Изобретение относится к области молекулярной биотехнологии и генной инженерии и касается нового штамма-продуцента рекомбинантного циклофилина А человека (рчЦфА).
Применение противоопухолевой химиотерапии ведет к подавлению гемопоэза и создает риск возникновения инфекционных осложнений, что препятствует проведению своевременного лечения. В связи с этим существует необходимость использования средств, поддерживающих кроветворную функцию костного мозга.
Циклофилин А (ЦфА) - белок с молекулярной массой 18 кД, находится во всех тканях млекопитающих, участвует в процессах внутриклеточного транспорта белков, регуляции клеточной пролиферации и проведении сигнала от рецептора в Т-лимфоцитах [Fischer G, Bang Н, Mech С. Determination of enzymatic catalysis for the cis-trans-isomerization of peptide binding in proline-containing peptides. Biomed. Biochim. Acta. 1984; 43(10): 1101-1111; Colgan J, Asmal M, Yu B, Luban J. Cyclophilin A-deficient mice are resistant to immunosuppression by cyclosporine. Journal of immunology. 2005; 174(10): 6030-6038]. Секреторный ЦфА формирует очаг воспаления, привлекая туда зрелые макрофаги, нейтрофилы и активированные Т-лимфоциты [Sherry В, Yarlett N, Strupp A, Cerami А. Identification of cyclophilin as a proinflammatory secretory product of lipopolysaccharide-activated macrophages. Proceedings of the National Academy of Sciences of the United States of America. 1992; 89(8): 3511-3515].
ЦфА усиливает миграцию стволовых клеток и предшественников из костного мозга на периферию, что позволяет рассматривать данный белок как фактор регенерации [Khromykh LM, Kulikova NL, Anfalova TV, et al. Cyclophilin A produced by thymocytes regulates the migration of murine bone marrow cells. Cell Immunol. 2007; 249 (1): 46-53].
ЦфА является фактором дифференцировки стволовых клеток миелоидного ряда и способствует созреванию дендритных клеток, что позволяет рассматривать его в качестве гемопоэтического фактора, способного участвовать в восстановлении иммунной системы.
Для всестороннего изучения биологических свойств ЦфА, особенно в системах in vivo, требуется значительное количество этого белка.
Известен штамм Escherichia coli (E.coli), продуцирующий рчЦфА в виде белка с полигистидиновой последовательностью [В. Sherry, G. Zybarth, М. Alfano, L. Dubrovsky, R. Mitchell, D. Rich, P. Ulrich, R. Bucala, A. Cerami, M. Bukrinsky. Role of cyclophilin A in the uptake of HTV-1 by macrophages and T lymphocytes. Proc. Natl. Acad. Sci. 1998, 95, pp.1758-1763].
Недостатки: рчЦфА продуцируется в виде белка, отличного от нативного, содержащего в своем составе полигистидиновую последовательность; невысокое содержание полученного конечного продукта (белка); необходимость дополнительных стадий очистки.
Известен штамм E.coli, продуцирующий рчЦфА без дополнительных полипептидных последовательностей [Liu J, Albers MW, Chen CM, Schreiber SL, Walsh CT. Cloning, expression, andpurification of human cyclophilin in Escherichia coli and assessment of the catalytic role of cysteines by site-directed mutagenesis. ProcNatlAcadSciUSA, 1990 Mar; 87(6): 2304-2308].
Недостатки: невысокое содержание конечного продукта и сложная схема очистки.
Задачей заявляемого изобретения является создание штамма E.coli - продуцента рчЦфА с высоким содержанием белка, структурно максимально приближенного к нативному и не требующего сложной системы очистки.
Поставленная задача решается путем трансформации компетентных клеток E.coli BL21(DE3)Gold плазмидой pETmin-CypA
Технический результат изобретения: получен штамм E.coli BL21(DE3)Gold/pETmin-CypA - продуцент рекомбинантного циклофилина А, структурно максимально приближенный к нативному и не требующий сложной системы очистки.
Для получения плазмиды pETmin-CypA проводили полимеразную цепную реакцию (ПЦР) с олигонуклеотидными праймерами (5′-TTATACATATGGTCAACCCGACCGTGTTCTTC и 5′-TTTCTCGAGTTATTCGAGTTGTCCACAGTCAGC), используя в качестве матрицы плазмиду pCyPAwt/pGEX-2TK. Полученный в результате реакции фрагмент длиной 515 п.н. и вектор рЕТ-22b(+) расщепляли эндонуклеазами рестрикции BamHI и NdeI. После этого ферменты инактивировали при t=65°C в течение 20 мин и проводили реакцию лигирования. Штамм E.coli ТОР10 трансформировали лигазной смесью (вода, трансформируемый вектор, буфер для лигазы, лигаза, полиэтиленгликоль). Отбирали устойчивые к ампициллину клоны, выделяли плазмиды и проводили их рестрикционный анализ.
