Настоящее изобретение относится к способу приготовления экспрессионного вектора, кодирующего адаптированную рекомбиназу, которая способна к рекомбинации асимметричных последовательностей-мишеней внутри длинного концевого повтора (LTR) провирусной ДНК множества штаммов ретровирусов, которая может быть встроена в геном клетки-хозяина, а также к полученному экспрессионному вектору, клеткам, трансфицированным этим, экспрессированной рекомбиназе и фармацевтическим композициям, включающим в экспрессионный вектор, клетки и/или рекомбиназу. Фармацевтические композиции полезны, например, при лечении и/или профилактике ретровирусной инфекции. В частности, раскрыты асимметричные последовательности-мишени, присутствующие во множестве штаммов ВИЧ-1, а также адаптированные рекомбиназы, способные комбинировать эти последовательности (Tre 3.0 и 4.0), и кодирующие их экспрессионные векторы.
ТЕХНИЧЕСКИЕ ПРЕДПОСЫЛКИ
Ретровирусные инфекции, такие как, например, инфекции вирусом иммунодефицита человека (ВИЧ), по-прежнему являются одними из важнейших и наиболее распространенных заболеваний человека.
Один из подходов к лечению ретровируса, например, ВИЧ, является нацеливание на провирус, встроенный в геном клетки-хозяина. Эксцизия провирусной ДНК из генома хозяина, например, могла бы предотвратить дальнейшую репликацию ВИЧ, и отличается от общепринятых методик тем, что она позволяет искоренить даже неактивные вирусы, присутствующие в геноме хозяина.
Один класс белков, который рассматривался для использования в этом альтернативном подходе, представляют собой сайт-специфические рекомбиназы (Flowers et al, 1997). Сайт-специфические рекомбиназы опосредуют множество функций в природе, от перестановки генов до геномной сегрегации, таких как, например, эксцизия, инверсия или интеграция определенных элементов ДНК (обзор в STARK et al, 1992).
Одной из самых простых и наиболее изученных рекомбиназ является рекомбиназа Cre бактериофага P1, которая разделяет геномные димеры до мономеров путем рекомбинации между двумя идентичными участками двухцепочечной ДНК определенной последовательности (Hoess & Abremski, 1985). Рекомбиназа Cre нашла широкое применение в генетике мыши (Nagy, 2000). Cre представляет собой белок размером 38 кДа, который был назван по его функции, поскольку он вызывает рекомбинацию (Sternberg & Hamilton, 1981). Предпосылкой для этой рекомбинации является выравнивание двух участков рекомбинации, распознаваемых Cre в антипараллельной ориентации, которые затем связываются четырьмя идентичными субъединицами Cre, соединяющимися с образованием кольца, в котором каждая субъединица контактирует с двумя соседними субъединицами и одной половиной участка одного участка рекомбинации (Hoess & Abremski, 1985). Участок рекомбинации, узнаваемый Cre, представляет собой последовательность двухцепочечной ДНК длиной 34 п.н., известную как loxP (от locus of crossing over (x), P1; Sternberg & Hamilton, 1981), которая является палиндромной, за исключением восьми его самых внутренних пар оснований (называемых спейсером), которые придают направление на участок.
Некоторые системы сайт-специфической рекомбинации, в том числе система Cre/loxP, функционируют без вспомогательных белков или кофакторов и функционируют в широком диапазоне состояний клетки. Однако, поскольку сайт-специфические рекомбиназы функционируют с помощью специфических взаимодействий субъединиц реконбиназного фермента с узнаваемыми ими последовательностями ДНК-мишенями, использование этих ферментов ограничено требованием, что области ДНК-мишени должны содержать соответствующим образом расположенные участки-мишени (Lewandoski, 2001). До настоящего времени, не выявлена рекомбиназа дикого типа, которая узнает нативные ретровирусные последовательности как их последовательности ДНК-мишени.
В последние годы были проведены обширные мутационные и структурные анализы сайт-специфических рекомбиназ, чтобы изменить их свойства и добиться лучшего понимания сложных механизмов этих ферментов (в качестве обзора см. van Duyne, 2001; и Coates et al., 2005). Много исследований было сфокусировано на рекомбиназе Cre, чтобы изучить ее способность к развитию. Несколько исследований показало, что специфичность Cre к мишени может изменяться в случае, когда были изменены несколько нуклеотидов в ее участке узнавания loxP (Buchholz & Stewart, 2001; Santoro & Schultz, 2002; Rufer & Sauer, 2002). Дальнейшие исследования направлены на конструирование мутантных участков-мишеней loxP, содержащих последовательности из LTR ВИЧ-1 для разработки возможных участков-мишеней для использования Cre в качестве противовирусной стратегии (Lee & Park, 1998; Lee et al, 2000).
Способ направленной эволюции является мощным способом отбора ферментов с модифицированными специфичностями (обобщено в Yuan et al., 2005; и Johannes & Zhao, 2006). Вначале этот способ был использован для изоляции улучшенных ферментов на основе РНК путем отбора молекул РНК с модифицированными субстратными участками. Применение способов на основе ПЦР позволяет проводить скрининг очень больших библиотек и отбирать удачные кодирующие области из числа кандидатов. В направленной эволюции белков, напротив, проведение скрининга и отбор улучшенных мутантов, которые идентифицируются путем изменений в свойствах белка, требует способа определения последовательности нуклеиновых кислот, кодирующей белок. Связь между белком и его кодирующей последовательностью часто поддерживалась компартментализацией. Соответственно, скрининг библиотек при направленной белковой эволюции был ограничен «последовательными» подходами, которые сохраняют компартменты, и преимущества, связанные со скринингом пулов кандидатов, не были достигнуты.
Это ограничение удалось преодолеть путем разработки способов, которые позволяют сшивать белки с соответствующими им матричными РНК (мРНК) с помощью слияний мРНК-белок и рибосомного дисплея. Поэтому функциональные скрининги для улучшения белковых свойств были связаны с прямым поиском соответствующих кодирующих молекул, и большой пул был скринирован in vitro (см., например, Buchholz et al, 1998). Дальнейшее улучшение направленной белковой эволюции было достигнуто при помощи так называемой субстрат-связанной белковой эволюции (SLiPE; Buchholz & Stewart, 2001), где субстрат рекомбиназы помещали на ту же молекулу ДНК, что и кодирующую белок область. Таким образом, когда рекомбиназу экспрессировали в компартменте, его действие изменяло ДНК-субстрат, следующий за его собственной кодирующей областью. Следовательно, библиотеку можно скринировать в качестве пула с помощью ПЦР, чтобы амплифицировать только кандидатные кодирующие области, которые находились рядом с модифицированным субстратом. Это позволяет удобно скринировать большие библиотеки для быстрого поиска удачных кодирующих областей. Этот способ был применен для изменения ДНК-специфичности рекомбиназы Cre и ее адаптации к новому узнаваемому участку-мишени (Buchholz & Stewart, 2001).
Принимая во внимание потенциал сайт-специфических рекомбиназ и необходимость поиска терапии против СПИДа, позволяющей искоренять провирус ВИЧ-1 из генома клетки-хозяина, WO 2008/083931 раскрыл образование адаптированной рекомбиназы (TRE), которая способна к рекомбинации асимметричных участков-мишеней внутри LTR провирусной ДНК ретровируса, встроенного в геном клетки-хозяина, тем самым удаляя провирус из генома клетки-хозяина. Сконструированная рекомбиназа Tre, раскрытая в примерах, узнает специфический loxP-подобный участок, присутствующий в определенном штамме ВИЧ-1. WO 2008/083931 принимает во внимание, что в связи с высокой вариабельностью последовательности ретровирусов, в частности ВИЧ, для лечения пациента с отличным штаммом ВИЧ, возможно, должна быть адаптирована отличная адаптированная рекомбиназа или коллекция подготовленных рекомбиназ, содержащих адаптированные рекомбиназы, специфические для множества последовательностей-мишеней.
В свете этого, авторы изобретения сейчас рассматривают проблему предоставления адаптированной рекомбиназы, способной удалять множество ретровирусов, например, штаммы ВИЧ. Соответственно, созданная рекомбиназа может быть использована для множества ВИЧ-инфекций, без образования новых рекомбиназ для каждого штамма. Эта проблема решена настоящим изобретением.
ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Авторы настоящего изобретения впервые предоставляют способ создания экспрессионного вектора, кодирующего рекомбиназу, способную к рекомбинации асимметричных последовательностей-мишеней внутри LTR провирусной ДНК множества ретровирусных штаммов одного вида, встроенных в геном клетки-хозяина. Рекомбиназы были адаптированы, чтобы узнавать асимметричные участки-мишени, отличающиеся от их нативных симметричных участков-мишеней, путем разбиения субстрата на ряд новых подгрупп с меньшими отличиями от первоначальной мишени, и постепенной адаптации рекомбиназ для узнавания этих подгрупп (WO 2008/083931). Комбинаторный подход позволяет отбирать функциональные молекулы, узнающие асимметричный участок-мишень внутри заданной последовательности. Таким образом, используя подход прохождения через субстратные интермедиаты во время направленной молекулярной эволюции, можно продуцировать ферменты с отличными новыми специфичностями к асимметричным мишеням. В настоящем изобретении такой подход используется в вопросе предоставления адаптированных рекомбиназ, способных к рекомбинации множества ретровирусных штаммов, предпочтительно, штаммов одного вида. Авторы обнаружили, что, несмотря на высокую вариабельность последовательностей ретровирусов, можно идентифицировать асимметричные последовательности-мишени, присутствующие в большей части вирусов определенного подтипа.
Изобретение предоставляет способ приготовления экспрессионного вектора, кодирующего адаптированную рекомбиназу, которая способна к рекомбинации асимметричных последовательностей-мишеней внутри LTR провирусных ДНК множества ретровирусных штаммов, которые могут быть встроены в геном клетки-хозяина, включающий стадии идентификации в последовательности LTR провирусной ДНК множества последовательностей ретровирусных штаммов с гомологий, по крайней мере, 30% к последовательности левой половины участка и последовательности правой половины участка, по крайней мере, одного известного рекомбиназного участка-мишени, где гомологичные последовательности разделены спейсером в 5-12 нуклеотидов, и где асимметричная последовательность-мишень обнаруживается во множестве ретровирусных штаммов; и генерация, путем повторяющихся стадий:
i) молекулярной направленной эволюции, по крайней мере, на одной рекомбиназе, узнающей известный гомологичный участок-мишень, с использованием в качестве субстрата модифицированных последовательностей-мишеней, основанных на последовательности асимметричной последовательности-мишени, но модифицированных так, чтобы содержать лишь ограниченное число вариаций известной последовательности-мишени, где, в каждом раунде, последовательность-мишень может отличаться от последовательности-мишени, на которую, как известно, действует рекомбиназа, на один, два или три нуклеотида; и
ii) перетасовки библиотек рекомбиназ для получения рекомбиназных библиотек, способных к рекомбинации последовательностей-мишеней, более гомологичных асимметричной последовательности-мишени;
до тех пор, пока не получится, по крайней мере, одна рекомбиназа, которая действует на асимметричную последовательность-мишень внутри LTR ретровирусной ДНК; выделение нуклеиновой кислоты, в меньшей мере, одной полученной рекомбиназы, и клонирование нуклеиновой кислоты, кодирующей рекомбиназу, в подходящем экспрессионном векторе.
Способ создания адаптированной рекомбиназы, раскрытый в WO 2008/083931, можно применять для создания адаптированной рекомбиназы, способной к рекомбинации асимметричной последовательности-мишени.
Изобретение также предоставляет способ приготовления экспрессионного вектора, кодирующего адаптированную рекомбиназу, которая способна к рекомбинации асимметричных последовательностей-мишеней внутри LTR провирусной ДНК множества ретровирусных штаммов, которые могут быть встроены в геном клетки-хозяина, включающий стадии
a) идентификация в последовательности LTR провирусной ДНК множества ретровирусных штаммов последовательностей с гомологий, по крайней мере, 30% к последовательности левой половины участка и последовательности правой половины участка, по крайней мере, одного известного рекомбиназного участка-мишени, где гомологичные последовательности разделены спейсером в 5-12 нуклеотидов, и где асимметричная последовательность-мишень обнаруживается в множестве ретровирусных штаммов;
(b) идентификация двух последовательностей, где первая последовательность соответствует последовательности асимметричной последовательности-мишени со стадии (а), гомологичной левой половине участка указанного известного участка-мишени, и называется «последовательность половины участка 1», и где вторая последовательность соответствует последовательности асимметричной последовательности-мишени со стадии (а), гомологичной правой половине участка, и называется «последовательность половины участка 2»;
(c) определение нуклеотидов внутри последовательности со стадии (b), отклоняющихся от соответствующих гомологичных последовательностей левой половины участка и правой половины участка известного гомологичного участка-мишени со стадии (a);
(d) создание первой подгруппы двух нуклеиновых кислот-мишеней, содержащих последовательности-мишени, где первая последовательность-мишень обозначена подгруппой 1 и включает в себя, рядом друг с другом и в направлении от 5’ к 3’, последовательность половины участка 1 со стадии (b), спейсерную последовательность асимметричной последовательности-мишени и инвертированный повтор последовательности половины участка 1, и где вторая последовательность-мишень обозначена подгруппой 2 и включает в себя, рядом друг с другом и в направлении от 5’ к 3’, последовательность половины участка 2, спейсерную последовательность асимметричной последовательности-мишени и последовательность половины участка 2 со стадии (b);
(е) создание второй подгруппы нуклеиновых кислот-мишеней, содержащих модифицированные последовательности-мишени на основе последовательностей-мишеней в первой подгруппе со стадии (d),
где, в последовательностях на основе подучастка 1, в последовательности левой половины участка, часть нуклеотидов, отклоняющихся от соответствующей гомологичной последовательности половины участка, по крайней мере, одного известного участка-мишени со стадии (а) заменяли нативными нуклеотидами, встречавшимися в указанном известном участке-мишени, до тех пор, пока указанная последовательность половины участка содержит один, два или три нуклеотида, отклоняющихся от указанного известного участка-мишени, где правая половина участка указанной модифицированной последовательности мишени формируется при помощи инвертированного повтора указанной модифицированной последовательности левой половины участка, которая отделена от указанной модифицированной последовательности левой половины участка при помощи спейсерной последовательности асимметричной последовательности-мишени, и
где, в последовательностях на основе подучастка 2, в последовательности правой половины участка, часть нуклеотидов, отклоняющихся от соответствующей гомологичной последовательности половины участка, по крайней мере, одного известного участка-мишени со стадии (а) заменяют нативными нуклеотидами, встречавшимися в указанном известном участке-мишени, до тех пор указанная последовательность половины участка содержит один, два или три нуклеотида, отклоняющихся от указанного известного участка-мишени, где левая половина участка указанной модифицированной последовательности-мишени формируется при помощи инвертированного повтора указанной модифицированной последовательности правой половины участка, которая отделена от указанной модифицированной последовательности правой половины участка при помощи спейсерной последовательности асимметричной последовательности-мишени,
так что во всех модифицированных последовательностях половин участка, происходящих из одной последовательности-мишени первой подгруппы со стадии (d), вместе взятых, можно обнаружить все отклоняющиеся нуклеотиды, принимая во внимание, что ни одна из указанных модифицированных последовательностей половин участка на всем протяжении не содержит все отклоняющиеся нуклеотиды,
(f) отдельно применение молекулярной направленной эволюции, по крайней мере, на одной рекомбиназе, узнающей известный гомологичный участок-мишень в соответствии со стадией (а), с использованием каждой нуклеиновой кислоты второй подгруппы, полученной на стадии (e), в качестве субстрата;
(g) перетасовывание рекомбиназных библиотек, образованных на стадии (f), где все рекомбиназные библиотеки, образованные на последовательностях на основе подучастка 1, объединяют и перетасовывают, и где все рекомбиназные библиотеки, образованные на последовательностях на основе подучастка 2, объединяют и перетасовывают;
(h) применение молекулярной направленной эволюции на перетасованных библиотеках, полученных на стадии (g), с использованием каждой нуклеиновой кислоты подгруппы в соответствии со стадией (d) в качестве субстрата;
(i) перетасовка рекомбиназных библиотек, образованных на стадии (h);
(j) применение молекулярной направленной эволюции на перетасованных библиотеках, полученных на стадии (g), с использованием нуклеиновой кислоты, содержащей асимметричную последовательность-мишень со стадии (а), в качестве субстрата, пока не получится, по крайней мере, одна рекомбиназа, которая действует на асимметричную последовательность-мишени внутри LTR ретровирусной ДНК со стадии (a);
(k) выделение нуклеиновой кислоты, в меньшей мере, одной рекомбиназы, полученной на стадии (j) из библиотеки; и
(l) клонирование нуклеиновой кислоты, полученной на стадии (k), в подходящем экспрессионном векторе.
На стадии (a) способа по настоящему изобретению последовательность LTR провирусной ДНК можно определить, например, путем секвенирования ДНК с использованием концевых ингибиторов (Sanger et al., 1977). Однако в случае, если последовательность LTR ретровирусной ДНК, встроенной в геном хозяина, уже была определена, последовательность может быть определена путем обращения к базе данных. На основе информации о последовательности выполняется компьютерный анализ информации о последовательности, чтобы определить в ней последовательности с гомологией, по крайней мере, 30% к последовательностям левой половины участка и правой половины участка известных участков-мишеней, соответственно, известных рекомбиназ, которые разделены подходящим спейсером из 5-12 нуклеотидов, где асимметричная последовательность-мишень обнаруживается во множестве ретровирусных штаммов. Предпочтительно, гомология с последовательностями левой половины участка и правой половины участка известных участков-мишеней составляет не менее 40% или, по крайней мере, 50%. Предпочтительно, чтобы эти ретровирусные штаммы относились к одному виду или одному подтипу этого. Предпочтительно, множество штаммов составляет более 10 штаммов, более предпочтительно, более 100 штаммов, более 130 штаммов, более 200 штаммов или более чем 300 штаммов, например, штаммов ВИЧ. Штаммы могут относиться к одному подтипу вируса, например, ВИЧ-1, ВИЧ-1 подтипа A и B или ВИЧ-1 подтипа В. Таким образом, полученная рекомбиназа или кодирующий ее экспрессионный вектор могут быть использованы для лечения инфекции множеством штаммов, например, более, чем 50%, более чем 70%, более чем 80%, более чем 90% или всеми известными штаммами ретровируса или подтипом этого.
Термин «рекомбиназа», используемый в настоящем документе, относится к белку, участвующему в рекомбинации. Как таковые, рекомбиназы распознают и связывают две специфические последовательности ДНК, называемые «участки рекомбинации» или «участки-мишени», и опосредуют рекомбинацию между этими двумя участками-мишенями. Соответственно, термин «рекомбиназа» относится к любому белковому компоненту любой системы рекомбинации, который опосредует перестройки ДНК в специфическом ДНК-локусе. Существующие в природе рекомбиназы узнают симметричные участки-мишени, состоящие из двух идентичных последовательностей, называемых «половина участка», размером примерно в 9-20 п.н., образующих инвертированный повтор, где последовательности половин участка разделены спейсерной последовательностью в 5-12 п.н. Рекомбиназы из семейства тирозиновых интеграз характеризуются наличием тирозина в качестве нуклеофила активного участка, который используется для расщепления ДНК, тогда как рекомбиназы из семейства сериновых интеграз используют серин вместо тирозина.
