ОБЛАСТЬ ПРИМЕНЕНИЯ ИЗОБРЕТЕНИЯ
В рамках настоящего изобретения истребуется приоритет заявки с серийным номером 61/116447, поданной 20 ноября 2008 года.
ОБЛАСТЬ ПРИМЕНЕНИЯ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к способам выращивания, размножения и дифференцирования плюрипотентных стволовых клеток на микроносителях.
ПРЕДПОСЫЛКИ СОЗДАНИЯ ИЗОБРЕТЕНИЯ
Плюрипотентные стволовые клетки, например эмбриональные стволовые клетки, имеют способность дифференцировать во все типы взрослых клеток. Таким образом, эмбриональные стволовые клетки могут являться источником резервных клеток и тканей для органов, поврежденных в результате заболевания, инфекции или врожденных аномалий. Реализация потенциальных возможностей использования эмбриональных стволовых клеток в качестве источника резервных клеток осложняется особенностями их размножения in vitro с сохранением плюрипотентности.
Существующие способы выращивания недифференцированных эмбриональных стволовых клеток требуют использования сложных условий культивирования, например культивирования эмбриональных стволовых клеток в присутствии слоя питающих клеток. В альтернативном варианте для культивирования эмбриональных стволовых клеток может использоваться среда, подвергнутая действию питающих клеток. В системах для культивирования, в которых реализованы указанные способы, нередко применяются клетки, полученные из организма другого вида по отношению к организму, чьи стволовые клетки культивируются (ксеногенные клетки). Кроме того, в эти культуральные среды может вводиться животная сыворотка.
Эмбриональные стволовые клетки обладают большим потенциалом для научно-исследовательской работы и скрининга лекарственных препаратов. В настоящее время крупномасштабное культивирование линий эмбриональных стволовых клеток человека представляет большие проблемы и связано со значительными трудностями. По существующим способам плюрипотентные стволовые клетки выращивают in vitro в культуральных флаконах, на плоских поверхностях, покрытых белками экстраклеточного матрикса (ЭКМ) или питающими клетками. При культивировании на плоскости необходимо часто пересевать клетки, поскольку ограниченная площадь не позволяет выращивать плюрипотентные стволовые клетки в течение долгого времени. Решением могут стать способы культивирования плюрипотентных стволовых клеток, основанные на использовании микроносителей. Микроносители имеют высокое отношение поверхности к объему, и, следовательно, снимается ограничение площади поверхности, характерное для выращивания плюрипотентных стволовых клеток на плоских поверхностях.
Например, в работе Fok et al. описаны культуральные системы для выращивания недифференцированных эмбриональных стволовых клеток (ЭСК) с перемешиванием суспензионных культур на микроносителе и агрегатных культур (Stem Cells 2005; 23: 1333-1342).
В другом примере Abranches et al. описал тестирование микропористого микроносителя Cytodex 3® (GE Healthcare Life Sciences, Нью Джерси, США), изготовленного из декстранового матрикса с коллагеновым слоем на поверхности, на предмет возможности выращивания мышиных ЭС клеток линии S25 в ротационных колбах (Biotechnol. Bioeng. 96 (2007), pp. 1211-1221).
В другом примере (патент № US20070264713) описан процесс культивирования недифференцированных стволовых клеток в суспензии и, в частности, способ культивирования стволовых клеток на микроносителях в сосудах.
В другом примере (заявка № WO2006137787) описано использование средства скрининга, состоящего из частиц или микроносителей, например капель, прикрепленных к твердой подложке, например планшете для микротитрования, для культивирования клеток на указанных микроносителях.
В другом примере (заявка № WO2008004990) описан способ стимулирования прикрепления, выживания и/или пролиферации стволовых клеток в культуре, заключающийся в культивировании стволовых клеток на опорной поверхности, которая имеет положительный заряд.
В другом примере (заявка № WO2007012144) описан биореактор, включающий опорную поверхность и синтетический прикрепляющий полипептид, связанный с опорной поверхностью, при этом синтетический прикрепляющий полипептид характеризуется высокой способностью к связыванию эмбриональных стволовых клеток или мультипотентных клеток.
КРАТКОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
В настоящем изобретении описаны способы выращивания, размножения и дифференцирования плюрипотентных стволовых клеток на микроносителях.
В одном из вариантов осуществления настоящего изобретения предлагается способ выращивания плюрипотентных стволовых клеток, включающий следующие этапы:
a) прикрепление популяции плюрипотентных стволовых клеток к первому объему микроносителей;
b) культивирование плюрипотентных стволовых клеток на первом объеме микроносителей;
c) отделение плюрипотентных стволовых клеток от первого объема микроносителей;
d) прикрепление популяции плюрипотентных стволовых клеток ко второму объему микроносителей.
КРАТКОЕ ОПИСАНИЕ ФИГУР
Фиг.1 - Ингибитор Rho-киназы стимулирует прикрепление человеческих эмбриональных стволовых клеток к микроносителям и их рост. Изображения клеток H9, выращенных в статичной культуре в течение двух суток на микроносителях HILLEX®II (Solohill, Мичиган, США). Клетки культивировали в среде, кондиционированной мышиными эмбриональными фибробластами (MEF-CM) в присутствии или отсутствие 10 мкМ ингибитора Rho-киназы, Y27632 ((Sigma-Aldrich, Миссури, США) A и B соответственно).
Фиг.2 - Клетки H9, выращенные на микроносителях. Клеткам H9 дали возможность прикрепиться к различным микроносителям и поместили их на качающуюся платформу при 37°C. Использовали микроносители Plastic, ProNectinF, HILLEX®II (Solohill, Мичиган, США) и микроносители Plastic Plus (A, B, C, D соответственно). Через 3 суток выращивания наилучшее прикрепление клеток наблюдалось на микроносителях HILLEX®II (Solohill, Мичиган, США). Стрелками показаны клетки, образующие агрегаты без прикрепления к микроносителям.
Фиг.3 - Пролиферация клеток H9 на микроносителях. Клетки H9 прикрепляли к микроносителям HILLEX®II, ProNectinF, Plastic Plus и Plastic (Solohill, Мичиган, США) и помещали в 6-луночном планшет на качающуюся платформу при 37°C в присутствии 10 мкМ Y27632 (Sigma-Aldrich, Миссури, США) и среды MEF-CM. Исходная плотность посева клеток - значение в день 0. Представлены количества клеток в день 3 и день 5.
Фиг.4 - Изображения клеток H1 после прикрепления к микроносителям. Представлены изображения клеток, прикрепленных к микроносителям ProNectinF, Plastic Plus и Plastic в дни 3, 5 и 7. Клетки выращивали в среде MEF-CM с добавлением 10 мкМ Y27632 (Sigma-Aldrich, Миссури, США) в 12-луночный планшет на качающейся платформе при 37°C. Клетки образовывали агрегаты независимо от прикрепления к микроносителям Plastic Plus и Plastic (стрелки на панелях G и H).
Фиг.5 - Изображения клеток H1 после прикрепления к микроносителям. Представлены изображения клеток, прикрепленных к микроносителям Cytodex 1®, Cytodex 3® (GE Healthcare Life Sciences, Нью Джерси, США) и HILLEX®II (Solohill, Мичиган, США). Клетки выращивали в среде MEF-CM с добавлением 10 мкМ Y27632 (Sigma-Aldrich, Миссури, США) в 12-луночный планшет на качающейся платформе при 37°C.
Фиг.6 - Пролиферация клеток H1 на микроносителях. Клеткам H1 дали возможность прикрепиться к микроносителям HILLEX®II (Solohill, Мичиган, США), Cytodex 1® (GE Healthcare Life Sciences, Нью-Джерси, США), Cytodex 3® (GE Healthcare Life Sciences, Нью-Джерси, США), ProNectinF (Solohill, Мичиган, США), Plastic Plus (Solohill, Мичиган, США) и Plastic (Solohill, Мичиган, США) и поместили в 12-луночный планшет на качающуюся платформу при 37°C в присутствии 10 мкМ Y27632 (Sigma-Aldrich, Миссури, США) и MEF-CM. Исходная плотность посева клеток - значение в день 0. Представлены количества клеток в дни 3, 5 и 7. Исходная плотность посева составляла 13333 клетки/см2, как показано линией.
Фиг.7 - Пролиферация клеток H9 на микроносителях при различных концентрациях ингибиторов Rho-киназы. Клетки выращивали в 12-луночном планшете на качающейся платформе и подсчитывали их количество в дни 4 и 7 для определения степени прикрепления и скорости пролиферации. А: Клетки H9 выращивали в среде MEF-CM с добавлением 1, 2,5, 5 или 10 мкМ Y27632 (Sigma-Aldrich, Миссури, США). B: Клетки H9 выращивали в среде MEF-CM с добавлением 0,5, 1, 2,5 или 5 мкМ глицил-H 1152 дигидрохлорида (Tocris, Миссури, США).
Фиг.8 - Клетки H1 выращивали в убывающих концентрациях ингибиторов Rho-киназы. Клетки H1p38 выращивали в присутствии Y27632 (Sigma-Aldrich, Миссури, США) или глицил-H 1152 дигидрохлорида (Tocris, Миссури, США) в течение двух дней в убывающих концентрациях (10 мкМ/5 мкМ, 2,5 мкМ/0,5 мкМ или 1,0 мкМ/0,5 мкМ) или при постоянной концентрации 0,25 мкМ глицил-H 1152 дигидрохлорида (Tocris, Миссури, США). Клеткам давали возможность прикрепиться к микроносителям HILLEX®II (Solohill, Мичиган, США), Cytodex 1® или Cytodex 3® ((GE Healthcare Life Sciences, Нью-Джерси, США) A, B, C соответственно). Количество клеток подсчитывали на 3, 5 и 7 день после посева.
Фиг.9 - Определение прикрепления клеток к микроносителям при разных плотностях посева в ротационные колбы. Клетки H1 сеяли на микроносители Cytodex 3® (GE Healthcare Life Sciences, Нью-Джерси, США) с уровнями плотности, указанными слева: низкой (0,4×104 клеток/см2), средней (1,2×104 клеток/см2) или высокой (3×104 клеток/см2). На 3, 5 и 7 дни клетки фотографировали и определяли процентную долю микроносителей с прикрепленными клетками (указано на изображении).
Фиг.10 - На рост клеток в ротационных пробирках влияет исходная плотность посева. Клетки H1 сеяли на микроносители Cytodex 3® (GE Healthcare Life Sciences, Нью-Джерси, США) с уровнями плотности, указанными слева: низкой (0,4×104 клеток/см2), средней (1,2×104 клеток/см2) или высокой (3×104 клеток/см2). На 3, 5 и 7 дни клетки отделяли от микроносителей и подсчитывали их количество.
Фиг.11 - Определение скорости роста клеток на микроносителях при разных плотностях посева в ротационные колбы. Клетки H1 сеяли на микроносители Cytodex 3® (GE Healthcare Life Sciences, Нью-Джерси, США) с разными уровнями плотности (день 0): низкой (0,4×104 клеток/см2), средней (1,2×104 клеток/см2) или высокой (3×104 клеток/см2). На 3, 5 и 7 дни клетки отделяли от микроносителей и подсчитывали их количество. Показана зависимость кратности увеличения количества клеток от исходной плотности посева.
Фиг.12 - Показан рост клеток H1 на микроносителях Cytodex 3® (GE Healthcare Life Sciences, Нью-Джерси, США) через 7 дней культивирования. Клетки получали среду MEF-CM без ингибитора Rho-киназы, начиная с 3 дня и далее. Клетки оставались прикрепленными к микроносителям.
Фиг.13 - Рост и диссоциация клеток H9 на микроносителях HILLEX®II (Solohill, Мичиган, США). А, B: 10- и 20-кратные изображения клеток H9, выращиваемых в течение шести дней на микроносителях HILLEX®II (Solohill, Мичиган, США). C: 20-кратное изображение клеток, диссоциировавшихся от микроносителей HILLEX®II (Solohill, Мичиган, США) за 10 минут воздействия 0,05% раствором Трипсин/EDTA. D: 20-кратное изображение клеток, диссоциировавшихся от микроносителей HILLEX®II (Solohill, Мичиган, США) за 10 минут воздействия раствором TrypLE™ Express.
Фиг.14 - Диссоциация клеток H9 от микроносителей. Клетки H9, выращенные на микроносителях HILLEX®II (Solohill, Мичиган, США) на качающейся платформе, диссоциировались при помощи раствора TrypLE™ Express или 0,05% Трипсин/EDTA. Представлено количество клеток и их жизнеспособность - A и B соответственно.
Фиг.15 - Диссоциация клеток H1 от микроносителей. Клетки H1, выращенные на микроносителях Cytodex 3® (GE Healthcare Life Sciences, Нью-Джерси, США) в ротационной колбе, диссоциировались при помощи реактива TrypLE™ Express (Invitrogen, CA), Accutase™ или коллагеназы (10 мг/мл). Представлено количество клеток и их жизнеспособность - A и B соответственно.
Фиг.16 - Клетки H9, выращенные на микроносителях HILLEX®II (Solohill, Мичиган, США), не переносятся между микроносителями.
Фиг.17 - Клетки H9, пассаж 43, выращивались на протяжении 5 пассажей на микроносителях HILLEX®II (Solohill, Мичиган, США) в ротационной колбе. Количество клеток подсчитывали каждые 2-3 дня и пересевали по достижении плотности 1-2×105 клеток/см2.
Фиг.18 - Клетки H9, пассаж 43, выращивались на протяжении 5 пассажей на микроносителях Cytodex 3® (GE Healthcare Life Sciences, Нью-Джерси, США) в ротационной колбе. Количество клеток подсчитывали каждые 2-3 дня и пересевали по достижении плотности 1-2×105 клеток/см2.
Фиг.19 - Путем сортировки флуоресцентно-активированных клеток (FACS) показана плюрипотентность клеток H9, выращенных в ротационных колбах. A: большинство клеток H9 p43, выращенных на микроносителях HILLEX®II (Solohill, Мичиган, США), экспрессируют белки-маркеры плюрипотентности. Для клеток пассажей 1 и 3 не проводился анализ на TRA-1-81. B: большинство клеток H9 p43, выращенных на микроносителях Cytodex 3® (GE Healthcare Life Sciences, Нью-Джерси, США), экспрессируют белки-маркеры плюрипотентности. Для клеток пассажа 1 не проводился анализ на TRA-1-81.
Фиг.20 - Клетки H1 p49 выращивались на протяжении 5 пассажей на микроносителях Cytodex 1® (GE Healthcare Life Sciences, Нью-Джерси, США) в ротационной колбе. Количество клеток подсчитывали каждые 2-3 дня и пересевали по достижении плотности 4-8×104 клеток/см2.
Фиг.21 - Клетки H1, пассаж 49, выращивались на протяжении 5 пассажей на микроносителях Cytodex 3® (GE Healthcare Life Sciences, Нью-Джерси, США) в ротационной колбе. Количество клеток подсчитывали каждые 2-3 дня и пересевали по достижении плотности 1-2×105 клеток/см2.
Фиг.22 - Путем сортировки флуоресцентно-активированных клеток (FACS) показана плюрипотентность клеток H1, выращенных в ротационных колбах.
Фиг.23 - Удвоение популяции клеток H1 и H9 на микроносителях. Сроки удвоения популяции клеток вычисляли от дня 3 до дня пересевания (день 5, 6 или 7).
Фиг.24 - Клетки H9, культивированные на микроносителях в определенной среде. Клетки культивировали на микроносителях HILLEX®II (HII, (Solohill, Мичиган, США)) или Cytodex 3® (C3, (GE Healthcare Life Sciences, Нью-Джерси, США)). Клетки культивировали на микроносителях в одной из следующих сред: mTESR (StemCell Technologies, Ванкувер, Канада), StemPro или MEF-CM. К среде добавляли 10 мкМ Y27632 (Y (Sigma-Aldrich, Миссури, США)) или 2,5 мкМ глицил-H 1152 дигидрохлорида (H (Tocris, Миссури, США)). Скорость роста определяли на 3, 5 и 7 день после посева.
Фиг.25 - Клетки H1, пассаж 38, культивировали на микроносителях в определенной среде. Клетки культивировали на микроносителях HILLEX®II (HII (Solohill, Мичиган, США)) или Cytodex 3® (C3 (GE Healthcare Life Sciences, Нью-Джерси, США)). Клетки культивировали на микроносителях в одной из следующих сред: mTESR (StemCell Technologies, Ванкувер, Канада), StemPro и MEF-CM. К среде добавляли 10 мкМ Y27632 (Y (Sigma-Aldrich, Миссури, США)) или 2,5 мкМ глицил-H 1152 дигидрохлорида (H (Tocris, Миссури, США)). Скорость роста определяли на 3, 5 и 7 день после посева.
Фиг.26 - Клетки H1, пассаж 50, культивировали на микроносителях HILLEX®II (Solohill, Мичиган, США) в определенной среде в ротационных колбах. A: изображения клеток H1 p50, выращенных в среде MEF-CM, через 3, 7 или 9 дней культивации в ротационной колбе. B: изображения клеток H1 p50, выращенных в среде mTESR (StemCell Technologies, Ванкувер, Канада), через 3, 7 или 9 дней культивации. Стрелками указаны кластеры клеток, не прикрепившихся к микроносителям.
Фиг.27 - Дифференцирование человеческих эмбриональных стволовых клеток, пересевавшихся пять раз в ротационных колбах. A: клетки H9, пассаж 43, пересевали пять раз на микроносители Cytodex 3® (GE Healthcare Life Sciences, Нью-Джерси, США). B: клетки H1, пассаж 49, пересевали пять раз на микроносители Cytodex 1® (GE Healthcare Life Sciences, Нью-Джерси, США). Оба типа клеток отделяли от микроносителей и сеяли на планшеты, покрытые материалом MATRIGEL (BD Biosciences, Калифорния, США). В отношении клеток, имевших конфлюэнтность 80-90%, реализовали протокол дифференцирования эмбриональных стволовых клеток в сформированную эндодерму. Анализ проводили методом FACS для определения процентной доли клеток, экспрессирующих CXCR4, маркер сформированной эндодермы. Процент CXCR4-положительных клеток указан в верхнем правом углу графика.
Фиг.28 - Дифференцирование клеток H1 в сформированную эндодерму на микроносителях. На графиках FACS представлена процентная доля клеток, экспрессирующих маркер сформированной эндодермы CXCR4. Процент клеток с положительным результатом указан в верхнем правом углу. Перед обработкой клетки выращивались на микроносителях в ротационных колбах. A: клетки H1, пассаж 40, перед дифференцированием выращивались на микроносителях Cytodex 1® (GE Healthcare Life Sciences, Нью-Джерси, США) в течение 6 дней после пассажа 5. B: клетки H1, пассаж 40, перед дифференцированием выращивались на микроносителях Cytodex 3® (GE Healthcare Life Sciences, Нью-Джерси, США) в течение 8 дней после пассажа 1. C: клетки H1, пассаж 50, перед дифференцированием выращивались на микроносителях HILLEX®II (Solohill, Мичиган, США) в течение 6 дней после пассажа 1.
Фиг.29 - Дифференцирование клеток H1 в сформированную эндодерму на микроносителях Cytodex 3® (GE Healthcare Life Sciences, Нью-Джерси, США). A: клетки H1, пассаж 40, выращивали на микроносителях 8 дней. B: клетки H1, пассаж 40, выращивали на микроносителях 11 дней. Затем обе популяции клеток дифференцировались в сформированную эндодерму на качающейся платформе при 37°C. На графиках FACS представлена процентная доля клеток, экспрессирующих маркер сформированной эндодермы CXCR4. Процент клеток с положительным результатом указан в верхнем правом углу.
Фиг.30 - Дифференцирование человеческих эмбриональных стволовых клеток линии H1, культивированных на микроносителях, в сформированную эндодерму. По оси Y представлены результаты FACS с процентной долей CXCR4-положительных клеток. Клетки H1 находились на микроносителях HILLEX®II, Cytodex 1® или Cytodex 3® как до, так и во время дифференцирования.
Фиг.31 - Дифференцирование человеческих эмбриональных стволовых клеток линии H1, культивированных на микроносителях, в клетки панкреатической эндодермы. По оси Y представлены значения CT для маркеров панкреатической эндодермы, Ngn3, Nkx6.1 и Pdx1. Клетки H1 дифференцировались на микроносителях HILLEX®II (HII), Cytodex 1® (C1) или Cytodex 3® (C3) в среде DMEM-High Glucose (HG) или DMEM-F12 (F12). Продолжительность протокола дифференцирования составила 13 дней.
Фиг.32 - Дифференцирование человеческих эмбриональных стволовых клеток линии H1, культивированных на микроносителях, в гормон-продуцирующие панкреатические клетки. По оси Y представлены результаты FACS с процентной долей клеток, положительных по маркерам гормон-продуцирующих панкреатических клеток, синаптофизину, глюкагону и инсулину. Клетки H1 сеяли в двух концентрациях - 10×105 (10) или 20×105 (20) - на микроносители Cytodex 3® (C-3). Клетки дифференцировались в среде DMEM-High Glucose (HG) с четвертого по девятый день, а затем подвергались дальнейшей дифференцировке в среде HG или DMEM-F12 (F12) с 10 по 24 день.
Фиг.33 - Дифференцирование клеток H1 в эндокринные клетки на микроносителях Cytodex 3® (GE Healthcare Life Sciences, Нью-Джерси, США). Клетки H1 дифференцировались в панкреатические эндокринные клетки через панкреатическую эндодерму (день 14), панкреатические эндокринные клетки (день 21) в инсулинэкспрессирующие клетки (день 28). Был определен уровень экспрессии генов Pdx1, глюкагона и инсулина (A, B, C соответственно). Клетки H1, выращенные и дифференцированные на микроносителях Cytodex 3® (GE Healthcare Life Sciences, Нью-Джерси, США) (C3), сравнивались с клетками, выращенными и дифференцированными на 6-луночных планшетах (плоских), покрытых материалом MATRIGEL (BD Biosciences, Калифорния, США). Значения уровня экспрессии генов для клеток, выращенных на микроносителях Cytodex 3® (GE Healthcare Life Sciences, Нью-Джерси, США), были получены в трех повторностях.
Фиг.34 - Клетки H9 дифференцировались в сформированную эндодерму (СЭ) на микроносителях Cytodex 3® (GE Healthcare Life Sciences, Нью-Джерси, США). Графики FACS по экспрессии CXCR4. Процентная доля клеток, положительных по маркеру сформированной эндодермы CXCR4, указана в верхнем правом углу. A: клетки H9, пассаж 39, росли на 6-луночных планшетах, покрытых материалом MATRIGEL (BD Biosciences, Калифорния, США), и дифференцировались в СЭ. B, C: дублированные образцы клеток H9 на микроносителях Cytodex 3® (GE Healthcare Life Sciences, Нью-Джерси, США) переносили из ротационных колб в 12-луночный планшет и инкубировали на качающейся платформе.
Фиг.35 - Дифференцирование клеток H9 в инсулин-экспрессирующие клетки на микроносителях Cytodex 3® (GE Healthcare Life Sciences, Нью-Джерси, США). Клетки H9 дифференцировались в панкреатические эндокринные клетки через панкреатическую эндодерму (день 14), эндокринные клетки (день 22) в инсулинэкспрессирующие клетки (день 29). Были определены уровни экспрессии генов Pdx1, глюкагона и инсулина (A, B, C соответственно). Клетки H9, выращенные и дифференцированные на микроносителях Cytodex 3® (GE Healthcare Life Sciences, Нью-Джерси, США) (C3), сравнивались с клетками, выращенными и дифференцированными на 6-луночных планшетах (плоских), покрытых материалом MATRIGEL (BD Biosciences, Калифорния, США).
Фиг.36 - Сохранение плюрипотентности человеческих эмбриональных стволовых клеток, культивированных в течение 5 пассажей на микроносителях Cytodex 3®, а затем перенесенных и культивированных на указанных плоских субстратах в присутствии ингибитора Rho-киназы. На панели A представлена экспрессия маркеров плюрипотентности CD9, SSEA3, SSEA4, Tra-160 и Tra-181, определенная проточной цитометрией. На панели B представлена экспрессия маркеров плюрипотентности Nanog, Pou5F1, SOX2 и ZFP42, а также маркеров дифференцирования FOXA2, FOXD3, GATA2, GATA4 и брахиурии, определенных методом ПЦР в реальном времени.
Фиг.37 - Образование сформированной эндодермы человеческими эмбриональными стволовыми клетками, культивированными в течение 5 пассажей на микроносителях Cytodex 3®, а затем перенесенными и культивированными на указанных плоских субстратах в присутствии ингибитора Rho-киназы. На панели А представлена экспрессия CXCR4, определенная проточной цитометрией. На панели B представлена экспрессия указанных маркеров, определенная методом ПЦР в реальном времени.
Фиг.38 - Образование сформированной эндодермы человеческими эмбриональными стволовыми клетками, культивированными в течение 5 пассажей на микроносителях Cytodex 3®, а затем перенесенными и культивированными на плоском субстрате PRIMARIA™. Экспрессия указанных генов определялась проточной цитометрией.
