Настоящее изобретение относится к способу выделения мочевины из проб крови.
Мочевина является органическим соединением, которое в организме человека представляет конечный продукт обмена азотных соединений. У человека мочевина выводится с мочой.
Образование мочевины протекает преимущественно в клетках печени и частично в почках. С образованием мочевины в организме связаны различные заболевания, частично наследственные, которые могут нанести значительный вред здоровью. Определение образования мочевины является индикатором функции печени, например, при пересадках печени или пересадке гепатоцитов.
Tuchman и другие в Pediatric Research 2008 (64), p. 213 описывают, например, дефект N-ацетилглутаматсинтазы и анализ образования мочевины.
Для измерения образования мочевины пациенты получали перорально меченный 13C ацетат натрия, который ведет в организме к образованию меченной 13C мочевины; из меченного 13C ацетата образуется 13CO2, который переходит в 13C-карбамоилфосфат и затем в 13C-мочевину.
Большинство химических элементов существуют в природе как смеси нескольких стабильных или радиоактивных изотопов. Распространенность изотопов обычно указывают также при радиоизотопных исследованиях с обогащенными соединения в единице "атомные проценты" (ат.%) или ppm. (части на млн). Для описания изменений в диапазоне естественной распространенности имеется относительная дельта-шкала в промилле (‰). Показатели δ (например, δ13C, δ15Ν, δ18O) определяются как разница соответствующих отношений изотопов R ([тяжелый изотоп]/[легкий изотоп], например, R13C=[13C]/[12C]) в пробе по сравнению со стандартом, отнесенная к этому стандарту.
Например, показатель δ13C рассчитывается согласно:
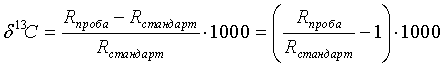
Стандартом для углерода является известь, PDB (Pee Dee Belemnite). Углерод, связанный в результате введения CO2 в фотосинтез, обычно обогащен 13C. Большинство растений восстанавливают CO2 в углеводы по циклу Кельвина-Бенсона или циклу C3. Это приводит к тому, что биомасса C3-растений (к ним относятся технические культуры: рис, картофель, соя, сахарная свекла и зерновые) имеет значения δ13C в диапазоне от -24 до -32‰. Другие растения связывают CO2 по циклу Хэтча-Слэка, или C4. Значения δ13C продуктов из C4-растений (кукуруза, просо, сахарный тростник) имеют δ13C в диапазоне от -10 до -16‰. Поэтому показатель δ13C может применяться для проверки происхождения и подлинности органических веществ.
Определение величины δ13C в мочевине из плазмы проводится обычно путем преобразования мочевины в CO2 с помощью фермента. Поэтому важно, чтобы выделенный из плазмы раствор мочевины не содержал постороннего CO2. Однако, как правило, в плазме всегда имеется CO2, либо как растворенный свободный CO2, либо как CO2, связанный в форме бикарбоната. Полностью освободить плазму от CO2 непросто, кроме того, требуется предотвращать внесение CO2 в пробу при выделении.
Для выделения мочевины из плазмы крови Tuchman и другие применяют способ со следующими этапами:
пробу плазмы объемом 0,5 мл соединяли с 0,5 мл H2O и 40 мкл 60%-ной перхлорной кислоты и удаляли осаждающийся белок. Затем сосуд перемешивали 30 минут, чтобы позволить выделиться CO2. После перемещения в новый сосуд и установления pH в диапазоне 6-7 с помощью 300 мкл KOH 1 М удаляли выпавший в осадок перхлорат калия. Остальной бикарбонат удаляли с помощью ионообменной колонки.
Колонку промывали 1 мл HCl 10 мМ и элюат сушили в стеклянной емкости при 80°C. Пробу перемешивали в течение ночи в закрытом сосуде, в котором был также заключен кусок марли, пропитанный гидроксидом натрия, чтобы уловить остатки CO2.
