Область техники, к которой относится настоящее изобретение
Настоящее изобретение относится к области биохимии, молекулярной биологии, медицины и может быть использовано в исследованиях функции сфингомиелинсинтазы человека.
Предшествующий уровень техники настоящего изобретения
В исследованиях биологической роли сфингомиелинсинтаз 1 и 2 (SMS1 и SMS2) уровень их экспрессии, как правило, определяют на уровне мРНК и суммарной ферментативной сфингомиелинсинтазной активности лизатов клеток и тканей [Qureshi A, Subathra M, Grey A, Schey K, Del Poeta M, Luberto C. Role of sphingomyelin synthase in controlling the antimicrobial activity of neutrophils against Cryptococcus neoformans. PLoS One. 2010 Dec 28; 5 (12): e15587, Hsiao JH, Fu Y, Hill AF, Halliday GM, Kim WS. Elevation in sphingomyelin synthase activity is associated with increases in amyloid-beta peptide generation. PLoS One. 2013 Aug 20; 8 (8): e74016, Giussani P, Tringali C, Riboni L, Viani P, Venerando B. Sphingolipids: key regulators of apoptosis and pivotal players in cancer drug resistance. Int J Mol Sci. 2014 Mar 12; 15 (3): 4356-92, Tafesse FG, Huitema K, Hermansson M, van der Poel S, van den Dikkenberg J, Uphoff A, Somerharju P, Holthuis JC. Both sphingomyelin synthases SMS1 and SMS2 are required for sphingomyelin homeostasis and growth in human HeLa cells. J Biol Chem. 2007 Jun 15; 282 (24): 17537-47, Ding T, Li Z, Hailemariam T, Mukherjee S, Maxfield FR, Wu MP, Jiang XC. SMS overexpression and knockdown: impact on cellular sphingomyelin and diacylglycerol metabolism, and cell apoptosis. J Lipid Res. 2008 Feb; 49 (2): 376-85, Separovic D, Semaan L, Tarca AL, Awad Maitah MY, Hanada K, Bielawski J, Villani M, Luberto C. Suppression of sphingomyelin synthase 1 by small interference RNA is associated with enhanced ceramide production and apoptosis after photodamage. Exp Cell Res. 2008 May 1; 314 (8): 1860-8]. Оценка содержания любого фермента необходима для выяснения его специфической активности. Определение уровня экспрессии SMS1 или SMS2 необходимо для оценки роли двух изоформ SMS и механизмов регуляции их активности. Обычным методом детекции и количественной оценки уровня экспрессии белка в клетках и тканях является иммуноблоттинг. Однако в исследованиях функции сфингомиелинсинтаз он практически не используется. Исключением являются работы, при которых клетки трансфицировали генноинженерными конструкциями, кодирующими белок, сшитый с антигенными пептидами, такими как V5 или flag, антитела к которым используются для детекции белка [Huitema K, van den Dikkenberg J, Brouwers JF, Holthuis JC. Identification of a family of animal sphingomyelin synthases. EMBO J. 2004 Jan 14; 23 (1): 33-44, Yeang C, Varshney S, Wang R, Zhang Y, Ye D, Jiang XC. The domain responsible for sphingomyelin synthase (SMS) activity. Biochim Biophys Acta. 2008 Oct; 1781 (10): 610-7]. Обычный способ оценки уровня экспрессии белка SMS1 в клетках и тканях путем иммуноблоттинга может быть затруднен из-за относительно низкого уровня его экспрессии. Например, в обогащенной фракции мембран аппарата Гольджи из тканей тонкой и толстой кишки свиньи [Guillén N, Navarro MA, Surra JC, Arnal C, Fernández-Juan M, Cebrián-Pérez JA, Osada J. Cloning, characterization, expression and comparative analysis of pig Golgi membrane sphingomyelin synthase 1. Gene. 2007 Feb 15; 388 (1-2): 117-24] и в суммарной фракции мембранных белков клеток HeLa [Burns ТА, Subathra М, Signorelli Р, Choi Y, Yang X, Wang Y, Villani M, Bhalla K, Zhou D, Luberto C. Sphingomyelin synthase 1 activity is regulated by the BCR-ABL oncogene. J Lipid Res. 2013 Mar; 54 (3): 794-805] SMS1 детектировали как слабую полосу, а в суммарной фракции мембранных белков клеток линий U937 и HL-60 SMS1 вообще не детектировали [Burns ТА, Subathra М, Signorelli Р, Choi Y, Yang X, Wang Y, Villani M, Bhalla K, Zhou D, Luberto C. Sphingomyelin synthase 1 activity is regulated by the BCR-ABL oncogene. J Lipid Res. 2013 Mar; 54 (3): 794-805].