В одном варианте осуществления настоящего изобретения, по крайней мере, одна известная рекомбиназа, чья последовательность-мишень используется на стадии (a) и в отношении которой применяется молекулярная направленная эволюция на стадиях (h) и (j), принадлежит к семейству сериновых интеграз. Предпочтительные рекомбиназы, принадлежащие семейству сериновых интеграз, выбирают из группы, состоящей из интегразы phiC31 (COMBES et al., 2002), любого компонента систем рекомбинации Gin или Hin, Tn3-резолвазы (Krasnow & Cozzarelli, 1983) или любого другого члена больших сериновых рекомбиназ, Rag1 Rag2 или любого другого компонента системы рекомбинации VDJ или вариантов этого.
В другом варианте осуществления, указанная рекомбиназа принадлежит к семейству тирозиновых интеграз. Предпочтительные рекомбиназы, принадлежащие семейству тирозиновых интеграз, выбирают из группы, состоящей из Cre фага P1 (Abremski et al., 1983, 1984), рекомбиназы FLP дрожжей (Volkert & Broach, 1986), Dre фага D6 (Sauer & McDermott, 2004), рекомбиназы R плазмиды pSR1 Zygosaccharomyces rouxii, рекомбиназы А плазмиды pKD1 Kluveromyces drosophilarium, рекомбиназы плазмиды pKW1 Kluveromyces waltii, Tnp1 транспозона Tn4430 Bacillus, любого компонента системы рекомбинации λ Int или вариантов этого. Предпочтительно, указанная рекомбиназа представляет собой рекомбиназу Cre или вариант этого. Например, можно использовать адаптированную рекомбиназу, раскрытую в WO 2008/083931 (Tre).
Термин «вариант» в данном контексте относится к белкам, которые образованы из вышеуказанных белков путем удаления, замены и/или добавления аминокислот и которые сохраняют некоторые или все функции, присущие белку, из которого они образованы.
В предпочтительном варианте осуществления, известная рекомбиназа представляет собой химерную рекомбиназу, полученную, например, «перетасовкой семейства», как описано в Crameri et al. (1998). Предпосылкой применения перетасовки семейства является значительная гомология между рекомбиназами, использованными для образования химерных рекомбиназ. Пример химерной рекомбиназы, который можно использовать в настоящем изобретении, представляет собой химерную рекомбиназу, состоящую из последовательностей рекомбиназы Cre и рекомбиназы Dre, соответственно.
В более предпочтительном варианте осуществления рекомбиназа представляет собой рекомбиназу Cre, узнающую симметричный участок-мишень в 34 п.н., известный как loxP (SEQ ID NO:3). Участок loxP (а также другие участки рекомбинации рекомбиназ дикого типа) представляет собой палиндром с двумя повторами по 13 п.н., разделенных при помощи восьми расположенных внутри пар оснований, которые представляют так называемый спейсер, который придает направленность участку. Рекомбинация происходит путем расщепления внутри спейсерной последовательности. В зависимости от относительного расположения и ориентации двух участвующих loxP-участков, Cre катализирует интеграцию, эксцизию или перестройку ДНК (Hoess & Abremski, 1985).
Одной из полезных рекомбиназ является Zre (SEQ ID NO:26), выделенная из Salmonella enterica, или варианты, фрагменты и гомологи этого, например, имеющие гомологию, по крайней мере, около 70%, по крайней мере, около 80%, по крайней мере, около 90% или, по крайней мере, около 95% с последовательностью дикого типа, и обладающие рекомбиназной функцией. Рекомбиназы Zre обеспечивают рекомбинацию ДНК по участкам zox (Фиг. 1). Они могут быть использованы на начальной стадии способа по изобретению, либо сами по себе, либо в контексте библиотеки.
В одном варианте осуществления, рекомбиназная библиотека используется в качестве отправной точки для молекулярной эволюции, например, рекомбиназная библиотека, включающая различные рекомбиназы дикого типа и/или адаптированные/перетасованные рекомбиназы, например, как описано ниже, или в Примере 2. Такая библиотека используется преимущественно в качестве отправной точки при создания адаптированных рекомбиназ, способных узнавать SEQ ID NO:1 и SEQ ID NO:2.
Адаптированная рекомбиназа способна к рекомбинации асимметричных последовательностей-мишеней внутри LTR провирусной ДНК множества ретровирусных штаммов. Провирусная ДНК, направляемая рекомбиназой, может быть встроена в геном клетки-хозяина. Альтернативно, адаптированная рекомбиназа по изобретению может обеспечивать рекомбинацию асимметричных последовательностей-мишеней внутри LTR провирусной ДНК множества ретровирусных штаммов, которые (пока) не интегрированы в геном клетки-хозяина, то есть которые присутствуют в качестве неинтегрированного преинтеграционного комплекса (PIC). Таким образом, ВИЧ, который еще не интегрирован в геном клетки-хозяина, а также ВИЧ, который уже интегрирован, могут быть инактивированы при помощи адаптированных рекомбиназ по изобретению.
Следует отметить, что в настоящем изобретении, а также в рассматриваемой области термины «последовательность-мишень», «участок-мишень» и «участок рекомбинации» используются как взаимозаменяемые.
В отличие от существующих в природе рекомбиназ, узнающих симметричный участок-мишень, способ по настоящему изобретению предоставляет адаптированные рекомбиназы, узнающие участки-мишени, которые не состоят из палиндромных последовательностей, разделенных спейсером. Вместо этого, в асимметричных участках-мишенях последовательности не образуют симметричный инвертированный повтор. Соответственно, адаптированная рекомбиназа, способная узнавать асимметричный участок-мишень, должна распознавать и обеспечивать рекомбинацию участков-мишеней, состоящих из половин участков варьирующей последовательности.
В асимметричном участке-мишени последовательности, называемые как «левая половина участка» и «правая половина участка», соответственно, определяются по их гомологии с левой и правой половиной участка известного участка-мишени. Последовательность, расположенная между последовательностями, гомологичными левой и правой половинам участка известного участка-мишени, называется спейсером.
При этом, если внутри LTR обнаруживаются последовательности, которые обладают гомологией лишь с последовательностью либо левой, либо правой половины участка известного участка-мишени, эти последовательности, тем не менее, могут использоваться в практике по настоящему изобретению. Размер участка-мишени, относящийся к рекомбиназе, чья нативная последовательность-мишень демонстрирует гомологию с последовательностями внутри LTR, известен специалистам в рассматриваемой области. Например, если гомология обнаруживается в последовательности LTR на последовательности-мишени, узнаваемой рекомбиназой Cre, асимметричный участок-мишень, чтобы узнаваться рекомбиназой Cre, должен состоять из 34 нуклеотидов с двумя последовательностями половин участков в 13 нуклеотидов, разделенных спейсером из 8 нуклеотидов. Соответственно, гомологичная последовательность внутри LTR определяется как-либо левой, либо правой половиной участка или спейсером асимметричного участка-мишени, в зависимости от гомологии с последовательностью известного участка-мишени. Таким образом, последовательности с гомологией к левой половине участка известной последовательности-мишени определяются как левая половина участка, последовательности с гомологией к правой половине участка известной последовательности-мишени определяются как правая половина участка. Исходя из этого определения, другие части асимметричных участков-мишеней определяются с учетом структуры известного участка-мишени. Таким образом, определив, например, последовательность правой половины участка внутри LTR по гомологии с участком loxP (узнаваемым рекомбиназой Cre), можно легко определить другие последовательности, соответствующие спейсеру и левой половине участка асимметричной последовательности-мишени. Спейсерная последовательность, например, определяется путем отсчета 8 нуклеотидов вверх по течению от 5’-конца последовательности, определенной как последовательность правой половины участка, тогда как последовательность левой половины участка определяется аналогичным образом путем отсчета 13 нуклеотидов вверх по течению от 5’-конца ранее определенной спейсерной последовательности.
Гомология в этом контексте, а также во всей заявке означает идентичность последовательностей. Предпочтительным сравнением в целях выявления гомологии является сравнение, по крайней мере, двух последовательностей с использованием стандартных способов, известных в рассматриваемой области, включая, в качестве неограничивающих примеров, алгоритм локальных гомологий Smith & Waterman (1981), алгоритм выравнивания гомологий Needleman & Wunsch (1970) или способ поиска сходства Pearson & Lipman (1988). Для целей настоящего приложения, гомологию последовательностей предпочтительно определять, используя компьютерные программы ClustalW, доступные из Европейского института биоинформатики (EBI), если не указано иное.
В связи с необходимостью наличия двух идентичных участков-мишеней, которые должны присутствовать в геноме провируса, чтобы дать возможность рекомбиназе удалить последовательность между этими двумя участками-мишенями, последовательности провирусной ДНК, которые присутствуют в геноме, по крайней мере, дважды, проверяют на стадии (a) способа по настоящему изобретению. Такие последовательности представляют собой, например, последовательности LTR провирусной ДНК. Соответственно, последовательность LTR сканируют предпочтительно, поскольку 5’-LTR и 3’-LTR провирусной ДНК являются идентичными. Асимметричный участок-мишень, присутствующий в 5’-LTR, также присутствует и в 3’-LTR и, таким образом, дает возможность эксцизии провирусной ДНК, расположенные между LTR.
Из последовательностей, идентифицированных в последовательности LTR, имеющих достаточную гомологию с известными участками-мишенями, предпочтительно выбирают последовательности, которые имеют наибольшую степень гомологии с последовательностью участка-мишени известных рекомбиназ. Однако можно также выбирать последовательности, отличные от тех, которые имеют наибольшую степень гомологии, например, такие, которые присутствуют в наибольшем числе штаммов ретровирусов, или в представляющих интерес штаммах ретровирусов, например, в случае, если пациент инфицирован конкретным штаммом.
Следует отметить, что потенциал способа по настоящему изобретению даже позволяет адаптировать рекомбиназы, которые узнают асимметричные участки-мишени с менее чем 30%-ой гомологией с известными участками-мишенями, например, по крайней мере, с 11%-ой или, по крайней мере, с 20%-ой гомологией. При этом, для уверенности в наличии рекомбинационной активности остатков для соответствующего асимметричного участка-мишени, предпочтительно проверяют последовательности, имеющие гомологию, по крайней мере, 30% с последовательностями левой половины участка и правой половины участка известных участков-мишеней известных рекомбиназ. В дополнительных предпочтительных вариантах осуществления выбраны асимметричные последовательности-мишени, имеющие гомологии, по крайней мере, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49, 50, 51, 52, 53, 54, 55, 56, 57, 58, 59, 60, 61, 62, 63, 64, 65, 66, 67, 68, 69, 70, 71 , 72, 73, 74, 75, 76, 77, 78, 79, 80%, более предпочтительно 85%, особенно предпочтительно 90% и наиболее предпочтительно 95% с последовательностями левой половины участка и правой половины участка известных участков-мишеней известных рекомбиназ.
В одном варианте осуществления по настоящему изобретению последовательность, выбранная внутри LTR, имеет гомологию с симметричными участками-мишенями loxP, узнаваемыми сайт-специфической рекомбиназой Cre.
В одном варианте осуществления, рекомбиназная библиотека используется в качестве отправной точки для молекулярной эволюции, например, рекомбиназная библиотека, содержащая различные рекомбиназы дикого типа и/или адаптированные/«перетасованные» рекомбиназы, такая как библиотека, описанная в Примере 2. Типичная библиотека состоит из рекомбиназы Cre и рекомбиназ, полученных на основе этого. Она может содержать Tre, Dre, рекомбиназы из Salmonella и Shewanella и/или рекомбиназы, полученные на основе этого. Библиотека может включать, например, Cre, Dre, Dre «Cre-ed» (SEQ ID NO: 24), рекомбиназу Shewanella (Shew), Shew «Cre-ed» (SEQ ID NO:25) и/или Zre (SEQ ID NO:26). Одна библиотека описана на Фиг. 3A. Tre представляет собой адаптированную рекомбиназу, как раскрыто в WO 2008/083931, которая в дальнейшем также называется Tre 1.0.
В одном варианте осуществления, все рекомбиназы в библиотеке узнают последовательность-мишень с такой же длиной спейсера. Общая длина последовательностей полуучастков 1 и 2, включая спейсер, предпочтительно составляет 34 нуклеотида.
В случае если, по крайней мере, одна рекомбиназа представляет собой рекомбиназную библиотеку, гомология представляет собой гомологию с пулом известных рекомбиназных участков-мишеней (т.е. гомология в заданной позиции, по крайней мере, с одной из последовательностей-мишеней определяется как гомология). Следовательно, на стадии (c), только те нуклеотиды, которые не соответствуют нуклеотиду, по крайней мере, в одной из известных последовательностей-мишеней, определяются как отклоняющиеся нуклеотиды. В случае рекомбиназной библиотеки «нативный нуклеотид» на стадии (e) может представлять собой нуклеотид, присутствующий в этом положении в любой из известных последовательностей-мишеней, предпочтительно, он является нуклеотидом, присутствующим в этом положении в нескольких или большинстве из известных последовательностей-мишеней.
Для идентификации последовательностей-мишеней, присутствующих во множестве ретровирусных штаммов, известные участки узнавания рекомбиназами, которые были описаны в литературе, могут быть использованы в качестве запроса для поиска консервативных асимметричных последовательностей-мишеней в отношении геномного фрагмента. При этом, учитывая повторяющийся характер областей, исключается использование стандартных инструментов поиска сходства последовательностей. В работе Sarkar et al., 2007, для нахождения Lox-подобного участка связывания среди штаммов ВИЧ используется BLAST (Altschul et al., 1997). Поиск BLAST для Lox-подобного участка при проведении среди последовательностей LTR ВИЧ-1 привел к открытию только одного участка, присутствующего в единственном штамме. Однако в случае, если рекомбиназы предполагается использовать в качестве терапевтических средств в отношении ретровирусных геномов, очень важно конструировать рекомбиназы для распознавания участков-мишеней, присутствующих у максимально большого количества штаммов ретровирусов.
Поскольку BLAST не выполняется хорошо с такими короткими избыточными последовательностями, было решено использовать HMMER (Eddy et al, 1998), RepeatMasker или программу палиндромов из набора пакетов Emboss. HMMER была разработана для нахождения отдаленных гомологий на основе вероятностной модели искомой структуры последовательности, которая не является содержанием поиска, предназначенного для проведения. HMMER может пытаться решить этот поисковый вопрос, но количество данных и параметров до и после обработки могут сделать его крайне ошибочным и неэффективным. RepeatMasker только проводит поиск повторов и областей с низкой сложностью, которые уже хорошо охарактеризованы, что опять-таки не является содержанием этого поиска. Программа палиндромов Emboss подходит ближе к решению проблемы поиска, но не дает возможность поиску быть определенным, а, скорее, дает в результате перечень возможных Lox-подобных участков. Затем они могут быть сопоставлены с представляющим интерес свойством lox-участка. Ясно, что такая стратегия могла бы только сделать поиск запутанным и усложненным. Для решения проблемы нехватки программы и способа нахождения Lox-подобных участков должна быть разработана программа, дающая возможность осуществлять поиск вырожденных Lox-подобных участков в геномных последовательностях.
Асимметричная последовательность-мишень, встречающаяся во множестве ретровирусных штаммов, может быть идентифицирована в соответствии с этой программой с помощью алгоритма, основанного на положении весовой матрицы для фланговых областей, на основании известного участка узнавания рекомбиназы. Предпочтительно, чтобы сделать поиск эффективным в вычислительном отношении, бинарные операции применяются к последовательностям после того, как они преобразованы в битовые строки.
В одном предпочтительном варианте осуществления изобретения, ретровирус представляет собой ВИЧ, в частности ВИЧ-1. Для ВИЧ-1 были определены подходящие асимметричные последовательности-мишени, имеющие последовательность, изложенную как SEQ ID NO:1 или SEQ ID NO:2, ниже.
Последовательности левой половины участка и правой половины участка подчеркнуты, а спейсерная последовательность набрана жирным шрифтом:
SEQ ID NO:1 AACCCACTGCTTAAGCCTCAATAAAGCTTGCCTT
SEQ ID NO:2 CTGGGCGGGACTGGGGAGTGGCGAGCCCTCAGAT
SEQ ID NO:1 идентична на 92% найденному в результате поиска штамму ВИЧ-1 подтипа B (348/379) и на 80% найденному в результате поиска штамму ВИЧ-1 подтипа А (32/40). SEQ ID NO:2 идентична на 76% найденному в результате поиска штамму подтипа B (288/379). 82% штаммов подтипа C также содержат эту последовательность. SEQ ID NO:2 не присутствует в каком-либо из найденных в результате поиска штаммов подтипа А.
Как показано на Фиг. 1, SEQ ID NO:1 имеет 54% гомологии с пулом известных участков-мишеней для рекомбиназ, и SEQ ID NO:2 имеет 42% гомологии с пулом из этих последовательностей (относительно левой и правой половин участков, соответственно). Гомологии для индивидуальных известных участков-мишеней являются более низкими, например, по крайней мере, 30% для SEQ ID NO:1 и, по крайней мере, 11% для SEQ ID NO:2. В частности, в случае низкой индивидуальной гомологий с известными участками-мишенями может быть выгодным использовать библиотеку рекомбиназ в качестве исходного материала, например, для создания адаптированной рекомбиназы, способной к рекомбинации SEQ ID NO:1 или SEQ ID NO:2, библиотеку, включающую Cre, Fre, Dre, Zre и Tre.
На стадии (b) способа по изобретению, последовательность асимметричного участка-мишени внутри LTR провируса, которая гомологична левой половине участка известного участка-мишени, определяется как «последовательность половины участка 1». Последовательность асимметричного участка-мишени внутри LTR провируса, которая гомологична правой половине участка известного участка-мишени, определяется как «последовательность половины участка 2. Последовательность между последовательностями, представляющими левую и правую половины участка, называется спейсером.
На стадии (c), нуклеотиды в пределах «последовательности половины участка 1» и «последовательности половины участка 2», соответственно, последовательностей стадии (b), отклоняющиеся от последовательностей соответствующих гомологичных последовательностей левой половины участка и правой половины участка известной мишени, определяются путем выравнивания последовательностей и сравнения последовательностей. В этом контексте, последовательность «последовательность половины участка 1» сравнивают с соответствующей нативной половиной участка, которая является предпочтительно последовательностью левой половины участка, а последовательность «последовательность половины участка 2» сравнивают с другой половиной участка, образующей палиндромный нативный участок-мишень, которая является предпочтительно последовательностью правой половины участка.
