ПЕРЕКРЕСТНАЯ ССЫЛКА НА РОДСТВЕННЫЕ ЗАЯВКИ
По настоящей заявке испрашивается приоритет временной заявки на патент США №61/641776, поданной 2 мая 2012 г., и временной заявки на патент США №61/780512, поданной 13 марта 2013 г., описания которых включены таким образом посредством ссылки в их полном объеме.
ЗАЯВЛЕНИЕ ПРАВ НА ИЗОБРЕТЕНИЯ, СДЕЛАННЫЕ В ПРОЦЕССЕ ИССЛЕДОВАНИЯ, ФИНАНСИРУЕМОГО ИЗ ФЕДЕРАЛЬНОГО БЮДЖЕТА
Не применимо.
ОБЛАСТЬ ТЕХНИКИ
Настоящее описание относится к области геномной инженерии, в частности, измененной экспрессии и/или направленной модификации эндогенного гена малатдегидрогеназы (MDH) растений.
ПРЕДПОСЫЛКИ СОЗДАНИЯ ИЗОБРЕТЕНИЯ
Биотехнология возникла, как важное средство, в попытках решить проблему увеличивающейся мировой потребности в производстве пищевых продуктов. Традиционные подходы к увеличению продуктивности сельского хозяйства, например, увеличению урожая или генно-инженерной устойчивости к вредителям, основаны или на мутационной селекции, или на введении новых генов в геномы видов культур с помощью трансформации. Оба процесса по своему существу являются неспецифическими и относительно неэффективными. Например, с помощью традиционных способов трансформации растений доставляется экзогенная ДНК, которая интегрируется в геном в произвольных положениях. Таким образом, для идентификации и изоляции трансгенных линий с желаемыми свойствами необходимо создание тысяч уникальных событий произвольной интеграции и последующий скрининг на предмет желаемого события. В результате, традиционная инженерия особенности растения является трудным, трудоемким и непредсказуемым предприятием. Кроме того, неспецифическая природа этих интеграций затрудняет предсказание того, возникли ли плейотропные эффекты вследствие непреднамеренного разрушения генома. В результате, создание, изоляция или определение характеристик линий растений с конструированными трансгенами или генно-инженерными свойствами было очень трудным и затратным процессом с низкой вероятностью успеха.
Направленная модификация генов позволяет преодолеть логистические проблемы традиционных технологических приемов в случае растительных систем и как таковая была многолетней, но труднодостижимой целью и в основном биологическом исследовании растений, и сельскохозяйственной биотехнологии. Однако, за исключением «направленного воздействия на ген» через позитивно-негативную селекцию с использованием лекарственных средств в случае риса или использование заранее сконструированных рестрикционных сайтов, направленная модификация генома во всех видах растений, и модельных растениях, и культурных растениях, оказывалась до недавнего времени очень трудной. Terada et al. (2002) Nat Biotechnol 20(10):1030; Terada et al. (2007) Plant Physiol 144(2):846; D'Halluin et al. (2008) Plant Biotechnology J. 6(1):93.
Недавно были описаны способы и композиции для направленного расщепления геномной ДНК. Такие события направленного расщепления могут использоваться, например, для индукции направленного мутагенеза, для индукции направленных мутаций (например, делеций, заменен и/или вставок) клеточных последовательностей ДНК и содействия направленной рекомбинации и интеграции в заданный локус хромосомы. См., например, Urnov et al. (2010) Nature 435(7042):646-51; публикации заявок на патенты США №20030232410; 20050208489; 20050026157; 20050064474; 20060188987; 20090263900; 20090117617; 20100047805; 20110207221; 20110301073; 2011089775 и публикацию международной заявки WO 2007/014275, описания которых включены посредством ссылки в их полном объеме для всех целей. Расщепление может происходить благодаря использованию специфических нуклеаз, таких как сконструированные нуклеазы с доменами «цинковые пальцы» (ZFN), активатора транскрипции вроде эффекторных нуклеаз (TALEN), хоминг-эндонуклеаз, или используя систему CRISPR/Cas вместе со сконструированной CRISPR РНК (РНК в виде коротких палиндромных повторов, регулярно расположенных группами)/трансактивирующей CRISPR РНК («единой направляющей РНК») для проведения специфического расщепления. В публикации заявки на патент США №20080182332 описывается использование нуклеаз с доменами «неканонические цинковые пальцы» (ZFN) для направленной модификации геномов растений; в публикации заявки на патент США №20090205083 описывается ZFN-опосредованная направленная модификация локуса EPSPS у растений; в публикации заявки на патент США №20100199389 описывается направленная модификация локуса Zp15 у растений, а в публикации заявки на патент США №20110167521 описывается направленная модификация генов, участвующих в биосинтезе жирных кислот. Кроме того, в Moehle et al. (2007) Proc. Natl. Acad, Sci. USA 104(9):3055-3060 описывается использование сконструированных ZFN для направленного добавления гена в конкретный локус.
Ассимиляция углерода является основной в метаболическом функционировании всех живых организмов. Способность синтезировать АТФ и использовать его энергию для гомеостаза, роста и размножения сохраняется во всех царствах и оказывает влияние на большинство известных биологических процессов. Основным компонентом синтеза АТФ у эукариот является цикл трикарбоновых кислот (ТКК), также известный как цикл лимонной кислоты или цикл Кребса, в ходе которого электроны с органических кислот переходят к окисленным редокс-кофакторам НАД+ и ФАД, образуя НАДН, ФАДН2 и углекислый газ. Цикл ТКК протекает в митохондриях; в растениях промежуточные продукты, образуемые в ходе его реакций, служат в качестве субстратов для многочисленных путей биосинтеза; основные поставки для образования аспартата, глютамата, нуклеиновых кислот, порфиринов и жирных кислот идут из цикла ТКК. Кроме того, промежуточные продукты цикла ТКК играют ключевую роль в энергетических процессах фотодыхания и фотосинтеза. По этой причине полагают, что цикл ТКК действуют в качестве связующего звена между окислительно-восстановительными функциями хлоропластов, митохондрий и цитозоля.
Малат является одним из промежуточных продуктов цикла ТКК и функционирует в качестве субстрата и для декарбоксилирующей малатдегидрогеназы, которая порождает пируват, и для малатдегидрогеназы (MDH). MDH катализирует обратимое восстановление оксалоацетата (OAA) до малата через НАДН и включена в малат-аспартатную челночную систему. Большинство растений содержат множество изоформ MDH, в том числе митохондриальные и цитозольные ферменты, которые кодируются ядерными генами. Митохондриальная MDH (mMDH) растений участвует в 3 типах реакций: превращении малата в OAA, восстановлении OAA до малата и восстановлении OAA в С4-пути фотосинтеза. У кукурузы (C4 злаковой травы) существует 5 различных локусов MDH в 5 независимых хромосомах, 2 из которых кодируют цитозольные изоформы, в то время как другие 3 кодируют митохондриальные ферменты. Используя классические анализы мутаций, было показано, что полная утрата функции 2 цитозольных форм MDH не оказывала вредное влияние на рост и размножение растений - функция в цитозоле, по-видимому, была несущественной. Напротив, полная утрата 3 митохондриальных ферментов приводила к летальности - растениям нужен был по крайней мере один функциональнй аллель, чтобы быть жизнеспособными (Goodman et al. (1981) Proc. Nat. Acad. Sci. USA 78:1783-1785). Так же данные наблюдений встречающихся в природе, самопроизвольных молчащих аллелей митохондриальной MDH-1 (Mdh1-n) у сои показали, что не было заметного изменения фенотипа растения до тех пор, пока ген митохондриальной Mdh2 оставался стабильным (Imsande et al. (2001) J. Heredity 92:333-338).
Несмотря на основную роль малата в метаболизме растений, его функции в цикле ТКК все еще не полностью поняты. Мутантные по MDH растения демонстрируют более медленные скорости роста и измененные характеристики фотодыхания. См., например, Tomaz et al. (2010) Plant Physiol. 154(3):1143-1157. Исследования антисмысловых олигонуклеотидов и РНК-интерференции в целых растениях или плоде дали противоречивые результаты, включающие растения с увеличенным весом сухого (не сырого) плода, а также растения с более высокими уровнями аскорбата в их листьях, чем в контролях дикого типа, но при выращивании в световых условиях короткого дня (которые способствуют фотодыханию) растения продемонстрировали фенотип карликовости и имели уменьшенную биомассу листьев, стеблей и корней. Nunes-Nesi et al. (2005) Plant Physiol. 137:611-622; Nunes-Nesi et al. (2007) Physiol. Plant. 129:45-56; Nunes-Nesi (2008) J. Exp. Bot. 59:1675-1684; Finkmeier and Sweetlove (2009) F1000 Biology Reports I:47; doi: 10.3410/B1-47. Кроме того, подвергнутые воздействию антисмысловых олигонуклеотидов для гена mMDH линии с уменьшенной экспрессией mMDH продемонстрировали уменьшенную активность (39% от таковой дикого типа) этого фермента, приведшую к уменьшению корневой зоны и остановке роста корней. Van der Merwe et al. (2009) Plant Physiol. 149:653-669; Van Der Merwe et al. (2010) Plant Physiol. 153:611-621. Кроме того, подвергнутые воздействию антисмысловых олигонуклеотидов для гена mMDH линии продемонстрировали увеличение высыхания плода (большую потерю H2О) и увеличенную подверженность заражения грибами. Centeno et al. (2011) Plant Cell 23:162-184. В публикации заявки на патент США №20090123626 описывается использование РНК-интерференции для MDH для уменьшения уровня аспарагина, что в свою очередь приводит к снижению уровня акриламида, который накапливается в результате связанного с обработкой нагревания растений и растительных продуктов.
Таким образом, остаются потребности в композициях и способах для изменения экспрессии генов MDH, например, с помощью направленной геномной модификации генов MDH, в растениях для создания стабильных, наследуемых генетических модификаций в растении и его потомстве.
КРАТКОЕ ИЗЛОЖЕНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
В настоящем описании представлены способы и композиции для направленной модификации гена(ов) MDH, а также клетки (например, семена), линии клеток, организмы (например, растения) и т.д., включающие одну или более направленных мутаций в MDH. Геном MDH может быть, например, ген митохондриальной MDH (mMDH). Как отмечено выше, исследования, демонстрирующие уменьшение ферментативной функции MDH с помощью технологии антисмысловых олигонуклеотидов и РНК-интерференции, дают противоречивые результаты, касающиеся эффекта(ов) ингибирования MDH, например, на урожай плодов. Основываясь на этих исследованиях можно было ожидать, что ингибирование протекания реакций цикла ТКК будет оказывать отрицательный эффект на фотосинтез. Таким образом, является внезапным и неожиданным, что авторы настоящего изобретения показали, что растения (и клетки растений), включающие направленные мутации в MDH, которые уменьшают функцию (активность) MDH, приводят к увеличенной урожайности от растений, включающих MDH-модифицированные клетки. Увеличенный урожай может включать, например, увеличенную величину урожая плодов, увеличенную биомассу растения (или плода растения), большее содержание мякоти плода, растения большего размера, увеличенный сухой вес, увеличенное твердое содержимое, более высокий общий вес при сборе урожая, увеличенную интенсивность и/или однородность цвета культуры, измененные химические характеристики (например, имеющие отношение к маслам, жирным кислотам, углеводам, белкам) и т.д.
Таким образом, в одном аспекте, здесь описываются растения, включающие клетки растений, в которых экспрессия эндогенного гена MDH модифицирована так, что экспрессия MDH является уменьшенной, и которые демонстрируют увеличенную урожайность. В некоторых вариантах осуществления экспрессию эндогенного гена MDH изменяют, используя составной белок, включающий ДНК-связывающий белок (например, белок с цинковыми пальцами, эффекторный домен TAL) и функциональный домен. В некоторых вариантах осуществления клетки растений содержат направленную модификацию гена MDH (например, mMDH), причем направленную модификацию, которая уменьшает экспрессию MDH, индуцирует нуклеазу, например, составной белок, включающий ДНК-связывающий домен и функциональный домен (например, нуклеаза с доменами «цинковые пальцы»), которая расщепляет эндогенный ген и уменьшает его экспрессию. Модификация (например, делеция, замена и/или вставка) может быть, например, модификацией одной или более аминокислот в НАДН-связывающем участке гена MDH (например, первом и/или втором НАДН-связывающем участке гена). В некоторых вариантах осуществления модификация включает изменение одной или более аминокислот в первом и/или втором НАДН-связывающем участке эндогенного гена MDH в клетке растения, например, одной или более аминокислот в положениях 104-136 и/или 171-220, пронумерованных относительно аминокислотной последовательности MDH дикого типа (например, SEQ ID NO: 1 (последовательности MDH помидора дикого типа), SEQ ID NO: 126 (последовательности MDH кукурузы дикого типа) и/или SEQ ID NO: 125 (последовательности MDH сои дикого типа)) и совмещенных с ней (например, фиг. 10).
Белок с цинковыми пальцами может включать распознающие спиральные участки, продемонстрированные в одном ряду таблицы 1A, и/или связываться с последовательностью-мишенью, продемонстрированной в таблице 1B. В других вариантах осуществления нуклеаза включает эффекторный домен TAL, хоминг-нуклеазу и/или Crispr/Cas единую направляющую РНК. Направленное изменение экспрессии MDH (например, направленная геномная модификация) может увеличить или уменьшить активность MDH, например, уменьшение активности MDH может быть в результате создания мутации, которая приводит к аберрантной транскрипции продукта гена (например, через сдвиг рамки считывания, новый стоп-кодон или другую мутацию). В некоторых вариантах осуществления направленная модификация, используя нуклеазу, включает небольшую вставку и/или делецию, также известную как вставка или делеция, например, вставка или делеция, продемонстрированная в таблице 4. Модификация в клетке может быть модификацией одного или более аллелей (например, в гомозиготах, гетерозиготах, в паралогичных генах). Любая из клеток растений, описываемых здесь, может находиться в растении или части растения (например, семенах, цветке, плоде), например, любого сорта: помидора (например, M82 или Moneymaker), сои, кукурузы, картофеля, люцерны или т.п.
