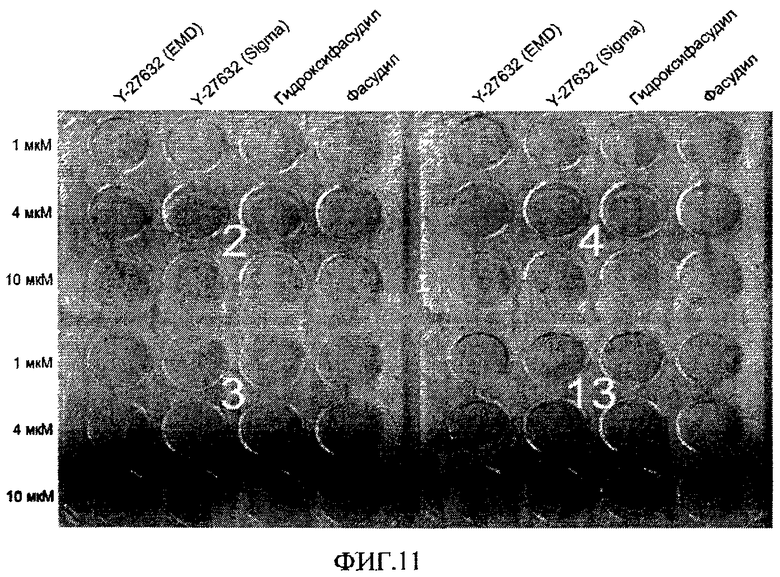


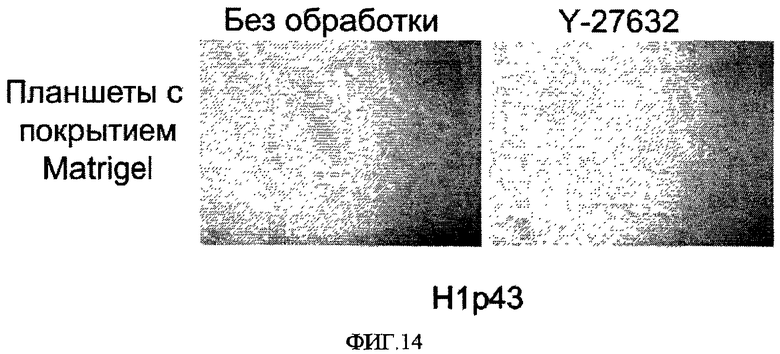
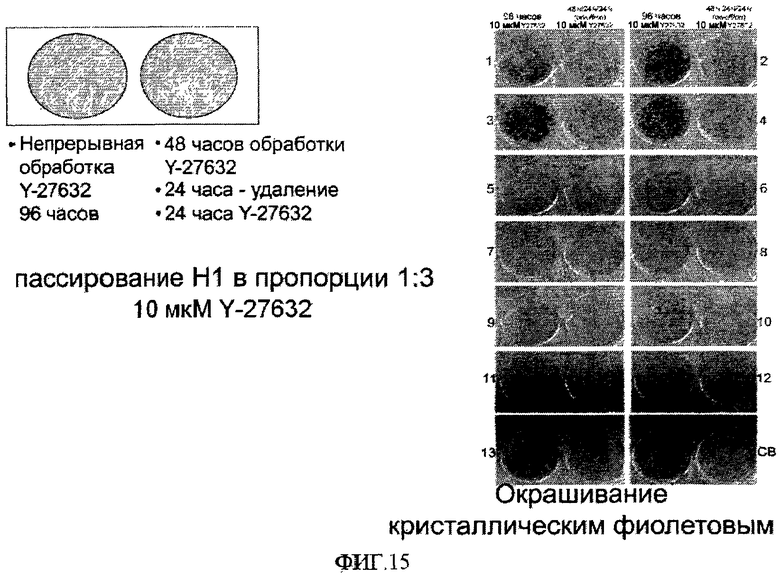

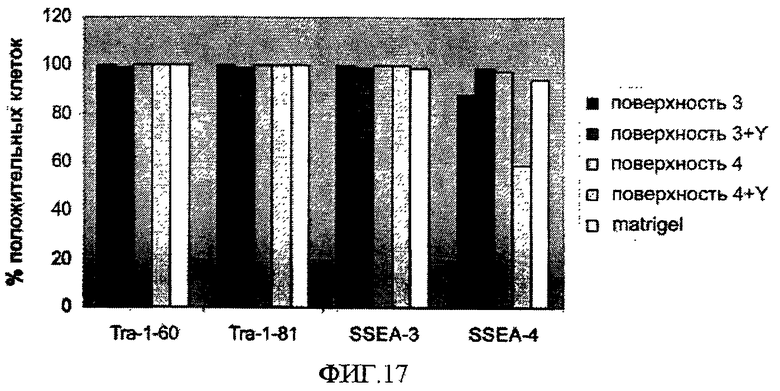
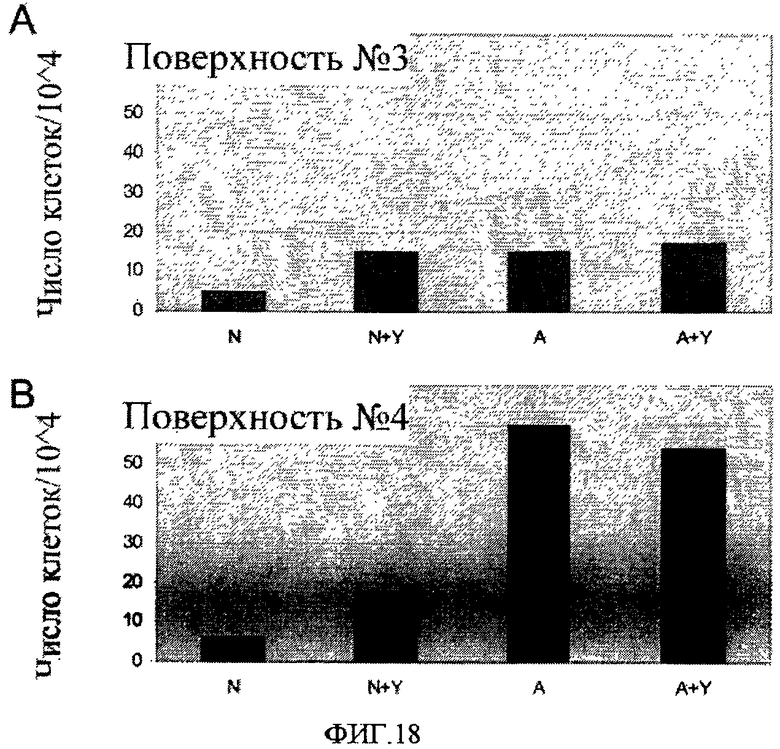
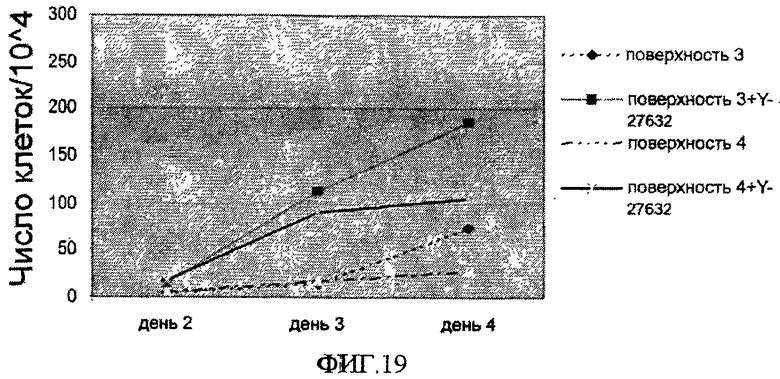
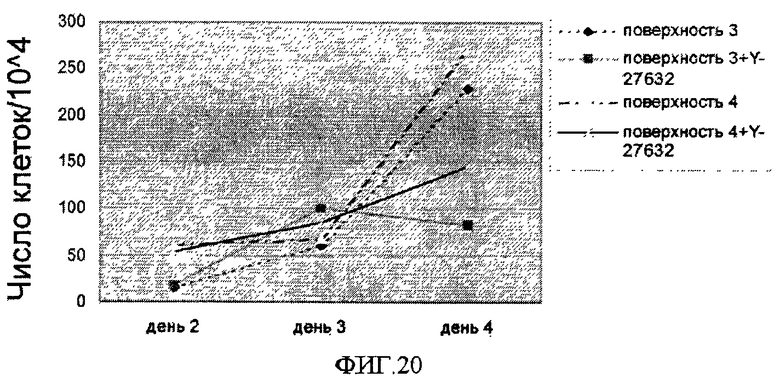


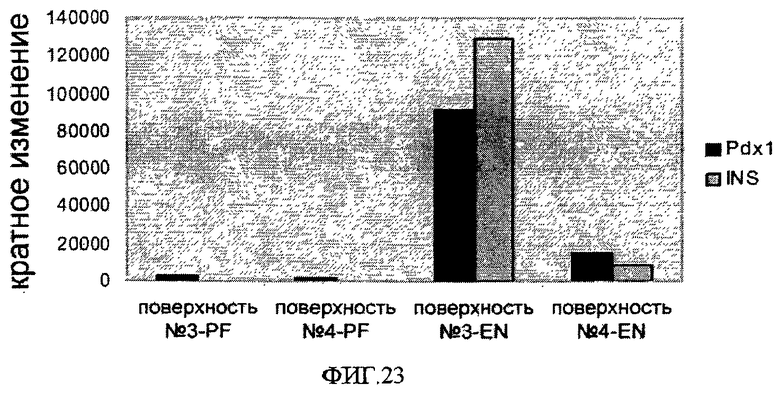

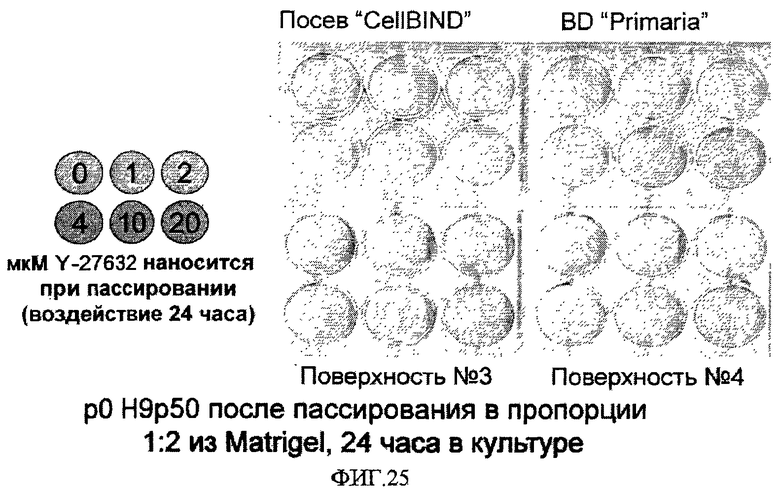
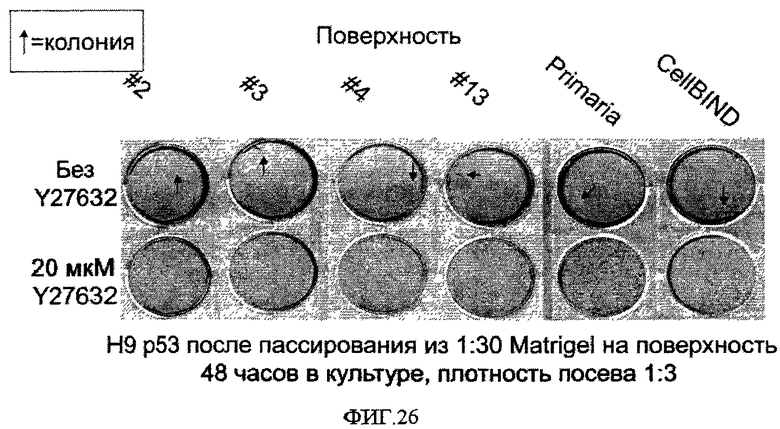
