Область изобретения
Настоящее изобретение относится к клеткам и нуклеиновым кислотам, а также к их применению для получения рамнолипидов, и к способу получения рамнолипидов.
Уровень техники
Поверхностно-активные вещества в настоящее время по существу получают на основе базы из петрохимического сырья. Применение поверхностно активных веществ на основе возобновляемого сырья является подходящей альтернативой по причине предсказуемой нехватки петрохимического сырья и возрастающей потребности в продуктах на основе возобновляемого сырья или биоразлагаемых продуктах.
Рамнолипиды состоят из одного (монорамнозиллипид) или двух остатков рамнозы (дирамнозиллипид) и одного или двух остатков 3-гидрокси жирной кислоты (смотрите Handbook of Hydrocarbon and Lipid Microbiology, 2010, Seiten 3037-51). Они имеют поверхностно-активные свойства, которые необходимы для всех видов применения в качестве поверхностно-активного вещества (siehe Leitermann et al., 2009).
Эти липиды в настоящее время получают, применяя изоляты различных патогенных бактерий немутантного типа человека и животных, в частности представителей рода (смотрите Handbook of Hydrocarbon and Lipid Microbiology, 2010, Seiten 3037-51). На самом деле эти организмы для получения способны вызывать заболевания, уменьшающие приемлемость для потребителя получаемых обычным образом рамнолипидов в значительной степени. Более того, более высокие требования безопасности приводят также к повышению затрат на производство из-за увеличения капитальных расходов и возможности дополнительных стадий обработки.
Хотя в некоторой степени высокие титры продукта, а также пространство-время и/или выходы углерода могут быть достигнуты с помощью этих организмов для получения, это требует применение растительных масел в качестве единственного или сосубстратов (смотрите Handbook of Hydrocarbon and Lipid Microbiology, 2010, Seiten 3037-51). Растительные масла, однако, являются сравнительно дорогостоящим сырьем по сравнению с другими источниками углерода, таким как, например, глюкоза, сахароза или полисахариды, такие как, например, крахмал, целлюлоза и гем и целлюлоза, глицерин, СО, CO2 или CH4. Более того, рамнолипиды сами по себе отличаются их поверхностно-активным характером, состоящим в том, что они склонны к бурному пенообразованию в ходе ферментационных процессов. Это происходит, в частности, когда применяются липофильные субстраты. Эта проблема заметно уменьшается при применении водорастворимых субстратов, таких как например, глюкоза, сахароза, полисахариды (крахмал, целлюлоза, гем и целлюлоза) или глицерин.
Наконец, на свойства рамнолипидов, получаемых из изолятов немутантного типа, можно влиять лишь в ограниченной степени. До настоящего времени влияние осуществлялось исключительно через оптимизацию управления процессом (значение рН, подача кислорода, состав среды, стратегия подпитки, подача азота, температура, выбор субстрата и т.д.). Однако было бы желательно очень специфично влиять на определенные свойства продукта, такие как, например, соотношение различных видов рамнолипидов (число остатков рамнозы и 3-гидроксижирной кислоты) или длина цепи и степень насыщенности остатков 3-гидроксижирной кислоты, чтобы контролировать свойства продукта, важные для применения.
Рамнолипиды, если они применяются в большей степени в качестве поверхностно-активных веществ для ухода за домом, очищения, косметических средств, технологии производства пищевых продуктов, фармацевтических средств, защиты растений и других применений, должны применяться конкурентоспособным образом с применяемыми на сегодняшний день поверхностно-активными веществами. Имеется большой объем химических средств, которые могут быть получены с очень низкими затратами, без очевидного риска для здоровья потребителя и с ясно определенными и модулируемыми характеристиками продукта. Поэтом должна быть возможность получения рамнолипидов с таким низкими затратами, как только это возможно, без риска для здоровья потребителя и с определенными свойствами, настолько, насколько это возможно.
Хотя рамнолипиды уже были получены в организмах GRAS (в общем безопасные) с применением обычных источников углерода, таких как, например, глюкоза или глицерин, в этом случае получали исключительно монорамнозиллипиды (Ochsner et at. Appl. Environ. Microbiol. 1995. 61(9):3503-3506).
С другой стороны Cha et al. в Bioresour Technol. 2008. 99(7):2192-9 описали получение монорамнозиллипидов из соевого масла в Р. putida путем введения генов Gene rhIA и rhIB из Pseudomonas aeruginosa.
Поэтому возросла потребность в не дорогостоящем и безопасном с точки зрения здоровья получении моно- и дирамнозиллипидов, имеющих определенные и модулируемые свойства. Эта модуляция может осуществляться, например, посредством сбалансированной подачи отдельных ферментных активностей, которые уменьшают обогащение монорамнозиллипидами. Такая модуляция, однако, может также осуществляться, например, посредством применения ферментов, имеющих определенные свойства, например, в отношении специфичности к субстрату, и, таким образом, например, меняется длина цепи гидроксижирных кислот, включенных в рамнолипиды. Поэтому задачей настоящего изобретения является обеспечение возможности получения рамнолипидов из легко доступных источников углерода, с применением безопасных хозяев для получения.
Описание изобретения
Неожиданно было обнаружено, что клетки и способы, описанные ниже, в которых эти клетки применяются, вносят вклад в решение поставленной задачи настоящего изобретения.
Поэтому настоящее изобретение относится к клеткам, которые способны формировать рамнолипиды и по сравнению с их немутированным типом имеют по меньшей мере одну повышенную активность генного продукта гомологов генных продуктов rhIA, rhIB и rhIC.
Настоящее изобретение, кроме того, относится к способу получения рамнолипидов с применением вышеупомянутых клеток в качества биокатализатора и простых источников углерода.
Преимуществом настоящего изобретения является то, что могут применяться организмы, которые являются непатогенными и простыми для применения.
Другим преимуществом настоящего изобретения является то, что применение масла или сосубстрата является необязательным.
Другим преимуществом является то, что с помощью настоящего изобретения могут быть получены рамнолипиды, имеющие определенные и модулируемые свойства.
Другое преимущество настоящего изобретения состоит в том, что могут быть получены дирамнозиллипиды.
Другое преимущество состоит в том, что могут быть получены рамнолипиды с более высоким пространство-временем и выходом углерода, чем при применении клеток без повышения их активностей.
Вклад в достижение поставленной в начале задачи, может быть сделан клеткой, предпочтительно выделенной клеткой, которая способна образовывать по меньшей мере один рамнолипид общей формулы (I) или его соли,
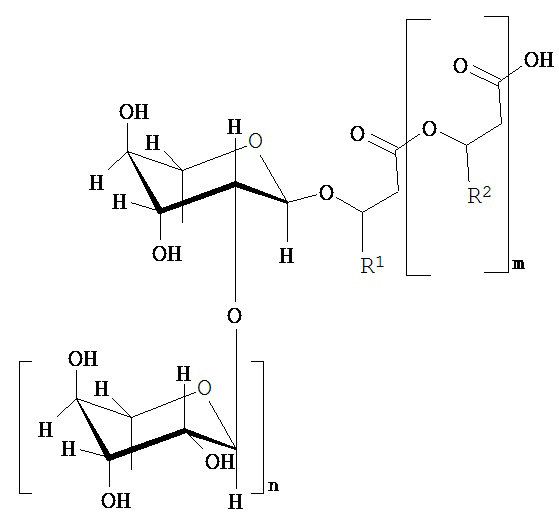
где
m равно 2, 1 или 0, в частности 1 или 0,
n равно 1 или 0, в частности 1,
R1 и R2 независимо друг от друга представляют собой идентичные или различные органические радикалы, имеющие от 2 до 24, предпочтительно от 5 до 13 атомов углерода, в частности необязательно разветвленный, необязательно замещенный, в частности гидрокси-замещенный, необязательно ненасыщенный, в частности, моно-, ди- или триненасыщенный, алкильный радикал, предпочтительно выбранный из группы, состоящей из пентенила, гептенила, ноненила, ундеценила и тридеценила и (СН2)0-СН3со=1-23, предпочтительно 4-12,
отличающейся тем, что она была генетически модифицирована таким образом, что, по сравнению с немутированным типом, она имеет повышенную активность по меньшей мере одного из ферментов E1, E2 и Е3, где фермент E1 способен катализировать превращение 3-гидроксиалканоил-АСР через 3-гидроксиалканоил-3-гидроксиалкановую кислоту-АСР в гидроксиалканоил-3-гидроксиалкановую кислоту, фермент Е2 представляет собой рамнозилтрансферазу I и способен катализировать превращение dTDP-рамнозы и 3-гидроксиалканоил-3-гидроксиалканоата в α-L-рамнопиранозил-3-гидроксиалканоил-3-гидроксиалканоат, и фермент Е3 представляет собой рамнозилтрансферазу II и способен катализировать превращение dTDP-рамнозы и α-L-рамнопиранозил-3-гидроксиалканоил-3-гидроксиалканоат в α-L-рамнопиранозил-(1-2)-α-L-рамнопиранозил-3-гидроксиалканоил-3-гидроксиалканоат, где эти ферменты E1, Е2 и Е3 предпочтительно выбираются из группы, состоящей из
по меньшей мере одного фермента E1, выбранного из:
фермент E1a, имеющий полипептидную последовательность Seq ID №2 или имеющий полипептидную последовательность, в которой до 25%, предпочтительно до 20%, особенно предпочтительно до 15%, особенно до 10, 9, 8, 7, 6, 5, 4, 3, 2, 1% аминокислотных остатков модифицированы по сравнению со ссылочной последовательностью Seq ID №2 посредством делеции, инсерции, замещения или их комбинации, и который все еще имеет по меньшей мере 10%, предпочтительно 50%, особенно предпочтительно 80%, особенно более чем 90% от ферментативной активности фермента, имеющего ссылочную последовательность Seq ID №2, где ферментативная активность фермента E1a, как понимается, означает способность предпочтительно превращать 3-гидроксидеканоил-АСР через 3-гидроксидеканоил-3-гидроксидекановой кислоты-АСР в гидроксидеканоил-3-гидроксидекановую кислоту,
фермент E1b, имеющий полипептидную последовательность Seq ID №18 или имеющий полипептидную последовательность, в которой до 25%, предпочтительно до 20%, особенно предпочтительно до 15%, особенно до 10, 9, 8, 7, 6, 5, 4, 3, 2, 1% аминокислотных остатков модифицированы по сравнению со ссылочной последовательностью Seq ID №18 посредством делеции, инсерции, замещения или их комбинации, и который все еще имеет по меньшей мере 10%, предпочтительно 50%, особенно предпочтительно 80%, особенно более чем 90% от ферментативной активности фермента, имеющего ссылочную последовательность Seq ID №18, где ферментативная активность фермента E1b, как понимается, означает способность предпочтительно превращать 3-гидрокситетрадеканоил-АСР через 3-гидрокситетрадеканоил-3-гидрокситетрадекановую кислоту-АСР в гидрокситетрадеканоил-3-гидрокситетрадека новую кислоту,
фермент E1c, имеющий полипептидную последовательность Seq ID №78 или имеющий полипептидную последовательность, в которой до 25%, предпочтительно до 20%, особенно предпочтительно до 15%, особенно до 10, 9, 8, 7, 6, 5, 4, 3, 2, 1% аминокислотных остатков модифицированы по сравнению со ссылочной последовательностью Seq ID №78 посредством делеции, инсерции, замещения или их комбинации, и который все еще имеет по меньшей мере 10%, предпочтительно 50%, особенно предпочтительно 80%, особенно более чем 90% от ферментативной активности фермента, имеющего ссылочную последовательность Seq ID №78, где ферментативная активность фермента E1c, как понимается, означает способность предпочтительно превращать 3-гидрокситетрадеканоил-АСР через 3-гидрокситетрадеканоил-3-гидрокситетрадекановую кислоту-АСР в гидрокситетрадеканоил-3-гидрокситетрадекановую кислоту,
фермент E1d, имеющий полипептидную последовательность Seq ID №80 или имеющий полипептидную последовательность, в которой до 25%, предпочтительно до 20%, особенно предпочтительно до 15%, особенно до 10, 9, 8, 7, 6, 5, 4, 3, 2, 1% аминокислотных остатков модифицированы по сравнению со ссылочной последовательностью Seq ID №80 посредством делеции, инсерции, замещения или их комбинации, и который все еще имеет по меньшей мере 10%, предпочтительно 50%, особенно предпочтительно 80%, особенно более чем 90% от ферментативной активности фермента, имеющего ссылочную последовательность Seq ID №80, где ферментативная активность фермента Е1d, как понимается, означает способность предпочтительно превращать 3-гидрокситетрадеканоил-АСР через 3-гидрокситетрадеканоил-3-гидрокситетрадекановую кислоту-АСР в гидрокситетрадеканоил-3-гидрокситетрадекановую кислоту, и
фермент E1e, имеющий полипептидную последовательность Seq ID №82 или имеющий полипептидную последовательность, в которой до 25%, предпочтительно до 20%, особенно предпочтительно до 15%, особенно до 10, 9, 8, 7, 6, 5, 4, 3, 2, 1% аминокислотных остатков модифицированы по сравнению со ссылочной последовательностью Seq ID №82 посредством делеции, инсерции, замещения или их комбинации, и который все еще имеет по меньшей мере 10%, предпочтительно 50%, особенно предпочтительно 80%, особенно более чем 90% от ферментативной активности фермента, имеющего ссылочную последовательность Seq ID №82, где ферментативная активность фермента E1e, как понимается, означает способность предпочтительно превращать 3-гидрокситетрадеканоил-АСР через 3-гидрокситетрадеканоил-3-гидрокситетрадекановую кислоту-АСР в гидрокситетрадеканоил-3-гидрокситетрадекановую кислоту,
по меньшей мере одного фермента Е2 с полипептидной последовательностью, выбранного из:
фермент Е2а, имеющий полипептидную последовательность Seq ID №4 или имеющий полипептидную последовательность, в которой до 25%, предпочтительно до 20%, особенно предпочтительно до 15%, особенно до 10, 9, 8, 7, 6, 5, 4, 3, 2, 1% аминокислотных остатков модифицированы по сравнению со ссылочной последовательностью Seq ID №4 посредством делеции, инсерции, замещения или их комбинации, и который все еще имеет по меньшей мере 10%, предпочтительно 50%, особенно предпочтительно 80%, особенно более чем 90% от ферментативной активности фермента, имеющего ссылочную последовательность Seq ID №4, где ферментативная активность фермента Е2а, как понимается, означает способность предпочтительно превращать dTDP-рамнозу и 3-гидроксидеканоил-3-гидроксиддекановую кислоту в α-L-рамнопиранозил-3-гидроксидеканоил-3-гидроксидекановую кислоту,
фермент E2b, имеющий полипептидную последовательность Seq ID №20 или имеющий полипептидную последовательность, в которой до 25%, предпочтительно до 20%, особенно предпочтительно до 15%, особенно до 10, 9, 8, 7, 6, 5, 4, 3, 2, 1% аминокислотных остатков модифицированы по сравнению со ссылочной последовательностью Seq ID №20 посредством делеции, инсерции, замещения или их комбинации, и который все еще имеет по меньшей мере 10%, предпочтительно 50%, особенно предпочтительно 80%, особенно более чем 90% от ферментативной активности фермента, имеющего ссылочную последовательность Seq ID №20, где ферментативная активность фермента E2b, как понимается, означает способность предпочтительно превращать dTDP-рамнозу и 3-гидрокситетрадеканоил-3-гидрокситетрадекановую кислоту в α-L-рамнопиранозил-3-гидрокситетрадеканоил-3-гидрокситетрадекановую кислоту,
фермент Е2с, имеющий полипептидную последовательность Seq ID №84 или имеющий полипептидную последовательность, в которой до 25%, предпочтительно до 20%, особенно предпочтительно до 15%, особенно до 10, 9, 8, 7, 6, 5, 4, 3, 2, 1% аминокислотных остатков модифицированы по сравнению со ссылочной последовательностью Seq ID №84 посредством делеции, инсерции, замещения или их комбинации, и который все еще имеет по меньшей мере 10%, предпочтительно 50%, особенно предпочтительно 80%, особенно более чем 90% от ферментативной активности фермента, имеющего ссылочную последовательность Seq ID №84, где ферментативная активность фермента Е2с, как понимается, означает способность предпочтительно превращать dTDP-рамнозу и 3-гидрокситетрадеканоил-3-гидрокситетрадекановую кислоту в α-L-рамнопиранозил-3-гидрокситетрадеканоил-3-гидрокситетрадекановую кислоту,
фермент E2d, имеющий полипептидную последовательность Seq ID №86 или имеющий полипептидную последовательность, в которой до 25%, предпочтительно до 20%, особенно предпочтительно до 15%, особенно до 10, 9, 8, 7, 6, 5, 4, 3, 2, 1% аминокислотных остатков модифицированы по сравнению со ссылочной последовательностью Seq ID №86 посредством делеции, инсерции, замещения или их комбинации, и который все еще имеет по меньшей мере 10%, предпочтительно 50%, особенно предпочтительно 80%, особенно более чем 90% от ферментативной активности фермента, имеющего ссылочную последовательность Seq ID №86, где ферментативная активность фермента Е2d, как понимается, означает способность предпочтительно превращать dTDP-рамнозу и 3-гидрокситетрадеканоил-3-гидрокситетрадекановую кислоту в α-L-рамнопиранозил-3-гидрокситетрадеканоил-3-гидрокситетрадекановую кислоту, и фермент Е2е, имеющий полипептидную последовательность Seq ID №88 или имеющий полипептидную последовательность, в которой до 25%, предпочтительно до 20%, особенно предпочтительно до 15%, особенно до 10, 9, 8, 7, 6, 5, 4, 3, 2, 1% аминокислотных остатков модифицированы по сравнению со ссылочной последовательностью Seq ID №88 посредством делеции, инсерции, замещения или их комбинации, и который все еще имеет по меньшей мере 10%, предпочтительно 50%, особенно предпочтительно 80%, особенно более чем 90% от ферментативной активности фермента, имеющего ссылочную последовательность Seq ID №88, где ферментативная активность фермента Е2е, как понимается, означает способность предпочтительно превращать dTDP-рамнозу и 3-гидрокситетрадеканоил-3-гидрокситетрадекановую кислоту в α-L-рамнопиранозил-3-гидрокситетрадеканоил-3-гидрокситетрадекановую кислоту, и
по меньшей мере одного фермента Е3, выбранного из:
фермент Е3а, имеющий полипептидную последовательность Seq ID №6 или имеющий полипептидную последовательность, в которой до 25%, предпочтительно до 20%, особенно предпочтительно до 15%, особенно до 10, 9, 8, 7, 6, 5, 4, 3, 2, 1% аминокислотных остатков модифицированы по сравнению со ссылочной последовательностью Seq ID №6 посредством делеции, инсерции, замещения или их комбинации, и который все еще имеет по меньшей мере 10%, предпочтительно 50%, особенно предпочтительно 80%, особенно более чем 90% от ферментативной активности фермента, имеющего ссылочную последовательность Seq ID №6, где ферментативная активность фермента Е3а, как понимается, означает способность предпочтительно превращать dTDP-рамнозу и α-L-рамнопиранозил-3-гидроксидеканоил-3-гидроксидекановую кислоту в α-L-рамнопиранозил-(1-2)-α-L-рамнопиранозил-3-гидроксидеканоил-3-гидроксидекановую кислоту, фермент E3b, имеющий полипептидную последовательность Seq ID №22 или имеющий полипептидную последовательность, в которой до 25%, предпочтительно до 20%, особенно предпочтительно до 15%, особенно до 10, 9, 8, 7, 6, 5, 4, 3, 2, 1% аминокислотных остатков модифицированы по сравнению со ссылочной последовательностью Seq ID №22 посредством делеции, инсерции, замещения или их комбинации, и который все еще имеет по меньшей мере 10%, предпочтительно 50%, особенно предпочтительно 80%, особенно более чем 90% от ферментативной активности фермента, имеющего ссылочную последовательность Seq ID №22, где ферментативная активность фермента E3b, как понимается, означает способность предпочтительно превращать dTDP-рамнозу и α-L-рамнопиранозил-3-гидрокситетрадеканоил-3-гидрокситетрадекановую кислоту в α-L-рамнопиранозил-(1-2)-α-L-рамнопиранозил-3-гидрокситетрадеканоил-3-гидрокситетрадекановую кислоту,
фермент Е3с, имеющий полипептидную последовательность Seq ID №90 или имеющий полипептидную последовательность, в которой до 25%, предпочтительно до 20%, особенно предпочтительно до 15%, особенно до 10, 9, 8, 7, 6, 5, 4, 3, 2, 1% аминокислотных остатков модифицированы по сравнению со ссылочной последовательностью Seq ID №90 посредством делеции, инсерции, замещения или их комбинации, и который все еще имеет по меньшей мере 10%, предпочтительно 50%, особенно предпочтительно 80%, особенно более чем 90% от ферментативной активности фермента, имеющего ссылочную последовательность Seq ID №90, где ферментативная активность фермента Е3с, как понимается, означает способность предпочтительно превращать dTDP-рамнозу и α-L-рамнопиранозил-3-гидрокситетрадеканоил-3-гидрокситетрадекановую кислоту в α-L-рамнопиранозил-(1-2)-α-L-рамнопиранозил-3-гидрокситетрадеканоил-3-гидрокситетрадекановую кислоту, и
фермент E3d, имеющий полипептидную последовательность Seq ID №92 или имеющий полипептидную последовательность, в которой до 25%, предпочтительно до 20%, особенно предпочтительно до 15%, особенно до 10, 9, 8, 7, 6, 5, 4, 3, 2, 1% аминокислотных остатков модифицированы по сравнению со ссылочной последовательностью Seq ID №92 посредством делеции, инсерции, замещения или их комбинации, и который все еще имеет по меньшей мере 10%, предпочтительно 50%, особенно предпочтительно 80%, особенно более чем 92% от ферментативной активности фермента, имеющего ссылочную последовательность Seq ID №92, где ферментативная активность фермента E3, как понимается, означает способность предпочтительно превращать dTDP-рамнозу и α-L-рамнопиранозил-3-гидрокситетрадеканоил-3-гидрокситетрадекановую кислоту в α-L-рамнопиранозил-(1-2)-α-L-рамнопиранозил-3-гидрокситетрадеканоил-3-гидрокситетрадекановую кислоту.
Для общего сведения смотрите Фиг.1.
Термин "немутантный тип" клетки согласно настоящему изобретению означает клетку, геном которой находится в том состоянии, в котором он сформировался естественным образом в ходе эволюции. Этот термин применяется как в отношении клетки в целом, так и в отношении отдельных генов. Термин "немутантный тип" поэтому в частности не включает те клетки или те гены, генные последовательности которых были модифицированы по меньшей мере частично с помощью рекомбинантных способов.
Термин "рамнолипид", как понимается, в контексте настоящего изобретения означает соединение общей формулы (I) или его соли.
Очевидно, что активности, указанные выше для ферментов E1a-E3b являются лишь конкретными примерами из широкого спектра активностей вышеуказанных ферментов; соответствующая упомянутая активность является такой, для которой надежный способ измерения доступен в случае данного фермента. Таким образом, очевидно, что фермент, который имеет субстрат с неразветвленными насыщенными С10-алкильными радикалами, даже, хотя необязательно, при пониженной активности будет превращать эти субстраты, которые содержат С6- или С16-алкильные радикалы, которые при необходимости могут быть разветвленными или ненасыщенными.
Термин "повышенная активность фермента", как понимается, предпочтительно означает повышенную внутриклеточную активность.
Описанные варианты выполнения настоящего изобретения для повышения ферментативной активности в летках, применяются как для повышения активности ферментов E1-Е3, так и для всех далее упомянутых ферментов, активность которых может при необходимости быть повышена.
На самом деле, повышение ферментативной активности может быть достигнуто повышением числа копий генной последовательности или генных последовательностей, которые кодируют фермент, с применением сильного промотора или улучшенного сайта связывания рибосом, уменьшая негативную регуляцию генной экспрессии, например, посредством регуляторов транскрипции, или усиливая положительную регуляцию генной экспрессии, модифицируя использование кодона гена, различными путями повышая период полураспада мРНК или фермента, модифицируя регуляцию экспрессии гена или применяя ген или аллель, которые кодируют соответствующий фермент, имеющий повышенную активность, или при необходимости комбинируя эти средства. Согласно настоящему изобретению получают генетически модифицированные клетки, например, посредством трансформации, трансдукции, конъюгации или комбинации этих способов, применяя вектор, который содержит желательный ген, аллель этого гена или их части и при необходимости содержит промотор, делающий возможной экспрессию гена. Гетерологичная экспрессия, в частности, достигается интеграцией гена или аллелей в хромосому клетки или экстрахромосомно реплицирующийся вектор.
B DE-A-10031999 приводится общий обзор возможностей повышения ферментативной активности в клетках, как показано в качестве примера на пируват карбоксилазе, указанный документ включен в настоящую заявку в качестве ссылки, и его содержание в отношении возможностей повышения ферментативной активности в клетках, формирует часть описания настоящего изобретения.
Экспрессия ферментов или генов, указанных выше и далее, определяется с помощью одно- и двухмерного гелевого разделения белков и последующей оптической идентификации концентрации белка в геле, с применением соответствующего аналитического программного обеспечения. Если повышение ферментативной активности основывается исключительно на повышении экспрессии соответствующего гена, количественное определение повышения ферментативной активности может быть определено простым образом путем сравнения одно- или двухмерных разделений белков между клеткой немутантного типа и генетически модифицированной клеткой. Обычный способ получения белковых гелей в случае коринеформных бактерий и для идентификации белков представляет собой методику, описанную Hermann et al. (Electrophoresis, 22: 1712.23 (2001)). Концентрация белка может быть проанализирована подобным образом с помощью Вестерн-блот гибридизации с применением антитела, специфичного для определяемого белка (Sambrook et al., Molecular Cloning: a laboratory manual, 2nd Ed. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y. USA, 1989) и последующего оптического анализа с применением соответствующего программного обеспечения для определения концентрации (Lohaus und Meyer (1989) Biospektrum, 5: 32-39; Lottspeich (1999) Angewandte Chemie 111: 2630-2647). Активность ДНК-связывающих белков может быть измерена посредством анализа сдвига полосы ДНК (также называется задержкой в геле) (Wilson et al. (2001) Journal of Bacteriology, 183: 2151-2155). Действие ДНК-связывающих белков на экспрессию других генов может быть обнаружено различными хорошо описанными способами анализа репортерного гена (Sambrook et al., Molecular Cloning: a laboratory manual, 2nd Ed. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y. USA, 1989). Внутриклеточные ферментативные активности могут быть определены в соответствии с различными описанными способами (Donahue et al. (2000) Journal of Bacteriology 182 (19): 5624-5627; Ray et al. (2000) Journal of Bacteriology 182 (8): 2277-2284; Freedberg et al. (1973) Journal of Bacteriology 115 (3): 816-823). Если в следующих вариантах выполнения настоящего изобретения не указано практических способов определения активности определенного фермента, определения повышения ферментативной активности, а также определения уменьшения ферментативной активности, они предпочтительно осуществляются с помощью способов, описанных в Hermann et al., Electophoresis, 22: 1712-23 (2001), Lohaus et al., Biospektrum 5 32-39 (1998), Lottspeich, Angewandte Chemie 111: 2630-2647 (1999) и Wilson et al., Journal of Bacteriology 183: 2151-2155 (2001).
Если повышение ферментативной активности сопровождается мутацией эндогенного гена, такие мутации могут быть получены случайным образом, либо обычными способами, такими как, например, УФ-облучение, или с помощью мутагенных химических веществ, или селективно посредством способов генной инженерии, как например делеция (делеции), инсерция (инсерции) и/или нуклеотидный обмен (обмены). Модифицированные клетки получают с помощью этих мутаций. Особенно предпочтительными мутантами ферментов являются, в частности, также те ферменты, которые более не ингибируются обратной связью, продуктом или субстратом, или ингибируются по меньшей мере в меньшей степени по сравнению с ферментом немутантного типа.
Если повышение ферментативной активности сопровождается повышением синтеза фермента, число копий соответствующих генов увеличивается, или промотор и область регуляции или сайт связывания рибосом, который расположен в 3’-5’ направлении выше структурного гена, являются мутированными. Экспрессионные кассеты, которые встраиваются в 3’-5’ направлении выше структурного гена действуют таким же образом. Кроме того, посредством индуцируемых промоторов возможно повысить экспрессию в любой желательный момент времени. Кроме того, однако, также "энхансеры" могут играть роль регуляторных последовательностей для гена фермента, которые подобным образом приводят к повышенной генной экспрессии посредством улучшенного взаимодействия между РНК-полимеразой и ДНК. В результате пролонгации периода полураспада мРНК экспрессия подобным образом улучшается. Кроме того, путем предотвращения разложения ферментного белка ферментативная активность подобным образом повышается. Гены или генные конструкции присутствуют либо в плазмидах, имеющих различное число копий или интегрированных и амплифицированных в хромосоме. Кроме того, сверхэкспрессия упомянутых генов может быть достигнута посредством модификации состава среды и управления культурой. Специалист в данной области техники найдет руководство для этого, среди прочего, в Martin et al. (Bio/Technology 5, 137-146 (1987)), в Guerrero et al. (Gene 138, 35-41 (1994)), Tsuchiya und Morinaga (Bio/Technology 6, 428-430 (1988)), в Eikmanns et al. (Gene 102, 93-98 (1991)), в ЕР-А-0 472 869, в US 4,601,893, в Schwarzer und Puhler (Bionechnology 9, 84-87 (1991), в Reinscheid et al. (Applied and Environmental Microbiology 60, 126-132 (1994)), в LaBarre et al. (Journal of Bacteriology 175, 1001-1007 (1993)), в WO-A-96/15246, в Malumbres et al. (Gene 134, 15-24 (1993)), в JP-A-10-229891, в Jensen und Hammer (Biotechnology and Bioengineering 58, 191-195 (1998)) и в известных учебниках по генетике и молекулярной биологии. Средства, описанные выше, образом, подобным мутациям, приводят к генетически модифицированным клеткам.
Эписомальные плазмиды, например, применяются для повышения экспрессии соответствующих генов. Подходящие плазмиды или вектора, в принципе, представляют собой все варианты, доступные для этой цели специалистам в данной области техники. Такие плазмиды и вектора могут быть получены, например, у компаний Novagen, Promega, New England Biolabs, Clontech или Gibco BRL Другие предпочтительные плазмиды и вектора могут быть обнаружены в: Glover, D. М. (1985) DNA cloning: a practical approach, Vol.l-lll, IRL Press Ltd., Oxford; Rodriguez, R.L. und Denhardt, D. Т (eds) (1988) Vectors: a survey of molecular cloning vectors and their uses, 179-204, Butterworth, Stoneham; Goeddel, D. V. (1990) Systems for heterologous gene expression, Methods Enzymol. 185, 3-7; Sambrook, J.; Fritsch, E.F. und Maniatis, T. (1989), Molecular cloning: a laboratory manual, 2nd ed., Cold Spring Harbor Laboratory Press, New York.