Фиг.39 - Человеческие эмбриональные стволовые клетки, культивированные на плоских субстратах, сохраняют плюрипотентность. Отбирались образцы мРНК, пересеянные с помощью TrypLE™, Accutase™ или коллагеназы человеческих эмбриональных стволовых клеток, и проводился анализ на экспрессию в мРНК гена плюрипотентности. Клетки выращивались в течение 4 дней (один пассаж) либо в культуре на материале MATRIGEL в среде, кондиционированной мышиными эмбриональными фибробластами (МЭФ) (A), либо в течение одного пассажа на материале Primaria™ в среде, кондиционированной МЭФ, с добавлением ингибитора Rock (B), либо в течение двух пассажей на материале Primaria™ в среде, кондиционированной МЭФ, с добавлением ингибитора Rock (C).
Фиг.40 - Человеческие эмбриональные стволовые клетки H1, выращиваемые в течение свыше 7 пассажей (больше p45) на материале PRIMARIA, пересеянные с использованием реактива AccutaseTM или TrypLETM с разведением 1:4, 1:8 или 1:16 на материал PRIMARIA в присутствии ингибитора Rho-киназы глицил-H 1152 дигидрохлорида, были проверены на плюрипотентность (A) и на способность дифференцироваться в сформированную эндодерму (B). Контролем служат человеческие эмбриональные стволовые клетки H1 p48, выращенные на 1:30 MATRIGEL, пересеянные с использованием коллагеназы. 10mA = пересев с 10-минутным воздействием Accutase™. 10mT = пересев с 10-минутным воздействием TrypLE™. Указана информация о соотношении при пересеве 1:4, 1:8 или 1:16. Значение P(X) показывает номер пассажа с момента переноса с питающих клеток МЭФ на пластик Primaria™.
Фиг.41 - Человеческие эмбриональные стволовые клетки H1, выращиваемые в течение свыше 7 пассажей (больше p45) на материале PRIMARIA, пересеянные с использованием реактива Accutase™ или TrypLE™ с разведением 1:4 на PRIMARIA в присутствии ингибитора Rho-киназы глицил-H 1152 дигидрохлорида, были проверены на экспрессию мРНК маркеров плюрипотентности и дифференцирования. Контролем является исходная популяция клеток пассажа - 37. 10min Accutase™ = пересев с 10-минутным воздействием Accutase™. Значение P(X) показывает номер пассажа с момента переноса с питающих клеток МЭФ на пластик PRIMARIA™.
Фиг.42 - Человеческие эмбриональные стволовые клетки H1, выращиваемые в течение свыше 7 пассажей (больше p45) на материале PRIMARIA™, пересеянные с использованием реактива Accutase™ или TrypLE™ с разведением 1:8 на PRIMARIA в присутствии ингибитора Rho-киназы глицил-H 1152 дигидрохлорида, были проверены на экспрессию мРНК маркеров плюрипотентности и дифференцирования. Контролем является исходная популяция клеток пассажа - 37. 10min Accutase™ = пересев с 10-минутным воздействием Accutase™. Значение P(X) показывает номер пассажа с момента переноса с питающих клеток МЭФ на пластик PRIMARIA™.
Фиг.43 - Человеческие эмбриональные стволовые клетки H1, выращиваемые в течение свыше 7 пассажей (больше p45) на материале PRIMARIA, пересеянные с использованием реактива Accutase™ или TrypLE™ с разведением 1:16 на PRIMARIA в присутствии ингибитора Rho-киназы глицил-H 1152 дигидрохлорида, были проверены на экспрессию мРНК маркеров плюрипотентности и дифференцирования. Контролем является исходная популяция клеток пассажа - 37. 10min Accutase™ = пересев с 10-минутным воздействием Accutase™. Значение P(X) показывает номер пассажа с момента переноса с питающих клеток МЭФ на пластик PRIMARIA™.
Фиг.44 - Изображения клеток H1, выращенных на плоских субстратах Primaria™ (№ по каталогу 353846, Becton Dickinson, Franklin Lakes, Нью-Джерси, США), а затем перенесенных на микроносители через 3 дня после посева. A-C: клетки H1 сеяли на микроносители Cytodex 3® (GE Healthcare Life Sciences, Нью-Джерси, США). D-F: клетки сеяли на микроносители HILLEX®II (Solohill, Мичиган, США). A, D: клетки H1 сеяли на плоские планшеты Primaria™ (№ по каталогу 353846, Becton Dickinson, Franklin Lakes, Нью-Джерси, США) и проходили 10-минутную обработку реактивом TrypLE™ Express (Invitrogen, Калифорния, США) перед переносом на микроносители. A, E: клетки H1 сеяли на плоские планшеты Primaria™ (№ по каталогу 353846, Becton Dickinson, Franklin Lakes, Нью-Джерси, США) и проходили 10-минутную обработку реактивом Accutase™ перед переносом на микроносители. C, F: клетки H1, пассаж 46, сеяли на планшеты с покрытием MATRIGEL (BD Biosciences, Калифорния, США) и обрабатывали коллагеназой (1 мг/мл) перед переносом на микроносители.
Фиг.45 - Плюрипотентность клеток H1, выращенных на плоских субстратах Primaria™ (№ по каталогу 353846, Becton Dickinson, Franklin Lakes, Нью-Джерси, США), а затем перенесенных на микроносители Cytodex 3® (GE Healthcare Life Sciences, Нью-Джерси, США) и HILLEX®II. Результаты FACS-анализа на белки клеточной поверхности, являющиеся маркерами плюрипотентности. Клетки при пересеве на планшеты Primaria™ (№ по каталогу 353846, Becton Dickinson, Franklin Lakes, Нью-Джерси, США) проходили обработку реактивом Accutase™ или TrypLE™ Express (Invitrogen, Калифорния, США) в течение 3-10 минут.
Фиг.46 - Дифференцирование клеток H1, выращенных на планшетах Primaria™ (№ по каталогу 353846, Becton Dickinson, Franklin Lakes, Нью-Джерси, США), а затем перенесенных на микроносители Cytodex 3® (GE Healthcare Life Sciences, Нью-Джерси, США). Результаты FACS-анализа на экспрессию CXCR4, маркера сформированной эндодермы, на поверхности клеток. Клетки при пересеве на планшеты Primaria™ (№ по каталогу 353846, Becton Dickinson, Franklin Lakes, Нью-Джерси, США) проходили обработку реактивом Accutase™ или TrypLE™ Express (Invitrogen, Калифорния, США) в течение 3-10 минут.
Фиг.47 - Результаты FACS-анализа для человеческих эмбриональных стволовых клеток, которые перед культивированием на микроносителях культивировали на плоских субстратах, состоящих из смеси эфиров целлюлозы.
Фиг.48 - Результаты FACS-анализа по экспрессии маркеров, характерных для линии сформированной эндодермы, в эмбриональных стволовых клетках, которые перед культивированием и дифференцированием на микроносителях культивировали на плоских субстратах из смеси эфиров целлюлозы.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Для ясности описания, а не для ограничения изобретения, подробное описание изобретения разделено на следующие подразделы, описывающие или иллюстрирующие определенные особенности, варианты осуществления или области применения настоящего изобретения.
Определения
Стволовые клетки представляют собой недифференцированные клетки, определяемые по способности на уровне единичной клетки как самообновляться, так и дифференцироваться с образованием клеток-потомков, таких как самообновляющиеся клетки-предшественники, необновляющиеся клетки-предшественники и окончательно дифференцированные клетки. Стволовые клетки также характеризуются способностью дифференцироваться in vitro в функциональные клетки различных клеточных линий дифференцирования из нескольких зародышевых листков (эндодермы, мезодермы и эктодермы), а также после трансплантации давать начало тканям, происходящим от нескольких зародышевых листков, и вносить существенный вклад в формирование большинства, если не всех, тканей после инъекции в бластоцисты.
По потенциалу развития стволовые клетки классифицируются следующим образом: (1) тотипотентные, т.е. способные давать начало всем эмбриональным и внеэмбриональным типам клеток; (2) плюрипотентные, т.е. способные давать начало всем эмбриональным типам клеток; (3) мультипотентные, т.е. способные давать начало группе клеточных линий дифференцирования в пределах конкретной ткани, органа или физиологической системы (например, гематопоэтические стволовые клетки (HSC) могут давать таких потомков, как HSC (самообновление), олигопотентные предшественники, ограниченные клетками крови, и все типы клеток и клеточных элементов (таких как тромбоциты), являющиеся нормальными компонентами крови); (4) олигопотентные, т.е. способные давать начало более ограниченному набору клеточных линий дифференцировки, чем мультипотентные стволовые клетки; (5) унипотентные, т.е. способные давать начало единственной клеточной линии дифференцировки (например, сперматогенные стволовые клетки).
Дифференцирование представляет собой процесс, при помощи которого неспециализированная («некоммитированная») или менее специализированная клетка приобретает свойства специализированной клетки, например нервной или мышечной клетки. Дифференцированная клетка или клетка с индуцированным дифференцированием представляет собой клетку, занявшую более специализированное («коммитированное») положение в линии дифференцирования клетки. Термин «коммитированная» применительно к процессу дифференцирования обозначает клетку, дошедшую в ходе процесса дифференцирования до стадии, от которой в нормальных условиях она продолжит дифференцироваться до определенного типа клеток или набора типов клеток и не сможет в нормальных условиях дифференцироваться в иной тип клеток или вернуться обратно к менее дифференцированному типу. Дедифференцированием называется процесс, в ходе которого клетка возвращается к менее специализированному (или коммитированному) положению в линии дифференцирования. Используемый в настоящем документе термин «линия дифференцирования клетки» определяет наследственность клетки, то есть определяет, из какой клетки произошла данная клетка и каким клеткам она может дать начало. В линии дифференцирования клетка помещается в наследственную схему развития и дифференцирования. Маркером, специфичным для линии дифференцирования, называется характерная особенность, специфически ассоциированная с фенотипом клеток конкретной линии дифференцирования, которая может использоваться для оценки дифференцирования некоммитированных клеток в клетки данной линии дифференцирования.
Для описания клеток в процессе культивирования используются различные термины. Термин «поддержание» относится в целом к клеткам, помещенным в среду роста в условиях, которые способствуют росту и/или делению клеток, в результате чего популяция клеток может увеличиваться или не увеличиваться. Термин «пересевание» означает процесс изъятия клеток из одного культурального сосуда и переноса их в другой культуральный сосуд в условия, которые способствуют росту и/или делению клеток.
Конкретная популяция клеток или клеточная линия иногда описывается или характеризуется количеством выполненных с ней пересеваний (пассажей). Например, пересеянная 10 раз культивируемая популяция клеток может описываться как культура десятого пассажа, или культура P10. Первичная культура, т.е. первая культура после выделения клеток из ткани, получает обозначение P0. После первого пересева клетки описываются как вторичная культура (P1, или культура первого пассажа). После второго пересева клетки превращаются в третичную культуру (P2, или культура второго пассажа) и т.д. Специалистам в данной области будет понятно, что за период пассажа численность популяции может многократно удваиваться. Следовательно, количество удвоений численности популяции превышает номер пассажа. Уровень размножения клеток (то есть количество удваиваний популяции) за промежуток времени между последовательными пересевами зависит от многих факторов, включая, помимо прочего, плотность посева, тип субстрата, тип среды, условия роста и время между пересевами.
Термин «β-клеточная линия дифференцирования» относится к клеткам, положительным по экспрессии гена транскрипционного фактора PDX-1 и по меньшей мере одного из следующих транскрипционных факторов: NGN-3, Nkx2.2, Nkx6.1, NeuroD, Isl-1, HNF-3 beta, MAFA, Pax4 или Pax6. К клеткам, экспрессирующим маркеры, характерные для β-клеточной линии дифференцирования, относятся β-клетки.
Термин «клетки, экспрессирующие маркеры, характерные для линии сформированной эндодермы» в настоящем документе означает клетки, экспрессирующие по меньшей мере один из следующих маркеров: SOX-17, GATA-4, HNF-3 beta, GSC, Cer1, Nodal, FGF8, Brachyury, гомеобоксный белок Mix-like, FGF4, CD48, эомезодермин (EOMES), DKK4, FGF17, GATA-6, CXCR4, C-Kit, CD99 или OTX2. К клеткам, экспрессирующим маркеры, характерные для линии сформированной эндодермы, относятся клетки-предшественники первичной полоски, клетки первичной полоски, клетки мезоэндодермы и клетки сформированной эндодермы.
Термин «клетки, экспрессирующие маркеры, характерные для линии панкреатической эндодермы» в настоящем документе означает клетки, экспрессирующие по меньшей мере один из следующих маркеров: PDX-1, HNF-1 beta, PTF-1 alpha, HNF-6 или HB9. К клеткам, экспрессирующим маркеры, характерные для линии панкреатической эндодермы, относятся клетки панкреатической эндодермы.
Используемый в настоящей заявке термин «клетки с экспрессией маркеров, характерных для линии дифференцирования в панкреатические эндокринные клетки» относится к клеткам с экспрессией по меньшей мере одного из следующих маркеров: NGN-3, NeuroD, Islet-1, PDX-1, NKX6.1, Pax-4, Ngn-3 и PTF-1 alpha. Клетки, экспрессирующие маркеры, характерные для линии дифференцирования в панкреатические эндокринные клетки, включают панкреатические эндокринные клетки, панкреатические экспрессирующие гормоны клетки и панкреатические секретирующие гормоны клетки, а также клетки β-клеточной линии дифференцирования.
Используемый в настоящем документе термин «сформированная эндодерма» относится к клеткам, обладающим характерными особенностями клеток, происходящих в ходе гаструляции от эпибласта и формирующих желудочно-кишечный тракт и его производные. Клетки сформированной эндодермы экспрессируют следующие маркеры: CXCR4, HNF-3 beta, GATA-4, SOX-17, Cerberus, OTX2, goosecoid, c-Kit, CD99 и Mixl1.
Термином «внеэмбриональная эндодерма» в настоящем документе означает популяцию клеток, экспрессирующих по меньшей мере один из следующих маркеров: SOX-7, AFP или SPARC.
Используемый в настоящей заявке термин «маркеры» означает молекулы нуклеиновых кислот или полипептидов с дифференциальной экспрессией в интересующих клетках. В данном контексте под дифференциальной экспрессией подразумевается повышение уровня экспрессии для положительного маркера и понижение уровня экспрессии для отрицательного маркера. Поддающийся обнаружению уровень маркерной нуклеиновой кислоты или полипептида в интересующих клетках оказывается значительно выше или ниже по сравнению с другими клетками, что позволяет идентифицировать интересующую клетку и отличить ее от других клеток с помощью любого из множества известных в данной области способов.
Термин «мезэндодермальная клетка» в настоящем документе означает клетку, экспрессирующую по меньшей мере один из следующих маркеров: CD48, эомезодермин (EOMES), SOX-17, DKK4, HNF-3 beta, GSC, FGF17 и GATA-6.
Термин «панкреатическая эндокринная клетка» или «клетка, экспрессирующая панкреатические гормоны» в настоящем документе относится к клеткам, способным к экспрессии по меньшей мере одного из следующих гормонов: инсулина, глюкагона, соматостатина и панкреатического полипептида.
Термин «панкреатическая секретирующая гормоны клетка» в настоящем документе относится к клеткам, способным к секреции по меньшей мере одного из следующих гормонов: инсулин, глюкагон, соматостатин и панкреатический полипептид.
Термин «клетка-предшественник клетки первичной полоски» в настоящем документе относится к клеткам, экспрессирующим по меньшей мере один из следующих маркеров: Nodal и FGF8.
Термин «клетка первичной полоски» в настоящем документе относится к клеткам, экспрессирующим по меньшей мере один из следующих маркеров: брахиурия, гомеобоксный белок Mix-like или FGF4.
Микроносители
Микроносителями называются частицы, сферы или гранулы, которые могут использоваться для связывания и культивирования клеток, зависимых от культуральной подложки. Микроносители обладают следующими свойствами: (a) они имеют достаточно малые размеры, чтобы их можно было использовать в суспензионных культурах (при скорости перемешивания, не приводящей к сдвиговым повреждениям микроносителей или клеток); (b) они являются цельными или имеют цельное ядро и пористое покрытие на поверхности; (c) их поверхности (внешняя и внутренняя, в случае пористых носителей) могут иметь положительный или отрицательный заряд. В одном аспекте изобретения общий диаметр частицы находится в диапазоне от приблизительно 150 до 350 мкм, а плотность заряда - в диапазоне от приблизительно 0,8 до 2,0 мг-экв./г. К применимым микроносителям относятся, помимо прочего, Cytodex 1®, Cytodex 2® или Cytodex 3® (GE Healthcare Life Sciences, Нью-Джерси, США).
В другом аспекте микроноситель представляет собой цельный носитель. Цельные носители хорошо подходят для адгезионных клеток, например для клеток, зависимых от культуральной подложки. Частица носителя может также представлять собой пористый микроноситель.
Термин «пористые микроносители» относится к частицам, которые могут использоваться для связывания и культивирования клеток, зависимых от культуральной подложки. Пористые микроносители обладают следующими свойствами: (a) они имеют достаточно малые размеры, чтобы их можно было использовать в суспензионных культурах (при скорости перемешивания, не приводящей к сдвиговым повреждениям микроносителей или клеток); (b) они имеют поры и внутренние лакуны, размеры которых достаточны для миграции клеток в лакуны; (c) их поверхности (внешняя и внутренняя) могут иметь положительный или отрицательный заряд. В одной серии вариантов осуществления носители: (a) имеют общий диаметр частицы от приблизительно 150 до 350 мкм; (b) имеют поры с диаметром отверстия от приблизительно 15 до приблизительно 40 мкм; (c) имеют плотность положительного заряда от приблизительно 0,8 до 2,0 мг-экв./г. В некоторых вариантах осуществления положительный заряд придается группами DEAE (N,N,-диэтиламиноэтил). К применимым пористым микроносителям относятся, помимо прочего, Cytopore 1® и Cytopore 2® (GE Healthcare Life Sciences, Piscataway, Нью-Джерси, США). Микроносители могут иметь любую форму, но, как правило, они имеют приблизительно сферическую форму, могут содержать макро- или микропоры или могут быть цельными.
В продаже имеются как цельные, так и пористые типы микроносителей различных производителей. К примерам имеющихся в продаже микроносителей относятся Cytodex 1® и Cytodex 3® (GE Healthcare Life Sciences, Нью-Джерси, США), представляющие собой декстрановые микроносители производства GE Healthcare Life Sciences. К пористым микроносителям, имеющимся на рынке, относятся продукты Cytoline и Cytopore, также производства GE Healthcare Life Sciences. Кроме того, в продаже представлены продукты Biosilon (NUNC) и Cultispher (Percell Biolytica). В другом аспекте микроносители могут состоять или иметь покрытие из поликарбоната или смеси эфиров целлюлозы.
Микроносители, которые используются в рамках настоящего изобретения, могут состоять из материалов природного или синтетического происхождения. Примерами являются микроносители на основе коллагена, целлюлозы, а также стекла, керамики, полимеров и металлов. Микроноситель может не содержать белков или иметь белковое покрытие, например, из коллагена. В дополнительном аспекте микроноситель может состоять или иметь покрытие из соединений, улучшающих связывание клетки с микроносителем и улучшающих отделение клетки от микроносителя, включая, помимо прочего, такие соединения, как поли(моностеароилглицерид-ко-янтарная кислота), поли-D,L-лактид-ко-гликолид, натрия гиалуронат, коллаген, фибронектин, ламинин, эластин, лизин, n-изопропилакриламид, витронектин.
Микроносители для культивирования клеток
Культивирование на микроносителях представляет собой технологию, которая обеспечивает возможность получения высокого практического выхода в культуре клеток, зависимых от культуральной подложки, например человеческих эмбриональных стволовых клеток. Микроносители специально разработаны для культивирования клеток, например человеческих эмбриональных стволовых клеток, при объемах культуры в диапазоне от нескольких миллилитров до более тысячи литров. Микроноситель является биологически инертным. Он образует надежный, но не жесткий субстрат для смешиваемых культур на микроносителях. Микроносители могут быть прозрачными, что позволяет проводить микроскопическое исследование прикрепленных клеток. Микроноситель Cytodex 3® (GE Healthcare Life Sciences, Нью-Джерси, США) состоит из тонкого слоя денатурированного коллагена, химически связанного с матриксом из сшитого декстрана. Слой денатурированного коллагена на носителях Cytodex 3® (GE Healthcare Life Sciences, Нью-Джерси, США) может разлагаться различными протеазами, включая трипсин и коллагеназу, что дает возможность извлекать клетки из микроносителя, сохраняя максимальную жизнеспособность, функциональность и целостность клеток.
Для культивирования человеческих эмбриональных стволовых клеток могут использоваться микроносители, не содержащие белка. Например, микроносители, предназначенные для производственного, лабораторного или исследовательского применения, предлагаемые под торговым наименованием HILLEX® (SoloHill Engineering, Inc., Мичиган, США), представляют собой модифицированные полистироловые сферы с закрепленными на поверхности катионами триметиламмония для придания поверхности микроносителя положительного заряда. Диаметр сфер находится в диапазоне от приблизительно 90 до приблизительно 200 микрон.
Способы культивирования клеток с использованием микроносителей дают много преимуществ, включая простоту дальнейшей технологической обработки во многих областях применения. Микроносители, как правило, имеют приблизительно сферическую форму и могут быть пористыми или цельными. Использование микроносителей для прикрепления к ним клеток упрощает использование смесительных сосудов и вспомогательных реакторов для выращивания клеток, зависимых от культуральной подложки. Клетки прикрепляются к микроносителям, легко образующим суспензию. Необходимость образования суспензии ограничивает физические характеристики микроносителей. Вследствие этого микроносители, как правило, имеют средний диаметр в диапазоне 50-2000 микрон. В некоторых областях применения цельные микроносители имеют диаметр от приблизительно 100 до приблизительно 250 микрон, а пористые микроносители - от приблизительно 250 до приблизительно 2500 микрон. Диапазоны размеров позволяют выбирать микроносители, достаточно крупные, чтобы на них прикреплялось много клеток, зависимых от культуральной подложки, и достаточно мелкие, чтобы образовывать суспензии со свойствами, подходящими для использования в смесительных реакторах.
При использовании микроносителей и аналогичных средств учитываются, помимо прочего, следующие факторы: эффективность прикрепления, иммуногенность, биосовместимость, возможность биологической деградации, время достижения конфлюэнтности, параметры роста прикрепившихся клеток, в том числе максимально достижимая плотность на единице поверхности, технологии отделения клеток (при необходимости) и эффективность отделения, масштабируемость условий культивирования, а также гомогенность культуры при увеличении масштабов производства, возможность успешного масштабирования процедур отделения клеток, а также будут ли микроносители использоваться для имплантации. На эти параметры могут влиять характеристики поверхности микроносителя, а также пористость, диаметр, плотность и свойства микроносителя при обработке.
Например, важным свойством является плотность микроносителя. Слишком высокая плотность может приводить к оседанию микроносителей из суспензии или к скапливанию в нижней части сосуда для культивирования, что может приводить к плохому перемешиванию массы клеток, культуральной среды и газовых фаз в реакторе. С другой стороны, слишком низкая плотность может приводить к излишней плавучести микроносителя. Стандартной для многих микроносителей является плотность 1,02-1,15 г/см3.
Малый диаметр микроносителей и большое количество частиц, добавляемых в реактор, позволяют получить значительную площадь поверхности, значительно превышающую площадь роллер-флаконов и другого оборудования для культивирования клеток, зависимых от культуральной подложки, например планшет. Пористые микроносители обеспечивают еще большую площадь на единицу объема или массы. Пористые микроносители имеют большие лакуны, в которых могут расти клетки, зависимые от культуральной подложки. Такие лакуны значительно увеличивают площадь поверхности и могут защищать клетки от разрушительных механических влияний, например, при перемешивании или пропускании газа.
Поверхность микроносителя может быть рельефной для обеспечения лучшего прикрепления и пролиферации клеток. Рельеф на поверхности микроносителей может быть сформирован различными способами, включая, помимо прочего, прессовку, отливку, выщелачивание и травление. Размер элементов рельефа может измеряться нанометрами. Рельеф поверхности может использоваться для обеспечения точного выравнивания клеток на поверхности микроносителя. Поверхность пор в пористых микроносителях также может быть рельефной для облегчения прикрепления и пролиферации клеток. Рельеф на поверхность пор может быть нанесен различными способами, включая, помимо прочего, прессовку, отливку, выщелачивание и травление.
Поверхность микроносителя может быть покрыта плазмой для придания ей заряда. Заряд может улучшать прикрепление и пролиферацию клеток.
В других вариантах осуществления микроносители состоят или имеют покрытие из термочувствительных полимеров, например поли-N-изопропилакриламида, или имеют электромеханические свойства.
В продаже есть как цельные, так и пористые типы микроносителей разных производителей. К примерам доступных в продаже цельных микроносителей относятся Cytodex 1® и Cytodex 3® (GE Healthcare Life Sciences, Нью-Джерси, США), представляющие собой декстрановые микроносители производства GE Healthcare Life Sciences. К пористым микроносителям, представленным на рынке, относятся продукты Cytoline и Cytopore также производства GE Healthcare Life Sciences. Кроме того, в продаже представлены продукты Biosilon (NUNC) и Cultispher (Percell Biolytica).
