Способ получения протеолитического препарата для медицинского применения
Изобретение относится к биотехнологии, а именно к получению комплекса протеолитических ферментов, обладающих широким спектром протеолитической активности, способных гидролизовать белковые субстраты. Изобретение может найти применение в производстве медицинских препаратов.
Известно использование в медицине для очищения ран препаратов на основе микробной коллагеназы, выделяемой штаммами-продуцентами Clostridium histolyticum. Микробная коллагеназа обладает высокой специфичностью в отношении основного белка соединительной ткани коллагена. Однако, высокая специфичность, для целей очищения ран и лечения келоидных рубцов является недостатком. Это связано с тем, что межклеточный матрикс и соединительная ткань содержит не только коллагеновые волокна, она представляет собой сложную сеть, включающую помимо коллагена эластин и адгезивные белки. Другой важной составляющей межклеточного матрикса являются протеогликаны, осевой молекулой которых служит гиалуроновая кислота, к которой как щетинки к щетке присоединены белки, в свою очередь соединенные с множеством гликозаминогликанов.
Омертвевшие ткани в ожоговой ране представлены, главным образом, коагулированными белками дермы (коллагена, эластина и др.), а также поврежденными компонентами внеклеточного матрикса. На поверхности ран может находиться фибринозный налет. Видимые эффекты очищения ран под действием протеолитических ферментов чаще являются результатом расщепления фибрина, а не коллагена. При лечении келоидных рубцов основной задачей ферментных препаратов является разрушение неправильно собранных коллагенновых фибрилл и протеогликанов до олигомеров и индивидуальных мономеров, которые частично используются для сборки новой более упорядоченной соединительной ткани.
На основании этого можно полагать, что в медицинском применении могут оказаться более эффективными не препараты с узкоспецифичным действием (микробные коллагеназы), а комплексы протеолитических ферментов с более широким спектром субстратов.
Под комплексом протеолитических ферментов мы понимаем совокупность индивидуальных ферментов пищеварительной системы, обладающих протеолитическим действием, которые выделяются совместно в технологическом процессе.
К таким ферментным комплексам можно отнести сериновые протеазы, выделяемые из морских беспозвоночных (крабовая коллагеназа) и пищеварительные сериновые протеазы рыб.
В отличие от истинных коллагеназ сериновые протеазы беспозвоночных и рыб расщепляют все три полипептидных цепи тропоколлагена, причем образующиеся пептиды подвергаются дальнейшему гидролизу вплоть до индивидуальных аминокислот, которые либо включаются в процессы анаболизма, либо быстро выводятся из организма, не вызывая интоксикации.
Наиболее широко разработаны и являются аналогами заявляемого способа методы получения крабовой коллагеназы из гепатопанкреаса крабов. Эффективность крабовой коллагеназы для очищения ран и лечения келоидных рубцов установлена и описана в патентной литературе (US 4801451, US 4963491, US 5945102 и US 6030612, а также RU 2008353, RU 2039819, RU 226505, RU 2365623).
Основной проблемой при промышленном получении препаратов на основе коллагеназы краба является нестабильность поставок сырья и длительный срок хранения сырья до доставки на перерабатывающее предприятие, что часто приводит обильному обсеменению сырья микроорганизмами и накоплению микробных эндотоксинов (по химической структуре представляющих собой липополисахариды), которые способны вызывать пирогенный эффект и аллергию. Это затрудняет промышленное выделение комплекса протеолитических ферментов и снижает его качество, а применение таких препаратов у пациентов сопровождается частыми аллергическими реакциями.
Как известно из уровня техники, комплексы протеолитических ферментов со сходными с крабовыми N-концевыми аминокислотными последовательностями можно выделить из пищеварительных органов рыб Изучение субстратной специфичности этих комплексов протеолитических ферментов показывает, что, в отличие от пищеварительных протеаз млекопитающих, они способны гидролизовать более широкий спектр субстратов, в том числе и коллагены различных типов, которые, вследствие особенностей аминокислотной последовательности и пространственной структуры, устойчивы к действию большинства протеаз и доступны для гидролиза только специфическими ферментами - коллагеназами. Источником комплекса протеолитических ферментов служат отходы пищевого производства рыбы - потроха (пищеварительные органы), которые обычно идут в отход и на утилизацию. По сравнению с крабовой коллагеназой этот источник гораздо более доступен и дешев (Zwilling R., et al., 1975 г, FEBS Lett. 60, 247-9; Gran G.A., et al., 1981 г., Methods Enzymol 80, 722-734; Reeck G.R. and H. Neurath. 1972 г., Biochem. 11: 503-510, O.A. Klimova, et al., 1990 г., BBRC, v. 166, N3, 1411-1420).
Для медицинского использования наиболее предпочтительно использование комплексов протеолитических ферментов, выделяемых из пищеварительных органов рыб, так как они имеют сериновые протеазы, активные в отношении глубокого гидролиза коллагена и других белков соединительной ткани. Как показано в диссертационной работе «Соловьев М.М. Характеристика пищеварительных ферментов рыб озера Чаны на ранних этапов онтогенеза. / Диссертация на соискание ученой степени к.б.н.: Новосибирск. - 2011.» на всех этапах развития мирных и хищных рыб наибольший вклад в активность щелочных протеиназ вносит группа сериновых протеиназ (до 70-80%). Доля цистеиновых протеиназ и металлопротеаз значительно меньше и не превышает 10-12%. У нехищных рыб выявлено 4-6 изоформ, у хищных 7-8 изоформ щелочных протеаз.
Известен «Способ выделения комплекса протеолитических ферментов из морепродуктов» путем гомогенизации сырья и осаждения ферментов, отличающийся тем, что, с целью расширения сырьевой базы и повышения выхода целевого продукта, в качестве сырья используют пилорические придатки камбалы, а осаждение ферментов ведут ацетоном при соотношении гомогенат:ацетон 1:2 (2,3). При этом выделяемый комплекс представляет собой ценный набор ферментов, в частности щелочную, нейтральную и кислые протеиназы, аминопепсидазы и карбоксипептидазы. Способ выделения заключается в том, что выделенные из камбалы придатки гомогенизируют и быстро смешивают с двумя объемами, охлажденного до -15°С ацетона. Через 20 мин материал отжимают от ацетона и высушивают при +20°С. Удельная активность равна 0,76 по субстрату казеину. Протеолитическая активность выделенных ферментов в нейтральной и слабокислой области превосходит активность фермента, выделяемого из гепатопанкреаса краба (SU 1030409 C12N 9/64. «Способ выделения комплекса протеолитических ферментов из морепродуктов». М.В. Колодзейская, С.А. Кудинов, Л.М. Эпштейн, В.Н. Акулин. Заявл. 12.03.82, опубл. 23.07.83).
Основным недостатком данного метода является использование больших количеств органического растворителя - ацетона, что требует соблюдения особых условий на производстве. Другим недостатком является присутствие в препарате балластных белков и липополисахаридов, что препятствует применению такого фермента в медицинских целях.
Известен ряд технических решений, в который описывается выделение протеолитических ферментов из рыб, но получаемый продукт также характеризуется наличием балластных белков и липополисахаридов, поэтому выделенные протеолитические ферменты пригодны только для использования в пищевой и кожевной промышленности.
В частности, известен «Способ получения протеолитического ферментного препарата из внутренних органов». Это изобретение относится к биотехнологии, в частности к получению ферментного препарата, и использовано в производстве пищевых продуктов из слабосозревающих объектов промысла Волго-Каспийского района. Способ предусматривает измельчение сырья. Измельченное сырье подвергают автолизу в присутствии воды при гидромодуле 1:0,5 при рН 6,2±0,2 в течение 1-5 часов. После проведения автолиза жидкий автолизат, содержащий протеолитические ферменты, отделяют центрифугированием и подвергают очистке изопропиловым спиртом при гидромодуле 3:1 в течение 30-40 мин. Полученный в результате очистки жидкий ферментный препарат высушивают распылительно при температуре 150-180°С в течение 5-8 с. Изобретение позволяет повысить выход протеолитического препарата и повысить его активность (RU №2288951 C12N 9/64 «Способ получения протеолитического ферментного препарата из внутренних органов рыб», опубл. 10.12.2006).