Плазмидный вектор, который содержит ген, подлежащий амплификации, затем превращается в желательный штамм посредством конъюгации или трансформации. Способ конъюгации описывается, например, в Schafer et al., Applied and Environmental Microbiology 60: 756-759 (1994). Способ трансформации описывается, например, в Thierbach et al., Applied Microbiology and Biotechnology 29: 356-362 (1988), Dunican und Shivnan, Bio/Technology 7: 1067-1070 (1989) и Tauch et al., FEMS Microbiology Letters 123: 343-347 (1994). После гомологической рекомбинации посредством события „cross-over", полученный штамм содержит по меньшей мере две копии рассматриваемого.
Под выражением, применяемым выше и в следующих вариантах выполнения настоящего изобретения "активность фермента Ех повышенная по сравнению с его немутантным типом" предпочтительно всегда понимается активность соответствующего фермента Ех, повышенная по меньшей мере в 2 раза, особенно предпочтительно по меньшей мере в 10 раз, более предпочтительно по меньшей мере в 100 раз, еще более предпочтительно в 1.000 раз и по меньшей мере предпочтительно в 10.000 раз. Кроме того, клетка согласно настоящему изобретению, которая имеет "повышенную активность фермента Ех по сравнению с его немутантным типом", в частности также содержит клетку, немутантный тип которой не имеет активности или имеет по меньшей мере не обнаруживаемую активность этого фермента Ех и который показывает обнаруживаемую активность этого фермента Ех только после повышения ферментативной активности, например, посредством сверхэкспрессии. В этом контексте термин "сверхэкспрессия" или выражение, применяемое в следующих вариантах выполнения настоящего изобретения "повышенная экспрессия", также охватывает случай, когда сходная клетка, например, клетка немутантного типа, не показывает экспрессию или по меньшей мере не показывает обнаруживаемую экспрессию, и обнаруживаемый синтез фермента Ех индуцируется только рекомбинантными способами.
Изменения аминокислотных остатков данной полипептидной последовательности, которые не приводят к существенным изменениям свойств и функции данного полипептида, известны специалистам в данной области техники. Таким образом, например, консервативные аминокислоты могут взаимно заменяться; примерами таких подходящих аминокислотных замещений являются: Ala на Ser; Arg на Lys; Asn на Gln или His; Asp на Glu; Cys на Ser; Gln на Asn; Glu на Asp; Gly на Pro; His на Asn или Gln; Ile на Leu или Val; Leu на Met или Val; Lys на Arg или Gln или Glu; Met на Leu или Ile; Phe на Met или Leu или Tyr; Ser на Thr; Thr на Ser; Trp на Tyr; Tyr на Trp или Phe; Val на Ile или Leu. Подобным образом известно, что изменения, особенно на N- или С-концах полипептида, в виде, например, аминокислотных вставок или делеций, часто не оказывают существенного влияния на функцию полипептида.
Активность фермента может быть определена путем разрушения клеток, которые содержат эту активность, образом, известным специалистам в данной области техники, например, с помощью шаровой мельницы, френч-пресса или ультразвукового дезинтегратора, и последующего разделения клеток, остатков клеток и средств разделения, как например, стеклянные шарики, путем центрифугирования в течение 10 минут при 13.000 оборотах в минуту и 4°С. Применяя полученный свободный от клеток неочищенный экстракт затем может быть осуществлен ферментный анализ с последующим LC-ESI-MS обнаружением продуктов. Альтернативно, фермент может быть обогащен образом, известным специалистам в данной области техники, хроматографическими способами (как например, аффинная хроматография на основе никель-нитрилтриуксусной кислоты, аффинной хроматографии на основе стрептавидина, хроматографии на основе гель-фильтрации и ион-обменной хроматографии), или еще очищен до гомогенности.
Активность фермента Е1 затем определяется с применением образцов, описанных как получено выше, следующим образом: Стандартный образец содержит 100 мкМ Е. coli АСР, 1 мМ β-Меркаптоэтанола, 200 мкМ малонил-коэнзима А, 40 мкМ октанил-коэнзима А (для E1a) или додеканоил-коэнзима А (для E1b), 100 мкМ NADPH, 2 мкг Е. coli FabD, 2 мкг Mycobacterium tuberculosis FabH, 1 мкг Е. coli FabG, 0,1 М натрий-фосфатного буфера, рН 7,0, и 5 мкг фермента E1 с конечным объемом 120 мкл. АСР, β-меркаптоэтаноли натрий-фосфатный буфер проинкубировали в течение 30 минут при 37°С, чтобы полностью восстановить АСР. Реакцию запустили добавлением фермента E1. Реакцию остановили, применяя 2 мл воды, которая была подкислена с помощью HCl до рН 2,0, и затем дважды экстрагировали 2 мл хлороформ/метанол (2:1 (об.:об.)). Разделение фаз осуществили с помощью центрифугирования (16.100 g, 5 минут, комнатная температура). Нижнюю органическую фазу удалили, испарение завершили в вакуумной центрифуге, и остаток растворили в 50 мкл этанола. Нерастворившиеся составляющие осадили с помощью центрифугирования (16.100 g, 5 минут, комнатная температура), и образец проанализировали посредством LC-ESI-MS. Идентификацию продуктов осуществили с помощью анализа соответствующих массовых следов и MS2-спектра.
Активность фермента Е2 затем определяется с применением образцов, описанных как получено выше, следующим образом: стандартный образец может состоять из 185 мкл 10 мМ Tris-HCl (рН 7,5), 10 мкл 125 мМ dTDP-рамнозы и 50 мкл неочищенного экстракта белка (около 1 мг общего белка) или очищенного белка в растворе (5 мкг очищенного белка). Реакцию запустили добавлением 10 мкл 10 мМ этанольного раствора 3-гидроксидеканоил-3-гидроксидекановой кислоты (для Е2а) или 3-гидрокситетрадеканоил-3-гидрокситетрадекановой кислоты (для E2b) и инкубировали в течение 1 часа при 30°С при перемешивании (600 оборотов в минуту). Затем реакционную смесь обработали 1 мл ацетона. Нерастворившиеся составляющие осадили с помощью центрифугирования (16.100 g, 5 минут, комнатная температура), и образец проанализировали посредством LC-ESI-MS. Идентификацию продуктов осуществили с помощью анализа соответствующих массовых следов и MS2-спектра.
Активность фермента Е3 затем определяется с применением образцов, описанных как получено выше, следующим образом: стандартный образец может состоять из 185 мкл 10 мМ Tris-HCl (pH 7,5), 10 мкл 125 мМ dTDP-рамнозы и 50 мкл неочищенного экстракта белка (около 1 мг общего белка) или очищенного белка в растворе (5 мкг очищенного белка). Реакцию запустили добавлением 10 мкл 10 мМ этанольного раствора α-L-рамнопиранозил-3-гидроксидеканоил-3-гидрокси декановой кислоты (для Е3а) или α-L-рамнопиранозил-3-гидрокситетрадеканоил-3-гидрокси тетрадекановой кислоты (для E3b) и инкубировали в течение 1 часа при 30°С при перемешивании (600 оборотов в минуту). Затем реакционную смесь обработали 1 мл ацетона. Нерастворившиеся составляющие осадили с помощью центрифугирования (16.100 g, 5 минут, комнатная температура), и образец проанализировали посредством LC-ESI-MS. Идентификацию продуктов осуществили с помощью анализа соответствующих массовых следов и MS2-спектра.
Клетками согласно настоящему изобретению предпочтительно являются те клетки, которые имеют повышенные активности следующих комбинаций ферментов:
Е1, Е2, Е3, Е1Е2, E1E3, Е2Е3 И E1E2E3,
где комбинация
Е2, Е2Е3 и E1E2E3, особенно E1E2E3
особенно предпочтительна.
В предпочтительном варианте выполнения настоящего изобретения клетка согласно настоящему изобретению представляет собой клетку, имеющую повышенную активность комбинации ферментов E1E2E3, n предпочтительно =1.
Клетки согласно настоящему изобретению могут быть прокариотическими или эукариотическими. Они могут представлять собой клетки млекопитающих (как например, клетки человека), растительные клетки и микроорганизмы, как например, дрожжи, грибы или бактерии, где микроорганизмы являются предпочтительными, и бактерии и дрожжи являются наиболее предпочтительными.
Подходящими бактериями, дрожжами или грибами являются в частности те бактерии, дрожжи или грибы, которые депонированы в Deutschen Sammlung von Mikroorganismen und Zeilkulturen GmbH (DSMZ), Braunschweig, Германия, в виде штаммов бактерий, грибов или дрожжей. Подходящие бактерии согласно настоящему изобретению принадлежат родам, перечисленным в:
http://www.dsmz.de/species/bacteria.htm;
подходящие дрожжи согласно настоящему изобретению принадлежат родам, перечисленным в:
http://www.dsmz.de/species/yeasts.htm;
подходящие грибы согласно настоящему изобретению принадлежат родам, перечисленным в:
http://www.dsmz.de/species/fungi.htm.
Предпочтительные клетки согласно настоящему изобретению принадлежат родам Aspergillus, Corynebacterium, Brevibactehum, Bacillus, Acinetobacter, Alcaligenes, Lactobacillus, Paracoccus, Lactococcus, Candida, Pichia, Hansenula, Kluyveromyces, Saccharomyces, Escherichia, Zymomonas, Yarrowia, Methylobacterium, Ralstonia, Pseudomonas, Rhodospirillum, Rhodobacter, Burkholderia, Clostridium и Cupriavidus, где Aspergillus nidulans, Aspergillus niger, Alcaligenes latus, Bacillus megaterium, Bacillus subtilis, Brevibacterium flavum, Brevibactehum lactofermentum, Burkholderia andropogonis, B. brasilensis, B. caledonica, B. caribensis, B. caryophylli, B. fungorum, B. gladioli, B. glathei, B. glumae, B. graminis, B. hospita, B. kururiensis, B. phenazinium, B. phymatum, B. phytofirmans, B. plantani, B. sacchari, B. singaporensis, B. sordidicola, B. terricola, B. tropica, B. tuberum, B. ubonensis, B. unamae, B. xenovorans, B. anthina, B. pyrrocinia, B. thailandensis, Candida blankii, Candida rugosa, Corynebacterium glutamicum, Corynebacterium efficiens, Escherichia coli, Hansenula polymorpha, Kluveromyces lactis, Methylobacterium extorquens, Paracoccus versutus, Pseudomonas argentinensis, P. borbori, P. citronellolis, P. flavescens, P. mendocina, P. nitroreducens, P. oleovorans, P. pseudoalcaligenes, P. resinovorans, P. straminea, P. aurantiaca, P. aureofaciens, P. chlororaphis, P. fragi, P. lundensis, P. taetrolens, P. antarctica, P. azotoformans, ‘P. blatchfordae’, P. brassicacearum, P. foremen, P. cedrina, P. corrugata, P. fluorescens, P. gessardii, P. libanensis, P. mandelii, P. marginalis, P. mediterranea, P. meridiana, P. migulae, P. mucidolens, P. orientalis, P. panacis, P. proteolytica, P. rhodesiae, P. synxantha, P. thivervalensis, P. tolaasii, P. veronii, P. denitrificans, P. pertucinogena, P. cremoricolorata fulva, P. mosselii, P. parafulva, P. putida, P. balearica, P. stutzeri, P. amygdali, P. avellanae, P. caricapapayae, P. cichorii, P. coronafaciens, P. ficuserectae, ‘P. helianthi’, P. meliae, P. savastanoi, P. syringae, P. tomato, P. viridiflava, P. abietaniphila, P. acidophila, P. agarici, P. alcaliphila, P. alkanolytica, P. amyloderamosa, P. asplenii, P. azotifigens, P. cannabina, P. coenobios, P. congelans, P. costantinii, P. cruciviae, P. delhiensis, P. excibis, P. extremorientalis, P. frederiksbergensis, P. fuscovaginae, P. gelidicola, P. grimontii, P. indica, P. jessenii, P. jinjuensis, P. kilonensis, P. knackmussii, P. koreensis, P. lini, P. lutea, P. moraviensis, P. otitidis, P. pachastrellae, P. palleroniana, P. papaveris, P. peli, P. perolens, P. poae, P. pohangensis, P. psychrophila, P. psychrotolerans, P. rathonis, P. reptilivora, P. resiniphila, P. rhizosphaerae, P. rubescens, P. salomonii, P. segitis, P. septica, P. simiae, P. suis, P. thermotolerans, P. aeruginosa, P. tremae, P. trivialis, P. turbinellae, P. tuticorinensis, P. umsongensis, P. vancouverensis, P. vranovensis, P. xanthomaiina, Ralstonia eutropha, Rhodospirillum rubrum, Rhodobacter sphaeroides, Saccharomyces cerevisiae, Yarrowia lipolytica и Zymomonas mobilis, особенно Pseudomonas putida, Escherichia coli и Burkholderia thailandensis являются особенно предпочтительными.
Предпочтительные клетки согласно настоящему изобретению, в виде их немутантного типа, не образуют или образуют необнаруживаемые количества рамнолипидов и, более того, в виде немутантного типа предпочтительно не показывают или показывают необнаруживаемую активность ферментов E1, E2 и Е3.
Предпочтительно в соответствии с настоящим изобретением клетка согласно настоящему изобретению представляет собой клетку, которая способна в форме немутантного типа образовывать полигидроксиалканоаты, с длиной цепи моноалканоата, составляющей от 6 до 16 атомов углерода. Такие клетки предпочтительно представляют собой, например, Burkholderia sp., Burkholderia thailandensis, Pseudomonas sp., Pseudomonas putida, Pseudomonas aeruginosa, Pseudomonas oleovorans, Pseudomonas stutzeri, Pseudomonas fluorescens, Pseudomonas citronellolis, Pseudomonas resinovorans, Comamonas testosteroni, Aeromonas hydrophila, Cupriavidus necator, Alcaligenes latus и Ralstonia eutropha. В этом контексте предпочтительные клетки согласно настоящему изобретению являются генетически модифицированными, таким образом, что по сравнению с их немутантным типом они способны формировать меньше полигидроксиалканоатов.
Такие клетки описываются, например, в De Eugenio et al., Environ Microbiol. 2010. 12(1):207-21 и Rehm et al., AppI Environ Microbiol. 2001. 67(7):3102-9.
Такая клетка, способная образовывать меньше полигидроксиалканоатов, по сравнению с ее немутантным типом, в частности характеризуются тем, что по сравнению с ей немутантным типом она имеет пониженную активность по меньшей мере одного фермента E9 или Е10,
где E9 представляет собой полигидроксиалканоат-синтазу, ЕС:2.3.1., в частности имеющую полипептидную последовательность Seq ID №30 или Seq ID №32 или имеющую полипептидную последовательность, в которой до 25%, предпочтительно до 20%, особенно предпочтительно до 15%, особенно до 10, 9, 8, 7, 6, 5, 4, 3, 2, 1% аминокислотных остатков по сравнению с соответствующей ссылочной последовательностью Seq ID №30 или Seq ID №32 модифицированы посредством делеции, инсерции, замещения или их комбинации, и которая все еще имеет по меньшей мере 10%, предпочтительно 50%, особенно предпочтительно 80%, особенно более чем 90% ферментативной активности фермента, имеющего соответствующую ссылочную последовательность Seq ID №30 или Seq ID №32, где ферментативная активность для фермента E9, как понимается, означает способность превращать 3-гидроксиалканоил-коэнзим А в поли-3-гидроксиалкановую кислоту, особенно 3-гидрокситетрадеканоил-коэнзим А в поли-3-гидрокситетрадекановую кислоту, и
Е10 представляет собой 3-гидроксиалканоил-АСР:Коэнзим А трансферазу, особенно имеющую полипептидную последовательность Seq ID №34 или Seq ID №36 или имеющую полипептидную последовательность, в которой до 25%, предпочтительно до 20%, особенно предпочтительно до 15%, особенно до 10, 9, 8, 7, 6, 5, 4, 3, 2, 1% аминокислотных остатков по сравнению с соответствующей ссылочной последовательностью Seq ID №34 или Seq ID №36 модифицированы посредством делеции, инсерции, замещения или их комбинации, и которая все еще имеет по меньшей мере 10%, предпочтительно 50%, особенно предпочтительно 80%, особенно более чем 90% ферментативной активности фермента, имеющего соответствующую ссылочную последовательность Seq ID №34 или Seq ID №36, где ферментативная активность для фермента Е10, как понимается, означает способность превращать 3-гидроксиалканоил-АСР в 3-гидроксиалканоил-коэнзим А, особенно 3-гидроксиалканоил-АСР в 3-гидрокситетрадеканоил-коэнзим А.
Для общего сведения смотрите Фиг.1.
Активность фермента E9 затем определяется с применением образцов, полученных как описано выше для ферментов E1-Е3, посредством сначала смешивания 560 мкл 100 мМ Tris/HCl, pH 7,5, 20 мкл 35 мМ DTNB в DMSO и 20 мкл 41 мМ 3-гидроксидеканоил-коэнзим А. Затем добавляют 5 мкг очищенного фермента E9 в 100 мкл Tris/HCl, pH 7,5, и затем повышение экстинкции при 412 нм (вызванное добавлением 5,5’-дитиобис(2-нитробензоата) (DTNB) к свободным SH-группам) со временем (АЕ/мин) непрерывно регистрируется в течение 1 минуты в спектрофотометре.
Активность фермента Е10 затем определяется с применением образцов, полученных как описано выше для ферментов E1-Е3. Стандартный образец содержит 3 мМ MgCl2, 40 мкМ гидроксидеканоил-коэнзим А и 20 мкМ Е. coli ACP в 50 мМ Tris-HCl, pH 7,5, при общем объеме 200 мкл. Реакцию запускают добавлением 5 мкг очищенного фермента Е10 в 50 мкл Tris/HCl, pH 7,5 и инкубируют в течение 1 часа при 30°С. Реакцию останавливают добавлением 50% (масс./об.) трихлоруксусной кислоты и 10 мг/мл BSA (30 мкл). Выделившийся Коэнзим А определяют спектрофотометрически путем регистрации повышения экстинкции при 412 нм, вызванного добавлением 5,5’-дитиобис(2-нитробензоата) (DTNB) к свободным SH-группам), со временем.
Выражение "пониженная активность фермента Ех", применяемое в соответствии с настоящим изобретением, как понимается, означает активность, пониженную на по меньшей мере 0,5, особенно предпочтительно на по меньшей мере 0,1, особенно предпочтительно на по меньшей мере 0,01, еще более предпочтительно на по меньшей мере 0,001 и наиболее предпочтительно на по меньшей мере 0,0001. Выражение "пониженная активность" также охватывает не обнаруживаемую активность ("нулевая активность"). Понижение активности определенного фермента может быть вызвано, например. Селективной мутацией или другими средствами, хорошо известными специалистам в данной области техники, для понижения активности определенного фермента.
Способы понижения ферментативных активностей микроорганизмов известны специалистам в данной области техники.
В частности, здесь предлагаются молекулярно-биологические методики. Специалист в данной области техники найдет инструкции по модификации и понижению экспрессии белка и сопутствующему понижению ферментативной активности, особенно для Pseudomonas и Burkholderia, в частности для прерывающихся специфических генов, например в Dubeau et al. 2009. BMC Microbiology 9:263; Singh & Rohm. Microbiology. 2008. 154:797-809 или Lee et al. FEMS Microbiol Lett. 2009. 297(1):38-48.
Клетки, предпочтительные в соответствии с настоящим изобретением, характеризуются тем, что уменьшение ферментативной активности достигается посредством модификации гена, содержащего одну из указанных последовательностей нуклеиновых кислот, где модификация выбирается из группы, предпочтительно состоящей из вставки чужеродной ЛНК в ген, делеции по меньшей мере частей гена, точечных мутаций в генной последовательности, интерференции РНК (siRNA), антисмысловой РНК или модификации (вставки, делеции или точечных мутаций) регуляторных последовательностей, как например, промоторы и терминаторы или сайты связывания рибосом, которые фланкируют ген.
Чужеродная ДНК, как понимается в этом контексте, означает любую последовательность ДНК, которая является "чужеродной" для гена (но не для организма), то есть эндогенные последовательности ДНК в этом контексте могут также выступать в качестве "чужеродной ДНК".
В этом контексте особенно предпочтительно, чтобы ген прерывался вставкой селекционного маркерного гена, таким образом, чужеродная ДНК представляет собой селекционный маркерный ген, где инсерция предпочтительно осуществляется посредством гомологической рекомбинации в локус гена.
В предпочтительном варианте выполнения настоящего изобретения клетки представляют собой клетки Pseudomonas putida, которые показывают пониженный синтез полигидроксиалканоатов, по сравнению с их немутантным типом. Такие клетки описываются, Например, в Ren et al., Journal Applied Microbiology and Biotechnology 1998 Jun, 49(6):743-50 как GPp121, GPp122, GPp123 и GPp124, в Huisman et al., J Biol Chem. 1991 Feb 5;266(4):2191-8 как GPp104, а также в De Eugenio et al., Environ Microbiol. 2010. 12(1):207-21 как KT42C1 и в Ouyang et al. Macromol Biosci. 2007. 7(2):227-33 как KTOY01 и KTOY02.
Когда клетка согласно настоящему изобретению способна образовывать рамнолипид, имеющий m=1, радикал, определяемый R1 и R2, предпочтительно получают из
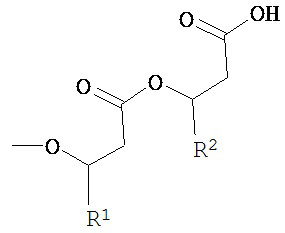
3-гидроксиоктаноил-3-гидроксиоктановой кислоты, 3-гидроксиоктаноил-3-гидроксидекановой кислоты, 3-гидроксидеканоил-3-гидроксиоктановой кислоты, 3-гидроксиоктаноил-3-гидроксидеценовой кислоты, 3-гидроксидеценоил-3-гидроксиоктановой кислоты, 3-гидроксиоктаноил-3-гидроксидодекановой кислоты, 3-гидроксидодеканоил-3-гидроксиоктановой кислоты, 3-гидроксиоктаноил-3-гидроксидодеценовой кислоты, 3-гидроксидодеценоил-3-гидроксиоктановой кислоты, 3-гидроксидеканоил-3-гидроксидекановой кислоты, 3-гидроксидеканоил-3-гидроксидеценовой кислоты, 3-гидроксидеценоил-3-гидроксидекановой кислоты, 3-гидроксидеценоил-3-гидроксидеценовой кислоты, 3-гидроксидеканоил-3-гидроксидодекановой кислоты, 3-гидроксидодеканоил-3-гидроксидекановой кислоты, 3-гидроксидеканоил-3-гидроксидодеценовой кислоты, 3-гидроксидеканоил-3-гидрокситетрадеценовой кислоты, 3-гидрокситетрадеканоил-3-гидроксидеценовой кислоты, 3-гидроксидодеценоил-3-гидроксидекановой кислоты, 3-гидроксидеканоил-3-гидрокситетрадекановой кислоты, 3-гидрокситетрадеканоил-3-гидроксидекановой кислоты, 3-гидроксидеканоил-3-гидрокситетрадеценовой кислоты, 3-гидрокситетрадеценоил-3-гидроксидекановой кислоты, 3-гидроксидодеканоил-3-гидроксидодекановой кислоты, 3-гидроксидодеценоил-3-гидроксидодекановой кислоты, 3-гидроксидодеканоил-3-гидроксидодеценовой кислоты, 3-гидроксидодеканоил-3-гидрокситетрадекановой кислоты, 3-гидрокситетрадеканоил-3-гидроксидодекановой кислоты, 3-гидрокситетрадеканоил-3-гидрокситетрадекановой кислоты, 3-гидроксигексадеканоил-3-гидрокситетрадекановой кислоты, 3-гидрокситетрадеканоил-3-гидроксигексадекановой кислоты или 3-гидроксигексадеканоил-3-гидроксигексадекановой кислоты.
Специалисту в данной области техники очевидно, что клетка согласно настоящему изобретению также способна образовывать смеси различных рамнолипидов общей формулы (I).
В этом контексте клетки согласно настоящему изобретению предпочтительно способны образовывать смеси различных рамнолипидов общей формулы (I), которые характеризуются тем, что в более 80 мас.%, предпочтительно более 90 мас.%, особенно предпочтительно более 95 мас.% сформированных рамнолипидов n=1, и радикал, определенный R1 и R2, происходит в менее 10 мас.%, предпочтительно менее 5 мас.%, особенно предпочтительно менее 2 мас.% сформированных рамнолипидов из 3-гидроксидеканоил-3-гидроксиоктановой кислоты или 3-гидроксиоктаноил-3-гидроксидекановой кислоты, где масс.% относятся к сумме всех сформированных рамнолипидов общей формулы (I).
Клетка согласно настоящему изобретению предпочтительно была дополнительно генетически модифицирована в отношении E1-Е3, так что по сравнению с ее немутантным типом она имеет повышенную активность, как в каждом из приведенных далее случаев, по меньшей мере одного из ферментов, выбранных из группы, состоящей из:
по меньшей мере одного фермента Е4, dTTP:α-D-глюкоза-1-фосфат тимидилилтрансфераза, ЕС 2.7.7.24, в частности имеющего полипептидную последовательность Seq ID №10 или имеющего полипептидную последовательность, в которой до 25%, предпочтительно до 20%, особенно предпочтительно до 15%, особенно до 10, 9, 8, 7, 6, 5, 4, 3, 2, 1% аминокислотных остатков модифицированы по сравнению со ссылочной последовательностью Seq ID №10 посредством делеции, инсерции, замещения или их комбинации, и который все еще имеет по меньшей мере 10%, предпочтительно 50%, особенно предпочтительно 80%, особенно более чем 90% от ферментативной активности фермента, имеющего ссылочную последовательность Seq ID №10, где ферментативная активность фермента Е4 понимается как способность превращать α-D-глюкоза-1-фосфат и dTTP в dTDP-глюкозу,
по меньшей мере одного фермента Е5, dTTP-глюкоза-4,6-гидролаза, ЕС 4.2.1.46, в частности имеющего полипептидную последовательность Seq ID №12 или имеющего полипептидную последовательность, в которой до 25%, предпочтительно до 20%, особенно предпочтительно до 15%, особенно до 10, 9, 8, 7, 6, 5, 4, 3, 2, 1% аминокислотных остатков модифицированы по сравнению со ссылочной последовательностью Seq ID №12 посредством делеции, инсерции, замещения или их комбинации, и который все еще имеет по меньшей мере 10%, предпочтительно 50%, особенно предпочтительно 80%, особенно более чем 90% от ферментативной активности фермента, имеющего ссылочную последовательность Seq ID №12, где ферментативная активность фермента Е5 понимается как способность превращать dTDP-глюкозу в dTDP-4-дегидро-6-деокси-D-глюкозу,
по меньшей мере одного фермента Е6, dTDP-4-дегидрорамноза-3,5-эпимераза, ЕС 5.1.3.13, в частности имеющего полипептидную последовательность Seq ID №14 или имеющего полипептидную последовательность, в которой до 25%, предпочтительно до 20%, особенно предпочтительно до 15%, особенно до 10, 9, 8, 7, 6, 5, 4, 3, 2, 1% аминокислотных остатков модифицированы по сравнению со ссылочной последовательностью Seq ID №14 посредством делеции, инсерции, замещения или их комбинации, и который все еще имеет по меньшей мере 10%, предпочтительно 50%, особенно предпочтительно 80%, особенно более чем 90% от ферментативной активности фермента, имеющего ссылочную последовательность Seq ID №14, где ферментативная активность фермента Е6 понимается как способность превращать dTDP-4-дегидро-6-деокси-D-глюкозу в dTDP-4-дегидро-6-деокси-L-маннозу,
по меньшей мере одного фермента Е7, dTDP-4-дегидрорамноза-редуктаза, ЕС 1.1.1.133, в частности имеющего полипептидную последовательность Seq ID №16 или имеющего полипептидную последовательность, в которой до 25%, предпочтительно до 20%, особенно предпочтительно до 15%, особенно до 10, 9, 8, 7, 6, 5, 4, 3, 2, 1% аминокислотных остатков модифицированы по сравнению со ссылочной последовательностью Seq ID №16 посредством делеции, инсерции, замещения или их комбинации, и который все еще имеет по меньшей мере 10%, предпочтительно 50%, особенно предпочтительно 80%, особенно более чем 90% от ферментативной активности фермента, имеющего ссылочную последовательность Seq ID №16, где ферментативная активность фермента Е7 понимается как способность превращать dTDP-4-дегидро-6-деокси-L-маннозу в dTDP-6-деокси-L-маннозу.
Активность фермента Е4 определяется с применением образцов, полученных, как описано выше для ферментов E1-Е3, путем инкубации α-D-глюкоза-1-фосфата (1,3 мМ) с dTTP (5 мМ) и 5 мкг очищенного фермента Е4 в 50 мкл натрий-фосфатного буфера, рН 8,5, и остановки реакции через 5, 10 и 20 минут инкубации при 30°С путем добавления 20 мкл хлороформа. Смесь вортексировали и центрифугировали в течение 5 минут при 16.000 g и комнатной температуре. Водную фазу перенесли в новый реакционный сосуд, и органическую фазу снова экстрагировали с помощью 80 мкл воды. Обе водные фазы объединили и проанализировали посредством ВЭЖХ. Применяли колонку Phenosphere ODS2 (250×4,6 мм; Phenomenex, Torrance, USA) или колонку Spheresorb ODS2 (250×4,6 мм; Waters, Milford, USA). Элюацию аналитов осуществляли при скорости потока 1 мл мин-1, применяя 0,5 М KH2PO4 (элюент А) в течение 15 минут, с последующим линейным градиентом до 80% элюента А и 20% метанола в течение 14 минут при скорости потока 0,7 мл мин-1. Аналиты, которые элюировали с колонок ODS2 затем впрыскивали в ион-обменную колонку Phenosphere-SAX (250×4,6 мм; Phenomenex, Torrance, USA), и аналиты элюировали, применяя скорость потока 1 мл мин-1 и линейный градиент формиата аммония (2-600 мМ в течение 25 минут). Количественное определение dTDP-глюкозы осуществляли с помощью УФ-поглощения, применяя матричный фотодиодный детектор (DAD). Максимум поглощения тимидина находится при 267 нм. Калибровка осуществляется посредством аутентичного нуклеотидного сахара (Sigma-Aldrich, München, USA).
Активность фермента E5 определяется с применением образцов, полученных, как описано выше для ферментов E1-Е3, путем инкубации dTDP-α-D-глюкозы (1,3 мМ) с 5 мкг очищенного фермента E5 в 50 мкл натрий-фосфатного буфера, рН 8,5, и остановки реакции через 5, 10 и 20 минут инкубации при 30°С путем добавления 20 мкл хлороформа. Смесь вортексировали и центрифугировали в течение 5 минут при 16.000 g и комнатной температуре. Водную фазу перенесли в новый реакционный сосуд, и органическую фазу снова экстрагировали с помощью 80 мкл Воды. Обе водные фазы объединили и проанализировали посредством ВЭЖХ. Применяли колонку Phenosphere ODS2 (250×4,6 мм; Phenomenex, Torrance, USA) или колонку Spheresorb ODS2 (250×4,6 мм; Waters, Milford, USA). Элюацию аналитов осуществляли при скорости потока 1 мл мин-1, применяя 0,5 М KH2PO4 (элюент А) в течение 15 минут, с последующим линейным градиентом до 80% элюента А и 20% метанола в течение 14 минут при скорости потока 0,7 мл мин-1. Аналиты, которые элюировали с колонок ODS2 затем впрыскивали в ион-обменную колонку Phenosphere-SAX (250×4,6 мм; Phenomenex, Torrance, USA), и аналиты элюировали, применяя скорость потока 1 мл мин-1 и линейный градиент формиата аммония (2-600 мМ в течение 25 минут). Количественное определение dTDP-глюкозы и dTDP-4-дегидро-6-деокси-D-глюкозы осуществляли с помощью УФ-поглощения, применяя матричный фотодиодный детектор (DAD). Максимум поглощения тимидина находится при 267 нм. Калибровка осуществляется посредством аутентичного нуклеотидного сахара (Sigma-Aldrich, München, USA).