Затем сосуд продували гелием и воздухонепроницаемо закрывали резиновой пробкой. Через резиновую пробку впрыскивали 400 мкл калийфосфатного буфера (0,5 M pH 6,0) с 3 мг фермента уреазы (400 мкл). Через час добавляли 100 мкл 20%-ной фосфорной кислоты, чтобы высвободить CO2 и остановить реакцию уреазы. Выделившийся 13CO2 измеряли с помощью изотопного масс-спектрометра (IRMS).
Исследования авторов настоящей заявки дали, что вышеописанный способ является очень чувствительным. Некоторые источники постороннего CO2 могут при этом способе искажать измеренные значения δ для CO2, происходящего из мочевины. Кроме того, этот способ является очень трудоемким и длительным. Из-за низкого выхода по мочевине требуется больший объем (0,5 мл) плазмы. В случае детей это может вызывать проблемы. Также, при перекрестной проверке на достоверность в США и Европе обнаружилась необъяснимая разница в результатах.
То, что в способе выделения мочевины из плазмы, описанном Tuchman и другие, не удается полностью удалить мешающий CO2, можно доказать путем сравнения значений δ13C для CO2, полученного из мочевины. Наблюдаемые Tuchmann et al. значения δ13C являются очень низкими, от -25 до -26‰.
Естественная мочевина в плазме имеет значение δ13C от -19 до -23‰, в зависимости от питания.
Задачей изобретения было разработать способ, который устраняет по меньшей мере некоторые из недостатков известного способа.
Эта задача решена способом выделения мочевины и удаления CO2 из пробы плазмы, содержащим следующие этапы:
a) получение пробы плазмы, содержащей мочевину,
b) добавление кислоты, чтобы частично удалить CO2,
c) лиофилизация пробы, чтобы дополнительно удалить CO2 и получить сухую пробу,
d) повторное растворение высушенной пробы и нейтрализация буферным раствором до значения pH от 4 до 7, предпочтительно от 4 до 6,9, в частности от 4 до 6.
Исходным пунктом для выделения мочевины из пробы плазмы является проба плазмы, которая содержит мочевину. Пробы плазмы могут быть получены известными способами из проб крови. Благодаря высокой воспроизводимости способа согласно изобретению достаточно иметь пробы плазмы объемом в диапазоне от 0,2 до 0,3 мл, но могут также использоваться и более значительные количества.
Проба плазмы содержит по меньшей мере мочевину с естественным изотопным составом. Дополнительно к ней может быть добавлена мочевина, обогащенная 13C. Но она может быть также обогащена изотопом 13C путем введения меченных 13C предшественников, например ацетата или бикарбоната. Для этого могут вводиться совместимые соли, например соли Na или K.
Согласно изобретению мочевину определяют путем реакции мочевины с уреазой для выделения CO2, поэтому сначала нужно удалить имеющийся CO2.
В одном варианте осуществления изобретения сначала проводят фильтрацию. Для этого для осаждения растворителя добавляют, например, ацетонитрил и/или муравьиную кислоту. В результате этой фильтрации можно удалить из плазмы белки и липиды. Особенно подходят для этого так называемые колонки HybridSPE™, выпускаемые фирмой SUPELCO. Они годятся также, в частности, для удаления фосфолипидов.
Однако оказалось, что от этапа отделения белков и липидов из плазмы также можно отказаться и, тем не менее, получить очень хорошо воспроизводимые значения. Отказ от этапа фильтрации, во-первых, экономит время, а во-вторых, также издержки на приобретение соответствующего фильтра.
Согласно изобретению добавляют кислоту. В результате добавления кислоты часть CO2 из пробы плазмы удаляется. Подходит, например, фосфорная кислота в концентрации примерно 20%. В расчете на пробу плазмы объемом 0,3 мл достаточно кислоты (например, фосфорной кислоты) в количестве около 50 мкл. Разумеется, могут также использоваться и другие кислоты.
Существенным этапом способа по изобретению является последующая лиофилизация пробы. Лиофилизация есть способ, при котором пробу замораживают и в вакууме сублимируют содержащуюся воду. Согласно изобретению этот способ особенно хорошо подходит для удаления остаточных количеств CO2 из пробы.