Известен также способ получения ферментного препарата из рыбного сырья. Это изобретение относится к биотехнологии, в частности к способам извлечения протеолитических ферментов из рыбного сырья, и предназначено для использования в пищевой промышленности. Способ предусматривает измельчение сырья, выделение фермента экстракцией в присутствии раствора хлорида щелочного металла, отделение осадка, две стадии ультрафильтрации с добавлением в жидкую фракцию дробленого льда и лиофильную сушку концентрата. На первой стадии ультрафильтрацию осуществляют на мембране с лимитом пропускания 50 кДа, а на второй стадии на мембране с лимитом пропускания 15 кДа. Изобретение позволяет повысить выход протеолитических ферментов и их активность в 3-5 раз по сравнению с известными аналогами (RU №2352634 C12N9/00 «Способ получения ферментного препарата из рыбного сырья», опубл. 20.04.2009).
Наиболее близким техническим решением к заявляемому изобретению является следующая последовательность действий для получения комплекса протеолитических ферментов из тканей рыб (Daboor S.M., Budge S.M., Chaly А.Е., et al. Extraction and Purification of Collagenase Enzymes: A Critical Review. // American Journal of Biochemistry and Biotechnology 6 (4): 239-263,2010.):
- Гомогенизация сырья в 0,05 M Tris-HCl буфере с добавкой 0,005 М хлористого кальция при рН 7,4;
- Центрифугирование гомогената при 25000×g в течение 30 мин при +4°С и сбор супернатанта;
- Повторная экстракция осадка в 0,05 М Tris-HCl буфере с добавкой 0,005 М хлористого кальция при рН 7,4 с последующим центрифугированием гомогената при 25000×g в течение 30 мин и сбором супернатанта;
- Объединение супернатантов и центрифугирование при 40000×g в течение 30 мин при +4°С;
- Фильтрование полученного супернатанта через мембрану с порами 0,2 мкм;
- Фракционирование белка переосаждением в растворе сульфата аммония при насыщении в пределах 40-80%;
- Выделение сульфатаммонийного осадка фракционированного фермента центрифугированием;
- Растворение осадка в буфере;
- Обессоливание путем пропускания раствора через полые волокна с размером пор 10 кДа;
- Вторичная очистка комплекса протеолитических ферментов с помощью ионообменной хроматографии.
Большая часть протеолитических ферментов находятся в клетках тканей либо в неактивной форме, либо в лизосомах, иначе бы они переваривали сами клетки. Поэтому, чтобы их экстрагировать необходимо разрушить клетки. Гомогенизация это процедура разрушения клеток и лизосом под действием механических, температурных или химических факторов. От степени разрушения (измельчения клеток) зависит скорость и количество выхода ферментов в экстрагирующий раствор. Обычно продолжительность экстракции составляет от нескольких часов до 2-3 суток. Другой процедурой извлечения протеолитических ферментов является автолиз, используемый в большинстве вышеприведенных патентов-аналогов. Автолиз тканей протекает при повышенных температурах. В ходе него наблюдается ускоренный транспорт протеаз через лизосомальные барьеры с последующей их активацией и разрушением клеточных и лизосомальных барьеров под действием активных протеаз. Также автолиз сопровождается побочными явлениями, к которым можно отнести автогидролиз самих ферментов и накопление в автолизированных тканях микробной биомассы, что приводит к резкому повышению содержания ЛПС и других микробных токсинов.
Центрифугирование и мембранная фильтрация это технологические процедуры, направленные на удаление из гомогенизата нерастворимых частиц тканей, нерастворимых липидно-белковых комплексов и других нерастворимых частиц.
Фракционирование белка в сульфате аммония это процедура первичной очистки при выделении белков. Высаливание белков идет по двум механизмам: сульфат-ионы делают молекулу белка более компактной, а следовательно, менее растворимой, сульфат-ионы связывают свободную воду. Один ион SO42- имеет 13-14 молекул H2O только в первом гидратном слое и, возможно, больше - во втором. Считается, что при концентрации 5,4 Моля сульфат аммония связывает всю свободную воду - это 100% насыщение. Достоинствами сульфата аммония является то, что он имеет примерно одинаковую растворимость в диапазоне температур 0-+30 градусов Цельсия и то, что в концентрации выше 0,5 М он стабилизирует белки против денатурации. Обычно, индивидуальные белки теряют растворимость и выпадают в осадок при определенной степени насыщения раствора сульфатом аммония. При выделении сразу нескольких индивидуальных белков, как в нашем случае, происходит их осаждение в достаточно широком диапазоне насыщения сульфата аммония. Очищенный комплекс протеолитических ферментов, выделенный из рыб обычно высаждается в области насыщения сульфата аммония от 40 до 80%. Осадок отделяют от раствора сульфата аммония осаждением в центрифуге. Однако наличие в растворе нуклеиновых кислот и липидно-белковых комплексов ухудшает селективность фракционирования и приводит к соосаждению целевого комплекса протеолитических ферментов во всех фракциях сульфатаммонийного осадка.
После растворения осадка фракции белков в буфере, раствор содержит значительное количество сульфата аммония, который будет мешать дальнейшей очистке комплекса протеолитических ферментов. Раствор подвергают обессаливанию. Обычно, эту процедуру проводят путем ультрафильтрации через мембрану с определенным размером пор. Однако, в нашем случае появление тонкодисперсного осадка липидно-белкового комплекса приводит к быстрой закупорке пор и резкому торможению процесса. Поэтому для обессаливания необходимо использовать тангенциальную фильтрацию. При тангенциальной фильтрации поток раствора направляют параллельно поверхности мембраны, что позволяет непрерывно смывать, забивающий поры осадок. Разновидностью тангенциальной фильтрации являются аппараты с полыми волокнами и различные мембранные модули для тангенциальной фильтрации.
Ионообменная хроматография эффективный способ разделения индивидуальных белков. Белки связываются с сорбентом и элюируются с него в соответствии со своим зарядом. Ионообменная хроматография позволяет отделить целевые белки от сопутствующих. Однако при выделении комплекса протеолитических ферментов из пищеварительных органов рыб ионообменная хроматография становится неэффективна, опять же из-за присутствия белково-липидного комплекса. Белково-липидный комплекс необратимо садится на сорбент и связывает значительное количество белков комплекса протеолитических ферментов. В результате их выход с ионообменной хроматографии не превышает 10%.
Недостатком данного известного способа является то, что предложенная в нем методика не учитывает особенностей тканей рыб. В них присутствует большое количество липидов, балластных и гидрофобных белков, а также нуклеиновых кислот и других полианионных молекул, которые склонны к образованию липидно-белковых комплексов, препятствующих выделению ферментов высокой степени очистки. Липиды и нуклеиновые кислоты способны соосаждаться с ферментами. Образование липидно-белковых и полиэлектролитных комплексов затрудняет ультрафильтрацию и мембранную фильтрацию, а также приводит к неспецифическому необратимому связыванию больших количеств целевых ферментов с сорбентом на технологических стадиях ионообменной хроматографии. В результате существенными недостатками взятой в качестве прототипа схемы являются низкий выход конечной продукции - (менее 10% от содержания в сырье) и низкая коллагенолитическая активность.
Под липидно-белковым комплексом понимаются комплексы, образующиеся в экстракте в результате связывания фосфолипидов с белками за счет ионных и гидрофобных взаимодействий, которые в процессе выделения целевых ферментов теряют растворимость и выпадают в виде нерастворимых комплексов и мицелл.
Они способны захватывать и осаждать целевые ферменты, что приводит к снижению выхода целевого продукта.
Технической задачей изобретения является создание эффективного способа получения очищенного комплекса протеолитических ферментов медицинского применения и расширение арсенала способов получения очищенного комплекса протеолитических ферментов.
Технический результат, обеспечивающий решение поставленной задачи, состоит в получении комплекса протеолитических ферментов, содержащего щелочные, нейтральные и кислые протеиназы, с повышенной коллагенолитической активностью за счет повышения специфической активности в отношении глубокого гидролиза коллагена и желатины, а также расщепления гемоглобина, казеина, снижении пирогенности (способность вещества вызывать повышение температуры тела), одновременно с повышением количественного выхода целевого продукты. Обнаружено, что помимо вышеназванных субстратов комплекс протеолитических ферментов способен в меньшей степени расщеплять хитозан, аналог гликозоаминогликанов соединительной ткани.