В другом аспекте здесь описывается ДНК-связывающий домен (например, белок с цинковыми пальцами (ZFP)), который специфически связывается с геном MDH. Белок с цинковыми пальцами может включать один или более цинковых пальцев (например, 2, 3, 4, 5, 6, 7, 8, 9 или более цинковых пальцев), и может быть сконструирован белок с цинковыми пальцами, который связывается с любой последовательностью в любом гене MDH. Любой из белков с цинковыми пальцами, описываемый здесь, может связываться с сайтом-мишенью в кодирующей последовательности MDH или в примыкающих последовательностях (например, промоторе или других контролирующих экспрессию элементах) при условии, что достигается модификация экспрессии MDH. В некоторых вариантах осуществления белок с цинковыми пальцами связывается с сайтом-мишенью в гене mMDH, например, являющейся мишенью последовательности, продемонстрированной в таблице 1B. В других вариантах осуществления распознающие спиральные участки компонентных цинковых пальцев расположены в порядке от пальца 1 к пальцу 5 (от F1 к F5) или от пальца 1 к пальцу 6 (от F1 к F6), как продемонстрировано в одном ряду таблицы 1A. Один или более компонентных связывающих доменов «цинковые пальцы» белка с цинковыми пальцами может быть каноническим (C2H2) цинковым пальцем или неканоническим (например, C3H) цинковым пальцем (например, N-концевой и/или C-концевой цинковый палец может быть неканоническим пальцем).
В другом аспекте здесь описываются составные белки, при этом каждый составной белок включает ДНК-связывающий домен (например, белок с цинковыми пальцами), который специфически связывается с одним или более генов MDH. В некоторых вариантах осуществления белками являются составные белки, включающие белок с MDH-связывающимися цинковыми пальцами и функциональный домен, например, активирующий транскрипцию домен, подавляющий транскрипцию домен и/или расщепляющий домен (или полдомена для расщепления). В некоторых вариантах осуществления составным белком является нуклеаза с доменами «цинковые пальцы» (ZFN). Расщепляющие домены и половины доменов для расщепления можно получить, например, из различных эндонуклеаз рестрикции и/или хоминг-эндонуклеаз. В одном варианте осуществления половины доменов для расщепления происходят из эндонуклеазы рестрикции типа IIS (например, Fok I).
В других аспектах здесь обеспечиваются полинуклеотиды, кодирующие любой из ДНК-связывающих доменов (например, белки с цинковыми пальцами) и/или составных белков, описываемых здесь. В некоторых вариантах осуществления здесь описывается вектор для экспрессии ZFP, включающий полинуклеотид, кодирующий один или более ZFP, описываемых здесь, функционально связанный с промотором. В одном варианте осуществления одним или более ZFP являются ZFN.
ZFP и составные белки, включающие эти ZFP, могут связывать и/или расщеплять один или более генов MDH (например, ген mMDH) в кодирующей области гена или в некодирующей последовательности в гене или рядом с ним, такой как, например, лидерная последовательность, трейлерная последовательность или интрон, или промоторная последовательность, или в нетранскрибируемой области, или 5', или 3' от кодирующей области. В некоторых вариантах осуществления ZFP или ZFN связываются с кодирующей последовательностью или регуляторной последовательностью гена MDH и/или расщепляют ее.
В другом аспекте здесь описываются композиции, включающие один или более белков, составных белков и/или полинуклеотидов, описываемых здесь. Клетки растений могут содержать один уникальный ген MDH-мишень или множество паралогичных генов MDH-мишеней. Таким образом, описываемые здесь композиции могут включать один или более ZFP-содержащих белков (и полинуклеотидов, кодирующих их), мишенью которых является один или более генов MDH в клетке растения. Мишенями ZFP могут быть любые паралогичные или гомологичные гены и избранные конкретные паралогичные или гомологичные гены в клетке растения или комбинация некоторых паралогичных и некоторых гомологичных генов.
В другом аспекте здесь обеспечивается способ изменения экспрессии одного или более генов MDH (например, эндогенного гена mMDH) в клетке растения, при этом способ включает экспрессию одного или более содержащих ДНК-связывающий домен белков (например, белков с цинковыми пальцами) в клетке из условия, чтобы экспрессия MDH изменялась. В некоторых вариантах осуществления способы включают использование пары нуклеаз с доменами «цинковые пальцы» (белков и/или полинуклеотидов, кодирующих эти белки) для создания небольшой вставки и/или делеции («вставки или делеции»), которая нарушает экспрессию MDH. В других вариантах осуществления способы включают использование пары нуклеаз с доменами «цинковые пальцы» для увеличения экспрессии MDH, например, посредством направленной вставки трансгена или увеличивающего экспрессию элемента. В других вариантах осуществления способы изменения экспрессии MDH включают использование одного или более факторов транскрипции с доменами «цинковые пальцы» (составных белков, включающих белки с MDH-связывающимися цинковыми пальцами и функциональный домен, который является регулирующим транскрипцию доменом, таким как домен активации или репрессии). В некоторых вариантах осуществления измененная экспрессия/функция MDH приводит к увеличению фотосинтеза в клетках растений. В некоторых вариантах осуществления измененная экспрессия/функция MDH приводит к модификациям в цикле лимонной кислоты в клетках растений. В некоторых вариантах осуществления измененная экспрессия/функция MDH приводит к более высоким уровням малата в клетке растения. В других вариантах осуществления измененная экспрессия/функция MDH приводит к снижению уровней OAA в клетке. В одном варианте осуществления измененная экспрессия/функция MDH в клетках растений имеет следствием растения с повышенной урожайностью. В некоторых вариантах осуществления увеличение урожайности приводит к большему сырому весу полученного плода и общему сырому весу всех плодов, собранных с первой кисти мутантных растений.
В другом аспекте здесь обеспечиваются нуклеиновые кислоты и антитела, и способы их применения для обнаружения и/или измерения измененной экспрессии генов MDH и их модификаций.
В другом аспекте здесь описывается способ модификации одного или более генов MDH в клетке. В некоторых вариантах осуществления способ включает: (a) введение в клетку растения одной или более нуклеаз в форме белка и/или одного или более экспрессионных векторов, кодирующих одну или более нуклеаз (например, ZFN), которые связываются с сайтом-мишенью в одном или более генов MDH, в условиях, чтобы нуклеазы (например, ZFN) экспрессировались и один или более генов MDH расщеплялись, в силу чего осуществляется модификация одного или более генов MDH. В некоторых вариантах осуществления по крайней мере один сайт-мишень находится в гене mMDH. В других вариантах осуществления расщепляется более чем один ген MDH. Более того, в любом из описываемых здесь способах расщепление одного или более генов может приводить к делеции, добавлению и/или замене нуклеотидов в расщепленном участке, например, так, что активность MDH изменяется (например, увеличивается или уменьшается).
Тем не менее, в другом аспекте здесь описывается способ введения экзогенной последовательности (трансгена) в геном клетки растения из условия, чтобы активность MDH в клетке растения изменялась, при этом способ включает стадии: (a) приведения клетки в контакт с экзогенной последовательностью (вектором-донором); и (b) экспрессии одной или более нуклеаз (например, нуклеаз с доменами «цинковые пальцы»), описываемых здесь, в клетке, причем одна или боле нуклеаз расщепляет хромосомную ДНК; так что расщепление хромосомной ДНК на стадии (b) стимулирует включение вектора-донора в геном в результате гомологичной рекомбинации. В некоторых вариантах осуществления экзогенная последовательность встраивается в ген MDH. В других вариантах осуществления экзогенная последовательность встраивается близко от гена MDH. Активность MDH может увеличиваться или уменьшаться. В любом из описываемых здесь способов одна или более нуклеаз могут представлять собой объединения расщепляющего домена эндонуклеазы рестрикции типа IIs с сконструированным связывающим доменом «цинковые пальцы». В других вариантах осуществления нуклеаза включает хоминг-нуклеазу, например, хоминг-нуклеазу с модифицированным ДНК-связывающим доменом. В любом из описываемых здесь способов экзогенная последовательность может кодировать белковый продукт.
Тем не менее, в дальнейшем аспекте также обеспечивается клетка растения, полученная в соответствии с любым из способов, описываемых здесь.
В другом аспекте здесь обеспечивается растение, включающее клетку растения, описываемую здесь.
В другом аспекте здесь обеспечивается семя растения, включающего клетку растения, которая получена как описывается здесь.
В другом аспекте здесь обеспечивается плод, полученный от растения, включающего клетку растения, полученную как описывается здесь.
В любой из композиций (клеток или растений) или способов, описываемых здесь, клетка растения может включать клетку однодольного или двудольного растения. В некоторых вариантах осуществления клеткой растения является таковая культурного растения, например, помидора (или другой плодовой культуры), картофеля, кукурузы, сои, люцерны и т.д.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
Фиг. 1 представляет собой схему, на которой изображены различные структурные элементы гена митохондриальной MDH (mMDH) из Solanum lyocpersicum (v. M82).
На фиг. 2, панели A и B, изображена геномная организация и последовательность гена митохондриальной малатдегидрогеназы (mMDH) помидора. На фиг. 2A продемонстрированы сайты-мишени для ZFN (короткие указывающие налево и направо стрелки над экзонами) в экзонах 1, 3, 4 и 6. Обозначенный на фиг. 2А номер связывающей последовательности ZFN соответствует номеру ZFN, описанному в таблице 1; 107830L в таблице 1 описан на фиг. 2A как 830L, 107830R в таблице 1 описан на фиг. 2A как 830R, 107832L в таблице 1 описан на фиг. 2A как 832L, 107832R в таблице 1 описан на фиг. 2A как 832R, 107833L в таблице 1 описан на фиг. 2A как 833L, 107833R в таблице 1 описан на фиг. 2A как 833R, 107835L в таблице 1 описан на фиг. 2A как 835L, 107835R в таблице 1 описан на фиг. 2A как 835R. На фиг. 2B (SEQ ID NO: 2) представлена последовательность локуса mMDH; экзоны подчеркнуты, и сайты-мишени ZFN указаны жирным шрифтом.
Фиг. 3 представляет собой схему, на которой представлена карта плазмиды pKG7479.
Фиг. 4 представляет собой схему, на которой представлена карта плазмиды pKG7480.
Фиг. 5 представляет собой схему, на которой представлена карта плазмиды pKG7481.
Фиг. 6 представляет собой схему, на которой представлена карта плазмиды pKG7482.
На фиг. 7 изображен анализ последовательностей небольших вставок или делеций («вставок или делеций»), индуцированных в гене mMDH помидора в результате активности ZFN в протопластах. ZFN были транзиторно экспрессированы в протопластах клеток помидора, и вставки или делеции детектировали, используя анализ HRM. Сайты-мишени в mMDH для каждой ZFN представлены вместе с подчеркнутыми сайтами связывания. Амплифицированные продукты, содержащие делеции (представленные как -) или вставки (жирный шрифт), представлены под каждой последовательностью-мишенью.
Фиг. 8, панели A и B, представляет собой графики, на которых представлена активность mMDH в растениях F2. Фиг. 8A представляет собой график, на котором представлено определение активности mMDH в растениях F2, происходящих от линии 107832_9-6 (с делецией -3 п.о.). «126 9-6 WT» обозначает результаты биохимического анализа растения F2, в котором отсутствует мутация в виде вставки или делеции, «115 9-6 M» обозначает растения F2, гомозиготные по мутации в виде вставки или делеции, и «132 9-6 H» обозначает растение F2, гетерозиготное по мутации в виде вставки или делеции. Фиг. 8В представляет собой график, на котором представлено определение активности mMDH в растениях F2, происходящих от линии 107832_10-2 (с делецией -2 п.о.). WT48 обозначает растения F2, в которых отсутствует мутация в виде вставки или делеции, «M60» обозначает растения F2, гомозиготные по мутации в виде вставки или делеции, и «H32» обозначает растение F2, гетерозиготное по мутации в виде вставки или делеции.
Фиг. 9 представляет собой диаграмму, на которой представлен урожай плодов помидора линии 107832 9-6. Средний вес помидора (г) представлен по оси Y для 3 классов растений F2, выделяющихся по мутации в виде делеции -3 п.о. в локусе mMDH. «WT» означает растения F2, в которых отсутствует делеция, «Het» означает растения F2, гетерозиготные по делеции, и «Homo» означает растения F2, гомозиготные по делеции.
Фиг. 10 представляет собой совмещение последовательностей ферментов mMDH сои (SEQ ID NO: 125), кукурузы (SEQ ID NO: 126) и помидора (SEQ ID NO: 1).
Фиг. 11 представляет собой совмещение последовательностей мутаций mMDH, возникающих в геноме помидора.
Фиг. 12 представляет собой схему, на которой представлена биохимическая реакция, катализируемая ферментом mMDH.
Фиг. 13 представляет собой таблицу, в которой представлены специфические активности mMDH помидора дикого типа и двух мутантных ферментов mMDH помидора, которые определяли, спектрофотометрически следя за окислением НАДН. Мутация «mMDH del 3» сохраняет приблизительно 23% активности фермента дикого типа, а активность фермента с мутацией «mMDH del3 NADH BS1» является значительно уменьшенной на уровне приблизительно 1,5% от активности фермента дикого типа.