Затем полученную пробу снова растворяют и устанавливают ее pH в диапазоне значений от 4 до 7, предпочтительно от 4 до 6,9, в частности от 4 до 6, предпочтительно от 5 до 6. Для установления подходят, в частности, буферные растворы, например фосфатные буферные растворы с pH 9,0. Могут применяться и другие буферные растворы. Раствор едкого калия, используемый Tuchman et al., особенно неблагоприятен, так как он иногда содержит повышенные количества CO2.
В одном варианте осуществления изобретения после повторного растворения высушенной пробы ее дегазируют, чтобы удалить остатки постороннего CO2, в частности, из-за добавления буфера. Оказалось подходящим приложение вакуума в диапазоне от 1 до 10 мбар на 2-3 часа.
Результатом этого процесса является проба, которая еще содержит мочевину из плазмы, но по существу не содержит CO2 из плазмы.
Теперь для определения изотопного состава мочевины можно прибегнуть к известному способу реакции с уреазой:
вновь растворенную пробу продувают защитным газом, например гелием, азотом или аргоном, и герметично закрывают. Затем проводят добавление уреазы, чтобы образовать из имеющейся мочевины CO2. Подходит, например, уреаза, какую можно приобрести у фирмы Sigma. Особенно подходящим показало себя количество от 20 до 100 единиц уреазы на пробу плазмы объемом 0,3 мл.
Затем проводят инкубацию раствора. Оправдала себя инкубация при температуре примерно 36°C в течение периода примерно 60 минут.
Затем добавляют кислоту, чтобы остановить дальнейшую реакцию уреазы. Кроме того, в результате добавления кислоты высвобождается образованный CO2. Для объема пробы плазмы 0,3 мл подходящим оказалось использование 100 мкл 20%-ой фосфорной кислоты. Теперь в газонепроницаемом сосуде выделился CO2, который образован при конверсии мочевины в плазме. Теперь можно определить изотопный состав CO2. Для этого определения подходит, в частности, масс-спектрометрия изотопных соотношений (IRMS). При IRMS измеряется соотношение между 13C и 12C относительно стандарта, как описано выше.
Для контроля процесса может иметь смысл проверить перед добавлением уреазы, что раствор не содержит CO2. Это можно осуществить, например, посредством спектроскопии ИК/МС.
В одном применении изобретения для измерения кинетики образования мочевины пациенту вводятся меченные 13C вещества, чтобы определить образование меченной 13C мочевины. В одном предпочтительном варианте осуществления изобретения кинетику определяют тем, что сначала, прежде чем пациенту введут меченный 13C предшественник мочевины (базовое значение), отбирают пробу крови, затем, после того как пациенту ввели меченный 13C предшественник мочевины, проводят отбор одной или, предпочтительно, нескольких проб крови. Так можно проверить образование мочевины, меченной 13C.
Согласно изобретению можно использовать количества плазмы примерно 100-200 или 200-300 мл, т.е. достаточно половинного количества пробы крови по сравнению со способом Tuchman. В частности, когда кинетика определена, и способ применяется на детях, выгодно, если количество взятой крови будет особенно малым. По сравнению со способом Tuchman и другие, способ по изобретению имеет лучшую воспроизводимость и менее трудоемкий.
Настоящее изобретение относится к следующим аспектам в особенно предпочтительных вариантах осуществления:
аспект 1: способ выделения мочевины и удаления CO2 из пробы плазмы, содержащий следующие этапы: a) получение пробы плазмы, b) добавление кислоты, чтобы частично удалить CO2, c) лиофилизация пробы, чтобы дополнительно удалить CO2 и получить сухую пробу, d) повторное растворение высушенной пробы и нейтрализация буферным раствором до значения pH 4-7, предпочтительно 4-6,9, в частности 4-6.
Аспект 2: способ согласно аспекту 1, отличающийся тем, что перед добавлением кислоты на этапе b) проводят фильтрацию.
Аспект 3: способ согласно аспекту 1 или 2, отличающийся тем, что после этапа d) из пробы удаляют газ при пониженном давлении.
Аспект 4: способ по меньшей мере по одному из аспектов 1-3, отличающийся тем, что pH пробы на этапе d) устанавливают на значение от 5 до 6.
