Изобретение относится к плазмидной генетической конструкции pVEAL3-10H10ch, штамму рекомбинантной клеточной линии СНО-K1-10H10ch и химерному антителу 10H10ch против вируса клещевого энцефалита, продуцируемое указанным штаммом клеточной линии СНО-K1-10H10ch и может быть использовано в генной инженерии, биотехнологии и медицине.
Уровень техники.
Семейство Flaviviridae представлено четырьмя родами объединяющих 89 видов вирусов. Наиболее изученным является род Flavivirus насчитывающий 53 вида, Hepacivirus - 14 видов, Pegivirus - 11 видов, и Pestivirus - 11 видов, а также более 51 некласифицированных вирусов. Некоторые представители рода Flavivirus имеют ареал распространения охватывающий 5 континентов. Один из таких вирусов, вирус денге вызывающий геморрагическую лихорадку денге или шоковый синдром денге (Neufeldt CJ., et al., 2018, Begum F, et al., 2019). Тяжелое течение этого заболевания характеризуется повреждением тканей и капилляров, что приводит к 500 тыс. ежегодных госпитализаций с летальностью около 5% (Kyle JL., et al. 2008). Кроме вируса денге опасение вызывает ежегодный рост заболеваемости обусловленный другими представителями рода Flavivirus такими как вирус Западного Нила (WNV), вирус клещевого энцефалита (TBEV), вирус японского энцефалита (JEV) (Neufeldt CJ., et al., 2018), вирус Зика (ZIKV) (Basarab M, et al 2016, Aubry F, et al., 2021). Известны примеры создания высокоэффективных вакцин против флавивирусов. Это инактивированные вакцины против JEV и TBEV, аттенуированная вакцина против YFV (Fox J. P. et al. 1948, Durbin A., Wilder-Smith A, 2017, Monath TP 2005). Многочисленные попытки создания вакцины против лихорадки денге наткнулись на существенные трудности, включая феномен антитело-зависимого усиления (antybody-dependent enhancement, ADE) (Baykov IK, et al., 2014, Yu L, et al. 2017, Priyamvada L, et al., 2016).
Внимание к моноклональным антителам резко возрастает с каждым годом. Их высокоспецифичное нацеливание на антигены может обеспечить очень эффективное лечение, а появление адресных агентов позволяет разрабатывать терапевтические агенты нового поколения. За последние годы наблюдается увеличения количество моноклональных антител на фармакологическом рынке, а также увеличение доли человеческих и гуманизированных антител в общем количестве антител. Биспецифические антитела (Asano R, et al., 2012, Carlring J, et al 2011, Satt A, et al., 2018, Asano R, et al. 2018, Li J, et al., 2018), конъюгаты антитело-лекарственное средство (Smith I, et al 2007), антитела с модифицированными гликозилированием (Yamane-Ohnuki N, et al., 2004, Busse WW, et a., 2010, Yamamoto K, et al., 2010, Ishida T, et al., 2012) и фрагменты антител (Asano R, et al 2012, Ferrara N, et al 2006) в настоящее время рассматриваются как продукты следующего поколения. Некоторые биспецифические антитела и конъюгаты антитело-лекарственное средство уже представлены на рынке, например, эмицизумаб (Oldenburg J, et al 2017) гемтузумаб озогамицин (Jen EY, et al., 2018), инотузумаб озогамицин (Choudhry A, et al., 2017) и брентуксимаб ведотин (de Claro RA, et al., 2012).
Одни из феноменов связанных с гуморальным иммунитетом, находящихся в центре внимание, в настоящее время является феномен широкой реактивности. Это явление, было подробно изучено на примере ВИЧ-1. (Muster Т. et al. 1993, Buchacher A. et al. 1994, Montefiori D.C. et al. 1996), Широкореактивные антитела помимо ВИЧ-инфицированных также были обнаружены у больных с другими вирусными заболеваниями такие как вирусы гриппа, Эбола, ласса, флавивирусы, метапневмовирус и другие (Walker L.M.,et al 2018). Найдены они была так же у людей перенесших флавивирусные инфекции, подобные антитела способны связываться не только с разными изолятами одного вируса, но и с родственными вирусами (Barba-Spaeth G. et al. 2016). Использование подобных антител в качестве терапевтических препаратов в случае флавивирусов, однако, может быть осложнено феноменом антитело-зависимого усиления инфекции (ADE) (Миронов А. Н., и др., 2013, Dejnirattisai W. et al. 2010, Dejnirattisai W. et al. 2016, Demina A.V. et al. 2018). В качестве примера можно рассмотреть лихорадки денге (DENV1-4) и Зика (ZIKV), которые являются флавивирусами, переносимыми комарами. Моноклональные антитела и плазма от DENV-иммунных доноров способны как нейтрализовать, так и усиливать ZIKV, и наоборот (Montoya M. et al. 2018, Fernandez E. et al. 2017). Поэтому в случае флавивирусов, создание эффективных и безопасных препаратов требует не только наличия антиген-распознающей области направленной на консервативный участок поверхностных белков, но специальных усилий по предотвращению возможного ADE-эффекта.
Ближайшие аналоги.
Известно получение (VanBlargan L. A. et al. Broadly neutralizing monoclonal antibodies protect against multiple tick-borne flaviviruses // Journal of Experimental Medicine. - 2021. - T. 218. - №. 5. - C. e20210174.) панели из 21 антитела против вируса Повассан. Панель антител получали при помощи инфицирования мышей POWV SPO или получавших мРНК-вакцина POWV, и в дальнейшем выделяли 84, гибридомы продуцирующие антитела против вируса Повассан, при помощи проточной цитометрии и скрининга на основе ИФА с инфицированными клетками и рекомбинантным оболочечным белком вируса Повассан. Однако при проверке на перекрестную реактивность этих антител было обнаружено, что антитела способны взаимодействовать с родственными флавивирусами, а также некоторые из них взаимодействуют с петлей слияния.
Известна разработка Yong-Qiang Deng с коллегами по получению человеческого антитела против вируса клещевого энцефалита с помощью метода гибдридомных технологий, а эпитоп охарактеризован при помощи фаговой пептидной библиотеки (Deng Y. Q. et al. A broadly flavivirus cross-neutralizing monoclonal antibody that recognizes a novel epitope within the fusion loop of E protein // PloS one. - 2011. - T. 6. - №. 1. - с. e16059.). В этом исследовании было установлено, что новое МКА, обозначенное как МКА 2A10G6, обладает широкой перекрестной реактивностью с DENV 1-4, YFV, WNV, JEV и TBEV. Широкая кроссреактивность достигалась за счет взаимодействия с эпитопом в высококонсервативном пептиде петли слияния флавивируса, мотиве 98DRXW101. Функциональные исследования показали, что 2A10G6 блокирует инфекцию на этапе после прикрепления вируса (Deng Y. Q. et al. 2011).
