Изобретение относится к медицине, в частности наркологии и неврологии, а также к химико-фармацевтической отрасли и касается применения биологически активного флавоноидного соединения, 7-O-[6-O-(4-ацетил-α-L-рамнопиранозил)-β-D-глюкопиранозидо]-5-гидрокси-6-метокси-2-(4-метокси-фенил)-4Н-хромон-4-он (ацетилпектолинарин - АЦП) формулы,
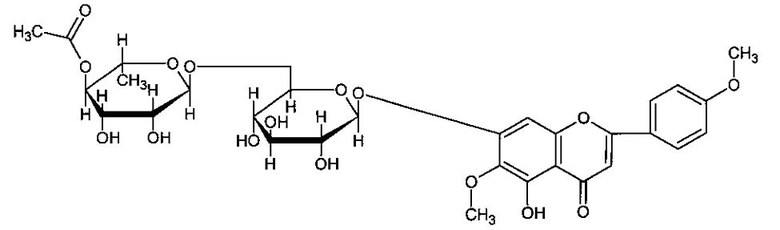
выделенного из Linaria vulgaris Mill. и обладающего выраженным антиалкогольным действием на нервную систему, что позволяет предположить возможность его использования в качестве лекарственного средства для лечения алкогольной зависимости, а также для коррекции нарушений таких ключевых параметров поведения, как координация движений и внимания, трезвая оценка ситуации, память и т.д., вызванных как кратковременным алкогольным опьянением, так и хроническим употреблением алкоголя.
Пристрастие к алкоголю является существенной социальной и медицинской проблемой во всем мире. Алкоголизм - одна из важных причин серьезных экономических и социальных проблем, начиная с нарушения производственного процесса и до ущерба трудоспособности населения планеты. Затраты на лечение больных, снижение производительности труда, повышенный риск развития соматических и психических заболеваний, ранняя смертность - все это последствия острого и хронического злоупотребления алкоголем.
Мозг является одной из основных мишеней алкогольной интоксикации. Потенциальные расходы, связанные с повреждением мозга алкоголем, огромны. У 7-10% населения развитых стран диагностируется алкогольная зависимость; у 9% из них выявлены клинические повреждения мозга. При этом показано, что мозг сохраняет накопленные в прошлом дисфункции, даже если интоксикация прекращена [Special Report to the US Congress: Alcohol and Health, Chapter 2: Alcohol and the Brain: Neuroscience and Neurobehavior; The Neurotoxicity of Alcohol].
Интенсивные и экстенсивные нейробиологические исследования выявили целый ряд синаптических и внесинаптических механизмов, в той или иной степени затрагиваемых воздействием этанола. Известно, что этанол способен вступать во взаимодействие с липидами и тем самым влиять на вязкость клеточных мембран [Electron paramagnetic resonance studies of ethanol on membrane fluidity / JH Chin, DB Goldstein // Adv Exp Med Biol. 1977; 85А: 111-22]. Среди исследователей существует консенсус, что мишенью этанола является ГАМК-эргическая передача: либо напрямую путем влияния на синаптические и экстрасинаптические ГАМК рецепторы, либо посредством эффекта на нейростероиды [Ethanol potentiation of GABAergic synaptic transmission may be self-limiting: role of presynaptic GABA(B) receptors / OJ Ariwodola, JL Weiner // J Neurosci. 2004 Nov 24; 24(47): 10679-86, The effects of acute and chronic ethanol exposure on presynaptic and postsynaptic gamma-aminobutyric acid (GABA) neurotransmission in cultured cortical and hippocampal neurons / RL Fleming, PB Morris, AL Morrow // Alcohol. 2009 Dec; 43(8): 603-18. doi: 10.1016 /j. alcohol.2009.10.006, Brain steroidogenesis mediates ethanol modulation of GABAA receptor activity in rat hippocampus / E Sanna, G Talani, F Busonero, MG Pisu, RH Purdy M Serra G Biggio // J Neurosci. 2004 Jul 21; 24(29): 6521-30, Effects of ethanol on GABA(A) receptors in GABAergic and glutamatergic presynaptic nerve terminals / M Wakita, MC Shin, S Iwata, N Akaike // J Pharmacol Exp Ther. 2012 Jun; 341(3): 809-19. doi: 10.1124/ jpet. 111.189126. Epub 2012 Mar 20]. При этом удовлетворительного объяснения, возбуждающего и растормаживающего действия алкоголя на психику не получено.
Несмотря на интенсивные исследования возможных молекулярных механизмов, поведенческих эффектов, до сих пор отсутствуют медикаментозные средства, оказывающие терапевтическое влияние на алкогольную интоксикацию головного мозга и нервной системы. Недостаточно информации по влиянию этанола на поведение малых нейронных сетей, изолированных от остального мозга. Данная тестовая система демонстрирует действие вещества на локальную сеть без привлечения сопутствующих эффектов внешних афферентов или действия нейронов, внешних по отношению к данной сети.
Ученые Калифорнийского университета в исследованиях на животных обнаружили, что Dihydromyricetin (DHM), выделенный из Hovenia dulcis Thub., противодействует острой алкогольной интоксикации, блокируя действие алкоголя на мозг, снижает пристрастие к его добровольному употреблению животными и способствует снятию симптомов похмелья. Установлено, что DHM устраняет эффект алкоголя на тормозные ГАМК(А) рецепторы мозга [Dihydromyricetin As a Novel Anti-Alcohol Intoxication Medication / Yi Shen, A. Kerstin Lindemeyer, Claudia Gonzalez, Xuesi M. Shao, Igor Spigelman, Richard W. Olsen, Jing Liang // The Journal of Neuroscience, January 4, 2012 - 32(1) - S. 390-401].
Это исследование целесообразно принять за прототип вследствие сходного класса химического вещества и вида биологической активности. Недостатком прототипа являются следующие положения.
1. Установленная анти-ГАМКэргическая активность DHM характеризует его действие только при интоксикации высокими дозами алкоголя.
2. В исследовании отсутствуют данные по влиянию алкоголя на нейронную активность сетей в области физиологических концентраций 0.09-0.5%. Содержание алкоголя в крови человека и экспериментальных животных в концентрации порядка 0.5% и выше являются летальными и не представляют практического интереса.
3. Отсутствуют исследования по установлению возможного механизма действия физиологических концентраций алкоголя на активность нейронных сетей.
4. Отсутствуют исследования по влиянию DHM на хроническую алкогольную интоксикацию.
5. Ограниченное распространение производящего растения. Родина Hovenia dulcis Thub. - Китай; разводят в Индии (Гималаи), Японии. В России и сопредельных государствах культивируют в теплых районах Кавказа, Средней Азии и Крыма (иногда побеги подмерзают) [Официальный сайт словаря «Большая советская энциклопедия», http://www.big-soviet.ru/ (дата обращения 22.04.2014)].
Предложенное изобретение отличается от прототипа тем, что биологически активное вещество АЦП, выделенное из Linaria vulgaris Mill, начинает действовать уже при низком и среднем уровне алкогольной интоксикации. Флавоноидное соединение АЦП является «мягким» избирательным ингибитором SK-каналов, устраняет прямое нейротоксическое действие алкоголя на мозг и корректирует аномальное поведение, вызванное как кратковременным алкогольным опьянением, так и его хроническим употреблением. Установлено, что малые калиевые {small potassium, SK) каналы являются мишенью всего спектра физиологических концентраций этанола. Кроме того, производящее растение Linaria vulgaris Mill. широко распространено на территории России и сопредельных государств. Сырьевая база растения достаточна для заготовки сырья в промышленных масштабах.
