(54) СПОСОБ ПОЛУЧЕНИЯ РНК-ЛИГАЗЫ
I
Изобретение относится к биохимии и может быть использовано в процессах получения полирибонуклеотидов с определемис последовательностью гетеро- циклических оснований.
Пригодность препаратов индуцируемой бактери.офагом Т4 поли (рибонуклеотид) t поли(рибонуклеотид) лигаэы (образующей АНФ) или РНК-лигазы для Q целей полирибонуклеотидного Синтеза определяется в первую очередь уровнем активности сопутствующей 3 - экзойуклеазы.
Известен способ получения РНК-лигазы, включающий разрушение клеток E.coli, инфицированных бактериофагом Т4, осгикдение нуклеиновых кислот стрептомидинсульфатом, фракционирование белков сульфатом аммония, последо- 0 вательные хроматографические стадии на традиционных хроматографических носителях (ЦЭ)-целлюлоз а, сефадекс G-,100, гидроксилапатит, ДЭАЭ-сефадекс, А-25, ОАЕ-сефадекс А-25) и за- js ключительную аффинную хроматографию 1 .
Известны и другие аналогичные способы получения РНК-лигаэы, незначительно отличающиеся от указанного JQ способа и друг от друга порядком
проведения и числом хроматографических стадий, а также выбором носителя для аффинной хроматографии 21 и 3.
Однако эти способы являются многостадийными, трудоемки, выход фермента уменьшается пропорционально увелнчению числа хроматографических стадий, носители для аффинной хроматографии дороги и труднодоступны, к тому же они часто нестабильны и загрязняют препараты РНК-лигазы на последней стадии очистки. Кроме того, во всех случаях получения РНК -лигазы в качестве исходного материала ислользованы специально сконструирован ные двойные мутанты бактериофага Т4 Ш и 21 или мутантный штамм Е.соИ В 3.
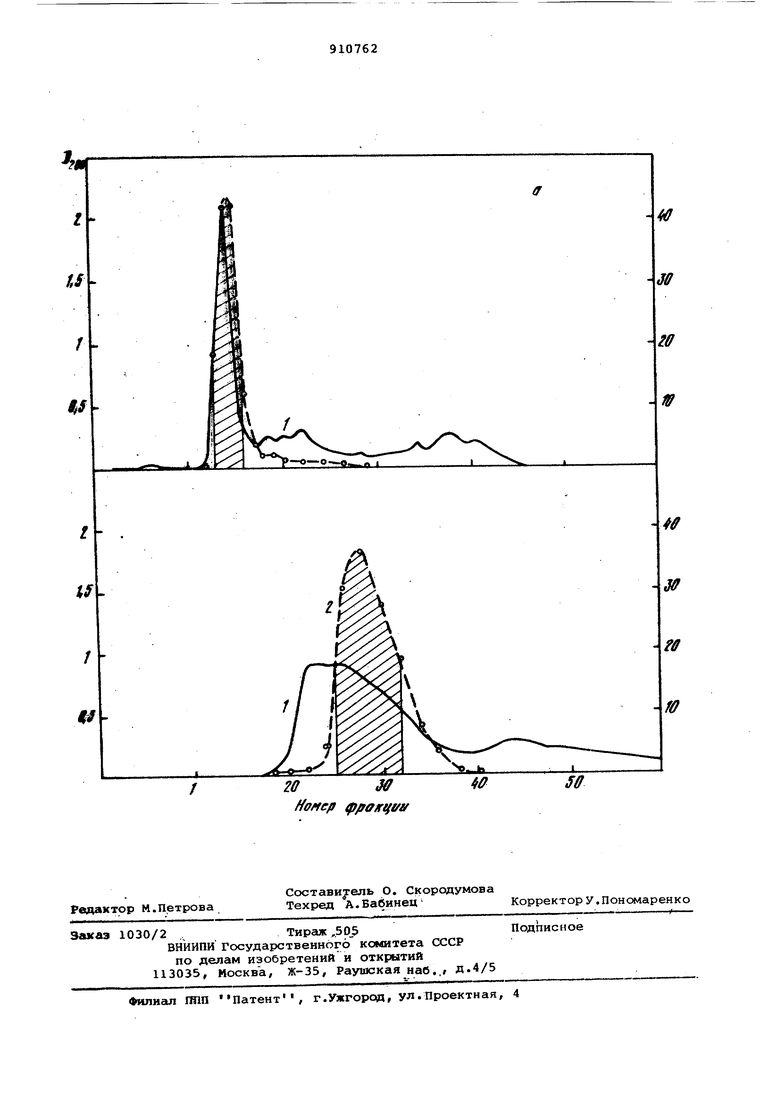
Наиболее близким по технической сущности к предлагаемому является способ получения РНК-лигазы, пригодной для целей полирибонуклеотидного синтеза, заключающийся в инкубации клеток E.coli с бактериофагом Т4 в течение 3 ч, разрушении клеток ультразвуковом с отделением экстракта и последующей обработке экстракта стрептомицинсульфатом и сульфатом а()мсния, после чего для выделения целевого продукта полученный ферментный раствор подвергают последователь но проводимой хроматографии в услови ях буфера рН 7,5 ± 0,1 на следуквдих сорбентах в следующих условиях: а)хроматография на диэтиламиноэтилцеллюлозе с элюцией 0,3 М раство ром Nad в исходном буфере (в качест ве исходного обычно используют 0,01 трис-НС буфер, содержащий 10 мМ 2меркаптоэтанола, рН 7,5); б)повторная хроматография получе него на стадии а элюата,на диэтилами ноэтилцеллюлозев условиях того же исходного буфера с элюцией линейным градиентом NaC 0-0,3 М в исходном буфере; в)хроматография полученного на стадии б.