Изобретение относится к области микробиологии и биотехнологии и касается оптимизации процессов культивирования штамма-продуцента Е.coli, а точнее способа получения культуры клеток штаммов Е.coli, полученных на основе штамма BL21(DE3). Изобретение направлено как на повышение выхода биомассы с единицы объема культуральной среды, так и на повышение выхода целевого белка путем оптимизации процесса культивирования культуры клеток различных штаммов Е. coli.
Под различными штаммами Е.coli следует понимать штаммы, полученные на основе штамма BL21(DE3), который широко используется для экспрессии нетоксичных для клетки белков в системе Т7 промотора под контролем lacUV5 с использованием векторов рЕТ, pCAL и др.
Способ культивирования основан на использовании созданных на основе штамма BL21(DE3) рекомбинантных штаммов-продуцентов и предусматривает их глубинное культивирование на питательной среде при дробном добавлении питательных субстратов, являющихся ингибитором, нейтральным веществом и индуктором по отношению к lacUV5 промотору, в процессе биосинтеза.
Техническим результатом, достигаемом при использовании заявляемого способа, является обеспечение оптимальных условия культивирования и, как следствие, максимальный рост биомассы и накопление целевых продуктов в клетках бактерий при одновременном снижении энергетических затрат и трудоемкости процесса.
Штамм BL21(DE3) известен способностью образовывать, так называемые, тельца включения, представляющие собой частицы, состоящие из агрегатов рекомбинантного белка и ряда бактериальных белков. Тельца включения образуются при высоком уровне биосинтеза и некоторых специфических особенностях белка. Рекомбинантные белки в тельцах включения находятся в денатурированном неактивном состоянии. Малая растворимость телец включений упрощает и удешевляет очистку рекомбинантного белка.
Предложенный способ культивирования позволяет получать продукт в виде нерастворимых телец включений с высоким выходом как по продукту, так и по выходу биомассы с объема культуральной среды.
Известен способ получения человеческого рекомбинантного интерферона гамма в тельцах включения с использованием рекомбинантного штамма Escherichia BL21 (DE3)[pET3a-hifn-γ], модифицированного плазмидой рЕТЗа-hifh-γ, содержащей структурный ген интерферона-γ. Способ заключается в культивировании клеток в 2-литровом ферментере. Клетки культивируют в 1 л среды, содержащей 10,0 г/л глюкозы, 12,8 г/л Na2HPO4·7H2O, 3 г/л KH2PO4, 0,5 г/л NaCl, 1 г/л NH4Cl, 0,24 г/л MgSO4, 1,0 мл/л раствора металлов (2,8 г FeSO4·7H2O, 2,0 г MnCl2 · 4Н2О, 2,8 г CoSO4·7Н2О, 1,5 г CaCl2·2Н2О, 0,2 г CuCl2·2Н2О и 0,3 г ZnSO4·7H2O на литр 1 М HCl). В процессе культивирования ведется экспоненциальная подпитка растворами глюкозы, сульфата магния и раствором солей металлов. На 18 часу культивирования при плотности клеток 50 г/л культура индуцируется внесением раствора IPTG до конечной концентрации 3 мM, индукция продолжается до 24 часа культивирования. Выход в культивировании составляет 59 г биомассы /л с содержанием интерферона 35% [1].
Недостатком данного способа являются:
- большая продолжительность культивирования, являющаяся следствием низкой скорости роста культуры в бедной питательной среде,
- использование дорогостоящего индуктора IPTG,
- внесение в среду культивирования высоких концентраций металлов, чьи токсические свойства потенциально опасны для человека при подкожном или внутримышечном введении препарата [2],
- низкий выход биомассы с объема культуральной жидкости.