Фиг. 1 показывает результат такого сравнения для SEQ ID NO:1 и 2, по сравнению с библиотекой рекомбиназ. Отклоняющиеся нуклеотиды показаны перед темным фоном.
Это сравнение не обязательно выполнять после стадии (b) и перед стадией (d) способа по изобретению, но также можно проводить на другой стадии способа после стадии (a) и до стадии (e).
На стадии (d), генерируется первая подгруппа двух нуклеиновых кислот-мишеней, содержащих последовательности-мишени, где первая последовательность-мишень обозначается подучастком 1 и включает в себя, рядом друг с другом и в направлении 5’-3’, последовательность половины участка 1 со стадии (b), спейсерную последовательность асимметричной последовательности-мишени и инвертированный повтор последовательности половины участка 1, и где вторая последовательность-мишень обозначается подучастком 2 и включает в себя, рядом друг с другом и в направлении 5’-3’, инвертированный повтор последовательности половины участка 2, спейсерную последовательность асимметричной последовательности-мишени и последовательность половины участка 2 со стадии (b). Последовательность-мишень первой подгруппы представляет собой палиндромные олигонуклеотидные последовательности, имеющие структуру с симметричного участка-мишени. Эти искусственные симметричные участки-мишени синтезируют на основе последовательностей половины участка со стадии (b) путем дополнения недостающей последовательности половины участка в каждой олигонуклеотидной последовательности в качестве инвертированного повтора, где последовательность «последовательность половины участка 1» и «последовательность половины участка 2», соответственно, используется для дополнения второй последовательности половины участка на противоположном конце спейсерной последовательности. Соответственно, первая последовательность-мишень в первой подгруппе (названная как «подучасток 1») состоит из инвертированного повтора, состоящего из «последовательности половины участка 1» и инвертированно повторенной «последовательности половины участка 1», разделенных спейсерной последовательностью, принимая во внимание, что вторая последовательность-мишень в первой подгруппе (именуемая «подучасток 2») содержит инвертированный повтор, состоящий из инвертированно повторенной «последовательности половины участка 2» и «последовательности половины участка 2», разделенных спейсерной последовательностью. В «подучастке 1» последовательности располагаются следующим образом: 5’-«последовательность половины участка 1»-спейсер-«инвертированный повтор последовательности половины участка 1»-3’, в «подучастке 2» последовательности располагаются следующим образом: 5’-«инвертированный повтор последовательности половины участка 2»-спейсер-«последовательность половины участка 2»-3’.
Спейсерные последовательности в пределах каждой из двух синтетических последовательностей-мишеней первой подгруппы, предпочтительно, являются идентичными и соответствуют последовательности LTR, представленной или определенной как спейсерная последовательность асимметричного участка-мишени. При этом в дополнительном варианте осуществления, спейсерные последовательности могут содержать одно или два отклонения в последовательности, происходящие от нуклеотидных замен.
Как правило, эта стадия представляет первое расхождение последовательностей асимметричного участка-мишени, выбранного для адаптации конкретной рекомбиназы (см. Фиг. 1 в WO 2008/083931, который включен в настоящем документе в полном объеме в качестве ссылки, и Фиг. 2 настоящего приложения). На этой стадии образуются последовательности, несущие симметричные участки-мишени, полученные от полуучастков асимметричного участка-мишени, выбранного для адаптирования специфической рекомбиназы. Как следствие, каждая мутация (т.е. отличие от участка-мишени, узнаваемого рекомбиназой дикого типа), присутствующая в одном полуучастке указанного асимметричного участка-мишени, теперь распределена между симметричными последовательностями-мишенями в первой подгруппе.
На стадии (e) способа по изобретению, генерируют вторую подгруппу нуклеиновых кислот-мишеней, содержащих модифицированные последовательности-мишени, на основе последовательностей-мишеней в первой подгруппе со стадии (d). В последовательностях, основанных на подучастке 1, в последовательности левой половины участка, часть нуклеотидов, отклоняющихся от соответствующей гомологичной последовательности половины участка, по крайней мере, одного известного участка-мишени со стадии (а), заменяется нативными нуклеотидами, обнаруживаемыми в указанном известном участке-мишени, до тех пор, пока указанная последовательность половины участка содержит один, два или три (предпочтительно два) нуклеотида, отклоняющихся от указанного известного участка-мишени, где правая половина участка указанной модифицированной последовательности-мишени формируется инвертированным повтором указанной модифицированной последовательности левой половины участка, которая отделена от указанной модифицированной последовательности левой половины участка спейсерной последовательностью асимметричной последовательности-мишени.
В последовательностях на основе подучастка 2, в последовательности правой половины участка, часть нуклеотидов, остлоняющихся от соответствующей гомологичной последовательности половины участка, по крайней мере, одного известного участка-мишени со стадии (а), заменяется на нативные нуклеотиды, обнаруживаемые в указанном известном участке-мишени, до тех пор, пока указанная последовательность половины участка содержит один, два или три (предпочтительно два) нуклеотида, остлоняющихся от указанного известного участка-мишени, где левая половина участка указанной модифицированной последовательности-мишени формируется путем инвертированного повтора указанной модифицированной последовательности правой половины участка, которая отделена от указанной модифицированной последовательности правой половины участка спейсерной последовательностью асимметричной последовательности-мишени.
Например, если один подучасток содержит шесть отклоняющихся нуклеотидов, такой как оба подучастка на основе SEQ ID NO:1 или подучастка 2 SEQ ID NO:2 в отношении библиотеки рекомбиназ, показанной на Фиг. 1, на основе подучастка могут быть созданы три модифицированные последовательности-мишени, каждая из которых содержит два (различных) отклоняющихся нуклеотида в левой половине участка (если на основе подучастка 1) или правой половине участка (если на основе подучастка 2). Следовательно, в каждой модифицированной последовательности-мишени, последовательность соответствующего подучастка модифицируют для соответствия с последовательностью известной последовательности-мишени (или, по крайней мере, одной известной последовательностью-мишенью) по четырем нуклеотидам (Фиг. 2). Разумеется, также можно генерировать шесть модифицированных последовательностей-мишеней, каждая из которых содержит один из отклоняющихся нуклеотидов, или две последовательности-мишени, каждая из которых содержит три отклоняющихся нуклеотида.
В другом примере, если один подучасток включает в себя девять отклоняющихся нуклеотидов, как, например, подучасток 1 SEQ ID NO:2 в отношении библиотеки рекомбиназ, показанной на Фиг. 1, на основе подучастка могут быть созданы три модифицированных последовательности-мишени, каждая из которых содержит три (различных) отклоняющихся нуклеотида в половине участка.
Как следствие, во всех модифицированных последовательностях половин участков, происходящих из одной последовательности-мишени первой подгруппы со стадии (d), вместе взятых, можно найти все отклоняющиеся нуклеотиды, принимая во внимание, что ни одна из указанных модифицированных последовательностей половин участков сама по себе не содержит все отклоняющиеся нуклеотиды.
Опять же, инвертированный повтор генерируют на основе модифицированной последовательности половины участка, так, что спейсерная последовательность разделяет обе последовательности, образующие инвертированный повтор (см. Фиг. 2). Спейсерные последовательности в пределах каждой модифицированной последовательности-мишени новой подгруппы, будучи выведенными из последовательности-мишени более высокой подгруппы, предпочтительно являются идентичными и соответствуют последовательности LTR, представленной или определенной в качестве спейсерной последовательности асимметричного участка-мишени. При этом в дополнительном варианте осуществления спейсерные последовательности могут включать в себя одно или два отклонения последовательности, происходящих от нуклеотидных замен. С использованием этого подхода, количество мутаций (т.е. отличия в участке-мишени, узнаваемом рекомбиназой дикого типа) в последовательностях-мишенях, представляющих каждую подгруппу, является меньшим, чем в начальной асимметричной последовательности-мишени, но все мутации по-прежнему представлены в одной из последовательностей-мишеней (см. Фиг. 1 в WO 2008/083931, Фиг. 2 настоящего приложения).
Термин «отклоняющийся нуклеотид», используемый в настоящем документе, относится к нуклеотиду в пределах асимметричной последовательности-мишени, идентифицированной или определенной внутри LTR или в пределах последовательности-мишени подгруппы, полученной в соответствии с настоящим изобретением, который отклоняется (т.е. отличается) от нуклеотида, присутствующего в том же положении в соответствующей гомологичной последовательности известной гомологичной симметричной последовательности-мишени известной рекомбиназы, отобранной на стадии (a) способа по настоящему изобретению. В данном контексте термины «отклоняющиеся нуклеотиды» и «мутации» используются как взаимозаменяемые.
WO 2008/083931 наставляет, что рекомбиназы можно адаптировать с помощью молекулярной направленной эволюции с использованием последовательностей-мишеней в качестве субстрата, если последовательность-мишень, используемая в качестве субстрата, отличается не более чем 3 нуклеотидами от нативной последовательности-мишени. Таким образом, создание подгрупп различных порядков, описанных выше, служит для того, чтобы сократить число отклоняющихся нуклеотидов в последовательности-мишени до 3 или меньше (см. Фиг. 1 в WO 2008/083931). Постепенное сокращение числа отклоняющихся нуклеотидов, наконец, дает число подгрупп последовательностей-мишеней разных порядков с уменьшенным числом отклоняющихся нуклеотидов до того, как окончательно создается подгруппа, которая может быть использована в качестве субстрата для молекулярной направленной эволюции. При создании различных подгрупп и, вследствие этого, сокращения числа отклоняющихся нуклеотидов, различия с участком-мишенью, узнаваемым рекомбиназой дикого типа, распределены между несколькими последовательностями-мишенями, каждый из которых не содержит более чем 3 из этих отклоняющихся нуклеотидов, в то время как последовательности-мишени конечного порядка в целом по-прежнему представляют все отклоняющиеся нуклеотиды.
При желании, в способе по изобретению можно создать дополнительные подгруппы последовательностей-мишеней, начиная с последовательностей-мишеней второй подгруппы, путем поэтапного повторения процесса со стадии (e), т.е. разбиение последовательностей-мишеней на соответствующие последовательности половин участков и создание новых палиндромных структур на основе этих последовательностей половин участков после изменения последовательности половины участка, полученного из последовательности-мишени второй подгруппы, каждый раз создавая новую подгруппу последовательностей-мишеней, где последовательности половин участков, используемые для образования инвертированных повторов, содержат меньше нуклеотидов, отклоняющихся от соответствующих гомологичных последовательностей половин участков, по крайней мере, одного известного участка-мишени. Эти дополнительные последовательности-мишени можно использовать для дополнительных стадий направленной молекулярной эволюции и перетасовки рекомбиназных библиотек. Конечно, такая дополнительная стадия также может быть выполнена только для некоторых из последовательностей, например, для последовательностей, при которых получаются рекомбиназы с низкой эффективностью рекомбинации. Если генерируются дополнительные подгруппы и рекомбиназы, образованные на них, в процессе, эволюционная библиотека рекомбиназ используется на стадии (f) способа по изобретению.
Начиная со второй подгруппы последовательностей-мишеней, полученной на стадии (e), может быть создана третья подгруппа, за которой следует четвертая, пятая, шестая и т.д. подгруппа, если это необходимо. При этом, создание третьей подгруппы, как правило, необходимо только в том случае, если последовательности-мишени второй подгруппы все еще содержат более трех отклоняющихся нуклеотидов. То же самое относится и к созданию следующих подгрупп, которые необходимы только в том случае, если последовательности-мишени предыдущей подгруппы все еще содержат более трех отклоняющихся нуклеотидов. Следует отметить, что в одном варианте осуществления подгруппы последовательностей-мишеней будут генерироваться до тех пор, пока последовательности-мишени конечной подгруппы содержат только один отклоняющийся нуклеотид. Соответственно, в зависимости от числа отклоняющихся нуклеотидов в последовательности каждой половины участка, количество подгрупп, образованных для последовательности каждой половины участка асимметричного участка-мишени, может отличаться. Например, может оказаться необходимым генерировать только две подгруппы для последовательности левой половины участка, принимая во внимание, что три или четыре подгруппы должно быть создано для правой половины участка для правильного распределения отклоняющихся нуклеотидов между несколькими последовательностями-мишенями, так что единая последовательность-мишень не содержит более 3 из этих отклоняющихся нуклеотидов.
Принцип создания дополнительных подгрупп последовательностей-мишеней для уменьшения числа отклоняющихся нуклеотидов до числа меньше трех, иллюстрируемый на Фиг. 1 в WO 2008/083931 и Фиг. 2 настоящего приложения, предоставляет конкретные примеры модифицированных последовательностей-мишеней.
На стадии (f), способ молекулярной направленной эволюции применяется, по крайней мере, на одной рекомбиназе, узнающей известный гомологичный участок-мишень со стадии (а), используя последовательность-мишень конечной или второй подгруппы, полученной на стадии (e), содержащую один, два или три нуклеотида, отклоняющиеся от соответствующей гомологичной последовательности половины участка указанного известного гомологичного участка-мишени в качестве субстрата.
Термин «конечная подгруппа», используемый в настоящем документе, относится к последней подгруппе, образованной на стадии (e), т.е. ко второй подгруппе, если никакие дополнительные подгруппы не образованы. В зависимости от числа отклоняющихся нуклеотидов в асимметричном участке-мишени и числа подгрупп, которые должны были быть получены, чтобы уменьшить число отклоняющихся нуклеотидов в последовательности-мишени ниже 3, «конечная подгруппа» может соответствовать любой подгруппе, например, второй, третьей, четвертой или более поздней подгруппе, и может быть разной для последовательностей половины участка асимметричной последовательности-мишени внутри LTR. Если рекомбиназы предварительно были образованы на дополнительных подгруппах модифицированных последовательностей-мишеней, имеющих меньше нуклеотидов, отклоняющихся от соответствующей гомологичной последовательности половины участка указанного известного гомологичного участка-мишени, используется рекомбиназа, полученная на этой стадии.
Конечно, можно начинать процесс изобретения со специфической рекомбиназы для специфической модифицированной последовательности-мишени, и с другой рекомбиназы (или библиотеки) для другой специфической модифицированной последовательности-мишени.
Способы молекулярной направленной эволюции, также именуемой как лабораторная эволюция или in vitro-эволюция, известны в рассматриваемой области (см. обзор Yuan et al, 2005 и ссылки в нем; Johannes & Zhao, 2006).
На первой стадии молекулярной направленной эволюции, библиотеки случайно мутировавших рекомбиназных последовательностей создают при помощи способов, известных в рассматриваемой области, например, используя подверженный ошибкам ПЦР и перетасовку ДНК, (обзор, например, в Yuan et al., 2005), либо способов, раскрытых в Международной патентной заявке WO 2002/44409. Плазмиды каждой библиотеки, содержащей мутированные рекомбиназы, также содержат одну из последовательностей-мишеней конечной подгруппы, полученной на стадии (f). После трансфекции созданной плазмидной библиотеки в соответствующие клетки, обеспечивается возможность экспрессии рекомбиназы и осуществляется молекулярная направленная эволюция, как известно специалистам в рассматриваемой области.
В предпочтительном варианте осуществления, молекулярная направленная эволюция, применяемая на стадии (f) способа по настоящему изобретению, представляет собой субстрат-связанную белковую эволюцию (SLiPE; Buckholz & Stewart, 2001; Международная патентная заявка WO 02/44409). Субстрат-связанная белковая эволюция может осуществляться, как подробно описано в примерах в WO 2008/083931. Вкратце, последовательности-мишени, полученные на стадии (e), клонируют в плазмиду (так называемый эволюционный вектор) вместе со случайным образом мутированной кодирующей последовательностью для рекомбиназы. Мутитование случайным образом осуществляется при помощи подверженного ошибкам ПЦР (см. Buckholz & Buckholz, 2001). Созданной плазмидной библиотекой затем трансфицируют клетки E. coli для обеспечения возможности экспрессии рекомбиназы. С помощью индуцируемого промотора, управляющего экспрессией рекомбиназы, можно регулировать уровень экспрессии. После инкубации в течение ночи, из клеток выделяют плазмидную ДНК и расщепляют Ndel, чтобы разрезать плазмиды, которые не рекомбинировали, и только рекомбинировавшие плазмиды впоследствии амплифицировали при помощи праймеров. ПЦР-продукт рекомбинантной формы плазмиды образует полосу в 1,7 т.п.н. ПЦР-продукт расщепляют BsrGI и XbaI и субклонируют обратно в аналогичным образом расщепленный эволюционный вектор для следующего эволюционного цикла.
На стадии (g), рекомбиназные библиотеки, образованные на стадии (f), объединяют и перетасовывают. Технология перетасовки ДНК известна в рассматриваемой области (в качестве обзора см. Minshull & Stemmer, 1999; Stemmer, 1994). Рекомбиназные библиотеки, образованные на модифицированных последовательностях-мишенях на основе подучастка 1, объединяют и перетасовывают, и, отдельно, рекомбиназные библиотеки, образованные на модифицированных последовательностях-мишенях на основе подучастка 2, объединяют и перетасовывают.
Объединенные и перетасованные библиотеки затем клонируют в новое поколение векторов, содержащих последовательности-мишени следующей, более высокой подгруппы, т.е. если генерируется две подгруппы, подгруппа генерируется на стадии (d). Например, вектор для библиотеки, образованной на последовательностях на основе подучастка 1, включает в себя последовательность подучастка 1 в качестве последовательности-мишени, и вектор для библиотеки, образованной на последовательностях на основе подучастка 2, включает последовательность подучастка 2 в качестве последовательности-мишени.
На стадии (h), применяется способ молекулярной направленной эволюции на перетасованных библиотеках, полученных на стадии (g) с использованием последовательности-мишени следующей, более высокой подгруппы, которая, как обсуждалась, может быть подгруппой в соответствии со стадией (d). На этой стадии можно использовать тот же способ молекулярной направленной эволюции, что и способ, который применялся ранее на стадии (f), но также можно использовать другой способ молекулярной направленной эволюции на этой стадии способа по настоящему изобретению. Примеры различных способов молекулярной направленной эволюции были описаны, например, в Yuan et al. (2005). На объединенных и перетасованных библиотеках, предпочтительно, также применяется способ субстрат-связанной белковой эволюции.
Эта стадия дает рекомбиназы, узнающие и обеспечивающие рекомбинацию последовательностей-мишеней, несущих комбинацию (и, таким образом, увеличенное число) мутаций от различных последовательностей-мишеней более низкой подгруппы. Сочетание мутаций из различных библиотек более низкой подгруппы последовательностей-мишеней приводит к синергическому эффекту и приводит к образованию рекомбиназ, которые теперь обеспечивают рекомбинацию последовательностей-мишеней более высокой подгруппы, демонстрируя, что эволюционная стратегия, проходя через интермедиаты, может быть использована для достижения желаемой активности.