Микроносители могут содержать биологически активный агент. Микроноситель может содержать биологически активный агент, который регулирует рост или функциональность клеток или тканевого окружения. К этим факторам могут относиться, помимо прочего, факторы роста фибробластов, эритропоэтин, факторы роста клеток эндотелия сосудов, тромбоцитарные факторы роста, костные морфогенетические белки, трансформирующие факторы роста, факторы роста опухолей, эпидермальные факторы роста и инсулинподобные факторы. Могут использоваться как полные факторы, так и миметики или активные фрагменты этих факторов.
На микроносителях может быть высеян второй тип клеток, культивируемый вместе с плюрипотентными стволовыми клетками. В одном из вариантов осуществления два (или несколько) типа клеток могут прикрепляться к микроносителю в равных или неравных соотношениях. Два или несколько типов клеток могут высевать на микроноситель одновременно или в разное время. Микроносители могут быть обработаны таким образом, чтобы конкретные типы клеток предпочтительно прикреплялись к конкретным зонам микроносителя. В дополнительном варианте осуществления микроноситель с прикрепившимися клетками одного или нескольких типов может культивироваться в сосуде для культивирования совместно со вторым типом клеток, выращиваемым в суспензии.
Ко второму типу клеток могут относиться, например, эпителиальные клетки (например, клетки слизистой полости рта, желудочно-кишечного тракта, назального эпителия, эпителия дыхательных путей, вагинального эпителия, эпителия роговицы), клетки костного мозга, адипоциты, стволовые клетки, кератиноциты, меланоциты, кожные фибробласты, кератиноциты, клетки сосудистого эпителия (например, клетки эндотелия аорты, клетки эндотелия коронарной артерии, клетки эндотелия легочной артерии, клетки эндотелия подвздошной артерии, клетки капиллярного эндотелия, клетки эндотелия пупочной артерии, клетки эндотелия пупочной вены и предшественники эндотелиальных клеток (например, клетки CD34+, CD34+/CD117+)), миобласты, миоциты, гепатоциты, клетки гладкой мускулатуры, клетки поперечнополосатой мускулатуры, стромальные клетки и другие клетки мягких тканей и клетки-предшественники, хондроциты, остеобласты, островковые клетки, нервные клетки, включая, помимо прочего, нейроны, астроциты, шванновские клетки, энтерические глиальные клетки, олигодендроциты.
Плюрипотентные стволовые клетки
Характеристика плюрипотентных стволовых клеток
Плюрипотентные стволовые клетки могут экспрессировать один или несколько стадийно-специфичных эмбриональных антигенов (SSEA) 3 и 4, а также маркеры, определяемые антителами, обозначенными Tra-1-60 и Tra-1-81 (Thomson et al., Science 282:1145, 1998). Дифференцирование плюрипотентных стволовых клеток in vitro приводит к потере экспрессии SSEA-4, Tra-1-60 и Tra-1-81 (при наличии) и повышению экспрессии SSEA-1. Недифференцированные плюрипотентные стволовые клетки, как правило, обладают щелочно-фосфатазной активностью, которую можно определить путем фиксирования клеток 4% раствором параформальдегида с последующим проявлением с помощью красителя Vector Red в качестве субстрата, следуя рекомендациям производителя (компания Vector Laboratories, Берлингейм, Калифорния, США). Недифференцированные плюрипотентные стволовые клетки также, как правило, экспрессируют OCT4 и TERT, определяемые с помощью ПЦР в реальном времени.
Другим желательным фенотипическим свойством выращенных плюрипотентных клеток является потенциал дифференцирования в клетки всех трех зародышевых листков: эндодермальные, мезодермальные и эктодермальные ткани. Плюрипотентность стволовых клеток может быть подтверждена, например, путем инъекции клеток мышам с тяжелым комбинированным иммунодефицитом (SCID), фиксирования образующихся тератом с помощью 4% параформальдегида и их гистологического исследования на предмет наличия клеточных типов, происходящих от трех зародышевых листков. В альтернативном варианте плюрипотентность можно определить по созданию эмбриоидных телец и анализу их на предмет присутствия маркеров, ассоциирующихся с тремя зародышевыми листками.
Выращенные линии плюрипотентных стволовых клеток могут быть кариотипированы с применением стандартного способа окрашивания с использованием красителя Гимза (G-banding) и сравнения с опубликованными кариотипами соответствующих видов приматов. Желательно получить клетки, имеющие «нормальный кариотип», т.е. эуплоидные клетки, в которых все человеческие хромосомы присутствуют и не имеют видимых изменений.
Источники плюрипотентных стволовых клеток
К типам плюрипотентных стволовых клеток, которые можно использовать, относятся стабильные линии плюрипотентных клеток, получаемых из ткани, формирующейся после наступления беременности, в том числе из преэмбриональной ткани (например, бластоциста), эмбриональной ткани или ткани плода, взятой в любой момент в процессе беременности, как правило, но не обязательно, до срока приблизительно 10-12 недель беременности. Неограничивающими настоящее изобретение примерами являются устойчивые линии эмбриональных стволовых клеток человека или эмбриональных зародышевых клеток человека, например линии эмбриональных стволовых клеток человека H1, H7 и H9 (WiCell). Также возможно использование описываемых в настоящем документе составов во время первоначального установления или стабилизации таких клеток. В этом случае исходными клетками являются первичные плюрипотентные клетки, взятые напрямую из тканей-источников. Также целям настоящего изобретения соответствуют клетки, взятые из популяции плюрипотентных стволовых клеток, уже культивированных в отсутствие питающих клеток. Также целям настоящего изобретения соответствуют клетки мутантных линий эмбриональных стволовых клеток человека, таких как, например, BG01v (BresaGen, Атенс, Джорджия, США). Также целям настоящего изобретения соответствуют плюрипотентные стволовые клетки, происходящие от неплюрипотентных клеток, например соматических клеток взрослого организма.
Прикрепление плюрипотентных стволовых клеток к микроносителям, подходящим для использования в рамках настоящего изобретения
Плюрипотентные стволовые клетки до прикрепления к микроносителям могут культивироваться на плоском субстрате любым известным специалистам способом. Например, плюрипотентные стволовые клетки могут культивироваться на плоских субстратах, обработанных белком экстраклеточного матрикса (например, MATRIGEL). В альтернативном варианте плюрипотентные стволовые клетки могут культивироваться на плоских субстратах, на которых высеян слой питающих клеток.
В одном из вариантов осуществления настоящего изобретения указанные плюрипотентные стволовые клетки представляют собой эмбриональные стволовые клетки. В альтернативном варианте осуществления эмбриональные стволовые клетки являются человеческими.
В одном аспекте настоящего изобретения плюрипотентные стволовые клетки отделяются от плоского субстрата путем обработки плюрипотентных стволовых клеток протеазой, которая отделяет клетки от плоского субстрата. Протеаза может представлять собой, например, коллагеназу, TrypLE™ Express, Accutase™, трипсин и т.п.
В одном из вариантов осуществления плюрипотентные стволовые клетки отделяются от субстрата-микроносителя путем обработки клеток реактивом Accutase™ в течение приблизительно 5-10 минут.
В одном из вариантов осуществления плюрипотентные стволовые клетки отделяются от субстрата-микроносителя путем обработки клеток 0,05% раствором трипсин/EDTA в течение от приблизительно 10 до приблизительно 20 минут.
В одном из вариантов осуществления плюрипотентные стволовые клетки отделяются от субстрата-микроносителя путем обработки клеток реактивом TrypLE™ Express в течение от приблизительно 5 до приблизительно 20 минут.
В одном из вариантов осуществления плюрипотентные стволовые клетки отделяются от субстрата путем обработки клеток раствором коллагеназы 10 мг/мл в течение от приблизительно 5 до приблизительно 10 минут.
Отделенные плюрипотентные клетки добавляют в среду, содержащую микроносители с конкретной плотностью. В одном из вариантов осуществления плюрипотентные стволовые клетки высеивали с плотностью от приблизительно 4000 до приблизительно 30000 клеток на см2 микроносителей.
Отделенные плюрипотентные клетки добавляют в среду, содержащую микроносители. В одном из вариантов осуществления прикрепление плюрипотентных стволовых клеток ускоряется путем обработки плюрипотентных стволовых клеток ингибитором Rho-киназы. Ингибитор Rho-киназы может представлять собой Y27632 (Sigma-Aldrich, Миссури, США). В альтернативном варианте ингибитор Rho-киназы представляет собой глицил-H 1152 дигидрохлорид.
В одном из вариантов осуществления плюрипотентные стволовые клетки обрабатывают Y27632 с концентрацией от приблизительно 1 мкМ до приблизительно 10 мкМ. В одном из вариантов осуществления плюрипотентные стволовые клетки обрабатывают Y27632 с концентрацией приблизительно 10 мкМ.
В одном из вариантов осуществления плюрипотентные стволовые клетки обрабатывают глицил-H 1152 дигидрохлоридом с концентрацией от приблизительно 0,25 мкМ до приблизительно 5 мкМ. В одном из вариантов осуществления плюрипотентные стволовые клетки обрабатывают глицил-H 1152 дигидрохлоридом с концентрацией приблизительно 2,5 мкМ.
Среда, содержащая микроносители, может подвергаться непрерывному перемешиванию. Перемешивание в рамках настоящего изобретения может означать движение культуральной среды. Такое перемешивание может осуществляться вручную или, в альтернативном варианте, с использованием оборудования, например качающейся платформы, ротационной колбы и т.п. В одном из вариантов осуществления среда, содержащая микроносители, перемешивается вручную. Планшет, содержащий микроносители и клетки, движется вперед и назад менее 30 секунд.
Среда, содержащая микроносители, может подвергаться непрерывному перемешиванию. В одном из вариантов осуществления среда, содержащая микроносители, непрерывно перемешивается в ротационной колбе. Ротационная колба (Corning, Lowell, Массачусетс, США) помещается на пластину для смешивания при 30-70 об/мин, в зависимости от типа сфер.
В альтернативном варианте осуществления среда, содержащая микроносители, непрерывно перемешивается с помощью качающейся платформы (Vari-mix, Barnstead, Dubuque, Айова, США). Скорость качающейся платформы составляет приблизительно 1 оборот в 2 секунды.
Дифференцирование плюрипотентных стволовых клеток на микроносителях
В одном из вариантов осуществления плюрипотентные стволовые клетки могут дифференцироваться в клетки, экспрессирующие маркеры, характерные для линии сформированной эндодермы, на микроносителях. В альтернативном варианте плюрипотентные стволовые клетки могут дифференцироваться в клетки, экспрессирующие маркеры, характерные для линии панкреатической эндодермы, на микроносителях. В альтернативном варианте плюрипотентные стволовые клетки могут дифференцироваться в клетки, экспрессирующие маркеры, характерные для линии панкреатических эндокринных клеток, на микроносителях.
В альтернативном варианте осуществления плюрипотентные стволовые клетки могут выращиваться на микроносителях, а затем дифференцироваться в клетки, экспрессирующие маркеры, характерные для линии сформированной эндодермы, на плоских поверхностях. В альтернативном варианте плюрипотентные стволовые клетки могут выращиваться на микроносителях, а затем дифференцироваться в клетки, экспрессирующие маркеры, характерные для линии панкреатической эндодермы, на плоских поверхностях. В альтернативном варианте плюрипотентные стволовые клетки могут выращиваться на микроносителях, а затем дифференцироваться в клетки, экспрессирующие маркеры, характерные для линии панкреатических эндокринных клеток, на плоских поверхностях.
Плюрипотентные стволовые клетки, обработанные в соответствии со способами, составляющими предмет настоящего изобретения, могут быть дифференцированы в различные клетки иных типов с использованием любых соответствующих способов, известных специалистам в данной области. Например, плюрипотентные стволовые клетки, обработанные в соответствии со способами, составляющими предмет настоящего изобретения, могут быть дифференцированы в нервные клетки, сердечные клетки, гепатоциты и т.д.
Например, плюрипотентные стволовые клетки, обработанные в соответствии со способами, составляющими предмет настоящего изобретения, могут быть дифференцированы в нервные клетки-предшественники и кардиомиоциты в соответствии со способами, описанными в заявке № WO2007030870.
В другом примере плюрипотентные стволовые клетки, обработанные в соответствии со способами, составляющими предмет настоящего изобретения, могут быть дифференцированы в гепатоциты в соответствии со способами, описанными в заявке на патент США № 6458589.
Образование клеток, экспрессирующих маркеры, характерные для линии сформированной эндодермы
Плюрипотентные стволовые клетки могут быть дифференцированы в клетки, экспрессирующие маркеры, характерные для линии сформированной эндодермы, любым способом, известным специалистам в данной области.
Например, плюрипотентные стволовые клетки могут быть дифференцированы в клетки, экспрессирующие маркеры, характерные для линии сформированной эндодермы, способами, описанными в публикации D'Amour et al., Nature Biotechnology 23, 1534-1541 (2005).
Например, плюрипотентные стволовые клетки могут быть дифференцированы в клетки, экспрессирующие маркеры, характерные для линии сформированной эндодермы, способами, описанными в публикации Shinozaki et al., Development 131, 1651-1662 (2004).
Например, плюрипотентные стволовые клетки могут быть дифференцированы в клетки, экспрессирующие маркеры, характерные для линии сформированной эндодермы, способами, описанными в публикации McLean et al., Stem Cells 25, 29-38 (2007).
Например, плюрипотентные стволовые клетки могут быть дифференцированы в клетки, экспрессирующие маркеры, характерные для линии сформированной эндодермы, способами, описанными в публикации D'Amour et al., Nature Biotechnology 24, 1392-1401 (2006).
Маркеры, характерные для линии сформированной эндодермы, выбраны из группы, включающей следующие маркеры: SOX17, GATA4, HNF-3 beta, GSC, CER1, Nodal, FGF8, брахиурия, гомеобоксный белок Mix-like, FGF4, CD48, эомезодермин (EOMES), DKK4, FGF17, GATA6, CXCR4, C-Kit, CD99 и OTX2. Клетка, соответствующая целям настоящего изобретения, экспрессирует по меньшей мере один из маркеров, характерных для линии сформированной эндодермы. В одном из аспектов настоящего изобретения клетка, экспрессирующая маркеры, характерные для линии сформированной эндодермы, представляет собой клетку-предшественника первичной полоски. В другом аспекте настоящего изобретения клетка, экспрессирующая маркеры, характерные для линии сформированной эндодермы, представляет собой мезэндодермальную клетку. В другом аспекте настоящего изобретения клетка, экспрессирующая маркеры, характерные для линии сформированной эндодермы, представляет собой клетку сформированной эндодермы.
В другом примере плюрипотентные стволовые клетки, обработанные способами, составляющими предмет настоящего изобретения, могут быть дифференцированы в клетки, экспрессирующие маркеры, характерные для линии сформированной эндодермы, путем культивирования плюрипотентных стволовых клеток в среде, содержащей активин A в отсутствие сыворотки, затем культивирования клеток с активином A и сывороткой и культивирования клеток с активином A и сывороткой в другой концентрации. Пример данного способа описан в публикации Nature Biotechnology 23, 1534-1541 (2005).
В другом примере плюрипотентные стволовые клетки, обработанные способами, составляющими предмет настоящего изобретения, могут быть дифференцированы в клетки, экспрессирующие маркеры, характерные для линии сформированной эндодермы, путем культивирования плюрипотентных стволовых клеток в среде, содержащей активин А в отсутствие сыворотки, затем культивирования клеток с активином A и сывороткой в другой концентрации. Пример данного способа описан в публикации D'Amour et al., Nature Biotechnology, 2005.
В другом примере плюрипотентные стволовые клетки, обработанные способами, составляющими предмет настоящего изобретения, могут быть дифференцированы в клетки, экспрессирующие маркеры, характерные для линии сформированной эндодермы, путем культивирования плюрипотентных стволовых клеток в среде, содержащей активин А и лиганд Wnt в отсутствие сыворотки, затем удаления лиганда Wnt и культивирования клеток с активином A и сывороткой. Пример использования данного способа приведен в публикации Nature Biotechnology 24, 1392-1401 (2006).
В другом примере плюрипотентные стволовые клетки, обработанные способами, составляющими предмет настоящего изобретения, могут быть дифференцированы в клетки, экспрессирующие маркеры, характерные для линии сформированной эндодермы, способами, изложенными в заявке на патент США с серийным № 11/736908, принадлежащей LifeScan, Inc.
В другом примере плюрипотентные стволовые клетки, обработанные способами, составляющими предмет настоящего изобретения, могут быть дифференцированы в клетки, экспрессирующие маркеры, характерные для линии сформированной эндодермы, способами, изложенными в заявке на патент США с серийным № 11/779311, принадлежащей LifeScan, Inc.
Образование клеток, экспрессирующих маркеры, характерные для линии панкреатической эндодермы
Плюрипотентные стволовые клетки могут быть дифференцированы в клетки, экспрессирующие маркеры, характерные для линии панкреатической эндодермы, любым способом, известным специалистам в данной области.
Например, плюрипотентные стволовые клетки могут быть дифференцированы в клетки, экспрессирующие маркеры, характерные для линии панкреатической эндодермы, способами, описанными в публикации D'Amour et al., Nature Biotechnology 24, 1392-1401 (2006).
Например, клетки, экспрессирующие маркеры, характерные для линии сформированной эндодермы, полученные способами, составляющими предмет настоящего изобретения, далее дифференцируются в клетки, экспрессирующие маркеры, характерные для линии панкреатической эндодермы, путем обработки клеток, экспрессирующих маркеры, характерные для линии сформированной эндодермы, фактором роста фибробластов и ингибитором сигнального пути Hedgehog KAAD-циклопамином с последующим удалением содержащей фактор роста фибробластов и KAAD-циклопамин среды и культивированием клеток в среде, содержащей ретиноевую кислоту, фактор роста фибробластов и KAAD-циклопамин. Пример использования данного способа приведен в публикации Nature Biotechnology 24, 1392-1401 (2006).
Например, клетки, экспрессирующие маркеры, характерные для линии сформированной эндодермы, полученные способами, являющимися предметом настоящего изобретения, дифференцируются в клетки, экспрессирующие маркеры, характерные для линии панкреатической эндодермы, путем обработки клеток, экспрессирующих маркеры, характерные для линии сформированной эндодермы, ретиноевой кислотой и одним фактором роста фибробластов в течение периода времени, соответствующего способам, описанным в заявке на патент США с серийным № 11/736908, принадлежащей LifeScan, Inc.
Например, клетки, экспрессирующие маркеры, характерные для линии сформированной эндодермы, полученные способами, составляющими предмет настоящего изобретения, далее дифференцируются в клетки, экспрессирующие маркеры, характерные для линии панкреатической эндодермы, путем обработки клеток, экспрессирующих маркеры, характерные для линии сформированной эндодермы, ретиноевой кислотой (Sigma-Aldrich, Миссури, США) и экзендином 4 и последующего удаления среды, содержащей DAPT (Sigma-Aldrich, Миссури, США) и экзендин 4, с дальнейшим культивированием клеток в среде, содержащей экзендин 1, IGF-1 и HGF. Пример использования данного способа приведен в публикации Nature Biotechnology 24, 1392-1401 (2006).
Например, клетки, экспрессирующие маркеры, характерные для линии панкреатической эндодермы, полученные способами, составляющими предмет настоящего изобретения, далее дифференцируются в клетки, экспрессирующие маркеры, характерные для линии панкреатической эндокринной линии, путем обработки клеток, экспрессирующих маркеры, характерные для линии панкреатической эндодермы, средой, содержащей экзендин 4, затем удаления среды, содержащей экзендин 4, с последующим культивированием клеток в среде, содержащей экзендин 1, IGF-1 и HGF. Пример данного способа описан в публикации D'Amour et al., Nature Biotechnology, 2006.
Например, клетки, экспрессирующие маркеры, характерные для линии панкреатической эндодермы, полученные способами, составляющими предмет настоящего изобретения, далее дифференцируются в клетки, экспрессирующие маркеры, характерные для линии панкреатической эндокринной линии, путем обработки клеток, экспрессирующих маркеры, характерные для линии панкреатической эндодермы, средой, содержащей DAPT (Sigma-Aldrich, Миссури, США) и экзендин 4. Пример данного способа описан в публикации D'Amour et al., Nature Biotechnology, 2006.
Например, клетки, экспрессирующие маркеры, характерные для линии панкреатической эндодермы, полученные способами, составляющими предмет настоящего изобретения, далее дифференцируются в клетки, экспрессирующие маркеры, характерные для линии панкреатической эндокринной линии, путем обработки клеток, экспрессирующих маркеры, характерные для линии панкреатической эндодермы, средой, содержащей экзендин 4. Пример данного способа описан в публикации D' Amour et al., Nature Biotechnology, 2006.
Например, клетки, экспрессирующие маркеры, характерные для линии панкреатической эндодермы, полученные способами, составляющими предмет настоящего изобретения, далее дифференцируются в клетки, экспрессирующие маркеры, характерные для линии панкреатических эндокринных клеток, путем обработки клеток, экспрессирующих маркеры, характерные для линии панкреатической эндодермы, фактором, ингибирующим сигнальный путь Notch, способами, описанными в заявке на патент США с серийным № 11/736908, принадлежащей LifeScan, Inc.
Например, клетки, экспрессирующие маркеры, характерные для линии панкреатической эндодермы, полученные способами, составляющими предмет настоящего изобретения, далее дифференцируются в клетки, экспрессирующие маркеры, характерные для линии панкреатических эндокринных клеток, путем обработки клеток, экспрессирующих маркеры, характерные для линии панкреатической эндодермы, фактором, ингибирующим сигнальный путь Notch, способами, описанными в заявке на патент США с серийным № 11/779311, принадлежащей LifeScan, Inc.
Например, клетки, экспрессирующие маркеры, характерные для линии панкреатической эндодермы, полученные способами, составляющими предмет настоящего изобретения, далее дифференцируются в клетки, экспрессирующие маркеры, характерные для линии панкреатических эндокринных клеток, путем обработки клеток, экспрессирующих маркеры, характерные для линии панкреатической эндодермы, фактором, ингибирующим сигнальный путь Notch, способами, описанными в заявке на патент США с серийным № 11/736908, принадлежащей LifeScan, Inc.
Например, клетки, экспрессирующие маркеры, характерные для линии панкреатической эндодермы, полученные способами, составляющими предмет настоящего изобретения, далее дифференцируются в клетки, экспрессирующие маркеры, характерные для линии панкреатических эндокринных клеток, путем обработки клеток, экспрессирующих маркеры, характерные для линии панкреатической эндодермы, фактором, ингибирующим сигнальный путь Notch, способами, описанными в заявке на патент США с серийным № 11/779311, принадлежащей LifeScan, Inc.
Маркеры, характерные для линии панкреатических эндокринных клеток, выбраны из группы, включающей следующие маркеры: NGN3, NEUROD, ISL1, PDX1, NKX6.1, PAX4, NGN3 и PTF-1 alpha. В одном из вариантов осуществления панкреатическая эндокринная клетка способна экспрессировать по меньшей мере один из следующих гормонов: инсулин, глюкагон, соматостатин и панкреатический полипептид. Соответствующей целям настоящего изобретения является клетка, экспрессирующая по меньшей мере один из маркеров, характерных для линии панкреатических эндокринных клеток. В одном из аспектов настоящего изобретения клетка, экспрессирующая маркеры, характерные для линии панкреатических эндокринных клеток, представляет собой панкреатическую эндокринную клетку. Указанная панкреатическая эндокринная клетка может представлять собой панкреатическую клетку, экспрессирующую гормоны. В альтернативном варианте указанная панкреатическая эндокринная клетка может представлять собой панкреатическую клетку, секретирующую гормоны.
В одном из аспектов настоящего изобретения указанная панкреатическая эндокринная клетка представляет собой клетку, экспрессирующую маркеры, характерные для β-клеточной линии дифференцирования. Клетка, экспрессирующая маркеры, характерные для β-клеточной линии дифференцирования, экспрессирует PDX1 и по меньшей мере один из следующих факторов транскрипции: NGN3, NKX2.2, NKX6.1, NEUROD, ISL1, HNF-3 beta, MAFA, PAX4 или PAX6. В одном из аспектов настоящего изобретения клетка, экспрессирующая маркеры, характерные для β-клеточной линии дифференцирования, представляет собой β-клетку.
Настоящее изобретение далее иллюстрируется, помимо прочего, следующими примерами.
ПРИМЕРЫ
Пример 1. Прикрепление и пролиферация человеческих эмбриональных стволовых клеток на микроносителях
Для выяснения того, могут ли человеческие эмбриональные стволовые клетки прикрепляться и пролиферировать на микроносителях, клетки H9, пассаж 52, были отделены от планшетов, покрытых материалом MATRIGELTM (BD Biosciences, Калифорния, США), с помощью реактива TrypLE™ Express. Затем их инкубировали с микроносителями в среде MEF-CM. Суспензии микроносителей ProNectinF (PN), Plastic (P), PlasticPlus (PP), HILLEX®II (H), коллаген (Col) и FACT III (SoloHill, Мичиган, США) приготовили в соответствии с инструкциями производителей. В таблице 1 описаны прикрепление и рост клеток H9 на микроносителях через 2 дня после инкубации при 37°C, определяемые по ежедневно генерируемым изображениям. На большинстве протестированных микроносителей прикрепилось и/или пролиферировало небольшое количество клеток. На микроносителях HILLEX®II (Solohill, Мичиган, США) клетки H9 прикрепились и пролиферировали, но изображения показывают уменьшение количества агрегатов клетка-сфера через 2 дня статичного культивирования (фиг.1,B).
