ПЕРЕКРЕСТНАЯ ССЫЛКА НА СВЯЗАННЫЕ ЗАЯВКИ
По данной заявке испрашивается приоритет по предварительным заявкам США под номерами 61/072572, поданной 31 марта 2008 года, 61/041480, поданной 1 апреля 2008 года, 61/128383, поданной 20 мая 2008 года, и 61/205392, поданной 16 января 2009 года.
ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Предоставляются композиции и способы для лечения и диагностики подтипов астмы у пациентов. Также предоставляются способы идентификации эффективных лекарственных средств против астмы и предсказания способности отвечать на лекарственные средства против астмы.
УРОВЕНЬ ТЕХНИКИ
Традиционно полагают, что астма является следствием индуцированного аллергенами воздуха воспаления, запускаемого процессами T-хелперов 2 типа (Th2) и опосредуемого цитокинами, включающими интерлейкин (IL)-4, IL-5 и IL-13. IL-13 представляет собой плейотропный Th2-цитокин, продуцируемый активированными T-клетками, базофилами, эозинофилами и тучными клетками, и он строго вовлечен в патогенез астмы в доклинических моделях [2]. В дыхательных путях пациентов-людей с астмой были выявлены повышенные уровни IL-13; однако это повышение наблюдается только у подгруппы астматиков [3-6]. Недавние исследования были направлены на понимание того, как Th2-цитокины вызывают подобную астме патологию и физиологию [49, 50].
Хотя астма часто характеризуется эозинофильной инфильтрацией дыхательных путей, имеются накапливающиеся данные о том, что существуют другие подтипы этого заболевания, запускаемые альтернативными формами воспаления [1, 39, 48]. Например, исследования клеточных компонентов воспаления дыхательных путей при астме обеспечивают данные об отличающихся эозинофильном и неэозинофильном фенотипах астмы [1, 39, 48]. Неизвестно, отличаются ли молекулярные механизмы, лежащие в основе этих клинических и клеточных фенотипов астмы. Идентификация и изучение биомаркеров различных молекулярных фенотипов астмы может дать направление для фундаментального исследования и клинического применения развивающихся способов лечения астмы, которые специфично нацелены на Th2-ответы в легком.
Периостин представляет собой секретируемый белок, ассоциированный с фиброзом, экспрессия которого повышается рекомбинантным IL-4 и IL-13 в эпителиальных клетках бронхов [7, 8] и фибробластах бронхов [9]. Он экспрессируется на повышенных уровнях in vivo в эпителиальных клетках бронхов [8] и в субэпителиальном слое бронхов [9] людей с астмой, а также в модели астмы на мышах [10]. Также он на повышенных уровнях экспрессируется в эпителии пищевода пациентов с эозинофильным эзофагитом зависимым от IL-13 образом [11]. Повышенную экспрессию периостина наблюдали в нескольких типах происходящих из эпителия злокачественных опухолей [64-67], и в сыворотке некоторых пациентов со злокачественной опухолью наблюдали повышенные уровни растворимого периостина [64, 68-70].
Был проведен полногеномный анализ на микрочипах экспрессии эпителиальных клеток бронхов от 42 не принимавших стероиды астматиков с астмой от мягкой до умеренной, и от 28 здоровых контрольных субъектов [8]. В этих исследованиях тремя генами с наиболее отличающейся экспрессией между всеми астматиками и всеми здоровыми контролями были ген периостина, CLCA1 и serpinB2 [8]. Более того, эти гены значительно подавлялись в эпителиальных клетках бронхов астматиков после лечения в течение 7 суток ингалируемым кортикостероидом (ICS) [8]. Все три из этих генов индуцировались в эпителиальных клетках бронхов обработкой рекомбинантным IL-13 in vitro и их экспрессия значительно ослаблялась добавлением в культуральную среду кортикостероидов [8].
На сегодняшний день такие полногеномные анализы экспрессии не идентифицировали генетические биомаркеры, которые являются прогностическими или предсказывают терапевтический ответ на лечение отдельных пациентов с астмой, а также они не идентифицировали генетические маркеры, которые характеризуют подтипы пациентов с астмой. Кроме того, не было идентифицировано надежных негенетических биомаркеров с широкой клинической применимостью для прогноза или предсказания ответов на медикаментозное лечение, или для диагностики подтипов астмы. Таким образом, поскольку пациентам с астмой требуется лечение, поиск лекарственного средства(средств), эффективного для конкретного пациента, осуществляют по существу методом проб и ошибок. Такой метод проб и ошибок при поиске наиболее эффективного лекарственного средства часто вовлекает существенный риск и дискомфорт у пациента.
Таким образом, существует потребность в более эффективных средствах для определения того, какие пациенты будут отвечать на какое лечение, и во включении такого определения в более эффективные режимы лечения пациентов с астмой.
Изобретение, описанное в настоящем документе, удовлетворяет описанные выше потребности и обеспечивает другие преимущества.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
С использованием профилей экспрессии генов в эпителии бронхов, авторы настоящего изобретения определили молекулярные подтипы астмы. Удивительно, что координированная кластеризация данных на основе набора генов, экспрессия которых высоко коррелировала с генами, которые, как известно, активируются при стимуляции IL-4 или IL-13, выявила не один, а два различных кластера пациентов с астмой. Более того, анализ этих дихотомических подгрупп астматиков выявил значимые ассоциации между статусом "профиля IL-4/13" и уровнями общих IgE в сыворотке, уровнями CEA в сыворотке, уровнями периостина в сыворотке, эозинофилией периферической крови, эозинофилией BAL (бронхоальвеолярного лаважа), и способностью отвечать на ингалируемые кортикостероиды (в каждом случае p<0,05 при помощи критерия суммы рангов Вилкоксона).
Таким образом, настоящее изобретение относится к способам диагностики субпопуляции пациентов с астмой, включающим определение экспрессии любого или комбинации генов, выбранных из POSTN, CST1, CCL26, CLCA1, CST2, PRR4, SERPINB2, CEACAM5, iNOS, SERPINB4, CST4, PRB4, TPSD1, TPSG1, MFSD2, CPA3, GPR105, CDH26, GSN, C2ORF32, TRACH2000196 (TMEM71), DNAJC12, RGS13, SLC18A2, SERPINB10, SH3RF2, FCER1B, RUNX2, PTGS1 и ALOX15. В одном варианте осуществления, определяют экспрессию любого или комбинации генов, выбранных из группы, состоящей из POSTN, CST1, CST2, CCL26, CLCA1, PRR4, PRB4, SERPINB2, CEACAM5, iNOS, SERPINB4, CST4 и SERPINB10. Согласно одному варианту осуществления, экспрессию генов определяют с помощью микрочипа. Согласно другому варианту осуществления, экспрессию генов определяют путем определения уровней экспрессии белка указанного выше гена. Согласно другому варианту осуществления, экспрессию гена считают повышенной по сравнению со здоровым контролем, если относительный уровень мРНК представляющего интерес гена более чем в 2,5 раза превышает уровень мРНК контрольного гена. Согласно другому варианту осуществления, относительный уровень мРНК представляющего интерес гена более чем в 3 раза, 5 раз, 10 раз, 15 раз, 25 раз или 30 раз превышает уровень экспрессии гена здорового контроля. Согласно одному варианту осуществления, экспрессию гена определяют способом, выбранным из группы, состоящей из способа ПЦР, способа микрочипов или способа иммунологического анализа. В одном варианте осуществления, способ микрочипов включает применение микрочипа, имеющего одну или несколько молекул нуклеиновых кислот, которые могут гибридизоваться в строгих условиях с молекулой нуклеиновой кислоты, кодирующей ген, упомянутый выше, или имеющего один или несколько полипептидов (таких как пептиды или антитела), которые могут связываться с одним или несколькими из белков, кодируемых упомянутыми выше генами. В одном варианте осуществления, способ ПЦР представляет собой qPCR. Согласно одному варианту осуществления, способ иммунологического анализа включает стадии связывания антитела с белком, экспрессируемым с указанного выше гена, в образце указанного выше пациента и определения того, является ли уровень белка в образце от пациента повышенным. Согласно одному варианту осуществления, контрольный ген представляет собой ген домашнего хозяйства, выбранный из группы, состоящей из гена актина, GAPDH, GASB и GUSB.
Настоящее изобретение относится к микрочипу, содержащему последовательности нуклеиновых кислот, кодирующие следующие гены: POSTN, CST1, CST2, CCL26, CLCA1, PRR4, SERPINB2, CEACAM5, iNOS, SERPINB4, CST4 и SERPINB10 или их фрагменты. Настоящее изобретение относится к микрочипу, содержащему последовательности нуклеиновых кислот, кодирующие следующие гены: POSTN, CST1, CCL26, CLCA1, CST2, PRR4, SERPINB2, CEACAM5, iNOS, SERPINB4, CST4, PRB4, TPSD1, TPSG1, MFSD2, CPA3, GPR105, CDH26, GSN, C2ORF32, TRACH2000196 (TMEM71), DNAJC12, RGS13, SLC18A2, SERPINB10, SH3RF2, FCER1B, RUNX2, PTGS1 и ALOX1 или их фрагменты.
Настоящее изобретение относится к субпопуляции пациентов с астмой, подлежащих лечению лекарственными средствами по этому изобретению, где соотношение уровней белка или мРНК Muc5AC:MUC5B в эпителиальных клетках дыхательных путей пациентов с астмой превышает 25.
Настоящее изобретение также относится к способам диагностики субпопуляции пациентов с астмой путем проведения однократных или комбинированных измерений системных биомаркеров, выбранных из уровней CEA в сыворотке, уровней IgE в сыворотке, уровней периостина в сыворотке, количества эозинофилов в периферической крови и процента эозинофилов в жидкости бронхоальвеолярного лаважа (BAL). Системные биомаркеры, как правило, представляют собой негенетические биомаркеры и, как правило, их измеряют в образцах, полученных неинвазивными процедурами, например, но не ограничиваясь ими, взятием крови или компонентов крови, например, сыворотки или плазмы. Согласно одному варианту осуществления, уровни IgE более 100 МЕ/мл и/или эозинофилов более 0,14×10e9/л предсказывают группу пациентов, подлежащую лечению лекарственным средством по этому изобретению.
Настоящее изобретение относится к способам лечения астмы, включающим введение лекарственного средства пациенту с повышенными уровнями экспрессии одного или комбинации генов, выбранных из POSTN, CST1, CCL26, CLCA1, CST2, PRR4, SERPINB2, CEACAM5, iNOS, SERPINB4, CST4, PRB4, TPSD1, TPSG1, MFSD2, CPA3, GPR105, CDH26, GSN, C2ORF32, TRACH2000196 (TMEM71), DNAJC12, RGS13, SLC18A2, SERPINB10, SH3RF2, FCER1B, RUNX2, PTGS1, ALOX15. Согласно одному варианту осуществления, у пациента экспрессируются повышенные уровни одного или комбинации генов, выбранных из группы, состоящей из гена периостина, CST1, CST2, CCL26, CLCA1, PRR4, SerpinB2, CEACAM5, iNOS, PRB4, SerpinB4, SERPINB10 и CST4. Согласно одному варианту осуществления, пациент представляет собой стероид-наивного (никогда не подвергавшегося лечению стероидами) пациента с астмой от мягкой до умеренной. Согласно другому варианту осуществления, пациент, подлежащий лечению, представляет собой устойчивого к стероидам (не отвечающего на стероиды) пациента с астмой от мягкой до умеренной. Таких пациентов лечат терапевтически эффективным количеством лекарственного средства. В одном варианте осуществления, пациент имеет астму, индуцированную каскадом TH2.
Согласно одному варианту осуществления, лекарственное средство представляет собой ингибитор каскада IL13/IL4. Согласно другому варианту осуществления, лекарственное средство нацелено на индуцируемый TH2 каскад астмы. Иллюстративные мишени включают, но не ограничиваются ими, цитокины или лиганды, такие как: IL-9, IL-5, IL-13, IL-4, OX40L, TSLP, IL-25, IL-33 и IgE; и рецепторы, такие как: рецептор IL-9, рецептор IL-5, рецептор-альфа для IL-4, рецептор-альфа 1 для IL-13 и рецептор-альфа-2 для IL-13, OX40, TSLP-R, IL-7R-альфа (корецептор для TSLP), IL-17RB (рецептор для IL-25), ST2 (рецептор для IL-33), CCR3, CCR4, CRTH2, Fc-эпсилон-RI и Fc-эпсилон-RII/CD23 (рецепторы для IgE). Таким образом, лекарственное средство согласно этому изобретению включает средство, которое может связываться с указанной выше мишенью, такое как полипептид(ы) (например, антитело, иммуноадгезин или пептидное антитело), аптамер или низкомолекулярное соединение.
Согласно одному варианту осуществления, лекарственное средство представляет собой антитело против IL-13. Согласно другому варианту осуществления, антитело против IL-13 содержит последовательность VH, содержащую SEQ ID NO:193, и последовательность VL, содержащую SEQ ID NO:194. Согласно другому варианту осуществления, антитело против IL13 содержит: (a) HVR-L1, содержащую аминокислотную последовательность RASKSVDSYGNSFMH (SEQ ID NO:195); (b) HVR-L2, содержащую аминокислотную последовательность LASNLES (SEQ ID NO:196); (c) HVR-L3, содержащую аминокислотную последовательность QQNNEDPRT (SEQ ID NO:197); (d) HVR-H1, содержащую аминокислотную последовательность AYSVN (SEQ ID NO:198); (e) HVR-H2, содержащую аминокислотную последовательность MIWGDGKIVYNSALKS (SEQ ID NO:199); и (f) HVR-H3, содержащую аминокислотную последовательность DGYYPYAMDN (SEQ ID NO:200). Согласно другому варианту осуществления, лекарственное средство представляет собой антитело против лиганда OX40 (OX40L). Согласно другому варианту осуществления, лекарственное средство представляет собой биспецифическое антитело против IL13/против IL4. Согласно другому варианту осуществления, лекарственное средство представляет собой антитело против IgE. Согласно другому варианту осуществления, лекарственное средство представляет собой антитело, направленное против мембранной проксимальной М1'-области IgE, экспрессированного на поверхности B-клеток. Согласно другому варианту осуществления, лекарственное средство представляет собой ингалируемый кортикостероид. В определенных вариантах осуществления, ингалируемый кортикостероид выбран из беклометазона дипропионата, будезонида, флунизолида, флутиказона пропионата, мометазона и триамцинолона ацетонида.
Согласно одному варианту осуществления, антитело против OX40L содержит: (a) HVR-L1, содержащую последовательность RSSQSPVHSNGNTYLH (SEQ ID NO:201); (b) HVR-L2, содержащую последовательность KVSNRFS (SEQ ID NO:202); (c) HVR-L3, содержащую последовательность SQSTHIPWT (SEQ ID NO:203); (d) HVR-H1, содержащую последовательность SYWMH (SEQ ID NO:204); (e) HVR-H2, содержащую последовательность EIDPSNGRTNYNEKFKS (SEQ ID NO:205); и (f) HVR-H3, содержащую последовательность ERSPRYFDV (SEQ ID NO:206). Согласно другому варианту осуществления, антитело против OX40L содержит: (a) HVR-L1, содержащую последовательность RSSQSIVHGNGNTYLE (SEQ ID NO:207); (b) HVR-L2, содержащую последовательность RVSNRFS (SEQ ID NO:208); (c) HVR-L3, содержащую последовательность FQGSHVPYT (SEQ ID NO:209); (d) HVR-H1, содержащую последовательность SYWLN (SEQ ID NO:210); (e) HVR-H2, содержащую последовательность MIDPSDSETHYNQVFKD (SEQ ID NO:211); и (f) HVR-H3, содержащую последовательность GRGNFYGGSHAMEY (SEQ ID NO:212). Согласно другому варианту осуществления, антитело против OX40L содержит (a) HVR-H1, содержащую последовательность SYTMH (SEQ ID NO:215), SYAMS (SEQ ID NO:216), NFGMH (SEQ ID NO:217) или NYGMH (SEQ ID NO:218), (b) HVR-H2, содержащую последовательность IISGSGGFTYYADSVKG (SEQ ID NO:219), AIWYDGHDKYYSYYVKG (SEQ ID NO:220), AIWYDGHDKYYAYYVKG (SEQ ID NO:221), VIWYDGSNKYYVDSVKG (SEQ ID NO:222) или VIWNDGSNKYYVDSVKG (SEQ ID NO:223), (c) HVR-H3, содержащую последовательность DSSSWYRYFDY (SEQ ID NO:224), DRLVAPGTFDY (SEQ ID NO:225), KNWSFDF (SEQ ID NO:226) или DRMGIYYYGMDV (SEQ ID NO:227), (d) HVR-L1, содержащую последовательность RASQGISSWLA (SEQ ID NO:228), RASQSVSSSYLA (SEQ ID NO:229), RASQSVSSNYLA (SEQ ID NO:230), RASQGVSRYLA (SEQ ID NO:231) или RASQSVSSYLA (SEQ ID NO:232), (e) HVR-L2, содержащую последовательность GASSRAT (SEQ ID NO:233), AASSLQS (SEQ ID NO:234), MPPVWKV (SEQ ID NO:235), DASNRAT (SEQ ID NO:236) или LHPLCKV (SEQ ID NO:237); и (f) HVR-L3, содержащую последовательность NSLIVTLT (SEQ ID NO:238), QQYNSYPYT (SEQ ID NO:239), QQYGSSFT (SEQ ID NO:240), QQRSNWQYT (SEQ ID NO:241), QQRSNWT (SEQ ID NO:242) или NSIIVSLT (SEQ ID NO:243), где антитело против OX40L связывает OX40L. Согласно одному варианту осуществления, антитело против IgE содержит последовательность VL, содержащую SEQ ID NO:213, и последовательность VH, содержащую SEQ ID NO:214. Согласно другому варианту осуществления, антитело против IgE содержит: (a) HVR-L1, содержащую последовательность RSSQSLVHNNANTYLH (SEQ ID NO:244); (b) HVR-L2, содержащую последовательность KVSNRFS (SEQ ID NO:245); (c) HVR-L3, содержащую последовательность SQNTLVPWT (SEQ ID NO:246); (d) HVR-H1, содержащую последовательность GFTFSDYGIA (SEQ ID NO:247); (e) HVR-H2, содержащую последовательность AFISDLAYTIYYADTVTG (SEQ ID NO:248); и (f) HVR-H3, содержащую последовательность ARDNWDAMDY (SEQ ID NO:249). Согласно одному варианту осуществления, антитело против IgE содержит последовательность VH, содержащую SEQ ID NO:250, и последовательность VL, содержащую SEQ ID NO:251. Согласно одному варианту осуществления, антитело против IgE содержит последовательность VH, содержащую SEQ ID NO:252, и последовательность VL, содержащую SEQ ID NO:253. Согласно другому варианту осуществления, антитело против IgE содержит: (a) HVR-L1, содержащую последовательность RSSQDISNSLN (SEQ ID NO:254); (b) HVR-L2, содержащую последовательность STSRLHS (SEQ ID NO:255); (c) HVR-L3, содержащую последовательность QQGHTLPWT (SEQ ID NO:256); (d) HVR-H1, содержащую последовательность GYTFTDYYMM (SEQ ID NO:257); (e) HVR-H2, содержащую последовательность GDNIDPNNYDTSYNQKFKG (SEQ ID NO:258); и (f) HVR-H3, содержащую последовательность ASKAY (SEQ ID NO:259). Согласно другому варианту осуществления, антитело против IgE содержит: (a) HVR-L1, содержащую последовательность RSSQDISNALN (SEQ ID NO:260); (b) HVR-L2, содержащую последовательность STSRLHS (SEQ ID NO:255); (c) HVR-L3, содержащую последовательность QQGHTLPWT (SEQ ID NO:256); (d) HVR-H1, содержащую последовательность GYTFTDYYMM (SEQ ID NO:257); (e) HVR-H2, содержащую последовательность GDNIDPNNYDTSYNQKFKG (SEQ ID NO:258); и (f) HVR-H3, содержащую последовательность ASKAY (SEQ ID NO:259). Согласно другому варианту осуществления, антитело против IgE содержит: (a) HVR-L1, содержащую последовательность RSSQDISNALN (SEQ ID NO:260); (b) HVR-L2, содержащую последовательность STSRLHS (SEQ ID NO:255); (c) HVR-L3, содержащую последовательность QQGHTLPWT (SEQ ID NO:256); (d) HVR-H1, содержащую последовательность GYTFTDYYIM (SEQ ID NO:261); (e) HVR-H2, содержащую последовательность GDNIDPNNYDTSYNQKFKG (SEQ ID NO:258); и (f) HVR-H3, содержащую последовательность ASKAY (SEQ ID NO:259).
Согласно одному варианту осуществления, пациент имеет астму, которая не вовлекает каскад TH2 (не-TH2 астма). В одном варианте осуществления, лекарственное средство нацелено на не-TH2 астму. Согласно одному варианту осуществления, лекарственное средство представляет собой ингибитор каскада IL-17. В одном варианте осуществления, лекарственное средство представляет собой антитело против IL-17. В одном варианте осуществления, лекарственное средство представляет собой антитело, перекрестно реагирующее как с IL-17A, так и с IL-17F. В одном варианте осуществления, лекарственное средство представляет собой биспецифическое антитело, способное связываться как с IL-17A, так и с IL-17F. В одном варианте осуществления, лекарственное средство представляет собой антитело против IL-17A/F.
Настоящее изобретение относится к набору для диагностики подтипа астмы у пациента, содержащему (1) одну или несколько молекул нуклеиновых кислот, которые гибридизуются с геном, где ген выбран из группы, состоящей из POSTN, CST1, CST2, CCL26, CLCA1, PRR4, PRB4, SERPINB2, CEACAM5, iNOS, SERPINB4, CST4 и SERPINB10 и (2) инструкции по измерению уровней экспрессии гена в образце от пациента с астмой, где повышенные уровни экспрессии любого, комбинации или всех из указанных генов указывают на подтип астмы. Согласно одному варианту осуществления, набор дополнительно содержит ген, выбранный из группы, состоящей из: PRB4, TPSD1, TPSG1, MFSD2, CPA3, GPR105, CDH26, GSN, C2ORF32, TRACH2000196 (TMEM71), DNAJC12, RGS13, SLC18A2, SH3RF2, FCER1B, RUNX2, PTGS1 и ALOX15. В одном дополнительном варианте осуществления, уровень экспрессии гена измеряют путем анализа уровней мРНК. В другом дополнительном варианте осуществления, анализ включает способ ПЦР или применение микрочипа. В другом дополнительном варианте осуществления, способ ПЦР представляет собой qPCR. В одном варианте осуществления, уровни мРНК представляющего интерес гена, более чем в 2,5 раза превышающие уровни мРНК контрольного гена, указывают на подтип астмы.
Изобретение относится к набору для диагностики подтипа астмы у пациента, содержащему (1) одну или несколько молекул белка, которые связываются с белком, выбранным из группы, состоящей из POSTN, CST1, CST2, CCL26, CLCA1, PRR4, PRB4, SERPINB2, CEACAM5, iNOS, SERPINB4, CST4 и SERPINB10, и (2) инструкции по измерению уровней экспрессии белка в образце от пациента, где повышенные уровни экспрессии любого, комбинации или всех из указанных белков указывают на подтип астмы. В одном варианте осуществления, набор дополнительно содержит молекулу белка, которая связывается с белком, выбранным из группы, состоящей из: PRB4, TPSD1, TPSG1, MFSD2, CPA3, GPR105, CDH26, GSN, C2ORF32, TRACH2000196 (TMEM71), DNAJC12, RGS13, SLC18A2, SH3RF2, FCER1B, RUNX2, PTGS1 и ALOX15. В одном варианте осуществления молекула белка представляет собой антитело, пептид или пептидное антитело. В следующем варианте осуществления, набор содержит микрочип, содержащий молекулу(ы) белка.
Настоящее изобретение относится к набору для диагностики подтипа астмы у пациента, включающему инструкции по определению одного из биомаркеров в образце от пациента, выбранного из группы, состоящей из: уровней общих IgE в сыворотке, уровней CEA в сыворотке, уровней периостина в сыворотке, эозинофилов периферической крови и эозинофилов бронхоальвеолярного лаважа (BAL), где уровни CEA, периостина в сыворотке, эозинофилов периферической крови и эозинофилов бронхоальвеолярного лаважа (BAL) являются повышенными. Согласно одному варианту осуществления, в наборе предоставляются инструкции, где уровень IgE, превышающий 100 МЕ/мл, указывает на подтип астмы. Согласно другому варианту осуществления, в наборе предоставляются инструкции, где уровень эозинофилов в периферической крови, превышающий 0,14×10e9/л, указывает на подтип астмы.
Настоящее изобретение относится к набору для диагностики подтипа астмы у пациента, содержащему инструкции по определению соотношения мРНК или белка Muc5AC:MUC5B в образце от пациента с астмой, где соотношение более 25 указывает на подтип астмы. В одном варианте осуществления, образец получают из эпителиального соскоба. В другом варианте осуществления, образец содержит эпителиальные клетки дыхательных путей. В одном варианте осуществления, в наборе предоставляется молекула нуклеиновой кислоты, которая гибридизуется в строгих условиях с Muc5AC, и молекула нуклеиновой кислоты, которая гибридизуется в строгих условиях с MUC5B. В одном варианте осуществления, в наборе предоставляется молекула белка, которая связывается с Muc5AC, и молекула белка, которая связывается с MUC5B. В одном варианте осуществления, молекула белка представляет собой антитело.
КРАТКОЕ ОПИСАНИЕ РИСУНКОВ
На фигуре 1 представлены уровни экспрессии генов в эпителии дыхательных путей, как описано в примерах 1 и 2. (A) Представлены относительные уровни экспрессии периостина (левая панель), CLCA1 (средняя панель) и serpinB2 (правая панель) у здоровых контролей (N=27) и у астматиков (N=42). На левой оси каждого графика указаны нормализованные единицы флуоресценции. (B) Представлены двухсторонние сравнения уровней экспрессии периостина и CLCA1 (левая панель), периостина и serpinB2 (средняя панель) и CLCA1 и serpinB2 (правая панель) у 42 астматиков. На каждой панели указаны коэффициент ранговой корреляции Спирмана (ρ) и значения p. (C) Анализ на микрочипах экспрессии генов у здоровых контролей и астматиков, идентифицирующий уровни экспрессии периостина и корегулируемых генов; кластер с профилем "IL-4/13 высокий" (кластер 1); кластер с профилем "IL-4/13 низкий" (кластер 2); здоровые контроли. (D) Теплокарта, на которой представлена неконтролируемая иерархическая кластеризация (евклидова полная) уровней экспрессии периостина, CLCA1 и serpinB2 в эпителии бронхов у всех субъектов на исходном уровне. (E) Средние (+SEM) уровни экспрессии IL-4, IL-5 и IL-13 в гомогенатах биоптатов бронхов, полученных одновременно с соскобами бронхов от подгруппы субъектов, представленных на фигурах 1A-D (кластер 1: 18 астматиков с "IL-13 высоким"; кластер 2: 16 здоровых контролей и 14 астматиков с "IL-13 низким"). Двухсторонние корреляции IL-4, IL-5 и IL-13 для всех субъектов, указанных справа (коэффициент ранговой корреляции Спирмана, ρ, и значения p).
На фигуре 2 представлены семейства генов для серпинов, цистатинов и PRR, и уровни экспрессии этих генов, как описано в примере 3. (A) Геномные локусы и организация серпинов (сверху), цистатинов (середина) и PRR (низ), как представлено на браузере для просмотра геномов University of California Santa Cruz, доступном на сайте http://genome.ucsc.edu. (B) Иерархическая кластеризация всех зондов, кодирующих гены цистатинов и серпинов, как представлено на панели A. (C) Представлены относительные уровни экспрессии генов в эпителии дыхательных путей PRR4 (левая панель), PRB4 (средняя панель) и CEACAM5 (правая панель) у здоровых контролей (N=27) и у астматиков (N=42). На левой оси каждого графика указаны нормализованные единицы флуоресценции.
На фигуре 3 представлен анализ на микрочипах соскобов эпителия бронхов на исходном уровне и после введения ингалируемого флукатизона пропионата (ICS) в течение одной недели, как описано в примере 6. (A) Экспрессия периостина; (B) экспрессия PRR4; (C) экспрессия RUNX2.
На фигуре 4 представлен объединенный график для IgE в сыворотке и эозинофилов периферической крови у пациентов с астмой, как описано в примерах 7 и 9.
На фигуре 5 представлены различные клинические признаки субфенотипов астмы "IL-13 высокий" и "IL-13 низкий", как описано в примере 8. (A) Объем воздуха, выдыхаемого за первую секунду форсированного выдоха (FEV1), показатель обструкции дыхательных путей. (B) Улучшение FEV1 после 4 вдыханий (360 мкг) альбутерола (тестирование обратимости с помощью бронхорасширяющего средства). (C) Провокационная концентрация метахолина, требуемая для индукции 20% снижения FEV1 (PC20), показатель гиперреактивности дыхательных путей.
На фигуре 6 представлены различные маркеры аллергии, эозинофильного воспаления и ремоделирования дыхательных путей для субфенотипов астмы "IL-13 высокий" и "IL-13 низкий", как описано в примере 8. (A) Результаты инъекционной кожной пробы с аллергенами (SPT) с использованием панели из 12 аллергенов воздуха. (B) Концентрация IgE в сыворотке. (C) Количество эозинофилов в периферической крови. (D) Эозинофилы в качестве процента от всех клеток жидкости бронхоальвеолярного лаважа (BAL). (E) Стереологическое измерение толщины ретикулярной базальной мембраны (RBM) в эндобронхиальном биоптате, показателя субэпителиального фиброза. (F) Соотношение экспрессии MUC5AC и MUC5B в эпителиальных соскобах при определении посредством qPCR.
На фигуре 7 представлены различные клинические признаки субфенотипов астмы "IL-13 высокий" и "IL-13 низкий", как описано в примере 8. (A) Процент субъектов, отвечающих на конкретные аллергены воздуха, как указано вдоль нижней оси. Субфенотип астмы "IL-13 низкий"; субфенотип астмы "IL-13 высокий" (*, p<0,05). (B) Количество положительных реакций SPT против процента эозинофилов BAL; субфенотип астмы по IL-13, как указано (высокий, незакрашенные квадраты; низкий, закрашенные круги). (C) Количество положительных реакций SPT против IgE в сыворотке; субфенотип астмы по IL-13, как указано (высокий, незакрашенные квадраты; низкий, закрашенные круги). (D) Количество положительных реакций SPT против количества эозинофилов периферической крови; субфенотип астмы IL-13, как указано (высокий, незакрашенные квадраты; низкий, закрашенные круги). Коэффициент ранговой корреляции Спирмана (ρ) и значения p указаны на каждом графике для B-D.
На фигуре 8 представлены содержание и состав муцина в эпителии дыхательных путей у субъектов с субфенотипами астмы "IL-13 высокий" и "IL-13 низкий" и у здоровых контролей, как описано в примере 8. (A) Объем муцина на объем эпителия, показатель содержания муцина в эпителии дыхательных путей. (B) Экспрессия муцина MUC2 при определении qPCR. (C) Экспрессия муцина MUC5AC при определении qPCR. (D) Экспрессия муцина MUC5B при определении qPCR.
На фигуре 9 представлены ответы субъектов с субфенотипами астмы "IL-13 высокий" и "IL-13 низкий" на ингалируемые кортикостероиды. (A) FEV1, измеренная на исходном уровне (0 неделя), после 4 и 8 недель введения флукатизона раз в сутки, и через одну неделю после прекращения введения флукатизона (неделя 9). (*): см. таблицу 5 для количества субъектов в каждой группе и значений p. (B) Теплокарта, на которой представлена неконтролируемая иерархическая кластеризация периостина, CLCA1 и serpinB2 (как на фиг. 1D) в эпителии бронхов астматиков через одну неделю после начала введения либо флукатизона (N=19), либо плацебо (N=13). Обозначение кластера на исходном уровне для отдельных субъектов и введение указаны ниже теплокарты (кластер 1: астматики с "IL-13 высоким"; кластер 2: астматики с "IL-13 низким").
На фигуре 10 представлена экспрессия генов альвеолярных макрофагов у субъектов с субфенотипами астмы "IL-13 высокий" и "IL-13 низкий", как описано в примере 8. Указаны здоровые контроли (N=15); субфенотип астмы "IL-13 низкий" (N=5); субфенотип астмы "IL-13 высокий" (N=9). На фигуре представлены средние (+SEM) уровни экспрессии 15-липоксигеназы (ALOX15) и фактора некроза опухоли-α (TNF-α) при определении посредством qPCR. (*): p<0,03.
На фигуре 11 представлен анализ на микрочипах экспрессии генов с использованием 35 зондов, охватывающих 28 генов, образцов от здоровых контролей и астматиков, как описано в примере 9.
