ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к антителам и, в частности, антителам, связывающимся с человеческим 4-1ВВ.
ПРЕДШЕСТВУЮЩИЙ УРОВЕНЬ ТЕХНИКИ
4-1ВВ (также называемый CD137, TNFRSF9 и так далее) представляет собой трансмембранный белок суперсемейства рецептора фактора некроза опухоли (TNFRS). Согласно современному пониманию 4-1ВВ, его экспрессия обычно зависит от активации и происходит в широком спектре иммунокомпетентных клеток, включая активированные NK- и МКТ-клетки, регуляторные Т-клетки, дендритные клетки (DC), стимулированные тучные клетки, дифференцирующиеся миелоидные клетки, моноциты, нейтрофилы и эозинофилы (Wang, 2009, Immunological Reviews 229: 192-215). Также была показана экспрессия 4-1ВВ в сосудистой сети опухолей (Broil, 2001, Amer. J Clin. Pathol. 115(4):543-549; Seaman, 2007, Cancer Cell 11: 539-554) и в областях воспаленного или атеросклеротического эндотелия (Drenkard, 2007 FASEB J. 21: 456-463; Olofsson, 2008, Circulation 117: 1292-1301). Лиганд, стимулирующий 4-1ВВ, то есть лиганд 4-1ВВ (4-1 BBL), экспрессирован на активированных антигенпрезентирующих клетках (АРС), миелоидных клетках-предшественниках и гемопоэтических стволовых клетках.
Человеческий 4-1ВВ представляет собой белок из 255 аминокислот (регистрационный номер NM_001561; NP_001552). Аминокислотная последовательность полноразмерного человеческого 4-1ВВ представлена в SEQ ID NO:68. Данный белок содержит сигнальную последовательность (аминокислотные остатки 1-17) с последующим внеклеточным доменом (169 аминокислот), трансмембранной областью (27 аминокислот) и внутриклеточным доменом (42 аминокислоты) (Cheuk АТС et al. 2004 Cancer Gene Therapy 11: 215-226). Данный рецептор экспрессирован на поверхности клеток в мономерной и димерной формах и, вероятно, тримеризуется с лигандом 4-1ВВ при передаче сигнала.
Многочисленные исследования мышиных и человеческих Т-клеток показывают, что 4-1ВВ стимулирует усиленную пролиферацию клеток, их выживание и образование цитокинов (Croft, 2009, Nat Rev Immunol 9:271-285). Исследования продемонстрировали, что некоторые агонисты моноклональных антител (mAb) 4-1ВВ повышают экспрессию костимулирующих молекул и заметно усиливают цитолитические Т-лимфоцитарные ответы, приводя к противоопухолевым эффектам во многих моделях. mAb-агонисты 4-1ВВ показали эффективность в профилактических и терапевтических условиях. Кроме того, в моделях опухолей с монотерапией и комбинированной терапией 4-1ВВ были получены продолжительные противоопухолевые защитные Т-клеточные ответы (Lynch, 2008, Immunol Rev. 22: 277-286). Также было показано, что агонисты 4-1ВВ ингибируют аутоиммунные реакции во многих моделях аутоиммунитета, известных в данной области техники (Vinay, 2006, J Mol Med 84:726-736). Эта двойная активность 4-1ВВ приводит к возможности обеспечения противоопухолевой активности с одновременным смягчением аутоиммунных побочных эффектов, которые могут быть связаны со способами иммунотерапии, приводящими к нарушению иммунологической толерантности.
Существует назревшая неудовлетворенная потребность в антителах, связывающихся с человеческим 4-1ВВ, усиливающих 4-1ВВ-опосредованный ответ и обеспечивающих посредством этого потенциальное лекарственное средство для лечения различных заболеваний и состояний, включая рак.
КРАТКОЕ ИЗЛОЖЕНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
Задачей изобретения является обеспечение выделенной связывающей молекулы, связывающейся с человеческим 4-1ВВ, такой как антитело, или его антигенсвязывающий фрагмент, или его производное. Другой задачей изобретения является обеспечение композиции, содержащей связывающую молекулу, связывающуюся с 4-1ВВ. Задачей настоящего изобретения также является обеспечение способов лечения заболевания и/или состояния, связанного с или опосредованного передачей сигнала 4-1ВВ, с использованием одной или более связывающих молекул по изобретению. Эти и другие задачи изобретения описаны здесь более подробно.
В некоторых аспектах согласно изобретению предложены выделенные антитела, связывающиеся с человеческим 4-1ВВ.
В одном конкретном аспекте выделенное антитело связывается с человеческим 4-1ВВ в эпитопе, содержащем аминокислотные остатки 115-156 SEQ ID NO:68. В некоторых конкретных воплощениях антитело содержит аминокислотную последовательность гипервариабельного участка 1 тяжелой цепи (H-CDR1) по SEQ ID NO:29, аминокислотную последовательность гипервариабельного участка 2 тяжелой цепи (H-CDR2) по SEQ ID NO:30 и аминокислотную последовательность гипервариабельного участка 3 тяжелой цепи (H-CDR3) по SEQ ID NO:31. В других определенных воплощениях антитело содержит аминокислотную последовательность гипервариабельного участка 1 легкой цепи (L-CDR1) по SEQ ID NO:34, аминокислотную последовательность гипервариабельного участка 2 легкой цепи (L-CDR2) по SEQ ID NO:35 и аминокислотную последовательность гипервариабельного участка 3 легкой цепи (L-CDR3) по SEQ ID NO:36.
В другом определенном аспекте выделенное антитело связывается с человеческим 4-1ВВ с константной диссоциации (Ко) 600 нМ или менее, 100 нМ или менее, 50 нМ или менее, 10 нМ или менее, 5 нМ или менее, или 1 нМ или менее, как измерено для внеклеточного домена человеческого 4-1ВВ анализом BIACore, описанным в данном изобретении.
В другом определенном аспекте выделенное антитело содержит: (а) Н-CDR1, как изложено в SEQ ID NO:1, SEQ ID NO:15 или SEQ ID NO:29; (б) Н-CDR2, как изложено в SEQ ID NO:2, SEQ ID NO:16 или SEQ ID NO:30; и (в) Н-CDR3, как изложено в SEQ ID NO:3, SEQ ID NO:17 или SEQ ID NO:31.
В другом определенном аспекте выделенное антитело содержит: (a) L-CDR1, как изложено в SEQ ID NO:6, SEQ ID NO:20 или SEQ ID NO:34; (б) L-CDR2, как изложено в SEQ ID NO:7, SEQ ID NO:21 или SEQ ID NO:35; и (в) L-CDR3, как изложено в SEQ ID NO:8, SEQ ID NO:22, SEQ ID NO:36 или SEQ ID NO:55.
В другом аспекте выделенное антитело содержит: (а) H-CDR1, как изложено в SEQ ID NO:1, SEQ ID NO:15 или SEQ ID NO:29; (б) H-CDR2, как изложено в SEQ ID NO:2, SEQ ID NO:16 или SEQ ID NO:30; и (в) H-CDR3, как изложено в SEQ ID NO:3, SEQ ID NO:17 или SEQ ID NO:31; и дополнительно содержит: (г) L-CDR1, как изложено в SEQ ID NO:6, SEQ ID NO:20 или SEQ ID NO:34; (д) L-CDR2, как изложено в SEQ ID NO:7, SEQ ID NO:21 или SEQ ID NO:35; и (e) L-CDR3, как изложено в SEQ ID NO:8, SEQ ID NO:22, SEQ ID NO:36 или SEQ ID NO:55.
В некоторых других определенных аспектах выделенное антитело выбрано из группы, состоящей из:
а) антитела или его антигенсвязывающего фрагмента, содержащих: Н-CDR1, как изложено в SEQ ID NO:1, H-CDR2, как изложено в SEQ ID NO:2, и Н-CDR3, как изложено в SEQ ID NO:3;
б) антитела или его антигенсвязывающего фрагмента, содержащих Н-CDR1, как изложено в SEQ ID NO:15, H-CDR2, как изложено в SEQ ID NO:16, и H-CDR3, как изложено в SEQ ID NO:17, и
в) антитела или его антигенсвязывающего фрагмента, содержащих Н-CDR1, как изложено в SEQ ID NO:29, H-CDR2, как изложено в SEQ ID NO:30, и H-CDR3, как изложено в SEQ ID NO:31.
В некоторых других аспектах согласно изобретению предложены выделенное антитело или его антигенсвязывающий фрагмент, специфично связывающиеся с человеческим 4-1ВВ, выбранные из группы, состоящей из:
а) антитела или его антигенсвязывающего фрагмента, содержащих L-CDR1, как изложено в SEQ ID NO:6, L-CDR2, как изложено в SEQ ID NO:7, и L-CDR3, как изложено в SEQ ID NO:8;
б) антитела или его антигенсвязывающего фрагмента, содержащих L-CDR1, как изложено в SEQ ID NO:20, L-CDR2, как изложено в SEQ ID NO:21, и L-CDR3, как изложено в SEQ ID NO:22;
в) антитела или его антигенсвязывающего фрагмента, содержащих L-CDR1, как изложено в SEQ ID NO:34, L-CDR2, как изложено в SEQ ID NO:35, и L-CDR3, как изложено в SEQ ID NO:36; и
г) антитела или его антигенсвязывающего фрагмента, содержащих L-CDR1, как изложено в SEQ ID NO:34, L-CDR2, как изложено в SEQ ID NO:35, и L-CDR3, как изложено в SEQ ID NO:55.
В некоторых других определенных аспектах выделенное антитело выбрано из группы, состоящей из:
а) антитела или его антигенсвязывающего фрагмента, содержащих: Н-CDR1, как изложено в SEQ ID NO:1, H-CDR2, как изложено в SEQ ID NO:2, Н-CDR3, как изложено в SEQ ID NO:3; L-CDR1, как изложено в SEQ ID NO:6, L-CDR2, как изложено в SEQ ID NO:7, и L-CDR3, как изложено в SEQ ID NO:8;
б) антитела или его антигенсвязывающего фрагмента, содержащих Н-CDR1, как изложено в SEQ ID NO:15, H-CDR2, как изложено в SEQ ID NO:16, Н-CDR3, как изложено в SEQ ID NO:17; L-CDR1, как изложено в SEQ ID NO:20, L-CDR2, как изложено в SEQ ID NO:21, и L-CDR3, как изложено в SEQ ID NO:22;
в) антитела или его антигенсвязывающего фрагмента, содержащих Н-CDR1, как изложено в SEQ ID NO:29, H-CDR2, как изложено в SEQ ID NO:30, Н-CDR3, как изложено в SEQ ID NO:31; L-CDR1, как изложено в SEQ ID NO:34, L-CDR2, как изложено в SEQ ID NO:35, и L-CDR3, как изложено в SEQ ID NO:36; и
г) антитела или его антигенсвязывающего фрагмента, содержащих Н-CDR1, как изложено в SEQ ID NO:29, H-CDR2, как изложено в SEQ ID NO:30, Н-CDR3, как изложено в SEQ ID NO:31; L-CDR1, как изложено в SEQ ID NO:34, L-CDR2, как изложено в SEQ ID NO:35, и L-CDR3, как изложено в SEQ ID NO:55.
В другом определенном аспекте выделенное антитело содержит аминокислотную последовательность цепи вариабельного домена (вариабельной области) тяжелой цепи (VH), как изложено в SEQ ID NO:4, SEQ ID NO:18, SEQ ID NO:32 и SEQ ID NO:43.
В другом определенном аспекте выделенное антитело содержит аминокислотную последовательность вариабельного домена (вариабельной области) легкой цепи (VL), как изложено в SEQ ID NO:9, SEQ ID NO:23, SEQ ID NO:37, SEQ ID NO:45, SEQ ID NO:51, SEQ ID NO:56, SEQ ID NO:60 или SEQ ID NO:64.
В другом определенном аспекте выделенное антитело содержит аминокислотную последовательность домена VH, как изложено в любой из SEQ ID NO:4, 18, 32 и 43, и дополнительно содержит аминокислотную последовательность домена VL, как изложено в любой из SEQ ID NO:9, SEQ ID NO:23, SEQ ID NO:37, SEQ ID NO:45, SEQ ID NO:51, SEQ ID NO:56, SEQ ID NO:60 и SEQ ID NO:64.
В другом определенном аспекте выделенное антитело выбрано из группы, состоящей из:
а) антитела, содержащего аминокислотную последовательность цепи VH, как изложено в SEQ ID NO:4, и аминокислотную последовательность цепи VL, как изложено в SEQ ID NO:9;
б) антитела, содержащего аминокислотную последовательность цепи VH, как изложено в SEQ ID NO:18, и аминокислотную последовательность цепи VL, как изложено в SEQ ID NO:23;
в) антитела, содержащего аминокислотную последовательность цепи VH, как изложено в SEQ ID NO:32, и аминокислотную последовательность цепи VL, как изложено в SEQ ID NO:37 или SEQ ID NO:56; и
г) антитела, содержащего аминокислотную последовательность цепи VH, как изложено в SEQ ID NO:43, и аминокислотную последовательность цепи VL, как изложено в SEQ ID NO:45, SEQ ID NO:51, SEQ ID NO:60 или SEQ ID NO:64.
В другом определенном аспекте выделенное антитело, предложенное согласно настоящему изобретению, содержит цепь VH, кодируемую (1) последовательностью нуклеиновой кислоты, содержащей SEQ ID NO:11, SEQ ID NO:25, SEQ ID NO:39, SEQ ID NO:47, или (2) последовательностями нуклеиновых кислот, гибридизующимися в строгих условиях с комплементарной цепью SEQ ID NO:11, SEQ ID NO:25, SEQ ID NO:39 или SEQ ID NO:47.
В другом определенном аспекте выделенное антитело содержит цепь VL, кодируемую (1) последовательностью нуклеиновой кислоты, содержащей SEQ ID NO:12, SEQ ID NO:26, SEQ ID NO:40, SEQ ID NO:48, SEQ ID NO:53, SEQ ID NO:58, SEQ ID NO:62 или SEQ ID NO:66, или (2) последовательностями нуклеиновых кислот, гибридизующимися в строгих условиях с комплементарной цепью SEQ ID NO:12, SEQ ID NO:26, SEQ ID NO:40, SEQ ID NO:48, SEQ ID NO:53, SEQ ID NO:58, SEQ ID NO:62 или SEQ ID NO:66.
В другом определенном аспекте предложено выделенное антитело, конкурирующее и/или перекрестно конкурирующее за связывание с человеческим 4-1ВВ с иллюстративным антителом, выбранным из MOR-6032, MOR-7361, MOR-7480, MOR-7480.1, MOR-7480.2, MOR 7483, MOR-7483.1 или MOR-7483.2.
В другом определенном аспекте предложено выделенное антитело, связывающееся с тем же эпитопом человеческого 4-1ВВ, что и любое из антител, описанных здесь. В некоторых воплощениях согласно изобретению предложено выделенное антитело, связывающееся с тем же эпитопом человеческого 4-1ВВ, что и иллюстративное антитело, выбранное из MOR-6032, MOR-7361, MOR-7480, MOR-7480.1, MOR-7480.2, MOR 7483, MOR-7483.1 или MOR-7483.2.
В другом определенном аспекте согласно настоящему изобретению предложено выделенное антитело, связывающееся с человеческим 4-1ВВ, содержащее вариабельную область тяжелой цепи, являющуюся продуктом или имеющую происхождение от человеческого гена VH 3-23, гена VH 1-69 или VH 5. В другом определенном аспекте согласно настоящему изобретению предложено выделенное антитело, связывающееся с человеческим 4-1ВВ, содержащее вариабельную область легкой цепи, являющуюся продуктом или имеющую происхождение от человеческого гена VL A3 или А1-13.
В некоторых воплощениях выделенные антитела, описанные здесь, имеют одно или более из следующих свойств или характеристик:
а) специфично связываются с человеческим 4-1ВВ;
б) связываются с человеческим 4-1ВВ и 4-1ВВ яванского макака;
в) связываются с человеческим 4-1ВВ или 4-1ВВ яванского макака, но не с крысиным или мышиным 4-1ВВ;
г) представляют собой IgG, такой как IgG1, IgG2, IgG3 или IgG4; и
д) представляют собой человеческие антитела или гуманизированные антитела.
В некоторых других аспектах согласно настоящему изобретению предложен антигенсвязывающий фрагмент любого из антител, предложенных согласно настоящему изобретению. В некоторых воплощениях антигенсвязывающий фрагмент представляет собой Fab- или scFv-фрагмент.
В некоторых других аспектах согласно настоящему изобретению предложено производное любого из антител, предложенных согласно настоящему изобретению.
В некоторых других аспектах согласно изобретению предложена выделенная нуклеиновая кислота, кодирующая цепь VH антитела или его антигенсвязывающего фрагмента, связывающегося с человеческим 4-1ВВ, выбранная из группы, состоящей из:
1) последовательности нуклеиновой кислоты, кодирующей аминокислотную последовательность цепи VH, как изложено в SEQ ID NO:4, SEQ ID NO:18. SEQ ID NO:32 или SEQ ID NO:43;
2) последовательности нуклеиновой кислоты, как изложено в SEQ ID NO:11, SEQ ID NO:25, SEQ ID NO:39 или SEQ ID NO:47; или
3) последовательности нуклеиновой кислоты, гибридизующейся в строгих условиях с комплементарной цепью последовательности нуклеиновой кислоты, как изложено в SEQ ID NO:11, SEQ ID NO:25, SEQ ID NO:39 или SEQ ID NO:47.
В некоторых других аспектах согласно изобретению предложена выделенная нуклеиновая кислота, кодирующая цепь VL антитела или его антигенсвязывающего фрагмента, связывающихся с человеческим 4-1ВВ, выбранная из группы, состоящей из:
1) последовательности нуклеиновой кислоты, кодирующей аминокислотную последовательность цепи VL, как изложено в SEQ ID NO:9, SEQ ID NO:23, SEQ ID NO:37, SEQ ID NO:45, SEQ ID NO:51, SEQ ID NO:56, SEQ ID NO:60, или SEQ ID NO:64;
2) последовательности нуклеиновой кислоты, как изложено в SEQ ID NO:12, SEQ ID NO:26, SEQ ID NO:40, SEQ ID NO:48, SEQ ID NO:53, SEQ ID NO:58, SEQ ID NO:62 или SEQ ID NO:66; или
3) последовательностей нуклеиновых кислот, гибридизующихся в строгих условиях с комплементарной цепью последовательности нуклеиновой кислоты, как изложено в SEQ ID NO:12, SEQ ID NO:26, SEQ ID NO:40, SEQ ID NO:48, SEQ ID NO:53, SEQ ID NO:58, SEQ ID NO:62 или SEQ ID NO:66.
В некоторых других аспектах согласно изобретению предложен вектор, содержащий любую из нуклеиновых кислот, описанных здесь. В другом аспекте согласно изобретению предложена клетка-хозяин, содержащая любой из векторов, описанных здесь. Такие клетки-хозяева могут представлять собой бактериальные клетки или клетки млекопитающих.
В некоторых других аспектах согласно изобретению предложена фармацевтическая композиция, содержащая любые из антител, их антигенсвязывающие фрагменты или их производные и фармацевтически приемлемый носитель.
Согласно изобретению также предложены способы лечения аномального клеточного роста у субъекта, нуждающегося в этом, включающие введение субъекту эффективного количества связывающей молекулы по изобретению или фармацевтической композиции, описанной здесь. Согласно изобретению также предложены способы уменьшения метастазирования опухолевых клеток у субъекта, включающие введение указанному субъекту эффективного количества связывающей молекулы или фармацевтических композиций, описанных здесь.
В другом аспекте согласно изобретению предложено применение любой из связывающих молекул или фармацевтической композиции, описанной здесь, для изготовления лекарственного средства для лечения аномального клеточного роста у субъекта, нуждающегося в этом. В другом аспекте согласно изобретению предложена связывающая молекула или фармацевтическая композиция, как описано здесь, для применения в лечении аномального клеточного роста у субъекта, нуждающегося в этом. В другом аспекте согласно изобретению предложена связывающая молекула или фармацевтическая композиция, как описано здесь, для применения в лечении метастазирования опухолевых клеток у субъекта, нуждающегося в этом. В другом аспекте согласно изобретению предложено применение любой из связывающих молекул или фармацевтической композиции, описанной здесь, для изготовления лекарственного средства для лечения метастазирования опухолевых клеток у субъекта, нуждающегося в этом.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
На Фиг.1 представлены четыре столбчатые диаграммы, на которых показана средняя интенсивность флуоресценции нестимулированных (черные столбцы) и стимулированных фитогемагглютинином (РНА) (светло-серые столбцы) первичных мононуклеарных клеток периферической крови (РМВС) человека (сверху слева), яванского макака (сверху справа), собаки (снизу слева) и крысы (снизу справа), инкубированных с указанным антителом против 4-1ВВ или контрольным антителом, конъюгированными с Alexafluor 647. Диаграммы демонстрируют связывание с человеческими РВМС и РВМС яванского макака, стимулированными РНА.
На Фиг.2 представлены две линейные диаграммы, на которых показана активность репортерного гена люциферазы в клетках 293Т, экспрессирующих 4-1ВВ, стимулированных несколькими концентрациями mAb, специфичных в отношении 4-1ВВ, или контрольным mAb того же изотипа. На левой диаграмме показана активность репортерного гена в клетках, экспрессирующих 4-1ВВ яванского макака. На правой диаграмме показана активность в клетках, экспрессирующих человеческий 4-1ВВ. Данные представлены как кратность стимуляции по сравнению с контрольным mAb того же изотипа.
На Фиг.3 (3А и 3Б) представлены линейные диаграммы, на которых показана концентрация человеческого интерлейкина-2 (IL-2), присутствующего в культуральной среде клеток через 72 часа стимуляции человеческих Т-клеток антителом против CD3 и несколькими концентрациями антител против 4-1ВВ. Каждая из диаграмм (А и Б) относится к отдельному донору.
На Фиг.4 представлена диаграмма рассеяния, на которой показана экспансия человеческих мононуклеарных клеток периферической крови у мышей, которым вводили mAb против 4-1ВВ или контрольное mAb того же изотипа. Данные представлены как процент клеток, экспрессирующих человеческий CD45, в периферической крови отдельных мышей NSG на 24-28 сутки исследования, которым были проведены инъекции шести миллионов человеческих мононуклеарных клеток периферической крови на 0 сутки и инъекции 1 мг/кг mAb против 4-1ВВ или контрольного mAb того же изотипа на 9 сутки. Статистическую значимость определяли двусторонним критерием Манна-Уитни, *р<0,05, **р<0,005. «Без НВРТ» относится к животным, которым не проводили инъекцию человеческих клеток.
На Фиг.5 представлены две столбчатые диаграммы, на которых показано изменение пролиферации CD8 центральных Т-клеток памяти в нескольких временных точках после введения mAb против 4-1ВВ у яванских макаков. Данные показаны в форме столбцов, соответствующих отдельным животным, обозначенным как (уровень дозы - номер животного), и представлены как изменение числа KJ-67+ клеток у животного по сравнению с их числом до исследования {[(число KJ-67+клеток на указанные сутки исследования - число KI-67+ клеток перед введением)/число KJ-67+ клеток перед введением]*100}. CD8 центральные клетки памяти определяли как CD3+, CD8+, CD28+ и CD95+.
На Фиг.6 представлены линейные диаграммы, на которых показан рост опухолей после подкожного введения опухолевых клеток (РС3, левая диаграмма; LOVO, правая диаграмма) и человеческих мононуклеарных клеток периферической крови на 0 сутки исследования. Мышам вводили 10 мг/кг указанных mAb против 4-1ВВ на 0 сутки.
Фиг.7. Слева представлена диаграмма рассеяния, на которой показан процент РВМС, положительных в отношении поверхностного маркера Т-клеток CD8+и содержащих нуклеозидный аналог бромдезоксиуридин (BrdU) после введения 4-1ВВ - «нок-ин» мышам mAb против 4-1ВВ или контроля наполнителем. Справа представлена линейная диаграмма, на которой показан рост опухолей мышиной меланомы, введенных подкожно 4-1ВВ «нок-ин» мышам, которым вводили mAb против 4-1ВВ в указанной концентрации.
На Фиг.8 показаны выравнивания аминокислотных последовательностей вариабельных областей тяжелой цепи и вариабельных областей легкой цепи (с подчеркнутыми гипервариабельными областями (CDR)) с соответствующими последовательностями эмбрионального типа. «GERMLINE VH1-69» - эмбриональный VH 1-69; «GERMLINE VH3-23» - эмбриональный VH 3-23; «GERMLINE VH5» - эмбриональный VH 5; «GERMLINE VL3-r» - эмбриональный VL 3-r.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
А. ОПРЕДЕЛЕНИЯ
Если здесь не определено иное, научные и технические термины, используемые в связи с настоящим изобретением, имеют значение, обычно подразумеваемое специалистами в данной области техники. Кроме того, если контекст не требует иного, термины в единственном числе включают множественное число, и термины во множественном числе включают единственное число. Обычно методики клеточных и тканевых культур, молекулярной биологии, иммунологии, микробиологии, генетики, химии белков и нуклеиновых кислот и гибридизации, описанные здесь, и используемые в связи с ними номенклатуры являются хорошо известными и широко применяемыми в данной области техники.
При использовании здесь каждый из следующих терминов имеет значение, связанное с ним в данном разделе.
Термин «антитело против 4-1ВВ» относится к антителу, как определено здесь, способному связываться с человеческим рецептором 4-1ВВ.
Термины «4-1ВВ» и «рецептор 4-1ВВ» использованы в настоящей заявке взаимозаменяемо и включают человеческий рецептор 4-1ВВ, а также его варианты, изоформы и видовые гомологи. Соответственно, связывающая молекула, как определено и раскрыто здесь, может также связываться с формами 4-1ВВ, отличными от человеческой. В других случаях связывающая молекула может быть полностью специфична в отношении человеческого 4-1ВВ и может не демонстрировать перекрестную реактивность с другими видами или другими типами.
Формы единственного числа относятся к одному или более (то есть по меньшей мере одному) грамматическим объектам. Например, «элемент» означает один элемент или более чем один элемент.
Термин «агонист» относится к связывающей молекуле, как определено здесь, которая при связывании с 4-1ВВ (1) стимулирует или активирует 4-1ВВ, (2) усиливает, повышает, стимулирует, индуцирует или продлевает активность, функцию или присутствие 4-1ВВ, или (3) усиливает, повышает, стимулирует или индуцирует экспрессию 4-1ВВ.
Термин «аминокислота» относится к встречающимся в природе и синтетическим аминокислотам, а также аналогам аминокислот и миметикам аминокислот, функционирующим сходным с встречающимися в природе аминокислотами образом. Встречающиеся в природе аминокислоты представляют собой аминокислоты, кодируемые генетическим кодом, а также аминокислоты, модифицируемые позднее, например, гидроксипролин, гамма-карбоксиглутамат и O-фосфосерин. Термин «аналоги аминокислот» относится к соединениям, имеющим такую же основную химическую структуру, как встречающаяся в природе аминокислота, но С-концевая карбоксильная группа, N-концевая аминогруппа или функциональная группа боковой цепи были химически модифицированы с образованием другой функциональной группы. Термин «миметики аминокислот» относится к химическим соединениям, имеющим структуру, отличную от обычной химической структуры аминокислоты, но функционирующим сходным образом с встречающейся в природе аминокислотой.
Термин «антитело» является термином, известным в данной области техники, и относится к антигенсвязывающему белку (то есть иммуноглобулину), имеющему основную структуру из четырех полипептидных цепей, состоящую из двух идентичных тяжелых (Н) цепей и двух идентичных легких (L) цепей. Каждая L-цепь связана с Н-цепью одной ковалентной дисульфидной связью, в то время как две Н-цепи связаны друг с другом одной или более дисульфидными связями, в зависимости от изотипа Н-цепей. Каждая тяжелая цепь имеет на N-конце вариабельную область (обозначаемую здесь аббревиатурой VH) с последующей константной областью. Константная область тяжелой цепи состоит из трех доменов: CH1, CH2 и CH3. Каждая легкая цепь имеет на N-конце вариабельную область (обозначаемую здесь аббревиатурой VL) с последующей константной областью на ее другом конце. Константная область легкой цепи состоит из одного домена CL. VL расположена напротив VH, и CL расположена напротив первого константного домена тяжелой цепи (СН1). Совместно пара VH и VL образует один антигенсвязывающий сайт. Антитело IgM состоит из 5 основных гетеротетра мерных единиц вместе с дополнительным полипептидом, называемым J-цепью, и, таким образом, содержит 10 антигенсвязывающих сайтов, в то время как секретируемые антитела IgA способны к полимеризации с образованием поливалентных структур, содержащих 2-5 основных 4-хцепочечных единиц с J-цепью.
Области VH и VL могут быть дополнительно разделены на гипервариабельные участки, называемые областями, определяющими комплементарность (CDR), разделенные более консервативными областями, называемыми каркасными областями (FR). CDR-участки могут быть определены с использованием систем нумерации Kabat или Chothia, обе из которых хорошо известны специалистам в данной области техники. См., например, Kabat, Е.A., et al. (1991) Sequences of Proteins of Immunological Interest, Fifth Edition, U.S. Department of Health and Human Services, NIH Publication No.91-3242; Chothia and Lesk, J. Mol. Biol. 196:901-917 (1987). Каждая VH и VL состоит из трех CDR и четырех FR, расположенных, от М-конца к С-концу, в следующем порядке: FR1, CDR1, FR2, CDR2, FR3, CDR3, FR4. В настоящем изобретении три CDR тяжелой цепи названы H-CDR1, H-CDR2 и Н-CDR3. Сходным образом, три CDR легкой цепи названы L-CDR1, L-CDR2 и L-CDR3. Вариабельные области тяжелой и легкой цепей содержат связывающий домен, взаимодействующий с антигеном. Константные области антител могут опосредовать связывание иммуноглобулина с тканями или факторами хозяина, включая различные клетки иммунной системы (например, эффекторные клетки) и первый компонент (C1q) классической системы комплемента. В легкой и тяжелой цепях вариабельные и константные области соединены «J»-областью из приблизительно 12 или более аминокислот, при этом тяжелая цепь также содержит «D»-o6nacTb из приблизительно 10 или более аминокислот. В целом, см. Fundamental Immunology Ch. 7 (Paul, W., ed., 2nd ed. Raven Press, N.Y. (1989)).
L-цепь любого вида позвоночных может быть отнесена к одному из двух четко разделенных типов, называемых каппа и лямбда, на основании аминокислотных последовательностей их константных доменов. В зависимости от аминокислотной последовательности константного домена их тяжелых цепей (СН), антитела могут быть отнесены к различным классам или изотипам. Существует пять классов антител: IgA, IgD, IgE, IgG и IgM, имеющие тяжелые цепи, обозначаемые а (альфа), 5 (дельта), £ (эпсилон), у (гамма) и р (мю), соответственно. Класс антител IgG может быть дополнительно разделен на четыре подкласса: IgG1, IgG2, IgG3 и IgG4, на основании тяжелых цепей гамма, Y1-Y4, соответственно.
Термин «производное антитела» или «производное» антитела относится к молекуле, способной связываться с тем же антигеном (то есть 4-1ВВ), с которым связывается антитело, и содержащей аминокислотную последовательность антитела, связанную с дополнительной молекулярной группировкой. Аминокислотная последовательность антитела, входящая в производное антитела, может представлять собой полноразмерную тяжелую цепь, полноразмерную легкую цепь, любую часть или части полноразмерной тяжелой цепи, любую часть или части полноразмерной легкой цепи антитела, любой(ые) другой(ие) фрагмент(ы) антитела или полноразмерное антитело. Дополнительная молекулярная группировка может представлять собой химическую или биологическую молекулу. Примеры дополнительных молекулярных группировок включают химические группы, аминокислоты, пептиды, белки (такие как ферменты, антитела) и химические соединения. Дополнительная молекулярная группировка может быть использована в качестве, например, агента для выявления, метки, маркера, фармацевтического или терапевтического агента. Аминокислотная последовательность антитела может быть присоединена или связана с дополнительной молекулярной группировкой химическим сочетанием, генетическим слиянием, нековалентным соединением или иным образом. Термин «производное антитела» также включает химерные антитела, гуманизированные антитела и молекулы, имеющие происхождение от модификаций аминокислотных последовательностей антитела против 4-1ВВ, таких как консервативные аминокислотные замены, присоединения и вставки.
Термин «антигенсвязывающий фрагмент» или «антигенсвязывающая часть» антитела относится к одной или более частям антитела, сохраняющим способность связываться с антигеном, с которым связывается антитело (например, 4-1ВВ). Примеры «антигенсвязывающего фрагмента» антитела включают: (1) Fab-фрагмент, моновалентный фрагмент, состоящий из доменов VL, VH, CL и CH1; (2) F(ab')2-фрагмент, бивалентный фрагмент, содержащий два Fab-фрагмента, связанных дисульфидной связью в шарнирной области; (3) Fd-фрагмент, состоящий из доменов VH и CH1; (4) Fv-фрагмент, состоящий из доменов VL и VH одной части антитела, (5) dAb-фрагмент (Ward et al., Nature 341:544-546 (1989)), состоящий из домена VH; и (6) выделенный гипервариабельный участок (CDR).
Термин «связывающая молекула» включает: (1) антитело; (2) антигенсвязывающий фрагмент антитела; и (3) производное антитела, как они определены здесь.
Термин «связывание 4-1ВВ», «связывает 4-1ВВ», «связывание с 4-1ВВ» или «связывается с 4-1ВВ» относится к связыванию связывающей молекулы, как определено здесь, с человеческим 4-1ВВ в in vitro-анализе, таком как анализ BIAcore, как описано в Примере 6, с аффинностью (KD) 500 нМ или менее.
Термин «химерное антитело» относится к антителу, содержащему аминокислотные последовательности, имеющие происхождение от разных видов животных, такому как антитела, имеющие вариабельную область, имеющую происхождение от человеческого антитела, и константную область мышиного иммуноглобулина.
Термин «конкурирует за связывание» относится к взаимодействию двух антител при их связывании с мишенью связывания. Первое антитело конкурирует за связывание со вторым антителом, если связывание первого антитела с его когнатным эпитопом выявляемо снижено в присутствии второго антитела по сравнению со связыванием первого антитела в отсутствие второго антитела. Возможен, но не обязателен, альтернативный вариант, где связывание второго антитела с его эпитопом также выявляемо снижено в присутствии первого антитела. То есть, первое антитело может ингибировать связывание второго антитела с его эпитопом без ингибирования вторым антителом связывания первого антитела с его соответствующим эпитопом. Тем не менее, если каждое антитело выявляемо ингибирует связывание другого антитела с его когнатным эпитопом, в такой же, большей или меньшей степени, говорят, что антитела «перекрестно конкурируют» друг с другом за связывание с их соответствующим(и) эпитопом(ами).