Наиболее близким аналогом (прототипом) является рекомбинантная плазмидная днк pclm4/hygro-14d5, кодирующая полипептид со свойствами легкой цепи химерного антитела против вируса клещевого энцефалита, и рекомбинантная плазмидная днк pchm2-14d5, кодирующая полипептид со свойствами тяжелой цепи химерного антитела против вируса клещевого энцефалита, а также химерное антитело, обеспечивающее экстренную профилактику клещевого энцефалита у мышей (патент РФ №2550252, МПК С 12N 15/63, опубл. 10.05.2015 г.). Рекомбинантная плазмидная ДНК pCLm4/hygro-14D5, кодирующая полипептид со свойствами легкой цепи химерного антитела против вируса клещевого энцефалита имеет размер 6232 п.н., молекулярную массу 4,11 МДа и фрагмент NheI/ApaI размером 741 п.н., содержащий искусственный ген, кодирующий гибридный белок, в котором вариабельный домен легкой цепи мышиного моноклонального антитела 14D5, осуществляющего протекцию против вируса клещевого энцефалита, соединен с константным доменом каппа-цепи антител человека. Рекомбинантная плазмидная ДНК pCHm2-14D5, кодирующая полипептид со свойствами тяжелой цепи химерного антитела против вируса клещевого энцефалита имеет размер 6765 п.н., молекулярную массу 4,47 МДа и фрагмент NheI/XbaI размером 1433 п.н., содержащий искусственный ген, кодирующий гибридный белок, в котором вариабельный домен тяжелой цепи мышиного моноклонального антитела 14D5, осуществляющего протекцию против вируса клещевого энцефалита, соединен с константными CH1-CH2-CH3-доменами IgG1 человека. Химерное антитело, способное связывать белок E вируса клещевого энцефалита и обеспечивающее экстренную профилактику клещевого энцефалита у мышей, включающее полипептиды со свойствами легкой и тяжелой цепей химерного антитела класса IgG1/каппа, полученное совместной трансфекцией клеток млекопитающих рекомбинантными плазмидными ДНК pCLm4/hygro-14D5 и pCHm2-14D5. Константа аффинности химерного антитела к рекомбинантному гликопротеину E составляет около 6,0⋅10-11 M. Полученное химерное антитело ch14D5, введенное в дозировке 1 мг на кг веса животного, обеспечивает экстренную профилактику мышей от вирусного клещевого энцефалита, что делает возможным использование химерного антитела ch14D5 в качестве основы для создания лекарственных препаратов для лечения вирусного клещевого энцефалита.
Однако в прототипе не исследована функциональная активность химерного антитела ch14D5 в отношении других флавивирусов (Зика, денге, Западного Нила, желтой лихорадки).
Техническим результатом заявляемого изобретения является получение химерного полноразмерного человеческого антитела 10H10ch против вируса клещевого энцефалита, продуцируемого рекомбинантным штаммом клеточной линии яичника китайского хомячка CHO-K1-10H10ch, и обладающего широкой перекрестной функциональной активностью в отношении вирусов клещевого энцефалита, Зика, денге, Западного Нила, желтой лихорадки.
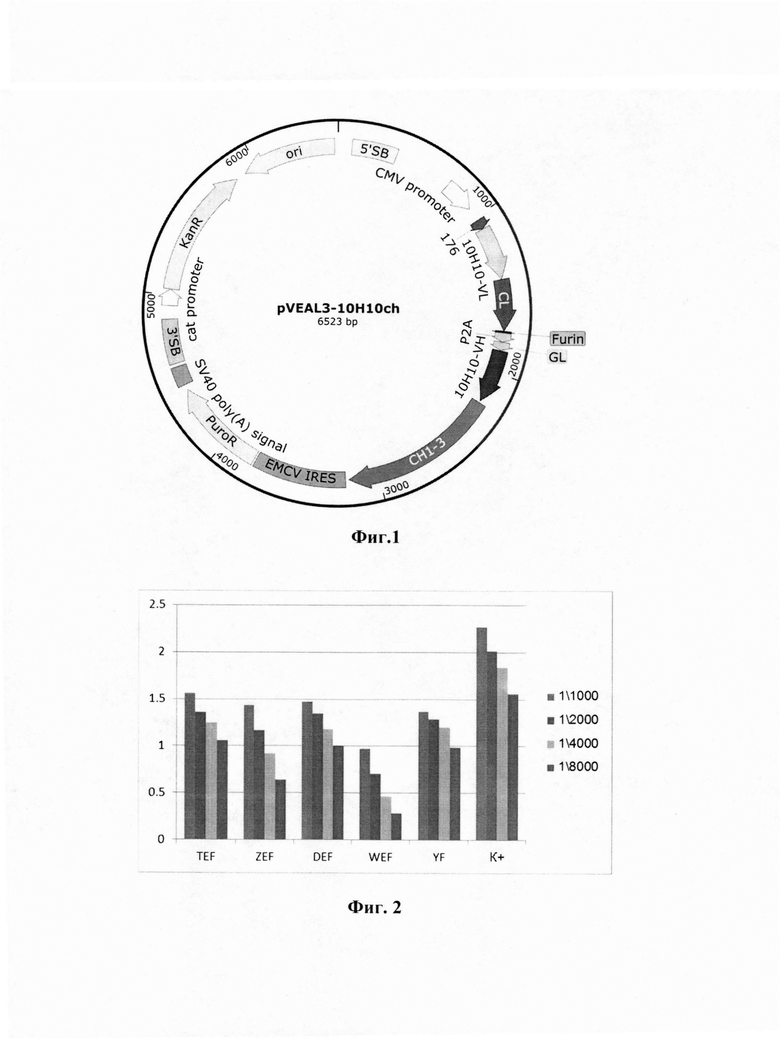
Технический результат достигается созданием интегративного плазмидного вектора pVEAL3-10H10ch, обеспечивающего экспрессию и секрецию антитела 10Н10 против вируса клещевого энцефалита в клетках млекопитающих, имеющего нуклеотидную последовательность SEQ ID NO:1, размер 6523 п.н. и содержащего в соответствии с физической и генетической картой, представленной на фиг. 1, следующие элементы:
- 5'SB с координатами 88-368 п.н. и 3'SB с координатами 4573-4850 п.н. - сайт связывания транспозазы SB100;
- CMV promoter, имеющего координаты с 734 по 937 п.н., - область промотора CMV;
- 176 - нуклеотидная последовательность, кодирующая гибридный лидерный пептид люциферазы (Cypridina noctiluca) и фиброина (Dendrolimus spectabilis), обеспечивающая экспорт белка из клетки и имеющая координаты с 1023 по 1085 п.н.;
- 10H10-VL - нуклеотидная последовательность, кодирующая вариабельный фрагмент легкой цепи антитела 10Н10 и имеющая координаты с 1086 по 1421 п.н.;
- CL - нуклеотидная последовательность, кодирующая константную часть легкой цепи антитела и имеющая координаты с 1422 по 1742 п.н.;
- Furin - нуклеотидная последовательность, кодирующая сайт протеолиза клеточной протеазы фурина и имеющая координаты с 1743 по 1754 п.н.;
- Р2А - нуклеотидная последовательность, кодирующая саморасщепляющийся пептид р2а и имеющая координаты с 1755 по 1811 пл.;
- GL - нуклеотидная последовательность, кодирующая лидерную последовательность gaussia luciferase и имеющая координаты с 1812 по 1862 п.н.;
- 10H10-VH - нуклеотидная последовательность, кодирующая вариабельный фрагмент тяжелой цепи антитела 10Н10 и имеющая координаты с 1866 по 2201 п.н.;
- СН1-3 - нуклеотидная последовательность, кодирующая константную цепь антитела человека IgG1 (СН1-СН2-СН3) и имеющая координаты с 2215 по 3191 п.н.;