ПОДРОБНОЕ ОПИСАНИЕ НАСТОЯЩЕГО ИЗОБРЕТЕНИЯ
Настоящее описание относится к способам и композициям для измененной экспрессии одного или более генов малатдегидрогеназы (MDH) в клетке растения или растении, например, направленной геномной модификации гена MDH, такого как ген митохондриальной малатдегидрогеназы (mMDH), в клетке растения (например, кукурузы, помидора, сои и т.д.). В частности, экспрессию MDH изменяют посредством использования составных белков, включающих ДНК-связывающий домен (например, белок с цинковыми пальцами) и функциональный домен (например, регулирующий транскрипцию домен и/или нуклеазу). В некоторых вариантах осуществления направленная модификация достигается при расщеплении гена MDH, используя одну или более нуклеаз (например, ZFN) для создания модификаций (например, мутаций) в локусе MDH. Расщепление является направленным благодаря использованию составных белков, включающих ДНК-связывающий домен, такой как ДНК-связывающий домен мегануклеазы, ДНК-связывающий домен «лейциновая молния», ДНК-связывающий домен TAL, белок с цинковыми пальцами (ZFP), Crispr/Cas систему или химерные комбинации вышеупомянутых доменов. В некоторых вариантах осуществления модификация включает мутацию (замены, делеции и/или вставки) гена MDH из условия, чтобы была изменена одна или более аминокислот в первом и/или втором НАДН-связывающем участке эндогенного гена MDH, например, одна или более аминокислот в положениях 104-136 и/или 171-220, пронумерованных относительно SEQ ID NO: 1 (последовательности MDH помидора дикого типа), SEQ ID NO: 125 (последовательности MDH кукурузы дикого типа) и/или SEQ ID NO: 126 (последовательности MDH сои дикого типа) и совмещенных с ней.
В некоторых вариантах осуществления нуклеазы(а) включают одну или более ZFN. ZFN типично включают расщепляющий домен (или полдомена для расщепления) и связывающий домен «цинковые пальцы», который связывается с сайтом-мишенью в эндогенном гене MDH. ZFN могут быть введены в виде белков, в виде полинуклеотидов, кодирующих эти белки, и/или в виде комбинаций полипептидов и кодирующих полипептиды полинуклеотидов. Нуклеазы с доменами «цинковые пальцы» типично функционируют в виде димерных белков после димеризации половин доменов для расщепления и могут образовывать гомодимеры и/или гетеродимеры. Были описаны облигатные гетеродимерные ZFN, в случае которых мономеры ZFN, связывающиеся с «левым» и «правым» доменами распознавания, могут объединяться с образованием активной нуклеазы. См., например, публикацию заявки на патент США №2008/0131962. Таким образом, если имеются соответствующие сайты-мишени, «левый» мономер мог бы образовывать активную ZFN с любым «правым» мономером. Это значительно увеличивает число пригодных для нуклеаз сайтов на основе опробованных «левых» и «правых» доменов, которые могут использоваться в различных комбинациях. Например, рекомбинирование связывающих центров 4 гомодимерных ZFN даст дополнительно 12 гетеродимерных ZFN. Что еще более важно, оно делает возможным системный подход к разработке трансгенов так, чтобы каждая новая введенная экзогенная последовательность (трансген) становилась фланкированной уникальным сайтом для ZFN, который может использоваться для обратного вырезания гена или для направления дополнительных генов рядом с ним. Кроме того, этот способ может упростить стратегии установки в один локус, которая обусловлена ZFN-зависимыми двухцепочечными разрывами.
Связывающий домен «цинковые пальцы» может быть каноническим (C2H2) цинковым пальцем или неканоническим (например, C3H) цинковым пальцем. Кроме того, связывающий домен «цинковые пальцы» может включать один или более цинковых пальцев (например, 2, 3, 4, 5, 6, 7, 8, 9 или более цинковых пальцев), и можно сконструировать такой домен, который связывается с любой последовательностью в любом гене MDH. Распознающие спиральные участки приводимых в качестве примера белков с MDH-связывающими цинковыми пальцами для применения для связывания с геном MDH продемонстрированы в таблице 1A, а приводимые в качестве примера сайты-мишени в гене MDH продемонстрированы в таблице 1B. Присутствие такого составного белка (или белков, и/или полинуклеотидов, кодирующих эти составные белки) в клетке приводит к связыванию составного белка(ов) со своим сайтом(ами) связывания и расщеплению в гене(ах) MDH.
Общие сведения
Для осуществления на практике способов, а также приготовления и применения композиций, описываемых здесь, используются, кроме особо оговоренных способов, традиционные методы в молекулярной биологии, биохимии, исследовании структуры хроматина, вычислительной химии, методы культивирования клеток, рекомбинантных ДНК и в связанных областях, которые находятся в пределах компетентности в данной области техники. Эти методы в полной мере объяснены в литературе. См., например, Sambrook et al. MOLECULAR CLONING: A LABORATORY MANUAL, Second edition, Cold Spring Harbor Laboratory Press, 1989 и третье издание, 2001; Ausubel et al, CURRENT PROTOCOLS IN MOLECULAR BIOLOGY, John Wiley & Sons, New York, 1987 и периодические обновления; выпуски METHODS IN ENZYMOLOGY, Academic Press, San Diego; Wolffe, CHROMATIN STRUCTURE AND FUNCTION, Third edition, Academic Press, San Diego, 1998; METHODS IN ENZYMOLOGY, Vol. 304, «Chromatin» (P.M. Wassarman and A.P. Wolffe, eds.), Academic Press, San Diego, 1999; и METHODS IN MOLECULAR BIOLOGY, Vol. 119, «Chromatin Protocols» (P.B. Becker, ed.) Humana Press, Totowa, 1999.
Определения
Термины «нуклеиновая кислота», «полинуклеотид» и «олигонуклеотид» используются взаимозаменяемо и относятся к полимеру дезоксирибонуклеотидов или рибонуклеотидов, в линейной или замкнутой в круг конформации, и в или одноцепочечной, или двухцепочечной форме. Применительно к целям настоящего описания, эти термины не должны рассматриваться как ограничение по длине полимера. Эти термины могут охватывать известные аналоги природных нуклеотидов, а также нуклеотиды с подвергнутыми модификациям основаниями, сахарными и/или фосфатными составляющими (например, фосфоротиоатными связями). Как правило, аналог конкретного нуклеотида обладает такой же специфичностью спаривания оснований; т.е. аналог A будет подвергаться спариванию с основанием T.
Термины «полипептид», «пептид» и «белок» используются взаимозаменяемо и относятся к полимеру аминокислотных остатков. Термин также применяется в отношении полимеров аминокислот, в которых одна или более аминокислот являются химическими аналогами или модифицированными производными соответствующих встречающихся в природе аминокислот.
«Связывание» относится к специфическому в отношении последовательности, нековалентному взаимодействию между макромолекулами (например, между белком и нуклеиновой кислотой). Не требуется, чтобы все компоненты взаимодействия-связывания были специфическими в отношении последовательности (например, контакты с фосфатными остатками в остове ДНК) при условии, что взаимодействие в целом является специфическим в отношении последовательности. Такие взаимодействия, как правило, характеризуются константой диссоциации (Kd), составляющей 10-6 M-1 или меньше. «Сродство» относится к силе связывания: при этом увеличенное сродство связывания коррелирует с меньшей Kd.
«Связывающий белок» является белком, который способен к связыванию с другой молекулой. Связывающий белок может связываться, например, с молекулой ДНК (ДНК-связывающий белок), молекулой РНК (РНК-связывающий белок) и/или молекулой белка (белок-связывающий белок). В случае белок-связывающего белка он может связываться с самим собой (с образованием гомодимеров, гомотримеров и т.д.), и/или он может связываться с одной или более молекулами отличного белка или белков. Связывающий белок может обладать более чем одним типом активности связывания. Например, белки с цинковыми пальцами обладают ДНК-связывающей, РНК-связывающей и белок-связывающей активностью.
«ДНК-связывающий белок с цинковыми пальцами» (или связывающий домен) является белком, или доменом в более большом белке, который связывается с ДНК специфическим в отношении последовательности образом благодаря одному или более цинковым пальцам, которые представляют собой участки аминокислотной последовательности внутри связывающего домена, структура которых стабилизирована благодаря координации иона цинка. Термин «ДНК-связывающий белок с цинковыми пальцами» часто сокращенно называют белком с цинковыми пальцами или ZFP.
Можно сконструировать связывающие домены «цинковые пальцы», которые связываются с заданной нуклеотидной последовательностью. Неограничивающими примерами способов конструирования белков с цинковыми пальцами являются разработка и отбор. Сконструированный белок с цинковыми пальцами представляет собой не встречающийся в природе белок, разработка/состав которого вытекают в основном из критериев рациональности. Критерии рациональности для разработки включают применение правил замещений и компьютерных алгоритмов для обработки информации в базе данных, хранящей информацию о существующих разработках ZFP и данные о связывании. См., например, патенты США №№6140081, 6453242 и 6534261; см. также WO 98/53058; WO 98/53059; WO 98/53060; WO 02/016536 и WO 03/016496.
«Отобранный» белок с цинковыми пальцами представляет собой не обнаруживаемый в природе белок, получение которого является в основном результатом эмпирического процесса, такого как фаговый дисплей, ловушка взаимодействия (двугибридная система) или отбор гибридов. См., например, патент США №5789538, патент США №5925523, патент США №6007988, патент США №6013453, патент США №6200759, WO 95/19431; WO 96/06166; WO 98/53057; WO 98/54311; WO 00/27878; WO 01/60970 WO 01/88197 и WO 02/099084.
Термин «последовательность» относится к нуклеотидной последовательности любой длины, которая может представлять собой ДНК или РНК; может быть линейной, замкнутой в круг или разветвленной и может быть или одноцепочечной, или двухцепочечной. Термин «последовательность-донор» относится к нуклеотидной последовательности, которая встраивается в геном. Последовательность-донор может быть любой длины, например, длиной между 2 и 10000 нуклеотидов (или любым целым числом между ними или больше значений этого диапазона), предпочтительно длиной между 100 и 1000 нуклеотидов (или любым целым числом между ними), более предпочтительно длиной между приблизительно 200 и 500 нуклеотидов.
«Гомологичная, неидентичная последовательность» относится к первой последовательности, которая обладает некоторой степенью идентичности последовательности со второй последовательностью, но чья последовательность не идентична таковой второй последовательности. Например, полинуклеотид, включающий последовательность дикого типа мутантного гена, гомологичен и не идентичен последовательности мутантного гена. В некоторых вариантах осуществления степень гомологии между двумя последовательностями является достаточной для допуска гомологичной рекомбинации между ними, используя нормальные клеточные механизмы. Две гомологичные, неидентичные последовательности могут быть любой длины, и степень их негомологии может быть столь мала, как один нуклеотид (например, для исправления геномной точечной мутации с помощью направленной гомологичной рекомбинации), или столь велика, как 10 или более тысяч оснований (например, для встраивания гена в заданный эктопический сайт в хромосоме). Не требуется, чтобы два полинуклеотида, включающие гомологичные, неидентичные последовательности, были одинаковой длины. Например, может использоваться экзогенный полинуклеотид (т.е. полинуклеотид-донор) длиной между 20 и 10000 нуклеотидов или пар нуклеотидов.
Методы определения идентичности нуклеотидных и аминокислотных последовательностей известны в данной области техники. Типично такие методы включают определение нуклеотидной последовательности мРНК для гена и/или определение аминокислотной последовательности, кодируемой им, и сравнение этих последовательностей со второй нуклеотидной или аминокислотной последовательностью. Таким образом могут быть также определены и сравнены геномные последовательности. В общем, идентичность относится к точному соответствию нуклеотида нуклеотиду или аминокислоты аминокислоте в двух последовательностях полинуклеотидов или полипептидов, соответственно. Две или более последовательностей (полинуклеотидных или аминокислотных) можно сравнить посредством определения процента их идентичности. Процент идентичности двух последовательностей, или нуклеотидных, или аминокислотных последовательностей, является числом точных соответствий между двумя совмещенными последовательностями, разделенным на длину более короткой последовательности и умноженным на 100. Аппроксимируемое совмещение нуклеотидных последовательностей обеспечивается за счет алгоритма поиска локальной гомологии Smith и Waterman, в Advances in Applied Mathematics 2:482-489 (1981). Этот алгоритм может применяться для аминокислотных последовательностей, используя таблицу замен, разработанную Dayhoff, Atlas of Protein Sequences and Structure, M.O. Dayhoff ed., 5 suppl. 3:353-358, National Biomedical Research Foundation, Washington, D.C., США, и упорядоченную Gribskov, Nucl. Acids Res. 14(6):6745-6763 (1986). Приводимое в качестве примера внедрение этого алгоритма для определения процента идентичности последовательности обеспечено Genetics Computer Group (Madison, WI) в прикладной программе «BestFit». Программы, подходящие для расчета процента идентичности или схожести между последовательностями, как правило, известны в данной области техники, например, другой программой для совмещения является BLAST, используемая с параметрами по умолчанию. Например, BLASTN и BLASTP могут использоваться, используя следующие параметры по умолчанию: генетический код = стандартный; фильтр = нет; цепь = обе цепи; порог = 60; ожидаемое значение = 10; таблица = BLOSUM62; описания = 50 последовательностей; сортировка с помощью = HIGH SCORE; базы данных = неизбыточные, GenBank + EMBL + DDBJ + PDB + трансляции с кодирующих последовательностей GenBank + белок Swiss + Spupdate + PIR. Детали этих программ можно найти в Интернете. Что касается описываемых здесь последовательностей, диапазоном желаемых степеней идентичности последовательностей является приблизительно 80-100% и любое целое число между ними. Типично проценты идентичностей между последовательностями составляют по крайней мере 70-75%, предпочтительно 80-82%, более предпочтительно 85-90%, даже более предпочтительно 92%, еще более предпочтительно 95% и наиболее предпочтительно 98%.
