Настоящее изобретение относится к области культивирования плюрипотентных стволовых клеток и улучшенным способам культивирования плюрипотентных стволовых клеток на промышленном уровне.
Уровень техники
Плюрипотентные стволовые клетки, такие как, например, эмбриональные стволовые клетки, способны дифференцироваться во все типы дифференцированных (зрелых) клеток. Как таковые, эмбриональные стволовые клетки могут быть источником пополнения клеток и ткани для органов, которые были повреждены в результате заболевания, инфекции или врожденных патологий. Возможности использовать эмбриональные стволовые клетки в качестве источника пополнения клеток препятствует трудность размножения этих клеток in vitro с сохранением их плюрипотентности.
Существующие способы культивирования недифференцированных эмбриональных стволовых клеток требуют сложных условий культивирования, таких как, например, культивирование эмбриональных стволовых клеток в присутствии слоя питающих клеток. Альтернативно, среды, полученные посредством воздействия на них культур питающих клеток, могут быть использованы для культивирования эмбриональных стволовых клеток. Системы культур, в которых используют эти способы, часто используют клетки, полученные из видов, отличных от вида, из которого получают культивируемые стволовые клетки (ксеногенные клетки). Кроме того, в эти системы культур может добавляться сыворотка животного.
Например, в статье Reubinoff et al. (Nature Biotechnology 18:399-404 (2000)) и Thompson et al. (Science 6 November 1998: Vol. 282. no. 5391, pp. 1145-1147) описана культура линий эмбриональных стволовых клеток из бластоцист человека, в которой используются эмбриональные фибробласты мыши в качестве слоя питающих клеток.
В другом примере, в документе WO2005014799, описывается кондиционированная среда для поддержания, пролиферации и дифференцировки клеток млекопитающих. В документе WO2005014799 указано: "Культуральная среда, полученная в соответствии с изобретением, обусловлена активностью секреции клеток мышиных клеток, в частности, клеток дифференцированных и иммортализованных трансгенных гепатоцитов, названных ММН (Met Murine Hepatocyte)".
Однако использование ксеногенных клеток или продуктов ксеногенных клеток повышает риск загрязнения полученных такими способами популяций эмбриональных стволовых клеток вирусными белками и/или ксенопротеинами иммуногенного характера.
В статье Richards et al. (Stem Cells 21: 546-556, 2003) оценивали набор из 11 различных слоев питающих клеток, состоящих из клеток взрослого человека, фетальных и неонатальных клеток, на их способность поддерживать культуру эмбриональных стволовых клеток человека. В статье Richards et al. указано: "Линии эмбриональных стволовых клеток человека, культивируемые на питающих фибробластах кожи взрослого человека, сохраняют морфологию эмбриональных стволовых клеток человека и остаются плюрипотентными".
В документе US6642048 описаны среды, которые поддерживают рост плюрипотентных стволовых (pPS) клеток в культуре без питающих клеток, и клеточные линии, подходящие для получения таких сред. В документе US6642048 указано: "Изобретение относится к мезенхимным и фибробластподобным клеточным линиям, полученным из эмбриональной ткани или дифференцированным из эмбриональных стволовых клеток. Способы получения клеточных линий, приготовления сред и выращивания стволовых клеток с использованием таких кондиционированных сред описаны и проиллюстрированы в предлагаемом описании".
В документе US20020072117 описаны клеточные линии, которые продуцируют среды, поддерживающие рост плюрипотентных стволовых клеток в культуре без питающих клеток. Такие используемые клеточные линии являются мезенхимными и фибробластподобными клеточными линиями, полученными из эмбриональной ткани или дифференцированными из эмбриональных стволовых клеток. В документе US20020072117 также описано применение этих клеточных линий в качестве первичного слоя питающих клеток.
В другом примере, в документе Wang et al. (Stem Cells 23: 1221-1227, 2005), описаны способы длительного выращивания эмбриональных стволовых клеток человека на слоях питающих клеток, полученных из эмбриональных стволовых клеток человека.
В другом примере, в документе Xu et al. (Stem Cells 22: 972-980, 2004) описана кондиционированная среда, полученная из производных эмбриональных стволовых клеток человека, которые были генетически модифицированы для сверхэкспрессии теломеразы, обратной транскриптазы человека.
В другом примере, в статье Stojkovic et al. (Stem Cells 23: 306-314, 2005), описана система питающих клеток, полученная путем спонтанной дифференцировки эмбриональных стволовых клеток человека.
В следующем примере, в статье Miyamoto et al. (Stem Cells 22: 433-440, 2004), описан источник питающих клеток, полученный из плаценты человека.
В статье Amit et al. (Biol. Reprod 68: 2150-2156, 2003) описан слой питающих клеток, полученный из крайней плоти человека.
В другом примере, в статье Inzunza et al. (Stem Cells 23: 544-549, 2005), описан слой питающих клеток из фибробластов постнатальной крайней плоти человека.
В альтернативной культуральной системе используется бессывороточная среда, дополненная факторами роста, способными стимулировать пролиферацию эмбриональных стволовых клеток. Например, в статье Cheon et al. (BioReprod DOI:10.1095/biolreprod.105.046870, October 19, 2005) описана не содержащая питающих клеток бессывороточная культуральная система, в которой эмбриональные стволовые клетки поддерживаются в некондиционированной среде с заменой сыворотки (SR), дополненной различными факторами роста, способная запускать самообновление эмбриональных стволовых клеток.
В другом примере, в статье Levenstein et al. (Stem Cells 24: 568-574, 2006), описаны способы длительного культивирования эмбриональных стволовых клеток человека в отсутствие фибробластов или кондиционированной среды, с использованием сред, дополненных bFGF.
В другом примере, в документе US20050148070, описан способ культивирования эмбриональных стволовых клеток человека в средах определенного состава без сыворотки и без фибробластных питающих клеток, причем этот способ включает культивирование стволовых клеток в культуральной среде, содержащей альбумин, аминокислоты, витамины, минеральные соединения, по меньшей мере трансферрин или заменитель трансферрина, по меньшей мере инсулин или заменитель инсулина, причем эта культуральная среда по существу не содержит фетальной сыворотки млекопитающего и содержит по меньшей мере приблизительно 100 нг/мл фактора роста фибробластов, способного активировать рецептор передачи сигнала фактора роста фибробластов, где фактор роста получают из источника, отличного от слоя питательных фибробластов, причем эта среда поддерживает пролиферацию стволовых клеток в недифференцированном состоянии без питающих клеток или кондиционированной среды.
В другом примере, в документе US20050233446, описаны среды определенного состава, которые можно использовать для культивирования стволовых клеток, в том числе недифференцированных примордиальных стволовых клеток приматов. В растворе эта среда является по существу изотоничной по сравнению с культивируемыми стволовыми клетками. В конкретной культуре эта конкретная среда содержит базальную среду и некоторое количество каждого bFGF, инсулина и аскорбиновой кислоты, необходимых для поддержания по существу недифференцированного роста примордиальных стволовых клеток.
В другом примере, в документе US6800480, указано: "В одном из вариантов осуществления описывается среда для культивирования клеток для выращивания полученных из приматов примордиальных стволовых клеток по существу в недифференцированном состоянии, которая включает базальную среду с низким осмотическим давлением, низким содержанием эндотоксина и которая эффективна для поддержания роста полученных из приматов примордиальных стволовых клеток. Эту базальную среду объединяют с питательной сывороткой, эффективной для поддержания роста полученных из приматов примордиальных стволовых клеток, и субстратом, выбранным из группы, состоящей из питающих клеток и компонента внеклеточного матрикса, происходящего из питающих клеток. Эта среда включает дополнительно аминокислоты, не являющиеся незаменимыми, антиоксидант и первый фактор роста, выбранный из группы, состоящей из нуклеозидов и соли пировиноградной кислоты”.
В другом примере, в документе US20050244962, указано: "В одном из аспектов изобретение относится к способу культивирования эмбриональных стволовых клеток приматов. Эти стволовые клетки культивируют в культуре, по существу не содержащей фетальной сыворотки млекопитающего (предпочтительно, по существу не содержащей любой сыворотки животного) и в присутствии фактора роста фибробластов, который получают из источника, отличного от слоя питательных фибробластов. В предпочтительном варианте этот слой питательных фибробластов, ранее необходимый для поддержания культуры стволовых клеток, снимает необходимость добавления достаточного количества фактора роста фибробластов".
В следующем примере, в документе WO2005065354, описана изотоническая культуральная среда определенного состава, которая является по существу не содержащей слоя питающих клеток и сыворотки, содержащая: a) базальную среду; b) количество bFGF, достаточное для поддержания роста по существу недифференцированных стволовых клеток млекопитающих; c) количество инсулина, достаточное для поддержания роста по существу недифференцированных стволовых клеток млекопитающих; и d) количество аскорбиновой кислоты, достаточное для поддержания по существу недифференцированных стволовых клеток млекопитающих.
В другом примере, в документе WO2005086845, описан способ поддержания недифференцированной стволовой клетки, причем указанный способ предусматривает обработку стволовой клетки представителем семейства белков трансформирующего фактора роста бета (TGFβ), представителем семейства белков фактора роста фибробластов (FGF) или никотинамида (NIC) в количестве, достаточном для поддержания этой клетки в недифференцированном состоянии в течение достаточного периода времени для достижения желаемого результата.
Эмбриональные стволовые клетки являются возможным источником исследования и скрининга лекарственных средств. В настоящее время промышленное культивирование линий ES-клеток человека затруднено и имеет значительные препятствия. Возможным решением этих задач является пассирование и культивирование эмбриональных стволовых клеток человека в виде отдельных клеток. Отдельные клетки являются более доступными для стандартных способов культивирования ткани, таких как, например, подсчет клеток, трансфекция и т.п.
Например, в статье Nicolas et al. описан способ получения и увеличения количества клеточных линий hES из отдельных клеток, которые были выделены сортингом клеток с возбуждением флуоресценции (FACS) после генетической модификации лентивирусными векторами. Stem Cells and Development (2007), 16(1), 109-118.
В другом примере, в заявке на патент США US2005158852, описан способ "улучшения роста и выживания отдельных эмбриональных стволовых клеток человека. Этот способ включает стадию получения отдельной недифференцированной клетки HES; смешивание этой единственной недифференцированной клетки с внеклеточным матриксом (ECM) для включения этой клетки; и инокуляцию этой смеси на питающие клетки с питательной средой в среде роста".
В другом примере, в статье Sidhu, KS et al. (Stem Cells Dev. 2006 Feb; 15(1):61-9), описано первое сообщение о трех клонах эмбриональных стволовых клеток человека (hESC), hES 3.1, 3.2 и 3.3, которые получены из родительской линии hES3 сортингом препаратов отдельных клеток проточной цитометрией. Значение жизнеспособности препаратов отдельных клеток до и после сортинга клеток сохранялось >98%.
При этом пассирование и культивирование эмбриональных стволовых клеток человека в виде отдельных клеток приводит к генетическим патологиям и потере плюрипотентности. Условия культивирования являются важными для поддержания плюрипотентности и генетической стабильности. Обычно пассирование клеточных линий hES проводят вручную или с ферментативными агентами, такими как коллагеназа, либераза или диспаза.
Например, в статье Draper JS et al. указано на наличие "кариотипических изменений, включающих конверсию хромосомы 17q в три независимые линии эмбриональных стволовых клеток человека в пяти независимых случаях" (Nat Biotechnol. 2004 Jan; 22(l):53-4. Epub 2003 Dec 7).
В другом примере, в статье Buzzard et al., указано: "Авторы всегда обнаруживали одно изменение кариотипа… используемые способы культивирования могли иметь отношение к таким результатам, при условии, что способы значительно отличаются от способов, используемых большинством других групп. Обычно авторы пассировали ES-клетки человека через 7 дней после начала иссечением колонии краем разбитой пипетки … В этот способ не включены никакие ферментативные или химические способы диссоциации клеток. Авторы предполагают, что это может объяснять относительную генетическую способность восстановления клеток ES в опытах". (Nat Biotechnol. 2004 Apr;22(4):381-2; author reply 382).
В другом примере, в статье Mitalipova MM et al., указано: "Способы пассирования основной массы материала... могут делать популяции анэуплоидных клеток бессмертными после продолжительного пассирования в культуре, но могут быть использованы в течение более коротких периодов (по меньшей мере, до 15 пассажей) без нарушения кариотипов... можно поддерживать нормальный кариотип в клетках hES при длительном размножении вручную с последующим пассированием основной массы в экспериментах, требующих более высоких количеств клеток hES, чем могут обеспечивать только способы ручного пассажа". (Nat Biotechnol. 2005 Jan;23(l):19-20).
В другом примере, в статье Heng BC et al., указано: "Эти результаты продемонстрировали, что второй протокол (трипсинизация с осторожным пипетированием) является гораздо менее вредным для клеточной жизнеспособности, чем первый протокол (обработка коллагеназой с экскориацией). Это, в свою очередь, способствует более высоким степеням выживания при замораживании-оттаивании" (Biotechnology and Applied Biochemistry (2007), 47(1), 33-37).
В другом примере, в статье Hasegawa K. et al., указано: "Авторы получили сублинии hESC, устойчивые к полной диссоциации. Эти клетки проявляют высокую эффективность реинокуляции, а также высокую эффективность клонирования, и они могут сохранять свою способность к дифференцировке в три зародышевых слоя" (Stem Cells. 2006 Dec;24(12):2649-60. Epub 2006 Aug 24).
Сущность изобретения
Настоящее изобретение относится к способам поддержания, пассажа и дифференцировки плюрипотентных стволовых клеток, которые были выделены как отдельные клетки с использованием ферментов. В частности, настоящее изобретение относится к способам поддержания, пассажа и дифференцировки плюрипотентных стволовых клеток, которые были выделены как отдельные клетки, без последующей потери плюрипотентности и без приобретения хромосомных аномалий.
В одном из вариантов осуществления настоящее изобретение относится к способу дифференцировки плюрипотентных стволовых клеток, включающему стадии:
a) культивирования плюрипотентных стволовых клеток в виде кластеров,
b) высвобождения плюрипотентных стволовых клеток в виде отдельных клеток,
c) посева отдельных плюрипотентных стволовых клеток на субстрате культуры ткани и
d) дифференцировки этих клеток.
В одном из вариантов осуществления настоящее изобретение относится к способу поддержания плюрипотентных стволовых клеток, включающему стадии:
a) получения плюрипотентных стволовых клеток,
b) высвобождения плюрипотентных стволовых клеток в виде отдельных клеток, и
c) посева отдельных плюрипотентных стволовых клеток на субстрате культуры ткани.
В одном из вариантов осуществления настоящее изобретение относится к способу пассирования плюрипотентных стволовых клеток, включающему стадии:
a) получения кластеров плюрипотентных стволовых клеток,
b) высвобождения плюрипотентных стволовых клеток в виде отдельных клеток,
c) посева отдельных плюрипотентных стволовых клеток на субстрате культуры ткани,
d) обеспечения возможности деления отдельных плюрипотентных клеткок,
e) высвобождения отдельных плюрипотентных стволовых клеток и
f) посева отдельных плюрипотентных стволовых клеток на новом субстрате культуры ткани.
Краткое описание фигур
Фигура 1: Изображение при 4-кратном увеличении клеток ES человека H9ccp33, выращенных на разведенном 1:30 фактором роста MATRIGEL™ в MEF-кондиционированной среде.
Фигура 2: Процент клеток, экспрессирующих CXCR4 после обработки, вызывающей дифференцировку для получения дефинитивной (развитой) эндодермы. Темно-серый столбец: среднее шести экспериментов по дифференцировке DE с кластерами клеток Н1 (H1 cc) между пассажами 45 и 55. Черный столбец: среднее двух экспериментов по дифференцировке DE с отдельными клетками Н1 (H1 sc) при пассаже 47 и 54. Белый столбец: среднее пяти экспериментов по дифференцировке DE с кластерами клеток Н9 (Н9 сс) между пассажами 37 и 55. Светло-серый столбец: среднее пяти экспериментов по дифференцировке DE с кластерами клеток Н9 (Н9 sc) между пассажами 36 и 48. Стержни ошибок представляют стандартное отклонение повторяемых экспериментов.
Фигура 3: Анализ экспрессии генов при помощи ПЦР реального времени после подвергания 14 или 17 дням протокола панкреатической эндокринной дифференцировки. A. Анализировали отдельные клетки Н9 и кластеры клеток при пассаже 37. B. Продолжение кластеров клеток и отдельных клеток H1 p47 и H9 p37 до стадии панкреатической эндодермы. Экспрессия Pdx1 после дней 14 и 17. Экспрессию генов для указанных маркеров для необработанных клеток принимали как величину 1 для каждого набора данных.
Фигура 4: Изображение, показанное при 4-кратном увеличении, отдельных клеток ES человека H9scp22, выращенных на разведенном 1:30 фактором роста MATRIGEL™ в MEF-кондиционированной среде.
Фигура 5: Оценивание при помощи FACS на экспрессию маркеров плюрипотентности клеток hES. Процент клеток, положительных в отношении указанных маркеров, показан на Х-оси.
