Область техники, к которой относится изобретение
Изобретение относится к области клинико-лабораторной диагностики, в частности к определению эффективности неспецифического иммунного ответа на бактериальную инфекцию пародонта по соотношению в пародонтальном смыве мРНК фактора некроза опухолей α (TNFα), металлопротеиназ ММР8 и ММР9 с помощью ПЦР в реальном времени. Предметом изобретения является метод количественного анализа мРНК TNFα, ММР8 и ММР9 с помощью ПЦР в реальном времени, а также принцип использования полученного значения для интерпретации результатов с целью измерения эффективности работы системы неспецифического иммунитета. Изобретение может быть использовано для оценки локального иммунного статуса пародонта у индивидуальных пациентов при профилактическом осмотре у пародонтолога, у больных с агрессивным и хроническим пародонтом, в том числе, после потери зуба, а также в тканях, соприкасающихся с имплантатами и другими ортодонтическими конструкциями. Использование метода обеспечивает высокочувствительную и объективную количественную характеристику иммунного статуса пародонта, что позволяет оценивать риск возникновения острого локального воспаления и разрушения пародонта в будущем, приживаемость имплантатов и других ортодонтических конструкций, оценивать эффективность лечебных мероприятий.
Уровень техники
Данные эпидемиологических исследований свидетельствуют, что распространенность воспалительных заболеваний пародонта среди взрослого населения составляет от 90 до 100% случаев [Грудянов А.И. и соавт., 1998; Дмитриева Л.А., 2001]. Тем не менее, в настоящее время прослеживается тенденция к увеличению частоты распространенности атипичных форм заболеваний пародонта, к числу которых относятся агрессивные формы пародонтита [Безрукова И.В., 2002; Choi Н.М. et al, 2015].
В качестве основной причины возникновения хронического пародонтита большинством авторов рассматривается инфицирование поверхности пародонта патогенными бактериями [Page R.C., 1995; Tanner A.C.R. et al., 2005]. В полости рта выявлено до 700 видов микроорганизмов, однако в качестве истинных пародонтопатогенов большинство авторов называют только виды A. actinomycetemcomitans, P. gingivalis, P. intermedia, Т. forsythensis, Т. denticola и С. albicans [Царев B.Н. и соавт., 2005; Atieh М.А., 2008; Cortelli J.R. et al, 2005; Hyvärinen K. et al, 2009; Finoti L.S. et al, 2013]. В результате реакции организма на бактериальную инфекцию происходит деструкция тканей, сопровождающаяся прогрессирующим разрушением кости альвеолярного отростка, и, в конечном итоге, потерей зуба [Бузрукова И.В., 2001; Dong Y. et al, 2015; Zhang Y & Li X., 2014; Behl Y. et al, 2008].
Возникновение и течение любого заболевания инфекционной природы определяется силой ответной реакции организма [Dosseva-Panova V.T. et al, 2014]. Характер этого ответа зависит от антигенной специфичности ранее приобретенного иммунитета и в значительной степени определяется генотипом человека [Dyer J.K. et al, 1997; Elamin A. et al, 2011; Gilowski L. et al, 2012]. При этом только часть наблюдаемых реакций организма на инфекцию можно рассматривать в качестве протективных [Gemmell Е. et al, 2007]. Помимо этого нередко возникают реакции нейтрального характера - бесполезные с точки зрения защиты организма, но оттягивающие не себя ресурсы иммунной системы [Graswinckel J.E.M. et al, 2004], а также аутотоксические реакции, вызывающие массовое повреждение и гибель собственных клеток и тканей [Graves D.T. & Cochran D., 2003; Godoy-Gallardo M. et al, 2015]. Нередко именно аутотоксические реакции определяют патофизиологическую картину заболевания в целом. Кроме того, при рассмотрении бактериальных инфекций, инициирующихся на слизистых оболочках человека, к которым относится и пародонт, необходимо принимать в расчет симбиотические и антагонистические отношения между бактериями [Haffajee A.D. et al, 2004; Gomez S.C., 2006].
Исследования в области этиологии как агрессивного, так и хронического пародонтита, привели к выявлению факта резкого подъема активности матриксинов (матриксных протеиназ, синтезируемых, в частности, фибробластами и макрофагами пародонта) при возникновении воспаления пародонта любой природы [Gürkan A. et al, 2007; Hayashi N. et al, 2007; Holla L.I. et al, 2004]. Особенно существенно - в сотни и тысячи раз, при этом поднимается активность нейтрофильной коллагеназы ММР8 и желатиназы ММР9: ферментов, участвующих в конечных стадиях деградации коллагена [Irshad М. et al, 2013; Javed F. et al, 2014 (1)]. На этом основании большинство исследователей делают вывод об аутопротеолитической природе деградации соединительнотканного матрикса пародонта, лежащего в основе пародонтита [Jayaprakash K. et al, 2014; Javed F. et al, 2014 (2)]. При этом роль бактериальных протеиназ в качестве разрушителей коллагена и других компонентов матрикса рассматривается как сигнальная, хотя иногда ей также приписывается существенное значение [Holt S.C. & Ebersole J.L., 2005]. Однако, этой точке зрения во многом противоречит тот факт, что подъем активности коллагеназ, непосредственно способных атаковать нативный коллаген, например, ММР1 и ММР13, ни в одном исследовании не удалось связать с поражением пародонта [Khalaf Н. et al, 2014; Kinney J.S., 2014 (1); Kinney J.S., 2014 (2)]. Между тем, активация одних только терминальных матриксинов, даже достигающая нескольких порядков по сравнению с базовым уровнем, не способна влиять на целостность коллагеновой основы пародонтальной связки [Leppilahti J.M. et al, 2014].
При рассмотрении влияния ММР8 и ММР9 на деградацию матрикса пародонта важным вопросом является регуляторный механизм подъема их активности [Lee W. et al, 2004; Loesche W.J. et al, 2001; Moutsopoulos N.M. et al, 2015]. Исследования, выполненные методами ПЦР в реальном времени и иммуноферментного анализа показывают существенное увеличение уровня мРНК ММР8 и ММР9 [Noack В. et al, 2009; Parra В. & Slots J., 1996], а также соответствующих белковых продуктов [Vandesompele J. et al, 2002; Ram V.S. et al, 2015]. Однако эти показатели лишь опосредованно отражают уровень активности соответствующих ферментов [Salminen A. et al, 2014]. Дело в том, что большая часть матриксинов любой нормальной ткани находится в форме предшественников, лишенных ферментативной активности [Savitri I.J. et al, 2014]. Активация предшественников происходит по каскадному механизму, причем активация терминальных ММР (желатиназ и стромелизинов) достигается преимущественно за счет протеолитической активности матриксинов, находящихся в начале сигнального пути [Schaumann Т. et al, 2013]. Таким образом, сама по себе гиперподукция предшественников ММР8 и ММР9, несомненно, имеющая место при воспалении пародонта, не является достаточным условием для подъема активности этих протеаз: для этого требуется еще предварительная активация ММР1, ММР2 и ММР3 [Pirhan D. et al, 2008; Trivedi S. et al, 2015]. Мало того, присутствие в тканях ингибиторов ММР - TIMP1, TIMP2, TIMP3 и TIMP4, общая концентрация которых в несколько раз превышает концентрацию любой ММР, позволяет полностью подавлять активность даже тех молекул ММР, которые подверглись протеолитической активации и не имеют в составе пропептида (как правило, пропептид проявляет высокую ингибирующую активность в отношении собственной ММР) [Tsilingaridis G. et al, 2013; Türkoğlu О. et al, 2014].
Но опубликованные экспериментальные данные не дают никаких свидетельств того, что подобные процессы имеют место при воспалении пародонта [Wohlfahrt J.C. et al, 2014; Wan С. et al, 2014]. При этом, однако, необходимо иметь в виду, что используемые в современных работах методы ПЦР-РВ, ИФА и ФА практически не позволяют отличать активную форму ММР от предшественника [Schaumann Т. et al, 2013]. Тем более трудной эта задача является в отношении первичных ММР (ММР1, ММР2 и ММР3), концентрация которых в тканях существенно ниже, чем терминальных ММР (ММР8 и ММР9) [Zhou Q. et al, 2005].
Резюмируя сказанное, необходимо отметить, что роль ММР8 и ММР9 в деградации пародонтальной связки и альвеолярной кости нуждается в конкретизации, хотя статистическая связь между гиперпродукцией предшественников этих матриксинов и тяжестью пародонтита можно считать доказанной [Miller N. et al, 2003].
