Настоящая группа изобретений относится к периодическим самособирающимся белкам-предшественникам, кодирующим их последовательностям нуклеиновых кислот и экспрессионным конструкциям, а также к способам рекомбинантного получения пептидов с применением таких белков-предшественников.
Уровень техники
Известны различные способы биотехнологического получения пептидов. Так как стабильность коротких полипептидных цепей в микробных клетках-хозяевах, как правило, является низкой, и так как свободные пептиды могут оказывать возможный токсичный эффект на организм хозяина (например, противомикробные пептиды), большинство способов включают продуцирование более крупных белков-предшественников, из которых пептид вырезается после того, как белок-предшественник был очищен.
Существует возможность получения стабильного белка-предшественника посредством экспрессии пептида вместе со стабильным белком в виде слитого белка. Свойства указанного слитого белка, которые сильно влияют на последующие стадии получения, определяются партнером слияния, в значительной степени независимо от пептидной последовательности, и поэтому являются легко контролируемыми и подходят для получения белков с различными последовательностями.
В WO 2008/085543 раскрывается конкретный способ поучения белков и пептидов с помощью слитого белка. Этот слитый белок содержит наряду с желаемой пептидной последовательностью партнер слияния, для которого характерен обратный фазовый переход. Эта характеристика, прежде всего, обеспечивает простую и недорогую очистку слитого белка от клеточных фрагментов. Во-вторых, партнер слияния также может быть выделен простым и недорогим образом, после того как пептид был удален посредством протеолитического расщепления. Тогда как слитый белок часто может быть получен с хорошим выходом, пептидная часть белка-предшественника, как правило, является маленькой, и поэтому эффективность способа не является оптимальной.
Другой подход относится к периодическим белкам-предшественникам, которые содержат множество копий целевого пептида, который подлежит рекомбинантному получению. В WO 03/089455 раскрывается получение мультимерных белков-предшественников, из которых целевые пептидные последовательности с противомикробными свойствами вырезаются посредством кислотного расщепления.
Существует ряд других концепций в опубликованном виде (например, Metlitskaya et al. Biotechnol Appl. Biochem 39; 339-345 (2004); Wang & Cai Appl. Biochem and Biotechnol. 141; 203-213 (2007)), которые показывают, что пептидные последовательности или семейства пептидных последовательностей могут быть получены установленным способом с помощью периодических белков-предшественников. До некоторой степени было описано применение особых вспомогательных последовательностей, которые локализованы между повторами целевых пептидных последовательностей. Более конкретно, были предложены анионные вспомогательные последовательности, которые очевидным образом уменьшают вредное воздействие катионных противомикробных пептидных последовательностей внутри периодического белка-предшественника на клетку-хозяина (смотрите, например, WO 00/31279 и US 2003/0219854). Так как белок-предшественник, согласно этой стереотипной концепции, имеет более значительную часть целевой пептидной последовательности, чем в случае со слитыми белками, последовательность целевого катионного пептида оказывает сильное влияние на свойства повторяющихся белков-предшественников.
Авторы не нашли каких-либо сведений о каком-либо ранее существующем способе получения какой-либо пептидной последовательности с помощью периодических белков-предшественников, который может быть осуществлен простым, выгодным и эффективным образом.
Различные противомикробные пептиды уже были описаны в литературе и рассмотрены в обзорах (Hancock, R.E.W. und Lehrer, R. 1998 in Trends in Biotechnology, 16: 82-88; Hancock, R.E.W. und Sahl, H.G. 2006 in Nature Biotechnology, 24: 1551-1557).
Слитые белки, в которых объединены два активных пептида, подобным образом описаны в литературе. Автор Wade с соавторами сообщают о противомикробном действии различных слияний цекропина А из Hyalophora cecropia и мелиттина пчелиного яда (Wade, D. et al., 1992, International Journal of Peptide and Protein Research, 40: 429-436). Автор Shin и его соавторы описывают противобактериальное действие слитого пептида цекропина А из Hyalophora cecropia и магаинина 2 из Xenopus laevis, содержащего 20 аминокислот. Цекропин А состоит из 37 аминокислот и проявляет активность в отношении грамотрицательных бактерий, но более низкую активность в отношении грамположительных бактерий. Магаинин 2 состоит из 23 аминокислот и проявляет активность в отношении бактерий, а также в отношении опухолевых клеточных линий. По сравнению со слитым пептидом цекропина А и мелиттина, этот слитый пептид проявляет намного более низкую гемолитическую активность при сопоставимом противобактериальном действии (Shin, S.Y. Kang, J.H., Lee, M.K., Kirn, S.Y., Kirn, Y., Hahm, K.S., 1998, Biochemistry and Molecular Biology International, 44: 1119-1126). В US 2003/0096745 A1 и в US 6,800,727 В2 раскрываются такие гибридные пептиды, состоящие из 20 аминокислот, а также раскрываются варианты такого слияния, которые являются более сильно положительно заряженными и являются более гидрофобными в результате замены аминокислот, особенно положительно заряженных аминокислот и гидрофобных аминокислот.
Автор Shin и его соавторы в 1999 г. описали другие разработки в отношении этого слитого пептида цекропина А и магаинина 2. Они продемонстрировали, что пептид, имеющий SEQ ID N0:6, обладает более низкой гомолитической активностью по сравнению с исходным слиянием, однако противобактериальная активность в отношении Escherichia coli и Bacillus sub-tilis не была понижена (Shin et al. 1999 Journal of Peptide Research, 53: 82-90).
Сущность изобретения
Таким образом, объектом настоящего изобретения является широко применяемый способ получения пептидов с помощью периодических белков-предшественников.
Данный объект обеспечивается благодаря новому подходу к биотехнологическому получению пептидов посредством получения периодических белков-предшественников, которые содержат высокую долю целевой пептидной последовательности и которые содержат вспомогательные последовательности, которые управляют свойствами белка-предшественника прогнозируемым образом. Указанный способ может применяться для получения различных пептидных последовательностей, без необходимости коренным образом заново устанавливать условия экспрессии молекулы-предшественника или процедуры последующей обработки для каждой из различных пептидных последовательностей. Более того, также возможно получать пептиды, для которых ранее применяемые способы не были эффективными.
Описание чертежей
На приведенных чертежах:
Фиг.1 показывает изображение проекции структуры альфа-спирали в виде диаграммы "зубчатое колесо" аминокислотных последовательностей. Аминокислотные последовательности, содержащиеся в периодическом белке-предшественнике, А1-А7 (А), изображены на круге (В). Это расположение позволяет визуализировать положение аминокислот в альфа-спирали;
Фиг.2 показывает обращение - фазовую хроматограмму для пептида "ZnO" после кислотного расщепления;
Фиг.3 показывает масс-спектр для пептида "ZnO" после кислотного расщепления и обращенно-фазовой ВЭЖХ; приведенные числа показывают величину m/z соответствующего моноизотопического пика;
Фиг.4 показывает обращено-фазовую хроматограмму для пептида "Р18" после кислотного расщепления и катионообменной хроматографии;
Фиг.5 показывает масс-спектр для пептида "Р18" после кислотного расщепления, катионообменной хроматографии и обращенно-фазовой ВЭЖХ;
приведенные числа показывают величину m/z конкретного моноизотопического пика;
Фиг.6 показывает обращенно-фазовую хроматограмму для пептида "Min" после кислотного расщепления;
Фиг.7 показывает масс-спектр для пептида "Min" после кислотного расщепления и обращенно-фазовой ВЭЖХ; приведенные числа показывают величину m/z конкретного моноизотопического пика;
Фиг.8 показывает обращенно-фазовую хроматограмму для пептида SEQ ID NO: 6 после кислотного расщепления и катионообменной хроматографии;
Фиг.9 показывает масс-спектр для пептида SEQ ID NO: 6 после кислотного расщепления, катионообменной хроматографии и обращено-фазовой ВЭЖХ; приведенные числа показывают величину m/z конкретного моноизотопического пика;
Фиг.10 показывает хроматограмму, полученную после ВЭЖХ и катионообменной хроматографии, для пептида "Р18" до и после амидирования согласно примеру 6; хроматограмма химически синтезированного и амидированного контрольного пептида с последовательностью пептида "Р18" показана для сравнения;
Фиг.11 показывает хроматограмму, полученную после ВЭЖХ и катионообменной хроматографии, для пептида "Р18" до и после амидирования согласно примеру 7; хроматограмма химически синтезированного и амидированного контрольного пептида с последовательностью пептида "Р18" показана для сравнения;
Предпочтительные варианты выполнения изобретения
Настоящая группа изобретений, в частности, относится к следующим вариантам выполнения изобретения:
1. Синтетический, в частности рекомбинантно полученный, белок-предшественник, содержащий ферментативно и/или химически расщепляемую повторяющуюся последовательность повторяющихся звеньев целевых пептидных элементов (Pep) и вспомогательных пептидных элементов (Aux) общей формулы:
(Pep-Aux)x или
(Aux-Pep)x,
где x>1, причем
элементы Aux являются идентичными или различными и содержат элементы аминокислотной последовательности, которые обеспечивают свойства самосборки указанного белка-предшественника; и
элементы Pep являются идентичными или различными и содержат аминокислотную последовательность идентичных или различных пептидных молекул.
2. Белок-предшественник согласно варианту выполнения изобретения 1, где элементы Pep и Aux пептидно связаны друг с другом напрямую или через расщепляемую пептидную последовательность, и пептидная связь специфически расщепляется химическим или ферментативным путем, то есть является исключительно или по существу расщепляемой по определенной аминокислоте или череде аминокислот последовательности.
3. Белок-предшественник согласно любому из предшествующих вариантов выполнения изобретения, который обладает свойствами самосборки, так чтобы образовываться самопроизвольно, то есть самостоятельно, или образует индуцируемые стабильные нековалентные ассоциаты, которые являются нерастворимыми в стандартных условиях, таких как, в частности, 0.2 М NaOH в течение одного часа или 2 М мочевина или 1 М гидрохлорид гуанидина, в каждом случае, в течение 10 минут, при комнатной температуре. Стабильный ассоциат согласно изобретению образуется в результате удовлетворения по меньшей мере одного из этих трех критериев.
4. Белок-предшественник согласно любому из предшествующих вариантов выполнения изобретения, где по меньшей мере один элемент Aux содержит самособирающийся пептидный элемент (SA), причем указанный элемент SA содержит по меньшей мере один мотив последовательности из по меньшей мере 8, как например, 8-10, 8-12, 8-14, 8-16, 8-18 или 8-20, примыкающих друг к другу аминокислот, которые содержат по меньшей мере 50%, например, 50-100%, 60-90% или 70-80%, остатков аланина, по меньшей мере 50%, например, 50-100%, 60-90% или 70-80% остатков валина, или по меньшей мере 50%, например, 50-100%, 60-90% или 70-80% остатков глутамина, или по меньшей мере 80% которых составляет по меньшей мере один из этих остатков; элемент SA может содержать, например, в частности, по меньшей мере один из следующих мотивов последовательности:
где А представляет собой аланин, G представляет собой глицин, V представляет собой валин, n представляет собой целое число от 2 до 12, m представляет собой целое число от 2 до 10, и о представляет собой целое число от 1 до 6, где, более конкретно, n=5-10, m=4-8 и о=2-4, например, n=7-9, m=6-7 и о=2-3.
Указанные SA последовательности могут быть продолжены на С- и/или N-концах, в каждом случае, на еще 1-3 случайных аминокислотных остатка. Примерами подходящих N-терминальных продолжений являются мотивы последовательностей "G-", "GS-", "GAG-", "GPG-", "GPS-", "GAS-", "GQQ-" и "GSS-"; примерами подходящих С-терминальных продолжений являются мотивы последовательностей "-SGP", "-GGA", "-GPG", "-SGA", "-GGQ", "-GGY" и "-GGL".
5. Белок-предшественник согласно варианту выполнения изобретения 4, где элемент SA содержит аминокислотную последовательность, выбранную из аминокислотных последовательностей SEQ ID NO: 1-SEQ ID NO: 5 или SED ID No: 73.
6. Белок-предшественник согласно любому из предшествующих вариантов выполнения изобретения, где по меньшей мере один пептид Aux содержит защитный пептидный элемент (SU).
7. Белок-предшественник согласно варианту выполнения изобретения 6, где элемент SU обладает "повышенной долей" заряженных аминокислотных остатков, то есть (например, при рН=7) общий заряд отличен от О, например, составляет от +20 до -20 или от +10 до -10, или от +5 до -5, в частности отрицательно заряженных аминокислотных остатков, когда, например, при рН=7 общий заряд отличен от 0 и, например, составляет от -1 до -20, в частности от -4 до -10.
8. Белок-предшественник согласно варианту выполнения изобретения 7, где элемент SU в белке-предшественнике способен образовывать амфифильную спиральную структуру.
9. Белок-предшественник согласно варианту выполнения изобретения 8, где элемент SU представляет собой амфифильный пептид, содержащий сегмент последовательности из по меньшей мере семи пептидно связанных аминокислот, способных образовывать амфифильную альфа-спираль, причем указанная спираль в ее вертикальной проекции показывает разделение аминокислотных остатков на гидрофобную половину и гидрофильную половину спирали, причем гидрофобная половина спирали в ее вертикальной проекции имеет по меньшей мере 3, например, 3 или 4 идентичных или различных смежных гидрофобных аминокислотных остатка, и гидрофильная половина спирали в ее вертикальной проекции имеет по меньшей мере 3, например, 3 или 4 идентичных или различных смежных гидрофильных аминокислотных остатка.
10. Белок-предшественник согласно варианту выполнения изобретения 7, 8 или 9, где доля заряженных аминокислотных остатков элемента SU выбирается таким образом, что общий суммарный заряд белка-предшественника при рН=7 больше -10 и меньше +10, например больше -8 и меньше +8; больше -5 и/или меньше +5, больше -2 и меньше +2.
11. Белок-предшественник согласно вариантам выполнения изобретения 7-10, где элемент SU содержит аминокислотную последовательность, выбранную из аминокислотных последовательностей SEQ ID NO: 16-SEQ ID NO: 19 и SEQ ID NO: 68.
12. Белок-предшественник согласно любому из предшествующих вариантов выполнения изобретения, где элемент Pep содержит противомикробную пептидную последовательность, имеющую общий катионный положительный заряд.
13. Белок-предшественник согласно варианту выполнения изобретения 12, где элемент Pep содержит аминокислотную последовательность, выбранную из катионных аминокислотных последовательностей SEQ ID NO: 6-SEQ ID NO: 15, SEQ ID NO: 23, SEQ ID NO: 26 и SEQ ID NO: 69 - SEQ ID NO: 72 или любой из их C-терминально и/или N-терминально модифицированных форм, указанных ниже.
14. Белок-предшественник согласно вариантам выполнения изобретения 1-5, где пептид Pep содержит аминокислотную последовательность, выбранную из аминокислотных последовательностей SEQ ID NO: 20 или SEQ ID NO: 29-67 или любой из их C-терминально и/или N-терминально модифицированных форм, указанных ниже.
15. Белок-предшественник согласно любому из предшествующих вариантов выполнения изобретения, где элементы Aux независимо друг от друга имеют одно из следующих значений:
SA,
SA-SU,
SU-SA,
SA-SU-SA,
SU-SA-SU,
где элементы SA и SU пептидно связаны друг с другом, и элементы Aux терминально пептидно связан с по меньшей мере одним элементом Pep, то есть напрямую или через расщепляемую пептидную последовательность, где по меньшей мере пептидная связь с элементами Pep специфически расщепляется химическим или ферментативным путем.
16. Последовательность нуклеиновой кислоты, кодирующая по меньшей мере один белок-предшественник согласно одному из предшествующих вариантов выполнения изобретения.
17. Последовательность нуклеиновой кислоты согласно варианту выполнения изобретения 16, содержащая по меньшей мере одну кодирующую последовательность SEQ ID NO: 21, 24, 27, 74 и 76.
18. Экспрессионная кассета, содержащая по меньшей мере одну последовательность нуклеиновой кислоты согласно варианту выполнения изобретения 16 или 17, оперативно связанную с по меньшей мере одной регуляторной последовательностью нуклеиновой кислоты.