Ранее, в единичных исследованиях, установлено, что АЦП обладает кардиотоническим действием, увеличивает амплитуду сердечных сокращений и замедляет их ритм, при увеличении дозы вызывает гипотензивный эффект, сопровождающийся учащением и углублением дыхания [Распространение льнянки обыкновенной в СССР и содержание в ней ацетилпектолинарина и пектолинарина / А.В. Патудин, Б.А. Кривут, Л.С. Демидова // Первая республиканская конференция по медицинской ботанике: тезисы докладов. - Киев, 1984.; К фармакологической характеристике ацетилпектолинарина / М.И. Рабинович, А.Р. Зельцер // Труды Троицкого ветеринарного института, 1970. - Т. 13, вып. 1. - С. 69-73.; Семейство норичниковых - источник получения новых растений сердечно-сосудистого действия / М.И. Рабинович, А.И. Шретер, А.В. Ермолин и др. // Труды Троицкого ветеринарного института, 1970. - Т. 13, вып. 1. - С. 74-81]. Данных об антиалкогольной активности АЦП в литературе не обнаружено.
Флавоноид АЦП, выделенный из Linaria vulgaris Mill., относится к малотоксичным соединениям. LD50 при подкожном введении составила 825 мг/кг, при длительном введении АЦП в течение 30 дней отклонений в состоянии подопытных животных не установлено, после вскрытия патологических изменений во внутренних органах не отмечено [К фармакологической характеристике ацетилпектолинарина / М.И. Рабинович, А.Р. Зельцер // Труды Троицкого ветеринарного института, 1970. - Т. 13, вып. 1. - С. 69-73].
Целью предлагаемого изобретения является установление и описание высокоактивного соединения растительного происхождения с достаточной сырьевой базой производящего растения, действующего на различном уровне и при различной длительности алкогольной интоксикации, устраняющего прямое нейротоксическое действие алкоголя на мозг и корректирующего аномальное поведение, вызванное его однократным и хроническим употреблением.
Поставленная цель достигается выделением АЦП из надземной части Linaria vulgaris Mill. [Смирнова, Л.П. Химическое изучение флавоноидов некоторых видов льнянок и шалфеев: дис. …канд. фармац. наук: 15.00.03: защищена 26.04.76 / Смирнова Лилия Порфирьевна. - Москва, 1976. - С. 53-64].
Методика получения АЦП. Растительный материал 50 г измельчают до размера частиц не более 2 мм, экстрагируют 95% этанолом по 400 мл при нагревании на водяной бане с обратным холодильником в течение 2 часов три раза. Полученные извлечения объединяют, высушивают под вакуумом. Сухой остаток растворяют в минимальном количестве смеси диоксан-вода (1:1) и смешивают с 10 г силикагеля. Полученный порошок высушивают при температуре 40°C, наносят на хроматографическую колонку диаметром 30 мм, приготовленную из 40 г силикагеля. Колонку промывают 1 л четыреххлористого углерода, а затем элюируют смесью хлороформ-метанол (96:4). Элюаты собирают порциями по 200 мл и исследуют методом ТСХ в системе растворителей этилацетат-этанол-вода (100:27:13). Элюаты, имеющие на хроматограммах после обработки 5%-ным спиртовым раствором алюминия хлорида одну зону адсорбции желтого цвета со значением Rf=0,68±0,01, объединяют. Элюент отгоняют под вакуумом, остаток высушивают при температуре 40°C, кристаллизуют из этанола. Выход готового продукта составляет 1.8% от массы растительного сырья. Тпл=242-244°C.
Заявляемое соединение представляет собой светло-желтое кристаллическое вещество, растворимое в этаноле, при нагревании на водяной бане до 30°C, легко растворимо в хлороформе. При растворении образует мутные растворы. Очень легко растворяется в диметилсульфоксиде и 1,4-диоксане с образованием прозрачного раствора. Практически нерастворимо в воде. В спектре ПМР ("Bruker-300" (300 МГц) в ДМСО-d6, внутренний стандарт - TMS) соединения I имеются: сигналы протонов -ОСН3 групп (3.78, 3.86 м.д.), сигналы протонов паразамещенного ароматического кольца (два дублета при δ=7.15 м.д. и δ=8.05 м.д. с J=8.9 Гц), сигнал протонов -COCH3 (синглет при δ=1.97 м.д.), сигнал протона ОН группы флавона (синглет при δ=12.94 м.д.), сигналы протонов при третьем и восьмом атомах углерода (синглеты при δ=6.99 м.д. и δ=6.41 м.д., соответственно), сигнал -СН3 группы (дублет при δ=0.86 с J=6.3 Гц), сигналы десяти метановых протонов, одной метиленовой группы и пяти гидроксильных групп в интервале от 5.50 м.д. до 3.20 м.д.
Методика приготовления нейронной культуры. Влияние АЦП на клеточную активность на фоне алкоголя проводили на культуре нейронов, изготовленной по общепринятому протоколу [Functional plasticity triggers formation and pruning of dendritic spines in cultured hippocampal networks / M Goldin, M Segal, E Avignone // J Neurosci. 2001 Jan 1; 21(1): 186-93]. Крысята подвергались быстрой декапитации непосредственно в день рождения (Р0), после чего их мозг быстро изымался и помещался в охлажденный (до 4°C), насыщенный кислородом раствор Лейбовица L15 (от компании Gibco), обогащенный глюкозой в количестве 0,6% и антибиотиком - гентамицином (Sigma, из расчета 20 мкг/мл). Ткань гиппокампа подвергалась механической диссоциации после инкубации в растворе с добавлением трипсина (0,25%) и дезоксирибонуклеазы (50 мкг/мл) и перемещалась в инкубационный раствор, содержащий 5% инактивированной лошадиной сыворотки (HS), 5% эмбриональной телячьей сыворотки и комплексного фактора роста В-27 (1 мкл/1 мл), на основе минимальной базовой среды (minimum essential medium, MEM), изготовленной по протоколу Эрла (Gibco), обогащенной 0.6% раствором глюкозы, гентамицином (20 мкг/мл), и 2 мМ GlutaMax (Gibco) (так называемый обогащенный MEM).
Примерно 105 клеток в 1 мл среды высевалось в каждую лунку 24-луночного планшета, поверх слоя гиппокампальных глиальных клеток, высеянных и выращенных на 13 мм покровном стекле за 2 недели до привнесения туда нейронов [Morphological analysis of dendritic spine development in primary cultures of hippocampal neurons / M Papa, MC Bundman, V Greenberg, M Segal // J Neurosci. 1995 Jan; 15 (l Pt 1): 1-11]. Клетки помещались для роста и развития в стерильный инкубатор при температуре 37°C в атмосфере, содержащей 5% углекислого газа, в течение четырех дней. После этого среду меняли на 10% HS в обогащенном MEM с добавлением смеси из 5-фтор-2′-дезоксиуридина (5-fluoro-2-deoxyuridine/uridine, FUDR) (Sigma; 20 мкг и 50 мкг/мл, соответственно) для блокирования пролиферации глии. Этот раствор через четыре дня заменялся на 10% HS в MEM и оставлялся без изменений до дня использования культуры в эксперименте.