Аспект 5: способ по меньшей мере по одному из аспектов 1-4, отличающийся тем, что проба плазмы берется у участника эксперимента или пациента, которому перед отбором вводился меченный 13C предшественник мочевины.
Аспект 6: способ определения доли изотопа 13C в мочевине в пробе плазмы, содержащий этапы: выделение мочевины способом по одному из аспектов 1-5, продувка защитным газом, добавление уреазы, чтобы образовать CO2, инкубация, добавление кислоты, чтобы высвободить образовавшийся CO2, и измерение доли изотопа 13C в выделившемся CO2.
Аспект 7: способ диагностики мочевинного обмена, содержащий этапы: получение первой пробы плазмы от пациента, определение доли изотопа 13C в мочевине из первой пробы плазмы, согласно аспекту 6, получение по меньшей мере одной следующей пробы плазмы от пациента, причем пациенту перед отбором вводился меченый 13C предшественник мочевины, определение доли изотопа 13C в мочевине по меньшей мере одной следующей пробы плазмы, согласно аспекту 6, и определение количества образованной мочевины по доле изотопа 13C в мочевине в первой и по меньшей мере одной следующей пробе плазмы.
Аспект 8: способ согласно аспекту 7, отличающийся тем, что применяются по меньшей мере две следующие пробы плазмы.
Аспект 9: способ согласно аспекту 8, отличающийся тем, что следующие пробы плазмы отбирались с промежутком от 15 до 240 мин после того, как пациенту был введен меченный 13C предшественник мочевины.
Аспект 10: способ по одному из аспектов 8 или 9, отличающийся тем, что следующие пробы плазмы отбирались через 15 мин после того как пациенту был введен меченный 13C предшественник мочевины.
Настоящее изобретение относится, в частности, также к следующим аспектам A), B), C), D), E), F), G), H), I):
аспект A): способ определения мочевины в пробе плазмы, содержащий следующие этапы: a) получение пробы плазмы, содержащей мочевину, b) добавление кислоты, c) лиофилизация пробы, чтобы удалить CO2 и получить сухую пробу, d) повторное растворение высушенной пробы и нейтрализация до значения pH 5-7 буферным раствором, e) продувка защитным газом, f) добавление уреазы, чтобы образовать CO2, g) инкубация, h) добавление кислоты, чтобы высвободить образованный CO2, и i) определение изотопного состава выделившегося CO2.
Аспект B): способ согласно аспекту A), отличающийся тем, что перед лиофилизацией проводится фильтрация.
Аспект C): способ согласно аспекту A) или B), отличающийся тем, что после этапа d) пробу нагревают до температуры от 40 до 70°C.
Аспект D): способ по меньшей мере по одному из аспектов A)-C), отличающийся тем, что для определения изотопного состава CO2 применяется IRMS.
Аспект E): способ по меньшей мере по одному из аспектов A)-D), отличающийся тем, что перед отбором пробы плазмы был введен ацетат, меченный 13C.
Аспект F): способ по меньшей мере по одному из аспектов A)-E), отличающийся тем, что pH пробы на этапе d) устанавливают на значение от 5 до 6.
Аспект G): способ диагностики мочевинного обмена, содержащий этапы: отбор пробы крови у пациента, получение пробы плазмы из пробы крови, определение мочевины в пробе плазмы способом согласно одному из аспектов A)-F), введение меченного 13C предшественника мочевины, отбор по меньшей мере двух проб крови с временным промежутком и определение мочевины после получения пробы плазмы из пробы крови, способом согласно одному из аспектов A)-F).
Аспект H): способ согласно аспекту G), отличающийся тем, что после введения предшественника мочевины отбираются по меньшей мере две пробы крови с промежутком по меньшей мере 120 минут, предпочтительно по меньшей мере 240 минут.
Аспект I): Способ согласно аспекту G) или H), отличающийся тем, что между двумя отборами проб крови имеется промежуток от 10 до 20 минут.
Описание фигур
Фиг. 1 показывает воспроизводимость способа согласно примеру 3.