Для улучшения прикрепления и пролиферации человеческих эмбриональных стволовых клеток на микроносителях в среду был добавлен низкомолекулярный ингибитор образующей двойную спираль серин/треониновой протеинкиназы, связанной с Rho, т.е. ингибитор Rho-киназы. В частности, использовалось соединение Y27632, Y (Sigma-Aldrich, Миссури, США). Среду MEF-CM с добавлением 10 мкМ Y27632 (Sigma-Aldrich, Миссури, США) меняли ежедневно. В присутствии 10 мкМ Y27632 (Sigma-Aldrich, Миссури, США) клетки H9 прикреплялись и образовывали агрегаты со всеми протестированными микроносителями (таблица 2). По результатам анализа изображений был сделан вывод, что человеческие эмбриональные стволовые клетки, выращенные на микроносителях HILLEX®II (Solohill, Мичиган, США), прикреплялись и пролиферировали лучше, чем на других испытанных микроносителях. Кроме того, клетки H9 лучше прикреплялись к микроносителям HILLEX®II (Solohill, Мичиган, США) в присутствии ингибитора Rho-киназы (фиг.1,A в сравнении с фиг.1,B).
Для того чтобы соблюсти требования, предъявляемые к продукции в сфере клеточной терапии, эмбриональные стволовые клетки должны размножаться. В настоящее время передовые технологии размножения клеток связаны с использованием ротационных колб и биореакторов. Обе технологии требуют, чтобы микроносители в суспензии находились в физическом движении. Для определения воздействия движения на рост человеческих эмбриональных стволовых клеток 6- или 12-луночные планшеты были помещены на качающейся платформе в инкубатор при 37°C. Через 3 суток инкубации агрегаты клеток начали отделяться от микроносителей. На фиг.2 A, B, D показано отделение агрегатов клеток от микроносителей Plastic Plus, Plastic и Pronectin. Напротив, на микроносителях HILLEX®II (Solohill, Мичиган, США) клетки оставались прикрепленными и пролиферирующими - фиг.2,C. В примере 4 описан способ отделения, использованный до подсчета количества клеток в приборе Guava PCA-96 с использованием реагента Viacount Flex (Guava Technologies, Hayward, Калифорния, США). При определении скорости роста клеток на микроносителях было обнаружено снижение количества клеток на 3 день относительно исходного количества при посеве. Вероятно, это объясняется плохим первоначальным прикреплением клеток к микроносителям, за которым последовал этап размножения, продолжавшийся до окончания эксперимента на 5 сутки. Клетки H9 на микроносителях HILLEX®II (Solohill, Мичиган, США) имели самую высокую скорость пролиферации в сравнении с другими типами сфер, вероятно, вследствие лучшего прикрепления клеток к микроносителям HILLEX®II (Solohill, Мичиган, США) (фиг.2, 3). Это показывает, что микроносители HILLEX®II (Solohill, Мичиган, США) могут обеспечивать рост клеток H9 в суспензии. Данный вывод в дальнейшем подтвердился при повторном посеве, см. пример 5.
Линия человеческих эмбриональных стволовых клеток H1 также была испытана выращиванием на микроносителях в условиях крупномасштабного производства. Поскольку ингибитор Rho-киназы Y27632 (Sigma-Aldrich, Миссури, США) был необходим для прикрепления клеток линии H9, предположили, что он также необходим для клеток H1. Микроносители Cytodex 1®, Cytodex 3® (GE Healthcare Life Sciences, Нью-Джерси, США), HILLEX®II, Plastic, ProNectinF, Plastic Plus (SoloHill Ann Arbor, Мичиган, США) были подготовлены в соответствии с инструкциями производителей. Человеческие эмбриональные стволовые клетки H1, пассаж 47, высеяли на микроносители с плотностью 13333 клеток/см2 в среде MEF-CM с добавлением 10 мкМ Y27632 (Sigma-Aldrich, Миссури, США). Клетки и микроносители переносили в 12-луночный планшет, не получавший обработку для тканевого культивирования, площадью 15 см2 на 12 лунок, и помещали на качающуюся платформу при 37°C, чтобы микроносители и среда находились в движении. Спустя 3, 5 и 7 суток с одной лунки получали изображение, клетки собирали и подсчитывали их количество. Способность клеток к прикреплению зависела от типа сфер. Результаты для клеточных линий H1 и H9 были аналогичными. В частности, для клеток, высеянных на микроносители Plastic, Plastic Plus или ProNectinF, не наблюдалось хорошего прикрепления и/или пролиферации (фиг.4). Клетки, высеянные на микроносители HILLEX®II (Solohill, Мичиган, США), Cytodex 1® или Cytodex 3® (GE Healthcare Life Sciences, Нью-Джерси, США), хорошо прикреплялись и пролиферировали (фиг.5). Клетки были отделены, как описано в примере 4, и их количество подсчитано для определения выхода. Клетки, выращиваемые на микроносителях Cytodex 3® (GE Healthcare Life Sciences, Нью-Джерси, США), через 7 дней культивирования давали наибольшее количество клеток (фиг.6).
Пример 2. Оптимальные концентрации Y27632 и других ингибиторов Rho-киназы для прикрепления и выращивания клеток
Для того чтобы определить наиболее эффективную концентрацию ингибитора Rho-киназы для обеспечения прикрепления и роста человеческих эмбриональных стволовых клеток на микроносителях, были проведены описанные ниже эксперименты.
Исходную аликвоту клеток H9, пассаж 44, 13333 клеток/см2, высеяли на микроносители площадью 15 см2 в одной лунке 12-луночного планшета, не обработанного для тканевого культивирования. Клетки поместили в температуру 37°C по меньшей мере на 60 минут, после чего поместили на качающуюся платформу при температуре 37°C. Перед добавлением к клеткам микроносители HILLEX®II (Solohill, Мичиган, США) и Cytodex 3® (GE Healthcare Life Sciences, Нью-Джерси, США) подготовили в соответствии с инструкциями производителя. Клетки выращивались в среде MEF-CM с добавлением ингибитора Rho-киназы Y27632 в концентрациях 10, 5, 2,5 или 1 мкМ или (S)-(+)-4-глицил-2-метил-1-[(4-метил-5-изохинолинил)сульфонил]-гексагидро-1H-1,4-диазепина дигидрохлорида (глицил-H 1152 дигидрохлорида (H), Tocris, Миссури, США) в концентрациях 5, 2,5, 1 или 0,5 мкМ. Среду меняли ежедневно и количество клеток в одной лунке подсчитывали на 4 и 7 сутки после посева, чтобы определить выход и жизнеспособность (фиг.7,A и B). В целом при концентрациях 10 или 5 мкМ Y27632 (Sigma-Aldrich, Миссури, США) наблюдалась наилучшая пролиферация клеток (7 день), а при концентрациях 2,5 и 1,0 мкМ наилучшим оказалось прикрепление (4 день). При концентрациях 1 и 0,5 мкМ глицил-H 1152 дигидрохлорида (Tocris, Миссури, США) наблюдалась наилучшая пролиферация клеток (7 день), а при 5 мкМ наилучшим оказалось прикрепление (4 день).
Также было проведено титрование доз ингибитора Rho-киназы, поскольку он является стимулятором апоптоза. Клетки H1, пассаж 48, были отделены от планшетов, покрытых материалом MATRIGELTM (BD Biosciences, Калифорния, США), с помощью реактива TrypLE™ Express. Затем клетки высевали на микроносители площадью 15 см2 в одной лунке 12-луночного планшета, не обработанного для тканевого культивирования. Микроносители HILLEX®II (Solohill, Мичиган, США), Cytodex 1® или Cytodex 3® (GE Healthcare Life Sciences, Нью-Джерси, США) испытывались с убывающими концентрациями ингибитора Rho-киназы: 10 мкМ Y27632 (Sigma-Aldrich, Миссури, США) в первый день, затем 0,5 мкМ во второй день (Y10/5 мкМ); 2,5 мкМ глицил-H 1152 дигидрохлорида (Tocris, Миссури, США) в первый день, затем 0,5 мкМ во второй день (H2,5/0,5 мкМ); 1 мкМ глицил-H 1152 дигидрохлорида (Tocris, Миссури, США) в первый день, затем 0,5 мкМ глицил-H 1152 дигидрохлорида (Tocris, Миссури, США) во второй день (H1/0,5 мкМ); или же применялось длительное добавление 0,25 мкМ глицил-H 1152 дигидрохлорида (Tocris, Миссури, США) ежедневно в среду MEF-CM (H0,25 мкМ). Клетки H1 и микроносители перемешивались каждые 45 минут в течение 3 часов при 37°C, после чего помещались на качающуюся платформу при 37°C. Количество клеток подсчитывали после 3, 5 и 7 дней инкубации на качающейся платформе при 37°C (фиг.8). В целом, наилучшей концентрацией глицил-H 1152 дигидрохлорида (Tocris, Миссури, США) была концентрация 1-2,5 мкМ в первый день и 0,5 мкМ во второй день с последующим удалением соединения. При данных концентрациях глицил-H 1152 дигидрохлорида (Tocris, Миссури, США) клетки имели скорость роста, аналогичную скорости при концентрации 10 мкМ Y27632 (Sigma-Aldrich, Миссури, США) на всех протестированных микроносителях. Выращивание клеток при постоянной концентрации 0,25 мкМ глицил-H 1152 дигидрохлорида (Tocris, Миссури, США) давало низкий выход клеток. Использование минимального количества ингибитора Rho-киназы также позволяет снизить технологические затраты и может способствовать пролиферации клеток. Данные также показывают, что человеческим эмбриональным стволовым клеткам не требуется ингибитор Rho-киназы для того, чтобы оставаться прикрепленными к микроносителям и для пролиферации.
Пример 3. Влияние плотности клеток на прикрепление и рост на микроносителях
Оптимизация плотности посева является способом снижения необходимого количества клеток. Для определения подходящей плотности посева было подсчитано количество микроносителей в поле зрения 4-кратного объектива. Клетки H1 высевали с плотностями 0,4×104 клеток/см2 (низкая), 1,2×104 клеток/см2 (средняя) или 3×104 клеток/см2 (высокая) в планшете на 10 см с микроносителями Cytodex 3 (GE Healthcare Life Sciences, Нью-Джерси, США) в среде MEF-CM с добавлением 10 мкМ Y27632 (Sigma-Aldrich, Миссури, США). Затем содержимое планшета перемешивали каждые 45 минут в течение 6 часов при 37°C. Клетки и микроносители были перенесены в ротационную колбу (описанную в примере 5) при 37°C и скорости 30 об/мин в 50 мл среды MEF-CM с добавлением 10 мкМ Y27632 (Sigma-Aldrich, Миссури, США). Через 24 часа добавили 25 мл среды MEF-CM с 5 мкМ Y27632 (Sigma-Aldrich, Миссури, США). Через 24 часа скорость вращения увеличили до 40 об/мин. На 3 и 5 сутки культивирования извлекли 50 мл среды из 75 мл, заменив ее средой MEF-CM. Получали изображения аликвот, отбираемых из ротационных колб через 6 часов, 3 суток, 5 суток и 7 суток. Процентная доля микроносителей с прикрепленными к ним клетками указана в нижнем правом углу изображений на фиг.9. На 3 сутки после посева количество микроносителей, покрытых клетками, соответствовало исходной плотности посева, но к 5 и 7 суткам количество покрытых клетками микроносителей при низкой плотности посева не увеличивалось. Это позволяет предположить, что 0,4×104 клеток/см2 не является достаточным количеством клеток для включения микроносителей в агрегаты. При плотности посева 3×104 клеток/см2 количество микроносителей с прикрепленными клетками на 5 и 7 сутки аналогично количеству при плотности посева 1,2×104 клеток/см2 (фиг. 9). Если рассматривать количество клеток, очевидно, что при более высокой плотности посева большее количество клеток прикрепляется к микроносителям (фиг.10). Анализ кратности увеличения количества клеток относительно исходного показывает, что в культурах с высокой плотностью посева на 3 и 5 сутки прикрепляется большее количество клеток (фиг.11). На 7 сутки контрольные культуры и культуры с высокой плотностью посева имели аналогичную кратность увеличения числа клеток от исходной плотности при посеве. На основании этих данных можно сделать вывод о том, что 1,2×104 клеток/см2 является минимально необходимым количеством для эффективного прикрепления и выращивания клеток H1 на микроносителях. Переход к более высоким плотностям посева поможет уменьшить количество дней, необходимое для размножения клеток.
Пример 4. Диссоциация клеток от микроносителей
Для определения скорости роста было необходимо отделить клетки от микроносителей. Отмена ингибитора Rho-киназы Y27632 (Sigma-Aldrich, Миссури, США) не приводила к диссоциации клеток H1 от микроносителей (пример 2, фиг.12). Изображения клеток H9 на микроносителях HILLEX®II (Solohill, Мичиган, США) получали при 10- и 20-кратном увеличении перед диссоциацией их от микроносителей (фиг.13,A, B соответственно). Ферментативная обработка клеток H9 на микроносителях HILLEX®II (Solohill, Мичиган, США) позволила отделить жизнеспособные клетки (фиг.13,C, D и фиг.14). Клетки H9 выращивались в течение 6 дней в 6-луночном планшете с микроносителями HILLEX®II (Solohill, Мичиган, США) на качающейся платформе при 37°C. Клетки, прикрепленные к микроносителям, помещались в коническую пробирку на 15 мл; после оседания микроносителей среду отсасывали. Осевшие микроносители троекратно промывали 4 мл PBS (не содержащим ионов кальция и магния), давая возможность микроносителям оседать под действием силы тяжести. PBS отсосали, после чего добавили 1 мл PBS. Микроносители с клетками были перенесены в одну лунку 12-луночного планшета, не обработанного для тканевого культивирования. Планшет установили под углом в состоянии покоя, чтобы микроносители осели. PBS отсосали и добавили в лунку 1 мл реактива TrypLE™ Express (Invitrogen, Калифорния, США) или 0,05% раствора трипсин/EDTA. Планшет поместили на качающуюся платформу при 37°C на 10 или 20 минут. Планшет сняли с платформы и добавили в лунку 3 мл среды DMEM/F12 или MEF-CM. Среду энергично пропипетировали, отделяя клетки (фиг.13,C, D). Наблюдение микроносителей в микроскоп позволило выявить отделение клеток от микроносителей. Клетки процентрифугировали при 200×g в течение 5 минут. Среду отсосали, а осадок ресуспендировали в 1 мл среды DMEM/F12 или MEF-CM. Затем количество клеток подсчитали с помощью прибора Guava PCA-96 (Guava Technologies, Hayward, Калифорния, США) с использованием красителя Viacount. В частности, 200 мкл разведенных в необходимом количестве среды клеток инкубировали с 2 мкл красителя Viacount в течение 10 минут. Определяли жизнеспособность и количество клеток (фиг.14). Оба реактива, TrypLE™ Express и трипсин/EDTA, эффективно отделяли клетки от микроносителей.
Поскольку реактив TrypLE™ Express отделял клетки от микроносителей и предлагался как продукт, соответствующий требованиям GMP, он был протестирован в сравнении с другими возможными агентами для диссоциирования, в частности коллагеназой и Accutase™ (Sigma-Aldrich, Миссури, США). Клетки H1 p48 выращивали в ротационной колбе (пример 5) в течение 10 дней. Затем микроносители отбирали и переносили в коническую пробирку объемом 50 мл. Клетки отмывали в PBS, как описано выше, и переносили в 12-луночный планшет. PBS отсосали, в лунку добавили 1 мл TrypLE™ Express, Accutase™ или коллагеназы (10 мг/мл) и поместили на качающуюся платформу при 37°C на 5 или 10 минут. Клетки/микроносители энергично ресуспендировали в среде DMEM/F12, после чего диссоциированные клетки и микроносители пропустили через клеточный фильтр на 40 мкм, установленный на коническую пробирку объемом 50 мл. Лунку промыли дополнительными 2 мл среды, которые затем также пропустили через фильтр, после чего провели центрифугирование при 200×g в течение 5 минут. Клетки ресуспендировали в 1 мл среды DMEM/F12 и развели для подсчета, как описано выше. Жизнеспособность клеток была аналогичной во всех испытаниях ферментов. Реактивы Accutase™ и TrypLE™ Express отделяли после 5- и 10-минутной инкубации аналогичные количества клеток (фиг.15). Это иллюстрирует возможность использования реагентов Accutase™ и TrypLE™ Express как средств диссоциации клеток при использовании человеческих эмбриональных стволовых клеток на микроносителях.
Пример 5. Выращивание недифференцированных плюрипотентных стволовых клеток на микроносителях
Для выращивания клеток на микроносителях должна существовать возможность отделять или ферментативно диссоциировать клетки от микроносителей и снова прикреплять их к новым микроносителям. Типичные методы выращивания клеток на микроносителях основываются на свойстве клеток прикрепляться и открепляться. Следующий эксперимент показал, что такое свойство не характерно для человеческих эмбриональных стволовых клеток. В частности, клетки H9 p43 были высеяны на микроносители HILLEX®II (Solohill, Мичиган, США) и инкубировались в ротационной колбе объемом 125 мл (см. ниже). Краситель феноловый красный присутствовал в среде и захватывался микроносителями HILLEX®II (Solohill, Мичиган, США). Через 8 дней инкубации аликвота клеток на микроносителях объемом 10 мл была помещена в новую ротационную колбу, где находились не содержащая фенолового красного среда MEF-CM, 440 мг микроносителей HILLEX®II и 5 мкМ Y27632 (Sigma-Aldrich, Миссури, США). Через 5 дней инкубации при 37°C со скоростью вращения 30 об/мин микроносители были извлечены, и были сделаны их изображения (фиг.16). Показанные на изображениях темные микроносители представляют собой микроносители, покрытые клетками H9, выращенными в среде, содержащей феноловый красный. Светлые микроносители представляют собой новые добавленные микроносители. Предполагалось, что клетки H9 будут отделяться и снова прикрепляться к новым микроносителям, однако вместо этого клетки образовывали агрегаты с новыми микроносителями. Отсутствуют такие светлые микроносители, на которых имелись бы прикрепленные клетки и которые при этом не создавали агрегаты с темными микроносителями. Это свидетельствует о том, что клетки не могли отделяться и снова прикрепляться к микроносителям. Для выращивания клеток на микроносителях должна существовать возможность отделять клетки от микроносителей с помощью ферментов (см. пример 4).
Поскольку было установлено, каким образом человеческие эмбриональные стволовые клетки можно выращивать на микроносителях, необходимо определить, как человеческие эмбриональные стволовые клетки растут в более крупных ротационных колбах. Ротационные колбы позволяют выращивать клетки в системах с высокой плотностью посева. Это позволяет экономить место, а также считается первым шагом к выращиванию клеток в биореакторах. Для проверки способности человеческих эмбриональных стволовых клеток пролиферировать в ротационных колбах, клетки H9, пассаж 43, были посеяны в ротационные колбы объемом 125 мл. Изначально, до переноса в ротационную колбу, клетки были прикреплены к микроносителям на планшете 10 см. В частности, клетки H9 были сняты с двух шестилуночных планшетов путем пятиминутной инкубации с реактивом TrypLE™ Express при 37°C. Перед пересеванием с использованием TrypLE™ Express клетки были пересеяны с помощью коллагеназы (1 мг/мл) и высеяны на планшеты, покрытые разведенным 1:30 материалом MATRIGELTM (BD Biosciences, Калифорния, США) с пониженным содержанием ростового фактора. Клетки ресуспендировали в среде DMEM/F12 и их количество подсчитывали с помощью прибора Guava с добавлением реактива Viacount. После центрифугирования 3×106 клеток посеяли на планшет 10 см, содержащий среду MEF-CM с добавлением 10 мкМ Y27632 (Sigma-Aldrich, Миссури, США) и 250 см2 микроносителей HILLEX®II (Solohill, Мичиган, США), подготовленных в соответствии с инструкциями производителя. Планшет инкубировали при 37°C, аккуратно вращали и перемешивали один раз в 45 минут в течение 4,5 часов. Затем клетки, микроносители и среду переносили в ротационную колбу объемом 125 мл. Ротационную колбу наполняли до 50 мл средой MEF-CM с добавлением 10 мкМ Y27632 (Sigma-Aldrich, Миссури, США) и устанавливали на качающуюся пластину при 37°C и скорости 40 об/мин. На следующий день среду меняли и доводили до объема 75 мл средой MEF-CM с добавлением 5 мкМ Y27632 (Sigma-Aldrich, Миссури, США). Скорость перемешивания увеличивали до 70 об/мин. Среду меняли через день, не добавляя соединение Y27632 (Sigma-Aldrich, Миссури, США). Клетки пересевали в соответствии со способами, изложенными в примере 4, и 3×106 клеток высеяли на 250 см2 новых микроносителей. Культуры пересевали по достижении конфлюэнтности 1-2×105 клеток/см2. Для этого требовалось 5 пассажей (фиг.17). При каждом пассаже оценивали экспрессию маркеров плюрипотентности. Анализ показал, что 80-95% клеток экспрессировали маркеры плюрипотентности CD9, SSEA4, SSEA3, TRA-1-60 и TRA-1-81 (фиг.19,A). Аналогичный эксперимент был проведен с использованием клеток H9 p43 на микроносителях Cytodex 3® ((GE Healthcare Life Sciences, Нью-Джерси, США), фиг.18, 19,B). В целом, клетки хорошо пролиферировали как на микроносителях HILLEX®II (Solohill, Мичиган, США), так и на микроносителях Cytodex 3® (GE Healthcare Life Sciences, Нью-Джерси, США), оставаясь плюрипотентными. Анализ кариотипа проводился после 5 пассажей в ротационных колбах; была обнаружена аномальная трисомия по 12 хромосоме в 1,5% клеток. Поскольку к концу эксперимента номер пассажа клеток приближался к 50, наблюдаемые аномалии, вероятно, являются типичными. Это предположение можно проверить, если начать культивирование с меньшего номера пассажа.
Аналогичные эксперименты были проведены на клетках линии H1, пассажи 48 и 49. Все параметры были такими же, за исключением скорости вращения и плотности посева. Скорость вращения в ротационной колбе составляла 30 об/мин в течение ночи дня 1 и была увеличена до 40 об/мин в последующие дни. Плотность посева для микроносителей Cytodex 3® (GE Healthcare Life Sciences, Нью-Джерси, США) составила приблизительно 11000 клеток/см2, а для микроносителей Cytodex 1® (GE Healthcare Life Sciences, Нью-Джерси, США) плотность посева составила приблизительно 7000 клеток/см2. Количество клеток при посеве оставалось постоянным - 3×106 клеток на ротационную колбу. Вес микроносителей оставался постоянным и составлял 100 мг для Cytodex 1® и Cytodex 3®. Одним из преимуществ микроносителей Cytodex 1® и Cytodex 3® перед микроносителями HILLEX®II является увеличенная площадь поверхности. На фиг.20 и 21 показано размножение клеток H1 на микроносителях Cytodex 1® и Cytodex 3® соответственно. На протяжении пяти пассажей клетки оставались плюрипотентными (фиг.22). Анализ кариотипа клеток H1 на микроносителях Cytodex 3® выявил дупликацию Y-хромосомы у 10% протестированных клеток. Эти клетки H1 высевали на микроносителях в пассаже 48 и анализировали через 5 пассажей. Клетки H1p55, выращенные на субстрате MATRIGELTM (BD Biosciences, Калифорния, США) на плоской поверхности, имели нормальный кариотип. Анализ скорости удвоения популяции этих клеток за период между 3 сутками и днем пересевания (день 5, 6 или 7) показал отсутствие общего изменения времени удвоения (фиг.23). Для клеток H1, выращенных на микроносителях Cytodex 1®, и клеток H9, выращенных на микроносителях HILLEX®II (Solohill, Мичиган, США), наблюдалось наиболее стабильное время удвоения численности (таблица 3).
Пример 6. Пролиферация человеческих эмбриональных стволовых клеток на микроносителях в определенной среде
Для производства терапевтического препарата желательно удалить из среды для культивации человеческих эмбриональных стволовых клеток любые животные компоненты. В настоящее время человеческие эмбриональные стволовые клетки культивируются на материале MATRIGELTM (BD Biosciences, Калифорния, США) в среде, кондиционированной мышиными эмбриональными фибробластами (MEF-CM). И материал MATRIGELTM (BD Biosciences, Калифорния, США), и среду MEF-CM получают из мышиных клеток. Кроме того, среда MEF-CM является дорогостоящей, и ее производство требует много времени. Для выяснения того, можно ли культивировать человеческие эмбриональные стволовые клетки на микроносителях в определенных средах, клетки H9 высеяли на микроносителях Cytodex 3® (GE Healthcare Life Sciences, Нью-Джерси, США) и HILLEX®II (Solohill, Мичиган, США) в присутствии ингибиторов Rho-киназы, 10 мкМ Y27632 (Sigma-Aldrich, Миссури, США) или 2,5 мкМ глицил-H 1152 дигидрохлорида (Tocris, Миссури, США) в среде Stem Pro (Invitrogen, Калифорния, США), mTESR (StemCell Technologies, Ванкувер, Канада) или MEF-CM. Клетки культивировали в 12-луночном планшете на качающейся платформе при 37°C. Подсчет количества клеток производили на 3, 5 и 7 сутки. Клетки H9 p39, выращенные в среде MEF-CM на обоих типах сфер, имели типичные характеристики роста (фиг.24). Подобные клетки, выращенные в среде mTESR (StemCell Technologies, Ванкувер, Канада), хорошо пролиферировали на микроносителях Cytodex 3® в присутствии 10 мкМ Y27632 (Sigma-Aldrich, Миссури, США), но демонстрировали более низкую скорость роста на микроносителях HILLEX®II. Человеческие эмбриональные стволовые клетки линии H9, пассаж 64 (H9p64), прошедшие акклиматизацию в среде StemPro на протяжении 20 пассажей, хорошо пролиферировали как на микроносителях HILLEX®II, так и на Cytodex 3® в присутствии 10 мкМ Y27632 (Sigma-Aldrich, Миссури, США). Удивительно, что эти клетки недостаточно хорошо пролиферировали в присутствии 2,5 мкМ глицил-H 1152 дигидрохлорида (Tocris, Миссури, США) на микроносителях Cytodex 3®. Таким образом, на способность человеческих эмбриональных стволовых клеток к пролиферации влияют, и тип микроносителя, и ингибитор Rho-киназы, и среда.