эЛюата на сшитом эпихлоргид рином дйэтил-2-оксипропиламиноэтилдекстране (QAE-сефадекс), в колонку которым предварительно вводят раство KC.I. в линейном градиенте концентраци 0,175-0,4 М в исходном буфере с элюцией 1 М раствором КС (в качестве исходного обычно используют 0,1 М трис-HCt буфер, содержащий 10 мМ 2меркаптоэтанола, 0,1 мМ этилендиамин тетрауксусной кислоты, рН 7,5), пол ченный на стсщии в элюат подвергают гель-фильтрации с помощью сшитого эпихлоргидрином декстрана ( сефадек са G-100) Б условиях буфера рН 7,5+ 0,1(обычно 0,02 М трис-HCt буфер, с держащий 0,1 М Кс1, 10 мМ 2-меркапт этанола , О, 1 мМ этилендиаминтетраук ной кислоты, рН 7,5); , заключительной стадией процесса является изоэлектрическое фокусиров ние на инертном носителе биогеле Рв присутствии амфолинов (LKB, Швеция) 4 . после проведения этой стадии из 100 г биомассы получают 2,35 нмоль препарата, РНК-лигазы, содержащего 4,38 ед. активности З-экзонуклеазы на 1 пмоль- РНК-лигазы, что в условиях синтеза полирибонуклеотидов соответствует в среднем разрыву одной фосфодиэфирной связи на каждые 250 вновь образованных (методы определения активности тестируемых ферментов и размерности единиц описаны 4 . Недостатком способа является нал чие в схеме очистки трудоемкого изо электрического фокусирования, для выполнения которого необходимы дефи цитные импортные амфолины, основные потери РНК-лигазы (до 75% фермента) связаны также с этой стадией. Крсиле того, этот способ характеризуется недостаточно высоким выходом целево го продукта, и Нсшичием высокого уровня примесной 3 -экзонуклеазы. Целью изобретения является упрсвд ние процесса получения РНК-лигазы, увеличение ее выхода и улучшение ка чества, определяемое снижением содержания 3 -экзонуклеаэы в целевом продукте, что делает его более пригодным для синтеза полирибонуклеотидов. Поставленная цель достигается спо собом получения РНК-лигаэы, который заключается в том, что проводят инкубацию клеток E.cbli с бактериофагом Т4 в течение 90±20 мин, после чего клетки разрушают ультразвуковом и отделяют экстракт;для выделения целевого продукта из экстракта последний обрабатывают стрептомицинсульфатом и сульфатом аммония и затем подвергают хроматографии в условиях буфера рН 7,5+0,1, проводимой последовательно на диэтиламиноэтилцеллюлозе с элюцией исходным буфером, содержащим 0,3 М NaCi, причем перед элюцией колонку с сорбентом промывают 0,12+0,01 М раствором NaCl в исходном буфере; повторно - на диэтиламиноэтилцеллюлозе с элюцией линейным градиентом NaCE от О до 0,3 М в исходном буфере; на сшитом эпихлоргидрином ДИЭТИЛ-2-ОКСИпропиламиноэтилдекстране { АЕ-сефадекс А-25) ,в колонку с которялл перед хроматографией предварительно вводят раствор KCi в линейном градиенте концентрации от 0,1+0,01 до 0,4+0,0-1 М в исходном буфере,с элюцией 1+0,01 М-, раствором КС1 и гель-фильтрацией полученного элюата с помощью сшитого эпихлоргидрином декстрана (сефадекс G-100) в условиях буфера рН 7,5+0,1. Обычно в качества исходного буфера при хроматографии на ДЭАЭ- целлюлозе, в обоих случаях используют 0,01 М трис-буфер, содержсцдий 10 мМ 2-меркаптоэтанола, рН 7,5. При хроматографии на QAE-сефалексе в качестве исходного буфера обычно используют 0,01 М трис-НС буфер, содержащий 10 мМ 2-меркаптоэтанола 0,1 мМ этилендиаминтетрауксусной кислоты, рН 7,5. При проведении г.