Для анализа экспрессии мембранных белков, к которым относятся сфингомиелинсинтазы, в клетках и тканях перед иммунодетекцией проводят экстракцию суммарного белка с помощью буферов, содержащих такие детергенты, как SDS, тритон Х-100 или NP-40 [Weissman AM. Solubilization of lymphocytes. Curr Protoc Immunol. 2003 Nov; Chapter 8: Unit 8.1A, Bonifacino JS, Dell′Angelica EC, Springer ТА. Immunoprecipitation. Curr Protoc Immunol. 2001 May; Chapter 8: Unit 8.3, Abeam, Western blotting - a beginner′s guide, http://www.abcam.com/ps/pdf/protocols/WB-beginner.pdf]. К числу часто используемых растворов относится буфер RIPA (Radio Immuno Precipitation Assay), содержащий 50 мМ трис-HCl, [pH 7,5; 150 мМ NaCl; 1% NP-40; 0,5% дезоксихолат натрия; 0,1% SDS; 5 мМ EDTA) [MacPhee DJ. Methodological considerations for improving Western blot analysis. J Pharmacol Toxicol Methods. 2010 Mar-Apr; 61 (2): 171-7, Ngoka LC. Sample prep for proteomics of breast cancer: proteomics and gene ontology reveal dramatic differences in protein solubilization preferences of radioimmunoprecipitation assay and urea lysis buffers. Proteome Sci. 2008 Oct 24; 6: 30]. Однако получение белкового экстракта с помощью RIPA буфера или полный лизис ткани буфером, содержащим 4% SDS, не позволили авторам настоящего изобретения исследовать экспрессию SMS1 в тканях человека с помощью иммуноблоттинга, так как детектируемые полосы в нанесенных на форез образцах были очень слабыми.
Ранее Wassler M. и сотр. показали, что при экстракции культуры клеток гепатоцитов крысы растворами с разными концентрациями сапонина Gypsophila paniculata происходит дифференциальная пермеабилизация (достижение проницаемости) клеточных мембран [Wassler M, Jonasson I, Persson R, Fries E. Differential permeabilization of membranes by saponin treatment of isolated rat hepatocytes. Release of secretory proteins. Biochem J. 1987 Oct 15; 247 (2): 407-15]. При концентрации сапонина 0,04 мг/мл происходит пермеабилизация плазматической мембраны и экстракция цитозоля, при 0,2 мг/мл происходит пермеабилизация аппарата Гольджи и эндоплазматического ретикулума, а также экстракция люменального содержимого, в частности, секреторных белков. При этом даже большие концентрации сапонина (5 мг/мл) солюбилизировали лишь небольшую часть, менее 10%, мембраны эндоплазматического ретикулума [Wassler M, Jonasson I, Persson R, Fries E. Differential permeabilization of membranes by saponin treatment of isolated rat hepatocytes. Release of secretory proteins. Biochem J. 1987 Oct 15; 247 (2): 407-15], в то время как при экстракции клеток 0,5% тритоном Х-100 клеточные мембраны почти полностью растворялись [Fey EG, Wan KM, Penman S. Epithelial cytoskeletal framework and nuclear matrix-intermediate filament scaffold: three-dimensional organization and protein composition. J Cell Biol. 1984 Jun; 98 (6): 1973-84].
Методы обработки биологических образцов лизирующими растворами, содержащими сапонин различного происхождения и в различной концентрации, для анализа белков и других биологических молекул широко известны из уровня техники. Например, в международной патентной публикации WO 2006/117557 (дата публикации 09.11.2006 г.) раскрыт способ выявления представляющих интерес клеток в образце путем обработки его лизирующим реагентом, которым предпочтительно является сапонин, с последующим измерением активности внутриклеточных ферментов. В международной патентной публикации WO 2012/162133 (дата публикации 29.11.2012 г.) раскрыты способ селективного лизиса эндогенных клеток в биологическом образце, предусматривающий контакт биологического образца с лизирующим раствором и воздействие на смесь ультразвуком; а также лизирующий раствор, содержащий сапонин и протеиназу. Из патента США №8617839, опубл. 31.12.2013 г., известен способ обработки биологического образца сапонином с метил-6-O-CN-гептилкарбамоил)-альфа-D-глюкопиранозидом.
Наиболее близкие к настоящему изобретению технические решения изложены в патенте Великобритании №1533835, опубл. 29.11.1978 г., в котором раскрыт способ получения образца из дыхательных путей пациента для выявления патогенов, предусматривающий смешивание образца с сапонином в качестве лизирующего средства, при этом концентрация сапонина в растворе с различными реагентами составляет от 1% до 5%; а также в международной патентной публикации WO 2007/090862 (дата публикации 16.08.2007 г.), касающейся способа диагностики и/или контроля заболевания по изменению экспрессии сфингомиелинсинтазы 3 (SMS3), для чего предлагается производить получение биологического образца от субъекта и выявление в нем количества и/или активности SMS3 или кодирующей ее нуклеиновой кислоты, при этом для лизиса клеток используют 0,1% сапонин в PBS.