На фигуре 12 представлен анализ на микрочипах экспрессии генов и анализ посредством qPCR периостина и CEACAM5, как описано в примере 9. (A) Экспрессия периостина у здоровых контролей, астматики с кластером 2 ("IL-13 низкий"), и астматики с кластером 1 ("IL-13 высокий"); (B) экспрессия CEACAM5 у здоровых контролей, астматики с кластером 2 ("IL-13 низкий"), и астматики с кластером 1 ("IL-13 высокий"); (C) объединенный график CEACAM5 и периостина у астматиков с "IL-13 высоким" (квадраты) и у астматиков с "IL-13 низким" (круги); (D) Анализ операционных характеристик приемника (ROC) в отношении оптимизированного алгоритма для уровней экспрессии периостина CEACAM5 на основе qPCR, показывающий чувствительность и специфичность для здоровых контролей, астматиков с "IL-13 высоким", и астматиков с "IL-13 низким".
На фигуре 13 представлены уровни в сыворотке сывороточных белков у астматиков и здоровых контролей, как описано в примере 9. (A) уровни IgE в сыворотке; (B) уровни периостина в сыворотке; (C) уровни CEA в сыворотке; (D) уровни YKL-40 в сыворотке; (E) уровни IgE в сыворотке у астматиков, которым вводили ингалируемые кортикостероиды (ICS) (+) или не вводили их (-); (F) уровни периостина в сыворотке у астматиков, которым вводили ингалируемые кортикостероиды (ICS) (+) или не вводили их (-); (G) уровни CEA в сыворотке у астматиков, которым вводили ингалируемые кортикостероиды (ICS) (+) или не вводили их (-); (H) уровни YKL-40 в сыворотке астматиков, которым вводили ингалируемые кортикостероиды (ICS) (+) или не вводили их (-); (I) объединенный график уровней периостина в сыворотке у астматиков, имеющих <100 МЕ/мл IgE в сыворотке (<100) и у астматиков, имеющих ≥100 МЕ/мл IgE в сыворотке (≥100); (J) объединенный график уровней CEA в сыворотке, имеющих <100 МЕ/мл IgE в сыворотке (<100) и у астматиков, имеющих ≥100 МЕ/мл IgE в сыворотке (≥100); (K) объединенный график уровней YKL-40 в сыворотке у астматиков, имеющих <100 МЕ/мл IgE в сыворотке (<100) и у астматиков, имеющих ≥100 МЕ/мл IgE в сыворотке (≥100); (L) объединенный график уровней периостина в сыворотке и CEA у астматиков, имеющих <100 МЕ/мл IgE в сыворотке (круги) и у астматиков, имеющих ≥100 МЕ/мл IgE в сыворотке (квадраты).
ПОДРОБНОЕ ОПИСАНИЕ
ОПРЕДЕЛЕНИЯ
Если не определено иначе, подразумевается, что все термины данной области, обозначения и другая научная терминология, используемые в настоящем документе, имеют значения, обычно понимаемые специалистами в области, к которой это изобретение относится. В некоторых случаях, термины с обычно понимаемыми значениями определены в настоящем документе для ясности и/или для простоты отсылки, и включение таких определений в настоящий документ не обязательно следует истолковывать как существенно отличающееся от того, что обычно понимают в данной области. Способы и процедуры, которые описаны и которые процитированы в настоящем документе, в общем, являются хорошо понятными и обычно применяемыми специалистами в данной области при использовании общепринятой методологии, например, такой как широко используемые методологии молекулярного клонирования, описанные в Sambrook et al., Molecular Cloning: A Laboratory Manual 2nd. edition (1989) Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y. В соответствующих случаях, способы, вовлекающие применение коммерчески доступных наборов и реагентов, обычно проводят согласно установленным изготовителями протоколам и/или параметрам, если нет иных указаний.
Термины "профиль генов IL-4/IL-13", "профиль IL-4/IL-13", "профиль генов IL-13" и "профиль IL-13" используют в настоящем документе взаимозаменяемо и они относятся к комбинации из 30 генов, как указано в таблице 4, или подкомбинации этих 30 генов, как указано в таблице 9, паттерн экспрессии которых коррелирует с определенными пациентами с астмой. Эти 30 генов включают POSTN, CST1, CCL26, CLCA1, CST2, PRR4, SERPINB2, CEACAM5, iNOS, SERPINB4, CST4, PRB4, TPSD1, TPSG1, MFSD2, CPA3, GPR105, CDH26, GSN, C2ORF32, TRACH2000196 (TMEM71), DNAJC12, RGS13, SLC18A2, SERPINB10, SH3RF2, FCER1B, RUNX2, PTGS1, ALOX15. Полипептиды профиля генов IL-4/IL13 представляют собой "полипептиды-мишени" по этому изобретению.
Термин "полипептид-мишень", когда его используют в настоящем документе, относится к полипептидам с "нативной последовательностью" и вариантам (которые определены в настоящем документе далее).
Полипептид с "нативной последовательностью" включает полипептид, имеющий ту же аминокислотную последовательность, что и соответствующий полипептид, встречающийся в природе. Таким образом, термин "полипептид с нативной последовательностью" включает встречающиеся в природе укороченные, удлиненные формы полипептида и формы полипептида со сдвигом рамки считывания, включая, но не ограничиваясь ими альтернативно сплайсированные формы, изоформы и полиморфизмы.
"Встречающийся в природе вариант" означает полипептид, имеющий по меньшей мере приблизительно 60% идентичность аминокислотной последовательности с эталонным полипептидом и сохраняющий по меньшей мере один вид биологической активности встречающегося в природе эталонного полипептида. Встречающиеся в природе варианты могут включать полипептидные варианты, обладающие по меньшей мере приблизительно 65% идентичностью аминокислотной последовательности, по меньшей мере приблизительно 70% идентичностью аминокислотной последовательности, по меньшей мере приблизительно 75% идентичностью аминокислотной последовательности, по меньшей мере приблизительно 80% идентичностью аминокислотной последовательности, по меньшей мере приблизительно 80% идентичностью аминокислотной последовательности, по меньшей мере приблизительно 85% идентичностью аминокислотной последовательности, по меньшей мере приблизительно 90% идентичностью аминокислотной последовательности, по меньшей мере приблизительно 95% идентичностью аминокислотной последовательности, по меньшей мере приблизительно 98% идентичностью аминокислотной последовательности или по меньшей мере приблизительно 99% идентичностью аминокислотной последовательности с эталонным полипептидом.
Примеры POSTN включают полипептид, содержащий SEQ ID NO:1, и другие полипептиды POSTN с нативной последовательностью, такие как встречающиеся в природе варианты и полипептиды с нативной последовательностью, кодируемые последовательностью нуклеиновой кислоты, которая может гибридизоваться в строгих условиях с SEQ ID NO:31 и/или 32.
Примеры CST1 включают полипептид, содержащий SEQ ID NO:2, и другие полипептиды CST1 с нативной последовательностью, такие как встречающиеся в природе варианты и полипептиды с нативной последовательностью, кодируемые последовательностью нуклеиновой кислоты, которая может гибридизоваться в строгих условиях с SEQ ID NO:33.
Примеры CCL26 включают полипептид, содержащий SEQ ID NO:3, и другие полипептиды CCL26 с нативной последовательностью, такие как встречающиеся в природе варианты и полипептиды с нативной последовательностью, кодируемые последовательностью нуклеиновой кислоты, которая может гибридизоваться в строгих условиях с SEQ ID NO:34.
Примеры CLCA1 включают полипептид, содержащий SEQ ID NO:4, и другие полипептиды CLCA1 с нативной последовательностью, такие как встречающиеся в природе варианты и полипептиды с нативной последовательностью, кодируемые последовательностью нуклеиновой кислоты, которая может гибридизоваться в строгих условиях с SEQ ID NO:35.
Примеры CST2 включают полипептид, содержащий SEQ ID NO:5, и другие полипептиды CST с нативной последовательностью, такие как встречающиеся в природе варианты и полипептиды с нативной последовательностью, кодируемые последовательностью нуклеиновой кислоты, которая может гибридизоваться в строгих условиях с SEQ ID NO:36.
Примеры PRR4 включают полипептид, содержащий SEQ ID NO:6, и другие полипептиды PRR4 с нативной последовательностью, такие как встречающиеся в природе варианты и полипептиды с нативной последовательностью, кодируемые последовательностью нуклеиновой кислоты, которая может гибридизоваться в строгих условиях с SEQ ID NO:37.
Примеры SERPINB2 включают полипептид, содержащий SEQ ID NO:7, и другие полипептиды SERPINB2 с нативной последовательностью, такие как встречающиеся в природе варианты и полипептиды с нативной последовательностью, кодируемые последовательностью нуклеиновой кислоты, которая может гибридизоваться в строгих условиях с SEQ ID NO:38.
Примеры CEACAM5 включают полипептид, содержащий SEQ ID NO:8, и другие полипептиды CEACAM5 с нативной последовательностью, такие как встречающиеся в природе варианты и полипептиды с нативной последовательностью, кодируемые последовательностью нуклеиновой кислоты, которая может гибридизоваться в строгих условиях с SEQ ID NO:39.
Примеры iNOS включают полипептид, содержащий SEQ ID NO:9 и другие iNOS полипептиды с нативной последовательностью, такие как встречающиеся в природе варианты и полипептиды с нативной последовательностью, кодируемые последовательностью нуклеиновой кислоты, которая может гибридизоваться в строгих условиях с SEQ ID NO:40.
Примеры SERPINB4 включают полипептид, содержащий SEQ ID NO:10, и другие полипептиды SERPINB4 с нативной последовательностью, такие как встречающиеся в природе варианты и полипептиды с нативной последовательностью, кодируемые последовательностью нуклеиновой кислоты, которая может гибридизоваться в строгих условиях с SEQ ID NO:41 и/или 42.
Примеры CST4 включают полипептид, содержащий SEQ ID NO:11, и другие полипептиды CST4 с нативной последовательностью, такие как встречающиеся в природе варианты и полипептиды с нативной последовательностью, кодируемые последовательностью нуклеиновой кислоты, которая может гибридизоваться в строгих условиях с SEQ ID NO:43.
Примеры PRB4 включают полипептид, содержащий SEQ ID NO:12, и другие полипептиды PRB4 с нативной последовательностью, такие как встречающиеся в природе варианты и полипептиды с нативной последовательностью, кодируемые последовательностью нуклеиновой кислоты, которая может гибридизоваться в строгих условиях с SEQ ID NO:44.
Примеры TPSD1 включают полипептид, содержащий SEQ ID NO:13, и другие полипептиды TPSD1 с нативной последовательностью, такие как встречающиеся в природе варианты и полипептиды с нативной последовательностью, кодируемые последовательностью нуклеиновой кислоты, которая может гибридизоваться в строгих условиях с последовательностью, выбранной из группы, состоящей из SEQ ID NO:45-51.
Примеры TPSG1 включают полипептид, содержащий SEQ ID NO:14, и другие полипептиды TPSG1 с нативной последовательностью, такие как встречающиеся в природе варианты и полипептиды с нативной последовательностью, кодируемые последовательностью нуклеиновой кислоты, которая может гибридизоваться в строгих условиях с последовательностью, выбранной из группы, состоящей из SEQ ID NO:52-55.
Примеры MFSD2 включают полипептид, содержащий SEQ ID NO:15, и другие полипептиды MFSD2 с нативной последовательностью, такие как встречающиеся в природе варианты и полипептиды с нативной последовательностью, кодируемые последовательностью нуклеиновой кислоты, которая может гибридизоваться в строгих условиях с SEQ ID NO:56.
Примеры CPA3 включают полипептид, содержащий SEQ ID NO:16, и другие полипептиды CPA3 с нативной последовательностью, такие как встречающиеся в природе варианты и полипептиды с нативной последовательностью, кодируемые последовательностью нуклеиновой кислоты, которая может гибридизоваться в строгих условиях с SEQ ID NO:57.
Примеры GPR105 включают полипептид, содержащий SEQ ID NO:17, и другие полипептиды GPR105 с нативной последовательностью, такие как встречающиеся в природе варианты и полипептиды с нативной последовательностью, кодируемые последовательностью нуклеиновой кислоты, которая может гибридизоваться в строгих условиях с SEQ ID NO:58.
Примеры CDH26 включают полипептид, содержащий SEQ ID NO:18, и другие полипептиды CDH26 с нативной последовательностью, такие как встречающиеся в природе варианты и полипептиды с нативной последовательностью, кодируемые последовательностью нуклеиновой кислоты, которая может гибридизоваться в строгих условиях с SEQ ID NO:59.
Примеры GSN включают полипептид, содержащий SEQ ID NO:19, и другие полипептиды GSN с нативной последовательностью, такие как встречающиеся в природе варианты и полипептиды с нативной последовательностью, кодируемые последовательностью нуклеиновой кислоты, которая может гибридизоваться в строгих условиях с SEQ ID NO:60.
Примеры C2ORF32 включают полипептид, содержащий SEQ ID NO:20, и другие полипептиды C2ORF32 с нативной последовательностью, такие как встречающиеся в природе варианты и полипептиды с нативной последовательностью, кодируемые последовательностью нуклеиновой кислоты, которая может гибридизоваться в строгих условиях с SEQ ID NO:61.
Примеры TRACH2000196 (TMEM71) включают полипептид, содержащий SEQ ID NO:21, и другие полипептиды TRACH2000196 (TMEM71) с нативной последовательностью, такие как встречающиеся в природе варианты и полипептиды с нативной последовательностью, кодируемые последовательностью нуклеиновой кислоты, которая может гибридизоваться в строгих условиях с SEQ ID NO:62.
Примеры DNAJC12 включают полипептид, содержащий SEQ ID NO:22, и другие полипептиды DNAJC12 с нативной последовательностью, такие как встречающиеся в природе варианты и полипептиды с нативной последовательностью, кодируемые последовательностью нуклеиновой кислоты, которая может гибридизоваться в строгих условиях с SEQ ID NO:63.
Примеры RGS13 включают полипептид, содержащий SEQ ID NO:23, и другие полипептиды RGS13 с нативной последовательностью, такие как встречающиеся в природе варианты и полипептиды с нативной последовательностью, кодируемые последовательностью нуклеиновой кислоты, которая может гибридизоваться в строгих условиях с SEQ ID NO:64.
Примеры SLC18A2 включают полипептид, содержащий SEQ ID NO:24, и другие полипептиды SLC18A2 с нативной последовательностью, такие как встречающиеся в природе варианты и полипептиды с нативной последовательностью, кодируемые последовательностью нуклеиновой кислоты, которая может гибридизоваться в строгих условиях с SEQ ID NO:65.
Примеры SERPINB10 включают полипептид, содержащий SEQ ID NO:25, и другие полипептиды SERPINB10 с нативной последовательностью, такие как встречающиеся в природе варианты и полипептиды с нативной последовательностью, кодируемые последовательностью нуклеиновой кислоты, которая может гибридизоваться в строгих условиях с SEQ ID NO:66.
Примеры SH3RF2 включают полипептид, содержащий SEQ ID NO:26, и другие полипептиды SH3RF2 с нативной последовательностью, такие как встречающиеся в природе варианты и полипептиды с нативной последовательностью, кодируемые последовательностью нуклеиновой кислоты, которая может гибридизоваться в строгих условиях с SEQ ID NO:67.
Примеры FCER1B включают полипептид, содержащий SEQ ID NO:27 и другие полипептиды FCER1B с нативной последовательностью, такие как встречающиеся в природе варианты и полипептиды с нативной последовательностью, кодируемые последовательностью нуклеиновой кислоты, которая может гибридизоваться в строгих условиях с SEQ ID NO:68.
Примеры RUNX2 включают полипептид, содержащий SEQ ID NO:28, и другие полипептиды RUNX2 с нативной последовательностью, такие как встречающиеся в природе варианты и полипептиды с нативной последовательностью, кодируемые последовательностью нуклеиновой кислоты, которая может гибридизоваться в строгих условиях с SEQ ID NO:69.
Примеры PTGS1 включают полипептид, содержащий SEQ ID NO:29 и другие полипептиды PTGS1 с нативной последовательностью, такие как встречающиеся в природе варианты и полипептиды с нативной последовательностью, кодируемые последовательностью нуклеиновой кислоты, которая может гибридизоваться в строгих условиях с SEQ ID NO:70.
Примеры ALOX15 включают полипептид, содержащий SEQ ID NO:30 и другие полипептиды ALOX15 с нативной последовательностью, такие как встречающиеся в природе варианты и полипептиды с нативной последовательностью, кодируемые последовательностью нуклеиновой кислоты, которая может гибридизоваться в строгих условиях с SEQ ID NO:71.
"Ингибитор каскада IL13/IL4" относится к средству, которое блокирует передачу сигнала IL-13 и/или IL-4. Примеры ингибиторов IL13, IL4 или IL13/IL4 включают, но не ограничиваются ими, связывающие IL13 соединения, связывающие IL4 соединения, связывающие рецептор-альфа для IL4 соединения, связывающие рецептор-альфа-1 для IL13 соединения и связывающие рецептор-альфа-2 для IL13 соединения. Конкретно, в ингибиторы включены однодоменные антитела, которые связывают IL-13, IL-4, IL-13R-альфа-1, IL-13R-альфа-2 или IL-4R-альфа. Следует отметить, что молекулы, которые могу связывать более чем одну мишень, также включены в изобретение.
"Связывающие IL4 соединения" относятся к средству, которое специфично связывается с IL-4 человека. Такие связывающие соединения могут включать низкомолекулярное соединение, аптамер или полипептид. Такой полипептид может включать, но не ограничиваться ими, полипептид(ы), выбранный из группы, состоящей из иммуноадгезина, антитела, пептидного антитела и пептида. Согласно одному варианту осуществления, связывающее соединение связывается с последовательностью IL-4 человека с аффинностью между 1 мкМ-1 пМ. Конкретные примеры связывающих IL4 соединений могут включать растворимый рецептор-альфа для IL4 (например, внеклеточный домен рецептора для IL4, слитый с Fc-областью человека), антитело против IL4 и растворимый рецептор-альфа-1 для IL13 (например, внеклеточный домен рецептора-альфа-1 для IL13, слитый с Fc-областью человека).
"Связывающие рецептор-альфа для IL4 соединения" относится к средству, которое специфично связывается с рецептором-альфа для IL4 человека. Такие связывающие соединения могут включать низкомолекулярное соединение, аптамер или полипептид. Такой полипептид может включать, но не ограничиваться ими, полипептид(ы), выбранный из группы, состоящей из иммуноадгезина, антитела, пептидного антитела и пептида. Согласно одному варианту осуществления, связывающее соединение связывается с последовательностью рецептора-альфа для IL-4 человека с аффинностью между 1 мкМ-1 пМ. Конкретные примеры связывающих рецептор-альфа для IL4 соединений могут включать антитела против рецептора-альфа для IL4.
"Связывающее IL13 соединение" относится к средству, которое специфично связывается с IL-13 человека. Такие связывающие соединения могут включать низкомолекулярное соединение, аптамер или полипептид. Такой полипептид может включать, но не ограничиваться ими, полипептид(ы), выбранный из группы, состоящей из иммуноадгезина, антитела, пептидного антитела и пептида. Согласно одному варианту осуществления, связывающее соединение связывается с последовательностью IL-13 человека с аффинностью между 1 мкМ-1 пМ. Конкретные примеры связывающих IL13 соединений могут включать антитела против IL13, растворимый рецептор-альфа-2 для IL13, слитый с Fc человека, растворимый рецептор-альфа для IL4, слитый с Fc человека, растворимый рецептор-альфа для IL-13, слитый с Fc человека. Согласно одному варианту осуществления, антитело против IL13 содержит вариабельные домены антитела TNX-650 (WO 2005/062972). Вариабельные домены антитела TNX-650 содержат (1) VH, содержащую QVTLRESGPALVKPTQTLTLTCTVSGFSLSAYSVNWIRQPPGKALEWLAMIWGDGKIVYNSALKSRLTISKDTSKNQVVLTMTNMDPVDTATYYCAGDGYYPYAMDNWGQGSLVTVSS (SEQ ID NO:193), и (2) VL, содержащую: DIVMTQSPDSLSVSLGERATINCRASKSVDSYGNSFMHWYQQKPGQPPKLLIYLASNLESGVPDRFSGSGSGTDFTLTISSLQAEDVAVYYCQQNNEDPRTFGGGTKVEIK (SEQ ID NO:194). Другие примеры антител против IL13 описаны в WO 2008/083695 (например, IMA-638 и IMA-026), US 2008/0267959, US 2008/0044420 и US 2008/0248048.
"Связывающие рецептор-альфа-1 для IL13 соединения" относятся к средству, которое специфично связывается с рецептором-альфа-1 для IL13 человека. Такие связывающие соединения могут включать низкомолекулярное соединение, аптамер или полипептид. Такой полипептид может включать, но не ограничиваться ими, полипептид(ы), выбранный из группы, состоящей из иммуноадгезина, антитела, пептидного антитела и пептида. Согласно одному варианту осуществления, связывающее соединение связывается с последовательностью рецептора-альфа-1 для IL13 человека с аффинностью между 1 мкМ-1 пМ. Конкретные примеры связывающих рецептор-альфа-1 для IL13 соединений могут включать антитела против рецептора-альфа-1 для IL13.
"Связывающие рецептор-альфа-2 для IL13 соединения" относятся к средству, которое специфично связывается с рецептором-альфа-2 для IL13 человека. Такие связывающие соединения могут включать низкомолекулярное соединение, аптамер или полипептид. Такой полипептид может включать, но не ограничиваться ими, полипептид(ы), выбранный из группы, состоящей из иммуноадгезина, антитела, пептидного антитела и пептида. Согласно одному варианту осуществления, связывающее соединение связывается с последовательностью рецептора-альфа-2 для IL13 человека с аффинностью между 1 мкМ-1 пМ. Конкретные примеры связывающих рецептор-альфа-2 для IL13 соединений могут включать антитела против рецептора-альфа-2 для IL13.
"Связывающие IgE соединения" относятся к средству, которое специфично связывается с IgE человека. Такие связывающие соединения могут включать низкомолекулярное соединение, аптамер или полипептид. Такой полипептид может включать, но не ограничиваться ими, полипептид(ы), выбранный из группы, состоящей из иммуноадгезина, антитела, пептидного антитела и пептида. Согласно одному варианту осуществления, антитело против IgE содержит последовательность VL, содержащую Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Val Asp Tyr Asp Gly Asp Ser Tyr Met Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile Tyr Ala Ala Ser Tyr Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Ser His Glu Asp Pro Tyr Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val (SEQ ID NO:213), и последовательность VH, содержащую Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Val Ser Gly Tyr Ser Ile Thr Ser Gly Tyr Ser Trp Asn Trp Ile Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ala Ser Ile Thr Tyr Asp Gly Ser Thr Asn Tyr Asn Pro Ser Val Lys Gly Arg Ile Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr Phe Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg Gly Ser His Tyr Phe Gly His Trp His Phe Ala Val Trp Gly Gln Gly (SEQ ID NO:214).
"Связывающие M1' соединения" относятся к средству, которое специфично связывается с мембранной проксимальной М1'-областью IgE, экспрессируемого на поверхности B-клеток. Такие связывающие соединения могут включать низкомолекулярное соединение, аптамер или полипептид. Такой полипептид может включать, но не ограничиваться ими, полипептид(ы), выбранный из группы, состоящей из иммуноадгезина, антитела, пептидного антитела и пептида. Согласно одному варианту осуществления, антитело против IgE включает антитело, описанное в WO2008/116149, или его вариант.
Термин "низкомолекулярное соединение" относится к органической молекуле, имеющей молекулярную массу между 50 дальтон и 2500 дальтон.
Термин "антитело" используют в наиболее широком значении и, конкретно, он охватывает, например, моноклональные антитела, поликлональные антитела, антитела с полиэпитопной специфичностью, одноцепочечные антитела, полиспецифические антитела и фрагменты антител. Такие антитела могут быть химерными, гуманизированными, человеческими и синтетическими. Такие антитела и способы их получения более подробно описаны ниже.
Термин "вариабельный" относится к тому факту, что последовательности определенных сегментов вариабельных доменов значительно отличаются среди антител. V-области опосредуют связывание антигена и определяют специфичность конкретного антитела к его конкретному антигену. Однако вариабельность не является равномерной на протяжении участка вариабельных доменов из 110 аминокислот. Вместо этого, V-участки состоят из относительно инвариантных участков, называемых каркасными областями (FR), из 15-30 аминокислот, разделенных более короткими участками высокой вариабельности, называемыми "гипервариабельными областями", длина которых составляет 9-12 аминокислот. Вариабельные домены нативных тяжелых и легких цепей содержат четыре FR, главным образом, принимающих конфигурацию бета-слоев, соединенных тремя гипервариабельными участками, которые формируют петли, объединяющие структуру бета-слоев, и, в некоторых случаях, формирующие ее часть. Гипервариабельные области в каждой цепи расположены вместе в непосредственной близости от FR и, совместно с гипервариабельными областями другой цепи, участвуют в формировании антигенсвязывающего центра антител (см. Kabat et al., Последовательности of Белок of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, MD. (1991)). Константные домены не вовлечены непосредственно в связывание антитела с антигеном, но они проявляют различные эффекторные функции, такие как участие антитела в антителозависимой клеточной цитотоксичности (ADCC).
Термины "гипервариабельная область" (или "HVR"), как используют в настоящем документе, относится к аминокислотным остаткам антитела, которые отвечают за связывание антигена. Гипервариабельная область, как правило, содержит аминокислотные остатки из "определяющей комплементарность области" или "CDR" (например, приблизительно в области остатков 24-34 (L1), 50-56 (L2) и 89-97 (L3) в VL, и приблизительно в области 31-35B (H1), 50-65 (H2) и 95-102 (H3) в VH (Kabat et al., Sequences of Protein of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, Md. (1991)), и/или эти остатки из "гипервариабельной петли" (например остатки 26-32 (L1), 50-52 (L2) и 91-96 (L3) в VL, и 26-32 (H1), 52A-55 (H2) и 96-101 (H3) в VH (Chothia and Lesk J. Mol. Biol. 196:901-917 (1987)).
Гипервариабельные области могут включать следующие "удлиненные гипервариабельные области": 24-36 (L1), 46-56 (L2) и 89-97 (L3) в VL и 26-35B (H1), 47-65 (H2) и 93-102 (H3) в VH. Остатки вариабельных доменов пронумерованы согласно Kabat et al., выше, для каждого из этих определений.
"Каркасные" или "FR" остатки представляют собой остатки вариабельного домена, отличные от остатков гипервариабельной области, как определено в настоящем документе. Например, каркасная область 1 (LC-FR1), каркасная область 2 (LC-FR2), каркасная область 3 (LC-FR3) и каркасная область 4 (LC-FR4) легкой цепи может содержать остатки антитела под номерами 1-23, 35-49, 57-88 и 98-107 (система нумерации по Kabat), соответственно. В другом примере, каркасная область 1 тяжелой цепи (HC-FR1), каркасная область 2 тяжелой цепи (HC-FR2), каркасная область 3 тяжелой цепи (HC-FR3) и каркасная область 4 тяжелой цепи (HC-FR4) может содержать остатки антитела 1-25, 36-48, 66-92 и 103-113, соответственно (система нумерации по Kabat).
Как обозначают в настоящем документе, "консенсусная последовательность" или консенсусная последовательность V-домена представляет собой искусственную последовательность, полученную при сравнении аминокислотных последовательностей известных последовательностей вариабельных областей иммуноглобулинов человека.
Как используют в настоящем документе, термин "моноклональное антитело" относится к антителу из совокупности по существу однородных антител, т.е. отдельные антитела, составляющие совокупность, являются идентичными и/или связывают один эпитоп(ы), за исключением возможных вариантов, которые могут возникать в процессе продукции моноклонального антитела, причем такие варианты обычно представлены в небольших количествах. Такое моноклональное антитело, как правило, включает антитело, содержащее полипептидную последовательность, которая связывает мишень, где связывающая мишень полипептидная последовательность получена способом, который включает селекцию единичной связывающей мишень полипептидной последовательности из множества полипептидных последовательностей. Например, способ селекции может представлять собой селекцию единичного клона из множества клонов, таких как пул гибридомных клонов, фаговых клонов или клонов рекомбинантных ДНК. Следует понимать, что отобранную связывающую мишень последовательность можно дополнительно изменять, например, для повышения аффинности к мишени, для гуманизации связывающей мишень последовательности, для повышения ее продукции в клеточной культуре, для снижения ее иммуногенности in vivo, для создания полиспецифичного антитела, и т.д., и что антитело, содержащее измененную связывающую мишень последовательность, также является моноклональным антителом по этому изобретению. В противоположность препаратам поликлональных антител, которые как правило, включают различные антитела, направленные против различных детерминант (эпитопов), каждое моноклональное антитело в препарате моноклонального антитела направлено против одной детерминанты на антигене. В дополнение к их специфичности, препараты моноклонального антитела является преимущественными в том, что они, как правило, не содержат примесей других иммуноглобулинов. Определение "моноклональный" указывает на тот признак антитела, что его получают из по существу гомогенной совокупности антител, и не подразумевает того, что антитело должно быть получено каким-либо конкретным способом. Например, моноклональные антитела, предназначенные для применения в соответствии с настоящим изобретением, можно получать множеством способов, включая, например, способ гибридом (например, Kohler et al., Nature, 256:495 (1975); Harlow et al., Antibodies: A Laboratory Manual, (Cold Spring Harbor Laboratory Press, 2nd ed. 1988); Hammerling et al., Monoclonal Antibodies and T-Cell Hybrodimas 563-681 (Elsevier, N.Y., 1981)), способы рекомбинантных ДНК (см., например, патент США No. 4816567), технологии фагового дисплея (см., например, Clackson et al., Nature, 352:624-628 (1991); Marks et al., J. Mol. Biol., 222:581-597 (1991); Sidhu et al., J. Mol. Biol. 338(2):299-310 (2004); Lee et al., J.Mol.Biol.340(5):1073-1093 (2004); Fellouse, Proc. Nat. Acad. Sci. USA 101(34):12467-12472 (2004); и Lee et al., J. Immunol. Methods 284(1-2):119-132 (2004), и технологии продуцирования антитела человека или антител, подобных антителам человека, у животных, которые имеют части локусов или генов иммуноглобулинов человека, кодирующие последовательности иммуноглобулинов, или все эти локусы или гены (см., например, WO 1998/24893; WO 1996/34096; WO 1996/33735; WO 1991/10741; Jakobovits et al., Proc. Natl. Acad. Sci. USA, 90:2551 (1993); Jakobovits et al., Nature, 362:255-258 (1993); Bruggemann et al., Year in Immuno., 7:33 (1993); патенты США No. 5545806; 5569825; 5591669 (all of GenPharm); 5545807; WO 1997/17852; патенты США No. 5545807; 5545806; 5569825; 5625126; 5633425; и 5661016; Marks et al., Bio/Technology, 10: 779-783 (1992); Lonberg et al., Nature, 368: 856-859 (1994); Morrison, Nature, 368: 812-813 (1994); Fishwild et al., Nature Biotechnology, 14: 845-851 (1996); Neuberger, Nature Biotechnology, 14: 826 (1996); и Lonberg and Huszar, Intern. Rev. Immunol., 13: 65-93 (1995).
Моноклональные антитела в настоящем документе конкретно включают "химерные" антитела (иммуноглобулины), в которых участок тяжелой и/или легкой цепи идентичен или гомологичен соответствующим последовательностям в антителах, полученных из конкретного вида или принадлежащих к конкретному классу или подклассу антител, в то время как оставшаяся часть цепи(ей) идентична или гомологична соответствующим последовательностям в антителах, полученных из другого вида или принадлежащих другому классу или подклассу антител, а также фрагменты таких антител, при условии, что они проявляют требуемую биологическую активность (патент США No 4816567; и Morrison et al., Proc. Natl. Acad. Sci USA, 81:6851-6855 (1984)). Способы получения химерных антител известны в данной области.
"Гуманизированные" формы не являющихся человеческими антител (например, грызунов) представляют собой химерные иммуноглобулины, цепи иммуноглобулинов или их фрагменты (такие как Fv, Fab, Fab', F(ab')2 или другие антигенсвязывающие подпоследовательности антител), которые содержат минимальную последовательность из иммуноглобулина, не являющегося человеческим. В некоторых вариантах осуществления, гуманизированные антитела представляют собой иммуноглобулины человека (реципиентное антитело), в которых остатки определяющей комплементарной области (CDR) реципиента заменены остатками из CDR видов, не являющихся человеком, (донорное антитело), таких как мышь, крыса или кролик, которые обладают требуемой специфичностью, аффинностью и емкостью. В некоторых случаях, остатки каркасной области (FR) иммуноглобулина человека заменяют соответствующими остатками, не являющиеся человеческими. Более того, гуманизированные антитела могут содержать остатки, которые отсутствуют как в реципиентном антителе, так и в донорном антителе. Эти модификации обычно проводят для дополнительного улучшения или максимизации параметров антитела. Как правило, гуманизированное антитело содержит по существу все из по меньшей мере одного вариабельного домена, в котором все или по существу все из гипервариабельных петель происходят из иммуноглобулина, не являющегося человеческим, и все или по существу все из FR-областей происходят из последовательности иммуноглобулина человека, хотя FR-области могут включать одну или несколько аминокислотных замен, например, для повышения аффинности связывания. В одном предпочтительном варианте осуществления, гуманизированное антитело необязательно содержит по меньшей мере часть константного домена (Fc) иммуноглобулина, как правило, константного домена иммуноглобулина человека или консенсусной константной последовательности человека. Для более подробной информации см. Jones et al., Nature 321:522-525 (1986); Riechmann et al., Nature 332:323-329 (1988); и Presta, Curr. Op. Struct. Biol. 2:593-596 (1992). Гуманизированное антитело включает антитело PRIMATIZED®, где антигенсвязывающая область антитела происходит из антитела, полученного, например, иммунизацией макак представляющим интерес антигеном. Способы получения гуманизированных антител известны в данной области.
