ОБЛАСТЬ ТЕХНИКИ, К КОТОРОЙ ОТНОСИТСЯ ИЗОБРЕТЕНИЕ
Изобретение относится к антитело-подобным связывающим белкам, содержащим четыре полипептидных цепи, которые образуют четыре антигенсвязывающих участка, при этом каждая пара полипептидов, образующих антитело-подобный связывающий белок, имеет двойные вариабельные домены, имеющие ориентацию «крест-накрест». Изобретение также относится к способам получения таких антиген-подобных связывающих белков.
УРОВЕНЬ ТЕХНИКИ
Встречающиеся в природе IgG-антитела являются бивалентными и моноспецифичными. Биспецифичные антитела, обладающие специфичностями связывания с двумя разными антигенами, могут быть получены с использованием способов рекомбинации и могут быть предназначены для широкого клинического применения. Хорошо известно, что полные молекулы IgG-антител представляют собой Y-образные молекулы, содержащие четыре полипептидных цепи: две тяжелых цепи и две легких цепи. Каждая легкая цепь содержит два домена, при этом N-концевой домен назван вариабельным или VL-доменом (или областью), и C-концевой домен называют константным (или CL) доменом (константный каппа (Cκ) или константный лямбда (Cλ) домен). Каждая тяжелая цепь состоит из четырех или пяти доменов, в зависимости от класса антитела. N-концевой домен называют вариабельным (или VH) доменом (или областью), за которым следует первый константный (или CH1) домен, шарнирная область и затем второй и третий константные (или CH2 и CH3) домены. В собранном антителе домены VL и VH ассоциированы вместе с образованием антигенсвязывающего участка. Также домены CL и CH1 ассоциированы вместе, удерживая одну тяжелую цепь в ассоциации с одной легкой цепью. Два гетеродимера тяжелой - легкой цепи ассоциированы вместе за счет взаимодействия доменов CH2 и CH3 и взаимодействия между шарнирными областями двух тяжелых цепей.
Известно, что протеолитическое расщепление антитела может приводить к образованию фрагментов антител (Fab и Fab2). Такие фрагменты целых антител могут проявлять антигенсвязывающую активность. Фрагменты антител также могут быть получены рекомбинантно. Могут быть получены Fv-фрагменты, состоящие только из вариабельных доменов тяжелой и легкой цепей, ассоциированных друг с другом. Такие Fv-фрагменты являются моновалентными в отношении связывания антигена. Также было показано, что более мелкие фрагменты, такие как отдельные вариабельные домены (доменные антитела или дАт (dAB); Ward с соавторами, 1989, Nature 341(6242): 544-46), и отдельные определяющие комплементарность области CDR (Williams с соавторами, 1989, Proc. Natl. Acad. Sсi. U.S.A. 86(14): 5537-41) сохраняют характеристики связывания родительского антитела, хотя для большинства встречающихся в природе антител необходимо, чтобы и VH и VL полностью сохраняли эффективность связывания.
Конструкции одноцепочечного вариабельного фрагмента (scFv) содержат VH- и VL-домены антитела, находящиеся в одной полипептидной цепи, при этом домены разделены гибким линкером достаточной длины (более 12 аминокислотных остатков), который способствует вынужденному внутримолекулярному взаимодействию, обеспечивающему самосборку двух доменов в функциональный эпитопсвязывающий участок (Bird с соавторами, 1988, Science 242(4877): 423-26). Такие небольшие белки (М.м. ~ 25000 Да) обычно сохраняют специфичность и аффинность по отношению своему антигену в одном полипептиде и могут служить удобным строительным блоком для более крупных антигенспецифичных молекул.
Преимущество применения фрагментов антител, а не целых антител в диагностике и терапии, заключается в их небольшом размере. Очевидно, они являются менее иммуногенными, чем целые антитела и обладают большей способностью проникать в ткани. Недостаток, связанный с применением таких фрагментов состоит в том, что они имеют только один антигенсвязывающий участок, что приводит к пониженной авидности. Кроме того, благодаря их небольшому размеру, они очень быстро выводятся из сыворотки и, следовательно, имеют короткое время полужизни.
Интерес представляло получение биспецифичных антител (BsAb), которые объединяют антигенсвязывающие участки двух антител в одной молекуле, и поэтому могут быть способны связывать два разных антигена одновременно. Кроме применений для диагностических целей такие молекулы открывают путь для новых терапевтических применений, например, благодаря перенаправлению мощных эффекторных систем на пораженные заболеванием области (в которых злокачественные клетки часто развивают механизмы подавления нормальных иммунных ответов, запускаемых моноклональными антителами, подобных зависимой от антител клеточной цитотоксичности (ADCC) или зависимой от комплемента цитотоксичности (CDC)), или, благодаря повышению нейтрализующей или стимулирующей активности антител. Такая способность была известна раньше, что привело к разработке ряда способов получения таких биспецифичных антител. В ходе начальных попыток соединить специфичности связывания двух целых антител против разных антигенов-мишеней для терапевтических целей использовали химически слитые молекулы гетероконъюгатов (Staerz с соавторами, 1985, Nature 314(6012): 628-31).
Биспецифичные антитела исходно получали слиянием двух гибридом, которые способны продуцировать разные иммуноглобулины (Milstein с соавторами, 1983, Nature 305(5934): 537-40), но сложность видового состава (до десяти разных видов), продуцируемого в клеточной культуре, делает очистку трудной и дорогостоящей (George с соавторами, 1997, The antibodies 4: 99-141 (Capra et al., ed., Harwood Academic Publishers)). С использованием такого формата получили мышиное IgG2a-антитело и крысиное IgG2b-антитело в одной и той же клетке (например, либо в виде квадромного слияния двух гибридом, либо в сконструированных клетках CHO). Поскольку легкие цепи каждого антитела преимущественно связываются с тяжелыми цепями своего родственного им вида, собираются три основных вида антитела: два исходных антитела и гетеродимер из двух антител, содержащий по одной паре тяжелой/легкой цепи каждого антитела, которые ассоциированы своими Fc-частями. Требуемый гетеродимер может быть очищен из такой смеси, поскольку его свойства связывания с белком A отличаются от свойств исходных антител: IgG2b крысы не связывается с белком A, тогда как IgG2a мыши связывается с таким белком. Поэтому гетеродимер мыши-крысы связывается с белком A, но элюируется при более высоком значении pH, чем гомодимер IgG2a мыши, и это делает возможной избирательную очистку биспецифичного гетеродимера (Lindhofer с соавторами, 1995, J. Immunol. 155(1): 219-25). Полученный в результате биспецифичный гетеродимер является полностью нечеловеческим, следовательно, высоко иммуногенным, что может иметь вредные побочные эффекты (например, реакции «HAMA» или «HARA»), и/или нейтрализовать терапевтическое средство. Сохраняется потребность в сконструированных биспецифичных средствах с превосходящими свойствами, которые можно легко получить с высоким выходом из культуры клеток млекопитающих.
Несмотря на многообещающие результаты, полученные с использованием гетероконъюгатов или биспецифичных антител, полученных из слияний клеток, которые описаны выше, несколько факторов делают их непригодными для крупномасштабных терапевтических применений. Такие факторы включают: быстрый клиренс гетероконъюгатов in vivo, трудоемкие лабораторные методики, необходимые для создания любого типа молекулы, необходимость тщательной очистки гетероконъюгатов от гомоконъюгатов или моноспецифичных антител, и обычно низкие получаемые выходы.
Все чаще применяли генетическую инженерию для конструирования, модификации и получения антител или производных антител с требуемым набором связывающих свойств и эффекторных функций. Было разработано несколько основанных на рекомбинации способов для эффективного получения BsAb, как в форме фрагментов антител (Carter с соавторами, 1995, J. Hematother. 4(5): 463-70; Pluckthun с соавторами, 1997, Immunotechnology 3(2): 83-105; Todorovska с соавторами, 2001, J. Immunol. Methods 248(1-2): 47-66), так и в форме полноразмерных IgG (Carter, 2001, J. Immunol. Methods 248(1-2): 7-15).
Объединение двух разных scFv приводит к получению форм BsAb с минимальной молекулярной массой, называемых sc-BsAb или Ta-scFv (Mack с соавторами, 1995, Proc. Natl. Acad. Sci. U.S.A. 92(15): 7021-25; Mallender с соавторами, 1994, J. Biol. Chem. 269(1): 199-206). BsAb были сконструированы с использованием генетического слияния двух scFv с использованием для димеризации функциональной группы, такой как лейциновая молния (Kostelny с соавторами, 1992, J. Immunol. 148(5): 1547-53; de Kruif с соавторами, 1996, J. Biol. Chem. 271(13): 7630-34).
Диантитела представляют собой небольшие бивалентные и биспецифичные фрагменты антител. Фрагменты содержат VH, связанный с VL в одной и той же полипептидной цепи с использованием линкера, который является слишком коротким (менее 12 аминокислотных остатков), чтобы обеспечить возможность спаривания между двумя доменами в одной и той же цепи. Домены вынуждены спариваться межмолекулярно с комплементарными доменами в другой цепи и создавать два антигенсвязывающих участка. Такие димерные фрагменты антител или «диантитела» являются бивалентными и биспецифичными (Holliger с соавторами, 1993, Proc. Natl. Acad. Sci. U.S.A. 90(14): 6444-48). Диантитела близки по размеру к Fab-фрагменту. Полипептидные цепи доменов VH и VL связаны линкером длиной от 3 до 12 аминокислотных остатков преимущественно с образованием димеров (диантител), тогда как в случае связывания линкером длиной от 0 до 2 аминокислотных остатков преобладают тримеры (триантитела) и тетрамеры (тетраантитела). Кроме зависимости от длины линкера точная картина олигомеризации, по-видимому, зависит от состава, а также от ориентации вариабельных доменов (Hudson с соавторами, 1999, J. Immunol. Methods 231(1-2): 177-89). Вероятность прогнозирования конечной структуры молекул диантител очень низка.
Хотя конструкции, основанные на sc-BsAb и диантителах, проявляют интересные клинические возможности, было показано, что такие нековалентно ассоциированные молекулы недостаточно стабильны в физиологических условиях. Общая стабильность scFv-фрагмента зависит от стабильности, присущей доменам VL и VH, а также от стабильности зоны контакта доменов. Часто недостаточную стабильность зоны контакта VH-VL scFv-фрагментов считали основной причиной необратимой инактивации scFv, так как временное раскрывание зоны контакта, которое может позволять пептидный линкер, открывает гидрофобные участки, которые способствуют агрегации и, следовательно, нестабильности и низкому выходу продукта (Worn с соавторами, 2001, J. Mol. Biol. 305(5): 989-1010).
Альтернативный способ производства биспецифичных бивалентных антигенсвязывающих белков из доменов VH и VL описан в патенте США № 5989830. Такие конфигурации, имеющие две головки и двойной Fv, получают в результате экспрессии бицистронного вектора, который кодирует две полипептидных цепи. В конфигурации с двойным Fv вариабельные домены двух разных антител экспрессируются в тандемной ориентации на двух разных цепях (одна тяжелая цепь и одна легкая цепь), при этом одна полипептидная цепь имеет два VH, расположенных последовательно и разделенных пептидным линкером (VH1-линкер-VH2), а другая полипептидная цепь состоит их комплементарных доменов VL, соединенных последовательно пептидным линкером (VL1-линкер-VL2). В конфигурации с двумя головками крест-накрест вариабельные домены двух разных антител экспрессируются в тандемной ориентации в двух отдельных полипептидных цепях (одна тяжелая цепь и одна легкая цепь), при этом одна полипептидная цепь имеет два VH, расположенных последовательно и разделенных пептидным линкером (VH1-линкер-VH2), а другая полипептидная цепь состоит из комплементарных доменов VL, соединенных последовательно пептидным линкером в противоположной ориентации (VL2-линкер-VL1). Молекулярное моделирование конструкций предполагает, чтобы размер линкера был достаточно длинным и простирался на 30-40 Å (15-20 аминокислотных остатков).
Возрастание валентности антитела представляет интерес, так как это усиливает функциональную аффинность такого антитела, вследствие эффекта авидности. Поливалентные белковые комплексы (PPC) с увеличенной валентностью описаны в публикации заявки на выдачу патента США № US 2005/0003403 A1. PPC содержат две полипептидных цепи, обычно располагаются латерально друг к другу. Каждая полипептидная цепь обычно содержит три или четыре «v-области», которые содержат аминокислотные последовательности, способные образовывать антигенсвязывающий участок при спаривании с соответствующей v-областью на противоположной полипептидной цепи. Можно использовать примерно до шести «v-областей» на каждой полипептидной цепи. V-области каждой полипептидной цепи соединены друг с другом линейно и могут быть соединены промежуточными связывающими областями. В случае расположения в форме PPC v-области на каждой полипептидной цепи образуют антигенсвязывающие участки. Комплекс может иметь одну или несколько специфичностей связывания.
Carter с соавторами (Ridgway с соавторами, 1996, Protein Eng. 9(7): 617-21; Carter, 2011, J. Immunol. Methods 248(1-2): 7-15) предложили методику получения Fc-гетеродимера с использованием набора мутаций «выступ во впадину» в домене CH3 Fc. Такие мутации приводят к изменению комплементарности при упаковке остатков между зоной контакта домена CH3 и структурно консервативной гидрофобной сердцевиной так, чтобы предпочтительно происходило образование гетеродимера по сравнению с гомодимерами, что приводит к хорошей экспрессии гетеродимеров в культуре клеток млекопитающих. Хотя методика приводит к более высокому выходу гетеродимеров, образование гомодимеров не было полностью подавлено (Merchant с соавторами, 1998, Nat. Biotechnol. 16(7): 677-81).
Gunasekaran с соавторами исследовали возможность сохранения целостности гидрофобной сердцевины при приведении в действие образования Fc-гетеродимера, за счет изменения комплементарности зарядов в зоне контакта домена CH3 (Gunasekaran с соавторами, 2010, J. Biol. Chem. 285(25): 19637-46). Используя преимущество механизма ориентирования в электростатическом поле, такие конструкции эффективно стимулируют образование Fc-гетеродимеров с минимальным загрязнением гомодимерами в результате мутации двух пар периферически расположенных заряженных остатков. В противоположность конструкции «выступ во впадину» образование гомодимеров в одинаковой мере подавляется, вследствие природы механизма электростатического отталкивания, но его не избегают полностью.
Davis с соавторами описывают способ конструирования антител с целью превращения Fc-гомодимеров в гетеродимеры посредством переплетения β-ничтатых участков CH3-доменов IgG и IgA человека без введения дополнительных дисульфидных связей между цепями (Davis с соавторами, 2010, Protein Eng. Des. Sel. 23(4): 195-202). Экспрессия слитых белков SEEDbody (Sb) клетками млекопитающих дает гетеродимеры Sb с высоким выходом, которые легко очищают, исключая минорные побочные продукты.
В публикации заявки на выдачу патента США № US 2010/331527 A1 описано биспецифичное антитело на основе гетеродимеризации домена CH3, введении в одну тяжелую цепь мутаций H95R и Y96F в домене CH3. Такие аминокислотные замены происходят из домена CH3 подтипа IgG3, и будут гетеродимеризоваться с остовом IgG1. Типичная легкая цепь, склонная спариваться с любой тяжелой цепью, является предпосылкой для всех форм, основанных на гетеродимеризации посредством домена CH3. Поэтому получают всего три типа антител: 50% имеющих чистый остов IgG1, одну треть имеющих чистый остов с мутациями H95R и Y96F, и одну треть имеющих две разных тяжелых цепи (биспецифичные). Требуемый гетеродимер может быть очищен из такой смеси, так как его связывающие свойства по отношению к белку A отличаются от свойств исходных антител: полученные из IgG3 домены CH3 не связываются с белком A, тогда как IgG1 связывается. Следовательно, гетеродимер связывается с белком A, но элюируется при более высоком значении pH, чем чистый гомодимер IgG1, и это делает возможным избирательную очистку биспецифичного гетеродимера.
В патенте США № 7612181 описаны биспецифичные антитела с двойными вариабельными доменами IgG (DVD-IgG), которые основаны на формате двойных Fv, описанном в патенте США № 5989830. Сходная биспецифичная форма также описана в публикации заявки на выдачу патента США № US 2010/0226923 A1. Добавление константных доменов к соответствующим цепям двойного Fv (CH1-Fc к тяжелой цепи и константного каппа- или лямбда-домена к легкой цепи) приводит к образованию функциональных биспецифичных антител без какой-либо необходимости в дополнительных модификациях (т.е., очевидное добавление константных доменов для повышения стабильности). В случае некоторых из антител, экспрессируемых в форме DVD-Ig/TBTI, наблюдают эффект положения по отношению ко второму (или находящемуся глубже внутри) антигенсвязывающему положению (Fv2). В зависимости от последовательности и природы антигена, узнаваемого положением Fv2, такой домен антитела проявляет пониженную аффинность к своему антигену (т.е., уменьшение скорости ассоциации по сравнению с исходным антителом). Одним возможным объяснением такого наблюдения является то, что линкер между VL1 и VL2 выступает в CDR-область Fv2, делая Fv2 в определенной степени недоступным для более крупных антигенов.
Второй конфигурацией биспецифичного фрагмента антитела, описанного в патенте США № 5989830, является конфигурация с двумя головками крест-накрест (CODH), имеющая следующую ориентацию вариабельных доменов, экспрессируемых в двух цепях:
VL1-линкер-VL2 в случае легкой цепи и
VH2-линкер-VH1 в случае тяжелой цепи.
В патенте ’830 описан тот факт, что биспецифичный фрагмент антитела с двумя головками крест-накрест (конструкция GOSA.E) сохраняет более высокую активность связывания, чем двойной Fv (см. страницу 20, строки 20-50 в патенте ’830), и дополнительно раскрыт тот факт, что такая форма менее подвержена влиянию линкеров, которые используют между вариабельными доменами (см. страницы 20-21 патента ’830).
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Изобретение относится к антитело-подобному связывающему белку, содержащему четыре полипептидных цепи, которые образуют четыре антигенсвязывающих участка, при этом две полипептидные цепи имеют структуру, представленную формулой:
и две полипептидные цепи имеют структуру, представленную формулой:
где:
VL1 означает первый вариабельный домен легкой цепи иммуноглобулина;
VL2 означает второй вариабельный домен легкой цепи иммуноглобулина;
VH1 означает первый вариабельный домен тяжелой цепи иммуноглобулина;
VH2 означает второй вариабельный домен тяжелой цепи иммуноглобулина;
CL означает константный домен легкой цепи иммуноглобулина;
CH1 означает константный домен тяжелой цепи иммуноглобулина CH1;
Fc означает шарнирную область иммуноглобулина и константные домены тяжелой цепи иммуноглобулина CH2, CH3;
L1, L2, L3 и L4 означают аминокислотные линкеры;
и где полипептиды формулы I и полипептиды формулы II образуют пару легкая цепь - тяжелая цепь в ориентации крест-накрест.
Изобретение также относится к антитело-подобному связывающему белку, содержащему две полипептидных цепи, которые образуют два антигенсвязывающих участка, при этом первая полипептидная цепь имеет структуру, представленную формулой:
и вторая полипептидная цепь имеет структуру, представленную формулой:
где:
VL1 означает первый вариабельный домен легкой цепи иммуноглобулина;
VL2 означает второй вариабельный домен легкой цепи иммуноглобулина;
VH1 означает первый вариабельный домен тяжелой цепи иммуноглобулина;
VH2 означает второй вариабельный домен тяжелой цепи иммуноглобулина;
CL означает константный домен легкой цепи иммуноглобулина;
CH1 означает константный домен тяжелой цепи иммуноглобулина CH1;
L1, L2, L3 и L4 означают аминокислотные линкеры;
и при этом первый и второй полипептиды образуют пару легкая цепь - тяжелая цепь в ориентации крест-накрест.
Изобретение, кроме того, относится к способу получения антитело-подобного связывающего белка, содержащего четыре полипептидных цепи, которые образуют четыре антигенсвязывающих участка, включающему в себя идентификацию первого вариабельного домена антитела, который связывает первый антиген-мишень, и второго вариабельного домена антитела, который связывает второй антиген-мишень, при этом каждый содержит VL и VH; определение либо легкой цепи, либо тяжелой цепи в качестве матричной цепи; определение VL первого вариабельного домена антитела или второго вариабельного домена антитела в качестве VL1; определение VL2, VH1 и VH2 согласно формулам [I] и [II], приведенным ниже:
определение максимальной и минимальной длины L1, L2, L3 и L4; создание полипептидных структур формул I и II; отбор полипептидных структур формул I и II, которые связывают первый антиген-мишень и второй антиген-мишень при объединении с образованием антитело-подобного связывающего белка;
где:
VL1 означает первый вариабельный домен легкой цепи иммуноглобулина;
VL2 означает второй вариабельный домен легкой цепи иммуноглобулина;
VH1 означает первый вариабельный домен тяжелой цепи иммуноглобулина;
VH2 означает второй вариабельный домен тяжелой цепи иммуноглобулина;
CL означает константный домен легкой цепи иммуноглобулина;
CH1 означает константный домен тяжелой цепи иммуноглобулина CH1;
Fc означает шарнирную область иммуноглобулина и константные домены тяжелой цепи иммуноглобулина CH2, CH3; и
L1, L2, L3 и L4 означают аминокислотные линкеры;
и при этом полипептиды формулы I и полипептиды формулы II образуют пару легкая цепь - тяжелая цепь в ориентации крест-накрест.
Изобретение, кроме того, относится к способу получения антитело-подобного связывающего белка, содержащего четыре полипептидных цепи, которые образуют четыре антигенсвязывающих участка, включающему в себя идентификацию первого вариабельного домена антитела, который связывает первый антиген-мишень, и второго вариабельного домена антитела, который связывает второй антиген-мишень, при этом каждый содержит VL и VH; определение либо легкой цепи, либо тяжелой цепи в качестве матричной цепи; определение VL первого вариабельного домена антитела или второго вариабельного домена антитела в качестве VL1; определение VL2, VH1 и VH2 согласно формулам [I] и [II], приведенным ниже:
определение максимальной и минимальной длины L1, L2, L3 и L4; создание полипептидных структур формулы I и II; отбор полипептидных структур формул I и II, которые связывают первый антиген-мишень и второй антиген-мишень при объединении с образованием антитело-подобного связывающего белка;
где:
VL1 означает первый вариабельный домен легкой цепи иммуноглобулина;
VL2 означает второй вариабельный домен легкой цепи иммуноглобулина;
VH1 означает первый вариабельный домен тяжелой цепи иммуноглобулина;
VH2 означает второй вариабельный домен тяжелой цепи иммуноглобулина;
CL означает константный домен легкой цепи иммуноглобулина;
CH1 означает константный домен тяжелой цепи иммуноглобулина CH1; и
L1, L2, L3 и L4 означают аминокислотные линкеры;
и при этом полипептиды формулы I и полипептиды формулы II образуют пару легкая цепь - тяжелая цепь в ориентации крест- накрест.
Конкретные варианты осуществления изобретения станут очевидными на основании следующего подробного описания некоторых вариантов и формулы изобретения.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
Фигура 1. Схематичное представление антигенсвязывающих доменов Fv1 и Fv2 в конфигурации двойной V-области и расположение соответствующих им пептидных линкеров LL и LH в форме TBTI.
Фигура 2. Схематичная диаграмма (2D) антигенсвязывающих доменов Fv1 (против IL4) и Fv2 (против IL13) в конфигурации двойной вариабельной области крест-накрест (CODV) и расположение соответствующих им пептидных линкеров.
Фигура 3. Схематичное представление Fv против IL4 и Fab против IL13, показывающее одно возможное пространственное расположение, получаемое в результате белок-белкового докинга Fv против IL4 и Fv против IL13.
Фигура 4. Оценка тетравалентной и биспецифичной способности связывания белка CODV в анализе BIACORE при инъекции двух антигенов последовательно или одновременно на чип, покрытый белком DVD-Ig. Максимальный сигнал, наблюдаемый при последовательной инъекции, может быть получен при совместной инъекции двух антигенов, что свидетельствует о насыщении всех участков связывания.
Фигура 5. Схематичная диаграмма (2D) антигенсвязывающих доменов в конфигурации CODV и расположение соответствующего им пептидного линкера LL (L1 и L2) и LH (L3 и L4). На панели A легкая цепь поддерживается в «линейном или матричном» расположении, тогда как тяжелая цепь находится в конфигурации «крест-накрест». На панели B тяжелая цепь поддерживается в «линейном или матричном» расположении, а легкая цепь находится в конфигурации «крест-накрест».
Фигура 6. Схематичное представление конструкции CODV-Ig на основе того, какую - легкую цепь или тяжелую цепь, используют в качестве «матричной».
Фигура 7. Сравнение молекул TBTI/DVD-Ig или CODV-Ig, включающих последовательности против IL4 и против IL13.