- EMCV IRES - участок внутренней посадки рибосомы, имеющий координаты с 3210 по 3784 п.н.;
- PuroR - нуклеотидная последовательность, кодирующая фактор устойчивости к антибиотику пуромицину и имеющая координаты с 3797 по 4396 п.н.;
- SV40 poly (A) signal - последовательность для стабилизации мРНК-транскриптов за счет полиаденилирования, имеющая координаты с 4431 по 4552 п.н.;
- cat promoter - синтетический бактериальный промотер, имеющий координаты с 4934-5036 п.н.;
- KanR - нуклеотидная последовательность, кодирующая фактор устойчивости к антибиотику канамицину и имеющая координаты с 5037 по 5852 п.н.;
- ori - участок начала репликации, имеющий координаты с 5915 по 6502 п.н.
Интегративный плазмидный вектор pVEAL3-10H10ch позволяет осуществить высокую и стабильную экспрессию химерного рекомбинантного полноразмерного антитела.
Технический результат достигается также созданием рекомбинантного штамма клеточной линии яичника китайского хомячка CHO-K1-10H10ch продуцента рекомбинантного химерного антитела 10H10ch против вируса клещевого энцефалита, содержащего интегративный плазмидный вектор pVEAL3-10H10ch, имеющий нуклеотидную последовательность SEQ ID NO: 1, обеспечивающий экспрессию и секрецию полноразмерного человеческого антитела 10H10ch и депонированный под номером 325 в коллекции культур клеток ФБУН ГНЦ ВБ «Вектор» Роспотребнадзора.
Указанный технический результат достигается также получением химерного антитела 10H10ch против вируса клещевого энцефалита, продуцируемого рекомбинантным штаммом клеточной линии яичника китайского хомячка CHO-K1-10H10ch по п. 2, имеющего аминокислотные последовательности вариабельных фрагментов тяжелой цепи SEQ ID NO: 2 и легкой цепи SEQ ID NO: 3, и предназначенного для создания терапевтических препаратов против вируса клещевого энцефалита.
Изобретение поясняется графическими материалами, представленными на фиг. 1, 2. На фиг. 1 изображена физическая и генетическая карта универсального рекомбинантного вектора pVEAL3-10H10ch. На фиг. 2. представлены результаты иммуноферментного анализа взаимодействия моноклонального химерного антитела 10H10ch с рекомбинантным фрагментом белка вируса клещевого энцефалита.
Для лучшего понимания сущности предлагаемого изобретения ниже приведены примеры его осуществления. Все стандартные генно-инженерные и микробиологические манипуляции, а также амплификацию и секвенирование ДНК проводили по известным методикам (Маниатис Т., и др., 1984, Гловер Д. 1988, Saiki R.K. et al. 1988, Sanger F. et al. 1977).
Пример 1. Конструирование рекомбинантной плазмиды pVEAL3-10H10ch, обеспечивающей синтез и секрецию химерного антитела 10Н10 против вируса клещевого энцефалита.
На основе вектора pVEAL3 получали интеграционный вектор pVEAL3-10H10ch (фиг. 1), содержащий последовательности вариабельных легкой (SEQ ID NO: 3) и тяжелой (SEQ ID NO: 2) цепи антитела 10H10ch. Последовательность гена, кодирующего тяжелую цепь 10H10ch (GenBank OK483332) и последовательность гена, кодирующего легкую цепь 10H10ch (GenBank OL448869) извлекали из базы данных GenBank и проводили оптимизацию кодонного состава для клеток СНО при помощи инструмента Gene Optimizer (https://www.thermofisher.com/ru/en/home/life-science/cloning/gene-synthesis/geneart-gene-synthesis/geneoptimizer.html).
Итоговая нуклеотидная последовательность (SEQ ID NO:1) содержит последовательности, кодирующие сигнальный пептид 176, константную часть легкой цепи антитела, сайт протеолиза клеточной протеазы фурина, саморасщепляющийся пептид р2а, лидерную последовательность gaussia luciferase, и константную часть тяжелой цепи антитела (СН1), а также сайты гидролиза рестриктаз NheI и BstX I и синтезировали на заказ в ООО «ДНК-синтез».

Для получения рекомбинантной плазмиды pVEAL3-10H10ch в вектор pVEAL3 встраивали синтезированную нуклеотидную последовательность 10H10ch. Последовательности олигонуклеотидных праймеров, использованных для сборки плазмиды pVEAL3-10H10ch, представлены в таблице 1.
С помощью ПЦР была амплифицирована нуклеотидная последовательность 10H10ch. ПЦР проводили с использованием ПЦР-амплификатора «БИС» фирмы ООО БИС-Н (Россия). Реакционная смесь объемом 50 мкл, содержащая 2 мкг ДНК (плазмида pGH-10H10), 10 пкМ каждого праймера (10H10-pVL3_F и 10H10-BstXI_R) (таблица 1), 10 мкл 5х Q5 реакционного буфера, 10 мкл 5xQ5 High GC Enhancer, смесь дезоксинуклеотидтрифосфатов (дАТФ, дЦТФ, дТТФ, дГТФ по 2,5 мМ) и 0.5 ед. Q5 High-Fidelity ДНК-полимеразы, реакцию осуществляли при следующих параметрах: 10 с - 98°С, 20 с - 61°С, 30 с - 72°С (30 циклов). Готовые ПЦР-продукты были выделены и очищены из геля с помощью набора «Gel Extraction Kit» фирмы Qiagen (Германия) в соответствии с инструкцией производителя.