Человеческие эмбриональные стволовые клетки H1, пассаж 38, высевали на микроносители Cytodex 3® или HILLEX®II в присутствии ингибиторов Rho-киназы, 10 мкМ Y27632 (Sigma-Aldrich, Миссури, США) или 2,5 мкМ глицил-H 1152 дигидрохлорида в среде mTESR (StemCell Technologies, Ванкувер, Канада) или MEF-CM в 12-луночный планшет. Клетки помещали на качающуюся платформу при 37°C. Подсчет количества клеток проводили на 3, 5 и 7 сутки. Клетки, выращенные в среде MEF-CM на обоих типах шариков, имели типичные характеристики роста в присутствии Y27632 (Sigma-Aldrich, Миссури, США), но плохо росли в присутствии глицил-H 1152 дигидрохлорида (Tocris, Миссури, США) на микроносителях Cytodex 3® (фиг.25). Среда mTESR (StemCell Technologies, Ванкувер, Канада) позволяла клеткам H1 пролиферировать на микроносителях HILLEX®II в присутствии обоих ингибиторов Rho-киназы, но на микроносителях Cytodex 3® скорость роста была низкой.
Принимая во внимание, что клетки H1 p50 хорошо пролиферировали в среде mTESR (StemCell Technologies, Ванкувер, Канада) на микроносителях HILLEX®II, был произведен посев 3×106 клеток на микроносители HILLEX®II площадью 250 см2. Клетки инкубировали при 37°C в планшете площадью 10 см2 в течение 5 часов при непрерывном перемешивании вручную каждые 45 минут. Среду mTESR (StemCell Technologies, Ванкувер, Канада) с добавлением 10 мкМ Y27632 (Sigma-Aldrich, Миссури, США) меняли через день. Параллельно выращивали клетки в среде MEF-CM (пример 5). В отличие от клеток, выращенных в MEF-CM, клетки, выращенные в среде mTESR (StemCell Technologies, Ванкувер, Канада), начали отделяться от микроносителей HILLEX®II через 7 суток инкубации (фиг.26,A в сравнении с фиг.26,B). Это показывает, что в среду mTESR (StemCell Technologies, Ванкувер, Канада) необходимо добавить дополнительные ингредиенты, чтобы человеческие эмбриональные стволовые клетки оставались прикрепленными и пролиферировали на микроносителях HILLEX®II (Solohill, Мичиган, США).
Пример 7. Дифференцирование человеческих эмбриональных стволовых клеток на микроносителях
Поскольку человеческие эмбриональные стволовые клетки могут выращиваться на микроносителях, необходимо определить потенциал данных клеток к дифференцированию. Человеческие эмбриональные стволовые клетки линии H9, пассаж 43, пятикратно пересевали на микроносители Cytodex 3® (GE Healthcare Life Sciences, Нью-Джерси, США). Клетки 5 пассажа выращивали в течение 6 суток, после чего диссоциировали от микроносителей с помощью реактива TrypLE™ Express (см. пример 4). Затем клетки высеяли на планшеты, покрытые 1:30 MATRIGELTM:DMEM/F12. После достижения 80-90% конфлюэнтности на планшетах на клетки воздействовали дифференцирующими реагентами. Дифференцирование человеческих эмбриональных стволовых клеток в сформированную эндодерму производили путем обработки клеток в течение 2 суток 2% раствором фракции V бычьего альбумина, не содержащего жирных кислот (FAF BSA, MP Biomedicals, Огайо, США) в среде RPMI с добавлением 100 нг/мл активина A (PeproTech, Нью-Джерси, США), 20 нг/мл Wnt3a (R&D Biosciences, Миннесота, США) и 8 нг/мл bFGF (PeproTech, Нью-Джерси, США). В течение дополнительных двух дней клетки обрабатывали 2% FAF BSA в RPMI с добавлением 100 нг/мл активина A (PeproTech, Нью-Джерси, США) и 8 нг/мл bFGF (PeproTech, Нью-Джерси, США). Среду заменяли ежедневно. Анализ методом FACS на наличие маркера сформированной эндодермы CXCR4 на поверхности клетки показал, что 87% клеток экспрессировали данный белок (фиг.27,A). Подобный эксперимент был проведен на человеческих эмбриональных стволовых клетках линии H1, пассаж 49, выращенных на микроносителях Cytodex 1® (GE Healthcare Life Sciences, Нью-Джерси, США) в течение 5 пассажей. Результаты показали, что 91% клеток, дифференцированных на микроносителях, экспрессировали CXCR4 (фиг.27,B). Данные показывают, что клетки, выращенные на микроносителях, могут дифференцироваться в сформированную эндодерму, что является первым этапом превращения их в инсулинпродуцирующие клетки.
Три типа микроносителей, Cytodex 1®, Cytodex 3® (GE Healthcare Life Sciences, Нью-Джерси, США) и HILLEX®II (Solohill, Мичиган, США), обеспечивают прикрепление и рост клеток H1. Было проведено дифференцирование клеток H1 на этих трех микроносителях. Клетки выращивали на трех типах микроносителей в ротационных колбах (пример 5) в течение разного количества пассажей (от 1 до 5). Через 6-8 дней после последнего пересевания аликвоты суспензии микроносителей с клетками переносились в 6- или 12-луночные планшеты. Всего было перенесено 15 см2 микроносителей с клетками на лунку 12-луночного планшета или 30 см2 микроносителей с клетками в 6-луночный планшет. В лунки планшета добавили среду для дифференцирования и поместили планшет на качающуюся платформу при 37°C. Дифференцирование человеческих эмбриональных стволовых клеток в сформированную эндодерму производили путем обработки клеток в течение 2 суток 2% раствором фракции V бычьего альбумина, не содержащего жирных кислот (MP Biomedicals, Огайо, США), в среде RPMI с добавлением 100 нг/мл активина A (PeproTech, Нью-Джерси, США), 20 нг/мл Wnt3a (R&D Biosciences, Миннесота, США) и 8 нг/мл bFGF (PeproTech, Нью-Джерси, США). В течение дополнительных двух дней клетки обрабатывали 2% FAF BSA в RPMI с добавлением 100 нг/мл активина A (PeproTech, Нью-Джерси, США) и 8 нг/мл bFGF (PeproTech, Нью-Джерси, США). Среду заменяли ежедневно. Методом FACS был проведен анализ на наличие маркера сформированной эндодермы CXCR4 на поверхности клетки (фиг.28). Клетки, выращенные на микроносителях Cytodex 1® и Cytodex 3®, могли дифференцироваться в сформированную эндодерму (87% и 92% соответственно), тогда как микроносители HILLEX®II не обеспечивали дифференцирование в той мере, как другие микроносители, протестированные в данном эксперименте (42%).
Для определения того, влияет ли плотность клеток на их дифференцирование на микроносителях, человеческие эмбриональные стволовые клетки линии H1, пассаж 40, выращивались на микроносителях Cytodex 3® в ротационной колбе в течение 8 или 11 суток. Затем эквивалент приблизительно 15 см2 площади микроносителей с клетками помещали в 6-луночный планшет на качающуюся платформу. Далее клетки инкубировали в среде для дифференцирования в сформированную эндодерму, как описано выше. Через 4 дня клетки анализировали на экспрессию CXCR4 методом FACS. 87% клеток, выращенных в течение 6 суток в ротационной колбе, экспрессировали CXCR4, тогда как среди клеток, выращенных в течение 11 суток в ротационной колбе, CXCR4 экспрессировали 56% клеток (фиг.29). Данные показывают, что количество дней культивирования клеток важно для дифференцирования, в частности, если плотность клеток слишком высока, это может помешать клеткам дифференцироваться эффективно.
Для определения того, могут ли человеческие эмбриональные стволовые клетки дифференцироваться в клетки панкреатической эндодермы на трех типах микроносителей, которые имели подходящие характеристики для прикрепления и роста, человеческие эмбриональные стволовые клетки линии H1, пассаж 41 (H1p41), были посеяны на микроносители Cytodex 1®, Cytodex 3® (GE Healthcare Life Sciences, Нью-Джерси, США) и HILLEX®II (Solohill, Мичиган, США) (см. пример 1). Микроносители были подготовлены в соответствии с инструкциями производителей. 30 см2 микроносителей были перенесены в 6-луночные планшеты, имеющие низкую способность к прикреплению клеток. Клетки H1 были диссоциированы от двух планшетов площадью 10 см2 с помощью реактива TrypLE™ Express в соответствии с инструкциями производителя. Клетки высеяли в количестве 5×105 клеток в лунку. Прикрепление клеток к сферам осуществляли способами, описанными в примере 3. Вкратце, клетки и микроносители инкубировали в среде, кондиционированной мышиными эмбриональными фибробластами, с добавлением 10 мкМ Y27632 при 37°C в течение 4 часов с кратковременным перемешиванием каждый час. Клетки на микроносителях HILLEX®II и Cytodex 1® помещали на качающуюся платформу. Клеткам на микроносителях Cytodex 3® давали возможность осесть в течение ночи в спокойном состоянии. Среду меняли ежедневно; в ней отсутствовал реагент Y27632.
Вследствие плохого прикрепления большинство клеток в планшете с Cytodex 1® оказались не прикрепленными к микроносителям. Однако увеличение срока прикрепления и/или уменьшение скорости качания платформы может повысить прикрепление клеток. Через 7 дней провели дифференцирование клеток в сформированную эндодерму с помощью 2-процентного раствора, не содержащего жирных кислот (FAF) BSA (Proliant, Айова, США), в среде RPMI со следующими ростовыми факторами: bFGF (8 нг/мл, (PeproTech, Нью-Джерси, США)), активин A (100 нг/мл, (PeproTech, Нью-Джерси, США)), Wnt3a (20 нг/мл, (R&D Biosciences, Миннесота, США)). Со второго по четвертый день дифференцирования клетки обрабатывали той же средой, но без Wnt3a. Анализ методом FACS, проведенный в двукратно повторенных образцах через 4 суток, показал 77-83% положительных по экспрессии CXCR4 клеток. Экспрессия маркера сформированной эндодермы в клетках, выращенных на разных микроносителях, была одинаковой (см. фиг.30).
Затем клетки дифференцировались в течение дополнительных 2 дней с использованием FGF7 (50 нг/мл, (R&D Systems, Миннесота, США)), KAAD-Cyclopamine (0,25 мкМ, (Calbiochem, Нью-Джерси, США)) в среде DMEM/F12 или DMEM-HG с добавлением 2% FAF BSA (Proliant, Айова, США). После этого была проведена 4-дневная обработка фактором Noggin (100 нг/мл, (R&D Biosciences, Миннесота, США)), FGF7 (50 нг/мл, (R&D Systems, Миннесота, США)), ретиноевой кислотой (2 мкМ, (Sigma-Aldrich, Миссури, США)) и KAAD-Cyclopamine (0,25 мкМ, (Calbiochem, Нью-Джерси, США)) в среде DMEM/F12 или DMEM-HG с добавлением 1% B-27 (Invitrogen, Калифорния, США). Затем клетки дифференцировались в течение трех дней с помощью Noggin (100 нг/мл, (R&D Biosciences, Миннесота, США)), DAPT (1 мкМ, (Sigma-Aldrich, Миссури, США)), Alk5 ингибитора II (1 мкМ, (Axxora, Калифорния, США)) в среде DMEM/F12 или DMEM-HG с добавлением 1% B-27 (день 13, панкреатическая эндодерма, (Invitrogen, Калифорния, США)). На фиг.31 показан измеренный путем количественной ПЦР уровень экспрессии специфических панкреатических генов NKX6.1, PDX1 и NGN3. Значения CT четко показывают, что клетки на микроносителях HILLEX®II не дифференцировались эффективно до экспрессии маркеров клеток-предшественников бета-клеток. Хотя на всех трех типах микроносителей клетки эффективно дифференцировались до сформированной эндодермы, дальнейшее дифференцирование в панкреатические клетки-предшественники на микроносителях HILLEX®II эффективно не происходит.
Для определения того, могут ли человеческие эмбриональные стволовые клетки далее дифференцироваться в инсулинпродуцирующие клетки, клетки H1 p45 выращивали на микроносителях Cytodex 3® (GE Healthcare Life Sciences, Нью-Джерси, США) и дифференцировали способом, аналогичным описанному выше. Коротко говоря, клетки H1 диссоциировали от планшеты площадью 10 см2 при помощи реактива TrypLE™ Express в соответствии с инструкциями производителя. Клетки высеяли в концентрации 1 или 2×106 клеток на одну лунку 6-луночного планшета. Прикрепление клеток к сферам описано в примере 3. Коротко говоря, клетки и микроносители инкубировали в среде, кондиционированной мышиными эмбриональными фибробластами, с добавлением 10 мкМ Y27632 при 37°C в течение 4 часов с кратковременным перемешиванием каждый час. Клетки инкубировали в течение ночи в спокойном состоянии. В день 2 среду заменяли средой, кондиционированной мышиными эмбриональными фибробластами, с добавлением 5 мкМ Y27632 и помещали на качающуюся платформу. Среду меняли через день, не добавляя Y27632. На пятые сутки среду заменяли средой для дифференцирования в сформированную эндодерму, 2% не содержащей жирных кислот (FAF) BSA (Proliant, Айова, США) в среде RPMI, с добавлением следующих ростовых факторов: bFGF (8 нг/мл, (PeproTech, Нью-Джерси, США)), активин А (100 нг/мл, (PeproTech, Нью-Джерси, США)), Wnt3a (20 нг/мл (R&D Biosciences, Миннесота, США)). На второй и третий дни дифференцирования клетки обрабатывали той же средой, но без добавления Wnt3a. Анализ методом FACS, проведенный в двукратно повторенных образцах через 3 дня, показал 97-98% положительных по экспрессии CXCR4 клеток. Затем клетки дифференцировали в течение дополнительных двух дней с добавлением FGF7 (50 нг/мл, (R&D Systems, Миннесота, США)), KAAD-Cyclopamine (0,25 мкМ, (Calbiochem, Нью-Джерси, США)) в среде DMEM-High Glucose (HG) с добавлением 2% FAF BSA (Proliant, Айова, США). После этого была проведена четырехдневная обработка фактором Noggin (100 нг/мл, (R&D Biosciences, Миннесота, США)), FGF7 (50 нг/мл, (R&D Systems, Миннесота, США)), ретиноевой кислотой (2 мкМ, (Sigma-Aldrich, Миссури, США)) и KAAD-Cyclopamine (0,25 мкМ, (Calbiochem, Нью-Джерси, США)) в среде DMEM-HG с добавлением 1% B-27 (Invitrogen, Калифорния, США). Затем клетки дифференцировали в течение трех дней с использованием Noggin (100 нг/мл, (R&D Biosciences, Миннесота, США)), DAPT (1 мкМ, (Sigma-Aldrich, Миссури, США)), Alk5-ингибитора II (1 мкМ, (Axxora, Калифорния, США)) в среде DMEM-HG или DMEM-F12 с добавлением 1% B-27 (Invitrogen, Калифорния, США). Далее выполнялось дифференцирование в среде DMEM-HG или DMEM-F12 с добавлением Alk5-ингибитора II (1 мкМ, (Axxora, Калифорния, США)) в течение семи дней. Заключительное дифференцирование проводилось в среде DMEM-HG или DMEM-F12 соответственно. В целом процесс дифференцирования занимал 24 дня и приводил к экспрессии панкреатических эндокринных гормонов. На фиг.32 показаны данные анализа методом FACS для клеток на заключительной стадии. Клетки с наибольшей плотностью посева, дифференцированные в среде DMEM-HG с 6 по 24 сутки, имели наивысший уровень экспрессии инсулина (фиг.32).
В альтернативном варианте с целью определения того, могут ли человеческие эмбриональные стволовые клетки быть дифференцированы в инсулинпродуцирующие клетки, клетки H1 p44 выращивались в ротационной колбе в течение 7 дней на микроносителях Cytodex 3® (см. пример 5). Клетки с микроносителями были перенесены в 12-луночный планшет площадью 15 см2/лунка и поставлены на качающуюся платформу при 37°C. Клетки дифференцировали в сформированную эндодерму, как описано выше, но вместо DMEM/F12 использовалась среда RMPI. Анализ методом FACS, проведенный через 4 дня в трех повторностях, показал 75-77% положительных по экспрессии CXCR4 клеток. Далее клетки дифференцировали в течение дополнительных трех дней путем обработки FGF7 (50 нг/мл, (R&D Systems, Миннесота, США)), KAAD-Cyclopamine (0,25 мкМ, (Calbiochem, Нью-Джерси, США)) в среде DMEM/F12 с добавлением 2% раствора фракции V бычьего альбумина, не содержащего жирных кислот. После этого была проведена четырехдневная обработка фактором Noggin (100 нг/мл, (R&D Biosciences, Миннесота, США)), FGF7 (50 нг/мл, (R&D Systems, Миннесота, США)), ретиноевой кислотой (2 мкМ, (Sigma-Aldrich, Миссури, США)) и KAAD-Cyclopamine (0,25 мкМ, (Calbiochem, Нью-Джерси, США)) в среде DMEM/F12 с добавлением 1% B-27 (Invitrogen, Калифорния, США). Затем клетки дифференцировали в течение трех дней с использованием Noggin (100 нг/мл, (R&D Biosciences, Миннесота, США)), Netrin4 (100 нг/мл, (R&D Biosciences, Миннесота, США)), DAPT (1 мкМ, (Sigma-Aldrich, Миссури, США)), Alk5-ингибитора II (1 мкМ, (Axxora, Калифорния, США)) в среде DMEM/F12 с добавлением 1% B-27 (день 15, панкреатическая эндодерма (Invitrogen, Калифорния, США)). После этого была проведена шестидневная обработка Alk5-ингибитором II (1 мкМ, (Axxora, Калифорния, США)) в среде DMEM/F12 с добавлением 1% B-27 (день 21, панкреатические эндокринные клетки (Invitrogen, Калифорния, США)). В качестве заключительного этапа проводилась семидневная обработка средой DMEM/F12 с добавлением 1% B-27 (день 28, инсулинэкспрессирующие клетки (Invitrogen, Калифорния, США)). На фиг.33 представлен измеренный методом количественной ПЦР уровень экспрессии специфических панкреатических генов, инсулина, Pdx1 и глюкагона. Данные по микроносителям сравнивали с ранее полученными результатами для клеток H1 p42, дифференцированных на плоской поверхности, покрытой материалом MATRIGELTM (BD Biosciences, Калифорния, США). Уровень экспрессии генов, специфичных для поджелудочной железы, в клетках, дифференцированных на микроносителях, был аналогичным или лучшим, чем в клетках, дифференцированных на плоских поверхностях.
Подобные эксперименты были проведены на клетках H9p38, посеянных на микроносители Cytodex 3® и выращенных в ротационных колбах. Аликвоту микроносителей площадью 15 см2 с клетками помещали в 12-луночный планшет со средой для дифференцирования и устанавливали на качающуюся платформу. Эти клетки сравнивались с клетками, посеянными на 6-луночные планшеты, покрытые материалом MATRIGELTM (BD Biosciences, Калифорния, США). Дифференцирование клеток в сформированную эндодерму осуществляли в среде RPMI с добавлением реактивов. В среднем CXCR4 экспрессировали 83% клеток (образцы имели две повторности). Для сравнения, на плоском субстрате 72% клеток экспрессировали CXCR4 (фиг.34). Во время дальнейшего дифференцирования в панкреатическую эндодерму (день 15) панкреатические эндокринные клетки (день 22) и инсулинэкспрессирующие клетки (день 29) имели похожие уровни экспрессии инсулина клетками, выращенными на микроносителях и на плоском субстрате (фиг.35). Компоненты среды были идентичны перечисленным выше в эксперименте с дифференцированием клеток H1, но эксперимент на этапе дифференцирования в эндокринные клетки длился на день больше. На этапе экспрессии инсулина в клетках наблюдали неожиданное падение экспрессии инсулина в сравнении с днем 22. Поскольку это падение наблюдалось как на микроносителях, так на плоском субстрате, вероятно, оно не связано с субстратом для прикрепления. Таким образом, было показано, что клетки H9 также могут быть успешно дифференцированы на микроносителях, по меньшей мере, в панкреатические эндокринные клетки.
В целом, две линии человеческих эмбриональных стволовых клеток, H1 и H9, могут дифференцировать в панкреатические эндокринные клетки на микроносителях Cytodex 3®, что иллюстрирует потенциал выращивания и дифференцирования этих клеток в крупномасштабной системе культивирования (фиг.17, 21, 33 и 35). Человеческие эмбриональные стволовые клетки могли прикрепляться и пролиферировать по меньшей мере на трех типах сферических микроносителей. Клетки могли дифференцировать, по меньшей мере, до этапа сформированной эндодермы (фиг.28). Результаты иллюстрируют способ, с помощью которого человеческие эмбриональные стволовые клетки можно выращивать и дифференцировать в терапевтических целях.
Пример 8. Человеческие эмбриональные стволовые клетки, посеянные в виде одиночных клеток в культуру на основе трехмерного микроносителя, могут быть перенесены в культуру на поверхности, не содержащей экстраклеточного матрикса (ЭКМ), с сохранением плюрипотентности
Человеческие эмбриональные стволовые клетки H1 культивировали на микроносителях способами, описанными в примере 5. Клетки отделяли от микроносителей и высевали на плоские поверхности из полистирола для тканевого культивирования (TCPS) Nunc4, Nunc13, CELLBIND™ или PRIMARIATM в среде MEFCM16 с добавлением 3 мкМ глицил-H 1152 дигидрохлорида. Клетки высевали с плотностью 100000 клеток/см2 в шестилуночные планшеты, а затем культивировали на соответствующей поверхности в течение одного дополнительного пассажа. Затем клетки отделяли с помощью TrypLE и анализировали проточной цитометрией на маркеры плюрипотентности, либо лизировали в лунке с помощью RLT для очистки мРНК для выполнения количественной ОТ-ПЦР, либо дифференцировали в сформированную эндодерму. Дифференцирование инициировали путем обработки клеток средой RPMI с добавлением 2% BSA, 100 нг/мл активина А, 20 нг/мл Wnt3a, 8 нг/мл bFGF и 3 мкМ глицил-H 1152 дигидрохлорида в течение 24 часов. Затем среду меняли на среду RPMI с добавлением 2% BSA, 100 нг/мл активина А, 8 нг/мл bFGF и 3 мкМ глицил-H 1152 дигидрохлорида. Обработка велась в течение дополнительных 48 часов при ежедневной замене среды.
Как показывает анализ маркеров плюрипотентности, выполненный проточной цитометрией или количественной ОТ-ПЦР, клетки, культивированные на микроносителях и перенесенные в культуру на плоском полистироле для культивирования тканей Nunc4, Nunc13, CellBIND или Primaria, сохраняли плюрипотентность после двух пассажей на соответствующей плоской поверхности (фиг.36). Более того, как показывают данные проточной цитометрии и количественной ОТ-ПЦР, клетки сохраняли способность к дифференцированию в сформированную эндодерму (фиг.37). Аналогичные результаты были получены в выполняемых параллельно тестах с человеческими эмбриональными стволовыми клетками H1 и H9, которые высевались на микроносители Cytodex 3® и дифференцировались в сформированную эндодерму (фиг.38).
Результаты тестов показывают, что человеческие эмбриональные стволовые клетки могут высеваться на микроносители, а затем культивироваться на другой поверхности, сохраняя плюрипотентность. Клетки также могут быть перенесены на другую поверхность, после чего возможно инициировать дифференцирование.