Известен способ получения рекомбинантного металлопротеина Δ-9-Десатуразы в тельцах включения с использованием рекомбинантного штамма Escherichia BL21(DE3), модифицированного плазмидой pRCMD9, содержащей структурный ген десатуразы. Культивирование проводили в ферментере объемом 80 л. Культуру растили в минимальной питательной среде, содержащей 6,0 г/л Na2HPO4, 3 г/л KH2PO4, 0,5 г/л NaCl, 0,5 г/л NH4Cl, 0,5 г/л MgSO4, 0,025 г/л CaCl2·2Н2О и 2 мл/л раствора солей металлов (0,288 г/л ZnSO4·7H2O, 0,142 г/л MnSO4·2H2O, 0,062 г/л H3BO3, 0,048 г/л CoCl2·6H2O, 0,083 г/л KI, 0,125 CuSO4·5H2O и 1 мл 0,25 М H2SO4) и 1 мл/л железосодержащего раствора (1,12 г FeSO4 на 95 мл Н2О и 5 мл 0,25М H2SO4) с подпиткой глюкозой. При оптике культуры 32 о.е. проводили индукцию лактозой. Время индукции составляло 12 ч, конечная оптика бактериальной культуры 40 о.е., выход по биомассе 12 г сухого веса/л, выход по белку 4,5% [3].
Недостатком данного способа являются:
- большая продолжительность культивирования, являющаяся следствием низкой скорости роста культуры в бедной питательной среде,
- использование в качестве основного углеродсодержащего субстрата глюкозы, которая является ингибитором lacUV5 промотора,
- низкий выход биомассы и белка с объема культуральной жидкости.
В качестве наиболее близкого аналога может быть указан способ получения человеческого рекомбинантного интерферона альфа 2b с использованием рекомбинантного штамма Escherichia coli BL21(DE3), модифицированного плазмидой [pET22(b+)-IFN-2T], содержащей структурный ген интерферона альфа 2b. Способ состоит в культивировании штамма BL21(DE3)[pET22(b+)-IFN-2Tφ] в ферментере в объеме 30 л в синтетической среде М9, содержащей 10 г/л триптона, 5 г/л дрожжевого экстракта при температуре 37°С и рН 7. Индукция проводится лактозой на 7-ом часу роста. Конечная оптическая плотность культуры равна 60 о.е. при содержании целевого белка 27% [3].
Недостатком данного способа является:
- невысокий выход биомассы в процессе культивирования, связанный с использованием индуктора лактозы как субстрата, который является малоэффективным для роста клеток, т.к. продуктами катаболизма лактозы являются глюкоза и галактоза. BL21(DE3) не способен утилизировать галактозу, таким образом, половина углерода и энергии, содержащейся в лактозе, недоступна.
Согласно изобретению предлагается способ культивирования штаммов, созданных на основе штамма BL21(DE3), на среде, содержащей 10 г/л триптона, 5 г/л дрожжевого экстракта, 7,05 г/л Na2HPO4, 6,8 г/л KH2PO4, 3,3 г/л (NH4)2SO4, 0,24 г/л MgSO4, 40 мкг/мл канамицина сульфат, при температуре 38°С и рН 7,0. В течение культивирования максимальный уровень аэрации 1 объем воздуха /мин: 1,2 объема культуральной жидкости, перемешивание 1200 об/мин, концентрация растворенного кислорода изменяется в пределах от 80 до 10% от максимально возможной при данных условиях.
Засев ферментера производится инокулятом со стандартными ростовыми свойствами (середина логарифмической фазы роста) в объеме 10% от объема ферментера.
В процессе роста используется три вида углеродсодержащего субстрата, который является основным для клетки источником энергии. На раннем этапе роста в лаг-фазе и до середины логарифмической фазы роста в качестве субстрата используется глюкоза, которая является приоритетным для клетки субстратом и ингибитором lacUV5 промотора. Культура при этом растет активно, глюкоза подавляет индукцию синтеза целевого белка, которая может возникнуть из-за наличия лактозы в компонентах питательной среды, таких как пептон, и сильной активности промотора. Вся энергия в клетке тратится на прирост биомассы, синтез целевого белка блокируется.
В середине логарифмической фазы роста вносится субстрат глицерин - нейтральный по отношению к лактозному оперону. После утилизации глюкозы в среде культура начинает использование глицерина. При этом переход на утилизацию глицерина занимает около 20 минут. Культура на нем растет активно и при этом индукция целевого белка не ингибируется.
К окончанию логарифмической фазы вносится лактоза, которая является одновременно и индуктором, и субстратом. Продуктами катаболизма лактозы являются глюкоза и галактоза. BL21(DE3) не способен утилизировать галактозу, таким образом, половина углерода и энергии, содержащейся в лактозе, недоступна. Процесс прямой перестройки культуры с утилизации глюкозы на утилизацию лактозы занимает 1,5-2 часа, культура при этом тормозится в росте, кроме того, пока ингибитор индукции глюкоза не будет до конца утилизирован, синтез продукта не начнется. При использовании глицерина культура утилизирует его, активно растет, при этом идет синтез белка.
