Изобретение относится к микробиологии, биотехнологии, фармакологии и медицине и может быть использовано для дифференцировки промоноцитов и моноцитов в клинической и экспериментальной медицине.
В качестве модельных систем для исследований механизмов дифференциации широко используются промоноцитарные клеточные линии лейкемии человека (U937 и ТНР-1). Дифференциация характеризуется изменением различных клеточных параметров клеток по сравнению с соответствующим недифференцированным фенотипом. Такие изменения касаются морфологии, метаболических процессов и роста клеток. Клетки U937 и ТНР-1 представляют собой различные этапы развития моноцитов [14].
Существуют различные агенты, способные индуцировать дифференциацию клеток этих линий в зрелые моноциты. Такие агенты, как диметилсульфоксид (ДМСО) [7], ретиноевая кислота [9], витамин D3 [10], цитокины [3, 13] и сложные форболовые эфиры, в частности 12-O-тетрадеканоилфорбол-13-ацетат (TPA) [4], могут способствовать дифференциации незрелых моноцитов в моноциты/макрофаги.
Условием дифференциации в зрелые макрофаги является рост адгезии к поверхности культуральных плашек и, как следствие, резкое изменение формы клетки. Недифференцированные U937 и ТНР-1 клетки имеют округлую форму и растут в суспензии. Оба типа клеток имеют небольшую цитоплазму и крупное ядро. После дифференциации с ТРА большинство клеток прикрепляются к культуральным плашкам. Для ТНР-1 клеток особенно характерно образование агрегатов за счет развития межклеточных псевдоподий [11].
CD14, первоначально описанный как маркер дифференцировки моноцитов, представляет собой гликопротеин массой 55 кДа. Обширные исследования показали, что CD14 вовлекается в опосредование ответов на несколько продуктов бактериальной мембраны, особенно на бактериальные ЛПС. CD14 не обнаруживается на поверхности моноцитов-предшественников и резко возрастает во время их дифференцировки в моноциты. Таким образом, поверхностная экспрессия CD14 является отличной моделью для изучения механизмов миелоидного созревания клеток [12].
CR3 (β2-интегрин) - еще один классический маркер моноцитарной дифференциации, участвующий в клеточной адгезии и также функционирующий как рецептор комплемента. CD11b, субъединица CR3, представляет собой гликопротеин (масса 160 кДа), который связывается нековалентно с β2-субъединицей партнера CD18. Комплекс CD11b/CD18 экспрессируется на зрелых миелоидных клетках и широко используется в качестве раннего маркера дифференцировки моноцитов [5].
Ретиноевая кислота (RA) представляет собой активную форму ретинола (витамина A). RA, являясь физиологическим агентом, способна образовывать связи с ядерными рецепторами, при помощи которых запускает ответ клетки. Следует отметить, что к ретинолу проявляют чувствительность практически все клетки организма.
Рост ТНР-1 клеток ингибируется после добавления RA в концентрации 10-6 М. Помимо остановки роста происходят морфологические изменения: клетки приобретают неправильную форму и дольчатые ядра. Помимо морфологии дифференциация ТНР-1 клеток с помощью RA приводит и к другим изменениям. Заметно увеличивается фагоцитирующая активность. Однако RA не вызывает увеличения адгезивной способности [6].
1α, 25-дигидроксивитамин D3 (VD3), как и ретиноевая кислота, является физиологическим агентом и лигандом суперсемейства ядерных рецепторов. Легко проникая через клеточную и ядерную мембраны (по такому же типу, как и RA), рецепторы такого типа воздействуют непосредственно на гены, играя роль факторов транскрипции, контролирующих экспрессию генов.
Плейотропные эффекты VD3 опосредуются, главным образом, через внутриклеточный рецептор витамина D (VDR). VDR является членом надсемейства ядерных стероидов и рецепторов ретиноевой кислоты. Таким образом, он функционирует как лиганд-зависимый фактор транскрипции, регулирующий активацию генов-мишеней витамина D. На молекулярном уровне VDR активизирует свои гены-мишени, взаимодействуя со специфическими последовательностями ДНК, существующими в качестве элементов витамин-D-реагирования (VDR), называемыми "genomicaction" [5].