Указанный технический результат достигается за счет включения в процесс получения комплекса протеолитических ферментов на оптимальных стадиях операций отделения нуклеиновых кислот и липидно-белковых комплексов путем обработки экстракта полиэтиленимином, а также операции хроматографической очистки на гидрофобном сорбенте, что позволяет своевременно удалить из выделяемого фермента липополисахариды, ответственные за пирогенность. Отсутствие пирогености значительно повышает качество препарата по сравнению с аналогами при медицинском применении.
Сущность изобретения состоит в том, что способ выделения комплекса протеолитических ферментов из пищеварительных органов рыб включает … последовательно выполняемые гомогенизацию тканей в буфере, отделение измельченных тканей центрифугированием, фильтрование супернатанта через мембрану, высаливание белков в сульфате аммония, выделение сульфатаммонийного осадка центрифугированием, растворение осадка в буфере, обессоливание экстракта диафильтрацией, очистку ионообменной хроматографией на DEAE-целлюлозе, отличающийся тем, что до осаждения белков в сульфате аммония нуклеиновые кислоты и липидно-белковые комплексы удаляют из экстракта обработкой раствором полиэтиленимина, который добавляют до конечной концентрации полиэтиленимина в экстракте от 0,1 до 0,01 вес. %, а после ионообменной хроматографии осуществляют отделение остаточных пирогенов на гидрофобном сорбенте Сфероцелл С-80, стерилизующую фильтрацию через мембрану с размером пор 0,2 мкм и повторное обессоливание экстракта диафильтрацией с последующим приготовлением готовой формы лиофилизацией.
Предпочтительно, для получения готовой дозированной формы после отделения остаточных пирогенов вводят криопротектор, например, физиологически приемлемый буфер, моно- или дисахарид, в количестве от 1 до 10% вес. %, и/или медицинский биополимер, в количестве от 0,1 до 10% вес. %.
Предпочтительно, гомогенизацию производят постепенно добавляя 0,05 М Трис-буфер с добавкой 0,005М хлористого кальция CaCl2 рН 7,4. с последующей экстракцией на холоду.
В одних частных случаях реализации способа обессоливание экстракта диафильтрацией осуществляют путем пропускания раствора через полые волокна с размером пор 10 кДа, в иных случаях обессоливание экстракта диафильтрацией осуществляют на модуле тангенциальной ультрафильтрации.
Предпочтительно, моно- или дисахарид, выбирают из группы: маннитол, лактоза или трегалоза, а медицинский биополимер - из группы декстранов, поливинилпирралидонов, поливиниловых спиртов.
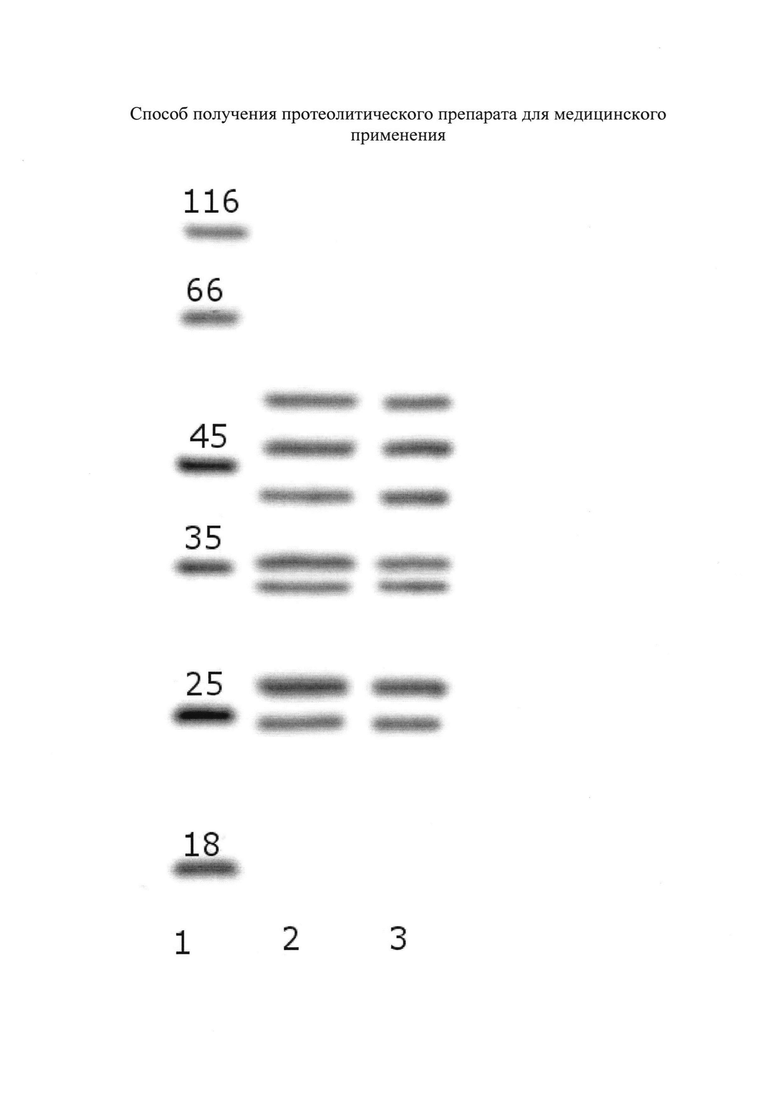
На чертеже представлен рисунок электрофореза.
ПРИМЕРЫ ПОЛУЧЕНИЯ КОМПЛЕКСА ПРОТЕОЛИТИЧЕСКИХ ФЕРМЕНТОВ
Пример 1.
В качестве исходного сырья использовали 400 г пищеварительных органов африканского сома. Органы отмыли от крови водой и провели гомогенизацию тканей в буфере, для чего дезинтегрировали в гомогенизаторе (10 мин), постепенно добавляя 0,05 М Трис-буфер с добавкой 0,005М хлористого кальция CaCl2 рН 7,4 (800 мл). Для отделения измельченных тканей (гомогената) центрифугированием предварительно провели экстракцию на холоду в течение суток. Общий объем экстракта составил 1200 мл. Гомогенат центрифугировали при 25000 об/мин, +4°С 30 мин. Верхний слой липидов откинули.
Осадок повторно гомогенизировали в 0,05 М Трис-буфере с добавкой 0,005М хлористого кальция CaCl2 рН 7,4 (200 мл), провели повторную экстракцию на холоду в течение суток. Общий объем экстракта составил 300 мл. Экстракт (Гомогенат) для отделение измельченных тканей центрифугировали при 25000 об/мин, +4°С 30 мин. Верхний слой липидов и осадок откинули.
Оба супернатанта объединили и получили 1200 мл раствора. Полученный раствор профильтровали через мембрану с размерами пор 0,2 мкм.
Раствор подтитровали 0,1 М раствором гидроокиси натрия до рН 8,0. Отдельно приготовили раствор, содержащий 5 вес. % полиэтиленимина. При перемешивании в 1200 мл раствора супернатанта добавили 24 мл раствора полиэтиленимина (ПЭИ) до конечной концентрации полиэтиленимина в растворе 0,1 вес. %. Конечную концентрацию полиэтиленимина в растворе (СПЭИ кон) рассчитываем по формуле 1:
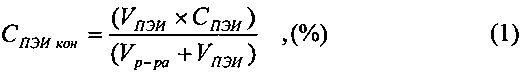
где VПЭИ - объем добавляемого раствора полиэтиленимина в миллилитрах,
СПЭИ - концентрация взятого раствора ПЭИ в весовых процентах,
Vp-pa - объем раствора супернатанта в миллилитрах.
Конечная концентрация полиэтиленимина составит (24*5)/1224≈0,1 (весовой процент полиэтиленимина в экстракте).
Образовался осадок нуклеиновых кислот с ПЭИ. Раствор перемешивали еще 40 минут при комнатной температуре для укрупнения осадка.
Осадок (нуклеиновые кислоты) выделили на центрифуге 12000 об/мин в течение 25 мин. Осадок ПЭИ и нуклеиновых кислот практически не имел протеолитической активности. Его отбросили.