Еще одним хорошо освещенным в литературе моментом, касающимся роли ММР8 и ММР9 в развитии пародонтита, является механизм активации их синтеза лимфокинами [Sakalauskiene J. et al, 2005]. Существуют неопровержимые доказательства того, что продукция ММР8 и ММР9 на пораженных участках пародонта коррелирует с увеличением концентрации там лимфокинов острого неспецифического ответа на инвазию (Th1-тип): IL1β, IL12, IL18, TNFα, лиганда-активатора NF-κВ (RANKL) и остеопротегерина (OPG) [Livingstone D. et al, 2015]. Напротив, повышение концентрации лимфокинов, стимулирующих антиген-зависимый ответ (Th2-тип): IL10 и IL4 коррелирует с падением продукции ММР8 и ММР9 [Lee S.I. et al, 2012]. В этот же момент наблюдается повышение продукции IL17 - фактора, стимулирующего заживление раневой поверхности после устранения инфекции [Finoti L.S. et al, 2013]. При исследовании смывов пародонта этот процесс удается наблюдать в основном на стадии ремиссии пародонтита [Folwaczny М. et al, 2005]. Имеются и прямые доказательства того, что стимуляция культивируемых in vitro фибробластов Th1-лимфокинами: IL1β, IL12, IL18, TNFα и RANKL на 2-3 порядка повышает уровень накопления предшественников металлопротеиназ, прежде всего, ММР8 и ММР9 (результаты определения мРНК и кодируемых ими белков-предшественников совпадают) [Birkedal-Hansen Н., 1993; Finoti L.S. et al, 2013]. Таким образом, лимфокины должны рассматриваться в качестве одного из ключевых факторов активации продукции предшественников ММР в ответ на инвазию [Galbraith G.M.P. et al, 1999; Emingil G. et al, 2006]. При обобщении изложенных фактов вырисовывается следующая сигнальная цепь ответа на бактериальную инвазию, при определенных обстоятельствах способная вызывать распад пародонтальной связки [Fine D.H. et al, 2000]:
1) Бактерии, их специфические сигнальные факторы или продукты распада (липополисахарид и ДНК) воздействуют на TLR-рецепторы (TLR1, TLR4, TLR9) поверхности резидентных клеток пародонта (фибробластов или макрофагов);
2) Клетки пародонта, получившие стимул, выбрасывают лимфокины острого неспецифического ответа (Th1): IL1β, IL12, IL18, TNFα и RANKL.
3) Лимфокины Th1-ответа вызывают гиперпродукцию металлопротеиназ ММР8 и ММР9. Эта реакция, вероятно, необходима для облегчения миграции в очаг инфекции лимфоцитов из циркуляции, а также облегчения проникновения компонентов комплемента и других бактерицидных факторов крови в очаг инфекции. За повышение проницаемости стенок сосудистых капилляров отвечает IL6, формально относимый к лимфокинам Th2-ответа. Одновременно все перечисленные факторы стимулируют положительный хемотаксис Т-лимфоцитов и NK-клеток из периферической крови, которые, в свою очередь, мобилизуются из костного мозга и региональных лимфоузлов.
4) В случае быстрого подавления развития инфекции за счет факторов Th1-ответа вызвавший его стимул исчезает, и все перечисленные реакции угасают.
5) В случае нечувствительности инфекции к Th1-ответу за счет IL12-зависимой выработки тормозного медиатора IL10 и активности привлеченных в очаг NK и Т-клеток через небольшое время (1-2 суток или менее) происходит подавление Th1-ответа, и стимулируется Th2-ответ, опосредующий выработку специфических антител к антигенам проникших во внутреннюю среду бактерий. Накопление антител стимулирует резкое усиление ударной мощи комплемента и цитотоксических клеток в отношении инфекционного агента, опсонизацию макрофагов, что приводит к подавлению инфекции. В качестве маркера накопления на пародонте лимфоцитов целесообразно рассматривать IL2, который не может вырабатываться другими типами клеток.
6) В случае нарушения переключения Th1/Th2 ответа происходит неконтролируемое накопление ММР, приводящее к хроническому воспалению, распаду коллагенового матрикса пародонтальной связки и постепенной резорбции кости. Эти явления и лежат в основе известной клинической картины как агрессивного, так и хронического пародонтита.
Существенным элементом антибактериального ответа с участием факторов, рекрутируемых из центральной циркуляции, являются нейтрофилы - самая многочисленная популяция клеток крови [Hajishengallis G. et al, 2015]. Функция нейтрофилов заключается в доставке в очаг инфекции окислителей - активных форм кислорода, обладающих высокой бактерицидной активностью [Орехова Л.Ю. и соавт., 2000]. При этом нейтрофилы погибают, формируя гной [Левин М.Я. и соавт, 1999].
Основным хемотактическим фактором привлечения нейтрофилов в очаг инфекции является IL8 [Anovazzi G. et al, 2013]. Имеются многочисленные экспериментальные данные о локальном подъеме уровня IL8 в очагах воспаления, в том числе, при хроническом пародонтите [Микробиология и иммунология для стоматологов], что заставляет некоторых авторов причислять его к факторам Th1-ответа. В случае пародонтита любой формы гиперпродукцию IL8 можно рассматривать как маркер, свидетельствующий о неудаче попыток иммунной системы сдержать инфекцию за счет Th1-ответа [Berglundh Т. et al, 2003]. При этом тип клеток, являющийся основным источником IL8, остается неизвестным [Borilova-Linhartova P. et al, 2013]. Несомненно также, что длительная и массивная инфильтрация избытка нейтрофилов в ткань пародонта не может обеспечить протективности иммунного ответа [Berker Е. et al, 2005], хотя не исключено, что при первом контакте нейтрофилов с бактериями они способны в течение определенного срока сдерживать развитие инфекции и даже полностью стерилизовать небольшие очаги инвазии [de Goncalves L.S. et al, 2006].
В работе бразильских авторов Finoti et al, 2013 «Обсемененность патогенами и клиническая эффективность ответа на лечение пародонтита у пациентов с различным гаплотипом по гену интерлейкина 8» с помощью ПЦР в реальном времени исследовали обсемененность пародонта у пациентов с различным генотипом по позициям АТС/ТТС и AGT/TTC в гене IL8 тремя пародонтопатогенами: P. gingivalis, Т. forsythensis и Treponema denticola. Было обследовано 240 образцов пародонтальных смывов, причем их отбирали как из пораженных участков пародонта, так и из здоровых участков пародонта тех же больных. Было установлено, что ни один из гаплотипов не приводит к изменению обсемененности пародонта в целом. Однако, при анализе только пораженных участков пародонта наблюдалась тенденция к вредному влиянию генотипа AGT/TTC по сравнению с АТС/ТТС: у этих больных пораженные участки содержали большее количество P. gingivalis, Т. denticola, Т. forsythensis и красного комплекса в целом. В остальном генотип по гену IL8 не сказывался на ходе течения пародонтита и эффективности его хирургического и терапевтического лечения.
Большинство работ по экспериментальному исследованию механизмов индукции синтеза IL8 при пародонтите выполнено на модели клеток гингивиального эпителия человека, в частности, на иммортализованной клеточной линии ОВА-9 или на первичных гингивиальных фибробластах. Примером такой работы является статья Savitri et al, 2014 «Малеат ирзогладина ингибирует индукцию синтеза toll-рецепторов 2 типа и IL8, индуцированную Porphyromonas gingivalis, в клетках гингивиального эпителия человека». В работе постулируется, что малеат ирзогладина (ИМ) является общеизвестным регулятором воспаления и состояния плотных межклеточных контактов. Авторы поставили целью исследовать влияние этого биологически активного вещества на взаимодействие эпиталиальных клеток десны с пародонтопатогеном P. gingivalis. Выбор этой бактерии, входящей в красный комплекс пародонтопатогенов по Сокранскому, в качестве модели авторами не аргументировался, но можно предполагать, что причиной этого является относительная легкость культивирования этого вида по сравнению с другими известными пародонтпатогенами. В качестве технического средства авторы использовали стимуляцию клеток человека in vitro цельными клетками P. gingivalis или ее очищенным полисахаридом. Уровень продукции клетками IL8 и TLR2 (компонент системы антиген-независимого (врожденного) иммунитета) определяли с помощью ПЦР в реальном времени. Оценивая зависимость между уровнем синтеза IL8 и TLR2, экспрессию гена TLR2 в культуре подавления путем трансфекции малыми интерферирующими РНК (siRNA). Было показано, что ИМ снимает эффект индукции синтеза IL8 P. gingivalis или ее очищенным полисахаридом на клетках ОВА-9 (Р<0,01). Точно такой эффект наблюдается и в отношении TLR2, причем как на культуре ОВА-9, так и на первичных фибробластах (Р<0,01). Результаты ПЦР-РВ были подтверждены иммуногистохимическим окрашиванием. Показано, что стимуляция пептидогликаном (типовым лигандом TLR2) в сочетании с ИМ приводит к снижению уровня продукции другого toll-рецептора TLR4 (Р<0,01). Напротив, липополисахарид Escherichia coli, основной лиганд TLR4, не влиял на уровень синтеза мРНК TLR2. Индуцирующий эффект полисахарида P. gingivalis на продукцию мРНК TLR4 снимался добавлением ИМ. Нарушение синтеза TLR2 с помощью siRNA приводило к снижению уровня синтеза мРНК IL8 под действием P. gingivalis или ее полисахарида на клетках ОВА-9.