19. Рекомбинантный вектор для трансформации эукариотического или прокариотического хозяина, содержащий последовательность нуклеиновой кислоты согласно любому из вариантов выполнения изобретения 16 и 17, или экспрессионную кассету согласно варианту выполнения изобретения 18.
20. Способ получения целевого пептида (Pep), который содержит
a) получения белка-предшественника согласно одному из вариантов выполнения изобретения 1-15;
b) удаление пептидов Pep из белка-предшественника; и
с) при необходимости ферментативную или химическую модификацию, как например, амидирование, этерифицирование с образованием сложных эфиров, окисление, алкилирование пептида или связывание его с другой молекулой (например, с помощью естественного химического лигирования или посредством добавления по Михаэлю); где, например, пептид модифицируют молекулой, которая повышает гидрофобность указанного пептида, например, модифицируют молекулой, содержащей алкильный радикал; где указанную модификацию возможно осуществлять перед или после необязательной очистки пептида, далее приводятся соответствующие примеры в целях иллюстрации.
Примерами подходящих алкильных радикалов являются С2-С16-алкильные радикалы, такие как этил, изопропил или н-пропил, н-бутил, изобутил, втор-бутил или трет-бутил, н-пентил или изопентил; а также н-гексил, н-гептил, н-октил, н-нонил, н-децил, н-ундецил, н-додецил, н-тридецил, н-тетрадецил, н-пентадецил и н-гексадецил, или их однократно или многократно разветвленные аналоги, и их при необходимости замещенные модификации, которые могут иметь один или более заместителей, как, например, 1, 2 или 3 галогена (как, например, F, Cl, Br), гидрокси, меркапто, амино, C1-C4-алкиламино, или могут прерываться одним или более, например, 1, 2 или 3, гетероатомами, такими как О или N, в алкильной цепи. Более конкретно, C1-C4-алкил представляет собой метил, этил, изопропил или н-пропил, н-бутил, изобутил, втор-бутил или трет-бутил.
21. Способ согласно варианту выполнения изобретения 20, где белок-предшественник продуцируется в рекомбинантном микроорганизме, несущем по меньшей мере один вектор согласно варианту выполнения изобретения 19.
22. Способ согласно варианту выполнения изобретения 21, где белок-предшественник продуцируется в рекомбинантном штамме Е.coli.
23. Способ согласно любому из вариантов выполнения изобретения 20-22, где экспрессированный белок-предшественник, при необходимости, после того как был превращен в стабильную ассоциированную форму, подвергают очистке и химическому или ферментативному расщеплению с высвобождением целевого пептида (Pep).
24. Белок-предшественник, содержащий расщепляемую последовательность целевых пептидных элементов (Pep) и вспомогательных пептидных элементов (Aux') общей формулы:
(Pep-Aux')x или
(Aux'-Pep)x,
где x>1, причем
элементы Aux' являются идентичными или различными и содержат амфифильный пептид, образующий альфа-спираль, причем указанный амфифильный пептид содержит сегмент последовательности из по меньшей мере семи пептидно связанных аминокислот, способных образовывать амфифильную альфа-спираль, причем указанная спираль в ее вертикальной проекции показывает разделение аминокислотных остатков на гидрофобную половину и гидрофильную половину спирали, причем гидрофобная половина спирали в ее вертикальной проекции имеет по меньшей мере 3, например, 3 или 4 смежных идентичных или различных гидрофобных аминокислотных остатка, и гидрофильная половина спирали в ее вертикальной проекции имеет по меньшей мере 3, например, 3 или 4 смежных идентичных или различных гидрофильных аминокислотных остатка;
и элементы Pep являются идентичными или различными и содержат аминокислотную последовательность идентичных или различных пептидных молекул.
25. Белок-предшественник согласно варианту выполнения изобретения 24, где элементы Aux' содержат по меньшей мере один самособирающийся пептидный элемент (SA), как определено в любом из вариантов выполнения изобретения 4 и 5.
26. Белок-предшественник согласно варианту выполнения изобретения 24 или 25, где целевым пептидом (Pep) является катионный противомикробный пептид, и элементом Aux' является анионный пептид, формирующий амфифильную альфа-спираль.
27. Применение амфифильного пептида в качестве защитного пептида для рекомбинантного получения противомикробного целевого пептида, отличного от него; где указанный амфифильный пептид содержит участок последовательности из по меньшей мере семи пептидно связанных аминокислот, способных образовывать амфифильную альфа-спираль, причем указанная спираль в ее вертикальной проекции показывает разделение аминокислотных остатков на гидрофобную половину и гидрофильную половину спирали, причем гидрофобная половина спирали (в ее вертикальной проекции) имеет по меньшей мере 3 смежных идентичных или различных гидрофобных аминокислотных остатка, и гидрофильная половина спирали (в ее вертикальной проекции) имеет по меньшей мере 3 смежных идентичных или различных гидрофильных аминокислотных остатка;
28. Применение согласно варианту выполнения изобретения 27, где целевой пептид (Pep) представляет собой катионный противомикробный пептид, и элемент Aux' представляет собой анионный пептид, формирующий амфифильную альфа-спираль.
29. Способ согласно любому из вариантов выполнения изобретения 20-22, где получают белок-предшественник согласно варианту выполнения изобретения 12 или 13, как, например, белок-предшественник, содержащий Р18 пептидные строительные блоки согласно SEQ ID NO: 23 или SEQ ID NO: 6.
30. Способ согласно варианту выполнения изобретения 29, который включает следующие стадии:
- промывание ассоциатов белка-предшественника растворителем, который растворяет содержащиеся белки, но не растворяет или по существу не растворяет указанные ассоциаты, как, например, 0.1 М-1.0 М NaOH;
- расщепление белков-предшественников, например, кислотой, если целевой пептид, например, Р18, включен в белок-предшественник посредством расщепляемых кислотой групп.
31. Способ согласно варианту выполнения изобретения 30, который включает по меньшей мере одну из следующих дополнительных стадий:
- обработка ассоциата белка-предшественника осаждающим агентом, таким как, например, фосфорная кислота, после разрушения клетки;
- очистка пептида от продуктов расщепления с помощью хроматографического способа;
- промывание очищенного и высушенного пептида кислотным растворителем или смесью растворителей.
32. Способ согласно любому из вариантов выполнения изобретения 20-23 и 29-31 для получения пептида SEQ ID NO: 23, где способ включает следующие стадии:
- обработка ассоциатов белка-предшественника после разрушения клетки путем добавления 85% сильной фосфорной кислоты до рН=3;
- промывание ассоциатов белка-предшественника раствором гидроксида натрия, например 0.4 М NaOH;
- расщепление белка-предшественника фосфорной кислотой или муравьиной кислотой, например, 2% фосфорной кислотой;
- при необходимости промывание высушенного пептида капроновой кислотой или смесью из 99 частей гексана и одной части уксусной кислоты.
33. Способ согласно любому из вариантов выполнения изобретения 20-23 и 29-31 для получения пептида SEQ ID NO: 6, где способ включает следующие стадии:
- гидролиз или расщепление гранул, например, посредством 5% H3PO4;
- центрифугирование;
- доведение pH супернатанта до около 4.0, например, с помощью 25% NaOH;
- очищение супернатанта посредством катионообменной хроматографии;
- осаждение целевого пептида, например, путем добавления NaOH к элюату;
- центрифугирование;
- ресуспендирование гранул в воде;
- растворение пептида, например, путем добавления уксусной кислоты;
- лиофилизация.
34. Изобретение, кроме того, относится к пептиду Р18 (SEQ ID NO: 23) и пептиду SEQ ID NO: 6, а также к их получению согласно изобретению, и к их применению в косметических или фармацевтических средствах для лечения или предотвращения шелушения, в частности, перхоти; или для ингибирования роста и/или активности липофильных грибов, в частности Malassezia ssp., конкретно Malassezia furfur. Это также описывается, например, в другой международной заявке РСТ/ЕР2008/010912, поданной 19 декабря 2008 г., описание которой включено в настоящую заявку в полном объеме посредством ссылки.
Подробное описание отдельных объектов изобретения
1. Пептиды
Пептиды (Pep) согласно настоящему изобретению, которые также могут упоминаться как "желаемые пептиды" или "целевые пептиды", представляют собой аминокислотные цепочки, в которых от 2 до 100, например, 5-70 и, в частности, 7-50, например, 10-40, 12-35 или 15-25, аминокислот связаны пептидными связями. Пептиды могут состоять из любых α-аминокислот, в частности протеиногенных аминокислот.
Пептиды могут иметь конкретные желательные биологические или химические и, в частности, также фармацевтические свойства. Примерами таких свойств являются: противомикробная активность, специфическое связывание с конкретными поверхностями, свойства образовывать зародыш в процессах кристаллизации и при образовании частицы, контроль кристаллических структур, связывание металлов или металлических ионов, свойства поверхностно-активных веществ, эмульгирующие свойства, свойства стабилизации пены, влияние на клеточную абсорбцию.
Указанные пептиды могут иметь одно или более из этих свойств.
В одном варианте выполнения изобретения наше изобретение относится к способу получения противомикробных пептидов. Такие "противомикробные пептиды" характеризуются способностью ингибировать рост и/или размножение по меньшей мере одного вида грамположительных или грамотрицательных бактерий и/или по меньшей мере одного вида дрожжей и/или по меньшей мере одного вида мицелиальных грибов и/или по меньшей мере одного вида водорослей, и и/или разрушать клетки соответствующих организмов, при концентрациях противомикробного пептида ≤100 мкМ.
В одном варианте выполнения изобретения, изобретение обеспечивает катионные противомикробные пептиды. Катионные противомикробные пептиды отличаются противомикробном действием, как указано выше, и имеют суммарный заряд более 0 при рН 7.
Катионные пептиды этого вида содержат, например, следующую последовательность:
X1X2KX3X4X5KIPX10KFX6X7X8AX9KF (SEQ ID NO: 7), где
Х10 представляет собой пептидную связь или любой один или два основных или гидрофобных аминокислотных остатка, или один или два остатка пролина, и
Х1-Х9 представляют собой любой из основных или отличных от пролина гидрофобных аминокислотных остатков;
и/или ее мутанты или производные,
где соответствующие мотивы последовательности, присутствующие в белке-предшественнике, могут быть идентичными или различными.
В следующем конкретном варианте выполнения изобретения, изобретение относится к получению пептидов, содержащих следующую последовательность
X1X2KX3X4X5KIPX11X12KFX6X7X8AX9KF (SEQ ID NO: 8),
где
X1 представляет собой лизин, аргинин или фенилаланин,
Х2 представляет собой лизин или триптофан,
X3 представляет собой лейцин или лизин,
Х4 представляет собой фенилаланин или лейцин,
Х5 представляет собой лейцин или лизин,
X6 представляет собой лейцин или лизин,
Х7 представляет собой гистидин или лизин,
Х8 представляет собой аланин, лейцин, валин или серин,
Х9 представляет собой лейцин или лизин,
X11 представляет собой пролин или химическую связь, и
Х12 представляет собой пролин или химическую связь,
и/или ее мутанты или производные;
где соответствующие мотивы последовательности, присутствующие в белке-предшественнике, могут быть идентичными или различными.
Неограничивающими примерами последовательностей или повторяющихся мотивов последовательностей являются SEQ ID NO: 6, SEQ ID NO: 9-SEQ ID NO: 15, SEQ ID NO: 23, SEQ ID NO: 69, SEQ ID NO: 71, и/или их мутанты или производные.
Другие подходящие пептиды описываются, например, в международной заявке того же заявителя, РСТ/ЕР2008/010912, поданной 19 декабря 2008, которая включена в настоящую заявку в полном объеме посредством ссылки.
2. Периодические белки-предшественники
Периодические белки-предшественники согласно настоящему изобретению отличаются тем, что по меньшей мере 60%, в частности по меньшей мере 80% их аминокислотной последовательности, например 60-99%, 70-95%, 75-85%, в каждом случае от общей длины последовательности, состоит из повторяющихся пептидных звеньев (как определено далее). Оставшаяся часть может содержать, например, неповторяющиеся пептиды, такие как, например, сигнальные пептиды, метки и тому подобное.
3. Повторяющиеся звенья
Повторяющиеся пептидные звенья содержат по меньшей мере один пептид, предпочтительно полученный согласно настоящему изобретению предпочтительным образом, и, фактически, сконструированы следующим образом:
(Pep-Aux)x или
(Aux-Pep)x,
где x>1, и Pep представляет собой пептид, отмеченный выше, и Aux является таким, как определено выше.
Повторяющееся звено (Pep-Aux или Aux-Pep) согласно настоящему изобретению представляет собой аминокислотную последовательность, имеющую 10-200, например, 20-130 и/или 30-80 аминокислот в длину, которая присутствует в белке-предшественнике множество раз, либо в виде идентичных последовательностей, либо в виде вариантов конкретной последовательности, обладающих по меньшей мере 70%, например, по меньшей мере 80% и, в частности, по меньшей мере около 90% идентичностью, например, 91, 92, 93, 94, 95, 96, 97, 98 или 99% идентичности. Периодические белки-предшественники согласно настоящему изобретению могут, таким образом, содержать, например, идентичные копии или варианты одной аминокислотной последовательности или множества различных аминокислотных последовательностей, например Pep и/или Aux строительные блоки.
Более того, многие из указанные повторяющихся звеньев, например, 1-100, 1-50 или 2-32 и, в частности, 4-16 могут быть соединены вместе в периодическом белке-предшественнике.
Доля пептида согласно настоящему изобретению в повторяющемся звене, на основе молярной массы, составляет 20%-80%, например 30%-70% или 40-60%. Оставшаяся часть повторяющегося звена состоит из Aux последовательностей, в частности SA и SU последовательностей, определенных выше, и при необходимости специфически расщепляемых последовательностей для селективного удаления Pep строительного блока.
4. Вспомогательные последовательности
Вспомогательные последовательности в самом широком смысле представляют собой аминокислотные последовательности в белке-предшественнике согласно изобретению, которые влияют на свойства белка-предшественника, улучшая экспрессию, стабильность и/или обработку белка-предшественника. Вспомогательные последовательности в периодическом белке-предшественнике могут быть частью повторяющегося звена (Aux строительные блоки указаны выше) или могут быть присоединены к амино-концу или карбокси-концу белка-предшественника, как, например, 6 × His-Tag (HHHHHH), Т7-Tag (MASMTGGQQMG), S-Tag (KETAAAKFERQHMDS), c-Myc-Tag (EQKLISEEDL), Strep-Tag (WSHPQFEK) или НА-Tag (YPYDVPDYA), глутатион S-трансфераза, связывающий мальтозу белок, связывающий целлюлозу белок. Эти и другие вспомогательные последовательности описываются в Terpe; AppI Microbiol Biotechnol; 60(5): 523-33 (2003). Корме того, вспомогательная последовательность CanA (Maijn Vitro Untersuchungen zum extrazellularen Netzwerk von Pyrodictium abyssi TAG11" Dissertation, Universitat Regensburg (1998)) и yaaD (Wohlleben Eur Biophys J, (2009) онлайн публикация) полезны для присоединения к амино-концу или карбокси-концу белка-предшественника.
В одном варианте выполнения изобретения белок-предшественник содержит вспомогательные последовательности, которые влияют на стабильность указанного белка-предшественника.
В предпочтительном варианте выполнения изобретения вспомогательные последовательности влияют на свойства "самосборки" белка-предшественника. Указанные свойства самосборки белка-предшественника отличаются тем, что указанный белок-предшественник уже в ходе экспрессии "самопроизвольно" формирует стабильные ассоциаты, то есть самостоятельно, без дополнительно требуемых средств, или образование таких стабильных ассоциатов из растворимых белков-предшественников может быть запущено "индуцируемым образом", то есть через «запускающие» агенты. Белки-предшественники, обладающие свойствами самосборки, имеют преимущества по сравнению с другими белками-предшественниками, благодаря тому, что они могут быть очищены простым и эффективным образом. Ассоциаты такого вида, как правило, содержат исключительно или по существу нековалентные связи, такие как, например, водородные связи, ионные и/или гидрофобные взаимодействия.