Фигура 6: Хромосомный препарат отдельных клеток H9 hES при пассажах 38 раз в виде кластеров с последующими 20 пассажами в виде отдельных клеток.
Фигура 7: Сравнение отдельных клеток Н9 и кластеров клеток во время дифференцировки дефинитивной (развитой) эндодермы. Процент клеток, положительных в отношении CXCR4, показан после подвергания этих клеток протоколу дифференцировки дефинитивной эндодермы. N=2 для H9sc-p и N=5 для H9cc. Стержни ошибок представляют стандартное отклонение повторяемых экспериментов.
Фигура 8: Увеличение маркеров панкреатической эндодермы после дифференцировки отдельных клеток Н9 (пассаж 22). Анализ экспрессии генов при помощи ПЦР реального времени показан после 11, 14 или 17 дней панкреатической эндокринной дифференцировки. Величины для дней 14 и 17 являются средними величинами двух лунок из 6-луночной чашки.
Фигура 9: Отдельные клетки hES могут дифференцироваться на MEF. Результаты FACS из клеток H1scp4, выращенных на питательных MEF-клетках и дифференцированных на дефинитивной (развитой) эндодерме. Маркер дефинитивной эндодермы CXCR4 (CD184) экспрессируется в 89% этих клеток по сравнению с 0% в недифференцированных клетках (см. фигуру 5).
Фигура 10: Отдельные клетки hES (H9scpl8) могут дифференцироваться в 96-луночном формате в дефинитивную эндодерму. Показаны данные иммунофлуоресценции для Sox17-положительного детектирования. Восемь лунок обрабатывали MEF-кондиционированной средой на протяжении эксперимента: MEF-среда. Восемь лунок обрабатывали базальной средой для дифференцировки без компонентов: без соединений. Наборы повторяемых данных восьми лунок усредняли для каждой диаграммы данных. В целом, каждую из 40 лунок обрабатывали ингибитором Wnt3a или Gsk3b и определяли среднее для каждого набора данных. Стержни ошибок представляют стандартное отклонение для каждого повторяемого набора данных.
Фигура 11: Скрининг активных компонентов лекарственных средств с отдельными клетками hES (H9scp19). Всего 13 экспериментальных соединений с малыми молекулами испытывали на их способность заменять Wnt3a в протоколе дифференцировки дефинитивной эндодермы. Показаны три эффективных соединения. Эти наборы данных представляют среднее Sox 17-положительных клеток в двух или более лунках. Клетки, обработанные MEF-кондиционированной средой или базальной средой, использовали в качестве отрицательных контролей.
Фигура 12: Оценивание эффективности трансфекции между кластерами клеток и отдельными клетками H9p33 hES. CMV-GFP трансфицировали в клетки с использованием 8 мкл липофектамина 2000 (Invitrogen, Carlesbad, CA) и 4 или 8 мкг ДНК, черный и серый столбцы, соответственно.
Фигура 13: Фазово-контрастные микрофотографии клеток H1 hES, выращенных на MEF и затем пассированных на MATRIGEL™, в виде кластеров или отдельных клеток. Клетки H1 hES пассажа 37, пассированные из MEF на MATRIGEL™ с коллагеназой, образуют дискретные, плотно упакованные колонии с некоторым количеством рыхлых дифференцированных клеток. Клетки H1 hES, пассированные один раз с Accutase™ или TrypLE™, образуют монослойную культуру с очагами плотно упакованных дифференцированных клеток.
Фигура 14: Клетки H1 hES, пассированные непосредственно на MATRIGEL™ из MEF в виде отдельных клеток, дифференцируются спонтанно. Панель A: процент клеток, остающихся в этой популяции после двух пассажей с Accutase™ из MEF на MATRIGEL™. Панель В: экспрессия маркеров плюрипотентности и дифференцировки в клетках hES после двух пассажей с Accutase™ из MEF на MATRIGEL™. Панель С: фазовые микрофотографии клеток H1 hES после двух пассажей с Accutase™ из MEF на MATRIGEL™.
Сведения, подтверждающие возможность
осуществления изобретения
Для большей ясности, но при этом не ограничивая объем притязаний, подробное описание этого изобретения разделено на следующие подразделы, которые описывают или иллюстрируют определенные признаки, варианты осуществления или варианты применения изобретения.
Определения
Стволовые клетки являются недифференцированными клетками, определяемыми по их способности на уровне этих отдельных клеток как к самообновлению, так и к дифференцировке с образованием клеток-предшественников, в том числе самообновляющихся предшественников, несамообновляющихся предшественников и терминально дифференцированных клеток. Стволовые клетки отличаются также их способностью дифференцироваться in vitro в функциональные клетки различных линий (направлений) дифференцировки из множественных слоев зародышевых клеток (эндодермы, мезодермы и эктодермы), а также давать начало тканям множественных зародышевых слоев после трансплантации и вносить вклад по существу в большинство, если не все, ткани после инъекции в бластоцисты.
Стволовые клетки классифицируют по возможности их развития как: (1) тотипотентные, т.е. способные давать начало всем зародышевым и внезародышевым типам клеток; (2) плюрипотентные, т.е. способные давать начало всем зародышевым типам клеток; (3) мультипотентные, т.е. способные давать начало субпополяции линий (направлений) дифференцировки, но все в пределах конкретных ткани, органа или физиологической системы (например, гемопоэтические стволовые клетки (HSC могут продуцировать потомство, которое включает HSC (самообновляющиеся), ограниченные клетками крови олигопотентные предшественники и все типы клеток и элементы (например, тромбоциты), которые являются нормальными компонентами крови); (4) олигопотентные, т.е. способные давать начало более ограниченной субпопуляции линий (направлений) дифференцировки, т.е. способные давать начало более ограниченной субпопуляции линий (направлений) дифференцировки, чем мультипотентные стволовые клетки; и (5) унипотентные, т.е. способные давать начало линии (направлению) дифференцировки отдельных клеток (например, сперматогенные стволовые клетки).
Дифференцировка является процессом, при котором неспециализированная ("некоммитированная") или менее специализированная клетка приобретает признаки специализированной клетки, такой как, например, нервная клетка или мышечная клетка. Дифференцированной или индуцированной к дифференцировке клеткой является клетка, которая взята на более специализированном ("коммитированном") положении линии дифференцировки клетки. Термин "коммитированная", при применении к процессу дифференцировки, относится к клетке, которая перешла в пути дифференцировки к точке, в которой, при нормальных обстоятельствах, она будет продолжать дифференцироваться в конкретный тип клеток или субпопуляцию типов клеток и не может, при нормальных обстоятельствах, дифференцироваться в другой тип клеток или вернуться к менее дифференцированному типу клеток. Дедифференцировкой называют процесс, при помощи которого клетка возвращается в менее специализированное (или коммитированное) положение в пределах линии дифференцировки клетки. В контексте настоящего изобретения линия (направление) дифференцировки клетки определяет наследственность этой клетки, т.е. из каких клеток она происходит и каким клеткам она может дать начало. Эта линия дифференцировки клетки помещает эту клетку в схему наследственности развития и дифференцировки. Специфическим для линии дифференцировки маркером называют признак, специфически ассоциированный с фенотипом клеток представляющей интерес линии дифференцировки, и он может быть использован для оценки дифференцировки некоммитированной клетки в представляющую интерес линию дифференцировки.
Для описания клеток в культуре могут быть использованы различные термины. "Поддержание" относится обычно к клеткам, помещенным в среду для выращивания при условиях, которые облегчают рост и/или деление клеток, которые могут приводить или могут не приводить к большей популяции этих клеток. "Пассирование" относится к процессу удаления клеток из одного культурального сосуда и помещения их во второй культуральный сосуд при условиях, которые облегчают рост и/или деление клеток.
Конкретную популяцию клеток, или линию клеток, называют или характеризуют иногда количеством проведенных пассажей. Например, популяция культивируемых клеток, которую пассировали десять раз, может называться культурой Р10. Первичную культуру, т.е. первую культуру после выделения клеток из ткани, обозначают Р0. После первой субкультуры клетки описывают как вторичную культуру (Р1 или пассаж 1). После второй субкультуры эти клетки становятся третичной культурой (Р2 или пассаж 2) и т.д. Квалифицированному в данной области специалисту будет понятно, что во время периода пассирования могут быть многочисленные удвоения популяции; таким образом, количество удвоений популяции культуры является большим, чем количество пассажей. Размножение клеток (т.е. количество удвоений популяции) во время периода между пассажами зависит от многих факторов, включающих в себя, но не ограничивающихся ими, плотность посева, субстрат, среду, условия роста и время между проведением пассажей.
"AFP", или "белок альфа-фетопротеин", относится в контексте настоящего изобретения к антигену, продуцируемому в начале развития печени. AFP может также экспрессироваться во внезародышевых клетках.
"β-клеточная линия дифференцировки" относится к клеткам с положительной экспрессией гена для фактора транскрипции PDX-1 и по меньшей мере одного из следующих факторов транскрипции: NGN-3, Nkx2.2, Nkx6.1, NeuroD, Is1-1, HNF-3 бета, MAFA, Pax4 и Pax6. Клетки, экспрессирующие маркеры, характерные для β-клеточной линии дифференцировки, включают β-клетки.
"Brachyury", в контексте настоящего изобретения, является членом семейства T-бокс-генов. Он является маркером клеток первичной полоски и мезодермы.
"Клетки, экспрессирующие маркеры, характерные для линии дифференцировки дефинитивной (развитой) эндодермы", обозначают в контексте настоящего изобретения клетки, экспрессирующие по меньшей мере один из следующих маркеров: SOX-17, GATA-4, HNF-3 бета, GSC, Cer1, Nodal, FGF8, Brachyury, Mix-подобный белок гомеобокса, FGF4, CD48, эомезодермин (EOMES), DKK4, FGF 17, GATA-6, CXCR4, C-Kit, CD99 или OTX2. Клетки, экспрессирующие маркеры, характерные для линии дифференцировки дефинитивной эндодермы, включают клетки-предшественники первичной полоски, клетки первичной полоски, клетки мезодермы и клетки дефинитивной (развитой) эндодермы.
"CD99" относится в контексте настоящего изобретения к белку, кодируемому геном с номером доступа NM_002414.
"Клетки, экспрессирующие маркеры, характерные для линии дифференцировки панкреатической эндодермы", обозначают в контексте настоящего изобретения клетки, экспрессирующие по меньшей мере один из следующих маркеров: PDX-1, HNF-1 бета, PTF-1 альфа, HNF-6 или HB9. Клетки, экспрессирующие маркеры, характерные для линии дифференцировки панкреатической эндодермы, включают клетки панкреатической эндодермы.
“Клетки, экспрессирующие маркеры, характерные для линии панкреатической эндокринной дифференцировки", обозначают в контексте настоящего изобретения клетки, экспрессирующие по меньшей мере один из следующих маркеров: NGN-3, NeuroD, Islet-1, PDX-1, NKX6.1, Pax-4, Ngn-3 или PTF-1 альфа. Клетки, экспрессирующие маркеры, характерные для линии панкреатической эндокринной дифференцировки, включают панкреатические эндокринные клетки, панкреатические экспрессирующие гормоны клетки и панкреатические секретирующие гормоны клетки и клетки β-клеточной линии дифференцировки.
"CXCR4" относится в контексте настоящего изобретения к рецептору происходящего из стромальных клеток фактора 1 (SDF-1), также известному как "LESTR" или "фузин". В гаструлирующем зародыше мыши CXCR4 экспрессируется в дефинитивной эндодерме и мезодерме, но не во внезародышевой эндодерме.
"Дефинитивная (развитая) эндодерма" относится в контексте настоящего изобретения к клеткам, которые имеют характеристики клеток, возникающих из эпибласта во время гаструляции и которые образуют желудочно-кишечный тракт и его производные. Клетки дефинитивной эндодермы экспрессируют следующие маркеры: CXCR4, HNF-3 бета, GATA-4, SOX-17, Cerberus, OTX2, goosecoid, c-Kit, CD99 и Mix11.
"Внезародышевой эндодермой" называют в контексте настоящего изобретения популяцию клеток, экспрессирующих по меньшей мере один из следующих маркеров: SOX-7, AFP и SPARC.
"GATA-4" и "GATA-6" являются членами семейства факторов транскрипции GATA. Это семейство факторов транскрипции индуцируется лигандом передачи сигнала TGF-β и способствует сохранению ранних маркеров эндодермы.
"GLUT-2" в контексте настоящего изобретения относится к молекуле-транспортеру глюкозы, которая экспрессируется в многочисленных фетальных и дифференцированных (зрелых) тканях, включающих поджелудочную железу, печень, кишечник, головной мозг и почки.
"Goosecoid", или "GSC", относится в контексте настоящего изобретения к имеющему гомеодомен фактору транскрипции, экспрессируемому в дорсальной губе бластопора.
"Islet-1" или "Isl-1" является в контексте настоящего изобретения членом LIM/гомеодоменного семейства факторов транскрипции и экспрессируется в развивающихся поджелудочных железах.
"MafA" является в контексте настоящего изобретения фактором транскрипции, экспрессируемым в поджелудочной железе, и регулирует экспрессию генов, участвующих в биосинтезе и секреции инсулина.
"Маркеры" являются в контексте настоящего изобретения молекулами нуклеиновых кислот или полипептидов, которые дифференциально экспрессируются в представляющей интерес клетке. В контексте настоящего изобретения дифференциальная экспрессия означает увеличенный уровень в отношении положительного маркера и уменьшенный уровень в отношении отрицательного маркера. Детектируемый уровень маркерной нуклеиновой кислоты или полипептида является достаточно более высоким или более низким в представляющих интерес клетках по сравнению с другими клетками, так что представляющие интерес клетки могут быть идентифицированы и отличены от других клеток с использованием любого из различных способов, известных в данной области.
"Клетка мезоэндодермы" относится в контексте настоящего изобретения к клетке, экспрессирующей по меньшей мере один из следующих маркеров: CD48, эомезодермин (EOMES), SOX-17, DKK4, HNF-3 бета, GSC, FGF 17, GATA-6.
"Nodal" является в контексте настоящего изобретения членом суперсемейства белков TGF бета.
"Oct-4" является членом фактора транскрипции с POU-доменом и является широко признаваемым отличительным признаком плюрипотентных стволовых клеток. Взаимосвязь Oct-4 с плюрипотентными стволовыми клетками демонстрируется по его прочно ограниченной экспрессии в недифференцированных плюрипотентных стволовых клетках. После дифференцировки в соматические линии дифференцировки экспрессия Oct-4 быстро исчезает.
"Панкреатическая эндокринная клетка" или "панкреатическая экспрессирующая гормоны клетка" относится в контексте настоящего изобретения к клетке, способной экспрессировать по меньшей мере один из следующих гормонов: инсулин, глюкагон, соматостатин и панкреатический полипептид.
"Панкреатическая секретирующая гормоны клетка" относится в контексте настоящего изобретения к клетке, способной секретировать по меньшей мере один из следующих гормонов: инсулин, глюкагон, соматостатин и панкреатический полипептид.
"Pax-4" и "Pax-6" являются в контексте настоящего изобретения факторами транскрипции, специфическими в отношении панкреатических β-клеток, которые участвуют в развитии островков.
"PDX-1" относится в контексте настоящего изобретения к имеющему гомеодомен фактору транскрипции, участвующему в развитии поджелудочной железы.
"Клетка-предшественник клеток первичной полоски" относится в контексте настоящего изобретения к клетке, экспрессирующей по меньшей мере один из следующих маркеров: Nodal или FGF8.
"Клетка первичной полоски" относится в контексте настоящего изобретения к клетке, экспрессирующей один из следующих маркеров: Brachyury, Mix-подобный белок гомеобокса или FGF4.
"PTF-1 альфа" относится в контексте настоящего изобретения к основному белку спираль-петля-спираль 48 кД, который является последовательность-специфической ДНК-связывающей субъединицей тримерного фактора транскрипции 1 поджелудочной железы (PTF1).
"SPARC" в контексте настоящего изобретения известен также как "секретируемый кислый белок, богатый цистеином".
"SSEA-1" (стадиеспецифический зародышевый антиген-1) является гликолипидным поверхностным антигеном на поверхности стволовых клеток (ES) тератокарциномы человека, зародышевых клеток (половых клеток) (EG) мышиных и человека и мышиных эмбриональных стволовых клеток (ES).
"SSEA-3" (стадиеспецифический зародышевый антиген-3) является гликолипидным поверхностным антигеном на поверхности стволовых клеток (ES) тератокарциномы человека, зародышевых клеток (половых клеток) (EG) человека и эмбриональных стволовых клеток (ES) человека.
"SSEA-4" (стадиеспецифический зародышевый антиген-4) является гликолипидным поверхностным антигеном на поверхности стволовых клеток (ES) тератокарциномы человека, зародышевых клеток (половых клеток) (EG) человека и эмбриональных стволовых клеток (ES) человека.
"TRA1-60" является родственным кератинсульфату антигеном, который экспрессируется на поверхности стволовых клеток (ES) тератокарциномы человека и зародышевых клеток (половых клеток) (EG) человека и эмбриональных стволовых клеток (ES) человека.