Пример 9. Человеческие эмбриональные стволовые клетки могут быть прямо перенесены из культуры кластерного/колониального типа на слое митотически инактивированных питающих фибробластов в культуру одиночных клеток на поверхностях, не имеющих экстраклеточного матрикса, на срок по меньшей мере 10 пассажей, без потери плюрипотентности и без необходимости удаления питающих фибробластов вручную
Линии человеческих эмбриональных стволовых клеток в настоящее время получают способом, состоящим в стимулировании роста колонии из одиночной клетки или кластера нескольких клеток из бластоцисты. Затем разросшуюся колонию последовательно пересевают и растят до тех пор, пока не получат достаточное количество кластеров/колоний клеток, которые могут составить клеточную линию. После получения клеточной линии для сохранения плюрипотентности и кариотипической стабильности человеческих эмбриональных стволовых клеток текущий стандарт высококачественной воспроизводимой культуры человеческих эмбриональных стволовых клеток требует поддерживать человеческие эмбриональные стволовые клетки на питающем слое из митотически инактивированных фибробластов и пересевать их путем разрушения питающих клеток вручную или, в случае больших партий, путем мягкой ферментативной обработки коллагеназой, нейтральной протеазой или смесью веществ. Такие способы пересевания сохраняют кластеры человеческих эмбриональных стволовых клеток и способствуют росту таких клеток по типу колоний. После образования стабильной линии человеческих эмбриональных стволовых клеток клетки могут быть перенесены на субстрат с экстраклеточным матриксом (ЭКМ), например MATRIGEL™. Однако независимо от того, выращиваются клетки на питающих фибробластах или на субстрате ЭКМ, в рекомендуемом способе пересевания человеческих эмбриональных стволовых клеток представлены особые указания для специалистов о недопущении полной диссоциации колоний человеческих эмбриональных стволовых клеток.
Передовая практика крупномасштабного культивирования клеток млекопитающих включает использование культурального сосуда с трехмерной структурой, в котором строго соблюдаются постоянные гомеостатические условия и в который могут внедряться микроносители для клеток, зависимых от культуральной подложки. Однако современные стандартные способы, применяемые для выращивания человеческих эмбриональных стволовых клеток на питающих фибробластах или на субстрате с ЭКМ и в культуре кластерного/колониального типа, налагают технические ограничения на успешное выращивание культуры человеческих эмбриональных стволовых клеток на микроносителях, поскольку такие способы нелегко перенести на условия крупномасштабного культивирования на микроносителях. Для того чтобы эффективно выращивать человеческие эмбриональные стволовые клетки на микроносителях, необходима возможность пересевать культуру человеческих эмбриональных стволовых клеток в виде одиночных клеток, а не колоний и кластеров, как требует действующий стандарт в данной сфере. Более того, должна существовать возможность выращивания человеческих эмбриональных стволовых клеток без слоя питающих клеток или ЭКМ-субстрата.
Ниже описан способ, позволяющий преодолеть эти технические ограничения. Мы показываем, как можно преобразовать культуру человеческих эмбриональных стволовых клеток из колоний/кластеров на слое митотически инактивированных фибробластов непосредственно в культуральную систему одиночных клеток, не требующую слоя питающих фибробластов или поверхности, покрытой материалом MATRIGEL, или иного субстрата с экстраклеточным матриксом. В данном способе используется крупномасштабное пересевание человеческих эмбриональных стволовых клеток без удаления питающих фибробластов вручную или отбора плюрипотентных клеток из клеточной популяции и производится прямое преобразование из культуры колониального типа с питающими клетками в культуру без питающих клеток и матрикса на субстрате PRIMARIA в присутствии ингибитора Rho-киназы (ROCK), глицил-H 1152 дигидрохлорида. Данный способ может быть реализован в герметичном сосуде в соответствии с нормативными требованиями. Он позволяет получить однородную культуру человеческих эмбриональных стволовых клеток, сохраняющих плюрипотентность и способность дифференцироваться в сформированную эндодерму, не содержащую популяции фибробластов.
Описание способа. Клетки регулярно пересевали путем отсасывания среды, промывания PBS и последующей обработки клеток ферментом, способствующим диссоциации (коллагеназой, реактивом Accutase™ или TrypLE). Коллагеназу использовали в концентрации 1 мг/мл. Accutase™ или TrypLE применяли в исходной концентрации (1Х). Все ферменты использовались после нагрева до комнатной температуры. В каждую лунку добавляли 2% раствор BSA в DMEM/F12, клетки равномерно суспендировали в растворе после обработки ферментом. Затем клетки центрифугировали 5 минут при 200×g, к осадку снова добавляли 2% BSA в DMEM/F12 для ресуспендирования клеток, суспензию клеток распределяли по трем коническим пробиркам объемом 50 мл и центрифугировали 5 мин при 200×g.
В соответствии с последовательным методом мы удаляли питающие фибробласты путем высокоплотного пересевания кластерной/колониальной культуры человеческих эмбриональных стволовых клеток на субстрат Primaria с обработкой культуры на основе мышиных эмбриональных фибробластов (МЭФ) реактивом Accutase™, TrypLE™ или коллагеназой. При первом пересевании клетки высевали во флаконы T-25, покрытые материалом MATRIGELTM, разведенным 1 к 30, в среду, кондиционированную мышиными эмбриональными фибробластами (MEF-CM), или клетки высевали в культуральные флаконы T-25 PRIMARIA™ в среду MEF-CM с добавлением 3 мкМ глицил-H 1152 дигидрохлорида. Все клетки пересевали с показателем разведения от 1 до 3,5; срок воздействия фермента составлял 10 минут. Количество клеток, отделенных реактивом TrypLETM или AccutaseTM, определяли путем подсчета количества клеток, окрашенных трипановым синим, при помощи гемоцитометра. После высевания клеток среду меняли ежедневно, а к клеткам, которые были высеяны в среду MEF-CM+3 мкМ глицил-H 1152 дигидрохлорида, ежедневно добавляли MEF-CM+1 мкМ глицил-H 1152 дигидрохлорида. Образцы анализировали на экспрессию мРНК маркеров плюрипотентности и дифференцирования. В человеческих эмбриональных стволовых клетках (чЭСК), дважды пересеянных в виде одиночных клеток без матрикса, сохранялась экспрессия генов плюрипотентности, а гены дифференцирования были заторможены (фиг.39).
2-й пассаж. Клетки пересевали в соотношении 1 к 4 с 10-минутным воздействием реагента TrypLETM или AccutaseTM. Мы также использовали более кратковременное воздействие фермента, определяемое эмпирическим способом путем обработки клеток и наблюдения за их отделением. Мы наблюдали, что 3-минутного воздействия TrypLETM и 5-минутного воздействия AccutaseTM достаточно, чтобы отделить клетки. После обработки клеток ферментом осуществлялось пересевание, как описано выше. В момент пересевания отбирались аликвоты мРНК клеток для проведения количественной ОТ-ПЦР.