В раствор, собранный после обработки ПЭИ, ввели сульфат аммония до 80% насыщения. Осадок формировали в холодильнике в течение суток. Осадок выделили центрифугированием при 12000 об/мин, +4°С 20 мин. Осадок белков растворили в 0,05М Трис-буфере. Раствор обессолили диафильтрацией на полых волокнах с размерами пор 10 кДа.
Провели ионообменную хроматографией на DEAE-целлюлозе. Колонку, содержащую 80 мл суспензии сорбента DEAE-целлюлозы, предварительно уравновесили 0,01 М Трис-буфером. Обезсоленый раствор белков нанесли на колонку. Элюцию сорбированных белков провели градиентом натрия хлористого 0,05-1,0 М NaCl в 0,01 М Трис буфере, рН 8,0. Скорость элюции 20-40 мл/час. Контроль содержания белка провели с помощью УФ-детектора при длине волны 280 нм. При элюции собрали три фракции белка, имеющего активность. Все активные фракции объединили. Объединенный раствор обессолили диафильтрацией на установке для тангенциальной ультрафильтрации.
Провели хроматографию полученного раствора на гидрофобном сорбенте для отделения остаточных пирогенов. В колонку поместили 50 мл гидрофобного сорбента Сфероцелл С-80, содержащего гидрофобные лиганды С8-С10 (сорбент на основе макропористой целлюлозной матрицы, модифицированной октил/децилглицидиловым эфиром). Колонку уравновесили 0,01 М фосфатным буфером с добавкой 0,05М NaCl, рН 8,0. Раствор пропускали через колонку с сорбентом со скоростью 20-40 мл/час. Выход фермента контролировали УФ-детектором по оптической плотности раствора. Собранный раствор обессолили с помощью диафильтрации на установке для тангенциальной ультрафильтрации и сконцентрировали до содержания белка 4 мг/мл (по Лоури). Довели рН полученного раствора до 7,4. Далее провели стерилизующую фильтрацию через мембрану 0,2 мкм.
В полученный раствор ввели моносахарид маннитол до концентрации 3,2% и полимер полиглюкин (декстран) до концентрации 0,1%. Раствор стерилизовали фильтрацией через мембрану с порами 0,2 мкм. Раствор разлили по стерильным флаконам по 1,25 мл/флакон.
Флаконы заморозили до -40°С в течение 2-10 часов и лиофилизовали в течение суток по стандартной программе. После лиофилизации флаконы герметично укупорили.
Получили очищенный комплекс протеолитических ферментов с активностью по коллагену 1500-2500 мкг субстрата/мин×мг фермента, который может быть использован для медицинских целей. В 1 флаконе 5 мг очищенного комплекса протеолитических ферментов. Всего получили 120 флаконов.
Выполнен электрофорез выделенного комплекса протеолитических ферментов в полиакриламидном геле. После окрашивания пластины геля красителем кумасси синим виден большой набор полос белков с различными массами. Наиболее интенсивно окрашены полосы соответствующие массам 25±2 и 36±3 кДа и мене интенсивно в области 45 кДа.
Специфическую коллагенолитическую активность выделенного в примерах комплекса протеолитических ферментов определялась по скорости ферментативного гидролиза коллагена методом окрашивания освобождающихся аминогрупп тринитробензолсульфокислотой (ТНБС). Активность определяли в микрограммах гидролизованного субстрата 1 миллиграммом фермента в минуту.
Метод ТНБС известен из технической литературы: Практическая химия белка: Пер. с англ. / Под ред. А. Дарбре. - М.: Мир, 1989. - 623 с.
Пример 2.
В качестве исходного сырья использовали 400 г пищеварительных органов щук. Органы отмыли от крови водой и провели гомогенизацию тканей в буфере, для чего дезинтегрировали в гомогенизаторе (10 мин), постепенно добавляя 0,05 М Трис-буфер включающий 1 М хлористого натрия NaCl и 0,005М хлористого кальция CaCl2 рН 7,4 (800 мл). Для отделения измельченных тканей (гомогената) центрифугированием предварительно провели экстракцию на холоду в течение суток. Общий объем экстракта составил 1200 мл. Гомогенат центрифугировали при 25000 об/мин, +4°С 30 мин. Верхний слой липидов откинули.
Осадок повторно гомогенизировали в 0,05 М Трис-буфере с добавкой 0,005М хлористого кальция CaCl2 рН 7,4 (200 мл). провели повторную Экстракцию провели на холоду в течение суток. Общий объем экстракта составил 300 мл. Экстракт (Гомогенат) для отделение измельченных тканей центрифугировали при 25000 об/мин, +4°С 30 мин. Верхний слой липидов и осадок откинули.
Оба супернатанта объединили и получили 1200 мл раствора. Полученный раствор профильтровали через мембрану с размерами пор 0,2 мкм.
Раствор подтитровали 0,1 М раствором гидроокиси натрия до рН 8,0. При перемешивании в 1200 мл раствора супернатанта добавили 6 мл раствора полиэтиленимина (ПЭИ) до конечной концентрации полиэтиленимина в растворе 0,025 вес. %.
Конечная концентрация полиэтиленимина составила (6*5)/1206≈0,025 (весовой процент полиэтиленимина в экстракте).
Образовался осадок нуклеиновых кислот с ПЭИ. Раствор перемешивали еще 40 минут при комнатной температуре для укрупнения осадка.
Осадок (нуклеиновые кислоты) выделили на центрифуге 12000 об/мин в течение 25 мин. Осадок ПЭИ и нуклеиновых кислот практически не имел протеолитической активности. Его отбросили.
В раствор, собранный обработки ПЭИ, ввели сульфат аммония до 80% насыщения. Осадок формировали в холодильнике в течение суток. Осадок выделили центрифугированием при 12000 об/мин, +4°С 20 мин. Осадок белков растворили в 0,05М Трис-буфере. Раствор обессолили диафильтрацией на полых волокнах с размерами пор 10 кДа.
Провели ионообменную хроматографией на DEAE-целлюлозе. Колонку, содержащую 80 мл суспензии сорбента DEAE-целлюлозы, предварительно уравновесили 0,01 М Трис-буфером. Обезсоленый раствор белков нанесли на колонку. Элюцию сорбированных белков провели градиентом натрия хлористого 0,05-1,0 М NaCl в 0,01 М Трис буфере, рН 8,0. Скорость элюции 20-40 мл/час. Контроль содержания белка провели с помощью УФ-детектора при длине волны 280 нм. При элюции собрали три фракции белка, имеющего активность. Все активные фракции объединили. Объединенный раствор обессолили диафильтрацией на установке для тангенциальной ультрафильтрации.
Провели хроматографию полученного раствора на гидрофобном сорбенте для отделения остаточных пирогенов. В колонку поместили 50 мл гидрофобного сорбента Сфероцелл С-80, содержащего гидрофобные лиганды С8-С10. Колонку уравновесили 0,01 М фосфатным буфером с добавкой 0,05М NaCl, рН 8,0. Раствор пропускали через колонку с сорбентом со скоростью 20-40 мл/час. Выход фермента контролировали УФ-детектором по оптической плотности раствора. Собранный раствор обессолили с помощью диафильтрации на установке для тангенциальной ультрафильтрации и сконцентрировали до содержания белка 2 мг/мл (по Лоури). Довели рН полученного раствора до 7,4. Далее провели стерилизующую фильтрацию через мембрану 0,2 мкм.
В полученный раствор ввели дисахарид лактозу до концентрации 10% и г полимер поливинилпирралидон (марки коллидон 17 PF) до концентрации 2,5%. Раствор стерилизовали фильтрацией через мембрану с порами 0,2 мкм. Раствор разлили по стерильным флаконам по 2 мл/флакон.
Флаконы заморозили до -40°С в течение 2-10 часов и лиофилизовали в течение суток по стандартной программе. После лиофилизации флаконы герметично укупорили.
Получили очищенный комплекс протеолитических ферментов с активностью по коллагену 1200-2000 мкг субстрата/мин×мг фермента, который может быть использован для медицинских целей. В 1 флаконе 4 мг очищенного комплекса протеолитических ферментов. Всего получили 125 флаконов.