Предварительно наработанную и очищенную плазмиду pVEAL3 и готовый ПЦР-продукт обрабатывали эндонуклеазами рестрикции Nhe I, BstX I. Реакцию гидролиза проводили в условиях, рекомендованных производителем. С целью очистки линеаризованного вектора, плазмидную ДНК наносили на 1%-й агарозный гель и выделяли из геля с использованием набора «Gel Extraction Kit» фирмы Qiagen (Германия). Реакцию лигирования проводили с использованием ДНК-лигазы бактериофага Т4 («СибЭнзим», г. Новосибирск). Реакция проводилась при +4°С в течение ночи. Полученной лигазной смесью трансформировали компетентные клетки Е. coli штамм NebStable.
Первичную проверку на наличие вставки проводили при помощи ПЦР с колонии. Разделение продуктов амплификации проводили в 1%-м агарозном геле с последующим окрашиванием бромистым этидием (0,5 мкг/мл).
Положительные колонии, вносили в 5 мл среды LB с ампициллином (50 мкг/мл) и растили в течение ночи при 37°С при 170 об/мин. Затем плазмидную ДНК выделяли из бактериальных клеток с помощью коммерческих наборов DNAminikit фирмы «Qiagen» согласно рекомендациям производителя. Первичную структуру экспрессионного вектора подтверждали секвенированием. Секвенирование проводили по методу Сэнгера в ЦКП «Геномика» СО РАН (г. Новосибирск). В результате была получена рекомбинантная плазмида pVEAL3-10H10ch, обеспечивающей синтез химерного антитела 10Н10 против вируса клещевого энцефалита в одноцепочечном формате.
Секвенирование плазмидной ДНК положительных клонов в районе встройки позволило отобрать клоны с отсутствием дефектов встраиваемых генов (вставки, делеции, замены), после чего из отобранных клонов была наработана и выделена целевая плазмидная ДНК.
Пример 2. Трансфекция клеток СНО-K1 плазмидой pVEAL3-10H10ch и получение штамма-продуцента клеток яичника китайского хомячка CHO-K1-10H10ch, продуцирующего химерное антитело 10H10ch против вируса клещевого энцефалита
Штамм клеток яичника китайского хомячка CHO-K1-10H10ch получен на основе клеточной линии яичников китайского хомячка СНО-K1 с использованием разработанной конструкции pVEAL3-10H10ch. Клетки СНО-K1, растили в инкубаторе при 5% содержания СО2, 80%-ной влажности. При достижении 80% плотности монослоя проводили трансфекцию клеток с помощью Lipofectamine 3000 (ThermoFisher, США) в соответствии с инструкцией производителя. Трансфецировали клетки смесью плазмид pVEAL3-10H10ch и pCMV(CAT)T7-SB100, взятых в соотношении 1:10. Через 48 часов после трансфекции добавляли селективный антибиотик пуромицин в концентрации 2 мкг/мл. В течении 3-х дней наблюдалось активное отмирание основной массы клеток. На четвертый день клеточный пул был перенесен в 6-луночный культуральный планшет, так же с добавление селективного антибиотика. Через 2 дня достигался монослой. Полученный пул клеток был криоконсервирован, после чего было проведено клонирование с целью отбора индивидуальных клонов с высокой продукцией рекомбинантных антител. Клонирование выполнялось методом лимитирующих разведений в 96-луночных планшетах. Через 14 суток визуально отбирали лунки, содержащие единственный клон, для которых проводили отбор культуральной жидкости с целью оценки продуктивности клонов при помощи ИФА, из которых и получен заявляемый рекомбинантный штамм CHO-K1-10H10ch.
Штамм депонирован 02.09.2022 г. в коллекции ФБУН ГНЦ ВБ «Вектор» Роспотребнадзора под номером 325 (копия справки о депонировании прилагается).
Характеристика рекомбинантного штамма CHO-K1-10H10ch.
Морфология: веретеновидные и эпителиоподобные клетки с круглыми ядрами, содержащими от 1 до 2 ядрышек.
Способ культивирования: монослойный.
Среда для культивирования: питательная среда DMEM/F-12 (1:1) - 90%, сыворотка крови плодов коровы - 10%.
Температура культивирования: 37°С.
Посевная концентрация: 100 тыс. клеток в 1 мл.
Метод снятия: 0,25% трипсин (1/3) и 0,02% версен (2/3).
Кратность рассева: 1:3.
Частота пассирования: 3-4 суток.
Условия криоконсервации: питательная среда DMEM/F-12 (1:1) - 50%, сыворотка крови плодов коровы - 40%, ДМСО - 10%.
Режим замораживания: при температуре 4°С - 1 ч, минус 80°С - 12 ч, минус 196°С.
Условия хранения: в криопробирках в количестве 5 шт. хранится в жидком азоте при температуре минус 196°С.
Номер пассажа в жидком азоте: 3.
Жизнеспособность после криоконсервации: 85-90%.
Маркерый признак: наличие в культуральной среде рекомбинантного антитела 10Н10 размером ~55 кДа, подтверждается с помощью белкового электрофореза и иммуноблотинга. Область применения: биотехнология.
Пример 3. Иммуноферментный анализ взаимодействия моноклонального химерного антитела 10H10ch с рекомбинантными фрагментами оболочечных белков флавивирусов
В качестве антигена использовали рекомбинантные фрагменты оболочечных белков вирусов клещевого энцефалита, Зика, денге, Западного Нила, желтой лихорадки (TEF, ZEF, DEF, WEF, YF соответственно) в концентрации 200 нг/лунка, его сорбировали в лунках 96-луночного планшета в фосфатно-солевом буфере (PBS) (Greiner bio one, Германия) при 4°С в течение ночи. Затем отмывали буфером PBST (0.1% р-р Tween-20 в PBS) и блокировали 1% раствором казеина в буфере PBST в течение 1 часа при комнатной температуре. После этого образцы вносили в последовательном разведении, начиная с 1/1000 до 1/8000, и инкубировали в течение 1 часа при 37°С. Затем добавляли для рекомбинантного химерного антитела 10Н10 - конъюгат (Goat a Hum IgG-per Sigma - 1/5000), для нативного антитела (положительного контроля) - конъюгат (Goat a mouse IgG-per Sigma - 1/5000) разбавленные в 1% блокирующем растворе казеина, с пероксидазой хрена (США). После каждого этапа проводили трехкратную отмывку несвязавшихся белков буфером PBST 0,1%. Затем вносили раствор субстрата ТМВ (Amresco, США) и инкубировали в течение 10 минут, реакцию терминировали раствором 1Н HCl и сразу измеряли оптическую плотность при длине волны 450 нм на ИФА-ридере (Varioska LUX, Thermo Scientific, США).