В работе шведских авторов Palm et al, 2013 «Porphyromonas gingivalis снижает уровень иммунного ответа фибробластов» утверждается, что P. gingivalis является основной причиной развития хронического пародонтита. При этом в качестве основного фактора патогенности этой бактерии, обеспечивающей также ее защиту и выживание на пародонте, называют гингвипаины - протеиназы семейства папаина. Отталкиваясь от этого, авторы исследовали поведение P. gingivalis в культуре первичных фибробластов слизистой десны и кожи человека. В качестве определяемого параметра служил уровень накопления хемокинов и цитокинов, в частности, TNFα и CXCL8 (IL8). Измерение уровня продукции этих факторов проводили с помощью ИФА и цитокинового теста. Авторы сделали вывод, что TNFα-зависимый синтез CXCL8 полностью подавляется живыми клетками P. gingivalis, тогда как клетки Р. gingivalis, убитые нагреванием, не оказывают влияния на скорость продукции фибробластами CXCL8. Добавление ингибиторов аргинин-специфичной протеазы частично снимало эффект супрессии стимулирующего действия TNFα на синтез CXCL8. Кроме того, в присутствии ингибитора аргинин-специфичной протеазы восстанавливался синтез ряда других провоспалительных медиаторов, подавленный живой культурой P. gingivalis. На этом основании авторы делают предположение о ключевой роли аргинин-специфичной протеазы P. gingivalis в подавлении иммунного ответа на микрофлору пародонта.
В еще одной работе тех же авторов Palm et al, 2014 «Подавление воспалительной реакции гингивиальных фибробластов человека гингвипаинами Porphyromonas gingivalis» описываются углубленные исследования механизма действия гингвипаинов Rgp (Arg-специфичный) и Kgp (Lys-специфичный) в отношении факторов, продуцируемых фиброобластами эпителия десны человека: фактор β1 (TGFβ1), CXCL8 (IL8), секреторный ингибитор протеазы лейкоцитов (SLPI), c-Jun и индоламин-2,3-диоксигеназы (IDO). Первичную культуру фибробластов в течение 6 и 24 часов инкубировали с культурами P. gingivalis дикого типа АТСС 33277 и W50, а также двух мутантов W50 по генам гингвипаинов или прогретой культурой P. gingivalis АТСС 33277. Было показано, что дикие штаммы P. gingivalis влияют на уровень продукции фибробластами CXCL8 и TGFβ1, причем мутант по гену kgp существенно в большей мере стимулирует продукцию CXCL8 (Р<0,001) и снижение уровня TGFβ1, чем дикие штаммы. Продукция SLPI подавляется P. gingivalis (Р<0,05). Таким образом, подавляя активность SLPI, P. gingivalis может непосредственно влиять на скорость деградации пародонта. Обнаружено также, что P. gingivalis подавляет продукцию такого антимикробного фактора, как IDO, причем этот механизм зависим от уровня внутриклеточного c-Jun (Р<0,05). Все эти наблюдения свидетельствуют о наличии у P. gingivalis целого ряда механизмов непосредственного влияния на работу местного иммунитета при пародонтите.
В работе Schaumann et al, 2013 «Возможная иммуномодулирующая активность глицина при воспалении десны» описаны эксперименты по воздействию глицина на клетки эпителия десны. Авторы предположили существование специфических рецепторов глицина и выполнили эксперименты по их поиску гистохимическими методами. Первичные фибробласты эпителия десны в культуре обрабатывали IL1β в присутствии или в отсутствие глицина, после чего с помощью ПЦР в реальном времени исследовалась динамика синтеза IL6 и IL8. Кроме того, с помощью иммунофлуоресцентной микроскопии исследовался транспорт регуляторного фактора NFκB в ядро. Было показано, что стимуляция глицином усиливает эффективность действия IL1β в качестве индуктора синтеза мРНК IL6 и IL8, а также интенсифицирует транспорт в ядро NFκB. Таким образом, глицин способствует ослаблению воспаления эпителия десны. По мнению авторов, существует перспектива применения глицина в качестве антивоспалительного средства при пародонтите.
В работе шведских авторов Khalaf et al, 2014 «Цитокины и хемокины по-разному экспрессируются у больных пародонтитом: возможная роль TGF1β в качестве маркера заболевания» утверждается, что тяжелая форма пародонтита повышает риск развития системных заболеваний воспалительной природы, прежде всего, атеросклероза. С учетом этого они провели комплексное исследование представленности различных цитокинов и лимфокинов в сыворотке крови, слюне и пародонтальных смывах больных пародонтитом и контрольной выборки лиц со здоровым пародонтом. Наиболее значимым отличием выборки больных оказался сниженный уровень IL6 в крови и слюне. Напротив, уровень CXCL8 (IL8) в крови не отличался, в слюне он был снижен, а в пародонтальных смывах - повышен. Лимфокин Т-клеточного происхождения IL2 в крови и слюне пациентов обеих выборок содержался в одинаковой концентрации, но его содержание в пародонтальных смывах больных было существенно снижено. Неожиданным результатом оказалось существенное повышение уровня TGFβ1 в крови, слюне и пародонтальных смывах больных ХП. Исследуя это наблюдение, авторы обнаружили, что стимуляция фибробластов in vitro живыми клетками P. gingivalis дикого типа или их беспротеазных мутантов приводит к снижению уровня продукции CXCL8 и IL6 и повышению экспрессии TGFβ1. По мнению авторов, основным фактором, ответственным за этот эффект, является лизин-специфичная цистеиновая протеиназа P. gingivalis гингвипаин Kgp. Авторы считают TGFβ1 одним из основных кандидатов на роль фактора, опосредующего системные эффекты воспаления пародонта.
Известен патент РФ №2381746 «Способ диагностики степени тяжести пародонтита». Изобретение относится к стоматологии и применяется для диагностики степени тяжести пародонтита. Метод состоит в том, что измеряют длины корней зубов на нижней и на верхней челюсти, В результате авторами установлена корреляция между степенью тяжести воспалительного процесса в пародонте, степенью деструкции костной ткани межальвеолярной перегородки и анатомическим строением зубов, а именно длиной корней зубов. На основании статистических данных, полученных при обследовании больных, авторами доказано, что метрическая характеристика зубов, а именно длина корня, может использоваться предсказания тяжести протекания пародонтита даже в случаях, когда обострение заболевания в явном виде себя не проявляет, а клиническая картина сглажена.
Известен патент РФ №2451937 «Способ диагностики рака молочной железы по уровню РНК интерлейкинов IL8 и/или IL18 в плазме крови». Изобретение относится к области медицины, онкологии и молекулярной биологии и может быть использовано для диагностики рака молочной железы. Способ основан на измерении в плазме крови уровня РНК интерлейкинов IL8 и/или IL18 и сравнении уровня представленности РНК IL8 и/или IL18 с уровнем представленности референсных транскриптов. Для осуществления способа используют обратную транскрипцию и полимеразную цепную реакцию в «реальном времени». В качестве референсных транскриптов используют РНК ABL, и/или HPRT, и/или IL lb. Результат исследования считают положительным при разнице уровней представленности РНК IL8 и HPRT более десяти раз. Использование заявленного способа позволяет осуществлять раннюю диагностику рака молочной железы. Патент не описывает применения ПЦР в реальном времени для диагностики пародонтита, не содержит описания метода определения мРНК IL8 в смывах пародонта и последовательностей праймеров и зондов. Таким образом, заявляемое изобретение не нарушает прав интеллектуальной собственности, описанной в патенте РФ №2451937.