Активность фермента Е6 определяется с применением образцов, полученных, как описано выше для ферментов E1-Е3, путем инкубации dTDP-a-D-глюкозы (1,3 мМ) с 5 мкг очищенного фермента Е5 в 50 мкл натрий-фосфатного буфера, рН 8,5, в течение 10 минут при 30°С. Затем добавили 0,5 мкг очищенного фермента Е6, и реакцию остановили через 5, 10 и 20 минут инкубации при 30°С путем добавления 20 мкл хлороформа. Смесь вортексировали и центрифугировали в течение 5 минут при 16.000 g и комнатной температуре. Водную фазу перенесли в новый реакционный сосуд, и органическую фазу снова экстрагировали с помощью 80 мкл Воды. Обе водные фазы объединили и проанализировали посредством ВЭЖХ. Применяли колонку Phenosphere ODS2 (250×4,6 мм; Phenomenex, Torrance, USA) или колонку Spheresorb ODS2 (250×4,6 мм; Waters, Milford, USA). Элюацию аналитов осуществляли при скорости потока 1 мл мин-1, применяя 0,5 М КН2РО4 (элюент А) в течение 15 минут, с последующим линейным градиентом до 80% элюента А и 20% метанола в течение 14 минут при скорости потока 0,7 мл мин-1. Аналиты, которые элюировали с колонок ODS2 затем впрыскивали в ион-обменную колонку Phenosphere-SAX (250×4,6 мм; Phenomenex, Torrance, USA), и аналиты элюировали, применяя скорость потока 1 мл мин-1 и линейный градиент формиата аммония (2-600 мМ в течение 25 минут). Количественное определение dTDP-глюкозы, dTDP-4-дегидро-6-деокси-D-глюкозы и dTDP-6-деокси-L-маннозы осуществляли с помощью УФ-поглощения, применяя матричный фотодиодный детектор (DAD). Максимум поглощения тимидина находится при 267 нм. Калибровка осуществляется посредством аутентичного нуклеотидного сахара (Sigma-Aldrich, München, USA).
Активность фермента Е7 определяется с применением образцов, полученных, как описано выше для ферментов E1-Е3, путем инкубации dTDP-a-D-глюкозы (1,3 мМ) с 5 мкг очищенного фермента Е5 в 50 мкл натрий-фосфатного буфера, рН 8,5, в течение 10 минут при 30°С. Затем добавили 5 мкг очищенного фермента Е6 и 0,5 мкг очищенного фермента Е7, а также NADPH (10 мМ), и реакцию остановили через 5, 10 и 20 минут инкубации при 30°С путем добавления 20 мкл хлороформа. Смесь вортексировали и центрифугировали в течение 5 минут при 16.000 g и комнатной температуре. Водную фазу перенесли в новый реакционный сосуд, и органическую фазу снова экстрагировали с помощью 80 мкл Воды. Обе водные фазы объединили и проанализировали посредством ВЭЖХ. Применяли колонку Phenosphere ODS2 (250×4,6 мм; Phenomenex, Torrance, USA) или колонку Spheresorb ODS2 (250×4,6 мм; Waters, Milford, USA). Элюацию аналитов осуществляли при скорости потока 1 мл мин-1, применяя 0,5 М KH2PO4 (элюент А) в течение 15 минут, с последующим линейным градиентом до 80% элюента А и 20% метанола в течение 14 минут при скорости потока 0,7 мл мин-1. Аналиты, которые элюировали с колонок ODS2 затем впрыскивали в ион-обменную колонку Phenosphere-SAX (250×4,6 мм; Phenomenex, Torrance, USA), и аналиты элюировали, применяя скорость потока 1 мл мин-1 и линейный градиент формиата аммония (2-600 мМ в течение 25 минут). Количественное определение dTDP-глюкозы, dTDP-4-дегидро-6-деокси-D-глюкозы, dTDP-6-деокси-L-маннозы и dTDP-4-дегидро-6-деокси-L-маннозы осуществляли с помощью УФ-поглощения, применяя матричный фотодиодный детектор (DAD). Максимум поглощения тимидина находится при 267 нм. Калибровка осуществляется посредством аутентичного нуклеотидного сахара (Sigma-Aldrich, München, USA).
Предпочтительными согласно настоящему изобретению являются те клетки, которые имеют повышенные активности следующих комбинаций ферментов:
Е4Е5, Е4Е6, Е4Е7, E5E6, E5E7, E6E7, Е4Е5Е6, Е4Е5Е7, E5E6E7, Е4Е6Е7, E4E5E6E7, особенно предпочтительно Е4Е5Е5Е7.
Предпочтительно согласно настоящему изобретению клетка согласно настоящему изобретению была генетически модифицирована в отношении биосинтеза жирных кислот, так что ферментативные реакции, которые приводят к превращению ацил-АСР и малонил-коэнзим А в 3-кетоацил-АСР и/или превращению 3-кетоацил-АСР в (R)-3-гидроксиалканоил-АСР, усиливаются. Дополнительно или альтернативно предпочтительно согласно настоящему изобретению, клетка согласно настоящему изобретению была генетически модифицирована в отношении биосинтеза жирных кислот, так что ферментативные реакции, которые приводят к превращению (R)-3-гидроксиалканоил-АСР в транс-2-еноил-АСР и/или к превращению транс-2-еноил-АСР в ацил-АСР уменьшаются.
Также предпочтительно согласно настоящему изобретению, когда клетка согласно настоящему изобретению была генетически модифицирована в отношении β-окисления жирных кислот, так что ферментативные реакции, которые приводят к превращению ацил-коэнзима А в транс-2-еноил-коэнзим А и/или к превращению транс-2-еноил-коэнзима А в (S)-3-гидроксиалканоил-коэнзим А, усиливаются. Дополнительно или альтернативно предпочтительно согласно настоящему изобретению, клетка согласно настоящему изобретению была генетически модифицирована в отношении β-окисления жирных кислот, так что ферментативные реакции, которые приводят к превращению (S)-3-гидроксиалканоил-коэнзима А в 3-кетоацил-коэнзим А и/или к превращению 3-кетоацил-коэнзима А в ацил-коэнзим А и ацетил-коэнзим А, уменьшаются.
Для общего сведения смотрите Фиг.1.
Так как клетки согласно настоящему изобретению могут применяться предпочтительно для получения рамнолипидов, и так как эти липиды затем при необходимости очищаются, предпочтительно, если клетки согласно настоящему изобретению имеют повышенную активность по сравнению с их немутантным типом по меньшей мере фермента E8, который катализирует экспорт рамнолипидов общей формулы (I) из клетки в окружающую среду.
Предпочтительно в этом отношении белки Е8, выбранные из группы, состоящей из:
фермента Е8, имеющего полипептидную последовательность Seq ID №8, Seq ID №24, Seq ID №26 или Seq ID №28 или имеющего полипептидную последовательность, в которой до 25%, предпочтительно до 20%, особенно предпочтительно до 15%, особенно до 10, 9, 8, 7, 6, 5, 4, 3, 2, 1% аминокислотных остатком модифицированы путем делеции, инсерции, замещения или их комбинации по сравнению с соответствующей ссылочной последовательностью Seq ID №8, Seq ID №24, Seq ID №26 или Seq ID №28, и который все еще имеет по меньшей мере 50%, предпочтительно 65%, особенно предпочтительно 80%, особенно более чем 90% от ферментативной активности фермента, имеющего соответствующую ссылочную последовательность Seq ID №8, Seq ID №24, Seq ID №26 или Seq ID №28, где ферментативная активность фермента Е8, как понимается, означает способность экспортировать рамнолипид общей формулы (I) из клетки в окружающую среду.
Другой предпочтительный вариант клеток согласно настоящему изобретению характеризуется тем, что он содержит по меньшей мере одну из нуклеиновых кислот или векторов согласно настоящему изобретению.
Клетки согласно настоящему изобретению могут предпочтительно применяться для производства рамнолипидов.
Таким образом, другим объектом настоящего изобретения является применение клеток согласно настоящему изобретению для получения соединений общей формулы (I).
Другим объектом настоящего изобретения является способ получения рамнолипидов общей формулы (I),
где
m равно 2, 1 или 0, в частности 1 или 0,
n равно 1 или 0, в частности 1,
R1 и R2 независимо друг от друга представляют собой идентичные или различные органические радикалы, имеющие от 2 до 24, предпочтительно от 5 до 13 атомов углерода, в частности необязательно разветвленный, необязательно замещенный, в частности гидрокси-замещенный, необязательно ненасыщенный, в частности, моно-, ди- или триненасыщенный, алкильный радикал, предпочтительно выбранный из группы, состоящей из пентенила, гептенила, ноненила, ундеценила и тридеценила и (СН2)0-СН3со=1-23, предпочтительно 4-12,
содержащий следующие стадии:
I) введение клетки согласно настоящему изобретению в контакт со средой, содержащей источник углерода; и
II) культивирование клетки при условиях, при которых клетка образует рамнолипиды из источника углерода,
III) при необходимости выделение полученных рамнолипидов.
Генетически модифицированные клетки согласно настоящему изобретению могут вводиться в контакт с питательной средой непрерывно или периодически в периодическом процессе (периодическая культура) или в процессе с подпиткой (подпитываемая культура) или в процессе с периодической подпиткой (периодически подпитываемая культура) с целью получения вышеупомянутых продуктов и, таким образом, культивироваться. Полунепрерывный процесс также возможен, как описывается в GB-A-1009370. Сведения об известных сведениях культивирования описываются в учебнике Chmiel ("Bioprozesstechnik 1. Einfuhrung in die Bioverfahrenstechnik" (Gustav Fischer Verlag, Stuttgart, 1991)) или в учебнике Storhas ("Bioreaktoren und periphere Einrichtungen", Vieweg Verlag, Braunschweig/Wiesbaden, 1994).
Культуральная среда, которая подлежит применению, должна соответствующим образом удовлетворять требованиям соответствующих штаммов. Описания культуральных сред различных штаммов дрожжей приводятся, например, в "Nonconventional yeast in biotechnology" (Hrsg. KlausWolf, Springer-Verlag Berlin, 1996).
Применяемым источником углерода могут быть углеводы, такие как, например, глюкоза, сахароза, арабиноза, ксилоза, лактоза, фруктоза, мальтоза, меласса, крахмал, целлюлоза и гемицеллюлоза, растительные и животные масла и жиры, как например соевое масло, сафлоровое масло, арахисовое масло, конопляное масло, масло зародышей пшеницы, кокосовое масло, масло из тыквенного семени, льняное масло, кукурузное масло, маковое масло, масло ослинника, оливковое масло, пальмоядровое масло, пальмовое масло, рапсовое масло, кунжутное масло, подсолнечное масло, виноградное масло, масло грецкого ореха, масло из зародышей пшеничного зерна и кокосовое масло, жирные кислоты, такие как, например, каприловая кислота, каприновая кислота, лауриновая кислота, миристиновая кислота, пальмитиновая кислота, пальмитолеиновая кислота, стеариновая кислота, арахидоновая кислота, бегеновая кислота, олеиновая кислота, линолевая кислота, линоленовая кислота, гамма-линоленовая кислота и их метиловый или этиловый сложные эфиры, а также смеси жирных кислот, моно-, ди- и триглицериды содержащие только что упомянутые жирные кислоты, спирты, такие как, например, глицерин, этанол и метанол, углеводороды, такие как метан, содержащие углерод газы и смеси газов, как например СО, CO2, синтез-газ или дымовой газ, аминокислоты, такие как L-глутамат или L-валин, или органические кислоты, такие как, например уксусная кислота. Эти вещества могут применяться по отдельности или в виде смеси. Применение углеводов, особенно предпочтительно моносахаридов, олигосахаридов или полисахаридов, в качестве источника углерода US 6,01,494 и US 6,136,576, а также углеводородов, особенно алканов, алкенов и алкинов, а также полученных из них монокарбоновых кислот и полученных из этих монокарбоновых кислот моно-, ди- и триглицеридов, а также глицерина и ацетата, является особенно предпочтительным. Наиболее предпочтительными являются моно-, ди- и триглицериды, содержащие продукты эстерификации глицерина каприловой кислотой, каприновой кислотой, лауриновой кислотой, миристиновой кислотой, пальмитиновой кислотой, пальмитолеиновой кислотой, стеариновой кислотой, арахидоновой кислотой, бегеновой кислотой, олеиновой кислотой, линолевой кислотой, линоленовой кислотой и/или гамма-линоленовой кислотой.
Значительное преимущество настоящего изобретения состоит в том, что клетки согласно настоящему изобретению способны образовывать рамнолипиды из самых простых источников углерода, таких как, например, глюкоза, сахароза или глицерин, так что обеспечение длинноцепочечных источников углерода в среде в ходе способа согласно настоящему изобретению не является необходимым. Таким образом, преимущество состоит в том, что в случае отсутствия доступности среда на стадии I) способа согласно настоящему изобретению может не содержать или содержать необнаруживаемые количества карбоновых кислот с длиной цепи более шести атомов углерода или полученных из них сложных эфиров или глицеридов.
Применяемыми источниками азота могут быть содержащие азот органические соединения, такие как пептоны, дрожжевой экстракт, мясной экстракт, солодовый экстракт, кукурузный экстракт, мука из соевых бобов и мочевина, или неорганические соединения, такие как сульфат аммония, хлорид аммония, фосфат аммония, карбонат аммония нитрат аммония, аммиак, гидроксид аммония или водный раствор аммиака. Источники азота могут применяться по отдельности или в виде смеси.
Применяемым источником фосфора может быть фосфорная кислота, дигидрофосфат калия или гидрофосфат дикалия или соответствующие натрийсодержащие соли. Культуральная среда должна, кроме того, содержать соли металлов, такие как, например, сульфат магния или сульфат железа, которые необходимы для роста. Наконец, необходимые промоторы роста, такие как аминокислоты и витамины, могут применяться в дополнение к вышеупомянутым веществам. Кроме того, в культуральную среду могут быть добавлены подходящие предшественники. Указанное сырье может быть добавлено в культуры в форме одной порции или посредством подпитки подходящим образом в ходе культивирования.
Основные соединения, такие как гидроксид натрия, гидроксид калия, аммиак или водный раствор аммиака, или кислотные соединения, такие как фосфорная кислота или серная кислота, подходящим образом применяются для контроля значения рН культуры. Пеногасители, такие как, например, полигликолевые сложные эфиры жирных кислот, могут применяться для контроля пенообразования. Подходящие селективно действующие вещества, такие как, например, антибиотики, могут добавляться в среду для поддержания стабильности плазмид. Для поддержания аэробных условий в культуру включаются кислород и содержащие кислород газовые смеси.
Температура культуры, как правило, составляет более 20°С, предпочтительно более 25°С, она также может быть более 40°С, причем температура культивирования предпочтительно не превышает 95°С, особенно предпочтительно 90°С и наиболее предпочтительно 80°С.
На стадии III) способа согласно настоящему изобретению, рамнолипиды, образованные клетками, могут при необходимости быть выделены из клеток и/или питательной среды, где для выделения могут применяться все способы, известные специалистам в данной области техники для выделения низкомолекулярных веществ из сложных композиций, такие как, например, фильтрация, экстракция, адсорбция (хроматография) или кристаллизация.
Более того, фаза продукта содержит остатки биомассы и различные примеси, такие как масла, жирные кислоты и другие компоненты питательной среды. Отделение примесей предпочтительно осуществляется свободным от растворителей способом. Так, например, фаза продукта может быть разбавлена водой, чтобы установить рН. Фаза продукта и водная фаза могут затем быть гомогенизированы путем превращения рамнолипидов в растворимую в воде форму путем понижения или повышения рН с помощью кислот или щелочей. Потенциально, солюбилизации рамнолипидов в водной фазе может способствовать инкубация при более высоких температурах, например, от 60 до 90°С, и непрерывное перемешивание. Посредством последующего повышения или понижения рН с помощью щелочей или кислот рамнолипиды могут затем снова превращаться в нерастворимую в воде форму, так что они могут быть легко отделены от водной фазы. Фаза продукта может затем быть промыта один или несколько раз водой для удаления растворимых в воде примесей.
Остатки масла могут быть отделены, например, экстракцией с помощью подходящих растворителей, предпочтительно посредством органических растворителей. Предпочтительно в качестве растворителя применяется алкан, например, н-гексан.
Отделение продукта от водной фазы альтернативно может осуществляться свободным от растворителя способом, описанным выше, с применением подходящего растворителя, например, сложного эфира, такого как, например, этилацетат или бутилацетат. Указанные стадии экстракции могут осуществляться в любой желательной последовательности.
В этом случае предпочтительно применяются растворители, в частности органические растворители. Особенно предпочтительно в качестве растворителя применяется н-пентанол. Для удаления растворителя применяется, например, дистилляция. Затем лиофилизированный продукт может быть далее очищен, например, с помощью хроматографических способов. Здесь, например, можно упомянуть осаждение посредством подходящих растворителей, экстракцию посредством подходящих растворителей, комплексообразование, например, с помощью циклодекстринов или производных циклодекстринов, кристаллизацию, очищение или выделение с помощью хроматографических способов или превращение рамнолипидов в легко отделяемые производные.
Рамнолипиды, которые могут быть получены с применением способа согласно настоящему изобретению, подобным образом являются объектом настоящего изобретения, в частности также смеси рамнолипидов, описанные выше, которые могут быть получены с применением способа согласно настоящему изобретению.
Рамнолипиды и смеси, которые могут быть получены с применением способа согласно настоящему изобретению, могут предпочтительно применяться в очищающих средствах, в косметических или фармацевтических композициях, а также в композициях для защиты растений.
Таким образом, другим объектом настоящего изобретения является применение рамнолипидов, полученных способом по настоящему изобретению, для получения косметических, дерматологических или фармацевтических композиций, композиций для защиты растений, а также средств для ухода и защиты, а также концентратов поверхностно-активных веществ.
Термин "средство для ухода", как понимается, означает композицию, назначение которой состоит в сохранении изделия в его первоначальной форме, уменьшение или исключение эффектов внешних воздействий (например, время, свет, температура, давление, загрязнения окружающей среды, химическая реакция с другими реакционноспособными соединениями, вступающими в контакт с изделием), как например, старение, загрязнение, усталость материала или даже улучшение желательных положительных свойств изделия. Для последнего необходимо отметить, например, улучшенный блеск волос или большую эластичность изделия.
Термин "композиции для защиты растения", как понимается, означает те композиции, которые по природе их получения очевидным образом применяются для защиты растений; то есть, в частности, когда по меньшей мере одно соединение из классов, состоящих из гербицидов, фунгицидов, инсектицидов, акарицидов, нематоцидов, защитных веществ от повреждения птицами, питательных веществ для растений и средств для улучшения структуры почвы, содержится в композиции.
Согласно настоящему изобретению, рамнолипиды, полученные с применением способа согласно настоящему изобретению, предпочтительно применяются в средствах для ухода и защиты, предназначенных для домашнего хозяйства, промышленности, в частности для твердых поверхностей, кожи или текстиля.
Вклад в достижение поставленной задачи обеспечивается выделенной нуклеиновой кислотой, которая содержит в каждом случае по меньшей мере одну последовательность, выбранную из трех групп [А1-G1], [А2-G2] и [A3-G3], где
группа [А1-G1] состоит из следующих последовательностей:
А1а) последовательность согласно Seq ID №1, где эта последовательность кодирует белок, который способен превращать 3-гидроксидеканоил-АСР через 3-гидроксидеканоил-3-гидроксидеканоил-АСР в 3-гидроксидеканоил-3-гидроксидекановую кислоту,
В1а) свободная от интронов последовательность, которая происходит из последовательности согласно А1а) и которая кодирует такой же белок или пептид, что и последовательность согласно Seq ID №1,
С1а) последовательность, которая кодирует белок или пептид, который содержит аминокислотную последовательность согласно Seq ID №2 и который предпочтительно способен превращать 3-гидроксидеканоил-АСР через 3-гидроксидеканоил-3-гидроксидеканоил-АСР в 3-гидроксидеканоил-3-гидроксидекановую кислоту,
D1a) последовательность, которая идентична последовательности согласно одной из групп А1а)-С1а), особенно предпочтительно согласно группе А1а), на по меньшей мере 70%, особенно предпочтительно на по меньшей мере 90%, более предпочтительно на по меньшей мере 95% и наиболее предпочтительно на по меньшей мере 99%, где эта последовательность предпочтительно кодирует белок или пептид, который способен превращать 3-гидроксидеканоил-АСР через 3-гидроксидеканоил-3-гидроксидеканоил-АСР в 3-гидроксидеканоил-3-гидроксидекановую кислоту,
Е1а) последовательность, которая гибридизуется или, принимая во внимание вырожденность генетического кода, будет гибридизоваться с комплементарной нитью последовательности согласно одной из групп А1а)-D1a), особенно предпочтительно согласно группе А1а), где эта последовательность предпочтительно кодирует белок или пептид, который способен превращать 3-гидроксидеканоил-АСР через 3-гидроксидеканоил-3-гидроксидеканоил-АСР в 3-гидроксидеканоил-3-гидроксидекановую кислоту,
F1a) производная последовательности согласно одной из групп А1а)-Е1а), особенно предпочтительно согласно группе А1а), полученная замещением, добавлением, инверсией и/или делецией по меньшей мере одного основания, предпочтительно по меньшей мере 2 оснований, более предпочтительно по меньшей мере 5 оснований и наиболее предпочтительно по меньшей мере 10 оснований, но предпочтительно не более 100 оснований, особенно предпочтительно не более 50 оснований и наиболее предпочтительно не более 25 оснований, где эта производная предпочтительно кодирует белок или пептид, который способен превращать 3-гидроксидеканоил-АСР через 3-гидроксидеканоил-3-гидроксидеканоил-АСР в 3-гидроксидеканоил-3-гидроксидекановую кислоту,
G1a) последовательность, комплементарная последовательности согласно одной из групп А1а)-F1a), особенно предпочтительно согласно группе А1а),
A1b) последовательность согласно Seq ID №17, где эта последовательность кодирует белок, который способен превращать 3-гидрокситетрадеканоил-АСР через 3-гидрокситетрадеканоил-3-гидрокситетрадеканоил-АСР в 3-гидрокситетрадеканоил-3-гидрокситетрадекановую кислоту,
B1b) свободная от интронов последовательность, которая происходит из последовательности согласно A1b) и которая кодирует такой же белок или пептид, что и последовательность согласно Seq ID №17,
C1b) последовательность, которая кодирует белок или пептид, который содержит аминокислотную последовательность согласно Seq ID №18 и который предпочтительно способен превращать 3-гидрокситетрадеканоил-АСР через 3-гидрокситетрадеканоил-3-гидрокситетрадеканоил-АСР в 3-гидрокситетрадеканоил-3-гидрокситетрадекановую кислоту,
D1b) последовательность, которая идентична последовательности согласно одной из групп A1b)-C1b), особенно предпочтительно согласно группе A1b), на по меньшей мере 70%, особенно предпочтительно на по меньшей мере 90%, более предпочтительно на по меньшей мере 95% и наиболее предпочтительно на по меньшей мере 99%, где эта последовательность предпочтительно кодирует белок или пептид, который способен превращать 3-гидрокситетрадеканоил-АСР через 3-гидрокситетрадеканоил-3-гидрокситетрадеканоил-АСР в 3-гидрокситетрадеканоил-3-гидрокситетрадекановую кислоту,
E1b) последовательность, которая гибридизуется или, принимая во внимание вырожденность генетического кода, будет гибридизоваться с комплементарной нитью последовательности согласно одной из групп A1b)-D1b), особенно предпочтительно согласно группе A1b), где эта последовательность предпочтительно кодирует белок или пептид, который способен превращать 3-гидрокситетрадеканоил-АСР через 3-гидрокситетрадеканоил-3-гидрокситетрадеканоил-АСР в 3-гидрокситетрадеканоил-3-гидрокситетрадекановую кислоту, F1b) производная последовательности согласно одной из групп A1b)-E1b), особенно предпочтительно согласно группе A1b), полученная замещением, добавлением, инверсией и/или делецией по меньшей мере одного основания, предпочтительно по меньшей мере 2 оснований, более предпочтительно по меньшей мере 5 оснований и наиболее предпочтительно по меньшей мере 10 оснований, но предпочтительно не более 100 оснований, особенно предпочтительно не более 50 оснований и наиболее предпочтительно не более 25 оснований, где эта производная предпочтительно кодирует белок или пептид, который способен превращать 3-гидрокситетрадеканоил-АСР через 3-гидрокситетрадеканоил-3-гидрокситетрадеканоил-АСР в 3-гидрокситетрадеканоил-3-гидрокситетрадекановую кислоту, и
G1b) последовательность, комплементарная последовательности согласно одной из групп A1b)-F1b), особенно предпочтительно согласно группе A1b), и
А1с) последовательность согласно Seq ID №77, где эта последовательность кодирует белок, который способен превращать 3-гидрокситетрадеканоил-АСР через 3-гидрокситетрадеканоил-3-гидрокситетрадеканоил-АСР в 3-гидрокситетрадеканоил-3-гидрокситетрадекановую кислоту,
В1с) свободная от интронов последовательность, которая происходит из последовательности согласно А1с) и которая кодирует такой же белок или пептид, что и последовательность согласно Seq ID №77,
С1с) последовательность, которая кодирует белок или пептид, который содержит аминокислотную последовательность согласно Seq ID №78 и который предпочтительно способен превращать 3-гидрокситетрадеканоил-АСР через 3-гидрокситетрадеканоил-3-гидрокситетрадеканоил-АСР в 3-гидрокситетрадеканоил-3-гидрокситетрадекановую кислоту,
D1c) последовательность, которая идентична последовательности согласно одной из групп А1с)-С1с), особенно предпочтительно согласно группе А1с), на по меньшей мере 70%, особенно предпочтительно на по меньшей мере 90%, более предпочтительно на по меньшей мере 95% и наиболее предпочтительно на по меньшей мере 99%, где эта последовательность предпочтительно кодирует белок или пептид, который способен превращать 3-гидрокситетрадеканоил-АСР через 3-гидрокситетрадеканоил-3-гидрокситетрадеканоил-АСР в 3-гидрокситетрадеканоил-3-гидрокситетрадекановую кислоту,
Е1с) последовательность, которая гибридизуется или, принимая во внимание вырожденность генетического кода, будет гибридизоваться с комплементарной нитью последовательности согласно одной из групп А1с)-D1c), особенно предпочтительно согласно группе А1с), где эта последовательность предпочтительно кодирует белок или пептид, который способен превращать 3-гидрокситетрадеканоил-АСР через 3-гидрокситетрадеканоил-3-гидрокситетрадеканоил-АСР в 3-гидрокситетрадеканоил-3-гидрокситетрадекановую кислоту,
F1c) производная последовательности согласно одной из групп А1с)-Е1с), особенно предпочтительно согласно группе А1с), полученная замещением, добавлением, инверсией и/или делецией по меньшей мере одного основания, предпочтительно по меньшей мере 2 оснований, более предпочтительно по меньшей мере 5 оснований и наиболее предпочтительно по меньшей мере 10 оснований, но предпочтительно не более 100 оснований, особенно предпочтительно не более 50 оснований и наиболее предпочтительно не более 25 оснований, где эта производная предпочтительно кодирует белок или пептид, который способен превращать 3-гидрокситетрадеканоил-АСР через 3-гидрокситетрадеканоил-3-гидрокситетрадеканоил-АСР в 3-гидрокситетрадеканоил-3-гидрокситетрадекановую кислоту, и
G1c) последовательность, комплементарная последовательности согласно одной из групп А1с)-F1c), особенно предпочтительно согласно группе А1с), и
A1d) последовательность согласно Seq ID №79, где эта последовательность кодирует белок, который способен превращать 3-гидрокситетрадеканоил-АСР через 3-гидрокситетрадеканоил-3-гидрокситетрадеканоил-АСР в 3-гидрокситетрадеканоил-3-гидрокситетрадекановую кислоту,
B1d) свободная от интронов последовательность, которая происходит из последовательности согласно A1d) и которая кодирует такой же белок или пептид, что и последовательность согласно Seq ID №79,
C1d) последовательность, которая кодирует белок или пептид, который содержит аминокислотную последовательность согласно Seq ID №80 и который предпочтительно способен превращать 3-гидрокситетрадеканоил-АСР через 3-гидрокситетрадеканоил-3-гидрокситетрадеканоил-АСР в 3-гидрокситетрадеканоил-3-гидрокситетрадекановую кислоту,
D1d) последовательность, которая идентична последовательности согласно одной из групп A1d)-C1d), особенно предпочтительно согласно группе A1d), на по меньшей мере 70%, особенно предпочтительно на по меньшей мере 90%, более предпочтительно на по меньшей мере 95% и наиболее предпочтительно на по меньшей мере 99%, где эта последовательность предпочтительно кодирует белок или пептид, который способен превращать 3-гидрокситетрадеканоил-АСР через 3-гидрокситетрадеканоил-3-гидрокситетрадеканоил-АСР в 3-гидрокситетрадеканоил-3-гидрокситетрадекановую кислоту,
E1d) последовательность, которая гибридизуется или, принимая во внимание вырожденность генетического кода, будет гибридизоваться с комплементарной нитью последовательности согласно одной из групп A1d)-D1d), особенно предпочтительно согласно группе A1d), где эта последовательность предпочтительно кодирует белок или пептид, который способен превращать 3-гидрокситетрадеканоил-АСР через 3-гидрокситетрадеканоил-3-гидрокситетрадеканоил-АСР в 3-гидрокситетрадеканоил-3-гидрокситетрадекановую кислоту,
F1d) производная последовательности согласно одной из групп A1d)-E1d), особенно предпочтительно согласно группе A1d), полученная замещением, добавлением, инверсией и/или делецией по меньшей мере одного основания, предпочтительно по меньшей мере 2 оснований, более предпочтительно по меньшей мере 5 оснований и наиболее предпочтительно по меньшей мере 10 оснований, но предпочтительно не более 100 оснований, особенно предпочтительно не более 50 оснований и наиболее предпочтительно не более 25 оснований, где эта производная предпочтительно кодирует белок или пептид, который способен превращать 3-гидрокситетрадеканоил-АСР через 3-гидрокситетрадеканоил-3-гидрокситетрадеканоил-АСР в 3-гидрокситетрадеканоил-3-гидрокситетрадекановую кислоту, и