ель-фильтрации обычно используют 0,02 М трис-НС1 буфер, содержащий 0,1 М КСЕ, 10 мМ 2-меркаптоэтанола, 0,1 мМ этилендиаминтетрауксусной кислоты, рН 7,5. Существенными отличиями способа являются: продолжительность инкубации клеток E.coli с бактериофагом Г4 90+20 мин, промывка колонки с ДЭАЭ-целлкшозой (первая xpcwaTorpaФическая стадия) перед элюцией 0,12+ „ „, „ ,, 0,01 М раствором NaCi в исходном буфере, а также предварительное введение в колонку с QAE-сефадексом перед хроматографией раствора KCI в измененном интервале линейного градиента концентрации данной соли в исходном буфере от 0,1+0,01 до 0,4 0,01 М. Таким образом, установление оптимального времени инкубации E.coli с бактериофагом Т4, введение дополнительной ступени при хроматографии фермента на ДЭАЭ-целлюлозе и снижение ионной силы раствора преформируемого градиента концентрации соли КС2 в колонне при хроматографии на QAE-сефадексе А-25 позволяет из 100 биомассы получать 10-12 нмолей РНКлигазы, содержащей 1-2 ед. активности З -экзонуклеазы на 1 пмоль РНКлигазы, препарат РНК-лигазы получают необходимой степени чистоты при одновременном 4-кратном увеличении выхода фермента и снижении трудоемко ти процесса (исключена стадия изоэлектрического фокусирования, на которой происходят основные потери продукта). На чертеже изображена хроматография РНК-лигазы на (}АЕ-сефалексе,где i - поглощение при 280 нм; 2 - радио активность 8- И АМР-лигазного ксяч плекса; 3 - преформируемый градиент ионной силы в колонке от 0,275 до 0,5; б - то же, от 0,11 до 0,41, заштрихован объединяекый для дальней шей очистки пул РНК-лигазы. Способ осуществляют следукидим образом. РНК-лигазу выделяют из биомассы E.coli, инфицированной бактериофагом Т4 am № 82, клетки разрушают ультразвуком, экстракт обрабатывают стрептомицином и сульфатом аммония, далее проводят фракционирование и хроматографию на ДЭАЭ-целлюлозе, гел фильтрационно-ионообменную хроматографию на (1АЕ-сефадексе А-25 и гельфильтрацию не сефадексе G-100. При этом E.coli инкубируют с фагом 90 мин i 20 мин {экспериментально ус тановленное время, при которся в клетках E.coli, инфицированный бакте риофагом Т4 am 82, на единицу РНКлигазы продуцируется минимальное количество з-экзонуклеазы), при фракционировании на ДЭАЭ-целлюлозе перед элюцией РНК-лигазы колонку про мывают раствором NaCt с ионной силой 0,13+0,01, перед хроматографией на QAE-сефадексе А-25 в колонку вводят раствор KCt в линейном градиенте ионной силы от 0,11+0,01 до 0,41+0,01 (экспериментально установленные значе ния ионной силы растворов, обеспечива ющие достижение максимгшьной степени очистки РНК-лигазы на соответствующей стадии при сохранении высокого (более 80%) выхода фермента). Пример 1. Все операции прово дят при . 