Методика электрофизиологической регистрации. Нейронная культура помещалась для физиологической регистрации в проточную камеру. Позиционирование ее относительно микроскопа в плоскостях X/Y осуществлялось моторизированной платформой производства фирмы Luigs & Neuman (Германия). Клетки помещались на предметный столик конфокального микроскопа фирмы Zeiss - PASCAL LSM-5. Использовались водно-иммерсионные объективы: Olympus х63 (0.9NA) или Zeiss х40 (0.8NA). Для обзора клеток и их сканирования один из объективов погружался непосредственно в регистрационный омывающий раствор. Состав стандартного раствора для регистрации: NaCl (129 мМ), KCl (4 мм), MgCl2 (1 мМ), CaCl2 (2 мМ), глюкоза (10 мМ), HEPES (10 мМ). pH=7.4 устанавливают добавлением NaOH, осмолярность на уровне около 320 мОсм - с помощью сахарозы.
Нейронная активность записывалась методом патч-кламп при помощи стеклянных микропипеток, содержащих раствор для внутриклеточной регистрации: K-глюконат или CsCl (140 мМ), NaCl (2 мМ), HEPES (10 мМ), Na-GTP (0,3 мМ), Mg-АТФ (2 мМ), фосфокреатин (10 мМ), pH 7.4. Сопротивление току для пипеток подбиралось на уровне 6-10 мОм. Сигналы усиливались при помощи усилителя Axopatch 200А (Axon Instruments), и сохранялись для компьютерной обработки. Данные анализировались с помощью программного обеспечения pCLAMP (MolecularDevices).
Метод визуализации внутриклеточного кальция. Культура инкубировалась в течение одного часа при комнатной температуре и в указанном выше регистрационном растворе, содержащем 3 мкМ кальциевого сенсора Fluo-2HA (AM). Вследствие пассивного одностороннего проникновения этого сенсора внутрь клеток его концентрация там достигала значений около 50 мкМ, что достаточно для надежной флуоресцентной записи внутриклеточных кальциевых сигналов. После завершения инкубационного периода клетки на стекле переводились в идентичный раствор, не содержащий краситель, и закреплялись в записывающей проточной камере на предметном столике одного из конфокальных микроскопов: прямого Zeiss PASCAL LSM-5 (Германия), оснащенного водно-иммерсионными объективами Olympus х63 (0.9NA) или инвертированного Zeiss LSM-510 (Германия) с масляно-иммерсионным объективом Zeiss х40 (1.3NA). Во втором случае моторизованный столик позволял фиксировать, заносить в память компьютера и воспроизводить местоположение нескольких полей записи в пределах одного препарата. Проточная перфузионная система была построена на основе пассивного притока из источника раствора на возвышенной платформе - стерильных 30 мл одноразовых шприцев. Отток осуществлялся принудительным избыточным отсосом с регулируемым уровнем по высоте на базе перистальтического насоса с переменной скоростью вращения. Пинхол устанавливался в закрытое положение, позволяющее получать оптические срезы толщиной 3-5 мкм с применением акустооптического модулятора. Данная экспериментальная система делала возможным двукратное уменьшение концентрации кальциевого сенсора (что снижало проявления артефактов) с сохранением качества визуализации. Изображения формировались путем сканирования высокой скорости и разрешающей способности в возбуждающем флуоресценцию свете аргонового лазера при длине волны света 488 нм и в спектре излучения около 505-550 нм. Высокая скорость сканирования достигалась методом двунаправленного движения зеркала, а также сканирования ограниченных зон поля зрения (так зазываемых «регионов интереса») при уменьшенном суммарном числе пикселей. Общая скорость сканирования достигала 50 мс/кадр и менее, что составляет 20 и более Гц. Серия изображений длительностью от несколько минут до нескольких десятков минут (обычно от 5 до 40 минут), в зависимость от размера текущего кадра в пикселях и от характера эксперимента, записывалась в стек. Серии изображений предварительно обрабатывались с применением программного обеспечения Zeiss LSM, Image J и Kaleida Graph.
Исследование взаимодействия АЦП и этанола in vivo и in vitro. Влияние АЦП in vitro на клеточную активность на фоне широкого спектра концентраций алкоголя (от 0.1 до 4% в омывающем солевом растворе) проводили в примарной культуре нейронов гиппокампа крысы; влияние in vivo на коррекцию поведения у животных, находящихся под воздействием физиологических концентраций алкоголя (при концентрации в крови в пределах 0.01%-0.5%), проводили на лабораторных мышах и крысах.
Оценка концентрации алкоголя в эксперименте. Следует отметить, что концентрации алкоголя в крови, после его употребления в виде спиртных напитков, достигает значений в 2-3 промилле (0.2-0.3%) при легких и средних уровнях опьянения и порядка 5 промилле (0.5%) при сильном опьянении. Дальнейшее повышение алкоголя в крови считается несовместимым с жизнью [A hybridizing of effects as described at Alcohol′s Effects from Virginia Tech and Federal Aviation Regulation (CFR) 91.17: Alcohol and Flying]. Для человека (мужчин и женщин весом около 70 кг), а также для лабораторных крыс (породы Wistar весом около 100 г) существует следующее соотношения количества выпитого этанола (EtOH) и его концентрации в крови, измеренной с интервалом в полчаса:

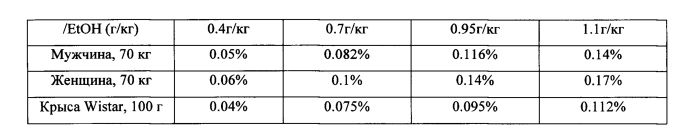
Количество этанола в крови связано с его концентрацией в мозге. Для человека не существует формулы простой корреляции между содержанием этанола в крови и в мозговой ткани. Однако, используя экспериментальные данные с крысами различных пород, при параллельном замере уровня EtOH в крови и в мозге [М. Nurmi, K. Kllanmaa, J.D. Sinklair, Brain ethanol levels after voluntary ethanol drinking in AA and Wistar rats, Alcohol, 19, 2, 113-118, 1999], установили, что концентрации этанола в мозге в течение последующих 5-15 минут после приема превышают таковые в крови в 1.5 раза, а при быстром его поступлении - в 3 раза.
Таким образом, физиологические концентрации этанола в мозге не могут превышать 1.5-2%. Предполагается, что этанол свободно преодолевает гематоэнцефалический барьер и поступает в мозг, где накапливается до указанного предела в силу аффинных свойств. Таким образом, концентрацию этанола в 4%, использованную в экспериментах in vitro, когда наблюдается полное угнетение активности нейронов, нельзя признать физиологически значимой, в то время как область 0.25-1.5% в ткани мозга является физиологической и составляет практический интерес.
Статистический анализ. Зоны, соответствующие отдельным нейронам в пределах поля, усреднялись по уровню серого и трансформировались в цифровые матрицы. Записи присваивался уникальный кодовый номер, и данные всего оптического поля сохранялись для дальнейшей обработки. Анализ цифровых данных, полученных из оптических стеков, производился с помощью созданной авторами системы программ в среде программирования высокого уровня MatLab для решения ряда задач1 (1 Компенсации эффекта выцветания красителя, заметного при длительных записях. Разделения записи на отдельные участки в соответствии с фармакологическим воздействием. Нормализации различных записей по частоте сканирования, зависящей от размера оптического поля. Автоматического выявление спайков по разработанному авторами критерию, в сравнении с динамическим значением стандартного отклонения уровня шума. Подсчета числа спайков за единицу времени. Измерения средней амплитуды спайков. Преобразования записей по принципу ΔF/F (вычитание среднего значения «фона» и деление на это значение). Вычисления усредненного спайка на основе всех записанных спайков. Определения коэффициента синхронизации между отдельными клетками в исследуемых полях (1 - максимум; 0 - минимум). Все виды вычислений сопровождались статистическим анализом: определением дисперсии, стандартного отклонения, дисперсионным анализом ANOVA, а также ретроспективным или post-hoc.).