На стадии (i), стадию (g), т.е. объединение и перетасовка рекомбиназных библиотек, и (j), т.е. применения молекулярной направленной эволюции на объединенных и перетасованных библиотеках, повторяют до тех пор, пока не получится, по крайней мере, одна рекомбиназа, которая активна на асимметричной последовательности-мишени, присутствующей внутри LTR провирусной ДНК.
Например, в способе, при котором образование двух подгрупп последовательностей-мишеней было необходимо для образования последовательностей-мишеней только с одним, двумя или тремя нуклеотидными отклонениями, рекомбиназные библиотеки, образованные, например, для второй подгруппы последовательностей-мишеней, объединяют и перетасовывают, и молекулярную направленную эволюцию применяют на этой перетасованной библиотеке с использованием последовательности-мишени первой подгруппы. На следующей (и конечной) стадии асимметричная последовательность-мишень со стадии (а) внутри LTR провирусной ДНК используется для создания рекомбиназной библиотеки, включающей рекомбиназы, узнающие последовательности-мишени первой подгруппы, путем молекулярной направленной эволюции, чтобы получить, по крайней мере, одну рекомбиназу, которая активна на асимметричной последовательности-мишени внутри LTR ретровирусной ДНК. На этой стадии, способ молекулярной направленной эволюции, предпочтительно, является способом субстрат-связанной белковой эволюции.
На стадии (k), из библиотеки выделяют нуклеиновую кислоту рекомбиназы, обладающей активностью на асимметричной последовательность-мишени со стадии (а) внутри LTR ретровирусной ДНК. Нуклеиновую кислоту выделяют из соответствующей плазмиды из библиотеки, используя соответствующие рестрикционные ферменты. Способ расщепления рестрикционными эндонуклеазами известен в рассматриваемой области. Нуклеиновую кислоту, кодирующую рекомбиназу, можно затем извлечь с помощью известных способов, таких как, например, гель-электрофорез.
Нуклеиновую кислоту можно хранить (предпочтительно при температурах ниже -80 °C) или, при желании, можно клонировать на стадии (l) в экспрессионный вектор для использования в дальнейшем анализе, в способах экспрессии белка, или для введения субъекту для лечения и/или профилактики ретровирусной инфекции, в частности, ВИЧ-инфекции и/или СПИДа. Подходящие экспрессионные векторы известны в рассматриваемой области или раскрыты ниже.
Предпочтительно, асимметричная последовательность-мишень, идентифицированная на стадии (a), локализована и в 5’-LTR, и в 3’-LTR провируса, чтобы дать возможность эксцизии провирусной ДНК из генома клетки-хозяина.
Используя субстрат-связанную направленную эволюцию и последовательности, идентифицированные в SEQ ID NO:1 и SEQ ID NO:2 в качестве субстрата, авторы настоящего изобретения продуцируют адаптированную рекомбиназу, которая обеспечивает рекомбинацию этой асимметричной последовательности ДНК-мишени, присутствующей в длинных концевых повторах ВИЧ-1 множества штаммов ВИЧ-1. Разработка таких адаптированных рекомбиназ, которые специфически нацелены на асимметричные последовательности в пределах множества LTR ВИЧ-1, позволяет осуществлять эксцизию соответствующего провируса из участков его хромосомной интеграции для большинства субъектов, инфицированных ВИЧ-1. Таким образом, в одном варианте осуществления, адаптированную рекомбиназу получают из библиотеки рекомбиназ, содержащей рекомбиназу Cre, например, библиотеки, как показано на Фиг. 3, и она нацелена на множество штаммов ВИЧ-1, узнавая последовательности-мишени, имеющие последовательность, приведенную как SEQ ID NO:1 или SEQ ID NO:2. Предпочтительно, молекулярная направленная эволюция представляет собой субстрат-связанную белковую эволюцию (SLiPE). Экспрессионный вектор, кодирующий адаптированную рекомбиназу, предпочтительно получают из лентивирусного вектора. Этот экспрессионный вектор, трансфицированные им клетки и/или рекомбиназный белок, полученные из этого, имеют медицинские применения, например, при лечении и/или профилактике ВИЧ-1-инфекции.
В одном варианте осуществления, изобретение предоставляет способ приготовления экспрессионного вектора, кодирующего адаптированную рекомбиназу, которая способна к рекомбинации асимметричных последовательностей-мишеней внутри LTR провирусной ДНК множества штаммов ВИЧ-1, которая может быть встроена в геном клетки-хозяина, включающий стадии
(a) идентификация SEQ ID NO:1 или SEQ ID NO:2, имеющих гомологию с последовательностью левой половины участка и последовательностью правой половины участки рекомбиназного участка-мишени рекомбиназной библиотеки, содержащей рекомбиназы, указанные на Фиг. 1;
(b) идентификация двух последовательностей, где первая последовательность соответствует последовательности асимметричной последовательности-мишени со стадии (а), гомологичной левой половине участка указанного известного участка-мишени, и называется «последовательность половины участка 1», и где вторая последовательность соответствует последовательности асимметричной последовательности-мишени со стадии (а), гомологичной правой половине участка, и называется «последовательность половины участка 2»;
(c) определение нуклеотидов в пределах последовательностей со стадии (b), отклоняющихся от соответствующих гомологичных последовательностей левой половины участка и правой половины участка известного гомологичного участка-мишени со стадии (а), как показано на Фиг. 1;
(d) образование первой подгруппы двух нуклеиновых кислот-мишеней, содержащих последовательности-мишени, где первая последовательность-мишень обозначает подучасток 1 и включает SEQ ID NO: 8 (для SEQ ID NO:1) или SEQ ID: 16 (для SEQ ID NO:2), и где вторая последовательность-мишень обозначает подучасток 2 и включает SEQ ID NO:12 (для SEQ ID NO:1) или SEQ ID NO:20 (SEQ ID NO:2);
е) образование второй подгруппы нуклеиновых кислот-мишеней, содержащих модифицированные последовательности-мишени на основе последовательностей-мишеней в первой подгруппе со стадии (d), включающей SEQ ID NO:9-11 и 13-15 (для SEQ ID NO:1), или SEQ ID NO:17-19 и 21-23 (для SEQ ID NO:2), соответственно,
(f) раздельное применение молекулярной направленной эволюции на указанной рекомбиназной библиотеке с использованием каждой нуклеиновой кислоты второй подгруппы, полученной на стадии (e), в качестве субстрата;
(g) перетасовка рекомбиназных библиотек, образованных на стадии (f), где все рекомбиназные библиотеки, образованные на последовательностях на основе подучастка 1, объединяют и перетасовывают, и где все рекомбиназные библиотеки, образованные на последовательностях на основе подучастка 2, объединяют и перетасовывают;
(h) применение молекулярной направленной эволюции на перетасованных библиотеках, полученных на стадии (g), с использованием каждой нуклеиновой кислоты подгруппы, в соответствии со стадией (d), в качестве субстрата;
(i) перетасовка рекомбиназных библиотек, образованных на стадии (h);
(j) применение молекулярной направленной эволюции на перетасованной библиотеке, полученной на стадии (g), с использованием нуклеиновых кислот, содержащих асимметричную последовательность-мишень со стадии (a) в качестве субстрата, до тех пор, пока не получится, по крайней мере, одна рекомбиназа, которая действует на асимметричную последовательность-мишень внутри LTR ретровирусной ДНК со стадии (a);
(k) выделение нуклеиновой кислоты, в меньшей мере, одной рекомбиназы, полученной на стадии (j), из библиотеки; и
(l) клонирование нуклеиновой кислоты, полученной на стадии (k), в подходящий экспрессионный вектор,
где способ молекулярной направленной эволюции, предпочтительно, представляет собой субстрат-связанную белковую эволюцию.
Рекомбиназа TRE, адаптированная для узнавания SEQ ID NO:1, обозначена TRE 3.0. Рекомбиназа TRE, адаптированная для узнавания SEQ ID NO:2, обозначена TRE 4.0.
При этом для специалиста в рассматриваемой области очевидно, что можно создать другие адаптированные сайтспецифические рекомбиназы, которые обеспечивают рекомбинацию дивергентных участков-мишеней, встречающихся в геноме множества ретровирусных провирусов, встроенных в геном клетки-хозяина. Например, могут быть определены другие кандидатуры последовательностей-мишеней, которые присутствуют в геноме множества ретровирусных штаммов.
Провирусная ДНК, которая может быть встроена в геном клетки-хозяина, или которая может еще не быть встроена, предпочтительно представляет собой ДНК ретровируса. Ретровирусы включают большое и разнообразное семейство оболочечных РНК-вирусов. Отличительной особенностью семейства является его репликативная стратегия, которая включает существенные стадии обратной транскрипции вирусной РНК в линейную двухцепочечную ДНК и последующей интеграции этой ДНК (провирусной ДНК) в геном клетки-хозяина. Ретровирусы подразделяются на семь групп, определенных по их близости в эволюционном плане. Пять из этих групп (альфа-, бета-, дельта-, эпсилон- и гамма-ретровирусов) представляют ретровирусы с онкогенным потенциалом, а другие две группы представляют собой лентивирусы и спумавирусы. Патогенные для человека вирусы Т-клеточного лейкоза человека типа I и типа II (HTLV-I и HTLV-II) относятся к группе дельта-ретровирусов, в то время как вирусы СПИДа, вирус иммунодефицита человека типа 1 и типа 2 (ВИЧ-1 и ВИЧ-2), относятся к группе лентивирусов (в качестве обзора см. стандартный учебник «Retroviruses» of Coffin JM, Hughes SH, Varus HE (Eds.) 1997, Cold Spring Harbor Laboratory Press, New York).
В одном варианте осуществления, провирусная ДНК, которая может быть встроена в геном клетки-хозяина, представляет собой ДНК ретровируса, выбранного из группы, состоящей из вируса опухоли молочной железы мышей (MMTV), обезьяньего вируса Мейсон-Пфайзера (MPMV), вируса Т-клеточного лейкоза человека типа I (HTLV-I), вируса Т-клеточного лейкоза человека типа II (HTLV-II), обезьяньего вируса Т-клеточного лейкоза типа I (STLV-I), обезьяньего вируса Т-клеточного лейкоза типа II (STLV-II), вируса лейкоза крупного рогатого скота (BLV), вируса лейкоза кошек (FeLV) и вируса мышиной лейкемии Молони (MoMLV).
В другом варианте осуществления, ретровирус представляет собой лентивирус, выбранный из группы, состоящей из вируса иммунодефицита человека типа 1 (ВИЧ-1), вируса иммунодефицита человека типа 2 (ВИЧ-2), обезьяньего вируса иммунодефицита (SIV), вируса кошачьего иммунодефицита (FIV), вируса иммунодефицита крупного рогатого скота (BIV), вируса Маеди-Висна (MVV), вируса инфекционной анемии у лошадей (EIAV) и вируса козьего артрита-энцефалита (CAEV). Как указано выше, ВИЧ, в частности, ВИЧ-1 является предпочтительным.
В более предпочтительном варианте осуществления, асимметричная последовательность-мишень, идентифицированная на стадии (a) способа по настоящему изобретению, локализована как в 5’-LTR, так и в 3’-LTR провируса ВИЧ. Предпочтительно, указанная асимметричная последовательность-мишень, локализованная как в 5’-LTR, так и в 3’-LTR провируса ВИЧ, имеет последовательность, представленную в SEQ ID NO: 1 или SEQ ID NO:2.
В предпочтительном варианте осуществления, способ молекулярной направленной эволюции, применяемый в способе по настоящему изобретению, представляет собой способ субстрат-связанной белковой эволюции (SLiPE; Buckholz & Stewart, 2001; см. также WO 02/44409).
При помощи проведения способа по изобретению, описанного в настоящем документе, авторы изобретения создали несколько нуклеиновых кислот, кодирующих адаптированную рекомбиназу, и собственно адаптированные рекомбиназы. Изобретение, таким образом, предоставляет адаптированную рекомбиназу, включающую последовательность в соответствии с любой из SEQ ID NO: 40-64 или состоящую из этого, или нуклеиновые кислоты, кодирующие это.
Было с удивлением обнаружено, что эти адаптированные рекомбиназы предпочтительно содержат определенные консенсусные последовательности, как показано на Фиг. 5. В частности, все проанализированные адаптированные рекомбиназы, способные к рекомбинации асимметричной последовательности-мишени, включают последовательность SEQ ID NO: 37 («общая для Tre» консенсусная последовательность). Далее было обнаружено, что все проанализированные адаптированные рекомбиназы, способные к рекомбинации последовательности-мишени SEQ ID NO: 1, включают последовательность SEQ ID NO: 38. 95% всех проанализированных адаптированных рекомбиназ, способных к рекомбинации последовательности-мишени SEQ ID NO: 1, включают последовательность SEQ ID NO: 39. В SEQ ID NO: 37, 38 и 39 вариабельные аминокислоты представлены как X (см. Фиг. 5).
Изобретение, таким образом, предоставляет адаптированную рекомбиназу, способную к рекомбинации асимметричных последовательностей-мишеней внутри LTR провирусной ДНК ретровирусного штамма, который может быть встроена в геном клетки-хозяина (т.е. функциональная адаптированная рекомбиназа), которые предпочтительно включают SEQ ID NO: 37. Предпочтительно, указанная адаптированная рекомбиназа способна к рекомбинации асимметричных последовательностей-мишеней внутри LTR провирусной ДНК множества ретровирусных штаммов, которая может быть встроена в геном клетки-хозяина. Такая адаптированная рекомбиназа является доступной, например, в соответствии со способом по изобретению. Предпочтительно, указанная адаптированная рекомбиназа содержит SEQ ID NO: 37 и включает в себя, по крайней мере, одну, предпочтительно, 2, 3, 4, 5, 6, или 7 следующих определенных аминокислотных замен, по сравнению с последовательностью Cre (SEQ ID NO: 36): P12S, P15L, M44V, K86N, G93A, A175S, P307A. Эти замены представляют фермент, особенно подходящий для рекомбинации в последовательности-мишени SEQ ID NO: 1 или последовательности-мишени, обладающей высокий идентичностью с последовательностью SEQ ID NO: 1 (например, по крайней мере, 80%, по крайней мере, 90% или, по крайней мере, 95% идентичности с последовательностью SEQ ID NO: 1).
В одном варианте осуществления, последовательность адаптированной рекомбиназы по изобретению, в частности, адаптированная рекомбиназа, содержащая SEQ ID NO: 37, не раскрывается в WO 2008/083931. Предпочтительно, последовательность не включает SEQ ID NO: 65 настоящего приложения (которая идентична SEQ ID NO: 3 WO 2008/083931), или положения 11-351 из SEQ ID NO: 65, и последовательность нуклеиновой кислоты, кодирующая адаптированную рекомбиназу по изобретению, не кодирует белок, включающий положения 11-351 из SEQ ID NO: 65. Последовательность адаптированной рекомбиназы по изобретению, включающая SEQ ID NO: 37, также отличается от существующих в природе рекомбиназ, таких, как Cre, Dre, Fre или Zre, что видно из той особенности, что она способна к рекомбинации асимметричных последовательностей-мишеней внутри LTR провирусной ДНК ретровирусного штамма, который может быть встроен в геном клетки-хозяина, и, предпочтительно, что она способна к рекомбинации асимметричных последовательностей-мишеней внутри LTR провирусной ДНК множества ретровирусных штаммов, которые могут быть встроены в геном клетки-хозяина.
Если адаптированная рекомбиназа, способная к рекомбинации асимметричных последовательностей-мишеней внутри LTR провирусной ДНК множества ретровирусных штаммов, встроенных в геном клетки-хозяина, предназначена для обеспечения рекомбинации последовательности-мишени с SEQ ID NO: 1, то она предпочтительно включает SEQ ID NO: 38 или SEQ ID NO: 39.
Функциональные адаптированные рекомбиназы, способные к рекомбинации асимметричных последовательностей-мишеней внутри LTR провирусной ДНК множества ретровирусных штаммов, которые могут быть встроены в геном клетки-хозяина, которые могут быть получены, например, при помощи способа по изобретению, могут отличаться от указанной последовательности, однако последовательности служат ценным руководством для специалиста в рассматриваемой области, чтобы продуцировать адаптированную рекомбиназу, способную к рекомбинации асимметричных участков-мишеней, таких как SEQ ID NO: 1, даже без проведения способа по изобретению. Изобретение также предоставляет адаптированную рекомбиназу, включающую последовательности, имеющие, по меньшей мере, 97% идентичностью последовательности с SEQ ID NO: 38, и/или имеющие, по крайней мере, 98% идентичности последовательности с SEQ ID NO: 38 и/или имеющие, по крайней мере, 99% идентичности последовательности с SEQ ID NO: 38. Предпочтительно, адаптированная рекомбиназа имеет, по крайней мере, 97% идентичности последовательности с SEQ ID NO: 39, и/или, по крайней мере, 98% идентичности последовательности SEQ ID NO: 39 и/или, по крайней мере, 99% идентичности последовательности с SEQ ID NO: 39. Наличие определенного процента идентичности последовательности с последовательностью SEQ ID NO: 37, 38 или 39 в контексте настоящего изобретения означает идентичность последовательности в положениях, определенных в соответствующей последовательности, например, при 97% идентичности последовательности, 3% положений определенных аминокислот указанных последовательностей могут варьировать. Предпочтительно, аминокислотные замены в отношении референсных последовательностей являются консервативными заменами, которые известны специалистам в рассматриваемой области (например, Creighton (1984), Proteins. W. H. Freeman and Company (Ed.)). Например, консервативные замены замещают одну аминокислоту из группы отрицательно заряженных аминокислот на другую.
Адаптированные рекомбиназы, способные к рекомбинации асимметричных последовательностей-мишеней внутри LTR провирусной ДНК множества ретровирусных штаммов, которые могут быть встроены в геном клетки-хозяина, также могут содержать комбинации из 2, 3, 4 или более последовательностей, выбранных из группы, состоящей из SEQ ID NO: 40-64, например, C-концевую часть любой из этих последовательностей, например, SEQ ID NO: 40, и N-концевую частью любой другой из этих последовательностей, например, SEQ ID NO: 64. C-концевая часть может иметь длину в 1-342 аминокислот. В объединении двух последовательностей, N-концевая часть может иметь длину в 1-342 аминокислот. Объединение в любом случае содержит консенсусный мотив TRE, например, SEQ ID NO: 37, или, предпочтительно, SEQ ID NO: 38 или SEQ ID NO: 39.
Изобретение дополнительно предоставляет композицию, включающую две или более адаптированных рекомбиназ, содержащих различные последовательности SEQ ID NO: 37, предпочтительно, SEQ ID NO: 38 или SEQ ID NO: 39, или нуклеиновые кислоты, кодирующие их. В одном варианте осуществления, композиция включает две или более, три или более, четырех или более, пять или более, десять и более, 20 или более, или 25 рекомбиназ, содержащих последовательности в соответствии с любой из SEQ ID NO: 40-64 или нуклеиновых кислот, кодирующих их. Такие композиции могут представлять собой фармацевтические композиции, как описано ниже.