Однако несмотря на наличие многочисленных способов получения белков и их экстрактов, в том числе и с использованием сапонина, остается нерешенной проблема анализа в биологических образцах количества и/или активности сфингомиелинсинтаз 1 и 2 для дальнейшего изучения патогенеза ряда заболеваний, а также для их диагностики. В преодолении этой проблемы и состоит цель настоящего изобретения.
Краткое описание чертежей
В прилагаемых чертежах представлено сравнение эффективности экстракции SMS1 из тканей с использованием возрастающих концентраций сапонина и RIPA буфера.
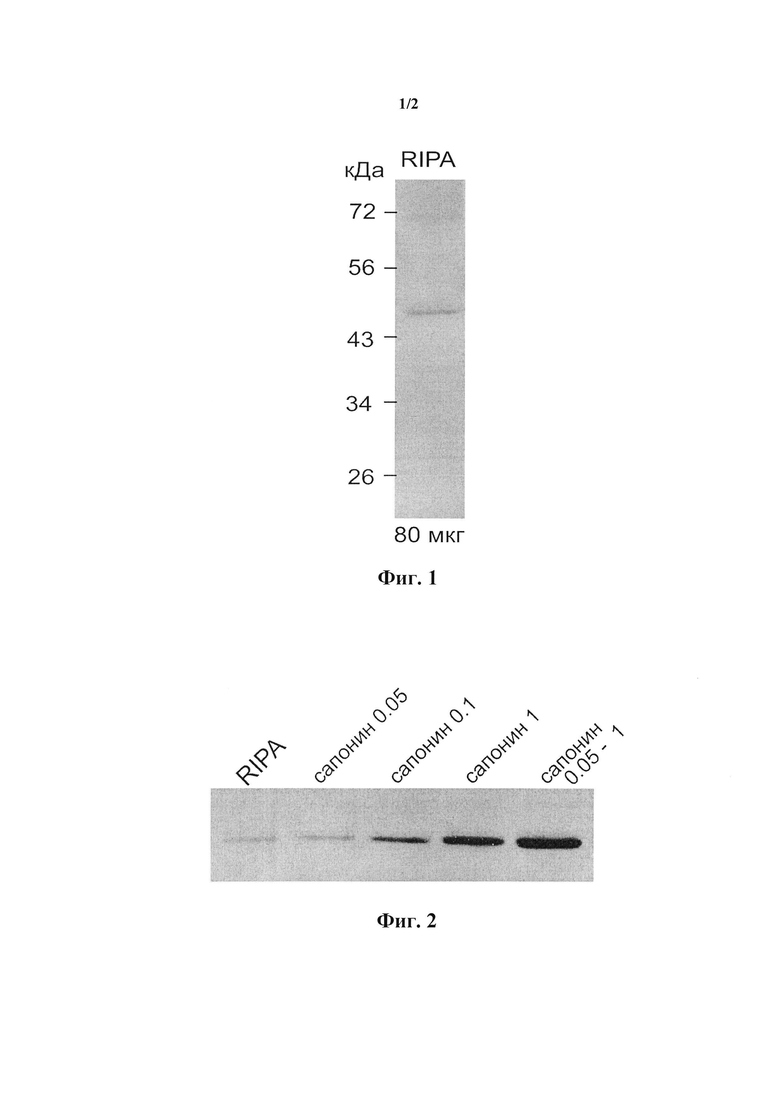
На фиг. 1 представлены результаты иммуноблоттинга 80 мкг белка, полученного из коры почки человека с использованием RIPA буфера, с поликлональным кроличьим антителом РАВ10379 к полноразмерному белку SMS1. При инкубации мембран с антителом в течение ночи наблюдали слабую полосу ожидаемого размера (молекулярная масса, рассчитанная по подвижности полосы, составила около 48 кДа).