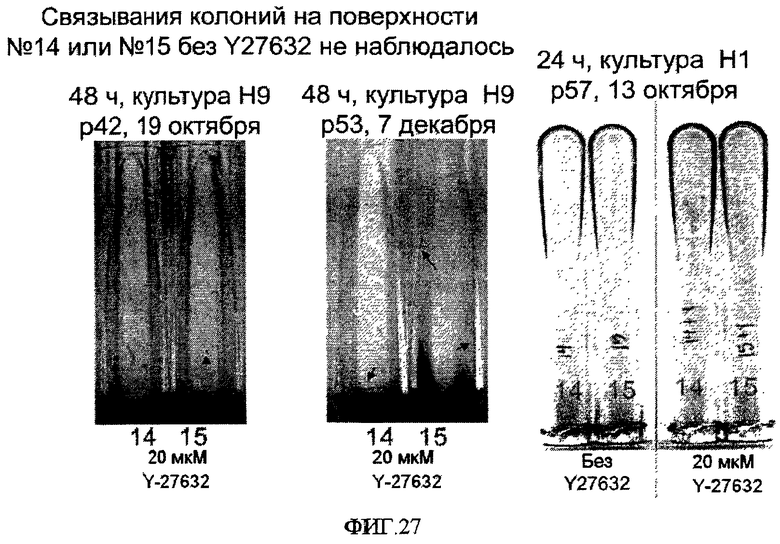
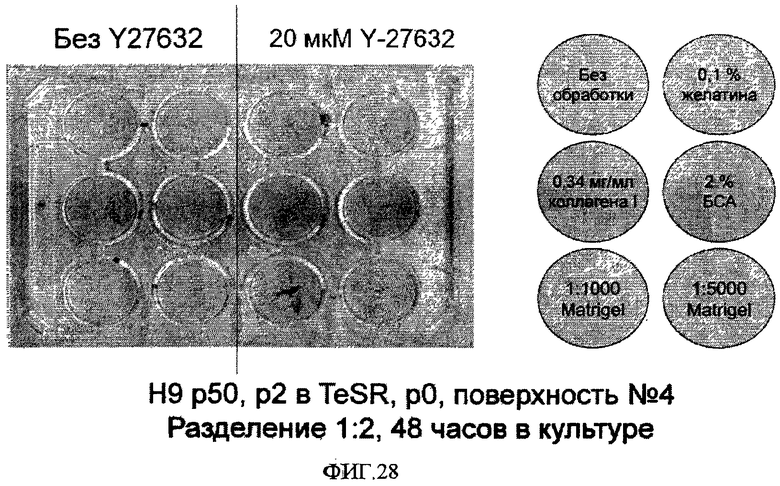
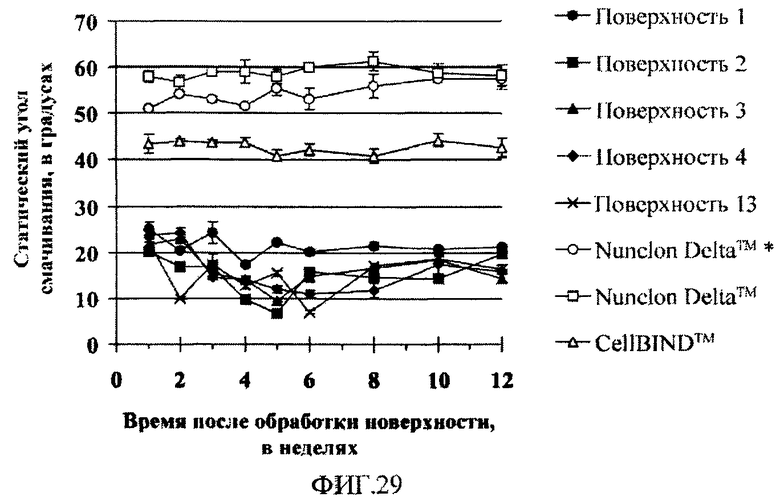
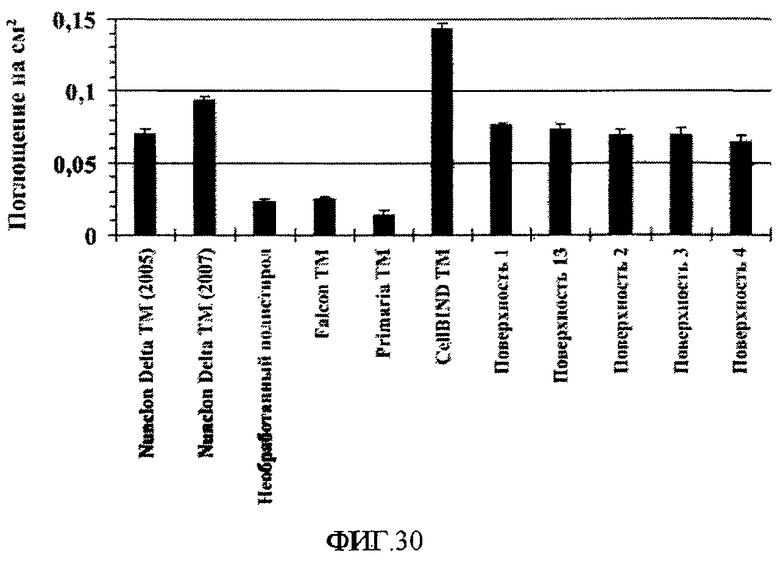
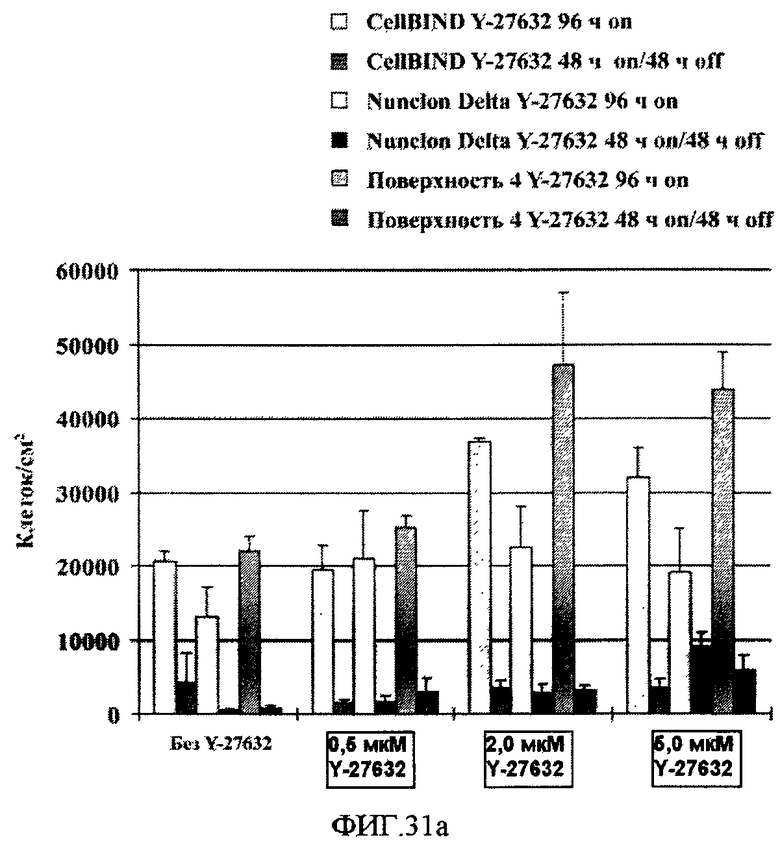

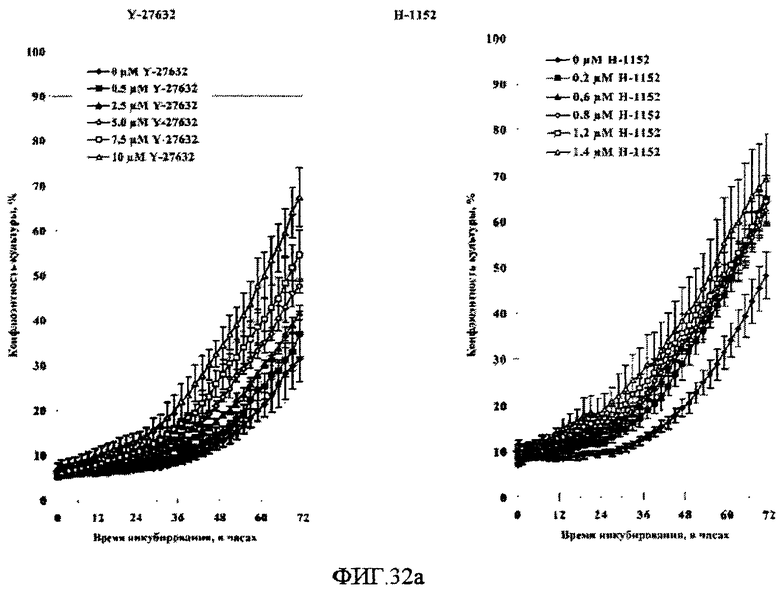
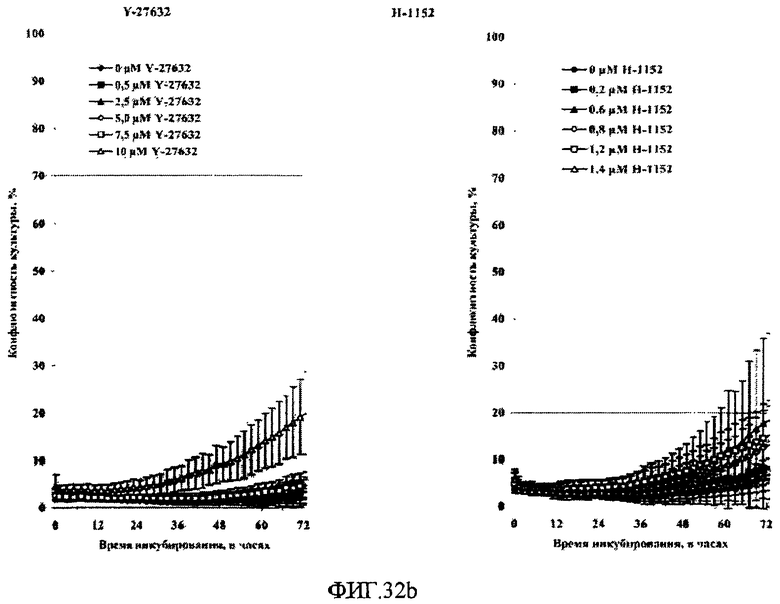
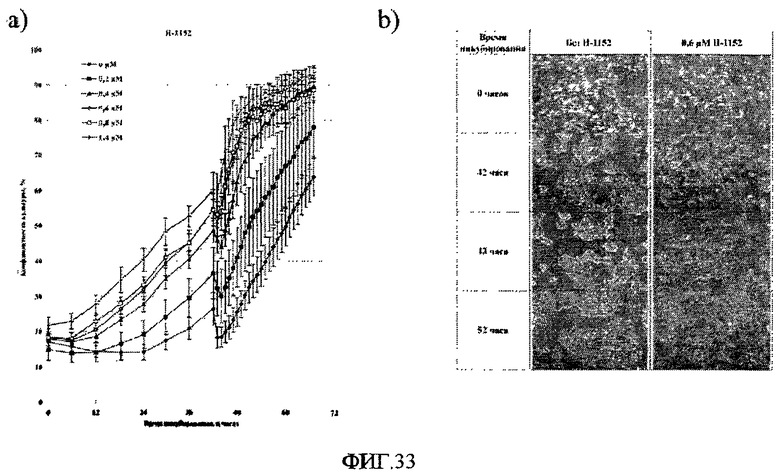
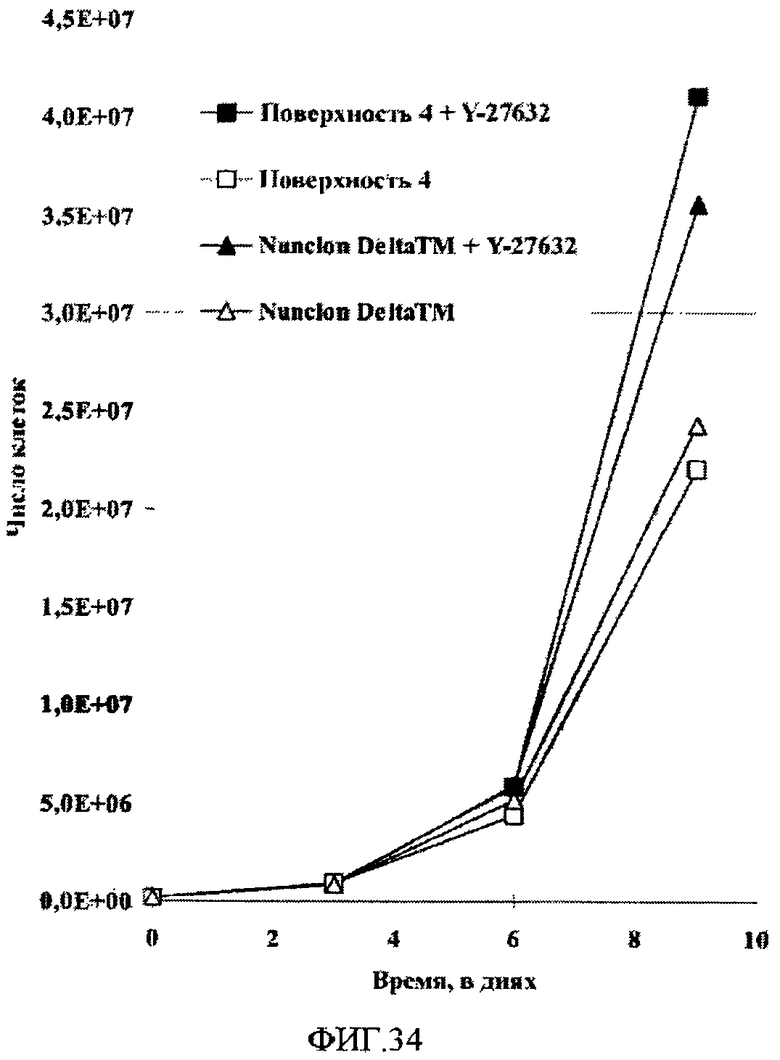