Результаты ИФА представлены на фиг. 2, из которых видно, что новый штамм-продуцент клеток яичника китайского хомячка CHO-K1-10H10ch, продуцирует химерное полноразмерное человеческое антитело 10Н10 против вируса клещевого энцефалита, которое обладает широкой функциональной перекрестной активностью (в отношении вирусов клещевого энцефалита, Зика, денге, Западного Нила, желтой лихорадки).
Источники научно-технической и патентной информации
1. Гловер Д. Клонирование ДНК. Методы. - 1988.
2. Маниатис Т., Фрич Э., Сэмбрук Д. Молекулярное клонирование: Пер. с англ. - Мир, 1984.
3. Миронов А.Н., Супотницкий М.В., Лебединская Е.В. Феномен антитело-зависимого усиления инфекции у вакцинированных и переболевших // БИОпрепараты. Профилактика, диагностика, лечение. - 2013. - №. 3 (47). - С. 12-25.
4. Asano R. et al. Comprehensive study of domain rearrangements of single-chain bispecific antibodies to determine the best combination of configurations and microbial host cells // MAbs. - Taylor & Francis, 2018. - T. 10. - №. 6. - C. 854-863.
5. Asano R. et al. Construction and humanization of a functional bispecific EGFR× CD16 diabody using a refolding system // The FEBS Journal. -2012. - T. 279. - №. 2. - C. 223-233.
6. Aubry F. et al. Recent African strains of Zika virus display higher transmissibility and fetal pathogenicity than Asian strains //Nature communications. -2021.-Т. 12.-№. 1.-C. 1-14.
7. Barba-Spaeth G. et al. Structural basis of potent Zika-dengue virus antibody cross-neutralization // Nature. - 2016. - T. 536. - №. 7614. - C. 48-53.
8. Basarab M. et al. Zika virus // Bmj. - 2016. - T. 352.
9. Baykov I.K. et al. A protective chimeric antibody to tick-borne encephalitis virus // Vaccine. - 2014. - T. 32. - №. 29. - C. 3589-3594.
10. Begum F. et al. Insight into the tropism of dengue virus in humans //Viruses. - 2019.-Т. 11. - №. 12. - С.1136.
11. Buchacher A. et al. Generation of human monoclonal antibodies against HIV-1 proteins; electrofusion and Epstein-Barr virus transformation for peripheral blood lymphocyte immortalization // AIDS research and human retroviruses. - 1994. - T. 10. -№. 4. - C. 359-369.
12. Busse W. W. et al. Safety profile, pharmacokinetics, and biologic activity of MEDI-563, an anti-IL-5 receptor α antibody, in a phase I study of subjects with mild asthma // Journal of Allergy and Clinical Immunology. - 2010. - T. 125. - №. 6. - C. 1237-1244. e2.
13. Carlring J., De Leenheer E., Heath A. W. A novel redox method for rapid production of functional bi-specific antibodies for use in early pilot studies // PLoS One. - 2011. - T. 6. - №. 7. - C. e22533.
14. Choudhry A., O'Brien S.M. Inotuzumab ozogamicin for the treatment of patients with acute lymphocytic leukemia // Drugs of Today (Barcelona, Spain: 1998). - 2017. - T. 53. - №. 12. - C. 653-665.
15. de Claro R.A. et al. US Food and Drug Administration Approval Summary: Brentuximab Vedotin for the Treatment of Relapsed Hodgkin Lymphoma or Relapsed Systemic Anaplastic Large-Cell LymphomaBrentuximab Vedotin for HL and sALCL // Clinical Cancer Research. - 2012. - T. 18. - №. 21. - C. 5845-5849.
16. Dejnirattisai W. et al. Cross-reacting antibodies enhance dengue virus infection in humans // Science. - 2010. - T. 328. - №. 5979. - C. 745-748.
17. Dejnirattisai W. et al. Dengue virus sero-cross-reactivity drives antibody-dependent enhancement of infection with zika virus //Nature immunology. - 2016. - Т. 17. - №. 9. - С.1102-1108.
18. Demina A.V. et al. TBE vaccine and post TBE disease Abs drive antibody dependent enhancement of Zika infection. - 2018.
19. Deng Y.Q. et al. A broadly flavivirus cross-neutralizing monoclonal antibody that recognizes a novel epitope within the fusion loop of E protein // PloS one. - 2011. - T. 6. - №. 1. - C. e16059.
20. Durbin A., Wilder-Smith A. An update on Zika vaccine developments // Expert review of vaccines. - 2017. - T. 16. - №. 8. - C. 781-787.
21. Fernandez E. et al. Human antibodies to the dengue virus E-dimer epitope have therapeutic activity against Zika virus infection // Nature immunology. - 2017. - T. 18. - №. 11. - C. 1261-1269.
22. Ferrara N. et al. Development of ranibizumab, an anti-vascular endothelial growth factor antigen binding fragment, as therapy for neovascular age-related macular degeneration //Retina. - 2006. - T. 26. - №. 8. - C. 859-870.
23. Fox J. P. et al. Additional observations on the duration of humoral immunity following vaccination with the 17D strain of yellow fever virus //American journal of hygiene. - 1948. - T. 47. -№. 1. - C. 64-70.
24. Ishida T. et al. Defucosylated anti-CCR4 monoclonal antibody (KW-0761) for relapsed adult T-cell leukemia-lymphoma: a multicenter phase II study // J Clin Oncol. - 2012. - T. 30. - №. 8. - C. 837-842.
25. Jen E.Y. et al. FDA Approval: Gemtuzumab Ozogamicin for the Treatment of Adults with Newly Diagnosed CD33-Positive Acute Myeloid LeukemiaFDA Approval Summary: Gemtuzumab Ozogamicin // Clinical cancer research. - 2018. - T. 24. - №. 14. - C. 3242-3246.
26. Kyle J. L., Harris E. Global spread and persistence of dengue //Annual review of microbiology. - 2008. - T. 62. - №. 1. - C. 71-92.
27. Li J. et al. IFNγ-induced Chemokines Are Required for CXCR3-mediated T-Cell Recruitment and Antitumor Efficacy of Anti-HER2/CD3 Bispecific AntibodyT-Cell Recruitment By Anti-HER2/CD3 // Clinical Cancer Research. - 2018. - T. 24. - №. 24. - C. 6447-6458.