Выполнен электрофорез выделенного комплекса протеолитических ферментов в полиакриламидном геле. После окрашивания пластины геля красителем кумасси синим виден большой набор полос белков с различными массами. Наиболее интенсивно окрашены полосы соответствующие массам 25±2 и 36±3 кДа и мене интенсивно в области 45 кДа.
Пример 3
400 г пищеварительных органов камбалы отмыли от крови водой. Провели гомогенизацию тканей в буфере, для чего ткани дезинтегрировали в гомогенизаторе (10 мин за 5 раз), постепенно добавляя 0,05 М Трис буфер, включающий 3М хлористого натрия NaCl, 0,005М хлористого кальция CaCl2, рН 7,4, 0,02% азида натрия NaN3. (800 мл). Экстракцию провели в холодильнике в течение трех суток. Общий объем экстракта около 1200 мл. Провели отделение измельченных тканей центрифугированием. Экстракт центрифугировали при 14000 Об/мин, +4°С 20 мин. Верхний слой липидов и осадок откинули.
Получили 800 мл раствора. Полученный раствор профильтровали через мембрану с размерами пор 0,2 мкм.
Раствор подтитровали 0,1 М раствором гидроокиси натрия до рН 8,0. При перемешивании в 800 мл раствора супернатанта добавили 1,6 мл раствора полиэтиленимина (ПЭИ) до конечной концентрации полиэтиленимина в растворе 0,01 вес. %.
Конечная концентрация полиэтиленимина составила (1,6*5)/801,6≈0,01 (весовой процент полиэтиленимина в экстракте).
Образовался осадок нуклеиновых кислот с ПЭИ. Раствор перемешивали еще 25 минут при комнатной температуре для укрупнения осадка.
Осадок (нуклеиновые кислоты) выделили на центрифуге 12000 об/мин в течение 25 мин. Осадок ПЭИ и нуклеиновых кислот практически не имел протеолитической активности. Его отбросили.
В раствор, собранный обработки ПЭИ, ввели сульфат аммония до 80% насыщения. Осадок формировали в холодильнике в течение суток. Осадок выделили центрифугированием при 12000 об/мин, +4°С 20 мин. Осадок белков растворили в 0,05М Трис-буфере. Раствор обессолили диафильтрацией на полых волокнах с размерами пор 10 кДа.
Провели ионообменную хроматографией на DEAE-целлюлозе. Колонку, содержащую 80 мл суспензии сорбента DEAE-целлюлозы, предварительно уравновесили 0,01 М Трис-буфером. Обезсоленый раствор белков нанесли на колонку. Элюцию сорбированных белков провели градиентом натрия хлористого 0,05-1,0 М NaCl в 0,01 М Трис буфере, рН 8,0. Скорость элюции 20-40 мл/час. Контроль содержания белка провели с помощью УФ-детектора при длине волны 280 нм. При элюции собрали три фракции белка, имеющего активность. Все активные фракции объединили. Объединенный раствор обессолили диафильтрацией на установке для тангенциальной ультрафильтрации.
Провели хроматографию полученного раствора на гидрофобном сорбенте для отделения остаточных пирогенов. В колонку поместили 50 мл гидрофобного сорбента Сфероцелл С-80, содержащего гидрофобные лиганды С8-С10. Колонку уравновесили 0,01 М фосфатным буфером с добавкой 0,05М NaCl, рН 8,0. Раствор пропускали через колонку с сорбентом со скоростью 20-40 мл/час. Выход фермента контролировали УФ-детектором по оптической плотности раствора. Собранный раствор обессолили с помощью диафильтрации на установке для тангенциальной ультрафильтрации и сконцентрировали до содержания белка 2 мг/мл (по Лоури). Довели рН полученного раствора до 7,4. Далее провели стерилизующую фильтрацию через мембрану 0,2 мкм.
В полученный раствор ввели фосфатный буфер до 0,1 М и полимер поливинилпирралидон (марки коллидон 17 PF) до концентрации 10%. Раствор стерилизовали фильтрацией через мембрану с порами 0,2 мкм. Раствор разлили по стерильным флаконам по 2 мл/флакон.
Флаконы заморозили до -40°С в течение 2-10 часов и лиофилизовали в течение суток по стандартной программе. После лиофилизации флаконы герметично укупорили.
Получили очищенный комплекс протеолитических ферментов с активностью по коллагену 1500-2000 мкг субстрата/мин×мг фермента, который может быть использован для медицинских целей. В 1 флаконе 4 мг очищенного комплекса протеолитических ферментов. Всего получили 150 флаконов.
Выполнен электрофорез в полиакриламидном геле выделенного комплекса протеолитических ферментов. После окрашивания пластины геля красителем кумасси синим наиболее интенсивно окрашены полосы соответствующие массам 25 и 36 кДа и мене интенсивно в области 45 кДа (фиг. 1).
Пример 4.
400 г внутренних органов пираньи отмыли от крови водой. Провели гомогенизацию тканей в буфере. Ткани дезинтегрировали в гомогенизаторе, постепенно добавляя 0,05 М Трис буфер, 0,005М CaCl2, рН 7,4 с 0,02% Азидом натрия NaN3. (800 мл). Экстракцию провели в холодильнике в течение суток. Общий объем экстракта около 1200 мл. Отделение измельченных тканей центрифугированием Экстракт центрифугировали при 14000 Об/мин, +4°С 20 мин. Верхний слой липидов и осадок откинули.
Получили 600 мл раствора. Полученный раствор профильтровали через мембрану с размерами пор 0,2 мкм.
Раствор подтитровали 0,1 М раствором гидроокиси натрия до рН 8,0. При перемешивании в 600 мл раствора супернатанта добавили 3 мл раствора полиэтиленимина (ПЭИ) до конечной концентрации полиэтиленимина в растворе 0,025 вес. %.
Конечная концентрация полиэтиленимина составила (3*5)/603≈0,025 (весовой процент полиэтиленимина в экстракте).
Образовался осадок нуклеиновых кислот с ПЭИ. Раствор перемешивали еще 30 минут при комнатной температуре для укрупнения осадка.
Осадок (нуклеиновые кислоты) выделили на центрифуге 12000 об/мин в течение 25 мин. Осадок ПЭИ и нуклеиновых кислот практически не имел протеолитической активности. Его отбросили.
В раствор, собранный обработки ПЭИ, ввели сульфат аммония до 80% насыщения. Осадок формировали в холодильнике в течение суток. Осадок выделили центрифугированием при 12000 об/мин, +4°С 20 мин. Осадок белков растворили в 0,05М Трис-буфере. Раствор обессолили диафильтрацией на полых волокнах с размерами пор 10 кДа.
Провели ионообменную хроматографией на DEAE-целлюлозе. Колонку, содержащую 80 мл суспензии сорбента DEAE-целлюлозы, предварительно уравновесили 0,01 М Трис-буфером. Обезсоленый раствор белков нанесли на колонку. Элюцию сорбированных белков провели градиентом натрия хлористого 0,05-1,0 М NaCl в 0,01 М Трис буфере, рН 8,0. Скорость элюции 20-40 мл/час. Контроль содержания белка провели с помощью УФ-детектора при длине волны 280 нм. При элюции собрали три фракции белка, имеющего активность. Все активные фракции объединили. Объединенный раствор обессолили диафильтрацией на установке для тангенциальной ультрафильтрации.
Провели хроматографию полученного раствора на гидрофобном сорбенте для отделения остаточных пирогенов. В колонку поместили 50 мл гидрофобного сорбента Сфероцелл С-80, содержащего гидрофобные лиганды С8-С10. Колонку уравновесили 0,01 М фосфатным буфером с добавкой 0,05М NaCl, рН 8,0. Раствор пропускали через колонку с сорбентом со скоростью 20-40 мл/час. Выход фермента контролировали УФ-детектором по оптической плотности раствора. Собранный раствор обессолили с помощью диафильтрации на установке для тангенциальной ультрафильтрации и сконцентрировали до содержания белка 2 мг/мл (по Лоури). Довели рН полученного раствора до 7,4. Далее провели стерилизующую фильтрацию через мембрану 0,2 мкм.
В полученный раствор ввели Трис-буфер до 0,025 М и полимер поливиниловый спирт до концентрации 0,1%, и дисахарид трегалозу до концентрации 4%. Раствор стерилизовали фильтрацией через мембрану с порами 0,2 мкм. Раствор разлили по стерильным флаконам по 2 мл/флакон.