Изобретение также предоставляет нуклеиновую кислоту, кодирующую адаптированную рекомбиназу, способную к рекомбинации асимметричных последовательностей-мишеней внутри LTR провирусной ДНК множества ретровирусных штаммов, которые могут быть встроены в геном клетки-хозяина, адаптированную рекомбиназу, содержащую аминокислотную последовательность, как определено выше. В предпочтительных вариантах изобретения, указанная аминокислотная последовательность имеет, по крайней мере, 97% идентичности последовательности с последовательностью SEQ ID NO: 38, по крайней мере, 98% идентичности последовательности с последовательностью SEQ ID NO: 38 или, по крайней мере, 99% идентичности последовательности с SEQ ID N: 38, или включает SEQ ID NO: 38. Изобретение также предоставляет нуклеиновую кислоту, кодирующую адаптированную рекомбиназу, содержащую аминокислотную последовательность, имеющую, по меньшей мере, 97% идентичности последовательности с последовательностью SEQ ID NO: 39, по крайней мере, 98% идентичности последовательности с последовательностью SEQ ID NO: 39 или, по крайней мере, 99% идентичности последовательности с SEQ ID NO: 38, или аминокислотную последовательность, включающую SEQ ID NO: 39. Изобретение также предоставляет нуклеиновую кислоту, кодирующую адаптированную рекомбиназу, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 40-64 или комбинации указанных последовательностей.
В контексте изобретения, нуклеиновая кислота и белок, включающий последовательность, может также состоять из этой последовательности.
Она также может содержать дополнительные последовательности, например, сигнальную последовательность, обеспечивающая экспрессию/локализацию в специфическом клеточном компартменте, такую как сигнал ядерной локализации (например, сигнал в положениях 2-9 SEQ ID NO: 65). Если белок предназначен для использования в фармацевтической композиции, особенно предпочтительным является его экспрессия в качестве слитого белка с доменом трансдукции белка, например, доменом трансдукции белка tat, что делает возможным трансдукцию белка в клетки-мишени. Предпочтительно, адаптированную рекомбиназу по изобретению, которая предназначена для использования в фармацевтической композиции, подготавливают в качестве слитого белка с последовательностью ядерной локализации и доменом трансдукции белка, например, из tat, и нуклеиновая кислота, кодирующая адаптированную рекомбиназу по изобретению, может кодировать такой слитый белок. Например, можно использовать следующие домены трансдукции белка в слитом белке с адаптированной рекомбиназой по изобретению, которые предпочтительно также включают сигнал ядерной локализации:
- Основной домен Tat-трансактиватора ВИЧ-1 (Fawell S, Seery J, Daikh Y, Moore C, Chen LL, Pepinsky B, Barsoum J., Tat-mediated delivery of heterologous proteins into cells., Proc Natl Acad Sci USA. 1994 Jan 18; 91(2):664-8).
- Гомеодомен антеннопедии Drosohila (Antp) (Derossi D, Joliot AH, Chassaing G, Pro-chiantz A., The third helix of the Antennapedia homeodomain translocates through biological membranes., J Biol Chem. 1994 Apr 8; 269(14): 10444-50).
- Транскрипционный фактор VP22 HVS (Elliott G, O’Hare P., Intercellular trafficking and protein delivery by a herpesvirus structural protein., Cell. 1997 Jan 24; 88(2):223-33).
- Транслокационный мотив клеточной проницаемости (TLM) поверхностного антигена PreS2 вируса гепатита В (HBV) (Oess S, Hildt E., Novel cell permeable motif derived from the PreS2-domain of hepatitis-B virus surface antigens., Gene Ther. 2000 May;7(9):750-8).
В случае если белок будет подвергаться очистке, можно также добавлять тег, облегчающий очистку белка, такой как His-tag.
Использование кодонов в нуклеиновой кислоте по изобретению, кодирующей рекомбиназу Tre, как определено выше, может быть выбрано специалистом в рассматриваемой области. Например, можно выбрать использование кодонов, подходящее для экспрессии в клетке человека, в частности, если предполагается экспрессия в клетках человека, например, для терапевтических целей. Использование кодонов также может основываться на использовании кодонов, например, для рекомбиназы Cre.
Адаптированную рекомбиназу или нуклеиновую кислоту, кодирующую указанную адаптированную рекомбиназу, можно получить способом по изобретению, как описано в настоящем документе, или ее можно получить с помощью этого способа. Она также может быть получена на основе раскрытых в настоящем документе последовательностей, необязательно, путем комбинирования и/или дальнейшего изменения этих последовательностей, при необходимости, тестирования на активность в рекомбинации асимметричных участков-мишеней, таких как SEQ ID NO: 1 или SEQ ID NO: 2.
Изобретение также предоставляет композицию, например, библиотеку, состоящую из двух или более нуклеиновых кислот, кодирующих адаптированную рекомбиназу, как определено выше, например, кодирующих две или более адаптированных рекомбиназ, включающих различные последовательности SEQ ID NO: 37, предпочтительно, различные последовательности SEQ ID NO: 38 или SEQ ID NO: 39. В одном варианте осуществления, композиция включает нуклеиновые кислоты, кодирующие адаптированные рекомбиназы, состоящие из двух или более, трех или более, четырех или более, пяти или более, десяти и более, 20 или более, или 25 или более рекомбиназ, включающих последовательности в соответствии с любой из SEQ ID NO: 40-64 или комбинаций этих последовательностей. Такие композиции, в частности, композиции, в которой нуклеиновая кислота представляет собой экспрессионный вектор, могут быть особенно подходящими в качестве фармацевтических композиций, как описано ниже.
В способе по настоящему изобретению нуклеиновую кислоту, кодирующую, по крайней мере, одну адаптированную рекомбиназу, которая действует на асимметричную последовательность-мишень внутри LTR ретровирусной ДНК, клонируют в экспрессионный вектор. Экспрессионные векторы представляют собой генетические конструкции для экспрессии белков, кодируемых нуклеиновыми кислотами в составе вектора. Такие экспрессионные векторы могут представлять собой либо самовоспроизводящиеся экстрахромосомные векторы, либо векторы, которые встраиваются в геном хозяина. Как правило, эти экспрессионные векторы включают транскрипционно и трансляционно регулируемую нуклеиновую кислоту, функционально связанную с нуклеиновой кислоты, кодирующей адаптированную рекомбиназу по настоящему изобретению.
Термин «контролирующие последовательности» относится к последовательностям ДНК, необходимым для экспрессии функционально связанной кодирующей последовательности в конкретном организме-хозяине. Контролирующие последовательности, которые подходят для прокариот, включают, например, промотор, необязательно операторную последовательность и участок связывания с рибосомой. Эукариотические клетки, как известно, используют промоторы, сигналы полиаденилирования и энхансеры.
Нуклеиновая кислота является «функционально связанной», когда она находится в функциональной взаимосвязи с другой последовательностью нуклеиновых кислот. Например, промотор или энхансер является функционально связанным с кодирующей последовательностью, если он действуют на транскрипцию последовательности; или участок связывания с рибосомой является функционально связанным с кодирующей последовательностью, если оно расположен таким образом, чтобы способствовать трансляцию. Связывание осуществляется путем лигирования ковалентных участков рестрикции. Если таких участков не существует, в соответствии с обычной практикой используют синтетические олигонуклеотидные адапторы или линкеры. Нуклеиновые кислоты, регулирующие транскрипцию и трансляцию, как правило, должны соответствовать клетке-хозяину, используемой для экспрессии адаптированной рекомбиназы. В рассматриваемой области известны многочисленные типы соответствующих экспрессионных векторов и подходящих регуляторных последовательностей для различных клеток-хозяев.
Экспрессионный вектор, используемый в настоящем изобретении, может представлять собой ретровирусный вектор, лентивирусный вектор, спумавирусный вектор или аденовирусный вектор. Однако в предпочтительном варианте осуществления экспрессионный вектор представляет собой лентивирусный вектор, выбранный из группы, состоящей из лентивирусных векторов, полученных из ВИЧ-1, SIV, FIV или EIAV. Лентивирусные векторы, например, описывается в Schambach et al. (2006) или в Европейской патентной заявке No 11000751.5.
В предпочтительном варианте настоящего изобретения экспрессионный вектор содержит клеточный, бактериальный, вирусный или гибридный промотор.
В большинстве случаев, для целей настоящего изобретения, промотор может представлять собой конститутивный или индуцируемый промотор. Далее, промоторы могут представлять собой либо промоторы, встречающиеся в природе, например, клеточный, бактериальный или вирусный промотор, или гибридный промотор. Гибридные промоторы, которые сочетают элементы более чем одного промотора, известны в рассматриваемой области и пригодны для настоящего изобретения. Далее, промотор, используемый в настоящем изобретении, также может быть производным от промотора, существующего в природе. «Производное» от промотора, существующего в природе, как используется в настоящем документе, может представлять собой комбинацию из цис-активных элементов, полученных от промоторов или последовательностей различного происхождения, или, альтернативно, может быть получено путем делеции или мутации цис-активных элементов в пределах определенного промотора, существующего в природе (Edelman et al., 2000; Alper et al., 2006; Hartenbach & Fussenegger, 2006).
В варианте осуществления настоящего изобретения, конститутивный промотор или производное этого выбраны или получены из группы, состоящей из промоторов цитомегаловируса, вируса саркомы Рауса, ретровирусов, родственных вирусу мышиной лейкемии, фосфоглицерокиназного гена, очаг-образующего вируса мышиный селезенки или фактора элонгации 1-альфа человека.
В дополнительном варианте осуществления настоящего изобретения, индуцируемый промотор или производное этого выбран или получен из группы, состоящей из LTR или производных этого, полученных из лентивирусов, спумавирусов и дельта-ретровирусов.
В данном контексте, термин «LTR» относится как к 5’-, так и к 3’- длинным концевым повторам провируса, обладающим промоторной функцией (в качестве обзора см. стандартный учебник «Retroviruses» (Coffin JM, Hughes SH, Varmus ОН (Eds.) 1997, Cold Spring Harbor Laboratory Press, New York).
Предпочтительно, индуцируемый промотор или производное этого выбирают или получают из LTR или производного этого, полученного из ВИЧ-1, ВИЧ-2, MVV, EIAV, CAEV, SIV, FIV, BIV, HTLV-I и HTLV-II.
Настоящее изобретение также предоставляет способ приготовления адаптированной рекомбиназы, где указанный способ включает вышеизложенный способ приготовления экспрессионного вектора, кодирующего адаптированную рекомбиназу, и дополнительную стадию экспрессии адаптированной рекомбиназы или слитого полипептида, содержащего аминокислотную последовательность указанной адаптированной рекомбиназы из нуклеиновой кислоты, кодирующей рекомбиназу, встроенной в экспрессионный вектор, полученный вышеупомянутым способом приготовления экспрессионного вектора, кодирующего адаптированную рекомбиназу, в подходящей клетке-хозяине.
Предпочтительно, полученные в результате рекомбиназы тестируют в клетках млекопитающих, чтобы убедиться, что они функционируют в окружающей среде клеток млекопитающих. Далее, чтобы получить хорошую экспрессию в клетках млекопитающих, рекомбиназы могут быть оптимизированы для экспрессии в этих клетках (например, оптимизация использования кодонов при помощи способов, известных в рассматриваемой области. См., например, Shimshek et al., 2002), или к нуклеиновой кислоте адаптированной рекомбиназы можно добавить сигнальные последовательности, необходимые для направления белка в ядро клетки млекопитающего, такие, как NLS-последовательности (Macara, 2001).
Экспрессию кодирующей адаптированную рекомбиназу нуклеиновой кислоты, клонированной в экспрессионный вектор, например, в соответствии со стадией (l) способа приготовления экспрессионного вектора, кодирующего адаптированную рекомбиназу, можно проводить с использованием, например, экспрессионных систем бактерий, насекомых и млекопитающих. При этом также можно использовать другие экспрессионные системы, известные в рассматриваемой области. Способы введения экзогенных нуклеиновых кислот в организмы-хозяева млекопитающих, насекомых или бактерий, равно как и в другие организмы-хозяева, также известны в рассматриваемой области, и меняются в зависимости от используемой клетки-хозяина. Способы включают в себя декстран-опосредованную трансфекцию, осаждение фосфатом кальция, полибрен-опосредованная трансфекция, слияние протопластов, электропорацию, инфицирование вирусами, инкапсуляции полинуклеотид(ов) в липосомы и прямую микроинъекцию ДНК в ядро.
Слитые белки подготовливают при помощи способов, известных в рассматриваемой области. Например, нуклеиновую кислоту, кодирующую адаптированную рекомбиназу, клонируют в экспрессионный вектор, который уже содержит нуклеотидную последовательность, кодирующую второй полипептид или белок. При клонировании нуклеиновой кислоты, кодирующей адаптированную рекомбиназу, в рамке с последовательностью второго полипептида или белка, обе последовательности будут экспрессироваться в качестве слитого белка.
Клетки-хозяева, используемые для экспрессии адаптированной рекомбиназы с экспрессионного вектора, представляют собой, предпочтительно, клетки-хозяева, включающие прокариотические клетки, такие как, например, клетки бактерий и клетки дрожжей, или эукариотические клетки, такие как, например, клетки насекомых или клетки млекопитающих. Клетка-хозяин может представлять собой кроветворную клетку, например, взрослую гемопоэтическую стволовую клетку и Т-клетку, например, клетку CD4+. Клетки могут быть получены от субъекта, инфицированного ретровирусом, и клетка может быть введена обратно субъекту после трансформации, и, необязательно, культивирования и/или размножения.
Настоящее изобретение также предоставляет способ приготовления трансформированной взрослой стволовой клетки, где указанный способ включает вышеизложенный способ приготовления экспрессионного вектора, кодирующего адаптированную рекомбиназу, и дополнительную стадию введения экспрессионного вектора, полученного в вышеупомянутом способе приготовления экспрессионного вектора, кодирующего адаптированную рекомбиназу, in vitro в подходящую взрослую стволовую клетку.
В еще одном аспекте, настоящее изобретение направлено на нуклеиновую кислоту, получаемую при помощи вышеизложенного способа по настоящему изобретению. В настоящем документе также предоставлены нуклеиновые кислоты, кодирующие адаптированную рекомбиназу, определенную последовательностью.
«Нуклеиновая кислота», как используется в настоящем документе, представляет собой полимерное соединение, состоящее из ковалентно связанных субъединиц, называемых нуклеотидами. Нуклеиновая кислота включает полирибонуклеиновую кислоту (РНК) и полидезоксирибонуклеиновую кислоту (ДНК), каждая из которых может быть одноцепочечной или двухцепочечной. ДНК включает кДНК, геномную ДНК, синтетическую ДНК и полусинтетическую ДНК.
В самом широком аспекте, нуклеиновая кислота, полученная при помощи способа по настоящему изобретению или определенная в этом документе, представляет собой нуклеиновую кислоту, кодирующую адаптированную рекомбиназу, где адаптированная рекомбиназа обеспечивает рекомбинацию асимметричных последовательностей-мишеней в пределах ДНК провируса из множества вирусных штаммов, которые могут быть встроены в геном клетки-хозяина, приводящую к эксцизии провируса из генома клетки-хозяина, где асимметричные участки-мишени отличаются от участков-мишеней рекомбиназы дикого типа.
В еще одном аспекте, настоящее изобретение направлено также на экспрессионный вектор, как получаемый вышеизложенным способом по настоящему изобретению, и экспрессионный вектор, содержащий нуклеиновую кислоту, кодирующую адаптированную рекомбиназу, как определено в настоящем документе.
Термин «белок», используемый в настоящем документе, включает в себя белки, полипептиды и пептиды. Как могут оценить специалисты в рассматриваемой области, последовательности нуклеиновых кислот по изобретению могут быть использованы для получения белковых последовательностей. Еще один аспект изобретения представляет собой белок адаптированной рекомбиназы, получаемый, например, вышеизложенным способом по настоящему изобретению, который необязательно может представлять собой слитый белок, включающий функциональную рекомбиназу. В одном варианте осуществления, белок адаптированной рекомбиназы можно подготовить как слитый полипептид, используя приемы, известные в рассматриваемой области. В предпочтительном варианте осуществления, белок адаптированной рекомбиназы связан со вторым полипептидом. Предпочтительно, слитый полипептид получается вышеизложенным способом по настоящему изобретению, при котором адаптированная рекомбиназа связывается со вторым полипептидом.
В одном варианте осуществления, белок адаптированной рекомбиназы подготавливают в качестве слитого полипептида для увеличения экспрессии. В дополнительном варианте осуществления, белок адаптированной рекомбиназы выполняют в виде слитого полипептида, чтобы дать возможность вводить полипептид в живые клетки. Как правило, очищенные белки не могут попадать в клетки, потому что они не способны проходить через клеточную мембрану из-за своих размеров. При этом, слияние специфических белковых последовательностей с белками может привести к поглощению этих слитых белков клетками. В клетке белок может затем выполнить свою функцию. Сайтспецифические рекомбиназы, в том числе рекомбиназа Cre, были успешно доставлены в клетки при помощи этого подхода (Peltz et al., 2002). Проникающая в клетку рекомбиназа также была описана Nolden et al. (2006) и Lin et al. (2004). Следовательно, эта стратегия может быть использована для доставки адаптированной рекомбиназы в клетки для удаления провируса из инфицированных клеток. Так, второй полипептид в слитом полипептиде может содержать сигнальный пептид. Сигнальный пептид может представлять собой домен трансдукции белка, такой как ТАТ-пептид или пептид третьей спирали гомеодомена антеннапедии (Derossi et al, 1994, 1996; Vives et al., 1997; Vives, 2003; Richard et al., 2005) или NLS (последовательность ядерной локализации) для доставки слитого полипептида в ядро эукариотической клетки (Macara, 2001).
Еще один аспект настоящего изобретения направлен на взрослую стволовую клетку, получаемую вышеизложенным способом, для приготовления трансформированной взрослой стволовой клетки по настоящему изобретению. Стволовые клетки, предпочтительно, инфицируют или трансфицируют экспрессионным вектором в соответствии с изобретением. В предпочтительном варианте осуществления, взрослая стволовая клетка представляет собой стволовую клетку из кроветворной линии, экспрессирующей адаптированную рекомбиназу, вышеупомянутый слитый полипептид или включающей вышеуказанный экспрессионный вектор. Гемопоэтические стволовые клетки (ГСК) представляют собой полученные из костного мозга клетки CD34+, которые могут, например, быть очищены от G-CSF-мобилизованной периферической крови доноров (например, ВИЧ-инфицированных пациентов) при помощи обычного лейкафереза (Scherr & Eder, 2002). Генетически модифицированные in vitro клетки можно затем составлять для реинфузии пациентам.