"TRA1-81" является родственным кератинсульфату антигеном, который экспрессируется на поверхности стволовых клеток (ES) тератокарциномы человека и зародышевых клеток (половых клеток) (EG) человека и эмбриональных стволовых клеток (ES) человека.
"TRA2-49" является изоферментом щелочной фосфатазы, экспрессируемым на поверхности стволовых клеток (ES) тератокарциномы человека и зародышевых клеток (половых клеток) (EG) человека.
Настоящее изобретение относится к способам поддержания, пассирования и дифференцировки плюрипотентных стволовых клеток, которые были выделены в виде отдельных клеток с использованием ферментов. В частности, изобретение относится к способам поддержания, пассирования и дифференцировки плюрипотентных стволовых клеток, которые были выделены в виде отдельных клеток, без последующей потери плюрипотентности и без приобретения хромосомных аномалий.
В одном из вариантов осуществления настоящее изобретение относится к способу дифференцировки плюрипотентных стволовых клеток, включающему стадии:
a) культивирования плюрипотентных стволовых клеток в виде кластеров,
b) высвобождения плюрипотентных стволовых клеток в виде отдельных клеток,
c) посева отдельных плюрипотентных стволовых клеток на субстрате культуры ткани и
d) дифференцировки отдельных плюрипотентных стволовых клеток.
Кластеры плюрипотентных стволовых клеток могут быть выделены в виде отдельных клеток ферментативной обработкой. Эта ферментативная обработка может выполняться с использованием TrypLE™Express, альтернативно, с использованием TrypLE™Select или, альтернативно, с использованием Трипсин/ЭДТА.
Ферментативная обработка может выполняться в течение приблизительно двух - приблизительно пяти минут. Альтернативно, ферментативная обработка может выполняться в течение приблизительно пяти минут.
Ферменты могут быть использованы в концентрации от приблизительно 0,5 г/л до приблизительно 2,5 г/л.
В одном из вариантов осуществления кластеры плюрипотентных стволовых клеток выделяют в виде отдельных клеток с использованием TrypLE™EXPRESS.
В одном из вариантов осуществления плюрипотентные стволовые клетки являются эмбриональными стволовыми клетками. В альтернативном варианте осуществления эти эмбриональные стволовые клетки является клетками человека.
В одном из вариантов осуществления высвобожденные отдельные плюрипотентные стволовые клетки высевают на субстрате для культуры ткани. Этим субстратом может быть MATRIGEL™, альтернативно, этим субстратом может быть фибронектин, альтернативно, этим субстратом может быть ламинин, альтернативно, этим субстратом может быть сыворотка человека или, альтернативно, этим субстратом может быть коллаген.
В одном из вариантов осуществления высвобожденные отдельные плюрипотентные клетки высевают на трехмерный носитель. В этот носитель может быть включен по меньшей мере один фармацевтический агент, который облегчает выживаемость и функцию выделенных отдельных плюрипотентных клеток. Материалы-носители, подходящие для применения для целей изобретения, включают синтетические и природные материалы в форме пен, губок, гелей, гидрогелей, текстильных материалов и нетканых структур.
В одном из вариантов осуществления субстратом культуры ткани является MATRIGEL™. MATRIGEL™ может быть использован при разведении от приблизительно 1:30 до приблизительно 1:10. В одном из вариантов осуществления MATRIGEL™ используют при разведении 1:10.
Отдельные плюрипотентные стволовые клетки могут быть дифференцированы в клетки, экспрессирующие маркеры, характерные для линии дифференцировки дефинитивной (развитой) эндодермы. Альтернативно, эти отдельные плюрипотентные стволовые клетки могут быть дифференцированы в клетки, экспрессирующие маркеры, характерные для линии дифференцировки панкреатической эндодермы. Альтернативно, эти отдельные плюрипотентные стволовые клетки могут быть дифференцированы в клетки, экспрессирующие маркеры, характерные для линии панкреатической эндокринной дифференцировки.
В одном из вариантов осуществления настоящее изобретение относится к способу поддержания плюрипотентных стволовых клеток, включающему стадии:
a) получения кластеров плюрипотентных стволовых клеток,
b) высвобождения плюрипотентных стволовых клеток в виде отдельных клеток, и
c) посева отдельных плюрипотентных стволовых клеток на субстрате культуры ткани.
Кластеры плюрипотентных стволовых клеток могут быть выделены в виде отдельных клеток ферментативной обработкой. Ферментативная обработка может выполняться с использованием TrypLE™Express, альтернативно, с использованием TrypLE™Select или, альтернативно, с использованием Трипсин/ЭДТА.
Ферментативная обработка может выполняться в течение приблизительно двух - приблизительно пяти минут. Альтернативно, ферментативная обработка может выполняться в течение приблизительно пяти минут.
Ферменты могут быть использованы в концентрации от приблизительно 0,5 г/л до приблизительно 2,5 г/л.
В одном из вариантов осуществления кластеры плюрипотентных стволовых клеток выделяют в виде отдельных клеток с использованием TrypLE™EXPRESS.
В одном из вариантов осуществления плюрипотентные стволовые клетки являются эмбриональными стволовыми клетками. В альтернативном варианте осуществления эти эмбриональные стволовые клетки является клетками человека.
В одном из вариантов осуществления высвобожденные отдельные плюрипотентные стволовые клетки высевают на субстрате для культуры ткани. Этим субстратом может быть MATRIGEL™, альтернативно, этим субстратом может быть разведенный фактором роста MATRIGEL™, альтернативно, этим субстратом может быть фибронектин, альтернативно, этим субстратом может быть ламинин, альтернативно, этим субстратом может быть сыворотка человека или, альтернативно, этим субстратом может быть коллаген.
В одном из вариантов осуществления высвобожденные отдельные плюрипотентные клетки высевают на трехмерный носитель. В носитель может быть включен по меньшей мере один фармацевтический агент, который облегчает выживаемость и функцию выделенных отдельных плюрипотентных клеток. Материалы-носители, подходящие для применения для целей изобретения, включают синтетические и природные материалы в форме пен, губок, гелей, гидрогелей, текстильных материалов и нетканых структур.
В одном из вариантов осуществления субстратом культуры ткани является разведенный фактором роста MATRIGEL™. Этот разведенный фактором роста MATRIGEL™ может быть использован при разведении от приблизительно 1:30 до приблизительно 1:10. В одном из вариантов осуществления MATRIGEL™ используют при разведении 1:30.
Отдельные плюрипотентные стволовые клетки могут быть дифференцированы в клетки, экспрессирующие маркеры, характерные для линии дифференцировки дефинитивной (развитой) эндодермы. Альтернативно, отдельные плюрипотентные стволовые клетки могут быть дифференцированы в клетки, экспрессирующие маркеры, характерные для линии дифференцировки панкреатической эндодермы. Альтернативно, отдельные плюрипотентные стволовые клетки могут быть дифференцированы в клетки, экспрессирующие маркеры, характерные для линии панкреатической эндокринной дифференцировки.
В одном из вариантов осуществления настоящее изобретение относится к способу пассирования плюрипотентных стволовых клеток, включающему стадии:
a) получения кластеров плюрипотентных стволовых клеток,
b) высвобождения плюрипотентных стволовых клеток в виде отдельных клеток,
c) посева отдельных плюрипотентных стволовых клеток на субстрате культуры ткани,
d) предоставления возможности размножения отдельным плюрипотентным клеткам,
e) высвобождения отдельных плюрипотентных стволовых клеток и
f) посева отдельных плюрипотентных стволовых клеток на новом субстрате культуры ткани.
Кластеры плюрипотентных стволовых клеток могут быть выделены в виде отдельных клеток ферментативной обработкой. Эта ферментативная обработка может выполняться с использованием TrypLE™Express, альтернативно, с использованием TrypLE™Select или, альтернативно, с использованием Трипсин/ЭДТА.
Ферментативная обработка может выполняться в течение приблизительно двух - приблизительно пяти минут. Альтернативно, ферментативная обработка может выполняться в течение приблизительно пяти минут.
Ферменты могут быть использованы в концентрации от приблизительно 0,5 г/л до приблизительно 2,5 г/л.
В одном из вариантов осуществления кластеры плюрипотентных стволовых клеток выделяют в виде отдельных клеток с использованием TrypLE™EXPRESS.
В одном из вариантов осуществления отдельные плюрипотентные клетки выращивают до плотности приблизительно 70%-80% перед повторной обработкой клеток ферментативным пассированием на новый субстрат культуры ткани. Отдельные плюрипотентные клетки могут быть пассированы один раз, или они могут быть пассированы более одного раза с использованием способов изобретения.
В одном из вариантов осуществления плюрипотентные стволовые клетки являются эмбриональными стволовыми клетками. В альтернативном варианте осуществления эмбриональные стволовые клетки является клетками человека.
В одном из вариантов осуществления высвобожденные отдельные плюрипотентные стволовые клетки высевают на субстрате для культуры ткани. Этим субстратом может быть MATRIGEL™, альтернативно, этим субстратом может быть разведенный фактором роста MATRIGEL™, альтернативно, этим субстратом может быть фибронектин, альтернативно, этим субстратом может быть ламинин, альтернативно, этим субстратом может быть сыворотка человека или, альтернативно, этим субстратом может быть коллаген.
В одном из вариантов осуществления высвобожденные отдельные плюрипотентные клетки высевают на трехмерный носитель. В этот носитель может быть включен по меньшей мере один фармацевтический агент, который облегчает выживаемость и функцию выделенных отдельных плюрипотентных клеток. Материалы-носители, подходящие для применения для целей изобретения, включают синтетические и природные материалы в форме пен, губок, гелей, гидрогелей, текстильных материалов и нетканых структур.
В одном из вариантов осуществления субстратом культуры ткани является разведенный фактором роста MATRIGEL™. Разведенный фактором роста MATRIGEL™ может быть использован при разведении от приблизительно 1:30 до приблизительно 1:10. В одном из вариантов осуществления MATRIGEL™ используют при разведении 1:30.
Другие способы выделения, размножения и культивирования плюрипотентных стволовых клеток
Характеристика плюрипотентных стволовых клеток
Плюрипотентные стволовые клетки могут экспрессировать один или несколько эмбриональных антигенов (SSEA) 3 и 4, и маркеры, детектируемые с использованием антител, названы Tra-1-60 и Tra-1-81 (Thomson et al, Science 282: 1145, 1998). Дифференцировка плюрипотентных стволовых клеток in vitro приводит к потере экспрессии SSEA-4, Tra-1-60 и Tra-1-81 (если она присутствует) и увеличенной экспрессии SSEA-1. Недифференцированные плюрипотентные стволовые клетки обычно имеют активность щелочной фосфатазы, которая может быть детектирована фиксацией этих клеток 4% параформальдегидом и затем проявлением с использованием Vector Red в качестве субстрата, как описано изготовителем (Vector Laboratories, Burlingame Calif.) Недифференцированные стволовые клетки обычно также экспрессируют Oct-4 и TERT, детектированные при помощи ОТ-ПЦР.
Другим желаемым фенотипом размноженных плюрипотентных стволовых клеток является потенциал дифференцировки в клетки всех трех зародышевых слоев: тканей эндодермы, мезодермы и эктодермы. Плюрипотентность стволовых клеток может быть подтверждена, например, инъекцией клеток в мышей с тяжелым комбинированным иммунодефицитом (SCID), фиксацией тератом, которые образуются, с использованием 4% параформальдегида и затем исследованием их гистологически для доказательства типов клеток из этих трех зародышевых слоев. Альтернативно, плюрипотентность может быть определена созданием эмбриоидных телец и оцениванием этих эмбриоидных телец на присутствие маркеров, ассоциированных с этими тремя зародышевыми слоями.
Размноженные линии плюрипотентных стволовых клеток могут быть кариотипированы с использованием стандартного способа G-бэндинга и сравнены с опубликованными кариотипами соответствующих видов приматов. Желательным является получение клеток, которые имеют "нормальный кариотип", что означает, что эти клетки являются эуплоидными клетками, в которых все хромосомы человека присутствуют и не имеют заметных изменений.
Источники плюрипотентных стволовых клеток
Типы плюрипотентных стволовых клеток, которые могут использоваться, включают установленные линии плюрипотентных стволовых клеток, полученные из ткани, образованной после гестации, включающие преэмбриональную (зародышевую) ткань (такую как, например, бластоциста), эмбриональную (зародышевую) ткань или фетальную ткань, взятую в любое время во время гестации, обычно, но необязательно, перед приблизительно 10-12 неделями гестации. Не ограничивающими примерами являются установленные линии эмбриональных стволовых клеток человека или эмбриональные зародышевые клетки человека, такие как, например, линии эмбриональных стволовых клеток человека Н1, Н7 и Н9 (WiCell). Обсуждается также применение композиций этого описания во время начального установления или стабилизации таких клеток, и в этом случае клетками-источником могут быть первичные плюрипотентные клетки, взятые непосредственно из этого источника ткани. Подходящими являются также клетки, взятые из популяции плюрипотентных стволовых клеток, уже культивируемой в отсутствие питающих клеток. Подходящими являются также мутантные линии эмбриональных стволовых клеток человека, такие как, например, BGO1v (BresaGen, Athens, GA).
В одном из вариантов осуществления эмбриональные стволовые клетки человека получают, как описано Thomson et al. (U.S. Pat. No. 5843780; Science 282: 1145, 1998; Curr. Top. Dev. Biol. 38:
133 ff, 1998; Proc. Natl. Acad. Sci. U.S.A. 92:7844, 1995).
Культивирование плюрипотентных стволовых клеток
В одном из вариантов осуществления плюрипотентные стволовые клетки обычно культивируют на слое питающих клеток, которые поддерживают плюрипотентные стволовые клетки различными путями. Альтернативно, плюрипотентные стволовые клетки культивируют в культуральной системе, которая по существу не содержит питающих клеток, но, тем не менее, поддерживает пролиферацию плюрипотентных стволовых клеток, не подвергая их существенной дифференцировке. Рост плюрипотентных стволовых клеток в не содержащей питающих клеток культуре без дифференцировки поддерживают с использованием среды, кондиционированной культивированием предварительно с другим типом клеток. Альтернативно, рост плюрипотентных стволовых клеток в не содержащей питающих клеток культуре без дифференцировки поддерживают с использованием среды определенного химического состава.
Например, в статье Reubinoff et al. (Nature Biotechnology 18: 399-404 (2000)) и в статье Thompson et al. (Science 6 November 1998: Vol. 282. no. 5391, pp. 1145-1147), описана культура линий плюрипотентных стволовых клеток из бластоцист человека, в которой используется слой эмбриональных фибробластных питающих клеток мыши.
Richards et al. (Stem Cells 21: 546-556, 2003) оценивали набор из 11 различных слоев дифференцированных, фетальных и неонатальных питающих клеток человека на их способность поддерживать культуру плюрипотентных стволовых клеток человека. В статье Richards et al. указано: "Линии эмбриональных стволовых клеток человека, культивируемые на дифференцированных фибробластах кожи в качестве питающих клеток, сохраняют морфологию эмбриональных стволовых клеток человека и остаются плюрипотентными".
В документе US20020072117 описаны клеточные линии, которые продуцируют среды, поддерживающие рост плюрипотентных стволовых клеток приматов в не содержащей питающих клеток культуре. Используемые клеточные линии являются мезенхимными и фибробластподобными клеточными линиями, полученными из эмбриональной ткани или дифференцированные из эмбриональных стволовых клеток. В документе US20020072117 также описано применение этих клеточных линий в качестве первичного слоя питающих клеток.
В другом примере, в статье Wang et al. (Stem Cells 23: 1221-1227, 2005), описаны способы длительного выращивания плюрипотентных стволовых клеток человека на слоях питающих клеток, полученных из эмбриональных стволовых клеток человека.
В другом примере, в статье Stojkovic et al. (Stem Cells 23: 306-314, 2005), описана система питающих клеток, полученная путем спонтанной дифференцировки эмбриональных стволовых клеток человека.
В следующем примере, в статье Miyamoto et al. (Stem Cells 22: 433-440, 2004), описан источник питающих клеток, полученный из плаценты человека.
В статье Amit et al. (Biol. Reprod 68: 2150-2156, 2003) описан слой питающих клеток, полученный из крайней плоти человека.
В другом примере, в статье Inzunza et al. (Stem Cells 23: 544-549, 2005), описан слой питающих клеток из постнатальных фибробластов крайней плоти человека.
В документе US6642048 описаны среды, которые поддерживают рост плюрипотентных стволовых клеток (pPS) приматов в не содержащей питающих клеток культуре, и клеточные линии, подходящие для получения таких сред. В документе US6642048 указано: "Изобретение включает мезенхимные и фибробластподобные клеточные линии, полученные из эмбриональной ткани или дифференцированные из эмбриональных стволовых клеток. Способы получения таких клеточных линий, рабочие среды и выращивание стволовых клеток с использованием кондиционированных сред описаны и иллюстрированы в изобретении".
В другом примере, в документе WO2005014799, описана кондиционированная среда для поддержания, пролиферации и дифференцировки клеток млекопитающих. В документе WO2005014799 указано: "Культуральная среда, полученная в соответствии с изобретением, является кондиционированной активностью секреции клеток мышиных клеток, в частности, дифференцированных и иммортализованных трансгенных гепатоцитов, названных MMH (Met Murine Hepatocyte)".