В попытках определения молекулярных механизмов регулирования клеточных ответов на VD3 рассматривалась возможность существования альтернативных путей передачи сигналов к классической программе действия генома. Действительно, в течение последнего десятилетия экспериментальные данные для "негеномной" передачи сигналов имеют концепцию эксклюзивного VDR-опосредованного геномного действия. Например, VD3 стимулирует быстрое образование вторичных мессенджеров, в том числе керамидов, цАМФ, инозитов, высвобождение катионов кальция, и активирует различные протеинкиназы, включая протеинкиназу С, Raf, митоген-активированный белок (МАР-киназу) и Src-семейство киназ. Тем не менее, физиологическое значение негеномной передачи сигналов само по себе или относительно VDR-опосредованного геномного действия до сих пор неясно. Кроме того, неизвестно, взаимодействует ли VD3 с классическим VDR для негеномной передачи сигналов или участвует система альтернативных рецепторов.
Обработка с помощью VD3 увеличивает поверхностную экспрессию CD11b и CD14 на HL-60, U937 и ТНР-1 клетках [5]. VD3 вызывает усиление фагоцитарной активности, регистрируемое по флуоресценции латексных шариков.
Форболовый эфир РМА (phorbol 12-myristate 13-acetate) - один из самых сильных нефизиологических агентов дифференциации. РМА легко проникает в клетку и, благодаря структурному сходству с диацилглицерином (DAG), взаимодействует с участком его связывания на молекуле протеинкиназы С (РКС) [8].
Экспрессия поверхностных M-CSF рецепторов является одним из первых событий, связанных с моноцитарной дифференцировкой лейкозных клеток в моноциты. Недавние исследования, использующие несколько лейкозных клеток-моделей, показали, что уровень c-fms транскриптов сильно возрастает, когда они начинают проходить моноцитарную дифференцировку под действием РМА. Считается, что активация протеинкиназы С (РКС) играет главную регуляторную роль в начале дифференцировки. В нормальном физиологическом процессе РКС активируется эндогенным диацилглицерином (DAG), продуктом гидролиза мембранных фосфолипидов под действием фосфолипазы С (PLC).
Эффекты, вызываемые в ТНР-1 клетках дифференцировкой, начинают проявляться при обработке РМА концентрацией 10 нМ: ингибируется рост клеток, в течение 24 часов после воздействия РМА более 80% клеток становились адгезивными. После РМА-обработки наблюдается увеличение количества вакуолей, а дальнейшая инкубация в присутствии этого агента приводит к увеличению размеров и удлинению формы, как у нормальных моноцитов крови в культуре. Кроме того, когда ТНР-1 клетки подвергаются обработке РМА, через 6-12 часов в них обнаруживается повышенный уровень транскриптов c-fms.
Молекулы РМА относительно стабильны и оказывают длительное воздействие на ТНР-1 клетки, что обусловливает их длительный эффект относительно воздействия физиологических агентов. Однако РМА настолько сильно активирует клетки, что делает невозможным их дальнейшее использование в качестве объекта исследования влияния липополисахаридов (эндотоксинов) [12].
Задача изобретения - физиологическое, практически нетоксичное средство для дифференцировки моноцитоподобных клеток.
Поставленная задача решается нетоксичным липополисахаридом из фототрофной бактерии Rhodobacter capsulatus PG.
Ранее известно использование штамма Rhodobacter capsulatus PG в качестве продуцента липополисахарида, антагониста эндотоксинов [1].
Данный штамм Rhodobacter capsulatus PG депонирован Всероссийской коллекцией микроорганизмов ИБФМ РАН [2].
Исследовалась способность нетоксичного ЛПС Rhodobacter capsulatus PG дифференцировать клетки моноцитарной линии ТНР-1 в моноциты и макрофаги. Дифференцирующую активность ЛПС исследовали по экспрессии поверхностных рецепторов (TLR4, CD11b и CD14) и изменению морфологических характеристик ТНР-1 клеток.
1. Исследование влияния ЛПС Rhodobacter capsulatus PG в диапазоне 100-1000 нг/мл на морфологию ТНР-1 клеток.