Флаконы заморозили до -40°С в течение 2-10 часов и лиофилизовали в течение суток по стандартной программе. После лиофилизации флаконы герметично укупорили.
Получили очищенный комплекс протеолитических ферментов с активностью по коллагену 1500-2200 мкг субстрата/мин×мг фермента, который может быть использован для медицинских целей. В 1 флаконе 4 мг очищенного комплекса протеолитических ферментов. Всего получили 100 флаконов.
Выполнен электрофорез в полиакриламидном геле выделенного комплекса протеолитических ферментов. После окрашивания пластины геля красителем кумасси синим наиболее интенсивно окрашены полосы соответствующие массам 25 и 36 кДа и мене интенсивно в области 45 кДа.
Пример 5
В качестве исходного сырья использовали 500 г пищеварительных органов катрановой акулы (Squalus acanthias). Органы отмыли от крови очищенной водой и дезинтегрировали в гомогенизаторе (10 мин), постепенно добавляя 0,05 М Трис-буфер с добавкой 1М NaCl и 0,005М хлористого кальция CaCl2 рН 7,4 (1000 мл). Экстракцию провели на холоду. Общий объем экстракта составил 1500 мл. Гомогенат центрифугировали при 25000 об/мин, +4°С 30 мин. Верхний слой липидов откинули.
Получили 1200 мл раствора. Полученный раствор профильтровали через мембрану с размерами пор 0,2 мкм.
Раствор подтитровали 0,1 М раствором гидроокиси натрия до рН 8,0. При перемешивании в 1200 мл раствора супернатанта добавили 12 мл раствора полиэтиленимина (ПЭИ) до конечной концентрации полиэтиленимина в растворе 0,05 вес. %.
Конечная концентрация полиэтиленимина составила (12*5)/1212≈0,05 (весовой процент полиэтиленимина в экстракте).
Образовался осадок нуклеиновых кислот с ПЭИ. Раствор перемешивали еще 30 минут при комнатной температуре для укрупнения осадка.
Осадок (нуклеиновые кислоты) выделили на центрифуге 12000 об/мин в течение 25 мин. Осадок ПЭИ и нуклеиновых кислот практически не имел протеолитической активности. Его отбросили.
В раствор, собранный обработки ПЭИ, ввели сульфат аммония до 80% насыщения. Осадок формировали в холодильнике в течение суток. Осадок выделили центрифугированием при 12000 об/мин, +4°С 20 мин. Осадок белков растворили в 0,05М Трис-буфере. Раствор обессолили диафильтрацией на полых волокнах с размерами пор 10 кДа.
Провели ионообменную хроматографией на DEAE-целлюлозе. Колонку, содержащую 80 мл суспензии сорбента DEAE-целлюлозы, предварительно уравновесили 0,01 М Трис-буфером. Обезсоленый раствор белков нанесли на колонку. Элюцию сорбированных белков провели градиентом натрия хлористого 0,05-1,0 М NaCl в 0,01 М Трис буфере, рН 8,0. Скорость элюции 20-40 мл/час. Контроль содержания белка провели с помощью УФ-детектора при длине волны 280 нм. При элюции собрали три фракции белка, имеющего активность. Все активные фракции объединили. Объединенный раствор обессолили диафильтрацией на установке для тангенциальной ультрафильтрации.
Провели хроматографию полученного раствора на гидрофобном сорбенте для отделения остаточных пирогенов. В колонку поместили 50 мл гидрофобного сорбента Сфероцелл С-80, содержащего гидрофобные лиганды С8-С10. Колонку уравновесили 0,01 М фосфатным буфером с добавкой 0,05М NaCl, рН 8,0. Раствор пропускали через колонку с сорбентом со скоростью 20-40 мл/час. Выход фермента контролировали УФ-детектором по оптической плотности раствора. Собранный раствор обессолили с помощью диафильтрации на установке для тангенциальной ультрафильтрации и сконцентрировали до содержания белка 2,5 мг/мл (по Лоури). Довели рН полученного раствора до 7,4. Далее провели стерилизующую фильтрацию через мембрану 0,2 мкм.
В полученный раствор ввели Трис-буфер до 0,025 М моносахарид маннитол до концентрации 4%. Раствор простерилизовали фильтрацией через мембрану с порами 0,2 мкм. Раствор разлили по стерильным флаконам по 2 мл/флакон.
Флаконы заморозили до -40°С в течение 2-10 часов и лиофилизовали в течение суток по стандартной программе. После лиофилизации флаконы герметично укупорили.
Получили очищенный комплекс протеолитических ферментов с активностью по коллагену 1800-2600 мкг субстрата/мин×мг фермента, который может быть использован для медицинских целей. В 1 флаконе 5 мг очищенного комплекса протеолитических ферментов. Всего получили 130 флаконов.
Выполнен электрофорез в полиакриламидном геле выделенного комплекса протеолитических ферментов. После окрашивания пластины геля красителем кумасси синим наиболее интенсивно окрашены полосы соответствующие массам 25 и 36 кДа и мене интенсивно в области 45 кДа.
Пример 6
В качестве исходного сырья использовали 500 г пищеварительных органов ската морская лисица (Raja clavata). Органы отмыли от крови очищенной водой и дезинтегрировали в гомогенизаторе (10 мин), постепенно добавляя 1М NaCl и 0,005М хлористого кальция CaCl2, рН 7,4 (1000 мл). Экстракцию провели на холоду. Общий объем экстракта составил 1500 мл. Гомогенат центрифугировали при 25000 об/мин, +4°С 30 мин. Верхний слой липидов откинули.
Получили 1200 мл раствора. Полученный раствор профильтровали через мембрану с размерами пор 0,2 мкм.
Раствор подтитровали 0,1 М раствором гидроокиси натрия до рН 8,0. При перемешивании в 1200 мл раствора супернатанта добавили 4 мл раствора полиэтиленимина (ПЭИ) до конечной концентрации полиэтиленимина в растворе 0,017 вес. %.
Конечная концентрация полиэтиленимина составила (4*5)/1204≈0,017 (весовой процент полиэтиленимина в экстракте).
Образовался осадок нуклеиновых кислот с ПЭИ. Раствор перемешивали еще 40 минут при комнатной температуре для укрупнения осадка.
Осадок (нуклеиновые кислоты) выделили на центрифуге 12000 об/мин в течение 25 мин. Осадок ПЭИ и нуклеиновых кислот практически не имел протеолитической активности. Его отбросили.
В раствор, собранный обработки ПЭИ, ввели сульфат аммония до 80% насыщения. Осадок формировали в холодильнике в течение суток. Осадок выделили центрифугированием при 12000 об/мин, +4°С 20 мин. Осадок белков растворили в 0,05М Трис-буфере. Раствор обессолили диафильтрацией на полых волокнах с размерами пор 10 кДа.
Провели ионообменную хроматографией на DEAE-целлюлозе. Колонку, содержащую 80 мл суспензии сорбента DEAE-целлюлозы, предварительно уравновесили 0,01 М Трис-буфером. Обезсоленый раствор белков нанесли на колонку. Элюцию сорбированных белков провели градиентом натрия хлористого 0,05-1,0 М NaCl в 0,01 М Трис буфере, рН 8,0. Скорость элюции 20-40 мл/час. Контроль содержания белка провели с помощью УФ-детектора при длине волны 280 нм. При элюции собрали три фракции белка, имеющего активность. Все активные фракции объединили. Объединенный раствор обессолили диафильтрацией на установке для тангенциальной ультрафильтрации.
Провели хроматографию полученного раствора на гидрофобном сорбенте для отделения остаточных пирогенов. В колонку поместили 50 мл гидрофобного сорбента Сфероцелл С-80, содержащего гидрофобные лиганды С8-С10. Колонку уравновесили 0,01 М фосфатным буфером с добавкой 0,05М NaCl, рН 8,0. Раствор пропускали через колонку с сорбентом со скоростью 20-40 мл/час. Выход фермента контролировали УФ-детектором по оптической плотности раствора. Собранный раствор обессолили с помощью диафильтрации на установке для тангенциальной ультрафильтрации и сконцентрировали до содержания белка 2,5 мг/мл (по Лоури). Довели рН полученного раствора до 8,0. Далее провели стерилизующую фильтрацию через мембрану 0,2 мкм.