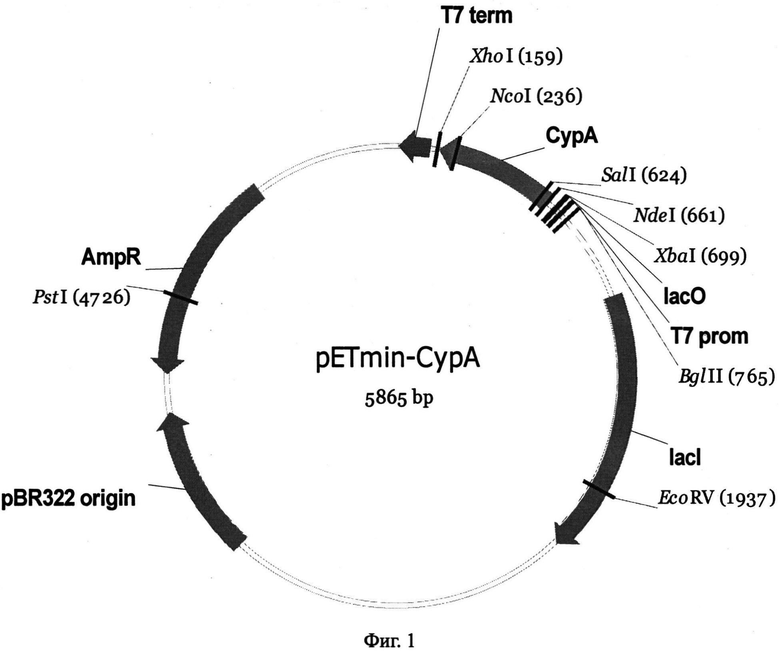
Физическая карта полученной плазмиды представлена на фиг.1.
Структуру клонированного гена в отобранных клонах подтверждали определением нуклеотидной последовательности с использованием набора Big Dye Terminator Cycle Sequencing Kit (v. 3.1). В результате получали экспрессионную плазмиду pETmin-CypA (фиг.1). Плазмида pETmin-CypA содержит уникальные сайты узнавания рестрикционными эндонуклеазами, имеющими следующие координаты: PstI - 4726, EcoRV - 1937, BgIII - 765.
Плазмида pETmin-CypA имеет размер 5865 пар нуклеотидов (п.н.) и содержит: фрагмент XhoI-NdeI вектора рЕТ-22b(+), несущего ген устойчивости к ампициллину ampR, промотор и терминатор РНК-полимеразы фага Т7 и полилинкер, в котором по сайтам XhoI-NdeI клонирован фрагмент, полученный с помощью ПЦР с использованием олигонуклеотидных праймеров 5′-TTATACATATGGTCAACCCGACCGTGTTCTTC и 5′-TTTCTCGAGTTATTCGAGTTGTCCACAGTCAGC плазмида pCyPAwt/pGEX-2TK с клонированным фрагментом гена рчЦфА размером 498 п.н. (М. Bukrinsky Albert Einstein College of Medicine of Yeshiva University, США):

Способ получения штамма E.coli BL21(DE3)Gold/pETmin-CypA.
Компетентные клетки E.coli BL21(DE3)Gold трансформировали плазмидой pETmin-CypA и после выращивания рекомбинантных клонов на агаризированной среде LB, содержащей 150 мг/л ампициллина при t=37°C, получали штамм-продуцент полипептида рчЦфА.
Выращивание заявляемого штамма в ферментере.
Единичную колонию заявляемого штамма инокулировали в 5 мл бульона ТВ, содержащего 150 мг/л ампициллина, и выращивали в течение 18 ч при перемешивании со скоростью 180 об/мин и t=37°C. Инокулят для ферментации готовили, пересевая адаптированную культуру в 500 мл вновь приготовленной среды, культивировали в условиях аэрации в течение 18 ч и вносили в ферментер.