100 г биомассы E.coli б, инфицированной бактериофагом. Т.4 am 82 (время инкубгщии E,colic фагом 90 мин), суспендируют в 200 мл бу фера А (0,05 М трис-HCt, 1 мМ глутати он, рН 7,5). Суспензию клеток обрабатывают ультразвуке (мощность 250-г 280 Вт, частота 20 кГц) 30 с, после чего выдерживают 10 мин при О С,Цикл разрушения (30 с, ультразвук, 10 мин охлаждение) повторяют 12 раз, затем суспензию центрифугируют (центрифуга а-21 Beckman, ротор ЭА-20, 20000 об/мин 1 ч) и осадок отбрасывают. К 240 мл супернатанта, полученного на предыдущей стадии, добавляют 560 кп буфера Д и по каплям при непрерсгвном перемеишвании на магнитной мешалке 60 мл 10%-ного стрептсжшцинсульфата, приготовленного на 0,5 М трис-НС буфере, рН 7,5. Смесь выдерживают 1 ч и затем центрифугируют (центрифуга 3-21, ротор аа-14, 14000 об/ми.н, 30 мин). Осадок собирают и замораживают (далее из него может быть выдр;лена полинуклеотидКиназа; „ К 805 мл супернатанта, полученного на предыдущей стадии, добавляют по порциям при непрерывном перемешивании на магнитной мешалке в течение 1ч 132 г сульфата аммония (до 30% от насыщения при ) . рН раствора при добавлении сульфата аммония контролируют универсальной индикаторной бумагой Реагент и по мере необходимости подтитровывают 0,2 М NaOH до рН 7-7,5. Раствор оставляют на 24 ч для формирования осадка и затем центрифугируют (центрифуга Q-21, ротор ОА-14, 14000 об/мин, 30 мин). Осадок отбрасывают, а к 870 мл супернатанта добавляют 129 г сульфата аммония (до 55% от насыщения при ) как и в предыдущем случае. Сформировавшийся в течение 24 ч осадок отделяют центрифугированием (центрифуга G-21, ротор GA-14, 14000 об/мин, 30 мин), растворяют в 100 мл буфера Б (0,01 М трис-KCi, 10 мМ 2-меркаптоэтанал, рН 7,5), содержащего NaCI в концентрации 0,1 М. Раствор переносят в целлофановый диализный мешок и диализуют против 2л буфера Б в течение 24 ч. Отдиализованный препарат РНК-лигазы (объем 120 мл) наносят на колонку (2,6x25 см) с ДЭАЭ-целлюлозой ДЕ-52, уравновешенную буфером Б. Колонку промывают последовательно со скоростью 50 мл/ч 100 мл буфера и 300 мл буфера Б, содержащего NaC2 в концентрации 0,12 М (0,12 М NaCt в буфере Б и числе прочих белков элюируется ДНКлигаза). РНК-лигаэу далее элюируют аналогичным буфером, содержащим NaCH. в концентрации 0,3 М. Скорость элюции 50 мл/ч. Фракции объемом 12,5 мл собирают на коллекторе, измеряют в них оптическую плотность при 280 им и содержание РНК-лигазы по-образованию кислотонерастворимого АМР-лигазного комплекса. Фракции, содержащие РНК-лигазу, объединяют .