G1d) последовательность, комплементарная последовательности согласно одной из групп A1d)-F1d), особенно предпочтительно согласно группе A1d), и
А1е) последовательность согласно Seq ID №81, где эта последовательность кодирует белок, который способен превращать 3-гидрокситетрадеканоил-АСР через 3-гидрокситетрадеканоил-3-гидрокситетрадеканоил-АСР в 3-гидрокситетрадеканоил-3-гидрокситетрадекановую кислоту,
В1е) свободная от интронов последовательность, которая происходит из последовательности согласно А1е) и которая кодирует такой же белок или пептид, что и последовательность согласно Seq ID №81,
С1е) последовательность, которая кодирует белок или пептид, который содержит аминокислотную последовательность согласно Seq ID №82 и который предпочтительно способен превращать 3-гидрокситетрадеканоил-АСР через 3-гидрокситетрадеканоил-3-гидрокситетрадеканоил-АСР в 3-гидрокситетрадеканоил-3-гидрокситетрадекановую кислоту,
D1e) последовательность, которая идентична последовательности согласно одной из групп А1е)-С1е), особенно предпочтительно согласно группе А1е), на по меньшей мере 70%, особенно предпочтительно на по меньшей мере 90%, более предпочтительно на по меньшей мере 95% и наиболее предпочтительно на по меньшей мере 99%, где эта последовательность предпочтительно кодирует белок или пептид, который способен превращать 3-гидрокситетрадеканоил-АСР через 3-гидрокситетрадеканоил-3-гидрокситетрадеканоил-АСР в 3-гидрокситетрадеканоил-3-гидрокситетрадекановую кислоту,
Е1е) последовательность, которая гибридизуется или, принимая во внимание вырожденность генетического кода, будет гибридизоваться с комплементарной нитью последовательности согласно одной из групп А1е)-D1e), особенно предпочтительно согласно группе А1е), где эта последовательность предпочтительно кодирует белок или пептид, который способен превращать 3-гидрокситетрадеканоил-АСР через 3-гидрокситетрадеканоил-3-гидрокситетрадеканоил-АСР в 3-гидрокситетрадеканоил-3-гидрокситетрадекановую кислоту,
F1e) производная последовательности согласно одной из групп А1е)-Е1е), особенно предпочтительно согласно группе А1е), полученная замещением, добавлением, инверсией и/или делецией по меньшей мере одного основания, предпочтительно по меньшей мере 2 оснований, более предпочтительно по меньшей мере 5 оснований и наиболее предпочтительно по меньшей мере 10 оснований, но предпочтительно не более 100 оснований, особенно предпочтительно не более 50 оснований и наиболее предпочтительно не более 25 оснований, где эта производная предпочтительно кодирует белок или пептид, который способен превращать 3-гидрокситетрадеканоил-АСР через 3-гидрокситетрадеканоил-3-гидрокситетрадеканоил-АСР в 3-гидрокситетрадеканоил-3-гидрокситетрадекановую кислоту, и
G1e) последовательность, комплементарная последовательности согласно одной из групп А1е)-F1e), особенно предпочтительно согласно группе А1е), и
группа [А2-G2] состоит из следующих последовательностей:
А2а) последовательность согласно Seq ID №3, где эта последовательность кодирует белок, который способен превращать dTDP-рамнозу и 3-гидроксидеканоил-3-гидроксидекановую кислоту в α-L-рамнопиранозил-3-гидроксидеканоил-3-гидроксидекановую кислоту,
В2а) свободная от интронов последовательность, которая происходит из последовательности согласно А2а) и которая кодирует такой же белок или пептид, что и последовательность согласно Seq ID №3,
С2а) последовательность, которая кодирует белок или пептид, который содержит аминокислотную последовательность согласно Seq ID №4 и который предпочтительно способен превращать dTDP-рамнозу и 3-гидроксидеканоил-3-гидроксидекановую кислоту в α-L-рамнопиранозил-3-гидроксидеканоил-3-гидроксидекановую кислоту,
D2a) последовательность, которая идентична последовательности согласно одной из групп А2а)-С2а), особенно предпочтительно согласно группе А2а), на по меньшей мере 80%, особенно предпочтительно на по меньшей мере 90%, более предпочтительно на по меньшей мере 95% и наиболее предпочтительно на по меньшей мере 99%, где эта последовательность предпочтительно кодирует белок или пептид, который способен превращать dTDP-рамнозу и 3-гидроксидеканоил-3-гидроксидекановую кислоту в α-L-рамнопиранозил-3-гидроксидеканоил-3-гидроксидекановую кислоту,
Е2а) последовательность, которая гибридизуется или, принимая во внимание вырожденность генетического кода, будет гибридизоваться с комплементарной нитью последовательности согласно одной из групп А2а)-D2a), особенно предпочтительно согласно группе А2а), где эта последовательность предпочтительно кодирует белок или пептид, который способен превращать dTDP-рамнозу и 3-гидроксидеканоил-3-гидроксидекановую кислоту в α-L-рамнопиранозил-3-гидроксидеканоил-3-гидроксидекановую кислоту,
F2a) производная последовательности согласно одной из групп А2а)-Е2а), особенно предпочтительно согласно группе А2а), полученная замещением, добавлением, инверсией и/или делецией по меньшей мере одного основания, предпочтительно по меньшей мере 2 оснований, более предпочтительно по меньшей мере 5 оснований и наиболее предпочтительно по меньшей мере 10 оснований, но предпочтительно не более 100 оснований, особенно предпочтительно не более 50 оснований и наиболее предпочтительно не более 25 оснований, где эта производная предпочтительно кодирует белок или пептид, который способен превращать dTDP-рамнозу и 3-гидроксидеканоил-3-гидроксидекановую кислоту в α-L-рамнопиранозил-3-гидроксидеканоил-3-гидроксидекановую кислоту,
G2a) последовательность, комплементарная последовательности согласно одной из групп А2а) - F2a), особенно предпочтительно согласно группе А2а),
A2b) последовательность согласно Seq ID №19, где эта последовательность кодирует белок, который способен превращать dTDP-рамнозу и 3-гидрокситетрадеканоил-3-гидрокситетрадекановую кислоту в α-L-рамнопиранозил-3-гидрокситетрадеканоил-3-гидрокситетрадекановую кислоту,
B2b) свободная от интронов последовательность, которая происходит из последовательности согласно A2b) и которая кодирует такой же белок или пептид, что и последовательность согласно Seq ID №19,
C2b) последовательность, которая кодирует белок или пептид, который содержит аминокислотную последовательность согласно Seq ID №20 и который предпочтительно способен превращать dTDP-рамнозу и 3-гидрокситетрадеканоил-3-гидрокситетрадекановую кислоту в α-L-рамнопиранозил-3-гидрокситетрадеканоил-3-гидрокситетрадекановую кислоту,
D2b) последовательность, которая идентична последовательности согласно одной из групп A2b)-C2b), особенно предпочтительно согласно группе A2b), на по меньшей мере 70%, особенно предпочтительно на по меньшей мере 90%, более предпочтительно на по меньшей мере 95% и наиболее предпочтительно на по меньшей мере 99%, где эта последовательность предпочтительно кодирует белок или пептид, который способен превращать dTDP-рамнозу и 3-гидрокситетрадеканоил-3-гидрокситетрадекановую кислоту в α-L-рамнопиранозил-3-гидрокситетрадеканоил-3-гидрокситетрадекановую кислоту,
E2b) последовательность, которая гибридизуется или, принимая во внимание вырожденность генетического кода, будет гибридизоваться с комплементарной нитью последовательности согласно одной из групп A2b)-D2b), особенно предпочтительно согласно группе A2b), где эта последовательность предпочтительно кодирует белок или пептид, который способен превращать dTDP-рамнозу и 3-гидрокситетрадеканоил-3-гидрокситетрадекановую кислоту в α-L-рамнопиранозил-3-гидрокситетрадеканоил-3-гидрокситетрадекановую кислоту,
F2b) производная последовательности согласно одной из групп A2b)-E2b), особенно предпочтительно согласно группе A2b), полученная замещением, добавлением, инверсией и/или делецией по меньшей мере одного основания, предпочтительно по меньшей мере 2 оснований, более предпочтительно по меньшей мере 5 оснований и наиболее предпочтительно по меньшей мере 10 оснований, но предпочтительно не более 100 оснований, особенно предпочтительно не более 50 оснований и наиболее предпочтительно не более 25 оснований, где эта производная предпочтительно кодирует белок или пептид, который способен превращать dTDP-рамнозу и 3-гидрокситетрадеканоил-3-гидрокситетрадекановую кислоту в α-L-рамнопиранозил-3-гидрокситетрадеканоил-3-гидрокситетрадекановую кислоту, и
G2b) последовательность, комплементарная последовательности согласно одной из групп A2b)-F2b), особенно предпочтительно согласно группе A2b),
А2с) последовательность согласно Seq ID №83, где эта последовательность кодирует белок, который способен превращать dTDP-рамнозу и 3-гидрокситетрадеканоил-3-гидрокситетрадекановую кислоту в α-L-рамнопиранозил-3-гидрокситетрадеканоил-3-гидрокситетрадекановую кислоту,
В2с) свободная от интронов последовательность, которая происходит из последовательности согласно А2с) и которая кодирует такой же белок или пептид, что и последовательность согласно Seq ID №83,
С2с) последовательность, которая кодирует белок или пептид, который содержит аминокислотную последовательность согласно Seq ID №84 и который предпочтительно способен превращать dTDP-рамнозу и 3-гидрокситетрадеканоил-3-гидрокситетрадекановую кислоту в α-L-рамнопиранозил-3-гидрокситетрадеканоил-3-гидрокситетрадекановую кислоту,
D2c) последовательность, которая идентична последовательности согласно одной из групп А2с)-С2с), особенно предпочтительно согласно группе А2с), на по меньшей мере 70%, особенно предпочтительно на по меньшей мере 90%, более предпочтительно на по меньшей мере 95% и наиболее предпочтительно на по меньшей мере 99%, где эта последовательность предпочтительно кодирует белок или пептид, который способен превращать dTDP-рамнозу и 3-гидрокситетрадеканоил-3-гидрокситетрадекановую кислоту в α-L-рамнопиранозил-3-гидрокситетрадеканоил-3-гидрокситетрадекановую кислоту,
Е2с) последовательность, которая гибридизуется или, принимая во внимание вырожденность генетического кода, будет гибридизоваться с комплементарной нитью последовательности согласно одной из групп А2с)-D2c), особенно предпочтительно согласно группе А2с), где эта последовательность предпочтительно кодирует белок или пептид, который способен превращать dTDP-рамнозу и 3-гидрокситетрадеканоил-3-гидрокситетрадекановую кислоту в α-L-рамнопиранозил-3-гидрокситетрадеканоил-3-гидрокситетрадекановую кислоту,
F2c) производная последовательности согласно одной из групп А2с)-Е2с), особенно предпочтительно согласно группе А2с), полученная замещением, добавлением, инверсией и/или делецией по меньшей мере одного основания, предпочтительно по меньшей мере 2 оснований, более предпочтительно по меньшей мере 5 оснований и наиболее предпочтительно по меньшей мере 10 оснований, но предпочтительно не более 100 оснований, особенно предпочтительно не более 50 оснований и наиболее предпочтительно не более 25 оснований, где эта производная предпочтительно кодирует белок или пептид, который способен превращать dTDP-рамнозу и 3-гидрокситетрадеканоил-3-гидрокситетрадекановую кислоту в α-L-рамнопиранозил-3-гидрокситетрадеканоил-3-гидрокситетрадекановую кислоту, и
G2c) последовательность, комплементарная последовательности согласно одной из групп А2с)-F2c), особенно предпочтительно согласно группе А2с),
A2d) последовательность согласно Seq ID №85, где эта последовательность кодирует белок, который способен превращать dTDP-рамнозу и 3-гидрокситетрадеканоил-3-гидрокситетрадекановую кислоту в α-L-рамнопиранозил-3-гидрокситетрадеканоил-3-гидрокситетрадекановую кислоту,
B2d) свободная от интронов последовательность, которая происходит из последовательности согласно A2d) и которая кодирует такой же белок или пептид, что и последовательность согласно Seq ID №85,
C2d) последовательность, которая кодирует белок или пептид, который содержит аминокислотную последовательность согласно Seq ID №86 и который предпочтительно способен превращать dTDP-рамнозу и 3-гидрокситетрадеканоил-3-гидрокситетрадекановую кислоту в α-L-рамнопиранозил-3-гидрокситетрадеканоил-3-гидрокситетрадекановую кислоту,
D2d) последовательность, которая идентична последовательности согласно одной из групп A2d)-C2d), особенно предпочтительно согласно группе A2d), на по меньшей мере 70%, особенно предпочтительно на по меньшей мере 90%, более предпочтительно на по меньшей мере 95% и наиболее предпочтительно на по меньшей мере 99%, где эта последовательность предпочтительно кодирует белок или пептид, который способен превращать dTDP-рамнозу и 3-гидрокситетрадеканоил-3-гидрокситетрадекановую кислоту в α-L-рамнопиранозил-3-гидрокситетрадеканоил-3-гидрокситетрадекановую кислоту,
E2d) последовательность, которая гибридизуется или, принимая во внимание вырожденность генетического кода, будет гибридизоваться с комплементарной нитью последовательности согласно одной из групп A2d)-D2d), особенно предпочтительно согласно группе A2d), где эта последовательность предпочтительно кодирует белок или пептид, который способен превращать dTDP-рамнозу и 3-гидрокситетрадеканоил-3-гидрокситетрадекановую кислоту в α-L-рамнопиранозил-3-гидрокситетрадеканоил-3-гидрокситетрадекановую кислоту,
F2d) производная последовательности согласно одной из групп A2d)-E2d), особенно предпочтительно согласно группе A2d), полученная замещением, добавлением, инверсией и/или делецией по меньшей мере одного основания, предпочтительно по меньшей мере 2 оснований, более предпочтительно по меньшей мере 5 оснований и наиболее предпочтительно по меньшей мере 10 оснований, но предпочтительно не более 100 оснований, особенно предпочтительно не более 50 оснований и наиболее предпочтительно не более 25 оснований, где эта производная предпочтительно кодирует белок или пептид, который способен превращать dTDP-рамнозу и 3-гидрокситетрадеканоил-3-гидрокситетрадекановую кислоту в α-L-рамнопиранозил-3-гидрокситетрадеканоил-3-гидрокситетрадекановую кислоту, и
G2d) последовательность, комплементарная последовательности согласно одной из групп A2d)-F2d), особенно предпочтительно согласно группе A2d), и
А2е) последовательность согласно Seq ID №87, где эта последовательность кодирует белок, который способен превращать dTDP-рамнозу и 3-гидрокситетрадеканоил-3-гидрокситетрадекановую кислоту в α-L-рамнопиранозил-3-гидрокситетрадеканоил-3-гидрокситетрадекановую кислоту,
В2е) свободная от интронов последовательность, которая происходит из последовательности согласно А2е) и которая кодирует такой же белок или пептид, что и последовательность согласно Seq ID №87,
С2е) последовательность, которая кодирует белок или пептид, который содержит аминокислотную последовательность согласно Seq ID №88 и который предпочтительно способен превращать dTDP-рамнозу и 3-гидрокситетрадеканоил-3-гидрокситетрадекановую кислоту в α-L-рамнопиранозил-3-гидрокситетрадеканоил-3-гидрокситетрадекановую кислоту,
D2e) последовательность, которая идентична последовательности согласно одной из групп А2е)-С2е), особенно предпочтительно согласно группе А2е), на по меньшей мере 70%, особенно предпочтительно на по меньшей мере 90%, более предпочтительно на по меньшей мере 95% и наиболее предпочтительно на по меньшей мере 99%, где эта последовательность предпочтительно кодирует белок или пептид, который способен превращать dTDP-рамнозу и 3-гидрокситетрадеканоил-3-гидрокситетрадекановую кислоту в α-L-рамнопиранозил-3-гидрокситетрадеканоил-3-гидрокситетрадекановую кислоту,
Е2е) последовательность, которая гибридизуется или, принимая во внимание вырожденность генетического кода, будет гибридизоваться с комплементарной нитью последовательности согласно одной из групп А2е)-D2e), особенно предпочтительно согласно группе А2е), где эта последовательность предпочтительно кодирует белок или пептид, который способен превращать dTDP-рамнозу и 3-гидрокситетрадеканоил-3-гидрокситетрадекановую кислоту в α-L-рамнопиранозил-3-гидрокситетрадеканоил-3-гидрокситетрадекановую кислоту,
F2e) производная последовательности согласно одной из групп А2е)-Е2е), особенно предпочтительно согласно группе А2е), полученная замещением, добавлением, инверсией и/или делецией по меньшей мере одного основания, предпочтительно по меньшей мере 2 оснований, более предпочтительно по меньшей мере 5 оснований и наиболее предпочтительно по меньшей мере 10 оснований, но предпочтительно не более 100 оснований, особенно предпочтительно не более 50 оснований и наиболее предпочтительно не более 25 оснований, где эта производная предпочтительно кодирует белок или пептид, который способен превращать dTDP-рамнозу и 3-гидрокситетрадеканоил-3-гидрокситетрадекановую кислоту в α-L-рамнопиранозил-3-гидрокситетрадеканоил-3-гидрокситетрадекановую кислоту, и
G2e) последовательность, комплементарная последовательности согласно одной из групп А2е)-F2e), особенно предпочтительно согласно группе А2е), и
группа [A3-G3] состоит из следующих последовательностей:
А3а) последовательность согласно Seq ID №5, где эта последовательность кодирует белок, который способен превращать dTDP-рамнозу и α-L-рамнопиранозил-3-гидроксидеканоил-3-гидроксидекановую кислоту в α-L-рамнопиранозил-(1-2)-α-L-рамнопиранозил-3-гидроксидеканоил-3-гидроксидекановую кислоту,
В3а) свободная от интронов последовательность, которая происходит из последовательности согласно А3а) и которая кодирует такой же белок или пептид, что и последовательность согласно Seq ID №5,
С3а) последовательность, которая кодирует белок или пептид, который содержит аминокислотную последовательность согласно Seq ID №6 и который предпочтительно способен превращать dTDP-рамнозу и α-L-рамнопиранозил-3-гидроксидеканоил-3-гидроксидекановую кислоту в α-L-рамнопиранозил-(1-2)-α-L-рамнопиранозил-3-гидроксидеканоил-3-гидроксидекановую кислоту,
D3a) последовательность, которая идентична последовательности согласно одной из групп А3а)-С3а), особенно предпочтительно согласно группе А3а), на по меньшей мере 80%, особенно предпочтительно на по меньшей мере 90%, более предпочтительно на по меньшей мере 95% и наиболее предпочтительно на по меньшей мере 99%, где эта последовательность предпочтительно кодирует белок или пептид, который способен превращать dTDP-рамнозу и α-L-рамнопиранозил-3-гидроксидеканоил-3-гидроксидекановую кислоту в α-L-рамнопиранозил-(1-2)-α-L-рамнопиранозил-3-гидроксидеканоил-3-гидроксидекановую кислоту,
Е3а) последовательность, которая гибридизуется или, принимая во внимание вырожденность генетического кода, будет гибридизоваться с комплементарной нитью последовательности согласно одной из групп А3а)-D3a), особенно предпочтительно согласно группе А3а), где эта последовательность предпочтительно кодирует белок или пептид, который способен превращать dTDP-рамнозу и α-L-рамнопиранозил-3-гидроксидеканоил-3-гидроксидекановую кислоту в α-L-рамнопиранозил-(1-2)-α-L-рамнопиранозил-3-гидроксидеканоил-3-гидроксидекановую кислоту,
F3a) производная последовательности согласно одной из групп А3а)-Е3а), особенно предпочтительно согласно группе А3а), полученная замещением, добавлением, инверсией и/или делецией по меньшей мере одного основания, предпочтительно по меньшей мере 2 оснований, более предпочтительно по меньшей мере 5 оснований и наиболее предпочтительно по меньшей мере 10 оснований, но предпочтительно не более 100 оснований, особенно предпочтительно не более 50 оснований и наиболее предпочтительно не более 25 оснований, где эта производная предпочтительно кодирует белок или пептид, который способен превращать dTDP-рамнозу и α-L-рамнопиранозил-3-гидроксидеканоил-3-гидроксидекановую кислоту в α-L-рамнопиранозил-(1-2)-α-L-рамнопиранозил-3-гидроксидеканоил-3-гидроксидекановую кислоту,
G3a) последовательность, комплементарная последовательности согласно одной из групп А3а)-F3a), особенно предпочтительно согласно группе А3а),
A3b) последовательность согласно Seq ID №21, где эта последовательность кодирует белок, который способен превращать dTDP-рамнозу и α-L-рамнопиранозил-3-гидрокситетрадеканоил-3-гидрокситетрадекановую кислоту в α-L-рамнопиранозил-(1-2)-α-L-рамнопиранозил-3-гидрокситетрадеканоил-3-гидрокситетрадекановую кислоту,
B3b) свободная от интронов последовательность, которая происходит из последовательности согласно A3b) и которая кодирует такой же белок или пептид, что и последовательность согласно Seq ID №21,
C3b) последовательность, которая кодирует белок или пептид, который содержит аминокислотную последовательность согласно Seq ID №22 и который предпочтительно способен превращать dTDP-рамнозу и α-L-рамнопиранозил-3-гидрокситетрадеканоил-3-гидрокситетрадекановую кислоту в α-L-рамнопиранозил-(1-2)-α-L-рамнопиранозил-3-гидрокситетрадеканоил-3-гидрокситетрадекановую кислоту,
D3b) последовательность, которая идентична последовательности согласно одной из групп A3b)-C3b), особенно предпочтительно согласно группе A3b), на по меньшей мере 60%, особенно предпочтительно на по меньшей мере 90%, более предпочтительно на по меньшей мере 95% и наиболее предпочтительно на по меньшей мере 99%, где эта последовательность предпочтительно кодирует белок или пептид, который способен превращать dTDP-рамнозу и α-L-рамнопиранозил-3-гидрокситетрадеканоил-3-гидрокситетрадекановую кислоту в α-L-рамнопиранозил-(1-2)-α-L-рамнопиранозил-3-гидрокситетрадеканоил-3-гидрокситетрадекановую кислоту,
E3b) последовательность, которая гибридизуется или, принимая во внимание вырожденность генетического кода, будет гибридизоваться с комплементарной нитью последовательности согласно одной из групп A3b)-D3b), особенно предпочтительно согласно группе A3b), где эта последовательность предпочтительно кодирует белок или пептид, который способен превращать dTDP-рамнозу и α-L-рамнопиранозил-3-гидрокситетрадеканоил-3-гидрокситетрадекановую кислоту в α-L-рамнопиранозил-(1-2)-α-L-рамнопиранозил-3-гидрокситетрадеканоил-3-гидрокситетрадекановую кислоту,
F3b) производная последовательности согласно одной из групп A3b)-E3b), особенно предпочтительно согласно группе A3b), полученная замещением, добавлением, инверсией и/или делецией по меньшей мере одного основания, предпочтительно по меньшей мере 2 оснований, более предпочтительно по меньшей мере 5 оснований и наиболее предпочтительно по меньшей мере 10 оснований, но предпочтительно не более 100 оснований, особенно предпочтительно не более 50 оснований и наиболее предпочтительно не более 25 оснований, где эта производная предпочтительно кодирует белок или пептид, который способен превращать dTDP-рамнозу и α-L-рамнопиранозил-3-гидрокситетрадеканоил-3-гидрокситетрадекановую кислоту в α-L-рамнопиранозил-(1-2)-α-L-рамнопиранозил-3-гидрокситетрадеканоил-3-гидрокситетрадекановую кислоту, и
G3b) последовательность, комплементарная последовательности согласно одной из групп A3b)-F3b), особенно предпочтительно согласно группе A3b),
А3с) последовательность согласно Seq ID №89, где эта последовательность кодирует белок, который способен превращать dTDP-рамнозу и α-L-рамнопиранозил-3-гидрокситетрадеканоил-3-гидрокситетрадекановую кислоту в α-1-рамнопиранозил-(1-2)-α-L-рамнопиранозил-3-гидрокситетрадеканоил-3-гидрокситетрадекановую кислоту,
В3с) свободная от интронов последовательность, которая происходит из последовательности согласно А3с) и которая кодирует такой же белок или пептид, что и последовательность согласно Seq ID №89,
С3с) последовательность, которая кодирует белок или пептид, который содержит аминокислотную последовательность согласно Seq ID №90 и который предпочтительно способен превращать dTDP-рамнозу и α-L-рамнопиранозил-3-гидрокситетрадеканоил-3-гидрокситетрадекановую кислоту в α-L-рамнопиранозил-(1-2)-α-L-рамнопиранозил-3-гидрокситетрадеканоил-3-гидрокситетрадекановую кислоту,
D3c) последовательность, которая идентична последовательности согласно одной из групп А3с)-С3с), особенно предпочтительно согласно группе А3с), на по меньшей мере 60%, особенно предпочтительно на по меньшей мере 90%, более предпочтительно на по меньшей мере 95% и наиболее предпочтительно на по меньшей мере 99%, где эта последовательность предпочтительно кодирует белок или пептид, который способен превращать dTDP-рамнозу и α-L-рамнопиранозил-3-гидрокситетрадеканоил-3-гидрокситетрадекановую кислоту в α-L-рамнопиранозил-(1-2)-α-L-рамнопиранозил-3-гидрокситетрадеканоил-3-гидрокситетрадекановую кислоту,
Е3с) последовательность, которая гибридизуется или, принимая во внимание вырожденность генетического кода, будет гибридизоваться с комплементарной нитью последовательности согласно одной из групп А3с)-D3c), особенно предпочтительно согласно группе А3с), где эта последовательность предпочтительно кодирует белок или пептид, который способен превращать dTDP-рамнозу и α-L-рамнопиранозил-3-гидрокситетрадеканоил-3-гидрокситетрадекановую кислоту в α-L-рамнопиранозил-(1-2)-α-L-рамнопиранозил-3-гидрокситетрадеканоил-3-гидрокситетрадекановую кислоту,
F3c) производная последовательности согласно одной из групп А3с)-Е3с), особенно предпочтительно согласно группе А3с), полученная замещением, добавлением, инверсией и/или делецией по меньшей мере одного основания, предпочтительно по меньшей мере 2 оснований, более предпочтительно по меньшей мере 5 оснований и наиболее предпочтительно по меньшей мере 10 оснований, но предпочтительно не более 100 оснований, особенно предпочтительно не более 50 оснований и наиболее предпочтительно не более 25 оснований, где эта производная предпочтительно кодирует белок или пептид, который способен превращать dTDP-рамнозу и α-L-рамнопиранозил-3-гидрокситетрадеканоил-3-гидрокситетрадекановую кислоту в α-L-рамнопиранозил-(1-2)-α-L-рамнопиранозил-3-гидрокситетрадеканоил-3-гидрокситетрадекановую кислоту, и
G3c) последовательность, комплементарная последовательности согласно одной из групп А3с)-F3c), особенно предпочтительно согласно группе А3с), и
A3d) последовательность согласно Seq ID №91, где эта последовательность кодирует белок, который способен превращать dTDP-рамнозу и α-L-рамнопиранозил-3-гидрокситетрадеканоил-3-гидрокситетрадекановую кислоту в α-L-рамнопиранозил-(1-2)-α-L-рамнопиранозил-3-гидрокситетрадеканоил-3-гидрокситетрадекановую кислоту,
B3d) свободная от интронов последовательность, которая происходит из последовательности согласно A3d) и которая кодирует такой же белок или пептид, что и последовательность согласно Seq ID №91,
C3d) последовательность, которая кодирует белок или пептид, который содержит аминокислотную последовательность согласно Seq ID №92 и который предпочтительно способен превращать dTDP-рамнозу и α-L-рамнопиранозил-3-гидрокситетрадеканоил-3-гидрокситетрадекановую кислоту в α-L-рамнопиранозил-(1-2)-α-L-рамнопиранозил-3-гидрокситетрадеканоил-3-гидрокситетрадекановую кислоту,
D3d) последовательность, которая идентична последовательности согласно одной из групп A3d)-C3d), особенно предпочтительно согласно группе A3d), на по меньшей мере 60%, особенно предпочтительно на по меньшей мере 90%, более предпочтительно на по меньшей мере 95% и наиболее предпочтительно на по меньшей мере 99%, где эта последовательность предпочтительно кодирует белок или пептид, который способен превращать dTDP-рамнозу и α-L-рамнопиранозил-3-гидрокситетрадеканоил-3-гидрокситетрадекановую кислоту в α-L-рамнопиранозил-(1-2)-α-L-рамнопиранозил-3-гидрокситетрадеканоил-3-гидрокситетрадекановую кислоту,
E3d) последовательность, которая гибридизуется или, принимая во внимание вырожденность генетического кода, будет гибридизоваться с комплементарной нитью последовательности согласно одной из групп A3d)-D3d), особенно предпочтительно согласно группе A3d), где эта последовательность предпочтительно кодирует белок или пептид, который способен превращать dTDP-рамнозу и α-L-рамнопиранозил-3-гидрокситетрадеканоил-3-гидрокситетрадекановую кислоту в α-L-рамнопиранозил-(1-2)-α-L-рамнопиранозил-3-гидрокситетрадеканоил-3-гидрокситетрадекановую кислоту,
F3d) производная последовательности согласно одной из групп A3d)-E3d), особенно предпочтительно согласно группе A3d), полученная замещением, добавлением, инверсией и/или делецией по меньшей мере одного основания, предпочтительно по меньшей мере 2 оснований, более предпочтительно по меньшей мере 5 оснований и наиболее предпочтительно по меньшей мере 10 оснований, но предпочтительно не более 100 оснований, особенно предпочтительно не более 50 оснований и наиболее предпочтительно не более 25 оснований, где эта производная предпочтительно кодирует белок или пептид, который способен превращать dTDP-рамнозу и α-L-рамнопиранозил-3-гидрокситетрадеканоил-3-гидрокситетрадекановую кислоту в α-L-рамнопиранозил-(1-2)-α-L-рамнопиранозил-3-гидрокситетрадеканоил-3-гидрокситетрадекановую кислоту, и
G3d) последовательность, комплементарная последовательности согласно одной из групп A3d)-F3d), особенно предпочтительно согласно группе A3d).
"Нуклеотидная идентичность" или "аминокислотная идентичность" согласно настоящему изобретению определяются известными способами. Как правило, применяются конкретные компьютерные программы, в которых учитываются конкретные требования.
Предпочтительные способы определения идентичности согласно настоящему изобретению обеспечивают наилучшее сопоставление сравниваемых последовательностей. Компьютерные программы для определения идентичности содержат, но без ограничения к этому, GCG-программное обеспечение, включая GAP (Deveroy, J. et al., Nucleic Acid Research 12 (1984), Seite 387, Genetics Computer Group University of Wisconsin, Medicine (Wi), и BLASTP, BLASTN и FASTA (Altschul, S. et al., Journal of Molecular Biology 215 (1990), Seiten 403-410. BLAST программа может быть доступна от National Center For Biotechnology Information (NCBI) и из других источников (руководство BLAST, Altschul S. et al., NCBI NLM NIH Bethesda ND 22894; Altschul S. et al., выше).
Известный алгоритм Смита-Ватермана может подобным образом применяться для определения нуклеотидной идентичности.
Предпочтительные параметры для определения "нуклеотидной идентичности" при применении программы BLASTN (Altschul, S. et al., Journal of Molecular Biology 215 (1990), Seiten 403-410) представляют собой:
Вышеприведенные параметры являются параметрами по умолчанию для сравнения нуклеотидных последовательностей.
Программа GAP подобным образом подходит для применения с приведенными выше параметрами.