Самособирающиеся последовательности могут составлять, например, 8 последовательных аминокислот в длину. Подходящие последовательности могут быть локализованы, например, в известных белках, в которых ранее уже была обнаружена сборка в высокомолекулярные ассоциаты. Примерами таких ассоциатов являются амилоидные фибриллы, актиновая или миозиновая нить, белковые волокна, такие как волокна эластина, волокна коллагена, биссусные нити мидий, волокна кератина или нити шелка. Эти и другие белки, содержащие самособирающиеся последовательности, описываются в документе Scheibel, Current Opinion in Biotechnology 16; 1-7 (2005), который включен в настоящую заявку в полном объеме посредством ссылки.
Растворы космотропных солей могут применяться в качестве "запускающих агентов". В качестве примеров космотропных солей можно упомянуть, например, соли, содержащие по меньшей мере один тип иона, который имеет более выраженные космотропные свойства, чем ион натрия или хлоридный ион в соответствии с так называемым рядом Гофмейстера. Примерами таких солей являются фосфат калия и сульфат аммония. Примерами таких растворов солей являются 0.5 М раствор фосфата калия и 0.8 М раствор сульфата аммония.
Подходящие ассоциаты белков-предшественников по изобретению отличаются возможностью сохранения их ассоциированной формы в течение определенного периода времени, в ходе обработки растворами, как правило, способными солюбилизировать множество агрегированных белков, и, таким образом, могут быть отделены от белковых загрязнителей. Примерами таких растворов являются растворы оснований, кислот, мочевины, солей и детергентов. Более конкретно, стабильные ассоциаты согласно изобретению нерастворимы в течение определенного периода времени в растворах гидроксидов щелочных металлов, мочевины, солей гуанидина или заряженных детергентов, таких как, например, соли алкилтримети-ламмония или алкилсульфаты.
Более конкретно, стабильные ассоциаты нерастворимы в течение определенного периода времени в растворах ≥0.2 М гидроксида натрия, ≥2 М мочевины, ≥1 М гидрохлорида гуанидина, ≥1 М тиоцианата гуанидина или ≥0.1% додецилсульфата натрия, или ≥0.1% бромида цетилтриметиламмония. Более конкретно, стабильные ассоциаты белков-предшественников являются стабильными в вышеописанных растворах в течение ≥10 минут, например ≥30 минут и, в частности, ≥60 минут.
Стабильный ассоциат, в частности, не растворяется
a) 0.2 М раствором NaOH в течение одного часа, и/или
b) 2 М раствором мочевины и/или
c) 1 М раствором гидрохлорида гуанидина в течение 10 минут при комнатной температуре (то есть, около 20°C).
В следующем конкретном варианте выполнения изобретения, белок-предшественник содержит вспомогательные последовательности (SU), которые защищают клетку-хозяина от вредных воздействий периодического белка-предшественника.
В конкретном варианте выполнения изобретения белок-предшественник содержит вспомогательные последовательности SU, которые защищают клетку-хозяина от вредных влияний катионных противомикробных пептидных последовательностей, присутствующих в периодическом белке-предшественнике. Более конкретно, эти защитные последовательности содержат отрицательно заряженные аминокислоты (Asp, Glu). Более конкретно, вспомогательная последовательность содержит ряд отрицательно заряженных аминокислот глутамата и/или аспартата, что приводит к общему отрицательному заряду при рН=7 более -10 и менее +10, особенно более -5 и менее +5, например, более -2 и менее +2, внутри периодического белка-предшественника.
В следующем конкретном варианте выполнения изобретения, отрицательно заряженная защитная последовательность формирует амфипатическую спираль. Амфипатическая спираль согласно настоящему изобретению формируется, если при кругообразном расположении (то есть в ее аксиальной проекции (вдоль оси спирали) или при виде сверху) последовательности из 7 последовательных аминокислот в первичной структуре (А1-А7) имеет следующий порядок: А1-А5-А2-А6-А3-А7-А4 (Фиг.1), где по меньшей мере 3 соседние аминокислоты на указанном круге являются гидрофобными аминокислотами (Ala, Met, Cys, Phe, Leu, Val, IIe) или глицином, и 3 соседние аминокислоты на указанном круге являются гидрофильными аминокислотами (Thr, Ser, Trp, Tyr, Pro, His, Glu, Gin, Asp, Asn, Lys, Arg) или глицином. Это кругообразное расположение также обозначается как проекция "зубчатое колесо".
В предпочтительном варианте выполнения изобретения отрицательно заряженная защитная последовательность соответствует любой из последовательностей SEQ ID NO: 16-SEQ ID NO: 19.
5. Расщепляемые последовательности
Расщепляемыми последовательностями являются аминокислотные последовательности, которые располагаются выше и ниже в 3'-5' направлении от целевых пептидных последовательностей (Pep) согласно изобретению. Эти последовательности обеспечивают удаление Pep строительных блоков из периодического белка-предшественника посредством "специфического" расщепления. В этом контексте термин "специфическое" означает, что указанное расщепление происходит в белке-предшественнике по существу, в частности, исключительно, в одном или более конкретных положениях, где удаляется конкретный пептид или его предшественник.
"Предшественник" может, например, состоять из пептидной цепи, которая содержит на одном конце или на обоих концах аминокислотные остатки, которые не являются частью природной оригинальной пептидной последовательности, но которые не мешают ее дальнейшему применению или функциональности, или которые могут быть удалены путем расщепления, если требуется, применяя обычные химические или биохимические способы.
Расщепление последовательностей может осуществляться благодаря специфической последовательности, распознаваемой протеолитически активными ферментами, которые связываются с указанной последовательностью и расщепляют пептидную связь между двумя конкретными аминокислотами. Примерами являются последовательности, распознаваемые Arg-C протеиназой, Asp-N эндопептидазой, каспазами, химотрипсином, клострипаином, энтерокиназой, фактором Ха, глутамилэндопептидазой, гранзимом В, LysC лизилэндопептидазой (ахромобактерильная про-теиназа I), LysN пептидил-Lys металлоэндопептидазой, пепсином, пролин эндопептидазой, протеиназой К, стафилококковой протеиназой I, термолизином, тромбином, трипсином. Соответствующие распознаваемые последовательности описываются в литературе, например, в Keil, "Specificity of proteolysis" p.335 Springer-Verlag (1992).
Альтернативно, конкретные аминокислотные последовательности обеспечивают селективное расщепление полипептидной основной цепи конкретными химическими веществами, такими как, например, BNPS - скатол (2-(2'-нитрофенилсульфенил)-3-метил-3-броминдоленин), бромциан, кислоты, гидроксиламин, иодозобензойная кислота, NTCB (2-нитор-5-тиоцианобензойная кислота).
Более конкретно, применяемые расщепляемые последовательности обеспечивают расщепление периодических белков-предшественников химическими реагентами. Особенно подходящие расщепляемые последовательности содержат мотивы последовательности Asn-Gly, которые обеспечивают расщепление гидроксиламином, или Asp-Pro или Asp-Xxx, которые обеспечивают расщепление кислотой, причем Xxx представляет собой любую протеиногенную аминокислоту.
6. Последовательности согласно изобретению
6.1 Аминокислотные последовательности
Помимо последовательностей пептидов (Pep), конкретно описанных в настоящей заявке, и вспомогательных последовательностей (Aux, SA, SU), повторяющихся последовательностей, расщепляемых последовательностей и последовательностей периодических белков-предшественников, настоящее изобретение также относится к функциональным эквивалентам, функциональным производным и солям указанной последовательности.
Термин "функциональные эквиваленты", как понимается согласно настоящему изобретению, в частности, означает мутантов, которые в по меньшей мере одном положении вышеупомянутых аминокислотных последовательностей имеют аминокислоту, отличную от обозначенных, но тем не менее обладают желательными свойствами первоначального неизмененного пептида. "Функциональные эквиваленты", таким образом, включают мутантов, получаемых путем одного или более аминокислотных добавлений, замещений, делеций и/или инверсий, где упомянутые изменения могут происходить в любом положении последовательности, при условии, что они приводят к мутанту с профилем свойств согласно изобретению. Функциональные эквиваленты, в частности, также существуют, если реакционная способность мутанта качественно соответствует реакционной способности неизменного полипептида.
"Функциональные эквиваленты" в вышеописанном смысле также включают "предшественники" описанных полипептидов, и "функциональные производные" и "соли" полипептидов.
"Предшественники" представляют собой природные или синтетические предшественники полипептидов с или без желательной биологической активности.
Примеры подходящих аминокислотных замещений можно найти в следующей таблице:
Выражение "соли", как понимается, означает как соли карбоксильных групп, так и кислотные аддитивные соли аминогрупп молекул пептида по изобретению. Соли карбоксильных групп могут быть непосредственно получены известным образом и содержат неорганические соли, например соли натрия, кальция, аммония, железа и цинка, и соли с органическим основаниями, как, например, амины, такие как триэтаноламин, аргинин, лизин, пиперидин и тому подобное. Кислотные аддитивные соли, например соли с минеральными кислотами, такими как соляная кислота и серная кислота, и соли с органическими кислотами, такими как уксусная кислота и щавелевая кислота, подобным образом являются объектом настоящего изобретения.
"Функциональные производные" (или "производные") полипептидов по изобретению могут, подобным образом, быть получены при функциональных боковых группах аминокислот или при N- или C-терминальном конце, с помощью известных методик. Такие производные содержат, например, алифатические сложные эфиры групп карбоновой кислоты, амиды групп карбоновой кислоты, получаемые путем реакции с аммиаком или с первичным или вторичным амином; N-ацильные производные свободных аминогрупп, получаемые путем реакции с ацильными группами; или O-ацильные производные свободных гидроксильных групп, получаемые путем реакции с ацильными группами. Кроме того, также возможно ковалентное (пептидное) связывание любых 1-5, например, 2, 3 или 4, D- или L-аминокислотных остатков при N- и/или C-терминальном конце;
6.2 Нуклеиновые кислоты, экспрессионные конструкции, вектора и микроорганизмы для их получения
Нуклеиновые кислоты
Настоящая группа изобретений, кроме того, относится к молекулам нуклеиновой кислоты, которые кодируют пептиды и пептидные последовательности по изобретению.
Все последовательности нуклеиновых кислот, упомянутые в настоящем изобретении (одно- и двухцепочечные ДНК и РНК последовательности, например, кДНК и мРНК), могут быть непосредственно получены известным образом, путем химического синтеза из нуклеотидных звеньев, например, путем конденсации фрагментов отдельных перекрывающихся комплементарных звеньев нуклеиновых кислот двойной спирали. Химический синтез олигонуклеотидов может быть осуществлен, например, известным образом, с помощью фосфорамидитного способа (Voet, Voet, 2. Auflage, Wiley Press New York, Seiten 896-897). Присоединение синтетических олигонуклеотидов и заполнение гэпов с использованием фрагмента Кленова ДНК-полимеразы и реакций лигирования, а также общие способы кпонирования описаны в Sambrook et al. (1989), Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory Press.
Настоящая группа изобретений относится как к выделенным молекулам нуклеиновых кислот, которые кодируют полипептиды или белки согласно изобретению или их биологически активные сегменты, так и к фрагментам нуклеиновых кислот, которые, например, могут использоваться для применения в качестве гибридизационных зондов или праймеров для идентификации или амплификации кодирующих нуклеиновых кислот по изобретению.
Молекулы нуклеиновой кислоты по изобретению могут дополнительно содержать нетранслируемые последовательности у 3' и/или 5' конца кодирующей области гена.
"Выделенная" молекула нуклеиновой кислоты отделена от других молекул нуклеиновой кислоты, присутствующих в природном источнике нуклеиновой кислоты, и, более того, может быть по существу свободна от клеточного материала или культуральной среды, если ее получают рекомбинантными способами, или свободна от химических предшественников или других химических веществ, в случае синтеза химическим путем.
Молекула нуклеиновой кислоты по изобретению может быть выделена с помощью стандартных молекулярно-биологических способов и информации, раскрытой в настоящем изобретении. Например, кДНК может быть выделена из подходящей библиотеки кДНК, применяя одну из раскрытых конкретным образом полных последовательностей или ее часть в качестве зонда гибридизации, и стандартные методики гибридизации (как описывается, например, в Sambrook, J., Fritsch, E.F. und Maniatis, Т. Molecular Cloning: A Laboratory Manual. 2. Aufl., Cold Spring Harbor Laboratory, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY, 1989). Более того, возможно выделить молекулу нуклеиновой кислоты, содержащую одну из раскрытых последовательностей или ее часть с помощью полимеразной цепной реакции, применяя олигонуклеотидные праймеры, которые были установлены на основе этой последовательности. Нуклеиновая кислота, амплифицированная таким образом, может быть клонирована в подходящий вектор и охарактеризована с помощью анализа последовательности ДНК. Олигонуклеотиды по изобретению также могут быть получены с помощью стандартных способов синтеза, например, с помощью автоматизированного ДНК-синтезатора.
Настоящая группа изобретений, кроме того, относится к молекулам нуклеиновой кислоты, комплементарным конкретным описанным нуклеотидным последовательностям, или их части.
Нуклеотидные последовательности по изобретению обеспечивают возможность получения зондов или праймеров, которые могут применяться для идентификации и/или клонирования гомологических последовательностей в других типах клеток и организмах. Такие зонды или праймеры, как правило, содержат область нуклеотидной последовательности, которая при строгих условиях гибридизуется с по меньшей мере около 12, предпочтительно по меньшей мере около 25, например, около 40, 50 или 75, следующими друг за другом нуклеотидами смысловой нити последовательности нуклеиновой кислоты по изобретению или соответствующей антисмысловой нити.
Настоящая группа изобретений также относится к таким последовательностям нуклеиновых кислот, которые содержат так называемые молчащие мутации или были изменены в соответствии с использованием кодонов конкретного организма-источника или организма-хозяина, по сравнению с конкретной названной последовательностью, а также их вариантам природного происхождения, например, их сплайсинговым вариантам или аллельным вариантам. Подобным образом обеспечиваются последовательности, получаемые с помощью консервативных нуклеотидных замещений (то есть, рассматриваемые аминокислоты замещаются аминокислотой с таким же зарядом, размером, полярностью и/или растворимостью).
Настоящая группа изобретений также обеспечивает молекулы, происходящие из специфически описанных нуклеиновых кислот в результате полиморфизма последовательности. Эти генетические полиморфизмы могут существовать между индивидуумами внутри популяции на основе природной изменчивости. Эти природные вариации вызывают обычно дисперсию 1-5% в нуклеотидной последовательности гена.
Кроме того, изобретение также относится к последовательностям нуклеиновых кислот, которые гибридизуются с вышеупомянутыми кодирующими последовательностями или являются комплементарными им. Эти полинуклеотиды могут быть обнаружены при проведении поиска по геномным библиотекам или библиотекам кДНК, и при необходимости могут быть амплифицированы с помощью подходящих праймеров посредством ПЦР и затем выделены, например, с помощью подходящих зондов. Следующая возможность состоит в трансформации подходящих микроорганизмов полинуклеотидами или векторами по изобретению, размножении микроорганизмов и, следовательно, полинуклеотидов, и последующем их выделении. Кроме того, также возможно синтезировать полинуклеотиды по изобретению химическим путем.
Под свойством быть способным к гибридизации с полинуклеотидами имеют в виду способность поли- или олигонуклеотида связываться при строгих условиях с почти комплементарной последовательностью, в то время как при этих условиях не образуются неспецифические связи между некомплементарными партнерами. Для этого эти последовательности должны быть комплементарными до 70-100%, предпочтительно до 90-100%. Свойство комплементарных последовательностей специфически связываться друг с другом используют, например, в Нозерн- или Саузерн-блоттинге или при связывании праймера в ПЦР или ОТ-ПЦР. Для этого обычно используют олигонуклеотиды с длиной от 30 пар оснований. Строгие условия, например, при Нозерн-блоттинге означают применение промывочного раствора при температуре 50-70°С, предпочтительно при 60-65°С, например, 0.1*SSC буфер с 0.1% SDS (20*SSC: 3 М NaCl, 0.3 М цитрат натрия, pH 7.0), для элюирования неспецифически гибридизованных кДНК зондов или олигонуклеотидов. Как упоминается выше, только нуклеиновые кислоты, которые обладают высокой степенью комплементарности, остаются связанными друг с другом. Создание строгих условий известно специалистам в данной области техники и описывается, например, в Ausubel et al., Current Protocols in Molecular Biology, John Wiley & Sons, N.Y. (1989), 6.3.1-6.3.6.