Фигура 8. Сравнение форм CODV-Fab и B-Fab в анализе цитотоксичности с использованием клеток NALM-6.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Изобретение относится к антитело-подобным связывающим белкам, содержащим четыре полипептидных цепи, которые образуют четыре антигенсвязывающих участка, при этом каждая пара полипептидов, образующих антитело-подобный связывающий белок, обладает двойными вариабельными доменами, имеющими ориентацию крест-накрест. Изобретение также относится к способам получения таких антиген-подобных связывающих белков.
С использованием компьютерного моделирования было спрогнозировано, что дизайн с двумя головками крест-накрест (CODH), описанный в патенте США № 5989830, может давать комплекс, в котором оба связывающих участка находятся лицом к лицу в противоположном направлении без ограничений, предполагаемых в случае конфигурации с двойным Fv, описанной в патенте США № 7612181. В частности, компьютерное моделирование показало, что длина аминокислотных линкеров между вариабельными доменами не является критически важной в случае дизайна CODH, но важна для обеспечения полного доступа к обоим антигенсвязывающим участкам в случае дизайна с двойным Fv. Как в случае формы DVD-Ig/TBTI, были получены конструкции антитело-подобных связывающих белков, в которых константные домены были связаны с конфигурацией CODH с образованием антитело-подобных связывающих белков, содержащих четыре полипептидных цепи, которые образуют четыре антигенсвязывающих участка, при этом каждая пара полипептидов, образующая антитело-подобный связывающий белок, обладает двойными вариабельными доменами, имеющими ориентацию крест-накрест (т.е., CODH-Ig). Молекулы CODH-Ig предположительно обладают значительно повышенной стабильностью по сравнению с молекулами CODH (так как DVD-Ig/TBTI обладали повышенной стабильностью по сравнению с молекулами с двойным Fv).
Чтобы проверить указанные выше гипотезы, получали молекулу CODH-Ig, используя последовательности антител против IL4 и против IL13, описанные в публикации заявки на выдачу патента США № US 2010/0226923 A1. Молекула CODH-Ig отличалась от молекулы CODH, описанной в US 2010/0226923, в отношении длин аминокислотных линкеров, разделяющих вариабельные домены на соответствующих полипептидных цепях. Молекулы CODH-Ig экспрессировали в клетках после временной трансфекции и затем очищали хроматографией на белке A. Хотя на профилях эксклюзионной хроматографии по размеру (SEC) в них наблюдали уровни агрегации 5-10%, ни одна из молекул CODH-Ig не была функциональной и, следовательно, ни одна из молекул CODH-Ig не была способна связывать ни один из своих антигенов-мишеней. Отсутствие антигенсвязывающей активности могло быть следствием нарушенной димеризации Fv-областей тяжелой и легкой цепей из-за неподходящих длин линкеров, мешающих образованию правильного паратопа. В результате был разработан протокол для идентификации подходящих аминокислотных линкеров для инсерции между двумя вариабельными доменами и вторым вариабельным доменом и константным доменом на обеих тяжелой и легкой полипептидных цепях антитело-подобного связывающего белка. Такой протокол был основан на белок-белковом докинге по гомологии и экспериментальных моделях областей FvIL4 и FvIL13, соответственно, включении домена Fc1 в модель и конструировании подходящих линкеров между областями FvIL4 и FvIL13 и между Fv и константными областями Fc1.
Применяли стандартную методику рекомбинантной ДНК для конструирования полинуклеотидов, которые кодируют полипептиды, которые образуют антитело-подобные связывающие белки согласно изобретению, включали такие полинуклеотиды в рекомбинантные экспрессирующие векторы и вводили такие векторы в клетки-хозяева. См., например, публикацию Sambrook с соавторами, 2001, Molecular Cloning: A Laboratory Manual (Cold Spring Harbor Laboratory Press, 3rd ed.). Ферментативные реакции и способы очистки могут быть осуществлены согласно инструкциям производителя, как обычно делают в данной области, или как описано в настоящей публикации. Если не приведены специальные определения, номенклатура, используемая в связи с лабораторными способами и методиками аналитической химии, химии органического синтеза и медицинской и фармацевтической химии, описанными в настоящей публикации, представляет собой номенклатуру, хорошо известную и обычно используемую в данной области. Подобным образом, можно применять обычные методики химического синтеза, химического анализа, получения, приготовления, доставки фармацевтических препаратов и лечения пациентов.
1. Общие определения
В используемом в соответствии с настоящим описанием смысле следует понимать, что следующие далее термины, если не указано иное, имеют следующее значение. Если контекст не требует иное, термины, употребляемые в единственном числе должны включать и формы множественного числа, а термины, употребляемые во множественном числе должны включать и формы единственного числа.
Термин «полинуклеотид» в используемом в настоящем описании смысле относится к однонитевым или двунитевым полимерам нуклеиновых кислот длиной, по меньшей мере, 10 нуклеотидов. В некоторых вариантах нуклеотиды, составляющие полинуклеотид, могут представлять собой рибонуклеотиды или дезоксирибонуклеотиды или модифицированную форму любого типа нуклеотида. Такие модификации включают модификации оснований, такие как бромуридин, модификации рибозы, такие как арабинозид и 2’,3’-дидезоксирибоза, и модификации межнуклеотидных связей, такие как фосфоротиоат, фосфородитиоат, фосфороселеноат, фосфородиселеноат, фосфороанилотиоат, фосфораниладат и фосфороамидат. Термин «полинуклеотид» специально включает однонитевые и двунитевые формы ДНК.
Термин «изолированный полинуклеотид» означает полинуклеотид геномного, кДНК или синтетического происхождения, или их определенного сочетания, при этом в силу своего происхождения изолированный полинуклеотид: (1) не ассоциирован со всем или частью полинуклеотида, в составе которого изолированный полинуклеотид встречается в природе, (2) связан с полинуклеотидом, с которым он не связан в природе, или (3) не встречается в природе в виде части более крупной последовательности.
«Изолированный полипептид» представляет собой полипептид, который: (1) не содержит, по меньшей мере, некоторых других полипептидов, с которыми он может быть найден в природе, (2) по существу не содержит других полипептидов из того же источника, например, из того же самого вида, (3) экспрессируется клеткой другого вида, (4) был отделен, по меньшей мере, примерно от 50 процентов полинуклеотидов, липидов, углеводов или других веществ, с которыми он ассоциирован в природе, (5) не ассоциирован (в результате ковалентного или нековалентного взаимодействия) с частями полипептида, с которыми «изолированный полипептид» ассоциирован в природе, (6) функционально ассоциирован (в результате ковалентного или нековалентного взаимодействия) с полипептидом, с которым он ассоциирован в природе, или (7) не встречается в природе. Такой изолированный полипептид может быть закодирован в геномной ДНК, кДНК, мРНК или другой РНК синтетического происхождения или в любом их сочетании. Предпочтительно изолированный полипептид по существу не содержит полипептидов или других примесей, которые встречаются в его природном окружении, которые могут мешать его применению (терапевтическому, диагностическому, профилактическому, исследовательскому или иному).
Термин «антитело человека» в используемом в настоящем описании смысле включает антитела, имеющие вариабельные и константные области, по существу соответствующие последовательностям иммуноглобулина зародышевой линии человека. В некоторых вариантах антитела человека получают в организме млекопитающих, отличных от человека, включая без ограничения грызунов, таких как мыши и крысы, и зайцеобразных, таких как кролики. В других вариантах антитела человека получают в клетках гибридом. В следующих вариантах антитела человека получают рекомбинантно.
Встречающиеся в природе антитела обычно содержат тетрамер. Каждый такой тетрамер обычно состоит из двух идентичных пар полипептидных цепей, при этом каждая пара имеет одну полноразмерную «легкую» цепь (обычно имеющую молекулярную массу приблизительно 25 кДа) и одну полноразмерную «тяжелую» цепь (обычно имеющую молекулярную массу приблизительно 50-70 кДа). Термины «тяжелая цепь» и «легкая цепь» в используемом в настоящем описании смысле относятся к любому полипептиду иммуноглобулина, имеющему достаточную последовательность вариабельного домена, чтобы придать специфичность по отношению к антигену-мишени. Аминоконцевая часть каждой легкой и тяжелой цепи обычно содержит вариабельный домен длиной приблизительно от 100 до 110 или больше аминокислот, который обычно отвечает на узнавание антигена. Находящаяся на карбоксильном конце часть каждой цепи обычно определяет границы константного домена, ответственного за эффекторную функцию. Таким образом, во встречающемся в природе антителе полноразмерный полипептид тяжелой цепи иммуноглобулина содержит вариабельный домен (VH) и три константных домена (CH1, CH2 и CH3), при этом домен VH находится на амино-конце полипептида, а домен CH3 находится на карбоксильном конце, и полноразмерный полипептид легкой цепи иммуноглобулина содержит вариабельный домен (VL) и константный домен (CL), при этом домен VL находится на амино-конце полипептида, а домен CL находится на карбоксильном конце.
Легкие цепи человека обычно классифицируют как легкие цепи каппа и лямбда, и тяжелые цепи человека обычно классифицируют как мю, дельта, гамма, альфа или эпсилон и определяют изотип антитела как IgM, IgD, IgG, IgA и IgE, соответственно. IgG имеет несколько подклассов, включая без ограничения IgG1, IgG2, IgG3 и IgG4. IgM имеет подклассы, включая без ограничения IgM1 и IgM2. IgA подобным образом подразделяют на подклассы, включая без ограничения IgA1 и IgA2. В полноразмерных легких и тяжелых цепях вариабельные и константные домены обычно связаны областью «J», состоящей приблизительно из 12 или больше аминокислот, при этом тяжелая цепь также включает область «D» приблизительно еще из 10 аминокислот. См., например, публикацию Fundamental Immunology (Paul, W., ed., Raven Press, 2nd ed., 1989), которая включена в виде ссылки в полном объеме для всех целей. Вариабельные области каждой пары легкая/тяжелая цепи обычно образуют антигенсвязывающий участок. Вариабельные домены встречающихся в природе антител обычно имеют одну и ту же общую структуру относительно консервативных каркасных областей (FR), связанных тремя гипервариабельными областями, также называемыми определяющими комплементарность областями или CDR. CDR из двух цепей каждой пары обычно выстроены вряд с каркасными областями, которые могут обеспечивать связывание с конкретным эпитопом. Начиная с амино-конца и до карбоксильного конца вариабельные домены и легкой и тяжелой цепей обычно содержат домены FR1, CDR1, FR2, CDR2, FR3, CDR3 и FR4.
Термин «нативный Fc» в используемом в настоящем описании смысле относится к молекуле, содержащей последовательность не связывающего антиген фрагмента, получаемого в результате расщепления антитела или получаемого другими способами, либо в мономерной, либо в мультимерной форме, и может содержать шарнирную область. Исходный иммуноглобулиновый источник нативного Fc предпочтительно имеет человеческое происхождение и может представлять собой любой из иммуноглобулинов, хотя предпочтительными являются IgG1 и IgG2. Молекулы нативного Fc состоят из мономерных полипептидов, которые могут быть связаны в димерные или мультимерные формы в результате ковалентного (т.е., дисульфидными связями) и нековалентного связывания. Количество межмолекулярных дисульфидных связей между мономерными субъединицами нативных молекул Fc находится в диапазоне от 1 до 4, в зависимости от класса (например, IgG, IgA и IgE) или подкласса (например, IgG1, IgG2, IgG3, IgA1 и IgA2). Одним из примеров нативного Fc является связанный дисульфидными связями димер, получаемый в результате расщепления IgG папаином. Термин «нативный Fc» в используемом в настоящем описании смысле является родовым термином для мономерных, димерных и мультимерных форм.
Термин «вариант Fc» в используемом в настоящем описании смысле относится к молекуле или последовательности, которая модифицирована по сравнению с нативным Fc, но все еще содержит участок связывания для рецептора спасения, FcRn (неонатальный Fc-рецептор). Примеры вариантов Fc и их взаимодействия с рецептором спасения известны в данной области. Таким образом, термин «вариант Fc» может включать молекулу или последовательность, которая гуманизирована по сравнению с нативным Fc животного, отличного от человека. Кроме того, нативный Fc содержит области, которые могут быть удалены, поскольку они обеспечивают структурные признаки или биологическую активность, которая не требуется для антитело-подобных связывающих белков согласно изобретению. Таким образом, термин «вариант Fc» включает молекулу или последовательность, в которой отсутствует один или несколько участков или остатков нативного Fc, или в котором модифицированы один или несколько участков или остатков Fc, которые влияют или вовлечены в: (1) образование дисульфидных связей, (2) несовместимость с выбранной клеткой-хозяином, (3) N-концевую гетерогенность при экспрессии в выбранной клетке-хозяине, (4) гликозилирование, (5) взаимодействие с комплементом, (6) связывание с другим Fc-рецептором, отличным от рецептора спасения, или (7) зависимую от антител клеточную цитотоксичность (ADCC).
Термин «Fc-домен» в используемом в настоящем описании смысле охватывает нативный Fc и варианты Fc и последовательности, которые определены выше. Как в случае вариантов Fc и нативных молекул Fc, термин «Fc-домен» включает молекулы в мономерной или мультимерной форме, либо отщепляемый от целого антитела, либо получаемый другими способами.
Термин «антитело-подобный связывающий белок» в используемом в настоящем описании смысле, относится к не встречающейся в природе (или рекомбинантной) молекуле, которая специфично связывается, по меньшей мере, с одним антигеном-мишенью и которая содержит четыре полипептидных цепи, которые образуют четыре антигенсвязывающих участка, при этом две полипептидных цепи имеют структуру, представленную формулой:
и две полипептидные цепи имеют структуру, представленную формулой:
где:
VL1 означает первый вариабельный домен легкой цепи иммуноглобулина;
VL2 означает второй вариабельный домен легкой цепи иммуноглобулина;
VH1 означает первый вариабельный домен тяжелой цепи иммуноглобулина;
VH2 означает второй вариабельный домен тяжелой цепи иммуноглобулина;
CL означает константный домен легкой цепи иммуноглобулина;
CH1 означает константный домен тяжелой цепи иммуноглобулина CH1;
Fc означает шарнирную область иммуноглобулина и константные домены тяжелой цепи иммуноглобулина CH2, CH3;
L1, L2, L3 и L4 означают аминокислотные линкеры;
и где полипептиды формулы I и полипептиды формулы II образуют пару легкая цепь - тяжелая цепь в ориентации крест-накрест. Термин «антитело-подобный связывающий белок» в используемом в настоящем описании смысле, также относится к не встречающейся в природе (или рекомбинантной) молекуле, которая специфично связывается, по меньшей мере, с одним антигеном-мишенью и которая содержит две полипептидных цепи, которые образуют два антигенсвязывающих участка, при этом первая полипептидная цепь имеет структуру, представленную формулой:
и две полипептидные цепи имеют структуру, представленную формулой:
где:
VL1 означает первый вариабельный домен легкой цепи иммуноглобулина;
VL2 означает второй вариабельный домен легкой цепи иммуноглобулина;
VH1 означает первый вариабельный домен тяжелой цепи иммуноглобулина;
VH2 означает второй вариабельный домен тяжелой цепи иммуноглобулина;
CL означает константный домен легкой цепи иммуноглобулина;
CH1 означает константный домен тяжелой цепи иммуноглобулина CH1;
L1, L2, L3 и L4 означают аминокислотные линкеры;
и где первый и второй полипептиды образуют пару легкая цепь - тяжелая цепь в ориентации крест-накрест. «Рекомбинантная» молекула представляет собой молекулу, которая была получена, экспрессирована, создана или выделена способами рекомбинации.
Один вариант осуществления изобретения относится к антитело-подобным связывающим белкам, обладающим биологической и иммунологической специфичностью к одному-четырем антигенам-мишеням. Другой вариант осуществления изобретения относится к молекулам нуклеиновых кислот, содержащим нуклеотидные последовательности, кодирующие полипептидные цепи, которые образуют такие антитело-подобные связывающие белки. Другой вариант осуществления изобретения относится к экспрессирующим векторам, содержащим молекулы нуклеиновых кислот, содержащие нуклеотидные последовательности, кодирующие полипептидные цепи, которые образуют такие антитело-подобные связывающие белки. Еще один вариант осуществления изобретения относится к клеткам-хозяевам, которые экспрессируют такие антитело-подобные связывающие белки (т.е., содержащим молекулы нуклеиновых кислот или векторы, кодирующие полипептидные цепи, которые образуют такие антитело-подобные связывающие белки).
Термин «способность меняться местами» в используемом в настоящем описании смысле относится к взаимозаменяемости вариабельных доменов в форме CODV c сохранением укладки и конечной аффинности связывания. «Способность полностью меняться местами» относится к способности менять порядок доменов VH1 и VH2 и, следовательно, порядок доменов VL1 и VL2 в CODV-Ig (т.е., менять порядок на противоположный) или CODV-Fab, при сохранении полной функциональности антитело-подобного связывающего белка, о которой свидетельствует сохранение аффинности связывания. Кроме того, следует отметить, что указание VH и VL в конкретном CODV-Ig или CODV-Fab относится только к локализации домена в конкретной белковой цепи в конечной форме. Например, VH1 и VH2 могут быть получены из доменов VL1 и VL2 в исходных антителах и помещены в положения VH1 и VH2 в антитело-подобном связывающем белке. Подобным образом VL1 и VL2 могут быть получены из доменов VH1 и VH2 в исходных антителах и помещены в положения VH1 и VH2 в антитело-подобном связывающем белке. Таким образом, указания VH и VL относятся к настоящей локализации, а не к исходной локализации в исходном антителе. Поэтому домены VH и VL «могут меняться местами».
«Изолированный» антитело-подобный связывающий белок представляет собой белок, который был идентифицирован и отделен и/или извлечен из смеси с компонентом его природного окружения. Загрязняющими компонентами из его природного окружения являются вещества, которые могут мешать диагностическим или терапевтическим применениям антитело-подобного связывающего белка, и они могут включать ферменты, гормоны и другие белковые или небелковые растворимые вещества. В предпочтительных вариантах антитело-подобный связывающий белок может быть очищен: (1) до более чем 95% масс. антитела, судя по определению способом Лоури, и наиболее предпочтительно более чем на 99% масс., (2) в достаточной степени, чтобы получить, по меньшей мере, 15 остатков N-концевой или внутренней аминокислотной последовательности, с использованием секвенатора с вращающимся стаканом, или (3) до гомогенности в SDS-ПААГ в восстанавливающих или не восстанавливающих условиях, с использованием окрашивания Кумасси синим, или, предпочтительно, серебром. Изолированные антитело-подобные связывающие белки включают антитело-подобный связывающий белок in situ в рекомбинантных клетках, поскольку в этом случае отсутствует, по меньшей мере, один компонент природного окружения антитело-подобного связывающего белка.
Термины «по существу чистый» или «по существу очищенный» в используемом в настоящем описании смысле, относятся к соединению или виду соединения, который является преобладающим присутствующим видом (т.е., при расчете в молях он является более широко представленным, чем любой другой вид в композиции). В некоторых вариантах по существу очищенная фракция представляет собой композицию, в которой данный вид составляет, по меньшей мере, приблизительно 50% (при расчете в молях) от всех присутствующих видов макромолекул. В других вариантах по существу чистая композиция будет содержать более чем приблизительно 80%, 85%, 90%, 95% или 99% от всех видов макромолекул, присутствующих в композиции. В следующих вариантах данный вид очищают по существу до гомогенности (загрязняющие виды не могут быть выявлены в композиции обычными способами детекции), при этом композиция состоит по существу из одного вида макромолекул.
Термин «антиген» или «антиген-мишень» в используемом в настоящем описании смысле относится к молекуле или части молекулы, которая может быть связана антитело-подобным связывающим белком, и дополнительно может быть использована у животного для продуцирования антител, способных связываться с эпитопом такого антигена. Антиген-мишень может иметь один или несколько эпитопов. По отношению к каждому антигену-мишени, узнаваемому антитело-подобным связывающим белком, антитело-подобный связывающий белок способен конкурировать с интактным антителом, которое узнает данный антиген-мишень. Следует понимать, что «бивалентный» антитело-подобный связывающий белок, отличный от «полиспецифичного» или «полифункционального» антитело-подобного связывающего белка, содержит антигенсвязывающие участки, обладающие идентичными антигенными специфичностями.
Биспецифичное или бифункциональное антитело обычно представляет собой искусственное гибридное антитело, имеющее две разных пары тяжелая цепь/легкая цепь и два разных участка связывания или эпитопа. Биспецифичные антитела могут быть получены различными способами, включая без ограничения слияние гибридом или связывание F(ab’)-фрагментов.
F(ab)-фрагмент обычно содержит одну легкую цепь и домены VH и CH1 одной тяжелой цепи, при этом часть тяжелой цепи VH-CH1 F(ab)-фрагмента не может образовывать дисульфидную связь с другим полипептидом тяжелой цепи. В используемом в настоящем описании смысле, F(ab)-фрагмент также может включать в себя одну легкую цепь, содержащую два вариабельных домена, разделенных аминокислотным линкером, и одну тяжелую цепь, содержащую два вариабельных домена, разделенных аминокислотным линкером, и домен CH1.
F(ab’)-фрагмент обычно включает в себя одну легкую цепь и часть одной тяжелой цепи, которая содержит дополнительную константную область (между доменами CH1 и CH2), так что может быть образована межцепочечная дисульфидная связь между двумя тяжелыми цепями с образованием молекулы F(ab’)2.
Фразы «биологическое свойство», «биологическая характеристика» и термин «активность» в отношении антитело-подобного связывающего белка согласно изобретению используют в настоящем описании взаимозаменяемо, и они охватывают без ограничения аффинность и специфичность по отношению к эпитопу, способность антагонизировать активность антигена-мишени (или полипептида-мишени), стабильность антитело-подобного связывающего белка in vivo и иммуногенные свойства антитело-подобного связывающего белка. Другие идентифицируемые биологические свойства или характеристики антитело-подобного связывающего белка включают, например, перекрестную реактивность (т.е., с гомологами антигена-мишени животных, отличных от человека, или с другими антигенами-мишенями или тканями, в общем) и способность сохранять высокие уровни экспрессии белка в клетках млекопитающих. Указанные выше свойства или характеристики можно наблюдать и измерять, используя известные в данной области способы, включая без ограничения ELISA, конкурентный ELISA, анализ резонанса поверхностного плазмона, анализы нейтрализации in vitro и in vivo и иммуногистохимию с использованием срезов тканей из разных источников, включая человека, примата или любой другой источник, который может быть необходим.
Термин «иммунологически функциональный фрагмент иммуноглобулина» в используемом в настоящем описании смысле относится к полипептидному фрагменту, который содержит, по меньшей мере, CDR тяжелой и легкой цепей иммуноглобулина, из которого получен полипептидный фрагмент. Иммунологически функциональный фрагмент иммуноглобулина способен связываться с антигеном-мишенью.
«Нейтрализующий» антитело-подобный связывающий белок в используемом в настоящем описании смысле относится к молекуле, которая способна блокировать или значимо снижать эффекторную функцию антигена-мишени, с которым она связывается. В используемом в настоящем описании смысле «значимо уменьшать» означает, по меньшей мере, приблизительно 60%, предпочтительно, по меньшей мере, приблизительно 70%, более предпочтительно, по меньшей мере, приблизительно 75%, еще более предпочтительно, по меньшей мере, приблизительно 80%, еще более предпочтительно, по меньшей мере, приблизительно 85%, наиболее предпочтительно, по меньшей мере, приблизительно 90% снижение эффекторной функции антигена-мишени.
Термин «эпитоп» включает любую детерминанту, предпочтительно полипептидную детерминанту, способную специфично связываться с иммуноглобулином или T-клеточным рецептором. В некоторых вариантах эпитопные детерминанты включают химические активные поверхностные группы молекул, таких как аминокислоты, боковые цепи сахаров, фосфорильные группы или сульфонильные группы, и в некоторых вариантах могут имеет специфичные трехмерные структурные характеристики и/или специфичные характеристики заряда. Эпитоп представляет собой область антигена, которая связывается антителом или антитело-подобным связывающим белком. В некоторых вариантах говорят, что антитело-подобный связывающий белок специфично связывает антиген, когда он предпочтительно узнает свой антиген-мишень в сложной смеси белков и/или макромолекул. В предпочтительных вариантах говорят, что антитело-подобный связывающий белок специфично связывает антиген, когда равновесная константа диссоциации ≤ 10-8 М, более предпочтительно, когда равновесная константа диссоциации ≤ 10-9 М, и наиболее предпочтительно, когда константа диссоциации ≤ 10-10 М.