В полученный раствор ввели Трис-буфер до 0,025 М моносахарид маннитол до концентрации 4%. Раствор простерилизовали фильтрацией через мембрану с порами 0,2 мкм. Раствор разлили по стерильным флаконам по 2 мл/флакон.
Флаконы заморозили до -40°С в течение 2-10 часов и лиофилизовали в течение суток по стандартной программе. После лиофилизации флаконы герметично укупорили.
Получили очищенный комплекс протеолитических ферментов с активностью по коллагену 1800-2600 мкг субстрата/мин×мг фермента, который может быть использован для медицинских целей. В 1 флаконе 5 мг очищенного комплекса протеолитических ферментов. Всего получили 120 флаконов.
Выполнен электрофорез в полиакриламидном геле выделенного комплекса протеолитических ферментов. После окрашивания пластины геля красителем кумасси синим наиболее интенсивно окрашены полосы соответствующие массам 25 и 36 кДа и мене интенсивно в области 45 кДа.
СРАВНЕНИЕ ПОЛУЧЕННОГО КОМПЛЕКСА ПРОТЕОЛИТИЧЕСКИХ ФЕРМЕНТОВ С ИЗВЕСТНЫМИ АНАЛОГАМИ
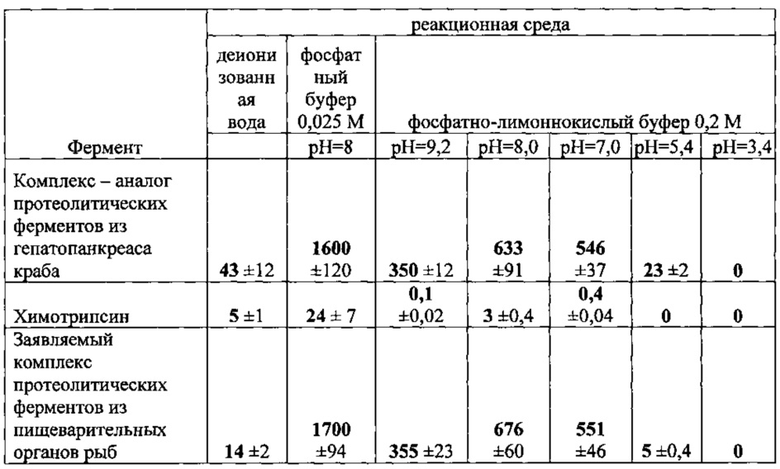
Сравнительную специфическую активность определяли для полученного комплекса протеолитических ферментов в сравнении с коммерческим препаратом коллагеназы краба и коммерческой сериновой протеазой химотрипсин. В качестве субстрата использовали высокомолекулярный фотожелатин. Реакцию ферментативного гидролиза провели при +37°С. Для получения сравнимых результатов испытуемые препараты растворили в одинаковой концентрации по белку. Концентрацию белка нормировали по калибровочным кривым по оптической плотности при 280 нм. Для определения активности использовали концентрации ферментов 0,25, 0,1, 0,05, 0,025 мг/мл. Активность фермента зависит от ионной силы, природы присутствующих солей и рН среды. В деионизированной воде протеолитические ферменты мало активны. Для создания различного кислотно-основного баланса среды использовали 0,2 М фосфатно-лимоннокислый буфер.
Полученные результаты представлены в таблице. Специфическая активность выделенного комплекса протеолитических ферментов в области рН 7,0-9,2 практически совпадает со специфической активностью комплекса ферментов, выделенных из гепатопанкреаса краба. Химотрипсин показал низкую активность в отношении к субстрату.
В таблице приведена специфическая активность протеолитических ферментов в мкг субстрата*мин/мг фермента.
Известно, что для каждого фермента существует оптимальная область рН, при котором его ферментативная активность максимальна. Для установления рН-оптимума действия обычно используют универсальные буферы, эффективные в широком диапазоне рН. Мы использовали фосфатно-лимоннокислый буфер. К сожалению, оказалось, что анион лимонной кислоты (цитрат), снижает активность протеолитических ферментов. Тем не менее, во-первых удалось установить, что рН-оптимум для комплекса протеолитических ферментов лежит в области рН 7,0-9,0, во вторых комплекс протеолитических ферментов, выделенных из пищеварительных органов рыб имеет близкую специфическую активность и рН-оптимум к характеристикам крабовой коллагеназы
Таблица
ПРИМЕР ПРИМЕНЕНИЯ ПОЛУЧЕННОГО КОМПЛЕКСА ПРОТЕОЛИТИЧЕСКИХ ФЕРМЕНТОВ
Терапевтическое действие препарата - эффект очищения ран и отторжение струпа исследовали на модели термического ожога на крысах самцах. Эксперименты выполнены на белых беспородных крысах, самцах - 60 штук. Животные были распределены на группы с помощью метода рандомизации с использованием в качестве основного критерия массы тела. При сравнительном изучении влияния комплекса на динамику отторжения некротического струпа было использовано по 5 особей крыс. При изучении специфической активности опытные и контрольные группы состояли из 10 крыс (ожоги). В группе изучения местно-раздражающего действия комплекса использовали группы по 5 крыс.
Животных из контрольной и опытных групп наркотизировали диэтиловым эфиром и наносили на депилированную кожу спины в грудино-поясничной области термический ожог раскаленной до 200°С металлической пластиной площадью 2 см2 с экспозицией 10 секунд. Экспериментальную терапию проводили комплексом протеолитических ферментов, полученным по примерам 1-6 и препаратом сравнения крабовой коллагеназой при разведении их водой.
На модели термических ожогов у крыс были установлены более ранние клинически значимые признаки коллагенолитических свойств исследуемого комплекса - краевое, неполное и полное отторжение некротического струпа, сопоставимое с таковыми показателями при терапии ожоговых ран кожи крыс крабовой коллагеназой. Положительное действие комплекса протеолитических ферментов отражалось в активной эпителизации поверхности раневого дефекта с менее выраженными реактивными изменениями многослойного плоского эпителия в краях раны по сравнению с контрольной группой, в которой отмечались менее выраженные процессы регенерации с тенденцией к псевдоэпителиоматозной гиперплазии в краях раны.
Комплекс протеолитических ферментов, полученный в соответствии с заявляемым способом, в частности, по примерам 1-6, не обладает местно-раздражающим действием на кожу крыс при 21-дневном нанесении.
Одновременно он обладает регенерирующим, анальгетическим и дезодорирующим действием.
Определение подлинности и чистоты комплекса протеолитических ферментов методом электрофореза
Для определения подлинности (соответствия молекулярной массе) и чистоты (отсутствие посторонних примесей) используют метод вертикального электрофореза в полиакриламидном геле в нативных и редуцирующих условиях с последующей визуализацией окрашивающим раствором Кумасси. Использовали систему для электрофореза "Mini-PROTEAN 3". Электрофорез проводили при следующих условиях: концентрация разделяющего геля 4-20%, концентрация концентрирующего геля 4%, сила тока 25 мА, напряжение при разделении белков 240 В, продолжительность 50 мин, температура среды +7 град. Цельсия. В лунки дорожек вносили исследуемый раствор препарата в 0,5 М трис-HCl буфере ((гидроксиметил)аминометан) рН 6,8 в количестве 2-7 мкл, из расчета чтобы нагрузка вещества на дорожку геля составляла около 2,5 мкг.
По окончании электрофореза гель фиксировали в спиртовом растворе уксусной кислоты, далее окрашивали коллоидным красителем Кумасси в течение 40-60 мин на шейкере, далее краситель сливали и отмывали гель водой до исчезновения фона. Для оценки результатов при выполнении электрофореза в одну из дорожек вносят смесь калибровочных белков с молекулярными массами 18 кДа, 25 кДа, 35 кДа, 45 кДа, 66 кДа, 116 кДа.
На чертеже дорожка 1 рисунка электрофореза содержит калибровочные белки. Дорожки 2 и 3 содержат характерные полосы белков комплекса протеолитических ферментов в редуцирующих условиях. Нагрузка на дорожку 2 составляла 3 мкл раствора, нагрузка на дорожку 3 5 мкл раствора.