"Идентичность" между двумя нуклеиновыми кислотами означает идентичность нуклеотидов, в каждом случае, по всей длине нуклеиновых кислот, в частности идентичность вычисляется посредством сравнения с помощью программного обеспечения Vector NTI Suite 7.1 Software от Informax (США) и применяя способ Clustal (Higgins D G, Sharp P M. Fast and sensitive multiple sequence alignments on a microcomputer. Comput Appl. Biosci. 1989 April; 5(2): 151-1), при следующих параметрах:
Параметры множественного выравнивания:
Параметры попарного выравнивания:
Экспрессионные конструкции и вектора
Настоящая группа изобретений также обеспечивает экспрессионные конструкции, содержащие, под генетическим контролем регуляторных последовательностей нуклеиновых кислот, последовательность нуклеиновой кислоты, кодирующую пептид или белок-предшественник по изобретению, а также вектора, содержащие по меньшей мере одну из этих экспрессионных конструкций. Такие конструкции по изобретению предпочтительно содержат промотор в направлении 5' (слева) от конкретной кодирующей последовательности, и терминаторную последовательность в направлении 3' (справа), и при необходимости другие обычные регуляторные элементы, каждый из которых оперативно связан с кодирующей последовательностью. Термин "оперативная связь", как понимается, означает последовательное расположение промотора, кодирующей последовательности, терминатора и при необходимости других регуляторных элементов, таким образом, что каждый из регуляторных элементов может выполнять свою функцию, как предназначено, при экспрессии кодирующей последовательности. Примерами оперативно связываемых последовательностей являются нацеливающие последовательности и энхансеры, сигналы полиаденилирования и тому подобное. Другие регуляторные элементы содержат селектируемые маркеры, сигналы амплификации, ориджины репликации и тому подобное. Подходящие регуляторные последовательности описываются, например, в Goeddel, Gene Expression Technology: Methods in Enzymology 185, Academic Press, San Diego, Calif (1990).
Помимо искусственных регуляторных последовательностей, природная регуляторная последовательность может все еще присутствовать перед собственно структурным геном. Путем генетической модификации эта природная регуляция при необходимости может быть выключена, и экспрессия генов может быть увеличена или уменьшена. Однако генная конструкция также может представлять собой более простую конструкцию, что означает, что никакие дополнительные регуляторные сигналы не устанавливаются перед структурным геном, и природный промотор с его регуляцией не удаляется. Напротив, природную регуляторную последовательность мутируют таким образом, что регуляция более не происходит и экспрессия гена повышается или уменьшается. Последовательности нуклеиновых кислот могут содержаться в данной конструкции гена в виде одной или в виде нескольких копий.
Примерами подходящих промоторов являются: cos, tac, trp, tet, trp-tet, lpp, lac, lpp-lac, laclq, T7, T5, T3, gal, trc, ara, SP6, лямбда-PR или лямбда-PL промотор, которые предпочтительно применяются в грамотрицательных бактериях; и грамположительные промоторы amy и SP02, промоторы дрожжей ADC1, MFa, AC, P-60, CYC1, GAPDH, или растительные промоторы CaMV/355, SSU, OCS, lib4, usp, STLS1, B33, not, или промотор убик-витина или фазеолина. Особенно предпочтительным является применение индуцируемых промоторов, таких как, например, индуцируемые светом и особенно индуцируемые температурой промоторы, такие как PrPl-промотор. В принципе могут использоваться все природные промоторы с их регуляторными последовательностями. Кроме того, могут также предпочтительно использоваться синтетические промоторы.
Упомянутые регуляторные последовательности, как предполагается, обеспечивают возможность контролируемой экспрессии последовательностей нуклеиновой кислоты и экспрессии белка. В отношении организма-хозяина это может означать, например, что ген экспрессируется или сверхэкспрессируется только после индукции, или что он экспрессируется и/или сверхэкспрессируется немедленно.
Регуляторные последовательности или факторы предпочтительно могут позитивно влиять на экспрессию и, в результате, повышать или понижать ее. Например, регуляторные элементы предпочтительно могут быть усилены на уровне транскрипции, путем применения сильных сигналов транскрипции, таких как промоторы и/или "энхансеры". Кроме того, однако, также возможно усиливать трансляцию, например, путем улучшения стабильности мРНК.
Экспрессионную кассету получают путем слияния подходящего промотора с подходящей кодирующей нуклеотидной последовательностью и терминирующим сигналом или сигналом полиаденилирования. Для этого применяются стандартные методики рекомбинации и клонирования, как описывается, например, в Т.Maniatis, E.F.Fritsch und J.Sambrook, Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory, Cold Spring Harbor, NY (1989), а также в T.J.Silhavy, M.L.Berman und L.W.Enquist, Experiments with Gene Fusions, Cold Spring Harbor Laboratory, Cold Spring Harbor, NY (1984) и в Ausubel, F.M. et al., Current Protocols in Molecular Biology, Greene Publishing Assoc. and Wiley Interscience (1987).
Для экспрессии в подходящем организме-хозяине, рекомбинантная конструкция нуклеиновой кислоты или генная конструкция вставляется в специфический по отношению к хозяину вектор, который обеспечивает возможность оптимальной экспрессии генов в хозяине. Вектора хорошо известны специалистам в данной области техники и их можно найти, например, в "Cloning Vectors" (Pouwels P.H. et al., eds., Elsevier, Amsterdam-New York-Oxford, 1985). Вектора, за исключением плазмид, также представляют собой все другие вектора, известные специалистам в данной области техники, например, фаги, вирусы, такие как SV40, CMV, бакуловирусы и аденовирусы, транспозоны, IS-элементы (мобильные нуклеотидные последовательности, встречающиеся в геномах бактериальных популяций), фагмиды, космиды и линейные или кольцевые ДНК. Эти векторы могут реплицироваться автономно в организме-хозяине или хромосомно.
В качестве примеров экспрессионных векторов могут быть названы:
обычные слитые экспрессионные векторы, такие как pGEX (Pharmacia Biotech Inc; Smith, D.B. and Johnson, K.S. (1988) Gene 67:31-40), pMAL (New England Biolabs, Beverly, MA) и pRIT 5 (Pharmacia, Piscataway, NJ), у которых глутатион-S-трансфераза, мальтозу E-связывающий белок или белок А слиты с рекомбинантным целевым белком;
экспрессионные векторы, не являющиеся слитыми белками, такие как pTrc (Amann et al., (1989) Gene 69:301-315) и рЕТ 11d (Studier et al. Gene Expression Technology: Methods in Enzymology 185, Academic Press, San Diego, Kalifornien (1990) 60-89);
дрожжевой экспрессионный вектор для экспрессии в дрожжах S. сеrе-visiae, такой как pYepSed (Baldari et al., (1987) Embo J. 6:229-234), pMFa (Kurjan und Herskowitz (1982) Cell 30:933-943), pJRY88 (Schultz et al. (1987) Gene 54:113-123), а также pYES2 (Invitrogen Corporation, San Diego, CA). Векторы и способы конструирования векторов, которые пригодны для применения в других грибах, таких как нитевидные грибы, включают в себя векторы и способы, которые подробно описаны в van den Hondel, C.A.M.J.J. & Punt, P.J. (1991) Gene transfer systems and vector development for filamentous fungi, in: Applied Molecular Genetics of Fungi, J.F. Peberdy et al., Hrsg., S.1-28, Cambridge University Press: Cambridge;
бакуловирусные векторы, которые пригодны для экспрессии белков в культивируемых клетках насекомых (например, Sf9-клетках), включают в себя рАс-серию (Smith et al., (1983) Mol. Cell Biol., 3: 2156-2165) и pVL-серию (Lucklow und Summers (1989) Virology 170: 31-39);
экспрессионные векторы растений, такие, какие подробно описаны в: Becker, D., Kemper, E., Schell, J. und Masterson, R. (1992) New plant binary vectors with selectable markers located proximal to the left border Plant Mol. Biol. 20:1195-1197; и Bevan, M.W. (1984) Binary Agrobacterium vectors for plant transformation, Nucl. Acids Res.12: 8711-8721;
экспрессионные векторы млекопитающих, такие как pCDM8 (Seed, В. (1987) Nature 329:840) и pMT2PC (Kaufman et al. (1987) EMBO J. 6:187-195);
другие подходящие Экспрессионные системы для эукариотических и прокариотических клеток описываются в главах 16 и 17 источника Sambrook, J., Fritsch, E.F. und Maniatis, Т., Molecular cloning: A Laboratory Manual, 2. Auflage, Cold Spring Harbor Laboratory, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY, 1989.
Рекомбинантные микроорганизмы:
При помощи векторов по изобретению могут быть получены рекомбинантные микроорганизмы, как, например, трансформированные по меньшей мере одним вектором по изобретению, и они могут быть использованы для получения полипептидов по изобретению. Предпочтительно, вышеописанные рекомбинантные конструкции по изобретению вводят в подходящую систему-хозяина и экспрессируют. При этом используют предпочтительно известные специалисту общепринятые способы клонирования и трансфекции, такие как, например, копреципитация, слияние протопластов, электропорация, ретровирусная трансфекция и т.п., чтобы заставить вышеупомянутые нуклеиновые кислоты экспрессироваться в соответствующей системе экспрессии. Подходящие системы описаны, например, в Current Protocols in Molecular Biology, F. Ausubel et al., Hrsg, Wiley Interscience, New York 1997 или Sambrook et al. Molecular Cloning: A Laboratory Manual, Second Edition, Cold Spring Harbor Laboratory, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY, 1989.
В соответствии с изобретением могут быть получены также гомологично рекомбинированные микроорганизмы. Для этого получают вектор, который содержит по меньшей мере один сегмент гена по изобретению или кодирующей последовательности, в который были введены в случае необходимости делеция, добавление или замена по меньшей мере одной аминокислоты для изменения последовательности по изобретению, например, функционального разрушения (нокаут-вектор). Введенная последовательность может быть, например, также гомологом из родственного микроорганизма или быть произведена из источника, являющегося млекопитающим, дрожжами или насекомым. Альтернативно, используемый для гомологичной рекомбинации вектор может быть построен таким образом, что эндогенный ген мутирован при гомологичной рекомбинации или изменен по-другому, однако все еще кодирует функциональный белок (например, лежащая слева (против хода транскрипции) регуляторная область может быть изменена таким образом, что посредством этого изменяется экспрессия эндогенного белка). Этот измененный сегмент гена по изобретению находится в векторе гомологичной рекомбинации. Конструирование подходящих векторов для гомологичной рекомбинации описано, например, в Thomas, K.R. und Capecchi, M.R. (1987) Cell 51:503.
Подходящими организмами-хозяевами, в принципе, являются любые организмы, которые способны экспрессировать нуклеиновые кислоты по изобретению, их аллельные варианты, их функциональные эквиваленты или производные. Организмы-хозяева означают, например, бактерии, грибы, дрожжи, растительные или животные клетки.
Неограничивающими примерами прокариотических экспрессионных организмов являются Escherichia coli, Bacillus subtilis, Bacillus megaterium, Cory-nebacterium glutamicum и другие. Неограничивающими примерами эукариотических экспрессионных организмов являются дрожжи, такие как Sac-charomyces cerevisiae, Pichia pastoris и другие, мицелиальные грибы, такие как Aspergillus niger, Aspergillus oryzae, Aspergillus nidulans, Trichoderma reesei, Acremonium chrysogenum и другие, клетки млекопитающих, такие как клетки Hela, клетки COS, клетки СНО и другие, клетки насекомых, такие как клетки Sf9, клетки MEL и другие, растения или растительные клетки, такие как Solarium tuberosum, Nicotiana и другие.
Отбор успешно трансформированных организмов может происходить с использованием маркерных генов, которые также содержатся в векторе или в экспрессионной кассете. Примерами таких маркерных генов являются гены устойчивости к антибиотикам и гены ферментов, катализирующих цветную реакцию, которая влияет на окраску трансформированных клеток. Они могут также отбираться при помощи автоматизированного сортинга клеток. Успешно трансформированные вектором микроорганизмы, которые несут соответствующий ген устойчивости к антибиотикам (например, к G418 или гигромицину), могут отбираться также с использованием соответствующих содержащих антибиотики сред или питательных сред. Маркерные белки, которые присутствуют на клеточной поверхности, могут быть использованы для отбора с использованием аффинной хроматографии.
7. Рекомбинантное получение белков-предшественников и пептидов
Пептиды и белки-предшественники, применяемые согласно настоящему изобретению, могут быть получены рекомбинантным путем согласно известному способу, например, посредством культивации микроорганизмов, которые продуцируют пептиды/белки-предшественники, при необходимости индуцируя экспрессию полипептидов и выделяя их из культуры. Пептиды и белки-предшественники могут, таким образом, продуцироваться также и в промышленном масштабе, если это желательно.
Рекомбинантные микроорганизмы могут культивироваться и ферментироваться согласно известным способам. Бактерии могут, например, размножаться в ТВ- или LB-среде и при температуре 20-40°С и показателе pH 6-9. В частности, подходящие условия культивирования описаны, например, в Т.Maniatis, E.F., Fritsch und J.Sambrook, Molecular Cloning: A Laboratory Manual, Second Edition, Cold Spring Harbor Laboratory, Cold Spring Harbor, NY (1989).
Если пептиды или белки-предшественники не секретируются в культуральную среду, клетки затем разрушают, и продукт получают согласно известным способам выделения белков из лизата. Клетки могут быть разрушены, по выбору, посредством высокочастотного ультразвука, посредством высокого давления, например, во French-прессе, посредством осмолиза, посредством обработки детергентами, литическими ферментами или органическими растворителями, посредством использования гомогенизаторов, или комбинация нескольких из названных способов.
Очистка пептидов или белков-предшественников может достигаться известными хроматографическими способами, такими как хроматография с молекулярными ситами (гель-фильтрация), такими как хроматография с использованием Q-сефарозы, ионообменная хроматография и гидрофобная хроматография, а также другими обычными способами, такими как ультрафильтрация, кристаллизация, высаливание, диализ и нативный гель-электрофорез. Подходящие способы описаны, например, в Cooper, F.G., Biochemische Arbeitsmethoden, Verlag Walter de Gruyter, Berlin, New York или в Scopes, R., Protein Purification, Springer Verlag, New York, Heidelberg, Berlin.
Особенно предпочтительно использовать для выделения рекомбинантных пептидов или белков-предшественников векторные системы или олигонуклеотиды, которые удлиняют кДНК на определенные нуклеотидные последовательности и тем самым кодируют удлиненные полипептиды или слитые белки, которые, например, служат для более простой очистки. Подобными подходящими модификациями являются, например, действующие в качестве якоря так называемые «метки», такие как, например, известная как гексагистидиновый якорь модификация или эпитопы, которые могут узнаваться в качестве антигенов антителами (описанные, например, в Har-low, Е. and Lane, D., 1988, Antibodies: A Laboratory Manual, Cold Spring Harbor (N.Y.) Press). Эти якоря могут использоваться для прикрепления белков к твердому носителю, такому как полимерный матрикс, который, например, может быть помещен в хроматографическую колонку, или к микротитрационному планшету или иному носителю.
Одновременно эти якоря могут использоваться также для узнавания белков. Кроме того, для узнавания белков могут также применяться обычные маркеры, такие как флуоресцентные красители, ферментные маркеры, которые после реакции с субстратом образуют детектируемый продукт реакции, или радиоактивные маркеры, по отдельности или в комбинации с якорями для дериватизации белков.
Более конкретно, периодические белки-предшественники получают путем экспрессии синтетически полученных генных последовательностей, которые кодируют периодические белки-предшественники по изобретению. Возможное получение синтетических генных последовательностей описывается в Hummerich et al. Biochemistry 43; 13604-13612 (2004).