Известен патент РФ №2180119 «Способ ранней диагностики сепсиса у новорожденных. Изобретение относится к методам ранней лабораторной диагностики септической инфекции у новорожденных. Способ заключается в исследовании уровня цитокинов: интерлейкина-8 (ИЛ-8) и фактора некроза опухоли (ФНО) и при концентрации ИЛ-8 более 5 пг/мл и ФНО более 2 пг/мл диагностируют развитие септической инфекции. Техническим результатом является получение высокочувствительного метода ранней диагностики септической инфекции у новорожденных, что позволяет поставить клинический диагноз с наибольшей степенью вероятности и в максимально ранние сроки. Способ не использует ПЦР, в том числе, ПЦР в реальном времени, и не применим в пародонтологии, так как не описывает калибровку и пробоподготовку к анализу пародонтальных смывов.
Таким образом, в настоящее время в РФ нет действующих аналогов заявляемого изобретения.
Раскрытие изобретения
Смывы пародонтальных карманов получают, отбирая содержимое из наиболее глубоких участков с помощью стерильных бумажных эндодонтических штифтов (размер №25), которые затем помещают в пробирку с 1,5 мл реактива РНК-латер (консервант для РНК) (Life technologies - Invitrogen, каталожный номер №7020) и транспортируют в лабораторию в охлажденном состоянии, в течение 6 час.
Забор проводят в двух повторностях от каждого пациента. На стадии выделения РНК и проведения ПЦР-анализа каждая повторность обрабатывается отдельно.
Для проведения обратной транскрипции используют наборы производства ООО «НПО ДНК-Технология» (Россия). Ингредиенты набора «ОТ-буфер», «Праймеры ОТ-TNFα», «Праймеры ОТ-ММР8» «Праймеры ОТ-ММР9» из комплекта реагентов для обратной транскрипции оттаивают при комнатной температуре в течение 10 мин.
Для получения реакционной смеси для проведения ПЦР используют следующие объемы ингредиентов:
1. Смесь готовят в стрипованных пробирках Scintific Specialties Inc. (SSI) Part # 3135-00 UltraFlux Flat Cap PCR Tubes 200 μL, unit 1×125 tube strips with caps.
2. Промаркировать ряды пробирок для каждого исследуемого образца с учетом отрицательного контрольного образца (К(-)) и положительного контрольного образца (К(+)) и 2 пробирки для контроля фона флуоресценции.
3. В пробирки внести буфер «Н» объемом 20 мкл, в состав которого входят все компоненты смеси для проведения ПЦР реакции за исключением ДНК-полимеразы и ДНК-матрицы.
4. В каждую пробирку внести каплю расплавленного парафина, отделив реакционную смесь.
5. Подготовить раствор Tag-полимеразы (Комплект реагентов для ПЦР амплификации, ООО «НПО ДНК-Технология», Россия), встряхнув пробирку на вортексе и сбросив капли со стенок.
6. Внести в каждую промаркированную пробирку (кроме пробирок контроля фона) по 10 мкл раствора Tag-полимеразы, не повреждая слой парафина.
7. Добавить в каждую пробирку по капле минерального масла, не повреждая слой парафина.
8. В пробирку с исследуемым образцом внести по 5 мкл препарата ДНК, не повреждая слой парафина.
9. В пробирку с положительным контролем (К(+)) внести 5 мкл положительного контрольного образца ДНК, в пробирки с отрицательным контролем (К(-)) и контролем фона внести 5 мкл отрицательного контрольного образца или воды, соответственно, не повреждая слой парафина.
10. Центрифугировать пробирки на микроцентрифуге или вортексе 3-5 сек.
11. Установить пробирки в термоциклер ДТ-322 (ООО «НПО ДНК-Технология», Россия), установив объем реакционной смеси «35 мкл».
Для каждого препарата ДНК, выделенного из смыва, проводят реакцию с шестью парами праймеров/зондом. Реакцию проводят со следующими параметрами циклирования:
Программа амплификации (50 циклов):
1. 94,0°C - 01'30ʺ
2. 94,0°C - 00'10ʺ
3. 64,0°C - 00'15ʺ
Настройки анализа, устанавливаемые на приборе:
Метод - Геометрический (Ср) (BF), cr=9, vt=10, tp=30, tv=5
Для каждой реакции программное обеспечение термоциклера автоматически вычисляет величину порогового цикла Ct. Полученные величины усредняют по двум образцам одной серии.
Анализировали содержание в пародонтальных смывах специфических транскриптов провоспалительного лимфокина TNFα и матриксинов ММР8 и ММР9. Кроме того, проводилась оценка содержания суммарной мРНК человека по маркеру КВМ (β2-микроглоублин человека).
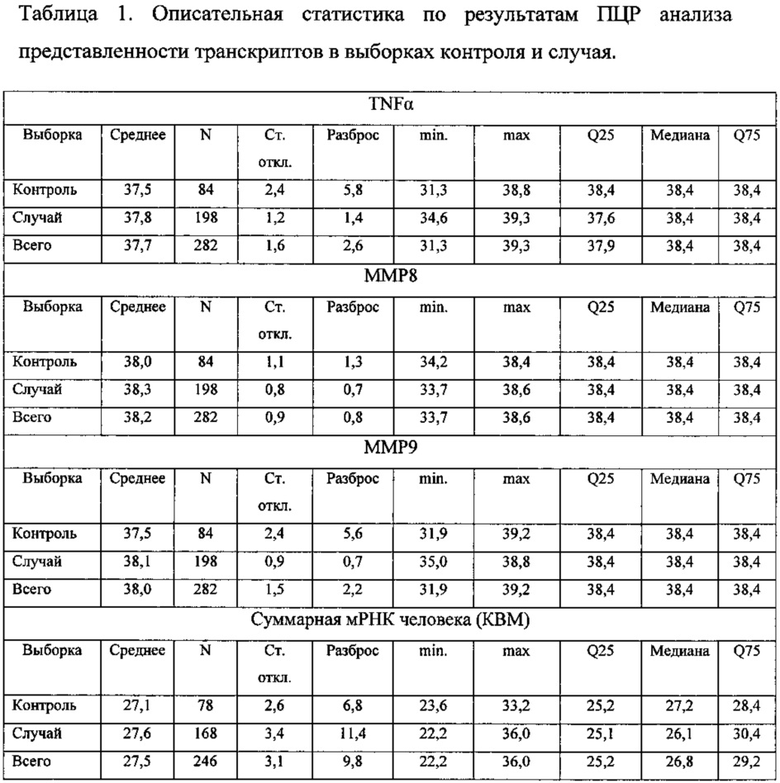
На основании полученных уровней генетических маркеров вычисляется их отношение. Если отношение уровней маркеров TNFα:MMP8:MMP9 находится в диапазоне 1:(1±0,2):(1±0,2), следует диагностировать нормальный статус локального неспецифичекого иммунитета в обследуемой точке пародонта, а при выходе отношении уровней маркеров за пределы указанного диапазона - наличие нарушения в работе неспецифического иммунитета, что увеличивает риск развития воспалительного поражения пародонта.
Осуществление изобретения:
Клинический материал был собран на базе ЦНИСиЧЛХ МЗ РФ. Было обследовано 288 пациентов в возрасте от 21 до 63 лет без тяжелой соматической патологии. В исследуемой выборке находилось 204 пациента с диагнозом «пародонтит» и 84 пациента без патологии пародонта.
Основным критерием для постановки диагноза хронический генерализованный пародонтит (ХГП) являлось разрушение зубодесневого прикрепления. Степень тяжести устанавливали на основании глубины пародонтальных карманов и степени деструкции костной ткани.
Смывы пародонтальных карманов извлекали из наиболее глубоких участков с помощью стерильных бумажных эндодонтических штифтов. Для выделения РНК штифты консервировали в пробирках типа Эппендорф объемом 1,5 мл, содержащих по 0,5 мл РНК-латера (консервант для РНК) (Life technologies - Invitrogen, каталожный номер №7020).