В рассматриваемой области, термин «стволовые клетки» обозначает клетки, которые (а) обладают способностью самообновления и (b) способностью образовывать, по крайней мере, одну, а часто несколько специальных типов клеток, из-за их способности асимметричного деления (Donovan & Gearhart, 2001). Взрослые стволовые клетки можно выделить из разных тканей взрослого, т.е. из дифференцированных индивидуумов. Такие стволовые клетки в рассматриваемой области называются «мультипотентными взрослыми стволовыми клетками». Существенная разница между эмбриональными плюрипотентными стволовыми клетками и взрослыми мультипотентными стволовыми клетками заключается в числе дифференцированных тканей, которые могут быть получены из соответствующих клеток.
В еще одном варианте осуществления, экспрессионный вектор по настоящему изобретению используется для трансформации T-клеток, например, первичных клеток CD4+ (клеток крови) пациентов, инфицированных ретровирусом (например, ВИЧ).
В дополнительной стадии способа по настоящему изобретению, экспрессионный вектор, содержащий последовательность нуклеиновых кислот, кодирующую адаптированную рекомбиназу по изобретению, белок рекомбиназы, слитый белок или взрослая стволовая клетка, полученные способами по настоящему изобретению, составляют в качестве фармацевтической композиции для применения при профилактике и/или лечении ретровирусной инфекции и/или для снижения вирусной нагрузки у субъекта, инфицированного ретровирусом, например, ВИЧ, в частности, ВИЧ-1.
Еще одним предметом изобретения является фармацевтическая композиция, полученная вышеизложенным способом. Фармацевтическая композиция предпочтительно присутствует в виде раствора, пригодного для внутривенного применения (инфузии).
Фармацевтический препарат может дополнительно включать в себя один или несколько фармацевтически приемлемых носителей, наполнителей и/или адъювантов. Носители, наполнители и адъюванты, пригодные для использования в фармацевтической композиции, известны в рассматриваемой области.
Фармацевтическая композиция по настоящему изобретению предпочтительно снижает вирусную нагрузку у субъекта, инфицированного ретровирусом, ниже 5,000 геномных эквивалентов/мл плазмы, предпочтительно ниже 500 геномных эквивалентов/мл плазмы и, более предпочтительно, ниже 50 геномных эквивалентов/мл плазмы при введении субъекту. Таким образом, фармацевтическая композиция по настоящему изобретению, включающая экспрессионный вектор, кодирующий адаптированную рекомбиназу (или адаптированную рекомбиназу в качестве белка или слитого полипептида или стволовую клетку, содержащую экспрессионный вектор), способна снизить вирусную нагрузку у субъекта, инфицированного ретровирусом, путем эрадикации генетического источника ретровирусов в клетках-хозяевах, тем самым предотвращая дальнейший жизненный цикл вируса.
Термин «вирусная нагрузка», используемый в настоящем документе, относится, например, на РНК-эквивалентам ВИЧ (т.е. геномам), которые ассоциированы с 1 мл плазмы пациента (Dybul et al., 2002). Таким образом, вирусная нагрузка определяется путем измерения содержания вирусной ДНК в образце, полученном от пациента. В настоящее время, существует три основных типа доступных тестов вирусной нагрузки:
1) Полимеразная цепная реакция обратно транскрибированной РНК ВИЧ (RT-PCR): AmplicorTM, HIV-1 Monitor Test; Roche Diagnostics
2) ДНК с разветвленной цепью (bDNA): VersantTM HIV RNA Assay; Bayer Diagnostics; и
3) Амплификация на основе последовательности нуклеиновых кислот (NASBA): NucliSensTM Assay; bioMerieux.
В предпочтительном варианте осуществления, фармацевтическая композиция по настоящему изобретению способны снизить вирусную нагрузку у субъекта, инфицированного ретровирусом, ниже 5,000 геномных эквивалентов/мл плазмы, предпочтительно ниже 500 геномных эквивалентов/мл плазмы и, более предпочтительно, ниже 50 геномных эквивалентов/мл плазмы. Пациент с вирусной нагрузкой ниже уровня 5000 геномных эквивалентов/мл плазмы считаются относительно хорошо приспособленным к медикаментозному лечению. Однако целью в современной терапии СПИДа является снижение вирусной нагрузки ниже предела обнаружения теста вирусной нагрузки, которые в настоящее время составляет ниже около 50 геномных эквивалентов/мл плазмы.
Фармацевтическая композиция предпочтительно снижает вирусную нагрузку ретровирусов, выбранных из группы, состоящей из вируса опухоли молочной железы мышей (MMTV), обезьяньего вируса Мейсон-Пфайзера (MPMV), вируса Т-клеточного лейкоза человека типа I (HTLV-I), вируса Т-клеточного лейкоза человека типа II (HTLV-II), обезьяньего вируса Т-клеточного лейкоза типа I (STLV-I), обезьяньего вируса Т-клеточного лейкоза типа II (STLV-II), вируса лейкоза крупного рогатого скота (BLV), вируса лейкоза кошек (FeLV) и вируса мышиной лейкемии Молони (MoMLV). В еще одном предпочтительном варианте осуществления ретровирус, подвергаемый лечению фармацевтической композицией по настоящему изобретению, является лентивирусом. Указанный лентивирус предпочтительно, выбирают из группы, состоящей из вируса иммунодефицита человека типа 1 (ВИЧ-1), вируса иммунодефицита человека типа 2 (ВИЧ-2), обезьяньего вируса иммунодефицита (SIV), вируса кошачьего иммунодефицита (FIV), вируса иммунодефицита крупного рогатого скота (BIV), вируса Маеди-Висна (MVV), вируса инфекционной анемии у лошадей (EIAV) и вируса козьего артрита-энцефалита (CAEV). Наиболее предпочтительно, ретровирус представляет собой ВИЧ, в частности ВИЧ-1. При этом для специалиста в рассматриваемой области является очевидным, что настоящее изобретение также применимо к ретровирусным инфекциям ретровирусами, отличными от тех, о которых упоминалось выше.
Субъект, инфицированный ретровирусом, которому должна быть введена фармацевтическая композиция, выбирается из группы, состоящей из людей, приматов, обезьян, крупного рогатого скота, лошадей, коз, овец и домашних кошек. Однако, субъектом, предпочтительно, является человек.
В большинстве случаев, к субъекту должно быть применено эффективное количество экспрессионного вектора, адаптированной рекомбиназы или трансформированной клетки по изобретению. Введение может быть, например, внутривенным или внутримышечным введением.
В одном варианте осуществления, фармацевтическую композицию составляют для сопутствующего введения с другими активными веществами высокоактивной антиретровирусной терапии (ВААРТ). Высокоактивная антиретровирусная терапия ВААРТ представляет собой комбинированную терапию, нацеленную на вирусную обратную транскриптазу, протеазу и слияние (Gulic et al., 1997; Lalezari et al., 2003).
В другом варианте осуществления, фармацевтическую композицию составляют для введения одновременно или после глобального терапии иммунной активации или специфической активации экспрессии генов провируса. Предположение о терапии иммунной активации основывается на гипотезе о том, что преднамеренная активация клеток, инфицированных латентным ВИЧ, может ускорить эрадикацию стойких вирусной источников. Эрадикация будет происходить через иммунную очистку путем запрограммированной гибели тех клеток, которые активно экспрессируют (проапоптотические) продукты ВИЧ-1 (Kulkovsky & Bray, 2006). Глобальная иммунная активация (активация иммунных клеток, в том числе покоящихся клеток) обычно достигается введением иммунотоксинов, цитокинов (например, ИЛ-2) или активирующих Т-клетки антител (например, OKT3).
В связи с тем, что иммунная активация, проведенная для намеренной активации ВААРТ-устойчивых латентных источников, к сожалению, не достигла успеха в окончательной элиминации ВИЧ-1 и в плане восстановления вируса после лечения (в качестве обзора см. Kulkovsky & Bray 2006; Marcello, 2006; Shehu-Xhilaga et al., 2005) в связи с тем, что глобальная Т-клеточная активация, по-видимому, также индуцирует вирусную репликацию и увеличивает количество потенциальных клеток-мишеней ВИЧ-1 сверх того уровня, который может удерживаться при помощи ВААРТ (Fraser et al., 2000), для лечения ВИЧ необходимы дополнительные специфические процедуры. Один из подходов заключается в активации транскрипции в ином случае латентных вирусных геномов. Специфическая активация экспрессии генов латентного провируса может быть достигнута путем введении форболового эфира простратина или цитокина IL-7 человека, каждый из которых, как представляется, реактивирует латентный ВИЧ-1 при отсутствии клеточной пролиферации (Marcello, 2006). Кроме того, селективной активации транскрипции ВИЧ-1 также можно достичь при помощи гистон-деацетилазных (HDACl)-ингибиторов, таких как, например, вальпроевая кислота, которая, в конечном счете, вызывает рост ВИЧ-1 из покоящихся клеток в отсутствие клеточной активации (Marcello, 2006; Lehrman et al., 2005).
Тем не менее, терапии глобальной иммунной активации или специфическая активация экспрессии генов провируса или аналогичные терапевтические стратегии значительно выигрывают от одновременного удаления провирусной ДНК, тем самым снижая у пациента пул инфицированных клеток.
Настоящее изобретение также предоставляет способ лечения и/или профилактики ретровирусной инфекции, в частности, ВИЧ-инфекции, у субъекта. В одном варианте осуществления, последовательность ретровируса, инфицирующего субъекта, анализируют в образце, полученном от субъекта, и, по крайней мере, один экспрессионный вектор, кодирующий адаптированную рекомбиназу, по крайней мере, одна адаптированная рекомбиназа или, по крайней мере, одна клетка, трансформированная указанным экспрессионным вектором, например, одна взрослая стволовая клетка, должна быть введена субъекту, если провирусная ДНК от субъекта содержит асимметричную последовательность-мишень, определенную на стадии (а), на которой была отобрана рекомбиназа. Образец, полученный от субъекта, может представлять собой образец крови, например, содержащий инфицированные клетки CD4+.
Краткое описание фигур
Фиг. 1: Фиг. 1 представляет участки-мишени специфических рекомбиназ, loxP (узнаваемый Cre), loxH (узнаваемый Fre), rox (узнаваемый Dre), zox (узнаваемый Zre) и loxLTR (узнаваемый Tre) (SEQ ID NO: 3-7), а также асимметричные участки-мишени, присутствующие во множестве штаммов ВИЧ-1 (SEQ ID NO: 1 и 2). Спейсерная последовательность представлена жирным шрифтом. Положения нуклеотидов в левой половине участка и правой половине участка SEQ ID NO: 1 или SEQ ID NO:2, которые не гомологичны одному из известных участков-мишеней, подчеркнуты. SEQ ID NO:1 имеет 54% гомологии с пулом известных рекомбиназных участков-мишеней; SEQ ID NO:2 имеет 42% гомологии с пулом известных рекомбиназных участков-мишеней.
Фиг. 2: Фиг. 2 представляет последовательность подучастков, использованных для создания адаптированных рекомбиназ TRE 3.0 и TRE 4.0. Положения нуклеотидов в левой половине участка и правой половине участка SEQ ID NO:1 или 2, которые не гомологичны одному из известных участков-мишеней, подчеркнуты. Подучастки 1 и 2 создают на стадии (d) способа по изобретению, и подучастки 1A-C и подучастки 2А-С создают на стадии (e) способа по изобретению. Подучастки 1a/b и подучастки 2a/b, а также промежуточные подучастки 1b+1, 1b+2, 2а+1 и 2a+2 создают на дальнейших стадиях способа по изобретению.
Фиг. 3: Фиг. 3 представляет схему эволюционной стратегии и показывает результаты первой стадии эволюции для SEQ ID NO:1. Начальная библиотека рекомбиназ была создана путем объединения и перетасовки семейства Cre и нескольких известных Cre-подобных рекомбиназ (А). Удачные рекомбиназы были обогащены путем проведения нескольких раундов субстрат-связанной белковой эволюции на подучастках 2-й подгруппы. В (В) рекомбинационную активность после 1-го эволюционного цикла анализируют, в (E) - после 6-го эволюционного цикла. Рекомбинированный продукт является более коротким, так как последовательность между участками-мишенями была эксцизирована, он отмечен одним треугольником. Нерекомбинированный продукт отмечен двумя треугольниками. Удачные рекомбиназы были амплифицированы при помощи ПЦР, в результате был получен продукт размером 1,7 т.п.н. (C и D). К одному из несоответствий в подучастках 1С и 2С, соответственно, был применен отдельный подход (через мутированный участок-мишень loxP, названный ΔloxP, чтобы получить более высокой рекомбинационной активности остатков на подучастках 1С и 2С (D).
Фиг. 4: Фиг. 4 представляет схему дополнительных стадий эволюционной стратегии и показывает результат конечный эволюционной стадии для SEQ ID NO:1. Часть (А) фигуры соответствует стадиям первых эволюционных циклов, подробно охарактеризованных на Фигуре 3.
(B) Удачные рекомбиназы были, кроме того, обогащены путем проведения нескольких раундов субстрат-связанной белковой эволюции на подучастках 3-й группы, за которым следуют несколько раундов субстрат-связанных эволюционных циклов на некоторых промежуточных субстратах (C) и, наконец, на 1-ых подучастках (D), чтобы получить рекомбиназы, специфичные для асимметричных участков-мишеней SEQ ID NO: 1. В (E) проверяют рекомбинационную активность после 8-го эволюционного цикла с относительно высоким количеством транскрипционного активатора L-арабинозы (L-ara; 200 мкг/мл), в (F) рекомбинационную активность определяли после 16-го эволюционного цикла с более низкой индукцией транскрипции только при 50 мкг/мл L-ara. Рекомбинированный продукт отмечен одним треугольником, нерекомбинированный - двумя треугольниками. (G) и (H) показывают соответствующую рекомбинационную активность после указанных эволюционных циклов, указанные концентрации L-ara и на указанных субучастках. После конечного эволюционного цикла (цикл 43) популяция рекомбиназ с наиболее высокой активностью была изолированы и обогащена, в результате образована активная библиотека. Отдельные клоны рекомбиназ были отобраны и подвергнуты дальнейшему анализу.
Фиг. 5: Фиг. 5 представляет выравнивание белковых последовательностей (а) рекомбиназы Cre SEQ ID NO:36, (b) общей консенсусной последовательности SEQ ID NO:37 Tre-рекомбиназ (специфической для асимметричного участка-мишени внутри LTR ВИЧ-1), (c) обязательной/жесткой консенсусной последовательности SEQ ID NO:38 Tre-рекомбиназ, специфичных для SEQ ID NO:1 и d) менее строгой консенсусной последовательности SEQ ID NO:39 (с гарантией 95%) Tre-рекомбиназ, специфичных для SEQ ID NO:1. Жирным шрифтом показаны консервативные аминокислоты, не точно определенные положения показаны как X.
ПРИМЕРЫ
Пример 1: Идентифицированные асимметричные последовательности-мишени, присутствующие во множестве штаммов
Асимметричные последовательности-мишени были идентифицированы способом, разделенным на 3 стадии: формирование весовой матрицы положений на основе известного участка узнавания рекомбиназой, предоставление геномных последовательностей, которые должны быть исследованы на участки узнавания, и бинарный поиск потенциальных участков-мишеней в предоставленных последовательностях и балльная оценка полученных в результате удачных вариантов на основании весовой матрицы исходных положений, где нуклеотиды преобразуются в двоичное пространство.
SEQ ID NO:1 обнаружена присутствующей в 348 из 379 штаммов ВИЧ-1 подтипа B. Кроме того, SEQ ID NO:1 обнаружена присутствующей в 32 из 40 штаммов ВИЧ-1 подтипа А. Она расположена в R-области LTR.
SEQ ID NO:2 обнаружена присутствующей в 288 из 379 штаммов ВИЧ-1 подтипа В. SEQ ID NO:2 не была обнаружена присутствующей ни в одном из исследованных 40 штаммов ВИЧ-1 подтипа А. Она расположена в области U3 LTR.
Пример 2: Создание адаптированной рекомбиназы, узнающей и обеспечивающей рекомбинацию асимметричный последовательности-мишени внутри LTR ВИЧ-1
Чтобы начать процесс эволюции, были выбраны LTR-последовательности ВИЧ-1, которые являются чрезвычайно консервативными среди штаммов ВИЧ-1.
SEQ ID NO:1 и SEQ ID NO:2, как было обнаружено, соответствуют этим критериям и представляют асимметричные последовательности-мишени, для которых может быть отобрана адаптированная рекомбиназа.
В дальнейшем подробно описывается создание адаптированной рекомбиназы, узнающей SEQ ID NO:1.
Упрощенная схема эволюционной стратегии и прогресс, достигнутый на сегодняшний день, представлен на Фиг. 2. В целом, эволюцию рекомбиназы Tre, специфически узнающей нужную последовательность-мишень, проводили, как раскрыто в WO 2008/083931 и описаны в Sarkar et al. (Sarkar et al., Science 2007).
Начальную библиотеку рекомбиназ генерировали путем объединения и перетасовки семейства Cre и нескольких известных Cre-подобных рекомбиназ, а именно библиотеки Cre-мутантов (Cre/Fre) (Buchholz & Stewart, Nature 2001), сформированной ранее Tre-библиотеки (Sarkar et al., Science 2007), Dre (Sauer et al., Nucl Acids Res 2004), Zre (выделенной из Salmonella enterica, инвентарный номер в Genbank NZ_ABEW01000015), и Shew (штамм Shewanella sp. АНА-3, инвентарный номер в Genbank CP000470) (Фиг.2а). В целях содействия эффективности перетасовки семейства, Dre и Shew были модифицированы («Cre-ed») путем изменения нуклеотидной последовательности путем, насколько это было возможно, аналогичным таковому для Cre, без изменения в результате белковой последовательности. Для выполнения субстрат-связанной белковой эволюции, начальная библиотека рекомбиназ была клонирована в эволюционные векторы (Sarkar et al., Science 2007), содержащие соответствующий подучасток-мишень.
Была создана первая подгруппа двух нуклеиновых кислот-мишеней, содержащих последовательности-мишени, где первая последовательность-мишень обозначена подучастком 1 и включает SEQ ID NO:8, и где вторая последовательность-мишень обозначена подучастком 2 и включает SEQ ID NO: 12.