Периодический белок-предшественник может присутствовать в клетке-хозяине в растворимой или нерастворимой форме. В обоих случаях клетки подвергаются разрушению. Более конкретно, разрушение осуществляется посредством гомогенизатора высокого давления при 1000-1500 бар. В случае растворимого периодического белка-предшественника, большая часть клеточных белков осаждается посредством нагревания лизата до 60-100°С, как, например, 70-90°С или 75-85°С, и отделяется от растворимого периодического белка-предшественника подходящим способом разделения (например, седиментация или фильтрация). Периодический белок-предшественник затем осаждают посредством добавления космотропной соли (как описано выше). При этом периодические белки-предшественники образуют стабильные ассоциаты. В зависимости от ассоциата, конечные концентрации добавленных космоторпных солей могут варьироваться и находятся в интервале около 0.2-3 М или, например, 0.8-2 М. Оптимальные концентрации могут быть определены простым образом, известным химику-специалисту в данной области техники.
Периодические белки-предшественники также могут собираться с помощью внешнего запускающего агента. В этом случае, белки-предшественники собираются уже в клетке-хозяине, с образованием соответствующих стабильных ассоциатов. После разрушения клеток, указанные ассоциаты отделяются от растворимых компонентов подходящим способом разделения (например, седиментация или фильтрация).
Отделение ассоциатов может быть усовершенствовано путем добавления вспомогательных осаждающих веществ после разрушения клеток. Указанные вспомогательные осаждающие вещества вызывают коагуляцию ассоциатов, в результате чего, например, для седиментации требуются более низкие ускорения, например, для того, чтобы отделить ассоциаты от водной среды. Вспомогательными осаждающими веществами, которые могут применяться, являются кислоты, щелочи, полимерные растворы, в частности водные растворы заряженных полимеров. Примерами вспомогательных заряженных веществ являются фосфорная кислота или растворы полиэтиленимина.
Стабильные ассоциаты белков-предшественников по настоящему изобретению могут быть затем очищены. С этой целью для очистки применяются растворы, в которых стабильные вещества нерастворимы, но растворимы другие загрязняющие вещества. Более конкретно, применяются водные растворы оснований, кислот, мочевины, солей и детергентов. Особенно подходящим является применение растворов гидроксидов щелочных металлов, мочевины, солей гуанидина или заряженных детергентов, как например, соли алкилтриметиламмония или алкилсульфаты. Более конкретно, применяются растворы ≥0.2 М гидроксида натрия, ≥2 М мочевины, ≥1 М гидорхлорида гуанидина, ≥1 М тиоцианата гуанидина или ≥0.1% доде-цилсульфата натрия, или ≥0.1% бромида цетилтри метила ммония. Для очищения стабильные ассоциаты ресуспендируют в соответствующих растворах и затем отделяют от раствора простым способом разделения (например, седиментацией или фильтрацией). Периодические белки-предшественники затем промывают водой и высушивают, применяя способы, известные специалистам в данной области техники.
Для извлечения пептидов из периодических белков-предшественников, эти последовательности должны быть подвергаемыми расщеплению для выделения из указанных белков-предшественников и отделения от вспомогательных последовательностей. Расщепление происходит в расщепляемых последовательностях, присутствующих в периодических белках-предшественниках. Способы специфического расщепления аминокислотных цепей описываются в литературе. Периодические белки-предшественники могут расщепляться ферментативным или химическим путем. Примерами ферментов, которые могут применяться для специфического расщепления аминокислотных цепей являются Arg-С протеиназа, Asp-N эндопептидаза, каспазы, химотрипсин, клострипаин, энтерокиназа, фактор Ха, глутамилэндопептидаза, гранзим В, LysC лизилэндопептидаза (ахромобактерильная протеиназа I), LysN пептидил-Lys металлоэндопептидаза, пепсин, пролин эндопептидаза, протеиназа K, стафилококковая протеиназа I, термолизин, тромбин, трипсин. Примерами химических веществ, которые могут применяться для специфического расщепления аминокислотных цепей, являются BNPS-скатол (2-(2'-нитрофенилсульфенил)-3-метил-3-броминдоленин), бромциан, кислоты, гидроксиламин, иодозобензойная кислота, NTCB (2-нитор-5-тиоцианобензойная кислота).
Более конкретно, периодические белки-предшественники расщепляются химическим образом, например, посредством расщепления гидроксиламином или кислотой. Любая неорганическая или органическая кислота, имеющая значение pKs менее 5 и более 0, предпочтительно менее 4 и более 1, подходит для кислотного расщепления. Более конкретно, 1-5% фосфорная кислота и 1-5% муравьиная кислота применяются для указанного расщепления. В зависимости от условий кислотного расщепления имеет место либо однократное кислотное расщепление между аминокислотами Asp и Pro или Asp и Xxx, причем Xxx является любая протеиногенная аминокислота, либо прежде всего происходит расщепление между аминокислотами Asp и Pro или Asp и Xxx, и затем аспартат полностью отщепляется от аминокислоты, лежащей у М-конца перед аспартатом в аминокислотной последовательности пептида.
Расщепление можно осуществлять, применяя очищенный белок-предшественник или применяя клеточную фракцию, содержащую периодический белок-предшественник (например, растворимые компоненты клетки-хозяина или нерастворимые компоненты клетки-хозяина), или применяя целые клетки-хозяева, содержащие периодические белки-предшественники. Расщепляющий агент может быть инактивирован после расщепления. Применяемые с этой целью способы известны специалистам в данной области техники.
После инактивации, смесь, полученная после расщепления, содержит, среди прочего, целевой пептид, высвобожденные вспомогательные последовательности и инактивированные расщепляющие агенты. В этом случае пептиды могут уже обладать их целевой активностью. Если требуется более высокая степень чистоты, пептиды, высвобожденные из периодических белков-предшественников, могут быть отделены от вспомогательных последовательностей после расщепления. Преимущество самособирающихся вспомогательных последовательностей состоит в том, что указанные вспомогательные последовательности собираются в ходе или после расщепления. Они могут либо собираться самопроизвольно в ходе расщепления при выбранных условиях расщепления, либо в результате добавления веществ, способствующих сборке указанных вспомогательных последовательностей. Такими веществами, способствующими сборке, являются, например, космотропные соли, содержащие по меньшей мере один тип иона, который обладает более космотропными свойствами, чем натриевый или хлоридный ионы согласно ряду Гофмейстера. Другими веществами, способствующими сборке, являются кислоты или щелочи, или органические растворители, смешиваемые с водой, такие как спирты, например. Собирающиеся вспомогательные вещества могут быть удалены из растворимого высвободившегося пептида путем седиментации или фильтрации. Последующие стадии очистки могут потребоваться для того, чтобы отделить от целевого пептида оставшиеся белковые или пептидные составляющие или соли, или другие вещества, добавленные в ходе или после расщепления. С этой целью могут применяться, например, хроматографические способы, преципитация, диализ, двухфазные экстракции и другие способы, известные специалистам в данной области техники.
Раствор, содержащий пептид, может затем непосредственно применяться в целях желательного применения, или раствор может быть высушен, применяя способы, известные специалистам (распылительная сушка или сушка вымораживанием), и применяется соответствующий сухой продукт.
После сушки можно удалить загрязняющие вещества, которые нельзя удалить из пептида, поскольку он растворим в воде, путем промывания растворами, в которых указанный пептид нерастворим. Подходящими для этого органическими растворителями являются, например, н-гексан, N-метилпирролидон или смеси растворителей и кислот, как например, смеси н-гексана и уксусной кислоты, или органические кислоты, такие как, например, уксусная кислота или капроновая кислота. Для этой стадии очистки, высушенный пептид ресуспендируют в соответствующем растворителе/смеси растворителей и затем снова удаляют путем седиментации или фильтрации. Остаточный растворитель/смесь растворителей удаляют путем сушки.
Целевые пептиды могут обладать целевой активностью в форме, полученной путем расщепления. Однако также может потребоваться дальнейшая модификация пептидов после расщепления. Например, пептид может быть амедирован, этерифицирован с образованием сложного эфира, окислен, алкилирован или химически связан с любыми молекулами. Примерами молекул, которые могут применяться для таких модификаций, являются спирты, сложные цистеиновые эфиры спиртов, карбоновые кислоты, сложные тиоэфиры или малеимиды. Более конкретно, молекулами, применяемыми для таких модификаций, являются те, которые повышают гидрофобность пептида. Такие молекулы могут содержать модифицированные или немодифицированные алкильные радикалы, как определено выше. Предпочтительно такие молекулы содержат С2-C16-алкильные остатки, особенно C6-C14-алкильные остатки. Соответствующие радикалы известны специалисту в данной области техники. Модификация может осуществляться в любой момент: например, непосредственно после разрушения клетки, после очищения белка-предшественника, после расщепления белка-предшественника или после очистки пептида.
Растворы пептида, имеющие желательную степень очистки, могут применяться непосредственным образом. Альтернативно, могут применяться различные способы консервации для продолжительного хранения. Примерами способов консервации являются охлаждение, замораживание, добавление консервантов. Альтернативно, пептиды могут быть высушены. Примерами способов сушки являются лиофилизация или сушка распылением. Высушенные пептиды можно затем хранить. Для того чтобы применять пептиды, высушенное вещество растворяют в подходящем растворителе, предпочтительно в водном растворе. Указанный водный раствор может содержать соли или буферные вещества или не содержать добавок.
8. Определение различных других общих терминов
Если иного не указано, последовательность, "полученная" из конкретно описанной последовательности или "гомологичная" ее, например, производная последовательности нуклеиновой кислоты или аминокислоты означает согласно настоящему изобретению последовательность, которая на по меньшей мере 80% или по меньшей мере 90%, в частности 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% и 99%, идентичная исходной последовательности.
Примеры
Описанные в настоящем изобретение общеприменимые способы получения любых пептидных последовательностей (Pep) продемонстрированы на основе получения трех пептидов с различными последовательностями и аминокислотными составами (ZnO, P18, Min).
Если иного не указано, применяются стандартные способы органического и биохимического анализа, а также рекомбинантного получения белков и культивации микроорганизмов.
Пример 1: Получение пептида ZnO (SEQ ID NO: 20)
Пептид ZnO представляет собой пептид, полученный из известной (опубликованной) последовательности, которая влияет на образование частиц оксида цинка (Umetsu et al. Adv. Mat. 17: 2571-75 (2005)). Синтетический ген ZnO4 (SEQ ID NO: 21) клонировали в векторе pAZL, описанном в Hum-merich et al. Biochemistry 43; 13604-13612 (2004), применяя эндонуклеазы рестрикции BamHI и Hindlll, и затем димеризовали, согласно протоколу, описанному там, и клонировали в вектор рЕТ21 (Novagen). Последовательность, впоследствии представленная в указанном векторе, кодирует периодический белок-предшественник ZnO8 (SEQ ID NO: 22). Указанный периодический белок-предшественник содержит 8 повторяющихся звеньев, каждый из которых содержит копию ZnO пептида и вспомогательную последовательность. Указанная вспомогательная последовательность содержит полиаланиновую последовательность и придает периодическому белку-предшественнику свойства самосборки. Аминокислоты Asp-Pro, которые, как предполагается, обеспечивают селективное выщепление ZnO пептида из белка-предшественника с применением кислоты, должны быть локализованы между вспомогательными последовательностями и пептидными последовательностями. Экспрессию осуществляли в штамме Е.coli BL21 [DE3] (Novagen).
Культивацию и белковый синтез осуществляли при pO2>20% и pH=6.8, применяя полупериодический процесс с подпиткой.
Среда:
Солевой раствор:
Подпитывающий раствор:
После того как глицерин, присутствующий в основной среде, был израсходован, начали непрерывную подпитку при скорости 100 мл/ч.
Белковый синтез индуцировали добавлением 1 мМ изопропил-β-D-тиогалактопиранозида, после того как бактериальная культура достигла оптической плотности OD600=60. В этой точке температуру культуры понизили от 37°С до 30°С. Клетки собрали через 5 часов после индукции.
ZnOs очистили согласно следующему протоколу:
- ресуспендирование осажденной клеточной массы в 5 мл 20 мМ MOPS (3-(N-морфолино)пропансульфоновой кислоты), pH 7.0, на грамм влажной массы
- разрушение клеток в гомогенизаторе высокого давления при 1400 бар
- центрифугирование 30 минут при 5000* g
- инкубация супернатанта 30 минут при 80°C
- центрифугирование 30 минут при 5000* g
- осаждение ZnOs из супернатанта путем добавления 1.8 М сульфата аммония (конечная концентрация) при 4°C всю ночь
- промывание осажденной массы 8 М мочевиной
- 2-х кратное промывание осажденной массы водой
- лиофилизация
- хранение лиофилизованного ZnOs при -20°C.
Из каждого литра культуральной среды получили 2.2 г чистого ZnO8.
Для расщепления 250 мг лиофилизованного ZnO8 белка-предшественника ресуспендировали в 5 мл 1% муравьиной кислоты и инкубировали при 90°С в течение 6 часов. В ходе этой инкубации лиофилизат растворился с образованием желеподобного вещества. После охлаждения до комнатной температуры, желеподобное вещество отделили от растворимых компонентов путем седиментации при 18000* g. Полученный раствор нейтрализовали, применяя 2 М NaOH. Затем раствор лиофилизовали. Лиофилизат содержал целевой продукт расщепления и формиат натрия от нейтрализации муравьиной кислотой.
Лиофилизованный продукт проанализировали посредством ВЭЖХ: для этого продукт растворили в воде до концентрации 1 мг/мл и проанализировали с помощью обращеннофазной хроматографической колонки (Jupiter Proteo 4.6*250 mm; Phenomenex). Применяемый элюат содержал 0.1% трифторуксусной кислоты в воде, которая затем замещалась на 0.1% трифторуксусной кислоты в ацетонитриле посредством линейного градиента. Обнаружение осуществляли при 206 нм (Фиг.2). Для дальнейшего анализа собирали фракции основного пика, и затем изучали присутствующее в них вещество. М-терминальное секвенирование подтвердило, что этим компонентом является пептид ZnO. Изучение посредством масс-спектрометрии (MALDI-TOF) позволило установить массу 2002, которая идентична теоретической массе пептида ZnO (Фиг.3). Анализ ВЭЖХ позволил определить чистоту 62% относительно УФ-активных компонентов.
Пример 2: Получение пептида Р18 (SEQ ID NO: 23)
Пептид Р18 представляет собой пептид, полученный на основе высокоактивной противомикробной последовательности, описанной Shin et al. J. Peptide Res. 58:504-14 (2001). Синтетический ген АНеАР182 (SEQ ID NO: 24) клонировали в вектор pAZL, описанный в Hummerich et al. Biochemistry 43; 13604-13612 (2004), применяя эндонуклеазы рестрикции BamHI и Hind I II, и димеризовали согласно протоколу, описанному в указанном источнике, и клонировали в вектор рЕТ21 (Novagen). Последовательность, затем представленная в указанном векторе, кодирует периодический белок-предшественник АНеАР184 (SEQ ID NO: 25). Указанный периодический белок-предшественник содержит 4 повторяющихся звена, каждый из которых содержит копию пептида Р18 и вспомогательную последовательность. Указанная вспомогательная последовательность содержит полиаланиновую последовательность и придает периодическому белку-предшественнику свойства самосборки. Кроме того, вспомогательная последовательность содержит отрицательно заряженную вспомогательную последовательность. Аминокислоты Asp-Pro, которые, как предполагается, обеспечивают селективное выщепление Р18 пептида из белка-предшественника с применением кислоты, должны быть локализованы между вспомогательными последовательностями и пептидными последовательностями. Экспрессию осуществляли в штамме Е.coli BL21 [DE3] (Novagen).
Культивацию и белковый синтез осуществили посредством периодического процесса с подпиткой при pO2>20% и pH=6.8. Среда, остаточный раствор соли и подпитывающий раствор имели состав, описанный в примере 1.
После того как глицерин, присутствующий в основной среде, был израсходован, начали непрерывную подпитку при скорости 100 мл/ч.
Белковый синтез индуцировали добавлением 1 мМ изопропил-β-D-тиогалактопиранозида после того, как бактериальная культура достигла оптической плотности OD600=60. Через 10 часов после индукции клетки собрали путем осаждения при 5000* g в течение 30 минут. Влажная биомасса составила 1932 г.