Предпочтительными параметрами для определения "аминокислотной идентичности" при применении программы BLASTP (Altschul, S. et al., Journal of Molecular Biology 215 (1990), Seiten 403-410), являются следующие:
Вышеприведенные параметры представляют собой параметры по умолчанию для сравнения аминокислотных последовательностей.
Программа GAP подобным образом подходит для применения с вышеприведенными параметрами.
Идентичность 60% согласно вышеприведенному алгоритму означает идентичность 60% в контексте настоящего изобретения. То же относится к большим идентичностям.
Выражение "последовательность, которая гибридизуется или, принимая во внимание вырожденность генетического кода, будет гибридизоваться с комплементарной нитью последовательности" означает последовательность, которая при предпочтительно жестких условиях гибридизации гибридизуется или, принимая во внимание вырожденность генетического кода, будет гибридизоваться с комлементарной нитью ссылочной последовательности. Например, гибридизация может осуществляться при 68°С в 2×SSC или согласно протоколу для Digoxigenin-Labelling-Kits компании Boehringer (Mannheim). Предпочтительными условиями гибридизации является, например, инкубация при б5°С всю ночь в 7% SDS, 1% BSA, 1 мМ EDTA, 250 мМ натрий-фосфатного буфера (рН 7,2) с последующим промыванием при 65°С с помощью 2×SSC; 0,1% SDS.
Производные выделенных ДНК согласно настоящему изобретению, которые согласно альтернативам F1), F2) или F3) могут быть получены замещением, добавлением, инверсией и/или делецией одного или более оснований последовательности согласно одной из групп А1)-Е1), А2)-Е2) и A3)-Е3), включают в частности те последовательности, которые приводят к консервативным аминокислотным заменам в белке, который они кодируют, как например, к замене глицина на аланин или аспарагиновой кислоты на глутаминовую кислоту. Такие функционально нейтральные мутации описываются как смысловые мутации (sense mutations) и не приводят к фундаментальной модификации активности полипептида. Кроме того, известно, что замены на N- и/или С-концах полипептида существенно не влияют на его функцию и могут даже стабилизировать его, так что настоящее изобретение также охватывает последовательности ДНК, в которых основания присоединяются на 3’-конце или 5’-конце последовательности, содержащей нуклеиновые кислоты согласно настоящему изобретению. Специалист в данной области техники может найти соответствующую информацию, среди прочего в Веп-Bassat et at. (Journal of Bacteriology 169:751-757 (1987)), в O’Regan et al. (Gene 77:237-251 (1989)), в Sahin-Toth et al. (Protein Sciences 3:240-247 (1994)), в Hochuli et al. (Bio/Technology 6:1321-1325 (1988)) и в известных учебниках по генетике и молекулярной биологии.
Нуклеиновая кислота согласно настоящему изобретению предпочтительно представляет собой вектор, в частности экспрессионный вектор или генную сверхэкспрессионную кассету. Подходящими векторами являются все вектора, известные специалистам в данной области техники, которые обычно применяются для включения ДНК в клетку-хозяин. Эти вектора могут как реплицироваться самостоятельно, так и иметь ориджины репликации, как например, в 2р плазмиде или ARS (самостоятельно реплицирующиеся последовательности) или интегрироваться в хромосомы (нерепликативные плазмиды). Вектора, как понимается, также означают линейные фрагменты ДНК, которые не имеют ориджинов репликации вообще, как например, кассеты генной инсерции или генной сверхэкспрессии. Кассеты генной сверхэкспрессии обычно состоят из маркера, генов, подлежащих сверхэкспрессии, а также регуляторных областей, ответственных за экспрессию генов, как например, промоторы и терминаторы. Предпочтительные вектора выбираются из группы, содержащей плазмиды и кассеты, как например, Е. Coli шатл-плазмиды дрожжей; экспрессионные вектора, кассеты генной инсерции или генной сверхэкспрессии особенно предпочтительны, в частности вектора Seq ID №38, Seq ID №40, Seq ID №42, Seq ID №45 и Seq ID №47.
Согласно предпочтительному варианту выполнения вектора согласно настоящему изобретению, последовательности групп [А1-G1], [А2-G2] и [A3-G3] находятся под контролем по меньшей мере одного конститутивного или регулируемого промотора, который подходит для экспрессии полипептида, кодируемого этими последовательностями ДНК в клетке микроорганизма, предпочтительно клетки бактерий, грибов или дрожжей, причем Aspergillus nidulans, Aspergillus niger, Alcaligenes latus, Bacillus megatehum, Bacillus subtilis, Brevibacterium flavum, Brevibacterium lactofermentum, Burkholderia andropogonis, B. brasilensis, B. caledonica, B. caribensis, B. caryophylli, B. fungorum, B. gladioli, B. glathei, B. glumae, B. graminis, B. hospita, B. kururiensis, B. phenazinium, B. phymatum, B. phytofirmans, B. plantarii, B. sacchari, B. singaporensis, B. sordidicola, B. temcola, B. tropica, B. tuberum, B. ubonensis, B. unamae, B. xenovorans, B. anthina, B. pyrrocinia, B. thailandensis, Candida blankii, Candida rugosa, Corynebacterium glutamicum, Corynebacterium efficiens, Escherichia coli, Hansenula polymorpha, Kluveromyces lactis, Methylobacterium extorquens, Paracoccus versutus, Pseudomonas argentinensis, P. borbori, P. citronellolis, P. flavescens, P. mendocina, P. nitroreducens, P. oleovorans, P. pseudoalcaligenes, P. resinovorans, P. straminea, P. aurantiaca, P. aureofaciens, P. chlororaphis, P. fragi, P. lundensis, P. taetrolens, P. antarctica, P. azotoformans, ‘P. blatchfordae’, P. brassicacearum, P. brenneri, P. cedrina, P. corrugate, P. fluorescens, P. gessardii, P. libanensis, P. mandelii, P. marginalis, P. mediterranea, P. meridiana, P. migulae, P. mucidolens, P. orientalis, P. panacis, P. proteolytica, P. rhodesiae, P. synxantha, P. thivervalensis, P. tolaasii, P. veronii, P. denitrificans, P. pertucinogena, P. cremoricolorata, P. fulva, P. monteilii, P. mosselii, P. parafulva, P. putida, P. balearica, P. stutzeri, P. amygdali, P. avellanae, P. caricapapayae, P. cichorii, P. coronafaciens, P. ficuserectae, ‘P. helianthi’, P. meliae, P. savastanoi, P. syringae, P. tomato, P. viridiflava, P. abietaniphila, P. acidophila, P. agahci, P. alcaliphila, P. alkanolytica, P. amyloderamosa, P. asplenii, P. azotifigens, P. cannabina, P. coenobios, P. congelans, P. costantinii, P. cruciviae, P. delhiensis, P. excibis, P. extremorientalis, P. frederiksbergensis, P. fuscovaginae, P. gelidicola, P. grimontii, P. indica, P. jessenii, P. jinjuensis, P. kilonensis, P. knackmussii, P. koreensis, P. lini, P. lutea, P. moraviensis, P. otitidis, P. pachastrellae, P. palleroniana, P. papaveris, P. peli, P. perolens, P. poae, P. pohangensis, P. psychrophila, P. psychrotolerans, P. rathonis, P. reptilivora, P. resiniphila, P. rhizosphaerae, P. rubescens, P. salomonii, P. segitis, P. septica, P. simiae, P. suis, P. thermotolerans, P. aeruginosa, P. tremae, P. trivialis, P. turbinellae, P. tuticorinensis, P. umsongensis, P. vancouverensis, P. vranovensis, P. xanthomarina, Ralstonia eutropha, Rhodospirillum rubrum, Rhodobacter sphaeroides, Saccharomyces cerevisiae, Yarrowia lipolytica, Zymomonas mobilis, в частности Pseudomonas putida, Escherichia coli и Burkholderia thailandensis, особенно предпочтительны.
Примерами конститутивных промоторов являются lac, lacUV5, tac, trc (в каждом случае в отсутствии Lad репрессора в клетках согласно настоящему изобретению), Ltet-O1 (в каждом случае в отсутствии TetR репрессора в клетках согласно настоящему изобретению), Т5 и дар. Примерами индуцируемых промоторов являются lac, lacUV5, tac, trc (в каждом случае в присутствии LacI репрессора в клетках согласно настоящему изобретению), Ltet-O1 (в каждом случае в присутствии TetR репрессора в клетках согласно настоящему изобретению), Т5 (в комбинации с lac оператором и в присутствии LacI репрессора в клетках согласно настоящему изобретению), SP6 и Т7 (в присутствии гена, кодирующего родственную РНК-полимеразу, экспрессия которой, в свою очередь, регулируется). Вектор согласно настоящему изобретению должен в дополнение к промотору предпочтительно содержать сайт связывания рибосом, а также терминатор. Особенно предпочтительно согласно настоящему изобретению, чтобы нуклеиновая кислота согласно настоящему изобретению встраивалась в экспрессионную кассету вектора, содержащего промотор, сайт связывания рибосом и терминатор. В дополнение к вышеупомянутым структурным элементам вектор может дополнительно содержать гены селекции, известные специалистам в данной области техники.
Все приведенные проценты (%) представляют собой проценты по массе, если иного не указано.
В приведенных далее примерах настоящее изобретение описывается примерным образом, без ограничения объема настоящего изобретения, который определяется приведенным описанием и приложенной формулой изобретения.
Краткое описание чертежей:
Фиг.1: показаны биосинтез жирных кислот, β-окисление жирных кислот и связь этих метаболических путей с биосинтезом рамнолипидов (ферменты E1, Е2 и Е3) и полигидроксиалканоатов (ферменты E9 и Е10). Подача углерода для биосинтеза жирных кислот, β-окисление жирных кислот, биосинтез рамнолипидов и биосинтез полигидроксиалканоатов. Потребление и формирование коэнзимов, окислительно-восстановительные равновесия и нуклеотида также показаны.
Фиг.2: образование дирамнозиллипида (мг/л/OD 600 нм) рекомбинантных штаммов Р. putida KT2440 pBBR1MCS-2 и pBBR1MCS-2::ABC, а также GPp104 pBBR1MCS-2 и pBBR1MCS-2::ABC после 48 часов, 72 часов и 96 часов культивирования в среде СМР. Анализ концентрации рамнолипидов осуществляется посредством ВЭЖХ.
Фиг.3: образование монорамнозиллипида (площадь пика/OD 600 нм) рекомбинантных штаммов Р. putida KT2440 pBBR1MCS-2, pBBRIMCS-2::АВ и pBBR1MCS-2::ABM, а также GPp104 pBBR1MCS-2, pBBRIMCS-2::АВ и pBBR1MCS-2::ABM после 72 часов, 96 часов и 120 часов культивирования в среде СМР. Анализ концентрации рамнолипидов осуществляется посредством ВЭЖХ.
Примеры:
1. Конструирование вектора pBBR1MCS-2::AB для гетерологичной экспрессии генов rhIA и rhIB Pseudomonas aeruginosa 1707 в Pseudomonas putida
Для гетерологичной экспрессии Pseudomonas aeruginosa DSM1707 генов rhlA и rhlB, сконструировали плазмиду pBBR1MCS-2::AB (Seq ID №38). Для этого синтетический оперон rhlAB (Seq ID №37) был синтезирован фирмой GeneArt AG (Regensburg) и клонирован в коммерческом векторе рМА (GeneArt AG). В качестве основы для синтеза применялась уже известная геномная последовательность Pseudomonas aeruginosa DSM1707. Исходя из вектора рМА::АВ синтетический оперон был выделен из вектора посредством BglII и XbaI и далее лигирован в подобным образом расщепленный с помощью BamHI и XbaI экспрессионный вектор pBBR1MCS-2 (Seq ID №49) (описано Kovach et al., 1995: Four new derivatives of the broad-host-range cloning vector pBBRIMCS carrying different antibiotic-resistance cassettes. Gene, 166:175-176). Полученная плазмида pBBR1MCS-2::AB (Seq ID №38) составляет 7422 пар оснований по размеру. Лигирование и трансформация химически компетентных клеток Е. coli DH5a (Gibco-BRL, Karlsruhe) осуществлялась образом, известным специалистам в данной области техники. Аутентичность вставки была проверена посредством анализа последовательности ДНК.
Трансформация Pseudomonas putida KT2440 und GPp104 с применением векторов pBBR1MCS-2 (Seq ID №49) и pBBR1MCS-2::AB осуществляется как описано ранее (Iwasaki et al. Biosci. Biotech. Biochem. 1994. 58(5):851-854). ДНК плазмида каждых десяти клонов была выделена и проанализирована. Полученные штаммы, несущие плазмиду, были названы Р. putida KT2440 pBBR1MCS-2, Р. putida GPp104 pBBR1MCS-2, Р. putida KT2440 pBBR1MCS-2::AB и Р. putida GPp104 pBBR1MCS-2::AB.
2. Конструирование вектора pBBR1MCS-2::ABC для гетерологичной экспрессии Pseudomonas aeruginosa DSM1707 генов rhlA, rhlB и rhlC в Pseudomonas putida
Для гетерологичной экспрессии Pseudomonas aeruginosa DSM1707 генов rhlA, rhlB и rhlC, сконструировали плазмиду pBBR1MCS-2::ABC (Seq ID №40). Для этого синтетический оперон rhlABC (Seq ID №39) был синтезирован фирмой GeneArt AG (Regensburg) и клонирован в коммерческом векторе рМА (GeneArt AG). В качестве основы для синтеза применялась уже известная геномная последовательность Pseudomonas aeruginosa DSM1707. Исходя из вектора рМА::АВС синтетический оперон был выделен из вектора посредством BglII и XbaI и далее лигирован в подобным образом расщепленный с помощью BamHI и XbaI экспрессионный вектор pBBR1MCS-2 (Seq ID №49) (Kovach et al., 1995: Four new derivatives of the broad-host-range cloning vector pBBRIMCS carrying different antibiotic-resistance cassettes. Gene, 166:175-176). Полученная плазмида pBBR1MCS-2::ABC (Seq ID №40) состояла из 8409 пар оснований. Лигирование и трансформация химически компетентных клеток Е. coli DH5α (Gibco-BRL, Karlsruhe) осуществлялась образом, известным специалистам в данной области техники. Аутентичность вставки была проверена посредством анализа последовательности ДНК.
Трансформация Pseudomonas putida KT2440 и GPp104 с применением вектора pBBR1MCS-2::ABC осуществляется как описано ранее (Iwasaki et al. Biosci. Biotech. Biochem. 1994. 58(5):851-854). ДНК плазмида каждых десяти клонов была выделена и проанализирована. Полученные штаммы, несущие плазмиду, были названы Р. putida KT2440 pBBR1MCS-2::ABC и Р. putida GPp104 pBBR1MCS-2::ABC.
3. Конструирование вектора pBBR1MCS-2::ABM для гетерологичной экспрессии Pseudomonas aeruginosa DSM1707 генов rhlA, rhlB и ра1131 в Pseudomonas putida
Для гетерологичной экспрессии Pseudomonas aeruginosa DSM1707 генов rhlA, rhlB и ра1131, сконструировали плазмиду pBBR1MCS-2::ABM (Seq ID №42). Для этого синтетический оперон rhlAB-pa1131 (Seq ID №41) был синтезирован фирмой GeneArt AG (Regensburg) и клонирован в коммерческом векторе рМА (GeneArt AG). В качестве основы для синтеза применялась уже известная геномная последовательность Pseudomonas aeruginosa DSM1707. Исходя из вектора рМА::АВМ синтетический оперон был выделен из вектора посредством BglII и XbaI и далее лигирован в подобным образом расщепленный с помощью BamHI и XbaI экспрессионный вектор pBBR1MCS-2 (Seq ID №49) (Kovach et al., 1995: Four new derivatives of the broad-host-range cloning vector pBBRIMCS carrying different antibiotic-resistance cassettes. Gene, 166:175-176). Полученная плазмида pBBR1MCS-2::ABM (Seq ID №42) состояла из 8702 пар оснований. Лигирование и трансформация химически компетентных клеток Е. coli DH5α (Gibco-BRL, Karlsruhe) осуществлялась образом, известным специалистам в данной области техники. Аутентичность вставки была проверена посредством анализа последовательности ДНК.
Трансформация Pseudomonas putida KT2440 и GPp104 с помощью вектора pBBR1MCS-2::ABM осуществляется как описано ранее (Iwasaki et al. Biosci. Biotech. Biochem. 1994. 58(5):851-854). ДНК плазмида каждых десяти клонов была выделена и проанализирована. Полученные штаммы, несущие плазмиду, были названы Р. putida KT2440 pBBR1MCS-2::ABM и Р. putida GPp104 pBBR1MCS-2::ABM.
4. Количественное определение рамнолипидов, продуцируемых рекомбинантными штаммами Р. putida
Рекомбинантные штаммы Р. putida KT2440 pBBR1MCS-2; Р. putida KT2440 pBBR1MCS-2::AB; P. putida KT2440 pBBR1MCS-2::ABC; P. putida KT2440 pBBR1MCS-2::ABM; Р. putida GPp104 pBBR1MCS-2; Р. putida GPp104 pBBR1MCS-2::AB, Р. putida GPp104 pBBR1MCS-2::ABC и Р. putida GPp104 pBBR1MCS-2::ABM были культивированы на чашках с LB-агар-канамицином (50 мкг/мл).
Для получения рамнолипидов применялась среда, обозначенная ниже как среда СМР. Она состоит из 2% (мас./об.) глюкозы, 0,007% (мас./об.) KH3PO4, 0,11% Na2HPO4×2H2O, 0,2% (мас./об.) NaNO3, 0,04% (мас./об.) MgSO4×H2O, 0,01% (мас./об.) CaCl2×2H2O и 0,2% (об./об.) раствора микроэлементов. Он состоит из 0,2% (мас./об.) FeSO4×7Н2О, 0,15% (мас./об.) MnSO4×H2O и 0,06% (мас./об.) (NH4)MO7O24×4H2O. Значение рН среды было доведено с помощью NaOH до 6,7, и среда затем была стерилизована с помощью автоклава (121°С, 20 минут). Регулировка рН в ходе культивации не является обязательной.
Для исследования продуцирования рамнолипидов сначала была получена прекультура во встряхиваемой колбе. Для этого применялся инокулят штамма, свежо нанесенный на LB-агаровую чашку, и 10 мл LB среды инокулировали в колбе Эрленмейера, объемом 100 мл. Все рекомбинантные штаммы Р. putida были культивированы в среде LB, в которую добавили 50 мкг/мл канамицина. Культивирование штаммов осуществляли всю ночь при 30°С и 200 оборотах в минуту.
Прекультуры применялись для инокулирования 50 мл среды СМР в колбе Эрленмейера, объемом 250 мл (старт OD600 0,1). Культуры культивировали при 200 оборотах в минуту и при 30°С в течение самое большое 120 часов. С интервалами, равными 24 часам, брали образцы бульона объемом 1 мл из колбы с культурой. Получение образца для последующих хроматографических анализов осуществляли следующим образом:
Применяя поршневую пипетку (Combitip) 1 мл ацетона ввели в реакционный сосуд, объемом 2 мл, и затем реакционный сосуд немедленно закрыли для минимизации испарения. Затем добавили 1 мл бульона. После вортексирования смеси бульон/ацетон, ее центрифугировали в течение 3 минут при 13.000 оборотах в минуту, и 800 мкл супернатанта перенесли в сосуд для ВЭЖХ.
Для обнаружения и количественного определения рамнолипидов применялся испарительный детектор светорассеяния (Sedex LT-ELSD Model 85LT). Точно измерение осуществляли с помощью Agilent Technologies 1200 Series (Santa Clara, Kalifornien) и Zorbax SB-C8 колонок быстрого разрешения (4,6×150 мм, 3,5 мкМ, Agilent). Впрыскиваемый объем составлял 5 мкл, и время удерживания способа составляло 20 минут. В качестве подвижной фазы применялась водная 0,1% TFA (трифторуксусная кислота, раствор А) и метанол (раствор В). Температура колонки составляла 40°С. В качестве детекторов применялись ELSD (температура детектора 60°С) и DAD (матричный диодный фотодетектор, 210 нм). В способе применялся следующий градиент:
В то время как Р. putida KT2440 pBBR1MCS-2 и GPp104 pBBR1MCS-2 не продуцировали рамнолипиды, в рекомбинантных штаммах Р. putida KT2440 pBBR1MCS-2::AB, P. putida KT2440 pBBR1MCS-2::ABC, P. putida KT2440 pBBR1MCS-2::ABM, P. putida GPp104 pBBR1MCS-2::AB, P. putida GPp104 pBBR1MCS-2::ABC и Р. putida GPp104 pBBR1MCS-2::ABM было обнаружено образование различных видов рамнолипидов (Фиг.2 и 3).
Путем встраивания pBBR1MCS-2::AB и pBBR1MCS-2::ABM в Р. putida было возможно получить монорамнозиллипиды (Фиг.3). Так как никакого контрольного материала для монорамнозиллипидов не было представлено, идентификация продуктов осуществлялась путем анализа соответствующих массовых следов и MS2-спектра в LC-MS.
Если rhlC (pBBR1MCS-2::ABC) был дополнительно включен в штаммы, были получены моно- и дирамнозиллипиды (Фиг.2).
Прямое сравнение образования рамнолипида с применением Р. putida pBBR1MCS-2::AB и Р. putida pBBR1MCS-2::ABM показывает, что соэкспрессия Р. aeruginosa p3111 с Р. aeruginosa rhIAB приводит к улучшению биосинтеза рамнолипидов (Фиг.3). Тогда как штаммы Р. putida КТ2440 pBBR1MCS-2::AB и Р. putida GPp104 pBBR1MCS-2::AB продуцировали около 39 (Р. putida КТ2440 pBBR1MCS-2::AB) и 23 площадей пиков рамнолипидов/OD 600 нм (Р. putida GPp104 pBBR1MCS-2::AB) через 120 часов, штаммы Р. putida KT2440 pBBR1MCS-2::ABM и Р. putida GPp104 pBBR1MCS-2::ABM формировали около 50 (Р. putida КТ2440 pBBR1MCS-2::ABM) и 62 площадей пиков рамнолипидов /OD 600 нм (Р. putida GPp104 pBBR1MCS-2::ABM) через 120 часов.
Если сравнить синтез монорамнозиллипидов штаммов Р. putida КТ2440 pBBR1MCS-2::ABM и Р. putida GPp104 pBBR1MCS-2::ABM, возможно обнаружить в РНА-негативном мутанте Р. putida GPp104 pBBR1MCS-2::ABM 62 площади пиков/OD 600 нм (120 часов культивирования) и для Р. putida КТ2440 pBBR1MCS-2::ABM - 50 площадей/OD 600 нм монорамнозиллипидов (Фиг.3).
Сравнительный анализ образования дирамнозиллипидов (мг/л/OD 600 нм) в штаммах Р. putida КТ2440 и GPp104 подобным образом показывает большее образование дирамнозиллипидов в РИА-негативного штамма Р. putida GPp104. Р. putida GPp104 pBBR1MCS-2::ABC, составляющее в среднем 113 мг/л/OD 600 нм дирамнозиллипидов (96 ч), где для Р. putida КТ2440 pBBR1MCS-2::ABC было обнаружено только 55 мг/л/OD 600 нм дирамнозиллипидов через 96 часов (Фиг.2).
Таким образом, возможно показать, что применение штамма с уменьшенным синтезом РНА приводит к улучшению биосинтеза рамнолипидов.
5. Конструирование вектора pBBR1MCS-2::ABMC для гетерологичной экспрессии Pseudomonas aeruginosa DSM1707 генов rhlA, rhLB, pa1131 и rhIC в Pseudomonas putida
Для гетерологичной экспрессии Pseudomonas aeruginosa DSM1707 генов rhlA, rhlB, pa1131 и rhlC, сконструировали плазмиду pBBR1MCS-2::ABMC (Seq ID №51). Для этого синтетический оперон rhlAB-pal131-rhIC (Seq ID №50) был синтезирован фирмой GeneArtAG (Regensburg) и клонирован в коммерческом векторе рМА (GeneArt AG). В качестве основы для синтеза применялась уже известная геномная последовательность Pseudomonas aeruginosa DSM1707. Исходя из вектора рМА::АВМС синтетический оперон был выделен из вектора посредством BglII и XbaI и далее лигирован в подобным образом расщепленный с помощью BamHI и XbaI экспрессионный вектор pBBR1MCS-2 (Seq ID №49) (Kovach et al., 1995: Four new derivatives of the broad-host-range cloning vector pBBRIMCS carrying different antibiotic-resistance cassettes. Gene, 166:175-176). Полученная плазмида pBBR1MCS-2::ABMC (Seq ID №51) состояла из 9663 пар оснований. Лигирование и трансформация химически компетентных клеток Е. coli DH5α (Gibco-BRL, Karlsruhe) осуществлялась образом, известным специалистам в данной области техники. Аутентичность вставки была проверена посредством анализа последовательности ДНК.
Трансформация Pseudomonas putida KT2440 и GPp104 с помощью вектора pBBR1MCS-2::ABMC осуществляется как описано ранее (Iwasaki et al. Biosci. Biotech. Biochem. 1994. 58(5):851-854). ДНК плазмида каждых десяти клонов была выделена и проанализирована. Полученные штаммы, несущие плазмиду, были названы Р. putida KT2440 pBBR1MCS-2::ABMC и Р. putida GPp104 pBBR1MCS-2::ABMC.
6. Количественное сравнение рамнолипидов, продуцируемых рекомбинантным штаммом Р. putida и штаммом Р. aeruginosa
Рекомбинантные штаммы Р. putida GPp104 pBBR1MCS-2 и Р. putida GPp104 pBBR1MCS-2::ABMC, а также Р. aeruginosa DSM 19880 были культивированы на чашках с LB-агар-канамицином (50 мкг/мл; Р. putida) и чашках с LB-агаром (Р. aeruginosa).
Для получения рамнолипидов применялась среда, обозначенная ниже как среда СМР. Она состоит из 2% (мас./об.) глюкозы, 0,007% (мас./об.) KH2PO4, 0,11% Na2HPO4×2H2O, 0,2% (мас./об.) NaNO3, 0,04% (мас./об.) MgSO4×H2O, 0,01% (мас./об.) CaCl2×2H2O и 0,2% (об./об.) раствора микроэлементов. Он состоит из 0,2% (мас./об.) FeSO4×7H2O, 0,15% (мас./об.) MnSO4×H2O и 0,06% (мас./об.) (NH4)MO7O24×4H2O. Значение рН среды было доведено с помощью NaOH до 6,7, и среда затем была стерилизована с помощью автоклава (121°С, 20 минут). Регулировка рН в ходе культивации не является обязательной.
Для исследования продуцирования рамнолипидов сначала была получена прекультура во встряхиваемой колбе. Для этого применялся инокулят штамма, свежо нанесенный на LB-агаровую чашку, и 10 мл LB среды инокулировали в колбе Эрленмейера, объемом 100 мл. Рекомбинантные штаммы Р. putida были культивированы в среде LB, в которую добавили 50 мкг/мл канамицина. Р. aeruginosa культивировали в среде LB. Культивирование штаммов осуществляли всю ночь при 30°С и 200 оборотах в минуту.
Прекультуры применялись для инокулирования 50 мл среды СМР в колбе Эрленмейера, объемом 250 мл (старт OD600 0,1). Культуры культивировали при 200 оборотах в минуту и при 30°С в течение самое большое 120 часов. С интервалами, равными 24 часам, брали образцы бульона объемом 1 мл из колбы с культурой. Получение образца для последующих хроматографических анализов осуществляли следующим образом:
Применяя поршневую пипетку (Combitip) 1 мл ацетона ввели в реакционный сосуд, объемом 2 мл, и затем реакционный сосуд немедленно закрыли для минимизации испарения. Затем добавили 1 мл бульона. После вортексирования смеси бульон/ацетон, ее центрифугировали в течение 3 минут при 13.000 оборотах в минуту, и 800 мкл супернатанта перенесли в сосуд для ВЭЖХ.
Для идентификации полученного продукта 5 мкл впрыскивали в UPLC-Aniage (Thermo Scientific, Dreieich). Исследуемые вещества анализировали с применением колонки semi UPLC "Pursuit XRs ULTRA" (C8, 2,8 мкМ, 2,1×100 мМ) (Varian, Darmstadt). Разделение осуществляли в течение 25 минут посредством градиента, состоящего из подвижной фазы А1 (H2O, 0,1% (об./об.) TFA) и подвижной фазы В1 (Methanol, 0,1% (об./об.) TFA) со скоростью потока 0,3 мл/мин при 40°С. Применялся следующий градиент:
Обнаружение осуществляли посредством детектора DAD в интервале длин волн 200-600 нм, масс-селективно с применением масс-спектрометра FT-ICR LTQ-FT высокого разрешения (Thermo Scientific, Dreieich) в диапазоне сканирования m/z 100-1000. Ионизация осуществлялась посредством ESI (электрораспылительная ионизация). Точные массы и эмпирические химические формулы определяли с помощью FT-ICR масс-анализатора, применяя разрешение R=100000, с точностью массы менее ≤2 частей на миллион. Идентификация продуктов осуществлялась путем анализа соответствующих массовых следов и MS2 спектра в LC-MS. Для возможности сравнения штаммов площади пиков соответствующих веществ были сделаны контрастными.
Как показано на Фиг.4, штамм Р. putida GPp104 pBBR1MCS-2 вообще не продуцирует рамнолипиды. Р. putida GPp104 pBBR1MCS-2::ABMC и Р. aeruginosa DSM 19880 образуют рамнолипиды, где отношение между ди- и монорамнозиллипидами, образованными Р. putida GPp104 pBBR1MCS-2::ABMC составляло, например, 4:1, а для Р. aeruginosa DSM 19880, например, 2:1. Более того, штамм Р. putida GPp104 pBBR1MCS-2::ABMC в отличие от Р. aeruginosa DSM 19880 не образовывал или образовывал незначительное количество рамнолипидов, имеющих радикал, определенный R1 и R2, полученный из 3-гидроксиоктаноил-3-гидроксидекановой кислоты или 3-гидроксидеканоил-3-гидроксиоктановой кислоты.
7. Конструирование вектора pBBR1MCS-2::rfbBDAC и pBBRIMCS-2::ABC_rfbBDAC для гетерологической экспрессии в Pseudomonas putida
Компанией Trenzyme GmbH (Konstanz) оперон биосинтеза рамнозы rfbBDAC был амплифицирован исходя из хромосомной ДНК Pseudomonas putida KT2440. Для этого применялся следующий праймер:
RL1: 5’-TATATATAGAATTCGCGTCATCTGTCTACGACAACAC-3’ (Seq ID №48)
RL2: 5’-TATATATAGAATTCGGCTGCGCTACCGCAGCCCTTC-3’ (Seq ID №43)
Полученный ПЦР продукт был клонирован в Trenzyme’s Alligator клонирующую систему и трансформирован в Е. coli DH5α (New England Biolabs; Frankfurt). Векторы различных кандидатов были проанализированы и секвенированы. После успешного и безошибочного секвенирования ДНК вектор был расщеплен с помощью EcoRI, и целевой фрагмент rfbBDAC был выделен. Для последующего клонирования вектор pBBR1MCS-2 (Kovach et al., 1995: Four new derivatives of the broad-host-range cloning vector pBBRIMCS carrying different antibiotic-resistance cassettes. Gene, 166:175-176) был расщеплен таким же образом. Расщепленный целевой фрагмент (rfbBDAC) и расщепленный вектор (pBBR1MCS-2) были объединены с помощью обычной лигации. Полученный вектор pBBR1MCS-2::rfbBDAC (Seq ID №45) был подобным образом трансформирован в Е. coli DH5α (New England Biolabs; Frankfurt). Некоторые кандидаты трансформантов были исследованы в отношении успешного включения плазмиды.