Константа диссоциации (KD) антитело-подобного связывающего белка может быть определена, например, с использованием резонанса поверхностного плазмона. В общем, в анализе резонанса поверхностного плазмона в реальном времени измеряют взаимодействия при связывании между лигандом (антиген-мишень на матриксе биосенсора) и аналитом (антитело-подобным связывающим белком в растворе) на основании резонанса поверхностного плазмона (SPR), используя систему BIAcore (Pharmacia Biosensor; Piscataway, NJ). Анализ поверхностного плазмона также можно осуществлять, используя иммобилизацию аналита (антитело-подобного связывающего белка на матриксе биосенсора) и презентируя лиганд (антиген-мишень). Термин «KD» в используемом в настоящем описании смысле относится к константе диссоциации для взаимодействия между конкретным антитело-подобным связывающим белком и антигеном-мишенью.
Термин «специфично связывается» в используемом в настоящем описании смысле относится к способности антитело-подобного белка или его антигенсвязывающего фрагмента связываться с антигеном, содержащим эпитоп, с Kd, составляющим, по меньшей мере, приблизительно 1×10-6 М, 1×10-7 М, 1×10-8 М, 1×10-9 М, 1×10-10 М, 1×10-11 М, 1×10-12 М или больше, и/или связываться с эпитопом с аффинностью, которая, по меньшей мере, в два-три раза выше, чем его аффинность по отношению к неспецифичному антигену.
Термин «линкер» в используемом в настоящем описании смысле относится к одному или нескольким аминокислотным остаткам, встроенным между доменами иммуноглобулина, чтобы обеспечить достаточную подвижность доменов легких и тяжелых цепей для укладки в иммуноглобулины с двойной вариабельной областью крест-накрест. Линкер встраивают в переходный участок между вариабельными доменами или между вариабельным и константным доменами, соответственно, на уровне последовательности. Переходный участок между доменами можно идентифицировать, поскольку примерный размер доменов иммуноглобулинов хорошо известен. Точное положение переходного участка между доменами можно определить по локализации пептидных участков, которые не образуют элементы вторичной структуры, такие как бета-слои или альфа-спирали, которые показаны на основании экспериментальных данных или которые можно предположить, используя способы моделирования или прогнозирования вторичной структуры. Линкеры, описанные в настоящей публикации, называют: L1, который локализован в легкой цепи между N-концевым доменом VL1 и доменом VL2; L2, который также находится в легкой цепи и локализован между доменом VL2 и C-концевым доменом CL. Линкеры тяжелой цепи названы: L3, который локализован между N-концевым доменом VH2 и VH1; и L4, который локализован между доменами VH1 и CH1-Fc. Линкеры L1, L2, L3 и L4 независимы, но в некоторых случаях они могут иметь одну и ту же последовательность и/или длину.
Термин «вектор» в используемом в настоящем описании смысле относится к любой молекуле (например, нуклеиновой кислоте, плазмиде или вирусу), которую используют для переноса информации о кодировании в клетку-хозяина. Термин «вектор» включает молекулу нуклеиновой кислоты, которая способна к транспорту другой нуклеиновой кислоты, с которой она связана. Одним из типов векторов является «плазмида», которая относится к кольцевой двунитевой молекуле ДНК, в которую могут быть встроены дополнительные участки ДНК. Другим типом вектора является вирусный вектор, в случае которого дополнительные фрагменты ДНК могут быть встроены в вирусный геном. Некоторые векторы способны к автономной репликации в клетке-хозяине, в которую они введены (например, бактериальные векторы, имеющие бактериальное начало репликации, и эписомные векторы млекопитающих). Другие векторы (например, неэписомные векторы млекопитающих) могут быть интегрированы в геном клетки-хозяина при введении в клетку-хозяина и при этом реплицируются вместе с геномом хозяина. Кроме того, некоторые векторы способны управлять экспрессией генов, с которыми они оперативно связаны. Такие векторы в настоящем описании называют «рекомбинантными экспрессирующими векторами» (или просто «экспрессирующими векторами»). В общем, экспрессирующие векторы, применимые в способах рекомбинации ДНК, часто имеют форму плазмид. В настоящем описании термины «плазмида» и «вектор» могут быть использованы взаимозаменяемо, так как плазмида является наиболее широко применяемой формой вектора. Однако подразумевается, что изобретение включает и другие формы экспрессирующих векторов, такие как вирусные векторы (например, дефицитные по репликации ретровирусы, аденовирусы и аденоассоциированные вирусы), которые выполняют эквивалентные функции.
Термин «оперативно связанный» используют в настоящем описании по отношению к расположению фланкирующих последовательностей, при этом фланкирующие последовательности, описываемые таким образом, сконфигурированы или собраны так, чтобы они выполняли свою обычную функцию. Таким образом, фланкирующая последовательность, оперативно связанная с кодирующей последовательностью, может быть способна влиять на репликацию, транскрипцию и/или трансляцию кодирующей последовательности. Например, кодирующая последовательность оперативно связана с промотором, если промотор способен управлять транскрипцией данной кодирующей последовательности. Фланкирующая последовательность не обязательно должна быть смежной с кодирующей последовательностью, при условии, что она правильно функционирует. Таким образом, например, промежуточные нетранслируемые, но еще транскрибируемые последовательности могут присутствовать между последовательностью промотора и кодирующей последовательностью, и последовательность промотора можно еще считать «оперативно связанной» с кодирующей последовательностью.
Фраза «рекомбинантная клетка-хозяин» (или «клетка-хозяин») в используемом в настоящем изобретении смысле относится к клетке, в которую был введен рекомбинантный экспрессирующий вектор. Подразумевают, что рекомбинантная клетка-хозяин или клетка-хозяин относится не только к конкретной клетке субъекта, но также к потомству такой клетки. Так как в последующих поколениях могут иметь место некоторые модификации либо вследствие мутации, либо вследствие влияния окружающей среды, такое потомство в действительности может быть не идентичным исходной клетке, но все еще будет входить в объем термина «клетка-хозяин» в используемом в настоящем описании смысле. Широкое множество систем экспрессии в клетках-хозяевах можно использовать для экспрессии антитело-подобных связывающих белков согласно изобретению, включая системы экспрессии бактерий, дрожжей, бакуловирусов и млекопитающих (а также системы экспрессии на основе фагового дисплея). Примером подходящего бактериального экспрессирующего вектора является pUC19. Чтобы экспрессировать антитело-подобный связывающий белок рекомбинантно, клетку-хозяина трансформируют или трансфицируют одним или несколькими рекомбинантными экспрессирующими векторами, несущими фрагменты ДНК, кодирующие полипептидные цепи антитело-подобного связывающего белка, так что полипептидные цепи экспрессируются в клетке-хозяине и предпочтительно секретируются в среду, в которой культивируют клетки-хозяева, и из такой среды антитело-подобный связывающий белок может быть извлечен.
Термин «трансформация» в используемом в настоящем описании смысле относится к изменению генетических характеристик клетки, и клетка является трансформированной, когда она модифицирована таким образом, что содержит новую ДНК. Например, клетка трансформирована, когда она генетически модифицирована по сравнению с нативным состоянием. После трансформации трансформирующая ДНК может рекомбинировать с ДНК клетки посредством физической интеграции в хромосому клетки или может сохраняться в виде эписомного элемента без репликации или может реплицироваться независимо, как плазмида. Клетку считают стабильно трансформированной, когда ДНК реплицируется с делением клетки. Термин «трансфекция» в используемом в настоящем описании смысле, относится к захвату чужеродной или экзогенной ДНК клеткой, и клетка является «трансфицированной», когда экзогенная ДНК была введена внутрь через клеточную мембрану. Несколько способов трансфекции хорошо известны в данной области. Такие способы можно применять для введения одной или нескольких экзогенных молекул ДНК в подходящие клетки-хозяева.
Термин «встречающийся в природе» в используемом в настоящем описании смысле и применимо к определенному объекту относится к тому факту, что объект может быть найден в природе, а не получен человеком. Например, полинуклеотид или полипептид, который присутствует в организме (включая вирусы), который может быть выделен из источника в природе и который не был преднамеренно модифицирован человеком, является встречающимся в природе. Подобным образом «не встречающийся в природе» в используемом в настоящем описании смысле относится к объекту, который не встречается в природе или который был структурно модифицирован или синтезирован человеком.
В используемом в настоящем описании смысле названия двадцати обычных аминокислот и их сокращения соответствуют традиционному использованию. Стереоизомеры (например, D-аминокислоты) двадцати обычных аминокислот, неприродные аминокислоты, такие как α-,α-дизамещенные аминокислоты, N-алкил-аминокислоты, молочная кислота и другие необычные аминокислоты также могут быть подходящими компонентами полипептидных цепей антитело-подобных связывающих белков согласно изобретению. Примеры необычных аминокислот включают: 4-гидроксипролин, γ-карбоксиглутамат, ε-N,N,N-триметиллизин, ε-N-ацетиллизин, O-фосфосерин, N-ацетилсерин, N-формилметионин, 3-метилгистидин, 5-гидроксилизин, σ-N-метиларгинин и другие подобные аминокислоты и иминокислоты (например, 4-гидроксипролин). В используемом в настоящем описании обозначении полипептида направление влево является направлением амино-конца, а направление вправо является направлением карбоксильного конца в соответствии со стандартным использованием и правилами.
Встречающиеся в природе остатки могут быть разделены на классы на основании общих свойств боковых цепей:
(1) гидрофобные: Met, Ala, Val, Leu, Ile, Phe, Trp, Tyr, Pro;
(2) полярные гидрофильные: Arg, Asn, Asp, Gln, Glu, His, Lys, Ser, Thr;
(3) алифатические: Ala, Gly, Ile, Leu, Val, Pro;
(4) алифатические гидрофобные: Ala, Ile, Leu, Val, Pro;
(5) нейтральные гидрофильные: Cys, Ser, Thr, Asn, Gin;
(6) кислые: Asp, Glu;
(7) основные: His, Lys, Arg;
(8) остатки, которые влияют на ориентацию цепи: Gly, Pro;
(9) ароматические: His, Trp, Tyr, Phe; и
(10) ароматические гидрофобные: Phe, Trp, Tyr.
Консервативные аминокислотные замены могут заключаться в замене представителя одного из таких классов другим представителем того же класса. Консервативные аминокислотные замены могут охватывать не встречающиеся в природе аминокислотные остатки, которые обычно включают при химическом синтезе пептидов, но которые не включаются при синтезе в биологических системах. К ним относятся пептидомиметики и другие обратные или инвертированные формы аминокислотных остатков. Неконсервативные замены могут включать замену представителя одного из таких классов представителем другого класса.
Специалист в данной области может определить подходящие варианты полипептидных цепей антитело-подобных связывающих белков согласно изобретению с использованием хорошо известных методик. Например, специалист в данной области может идентифицировать подходящие области полипептидной цепи, которые могут быть изменены без нарушения активности посредственно целенаправленного воздействия на области, которые, как полагают, не важны для активности. Альтернативно, специалист в данной области может идентифицировать остатки и части молекул, которые являются консервативными в сходных полипептидах. Кроме того, даже области, которые могут быть важны для биологической активности или для структуры, могут быть подвергнуты консервативным аминокислотным заменам без нарушения биологической активности или без неблагоприятного влияния на структуру полипептида.
Термин «пациент» в используемом в настоящем описании смысле включает человека и животных.
«Расстройство» представляет собой любое состояние, при котором может быть полезным лечение с использованием антитело-подобных связывающих белков согласно изобретению. Термины «расстройство» и «состояние» используют в настоящем описании взаимозаменяемо, и термины охватывают хронические и острые расстройства или заболевания, включая такие патологические состояния, которые обусловливают предрасположенность пациента к рассматриваемому расстройству.
Термины «лечение» или «лечить» в используемом в настоящем описании смысле относится как к терапевтическому лечению, так и к профилактическим или превентивным мерам. К субъектам, нуждающимся в лечении, относятся субъекты, уже имеющие расстройство, а также субъекты, которые склонны к развитию расстройства, или субъекты, у которых необходимо предотвратить расстройство.
Термины «фармацевтическая композиция» или «терапевтическая композиция» в используемом в настоящем описании смысле относятся к соединению или композиции, способной индуцировать требуемый терапевтический эффект при правильном введении пациенту.
Термин «фармацевтически приемлемый носитель» или «физиологически приемлемый носитель» в используемом в настоящем описании смысле относится к одному или нескольким веществам в препарате, подходящим для осуществления или усиления доставки антитело-подобного связывающего белка.
Термины «эффективное количество» и «терапевтически эффективное количество» при использовании в отношении фармацевтической композиции, содержащей один или несколько антитело-подобных связывающих белков, относятся к количеству или дозе, достаточной для получения требуемого терапевтического результата. Более конкретно, терапевтически эффективное количество представляет собой количество антитело-подобного связывающего белка, достаточное для ингибирования в течение определенного периода времени одного или нескольких клинически определенных патологических процессов, ассоциированных с состоянием, подвергаемым лечению. Эффективное количество может варьировать в зависимости от конкретного используемого антитело-подобного связывающего белка, и также зависит от множества факторов и условий, связанных с пациентом, подвергаемым лечению, и тяжестью расстройства. Например, если антитело-подобный связывающий белок необходимо вводить in vivo, среди учитываемых факторов могут быть такие факторы, как возраст, масса и состояние здоровья пациента, а также кривые «доза-ответ» и данные о токсичности. Специалисты в данной области могут определить эффективное количество или терапевтически эффективное количество данной фармацевтической композиции.
Один вариант осуществления изобретения относится к фармацевтической композиции, содержащей фармацевтически приемлемый носитель и терапевтически эффективное количество антитело-подобного связывающего белка.
2. Антитело-подобные связывающие белки
В одном варианте осуществления изобретения антитело-подобные связывающие белки содержат четыре полипептидных цепи, которые образуют четыре антигенсвязывающих участка, при этом две полипептидных цепи имеют структуру, представленную формулой:
и две полипептидные цепи имеют структуру, представленную формулой:
где:
VL1 означает первый вариабельный домен легкой цепи иммуноглобулина;
VL2 означает второй вариабельный домен легкой цепи иммуноглобулина;
VH1 означает первый вариабельный домен тяжелой цепи иммуноглобулина;
VH2 означает второй вариабельный домен тяжелой цепи иммуноглобулина;
CL означает константный домен легкой цепи иммуноглобулина;
CH1 означает константный домен тяжелой цепи иммуноглобулина CH1;
Fc означает шарнирную область иммуноглобулина и константные домены тяжелой цепи иммуноглобулина CH2, CH3;
L1, L2, L3 и L4 означают аминокислотные линкеры;
и где полипептиды формулы I и полипептиды формулы II образуют пару легкая цепь - тяжелая цепь в ориентации крест-накрест.
В другом варианте осуществления изобретения антитело-подобные связывающие белки содержат две полипептидных цепи, которые образуют два антигенсвязывающих участка, при этом первая полипептидная цепь имеет структуру, представленную формулой:
и вторая полипептидная цепь имеет структуру, представленную формулой:
где:
VL1 означает первый вариабельный домен легкой цепи иммуноглобулина;
VL2 означает второй вариабельный домен легкой цепи иммуноглобулина;
VH1 означает первый вариабельный домен тяжелой цепи иммуноглобулина;
VH2 означает второй вариабельный домен тяжелой цепи иммуноглобулина;
CL означает константный домен легкой цепи иммуноглобулина;
CH1 означает константный домен тяжелой цепи иммуноглобулина CH1;
L1, L2, L3 и L4 означают аминокислотные линкеры;
и при этом первый и второй полипептиды образуют пару легкая цепь - тяжелая цепь в ориентации крест-накрест.
Антитело-подобные связывающие белки согласно изобретению могут быть получены с использованием доменов или последовательностей, полученных или производных из любого антитела человека или животного, отличного от человека, включая, например, антитела человека, мыши или гуманизированные антитела.
В некоторых антитело-подобных связывающих белках согласно изобретению длина L3, по меньшей мере, в два раза больше длины L1. В других антитело-подобных связывающих белках согласно изобретению длина L4, по меньшей мере, в два раза больше длины L2. В некоторых антитело-подобных связывающих белках согласно изобретению длина L1, по меньшей мере, в два раза больше длины L3. В других антитело-подобных связывающих белках согласно изобретению длина L2, по меньшей мере, в два раза больше длины L4.
В некоторых антитело-подобных связывающих белках согласно изобретению L1 имеет длину от 3 до 12 аминокислотных остатков, L2 имеет длину от 3 до 14 аминокислотных остатков, L3 имеет длину от 1 до 8 аминокислотных остатков, и L4 имеет длину от 1 до 3 аминокислотных остатков. В других антитело-подобных связывающих белках L1 имеет длину от 5 до 10 аминокислотных остатков, L2 имеет длину от 5 до 8 аминокислотных остатков, L3 имеет длину от 1 до 5 аминокислотных остатков, и L4 имеет длину от 1 до 2 аминокислотных остатков. В предпочтительном антитело-подобном связывающем белке L1 имеет длину 7 аминокислотных остатков, L2 имеет длину 5 аминокислотных остатков, L3 имеет длину 1 аминокислотный остаток, и L4 имеет длину 2 аминокислотных остатка.
В некоторых антитело-подобных связывающих белках согласно изобретению L1 имеет длину от 1 до 3 аминокислотных остатков, L2 имеет длину от 1 до 4 аминокислотных остатков, L3 имеет длину от 2 до 15 аминокислотных остатков, и L4 имеет длину от 2 до 15 аминокислотных остатков. В других антитело-подобных связывающих белках L1 имеет длину от 1 до 2 аминокислотных остатков, L2 имеет длину от 1 до 2 аминокислотных остатков, L3 имеет длину от 4 до 12 аминокислотных остатков, и L4 имеет длину от 2 до 12 аминокислотных остатков. В предпочтительном антитело-подобном связывающем белке L1 имеет длину 1 аминокислотный остаток, L2 имеет длину 2 аминокислотных остатка, L3 имеет длину 7 аминокислотных остатков, и L4 имеет длину 5 аминокислотных остатков.
В некоторых антитело-подобных связывающих белках согласно изобретению L1, L3 или L4 могут быть равны нулю. Однако антитело-подобные связывающие белки, в которых L1, L3 или L4 равны нулю, соответствующий переходный линкер между вариабельной областью и константной областью или между двумя вариабельными доменами на другой цепи не могут быть равны нулю. В некоторых вариантах L1 равен нулю, а L3 состоит из 2 или больше аминокислотных остатков, L3 равен нулю, а L1 состоит из 1 или больше аминокислотных остатков, или L4 равен 0, а L2 состоит из 3 или больше аминокислотных остатков.
В некоторых антитело-подобных связывающих белках согласно изобретению, по меньшей мере, один из линкеров, выбранных из группы, состоящей из L1, L2, L3 и L4, содержит, по меньшей мере, один остаток цистеина.
Примеры подходящих линкеров включают один остаток глицина (Gly); диглициновый пептид (Gly-Gly); трипептид (Gly-Gly-Gly); пептид с четырьмя остатками глицина (Gly-Gly-Gly-Gly; SEQ ID NO: 25); пептид с пятью остатками глицина (Gly-Gly-Gly-Gly-Gly; SEQ ID NO: 26); пептид с шестью остатками глицина (Gly-Gly-Gly-Gly-Gly-Gly; SEQ ID NO: 27); пептид с семью остатками глицина (Gly- Gly-Gly-Gly-Gly-Gly-Gly; SEQ ID NO: 28); пептид с восьмью остатками глицина (Gly-Gly-Gly-Gly-Gly-Gly-Gly-Gly; SEQ ID NO: 29). Также можно использовать другие сочетания аминокислотных остатков, такие как пептид Gly-Gly-Gly-Gly-Ser (SEQ ID NO: 30) и пептид Gly-Gly-Gly-Gly-Ser-Gly-Gly-Gly-Gly-Ser (SEQ ID NO: 31). Другие подходящие линкеры включают один остаток Ser и остаток Val; дипептид Arg-Thr, Gln-Pro, Ser-Ser, Thr-Lys и Ser-Leu; Thr-Lys-Gly-Pro-Ser (SEQ ID NO: 52), Thr-Val-Ala-Ala-Pro (SEQ ID NO: 53), Gln-Pro-Lys-Ala-Ala (SEQ ID NO: 54), Gln-Arg-Ile-Glu-Gly (SEQ ID NO: 55); Ala-Ser-Thr-Lys-Gly-Pro-Ser (SEQ ID NO: 48), Arg-Thr-Val-Ala-Ala-Pro-Ser (SEQ ID NO: 49), Gly-Gln-Pro-Lys-Ala-Ala-Pro (SEQ ID NO: 50) и His-Ile-Asp-Ser-Pro-Asn-Lys (SEQ ID NO: 51). Перечисленные выше примеры никоим образом не предназначены для ограничения объема изобретения, и было показано, что линкеры, содержащие аминокислоты, случайно выбранные из группы, состоящей из валина, лейцина, изолейцина, серина, треонина, лизина, аргинина, гистидина, аспартата, глутамата, аспарагина, глутамина, глицина и пролина, являются подходящими для антитело-подобных связывающих белков согласно изобретению (см. пример 12).
Индивидуальный состав и последовательность аминокислотных остатков в линкере могут варьировать в зависимости от типа элемента вторичной структуры, которую необходимо получить в линкере. Например, глицин, серин и аланин являются наилучшими для линкеров, имеющих максимальную гибкость. Определенное сочетание глицина, пролина, треонина и серина применимо в случае, если необходим более жесткий и удлиненный линкер. Любой аминокислотный остаток можно рассматривать в качестве линкера в сочетании с другими аминокислотными остатками, чтобы при необходимости сконструировать более крупные пептидные линкеры, в зависимости от требуемых свойств.
В некоторых антитело-подобных связывающих белках согласно изобретению VL1 содержит аминокислотную последовательность SEQ ID NO: 1; VL2 содержит аминокислотную последовательность SEQ ID NO: 3; VH1 содержит аминокислотную последовательность SEQ ID NO: 2; и VH2 содержит аминокислотную последовательность SEQ ID NO: 4.
В некоторых вариантах осуществления изобретения антитело-подобный связывающий белок способен специфично связывать один или несколько антигенов-мишеней. В предпочтительных вариантах осуществления изобретения антитело-подобный связывающий белок способен специфично связывать, по меньшей мере, один антиген-мишень, выбранный из группы, состоящей из B7.1, B7.2, BAFF, BlyS, C3, C5, CCL11 (эотаксин), CCL15 (MIP-1d), CCL17 (TARC), CCL19 (MIP-3b), CCL2 (MCP-1), CCL20 (MIP-3a), CCL21 (MIP-2), SLC, CCL24 (MPIF-2/эотаксин-2), CCL25 (TECK), CCL26 (эотаксин-3), CCL3 (MIP-1a), CCL4 (MIP-1b), CCL5 (RANTES), CCL7 (MCP-3), CCL8 (mcp-2), CD3, CD19, CD20, CD24, CD40, CD40L, CD80, CD86, CDH1 (E-кадгерин), хитиназы, CSF1 (M-CSF), CSF2 (GM-CSF), CSF3 (GCSF), CTLA4, CX3CL1 (SCYD1), CXCL12 (SDF1), CXCL13, EGFR, FCER1A, FCER2, HER2, IGF1R, IL-1, IL-12, IL13, IL15, IL17, IL18, ILIA, IL1B, IL1F10, IL1β, IL2, IL4, IL6, IL7, IL8, IL9, IL12/23, IL22, IL23, IL25, IL27, IL35, ITGB4 (интегрин b4), LEP (лептин), MHC класса II, TLR2, TLR4, TLR5, TNF, TNF-α, TNFSF4 (лиганд OX40), TNFSF5 (лиганд CD40), Toll-подобных рецепторов, TREM1, TSLP, TWEAK, XCR1 (GPR5/CCXCR1), DNGR-1(CLEC91) и HMGB1. В других вариантах осуществления изобретения антитело-подобный связывающий белок способен ингибировать функцию одного или нескольких антигенов-мишеней.
В некоторых вариантах осуществления изобретения антитело-подобный связывающий белок является биспецифичным и способен связывать два разных антигена-мишени или эпитопа. В предпочтительном варианте осуществления изобретения антитело-подобный связывающий белок является биспецифичным и каждая пара легкая цепь - тяжелая цепь способна связывать два разных антигена-мишени или эпитопа. В более предпочтительном варианте антитело-подобный связывающий белок способен связывать два разных антигена-мишени, которые выбраны из группы, состоящей из IL4 и IL13, IGF1R и HER2, IGF1R и EGFR, EGFR и HER2, BK и IL13, PDL-1 и CTLA-4, CTLA4 и MHC класса II, IL-12 и IL-18, IL-1α и IL-1β, TNFα и IL12/23, TNFα и IL-12p40, TNFα и IL-1β, TNFα и IL-23, и IL17 и IL23. В еще более предпочтительном варианте антитело-подобный связывающий белок способен связывать антигены-мишени IL4 и IL13.