После исчерпания в среде глицерина культура переходит на использование лактозы и до конца процесса культивирования лактоза дробно вносится в среду, чтобы избежать исчерпания индуктора в среде.
Продолжительность культивирования составляет 12 часов.
Существенными отличиями заявляемого способа от ближайшего аналога являются:
- использование эффективной системы контроля уровня растворенного кислорода, что позволяет управлять метаболизмом культуры,
- использование 3-компонентной подпитки углеродсодержащими субстратами, что создает оптимальные условия для максимального роста культуры в первой фазе процесса и максимального синтеза целевого белка во второй,
- индукция синтеза белка на ранней стадии роста, что позволяет увеличить время экспрессии при равной продолжительности культивирования,
- высокий выход и биомассы, и целевого белка.
Способ иллюстрируется примерами.
Пример 1
Штамм-продуцент гибридного белка интерферона альфа-2b человеческого рекомбинантного и фрагмента дрожжевого белка SUMO (используется для дальнейшей модификации - удаления концевого метионина и получения безметионинового интерферона альфа 2b человеческого рекомбинантного).
Предложенная схема была реализована для штамма-продуцента SUMO-IFN-α-2b при культивировании в ферментере объемом 42 л. Объем среды в ферментере 30 л, культивирование ведется при температуре 38°С, концентрации растворенного кислорода 80% (концентрация растворенного кислорода в процессе культивирования изменяется в пределах от 80 до 10%) при максимальной скорости перемешивания 1200 об/мин, максимальном уровне аэрации 25 л/мин, что соответствует отношению ~1 объем воздуха/мин: 1,2 объема культуральной жидкости. рН 7,0.
Инокулят в объеме 3 л был выращен до середины логарифмической фазы роста. Выращивание инокулята осуществляли на питательной среде, где в качестве источника углерода используют глюкозу в концентрации 10 г/л, при температуре 38°С, перемешивании 200 об/мин, времени культивирования 3 ч. Инокулят был внесен в ферментер со стерильной питательной средой, засев ферментера осуществляли инокулятом с посевной дозой 10%.
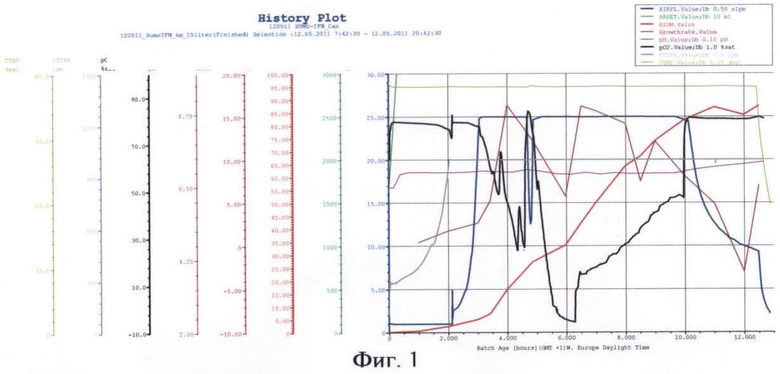
В качестве стартового углеродсодержащего субстрата в среду в начале культивирования добавляют 50%-ный (мас.%) раствор глюкозы. К середине логарифмической фазы роста, когда глюкоза была утилизирована, производится порционная добавка 80%-ного (мас.%) раствора глицерина. После перехода культуры на утилизацию глицерина была начата дробная подача 25%-ного (мас.%) раствора лактозы и началась индукция белка. После исчерпания глицерина в среде культура начала утилизировать лактозу как субстрат. Как видно из почасового электрофореза культуральной жидкости, синтез целевого белка начался на 5 ч культивирования. Окончание ферментации происходит на 12 ч культивирования. Конечная оптическая плотность - 85 о.е., выход биомассы - 73 г/л. Содержание целевого белка по отношению к общему клеточному белку - 35%.
На фиг.1 представлены параметры культивирования штамма-продуцента гибридного белка SUMO-IFN-α-2b по предлагаемой схеме.
На фиг.2 представлен анализ содержания белка в культуральной жидкости штамма-продуцента гибридного белка SUMO-IFN-α-2b методом электрофореза в ПААГ в ходе процесса культивирования по предлагаемой схеме.