В другом примере, в статье Xu et al. (Stem Cells 22: 972-980, 2004), описана кондиционированная среда, полученная из производных эмбриональных стволовых клеток человека, которые были генетически модифицированы для сверхэкспрессии теломеразы, обратной транскриптазы человека.
В другом примере, в документе US20070010011, описана культуральная среда определенного химического состава для поддержания плюрипотентных стволовых клеток.
Альтернативная культуральная система использует бессывороточную среду, дополненную факторами роста, способными стимулировать пролиферацию эмбриональных стволовых клеток. Например, в статье Cheon et al. (BioReprod DOI:10.1095/biolreprod.105.046870, October 19, 2005) описана не содержащая питающих клеток, бессывороточная культуральная система, в которой эмбриональные стволовые клетки поддерживаются в некондиционированной среде с заменой сыворотки (SR), дополненной различными факторами роста, способная запускать самообновление эмбриональных стволовых клеток.
В другом примере, в статье Levenstein et al. (Stem Cells 24: 568-574, 2006) описаны способы для долгосрочной культуры эмбриональных стволовых клеток человека в отсутствие фибробластов или кондиционированной среды, с использованием сред, дополненных bFGF.
В другом примере, в документе US20050148070 описан способ культивирования эмбриональных стволовых клеток человека в средах определенного состава без сыворотки и без фибробластных питающих клеток, причем этот способ включает: культивирование стволовых клеток в культуральной среде, содержащей альбумин, аминокислоты, витамины, минеральные соединения, по меньшей мере трансферрин или заменитель трансферрина, по меньшей мере инсулин или заменитель инсулина, причем эта культуральная среда по существу не содержит фетальной сыворотки млекопитающего и содержит по меньшей мере приблизительно 100 нг/мл фактора роста фибробластов, способного активировать рецептор передачи сигнала фактора роста фибробластов, где фактор роста подается из источника, отличного от слоя питательных фибробластов, причем эта среда поддерживает пролиферацию стволовых клеток в недифференцированном состоянии без питающих клеток или кондиционированной среды.
В другом примере, в документе US20050233446, описаны среды определенного состава, которые можно использовать для культивирования стволовых клеток, в том числе недифференцированных примордиальных стволовых клеток приматов. В растворе эта среда является по существу изотоничной по сравнению с культивируемыми стволовыми клетками. В конкретной культуре конкретная среда содержит базальную среду и некоторое количество каждого bFGF, инсулина и аскорбиновой кислоты, необходимых для поддержания по существу недифференцированного роста примордиальных стволовых клеток.
В другом примере, в документе US6800480, указано: "В одном из вариантов осуществления описывается среда для культивирования клеток для выращивания полученных из приматов примордиальных стволовых клеток по существу в недифференцированном состоянии, которая включает базальную среду с низким осмотическим давлением, низким содержанием эндотоксина, которая является эффективной для поддержания роста полученных из приматов примордиальных стволовых клеток. Эту базальную среду объединяют с питательной сывороткой, эффективной для поддержания роста полученных из приматов примордиальных стволовых клеток, и субстратом, выбранным из группы, состоящей из питающих клеток и компонента внеклеточного матрикса, происходящего из питающих клеток. Эта среда включает дополнительно аминокислоты, не являющиеся незаменимыми, антиоксидант и первый фактор роста, выбранный из группы, состоящей из нуклеозидов и соли пировиноградной кислоты”.
В другом примере, в документе US20050244962, указано: "В одном аспекте изобретение относится к способу культивирования эмбриональных стволовых клеток приматов. Эти стволовые клетки культивируют в культуре, по существу не содержащей фетальной сыворотки млекопитающего (предпочтительно, по существу не содержащей любой сыворотки животного), и в присутствии фактора роста фибробластов, который добавляют из источника, отличного от слоя питательных фибробластов. В предпочтительном варианте слой питательных фибробластов, ранее необходимый для поддержания культуры стволовых клеток, снимает необходимость добавления достаточного количества фактора роста фибробластов".
В следующем примере, в документе WO2005065354, описана изотоническая культуральная среда определенного состава, которая является по существу не содержащей слоя питающих клеток и сыворотки, содержащая: a) базальную среду; b) количество bFGF, достаточное для поддержания роста по существу недифференцированных стволовых клеток млекопитающих; c) количество инсулина, достаточное для поддержания роста по существу недифференцированных стволовых клеток млекопитающих; и d) количество аскорбиновой кислоты, достаточное для поддержания по существу недифференцированных стволовых клеток млекопитающих.
В другом примере, в документе WO2005086845, описан способ поддержания недифференцированной стволовой клетки, причем указанный способ предусматривает обработку стволовой клетки представителем семейства белков трансформирующего фактора роста бета (TGFβ), представителем семейства белков фактора роста фибробластов (FGF) или никотинамида (NIC) в количестве, достаточном для поддержания этой клетки в недифференцированном состоянии в течение достаточного периода времени для достижения желаемого результата.
Плюрипотентные стволовые клетки могут быть посеяны на подходящий культуральный субстрат. В одном из вариантов осуществления подходящим культуральным субстратом является компонент внеклеточного матрикса, такой как, например, компоненты, полученные из базальной мембраны, или который может образовывать часть сопряжений рецептор-лиганд молекулы адгезии. В одном из вариантов осуществления подходящим культуральным субстратом является MATRIGEL™ (Becton Dickenson). MATRIGEL™ является растворимым препаратом из опухолевых клеток Энгельбрета-Хольма-Сварма, который превращается в гель при комнатной температуре с образованием воспроизведенной базальной мембраны.
В качестве альтернативы пригодны другие компоненты и смеси компонентов внеклеточного матрикса. В зависимости от типа пролиферирующих клеток они могут включать в себя ламинин, фибронектин, протеогликан, энтактин, гепаринсульфат и тому подобное, по отдельности или в различных комбинациях.
Плюрипотентные стволовые клетки могут быть посеяны на субстрат в подходящем распределении и в присутствии среды, которая стимулирует выживаемость, размножение клеток и сохранение желаемых характеристик. Для всех этих характеристик важным является внимание и осторожность в отношении распределения посева, и это распределение может быть легко определено квалифицированным в данной области специалистом.
Подходящие культуральные среды могут быть приготовлены из следующих компонентов, таких как, например, модифицированная по способу Дульбекко среда Игла (DMEM), Gibco #11965-092; модифицированная по способу Дульбекко среда Игла для нокаута (KO DMEM), Gibco #10829-018; Ham F12/50% базальная среда DMEM; 200 мМ L-глутамин, Gibco #15039-027; раствор заменимых аминокислот, Gibco 11140-050; β-меркаптоэтанол, Sigma #M7522; рекомбинантный основный фактор роста фибробластов человека (bFGF), Gibco #13256-029.
Дифференцировка плюрипотентных стволовых клеток
В одном из вариантов осуществления изобретения плюрипотентные стволовые клетки размножают в культуре при поддержании их плюрипотентности. Изменения плюрипотентности клеток со временем могут быть определены детектированием изменений в уровнях экспрессии маркеров, ассоциированных с плюрипотентностью. Альтернативно, изменения плюрипотентности могут быть подвергнуты мониторингу детектированием изменений в уровнях экспрессии маркеров, ассоциированных с дифференцировкой, или маркеров, ассоциированных с другим типом клеток.
В альтернативном варианте осуществления плюрипотентные стволовые клетки размножают в культуре и затем обрабатывают способом, который стимулирует их дифференцировку в другой тип клеток. Другим типом клеток может быть клетка, экспрессирующая маркеры, характерные для линии дифференцировки дефинитивной эндодермы. Альтернативно, этим типом клеток может быть клетка, экспрессирующая маркеры, характерные для линии дифференцировки панкреатической эндодермы. Альтернативно, этим типом клеток может быть клетка, экспрессирующая маркеры, характерные для β-клеточной линии дифференцировки.
Плюрипотентные стволовые клетки, обработанные в соответствии со способами изобретения, могут быть дифференцированы в различные другие типы клеток любым подходящим способом, известным в данной области. Например, плюрипотентные стволовые клетки, обработанные в соответствии со способами изобретения, могут дифференцироваться в нервные клетки, сердечные клетки, гепатоциты и т.п.
Например, плюрипотентные стволовые клетки, обработанные в соответствии со способами изобретения, могут быть дифференцированы в предшественники нервных клеток и кардиомиоциты в соответствии со способами, описанными в документе WO2007030870.
В другом примере плюрипотентные стволовые клетки, обработанные в соответствии со способами изобретения, могут быть дифференцированы в гепатоциты в соответствии со способами, описанными в патенте США 6458589.
Образование клеток, экспрессирующих маркеры, характерные для линии дифференцировки дефинитивной эндодермы
Плюрипотентные стволовые клетки могут быть дифференцированы в клетки, экспрессирующие маркеры, характерные для линии дифференцировки дефинитивной эндодермы, любым способом, используемым в данной области.
Например, плюрипотентные стволовые клетки могут быть дифференцированы в клетки, экспрессирующие маркеры, характерные для линии дифференцировки дефинитивной эндодермы, в соответствии со способами, описанными в статье D'Amour et al. Nature Biotechnology 23, 1534-1541 (2005).
Например, плюрипотентные стволовые клетки могут быть дифференцированы в клетки, экспрессирующие маркеры, характерные для линии дифференцировки дефинитивной эндодермы, в соответствии со способами, описанными в статье Shinozaki et al. Development 131, 1651-1662 (2004).
Например, плюрипотентные стволовые клетки могут быть дифференцированы в клетки, экспрессирующие маркеры, характерные для линии дифференцировки дефинитивной эндодермы, в соответствии со способами, описанными в статье McLean et al. Stem Cells 25, 29-38 (2007).
Например, плюрипотентные стволовые клетки могут быть дифференцированы в клетки, экспрессирующие маркеры, характерные для линии дифференцировки дефинитивной эндодермы, в соответствии со способами, описанными в статье D'Amour et al. Nature Biotechnology 24, 1392-1401 (2006).
Маркеры, характерные для линии дифференцировки дефинитивной эндодермы, выбраны из группы, состоящей из SOX17, GATA4, Hnf-3beta, GSC, Cer1, Nodal. FGF8, Brachyury, Mix-подобного белка гомеобокса, FGF4, CD48, эомезодермина (EOMES), DKK4, FGF17, GATA6, CXCR4, C-Kit, CD99 и OTX2. Подходящей для применения в настоящем изобретении является клетка, которая экспрессирует по меньшей мере один из маркеров, характерных для линии дифференцировки дефинитивной эндодермы. В одном аспекте изобретения клеткой, экспрессирующей маркеры, характерные для линии дифференцировки дефинитивной эндодермы, является клетка-предшественник первичной полоски. В альтернативном аспекте клеткой, экспрессирующей маркеры, характерные для линии дифференцировки дефинитивной эндодермы, является мезоэндодермальная клетка. В альтернативном аспекте клеткой, экспрессирующей маркеры, характерные для линии дифференцировки дефинитивной эндодермы, является клетка дефинитивной эндодермы.
Образование клеток, экспрессирующих маркеры, характерные для линии дифференцировки панкреатической эндодермы
Плюрипотентные стволовые клетки могут быть дифференцированы в клетки, экспрессирующие маркеры, характерные для линии дифференцировки панкреатической эндодермы, любым способом, используемым в данной области.
Например, плюрипотентные стволовые клетки могут быть дифференцированы в клетки, экспрессирующие маркеры, характерные для линии дифференцировки панкреатической эндодермы, в соответствии со способами, описанными в статье D'Amour et al. Nature Biotechnology 23, 1534-1541 (2005).
Маркеры, характерные для линии дифференцировки панкреатической эндодермы, выбраны из группы, состоящей из Pdx1, HNF-1бета, PTF1a, HNF-6, HB9 и PROX1. Для применения в изобретения является подходящей клетка, которая экспрессирует по меньшей мере один из маркеров, характерных для линии дифференцировки панкреатической эндодермы. В одном аспекте изобретения клеткой, экспрессирующей маркеры, характерные для линии дифференцировки панкреатической эндодермы, является клетка панкреатической эндодермы.
Образование клеток, экспрессирующих маркеры линии панкреатической эндокринной дифференцировки
Плюрипотентные стволовые клетки могут быть дифференцированы в клетки, экспрессирующие маркеры, характерные для линии панкреатической эндокринной дифференцировки, любым способом, используемым в данной области.
Например, плюрипотентные стволовые клетки могут быть дифференцированы в клетки, экспрессирующие маркеры, характерные для линии панкреатической эндокринной дифференцировки, в соответствии со способами, описанными в статье D'Amour et al. Nature Biotechnology 23, 1534-1541 (2006).
Например, плюрипотентные стволовые клетки могут быть дифференцированы в клетки, экспрессирующие маркеры, характерные для линии панкреатической эндокринной дифференцировки, в соответствии со способами, описанными в документе Nature Biotechnology 24, 1392-1401 (2006).
Например, плюрипотентные стволовые клетки могут быть дифференцированы в клетки, экспрессирующие маркеры, характерные для линии панкреатической эндокринной дифференцировки, в соответствии со способами, описанными в статье D'Amour et al. Nature Biotechnology 23, 1534-1541 (2006).
Маркеры, характерные для линии панкреатической эндокринной дифференцировки, выбраны из группы, состоящей из NGN-3, NeuroD, Islet-1, Pdx-1, NKX6.1, Pax-4, Ngn-3 и PTF-1 альфа. В одном из вариантов осуществления панкреатическая эндокринная клетка способна экспрессировать по меньшей мере один из следующих гормонов: инсулин, глюкагон, соматостатин и панкреатический полипептид. Подходящей для применения в изобретении является клетка, которая экспрессирует по меньшей мере один из маркеров, характерных для линии панкреатической эндогенной дифференцировки. В одном аспекте изобретения клеткой, экспрессирующей маркеры, характерные для линии панкреатической эндокринной дифференцировки, является панкреатическая эндокринная клетка. Панкреатическая эндокринная клетка может быть панкреатической экспрессирующей гормон клеткой. Альтернативно, панкреатической эндокринной клеткой является панкреатическая секретирующая гормон клетка.
В одном из аспектов изобретения панкреатической эндокринной клеткой является клетка, экспрессирующая маркеры, характерные для β-клеточной линии дифференцировки. Клетка, экспрессирующая маркеры, характерные для β-клеточной линии дифференцировки, экспрессирует по меньшей мере один из следующих факторов транскрипции: NGN-3, Nkx2.2, Nkx6.1, NeuroD, Isl-1, HNF-3 бета, MAFA, Pax4 и Pax6. В одном из аспектов изобретения клеткой, экспрессирующей маркеры, характерные для β-клеточной линии дифференцировки, является β-клетка.
Трехмерные носители
Материалы-носители, подходящие для применения для целей изобретения, включают синтетические и природные материалы в форме пен, губок, гелей, гидрогелей, текстильных материалов и нетканых структур, которые были использованы in vitro и in vivo для реконструирования или регенерации биологической ткани, а также для доставки хемотаксических агентов для индукции роста ткани, которые являются подходящими для применения в практике способов изобретения. Смотрите, например, материалы, описанные в патенте США 5770417, патенте США 6022743, патенте США 5567612, патенте США 5759830, патенте США 6626950, патенте США 6534084, патенте США 6306424, патенте США 6365149, патенте США 6599323, патенте США 6656488, опубликованной заявке на патент США 2004/0062753 A1, патенте США 4557264 и патенте США 6333029.
Для получения носителя с включенным фармацевтическим агентом фармацевтический агент может быть смешан с раствором полимера перед образованием носителя. Альтернативно, фармацевтический агент может быть нанесен в виде покрытия на полученный носитель, предпочтительно в присутствии фармацевтического носителя. Фармацевтический агент может присутствовать в виде жидкости, мелкоизмельченного твердого вещества или в любой другой подходящей физической форме. Альтернативно, к носителю могут быть добавлены эксципиенты для изменения скорости высвобождения фармацевтического агента. В альтернативном варианте осуществления в носитель включают по меньшей мере одно фармацевтическое соединение, которое является противовоспалительным соединением, таким как, например, соединения, описанные в патенте США 6509369.
В носитель может быть включено по меньшей мере одно фармацевтическое соединение, которое является антиапоптотическим соединением, таким как, например, соединения, описанные в патенте США 6793945.
В носитель может быть включено по меньшей мере одно фармацевтическое соединение, которое является ингибитором фиброза, таким как, например, соединения, описанные в патенте США 6331298.
В носитель может быть включено по меньшей мере одно фармацевтическое соединение, которое способно усиливать ангиогенез, такое как, например, соединения, описанные в опубликованной заявке на патент США 2004/0220393 и опубликованной заявке на патент США 2004/0209901.
В носитель может быть также включено по меньшей мере одно фармацевтическое соединение, которое является иммуносупрессивным соединением, таким как, например, соединения, описанные в опубликованной заявке на патент США 2004/0171623.
Далее настоящее изобретение иллюстрируется, но не ограничивается следующими примерами.