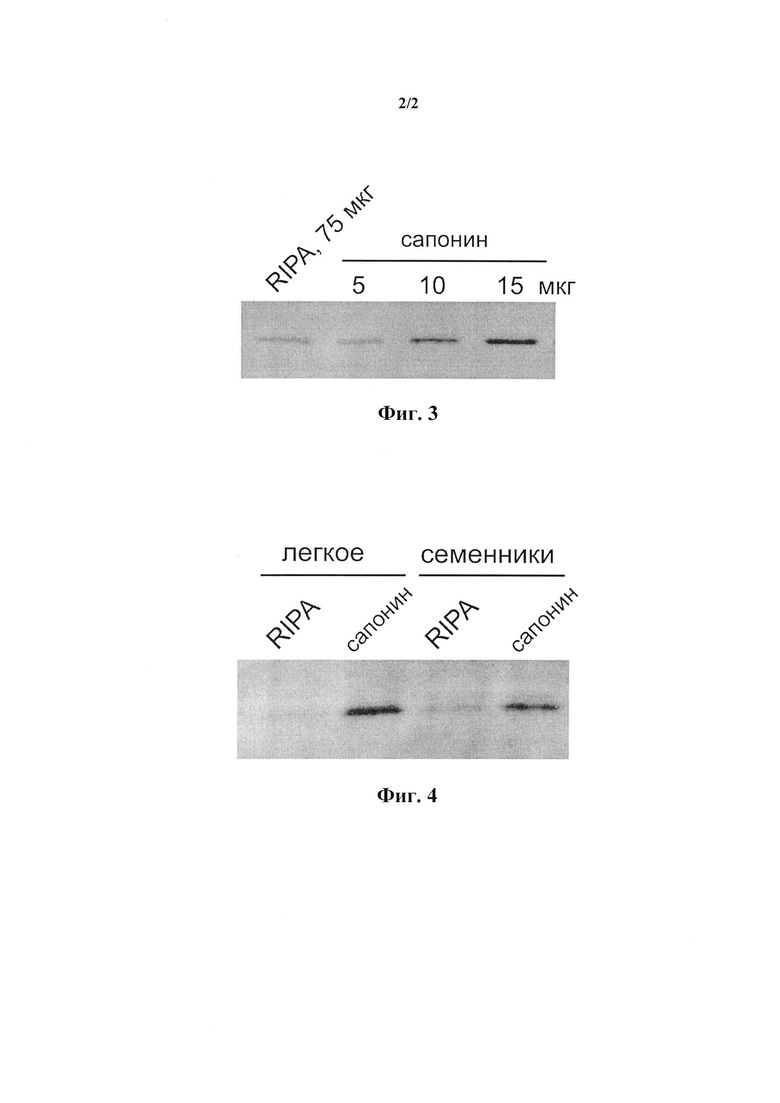
На фиг. 2 представлены результаты иммуноблоттинга 50 мкг белка, полученного из коры почки человека с использованием буфера, содержащего возрастающие концентрации сапонина 0,05 мг/мл, 0,1 мг/мл и 1 мг/мл, а также с использованием RIPA буфера.
На фиг. 3 представлено сравнение содержания SMS1 в 75 мг белка в экстрактах коры почки, полученных с использованием RIPA буфера, и в 5 мг, 10 мг и 15 мг белка этой же ткани, полученного последовательной экстракцией буфером, содержащим сапонин в концентрациях 0,05 мг/мл и 1 мг/мл.
На фиг. 4 представлены результаты анализа содержания SMS1 в легком и семенниках с использованием экстракции 50 мкг белка с помощью RIPA буфера и содержащего сапонин буфера.
Раскрытие настоящего изобретения
Сфингомиелинсинтазы 1 и 2 катализируют биосинтез мембранного фосфолипида сфингомиелина, который составляет 2-15% фосфолипидов клеток млекопитающих [Tafesse FG, Ternes Р, Holthuis JC. The multigenic sphingomyelin synthase family. J Biol Chem. 2006 Oct 6; 281 (40): 29421-5, M. Koval, R.E. Pagano, Intracellular transport and metabolism of sphingomyelin. Biochim. Biophys. Acta 1082 (1991) 113-125]. Синтез происходит путем обратимого переноса фосфохолина с фосфатидилхолина на церамид с образованием сфингомиелина и диацилглицерола [Tafesse FG, Ternes Р, Holthuis JC. The multigenic sphingomyelin synthase family. J Biol Chem. 2006 Oct 6; 281 (40): 29421-5]. Сфингомиелинсинтаза 1 (SMS1) локализуется в мембране аппарата Гольджи и синтезирует большую часть (до 90%) сфингомиелина клетки [Holthuis JC, Luberto С. Tales and mysteries of the enigmatic sphingomyelin synthase family. Adv Exp Med Biol. 2010; 688: 72-85]. Сфингомиелинсинтаза 2 локализуется преимущественно в плазматической мембране и в меньшей степени в аппарате Гольджи [Huitema K, van den Dikkenberg J, Brouwers JF, Holthuis JC. Identification of a family of animal sphingomyelin synthases. EMBO J. 2004 Jan 14; 23 (1): 33-44, Tani M, Kuge O. Sphingomyelin synthase 2 is palmitoylated at the COOH-terminal tail, which is involved in its localization in plasma membranes. Biochem Biophys Res Commun. 2009 Apr 10; 381 (3): 328-32]. Сфингомиелин формирует мембранные микродомены, и предполагается, что его синтез в аппарате Гольджи и дальнейший транспорт в плазматическую мембрану, где он концентрируется в экзоплазматическом листке, обеспечивает формирование композиционных различий между эндоплазматическим ретикулумом, аппаратом Гольджи и плазматической мембраной [Tafesse FG, Ternes Р, Holthuis JC. The multigenic sphingomyelin synthase family. J Biol Chem. 2006 Oct 6; 281 (40): 29421-5, Holthuis JC, Pomorski T, Raggers RJ, Sprang H, Van Meer G. The organizing potential of sphingolipids in intracellular membrane transport. Physiol Rev. 2001 Oct; 81 (4): 1689-723]. Синтез сфингомиелина сопровождается расходованием и образованием важных регуляторов клеточных процессов церамида и диацилглицерола, соответственно, что связывает функцию SMS1 с такими процессами, как мембранный везикулярный транспорт, передача сигнала рецепторов, пролиферация клеток и апоптоз [Tafesse FG, Ternes Р, Holthuis JC. The multigenic sphingomyelin synthase family. J Biol Chem. 2006 Oct 6; 281 (40): 29421-5, Holthuis JC, Luberto C. Tales and mysteries of the enigmatic sphingomyelin synthase family. Adv Exp Med Biol. 2010; 688: 72-85].