Антитела человека также можно получать с использованием различных способов, известных в данной области, включая библиотеки фагового дисплея. Hoogenboom and Winter, J. Mol. Biol., 227:381 (1991); Marks et al., J. Mol. Biol, 222:581 (1991). Также для получения моноклональных антител человека доступны способы Cole et al. и Boerner et al. Cole et al., Monoclonal Anribodies and Cancer Therapy, Alan R. Liss, p. 77 (1985); Boerner et al., J. Immunol, 147(l):86-95 (1991). См. также, Lonberg and Huszar, Int. Rev. Immunol. 13:65-93 (1995). Публикации PCT WO 98/24893; WO 92/01047; WO 96/34096; WO 96/33735; патент Европы No. 0598877; патенты США No. 5413923; 5625126; 5633425; 5569825; 5661016; 5545806; 5814318; 5885793; 5916771 и 5939598.
"Фрагменты антител" содержат часть полноразмерного антитела, как правило, его антигенсвязывающую или вариабельную область. Примеры фрагментов антител включают Fab, Fab', F(ab')2 и Fv-фрагменты; антитела-димеры; линейные антитела; молекулы одноцепочечных антител; и полиспецифические антитела, образованные из фрагментов антител.
"Fv" представляет собой минимальный фрагмент антитела, который содержит полный антигенраспознающий и антигенсвязывающий центр. Этот фрагмент состоит из димера, состоящего из одного вариабельного домена тяжелой цепи и одного вариабельного домена легкой цепи, связанных прочной нековалентной связью. Сворачивание этих двух доменов образует шесть гипервариабельных петель (3 петли в каждой из H- и L-цепи), которые предоставляют аминокислотные остатки для связывания антигена и обеспечивают специфичность связывания антигена антителом. Однако даже один вариабельный домен (или половина Fv, содержащая только три CDR, специфичных к антигену) обладает способностью распознавать и связывать антиген, хотя и с более низкой аффинностью, чем целый участок связывания.
"Функциональные фрагменты" антител по изобретению представляют собой фрагменты, которые сохраняют связывание с полипептидом по существу с той же аффинностью, что и интактная молекула с полной цепью, из которой они происходят, и являются активными по меньшей мере в одном анализе (например, ингибирование TH2-индуцированного каскада астмы, например, в моделях на мышах, или ингибирование биологической активности антигена, который связывается с фрагментом антитела in vitro).
"Эффекторные функции" антитела относятся к таким видам биологической активности, которые являются свойственными Fc-области (Fc-области с нативной последовательностью или варианту Fc-области по аминокислотной последовательности) антитела, и варьируют в зависимости от изотипа антитела. Примеры эффекторных функций антитела включают: связывание C1q и комплементзависимую цитотоксичность; связывание Fc-рецептора; антителозависимую клеточно-опосредуемую цитотоксичность (ADCC); фагоцитоз; отрицательную регуляцию рецепторов клеточной поверхности (например, B-клеточного рецептора) и активацию B-клеток. "Fc-область с нативной последовательностью" содержит аминокислотную последовательность, идентичную аминокислотной последовательности Fc-области, встречающейся в природе.
"Процентную (%) идентичность аминокислотных последовательностей" или "гомологию" в отношении последовательностей полипептидов антител, идентифицированных в настоящем документе, определяют как процент аминокислотных остатков в последовательности-кандидате, которые идентичны аминокислотным остаткам в подвергаемом сравнению полипептиде, после выравнивания последовательностей, считая какие-либо консервативные замены частью идентичности последовательностей. Выравнивание для целей определения процентной идентичности аминокислотных последовательностей можно проводить различными способами, которые находятся в пределах специальных знаний, например, с использованием общедоступного компьютерного программного обеспечения, такого как программное обеспечение BLAST, BLAST-2, ALIGN или Megalign (DNASTAR). Специалисты в данной области могут определить соответствующие параметры для проведения выравнивания, включая любые алгоритмы, требуемые для достижения максимального выравнивания на протяжении всей длины сравниваемых последовательностей. Однако для целей, представленных в настоящем документе, значения % идентичности аминокислотных последовательностей получают с использованием компьютерной программы для сравнения последовательностей ALIGN-2. Компьютерная программа для сравнения последовательностей ALIGN-2 составлена в Genentech, Inc. и исходный текст был представлен с пользовательской документацией в U.S. Copyright Office, Washington D.C., 20559, где он зарегистрирован под U.S. Copyright Registration No. TXU510087. Программа ALIGN-2 является общедоступной через Genentech, Inc., South San Francisco, California. Программа ALIGN-2 должна быть составлена для применения на операционной системе UNIX, предпочтительно цифровой UNIX V4.0D. Все параметры сравнения последовательностей установлены программой ALIGN-2 и не изменяются.
Термин "содержащий Fc-область полипептид" относится к полипептиду, такому как антитело или иммуноадгезин (см. определения ниже), который содержит Fc-область. C-концевой лизин (остаток 447 согласно системе нумерации EU) Fc-области может быть удален, например, в процессе очистки полипептида или путем рекомбинантной инженерии нуклеиновой кислоты, кодирующей полипептид. Таким образом, композиция, содержащая полипептиды, включая антитела, имеющие Fc-область по этому изобретению, может содержать совокупности полипептидов, в которых все остатки K447 удалены, совокупности полипептидов, в которых остатки K447 не удалены, или совокупности полипептидов, имеющие смесь полипептидов с остатком K447 или без него.
На протяжении настоящего описания и формулы изобретения, систему нумерации Kabat обычно используют при указании на остаток в вариабельном домене (приблизительно остатки 1-107 легкой цепи и остатки 1-113 тяжелой цепи) (например, Kabat et al., Sequences of Immunological Interest. 5th Ed. Public Health Service, National Institutes of Health, Bethesda, Md. (1991)). "Систему нумерации EU" или "индекс EU" обычно используют при указании на остаток в константной области тяжелой цепи иммуноглобулина (например, индекс EU, указанный в Kabat et al., Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, MD (1991), включенная в настоящий документ в качестве ссылки в полном объеме). Если в настоящем документе нет иных указаний, указания на номера остатков в вариабельных доменах антител означают нумерацию остатков в соответствии с системой нумерации Kabat. Если в настоящем документе нет иных указаний, указания на номера остатков в константном домене антител означают нумерацию остатков в соответствии с системой нумерации EU (например, см. предварительную заявку США No. 60/640323, фигуры для нумерации EU).
"Строгость" реакций гибридизации легко определяет специалист в данной области, и, как правило, она представляет собой эмпирическое вычисление, зависящее от длины зонда, температуры промывания и концентрации соли. Как правило, для надлежащего отжига более длинные зонды требуют более высоких температур, в то время как более короткие зонды требуют более низких температур. Гибридизация, как правило, зависит от способности денатурированной ДНК к повторному отжигу, когда комплементарные цепи присутствуют в окружающей среде при температуре, ниже их температуры отжига. Чем более высокой является степень требуемой гомологии между зондом и гибридизуемой последовательностью, тем более высокой является относительная температура, которую можно использовать. В результате, из этого следует, что более высокие относительные температуры будут обеспечивать тенденцию к более строгим условиям реакции, в то время как более низкие температуры снижают строгость. Для дополнительных деталей и разъяснения строгости реакций гибридизации, см. Ausubel et al., Current Protocols in Molecular Biology, Wiley Interscience Publishers, (1995).
"Строгие условия" или "условия высокой строгости", как определено в настоящем документе, можно определить как условия, в которых: (1) используют низкую ионную силу и высокую температуру промывания, например 0,015 M хлорид натрия/0,0015 M цитрат натрия/0,1% додецилсульфат натрия при 50°С; (2) используют в ходе гибридизации денатурирующее средство, такое как формамид, например, 50% (об./об.) формамид с 0,1% бычьим сывороточным альбумином/0,1% Ficoll/0,1% поливинилпирролидоном/50 мМ натрий-фосфатным буфером при pH 6,5 с 750 мМ хлоридом натрия, 75 мМ цитратом натрия при 42°C; или (3) проводят гибридизацию в течение ночи в растворе, в котором используется 50% формамид, 5x SSC (0,75 M NaCl, 0,075 M цитрат натрия), 50 мМ фосфат натрия (pH 6,8), 0,1% пирофосфат натрия, 5×раствор Денхардта, облученная ультразвуковым облучением ДНК спермы лосося (50 мкг/мл), 0,1% SDS и 10% сульфат декстрана при 42°C, с промыванием в течение 10 минут при 42°C в 0,2x SSC (хлорид натрия/цитрат натрия) с последующим промыванием в условиях высокой строгости в течение 10 минут, состоящим из 0,1x SSC, содержащего EDTA, при 55°C.
"Условия умеренной строгости" можно определить, как описано в Sambrook et al., Molecular Cloning: A Laboratory Manual, New York: Cold Spring Harbor Press, 1989, и они включают применение менее строгого раствора для промывания и условий гибридизации (например, температуры, ионной силы и %SDS), чем раствор для промывания и условия гибридизации, описанные выше. Примером умеренно строгих условий является инкубация в течение ночи при 37°C в растворе, содержащем: 20% формамид, 5x SSC (150 мМ NaCl, 15 мМ трицитрат натрия), 50 мМ фосфат натрия (pH 7,6), 5x раствор Денхардта, 10% сульфат декстрана и 20 мг/мл денатурированной расщепленной ДНК спермы лосося, с последующим промыванием фильтров в 1x SSC при приблизительно 37-50°С. Специалисту будет понятно, каким образом корректировать температуру, ионную силу и т.д., в случае необходимости приспосабливания к таким факторам, как длина зонда и т.п.
Как используют в настоящем документе, субъектом, подлежащим лечению, является млекопитающее (например, человек, не являющийся человеком примат, крыса, мышь, корова, лошадь, свинья, овца, коза, собака, кошка и т.д.). Субъектом может быть пациент в клинике, доброволец для клинического испытания, экспериментальное животное и т.д. У субъекта могут предполагать наличие риска астмы или он может иметь риск астмы или у него может быть диагностирована астма. Согласно одному предпочтительному варианту осуществления, субъектом, подлежащим лечению согласно этому изобретению, является человек.
"Проведение лечения" или "лечение" или "смягчение течения" относится как к терапевтическим, так и к профилактическим или превентивным мерам, где целью является предотвратить или замедлить (уменьшить) конкретное патологическое состояние или нарушение. Субъекты, нуждающиеся в лечении, включают субъектов, уже имеющих нарушение, а также субъектов, имеющих предрасположенность к нарушению, или индивидуумов у которых нарушение подлежит профилактике. Субъекта или млекопитающее успешно "лечат" от астмы, если после получения терапевтического количества средства по настоящему изобретению, у пациента наблюдают заметное и/или поддающееся измерению снижение или отсутствие одного или нескольких из следующих: рецидивирующее свистящее дыхание, кашель, затрудненное дыхание, сжимание в области груди, симптомы, которые возникают или ухудшаются ночью, симптомы, которые стимулируются холодным воздухом, физической нагрузкой или воздействием аллергенов.
Термин "терапевтически эффективное количество" относится к количеству полипептида по этому изобретению, эффективному для "смягчения течения" или "лечения" заболевания или нарушения у субъекта.
"Длительное" введение относится к введению средства(средств) постоянно в противоположность кратковременному режиму, чтобы поддерживать исходный терапевтический эффект (активность) в течение длительного периода времени. "Прерывающееся" введение представляет собой лечение, которое не проводится без прерывания, а вместо этого является циклическим.
"Объем форсированного выдоха (FEV1)" относится к стандартному тесту, который измеряет объем воздуха, выдыхаемого в течение первой секунды форсированного выдоха. FEV1 измеряют с помощью спирометра, который состоит из наконечника и одноразовой трубки, присоединенной к устройству, которое регистрирует результаты и отображает их на графике. Для проведения спирометрии, человек глубоко вдыхает, смыкает плотно губы вокруг трубки, а затем выдыхает через трубку, в то время как проводят измерения. Объем выдыхаемого воздуха, и продолжительность времени, которое занимает каждый акт дыхания, записывают и анализируют. Результаты спирометрии выражают в процентах. Примеры нормальных результатов спирометрии включают FEV1, составляющую 75 процентов жизненной емкости после одной секунды. Пример ненормальных результатов спирометрии включают показатели менее чем 80 процентов от нормального предсказанного значения. Ненормальный результат обычно указывает на наличие некоторой степени обструктивного заболевания легких, такого как астма, эмфизема или хронический бронхит, или рестриктивного заболевания легких, такого как легочный фиброз. Например, величины FEV1 (процент от предсказанного) можно использовать для классификации обструкции, которая может происходить при астме и других обструктивных заболеваниях легких, таких как эмфизема или хронический бронхит: FEV1 от 65 процентов до 79 процентов от предсказанного значения=мягкая обструкция, FEV1 от 40 процентов до 59 процентов от предсказанного значения=умеренная обструкция, и FEV1 менее 40 процентов от предсказанного значения=тяжелая обструкция.
Примеры зондов нуклеиновых кислот, которые можно использовать для идентификации белков, описанных в настоящем документе (например, посредством анализа на микрочипах), включают, но не ограничиваются ими, зонды, описанные в таблице 4.
"Повышенный уровень экспрессии" или "повышенные уровни" относится к повышенной экспрессии мРНК или белка у пациента относительно контроля, такого как индивидуум или индивидуумы, которые не страдают астмой.
Все публикации (включая патенты и патентные заявки), цитированные в настоящем документе, включены в настоящий документ в качестве ссылок в полном объеме.
На протяжении этого описания и формулы изобретения, слово "содержат" или его варианты, такие как "содержит" или "содержащий", следует понимать, как подразумевающие включение указанного числа или группы чисел, но исключение любого другого числа или группы чисел.
Представленное выше письменное описание считается достаточным, чтобы специалист в данной области мог осуществлять на практике это изобретение. Следующие примеры предназначены только для иллюстративных целей и не предназначены для ограничения объема настоящего изобретения никоим образом. Действительно, различные модификации изобретения в дополнение к модификациям, показанным и описанным в настоящем документе, будут очевидны специалистам в данной области из представленного выше описания, и они относятся к объему прилагаемой формулы изобретения.
Неполный список ссылок:
1. Haldar, P. and I.D. Pavord, Noneosinophilic asthma: a distinct clinical and pathologic phenotype. J Allergy Clin Immunol, 2007. 119(5): p. 1043-52; quiz 1053-4.
2. Hershey, G.K., IL-13 receptors and signaling pathways: an evolving web. J Allergy Clin Immunol, 2003. 111(4): p. 677-90; quiz 691.
3. Berry, M.A., et al., Sputum and bronchial submucosal IL-13 expression in asthma and eosinophilic bronchitis. J Allergy Clin Immunol, 2004. 114(5): p. 1106-9.
4. Humbert, M., et al., Elevated expression of messenger ribonucleic acid encoding IL-13 in the bronchial mucosa of atopic and nonatopic subjects with asthma. J Allergy Clin Immunol, 1997. 99(5): p. 657-65.
5. Truyen, E., et al., Evaluation of airway inflammation by quantitative Th1/Th2 cytokine mRNA measurement in sputum of asthma patients. Thorax, 2006. 61(3): p. 202-8.
6. Saha, S.K., et al., Increased sputum and bronchial biopsy 1L-13 expression in severe asthma. J Allergy Clin Immunol, 2008. 121(3): p. 685-91.
7. Yuyama, N., et al., Analysis of novel disease-related genes in bronchial asthma. Cytokine, 2002. 19(6): p. 287-96.
8. Woodruff, P.G., et al., Genome-wide profiling identifies epithelial cell genes associated with asthma and with treatment response to corticosteroids. Proc Natl Acad Sci USA, 2007. 104(40): p. 15858-63.
9. Takayama, G., et al., Periostin: a novel component of subepithelial fibrosis of bronchial asthma downstream of IL-4 and 1L-13 signals. J Allergy Clin Immunol, 2006. 118(1): p. 98-104.
10. Hayashi, N., et al., T helper 1 cells stimulated with ovalbumin and IL-18 induce airway hyperresponsiveness and lung fibrosis by IFN-gamma and IL-13 production. Proc Natl Acad Sci USA, 2007. 104(37): p. 14765-70.
11. Blanchard, C., et al., IL-13 involvement in eosinophilic esophagitis: transcriptome analysis and reversibility with glucocorticoids. J Allergy Clin Immunol, 2007. 120(6): p. 1292-300.
12. Nakanishi, A., et al., Role of gob-5 in mucus overproduction and airway hyperresponsiveness in asthma. Proc Natl Acad Sci USA, 2001. 98(9): p. 5175-80.
13. Zhou, Y., et al., Characterization of a calcium-activated chloride channel as a shared target of Th2 cytokine pathways and its potential involvement in asthma. Am J Respir Cell Mol Biol, 2001. 25(4): p. 486-91.
14. Kuperman, D.A., et al., Dissecting asthma using focused transgenic modeling and functional genomics. J Allergy Clin Immunol, 2005. 116(2): p. 305-11.
15. Ray, R., et al., Uteroglobin suppresses SCCA gene expression associated with allergic asthma. J Biol Chem, 2005. 280(11): p. 9761-4.
16. Sabatini, L.M., et al., Tissue distribution of RNAs for cystatins, histatins, statherin, and proline-rich salivary proteins in humans and macaques. J Dent Res, 1989. 68(7): p. 1138-45.
17. Lindahl, M., B. Stahlbom, and C. Tagesson, Newly identified proteins in human nasal and bronchoalveolar lavage fluids: potential biomedical and clinical applications. Electrophoresis, 1999. 20(18): p. 3670-6.
18. Cimerman, N., et al., Serum cystatin C, a potent inhibitor of cysteine proteinases, is elevated in asthmatic patients. Clin Chim Acta, 2000. 300(1-2): p. 83-95.
19. Finkelman, F.D., et al., Suppressive effect of IL-4 on IL-13-induced genes in mouse lung. J Immunol, 2005. 174(8): p. 4630-8.
20. Zimmermann, N., et al., Expression and regulation of small proline-rich protein 2 in allergic inflammation. Am J Respir Cell Mol Biol, 2005. 32(5): p. 428-35.
21. Warner, T.F. and E.A. Azen, Proline-rich proteins are present in serous cells of submucosal glands in the respiratory tract. Am Rev Respir Dis, 1984. 130(1): p. 115-8.
22. Maeda, Y., et al., [Concentrations of carcinoembryonic antigen in serum and bronchoalveolar lavage fluid of asthmatic patients with mucoid impaction], Nihon Kokyuki Gakkai Zasshi, 2004. 42(12): p. 988-93.
23. Suresh, V., J.D. Mih, and S.C. George, Measurement of IL-13-induced iNOS-derived gas phase nitric oxide in human bronchial epithelial cells. Am J Respir Cell Mol Biol, 2007. 37(1): p. 97-104.
24. Storey, J.D. and R. Tibshirani, Statistical significance for genomewide studies. Proc Natl Acad Sci USA, 2003. 100(16): p. 9440-5.
25. Eisen, M.B., et al., Cluster analysis and display of genome-wide expression patterns. Proc Natl Acad Sci USA, 1998. 95(25): p. 14863-8.
26. Saldanha, A.J., Java Treeview-extensible visualization of microarray data. Bioinformatics, 2004. 20(17): p. 3246-8.
27. Kelly-Welch, A. E., Hanson, E. M., Boothby, M. R. & Keegan, A. D. Interleukin-4 and interleukin-13 signaling connections maps. Science 300, 1527-8 (2003).
28. Fixman, E. D., Stewart, A. & Martin, J. G. Basic mechanisms of development of airway structural changes in asthma. Eur Respir J 29, 379-89 (2007).
29. Holgate, S. T. Pathogenesis of asthma. Clin Exp Allergy 38, 872-97 (2008).
30. Roche, W. R., Beasley, R., Williams, J. H. & Holgate, S. T. Subepithelial fibrosis in the bronchi of asthmatics. Lancet 1, 520-4 (1989).
31. Ferrando, R. E., Nyengaard, J. R., Hays, S. R., Fahy, J. V. & Woodruff, P. G. Applying stereology to measure thickness of the basement membrane zone in bronchial biopsy specimens. J Allergy Clin Immunol 112, 1243-5 (2003).
32. Ordonez, C. L. et al. Mild and moderate asthma is associated with airway goblet cell hyperplasia and abnormalities in mucin gene expression. Am J Respir Crit Care Med 163, 517-23 (2001).
33. Martin, R. J. et al. The Predicting Response to Inhaled Corticosteroid Efficacy (PRICE) trial. J Allergy Clin Immunol 119, 73-80(2007).
34. Wenzel, S., Wilbraham, D., Fuller, R., Getz, E. B. & Longphre, M. Effect of an interleukin-4 variant on late phase asthmatic response to allergen challenge in asthmatic patients: results of two phase 2a studies. Lancet 370, 1422-31 (2007).
35. Grunig, G. et al. Requirement for IL-13 independently of IL-4 in experimental asthma. Science 282, 2261-3 (1998).
36. Wills-Karp, M. et al. Interleukin-13: central mediator of allergic asthma. Science 282, 2258-61 (1998).
37. Simpson, J. L., Scott, R., Boyle, M. J. & Gibson, P. G. Inflammatory subtypes in asthma: assessment and identification using induced sputum. Respirology 11, 54-61 (2006).
38. Wenzel, S. E. et al. Evidence that severe asthma can be divided pathologically into two inflammatory subtypes with distinct physiologic and clinical characteristics. Am J Respir Crit Care Med 160, 1001-8 (1999).
39. Wenzel, S. E. Asthma: defining of the persistent adult phenotypes. Lancet 368, 804-13 (2006).
40. Berry, M. et al. Pathological features and inhaled corticosteroid response of eosinophilic and non-eosinophilic asthma. Thorax 62, 1043-9 (2007).
41. Pavord, I. D., Brightling, C. E., Woltmann, G. & Wardlaw, A. J. Non-eosinophilic corticosteroid unresponsive asthma. Lancet 353, 2213-4 (1999).
42. McKinley, L. et al. TH17 cells mediate steroid-resistant airway inflammation and airway hyperresponsiveness in mice. J Immunol 181, 4089-97 (2008).
43. Holgate, S. T. Epithelium dysfunction in asthma. J Allergy Clin Immunol 120, 1233-44; quiz 1245-6 (2007).
44. Martin, R. J., Kraft, M., Chu, H. W., Berns, E. A. & Cassell, G. H. A link between chronic asthma and chronic infection. J Allergy Clin Immunol 107, 595-601 (2001).
45. Dolganov, G. M. et al. A novel method of gene transcript profiling in airway biopsy homogenates reveals increased expression of a Na+-K+-Cl- cotransporter (NKCC1) in asthmatic subjects. Genome Res 11, 1473-83 (2001).
46. Innes, A. L. et al. Epithelial mucin stores are increased in the large airways of smokers with airflow obstruction. Chest 130, 1102-8 (2006).
47. Gentleman, R. C. et al. Bioconductor: open software development for computational biology and bioinformatics. Genome Biol 5, R80 (2004).
48. Douwes J, Gibson P, Pekkanen J, Pearce N. Non-eosinophilic asthma: importance and possible mechanisms. Thorax 2002;57(7):643-8.
49. Galli SJ, Tsai M, Piliponsky AM. The development of allergic inflammation. Nature 2008;454(7203):445-54.
50. Barnes PJ. The cytokine network in asthma and chronic obstructive pulmonary disease. J Clin Invest 2008; 118(11):3546-56.
51. Woodruff PG, Koth LL, Yang YH, et al. A distinctive alveolar macrophage activation state induced by cigarette smoking. Am J Respir Crit Care Med 2005; 172(11): 1383-92.
52. Weibel ER, Hsia CC, Ochs M. How much is there really? Why stereology is essential in lung morphometry. J Appl Physiol 2007;102(l):459-67.
53. Seibold MA, Donnelly S, Solon M, et al. Chitotriosidase is the primary active chitinase in the human lung and is modulated by genotype and smoking habit. J Allergy Clin Immunol 2008;122(5):944-50 e3.
54. Kim EY, Battaile JT, Patel AC, et al. Persistent activation of an innate immune response translates respiratory viral infection into chronic lung disease. Nat Med 2008;14(6):633-40.
55. Peters-Golden M. The alveolar macrophage: the forgotten cell in asthma. Am J Respir Cell Mol Biol 2004;31(l):3-7.
56. Chu HW, Balzar S, Westcott JY, et al. Expression and activation of 15-lipoxygenase pathway in severe asthma: relationship to eosinophilic phenotype and collagen deposition. Clin Exp Allergy 2002;32(11): 1558-65.
57. Gordon S. Alternative activation of macrophages. Nat Rev Immunol 2003;3(l):23-35.
58. Anderson GP. Endotyping asthma: new insights into key pathogenic mechanisms in a complex, heterogeneous disease. Lancet 2008;372(9643): 1107-19.
59. Berry MA, Hargadon B, Shelley M, et al. Evidence of a role of tumor necrosis factor alpha in refractory asthma. N Engl J Med 2006;354(7):697-708.
60. Takatsu K, Nakajima H. IL-5 and eosinophilia. Curr Opin Immunol 2008;20(3):288-94.
61. Zimmermann N, Hershey GK, Foster PS, Rothenberg ME. Chemokines in asthma: cooperative interaction between chemokines and IL-13. J Allergy Clin Immunol 2003; 111(2):227-42; quiz 43.
62. Flood-Page PT, Menzies-Gow AN, Kay AB, Robinson DS. Eosinophil's role remains uncertain as anti-interleukin-5 only partially depletes numbers in asthmatic airway. Am J Respir Crit Care Med 2003; 167(2): 199-204.
63. Flood-Page P, Swenson C, Faiferman I, et al. A study to evaluate safety and efficacy of mepolizumab in patients with moderate persistent asthma. Am J Respir Crit Care Med 2007; 176(11): 1062-71.
64. Baril, P., et al., Periostin promotes invasiveness and resistance of pancreatic cancer cells to hypoxia-induced cell death: role of the beta4 integrin and the PI3k pathway. Oncogene, 2007: 26(14): p. 2082-94.
65. Kudo, Y., et al., Periostin promotes invasion and anchorage-independent growth in the metastatic process of head and neck cancer. Cancer Res, 2006:66(14): p. 6928-35.
66. Puglisi, F., et al., Expression of periostin in human breast cancer. J Clin Pathol, 2008:61(4): p. 494-8. 15.
67. Siriwardena, B.S., et al., Periostin is frequently overexpressed and enhances invasion and angiogenesis in oral cancer. Br J Cancer, 2006:95(10): p. 1396-403.
68. Sasaki, H., et al., Serum level of the periostin, a homologue of an insect cell adhesion molecule, as a prognostic marker in nonsmall cell lung carcinomas. Cancer, 2001:92(4): p. 843-8.
69. Sasaki, H., et al., Expression of Periostin, homologous with an insect cell adhesion molecule, as a prognostic marker in non-small cell lung cancers. Jpn J Cancer Res, 2001:92(8): p. 869-73.
70. Sasaki, H., et al., Elevated serum periostin levels in patients with bone metastases from breast but not lung cancer. Breast Cancer Res Treat, 2003:77(3): p. 245-52.
71. Komiya, A., et al., Concerted expression of eotaxin-1, eotaxin-2, and eotaxin-3 in human bronchial epithelial cells. Cell Immunol, 2003:225(2): p. 91-100.
72. Chupp, G.L., et al., A chitinase-like protein in the lung and circulation of patients with severe asthma. N Engl J Med, 2007:357(20): p. 2016-27.
73. Liu, S.M., et al., Immune cell transcriptome datasets reveal novel leukocyte subset-specific genes and genes associated with allergic processes. J Allergy Clin Immunol, 2006:118(2): p. 496-503.
74. Nakajima, T., et al., Identification of granulocyte subtype-selective receptors and ion channels by using a high-density oligonucleotide probe array. J Allergy Clin Immunol, 2004:113(3): p. 528-35.
75. De Filippis, D., A. D'Amico, and T. luvone, Cannabinomimetic control of mast cell mediator release: new perspective in chronic inflammation. J Neuroendocrinol, 2008:20 Suppl 1: p. 20-5.
76. Reed, C.E. and H. Kita, The role of protease activation of inflammation in allergic respiratory diseases. J Allergy Clin Immunol, 2004:114(5): p. 997-1008; quiz 1009.
77. Blanchard, C., et al., Periostin facilitates eosinophil tissue infiltration in allergic lung and esophageal responses. Mucosal Immunol, 2008:1(4): p. 289-96.
78. Wenzel, S.E., et al., Bronchoscopic evaluation of severe asthma. Persistent inflammation associated with high dose glucocorticoids. Am J Respir Crit Care Med, 1997:156(3 Pt 1): p. 737-43.
79. Leung, D.Y., et al., Dysregulation of interleukin 4, interleukin 5, and interferon gamma gene expression in steroid-resistant asthma. J Exp Med, 1995:181(1): p. 33-40.
80. Walsh, G.M., Emerging drugs for asthma. Expert Opin Emerg Drugs, 2008:13(4): p. 643-53.
81. Ballesta, A.M., et al., Carcinoembryonic antigen in staging and follow-up of patients with solid tumors. Tumour Biol, 1995:16(1): p. 32-41.
Все ссылки, цитированные в настоящем документе, включая патентные заявки и публикации, включены в качестве ссылок в полном объеме для любой цели. Кроме того, предварительные заявки США US с серийным номером No. 61/072572, поданная 3/31/2008, US с серийным номером No. 61/041480, поданная 4/1/2008, US с серийным номером No.: 61/128383, поданная 5/20/2008, US с серийным номером No.: 61/205392, поданная 1/16/2009, включены в качестве ссылок в полном объеме. Также конкретно публикации PCT WO 2005/062972 и WO 2008/116149 включены в качестве ссылок в полном объеме.
ПРИМЕРЫ
Пример 1 - Способы
Банк тканей дыхательных путей
Авторы настоящего изобретения исследовали биологические образцы, хранившиеся в банке тканей дыхательных путей в University of California, San Francisco (UCSF), которые были собраны в ходе бронхоскопии, проведенной для исследовательских целей у здоровых добровольцев и добровольцев с астмой. Бронхоскопия для исследования включала сбор эпителиальных соскобов, бронхоальвеолярного лаважа (BAL) и биоптатов бронхов с использованием определенных способов, описанных ранее [8, 46]. Проводили подсчет клеток в BAL и определение состава клеток и результаты вносили в базу данных, и от жидкости BAL отсортировывали макрофаги с использованием проточной цитометрии [51]. От четырех до шести биоптатов бронхов было получено из киля трахеи от 2 до 5 порядка (контрлатерально области соскоба), их фиксировали в формалине, а затем погружали в парафин в изотропной единообразной случайной ориентации [31] для количественного определения воспаления и ремоделирования с использованием способов стереологии на основе изображений [52]. Дополнительные 2 биоптата бронхов гомогенизировали и обрабатывали для получения РНК с использованием набора Qiagen RNeasy minikit (Qiagen Inc., Valencia, CA). РНК, экстрагированную из эпителиальных соскобов, гомогенатов биоптатов бронхов, и макрофагов лаважа, подвергали проверке качества и разделяли на аликвоты для последующего профилирования генов, основанном на микрочипах и ПЦР. Все научные исследования бронхоскопии были одобрены UCSF Committee on Human Research (CHR), от всех субъектов было получено письменное информированное согласие, и все исследования проводили в соответствии с принципами, выраженными в декларации Хельсинки. Процедуры банка тканей дыхательных путей также рассмотрены и одобрены UCSF CHR. Образцы эпителиальных соскобов и макрофагов из этого банка тканей использовали в ранее описанных исследованиях [8, 14, 46, 51, 53]. Позднее, авторами настоящего изобретения были описаны анализы на микрочипах дифференциально экспрессированных генов в эпителиальных соскобах у субъектов-астматиков [8].
Для идентификации подгрупп пациентов с астмой, которые отличаются с точки зрения молекулярного механизма, лежащего в основе воспаления их дыхательных путей, и особых воспалительных, патологических и клинических фенотипических характеристик этих подгрупп, авторы настоящего изобретения сначала провели новые анализы на их ранее полученных с помощью микрочипов данных для эпителиальных клеток, а затем авторы настоящего изобретения дополнили эти новые анализы обзором дополнительных и детальных данных клинической охарактеризации (включая данные по обратимости после введения бронходилятатора и реактивности в кожных тестах с аллергенами) этих субъектов и вновь полученными данными, включающими: (i) профили экспрессии генов в гомогенатах биоптатов бронхов и альвеолярных макрофагах; (ii) количественные показатели субэпителиального коллагена и муцина эпителия дыхательных путей в биоптатах бронхов; (iii) количества всех и конкретных клеток в BAL.