Непосредственно перед выделением материал смыва осаждали центрифугированием на настольной центрифуге в течение 5 мин. Из пробирки тщательно удаляли супернатант, представляющий собой раствор фиксатора IntactRNA (латер) и вносили 200 мкл лизирующего раствора (из набора «Проба-ГС»), образец тщательно перемешивали и выдерживали в течение 1 мин при комнатной температуре. Далее в пробирку с образцом вносили 100 мкл водонасыщенного фенола (органическая фаза двухфазной системы фенол - буфер Трис-HCl 40 мМ, рН 8,0) и 100 мкл смеси хлороформа и изоамилового спирта (24:1). Содержимое тщательно перемешивали на вортексе и инкубировали в течение 5 мин при комнатной температуре, периодически повторяя перемешивание. Пробирки центрифугировали на настольной центрифуге при 13000 G в течение 10 мин. Тщательно избегая захвата органической фазы и интерфазы на поверхности раздела фаз, отбирали верхнюю (водную) фазу и переносили в чистую пробирку, к ней добавляли 200 мкл смеси хлороформа и изоамилового спирта (24:1). Центрифугировали на настольной центрифуге при 13000 G в течение 5 мин. Отбирали верхнюю (водную) фазу и переносили в чистую полипропиленовую пробирку объемом 1,5 мл. К водной фазе добавляли чистый изопропиловый спирт объемом 200 мкл (или в пропорции 1:1 по отношению к собранной водной фазе), перемешивали содержимое 5-6-кратным переворачиванием пробирки. Пробирки центрифугировали на настольной центрифуге при 13000 G в течение 10 мин. Супернатант тщательно удаляли с помощью водоструйного насоса, касаясь осадка, содержащего нуклеиновые кислоты. Осадок промывали, внося в пробирку 100 мкл 70% водного этилового спирта и переворачивая пробирку 5-6 раз.
Пробирки центрифугировали на настольной центрифуге при 13000 G в течение 10 мин. Супернатант тщательно удаляли с помощью водоструйного насоса, касаясь осадка, содержащего нуклеиновые кислоты. Осадок высушивали при температуре 50°C при открытой крышке пробирки в течение 5 мин.
К осадку добавляли 16,5 мкл дистиллированной воды, закрывали крышки пробирок и инкубировали при температуре 50°C в течение 5 мин. Пробирки встряхивали на настольном вортексе, добиваясь полного растворения. Препараты РНК хранили при температуре -20°C не свыше 2 месяцев.
Препараты, выделенные по методике, описанной выше, очищали от примеси геномной ДНК обработкой ДНКазой, свободной от РНКаз. Для этого к препарату суммарных нуклеиновых кислот объемом 15,5 мкл добавляли препарат ДНКазы, свободной от РНКаз, объемом 1 мкл, содержащий 1-5 ед. ферментативной активности. Смесь инкубировали при 37°C в течение 30 мин. После этого фермент инактивировали прогреванием смеси при 95°C в течение 5 мин. Капли со стенок пробирок собирали спин-центрифугированием в течение 3-5 сек.
Для проведения обратной транскрипции использовали наборы производства ООО «НПО ДНК-Технология» (Россия)
Ингредиенты набора «ОТ-буфер», «Праймеры ОТ-TNFα», «Праймеры ОТ-ММР8» и «Праймеры ОТ-ММР9» из комплекта реагентов для обратной транскрипции оттаивали при комнатной температуре в течение 10 мин. Капли со стенок пробирок собирали спин-центрифугированием в течение 3-5 сек.
Смесь ОТ готовили по следующей прописи в стерильной пробирке объемом 1,5 мл:
1. ОТ-буфер - 2×(N+1) мкл;
2. «Праймеры + дНТФ» - (N+1) мкл;
3. Обратная транскриптаза 2 ед./мкл - 0,5×(N+1) мкл,
где N - количество анализируемых образцов с учетом отрицательного контроля. Капли со стенок пробирок собирали спин-центрифугированием в течение 3-5 сек.
В пробирки, содержащие по 16,5 мкл препарата подготовленной РНК, добавляли по 3,5 мкл смеси ОТ. При постановке отрицательного контроля использовали пробирку, содержащую 16,5 мкл очищенной воды. Перемешивали реакционные смеси 5-7-кратным пипетированием. Пробирки инкубировали при 40°C в течение 30 мин, затем останавливали реакцию прогреванием при 95°C в течение 5 мин. Капли со стенок пробирок собирали спин-центрифугированием в течение 3-5 сек. Полученный препарат кДНК использовали для проведения ПЦР в реальном времени.
На основании полученных уровней генетических маркеров вычисляется их отношение, при этом если отношение уровней маркеров TNFα:MMP8:MMP9 находится в диапазоне 1:(1±0,2):(1±0,2), диагностировали нормальный статус локального неспецифического иммунитета в обследуемой точке пародонта, а при выходе отношении уровней маркеров за пределы указанного диапазона - наличие нарушения в работе неспецифического иммунитета, что увеличивает риск развития воспалительного поражения пародонта.
Список использованных источников
1. Anovazzi G, Finoti LS, Corbi SC, Kim YJ, Marcaccini AM, Gerlach RF, Capela MV, Orrico SR, Cirelli JA, Scarel-Caminaga RM. Interleukin 4 haplotypes of susceptibility to chronic periodontitis are associated with IL-4 protein levels but not with clinical outcomes of periodontal therapy. // Hum Immunol. - 2013 - Vol. 74, №12, doi: 10.1016/j.humimm.2013.08.286.
2. Atieh M.A. Accuracy of real-time polymerase chain reaction versus anaerobic culture in detection of Aggregatibacter actinomycetemcomitans and Porphyromonas gingivalis: a meta-analysis. // J. Periodontol. - 2008. - Vol. 79, №9. - P. 1620-1629.
3. Bai D, Nakao R, Ito A, Uematsu H, Senpuku H. Immunoreactive antigens recognized in serum samples from mice intranasally immunized with Porphyromonas gingivalis outer membrane vesicles. // Pathog Dis. - 2015. Vol. 73. №3.
4. Behl Y., Siqueira M., Ortiz J. et al. Activation of the acquired immune response reduces coupled bone formation in response to a periodontal pathogen. // J. Immunol. - 2008. - Vol. 181. - №12. - P. 8711-8718.
5. Berdeli A., Emingil G., Gurkan A. et al. Association of the IL-1RN2 allele with periodontal diseases. // Clin. Biochem. - 2006. - Vol. 39. - P. 357-362.
6. Berdeli A., Emingil G., Gurkan A. et al. Association of the IL-1RN2 allele with periodontal diseases. // Clin. Biochem. - 2006. - Vol. 39. - P. 357-362.
7. Berglundh Т., Donati M., Hahn-Zoric M. et al. Association of the -1087 IL-10 gene polymorphism with severe chronic periodontitis in Swedish Caucasians. // J. Clin. Periodontol. - 2003. - Vol. 30. - P. 249-254.
8. Berker E., Kantarci A., Hasturk H. et al. Effect of neutrophil apoptosis on monocytic cytokine response to Porphyromonas gingivalis lipopolyssaccharide. // J. Periodontol. - 2005. - Vol. 76. - P. 964-971.
9. Birkedal-Hansen H. Role of matrix metalloproteinases in human periodontal diseases. // J. Periodontol. - 1993. - Vol. 64, №5 (Suppl.). - P. 474-484.
10. Borilova-Linhartova P, Vokurka J, Poskerova H, Fassmann A, Izakovicova Holla L. Haplotype analysis of interleukin-8 gene polymorphisms in chronic and aggressive periodontitis. // Mediators Inflamm. - 2013; 2013:342351.
11. Borilova-Linhartova P, Vokurka J, Poskerova H, Fassmann A, Izakovicova Holla L. Haplotype analysis of interleukin-8 gene polymorphisms in chronic and aggressive periodontitis. // Mediators Inflamm. - 2013; 2013. P. 342351.
12. Choi НМ, Han K, Park YG, Park JB. Associations Among Oral Hygiene Behavior and Hypertension Prevalence and Control: The 2008-2010 Korea National Health and Nutrition Examination Survey. J Periodontol. 2015. V. 5. P. 1-19.
13. Corbi SC, Anovazzi G, Finoti LS, Kim YJ, Capela MV, Secolin R, Marcaccini AM, Gerlach RF, Orrico SR, Cirelli JA, Scarel-Caminaga RM. Haplotypes of susceptibility to chronic periodontitis in the Interleukin 8 gene do not influence protein level in the gingival crevicular fluid. // Arch Oral Biol. - 2012 Vol. 57, №10. P. 1355-61.