Биологическая роль сфингомиелинсинтаз в клетках и организме животных и человека активно изучается в последние годы. Результаты исследований показывают, что нарушения их функции могут быть вовлечены в патогенез ряда заболеваний. В частности, снижение активности или экспрессии SMS1 или SMS2 в клетках HL-60 промиелоцитарного лейкоза человека, дифференцированных по нейтрофильному пути, нарушало их киллерную активность в отношении криптококков, осуществляемую путем секреции антимикробных факторов, таких как дефенсины [Qureshi A, Subathra М, Grey A, Schey K, Del Poeta М, Luberto С. Role of sphingomyelin synthase in controlling the antimicrobial activity of neutrophils against Cryptococcus neoformans. PLoS One. 2010 Dec 28; 5 (12): е15587]. Увеличение экспрессии SMS1 или SMS2 в клетках гепатомы человека Huh7 повышало в них уровень сфингомиелина, холестерина и алипопротеина АроА-1, что свидетельствует о влиянии синтеза сфингомиелина на метаболизм холестерина [Yan N, Ding Т, Dong J, Li Y, Wu M. Sphingomyelin synthase overexpression increases cholesterol accumulation and decreases cholesterol secretion in liver cells. Lipids Health Dis. 2011 Mar 21; 10: 46]. У человека повышенный уровень сфингомиелина в плазме крови является фактором риска развития атеросклероза коронарных артерий [Jiang ХС, Paultre F, Pearson ТА, Reed RG, Francis CK, Lin M, Berglund L, Tall AR. Plasma sphingomyelin level as a risk factor for coronary artery disease. Arterioscler Thromb Vase Biol. 2000 Dec; 20 (12): 2614-8, Slotte JP. Biological functions of sphingomyelins. Prog Lipid Res. 2013 Oct; 52 (4): 424-37]. Болезнь Альцгеймера, для которой характерно образование в определенных структурах головного мозга бляшек, содержащих β-амилоидный пептид, сопровождается также нарушением баланса фосфолипидов в тканях мозга, в частности снижением уровня сфингомиелина и повышением уровня церамида [Haughey NJ, Bandaru VV, Bae M, Mattson MP. Roles for dysfunctional sphingolipid metabolism in Alzheimer′s disease neuropathogenesis. Biochim Biophys Acta. 2010 Aug; 1801 (8): 878-86]. Было показано, что при болезни Альцгеймера в гиппокампе человека значительно повышается экспрессия гена SMS1. Кроме того, ингибирование сфингомиелинсинтазной активности в клетках линии СНО-АРР с помощью D609 снижало секрецию этими клетками β-амилоидного пептида [Hsiao JH, Fu Y, Hill AF, Halliday GM, Kim WS. Elevation in sphingomyelin synthase activity is associated with increases in amyloid-beta peptide generation. PLoS One. 2013 Aug 20; 8(8): e74016]. Повышение уровня церамида и запуск апоптоза опосредуют действие многих химиотерапевтических препаратов. Различные пути снижения уровня церамида в клетках могут обусловливать устойчивость опухолей к химиотерапии. Одним из таких путей может быть повышение сфингомиелинсинтазной активности [Taniguchi М, Okazaki Т. The role of sphingomyelin and sphingomyelin synthases in cell death, proliferation and migration-from cell and animal models to human disorders. Biochim Biophys Acta. 2014 May; 1841 (5): 692-703, Itoh M, Kitano T, Watanabe M, Kondo T, Yabu T, Taguchi Y, Iwai K, Tashima M, Uchiyama T, Okazaki T. Possible role of ceramide as an indicator of chemoresistance: decrease of the ceramide content via activation of glucosylceramide synthase and sphingomyelin synthase in chemoresistant leukemia. Clin Cancer Res. 2003 Jan; 9 (1): 415-23]. С другой стороны, снижение сфингомиелинсинтазной активности в опухолевых клетках может обусловливать их устойчивость к апоптозу, который индуцируют клетки иммунной системы через Fas-, TNF- и TRAIL-рецепторы [Taniguchi М, Okazaki Т. The role of sphingomyelin and sphingomyelin synthases in cell death, proliferation and migration-from cell and animal models to human disorders. Biochim Biophys Acta. 2014 May; 1841 (5): 692-703, Morad SA, Cabot MC. Ceramide-orchestrated signalling in cancer cells. Nat Rev Cancer. 2013 Jan; 13 (1): 51-65].