Термин «эпитоп» относится к части антигена, с которой связывается антитело (или его антигенсвязывающий фрагмент). Эпитопы могут быть образованы смежными аминокислотами или несмежными аминокислотами, расположенными рядом друг с другом при формировании третичной структуры белка. Эпитопы, образованные смежными аминокислотами, обычно сохраняются при обработке денатурирующими растворителями, тогда как эпитопы, образованные при формировании третичной структуры, обычно не сохраняются при обработке денатурирующими растворителями. Эпитоп может содержать различные количества аминокислот в уникальной пространственной конформации. Способы определения пространственной конформации эпитопов включают, например, рентгеновскую кристаллографию и 2-мерный ядерный магнитный резонанс.См., например, Epitope Mapping Protocols in Methods in Molecular Biology, Vol.66, G.E. Morris, Ed. (1996). После определения желаемого эпитопа на антигене могут быть получены антитела к данному эпитопу, например, с применением методик, описанных здесь. При получении и определении свойств антител можно также получить информацию о желаемых эпитопах. На основании этой информации затем можно проводить конкурентный скрининг антител на связывание с одним и тем же эпитопом. Одним способом достижения описанного выше является проведение перекрестие-конкурентных исследований для обнаружения антител с конкурентным связыванием друг относительно друга, то есть антител, конкурирующих за связывание с антигеном. Высокоэффективный способ «селекции» антител на основании их перекрестной конкуренции описан в РСТ-публикации № WO 03/48731.
Термин «эмбриональный» относится к нуклеотидным последовательностям генов и сегментов генов антител при их передаче от родителей потомству через половые клетки. Эмбриональная последовательность отличается от нуклеотидных последовательностей, кодирующих антитела в зрелых В-клетках, измененных рекомбинацией и гипермутацией при созревании В-клеток.
Термин «сайты гликозилирования» относится к аминокислотным остаткам, распознаваемым эукариотической клеткой как места присоединения сахарных остатков. Аминокислоты, к которым происходит присоединение углевода, такого как олигосахарид, обычно представляют собой остатки аспарагина (N-связь), серина (O-связь) и треонина (O-связь). На точное место присоединения обычно указывает последовательность аминокислот, называемая здесь «последовательностью сайта гликозилирования». Последовательность сайта гликозилирования для N-гликозилирования представляет собой -Asn-X-Ser- или -Asn-X-Thr-, где Х может представлять собой любую обычную аминокислоту, не являющуюся пролином. Термины «N-гликозилирование» и «O-гликозилирование» относятся к химической группе, используемой в качестве сайта присоединения между молекулой сахара и аминокислотным остатком. Присоединение N-связанных Сахаров происходит через аминогруппу; присоединение O-связанных Сахаров происходит через гидроксильную группу. Термин «степень гликозилирования» относится к присутствию углеводной группировки, связанной с сайтом гликозилирования (то есть сайт гликозилирования занят). Там, где на пептиде присутствуют по меньшей мере два возможных сайта гликозилирования, углеводными группировками может не быть занят ни один сайт (степень гликозилирования 0), может быть занят один сайт (степень гликозилирования 1) или оба сайта (степень гликозилирования 2).
Термин «клетка-хозяин» относится к клеточной системе, которая может быть сконструирована для получения белков, фрагментов белков или интересующих пептидов. Клетки-хозяева включают, без ограничения, культивируемые клетки, например, культивируемые клетки млекопитающих, имеющие происхождение от грызунов (крыс, мышей, морских свинок или хомяков), такие как СНО, ВНК, NSO, SP2/0, YB2/0, или человеческие ткани или клетки гибридом, дрожжевые клетки, клетки насекомых и клетки трансгенного животного или культивированной ткани. Данный термин включает не только конкретную рассматриваемую клетку, но также потомство такой клетки. Поскольку в последующих поколениях возможны определенные модификации, обусловленные мутацией или влияниями среды, такое потомство может не быть идентичным исходной клетке, но оно все равно включено в объем термина «клетка-хозяин».
Термин «человеческое антитело» относится к антителу, в котором все аминокислотные последовательности легких цепей и тяжелых цепей имеют происхождение от генов человеческих иммуноглобулинов. Человеческое антитело может содержать мышиные углеводные цепи, если оно получено в мыши, мышиной клетке или в гибридоме, имеющей происхождение от мышиной клетки. Человеческие антитела могут быть получены множеством способов, известных в данной области техники.
Термин «гуманизированное антитело» относится к химерному антителу, содержащему аминокислотные остатки, имеющие происхождение от последовательностей человеческих антител. Гуманизированное антитело может содержать некоторые или все CDR антитела животного, не являющегося человеком, в то время как каркасные и константные области антитела содержат аминокислотные остатки, имеющие происхождение от последовательностей человеческих антител.
Термин «иллюстративное антитело» относится к любому из антител, описанных в изобретении и обозначенных MOR-6032, MOR-7361, MOR-7480, MOR-7480.1, MOR-7480.2, MOR-7483, MOR-7483.1 и MOR-7483.2. Эти антитела могут представлять собой антитела любого класса (например, IgA, IgD, IgE, IgG и IgM). Таким образом, каждое антитело, определенное выше, включает антитела всех пяти классов, имеющие одинаковые аминокислотные последовательности областей VL и VH. Кроме того, антитела класса IgG могут представлять собой антитела любого подкласса (например, IgG1, IgG2, IgG3 и IgG4). Таким образом, каждое антитело подкласса IgG, определенное выше, включает антитела всех четырех подклассов, имеющие одинаковые аминокислотные последовательности областей VL и VH. Аминокислотные последовательности константных областей тяжелой цепи человеческих антител пяти классов, а также четырех подклассов IgG, известны в данной области техники. Примеры аминокислотных последовательностей константных областей человеческих IgG1 и IgG2 приведены в SEQ ID NO:69 и 71, соответственно. Аминокислотная последовательность полноразмерной тяжелой цепи подкласса IgG2 каждого из иллюстративных антител приведена в данном описании.
Термин «выделенное антитело» или «выделенная связывающая молекула» относится к антителу или связывающей молекуле, как определено здесь, которые: (1) не связаны с природными компонентами, с которыми они связаны в их естественном состоянии; (2) свободны от других белков того же вида; (3) экспрессированы клеткой другого вида; или (4) не встречаются в природе. Примеры выделенных антител включают антитело против 4-1ВВ, аффинно очищенное с использованием 4-1ВВ, антитело против 4-1ВВ, полученное в гибридомах или другой клеточной линии in vitro, и антитело против 4-1ВВ, имеющее происхождение от трансгенного животного.
Термин «выделенная нуклеиновая кислота» относится к молекуле нуклеиновой кислоты, имеющей происхождение от генома, комплементарной ДНК (кДНК), или синтетического происхождения, или их комбинации, отделенной от других молекул нуклеиновых кислот, присутствующих в природном источнике нуклеиновой кислоты. Например, относительно геномной ДНК термин «выделенная» включает молекулы нуклеиновых кислот, отделенные от хромосомы, с которой геномная ДНК связана естественным образом. Предпочтительно, «выделенная» нуклеиновая кислота свободна от последовательностей, естественным образом окружающих нуклеиновую кислоту (то есть последовательностей, расположенных на 5'- и 3'-концах рассматриваемой нуклеиновой кислоты).
Термин «ka» относится к константе скорости ассоциации при взаимодействии определенного антитела с определенным антигеном, в то время как термин «kd» относится к константе скорости диссоциации при взаимодействии определенного антитела с определенным антигеном.
Термин «KD» относится к равновесной константе диссоциации при взаимодействии определенного антитела с определенным антигеном. Ее получают как отношение kd к ka (то есть kd/ka) и выражают как молярную концентрацию (М). KD используют как меру аффинности связывания антитела с его партнером по связыванию. Чем меньше KD, тем прочнее связывание антитела или тем выше аффинность антитела в отношении антигена. Например, антитело с наномолярной (нМ) константной диссоциации связывается с определенным антигеном прочнее антитела с микромолярной (мкМ) константной диссоциации. Значения Ко для антител могут быть получены с применением способов, хорошо известных в данной области техники. Одним способом определения Ко антитела является применение поверхностного плазменного резонанса, обычно с использованием биосенсорной системы, такой как система Biacore®. Способ анализа с использованием системы BIACORE™ (анализ BIAcore) описан в разделе «Примеры» данного описания.
Термин «млекопитающее» относится к любому виду животных класса Млекопитающие. Примеры млекопитающих включают: людей; лабораторных животных, таких как крысы, мыши, обезьяны и морские свинки; домашних животных, таких как кошки, собаки, кролики, крупный рогатый скот, овцы, козы, лошади и свиньи; и находящихся в неволе диких животных, таких как львы, тигры, слоны и тому подобное.
Термин «моноклональное антитело» относится к антителу, полученному из однородной по существу совокупности антител, то есть, отдельные антитела, составляющие совокупность, идентичны, за исключением возможных происходящих естественным образом мутаций, которые могут присутствовать в небольших количествах. Моноклональные антитела высокоспецифичны и направлены против одного антигенного сайта. Кроме того, в отличие от препаратов поликлональных антител, содержащих разные антитела, направленные против разных детерминант (эпитопов), каждое моноклональное антитело направлено против одной детерминанты на антигене. Помимо их специфичности, преимущества моноклональных антител состоят в возможности их синтеза без примесей других антител. Термин «моноклональное» не следует толковать как необходимость получения антитела каким-либо конкретным способом. Например, моноклональные антитела могут быть получены гибридомным способом или могут быть получены с применением способов рекомбинантных ДНК в клетках бактерий, животных-эукариот или растений (см., например, патент США №4816567). Моноклональные антитела могут также быть выделены из фаговых библиотек антител с применением методик, описанных, например, в Clackson et al., Nature, 352:624-628 (1991) и Marks et al., J. Mol. Biol., 222:581-597 (1991).
Термин «предотвращать» или «предотвращение» в связи с определенным болезненным состоянием у млекопитающего относится к предотвращению или задержке начала заболевания или предотвращению проявления его клинических или субклинических симптомов.
Термин «рекомбинантное антитело» относится к антителу, полученному, экспрессированному, созданному или выделенному рекомбинантным способом, такому как антитела, выделенные из трансгенного животного с генами иммуноглобулинов другого вида, антитела, экспрессированные с использованием клетки-хозяина, трансфицированной рекомбинантным вектором экспрессии, антитела, выделенные из рекомбинантной комбинаторной библиотеки антител, или антитела, полученные, экспрессированные, созданные или выделенные любым другим способом, включающим сплайсинг последовательностей генов иммуноглобулинов с другими последовательностями ДНК.
При использовании здесь «идентичность последовательности» двух полипептидных последовательностей указывает процент аминокислот, идентичных в данных последовательностях. Идентичность аминокислотных последовательностей полипептидов может быть определена обычными способами с использованием известных компьютерных программ, таких как Bestfit, FASTA или BLAST (см., например, Pearson, Methods Enzymol. 183:63-98 (1990); Pearson, Methods Mol. Biol. 132:185-219 (2000); Altschul et al, J. Mol. Biol. 215:403-410 (1990); Altschul et al., Nucelic Acids Res. 25:3389-3402 (1997)). При использовании Bestfit или любой другой программы выравнивания последовательностей для определения, является ли определенная последовательность, например, на 95% идентичной эталонной аминокислотной последовательности, параметры устанавливают таким образом, что происходит вычисление процента идентичности по всей длине эталонной аминокислотной последовательности при допустимом нарушении гомологии до 5% от общего числа аминокислотных остатков эталонной последовательности. Этот указанный выше способ определения процента идентичности полипептидов применим для всех белков, их фрагментов или вариантов, раскрытых здесь.
Термин «специфично связывает» или «специфично связывается с» в связи с взаимодействием связывающей молекулы, как определено здесь (например, антитела), с ее партнером по связыванию (например, антигеном), относится к способности связывающей молекулы различать интересующий антиген животного одного вида и ортолог антигена животного другого вида при данном наборе условий. Говорят, что связывающая молекула против 4-1ВВ специфично связывается с человеческим 4-1ВВ, если она связывается с человеческим 4-1ВВ со средней эффективной концентрацией (ЕС50), составляющей менее 50 процентов ЕС50, с которой она связывается с 4-1ВВ крысы или мыши, как определено в анализе in vitro. Специфичность связывания антитела может быть определена способами, известными в данной области техники. Примеры таких способов включают сортировку клеток с возбуждением флуоресценции (FACS) с использованием первичных клеток, стимулированных фитогемагглютинином (РНА), вестерн-блоттинг, твердофазный иммуноферментный анализ (ELISA), радиоиммунный анализ (RIA), электрохемилюминесцентный анализ (ECL), иммунорадиометрический анализ (IRMA) и пептидное сканирование.
Термин «селективно связывает» или «селективно связывается с» в связи с взаимодействием связывающей молекулы, как определено здесь (например, антитела), с ее партнером по связыванию (например, антигеном), относится к способности связывающей молекулы различать интересующий антиген животного одного вида (такой как человеческий 4-1ВВ) и другой антиген животного того же вида (такой как человеческий CD40) при данном наборе условий. Говорят, что связывающая молекула против 4-1ВВ селективно связывается с человеческим 4-1ВВ, если она связывается с человеческим 4-1ВВ с ЕС50, составляющей менее 10 процентов ЕС50, с которой она связывается с человеческим CD40 или человеческим CD134, как определено в анализе in vitro.
Термин «лечить» или «лечение» в связи с определенным болезненным состоянием у млекопитающего относится к оказанию желаемого или полезного эффекта у млекопитающего, имеющего болезненное состояние. Желаемый или полезный эффект может включать сниженную частоту или тяжесть одного или более чем одного симптома заболевания (то есть роста и/или метастазирования опухоли, или другой эффект, опосредованный количеством и/или активностью иммунокомпетентных клеток и тому подобным), или остановку или ингибирование дальнейшего развития заболевания, состояния или расстройства. В контексте лечения рака у млекопитающего желаемый или полезный эффект может включать ингибирование дальнейшего роста или распространения раковых клеток, гибель раковых клеток, ингибирование рецидивирования рака, ослабление боли, связанной с раком или улучшенную выживаемость млекопитающего. Эффект может быть субъективным или объективным. Например, если млекопитающее представляет собой человека, человек может отметить прилив сил или энергии или ослабление боли как субъективные симптомы улучшения или ответа на терапию. Альтернативно, врач может отметить уменьшение размера опухоли или опухолевой массы на основании медицинского осмотра, лабораторных параметров, опухолевых маркеров или рентгенологических данных. Некоторые лабораторные признаки, которые врач может наблюдать на предмет ответа на лечение, включают нормализацию таких показателей, как количество лейкоцитов, количество эритроцитов, количество тромбоцитов, скорость оседания эритроцитов и уровни различных ферментов. Кроме того, врач может наблюдать снижение уровня выявляемого опухолевого маркера. Альтернативно, для оценки объективного улучшения могут быть применены другие исследования, такие как ультразвуковое исследование, магнитно-резонансное исследование и позитронно-эмиссионное исследование.
Термин «вектор» относится к молекуле нуклеиновой кислоты, способной переносить чужеродную молекулу нуклеиновой кислоты. Чужеродную молекулу нуклеиновой кислоты соединяют с векторной молекулой нуклеиновой кислоты рекомбинантной методикой, такой как лигирование или рекомбинация. Это обеспечивает возможность увеличения количества, селекции, других манипуляций или экспрессии чужеродной молекулы нуклеиновой кислоты в клетке-хозяине или организме-хозяине. Вектор может представлять собой плазмиду, фаг, транспозон, космиду, хромосому, вирус или вирион. Векторы одного типа способны к интеграции в геном клетки-хозяина при введении в клетку-хозяина и, таким образом, проходят репликацию вместе с геномом хозяина (например, неэписомные векторы млекопитающих). Векторы другого типа способны к автономной репликации в клетке-хозяине, в которую ее вводят (например, бактериальные векторы, имеющие бактериальный репликатор, и эписомные векторы млекопитающих). Векторы еще одного определенного типа, способные управлять экспрессией экспрессируемых чужеродных нуклеиновых кислот, с которыми они функционально связаны, обычно называют «векторами экспрессии». Векторы экспрессии обычно имеют контрольные последовательности, управляющие экспрессией экспрессируемых чужеродных нуклеиновых кислот.Более простые векторы, известные как «транскрипционные векторы», способны только к транскрипции, но не к трансляции: в клетке-мишени возможна их репликация, но не экспрессия. Термин «вектор» включает векторы всех типов, независимо от их функции. Векторы, способные управлять экспрессией экспрессируемых нуклеиновых кислот, с которыми они функционально связаны, обычно называют «векторами экспрессии».
Все способы и методики по настоящему изобретению обычно осуществляют в соответствии со способами, хорошо известными в данной области техники, как описано в различных общих и более конкретных ссылках, упомянутых и обсужденных в настоящем описании, если не указано иное. Такие ссылки включают, например, Sambrook and Russell, Molecular Cloning, A Laboratory Approach, Cold Spring Harbor Press, Cold Spring Harbor, NY (2001), Ausubel et al., Current Protocols in Molecular Biology, John Wiley & Sons, NY (2002), и Harlow and Lane Antibodies: A Laboratory Manual, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY (1990). Ферментативные реакции и методики очистки осуществляют в соответствии с описаниями изготовителей, как это обычно делают в данной области техники или как описано здесь. Лабораторные способы и методики аналитической химии, синтетической органической химии и медицинской и фармацевтической химии, описанные здесь, и используемые в связи с ними номенклатуры являются хорошо известными и широко применяемыми в данной области техники. Для химических синтезов, химических анализов, фармацевтического изготовления, приготовления лекарственного средства, доставки и лечения пациентов применяют стандартные методики.
При использовании здесь двадцать обычных аминокислот с аббревиатурами соответствуют их обычному применению. См. Immunology-A Synthesis (2nd Edition, E. S. Golub and D.R. Gren, Eds., Sinauer Associates, Sunderiand, Mass. (1991)).
Б. СВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ, СВЯЗЫВАЮЩИЕСЯ С ЧЕЛОВЕЧЕСКИМ 4-1ВВ
Согласно настоящему изобретению предложены выделенные связывающие молекулы, связывающиеся с человеческим 4-1ВВ, включая антитела против 4-1ВВ, антигенсвязывающие фрагменты антител против 4-1ВВ и производные антител против 4-1ВВ.
Б-1. Антитела против 4-1ВВ
В некоторых аспектах согласно настоящему изобретению предложено выделенное антитело, связывающееся с человеческим 4-1ВВ в эпитопе в пределах аминокислотных остатков 115-156 SEQ ID NO:68. В некоторых воплощениях выделенное антитело содержит аминокислотную последовательность H-CDR1 по SEQ ID NO:29, аминокислотную последовательность H-CDR2 по SEQ ID NO:30 и аминокислотную последовательность H-CDR3 по SEQ ID NO:31. В некоторых других воплощениях выделенное антитело содержит аминокислотную последовательность L-CDR1 по SEQ ID NO:34, аминокислотную последовательность L-CDR2 по SEQ ID NO:35 и аминокислотную последовательность L-CDR3 по SEQ ID NO:36. В некоторых воплощениях антитела, описанные здесь выше, имеют одно или более биологических свойств, описанных здесь ниже.
В других аспектах согласно изобретению предложено выделенное антитело, связывающееся с человеческим 4-1ВВ, содержащее: (a) H-CDR1, как изложено в SEQ ID NO:1, SEQ ID NO:15 или SEQ ID NO:29; (б) H-CDR2, как изложено в SEQ ID NO:2, SEQ ID NO:16 или SEQ ID NO:30; и (в) H-CDR3, как изложено в SEQ ID NO:3, SEQ ID NO:17 или SEQ ID NO:31.
В другом аспекте согласно изобретению предложено выделенное антитело, связывающееся с человеческим 4-1ВВ, содержащее: (а) L-CDR1, как изложено в SEQ ID NO:6, SEQ ID NO:20 или SEQ ID NO:34; (б) L-CDR2, как изложено в SEQ ID NO:7, SEQ ID NO:21 или SEQ ID NO:35; и (в) L-CDR3, как изложено в SEQ ID NO:8, SEQ ID NO:22, SEQ ID NO:36 или SEQ ID NO:55.
В другом аспекте согласно изобретению предложено выделенное антитело, связывающееся с человеческим 4-1ВВ, содержащее: (а) H-CDR1, как изложено в SEQ ID NO:1, SEQ ID NO:15 или SEQ ID NO:29; (б) H-CDR2, как изложено в SEQ ID NO:2, SEQ ID NO:16 или SEQ ID NO:30; и (в) H-CDR3, как изложено в SEQ ID NO:3, SEQ ID NO:17 или SEQ ID NO:31; и дополнительно содержащее: (г) L-CDR1, как изложено в SEQ ID NO:6, SEQ ID NO:20 или SEQ ID NO:34; (д) L-CDR2, как изложено в SEQ ID NO:7, SEQ ID NO:21 или SEQ ID NO:35; и (е) L-CDR3, как изложено в SEQ ID NO:8, SEQ ID NO:22, SEQ ID NO:36 или SEQ ID NO:55.
В некоторых других аспектах согласно изобретению предложено выделенное антитело, связывающееся с человеческим 4-1ВВ, выбранное из группы, состоящей из:
а) антитела, содержащего H-CDR1, как изложено в SEQ ID NO:1, H-CDR2, как изложено в SEQ ID NO:2, и H-CDR3, как изложено в SEQ ID NO:3;
б) антитела, содержащего H-CDR1, как изложено в SEQ ID NO:15, Н-CDR2, как изложено в SEQ ID NO:16, и H-CDR3, как изложено в SEQ ID NO:17; и
в) антитела, содержащего H-CDR1, как изложено в SEQ ID NO:29, Н-CDR2, как изложено в SEQ ID NO:30, и H-CDR3, как изложено в SEQ ID NO:31.
В некоторых других аспектах согласно изобретению предложено выделенное антитело, связывающееся с человеческим 4-1ВВ, выбранное из группы, состоящей из:
а) антитела, содержащего L-CDR1, как изложено в SEQ ID NO:6, L-CDR2, как изложено в SEQ ID NO:7, и L-CDR3, как изложено в SEQ ID NO:8;
б) антитела, содержащего L-CDR1, как изложено в SEQ ID NO:20, L-CDR2, как изложено в SEQ ID NO:21, и L-CDR3, как изложено в SEQ ID NO:22;
в) антитела, содержащего L-CDR1, как изложено в SEQ ID NO:34, L-CDR2, как изложено в SEQ ID NO:35, и L-CDR3, как изложено в SEQ ID NO:36; и
г) антитела, содержащего L-CDR1, как изложено в SEQ ID NO:34, L-CDR2, как изложено в SEQ ID NO:35, и L-CDR3, как изложено в SEQ ID NO:55.
В некоторых других аспектах согласно изобретению предложено выделенное антитело, связывающееся с человеческим 4-1ВВ, выбранное из группы, состоящей из:
а) антитела, содержащего: H-CDR1, как изложено в SEQ ID NO:1, Н-CDR2, как изложено в SEQ ID NO:2, H-CDR3, как изложено в SEQ ID NO:3; L-CDR1, как изложено в SEQ ID NO:6, L-CDR2, как изложено в SEQ ID NO:7, и L-CDR3, как изложено в SEQ ID NO:8;
б) антитела, содержащего H-CDR1, как изложено в SEQ ID NO:15, Н-CDR2, как изложено в SEQ ID NO:16, H-CDR3, как изложено в SEQ ID NO:17; L-CDR1, как изложено в SEQ ID NO:20, L-CDR2, как изложено в SEQ ID NO:21, и L-CDR3, как изложено в SEQ ID NO:22;
в) антитела, содержащего H-CDR1, как изложено в SEQ ID NO:29, Н-CDR2, как изложено в SEQ ID NO:30, H-CDR3, как изложено в SEQ ID NO:31; L-CDR1, как изложено в SEQ ID NO:34, L-CDR2, как изложено в SEQ ID NO:35, и L-CDR3, как изложено в SEQ ID NO:36; и
г) антитела, содержащего H-CDR1, как изложено в SEQ ID NO:29, Н-CDR2, как изложено в SEQ ID NO:30, H-CDR3, как изложено в SEQ ID NO:31; L-CDR1, как изложено в SEQ ID NO:34, L-CDR2, как изложено в SEQ ID NO:35, и L-CDR3, как изложено в SEQ ID NO:55.
В другом аспекте согласно изобретению предложено выделенное антитело, связывающееся с человеческим 4-1ВВ, содержащее аминокислотную последовательность цепи VH, как изложено в SEQ ID NO:4, SEQ ID NO:18, SEQ ID NO:32 и SEQ ID NO:43.
В другом аспекте согласно изобретению предложено выделенное антитело, связывающееся с человеческим 4-1ВВ, содержащее аминокислотную последовательность цепи VL, как изложено в SEQ ID NO:9, SEQ ID NO:23, SEQ ID NO:37, SEQ ID NO:45, SEQ ID NO:51, SEQ ID NO:56, SEQ ID NO:60 или SEQ ID NO:64.
В другом аспекте согласно изобретению предложено выделенное антитело, связывающееся с человеческим 4-1ВВ, содержащее (1) аминокислотную последовательность цепи VH, как изложено в SEQ ID NO:4, SEQ ID NO:18, SEQ ID NO:32 или SEQ ID NO:43, и (2) аминокислотную последовательность цепи VL, как изложено в SEQ ID NO:9, SEQ ID NO:23, SEQ ID NO:37, SEQ ID NO:45, SEQ ID NO:51, SEQ ID NO:56, SEQ ID NO:60 или SEQ ID NO:64.
В другом аспекте согласно изобретению предложено выделенное антитело, связывающееся с человеческим 4-1ВВ, выбранное из группы, состоящей из:
а) антитела, содержащего аминокислотную последовательность цепи VH, как изложено в SEQ ID NO:4, и аминокислотную последовательность цепи VL, как изложено в SEQ ID NO:9;
б) антитела, содержащего аминокислотную последовательность цепи VH, как изложено в SEQ ID NO:18, и аминокислотную последовательность цепи VL, как изложено в SEQ ID NO:23;
в) антитела, содержащего аминокислотную последовательность цепи VH, как изложено в SEQ ID NO:32, и аминокислотную последовательность цепи VL, как изложено в SEQ ID NO:37 или SEQ ID NO 56; и
г) антитела, содержащего аминокислотную последовательность цепи VH, как изложено в SEQ ID NO:43, и аминокислотную последовательность цепи VL, как изложено в SEQ ID NO:45, SEQ ID NO:51, SEQ ID NO:60 или SEQ ID NO:64.
В некоторых воплощениях антитела, описанные здесь выше, включая антитела, описанные со ссылкой на связывание эпитопов, и антитела, описанные со ссылкой на конкретные аминокислотные последовательности CDR или вариабельных областей, имеют по меньшей мере одно из следующих функциональных свойств: (а) связываются с человеческим 4-1ВВ с Ко 500 нМ или менее; (б) имеют агонистическую активность в отношении человеческого 4-1ВВ; (в) не связываются с человеческим рецептором CD40 в концентрациях до 1000 нМ; (г) не связываются с человеческим рецептором CD134 в концентрациях до 1000 нМ; (д) не связываются с крысиным или мышиным 4-1ВВ в концентрациях до 100 нМ; (е) способны ингибировать рост опухолевых клеток; и (ж) оказывают терапевтический эффект при раке. В некоторых других воплощениях антитела специфично связываются с человеческим 4-1ВВ с kd 500 нМ или менее, 100 нМ или менее, 50 нМ или менее, 10 нМ или менее, 5 нМ или менее или 1 нМ или менее, как измерено для внеклеточного домена человеческого 4-1ВВ анализом BIACore, описанным в данном изобретении. В других воплощениях антитело представляет собой человеческое антитело или гуманизированное антитело, специфично связывающиеся с человеческим 4-1ВВ с KD 500 нМ или менее, 100 нМ или менее, 50 нМ или менее, 10 нМ или менее, 5 нМ или менее или 1 нМ или менее, как измерено для внеклеточного домена человеческого 4-1ВВ анализом BIACore, описанным в данном изобретении. В некоторых других воплощениях антитело представляет собой человеческое антитело, специфично и селективно связывающееся с человеческим 4-1ВВ.
В других воплощениях антитела, описанные здесь выше, содержат вариабельную область тяжелой цепи определенного эмбрионального гена тяжелой цепи иммуноглобулина и/или вариабельную область легкой цепи определенного эмбрионального гена легкой цепи иммуноглобулина, как, например, антитело, содержащее вариабельную область тяжелой цепи, являющуюся продуктом или имеющую происхождение от человеческих генов VH 1-69, VH 3-23 или VH 5. Типичные антитела включают MOR-7480.1, MOR-7480.2, MOR-7483.1 и MOR-7483.2, каждое из которых содержит аминокислоты, имеющие происхождение от человеческого эмбрионального гена VH5.
В других воплощениях антитела, описанные здесь выше, содержат вариабельную область легкой цепи, имеющую происхождение от человеческого гена VL λ3. В другом воплощении антитела, описанные здесь выше, содержат вариабельную область тяжелой цепи, являющуюся продуктом или имеющую происхождение от человеческих генов VH 1-69, VH 3-23 или VH 5, и дополнительно содержат вариабельную область легкой цепи, являющуюся продуктом или имеющую происхождение от человеческого гена VL λ3, где антитело или его часть специфично связывается с человеческим 4-1ВВ. Типичные антитела включают MOR-7480.1, MOR-7480.2, MOR-7483.1 и MOR-7483.2, каждое из которых содержит аминокислоты, имеющие происхождение от человеческих эмбриональных генов VH5 и VL λ3, соответственно.
При использовании здесь человеческое антитело содержит вариабельные области тяжелой или легкой цепи, «имеющие происхождение от» определенной эмбриональной последовательности, если вариабельные области антитела получены из системы, в которой использованы человеческие эмбриональные гены иммуноглобулинов. Такие системы включают иммунизацию трансгенной мыши, несущей человеческие гены иммуноглобулинов, интересующим антигеном или скрининг библиотеки человеческих генов иммуноглобулинов, представленной на фаге, с использованием интересующего антигена. Человеческое антитело, «имеющее происхождение от» человеческой эмбриональной последовательности иммуноглобулина, может быть определено как таковое сравнением аминокислотной последовательности человеческого антитела с аминокислотными последовательностями человеческих эмбриональных иммуноглобулинов и выбором человеческой эмбриональной последовательности иммуноглобулина, наиболее близкой (то есть с наибольшим % идентичности) к последовательности человеческого антитела. Человеческое антитело, «имеющее происхождение от» определенной человеческой эмбриональной последовательности иммуноглобулина, может иметь аминокислотные отличия по сравнению с эмбриональной последовательностью ввиду, например, естественных соматических мутаций или преднамеренного введения сайт-направленной мутации. Тем не менее, аминокислотная последовательность выбранного человеческого антитела обычно по меньшей мере на 90% идентична аминокислотной последовательности, кодируемой человеческим эмбриональным геном иммуноглобулина, и содержит аминокислотные остатки, определяющие человеческое антитело как человеческое при сравнении с аминокислотными последовательностями эмбриональных иммуноглобулинов другого вида (например, с мышиными эмбриональными последовательностями). В определенных случаях аминокислотная последовательность человеческого антитела может быть по меньшей мере на 95% или даже по меньшей мере на 96%, 97%, 98% или 99% идентична аминокислотной последовательности, кодируемой эмбриональным геном иммуноглобулина. В определенных случаях аминокислотная последовательность человеческого антитела идентична аминокислотной последовательности, кодируемой эмбриональным геном иммуноглобулина. Обычно человеческое антитело, имеющее происхождение от определенной человеческой эмбриональной последовательности, будет иметь не более 10 аминокислотных отличий от аминокислотной последовательности, кодируемой человеческим эмбриональным геном иммуноглобулина. В определенных случаях человеческое антитело может иметь не более 5 или даже не более 4, 3, 2 или 1 аминокислотного отличия от аминокислотной последовательности, кодируемой эмбриональным геном иммуноглобулина. Выравнивания аминокислотных последовательностей вариабельных областей иллюстративных антител и соответствующих эмбриональных последовательностей представлены на Фиг.6.
В другом аспекте согласно изобретению предложены выделенные антитела, конкурирующие или перекрестно конкурирующие за связывание с человеческим 4-1ВВ с любым из иллюстративных антител по изобретению, таким как MOR-6032, MOR-7361, MOR-7480, MOR-7480.1, MOR-7480.2, MOR-7483, MOR-7483.1 или MOR-7483.2. В определенном воплощении согласно изобретению предложены выделенные антитела, конкурирующие или перекрестно конкурирующие за связывание с одним и тем же эпитопом человеческого 4-1ВВ с любым из иллюстративных антител по изобретению. Способность антитела конкурировать или перекрестно конкурировать за связывание с другим антителом может быть определена с применением стандартных анализов связывания, известных в данной области техники, таких как анализ BIAcore, ELISA-анализы или проточная цитометрия. Например, возможно связывание иллюстративного антитела по изобретению с человеческим 4-1ВВ в насыщающих условиях с последующим измерением способности исследуемого антитела связываться с человеческим 4-1ВВ. Если исследуемое антитело способно связываться с 4-1ВВ одновременно с иллюстративным антителом, то исследуемое антитело и иллюстративное антитело связываются с разными эпитопами. Тем не менее, если исследуемое антитело не способно связываться с 4-1ВВ одновременно с иллюстративным антителом, то исследуемое антитело связывается с тем же эпитопом, перекрывающимся эпитопом или эпитопом, расположенным в непосредственной близости от эпитопа, с которым связывается иллюстративное антитело. Этот эксперимент может быть проведен с применением различных способов, таких как ELISA, RIA, FACS или поверхностный плазменный резонанс.
Антитела против 4-1ВВ, описанные здесь, могут представлять собой антитела любого класса, такого как IgG, IgM, IgE, IgA или IgD. Предпочтительно, антитела против 4-1ВВ представляют собой антитела класса IgG, например, подкласса IgG1, IgG2, IgG3 или IgG4, более предпочтительно подкласса IgG2. Антитело против 4-1ВВ может быть преобразовано из одного класса или подкласса в другой класс или подкласс способами, известными в данной области техники. Типичный способ получения антитела желаемого класса или подкласса включает стадии выделения нуклеиновой кислоты, кодирующей тяжелую цепь антитела против 4-1ВВ, и нуклеиновой кислоты, кодирующей легкую цепь антитела против 4-1ВВ, выделение последовательности, кодирующей область VH, лигирование последовательности VH с последовательностью, кодирующей константную область тяжелой цепи желаемого класса или подкласса, экспрессию гена легкой цепи и конструкции тяжелой цепи в клетке и выделение антитела против 4-1ВВ.