Пример 2.
Штамм-продуцент гибридного белка интерферона альфа-2а человеческого рекомбинантного и фрагмента дрожжевого белка SUMO (используется для дальнейшей модификации - удаления концевого метионина и получения безметионинового интерферона альфа-2а человеческого рекомбинантного).
Предложенная схема была реализована для штамма-продуцента SUMO-IFN-α-2a при культивировании в объеме 30 л. Культивирование ведется при температуре 38°С, концентрации растворенного кислорода 80% (концентрация растворенного кислорода в процессе культивирования изменяется в пределах от 80 до 10%) при максимальной скорости перемешивания 1200 об/мин, максимальном уровне аэрации 25 л/мин, что соответствует отношению 1 объем воздуха/мин: 1,2 объема культуральной жидкости. рН 7,0.
Инокулят в объеме 3 л был выращен до середины логарифмической фазы роста. Выращивание инокулята осуществляли на питательной среде, где в качестве источника углерода используют глюкозу в концентрации 10 г/л, при температуре 38°С, перемешивании 200 об/мин, времени культивирования 3 ч. Инокулят был внесен в ферментер со стерильной питательной средой, засев ферментера осуществляли инокулятом с посевной дозой 10%.
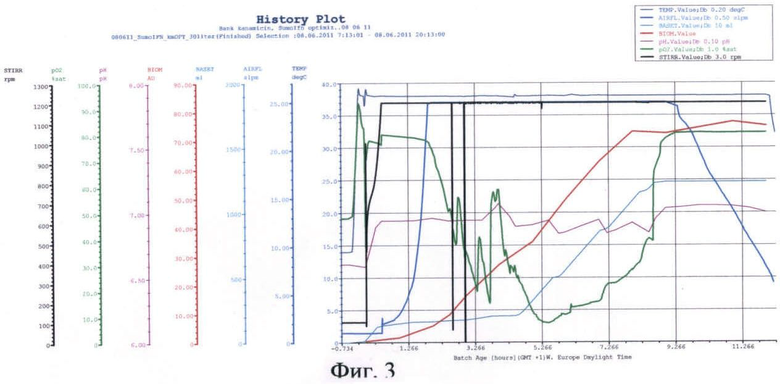
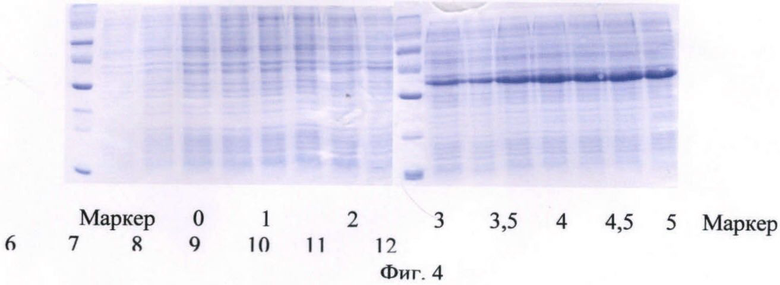
В качестве стартового углеродсодержащего субстрата в среду в начале культивирования добавляют 50%-ный (мас.%) раствор глюкозы. К середине логарифмической фазы роста, когда глюкоза была утилизирована, производится порционная добавка 80%-ного (мас.%) раствора глицерина. После перехода культуры на утилизацию глицерина была начата дробная подача 25%-ного (мас.%) раствора лактозы и началась индукция белка. После исчерпания глицерина в среде культура начала утилизировать лактозу как субстрат. Как видно из почасового электрофореза культуральной жидкости, синтез целевого белка начался на 6 ч культивирования. Окончание ферментации происходит на 12 ч культивирования. Конечная оптическая плотность - 85 о.е., выход биомассы - 72 г/л. Содержание целевого белка по отношению к общему клеточному белку - 35%.
На фиг.3 представлены параметры культивирования штамма-продуцента гибридного белка SUMO-IFN-α-2a по разработанной схеме.
На фиг.4 представлен анализ содержания белка в культуральной жидкости штамма-продуцента гибридного белка SUMO-IFN-α-2a методом электрофореза в ПААГ в ходе процесса культивирования по предлагаемой схеме.
Пример 3.
Штамм-продуцент интерферона-γ человеческого рекомбинантного.