ПРИМЕРЫ
Пример 1
Пассирование и поддержание hESC в виде кластеров клеток
Эмбриональные стволовые клетки человека Н1 и Н9 поддерживали на инактивированных митомицином С первичных эмбриональных фибробластах мыши (MEF). Клетки hES переключали из питающих MEF-клеток на MATRIGEL™ посредством повторяемых пассажей.
Покрытие MATRIGEL™ чашек для культуры ткани: разведенный фактором роста MATRIGEL™ (Becton-Dickinson, Bedford, Mass.) оттаивали при 4°С и затем разбавляли 1:30 в холодной среде DMEM/F12 (Invitrogen, Carlsbad, CA). Объемы, достаточные для покрытия, добавляли в каждую чашку диаметром 6 см (2 мл) или в каждую лунку 6-луночной чашки (1 мл) и инкубировали 1 час при комнатной температуре. Чашки использовали в течение нескольких часов или выдерживали при 4°C до двух недель.
Культура эмбриональных стволовых клеток человека: недифференцированные колонии эмбриональных стволовых клеток человека (Н9 и Н1) собирали из слоев питающих клеток инкубированием в 1 мг/мл коллагеназы IV (Sigma-Aldrich, St. Louis, MO) в DMEM/F12 в течение 10 минут, с последующим выскребанием пипеткой. Скопления клеток осаждали центрифугированием при 1000 об/мин в течение четырех минут, и осадок осторожно диспергировали пипеткой на 2 мл для разрушения колоний на малые кластеры клеток. Эти кластеры клеток высевали на покрытые MATRIGEL™ чашки в MEF-CM, дополненной bFGF (8 нг/мл; R&D Systems, Minneapolis, MN), 50-150 колоний на чашку диаметром 6 см в 5 мл среды для выращивания. Среду заменяли ежедневно. Колонии на MATRIGEL™ в MEF-CM становились большими, и их пассировали, когда они занимали 70-80% площади поверхности, приблизительно каждые 3-4 дня. Клетки hES в колониях имели высокое отношение ядра к цитоплазме и имели выступающие ядрышки, подобно клеткам hES, поддерживаемым на питающих клетках (фигура 1). Дифференцированные клетки представляли менее 5% общих клеток в культуре.
Для рутинного пассажа клеток в MEF-CM на MATRIGEL™ клетки инкубировали в 1 мг/мл коллагеназы IV в DMEM/F12 в течение до 60 минут и удаляли из чашек сильными струями DMEM/F12 с выскребанием. Клетки осаждали, диспергировали и высевали при отношении 1:3 или 1:4.
Пример 2
Пассирование эмбриональных стволовых клеток человека в виде отдельных клеток: оценивание ферментов
Для облегчения манипулирования клетками hES в способах пассирования могут использоваться другие ферментативные растворы, которые требуют более короткого времени инкубации и не требуют стадии выскребания. Кроме того, пассирование клеток в виде кластеров клеток коллагеназой не позволяет определить численное количество высеваемых клеток. Доступны многие ферментативные растворы для высвобождения отдельных клеток в одной быстрой стадии. Быстродействующий фермент, который вызывает минимальное повреждение клеток и не препятствует прикреплению клеток или росту клеток, был идентифицирован посредством следующего эксперимента.
Эмбриональные стволовые клетки H9p33 человека, выращенные в кластерах в чашке с 6 лунками, инкубировали со следующими ферментами: TrypLE™ Express, TrypLE™ Select, трипсин/ЭДТА (0,05%) или трипсином (0,25%) в течение двух минут при 36°C. Все ферменты высвобождали клетки в пределах двух минут, за исключением трипсина. Высвобождение трипсином достигалось после 5 минут при 36°C. Клетки повторно высевали из расчета 200000 клеток на лунку в 6-луночную чашку, покрытую MATRIGEL™, и им давали размножаться в течение трех дней. Эмбриональные стволовые клетки человека пассировали также в виде кластеров с использованием коллагеназы (30-минутная инкубация) и повторно высевали при разведении 1:5 на покрытые MATRIGEL™ лунки, подобно клеткам, число которых может быть определено с использованием TrypLE™ Express. Спустя три дня hESC высвобождали пятиминутной инкубацией с TrypLE™ Express. Клетки инкубировали с 0,01% трипановым синим и затем считали (таблица 1). Жизнеспособность клеток сразу после высвобождения была больше чем 98% для всех испытанных ферментов. Пассаж эмбриональных стволовых клеток человека с коллагеназой является стандартным способом пассажа. После трех дней культивирования как TrypLE™ Select, так и TrypLE™ Express давали числа извлеченных клеток, сходные с коллагеназой. Смесь трипсин/ЭДТА и трипсин были значимо менее эффективными в поддержании прикрепления/роста клеток. TrypLE™ Select и TrypLE™ Express были наилучшими оцениваемыми ферментами, и их дополнительно испытывали в эксперименте с временным ходом, в примере 3.
Пример 3
Пассирование эмбриональных стволовых клеток человека в виде отдельных клеток: оптимизация времени воздействия фермента
TrypLE™ Select и TrypLE™ Express оказались оптимальными из всех испытанных ферментов. Для определения идеального времени инкубации для ферментов с клетками hES TrypLE™ Select и TrypLE™ Express инкубировали с кластерами H9p34 hESC в течение двух минут или 10 минут при 37°С. Клетки удаляли из лунки, считали и осаждали центрифугированием. Аликвоты 200000 клеток на лунку высевали в 6-луночную чашку. Клетки выращивали в течение трех дней с последующим высвобождением с использованием TrypLE™ Express и счетом в присутствии 0,01% трипанового синего.
Жизнеспособность клеток была более 98% для обоих ферментов для обоих периодов инкубирования. Таблица III показывает количество извлеченных клеток на лунку спустя 36 часов после посева. Клетки, пассированные с TrypLE™ Express, достигали первоначальной плотности посева после трех дней. Это указывает на то, что большинство клеток не прикреплялось повторно после посева; однако клетки, которые действительно прикреплялись, способны пролиферировать и размножаться. При предположении, что клетки размножаются при той же скорости после прикрепления, эти данные демонстрируют, что обработка в течение двух минут TrypLE™ Express приводит к наивысшей степени прикрепления. Затем обработки TrypLE™ Express в течение двух минут использовали для получения отдельных клеток во всех последующих экспериментах.
Пример 4
Дифференцировка эмбриональных стволовых клеток человека в виде отдельных клеток и в виде кластеров клеток в дефинитивную эндодерму
Эмбриональные стволовые клетки могут дифференцироваться в множественные линии дифференцировки клеток. Клетки ES человека, пассированные в виде отдельных клеток, предоставляют значительно лучшие возможности определения количества введенных клеток и облегчения манипулирования. Определяли способность отдельных клеток hES дифференцироваться.
Посев кластеров клеток и отдельных клеток: чашку диаметром 6 см кластеров клеток Н9 или Н1 на разведенном фактором роста MATRIGEL™ инкубировали с 2 мл коллагеназы (1 мг/мл) в DMEM:F12 до 60 минут при 37°C. Клетки удаляли пипетированием и выскребанием и центрифугировали в течение 4 минут при 900 об/мин. Затем кластеры клеток высевали в 6-луночную чашку, покрытую разведенным 1:15 или 1:30 фактором роста MATRIGEL™. Этот способ пассирования приводил к посеву кластеров клеток (cc). Альтернативно, чашку диаметром 6 см кластеров клеток Н9 или Н1 инкубировали на разведенном фактором роста MATRIGEL™ с TrypLE™ Express (2 мл) в течение 5 минут при 37°C и диспергировали пипетированием. После центрифугирования при 900 об/мин в течение 4 минут клетки высевали в 6-луночную чашку, покрытую разведенным 1:10 фактором роста MATRIGEL™, и их назвали отдельными клетками (sc).
Дифференцировка в дефинитивную эндодерму: культуры H9 и H1 sc и cc при приблизительно 60-70% конфлюэнтности подвергали действию среды DMEM:F12, дополненной 0,5% ФТС, 10 нг/мл Wnt3a (R&D Systems) и 100 нг/мл активина А (AA; R&D Systems), в течение двух дней с последующей обработкой средой DMEM/F12, дополненной 2% ФТС и 100 нг/мл активина А, в течение дополнительных трех дней.
Эти культуры анализировали при помощи FACS на экспрессию CXCR4, CD99 и CD9 и при помощи ПЦР реального времени на SOX-17, SOX-7, фетальный белок альфа (AFP), CXCR4, Brychyury (Bry), goosecoid (GSC), HNF-3 бета и GATA4. AFP и SOX-7 считали маркерами висцеральной эндодермы, тогда как GATA4, HNF-3 бета и SOX-17 представляют маркеры дефинитивной эндодермы, а GSC, Bry и CXCR4 представляют маркеры первичной полоски. Отдельные клетки дифференцировались в DE в сходной с кластерами клеток степени, как наблюдали по проценту получения CXCR4-положительных клеток (фигура 2).
Пример 5
Дифференцировка отдельных клеток и кластеров hES человека в панкреатическую эндодерму
Дополнительную дифференцировку в соответствии с протоколом, опубликованным Novocell (Baetge, EE et al. Nature Biotechnology (2006), 24(11), 1392-1401), с модификациями испытывали также для определения способности к дифференцировке отдельных клеток hES. Эти клетки дополнительно дифференцировались в панкреатическую эндодерму в соответствии с протоколом DE, описанным в примере 4.
Дифференцировка в панкреатическую эндодерму: один из экспериментов с DE H9 (p37), использующих как кластеры клеток, так и отдельные клетки из примера 4, использовали дополнительно для дифференцировки кластеров и отдельных клеток в панкреатическую эндодерму. После завершения протокола дефинитивной эндодермы клетки инкубировали в течение 3 дней с FGF10 (50 нг/мл; R&D Systems), ингибитором sonic hedgehog, KAAD циклопамином (2,5 мкМ; Sigma-Aldrich) и 2% ФТС в среде DMEM:F12. Затем клетки инкубировали в течение трех дополнительных дней с FGF10 (50 нг/мл), KAAD циклопамином (2,5 мкМ), ретиноевой кислотой (1 мкМ; Sigma-Aldrich) и 1% B27 (Invitrogen) в DMEM с низким содержанием глюкозы. Затем клетки инкубировали в течение дополнительных трех дней в эксендине 4 (50 нг/мл; Sigma-Aldrich), DAPT (1 мкМ; Calbiochem) и 1% В27 в DMEM с низким содержанием глюкозы. Пробы РНК брали из одной лунки 6-луночной чашки для каждого типа клеток и затем анализировали при помощи ПЦР реального времени в этой стадии на панкреатические маркеры Pdx1, Nkx6.1, Nkx2.2, Pax4, NeuroD, HNF3b, Ptf1a, инсулин и AFP. Дифференцировку продолжали в течение трех дней со средой CMRL (Invitrogen), содержащей 50 нг/мл HGF, IGF (R&D Systems), и эксендин 4 (50 нг/мл), и 1% B27. Оценивание тех же маркеров панкреатической эндодермы повторяли на этой стадии. РНК-пробы из необработанных клеток hES той же самой линии подвергали ПЦР реального времени параллельно с обработанными пробами. Обработанные пробы нормализовали относительно необработанных контролей, установленных на кратное изменение 1. Экспрессию Pdx1 подвергали мониторингу и сравнивали между отдельными клетками и кластерами клеток. Индукция экспрессии маркера панкреатической эндодермы была эквивалентной между отдельными клетками и кластерами клеток (фигура 3). Таким образом, отдельные клетки hES имеют присущую им способность к дифференцировке, сходную со способностью кластеров клеток hES.
Пример 6
Пассирование клеток hES в виде отдельных клеток
Пассирование клеток hES в виде отдельных клеток могло бы способствовать возможности увеличения масштаба культур и облегчению процесса производства.
Описание генерирования и пассирования отдельных клеток: клетки H9 выращивали в виде кластеров на MATRIGEL™, как описано выше. При пассаже 38 чашку диаметром 6 см клеток Н9 инкубировали с 2 мл TrypLE™ Express в течение 5 минут при 37°C. Эти клетки ресуспендировали в DMEM:F12 и центрифугировали в течение 4 минут при 900 об/мин. Клетки повторно высевали при отношении 1:4 на чашки, покрытые разведенным 1:30 фактором роста MATRIGEL™. После приблизительно пяти пассажей эти клетки считали перед повторным посевом при плотности 14000 клетка/см2. Пассирование и повторный посев при этой плотности продолжали с интервалами каждые четыре дня. Клетки сохраняли компактную структуру при росте из отдельных клеток, но никогда не образовывали прочные кластеры (сравните фигуру 1 и фигуру 4).
Пример 7
Анализ плюрипотентности отдельных клеток hES
Поддержание плюрипотентности клеток hES является необходимым при любом способе пассирования. Таким образом, отдельные клетки оценивали на плюрипотентность после множественных пассажей с использованием TrypLE™ Express.
FACS-анализ плюрипотентности: отдельные клетки Н9 культивировали с 38 пассажами в виде кластеров с последующими 16 пассажами в виде отдельных клеток, включающими одно замораживание-оттаивание при криоконсервации. Затем клетки анализировали при помощи FACS на экспрессию маркеров плюрипотентности. Прикрепившиеся клетки удаляли из культуральных чашек с использованием 5-минутной инкубации с раствором TrypLE™ Express. Высвободившиеся клетки ресуспендировали в буфере для окрашивания, состоящем из 2% БСА (Sigma-Aldrich, St. Louis, MO), 0,05% азида натрия в ЗФР. При необходимости, эти клетки блокировали в отношении Fc-рецептора в течение 15 минут с использованием 0,1% раствора гамма-глобулина (Sigma). Аликвоты (приблизительно 105 клеток) инкубировали с конъюгированными либо с фикоэритрином (РЕ), либо с аллофикоцианином (АРС) моноклональными антителами (5 мкл антитела на 106 клеток), как показано в таблице III, или с неконъюгированным первичным антителом. Контроли включали подходящие изотипически сходные антитела, неокрашенные клетки и клетки, окрашенные только вторичным конъюгированным антителом. Все инкубации с антителами выполняли в течение 30 минут при 4°C, после чего клетки промывали буфером для окрашивания. Пробы, которые окрашивались неконъюгированными первичными антителами, инкубировали в течение дополнительных 30 минут при 4°C со вторичными конъюгированными меченными РЕ или АРС антителами. Смотрите таблицу III в отношении перечня использованных вторичных антител. Промытые клетки осаждали и ресуспендировали в буфере для окрашивания, и молекулы клеточной поверхности идентифицировали с использованием прибора FACS Array (BD Biosciences) со сбором по меньшей мере 10000 событий. Пассированные с использованием коллагеназы hESC H1p40 и H9scp16 имеют эквивалентные профили экспрессии белков плюрипотентности (фигура 5).
Пример 8
Анализ стабильности кариотипа эмбриональных стволовых клеток человека
Кариотип отдельных клеток hES должен оставаться стабильным на протяжении множественных пассажей. Отдельные клетки H9 культивировали с 38 пассажами в виде кластеров с последующими множественными пассажами в виде отдельных клеток. Кариотип клеток Н9 определяли стандартным анализом кариотипа с использованием G-бэндинга (Гимза-бэндинга) (Cell Line Genetics, Madison, WI). Оценивали всего 20 подвергнутых G-бэндингу клеток, и 200 интерфазных ядер анализировали при помощи FISH (флуоресцентной гибридизации in situ). В анализируемых клетках не обнаруживали аберрации хромосом. Цитогенетический анализ показал, что клетки имели нормальное число аутосом и модальное число хромосом 46. Кариотипирование отдельных клеток H9 проводили при пассаже 13 и пассаже 20. Клетки пассажа 20 подвергали одному событию замораживания-оттаивания криоконсервации. Клетки пассажа 13 и 20 были кариотипически нормальными (фигура 6).
Пример 9
Дифференцировка отдельных клеток hES и кластеров клеток hES в дефинитивную эндодерму
Для конечного применения клетки hES, пассированные в виде отдельных клеток, должны сохранять потенциал дифференцировки. Отдельные клетки подвергали протоколу дефинитивной эндодермы следующим образом. Клетки H9 после множественных пассажей с использованием TrypLE™ Express (sc-p) высвобождали с использованием TrypLE™ Express (sc) из пассированных отдельных клеток (пассаж 6 и 21) на MATRIGEL™ (разведение 1:10) для дифференцировки. H9 sc, при приблизительно 60-70% конфлюэнтных монослоев, подвергали действию среды DMEM:F12, дополненной 0,5% ФТС, 10 нг/мл Wnt3a и 100 нг/мл активина А, в течение двух дней, с последующей обработкой средой DMEM/F12, дополненной 2% ФТС и 100 нг/мл активина А, в течение дополнительных трех дней.
В день 5 клетки анализировали при помощи FACS на экспрессию CXCR4, CD99 и CD9 и при помощи ПЦР реального времени на SOX-17, SOX-7, фетальный белок альфа (AFP), CXCR4, Brychyury (Bry), goosecoid (GSC), HNF-3 бета и GATA4. AFP и SOX-7 считаются маркерами висцеральной эндодермы, тогда как GATA4, HNF-3 бета и SOX-17 представляют маркеры дефинитивной эндодермы, а GSC, Bry и CXCR4 представляют маркеры первичной полоски. Отдельные клетки, пассированные много раз (пассаж 6 и 21), сохраняют свою способность дифференцироваться в DE, аналогично кластерам клеток (пассажи 36-55) (фигура 7).