Кроме того, антитела, предложенные согласно настоящему изобретению, могут быть моноклональными или поликлональными, но предпочтительно моноклональными.
Примеры конкретных выделенных антител, предложенных согласно настоящему изобретению, включают следующие иллюстративные антитела:
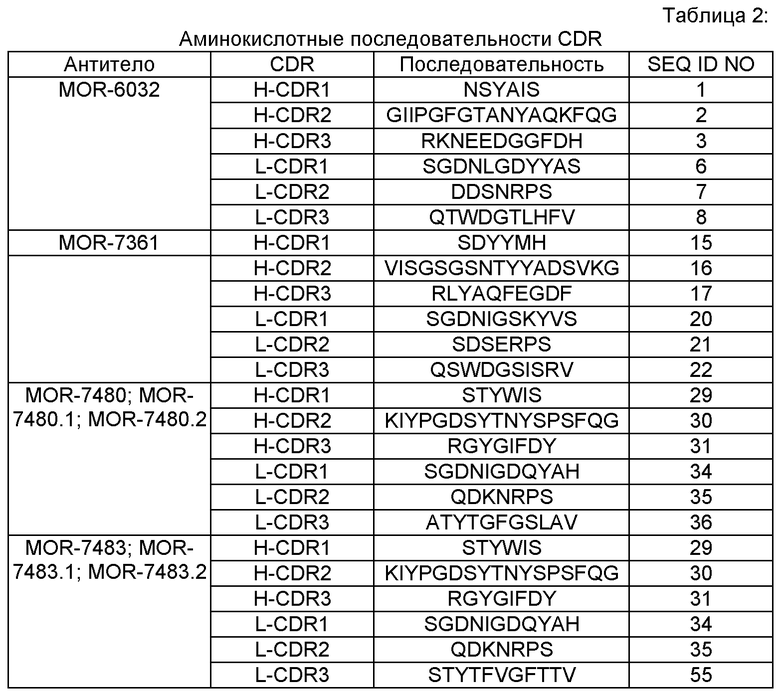
MOR-6032, MOR-7361, MOR-7480, MOR-7480.1, MOR-7480.1, MOR-7480.2, MOR-7483, MOR-7483, MOR-7483.1 и MOR-7483.2. Нуклеотидные и аминокислотные последовательности вариабельных областей тяжелой цепи, полноразмерных тяжелых цепей подкласса IgG2, вариабельных областей легкой цепи и полноразмерных легких цепей этих антител представлены в данном описании; указатель SEQ ID NO этих последовательностей приведен в Таблице 1. Аминокислотные последовательности CDR этих иллюстративных антител показаны в Таблице 2.
Антитела по настоящему изобретению могут быть получены методиками, известными в данной области техники, включая обычную методологию моноклональных антител, например, стандартную методику гибридизации соматических клеток (см., например, Kohler and Milstein, Nature 256:495 (1975)), вирусную или онкогенную трансформацию В-лимфоцитов или технологии рекомбинантных антител, как подробно описано здесь ниже.
Получение антител в гибридомах является общепризнанным способом. Обычной животной системой для получения гибридом является мышиная система. Протоколы иммунизации и методики выделения иммунизированных спленоцитов для слияния известны в данной области техники. Также известны партнеры слияния (например, клетки мышиной миеломы) и способы слияния. Один хорошо известный способ, который может быть применен для получения антител против человеческого 4-1ВВ, предложенных согласно настоящему изобретению, включает использование животной системы XenoMouse™. Мыши XenoMouse™ представляют собой искусственные мышиные штаммы, содержащие большие фрагменты локусов тяжелых цепей и легких цепей человеческих иммуноглобулинов, и не способные продуцировать мышиные антитела. См., например, Green et al., Nature Genetics 7:13-21 (1994) и WO 2003/040170. Животное иммунизируют антигеном 4-1ВВ. Антиген 4-1ВВ представляет собой выделенный и/или очищенный 4-1ВВ, предпочтительно 4-1ВВ. Он может представлять собой фрагмент 4-1ВВ, такой как внеклеточный домен 4-1ВВ, в частности фрагмент внеклеточного домена 4-1ВВ, содержащий аминокислотные остатки 115-156 SEQ ID NO:68. Иммунизация животных может быть проведена любым способом, известным в данной области техники. См., например, Harlow and Lane, Antibodies: A Laboratory Manual, New York: Cold Spring Harbor Press, 1990. Способы иммунизации животных, не являющихся людьми, таких как мыши, крысы, овцы, козы, свиньи, крупный рогатый скот и лошади, хорошо известны в данной области техники. См., например, Harlow and Lane, цит. выше, и патент США №5994619. Антиген 4-1ВВ может быть введен с адъювантом для стимуляции иммунного ответа. Типичные адъюванты включают полный или неполный адъювант Фрейнда, RIBI (мурамилдипептиды) или ISCOM (иммуностимулирующий комплекс). После иммунизации животного антигеном 4-1ВВ, из клеток, выделенных от иммунизированного животного, получают антителообразующие иммортализованные клеточные линии. После иммунизации животное умерщвляют и иммортализуют В-клетки лимфатических узлов и/или селезенки. Способы иммортализации клеток включают, без ограничения, введение в них онкогенов, инфицирование их онкогенным вирусом с их культивацией в условиях, приводящих к селекции иммортализованных клеток, воздействие канцерогенных или мутагенных соединений, их слияние с иммортализованной клеткой, например, клеткой миеломы, и инактивацию гена-супрессора опухоли. См., например, Harlow and Lane, цит. выше. При применении слияния с клетками миеломы клетки миеломы предпочтительно не секретируют иммуноглобулиновые полипептиды (несекреторная клеточная линия). Проводят скрининг иммортализованных клеток, применяя 4-1ВВ, его часть или клетку, экспрессирующую 4-1ВВ. Клетки, образующие антитела против 4-1ВВ, например, гибридомы, отбирают, клонируют и проводят их дальнейший скрининг на предмет желаемых свойств, включая активный рост, образование большого количества антител и желаемые свойства антител, как дополнительно обсуждено ниже. Гибридомы могут быть культивированы in vivo в сингенных животных, в животных с недостаточностью иммунной системы, например, «голых» мышах (бестимусных мышах), или в культуре клеток in vitro. Способы селекции, клонирования и культивирования гибридом хорошо известны специалистам в данной области техники.
Антитела по изобретению могут также быть получены с применением способов фагового дисплея. Такие способы фагового дисплея для выделения человеческих антител известны в данной области техники, как например библиотеки HuCAL®, как описано далее в Примере 1. См. также, например, Achim Knappik, et al: Fully Synthetic Human Combinatorial Antibody Libraries (HuCAL) Based on Modular Consensus Frameworks and CDRs Randomized with Trinucleotides. J. Mol. Biol. (2000) 296, 57-86.
Б-2. Антигенсвязывающие фрагменты
В некоторых других аспектах согласно настоящему изобретению предложены антигенсвязывающие фрагменты любого из антител против 4-1ВВ, предложенных согласно настоящему изобретению.
Антигенсвязывающий фрагмент может содержать любые последовательности антитела. В некоторых воплощениях антигенсвязывающий фрагмент содержит аминокислотную последовательность: (1) легкой цепи антитела против 4-1ВВ; (2) тяжелой цепи антитела против 4-1ВВ; (3) вариабельной области легкой цепи антитела против 4-1ВВ; (4) вариабельной области тяжелой цепи антитела против 4-1ВВ; (5) одного или более чем одного CDR (двух, трех, четырех, пяти или шести CDR) антитела против 4-1ВВ; или (6) трех CDR легкой цепи и трех CDR тяжелой цепи антитела против 4-1ВВ.
В некоторых определенных воплощениях согласно изобретению предложен антигенсвязывающий фрагмент антитела, выбранного из: MOR-6032, MOR-7361, MOR-7480, MOR-7480.1, MOR-7480.2, MOR-7483, MOR-7483.1 или MOR-7483.2.
В некоторых других определенных воплощениях антигенсвязывающие фрагменты антитела против 4-1ВВ включают: (1) Fab-фрагмент, представляющий собой моновалентный фрагмент, состоящий из доменов VL, VH, CL и CH1; (2) F(ab')2-фрагмент, представляющий собой бивалентный фрагмент, содержащий два Fab-фрагмента, связанных дисульфидной связью в шарнирной области; (3) Fd-фрагмент, состоящий из доменов VH и CH1; (4) Fv-фрагмент, состоящий из доменов VL и VH одной части антитела; (5) dAb-фрагмент (Ward et al., (1989) Nature 341:544-546), состоящий из домена VH; (6) выделенный CDR; и (7) одноцепочечное антитело (scFv), представляющее собой полипептид, содержащий область VL антитела, связанную с областью VH антитела. Bird et al., (1988) Science 242:423-426 и Huston et al., (1988) Proc. Natl. Acad. Sci. USA 85:5879-5883.
В некоторых определенных воплощениях антигенсвязывающий фрагмент представляет собой Fab-фрагмент, выбранный из группы, состоящей из Fab-6032, Fab-7361, Fab-7480 и Fab-7483.
Б-3. Производные антител
В некоторых других аспектах согласно настоящему изобретению предложены производные любого из антител против 4-1ВВ, предложенных согласно настоящему изобретению.
В одном аспекте производное антитела имеет происхождение от модификаций аминокислотных последовательностей иллюстративного антитела («исходного антитела») по изобретению при сохранении общей молекулярной структуры аминокислотной последовательности исходного антитела. Возможна модификация аминокислотных последовательностей любых областей исходного антитела, таких как каркасные области, CDR-участки или константные области. Типы модификаций включают замены, вставки, делеции одной или более аминокислот исходного антитела, или их комбинации. В некоторых воплощениях производное антитела содержит область VH, по меньшей мере на 65%, по меньшей мере на 75%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98% или по меньшей мере на 99% идентичную аминокислотной последовательности, как изложено в любой из SEQ ID NO:4, 18, 32 или 43. В некоторых других воплощениях производное антитела содержит область VL, по меньшей мере на 65%, по меньшей мере на 75%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98% или по меньшей мере на 99% идентичную аминокислотной последовательности, как изложено в любой из SEQ ID NO:9, 23, 37, 45, 51, 56, 60 или 64. В некоторых определенных воплощениях производное содержит 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14 или 15 консервативных или неконсервативных замен и/или 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14 или 15 присоединений или делеций, по сравнению с аминокислотной последовательностью, как изложено в любой из SEQ ID NO:4, 18, 32, 43, 9, 23, 37, 45, 51, 56, 60 или 64.
Аминокислотные замены включают как консервативные замены, так и неконсервативные замены. Термин «консервативная аминокислотная замена» обозначает замену одной аминокислоты другой аминокислотой, где две аминокислоты имеют сходство определенных физико-химических свойств, таких как полярность, заряд, растворимость, гидрофобность, гидрофильность и/или амфипатические свойства рассматриваемых остатков. Например, обычно возможны замены в пределах каждой из следующих групп: (а) неполярные (гидрофобные) аминокислоты, такие как аланин, лейцин, изолейцин, валин, пролин, фенилаланин, триптофан и метионин; (б) полярные нейтральные аминокислоты, такие как глицин, серин, треонин, цистеин, тирозин, аспарагин и глутамин; (в) положительно заряженные (основные) аминокислоты, такие как аргинин, лизин и гистидин; и (г) отрицательно заряженные (кислые) аминокислоты, такие как аспарагиновая кислота и глутаминовая кислота.
Модификации могут быть проведены в любых положениях аминокислотных последовательностей антитела, включая CDR, каркасные области или константные области. В одном воплощении согласно настоящему изобретению предложено производное антитела, содержащее последовательности CDR VH и VL иллюстративного антитела по данному изобретению, но содержащее последовательности каркасных областей, отличающиеся от соответствующих последовательностей иллюстративного антитела. Такие последовательности каркасных областей могут быть получены из общедоступных баз данных ДНК или опубликованных источников, содержащих последовательности эмбриональных генов антител. Например, эмбриональные последовательности ДНК человеческих генов вариабельных областей тяжелых и легких цепей могут быть обнаружены в базе данных Genbank или в базе данных человеческих эмбриональных последовательностей "VBase" (Kabat, E.A., et al., Sequences of Proteins of Immunological Interest, Fifth Edition, U.S. Department of Health and Human Services, NIH Publication No. 91-3242 (1991); Tomlinson, I.M., et al., J. Mot. Biol. 227:776-798 (1992); и Сох, J. P.L. et al., Eur. J. Immunol. 24:827-836 (1994)). Последовательности каркасных областей, которые могут быть использованы при конструировании производного антитела, включают последовательности, структурно сходные с последовательностями каркасных областей, использованными в иллюстративных антителах по изобретению, например, сходные с последовательностями каркасных областей VH 3-23 и/или последовательностями каркасных областей VL λ3 или λ1-13, использованными в иллюстративных антителах по изобретению. Например, последовательности H-CDR1, H-CDR2 и H-CDR3 и последовательности L-CDR1, L-CDR2 и L-CDR3 иллюстративного антитела могут быть перенесены на каркасные области, последовательность которых идентична последовательности, обнаруженной в эмбриональном гене иммуноглобулина, от которого происходит последовательность каркасной области, или последовательности CDR могут быть перенесены на каркасные области, содержащие одну или более мутаций по сравнению с эмбриональными последовательностями.
В определенном воплощении производное антитела представляет собой химерное антитело, содержащее аминокислотную последовательность иллюстративного антитела по изобретению. В одном примере один или более чем один CDR одного или более чем одного иллюстративного человеческого антитела сочетают с CDR антитела от животного, не являющегося человеком, такого как мышь или крыса. В другом примере все CDR химерного антитела имеют происхождение от одного или более чем одного иллюстративного антитела. В некоторых определенных воплощениях химерное антитело содержит один, два или три CDR вариабельной области тяжелой цепи или вариабельной области легкой цепи иллюстративного антитела. Химерные антитела могут быть получены с применением обычных способов, известных в данной области техники.
Модификация другого типа состоит в мутации аминокислотных остатков в пределах участков CDR1, CDR2 и/или CDR3 цепи VH и/или VL. Для введения мутации (мутаций) могут быть проведены сайт-направленный мутагенез или мутагенез, опосредованный полимеразной цепной реакцией (PCR, PCR-опосредованный мутагенез), и эффект на связывание антитела или другое интересующее функциональное свойство может быть оценен в анализах in vitro или in vivo, известных в данной области техники. Обычно вводят консервативные замены. Мутации могут представлять собой аминокислотные присоединения и/или делеции. Более того, обычно изменяют не более одного, двух, трех, четырех или пяти остатков в пределах CDR-участка. В некоторых воплощениях производное антитела содержит 1, 2, 3 или 4 аминокислотные замены в H-CDR и/или CDR легкой цепи. В другом воплощении аминокислотная замена состоит в замене одного или более чем одного цистеина антитела на другой аминокислотный остаток, такой как, без ограничения, аланин или серин. Цистеин может представлять собой канонический или неканонический цистеин. В одном воплощении производное антитела содержит 1, 2, 3 или 4 консервативные аминокислотные замены в H-CDR-участках, по сравнению с аминокислотными последовательностями иллюстративного антитела.
Также могут быть проведены модификации остатков каркасной области в пределах областей VH и/или VL. Обычно такие варианты каркасных областей получают для снижения иммуногенности антитела. Одним способом является «обратная мутация» одного или более чем одного остатка каркасных областей с восстановлением соответствующей эмбриональной последовательности. Антитело, прошедшее соматическую мутацию, может содержать остатки каркасных областей, отличающиеся от эмбриональной последовательности, от которой происходит антитело. Такие остатки могут быть определены сравнением последовательностей каркасных областей антитела с эмбриональными последовательностями, от которых происходит антитело. Для возврата последовательностей каркасных областей к их эмбриональной конфигурации возможна «обратная мутация» соматических мутаций с восстановлением эмбриональной последовательности, например, сайт-направленным мутагенезом или PCR-опосредованным мутагенезом. В некоторых иллюстративных антителах по настоящему изобретению были проведены такие «обратные мутации» с восстановлением определенных эмбриональных последовательностей, как описано более подробно в Примере 6.
Кроме того, могут также быть проведены модификации в пределах Fc-области иллюстративного антитела, обычно для изменения одного или более чем одного функционального свойства антитела, такого как период полувыведения из сыворотки, фиксация комлемента, связывание с Fc-рецепторами и/или антигензависимая клеточная цитотоксичность. В одном примере шарнирная область СН1 модифицирована таким образом, что число остатков цистеина в шарнирной области изменено, например, увеличено или уменьшено. Этот способ описан более подробно в патенте США №5677425. Число остатков цистеина в шарнирной области СН1 изменяют, например, для облегчения сборки тяжелых и легких цепей или для увеличения или уменьшения стабильности антитела. В другом случае Fc-шарнирную область антитела изменяют для уменьшения биологического периода полувыведения антитела.
Кроме того, антитело по изобретению может быть модифицировано для изменения его потенциального сайта или паттерна гликозилирования. Иллюстративные антитела MOR-7480 и MOR-7483, и их любые эмбриональные варианты, и антитела, содержащие аминокислотные последовательности вариабельной области тяжелой цепи MOR-7480 и MOR-7483, содержат потенциальный сайт N-гликозилирования (NYS) при аспарагине 59 в вариабельном домене тяжелой цепи. IgG-варианты этих антител дополнительно содержат второй сайт N-гликозилирования в Fc-домене тяжелой цепи. Конкретнее, в IgG2 варианте этих антител Fc-сайт N-гликозилирования (NST) расположен при аспарагине 292 в домене СН2 тяжелой цепи. То есть, степень гликозилирования каждой тяжелой цепи может составлять 0, 1 (в Fab или Fc) или 2, таким образом, что степень гликозилирования антитела, содержащего две тяжелые и две легкие цепи, может быть любой от 0 (то есть отсутствие гликозилирования во всех четырех потенциальных сайтах гликозилирования) до 4 (то есть, антитело гликозилировано в Fab- и Fc-сайтах каждой цепи). В другом аспекте согласно настоящему изобретению предложено производное антитела против 4-1ВВ, содержащее по меньшей мере одну мутацию в вариабельной области легкой цепи или тяжелой цепи, изменяющую паттерн гликозилирования вариабельной области. Такое производное антитела может иметь повышенную аффинность и/или измененную специфичность связывания с антигеном. Мутации могут приводить к появлению нового сайта гликозилирования в вариабельной области (V-области), изменению расположения одного или более чем одного сайта гликозилирования в V-области или удалению предсуществующего сайта гликозилирования в V-области. В одном воплощении согласно настоящему изобретению предложено производное антитела против 4-1ВВ, имеющее потенциальный сайт N-гликозилирования при аспарагине 59 в вариабельной области тяжелой цепи, где потенциальный сайт М-гликозилирования в одной вариабельной области тяжелой цепи удален. В другом воплощении согласно настоящему изобретению предложено производное антитела против 4-1ВВ, имеющее потенциальный сайт N-гликозилирования при аспарагине 59 в вариабельной области тяжелой цепи, где потенциальный сайт N-гликозилирования удален в вариабельных областях обеих тяжелых цепей. Способ изменения паттерна гликозилирования антитела известен в данной области техники, как, например, способы, описанные в патенте США №6933368, содержание которого полностью включено сюда посредством ссылки.
В другом аспекте согласно настоящему изобретению предложено производное антитела, содержащее антитело против 4-1ВВ или его антигенсвязывающий фрагмент, как описано здесь, связанное с дополнительной молекулярной группировкой. Примеры дополнительных молекулярных группировок включают фармацевтические агенты, пептиды или белки, агент для выявления, или метки, и антитела.
В некоторых воплощениях производное антитела содержит антитело по изобретению, связанное с фармацевтическим агентом. Примеры фармацевтических агентов включают цитотоксические агенты или другие противораковые терапевтические агенты и радиоактивные изотопы. Конкретные примеры цитотоксических агентов включают таксол, цитохалазин В, грамицидин D, бромид этидия, эметин, митомицин, этопозид, тенопозид, винкристин, винбластин, колхицин, доксорубицин, даунорубицин, дигидроксиантрациндион, митоксантрон, митрамицин, актиномицин D, 1-дегидротестостерон, глюкокортикоиды, прокаин, тетракаин, лидокаин, пропранолол, и пуромицин, и их аналоги или гомологи. Терапевтические агенты также включают, например, антиметаболиты (например, метотрексат, 6-меркаптопурин, 6-тиогуанин, цитарабин, 5-фторурацил, декарбазин), алкилирующие агенты (например, мехлорэтамин, тиотепу, хлорамбуцил, мелфалан, кармустин (BSNU) и ломустин (CCNU), циклофосфамид, бусульфан, дибромманнит, стрептозотоцин, митомицин С и цис-дихлордиаминплатину (II) (DDP) (цисплатин)), антрациклины (например, даунорубицин (ранее дауномицин) и доксорубицин), антибиотики (например, дактиномицин (ранее актиномицин), блеомицин, митрамицин и антрамицин (АМС)) и антимитотические агенты (например, винкристин и винбластин). Примеры радиоактивных изотопов, которые могут быть конъюгированы с антителами для диагностического или терапевтического применения, включают, без ограничения, йод131, индий111, иттрий90 и лютеций177. Способы связывания антитела с фармацевтическим агентом известны в данной области техники, как, например, применение различных линкерных технологий. Примеры типов линкеров включают гидразоны, тиоэфиры, эфиры, дисульфиды и пептидсодержащие линкеры. Более подробное обсуждение линкеров и способов связывания терапевтических агентов с антителами см. также в Saito et al., Adv. Drug Deliv. Rev. 55:199-215 (2003); Trail, et al., Cancer Immunol. Immunother. 52:328-337 (2003); Payne, Cancer Cell 3:207-212 (2003); Alien, Nat. Rev. Cancer 2:750-763 (2002); Pastan, I. and Kreitman, Curr. Opin. Investig. Drugs 3:1089-1091 (2002); Senter, P.D. and Springer, C.J. (2001) Adv. Drug Deliv. Rev. 53:247-264.
В определенном воплощении производное антитела представляет собой мультимер антитела против 4-1ВВ, являющийся мультимерной формой антитела против 4-1ВВ, такой как димеры, тримеры антител или мультимеры мономерных антител более высокого порядка. Отдельные мономеры мультимера антител могут быть одинаковыми или разными. Кроме того, отдельные антитела мультимера могут иметь одинаковые или разные специфичности связывания. Мультимеризация антител может быть проведена естественной агрегацией антител. Например, некоторый процент препаратов очищенных антител (например, очищенных молекул IgG1) спонтанно образует белковые агрегаты, содержащие гомодимеры антител и другие мультимеры антител более высокого порядка. Альтернативно, гомодимеры антител могут быть получены методиками химического связывания, известными в данной области техники, такими как использование поперечно сшивающих агентов. Подходящие поперечно сшивающие агенты включают гетеробифункциональные поперечно сшивающие агенты, имеющие две отдельные реакционноспособные группы, разделенные подходящим спейсером (такие как м-малеимидбензоил-N-гидроксисукцинимидный эфир, сукцинимидил-4-(малеимидметил)циклогексан-1-карбоксилат и N-сукцинимидил-S-ацетилтиоацетат), или гомобифункциональные поперечно сшивающие агенты (такие как дисукцинимидилсуберат). Такие линкеры имеются в продаже, например, от Pierce Chemical Company, Rockford, IL. Мультимеризация антител может также быть проведена методиками рекомбинантных ДНК, известными в данной области техники.
Примеры других производных антител, предложенных согласно настоящему изобретению, включают одноцепочечные антитела, диатела, однодоменные антитела, нанотела (nanobodies) и унитела (unibodies). «Одноцепочечное антитело» (scFv) состоит из одной полипептидной цепи, содержащей домен VL, связанный с доменом VH, где домен VL и домен VH спарены с образованием моновалентной молекулы. Одноцепочечное антитело может быть получено способом, известным в данной области техники (см., например, Bird et al., (1988) Science 242:423-426 и Huston et al., (1988) Proc. Natl. Acad. Sci. USA 85:5879-5883). «Диатело» состоит из двух цепей, каждая из которых содержит вариабельную область тяжелой цепи, связанную с вариабельной областью легкой цепи на одной полипептидной цепи, соединенные коротким пептидным линкером, где две области одной цепи не образуют пару друг с другом, но вместе с комплементарными доменами другой цепи образуют биспецифичную молекулу. Способы получения диател известны в данной области техники (см., например, Holliger Р. et al., (1993) Proc. Natl. Acad. Sci. USA 90:6444-6448, и Poljak R. J. et al., (1994) Structure 2:1121-1123). Однодоменные антитела (dAb) представляют собой малые функциональные связывающие единицы антител, соответствующие вариабельным областям тяжелых или легких цепей антител. Однодоменные антитела хорошо экспрессируются в бактериальных, дрожжевых клеточных системах и системах с клетками млекопитающих. Дополнительные подробности об однодоменных антителах и способы их получения известны в данной области техники (см., например, патенты США №№6291158; 6582915; 6593081; 6172197; 6696245; Европейские патенты 0368684 и 0616640; WO 05/035572, WO 04/101790, WO 04/081026, WO 04/058821, WO 04/003019 и WO 03/002609). Нанотела имеют происхождение от тяжелых цепей антитела. Нанотело обычно содержит один вариабельный домен и два константных домена (СН2 и СНЗ) и сохраняет антигенсвязывающую способность исходного антитела. Нанотела могут быть получены способами, известными в данной области техники (см., например, патент США №6765087, патент США №6838254, WO 06/079372). Унитела состоят из одной легкой цепи и одной тяжелой цепи антитела IgG4. Унитела могут быть получены удалением шарнирной области антител IgG4. Дополнительные подробности об унителах и способы их получения могут быть обнаружены в WO 2007/059782.
В. НУКЛЕИНОВЫЕ КИСЛОТЫ, ВЕКТОРЫ, КЛЕТКИ-ХОЗЯЕВА И РЕКОМБИНАНТНЫЕ СПОСОБЫ ПОЛУЧЕНИЯ АНТИТЕЛ ПРОТИВ 4-1ВВ
Согласно другому аспекту изобретения предложена выделенная молекула нуклеиновой кислоты, содержащая нуклеотидную последовательность, кодирующую аминокислотную последовательность связывающей молекулы, предложенной согласно настоящему изобретению. Аминокислотная последовательность, кодируемая нуклеотидной последовательностью, может представлять собой любую часть антитела, такую как CDR, последовательность, содержащую один, два или три CDR, вариабельную область тяжелой цепи, вариабельную область легкой цепи, или может представлять собой полноразмерную тяжелую цепь или полноразмерную легкую цепь. Нуклеиновая кислота по изобретению может представлять собой, например, ДНК или РНК и может содержать или не содержать интронные последовательности. Обычно нуклеиновая кислота представляет собой молекулу кДНК.
В некоторых воплощениях согласно изобретению предложена выделенная молекула нуклеиновой кислоты, содержащая или состоящая из нуклеотидной последовательности, кодирующей аминокислотную последовательность, выбранную из группы, состоящей из: (1) аминокислотной последовательности H-CDR3 или L-CRD3 иллюстративного антитела; (2) вариабельной области тяжелой цепи или вариабельной области легкой цепи иллюстративного антитела; или (3) полноразмерной тяжелой цепи или полноразмерной легкой цепи иллюстративного антитела.
В других воплощениях молекула нуклеиновой кислоты содержит или состоит из нуклеотидной последовательности, кодирующей аминокислотную последовательность, как изложено в любой из SEQ ID NO:1-10, 15-24, 29-38, 43, 44, 45, 46, 51, 52, 55-57, 60, 61, 64 и 65.
В других воплощениях молекула нуклеиновой кислоты содержит или состоит из нуклеотидной последовательности, выбранной из группы, состоящей из SEQ ID NO:11-14, 25-28, 39-42, 47-50, 53, 54, 58, 59, 62, 63, 66 и 67.
Нуклеиновые кислоты по изобретению могут быть получены с применением любых подходящих молекулярно-биологических методик. Для антител, экспрессируемых в гибридомах, кДНК, кодирующая легкие и тяжелые цепи антитела, получаемого в гибридоме, может быть получена методиками PCR-амплификации или клонирования кДНК. Для антител, получаемых из библиотеки генов иммуноглобулинов (например, с применением методик фагового дисплея), нуклеиновая кислота, кодирующая антитело, может быть получена из библиотеки.
Выделенная ДНК, кодирующая область VH, может быть превращена в ген полноразмерной тяжелой цепи функциональным связыванием ДНК, кодирующей VH, с другой молекулой ДНК, кодирующей константные области тяжелой цепи (СН1, СН2 и СН3). Последовательности человеческих генов константных областей тяжелой цепи известны в данной области техники (см., например, Kabat el al. (1991) Sequences of Proteins of Immunological Interest, Fifth Edition, U.S. Department of Health and Human Services, NIH Publication No. 91-3242), и фрагменты ДНК, содержащие эти области, могут быть получены стандартной PCR-амплификацией. Константная область тяжелой цепи может представлять собой константную область IgG1, IgG2, IgG3, IgG4, IgA, IgE, IgM или IgD, но наиболее предпочтительно представляет собой константную область IgG1 или IgG4. Последовательность константной области IgG1 может представлять собой последовательность любого из различных аллелей или аллотипов, встречающихся у разных индивидов, таких как Gm(1), Gm(2), Gm(3) и Gm(17). Эти аллотипы отражают встречающиеся в природе аминокислотные замены в константных областях IgG1. Для гена Fab-фрагмента тяжелой цепи, ДНК, кодирующая VH, может быть функционально связана с другой молекулой ДНК, кодирующей только константную область СН1 тяжелой цепи.
Выделенная ДНК, кодирующая область VL, может быть превращена в ген полноразмерной легкой цепи (а также в ген Fab-фрагмента легкой цепи) функциональным связыванием ДНК, кодирующей VL, с другой молекулой ДНК, кодирующей константную область легкой цепи CL. Последовательности человеческих генов константной области легкой цепи известны в данной области техники (см., например, Kabat et al. (1991) Sequences of Proteins of Immunological Interest, Fifth Edition, U.S. Department of Health and Human Services, NIH Publication No.91-3242), и фрагменты ДНК, содержащие эти области, могут быть получены стандартной PCR-амплификацией. Константная область легкой цепи может представлять собой константную область каппа или лямбда.
Для получения гена scFv фрагменты ДНК, кодирующие VH и VL, функционально связывают с другим фрагментом, кодирующим гибкий линкер, например, кодирующим аминокислотную последовательность (Gly4-Ser)3, таким образом, что последовательности VH и VL могут быть экспрессированы в форме непрерывного одноцепочечного белка, где области VL и VH соединены гибким линкером (см., например, Bird et al., Science 242:423-426 (1988); Huston et al., Proc. Natl. Acad. Sci. USA 85:5879-5883 (1988); и McCafferty et al., Nature 348:552-554(1990)).
Согласно настоящему изобретению также предложен вектор, содержащий молекулу нуклеиновой кислоты, предложенную согласно настоящему изобретению. Молекула нуклеиновой кислоты может кодировать часть легкой цепи или тяжелой цепи (такую как CDR или вариабельная область), полноразмерную легкую или тяжелую цепь, полипептид, содержащий полноразмерную тяжелую или легкую цепь или их часть, или аминокислотную последовательность производного или антигенсвязывающего фрагмента антитела. В некоторых воплощениях вектор представляет собой вектор экспрессии, применимый для экспрессии связывающей молекулы, такой как антитело или его антигенсвязывающий фрагмент.
Для экспрессии связывающей молекулы по изобретению, ДНК, кодирующие полноразмерные легкие и тяжелые цепи или их часть, вводят в векторы экспрессии таким образом, что молекулы ДНК функционально связаны с последовательностями контроля транскрипции и трансляции. В данном контексте термин «функционально связанный» означает, что ген антитела лигирован в вектор, таким образом, что последовательности контроля транскрипции и трансляции в векторе выполняют их предполагаемую функцию регуляции транскрипции и трансляции молекулы ДНК. Выбирают вектор экспрессии и последовательности контроля экспрессии, совместимые с клеткой-хозяином, используемой для экспрессии. Ген легкой цепи антитела и ген тяжелой цепи антитела могут быть введены в разные векторы, или, чаще, оба гена вводят в один и тот же вектор экспрессии. Гены антитела вводят в вектор экспрессии любыми подходящими способами (например, лигированием комплементарных сайтов рестрикции фрагмента гена антитела и вектора или лигированием тупых концов при отсутствии сайтов рестрикции). Вариабельные области легких и тяжелых цепей антител, описанных здесь, могут быть использованы для создания генов полноразмерных антител любого изотипа и подкласса введением их в векторы экспрессии, уже кодирующие константные области тяжелой цепи и легкой цепи желаемого изотипа и подкласса, таким образом, что сегмент VH функционально связан с сегментом(ами) СН вектора и сегмент VK функционально связан с сегментом CL вектора. Дополнительно или альтернативно, рекомбинантный вектор экспрессии может кодировать сигнальный пептид, способствующий секреции цепи антитела из клетки-хозяина. Ген цепи антитела может быть клонирован в вектор, таким образом, что сигнальный пептид связан в рамке считывания с N-концом гена цепи антитела. Сигнальный пептид может представлять собой сигнальный пептид иммуноглобулина или гетерологичный сигнальный пептид (то есть сигнальный пептид белка, не являющегося иммуноглобулином).
Помимо генов цепей антител векторы экспрессии по изобретению обычно содержат регуляторные последовательности, контролирующие экспрессию генов цепей антител в клетке-хозяине. Подразумевают, что термин «регуляторная последовательность» включает промоторы, энхансеры и другие элементы контроля экспрессии (например, сигналы полиаденилирования), контролирующие транскрипцию или трансляцию генов цепей антител. Такие регуляторные последовательности описаны, например, в Goeddel (Gene Expression Technology. Methods in Enzymology 185, Academic Press, San Diego, CA (1990)). Специалистам в данной области техники будет ясно, что конструкция вектора экспрессии, включая выбор регуляторных последовательностей, может зависеть от таких факторов, как выбор клетки-хозяина для трансформации, желаемый уровень экспрессии белка и так далее. Примеры регуляторных последовательностей для экспрессии в клетках-хозяевах, являющихся клетками млекопитающих, включают вирусные элементы, обеспечивающие высокие уровни экспрессии белков в клетках млекопитающих, такие как промоторы и/или энхансеры, имеющие происхождение от цитомегаловируса (CMV), вируса обезьян 40 (SV40), аденовируса (например, основной поздний промотор аденовируса (AdMLP)) и вируса полиомы. Альтернативно, могут быть использованы невирусные регуляторные последовательности, такие как промотор убиквитина или промотор β-глобина. Кроме того, могут быть использованы регуляторные элементы, состоящие из последовательностей от различных источников, такие как промоторная система SR, содержащая последовательности раннего промотора SV40 и длинный концевой повтор вируса Т-клеточного лейкоза человека 1 типа (Takebe, Y. et al. (1988) Mol. Cell. Biol. 8:466-472).