Предложенная схема была реализована для штамма-продуцента hrIFN-γ при культивировании в объеме 30 л. Культивирование ведется при температуре 38°С, концентрации растворенного кислорода 80% (концентрация растворенного кислорода в процессе культивирования изменяется в пределах от 80 до 10%) при максимальной скорости перемешивания 1200 об/мин, максимальном уровне аэрации 25 л/мин, что соответствует отношению - 1 объем воздуха/мин: 1,2 объема культуральной жидкости. рН 7,0. Инокулят в объеме 3 л был выращен до середины логарифмической фазы роста. Выращивание инокулята осуществляли на питательной среде, где в качестве источника углерода используют глюкозу в концентрации 10 г/л, при температуре 38°С, перемешивании 200 об/мин, времени культивирования 3 ч. Инокулят был внесен в ферментер со стерильной питательной средой, засев ферментера осуществляли инокулятом с посевной дозой 10%.
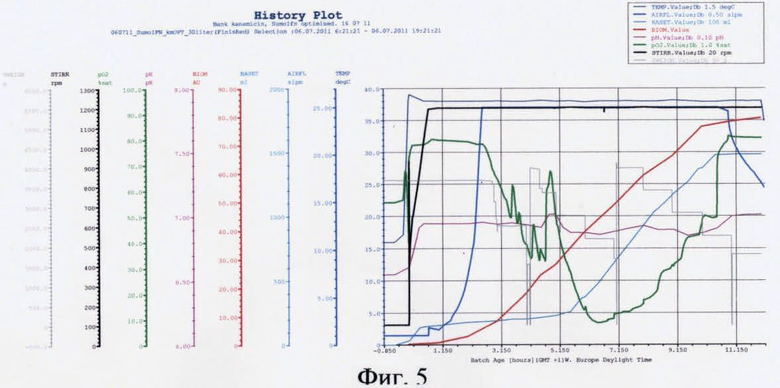
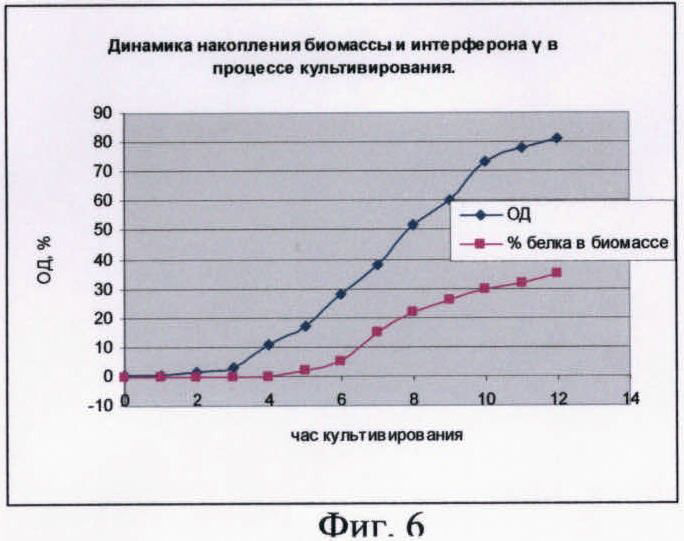
В качестве стартового углеродсодержащего субстрата в среду в начале культивирования добавляют 50%-ный (мас.%) раствор глюкозы. К середине логарифмической фазы роста, когда глюкоза была утилизирована, производится порционная добавка 80%-ного (мас.%) раствора глицерина. После перехода культуры на утилизацию глицерина была начата дробная подача 25%-ного (мас.%) раствора лактозы и началась индукция белка. После исчерпания глицерина в среде культура начала утилизировать лактозу как субстрат. Как видно из почасового электрофореза культуральной жидкости, синтез целевого белка начался на 6 ч культивирования. Окончание ферментации происходит на 13 ч культивирования. Конечная оптическая плотность - 80 о.е., выход биомассы - 68 г/л. Содержание hrIFN-γ по отношению к общему клеточному белку - 31%.
На фиг.5 представлены параметры культивирования штамма-продуцента hrIFN-γ по разработанной схеме.
На фиг.6 показана динамика накопления биомассы штамма-продуцента и hrIFN-γ в ходе процесса культивирования по предагаемой схеме.
Пример 4.