Объектом исследования служила моноцитарная линия клеток ТНР-1 из коллекции АТСС®TIB™202 (США). Клетки ТНР-1 культивировали в среде RPMI 1640 (Sigma), содержащей 2 ммоль/л L-глутамина, 100 Ед/мл пенициллина, 100 мкг/мл стрептомицина, (L-Glutamin-penicillin-streptomycin solution, Sigma) и 10% инактивированной бычьей сыворотки (Hyclon) во флаконах для культивирования объемом 25 см2, 50 мл (Orange Scientific) в CO2-инкубаторе (Jouan, Франция) при температуре 37°C и 5% CO2. Жизнеспособность, определяемая по окрашиванию клеток трипановым синим, составляла 88-95%.
К суспензии клеток ТНР-1 в среде RPMI 1640 (2×106 кл/мл) добавляли водный раствор ЛПС R. capsulatus PG в конечной концентрации 100 или 1000 нг/мл и инкубировали 24 ч в CO2-инкубаторе при температуре 37°C и 5% CO2.
Для оценки степени дифференцировки использовали визуальный анализ ТНР-1 клеток после 24-часовой инкубации с ЛПС R. capsulatus PG. С этой целью определялась доля (в % от общего числа клеток в 7-8 полных полях зрения) клеток с измененной морфологией, в частности имеющих заметные псевдоподии, увеличенные размеры, веретеновидную или вытянутую форму клеток. Нормальная морфология недифференцированных ТНР-1 клеток - округлые клетки без заметных псевдоподий. Результаты визуального анализа представлены в таблице 1 и на фиг. 1, 2а, 2б, 2в.


На фиг. 1. представлено изменение морфологии ТНР-1 клеток после 24 часовой экспозиции с ЛПС R. capsulatus PG (*- различия от контроля достоверны при p<0,05).
На фиг. 2а показано изменение морфологии ТНР-1 клеток под действием ЛПС R. capsulatus PG (увеличение 40×1,5×10, окрашивания нет), контроль.
На фиг. 2б представлено изменение морфологии ТНР-1 клеток под действием ЛПС R. capsulatus PG (увеличение 40×1,5×10, окрашивания нет), клетки после стимуляции ЛПС R. capsulatus PG 100 нг/мл.
На фиг. 2в показано изменение морфологии ТНР-1 клеток под действием ЛПС R. capsulatus PG (увеличение 40×1,5×10, окрашивания нет), клетки после стимуляции ЛПС R. capsulatus PG 1000 нг/мл.
Доля ТНР-1 клеток с измененной морфологией составляла в контрольном варианте (25,2±2,4)%. Из представленных в таблице 1 и на фиг. 1, 2 (а, б, в) результатов видно, что ЛПС R. capsulatus PG, внесенный в минимальной дозе (100 нг/мл), не вызывал достоверного изменение морфологии ТНР-1 клеток (доля клеток с измененной морфологией равна 23,9±1,8% по сравнению с контролем). Следовательно, внесенный в данной концентрации ЛПС R. capsulatus PG не оказывает биологического действия на изменение морфологии ТНР-1 клеток.
ЛПС R. capsulatus PG в концентрации 1000 нг/мл вызывал значительное увеличение доли измененных ТНР-1 клеток до (37,4±4,3) пг/мл. Данное значение достоверно отличается от контроля (25,2±2,4)% и свидетельствует о способности ЛПС из R. capsulatus PG вызывать изменение морфологии ТНР-1 клеток и, возможно, усиливать степень их дифференцировки.
2. Поиск оптимальной концентрации ЛПС Rhodobacter capsulatus PG в диапазоне 100-1000 нг/мл на изменение уровня поверхностных рецепторов (TLR4, CD11b и CD14) недифференцированных ТНР-1 клеток.
В работе использовали антитела к TLR4, CD14- и CD11b-рецепторам, меченные флуоресцентной меткой: Alexa Fluor 488 anti-human CD14 (Clone HCD14), Alexa Fluor 488 anti-human CD11b (Clone ICRF44) (BioLegend); Alexa Fluor 488 anti-human CD284 (TLR4) (Clone HTA 125) (eBioscience).
После инкубации клеток с ЛПС R. capsulatus PG в конечной концентрации 500 или 1000 нг/мл в течение 24 ч в CO2-инкубаторе при температуре 37°C и 5% CO2, ТНР-1 клетки отделяли от среды культивирования центрифугированием и ресуспендировали в буфере для окрашивания (Cell Staining Buffer, BioLegend) в расчете 105-106 клеток в 100 мкл буфера.