(объем 37,5 мл), разбавляют в. 3 раза буфером Б и наносят на колонку (5x21 см) с ДЭДЭ-целлюлозной ДЕ-52, уравновешенную буфером Б. Элюциюосуществляют 3 л раствора NaC в линейном градиенте концентрации от 0,1 до 0,3 М NaCl в буфере Б со скоростью 67 мл/ч. Фракции объемом 16,75 МП собирают на коллекторе, измеряют в них оптическую плотность при 280 нм и содержание РНК-лигазы, «Фракции, содержащие фермент, объединяют (объем 150 мл) и добавляют к ним по порциям при непрерывном перемешивании на магнитной мешгшке 1 ч 77,4 г сульфата аммония (до 80% от насьадения при 0°С) . рН раствора при добавлении сульфата аммония контролируют универсальной индикаторной бумагой и по мере необходимости подтитровывают твердым трис(оксиметил) гминометаном до рН 7-7,5. Раствор оставляют на 24 ч для формирования осадка, который Дсшее отделяют центрифугированием (центрифуга Q-21, ротор аА-14, 14000 об/мин, 1 ч) и растворяют в 4,5 мл буфера j (0,01 М трис-НС1, 10 мМ 2 меркаптоэтанол, 0,1 мМ ЭДТА, рН 7,5), содержащего КС1 в концентрации 0,75 М, Колонну SP 25/45 (Pharmacia, Швеция) , заполненную, на 35 см QAE-сефадёксом А-25, уравновоиивают 0,1 М КС1 в буфере . Снизу вверх в уравно вешенную колонку вводят со скоростью 40 мл/ч 50 мл раствора KUl в линейном градиенте концентрации от О,1 до 0,4 М КС1 в буфере . Дгшее аналогично-вводят в колонку 4,5 мл раствора фермента, полученного на предыдущей стадии, и следом 10 мл 1 М КС1 в 50%-ном глицерине. Элюцию осуществляют снизу вверх 1 М КС1 со скоростью 20 мл/ч. Фракции объемом 5 мл собирают на коллекторе, измеряют в них оптическую плотность при 280 нм и содержание РНК-лигазы. Фракции, содержащие фермент, объедин ют (объем 25 мл), переносят в центри фужный стакан (для ротора (ЗА-20) и добавляют по порциям при непрерывном перемешивании стеклянной палочкой 12,9 г сульфата аммония (до 80% от насыщения при 0° С) рН раствора при добавлении сульфата аммония контроли руют и подтитровывгиот твердым трис (оксиметил)аминометаном до 7-7,5. Сформировавшийся в течение 24 ч оса . док отделяют центрифугированием (центрифуга Q-21, ротор GA-20 20000 об/мин, 1 ч) и растворяют в .3 буфера Г, (0,02 М трис-НС1, 0,1 М КС 10 мМ 2 меркаптоэтанол,0,1 мМ ЭДТА, рН 7,5) . Колонку (2,5x60 см) с сефадексом G-100 (сверхмелким) уравновешивают буфере Г, наносят на нее 3 мл раст вора фермента, полученного на преды дущей стадии, и элюируют буфером Г со скоростью 5,2 мл/ч. Фракции объемом 2,6 мл собирают на коллекто измеряют в них оптическую плотность при 280 содержание РНК-лигаэы 1 акции, содержащие фермент, объедияют (объем 13 мл), переносят в целлофановый диализный мешок и концентрируют диализом против 0,5 л буфера 0,01 М трис-НС (рН 7,5) в 50%-ном глицерине с 10 мМ дитиотреитапом в течение 24 ч. Отдигипизованный препаат РНК-лигаэы (4,1 мл, 12 нмоль фер-. ента, 1,8 ед/пмоль РНК-лигазы - активность 3 -экзонуклеаэы в препарате) хранят при в герметично закрытом флаконе в течение года без потери активности. Пример 2. Поясняет выбор времени инкубации Е.