Альтернативно, степень схожести последовательностей полинуклеотидов можно определить с помощью гибридизации полинуклеотидов в условиях, допускающих образование стабильных дуплексов между гомологичными районами, с последующим расщеплением специфической в отношении одноцепочечной последовательности нуклеазой(ами), и определения размера расщепленных фрагментов. Две последовательности нуклеиновых кислот или полипептидов являются в значительной степени гомологичными друг другу, когда последовательности демонстрируют составляющую по крайней мере приблизительно 70-75%, предпочтительно 80-82%, более предпочтительно 85-90%, даже более предпочтительно 92%, еще более предпочтительно 95% и наиболее предпочтительно 98% идентичность последовательностей по всей определенной длине молекул, определенную с использованием вышеупомянутых способов. Используемый здесь термин «в значительной степени гомологичные» также относится к последовательностям, демонстрирующим полную идентичность с заданной последовательностью ДНК или полипептида. Последовательности ДНК, которые в значительной степени гомологичны, можно идентифицировать в эксперименте с использованием гибридизации по Саузерну, например, в жестких условиях, определенных для этой конкретной системы. Определение подходящих условий гибридизации известно квалифицированным в данной области техники специалистам. См., например, Sambrook et al., выше; Nucleic Acid Hybridization: A Practical Approach, editors B.D. Hames and S.J. Higgins, (1985) Oxford; Washington, DC; IRL Press).
Селективную гибридизацию двух фрагментов нуклеиновых кислот можно определить, как описано ниже. Степень идентичности последовательностей двух молекул нуклеиновых кислот влияет на эффективность и прочность гибридизационных событий между такими молекулами. Отчасти идентичная последовательность нуклеиновой кислоты будет по крайней мере частично ингибировать гибридизацию полностью идентичной последовательности с молекулой-мишенью. Ингибирование гибридизации полностью идентичной последовательности можно оценить, используя анализы гибридизации, которые хорошо известны в данной области техники (например, Саузерн (ДНК)-блот, Нозерн (РНК)-блот, гибридизация в растворе или т.п., см. Sambrook, et al., Molecular Cloning: A Laboratory Manual, Second Edition, (1989) Cold Spring Harbor, N.Y.). Такие анализы могут проводиться, используя варьирующие степени селективности, например, используя условия, меняющиеся от низкой жесткости к высокой жесткости. Если используются условия низкой жесткости, отсутствие неспецифического связывания можно определить, используя второй зонд без даже частичной степени идентичности последовательности (например, зонд с составляющей менее чем приблизительно 30% идентичностью последовательности с молекулой-мишенью), так что, в отсутствие событий неспецифического связывания, дополнительный зонд не будет гибридизоваться с мишенью.
Используя систему обнаружения на основе гибридизации, выбирают зонд в виде нуклеиновой кислоты, который комплементарен контрольной последовательности нуклеиновой кислоты, и затем в соответствии с выбором подходящих условий зонд и контрольная последовательность селективно гибридизуются, или связываются, друг с другом с образованием дуплексной молекулы. Молекула нуклеиновой кислоты, которая способна селективно гибридизоваться с контрольной последовательностью в умеренно жестких условиях гибридизации, типично гибридизуется в условиях, позволяющих обнаружить последовательность нуклеиновой кислоты-мишень длиной по крайней мере приблизительно 10-14 нуклеотидов, идентичную на по крайней мере приблизительно 70% последовательности выбранного зонда в виде нуклеиновой кислоты. Жесткие условия гибридизации типично позволяют обнаружить последовательности нуклеиновых кислот-мишеней длиной по крайней мере приблизительно 10-14 нуклеотидов, идентичные на более чем приблизительно 90-95% последовательности выбранного зонда в виде нуклеиновой кислоты. Условия гибридизации, применимые для гибридизации зонд/контрольная последовательность, причем зонд и контрольная последовательность имеют определенную степень идентичности последовательностей, можно определить, как это известно в данной области техники (см., например, Nucleic Acid Hybridization: A Practical Approach, editors B.D. Hames and S.J. Higgins, (1985) Oxford; Washington, DC; IRL Press).
Условия гибридизации хорошо известны квалифицированным в данной области техники специалистам. Жесткость гибридизации относится к степени, в которой условия гибридизации не содействуют образованию гибридов, содержащих несоответствующие нуклеотиды, при этом большая жесткость коррелирует с меньшим допуском несоответствующих гибридов. Факторы, которые влияют на жесткость гибридизации, хорошо известны квалифицированным в данной области техники специалистам и включают, но без ограничения, температуру, pH, ионную силу и концентрацию органических растворителей, таких как, например, формамид и диметилсульфоксид. Как это известно квалифицированным в данной области техники специалистам, жесткость гибридизации увеличивается при более высоких температурах, меньшей ионной силе и более низких концентрациях растворителя.
Что касается условий жесткости для гибридизации, в данной области техники хорошо известно, что многочисленные эквивалентные условия могут использоваться для установления конкретной жесткости, варьируя, например, следующие факторы: длину и природу последовательностей зондов, состав оснований различных последовательностей, концентрации солей и других компонентов раствора для гибридизации, присутствие или отсутствие блокирующих агентов в растворах для гибридизации (например, декстрана сульфата и полиэтиленгликоля), температуру реакции гибридизации и параметры времени, а также варьируя условия отмывки. Выбор конкретного множества условий гибридизации осуществляют, следуя стандартным методам в данной области техники (см., например, Sambrook, et al., Molecular Cloning: A Laboratory Manual, Second Edition, (1989) Cold Spring Harbor, N.Y.).
«Рекомбинация» относится к процессу обмена генетической информацией между двумя полинуклеотидами. Применительно к целям этого описания, «гомологичная рекомбинация (HR)» относится к специализированной форме такого обмена, которая имеет место, например, во время репарации двухцепочечных разрывов в клетках. Необходима гомология нуклеотидных последовательностей для этого процесса, в котором используется молекула «донора», служащая матрицей для репарации молекулы «мишени» (т.е. молекулы, в которой существует двухцепочечный разрыв), и который известен под разными названиями: «не являющаяся кроссинговером конверсия гена» или «генная конверсия, длина участка которой является короткой», поскольку он приводит к переносу генетической информации от донора к мишени. Без желания ограничиться какой-либо конкретной теорией, такой перенос может включать коррекцию несоответствия в гетеродуплексной ДНК, которая образуется между мишенью с разрывом и донором, и/или «синтез-зависимый отжиг цепей», при котором донор используется для ресинтеза генетической информации, которая будет становиться частью мишени, и/или связанные процессы. Такая специализированная HR часто приводит к изменению последовательности молекулы мишени, так что часть или вся последовательность полинуклеотида-донора включается в полинуклеотид-мишень.
«Расщепление» относится к разрыву ковалентного остова молекулы ДНК. Расщепление можно инициировать с помощью множества способов, включающих, но без ограничения, ферментативный или химический гидролиз фосфодиэфирной связи. Возможен как одноцепочечный разрыв, так и двухцепочечный разрыв, и двухцепочечный разрыв может происходить в результате двух отдельных событий одноцепочечного разрыва. Расщепление ДНК может приводить к созданию или тупых концов, или липких концов. В некоторых вариантах осуществления составные полипептиды используются для направленного расщепления двухцепочечной ДНК.
«Расщепляющий домен» включает одну или более полипептидных последовательностей, которые обладают каталитической активностью для расщепления ДНК. Расщепляющий домен может содержаться в одной полипептидной цепи, или активность расщепления может быть результатом ассоциации двух (или более) полипептидов.
«Полдомена для расщепления» представляет собой последовательность полипептида, которая, вместе со вторым полипептидом (или идентичным, или отличным), образует комплекс, обладающий активностью расщепления (предпочтительно активностью расщепления двухцепочечной ДНК).
«Сконструированный полдомена для расщепления» представляет собой половину домена для расщепления, которая была модифицирован, чтобы образовывать облигатные гетеродимеры с другой половиной домена для расщепления (например, другим сконструированным полдоменом для расщепления). См. также публикации заявок на патенты США №№2005/0064474, 20070218528 и 2008/0131962, включенные сюда посредством ссылки в их полном объеме.
«Хроматин» является нуклеопротеиновой структурой, включающей геном клетки. Хроматин клетки включает нуклеиновую кислоту, в основном ДНК, и белок, включая гистоны и не являющиеся гистонами хромосомные белки. Большая часть хроматина эукариотических клеток существует в форме нуклеосом, причем «ядро» нуклеосомы включает приблизительно 150 пар оснований ДНК, связанных с октамером, включающим два любых из гистонов H2A, H2B, H3 и H4; и линкерная ДНК (переменной длины в зависимости от организма) простирается между «ядрами» нуклеосом. С линкерной ДНК, как правило, связана молекула гистона H1. Применительно к целям настоящего описания, термин «хроматин», как подразумевается, охватывает все типы клеточного нуклеопротеина, как прокариотического, так эукариотического. Хроматин клетки включает как хромосомный, так и эписомный хроматин.
«Хромосома» представляет собой хроматиновый комплекс, включающий весь геном клетки или его часть. Геном клетки часто характеризуется своим кариотипом, который представляет собой совокупность всех хромосом, которые составляют геном клетки. Геном клетки может включать одну или более хромосом.
«Эписома» представляет собой реплицирующуюся нуклеиновую кислоту, нуклеопротеиновый комплекс или другую структуру, включающую нуклеиновую кислоту, которая не является частью хромосомного набора - кариотипа клетки. Примеры эписом включают плазмиды и некоторые вирусные геномы.
«Доступный участок» является местом в хроматине клетки, в котором сайт-мишень, присутствующий в нуклеиновой кислоте, может связываться экзогенной молекулой, которая распознает сайт-мишень. Без желания ограничиться какой-либо конкретной теорией, полагают, что доступным участком является участок, который не упакован в нуклеосомальную структуру. Отличную структуру доступного участка можно часто обнаружить по его чувствительности к химическим и ферментативным зондам, например, нуклеазам.
«Сайт-мишень» или «последовательность-мишень» является последовательностью нуклеиновой кислоты, которая определяет часть нуклеиновой кислоты, с которой будет связываться связывающая молекула, в том случае, если существуют достаточные условия для связывания. Например, последовательность 5'-GAATTC-3' является сайтом-мишенью для эндонуклеазы рестрикции EcoRI.
«Экзогенной молекулой» является молекула, которая обычно не присутствует в клетке, но может быть введена в клетку с помощью одного или более генетических, биохимических или других способов. «Обычное присутствие в клетке» определяется относительно конкретной стадии развития клетки и условий окружающей среды для клетки. Таким образом, например, молекула, которая присутствует в клетках только во время ранних стадий развития цветка, является экзогенной молекулой относительно клеток полностью сформировавшегося цветка. Так же молекула, индуцируемая тепловым шоком, является экзогенной молекулой относительно клетки, не подвергнутой тепловому шоку. Экзогенная молекула может включать, например, последовательность, кодирующую любой полипептид или его фрагмент, функционирующий вариант дисфункционирующей эндогенной молекулы или дисфункционирующий вариант нормально функционирующей эндогенной молекулы. Кроме того, экзогенная молекула может включать кодирующую последовательность от другого вида, которая является ортологом эндогенного гена в клетке-хозяине.
Экзогенной молекулой может быть, среди прочего, небольшая молекула, например, которая создается с помощью процесса комбинаторной химии, или макромолекула, такая как белок, нуклеиновая кислота, углевод, липид, гликопротеин, липопротеин, полисахарид, любое модифицированное производное вышеупомянутых молекул, или любой комплекс, включающий одну или более вышеупомянутых молекул. Нуклеиновые кислоты включают ДНК и РНК, могут быть одно- или двухцепочечными; могут быть линейными, разветвленными или замкнутыми в круг; и могут быть любой длины. Нуклеиновые кислоты включают те, которые способны образовывать дуплексы, а также образующие триплексы нуклеиновые кислоты. См., например, патенты США №№5176996 и 5422251. Белки включают, но без ограничения, ДНК-связывающие белки, факторы транскрипции, факторы реконструкции хроматина, связывающиеся с метилированной ДНК белки, полимеразы, метилазы, деметилазы, ацетилазы, деацетилазы, киназы, фосфатазы, интегразы, рекомбиназы, лигазы, топоизомеразы, гиразы и хеликазы. Таким образом, термин включает «трансгены» или «представляющие интерес гены», которые являются экзогенными последовательностями, введенными в клетку растения, например, в ген MDH в клетке растения.
Экзогенная молекула может быть молекулой того же типа, что и эндогенная молекула, например, экзогенным белком или нуклеиновой кислотой. Например, экзогенная нуклеиновая кислота может включать геном инфекционного вируса, плазмиду или эписому, введенный(ую) в клетку, или хромосому, которая обычно не присутствует в клетке. Способы введения экзогенных молекул в клетки известны квалифицированным в данной области техники специалистам и включают, но без ограничения, трансформацию протопластов, трансформацию с использованием карборунда (например, WHISKERS™), трансформацию с использованием Agrobacterium, липид-опосредованный перенос (т.е. липосомы, включающие нейтральные и катионные липиды), электропорацию, прямую инъекцию, слияние клеток, бомбардировку частицами (например, используя «генную пушку»), копреципитацию фосфатом кальция, перенос с использованием DEAE-декстрана и перенос с использованием вирусных векторов.
Напротив, «эндогенной» молекулой является молекула, которая обычно присутствует в конкретной клетке на конкретной стадии развития в конкретных условиях окружающей среды. Например, эндогенная нуклеиновая кислота может включать хромосому, геном митохондрии, хлоропласта или другой органеллы, или встречающуюся в природе эписомную нуклеиновую кислоту. Дополнительные эндогенные молекулы могут включать белки, например, факторы транскрипции и ферменты.