Для исключения неспецифического связывания антител с исследуемыми рецепторами в каждый образец добавляли по 5 мкл блокирующего буфера для Fc-рецепторов (Human Tru Stain FcX™, BioLegend) и инкубировали при комнатной температуре 10 минут. Затем в образцы добавляли по 5 мкл раствора соответствующих антител и инкубировали 30 мин при 4°C. Далее 2 раза отмывали Cell Staining Buffer (по 2 мл), ресуспендировали в 400 мкл этого же буфера и переносили в пробирку для цитофлуориметрии.
Анализ образцов проводили с помощью проточного цитофлуориметра EPICS XL-MCL (Beckman Coulter, США). Уровень поверхностной рецепции ТНР-1 клеток оценивали по величине средней интенсивности флуоресценции (MnIX) образца. В каждом образце просчитывалось 6000 клеток.
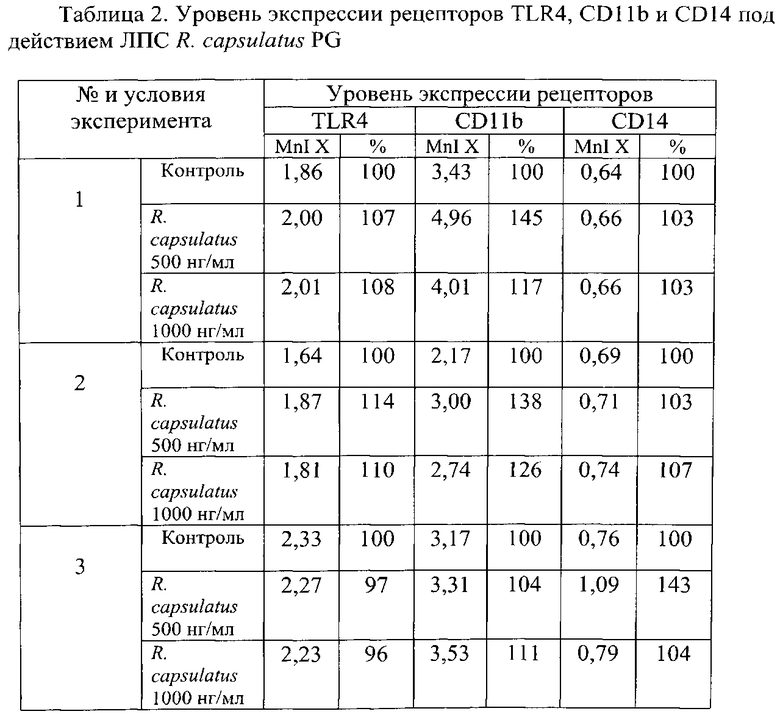

Фиг. 3. Изменение уровня экспрессии рецепторов TLR4, CD11b и CD14 под действием ЛПС R. capsulatus PG в %. Представлены средние значения изменения уровня экспрессии рецепторов (n=3) и стандартная ошибка в процентах (%) относительно контроля, принятого за 100%.
Из представленных в таблице 2 и на фиг. 3 результатов видно, что ЛПС R. capsulatus PG заметнее всего влияет на экспрессию CD11b-рецепторов. Из исследуемых концентраций наибольшим влиянием на изменение уровня поверхностных рецепторов обладает ЛПС R. capsulatus PG, внесенный в концентрации 500 нг/мл.
На основании результатов по изменению морфологии ТНР-1 клеток и экспрессии TLR4, CD11b и CD 14 рецепторов на их поверхности под воздействием нетоксичного ЛПС Rhodobacter capsulatus PG следует вывод, что нетоксичный ЛПС Rhodobacter capsulatus PG обладает максимальной дифференцирующей активностью в концентрации 500 нг/мл.
Предложенное средство может найти применение при создании лекарственных препаратов, в частности, для онкологии.
Список литературы
1. RU 2392309 С1, 24.11.2008. Прохоренко И.Р., Грачев С.В., Зубова С.В. Штамм Rhodobacter capsulatus PG - продуцент липополисахарида, антагониста эндотоксинов.
2. Свидетельство о депонировании штамма Rhodobacter capsulatus PG. Регистрационный номер ВКМ В-2381 Д, 21.12.2005.