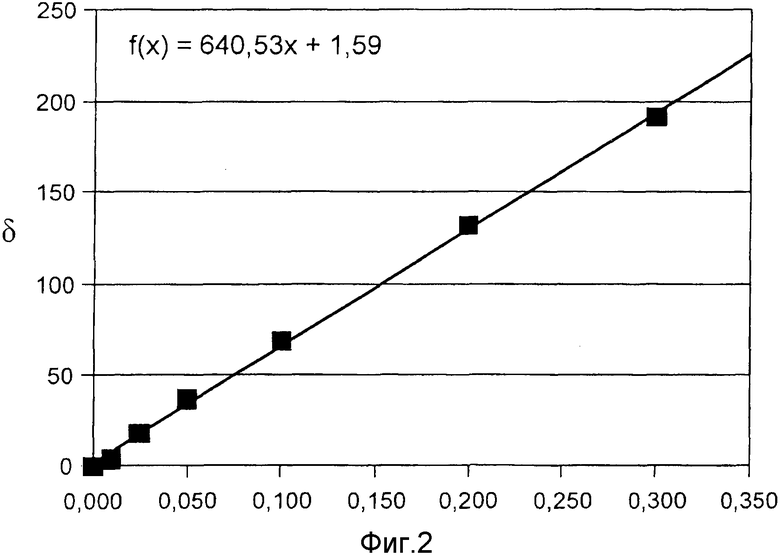
Фиг. 2 показывает калибровочную кривую в эксперименте с добавлением в образец активной субстанции для анализа, согласно примеру 4.
Фиг. 3 показывает воспроизводимость способа по примеру 5.
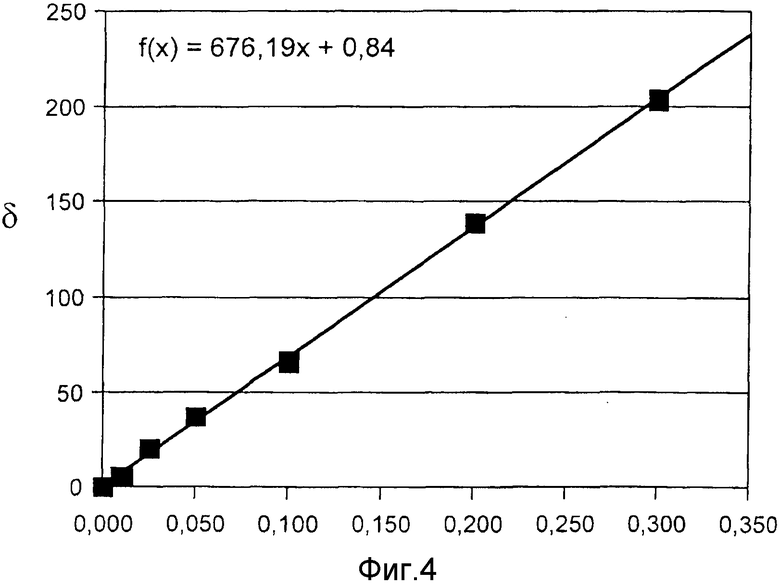
Фиг. 4 показывает калибровочную кривую в эксперименте с добавлением в образец активной субстанции для анализа, согласно примеру 6.