Результаты выводились в виде графиков, таблиц и отдельных цифровых матриц. Часть их них переводилась в программы KaleidaGraph и Photoshop для итоговой обработки и создания иллюстраций.
Электрофизиологические данные исследовались с помощью программного обеспечения pClamp 8. Статистический анализ производился методами Т-теста (критерий Стьюдента), ANOVA и Вилкоксона-Манна-Уитни. В ряде случаев применялась среда программирования MatLab.
1. Исследования in vitro
Описание фоновой импульсной активности нейронов. Спонтанная активность нейронных сетей в культуре отличалась стабильностью в течение 40 минут записи (Рис. 1А). Показаны примеры (кадры-врезки) межспайкового периода, пика, а также стадии развития спайка в трех временных точках (Рис. 1Б). Растры с пометкой net обозначают изображения с вычтенным уровнем фона. На рисунке видна различная динамика развития спайков в разных клетках сети при их общей синхронизации. Количество спайков, их динамика (форма), амплитуда и коэффициент синхронизации полей оставались неизменными (Рис. 1В-Е, соответственно).
Таким образом, фоновая активность достаточно стабильна для фармакологических исследований.
Влияние этанола на нейронную активность. Всего использованы усредненные результаты по 27 экспериментам, охватывающим 162 нейрона. Во время непрерывной записи уровня внутриклеточного кальция добавлялись возрастающие концентрации этанола (Рис. 2). Установлено, что низкие концентрации 0.5% (или ~85 мМ) и 1% (~170 мМ) вызывают существенное повышение частоты спайков в культивируемых нейронах (серия из 5 стекол, в общей сложности 34 клетки; 95% повышение частоты разрядов: с 13.5±1.45 до 26.4±1.84 спайков за 100 секунд, р<0.0004, в соответствии с парным Т-тестом). Данное учащение связано с укорочением длительности спайков (1/2 спада = 0.442±0.065 сек в контроле, против 0.319±0.041 сек в присутствии 0.5% этанола, р<0.0001) (Рис. 2А, примеры 1-3 и В). Кроме того, низкая концентрация этанола вызывала повышение коэффициента синхронизации между клетками (Рис. 2Г). Высокие концентрации этанола (2-4%), напротив, существенно снижали частоту разрядов до полного подавления спонтанной активности. В процессе отмывки этанола наблюдалось постепенное восстановление к норме.
В серии экспериментов (Рис. 2Б) при тестировании еще более низкой концентрации этанола (0.25% или около 40 мМ) установлено нарастание активности при низких концентрациях (0.25-0.5%) и постепенный ее спад при высоких концентрациях (1-4%). Минимальная концентрация этанола, способная вызвать статистически достоверное повышение импульсации, составила 0.1% (около 15 мМ). Отмывка этанола возвращала к исходному уровню как частоту спайковых разрядов, так и коэффициент синхронизации между ними.
Таким образом, во влиянии этанола на нейронную активность выделяется фаза активации (при низких и средних физиологических концентрациях по уровню в мозге: 0.1-1%) и фаза торможения (при высоких физиологических концентрациях более 1% по уровню в мозге, а также при запредельных концентрациях более 2%, которые могут создаваться лишь in vitro, но не in vivo, так как приводят к угнетению центра дыхания и смерти).
Хроническое действие этанола на культивируемые нейронные сети. Эффект этанола в концентрации 1% был изучен на гиппокампальной культуре нейронов через 0.5, 1, 2 и 5 суток его непрерывного воздействия. Стабильность концентрации этанола в растворе контролировалась. Спонтанная активность нейронов записывалась после удаления этанола и сравнивалась с контролем. Было установлено, что сравнительно кратковременное пребывание этанола в культуре нейронов (12-24 часа) достоверно увеличивает уровень спонтанной активности, измеренный за 100 секунд: 14.3±1.18 в контроле, 21.8±2.27 через 0.5 суток и 24.57±2.51 через 1 сутки (р<0.0001). Через 48 часов уровень активности приближался к контрольному и не отличался от него статистически (р>0.43), а через 5 суток снижался значительно ниже контрольного: 5.37±0.66 (р<0.0001).
Таким образом, как при остром, так и хроническом воздействии этанола, установлена фаза патологически высокой импульсации в нейронных сетях, которая постепенно переходит в фазу абнормального торможения.
Учитывая данный факт, терапевтическое воздействие на алкогольную интоксикацию мозга должно быть основано на предотвращении проявления данных симптомов, а также на устранении уже возникших симптомов.
Влияние АЦП на нейронную активность в контроле и на фоне этанола (EtOH). В целях противодействия патологическому эффекту этанола на импульсную активность нейронов, впервые протестировано биологически активное вещество флавоноидной природы АЦП. Установлено, что в дозе от 1 до 30 мкМ, АЦП практически не изменяет паттерн импульсации. Урежение активности наблюдалось при 50 мкМ и особенно 80 мкМ (Рис. 3А). Дозировка 60 мкМ и выше достоверно урежают импульсацию, частотные данные обобщают исследование на 134 клетках (Рис. 3Б). Аналогично меняется и амплитуда, она падает после добавления АЦП в дозе 60 мкМ и значительно снижается в дозе 120 мкМ (Рис. 3В). Отмыв эффективно возвращает активность к норме, указывая на нетоксический характер изменений. Особый интерес представляет динамический эффект флавоноидного соединения. Результаты, представленные на рисунке 3Г, Д, доказывают, что в начале наблюдается кратковременная, статистически достоверная фаза учащения, после чего активность нейронов возвращается к норме (Рис. 3Г); амплитуда спайков в период учащения незначительно и кратковременно падает (Д).
Воздействие АЦП в дозе 10 мкМ на нейронную культуру в течение 7 дней инкубации не приводило к уменьшению числа нейронов в сравнении с контролем (р>0.45), что указывает на отсутствие нейротоксичности АЦП при длительном (хроническом) введении.
Ключевым является влияние флавоноида на нейронную активность, изменившуюся под влиянием этанола. Установлено, что АЦП предотвращает возбудительную фазу реакции на этанол: 8.75±1.15 в контроле; 9.21±0.95 в 10 мкМ АЦП; 7.9±1.73 в АЦП + 0.5% EtOH; и 7.5±1.5 в АЦП + 1% EtOH (33 эксперимента, n=168 клеток). В том случае если эффект EtOH уже развился, введение АЦП возвращало импульсацию к норме: 8.46±0.98 в контроле; 14.31±1.71 в присутствии EtOH; и 9.34±1.12 после добавления АЦП (24 эксперимента, 122 клетки) (Рис. 4А, Б). Сравнительное влияние различных доз АЦП на фоне этанола приведено на рисунке 4. Показано, что интродукция 10 мкМ АЦП восстанавливает повышенный уровень импульсации до нормы, тогда как доза 100 мкМ снижает ее (Рис. 4Б), а также уменьшает амплитуду спайков (В). Указанные эффекты успешно устраняются отмыванием.
Таким образом, терапевтическими следует признать дозы АЦП, не превышающие 30-60 мкМ, так как АЦП в указанных дозировках достоверно устраняет или предотвращает патологический эффект этанола. Высокие дозы АЦП снижают импульсацию, вызванную этанолом, однако эффект обратим, что указывает на отсутствие нейротоксичности.