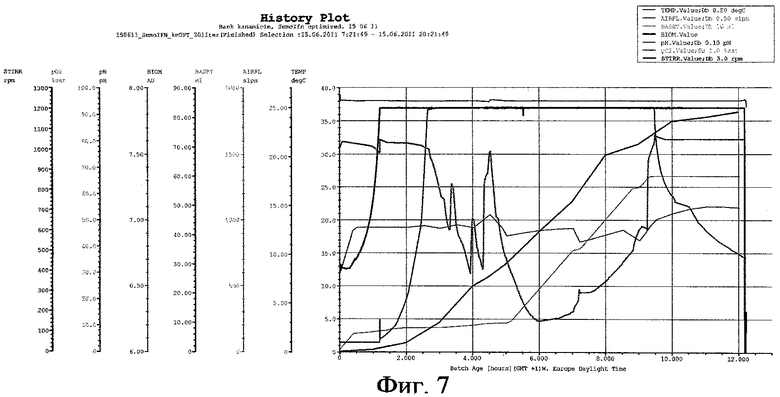
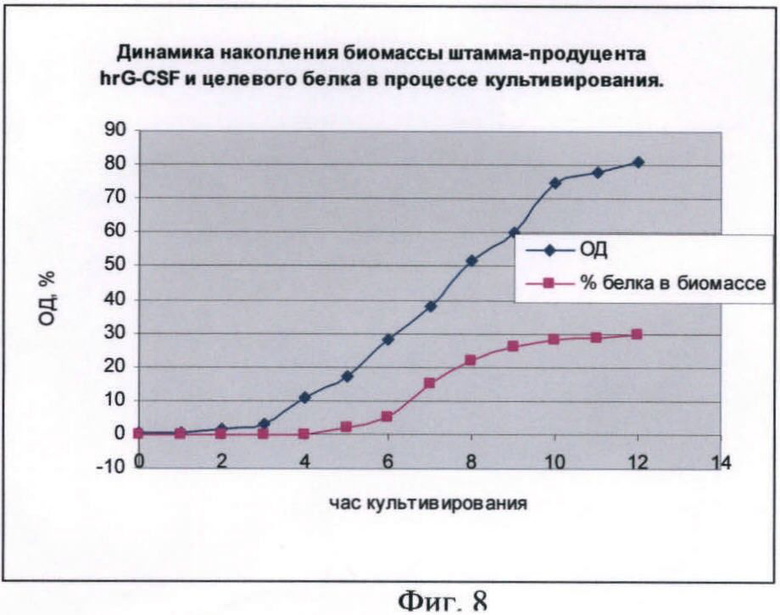
Штамм-продуцент G-CSF гранулоцитарнего колониестимулирующего фактора человеческого рекомбинантного. Предложенная схема была реализована для штамма-продуцента hrG-CSF при культивировании в объеме 30 л. Культивирование ведется при температуре 38°С, концентрации растворенного кислорода 80% (концентрация растворенного кислорода в процессе культивирования изменяется в пределах от 80 до 10%) при максимальной скорости перемешивания 1200 об/мин, максимальном уровне аэрации 25 л/мин, что соответствует отношению - 1 объем воздуха/мин: 1,2 объема культуральной жидкости. рН 7,0. Инокулят в объеме 3 л был выращен до середины логарифмической фазы роста. Выращивание инокулята осуществляли на питательной среде, где в качестве источника углерода используют глюкозу в концентрации 10 г/л, при температуре 38°С, перемешивании 200 об/мин, времени культивирования 3 ч. Инокулят был внесен в ферментер со стерильной питательной средой, засев ферментера осуществляли инокулятом с посевной дозой 10%.
В качестве стартового углеродсодержащего субстрата в среду в начале культивирования добавляют 50%-ный (мас.%) раствор глюкозы. К середине логарифмической фазы роста, когда глюкоза была утилизирована, производится порционная добавка 80%-ного (мас.%) раствора глицерина. После перехода культуры на утилизацию глицерина была начата дробная подача 25%-ного (мас.%) раствора лактозы и началась индукция белка. После исчерпания глицерина в среде культура начала утилизировать лактозу как субстрат. Как видно из почасового электрофореза культуральной жидкости, синтез целевого белка начался на 6 ч культивирования. Окончание ферментации происходит на 12 ч культивирования. Конечная оптическая плотность - 80 о.е., выход биомассы - 70 г/л. Содержание целевого белка по отношению к общему клеточному белку - 30%.