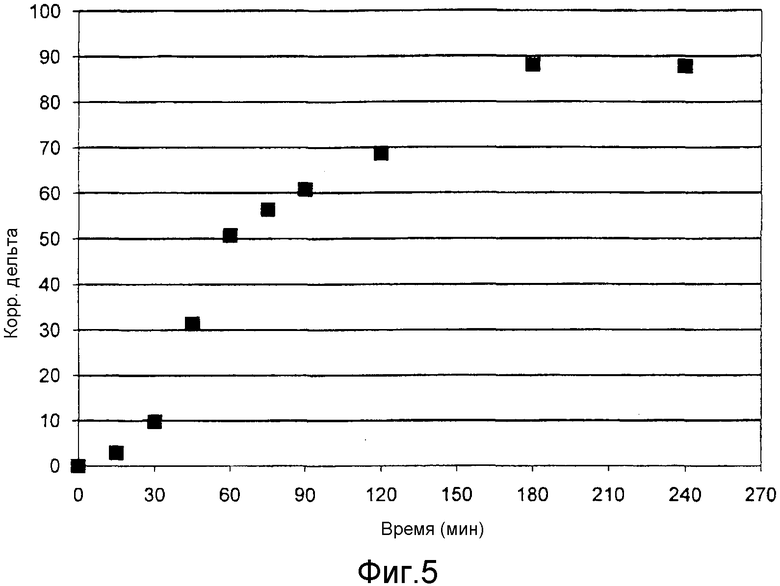
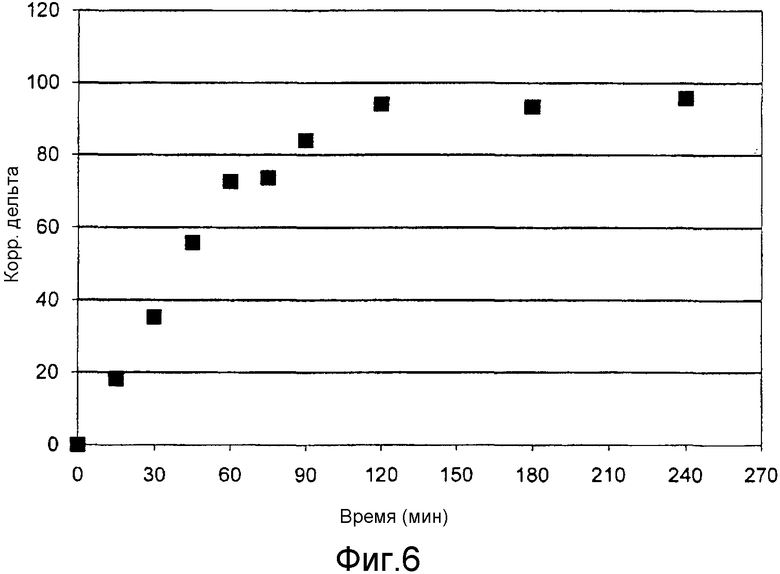
Фиг. 5 и 6 показывают результаты измерения образования мочевины согласно примеру 7.
Способ подробнее поясняется на следующих примерах.
Пример 1: Выделение мочевины с фильтрацией
Из пробы крови получали плазму. 300 мкл плазмы разбавляли 200 мкл деионизированной воды. К пробе добавляли 100 мкл ацетонитрила с 1% муравьиной кислоты. В сосуде образовывался осадок. Пробы фильтровали через колонку HybridSPE.
Фильтрат смешивали с 50 мкл фосфорной кислоты 1 М, замораживали и лиофилизовали. Дегазованным фосфатным буферным раствором (0,5 М, pH 9) устанавливали pH пробы на 5,5. Резервуар с пробой (Vacutainer) продували газообразным гелием. Затем добавляли 70 мкл раствора, содержащего 15 мг/мл уреазы канавалии мечевидной, тип III от фирмы Sigma, в фосфатном буфере. Пробу инкубировали один час при 36°C. Затем через мембрану впрыскивали 60 мкл 20%-ной фосфорной кислоты, чтобы остановить реакцию уреазы и высвободить CO2. На выделенном CO2 проводили определение изотопного состава методом IRMS.
Пример 2: Выделение мочевины без фильтрации
Способ осуществляли как в примере 1, но без добавления ацетонитрила и муравьиной кислоты, а также без фильтрации, т.е. пробу плазмы лиофилизовали прямо после добавления кислоты. Далее способ проводили идентично.
Пример 3: Воспроизводимость способа
Пробу плазмы разделяли на пять проб, которые по отдельности друг от друга подвергали способу согласно примеру 1. Измерение на каждой содержащей CO2 пробе проводили пять раз. Разница является минимальной, ср. фиг. 1.
Пример 4: Эксперимент с добавлением в образец активной субстанции для анализа
В пробы плазмы из примера 1 добавляли 99%-ую мочевину, меченную 13C, в количествах 0,01 мг, 0,25 мг, 0,5 мг, 0,1 мг, 0,2 мг, 0,3 мг, и пробу обрабатывали согласно способу 1. Фиг. 2 показывает соответствующие измеренные значения. Калибровочная кривая представляет собой прямую с коэффициентом корреляции 0,99923.