Влияние АЦП на культивируемые нейронные сети при хроническом действии этанола. Присутствие 10 мкМ АЦП в питательном растворе эффективно предотвращало патологическое влияние этанола, вызванное его длительным введением. Установлено выраженное увеличение (до 2-х суток инкубации) и уменьшение (5 суток инкубации) импульсной активности нейронов. Спонтанная активность клеток на всех сроках тестирования, измеренная после устранения как АЦП, так и этанола, статистически не отличалась от таковой в контрольной группе (14.3±1.18 в контроле; 15.9±2.56 через сутки; и 12.6±3.08 через 5 суток после хронического тестирования, р>0.72).
Таким образом, АЦП блокирует патологическое влияние этанола, в нейронной культуре гиппокампа, вызванное его длительным (хроническим) введением.
Анализ механизмов действия этанола in vitro на активность нейронных сетей
Исследование синаптических токов на фоне этанола. Исследование проведено по методике электрофизиологической фиксации локуса мембраны. Для блокады спайков применен тетродотоксин (1 мкМ), а для подавления ГАМК-эргической активности - бикукуллин (10 мкМ). В этих условиях велась запись миниатюрных постсинаптических токов (мВПСТ) в контроле и после добавления этанола в концентрации 0.5 и 3% (Рис. 5).
Анализ полученных данных показал, что в последнем случае достоверно снижалась амплитуда мВПСТ по сравнению с контролем (Рис. 5В). Число спонтанных событий нарастало с добавлением этанола (Рис. 5Г, Д). Достоверное увеличение частоты (р<0.05). Необходимо отметить, что присутствие бикукуллина не позволяло развиться тормозным постсинаптическим потенциалам, тогда как при его отсутствии этанол в концентрации 3% вызывал подавление активности (см. Рис. 2). Установлено, что фаза нарастания мВПСТ была достоверно короче в присутствии этанола (Рис. 5Е). Изменение динамики было количественно небольшим, однако отличия являются статистически значимыми (р<0.01).
Совокупность полученных данных указывает на наличие двух фаз, а именно возбудительной и тормозной, при действии этанола. Причем торможение наблюдается при высокой физиологически не релевантной концентрации (3%) и «маскирует» возбудительную компоненту. Кроме того, укорочение фазы нарастания мВПСТ возможно связано с вовлечением вольтажзависимых калиевых каналов в возбудительные эффекты этанола.
Таким образом, влияние этанола на нейронную активность является смешанным - возбудительным и тормозным (при этом возбудительный эффект, вероятно, связан с действием потенциалзависимых каналов).
Эффект торможения исследован в следующей серии экспериментов. Действие этанола на нейроактивность при блокаде торможения. В соответствии с ранее установленной парадигмой после добавления 0.5% этанола наблюдается фаза возбуждения, а после 4% - фаза торможения активности (Рис. 6А). Под влиянием бикукуллина наступает блокада ГАМК-эргической составляющей; спайки становятся длиннее и выше по амплитуде (Рис. 6А, примеры 1 и 2). На этом фоне добавление 0.5% этанола учащает, а 4% - более не урежает спонтанную активность, планомерно сужая ширину спайков (Рис. 6А, примеры 3 и 4). Общая картина, суммирующая пять экспериментов, частоты и амплитуды приведена на рисунке 6Б, В. Панель Г позволяет сравнить эффекты 4% этанола в присутствии бикукуллина и при его отсутствии. Этанол в концентрации 0.5% в обоих случаях усиливал активность, однако в концентрации 4% более не подавлял импульсацию в присутствии ГАМК-антагониста.
Таким образом, эффект торможения этанола обусловлен усилением ГАМК-эргической активности. В области физиологических концентраций (0.5-1.5%) эффекты возбуждения и торможения этанола перекрываются.
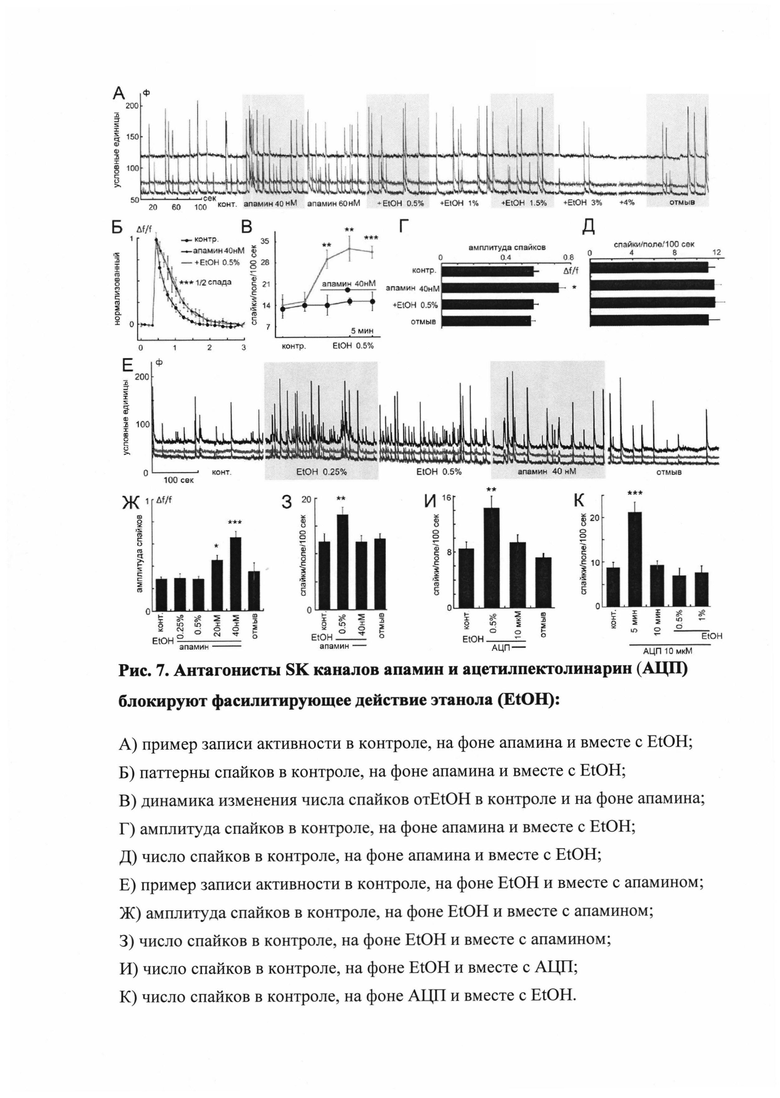
Канал возбудительного влияния этанола на нейронную активность. Вопрос о путях воздействия этанола на нервную систему имеет принципиальное значение. Наличие связи между этанолом и калиевыми каналами обнаружено в исследовании с применением специфического блокатора SK-каналов - токсина апамина. На рисунке 7А показана динамика влияния апамина (40-60 нМ) на импульсацию. За первой фазой кратковременной активации следует стабилизация нейронной активности на несколько повышенном или нормальном уровне при повышенной амплитуде разрядов (Рис. 7А, Д). Добавление этанола (0.5-1.5%) на фоне апамина не приводило к дальнейшему повышению импульсации, однако этанол в более высоких концентрациях по-прежнему угнетал ее, а отмыв - возвращал к норме. Также укороченная динамика спайков, вызванная этанолом, восстанавливалась апамином до нормы (Рис. 7Б). Сравнение эффекта этанола в концентрации 0,5% с апамином и без него приведено на рисунке 7Г. Апамин вызывает кратковременное увеличение частот, но блокирует эффект этанола (обобщены данные девяти экспериментов - 44 клетки).