Субъекты-люди и образцы
Субъекты с астмой (N=42) имели предшествующий поставленный врачом диагноз астмы, симптомы, согласующиеся с астмой, подтвержденные исследующим врачом, гиперреактивность дыхательных путей (определенную как снижение объема форсированного выдоха за первую секунду (FEV1), составляющее 20% или более, при ингаляции <8 мг/мл метахолина [PC20 метахолина]) и любое из следующих: 1) симптомы в течение 2 или более суток в неделю, 2) применение β-агониста в течение 2 или более суток в неделю, или 3) FEV1<85% от предсказанной величины. Они не принимали ингалируемые или пероральные кортикостероиды в течение 4 недель перед включением в исследование. У здоровых контролей (N=27) не было легочных заболеваний в анамнезе, и у них отсутствовала гиперреактивность дыхательных путей (PC20 метахолина >16 мг/мл). В некоторых исследованиях включали курящих в текущий момент людей без астмы (N=16). Критерии исключения для всех субъектов включали инфекцию верхних дыхательных путей в предшествующие 4 недели, обострение астмы в течение 6 недель и применение на текущий момент сальметерола, астемизола, недокромила натрия, кромогликата натрия, метилксантинов, монтелукаста или зафирлукаста. Субъектов подвергали исходной оценке исследующими врачами (включая спирометрию и тестирование с нагрузкой метахолином, как описано ранее [8]). Субъектов также подвергали инъекционной кожной пробе с аллергенами (ASPT) с панелью из 12 аллергенов воздуха, положительным контролем и отрицательным контролем (таблица 6).
Тридцать два из субъектов с астмой также были включены в двойное слепое рандомизированное контролируемое клиническое испытание ингалируемого флукатизона (500 мкг, два раза в сутки, N=19) или соответствующего плацебо (N=13) (идентификатор ClinicalTrials.gov: NCT00187499). Испытание было предназначено для определения эффекта ингалируемого стероида (флукатизона) на экспрессию генов в дыхательных путях и для сопоставления изменений в экспрессии генов с улучшениями функции легких. Субъектов с астмой в клиническом испытании подвергали бронхоскопии на исходном уровне и случайным образом распределяли для исследуемого лечения, а затем проводили повторную бронхоскопию через одну неделю после начала введения исследуемого лекарственного средства. Субъекты с астмой продолжали исследуемое лечение в течение всего 8 недель. Здоровых контрольных субъектов и курящих людей включали в одно из трех единовременных исследований, каждое из которых включало два посещения, первое для охарактеризации и второе для бронхоскопии через 1 неделю. У тридцати пяти субъектов бронхоскопия на исходном уровне была надлежащей, и у 32 имелась РНК, доступная из эпителиальных соскобов при обеих бронхоскопиях. Функцию легких измеряли (путем спирометрии) через 4 недели и 8 недель после исследуемого лечения, и конечную спирометрию проводили через одну неделю после его завершения. Способы бронхоскопии, эпителиального соскоба, бронхоальвеолярного лаважа, спирометрии, и обработки образцов были идентичны во всех исследованиях.
Бронхоальвеолярный лаваж (BAL) проводили инстилляцией 4 аликвот по 50 мл стерильного физиологического раствора либо в язычок, либо в правую среднюю долю, с извлечением отсасыванием. Подсчет числа клеток проводили с использованием гемоцитометра и раствора Тэрка (1% ледяная уксусная кислота и 0,01% генциановый фиолетовый в дистиллированной H2O). Затем на цитоцентрифугированных препаратах проводили подсчет в BAL конкретных типов клеток с использованием набора Shandon Kwik-Diff stain kit (Thermo Fisher Scientific, Waltham MA). Тридцать два субъекта с астмой также были включены в двойное слепое рандомизированное контролируемое клиническое испытание ингалируемого флукатизона (500 мг BID) или соответствующего плацебо. В дополнение к критериям включения, указанным выше, также требовалось, чтобы эти субъекты либо имели симптомы астмы в течение 2 или более суток в неделю, либо применяли β-агонист в течение 2 или более суток в неделю, либо имели FEV1<85% от предсказанного значения. Субъектов, проходивших в клиническом испытании исходное посещение и подвергавшихся бронхоскопии на исходном уровне, как описано выше, случайным образом разделяли для получения исследуемого лечения и подвергали повторной бронхоскопии через одну неделю. Затем они продолжали исследуемое лечение всего в течение 8 недель повторной оценкой с помощью спирометрии и тестирования с нагрузкой метахолином по расписанию. Все клинические испытания были одобрены University of California at San Francisco Committee on Human Research, от всех субъектов было получено письменное информированное согласие, и все исследования проводили в соответствии с принципами, указанными в декларации Хельсинки.
Анализы на микрочипах и морфометрия
Данные микрочипов для некурящих пациентов с мягкой-умеренной астмой и здоровых некурящих субъектов получали от предшествующего исследования, как описано [8]. Методологические детали и данные микрочипов также были доступны из общедоступной базы данных Gene Expression Omnibus, которая может быть доступна онлайн в National Center for Biotechnology Information, регистрационный номер GSE4302. Данные микрочипов в настоящем исследовании анализировали для определения того, отличается ли регуляция генов в группе астматиков. Также данные микрочипов анализировали для определения того, корегулируются ли со связанными с астмой индуцируемыми IL-13 генами другие гены. Двухстадийную ПЦР в реальном времени (qPCR) проводили, как описано ранее [45] с использованием праймеров и зондов, указанных в таблице 1 (т.е. мультиплексная ПЦР с последующей ПЦР в реальном времени на образованных из кДНК продуктах).
Морфометрические анализы проводили с использованием стереологии на основе изображений для 4-6 эндобронхиальных биоптатов от каждого субъекта, как описано ранее. Конкретно, анализ толщины ретикулярной базальной мембраны проводили в трихроматических 3-мкм срезах с использованием способа перпендикулярного сечения [31]. Содержание муцина в дыхательных путях измеряли в 3-мкм срезах, окрашенных альциановым синим/перйодной кислотой Шифа, с использованием способов подсчета с точечным и линейным пересечением [46].
Статистические способы
Предварительную обработку микрочипа проводили с использованием RMA с помощью программного обеспечения с открытым исходным кодом Bioconductor [47] в статистической среде R. Неконтролируемую иерархическую кластеризацию проводили с использованием евклидовой метрики с полным сцеплением. Все другие статистические анализы проводи с использованием пакета программ для статистического анализа JMP (SAS Institute, Cary, NC). Величины представлены в качестве среднего значения± стандартное отклонение или медианы (диапазона), если нет иных указаний. Корреляцию проводили с использованием коэффициента ранговой корреляции Спирмана. Для тестирования значимости PC20 и уровней IgE в сыворотке, данные подвергали логарифмической трансформации к нормальному распределению. В качестве статистически значимого было взято p<0,05 и после исходных сравнений трех групп с помощью ANOVA использовали коррекцию Сидака для множественных сравнений.

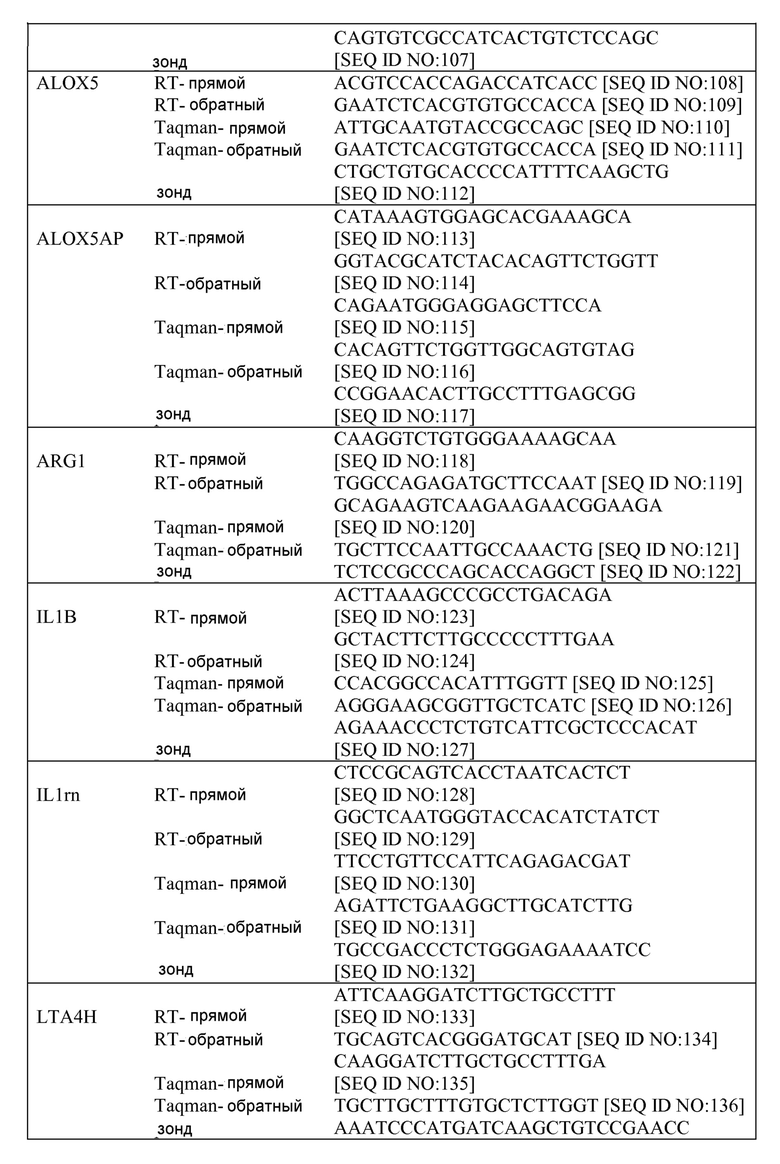

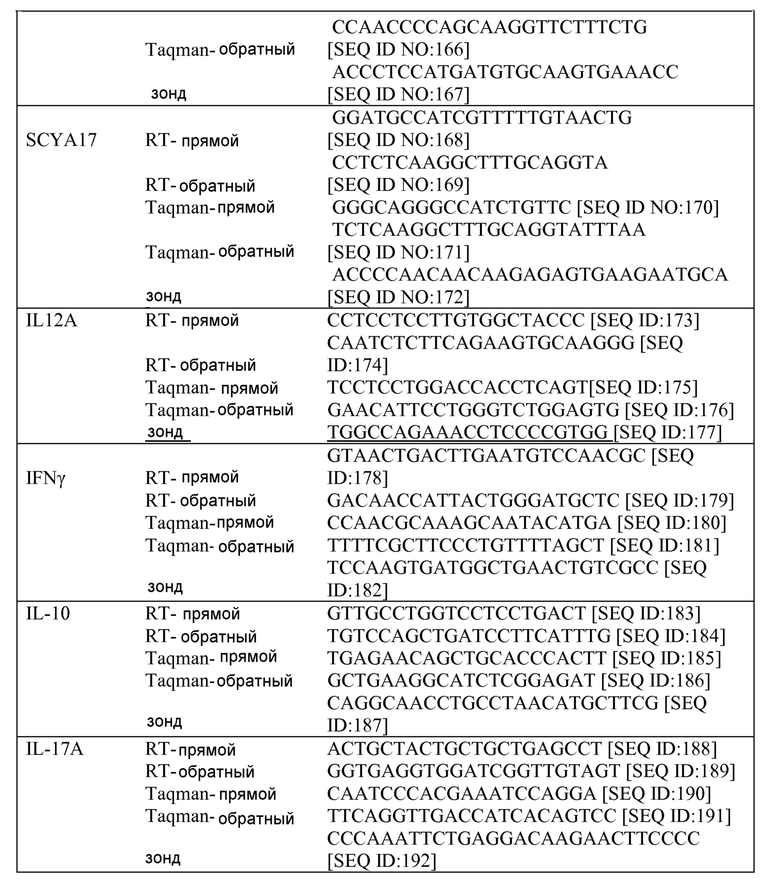
Для qPCR для периостина и CEACAM5, определяли относительное число копий для экспрессии периостина и CEACAM5 в образцах соскоба бронхиального эпителия на исходном уровне в соответствии с описанным ранее способом [45] и подвергали преобразованию log10. В качестве показателя ответа использовали профиль IL13 для 35 зондов, описанный в примере 9 (см. также фиг. 11). Все модели получали несколько раз с использованием платформы Fit Model в JMP 7.0. Ординальную логистическую регрессию проводили для предсказания ответа (статус IL-13 для 35 зондов), имеющего уровни (здоровый контроль; HC)<(IL-13 низкий)<(IL 13 высокий). Генерализованная предсказательная модель для вероятности для каждого уровня описана следующим образом:
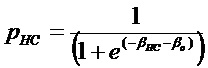
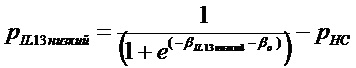
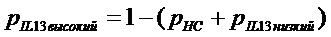
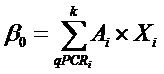
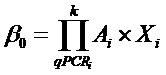
βх= оценка пересечения параметра x qPCR
Ординальную логистическую регрессию проводили для следующей модели: (статус IL-13 для 35 зондов)~(POSTN)+(CEACAM5). Значение p для полной модели, составляющее <0,0001, определяли из набора данных на основе многократной подгонки.
Отвечающие на IL-13 гены
Взаимосвязь между уровнями экспрессии периостина (также известного как специфический фактор остеобластов) (POSTN: 210809_s_at), CLCA1 (также известного как активируемый кальцием хлоридный канал, представитель 1 семейства) (CLCA1: 210107_at), и SERPINB2 (также известного как ингибитор серпиновой пептидазы, клад B (овальбумин), представитель 2) (SERPINB2: 204614_at) подтверждали с использованием критерия суммы рангов Вилкоксона. Уровень экспрессии POSTN использовали для разделения на категории исходных образцов от больных астмой. Использовали предел 800 единиц, получив в результате 21 образец от больных астмой на исходном уровне, классифицированный как "IL-13 низкий" (POSTN < 800 единиц) и остальные 21 образец как "IL13 высокий" (POSTN>800). Критерий суммы рангов Вилкоксона с последующим анализом уровня ложного открытия (величина q<0,05) [24] идентифицировали 35 зондов, дифференциально экспрессированных среди двух групп. Провели иерархическую кластеризацию с использованием этих зондов. Вследствие присутствия многих генов семейства цистатинов и серпинов в перечне дифференциально регулируемых зондов, были идентифицированы дополнительные зонды семейства цистатинов и серпинов и использованы в дополнительном кластерном анализе. Все статистические анализы проводили с использованием R. Кластерный анализ микрочипа проводили с использованием Cluster и визуализировали с использованием Java Treeview [25, 26].
Анализ анализируемых соединений в сыворотке
IgE в сыворотке определяли в клинических лабораториях UCSF или с помощью ELISA с использованием набора ELISA для IgE в сыворотке человека в соответствии с инструкциями изготовителя (Bethyl Laboratories). CEA в сыворотке измеряли с использованием набора ELISA для CEA в сыворотке человека в соответствии с инструкциями изготовителя (Alpco Diagnostics). Авторы настоящего изобретения разработали электрохемилюминесцентный анализ (ECLA) для измерения уровня периостина в сыворотке с использованием антител против периостина (R&D systems).
В кратком изложении, моноклональное антитело против периостина наносили на планшеты в количестве 1,5 микрограмм/мл в натрий-карбонатном буфере, pH 9,6 в течение ночи при 4°C. Планшеты блокировали в буфере для анализа (1X PBS pH 7,4, 0,35 M NaCl, 0,5% BSA, 0,05% Tween 20, 0,25% CHAPS, 5 мМ ЭДТА, 15 PPM Proclin)+3% BSA в течение 2 часов при комнатной температуре, затем промывали 4x TBST (Tris-буферный физиологический раствор+0,1% Tween-20). Сыворотку разбавляли 1:5 в буфере для анализа и инкубировали при встряхивании при комнатной температуре в течение 2 ч, затем промывали 4x с помощью TBST. Для установления стандартного диапазона использовали рекомбинантный периостин (R&D Systems). Биотинилированное поликлональное антитело против периостина человека (1,5 микрограмм/мл) (R&D Systems; биотинилированное in vitro стандартными способами, известными в данной области) и рутений-стрептавидин (0,75 микрограмм/мл) (Meso Scale Devices) добавляли в буфер для анализа+5% сыворотка козы и инкубировали в течение 90 минут при комнатной температуре. Добавляли буфер для считывания (Meso Scale Devices) и считывали электрохемилюминесценцию (Meso Scale Devices). Динамический диапазон составил 5-2000 нг/мл.
Пример 2 - Профиль IL-4/13 и подгруппы астматиков
Для определения того, отражают ли три индуцируемых IL-13 гена (ген периостина, CLCA1 и serpinB2) более широкий паттерн экспрессии генов в эпителии дыхательных путей астматиков, авторы настоящего изобретения исследовали, корегулируется ли их экспрессия на исходном уровне у отдельных субъектов среди 42 исследованных астматиков. При попарных сравнениях уровни экспрессии периостина, CLCA1 и serpinB2 в значительной степени коррелировали у отдельных астматиков. Более того, эти гены высоко экспрессировались у некоторых, но не у всех, из субъектов с астмой (фиг. 1A и 1B). Кроме того, уровни экспрессии этих трех генов высоко коррелировали у отдельных субъектов с астмой (фиг. 1B). Эти данные указывают на то, что определенные маркеры IL-13 сверхэкспрессируются у конкретной подгруппы пациентов с астмой. В дальнейших экспериментах авторы настоящего изобретения стремились идентифицировать дополнительные гены или маркеры, которые могут прямо или непрямо регулироваться IL-13, и они стремились охарактеризовать подгруппы пациентов с астмой на основе экспрессии маркеров IL-13.
Для идентификации других генов или маркеров, которые потенциально могут прямо или непрямо регулироваться IL-13 в эпителии дыхательных путей астматиков, авторы настоящего изобретения исследовали полный набор данных от микрочипов для 42 субъектов с астмой в отношении генов, экспрессия которых значимо коррелировала с экспрессией периостина. Авторы настоящего изобретения идентифицировали кластер из 653 зондов, экспрессия которых корегулировалась с периостином у отдельных субъектов на уровне ниже порогового значения q, равного 0,05. Неконтролируемая кластеризация всех субъектов, включая здоровых контролей и астматиков, на основе уровней экспрессии этих 653 зондов выявила два основных кластера: кластер с высокими уровнями экспрессии периостина и корегулируемых генов и кластер с низкими уровнями экспрессии периостина и корегулируемых генов. Центр этого кластера генов (фиг. 1C, правая панель) содержит подгруппу из 35 зондов, представляющих собой гены, представленные на фигуре 13, которые авторы настоящего изобретения относят в настоящем документе к "профилю IL-4/13", "генному профилю IL-4/13", "профилю IL-13", или "генному профилю IL-13". Как указано ранее, эти термины используют в настоящем документе в качестве синонимов. Кластер с высокой экспрессией периостина и корегулируемых генов включал 21 субъекта с астмой и не содержал здоровых контролей (фиг. 1C, правая панель, обозначен "профиль IL-4/13 высокий"), а кластер с низкой экспрессией периостина и корегулируемых генов включал остальных 21 астматиков (фиг. 1C, правая панель, обозначен "профиль IL-4/13 низкий"), вместе со всеми 27 здоровыми контролями (фиг. 1C, правая панель).
Кластер 1 ("профиль IL-4/13 высокий") характеризуется высокими уровнями экспрессии генов, соответствующих зондам для генов периостина, CST1, CST2, CST4, CCL26, CLCA1, CDH26, PRR4, serpinB2, SERPINB10, CEACAM5, iNOS, C2ORF32, PTGS1, P2RY14, RUNX2, SH3RF2, WLRW300, DNAJC12, ALOX15, GSN, RGS13, TGSAB1, PTSG1, FCER1B и CPA3, и он включает приблизительно половину астматиков в исследовании (N=23 из 42 астматиков) и один здоровый контроль всего из 27 здоровых контролей. Кластер 2 (здоровые контроли и "профиль IL-4/13 низкий") характеризуется низкими уровнями экспрессии генов, соответствующих указанным зондам, и он состоит из остальных 19 астматиков и 26/27 здоровых контролей. Зонды, соответствующие генам, преимущественно экспрессированным в тучных клетках, включая RGS13, TPSG1, TPSAB1, FCER1B, CPA3 и SLC18A2, указаны синим цветом в таблице 2, и зонды, соответствующие генам, преимущественно экспрессированным в эозинофилах, включая P2RY14 и ALOX15, указаны оранжевым цветом. Несмотря на то, что эпителиальные соскобы состояли преимущественно из эпителиальных клеток и бокаловидных клеток (среднее значение 97%, медиана 98%, минимум 91%), в соскобах астматиков из 1 кластера наблюдали небольшие количества тучных клеток и эозинофилов, и наличие генов тучных клеток и эозинофилов в профиле, вероятно, отражает эту инфильтрацию.
Для охарактеризации подгрупп субъектов с астмой на основе экспрессии маркеров IL-13, авторы настоящего изобретения провели неконтролируемую иерархическую кластеризацию всех 70 субъектов (42 астматиков и 27 здоровых контролей), на основе уровней экспрессии на микрочипах генов периостина, CLCA1 и serpinB2 (фигура 1D). В этом анализе, приблизительно половина субъектов с астмой (N=22) продемонстрировала постоянно высокие уровни экспрессии индуцируемых IL-13 генов, и они объединились в основную ветвь кластерной дендрограммы (кластер 1, подгрупп "IL-13 высокий"). Примечательно, что хотя гены периостина, CLCA1 и serpinB2 в значительной степени сверхэкспрессировались при сравнении всех 42 астматиков со всеми 27 здоровыми контролями [8], приблизительно половина астматиков, исследованных в этом исследовании (N=20), были неотличимы от здоровых контролей, исходя из экспрессии этих трех генов. Эта подгруппа астматиков (подгруппа "IL-13 низкий") и все здоровые контроли объединялись в группу второй главной ветви в дендрограмме (фигура 1D, кластер 2). Таким образом, иерархическая кластеризация на основе экспрессии генов в эпителии идентифицировала две различных подгруппы пациентов с астмой, обозначаемых в настоящем документе как подгруппа "IL-13 высокий" и подгруппа "IL-13 низкий".
Для подтверждения достоверности этих подгрупп пациентов с астмой, идентифицированных с использованием экспрессии индуцибельного маркера IL-13 в эпителиальных клетках, авторы настоящего изобретения измерили уровень экспрессии IL-13 и некоторых других Th2-цитокинов (т.е. IL-4 и IL-5) в биоптатах бронхов, полученных одновременно от 48 субъектов (14 здоровых контролей, 18 астматиков кластера 1, и 16 астматиков кластера 2). С использованием qPCR, авторы настоящего изобретения выявили, что экспрессия IL-13, IL-5 и IL-4 поддавалась детекции в гомогенатах биоптатов бронхов. Примечательно, что экспрессия IL-13 и IL-5, но не IL-4, была значительно более высокой (фиг. 1E, *, p<0,002) у астматиков кластера 1 по сравнению с астматиками кластера 2 или здоровыми контролями. Однако не было значимых отличий в экспрессии IL-4, IL-5 или IL-13 между астматиками в кластере 2 и здоровыми контролями (фиг. 1E). Кроме того, авторы настоящего изобретения выявили, что уровни экспрессии IL-13 и IL-5 в высокой степени коррелировали среди всех субъектов с астмой (коэффициент ранговой корреляции Спирмана ρ=0,58, p<0,0001; фиг. 1E). IL-4 имеет общий доминирующий каскад передачи сигнала с IL-13 и, как было показано, индуцирует экспрессию периостина [7, 9] и CLCA1 [12] аналогично IL-13. Поскольку были описаны повышенные уровни экспрессирующих IL-4 T-клеток в жидкости бронхоальвеолярного лаважа (BAL) [79] от астматиков, и авторы настоящего изобретения не исследовали конкретно экспрессию генов цитокинов в T-клетках BAL или уровни белков цитокинов в BAL или ткани бронхов в этом исследовании, авторы настоящего изобретения не могут определить вероятность того, что наблюдаемая индукция периостина, CLCA1, и serpinB2 частично является следствием IL-4, также как и IL-13. Исходя из данных, представленных в настоящем документе, авторы настоящего изобретения могут уверенно установить корреляцию между экспрессией IL-13 в бронхах и экспрессией периостина, CLCA1 и serpinB2 в эпителии. Таким образом, авторы настоящего изобретения используют термины "IL-4/13 высокий" и "IL-13 высокий" в качестве синонимов для обозначения астматиков кластера 1, и авторы настоящего изобретения используют термины "IL-4/13 низкий" и "IL-13 низкий" в качестве синонимов для обозначения астматиков кластера 2. Понятно, что когда используют термины "IL-13 высокий" и "IL-13 низкий", IL-4 и/или другие пока не идентифицированные факторы также могут частично вносить вклад в наблюдаемые паттерны экспрессии генов.
Пример 3 - Гены, составляющие профиль IL-4/13
В профиле IL-4/13 существует две основных группы генов: гены, экспрессируемые в эпителиальных и бокаловидных клетках, и гены, экспрессируемые в тучных клетках. Более 90% клеток в каждом образце бронхиального соскоба представляли собой клетки эпителия бронхов или бокаловидные клетки (среднее значение 97%, медиана 98%, минимум 91%). Уровни экспрессии зондов, соответствующих следующим генам эпителиальных и бокаловидных клеток, в наиболее значительной степени корегулировались с геном периостина: CST1, CST2, CCL26, CLCA1, PRR4, serpinB2, CEACAM5 и iNOS (таблица 2, указаны звездочками; экспрессия у субъектов с профилем "IL-4/13 высокий" >3 раз превышала экспрессию у субъектов с профилем "IL-4/13 низкий"). Ортолог CLCA1 мыши, mCLCA3 (также известный как gob-5) был ранее идентифицирован в качестве гена, ассоциированного с метаплазией бокаловидных клеток эпителия дыхательных путей и продукцией слизи; оба из них индуцируются Th2-цитокинами, включая IL-9 и IL-13 [12-14].
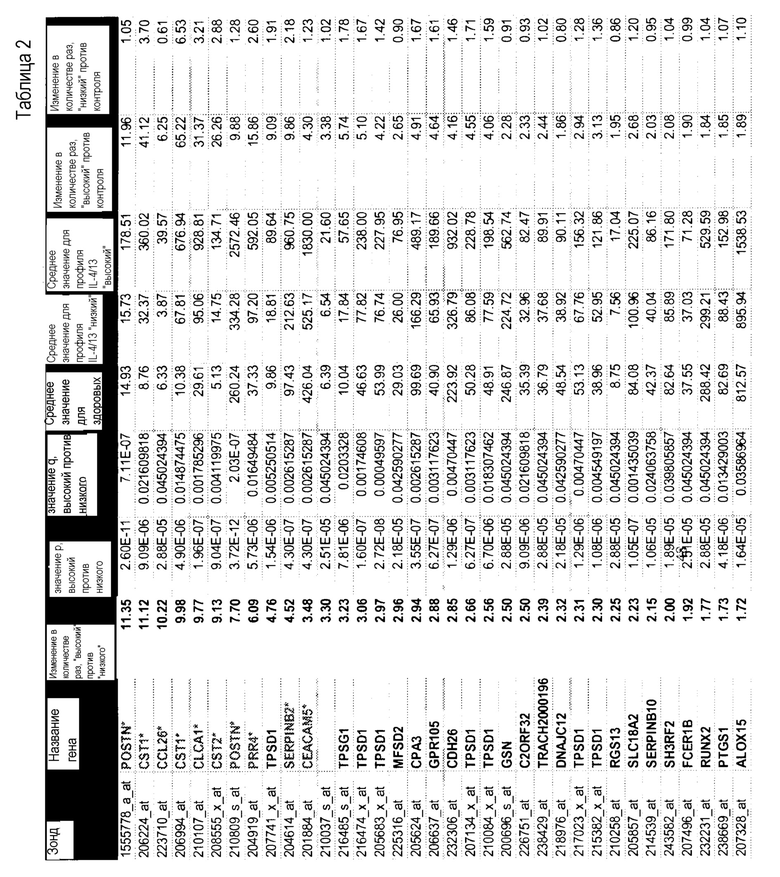
SerpinB2 является представителем большого семейства ингибиторов сериновых протеаз, кодируемых в генном кластере на хромосоме 18q21 (фиг. 2A, верх; фотография экрана с UCSC Genome Browser на http://genome.ucsc.edu). Уровни экспрессии серпинов B2 [8], B3 и B4 индуцируются в эпителиальных клетках при стимуляции рекомбинантным IL-4 и IL-13 [7, 15].
Цистатины (CST) 1 и 2 являются представителями большого семейства ингибиторов цистеиновых протеаз, кодируемых в генном кластере на хромосоме 20p11 (фиг. 2A, середина; фотография экрана с UCSC Genome Browser на http://genome.ucsc.edu). Несколько цистатинов экспрессируются в эпителии бронхов [16]; CST4 идентифицирован на повышенных уровнях в жидкости бронхоальвеолярного лаважа (BAL) астматиков [17]; сывороточный CST3 повышен у астматиков относительно здоровых контролей и его уровни снижаются при лечении ICS [18]. Поскольку семейства генов серпинов и CST колокализованы на хромосоме, авторы настоящего изобретения исследовали, существуют ли какие-либо дополнительные представители семейств генов серпинов и цистатинов, корегулируемые с уже идентифицированными представителями. Авторы настоящего изобретения провели неконтролируемую кластеризацию данных микрочипов, ограниченных генными семействами серпинов и цистатинов. Авторы настоящего изобретения выявили, что серпины B2, B4 и B10; и цистатины 1, 2 и 4 в значительной степени корегулировались, причем наиболее высокие уровни экспрессии встречались у астматиков, положительных по "профилю IL-4/13" (фиг. 2B).
PRR4 является представителем большого семейства белков, кодируемых в генном кластере на хромосоме 12p13 (фиг. 2A, низ; фотография экрана с UCSC Genome Browser на http://genome.ucsc.edu). Эти обогащенные пролином белки выявляются в слизистых секретах, включая слюну и слезы. Родственные, но неортологичные белки SPRR1a, 2a и 2b, были идентифицированы в эпителии бронхов в модели астмы на мышах и они индуцируются IL-13 [19, 20]. Обогащенные пролином белки из семейства PRR/PRB были идентифицированы в секретах бронхов [21] и их экспрессия была зарегистрирована в эпителии бронхов [16]. Среди семейства PRR/PRB, PRR4 и PRB4 в значительной степени активировались у астматиков с высокой экспрессией генного профиля IL-4/13 (фиг. 2C, слева и в середине).
CCL26 (эотаксин-3) представляет собой индуцируемый IL-4 и IL-13 хемокин в эпителии дыхательных путей астматиков.
CEACAM5 кодирует гликопротеин клеточной поверхности, встречающийся во многих эпителиальных тканях и на повышенных уровнях в сыворотке. CEACAM5 (карциноэмбриональный антиген; CEA) представляет собой документированный системный биомаркер злокачественных опухолей эпителия и метастатического заболевания. Повышенные уровни CEA были описаны у подгруппы астматиков, причем особенно высокие уровни в сыворотке наблюдали у астматиков с закупоркой мукоидом [22]. CEACAM5 в значительной степени активирован в эпителии дыхательных путей астматиков с профиле IL-4/13 "высокий" по сравнению с эпителием дыхательных путей у астматиков с профилем IL-4/13 "низкий" и здоровых контролей (фиг. 2C, справа), что указывает на то, что уровни CEA в сыворотке можно использовать для того, чтобы отличить эти два субфенотипа астматиков.
Индуцибельная синтаза оксида азота (iNOS) ассоциирована с воспалением дыхательных путей и индуцируется IL-13 в первичных культурах эпителиальных клеток бронхов человека [23]. Измерение выдыхаемого оксида азота (eNO), продукта ферментативной активности iNOS, широко используют при диагностике и мониторинге астмы.
Пример 4 - Тучные клетки
Несмотря на то, что соскобы дыхательных путей, используемые в этом исследовании, содержали преимущественно эпителиальные и бокаловидные клетки, во многих из образцов существовали небольшие, но существенные проценты инфильтрирующих лейкоцитов. Гены, экспрессия которых является специфичной для тучных клеток, включая гены триптазы (TPSD1, TPSG1), карбопептидазы A3 (CPA3), и Fc-эпсилон-RI-бета, в значительной степени коррелировали с генным профилем IL-4/13 (таблица 2 и таблица 4, гены тучных клеток, отмеченные двойными звездочками в таблице 4). С учетом значительной роли оседлых тканевых тучных клеток в аллергическом заболевании и недавнего наблюдения, что наличие экспрессирующих IL-13 тучных клеток в образцах эндобронхиальных биоптатов астматиков положительно коррелирует с поддающимися детекции уровнями IL-13 в мокроте [6], высокая корреляция между специфичными для тучных клеток генами и профилем IL-4/13 указывает на то, что: 1) тучные клетки могут быть существенным источником IL-13 в эпителии дыхательных путей и 2) инфильтрация тучных клеток в эпителий дыхательных путей может быть уникальной чертой подгруппы астматиков с профилем IL-4/13 "высокий".