Влажную биомассу очистили согласно следующему протоколу:
- ресуспендирование осажденной клеточной массы: на каждый грамм биомассы добавили 6 г 20 мМ натрий-фосфатного буфера и тщательно перемешали
- разрушение клеток в гомогенизаторе высокого давления при 1500 бар
- добавление фосфорной кислоты до pH=3±0.5
- 10 минут инкубации при 23°C с перемешиванием
- центрифугирование 20 минут при по меньшей мере 5000* g
- ресуспендирование осажденной массы: на каждый грамм влажной массы добавили 25 мл 0.2 М NaOH, гомогенизировали и инкубировали при перемешивании и при 23°C в течение 4 часов
- нейтрализация: установили pH 8.5+-0.5 с помощью 85% H3PO4
- центрифугирование: 20 минут при по меньшей мере 5000* g
- осажденную массу, состоящую из вымытых телец-включений, содержащих Р18 белок-предшественник, подвергли гидролизации или расщеплению посредством 2% Н3РО4.
Условия расщепления:
i. применение 5 мл H3PO4 на каждый грамм осажденной массы
ii. гомогенизация
iii. инкубация со встряхиванием при 90°C в течение 16 часов
- охлаждение реакционной смеси, полученной после расщепления
- центрифугирование: 20 минут при по меньшей мере 5000* g
- нейтрализация: установили pH 5.5±0.5 с помощью 10 М NaOH
- центрифугирование: 20 минут при по меньшей мере 5000* g
- разбавление супернатант водой до проводимости менее 10 мС/см
- очищение Р18 от супернатанта путем катионообменной хроматографии (Fractogel COO; Merck); элюирование с применением 450 мМ NaCl
- объединение фракций, содержащих пептид, и разбавление водой до проводимости менее 10 мС/см
- очищение Р18 от объединенных фракций посредством катионообменной хроматографии (Fractogel COO; Merck); элюирование с применением 50 мМ HCl
- нейтрализация элюата с применением 2 М NaOH
- лиофилизация нейтрализованного раствора.
Лиофилизованный продукт проанализировали с помощью ВЭЖХ: для этого продукт растворили до концентрации 1 мг/мл в воде и проанализировали, применяя обращеннофазную хроматографическую колонку (Jupiter Pro-teo 4.6*250 mm; Phenomenex). Применяемый элюат содержал 0.1% трифторуксусную кислоту в воде, которая затем была замещена на 0.1% трифторуксусную кислоту в ацетонитриле посредством линейного градиента. Обнаружение осуществляли при 280 нм (Фиг.4). Для дальнейшего анализа собрали фракции основного пика, и затем проанализировали вещество, содержащееся в них. N-терминальное секвенирование подтвердило, что этим компонентом является Р18 пептид.
Исследование посредством масс-спектрометрии (MALDI-TOF) позволило установить массу 2512 для пептида, которая идентична теоретической массе пептида Р18 (Фиг.5). С помощью анализа ВЭЖХ определили чистоту 85% относительно УФ-активных компонентов. Из 30 г влажной биомассы получили 52 мг Р18 пептида. Таким образом, около 330 мг чистого пептида Р18 получили из каждого литра ферментационной культуры.
Для того чтобы исследовать активность Р18 пептида, Е.coli В культуры в LB среде (5 г/л дрожжевого экстракта; 10 г/л триптона, 5 г/л хлорида натрия), где культура имеет оптическую плотность 0.1, как измерено при 600 нм, инкубировали со встряхиванием при 37°C с различными концентрациями пептида Р18. Бактериальный рост контролировали путем измерения оптической плотности через 24 часа. Полное ингибирование роста (оптическая плотность при 600 нм через 24 часа < 0.15) было достигнуто при концентрации пептида от 31 частей на миллион. Существует возможность дальнейшего улучшения противомикробной активности путем амидировния C-терминальных карбоксильных групп. Для этого указанные карбоксильные группы активировали с помощью гидрохлорида 1-этил-3-(3-диметиламинопропил)карбодиимида и N-гидрокси-сульфосукцинимида, и затем амидировли путем добавления аммиака.
Пример 3: Получение пептида Min (SEQ ID NO: 26)
Синтетический ген, AEMin4 (SEQ ID N0:27), клонировали в вектор pAZL, описанный в Hummerich et al. Biochemistry 43; 13604-13612 (2004), применяя эндонуклеазы рестрикции BamHI и Hindlll, и димеризовали, согласно протоколу, описанному в этом источнике, и клонировали в вектор рЕТ21 (Novagen). Последовательность, затем представленная в указанном векторе, кодирует периодический белок-предшественник AEMiris (SEQ ID NO: 28). Указанный периодический белок-предшественник содержит 8 повторяющихся звеньев, каждый из которых содержит копию пептида Min и вспомогательную последовательность. Указанная вспомогательная последовательность содержит полиаланиновую последовательность и придает периодическому белку-предшественнику свойства самосборки. Кроме того, вспомогательная последовательность содержит отрицательно заряженную вспомогательную последовательность. Аминокислоты Asp-Pro, которые, как предполагается, обеспечивают селективное выщепление Min пептида из белка-предшественника с применением кислоты, должны быть локализованы между вспомогательными последовательностями и пептидными последовательностями. Экспрессию осуществляли в штамме Е.coli BL21 [DE3] (Novagen).
Культивацию и белковый синтез осуществили посредством периодического процесса с подпиткой при pO2>20% и pH=6.8. Среда, остаточный раствор соли и подпитывающий раствор имели состав, описанный в примере 1.
После того как глицерин, присутствующий в основной среде, был израсходован, начали непрерывную подпитку при скорости 100 мл/ч.
Белковый синтез индуцировали добавлением 1 мМ изопропил-β-D-тиогалактопиранозида, после того как бактериальная культура достигла оптической плотности OD600=60. В этой точке температуру культуры понизили от 37°C до 30°C. Клетки собрали через 5 часов после индукции.
AEMin8 очистили согласно следующему протоколу:
- ресуспендирование осажденной клеточной массы в 5 мл 20 мМ MOPS (3-(N-морфолино)пропансульфоновая кислота), pH 7.0, на грамм влажной массы - разрушение клеток в гомогенизаторе высокого давления при 1400 бар
- центрифугирование 30 минут при 5000* g
- инкубация супернатанта 20 минут при 80°C
- центрифугирование 30 минут при 5000* g
- осаждение AEMin8 из супернатанта путем добавления 2 М сульфата аммония (конечная концентрация) при 4°C всю ночь
- промывание осажденной массы 8 М мочевиной
- 2-х кратное промывание осажденной массы водой
- лиофилизация
- хранение лиофилизованного AEMin8 при -20°C.
Из каждого литра культуральной среды получили 0.4 г чистого AEMirig.
Для расщепления 250 мг лиофилизованного AEMin8 белка-предшественника ресуспендировали в 12.5 мл 1% фосфорной кислоты и инкубировали 8 часов при 90°C. После охлаждения до комнатной температуры нерастворимые вещества отделили от растворимых компонентов путем седиментации при 18000* g. Оставшийся раствор нейтрализовали 2 М NaOH. Затем раствор лиофилизовали. Лиофилизат содержал целевой продукт расщепления и гидрофосфат натрия от нейтрализации фосфорной кислотой.
Лиофилизованый продукт проанализировали посредством ВЭЖХ: для этого продукт растворили до концентрации 1 мг/мл в воде и проанализировали, применяя обращеннофазную хроматографическую колонку (Jupiter Proteo 4.6*250 mm; Phenomenex). Применяемый элюат содержал 0,1% трифторуксусную кислоту в воде, которая затем замещалась на 0,1% трифторуксусную кислоту в ацетонитриле посредством линейного градиента. Обнаружение осуществляли при 206 нм (Фиг.6). Для дальнейшего анализа собрали фракции основного пика и затем изучили присутствующее в них вещество. N-терминальное секвенирование подтвердило, что этим компонентом является пептид Min. Исследование посредством масс-спектрометрии (MALDI-TOF) позволило установить массу 1900 для пептида, которая идентична теоретической массе пептида Min (Фиг.7). С помощью анализа ВЭЖХ определили чистоту 68% относительно УФ-активных компонентов.
Пример 4: Оптимизированное получение пептида Р18 (SEQ ID NO: 23)
Для того чтобы повысить выход пептида Р18 согласно примеру 2, изучили влияние различных последовательностей Aux на выход пептид. Экспрессия и общий выход заметно увеличились при применении синтетического гена АНе2АР182 (SEQ ID NO: 74), который после клонирования в вектор рЕТ21 согласно примеру 2 кодирует белок-предшественник, имеющий последовательность SEQ ID NO: 75. Ферментацию проводили в условиях, описанных в примере 2.
Влажную биомассу очистили согласно следующему протоколу:
- ресуспендирование осажденной клеточной массы: на каждый грамм биомассы было добавлено 6 г воды и тщательно перемешено
- разрушение клеток в гомогенизаторе высокого давления при 1500 бар
- добавление фосфорной кислоты до pH=3±0.5
-10 минут инкубации с перемешиванием при 23°C
- центрифугирование: 20 минут при по меньшей мере 5000* g
- ресуспендирование осажденной массы: на каждый грамм влажной биомассы добавили 20 мл 0.4 М NaOH, гомогенизировали и инкубировали при перемешивании при 23°C в течение одного часа
- нейтрализация: установили pH 8.5±0.5 с помощью 1 М калийфосфатного буфера pH 6.0
- центрифугирование: 20 минут при по меньшей мере 5000* g
- осажденную массу, состоящую из вымытых телец-включений, содержащих Р18 белок-предшественник, подвергли гидролизации или расщеплению посредством 2% H3PO4.
Условия расщепления:
i. применение 7 мл H3PO4 на каждый грамм осажденной массы
ii. гомогенизация
iii. инкубация со встряхиванием при 90°C в течение 16 часов
- охлаждение реакционной смеси, полученной после расщепления
- центрифугирование: 20 минут при по меньшей мере 5000* g
- установление pH 4.0+0.5 с помощью 25% NaOH
- центрифугирование: 20 минут при по меньшей мере 5000* g
- разбавление супернатанта водой до проводимости менее 30 мС/см
- очищение Р18 от супернатанта посредством катионообменной хроматографии (SP-Sepharose High Performance; GE Healthcare); промывочный буфер: 10 мМ натрий-ацетатный буфер pH 4+450 мМ NaCl; элюционный буфер: 10 мМ натрий-ацетатный буфер pH 4+1100 мМ NaCl
- осаждение пептида посредством добавления 25% NaOH к элюату до достижения pH=10.5±0.3
- центрифугирование: 20 минут при по меньшей мере 5000* g
- ресуспендирование осажденной массы в 5 мл воды на каждый грамм влажной массы
- растворение пептида: добавление уксусной кислоты до pH 6.0 pH=10.5±0.5
- лиофилизация.
Из каждого литра ферментационной культуры описанным образом можно получить около 1 г чистого пептида Р18.
Пример 5: Получение пептида SEQ ID NO: 6
Пептиды, перечисленные в примерах 1-4, были выделены из периодического белка-предшественника путем кислотного расщепления. При этом происходит гидролиз пептидной связи между аспартатом и пролином. Соответственно, как показано в примере 1, пептидная последовательность начинается с пролина на N-конце и заканчивается аспартатом на С-конце. В определенных условиях, C-терминальный аспартат, как показано в примерах 2 и 3, подобным образом может быть отщеплен, таким образом, позволяя получать пептидные последовательности со свободным выбором C-терминальной последовательности.
Пептид, имеющий последовательность SEQ ID NO: 6, был получен для того, чтобы продемонстрировать, что способ по изобретению может также применяться для получения пептидов, чьи N-концы не начинаются с пролина, и первая N-терминальная аминокислота которых может быть выбрана свободным образом. Для этого белок-предшественник, имеющий последовательность SEQ ID NO: 77, получили, применяя синтетический ген AHe2AP182-P-G (SEQ ID NO: 76), согласно примеру 4. Этот белок-предшественник отличается от белка-предшественника SEQ ID NO: 75 по примеру 4 только тем, что N-терминальные аминокислоты пептида Р18, Pro-Gly, и C-терминальный Gly были удалены. Клонирование и ферментацию осуществили в условиях, описанных в примере 4.
Влажную биомассу очистили согласно следующему протоколу:
- ресуспендирование осажденной клеточной массы: на каждый грамм биомассы было добавлено 6 г воды и тщательно перемешено
- разрушение клеток в гомогенизаторе высокого давления при 1500 бар
- центрифугирование: 20 минут при по меньшей мере 5000* g
- осажденную массу, состоящую из вымытых телец-включений, содержащих Р18 белок-предшественник, подвергли гидролизации или расщеплению посредством 5% H3PO4.
Условия расщепления:
iv. применение 5 мл H3PO4 на каждый грамм осажденной массы
v. гомогенизация
vi. инкубация со встряхиванием при 90°C в течение 16 часов
- охлаждение реакционной смеси, полученной после расщепления
- центрифугирование: 20 минут при по меньшей мере 5000* g
- установление pH 4.0±0.5 с помощью 25% NaOH
- центрифугирование: 20 минут при по меньшей мере 5000* g
- разбавление супернатанта водой до проводимости менее 30 мС/см
- очищение Р18 от супернатанта посредством катионообменной хроматографии (SP-Sepharose High Performance; GE Healthcare); промывочный буфер: 10 мМ натрий-ацетатный буфер pH 4+450 мМ NaCl; элюционный буфер: 10 мМ натрий-ацетатный буфер pH 4+1100 мМ NaCl
- осаждение пептида посредством добавления 25% NaOH к элюату до достижения pH=10.5±0.3
- центрифугирование: 20 минут при по меньшей мере 5000* g
- ресуспендирование осажденной массы в 5 мл воды на каждый грамм влажной массы
- растворение пептида: добавление уксусной кислоты до pH=6.0±0.5
- лиофилизация.
Лиофилизованый продукт проанализировали посредством ВЭЖХ: для этого продукт растворили до концентрации 1 мг/мл в воде и проанализировали, применяя обращеннофазную хроматографическую колонку (Jupiter Рго-teo 4.6*250 mm; Phenomenex). Применяемый элюат содержал 0,1% трифторуксусную кислоту в воде, которая затем замещалась на 0,1% трифторуксусную кислоту в ацетонитриле посредством линейного градиента. Обнаружение осуществляли при 280 нм (Фиг.8). Для дальнейшего анализа собрали фракции основного пика и затем изучили присутствующее в них вещество. С помощью масс-спектрометрии (MALDI-TOF) определили массу пептида 2300.6, которая равна теоретической массе пептида SEQ ID NO: 6 (Фиг.9).
Пример 6: Амидирование лиофилизованного пептида Р18 (SEQ ID NO: 23)
В некоторых случаях может быть предпочтительно с точки зрения активности пептида, если С-конец скорее амидирован, чем является свободной карбоксильной группой. Для того чтобы продемонстрировать это, лиофилизованный пептид Р18 по примеру 4 был амидирован согласно протоколу, приведенному ниже:
- 10 мг/мл Р18 пептида
- 30% EtOH
- 10 мМ 2-(N-морфолино)этансульфоновая кислота pH 5.0
- 3 М хлорид аммония
- 2.5 мМ N-гидроксисукцинимид
- 50 мМ 1-этил-3-(3-диметиламинопропил)карбодиимид
- инкубация в течение двух часов при комнатной температуре
- нейтрализация с помощью NaOH, pH 7.0.
Амидированный образец проанализировали посредством ВЭЖХ, применяя хроматографическую колонку Luna SCX 5р 100А (Phenomenex, Torrance, Calif., USA): применяемый элюент содержал 20 мМ KH2PO4 pH 2.5 с 25% ацетонитрила, который был замещен на 20 мМ KH2PO4 pH 2.5, 25% ацетонитрила и 1 М KCl посредством линейного градиента. Обнаружение осуществляли при 280 нм (Фиг.10). Фиг.10 показывает, для сравнения, хроматограмму синтезированного химическим путем и хроматограмму амидированного контрольного пептида с последовательностью "Р18" (полученного по заказу у Bachem A G, Бубендорф, Швейцария).