Вторая подгруппа нуклеиновых кислот-мишеней, содержащих модифицированные последовательности-мишени, была затем создана на основе подучастков 1 и 2. Как показано на Фиг. 2, они содержат SEQ ID NO:9-11 и 13-15, соответственно. Выделенные серым цветом представляют собой несоответствия последовательностей в контроле с пулом известных последовательностей-мишеней, узнаваемых рекомбиназами, содержащимися в начальной библиотеке. Библиотеки подучастков независимо наращивали в E. coli. В первые несколько эволюционных циклов (1-4) экспрессию рекомбиназ индуцировали 200 мкг/мл L-арабинозы, при следующих циклах - 100 мкг/мл (5-6 циклов). Культуры выращивали в течение 13 часов при 37°C с встряхиванием. Для проверки рекомбинационной активности, плазмидную ДНК, выделенную из культур, расщепляли BsrGI и XbaI. Смесь рекомбинированной плазмидной ДНК дает в результате более короткий фрагмент, чем таковой нерекомбинированной плазмиды (Sarkar et al., Science 2007). После первого эволюционного цикла, для всех библиотек подучастков 1A, 1B, 1C, 2A, 2B и 2C можно было регистрировать отсутствие или очень слабые полосы рекомбинированного продукта (Фиг. 3B). Чтобы изолировать удачные рекомбиназы, плазмидную ДНК расщепляли Ndel и амплифицировали при помощи ПЦР до клонирования их обратно в соответствующие эволюционные векторы для следующего эволюционного цикла (Sarkar et al., Science 2007). После первого эволюционного цикла была получена достаточная активность остатков для библиотек подучастков 1A, 1B, 2A, 2B, но не для библиотек подучастков 1С и 2С (Фиг. 3C). Было обнаружено, что несоответствие в положении 11 последовательности-мишени размером 34 п.н. было наиболее критичным для рекомбинационной активности. В результате, был создан модифицированный-участок loxP, названный ΔloxP, содержащий только эту единственную мутацию (T) и соответствующий инвертированный повтор в положении 24 (A). При тестировании начальной библиотеки рекомбиназ в эволюционном векторе, содержащем участок-мишен ΔloxP, была получена достаточная активность остатков (Фиг. 3D). Амплифицированная библиотека удачных рекомбиназ была клонирована обратно в эволюциионные векторы с подучастками 1С и 2С, соответственно, и проверены на рекомбинационную активность. Теперь достаточная активность остатков была также получена для библиотек подучастков 1С и 2С (Фиг. 3D), так что эволюционный процесс мог быть запущен на всех подучастках. 6 эволюционных циклов было выполнено на каждом из подучастков 1A IB, 1C, 2 A, 2B и 2C, что существенно обогащает библиотеки удачными рекомбиназами, как можно увидеть на проверочном геле, показанном на Фиг. 3Е (ср. с Фиг. 3B).
Обогащенные библиотеки подучастков объединяют и перетасовывают, чтобы получить библиотеки для продолжения эволюционного циклического процесса 2-ой подгруппы последовательностей-мишеней, а именно подучастка 1 и подучастка 2. Библиотеки удачных рекомбиназ затем обогащают, как описано выше, до их объединения и перетасовывания на заключительной стадии эволюционного циклирования, которая представляет собой эволюцию на участке-мишени, специфически узнаваемом рекомбиназой Tre. Нуклеиновая кислота, кодирующая рекомбиназу, может быть получена из библиотеки и клонирована в экспрессионный вектор.
Используются материалы и способы, как описано в WO 2008/083931 и Buchholz & Stewart, 2001 г., если не указано иное.
Полученные TRE-рекомбиназы, адаптированные для узнавания и обеспечения рекомбинации асимметричных последовательностей-мишеней SEQ ID NO:1 и SEQ ID NO:2, секвенируют и подтверждают их способность к рекомбинации провирусной ДНК различных штаммов-мишеней ВИЧ-1.
Список литературы
Abremski K, Hoess RH, Sternberg N (1983) “Studies on the properties of P1 site-specific recombination: evidence for topologically unlinked products following recombination.” Cell 32, 1301-1311.
Abremski K, Hoess R (1983) “Bacteriophage PI site-specific recombination. Purification and properties of the Cre recombinase protein.” J. Biol. Chem. 259, 1509-1514.
Adachi A, Gendelman HE, Koenig S, Folks T, Willey R, Rabson A, Martin MA (1986) “Production of acquired immunodeficiency syndrome-associated retrovirus in human and nonhuman cells transfected with an infectious molecular clone.” J. Virol. 59, 284-291.
Alper H, Fischer C, Nevoigt E, Stephanopoulos G (2006) “Tuning genetic control through promoter engineering” Proc. Natl. Acad. Sci. USA 102, 12678-12683.
Altschul, S.F., Madden, T.L., Schaffer, A.A., Zhang, J., Zhang, Z., Miller, W. and Lipman, D.J. (1997) Gapped BLAST and PSI-BLAST: a new generation of protein database search programs. Nucleic acids research, 25, 3389-3402.
Beyer WR, Westphal M, Ostertag W, von Laer D (2002) „Oncoretrovirus and lentivirus vectors pseudotyped with lymphocytic choriomeningitis virus glycoprotein: generation, concentration and broad host range.41 J. Virol. 76, 1488-1495.
Blackard JT, Renjifo BR, Mwakagile D, Montano MA, Fawzi WW, Essex M (1999) “Transmission of human immunodeficiency type 1 viruses with intersubtype recombinant long terminal repeat sequences.” Virology ISA, 220-225.
Bloom JD, Meyer MM, Meinhold P, Otey CR, MacMillan D, Arnold FH (2005) “Evolving strategies for enzyme engineering.” Curr. Opin. Struct. Biol. 15, 447-452.
Buchholz F, Ringrose L, Angrand PO, Rossi F, Stewart AF (1996) “Different thermostabilities of FLP and Cre recombinases: implications for applied site-specific recombination.” Nucl. Acids Res. 24, 4256-4262.
Buchholz F, Angrand PO, Stewart AF (1998) “Improved properties of FLP recombinase evolved by cycling mutagenesis.” Nat. Biotechnol. 16, 657-662.
Buchholz F, Stewart AF (2001) “Alteration of Cre recombinase site specificity by substrate-linked protein evolution”. Nat. Biotechnol. 19, 1047-1052.
Chiu YL, Soros VB, Kreisberg JF, Stopak K, Yonemoto W, Greene WC (2005) “Cellular APOBEC3G restricts HIV-1 infection in resting CD4+ T cells”. Nature 435, 108-114
Chun T-W, Engel D, Berrey MM, Shea T, Corey L, Fauci AS (1998) “Early establishment of a pool of latently infected, resting CD4+ T cells during primary HIV-1 infection”. Proc. Natl. Acad. Sci. USA 95, 8869-8873.
Coates CJ, Kaminski JM, Summers JB, Segal DJ, Miller AD, Kolb AF (2005) “Site-directed genome modification: derivatives of DNA-modifying enzymes as targeting tools”. Trends Biotechnol. 23, 407-419.
Collins CH, Yokobayashi Y, Umeno D, Arnold FH, (2003) “Engineering proteins that bind, move, make and break DNA.” Curr. Opin. Biotechnol. 14, 665.
Combes P, Till R, Bee S, Smith MC (2002) “The streptomyces genome contains multiple pseudo-attB sites for the (phi)C31-encoded site-specific recombination system.” J. Bacteriol. 184, 5746-5752.
Crameri A, Raillard SA, Bermudez E, Stemmer WP (1998) “DNA shuffling of a family of genes from diverse species accelerates directed evolution”. Nature 391, 288-291.
Derossi D, Joliot AH, Chassaing G, Prochiantz A (1994) “The thrid helix of the Antennapedia homeodomain translocates through biological membranes”. J. Biol. Chem. 269, 10444-10450.
Derossi D, Calvet S, Trembleau A, Chassaing G, Prochiantz A (1996) “Cell internalization of the third helix of the Antennapedia homeodomain is receptor-independent”. J Biol Chem 271, 18188-18193.
Donovan, P.J., Gearhart, J. (2001) “The end of the beginning for pluripotent stem cells”. Nature 414, 92-97.
Donzella GA, Schols D, Lin SW, Este JA, Nagashima KA, Maddon PJ, Allaway GP, Sakmar TP, Henson G, De Clercq E, Moore JP (1998) “AMD3100, a small molecule inhibitor of HIV-1 entry via the CXCR4 co-receptor.” Nature Medicine 4, 72-77.
Dybul M, Fauci AS, Bartlett JG, Kaplan JE, Pau AK (2002) “Guidelines for using antiretroviral agents among HIV-infected adults and adolescents.” Annals of Internal Medicine 137, 381-433.
Eddy, S.R. (1998) Profile hidden Markov models. Bioinformatics (Oxford, England), 14, 755-763.
Edelman GM, Meech R, Owens GC, Jones FS (2000) “Synthetic promoter elements obtained by nucleotide sequence variation and selection for activity”. Proc. Natl. Acad. Sci. USA 97, 3038-3043.
Emerman M, Malim MH (1998) “HIV-1 regulatory/accessory genes: keys to unraveling viral and host cell biology”. Science 280, 1880-1884.
Finzi D, Hemankova M, Pierson T, Carruth LM, Buck C, Chaisson RE, Quinn TC, Chadwick K, Margolick J, Brookmeyer R, Gallant J, Markowitz M, Ho DD, Richman DD, Siliciano RF (1997) “Identification of a reservoir for HIV-1 in patients on highly active antiretroviral therapy”. Science 278, 1295-1300.
Flowers CC, Woffendin C, Petryniak J, Yang S, Nabel GJ (1997) “Inhibition of recombinant human immunodeficiency virus type 1 replication by a site-specific recombinase.” J. Virol. 71, 2685-2692.
Gulick RM, Mellors JW, Havlir D, Eron JJ, Gonzalez C, McMahon D, Richman DD, Valentine FT, Jonas L, Meibohm A, Emini EA, Chodakewitz JA (1997) “Treatment with indinavir, zidovudine, and lamivudine in adults with human immunodeficiency virus infection and prior antiretroviral therapy.” N. Engl. J. Med. 337, 734-739.
Guzman LM, Belin D, Carson MJ, Beckwith J (1995) “Tight regulation, modulation, and high-level expression by vectors containing the arabinose PBAD promoter”. J. Bacteriol. 177, 4121-4130.
Hartenbach S, Fussenegger M (2006) “A novel synthetic mammalian promoter derived from an internal ribosome entry site”. Biotechnology and Bioengineering 95, 547-559.
Hauber I, Bevec D, Heukeshoven J, Kratzer F, Horn F, Choidas A, Harrer T, Hauber J (2005) “Identification of cellular deoxyhypusine synthase as a novel target for antiretroviral therapy”. J. Clin. Invest. 115, 76-85.
Hazuda DJ, Young SD, Guare JP, Anthony NJ, Gomez RP, Wai JS, Vacca JP, Handt L, Motzel SL, Klein HJ, Domadula G, Danovich RM, Witmer MV, Wilson KA, Tussey L, Schleif WA, Gabryelski LS, Jin L, Miller MD, Casimiro DR, Emini EA, Shiver JW (2004) “Integrase inhibitors and cellular immunity suppress retroviral replication in rhesus macaques”. Science 305, 528-532.
Hoess RH, Abremski К (1985) “ Mechanism of strand cleavage and exchange in the Cre-lox site-specific recombination system”. J! Mol. Biol. 181, 351-362.
Johannes TW, Zhao H (2006) “Directed evolution of enzymes and biosynthetic pathways”. Curr. Opin. Microbiol. 9, 261-267.
Krasnow MA, Cozzarelli NR (1983) “Site-specific relaxation and recombination by the Tn3 resolvase: Recognition of the DNA path between oriented res sites”. Cell 32, 1313-1324.
Kulkosky J, Bray S (2006) “HAART-persistent HIV-1 latent reservoirs: their origin, mechanisms of stability and potential strategies for eradication”. Curr. HIV Res. 4, 199-208.
Lalezari JP, Henry K, O’Heam M, Montaner JS, Piliero PJ, Trottier B, Walmsley S, Cohen C, Kuritzkes DR, Eron Jr. JJ, Chung J, DeMasi R, Donatacci L, Drobnes C, Delehanty J, Salgo M (2003) “Enfuvirtide, an HIV-1 fusion inhibitor, for drug-resistant HIV infection in North and South America”. N. Engl. J. Med. 348, 2175-2185.
Lee YS, Park JS (1998) “A novel mutant loxP containing part of long terminal repeat of HIV-1 in spacer region: presentation of possible target site for antiviral strategy using site-specific recombinase”. Biochem. Biophys. Res. Comm. 253, 588-593.
Lee YS, Kim ST, Kim GW, Lee M, Park JS (2000) “An engineered lox sequence containing part of a long terminal repeat of HIV-1 permits Cre recombinase-mediated DNA excision.” Biochem. Cell Biol. 78, 653-658.
Lehrman G, Hogue IB, Palmer S, Jennings C, Spina CA, Wiegand A, Landay AL, Coombs RW, Richman DD, Mellors JW, Coffin JM, Bosch RJ, Margolis DM (2005) “Depletion of latent HIV-1 infection in vivo: a proof-of-concept study” Lancet 366, 549-555.
Lewandoski, M. (2001) “Conditional control of gene expression in the mouse”. Nat. Rev. Genet. 2, 743-755.
Lin Q, Jo D, Gebre-Amlak KD, Ruley HE (2004) “Enhanced cell-permeant Cre protein for site-specific recombination in cultured cells”. BMC Biotechnol. 4, 25.
Little SJ, Holte S, Routy JP, Daar ES, Markowitz M, Collier AC, Koup RA, Mellors JW, Connick E, Conway B, Kilby M, Wang L, Whitcomb JM, Hellmann NS, Richman DD (2002) “Antiretroviral-drug resistance among patients recently infected with HIV”. N. Engl. J. Med. 347, 385-394.
Macara IG (2001) “Transport into and out of the nucleus”. Microbiology and molecular biology reviews 65, 570-594.
Malim MH, Hauber J, Fenrick R, Cullen BR (1988) “Immunodeficiency virus rev transactivator modulates the expression of the viral regulatory genes.” Nature 335, 181-183.
Marcello A (2006) “Latency: the hidden HIV-1 challenge”. Retrovirology 3, 7.
Matsumura I, Ellington AD (2001) “In vitro evolution of beta-glucuronidase into a beta- galactosidase proceeds through non-specific intermediates”. J. Mol. Biol. 305, 331-339.
Minshull J, Stemmer WP. (1999) “Protein evolution by molecular breeding”. Curr. Opin. Chem. Biol. 3, 284-290.
Nagy A (2000) “Cre recombinase: the universal reagent for genome tailoring”. Genesis 26, 99-109.
Needleman SB, Wunsch CD (1970) “A general method applicable to the search for similarities in the amino acid sequence of two proteins”. J. Mol. Biol. 48, 443-453.
Nolden L, Edenhofer F, Haupt S, Koch P, Wunderlich FT, Siemen H, Brustle O. (2006) “Site- specific recombination in human embryonic stem cells induced by cell-permeant Cre recombinase”. Nat. Methods 3, 461-467.
O’Doherty U, Swiggard WJ, Malim MH (2000) “Human immunodeficiency virus type 1 spinoculation enhances infection through virus binding.” J. Virol. 74, 10074-10080.
Pearson WR, Lipman DJ (1988) “Improved tools for biological sequence comparison”. Proc Natl Acad Sci USA 85, 2444-2448.
Peitz M, Pfannkuche K, Rajewsky K, Edenhofer F. (2002) “Ability of the hydrophobic FGF and basic TAT peptides to promote cellular uptake of recombinant Cre recombinase: A tool for efficient genetic engineering of mammalian genomes”. Proc. Natl. Acad. Sci. USA 99, 4489-4494.
Ratner L, Starcich B, Josephs SF, Hahn BH, Reddy EP, Livak KJ, Petteway SR, Jr., Pearson ML, Haseltine WA, Arya SK, (1985) “Polymorphism of the 3’ open reading frame of the virus associated with the acquired immune deficiency syndrome, human T-lymphotropic virus type III”. Nucl. Acids Res. 13, 8219-8229.
Richard JP, Melikov K, Brooks H, Prevot P, Lebleu B, Chemomordik LV (2005) “Cellular uptake of the unconjugated TAT peptide involves clathrin-dependent endocytosis and heparin sulfate receptors”. J. Biol. Chem. 280, 15300-15306.
Rufer AW, Sauer В (2002) “Non-contact positions impose site selectivity on Cre recombinase.” Nucl. Acids Res. 30, 2764-2771.
Ruhl M, Himmelspach M, Bahr GM, Hammerschmid F, Jaksche H, Wolff B, Aschauer H, Farrington GK, Probst H, Bevec D, Hauber J (1993) “Eukaryotic initiation factor 5A is a cellular target of the human immunodeficiency virus type 1 Rev activation domain mediating trans-activation” J. Cell Biol. 123, 1309-1320.
Sanger F, Nickler S, Coulson AR (1977) “DNA sequencing with chain-terminating inhibitors”. Proc. Natl. Acad. Sci. USA 74, 5463-5467.
Santoro SW, Schultz PG (2002) “Directed evolution of the site specificity of Cre recombinase”. Proc. Natl. Acad. Sci. USA 99, 4185-4190.
Saraf-Levy T, Santoro SW, Volpin H, Kushnirsky T, Eyal Y, Schultz PG, Gidoni D, Carmi N (2006) “Site-specific recombination of asymmetric lox sites mediated by a heterotetrameric Cre recombinase complex”. Bioorg. Med. Chem. 14, 3081-3089.
Sauer B, McDermott J (2004) “DNA recombination with a heterospecific Cre homolog identified from comparison of the pac-cl regions of PI-related phages.” Nucl. Acids. Res. 32, 6086- 6095.
Schambach A, Bohne J, Chandra S, Will E, Margison GP, Williams DA, Baum С (2006) “Equal potency of gammaretroviral and lentiviral SIN vectors for expression of O6- methylguanine-DNA methyltransferase in hematoietic cells”. Molecular Therapy 13, 391- 400.
Scherr M, Eder M (2002) “Gene Transfer into Hematopoietic Stem Cells Using Lentiviral Vectors”. Current Gene Therapy 2, 45-55.
Shehu-Xhilaga M, Tachedjian G, Crowe SM, Kedzierska K. (2005) “Antiretroviral compounds: mechanisms underlying failure of HAART to eradicate HIV-1”. Curr. Med. Chem. 12, 1705-1719.
Shimshek DR, Kim J, Hubner MR, Spergel DJ, Buchholz F, Casanova E, Stewart AF, See- burg PH, Sprengel R (2002) “Codon-improved Cre recombinase (iCre) expression in the mouse”. Genesis 32(1), 19-26.
Smith Tf, Waterman MS (1981) “Overlapping genes and information theory”. J. Theor. Biol. 91, 379-380.
Stark WM, Boocock MR, Sherratt DJ (1992) “Catalysis by site-specific recombinases”. Trends Genet. 8, 432-439.
Stemmer WPC (1994) “Rapid evolution of a protein in vitro by DNA shuffling”. Nature 370, 389-391.
Sternberg N, Hamilton D (1981) “Bacteriophage PI site-specific recombination. I. Recombination between loxP sites”. J. Mol. Biol. 150, 467-486.
Van Duyne GD (2001) “A structural view of cre-loxp site-specific recombination”. Annu. Rev. Biophys. Biomol. Struct. 30, 87-104.
Vives E, Brodin P, Lebleu В (1997) “A truncated HIV-1 Tat protein basic domain rapidly translocates through the plasma membrane and accumulates in the cell nucleus”. J. Biol. Chem. 272, 16010-16017.