Пример 10
Дифференцировка отдельных клеток hES
в панкреатическую эндодерму
Клетки дополнительно дифференцировались в панкреатическую эндодерму при следующих стадиях: три дня инкубации с FGF10 (50 нг/мл), ингибитором sonic hedgehog, KAAD циклопамином (2,5 мкМ) и 2% ФТС в среде DMEM:F12, с последующими тремя днями с FGF10 (50 нг/мл), KAAD циклопамином (2,5 мкМ), ретиноевой кислотой (1 мкМ; Sigma-Aldrich) и 1% B27 (Invitrogen) в DMEM с низким содержанием глюкозы. Оценивание этих клеток выполняли в день 11. Некоторые культуры продолжали обрабатывать в течение трех дней в эксендине 4 (50 нг/мл), DAPT (1 мкМ) и 1% В27 в DMEM с низким содержанием глюкозы. В день 14 пробы анализировали на панкреатические маркеры Pdx1, Nkx6.1, Nkx2.2, Pax4, NeuroD, HNF3b, Ptf1a, инсулин и AFP при помощи ПЦР реального времени. Некоторые культуры продолжали дифференцировку в течение дополнительных трех дней со средой CMRL, содержащей 50 нг/мл HGF, IGF и эксендин 4, и 1% B27. Оценивание некоторых маркеров панкреатической эндодермы повторяли в конце дня 17. Маркеры панкератической эндодермы, Nkx2.2, NeuroD, HNF6, HNF3b, преимущественно экспрессировались в конце дней 14 и 17. Экспрессия Pdx1 увеличивалась ступенчато на каждой стадии обработки (фигура 8).
Пример 11
Дифференцировка отдельных клеток hES на питающих MEF-клетках
Для достижения оптимальной дифференцировки клеток hES в панкреатическую эндодерму популяция дефинитивной эндодермы должна быть максимальной. В настоящее время выращивание hES на питающих MEF-клетках приводит к наивысшим достигаемым уровням дефинитивной эндодермы и панкреатической эндодермы. Для определения, способны ли отдельные клетки hES достигать этой цели, H1scp4 высевали на питающие MEF-клетки при плотности 14000 клетка/см2. Эти клетки выращивали в среде для клеток ES, содержащей раствор 20% заменителя сыворотки для нокаута (Invitrogen), 1× заменимых аминокислот (Invitrogen), 8 нг/мл bFGF, 1 мМ L-глутамина и 1 мМ 2-меркаптоэтанола в DMEM-F12. После 7 дней клетки были на 60-70% конфлюэнтными, и к этим клеткам применяли протокол дефинитивной эндодермы. Конкретно, два дня добавляли 100 нг/мл активина А, 10 нг/мл Wnt3a и 0,5% ФТС в среде DMEM:F12, 100 мкл на лунку, с последующей трехдневной обработкой 100 нг/мл активина А и 2% ФТС в среде DMEM:F12. Затем клетки удаляли с использованием TrypLE™ Express и анализировали при помощи FACS на экспрессию CXCR4, CD99 и CD9 (фиг. 9). Более 90% этих клеток экспрессировали CXCR4 и CD99. Менее 8% этих клеток экспрессировали CD9, как и ожидалось. Посев отдельных клеток Н1 на питающих MEF-клетках улучшает дифференцировку в дефинитивную эндодерму, как было определено по количествам CXCR4-положительных клеток.
Пример 12
Дифференцировка отдельных клеток hES в 96-луночных планшетах
Одной обычной трудностью при пассаже клеток hES в виде кластеров является то, что их трудно определять количественно, и то, что они не растут при одинаковых скоростях. Отдельные клетки имеют то преимущество, что они могут расти до конфлюэнтного монослоя, и их можно подсчитывать перед пассажем для гарантии равного посева для экспериментальных целей. Эти характерные черты являются предпосылкой успешной валидизации скрининга.
hES H9scp18 высевали в 96-луночные планшеты Packard View (Perkin-Elmer), покрытые разведенным 1:30 фактором роста MATRIGEL™, при плотности 14000 клетка/см2. Эти клетки выращивали в течение трех-четырех дней в MEF-кондиционированной среде и затем обрабатывали с использованием протокола дифференцировки DE. Одну субпопуляцию 40 лунок обрабатывали с использованием стандартного протокола (10 нг/мл Wnt3a, 100 нг/мл активина А и 0,5% ФТС в DMEM:F12 в течение двух дней). Вторую субпопуляцию 40 лунок обрабатывали ингибитором IX GSK3b (1000 нМ; EMD Chemicals, La Jolla, CA) вместо Wnt3a. Затем выполняли обработку для обеих популяций в течение трех дней с использованием 100 нг/мл активина А и 2% ФТС в DMEM:F12. В конце культивирования клетки фиксировали 4% параформальдегидом при комнатной температуре в течение 20 минут, промывали три раза ЗФР и выдерживали в 100 мкл ЗФР в течение ночи. Клетки обрабатывали для увеличения проницаемости 0,5% Тритоном Х-100 (Sigma-Aldrich), либо при комнатной температуре в течение 20 минут, либо при 4°С в течение пяти минут, затем промывали три раза ЗФР и блокировали 4% куриной сывороткой (Invitrogen-Gibco, Carlesbad, CA) в ЗФР в течение 30 минут при комнатной температуре. Первичные антитела (козьи анти-hSoxl7 (R&D Systems)) разбавляли и добавляли в 4% куриную сыворотку при разведении 1:100 в течение 1 часа при комнатной температуре. Вторичное антитело, Alexa Fluor 488, куриное антитело против козьего IgG (Invitrogen-Molecular Probes, Carlesbad, CA), разбавляли 1:200 в ЗФР и добавляли к клеткам после 3-кратного промывания ЗФР. Для контрастного окрашивания ядер к клеткам добавляли 5 мкМ Draq5 (Alexis Platform, Laufelfingen, Switzerland) в течение 5 минут при комнатной температуре. Клетки промывали один раз ЗФР и оставляли в 100 мкл на лунку ЗФР для визуализации для определения количества дифференцированных клеток DE. Этот планшет считывали на анализаторе IN Cell 1000 (GE Healthcare; Piscataway, NJ) для определения от лунки к лунке количества клеток и окрашивания Sox17. Обработка Wnt3a приводила к наивысшему количеству ядер клеток на лунку с небольшой вариабельностью между лунками (фигура 10). Обработка ингибитором Gsk3β обнаруживала плохую жизнеспособность клеток. Ясно, что обработка Wnt3a позволяет отдельным клеткам ES образовывать DE устойчивым и воспроизводимым образом в 96-луночном формате.
Пример 13
Анализы скрининга с отдельными клетками hES
После демонстрации того, что отдельные клетки пригодны для посева и дифференцировки в 96-луночном формате, определяли чувствительность к обработке с новыми соединениями. hES H9scp19 высевали в 96-луночные планшеты, покрытые разведенным 1:30 фактором роста MATRIGEL™, при плотности 14000 клетка/см2. Эти клетки выращивали в течение 3-4 дней в MEF-кондиционированной среде и затем обрабатывали в эксперименте дифференцировки DE. В целом, испытывали 13 новых соединений в трех повторностях при концентрации 1 или 3 мкМ, с заменой Wnt3a ингибитором с малой молекулой в протоколе дифференцировки DE. В качестве положительных контролей использовали лунки, содержащие Wnt3a (10 нг/мл) или ингибитор IX Gsk3b (100 нМ). Все лунки с дифференцировкой обрабатывали одним соединением плюс 100 нг/мл активина А и 0,5% ФТС в DMEM:F12 в течение двух дней с последующей обработкой в течение двух дней в отсутствие соединения 100 нг/мл активина А и 2% ФТС в DMEM:F12. В конце этого протокола клетки фиксировали, делали проницаемыми, блокировали и окрашивали, как описано в примере 11 выше, для определения экспрессии Sox17 и ядер. Затем планшет считывали на анализаторе IN Cell 1000 Analyzer. Этот скрининг показал, что H9scp21 отвечали на 3 из 13 соединений подобно Wnt3a в протоколе дифференцировки DE (фигура 11). Соединение А было эффективным при более низкой дозе 1 мкМ, тогда как соединения В и С были эффективны как при 1 мкМ, так и при 3 мкМ. В этом диапазоне эффективной обработки (более 60% Sox17-положительных клеток) стандартное отклонение между лунками было низким. Таким образом, отдельные клетки являются стойкими в их способности идентифицировать новые соединения посредством способов скрининга.
Пример 14
Размножение отдельных клеток hES в роллерных флаконах
Для облегчения увеличения масштаба для производственных задач клетки hES должны легко размножаться в больших количествах. В настоящее время обработка коллагеназой для пассирования клеток hES не поддается увеличению масштаба в роллерных флаконах или встряхиваемых колбах. Клетки hES, повторяемо размножаемые и выращиваемые в виде отдельных клеток, способны одинаково прикрепляться к различным поверхностям. В этом примере роллерные флаконы испытывали в качестве потенциальных увеличивающих масштаб производства сосудов. H9scp19 высевали при плотности 14000 клетка/см2 в роллерный флакон на 480 см2 (6,7×106 клеток; Corning, Acton, MA), покрытый разведенным 1:30 фактором роста MATRIGEL™. Объемы 100 мл MEF-кондиционированной среды на флакон использовали для поддержания этих клеток и сменяли каждые два дня. Эти флаконы устанавливали при 20 об/час в течение 24 часов и увеличивали до 60 об/час для остального времени. Клетки прикреплялись равномерно к роллерному флакону и явно размножались на протяжении четырехдневного периода оценивания. Эти клетки удаляли при помощи TrypLE™ Express и считали. Получаемые количества показали 13×106 клеток на флакон, что свидетельствует о минимальном удвоении встречающихся клеток по сравнению с первоначальным посевом. Авторы изобретения приближенно оценивают, что менее половины первоначально посеянных клеток прикреплялись к роллерному флакону, что позволяет предположить, что в роллерных флаконах имеет место большее фактическое размножение клеток. Это демонстрирует, что клетки hES, пассированные в виде отдельных клеток, могут размножаться в автоматизированной системе с роллерными флаконами. Будут проводиться дальнейшие эксперименты для определения скорости размножения вместе с сохранением плюрипотентности.
Пример 15
Трансфекция отдельных клеток hES
Введение ДНК в клетки для получения трансгенных типов клеток является полезным признаком. ДНК может содержать экспрессирующий вектор или репортерный ген для облегчения дифференцировки или для обеспечения возможности отбора клеток. Клетки hES оказались трудными для трансфекции в виде кластеров. Однако отдельные клетки hES могут быть более поддающимися способам трансфекции.
Клетки H9p33 пассировали с использованием TrypLE™ Express и высевали при плотности 20000 клетка/см2 на покрытый разведенным 1:30 фактором роста MATRIGEL™ планшет. Клетки H9p33 пассировали с использованием коллагеназы и высевали из чашки диаметром 6 см в одну лунку 6-луночной чашки, покрытой разведенным 1:30 фактором роста MATRIGEL™. Спустя 3 дня эти клетки трансфицировали CMV-GFP, где GFP экспрессируется в каждой клетке под контролем вирусного промотора CMV. Конкретно, 2 или 4 мкг ДНК разводили в 250 мкл среды Opti-MEM и 8 мкл липофектамина 2000 (Invitrogen) в 250 мкл Opti-MEM. Эти смеси объединяли после пяти минут при комнатной температуре. Объединенные компоненты инкубировали в течение 20 минут при комнатной температуре перед добавлением к клеткам, культивируемым в двух мл MEF-кондиционированной среды. После этого среду заменяли MEF-кондиционированной средой каждые 24 часа. Спустя три дня клетки фиксировали 4% параформальдегидом при комнатной температуре в
течение 20 минут, затем промывали два раза ЗФР и оставляли в ЗФР при 4°С на протяжении ночи. Для контрастного окрашивания ядер к клеткам добавляли 5 мкМ Draq5 (Alexis Platform) в течение пяти минут при комнатной температуре. Клетки промывали один раз ЗФР и оставляли в 1 мл на лунку ЗФР для визуализации на анализаторе IN Cell 1000 для определения эффективности трансфекции (фигура 12). Отдельные клетки hES обнаруживают двукратное улучшение эффективности трансфекции по сравнению с кластерами hES. Таким образом, отдельные клетки hES имеют улучшенную способность трансфекции и являются более легкими для генетической модификации. Посредством оптимизации, возможно с использованием других способов трансфекции или реагентов, может быть станет возможным достижение более разительного улучшения трансфекции отдельных клеток hES.
Пример 16
Превращение кластеров клеток hES, культивируемых на слое питающих клеток, в отдельные клетки hES, культивируемые на внеклеточном матриксе, требует удаления питающих клеток
Пассирование клеток млекопитающих с использованием ферментов, таких как трипсин, рекомбинантный трипсин (TrypLE™) или Accutase™ (полученный из беспозвоночных трипсин-подобный фермент), является существующим стандартом для культуры клеток как первичных, так и иммортализованных клеточных линий в исследовательской и производственной обстановках. Эти ферменты образуют гомогенную суспензию отдельных клеток, которые могут быть сосчитаны, проанализированы, подвергнуты манипуляциям и пассированы воспроизводимым образом для посева в таких различных условиях, как высокопроизводительный формат 384-луночного планшета или многолитровый масштабируемый культуральный сосуд.
При условии очевидных преимуществ пассирования отдельных клеток существует значительный интерес в развитии способа пассирования эмбриональных стволовых клеток человека в виде отдельных клеток. Однако существующие наилучшие применения на практике эмбриональных стволовых клеток человека требуют, чтобы эти клетки пассировали в виде кластеров либо разрушением вручную, либо пассированием с ферментом (коллагеназой или нейтральной протеазой), которое сохраняет кластеры эмбриональных клеток. Прошлые исследования в этой области приводили к отведению клеток, которые могут быть пассированы в виде отдельных клеток, на питающие клетки (Cellartis SCEDTM461 line) или отведению отдельных клеток из пассируемых коллагеназой кластеров в отдельные клетки на MATRIGEL™ с использованием Accutase™ (R. Bajpai et al. 2008).
В настоящем изобретении предпринята попытка разработки способа пассирования отдельных клеток для получения клеток hES непосредственно из питающих клеток в матригель в способе пассажа больших количеств с использованием либо TrypLE™, либо Accutase™. Результаты показывают, что применение пассирования на основе питающей клетки кластерного типа клеток hES непосредственно на MATRIGEL™ с использованием пассирования в больших количествах при помощи либо TrypLE™, либо Accutase™ без промежуточной стадии коллагеназного пассирования на основе кластеров на MATRIGEL™ является неэффективным вследствие появления популяции дифференцированных клеток и, следовательно, с меньшей вероятностью приведет к гомогенной и устойчивой культуре hES человека.
Результаты
В попытке продвижения к не содержащей питающих клеток культуре клеток hES от культуры на основе питающих клеток авторы изобретения пассировали клетки hES Н1 пассажа 37, которые поддерживали слоем питающих MEF-клеток в среде hESC и выращивали в течение 5 дней в лунках 10 см2 6-луночной чашки. Эти клетки пассировали с использованием TrypLE™, Accutase™ или 1 мг/мл коллагеназы.
Перед добавлением фермента к этим клеткам авторы изобретения аспирировали использованную среду, добавляли 1 мл ЗФР (без Ca2+ или Mg2+) в каждую лунку, аспирировали и затем добавляли 1 мл фермента при комнатной температуре (коллагеназу, Accutase™ или TrypLE™). Accutase™ или TrypLE™ использовали в виде исходной концентрации после достижения комнатной температуры. Коллагеназу вынимали из морозильника -80°C, оттаивали, смешивали с 9 мл DMEM/F12, стерильно фильтровали и давали нагреться до комнатной температуры перед использованием.
Клетки инкубировали в ферменте при 37°С в течение 10 минут. Accutase™ или TrypLE™ вызывали полное поднятие всех клеток из чашки после 10-минутной обработки. Коллагеназа не поднимала клетки после 10-минутной инкубации, так что эти клетки соскребали стеклянной пипеткой на 10 мл после 10-минутной инкубации. Один мл 2% пробумина в DMEM-F12 добавляли в каждую лунку и объединенный общий объем в лунке переносили в стерильную коническую пробирку, стараясь поднять и суспендировать как можно больше клеток.
Эти клетки центрифугировали в течение 5 минут при 200 × g с последующей дополнительной промывкой 2% пробумином и центрифугированием в течение 5 минут при 200 × g. Затем клетки ресуспендировали в MEF-кондиционированной среде и высевали при отношении 1:3,5 в покрытые MATRIGEL™ (разведение 1:30 в DMEM) колбы T25 и помещали в увлажненный термостат с температурой 37°С, 5% СО2.