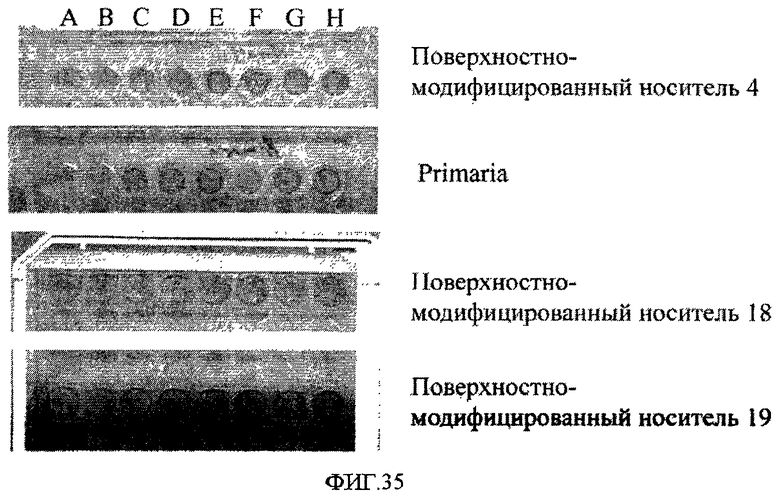
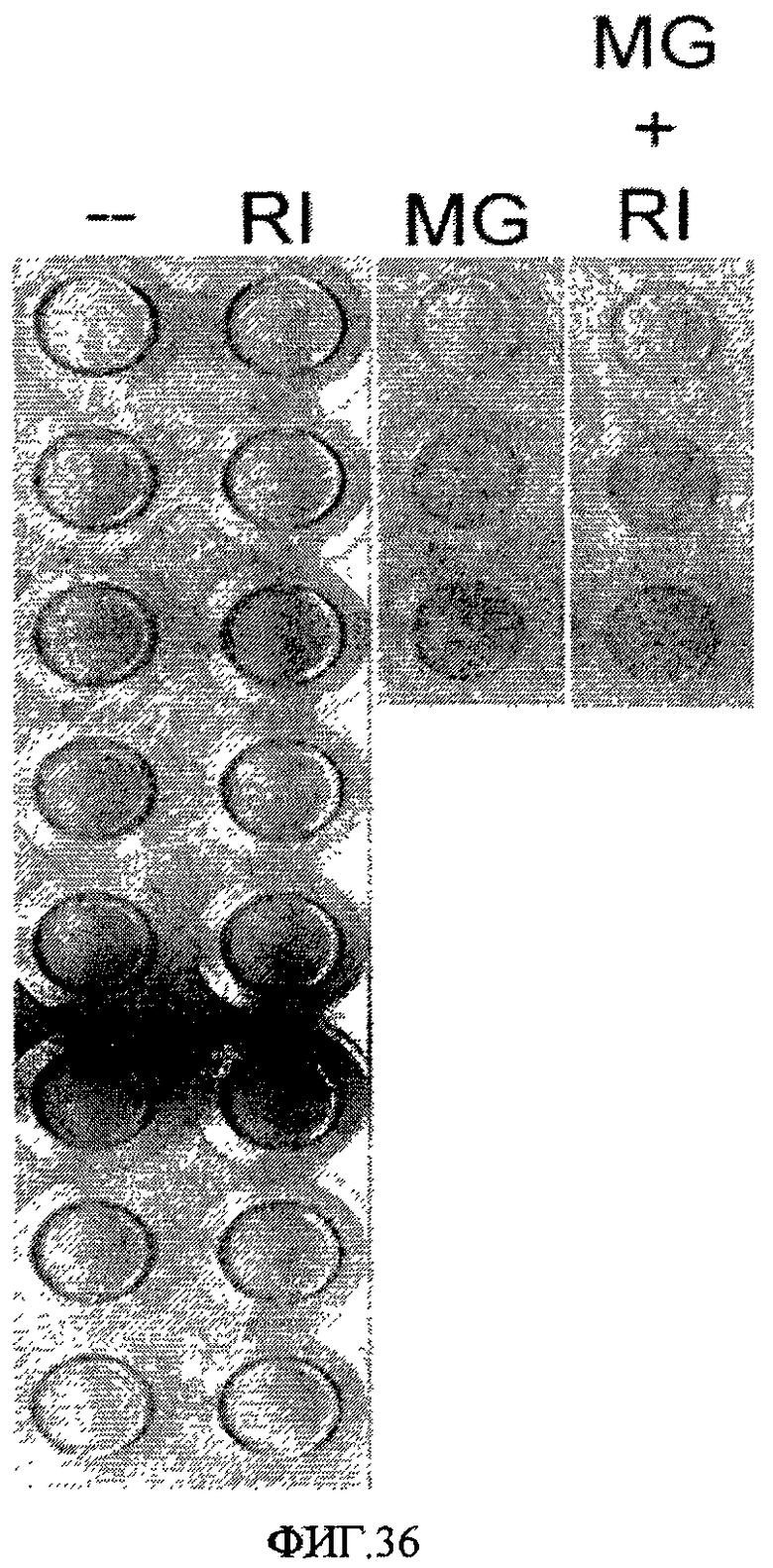



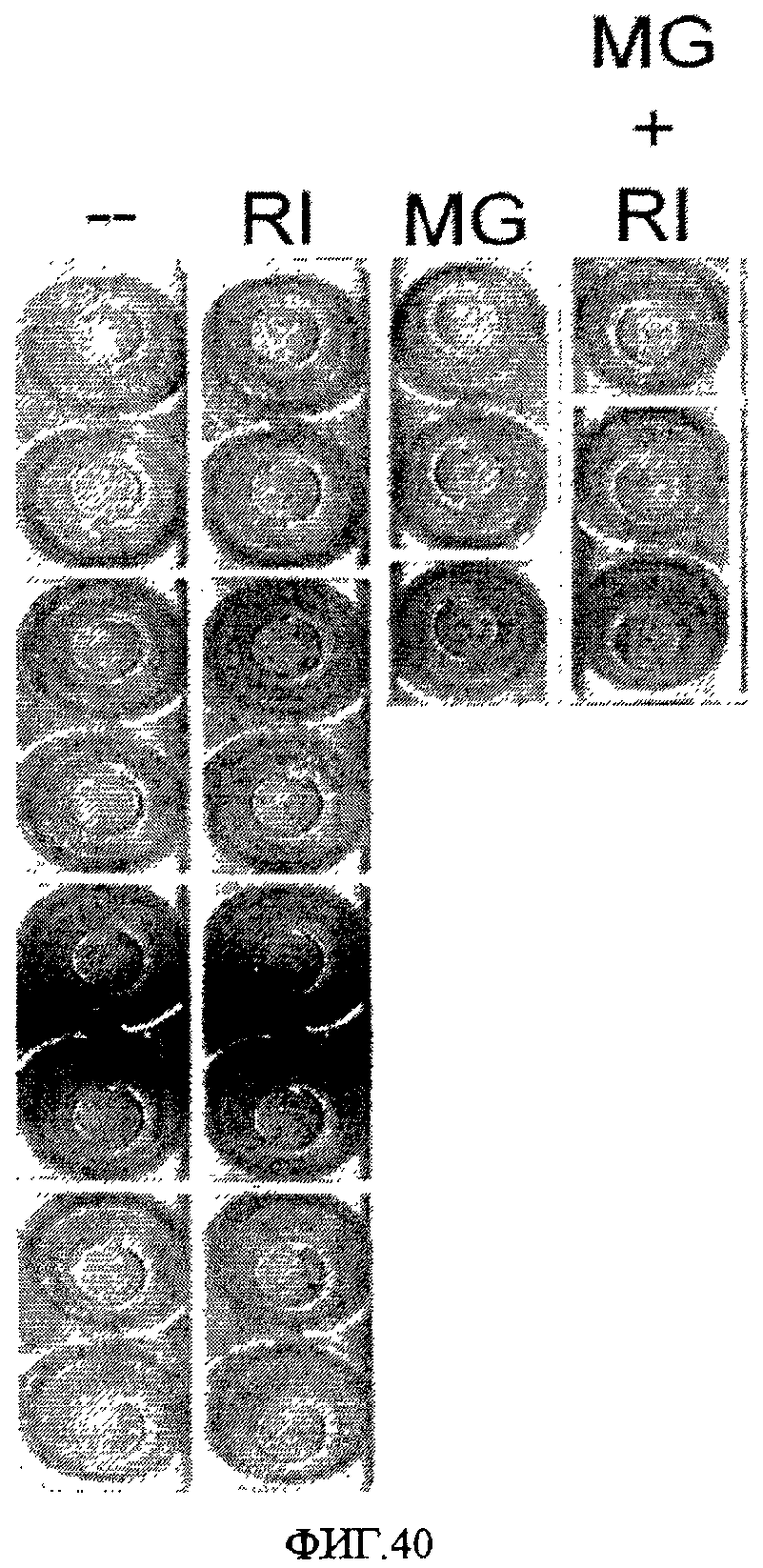

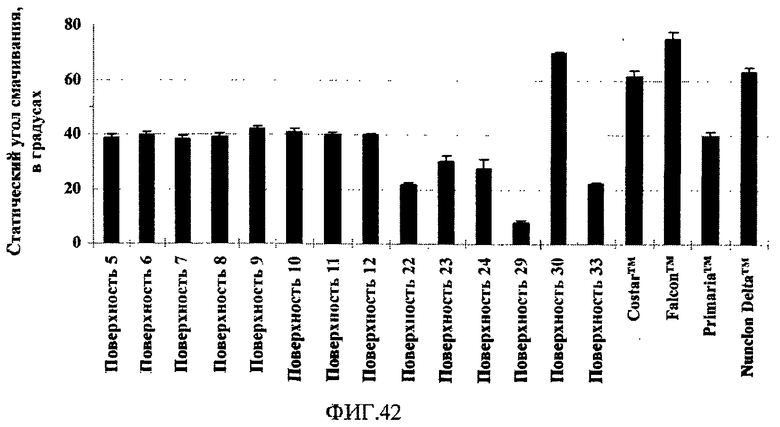
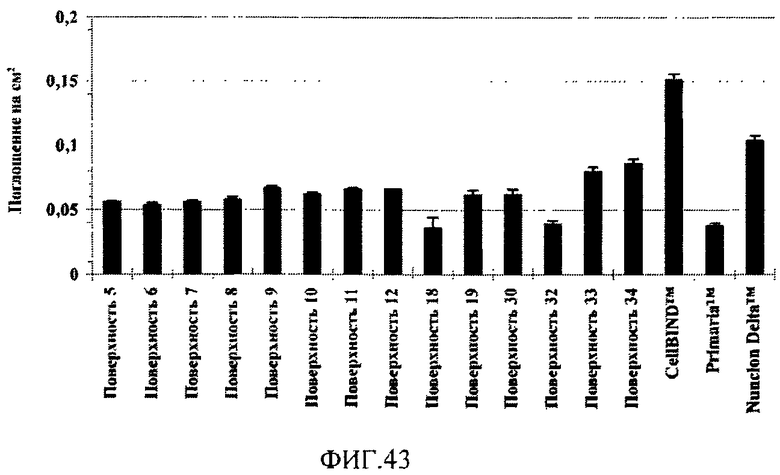
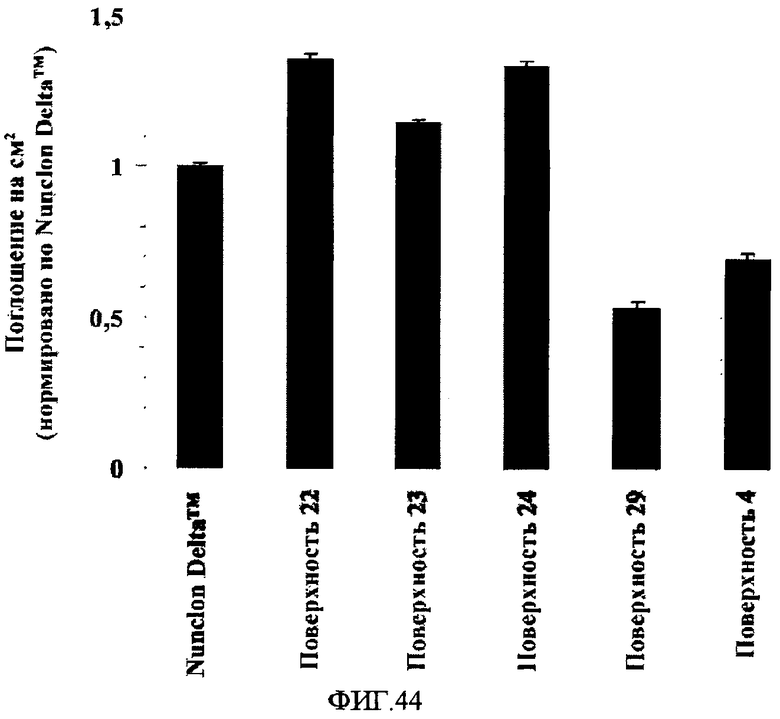
Изобретение относится к области биотехнологии. Описан способ закрепления плюрипотентных стволовых клеток человека на плоском носителе. Способ предусматривает получение