Помимо генов цепей антител и регуляторных последовательностей векторы экспрессии могут содержать дополнительные последовательности, такие как последовательности, регулирующие репликацию вектора в клетках-хозяевах (например, репликаторы), и селектируемые маркерные гены. Селектируемый маркерный ген облегчает селекцию клеток-хозяев, в которые был введен вектор (см., например, патенты США №№4399216, 4634665 и 5179017, авторами которых являются Axel et al.). Например, обычно селектируемый маркерный ген придает устойчивость клетке-хозяину, в которую был введен вектор, к лекарственным средствам, таким как G418, гигромицин или метотрексат.Селектируемые маркерные гены включают ген дигидрофолатредуктазы (DHFR) (для использования в dhfr- клетках-хозяевах с селекцией/амплификацией с метотрексатом) и ген neo (для селекции с использованием G418).
Для экспрессии легких и тяжелых цепей клетку-хозяина трансфицируют вектором(ами) экспрессии, кодирующим(ими) тяжелые и легкие цепи, с применением любых подходящих методик. Подразумевают, что различные формы термина «трансфекция» включают широкий спектр методик, обычно применяемых для введения экзогенной ДНК в прокариотическую или эукариотическую клетку-хозяина, например, электропорацию, преципитацию с фосфатом кальция, трансфекцию с использованием диэтиламиноэтилдекстрана (DEAE-декстрана) и тому подобное. Несмотря на возможность экспрессии антител по изобретению в прокариотических или эукариотических клетках-хозяевах, чаще всего экспрессию антител проводят в эукариотических клетках и обычно в клетках-хозяевах, являющихся клетками млекопитающих.
Согласно настоящему изобретению также предложена клетка-хозяин, содержащая молекулу нуклеиновой кислоты, предложенную согласно настоящему изобретению. Клетка-хозяин может представлять собой практически любую клетку, для которой доступны векторы экспрессии. Она может, например, представлять собой, клетку высших эукариот, такую как клетка млекопитающих, клетку низших эукариот, такую как дрожжевая клетка, и может представлять собой прокариотическую клетку, такую как бактериальная клетка. Введение рекомбинантной конструкции нуклеиновой кислоты в клетку-хозяина может быть проведено трансфекцией с использованием фосфата кальция, трансфекцией, опосредованной DEAE-декстраном, электропорацией или фаговой инфекцией.
Подходящие прокариотические хозяева для трансформации включают Е. coli, Bacillus subtilis, Salmonella typhimurium и различные виды родов Pseudomonas, Streptomyces и Staphylococcus.
Клетки-хозяева, являющиеся клетками млекопитающих, для экспрессии связывающей молекулы по изобретению включают, например, клетки яичника китайского хомячка (СНО) (включая dhfr-клетки СНО, описанные в Uriaub and Chasin, Proc. Natl. Acad. Sci. USA 77:4216-4220 (1980), используемые с селектируемым маркером DHFR, например, как описано в Kaufman and Sharp, J. Mol. Biol. 159:601-621 (1982)), клетки миеломы NSO, клетки COS и клетки Sp2. В частности, для применения с клетками миеломы NSO или клетками СНО, еще одной системой экспрессии является система экспрессии гена GS (глутаминсинтетазы), раскрытая в WO 87/04462, WO 89/01036 и ЕР 338841. Когда векторы экспрессии, кодирующие гены антител, вводят в клетки-хозяева, являющиеся клетками млекопитающих, антитела получают культивированием клеток-хозяев на протяжении периода времени, достаточного для экспрессии антитела в клетках-хозяевах или секреции антитела в культуральную среду, в которой проводят культивирование клеток-хозяев. Антитела могут быть выделены из культуральной среды с применением любых подходящих способов очистки белков.
Г. КОМПОЗИЦИИ
В других аспектах согласно настоящему изобретению предложена композиция, содержащая связывающую молекулу, предложенную согласно изобретению. В одном аспекте композиция представляет собой фармацевтическую композицию, содержащую связывающую молекулу и фармацевтически приемлемый носитель. Композиции могут быть изготовлены обычными способами, известными в данной области техники.
Термин «фармацевтическая композиция» относится к композициям, содержащим любые из связывающих молекул, любые из антител, любые из их антигенсвязывающих фрагментов или их производные, как описано здесь, и один или более чем один фармацевтически приемлемый эксципиент, разбавители или носители, необходимые для изготовления лекарственных форм для эффективной доставки связывающей молекулы.
В некоторых воплощениях согласно настоящему изобретению предложена композиция, содержащая антитело или его антигенсвязывающий фрагмент, предложенные согласно настоящему изобретению, и фармацевтически приемлемый носитель, где указанные антитело или антигенсвязывающий фрагмент содержат вариабельный домен, содержащий аминокислотную последовательность CDR, изложенную в SEQ ID NO:30, и где указанная композиция содержит не более приблизительно 11%, 10%, 8%, 5%, 3% или 2% указанного антитела или антигенсвязывающего фрагмента, гликозилированных при аспарагине указанной аминокислотной последовательности, по сравнению с общим количеством антитела или его антигенсвязывающего фрагмента, присутствующих в указанной композиции. В другом воплощении композиция содержит по меньшей мере приблизительно 2% указанного антитела или антигенсвязывающего фрагмента, гликозилированных при аспарагине указанной аминокислотной последовательности SEQ ID NO:30, по сравнению с общим количеством антитела или его антигенсвязывающего фрагмента, присутствующих в указанной композиции.
При использовании здесь термин «эксципиент» обозначает вещества, используемые для включения активных фармацевтических ингредиентов в фармацевтические композиции; в предпочтительном воплощении эксципиент не уменьшает или не препятствует первичному терапевтическому эффекту активных фармацевтических ингредиентов. Предпочтительно, эксципиент фармацевтически инертен. Термин «эксципиент» включает носители, разбавители, наполнители, растворители, стабилизаторы, наполнители, кислые или основные агенты для коррекции рН и связывающие агенты. Эксципиенты могут также представлять собой вещества, присутствующие в фармацевтической композиции как косвенный или непреднамеренный результат процесса изготовления. Предпочтительно, эксципиенты утверждены для введения человеку и животным, или их считают безопасными для введения человеку и животным, то есть они представляют собой вещества, признанные безвредными (GRAS(generally regarded as safe)-Beii4ecTBa). Список GRAS-веществ приведен Управлением по контролю за продуктами и лекарственными средствами (Food and Drug Administration) в Своде федеральных нормативных актов (Code of Federal Regulations (CFR)) в разделе 21, части 182 (21 CFR 182) и разделе 21, части 184 (21 CFR 184), включенных сюда посредством ссылки.
Термин «фармацевтически приемлемый носитель» относится к любому неактивному веществу, подходящему для применения в композиции для доставки связывающей молекулы. Носитель может представлять собой антиадгезионный агент, связывающий агент, оболочку, разрыхлитель, наполнитель или разбавитель, консервант (такой как антиоксидант, антибактериальный или противогрибковый агент), подсластитель, агент, замедляющий всасывание, смачивающий агент, эмульгатор, буфер и тому подобное. Примеры подходящих фармацевтически приемлемых носителей включают воду, этанол, полиолы (такие как глицерин, пропиленгликоль, полиэтиленгликоль и тому подобное), декстрозу, растительные масла (такие как оливковое масло), физиологический раствор, буфер, забуференный физиологический раствор и изотонические агенты, такие как сахара, полиспирты, сорбит и хлорид натрия.
Термин «буферный агент» относится к фармацевтически приемлемому эксципиенту, стабилизирующему рН фармацевтической композиции. Подходящие буферы хорошо известны в данной области техники и могут быть обнаружены в литературе. Например, могут быть использованы гистидиновые буферы, глициновые буферы, трис- или ацетатные буферы и/или их соответствующие свободные кислоты или основания, а также смеси различных солей и/или соответствующих кислот и оснований. В определенном воплощении буферы представляют собой гистидиновые буферы, то есть буферы, имеющие гистидин, обычно L-гистидин, в качестве буферного агента. В частности, буфер может представлять собой L-гистидиновый/НС1-буфер, содержащий L-гистидин, моногидрохлорид L-гистидина или смеси L-гистидина и моногидрохлорида L-гистидина. L-гистидиновые буферы обычно используют в концентрации от приблизительно 0,05 мг/мл до приблизительно 10 мг/мл, от приблизительно 0,1 мг/мл до приблизительно 5 мг/мл или от приблизительно 0,5 мг/мл до приблизительно 1 мг/мл. Буферы с моногидрохлоридом L-гистидина обычно используют в концентрации от приблизительно 0,1 мг/мл до приблизительно 10 мг/мл, от приблизительно 1 мг/мл до приблизительно 5 мг/мл или от приблизительно 2 мг/мл до приблизительно 4 мг/мл.
Независимо от того, какой буфер применяется, рН может быть скорректирован до значения в диапазоне от приблизительно 4,0 до приблизительно 7,0, или от приблизительно 5,0 до приблизительно 6,0, с использованием кислоты или основания, известных в данной области техники, например, соляной кислоты, уксусной кислоты, фосфорной кислоты, серной кислоты и лимонной кислоты, гидроксида натрия и гидроксида калия.
В одном аспекте настоящее изобретение относится к фармацевтической композиции, содержащей: от 0,1 до 1 мг/мл L-гистидина; от 1 до 5 мг/мл моногидрохлорида L-гистидина; от 50 до 100 мг/мл дигидрата трегалозы; от 0,01 до 0,1 мг/мл дигидрата двунатриевой соли этилендиаминтетрауксусной кислоты (EDTA); и от 0,05 до 1 мг/мл полисорбата 80; при рН в диапазоне от 4,0 до 7,0.
В определенном воплощении концентрация связывающей молекулы, входящей в композицию по изобретению, находится в диапазоне от приблизительно 10 до приблизительно 22 мг/мл.
В другом определенном воплощении буферный агент, входящий в композицию по изобретению, представляет собой гистидиновый буфер, например, L-гистидиновый/HCl-буфер, или ацетатный буфер, или буфер с ацетатом натрия. В определенном воплощении буферный агент представляет собой L-гистидин/HCl.
В определенном воплощении концентрация буфера составляет от приблизительно 0,1 мМ до приблизительно 50 мМ или от 1 мМ до приблизительно 25 мМ.
рН композиции по изобретению может быть выбран из следующих диапазонов: от 3 до 10 или от 4 до 9. В определенном воплощении буферный агент обеспечивает рН от 5,0 до 6,0 или 5,5±0.3. Могут быть необходимы рутинные коррекции рН в пределах или за пределами этих диапазонов для улучшения растворимости или стабильности полипептида или других компонентов композиции.
В одном воплощении композиция содержит полисорбат, например, полисорбат 20 или полисорбат 80. В определенном воплощении концентрация полисорбата 80 составляет от 0,01 до 10 мг/мл, от 0,5 до 5 мг/мл или от 0,1 до 0,5 мг/мл.
В одном воплощении композиция по изобретению содержит по меньшей мере один стабилизатор, выбранный из группы солей, например, хлорида натрия, сахаридов, дигидрата трегалозы или сахарозы и аминокислот, таких как гидрохлорид аргинина. В определенном воплощении концентрация по меньшей мере одного стабилизатора составляет от 10 до 200 мг/мл, от 20 до 150 мг/мл или от 50 до 100 мг/мл.
При использовании здесь термин «хелатирующий агент» обычно относится к эксципиенту, способному образовывать по меньшей мере одну связь (например, ковалентную, ионную или иную) с ионом металла. Хелатирующий агент обычно представляет собой мультидентатный лиганд, который может быть использован в выбранных жидких композициях в качестве стабилизатора для образования комплексов с частицами, которые могут приводить к нестабильности. Часто соединения, способные действовать как хелатирующие агенты, будут иметь богатые электронами функциональные группы. Подходящие богатые электронами функциональные группы включают карбоксильные группы, гидроксигруппы и аминогруппы. Расположение этих групп в аминополикарбоновых кислотах, гидроксиполикарбоновых кислотах, гидроксиаминокарбоновых кислотах и тому подобном приводит к образованию группировок, способных связываться с металлом.
Тем не менее, подразумевают, что настоящее изобретение не ограничено хелатирующими агентами, повышающими стабильность антител главным образом благодаря способности хелатирующего агента к образованию связей с ионом металла. Таким образом, подразумевают, что настоящее изобретение не ограничено конкретным механизмом, посредством которого хелатирующий агент стабилизирует композиции по настоящему изобретению, и свойства эксципиентов, называемых здесь хелатирующими агентами, обеспечивающие повышение стабильности антител, могут быть обусловлены главным образом механизмами, никак не связанными со способностью хелатирующего агента к образованию связей с ионом металла.
Хелатирующие агенты, подходящие для применения в настоящем изобретении, включают, без ограничения, аминополикарбоновые кислоты, гидроксиаминокарбоновые кислоты, N-замещенные глицины, 2-(2-амино-2-оксоэтил)-аминоэтансульфоновую кислоту (ВЕЗ), дефероксамин (DEF), лимонную кислоту, никотинамид и дезоксихолаты. Примеры подходящих аминополикарбоновых кислот включают этилендиаминтетрауксусную кислоту (EDTA), диэтилентриаминпентауксусную кислоту (DTPA), нитрилотриуксусную кислоту (NTA), N-2-ацетамидо-2-иминодиуксусную кислоту (ADA), бис(аминоэтил)гликолевый эфир, N,N,N',N'-тетрауксусную кислоту (EGTA), транс-диаминоциклогескантетрауксусную кислоту (DCTA), глутаминовую кислоту и аспарагиновую кислоту. Примеры подходящих гидроксиаминокарбоновых кислот включают N-гидроксиэтилиминодиуксусную кислоту (HIMDA), N,N-бис-гидроксиэтилглицин (бицин) и N-(трисгидроксиметилметил)-глицин (трицин). Примером подходящего N-замещенного глицина является глицилглицин. Примером подходящего дезоксихолата является дезоксихолат натрия.
Хелатирующие агенты, используемые в изобретении, могут присутствовать, по возможности, в форме свободной кислоты или свободного основания (например, называемых здесь взаимозаменяемо «EDTA» или «соль EDTA») или в форме соответствующей соли (например, соответствующей соли присоединения кислоты или соли присоединения основания, такой как динатриевая соль EDTA). Подходящие соли присоединения кислот включают, например, соли щелочных металлов (например, соли натрия или калия), соли щелочноземельных металлов (например, кальция), и соли могут быть получены с использованием других слабо связанных ионов металлов. Как известно в данной области техники, природа соли и число зарядов, подлежащих нейтрализации будут зависеть от числа присутствующих карбоксильных групп и рН, при котором используют стабилизирующий хелатирующий агент. Как также известно в данной области техники, сила связывания хелатирующих агентов с определенными связываемыми ионами-мишенями варьирует. В качестве дополнительной иллюстрации подходящие соли EDTA включают дикалиевую соль EDTA, динатриевую соль EDTA, кальциевую-динатриевую соль EDTA, натриевую соль EDTA, тринатриевую соль EDTA и калиевую соль EDTA; и подходящей солью дефероксамина (DEF) является мезилат дефероксамина (DFM).
Хелатирующие агенты, используемые в изобретении, могут присутствовать в безводной, сольватированной или гидратированной форме соединения или соответствующей соли. Если хелатирующий агент присутствует в сольватированной или гидратированной форме, он может быть представлен в различных состояниях сольватации или гидратации (включая, например, безводную, гидратированную, дигидратированную и тригидратированную формы). В качестве дополнительной иллюстрации подходящим гидратом EDTA является дигидрат динатриевой соли EDTA. В одном воплощении дигидрат динатриевой соли EDTA присутствует в концентрации от 0,001 до 5 мг/мл, от 0,01 до 2 мг/мл и от 0,02 до 0,5 мг/мл. В другом воплощении хелатирующий агент может уменьшать или предотвращать агрегацию антител в композициях, описанных здесь. Такие хелатирующие агенты могут уменьшать или предотвращать деградацию антитела, включенного в композицию без защиты хелатирующим агентом.
Фармацевтические композиции могут обычно быть приспособлены для определенного предполагаемого способа введения. В данной области техники существует множество методик введения соединения, включая, без ограничения, пероральное, аэрозольное, парентеральное и местное введение. Композиции могут быть представлены в любых подходящих формах, таких как жидкие, полутвердые и твердые лекарственные формы. Примеры жидких лекарственных форм включают раствор (например, растворы для инъекций и инфузий), микроэмульсию, липосому, дисперсию или суспензию. Примеры твердых лекарственных форм включают таблетку, драже, капсулу, микрокапсулу и порошок. В частности, формой композиции, подходящей для доставки связывающей молекулы, является стерильная жидкость, такая как раствор, суспензия или дисперсия, для инъекции или инфузий. Стерильные растворы могут быть получены включением антитела в необходимом количестве в подходящий носитель с последующей стерилизацией микрофильтрацией. Обычно дисперсии получают включением антитела в стерильный наполнитель, содержащий основную дисперсионную среду и другие носители. В случае стерильных порошков для приготовления стерильной жидкости, способы получения включают вакуумную сушку и сублимационную сушку (лиофилизацию) с получением порошка активного ингредиента с любым желаемым дополнительным ингредиентом из их раствора, предварительно стерилизованного фильтрацией. Различные лекарственные формы композиций могут быть изготовлены обычными методиками, известными в данной области техники.
Относительное количество связывающей молекулы, включенной в композицию, будет варьировать в зависимости от ряда факторов, таких как конкретные используемые связывающая молекула и носители, лекарственная форма и желаемые высвобождение и фармакодинамические свойства. В одном воплощении композиция содержит связывающую молекулу в концентрации от 1 до 100 мг/мл, от 5 до 50 мг/мл или от 10 до 22 мг/мл. Количество связывающей молекулы в одной лекарственной форме будет обычно представлять собой количество, оказывающее терапевтический эффект, но может также представлять собой меньшее количество. Обычно это количество будет варьировать от приблизительно 0,01 процента до приблизительно 99 процентов, от приблизительно 0,1 процента до приблизительно 70 процентов, от приблизительно 1 процента до приблизительно 30 процентов относительно общей массы лекарственной формы.
Помимо связывающей молекулы в композицию может быть включен один или более чем один дополнительный терапевтический агент. Примеры дополнительных терапевтических агентов описаны здесь ниже. Специалист в данной области техники легко выберет подходящее количество дополнительного терапевтического агента для включения в композицию, и оно будет варьировать в зависимости от ряда факторов, таких как конкретные используемые агент и носители, лекарственная форма и желаемые высвобождение и фармакодинамические свойства. Количество дополнительного терапевтического агента, включенного в одну лекарственную форму, будет обычно представлять собой количество агента, оказывающее терапевтический эффект, но может также представлять собой меньшее количество.
Д. ПРИМЕНЕНИЕ СВЯЗЫВАЮЩИХ МОЛЕКУЛ И ФАРМАЦЕВТИЧЕСКИХ КОМПОЗИЦИЙ
Связывающие молекулы и фармацевтические композиции, предложенные согласно настоящему изобретению, полезны для терапевтических, диагностических или других целей, таких как усиление иммунного ответа, лечение рака, повышение эффективности другой терапии рака, повышение эффективности вакцин или лечение аутоиммунных заболеваний. Таким образом, в других аспектах согласно настоящему изобретению предложены способы применения связывающих молекул или фармацевтических композиций. В одном аспекте согласно настоящему изобретению предложен способ лечения расстройства у млекопитающего, включающий введение млекопитающему, нуждающемуся в лечении, терапевтически эффективного количества связывающей молекулы, предложенной согласно изобретению. Связывающая молекула может представлять собой агонист или антагонист 4-1ВВ. В некоторых воплощениях связывающая молекула представляет собой агонист 4-1ВВ. В некоторых воплощениях млекопитающее представляет собой человека.
В некоторых воплощениях расстройство представляет собой рак. С 4-1ВВ связано множество опухолей, злокачественных или доброкачественных, первичных или вторичных, и их можно лечить или предотвращать способом, предложенным согласно изобретению. Примеры таких опухолей включают опухоли легкого, такие как бронхогенный рак (например, плоскоклеточный рак, мелкоклеточный рак, крупноклеточный рак и аденокарцинома), альвеолярно-клеточный рак, аденома бронха, хондроматозная гамартома (доброкачественная) и саркома (злокачественная); опухоли сердца, такие как миксомы, фибромы и рабдомиомы; опухоли кости, такие как остеохондромы, хондромы, хондробластомы, хондромиксоидные фибромы, остеоидные остеомы, гигантоклеточные опухоли, хондросаркома, множественная миелома, остеосаркома, фибросаркомы, злокачественные фиброзные гистиоцитомы, опухоль Юинга (саркома Юинга) и ретикулоклеточная саркома; опухоли головного мозга, такие как глиомы (например, мультиформная глиобластома), анапластические астроцитомы, астроцитомы, олигодендроглиомы, медуллобластомы, хордома, шванномы, эпендимомы, менингиомы, аденома гипофиза, пинеалома, остеомы, гемангиобластомы, краниофарингиомы, хордомы, герминомы, тератомы, дермоидные кисты и ангиомы; опухоли пищеварительной системы, такие как лейомиомы, плоскоклеточный рак, аденокарцинома, лейомиосаркома, аденокарциномы желудка, липомы кишечника, нейрофибромы кишечника, фибромы кишечника, полипы толстой кишки и колоректальные раки; опухоли печени, такие как печеночно-клеточные аденомы, гемангиома, печеночно-клеточный рак, фиброламеллярный рак, холангиокарцинома, гепатобластома и ангиосаркома; опухоли почек, такие как аденокарцинома почек, почечно-клеточный рак, гипернефрома и переходно-клеточный рак почечной лоханки; раки мочевого пузыря; гематологические злокачественные опухоли, такие как острый лимфоцитарный (лимфобластный) лейкоз, острый миелоидный (миелоцитарный, миелогенный, миелобластный, миеломоноцитарный) лейкоз, хронический лимфолейкоз (например, синдром Сезари и волосатоклеточный лейкоз), хронический миелоцитарный (миелоидный, миелогенный, гранулоцитарный) лейкоз, лимфома Ходжкина, неходжкинская лимфома, В-клеточная лимфома, грибовидный микоз и миелопролиферативные расстройства (включая такие миелопролиферативные расстройства, как истинная полицитемия, миелофиброз, тромбоцитемия и хронический миелоцитарный лейкоз); опухоли кожи, такие как базально-клеточный рак, плоскоклеточный рак, меланома, саркома Калоши и болезнь Педжета; раки головы и шеи; раки, связанные с глазом, такие как ретинобластома и внутриглазная меланома; опухоли мужской половой системы, такие как доброкачественная гиперплазия предстательной железы, рак предстательной железы и опухоли яичка (например, семинома, тератома, эмбриональная карцинома и хориокарцинома); рак молочной железы; опухоли женской половой системы, такие как рак матки (рак эндометрия), рак шейки матки (карцинома шейки матки), рак яичников (карцинома яичников), рак наружных женских половых органов, рак влагалища, рак фаллопиевых труб и пузырный занос; рак щитовидной железы (включая папиллярный, фолликулярный, анапластический или медуллярный рак); феохромоцитомы (надпочечника); доброкачественные гиперплазии паращитовидных желез; раки поджелудочной железы; и гематологические злокачественные опухоли, такие как лейкозы, миеломы, неходжкинские лимфомы и лимфомы Ходжкина.
В некоторых других воплощениях расстройство представляет собой аутоиммунное заболевание. Примеры аутоиммунных заболеваний, которые можно лечить связывающими молекулами, включают аутоиммунный энцефаломиелит, красную волчанку и ревматоидный артрит.Связывающая молекула может также быть использована для лечения воспаления (такого как аллергическая астма) и хронической болезни «трансплантат против хозяина».
В другом аспекте согласно настоящему изобретению предложен способ усиления иммунного ответа у млекопитающего, включающий введение млекопитающему терапевтически эффективного количества связывающей молекулы, предложенной согласно изобретению. В некоторых воплощениях связывающая молекула представляет собой антитело против 4-1ВВ или его антигенсвязывающий фрагмент, и млекопитающее представляет собой человека. В другом воплощении связывающая молекула представляет собой антитело-агонист 4-1ВВ или его антигенсвязывающий фрагмент.Термин «усиление иммунного ответа» или его грамматические варианты означают стимуляцию, индукцию, повышение, улучшение или усиление любого ответа иммунной системы млекопитающего. Иммунный ответ может представлять собой клеточный ответ (то есть клеточно-опосредованный, такой как, опосредованный цитотоксическими Т-лимфоцитами) или гуморальный ответ (то есть антителоопосредованный ответ) и может представлять собой первичный или вторичный иммунный ответ. Примеры усиления иммунного ответа включают повышение активности CD4+ Т-клеток-хелперов и образование цитолитических Т-клеток. Усиление иммунного ответа можно оценивать с применением ряда измерений in vitro или in vivo, известных специалистам в данной области техники, включая, без ограничения, анализы цитотоксических Т-лимфоцитов, высвобождение цитокинов (например, образование интерлейкина-2 (IL-2)), регрессию опухолей, выживаемость животных с опухолями, образование антител, пролиферацию иммунокомпетентных клеток, экспрессию маркеров клеточной поверхности и цитотоксичность. Обычно способы по изобретению усиливают иммунный ответ млекопитающего по сравнению с иммунным ответом млекопитающего, не получающего лечение, или млекопитающего, не получающего лечение с применением заявленных способов. В одном воплощении связывающую молекулу используют для усиления иммунного ответа человека на микробный патоген (такой как вирус). В другом воплощении связывающую молекулу используют для усиления иммунного ответа человека на вакцину. Связывающая молекула может представлять собой агонист или антагонист 4-1ВВ. В некоторых воплощениях связывающая молекула представляет собой агонист 4-1ВВ. В одном воплощении способ усиливает клеточный иммунный ответ, в частности цитотоксический Т-клеточный ответ.В другом воплощении клеточный иммунный ответ представляет собой Т-хелперный клеточный ответ. В другом воплощении иммунный ответ представляет собой образование цитокинов, в частности образование IL-2. Связывающая молекула может быть использована для усиления иммунного ответа человека на микробный патоген (такой как вирус) или на вакцину. Связывающая молекула может представлять собой агонист или антагонист 4-1ВВ. В некоторых воплощениях связывающая молекула представляет собой агонист 4-1ВВ.
При практическом применении способов лечения связывающие молекулы могут быть введены сами по себе в форме монотерапии или введены в комбинации с одним или более дополнительным терапевтическим агентом или способами лечения. Таким образом, в другом аспекте согласно настоящему изобретению предложена комбинированная терапия, включающая связывающую молекулу в комбинации с одним или более дополнительными способами лечения или терапевтическими агентами для раздельного, последовательного или одновременного введения. Термин «дополнительный способ лечения» относится к лечению без использования связывающей молекулы, предложенной согласно изобретению, в качестве терапевтического агента. Термин «дополнительный терапевтический агент» относится к любому терапевтическому агенту, не являющемуся связывающей молекулой, предложенной согласно изобретению. В одном определенном аспекте согласно настоящему изобретению предложена комбинированная терапия для лечения рака у млекопитающего, включающая введение млекопитающему терапевтически эффективного количества связывающей молекулы, предложенной согласно изобретению, в комбинации с одним или более дополнительным терапевтическим агентом. В другом воплощении млекопитающее представляет собой человека.
В комбинации со связывающей молекулой, предложенной согласно настоящему изобретению, может быть использован широкий спектр противораковых терапевтических агентов. Специалист в данной области техники будет осведомлен о существовании и разработке других способов лечения рака, которые могут быть применены в комбинации со способами и связывающими молекулами по настоящему изобретению, и не будет ограничен формами лечения, изложенными здесь. Примеры категорий дополнительных терапевтических агентов, которые могут быть использованы в комбинированной терапии для лечения рака, включают (1) химиотерапевтические агенты, (2) иммунотерапевтические агенты и (3) агенты для гормональной терапии.
Термин «химиотерапевтический агент» относится к химическому или биологическому веществу, способному приводить к гибели раковых клеток или препятствовать росту, делению, восстановлению и/или функционированию раковых клеток. Примеры химиотерапевтических агентов включают агенты, раскрытые в WO 2006/088639, WO 2006/129163 и US 20060153808, содержание которых включено сюда посредством ссылки. Примеры конкретных химиотерапевтических агентов включают: (1) алкилирующие агенты, такие как хлорамбуцил (LEUKERAN), циклофосфамид (CYTOXAN), ифосфамид (IFEX), гидрохлорид мехлорэтамина (MUSTARGEN), тиотепа (THIOPLEX), стрептозоцин (ZANOSAR), кармустин (BICNU, GLIADEL WAFER), ломустин (CEENU) и дакарбазин (DTIC-DOME); (2) алкалоиды или алкалоиды барвинка, включая цитотоксические антибиотики, такие как доксорубицин (ADRIAMYCIN), эпирубицин (ELLENCE, PHARMORUBICIN), даунорубицин (CERUBIDINE, DAUNOXOME), неморубицин, идарубицин (IDAMYCIN PFS, ZAVEDOS), митоксантрон (DHAD, NOVANTRONE), дактиномицин (актиномицин D, COSMEGEN), пликамицин (MITHRACIN), митомицин (MUTAMYCIN) и блеомицин (BLENOXANE), тартрат винорелбина (NAVELBINE), винбластин (VELBAN), винкристин (ONCOVIN) и виндезин (ELDIS1NE); (3) антиметаболиты, такие как капецитабин (XELODA), цитарабин (CYTOSAR-U), флударабин (FLUDARA), гемцитабин (GEMZAR), гидроксимочевина (HYDRA), метотексат (FOLEX, МЕХАТЕ, TREXALL), неларабин (ARRANON), триметрексат (NEUTREXIN) и пеметрексед (ALIMTA); (4) антагонисты пиримидина, такие как 5-фторурацил (5-FU); капецитабин (XELODA), ралтитрексид (TOMUDEX), тегафур-урацил (UFTORAL) и гемцитабин (GEMZAR); (5) таксаны, такие как доцетаксел (TAXOTERE), паклитаксел (TAXOL); (6) платиновые лекарственные средства, такие как цисплатин (PLATINOL), карбоплатин (PARAPLATIN) и оксалиплатин (ELOXATIN); (7) ингибиторы топоизомеразы, такие как иринотекан (CAMPTOSAR), топотекан (HYCAMTIN), этопозид (ETOPOPHOS, VEPESSID, TOPOSAR) и тенипозид (VUMON); (8) эпиподофиллотоксины (производные подофиллотоксина), такие как этопозид (ETOPOPHOS, VEPESSID, TOPOSAR); (9) производные фолиевой кислоты, такие как лейковорин (WELLCOVORIN); (10) нитрозомочевины, такие как камустин (BiCNU), ломустин (CeeNU); (11) ингибиторы рецепторных тирозинкиназ, включая рецептор эпидермального фактора роста (EGFR), фактор роста эндотелия сосудов (VEGF), рецептор инсулина, рецептор инсулиноподобного фактора роста (IGFR), рецептор фактора роста гепатоцитов (HGFR) и рецептор тромбоцитарного фактора роста (PDGFR), такие как гефитиниб (IRESSA), эрлотиниб (TARCEVA), бортезомиб (VELCADE), мезилат иматиниба (GLEEVEC), генефитиниб (genefitinib), лапатиниб, сорафениб, талидомид, сунитиниб (SUTENT), акситиниб, ритуксимаб (RITUXAN, MABTHERA), трастузумаб (HERCEPTIN), цетуксимаб (ERBITUX), бевацизумаб (AVASTIN) и ранибизумаб (LUCENTIS), lym-1 (ONCOLYM), антитела к рецептору инсулиноподобного фактора роста-1 (IGF-1R), раскрытые в WO 2002/053596; (12) ингибиторы ангиогенеза, такие как бевацизумаб (AVASTIN), сурамин (GERMANIN), ангиостатин, SU5416, талидомид, ингибиторы матриксных металлопротеиназ (такие как батимастат и маримастат) и агенты, раскрытые в WO 2002055106; и (13) ингибиторы протеасом, такие как бортезомиб (VELCADE).
Термин «иммунотерапевтические агенты» относится к химическому или биологическому веществу, способному усиливать иммунный ответ млекопитающего. Примеры иммунотерапевтических агентов включают: бациллу Кальметта-Герена (BCG); цитокины, такие как интерфероны; вакцины, такие как персонализированная иммунотерапия MyVax, Onyvax-P, Oncophage, GRNVAC1, Favid, Provenge, GVAX, Lovaxin C, BiovaxlD, GMXX и NeuVax; и антитела, такие как алемтузумаб (САМРАТН), бевацизумаб (AVASTIN), цетуксимаб (ERBITUX), гемтузумаб озогамицин (MYLOTARG), ибритумомаб тиуксетан (ZEVALIN), панитумумаб (VECTIBIX), ритуксимаб (RITUXAN, MABTHERA), трастузумаб (HERCEPTIN), тоситумомаб (BEXXAR), ипилимумаб (YERVOY), тремелимумаб, CAT-3888, антитела-агонисты рецептора 0Х40 (такие как описанные в WO 2009/079335), антитела-агонисты рецептора CD40 (такие как описанные в WO 2003/040170) и агонисты Toll-подобного рецептора-9 (TLR-9) (такие как описанные в WO 2003/015711, WO 2004/016805 и WO 2009/022215).
Термин «агент для гормональной терапии» относится к химическому или биологическому веществу, ингибирующему или подавляющему образование гормона, или ингибирующему или противодействующему эффекту гормона на рост и/или выживание раковых клеток. Примеры таких агентов, подходящих для способов, описанных здесь, включают агенты, раскрытые в US 20070117809. Примеры конкретных агентов для гормональной терапии включают тамоксифен (NOLVADEX), торемифен (Fareston), фулвестрант (FASLODEX), анастрозол (ARIMIDEX), эксеместан (AROMASIN), летрозол (FEMARA), ацетат мегестрола (MEGACE), госерелин (ZOLADEX) и леупролид (LUPRON). Связывающие молекулы по данному изобретению могут также быть использованы в комбинации с нелекарственными способами гормональной терапии, такими как (1) хирургические способы полного или частичного удаления органов или желез, участвующих в образовании гормона, таких как яичники, яички, надпочечник и гипофиз, и (2) лучевая терапия, при которой на органы или железы пациента воздействуют излучением в количестве, достаточном для ингибирования или подавления образования целевого гормона.