Сфингомиелинсинтаза в клетках представлена на очень низком уровне, и поэтому долгие годы этот белок не могли выделить, хотя определяли активность этого фермента. Попытки в получении белкового экстракта тканей известными методами и последующий иммуноблоттинг с антителами к SMS давали слабую полосу, да и то и не во всех тканях. Однако авторами настоящего изобретения было отмечено, что в присутствии сапонина белок выходит из мембранных структур в лизат. Поэтому для получения обогащенного SMS экстракта биологических тканей был использован содержащий сапонин буфер.
Сапонины, называемые еще сапонозидами, представляют собой гликозиды растительного происхождения с поверхностно-активными свойствами. Сапонины широко распространены в природе, встречаются в различных частях растений - листьях, стеблях, корнях, цветках, плодах. Как известно, сапонины обладают способностью вызывать лизис клеток.
Таким образом, целью настоящего изобретения является разработка способа получения экстракта тканей человека, обогащенного SMS, путем последовательной экстракции биологического образца содержащим сапонин буфером для последующего изучения уровня экспрессии белка SMS в тканях человека.
Указанная цель достигается тем, что в буфер включают сапонин, подбирают концентрацию и такую последовательность обработки биологических образцов, при которой SMS эффективно экстрагируется из тканей.
В соответствии с настоящим изобретением способ предусматривает несколько стадий. На первой стадии ткань механически измельчают в жидком азоте до консистенции тонкого порошка и гомогенизируют на льду с 10 объемами (1 мл на 100 мг ткани) буфера, содержащего 50 мМ трис-HCl, pH 7,5,150 мМ NaCl, 2 мМ EDTA, 2,4 мг/мл смеси ингибиторов протеаз (Complete protease inhibitor cocktail, Santa Cruz Biotechnology, США) и 0,05 мг/мл сапонина (BioClot GmbH, Германия) в гомогенизаторе Даунса со слабо притертым поршнем. Гомогенат центрифугируют при 4000 g, 4°C в течение 10 минут, супернатант удаляют, а осадок суспендируют в таком же объеме буфера с 0,05 мг/мл сапонином, перемешивают вращением (14 об/мин, 4°C, 30 минут) и центрифугируют при 13000 g, 4°C в течение 10 минут. Затем полученный осадок суспендируют в таком же объеме указанного выше раствора с увеличением концентрации сапонина до 1 мг/мл. После перемешивания вращением в указанном выше режиме гомогенат центрифугируют в течение 15 минут при 13000 g, 4°C, супернатант отбирают, не задевая осадок. Все манипуляции проводят на льду с охлажденными до 0°C растворами. Полученный супернатант представляет собой белковый экстракт ткани, обогащенный SMS. Полученные таким образом экстракты разных тканей содержат от около 1,5 до 3 мкг/мкл суммарного белка. Для дальнейшего иммуноблоттинга удобно получить более концентрированный раствор белка. Для этого можно осадить белок смесью хлороформа и метанола по методу Wessel-Flugge [Wessel D, Flügge UI. A method for the quantitative recovery of protein in dilute solution in the presence of detergents and lipids. Anal Biochem. 1984 Apr; 138 (1): 141-3], осадок промыть ацетоном, высушить и хранить при -20°C до использования. Полученный осадок промывают ацетоном, высушивают и хранят при -20°C. На следующем этапе проводят детекцию SMS путем иммуноблоттинга стандартным методом. Образцы белка, экстрагированного из тканей, растворяют в небольшом количестве раствора 2% SDS, 15 мМ NaOH при температуре 37-45°C, нейтрализуют добавлением 1/10 объема 150 мМ HCl (контролируя pH раствора с помощью индикаторной бумаги). После определения концентрации белка с помощью набора RC DC protein Assay (Bio-Rad, США) образцы разделяют в денатурирующем 10% полиакриламидном геле по методу Laemmli [Gallagher SR. One-dimensional SDS gel electrophoresis of proteins. Curr Protoc Mol Biol. 2012 Jan; Chapter 10: Unit 10.2A] и переносят путем электропереноса на PVDF-мембрану (0,2 мкм, Bio-Rad, США).