В некоторых вариантах осуществления изобретения антитело-подобный связывающий белок специфично связывает IL4 со скоростью ассоциации 2,97 E+07 и со скоростью диссоциации 3,30 E-04 и специфично связывает IL13 со скоростью ассоциации 1,39 E+06 и скоростью диссоциации 1,63 E-04. В других вариантах осуществления изобретения антитело-подобный связывающий белок специфично связывает IL4 со скоростью ассоциации 3,16 E+07 и скоростью диссоциации 2,89 E-04 и специфично связывает IL13 со скоростью ассоциации 1,20 E+06 и скоростью диссоциации 1,12 E-04.
В одном варианте осуществления изобретения антитело-подобный связывающий белок, содержащий четыре полипептидных цепи, которые образуют четыре антигенсвязывающих участка, получают посредством идентификации первого вариабельного домена антитела, который связывает первый антиген-мишень, и второго вариабельного домена антитела, который связывает второй антиген-мишень, при этом каждый содержит VL и VH; определение либо легкой цепи, либо тяжелой цепи в качестве матричной цепи; определение VL первого вариабельного домена антитела или второго вариабельного домена антитела в качестве VL1; определение VL2, VH1 и VH2 согласно формулам [I] и [II], приведенным ниже:
определения максимальной и минимальной длины L1, L2, L3 и L4; создания полипептидных структур формул I и II; отбора полипептидных структур формул I и II, которые связывают первый антиген-мишень и второй антиген-мишень при объединении с образованием антитело-подобного связывающего белка;
где:
VL1 означает первый вариабельный домен легкой цепи иммуноглобулина;
VL2 означает второй вариабельный домен легкой цепи иммуноглобулина;
VH1 означает первый вариабельный домен тяжелой цепи иммуноглобулина;
VH2 означает второй вариабельный домен тяжелой цепи иммуноглобулина;
CL означает константный домен легкой цепи иммуноглобулина;
CH1 означает константный домен тяжелой цепи иммуноглобулина CH1;
Fc означает шарнирную область иммуноглобулина и константные домены тяжелой цепи иммуноглобулина CH2, CH3; и
L1, L2, L3 и L4 означают аминокислотные линкеры;
и при этом полипептиды формулы I и полипептиды формулы II образуют пару легкая цепь - тяжелая цепь в ориентации крест-накрест.
В другом варианте осуществления изобретения антитело-подобный связывающий белок, содержащий четыре полипептидных цепи, которые образуют четыре антигенсвязывающих участка, получают посредством идентификации первого вариабельного домена антитела, который связывает первый антиген-мишень, и второго вариабельного домена антитела, который связывает второй антиген-мишень, при этом каждый содержит VL и VH; определение либо легкой цепи, либо тяжелой цепи в качестве матричной цепи; определение VL первого вариабельного домена антитела или второго вариабельного домена антитела в качестве VL1; определение VL2, VH1 и VH2 согласно формулам [I] и [II], приведенным ниже:
определения максимальной и минимальной длины L1, L2, L3 и L4; создания полипептидных структур формул I и II; отбор полипептидных структур формул I и II, которые связывают первый антиген-мишень и второй антиген-мишень при объединении с образованием антитело-подобного связывающего белка;
где:
VL1 означает первый вариабельный домен легкой цепи иммуноглобулина;
VL2 означает второй вариабельный домен легкой цепи иммуноглобулина;
VH1 означает первый вариабельный домен тяжелой цепи иммуноглобулина;
VH2 означает второй вариабельный домен тяжелой цепи иммуноглобулина;
CL означает константный домен легкой цепи иммуноглобулина;
CH1 означает константный домен тяжелой цепи иммуноглобулина CH1; и
L1, L2, L3 и L4 означают аминокислотные линкеры;
и при этом полипептиды формулы I и полипептиды формулы II образуют пару легкая цепь - тяжелая цепь в ориентации крест-накрест.
В других вариантах осуществления изобретения получают антитело-подобный связывающий белок, в котором первый вариабельный домен антитела и второй вариабельный домен антитела являются одинаковыми.
Один вариант осуществления изобретения относится к способу получения антитело-подобного связывающего белка, включающему в себя экспрессию в клетке одной или нескольких молекул нуклеиновых кислот, кодирующих полипептиды, имеющие структуры, представленные формулами [I] и [II], приведенными ниже:
где:
VL1 означает первый вариабельный домен легкой цепи иммуноглобулина;
VL2 означает второй вариабельный домен легкой цепи иммуноглобулина;
VH1 означает первый вариабельный домен тяжелой цепи иммуноглобулина;
VH2 означает второй вариабельный домен тяжелой цепи иммуноглобулина;
CL означает константный домен легкой цепи иммуноглобулина;
CH1 означает константный домен тяжелой цепи иммуноглобулина CH1;
Fc означает шарнирную область иммуноглобулина и константные домены тяжелой цепи иммуноглобулина CH2, CH3; и
L1, L2, L3 и L4 означают аминокислотные линкеры;
и при этом полипептиды формулы I и полипептиды формулы II образуют пару легкая цепь - тяжелая в ориентации крест-накрест.
Другой вариант осуществления изобретения относится к способу получения антитело-подобного связывающего белка, включающему в себя экспрессию в клетке одной или нескольких молекул нуклеиновых кислот, кодирующих полипептиды, имеющие структуры, представленные формулами [I] и [II], приведенными ниже:
где:
VL1 означает первый вариабельный домен легкой цепи иммуноглобулина;
VL2 означает второй вариабельный домен легкой цепи иммуноглобулина;
VH1 означает первый вариабельный домен тяжелой цепи иммуноглобулина;
VH2 означает второй вариабельный домен тяжелой цепи иммуноглобулина;
CL означает константный домен легкой цепи иммуноглобулина;
CH1 означает константный домен тяжелой цепи иммуноглобулина CH1; и
L1, L2, L3 и L4 означают аминокислотные линкеры;
и при этом полипептид формулы I и полипептид формулы II образуют пару легкая цепь - тяжелая цепь в ориентации крест-накрест.
3. Применения антитело-подобных связывающих белков
Антитело-подобные связывающие белки согласно изобретению можно применять в любом известном способе анализа, таком как анализы конкурентного связывания, прямые и непрямые анализы по типу «сэндвича» и анализы иммунопреципитации для выявления и количественной оценки одного или нескольких антигенов-мишеней. Антитело-подобные связывающие белки будут связывать один или несколько антигенов-мишеней с аффинностью, которая подходит для используемого способа анализа.
Для диагностических применений в некоторых вариантах антитело-подобные связывающие белки можно метить регистрируемым остатком. Регистрируемым остатком может быть любой остаток, который прямо или опосредованно способен генерировать регистрируемый сигнал. Например, регистрируемым остатком может быть радиоактивный изотоп, такой как 3H, 14C, 32P, 35S, 125I, 99Tc, 111In или 67Ga; флуоресцирующее или хемилюминесцирующее соединение, такое как изотиоцианат флуоресцеина, родамин или люциферин; или фермент, такой как щелочная фосфатаза, β-галактозидаза или пероксидаза хрена.
Антитело-подобные связывающие белки согласно изобретению также применимы для визуализации in vivo. Антитело-подобный связывающий белок, меченый регистрируемой меткой, можно вводить животному, предпочтительно в кровяное русло, и анализировать присутствие и положение меченого антитела в организме хозяина. Антитело-подобный связывающий белок можно метить любым остатком, который можно регистрировать у животного либо с использованием ядерного магнитного резонанса, либо радиологии, либо других способов детекции, известных в данной области.
Изобретение также относится к набору, содержащему антитело-подобный связывающий белок и другие реагенты, применимые для регистрации уровней антигена-мишени в биологических образцах. Такие реагенты могут включать регистрируемую метку, блокирующую сыворотку, образцы позитивного и негативного контроля и реагенты для регистрации.
4. Композиции антитело-подобных связывающих белков и их введение
Терапевтические или фармацевтические композиции, содержащие антитело-подобные связывающие белки, входят в объем изобретения. Такие терапевтические или фармацевтические композиции могут содержать терапевтически эффективное количество антитело-подобного связывающего белка или конъюгата антитело-подобный связывающий белок - лекарственное средство в смеси с фармацевтически или физиологически приемлемым средством для приготовления препарата, выбранного в соответствии с его пригодностью для способа введения.
Приемлемые вещества для приготовления препарата предпочтительно являются нетоксичными для реципиентов в используемых дозах и концентрациях.
Фармацевтическая композиция может содержать вещества для приготовления композиции с целью модификации, поддержания или сохранения, например, pH, осмолярности, вязкости, прозрачности, окраски, изотоничности, запаха, стерильности, стабильности, скорости растворения или высвобождения, всасывания или проникновения композиции. Подходящие вещества для приготовления композиции включают без ограничения аминокислоты (такие как глицин, глутамин, аспарагин, аргинин или лизин), противомикробные средства, антиоксиданты (такие как аскорбиновая кислота, сульфит натрия или гидросульфит натрия), буферы (такие как борат, бикарбонат, трис-HCl, цитраты, фосфаты или другие органические кислоты), объемообразующие средства (такие как маннит или глицин), хелаторы (такие как этилендиаминтетрауксусная кислота (EDTA)), комплексообразующие агенты (такие как кофеин, поливинилпирролидон, бета-циклодекстрин или гидроксипропил-бета-циклодекстрин), наполнители, моносахариды, дисахариды и другие углеводы (такие как глюкоза, манноза или декстрины), белки (такие как сывороточный альбумин, желатин или иммуноглобулины), красители, ароматизаторы и разбавители, эмульгаторы, гидрофильные полимеры (такие как поливинилпирролидон), низкомолекулярные полипептиды, солеобразующие противоионы (такие как натрий), консерванты (такие как хлорид бензалкония, бензойная кислота, салициловая кислота, тимеросал, фенетиловый спирт, метилпарабен, пропилпарабен, хлоргексидин, сорбиновая кислота или пероксид водорода), растворители (такие как глицерин, пропиленгликоль или полиэтиленгликоль), сахарные спирты (такие как маннит или сорбит), суспендирующие средства, поверхностно-активные вещества или увлажнители (такие как плюроники; ПЭГ; сложные эфиры сорбита; полисорбаты, такие как полисорбат 20 или полисорбат 80; тритон; трометамин; лецитин; холестерин или тилоксапол), агенты, повышающие стабильность (такие как сахароза или сорбит), агенты, повышающие тоничность (такие как галогениды щелочных металлов - предпочтительно хлорид натрия или калия, или маннит, сорбит), носители для доставки, разбавители, эксципиенты и/или фармацевтические адъюванты (см., например, Remington's Pharmaceutical Sciences (18th Ed., A.R. Gennaro, ed., Mack Publishing Company 1990), и последующие издания, включенные в настоящее описание в виде ссылки для любой цели).
Оптимальная фармацевтическая композиция может быть определена специалистом, например, в зависимости от предполагаемого пути введения, формы доставки и необходимой дозы. Такие композиции могут влиять на физическое состояние, стабильность, скорость высвобождения in vivo и скорость клиренса in vivo антитело-подобного связывающего белка.
Основной наполнитель или носитель в фармацевтической композиции может быть водным или неводным по своей природе. Например, подходящим наполнителем или носителем для инъекции может быть вода, физиологический раствор соли или искусственная спинномозговая жидкость, возможно с добавлением других веществ, обычных в случае композиций для парентерального введения. Нейтральный забуференный физиологический раствор или физиологический раствор, смешанный с сывороточным альбумином являются дополнительными примерами наполнителей. Другие примеры фармацевтических композиций содержат трис-буфер с pH приблизительно 7,0-8,5 или ацетатный буфер с pH приблизительно 4,0-5,5, которые могут дополнительно содержать сорбит или подходящий заменитель. В одном варианте осуществления изобретения композиции антитело-подобных связывающих белков могут быть приготовлены для хранения смешиванием выбранной композиции, имеющей необходимую степень чистоты с необязательными средствами для приготовления композиций в форме лиофилизированного остатка или водного раствора. Кроме того, антитело-подобный связывающий белок может быть приготовлен в виде лиофилизата с использованием подходящих эксципиентов, таких как сахароза.
Могут быть выбраны фармацевтические композиции согласно изобретению для парентеральной доставки. Альтернативно композиции могут быть выбраны для ингаляции или для доставки через пищеварительный тракт, такой как пероральная доставка. Приготовление таких фармацевтически приемлемых композиций входит в компетенции специалиста в данной области.
Компоненты препарата присутствуют в концентрациях, которые приемлемы для места введения. Например, буферы используют для поддержания значения pH композиции на уровне физиологического pH или на немного более низком уровне pH, обычно в диапазоне pH приблизительно от 5 до приблизительно 8.
Если предполагается парентеральное введение, то терапевтические композиции для применения в настоящем изобретении могут быть в форме апирогенного парентерально приемлемого водного раствора, содержащего требуемый антитело-подобный связывающий белок в фармацевтически приемлемом наполнителе. Особенно подходящим наполнителем для парентеральной инъекции является стерильная дистиллированная вода, в которой антитело-подобный связывающий белок готовят в виде стерильного изотонического раствора с соответствующими консервантами. Еще один способ приготовления включает в себя получение композиции требуемой молекулы с агентом, например, в виде микросфер, биологически разрушаемых частиц, полимерных соединений (таких как полимолочная кислота или полигликолевая кислота), шариков или липосом, которые обеспечивают контролируемое или длительное высвобождение продукта, которая затем может быть доставлена посредством инъекции депонируемого средства. Также можно использовать гиалуроновую кислоту, и она может оказывать влияние, увеличивая продолжительность циркуляции. Другие подходящие способы введения требуемой молекулы включают имплантируемые устройства для доставки лекарственных средств.
В одном варианте может быть приготовлена фармацевтическая композиция для ингаляции. Например, антитело-подобный связывающий белок может быть приготовлен в виде сухого порошка для ингаляции. Также могут быть приготовлены растворы антитело-подобных связывающих белков для ингаляции с использованием газа-вытеснителя для аэрозольной доставки. В еще одном варианте растворы можно распылять.
Также предполагается, что некоторые препараты могут быть введены перорально. В одном варианте осуществления изобретения антитело-подобные связывающие белки, которые вводят таким образом, могут быть приготовлены с носителями или без носителей, обычно используемых в приготовлении композиций в твердых дозированных формах, таких как таблетки и капсулы. Например, может быть приготовлена капсула для высвобождения активной части препарата в определенной точке желудочно-кишечного тракта, когда биодоступность максимальна, а пресистемное разложение минимально. Могут быть включены дополнительные средства для облегчения всасывания антитело-подобного связывающего белка. Также можно использовать разбавители, корригенты, воски с низкой точкой плавления, растительные масла, скользящие вещества, суспендирующие средства, дезинтегрирующие средства для таблеток и связывающие вещества.
Другая фармацевтическая композиция может включать в себя эффективное количество антитело-подобных связывающих белков в смеси с нетоксичными эксципиентами, которые подходят для производства таблеток. При растворении таблеток в стерильной воде или другом подходящем наполнителе могут быть получены растворы в стандартной лекарственной форме. Подходящие эксципиенты включают без ограничения инертные разбавители, такие как карбонат кальция, карбонат или бикарбонат натрия, лактоза или фосфат кальция; или связывающие средства, такие как крахмал, желатин или аравийская камедь; или скользящие средства, такие как стеарат магния, стеариновая кислота или тальк.
Дополнительные фармацевтические композиции согласно изобретению будут очевидны для специалистов в данной области, включая препараты, содержащие антитело-подобные связывающие белки, в виде препаратов замедленного или контролируемого высвобождения. Способы приготовления множества других средств длительной или контролируемой доставки, таких как липосомные носители, биологически разрушаемые микрочастицы или пористые шарики и инъекционные депонируемые средства также известны специалистам в данной области. Дополнительные примеры препаратов длительного высвобождения включают полупроницаемые полимерные матрицы в виде формованных изделий, например, пленок или микрокапсул. Матрицы для длительного высвобождения могут содержать сложные полиэфиры, гидрогели, полилактиды, сополимеры L-глутаминовой кислоты и гамма-этил-L-глутамата, поли(2-гидроксиэтилметакрилат), этиленвинилацетат или поли-D-(-)-3-гидроксимасляную кислоту. Композиции длительного высвобождения также могут включать липосомы, которые могут быть получены любым из нескольких способов, известных в данной области.
Фармацевтические композиции согласно изобретению, которые необходимо применять для введения in vivo, обычно должны быть стерильными. Это можно осуществить фильтрованием через стерильные мембраны для фильтрации. В том случае, когда композицию подвергают лиофилизации, стерилизация с использованием такого способа может быть проведена до или после лиофилизации и перерастворения. Композицию для парентерального введения можно хранить в лиофилизированной форме или в растворе. Кроме того, парентеральные композиции обычно помещают в емкость, имеющую стерильное входное отверстие, например, мешок для внутривенного раствора или флакон, имеющий пробку, прокалываемую иглой для подкожной инъекции.
После того, как фармацевтическая композиция приготовлена, ее можно хранить в стерильных флаконах в виде раствора, суспензии, геля, эмульсии, твердого вещества или в виде дегидратированного или лиофилизированного порошка. Такие препараты можно хранить либо в готовой для применения форме, либо в форме (например, лиофилизированной), требующей перерастворения перед введением.
Изобретение также охватывает наборы для получения стандартных единичных доз для введения. Каждый из наборов может содержать первую емкость, в которой находится высушенный белок, и вторую емкость, содержащую водный препарат. Также в объем настоящего изобретения включены наборы, содержащие однокамерные и многокамерные предварительно заполняемые шприцы (например, шприцы с жидкостью и шприцы с лиофилизатом).
Эффективное количество фармацевтической композиции антитело-подобного связывающего белка, которое необходимо использовать терапевтически, будет зависеть, например, от терапевтического контекста и целей. Специалисту в данной области будет понятно, что соответствующие уровни доз для лечения будут, поэтому варьировать, отчасти, в зависимости от доставляемой молекулы, показания, для которого применяют антитело-подобный связывающий белок, пути введения и размера (массы тела, поверхности тела или размера органа) и состояния (возраста и общего состояния здоровья) пациента. Соответственно, лечащий врач может титровать дозу и модифицировать путь введения, чтобы получить оптимальный терапевтический эффект. Типичная доза может быть в диапазоне приблизительно от 0,1 мкг/кг до приблизительно 100 мг/кг или больше, в зависимости от факторов, указанных выше. В других вариантах доза может быть в диапазоне от 0,1 мкг/кг до приблизительно 100 мг/кг; или от 1 мкг/кг до приблизительно 100 мг/кг; или 5 мкг/кг, 10 мкг/кг, 15 мкг/кг, 20 мкг/кг, 25 мкг/кг, 30 мкг/кг, 35 мкг/кг, 40 мкг/кг, 45 мкг/кг, 50 мкг/кг, 55 мкг/кг, 60 мкг/кг, 65 мкг/кг, 70 мкг/кг, 75 мкг/кг, приблизительно до 100 мг/кг.
Частота внесения доз будет зависеть от фармакокинетических параметров антитело-подобного связывающего белка в используемом препарате. Обычно лечащий врач будет вводить композицию вплоть до достижения дозы, которая приводит к требуемому эффекту. Поэтому композицию можно вводить в виде одной дозы, в виде двух или больше доз (которые могут содержать одинаковое или неодинаковое количество требуемой молекулы) в течение определенного периода времени, или в виде непрерывной инфузии с использованием имплантируемого устройства или катетера. Специалисты в данной области обычно проводят дополнительное уточнение подходящей дозы, и такое уточнение входит в объем задач, которые они обычно решают. Подходящие дозы могут быть установлены с использованием соответствующих данных исследования зависимости доза-ответ.
Путь введения фармацевтической композиции соответствует известным способам, например, введение перорально; посредством инъекции внутривенным, внутрибрюшинным, внутрицеребральным (интрапаренхимальным), интрацеребровентрикулярным, внутримышечным, внутриглазным, внутриартериальным, интрапортальным или внутриочаговым путями; с использованием систем длительного высвобождения или с использованием имплантируемых устройств. При необходимости композиции можно вводить с помощью болюсной инъекции или непрерывно посредством инфузии или с использованием имплантируемого устройства.
Композицию также можно вводить местно путем имплантации мембраны, губки или другого подходящего материала, на котором требуемая молекула была абсорбирована или в который она была инкапсулирована. В случае применения имплантируемого устройства такое устройство можно имплантировать в любую подходящую ткань или орган, и доставка требуемой молекулы может осуществляться посредством диффузии, высвобождаемого в определенное время болюса или непрерывного введения.
5. Примеры
Примеры, которые следуют далее, являются иллюстративными примерами конкретных вариантов осуществления изобретения и его различных применений. Примеры приведены только с целью пояснения, и их никоим образом не следует интерпретировать как ограничивающие объем изобретения.
Пример 1. Дизайн и конструирование биспецифичных антитело- подобных связывающих белков с двойной вариабельной областью в ориентации «крест-накрест»
Двойная вариабельная область в ориентации крест-накрест в форме Fv описана в патенте США № 5989830 и была названа конфигурацией с двумя головками, расположенными крест-накрест (CODH). С использованием молекулярного моделирования спрогнозировано, что дизайн с двумя головками крест-накрест (CODH) приводит к образованию комплекса, в котором оба связывающих участка находятся лицом к лицу в противоположном направлении без ограничений, предполагаемых в случае конфигурации с двойным Fv. Исследовали Fv-форму CODH, чтобы определить, можно ли ее превратить в полные антитело-подобные молекулы, благодаря добавлению домена CL к легкой цепи и Fc-области к тяжелой цепи. Сходное превращение было успешным в случае соответствующих двойных вариабельных доменов (DVD-Ig) и TBTI, которые описаны в патенте США № 7612181 и международной публикации № WO 2009/052081. Расположение вариабельных областей в форме CODH показано в изображенных ниже структурах, где указана ориентация пептидных цепей от амино-конца к карбоксильному концу:
(a) легкая цепь: NH2-VL1-линкер-VL2-COOH
(b) тяжелая цепь: NH2-VH2-линкер-VH1-COOH
Расположение вариабельных областей от амино-конца к карбоксильному концу в пунктах (a) и (b) можно отличить от расположения в случае двойной Fv-конфигурации, показанной в пунктах (c) и (d) ниже:
(c) легкая цепь: NH2-VL1-линкер-VL2-COOH
(d) тяжелая цепь: NH2-VH1-линкер-VH2-COOH
Главным отличием, которое необходимо отметить, является разное расположение соответствующих вариабельных областей легкой цепи и тяжелой цепи (VH1/VL1 и VH2/VL2) по отношению друг к другу в двух конфигурациях с двойными вариабельными областями. Оба соответствующих домена VL1 и VH1 были на N-конце легкой и тяжелой цепей в конфигурации с двойными вариабельными областями. Напротив, в конфигурации крест-накрест одна половина пары вариабельной области антитела была пространственно отделена в белковой цепи в конфигурации крест-накрест. В конфигурации крест-накрест домен VL1 может быть на N-конце легкой цепи белка, но составляющий пару домен VH1 находится на C-конце тяжелой цепи в конфигурации крест-накрест. Пространственная взаимосвязь между VL1 и VH1, выявляемая в конфигурации с двойной вариабельной областью, представляет сбой расположение, встречающееся в природных антителах.
Одним возможным недостатком конфигурации с двойным Fv является то, что линкер LL, разделяющий две вариабельные области, выпячивается в антигенсвязывающий участок домена Fv2 (см. фигуру 1). Такое выпячивание может мешать связыванию антигена и приводить к нарушению доступности антигена 2 для Fv2. Такая нарушенная доступность или препятствие могут предотвращать связывание антигена. Кроме того, такое препятствие может быть тем более выраженным, чем больше размер антигена 2. Действительно, в патенте № 7612181 документально показано, что аффинность связывания и способность к нейтрализации молекулы DVD-Ig зависит от того, какая антигенная специфичность присутствует на N-конце или C-конце. См. патент США № 7612181, пример 2.