Используемый здесь термин «продукт экзогенной нуклеиновой кислоты» включает продукты и в виде полинуклеотидов, и в виде полипептидов, например, продукты транскрипции (полинуклеотиды, такие как РНК) и продукты трансляции (полипептиды).
«Составной» молекулой является молекула, в которой две или более молекул субъединиц связаны, предпочтительно ковалентно. Молекулы субъединиц могут быть молекулами одинакового химического типа или могут быть молекулами различных химических типов. Примеры первого типа составной молекулы включают, но без ограничения, составные белки (например, объединение ДНК-связывающего домена ZFP с расщепляющим доменом) и составные нуклеиновые кислоты (например, нуклеиновая кислота, кодирующая составной белок, описанный выше). Примеры второго типа составной молекулы включают, но без ограничения, объединение образующей триплекс нуклеиновой кислоты с полипептидом и объединение образующей связи в малой бороздке молекулы с нуклеиновой кислотой.
Экспрессия составного белка в клетке может быть следствием доставки составного белка в клетку или в результате доставки полинуклеотида, кодирующего составной белок, в клетку, в которой полинуклеотид транскрибируется, и транскрипт транслируется, с порождением составного белка. Транс-сплайсинг, расщепление полипептида и лигирование полипептидов могут быть также вовлечены в экспрессию белка в клетке. Способы доставки в клетки полинуклеотидов и полипептидов представлены в этом описании в другом месте.
Применительно к целям настоящего описания, ген включает участок ДНК, кодирующий продукт гена (см. ниже), а также все участки ДНК, которые регулируют образование продукта гена, независимо от того, находятся ли регуляторные последовательности рядом с кодирующими и/или транскрибируемыми последовательностями или нет. Соответственно, ген включает, но без обязательного ограничения ими, промоторные последовательности, терминаторы, регулирующие трансляцию последовательности, такие как участки связывания рибосомы и участки внутренней посадки рибосомы, энхансеры, сайленсеры, инсуляторы, пограничные элементы, начала репликации, сайты присоединения к матриксу и локус-контролирующие элементы.
«Экспрессия гена» относится к превращению информации, содержащейся в гене, в продукт гена. Продуктом гена может быть непосредственный продукт транскрипции гена (например, мРНК, тРНК, рРНК, антисмысловая РНК, рибозим, структурная РНК или любой другой тип РНК) или белок, продуцируемый в результате трансляции с мРНК. Продукты гена также включают РНК, которые модифицированы в результате таких процессов, как кэппирование, полиаденилирование, метилирование и корректировка, и белки, модифицированные, например, в результате метилирования, ацетилирования, фосфорилирования, убиквитинирования, АДФ-рибозилирования, миристилирования и гликозилирования.
«Модуляция» экспрессии гена относится к изменению активности гена. Модуляция экспрессии может включать, но без ограничения, активацию гена и репрессию гена.
Клетки «растений» включают, но без ограничения, клетки однодольных или двудольных растений. Неограничивающие примеры однодольных растений включают хлебные злаки, такие как кукуруза, рис, ячмень, овес, пшеница, сорго, рожь, сахарный тростник, ананас, лук, банановое дерево и кокосовую пальму. Неограничивающие примеры двудольных растений включают табак, помидор, подсолнечник, хлопчатник, сахарную свеклу, картофель, салат-латук, дыню, сою, канолу (рапс) и люцерну. Клетки растений могут происходить из любой части растения и/или из любой стадии развития растения.
«Представляющим интерес участком» является любой участок хроматина клетки, такой как, например, ген или некодирующая последовательность в гене или рядом с ним, в котором желательно связывание экзогенной молекулы. Связывание может быть с целью направленного расщепления ДНК и/или направленной рекомбинации. Представляющий интерес участок может присутствовать в хромосоме, эписоме, геноме органеллы (например, митохондрии, хлоропласта) или геноме инфекционного вируса, например. Представляющий интерес участок может находиться в кодирующей области гена, в транскрибируемых некодирующих областях, таких как, например, лидерные последовательности, трейлерные последовательности или интроны, или в нетранскрибируемых областях, или 5', или 3' от кодирующей области. Представляющий интерес участок может быть настолько мал, как одна пара нуклеотидов, или составлять вплоть до 2000 пар нуклеотидов в длину, или любое целое число пар нуклеотидов.
Термины «функциональная связь» и «функционально связанный» используются взаимозаменяемо, что касается непосредственного соседства двух или более компонентов (таких как элементы последовательности), при котором компоненты расположены так, что оба компонента функционируют нормально, и предоставляется возможность опосредования по крайней мере одним из компонентов функции, которую проявляет по крайней мере один из других компонентов. В качестве иллюстрации регулирующая транскрипцию последовательность, такая как промотор, функционально связана с кодирующей последовательностью, если регулирующая транскрипцию последовательность контролирует уровень транскрипции с кодирующей последовательности в ответ на присутствие или отсутствие одного или более регулирующих транскрипцию факторов. Регулирующая транскрипцию последовательность, как правило, функционально связана в цис-положении с кодирующей последовательностью, но не требуется, чтобы она находилась рядом с кодирующей последовательностью. Например, энхансер представляет собой регулирующую транскрипцию последовательность, которая функционально связана с кодирующей последовательностью, даже если они не являются соприкасающимися.
Что касается составных полипептидов, термин «функционально связанный» может относиться к тому факту, что каждый из компонентов выполняет ту же функцию при нахождении в связи с другим компонентом, как было бы, если бы он не был связан подобным образом. Например, что касается составного полипептида, в котором ДНК-связывающий домен ZFP объединен с расщепляющим доменом, ДНК-связывающий домен ZFP и расщепляющий домен находятся в функциональной связи, если, в случае составного полипептида, часть в виде ДНК-связывающего домена ZFP способна к связыванию с своим сайтом-мишенью и/или со своим сайтом связывания, в то время как расщепляющий домен способен расщеплять ДНК вблизи сайта-мишени.
«Функциональным фрагментом» белка, полипептида или нуклеиновой кислоты является белок, полипептид или нуклеиновая кислота, последовательность которого(й) не идентична полноразмерному белку, полипептиду или нуклеиновой кислоте, при этом у него сохраняется та же функция, что у полноразмерного белка, полипептида или нуклеиновой кислоты. Функциональный фрагмент может обладать большим, меньшим или тем же количеством остатков, что и (чем) соответствующая природная молекула, и/или может содержать одну или более аминокислотных или нуклеотидных замен. Способы определения функции нуклеиновой кислоты (например, функции кодирования, способности к гибридизации с другой нуклеиновой кислотой) широко известны в данной области техники. Так же способы определения функции белков широко известны. Например, функцию связывания с ДНК полипептида можно определить, например, с помощью связывания на фильтрах, сдвига электрофоретической подвижности или анализов с использованием иммунопреципитации. Расщепление ДНК можно исследовать с помощью гель-электрофореза. См. Ausubel et al., выше. Способность белка к взаимодействию с другим белком можно определить, например, с помощью коиммунопреципитации, анализов с использованием двугибридной системы или комплементации, как генетической, так и биохимической. См., например, Fields et al. (1989) Nature 340:245-246; патент США №5585245 и PCT WO 98/44350.
ДНК-связывающие домены
В описываемых здесь способах может использоваться любой ДНК-связывающий домен. В некоторых вариантах осуществления ДНК-связывающий домен включает белок с цинковыми пальцами. Связывающий домен «цинковые пальцы» включает один или более цинковых пальцев. Miller et al. (1985) EMBO J. 4:1609-1614; Rhodes (1993) Scientific American Feb.: 56-65; патент США №6453242. Связывающие домены «цинковые пальцы», описываемые здесь, как правило, включают 2, 3, 4, 5, 6 или даже более цинковых пальцев.
Типично длина одного домена «цинковый палец» составляет приблизительно 30 аминокислот. Структурные исследования показали, что каждый домен (мотив) «цинковый палец» содержит два бета-складчатых слоя (удерживаемых в бета-изгибе, который содержит два инвариантных остатка цистеина) и альфа-спираль (содержащую два инвариантных остатка гистидина), которые удерживаются в особой конформации благодаря координации атома цинка двумя цистеинами и двумя гистидинами.
Цинковые пальцы включают как канонические C2H2 цинковые пальцы (т.е. те, в которых ион цинка координируется двумя остатками цистеина и двумя остатками гистидина), так и неканонические цинковые пальцы, такие как, например, C3H цинковые пальцы (те, в которых ион цинка координируется тремя остатками цистеина и одним остатком гистидина) и C4 цинковые пальцы (те, в которых ион цинка координируется четырьмя остатками цистеина). См. также WO 02/057293, а также публикацию заявки на патент США №20080182332 касательно неканонических ZFP для применения в растениях.
Сконструированный связывающий домен «цинковые пальцы» может обладать новой специфичностью связывания по сравнению со встречающимися в природе белком с цинковыми пальцами. Методы конструирования включают, но без ограничения, разработку на основе критериев рациональности и различные типы отбора. Разработка на основе критериев рациональности включает, например, использование баз данных, включающих последовательности нуклеотидов в триплетах (или квадруплетах) и аминокислотные последовательности отдельных цинковых пальцев, в которых последовательность нуклеотидов в триплетах или квадруплетах связана с одной или более аминокислотными последовательностями цинковых пальцев, которые связываются с конкретной последовательностью в виде триплета или квадруплета.
Приводимые в качестве примера методы отбора, включающие фаговый дисплей и двугибридные системы, описаны в патентах США №№5789538, 5925523, 6007988, 6013453, 6410248, 6140466, 6200759 и 6242568, а также WO 98/37186, WO 98/53057, WO 00/27878, WO 01/88197 и GB 2338237.
Увеличение специфичности связывания для связывающих доменов «цинковые пальцы» было описано, например, в WO 02/077227.
Поскольку отдельный цинковый палец связывается с последовательностью трех нуклеотидов (т.е. триплетом) (или четырехнуклеотидной последовательностью, которая может перекрываться, по одному нуклеотиду, с четырехнуклеотидным сайтом связывания для соседнего цинкового пальца), длина последовательности, с которой связывающий домен «цинковый палец» в соответствии с разработкой связывается (например, последовательности-мишени), будет определять число цинковых пальцев в конструируемом связывающем домене «цинковый палец». Например, в случае ZFP, в которых мотивы «цинковые пальцы» не связываются с перекрывающимися подсайтами, шестинуклеотидная последовательность-мишень связывается связывающим доменом, включающим два пальца; девятинуклеотидная последовательность-мишень связывается связывающим доменом, включающим три пальца, и т.д. Как здесь отмечается, не требуется, чтобы сайты связывания для отдельных цинковых пальцев (т.е. подсайты) в сайте-мишени были соприкасающимися, но они могут быть разделены одним или несколькими нуклеотидами, в зависимости от длины и природы аминокислотных последовательностей между цинковыми пальцами (т.е. линкеров между пальцами) в связывающем домене, включающем множество пальцев.
В связывающем домене, включающем множество пальцев, соседние цинковые пальцы могут быть разделены аминокислотными последовательностями-линкерами длиной приблизительно 5 аминокислот (так называемыми «каноническими» линкерами между пальцами) или, альтернативно, одним или более неканоническими линкерами. См., например, патенты США №№6453242 и 6534261. В случае сконструированных связывающих доменов «цинковые пальцы», включающих более трех пальцев, вставка более длинных («неканонических») линкеров между некоторыми из цинковых пальцев может быть желательной в некоторых случаях, поскольку она может увеличить сродство и/или специфичность связывания связывающим доменом. См., например, патент США №6479626 и WO 01/53480. Соответственно, связывающие домены, включающие множество цинковых пальцев, можно также охарактеризовать в отношении присутствия и локализации неканонических линкеров между пальцами. Например, связывающий домен с шестью цинковыми пальцами, включающий три пальца (соединенных с помощью двух канонических линкеров между пальцами), длинный линкер и три дополнительных пальца (соединенных с помощью двух канонических линкеров между пальцами), именуют конфигурацией 2×3. Аналогично, связывающий домен, включающий два пальца (с каноническим линкером между ними), длинный линкер и два дополнительных пальца (соединенных с помощью канонического линкера), именуют конфигурацией 2×2. Белок, который включает три единицы, включающие два пальца (в каждой из которых два пальца соединены с помощью канонического линкера), и в котором каждая единица, включающая два пальца, соединена с соседней единицей, включающей два пальца, с помощью длинного линкера, называют конфигурацией 3×2.
Присутствие длинного или неканонического линкерами между двумя соседними цинковыми пальцами в связывающем домене, включающем множество цинковых пальцев, позволяет двум пальцам связываться с подсайтами, которые не являются непосредственно соприкасающимися в последовательности-мишени. Соответственно, могут существовать промежутки из одного или более нуклеотидов между подсайтами в сайте-мишени; т.е. сайт-мишень может содержать один или более нуклеотидов, с которыми не контактирует цинковый палец. Например, связывающий домен «цинковые пальцы» 2×2 может связываться с двумя шестинуклеотидными последовательностями, разделенными одним нуклеотидом, т.е. он связывается с 13-нуклеотидным сайтом-мишенью. См. также Moore et al. (2001a) Proc. Natl. Acad. Sci. USA 98:1432-1436; Moore et al. (2001b) Proc. Natl. Acad. Sci. USA 98:1437-1441 и WO 01/53480.
Как обсуждалось ранее, подсайтом-мишенью является трех- или четырехнуклеотидная последовательность, которая связывается одним цинковым пальцем. В некоторых целях, единицу, включающую два пальца, именуют «связывающим модулем». Связывающий модуль можно получить, например, посредством отбора двух соседних пальцев в рамках белка с множеством пальцев (как правило, трех пальцев), которые связываются с конкретной шестинуклеотидной последовательностью-мишенью. Альтернативно, модули можно сконструировать с помощью сборки отдельных цинковых пальцев. См. также WO 98/53057 и WO 01/53480.