суспензии плюрипотентных стволовых клеток и добавление к суспензии соединения, способного ингибировать активность Rho-киназы. Изобретение может быть использовано в медицине. 7 з.п.ф-лы, 46 ил., 14 табл., 26 пр.
1. Способ улучшения закрепления эмбриональных стволовых клеток человека на поверхности, содержащей от по меньшей мере 0,5% N, с суммарным содержанием О и N не ниже 17,2% и с углом смачивания по меньшей мере 13,9 градусов, не имеющей слоя питающих клеток и адгезивного слоя, включающий стадии:
a. обработки суспензии указанных клеток по меньшей мере одним соединением, выбранным из группы, состоящей из: соединения, способного ингибировать активность Rho-киназы, и соединения, способного ингибировать активность Rho, и
b. нанесения указанной суспензии клеток на поверхность, где они закрепляются.
2. Способ по п. 1, в котором суммарное содержание О и N выше или равно 19,5%.
3. Способ по п. 1, в котором клетки поддерживают в культуре после закрепления клеток на поверхности.
4. Способ по п. 3, в котором по меньшей мере одно соединение удаляют после закрепления клеток на поверхности.
5. Способ по п. 1, в котором клетки открепляются от поверхности удалением по меньшей мере одного соединения.
6. Способ по п. 1, в котором суспензия клеток представляет собой суспензию кластеров клеток.
7. Способ по п. 1, в котором суспензия клеток представляет собой суспензию одиночных клеток.
8. Способ по п. 1, в котором поверхность является частью сосуда или матрицы.
| VAN KOOTEN T G ET AL: "Plasma-treated polystyrene surfaces: model surfaces for studying cell-biomaterial interactions" BIOMATERIALS, ELSEVIER SCIENCE PUBLISHERS BV., BARKING, GB, vol | |||
| Видоизменение пишущей машины для тюркско-арабского шрифта | 1923 |
|
SU25A1 |
| Печь-кухня, могущая работать, как самостоятельно, так и в комбинации с разного рода нагревательными приборами | 1921 |
|
SU10A1 |
| PARDO, A.M | |||
| ET AL: "Corning TM CellBIND TM Surface: An Improved Surface for Enhanced Cell Attachment" CORNING | |||