Пептид далее очистили с помощью катионообменной хроматографии, как описано в примере 4.
Пример 7: Получение пептида Р18 (SEQ ID NO: 23) с включенным амидированием
Для повышения экономической эффективности получения амидированного пептида Р18, амидирование скорее включают в процедуру получения, а не осуществляют после очищения пептида, как описано в примере 6. Для этого белок-предшественник SEQ ID NO: 75 получили путем ферментации согласно примеру 4, и пептид выделили из белка-предшественника путем кислотного расщепления.
После чего осуществили следующие стадии:
- охлаждение реакционной смеси, полученной после расщепления
- центрифугирование: 20 минут при по меньшей мере 5000* g
- установление pH 10.5±0.5 с 25% NaOH
- центрифугирование: 20 минут при по меньшей мере 5000* g
- растворение осажденной массы в 3 мл этанола на каждый грамм влажной массы
- центрифугирование; определение пептида в супернатанте (разбавление)
- смешивание следующих компонентов:
а. 12.5 мл растворенного пептида в этаноле
b. 4.2 мл воды
с. добавление 400 мкл 500 мМ 2-(N-морфолино)этансульфоновой кислоты
d. установление pH 5.0 с помощью HCl
е. добавление 2.14 г хлорида аммония
f. 200 мкл 500 мМ N-гидроксисукцинимида
g. 1 мл 1 М 1-этил-3-(3-диметиламинопропил)карбодиимида
- инкубирование в течение двух часов при комнатной температуре
- разбавление супернатанта водой до проводимости менее 30 мС/см
- очищение модифицированного пептида посредством катионообменной хроматографии (SP-Sepharose High Performance; GE Healthcare); промывочный буфер: 10 мМ натрий-ацетатный буфер pH 4+450 мМ NaCl; элюционный буфер: 10 мМ натрий-ацетатный буфер pH 4+1100 мМ NaCl
- осаждение пептида посредством добавления 25% NaOH к элюату до достижения pH=10.5±0.3
- центрифугирование: 20 минут при по меньшей мере 5000* g
- ресуспендирование осажденной массы в 5 мл воды на каждый грамм влажной массы
- растворение пептида: добавление уксусной кислоты до pH=6.0+-0.5
- лиофилизация.
Амидированный образец проанализировали с помощью ВЭЖХ, применяя хроматографическую колонку Luna SCX 5u 100А (Phenomenex, Torrance, Calif., USA)): применяемый элюент содержал 20 мМ KH2PO4 pH 2.5 с 25% ацетонитрила, который был замещен на 20 мМ KH2PO4 pH 2.5, 25% ацетонитрила и 1 М KCl посредством линейного градиента. Обнаружение осуществляли при 280 нм (Фиг.11). Фиг.11 показывает, для сравнения, хроматограмму синтезированного химическим путем и амидированного контрольного пептида с последовательностью "Р18" (полученного по заказу у Bachem A G, Бубендорф, Швейцария).
Последовательности по изобретению














спиральный
спиральный
спиральный
спиральный
















































неспиральный



(Min короткий)





SA=самособирающаяся последовательность
SU=защитный пептид
Pep=пептид, который должен быть получен.
В дополнение к вышеописанным конкретным Pep аминокислотным последовательностям, перечисленные последовательности могут быть изменены на С-конце и/или М-конце посредством добавления конкретных расщепляемых последовательностей (например, между остатками "DP" для кислотного расщепления; или между остатками "NG" для гидроксиламинного расщепления) или посредством аминокислотных остатков, оставшихся в результате такого расщепления. При необходимости, спейсерные остатки, такие как, например, остаток G, также могут быть дополнительно вставлены между последовательностью расщепления и последовательностями Pep. Необходимо также конкретно упомянуть следующие изменения вышеперечисленных Pep последовательностей, которые могут быть результатом кислотного или гидроксиламинного расщепления Pep последовательностей, получаемых согласно изобретению:
N-конец: добавление остатка PG-, Р- или G-;
С-конец: добавление остатка GD-; GN-; G-; N- или D-.
Такие изменения, в частности, осуществляются в отношении каждой из вышеперечисленных Pep последовательностей, в частности последовательностей SEQ ID NO: 6-15, 29-67 и 72.
Приводится ссылка на описание всех документов, процитированных в настоящей заявке.
| название | год | авторы | номер документа |
|---|---|---|---|
| ХИМЕРНЫЙ БЕЛОК, ОБЕСПЕЧИВАЮЩИЙ ОБРАЗОВАНИЕ БИОАКТИВНОЙ КОНФОРМАЦИИ, СПОСОБ ПОЛУЧЕНИЯ БЕЛКА С ИНСУЛИНОВОЙ АКТИВНОСТЬЮ, СПОСОБ ИДЕНТИФИКАЦИИ ПЕПТИДИЛЬНОГО ФРАГМЕНТА | 1998 |
|
RU2238951C2 |
| ВЫДЕЛЕННЫЙ ПЕПТИД, ОБЛАДАЮЩИЙ СПЕЦИФИЧЕСКОЙ СВЯЗЫВАЮЩЕЙ АНТИ-GDF-8 АНТИТЕЛО АКТИВНОСТЬЮ, ВЫДЕЛЕННАЯ МОЛЕКУЛА НУКЛЕИНОВОЙ КИСЛОТЫ, ВЕКТОР ЭКСПРЕССИИ, КЛЕТКА-ХОЗЯИН, СПОСОБ ПОЛУЧЕНИЯ ПЕПТИДА, ВАКЦИННАЯ КОМПОЗИЦИЯ И СПОСОБ ВЫЗЫВАНИЯ ИММУННОГО ОТВЕТА АНТИ-GDF-8, СПОСОБ СКРИНИНГА ДЛЯ ОТБОРА АНТИ-GDF-8 АНТИТЕЛА И СПОСОБ ПОНИЖАЮЩЕГО РЕГУЛИРОВАНИЯ АКТИВНОСТИ GDF-8 У ЖИВОТНОГО | 2004 |
|
RU2422460C2 |
| НОВЫЕ ГЕНЫ, КОДИРУЮЩИЕ НОВЫЕ ПРОТЕОЛИТИЧЕСКИЕ ФЕРМЕНТЫ | 2002 |
|
RU2423525C2 |
| НОВЫЕ ГЕНЫ, КОДИРУЮЩИЕ НОВЫЕ ПРОТЕОЛИТИЧЕСКИЕ ФЕРМЕНТЫ | 2002 |
|
RU2296160C2 |
| ГЕНЫ И БЕЛКИ ДЛЯ БИОСИНТЕЗА ЛАНТИБИОТИКА 107891 | 2007 |
|
RU2441915C9 |
| НЕОАНТИГЕННЫЕ КОМПОЗИЦИИ И ИХ ИСПОЛЬЗОВАНИЕ | 2020 |
|
RU2832574C2 |
| СПОСОБ СНИЖЕНИЯ СТЕПЕНИ ГЛИКОЗИЛИРОВАНИЯ БЕЛКОВ, СПОСОБЫ И БЕЛКИ | 2010 |
|
RU2575607C2 |
| МУТАНТ AAV, ОБЛАДАЮЩИЙ СПОСОБНОСТЬЮ НАЦЕЛИВАТЬСЯ НА ГОЛОВНОЙ МОЗГ | 2020 |
|
RU2809389C2 |
| Новые способы дисплея циклических пептидов на частицах бактериофага | 2015 |
|
RU2702087C2 |
| БИОЛОГИЧЕСКИ АКТИВИРУЕМЫЕ ЛЕКАРСТВЕННЫЕ СРЕДСТВА НА ОСНОВЕ ЦИТОКИНОВ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2019 |
|
RU2819307C2 |



























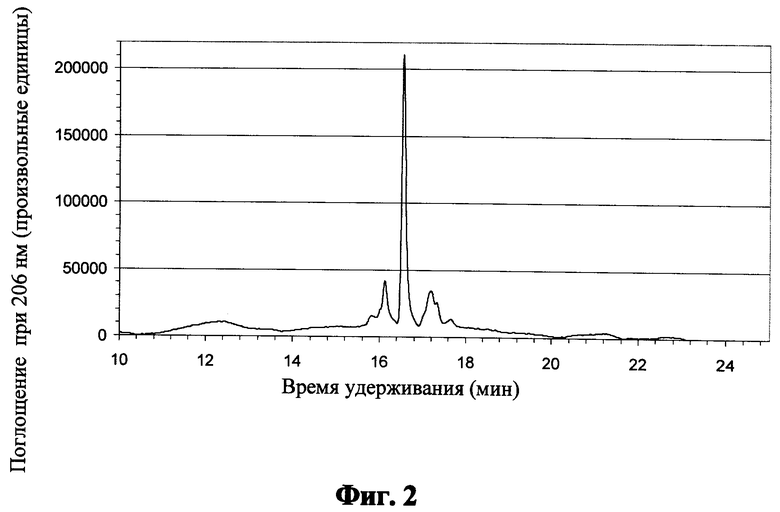
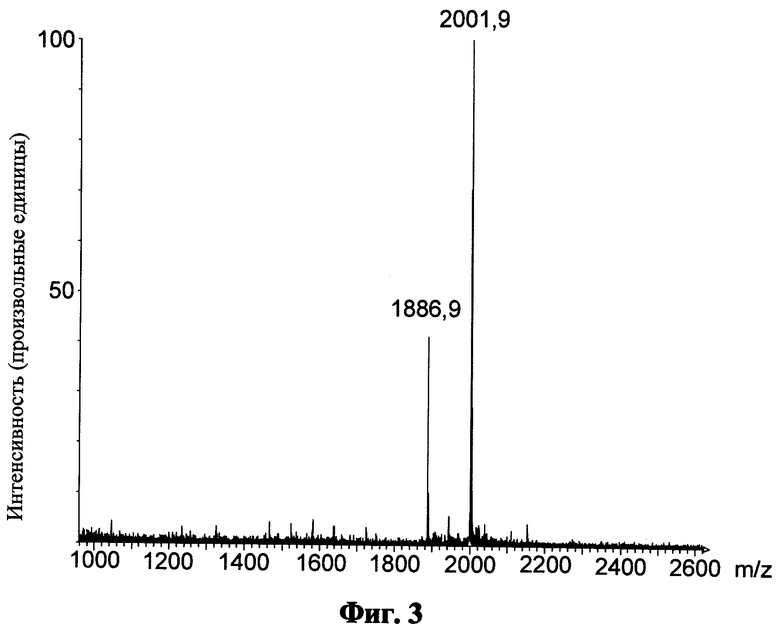
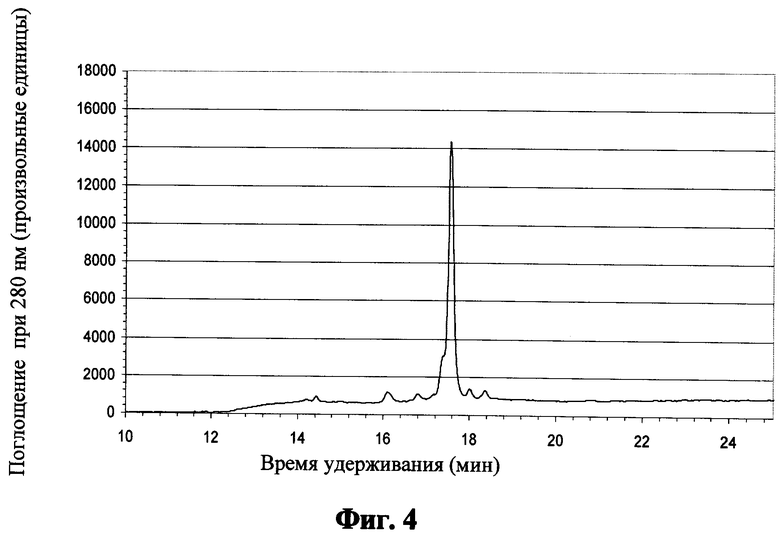
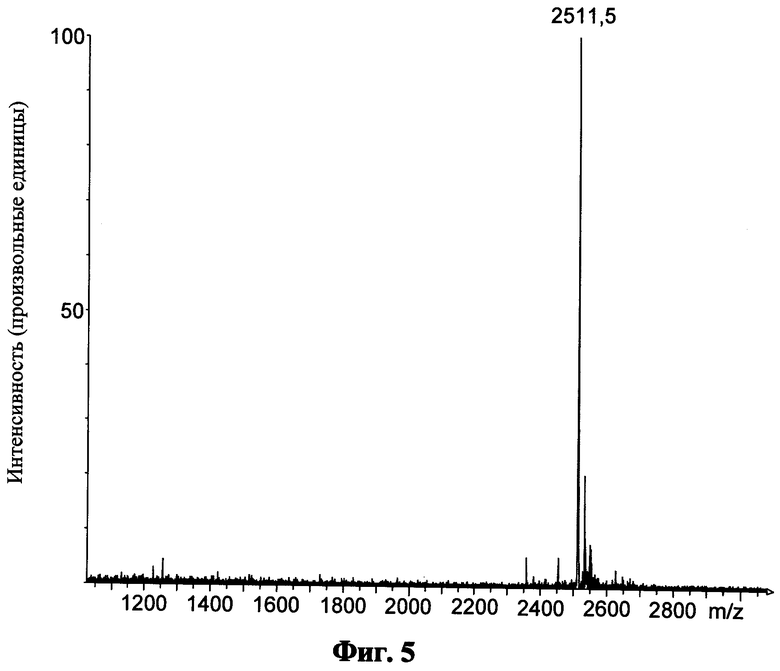
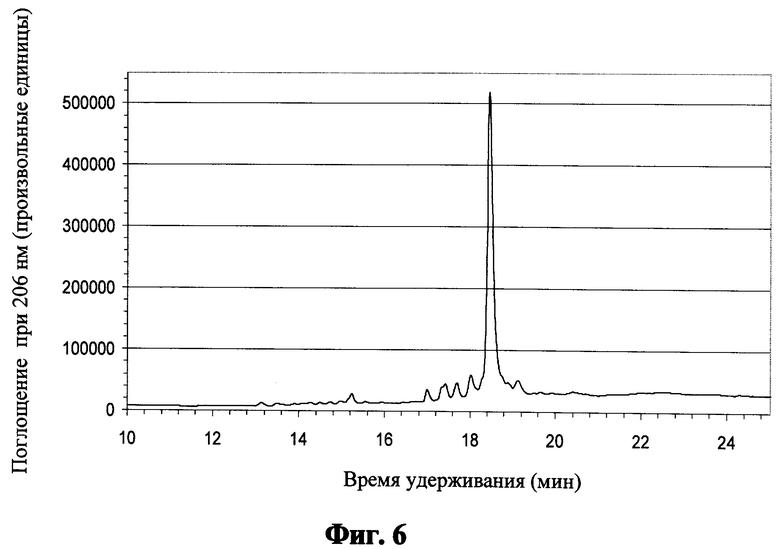
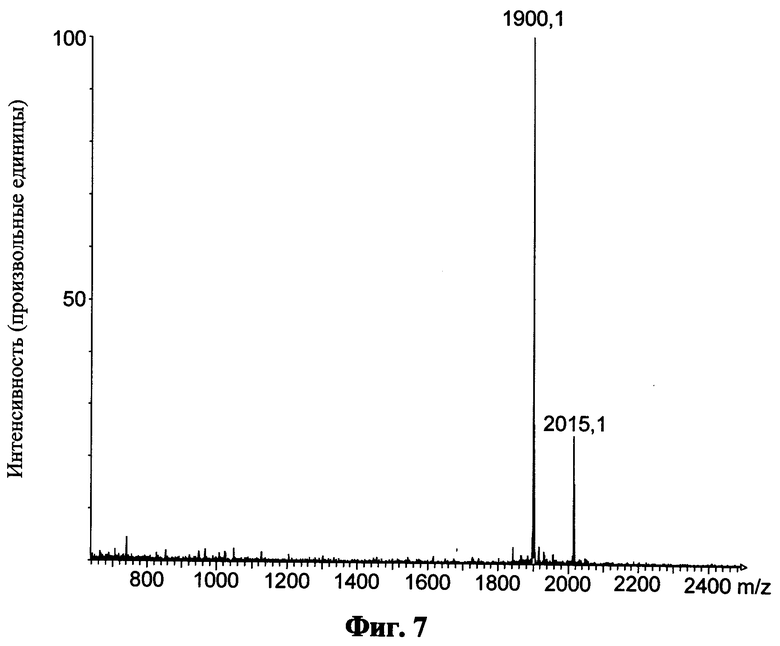
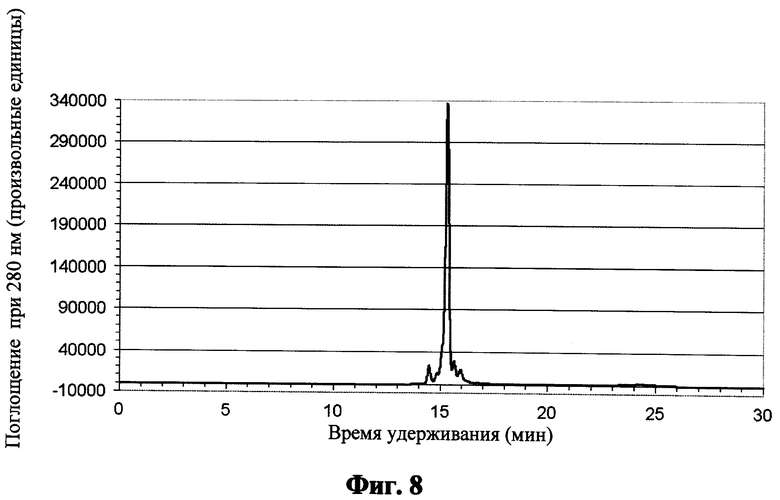
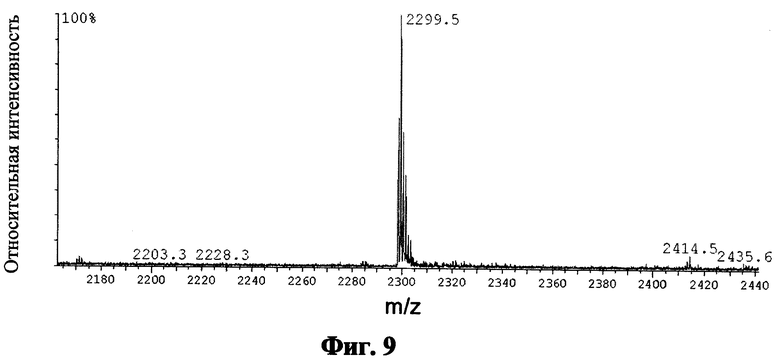
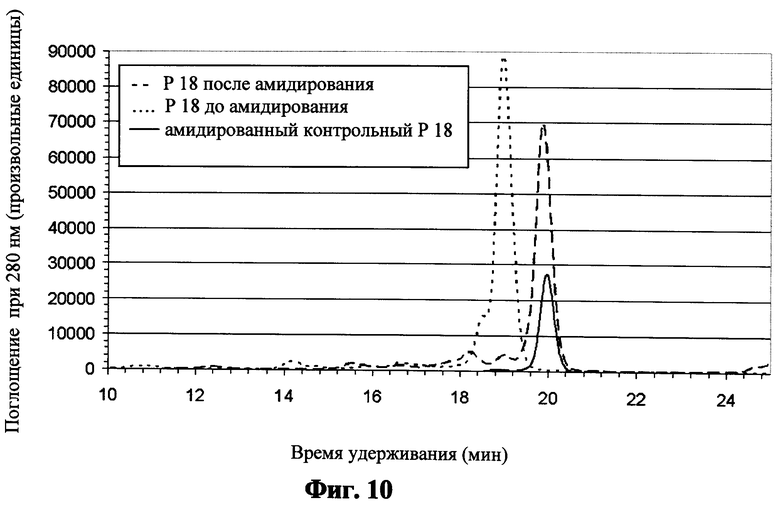

Группа изобретений относится к биотехнологии. Предложен белок-предшественник для рекомбинантного получения пептидов с длиной последовательности от 5 до 70 аминокислотных остатков. Указанный белок содержит расщепляемую повторяющуюся последовательность целевых пептидных элементов (Pep) и вспомогательных пептидных элементов (Aux) общей формулы: (Pep-Aux)x или (Aux-Pep)x, где x>1. Элементы Aux содержат самособирающийся пептидный элемент (SA). Элементы Pep содержат аминокислотную последовательность с длиной последовательности от 5 до 70 аминокислотных остатков, и на концах элементов Pep находятся расщепляемые последовательности, которые обеспечивают возможность высвобождения Pep элементов из белка-предшественника посредством специфического расщепления. Предложены также молекула нуклеиновой кислоты с последовательностью нуклеиновой кислоты, кодирующей указанный белок-предшественник, экспрессионная кассета и рекомбинантный вектор экспрессии, содержащие последовательность указанной нуклеиновой кислоты. Предложены также вариант белка-предшественника, способ получения целевого пептида Рер, а также применение амфифильного пептида для рекомбинантного получения противомикробного целевого пептида. Группа изобретений обеспечивает повышение выхода целевых пептидов. 7 н. и 17 з.п. ф-лы, 11 ил., 1 табл., 7 пр.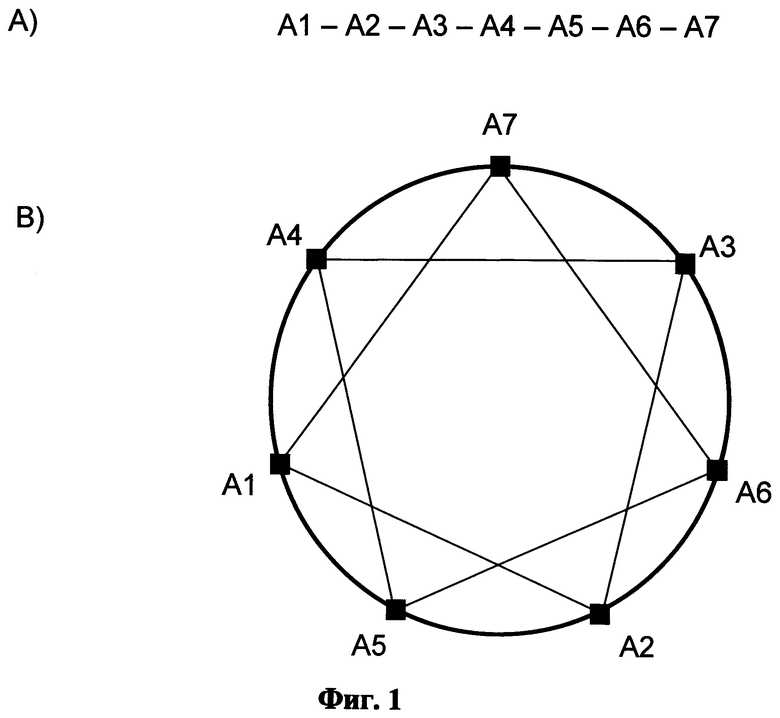
1. Белок-предшественник для рекомбинантного получения пептидов с длиной последовательности от 5 до 70 аминокислотных остатков, содержащий расщепляемую повторяющуюся последовательность целевых пептидных элементов (Pep) и вспомогательных пептидных элементов (Aux) общей формулы:
(Pep-Aux)x или
(Aux-Pep)x,
где x>1, причем
элементы Aux являются идентичными или различными и содержат элементы аминокислотной последовательности, которые обеспечивают свойства самосборки указанного белка-предшественника, причем элемент Aux содержит самособирающийся пептидный элемент (SA), причем элемент SA содержит по меньшей мере один из следующих мотивов последовательности:
где A представляет собой аланин, G представляет собой глицин, V представляет собой валин, n представляет собой целое число от 8 до 12, m представляет собой целое число от 4 до 10, и о представляет собой целое число от 2 до 6, и