Ферментацию культуры проводили после добавления в ферментер инокулята в соотношении 1:10 к объему среды при pH 7,0-7,4; t=37°C и аэрации 150 об/мин. Культуру выращивали в течении 2 ч, затем вносили индуктор изопропил-β-D-1-тиогалактопиранозид (ИПТГ) до конечной концентрации 0,5 мМ, культивировали в течение 4 ч. Клетки собирали центрифугированием 4000 об/мин в течение 10 мин и хранили при t=-20°C. Содержание рчЦфА в биомассе рекомбинантного штамма-продуцента составляло до 20% от суммарного содержания белка штамма-продуцента или до достижения 100 мг/литр бактериальной культуры.
Полученный штамм E.coli BL21(DE3)Gold/ pETmin-CypA характеризуется следующими признаками.
Морфологические признаки: клетки палочковидной формы, грамотрицательные, неспороносные.
Культуральные признаки: клетки хорошо растут на стандартных питательных средах. Время генерации составляет до 30 мин в жидкой LB-среде. При росте на агарезированной среде LB колонии круглые, гладкие, полупрозрачные, блестящие, серые, край ровный; диаметр колоний составляет 2-3 мм; консистенция пастообразная. Рост в жидкой среде LB характеризуется равномерным помутнением. Культуральную жидкость после охлаждения подвергали центрифугированию.
Физиолого-биохимические признаки: клетки росли в диапазоне температур 20-42°C, при значении pH 6,8-7,2.
Устойчивость штамма к антибиотикам: клетки устойчивы к ампициллину, что обусловлено наличием в плазмиде pETmin-CypA гена ampR.
Условия хранения штамма: среда LB с 15% содержанием глицерина, t=-70°C в криовиалах.
Штамм E.coli BL21(DE3)Gold/pETmin-CypA депонирован во Всероссийской Коллекции Промышленных Микроорганизмов ФГУП ГосНИИИ Генетика, регистрационный номер ВКПМ В-11722.
Способ получения рчЦфА осуществляли следующим образом. Культивирование штамма E.coli BL21(DE3)Gold/pETmin-CypA осуществляли при t=37°C в питательной среде на основе бульона ТВ (Terrific Broth) при pH 7,2; культуральную жидкость после охлаждения центрифугировали. Полученную биомассу разрушали ультразвуковым дезинтегратором. Растворимую фракцию подвергали ионообменной хроматографии на колонке с Fractogel TSKDEAE-650(S); несвязавшуюся фракцию пропускали через ионообменную колонку с CM Sepharose Fast Flow. Очистку конечного продукта от эндотоксинов проводили на колонке с Detoxi-Gel Endotoxin Removing Gel.
Разрушение биомассы проводили с помощью ультразвукового дезинтегратора Branson Sonifier 250 в лизирующем буфере 20 мМ TrisCl, pH 8,0 в течение 5 мин. Температуру суспензии поддерживали в пределах от 2 до 8°C при частоте колебаний наконечника, составляющей 22 КГц, амплитуде 14-16 мкм и мощности дезинтеграции 100 Вт. Полученный лизат центрифугировали при 50000 g в течение 15 мин.
Растворимую фракцию клеток наносили на колонку, содержащую Fractogel TSKDEAE-650(S) и уравновешенную буфером 20 мМ TrisCl pH 8,0 со скоростью 3 мл/мин. В данных условиях рчЦфА не связывался с сорбентом. Фракцию, содержащую рчЦфА, доводили до значения pH 6,2, добавляя 10% уксусную кислоту. Разделение фракции проводили на колонке, заполненной катионообменным сорбентом CM Sepharose Fast Flow и уравновешенной 20 мМ натрий-фосфатный буфером, pH 6,2 (буфер С) при скорости протока 5 мл/мин. После нанесения фракции колонку промывали буфером С до выхода оптической плотности протекающего раствора при 280 нм к значению, близкому к нулю. Элюцию проводили линейным градиентом от буфера С до буфера 20 мМ Na2CO3, 250 мМ NaCl, pH>10, объемом 150 мл.
Для удаления липополисахаридов использовали сорбент Detoxi-Gel Endotoxin Removing Gel (Thermo Scientific), который помещали в хроматографическую колонку ХК16/20 и уравновешивали пропусканием 50 мл фосфатно-солевого буфера. Далее через колонку пропускали раствор очищенного рчЦфА в фосфатно-солевом буфере при скорости потока 2 мл/мин. Максимальное суммарное количество белка, наносимого за один раз, составляло 200 мг, а объем раствора - 50 мл. Контроль процесса осуществляли путем проточного измерения оптической плотности раствора при длине волны 280 нм. Полученный продукт полностью удовлетворяет требованиям, предъявляемым к иммунобиологическим рекомбинантным препаратам по содержанию примесных бактериальных белков E.coli и эндотоксинов.