Пример 5 - Комбинации, которые предсказывают профиль IL-4/13
Уровни экспрессии отдельных генов в профиле IL-4/13 могут предсказывать статус профиля IL-4/13 у отдельных субъектов с переменной точностью; однако комбинации этих генов можно использовать для отнесения отдельных субъектов к категории профиля IL-4/13 "высокий" или "низкий" с повышенной чувствительностью и специфичностью.
Пример 6 - Эффект стероидов
Стандартом лечения при бронхиальной астме, которая не является хорошо контролируемой симптоматической терапией (т.е. бета-адренергическими агонистами), являются ингалируемые кортикостероиды (ICS). У астматиков с астмой от мягкой до умеренной с повышенными уровнями IL-13 в дыхательных путях [6] и у пациентов с эозинофильным эхофагитом с повышенными уровнями экспрессии IL-13 в ткани пищевода [11], лечение ICS по существу снижает уровень IL-13 и индуцируемых IL-13 генов в пораженных тканях. В эпителии дыхательных путей астматиков после лечения в течение одной недели посредством ICS и в культивированных эпителиальных клетках бронхов авторы настоящего изобретения показали, что лечение кортикостероидами по существу снижает индуцируемые IL-13 уровни экспрессии периостина, serpinB2 и CLCA1 [8]. Дальнейшее изучение генов, приведенных в таблице 2, показало, что в исследовании авторов настоящего изобретения у 19 субъектов, которые получали лечение ICS в течение одной недели перед второй бронхоскопией, большинство генов профиля IL-4/13 в значительной степени подавлялось посредством лечения ICS в эпителии дыхательных путей бронхов астматиков (периостин, показанный в качестве примера, фиг. 3A). Это подавление может быть результатом опосредуемого ICS снижения уровней IL-13, опосредуемого ICS снижения экспрессии гена-мишени, или комбинации этих двух случаев. Однако два гена в профиле IL-4/13, PRR4 (фиг. 3B) и RUNX2 (фиг. 3C), по существу не подавлялись у отдельных субъектов после лечения ICS в течение одной недели. Это указывает на то, что PRR4 и RUNX2 могут быть нечувствительными к стероидам маркерами профиля IL-4/13 в эпителии дыхательных путей астматиков. Другая возможность состоит в том, что PRR4 и RUNX2, только непрямо регулируются IL-4 и/или IL-13; например, поскольку PRR4 выявляется во многих секретах, он может представлять собой ген, специфичный для бокаловидных клеток. Поскольку дифференцировка бокаловидных клеток из эпителиальных клеток индуцируется IL-13, опосредуемое ICS ингибирование IL-13 и зависимых от IL-13 процессов может по существу не влиять на число бокаловидных клеток после только 7 суток лечения, однако после длительного лечения ICS, можно ожидать, что количества бокаловидных клеток (и, таким образом, экспрессия PRR4 в эндобронхиальных соскобах) снизятся. У тяжелых астматиков, рефрактерных к лечению ICS, было выявлено, что сходная часть субъектов (приблизительно 40%) имеет поддающиеся детекции уровни IL-13 в мокроте, по сравнению с уровнями, наблюдаемыми у астматиков с мягкой астмой, не подвергавшихся лечению ICS [6], что соответствует части субъектов с профилем IL-4/13, наблюдаемой в этом исследовании. Это наблюдение указывает на то, что, хотя профиль IL-4/13 в значительной степени подавляется лечением ICS у отвечающих на ICS астматиков с мягкой-умеренной астмой, исследованных в настоящем исследовании, он все еще может присутствовать у астматиков с тяжелой устойчивой к стероидам астмой.
Пример 7 - Взаимосвязь профиля IL-4/13 с клиническими признаками и другими биомаркерами
- Демографические данные
Эозинофильная астма, определяемая по повышенным уровням эозинофилов в дыхательных путях, ассоциирована с атопией и возникает приблизительно с равной распространенностью среди мужчин и женщин, в то время как неэозинофильный фенотип, определяемый по относительному отсутствию эозинофилов в дыхательных путях и ассоциированный с отсутствием атопии, демонстрирует выраженное преобладание у женщин [1]. Среди субъектов, классифицированных согласно генному профилю IL-4/13 в эпителии дыхательных путей, женщинами были 10/21 (48%) субъектов с профилем IL-4/13 "высокий", в то время как женщинами были 15/21 (71%) субъектов с профилем IL-4/13 "низкий" (таблица 3). Не было значимого отклонения в описанной самими субъектами этнической принадлежности в группах профилей IL-4/13 "низкий" и "высокий".
Распределение полов для профиля IL-4/13, N (%)
- FEV1 и способность отвечать на метахолин
В то время как отклонение полов между группами IL-4/13 "низкий" и "высокий" указывает на то, что наблюдаемые паттерны экспрессии генов в эпителии дыхательных путей астматиков отражают стабильные лежащие в их основе фенотипы, возможно, что наблюдаемые паттерны экспрессии генов отражают только тяжесть или активность заболевания во время бронхоскопии. Для определения того, коррелирует ли профиль IL-4/13 с тяжестью астмы, авторы настоящего изобретения сравнили объем форсированного выдоха за одну секунду (FEV1, в качестве процента от предсказанной величины, исходя из массы пациента, определенной при скрининговом посещении за одну неделю до бронхоскопии) между группами и выявили, что, хотя обе группы профилей IL-4/13 "высокий" и "низкий" имели значительно более низкую FEV1, чем здоровые контроли, не было статистически значимых отличий между группами (см. фиг. 5A), хотя в группе профиля IL-4/13 "высокий" было больше субъектов, которых можно было классифицировать как "умеренные" (т.е. FEV1 60-80% от предсказанной величины) чем группе профиля "низкий". Минимальная концентрация метахолина в мг/мл, требуемая для индукции снижения FEV1 20% (PC20, измеренная при скрининговом посещении перед бронхоскопией) является показателем гиперотвечаемости бронхов. Этот показатель является показателем гиперреактивности бронхов (BHR). Как группа профиля IL-4/13 "высокий" и так и группа профиля IL-4/13 "низкий" имели значимо более низкие значения PC20, чем здоровые контроли; хотя в группе профиля IL-4/13 "высокий" существовала тенденция в сторону более низких значений PC20, чем в группе профиля "низкий", это отличие не достигало статистической значимости (см. фиг. 5C).
- IgE и эозинофилы (периферические и дыхательных путей)
Для определения того, можно ли предсказать статус профиля IL-4/13 отдельного субъекта по стандартным показателям атопии, авторы настоящего исследовали уровни IgE в сыворотке (международные единицы на миллилитр; 1 МЕ=2,4 нг), количество эозинофилов в периферической крови (абсолютное число эозинофилов ×10^9 на литр крови), и процент эозинофилов в жидкости бронхоальвеолярного лаважа (BAL) (процент эозинофилов относительно общего количество нечешуйчатых клеток в жидкости бронхоальвеолярного лаважа) с использованием стандартных клинических лабораторных тестов, полученного во время бронхоскопии. Когда субъектов стратифицировали по статусу профиля IL-4/13, были выявлены значимые отличия в уровне IgE в сыворотке (см. фиг. 6B), количестве эозинофилов в периферической крови (см. фиг. 6C), и проценте эозинофилов в BAL (см. фиг. 6D), со значимо более высокими значениями для каждого анализируемого образца, наблюдаемыми в группе профиля IL-4/13 "высокий" относительно группы профиля "низкий". Взятые по отдельности, ни уровень IgE, ни количество эозинофилов в периферической крови не предсказывают статус профиля IL-4/13 в эпителии дыхательных путей у любого отдельного субъекта с высокой чувствительностью и специфичностью одновременно. Однако среди отдельных астматиков, уровень IgE и количество эозинофилов в периферической крови слабо, но значимо коррелировали (rho=0,44, p=3,4×10-3). При рассмотрении совместно, эмпирически установленные значения предела как 100 МЕ/мл IgE, так и 0,14×109/л, предсказывают статус профиля IL-4/13 в эпителии дыхательных путей отдельных субъектов с высокой чувствительностью и специфичностью (фиг. 4; 18/21 коррекция по профилю IL-4/13 "низкий", так и по профилю IL-4/13 "высокий"; чувствительность=86%, специфичность=86%).















Пример 8 - Взаимосвязь субфенотипов астмы "IL-13 высокий" и "IL-13 низкий" и клинических признаков
Субъектов с астмой далее анализировали в отношении дополнительных демографических показателей и клинических признаков, как описано в примере 7. Результаты представлены в таблице 5 и на фиг. 5 и 6. Хотя субъектов с субфенотипом астмы "IL-13 высокий" нельзя было отличить от субъектов с субфенотипом астмы "IL-13 низкий", исходя из демографических показателей, функции легких, или способности отвечать на бронходилятатор (дельта-FEV1 с альбутеролом) (таблица 5, фигуры 5 A-B), эти группы значимо отличались в отношении степени гиперреактивности дыхательных путей (AHR, PC20 метахолина, определенная как минимальная концентрация метахолина, требуемая для индукции 20%-снижения экспираторного потока воздуха, фигура 5C). Это отличие в AHR было очевидным, несмотря на критерии включения, которые требовали, чтобы все астматики имели значительную AHR (все астматики <8 мг/мл, все здоровые контроли >20 мг/мл).
Характеристики субъектов по фенотипу астмы
(22-64)
(0,06-7,3)
(0,05-1,9)
(19-1194)
(32-2627)
Для определения того, коррелирует ли субфенотип IL-13 отдельного субъекта с показателями аллергического воспаления, авторы настоящего изобретения исследовали инъекционные кожные пробы (SPT) с панелью из 12 аллергенов воздуха (таблица 6), уровни IgE в сыворотке, количество эозинофилов в периферической крови, и процент эозинофилов в жидкости бронхоальвеолярного лаважа (BAL). Результаты представлены на фигурах 6A-D и 7A-B. Как субфенотип астмы IL-13 "высокий", так и субфенотип астмы IL-13 "низкий" имели повышенную чувствительность в SPT на аллергены воздуха по сравнению со здоровыми контролями (фиг. 6A), хотя субфенотип астмы IL-13 "низкий" имел тенденцию к меньшему количеству положительных кожных проб, чем субфенотип астмы IL-13 "высокий" и к менее частой сенсибилизации аллергенами, такими как аллергены собаки и клещей домашней пыли (фиг. 7A). Субъекты с субфенотипом астмы IL-13 "высокий" имели более высокие уровни IgE в сыворотке и более высокое количество эозинофилов в периферической крови, чем субъекты с субфенотипом астмы IL-13 "низкий", хотя субфенотип астмы IL-13 "низкий" отличался от здоровых контролей с точки зрения этих признаков аллергического воспаления (фиг. 6B-C). Кроме того, субъекты с субфенотипом астмы IL-13 "высокий" имели повышенное количество эозинофилов в легком согласно оценке в BAL (фиг. 6D), в то время как астматики с IL-13 "низкий" не отличались от здоровых контролей по проценту эозинофилов в BAL. Эти данные демонстрируют увеличение AHR, уровней IgE, и эозинофильного воспаления у субъектов с субфенотипом астмы IL-13 "высокий", но чувствительность к аллергенам воздуха в SPT не ограничивалась этой подгруппой. Таким образом, вероятно, что у субъектов с субфенотипом астмы IL-13 "низкий" действуют альтернативные не-Th2 механизмы сенсибилизации аллергенами воздуха.
Панель аллергенов в инъекционной кожной пробе
Johnson
Для определения того, является ли субфенотип астмы IL-13 "высокий" длительным или временным проявлением запускаемого Th2 воспаления вследствие недавнего воздействия аллергена, авторы настоящего изобретения определили патологические изменения в биоптатах бронхов от тех же субъектов. Авторы настоящего изобретения и другие исследователи ранее продемонстрировали, что астма ассоциирована с патологическими изменениями, известными как ремоделирование дыхательных путей и отражающими либо продолжительное воспаление, либо эффекты повреждения и репарации с течением времени [28, 29]. Два специфических исхода ремоделирования при астме представляют собой фиброз дыхательных путей, проявляющийся как утолщение субэпителиальной ретикулярной базальной мембраны (RBM) [30, 31], и повышенное накопление муцина в эпителии дыхательных путей [32]. Авторы настоящего изобретения открыли, что толщина RBM была большей у субъектов с субфенотипом астмы IL-13 "высокий", чем субъектов субфенотипом астмы IL-13 "низкий" или здоровых контролей, и что толщина RBM была нормальной в случае субфенотипа астмы IL-13 "низкий" (фиг. 6E). Кроме того, несмотря на то, что авторы настоящего изобретения наблюдали тенденцию в сторону повышенного накопления эпителиального муцина у субъектов с астмой с обоими фенотипами, это повышение было значимым только у субъектов с субфенотипом астмы IL-13 "высокий" (фиг. 8A). Несмотря на то, что эти различия в общих накоплениях муцина были небольшими, qPCR выявила выраженное отличие в уровнях экспрессии основных гель-образующих муцинов в эпителиальных клетках дыхательных путей в случае субфенотипа астмы IL-13 "высокий" по сравнению с как с субфенотипом астмы IL-13 "низкий", так и со здоровыми контролями (фиг. 8B-D). Конкретно, субфенотип астмы IL-13 "высокий" отличался от субфенотипа астмы IL-13 "низкий" и здоровых контролей индукцией экспрессии MUC5AC и MUC2 и подавлением экспрессии MUC5B. Это изменение экспрессии конкретных генов муцина в случае субфенотипа астмы IL-13 "высокий" является наиболее очевидным из соотношения экспрессии MUC5AC и MUC5B (фиг. 6F). Без связи с теорией, авторы настоящего изобретения предполагают, что сосуществующие индукция и подавление конкретных гель-образующих муцинов могут объяснить относительно небольшое повышение накопления эпителиального муцина в случае субфенотипа астмы IL-13 "высокий" по сравнению с субфенотипом астмы IL-13 "низкий" и здоровыми контролями. Взятые вместе, эти данные указывают на то, что субфенотип астмы IL-13 "высокий" ассоциирован с изменениями при ремоделировании дыхательных путей, которые идентифицируют этот субфенотип как продолжающийся в течение длительного времени. Эти результаты также демонстрируют важность каскада IL-13 для ремоделирования дыхательных путей у человека.
Альвеолярные макрофаги могут модулировать аллергическое воспаление дыхательных путей при астме в качестве источника IL-13 [54] и лейкотриенов или эйкозаноидных липидов [55, 56] или через "альтернативную активацию" под влиянием IL-13 [57]. Для определения того, проявляют ли альвеолярные макрофаги от субъектов с астмой "IL-13 высокий" какие-либо из этих признаков, авторы настоящего изобретения определили экспрессию соответствующих генов с использованием qPCR у 14 субъектов с астмой и 15 здоровых контролей (таблица 7). Авторы настоящего изобретения не выявили доказательств индукции Th2-цитокинов или альтернативных маркеров активации при астмы в общем или в подгруппе "IL-13 высокий" в частности. Уровни экспрессии IL-13 были ниже предела детекции (порог циклов >40) у 26 из 29 субъектов, и IL-4 был ниже предела детекции у 20 из 29 субъектов (нет различий между тремя группами по какому-либо из цитокинов, все p>0,35). Все другие гены находились в пределах детекции для образцов. В этих анализах авторы настоящего изобретения выявили повышенную экспрессию 15-липоксигеназы при астме "IL-13 высокий" (фигура 10, таблица 8), что согласуется с предшествующими данными о повышенных продуктах 15-липоксигеназы в дыхательных путях при тяжелой эозинофильной астме [56]. Авторы настоящего изобретения также выявили повышение экспрессии TNFα, которое было ограничено подгруппой "IL-13 высокий" (фигура 10, таблица 8).
Только подгруппа астматиков проявляет улучшение функции легких при лечении ингалируемыми кортикостероидами (ICS) [33]. Для идентификации маркеров экспрессии генов, обеспечивающих способность к ответу на кортикостероиды, авторы настоящего изобретения измеряли FEV1 у подгруппы их субъектов с астмой в течение 8-недельного рандомизированного контролируемого испытания ингалируемого флукатизона или плацебо, как описано ранее [8]. Когда авторы настоящего изобретения повторно анализировали эти данные при стратификации субъектов по субфенотипу IL-13, авторы настоящего изобретения выявили, что улучшения FEV1 были ограничены субфенотипом IL-13 "высокий". Конкретно, субъекты с субфенотипом астмы IL-13 "высокий", которых лечили ингалируемым флукатизоном, имели значимые улучшения FEV1 как через 4, так и через 8 недель, по сравнению с субъектами, которым вводили плацебо, в то время как субъекты с субфенотипом астмы IL-13 "низкий" не имели улучшений (фиг. 9A). Эти улучшения FEV1 в группе IL-13 "высокий" утрачивались через одну неделю после отмены лекарственного средства. Ни в одной группе не было значимых изменений в FEV1 в ответ на плацебо в какой-либо момент времени (данные не представлены, N=5 "IL-13 высокий", N=6 "IL-13 низкий"). Как описано ранее [8], авторы настоящего изобретения проводили вторую бронхоскопию через одну неделю после начала лечения и анализировали экспрессию генов в эпителии бронхов с помощью микрочипа, как и на исходном уровне. В повторном анализе этих данных, при стратификации субъектов по субфенотипу IL-13, субъекты с астмой "IL-13 высокий" продолжали проявлять выраженный субфенотип IL-13 после введения плацебо в течение одной недели, что демонстрирует кратковременную стабильность этого субфенотипа в отсутствии лечения. Однако после введения в течение одной недели флукатизона, субъекты с астмой "IL-13 высокий" объединялись в кластер с субъектами, которые имели "IL-13 низкий" на исходном уровне, независимо от лечения (фиг. 9B). Таким образом, фенотипическая классификация астмы на основе профиля IL-13, описанного в настоящем документе, предсказывает ответ на ICS. Эти данные указывают на то, что общая польза лечения астмы с помощью ICS определяется субфенотипом "IL-13 высокий".
Результаты авторов настоящего изобретения обеспечивают новое понимание молекулярных механизмов, которые лежат в основе клинической гетерогенности при астме. Базовые исследования ранее установили IL-13 и родственные Th2-цитокины в качестве центральных регуляторов аллергического воспаления и многих патофизиологических изменений, ассоциированных с астмой [35, 36]. Здесь, с использованием профилирования экспрессии генов, авторы настоящего изобретения идентифицировали субфенотип "IL-13 высокий" у пациентов с астмой. С использованием строгих клинических критериев и тестирования с нагрузкой метахолином, авторы настоящего изобретения выявили, что этот субфенотип включает только ~50%, у которых диагностирована астма. Этот субфенотип "IL-13 высокий" также проявлял повышенные уровни экспрессии IL-5 и показал определенные отличительные клинические характеристики, включая повышенную гиперреактивность дыхательных путей, повышенные уровни IgE в сыворотке и эозинофильное воспаление, субэпителиальный фиброз, и измененную экспрессию гель-образующих муцинов по сравнению с субфенотипом "IL-13 низкий" и здоровыми контролями.
Работа авторов настоящего изобретения оспаривает определенные современные концепции патогенеза астмы, показывая, что генный профиль для запускаемого IL-13 воспаления в эпителиальных клетках дыхательных путей является выраженным только у половины астматиков; таким образом, у оставшейся половины должны действовать запускаемые не IL-13 механизмы. Данные, рассмотренные в настоящем документе, ведут авторов настоящего изобретения к предположению, что астма может быть подразделена на различные молекулярные субфенотипы, такие как субфенотипы "IL-13 высокий" и "IL-13 низкий", указанные в настоящем документе. Авторы настоящего изобретения подтвердили схему классификации IL-13 высокий/IL-13 низкий с помощью подтверждающих анализов экспрессии генов в биоптатов бронхов, анализа воспроизводимости при повторном исследовании, и исчерпывающей охарактеризации различных клинических, воспалительных, патологических и связанных с лечением характеристик этих двух молекулярных субфенотипов астмы. Эти данные обеспечивают механистическую основу для развивающегося клинического наблюдения, что астма представляет собой комплексное и гетерогенное заболевание [58].
Молекулярное фенотипирование астмы на основе Th2-воспаления имеет важное терапевтическое значение. Во-первых, обструкция дыхательных путей в случае субфенотипа "IL-13 высокий" улучшается с помощью ингалируемых стероидов, в то время как субфенотип "IL-13 низкий" демонстрирует от небольшого улучшения до его отсутствия. Th2-маркеры, которые идентифицировали авторы настоящего изобретения, можно использовать для направления развития клинических тестов способности отвечать на стероиды путем предоставления маркеров-заместителей отвечающего на стероиды фенотипа. Во-вторых, блокада IL-13 и родственных Th2-цитокинов находится в активной клинической разработке в качестве терапевтической стратегии при астме [34]. Данные авторов настоящего изобретения указывают на то, что клинический ответ на эти способы лечения может быть ограничен конкретным субфенотипом пациентов с астмой "IL-13 высокий". Таким образом, маркеры этого молекулярного фенотипа имеют прямое применение в клинических испытаниях.
Предшествующие исследования с использованием индуцируемого анализа мокроты указывают на то, что "эозинофильная астма" представляет собой отличающийся клеточный фенотип астмы, но молекулярные механизмы, лежащие в основе этого клеточного фенотипа не определены. Данные авторов настоящего изобретения указывают на то, что запускаемое IL-13 воспаление представляет собой молекулярный механизм, лежащий в основе "эозинофильной астмы" [37] вследствие эозинофилии дыхательных путей, которую авторы настоящего изобретения продемонстрировали в астме "IL-13 высокий". Кроме того, авторы настоящего изобретения продемонстрировали, что как "эозинофильная астма", так и астма "IL-13 высокий" характеризуются субэпителиальным фиброзом [38, 39], продукцией ALOX 15 альвеолярными макрофагами [55] и ответами функции легких на ингалируемые кортикостероиды [40, 41]. В дополнение к этим известным признакам эозинофильной астмы, авторы настоящего изобретения идентифицировали дополнительные клинические признаки астмы "IL-13 высокий", включая измененную экспрессию генов муцина в дыхательных путях и индукцию TNFα, медиатора, который не считается Th2-цитокином, но который ранее ассоциировали с тяжелой астмой [59]. Авторы настоящего изобретения предположили, что эти признаки также встречаются при эозинофильной астме. Кроме того, вероятно, что IL-5 является главным участником эозинофилии дыхательных путей и системной эозинофилии, которую авторы настоящего изобретения наблюдали при астме "IL-13 высокий", поскольку авторы настоящего изобретения выявили, что экспрессия IL-5 в значительной степени корегулируется с экспрессией IL-13 (фиг. 1E). IL-5 представляет собой главный стимул дифференцировки, привлечения, активации и выживания эозинофилов [60], но IL-13 может сильно индуцировать экспрессию хемоаттрактантов эозинофилов, таких как CCL11, CCL22 и CCL26 в дыхательных путях [61] и может, таким образом, действовать кооперативно с IL-5 для обеспечения инфильтрации, активации и выживания эозинофилов в дыхательных путях. Остаточная активность IL-13, таким образом, может объяснить неполное истощение эозинофилов в ткани, наблюдаемое в клинических испытаниях блокады IL-5 при астме [62, 63].
Кроме того, эти данные показывают, что значительный процент пациентов с астмой имели фенотип "IL-13 низкий", который проявляет такие клинические признаки астмы, как обструкция дыхательных путей, гиперреактивность дыхательных путей и обратимость после введения бронходилятатора, несмотря на слабо выраженное Th2-запускаемое воспаление. Причины астмы "IL-13 низкий" остаются неясными, однако возможные причины включают нейтрофильное воспаление [37], запускаемое IL-17 воспаление [42], внутренние дефекты барьерной функции [43] и хроническую субклиническую инфекцию атипичными внутриклеточными бактериями [44].
Гены, используемые в альвеолярных макрофагах при qPCR
Экспрессия генов альвеолярных макрофагов при qPCR
N=15
N=5
N=9
2343659
2235805
2211337
949034
2179904
2679369
163171
37416
2754206
2851849
1397415
3209466
2864216
2812301
1944551
1191877
1647800
Пример 9 - Взаимосвязь субфенотипов астмы "IL-13 высокий" и "IL-13 низкий" с сывороточными белковыми биомаркерами
Дальнейший анализ на микрочипах привел к идентификации авторами настоящего изобретения из набора генов и зондов, приведенных в таблице 4, группы из 35 зондов, соответствующей 28 генам, экспрессия которых корегулировалась с периостином у отдельных субъектов ниже значения q порогового уровня ложного открытия (FDR), равного 0,05. Эти гены и зонды и ассоциированные с ними данные представлены в таблице 9. Иерархический кластерный анализ всех субъектов, включая здоровых контролей и астматиков, на основе уровней экспрессии этих зондов подтвердил и далее определил два основных кластера, описанных выше, представляющих собой (1) кластер с высокими уровнями экспрессии периостина и корегулируемых генов и (2) кластер с низкими уровнями экспрессии периостина и корегулируемых генов (фигура 11). Гены тучных клеток включают RGS13, TPSG1, TPSAB1, FCER1B, CPA3 и SLC18A2. Гены эозинофилов включают P2RY14 и ALOX15.
Кластер с высокой экспрессией периостина и корегулируемых генов включал 23 субъекта с астмой и 1 здоровый контроль (фигура 11, кластер 1, указанный красным цветом), в то время как кластер с низкой экспрессией периостина и корегулируемых генов включал остальных 19 астматиков вместе с 26 здоровыми контролями (фигура 11, кластер 2, указанный зеленым цветом). В примере 8 авторы настоящего изобретения описали кластеризацию субъектов в этом наборе данных на основе определенных с помощью микрочипов уровней экспрессии трех из этих зондов: 210809_s_at (периостин), 210107_at (CLCA1) и 204614_at (serpinB2). Профиль из трех зондов, описанный в примере 8, высоко коррелирует с этим полным профилем из 35 зондов, отличаясь для семи астматиков и одного здорового контроля (отличающиеся сигналы указаны на фигуре 11 посредством *).
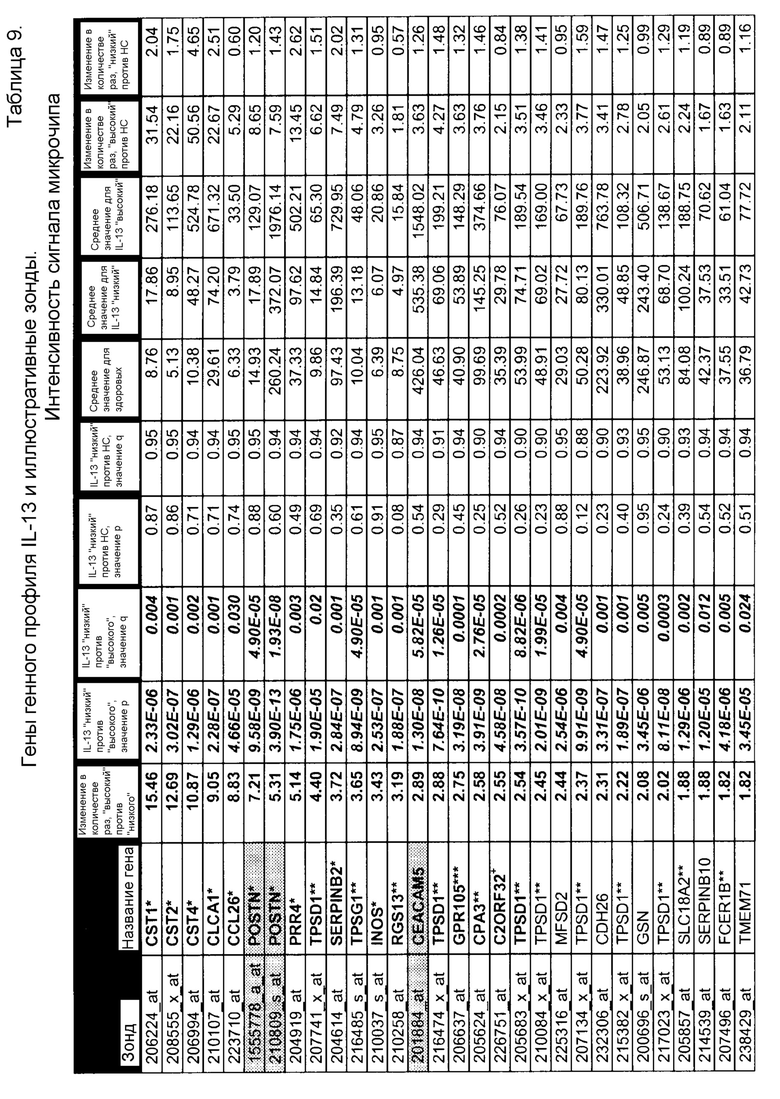

С использованием профиля IL-13 из трех генов (ген периостина, CLCA1 и serpinB2), авторы настоящего изобретения показали в примере 8, что системные маркеры аллергического воспаления, включая уровни IgE в сыворотке и эозинофилов в периферической крови, были значительно повышены у астматиков с субфенотипом "IL-13 высокий" относительно астматиков с субфенотипом "IL-13 низкий". Однако существовало значительное перекрывание между группами астматиков для каждого из этих показателей, взятых по отдельности. Кроме того, ни IgE в сыворотке, ни уровни эозинофилов в периферической крови отдельно, не являются неинавзивным показателем для предсказания профиля IL-13 и ассоциированного с ним субфенотипа "IL-13 высокий" или "IL-13 низкий" с высокой чувствительностью и специфичностью одновременно.
Для определения того, может ли совмещение уровней IgE и эозинофилов в периферической крови предсказать паттерны воспаления дыхательных путей с большей точностью, чем каждый из этих показателей отдельно, авторы настоящего изобретения оценили уровень IgE в сыворотке и количество эозинофилов в периферической крови вместе против статуса профиля IL-13 в дыхательных путях. Авторы настоящего изобретения выявили, что среди 42 астматиков, уровень IgE в сыворотке и количество эозинофилов в периферической крови коррелировали, хотя и слабо (фигура 4; данные показаны для профиля IL-4/13; сходные результаты были получены для профиля IL-13 [см. таблицу 10]). Для профиля IL-13, все из астматиков с "IL-13 высоким" имели количество эозинофилов более 0,14×109/л, но многие из астматиков с "IL-13 низким" имели более низкие количества эозинофилов. Все кроме двух астматиков с "IL-13 высоким" имели уровни IgE в сыворотке более 100 МЕ/мл, но многие из астматиков c "IL-13 низким" не имели их. Два показателя из (1) IgE в сыворотке ≥100 МЕ/мл и (2) количество эозинофилов ≥0,14×109/л вместе приводили к повышенной чувствительности и специфичности профиля IL-13 в дыхательных путях (таблица 10). Таким образом, объединение двух обычно используемых показателей периферической крови для аллергического воспаления может быть эффективным неинвазивным биомаркером для запускаемого IL-13 воспаления дыхательных путей.
Чувствительность, специфичность, положительные и отрицательные предсказательные значения показателей IgE и эозинофилов в периферической крови для профиля IL-13.
IgE в сыворотке>100 МЕ/мл
эозинофилы≥0,14×109/л
IgE>100 ME/мл и эозинофилы≥0,14×109/л
Для идентификации дополнительных системных (неинвазивных) биомаркеров-кандидатов профиля IL-13 в эпителии бронхов, авторы настоящего изобретения исследовали профиль генов, кодирующих внеклеточные или секретируемые белки, которые могут быть выявлены в периферической крови. Тремя представляющими особый интерес кандидатами были CCL26, периостин и CEACAM5. Поскольку CCL26 ранее был описан как индуцируемый Th2-цитокинами хемокин в эпителии бронхов [71], авторы настоящего изобретения сфокусировались на охарактеризации периостина и CEACAM5, которые ранее не были описаны в качестве сывороточных биомаркеров Th2-воспаления. CEACAM5 кодирует карциноэмбриональный антиген (CEA), который представляет собой часто используемый прогностический сывороточный биомаркер в происходящих из эпителия злокачественных опухолях. Периостин также описан в ограниченном количестве исследований как сывороточный биомаркер некоторых злокачественных опухолей и, что интересно, он поддавался детекции на уровне в диапазоне 10s-100s нг/мл сыворотки у большинства субъектов, что является привлекательной характеристикой сывороточного маркера для легкой детекции в иммунологических анализах.