Комбинированная терапия для лечения рака также включает комбинацию связывающей молекулы с хирургическим удалением опухоли. Связывающая молекула может быть введена млекопитающему до, во время или после хирургического вмешательства.
Комбинированная терапия для лечения рака также включает комбинацию связывающей молекулы с лучевой терапией, такой как ионизирующая (электромагнитная) лучевая терапия (например, рентгеновским или гамма-излучением) и лучевая терапия пучком частиц (например, излучением с высокой линейной энергией). Источник излучения может быть расположен вне или внутри млекопитающего. Связывающая молекула может быть введена млекопитающему до, во время или после лучевой терапии.
Связывающие молекулы и композиции, предложенные согласно настоящему изобретению, могут быть введены любым подходящим энтеральным или парентеральным путем введения. Термин «энтеральный путь» введения относится к введению через любую часть желудочно-кишечного тракта. Примеры энтеральных путей введения включают пероральный, трансмукозальный, трансбуккальный и ректальный путь введения или внутрижелудочный путь введения. «Парентеральный путь» введения относится к пути введения, отличному от энтерального пути введения. Примеры парентеральных путей введения включают внутривенное, внутримышечное, внутрикожное, внутрибрюшинное. внуриопухолевое, интравезикальное, внутриартериальное, интратекальное, внутрикапсулярное, интраорбитальное, интракардиальное, транстрахеальное, внутрисуставное, подкапсулярное, субарахноидальное, интраспинальное, эпидуральное и интрастернальное, подкожное или местное введение. Антитела и композиции по изобретению могут быть введены с применением любого подходящего способа, такого как пероральное введение, введение через назогастральный зонд, гастростомическую трубку, введение инъекцией, инфузией, имплантируемым инфузионным насосом и осмотическим насосом. Подходящий путь и способ введения могут варьировать в зависимости от ряда факторов, таких как конкретное используемое антитело, желаемая скорость всасывания, конкретная используемая композиция или лекарственная форма, тип или тяжесть расстройства, подлежащего лечению, конкретное место действия и состояние пациента, и могут быть легко выбраны специалистом в данной области техники.
Термин «терапевтически эффективное количество» связывающей молекулы относится к количеству, эффективному для предполагаемой терапевтической цели. Например, в контексте усиления иммунного ответа «терапевтически эффективное количество» представляет собой любое количество, эффективно стимулирующее, индуцирующее, повышающее, улучшающее или усиливающее любой ответ иммунной системы млекопитающего. В контексте лечения заболевания «терапевтически эффективное количество» представляет собой любое количество, достаточное для оказания любого желаемого или полезного эффекта у млекопитающего, получающего лечение. Конкретно, при лечении рака примеры желаемых или полезных эффектов включают ингибирование дальнейшего роста или распространения раковых клеток, гибель раковых клеток, ингибирование рецидивирования рака, ослабление боли, связанной с раком, или улучшенную выживаемость млекопитающего. Терапевтически эффективное количество антитела против 4-1ВВ обычно варьирует от приблизительно 0,001 до приблизительно 500 мг/кг и чаще от приблизительно 0,01 до приблизительно 100 мг/кг массы тела млекопитающего. Например, количество может составлять приблизительно 0,3 мг/кг, 1 мг/кг, 3 мг/кг, 5 мг/кг, 10 мг/кг, 50 мг/кг или 100 мг/кг массы тела млекопитающего. В некоторых воплощениях терапевтически эффективное количество антитела против 4-1ВВ находится в диапазоне приблизительно 0,01-30 мг/кг массы тела млекопитающего. В некоторых воплощениях терапевтически эффективное количество антитела против 4-1ВВ находится в диапазоне приблизительно 0,05-15 мг/кг массы тела млекопитающего. Конкретный уровень дозы для введения может быть легко определен специалистом в данной области техники и будет зависеть от ряда факторов, таких как тип и тяжесть расстройства, подлежащего лечению, конкретная используемая связывающая молекула, путь введения, время введения, продолжительность лечения, конкретный применяемый дополнительный способ лечения, возраст, пол, масса тела, состояние, общее состояние здоровья и предшествующий анамнез пациента, получающего лечение, и подобные факторы, хорошо известные в медицине.
Связывающую молекулу или композицию обычно вводят несколько раз. Интервалы между отдельными дозами могут составлять, например, неделю, месяц, три месяца или год. Типичная схема лечения включает введение один раз в неделю, один раз каждые две недели, один раз каждые три недели, один раз каждые четыре недели, один раз в месяц, один раз каждые три месяца или один раз каждые три-шесть месяцев. Типичные схемы введения для антитела против 4-1ВВ включают 1 мг/кг массы тела или 3 мг/кг массы тела внутривенным введением с применением одной из следующих схем введения: (1) каждые четыре недели для шести введений, затем каждые три месяца; (2) каждые три недели; (3) 3 мг/кг массы тела однократно с последующим введением 1 мг/кг массы тела каждые три недели.
Настоящее изобретение дополнительно проиллюстрировано следующими примерами, которые не следует толковать как дополнительное ограничение. Содержимое всех графических материалов и всех ссылок, патентов и опубликованных заявок на патенты, указанных в данном описании, прямо включено сюда посредством ссылки во всей их полноте.
ПРИМЕРЫ
ПРИМЕР 1: ПОЛУЧЕНИЕ Fab-ФРАГМЕНТОВ, СВЯЗЫВАЮЩИХСЯ С 4-1ВВ
Определенные антитела, предложенные согласно настоящему изобретению, были исходно получены из Fab-фрагментов, связывающихся с человеческим 4-1ВВ. Fab-фрагменты были отобраны из фаговой дисплейной библиотеки, фагмидной библиотеки MorphoSys HuCAL GOLD®, с последующим поочередным пэннингом на 4-1ВВ FC и клетки, экспрессирующие человеческий 4-1ВВ. Эти Fab-фрагменты включают Fab-фрагменты, обозначенные как «Fab-6032», «Fab-7361», «Fab-7480» и «Fab-7483». Антитела против 4-1 BB MOR-6032, MOR-7361, MOR-7480 и MOR-7483, описанные в данной заявке, были получены из «Fab-6032», «Fab-7361», «Fab-7480» и «Fab-7483», соответственно. Аминокислотная последовательность вариабельной области легкой цепи и вариабельной области тяжелой цепи данного Fab-фрагмента идентична аминокислотной последовательности вариабельной области легкой цепи и вариабельной области тяжелой цепи, соответственно, иллюстративного антитела, обозначение которого содержит тот же номер, что и обозначение Fab-фрагмента. Например, Fab-7480 и антитело MOR-7480 имеют идентичные аминокислотные последовательности их вариабельных областей легкой цепи и вариабельных областей тяжелой цепи, соответственно.
Фагмидная библиотека основана на концепции HuCAL® (Knappik et al., 2000, J. Mol. Biol. 296(1):57-86), и в ней применена технология CysDisplay™ для дисплея Fab-фрагментов на поверхности фагов (Lohning, WO 01/05950). HuCAL GOLD® обеспечивает возможность проведения селекции с шестью отдельными подбиблиотеками, каждая из которых содержит один основной ген VH (VH1, VH2, VH3, VH4, VH5, VH6) в сочетании со всеми семью основными генами VL, или проведение селекции с использованием объединенных совокупностей фагов. Фаги для первого раунда пэннинга получали посредством Hyperphage (M13KO7ΔpIII, полученного от Progen, Heidelberg, Germany). HuCAL GOLD® подробно описана в Christine Rothe, et. al, J. Mol. Biol. (2008) 376, 1182-1200.
Твердофазный пэннинг проводили с использованием рекомбинантного человеческого 4-1BB-Fc (R&D Systems, Cat. No.838-4B; Minneapolis, MN).
ПРИМЕР 2. ОПИСАНИЕ Fab-ФРАГМЕНТОВ
Свойства четырех Fab-фрагментов, описанных в Примере 1, определяли в анализах, описанных ниже, с использованием моновалентного Fab-формата, содержащего Fab-фрагмент, имеющий Flag/His-метку.
2А. Аффинность, определенная способом равновесного титрования в растворе (SET-способом)
Аффинность (выраженную как KD) четырех Fab-фрагментов определяли с применением SET-способа с использованием оборудования от Meso Scale Discovery («MSD»). Использовали мономерные белковые фракции антител (содержание мономера по меньшей мере 90%, определено аналитической гель-хроматографией (SEC); Superdex75 (Amersham Pharmacia) для Fab-фрагментов или Tosoh G3000SWXL (Tosoh Bioscience) для IgG, соответственно).
В целом, определение аффинности в растворе проводили, как описано в литературе (Friguet et al. 1985). Для улучшения чувствительности и точности SET-способа его проводили с применением технологии, основанной не на классическом ELISA, а на ECL (Haenel et al. 2005).
Проводили мечение козьих противочеловеческих антител, специфичных в отношении (Fab)2-фрагмента (Dianova), в концентрации 1 мг/мл меткой MSD Sulfo-TAG™ NHS-Ester (Meso Scale Discovery, Gaithersburg, MD, USA), следуя инструкциям изготовителя.
Эксперименты проводили в полипропиленовых титрационных микропланшетах с PBS (фосфатно-солевой буферный раствор), рН 7,4, с 0,5% BSA (бычий сывороточный альбумин) и 0,02% Tween 20 в качестве аналитического буфера. Проводили серийное разведение немеченого человеческого 4-1ВВ с кратностью 2n, начиная с концентрации, по меньшей мере в 10 раз превышающей предполагаемую KD. Лунки без антигена использовали для определения значений Bmax; лунки с аналитическим буфером использовали для определения фона. После добавления, например, 30 пМ Fab-фрагмента (конечная концентрация в конечном объеме 60 мкл) смесь инкубировали в течение ночи при комнатной температуре. Используемая концентрация Fab-фрагмента была близка к или ниже предполагаемой KD.
На стандартные MSD-планшеты сорбировали человеческий 4-1ВВ в концентрации 0,05 мкг/мл в PBS (30 мкл на лунку), планшеты инкубировали в течение ночи и блокировали 3% BSA в PBS в течение 1 часа. После отмывки планшетов аналитическим буфером в эти планшеты переносили уравновешенные образцы (30 мкл на лунку) и инкубировали в течение 20 минут.После отмывки в MSD-планшет добавляли меченное меткой MSD Sulfo-tag антитело для выявления (козье антитело против человеческого (Fab)2-фрагмента) в конечном разведении 1:1500 в количестве 30 мкл на лунку и инкубировали в течение 30 минут на качалке Eppendorf (700 об/мин).
После отмывки планшета и добавления буфера MSD Read Buffer Т с поверхностно-активным веществом в количестве 30 мкл на лунку электрохемилюминесцентные сигналы детектировали с использованием Sector Imager 6000 (Meso Scale Discovery, Gaithersburg, MD, USA).
Данные анализировали с применением программного обеспечения XLfit (IDBS), используя индивидуальные модели соответствия. Для определения Ко Fab-молекул использовали следующую модель соответствия (Haenel et al., 2005), модифицированную согласно Abraham et al. (1996, J. Molec. Recog. 9(5-6):456-461):
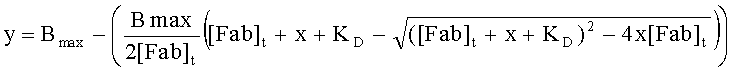
[Fab]t - использованная общая концентрация Fab-фрагмента;
x - использованная общая концентрация растворимого антигена (сайтов связывания);
Bmax - максимальный сигнал Fab-фрагмента без антигена;
KD - аффинность.
Результаты представлены в Таблице 3.
2Б. Определение KD на иммобилизованном прямым способом антигене с применением Biacore
Для определения KD в качестве аналита использовали мономерные фракции Fab-фрагментов (содержание мономера по меньшей мере 90%, определено аналитической SEC; Superdex75, Amersham Pharmacia). Связывание с иммобилизованным антигеном анализировали с использованием оборудования BIAcore3000 (Biacore, Sweden).
Кинетические константы скорости kon и koff определяли по связыванию серийных разведении соответствующих Fab-фрагментов с ковалентно иммобилизованным антигеном CD137/человеческим 4-1ВВ с использованием оборудования Biacore 3000 (Biacore, Uppsala, Sweden). Для ковалентной иммобилизации антигена применяли стандартную химию аминного сочетания EDC-NHS (1-этил-3-(3-диметиламинопропил)карбодиимид - N-гидроксисукцинимид). Кинетические измерения проводили в HBS-EP (10 мМ HEPES (N-2-гидроксиэтилпиперазин-N'-2-этансульфоновая кислота); рН 7,4; 150 мМ NaCl; 3 мМ EDTA (этилендиаминтетрауксусная кислота); Tween 20 0,005%) при скорости потока 20 мкл/мин с использованием концентраций Fab-фрагментов, варьирующих от приблизительно 16 до 500 нМ. Время инъекции для каждой концентрации составляло 1 минуту с последующей фазой диссоциации продолжительностью по меньшей мере 3 минуты. Для восстановления использовали одну или более инъекций 5 мкл глицина/HCl, рН 2.
Для оценки KD полноразмерных молекул IgG проводили инъекции образцов IgG на сенсорный чип F1 с низкой плотностью ковалентно иммобилизованного человеческого 4-1ВВ (приблизительно 130 RU (резонансных единиц)) с использованием серийного разведения с кратностью 2n и концентрациями, варьирующими от 16 до 500 нМ. Сенсограммы оценивали с применением бивалентной модели соответствия и проводили их количественное сравнение для ранжирования соответствующих значений Ко.
Анализ всех сенсограмм проводили с использованием программного обеспечения BIA evaluation 3.1 (Biacore). Результаты представлены в Таблице 3.
2В. Связывание Fab-фрагментов в ELISA-анализе
Связывание четырех Fab-фрагментов определяли с применением стандартных ELISA-методик на иммобилизованном прямым способом 4-1BB/Fc. Результаты представлены в Таблице 3.
2В. Связывание Fab-фрагментов в FACS-анализе
Связывание четырех Fab-фрагментов определяли с применением стандартных методик FACS-анализа на клетках HEK293, стабильно трансфицированных и экспрессирующих человеческий 4-1ВВ, а также клетках отрицательного контроля 300.19 (линии мышиных В-клеток). Результаты представлены в Таблице 3.
ПРИМЕР 3: ОПИСАНИЕ IgG
Несколько Fab-фрагментов, полученных пэннингом, как описано здесь, включая Fab-6032, Fab-7361, Fab-7480 и Fab-7483, были выбраны для превращения в полноразмерные антитела форматов IgG1 и IgG4 для дальнейшего описания их свойств, как описано в данном примере. Четыре иллюстративных антитела, описанных в данном примере, то есть MOR-6032, MOR-7361, MOR-7480 и MOR-7483, были получены из Fab-6032, Fab-7361, Fab-7480 и Fab-7483, соответственно. Антитела формата IgG экспрессировали и очищали, и затем определяли их свойства в ELISA, FACS-анализе и анализе с репортерным геном люциферазы.
3А. Превращение в IgG
Для экспрессии полноразмерного IgG, фрагменты вариабельных доменов тяжелой (VH) и легкой цепей (VL) субклонировали из векторов экспрессии Fab-фрагментов в подходящие векторы pMorph®_hIgG для человеческого IgG1 и человеческого IgG4.
3Б. Транзиторная экспрессия и очистка человеческого IgG
Транзиторную экспрессию полноразмерного человеческого IgG проводили в клетках HKB11, трансфицированных векторами экспрессии тяжелых и легких цепей IgG в соотношении 1:1. После трансфекции собирали супернатант клеточных культур и доводили его объем до трехкратного объема при трансфекции, соответственно. Супернатант очищали центрифугированием и фильтрацией и затем проводили стандартную аффинную хроматографию с белком A (MabSelect SURE, GE Healthcare). Белки элюировали и нейтрализовали. Последующая обработка включала замену буфера и стерилизацию фильтрацией. Концентрации белков определяли UV-спектрофотометрией. Степень очистки IgG анализировали в денатурирующих восстанавливающих и денатурирующих невосстанавливающих условиях при электрофорезе в полиакриламидном геле с додецилсульфатом натрия (SDS-PAGE) или с использованием Agilent BioAnalyzer. HP-SEC (высокоэффективную эксклюзионную хроматографию) проводили для анализа препаратов IgG в нативном состоянии.
3В. Описание IgG в ELISA-анализе
IgG использовали для описания связывания с применением ELISA с человеческим 4-1BB/Fc и мышиным 4-1BB/Fc при их прямой иммобилизации. В Таблице 4 ниже показаны результаты ELISA связывания для антител MOR-6032, MOR-7361, MOR-7480 и MOR-7483, все антитела формата IgG1.
3Г. Селективность связывания антител (FACS-анализ)
Селективность антител в отношении 4-1ВВ оценивали против внеклеточного домена белка 4-1ВВ и других представителей суперсемейства TNFR. Эти рецепторы включали CD40 (TNFRSF5) и OX-40 (CD134, TNFRSF4). IgG использовали для описания связывания с применением FACS на клетках отрицательного контроля HEK293, а также клетках НЕК293Т-h4-1ВВ, стабильно трансфицированных и экспрессирующих человеческий 4-1ВВ, клетках 300.19, стабильно трансфицированных и экспрессирующих ОХ-40, и клетках 300.19, стабильно трансфицированных и экспрессирующих CD40. Результаты FACS связывания для антител MOR-6032, MOR-7361, MOR-7480 и MOR-7483, все антитела формата IgG1, представлены в Таблице 5. В концентрациях до 1000 нМ не наблюдали значимого связывания с ОХ-40 или CD40; это показывает, что антитела по меньшей мере в 100 раз более селективны в отношении 4-1ВВ против других включенных в анализ родственных представителей семейства.
3Д. Описание IgG в анализе с репортерным геном люциферазы
Связывание IgG также описывали в анализе с репортерным геном люциферазы с использованием клеток HEK293T-h4-1ВВ с применением анализа связывания на планшете, анализа связывания в растворе и перекрестного анализа связывания. В Таблице 6 показаны результаты анализа с репортерным геном люциферазы для антител MOR-6032, MOR-7361, MOR-7480 и MOR-7483, все антитела формата IgG1.
ПРИМЕР 4. ОПИСАНИЕ СТРУКТУРЫ АНТИТЕЛ MOR-6032, MOR-7361, MOR-7480 И MOR-7483
Способы, описанные выше в Примерах 1-3, были применены для получения нескольких полностью человеческих антител IgG2 против 4-1ВВ, включая антитела, обозначенные «MOR-6032», «MOR-7361», «MOR-7480» и «MOR-7483». Последовательности кДНК, кодирующие вариабельные области тяжелых и легких цепей моноклональных антител MOR-6032, MOR-7361, MOR-7480 и MOR-7483, были получены с применением стандартных PCR-методик и были секвенированы с применением стандартных методик секвенирования ДНК.
Нуклеотидные и аминокислотные последовательности вариабельных областей тяжелой цепи, полноразмерных тяжелых цепей подкласса IgG2, вариабельных областей легкой цепи и полноразмерных легких цепей антител MOR-6032, MOR-7361, MOR-7480, MOR-7480.1, MOR-7480.2, MOR-7483, MOR-7483.1 и MOR-7483.2 представлены в данном описании; указатель SEQ ID NO этих последовательностей приведен в Таблице 1.
Сравнение последовательности тяжелой цепи иммуноглобулина MOR-6032 с известными человеческими эмбриональными последовательностями тяжелых цепей иммуноглобулинов продемонстрировало, что в тяжелой цепи MOR-6032 использован VH-сегмент человеческого эмбрионального VH 1-69, D-сегмент человеческого эмбрионального 4-23 и JH-сегмент человеческого эмбрионального JH 4а.
Дальнейший анализ последовательности VH MOR-6032 с применением системы определения CDR-участков по Kabat привел к определению участков H-CDR1, H-CDR2 и H-CDR3, как показано в SEQ ID NO:1, 2 и 3, соответственно.
Сравнение последовательности тяжелой цепи иммуноглобулина MOR-7361 с известными человеческими эмбриональными последовательностями тяжелых цепей иммуноглобулинов продемонстрировало, что в тяжелой цепи 7361 использован VH-сегмент человеческого эмбрионального VH 3-23, D-сегмент человеческого эмбрионального 2-8 и JH-сегмент человеческого эмбрионального JH 4а.
Дальнейший анализ последовательности VH MOR-7361 с применением системы определения CDR-участков по Kabat привел к определению участков H-CDR1, H-CDR2 и H-CDR3 тяжелой цепи, как показано в SEQ ID NO:15, 16 и 17, соответственно.
Сравнение последовательностей тяжелых цепей иммуноглобулинов MOR-7480 и MOR-7483 с известными человеческими эмбриональными последовательностями тяжелых цепей иммуноглобулинов продемонстрировало, что в тяжелых цепях 7480 и 7483 использован VH-сегмент человеческого эмбрионального VH 5, D-сегмент человеческого эмбрионального 5-18 и JH-сегмент человеческого эмбрионального JH 4а.
Дальнейший анализ последовательностей VH 7480 и 7483 с применением системы определения CDR-участков по Kabat привел к определению участков H-CDR1, H-CDR2 и H-CDR3, как показано в SEQ ID NO:29, 30 и 31, соответственно.
Сравнение последовательностей легких цепей иммуноглобулинов MOR-6032, MOR-7361, MOR-7480 и MOR-7483 с известными человеческими эмбриональными последовательностями легких цепей иммуноглобулинов продемонстрировало, что в легких цепях 6032, 7361, 7480 и 7483 использован VL-сегмент человеческого эмбрионального λ3-r и JL-сегмент человеческого эмбрионального JL 3b.
Дальнейший анализ последовательности VL MOR-6032 с применением системы определения CDR-участков по Kabat привел к определению участков CDR1, CDR2 и CDR3 легкой цепи, как показано в SEQ ID NO:6, 7 и 8, соответственно.
Дальнейший анализ последовательности VL MOR-7361 с применением системы определения CDR-участков по Kabat привел к определению участков L-CDR1, L-CDR2 и L-CDR3, как показано в SEQ ID NO:20, 21 и 22, соответственно.
Дальнейший анализ последовательности VL MOR-7480 с применением системы определения CDR-участков по Kabat привел к определению участков L-CDR1, L-CDR2 и L-CDR3, как показано в SEQ ID NO:34, 35 и 36, соответственно.
Дальнейший анализ последовательности VL MOR-7483 с применением системы определения CDR-участков по Kabat привел к определению участков L-CDR1, L-CDR2 и L-CDR3, как показано в SEQ ID NO:34, 35 и 55, соответственно.
ПРИМЕР 5: ЭМБРИОНАЛЬНЫЕ ВАРИАНТЫ АНТИТЕЛ MOR-7480 И MOR-7483
Для минимизации иммуногенности антител MOR-7480 и MOR-7483 проводили обратную мутацию нескольких аминокислотных остатков с восстановлением эмбриональной последовательности следующим образом. Эмбриональный вариант MOR-7480, обозначенный MOR-7480.1, получали заменой двух аминокислот в FR1-области вариабельной области тяжелой цепи на аминокислоты эмбриональной последовательности. Конкретнее, Q в аминокислотном положении номер 1 заменяли на Е эмбриональной последовательности и К в аминокислотном положении номер 19 заменяли на R. Два измененных аминокислотных остатка вариабельной области тяжелой цепи можно увидеть при сравнении аминокислотной последовательности MOR-7480 (SEQ ID NO:32) и MOR-7480.1 (SEQ ID NO:43). В вариабельной области легкой цепи MOR-7480 аминокислотами эмбриональной последовательности заменяли пять аминокислот в FRI-области (D1S, I2Y, A13S, R19S, S21T), две аминокислоты в РР2-области (A42S, V45L) и одну - в РР3-области (Е80М). Восемь измененных аминокислот вариабельной области легкой цепи можно увидеть при сравнении аминокислотной последовательности MOR-7480 (SEQ ID NO:37) и аминокислотной последовательности MOR-7480.1 (SEQ ID NO:45).
Более того, получали третий вариант MOR-7480, начиная с последовательности вариабельной области легкой цепи MOR-7480.1 (SEQ ID NO:45) и обратной замены L45 на V с получением MOR-7480.2 (SEQ ID NO:51).
«Эмбриональный» вариант MOR-7483, обозначенный MOR-7483.1, получали обратной мутацией двух аминокислот в FR1-области вариабельной области тяжелой цепи на аминокислоты эмбриональной последовательности. Эмбриональные варианты могут быть получены, начиная с эмбрионального варианта цепи антитела с последующим изменением желаемых аминокислот в CDR, или любой комбинацией мутаций, начиная с любого варианта. Для получения MOR-7483.1 Q в аминокислотном положении номер 1 заменяли на Е эмбриональной последовательности и К в аминокислотном положении номер 19 заменяли на R. Два измененных аминокислотных остатка вариабельной области тяжелой цепи можно увидеть при сравнении последовательности MOR-7483 (SEQ ID NO:32) и MOR-7483.1 (SEQ ID NO:43). В вариабельной области легкой цепи MOR-7483 аминокислотами эмбриональной последовательности заменяли пять аминокислот в FR1-области (D1S, I2Y, A13S, R19S, S21T), две аминокислоты в РР2-области (A42S, V45L) и одну - в РР3-области (Е80М). Восемь измененных аминокислот вариабельной области легкой цепи можно увидеть при сравнении аминокислотной последовательности MOR-7483 (SEQ ID NO:56) и аминокислотной последовательности MOR-7483.1 (SEQ ID NO:60).
Более того, получали третий вариант MOR-7483 обратной мутацией L45 последовательности вариабельной области легкой цепи MOR-7483.1 (SEQ ID NO:60) на эмбриональный V45 с получением MOR-7483.2 (SEQ ID NO:64).
ПРИМЕР 6: СВОЙСТВА АНТИТЕЛ IN VITRO, ВКЛЮЧАЯ ЭМБРИОНАЛЬНЫЕ
ВАРИАНТЫ
Аффинности связывания антител (анализ BIAcore)
Кинетику связывания определенных антител, связывающихся с человеческим 4-1ВВ, измеряли технологией поверхностного плазменного резонанса (SPR) с использованием оборудования Biacore 3000 (GE Healthcare). Химерный белок «рекомбинантный человеческий 4-IBB/Fc», содержащий аминокислоты 24-186 SEQ ID NO:68, приобретали от R&D Systems Inc. (#838-4 В). Лиофилизированный белок растворяли в буфере, содержащем 150 мМ NaCl, 25 мМ HEPES, рН 8,0, 6 мМ MgCl2, 0,005% полисорбата 20 и 0,5 мМ азида натрия в конечной концентрации 80 нМ на основании расчетной молекулярной массы (44,8 кДа), предоставленной R&D Systems. Fc-часть отщепляли обработкой бычьим фактором Ха (Pierce, #32521) в 150 мМ NaCl, 25 мМ HEPES, рН 8,0, 6 мМ MgCl2, 0,005% полисорбата 20, 0,5 мМ азида натрия с применением инкубации продолжительностью 20 часов при 22°С с 3% фактором Ха (3 мкг фактора Ха на 100 мкг химерного белка 4-1ВВ). 4-1ВВ-часть молекулы содержит аминокислотные остатки с 24 по 186 человеческого белка 4-1ВВ. Эксперименты связывания проводили при 25°С в аналитическом буфере, содержащем 150 мМ NaCl, 25 мМ HEPES, рН 8,0, 6 мМ MgCl2, 0,005% полисорбата 20 и 0,5 мМ азида натрия. Антитела иммобилизовали стандартным аминным сочетанием на сенсорном чипе СМ5 (GE Healthcare) с использованием 0,1 мг/мл раствора антитела в 10 мМ ацетате натрия при рН 5,0. 4-1ВВ вводили в диапазоне концентраций от 80 нМ до 0,16 нМ при скорости потока 50 мкл/мин в течение 3,6 минуты с последующим периодом диссоциации продолжительностью 26 минут с использованием функции Kinject оборудования Biacore 3000. Связанный комплекс восстанавливали введением 10 мМ фосфорной кислоты в воде в течение 1 минуты. Анализ данных проводили с использованием программного обеспечения Scrubber2 (BioLogic Software). Сенсограммы приводили к простой 1:1-модели связывания Лангмюра (Langmuir). Было показано, что антитела обратимо связываются с рекомбинантным человеческим 4-1ВВ. Результаты (средние значения) представлены в Таблице 7.
Связывание с внеклеточным доменом 4-1ВВ (ELISA-анализ)
Химерный белок «человеческий 4-1ВВ IgGIFc» (R&D Systems, Minneapolis, MN) ресуспендировали в забуференном фосфатом физиологическом растворе Дульбекко (DPBS), содержащем 0,1% бычьего сывороточного альбумина (BSA), в концентрации 0,2 мг/мл и разводили, применяя DPBS, до конечной концентрации 0,03 мкг/мл. На 96-луночные планшеты Nunc-Immuno Maxisorp сорбировали рекомбинантный химерный белок 4-1ВВ в количестве 0,1 мл на лунку, оставляя пустые лунки для контроля неспецифического связывания, и инкубировали при 4°С в течение ночи. Раствор 4-1ВВ удаляли, и планшеты отмывали три раза с использованием 0,2 мл отмывочного буфера (0,05% Tween-20 в DPBS). Во все лунки добавляли 0,2 мл блокирующего буфера (5% BSA, 0,05% Tween-20 в DPBS) и инкубировали при 4°С в течение 1 часа с перемешиванием. Блокирующий буфер удаляли, и планшеты отмывали три раза с использованием 0,2 мл отмывочного буфера. Проводили серийные разведения анализируемых антител против 4-1ВВ в DPBS и в каждую лунку добавляли 0,1 мл разведенного антитела. Планшеты инкубировали в течение 1,5 часов при комнатной температуре. Раствор антитела удаляли, и планшеты отмывали три раза с использованием 0,2 мл отмывочного буфера на лунку. Козье антитело против человеческого IgG, F(ab')2-антитело, специфичное в отношении F(ab')2-фрагмента, меченное пероксидазой хрена (Jackson Immunoresearch #109-036-097, West Grove, PA), разводили в соотношении 1:5000 в DPBS и добавляли в количестве 0,1 мл на лунку. Планшеты инкубировали в течение 1 часа при комнатной температуре и отмывали с использованием 0,2 мл отмывочного буфера на лунку. Добавляли 0,1 мл пероксидазного субстрата SureBlue TMB Microwell (Kirkegaard & Perry Labs, Gaithersburg, MD) и инкубировали в течение 20 минут при комнатной температуре. Реакцию останавливали добавлением равного объема 2 М H2S04 и оптическую плотность при 450 нм считывали спектрофотометром Molecular Devices Spectra Max 340 (Molecular Devices, Sunnyvale, CA). Результаты представлены в Таблице 8.
Лиганд-конкурентное связывание (ELISA-анализ)
Анализировали способность антител блокировать связывание химерного белка «человеческий 4-1BB_IgG1Fc» с сорбированным на планшет лигандом рекомбинантного 4-1ВВ (4-1 BBL). Лиганд рекомбинантного человеческого 4-1ВВ (Biosource/lnvitrogen, Carlsbad, CA) ресуспендировали в концентрации 0,2 мг/мл в DPBS+0,1% бычьего сывороточного альбумина и затем разводили до 1 мкг/мл в DPBS. На поверхность 96-луночных планшетов Nunc-Immuno Maxisorp сорбировали 4-1 BBL в количестве 0,1 мл раствора на лунку в течение ночи при 4°С. На следующий день раствор 4-1 BBL удаляли, добавляли 0,2 мл блокирующего буфера (1% бычий сывороточный альбумин, 0,05% Tween-20 в DPBS) и инкубировали при комнатной температуре в течение 2 часов. На стадии блокировки исходные растворы антител разводили в диапазоне от 8 нг/мл до 6 мкг/мл в DPBS. Рекомбинантный человеческий 4-1BB_IgG1Fc (R&D Systems, Minneapolis, MN) ресуспендировали 0,2 мг/мл в DPBS+0,1% бычьего сывороточного альбумина и затем разводили до 0,02 мкг/мл в DPBS. Блокированные планшеты с сорбированным 4-1 BBL отмывали три раза с использованием 0,2 мл отмывочного буфера (0,05% Tween 20 в DPBS). Добавляли 60 мкл разведении антител вместе с 60 мкл химерного белка 4-1BB_IgG1Fc и инкубировали при комнатной температуре в течение 1,5 часов. Планшеты отмывали, как описано выше. Антитело против 6х-гистидиновой метки, конъюгированное с пероксидазой хрена (R&D Systems, Minneapolis MN #МАВ050Н), разводили в соотношении 1:1000 в DPBS, 50 мкл полученного раствора добавляли в лунки отмытых планшетов и инкубировали их при комнатной температуре в течение 1 часа. Планшеты отмывали, как описано выше, в каждую лунку добавляли 50 мкл раствора тетраметилбензидинового (ТМВ) субстрата и инкубировали при комнатной температуре в течение 20 минут. Реакцию останавливали с использованием 50 мкл 0,2 н. H2SO4 и оптическую плотность при 450 нм считывали спектрофотометром для прочтения планшетов от Molecular Devices. Результаты представлены в Таблице 8.
Межвидовая перекрестная реактивность антител
Межвидовую перекрестную реактивность иллюстративных антител измеряли с использованием стимулированных фитогемагглютинином (РНА) первичных мононуклеарных клеток периферической крови (РМВС) человека, яванского макака (супо), собаки и крысы. Клетки выделяли способом, описанным ниже. Клетки (приблизительно 5,0·105 клеток на пробирку) отмывали один раз холодным отмывочным буфером (PBS, 2% фетальной бычьей сыворотки (FBS) и 0,02% азида натрия) и к каждому образцу добавляли конъюгированные с Alexa Fluor 647 контрольные антитела или антитела, реактивные в отношении 4-1ВВ, 100 мкл на пробирку, 15,5 мкг/мл (100 нМ), вместе с мечеными антителами к видоспецифичному Т-клеточному маркеру. Использованные антитела к Т-клеточному маркеру представляли собой следующее: конъюгированное с изотиоцианатом флуоресцеина (FITC) антитело против человеческого CD3e (BD Pharmingen, #555332), конъюгированное с FITC антитело против крысиного CD3e (BD Pharmingen, #559975), конъюгированное с FITC антитело против кроличьего CD4+конъюгированное с FITC антитело против кроличьего CDS (AbD Serotec, #MCA799F и #MCA1576F), конъюгированное с FITC антитело против CD3e собаки (AbD Serotec, #MCA1774F) и конъюгированное с перидинин-хлорофилл протеином (РегСР) антитело против CD3e человека/яванского макака (BD Pharmingen, #552851). Клетки инкубировали в темноте с антителами, конъюгированными с флуорохромами, на льду в течение 30 минут, отмывали три раза и ресуспендировали в 0,3 мл отмывочного буфера для анализа. Окрашивание антителами измеряли и анализировали с использованием Becton Dickinson FACS Calibur и программного обеспечения FlowJo 8.8.2.