Вектор pBBR1MCS-2::rfbBDAC служил в качестве матрицы для ПЦР. Применялись следующие олигонуклеотиды:
RL_Xbal-fw: 5’-TATATATATCTAGAATTAATGCAGCTGGCACGAC-3’ (Seq ID №44)
RL_Xba_rev: 5’-GGCCGCTCTAGAACTAGTGGA-3’ (Seq ID №46)
ПЦР проводили с применением полимеразы Phusion™ High-Fidelity Master Mix фирмы New England Biolabs (Frankfurt). ПЦР осуществлялась образом, известным специалистам в данной области техники. Целевая последовательность (lac промотор и rfbBDAC) клонировали в Trenzyme Alligator клонирующую систему. Е. coli DH5α (New England Biolabs; Frankfurt) трансформанты отобрали, и ДНК плазмиду различных кандидатов выделили и секвенировали. После того, как последовательность была проверена и исследована в отношении неточностей, вектор был расщеплен с применением XbaI. Целевой фрагмент был лигирован в подобным образом расщепленный с помощью XbaI pBBR1MCS-2::ABC (смотрите выше) с помощью обычных способов лигации. Полученный целевой вектор pBBR1MCS-2::ABC_rfbBDAC (Seq ID №47) имел размер 12249 пар оснований. Вставку вектора подвергли секвенированию. Осуществление ПЦР, проверку успешности амплификации ПЦР посредством электрофореза на агарозном геле, окрашивание ДНК бромидом этидия, определение размера фрагмента ПЦР, очищение продуктов ПЦР и определение концентрации ДНК осуществлялись образом, известным специалистам в данной области техники.
Трансформация Pseudomonas putida KT2440 и GPp104 с помощью вектора pBBR1MCS-2::ABC_rfbBDAC осуществляется как описано ранее (Iwasaki et al. Biosci. Biotech. Biochem. 1994. 58(5):851-854). ДНК плазмида каждых десяти клонов была выделена и проанализирована. Полученные штаммы, несущие плазмиду, были названы Р. putida KT2440 pBBRIMCS-2::ABC_rfbBDAC и Р. putida GPp104 pBBR1MCS-2::ABC_rfbBDAC.
8. Количественное определение рамнолипидов, продуцируемых рекомбинантными штаммами Р. putida с и без сверхэкспрессии оперона rfbBDAC
Рекомбинантные штаммы Р. putida KT2440 pBBR1MCS-2; P. putida KT2440 pBBR1MCS-2::ABC, P. putida KT2440 pBBR1MCS-2::ABC_rfbBDAC, P. putida GPp104 pBBR1MCS-2, P. putida GPp104 pBBR1MCS-2::ABC и Р. putida GPp104 pBBR1MCS-2::ABC_rfbBDAC были культивированы на чашках с LB-агар-канамицином (50 мкг/мл).
Для получения рамнолипидов применялась среда, обозначенная ниже как среда СМР. Она состоит из 2% (мас./об.) глюкозы, 0,007% (мас./об.) KH2PO4, 0,11% Na2HPO4×2H2O, 0,2% (мас./об.) NaNO3, 0,04% (мас./об.) MgSO4×H2O, 0,01% (мас./об.) CaCl2×2Н2О и 0,2% (об./об.) раствора микроэлементов. Он состоит из 0,2% (мас./об.) FeSO4×7H2O, 0,15% (мас./об.) MnSO4×H2O и 0,06% (мас./об.) (NH4)MO7O24×4H2O. Значение рН среды было доведено с помощью NaOH до 6,7, и среда затем была стерилизована с помощью автоклава (121°С, 20 минут). Регулировка рН в ходе культивации не является обязательной.
Для исследования продуцирования рамнолипидов сначала была получена прекультура во встряхиваемой колбе. Для этого применялся инокулят штамма, свежо нанесенный на LB-агаровую чашку, и 10 мл LB среды инокулировали в колбе Эрленмейера, объемом 100 мл. Все рекомбинантные штаммы Р. putida были культивированы в среде LB, в которую добавили 50 мкг/мл канамицина. Культивирование штаммов Р. putida осуществляли всю ночь при 30°С и 200 оборотах в минуту.
Прекультуры применялись для инокулирования 50 мл среды СМР в колбе Эрленмейера, объемом 250 мл (старт OD600 0,1). Культуры культивировали при 200 оборотах в минуту и при 30°С в течение самое большое 120 часов. С интервалами, равными 24 часам, брали образцы бульона объемом 1 мл из колбы с культурой. Получение образца для последующих хроматографических анализов осуществляли следующим образом:
Применяя поршневую пипетку (Combitip) 1 мл ацетона ввели в реакционный сосуд, объемом 2 мл, и затем реакционный сосуд немедленно закрыли для минимизации испарения. Затем добавили 1 мл бульона. После вортексирования смеси бульон/ацетон, ее центрифугировали в течение 3 минут при 13.000 оборотах в минуту, и 800 мкл супернатанта перенесли в сосуд для ВЭЖХ.
Для обнаружения и количественного определения рамнолипидов применялся испарительный детектор светорассеяния (Sedex LT-ELSD Model 85LT). Точно измерение осуществляли с помощью Agilent Technologies 1200 Series (Santa Clara, Kalifornien) и Zorbax SB-C8 колонок быстрого разрешения (4,6×150 мм, 3,5 мкМ, Agilent). Впрыскиваемый объем составлял 5 мкл, и время удерживания способа составляло 20 минут. В качестве подвижной фазы применялась водная 0,1% TFA (трифторуксусная кислота, раствор А) и метанол (раствор В). Температура колонки составляла 40°С. В качестве детекторов применялись ELSD (температура детектора 60°С) и DAD (матричный диодный фотодетектор, 210 нм). В способе применялся следующий градиент:
В то время как Р. putida KT2440 pBBR1MCS-2 и GPp104 pBBR1MCS-2 не продуцировали рамнолипиды, в рекомбинантных штаммах Р. putida KT2440 pBBR1MCS-2::ABC, P. putida KT2440 pBBR1MCS-2::ABC_rfbBDAC; Р. putida GPp104 pBBR1MCS-2::ABC и Р. putida GPp104 pBBRIMCS-2::ABC_rfbBDAC было обнаружено образование различных видов рамнолипидов.
Р. putida KT2440 pBBR1MCS-2::ABC_rfbBDAC по сравнению с Р. putida KT2440 pBBR1MCS-2::ABC и Р. putida GPp104 pBBR1MCS-2::ABC_rfbBDAC по сравнению с Р. putida GPp104 pBBR1MCS-2::ABC показывают повышенное образование ди- и монорамнозиллипидов. Это ясно показывает положительное влияние амплификации экспрессии rfbBDAC на образование моно- и дирамнозиллипидов.
Если сравнивать биосинтез моно- и дирамнозиллипидов штаммов Р. putida KT2440 pBBR1MCS-2::ABC_rfbBDAC и Р. putida GPp104 pBBRIMCS-2::ABC_rfbBDAC, повышенный синтез моно- и дирамнозиллипидов обнаруживается в РНА-негативом мутанте Р. putida GPp104 pBBRIMCS-2::ABC_rfbBDAC.
Как уже описано выше, биосинтез рамнолипидов повышается при применении штамма, инактивированного в отношении синтеза РНА.
9. Создание рекомбинантной Е. coli W3110 pBBR1MCS-2::ABC и Е. coli W3110 pBBR1MCS-2::ABC_rfbBDAC
Трансформация Е. coli W3110 осуществляется как описано ранее (Miller JH. A Short Course in Bacterial Genetics: A Laboratory Manual and Handbook for Escherichia coli and Related Bacteria. Plainview, NY: Cold Spring Harbor Lab. Press; 1992) с помощью электропорации. ДНК плазмида каждых десяти клонов была выделена и проанализирована. Полученные штаммы, несущие плазмиду, были названы Е. coli W3110 pBBRIMCS-2::ABC_rfbBDAC и Е. coli W3110 pBBR1MCS-2::ABC_rfbBDAC.
10. Количественное определение рамнолипидов, продуцируемых рекомбинантными штаммами Е. coli с и без сверхэкспрессии оперона rfbBDAC
Рекомбинантные штаммы Е. coli W3110 pBBR1MCS-2; Е. coli W3110 pBBR1MCS-2::ABC и Е. coli W3110 pBBR1MCS-2::ABC_rfbBDAC были культивированы на чашках с LB-агар-канамицином (50 мкг/мл).
Для получения рамнолипидов применялась среда, обозначенная ниже как среда СМР. Она состоит из 2% (мас./об.) глюкозы, 0,007% (мас./об.) KH2PO4, 0,11% Na2HPO4×2H2O, 0,2% (мас./об.) NaNO3, 0,04% (мас./об.) MgSO4×Н2О, 0,01% (мас./об.) CaCl2×2Н2О и 0,2% (об./об.) раствора микроэлементов. Он состоит из 0,2% (мас./об.) FeSO4×7Н2О, 0,15% (мас./об.) MnSO4×H2O и 0,06% (мас./об.) (NH4)MO7O24×4H2O. Значение рН среды было доведено с помощью NaOH до 6,7, и среда затем была стерилизована с помощью автоклава (121°С, 20 минут). Регулировка рН в ходе культивации не является обязательной.
Для исследования продуцирования рамнолипидов сначала была получена прекультура во встряхиваемой колбе. Для этого применялся инокулят штамма, свежо нанесенный на LB-агаровую чашку, и 10 мл LB среды инокулировали в колбе Эрленмейера, объемом 100 мл. Все рекомбинантные штаммы Е. coli были культивированы в среде LB, в которую добавили 50 мкг/мл канамицина. Культивирование штаммов Е. coli осуществляли всю ночь при 37°С и 200 оборотах в минуту.
Прекультуры применялись для инокулирования 50 мл среды СМР в колбе Эрленмейера, объемом 250 мл (старт OD600 0,1). Культуры культивировали при 200 оборотах в минуту и при 30°С в течение самое большое 120 часов. С интервалами, равными 24 часам, брали образцы бульона объемом 1 мл из колбы с культурой. Получение образца для последующих хроматографических анализов осуществляли следующим образом:
Применяя поршневую пипетку (Combitip) 1 мл ацетона ввели в реакционный сосуд, объемом 2 мл, и затем реакционный сосуд немедленно закрыли для минимизации испарения. Затем добавили 1 мл бульона. После вортексирования смеси бульон/ацетон, ее центрифугировали в течение 3 минут при 13.000 оборотах в минуту, и 800 мкл супернатанта перенесли в сосуд для ВЭЖХ.
Для обнаружения и количественного определения рамнолипидов применялся испарительный детектор светорассеяния (Sedex LT-ELSD Model 85LT). Точно измерение осуществляли с помощью Agilent Technologies 1200 Series (Santa Clara, Kalifornien) и Zorbax SB-C8 колонок быстрого разрешения (4,6×150 мм, 3,5 мкМ, Agilent). Впрыскиваемый объем составлял 5 мкл, и время удерживания способа составляло 20 минут. В качестве подвижной фазы применялась водная 0,1% TFA (трифторуксусная кислота, раствор А) и метанол (раствор В). Температура колонки составляла 40°С. В качестве детекторов применялись ELSD (температура детектора 60°С) и DAD (матричный диодный фотодетектор, 210 нм). В способе применялся следующий градиент:
В то время, как Е. coli W3110 pBBR1MCS-2 не продуцировали рамнолипидов, образование моно- и дирамнозиллипидов обнаруживалось в рекомбинантных штаммах Е. coli W3110 pBBR1MCS-2::ABC и Е. coli W3110 pBBR1MCS-2::ABC_rfbBDAC, где Е. coli W3110 pBBRIMCS-2::ABC_rfbBDAC образует значительно больше моно- и дирамнозиллипидов по сравнению с Е. coli W3110 pBBR1MCS-2::ABC. Это показывает, что гетерологическая экспрессия rhIABC из Pseudomonas aeruginosa DSM1707 приводит к образованию моно- и дирамнозиллипидов в Е. coli. Это, кроме того, показывает положительное влияние усиления экспрессии rfbBDAC на образование моно- и дирамнозиллипидов.
11. Конструирование вектора pBBR1MCS-2::ABC-BTH_II1077-II1080-II1081 для гетерологичной экспрессии Pseudomonas aeruginosa DSM1707 генов rhlA, rhlB и rhLC, а также Burkholderia thailandensis E264 генов BTH_II1077, BT_II1080 и ВТ_II1081 в Pseudomonas putida
Для гетерологичной экспрессии Pseudomonas aeruginosa DSM1707 генов rhlA, rhlB и rhlC, а также В. thailandensis E264 генов BT_II1077, BT_II1080 и BT_II1081 в Pseudomonas putida конструируется плазмида pBBR1MCS-2::АВС-ВТН_II1077-II1080-II1081 (Seq ID №69). Для этого синтетический оперон BTH_II1077, BT_II1080 и BTJI1081 (Seq ID №70) синтезируется компанией DNA 2.0 (Menio Park, CA, USA), и клонируется в коммерческом векторе pJ294 (DNA 2.0; Menio Park, CA, USA). В качестве основы для синтеза применялась уже известная геномная последовательность штамма S. thailandensis E264. Исходя из вектора pJ294-BTH_II1077-II1080-II1081 синтетический оперон был выделен из вектора посредством XbaI и затем лигирован в расщепленный подобным образом с помощью XbaI вектор pBBR1MCS-2::ABC (Seq ID №40). Целевой вектор pBBR1MCS-2::АВС-ВТН_II1077-II1080-II1081 (Seq ID №69) состоял из 13768 пар оснований. Вставку вектора подвергли секвенированию. Осуществление ПЦР, проверку успешности амплификации ПЦР посредством электрофореза на агарозном геле, окрашивание ДНК бромидом этидия, определение размера фрагмента ПЦР, очищение продуктов ПЦР и определение концентрации ДНК осуществлялись образом, известным специалистам в данной области техники.
Трансформация Pseudomonas putida KT2440 и GPp104 с помощью вектора pBBR1MCS-2::ABC-BTH_II1077-II1080-II1081 (Seq ID №69) осуществляется как описано ранее (Iwasaki et al. Biosci. Biotech. Biochem. 1994. 58(5):851-854). ДНК плазмида каждых десяти клонов была выделена и проанализирована. Полученные штаммы, несущие плазмиду, были названы Р. putida KT2440 pBBR1MCS-2::ABC-BTH_II1077-II1080-II1081 и Р. putida GPp104 pBBR1MCS-2::ABC-BTH_II1077-II1080-II1081.
12. Количественное определение рамнолипидов, продуцируемых рекомбинантными штаммами Р. putida с и без сверхэкспрессии В. thailandensis E264 генов BTH_II1077, BT_II1080 и BT_II1081
Рекомбинантные штаммы Р. putida P. putida KT2440 pBBR1MCS-2::AB, P. putida KT2440 pBBR1MCS-2::AB-BTH_II1077-II1080-II1081, P. putida GPp104 pBBR1MCS-2::AB, P. putida GPp104 pBBR1MCS-2::AB-BTH_II1077-II1080-II1081, P. putida KT2440 pBBR1MCS-2::ABC, P. putida KT2440 pBBR1MCS-2::ABC-BTH_II1077-II1080-II1081 P. putida GPp104 pBBR1MCS-2::ABC и Р. putida GPp104 pBBR1MCS-2::ABC-BTH_II1077-II1080-II1081, полученные в примерах 1, 2 и 11 были культивированы на чашках с LB-агар-канамицином (50 мкг/мл).
Для получения рамнолипидов применялась среда, обозначенная ниже как среда М9. Она состоит из 2% (мас./об.) глюкозы, 0,3% (мас./об.) KH2PO4, 0,679% Na2HPO4, 0,05% (мас./об.) NaCl, 0,2% (мас./об.) NH4Cl, 0,049% (мас./об.) MgSO4×7H2O и 0,1% (об./об.) раствора микроэлементов. Он состоит из 1,78% (мас./об.) FeSO4×7Н2О, 0,191% (мас./об.) MnCl2×7H2O, 3,65% (мас./об.) HCl, 0,187% (мас./об.) ZnSO4×7H2O, 0,084% (об./об.) Na-EDTA×2H2O, 0,03% (об./об.) Н3ВО3, 0,025% (мас./об.) Na2MoO4×2H2O и 0,47% (мас./об.) CaCl2×2Н2О. Значение рН среды было доведено с помощью NH4OH до 7,4, и среда затем была стерилизована с помощью автоклава (121°С, 20 минут). Регулировка рН в ходе культивации не является обязательной.
Для исследования продуцирования рамнолипидов сначала была получена прекультура во встряхиваемой колбе. Для этого применялся инокулят штамма, свежо нанесенный на LB-агаровую чашку, и 10 мл LB среды инокулировали в колбе Эрленмейера, объемом 100 мл. Все рекомбинантные штаммы Р. putida были с среде LB, в которую добавили 50 мкг/мл канамицина. Культивирование всех штаммов Р. putida осуществляли в среде LB, 50 мкг/мл, в которую добавили 50 мкг/мл канамицина. Культивирование штаммов Р. putida осуществляли при 37°С и 200 оборотах в минуту всю ночь.
Прекультуры применялись для инокулирования 50 мл среды М9 (+50 мкг/мл канамицина) в колбе Эрленмейера, объемом 250 мл (старт OD600 0,1). Культуры культивировали при 200 оборотах в минуту и при 30°С в течение самое большое 120 часов. С интервалами, равными 24 часам, брали образцы бульона объемом 1 мл из колбы с культурой. Получение образца для последующих хроматографических анализов и хроматографических анализов самих по себе осуществляли как описано в примере 4.
Это показывает, что рекомбинантные штаммы Р. putida KT2440 pBBR1MCS-2::AB-BTH_II1077-II1080-II1081 и Р. putida GPp104 pBBRIMCS-2::AB-BTH_II1077-II1080-II1081 образуют значительно больше монорамнозиллипидов по сравнению со штаммами Р. putida KT2440 pBBR1MCS-2::AB и Р. putida GPp104 pBBR1MCS-2::AB. Это показывает, что амплификация BTH_II1077-II1080-II1081 из В. thailandensis E264 повышает образование монорамнозиллипидов в штаммах Р. Putida, содержащих Pseudomonas aeruginosa DSM1707 ген rhIAB.
Кроме того, показано, что рекомбинантные штаммы Р. putida KT2440 pBBR1MCS-2::ABC-BTH_II1077-II1080-II1081 и Р. putida GPp104 pBBR1MCS-2::ABC-BTH_II1077-II1080-II1081 образуют значительно больше моно- и дирамнозиллипидов по сравнению со штаммами Р. putida KT2440 pBBR1MCS-2::ABC и Р. putida GPp104 pBBR1MCS-2::ABC. Это доказывает, что амплификация ВТН_II1077-II1080-II1081 из В. thailandensis E264 повышает образование моно- и дирамнозиллипидов в штаммах Р. Putida, содержащих Pseudomonas aeruginosa DSM1707 ген rhlABC.
Наконец показано, что уменьшение образования полигидроксибутирата в штамме Р. putida GPp104 по сравнению со штаммом Р. putida KT2440 приводит к повышенному образованию рамнолипидов, так штаммы Р. putida KT2440 pBBR1MCS-2::AB, Р. putida KT2440 pBBR1MCS-2::ABC, Р. putida KT2440 pBBR1MCS-2::AB-BTH_II1077-II1080-II1081 и Р. putida KT2440 pBBR1MCS-2::ABC-BTH_II1077-II1080-II1081 способны образовывать значительно меньше моно- () и моно- и дирамнозиллипидов(), чем соответствующие контрольные штаммы Р. putida GPp104 pBBR1MCS-2::AB, Р. putida GPp104 pBBR1MCS-2::ABC, Р. putida GPp104 pBBR1MCS-2::AB-BTH_II1077-II1080-II1081 и Р. putida GPp104 pBBR1MCS-2::ABC-BTH_II1077-II1080-II1081.
13. Конструирование вектора pBBR1MCS-2::ABCM для гетерологичной экспрессии Pseudomonas aeruginosa DSM1707 генов rhlA, rhlB, pa1131 и rhIC в Pseudomonas putida
Для гетерологичной экспрессии Pseudomonas aeruginosa DSM1707 генов rhlA, rhlB, pa1131 и rhLC, сконструировали плазмиду pBBR1MCS-2::ABCM (Seq ID №58). Для этого ген ра1131 (Seq ID №59) амплифицировали исходя из геномной ДНК штамма Pseudomonas aeruginosa PA01 (DSM 1707), применяя олигонуклеотиды
MFS2.0_xbaI_fw: 5’-AGGAAATCTAGATGAGAGGCCGGCAAGGATAC-3’ (Seq ID №60)
MFS2.0_XbaI_rev:5’-CCAGGTTCTAGACGCCAGGATTGAACAGTACC-3’ (Seq ID №61)
Амплификация продукта ПЦР (1483 пар оснований) осуществлялась с применением полимеразы Phusion™ High-Fidelity Master Mix от New England Biolabs (Frankfurt). Продукт ПЦР очистили с применением XbaI и лигировали подобным образом с применением XbaI вектор pBBRIMCS-2::АВС (Seq ID №40) с помощью набора Fast Link Ligation Kit (Epicentre Technologies; Madison, WI, USA). Полученный целевой вектор pBBRIMCS-2::ABCM (Seq ID №58) имел размер 9892 пар оснований. Вставку вектора секвенировали. Хромосомальную ДНК выделили посредством набора DNeasy Blood and Tissue Kit (Qiagen; Hilden) в соответствии с инструкциями производителя. Осуществление ПЦР, проверку успешности амплификации ПЦР посредством электрофореза на агарозном геле, окрашивание ДНК бромидом этидия, определение размера фрагмента ПЦР, очищение продуктов ПЦР и определение концентрации ДНК осуществлялись образом, известным специалистам в данной области техники.
Трансформация Pseudomonas putida KT2440 и GPp104 с помощью вектора pBBR1MCS-2::ABCM осуществляется как описано ранее (Iwasaki et al. Biosci. Biotech. Biochem. 1994. 58(5):851-854). ДНК плазмида каждых десяти клонов была выделена и проанализирована. Полученные штаммы, несущие плазмиду, были названы Р. putida KT2440 pBBR1MCS-2::ABCM и Р. putida GPp104 pBBR1MCS-2::ABCM.
14. Количественное определение рамнолипидов, продуцируемых рекомбинантными штаммами Р. putida с и без сверхэкспрессии Pseudomonas aeruginosa DSM1707 ра1131 гена
Рекомбинантные штаммы Р. putida P. putida KT2440 pBBR1MCS-2::ABC, P. putida KT2440 pBBR1MCS-2::ABCM, P. putida KT2440 pBBR1MCS-2::ABC и Р. putida GPp104 pBBR1MCS-2::ABCM, полученные в примерах 2 и 13, были культивированы на чашках с LB-агар-канамицином (50 мкг/мл). Последующее культивирование для получения рамнолипидов осуществлялось как описано в примере 12.
Получение образца для последующих хроматографических анализов и хроматографических анализов самих по себе осуществляли как описано в примере 4.
Результаты показаны в следующей таблице.
Образование моно- и дирамнозиллипидов штаммами Р. putida с и без сверхэкспрессии Р. aeruginosa-Gens ра1131 через 48 часа инкубации
Результаты показывают, что сверхэкспрессия Р. aeruginosa гена ра1131 в обоих штаммах (KT2440: немутантный тип и GPp104 с инактивированным образованием полигидроксибутирата) приводит к повышенному образованию ди- и монорамнозиллипидов. Результаты, кроме того, показывают, что уменьшение образования полигидроксибутирата в GPp104 в общем приводит к повышенному образованию рамнозиллипидов.
15. Конструирование вектора рЕС-ХТ99А::АВ для гетерологичной экспрессии генов rhlA и rhlB из Pseudomonas aeruginosa DSM1707 в Corynebacterium glutamicum
Для гетерологичной экспрессии генов rhlA и rhlB из Pseudomonas aeruginosa DSM1707 в Corynebacterium glutamicum была сконструирована плазмида рЕС-ХТ99А::АВ (Seq ID №52). Для этого синтетический оперон rhIAB (Seq ID №37) был синтезирован фирмой GeneArt AG (Regensburg) и клонирован в коммерческом векторе рМА (GeneArt AG). В качестве основы для синтеза применялась уже известная геномная последовательность Pseudomonas aeruginosa DSM1707. Исходя из вектора рМА::АВ синтетический оперон был выделен из вектора с помощью BglII и XbaI и далее лигирован в подобным образом расщепленный с помощью BamHI и XbaI экспрессионный вектор рЕС-ХТ99А (патент США 7118904). Полученная плазмида рЕС-ХТ99А::АВ (Seq ID №52) состоит из 9793 пар оснований. Лигирование и трансформация химически компетентных клеток Е. coli DH5α (Gibco-BRL, Karlsruhe) осуществляется образом, известным специалистам в данной области техники. Аутентичность вставки была проверена посредством анализа последовательности ДНК.
Трансформация С. glutamicum ATCC 13032 с помощью вектора рЕС-ХТ99А::АВ осуществляется как описано ранее (Liebl et al., FEMS Microbiol. Lett. 53:299-303 (1989)). Отбор трансформантов осуществляется на чашках с агаром LBHIS (18,5 г/л сердечно-мозгового инфузионного бульона, 0.5 М сорбита, 5 г/л бакто триптона, 2,5 г/л дрожжевого экстракта бакто, 5 г/л NaCl и 18 г/л бакто-агара, с добавкой 5 г/л тетрациклина).
Чашки инкубировали при 33°С в течение двух дней. Полученный штамм, несущий плазмиду, назвали С. glutamicum pEC-XT99A::AB.
16. Конструирование вектора рЕС-ХТ99А::АВС для гетерологичной экспрессии генов rhIA, rhIB и rhIC из Pseudomonas aeruginosa DSM1707 в Corynebacterium glutamicum
Для гетерологичной экспрессии генов rhlA, rhlB и rhlC из Pseudomonas aeruginosa DSM1707 в Corynebacterium glutamicum сконструировали плазмиду рЕС-ХТ99А::АВС (Seq ID №53). Для этого синтетический оперон rhlABC (Seq ID №39) был синтезирован фирмой GeneArt AG (Regensburg) и клонирован в коммерческом векторе рМА (GeneArt AG). В качестве основы для синтеза применялась уже известная геномная последовательность Pseudomonas aeruginosa DSM1707. Исходя из вектора рМА::АВС синтетический оперон был выделен с помощью BglII и XbaI и далее лигирован в подобным образом расщепленный с помощью BamHI и XbaI экспрессионный вектор рЕС-ХТ99А (US-Patent 7118904). Полученная плазмида рЕС-ХТ99А::АВС (Seq ID №53) состоит из 10780 пар оснований. Лигирование и трансформация химически компетентных клеток Е. coli DH5α (Gibco-BRL, Karlsruhe) осуществлялась образом, известным специалистам в данной области техники. Аутентичность вставки была проверена посредством анализа последовательности ДНК.
Трансформация С. glutamicum ATCC13032 с помощью вектора рЕС-ХТ99А::АВС осуществляется как описано ранее (Liebl et al., FEMS Microbiol. Lett. 53:299-303 (1989)). Отбор трансформантов осуществляется на чашках с агаром LBHIS (18,5 г/л сердечно-мозгового инфузионного бульона, 0.5 М сорбита, 5 г/л бакто триптона, 2,5 г/л дрожжевого экстракта бакто, 5 г/л NaCl и 18 г/л бакто-агара, с добавкой 5 г/л тетрациклина). Чашки инкубировали при 33°С в течение двух дней. Полученный штамм, несущий плазмиду, назвали рЕС-ХТ99А::АВС.
17. Конструирование вектора Vektors рЕС-ХТ99А;;АВМ для гетерологичной экспрессии гена rhlA, rhlB и ра1131 из Pseudomonas aeruginosa DSM1707 в Corynebacterium glutamicum
Для гетерологичной экспрессии генов rhlA, rhlB и ра1131 из Pseudomonas aeruginosa DSM1707 в Corynebacterium glutamicum сконструировали плазмиду рЕС-ХТ99А::АВМ (Seq ID №54). Для этого синтетический оперон rhlABM (Seq ID №41) был синтезирован фирмой GeneArt AG (Regensburg) и клонирован в коммерческом векторе рМА (GeneArt AG). В качестве основы для синтеза применялась уже известная геномная последовательность Pseudomonas aeruginosa DSM1707. Исходя из вектора рМА::АВМ синтетический оперон был выделен с помощью BglII и XbaI и далее лигирован в подобным образом расщепленный с помощью BamHI и XbaI экспрессионный вектор рЕС-ХТ99А (US-Patent 7118904). Полученная плазмида рЕС-ХТ99А::АВМ (Seq ID №54) состоит из 11073 пар оснований. Лигирование и трансформация химически компетентных клеток Е. coli DH5α (Gibco-BRL, Karlsruhe) осуществлялась образом, известным специалистам в данной области техники. Аутентичность вставки была проверена посредством анализа последовательности ДНК.
Трансформация С. glutamicum ATCC13032 с помощью вектора рЕС-ХТ99А::АВМ осуществляется как описано ранее (Liebl et al., FEMS Microbiol. Lett. 53:299-303 (1989)). Отбор трансформантов осуществляется на чашках с агаром LBHIS (18,5 г/л сердечно-мозгового инфузионного бульона, 0.5 М сорбита, 5 г/л бакто триптона, 2,5 г/л дрожжевого экстракта бакто, 5 г/л NaCl и 18 г/л бакто-агара, с добавкой 5 г/л тетрациклина). Чашки инкубировали при 33°С в течение двух дней. Полученный штамм, несущий плазмиду, назвали С. glutamicum рЕС-ХТ99А::АВМ.
18. Конструирование вектора рЕС-ХТ99А::АВСМ для гетерологичной экспрессии генов rhlA, rhlB, ра1131 и rhlC из Pseudomonas aeruginosa DSM1707 в Corynebacterium glutamicum
Для гетерологичной экспрессии генов rhlA, rhlB, pa1131 и rhlC из Pseudomonas aeruginosa DSM1707 в Corynebacterium glutamicum была сконструирована плазмида рЕС-ХТ99А::АВСМ (Seq ID №55). Для этого ген pa1131 (Seq ID №59) амплифицировали исходя из геномной ДНК штамма Pseudomonas aeruginosa PA01 (DSM 1707), применяя олигонуклеотиды:
MFS2.0_xbaI_fw: 5’-AGGAAATCTAGATGAGAGGCCGGCAAGGATAC-3’ (Seq ID №60)
MFS2.0_XbaI_rev:5’-CCAGGTTCTAGACGCCAGGATTGAACAGTACC-3’ (Seq ID №61)
Амплификация продукта ПЦР (1483 пар оснований) осуществлялась с применением полимеразы Phusion™ High-Fidelity Master Mix от New England Biolabs (Frankfurt). Продукт ПЦР очистили с применением XbaI и лигировали подобным образом с применением XbaI вектор pBBRIMCS-2::АВС (Seq ID №40) с помощью набора Fast Link Ligation Kit (Epicentre Technologies; Madison, WI, USA). Полученный целевой вектор рЕС-ХТ99А::АВСМ (Seq ID №55) имел размер 12263 пар оснований. Вставку вектора секвенировали. Хромосомальную ДНК выделили посредством набора DNeasy Blood and Tissue Kit (Qiagen; Hilden) в соответствии с инструкциями производителя. Осуществление ПЦР, проверку успешности амплификации ПЦР посредством электрофореза на агарозном геле, окрашивание ДНК бромидом этидия, определение размера фрагмента ПЦР, очищение продуктов ПЦР и определение концентрации ДНК осуществлялись образом, известным специалистам в данной области техники.