Как представлено на фигуре 12A-B, каждый из периостина и CEACAM5 являются хорошими отдельными представителями профиля IL-13, проявляющими значимо более высокую экспрессию у астматиков с "IL-13 высоким" чем у астматиков с "IL-13 низким" или здоровых контролей. У отдельных астматиков существовала сильная корреляция между уровнями экспрессии на микрочипе периостина и CEACAM5 (фигура 12C). Для подтверждения этих паттернов экспрессии генов и для определения того, можно ли экспрессию периостина и CEACAM5 использовать в алгоритме для того, чтобы отличить астматиков "IL-13 высокий" от астматиков "IL-13 низкий" и здоровых контролей, авторы настоящего изобретения анализировали уровни экспрессии двух генов посредством qPCR в тех же образцах соскобов эпителия бронхов, что использовали для анализа на микрочипах. Существовала высокая степень соответствия между значениями с микрочипа и qPCR у отдельных субъектов (не представлено). Авторы настоящего изобретения использовали анализ ординальной логистической регрессии для получения предсказательной модели для полученного от микрочипа статуса IL-13 от 35 зондов с использованием значений qPCR для периостина и CEACAM5. Модельное предсказательное значение было высоко значимым (p<0,0001) и каждая из оценок параметров периостина и CEACAM5 имела значимый эффект в этой модели (p<0,02 для CEACAM5; p<0,0001 для периостина). Анализ кривой операционных характеристик приемника (ROC) продемонстрировал превосходную предсказывающую способность для здоровых контролей и очень высокую чувствительность и специфичность для астмы с "IL-13 высоким" и "IL-13 низким" (фигура 12D). Взятые вместе, эти данные показывают, что уровни экспрессии в эпителии бронхов периостина и CEACAM5 являются хорошими заместителями для всего профиля IL-13.
Для определения того, являются ли повышенные уровни растворимых белков периостина и CEA поддающимися детекции в периферической крови, авторы настоящего изобретения исследовали периостин и CEA в сыворотке от 100 астматиков и 48 здоровых контролей с использованием иммунологических анализов. Кроме того, авторы настоящего изобретения измерили IgE и YKL-40, сывороточный маркер, который, как описано ранее, повышен у некоторых астматиков [72], в этих же сыворотках. Авторы настоящего изобретения выявили значимо повышенные уровни IgE, периостина, CEA и YKL-40 у астматиков относительно здоровых контролей (фигура 13 A-D). Однако в этих случаях между группами существовало значительное перекрывание в уровнях каждого биомаркера в сыворотке. Как показано в примере 8, введение ингалируемого кортикостероида (ICS) снижает экспрессию в эпителии бронхов периостина у астматиков, которые имеют повышенный уровень периостина на исходном уровне (см. также [8]). Среди 100 астматиков, сыворотку которых исследовали авторы настоящего изобретения, 51 получали ингалируемые кортикостероиды (ICS) и 49 не получали их. При сравнении астматиков без ICS и астматиков с ICS, субъекты, которых лечили ICS, имели значимо более низкие срединные уровни в сыворотке IgE и CEA, и они показали тенденцию к более низким уровням периостина, в то время как уровни YKL-40 были неизмененными (фигура 13E-H). Тем не менее, астматики с ICS имели более высокие срединные уровни в сыворотке IgE, периостина и CEA, чем здоровые контроли (таблица 13). Как представлено на фигуре 4 и в таблице 10, 21/23 астматиков, положительных по профилю IL-13 в эпителии бронхов ("IL-13 высокий") имели уровни IgE в сыворотке более 100 МЕ/мл, хотя часть астматиков с профилем "IL-13 низкий" также имели повышенные IgE. Авторы настоящего изобретения выявили, что уровни периостина в сыворотке имели тенденцию к повышению, и уровни CEA были значимо более высокими у астматиков с IgE≥100 МЕ/мл (N=68), чем у астматиков с IgE<100 МЕ/мл (N=32; фигура 13I-J). Однако уровни YKL-40 в сыворотке были значимо более низкими в группе с высоким уровнем IgE (фигура 13K). Поскольку уровни экспрессии в дыхательных путях периостина и CEACAM5 в высокой степени коррелировали у астматиков "IL-13 высокий", авторы настоящего изобретения исследовали корреляцию между периостином в сыворотке и CEA у всех астматиков (фигура 13L). Авторы настоящего изобретения выявили, что уровни периостина и CEA в сыворотке в значительной степени коррелировали друг с другом в популяции астматиков, и у астматиков без ICS или астматиков с IgE≥100 МЕ/мл, но не у здоровых контролей, астматиков с ICS, или астматиков с IgE<100 МЕ/мл (таблица 11). Взятые вместе, эти данные указывают на то, что периостин и CEA могут представлять собой сывороточные биомаркеры индуцированного IL-13 генного профиля эпителия бронхов у астматиков.
Корреляция между сывороточными биомаркерами.
В профиле IL-13 авторы настоящего изобретения наблюдали несколько функциональных групп множества генов, включая гены, кодирующие ингибиторы протеазы, и гены, экспрессируемые в тучных клетках и эозинофилах, которые могут отражать инфильтрацию в эпителий бронхов и/или анатомическую локализацию этих клеток в эпителии бронхов. Более чем 90% клеток в каждом образце бронхиального соскоба представляли собой эпителиальные клетки бронхов или бокаловидные клетки (среднее значение 97%, медиана 98%, минимум 91%), но на микрочипах выявлялись очень небольшие количества инфильтрирующих "загрязняющих" клеток со специфичными для клеток паттернами экспрессии генов. Специфичные для тучных клеток гены включали гены триптаз (TPSAB1 [TPSD1] и TPSG1), CPA3, FCER1B, RGS13 и SLC18A2 [73, 74]. Также с генами тучных клеток тесно кластеризовался ген CNRIP1 (C2ORF32) взаимодействующей c рецептором каннабиноидов GTP-азы. С учетом хорошо установленной роли миметиков каннабиноидов в регуляции функции тучных клеток [75], вероятно, что CNRIP1 также представляет собой ген, специфичный для тучных клеток. С учетом значительной роли оседлых тканевых тучных клеток в аллергическом заболевании и недавнего наблюдения, что присутствие экспрессирующих IL-13 тучных клеток в образцах эндобронхиальных биоптатов у астматиков положительно коррелировало с поддающимися детекции уровнями IL-13 в мокроте [6], высокая корреляция между специфичными для тучных клеток генами и профилем IL-13 указывает на то, что: 1) тучные клетки могут быть существенным источником IL-13 в эпителии дыхательных путей и 2) инфильтрация тучных клеток в эпителий дыхательных путей может быть уникальным признаком подгруппы астматиков с профилем "IL-13 высокий". Специфичные для эозинофилов гены включают P2RY14 (GPR105) и ALOX15, хотя в примере 8 авторы настоящего изобретения описали экспрессию ALOX15 в альвеолярных макрофагах от астматиков.
В профиле IL-13 присутствовало множество зондов, соответствующих ингибиторам сериновых и цистеиновых протеаз, включая серпины B2 и B10, и цистатины (CST) 1, 2 и 4. SerpinB2 является представителем большого семейства ингибиторов сериновых протеаз, кодируемых кластером генов на хромосоме 18q21. Уровни экспрессии серпинов B2 [8], B3 и B4 индуцируются в эпителиальных клетках дыхательных путей при стимуляции рекомбинантным IL-4 и IL-13 [7, 15]. Цистатины (CST) 1, 2 и 4 являются представителями большого семейства ингибиторов цистеиновых протеаз, кодируемых кластером генов на хромосоме 20p11. В эпителии бронхов экспрессируется несколько цистатинов [16]; CST4 был идентифицирован на повышенных уровнях в жидкости бронхоальвеолярного лаважа (BAL) у астматиков [17]; сывороточный CST3 повышен у астматиков относительно здоровых контролей, и его уровни снижаются при лечении посредством ICS [18]. Поскольку семейства генов серпинов и CST колокализованы на хромосоме, авторы настоящего изобретения исследовали, корегулируются ли какие-либо дополнительные представители семейств генов серпинов и цистатинов с уже идентифицированными представителями. Авторы настоящего изобретения провели неконтролируемую кластеризацию данных микрочипов, ограниченных генными семействами серпинов и цистатинов. Авторы настоящего изобретения выявили, что из более чем 40 генов ингибиторов протеаз, представленных на чипе, только серпины B2, B4 и B10; и цистатины 1, 2 и 4 в значительной степени корегулировались, причем наиболее высокие уровни экспрессии встречались у астматиков, имеющих профиль "IL-13 высокий" (фиг. 2B и таблица 12). Поскольку многие аллергены воздуха обладают активностью протеаз и активируемые протеазами рецепторы (PAR) ассоциированы с активацией аллергических воспалительных каскадов [76], активация ингибиторов протеаз Th2-цитокины может отражать компенсаторный ответ на содержащие протеазу аллергены воздуха.