элементы Pep содержат аминокислотную последовательность с длиной последовательности от 5 до 70 аминокислотных остатков, и на концах элементов Pep находятся расщепляемые последовательности, которые обеспечивают возможность высвобождения Pep элементов из белка-предшественника посредством специфического расщепления, причем элементы Pep и Aux пептидно связаны друг с другом, и пептидная связь специфически расщепляется химическим или ферментативным путем.
2. Белок-предшественник по п.1, который образует стабильные ассоциаты, которые являются нерастворимыми при комнатной температуре в 0.2 М NaOH в течение одного часа, или в 2 М мочевине или 1 М гидрохлориде гуанидина в течение 10 минут.
3. Белок-предшественник по п.1, где элемент SA содержит аминокислотную последовательность, выбранную из аминокислотных последовательностей SEQ ID NO: 1 - SEQ ID NO: 5 или SED ID NO: 73.
4. Белок-предшественник по п.1, где по меньшей мере один пептид Aux, кроме того, содержит защитный пептидный элемент (SU), содержащий повышенную долю отрицательно заряженных аминокислотных остатков.
5. Белок-предшественник по п.4, где элемент SU в белке-предшественнике способен образовывать амфифильную спиральную структуру.
6. Белок-предшественник по п.5, где элемент SU представляет собой амфифильный пептид, содержащий сегмент последовательности из по меньшей мере семи пептидно связанных аминокислот, способных образовывать амфифильную альфа-спираль, причем указанная спираль в ее вертикальной проекции показывает разделение аминокислотных остатков на гидрофобную половину и гидрофильную половину спирали, причем гидрофобная половина спирали (в ее вертикальной проекции) имеет по меньшей мере 3 идентичных или различных смежных гидрофобных аминокислотных остатка, и гидрофильная половина спирали (в ее вертикальной проекции) имеет по меньшей мере 3 идентичных или различных смежных гидрофильных аминокислотных остатка.
7. Белок-предшественник по п.4, 5 или 6, где доля заряженных аминокислотных остатков элемента SU выбирается таким образом, что общий суммарный заряд белка-предшественника при pH=7 больше -10 и меньше +10.
8. Белок-предшественник по п.7, где элемент SU содержит аминокислотную последовательность, выбранную из аминокислотных последовательностей SEQ ID NO: 16 - SEQ ID NO: 19 и SEQ ID NO: 68.
9. Белок-предшественник по п.1, где элемент Pep содержит катионную противомикробную пептидную последовательность.
10. Белок-предшественник по п.9, где элемент Pep содержит аминокислотную последовательность, выбранную из катионных аминокислотных последовательностей SEQ ID NO: 6 - SEQ ID NO: 15, SEQ ID NO: 23, SEQ ID NO: 26 и SEQ ID NO: 69-SEQ ID NO: 72.
11. Белок-предшественник по п.1, где элемент Pep содержит аминокислотную последовательность, выбранную из аминокислотных последовательностей SEQ ID NO: 20 или SEQ ID NO: 29-67.
12. Белок-предшественник по одному из пп.1-11, где элементы Aux независимо друг от друга имеют одно из следующих значений:
SA,
SA-SU,
SU-SA,
SA-SU-SA,
SU-SA-SU,
где элементы SA и SU пептидно связаны друг с другом, и элемент Aux терминально пептидно связан с по меньшей мере одним элементом Pep, и эта пептидная связь с элементами Pep специфически расщепляется химическим или ферментативным путем.
13. Молекула нуклеиновой кислоты с последовательностью нуклеиновой кислоты, кодирующей по меньшей мере один белок-предшественник по пп.1-12.
14. Молекула нуклеиновой кислоты по п.13, содержащая по меньшей мере одну кодирующую последовательность согласно SEQ ID NO:21, 24, 27, 74 и 76.
15. Экспрессионная кассета, содержащая по меньшей мере одну последовательность нуклеиновой кислоты по п.13 или 14, оперативно связанную с по меньшей мере одной регуляторной последовательностью нуклеиновой кислоты.
16. Рекомбинантный вектор экспрессии для трансформации прокариотического хозяина, содержащий последовательность нуклеиновой кислоты по одному из пп.13 или 14, или экспрессионную кассету по п.15.
17. Способ получения целевого пептида Pep с длиной последовательности от 5 до 70 аминокислотных остатков, в котором осуществляют:
а) получение белка-предшественника по одному из пп.1-12, и
b) высвобождение пептидов Pep из белка-предшественника посредством расщепления.
18. Способ по п.17, где белок-предшественник продуцируется в рекомбинантном микроорганизме, несущем по меньшей мере один вектор по п.16.
19. Способ по п.18, где белок-предшественник продуцируется в рекомбинантном штамме E.coli.
20. Способ по одному из пп.17-19, где экспрессированный белок-предшественник, при необходимости, после того как был превращен в стабильную ассоциированную форму, подвергается очистке и химическому или ферментативному расщеплению с высвобождением целевого пептида (Pep).
21. Белок-предшественник для рекомбинантного получения пептидов с длиной последовательности от 5 до 70 аминокислотных остатков, содержащий расщепляемую последовательность целевых пептидных элементов (Pep) и вспомогательных пептидных элементов (Aux') общей формулы:
(Pep-Aux')x или
(Aux'-Pep)x,
где x>1, причем
элементы Aux' являются идентичными или различными и содержат амфифильный пептид, образующий альфа-спираль, причем указанный амфифильный пептид содержит сегмент последовательности из по меньшей мере семи пептидно связанных аминокислот, способных образовывать амфифильную альфа-спираль, причем указанная спираль в ее вертикальной проекции показывает разделение аминокислотных остатков на гидрофобную половину и гидрофильную половину спирали, причем гидрофобная половина спирали (в ее вертикальной проекции) имеет по меньшей мере 3 смежных идентичных или различных гидрофобных аминокислотных остатка, и гидрофильная половина спирали (в ее вертикальной проекции) имеет по меньшей мере 3 смежных идентичных или различных гидрофильных аминокислотных остатка; и
элементы Pep содержат аминокислотную последовательность с длиной последовательности от 5 до 70 аминокислотных остатков, и на концах элементов Pep находятся расщепляемые последовательности, которые обеспечивают возможность высвобождения Pep элементов из белка-предшественника посредством специфического расщепления, а целевым пептидом Pep является катионный противомикробный пептид, и элементом Aux' является анионный пептид, формирующий амфифильную альфа-спираль.
22. Белок-предшественник по п.21, где элементы Aux' содержат по меньшей мере один самособирающийся пептидный элемент SA, как определено в любом из пп. 1 или 3.
23. Применение амфифильного пептида согласно определению по п.6 в качестве защитного пептида для рекомбинантного получения противомикробного целевого пептида с длиной последовательности от 5 до 70 аминокислотных остатков, отличного от него, где указанный амфифильный пептид содержит участок последовательности из по меньшей мере семи пептидно связанных аминокислот, способных образовывать амфифильную альфа-спираль, причем указанная спираль в ее вертикальной проекции показывает разделение аминокислотных остатков на гидрофобную половину и гидрофильную половину спирали, причем гидрофобная половина спирали (в ее вертикальной проекции) имеет по меньшей мере 3 смежных идентичных или различных гидрофобных аминокислотных остатка, и гидрофильная половина спирали (в ее вертикальной проекции) имеет по меньшей мере 3 смежных идентичных или различных гидрофильных аминокислотных остатка.
24. Применение по п.23, где целевой пептид Pep представляет собой катионный противомикробный пептид, и элемент Aux' представляет собой анионный пептид, формирующий амфифильную альфа-спираль.
| LEE J.H | |||
| ET AL | |||
| Acidic peptide-mediated expression of the antimicrobialpeptide buforin II as tandem repeats in Escherichia coli // protein Expression and Purification, Academic PRESS, vol | |||
| Способ гальванического снятия позолоты с серебряных изделий без заметного изменения их формы | 1923 |
|
SU12A1 |
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| WO 9854336 A1, 03.12.1998 | |||
| KIM H.-K | |||
| ET AL | |||
| Expression of the cationic antimicrobial peptide Lactoferrin fused with the anionic peptide in | |||