Выделение человеческих Т-лимфоцитов. Человеческую цельную кровь отбирали в шприцы, содержащие 1 мл 0,5 М EDTA и затем переносили в пробирки Sigma Accuspin (Sigma, St. Louis, МО) для выделения мононуклеарных клеток периферической крови (РВМС), как описано изготовителем. РВМС два раза отмывали DPBS, содержащим 5 мМ EDTA, и выделяли Т-лимфоциты с использованием колонки для очистки Т-клеток, как описано изготовителем (R&D Systems, Minneapolis, MN). Кратко, РВМС ресуспендировали в 2 мл колоночного буфера и помещали в предварительно отмытую колонку для выделения Т-клеток. РМВС инкубировали в течение 10 минут при комнатной температуре и Т-клетки элюировали колоночным буфером, отмывали один раз и ресуспендировали в концентрации 2·106 клеток/мл в среде для тканевых культур (ТСМ), состоящей из RPMI 1640 (Sigma, St Louis, МО), дополненной 10% фетальной бычьей сывороткой (Sigma, St. Louis, МО) и L-глутамином (2 мМ), HEPES (10 мМ), пенициллином (100 ЕД/мл), стрептомицином (50 мкг/мл) (Gibco, Grand Island, NY.).
Выделение РМВС яванского макака. Цельную кровь яванского макака (Bioreclamation; Hicksville, NY) отбирали в СРТ-пробирки Vacutainer с цитратом натрия (BD; Franklin Lakes, NJ) и затем центрифугировали при 1500 g в течение 20 минут при комнатной температуре. Пробирки оставляли на ночь при 4°С. РВМС-фракцию отбирали из СРТ-пробирок и отмывали 2 раза PBS, содержащим 5 мМ EDTA. После стадии отмывки проводили подсчет РВМС и доводили их до концентрации 2·106 клеток/мл в среде для тканевых культур (ТСМ). ТСМ состояла из RPMI 1640 (Sigma, St Louis, МО), дополненной 10% фетальной бычьей сывороткой (Sigma, St. Louis, МО) и L-глутамином (2 мМ), HEPES (10 мМ), пенициллином (100 ЕД/мл), стрептомицином (50 мкг/мл) (Gibco, Grand Island, NY.), приобретена от Gibco (Grand Island, NY.). Клетки стимулировали 10 мкг/мл РНА в течение 2-3 суток для индукции экспрессии 4-1ВВ.
Выделение РМВС собаки. Цельную кровь собаки отбирали в гепаринизированные пробирки Vacutainer (BD; Franklin Lakes, NJ) и разводили в соотношении 1:2 в PBS, содержащем 5 мМ EDTA. После перемешивания 4 мл разведенной крови аккуратно наслаивали на 3 мл Lympholyte-Mammal (Cedarlane Laboratories, Westbury, NY) и центрифугировали при 800 g в течение 20 минут при 25°С. Промежуточный РВМС-слой отбирали, отмывали два раза с использованием PBS и ресуспендировали в концентрации 2·106 клеток/мл в ТСМ, содержащей РИА в концентрации 10 мкг/мл (Remel, Lenexa, KS). Клетки культивировали в течение 48-72 часов перед анализом их связывания с антителами проточной цитометрией.
Выделение крысиных РВМС. Цельную кровь крысы отбирали в гепаринизированные пробирки Vacutainer (BD; Franklin Lakes, NJ) и разводили в соотношении 1:3 в PBS, содержащем 5 мМ EDTA. После перемешивания 6 мл разведенной крови аккуратно наслаивали на 4,5 мл Lympholyte-Mammal (Cedarlane Laboratories, Westbury, NY) и центрифугировали при 800 g в течение 20 минут при 25°С. Промежуточный РВМС-слой отбирали, отмывали два раза с использованием PBS и ресуспендировали в концентрации 2·106 клеток/мл в ТСМ, содержащей РНА в концентрации 10 мкг/мл (Remel, Lenexa, KS). Клетки культивировали в течение 48-72 часов перед анализом их связывания с антителами проточной цитометрией.
Результаты анализов связывания показаны на Фиг.1. Было обнаружено, что антитела связывались с человеческим 4-1ВВ и 4-1ВВ яванского макака с высокой аффинностью, в то время как связывания с 4-1ВВ собаки и крысы в концентрации 100 нМ, наибольшей проанализированной концентрации, не наблюдали.
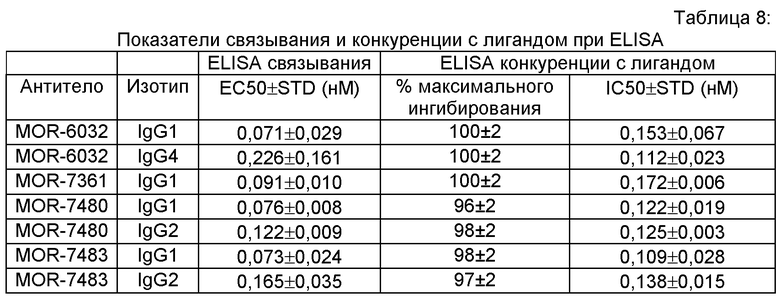
Картирование эпитопов
Для определения эпитопсвязывающей области антител-агонистов 4-1ВВ проводили ряд мутаций (Таблица 9) во внеклеточном домене человеческого 4-1ВВ с заменами на аминокислоты опубликованной последовательности 4-1ВВ собаки (Ref. Seq. XM_845243).
Все мутации «человек-собака» были проведены Gene Dynamics LLC, (Portland, OR) в ретровирусном векторе экспрессии pMSCVpuro (Clontech Laboratories Mountain View, CA). Кроме того, синтезом генов была получена полноразмерная последовательность кДНК собаки, соответствующая Ref. Seq. ХМ_845243.
Вирусные препараты получали транзиторной трансфекцией клеток 293Т с приблизительно 40-50% конфлюэнтностью в колбах Т-75. После культивирования вирусный супернатант стерилизовали фильтрацией и концентрировали. Концентрированный вирус собирали и хранили при -80°С.
Логарифмически растущие клетки 300-19 трансдуцировали ретровирусом с использованием концентрированного вируса, разведенного в соотношении 1:250, с 8 мкг/мл полибрена в полной модифицированной по способу Дульбекко среде Игла (DMEM). После 24 часов инкубации в культуры добавляли 2 мкг/мл пуромицин и поддерживали эту концентрацию в течение исследования.
Положительная экспрессия рецепторов 4-1ВВ селектированными пуромицином клетками была подтверждена окрашиванием поликлональным козьим антителом против человеческого 4-1ВВ (R&D Systems Inc.) в концентрации 1 мкг/мл с 1:200 разведением меченного фикоэритрином (РЕ) ослиного противокозьего IgG (H+L) F(ab')2 (Jackson Immunoresearch Inc.). Для определения распознавания мутантных рецепторов 4-1ВВ анализируемыми антителами селектированные пуромицином клетки окрашивали 100 нМ разведением немеченого первичного антитела на льду в течение 30 минут с последующими двумя отмывками FACS-буфером и добавлением 1:200 разведения видоспецифичного меченного РЕ ослиного антитела против IgG (H+L) F(ab')2. Клетки анализировали посредством FACS с использованием BD FACS Calibur и программного обеспечения FlowJo 8.8.6.
Относительное окрашивание каждой совокупности клеток показано в Таблице 10.
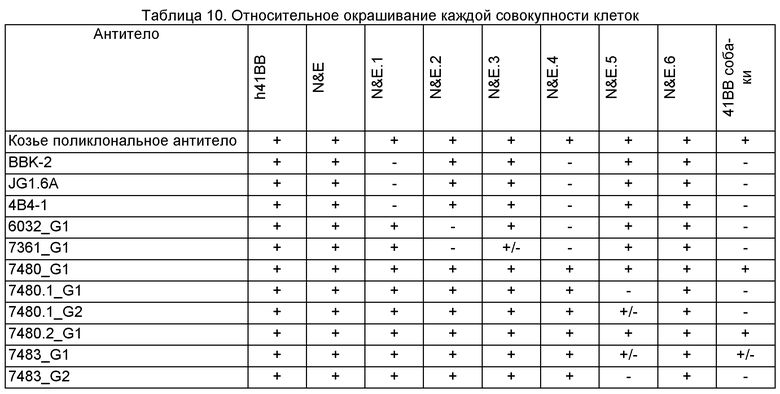
Различие связывания антител, имеющих сходные последовательности (MOR-7480, MOR-7480.1, MOR-7480.2, MOR-7483 и MOR-7483.1), было обнаружено с мутациями клона N&E.5, что указывает на расположение детерминант распознавания антитела в этой мутированной области.
Для определения относительной аффинности этих антител в отношении внеклеточного домена человеческого 4-1ВВ и мутантного внеклеточного домена человеческого 4-1ВВ, мутанта N&E.5, для каждого антитела строили FACS-кривую доза-ответ. MOR_7480, MOR_7480.1 и MOR_7480.2, меченные Alexa Fluor 647, разводили в FACS-буфере, начиная с 1 мкМ, с 8 серийными разведениями в соотношении 1:5 и использовали для окраски исходных клеток 300-19, клеток с hu4-1BB, hu4-1BB N&E.5 и клеток с 4-1ВВ собаки. Клетки анализировали посредством FACS с использованием BD FACS Calibur и программного обеспечения FlowJo 8.8.6. Среднее геометрическое флуоресценции рецептор-экспрессирующих клеток каждого типа нормализовали относительно окрашивания исходных клеток и выражали как кратность окрашивания и определяли ЕС50 ответа на дозу. ЕС50 представлены в Таблице 11. Для MORJ7480.2 и MOR_7480 было отмечено уменьшение связывания с мутантным человеческим 4-1ВВ N&E.5 более чем в 5 раз.
Агонистическая активность антител (люциферазный анализ активности)
Получали клетки 293Т, экспрессирующие человеческий 4-1ВВ вместе со стабильно интегрированным NFkB-люциферазным репортером. Клетки собирали, отмывали и ресуспендировали в полной среде, свободной от фенолового красного (DMEM, содержащей 10% фетальной бычьей сыворотки, HEPES-буфер, заменимые аминокислоты и L-глутамин) в концентрации 0,6·106 клеток/мл. 50 мкл клеток добавляли в каждую аналитическую лунку белого 96-луночного планшета (PerkinElmer, Waltham, MA). В каждую лунку добавляли анализируемые антитела в присутствии перекрестно связывающего козьего антитела Fab' против человеческого IgG Fc (Jackson ImmunoResearch, West Grove, PA) в соотношении 2,5:1. Планшет инкубировали в течение 5 часов при 37°С. Добавляли 75 мкл реагента Bright-Glo Luciferase (Promega, Madison Wl) и значение активности люциферазы измеряли с использованием сцинтилляционного счетчика Packard TopCount NXT.
Клетки 293Т, экспрессирующие 4-1ВВ яванского макака, получали вирусной трансдукцией и селекцией стабильных клеток с использованием 2 мкг/мл пуромицина. Клетки 293Т, экспрессирующие 4-1ВВ яванского макака, культивировали в колбе Т-75 до приблизительно 60-70% конфлюэнтности, затем трансфицировали их с использованием 10 мкг pLuc_6xNFkB и 0,1 мкг pRL-CMV в качестве контроля трансфекции. Трансфекцию проводили с использованием реагента для трансфекции Fugene 6 (Roche Indianapolis, IN) при соотношении 6 мкл Fugene к 1 мкг плазмидной ДНК, согласно инструкциям изготовителя. Клетки собирали на следующий день, отмывали и ресуспендировали в полной среде, свободной от фенолового красного (DMEM, содержащей 10% фетальной бычьей сыворотки, заменимые аминокислоты и L-глутамин) в концентрации 0,6·106 клеток/мл. 50 мкл клеток добавляли в каждую аналитическую лунку белого 96-луночного планшета (PerkinElmer, Waltham, MA). В каждую лунку добавляли анализируемые антитела в присутствии перекрестно связывающего козьего антитела Fab' против человеческого IgG Fc (Jackson ImmunoResearch, West Grove, PA) в соотношении 2,5:1. Планшет инкубировали в течение 5 часов при 37°С. Добавляли 75 мкл реагента для люциферазного анализа и значение активности люциферазы светлячка измеряли с использованием сцинтилляционного счетчика Packard TopCount NXT. Кроме того, добавляли 75 мкл реагента Stop & Glo для оценки активности люциферазы Renilla. Значение активности люциферазы Renilla измеряли с использованием сцинтилляционного счетчика Packard TopCount NXT. Результаты показаны на Фиг.2.
Агонистическая активность антител (анализ высвобождения IL-2 первичными Т-клетками)
96-луночные планшеты Nunc Maxisorp стерилизовали ультрафиолетом (UV) перед сорбцией на планшеты. Анализируемые антитела разводили в PBS до 60 мкг/мл. 0,2 мл разведенных антител добавляли в 2 лунки полипропиленового 96-луночного планшета и проводили серийное разведение в соотношении 1:3. В стерильный 96-луночный аналитический планшет Maxisorp добавляли 50 мкл разведенных антител и сразу добавляли 50 мкл 20 мкг/мл клона UCHT1 против человеческого CD3ε (Biolegend San Diego, CA). Затем все планшеты инкубировали в течение ночи при 4°С. На следующий день планшеты с сорбированными антителами отмывали 1 раз с использованием PBS и в лунки планшетов Nunc Maxisorp добавляли 0,15 мл полной среды RPMI. Человеческие Т-клетки выделяли, как описано выше, и в каждую лунку добавляли 50 мкл очищенных Т-клеток в количестве 2·106 клеток/мл (100000 клеток на лунку). Клетки инкубировали при 37°С, 5% CO2 в течение 3 суток. Из каждой лунки отбирали супернатант, анализируя его сразу или храня при -20°С до анализа. Перед ELISA-анализом на IL-2 (R&D Systems, Minneapolis, MN) супернатанты разводили полной средой. Результаты показаны на Фиг.3.
ПРИМЕР 7: РАЗМНОЖЕНИЕ ЧЕЛОВЕЧЕСКИХ ЛЕЙКОЦИТОВ, ИНДУЦИРОВАННОЕ АНТИТЕЛАМИ ПРОТИВ 4-1ВВ IN VIVO
Ввиду отсутствия выявляемой перекрестной реактивности антител против 4-1ВВ в отношении мышиного 4-1ВВ и необходимости присутствия человеческих иммунокомпетентных клеток было необходимо разработать модели для функциональной оценки антител против 4-1ВВ in vivo. Мыши с генетическим фоном NOD, мутацией, приводящей к тяжелому комбинированному иммунодефициту (SCID), и дефицитом общей гамма-цепи рецептора IL-2 (обычно называемые NSG) позволяют проводить перенос большого количества человеческих лейкоцитов периферической крови (huPBL) и сохранять их на протяжении по меньшей мере 30 суток (King, 2008, Clin. Immunol. 126:303-314). Эту модель на мышах, также известную как модель huPBL-NSG, использовали для оценки функционального эффекта системного введения антител на человеческие иммунокомпетентные клетки in vivo. Конкретно, проводили адоптивный перенос 6 миллионов свежевыделенных человеческих РВМС внутривенной инъекцией мышам-хозяевам NOD.Cg-Prkdcscid II2rgtmIWjI/SzJ (NSG). Через девять суток после инъекции РМВС животным вводили MOR7480, MOR7480.1 или контрольное антитело изотипа IgG2 в дозе 1 мг/кг однократно внутрибрюшинной инъекцией. На 24-28 сутки после введения РМВС их окрашивали антителами против человеческого и мышиного CD45 и анализировали проточной цитометрией. Для определения лимфоцитарного гейта использовали профили прямого и бокового рассеяния. Как показано на Фиг.4, MOR7480 и MOR7480.1 были способны усиливать размножение человеческих лейкоцитов, на что указывает повышенное содержание человеческих CD45+ клеток в периферической крови мышей с перенесенными РМВС. Для каждой группы n≥6 мышей.
Кроме того, введение MOR7480.1 яванским макакам усиливало пролиферацию центральных цитотоксических Т-клеток памяти (CD8 ТСМ) в образцах РВМС. Яванским макакам (2 животных на уровень дозы) проводили однократную внутривенную инъекцию MOR7480.1 в указанной дозе. РМВС получали на 7 сутки перед введением антитела (до введения) и на указанные сутки исследования после введения MOR7480.1 (на 1 сутки исследования). РВМС окрашивали антителами к CD3, CD4, CD8, CD28, CD95 и Ki-67 и анализировали проточной цитометрией. Данные получали на Canto II (Beckton Dickinson) и анализировали с использованием программного обеспечения DIVA (Becton Dickinson). CD8 центральные клетки памяти определяли как CD3+, CD8+, CD28+и CD95+. Показаны данные по отдельным животным, обозначенным как (уровень дозы-номер животного), и представлены как изменение числа Ki-67+клеток у животного по сравнению с их числом до исследования {[(число Ki-67+клеток на указанные сутки исследования - число Ki-67+клеток перед введением)/число Ki-67+клеток перед введением]*100}. Как показано на Фиг.5, усиление пролиферации центральных Т-клеток памяти в 1,5 раза или более на первые 7-13 суток исследования было отмечено у по меньшей мере одного животного из каждой группы, которым вводили антитело в дозе 0,3 мг/кг или более.
ПРИМЕР 8: ПРОТИВООПУХОЛЕВАЯ АКТИВНОСТЬ АНТИТЕЛ ПРОТИВ 4-1ВВ (МОДЕЛЬ IN VIVO)
Модель рака предстательной железы человека РС3
Отсутствие перекрестной реактивности антител против 4-1ВВ в отношении 4-1ВВ грызунов не позволяло использовать стандартные модели опухолей на сингенных мышах или с человеческим ксенотрансплантатом для оценки противоопухолевой эффективности антител. Соответственно, была разработана новая ксеногенная модель опухоли на мышах huPBL-SCID-Bg с использованием мышей SCID-Bg (CB.17/lcr.Cg PkrdscidLystbg/Crl), несущих Bg-мутацию дефицита мышиных Т- и В-лимфоцитов и функциональных NK-клеток. Противоопухолевую эффективность антител против 4-1ВВ оценивали с использованием этой модели, как описано ниже.
Клеточную линию предстательной железы человека РСЗ или клеточную линию толстой кишки человека LOVO получали из Американской коллекции типовых культур и культивировали в RPMI-1640 (Invitrogen), обогащенной следующими добавками Invitrogen: L-глутамином, пируватом натрия, заменимыми аминокислотами, пенициллином/стрептомицином, HEPES и 10% термоинактивированной фетальной бычьей сывороткой (FBS; Cat. No.F2442, Sigma Aldrich). Клетки выращивали до конфлюэнтности в колбах Falcon T-225. Затем клетки трипсинизировали (Trypsin 0,25%-EDTA; Cat. No. 2500-056, Invitrogen) и выращивали в большем объеме в колбах Hyperflask (Cat. No.3319 Corning Life Sciences) в течение трех суток. Для сбора клеточной линии использовали трипсин, отмывали ее 3 раза в ледяной PRMI, дополненной 10% FBS. От здоровых добровольцев получали не более 300 мл периферической крови. Лимфоциты периферической крови (РВМС) выделяли из гепаринизированной крови с использованием пробирок Accuspin, согласно протоколу изготовителя (Cat. No.A0561-100x 15 ml, Aldrich). Суспензии с подсчитанным количеством клеток смешивали, таким образом, что каждой мыши вводили 1,5·106 РВМС и 3·106 опухолевых клеток однократной болюсной инъекцией в 0,2 мл PBS. Смешанную суспензию клеток отмывали дважды холодным PBS, помещали на лед и сразу вводили подготовленным мышам.
Для каждой мыши смешанную суспензию клеток объемом 0,2 мл вводили подкожно в правый бок животного и однократно вводили антитело против 4-1ВВ или контрольное антитело (0,2 мл) подкожной инъекцией в левый бок. На протяжении экспериментов два раза в неделю измеряли опухоль штангенциркулем Pressier и также отмечали массу тела. Объем опухоли вычисляли по следующей формуле: длина×ширина2×0,44=объем (мм3). Мышей выводили из исследования в случае достижения опухолью объема 2000 мм3 или уменьшения массы тела животного на 20% до завершения эксперимента. На 23 сутки мышей из всех групп умерщвляли с применением процедур, изложенных Комитетом по содержанию и использованию лабораторных животных (IACUC) (Фиг.6). Процент ингибирования роста опухоли измеряли на последние сутки исследования и вычисляли как 100-{1-(введениепоследние сутки/контрольпоследние сутки)}. Сходные результаты наблюдали при измерении опухоли на 6 сутки после инъекции; животных рандомизировали по объему опухоли и однократно вводили им mAb против 4-1ВВ на 7 сутки после имплантации. Для большинства исследований каждая группа содержала 8 мышей.
ПРИМЕР 9: ОЦЕНКА АКТИВНОСТИ АНТИТЕЛ ПРОТИВ 4-1ВВ У «НОК-ИН»
МЫШЕЙ С ЧЕЛОВЕЧЕСКИМ 4-1ВВ IN VIVO
Получение «нок-ин» мышей с человеческим 4-1ВВ
Для лучшей оценки иммуномодулирующей активности моноклональных антител против человеческого 4-1ВВ, не обладающих перекрестной реактивностью в отношении мышиного 4-1ВВ, разрабатывали модель на мышах, у которых мышиный ген 4-1ВВ был заменен человеческим геном 4-1ВВ. Клоны бактериальной искусственной хромосомы (ВАС), несущие человеческий или мышиный геномный фрагмент 4-1ВВ, заказывали у Invitrogen (Carlsbad, СА.) и использовали для конструирования 4-1ВВ-направленного вектора на основе технологии рекомбинации Red/ET (Zhang, 1998, Nat Genet 20:123-128). Сначала проводили сборку вектора для удаления человеческого гена на каркасе pBR322, таким образом, что при расщеплении Xba1 два мышиных/человеческих гомологичных плеча (400 пар оснований (bp) каждое) будут удалять из ВАС-клона с человеческим 4-1ВВ 19994 bp человеческой геномной последовательности 4-1ВВ, начиная с кодона начала трансляции ATG, расположенного в экзоне 2, и заканчивая стоп-кодоном в TGA в экзоне 8. Затем проводили сборку неомициновой экспрессионной кассеты под контролем промоторов PGK/EM7 и фланкировали ее 100 парами оснований (bp) последовательностей, гомологичных последовательностям интрона 2 человеческого гена 4-1ВВ. Эту неомициновую экспрессионную кассету затем вводили в выделенный человеческий геномный фрагмент 4-1ВВ, полученный на первой стадии. В завершение, выделенный человеческий геномный фрагмент 4-1ВВ с неомициновой экспрессионной кассетой вводили в мышиный ВАС-клон, замещая мышиный ген 4-1ВВ модифицированным человеческим геномным фрагментом 4-1ВВ от кодона инициации ATG до стоп-кодона TGA.
Этот направленный ВАС-вектор вводили электропорацией в линию мышиных эмбриональных стволовых клеток с фоном C57BL/6NTAC (PRX-BL6N #1, Primogenix, Laurie, МО.), следуя стандартному протоколу, и проводили скрининг клонов, выживших при селекции G418 (также известным как генетицин), двумя Taqman-анализами на интрон 2 и экзон 8 мышиного гена 4-1ВВ для выявления клонов, модифицированных в мышином локусе 4-1ВВ механизмом гомологичной рекомбинации. Было обнаружено, что из 116 ES-клонов, использованных в скрининге, 7 клонов утратили один аллель мышиного локуса 4-1ВВ (эффективность введения 6%). Кариотипирование и гибридизация in situ (FISH) были проведены Coriell Institute for Medical Research (Camden, N.J.). Для клона LH15, 19 из 20 клеток имели кариотип 40XY, и для LH80, 20 из 20 клеток имели кариотип 40XY. В обоих клонах FISH с использованием мышиного ВАС-клона с геном 4-1ВВ в качестве зонда продемонстрировала один сигнал гибридизации с 4-1ВВ на каждой хромосоме 4 пары в области полосы Е2. Ни в каких других местах сигнала не наблюдали.
Оба клона LH15 и LH80 вводили в бластоцисты штамма BALB/c и эмбрионы имплантировали CD1-псевдобеременным самкам мышей для вынашивания. Химерных самцов скрещивали с мышами Ella-cre с фоном C57BL/6 для удаления кассеты устойчивости к неомицину и в исследовании использовали мышей, гомозиготных по человеческому гену 4-1ВВ.
Пролиферация лимфоцитов, опосредованная mAb-агонистами 4-1ВВ
Способность mAb-агонистов 4-1ВВ индуцировать пролиферацию лимфоцитов оценивали у 4-1ВВ-«нок-ин» мышей. 4-1ВВ-«нок-ин» мышам проводили внутрибрюшинную инъекцию 30 мг/кг MOR7480.1 на 0 сутки исследования (животным с еженедельным введением проводили инъекции mAb против 4-1ВВ на 0 сутки и 7 сутки). За 24 часа до забора образцов животным вводили внутрибрюшинно 2 мг BrdU. На указанные сутки после введения образцы периферической крови получали внутрисердечной пункцией. РВМС окрашивали антителами против CD3, 004, CD8 и BrdU и анализировали проточной цитометрией. Результаты показаны на Фиг.7, диаграмма А.
Противоопухолевая эффективность mAb-агонистов 4-1ВВ
Противоопухолевую эффективность MOR7480.1 оценивали у 4-1ВВ-«нок-ин» мышей с использованием B16-OVA/luc, линии меланомы, сконструированной для экспрессии модельного антигена овальбумина (OVA) и люциферазы (luc). В бок 4-1ВВ-«нок-ин» мышей имплантировали один миллион опухолевых клеток. Животных рандомизировали по размеру опухоли по достижении опухолью объема 50-100 мм3 (обычно на 7-10 сутки после имплантации опухоли) и проводили им однократную инъекцию указанной дозы mAb против 4-1ВВ. Размер опухоли оценивали измерением с использованием штангенциркуля два-три раза в неделю до прекращения исследования. Результаты показаны на Фиг.7, диаграмма Б.
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Пфайзер Инк.