соИв с бактериофагом Т4 am I 82. ,РНК-лигазу выделяют из 100 г биомассы E.coll В, инфицированной бакте риофагом Т4 am W 82, по примеру 1 до стадии хроматографии на ДЭАЭ-целлюлоэ е включительно. При этом E.coli инкубируют с фагом в течение 30 мин, 90 мин, и 3ч. В полученных препаратах определяют удельное содержание 3 -экзонуклеазы в ед. активности на 1 пмопь РНК-лигазы. Результаты приведены в табл.1. Таблица 1 Единица активности З Экспериментэкзонуклеазы на 1 пмоль РНК-лигазы Пример 3. Поясняет выбор ионной силы раствора вводимой ступени фракционирования на ДЭАЭ-целлюлозе. РНК-лигазу выделяют из 100 г биомассы E.coll В, инфицированной бактериофагом Т4 am № 82 по примеру 1 до стадии фракционирования на ДЭАЭ-целлюлозе включительно. При этсм перед эл1оцией РНК-лигазы колонку промывают раствором с ионной а - 0,11; б - 0,13) в - 0,16. РНК-лигазу далее элюируют раствором с ионной силой 0,31 аналогично примеру 1. в попученных препаратах определяют содержание РНК-лигаэы и белка, рассчитывают выход фермента (% к наиесенному на ко.лонку) и степень очистки. Результаты приведены в табл.2. Таблица 2 П р И м е р 4. Поясняет выбор ионной силы раствора преформируемого градиента концентрации соли в колонке при хроматографии на Ь-сефадексе А-25. РНК-лигазу вьщеляют из 100 г биомассы Б.coll В, иифи шрованной бакте риофагом Т4 am № 82, по примеру 1 до стс1лии хрс атографии на QAE-сефадексе А-25 включительно. При этсж пере хроматографией на (| АЕ-сефадексе А-25 в колонку вводят раствор в линейном градиенте ионной силы; а от 0,275 до 0,5, б от 0,11 до 0,41, в от 0,01 до 0,41. В полученных препаратах определяю содержание РНК-лигаэы и белка, рассч тывают выход .фермента (% к наиесеннсжу на колонку) и степень очис ки. Результаты приведены в табл.3. ТаблицаЗ Примечание. В случае JB проведение хроматографии осложнено резкой сменой ионной силы раствора колонке и вследствие этого значител ньм уменьшением объема хроматографи- ческого носителя при введении в колонну раствора в линейнс градиенте ионной силы от 0,01 до 0,41. формула изобретения 1. Способ получения РНК-лигазы, включающий инкубацию клеток E.coli с бактериофагом Т4, разрушение клеток ультразвуком с отделением экстракта и последующим выделением фермента из экстракта путем обработки его стрептомицинсульФатсж и сульфатом и хроматографией ферментного раствора в условиях буфера рН 7,5-0,1, проводимой последователь но на диэтиламиноэтилцеллюлозе с элюцйей исходным буфером с добавкой 0,3 М NaCl, повторной хрсматографией полученного элюата на. диэтиламиноэтилцеллюлозе с элюцйей линейньм Градиентом NaCl от О до 0,3 М в исходном буфере и хроматографией полученного элюата на сшитом эпихлоргидрином диэтил-2-оксипропиламиноэтилДекстране, в колонку с которым перед осуществлением хроматографии предцварительно вводят раствор КС1 в исходно буфере, с элюцйей 1+0,01 М раствором КС1 и гель-фильтрацией полученного элюата с помощью.