14. Corbi SC, Finoti LS, Anovazzi G, Tanaka MH, Kim YJ, Secolin R, Marcaccini AM, Gerlach RF, Orrico SR, Cirelli JA, Scarel-Caminaga RM. Clinical outcomes of periodontal therapy are not influenced by the ATC/TTC haplotype in the IL8 gene. J Periodontal Res. - 2014 Vol. 49, №4. P. 489-98.
15. Cortelli J.R., Cortelli S.C., Jordan S. et al. Prevalence of periodontal pathogens in Brazilians with aggressive or chronic periodontitis. // J. Clin. Periodontal. - 2005. - Vol. 32, №8. - P. 860-866.
16. de Goncalves L.S., Ferreira S.M., Souza C.O. et al. IL-1 gene polymorphism and periodontal status of HIV Brazilians on highly active antiretroviral therapy. // Acq. Immun. Defic. Syndr. - 2006. - Vol. 20. - P. 1779-1781.
17. Dong Y, Huihui Z, Li C. Pipeline inhibit inflammation, alveolar bone loss and collagen fibers breakdown in a rat periodontitis model. J Periodontal Res. 2015 Mar 2. doi: 10.1111/jre.l2262.
18. Dosseva-Panova VT, Popova CL, Panov VE Subgingival microbial profile and production of proinflammatory cytokines in chronic periodontitis. // Folia Med (Plovdiv). - 2014 - Jul - Vol. 56, №3 - P. 152-60.
19. Dyer J.K., Peck M.A., Reinhardt R.A. et al. HLA-D types and serum IgG responses to Capnocytophaga in diabetes and periodontitis. // J. Dent. Res. - 1997. - Vol. 76. - P. 1825-1832.
20. Elamin A., Albandar J.M., Poulsen K. et al. Prevalence of Aggregatibacter actinomycetemcomitans in Sudanese patients with aggressive periodontitis: a case-control study. // J. Periodontal. Res. - 2011. V. 46. №3. P. 285-91.
21. Fine D.H., Kaplan J.B., Kachlany S.C. et al. How we got attached to Actinobacillus actinomycetemcomitans: A model for infectious diseases. // Periodontol. - 2000. - 2006. - Vol. 42. - P. 114-157.
22. Finoti L.S., Anovazzi G., Pigossi S.C, Corbi S.C, Teixeira S.R., Braido G.V., Kim Y.J., Orrico S.R., Cirelli J.A., Mayer M.P., Scarel-Caminaga R.M. Periodontopathogens levels and clinical response to periodontal therapy in individuals with the interleukin-4 haplotype associated with susceptibility to chronic periodontitis. // Eur. J. Clin. Microbiol. Infect. Dis. - 2013. - Vol. 32, №12. - P. 1501-9.
23. Finoti L.S., Anovazzi G., Pigossi S.C, Corbi S.C, Teixeira S.R., Braido G.V., Kim Y.J., Orrico S.R., Cirelli J.A., Mayer M.P., Scarel-Caminaga R.M. Periodontopathogens levels and clinical response to periodontal therapy in individuals with the interleukin-4 haplotype associated with susceptibility to chronic periodontitis. // Eur. J. Clin. Microbiol. Infect. Dis. - 2013. - Vol. 32, №12. - P. 1501-9.
24. Finoti L.S., Corbi S.C, Anovazzi G., Teixeira S.R., Steffens J.P., Secolin R., Kim Y.J., Orrico S.R., Cirelli J.A., Mayer M.P., Scarel-Caminaga R.M. Association between IL8 haplotypes and pathogen levels in chronic periodontitis. // Eur. J. Clin. Microbiol. Infect. Dis. - 2013. - Vol. 32, №10. - P. 1333-40.
25. Folwaczny M., Glas J., Torok H.P. et al. Prevalence of the -295 T-to-C promoter polymorphism of the interleukin (IL)-16 gene in periodontitis. // Clin. Experiment. Immunol. - 2005. - Vol. 142. - P. 188-192.
26. Fujise O., Miura M., Hamachi T. et al. Risk of Porphyromonas gingivalis recolonization during the early period of periodontal maintenance in initially severe periodontitis sites. // J. Periodontol. - 2006. - Vol. 77, №8. - P. 1333-1339.
27. Galbraith G.M.P., Hendley T.M., Sanders J.J. et al. Polymorphic cytokine genotypes as markers of disease severity in adult periodontitis. // J. Clin. Periodontol. - 1999. - Vol. 26. - P. 705-709.
28. Gemmell E., Seymour G.J. Immunoregulatory control of Th1/Th2 cytokine profiles in periodontal disease // Periodontol. 2000. - 2004. - Vol. 35. - P. 21-41.
29. Gemmell E., Yamazaki K., Seymour G.J. The role of T cells in periodontal disease: homeostasis and autoimmunity. // Periodontol. - 2000. - 2007. - Vol. 43. - P. 14-40.
30. Gilowski L., Kondzielnik P., Wiench R., Płocica I., Strojek K., Krzemiński T.F. Efficacy of short-term adjunctive subantimicrobial dose doxycycline in diabetic patients-randomized study. // Oral. Dis. - 2012. - Vol. 18, №8. - P. 763-70.
31. Godoy-Gallardo M., Wang Z., Shen Y., Manero J.M., Gil F.J., Rodriguez D., Haapasalo M. Antibacterial coatings on titanium surfaces: a comparison study between in vitro single-species and multispecies biofilm. // ACS Appl. Mater. Interfaces. - 2015. - PMID. 25734758.
32. Gomez S.C. Periodontal status in smokers and never-smokers: clinical findings and real-time polymerase chain reaction quantification of putative periodontal pathogens. // J. Periodontol. - 2006. - Vol. 77, №9. - P. 1483-1490.
33. Goncalves P.F., Huang H., McAninley S., Alfant В., Harrison P., Aukhil I., Walker C, Shaddox L.M. Periodontal treatment reduces matrix metalloproteinase levels in localized aggressive periodontitis. // J. Periodontol. - 2013. - Vol. 84, №12. - P. 1801-8.
34. Graswinckel J.E.M., van der Velden U., van Winkelhoff A.J. et al. Plasma antibody levels in periodontitis patients and controls // J. Clin. Periodontol. - 2004. - Vol. 31. - P. 562-568.
35. Graves D.T., Cochran D. The contribution of interleukin-1 and tumor necrosis factor to periodontal tissue destruction. // J. Periodontol. - 2003. - Vol. 74. - P. 391-401.
36. Gürkan A., Emingil G., Saygan B.H. et al. Matrix metalloproteinase-2, -9, and -12 gene polymorphisms in generalized aggressive periodontitis. // J. Periodontol. - 2007. - Vol. 78, №12. - P. 2338-2347.
37. Gustafsson A., Ito H., Asman B. et al. Hyper-reactive mononuclear cells and neutrophils in chronic periodontitis. // J. Clin. Periodontol. - 2006. - Vol. 33. - P. 126-129.
38. Guzeldemir E., Gunhan M., Ozcelik O. et al. Interleukin-1 and tumor necrosis factoralpha gene polymorphisms in Turkish patients with localized aggressive periodontitis. // J. Oral Sci. - 2008. - Vol. 50. - P. 151-159.
39. Hagewald S., Bernimoulin J.P., Kottgen E. et al. Total IgA and Porphiromonas gingivalis - reactive IgA in the saliva of patients with generalized early - onset periodontitis. // Eur. J. Oral. Sci. - 2000. - Vol. 108, №2. - P. 147-153.
40. Hajishengallis G., Lamont R.J., Graves D.T. The enduring importance of animal models in understanding periodontal disease. // Virulence. - 2015. - Vol. 9. DOI: 10.4161/21505594.2014.990806.
41. Hayashi N., Kido J., Kido R. et al. Regulation of calprotectin expression by interleukin-1-alpha and transforming growth factor-beta in human gingival keratinocytes. // J. Periodontal. Res. - 2007. - Vol. 42, №1. - P. 1-7.
42. Holla L.I., Fassmann A., Stejskalova A. et al. Analysis of the interleukin-6 gene promoter polymorphisms in Czech patients with chronic periodontitis. // J. Periodontol. - 2004. - Vol. 75. - P. 30-36.
43. Holt S.C., Ebersole J.L. Porphyromonas gingivalis, Treponema denticola, and Tannerella forsythia: the "red complex", a prototype polybacterial pathogenic consortium in periodontitis. // Periodontol. - 2000. - 2005. - Vol. 38. - P. 72-122.
44. Hyvärinen K., Laitinen S., Paju S. et al. Detection and quantification of five major periodontal pathogens by single copy gene-based real-time PCR. // Innate Immun. - 2009. - Vol. 15, №4. - P. 195-204.