Для иммунодетекции можно использовать набор ECL Advance Western Blotting Detection Kit (GE Healthcare, США) и реагенты Lumigen TMA-6 (Beckman Coulter, США). Иммунодетекцию проводят согласно рекомендациям производителей с небольшими модификациями. Мембрану блокируют 5% обезжиренным молоком в TBS буфере (20 мМ трис-HCl, pH 7,6; 137 мМ NaCl), содержащем 0,1% Tween 20, в течение 1 часа при комнатной температуре. Затем мембрану ополаскивают TBS буфером, содержащим 0,06% Tween 20 (буфер TBS-T), и инкубируют с поликлональным кроличьим антителом к SMS РАВ10379 (Abnova, Тайвань) в разведении 1:30000 в TBS буфере, содержащем 0,05% Tween 20 и 0,5% обезжиренное молоко, в течение ночи при 4°C. Затем мембрану отмывают TBS-T буфером и инкубируют с конъюгированными с пероксидазой антителами козы к иммуноглобулинам кролика pGAR-Iss (ИМТЕК, Россия) в разведении 1:500000 в TBS буфере, содержащем 0,05% Tween 20 и 0,5% обезжиренное молоко, в течение 1,5 часа при 4°C. Мембрану отмывают, как описано выше, ополаскивают дважды TBS буфером и детектируют сигнал с использованием хемилюминесцентного субстрата Lumigen ТМА-6 и пленки Hyprefilm ECL (GE Healthcare, США).
Содержание SMS в экстрактах тканей оценивают путем денситометрического сканирования иммуноблотов с помощью программы ImageQuant 5.2 (Molecular Dynamics, США) в линейном диапазоне зависимости интегральной оптической плотности полос иммуноблотов от количества белка в образце, подвергнутом иммуноблоттингу. Количество SMS нормируют по количеству белка экстракта ткани. Результат выражают как среднее ±S.D по трем независимым экспериментам.
Итак, настоящее изобретение относится к способу получения белкового экстракта, обогащенного сфингомиелинсинтазой, с помощью содержащего сапонин буфера, отличающемуся тем, что биологический образец последовательно экстрагируют буфером с различной концентрацией сапонина.
В соответствии с одним из предпочтительных вариантов осуществления биологический образец последовательно экстрагируют буфером, содержащим сапонин в концентрациях 0,05 и 1 мг/мл.
В соответствии с одним из предпочтительных вариантов осуществления сфингомиелинсинтазой является сфингомиелинсинтаза 1.
В соответствии с одним из предпочтительных вариантов осуществления сапонином является сапонин, полученный из Quillaja saponaria.
Примеры осуществления настоящего изобретения
Пример 1
Согласно варианту осуществления настоящего изобретения детектировали сфингомиелинсинтазу 1 путем иммуноблоттинга в ткани коры почки человека с помощью поликлонального кроличьего антитела к полноразмерному белку (Abnova, Тайвань). С этой целью для обогащения лизата ткани мембранными белками экстрагировали ткань коры почки буфером, содержащим возрастающие концентрации сапонина Quillaja saponaria 0,05 и 1 мг/мл. Для уменьшения объема экстрагирующего буфера ткань предварительно гомогенизировали в буфере, содержащем 0,05 мг/мл сапонина, и осаждали гомогенат без инкубации с буфером. Эта отмывка в определенной мере уравновешивала образец ткани с буфером и удаляла значительную часть белка без заметных потерь SMS1. Экстракт коры почки, полученный этим методом, содержал SMS1 в 12 раз больше, чем экстракт, полученный в RIPA буфере (фиг. 1).
Пример 2
Из образца коры почки человека экстрагировали 50 мкг белка с использованием буфера, содержащего возрастающие концентрации сапонина 0,05, 0,1 и 1 мг/мл, а также с использованием RIPA буфера. Иммуноблоттинг экстрактов с поликлональным кроличьим антителом РАВ10379 на полноразмерный белок SMS1 показал, что буфер, содержащий 0,05 мг/мл сапонина, экстрагирует примерно столько же SMS1, сколько RIPA буфер. Буфер, содержащий 0,1 мг/мл сапонина, извлекал незначительно большее количество SMS1. Экстракция ткани буфером, содержащим 1 мг/мл сапонина, давала заметно больший выход SMS1. Экстракция ткани сапонином в концентрации 1 мг/мл после предварительной экстракции сапонином в концентрации 0,05 мг/мл давала самый больший выход SMS1 (фиг. 2).
Пример 3
Проводили сравнение содержания SMS1 в 75 мг белка в экстрактах коры почки, полученных с использованием RIPA буфера, и в 5 мг, 10 мг и 15 мг белка этой же ткани, полученного последовательной экстракцией сапонином в концентрациях 0,05 мг/мл и 1 мг/мл. Денситометрический анализ иммуноблотов RIPA и сапонинового экстрактов коры почки показал, что сапониновый экстракт содержал в 12,65±0,16 раза больше SMS1, чем RIPA экстракт. Из представленного примера видно, что заявляемый способ экстракции белка с использованием содержащего сапонин буфера более эффективен, чем экстракция RIPA буфером (фиг. 3).