Комплекс протеолитических ферментов дает специфические окрашенные полосы в области 23-28 кДа, соответствующие эластазам, в области 32-37 кДа, соответствующие трипсиноподобным ферментам, в области 40-50 кДа, соответствующие химотрипсиноподобным ферментам. В зависимости от процедуры исполнения и степени автолиза окрашенные полосы в этих регионах могут быть как одинарными, так и расщепляться на дублеты и триплеты изоферментов.
Характерная для комплекса протеолитических ферментов получаемая в электрофорезе последовательность полос отлична от последовательности полос при электрофорезе крабовой коллагеназе и индивидуальных препаратов протеолитических ферментов, таких как трипсин, химотрипсин и др. Для комплекса протеолитических ферментов не наблюдается различий в картине электрофореза в нередуцирующих и редуцирующих условиях.
Нативные ферменты и белки очень часто находятся в виде димеров, тетрамеров и октамеров. Для того чтобы их разделить в процедуре электрофореза до индивидуальных белков создают специальные условия, обычно, путем добавления восстановителя - меркаптоэтанола, который разрывает дисульфидные связи между белками.
Таким образом реализуется получение комплекса протеолитических ферментов, содержащего щелочные, нейтральные и кислые протеиназы, с повышенной коллагенолитической активностью за счет повышения специфической активности в отношении глубокого гидролиза коллагена и желатины, а также расщепления гемоглобина, казеина, снижении пирогенности (способность вещества вызывать повышение температуры тела), одновременно с повышением количественного выхода целевого продукты. Помимо вышеназванных субстратов комплекс протеолитических ферментов способен в меньшей степени расщеплять хитозан.
Преимущества заявляемого способа обусловлены включением в процесс получения комплекса протеолитических ферментов на оптимальных стадиях операций отделения нуклеиновых кислот и липидно-белковых комплексов путем обработки экстракта полиэтиленимином, а также операции хроматографической очистки на гидрофобном сорбенте, что позволяет своевременно удалить из выделяемого фермента липополисахариды, ответственные за пирогенность. Отсутствие пирогености значительно повышает качество препарата по сравнению с аналогами при медицинском применении
Изобретение позволяет получить протеолитический препарат, который может быть использован в медицине, в косметике и в биотехнологии, в медицине при очистке инфицированных ран от некротических тканей, ожоговых ран, отморожений, для ускорения отторжения струпов и некротических язв, при терапии грыжи интервертебрального диска позвоночника, для лечения келоидных рубцов, для лечения послеоперационных спаек в органах брюшной полости; в косметике для очистки проблемной кожи; в биотехнологии для получения и выделения клеточных культур человеческих и животных тканей.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ПОЛУЧЕНИЯ АЛЬФА-2 ИНТЕРФЕРОНА ЧЕЛОВЕКА | 1989 |
|
SU1640996A1 |
| СПОСОБ ПОЛУЧЕНИЯ РЕКОМБИНАНТНОГО АНТАГОНИСТА РЕЦЕПТОРА ИНТЕРЛЕЙКИНА-1 ЧЕЛОВЕКА "АРИЛ" | 2004 |
|
RU2326947C2 |
| Способ получения большого фрагмента ДНК-полимеразы I ЕSснеRIснIа coLI - фрагмента Кленова | 1988 |
|
SU1541256A1 |
| СПОСОБ ПОЛУЧЕНИЯ АКТИВАТОРА ПЛАЗМИНОГЕНА | 2007 |
|
RU2346983C1 |
| СПОСОБ ПОЛУЧЕНИЯ РЕКОМБИНАНТНОГО ИНГИБИТОРА СЕРИНОВЫХ ПРОТЕИНАЗ КАМЧАТСКОГО КРАБА | 2014 |
|
RU2560264C1 |
| СПОСОБ ПОЛУЧЕНИЯ ПРЕПАРАТА ИНТЕРЛЕЙКИНА-1-БЕТА | 2002 |
|
RU2234513C2 |
| Способ получения РНК-лигазы | 1979 |
|
SU910762A1 |
| Способ выделения полинуклеотидфосфорилазы | 1981 |
|
SU1076445A1 |
| СПОСОБ ПОЛУЧЕНИЯ ОВОИНГИБИТОРА | 1997 |
|
RU2129438C1 |
| Способ очистки трипсина | 1989 |
|
SU1742329A1 |
Изобретение относится к биотехнологии. Предложен способ извлечения протеолитических ферментов из пищеварительных органов рыб. Способ включает гомогенизацию тканей в буфере, выделение ферментов в присутствии сульфата аммония, причем до осаждения белков в сульфате аммония нуклеиновые кислоты и липидно-белковые комплексы удаляют из экстракта обработкой раствором полиэтиленимина, который добавляют до его конечной концентрации в экстракте от 0,1 до 0,01 вес.%. Отделение остаточных пирогенов осуществляют на гидрофобном сорбенте Сфероцелл С-80 с дальнейшим повторным обессоливанием экстракта. Раствор ферментов стерилизуют фильтрацией через мембрану с размером пор 0,2 мкм с последующим приготовлением лиофилизированной готовой формы ферментного препарата. Изобретение позволяет получить протеолитические ферменты с повышенной коллагенолитической активностью. 5 з.п. ф-лы, 1 ил., 1 табл., 7 пр.
1. Способ выделения комплекса протеолитических ферментов из пищеварительных органов рыб, включающий последовательно выполняемые гомогенизацию тканей в буфере, отделение измельченных тканей центрифугированием, фильтрование супернатанта через мембрану, высаливание белков в сульфате аммония, выделение сульфатаммонийного осадка центрифугированием, растворение осадка в буфере, обессоливание экстракта диафильтрацией, очистку ионообменной хроматографией на DEAE-целлюлозе, отличающийся тем, что до осаждения белков в сульфате аммония нуклеиновые кислоты и липидно-белковые комплексы удаляют из экстракта обработкой раствором полиэтиленимина, который добавляют до конечной концентрации полиэтиленимина в экстракте от 0,1 до 0,01 вес.%, а после ионообменной хроматографии осуществляют отделение остаточных пирогенов на гидрофобном сорбенте Сфероцелл С-80, стерилизующую фильтрацию через мембрану с размером пор 0,2 мкм и повторное обессоливание экстракта диафильтрацией с последующим приготовлением готовой формы лиофилизацией.
2. Способ по п. 1, отличающийся тем, что для получения готовой дозированной формы после отделения остаточных пирогенов вводят криопротектор, например физиологически приемлемый буфер, моно- или дисахарид, в количестве от 1 до 10 вес.%, и/или медицинский биополимер, в количестве от 0,1 до 10 вес.%.
3. Способ по п. 2, отличающийся тем, что гомогенизацию производят с медленным добавлением 0,05 М Трис-буфер с добавкой 0,005М хлористого кальция CaCl2 рН 7,4. с последующей экстракцией на холоду.
4. Способ по любому из пп. 1-3, отличающийся тем, что обессоливание экстракта диафильтрацией осуществляют путем пропускания раствора через полые волокна с размером пор 10 кДа.
5. Способ по любому из пп. 1-3, отличающийся тем, что обессоливание экстракта диафильтрацией осуществляют на модуле тангенциальной ультрафильтрации.
6. Способ по любому из пп. 1-3, отличающийся тем, что моно- или дисахарид выбирают из группы: маннитол, лактоза или трегалоза, а медицинский биополимер - из группы декстранов, поливинилпирралидонов, поливиниловых спиртов.
| TERUEL S.R.L., SIMPSON B.K | |||
| Characterization of the collagenolytic enzyme fraction from winter flouder (Pseudopleuronectes americanus).// Comp | |||
| Biochem | |||
| Physiol., 1995, v.112B, 131-136 | |||
| СПОСОБ ПОЛУЧЕНИЯ ПРОТЕОЛИТИЧЕСКОГО КОМПЛЕКСА | 1992 |
|
RU2034028C1 |
| СЕРИНОВЫЕ ПРОТЕИНАЗЫ РЫБ И ИХ ФАРМАЦЕВТИЧЕСКОЕ И КОСМЕТИЧЕСКОЕ ПРИМЕНЕНИЕ | 2000 |
|
RU2264824C2 |