45. Irshad M., Scheres N., Anssari Moin D., Crielaard W., Loos B.G., Wismeijer D., Laine M.L. Cytokine and matrix metalloproteinase expression in fibroblasts from peri-implantitis lesions in response to viable Porphyromonas gingivalis. // J. Periodontal. Res. - 2013. - Vol. 48, №5. - P. 647-56.
46. Javed F., Ahmed H.B., Mikami Т., Almas K., Romanos G.E., Al-Hezaimi K. Cytokine profile in the gingival crevicular fluid of rheumatoid arthritis patients with chronic periodontitis. // J. Investig. Clin. Dent. - 2014. - Vol. 5, №1. - P. 1-8.
47. Javed F., Ahmed H.B., Saeed A., Mehmood A., Bain C. Whole salivary interleukin-6 and matrix metalloproteinase-8 levels in patients with chronic periodontitis with and without prediabetes. // J. Periodontol. - 2014. - Vol. 85, №5. - P. 130-5.
48. Jayaprakash K., Khalaf H., Bengtsson T. Gingipains from Porphyromonas gingivalis play a significant role in induction and regulation of CXCL8 in THP-1 cells. // BMC Microbiol. - 2014. - Vol. 18. - P. 14-193.
49. Jayaprakash K., Khalaf H., Bengtsson T. Gingipains from Porphyromonas gingivalis play a significant role in induction and regulation of CXCL8 in THP-1 cells. // BMC Microbiol. - 2014. - Vol. 18. - P. 14-193.
50. Khalaf H., Lönn J., Bengtsson T. Cytokines and chemokines are differentially expressed in patients with periodontitis: possible role for TGF-β1 as a marker for disease progression. // Cytokine. - 2014. - Vol. 67(1). - P. 29-35.
51. Khosropanah H., Sarvestani E.K., Mahmoodi A., Golshah M. Association of IL-8 (-251A/T) gene polymorphism with clinical parameters and chronic periodontitis. // J. Dent (Tehran). - 2013. - Vol. 10(4). - P. 312-8.
52. Kinney J.S., Morelli Т., Oh M., Braun T.M., Ramseier C.A., Sugai J.V., Giannobile W.V. Crevicular fluid biomarkers and periodontal disease progression. // J. Clin. Periodontol. - 2014. - Vol. 41(2). - P. 113-20.
53. Kinney J.S., Morelli Т., Oh M., Braun T.M., Ramseier C.A., Sugai J.V., Giannobile W.V. Crevicular fluid biomarkers and periodontal disease progression. // J. Clin. Periodontol. - 2014. - Vol. 41(2). - P. 113-20.
54. Lee S.I., Kang K.L., Shin S.I., Herr Y., Lee Y.M., Kim E.C. Endoplasmic reticulum stress modulates nicotine-induced extracellular matrix degradation in human periodontal ligament cells. // J. Periodontal. Res. - 2012. - Vol. 47(3). - P. 299-308.
55. Lee W., Pankoski L., Zekavat A. et al. Treponema denticola immunoinhibitory protein induces irreversible G1 arrest in activated human lymphocytes. // Oral Microbiol. Immunol. - 2004. - Vol. 19. - P. 144-149.
56. Leppilahti J.M., Hernandez-Ríos P.A., Gamonal J.A., Tervahartiala Т., Brignardello-Petersen R., Mantyla P., Sorsa Т., Hernández M. Matrix metalloproteinases and myeloperoxidase in gingival crevicular fluid provide site-specific diagnostic value for chronic periodontitis. // J. Clin. Periodontol. - 2014. - Vol. 41(4). - P. 348-56.
57. Livingstone D, Murthy V, Reddy VK, Pillai A. Prosthodontic rehabilitation of a patient with aggressive periodontitis. // BMJ Case Rep. - 2015. - Vol. 5 - P. 2015.
58. Loesche W.J., Grossman N.S. Periodontal disease as a specific, albeit chronic, infection: diagnosis and treatment // Clin. Microbiol. Rev. - 2001. - Vol. 14, №10. - P. 727-752.
59. Miller N., Penaud J., Ambrosini P. et al. Analysis of etiologic factors and periodontal conditions involved with 309 abfractions. // J. Clin. Periodontal. - 2003. - Vol. 30. - P. 828-832.
60. Moutsopoulos NM, Chalmers NI, Barb JJ, Abusleme L, Greenwell-Wild T, Dutzan N, Paster BJ, Munson PJ, Fine DH, Uzel G, Holland SM. Subgingival microbial communities in leukocyte adhesion deficiency and their relationship with local immunopathology. // PLoS Pathog. - 2015. - Vol. 5. - P. 11(3).
61. Noack В., Görgens H., Lorenz K. et al. TLR4 and IL-18 gene variants in chronic periodontitis: impact on disease susceptibility and severity. // Immunol. Invest. - 2009. - Vol. 38, №3-4. - P. 297-310.
62. Page R.C. Critical issues in periodontal research. // J. Dent. Res. - 1995. - Vol. 74. - P. 1118-1128.
63. Palm E, Khalaf H, Bengtsson T. Porphyromonas gingivalis downregulates the immune response of fibroblasts. // BMC Microbiol. - 2013. - Vol. 10. - P. 13-155.
64. Palm E, Khalaf H, Bengtsson T. Suppression of inflammatory responses of human gingival fibroblasts by gingipains from Porphyromonas gingivalis. // Mol Oral Microbiol. - 2014. Vol. 23.
65. Parra В., Slots J. Detection of human viruses in periodontal pockets using Polymerase Chain Reaction. // Oral Microbiol. Immunol. - 1996. - Vol. 11, №5. - P. 289-293 Vandesompele J., De Preter K., Pattyn F. et al. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. // Genome Biol. - 2002. - Vol. 3. - P. 1796-1804.
66. Pirhan D., Atilla G., Emingil G. et al. Effect of MMP-1 promoter polymorphisms on GCF MMP-1 levels and outcome of periodontal therapy in patients with severe chronic periodontitis. // J. Clin. Periodontol. - 2008. - Vol. 35, №10. - P. 862-870.
67. Ram VS, Parthiban, Sudhakar U, Mithradas N, Prabhakar R. Bonebiomarkers in periodontal disease: a review article. // J Clin. Diagn Res. - 2015. Vol. 9(1). - P. 07-10.
68. Sakalauskiene J., Surna A., Ivanauskiene E. et al. Secretory function of neutrophilic leukocytes of the patients with periodontal diseases. // Stomatologija. - 2005. - Vol. 7, №3. - P. 90-94.
69. Salminen A., Gursoy U.K., Paju S., Hyvärinen K., Mäntylä P., Buhlin K., Könönen E., Nieminen MS, Sorsa T, Sinisalo J, Pussinen PJ. Salivary biomarkers of bacterial burden, inflammatory response, and tissue destruction in periodontitis. // J Clin Periodontol. - 2014. - Vol. 41(5). - P. 442-50.
70. Savitri IJ, Ouhara K, Fujita T, Kajiya M, Miyagawa T, Kittaka M, Yamakawa M, Shiba H, Kurihara H. Irsogladine maleate inhibits Porphyromonas gingivalis-mediated expression of toll-like receptor 2 and interleukin-8 in human gingival epithelial cells. // J Periodontal Res. - 2014. - Vol. 20.
71. Savitri IJ, Ouhara K, Fujita T, Kajiya M, Miyagawa T, Kittaka M, Yamakawa M, Shiba H, Kurihara H. Irsogladine maleate inhibits Porphyromonas gingivalis-mediated expression of toll-like receptor 2 and interleukin-8 in human gingival epithelial cells. // J Periodontal Res. - 2014. - Vol. 20.
72. Schaumann T, Kraus D, Winter J, Wolf M, Deschner J, Jäger A. Potential immune modularly role of glycine in oral gingival inflammation. // Clin Dev Immunol. - 2013. - PubMed PMID: 24348681.
73. Schaumann T, Kraus D, Winter J, Wolf M, Deschner J, Jäger A. Potential immune modularly role of glycine in oral gingival inflammation. // Clin Dev Immunol. - 2013. - PubMed PMID: 24348681.
74. Socransky S.S., Haffajee A.D. Periodontal microbial ecology. // Periodontol. 2000. - 2005. - Vol. 38. - С 135-187.
75. Tanner A.C.R., Kent R. Jr., Dyke Van T. et al. Clinical and other risk indicators for early periodontitis in adults. // J. Periodontal. - 2005. - Vol. 76, №4. - P. 573-581.