Vives E (2003) “Cellular uptake of the TAT peptide: an endocytosis mechanism following ionic interactions”. J. Mol. Recognit. 16, 265-271.
Volkert FC, Broach JR (1986) “Site-specific recombination promotes plasmid amplification in yeast”. Cell 46, 541-550.
Voziyanov Y, Konieczka JH, Stewart AF, Jayaram M (2003) “Stepwise manipulation of DNA specificity in Flp recombinase: progressively adapting Flp to individual and combinatorial mutations in its target site”. J. Mol. Biol. 326, 65-76.
Yuan L, Kurek I, English J, Keenan R (2005) “Laboratory-directed protein evolution” Microbiol. Mol. Biol. Rev. 69, 373-92.
WO 2002/44409
WO 2008/083931
Fawell S, Seery J, Daikh Y, Moore C, Chen LL, Pepinsky B, Barsoum J., Tat-mediated delivery of heterologous proteins into cells., Proc Natl Acad Sci USA. 1994 Jan 18;91(2):664-8.
Elliott G, O’Hare P., Intercellular trafficking and protein delivery by a herpesvirus structural protein., Cell. 1997 Jan 24;88(2):223-33
Oess S, Hildt E., Novel cell permeable motif derived from the PreS2-domain of hepatitis-B virus surface antigens., Gene Ther. 2000 May;7(9):750-8.































































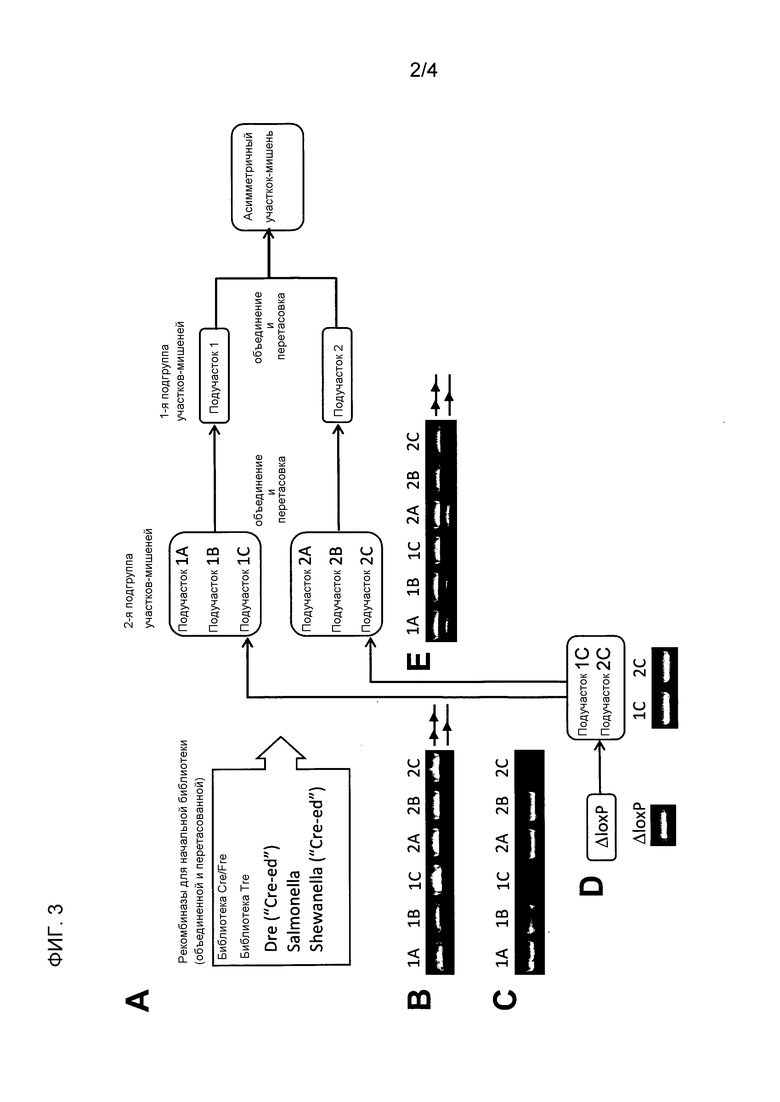
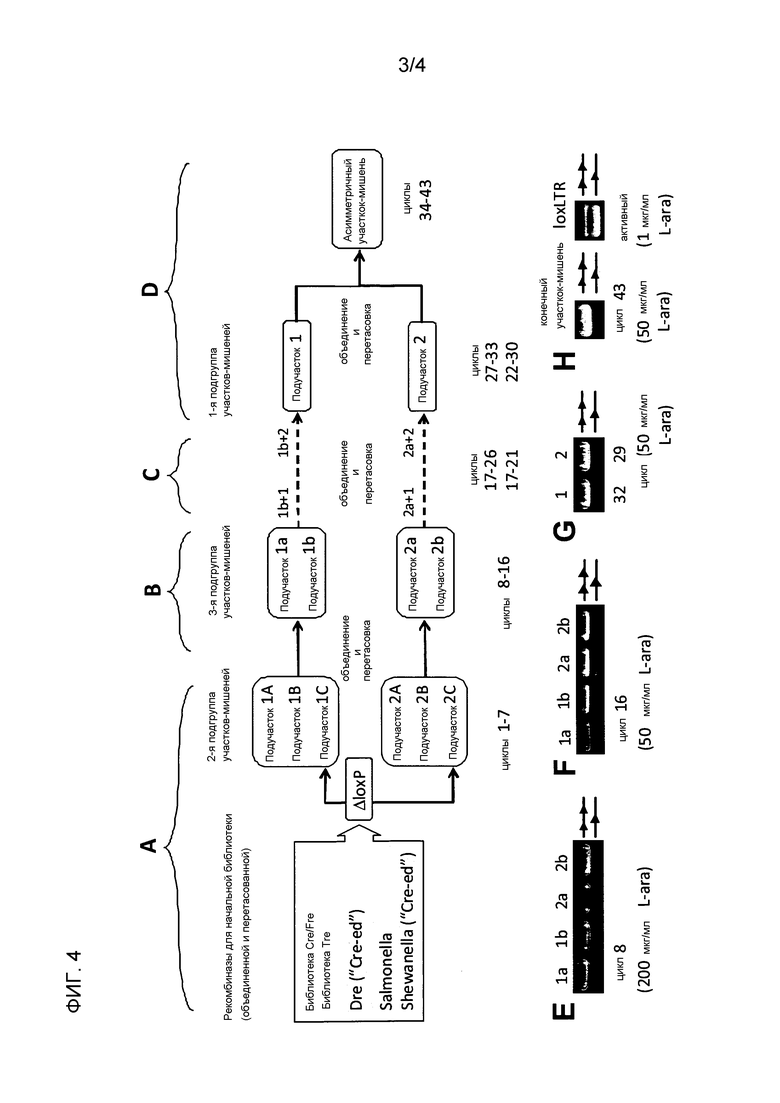

Изобретения касаются способа приготовления экспрессионного вектора, кодирующего адаптированную рекомбиназу, которая способна к рекомбинации асимметричных последовательностей-мишеней в пределах длинного концевого повтора (LTR) провирусной ДНК множества штаммов ВИЧ-1, встроенных в геном клетки-хозяина, а также к полученному экспрессионному вектору, клетке, трансфецированной этим вектором, экспрессированной рекомбиназе и фармацевтической композиции, включающей экспрессионный вектор, клетку и/или рекомбиназу. Представленные изобретения могут быть использованы при лечении или профилактике инфекции ВИЧ-1 у субъекта. 6 н и 22 з.п.ф-лы, 5 ил., 2 пр.
1. Способ приготовления экспрессионного вектора, кодирующего адаптированную рекомбиназу, которая способна к рекомбинации асимметричных последовательностей-мишеней внутри LTR провирусной ДНК множества штаммов ВИЧ-1, включающий стадии
а) идентификации последовательностей с гомологией, по крайней мере, 30% в отношении последовательности левой половины участка и последовательности правой половины участка, по крайней мере, одного известного рекомбиназного участка-мишени в последовательности LTR провирусной ДНК множества штаммов ВИЧ-1, где гомологичные последовательности разделены спейсером в 5-12 нуклеотидов, и где асимметричная последовательность-мишень, имеющая последовательность SEQ ID NO: 1, обнаруживается во множестве штаммов ВИЧ-1;
(b) идентификации двух последовательностей, где первая последовательность соответствует последовательности асимметричной последовательности-мишени стадии (а), гомологичной левой половине участка указанного известного участка-мишени, и называется «последовательность половины участка 1», и где вторая последовательность соответствует последовательности асимметричной последовательности-мишени стадии (а), гомологичной правой половине участка, и называется «последовательность половины участка 2»;
(c) определения нуклеотидов внутри последовательностей стадии (b), отклоняющихся от соответствующих гомологичных последовательностей левой половины участка и правой половины участка, по крайней мере, известного гомологичного участка-мишени стадии (а);
(d) генерации первой подгруппы двух нуклеиновых кислот-мишеней, содержащих последовательности-мишени, где первая последовательность-мишень обозначена «подучасток 1» и включает, друг рядом с другом и в направлении от 5' к 3', последовательность половины участка 1 стадии (b), спейсерную последовательность асимметричной последовательности-мишени и инвертированный повтор последовательности половины участка 1, и где вторая последовательность-мишень обозначена «подучасток 2» и включает, друг рядом с другом и в направлении от 5' к 3', инвертированный повтор последовательности половины участка 2, спейсерную последовательность асимметричной последовательности-мишени и последовательность половины участка 2 стадии (b);
(e) генерации второй подгруппы нуклеиновых кислот-мишеней, содержащих модифицированные последовательности-мишени на основе последовательностей-мишеней в первой подгруппе стадии (d),
где, в последовательностях на основе подучастка 1, в последовательности левой половины участка, часть нуклеотидов, отклоняющихся от соответствующей гомологичной последовательности половины участка, по крайней мере, одного известного участка-мишени стадии (а) заменяют нативными нуклеотидами, встречавшимися в указанном известном участке-мишени, до тех пор, пока указанная последовательность половины участка не будет содержать один, два или три нуклеотида, отклоняющихся от указанного известного участка-мишени, где правая половина участка указанной модифицированной последовательности-мишени образована инвертированным повтором указанной модифицированной последовательности левой половины участка, которая отделена от указанной модифицированной последовательности левой половины участка спейсерной последовательностью асимметричной последовательности-мишени, и
где, в последовательностях на основе подучастка 2, в последовательности правой половины участка, часть нуклеотидов, отклоняющихся от соответствующей гомологичной последовательности половины участка, по крайней мере, одного известного участка-мишени стадии (а) заменяют нативными нуклеотидами, встречавшимися в указанном известном участке-мишени, до тех пор, пока указанная последовательность половины участка не будет содержать один, два или три нуклеотида, отклоняющихся от указанного известного участка-мишени, где левая половина участка указанной модифицированной последовательности-мишени образована инвертированным повтором указанной модифицированной последовательности правой половины участка, которая отделена от указанной модифицированной последовательности правой половины участка спейсерной последовательностью асимметричной последовательности-мишени,
так что во всех модифицированных последовательностях половин участка, происходящих из одной последовательности-мишени первой подгруппы стадии (d), вместе взятых, можно обнаружить все отклоняющиеся нуклеотиды, тогда как ни одна из указанных модифицированных последовательностей половин участка на всем протяжении не содержит все отклоняющиеся нуклеотиды,
(f) применения отдельно молекулярной направленной эволюции,
по крайней мере, на одной рекомбиназе, узнающей известный гомологичный участок-мишень в соответствии со стадией (а), с использованием каждой нуклеиновой кислоты второй подгруппы, полученной на стадии (е), в качестве субстрата;
(q) перетасовывания рекомбиназных библиотек, образованных на стадии (f), где все рекомбиназные библиотеки, образованные на последовательностях на основе подучастка 1, объединяют и перетасовывают, и где все рекомбиназные библиотеки, образованные на последовательностях на основе подучастка 2, объединяют и перетасовывают;
(h) применения молекулярной направленной эволюции на перетасованных библиотеках, полученных на стадии (д), с использованием каждой нуклеиновой кислоты подгруппы в соответствии со стадией (d) в качестве субстрата;
(i) перетасовки рекомбиназных библиотек, образованных на стадии (h);
(j) применения молекулярной направленной эволюции на перетасованной библиотеке, полученной на стадии (д), с использованием нуклеиновой кислоты, содержащей асимметричную последовательность-мишень стадии (а), в качестве субстрата, до тех пор, пока не получится, по крайней мере, одна рекомбиназа, которая действует на асимметричную последовательность-мишени внутри LTR провирусной ДНК множества штаммов ВИЧ-1 стадии (а);
(k) выделения нуклеиновой кислоты, кодирующей, по меньшей мере, одну рекомбиназу, полученную на стадии (j) из библиотеки; и
(l) клонирования нуклеиновой кислоты, полученной на стадии (k), в подходящем экспрессионном векторе.
2. Способ по п. 1, где, по крайней мере, одна известная рекомбиназа, чья последовательность-мишень используется на стадии (а) и на которой молекулярная направленная эволюция применяется на стадии (f), является частью рекомбиназной библиотеки.
3. Способ по п. 1, где примененная молекулярная направленная эволюция представляет собой субстрат-связанную белковую эволюцию.
4. Способ по п. 1, где экспрессионный вектор на стадии (1) выбирают из группы, состоящей из векторов ретровирусов, векторов лентивирусов, векторов спумавирусов и векторов аденовирусов.
5. Способ приготовления адаптированной рекомбиназы, где указанный способ включает способ по любому из пп. 1-4 и дополнительную стадию экспрессии адаптированной рекомбиназы с нуклеиновой кислотой, кодирующей рекомбиназу, встроенную в экспрессионный вектор в подходящей клетке-хозяине.
6. Способ по п. 5, где рекомбиназа экспрессируется в качестве слитого полипептида, содержащего аминокислотную последовательность адаптированной рекомбиназы.
7. Способ по п. 6, где слитый белок содержит сигнал ядерной локализации.
8. Способ по п. 7, где сигнал ядерной локализации имеет последовательность, соответствующую положениям 2-9 SEQ ID NO: 65.
9. Способ по п. 7 или 8, где слитый белок дополнительно содержит домен трансдукции белка.
10. Способ по п. 9, где домен трансдукции белка происходит из tat.
11. Способ приготовления трансформированных клеток, где указанный способ включает способ по любому из пп. 1-4 и дополнительную стадию введения экспрессионного вектора in vitro в клетку, где клетка предпочтительно представляет собой взрослую стволовую клетку, где указанная клетка не является эмбриональной клеткой человека.
12. Способ по любому из пп. 1, 5 и 11, дополнительно включающий стадию приготовления экспрессионного вектора, рекомбиназы или клетки в качестве фармацевтической композиции с использованием по меньшей мере одного фармацевтически приемлемого носителя, наполнителя и/или адъюванта.
13. Нуклеиновая кислота, кодирующая адаптированную рекомбиназу, где указанная адаптированная рекомбиназа способна к рекомбинации асимметричных последовательностей-мишеней, содержащих SEQ ID NO: 1, внутри LTR провирусной ДНК множества штаммов ВИЧ-1, где аминокислотная последовательность адаптированной рекомбиназы имеет, по крайней мере, 97% идентичности последовательности SEQ ID NO: 38, где указанная адаптированная рекомбиназа включает аминокислотные замены P12S, P15L, M44V, K86N, G93A, A175S и Р307А по сравнению с последовательностью Cre SEQ ID NO: 36.
14. Нуклеиновая кислота по п. 13, где нуклеиновая кислота представляет собой экспрессионный вектор.
15. Нуклеиновая кислота по п. 13 или 14, полученная на стадии (k) способа по любому из пп. 1-4.
16. Нуклеиновая кислота по п. 13 или 14, где адаптированная рекомбиназа содержит аминокислотную последовательность SEQ ID NO: 3 7.
17. Нуклеиновая кислота по п. 13 или 14, где аминокислотная последовательность адаптированной рекомбиназы содержит последовательность SEQ ID NO: 38.
18. Нуклеиновая кислота по п. 13 или 14, где аминокислотная последовательность адаптированной рекомбиназы содержит последовательность SEQ ID NO: 39.
19. Нуклеиновая кислота по п. 13 или 14, где аминокислотная последовательность адаптированной рекомбиназы выбрана из группы, состоящей из SEQ ID NO: 40-64 и комбинации последовательностей из группы, состоящей из SEQ ID NO: 40-64.
20. Адаптированная рекомбиназа, кодируемая нуклеиновой кислотой по любому из пп. 13-19, где указанная адаптированная рекомбиназа способна к рекомбинации асимметричных последовательностей-мишеней, содержащих SEQ ID NO: 1, внутри LTR провирусной ДНК множества штаммов ВИЧ-1.
21. Рекомбиназа по п. 20, которую экспрессируют как слитый белок.
22. Рекомбиназа по п. 21, где слитый белок содержит сигнал ядерной локализации.
23. Рекомбиназа по п. 22, где сигнал ядерной локализации имеет последовательность, соответствующую положениям 2-9 SEQ ID NO: 65.
24. Рекомбиназа по п. 22 или 23, где слитый белок дополнительно содержит домен трансдукции белка.
25. Рекомбиназа по п. 24, где домен трансдукции белка происходит из tat.
26. Трансформированная клетка для экспрессии адаптированной рекомбиназы, содержащая нуклеиновую кислоту по любому из пп. 13-19, где указанная клетка предпочтительно представляет собой стволовую клетку из гематопоэтической линии, где указанная клетка не является эмбриональной клеткой человека.
27. Фармацевтическая композиция для применения при лечении или профилактике инфекции ВИЧ-1 у субъекта, содержащая эффективное количество нуклеиновой кислоты по любому из пп. 13-19, адаптированной рекомбиназы по п. 20 и/или трансформированной клетки по п. 26, и по меньшей мере один фармацевтически приемлемый носитель, наполнитель и/или адъювант.
28. Фармацевтическая композиция по п. 27, где провирусная ДНК, обнаруженная в образцах, полученных от субъекта, содержит асимметричную последовательность-мишень SEQ ID NO: 1, на которой рекомбиназа был отобрана.
| WO 2008083931 A1, 17.07.2008 | |||
| INDRANI SARKAR et al., HIV-1 Proviral DNA Excision Using an Envolved Recombinase, Science, 29 June 2007, Vol.316, pp.1912-1915 | |||
| WO 2005081632 A2, 09.09.2005 | |||
| ПРИМЕНЕНИЕ АДАПТИРОВАННЫХ РЕКОМБИНАЗ ДЛЯ ЛЕЧЕНИЯ РЕТРОВИРУСНЫХ ИНФЕКЦИЙ | 2008 |
|
RU2501860C2 |