Альтернативно, ДНК-связывающий домен может происходить из нуклеазы. Например, известны распознающие последовательности хоминг-нуклеаз и мегануклеаз, таких как I-SceI, I-CeuI, PI-PspI, PI-Sce, I-SceIV, I-CsmI, I-PanI, I-SceII, I-PpoI, I-SceIII, I-CreI, I-TevI, I-TevII и I-TevIII. См. также патент США №5420032; патент США №6833252; Belfort et al. (1997) Nucleic Acids Res. 25:3379-3388; Dujon et al. (1989) Gene 82:115-118; Perler et al. (1994) Nucleic Acids Res. 22, 1125-1127; Jasin (1996) Trends Genet. 12:224-228; Gimble et al. (1996) J Mol. Biol. 263:163-180; Argast et al. (1998) J. Mol. Biol. 280:345-353 и каталог New England Biolabs. Кроме того, можно создать специфичность связывания с ДНК хоминг-нуклеаз и мегануклеаз так, чтобы они связывались с неприродными сайтами-мишенями. См., например, Chevalier et al. (2002) Molec. Cell 10:895-905; Epinat et al. (2003) Nucleic Acids Res. 31:2952-2962; Ashworth et al. (2006) Nature 441:656-659; Paques et al. (2007) Current Gene Therapy 7:49-66; публикацию заявки на патент США №20070117128.
В качестве другой альтернативы, ДНК-связывающий домен может происходить из белка «лейциновая молния». Лейциновые молнии являются классом белков, которые вовлечены в белок-белковые взаимодействия во множестве регуляторных белков эукариотических клеток, которые являются важными факторами транскрипции, связанными с экспрессией генов. Лейциновая молния относится к общему структурному мотиву, совместно используемому этими факторами транскрипции, среди нескольких царств, включая животных, растения, дрожжи и т.д. Лейциновая молния образована двумя полипептидами (гомодимер или гетеродимер), которые связываются со специфическими последовательностями ДНК таким образом, что остатки лейцина равномерно распределяются по α-спирали, так что остатки лейцина двух полипептидов оказываются на одной и той же поверхности спирали. Специфичность связывания с ДНК лейциновых молний может использоваться в ДНК-связывающих доменах, описываемых здесь.
В некоторых вариантах осуществления ДНК-связывающим доменом является сконструированный домен эффектора TAL, происходящего из патогена растений Xanthomonas (см. Miller et al. (2011) Nature Biotechnology 29(2):143-8; Boch et al, (2009) Science 29 Oct 2009 (10.1126/science.117881) и Moscou and Bogdanove, (2009) Science 29 Oct 2009 (10.1126/science.l178817); и публикации заявок на патенты США №№20110239315, 20110145940 и 20110301073, которые включены сюда посредством ссылки в их полном объеме).
Нуклеазная система CRISPR (короткие палиндромные повторы, регулярно расположенные группами)/Cas (связываемая с CRISPR) является недавно разработанной нуклеазной системой на основе бактериальной системы, которая может использоваться для геномной инженерии. Она основана на части адаптивного иммунного ответа многих бактерий и Archea. Когда вирус или плазмида внедряется в бактерию, сегменты ДНК оккупанта превращаются в CRISPR РНК (crRNA) в результате «иммунной» реакции. Эта CRISPR РНК затем связывается, благодаря участку частичной комплементарности, с другим типом РНК, называемой трансактивирующей CRISPR РНК, для направления нуклеазы Cas9 на участок, гомологичный CRISPR РНК, в ДНК-мишени, называемый «протоспейсером». Cas9 расщепляет ДНК с порождением тупых концов в двухцепочечном разрыве в сайтах, заданных 20-нуклеотидной направляющей последовательностью, содержащейся в транскрипте CRISPR РНК. Для сайт-специфического распознавания ДНК и расщепления Cas9 требуется как CRISPR РНК, так и трансактивирующая CRISPR РНК. Теперь разработана такая система, что CRISPR РНК и трансактивирующая CRISPR РНК могут быть объединены в одну молекулу («единую направляющую РНК»), и можно сконструировать эквивалентную CRISPR РНК часть единой направляющей РНК, которая направляет нуклеазу Cas9 для воздействия на любую желаемую последовательность (см. Jinek et al (2012) Science 337, p. 816-821, Jinek et al, (2013), eLife 2:e00471, и David Segal, (2013) eLife 2:e00563). Таким образом, можно разработать систему CRISPR/Cas для создания двухцепочечного разрыва (DSB) в желательной мишени в геноме, и на репарацию DSB можно оказывать влияние с помощью использования ингибиторов репарации для вызова увеличения склонной к ошибкам репарации.
Расщепляющие домены
Как отмечено выше, ДНК-связывающий домен может быть ассоциирован с расщепляющим (нуклеазным) доменом. Например, может быть модифицирована специфичность связывания с ДНК хоминг-эндонуклеаз при сохранении их нуклеазной активности. Кроме того, белки с цинковыми пальцами могут быть также объединены с расщепляющим доменом с образованием нуклеазы с доменами «цинковые пальцы» (ZFN). Расщепляющую домен часть составных белков, описываемых здесь, можно получить из любой эндонуклеазы или экзонуклеазы. Приводимые в качестве примера эндонуклеазы, из которых может быть получен расщепляющий домен, включают, но без ограничения, эндонуклеазы рестрикции и хоминг-эндонуклеазы. См., например, 2002-2003 Catalogue, New England Biolabs, Beverly, MA; и Belfort et al. (1997) Nucleic Acids Res. 25:3379-3388. Известны дополнительные ферменты, которые расщепляют ДНК (например, нуклеаза S1; нуклеаза фасоли золотистой; ДНКаза I поджелудочной железы; нуклеаза микрококков; дрожжевая HO эндонуклеаза; см. также Linn et al. (eds.) Nucleases, Cold Spring Harbor Laboratory Press, 1993). Неограничивающие примеры хоминг-эндонуклеаз и мегануклеаз включают I-SceI, I-CeuI, PI-PspI, PI-Sce, I-SceIV, I-CsmI, I-PanI, I-SceII, I-PpoI, I-SceIII, I-CreI, I-TevI, I-TevII и I-TevIII. См. также патент США № 5420032, патент США №6833252; Belfort et al. (1997) Nucleic Acids Res. 25:3379-3388; Dujon et al. (1989) Gene 82:115-118; Perler et al. (1994) Nucleic Acids Res. 22, 1125-1127; Jasin (1996) Trends Genet. 12:224-228; Gimble et al. (1996) J Mol. Biol. 263:163-180; Argast et al. (1998) J Mol. Biol. 280:345-353 и каталог New England Biolabs. Один или более этих ферментов (или их функциональных фрагментов) могут использоваться в качестве источника расщепляющих доменов и половин доменов для расщепления.
Эндонуклеазы рестрикции (рестрикционные ферменты) присутствуют у многих видов и способны к специфическому в отношении последовательности связыванию с ДНК (в рестрикционном сайте) и расщеплению ДНК в или вблизи сайта(е) связывания. Некоторые рестрикционные ферменты (например, типа IIS) расщепляют ДНК в сайтах, удаленных от сайта распознавания, и имеют разделимые связывающий и расщепляющий домены. Например, фермент типа IIS FokI катализирует двухцепочечный разрыв ДНК на расстоянии 9 нуклеотидов от его сайта распознавания на одной цепи и на расстоянии 13 нуклеотидов от его сайта распознавания на другой цепи. См., например, патенты США №№5356802, 5436150 и 5487994; а также Li et al. (1992) Proc. Natl. Acad. Sci. USA 89:4275-4279; Li et al. (1993) Proc. Natl. Acad. Sci. USA 90:2764-2768; Kim et al. (1994a) Proc. Natl. Acad. Sci. USA 91:883-887; Kim et al. (1994b) J. Biol. Chem. 269:31978-31982. Таким образом, в одном варианте осуществления составные белки включают расщепляющий домен (или полдомена для расщепления) из по крайней мере одного рестрикционного фермента типа IIS и один или более связывающих доменов «цинковые пальцы», которые могут быть или могут не быть сконструированными.
Приводимым в качестве примера рестрикционным ферментом типа IIS, расщепляющий домен которого является отделимым от связывающего домена, является FokI. Этот конкретный фермент является активным в виде димера. Bitinaite et al. (1998) Proc. Natl. Acad. Sci. USA 95:10570-10575. Соответственно, применительно к целям настоящего описания, часть фермента FokI, используемая в описываемых составных белках, считается половиной домена для расщепления. Таким образом, для направленного двухцепочечного разрыва и/или направленного замещения клеточных последовательностей, используя объединения цинковый палец-FokI, два составных белка, каждый из которых включает полдомена для расщепления FokI, могут использоваться для воссоздания каталитически активного расщепляющего домена. Альтернативно, может также использоваться одна молекула полипептида, содержащая связывающий домен «цинковые пальцы» и две половины домена для расщепления FokI. Параметры для направленного расщепления и направленного изменения последовательностей, используя объединения цинковый палец-FokI, предоставлены в этом описании в другом месте.
Расщепляющий домен или полдомена для расщепления может быть любой частью белка, в которой сохраняется активность расщепления или в которой сохраняется способность к мультимеризации (например, димеризации) с образованием функционального расщепляющего домена.
Приводимые в качестве примера рестрикционные ферменты типа IIS описаны в публикации международной заявки WO 2007/014275, которая включена посредством ссылки в ее полном объеме.
Для увеличения специфичности расщепления расщепляющие домены можно также модифицировать. В некоторых вариантах осуществления используются варианты полдомена для расщепления, эти варианты минимизируют или предотвращают гомодимеризацию половин доменов для расщепления. Неограничивающие примеры таких модифицированных половин доменов для расщепления описаны подробно в WO 2007/014275, которая включена сюда посредством ссылки в ее полном объеме. См. также раздел «Примеры». В некоторых вариантах осуществления расщепляющий домен включает сконструированную половину домена для расщепления (также называемую мутантами домена, подвергаемого димеризации), которая минимизирует или предотвращает гомодимеризацию, эти мутанты известны квалифицированным в данной области техники специалистам и описаны, например, в публикациях заявок на патенты США №№20050064474; 20060188987; 20070305346 и 20080131962, описание каждой из которых включено сюда посредством ссылки в ее полном объеме. Все из аминокислотных остатков в положениях 446, 447, 479, 483, 484, 486, 487, 490, 491, 496, 498, 499, 500, 531, 534, 537 и 538 FokI являются мишенями для оказания влияния на димеризацию половин доменов для расщепления FokI.
Дополнительные сконструированные половины доменов для расщепления FokI, которые образуют облигатные гетеродимеры, могут также использоваться в ZFN, описываемых здесь. Приводимые в качестве примера сконструированные половины доменов для расщепления FokI, которые образуют облигатные гетеродимеры, включают пары, в которых первая половина домена для расщепления включает мутации в аминокислотных остатках в положениях 490 и 538 FokI, а вторая половина домена для расщепления включает мутации в аминокислотных остатках 486 и 499.
Таким образом, в одном варианте осуществления мутация в положении 490 приводит к замене Glu (E) на Lys (K); мутация в положении 538 приводит к замене Iso (I) на Lys (К); мутация в положении 486 приводит к замене Gln (Q) на Glu (E); и мутация в положении 499 приводит к замене Iso (I) на Lys (K). В частности, сконструированные половины доменов для расщепления, описываемые здесь, были приготовлены посредством вызова мутаций в положениях 490 (E→K) и 538 (I→K) в одной половине домена для расщепления с созданием сконструированного полдомена для расщепления, названного «E490K:I538K», и посредством вызова мутаций в положениях 486 (Q→E) и 499 (I→L) в другой половине домена для расщепления с созданием сконструированного полдомена для расщепления, названного «Q486E:I499L». Сконструированные половины доменов для расщепления, описываемые здесь, являются мутантами облигатных гетеродимеров, в которые аберрантное расщепление минимизировано или исключено. См., например, публикацию заявки на патент США №2008/0131962, описание которой включено посредством ссылки в ее полном объеме для всех целей. В некоторых вариантах осуществления сконструированная половина домена для расщепления включает мутации в положениях 486, 499 и 496 (пронумерованных относительно FokI дикого типа), например, мутации, которые приводят к замене остатка дикого типа Gln (Q) в положении 486 на остаток Glu (E), остатка дикого типа Iso (I) в положении 499 на остаток Leu (L) и остатка дикого типа Asn (N) в положении 496 на остаток Asp (D) или Glu (E) (также называемая доменами «ELD» и «ELE», соответственно). В других вариантах осуществления сконструированная половина домена для расщепления включает мутации в положениях 490, 538 и 537 (пронумерованных относительно FokI дикого типа), например, мутации, которые приводят к замене остатка дикого типа Glu (E) в положении 490 на остаток Lys (K), остатка дикого типа Iso (I) в положении 538 на остаток Lys (K) и остатка дикого типа His (H) в положении 537 на остаток Lys (K) или остаток Arg (R) (также называемая доменами «KKK» и «KKR», соответственно). В других вариантах осуществления сконструированная половина домена для расщепления включает мутации в положениях 490 и 537 (пронумерованных относительно FokI дикого типа), например, мутации, которые приводят к замене остатка дикого типа Glu (E) в положении 490 на остаток Lys (K) и остатка дикого типа His (H) в положении 537 на остаток Lys (K) или остаток Arg (R) (также называемая доменами «KIK» и «KIR», соответственно). (См. публикацию заявки на патент США №20110201055). В других вариантах осуществления сконструированная половина домена для расщепления включает «Sharkey»-мутации и/или мутации «Sharkey» (см. Guo et al, (2010) J. Mol. Biol. 400(1):96-107).