Количество клеток рассчитывали посредством подсчета с использованием улучшенного гемоцитометра Neubauer. Клетки, которые не поднимались ферментом Accutase™, имели плотность 3,8 миллиона клеток на лунку 10 см2. Клетки, которые поднимались TrypLE™, имели плотность 3,7 миллиона клеток на лунку 10 см2. Коллагеназа не могла считаться гемоцитометром вследствие типа (фасона) колоний. При условии индекса разведения 1:3,5 авторы изобретения высевали обработанные акутазой клетки при плотности посева 2,72 миллиона клеток на колбу Т25 или 105000 клетка/см2, обработанные TrypLE™ клетки при плотности посева 2,65 миллиона клеток на колбу Т25 или 105000 клетка/см2. Авторы изобретения предполагают, что обработанные коллагеназой лунки засевались при сходной плотности (105000-108000 клетка/см2), так как не было заметного различия в плотности hES от лунки к лунке перед обработкой ферментами. Дополнительную колбу засевали поднятыми коллагеназой клетками для целей счета в следующем пассаже.
Клетки поддерживали в течение 4 дней с ежедневными заменами MEF-кондиционированных сред, дополненных 16 мкг/мл bFGF. После 4 дней в культуре эти культуры были конфлюэнтными, и обработанные в виде отдельных клеток клетки hES образовывали монослои клеток hES со случайными очагами фибробластов, тогда как пассированные в виде кластеров клетки hES образовывали большие кластеры типа колоний клеток hES (фигура 13). Затем эти клетки снова пассировали.
Как описывалось ранее, клетки инкубировали в ферменте при 37°С в течение 10 минут. Accutase™ и TrypLE™ заставляли клетки подниматься от пластика и MATRIGEL™ после 10-минутной обработки. Коллагеназа не поднимала клетки, так что клетки инкубировали дополнительно в течение 45 минут в целом при 37°С. Дополнительную колбу с коллагеназой поднимали акутазой™. Затем отдельные клетки считали с использованием улучшенного гемоцитометра Neubauer.
Затем два мл 2% пробумина в DMEM-F12 добавляли в каждую колбу после инкубации с ферментом, общий объем (~4 мл) из колбы пипетировали вверх и вниз 5-10 раз для суспендирования как можно большего количества клеток. Затем суспензию переносили в коническую пробирку на 50 мл и центрифугировали в течение 5 минут при 200 × g. Затем клетки ресуспендировали в MEF-кондиционированной среде, дополненной 16 мкг/мл bFGF, и затем высевали в колбы Т25, предварительно покрытые 1:30 MATRIGEL™, при соотношении 1:4.
Пассированные с использованием Accutase™ клетки после четырех дней в культуре имели плотность 5,6 миллиона клеток в целом (223000 клетка/см2), пассированные с использованием TrypLE™ клетки после четырех дней в культуре имели плотность 5,05 миллиона клеток в целом (202000 клетка/см2), и пассированные с использованием коллагеназы клетки после четырех дней в культуре имели плотность 3,45 миллиона клеток в целом (138000 клетка/см2). При условии, что эти клетки пассировали при соотношении 1:4, клетки высевали при плотности 58000 клетка/см2 для пассажа с Accutase™, 51000 клетка/см2 для пассажа с TrypLE™ и 35000 клетка/см2 для пассажа с коллагеназой.
Затем клетки выращивали в течение дополнительных 6 дней с ежедневной заменой сред MEF-кондиционированными средами, дополненными 16 нг/мл bFGF. В то время как клетки, пассированные с коллагеназой, образуют большие колонии плотных и гомогенных клеток и дифференцированные/фибробластподобные колонии являются редкими, клетки, пассированные с Accutase™ или TrypLE™, росли очень медленно и со временем прекращали рост или зарастали фибробластподобными клетками, которые, вероятно, образовывались из дифференцирующихся клеток hES (фигура 14).
Эти результаты предполагают, что промежуточная стадия переноса из питающих клеток в не содержащую питающих клеток (MATRIGEL™) культуру при помощи пассирования вручную или с использованием фермента, которая поддерживает пассирование кластерного типа (коллагеназа или нейтральная протеаза), является наилучшей для акклиматизации клеток к не содержащей питающих клеток культуре на MATRIGEL™ перед началом пассирования отдельных клеток.
Публикации, цитируемые в настоящем документе, включены в настоящий документ посредством ссылки в их полном объеме. Хотя различные аспекты этого изобретения были иллюстрированы выше ссылкой на примеры и предпочтительные варианты осуществления, понятно, что объем настоящего изобретения определяется не предыдущим описанием, а следующей формулой изобретения, должным образом истолкованной в соответствии с нормами закона о патентах.

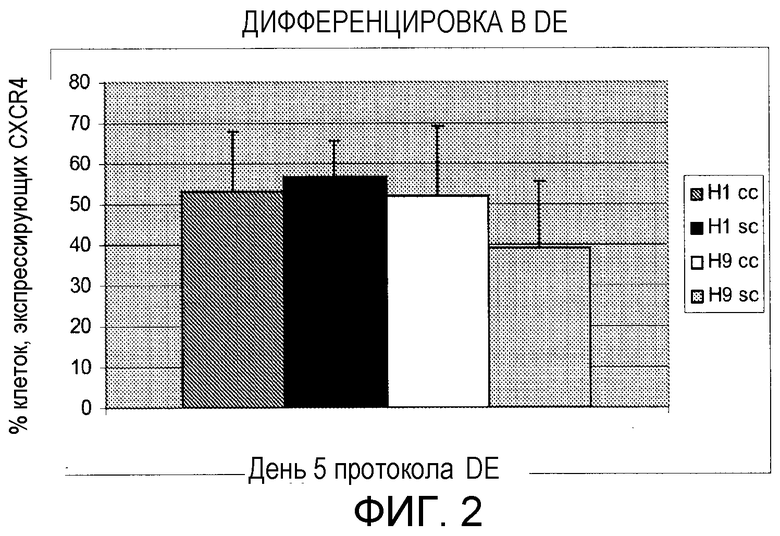
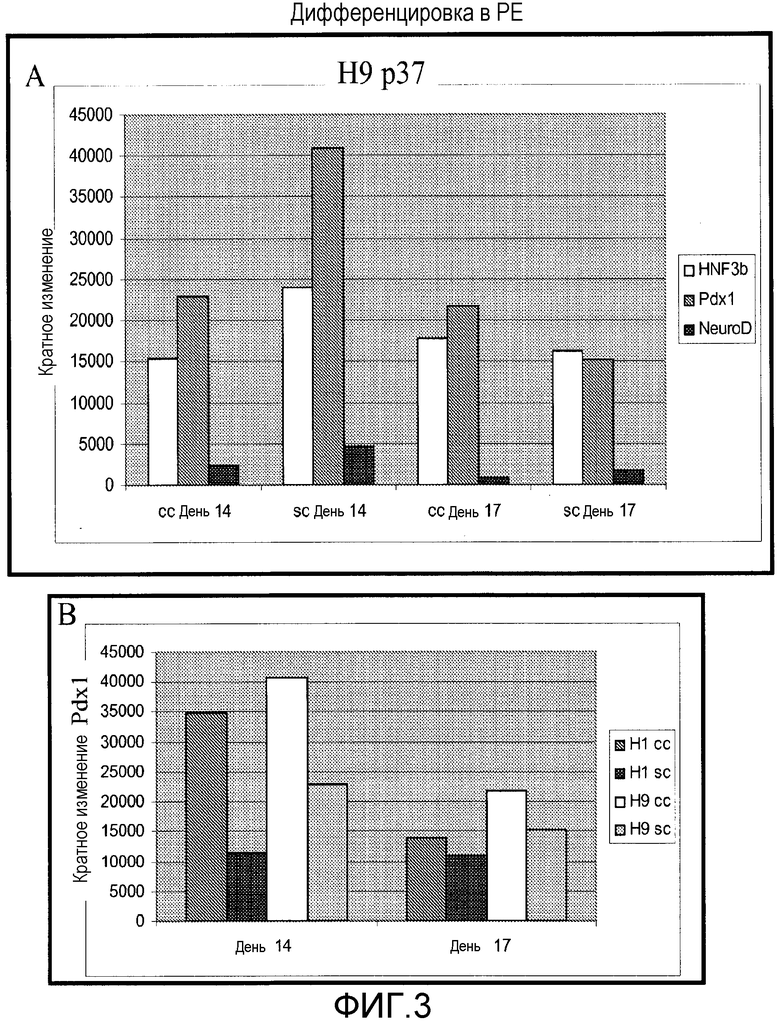
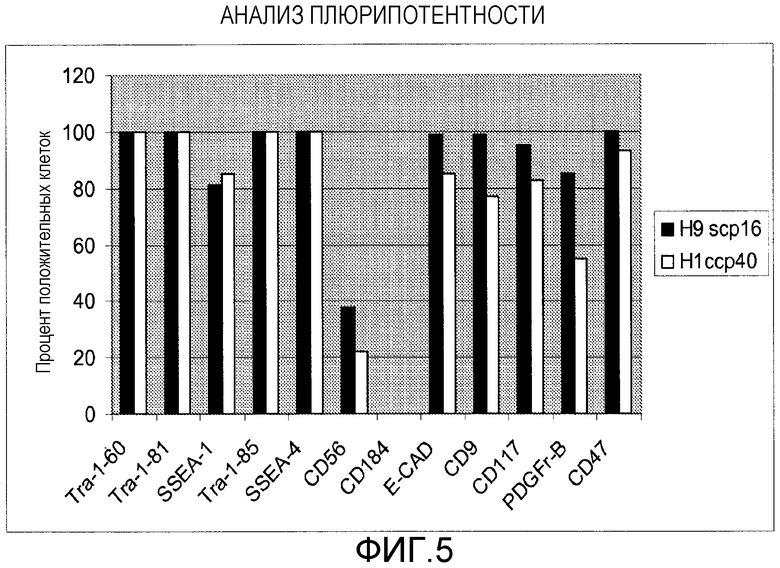

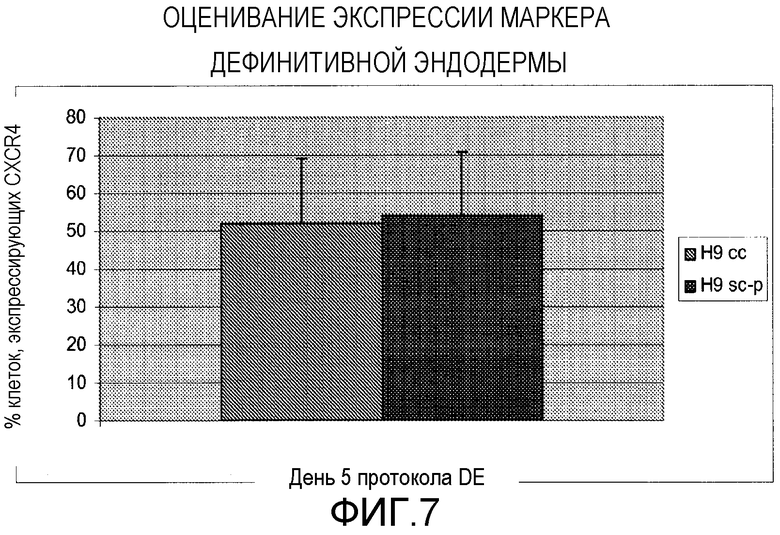
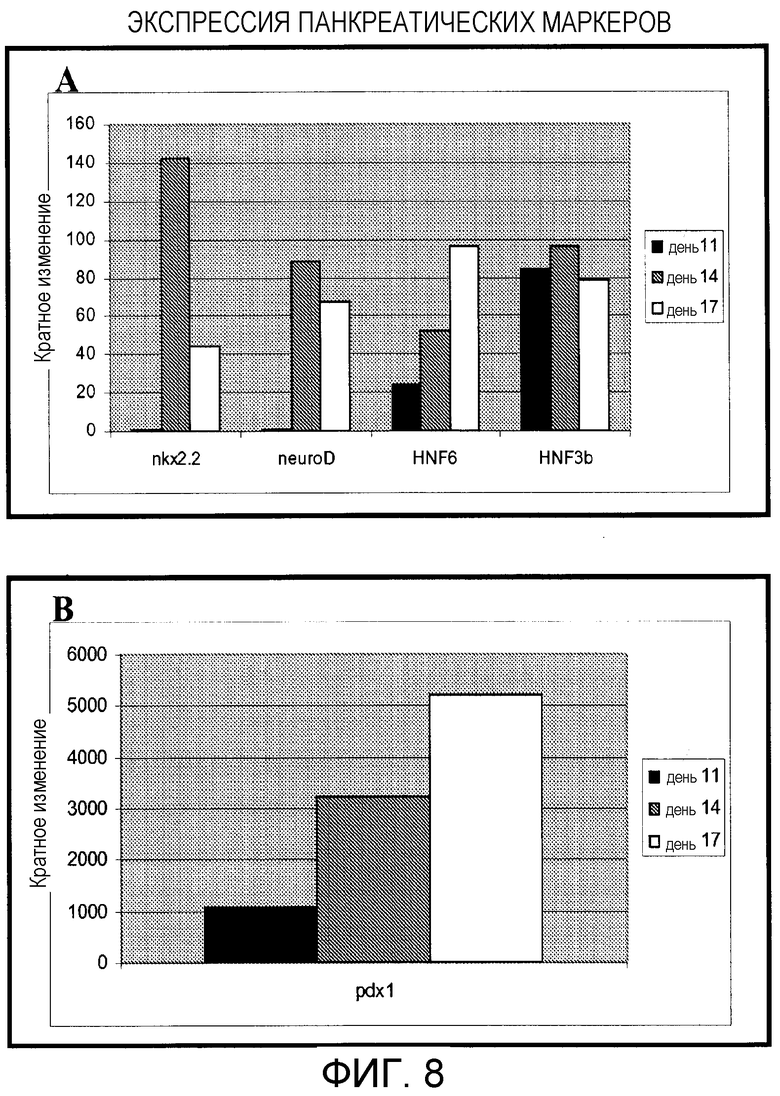

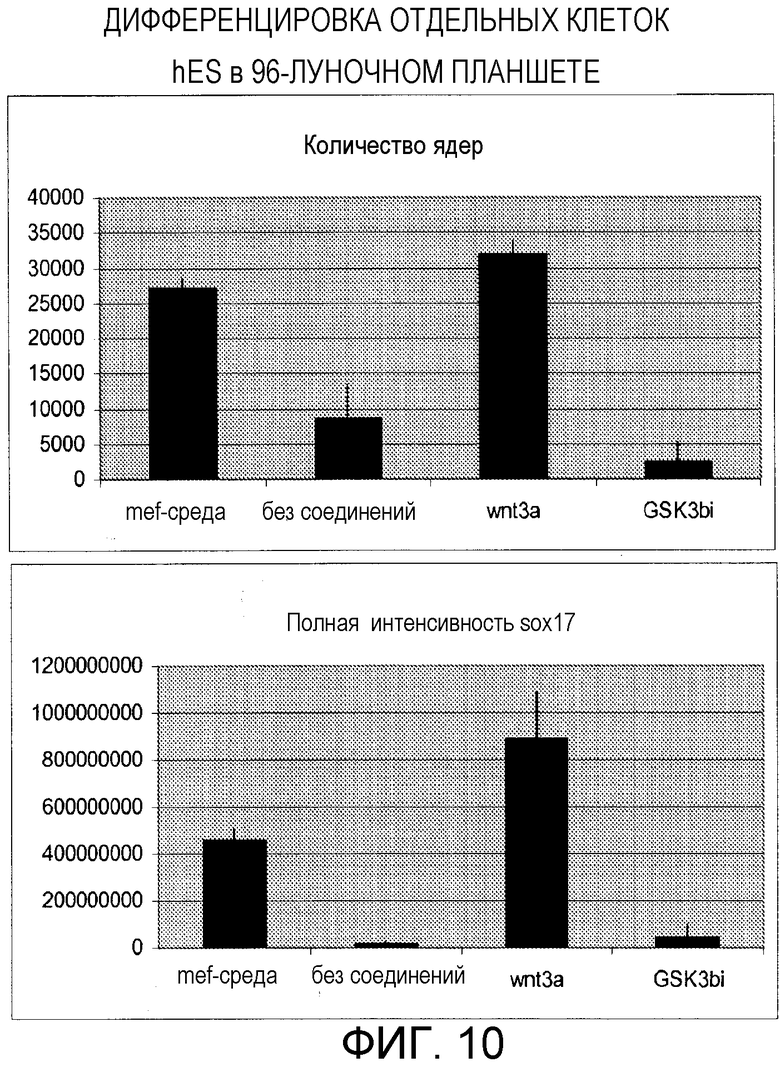
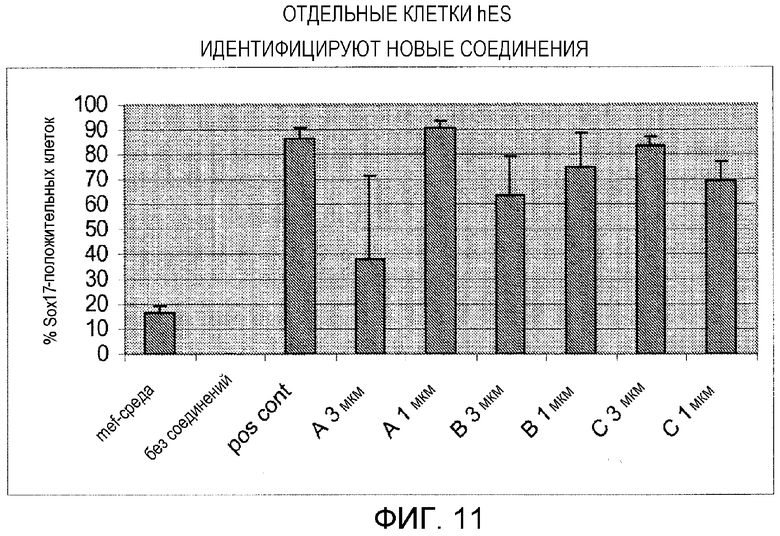
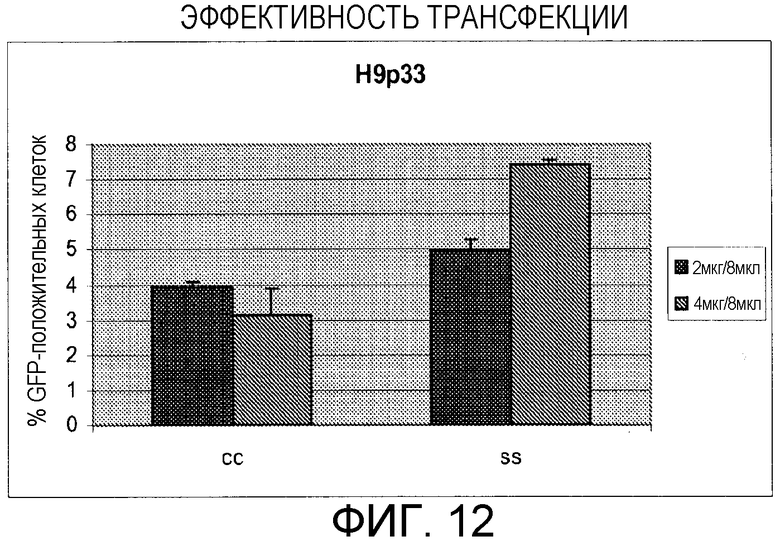
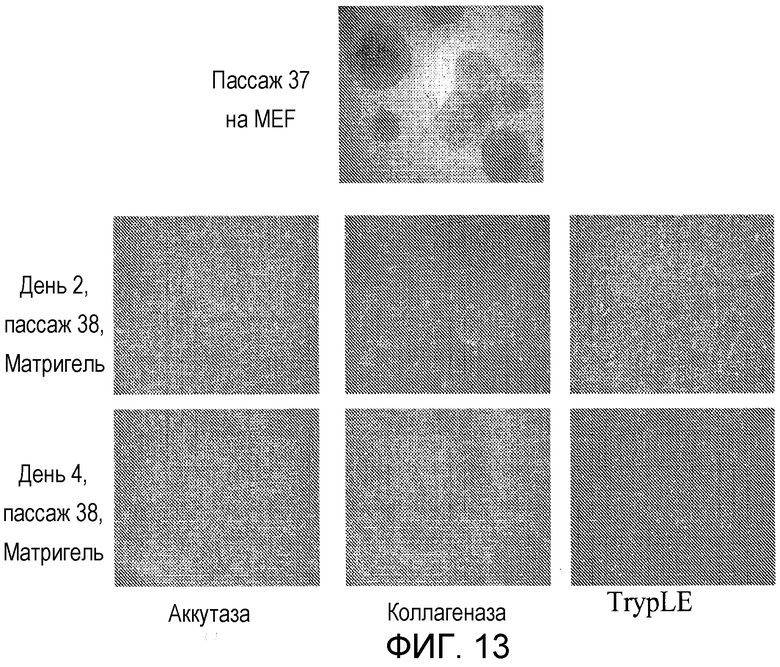
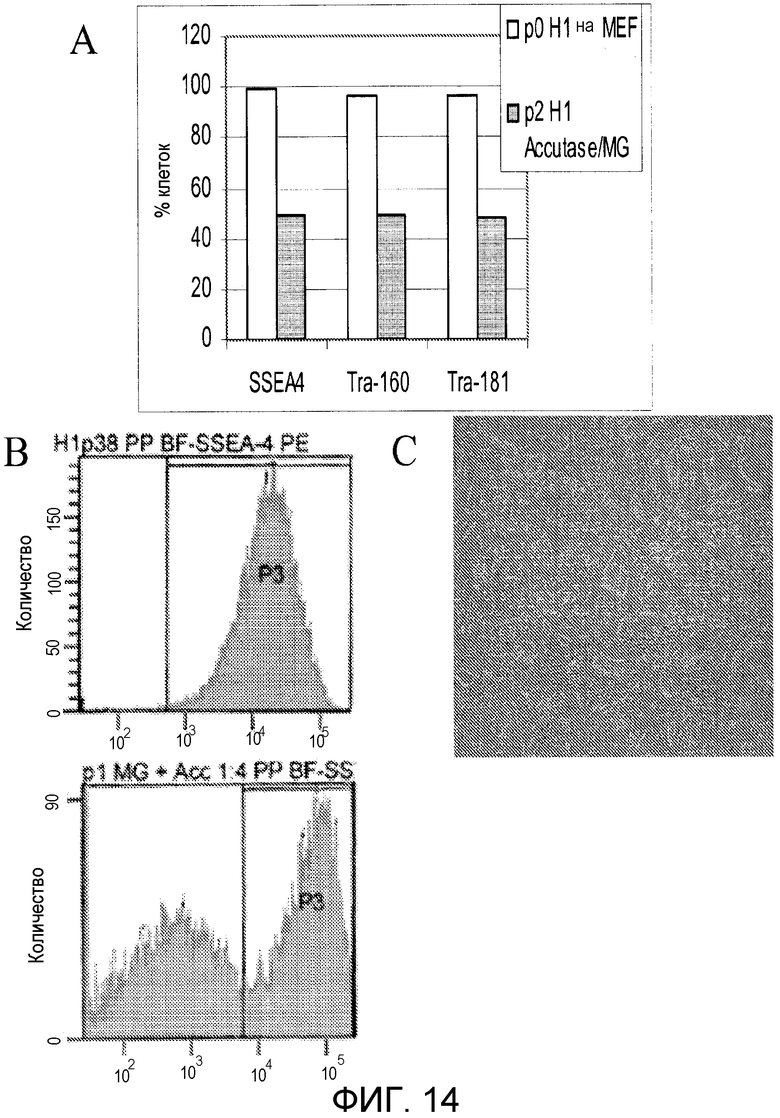
Изобретение относится к области биотехнологии и медицины. Раскрыт способ дифференцировки эмбриональных стволовых клеток человека в качестве единичных клеток в клетки, экспрессирующие маркеры, характерные для линии панкреотической эндодермы. Способ предусматривает культивирование эмбриональных стволовых клеток в виде кластеров и высвобождение эмбриональных стволовых клеток в виде отдельных клеток. Далее проводят посев отдельных эмбриональных стволовых клеток на субстрате культуры ткани и дифференцировку отдельных эмбриональных стволовых клеток. Также описаны способ поддержания эмбриональных стволовых клеток и способ пассирования эмбриональных стволовых клеток. Предложенная группа изобретений позволяет улучшить способы дифференцировки, поддержания и культивирования эмбриональных стволовых клеток. 3 н. и 46 з.п. ф-лы, 14 ил., 16 пр.
1. Способ дифференцировки эмбриональных стволовых клеток человека в качестве единичных клеток, полученных из существующих линий эмбриональных клеток в клетки, экспрессирующие маркеры, характерные для линии панкреатической эндодермы, включающий стадии:
a) культивирования эмбриональных стволовых клеток в виде кластеров,
b) высвобождения эмбриональных стволовых клеток в виде отдельных клеток,
c) посева отдельных эмбриональных стволовых клеток на субстрате культуры ткани и
d) дифференцировки отдельных эмбриональных стволовых клеток.
2. Способ по п.1, где эмбриональные стволовые клетки высвобождают в виде отдельных клеток обработкой ферментом, выбранным из группы, состоящей из трипсина, TrypLE™SELECT, TrypLE™EXPRESS и трипсин/ЭДТА.
3. Способ по п.2, где эмбриональные стволовые клетки высвобождают в виде отдельных клеток обработкой TrypLE™EXPRESS.
4. Способ по п.2, где эмбриональные стволовые клетки высвобождают в виде отдельных клеток обработкой ферментом при концентрации от приблизительно 0,5 г/л до приблизительно 2,5 г/л.
5. Способ по п.2, где эмбриональные стволовые клетки высвобождают в виде отдельных клеток обработкой ферментом при концентрации приблизительно 2,5 г/л.
6. Способ по п.2, где эмбриональные стволовые клетки высвобождают в виде отдельных клеток обработкой ферментом в течение приблизительно двух - приблизительно пяти минут.
7. Способ по п.2, где эмбриональные стволовые клетки высвобождают в виде отдельных клеток обработкой ферментом в течение приблизительно пяти минут.
8. Способ по п.3, где эмбриональные стволовые клетки высвобождают в виде отдельных клеток обработкой TrypLE™EXPRESS при концентрации от приблизительно 1X до приблизительно 0,001X.