Таким образом, чтобы создать более стабильные антитело-подобные связывающие белки, которые не подвержены потере аффинности по отношению к антигену по сравнению с исходным антителом, были спроектированы и сконструированы молекулы с двойными вариабельными областями в ориентации крест-накрест, имеющие домен CL в легкой цепи и Fc-область в тяжелой цепи. Полипептиды, которые образуют такие антитело-подобные белки, имеют структуры, показанные ниже, в которых имеет место указанная ниже ориентация полипептидных цепей от амино-конца к карбоксильному концу:
(e) легкая цепь: NH2-VL1-линкер-VL2-CL-COOH
(f) тяжелая цепь: NH2-VH2-линкер-VH1-CH1-Fc-COOH
Чтобы оценить, может ли биспецифичный антитело-подобный белок такого дизайна связывать два разных антигена, две ранее созданных и гуманизированных вариабельных области из антител, специфичных по отношению к IL4 (исходное гуманизированное против IL4) и IL13 (исходное гуманизированное против IL13), использовали для конструирования биспецифичных антитело-подообных молекул, показанных в таблице 1. Секвенирование антител мыши и способ гуманизации описаны в международной публикации № WO 2009/052081 (TBTI). Вкратце, аминокислотные последовательности вариабельных областей тяжелой и легкой цепей клона B-B13 антитела мыши против IL13 и клона 8D4-8 антитела мыши против IL4 определяли, используя секвенирование аминокислот. Последовательности мыши гуманизировали и затем обратно транслировали в нуклеотидные последовательности, как описано в примере 5 международной публикации № WO 2009/052081, которая включена в настоящее описание в виде ссылки в полном объеме. Последовательности VH и VL исходных гуманизированных антител мыши против IL4 и VH и VL исходных гуманизированных антител мыши против IL13 объединяли и располагали, как описано в таблице 1. Укороченные кодовые названия в колонке один в таблице 1 создавали для упрощения обсуждения таких антитело-подобных связывающих белков. Антитело-подобные связывающие белки отличаются размером линкера, встроенного между двумя вариабельными областями, как показано в таблице 1. Молекулы ДНК, кодирующие полипептиды, показанные в таблице 1, создавали на основе обратно транслированных исходных антител мыши против IL4- и антител мыши против IL13. Домены CH1, CL и Fc получали из IGHG1 (GenBank, номер доступа 569F4) и IGKC (GenBank, номер доступа Accession Q502W4).
Иммуноглобулины с двумя головками в ориентации крест-накрест
Сочетания белков, показанные в таблице 2, экспрессировали с использованием временной трансфекции и очищали хроматографией на белке A. В каждом случае эксклюзионная хроматография по размеру выявляла агрегацию, составляющую менее чем 12%, в наилучшем случае агрегацию менее 7%; но не обнаружено ни одного иммуноглобулина с двумя головками в ориентации крест-накрест, обладающего какой-либо способностью связывать либо IL4, либо IL13. Однако не могли выявить функционального антитело-подобного связывания, и не могли выяснить причины отсутствия такой активности. Ранее предполагали, что такое расположение может давать превосходную стабильность по сравнению с антителами с двойными доменами вариабельных областей, описанными в патенте США № 7612181 и в международной публикации № WO 2009/052081.
Связывание CODH-Ig с IL4 и IL13
против IL4 VL-(G4S)-против IL13 VL-CL
против IL4 VL-(G4S)2-против IL13 VL-CL
против IL4 VL-(G4S)-против IL13 VL-CL
против IL4 VL-(G4S)2-против IL13 VL-CL
против IL13 VL-(G4S)-против IL4 VL-CL
против IL13 VL-(G4S)2-против IL4 VL-CL
против IL13 VL-(G4S)-против IL4 VL-CL
против IL13 VL-(G4S)2-против IL4 VL-CL
Пример 2. Конструирование белков CODV-Ig с использованием молекулярного моделирования
Чтобы получить полностью функциональные антитело-подобные белки с использованием конфигурации с двумя головками в ориентации крест-накрест, в которые можно включить домены Fc и CL1, разработали протокол молекулярного моделирования для включения и оценки разных линкеров между константным и вариабельным доменами и между двумя вариабельными доменами на обеих, легкой и тяжелой цепях. Вопрос заключался в том, может ли добавление уникальных линкеров между участками контакта константного/вариабельного доменов и между участками контакта двух вариабельных доменов (вариабельный/вариабельный) в тяжелой и легкой цепях, обеспечить осуществление правильного фолдинга белка и получения функциональных антитело-подобных молекул в конфигурации с двумя вариабельными областями, расположенными крест-накрест (см. фигуру 2). Другими словами, оценивали всего четыре независимых и уникальных линкера (см. фигуру 2). Такой протокол молекулярного моделирования был основан на белок-белковом докинге в моделях гомологии и экспериментальных моделях областей FvIL4 и FvIL13, соответственно, в сочетании с подходящими линкерами между областями FVIL4 и FVIL13 и между Fv и константными или Fc-областями.
Независимым линкерам даны индивидуальные названия: L1 относится к линкеру между N-концевым VL и C-концевым VL в легкой цепи; L2 относится к линкеру между C-концевым VL и CL в легкой цепи; L3 относится к линкеру между N-концевым VH и C-концевым VH в тяжелой цепи; L4 относится к линкеру между C-концевым VH и CH1 (и Fc) в тяжелой цепи. Следует отметить, что обозначения VH и VL относятся только к положению доменов в конкретной белковой цепи в конечной форме. Например, VH1 и VH2 могут быть получены из доменов VL1 и VL2 исходных антител и помещены в положения VH1 и VH2 в CODV-Ig. Подобным образом, VL1 и VL2 могут быть получены из доменов VH1 и VH2 исходных антител и помещены в положения VH1 и VH2 в CODV-Ig. Таким образом, обозначения VH и VL относятся к данному положению, а не к исходному положению в исходном антителе.
Более подробно, модель гомологии FVIL4 конструировали на основе данных, введенных в базу PDB, 1YLD (легкая цепь) и 1IQW (тяжелая цепь). Димер FvIL4 реконструировали на полученной своими силами кристаллической структуре комплекса IL13/против IL13 FabIL13 и оптимизировали. Чтобы получить оценку объема, необходимого для IL4 в случае связывания с FvIL4, кристаллическую структуру IL4 (1RCB.pdb) стыковали с моделью гомологии для FvIL4. Затем создали двадцать две предполагаемых модели комплекса, которые заслуживали дальнейшего рассмотрения.
Параллельно модель гомологии FvIL4 стыковали с FvIL13, извлеченным из полученной собственными силами кристаллической структуры комплекса IL13/FabIL13. Было найдено одно прекрасное решение, которое позволяло конструировать относительно короткие линкеры без проявления при этом стерического влияния на связывание антигена и размещение константных доменов, как в случае иммуноглобулинов с двойными вариабельными областями (см. фигуру 3). При таком расположении FvIL4 (VL1) помещали на N-конце легкой цепи, затем следовал FvIL13 (VL2) и Fc (CL1) на C-конце легкой цепи. В тяжелой цепи FvIL13 (VH2) помещали на N-конце, затем следовал FvIL4 (VH1) и константные области (CH1-CH2-CH3).
Как показано в таблице 3, модели легкой цепи свидетельствуют о том, линкер L1 между доменами VL1 и VL2 и линкер L2 между доменами VL2 и CL1 должны иметь длину от одного до трех и от нуля до двух остатков глицина, соответственно. Модели тяжелой цепи свидетельствуют о том, что линкер L3 между доменами VH2 и VH1 и линкер L4 между доменами VH1 и CH1 должны иметь длину от двух до шести и от четырех до семи остатков глицина, соответственно (см. таблицу 3 и фигуру 2). В данном примере глицин использовали в качестве прототипной аминокислоты для линкеров, но другие аминокислотные остатки также могут служить в качестве линкеров. Стабильность структуры предложенных моделей подтверждали при использовании оптимизации конформации линкеров, минимизации и расчетов молекулярной динамики. Системное сочетание между четырьмя конструкциями легкой цепи и шестью конструкциями тяжелой цепи приводило к получению 24 возможных имеющих двойную вариабельную область в ориентации крест-накрест биспецифичных антитело-подобных связывающих белков против IL4 и против IL13 (см. таблицу 4).
Предлагаемые длины линкеров
CODV-Ig для экспрессии
В таблице 4, приставка «анти» не включена, но имеется в виду, что IL13 относится к против IL13, и IL4 относится к против IL4.
Пример 3. Создание плазмид для экспрессии CODV-Ig
Молекулы нуклеиновых кислот, кодирующие вариабельные области тяжелых и легких цепей для шести тяжелых цепей и четырех легких цепей, описанных в таблице 4, создавали посредством синтеза генов в Geneart (Regensburg, Germany). Вариабельные домены легкой цепи сливали с константными доменами легкой цепи (IGKC, GenBank, № доступа Q502W4), используя расщепление эндонуклеазами рестрикции ApaLI и BsiWI и затем лигирование в сайты ApaLI/BsiWI эписомного экспрессирующего вектора pFF, аналога вектора pTT, описанного Durocher с соавторами, (2002, Nuc. Acids Res. 30(2): E9), создавая экспрессирующую плазмиду млекопитающих для экспрессии легких цепей.
Вариабельные домены тяжелой цепи сливали с вариантом «Ted» константной области тяжелой цепи человека (IGHG1, GenBank, № доступа 569F4), или альтернативно с 6x His-меченым доменом CH1 из константной области человека IGHG1, чтобы создать биспецифичный Fab. Затем домен VH расщепляли эндонуклеазами рестрикции ApaLI и ApaI и затем сливали с IGHG1 или His-меченым доменом CH1, соответственно, с использованием лигирования в сайты ApaLI/ApaI эписомного экспрессирующего вектора pFF, создавая экспрессирующие плазмиды млекопитающих для экспрессии тяжелых цепей (IgG1 или Fab, соответственно).
Пример 4. Экспрессия CODV-Ig
Экспрессирующие плазмиды, кодирующие тяжелые и легкие цепи соответствующих конструкций, размножали в клетках E. coli DH5a. Плазмиды, используемые для трансфекции, получали из E. Coli, используя набор Qiagen EndoFree Plasmid Mega.
Клетки HEK 293-FS, растущие в среде Freestyle (Invitrogen), трансфицировали указанными LC- и HC-плазмидами, кодирующими тяжелые цепи и легкие цепи, показанные в таблице 4, используя реагент для трансфекции 293fectin (Invitrogen), как описано производителем. Через 7 дней клетки извлекали центрифугированием, и надосадок пропускали через фильтр 0,22 мкм, чтобы удалить частицы.
Конструкции CODV-IgG1 очищали аффинной хроматографией на колонках с белком A (колонки HiTrap Protein A HP, GE Life Sciences). После элюирования с колонки 100 мМ ацетатным буфером и 100 мМ NaCl, pH 3,5, конструкции CODV-IgG1 обессоливали, используя колонки для обессоливания HiPrep 26/10, готовили в виде препарата в PBS в концентрации 1 мг/мл и фильтровали, используя мембрану 0,22 мкм.
Биспецифичные конструкции CODV-Fab очищали, используя IMAC на колонках HiTrap IMAC HP (GE Life Sciences). После элюирования с колонки линейным градиентом (буфер для элюирования: 20 мМ фосфат натрия, 0,5 М NaCl, 50 - 500 мМ имидазол, pH 7,4), содержащие белок фракции объединяли и обессоливали, используя колонки для обессоливания HiPrep 26/10, готовили в PBS в концентрации 1 мг/мл и фильтровали, используя мембрану 0,22 мкм.
Концентрацию белка определяли посредством измерения поглощения при 280 нм. Каждую партию анализировали в SDS-ПААГ в восстанавливающих и невосстанавливающих условиях, чтобы определить чистоту и молекулярную массу каждой субъединицы и мономера.
Планшет Nunc F96-MaxiSorp-Immuno покрывали антителом козы против-IgG человека (Fc-специфичным) [NatuTec A80-104A]. Антитело разбавляли до концентрации 10 мкг/мл в карбонатном буфере для покрывания (50 мМ карбонат натрия, pH 9,6) и разносили по 50 мкл на лунку. Планшет герметично закрывали липкой лентой, и хранили в течение ночи при 4ºC. Планшет промывали три раза буфером для промывки (PBS, pH 7,4, и 0,1% твином 20). Разносили по 150 мкл блокирующего раствора (1% БСА/PBS) в каждую лунку, чтобы покрыть планшет. После инкубации в течение 1 часа при комнатной температуре планшет три раза промывали буфером для промывки. Добавляли 100 мкл образца или стандартов (в диапазоне от 1500 нг/мл до 120 нг/мл) и давали возможность отстояться в течение 1 часа при комнатной температуре. Планшет три раза промывали буфером для промывки. Добавляли 100 мкл конъюгата антитела козы против IgG-FC человека с HRP [NatuTec A80-104P-60], разбавленного 1:10000, используя раствор для инкубации (0,1% БСА, PBS, pH 7,4, и 0,05% твин 20). После инкубации в течение 1 часа при комнатной температуре планшет три раза промывали буфером для промывки. Добавляли 100 мкл субстрата ABTS (таблетка ABTS 10 мг (Pierce 34026) в 0,1 М Na2HPO4, 0,05 М раствор лимонной кислоты, pH 5,0). Добавляли 10 мкл 30% H2O2/10 мл буфера для субстрата перед использованием, разносили в каждую лунку, и давали возможность проявиться окраске. После проявления окраски (примерно 10-15 минут), добавляли 50 мкл 1% раствора SDS, чтобы остановить реакцию. Планшет считывали при A405.
Пример 5. Характеристика вариантов CODV-Ig
Чтобы определить могут ли тяжелые и легкие цепи антитело-подобного белка CODV-Ig образовывать пары и подвергаться правильному фолдингу, измеряли уровень агрегации, используя аналитическую эксклюзионную хроматографию по размеру (SEC). Аналитическую SEC осуществляли для собранных пар с использованием анализатора AKTA 10 (GE Healthcare), оборудованного колонкой TSKgel G3000SWXL (7,8 мм × 30 см) и предколонкой TSKgel SWXL (Tosoh Bioscience). Анализ проводили с расходом 1 мл/мин, используя 250 мМ NaCl, 100 мМ Na-фосфат, pH 6,7, с регистрацией при 280 нм. На колонку наносили 30 мкл образца белка (0,5-1 мг/мл). Для оценки размера молекул колонку калибровали, используя стандартную смесь для гель-фильтрации (MWGF-1000, SIGMA Aldrich). Оценку данных осуществляли, используя компьютерную программу UNICORN v5.11.
В таблице 5 показаны результаты для первого набора из 24 разных молекул CODV-Ig, полученных с использованием сочетаний вариабельных областей против IL4 и против IL13, описанных в таблице 4. Коды, присвоенные в таблице 4, представляют примыкающие структуры, показанные в таблице 4. Для пар легкой цепи и тяжелых цепей в случае продуцирования белка измеряли уровни агрегации, используя SEC. Результаты показаны в таблице 5, где LC4 (L1=1; L2=2) является наиболее успешным в отношении образования пар со всеми шестью тяжелыми цепями. LC4 соответствует структуре IL4 VL-(Gly)-IL13 VL-(Gly2)-CL1, имеющей линкер L1, равный 1, в которой один аминокислотный остаток разделял два домена VL легкой цепи с двойной вариабельной областью. Кроме того, LC4 имел L2, равный 2, который содержал дипептидный линкер Gly-Gly между центральным VL и C-концевым CH1.
Уровни агрегации среди пар цепей тяжелой и легкой цепей
В том случае, когда получали молекулы CODV-Ig, проводили эксперимент BIACORE с использованием одной концентрации, соответствующей промежуточным концентрациям IL13 и IL4, чтобы подтвердить связывание с антигенами-мишенями. Антитело-подобные молекулы CODV-Ig, соответствующие сочетаниям LC4:HC4 и LC4:HC6, описанные в таблице 4, выбирали для оценки полного кинетического анализа с использованием резонанса поверхностного плазмона.
Как показано в таблице 5, большинство молекул CODV-Ig нельзя было получить совсем или только в виде агрегатов (до 90%). Сочетания тяжелая цепь/легкая цепь, дающие приемлемые уровни агрегации (5-10%) после одной стадии хроматографии, представляли собой сочетания с легкой цепью IL4 VL-(Gly)-IL13 VL-(Gly2)-CL1. Указанная легкая цепь была наиболее тщательно разработанной цепью в таких вариантах CODV-Ig, и служила в качестве платформы для акцептирования разных тяжелых цепей с разными составами линкеров.
1. Кинетический анализ
Две пары тяжелой и легкой цепей были выбраны для полного кинетического анализа. Рекомбинантные IL13 и IL4 человека приобретали из Chemicon (USA). Кинетическую характеристику очищенных антител осуществляли с использованием методики, основанной на резонансе поверхностного плазмона, на BIACORE 3000 (GE Healthcare). Применяли анализ улавливания с использованием видоспецифичного антитела (например, специфичного для Fc человека MAB 1302, Chemicon) для улавливания и ориентации исследуемых антител. Улавливающее антитело иммобилизовали посредством первичных аминогрупп (11000 условных единиц) на чипе CM5 для исследования (GE Life Sciences), используя стандартные способы. Анализируемое антитело улавливали, используя скорость потока 10 мкл/мин, при этом скорректированное значение, выраженное в условных единицах, которое может приводить к максимальному связыванию аналита, составляло 30 условных единиц. Измеряли кинетику связывания IL4 и IL13 человека в диапазоне концентраций от 0 до 25 нМ в HBS EP (10 мМ HEPES, pH 7,4, 150 мМ NaCl, 3 мМ EDTA и 0,005% поверхностно-активное вещество P20) при скорости потока 30 мкл/мин. Поверхности чипов регенерировали, используя 10 мМ глицина, pH 2,5. Параметры кинетики анализировали и вычисляли с помощью пакета программ BIAevaluation v4.1, используя проточную ячейку без улавливающего антитела в качестве эталона.
В таблице 6 ниже показано сравнение кинетики исходных антител BB13 (против IL13) и 8D4 (против IL4) (экспрессированных в виде IgG) с соответствующими доменами в форме CODV-Ig (таблица 4, коды LC4:HC4 и LC4:HC6). Как показано в таблице 6, не наблюдали снижения связывающих свойств конструкций CODV-Ig по отношению к соответствующим антигенам по сравнению с исходными антителами против IL13 и против IL4. Снижения скорости ассоциации, которое наблюдали в случае формы DVD-Ig/TBTI, в которых использовали такие же последовательности Fv, не происходило в случае конфигурации CODV-Ig. Противоположно направленные участки связывания должны обеспечивать связывание крупных антигенов или образования мостиков между разными клетками с помощью биспецифичной антитело-подобной конфигурацией, а также могут подходить для более широкого отбора исходных антител. Следующим преимуществом CODV-Ig являлось то, что остатки линкера не выступали в антигенсвязывающий участок, и не уменьшали доступность антигена.
Кинетический анализ LC4:HC4 и LC4:HC6
2. Совместная инъекция IL4 и IL13 для демонстрации аддитивного связывания антигена молекулой CODV-Ig
Чтобы исследовать аддитивное связывание обоих антигенов, применяли способ управляемой программой-мастером совместной инъекции, в котором один антиген инъецировали непосредственно после другого антигена после лаг-периода (IL4 затем IL13 и наоборот). Получаемый в результате уровень связывания можно сравнивать с уровнем, достигаемым с использованием смеси 1:1 обоих антигенов в той же концентрации. Чтобы показать аддитивное связывание обоих антигенов IL4 и IL13 молекулами CODV-Ig, проводили эксперимент BIACORE с сочетанием CODV-Ig [HC4:LC4], используя совместную инъекцию обоих антигенов в трех разных циклах анализа (см. фигуру 4). Совместную инъекцию осуществляли, используя 3,125 нМ IL4/25 нМ IL13 (и наоборот) и используя смесь 1:1 3,125 нМ IL4 и 25 нМ IL13. Совместную инъекцию буфера HBS-EP осуществляли в качестве эталона. Во временной точке 800 секунд идентичный уровень связывания 63 условных единицы достигали после инъекции смеси антигенов или совместной инъекции антигенов, независимо от последовательности совместной инъекции. Когда белок CODV-Ig был насыщен первым антигеном (IL4), инъецировали второй антиген (IL13) и наблюдали второй сигнал связывания. Такое наблюдение было воспроизводимо в том случае, когда последовательность инъекции антигенов меняли на противоположную. Полученные данные демонстрируют аддитивное связывание и отсутствие ингибирования связывания обоих антигенов молекулами CODV-Ig. Следовательно, конструкция CODV-Ig способная связывать оба антигена одновременно (т.е., проявляет биспецифичность) с насыщением всех участков связывания (т.е., проявляет тетравалентность).
Пример 6. Допустимость длин линкеров для CODV-Ig
Допустимость линкеров разных длин оценивали, конструируя молекулы CODV-Ig, имеющие разные сочетания длин линкеров L1, L2 в легкой цепи и L3 и L4 в тяжелой цепи. Конструкции CODV-Ig создавали с линкерами тяжелой цепи L3 и L4 с варьирующей длиной от 1 до 8 остатков в случае L3 и либо 0, либо 1 остатком в случае L4. Тяжелая цепь содержала N-концевой связывающий домен против IL4 и C-концевой связывающий домен против IL13, за которым следовал CH1-Fc. Длины линкеров легкой цепи L1 и L2 варьировали от 3 до 12 остатков в случае L1 и от 3 до 14 остатков в случае L2. Легкая цепь содержала N-концевой связывающий домен против IL13 и C-концевой связывающий домен против IL4 с последующим CL1.
1. Характеристика вариантов CODV-Ig
Определение уровня агрегации с использованием аналитической эксклюзионной хроматографии по размеру (SEC). Аналитическую SEC осуществляли с использованием анализатора AKTA 10 (GE Healthcare), оборудованного колонкой TSKgel G3000SWXL (7,8 мм × 30 см) и предколонкой TSKgel SWXL (Tosoh Bioscience). Анализ проводили с расходом 1 мл/мин, используя 250 мМ NaCl, 100 мМ Na-фосфат, pH 6,7, с регистрацией при 280 нм. На колонку наносили 30 мкл образца белка (0,5-1 мг/мл). Для оценки размера молекул колонку калибровали, используя стандартную смесь для гель-фильтрации (MWGF-1000, SIGMA Aldrich). Оценку данных осуществляли, используя компьютерную программу UNICORN v5.11.
Рекомбинантные IL13 и IL4 человека приобретали из Chemicon (USA). Рекомбинантный TNF-α приобретали из Sigma Aldrich (H8916-10 мкг), рекомбинантный IL-1β человека (201-LB/CF), рекомбинантный IL-23 человека (1290-IL/CF), рекомбинантный EGFR человека (344 ER) и рекомбинантный HER2 человека (1129-ER-50) приобретали из R&D Systems.
Анализ кинетики связывания с использованием Biacore осуществляли следующим образом. Применяли методику резонанса поверхностного плазмона на Biacore 3000 (GE Healthcare) для подробной кинетической характеристики очищенных антител. Применяли анализ улавливания с использованием видоспецифичного антитела (например, специфичного для Fc человека MAB 1302, Chemicon) для улавливания и ориентации исследуемых антител. Для определения кинетики связывания IL4 и IL13 соответствующие Fab CODV, как описано в примере 10, таблица 12, улавливали, используя набор для улавливания антител против Fab человека (GE Healthcare). Улавливающее антитело иммобилизовали посредством первичных аминогрупп (11000 условных единиц) на чипе CM5 для исследования (GE Life Sciences), используя стандартные способы. Анализируемое антитело улавливали, используя скорость потока 10 мкл/мин, при этом скорректированное значение, выраженное в условных единицах, которое может приводить к максимальному связыванию аналита, составляло 30 условных единиц. Измеряли кинетику связывания рекомбинантных IL4 и IL13 человека в диапазоне концентраций от 0 до 25 нМ в HBS EP (10 мМ HEPES, pH 7,4, 150 мМ NaCl, 3 мМ EDTA и 0,005% поверхностно-активное вещество P20) при скорости потока 30 мкл/мин. Поверхности чипов регенерировали, используя 10 мМ глицина, pH 2,5. Параметры кинетики анализировали, и вычисляли с помощью пакета программ BIAevaluation v4.1, используя проточную ячейку без улавливающего антитела в качестве эталона.
Аффинности связывания CODV-Ig, CODV-Fab и TBTI по отношению к EGFR и HER2 измеряли, используя систему матриц для исследования взаимодействия белков Proteon XPR36 (Biorad). Антигены иммобилизовали за счет реактивного по отношению к аминам связывания на сенсорных чипах GLC (Biorad). Анализировали серию разбавлений вариантов биспецифичных антител в буфере PBSET (Biorad) в режиме исследования кинетики с использованием параллельной инъекции в одну стадию (one-shot kinetics) с двойными эталонами. Данные анализировали, используя компьютерную программу Proteon Manager v3.0 (Biorad) и либо модель 1:1 Ленгмюра с переносом массы, либо модель бивалентного аналита.
В таблице 7 суммированы результаты, относящиеся к выходу, агрегации (измеряемой с использованием эксклюзионной хроматографии по размеру), и аффинности связывания CODV-Ig, имеющих сочетания линкеров разных размеров. Результаты показали, что молекулы CODV-Ig, в которых L2 равен нулю, обычно не могут быть получены или в случае получения белка они давали высокий уровень агрегации (см. партии с идентификационными номерами 101, 102, 106-111 и 132-137 в таблице 7). Таким образом, в отличие от прогнозирования на основе молекулярного моделирования, описанного в примере 2, согласно которому L2, равный нулю, входил в приемлемый диапазон, полученные результаты показывают, что переходный участок VL2-CL (или L2) требует линкер длиной, по меньшей мере, в один остаток (см. таблицу 7).