Пример 5: Воспроизводимость
Измерение воспроизводимости согласно примеру 3 повторяли для способа без фильтра согласно примеру 2. Результаты показаны на фиг. 3. Здесь также имеется отличная воспроизводимость.
Пример 6: Эксперимент с добавлением в образец активной субстанции для анализа
Повторяли эксперимент согласно примеру 4, причем использовали способ согласно примеру 2. Соответствующую калибровочную прямую можно видеть на фиг. 4. Ее коэффициент корреляции равен R=0,99959.
Пример 7: Измерение образования мочевины
У двух участников эксперимента отбирали кровь и получали плазму, из каждых 300 мкл плазмы способом согласно изобретению выделяли мочевину и определяли отношение изотопов 13C/12C в мочевине.
При этом измерение на каждой пробе проводили 5 раз и по среднему значению определяли базовое значение. Затем участникам эксперимента вводилось 27 мг/кг 99%-ного Na-ацетата, меченного 13C.
Процедура отбора крови и определение отношения изотопов 13C/12C в выделенной мочевине проводили через 15, 30, 45, 60, 75, 90, 120, 180 и 240 минут. Меньшая доля меченного 13C ацетата превращалась в организме в мочевину и благодаря чувствительности измерения могла быть обнаружена. Кинетику вновь образованной мочевины определяли измерением значений дельта (повышение отношения изотопов 13C/12C). Кинетика образования мочевины представлена на фиг. 5 и 6.
| название | год | авторы | номер документа |
|---|---|---|---|
| Способ идентификации меда на основе изотопной масс-спектрометрии | 2022 |
|
RU2809285C1 |
| СПОСОБ НЕИНВАЗИВНОЙ ДИАГНОСТИКИ ХЕЛИКОБАКТЕРНОЙ ИНФЕКЦИИ | 2001 |
|
RU2229714C2 |
| СПОСОБ НЕИНВАЗИВНОЙ ДИАГНОСТИКИ ХЕЛИКОБАКТЕРИОЗА ИН ВИВО | 1996 |
|
RU2100010C1 |
| Способ синтеза линолевой и линоленовой кислот, меченных изотопами углерода С и С | 2016 |
|
RU2630691C1 |
| СПОСОБ ВЫЯВЛЕНИЯ РАННЕГО ПРОРЫВА НАГНЕТАЕМОЙ ВОДЫ В НЕФТЯНЫХ СКВАЖИНАХ, ИСПОЛЬЗУЮЩИЙ ПРИРОДНЫЙ ИЗОТОП УГЛЕРОДА-13 | 2007 |
|
RU2456448C2 |
| Способ неинвазивной диагностики инфекции Helicobacter pylori с помощью 13С-уреазного дыхательного теста | 2022 |
|
RU2790397C1 |
| Диагностическая тест-система для определения биомаркера, специфического для нейродегенеративных заболеваний или нейродегенеративных нарушений | 2024 |
|
RU2827060C1 |
| СПОСОБ ОПРЕДЕЛЕНИЯ ЭТАНОЛА НЕВИНОГРАДНОГО ПРОИСХОЖДЕНИЯ В ВИНОГРАДНЫХ ДИСТИЛЛЯТАХ И НАПИТКАХ НА ИХ ОСНОВЕ | 2009 |
|
RU2401428C1 |
| ПИЩЕВАЯ КОМПОЗИЦИЯ ДЛЯ УЛУЧШЕНИЯ НАКОПЛЕНИЯ БЕЛКА В МЫШЦАХ | 2001 |
|
RU2277355C2 |
| СПОСОБ ОПРЕДЕЛЕНИЯ ПРОИСХОЖДЕНИЯ ОРГАНИЧЕСКИХ ОКСИКИСЛОТ В ВИНАХ И СОКОСОДЕРЖАЩИХ НАПИТКАХ | 2012 |
|
RU2487348C1 |


Изобретение относится к медицине и описывает способ удаления CO2 из содержащей мочевину пробы плазмы, содержащий следующие стадии: a) получение пробы плазмы, b) добавление кислоты, чтобы частично удалить CO2, c) лиофилизация пробы, чтобы дополнительно удалить CO2 и получить сухую пробу, d) повторное растворение высушенной пробы и нейтрализация буферным раствором до значения pH 4-7. Данный способ позволяет более точно определить долю изотопа 13С в мочевине в первой пробе плазмы для дальнейшей диагностики мочевинного обмена. 3 н. и 7 з.п. ф-лы, 6 ил., 7 пр.