Аналогичным образом, на фоне возбудительного эффекта этанола интродукция апамина (40 нМ) возвращала импульсацию к норме (Рис. 7Е, 3). При этом, однако, как и в первом случае, возрастала амплитуда спайковых разрядов (Рис. 7Ж). Необходимо отметить, что длительное (хроническое) воздействие апамина на нейронную культуру сопровождалось нейротоксическим эффектом (27.3%±9.165 гибели клеток по сравнению с контролем), что, вероятно, связано с повышенной амплитудой импульсации.
Таким образом, возбудительный эффект этанола обусловлен участием потенциалзависимых калиевых каналов типа SK.
Механизм действия АЦП на нейронную активность. Биологически активное вещество АЦП, при идентичной стратегии эксперимента (см. выше), аналогично апамину воздействовало на патологическое возбуждение, провоцируемое 0.5% этанолом (Рис. 7И, К). АЦП в дозе 10 мкм блокирует возбудительное влияние этанола на импульсацию (114 клеток), при этом вызывает кратковременное повышение спонтанной активности нейронов. Через 10 минут она полностью возвращается к контрольному уровню, после чего влияние этанола оказывается заблокированным.
Очевидно практически полное совпадение картины как в случае с интродукцией АЦП на фоне этанола (И), так и при подаче этанола на фоне АЦП (К). В обоих случаях возбудительный эффект этанола был полностью заблокирован. Возбудительный эффект собственно АЦП был кратковременным и идентичным влиянию апамина. Однако принципиальным отличием эффекта АЦП от апамина явилось отсутствие повышения амплитуды спайков (Рис. 3) и признаков нейротоксичности.
Подобно апамину, АЦП не влиял на эффекты более высоких концентраций этанола. Данные приведены на рисунке 8. Как было показано выше, этанол в концентрации 3% существенно снижал импульсацию в нейронных сетях. Различные дозы АЦП от 5 до 100 мкМ не компенсировали данное тормозящее действие этанола, т.е. не повышали импульсацию (рис. 8А) и амплитуду спайков (рис. 8Б).
Доказательство работы АЦП через воздействие на SK-каналы было проведено с использованием избирательного агониста SK-каналов - 1-этил-1,3-дигидро-2Н-бензимидазола-2-один (1-EBIO) (рис. 9). Его воздействие на SK-каналы зависит от концентрации: в начале данный агонист, подобно этанолу, усиливает нейронную импульсацию (Рис. 9А). Затем по мере нарастания концентрации (с 200 нМ до 500 мкМ) импульсация начинает подавляться и окончательно исчезает. Данный эффект не связан с усилением ГАМК-эргической системы, так как не блокируется бикукуллином [Ethanol affects network activity in cultured rat hippocampus: mediation by potassium channels / E. Korkotian, T. Bombela; T. Odegova; P. Zubov; M. Segal / PLOS ONE, 2013. - N. 8, - Is. 11.]. Таким образом, действие 1-EBIO совпадало с действием этанола лишь в своей начальной фазе, но не в фазе торможения. Последняя связана с тем, что после открытия большей части или всех SK-каналов клетки теряли способность к деполяризации. Как и следовало ожидать, эффект 1-EBIO успешно устранялся специфическим блокатором SK-каналов - апамином (Рис. 9А, Б). Установлено, что аналогичным образом воздействовал и АЦП, в рамках терапевтических доз он восстанавливает спонтанную активность, что доказывает селективный и специфический характер его влияния на культуру нейронов (Рис. 9В).
Таким образом, биологически активное флавоноидное соединение АЦП в низких и средних дозах (от 1 до 30 мкМ) кратковременно усиливает импульсацию, не меняя общего ее паттерна. Высокие дозы АЦП (>80 мкМ) подавляют импульсацию. Эффекты АЦП обратимы, при кратковременном и длительном воздействии, что указывает на отсутствие нейротоксичности. АЦП не влияет на баланс торможения в нейронных сетях, но оказывает выраженное антиалкогольное воздействие в области физиологических концентраций этанола, устраняя фазу патологического учащения импульсации. Доказано, что действие АЦП опосредуется нетоксическим и обратимым воздействием на SK-каналы, которое выражается в избирательном снижении проводимости в данном типе калиевых каналов.
2. Исследования in vivo
1. Тест "крестообразный лабиринт". Крестообразный лабиринт состоит из 4 камер (пронумерованных 1, 2, 3, 4), соединяющихся между собой через такую же пятую центральную камеру. Тупиковые и центральная камера лабиринта для мышей представляют собой куб с ребром длиной 15 см и отверстием для входа сечением 7 × см. В лабиринте для крыс центральная камера представляет собой куб с ребром длиной 20 см, а тупики имеют размеры 30×20×20 см и соединяются с центральной камерой отверстием для входа сечением 12×12 см.
Животное помещали в центральную камеру, сверху лабиринт закрывали прозрачной крышкой и позволяли ему исследовать помещения аппарата до тех пор, пока животное не произведет 13 посещений его тупиковых камер. Заход в тупик считают состоявшимся, если животное переносит все 4 лапы в это отделение лабиринта.
Интерпретация результатов. Последовательность переходов и их продолжительность регистрируют с помощью программы персонального компьютера. Последующий анализ осуществляют с помощью программы, которая выделяет и оценивает следующие виды поведения.
- Общее время в лабиринте, затраченное на 13 заходов в его тупики, являющееся коррелятом исследовательской активности в тесте открытого поля.
- Латентный период начала исследования лабиринта - время между помещением животного в центральную камеру и его первым заходом в тупик. Этот показатель коррелирует с избеганием посещения открытых рукавов приподнятого плюсобразного лабиринта и предложен для отбора анксиолитического действия веществ.
- Полный обход лабиринта, продолжающийся до того момента, когда будут посещены все его отсеки. Это поведение отражает способность к пространственной ориентации и рассматривается как одна из форм элементарной рассудочной деятельности у животных. Чем более эффективна пространственная ориентация, тем меньше число визитов, затраченных на первый полный обход, и больше общее число совершенных полных обходов.
- Возврат в тупик, посещенный при предыдущем визите. Это поведение рассматривается как показатель ошибок краткосрочной памяти.
- Поочередное посещение двух из четырех тупиков, которое интерпретируется как поведенческая стереотипия. Такой эпизод поведения имеет место, если в течение трех или более визитов в тупики подряд посещались какие-либо 2 из 4 отсеков.
- Количество правых и левых поворотов при переходе из тупика в тупик через центральный отсек лабиринта, отражающие степень асимметрии локомоции.
Условия проведения эксперимента. Исследование проводили на нелинейных белых мышах обоего пола.
1. Контрольная группа: впрыскивался стерильный физиологический раствор, внутрибрюшинно.
2. Этанол-группа: впрыскивался спирт этиловый в количестве, необходимом для создания концентрации в крови в пределах 0.01%-0.5%.
3. Этанол-флавоноид-группа: впрыскивался раствор АЦП в количестве 10 мг/кг и через 10 мин впрыскивался спирт этиловый в количестве, необходимом для создания концентрации в пределах 0.01%-0.5% в крови.