Оптимизация размеров линкеров для CODV-Ig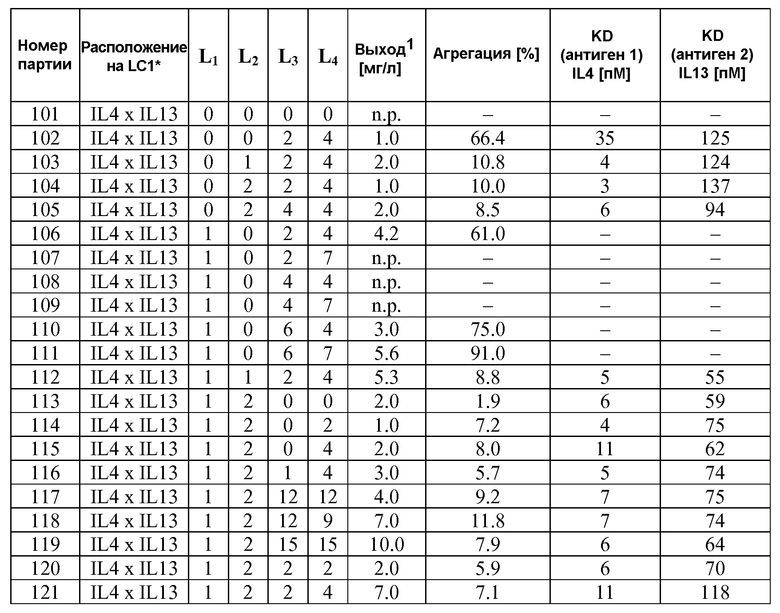
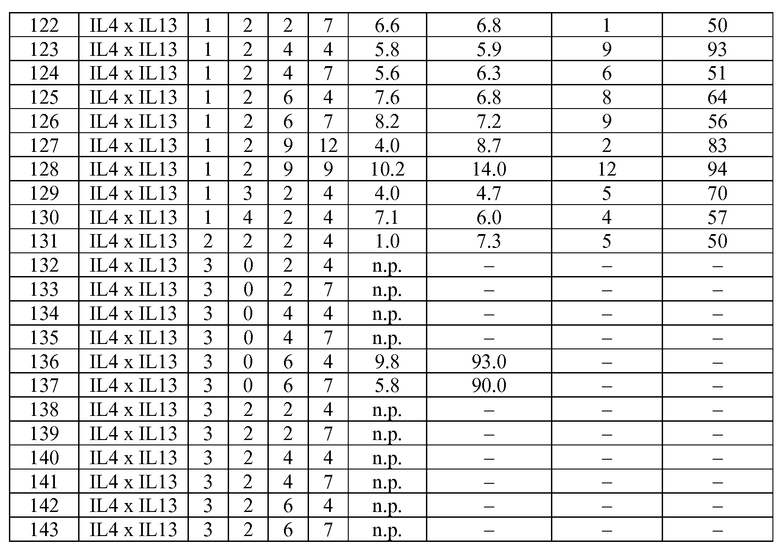
1n.p. означает, что конструкцию нельзя получить.
Кроме того, было обнаружено, длины линкеров CODV-Ig, описанные выше, являются более чувствительными к увеличению на 1 аминокислотный остаток, чем к увеличению на 2 аминокислотных остатков. Например, хотя партии № 103 и 104 отличаются на 1 аминокислотный остаток в L2, в партии № 103 наблюдали в 6 раз большую агрегацию, и в партии № 104 наблюдали меньше агрегации и в два раза более высокий выход. Напротив, в партиях № 104 и 105, которые отличаются на два остатка в L2, наблюдали сходные профили в отношении выхода, агрегации и связывания.
Пример 7. Тяжелая цепь в качестве матричной цепи для CODV-Ig
В примерах 1-5 оптимальные размеры коротких линкеров в легкой цепи свидетельствуют о том, что легкая цепь служила в качестве матрицы в случае сохранения линейного расположения, и что требовались более длинные линкеры в тяжелой цепи, чтобы тяжелая цепь правильно подвергалась фолдингу в конфигурацию крест-накрест, чтобы соответствовать матричной легкой цепи (см. фигуру 5, панель A). Затем оценивали, делали ли короткие линкеры в тяжелой цепи, специфично размещенные для поддержания линейной расположения в тяжелой цепи, такую тяжелую цепь «матричной» цепью, и могла ли картина повторяться сама по себе, и требовались ли более крупные линкеры для обеспечения возможности нематричной цепи правильно подвергаться фолдингу и вмещать новую матричную тяжелую цепь (см. фигуру 5, панель B).
Фигура 6 иллюстрирует такие принципы конструирования CODV-Ig на основе того, что в качестве «матрицы» принимают либо легкую цепь, либо тяжелую цепь. Чтобы оценить общую суть такой концепции, создавали конструкции CODV-Ig, имеющие линкеры тяжелой цепи L3 и L4, длина которых варьирует от 1 до 8 остатков в случае L3 и равна либо 0, либо 1 остатку в случае L4. Тяжелая цепь содержала N-концевой связывающий домен против IL4 и C-концевой связывающий домен против IL13, за которым следовал CH1-Fc. Длина линкеров легкой цепи L1 и L2 варьировала от 3 до 12 остатков в случае L1 и от 3 до 14 остатков в случае L2. Легкая цепь содержала N-концевой связывающий домен против IL13 и C-концевой связывающий домен против IL4 с последующим CL1.
В таблице 8 суммированы результаты, относящиеся к выходу, агрегации (измеряемой с использованием эксклюзионной хроматографии по размеру) и аффинности связывания CODV-Ig, имеющих сочетания линкеров разных размеров, и в которых тяжелую цепь сохраняли в линейном расположении в качестве матричной цепи, и легкой цепи давали возможность укладываться в конфигурацию крест-накрест. Результаты показали, что молекулы CODV-Ig, в которых L4 равен нулю, обычно не могут быть получены, или в случае получения белка они давали высокий уровень агрегации (сходный с молекулами, в которых L2 равен нулю) (см. партии с идентификационными номерами 207-209, 211-212, 219-224, 231-236, 243-252 и 263-266 в таблице 8). Одним исключением была партия № 210, в которой L1 был равен 7, L2 равен 5, L3 равен 2 и L4 равен нулю. Такое расположение давало достаточное количество белка и приемлемый уровень агрегации и связывания, что свидетельствовало о том, что в некоторых случаях можно обнаружить, что определенное сочетание размеров линкеров компенсирует линкер нулевой длины в положении L4.
Оптимизация размеров линкеров в случае тяжелой цепи в качестве матрицы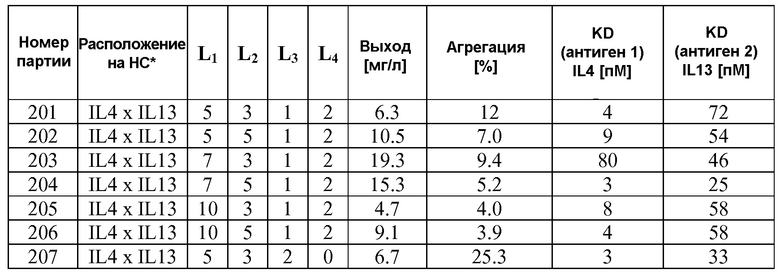
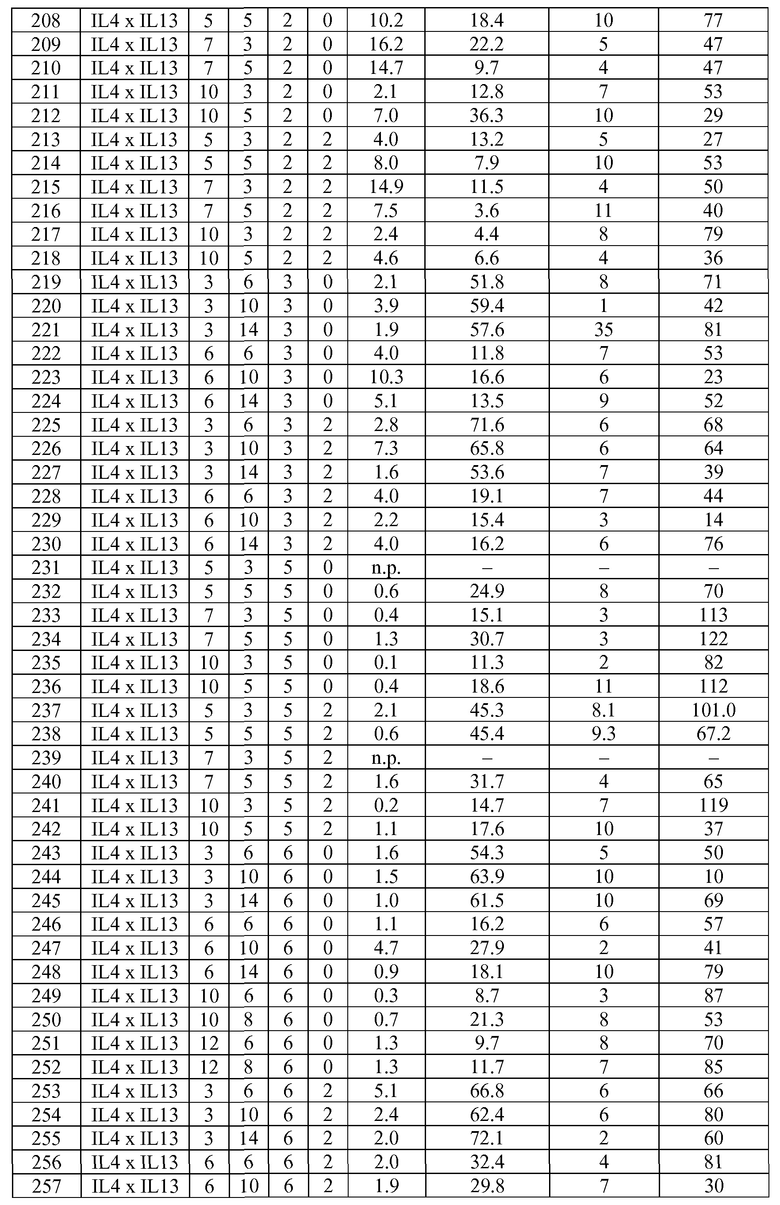
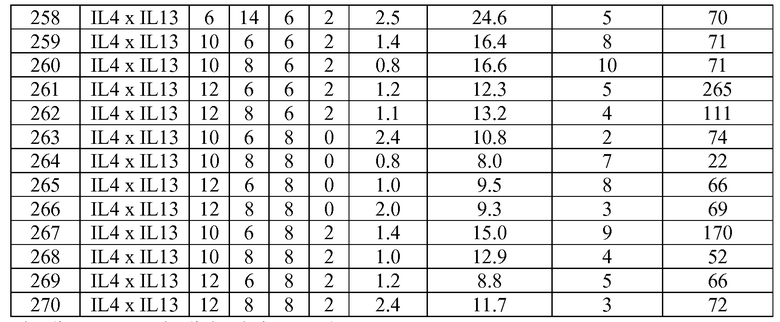
Результаты, приведенные в таблицах 7 и 8, ясно показывают, что необходимы линкеры между вариабельными и константными доменами, чтобы обеспечить оптимальный фолдинг. Только в редких схемах расположения допустим линкер, равный нулю (см. партии № 103-105, в которых L1 (LC) равен нулю, и партию № 210, в которой L4 равен нулю). Однако в каждом случае соответствующий переходный линкер между вариабельной областью и константной областью в другой цепи не может быть равен нулю.
Приведенные выше результаты показывают, что сочетания L1=7, L2=5, L3=1 и L4=2 были хорошей исходной точкой для оптимизации нового CODV-Ig, в котором тяжелая цепь является матрицей. Было показано, что диапазоны, представленные в таблице 9, являются приемлемыми диапазонами для успешного конструирования нового CODV-Ig из двух исходных антител.
Диапазоны размеров линкеров в случае либо LC, либо HC в качестве матрицы
Пример 8. Универсальная применимость формы CODV-Ig
Чтобы оценить приемлемость формы CODV-Ig для конструирования новых антитело-подобных связывающих белков, вариабельные области из многочисленных существующих антител человека и гуманизированных антител, обладающих специфичностью по отношению к рецептору инсулиноподобного фактора роста 1 (IGF1R(1)), второму антителу к рецептору инсулиноподобного фактора роста 1 (IGF1R(2)), рецептору эпидермального фактора роста человека 2 (HER2), рецептору эпидермального фактора роста (EGFR), фактору некроза опухоли - альфа (TNFα), интерлейкину 12 и 23 (IL-12/23) и интерлейкину 1 бета (IL-1β) включали в форму CODV-Ig (см. таблицу 10).
Описательные коды для тяжелых и легких цепей, используемых в биспецифичных CODV-Ig
Вариабельные области антител из известных антител человека и гуманизированных антител использовали для проверки универсальной применимости формы CODV-Ig в конструировании биспецифичных антитело-подобных связывающих белков. Кроме того, исследовали возможность эффектов положения в отношении размещения некоторых вариабельных областей антител либо на N-конце, либо на C-конце тяжелой цепи или легкой цепи. На основании дизайна молекул CODV-Ig, имеющих состав линкеров L1=7, L2=5, L3=1 и L4=2, вводили разные последовательности антител в форму CODV-Ig.
Активности биспецифичных антител или производных против IL1β и TNFα определяли, используя коммерчески доступные репортерные клетки HEK-Blue TNFα/IL1β (InvivoGen). Чтобы определить активности антител по отношению к TNFα и IL1β, цитокины предварительно инкубировали в течение 1 часа с разными концентрациями антител, и добавляли к 50000 клеток HEK Blue TNFα/IL1β. Опосредованную цитокинами индукцию SEAP измеряли через 24 часа в надосадке культуры, используя анализ QUANTI-Blue (InvivoGen).
Как показано в таблице 11, в случае всех конструкций наблюдали выход белка от хорошего до превосходного и приемлемые уровни агрегации (в частности, см. партии № 301 и 302 в таблице 11). Измеренная аффинность для каждого вариабельного домена антитела была в пределах опубликованных или ожидаемых значений аффинности. В случаях, когда оценивали аффинность, не выявляли эффектов положения. Таким образом, как показано в следующих таблицах, эффектов положения не наблюдали ни в одном случае использования какого-либо вариабельного домена антител, ни при использовании таких доменов в любой из цепей антитела.
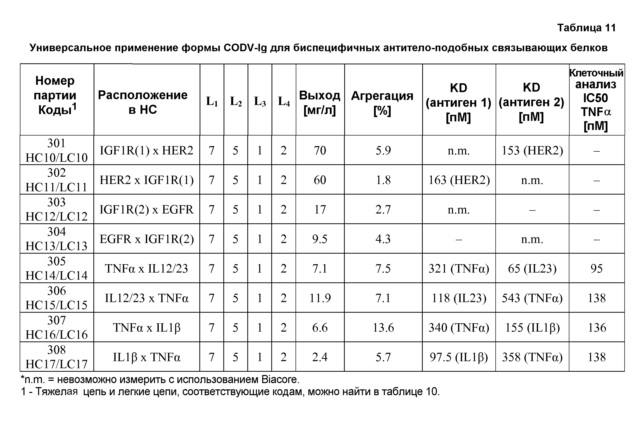
Пример 9. Сохранение аффинности исходного антитела в форме CODV-Ig
Идентичные последовательности антител против IL4 и против IL13 включали либо в форму TBTI/DVD-Ig, либо в форму CODV-Ig для прямого сравнения таких конфигураций, расположения линкеров и аффинностей получаемых в результате молекул. Как показано на фигуре 7, исходная аффинность каждого из антител сохранялась в форме CODV. Как показано на верхней панели фигуры 7, когда вариабельные области помещали в форму TBTI/DVD-Ig, уменьшение аффинности IL4-антитела, находящегося во внутреннем положении Fv2, проявлялось в виде снижения скорости ассоциации при связывании антитела с антигеном. Напротив, не было потери аффинности в случае формы CODV-Ig по сравнению с исходными антителами (см. фигуру 7, нижнюю панель).
Пример 10. Применимость CODV-Ig для формы Fab
Затем оценивали способность формы CODV-Ig давать такие фрагменты, как Fab-фрагменты. Две разных вариабельных области тяжелых цепей сливали друг с другом через линкер L3, и удлиняли на C-конце линкером L4. Затем такой комплекс VH сливали с доменом CH1 IGHG1 (GenBank, № доступа Q569F4), несущим на C-конце последовательность из пяти аминокислот DKTHT (SEQ ID NO: 60) из шарнирной области, за которыми следовали шесть остатков гистидина. Разные вариабельные области легких цепей сливали друг с другом в конфигурации крест-накрест с соответствующей тяжелой цепью через линкер L1 и удлиняли на C-конце линкером L2 и затем сливали с константной цепью каппа (IGKC, GenBank, № доступа Q502W4).
Fab-фрагменты экспрессировали в результате временной трансфекции, как описано ранее. Через семь дней после трансфекции клетки удаляли центрифугированием, добавляли 10% об./об. 1 М трис-HCl, pH 8,0, и надосадок пропускали через фильтр 0,22 мкм, чтобы удалить частицы. Fab-белки улавливали, используя высокоэффективные колонки HisTrap (GE Healthcare) и элюировали градиентом имидазола. Содержащие белок фракции объединяли и обессоливали, используя колонки PD-10 или колонки с сефадексом. Концентрированные и стерильно профильтрованные (0,22 мкм) растворы белков доводили до концентрации 1 мг/мл, и хранили при 4ºC вплоть до применения.
Непосредственные наблюдаемые преимущества заключались в том, что Fab-подобные молекулы в ориентации CODV не проявляли тенденции к агрегации, и сохраняли аффинности исходных антител (см. таблицу 12). Используя связывающие конструкции белков из партий № 401-421, непосредственно сравнивали антитело-подобные белки, в которых вариабельные области антител были расположены как в молекулах CODV-Ig с тяжелой цепью в качестве матрицы (401, 402, 406, и 407), CODV Fab-подобных фрагментах (402, 408, 413, 418 и 421), четырехдоменных антитело-подобных молекулах в форме TBTI/DVD-Ig (404, 409, 414, и 419) и CODV-Ig без линкеров (405, 410, 415 и 420). Как показано в таблице 12, результаты такого сравнения показали, что по всей вероятности происходит утрата аффинности по сравнению с исходными антителами, когда вариабельную область включают в форму TBTI или DVD-Ig. Напротив, CODV-Ig и CODV-Ig Fab-подобные формы были способны лучше сохранять исходные аффинности. Результаты дополнительно подтвердили, что в случае молекул CODV-Ig требуются линкеры между вариабельными областями и между вариабельными областями и константными доменами (см. таблицу 12).

Пример 11. Замена вариабельных доменов в CODV-Ig и CODV-Fab
Чтобы охарактеризовать форму CODV в способе привлечения T-клеток, создавали биспецифичные Fab CODV-подобные связывающие белки (CODV-Fab), имеющие участок связывания TCR (CD3epsilon) и участок связывания CD19, и сравнивали с биспецифичным Fab, полученным из формы TBTI/DVD-Ig (B-Fab). Чтобы исследовать важность ориентации участков связывания (TCR × CD19 по сравнению с CD19 × TCR), оценивали обе ориентации для каждого из связывающих белков.
Связывающие белки характеризовали в анализе цитотоксичности, используя клетки NALM-6 (экспрессирующие CD19) в качестве клеток-мишеней и первичные T-клетки человека в качестве эффекторных клеток. CD3-позитивные клетки выделяли из свежеполученных PBMC человека. Эффекторные клетки и клетки-мишени смешивали в соотношении 10:1 и инкубировали в течение 20 часов с указанными концентрациями биспецифичных связывающих белков (см. фигуру 8). Апоптозные клетки-мишени определяли в основанном на FACS анализе, используя окрашивание 7-аминоактиномицином.
Показано, что форма B-Fab в конфигурации CD3-CD19 (1060) является активной в индукции опосредованной T-клетками цитотоксичности по отношению к клеткам NALM-6 с EC50, составляющей 3,7 нг/мл. Сходную высокую активность наблюдали в случае CD19-CD3 CODV-Fab (1109) с EC50 3,2 нг/мл (см. фигуру 8).
Смена конфигурации молекулы B-Fab (Fab формы TBTI/DVD-Ig) на ориентацию CD19-CD3 приводила к значительной потере активности (см. фигуру 8). Молекула B-Fab после смены не проявляла активности в концентрациях, которые были максимальными для обеих ориентаций Fab CODV-Ig и другой ориентации B-Fab. В случае Fab CODV-Ig и одной ориентации B-Fab наблюдали максимальный ответ (в диапазоне от 1 до 100 нг/мл). В случае ориентации CD19-CD3 B-Fab, даже в максимальной концентрации (30 мкг/мл) не достигали оптимального цитотоксического ответа. И наоборот, изменение в ориентации доменов в CODV-Fab на CD3-CD19 (1108) приводило к получению молекулы, имеющей значительную активность в таком анализе (см. фигуру 8). Хотя смена доменов в CODV-Fab также снижала индукцию опосредованной T-клетками цитотоксичности (увеличение EC50 в 100 раз), такой эффект был намного менее выраженным, чем наблюдали в форме B-Fab, и молекула была способна индуцировать цитотоксичность до максимального уровня. Данные были репрезентативными и получены в трех независимых экспериментах.
Пример 12. Влияние индивидуального состава аминокислотной последовательности на линкеры CODV-Ig
Оптимизированную конструкцию, соответствующую партии № 204 (см. пример 7 и таблицу 8), выбирали для исследования влияния состава линкеров на линкеры L1-L4. Длины линкеров составляли 7, 5, 1 и 2 остатка в случае L1, L2, L3 и L4, соответственно (см. таблицу 13). Тестируемые последовательности получали из встречающихся в природе линкеров в участках переходов между доменами VH и CH1 природных антител или между доменами антител Fv и CL легких каппа- или лямбда-цепей. Выбранные для исследования последовательности представляли собой последовательности ASTKGPS (SEQ ID NO: 48), которая получена из переходного участка между доменами VH и CH1, RTVAAPS (SEQ ID NO: 49) и GQPKAAP (SEQ ID NO: 50), которые получены из переходных участков между доменами Fv и CL легких каппа- и лямбда-цепей, соответственно. Кроме того, создавали одну конструкцию с произвольным составом линкеров, чтобы показать, что потенциально можно использовать любую последовательность в линкерах L1-L4. Такой состав линкеров получали в результате случайного распределения аминокислот валина, лейцина, изолейцина, серина, треонина, лизина, аргинина, гистидина, аспартата, глутамата, аспарагина, глутамина, глицина и пролина в 15 положениях четырех линкеров. Ароматические аминокислоты фенилаланин, тирозин и триптофан, а также аминокислоты метионин и цистеин были предварительно исключены, чтобы избежать возможного повышения агрегации.
Трехмерную модель конструкции для партии № 204 создавали для того, чтобы убедиться в пригодности или уточнить варианты выбора состава линкеров. Таким образом, был выбран серин для линкера L3, так как в трехмерной модели вблизи наблюдали положительно и отрицательно заряженные остатки. Остатки в линкере L4 выбирали так, чтобы они были совместимы с влиянием растворителя в таких положениях, как предполагалось в модели. Подобным образом, не прогнозировали или не предполагали наличия проблем в отношении состава линкеров L1 и L2. Конструировали трехмерные модели выбранных предположений по составу линкеров.
Как показано в таблице 12, состав линкеров может оказывать сильное влияние на выход. Последовательности L1, которые были получены из лямбда цепи (сравните партии № 505-507 с партиями № 501-503) были более продуктивными генераторами белков (увеличение до 8 раз). Действительно, линкеры, основанные на случайной генерации, также давали хорошие выходы, как показано в таблице 13, партия № 508. Таким образом, состав линкеров должен быть одним из параметров, учитываемых при оптимизации CODV-Ig.