сшитого эпихлоргидрином декстрана с использованием буфера рН 7,5+0,1, о т л и ча1дщийся тем, .- что, с целью упрощения процесса и увеличения выхода целевого продукта, улучшения его качества, инкубацию E.coli с бактериофаге Т4 проводят 90+20 мин, при осуществлении первой хроматографии на диэтиламиноэтилцеллюлозе перед элюцйей И1К-лигазы колонку с сорбентом промавают 0,12+0,01 М раствором NaCl в исходном буфере, а перед хроматографией на сшитом эпихлоргидрином диэтил-2-оксипропиламиноэтилдекстране в колонку с сорбентом предварительно вводят раствор КС1 в линейном градиенте концентрации от 0,1± 0,01 до 0,,01 М в исходном буфере. 2.Способ поп.1,отличающ и и с я тем, что в качестве исходного буфера при двукратной хрсялатографии на диэтиламиноэтилцеллюлозе используют 0,4)1 М трис-HCl: буфер, содержащий 10 ыК 2-меркаптоэтанола, рН 7,5. 3.Способ по п.1, отличающий с я тем, что в качестве исходного буфера при хроматографии на сшитом эпихлоргидрином ДИЭТИЛ-2-ОКСИпропяламиноэтилдекстране иАпользуют 0,01 М трис-НС1 буфер, содер.жеидий 10 кй 2-меркаптоэтанола, 0,1 мМ этилендиаминтетрауксусной кислоты, рН 7,5. 4.Способ по П.1/ отличающийся тем, что при проведении гель-фильтрации используют 0,02 М трис-НС буфер, содержащий 0,1 М КС1, 10 мМ 2-меркаптоэтанола, 0,1 этилендиаминтетрауксусной кислоты, рН 7,5. Источники информации, принятые во внимание при экспертизе 1. Higgins N. Р., Geballe А.Р., Snopek T.I.eT al,Nucl. Acids, Res, 1977,4, 9, 3175-3186.. ohtsu2.Sugiura M., Suzuki M, Ohtsu jcgi E. ет al, FEBS Letters, 1979, 97, 1, 73-76. 3.Moseman McCoy M.I., Lubben Т.Н., Gumport P.I., Biodhim et. Biophys. Acta,1979, 562, 149-161. 4.Василенко С.К., Веньяминова А,Г. Ямковой В. И. и Майоров В.И. РНК-лигаза бактериофага Т4. .Биоорганическая химия, 1979, т.5, 4, 621-G27 (прототип).
t
ts
iiS
40
J0
Iff




| название | год | авторы | номер документа |
|---|---|---|---|
| Способ получения ферментов из биомассы BacILLUS амYLоLIQUеFасIеNS штамм ВКПМ В-3188 | 1989 |
|
SU1661211A1 |
| Способ получения рибонуклеаз из | 1978 |
|
SU787464A1 |
| Способ выделения полинуклеотидфосфорилазы | 1981 |
|
SU1076445A1 |
| Способ получения ДНК-полимеразы бактериофага Т4 в клетках ЕSснеRIснIа coLI | 1983 |
|
SU1147030A1 |
| Способ получения фосфорибулокиназы | 1983 |
|
SU1154329A1 |
| СПОСОБ ПОЛУЧЕНИЯ ЛАКТАТДЕГИДРОГЕНАЗЫ | 2001 |
|
RU2180688C1 |
| Способ выделения рибонуклеазы | 1977 |
|
SU734261A1 |
| Способ получения обратной транскриптазы из ЕSснеRIснIа coLI | 1989 |
|
SU1638162A1 |
| СПОСОБ ПОЛУЧЕНИЯ АКТИВАТОРА ПЛАЗМИНОГЕНА | 2007 |
|
RU2346983C1 |
| Способ очистки микробной эстеразы | 1991 |
|
SU1839190A1 |
t u
f/
4ff
Jff
fO
fO