Ортолог CLCA1 мыши, mCLCA3 (также известный как gob-5) был ранее идентифицирован в качестве гена, ассоциированного с метаплазией бокаловидных клеток эпителия дыхательных путей и продукцией слизи; оба из них индуцируются Th2-цитокинами, включая IL-9 и IL-13 [12-14]. PRR4 является представителем большого семейства генов, кодируемых в кластере на хромосоме 12p13. Эти гены кодируют обогащенные пролином белки, которые выявляются в слизистых секретах, включая слюну и слезы. Родственные, но неортологичные белки SPRR1a, 2a и 2b, были идентифицированы в эпителии бронхов в модели астмы на мышах и они индуцируются IL-13 [19, 20]. Обогащенные пролином белки из семейства PRR/PRB были идентифицированы в секретах бронхов [21] и их экспрессия была зарегистрирована в эпителии бронхов [16]. CCL26 (эотаксин-3) представляет собой документированный индуцируемый IL-4 и IL-13 привлекающий эозинофилы хемокин в эпителии дыхательных путей астматиков. CDH26 представляет собой кадгерин-подобную молекулу с неизвестной функцией, которую недавно идентифицировали в анализе на микрочипах эозинофильного эзофагита [11]. В этом исследовании было идентифицировано несколько дополнительных генов, перекрывающихся с профилем IL-13 эпителия бронхов, выявленным авторами настоящего изобретения, включая ген периостина, SerpinB4, и CCL26 [11]. Поскольку CDH26 корегулируется с эотаксинами и сверхэкспрессируется при заболеваниях, характеризующихся эозинофильным воспалением, можно предположить, что CDH26 участвует в эозинофильной инфильтрации в слизистые ткани. Индуцибельная синтаза оксида азота (iNOS) ассоциирована с воспалением дыхательных путей и индуцируется IL-13 в первичных культурах эпителиальных клеток бронхов [23]. Измерение выдыхаемого оксида азота (eNO) широко используют в диагностике и мониторинге астмы. Учитывая все это, многие из генов, описанные в настоящем документе в качестве компонентов профиля IL-13, в высокой степени согласуются моделями in vitro и моделями на животных Th2-воспаления и возможно участвуют в запускаемой Th2 патологии при астме у человека.
Уровни сывороточных биомаркеров.
Значения p представляют собой ранговую сумму Вилкоксона
* 99/100 астматиков имели значения CEA ≤7,5 нг/мл
CEACAM5 кодирует гликопротеин клеточной поверхности, встречающийся во многих эпителиальных тканях и на повышенных уровнях в сыворотке. CEACAM5 (карциноэмбриональный антиген; CEA) представляет собой документированный системный биомаркер злокачественных опухолей эпителия и метастатического заболевания. Повышенные уровни CEA были описаны у подгруппы астматиков, причем особенно высокие уровни в сыворотке наблюдали у астматиков с закупоркой мукоидом [75]. Интересно, что хотя верхняя граница нормы для сывороточного CEA находится в диапазоне 2,5-3 нг/мл, нижняя граница для предположения о злокачественной опухоли составляет 10 нг/мл. В анализах авторов настоящего изобретения, они выявили, что свыше 95% здоровых контролей имели уровни CEA ниже 3 нг/мл, в то время как 1/3 астматиков имели уровни CEA между 3 и 7,5 нг/мл, и из них, широкое большинство имели уровни IgE в сыворотке более 100 МЕ/мл. Это указывает на то, что у астматиков может присутствовать жесткое окно для детекции CEA с запускаемым Th2 воспалением дыхательных путей. Периостин описан в качестве индуцируемого IL-4 и IL-13 гена в дыхательных путях астматиков [7-9, 77], активируемого в злокачественных опухолях, происходящих из эпителия, который может быть ассоциирован с инвазивностью и изменением внеклеточного матрикса [64-67], и уровни белка которого в сыворотке поддаются детекции и повышены при некоторых злокачественных опухолях [68-70]. Поскольку он может участвовать в инфильтрации эозинофилов в ткани при эозинофильном эзофагите [11, 77], периостин может быть важным фактором при эозинофильных заболеваниях, таких как Th2-запускаемая астма, и он может быть их биомаркером. Стандартом лечения при бронхиальной астме, которая не является хорошо контролируемой симптоматической терапией (т.е. β-агонистами), являются ингалируемые кортикостероиды (ICS). У астматиков с астмой от мягкой до умеренной с повышенными уровнями IL-13 в дыхательных путях [6] и у пациентов с эозинофильным эхофагитом с повышенными уровнями экспрессии IL-13 в ткани пищевода [11], лечение ICS по существу снижает уровень IL-13 и индуцируемых IL-13 генов в пораженных тканях. В эпителии дыхательных путей астматиков после лечения в течение одной недели посредством ICS и в культивированных эпителиальных клетках бронхов, авторы настоящего изобретения показали, что лечение кортикостероидами по существу снижает индуцируемые IL-13 уровни экспрессии периостина, serpinB2 и CLCA1 [8]. Дальнейшее изучение генов, приведенных в таблице 9, показало, что у 19 субъектов в исследовании авторов настоящего изобретения, которые получали лечение ICS в течение одной недели перед второй бронхоскопией, большинство генов профиля IL-4/13 в значительной степени подавлялось посредством лечения ICS в эпителии дыхательных путей бронхов астматиков. Это подавление может быть результатом опосредуемого ICS снижения уровней IL-13, опосредуемого ICS снижения экспрессии гена-мишени, или комбинации этих двух случаев. У тяжелых астматиков, рефрактерных к лечению ICS, было выявлено, что сходная часть субъектов (приблизительно 40%) имеет поддающиеся детекции уровни IL-13 в мокроте, по сравнению с уровнями, наблюдаемые у астматиков с мягкой астмой, не подвергавшихся лечению ICS [6], что соответствует части субъектов с профилем IL-13, наблюдаемой в этом исследовании. Это наблюдение указывает на то, что, хотя профиль IL-13 в значительной степени подавляется лечением ICS у отвечающих на ICS астматиков с мягкой-умеренной астмой, исследованных в настоящем исследовании, он все еще может присутствовать у астматиков с тяжелой устойчивой к стероидам астмой. Сходные наблюдения были описаны для эозинофильного воспаления в биоптатах бронхов [78] и персистенции экспрессирующих IL-4 и IL-5 клеток в BAL [79] рефрактерных к стероидам астматиков. В настоящее время существует большое количество биологических лекарственных средств, находящихся в процессе клинической разработки, направленных против IL-13 или родственных факторов при Th2-воспалении [50, 80], включая, но не ограничиваясь этим, факторы, описанные в настоящем документе. Данные авторов настоящего изобретения указывают на то, что только у части астматиков с астмой от мягкой до умеренной, не принимавших стероиды, этот каскад может быть активным, и с учетом их чувствительности к лечению ICS, вероятно, что у меньшей части рефрактерных к стероидам астматиков с астмой от умеренной до тяжелой этот каска является активным. Таким образом, биомаркеры, которые идентифицируют астматиков, вероятно имеющих запускаемое IL-13 воспаление в их дыхательных путях, могут способствовать идентификации и селекции субъектов, наиболее вероятно отвечающих на эти экспериментальные способы нацеленной терапии.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБЫ ДИАГНОСТИКИ И ЛЕЧЕНИЯ, СВЯЗАННЫЕ С ИНГИБИРОВАНИЕМ TH2 | 2011 |
|
RU2578468C2 |
| СПОСОБЫ ДИАГНОСТИКИ И ЛЕЧЕНИЯ, СВЯЗАННЫЕ С ИНГИБИРОВАНИЕМ ТН2 | 2011 |
|
RU2737245C2 |
| СПОСОБЫ ДИАГНОСТИКИ И ЛЕЧЕНИЯ, СВЯЗАННЫЕ С ИНГИБИРОВАНИЕМ TH2 | 2020 |
|
RU2832013C2 |
| ТЕРАПЕВТИЧЕСКИЕ И ДИАГНОСТИЧЕСКИЕ СПОСОБЫ ДЛЯ ВОСПАЛИТЕЛЬНЫХ ЗАБОЛЕВАНИЙ, ОПОСРЕДОВАННЫХ ТУЧНЫМИ КЛЕТКАМИ | 2019 |
|
RU2795180C2 |
| СПОСОБЫ ЛЕЧЕНИЯ ИЛИ ПРЕДУПРЕЖДЕНИЯ АСТМЫ ПОСРЕДСТВОМ ВВЕДЕНИЯ АНТАГОНИСТА IL-33 | 2020 |
|
RU2838119C2 |
| СПОСОБЫ ЛЕЧЕНИЯ ИЛИ ПРЕДУПРЕЖДЕНИЯ АСТМЫ ПОСРЕДСТВОМ ВВЕДЕНИЯ АНТАГОНИСТА IL-4R | 2015 |
|
RU2713406C2 |
| СПОСОБЫ ЛЕЧЕНИЯ ИЛИ ПРЕДУПРЕЖДЕНИЯ АСТМЫ ПОСРЕДСТВОМ ВВЕДЕНИЯ АНТАГОНИСТА IL-4R | 2020 |
|
RU2832774C2 |
| СПОСОБЫ ЛЕЧЕНИЯ ИЛИ ПРЕДУПРЕЖДЕНИЯ АСТМЫ ПУТЕМ ВВЕДЕНИЯ АНТАГОНИСТА IL-4R | 2018 |
|