Пример 4
Согласно способу в соответствии с настоящим изобретением проводили анализ содержания SMS1 в легком и семенниках с использованием экстракции 50 мкг белка с помощью RIPA буфера и содержащего сапонин буфера. Результаты показали, что последовательная экстракция сапонином в концентрациях 0,05 мг/мл и 1 мг/мл позволила увеличить относительное содержание SMS1 в суммарном белке лизата исследуемых тканей (фиг. 4).
Таким образом, способ в соответствии с настоящим изобретением позволяет детектировать сфингомиелинсинтазу в биологических образцах различных тканей без предварительного выделения мембранной фракции, что делает его менее трудоемким и затратным, а также существенно повышает информативность и эффективность дальнейшего анализа, проводимого с использованием сфингомиелинсинтаз.
| название | год | авторы | номер документа |
|---|---|---|---|
| ЭНАНТИОМЕРЫ 2-ГИДРОКСИПРОИЗВОДНЫХ ЖИРНЫХ КИСЛОТ | 2012 |
|
RU2637937C2 |
| ЭНАНТИОМЕРЫ 2-ГИДРОКСИПРОИЗВОДНЫХ ЖИРНЫХ КИСЛОТ | 2012 |
|
RU2687967C2 |
| СПОСОБ ДИАГНОСТИКИ НАСЛЕДСТВЕННО ОБУСЛОВЛЕННОЙ ЭССЕНЦИАЛЬНОЙ ГИПЕРТЕНЗИИ | 2024 |
|
RU2841173C1 |
| ТЕСТ-СИСТЕМА ДЛЯ СКРИНИНГА ИНГИБИТОРОВ ПРОТЕИНКИНАЗЫ GSK3β; ЧЕЛОВЕКА | 2012 |
|
RU2528059C2 |
| ОДНОВРЕМЕННАЯ ХАРАКТЕРИСТИКА АНТИТЕЛ IgG И IgA ПРОТИВ НЕСКОЛЬКИХ ПИЩЕВЫХ АНТИГЕНОВ И ИММУННЫХ КОМПЛЕКСОВ С1q - ПИЩЕВОЙ БЕЛОК | 2015 |
|
RU2687976C2 |
| ТРИТЕРПЕНОВЫЕ КОМПОЗИЦИИ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 1999 |
|
RU2244547C2 |
| Способ получения рекомбинантного белка OMP25d-OMP19-OMP10His | 2024 |
|
RU2825400C1 |
| СПОСОБ ПОЛУЧЕНИЯ И ФАРМАКОЛОГИЧЕСКАЯ АКТИВНОСТЬ СРЕДСТВА АНТИОКСИДАНТНОГО ДЕЙСТВИЯ, ПОВЫШАЮЩЕГО ФИЗИЧЕСКУЮ РАБОТОСПОСОБНОСТЬ | 2021 |
|
RU2767675C1 |
| СПОСОБЫ ПОДДЕРЖАНИЯ ИЛИ УВЕЛИЧЕНИЯ РОСТА ИЛИ КОГНИТИВНОГО РАЗВИТИЯ | 2008 |
|
RU2523601C2 |
| НОВЫЙ АЛЛЕРГЕН-ПРОСТАТИЧЕСКИЙ КАЛЛИКРЕИН | 2007 |
|
RU2502074C2 |
Изобретение относится к фармацевтической промышленности, а именно к способу получения из тканей человека белкового экстракта, обогащенного сфингомиелинсинтазой 1. Способ заключается в последовательной экстракции ткани, выбранной из ткани коры почки человека, легкого или семенников, буфером, содержащим сапонин в концентрациях 0,05 и 1 мг/мл. Способ, описанный выше, позволяет получить экстракты с большим выходом сфингомиелинсинтазы 1. 4 ил., 4 пр.
Способ получения из тканей человека белкового экстракта, обогащенного сфингомиелинсинтазой 1, путем последовательной экстракции ткани, выбранной из ткани коры почки человека, легкого или семенников, буфером, содержащим сапонин в концентрациях 0,05 и 1 мг/мл, где сапонин получен из Quillaja saponaria.
| Способ прессования порошковых материалов | 1987 |
|
SU1533835A1 |
| Zhiqiang Li и др, Impact of Sphingomyelin Synthase 1 Deficiency on Sphingolipid Metabolism and Atherosclerosis in Mice, Arterioscler Thromb Vasc Biol, July 2012, С.1577-1584 | |||
| Н.И | |||
| Базыкина и др | |||
| Оптимизация условий экстрагирования природных антиоксидантов из растительного сырья | |||
| Химико-фармацевтический журнал | |||
| Коридорная многокамерная вагонеточная углевыжигательная печь | 1921 |
|
SU36A1 |
| WO | |||