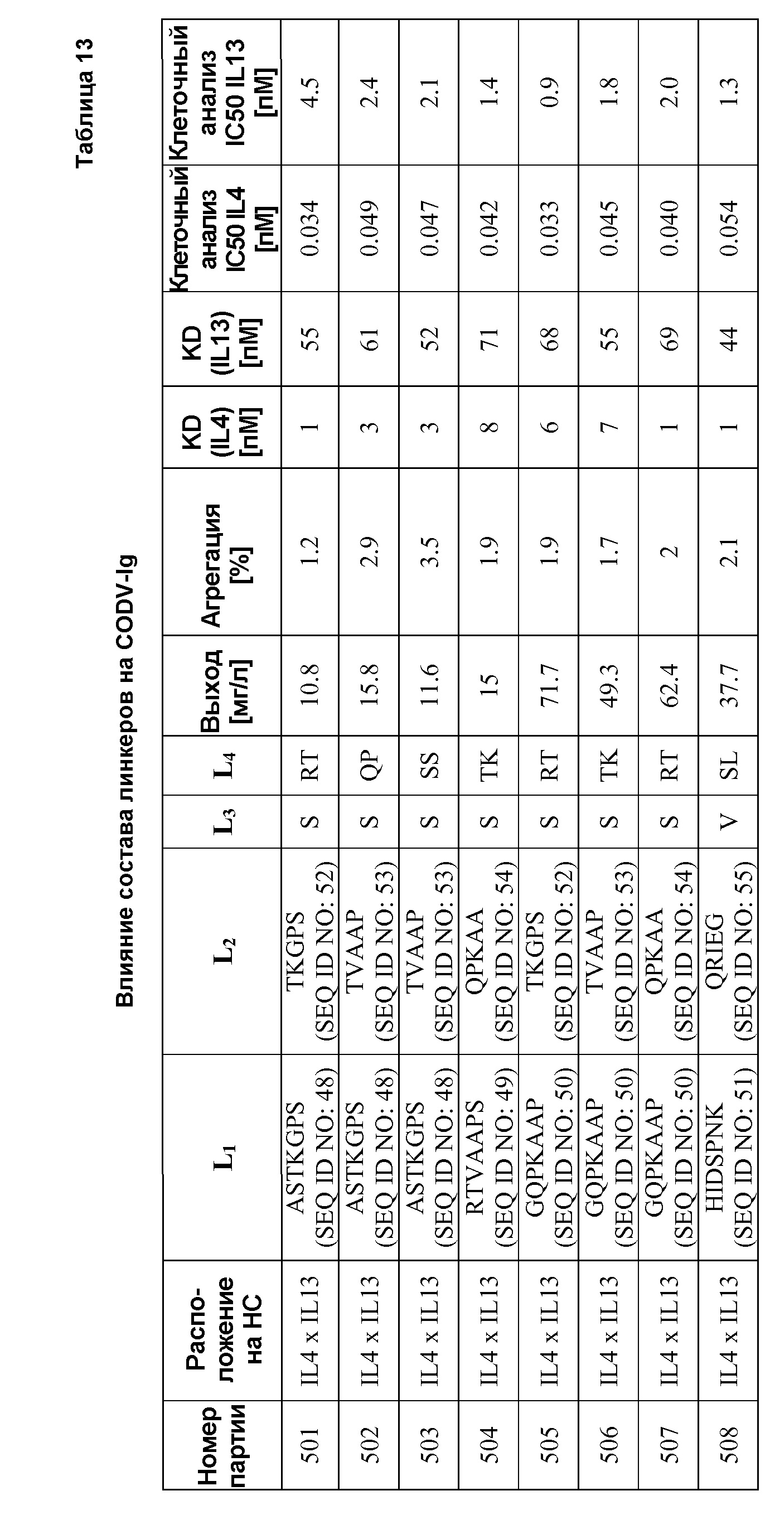
Активности биспецифичных антител или производных против цитокинов IL4 и IL13 определяли на коммерчески доступных репортерных клетках HEK-Blue IL-4/IL-13 (InvivoGen). Клетки HEK-Blue IL-4/IL-13 предназначены для наблюдения за активацией пути STAT6 под действием IL-4 или IL13. Стимуляция клеток любым цитокином приводит к продукции секретируемой эмбриональной щелочной фосфатазы (SEAP) репортерного гена, которую можно измерить в надосадке культуры с использованием анализа QUANTI-Blue. Чтобы тестировать активности антител против IL4 или IL13, цитокины предварительно инкубировали в течение 1 часа с разными концентрациями антител, и добавляли к 50000 клеток HEK-Blue IL-4/IL-13. Опосредованную цитокинами индукцию SEAP измеряли после 24-часовой инкубации в надосадке культуры клеток с использованием анализа QUANTI-Blue (InvivoGen).
Пример 13. Введение цистеинов а линкеры CODV-Ig
Опубликованные данные свидетельствуют, что стабильность антител и полученных из антител белков можно повысить введением, не встречающихся в природе дисульфидных мостиков (см. публикацию Wozniak-Knopp с соавторами, 2012, «Stabilisation of the Fc Fragment of Human IgG1 by Engineered Intradomain Disulfide Bonds», PLoS ONE 7(1): e30083). Чтобы исследовать, можно ли стабилизировать эквивалентный Fc-фрагмент, полученный из IgG1-антитела человека и сконструированный в виде молекулы CODV-Ig, путем введения межцепочечных и внутрицепочечных дисульфидных мостиков, эквивалентные положения Fc в конструкции CODV-Ig партии № 204 (из примера 7) подвергали мутации до остатков цистеина и мутантные белки сверхпродуцировали, очищали и характеризовали (см. таблицу 14).
Как показано в таблице 14, все мутантные молекулы CODV-Ig, содержащие дополнительные остатки цистеина, имели такие же температуры плавления, как и температура плавления конструкции CODV-Ig № 204.
Кроме того, были одновременно введены два цистеина в положения согласно нумерации Кабата 100 в случае легкой цепи и 44 в случае тяжелой цепи в каждом из вариабельных доменов, как описано Brinkmann с соавторами, 1993, Proc. Natl. Acad. Sci. U.S.A. 90: 7538-42. Было показано, что такие положения являются структурно консервативными в укладке антитела и поэтому допускают замену на цистеин, не мешающую целостности отдельных доменов.
Как показано в таблице 15, конструкции CODV и CODV-Ig, в которых были введены остатки цистеина в положениях согласно нумерации Кабата 100 в случае легкой цепи и 44 в случае тяжелой цепи в каждом из вариабельных доменов имели более высокие температуры плавления, чем конструкции CODV и CODV-Ig, в которых остатки цистеина не были введены в такие положения (см., например, партии № 704 и 706 и партии № 713 и 714).
1. Измерения термостабильности вариантов CODV и TBTI
Точки плавления (Tm) вариантов CODV и TBTI определяли, используя дифференциальную сканирующую флуориметрию (DSF). Образцы разбавляли в буфере D-PBS (Invitrogen) до конечной концентрации 0,2 мкг/мкл и добавляли 2 мкл концентрированного в 40 раз раствора красителя SYPRO-Orange (Invitrogen) в D-PBS в 96-луночные белые планшеты с полуокантовкой. Все измерения осуществляли в двух повторах, используя прибор для ПЦР в реальном времени MyiQ2 (Biorad). Значения Tm получали из отрицательной первой производной кривых плавления, используя компьютерную программу iQ5 v2.1.
Затем исследовали эффект введения остатков цистеина непосредственно в линкеры или в вариабельную область. В данном примере использовали партию № 204 (из примера 7 и таблицы 8) в качестве модельного связывающего белка CODV-Ig, и цистеином заменяли глицин в L1, L, или вариабельной области на основе трехмерной модели. Как показано в таблице 16 ниже, результаты показывают, как введение пары цистеинов может влиять на выход и агрегацию. Моделировали все предполагаемые мутации, чтобы убедиться, что дисульфидные связи были правильно образованы, и в моделях поддерживалась правильная геометрия линкеров и их окружения. При этом в партии № 808 наблюдали хороший выход и небольшую агрегацию, что свидетельствует о том, что может быть образован подходящий цистеиновый мостик.
Хотя изобретение описано на примерах различных вариантов осуществления, понятно, что специалисты в данной области могут осуществить изменения и модификации. Поэтому подразумевается, что прилагаемая формула изобретения охватывает все такие эквивалентные варианты, которые входят в объем заявленного изобретения. Кроме того, заголовки разделов, используемые в настоящем описании, предназначены только для организационных целей, и их не следует считать ограничивающими описанный объект изобретения.
Каждый вариант, описанный в настоящей публикации, можно сочетать с любым другим вариантом или вариантами, если ясно не указано иное. В частности, любой признак или вариант, указанный как предпочтительный или преимущественный, можно сочетать с любым другим признаком или признаками или вариантом или вариантами, указанными как предпочтительные или преимущественные, если ясно не указано иное.
Все ссылки, цитированные в настоящей заявке, специально включены в настоящее описание в виде ссылки.
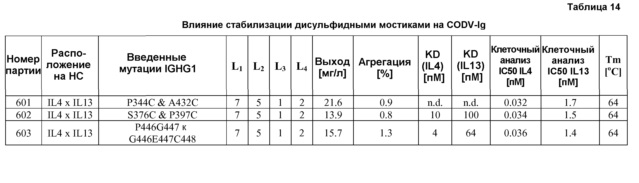
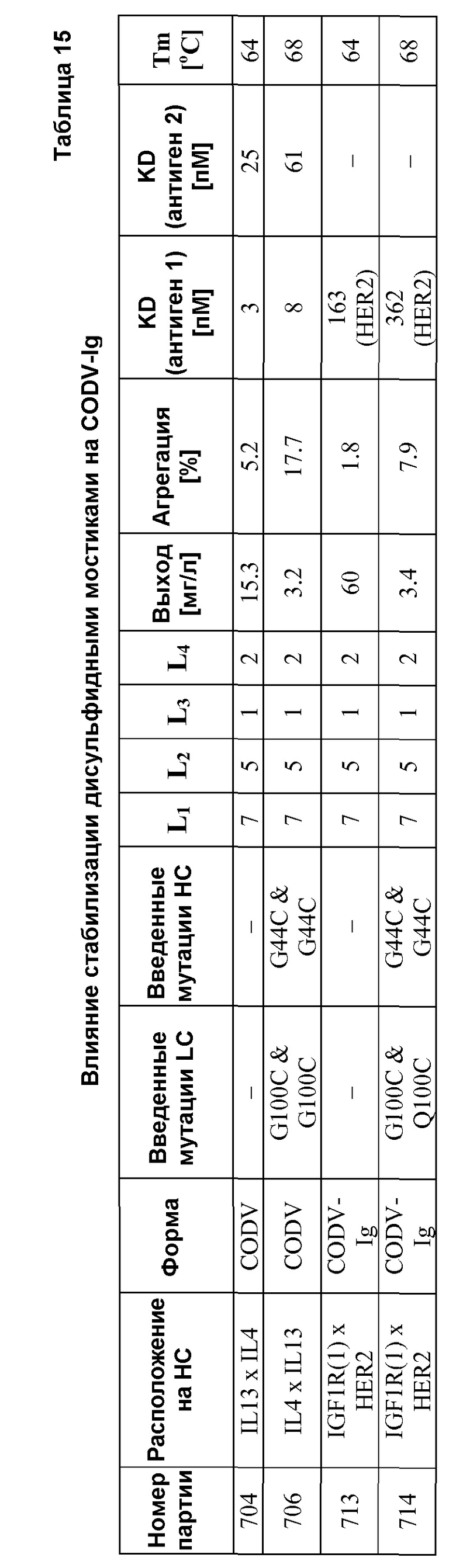
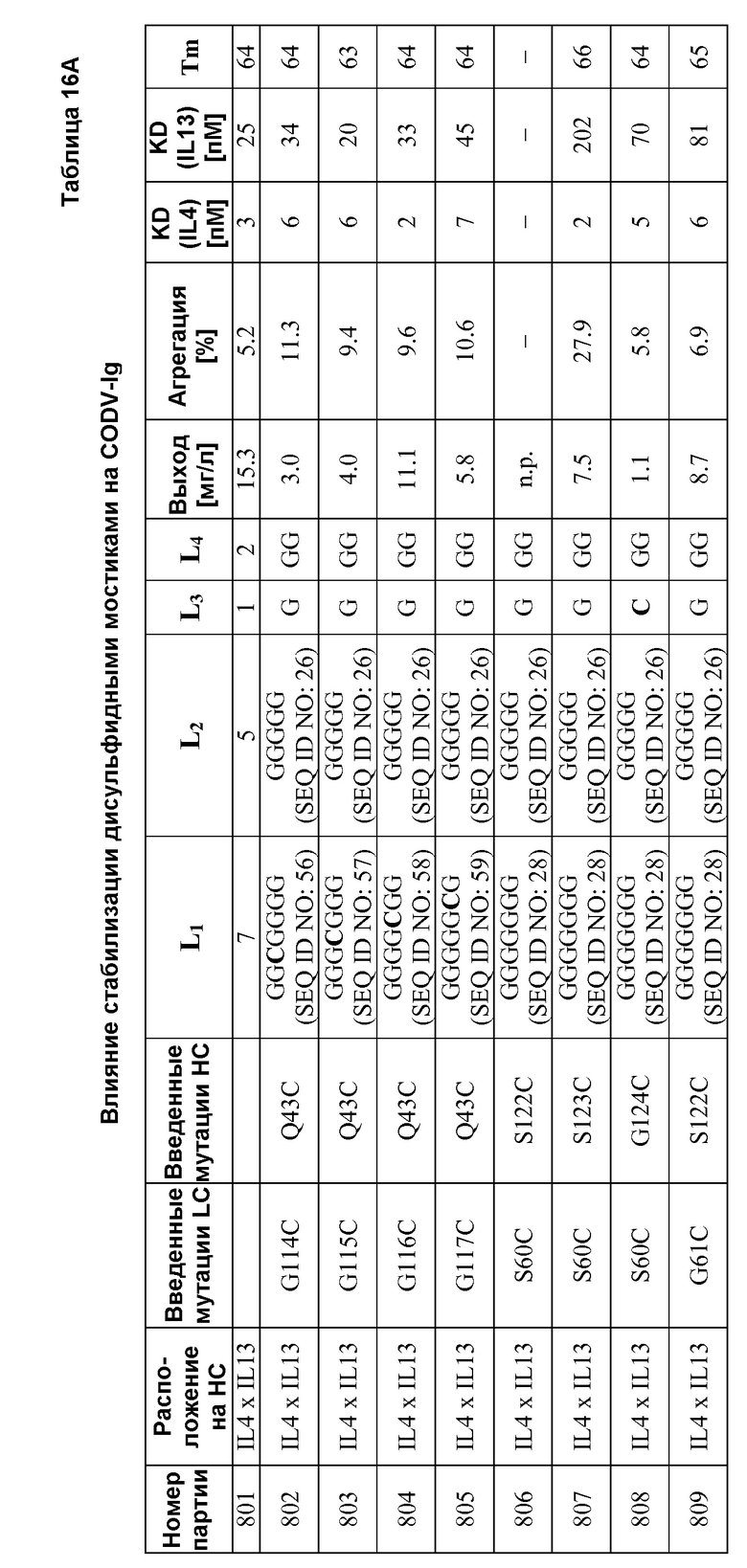

| название | год | авторы | номер документа |
|---|---|---|---|
| АНТИТЕЛО-ПОДОБНЫЕ СВЯЗЫВАЮЩИЕ БЕЛКИ С ДВОЙНЫМИ ВАРИАБЕЛЬНЫМИ ОБЛАСТЯМИ, ИМЕЮЩИЕ ОРИЕНТАЦИЮ СВЯЗЫВАЮЩИХ ОБЛАСТЕЙ КРЕСТ-НАКРЕСТ | 2012 |
|
RU2823693C2 |
| СПОСОБЫ АНАЛИЗА ОШИБОЧНОГО СПАРИВАНИЯ ЦЕПЕЙ В ПОЛИСПЕЦИФИЧЕСКИХ СВЯЗЫВАЮЩИХ БЕЛКАХ | 2020 |
|
RU2828030C1 |
| ТРИСПЕЦИФИЧЕСКИЕ СВЯЗЫВАЮЩИЕ БЕЛКИ, СВЯЗЫВАЮЩИЕ CD38, CD28 И CD3, А ТАКЖЕ СПОСОБЫ ПРИМЕНЕНИЯ ДЛЯ ЛЕЧЕНИЯ ВИРУСНОЙ ИНФЕКЦИИ | 2019 |
|
RU2820351C2 |
| АНТИТЕЛА К CD38 И КОМБИНАЦИИ С АНТИТЕЛАМИ К CD3 И CD28 | 2018 |
|
RU2812910C2 |
| СПОСОБЫ КОЛИЧЕСТВЕННОГО ОПРЕДЕЛЕНИЯ ПОЛИПЕПТИДОВ | 2019 |
|
RU2782464C2 |
| БИСПЕЦИФИЧЕСКИЕ СВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ, СПОСОБНЫЕ СВЯЗЫВАТЬ CD137 И ОПУХОЛЕВЫЕ АНТИГЕНЫ, И ВАРИАНТЫ ИХ ПРИМЕНЕНИЯ | 2018 |
|
RU2805648C2 |
| ГИПЕРГЛИКОЗИЛИРОВАННЫЕ СВЯЗЫВАЮЩИЕ ПОЛИПЕПТИДЫ | 2014 |
|
RU2708314C2 |
| МУЛЬТИСПЕЦИФИЧЕСКИЕ СВЯЗЫВАЮЩИЕ БЕЛКИ С МУТАНТНЫМИ ДОМЕНАМИ FAB | 2019 |
|
RU2827657C2 |
| ТРИСПЕЦИФИЧЕСКИЕ СВЯЗЫВАЮЩИЕ БЕЛКИ, ОТНОСЯЩИЕСЯ К НИМ СПОСОБЫ И ВАРИАНТЫ ИХ ПРИМЕНЕНИЯ | 2020 |
|
RU2822200C2 |
| БИСПЕЦИФИЧЕСКИЕ АНТИГЕНСВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ | 2012 |
|
RU2650775C2 |





































































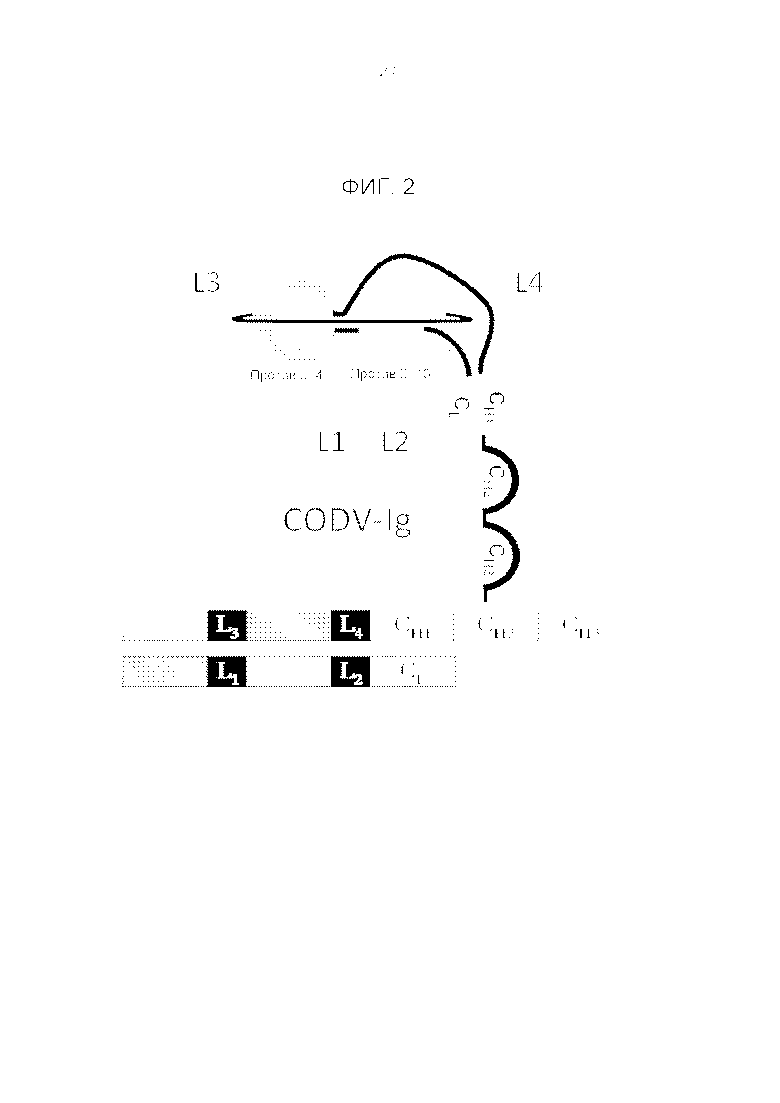
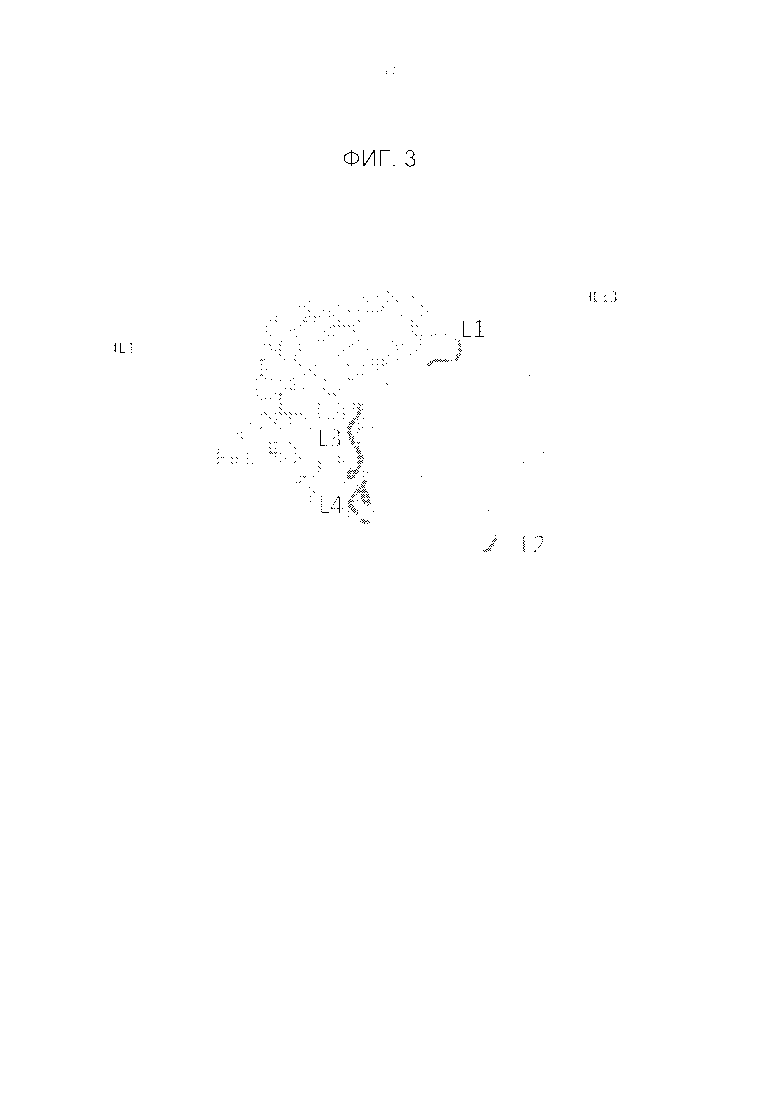

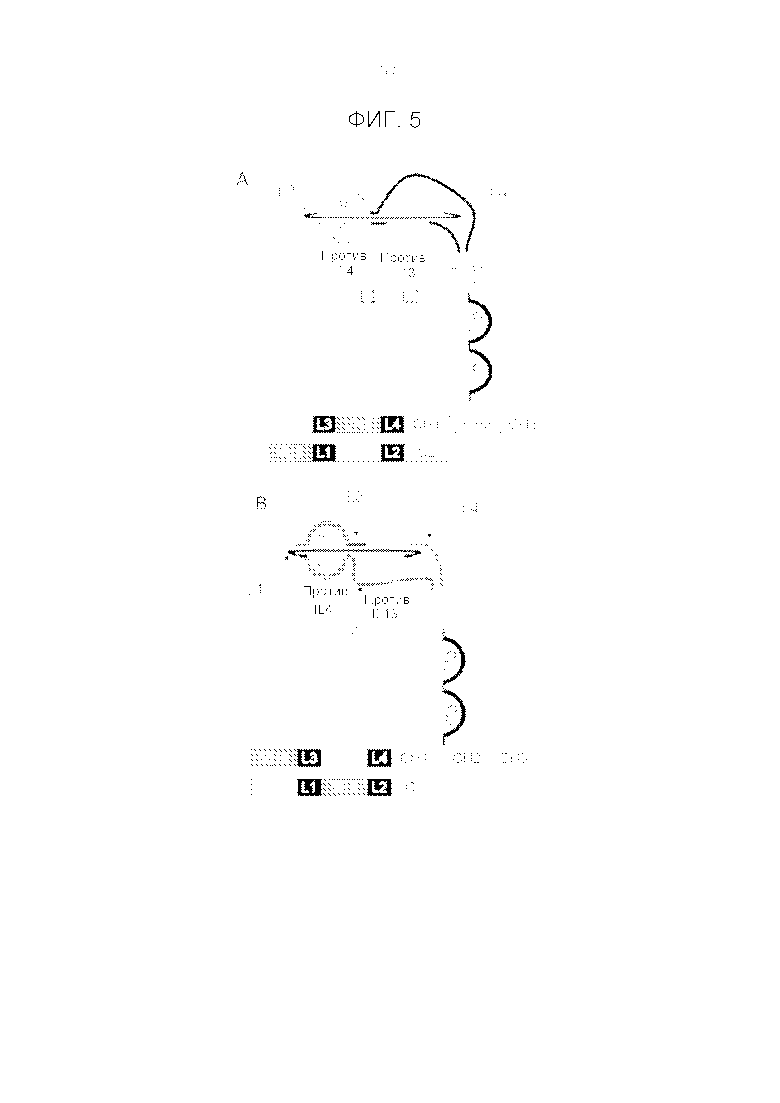

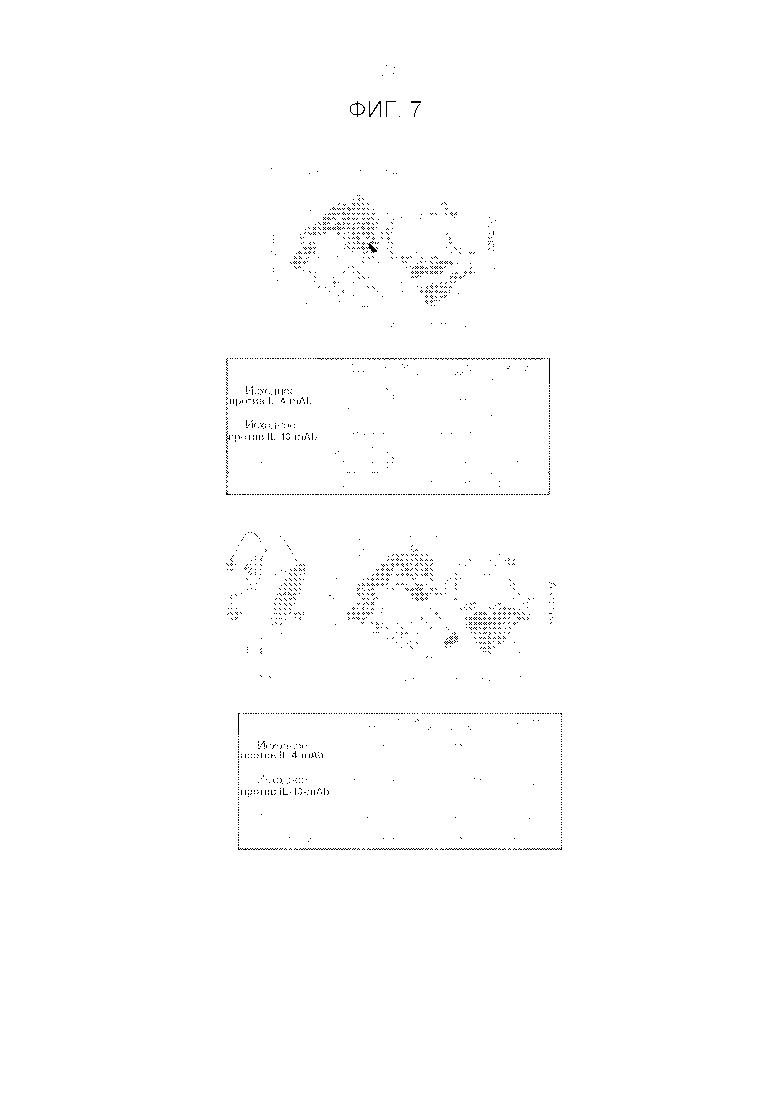
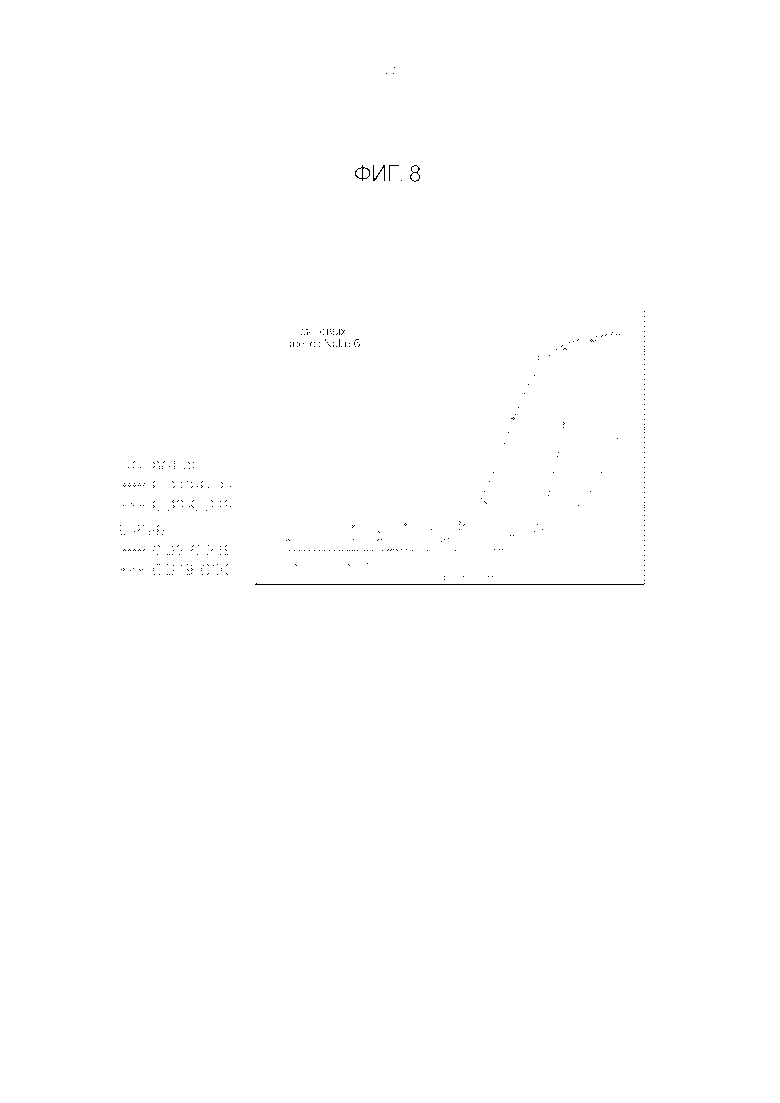
Изобретение относится к области биохимии, в частности к конструкции нуклеиновой кислоты для получения биспецифического антитело-подобного связывающего белка, связывающего пару антигенов. Указанная конструкция содержит четыре полипептидные цепи, которые образуют четыре антигенсвязывающих участка. Изобретение также относится к вектору экспрессии и выделенной клетке-хозяину, содержащим вышеуказанную конструкцию нуклеиновой кислоты. Изобретение позволяет эффективно получать биспецифический антитело-подобный связывающий белок, связывающий пару антигенов. 8 н. и 15 з.п. ф-лы, 8 ил., 16 табл., 13 пр.