Результаты. У алкоголизированных животных в тесте крестообразного лабиринта было отмечено сокращение латентного периода, общего времени в лабиринте и времени полного обхода в сравнении с контролем, что подтверждает установленное в опытах на культуре клеток гиппокампа повышение активности. Кроме того, наблюдалась четко выраженная стереотипность поведения и асимметрия локомоции в сравнении с контрольной группой животных.
У животных, которым предварительно внутрибрюшинно вводили раствор АЦП, наблюдалось увеличение общего времени нахождения в лабиринте в сравнении с алкоголизированными животными, а также времени, затраченного животными на совершение полного обхода, что свидетельствует о предотвращении возникновения спонтанной активности нервных клеток, вызываемой действием алкоголя.
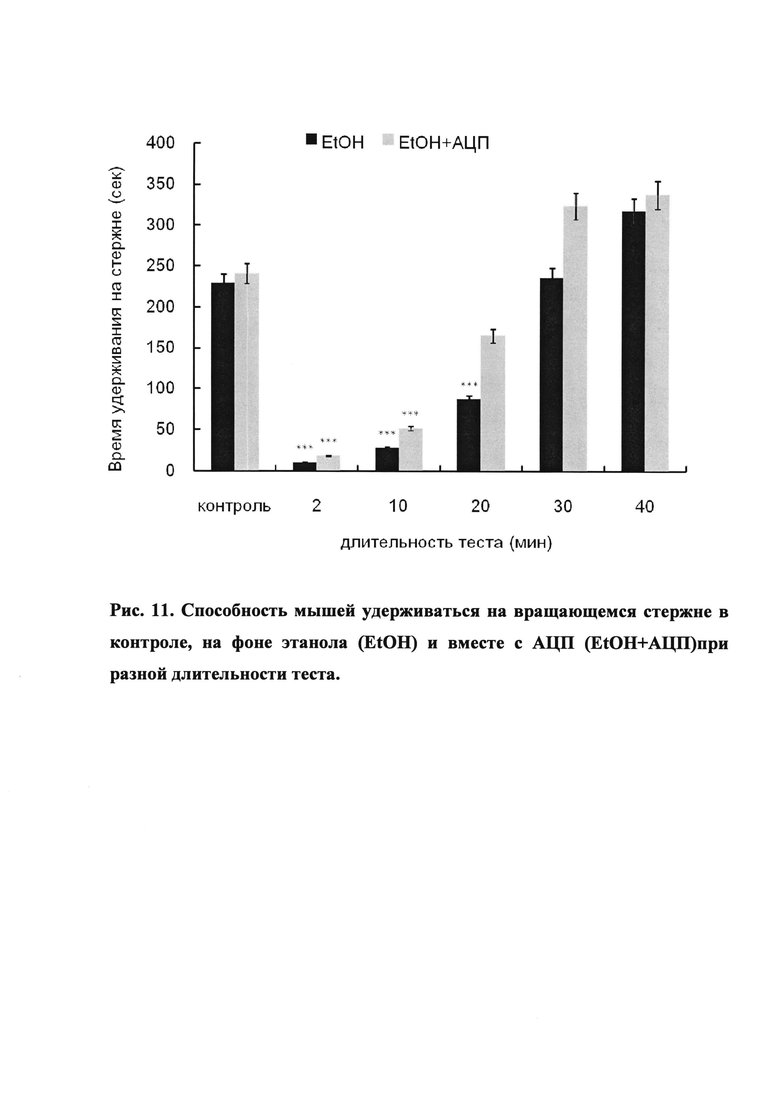
2. Тест «вращающийся стержень». Установка представляет собой приподнятый на высоту 60 см стержень диаметром 6 см с фиксированной скоростью вращения от 4,0 до 40,0 обротов/мин. При изучении влияния фармакологических веществ на алкогольную интоксикацию мышей предварительно тестируют до достижения стабильных показателей (время удерживания на стержне).
Интерпретация результатов. Способность животных сохранять равновесие и увеличивать время нахождения на вращающемся стержне под действием изучаемого вещества, вводимого на фоне этанола, рассматривают, как способность уменьшать токсическое действие этанола и восстанавливать координацию движения.
Условия проведения эксперимента. Скорость вращения стержня - 10 оборотов/минуту, мыши мужского пола массой 25-35 г.
1. Контрольная группа: впрыскивался стерильный физиологический раствор, внутрибрюшинно.
2. Этанол-группа: впрыскивался спирт этиловый 12% в количестве, необходимом для создания концентрации 0.05, 0.1, 0.2, 0.3, 0.5% в крови. Для белых мышей мужского пола массой около 25 г известно [ВАС Charts по Virginia Tech] следующее соотношение между количеством принятого 95% этанола и его содержанием в крови при внутрибрюшинном способе введения.
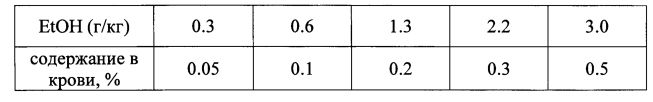
3. Этанол-флавоноид-группа: впрыскивался раствор АЦП в количестве 10 мг/кг, и через 10 мин впрыскивался спирт этиловый в количестве, необходимом для создания концентрации в крови 0.3%. Оценивали время удерживания мышей на вращающемся стержне.
Как следует из полученных данных (Рис. 10), поведение мышей при введении этилового спирта в концентрации 0.05% соответствует контрольной группе. При увеличении концентрации этанола в крови до 0.1 и 0.2% часть мышей становится более активна (перелезают через перегородки на стержне, в результате чего падают). Концентрация этанола 0.3% является наиболее приемлемой, у животных наблюдается состояние ажитации и мышечной слабости (мыши активны, но отсутствует координация движений). При введении этанола в дозе 0.5% мыши неподвижны (сон). При внутрибрюшинном введении АЦП поведение животных соответствует контрольной группе.
Отличие результатов контрольной группы от результатов этанол-группы статистически значимо (*, Т-тест).
Результаты. У алкоголизированных животных в тесте вращающегося стержня было отмечено состояние повышенной активности, а также наблюдалось изменение координации движений в сравнении с контрольной группой.
Установлено, что в интервале времени от 10 до 40 минут после внутрибрюшинного введения АЦП на фоне алкоголя значительно увеличивается время нахождения животных на стержне, что свидетельствует о восстановлении координации движений (Рис. 11).
3. Тест «открытое поле». Исследование проводилось на крысах разного возраста (от 5 до 9 недель). Отдельно исследовалось поведение самцов и самок. Открытое поле представляет собой белый ламинированный квадратный короб со стороной 1 метр и высотой 50 см, открытый сверху. Основание короба расчерчено на квадраты со стороной 25 см, таким образом, всего имеется 16 квадратов. Животное может свободно перемещаться по поверхности короба, пересекая линии внутренних квадратов. Эксперимент с каждым животным проводится в течение 3х минут. Животное помещается в один из угловых квадратов и одновременно с этим запускается таймер. Общее количество пересеченных линий за 3 минуты фиксируется автоматически.
Интерпретация результатов. Контрольные животные инстинктивно стараются избегать перемещений по открытому пространству, поэтому среднее количество пересеченных ими линий невелико. Эффект этанола на сознание животного может выразиться в расторможенном поведении, частичной или полной утрате страха перед перемещением по открытому пространству.
Условия проведения эксперимента.
1. Контрольная группа: впрыскивался стерильный физиологический раствор, внутрибрюшинно.
2. Этанол-группа: впрыскивался 30% раствор этанола в физиологическом растворе, внутрибрюшинно. Объем впрыснутого раствора вычислялся исходя из веса животного и контролировался по фактическому уровню в крови. Опыт проводился по истечении 30 минут после впрыскивания.