1. Способ удаления CO2 из содержащей мочевину пробы плазмы, содержащий следующие стадии:
a) получение пробы плазмы,
b) добавление кислоты, чтобы частично удалить CO2,
c) лиофилизация пробы, чтобы дополнительно удалить CO2 и получить сухую пробу,
d) повторное растворение высушенной пробы и нейтрализация буферным раствором до значения pH 4-7.
2. Способ по п. 1, отличающийся тем, что перед добавлением кислоты на стадии b) проводят фильтрацию.
3. Способ по п. 1 или 2, отличающийся тем, что после стадии d) из пробы удаляют газ при пониженном давлении.
4. Способ одному из пп. 1-3, отличающийся тем, что pH пробы на стадии d) устанавливают на значение от 4 до 6, предпочтительно от 5 до 6.
5. Способ по одному из пп. 1-4, отличающийся тем, что пробу плазмы берут у участника эксперимента или пациента, которому перед отбором вводился меченный 13С предшественник мочевины.
6. Способ определения доли изотопа 13С в мочевине в первой пробе плазмы, содержащий стадии:
- удаления CO2 из содержащей мочевину пробы плазмы способом по одному из пп. 1-5,
- продувка защитным газом,
- добавление уреазы, чтобы образовать CO2,
- инкубация,
- добавление кислоты, чтобы высвободить образовавшийся CO2,
- измерение доли изотопа 13С в выделившемся CO2.
7. Способ диагностики мочевинного обмена, содержащий стадии:
- взятие первой пробы плазмы у пациента,
- определение доли изотопа 13С в мочевине из первой пробы плазмы согласно п. 6,
- получение по меньшей мере одной следующей пробы плазмы от пациента, причем пациенту перед отбором вводился меченый 13С предшественник мочевины,
- определение доли изотопа 13С в мочевине из по меньшей мере одной следующей пробы плазмы согласно п. 6,
- определение количества образованной мочевины по доле изотопа 13С в мочевине из первой и по меньшей мере одной следующей пробы плазмы.
8. Способ по п. 7, отличающийся тем, что используются по меньшей мере две следующие пробы плазмы.
9. Способ по п. 8, отличающийся тем, что следующие пробы плазмы отбирались с промежутком от 15 до 240 мин после того, как пациенту был введен меченный 13С предшественник мочевины.
10. Способ по одному из пп. 8 или 9, отличающийся тем, что следующие пробы плазмы отбирались через 15 мин после того, как пациенту был введен меченный 13С предшественник мочевины.
| Tuchman Mendel et al | |||
| N-carbamylglutamate markedly enhances ureagenesis in N-acetylglutamate deficiency and propionic acidemia as measured by isotopic incorporation and blood biomarkers | |||
| Pediatric Research, Vol:64, No:2, Page(s):213 - 217 | |||
| US 5542419 A, 06.08.1996 | |||
| US 2008090268 A1, 17.04.2008 | |||
| Способ количественного определения новокаина | 1986 |
|
SU1415159A1 |
| СПОСОБ ОПРЕДЕЛЕНИЯ МОЧЕВИНЫ В БИОЛОГИЧЕСКИХ ЖИДКОСТЯХ И НАБОР РЕАКТИВОВ ДЛЯ ЕГО ОСУЩЕСТВЛЕНИЯ | 1995 |
|
RU2122740C1 |
| Стабилизированный состав для определения мочевины, содержащий уреазу | 1977 |
|
SU743600A3 |