<120> МОЛЕКУЛЫ, СВЯЗЫВАЮЩИЕСЯ С 4-1ВВ
<130> PC33845A
<160> 74
<170> PatentIn версия 3.5
<210> 1
<211> 6
<212> ПРТ
<213> Homo sapien
<400> 1
Asn Ser Tyr Ala Ile Ser
1 5
<210> 2
<211> 17
<212> ПРТ
<213> Homo sapien
<400> 2
Gly Ile Ile Pro Gly Phe Gly Thr Ala Asn Tyr Ala Gln Lys Phe Gln
1 5 10 15
Gly
<210> 3
<211> 11
<212> ПРТ
<213> Homo sapien
<400> 3
Arg Lys Asn Glu Glu Asp Gly Gly Phe Asp His
1 5 10
<210> 4
<211> 119
<212> ПРТ
<213> Homo sapien
<400> 4
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Asn Ser Tyr
20 25 30
Ala Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Ile Ile Pro Gly Phe Gly Thr Ala Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Glu Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Lys Asn Glu Glu Asp Gly Gly Phe Asp His Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 5
<211> 445
<212> ПРТ
<213> Homo sapien
<400> 5
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Asn Ser Tyr
20 25 30
Ala Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Ile Ile Pro Gly Phe Gly Thr Ala Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Glu Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Lys Asn Glu Glu Asp Gly Gly Phe Asp His Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Asn Phe Gly Thr Gln Thr Tyr Thr Cys Asn Val Asp His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Thr Val Glu Arg Lys Cys Cys Val Glu
210 215 220
Cys Pro Pro Cys Pro Ala Pro Pro Val Ala Gly Pro Ser Val Phe Leu
225 230 235 240
Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu
245 250 255
Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Gln
260 265 270
Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys
275 280 285
Pro Arg Glu Glu Gln Phe Asn Ser Thr Phe Arg Val Val Ser Val Leu
290 295 300
Thr Val Val His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys
305 310 315 320
Val Ser Asn Lys Gly Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys
325 330 335
Thr Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser
340 345 350
Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys
355 360 365
Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln
370 375 380
Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Met Leu Asp Ser Asp Gly
385 390 395 400
Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln
405 410 415
Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn
420 425 430
His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 6
<211> 11
<212> ПРТ
<213> Homo sapien
<400> 6
Ser Gly Asp Asn Leu Gly Asp Tyr Tyr Ala Ser
1 5 10
<210> 7
<211> 7
<212> ПРТ
<213> Homo sapien
<400> 7
Asp Asp Ser Asn Arg Pro Ser
1 5
<210> 8
<211> 10
<212> ПРТ
<213> Homo sapien
<400> 8
Gln Thr Trp Asp Gly Thr Leu His Phe Val
1 5 10
<210> 9
<211> 107
<212> ПРТ
<213> Homo sapien
<400> 9
Asp Ile Glu Leu Thr Gln Pro Pro Ser Val Ser Val Ala Pro Gly Gln
1 5 10 15
Thr Ala Arg Ile Ser Cys Ser Gly Asp Asn Leu Gly Asp Tyr Tyr Ala
20 25 30
Ser Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Ile Tyr
35 40 45
Asp Asp Ser Asn Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Gly Thr Gln Ala Glu
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Thr Trp Asp Gly Thr Leu His Phe
85 90 95
Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105
<210> 10
<211> 213
<212> ПРТ
<213> Homo sapien
<400> 10
Asp Ile Glu Leu Thr Gln Pro Pro Ser Val Ser Val Ala Pro Gly Gln
1 5 10 15
Thr Ala Arg Ile Ser Cys Ser Gly Asp Asn Leu Gly Asp Tyr Tyr Ala
20 25 30
Ser Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Ile Tyr
35 40 45
Asp Asp Ser Asn Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Gly Thr Gln Ala Glu
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Thr Trp Asp Gly Thr Leu His Phe
85 90 95
Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gln Pro Lys Ala
100 105 110
Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu Glu Leu Gln Ala
115 120 125
Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe Tyr Pro Gly Ala
130 135 140
Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val Lys Ala Gly Val
145 150 155 160
Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys Tyr Ala Ala Ser
165 170 175
Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser His Arg Ser Tyr
180 185 190
Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys Thr Val Ala
195 200 205
Pro Thr Glu Cys Ser
210
<210> 11
<211> 357
<212> ДНК
<213> Homo sapien
<400> 11
caggtgcaat tggttcagtc tggcgcggaa gtgaaaaaac cgggcagcag cgtgaaagtg 60
agctgcaaag cctccggagg cacttttaat tcttatgcta tttcttgggt gcgccaagcc 120
cctgggcagg gtctcgagtg gatgggcggt atcattccgg gttttggcac tgcgaattac 180
gcgcagaagt ttcagggccg ggtgaccatt accgcggatg aaagcaccag caccgcgtat 240
atggaactga gcagcctgcg tagcgaagat acggccgtgt attattgcgc gcgtaagaat 300
gaggaggatg gtggttttga tcattggggc caaggcaccc tggtgacggt tagctca 357
<210> 12
<211> 321
<212> ДНК
<213> Homo sapien
<400> 12
gatatcgaac tgacccagcc gccttcagtg agcgttgcac caggtcagac cgcgcgtatc 60
tcgtgtagcg gcgataatct tggtgattat tatgcttctt ggtaccagca gaaacccggg 120
caggcgccag ttcttgtgat ttatgatgat tctaatcgtc cctcaggcat cccggaacgc 180
tttagcggat ccaacagcgg caacaccgcg accctgacca ttagcggcac tcaggcggaa 240
gacgaagcgg attattattg ccagacttgg gatggtactc ttcattttgt gtttggcggc 300
ggcacgaagt taaccgttct t 321
<210> 13
<211> 1335
<212> ДНК
<213> Homo sapien
<400> 13
caggtgcaat tggttcagtc tggcgcggaa gtgaaaaaac cgggcagcag cgtgaaagtg 60
agctgcaaag cctccggagg cacttttaat tcttatgcta tttcttgggt gcgccaagcc 120
cctgggcagg gtctcgagtg gatgggcggt atcattccgg gttttggcac tgcgaattac 180
gcgcagaagt ttcagggccg ggtgaccatt accgcggatg aaagcaccag caccgcgtat 240
atggaactga gcagcctgcg tagcgaagat acggccgtgt attattgcgc gcgtaagaat 300
gaggaggatg gtggttttga tcattggggc caaggcaccc tggtgacggt tagctcagcc 360
tccaccaagg gcccatcggt cttccccctg gcgccctgct ccaggagcac ctccgagagc 420
acagcggccc tgggctgcct ggtcaaggac tacttccccg aaccggtgac ggtgtcgtgg 480
aactcaggcg ctctgaccag cggcgtgcac accttcccgg ctgtcctaca gtcctcagga 540
ctctactccc tcagcagcgt agtgaccgtg ccctccagca acttcggcac ccagacctac 600
acctgcaacg tagatcacaa gcccagcaac accaaggtgg acaagacagt tgagcgcaaa 660
tgttgtgtcg agtgcccacc gtgcccagca ccacctgtgg caggaccgtc agtcttcctc 720
ttccccccaa aacccaagga caccctcatg atctcccgga cccctgaggt cacgtgcgtg 780
gtggtggacg tgagccacga agaccccgag gtccagttca actggtacgt ggacggcgtg 840
gaggtgcata atgccaagac aaagccacgg gaggagcagt tcaacagcac gttccgtgtg 900
gtcagcgtcc tcaccgtcgt gcaccaggac tggctgaacg gcaaggagta caagtgcaag 960
gtctccaaca aaggcctccc agcccccatc gagaaaacca tctccaaaac caaagggcag 1020
ccccgagaac cacaggtgta caccctgccc ccatcccggg aggagatgac caagaaccag 1080
gtcagcctga cctgcctggt caaaggcttc taccccagcg acatcgccgt ggagtgggag 1140
agcaatgggc agccggagaa caactacaag accacacctc ccatgctgga ctccgacggc 1200
tccttcttcc tctacagcaa gctcaccgtg gacaagagca ggtggcagca ggggaacgtc 1260
ttctcatgct ccgtgatgca tgaggctctg cacaaccact acacacagaa gagcctctcc 1320
ctgtctccgg gtaaa 1335
<210> 14
<211> 639
<212> ДНК
<213> Homo sapien
<400> 14
gatatcgaac tgacccagcc gccttcagtg agcgttgcac caggtcagac cgcgcgtatc 60
tcgtgtagcg gcgataatct tggtgattat tatgcttctt ggtaccagca gaaacccggg 120
caggcgccag ttcttgtgat ttatgatgat tctaatcgtc cctcaggcat cccggaacgc 180
tttagcggat ccaacagcgg caacaccgcg accctgacca ttagcggcac tcaggcggaa 240
gacgaagcgg attattattg ccagacttgg gatggtactc ttcattttgt gtttggcggc 300
ggcacgaagt taaccgttct tggtcagccc aaggctgccc cctcggtcac tctgttccca 360
ccctcctctg aggagcttca agccaacaag gccacactgg tgtgtctcat aagtgacttc 420
tacccgggag ccgtgacagt ggcctggaag gcagatagca gccccgtcaa ggcgggagtg 480
gagaccacca caccctccaa acaaagcaac aacaagtacg cggccagcag ctacctgagc 540
ctgacgcctg agcagtggaa gtcccacaga agctacagct gccaggtcac gcatgaaggg 600
agcaccgtgg agaagacagt ggcccctaca gaatgttca 639
<210> 15
<211> 6
<212> ПРТ
<213> Homo sapien
<400> 15
Ser Asp Tyr Tyr Met His
1 5
<210> 16
<211> 17
<212> ПРТ
<213> Homo sapien
<400> 16
Val Ile Ser Gly Ser Gly Ser Asn Thr Tyr Tyr Ala Asp Ser Val Lys
1 5 10 15
Gly
<210> 17
<211> 10
<212> ПРТ
<213> Homo sapien
<400> 17
Arg Leu Tyr Ala Gln Phe Glu Gly Asp Phe
1 5 10
<210> 18
<211> 118
<212> ПРТ
<213> Homo sapien
<400> 18
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Tyr Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Val Ile Ser Gly Ser Gly Ser Asn Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Leu Tyr Ala Gln Phe Glu Gly Asp Phe Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ser
115
<210> 19
<211> 444
<212> ПРТ
<213> Homo sapien
<400> 19
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Tyr Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Val Ile Ser Gly Ser Gly Ser Asn Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Leu Tyr Ala Gln Phe Glu Gly Asp Phe Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Asn Phe Gly Thr Gln Thr Tyr Thr Cys Asn Val Asp His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Thr Val Glu Arg Lys Cys Cys Val Glu Cys
210 215 220
Pro Pro Cys Pro Ala Pro Pro Val Ala Gly Pro Ser Val Phe Leu Phe
225 230 235 240
Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val
245 250 255
Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Gln Phe
260 265 270
Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro
275 280 285
Arg Glu Glu Gln Phe Asn Ser Thr Phe Arg Val Val Ser Val Leu Thr
290 295 300
Val Val His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val
305 310 315 320
Ser Asn Lys Gly Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Thr
325 330 335
Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg
340 345 350
Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly
355 360 365
Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro
370 375 380
Glu Asn Asn Tyr Lys Thr Thr Pro Pro Met Leu Asp Ser Asp Gly Ser
385 390 395 400
Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln
405 410 415
Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His
420 425 430
Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440
<210> 20
<211> 11
<212> ПРТ
<213> Homo sapien
<400> 20
Ser Gly Asp Asn Ile Gly Ser Lys Tyr Val Ser
1 5 10
<210> 21
<211> 7
<212> ПРТ
<213> Homo sapien
<400> 21
Ser Asp Ser Glu Arg Pro Ser
1 5
<210> 22
<211> 10
<212> ПРТ
<213> Homo sapien
<400> 22
Gln Ser Trp Asp Gly Ser Ile Ser Arg Val
1 5 10
<210> 23
<211> 107
<212> ПРТ
<213> Homo sapien
<400> 23
Asp Ile Glu Leu Thr Gln Pro Pro Ser Val Ser Val Ala Pro Gly Gln
1 5 10 15
Thr Ala Arg Ile Ser Cys Ser Gly Asp Asn Ile Gly Ser Lys Tyr Val
20 25 30
Ser Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Ile Tyr
35 40 45
Ser Asp Ser Glu Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Gly Thr Gln Ala Glu
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Ser Trp Asp Gly Ser Ile Ser Arg
85 90 95
Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105
<210> 24
<211> 213
<212> ПРТ
<213> Homo sapien
<400> 24
Asp Ile Glu Leu Thr Gln Pro Pro Ser Val Ser Val Ala Pro Gly Gln
1 5 10 15
Thr Ala Arg Ile Ser Cys Ser Gly Asp Asn Ile Gly Ser Lys Tyr Val
20 25 30
Ser Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Ile Tyr
35 40 45
Ser Asp Ser Glu Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Gly Thr Gln Ala Glu
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Ser Trp Asp Gly Ser Ile Ser Arg
85 90 95
Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gln Pro Lys Ala
100 105 110
Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu Glu Leu Gln Ala
115 120 125
Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe Tyr Pro Gly Ala
130 135 140
Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val Lys Ala Gly Val
145 150 155 160
Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys Tyr Ala Ala Ser
165 170 175
Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser His Arg Ser Tyr
180 185 190
Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys Thr Val Ala
195 200 205
Pro Thr Glu Cys Ser
210
<210> 25
<211> 354
<212> ДНК
<213> Homo sapien
<400> 25
caggtgcaat tggtggaaag cggcggcggc ctggtgcaac cgggcggcag cctgcgtctg 60
agctgcgcgg cctccggatt taccttttct gattattata tgcattgggt gcgccaagcc 120
cctgggaagg gtctcgagtg ggtgagcgtt atctctggtt ctggtagcaa tacctattat 180
gcggatagcg tgaaaggccg ttttaccatt tcacgtgata attcgaaaaa caccctgtat 240
ctgcaaatga acagcctgcg tgcggaagat acggccgtgt attattgcgc gcgtctttat 300
gctcagtttg agggtgattt ttggggccaa ggcaccctgg tgacggttag ctca 354
<210> 26
<211> 327
<212> ДНК
<213> Homo sapien
<400> 26
gatatcgaac tgacccagcc gccttcagtg agcgttgcac caggtcagac cgcgcgtatc 60
tcgtgtagcg gcgataatat tggttctaag tatgtttctt ggtaccagca gaaacccggg 120
caggcgccag ttcttgtgat ttattctgat tctgagcgtc cctcaggcat cccggaacgc 180
tttagcggat ccaacagcgg caacaccgcg accctgacca ttagcggcac tcaggcggaa 240
gacgaagcgg attattattg ccagtcttgg gatggttcta tttctcgtgt gtttggcggc 300
ggcacgaagt taaccgtcct aggtcag 327
<210> 27
<211> 1332
<212> ДНК
<213> Homo sapien
<400> 27
caggtgcaat tggtggaaag cggcggcggc ctggtgcaac cgggcggcag cctgcgtctg 60
agctgcgcgg cctccggatt taccttttct gattattata tgcattgggt gcgccaagcc 120
cctgggaagg gtctcgagtg ggtgagcgtt atctctggtt ctggtagcaa tacctattat 180
gcggatagcg tgaaaggccg ttttaccatt tcacgtgata attcgaaaaa caccctgtat 240
ctgcaaatga acagcctgcg tgcggaagat acggccgtgt attattgcgc gcgtctttat 300
gctcagtttg agggtgattt ttggggccaa ggcaccctgg tgacggttag ctcagcctcc 360
accaagggcc catcggtctt ccccctggcg ccctgctcca ggagcacctc cgagagcaca 420
gcggccctgg gctgcctggt caaggactac ttccccgaac cggtgacggt gtcgtggaac 480
tcaggcgctc tgaccagcgg cgtgcacacc ttcccggctg tcctacagtc ctcaggactc 540
tactccctca gcagcgtagt gaccgtgccc tccagcaact tcggcaccca gacctacacc 600
tgcaacgtag atcacaagcc cagcaacacc aaggtggaca agacagttga gcgcaaatgt 660
tgtgtcgagt gcccaccgtg cccagcacca cctgtggcag gaccgtcagt cttcctcttc 720
cccccaaaac ccaaggacac cctcatgatc tcccggaccc ctgaggtcac gtgcgtggtg 780
gtggacgtga gccacgaaga ccccgaggtc cagttcaact ggtacgtgga cggcgtggag 840
gtgcataatg ccaagacaaa gccacgggag gagcagttca acagcacgtt ccgtgtggtc 900
agcgtcctca ccgtcgtgca ccaggactgg ctgaacggca aggagtacaa gtgcaaggtc 960
tccaacaaag gcctcccagc ccccatcgag aaaaccatct ccaaaaccaa agggcagccc 1020
cgagaaccac aggtgtacac cctgccccca tcccgggagg agatgaccaa gaaccaggtc 1080
agcctgacct gcctggtcaa aggcttctac cccagcgaca tcgccgtgga gtgggagagc 1140
aatgggcagc cggagaacaa ctacaagacc acacctccca tgctggactc cgacggctcc 1200
ttcttcctct acagcaagct caccgtggac aagagcaggt ggcagcaggg gaacgtcttc 1260
tcatgctccg tgatgcatga ggctctgcac aaccactaca cacagaagag cctctccctg 1320
tctccgggta aa 1332
<210> 28
<211> 327
<212> ДНК
<213> Homo sapien
<400> 28
gatatcgaac tgacccagcc gccttcagtg agcgttgcac caggtcagac cgcgcgtatc 60
tcgtgtagcg gcgataatat tggttctaag tatgtttctt ggtaccagca gaaacccggg 120
caggcgccag ttcttgtgat ttattctgat tctgagcgtc cctcaggcat cccggaacgc 180
tttagcggat ccaacagcgg caacaccgcg accctgacca ttagcggcac tcaggcggaa 240
gacgaagcgg attattattg ccagtcttgg gatggttcta tttctcgtgt gtttggcggc 300
ggcacgaagt taaccgtcct aggtcag 327
<210> 29
<211> 6
<212> ПРТ
<213> Homo sapien
<400> 29
Ser Thr Tyr Trp Ile Ser
1 5
<210> 30
<211> 17
<212> ПРТ
<213> Homo sapien
<400> 30
Lys Ile Tyr Pro Gly Asp Ser Tyr Thr Asn Tyr Ser Pro Ser Phe Gln
1 5 10 15
Gly
<210> 31
<211> 8
<212> ПРТ
<213> Homo sapien
<400> 31
Arg Gly Tyr Gly Ile Phe Asp Tyr
1 5
<210> 32
<211> 116
<212> ПРТ
<213> Homo sapien
<400> 32
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Glu
1 5 10 15
Ser Leu Lys Ile Ser Cys Lys Gly Ser Gly Tyr Ser Phe Ser Thr Tyr
20 25 30
Trp Ile Ser Trp Val Arg Gln Met Pro Gly Lys Gly Leu Glu Trp Met
35 40 45
Gly Lys Ile Tyr Pro Gly Asp Ser Tyr Thr Asn Tyr Ser Pro Ser Phe
50 55 60
Gln Gly Gln Val Thr Ile Ser Ala Asp Lys Ser Ile Ser Thr Ala Tyr
65 70 75 80
Leu Gln Trp Ser Ser Leu Lys Ala Ser Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Ala Arg Gly Tyr Gly Ile Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser
115
<210> 33
<211> 442
<212> ПРТ
<213> Homo sapien
<400> 33
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Glu
1 5 10 15
Ser Leu Lys Ile Ser Cys Lys Gly Ser Gly Tyr Ser Phe Ser Thr Tyr
20 25 30
Trp Ile Ser Trp Val Arg Gln Met Pro Gly Lys Gly Leu Glu Trp Met
35 40 45
Gly Lys Ile Tyr Pro Gly Asp Ser Tyr Thr Asn Tyr Ser Pro Ser Phe
50 55 60
Gln Gly Gln Val Thr Ile Ser Ala Asp Lys Ser Ile Ser Thr Ala Tyr
65 70 75 80
Leu Gln Trp Ser Ser Leu Lys Ala Ser Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Ala Arg Gly Tyr Gly Ile Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Asn Phe
180 185 190
Gly Thr Gln Thr Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Thr Val Glu Arg Lys Cys Cys Val Glu Cys Pro Pro
210 215 220
Cys Pro Ala Pro Pro Val Ala Gly Pro Ser Val Phe Leu Phe Pro Pro
225 230 235 240
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
245 250 255
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Gln Phe Asn Trp
260 265 270
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
275 280 285
Glu Gln Phe Asn Ser Thr Phe Arg Val Val Ser Val Leu Thr Val Val
290 295 300
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
305 310 315 320
Lys Gly Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Thr Lys Gly
325 330 335
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
340 345 350
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
355 360 365
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
370 375 380
Asn Tyr Lys Thr Thr Pro Pro Met Leu Asp Ser Asp Gly Ser Phe Phe
385 390 395 400
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
405 410 415
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
420 425 430
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440
<210> 34
<211> 11
<212> ПРТ
<213> Homo sapien
<400> 34
Ser Gly Asp Asn Ile Gly Asp Gln Tyr Ala His
1 5 10
<210> 35
<211> 7
<212> ПРТ
<213> Homo sapien
<400> 35
Gln Asp Lys Asn Arg Pro Ser
1 5
<210> 36
<211> 11
<212> ПРТ
<213> Homo sapien
<400> 36
Ala Thr Tyr Thr Gly Phe Gly Ser Leu Ala Val
1 5 10
<210> 37
<211> 108
<212> ПРТ
<213> Homo sapien
<400> 37
Asp Ile Glu Leu Thr Gln Pro Pro Ser Val Ser Val Ala Pro Gly Gln
1 5 10 15
Thr Ala Arg Ile Ser Cys Ser Gly Asp Asn Ile Gly Asp Gln Tyr Ala
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Val Val Ile Tyr
35 40 45
Gln Asp Lys Asn Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Gly Thr Gln Ala Glu
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Ala Thr Tyr Thr Gly Phe Gly Ser Leu
85 90 95
Ala Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105
<210> 38
<211> 214
<212> ПРТ
<213> Homo sapien
<400> 38
Asp Ile Glu Leu Thr Gln Pro Pro Ser Val Ser Val Ala Pro Gly Gln
1 5 10 15
Thr Ala Arg Ile Ser Cys Ser Gly Asp Asn Ile Gly Asp Gln Tyr Ala
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Val Val Ile Tyr
35 40 45
Gln Asp Lys Asn Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Gly Thr Gln Ala Glu
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Ala Thr Tyr Thr Gly Phe Gly Ser Leu
85 90 95
Ala Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gln Pro Lys
100 105 110
Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu Glu Leu Gln
115 120 125
Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe Tyr Pro Gly
130 135 140
Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val Lys Ala Gly
145 150 155 160
Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys Tyr Ala Ala
165 170 175
Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser His Arg Ser
180 185 190
Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys Thr Val
195 200 205
Ala Pro Thr Glu Cys Ser
210
<210> 39
<211> 348
<212> ДНК
<213> Homo sapien
<400> 39
caggtgcaat tggttcagag cggcgcggaa gtgaaaaaac cgggcgaaag cctgaaaatt 60
agctgcaaag gttccggata ttccttttct acttattgga tttcttgggt gcgccagatg 120
cctgggaagg gtctcgagtg gatgggcaag atctatccgg gtgatagcta taccaattat 180
tctccgagct ttcagggcca ggtgactatt agcgcggata aaagcattag caccgcgtat 240
cttcaatgga gcagcctgaa agcgagcgat acggccatgt attattgtgc gcgtggttat 300
ggtatttttg attattgggg ccaaggcacc ctggtcaccg tctcctca 348
<210> 40
<211> 324
<212> ДНК
<213> Homo sapien
<400> 40
gatatcgaac tgacccagcc gccttcagtg agcgttgcac caggtcagac cgcgcgtatc 60
tcgtgtagcg gcgataatat tggtgatcag tatgctcatt ggtaccagca gaaacccggg 120
caggcgccag ttgttgtgat ttatcaggat aagaatcgtc cctcaggcat cccggaacgc 180
tttagcggat ccaacagcgg caacaccgcg accctgacca ttagcggcac tcaggcggaa 240
gacgaagcgg attattattg cgctacttat actggttttg gttctcttgc tgtgtttggc 300
ggcggcacga agttaaccgt ccta 324
<210> 41
<211> 1326
<212> ДНК
<213> Homo sapien
<400> 41
caggtgcaat tggttcagag cggcgcggaa gtgaaaaaac cgggcgaaag cctgaaaatt 60
agctgcaaag gttccggata ttccttttct acttattgga tttcttgggt gcgccagatg 120
cctgggaagg gtctcgagtg gatgggcaag atctatccgg gtgatagcta taccaattat 180
tctccgagct ttcagggcca ggtgactatt agcgcggata aaagcattag caccgcgtat 240
cttcaatgga gcagcctgaa agcgagcgat acggccatgt attattgtgc gcgtggttat 300
ggtatttttg attattgggg ccaaggcacc ctggtcaccg tctcctcagc ctccaccaag 360
ggcccatcgg tcttccccct ggcgccctgc tccaggagca cctccgagag cacagcggcc 420
ctgggctgcc tggtcaagga ctacttcccc gaaccggtga cggtgtcgtg gaactcaggc 480
gctctgacca gcggcgtgca caccttcccg gctgtcctac agtcctcagg actctactcc 540
ctcagcagcg tagtgaccgt gccctccagc aacttcggca cccagaccta cacctgcaac 600
gtagatcaca agcccagcaa caccaaggtg gacaagacag ttgagcgcaa atgttgtgtc 660
gagtgcccac cgtgcccagc accacctgtg gcaggaccgt cagtcttcct cttcccccca 720
aaacccaagg acaccctcat gatctcccgg acccctgagg tcacgtgcgt ggtggtggac 780
gtgagccacg aagaccccga ggtccagttc aactggtacg tggacggcgt ggaggtgcat 840
aatgccaaga caaagccacg ggaggagcag ttcaacagca cgttccgtgt ggtcagcgtc 900
ctcaccgtcg tgcaccagga ctggctgaac ggcaaggagt acaagtgcaa ggtctccaac 960
aaaggcctcc cagcccccat cgagaaaacc atctccaaaa ccaaagggca gccccgagaa 1020
ccacaggtgt acaccctgcc cccatcccgg gaggagatga ccaagaacca ggtcagcctg 1080
acctgcctgg tcaaaggctt ctaccccagc gacatcgccg tggagtggga gagcaatggg 1140
cagccggaga acaactacaa gaccacacct cccatgctgg actccgacgg ctccttcttc 1200
ctctacagca agctcaccgt ggacaagagc aggtggcagc aggggaacgt cttctcatgc 1260
tccgtgatgc atgaggctct gcacaaccac tacacacaga agagcctctc cctgtctccg 1320
ggtaaa 1326
<210> 42
<211> 324
<212> ДНК
<213> Homo sapien
<400> 42
gatatcgaac tgacccagcc gccttcagtg agcgttgcac caggtcagac cgcgcgtatc 60
tcgtgtagcg gcgataatat tggtgatcag tatgctcatt ggtaccagca gaaacccggg 120
caggcgccag ttgttgtgat ttatcaggat aagaatcgtc cctcaggcat cccggaacgc 180
tttagcggat ccaacagcgg caacaccgcg accctgacca ttagcggcac tcaggcggaa 240
gacgaagcgg attattattg cgctacttat actggttttg gttctcttgc tgtgtttggc 300
ggcggcacga agttaaccgt ccta 324
<210> 43
<211> 116
<212> ПРТ
<213> Homo sapien
<400> 43
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Glu
1 5 10 15
Ser Leu Arg Ile Ser Cys Lys Gly Ser Gly Tyr Ser Phe Ser Thr Tyr
20 25 30
Trp Ile Ser Trp Val Arg Gln Met Pro Gly Lys Gly Leu Glu Trp Met
35 40 45
Gly Lys Ile Tyr Pro Gly Asp Ser Tyr Thr Asn Tyr Ser Pro Ser Phe
50 55 60
Gln Gly Gln Val Thr Ile Ser Ala Asp Lys Ser Ile Ser Thr Ala Tyr
65 70 75 80
Leu Gln Trp Ser Ser Leu Lys Ala Ser Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Ala Arg Gly Tyr Gly Ile Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser
115
<210> 44
<211> 442
<212> ПРТ
<213> Homo sapien
<400> 44
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Glu
1 5 10 15
Ser Leu Arg Ile Ser Cys Lys Gly Ser Gly Tyr Ser Phe Ser Thr Tyr
20 25 30
Trp Ile Ser Trp Val Arg Gln Met Pro Gly Lys Gly Leu Glu Trp Met
35 40 45
Gly Lys Ile Tyr Pro Gly Asp Ser Tyr Thr Asn Tyr Ser Pro Ser Phe
50 55 60
Gln Gly Gln Val Thr Ile Ser Ala Asp Lys Ser Ile Ser Thr Ala Tyr
65 70 75 80
Leu Gln Trp Ser Ser Leu Lys Ala Ser Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Ala Arg Gly Tyr Gly Ile Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Asn Phe
180 185 190
Gly Thr Gln Thr Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Thr Val Glu Arg Lys Cys Cys Val Glu Cys Pro Pro
210 215 220
Cys Pro Ala Pro Pro Val Ala Gly Pro Ser Val Phe Leu Phe Pro Pro
225 230 235 240
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
245 250 255
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Gln Phe Asn Trp
260 265 270
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
275 280 285
Glu Gln Phe Asn Ser Thr Phe Arg Val Val Ser Val Leu Thr Val Val
290 295 300
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
305 310 315 320
Lys Gly Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Thr Lys Gly
325 330 335
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
340 345 350
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
355 360 365
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
370 375 380
Asn Tyr Lys Thr Thr Pro Pro Met Leu Asp Ser Asp Gly Ser Phe Phe
385 390 395 400
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
405 410 415
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
420 425 430
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440
<210> 45
<211> 108
<212> ПРТ
<213> Homo sapien
<400> 45
Ser Tyr Glu Leu Thr Gln Pro Pro Ser Val Ser Val Ser Pro Gly Gln
1 5 10 15
Thr Ala Ser Ile Thr Cys Ser Gly Asp Asn Ile Gly Asp Gln Tyr Ala
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ser Pro Val Leu Val Ile Tyr
35 40 45
Gln Asp Lys Asn Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Gly Thr Gln Ala Met
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Ala Thr Tyr Thr Gly Phe Gly Ser Leu
85 90 95
Ala Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105
<210> 46
<211> 214
<212> ПРТ
<213> Homo sapien
<400> 46
Ser Tyr Glu Leu Thr Gln Pro Pro Ser Val Ser Val Ser Pro Gly Gln
1 5 10 15
Thr Ala Ser Ile Thr Cys Ser Gly Asp Asn Ile Gly Asp Gln Tyr Ala
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ser Pro Val Leu Val Ile Tyr
35 40 45
Gln Asp Lys Asn Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Gly Thr Gln Ala Met
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Ala Thr Tyr Thr Gly Phe Gly Ser Leu
85 90 95
Ala Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gln Pro Lys
100 105 110
Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu Glu Leu Gln
115 120 125
Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe Tyr Pro Gly
130 135 140
Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val Lys Ala Gly
145 150 155 160
Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys Tyr Ala Ala
165 170 175
Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser His Arg Ser
180 185 190
Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys Thr Val
195 200 205
Ala Pro Thr Glu Cys Ser
210
<210> 47
<211> 348
<212> ДНК
<213> Homo sapien
<400> 47
gaggtgcaat tggttcagag cggcgcggaa gtgaaaaaac cgggcgaaag cctgaggatt 60
agctgcaaag gttccggata ttccttttct acttattgga tttcttgggt gcgccagatg 120
cctgggaagg gtctcgagtg gatgggcaag atctatccgg gtgatagcta taccaattat 180
tctccgagct ttcagggcca ggtgactatt agcgcggata aaagcattag caccgcgtat 240
cttcaatgga gcagcctgaa agcgagcgat acggccatgt attattgtgc gcgtggttat 300
ggtatttttg attattgggg ccaaggcacc ctggtcaccg tctcctca 348
<210> 48
<211> 324
<212> ДНК
<213> Homo sapien
<400> 48
agctacgagc tgacccagcc ccccagcgtg tccgtgagcc ctggccagac cgccagcatc 60
acctgcagcg gcgacaacat cggcgaccag tacgcccact ggtatcagca gaagcccggc 120
cagagccccg tgctggtgat ctaccaggac aagaaccggc ccagcggcat ccccgagcgg 180
ttcagcggca gcaacagcgg caacaccgcc accctgacca tcagcggcac ccaggccatg 240