76. Trivedi S, Lal N, Mahdi AA, Singh B, Pandey S. Association of salivary lipid peroxidation levels, antioxidant enzymes, and chronic periodontitis. // Int J Periodontics Restorative Dent. - 2015. - Vol. 35(2). - P. 14-9.
77. Tsilingaridis G, Yucel-Lindberg T, Modéer T. Altered relationship between MMP-8 and TIMP-2 in gingival crevicular fluid in adolescents with Down's syndrome. // J Periodontal Res. - 2013. - Vol. 48(5). - P. 553-62.
78. Türkoğlu O, Becerik S, Tervahartiala T, Sorsa T, Atilla G, Emingil G. The effect of adjunctive chlorhexidine mouthrinse on GCF MMP-8 and TIMP-1 levels in gingivitis: a randomized placebo-controlled study. // BMC Oral Health. - 2014. - Vol. 20. - P. 14:55.
79. Wan C, Yuan G, Yang J, Sun Q, Zhang L, Zhang J, Zhang L, Chen Z. MMP9 deficiency increased the size of experimentally induced apical periodontitis. // J Endod. - 2014. Vol. 40(5). - P. 658-64.
80. Wohlfahrt JC, Aass AM, Granfeldt F, Lyngstadaas SP, Reseland JE. Sulcus fluid bone marker levels and the outcome of surgical treatment of peri-implantitis. // J Clin Periodontol. - 2014. Vol. 41(4). - P. 424-31.
81. Zhang Y, Li X. Lipopolysaccharide-regulated production of bone sialoprotein and interleukin-8 in human periodontal ligament fibroblasts: the role of toll-like receptors 2 and 4 and the МАРК pathway. // J Periodontal Res. - 2014. - Vol. 22.
82. Zhou Q., Daste Т., Fenton M. et al. Cytokine profiling of macrophages exposed to Porphyromonas gingivalis, its lipopolysaccharide, or its FimA protein. // Infect. Immun. - 2005. - Vol. 73. - P. 935-943
83. Безрукова И.В. Быстропрогрессирующий пародонтит. Этиология. Клиника. Лечение: дис.… д-ра мед. наук. - М., 2001. - 180 с.
84. Безрукова И.В., Грудянов А.И. Агрессивные формы пародонтита. // Руководство для врачей. - М: МИА 2002. - 126 с.
85. Грудянов А.И., Дмитриева Л.А., Максимовский Ю.М. Пародонтология: современное состояние, вопросы и направления научных разработок. // Пародонтология. - 1998. - №3(9). - С. 5-7.
86. Дмитриева Л.А. Современные аспекты клинической пародонтологии. / Л.А. Дмитриева. - М., МЕДпресс, 2001. - 128 с.
87. Иванов С.Ю., Царев В.Н., Чувилкин В.И. и др. Оценка эффективности антибактериальной санации пациентов от возбудителей периимплантитов с помощью молекулярно-генетических методов. // Мед. Вестн. МВД. - 2005. - №1. - С. 8-12.
88. Левин М.Я., Орехова Л.Ю., Антонова И.Н. и др. Иммунологические показатели слюны и крови при воспалительных заболеваниях тканей пародонта. // Пародонтология. - 1999. - №2(12). - С. 10-13.
89. Орехова Л.Ю., Левин М.Я., Антонова И.Н. и др. Местные проявления аутоиммунной реакции при воспалительных заболеваниях пародонта. // Ученые записки. - 2000. - Т. VII, №2. - С. 27-30.
90. Ребриков Д.В. Патент РФ №2451937. Способ диагностики рака молочной железы по уровню РНК интерлейкинов IL-8 и/или IL-18 в плазме крови. Заявка от 07.06.2010. Опубл. 27.05.20102.
91. Уварова Л.В., Еловикова Т.М. Патент РФ №2381746. Способ диагностики степени тяжести пародонтита. Заявка от 17.11.2008. Опубл. 20.02.2010 (прекратил действие).
92. Чуйко А.Н., Уварова Л.В. Метод конечных элементов. Пародонтология. 2008. Т. 46 №1. С. 25-40.
93. Эстрин В.В., Ефанова Е.А., Пухтинская М.Г. Патент РФ №2180119. Способ ранней диагностики сепсиса у новорожденных. Заявка от 21.12.1998. Опубл. 27.02.2002.
| название | год | авторы | номер документа |
|---|---|---|---|
| Способ определения степени гноетечения на пародонте по уровню мРНК гена интерлейкина-8 (IL-8) человека | 2015 |
|
RU2607041C2 |
| СПОСОБ ОПРЕДЕЛЕНИЯ ГЕНОТИПА ЧЕЛОВЕКА ПО ПОЛИМОРФИЗМУ В ГЕНЕ МАТРИКСНОЙ МЕТАЛЛОПРОТЕИНАЗЫ ММР9-1562 С>Т (rs3918242) | 2012 |
|
RU2548811C2 |
| Способ оценки обсемененности пародонта патогенными бактериями с применением полимеразной цепной реакции в реальном времени | 2015 |
|
RU2607046C2 |
| СПОСОБ ОПРЕДЕЛЕНИЯ ГЕНОТИПА ЧЕЛОВЕКА ПО ПОЛИМОРФИЗМУ В ГЕНЕ КОЛЛАГЕНА II ТИПА COL2A1 C>A (RS1635529) | 2012 |
|
RU2518301C1 |
| Способ оценки прогрессирования хронического пародонтита и набор реагентов для его осуществления | 2021 |
|
RU2777783C1 |
| Способ оценки состояния пародонта человека на устойчивость к развитию хронического генерализованного пародонтита на основании количественного определения бактерии-пародонтопротектора Streptococcus sanguinis методом ПЦР-РВ | 2015 |
|
RU2621858C2 |
| Способ оценки состояния пародонта человека на устойчивость к развитию хронического генерализованного пародонтита на основании количественного определения бактерии-пародонтопротектора Veillonella parvula методом ПЦР в реальном времени | 2015 |
|
RU2619172C1 |
| СПОСОБ ОЦЕНКИ ИНТЕНСИВНОСТИ РЕАКЦИИ ВОСПАЛЕНИЯ У БОЛЬНЫХ ХРОНИЧЕСКИМ ПАРОДОНТИТОМ | 2008 |
|
RU2397707C1 |
| Способ включения в группу риска по развитию сахарного диабета типа 2 у больных хроническим пародонтитом | 2017 |
|
RU2671569C1 |
| СПОСОБ ДИАГНОСТИКИ ПАТОЛОГИЧЕСКИХ ИЗМЕНЕНИЙ МИКРОФЛОРЫ ПОЛОСТИ РТА | 2006 |
|
RU2324182C1 |
Настоящее изобретение относится к медицине. Предложен способ диагностики локального иммунного статуса пародонта. Предложенный способ включает измерение уровня мРНК генов TNFα, MMP8 и MMP9 с использованием методов обратной транскрипции и полимеразной цепной реакции в “реальном времени”, с дальнейшим вычислением отношения полученных уровней генетических маркеров. В случае, если отношение уровней маркеров TNFα/MMP8/MMP9 находится в диапазоне 1/(1±0,2)/ (1±0,2), следует диагностировать нормальный статус локального неспецифического иммунитета в обследуемой точке пародонта, в случае, если отношение уровней маркеров выходит за пределы указанного диапазона, следует диагностировать наличие нарушения в работе неспецифического иммунитета. Предложенный метод обеспечивает чувствительную и объективную оценку локального иммунного статуса пародонта, что позволяет оценить риски локального воспаления, и может быть использован в клинико-лабораторной диагностике. 1 табл.
Способ диагностики локального иммунного статуса пародонта, отличающийся тем, что в соскобах из области поражения измеряется уровнь мРНК генов TNFα, ММР8 и ММР9, в качестве метода определения уровня мРНК используются обратная транскрипция и полимеразная цепная реакция «в реальном времени», на основании полученных уровней генетических маркеров вычисляется их отношение, при этом если отношение уровней маркеров TNFα : ММР8 : ММР9 находится в диапазоне 1:(1±0,2):(1±0,2), следует диагностировать нормальный статус локального неспецифичекого иммунитета в обследуемой точке пародонта, а при выходе отношения уровней маркеров за пределы указанного диапазона - наличие нарушения в работе неспецифического иммунитета, что увеличивает риск развития воспалительного поражения пародонта.