1. Конструкция нуклеиновой кислоты для получения биспецифического антитело-подобного связывающего белка, связывающего пару антигенов, содержащего четыре полипептидные цепи, которые образуют четыре антигенсвязывающих участка, где конструкция нуклеиновой кислоты содержит нуклеиновую кислоту, кодирующую две полипептидные цепи, содержащие структуру, представленную формулой:
VL1-L1-VL2-L2-CL [I]
и где конструкция нуклеиновой кислоты содержит нуклеиновую кислоту, кодирующую две полипептидные цепи, содержащие структуру, представленную формулой:
VH2-L3-VH1-L4-CH1-Fc [II]
где:
VL1 означает первый вариабельный домен легкой цепи иммуноглобулина;
VL2 означает второй вариабельный домен легкой цепи иммуноглобулина;
VH1 означает первый вариабельный домен тяжелой цепи иммуноглобулина;
VH2 означает второй вариабельный домен тяжелой цепи иммуноглобулина;
CL означает константный домен легкой цепи иммуноглобулина;
CH1 означает константный домен тяжелой цепи иммуноглобулина CH1;
Fc означает шарнирную область иммуноглобулина и константные домены тяжелой цепи иммуноглобулина CH2, CH3; и
L1, L2, L3 и L4 означают аминокислотные линкеры;
причем:
L1 имеет длину от 3 до 12 аминокислотных остатков;
L2 имеет длину от 3 до 14 аминокислотных остатков;
L3 имеет длину от 1 до 8 аминокислотных остатков; и
L4 имеет длину от 1 до 3 аминокислотных остатков;
и где полипептиды формулы I и полипептиды формулы II образуют пару легкая цепь - тяжелая цепь в ориентации крест-накрест, и
где антитело-подобный белок способен связываться с двумя различными антигенами-мишенями.
2. Конструкция нуклеиновой кислоты по п.1, где:
L1 имеет длину от 5 до 10 аминокислотных остатков;
L2 имеет длину от 5 до 8 аминокислотных остатков;
L3 имеет длину от 1 до 5 аминокислотных остатков; и
L4 имеет длину от 1 до 2 аминокислотных остатков.
3. Конструкция нуклеиновой кислоты по п.1, где:
L1 имеет длину 7 аминокислотных остатков;
L2 имеет длину 5 аминокислотных остатков;
L3 имеет длину 1 аминокислотный остаток; и
L4 имеет длину 2 аминокислотных остатка.
4. Конструкция нуклеиновой кислоты для получения биспецифического антитело-подобного связывающего белка, связывающего пару антигенов, содержащего четыре полипептидные цепи, которые формируют четыре антигенсвязывающих участка, где конструкция нуклеиновой кислоты содержит нуклеиновую кислоту, кодирующую две полипептидные цепи, содержащие структуру в соответствии со следующей формулой:
VL1-L1-VL2-L2-CL [I],
и где конструкция нуклеиновой кислоты содержит нуклеиновую кислоту, кодирующую две полипептидные цепи, содержащие структуру в соответствии со следующей формулой:
VH2-L3-VH1-L4-CH1-Fc [II],
где:
VL1 означает первый вариабельный домен легкой цепи иммуноглобулина;
VL2 означает второй вариабельный домен легкой цепи иммуноглобулина;
VH1 означает первый вариабельный домен тяжелой цепи иммуноглобулина;
VH2 означает второй вариабельный домен тяжелой цепи иммуноглобулина;
CL означает константный домен легкой цепи иммуноглобулина;
CH1 означает константный домен тяжелой цепи иммуноглобулина CH1;
Fc означает шарнирную область иммуноглобулина и константные домены тяжелой цепи иммуноглобулина CH2, CH3; и
L1, L2, L3 и L4 означают аминокислотные линкеры;
причем:
L1 имеет длину от 1 до 3 аминокислотных остатков;
L2 имеет длину от 1 до 4 аминокислотных остатков;
L3 имеет длину от 2 до 15 аминокислотных остатков; и
L4 имеет длину от 2 до 15 аминокислотных остатков;
и где полипептиды формулы I и полипептиды формулы II образуют пару легкая цепь - тяжелая цепь в ориентации крест-накрест, и
где антитело-подобный белок способен связываться с двумя различными антигенами-мишенями.
5. Конструкция нуклеиновой кислоты по п.4, где:
L1 имеет длину от 1 до 2 аминокислотных остатков;
L2 имеет длину от 1 до 2 аминокислотных остатков;
L3 имеет длину от 4 до 12 аминокислотных остатков; и
L4 имеет длину от 2 до 12 аминокислотных остатков.
6. Конструкция нуклеиновой кислоты по п.4, где:
L1 имеет длину 1 аминокислотный остаток;
L2 имеет длину 2 аминокислотных остатка;
L3 имеет длину 7 аминокислотных остатков; и
L4 имеет длину 5 аминокислотных остатков.
7. Конструкция нуклеиновой кислоты для получения биспецифического антитело-подобного связывающего белка, связывающего пару антигенов, содержащего четыре полипептидные цепи, которые формируют четыре антигенсвязывающих участка, где конструкция нуклеиновой кислоты содержит нуклеиновую кислоту, кодирующую две полипептидные цепи, содержащие структуру в соответствии со следующей формулой:
VL1-L1-VL2-L2-CL [I],
и где конструкция нуклеиновой кислоты содержит нуклеиновую кислоту, кодирующую две полипептидные цепи, содержащие структуру в соответствии со следующей формулой:
VH2-L3-VH1-L4-CH1-Fc [II],
где:
VL1 означает первый вариабельный домен легкой цепи иммуноглобулина;
VL2 означает второй вариабельный домен легкой цепи иммуноглобулина;
VH1 означает первый вариабельный домен тяжелой цепи иммуноглобулина;
VH2 означает второй вариабельный домен тяжелой цепи иммуноглобулина;
CL означает константный домен легкой цепи иммуноглобулина;
CH1 означает константный домен тяжелой цепи иммуноглобулина CH1;
Fc означает шарнирную область иммуноглобулина и константные домены тяжелой цепи иммуноглобулина CH2, CH3; и
L1, L2, L3 и L4 означают аминокислотные линкеры;
причем:
L1 имеет длину 0 аминокислотных остатков и L3 имеет длину от 2 до 15 аминокислотных остатков; или
L3 имеет длину 0 аминокислотных остатков и L1 имеет длину от 1 до 12 аминокислотных остатков; или
L4 имеет длину 0 аминокислотных остатков и L2 имеет длину от 3 до 14 аминокислотных остатков;
и где полипептиды формулы I и полипептиды формулы II образуют пару легкая цепь - тяжелая цепь в ориентации крест-накрест, и
где антитело-подобный белок способен связываться с двумя различными антигенами-мишенями.
8. Конструкция нуклеиновой кислоты для получения антитело-подобного связывающего белка по п.7, где:
L1 имеет длину 0 аминокислотных остатков и L3 имеет длину от 2 до 4 аминокислотных остатков; или
L3 имеет длину 0 аминокислотных остатков и L1 имеет длину от 1 до 2 аминокислотных остатков; или
L4 имеет длину 0 аминокислотных остатков и L2 имеет длину от 3 до 5 аминокислотных остатков.
9. Конструкция нуклеиновой кислоты для получения биспецифического антитело-подобного связывающего белка, связывающего пару антигенов, содержащего две полипептидные цепи, которые образуют два антигенсвязывающих участка, где конструкция нуклеиновой кислоты содержит нуклеиновую кислоту, кодирующую первую полипептидную цепь, содержащую структуру, представленную формулой:
VL1-L1-VL2-L2-CL [I]
и где конструкция нуклеиновой кислоты содержит
нуклеиновую кислоту, кодирующую вторую полипептидную цепь, содержащую структуру, представленную формулой:
VH2-L3-VH1-L4-CH1 [II]
где:
VL1 означает первый вариабельный домен легкой цепи иммуноглобулина;
VL2 означает второй вариабельный домен легкой цепи иммуноглобулина;
VH1 означает первый вариабельный домен тяжелой цепи иммуноглобулина;
VH2 означает второй вариабельный домен тяжелой цепи иммуноглобулина;
CL означает константный домен легкой цепи иммуноглобулина;
CH1 означает константный домен тяжелой цепи иммуноглобулина CH1; и
L1, L2, L3 и L4 означают аминокислотные линкеры;
причем:
L1 имеет длину от 3 до 12 аминокислотных остатков;
L2 имеет длину от 3 до 14 аминокислотных остатков;
L3 имеет длину от 1 до 8 аминокислотных остатков; и
L4 имеет длину от 1 до 3 аминокислотных остатков;
и где первый и второй полипептиды образуют пару легкая цепь - тяжелая цепь в ориентации крест-накрест, и
где антитело-подобный белок способен связываться с двумя различными антигенами-мишенями.
10. Конструкция нуклеиновой кислоты по п.9, где:
L1 имеет длину от 5 до 10 аминокислотных остатков;
L2 имеет длину от 5 до 8 аминокислотных остатков;
L3 имеет длину от 1 до 5 аминокислотных остатков; и
L4 имеет длину от 1 до 2 аминокислотных остатков.
11. Конструкция нуклеиновой кислоты по п.9, где:
L1 имеет длину 7 аминокислотных остатков;
L2 имеет длину 5 аминокислотных остатков;
L3 имеет длину 1 аминокислотный остаток; и
L4 имеет длину 2 аминокислотных остатка.
12. Конструкция нуклеиновой кислоты для получения биспецифическиого антитело-подобного связывающего белка, связывающего пару антигенов, содержащего две полипептидные цепи, которые образуют два антигенсвязывающих участка, где конструкция нуклеиновой кислоты содержит нуклеиновую кислоту, кодирующую первую полипептидную цепь, содержащую структуру, представленную формулой:
VL1-L1-VL2-L2-CL [I]
и где конструкция нуклеиновой кислоты содержит нуклеиновую кислоту, кодирующую вторую полипептидную цепь, содержащую структуру, представленную формулой:
VH2-L3-VH1-L4-CH1 [II]
где:
VL1 означает первый вариабельный домен легкой цепи иммуноглобулина;
VL2 означает второй вариабельный домен легкой цепи иммуноглобулина;
VH1 означает первый вариабельный домен тяжелой цепи иммуноглобулина;
VH2 означает второй вариабельный домен тяжелой цепи иммуноглобулина;
CL означает константный домен легкой цепи иммуноглобулина;
CH1 означает константный домен тяжелой цепи иммуноглобулина CH1; и
L1, L2, L3 и L4 означают аминокислотные линкеры;
причем:
L1 имеет длину от 1 до 3 аминокислотных остатков;
L2 имеет длину от 1 до 4 аминокислотных остатков;
L3 имеет длину от 2 до 15 аминокислотных остатков; и
L4 имеет длину от 2 до 15 аминокислотных остатков;
и где первый и второй полипептиды образуют пару легкая цепь - тяжелая цепь в ориентации крест-накрест, и
где антитело-подобный белок способен связываться с двумя различными антигенами-мишенями.
13. Конструкция нуклеиновой кислоты по п.12, где:
L1 имеет длину от 1 до 2 аминокислотных остатков;
L2 имеет длину от 1 до 2 аминокислотных остатков;
L3 имеет длину от 4 до 12 аминокислотных остатков; и
L4 имеет длину от 2 до 12 аминокислотных остатков.
14. Конструкция нуклеиновой кислоты по п.12, где:
L1 имеет длину 1 аминокислотный остаток;
L2 имеет длину 2 аминокислотных остатка;
L3 имеет длину 7 аминокислотных остатков; и
L4 имеет длину 5 аминокислотных остатков.
15. Конструкция нуклеиновой кислоты для получения биспецифического антитело-подобного связывающего белка, связывающего пару антигенов, содержащего две полипептидные цепи, которые образуют два антигенсвязывающих участка, где конструкция нуклеиновой кислоты содержит нуклеиновую кислоту, кодирующую первую полипептидную цепь, содержащую структуру, представленную формулой:
VL1-L1-VL2-L2-CL [I]
и где конструкция нуклеиновой кислоты содержит нуклеиновую кислоту, кодирующую вторую полипептидную цепь, содержащую структуру, представленную формулой:
VH2-L3-VH1-L4-CH1 [II]
где:
VL1 означает первый вариабельный домен легкой цепи иммуноглобулина;
VL2 означает второй вариабельный домен легкой цепи иммуноглобулина;
VH1 означает первый вариабельный домен тяжелой цепи иммуноглобулина;
VH2 означает второй вариабельный домен тяжелой цепи иммуноглобулина;
CL означает константный домен легкой цепи иммуноглобулина;
CH1 означает константный домен тяжелой цепи иммуноглобулина CH1; и
L1, L2, L3 и L4 означают аминокислотные линкеры;
причем:
L1 имеет длину 0 аминокислотных остатков и L3 имеет длину от 2 до 15 аминокислотных остатков; или
L3 имеет длину 0 аминокислотных остатков и L1 имеет длину от 1 до 12 аминокислотных остатков; или
L4 имеет длину 0 аминокислотных остатков и L2 имеет длину от 3 до 14 аминокислотных остатков;
и где полипептиды формулы I и полипептиды формулы II образуют пару легкая цепь - тяжелая цепь в ориентации крест-накрест, и
где антитело-подобный белок способен связываться с двумя различными антигенами-мишенями.
16. Конструкция нуклеиновой кислоты по п.15, где:
L1 имеет длину 0 аминокислотных остатков и L3 имеет длину от 2 до 4 аминокислотных остатков; или
L3 имеет длину 0 аминокислотных остатков и L1 имеет длину от 1 до 2 аминокислотных остатков; или
L4 имеет длину 0 аминокислотных остатков и L2 имеет длину от 3 до 5 аминокислотных остатков.
17. Конструкция нуклеиновой кислоты по любому из пп.1-16, где два разных антигена-мишени выбраны из группы, состоящей из B7.1, B7.2, BAFF, BlyS, C3, C5, CCL11 (эотаксина), CCL15 (MIP-1d), CCL17 (TARC), CCL19 (MIP-3b), CCL2 (MCP-1), CCL20 (MIP-3a), CCL21 (MIP-2), SLC, CCL24 (MPIF-2/эотаксина-2), CCL25 (TECK), CCL26 (эотаксина-3), CCL3 (MIP-1a), CCL4 (MIP-1b), CCL5 (RANTES), CCL7 (MCP-3), CCL8 (mcp-2), CD3, CD19, CD20, CD24, CD40, CD40L, CD80, CD86, CDH1 (E-кадгерина), хитиназы, CSF1 (M-CSF), CSF2 (GM-CSF), CSF3 (GCSF), CTLA4, CX3CL1 (SCYD1), CXCL12 (SDF1), CXCL13, EGFR, FCER1A, FCER2, HER2, IGF1R, IL-1, IL-12, IL13, IL15, IL17, IL18, IL1A, IL1B, IL1F10, IL1β, IL2, IL4, IL6, IL7, IL8, IL9, IL12/23, IL22, IL23, IL25, IL27, IL35, ITGB4 (интегрина b4), LEP (лептина), MHC класса II, TLR2, TLR4, TLR5, TNF, TNF-α, TNFSF4 (лиганда OX40), TNFSF5 (лиганда CD40), Toll-подобных рецепторов, TREM1, TSLP, TWEAK, XCR1 (GPR5/CCXCR1), DNGR-1(CLEC91) и HMGB1.
18. Конструкция нуклеиновой кислоты по любому из пп.1-16, где два разных антигена-мишени выбраны из группы, состоящей из IL4 и IL13, IGF1R и HER2, IGF1R и EGFR, EGFR и HER2, BK и IL13, PDL-1 и CTLA-4, CTLA4 и MHC класса II, IL-12 и IL-18, IL-1α и IL-1β, TNFα и IL12/23, TNFα и IL-12p40, TNFα и IL1β, TNFα и IL-23, и IL17 и IL23.
19. Конструкция нуклеиновой кислоты по любому из пп.1-18, в которой связывающий белок способен ингибировать функцию одного или нескольких антигенов-мишеней.
20. Конструкция нуклеиновой кислоты по любому из пп.1-19, где, по меньшей мере, один из линкеров, выбранный из группы, состоящей из L1, L2, L3 и L4, содержит, по меньшей мере, один остаток цистеина.
21. Вектор экспрессии, содержащий молекулу нуклеиновой кислоты по любому из пп.1-20.
22. Выделенная клетка-хозяин для получения антитело-подобного связывающего белка, связывающего пару антигенов, содержащая конструкцию нуклеиновой кислоты по любому из пп.1-20 или экспрессирующий вектор по п.21.
23. Клетка-хозяин по п.22, где клетка-хозяин является клеткой млекопитающего или клеткой насекомого.
| ВЫСЕВАЮЩИЙ АППАРАТ | 1992 |
|
RU2050764C1 |
| WO2011014659 A2, 03.02.2011 | |||
| WO9714719 A1, 24.04.1997 | |||
| CUESTA AM | |||
| et al., Multivalent antibodies: when design surpasses evolution, Trends Biotechnol, 2010, Vol.28, N.7, pp.355-362 | |||
| БИСПЕЦИФИЧЕСКАЯ МОЛЕКУЛА АНТИТЕЛА ДЛЯ ЛИЗИСА ОПУХОЛЕВЫХ КЛЕТОК, СПОСОБ ПОЛУЧЕНИЯ F(AB') 2 ФРАГМЕНТА БИСПЕЦИФИЧЕСКОЙ МОЛЕКУЛЫ АНТИТЕЛА, МОНОКЛОНАЛЬНОЕ АНТИТЕЛО (ВАРИАНТЫ), ФАРМАЦЕВТИЧЕСКИЙ ПРЕПАРАТ, ФАРМАЦЕВТИЧЕСКИЙ НАБОР ДЛЯ ЛИЗИСА ОПУХОЛЕВЫХ КЛЕТОК (ВАРИАНТЫ), СПОСОБ ЛИЗИСА ОПУХОЛЕВЫХ КЛЕТОК EX VIVO ПРИ АУТОГЕННОЙ ТРАНСПЛАНТАЦИИ КОСТНОГО МОЗГА | 1994 |
|
RU2203319C2 |