gacgaggccg actactactg cgccacctac accggcttcg gcagcctggc cgtgttcggc 300
ggagggacca agctgaccgt ccta 324
<210> 49
<211> 1326
<212> ДНК
<213> Homo sapien
<400> 49
gaggtgcaat tggttcagag cggcgcggaa gtgaaaaaac cgggcgaaag cctgaggatt 60
agctgcaaag gttccggata ttccttttct acttattgga tttcttgggt gcgccagatg 120
cctgggaagg gtctcgagtg gatgggcaag atctatccgg gtgatagcta taccaattat 180
tctccgagct ttcagggcca ggtgactatt agcgcggata aaagcattag caccgcgtat 240
cttcaatgga gcagcctgaa agcgagcgat acggccatgt attattgtgc gcgtggttat 300
ggtatttttg attattgggg ccaaggcacc ctggtcaccg tctcctcagc ctccaccaag 360
ggcccatcgg tcttccccct ggcgccctgc tccaggagca cctccgagag cacagcggcc 420
ctgggctgcc tggtcaagga ctacttcccc gaaccggtga cggtgtcgtg gaactcaggc 480
gctctgacca gcggcgtgca caccttcccg gctgtcctac agtcctcagg actctactcc 540
ctcagcagcg tagtgaccgt gccctccagc aacttcggca cccagaccta cacctgcaac 600
gtagatcaca agcccagcaa caccaaggtg gacaagacag ttgagcgcaa atgttgtgtc 660
gagtgcccac cgtgcccagc accacctgtg gcaggaccgt cagtcttcct cttcccccca 720
aaacccaagg acaccctcat gatctcccgg acccctgagg tcacgtgcgt ggtggtggac 780
gtgagccacg aagaccccga ggtccagttc aactggtacg tggacggcgt ggaggtgcat 840
aatgccaaga caaagccacg ggaggagcag ttcaacagca cgttccgtgt ggtcagcgtc 900
ctcaccgtcg tgcaccagga ctggctgaac ggcaaggagt acaagtgcaa ggtctccaac 960
aaaggcctcc cagcccccat cgagaaaacc atctccaaaa ccaaagggca gccccgagaa 1020
ccacaggtgt acaccctgcc cccatcccgg gaggagatga ccaagaacca ggtcagcctg 1080
acctgcctgg tcaaaggctt ctaccccagc gacatcgccg tggagtggga gagcaatggg 1140
cagccggaga acaactacaa gaccacacct cccatgctgg actccgacgg ctccttcttc 1200
ctctacagca agctcaccgt ggacaagagc aggtggcagc aggggaacgt cttctcatgc 1260
tccgtgatgc atgaggctct gcacaaccac tacacacaga agagcctctc cctgtctccg 1320
ggtaaa 1326
<210> 50
<211> 642
<212> ДНК
<213> Homo sapien
<400> 50
agctacgagc tgacccagcc ccccagcgtg tccgtgagcc ctggccagac cgccagcatc 60
acctgcagcg gcgacaacat cggcgaccag tacgcccact ggtatcagca gaagcccggc 120
cagagccccg tgctggtgat ctaccaggac aagaaccggc ccagcggcat ccccgagcgg 180
ttcagcggca gcaacagcgg caacaccgcc accctgacca tcagcggcac ccaggccatg 240
gacgaggccg actactactg cgccacctac accggcttcg gcagcctggc cgtgttcggc 300
ggagggacca agctgaccgt cctaggtcag cccaaggctg ccccctcggt cactctgttc 360
ccaccctcct ctgaggagct tcaagccaac aaggccacac tggtgtgtct cataagtgac 420
ttctacccgg gagccgtgac agtggcctgg aaggcagata gcagccccgt caaggcggga 480
gtggagacca ccacaccctc caaacaaagc aacaacaagt acgcggccag cagctacctg 540
agcctgacgc ctgagcagtg gaagtcccac agaagctaca gctgccaggt cacgcatgaa 600
gggagcaccg tggagaagac agtggcccct acagaatgtt ca 642
<210> 51
<211> 108
<212> ПРТ
<213> Homo sapien
<400> 51
Ser Tyr Glu Leu Thr Gln Pro Pro Ser Val Ser Val Ser Pro Gly Gln
1 5 10 15
Thr Ala Ser Ile Thr Cys Ser Gly Asp Asn Ile Gly Asp Gln Tyr Ala
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ser Pro Val Val Val Ile Tyr
35 40 45
Gln Asp Lys Asn Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Gly Thr Gln Ala Met
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Ala Thr Tyr Thr Gly Phe Gly Ser Leu
85 90 95
Ala Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105
<210> 52
<211> 214
<212> ПРТ
<213> Homo sapien
<400> 52
Ser Tyr Glu Leu Thr Gln Pro Pro Ser Val Ser Val Ser Pro Gly Gln
1 5 10 15
Thr Ala Ser Ile Thr Cys Ser Gly Asp Asn Ile Gly Asp Gln Tyr Ala
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ser Pro Val Val Val Ile Tyr
35 40 45
Gln Asp Lys Asn Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Gly Thr Gln Ala Met
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Ala Thr Tyr Thr Gly Phe Gly Ser Leu
85 90 95
Ala Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gln Pro Lys
100 105 110
Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu Glu Leu Gln
115 120 125
Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe Tyr Pro Gly
130 135 140
Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val Lys Ala Gly
145 150 155 160
Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys Tyr Ala Ala
165 170 175
Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser His Arg Ser
180 185 190
Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys Thr Val
195 200 205
Ala Pro Thr Glu Cys Ser
210
<210> 53
<211> 324
<212> ДНК
<213> Homo sapien
<400> 53
agctacgagc tgacccagcc ccccagcgtg tccgtgagcc ctggccagac cgccagcatc 60
acctgcagcg gcgacaacat cggcgaccag tacgcccact ggtatcagca gaagcccggc 120
cagagccccg tggtggtgat ctaccaggac aagaaccggc ccagcggcat ccccgagcgg 180
ttcagcggca gcaacagcgg caacaccgcc accctgacca tcagcggcac ccaggccatg 240
gacgaggccg actactactg cgccacctac accggcttcg gcagcctggc cgtgttcggc 300
ggagggacca agctgaccgt ccta 324
<210> 54
<211> 642
<212> ДНК
<213> Homo sapien
<400> 54
agctacgagc tgacccagcc ccccagcgtg tccgtgagcc ctggccagac cgccagcatc 60
acctgcagcg gcgacaacat cggcgaccag tacgcccact ggtatcagca gaagcccggc 120
cagagccccg tggtggtgat ctaccaggac aagaaccggc ccagcggcat ccccgagcgg 180
ttcagcggca gcaacagcgg caacaccgcc accctgacca tcagcggcac ccaggccatg 240
gacgaggccg actactactg cgccacctac accggcttcg gcagcctggc cgtgttcggc 300
ggagggacca agctgaccgt cctaggtcag cccaaggctg ccccctcggt cactctgttc 360
ccaccctcct ctgaggagct tcaagccaac aaggccacac tggtgtgtct cataagtgac 420
ttctacccgg gagccgtgac agtggcctgg aaggcagata gcagccccgt caaggcggga 480
gtggagacca ccacaccctc caaacaaagc aacaacaagt acgcggccag cagctacctg 540
agcctgacgc ctgagcagtg gaagtcccac agaagctaca gctgccaggt cacgcatgaa 600
gggagcaccg tggagaagac agtggcccct acagaatgtt ca 642
<210> 55
<211> 11
<212> ПРТ
<213> Homo sapien
<400> 55
Ser Thr Tyr Thr Phe Val Gly Phe Thr Thr Val
1 5 10
<210> 56
<211> 108
<212> ПРТ
<213> Homo sapien
<400> 56
Asp Ile Glu Leu Thr Gln Pro Pro Ser Val Ser Val Ala Pro Gly Gln
1 5 10 15
Thr Ala Arg Ile Ser Cys Ser Gly Asp Asn Ile Gly Asp Gln Tyr Ala
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Val Val Ile Tyr
35 40 45
Gln Asp Lys Asn Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Gly Thr Gln Ala Glu
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Ser Thr Tyr Thr Phe Val Gly Phe Thr
85 90 95
Thr Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105
<210> 57
<211> 214
<212> ПРТ
<213> Homo sapien
<400> 57
Asp Ile Glu Leu Thr Gln Pro Pro Ser Val Ser Val Ala Pro Gly Gln
1 5 10 15
Thr Ala Arg Ile Ser Cys Ser Gly Asp Asn Ile Gly Asp Gln Tyr Ala
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Val Val Ile Tyr
35 40 45
Gln Asp Lys Asn Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Gly Thr Gln Ala Glu
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Ser Thr Tyr Thr Phe Val Gly Phe Thr
85 90 95
Thr Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gln Pro Lys
100 105 110
Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu Glu Leu Gln
115 120 125
Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe Tyr Pro Gly
130 135 140
Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val Lys Ala Gly
145 150 155 160
Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys Tyr Ala Ala
165 170 175
Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser His Arg Ser
180 185 190
Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys Thr Val
195 200 205
Ala Pro Thr Glu Cys Ser
210
<210> 58
<211> 324
<212> ДНК
<213> Homo sapien
<400> 58
gatatcgaac tgacccagcc gccttcagtg agcgttgcac caggtcagac cgcgcgtatc 60
tcgtgtagcg gcgataatat tggtgatcag tatgctcatt ggtaccagca gaaacccggg 120
caggcgccag ttgttgtgat ttatcaggat aagaatcgtc cctcaggcat cccggaacgc 180
tttagcggat ccaacagcgg caacaccgcg accctgacca ttagcggcac tcaggcggaa 240
gacgaagcgg attattattg ctctacttat acttttgttg gttttactac tgtgtttggc 300
ggcggcacga agttaaccgt ccta 324
<210> 59
<211> 642
<212> ДНК
<213> Homo sapien
<400> 59
gatatcgaac tgacccagcc gccttcagtg agcgttgcac caggtcagac cgcgcgtatc 60
tcgtgtagcg gcgataatat tggtgatcag tatgctcatt ggtaccagca gaaacccggg 120
caggcgccag ttgttgtgat ttatcaggat aagaatcgtc cctcaggcat cccggaacgc 180
tttagcggat ccaacagcgg caacaccgcg accctgacca ttagcggcac tcaggcggaa 240
gacgaagcgg attattattg ctctacttat acttttgttg gttttactac tgtgtttggc 300
ggcggcacga agttaaccgt cctaggtcag cccaaggctg ccccctcggt cactctgttc 360
ccaccctcct ctgaggagct tcaagccaac aaggccacac tggtgtgtct cataagtgac 420
ttctacccgg gagccgtgac agtggcctgg aaggcagata gcagccccgt caaggcggga 480
gtggagacca ccacaccctc caaacaaagc aacaacaagt acgcggccag cagctacctg 540
agcctgacgc ctgagcagtg gaagtcccac agaagctaca gctgccaggt cacgcatgaa 600
gggagcaccg tggagaagac agtggcccct acagaatgtt ca 642
<210> 60
<211> 108
<212> ПРТ
<213> Homo sapien
<400> 60
Ser Tyr Glu Leu Thr Gln Pro Pro Ser Val Ser Val Ser Pro Gly Gln
1 5 10 15
Thr Ala Ser Ile Thr Cys Ser Gly Asp Asn Ile Gly Asp Gln Tyr Ala
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ser Pro Val Leu Val Ile Tyr
35 40 45
Gln Asp Lys Asn Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Gly Thr Gln Ala Met
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Ser Thr Tyr Thr Phe Val Gly Phe Thr
85 90 95
Thr Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105
<210> 61
<211> 214
<212> ПРТ
<213> Homo sapien
<400> 61
Ser Tyr Glu Leu Thr Gln Pro Pro Ser Val Ser Val Ser Pro Gly Gln
1 5 10 15
Thr Ala Ser Ile Thr Cys Ser Gly Asp Asn Ile Gly Asp Gln Tyr Ala
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ser Pro Val Leu Val Ile Tyr
35 40 45
Gln Asp Lys Asn Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Gly Thr Gln Ala Met
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Ser Thr Tyr Thr Phe Val Gly Phe Thr
85 90 95
Thr Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gln Pro Lys
100 105 110
Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu Glu Leu Gln
115 120 125
Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe Tyr Pro Gly
130 135 140
Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val Lys Ala Gly
145 150 155 160
Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys Tyr Ala Ala
165 170 175
Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser His Arg Ser
180 185 190
Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys Thr Val
195 200 205
Ala Pro Thr Glu Cys Ser
210
<210> 62
<211> 324
<212> ДНК
<213> Homo sapien
<400> 62
agctacgagc tgacccagcc ccccagcgtg tccgtgagcc ctggccagac cgccagcatc 60
acctgcagcg gcgacaacat cggcgaccag tacgcccact ggtatcagca gaagcccggc 120
cagagccccg tgctggtgat ctaccaggac aagaaccggc ccagcggcat ccccgagcgg 180
ttcagcggca gcaacagcgg caacaccgcc accctgacca tcagcggcac ccaggccatg 240
gacgaggccg actactactg ctctacttat acttttgttg gttttactac tgtgttcggc 300
ggagggacca agctgaccgt ccta 324
<210> 63
<211> 642
<212> ДНК
<213> Homo sapien
<400> 63
agctacgagc tgacccagcc ccccagcgtg tccgtgagcc ctggccagac cgccagcatc 60
acctgcagcg gcgacaacat cggcgaccag tacgcccact ggtatcagca gaagcccggc 120
cagagccccg tgctggtgat ctaccaggac aagaaccggc ccagcggcat ccccgagcgg 180
ttcagcggca gcaacagcgg caacaccgcc accctgacca tcagcggcac ccaggccatg 240
gacgaggccg actactactg ctctacttat acttttgttg gttttactac tgtgttcggc 300
ggagggacca agctgaccgt cctaggtcag cccaaggctg ccccctcggt cactctgttc 360
ccaccctcct ctgaggagct tcaagccaac aaggccacac tggtgtgtct cataagtgac 420
ttctacccgg gagccgtgac agtggcctgg aaggcagata gcagccccgt caaggcggga 480
gtggagacca ccacaccctc caaacaaagc aacaacaagt acgcggccag cagctacctg 540
agcctgacgc ctgagcagtg gaagtcccac agaagctaca gctgccaggt cacgcatgaa 600
gggagcaccg tggagaagac agtggcccct acagaatgtt ca 642
<210> 64
<211> 108
<212> ПРТ
<213> Homo sapien
<400> 64
Ser Tyr Glu Leu Thr Gln Pro Pro Ser Val Ser Val Ser Pro Gly Gln
1 5 10 15
Thr Ala Ser Ile Thr Cys Ser Gly Asp Asn Ile Gly Asp Gln Tyr Ala
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ser Pro Val Val Val Ile Tyr
35 40 45
Gln Asp Lys Asn Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Gly Thr Gln Ala Met
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Ser Thr Tyr Thr Phe Val Gly Phe Thr
85 90 95
Thr Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105
<210> 65
<211> 214
<212> ПРТ
<213> Homo sapien
<400> 65
Ser Tyr Glu Leu Thr Gln Pro Pro Ser Val Ser Val Ser Pro Gly Gln
1 5 10 15
Thr Ala Ser Ile Thr Cys Ser Gly Asp Asn Ile Gly Asp Gln Tyr Ala
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ser Pro Val Val Val Ile Tyr
35 40 45
Gln Asp Lys Asn Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Gly Thr Gln Ala Met
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Ser Thr Tyr Thr Phe Val Gly Phe Thr
85 90 95
Thr Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gln Pro Lys
100 105 110
Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu Glu Leu Gln
115 120 125
Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe Tyr Pro Gly
130 135 140
Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val Lys Ala Gly
145 150 155 160
Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys Tyr Ala Ala
165 170 175
Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser His Arg Ser
180 185 190
Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys Thr Val
195 200 205
Ala Pro Thr Glu Cys Ser
210
<210> 66
<211> 324
<212> ДНК
<213> Homo sapien
<400> 66
agctacgagc tgacccagcc ccccagcgtg tccgtgagcc ctggccagac cgccagcatc 60
acctgcagcg gcgacaacat cggcgaccag tacgcccact ggtatcagca gaagcccggc 120
cagagccccg tggtggtgat ctaccaggac aagaaccggc ccagcggcat ccccgagcgg 180
ttcagcggca gcaacagcgg caacaccgcc accctgacca tcagcggcac ccaggccatg 240
gacgaggccg actactactg ctctacttat acttttgttg gttttactac tgtgttcggc 300
ggagggacca agctgaccgt ccta 324
<210> 67
<211> 642
<212> ДНК
<213> Homo sapien
<400> 67
agctacgagc tgacccagcc ccccagcgtg tccgtgagcc ctggccagac cgccagcatc 60
acctgcagcg gcgacaacat cggcgaccag tacgcccact ggtatcagca gaagcccggc 120
cagagccccg tggtggtgat ctaccaggac aagaaccggc ccagcggcat ccccgagcgg 180
ttcagcggca gcaacagcgg caacaccgcc accctgacca tcagcggcac ccaggccatg 240
gacgaggccg actactactg ctctacttat acttttgttg gttttactac tgtgttcggc 300
ggagggacca agctgaccgt cctaggtcag cccaaggctg ccccctcggt cactctgttc 360
ccaccctcct ctgaggagct tcaagccaac aaggccacac tggtgtgtct cataagtgac 420
ttctacccgg gagccgtgac agtggcctgg aaggcagata gcagccccgt caaggcggga 480
gtggagacca ccacaccctc caaacaaagc aacaacaagt acgcggccag cagctacctg 540
agcctgacgc ctgagcagtg gaagtcccac agaagctaca gctgccaggt cacgcatgaa 600
gggagcaccg tggagaagac agtggcccct acagaatgtt ca 642
<210> 68
<211> 255
<212> ПРТ
<213> Homo sapien
<400> 68
Met Gly Asn Ser Cys Tyr Asn Ile Val Ala Thr Leu Leu Leu Val Leu
1 5 10 15
Asn Phe Glu Arg Thr Arg Ser Leu Gln Asp Pro Cys Ser Asn Cys Pro
20 25 30
Ala Gly Thr Phe Cys Asp Asn Asn Arg Asn Gln Ile Cys Ser Pro Cys
35 40 45
Pro Pro Asn Ser Phe Ser Ser Ala Gly Gly Gln Arg Thr Cys Asp Ile
50 55 60
Cys Arg Gln Cys Lys Gly Val Phe Arg Thr Arg Lys Glu Cys Ser Ser
65 70 75 80
Thr Ser Asn Ala Glu Cys Asp Cys Thr Pro Gly Phe His Cys Leu Gly
85 90 95
Ala Gly Cys Ser Met Cys Glu Gln Asp Cys Lys Gln Gly Gln Glu Leu
100 105 110
Thr Lys Lys Gly Cys Lys Asp Cys Cys Phe Gly Thr Phe Asn Asp Gln
115 120 125
Lys Arg Gly Ile Cys Arg Pro Trp Thr Asn Cys Ser Leu Asp Gly Lys
130 135 140
Ser Val Leu Val Asn Gly Thr Lys Glu Arg Asp Val Val Cys Gly Pro
145 150 155 160
Ser Pro Ala Asp Leu Ser Pro Gly Ala Ser Ser Val Thr Pro Pro Ala
165 170 175
Pro Ala Arg Glu Pro Gly His Ser Pro Gln Ile Ile Ser Phe Phe Leu
180 185 190
Ala Leu Thr Ser Thr Ala Leu Leu Phe Leu Leu Phe Phe Leu Thr Leu
195 200 205
Arg Phe Ser Val Val Lys Arg Gly Arg Lys Lys Leu Leu Tyr Ile Phe
210 215 220
Lys Gln Pro Phe Met Arg Pro Val Gln Thr Thr Gln Glu Glu Asp Gly
225 230 235 240
Cys Ser Cys Arg Phe Pro Glu Glu Glu Glu Gly Gly Cys Glu Leu
245 250 255
<210> 69
<211> 330
<212> ПРТ
<213> Homo sapien
<400> 69
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 70
<211> 990
<212> ДНК
<213> Homo sapien
<400> 70
gcctccacca agggcccatc ggtcttcccc ctggcaccct cctccaagag cacctctggg 60
ggcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtcg 120
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca 180
ggactctact ccctcagcag cgtagtgacc gtgccctcca gcagcttggg cacccagacc 240
tacatctgca acgtgaatca caagcccagc aacaccaagg tggacaagaa agttgagccc 300
aaatcttgtg acaaaactca cacatgccca ccgtgcccag cacctgaact cctgggggga 360
ccgtcagtct tcctcttccc cccaaaaccc aaggacaccc tcatgatctc ccggacccct 420
gaggtcacat gcgtggtggt ggacgtgagc cacgaagacc ctgaggtcaa gttcaactgg 480
tacgtggacg gcgtggaggt gcataatgcc aagacaaagc cgcgggagga gcagtacaac 540
agcacgtacc gtgtggtcag cgtcctcacc gtcctgcacc aggactggct gaatggcaag 600
gagtacaagt gcaaggtctc caacaaagcc ctcccagccc ccatcgagaa aaccatctcc 660
aaagccaaag ggcagccccg agaaccacag gtgtacaccc tgcccccatc ccgggaggag 720
atgaccaaga accaggtcag cctgacctgc ctggtcaaag gcttctatcc cagcgacatc 780
gccgtggagt gggagagcaa tgggcagccg gagaacaact acaagaccac gcctcccgtg 840
ctggactccg acggctcctt cttcctctat agcaagctca ccgtggacaa gagcaggtgg 900
cagcagggga acgtcttctc atgctccgtg atgcatgagg ctctgcacaa ccactacacg 960
cagaagagcc tctccctgtc tccgggtaaa 990
<210> 71
<211> 326
<212> ПРТ
<213> Homo sapien
<400> 71
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg
1 5 10 15
Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Asn Phe Gly Thr Gln Thr
65 70 75 80
Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Thr Val Glu Arg Lys Cys Cys Val Glu Cys Pro Pro Cys Pro Ala Pro
100 105 110
Pro Val Ala Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
115 120 125
Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp
130 135 140
Val Ser His Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly
145 150 155 160
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn
165 170 175
Ser Thr Phe Arg Val Val Ser Val Leu Thr Val Val His Gln Asp Trp
180 185 190
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro
195 200 205
Ala Pro Ile Glu Lys Thr Ile Ser Lys Thr Lys Gly Gln Pro Arg Glu
210 215 220
Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn
225 230 235 240
Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile
245 250 255
Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
260 265 270
Thr Pro Pro Met Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys
275 280 285
Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys
290 295 300
Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu
305 310 315 320
Ser Leu Ser Pro Gly Lys
325
<210> 72
<211> 978
<212> ДНК
<213> Homo sapien
<400> 72
gcctccacca agggcccatc ggtcttcccc ctggcgccct gctccaggag cacctccgag 60
agcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtcg 120
tggaactcag gcgctctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca 180
ggactctact ccctcagcag cgtagtgacc gtgccctcca gcaacttcgg cacccagacc 240
tacacctgca acgtagatca caagcccagc aacaccaagg tggacaagac agttgagcgc 300
aaatgttgtg tcgagtgccc accgtgccca gcaccacctg tggcaggacc gtcagtcttc 360
ctcttccccc caaaacccaa ggacaccctc atgatctccc ggacccctga ggtcacgtgc 420
gtggtggtgg acgtgagcca cgaagacccc gaggtccagt tcaactggta cgtggacggc 480
gtggaggtgc ataatgccaa gacaaagcca cgggaggagc agttcaacag cacgttccgt 540
gtggtcagcg tcctcaccgt cgtgcaccag gactggctga acggcaagga gtacaagtgc 600
aaggtctcca acaaaggcct cccagccccc atcgagaaaa ccatctccaa aaccaaaggg 660
cagccccgag aaccacaggt gtacaccctg cccccatccc gggaggagat gaccaagaac 720
caggtcagcc tgacctgcct ggtcaaaggc ttctacccca gcgacatcgc cgtggagtgg 780
gagagcaatg ggcagccgga gaacaactac aagaccacac ctcccatgct ggactccgac 840
ggctccttct tcctctacag caagctcacc gtggacaaga gcaggtggca gcaggggaac 900
gtcttctcat gctccgtgat gcatgaggct ctgcacaacc actacacaca gaagagcctc 960
tccctgtctc cgggtaaa 978
<210> 73
<211> 106
<212> ПРТ
<213> Homo sapien
<400> 73
Gly Gln Pro Lys Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser
1 5 10 15
Glu Glu Leu Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp
20 25 30
Phe Tyr Pro Gly Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro
35 40 45
Val Lys Ala Gly Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn
50 55 60
Lys Tyr Ala Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys
65 70 75 80
Ser His Arg Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val
85 90 95
Glu Lys Thr Val Ala Pro Thr Glu Cys Ser
100 105
<210> 74
<211> 318
<212> ДНК
<213> Homo sapien
<400> 74
ggtcagccca aggctgcccc ctcggtcact ctgttcccac cctcctctga ggagcttcaa 60
gccaacaagg ccacactggt gtgtctcata agtgacttct acccgggagc cgtgacagtg 120
gcctggaagg cagatagcag ccccgtcaag gcgggagtgg agaccaccac accctccaaa 180
caaagcaaca acaagtacgc ggccagcagc tacctgagcc tgacgcctga gcagtggaag 240
tcccacagaa gctacagctg ccaggtcacg catgaaggga gcaccgtgga gaagacagtg 300
gcccctacag aatgttca 318
| название | год | авторы | номер документа |
|---|---|---|---|
| Анти-PD-L1 и IL-2 цитокины | 2017 |
|
RU2769282C2 |
| PD-L1 специфические антитела | 2017 |
|
RU2756236C2 |
| БИСПЕЦИФИЧЕСКИЕ АНТИТЕЛА, СПЕЦИФИЧЕСКИЕ В ОТНОШЕНИИ КОСТИМУЛЯТОРНОГО TNF-РЕЦЕПТОРА | 2016 |
|
RU2761115C1 |
| Биспецифическое антитело анти-HER2/анти-4-1BB и его применение | 2020 |
|
RU2831838C2 |
| БИСПЕЦИФИЧЕСКОЕ АНТИТЕЛО АНТИ-EGFR/АНТИ-4-1ВВ И ЕГО ПРИМЕНЕНИЕ | 2020 |
|
RU2831836C2 |
| HER-2-НАПРАВЛЕННЫЕ АНТИГЕНСВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ, СОДЕРЖАЩИЕ 4-1BBL | 2019 |
|
RU2815451C2 |
| АНТАГОНИСТЫ | 2019 |
|
RU2839429C2 |
| ХИМЕРНЫЕ АНТИГЕННЫЕ РЕЦЕПТОРЫ, НАЦЕЛИВАЮЩИЕСЯ НА ВАРИАНТ III РЕЦЕПТОРА ЭПИДЕРМАЛЬНОГО ФАКТОРА РОСТА | 2017 |
|
RU2751662C2 |
| Т-КЛЕТКИ С КОСТИМУЛИРУЮЩИМ ХИМЕРНЫМ АНТИГЕННЫМ РЕЦЕПТОРОМ, НАЦЕЛЕННЫЕ НА IL13Rα2 | 2015 |
|
RU2749922C2 |
| T-КЛЕТОЧНЫЕ РЕЦЕПТОРЫ, РАСПОЗНАЮЩИЕ МУТАЦИЮ R175H ИЛИ Y220C В P53 | 2020 |
|
RU2830061C2 |
























































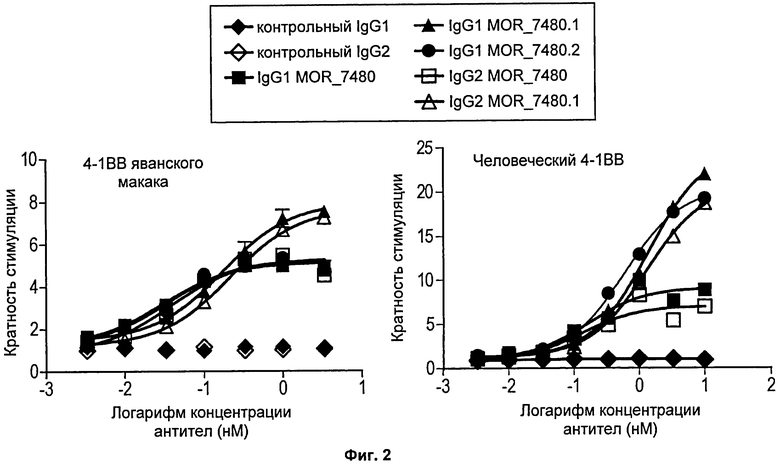
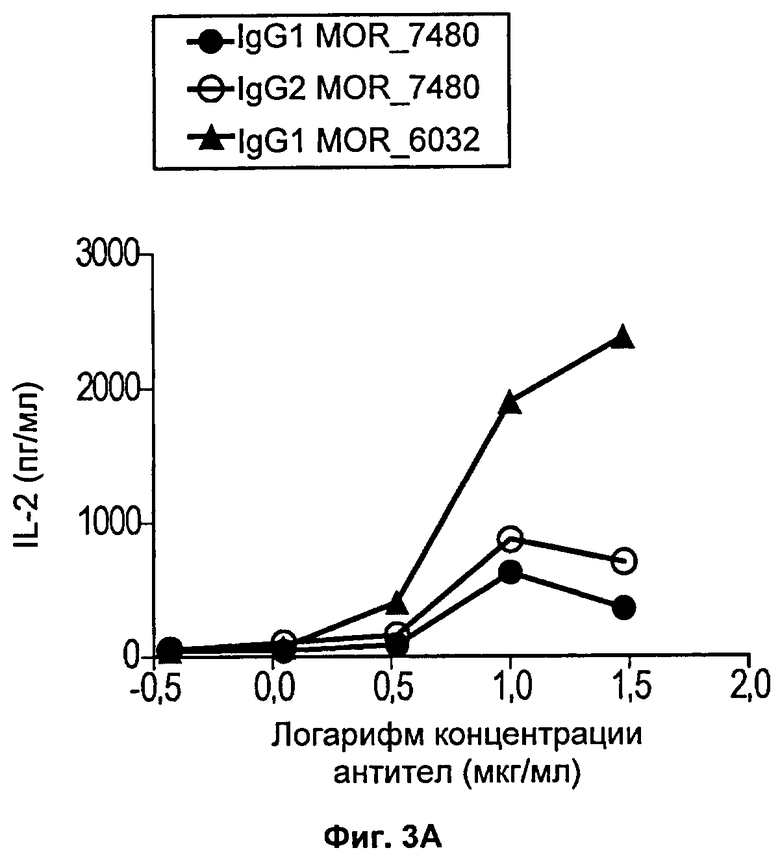
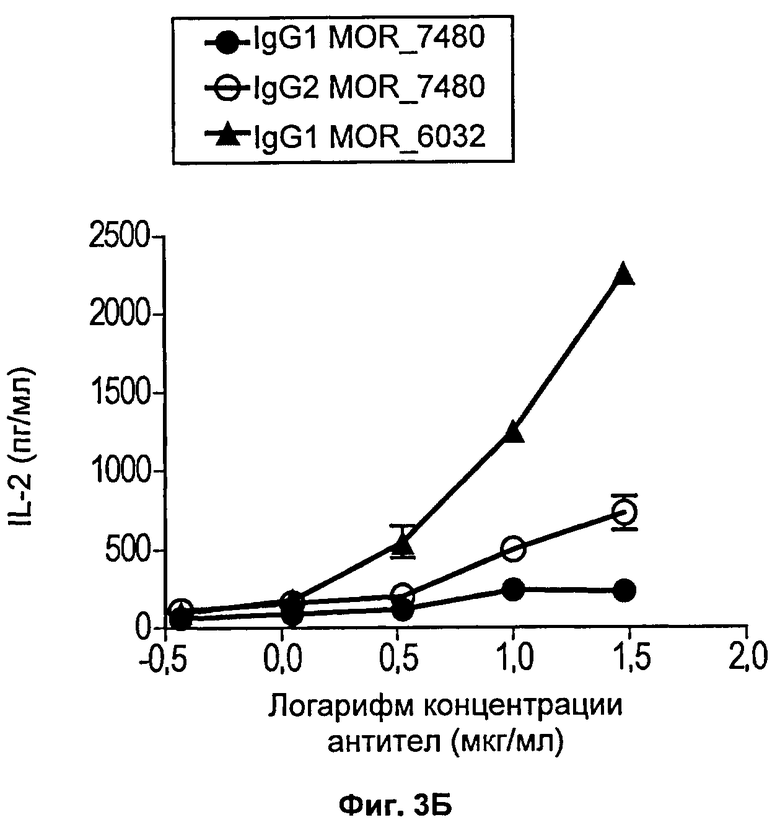
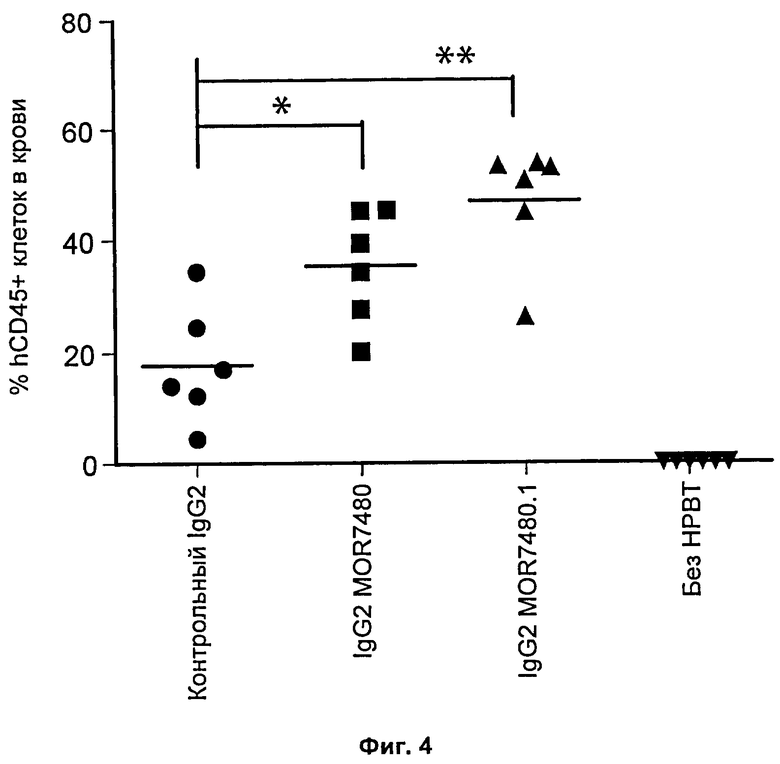
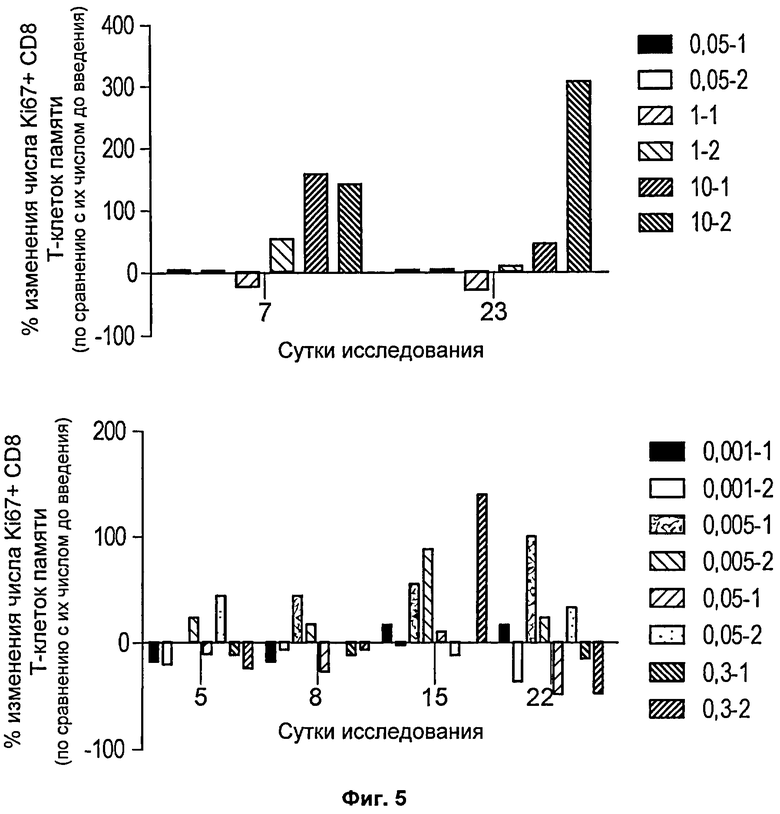
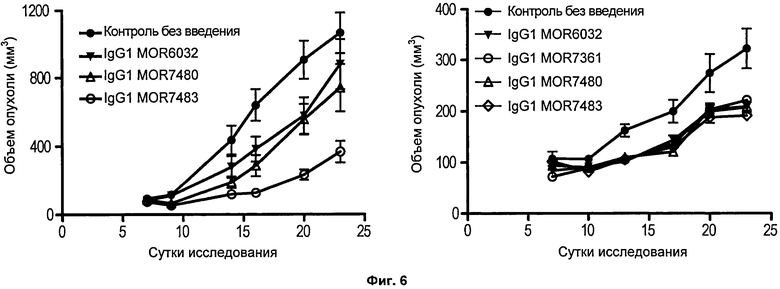
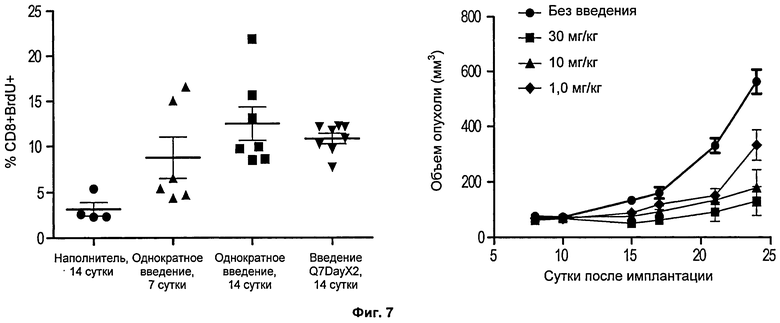

Предложенное изобретение относится к области иммунологии. Описаны варианты антител или антигенсвязывающих фрагментов, связывающиеся с человеческим 4-1ВВ. Один из вариантов характеризуется наличием соответствующих 3 CDR участков легкой цепи и 3 CDR участков тяжелой цепи. Другой вариант характеризуется наличием тяжелой и лёгкой цепи с соответствующими аминокислотными последовательностями. Описаны варианты фармацевтической композиции для снижения опухолевого роста или для лечения рака у субъекта, а также способы снижения опухолевого роста или лечения рака у субъекта, использующие варианты антител или антигенсвязывающих фрагментов в терапевтически эффективном количестве. Описан способ лечения рака с использованием комбинации антитела и иммунотерапевтического агента. Раскрыты: варианты кодирующих нуклеиновых кислот, вектор экспрессии и клетка-хозяин, содержащая вектор для экспрессии антитела. Описан способ получения антитела с использованием клетки. Изобретение обеспечивает новые агонистические антитела к 4-1ВВ (также называемому CD137 или TNFRSF9) человека, которые распознают эпитоп, расположенный в области аминокислотных остатков K115, C121, R134, R154, V156 антигена, имеют аффинность Kd, измеренную методом BIACORE, порядка нМ, например 0,4 нМ или 8 нМ (для формата антитела IgG1), что может найти применение в терапии рака и опухолевых заболеваний. 12 н. и 7 з.п. ф-лы, 8 ил., 11 табл., 9 пр.
1. Выделенное антитело или его антигенсвязывающий фрагмент, селективно связывающиеся с человеческим 4-1ВВ, содержащие:
а) гипервариабельный участок 1 тяжелой цепи (H-CDR1), как изложено в SEQ ID NO:29;
б) гипервариабельный участок 2 тяжелой цепи (H-CDR2), как изложено в SEQ ID NO:30;
в) гипервариабельный участок 3 тяжелой цепи (H-CDR3), как изложено в SEQ ID NO:31;
г) гипервариабельный участок 1 легкой цепи (L-CDR1), как изложено в SEQ ID NO:34;
д) гипервариабельный участок 2 легкой цепи (L-CDR2), как изложено в SEQ ID NO:35; и
е) гипервариабельный участок 3 легкой цепи (L-CDR3), как изложено в SEQ ID NO:36.
2. Антитело или антигенсвязывающий фрагмент по п. 1, содержащие аминокислотную последовательность вариабельной области тяжелой цепи (области VH), как изложено в SEQ ID NO:43.
3. Антитело или антигенсвязывающий фрагмент по п. 1, содержащие аминокислотную последовательность вариабельной области легкой цепи (области VL), как изложено в SEQ ID NO:45.
4. Антитело или антигенсвязывающий фрагмент по п. 1, содержащие аминокислотную последовательность области VH, как изложено в SEQ ID NO:43, и аминокислотную последовательность области VL, как изложено в SEQ ID NO:45.
5. Антитело по любому из пп. 1-4, где изотип антитела выбран из IgG1 или IgG2.
6. Выделенное антитело, селективно связывающееся с человеческим 4-1ВВ, имеющее аминокислотную последовательность тяжелой цепи, как изложено в SEQ ID NO:44, и аминокислотную последовательность легкой цепи, как изложено в SEQ ID NO:46, и где C-концевой остаток лизина SEQ ID NO:44 возможно отсутствует.
7. Выделенное антитело по п. 6, представляющее собой IgG2.
8. Фармацевтическая композиция для снижения опухолевого роста у нуждающегося в этом субъекта, содержащая антитело или его антигенсвязывающий фрагмент по любому из пп. 1-7 в терапевтически эффективном количестве и фармацевтически приемлемый носитель.
9. Фармацевтическая композиция для лечения рака у нуждающегося в этом субъекта, содержащая антитело или его антигенсвязывающий фрагмент по любому из пп. 1-7 в терапевтически эффективном количестве и фармацевтически приемлемый носитель.
10. Способ снижения опухолевого роста у нуждающегося в этом субъекта, включающий введение указанному субъекту эффективного количества антитела или его антигенсвязывающего фрагмента по любому из пп. 1-7.
11. Способ лечения рака у млекопитающего, включающий введение млекопитающему, нуждающемуся в лечении, терапевтически эффективного количества антитела или его антигенсвязывающего фрагмента по любому из пп. 1-7.
12. Способ по п. 11, где указанный рак выбран из группы, состоящей из колоректального рака, неходжкинской лимфомы, рака предстательной железы или меланомы.
13. Способ лечения рака у млекопитающего, включающий введение млекопитающему терапевтически эффективного количества антитела или его антигенсвязывающего фрагмента по любому из пп. 1-7 в комбинации с иммунотерапевтическим агентом.
14. Способ по п. 13, где указанный иммунотерапевтический агент представляет собой ритуксимаб.
15. Выделенная молекула нуклеиновой кислоты, кодирующая антитело или его антигенсвязывающий фрагмент по любому из пп. 1-7, которые содержат одну или более аминокислотных последовательностей, выбранных из SEQ ID NO:43, 44, 45 или 46.
16. Выделенная нуклеиновая кислота, кодирующая антитело или его антигенсвязывающий фрагмент по любому из пп. 1-7, содержащая одну или более нуклеотидных последовательностей, выбранных из SEQ ID NO:47, 48, 49 или 50.
17. Вектор экспрессии, содержащий выделенную нуклеиновую кислоту по п. 16.
18. Клетка-хозяин для экспрессии антитела, содержащая вектор по п. 17.
19. Способ получения антитела, которое содержит одну или более аминокислотных последовательностей, выбранных из SEQ ID NO:43, 44, 45 или 46, включающий экспрессирование антитела в клетке-хозяине по п. 18.
| ЧЕЛОВЕЧЕСКИЕ АНТИТЕЛА ПРОТИВ ЧЕЛОВЕЧЕСКОГО 4-1ВВ (CD137) | 2004 |
|
RU2376316C2 |
| WO 03049755 A1, 19.06.2003 | |||
| MURILLO et al | |||
| Способ приготовления строительного изолирующего материала | 1923 |
|
SU137A1 |