3-й пассаж. После достижения конфлюэнтности клетки отмывали PBS, отделяли путем воздействия фермента в течение 3 или 10 минут (TrypLETM) или 5 или 10 минут (AccutaseTM), суспендировали в 2% растворе BSA в DMEM/F12, центрифугировали, снова отмывали 2% раствором BSA в DMEM/F12, центрифугировали, а затем ресуспендировали и высевали в соответствующую среду. При данном пересевании клетки высеивали в соотношении 1:4, а также в двух других соотношениях - 1:8 и 1:16. При каждом пересевании отбирали аликвоты мРНК для количественной ОТ-ПЦР.
4-й и остальные пассажи. Условия, связанные со временем воздействия фермента и соотношением при пересевании, подобранные в пассажах 2 и 3, сохранялись в пассаже 4 и в последующих пассажах. Каждый раз, когда культура достигала конфлюэнтности, клетки отмывали PBS, отделяли путем воздействия фермента в течение заданного времени, суспендировали в 2% растворе BSA в DMEM/F12, центрифугировали, снова отмывали 2% раствором BSA в DMEM/F12, центрифугировали и ресуспендировали в соответствующей среде с соответствующим соотношением для пересевания. В среду для клеток, которые высевались на субстрат PRIMARIA, в момент посева добавляли 3мкМ глицил-H 1152 дигидрохлорида. После посева среду меняли ежедневно, и к клеткам, которые были высеяны в среду MEF-CM+3 мкМ глицил-H 1152 дигидрохлорида, ежедневно добавляли среду MEF-CM+1 мк глицил-H 1152 дигидрохлорида. В момент посева отбирали аликвоты клеточной мРНК для количественной ОТ-ПЦР.
После завершения более 8 пассажей клетки анализировали на плюрипотентность, для чего проточной цитометрией определяли поверхностные маркеры плюрипотентности (фиг.40), а количественной ОТ-ПЦР - маркеры плюрипотентности и дифференцирования (фиг.41, 42 и 43). Клетки также подвергались дифференцированию в сформированную эндодерму путем их обработки средой RPMI с добавлением 2% BSA, 100 нг/мл активина А, 20 нг/мл Wnt3a, 8 нг/мл bFGF и 3 мкМ глицил-H 1152 дигидрохлорида в течение 24 часов. После этого среду меняли на среду RPMI с добавлением 2% BSA, 100 нг/мл активина А, 8 нг/мл bFGF и 3 мкМ глицил-H 1152 дигидрохлорида. Обработка проводилась в течение дополнительных 48 часов при ежедневной замене среды. Образцы, дифференцированные в сформированную эндодерму, тестировали на наличие маркера сформированной эндодермы CXCR4 проточной цитометрией (фиг.40).
Результаты показывают, что крупномасштабное пересевание клеток из культуры колониального типа на основе питающих фибробластов в культуру без питающих клеток и матрикса на субстрате PRIMARIA в присутствии ингибитора Rho-киназы (ROCK) глицил-H 1152 дигидрохлорида позволяет получить высокогомогенную культуру человеческих эмбриональных стволовых клеток, сохраняющую плюрипотентность и способность к дифференцированию в сформированную эндодерму и не содержащую популяции фибробластов.
Пример 10. Человеческие эмбриональные стволовые клетки, перенесенные с пластика для тканевого культивирования на микроносителях
Клетки H1 культивировали на планшетах для тканевого культивирования PRIMARIA™ (№ по каталогу 353846, Becton Dickinson, Franklin Lakes, Нью-Джерси, США) (способ описан в примере 9), отделяли путем обработки реактивом TrypLE™ Express в течение 3-5 минут и высевали в 6-луночные планшеты, не обработанные для тканевого культивирования, с микроносителями Cytodex 3® (GE Healthcare Life Sciences, Нью-Джерси, США) или HILLEX®II (Solohill, Мичиган, США) в среде MEF-CM с добавлением 10 мкМ Y27632 (Sigma-Aldrich, Миссури, США). Для контроля клетки H1p46, выращенные на планшетах, покрытых материалом MATRIGEL (BD Biosciences, Калифорния, США) и пересеянных с использованием коллагеназы (1 мг/мл), отделяли от субстрата и высевали на микроносители аналогичным образом. Планшеты инкубировали при 37°C в течение 5 часов при перемешивании вручную каждые 45 минут. Далее планшеты устанавливались на качающуюся платформу при 37°C. Среду ежедневно меняли на MEF-CM с добавлением 10 мкМ Y27632 (Sigma-Aldrich, Миссури, США). Изображения показывают хорошее прикрепление клеток к микроносителям на 3 день (фиг.44). Через 7 дней клетки отделяли (как описано в примере 4) и анализировали методом FACS на маркеры плюрипотентности CD9, SSEA-4, SSEA-3, TRA-1-60, TRA-1-81 (фиг.45). Большая часть маркеров плюрипотентности была экспрессирована 90-100% клеток. Отсутствовали очевидные различия между клетками, пересеянными с использованием Accutase™ (Millipore, Массачусетс, США) и TrypLE™ Express (Invitrogen, Калифорния, США), а также между клетками, выращенными на микроносителях Cytodex 3® (GE Healthcare Life Sciences, Нью-Джерси, США) и HILLEX®II (Solohill, Мичиган, США). В целом, клетки оставались плюрипотентными после переноса с пластика для культивирования клеток PRIMARIA™ (№ по каталогу 353846, Becton Dickinson, Franklin Lakes, Нью-Джерси, США) на микроносители.
Затем клетки H1 на микроносителях были дифференцированы в сформированную эндодерму. Способ описан в примере 7. Через 4 дня дифференцирования клетки H1 отделяли от микроносителей и подвергали анализу методом FACS, который показал, что более 82% клеток экспрессировали CXCR4 (см. фиг.46). Клетки эффективно дифференцировались в сформированную эндодерму, независимо от типа микроносителя или использованного фермента при пересевании на пластик PRIMARIA™ (№ по каталогу 353846, Becton Dickinson, Franklin Lakes, Нью-Джерси, США). Это является доказательством гибкости системы культивирования, так как допустимо выращивание клеток на пластике и микроносителях.
Пример 11. Человеческие эмбриональные стволовые клетки, перенесенные с плоских субстратов, состоящих из смеси эфиров целлюлозы, на микроносители
Клетки H1 культивировали на плоских субстратах, состоящих из смеси эфиров целлюлозы, на протяжении 12 пассажей в соответствии со способами, изложенными в заявке на патент США № 61/116452. Клетки отделяли от плоского субстрата путем обработки реактивом TrypLE™ Express в течение 3-5 минут и высевали в 6-луночные планшеты, не обработанные для тканевого культивирования, с микроносителями CYTODEX 3® (GE Healthcare Life Sciences, Нью-Джерси, США) или HILLEX®II (Solohill, Мичиган, США) в среде MEF-CM с добавлением 10 мМ Y27632 (Sigma-Aldrich, Миссури, США). Для контроля клетки H1p44, выращенные на планшетах, покрытых материалом MATRIGELTM (BD Biosciences, Калифорния, США) и пересеянных с использованием коллагеназы (1 мг/мл), отделяли от субстрата и высевали на микроносители аналогичным образом. Планшеты инкубировали при 37°C в течение 5 часов при перемешивании вручную каждые 45 минут. Затем планшеты устанавливали на качающуюся платформу при 37°C. Среду меняли ежедневно. Через 7 дней клетки отделяли (как описано в примере 4) и анализировали методом FACS на маркеры плюрипотентности CD9, SSEA-4, SSEA-3, TRA-1-60, TRA-1-81 (фиг.47). Более 90% клеток экспрессировали большинство маркеров плюрипотентности. Не было очевидных различий между клетками, выращенными на микроносителях CYTODEX 3® и на микроносителях HILLEX®II. Контрольные клетки H1p44 не тестировали на плюрипотентность после выращивания на микроносителях HILLEX®II, поскольку плюрипотентность была подтверждена в других экспериментах (см. пример 5). В целом, клетки сохраняли плюрипотентность при переносе с плоских субстратов, состоящих из смеси эфиров целлюлозы, на микроносители.