RU2793745C2 |
| СПОСОБЫ ЛЕЧЕНИЯ ИЛИ ПРЕДОТВРАЩЕНИЯ АСТМЫ ПОСРЕДСТВОМ ВВЕДЕНИЯ АНТАГОНИСТА IL-4R | 2013 |
|
RU2690675C2 |
| СПОСОБ ДИАГНОСТИКИ МОЛЕКУЛЯРНОГО ФЕНОТИПА ПАЦИЕНТА, СТРАДАЮЩЕГО ЗАБОЛЕВАНИЕМ, СОПРОВОЖДАЮЩИМСЯ ХРОНИЧЕСКИМИ ВОСПАЛЕНИЯМИ | 2013 |
|
RU2672595C2 |


























































































































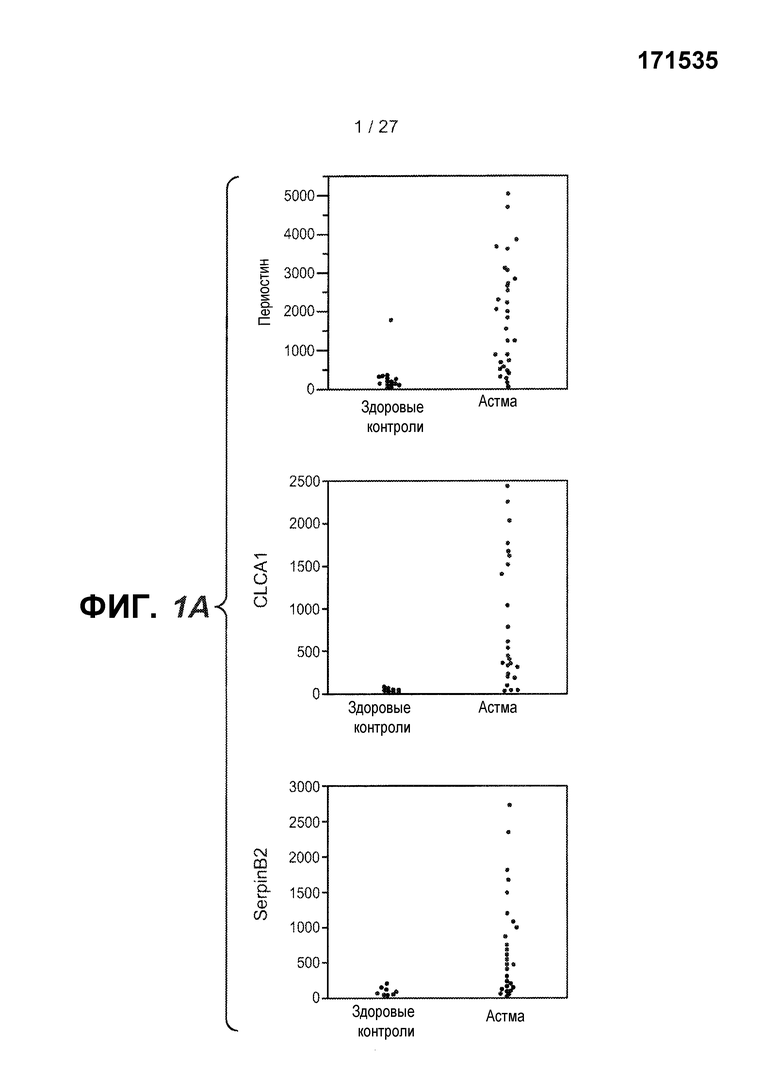
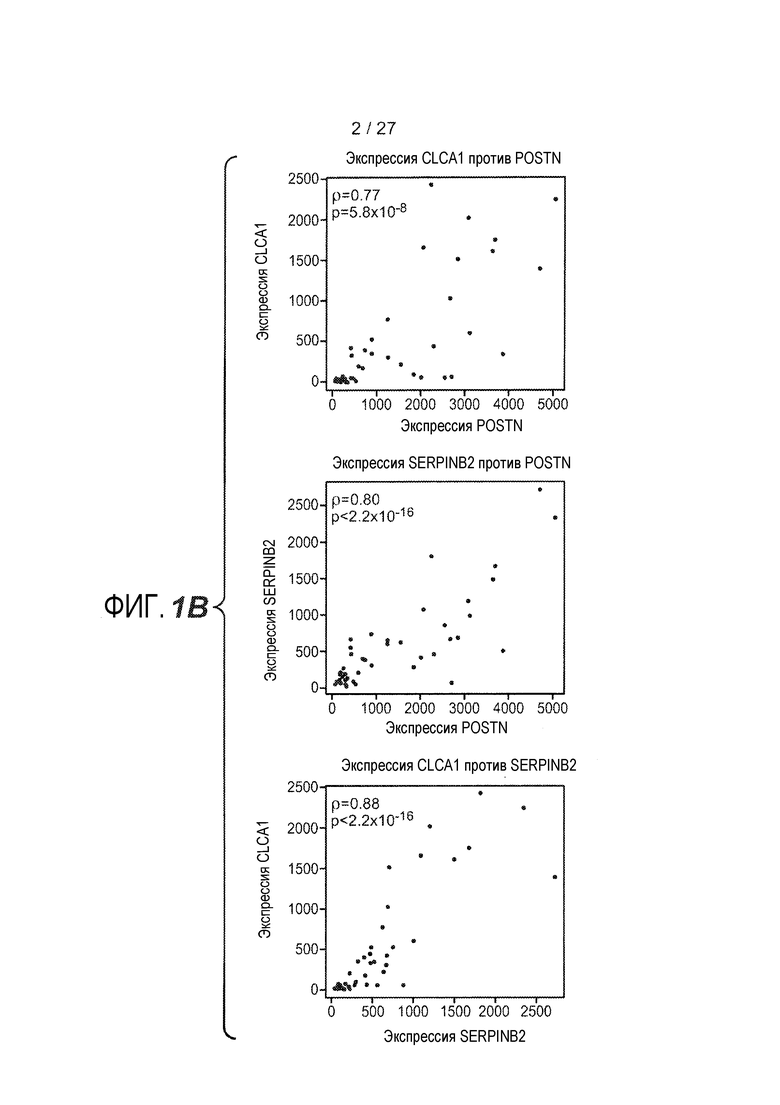
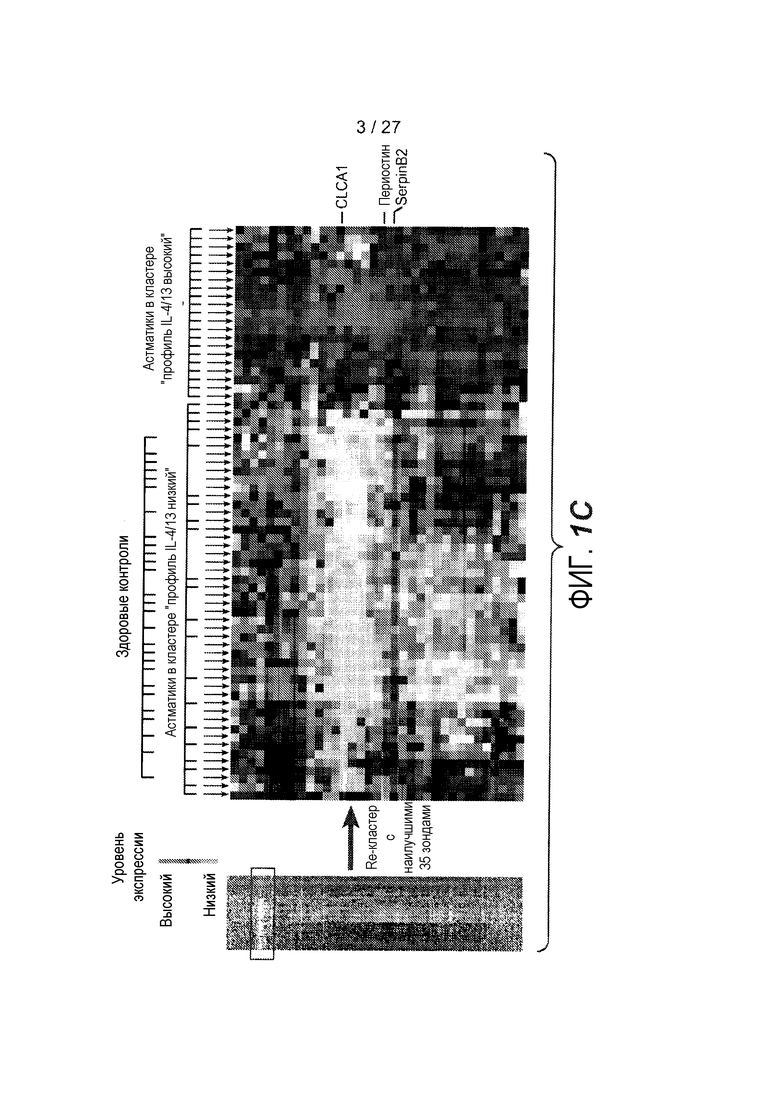
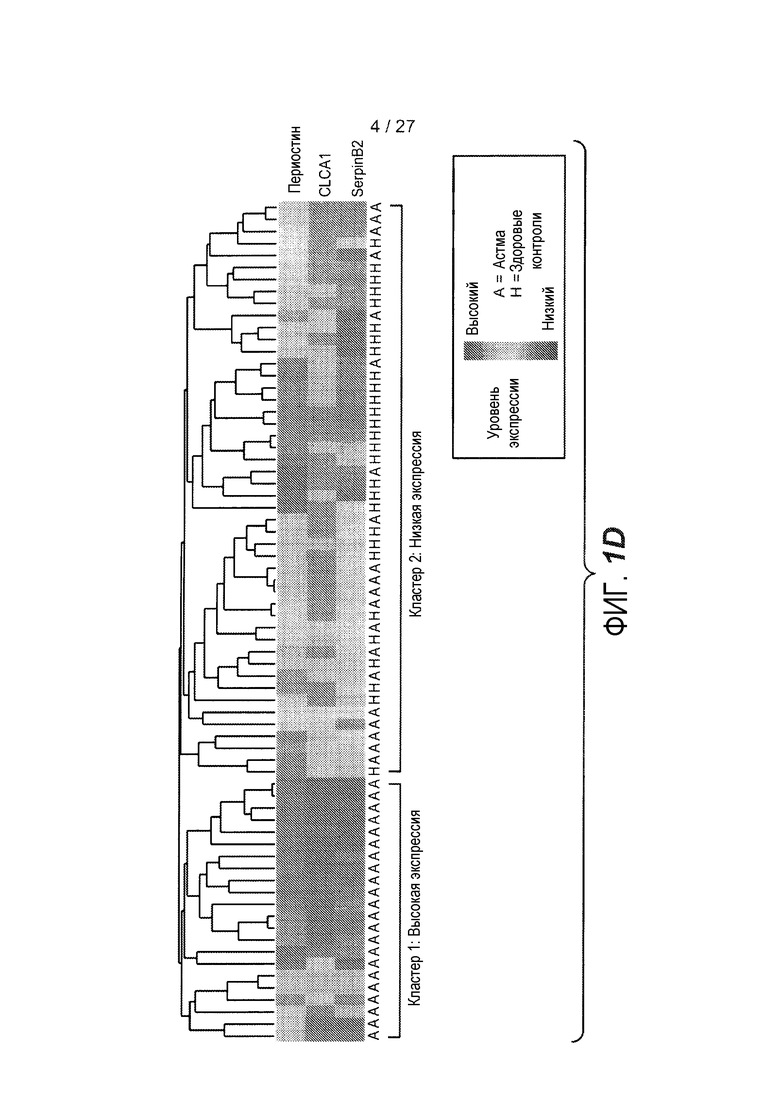
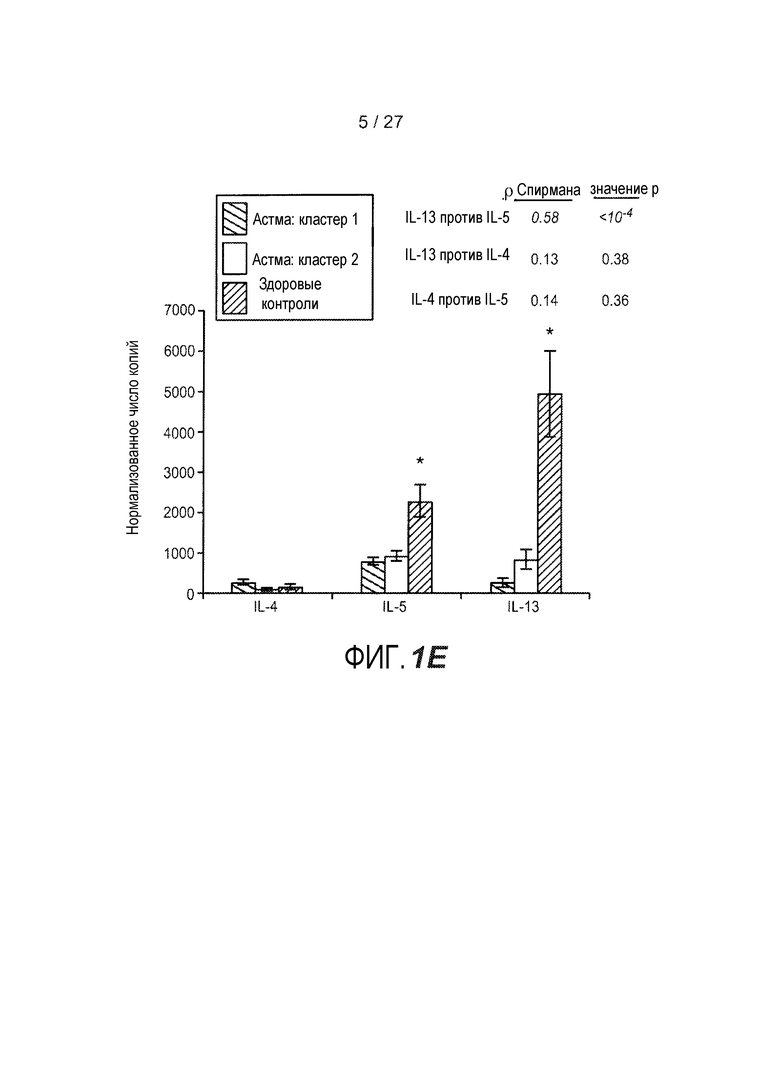
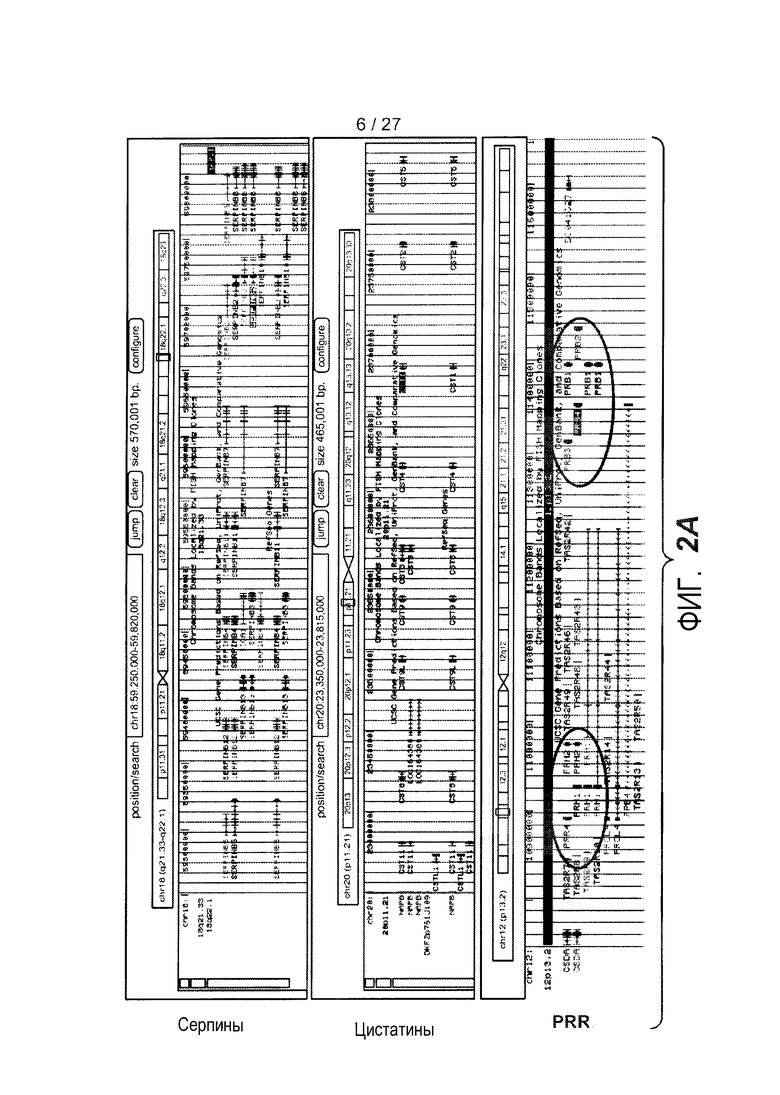
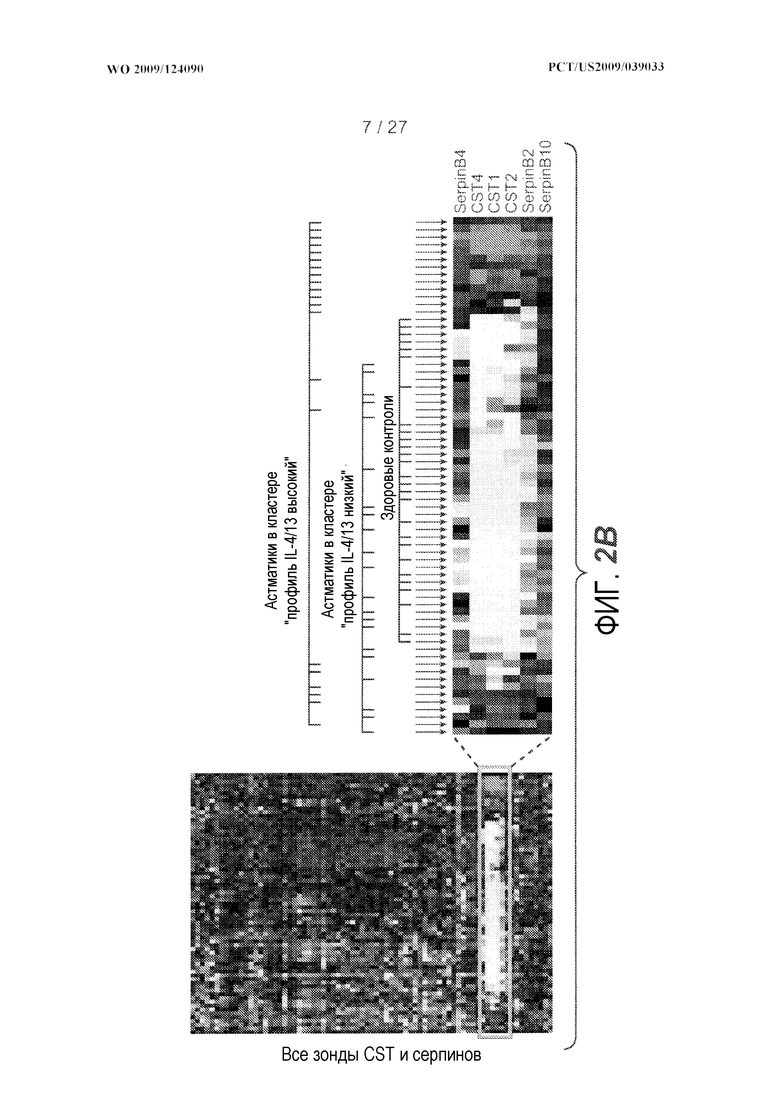
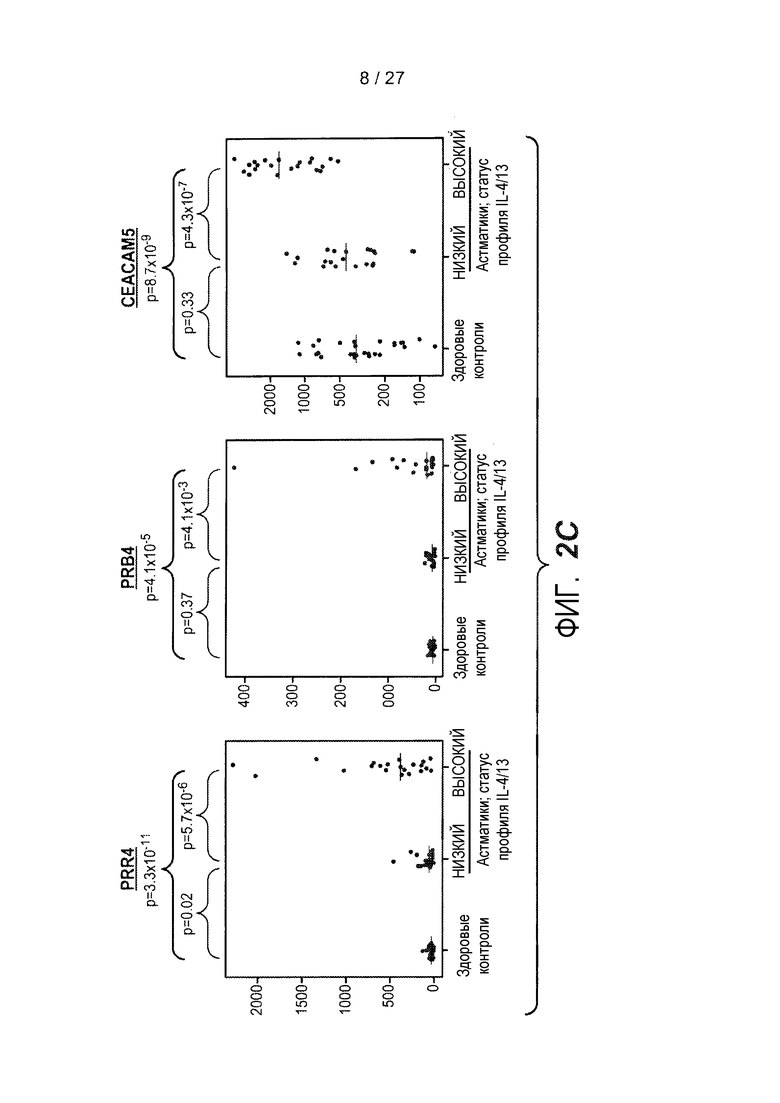
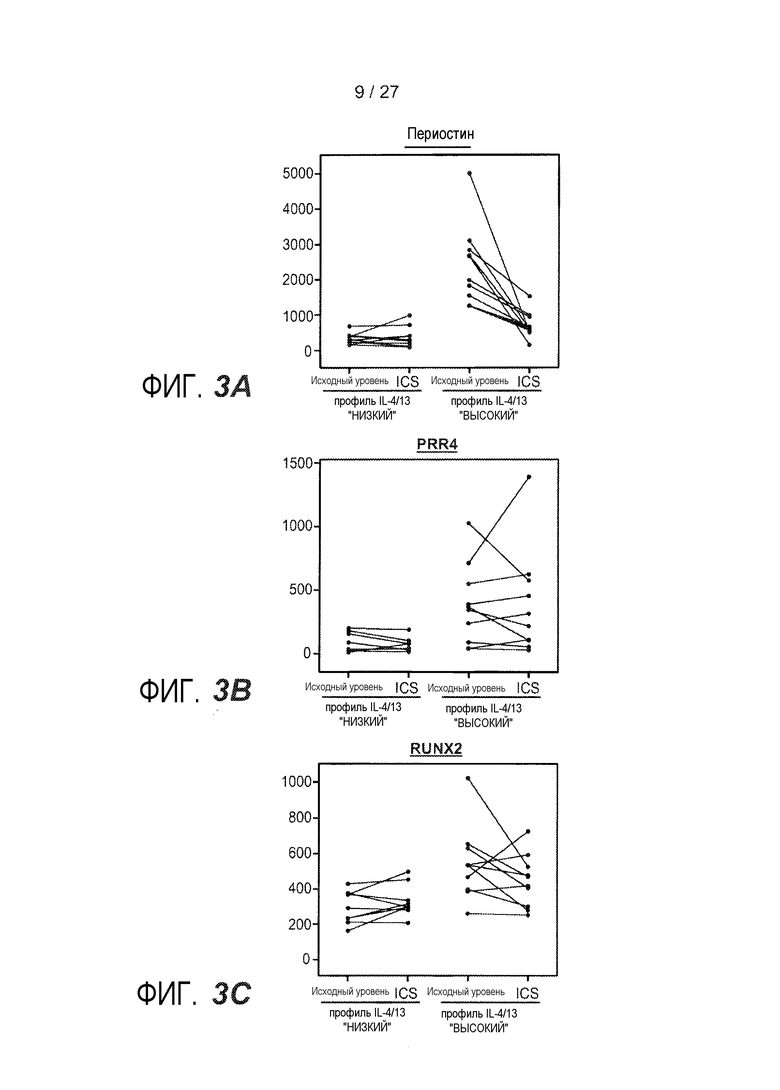
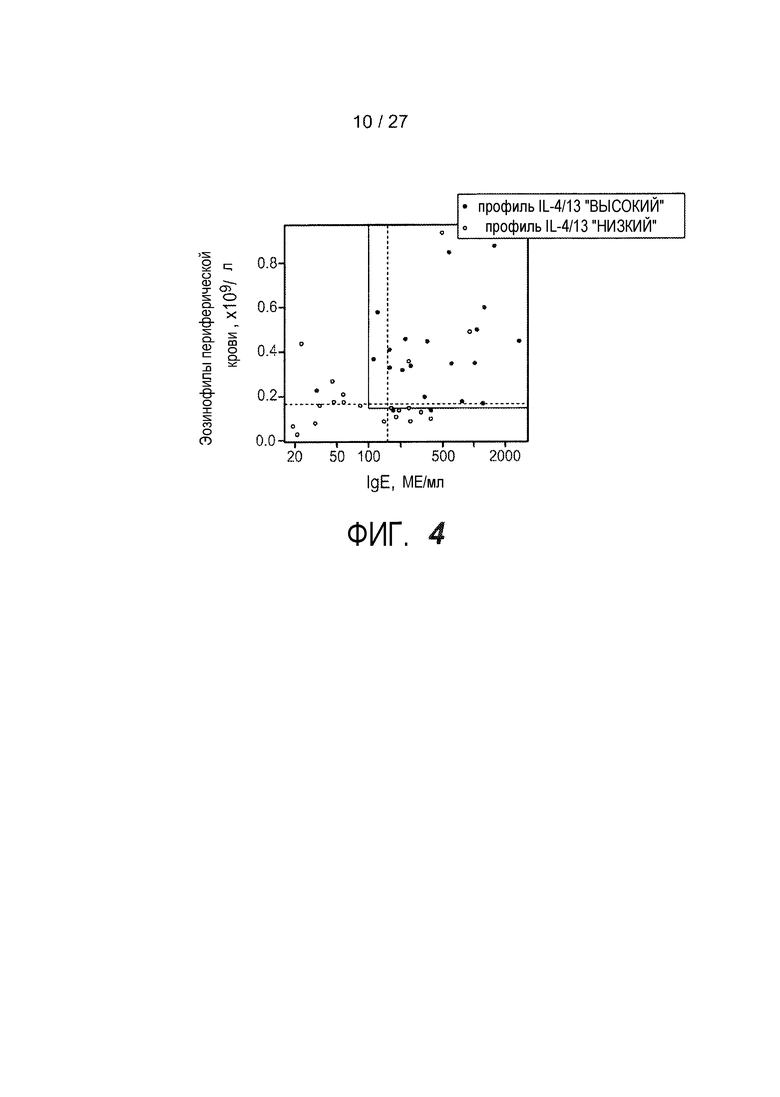
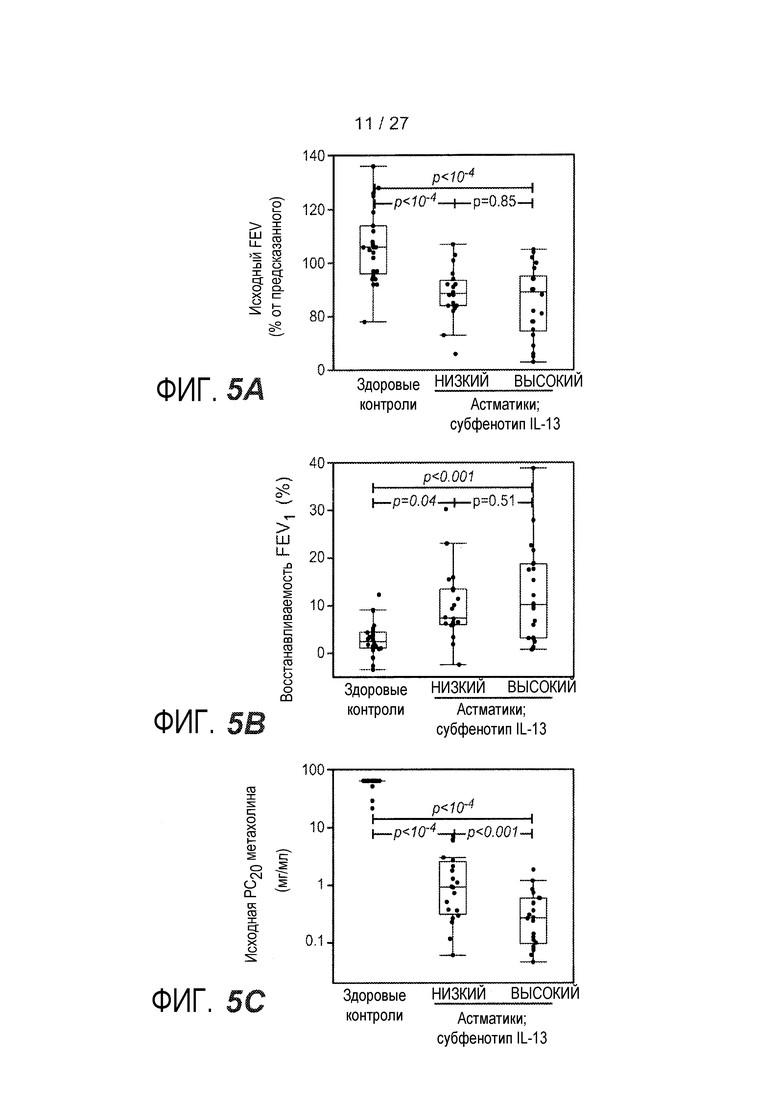
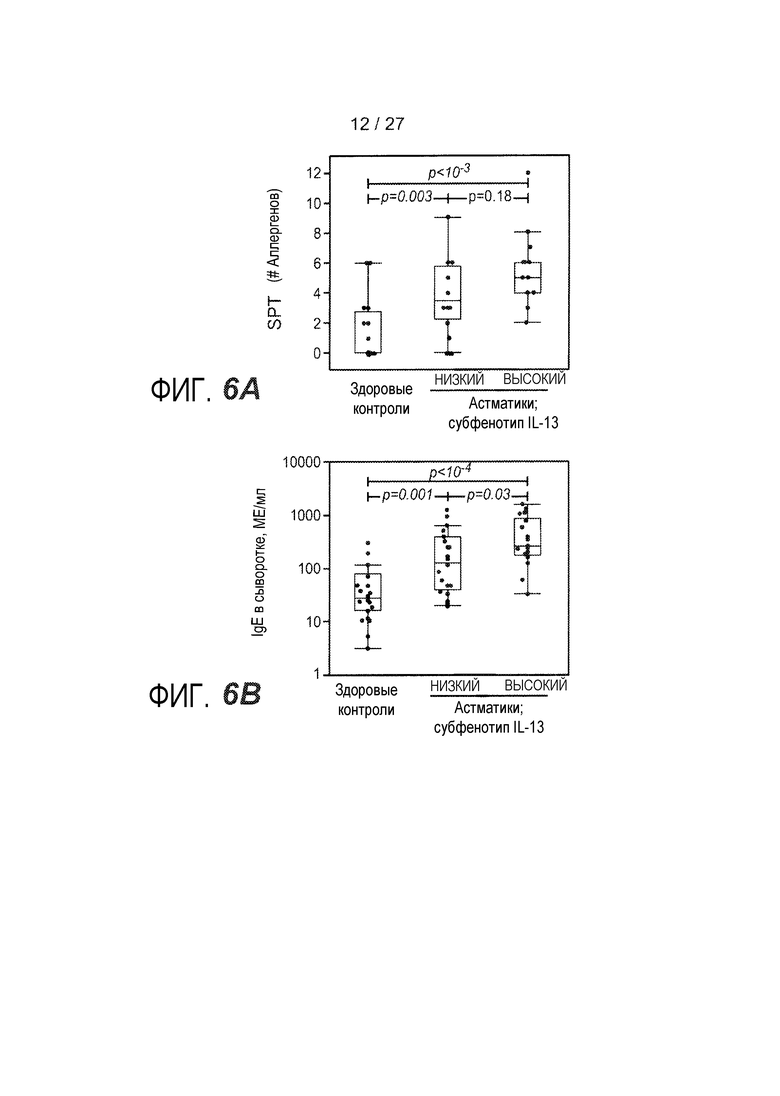
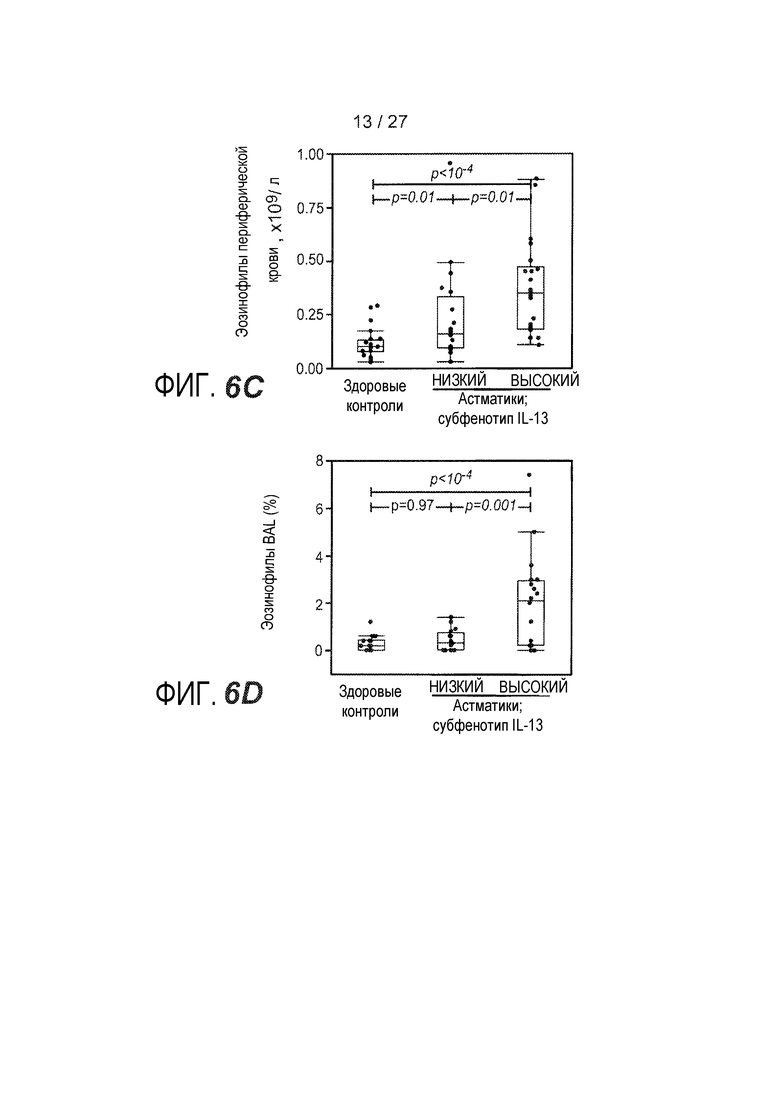
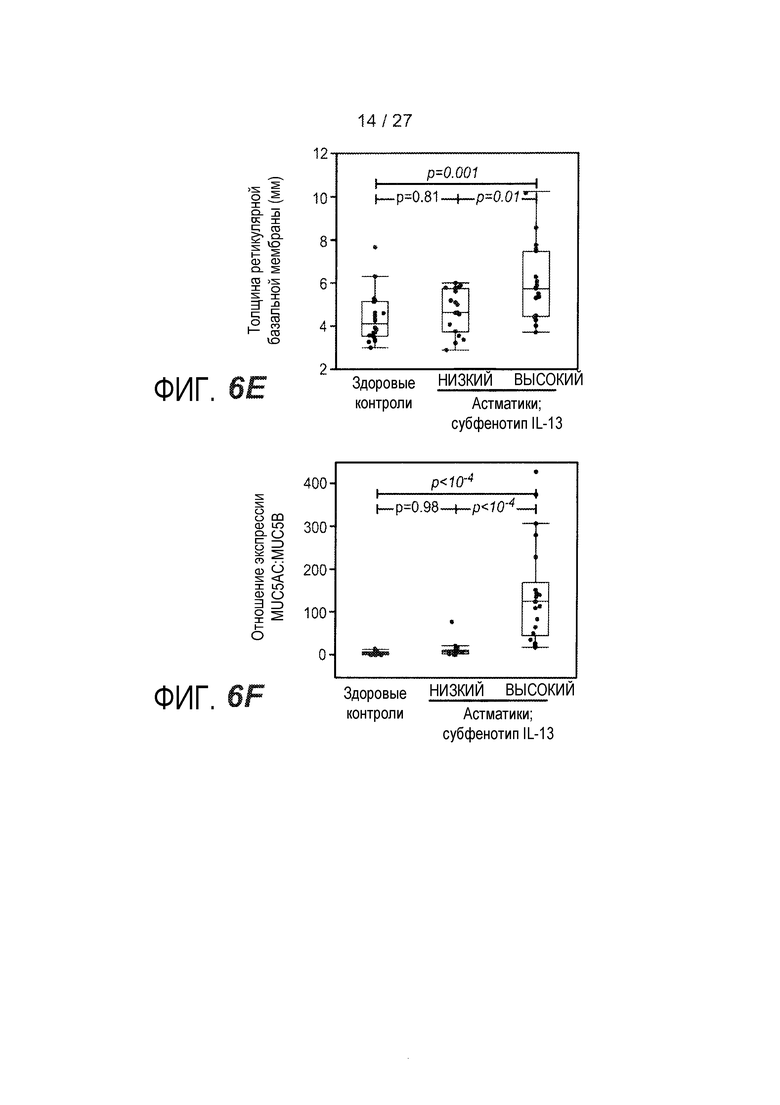
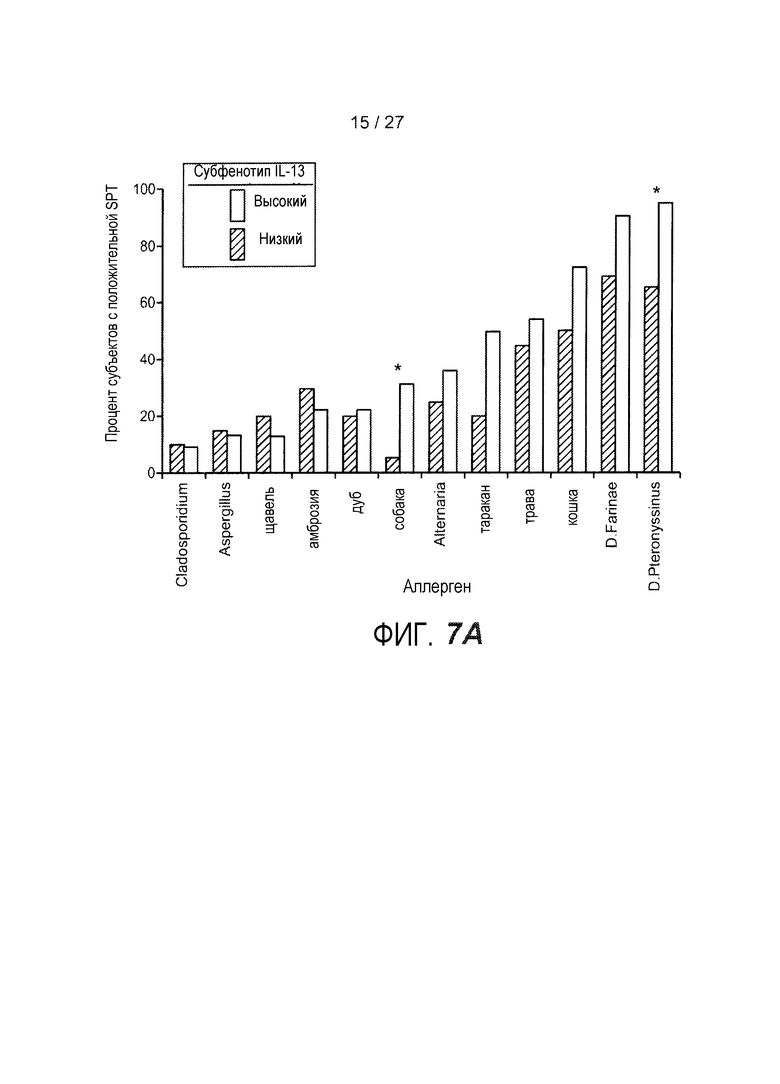
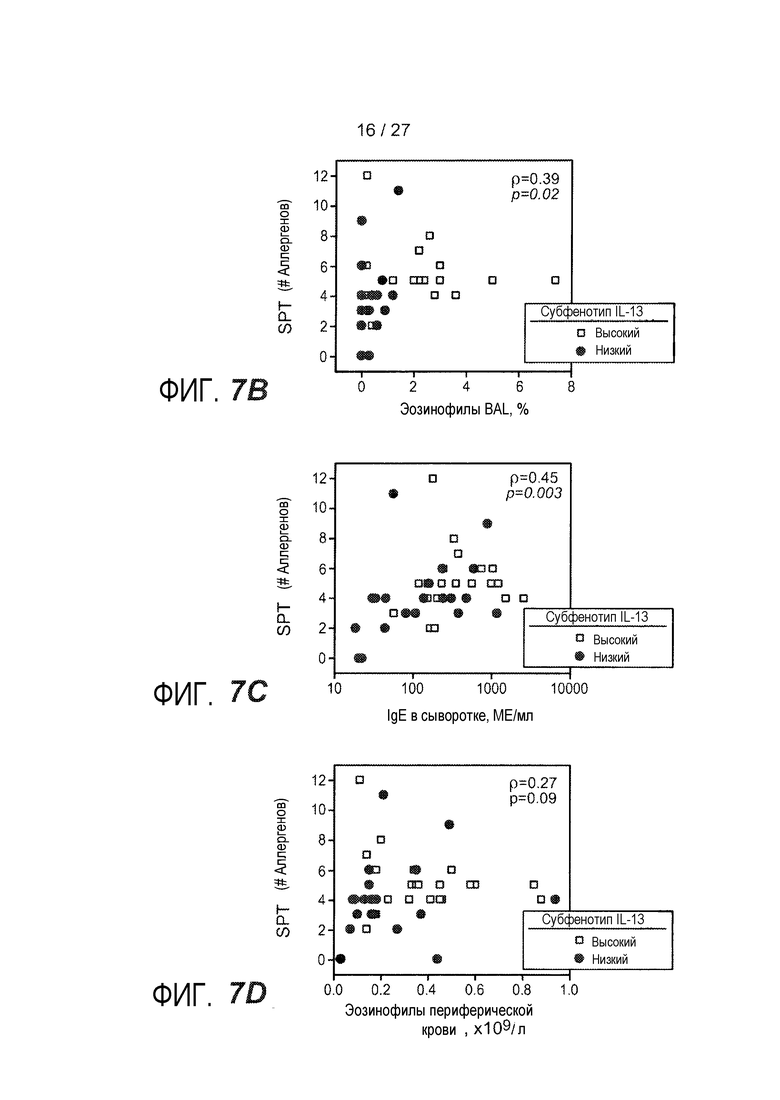
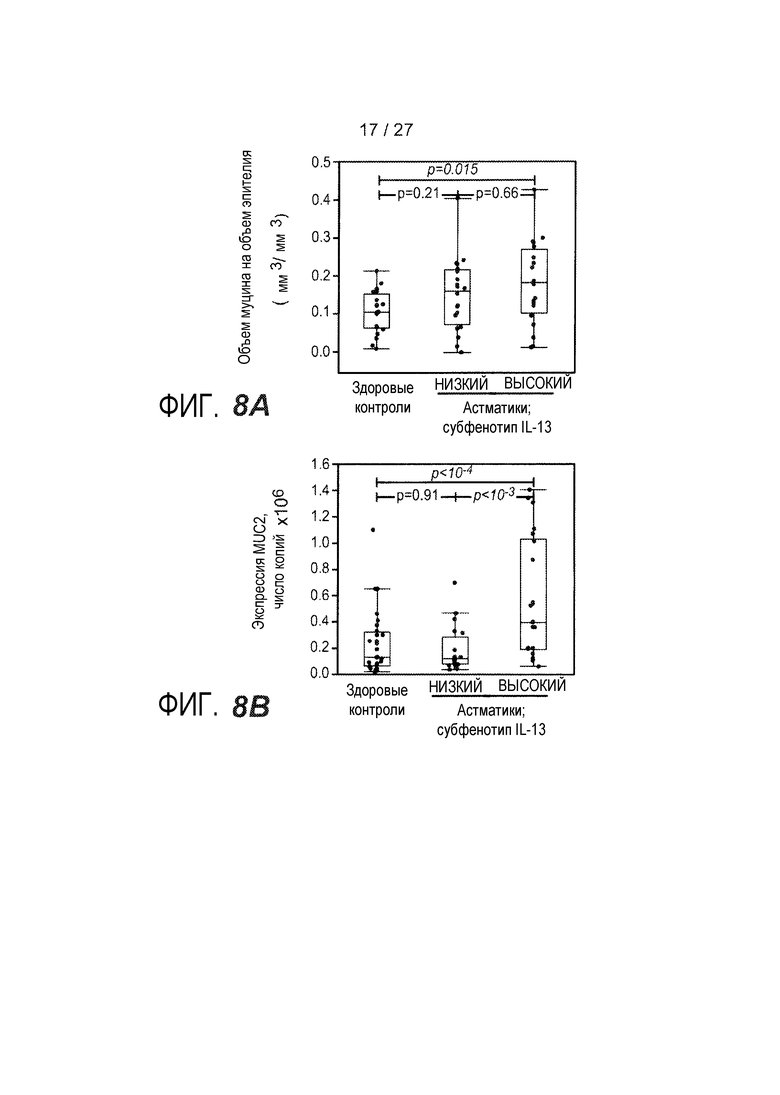
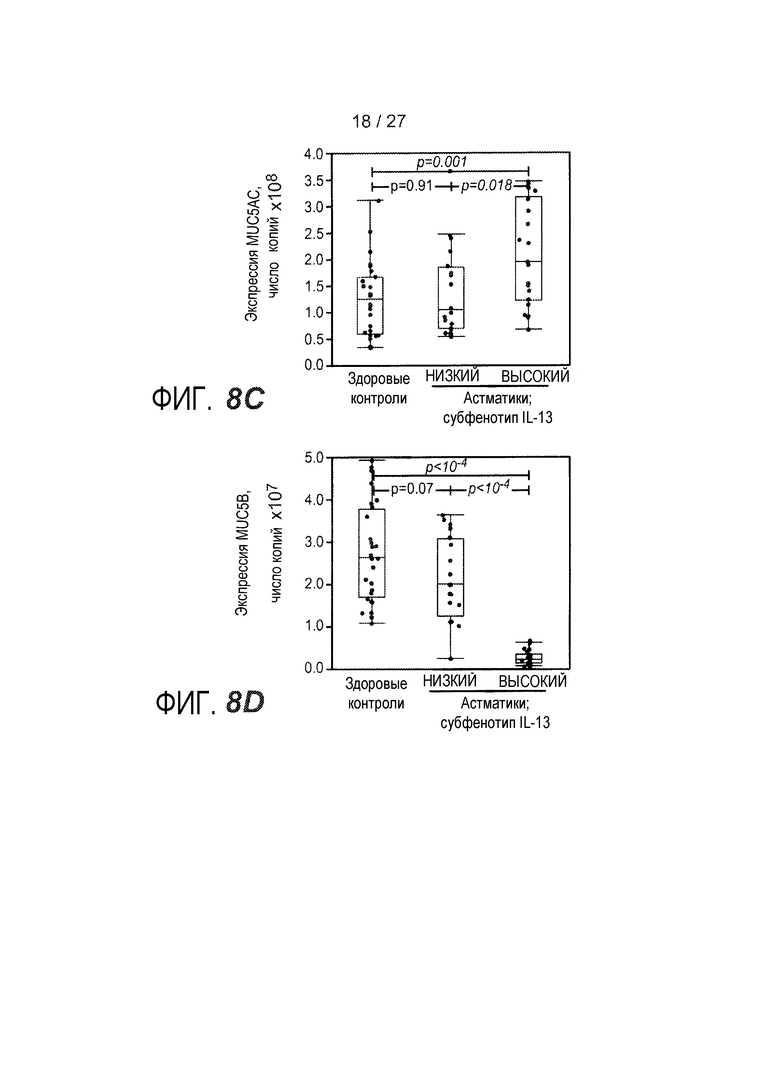
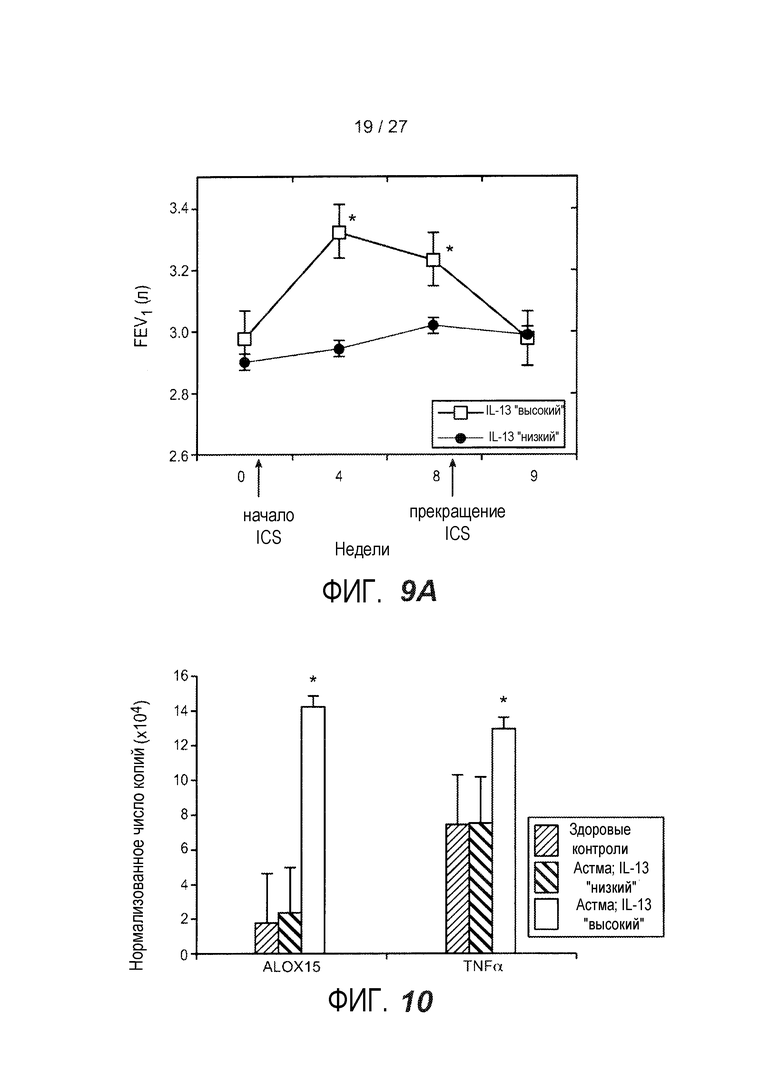
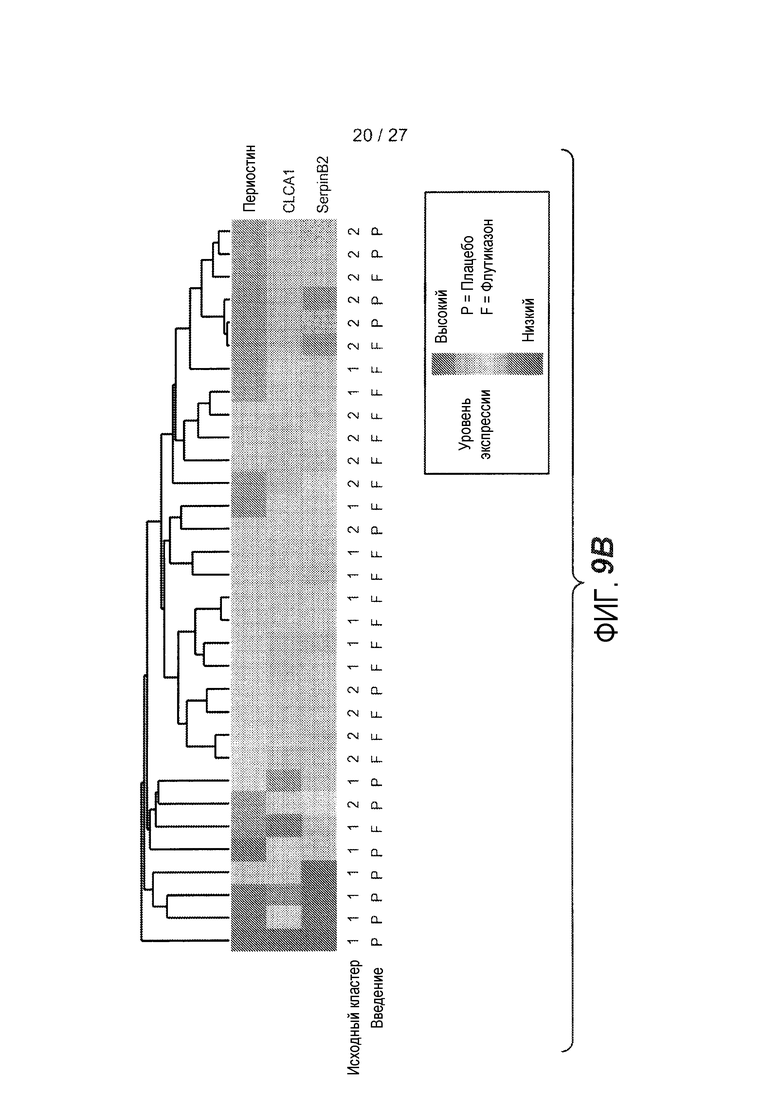
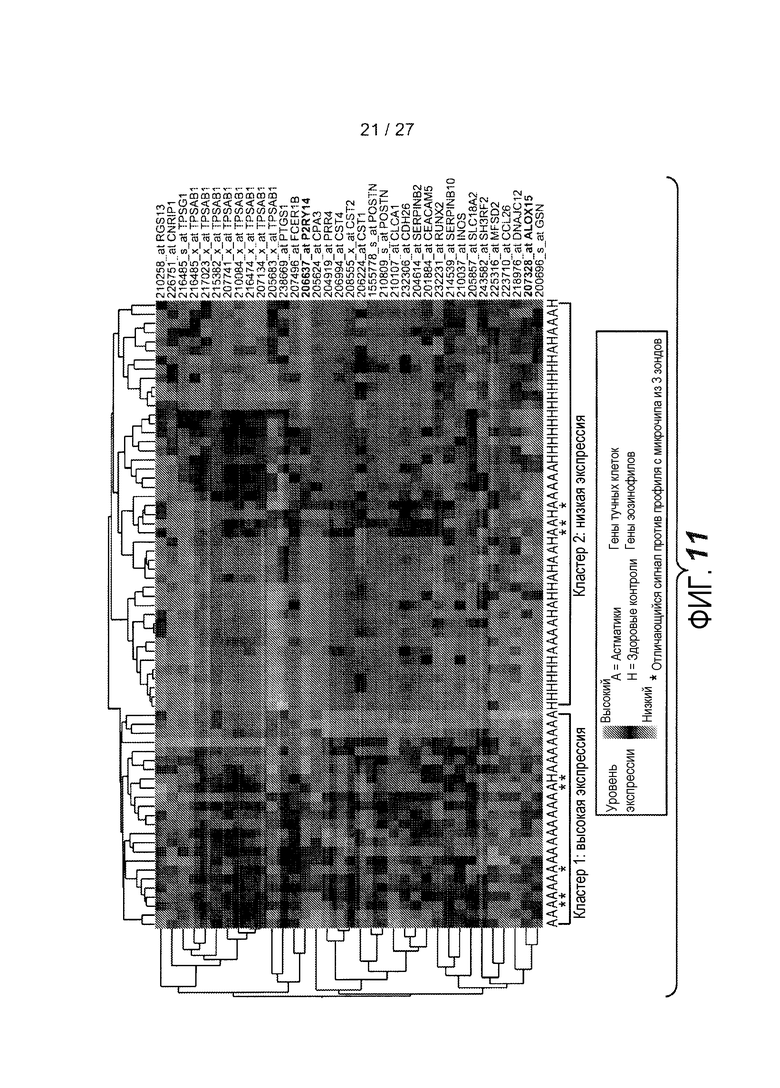
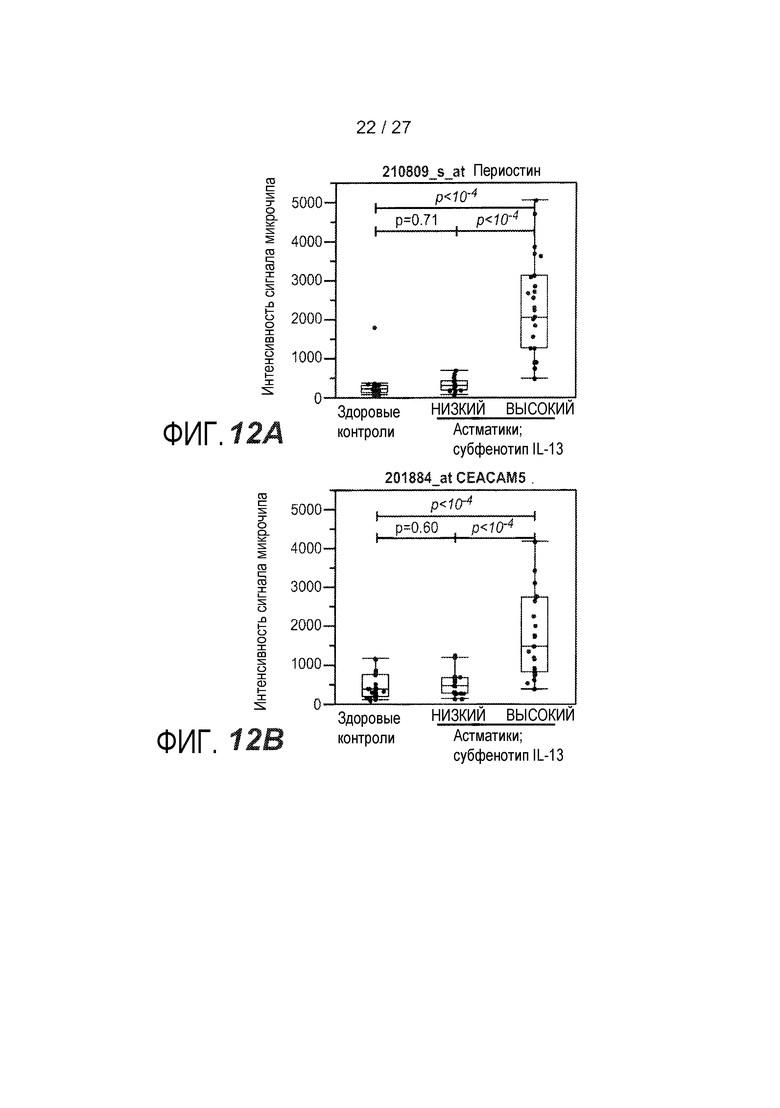
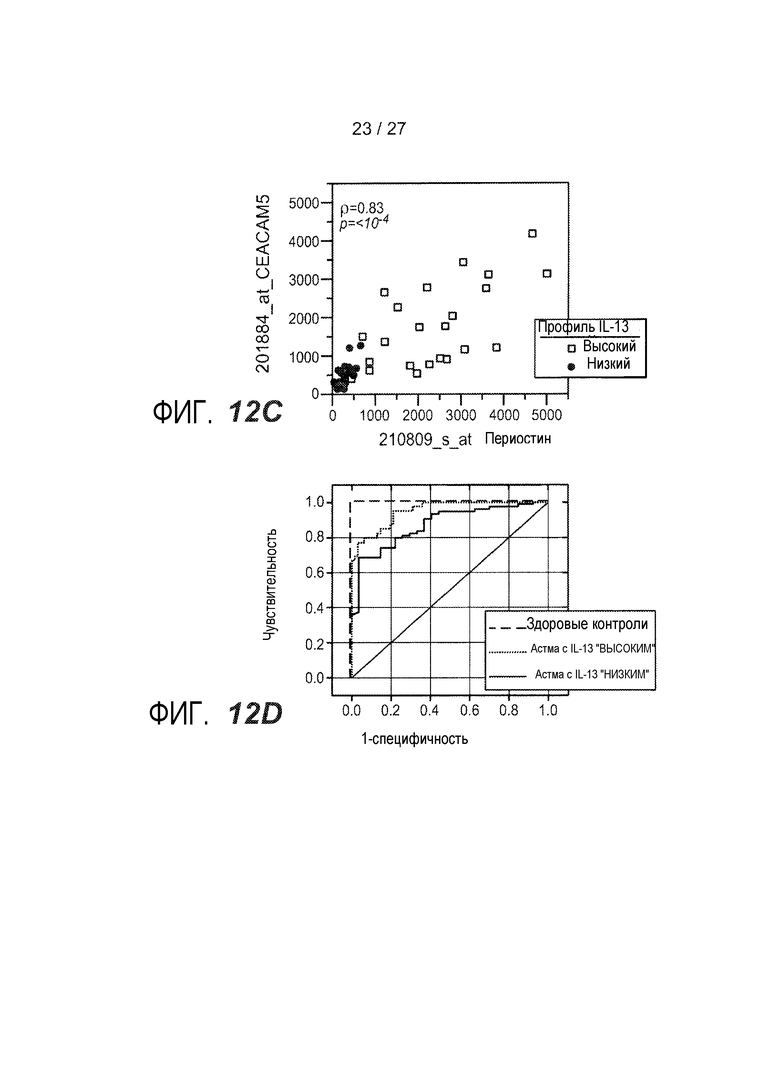
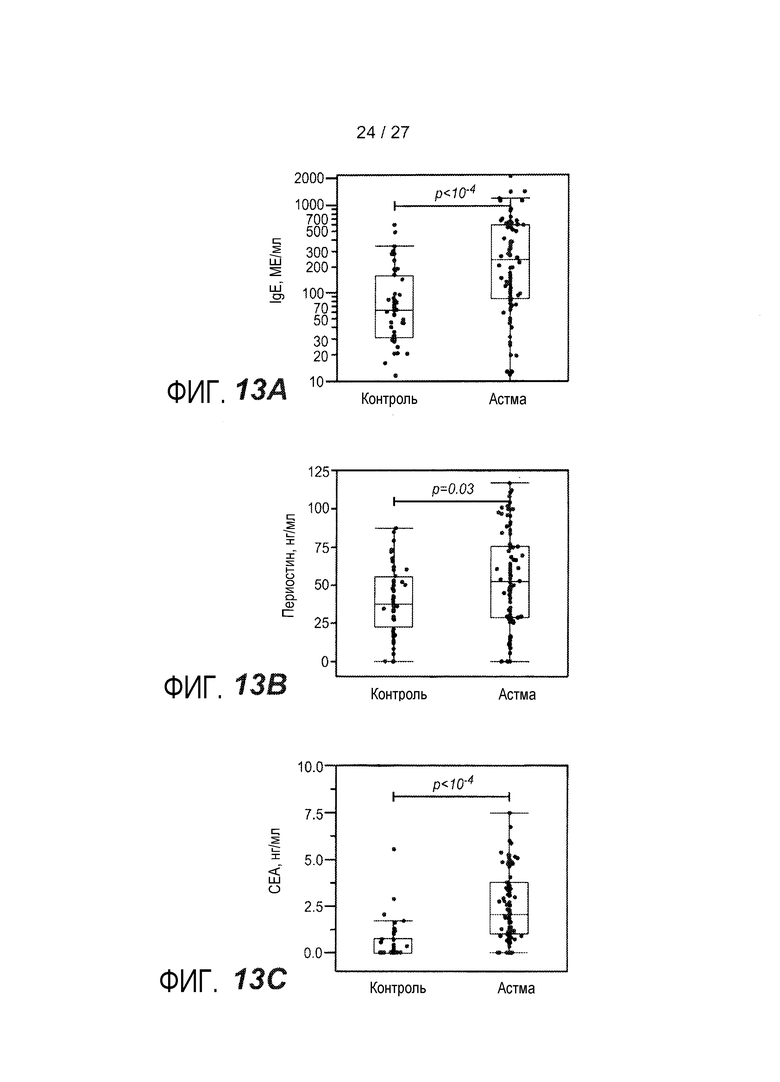
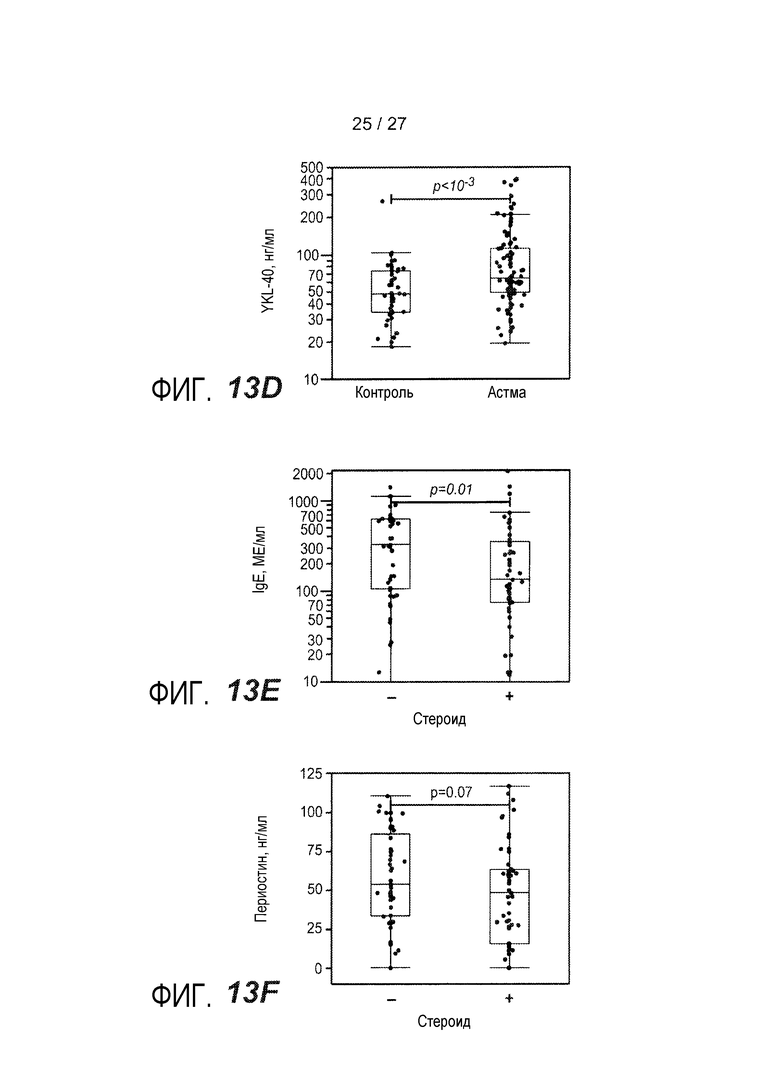
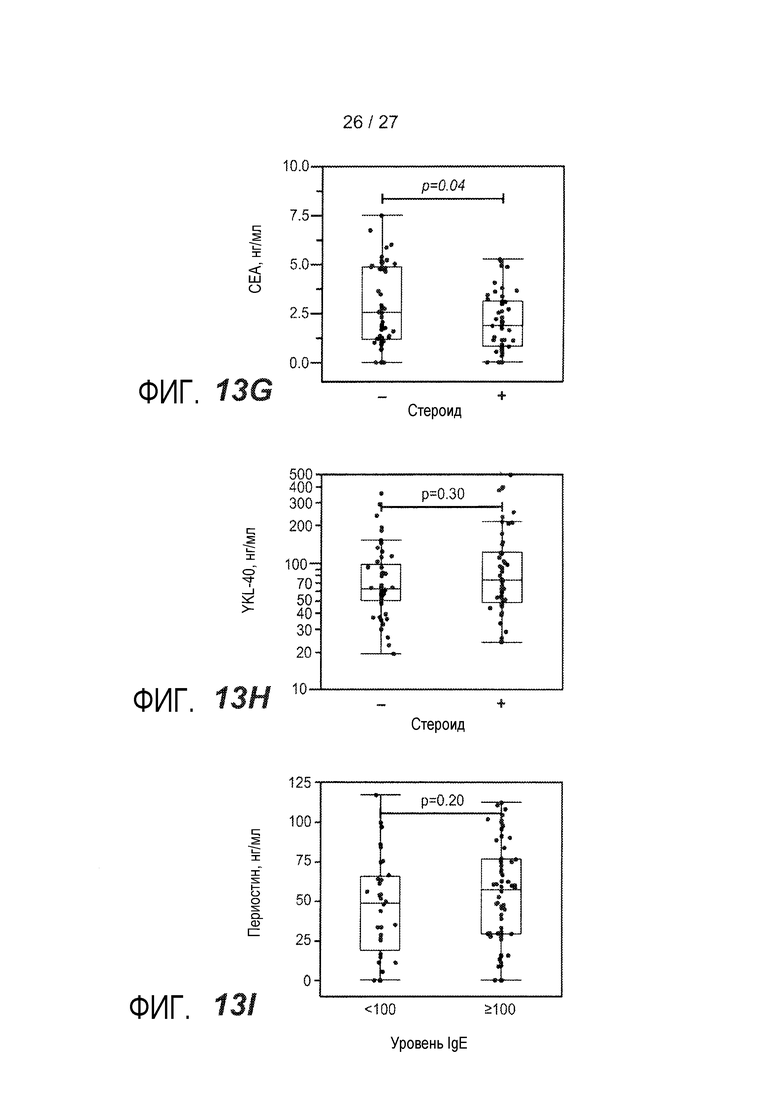
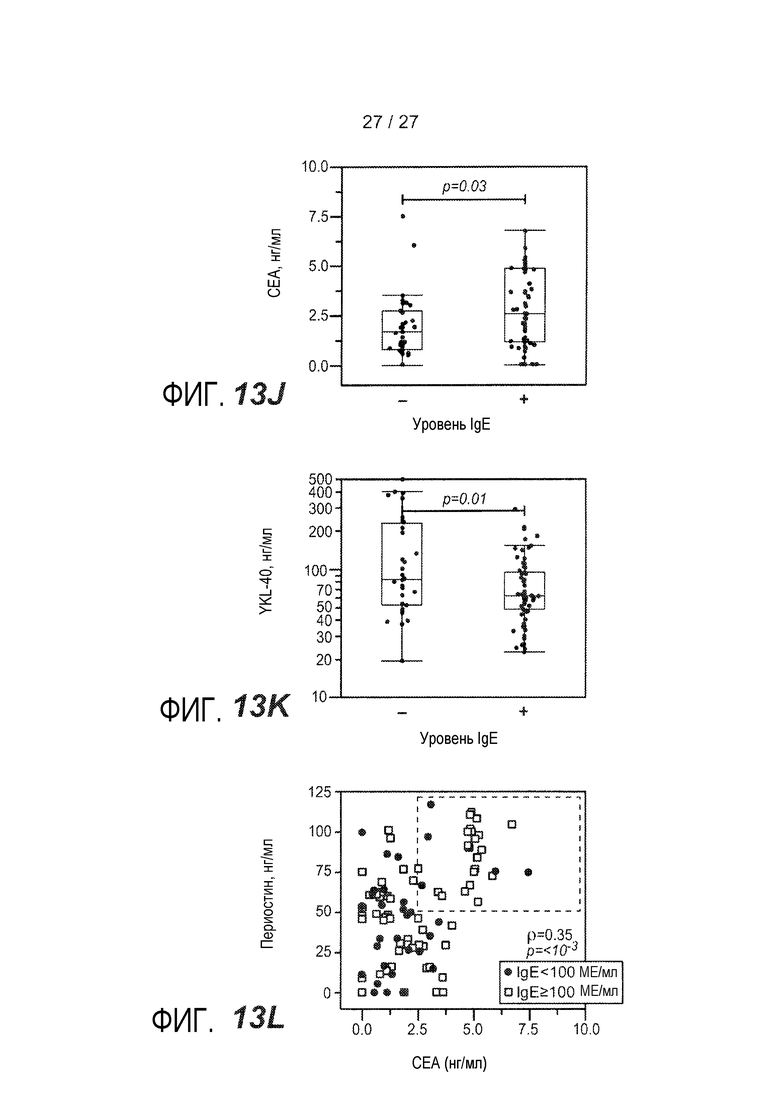
Изобретение относится к области биотехнологии, иммунологии и медицины. Предложено применение антитела, которое связывается с IL-13, в качестве лекарственного средства для лечения астмы у человека. При этом у пациента повышены уровни экспрессии POSTN по отношению к среднему значению или медиане уровня экспрессии POSTN. Также описан способ лечения подтипа астмы с использованием такого антитела. Предложенная группа изобретений может быть использована в медицине. 3 н. и 20 з.п. ф-лы, 49 ил., 13 табл., 9 пр.
1. Применение антитела, которое связывается с IL-13, в качестве лекарственного средства для лечения подтипа астмы у пациента с астмой, являющегося человеком, где у пациента были диагностированы повышенные уровни экспрессии POSTN по отношению к среднему значению или медиане уровня экспрессии POSTN популяции (а) здоровых индивидуумов, которые не страдают от астмы, или (б) пациентов с астмой.
2. Применение по п. 1, где у пациента дополнительно определены повышенные уровни экспрессии любого одного или комбинации генов, выбранных из группы, состоящей из CST1, CST2, CCL26, CLCA1, PRR4, PRB4, SERPINB2, СЕАСАМ5, iNOS, SERPINB4, CST4 и SERPINB10.
3. Применение по п. 2, где группа дополнительно содержит гены PRB4, TPSD1, TPSG1, MFSD2, CPA3, GPR105, CDH26, GSN, C2ORF32, TRACH2000196 (ТМЕМ71), DNAJC12, RGS13, SLC18A2, SH3RF2, FCER1B, RUNX2, PTGS1 и ALOX15.
4. Применение по п. 1, где антитело против IL-13 содержит вариабельный домен антитела TNX-650 или аминокислотные последовательности SEQ ID NO: 195-200, или аминокислотную последовательность SEQ ID NO: 193 и/или 194.
5. Применение по любому из пп. 1-4, где у пациента с астмой определены повышенные уровни периостина в сыворотке.
6. Применение по любому из пп. 1-4, где пациент, получающий лечение, представляет собой пациента с астмой в форме от мягкой до умеренной, не подвергавшейся лечению стероидами.
7. Применение по любому из пп. 1-4, где пациент, получающий лечение, представляет собой пациента с астмой в форме от умеренной до тяжелой, устойчивой к лечению стероидами.
8. Применение по любому из пп. 1-4, где у пациента измеряются уровни экспрессии гена POSTN, необязательно, при помощи иммуноанализа.
9. Применение лекарственного средства, которое связывается с IL-13, для лечения подтипа астмы у пациента с астмой, являющегося человеком, где лекарственное средство представляет собой антитело, и где у пациента были диагностированы повышенные уровни POSTN по отношению к среднему значению или медиане уровня экспрессии POSTN популяции (а) здоровых индивидуумов, которые не страдают от астмы, или (б) пациентов с астмой, а также повышенные уровни экспрессии любого одного или комбинации генов, выбранных из группы, состоящей из CST1, CST2, CLCA1, PRB4, SERPINB2, СЕАСАМ5, iNOS, SERPINB4, CST4 и SERPINB10.
10. Способ лечения подтипа астмы, включающий введение лекарственного средства пациенту с астмой, являющемуся человеком, у которого были диагностированы повышенные уровни POSTN по отношению к среднему значению или медиане уровня экспрессии POSTN популяции (а) здоровых индивидуумов, которые не страдают от астмы, или (б) пациентов с астмой, где лекарственное средство представляет собой антитело, которое связывается с IL-13.
11. Способ по п. 10, включающий стадию измерения у пациента экспрессии гена POSTN.
12. Способ по п. 10, где ингибитор, нацеленный на каскад IL13/IL4, представляет собой антитело против IL-13, содержащее
(i) аминокислотные последовательности SEQ ID NO: 193 и 194; или
(ii) аминокислотные последовательности SEQ ID NO: 195-200.
13. Способ по п. 10 или 11, где у пациента с астмой определены повышенные уровни периостина в сыворотке.
14. Способ по п. 10 или 11, где пациент, получающий лечение, представляет собой пациента с астмой в форме от мягкой до умеренной, не подвергавшейся лечению стероидами.
15. Способ по п. 10 или 11, где пациент, получающий лечение, представляет собой пациента с астмой в форме от умеренной до тяжелой, устойчивой к лечению стероидами.
16. Способ по п. 10 или 11, где у пациента дополнительно определены повышенные уровни экспрессии любого одного или комбинации генов, выбранных из группы, состоящей из POSTN, CST1, CST2, CCL26, CLCA1, PRR4, PRB4, SERPINB2, СЕАСАМ5, iNOS, SERPINB4, CST4 и SERPINB10.
17. Способ по п. 10, где группа дополнительно содержит гены PRB4, TPSD1, TPSG1, MFSD2, CPA3, GPR105, CDH26, GSN, C2ORF32, TRACH2000196 (ТМЕМ71), DNAJC12, RGS13, SLC18A2, SH3RF2, FCER1B, RUNX2, PTGS1 и ALOX15.
18. Способ по п. 10 или 11, где экспрессию гена измеряют анализом уровней белка или мРНК.
19. Способ по п. 18, где уровни мРНК измеряют с использованием метода ПЦР или с использованием микроматричного чипа.
20. Способ по п. 19, где метод ПЦР представляет собой двухстадийную ПЦР в реальном времени (qPCR).
21. Способ по п. 19, где уровни мРНК целевого гена, превышающие более чем в 2,5 раза уровни мРНК контрольного гена, указывают на подтип астмы.
22. Способ по п. 10 или 11, включающий измерение уровня периостина в образце сыворотки, полученном от пациента, являющегося человеком.
23. Способ по п. 22, где измерение уровня периостина производят при помощи иммуноанализа.
| WOODRUFF P.G | |||
| ET AL., Genome-wide profiling identifies elpithelial cell genes associated with asthma and with treatment response to corticosteroids, PROCEEDINGS OF THE NATIONAL ACADEMY OF SCIENCES OF USA, 2007, v | |||
| Счетная таблица | 1919 |
|
SU104A1 |
| Приспособление с иглой для прочистки кухонь типа "Примус" | 1923 |
|
SU40A1 |
| Приспособление для применения кинофильма в фильмовом канале в проекционных и съемочных киноаппаратах | 1929 |
|
SU15858A1 |
| TAKAYAMA G | |||
| ET AL., Periostin: A novel component of subepithelial fibrosis of bronchial asthma downstream of IL-4 and IL-13 signals, JOURNAL OF ALLERGY AND CLINICAL IMMUNOLOGY, 2006, v | |||
| Прибор для массовой выработки лекал | 1921 |
|
SU118A1 |
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| Дорожная спиртовая кухня | 1918 |
|
SU98A1 |
| IZUHARA K | |||
| ET AL., Clarification of the pathogenesis and development of clinical examination for allergic diseases, Rinsho Byori., 2007, v.55, no.4, p | |||
| Прибор для сжигания нефти | 1921 |
|
SU369A1 |
| IZUHARA K | |||
| ET AL., Microarray-based identification of novel biomarkers in asthma, ALLERGOLOGY INTERNATIONAL, 2006, v | |||
| Устройство двукратного усилителя с катодными лампами | 1920 |
|
SU55A1 |
| Очаг для массовой варки пищи, выпечки хлеба и кипячения воды | 1921 |
|
SU4A1 |
| Способ получения продуктов уплотнения фенолов с альдегидами | 1920 |
|
SU361A1 |
| WO 2007085815 A, 02.08.2007 | |||
| RU 2006104618 А, 27.08.2007. | |||