28. Monath T. P. Yellow fever vaccine //Expert review of vaccines. -2005. - T. 4. - №. 4. - C. 553-574.
29. Montefiori D. C. et al. Neutralizing and infection-enhancing antibody responses to human immunodeficiency virus type 1 in long-term nonprogressors //The Journal of infectious diseases. - 1996. - Т. 173. - №. 1.-C. 60-67.
30. Montoya M. et al. Longitudinal analysis of antibody cross-neutralization following Zika virus and dengue virus infection in Asia and the Americas // The Journal of infectious diseases. - 2018. - T. 218. - №. 4. - C. 536-545.
31. Muster T. et al. A conserved neutralizing epitope on gp41 of human immunodeficiency virus type 1 // Journal of virology. - 1993. - T. 67. - №. 11.-C. 6642-6647.
32. Neufeldt C. J. et al. Rewiring cellular networks by members of the Flaviviridae family // Nature Reviews Microbiology. - 2018. - T. 16. - №. 3. - C. 125-142.
33. Oldenburg J. et al. Emicizumab prophylaxis in hemophilia A with inhibitors // New England Journal of Medicine. - 2017. - T. 377. - №. 9. - C. 809-818.
34. Priyamvada L. et al. Human antibody responses after dengue virus infection are highly cross-reactive to Zika virus // Proceedings of the National Academy of Sciences.-2016.-T. 113.-№. 28. - C. 7852-7857.
35. Saiki R.K. et al. Primer-directed enzymatic amplification of DNA with a thermostable DNA polymerase // Science. - 1988. - T. 239. - №. 4839. - C. 487-491.
36. Sanger F., Nicklen S., Coulson A. R. DNA sequencing with chain-terminating inhibitors // Proceedings of the national academy of sciences. - 1977. -T. 74.-№. 12.-C. 5463-5467.
37. Satta A. et al. Design, selection and optimization of an anti-TRAIL-R2/anti-CD3 bispecific antibody able to educate T cells to recognize and destroy cancer cells // MAbs. - Taylor & Francis, 2018. - T. 10. - №. 7. - C. 1084-1097.
38. Smith I. et al. 2-year follow-up of trastuzumab after adjuvant chemotherapy in HER2-positive breast cancer: a randomised controlled trial //The lancet. - 2007. - T. 369. - №. 9555. - C. 29-36.
39. VanBlargan L. A. et al. Broadly neutralizing monoclonal antibodies protect against multiple tick-borne flaviviruses // Journal of Experimental Medicine. - 2021. - T. 218. - №. 5. - С.e20210174.
40. Walker L. M., Burton D. R. Passive immunotherapy of viral infections:'super-antibodies' enter the fray // Nature Reviews Immunology. - 2018. -Т. 18.-№. 5.-C. 297-308.
41. Yamamoto K. et al. Phase I study of KW-0761, a defucosylated humanized anti-CCR4 antibody, in relapsed patients with adult T-cell leukemia-lymphoma and peripheral T-cell lymphoma //Journal of Clinical Oncology. - 2010. -Т. 28.-№. 9.-C. 1591-1598.
42. Yamane-Ohnuki N. et al. Establishment of FUT8 knockout Chinese hamster ovary cells: an ideal host cell line for producing completely defucosylated antibodies with enhanced antibody-dependent cellular cytotoxicity //Biotechnology and bioengineering. - 2004. - T. 87. - №. 5. - C. 614-622.
43. Yu L. et al. Delineating antibody recognition against Zika virus during natural infection // JCI insight. - 2017. - T. 2. - №. 12.
44. Патент РФ №2550252, МПК C12N 15/63, опубл. 10.05.2015 г. (прототип).
Seq id no: 1
cgaagaaaggcccacccgtgaaggtgagccagtgagttgattgcagtccagttacgctggagtctgaggctcgtcctgaatggatccctatacagttgaagtcggaagtttacatacacttaagttggagtcattaaaactcgtttttcaactactccacaaatttcttgttaacaaacaatagttttggcaagtcagttaggacatctactttgtgcatgacacaagtcatttttccaacaattgtttacagacagattatttcacttataattcactgtatcacaattccagtgggtcagaagtttacatacactaagttcgactcctctgcagaatgcggcgatgtttcggtaaggggtccgctactagttattaatagtaatcaattacggggtcattagttcatagcccatatatggagttccgcgttacataacttacggtaaatggcccgcctggctgaccgcccaacgacccccgcccattgacgtcaataatgacgtatgttcccatagtaacgccaatagggactttccattgacgtcaatgggtggagtatttacggtaaactgcccacttggcagtacatcaagtgtatcatatgccaagtacgccccctattgacgtcaatgacggtaaatggcccgcctggcattatgcccagtacatgaccttatgggactttcctacttggcagtacatctacgtattagtcatcgctattaccatggtgatgcggttttggcagtacatcaatgggcgtggatagcggtttgactcacggggatttccaagtctccaccccattgacgtcaatgggagtttgttttggcaccaaaatcaacgggactttccaaaatgtcgtaacaactccgccccattgacgcaaatgggcggtaggcgtgtacggtgggaggtctatataagcagagctctctggctaactagagaacccactgcttactggcttatcgaaattaatacgactcactatagggagacccaagctggctagcaccatgatgcggaccctgatcctggctgtgctgctggtgtacttctgtgccaccgtgcactgctccgacattgtgctgacccagactccactcactttgtcggttaccattggacagccagcctccatctcttgcaagtcaagtcagagcctcttagatagtgatggaaagacatatttgaattggttgttacagaggccaggccagtctccaaagcgcctaatctatctggtgtctaaattggactctggagtccctgacaggttcactggcggtggatcagggacagatttcacactgaaaatcagcagagtggaggctgaggatttgggagtttattattgctggcaaggtacacattttcctcagacgttcggtggaggcaccaagctggaaatcaaacgtacggtggctgcaccatctgtcttcatcttcccgccatctgatgagcagttgaaatctggaactgcctctgttgtgtgcctgctgaataacttctatcccagagaggccaaagtacagtggaaggtggataacgccctccaatcgggtaactcccaggagagtgtcacagagcaggacagcaaggacagcacctacagcctcagcagcaccctgacgctgagcaaagcagactacgagaaacacaaagtctacgcctgcgaagtcacccatcagggcctgagctcgcccgtcacaaagagcttcaacaggggagagtgtagacggaagagagccaccaactttagtctgctgaagcaggccggcgacgtggaagagaatcctggacctatgggagtgaaggtgctgttcgccctgatctgtattgccgtggccgaagcttctgtgaagctggaggagtctgggggaggcttagtgaagcctggagggtccctgaaactctcctgtgcagcctctggattcagtttcagtgactattacatgtattgggttcgtcagactccggaaaagaggctggagtgggtcgcaaccattagtgatggtggtagtcacacctcctatcgagacagtgtgaaggggcgatttaccatctccagagacaatggcaagaacaacctgtacctgcaaatgagcagtctgaagtctgaggacacagccatgtattactgtgtaagaggtgcttactggggccaagggactctggtcactgtctcttctgccagcaccaaggggccaagcgtcttccccctggcaccctcctccaagagcacctctgggggcacagcggccctgggctgcctggtcaaggactacttccccgaaccggtgacggtgtcgtggaactcaggcgccctgaccagcggcgtgcacaccttcccggctgtcctacagtcctcaggactctactccctcagcagcgtggtgaccgtgccctccagcagcttgggcacccagacctacatctgcaacgtgaatcacaagcccagcaacaccaaggtggacaagaaagttgagcccaaatcttgtgacaaaactcacacatgcccaccgtgcccagcacctgaactcctggggggaccgtcagtcttcctcttccccccaaaacccaaggacaccctctacatcacccgggaacctgaggtcacatgcgtggtggtggacgtgagccacgaagaccctgaggtcaagttcaactggtacgtggacggcgtggaggtgcataatgccaagacaaagccgcgggaggagcagtacaacagcacgtaccgtgtggtcagcgtcctcaccgtcctgcaccaggactggctgaatggcaaggagtacaagtgcaaggtctccaacaaagccctcccagcccccatcgcgaaaaccatctccaaagccaaagggcagccccgagaaccacaggtgtacaccctgcccccatcccgggaggagatgaccaagaaccaggtcagcctgacctgcctggtcaaaggcttctatcccagcgacatcgccgtggagtgggagagcaatgggcagccggagaacaactacaagaccacgcctcccgtgctggactccgacggctccttcttcctctacagcaagctcaccgtggacaagagcaggtggcagcaggggaacgtcttctcatgctccgtgatgcatgaggctctgaagttccactacacgcagaagagcctctccctgtctccgggtaaatgagtcgaccgagcggttcccgcccctctccctcccccccccctaacgttactggccgaagccgcttggaataaggccggtgtgcgtttgtctatatgttattttccaccatattgccgtcttttggcaatgtgagggcccggaaacctggccctgtcttcttgacgagcattcctaggggtctttcccctctcgccaaaggaatgcaaggtctgttgaatgtcgtgaaggaagcagttcctctggaagcttcttgaagacaaacaacgtctgtagcgaccctttgcaggcagcggaaccccccacctggcgacaggtgcctctgcggccaaaagccacgtgtataagatacacctgcaaaggcggcacaaccccagtgccacgttgtgagttggatagttgtggaaagagtcaaatggctcacctcaagcgtattcaacaaggggctgaaggatgcccagaaggtaccccattgtatgggatctgatctggggcctcggtgcacatgctttacatgtgtttagtcgaggttaaaaaacgtctaggccccccgaaccacggggacgtggttttcctttgaaaaacacgatgataatatggccacaaccatgaccgagtacaagcccacggtgcgcctcgccacccgcgacgacgtccccagggccgtacgcaccctcgccgccgcgttcgccgactaccccgccacgcgccacaccgtcgatccggaccgccacatcgagcgggtcaccgagctgcaagaactcttcctcacgcgcgtcgggctcgacatcggcaaggtgtgggtcgcggacgacggcgccgcggtggcggtctggaccacgccggagagcgtcgaagcgggggcggtgttcgccgagatcggcccgcgcatggccgagttgagcggttcccggctggccgcgcagcaacagatggaaggcctcctggcgccgcaccggcccaaggagcccgcgtggttcctggccaccgtcggcgtctcgcccgaccaccagggcaagggtctgggcagcgccgtcgtgctccccggagtggaggcggccgagcgcgccggggtgcccgccttcctggagacctccgcgccccgcaacctccccttctacgagcggctcggcttcaccgtcaccgccgacgtcgaggtgcccgaaggaccgcgcacctggtgcatgacccgcaagcccggtgcctgattcgcatatgggttaatgcttcgagcagacatgataagatacattgatgagtttggacaaaccacaactagaatgcagtgaaaaaaatgctttatttgtgaaatttgtgatgctattgctttatttgtaaccattataagctgcaataaacaagttcctcgacctctagctagagctactcgggaccccttaccgaaacatcgccgcattctgcagaggagtcgagtgtatgtaaacttctgacccactgggaatgtgatgaaagaaataaaagctgaaatgaatcattctctctactattattctgatatttcacattcttaaaataaagtggtgatcctaactgacctaagacagggaatttttactaggattaaatgtcaggaattgtgaaaaagtgagtttaaatgtatttggctaaggtgtatgtaaacttccgacttcaactgtatagggatccgctcaatactgaccatttaaatcatacctgacctccatagcagaaagtcaaaagcctccgaccggaggcttttgacttgatcggcacgtaagaggttccaactttcaccataatgaaataagatcactaccgggcgtattttttgagttatcgagattttcaggagctaaggaagctaaaatgagccatattcaacgggaaacgtcttgctcgaggccgcgattaaattccaacatggatgctgatttatatgggtataaatgggctcgcgataatgtcgggcaatcaggtgcgacaatctatcgattgtatgggaagcccgatgcgccagagttgtttctgaaacatggcaaaggtagcgttgccaatgatgttacagatgagatggtcaggctaaactggctgacggaatttatgcctcttccgaccatcaagcattttatccgtactcctgatgatgcatggttactcaccactgcgatcccagggaaaacagcattccaggtattagaagaatatcctgattcaggtgaaaatattgttgatgcgctggcagtgttcctgcgccggttgcattcgattcctgtttgtaattgtccttttaacggcgatcgcgtatttcgtctggctcaggcgcaatcacgaatgaataacggtttggttggtgcgagtgattttgatgacgagcgtaatggctggcctgttgaacaagtctggaaagaaatgcataagcttttgccattctcaccggattcagtcgtcactcatggtgatttctcacttgataaccttatttttgacgaggggaaattaataggttgtattgatgttggacgagtcggaatcgcagaccgataccaggatcttgccatcctatggaactgcctcggtgagttttctccttcattacagaaacggctttttcaaaaatatggtattgataatcctgatatgaataaattgcagtttcacttgatgctcgatgagtttttctaatgagggcccaaatgtaatcacctggctcaccttcgggtgggcctttctgcgttgctggcgtttttccataggctccgcccccctgacgagcatcacaaaaatcgatgctcaagtcagaggtggcgaaacccgacaggactataaagataccaggcgtttccccctggaagctccctcgtgcgctctcctgttccgaccctgccgcttaccggatacctgtccgcctttctcccttcgggaagcgtggcgctttctcatagctcacgctgtaggtatctcagttcggtgtaggtcgttcgctccaagctgggctgtgtgcacgaaccccccgttcagcccgaccgctgcgccttatccggtaactatcgtcttgagtccaacccggtaagacacgacttatcgccactggcagcagccactggtaacaggattagcagagcgaggtatgtaggcggtgctacagagttcttgaagtggtggcctaactacggctacactagaagaacagtatttggtatctgcgctctgctgaagccagttacctcggaaaaagagttggtagctcttgatccggcaaacaaaccaccgctggtagcggtggtttttttgtttgcaagcagcagattacgcgcagaaaaaaaggatctcaagaagatcctttgattttctac