3. Harris Р.Е., Ralph P., Litcofsky P., Moore M.A.S. Distinct activities of interferon, lymphokine and cytokine differentiation-inducing factors acting on the human monoblastic leukemia cell line U937. Cancer Res. 1985. V. 45. P. 9-13.
4. Hass R., Bartels H., Topley M, Hadam M., Kohler L., Goppelt-Strube M., Resh K. TPA-induced differentiation and adhesion of U937 cells: changes in ultrastructure, cytoskeletal organization and expression of cell surface antigens. Eur. J. Cell Biol. 1989. V. 48. P. 282-293.
5. Hmama Z., Nandan D., Sly L., Knutson K.L., Herrera-Velit P., Reiner N.E. 1999. 1α, 25-Dihydroxyvitamin D3-induced myeloid cell differentiation is regulated by a vitamin D receptor-phosphatidylinositol 3-kinase signaling complex. J. Exp. Med. V. 190. P. 1583-1594.
6. Hyodoh F. Effects of Retinoic Acid on the Differentiation of THP-1 Cell Lines Containing Aneuploid or Diploid Chromosomes. Cell structure and function. 1987. №12. P. 225-242.
7. Nakamura Т., Kharbanda S., Spriggs D., Kufe D. Effect of dexamethasone of monocytic differentiation in human U937 cells by dimethylsulfoxide. J. Cell Physiol. 1990. V. 142. P. 261-267.
8. Nishizuka Y. Protein kinase С and lipid signaling for sustained cellular responses. FASEB. J. 1995. V. 9. P. 484-496.
9. Olsson I.L. and Breitman T.R. Induction of differentiation of the human histiocytic lymphoma cell line U937 by retinoic acid and cyclic adenosine 3′,5′-monophosphate-inducing agents. Cancer Res. 1982. V. 42. P. 3924-3927.
10. Olsson I.L., Gullberg U., Ivhed I., Nilsson K. Induction of differentiation of the human histiocytic lymphoma cell line U937 by 1α, 25-dihydroxycholecalciferol. Cancer Res. 1983. V. 43. P. 5862-5867.
11. Panzarini E., Ramires P.A., Miccoli M.A., Tenuzzo В., Scordari A., Dini L. Differentiation of THP-1 and U937 cells in presence of synthetic hydrogels. Caryologia. 2006. V. 59. №4. P. 395-402.
12.Schwende H., Fitzke E., Ambs P., Dieter P. Differences in the state of differentiation of THP-1 cells induced by phorbol ester and la, 25-dihydroxyvitamin D3. J. Leukocyte Biol. 1996. V. 59. V. 555.
13. Testa U., Ferbus D., Gabbianelli M., Pascucci В., Boccoli G., Louache F., Thang M.N. 1988 Effect of endogenous and exogenous interferons on the differentiation of human monocyte cell line U937. Cancer Res. 1988. V. 48. P. 82-88.
14. Tsuchiya S., Yamabe M., Yamaguchi Y., Kobayashi Y., Konno Т., Tada K. Establishment and characterization of a human acute monocytic leukemia cell line (THP-1). Int. J. Cancer. 1980. V. 26. №2. P. 171-176.