9. Способ по п.3, где эмбриональные стволовые клетки высвобождают в виде отдельных клеток обработкой TrypLE™EXPRESS при концентрации 1X.
10. Способ по п.3, где эмбриональные стволовые клетки высвобождают в виде отдельных клеток обработкой TrypLE™EXPRESS в течение приблизительно двух - приблизительно пяти минут.
11. Способ по п.3, где эмбриональные стволовые клетки высвобождают в виде отдельных клеток обработкой TrypLE™EXPRESS в течение приблизительно пяти минут.
12. Способ по п.1, где субстрат культуры ткани выбран из группы, состоящей из MATRIGEL™, фибронектина, ламинина, сыворотки человека и коллагена.
13. Способ по п.12, где субстратом культуры ткани является MATRIGEL™.
14. Способ по п.13, где MATRIGEL™ используют при разведении от приблизительно 1:30 до приблизительно 1:10.
15. Способ по п.14, где MATRIGEL™ используют при разведении 1:10.
16. Способ по п.1, где отдельные эмбриональные стволовые клетки впоследствии дифференцируются.
17. Способ поддержания эмбриональных стволовых клеток, полученных из существующих линий эмбриональных клеток, включающий стадии:
a) получения кластеров эмбриональных стволовых клеток,
b) высвобождения эмбриональных стволовых клеток в виде отдельных клеток и
c) посева отдельных эмбриональных стволовых клеток на субстрате культуры ткани.
18. Способ по п.17, где эмбриональные стволовые клетки высвобождают в виде отдельных клеток обработкой ферментом, выбранным из группы, состоящей из трипсина, TrypLE™SELECT, TrypLE™EXPRESS и трипсин/ЭДТА.
19. Способ по п.18, где эмбриональные стволовые клетки высвобождают в виде отдельных клеток обработкой TrypLE™EXPRESS.
20. Способ по п.18, где эмбриональные стволовые клетки высвобождают в виде отдельных клеток обработкой ферментом при концентрации от приблизительно 0,5 г/л до приблизительно 2,5 г/л.
21. Способ по п.18, где эмбриональные стволовые клетки высвобождают в виде отдельных клеток обработкой ферментом при концентрации приблизительно 2,5 г/л.
22. Способ по п.18, где эмбриональные стволовые клетки высвобождают в виде отдельных клеток обработкой ферментом в течение приблизительно двух - приблизительно пяти минут.
23. Способ по п.18, где эмбриональные стволовые клетки высвобождают в виде отдельных клеток обработкой ферментом в течение приблизительно пяти минут.
24. Способ по п.19, где эмбриональные стволовые клетки высвобождают в виде отдельных клеток обработкой TrypLE™EXPRESS при концентрации от приблизительно 1X до приблизительно 0,001X.
25. Способ по п.19, где эмбриональные стволовые клетки высвобождают в виде отдельных клеток обработкой TrypLE™EXPRESS при концентрации 1X.
26. Способ по п.19, где эмбриональные стволовые клетки высвобождают в виде отдельных клеток обработкой TrypLE™EXPRESS в течение приблизительно двух - приблизительно пяти минут.
27. Способ по п.19, где эмбриональные стволовые клетки высвобождают в виде отдельных клеток обработкой TrypLE™EXPRESS в течение приблизительно пяти минут.
28. Способ по п.17, где субстрат культуры ткани выбран из группы, состоящей из MATRIGEL™, разведенного фактором роста MATRIGEL™, фибронектина, ламинина, сыворотки человека и коллагена.
29. Способ по п.28, где субстратом культуры ткани является разведенный фактором роста MATRIGEL™.
30. Способ по п.29, где разведенный фактором роста MATRIGEL™ используют при разведении от приблизительно 1:30 до приблизительно 1:10.
31. Способ по п.30, где разведенный фактором роста MATRIGEL™ используют при разведении 1:30.
32. Способ по п.17, где отдельные эмбриональные стволовые клетки впоследствии дифференцируются.
33. Способ пассирования эмбриональных стволовых клеток, полученных из существующих линий эмбриональных клеток, включающий стадии:
a) получения кластеров эмбриональных стволовых клеток,
b) высвобождения эмбриональных стволовых клеток в виде отдельных клеток,
c) посева отдельных эмбриональных стволовых клеток на субстрате культуры ткани,
d) предоставления возможности размножения отдельным эмбриональным клеткам,
e) высвобождения отдельных эмбриональных стволовых клеток и
f) посева отдельных эмбриональных стволовых клеток на новом субстрате культуры ткани.
34. Способ по п.33, где эмбриональные стволовые клетки высвобождают в виде отдельных клеток обработкой ферментом, выбранным из группы, состоящей из трипсина, TrypLE™ SELECT, TrypLE™EXPRESS и трипсин/ЭДТА.
35. Способ по п.34, где эмбриональные стволовые клетки высвобождают в виде отдельных клеток обработкой TrypLE™EXPRESS.
36. Способ по п.34, где эмбриональные стволовые клетки высвобождают в виде отдельных клеток обработкой ферментом при концентрации от приблизительно 0,5 г/л до приблизительно 2,5 г/л.
37. Способ по п.34, где эмбриональные стволовые клетки высвобождают в виде отдельных клеток обработкой ферментом при концентрации приблизительно 2,5 г/л.
38. Способ по п.34, где эмбриональные стволовые клетки высвобождают в виде отдельных клеток обработкой ферментом в течение приблизительно двух - приблизительно пяти минут.
39. Способ по п.34, где эмбриональные стволовые клетки высвобождают в виде отдельных клеток обработкой ферментом в течение приблизительно пяти минут.
40. Способ по п.35, где эмбриональные стволовые клетки высвобождают в виде отдельных клеток обработкой TrypLE™EXPRESS при концентрации от приблизительно 1X до приблизительно 0,001X.
41. Способ по п.35, где эмбриональные стволовые клетки высвобождают в виде отдельных клеток обработкой TrypLE™EXPRESS при концентрации 1X.
42. Способ по п.35, где эмбриональные стволовые клетки высвобождают в виде отдельных клеток обработкой TrypLE™EXPRESS в течение приблизительно двух - приблизительно пяти минут.
43. Способ по п.35, где эмбриональные стволовые клетки высвобождают в виде отдельных клеток обработкой TrypLE™EXPRESS в течение приблизительно пяти минут.
44. Способ по п.33, где субстрат культуры ткани выбран из группы, состоящей из MATRIGEL™, разведенного фактором роста MATRIGEL™, фибронектина, ламинина, сыворотки человека и коллагена.
45. Способ по п.44, где субстратом культуры ткани является разведенный фактором роста MATRIGEL™.
46. Способ по п.45, где разведенный фактором роста MATRIGEL™ используют при разведении от приблизительно 1:30 до приблизительно 1:10.
47. Способ по п.46, где разведенный фактором роста MATRIGEL™ используют при разведении 1:30.
48. Способ по п.33, где отдельные эмбриональные стволовые клетки на новом субстрате культуры ткани пассируют на другой субстрат культуры ткани высвобождением клеток и посевом высвобожденных клеток на другой субстрат культуры ткани.
49. Способ по п.33, где отдельные эмбриональные стволовые клетки впоследствии дифференцируются.
| KLIMANSKAYA I | |||
| et al., "Human embryonic stem cells derived without feeder cells", Lancet | |||
| Способ обработки целлюлозных материалов, с целью тонкого измельчения или переведения в коллоидальный раствор | 1923 |
|
SU2005A1 |
| ПРИБОР ДЛЯ ЗАПИСИ УГЛОВОГО ПЕРЕМЕЩЕНИЯ НАВОЯ ТКАЦКОГО СТАНКА | 0 |
|
SU181549A1 |
| WANG G | |||
| et al., "Noggin and bFGF cooperate to maintain the pluripotency of human embryonic stem cells in the absence of feeder layers", Biochem Biophys Res Commun | |||
| Способ обработки целлюлозных материалов, с целью тонкого измельчения или переведения в коллоидальный раствор | 1923 |
|
SU2005A1 |
| XU C. | |||