После этого клетки H1 на микроносителях подвергались дифференцированию в сформированную эндодерму в соответствии со способами, описанными в примере 7. Через 4 дня дифференцирования клетки H1 отделяли от микроносителей и подвергали анализу методом FACS, который показал, что более 65% клеток экспрессировали CXCR4 (см. фиг.48). Клетки эффективно дифференцировались в сформированную эндодерму, независимо от типа микроносителя. Очевидно, на микроносителях HILLEX®II (Solohill, Мичиган, США) в сформированную эндодерму дифференцировалось несколько меньшее количество клеток. Способность клеток к дифференцированию является доказательством гибкости системы культивирования. Кроме того, клетки могут быть выращены и дифференцированы непосредственно на мембранах и микроносителях, что устраняет необходимость использования любых матриксов из животных компонентов.
Прикрепление клеток H9 к сферическим микроносителям в среде MEF-CM в статических культурах
полистирол
фибронектин
полистирол
полистирол
полистирол
триметиламмоний
полистирол
полистирол
коллаген
полистирол
стекло
коллаген
Прикрепление клеток H1 и H9 к сферическим микроносителям в среде MEF-CM с добавлением 10 мкМ ингибитора Rho-киназы Y27632
производитель
0-5*
полистирол
фибронектин
полистирол
полистирол
полистирол
триметиламмоний
полистирол
полистирол
коллаген
полистирол
стекло
коллаген
Удвоение популяции клеток H1 и H9, выращиваемых в течение 5 пассажей на Cytodex 1®, Cytodex 3® или HILLEX®II
Публикации, цитируемые в настоящем документе, полностью включены в настоящий документ путем ссылки. Хотя различные аспекты изобретения иллюстрируются ссылками на примеры и предпочтительные варианты осуществления, подразумевается, что область изобретения ограничивается не представленным выше описанием, а следующими пунктами формулы изобретения, составленными в соответствии с принципами патентного законодательства.
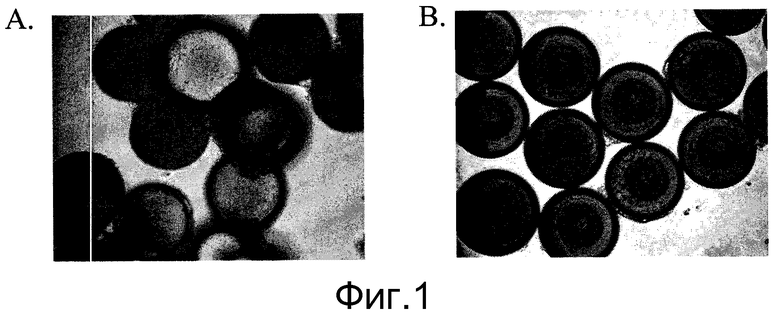

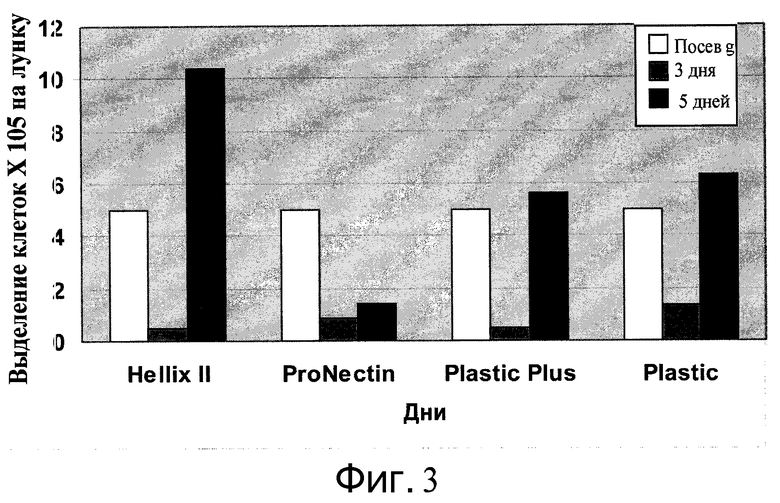
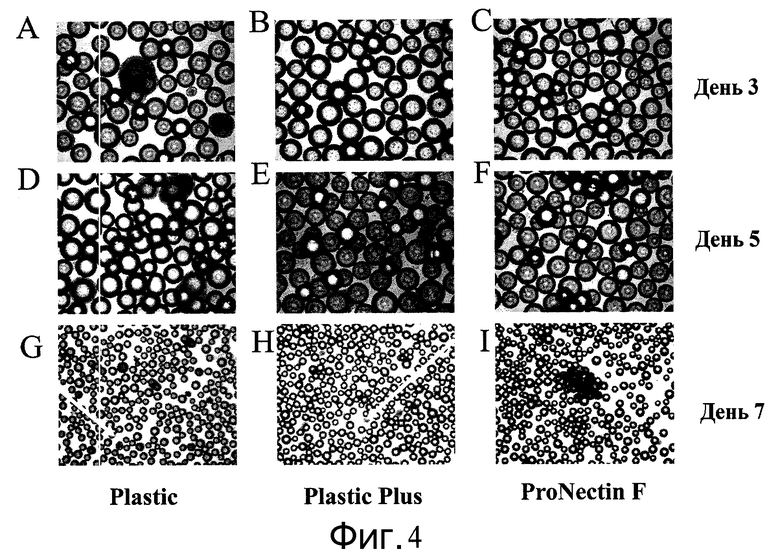

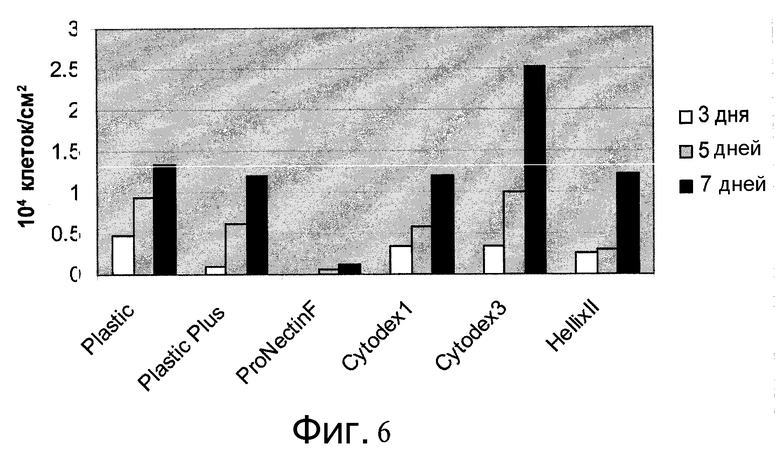
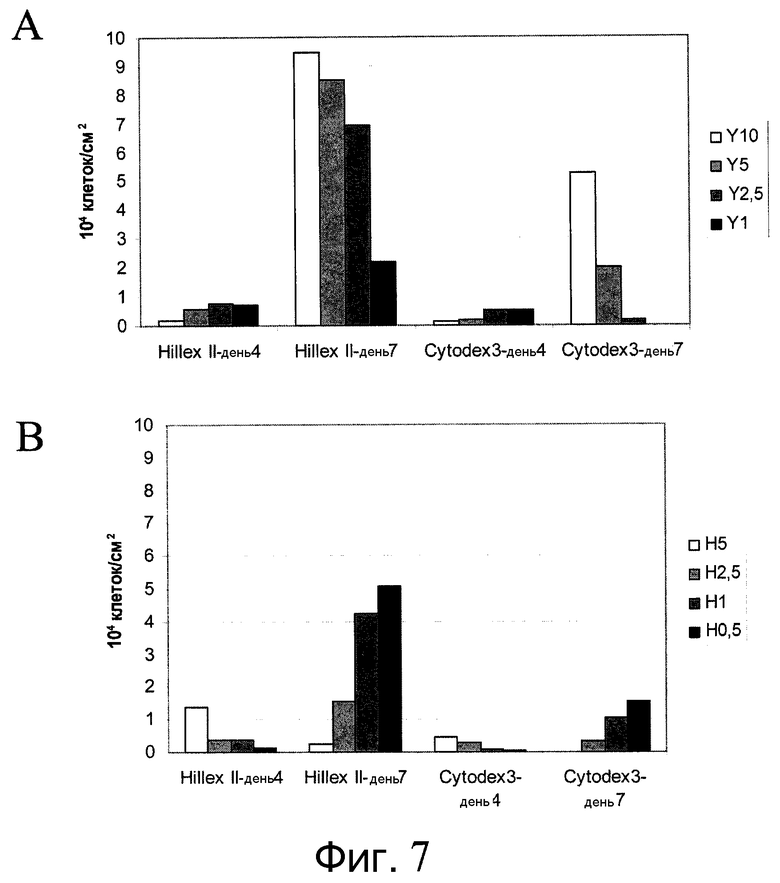
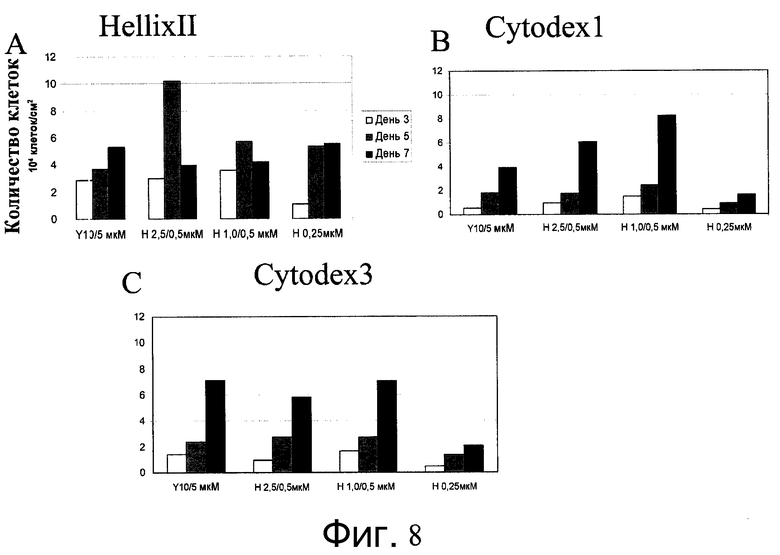

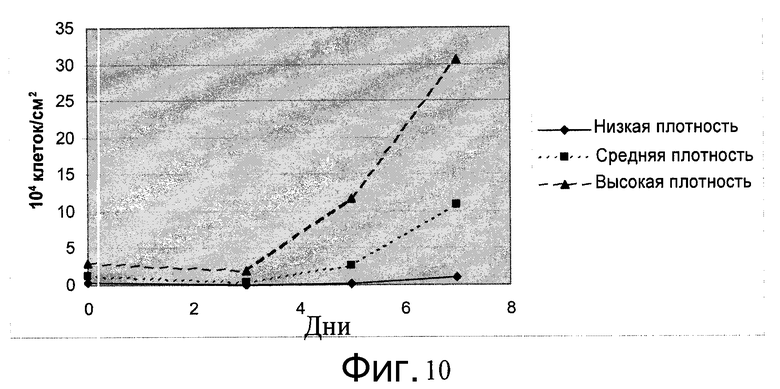
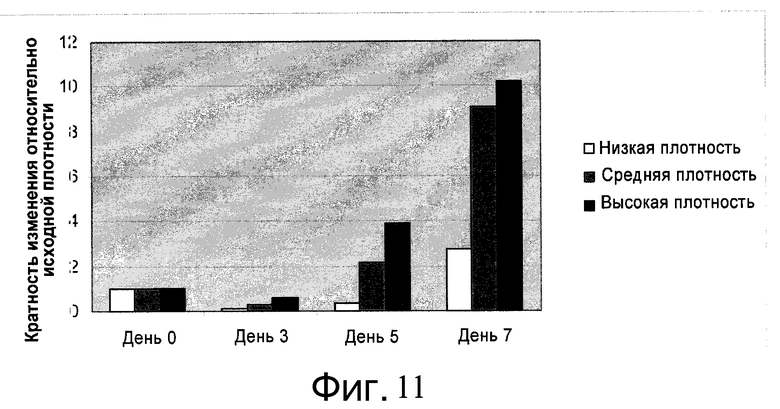
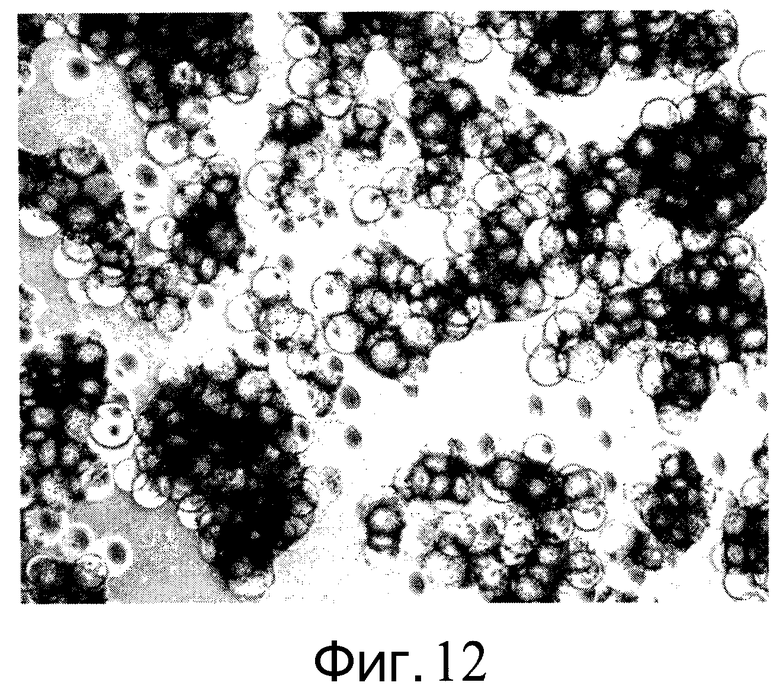

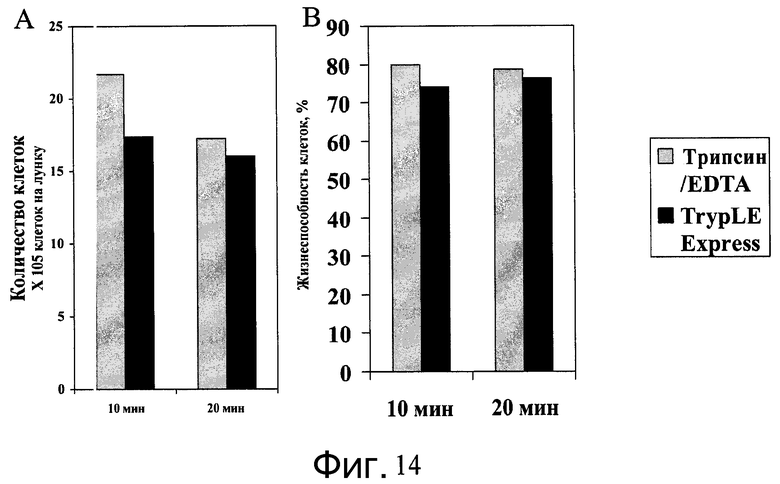
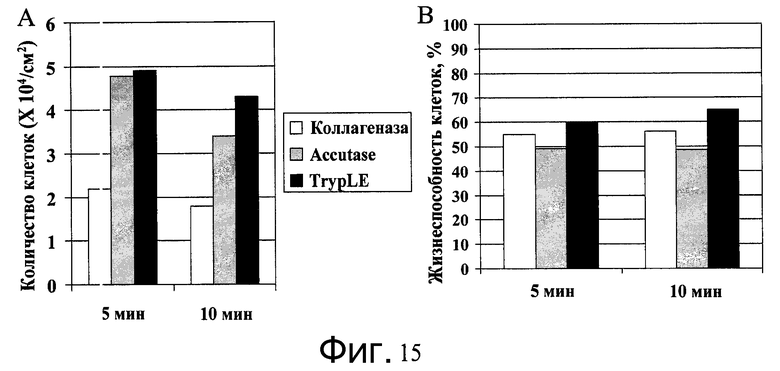
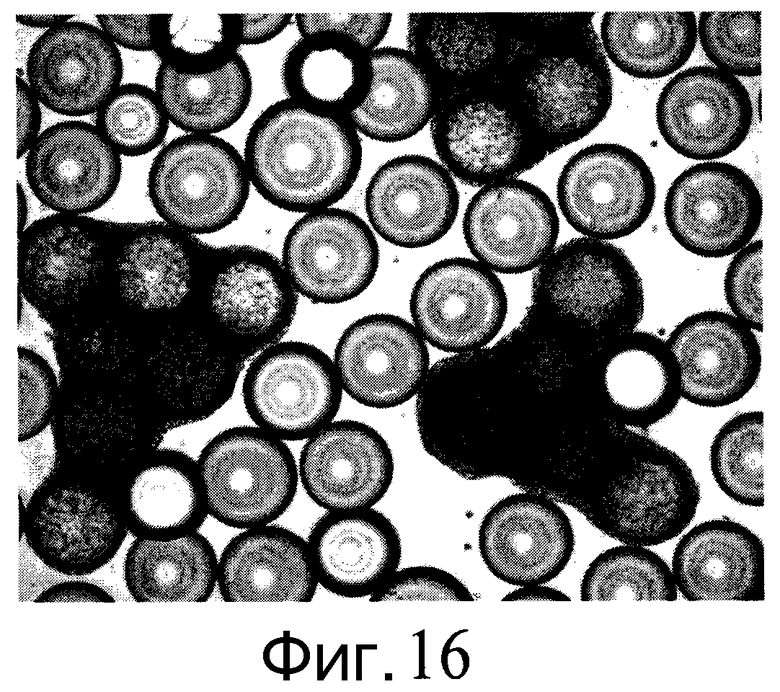
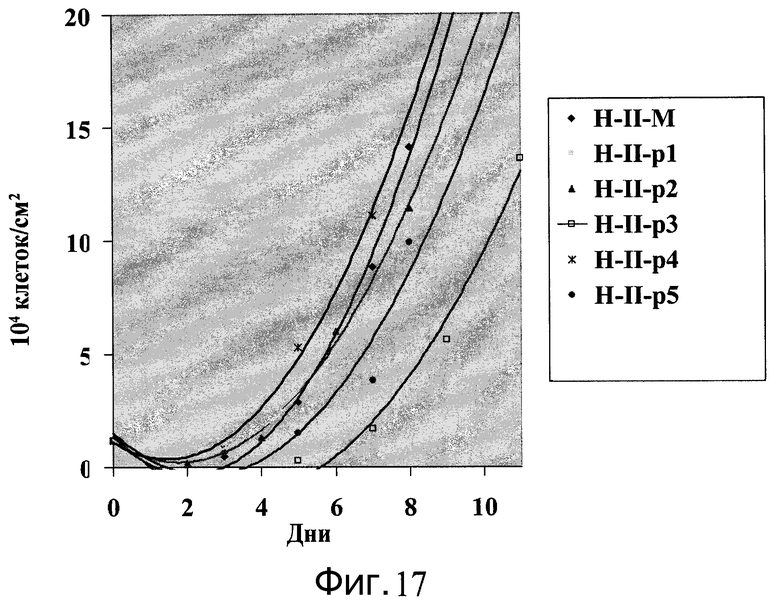
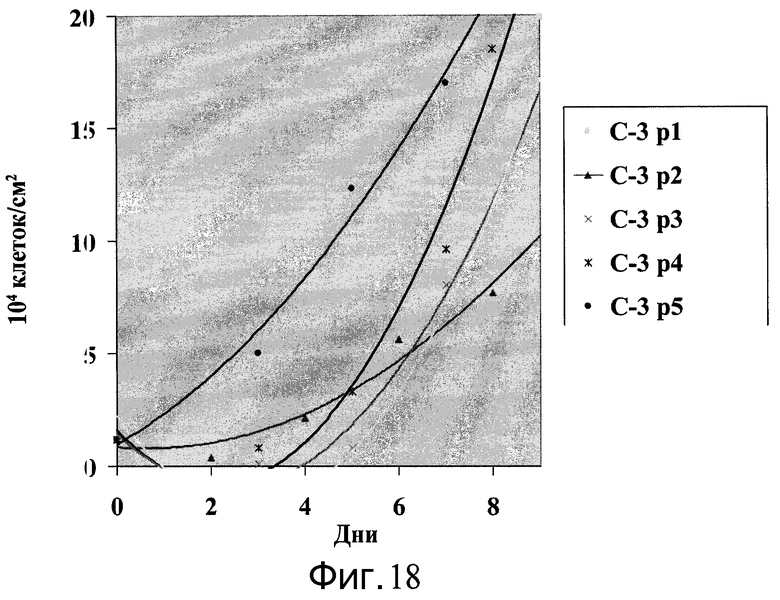
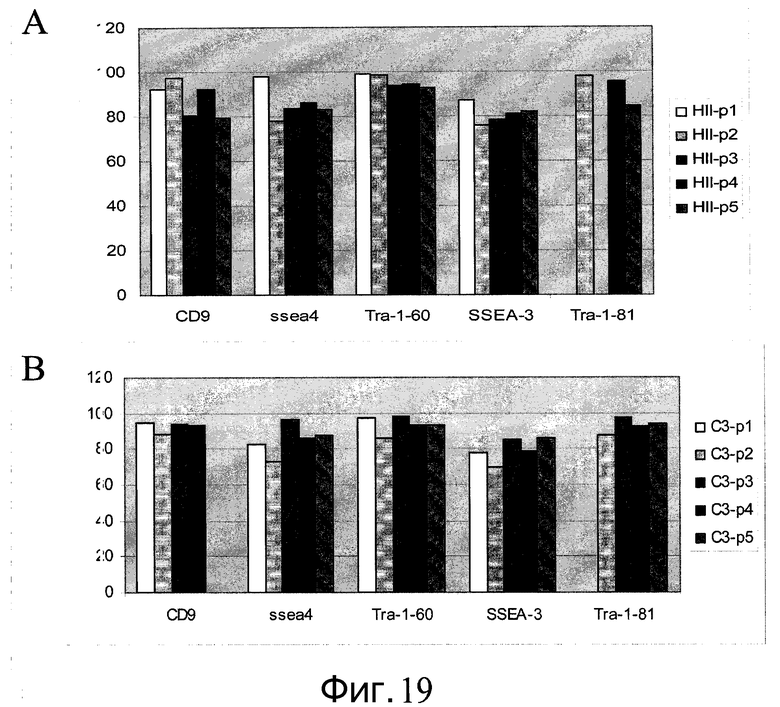

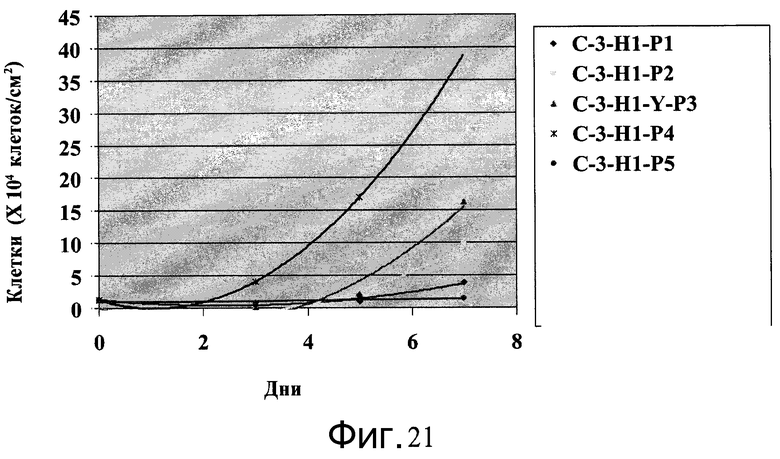
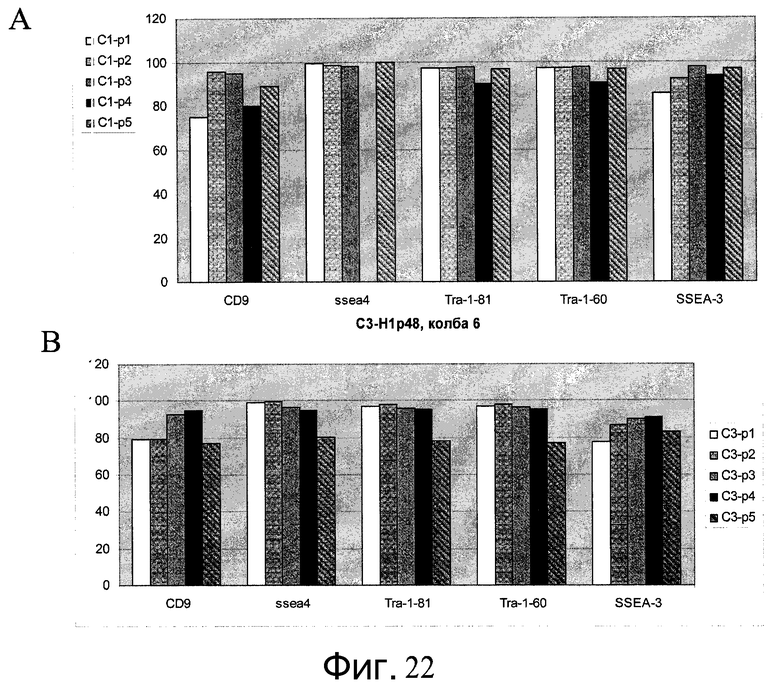
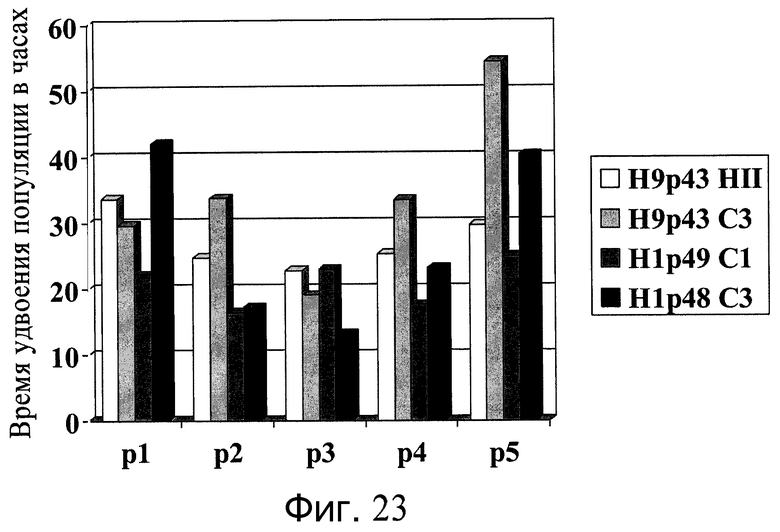
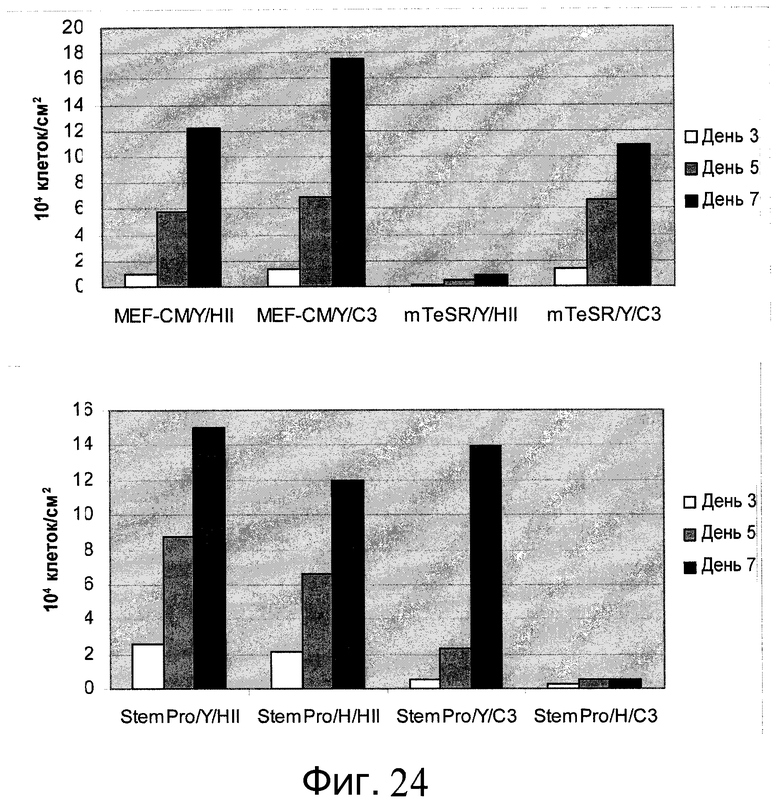
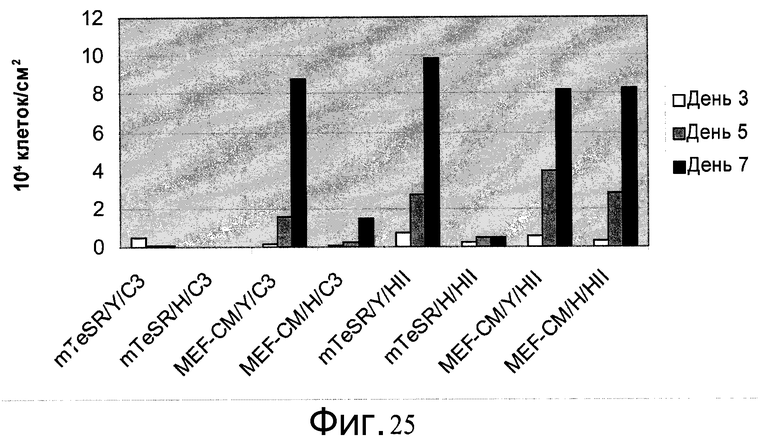

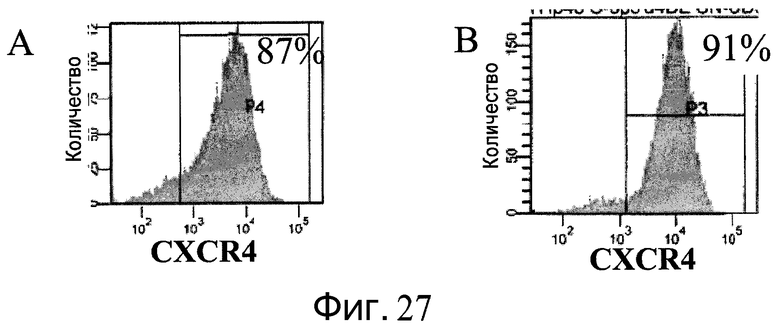
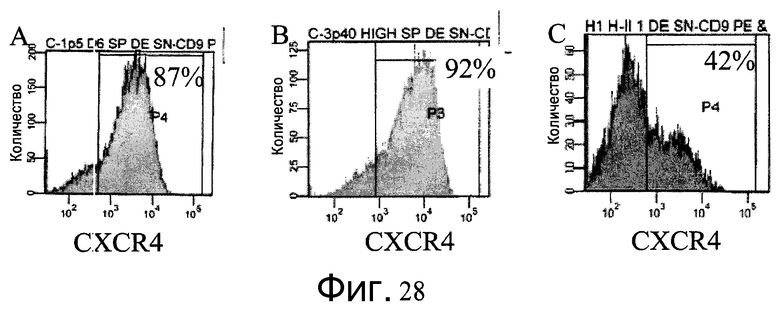
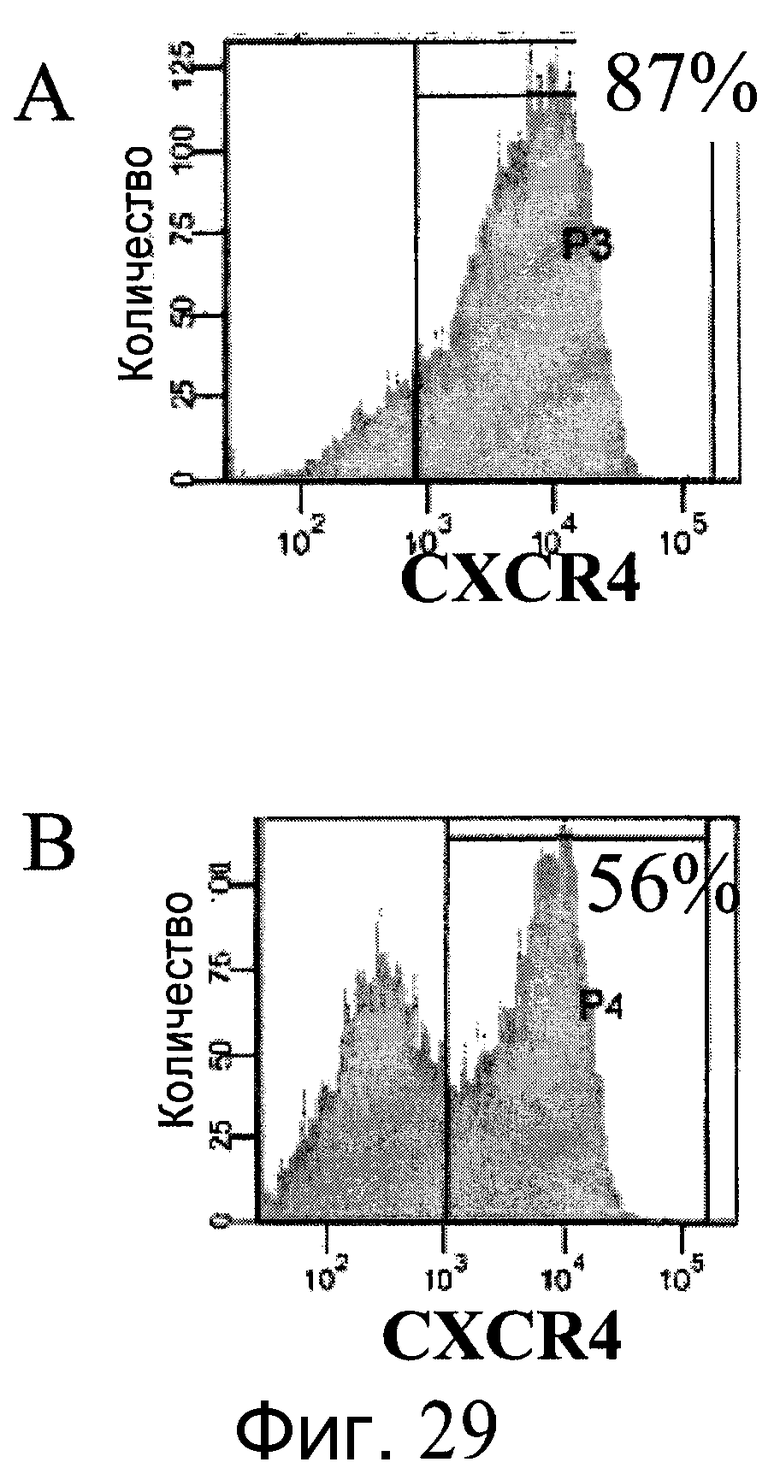
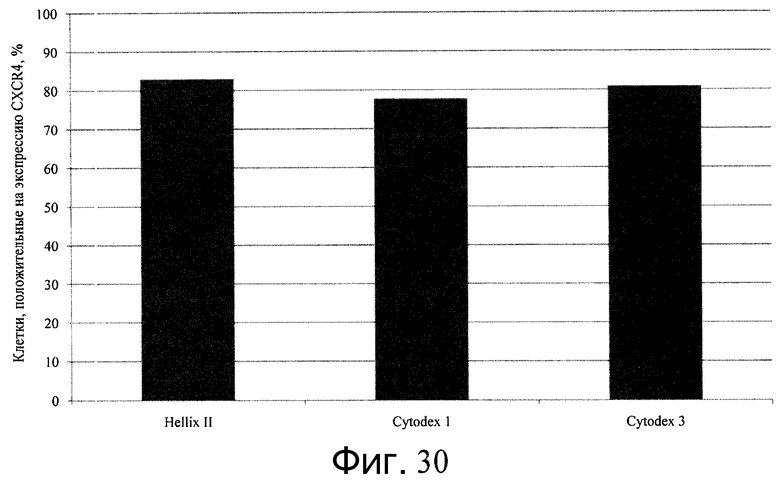
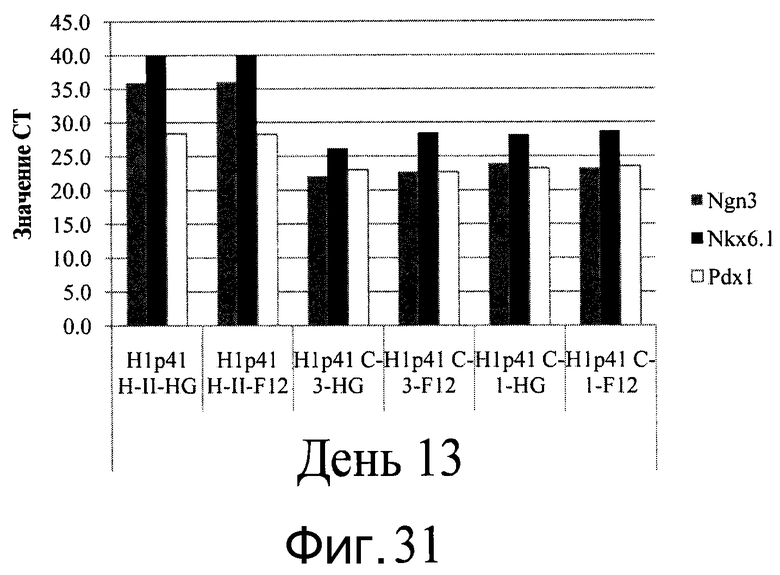
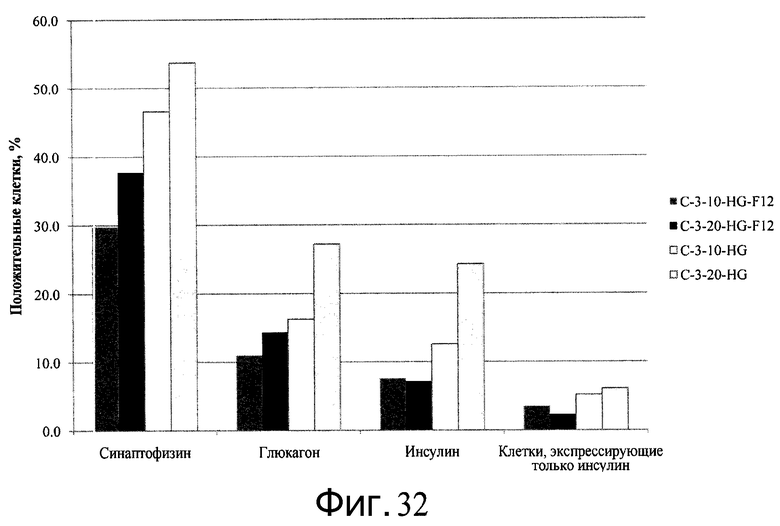
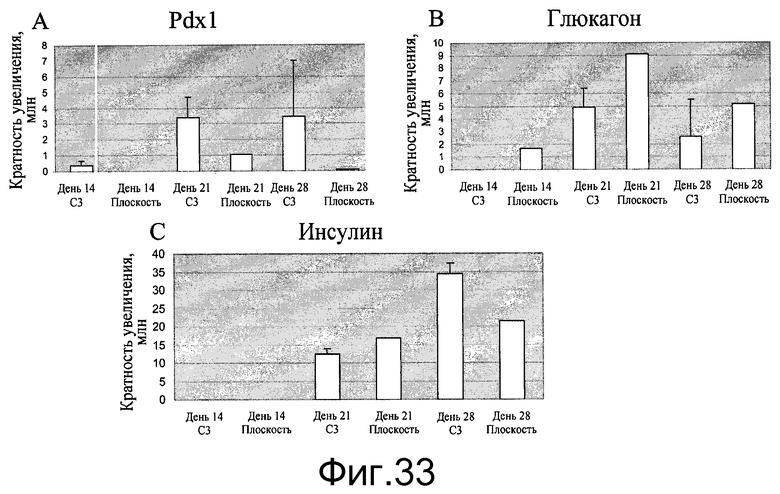
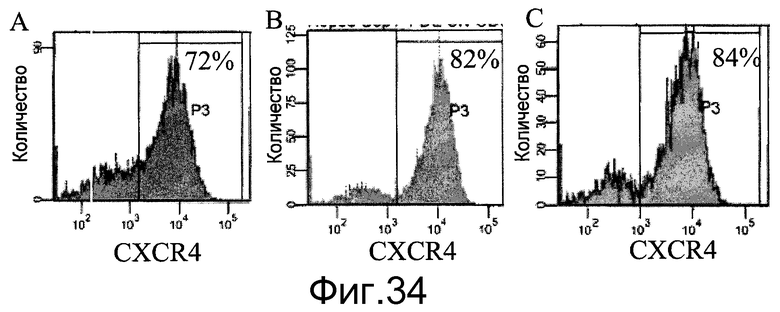
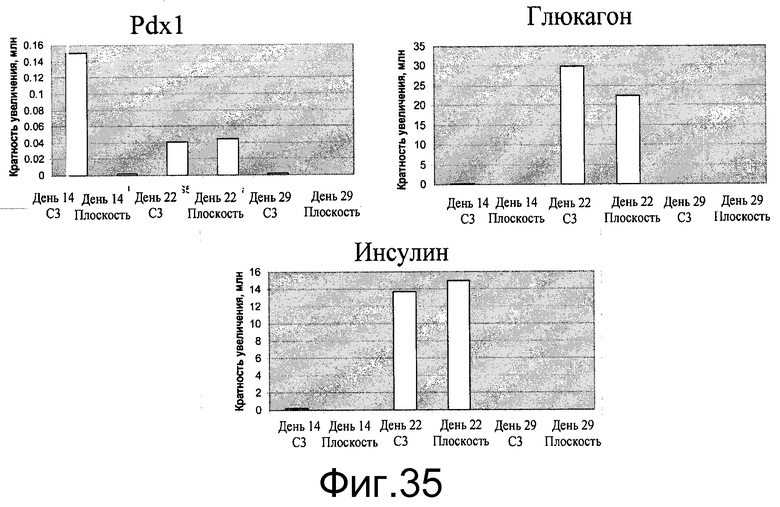
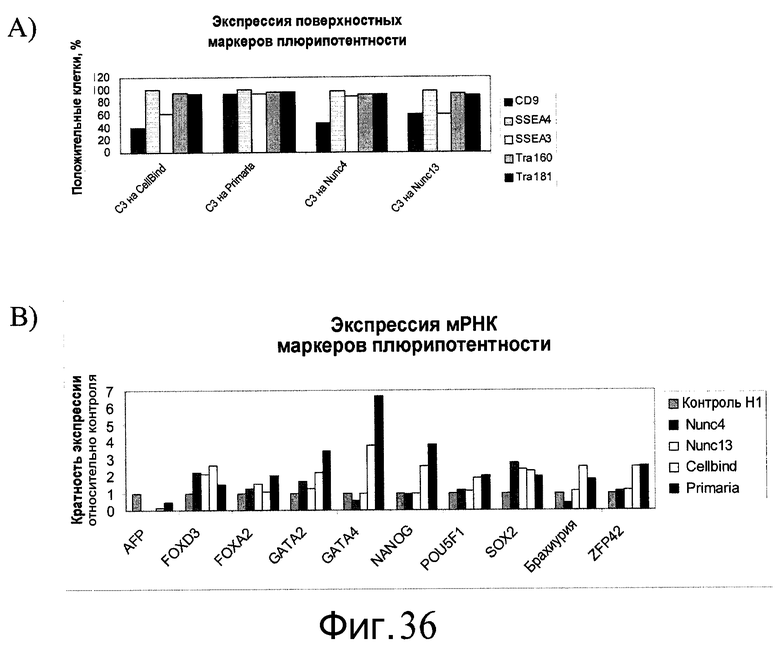
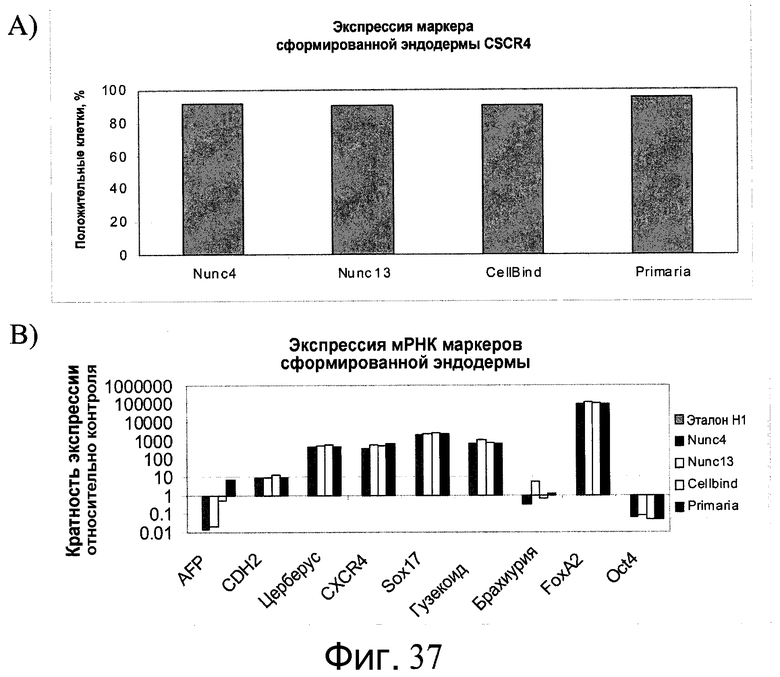
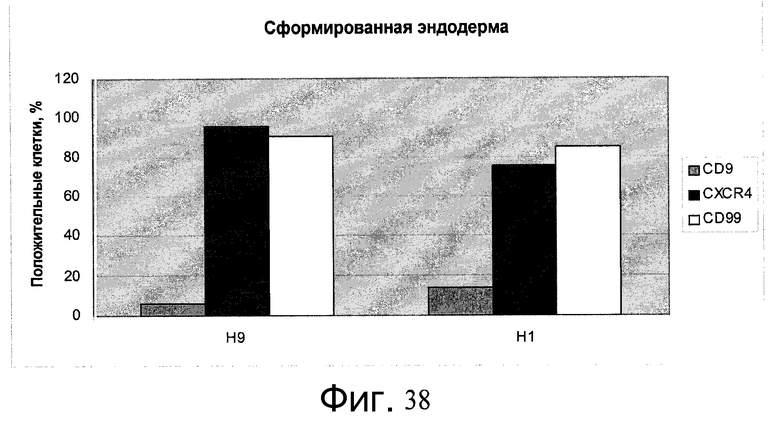
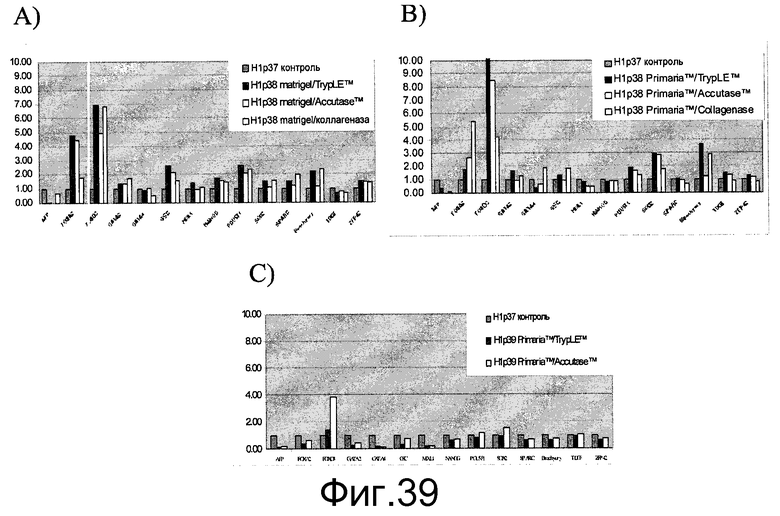
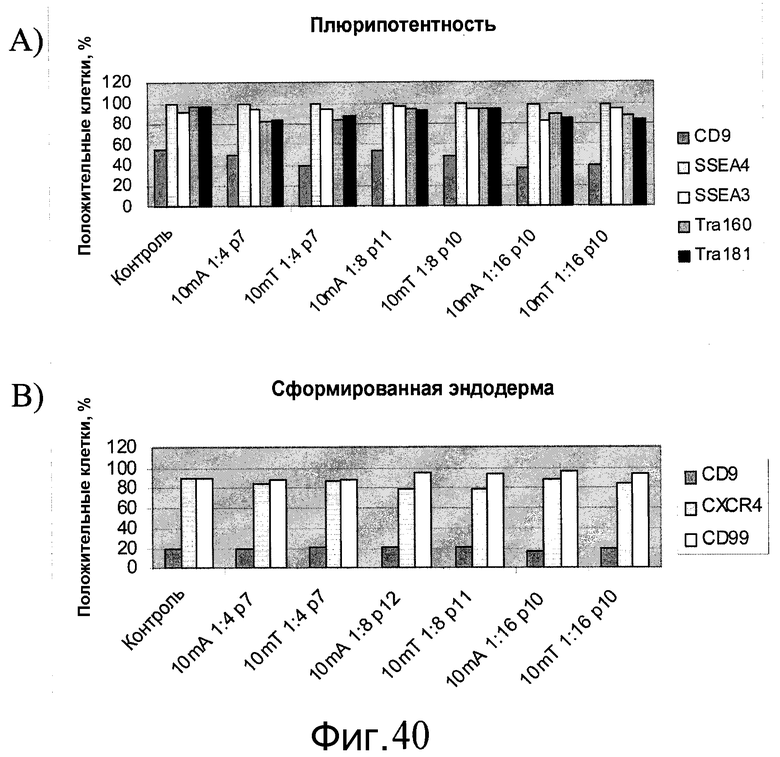
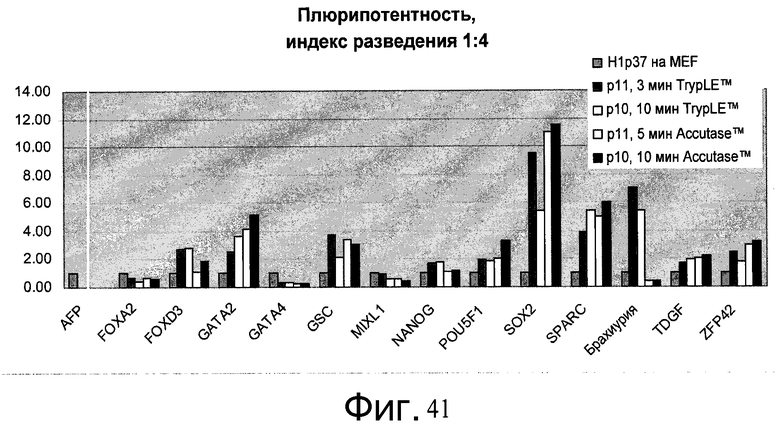
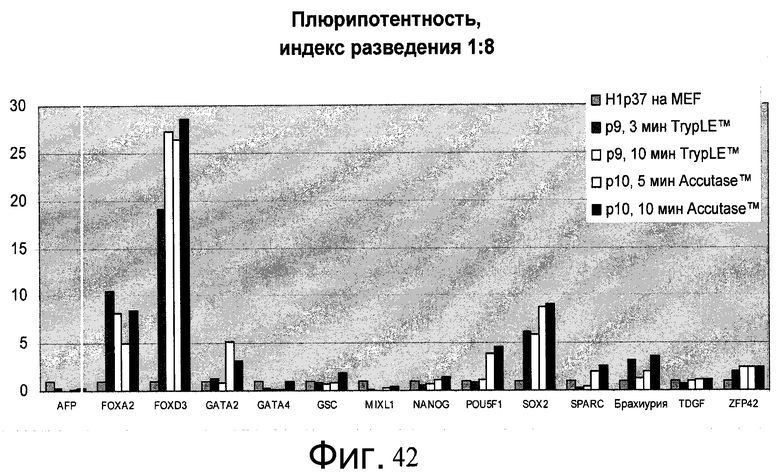
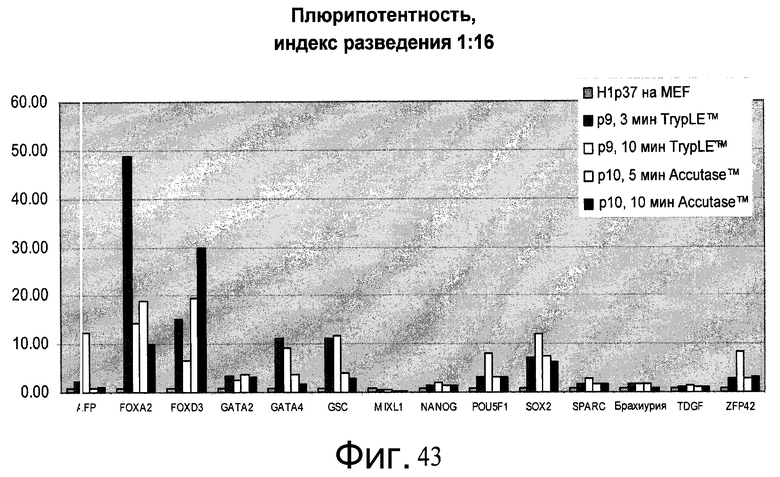
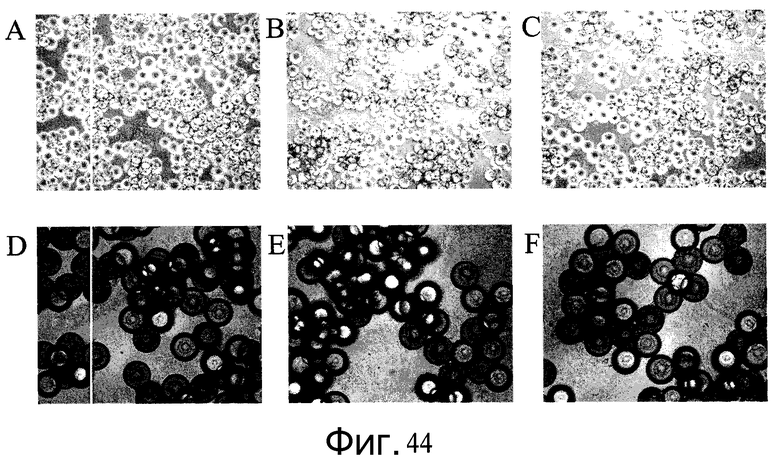
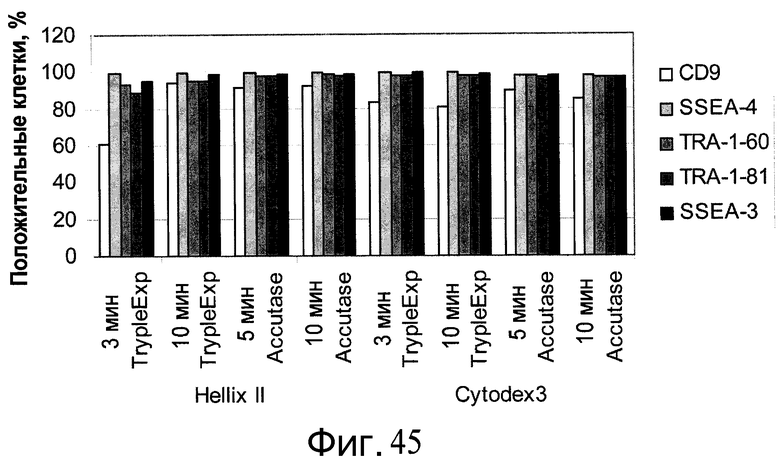
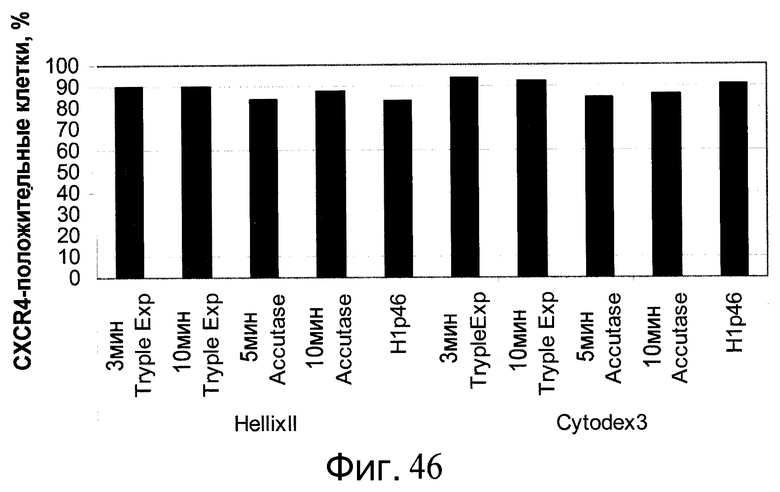
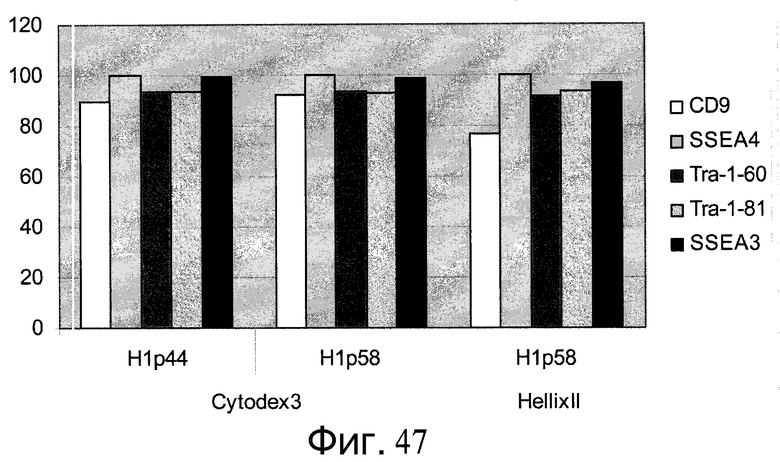
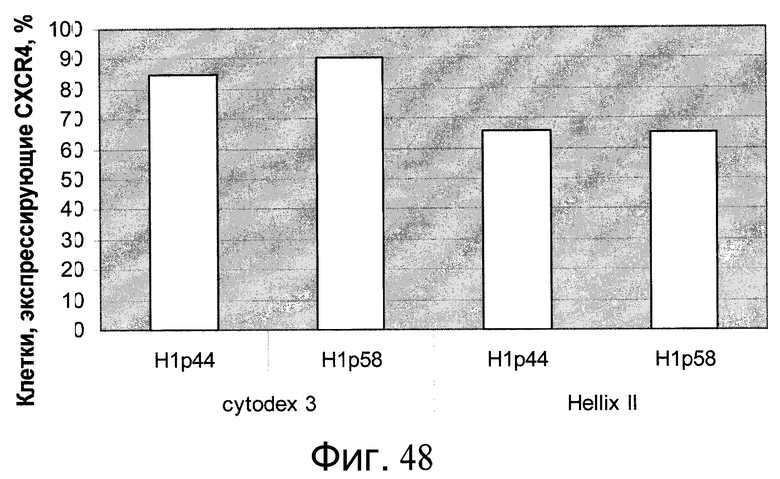
Изобретение относится к биотехнологии. Раскрыт способ выращивания эмбриональных стволовых клеток человека. Способ предусматривает прикрепление популяции клеток к микроносителю в среде, содержащей ингибитор Rho-киназы. Далее проводят культивирование клеток, отделение клеток от микроносителя и прикрепление полученных клеток ко второму микроносителю в среде, содержащей ингибитор Rho-киназы. Изобретение может быть использовано в медицине. 6 з.п. ф-лы, 48 ил., 3 табл., 11 пр.
1. Способ выращивания эмбриональных стволовых клеток человека, включающий стадии:
a) прикрепление популяции эмбриональных стволовых клеток человека к первому объему микроносителей в среде, содержащей ингибитор Rho-киназы, где эмбриональные стволовые клетки человека выращивают с плотностью по меньшей мере около 3,0×104 клеток/см2;
b) культивирование эмбриональных стволовых клеток человека на первом объеме микроносителей, где среда выбрана из среды, кондиционированной фибробластами, и среды, содержащей bFGF;
c) отделение эмбриональных стволовых клеток человека от первого объема микроносителей путем ферментативной обработки; и
d) прикрепление популяции эмбриональных стволовых клеток человека ко второму объему микроносителей в среде, содержащей ингибитор Rho-киназы.
2. Способ по п. 1, в котором стадии b), с) и d) успешно повторяют, используя следующие объемы свежих микроносителей.
3. Способ по п. 1, в котором первый объем микроносителей выбирается из группы, состоящей из декстрановых микроносителей и полистироловых микроносителей.
4. Способ по п. 1, в котором второй объем микроносителей выбирается из группы, состоящей из декстрановых микроносителей и полистироловых микроносителей.
5. Способ по п. 1, в котором эмбриональные стволовые клетки человека отделяют от первого объема микроносителей путем ферментативной обработки.
6. Способ по п. 1, в котором эмбриональные стволовые клетки человека отделяют от второго объема микроносителей путем ферментативной обработки.
7. Способ по п. 1, в котором первый объем микроносителей отделяется от эмбриональных стволовых клеток человека перед прикреплением клеток ко второму объему микроносителей.
| WO 2008004990 A2, 10.01.2008 | |||
| WO 2008102118 A1, 28.08.2008 | |||
| СПОСОБ КУЛЬТИВИРОВАНИЯ МЕЗЕНХИМАЛЬНЫХ СТВОЛОВЫХ КЛЕТОК ЧЕЛОВЕКА EX VIVO | 2006 |
|
RU2323252C1 |