3. Этанол-флавоноид-группа: впрыскивался 30% раствор этанола в физиологическом растворе (объем как в пункте 2) и по истечении 5 минут впрыскивался раствор АЦП в концентрации 3, 5 и 10 мг/кг веса животного. Опыт проводился по истечении 30 минут после впрыскивания этанола.
Результаты. Среднее число пересеченных линий в контрольной группе (6 животных в возрасте 4-х недель, 4 животных в возрасте 5-х недель, 4 животных в возрасте 9-и недель) не зависело от возраста животных и составило 8.2±3.6. После введения этанола низкой концентрации (0.1-0.15%) количество пересеченных линий резко возрастало - до 22.1±3.2. Кроме того, обнаружено более выраженное влияние этанола на группу молодых крыс (5 недель) в сравнении со старшей группой (9 недель). Высокие концентрации этанола подавляли двигательную активность животных, и число пересеченных линий снижалось по отношению к контролю вплоть до нуля. Установлено, что АЦП в дозе 5-10 мг/кг блокировал активирующий эффект только низких концентраций этанола, снижая число пересеченных линий до 10.7±4.6 (Рис. 12). При этом сам по себе АЦП не оказывал значимого влияния на поведение крыс в данном тесте, а также не влиял на эффекты высоких концентраций этанола.
Таким образом, установлено, что АЦП эффективно блокирует эффект низких концентраций этанола в крови крыс, ассоциирующийся с патологически расторможенным поведением и утратой естественной осторожности.
Отличие результатов контрольной группы от результатов этанол-группы статистически значимо (***, Т-тест). Отличие результатов этанол-флавоноид-группы от результатов этанол-группы статистически значимо (**, Т-тест), отличие результатов контрольной группы от результатов этанол-флавоноид-группы - статистически не значимо.
Таким образом, биологически активное флавоноидное соединение (АЦП) проявляет анти-SK-канальные свойства предотвращая патологическое усиление спонтанной активности нервных клеток центральной нервной системы, вызванное воздействием этанола. АЦП эффективно блокирует эффект физиологических концентраций этанола в крови животных, восстанавливает координацию движений, когнитивные способности, предотвращает возбуждение и патологические формы активности, вызванные воздействием этанола. Следовательно, заявляемое биологически активное флавоноидное соединение АЦП является перспективным для коррекции поведения в состоянии алкогольного опьянения, для применения в качестве лекарственного средства при лечении хронической алкогольной зависимости, а также широкого спектра неврологических и психиатрических заболеваний, вызванных злоупотреблением алкоголя.
| название | год | авторы | номер документа |
|---|---|---|---|
| ПРИМЕНЕНИЕ АСКОРБАТА ЛИТИЯ ДЛЯ ПРОФИЛАКТИКИ И ЛЕЧЕНИЯ ХРОНИЧЕСКОЙ АЛКОГОЛЬНОЙ ИНТОКСИКАЦИИ | 2017 |
|
RU2639496C1 |
| Низкомолекулярный миметик мозгового нейротрофического фактора с анальгетическим, анксиолитическим и антиаддиктивным действием | 2020 |
|
RU2759023C2 |
| СПОСОБ ПОЛУЧЕНИЯ СРЕДСТВА, ОБЛАДАЮЩЕГО СЕДАТИВНОЙ, ПРОТИВОСУДОРОЖНОЙ И НЕЙРОМОДУЛЯТОРНОЙ АНТИАЛКОГОЛЬНОЙ АКТИВНОСТЬЮ | 2014 |
|
RU2613312C2 |
| СРЕДСТВО, ОБЛАДАЮЩЕЕ АНКСИОЛИТИЧЕСКОЙ И ЦЕРЕБРОПРОТЕКТОРНОЙ АКТИВНОСТЬЮ, УМЕНЬШАЮЩЕЕ ВЛЕЧЕНИЕ К АЛКОГОЛЮ | 2008 |
|
RU2393855C1 |
| СРЕДСТВО ДЛЯ КУПИРОВАНИЯ АЛКОГОЛЬНОГО АБСТИНЕНТНОГО СИНДРОМА | 2012 |
|
RU2521226C2 |
| КОМПОЗИЦИЯ ДЛЯ ЛЕЧЕНИЯ ИЛИ ПРОФИЛАКТИКИ НЕЙРОДЕГЕНЕРАТИВНЫХ РАССТРОЙСТВ | 2012 |
|
RU2699038C2 |
| АНТИАЛКОГОЛЬНОЕ СРЕДСТВО | 2015 |
|
RU2624482C2 |
| СПОСОБ ПРОФИЛАКТИКИ И ЛЕЧЕНИЯ НЕЙРОДЕГЕНЕРАТИВНОЙ ПАТОЛОГИИ И СОСУДИСТОЙ ДЕМЕНЦИИ | 2014 |
|
RU2587617C1 |
| СПОСОБ ЛЕЧЕНИЯ ПАЦИЕНТОВ С ХРОНИЧЕСКОЙ АЛКОГОЛЬНОЙ ИНТОКСИКАЦИЕЙ | 2014 |
|
RU2566075C1 |
| 17α,21-ДИАЦЕТАТ 17α,21-ДИГИДРОКСИПРЕГН-4-ЕН-3,20-ДИОН, ОБЛАДАЮЩИЙ АНТИАЛКОГОЛЬНОЙ И АДАПТОГЕННОЙ АКТИВНОСТЬЮ | 1999 |
|
RU2172173C2 |










Изобретение относится к медицине и касается применения биологически активного флавоноида 7-O-[6-O-(4-ацетил-α-L-рамнопиранозил)-β-D-глюкопиранозидо]-5-гидрокси-6-метокси-2-(4-метокси-фенил)-4Н-хромон-4-он (ацетилпектолинарин - АЦП) формулы I, выделенного из Linaria vulgaris Mill. и обладающего выраженным антиалкогольным действием на нервную систему. Это позволяет предложить его к использованию в качестве лекарственного средства для коррекции нарушений параметров поведения: координация движений и внимания, трезвая оценка ситуации, память - вызванных как кратковременным алкогольным опьянением, так и хроническим употреблением алкоголя. Технический результат состоит в реализации указанного назначения. 12 ил., 2 табл.
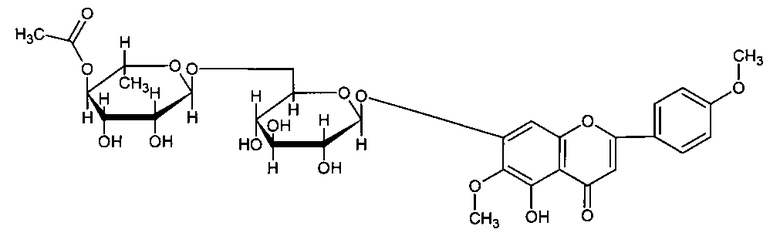
формула I
Применение флавоноидного соединения 7-O-[6-O-(4-ацетил-α-L-рамнопиранозил)-β-D-глюкопиранозидо]-5-гидрокси-6-метокси-2-(4-метокси-фенил)-4Н-хромон-4-он (ацетилпектолинарин - АЦП) в качестве лекарственного средства для коррекции нарушений параметров поведения: координации движений, внимания, трезвой оценки ситуации, памяти - вызванных как кратковременным алкогольным опьянением, так и хроническим употреблением алкоголя.