| название | год | авторы | номер документа |
|---|---|---|---|
| ПРИМЕНЕНИЕ ЛИПОПОЛИСАХАРИДА ФОТОТРОФНОЙ БАКТЕРИИ RHODOBACTER CAPSULATUS PG В КАЧЕСТВЕ ФАКТОРА, УСИЛИВАЮЩЕГО ДИФФЕРЕНЦИРУЮЩУЮ АКТИВНОСТЬ 1α,25-ДИГИДРОКСИВИТАМИНА D | 2016 |
|
RU2642309C1 |
| ПРОТИВОАЛЛЕРГИЧЕСКОЕ СРЕДСТВО | 2017 |
|
RU2690430C2 |
| Способ идентификации функционального М1 и М2 фенотипа макрофагов человека, генерированных in vitro из моноцитов крови | 2019 |
|
RU2717024C1 |
| СРЕДСТВО ДЛЯ БЛОКИРОВАНИЯ ЭФФЕКТОВ ЛИПОТЕЙХОЕВЫХ КИСЛОТ - АГОНИСТОВ ТОЛЛ-ПОДОБНОГО РЕЦЕПТОРА 2 | 2022 |
|
RU2781012C1 |
| СПОСОБ УСИЛЕНИЯ ИММУННОГО ОТВЕТА НА АНТИГЕН У МЛЕКОПИТАЮЩИХ | 2004 |
|
RU2341289C2 |
| ШТАММ RHODOBACTER CAPSULATUS PG - ПРОДУЦЕНТ ЛИПОПОЛИСАХАРИДА, АНТАГОНИСТА ЭНДОТОКСИНОВ | 2008 |
|
RU2392309C1 |
| ГОРМОН D (ВИТАМИН D) И ЕГО ПРОИЗВОДНЫЕ ДЛЯ ЛЕЧЕНИЯ И ПРЕДУПРЕЖДЕНИЯ РАКА | 2019 |
|
RU2794340C1 |
| СПОСОБ МОДИФИКАЦИИ МОНОЦИТОВ ПЕРИФЕРИЧЕСКОЙ КРОВИ ДЛЯ ПОВЫШЕНИЯ ИХ ПАРАКРИННОЙ АКТИВНОСТИ ПРИ АУТОЛОГИЧЕСКОЙ ТРАНСПЛАНТАЦИИ | 2012 |
|
RU2497947C1 |
| ПРИМЕНЕНИЕ БИОМАРКЕРОВ ДЛЯ ОЦЕНКИ ЭФФЕКТИВНОСТИ КОМПОЗИЦИИ В СНИЖЕНИИ ВОЗДЕЙСТВИЯ ПРОТИВОРАКОВЫХ ТЕРАПЕВТИЧЕСКИХ СРЕДСТВ НА КОЖУ | 2020 |
|
RU2836055C1 |
| СПОСОБ ПРОГНОЗИРОВАНИЯ ЭФФЕКТОРНОЙ НЕДОСТАТОЧНОСТИ ЦИТОТОКСИЧЕСКИХ КЛЕТОК ПО УВЕЛИЧЕНИЮ КОНЦЕНТРАЦИЙ В КРОВИ МОЛЕКУЛ SCD54, SCD56, SCD71 | 2023 |
|
RU2811001C1 |


Изобретение относится к биохимии. Описан липополисахарид, антагонист эндотоксинов, продуцируемый штаммом Rhodobacter capsulatus PG, депонированным в ВКМ ИБФМ РАН под номером В-2381 Д, в качестве средства для дифференцировки моноцитоподобных клеток. Представленный липополисахарид Rhodobacter capsulatus PG является физиологическим, практически нетоксичным. Изобретение может быть использовано в качестве средства для дифференцировки моноцитоподобных клеток. Предложенное средство может найти применение при создании лекарственных препаратов, в частности, для онкологии. 3 ил., 2 табл.
Липополисахарид, антагонист эндотоксинов, продуцируемый штаммом Rhodobacter capsulatus PG, депонированным в ВКМ ИБФМ РАН под номером В-2381 Д, в качестве средства для дифференцировки моноцитоподобных клеток.
| ГРАЧЕВ С.В | |||
| НЕЙТРАЛИЗАЦИЯ ЭНДОТОКСИНИНДУЦИРОВАННЫХ ОТВЕТОВ НЕЙТРОФИЛОВ И МОНОЦИТОВ ПЕРИФЕРИЧЕСКОЙ КРОВИ ЧЕЛОВЕКА ЛИПОПОЛИСАХАРИДОМ RHODOBACTER CAPSULATUS | |||
| Молекулярная медицина | |||
| Пломбировальные щипцы | 1923 |
|
SU2006A1 |
| Очаг для массовой варки пищи, выпечки хлеба и кипячения воды | 1921 |
|
SU4A1 |
| Приспособление для разматывания лент с семенами при укладке их в почву | 1922 |
|
SU56A1 |
| ШТАММ RHODOBACTER CAPSULATUS PG - ПРОДУЦЕНТ ЛИПОПОЛИСАХАРИДА, АНТАГОНИСТА ЭНДОТОКСИНОВ | 2008 |
|
RU2392309C1 |
| WO 2008080195 A1, 10.07.2008 | |||
| WO 2000062818 A1, 26.10.2000 | |||
| PIN-YU PERERA | |||
| Lipopolysaccharide and Its Analog Antagonists | |||