На фиг.7 представлены параметры культивирования штамма-продуцента hrG-CSF по предлагаемой схеме
На фиг.8 показана динамика накопления биомассы штамма-продуцента hrG-CSF в ходе процесса культивирования по предлагаемой схеме.
Изобретение позволяет повысить выход биомассы с единицы объема культуральной среды, так и повысить выход целевого белка, путем оптимизации процесса культивирования культуры клеток различных штаммов Е.coli.
Литература
1. Over-expression of recombinant human interferon-gamma in high eel density fermentation of Escherichia coli
R.Khalilzadeh, S.A.Shojaosadati, A.Bahrami & N.Maghsoudi
Biotechnology Group, Department of Chemical Engineering, Faculty of Engineering, Tarbiat Modarres University, Tehran, P.O. Box 14155-4838, Iran
Author for correspondence (Fax: +9821/8005040; E-mail: shoja_sa@modares.ac.ir)
Received 13 August 2003; Revisions requested 27 August 2003; Revisions received 30 September 2003; Accepted 2 October 2003.
2. GUIDELINE ON THE SPECIFICATION LIMITS FOR RESIDUES OF METAL CATALYSTS (Doc. Ref. CPMP/SWP/QWP/4446/00).
3. Lactose Fed-Batch Overexpression of Recombinant Metalloproteins inEscherichia coli BL21(DE3): Process Control Yielding High Levels of Metal-Incorporated, Soluble Protein B.J. Hoffman, J.A. Broadwater, P. Johnson, J. Harper, B.G. Fox, W.R.Kenealy.
4. Гавриков А.В. Оптимизация биотехнологического производства субстанций рекомбинантных интерферонов человека для создания на их основе препаратов ветеринарного назначения.
| название | год | авторы | номер документа |
|---|---|---|---|
| ПРОМЫШЛЕННЫЙ СПОСОБ МИКРОБИОЛОГИЧЕСКОГО СИНТЕЗА ФЕРМЕНТА ПЕНИЦИЛЛИН G АЦИЛАЗЫ ESCHERICHIA COLI | 2020 |
|
RU2729410C1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК pACYC-LANS(KM), ШТАММ Escherichia coli BL21(DE3), ТРАНСФОРМИРОВАННЫЙ РЕКОМБИНАНТНОЙ ДНК pACYC-LANS(KM), И СПОСОБ ПОЛУЧЕНИЯ РЕКОМБИНАНТНОЙ L-АСПАРАГИНАЗЫ Erwinia carotovora | 2010 |
|
RU2441916C1 |
| Рекомбинантная плазмидная ДНК pSMT3_HCRG21, кодирующая гибридный белок SMT3-HCRG21, штамм бактерий Escherichia coli BL21(DE3)/pSMT3_HCRG21 - продуцент анальгетического пептида HCRG21 и способ получения рекомбинантного анальгетического пептида HCRG21 | 2022 |
|
RU2798545C1 |
| ШТАММ Escherichia coli BL21 (DE3)[pAYC-ET-(hIFN-α2b)-IacI]-ПРОДУЦЕНТ РЕКОМБИНАНТНОГО ЧЕЛОВЕЧЕСКОГО АЛЬФА-2b ИНТЕРФЕРОНА И СПОСОБ ЕГО КУЛЬТИВИРОВАНИЯ | 2005 |
|
RU2303063C2 |
| Рекомбинантная плазмидная ДНК pET21-IFN-γ, содержащая ген, кодирующий гамма-интерферон человека, штамм E.coli BL21/pET21-IFN-γ - продуцент рекомбинантного гамма-интерферона человека и способ получения гамма-интерферона человека | 2023 |
|
RU2827613C1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК pET15/N-Dest+ И СПОСОБ ЕЕ ПОЛУЧЕНИЯ, ШТАММ Escherichia coli BL21(DE3)Gold, ТРАНСФОРМИРОВАННЫЙ РЕКОМБИНАНТНОЙ ДНК pET15/N-Dest+, И СПОСОБ ЕГО ПОЛУЧЕНИЯ, СПОСОБ ПОЛУЧЕНИЯ РЕКОМБИНАНТНОГО БИФУНКЦИОНАЛЬНОГО ФЕРМЕНТА - ДЕСТАБИЛАЗЫ МЕДИЦИНСКОЙ ПИЯВКИ | 2015 |
|
RU2658429C2 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДА pET32a-IFG144, ОБЕСПЕЧИВАЮЩАЯ СИНТЕЗ ИНТЕРФЕРОНА ГАММА, ШТАММ БАКТЕРИЙ ESCHERICHIA COLI JM109/pET32a-IFG144 ПРОДУЦЕНТ БЕЛКА ИНТЕРФЕРОН ГАММА И СПОСОБ ПОЛУЧЕНИЯ ИНТЕРФЕРОНА ГАММА | 2022 |