Способ позволяет получить 70-100 мг белка на 1 л бактериальной культуры.
Хроматографическое разделение рчЦфА методом электрофореза представлено на фиг.2.
Условные обозначения:
М - маркеры.
А - растворимая фракция лизата штамма E.coli BL21(DE3)Gold/pETmin-CypA.
В - фракция, полученная после флеш-хроматографии.
С - фракция, не связавшаяся с катионообменной колонкой.
1, 2, 3 - фракции, полученные при градиентной элюции катионообменной колонки.

Изобретение относится к области биотехнологии и касается штамма Escherichia coli BL21(DE3)Gold/pETmin-CypA - продуцента рекомбинантного циклофилина А человека. Охарактеризованный штамм получен путем трансформации клеток штамма BL21(DE3)Gold плазмидой pETmin-CypA. Плазмида имеет размер 5865 пар нуклеотидов и содержит фрагмент XhoI-NdeI вектора рЕТ-22b(+), несущего ген устойчивости к ампициллину ampR, промотор и терминатор РНК-полимеразы фага Т7 и полилинкер. По сайтам XhoI-NdeI клонирован фрагмент, полученный с помощью ПЦР с использованием олигонуклеотидных праймеров 5′-TTATACATATGGTCAACCCGACCGTGTTCTTC и 5′-TTTCTCGAGTTATTCGAGTTGTCCACAGTCAGC плазмиды pCyPAwt/pGEX-2TK с клонированным фрагментом гена рчЦфА размером 498 п.н. Предложенный штамм высокопродуктивен, не требует сложной системы очистки и может быть использован в противоопухолевой терапии. 2 ил.
Штамм Escherichia coli BL21(DE3)Gold/pETmin-CypA - продуцент рекомбинантного циклофилина А человека, полученный путем трансформации клеток штамма BL21(DE3)Gold плазмидой pETmin-CypA, имеющей размер 5865 пар нуклеотидов и содержащей фрагмент XhoI-NdeI вектора рЕТ-22b(+), несущего ген устойчивости к ампициллину ampR, промотор и терминатор РНК-полимеразы фага Т7 и полилинкер, в котором по сайтам XhoI-NdeI клонирован фрагмент, полученный с помощью ПЦР с использованием олигонуклеотидных праймеров 5′-TTATACATATGGTCAACCCGACCGTGTTCTTC и 5′-TTTCTCGAGTTATTCGAGTTGTCCACAGTCAGC плазмиды pCyPAwt/pGEX-2TK с клонированным фрагментом гена рчЦфА размером 498 п.н.:
1 atggtcaacc ccaccgtgtt cttcgacatt gccgtcgacg gcgagccctt
51 gggccgcgtc tcctttgagc tgtttgcaga caaggtccca aagacagcag
101 aaaattttcg tgctctgagc actggagaga aaggatttgg ttataagggt
151 tcctgctttc acagaattat tccagggttt atgtgtcagg gtggtgactt
201 cacacgccat aatggcactg gtggcaagtc catctatggg gagaaatttg
251 aagatgagaa cttcatccta aagcatacgg gtcctggcat cttatcgatg
301 gcaaatgctg gacccaacac aaatggttcc cagtttttca tctgcactgc
351 caagactgag tggttggatg gcaagcatgt ggtgtttggc aaagtgaaag
401 aaggcatgaa tattgtggag gccatggagc gctttgggtc caggaatggc
451 aagaccagca agaagatcac cattgctgac tgtggacaac tcgaataa
| Yu | |||
| A | |||
| Morozov et al., Recombinant Human Cyclophilin A in vitro Inhibits the Formation of Fibrin Clot, Acta naturae, 2012, VOL | |||
| Очаг для массовой варки пищи, выпечки хлеба и кипячения воды | 1921 |
|
SU4A1 |
| SEONMIN LEE et al., A new method for purification of functional recombinant GST-cyclophilin A protein from E.coli, Indian Journal of Biochemistry & Biophysics, December 2008, Vol.45, pp.374-378 | |||
| ЦИКЛОФИЛИН А - СРЕДСТВО, ОБЛАДАЮЩЕЕ ГЕПАРИНОПОДОБНЫМ ДЕЙСТВИЕМ | 2010 |
|
RU2422152C1 |