Трансформация С. glutamicum ATCC 13032 с помощью вектора рЕС-ХТ99А::АВСМ осуществляется как описано ранее (Liebl et al., FEMS Microbiol. Lett. 53:299-303 (1989)). Отбор трансформантов осуществляется на чашках с агаром LBHIS (18,5 г/л сердечно-мозгового инфузионного бульона, 0.5 М сорбита, 5 г/л бакто триптона, 2,5 г/л дрожжевого экстракта бакто, 5 г/л NaCl и 18 г/л бакто-агара, с добавкой 5 г/л тетрациклина).
Чашки инкубировали при 33°С в течение двух дней. Полученный штамм, несущий плазмиду, назвали С. glutamicum pEC-XT99A::ABCM.
19. Конструирование вектора pVWEX1::rfbBDAC для гетерологической экспрессии в С. glutamicum
Для гетерологичной экспрессии Gene rfbBDAC из Р. putida под контролем промотора lac в С. glutamicum сконструировали вектор pVWEX1::rfbBDAC (Seq ID №57). Для этого вектор pBBR1MCS-2::rfbBDAC (Seq ID №45) был расщеплен с помощью XbaI, и фрагмент (3840 пар оснований), содержащий ген rfbBDAC из Р. putida KT2440 и промотор lac, лигируется в расщепленный с помощью XbaI вектор pVWEXI (Seq ID №56). Полученная плазмида pVWEX1::rfbBDAC (Seq ID №57) состоит из 12311 пар оснований. Лигирование и трансформация химически компетентных клеток Е. coli DH5a (Gibco-BRL, Karlsruhe) осуществлялась образом, известным специалистам в данной области техники. Аутентичность вставки была проверена посредством анализа последовательности ДНК.
Трансформация С. glutamicum ATCC13032 рЕС-ХТ99А, АТСС13032 рЕС-ХТ99А::АВ, ATCC13032 рЕС-ХТ99А::АВМ, ATCC13032 рЕС-ХТ99А::АВС и ATCC13032 pEC-XT99A::ABCM с помощью вектора pVWEX1::rfbBDAC осуществлялась как описано ранее (Liebl et al., FEMS Microbiol. Lett. 53:299-303 (1989)). Отбор трансформантов осуществляется на чашках с агаром LBHIS (18,5 г/л сердечно-мозгового инфузионного бульона, 0.5 М сорбита, 5 г/л бакто триптона, 2,5 г/л дрожжевого экстракта бакто, 5 г/л NaCl и 18 г/л бакто-агара, с добавкой 5 г/л тетрациклина). Чашки инкубировали при 33°С в течение двух дней. Полученные штаммы, несущие плазмиду, назвали С. glutamicum рЕС-ХТ99А pVWEX1::rfbBDAC, С. glutamicum pEC-XT99A::AB pVWEX1::rfbBDAC, С. glutamicum pEC-ХТ99А::АВМ pVWEX1::rfbBDAC, С. glutamicum рЕС-ХТ99А::АВС pVWEX1::rfbBDAC и С. glutamicum pEC-XT99A::ABCM pVWEX1::rfbBDAC.
20. Количественное определение рамнолипидов, продуцируемых рекомбинантными штаммами С. glutamicum
Рекомбинантные штаммы С. glutamicum, получены в примерах 15 -19 С. glutamicum pEC-XT99A, С. glutamicum pEC-XT99A::AB, С. glutamicum pEC-ХТ99А::АВС, С. glutamicum pEC-XT99A::АВМ, С. glutamicum pEC-ХТ99А::АВСМ, С. glutamicum pEC-XT99A pVWEX1::rfbBDAC, С. glutamicum pEC-XT99A::AB pVWEX1::rfbBDAC, С. glutamicum pEC-XT99A::АВМ pVWEX1::rfbBDAC, С. glutamicum pEC-XT99A::АВС pVWEX1::rfbBDAC и С. glutamicum pEC-XT99A::АВСМ pVWEX1::rfbBDAC культивировали на агаровых чашках LBHIS, с применением 5 мг/л тетрациклина или 5 мг/л тетрациклина и 25 мг/л канамицина. Для исследования продуцирования рамнолипидов сначала была получена прекультура во встряхиваемой колбе. Для этого применялся инокулят штамма, свежо нанесенный на LBHIS агаровую тарелку, и 10 мл LBHIS среды (18,5 г/л сердечно-мозгового инфузионного бульона, 0.5 М сорбита, 5 г/л бакто триптона, 2,5 г/л дрожжевого экстракта бакто, 5 г/л NaCl и 18 г/л бакто-агара, с добавкой 5 г/л тетрациклина или 5 мг/л тетрациклина и 25 мг/л канамицина) инокулировали в колбе Эрленмейера, объемом 100 мл. Культивирование штаммов осуществляли при 33°С и 200 оборотах в минуту всю ночь. На следующее утро 50 мл среды CGXII (с применением 5 мг/л тетрациклина или 5 мг/л тетрациклина и 25 мг/л канамицина) инокулировали в колбе Эрленмейера, объемом 500 мл, содержащей перегородки, с 1 мл прекультуры (старт OD600 0,1).
CGXII среда:
- 20 г/л (NH4)2SO4 (Merck)
- 5 г/л мочевины (Merck)
- 1 г/л KH2PO4 (Merck)
- 1 г/л K2HPO4 (Merck)
- 0,25 г/л MgSO4⋅7H2O (Merck)
- 10 мг/л CaCl2 (Merck)
- 42 г/л MOPS (Roth)
- 0,2 мг/л биотин (Merck)
- 1 мл/л раствора микроэлементов
- Установка рН 7 с применением NaOH
- После автоклавирования добавили 1 мл/л протокатеховой кислоты (30 г/л растворенной в разбавленном NaOH, стерильная-отфильтрованная) и 40 г/л глюкозы (Merck)
Раствор микроэлементов:
- 10 г/л FeSO4⋅7H2O (Merck)
- 10 г/л MnSO4⋅H2O (Merck)
- 1 г/л ZnSO4⋅7H2O (Merck)
- 0,2 г/л CuSO4⋅5H2O(Merck)
- 20 мг/л NiCl2⋅6H2O (Merck)
- Для растворения подкислили до рН 1 с помощью HCl
Культуры культивировали при 200 оборотах в минуту и 33°С до оптической плотности (600 нм), равной 0,4-0,6. При этой оптической плотности культуры индуцировали добавлением IPTG (изопропил-β-D-тиогалактопиранозид; 1 мМ конечная концентрация). Затем экспрессия подобным образом осуществлялась при 33°С и 200 оборотах в минуту в течение 72 часов. С интервалами в 24 часа брали пробы объемом 1 мл бульона из колбы для культивирования. Получение образцов для последующих хроматографических анализов и хроматографические анализы сами по себе осуществляли как описано в примере 4.
Тогда как С. glutamicum pEC-XT99A не продуцирует рамнолипиды, рекомбинантные штаммы С. glutamicum pEC-XT99A::АВ, С. glutamicum pEC-XT99A::АВС, С. glutamicum pEC-XT99A::АВМ и С. glutamicum pEC-ХТ99А::АВСМ образуют обнаруживаемое количество рамнолипидов. С помощью контрольных материалов показано, что С. glutamicum pEC-ХТ99А::АВ и С. glutamicum pEC-XT99A::ABM образуют только монорамнозиллипиды, тогда как С. glutamicum pEC-XT99A::ABC, С. glutamicum pEC-XT99A::ABM и С. glutamicum рЕС-ХТ99А::АВСМ образуют дирамнозиллипиды и монорамнозиллипиды. Кроме того, показано, что С. glutamicum pEC-XT99A::ABM и С. glutamicum рЕС-ХТ99А::АВСМ способны образовывать больше монорамнозиллипидов или дирамнозиллипидов и монорамнозиллипидов, чем соответствующие ссылочные штаммы С. glutamicum pEC-XT99A::AB и С. glutamicum pEC-XT99A::ABC без амплификации ра1131 гена из Pseudomonas aeruginosa.
Более того, показано, что штаммы С. glutamicum pEC-XT99A::AB pVWEX1::rfbBDAC, С. glutamicum pEC-XT99A::ABM pVWEX1::rfbBDAC, С. glutamicum pEC-XT99A::ABC pVWEX1::rfbBDAC и С. glutamicum рЕС-ХТ99А::АВСМ pVWEX1::rfbBDAC образуют значительно больше моно- (С. glutamicum pEC-XT99A::AB pVWEX1::rfbBDAC и С. glutamicum pEC-XT99A::ABM pVWEX1::rfbBDAC) или моно- и дирамнозиллипидов (С. glutamicum pEC-XT99A::ABC pVWEX1::rfbBDAC и С. glutamicum рЕС-ХТ99А::АВСМ pVWEX1::rfbBDAC), чем штаммы С. glutamicum pEC-XT99A::ABM, С. glutamicum pEC-XT99A::ABC и С. glutamicum рЕС-ХТ99А::АВСМ без амплификации rfbBDA гена из Р. putida.
21. Конструкция штаммов Pseudomonas, которые несут плазмиду pBBR1MCS-2, pBBR1MCS-2::AB, pBBR1MCS-2::ABC, pBBR1MCS-2::ABM и pBBR1MCS-2::ABCM
Плазмиды pBBR1MCS-2, pBBR1MCS-2::AB, pBBR1MCS-2::ABC, pBBR1MCS-2::ABM и pBBR1MCS-2::ABCM встраиваются в Pseudomonas fluorescens DSM 50090, Pseudomonas fluorescens DSM 9958, Pseudomonas putida DSM 6899, Pseudomonas putida DSM 50204, Pseudomonas putida 50194, P. brassicacearum DSM 13227, P. stutzeri DSM 10701, Pseudomonas stutzeri DSM 4166 и Pseudomonas fulva DSM 17717 посредством электропорации. Трансформация штаммов Pseudomonas осуществляется как описано ранее (Iwasaki et al. Biosci. Biotech. Biochem. 1994. 58(5):851-854). Отбор трансформантов осуществляется на агаровых чашках с питательными элементами (5 г/л пептона; 3 г/л мясного экстракта; 15 г/л агара; рН 7; с добавлением 50 мг/л канамицина). Чашки инкубировали при 30°С или скорее при 28°С.Полученные штаммы, несущие плазмиды, назвали Pseudomonas fluorescens DSM 50090 pBBR1MCS-2, Pseudomonas fluorescens DSM 9958 pBBR1MCS-2, Pseudomonas putida DSM 6899 pBBR1MCS-2, Pseudomonas putida DSM 50204 pBBR1MCS-2, Pseudomonas putida 50194 pBBR1MCS-2, P. brassicacearum DSM 13227 pBBR1MCS-2, P. stutzeri DSM 10701 pBBR1MCS-2, Pseudomonas stutzeri DSM 4166 pBBR1MCS-2, Pseudomonas fulva DSM 17717 pBBR1MCS-2, Pseudomonas fluorescens DSM 50090 pBBR1MCS-2::AB, Pseudomonas fluorescens DSM 9958 pBBR1MCS-2::AB, Pseudomonas putida DSM 6899 pBBR1MCS-2::AB, Pseudomonas putida DSM 50204 pBBR1MCS-2::AB, Pseudomonas putida 50194 pBBR1MCS-2::AB, P. brassicacearum DSM 13227 pBBR1MCS-2::AB, P. stutzeri DSM 10701 pBBR1MCS-2::AB, Pseudomonas stutzeri DSM 4166 pBBR1MCS-2::AB, Pseudomonas fulva DSM 17717 pBBR1MCS-2::AB, Pseudomonas fluorescens DSM 50090 pBBR1MCS-2::ABC, Pseudomonas fluorescens DSM 9958 pBBR1MCS-2::ABC, Pseudomonas putida DSM 6899 pBBR1MCS-2::ABC, Pseudomonas putida DSM 50204 pBBR1MCS-2::ABC, Pseudomonas putida 50194 pBBR1MCS-2::ABC, P. brassicacearum DSM 13227 pBBR1MCS-2::ABC, P. stutzeri DSM 10701 pBBR1MCS-2::ABC, Pseudomonas stutzeri DSM 4166 pBBR1MCS-2::ABC, Pseudomonas fulva DSM 17717 pBBR1MCS-2::ABC, Pseudomonas fluorescens DSM 50090 pBBR1MCS-2::ABCM, Pseudomonas fluorescens DSM 9958 pBBRIMCS-2::ABCM, Pseudomonas putida DSM 6899 pBBR1MCS-2::ABCM, Pseudomonas putida DSM 50204 pBBR1MCS-2::ABCM, Pseudomonas putida 50194 pBBR1MCS-2::ABCM, P. brassicacearum DSM 13227 pBBR1MCS-2::ABCM, P. stutzeri DSM 10701 pBBR1MCS-2::ABCM, Pseudomonas stutzeri DSM 4166 pBBR1MCS-2::ABCM, Pseudomonas fulva DSM 17717 pBBR1MCS-2::ABCM, Pseudomonas fluorescens DSM 50090 pBBRIMCS-2::ABM, Pseudomonas fluorescens DSM 9958 pBBR1MCS-2::ABM, Pseudomonas putida DSM 6899 pBBR1MCS-2::ABM, Pseudomonas putidaDSM 50204 pBBR1MCS-2::ABM, Pseudomonas putida 50194 pBBRIMCS-2::ABM, P. brassicacearum DSM 13227 pBBR1MCS-2::ABM, P. stutzeri DSM 10701 pBBR1MCS-2::ABM, Pseudomonas stutzeri DSM 4166 pBBR1MCS-2::ABM и Pseudomonas fulva DSM 17717 pBBR1MCS-2::ABM.
22. Количественное определение рамнолипидов, продуцируемых рекомбинантными штаммами Pseudomonas
Рекомбинантные штаммы Pseudomonas Pseudomonas fluorescens DSM 50090, Pseudomonas fluorescens DSM 9958 pBBR1MCS-2, Pseudomonas putida DSM 6899 pBBR1MCS-2, Pseudomonas putida DSM 50204 pBBR1MCS-2, Pseudomonas putida 50194 pBBR1MCS-2, P. brassicacearum DSM 13227 pBBR1MCS-2, P. stutzeri DSM 10701 pBBR1MCS-2, Pseudomonas stutzeri DSM 4166 pBBR1MCS-2, Pseudomonas fulva DSM 17717 pBBR1MCS-2, Pseudomonas fluorescens DSM 50090 pBBRIMCS-2::AB, Pseudomonas fluorescens DSM 9958 pBBR1MCS-2::AB, Pseudomonas putida DSM 6899 pBBR1MCS-2::AB, Pseudomonas putida DSM 50204 pBBR1MCS-2::AB, Pseudomonas putida 50194 pBBR1MCS-2::AB, P. brassicacearum DSM 13227 pBBR1MCS-2::AB, P. stutzeri DSM 10701 pBBR1MCS-2::AB, Pseudomonas stutzeri DSM 4166 pBBR1MCS-2::AB, Pseudomonas fulva DSM 17717 pBBR1MCS-2::AB, Pseudomonas fluorescens DSM 50090 pBBR1MCS-2::ABC, Pseudomonas fluorescens DSM 9958 pBBR1MCS-2::ABC, Pseudomonas putida DSM 6899 pBBR1MCS-2::ABC, Pseudomonas putida DSM 50204 pBBR1MCS-2::ABC, Pseudomonas putida 50194 pBBR1MCS-2::ABC, P. brassicacearum DSM 13227 pBBR1MCS-2::ABC, P. stutzeri DSM 10701 pBBR1MCS-2::ABC, Pseudomonas stutzeri DSM 4166 pBBR1MCS-2::ABC, Pseudomonas fulva DSM 17717 pBBR1MCS-2::ABC, Pseudomonas fluorescens DSM 50090 pBBR1MCS-2::ABCM, Pseudomonas fluorescens DSM 9958 pBBR1MCS-2::ABCM, Pseudomonas putida DSM 6899 pBBR1MCS-2::ABCM, Pseudomonas putida DSM 50204 pBBR1MCS-2::ABCM, Pseudomonas putida 50194 pBBR1MCS-2::ABCM, P. brassicacearum DSM 13227 pBBR1MCS-2::ABCM, P. stutzeri DSM 10701 pBBR1MCS-2::ABCM, Pseudomonas stutzeri DSM 4166 pBBRIMCS-2::ABCM, Pseudomonas fulva DSM 17717 pBBR1MCS-2::ABCM, Pseudomonas fluorescens DSM 50090 pBBR1MCS-2::ABM, Pseudomonas fluorescens DSM 9958 pBBR1MCS-2::ABM, Pseudomonas putida DSM 6899 pBBR1MCS-2::ABM, Pseudomonas putida DSM 50204 pBBR1MCS-2::ABM, Pseudomonas putida 50194 pBBR1MCS-2::ABM, P. brassicacearum DSM 13227 pBBR1MCS-2::ABM, P. stutzeri DSM 10701 pBBR1MCS-2::ABM, Pseudomonas stutzeri DSM 4166 pBBR1MCS-2::ABM и Pseudomonas fulva DSM 17717 pBBR1MCS-2::ABM, полученные в примере 21, культивировали на чашках с LB-агар-канамицином (50 мкг/мл). Последующие культивирование для получения рамнолипидов осуществляли как описано в примере 12. Получение образцов для последующих хроматографических анализов и хроматографические анализы сами по себе осуществляли как описано в примере 4.
Тогда как Pseudomonas Pseudomonas fluorescens DSM 50090, Pseudomonas fluorescens DSM 9958 pBBR1MCS-2, Pseudomonas putida DSM 6899 pBBR1MCS-2, Pseudomonas putida DSM 50204 pBBR1MCS-2, Pseudomonas putida 50194 pBBR1MCS-2, P. brassicacearum DSM 13227 pBBR1MCS-2, P. stutzeri DSM 10701 pBBR1MCS-2, Pseudomonas stutzeri DSM 4166 pBBR1MCS-2, Pseudomonas fulva DSM 17717 pBBR1MCS-2 не продуцируют рамнолипиды, в рекомбинантных штаммах Stammen Pseudomonas fluorescens DSM 50090 pBBR1MCS-2::AB, Pseudomonas fluorescens DSM 9958 pBBR1MCS-2::AB, Pseudomonas putida DSM 6899 pBBR1MCS-2::AB, Pseudomonas putida DSM 50204 pBBR1MCS-2::AB, Pseudomonas putida 50194 pBBR1MCS-2::AB, P. brassicacearum DSM 13227 pBBR1MCS-2::AB, P. stutzeri DSM 10701 pBBR1MCS-2::AB, Pseudomonas stutzeri DSM 4166 pBBR1MCS-2::AB, Pseudomonas fulva DSM 17717 pBBR1MCS-2::AB, Pseudomonas fluorescens DSM 50090 pBBR1MCS-2::ABM, Pseudomonas fluorescens DSM 9958 pBBR1MCS-2::ABM, Pseudomonas putida DSM 6899 pBBR1MCS-2::ABM, Pseudomonas putida DSM 50204 pBBR1MCS-2::ABM, Pseudomonas putida 50194 pBBR1MCS-2::ABM, P. brassicacearum DSM 13227 pBBR1MCS-2::ABM, P. stutzeri DSM 10701 pBBR1MCS-2::ABM, Pseudomonas stutzeri DSM 4166 pBBRIMCS-2::АВМ и Pseudomonas fulva DSM 17717 pBBR1MCS-2::ABM образуются монорамнозиллипиды, а также в штаммах Pseudomonas fluorescens DSM 50090 pBBR1MCS-2::ABC, Pseudomonas fluorescens DSM 9958 pBBRIMCS-2::ABC, Pseudomonas putida DSM 6899 pBBR1MCS-2::ABC, Pseudomonas putida DSM 50204 pBBR1MCS-2::ABC, Pseudomonas putida 50194 pBBR1MCS-2::ABC, P. brassicacearum DSM 13227 pBBR1MCS-2::ABC, P. stutzeri DSM 10701 pBBR1MCS-2::ABC, Pseudomonas stutzeri DSM 4166 pBBR1MCS-2::ABC, Pseudomonas fulva DSM 17717 pBBR1MCS-2::ABC, Pseudomonas fluorescens DSM 50090 pBBR1MCS-2::ABCM, Pseudomonas fluorescens DSM 9958 pBBR1MCS-2::ABCM, Pseudomonas putida DSM 6899 pBBR1MCS-2::ABCM, Pseudomonas putida DSM 50204 pBBR1MCS-2::ABCM, Pseudomonas putida 50194 pBBR1MCS-2::ABCM, P. brassicacearum DSM 13227 pBBR1MCS-2::ABCM, P. stutzeri DSM 10701 pBBR1MCS-2::ABCM, Pseudomonas stutzeri DSM 4166 pBBR1MCS-2::ABCM и Pseudomonas fulva DSM 17717 pBBR1MCS-2::ABCM образуется обнаруживаемое количество моно- и дирамнозиллипидов.
Кроме того, рекомбинантными штаммами Pseudomonas Pseudomonas fluorescens DSM 50090 pBBR1MCS-2::ABM, Pseudomonas fluorescens DSM 9958 pBBR1MCS-2::ABM, Pseudomonas putida DSM 6899 pBBR1MCS-2::ABM, Pseudomonas putida DSM 50204 pBBR1MCS-2::ABM, Pseudomonas putida 50194 pBBR1MCS-2::ABM, P. brassicacearum DSM 13227 pBBR1MCS-2::ABM, P. stutzeri DSM 10701 pBBR1MCS-2::ABM, Pseudomonas stutzeri DSM 4166 pBBR1MCS-2::ABM Pseudomonas fulva DSM 17717 pBBR1MCS-2::ABM образуется меньше монорамнозиллипидов, и рекомбинантными штамма Pseudomonas Pseudomonas fluorescens DSM 50090 pBBR1MCS-2::ABCM, Pseudomonas fluorescens DSM 9958 pBBR1MCS-2::ABCM, Pseudomonas putida DSM 6899 pBBR1MCS-2::ABCM, Pseudomonas putida DSM 50204 pBBR1MCS-2::ABCM, Pseudomonas putida 50194 pBBR1MCS-2::ABCM, P. brassicacearum DSM 13227 pBBR1MCS-2::ABCM, P. stutzeri/DSM 10701 pBBR1MCS-2::ABCM, Pseudomonas stutzeri DSM 4166 pBBR1MCS-2::ABCM и Pseudomonas fulva DSM 17717 pBBR1MCS-2::ABCM образуется меньше моно- и дирамнозиллипидов, чем соответствующими контрольными штаммами без Р. Aeruginosa гена ра1131, а именно Pseudomonas fluorescens DSM 50090 pBBR1MCS-2::AB, Pseudomonas fluorescens DSM 9958 pBBR1MCS-2::AB, Pseudomonas putida DSM 6899 pBBR1MCS-2::AB, Pseudomonas putida DSM 50204 pBBR1MCS-2::AB, Pseudomonas putida 50194 pBBR1MCS-2::AB, P. brassicacearum DSM 13227 pBBR1MCS-2::AB, P. stutzeri DSM 10701 pBBR1MCS-2::AB, Pseudomonas stutzeri DSM 4166 pBBR1MCS-2::AB и Pseudomonas fulva DSM 17717 pBBR1MCS-2::AB или Pseudomonas fluorescens DSM 50090 pBBR1MCS-2::ABC, Pseudomonas fluorescens DSM 9958 pBBRIMCS-2::ABC, Pseudomonas putida DSM 6899 pBBR1MCS-2::ABC, Pseudomonas putida DSM 50204 pBBR1MCS-2::ABC, Pseudomonas putida 50194 pBBR1MCS-2::ABC, P. brassicacearum DSM 13227 pBBR1MCS-2::ABC, P. stutzeri DSM 10701 pBBR1MCS-2::ABC, Pseudomonas stutzeri DSM 4166 pBBR1MCS-2::ABC и Pseudomonas fulva DSM 17717 pBBR1MCS-2::ABC без амплификации ра1131 гена из Pseudomonas aeruginosa.
23. Конструирование векторов pBBR1MCS-2::ABPAO1-C1 и pBBRIMCS-2::АВРА 7-СЕ264 для гетерологической экспрессии альтернативных генов rhIA, rhIB и rhIC из Pseudomonas aeruginosa PAO1, Pseudomonas aeruginosa PAY, Pseudomonas aeruginosa 1 и Burkholderia thailandensis E264 в Р. putida
Для гетерологичной экспрессии генов rhlA, rhlB и rhlC из Pseudomonas aeruginosa PA01 или Pseudomonas aeruginosa PA7 плазмиды pBBRIMCS-2::ABPAO1 (Seq ID №62) и pBBR1MCS-2::ABPA7 (Seq ID №63) были сконструированы. Для этого синтетические опероны rhIABPAO1 (Seq ID Nr.64) и rhIABPAO1 (Seq ID №65) были синтезированы компанией DNA 2.0 (Menio Park, CA, U.S.A) и клонированы в коммерческом векторе pJ294 (DNA 2.0). Основой для синтеза является уже известная геномная последовательность штаммов Pseudomonas aeruginosa PA01 и Pseudomonas aeruginosa PA7. Исходя из векторов pJ294::ABPA01 и pJ294::АВРА7 синтетические опероны были выделены из векторов с помощью KpnI и XbaI и затем лигированы в расщепленный с помощью KPnI и XbaI экспрессионный вектор pBBR1MCS-2 (Seq ID №49) (Kovach et al., 1995: Four new derivatives of the broad-host-range cloning vector pBBRIMCS carrying different antibiotic-resistance cassettes. Gene, 166:175-176). Полученные плазмиды pBBR1MCS-2::ABPAO1 (Seq ID №62) и pBBR1MCS-2::ABPA7 (Seq ID №63) состояли из 7332и 7354 пар оснований. Лигирование и трансформация химически компетентных клеток Е. coli DH5a (Gibco-BRL, Karlsruhe) осуществлялась известным специалисту в данной области техники образом. Аутентичность вставки проверялась посредством анализа последовательности ДНК.
На второй стадии получили плазмиды pBBR1MCS-2::ABPAO1-C1 (Seq ID №66) и pBBR1MCS-2::ABPA7-CE264 (Seq ID №67). Для этого г/7/С-ген из Pseudomonas aeruginosa 1 (Seq ID №68) и Burkholderia thailandensis E264 (Seq ID №76) был синтезирован компанией DNA 2.0 (Menio Park, CA, U.S.A) и клонирован в коммерческом векторе pJ294 (DNA 2.0). Основой для синтеза являлась уже известная геномная последовательность штаммов Pseudomonas aeruginosa 1 и Burkholderia thailandensis E264. Исходя из векторов pJ294::С1 и pJ294::СЕ264 гены rhIC выделялись с помощью Хbа и Sad из векторов и затем лигировались в подобным образом расщепленные с помощью Хbа и Sad вектора pBBR1MCS-2::ABPAO1 (Seq ID №62) и pBBR1MCS-2::ABPA7 (Seq ID №63). Полученные плазмиды pBBR1MCS-2::ABPA01-C1 (Seq ID №66) и pBBR1MCS-2::ABPA7-CE264 (Seq ID №67) имели размер, равный 8325 и 8335 пар оснований. Лигирование и трансформация химически компетентных клеток Е. coli’ DH5a (Gibco-BRL, Karlsruhe) осуществлялась известным специалисту в данной области техники образом. Аутентичность вставки проверялась посредством анализа последовательности ДНК.
Трансформация Pseudomonas putida KT2440 и GPp104 с применением векторов pBBR1MCS-2, pBBR1MCS-2::ABPAO1-C1 и pBBR1MCS-2::ABPA7-CE264 осуществлялась как описано ранее (Iwasaki et al. Biosci. Biotech. Biochem. 1994. 58(5):851-854). ДНК плазмиду каждых десяти клонов выделили и проанализировали. Полученные штаммы, несущие плазмиду, были названы Р. putida KT2440 pBBR1MCS-2, Р. putida KT2440 pBBR1MCS-2::ABPAO1-C1, P. putida KT2440 pBBR1MCS-2::ABPA7-CE264, P. putida GPp104 pBBR1MCS-2, P. putida GPp104 pBBR1MCS-2::ABPA01-C1 и Р. putida GPp104 pBBR1MCS-2::ABPA7-CE264.
24. Количественное определение рамнолипидов, продуцируемых рекомбинантными штаммами Р. putida с альтернативными rhlA-, rhlB- и rhlC генами из Pseudomonas aeruginosa PA01, Pseudomonas aeruginosa PA7, Pseudomonas aeruginosa 1 и Burkholderia thailandensis E264
Рекомбинантные штаммы Р. putida, полученные в примере 23, культивировали на чашках с LB-агар-канамицином (50 мкг/мл). Последующие культивирование для получения рамнолипидов осуществляли как описано в примере 12. Получение образцов для последующих хроматографических анализов и хроматографические анализы сами по себе осуществляли как описано в примере 4.
Тогда как штаммы Р. putida KT2440 pBBR1MCS-2 и Р. putida GPp104 pBBR1MCS-2 не способны продуцировать моно- и дирамнозиллипиды, штаммы Р. putida KT2440 pBBR1MCS-2::ABPA01-C1, P. putida KT2440 pBBR1MCS-2::ABPA7-CE264, Р. putida GPp104 pBBR1MCS-2::ABPA01-C1 и Р. putida GPp104 pBBR1MCS-2::ABPA7-CE264 образуют как моно-, так и дирамнозиллипиды. Показано, что штаммы с уменьшенным образованием полигидроксибутирата способны продуцировать больше моно- и дирамнозиллипидов (Р. putida GPp104 pBBR1MCS-2::ABPA01-C1 и Р. putida GPp104 pBBR1MCS-2::ABPA7-CE264), чем штаммы без ослабления образования полигидроксибутирата (Р. putida KT2440 pBBR1MCS-2::ABPA01-C1 и Р. putida KT2440 pBBR1MCS-2::ABPA7-CE264).