|
RU2787131C1 |
| ШТАММ Escherichia coli BL21(DE3)Gold/pETCYPopti - ПРОДУЦЕНТ РЕКОМБИНАНТНОГО ЦИКЛОФИЛИНА А ЧЕЛОВЕКА | 2015 |
|
RU2603283C1 |
| ШТАММ Escherichia coli BL21(DE3)Gold/pETmin-CypA - ПРОДУЦЕНТ РЕКОМБИНАНТНОГО ЦИКЛОФИЛИНА А ЧЕЛОВЕКА | 2013 |
|
RU2557305C2 |
| ШТАММ-ПРОДУЦЕНТ БЕЗМЕТИОНИНОВОГО CRM197 НА ОСНОВЕ КЛЕТОК E. coli BL21 (DE3) | 2015 |
|
RU2575621C1 |

Способ по изобретению предусматривает выращивание инокулята на основе штамма BL21(DE3) E.coli, несущего ген Т7 RNA полимеразы под контролем lacUV5 промотора, до середины логарифмической фазы роста на питательной среде с использованием в качестве источника углерода глюкозы в концентрации 10 г/л при температуре 38°С, перемешивании 200 об/мин в течение 3 ч, засев ферментера инокулятом с посевной дозой 10%, культивирование микроорганизмов при 38°С, концентрации растворенного кислорода 80% при максимальной скорости перемешивания 1200 об/мин, максимальном уровне аэрации 1 объем воздуха/мин: 1,2 объема культуральной жидкости, рН 7,00; окончание ферментации на 12-13 ч культивирования; осаждение клеток штамма-продуцента Е.coli. Вначале культивирования добавляют 50%-ный (мас.%) раствор глюкозы, в середине логарифмической фазы роста производят порционную добавку 80%-го (мас.%) раствора глицерина и с конца логарифмической фазы роста культуры осуществляют дробные добавки 25%-ного (мас.%) раствора лактозы. После культивирования содержащую рекомбинантные целевые белки в виде нерастворимых тел включения биомассу осаждают. Изобретение позволяет повысить выход биомассы и целевого белка с единицы объема культуральной среды. 8 ил., 4 пр.
Способ промышленного культивирования штаммов трансформированных Escherichia coli, содержащих рекомбинантные целевые белки в виде нерастворимых тел включения, отличающийся тем, что в качестве продуцентов используют E.coli, полученных на основе штамма BL21(DE3), несущего ген Т7 RNA полимеразы под контролем lacUV5 промотора, и предусматривающий:
- выращивание инокулята до середины логарифмической фазы роста на питательной среде, где в качестве источника углерода используют глюкозу в концентрации 10 г/л, при температуре 38°С, перемешивании 200 об/мин, времени культивирования 3 ч;
- засев ферментера инокулятом с посевной дозой 10%;
- культивирование микроорганизмов при 38°С, концентрации растворенного кислорода 80% при максимальной скорости перемешивания 1200 об/мин, максимальном уровне аэрации 1 объем воздуха/мин: 1,2 объема культуральной жидкости, рН=7,00;
- окончание ферментации на 12-13 ч культивирования;
- осаждение клеток штамма-продуцента E.coli, при этом вначале культивирования добавляют 50%-ный (мас.%) раствор глюкозы, в середине логарифмической фазы роста производится порционная добавка 80%-ного (мас.%) раствора глицерина и с конца логарифмической фазы роста культуры осуществляются дробные добавки 25%-ного (мас.%) раствора лактозы.
| Способ культивирования дрожжей | 1983 |
|
SU1418337A1 |
| Аппарат для упаривания конфетных масс, овощных пюре, паст и т.п. масс | 1958 |
|
SU121775A1 |