Сконструированные половины доменов для расщепления, описываемые здесь, можно приготовить, используя любой подходящий способ, например, сайт-направленный мутагенез половин доменов для расщепления дикого типа (FokI), как описано в публикациях заявок на патенты США №№20050064474; 20080131962 и 20110201055.
Альтернативно, нуклеазы могут подвергаться сборке in vivo на сайте-мишени нуклеиновой кислоты, используя так называемую технологию «разделенных ферментов» (см., например, публикацию заявки на патент США №20090068164). Компоненты таких разделенных ферментов могут экспрессироваться или с отдельных экспрессионных конструкций, или могут быть соединены в одну открытую рамку считывания, в которой отдельные компоненты разделены, например, саморасщепляющимся пептидом 2A или последовательностью IRES. Компонентами могут быть отдельные связывающие домены «цинковые пальцы» или домены в виде связывающего нуклеиновую кислоту домена мегануклеазы.
Нуклеазы можно подвергнуть скринингу на предмет активности перед использованием, например, в хромосомной системе на основе дрожжей, описанной в WO 2009/042163 и 20090068164. Конструкции для экспрессии нуклеаз можно легко разработать, используя известные в данной области техники способы. См., например, публикации заявок на патенты США №№20030232410; 20050208489; 20050026157; 20050064474; 20060188987; 20060063231 и публикацию международной заявки WO 07/014275. Экспрессия нуклеазы может находиться под контролем конститутивного промотора или индуцируемого промотора, например, промотора галактокиназы, который активируется (подвергается дерепрессии) в присутствии раффинозы и/или галактозы и подвергается репрессии в присутствии глюкозы.
Составные белки
Способы разработки и конструирования составных белков (и кодирующих их полинуклеотидов) известны квалифицированным в данной области техники специалистам. Например, способы разработки и конструирования составных белков, включающих ДНК-связывающие домены (например, домены «цинковые пальцы») и регуляторные или расщепляющие домены (или половины доменов для расщепления), и полинуклеотидов, кодирующих такие составные белки, описаны в патентах США №№6453242 и 6534261 и публикациях заявок на патенты США №№2007/0134796; 2005/0064474; 20080182332; 20090205083; 20100199389; 20110167521, 20110239315, 20110145940 и 20110301073, которые включены сюда посредством ссылки в их полном объеме. В некоторых вариантах осуществления конструируют полинуклеотиды, кодирующие составные белки. Эти полинуклеотиды можно встроить в вектор, и вектор можно ввести в клетку (см. ниже ради дополнительного описания, касающегося векторов и способов введения полинуклеотидов в клетки).
В некоторых вариантах осуществления способов, описываемых здесь, нуклеаза с доменами «цинковые пальцы» включает составной белок, включающий связывающий домен «цинковые пальцы» и полдомена для расщепления из рестрикционного фермента FokI, и два таких составных белка экспрессируют в клетке. Экспрессия двух составных белков в клетке может быть результатом доставки двух белков в клетку; доставки одного белка и одной нуклеиновой кислоты, кодирующей один из белков, в клетку; доставки двух нуклеиновых кислот, каждая из которых кодирует один из белков, в клетку; или доставки одной нуклеиновой кислоты, кодирующей оба белка, в клетку. В дополнительных вариантах осуществления составной белок включает одну полипептидную цепь, включающую две половины домена для расщепления и связывающий домен «цинковые пальцы». В этом случае один составной белок экспрессируется в клетке и, как полагают, без желания ограничиться какой-либо теорией, расщепляет ДНК в результате образования внутримолекулярного димера половин домена для расщепления.
В некоторых вариантах осуществления компоненты составных белков (например, объединений ZFP-FokI) расположены так, что домен «цинковые пальцы» находится ближе всего к N-концу составного белка, а полдомена для расщепления находится ближе всего к С-концу. Это служит отображением относительной ориентации расщепляющего домена во встречающихся в природе, димеризующихся расщепляющих доменах, таких как те, которые происходят из фермента FokI, в котором ДНК-связывающий домен находится ближе всего к N-концу, а полдомена для расщепления находится ближе всего к С-концу. В этих вариантах осуществления димеризация половин домена для расщепления с образованием функциональной нуклеазы осуществляется при связывании составных белков с сайтами на противоположных цепях ДНК, при этом 5' концы сайтов связывания являются проксимальными по отношению друг к другу.
В дополнительных вариантах осуществления компоненты составных белков (например, объединений ZFP-FokI) расположены так, что полдомена для расщепления находится ближе всего к N-концу составного белка, а домен «цинковые пальцы» находится ближе всего к С-концу. В этих вариантах осуществления димеризация половин домена для расщепления с образованием функциональной нуклеазы осуществляется при связывании составных белков с сайтами на противоположных цепях ДНК, при этом 3' концы сайтов связывания являются проксимальными по отношению друг к другу.
Тем не менее, в дополнительных вариантах осуществления первый составной белок содержит полдомена для расщепления, который находится ближе всего к N-концу составного белка, и домен «цинковые пальцы», который находится ближе всего к С-концу, а второй составной белок организован так, что домен «цинковые пальцы» находится ближе всего к N-концу составного белка, а полдомена для расщепления находится ближе всего к С-концу. В этих вариантах осуществления оба составных белка связываются с одной и той же цепью ДНК, при этом сайт связывания для первого составного белка, содержащего домен «цинковые пальцы» ближе всего к С-концу, находится с 5' стороны от сайта связывания для второго составного белка, содержащего домен «цинковые пальцы» ближе всего к N-концу.
В некоторых вариантах осуществления описываемых составных белков аминокислотную последовательность между доменом «цинковые пальцы» и расщепляющим доменом (или половиной домена для расщепления) называют «ZC линкером». ZC линкер следует отличать от линкеров между пальцами, обсуждавшихся выше. См., например, публикации заявок на патенты США №№20050064474 и 20030232410 и публикацию международной заявки на патент WO 05/084190 ради деталей получения ZC линкеров, которые оптимизируют расщепление.
В одном варианте осуществления настоящим изобретением обеспечивается нуклеаза (например, ZFN), включающая белок с цинковыми пальцами, имеющий одну или более распознающих спиральных аминокислотных последовательностей, представленных в таблице 1A (например, белок с цинковыми пальцами, состоящий из компонентных доменов «цинковые пальцы» с распознающими спиральными участками, представленными в одном ряду таблицы 1A). В другом варианте осуществления здесь обеспечивается вектор для экспрессии ZFP, включающий нуклеотидную последовательность, кодирующую ZFP, содержащий один или более распознающих спиральных участков, представленных в таблице 1A, например, ZFP, содержащий распознающие спиральные участки, упорядоченные и представленные в одном ряду таблицы 1A. В другом варианте осуществления здесь обеспечивается ДНК-связывающий домен (например, ZFP), который связывается с сайтом-мишенью, представленным в таблице 1B, или полинуклеотид, кодирующий ДНК-связывающий домен (например, ZFP), который связывается с сайтом-мишенью, представленным в таблице 1B.
Сайты-мишени
Описываемые способы и композиции включают составные белки, включающие ДНК-связывающий домен (например, ZFP) и регуляторный домен или расщепляющий (например, нуклеазный) домен (или полдомена для расщепления), в которых ДНК-связывающий домен (например, домен «цинковые пальцы»), при связывании с последовательностью в хроматине клетки в одном или более генов MDH, направляет активность регулирующего транскрипцию домена или расщепляющего домена (или полдомена для расщепления) на окрестности последовательности и поэтому модулирует транскрипцию или индуцирует расщепление вблизи последовательности-мишени.
Как изложено в этом описании в другом месте, можно сконструировать ДНК-связывающий домен, такой как домен «цинковые пальцы», который связывается фактически с любой желаемой последовательностью. Соответственно, после идентификации представляющего интерес участка, содержащего последовательность, в (для) которой желательны регуляция гена, расщепление или рекомбинация, можно сконструировать один или более связывающих доменов «цинковые пальцы», которые связываются с одной или более последовательностями в представляющем интерес участке. В некоторых вариантах осуществления ZFP, описываемые здесь, связываются с сайтом-мишенью, продемонстрированным в таблице 1B.
Выбор сайта-мишени в представляющем интерес геномном участке в хроматине клетки любого гена MDH для связывания ДНК-связывающего домена (например, сайта-мишени) может быть осуществлен, например, в соответствии со способами, описанными в патенте США №6453242 и/или публикациях заявок на патенты США №№20110239315, 20110145940 и 20110301073, в которых также описаны способы разработки ZFP, которые связываются с выбранной последовательностью. Квалифицированным в данной области техники специалистам будет понятно, что простая визуальная проверка нуклеотидной последовательности может также использоваться для отбора сайта-мишени. Соответственно, любой способ отбора сайта-мишени может использоваться в заявленных способах.
Сайты-мишени, как правило, состоят из множества расположенных рядом подсайтов-мишеней. Подсайт-мишень относится к последовательности (обычно или триплету нуклеотидов, или квадруплету нуклеотидов, который может перекрываться по одному нуклеотиду с расположенным рядом квадруплетом), связываемой отдельным цинковым пальцем. См., например, WO 02/077227. Если цепь, с которой белок с цинковыми пальцами в основном контактирует, назвать цепью-мишенью, «основной цепью распознавания» или «основной цепью для осуществления контактов», некоторые белки с цинковыми пальцами связываются с трехосновным триплетом в цепи-мишени и четвертым основанием в не являющейся мишенью цепи. В случае белков с цинковыми пальцами длина сайта-мишени, как правило, составляет по крайней мере 9 нуклеотидов, и, соответственно, он связывается связывающим доменом «цинковые пальцы», включающим по крайней мере три цинковых пальца. Однако также возможно связывание, например, связывающего домена, включающего 4 пальца, с 12-нуклеотидным сайтом-мишенью, связывающего домена, включающего 5 пальцев, с 15-нуклеотидным сайтом-мишенью или связывающего домена, включающего 6 пальцев, с 18-нуклеотидным сайтом-мишенью. Как это и очевидно, также возможно связывание более больших связывающих доменов (например, включающих 7, 8, 9 пальцев или более) с более длинными сайтами-мишенями.
Не требуется, чтобы сайт-мишень был множеством трех нуклеотидов. Например, в тех случаях, когда происходят взаимодействия с перекрещиванием цепей (см., например, патент США №6453242 и WO 02/077227), один или более отдельных цинковых пальцев связывающего домена, включающего множество пальцев, может связываться с подсайтами в виде перекрывающихся квадруплетов. В результате, белок с тремя пальцами может связываться с 10-нуклеотидной последовательностью, в которой десятый нуклеотид является частью квадруплета, связываемого конечным пальцем, белок с четырьмя пальцами может связываться с 13-нуклеотидной последовательностью, в которой тринадцатый нуклеотид является частью квадруплета, связываемого конечным пальцем, и т.д.
Длина и природа аминокислотных последовательностей-линкеров между отдельными цинковыми пальцами в связывающем домене, включающем множество пальцев, также оказывает влияние на связывание с последовательностью-мишенью. Например, присутствие так называемого «неканонического» линкера, «длинного линкера» или «структурированного линкера» между соседними цинковыми пальцами в связывающем домене, включающем множество пальцев, может позволить этим пальцам связываться с подсайтами, которые не являются непосредственно примыкающими. Неограничивающие примеры таких линкеров описаны, например, в патенте США №6479626 и WO 01/53480. Соответственно, один или более подсайтов, в сайте-мишени для связывающего домена «цинковые пальцы», могут быть отделены друг от друга 1, 2, 3, 4, 5 или более нуклеотидами. Для предоставления всего только одного примера, связывающий домен, включающий четыре пальца, может связываться с 13-нуклеотидным сайтом-мишенью, включающим, один за другим, два расположенных рядом 3-нуклеотидных подсайта, промежуточный нуклеотид и два расположенных рядом подсайта в виде триплетов. См. также публикацию заявки на патент США №20090305419 ради составов и способов соединения искусственных нуклеаз, которые связываются с сайтами-мишенями, разделенными различным количеством нуклеотидов. Расстояние между последовательностями (например, сайтами-мишенями) относится к числу нуклеотидов или пар нуклеотидов, находящихся между двумя последовательностями, определяемому от краев последовательностей, ближайших друг к другу.
В некоторых вариантах осуществления создают ZFP с функцией фактора транскрипции, например, посредством конструирования составных белков, включающих ZFP и регулирующий транскрипцию домен (например, домен для активации или репрессии). Неограничивающие примеры подходящих регулирующих транскрипцию доменов, которые можно объединить с ДНК-связывающими доменами для модуляции экспрессии гена, описаны, например, в патентах США №№6534261, 6824978, 6933113 и 8399218 и публикации заявки на патент №20080182332. Для функционирования фактора транскрипции, простое связывание и достаточная близость к промотору - это все, что обычно требуется. Точное расположение относительно промотора, ориентация и, в определенных пределах, расстояние не имеют большое значение. Эта особенность делает возможной значительную гибкость при выборе сайтов-мишеней для конструирования искусственных факторов транскрипции. По этой причине сайтом-мишенью, распознаваемым ZFP, может быть любой подходящий сайт в гене-мишени, который сделает возможной активацию или подавление экспрессии гена с помощью ZFP, необязательно связанного с регуляторным доменом. Предпочтительные сайты-мишени включают участки, примыкающие к, находящиеся 3' или 5' от сайта(у) начала транскрипции. Кроме того, сайтами-мишенями могут быть сайты-мишени, которые расположены в энхансерных районах, сайтах-репрессорах, местах приостановки работы РНК-полимеразы и специфических регуляторных сайтах (например, сайтах для SP-1, чувствительных к гипоксии элементах, распознаваемых ядерными рецепторами элементах, сайтах связывания р53), сайтах в кодирующей области кДНК или в кодирующей маркер экспрессируемой последовательности (EST) области.