25. Конструирование векторов pBBR1MCS-2::AB_rfbBDAC, pBBRIMCS-2::ABM_rfbBDAC и pBBR1MCS-2::ABMC_rfbBDAC для сверхэкспрессии Р. putida rfbBDAC оперона в Р. putida и Е. coli
Для конструирования векторов pBBR1MCS-2::AB_rfbBDAC, pBBRIMCS-2::ABM_rfbBDAC и pBBR1MCS-2::ABMC_rfbBDAC для сверхэкспрессии Р. putida rfbBDAC оперона в Р. putida и Е. coli оперон Р. putida rfbBDAC подвергли сначала ПЦР амплификации. Вектор pBBR1MCS-2::rfbBDAC (Seq ID №45) служил в качестве матрицы для ПЦР. Применяли следующие олигонуклеотиды:
RL_Agel-fw: 5’-TATATATAACCGGTATTAATGCAGCTGGCACGAC-3’ (Seq ID №71)
RL_Agel_rev: 5’-GGCCGACCGGTACTAGTGGA-3’ (Seq ID №72)
ПЦР осуществляли с применением полимеразы Phusion™ High-Fidelity Master Mix от New England Biolabs (Frankfurt). Она осуществлялась образом, известным специалистам в данной области техники. Целевая последовательность (lac промотор и rfbBDAC) была клонирована в Trenzyme Alligator клонирующей системе. Е. coli DH5α (New England Biolabs; Frankfurt) трансформанты отобрали, и ДНК плазмиды различных кандидатов выделили и секвенировали. После того, как последовательно проверили и исследовали ошибки, вектор отщепили с помощью AgeI. Целевой фрагмент лигировали в расщепленные подобным образом с помощью AgeI вектора pBBR1MCS-2::AB (Seq ID №38), pBBRIMCS-2::ABM (Seq ID №42) и pBBR1MCS-2::ABMC (Seq ID №51). Полученные вектора pBBR1MCS-2::AB_rfbBDAC (Seq ID №73), pBBRIMCS-2::ABM_rfbBDAC (Seq ID №74) и pBBR1MCS-2::ABMC_rfbBDAC (Seq ID №75) состояли из 11960, 13289 и 14250 пар оснований. Вставку вектора подвергли секвенированию. Осуществление ПЦР, проверку успешности амплификации ПЦР посредством электрофореза на агарозном геле, окрашивание ДНК бромидом этидия, определение размера фрагмента ПЦР, очищение продуктов ПЦР и определение концентрации ДНК осуществлялись образом, известным специалистам в данной области техники.
Трансформация Pseudomonas putida KT2440 с применением векторов pBBR1MCS-2::AB_rfbBDAC, pBBR1MCS-2::ABM_rfbBDAC и pBBRIMCS-2::ABMC_rfbBDAC осуществляется как описано ранее (Iwasaki et al. Biosci. Biotech. Biochem. 1994. 58(5):851-854). ДНК плазмида каждых десяти клонов была выделена и проанализирована. Полученные штаммы, несущие плазмиду, были названы Р. putida KT2440 pBBRIMCS-2::AB_rfbBDAC, P. putida KT2440 pBBR1MCS-2::ABM_rfbBDAC и Р. putida KT2440 pBBR1MCS-2::ABMC_rfbBDAC.
26. Количественное определение рамнолипидов, продуцируемых рекомбинантными штаммами Р. putida KT2440 pBBR1MCS-2::AB_rfbBDAC, Р. putida KT2440 pBBR1MCS-2::ABM_rfbBDAC, P. putida KT2440 pBBR1MCS-2::ABC_rfbBDAC, P. putida KT2440 pBBRIMCS-2::ABMC_rfbBDAC, P. putida KT2440 pBBR1MCS-2::AB, P. putida KT2440 pBBR1MCS-2::ABM, P. putida KT2440 pBBR1MCS-2::ABC и Р. putida KT2440 pBBR1MCS-2::ABMC
Рекомбинантные штаммы Р. putida, полученные в примере 2, 7 и 25, культивировали на чашках с LB-агар-канамицином (50 мкг/мл). Последующие культивирование для получения рамнолипидов осуществляли как описано в примере 12. Получение образцов для последующих хроматографических анализов и хроматографические анализы сами по себе осуществляли как описано в примере 4.
Показано, что Р. putida KT2440 pBBR1MCS-2::AB_rfbBDAC, P. putida KT2440 pBBR1MCS-2::ABM_rfbBDAC, P. putida KT2440 pBBR1MCS-2::AB и Р. putida KT2440 pBBR1MCS-2::ABM способны образовывать монорамнозиллипиды, тогда как Р. putida KT2440 pBBRIMCS-2::ABMC_rfbBDAC, P. putida KT2440 pBBR1MCS-2::ABC_rfbBDAC, P. putida KT2440 pBBR1MCS-2::ABC и Р. putida KT2440 pBBR1MCS-2::ABMC способны образовывать моно- и дирамнозиллипиды.
Кроме того, показано, что Р. putida KT2440 pBBR1MCS-2::ABM_rfbBDAC, P. putida KT2440 pBBR1MCS-2::ABM, KT2440 pBBR1MCS-2::ABMC_rfbBDAC и KT2440 pBBR1MCS-2::ABMC способны образовывать больше моно- и дирамнозиллипидов, чем соответствующие контрольные штаммы Р. putida KT2440 pBBR1MCS-2::AB_rfbBDAC, P. putida KT2440 pBBR1MCS-2::AB, KT2440 pBBR1MCS-2::ABC_rfbBDAC и KT2440 pBBR1MCS-2::ABC без амплификации Pseudomonas aeruginosa гена ра1131.
Наконец, показано, что Р. putida KT2440 pBBR1MCS-2::AB_rfbBDAC, P. putida KT2440 pBBR1MCS-2::ABM_rfbBDAC, P. putida KT2440 pBBR1MCS-2::ABC_rfbBDAC, P. putida KT2440 pBBR1MCS-2::ABMC_rfbBDAC способны образовывать больше моно- (Р. putida KT2440 pBBR1MCS-2::AB_rfbBDAC и Р. putida KT2440 pBBR1MCS-2::ABM_rfbBDAC) или моно- и дирамнозиллипидов (Р. putida KT2440 pBBR1MCS-2::ABC_rfbBDAC и Р. putida KT2440 pBBR1MCS-2::ABMC_rfbBDAC), чем соответствующие контрольные штаммы Р. putida KT2440 pBBR1MCS-2::AB, Р. putida KT2440 pBBR1MCS-2::ABM, Р. putida KT2440 pBBR1MCS-2::ABC, Р. putida KT2440 pBBR1MCS-2::ABMC без амплификации Р. Putida гена rfbBDAC.
27. Образование рекомбинантного Е. coli W3110 pBBR1MCS-2::AB, Е. coli W3110 pBBR1MCS-2::ABM, Е. coli W3110 pBBR1MCS-2::ABC, Е. coli W3110 pBBR1MCS-2::ABCM, Е. coli W3110 pBBR1MCS-2::AB_rfbBDAC, Е. coli W3110 pBBR1MCS-2::ABM_rfbBDAC, Е. coli W3110 pBBRIMCS-2::ABC_rfbBDAC и Е. coli W3110 pBBR1MCS-2::ABCM_rfbBDAC
Трансформация Е. coli W3110 осуществляется как описано ранее (Miller JH. A Short Course in Bacterial Genetics: A Laboratory Manual and Handbook for Escherichia coli and Related Bacteria. Plainview, NY: Cold Spring Harbor Lab. Press; 1992) с помощью электропорации. ДНК плазмида каждых десяти клонов была выделена и проанализирована. Полученные штаммы, несущие плазмиду, были названы Е. coli W3110 pBBR1MCS-2::AB, Е. coli W3110 pBBR1MCS-2::ABM, Е. coli W3110 pBBR1MCS-2::ABC, Е. coli W3110 pBBR1MCS-2::ABCM, Е. coli W3110 pBBR1MCS-2::AB_rfbBDAC, Е. coli W3110 pBBR1MCS-2::ABM_rfbBDAC, E.coli W3110 pBBRIMCS-2::ABC_rfbBDAC и E.coli W3110 pBBR1MCS-2::ABCM_rfbBDAC.
28. Количественное определение рамнолипидов, продуцируемых рекомбинантными штаммами E.coli W3110 pBBR1MCS-2::AB, E.coli W3110pBBR1MCS-2::ABM, E.coli W3110 pBBR1MCS-2::ABC, E.coli W3110 pBBR1MCS-2::ABCM, E.coli W3110 pBBR1MCS-2::AB_rfbBDAC, E.coli W3110 pBBR1MCS-2::ABM_rfbBDAC, E.coli W3110 pBBRIMCS-2::ABC_rfbBDAC и E.coli W3110 pBBR1MCS-2::ABCM_rfbBDAC
Рекомбинантные штаммы E. Coli, полученные в примере 27, культивировали на чашках с LB-агар-канамицином (50 мкг/мл). Последующие культивирование для получения рамнолипидов осуществляли как описано в примере 10. Получение образцов для последующих хроматографических анализов и хроматографические анализы сами по себе осуществляли как описано в примере 4.
Показано, что E.coli W3110 pBBR1MCS-2::AB, E. coli W3110 pBBRIMCS-2::АВМ, E.coli W3110 pBBR1MCS-2::AB_rfbBDAC и E. coli W3110 pBBR1MCS-2::ABM_rfbBDAC способны формировать монорамнозиллипиды, тогда как E.coli W3110 pBBR1MCS-2::ABC, E.coli W3110 pBBR1MCS-2::ABCM, E.coli W3110 pBBR1MCS-2::ABC_rfbBDAC и E.coli W3110 pBBR1MCS-2::ABCM_rfbBDAC способны формировать моно- и дирамнозиллипиды. Кроме того, показано, что E.coli W3110 pBBRIMCS-2::АВМ и E.coli W3110 pBBR1MCS-2::ABM_rfbBDAC образуют больше монорамнозиллипидов, по сравнению с E.coli W3110 pBBR1MCS-2::AB и E.coli W3110 pBBR1MCS-2::AB_rfbBDAC без амплификации Pseudomonas aeruginosa гена ра1131.
Кроме того, показано, что E.coli W3110 pBBR1MCS-2::ABCM и E.coli W3110 pBBR1MCS-2::ABCM_rfbBDAC образуют больше моно- и дирамнозиллипидов, чем E.coli W3110 pBBR1MCS-2::ABC и E.coli W3110 pBBR1MCS-2::ABC_rfbBDAC без амплификации Pseudomonas aeruginosa гена ра1131. Кроме того, показано, что Е. coli W3110 pBBR1MCS-2::ABM и E. coli W3110 pBBR1MCS-2::ABM_rfbBDAC образуют больше монорамнозиллипидов, чем Е. coli W3110 pBBR1MCS-2::AB и Е. coli W3110 pBBR1MCS-2::AB_rfbBDAC без амплификации Pseudomonas aeruginosa гена ра1131.
Наконец показано, что Е. coli W3110 pBBR1MCS-2::AB_rfbBDAC, Е. coli W3110 pBBR1MCS-2::ABM_rfbBDAC, Е. coli W3110 pBBRIMCS-2::ABC_rfbBDAC и Е. coli W3110 pBBR1MCS-2::ABCM_rfbBDAC образуют больше моно- (Е. coli W3110 pBBR1MCS-2::AB_rfbBDAC, Е. coli W3110 pBBR1MCS-2::ABM_rfbBDAC) или моно- и дирамнозиллипидов (Е. coli W3110 pBBR1MCS-2::ABC_rfbBDAC и Е. coli W3110 pBBRIMCS-2::ABCM_rfbBDAC) по сравнению с соответствующими контрольными штаммами Е. coli W3110 pBBR1MCS-2::AB, Е. coli W3110 pBBR1MCS-2::ABM, Е. coli W3110 pBBR1MCS-2::ABC и Е. coli W3110 pBBRIMCS-2::АВСМ без амплификации Р. Putida гена rfbBDAC.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБЫ ПОЛУЧЕНИЯ РАМНОЛИПИДОВ | 2015 |
|
RU2694392C2 |
| Средство редактирования генома на основе белка LigD из бактерии Pseudomonas putida и Cas9 комплекса | 2022 |
|
RU2797049C1 |
| УСТОЙЧИВЫЕ К ЦИАНИДАМ НИТРИЛГИДРАТАЗЫ | 2005 |
|
RU2385876C2 |
| СПОСОБ БИОКАТАЛИТИЧЕСКОГО ОКИСЛЕНИЯ С ПРИМЕНЕНИЕМ ПРОДУКТА ГЕНА alkL | 2011 |
|
RU2573900C2 |
| Микроорганизм для продуцирования диамина и способ получения диамина с его использованием | 2015 |
|
RU2672323C2 |
| СПОСОБ ЭКСПРЕССИИ | 2013 |
|
RU2644199C2 |
| Микроорганизм для продуцирования диамина и способ получения диамина с его использованием | 2015 |
|
RU2696504C2 |
| СПОСОБ ПОЛУЧЕНИЯ ДИПЕПТИДА (ВАРИАНТЫ) | 2005 |
|
RU2316596C2 |
| ГЕН ПЕПТИДООБРАЗУЮЩЕГО ФЕРМЕНТА, ПЕПТИДООБРАЗУЮЩИЙ ФЕРМЕНТ И СПОСОБ ПОЛУЧЕНИЯ ДИПЕПТИДА | 2002 |
|
RU2280077C2 |
| 6-ФОСФОГЛЮКОНОЛАКТОНАЗА ИЗ ESCHERICHIA COLI, ФРАГМЕНТ ДНК, БАКТЕРИЯ, ПРИНАДЛЕЖАЩАЯ К РОДУ ESCHERICHIA - ПРОДУЦЕНТ L-АМИНОКИСЛОТЫ И СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТЫ | 2005 |
|
RU2288268C2 |



































































































































































Группа изобретений относится к области биотехнологии. Представлена бактериальная клетка, которая способна образовывать по меньшей мере один рамнолипид и генетически модифицированная таким образом, что по сравнению с ее диким типом она имеет повышенную активность двух ферментов Е1 и Е2 или трех ферментов Е1, Е2 и Е3 и по меньшей мере одного фермента Е8, который катализирует экспорт рамнолипидов из клетки в окружающую среду, при этом фермент Е1 способен катализировать превращение 3-гидроксиалканоил-АСР через 3-гидроксиалканоил-3-гидроксиалкановую кислоту АСР в гидроксиалканоил-3-гидроксиалкановую кислоту, фермент Е2 представляет собой рамнозилтрансферазу I, а фермент Е3 – рамнозилтрансферазу II. Представлен способ получения рамнолипидов с использованием указанной клетки. Группа изобретений позволяет повысить образование рамнолипидов. 2 н. и 10 з.п. ф-лы, 5 табл., 3 ил., 28 пр.
1. Бактериальная клетка, которая способна образовывать по меньшей мере один рамнолипид общей формулы (I),
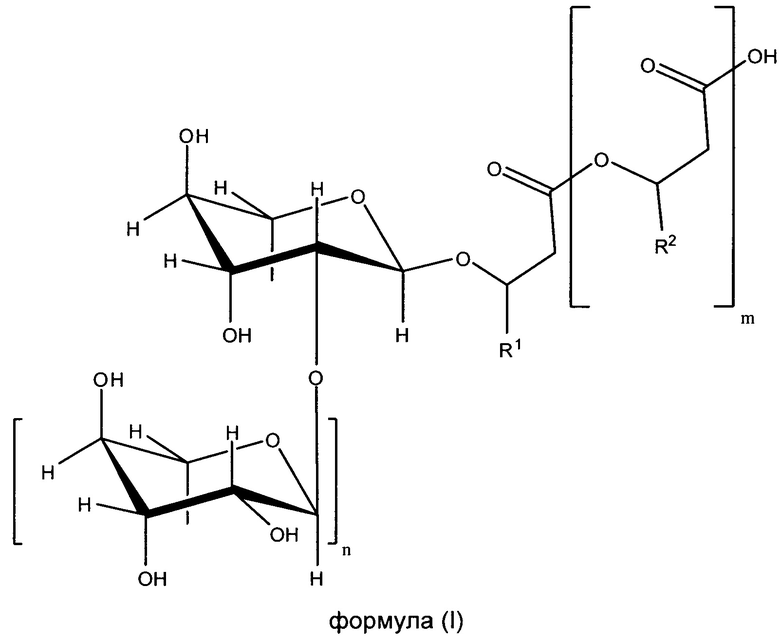 ,
,
где
m=2, 1 или 0,
n=1 или 0,
R1 и R2 независимо друг от друга означают идентичные или различные органические радикалы, имеющие от 2 до 24 атомов углерода,
отличающаяся тем, что она генетически модифицирована таким образом, что по сравнению с ее диким типом она имеет повышенную активность двух ферментов E1 и Е2 или трех ферментов E1, Е2 и Е3 и по меньшей мере одного фермента Е8, который катализирует экспорт рамнолипидов общей формулы (I) из клетки в окружающую среду, имеющего полипептидную последовательность Seq ID №8, Seq ID №24, Seq ID №26 или Seq ID №28 или имеющего полипептидную последовательность, в которой до 25% аминокислотных остатков модифицированы по сравнению с эталонной последовательностью Seq ID №8, Seq ID №24, Seq ID №26 или Seq ID №28 посредством делеции, инсерции, замещения или их комбинации, и которая все еще имеет по меньшей мере 10% от ферментативной активности фермента, имеющего эталонную последовательность Seq ID №8, Seq ID №24, Seq ID №26 или Seq ID №28.
причем
фермент E1 способен катализировать превращение 3-гидроксиалканоил-АСР через 3-гидроксиалканоил-3-гидроксиалкановую кислоту-АСР в гидроксиалканоил-3-гидроксиалкановую кислоту, фермент Е2 представляет собой рамнозилтрансферазу I и способен катализировать превращение dTDP-рамнозы и 3-гидроксиалканоил-3-гидроксиалканоата в α-L-рамнопиранозил-3-гидроксиалканоил-3-гидроксиалканоат, фермент Е3 представляет собой рамнозилтрансферазу II и способен катализировать превращение dTDP-рамнозы и α-L-рамнопиранозил-3-гидроксиалканоил-3-гидроксиалканоат в α-L-рамнопиранозил-(1-2)-α-L-рамнопиранозил-3-гидроксиалканоил-3-гидроксиалканоат.
2. Бактериальная клетка по п. 1, причем в формуле (I) m=1 или 0.
3. Бактериальная клетка по п. 1, причем в формуле (I) n=1.
4. Бактериальная клетка по п. 1, причем в формуле (I) R1 и R2 означают необязательно разветвленный, необязательно замещенный, в частности гидроксизамещенный, необязательно ненасыщенный, в частности необязательно моно-, ди- или триненасыщенный, алкильный радикал.
5. Бактериальная клетка по п. 1, отличающаяся тем, что она по сравнению с ее диким типом дополнительно имеет повышенную активность по меньшей мере одного из ферментов, выбранных из группы, состоящей из:
фермента Е4, dTTP: α-D-глюкоза-1-фосфат-тимидилилтрансфераза, ЕС 2.7.7.24, имеющего полипептидную последовательность, в которой до 15% аминокислотных остатков модифицированы по сравнению с эталонной последовательностью Seq ID №10 посредством делеции, инсерции, замещения или их комбинации, и которая все еще имеет по меньшей мере 10% от ферментативной активности фермента, имеющего эталонную последовательность Seq ID №10,
фермента Е5, dTTP-глюкоза-4,6-гидролаза, ЕС 4.2.1.46, имеющего полипептидную последовательность, в которой до 15% аминокислотных остатков модифицированы по сравнению с эталонной последовательностью Seq ID №12 посредством делеции, инсерции, замещения или их комбинации, и которая все еще имеет по меньшей мере 10% от ферментативной активности фермента, имеющего эталонную последовательность Seq ID №12,
фермента Е6, dTDP-4-дегидрорамноза-3,5-эпимераза, ЕС 5.1.3.13, имеющего полипептидную последовательность, в которой до 15% аминокислотных остатков модифицированы по сравнению с эталонной последовательностью Seq ID №14 посредством делеции, инсерции, замещения или их комбинации, и которая все еще имеет по меньшей мере 10% от ферментативной активности фермента, имеющего эталонную последовательность Seq ID №14, и
фермента Е7, dTDP-4-дегидрорамноза-редуктаза, ЕС 1.1.1.133, имеющего полипептидную последовательность, в которой до 15% аминокислотных остатков модифицированы по сравнению с эталонной последовательностью Seq ID №16 посредством делеции, инсерции, замещения или их комбинации, и которая все еще имеет по меньшей мере 10% от ферментативной активности фермента, имеющего эталонную последовательность Seq ID №16.
6. Бактериальная клетка по п. 5, отличающаяся тем, что фермент Е4, представляющий собой dTTP: α-D-глюкоза-1-фосфат-тимидилилтрансферазу, ЕС 2.7.7.24, имеет полипептидную последовательность Seq ID №10,
фермент Е5, представляющий собой dTTP-глюкоза-4,6-гидролазу, ЕС 4.2.1.46, имеет полипептидную последовательность Seq ID №12,
фермент Е6, представляющий собой dTDP-4-дегидрорамноза-3,5-эпимеразу, ЕС 5.1.3.13, имеет полипептидную последовательность Seq ID №14, и
фермент Е7, представляющий собой dTDP-4-дегидрорамноза-редуктазу, ЕС 1.1.1.133, имеет полипептидную последовательность Seq ID №16.
7. Бактериальная клетка по п. 1, отличающаяся тем, что ферменты Е1, Е2 и Е3 выбраны из группы, состоящей из:
по меньшей мере одного фермента E1, выбранного из следующего:
фермент E1a, имеющий полипептидную последовательность Seq ID №2 или имеющий полипептидную последовательность, в которой до 25% аминокислотных остатков модифицированы по сравнению с эталонной последовательностью Seq ID №2 посредством делеции, инсерции, замещения или их комбинации, и который все еще имеет по меньшей мере 10% от ферментативной активности фермента, имеющего эталонную последовательность Seq ID №2,
фермент E1b, имеющий полипептидную последовательность Seq ID №18 или имеющий полипептидную последовательность, в которой до 25% аминокислотных остатков модифицированы по сравнению с эталонной последовательностью Seq ID №18 посредством делеции, инсерции, замещения или их комбинации, и который все еще имеет по меньшей мере 10% от ферментативной активности фермента, имеющего эталонную последовательность Seq ID №18,
фермент E1d, имеющий полипептидную последовательность Seq ID №80 или имеющий полипептидную последовательность, в которой до 25% аминокислотных остатков модифицированы по сравнению с эталонной последовательностью Seq ID №80 посредством делеции, инсерции, замещения или их комбинации, и который все еще имеет по меньшей мере 10% от ферментативной активности фермента, имеющего эталонную последовательность Seq ID №80, и
фермент E1e, имеющий полипептидную последовательность Seq ID №82 или имеющий полипептидную последовательность, в которой до 25% аминокислотных остатков модифицированы по сравнению с эталонной последовательностью Seq ID №82 посредством делеции, инсерции, замещения или их комбинации, и который все еще имеет по меньшей мере 10% от ферментативной активности фермента, имеющего эталонную последовательность Seq ID №82,
по меньшей мере одного фермента Е2, выбранного из следующего:
фермент Е2а, имеющий полипептидную последовательность Seq ID №4 или имеющий полипептидную последовательность, в которой до 25% аминокислотных остатков модифицированы по сравнению с эталонной последовательностью Seq ID №4 посредством делеции, инсерции, замещения или их комбинации, и который все еще имеет по меньшей мере 10% от ферментативной активности фермента, имеющего эталонную последовательность Seq ID №4,
фермент E2b, имеющий полипептидную последовательность Seq ID №20 или имеющий полипептидную последовательность, в которой до 25% аминокислотных остатков модифицированы по сравнению с эталонной последовательностью Seq ID №20 посредством делеции, инсерции, замещения или их комбинации, и который все еще имеет по меньшей мере 10% от ферментативной активности фермента, имеющего эталонную последовательность Seq ID №20,
фермент Е2с, имеющий полипептидную последовательность Seq ID №84 или имеющий полипептидную последовательность, в которой до 25% аминокислотных остатков модифицированы по сравнению с эталонной последовательностью Seq ID №84 посредством делеции, инсерции, замещения или их комбинации, и который все еще имеет по меньшей мере 10% от ферментативной активности фермента, имеющего эталонную последовательность Seq ID №84,
фермент E2d, имеющий полипептидную последовательность Seq ID №86 или имеющий полипептидную последовательность, в которой до 25% аминокислотных остатков модифицированы по сравнению с эталонной последовательностью Seq ID №86 посредством делеции, инсерции, замещения или их комбинации, и который все еще имеет по меньшей мере 10% от ферментативной активности фермента, имеющего эталонную последовательность Seq ID №86, и
фермент Е2е, имеющий полипептидную последовательность Seq ID №88 или имеющий полипептидную последовательность, в которой до 25% аминокислотных остатков модифицированы по сравнению с эталонной последовательностью Seq ID №88 посредством делеции, инсерции, замещения или их комбинации, и который все еще имеет по меньшей мере 10% от ферментативной активности фермента, имеющего эталонную последовательность Seq ID №88, и
по меньшей мере одного фермента Е3, выбранного из следующего:
фермент Е3а, имеющий полипептидную последовательность Seq ID №6 или имеющий полипептидную последовательность, в которой до 25% аминокислотных остатков модифицированы по сравнению с эталонной последовательностью Seq ID №6 посредством делеции, инсерции, замещения или их комбинации, и который все еще имеет по меньшей мере 10% от ферментативной активности фермента, имеющего эталонную последовательность Seq ID №6,
фермент E3b, имеющий полипептидную последовательность Seq ID №22 или имеющий полипептидную последовательность, в которой до 25% аминокислотных остатков модифицированы по сравнению с эталонной последовательностью Seq ID №22 посредством делеции, инсерции, замещения или их комбинации, и который все еще имеет по меньшей мере 10% от ферментативной активности фермента, имеющего эталонную последовательность Seq ID №22,
фермент Е3с, имеющий полипептидную последовательность Seq ID №90 или имеющий полипептидную последовательность, в которой до 25% аминокислотных остатков модифицированы по сравнению с эталонной последовательностью Seq ID №90 посредством делеции, инсерции, замещения или их комбинации, и который все еще имеет по меньшей мере 10% от ферментативной активности фермента, имеющего эталонную последовательность Seq ID №90, и
фермент E3d, имеющий полипептидную последовательность Seq ID №92 или имеющий полипептидную последовательность, в которой до 25% аминокислотных остатков модифицированы по сравнению с эталонной последовательностью Seq ID №92 посредством делеции, инсерции, замещения или их комбинации, и который все еще имеет по меньшей мере 10% от ферментативной активности фермента, имеющего эталонную последовательность Seq ID №92.
8. Бактериальная клетка по п. 1, отличающаяся тем, что она имеет повышенную активность комбинации ферментов E1E2E3 и n=1.
9. Бактериальная клетка по п. 1, отличающаяся тем, что она, как и ее дикий тип, способна образовывать полигидроксиалканоаты, имеющие цепи длиной от 6 до 16 атомов углерода, в частности такая клетка, которая была генетически модифицирована таким образом, что способна образовывать меньше полигидроксиалканоатов по сравнению с ее диким типом.
10. Бактериальная клетка по п. 9, отличающаяся тем, что указанная клетка по сравнению с ее диким типом дополнительно имеет пониженную активность по меньшей мере одного из ферментов Е9 или Е10,
где Е9 представляет собой полигидроксиалканоат-синтазу, ЕС:2.3.1., имеющую способность превращать 3-гидроксиалканоил-коэнзим А в поли-3-гидроксиалкановую кислоту, в частности имеющую полипептидную последовательность Seq ID №30 или Seq ID №32 или имеющую полипептидную последовательность, в которой до 25% аминокислотных остатков по сравнению с соответствующей эталонной последовательностью Seq ID №30 или Seq ID №32 модифицированы посредством делеции, инсерции, замещения или их комбинации, и которая все еще имеет по меньшей мере 10% от ферментативной активности фермента, имеющего соответствующую эталонную последовательность Seq ID №30 или Seq ID №32, и
Е10 представляет собой 3-гидроксиалканоил-АСР: коэнзим А-трансферазу, имеющую способность превращать 3-гидроксиалканоил-АСР в 3-гидроксиалканоил-коэнзим А, особенно имеющую полипептидную последовательность Seq ID №34 или Seq ID №36 или имеющую полипептидную последовательность, в которой до 25% аминокислотных остатков по сравнению с соответствующей эталонной последовательностью Seq ID №34 или Seq ID №36 модифицированы посредством делеции, инсерции, замещения или их комбинации, и которая все еще имеет по меньшей мере 10% от ферментативной активности фермента, имеющего соответствующую эталонную последовательность Seq ID №34 или Seq ID №36.
11. Способ получения рамнолипидов общей формулы (I), содержащий стадии:
I) введения клетки по одному из пп. 1-10 в контакт со средой, содержащей источник углерода;
II) культивирования клетки при условиях, позволяющих клетке образовать рамнолипид из источника углерода.
12. Способ по п. 11, дополнительно содержащий стадию
III) выделения полученных рамнолипидов.
| X | |||
| Fang et al | |||
| Engineering Rhamnolipid Biosurfactants as Agents for Microbial Enhanced Oil Recovery, Society of Petroleum Engineers, Internacional Symposium on Olifield Chemistry, 28 February-2 March, Houston, Texas, U.S.A., 2007 | |||
| OCHSNER UA et al | |||
| Production of Pseudomonas aeruginosa Rhamnolipid Biosurfactants in Heterologous Hosts, Applied and Environmental Microbiology Vol | |||
| Устройство для сортировки каменного угля | 1921 |
|
SU61A1 |
| СТОЛ (ГЕРДА) ДЛЯ ОБОГАЩЕНИЯ ПОЛЕЗНЫХ ИСКОПАЕМЫХ | 1921 |
|
SU3503A1 |
| WO2004050882 A1, 17.06.2004 | |||
| S.KORNPELD et al | |||
| The Enzymatic Synthesis of Thymidine-linked Sugars I | |||
| THYMIDINE DIPHOSPHATE GLUCOSE, THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol | |||
| Стеклографический печатный станок с ножной педалью | 1922 |
|
SU236A1 |
| Приспособление для точного наложения листов бумаги при снятии оттисков | 1922 |
|
SU6A1 |
| Паровой двигатель с качающимся крыльчатым поршнем | 1925 |
|
SU1791A1 |
| R.RAHIM et al | |||
| Involvement of the rml locus in core oligosaccharide and O polysaccharide assembly in Pseudomonas aeruginosa, Microbiology Vol.146, pp | |||
| СПОСОБ ПОЛУЧЕНИЯ ФЕРРОВОЛЬФРАМА | 1925 |
|
SU2803A1 |
| Cabrera-Valladares et al | |||
| Переносная печь для варки пищи и отопления в окопах, походных помещениях и т.п. | 1921 |
|
SU3A1 |
| Способ подготовки рафинадного сахара к высушиванию | 0 |
|
SU73A1 |
| Индукционная катушка | 1920 |
|
SU187A1 |
| Шланговое соединение | 0 |
|
SU88A1 |
| Шланговое соединение | 0 |
|
SU88A1 |
| Шланговое соединение | 0 |
|
SU88A1 |
| Шланговое соединение | 0 |
|
SU88A1 |
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |
| Противоповторное устройство для рукояток ящика зависимости аппарата механической централизации и сопряженных сними клавиш блок-механизмов | 1936 |
|
SU51560A1 |
| Разборный с внутренней печью кипятильник | 1922 |
|
SU9A1 |
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |
| КАРТЕЛЬ Н.А | |||
| и др | |||
| Рамнолипиды: перспективы их использования для фиторемедиации, Биотехнология, N4, с.47-60, 2007. | |||