Seq id no: 2
VKLEESGGGLVKPGGSLKLSCAASGFSFSDYYMYWVRQTPEKRLEWVATISDGGSHTSYRDSVKGRFTISRDNGKNNLYLQMSSLKSEDTAMYYCVRGAYWGQGTLVTVSSA
Seq id no: 3
DIVLTQTPLTLSVTIGQPASISCKSSQSLLDSDGKTYLNWLLQRPGQSPKRLIYLVSKLDSGVPDRFTGGGSGTDFTLKISRVEAEDLGVYYCWQGTHFPQTFGGGTKLEIK
Изобретение относится к биотехнологии. Описана плазмида pVEAL3-10H10ch, штамм рекомбинантной клеточной линии CHO-K1-10H10ch и химерное антитело 10H10ch против вируса клещевого энцефалита. Создан интегративный плазмидный вектор pVEAL3-10H10ch, обеспечивающий экспрессию и секрецию антитела 10Н10 против вируса клещевого энцефалита в клетках млекопитающих. Вектор имеет нуклеотидную последовательность SEQ ID NO: 1, размер 6523 п.н. и содержит элементы в соответствии с физической и генетической картой, представленной на фиг. 1. Химерное антитело 10H10ch против вируса клещевого энцефалита имеет аминокислотные последовательности вариабельных фрагментов тяжелой цепи SEQ ID NO: 2 и легкой цепи SEQ ID NO: 3 и предназначено для создания терапевтических препаратов против вируса клещевого энцефалита. При этом антитело обладает широкой перекрестной активностью в отношении других флавивирусов. Изобретение может быть использовано в генной инженерии, биотехнологии и медицине. Техническим результатом заявляемого изобретения является получение химерного полноразмерного человеческого антитела 10H10ch против вируса клещевого энцефалита, продуцируемого рекомбинантным штаммом клеточной линии яичника китайского хомячка CHO-K1-10H10ch и обладающего широкой перекрестной функциональной активностью в отношении вирусов клещевого энцефалита, Зика, денге, Западного Нила, желтой лихорадки. 3 н.п. ф-лы, 2 ил., 1 табл., 3 пр.
1. Плазмидная генетическая конструкция pVEAL3-10H10ch, обеспечивающая экспрессию и секрецию антитела 10H10ch против вируса клещевого энцефалита в клетках млекопитающих, имеющая нуклеотидную последовательность SEQ ID NO: 1, размер 6523 п.н. и содержащая в соответствии с физической и генетической картой, представленной на фиг. 1, следующие элементы:
- 5'SB с координатами 88-368 п.н. и 3'SB с координатами 4573-4850 п.н. - сайт связывания транспозазы SB100;
- CMV promoter, имеющий координаты с 734 по 937 п.н. - область промотора CMV;
- 176 - нуклеотидная последовательность, кодирующая гибридный лидерный пептид люциферазы и фиброина, обеспечивающая экспорт белка из клетки и имеющая координаты с 1023 по 1085 п.н.;
- 10H10ch-VL - нуклеотидная последовательность, кодирующая вариабельный фрагмент легкой цепи антитела 10H10ch и имеющая координаты с 1086 по 1421 п.н.;
- CL - нуклеотидная последовательность, кодирующая константную часть легкой цепи антитела и имеющая координаты с 1422 по 1742 п.н.;
- Furin - нуклеотидная последовательность, кодирующая сайт протеолиза клеточной протеазы фурина и имеющая координаты с 1743 по 1754 п. н.;
- Р2А - нуклеотидная последовательность, кодирующая саморасщепляющийся пептид р2а и имеющая координаты с 1755 по 1811 п.н.;
- GL - нуклеотидная последовательность, кодирующая лидерную последовательность gaussia luciferase и имеющая координаты с 1812 по 1862 п.н.;
- 10H10ch-VH - нуклеотидная последовательность, кодирующая вариабельный фрагмент тяжелой цепи антитела 10H10ch и имеющая координаты с 1866 по 2201 п.н.;
- СН1-3 - нуклеотидная последовательность, кодирующая константную цепь антитела человека IgG1 СН1-СН2-СН3 и имеющая координаты с 2215 по 3191 п.н.;
- EMCV IRES - участок внутренней посадки рибосомы, имеющий координаты с 3210 по 3784 п.н.;
- PuroR - нуклеотидная последовательность, кодирующая фактор устойчивости к антибиотику пуромицину и имеющая координаты с 3797 по 4396 п.н.;
- SV40 poly (A) signal - последовательность для стабилизации мРНК-транскриптов за счет полиаденилирования, имеющая координаты с 4431 по 4552 п.н.;
- cat promoter - синтетический бактериальный промотер, имеющий координаты с 4934 по 5036 п.н.;
- KanR - нуклеотидная последовательность, кодирующая фактор устойчивости к антибиотику канамицину и имеющая координаты с 5037 по 5852 п.н.;
- ori - участок начала репликации, имеющий координаты с 5915 по 6502 п.н.
2. Рекомбинантный штамм клеточной линии яичника китайского хомячка CHO-K1-10H10ch - продуцент рекомбинантного химерного антитела 10H10ch против вируса клещевого энцефалита, полученный на основе штамма CHC-K1, содержащий интегративный плазмидный вектор pVEAL3-10H10ch по п. 1, обеспечивающий экспрессию и секрецию полноразмерного человеческого антитела 10H10ch.
3. Химерное антитело 10H10ch против вируса клещевого энцефалита, продуцируемое рекомбинантным штаммом клеточной линии яичника китайского хомячка CHO-K1-10H10ch по п. 2, имеющее аминокислотные последовательности вариабельных фрагментов тяжелой цепи SEQ ID NO: 2 и легкой цепи SEQ ID NO: 3, предназначенное для создания терапевтических препаратов против вируса клещевого энцефалита.