В других вариантах осуществления создают нуклеазы, например ZFN и/или TALEN. Экспрессия нуклеазы, включающей составной белок, включающий ДНК-связывающий домен и расщепляющий домен, (или двух составных белков, каждый из которых включает связывающий домен «цинковые пальцы» и полдомена для расщепления), в клетке вызывает расщепление вблизи последовательности-мишени. В некоторых вариантах осуществления расщепление зависит от связывания двух составных молекул домен «цинковые пальцы»/полдомена для расщепления с отдельными сайтами-мишенями. Два сайта-мишени могут находиться на противоположных цепях ДНК, или, альтернативно, оба сайта-мишени могут находиться на одной и той же цепи ДНК.
Регуляция экспрессии генов
Ряд анализов может использоваться для определения того, модулирует ли ДНК-связывающий белок, такой как ZFP, экспрессию гена. Активность конкретного белка можно оценить, используя множество in vitro и in vivo анализов, посредством определения, например, уровней белка или мРНК, уровней продуктов, ферментативной активности, активации или репрессии транскрипции гена-репортера, используя, например, иммуноанализы (например, ELISA и иммуногистохимические анализы с использованием антител), гибридизационные анализы (например, защиту от РНКазы, Нозерн-блоттинг, in situ гибридизацию, исследования с использованием ранжированного ряда олигонуклеотидов), колориметрические анализы, анализы амплификации, анализы ферментативной активности, фенотипические исследования и т.п.
Активность ДНК-связывающих белков типично сначала проверяют in vitro, используя анализы ELISA, а затем используя дрожжевую экспрессионную систему. Например, ZFP часто сначала проверяют, используя систему для транзиторной экспрессии с использованием гена-репортера, а затем проверяют регуляцию эндогенного гена-мишени в клетках или в целых растениях как in vivo, так и ex vivo. ДНК-связывающий белок можно рекомбинантно экспрессировать в клетке, рекомбинантно экспрессировать в клетках, трансплантируемых в растение, или рекомбинантно экспрессировать в трансгенном растении, а также ввести в виде белка в растение или клетку, используя средства для доставки, описанные ниже. Клетки могут быть иммобилизованными, находиться в растворе, быть подвергнутыми введению в растение или быть встречающимися в природе в трансгенном или нетрансгенном растении.
Модуляцию экспрессии гена проверяют, используя один из in vitro или in vivo анализов, описанных здесь. Образцы или пробы обрабатывают ДНК-связывающим белком (например, ZFP) и сравнивают с контрольными образцами без исследуемого соединения для исследования степени модуляции. Для регуляции экспрессии эндогенного гена ДНК-связывающий белок типично характеризуется Kd, составляющей 200 нМ или менее, более предпочтительно 100 нМ или менее, более предпочтительно 50 нМ, наиболее предпочтительно 25 нМ или менее.
Эффекты связывающих белков можно определить, исследуя любой из параметров, описанных выше. Экспрессия любого подходящего гена, фенотипическое или физиологическое изменение может использоваться для оценки влияния ДНК-связывающего белка (например, ZFP). При определении функциональных последствий, используя исходные клетки или растения, можно также определить ряд эффектов, таких как рост растений, изменения транскрипции и известных, и не охарактеризованных генетических маркеров (например, с помощью Нозерн-блотов или исследований с использованием ранжированного ряда олигонуклеотидов), изменения клеточного метаболизма, такие как изменения роста клеток или pH, и изменения внутриклеточных вторичных мессенджеров, таких как cGMP.
Предпочтительные исследования регуляции экспрессии эндогенного гена могут выполняться in vitro. В одном предпочтительном формате in vitro исследования регуляцию с помощью ZFP экспрессии эндогенного гена в культивируемых клетках определяют, исследуя продукцию белка с использованием анализа ELISA. Исследуемый образец сравнивают с контрольными клетками, подвергнутыми обработке пустым вектором или неродственным ZFP, мишенью которого является другой ген.
В другом варианте осуществления регуляцию экспрессии эндогенного гена определяют in vitro посредством измерения уровня экспрессии мРНК с гена-мишени. Уровень экспрессии гена измеряют, используя амплификацию, например, с использованием ПЦР (такой как ПЦР в режиме реального времени), LCR или гибридизационные исследования, например, Нозерн-гибридизацию, защиту от РНКазы, дот-блоттинг. В одном варианте осуществления используют защиту от РНКазы. Уровень белка или мРНК определяют, используя прямо или непрямо меченые агенты для обнаружения, например, флуоресцентно или радиоактивно меченые нуклеиновые кислоты, радиоактивно или ферментативно меченые антитела и т.п., как описано здесь.
Альтернативно, система с использованием гена-репортера может быть разработана, используя промотор гена-мишени, функционально связанный с геном-репортером, таким как люцифераза, зеленый флуоресцентный белок, CAT или β-gal. Конструкцию репортера типично совместно трансфецируют в культивируемые клетки. После обработки предпочтительным ДНК-связывающим белком (например, ZFP) величину транскрипции с гена-репортера, трансляции или его активности измеряют в соответствии со стандартными методами, известными квалифицированным в данной области техники специалистам.
Трансгенные и нетрансгенные растения также используются в качестве предпочтительного варианта осуществления для исследования регуляции экспрессии эндогенного гена in vivo. Трансгенные растения могут устойчиво экспрессировать предпочтительный ДНК-связывающий белок (например, ZFP). Альтернативно, могут использоваться растения, которые транзиторно экспрессируют предпочтительный ZFP, или в которые ZFP был введен в носителе для доставки. Регуляцию экспрессии эндогенного гена проверяют, используя любой из описанных здесь анализов.
Способы направленного расщепления
Описываемые способы и композиции могут использоваться для расщепления ДНК в представляющем интерес участке в хроматине клетки (например, в желаемом или заданном сайте в геноме, например, в гене MDH или вблизи него). Для такого направленного расщепления ДНК конструируют ДНК-связывающий домен, такой как связывающий домен «цинковые пальцы», который связывается с сайтом-мишенью в заданном сайте расщепления или вблизи него, и составной белок, включающий сконструированный ДНК-связывающий домен и расщепляющий домен, вводят в клетку и/или экспрессируют с полинуклеотида в клетке. После связывания части «цинковые пальцы» составного белка с сайтом-мишенью ДНК расщепляется вблизи сайта-мишени расщепляющим доменом.
Альтернативно, два составных белка, каждый из которых включает связывающий домен «цинковые пальцы» и полдомена для расщепления, экспрессируют в клетки, и они связываются с сайтами-мишенями, которые размещены рядом таким образом, что воссоздается функциональный расщепляющий домен, и ДНК расщепляется вблизи сайтов-мишеней. В одном варианте осуществления расщепление происходит между сайтами-мишенями двух связывающих доменов «цинковые пальцы». Один или оба связывающих домена «цинковые пальцы» могут быть сконструированными.
Для направленного расщепления, используя составной полипептид связывающий домен «цинковые пальцы» - расщепляющий домен, сайт связывания может охватывать сайт расщепления, или ближний край сайта связывания может находиться на расстоянии 1, 2, 3, 4, 5, 6, 10, 25, 50 или более нуклеотидов (или любого целого значения между 1 и 50 нуклеотидами) от сайта расщепления. Точное местоположение сайта связывания, относительно сайта расщепления, будет зависеть от конкретного расщепляющего домена и длины ZC линкера. В случае способов, в которых используются два составных белка, каждый из которых включает связывающий домен «цинковые пальцы» и полдомена для расщепления, сайты связывания, как правило, находятся по обе стороны сайта расщепления. Таким образом, ближний край первого сайта связывания может находиться на расстоянии 1, 2, 3, 4, 5, 6, 10, 25 или более нуклеотидов (или любого целого значения между 1 и 50 нуклеотидами) с одной стороны сайта расщепления, а ближний край второго сайта связывания может находиться на расстоянии 1, 2, 3, 4, 5, 6, 10, 25 или более нуклеотидов (или любого целого значения между 1 и 50 нуклеотидами) с другой стороны сайта расщепления. Способы картирования сайтов расщепления in vitro и in vivo известны квалифицированным в данной области техники специалистам.
Таким образом, в описываемых здесь способах может использоваться сконструированный домен «цинковые пальцы», объединенный с расщепляющим доменом. В этих случаях конструируют связывающий домен, который связывается с последовательностью-мишенью, в сайте или вблизи сайта, в котором желательно расщепление. Составной белок, или кодирующий его полинуклеотид, вводят в клетку растения. После введения в клетку, или экспрессии в ней, составной белок связывается с последовательностью-мишенью и осуществляет расщепление в последовательности-мишени или вблизи нее. Точный сайт расщепления зависит от природы расщепляющего домена и/или присутствия и/или природы последовательностей-линкеров между связывающим и расщепляющим доменами. В тех случаях, когда используются два составных белка, каждый из которых включает полдомена для расщепления, расстояние между ближними краями сайтов связывания может составлять 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 25 или более нуклеотидов (или любое целое значение между 1 и 50 нуклеотидами). Оптимальные уровни расщепления могут также зависеть и от расстояния между сайтами связывания двух составных белков (см., например, Smith et al. (2000) Nucleic Acids Res. 28:3361-3369; Bibikova et al. (2001) Mol. Cell. Biol. 21:289-297), и от длины ZC линкера в каждом составном белке. См. также патент США №7888121, публикацию заявки на патент США №20050064474 и публикации международных заявок на патенты WO 05/084190, WO 05/014791 и WO 03/080809.
В некоторых вариантах осуществления расщепляющий домен включает две половины домена для расщепления, обе из которых являются частью одного полипептида, включающего связывающий домен, первую половину домена для расщепления и вторую половину домена для расщепления. Половины домена для расщепления могут иметь одинаковую аминокислотную последовательность или различные аминокислотные последовательности при условии, что они функционируют с расщеплением ДНК.
Половины домена для расщепления могут быть также обеспечены в отдельных молекулах. Например, два составных белка могут вводиться в клетку, причем каждый полипептид включает связывающий домен и полдомена для расщепления. Половины домена для расщепления могут иметь одинаковую аминокислотную последовательность или различные аминокислотные последовательности при условии, что они функционируют с расщеплением ДНК. Кроме того, связывающие домены связываются с последовательностями-мишенями, которые типично расположены таким образом, что, после связывания составных полипептидов, две половины домена для расщепления представлены в пространственной ориентации друг относительно друга, которая делает возможным воссоздание расщепляющего домена (например, в результате димеризации половин домена), таким образом, две половины домена располагаются относительно друг друга с образованием функционального расщепляющего домена, приводящего к расщеплению хроматина клетки в представляющем интерес участке. Как правило, расщепление воссозданным расщепляющим доменом происходит в сайте, расположенном между двумя последовательностями-мишенями. Можно сконструировать один или оба белка, которые связываются со своим сайтом-мишенью.
Два составных белка могут связываться в представляющем интерес участке в одном и том же направлении или в противоположных направлениях, и сайты их связывания (т.е. сайты-мишени) могут быть разделены любым числом нуклеотидов, например, от 0 до 200 нуклеотидов или любым целым числом между ними. В некоторых вариантах осуществления сайты связывания для двух составных белков, каждый из которых включает связывающий домен «цинковые пальцы» и полдомена для расщепления, могут находиться на расстоянии 5-18 нуклеотидов, например, на расстоянии 5-8 нуклеотидов, или на расстоянии 15-18 нуклеотидов, или на расстоянии 6 нуклеотидов, или на расстоянии 16 нуклеотидов, измеряемом от края каждого сайта связывания, ближайшего к другому сайту связывания, и расщепление происходит между сайтами связывания.
Сайт, в котором расщепляется ДНК, как правило, находится между сайтами связывания для двух составных белков. Двухцепочечный разрыв ДНК часто является результатом двух одноцепочечных разрывов или «ников», сдвинутых на 1, 2, 3, 4, 5, 6 или более нуклеотидов (например, расщепление двухцепочечной ДНК природной FokI является результатом одноцепочечных разрывов, сдвинутых на 4 нуклеотида). Таким образом, расщепление не происходит обязательно в точно противоположных сайтах в каждой цепи ДНК. Кроме того, структура составных белков и расстояние между сайтами-мишенями может оказывать влияние на то, происходит ли расщепление около одной пары нуклеотидов или происходит ли расщепление в нескольких сайтах. Однако, в случае многих применений, включая направленную рекомбинацию или направленный мутагенез (см. ниже), расщепление в заданном диапазоне нуклеотидов, как правило, является достаточным, и расщепления между конкретными парами нуклеотидов не требуется.
Как отмечалось выше, описываемые здесь составные белки(ок) можно ввести в виде полипептидов и/или полинуклеотидов. Например, два полинуклеотида, каждый из которых включает последовательности, кодирующие один из вышеупомянутых полипептидов, можно ввести в клетку, и, когда полипептиды экспрессируются и каждый из них связывается со своей последовательностью-мишенью, происходит расщепление в последовательности-мишени или вблизи нее. Альтернативно, один полинуклеотид, включающий последовательности, кодирующие оба составных белка, вводят в клетку. Полинуклеотиды могут представлять собой ДНК, РНК или любые модифицированные формы или аналоги ДНК и/или РНК.
Для увеличения специфичности расщепления дополнительные композиции могут также использоваться в описываемых здесь способах. Например, взятые в отдельности половины доменов для расщепления могут демонстрировать ограниченную активность двухцепочечного расщепления. В способах, в которых два составных белка, каждый из которых содержит домен, включающий три цинковых пальца, и полдомена для расщепления, вводят в клетку, каждый из белков определяет сайт-мишень, составляющий приблизительно 9 нуклеотидов. Хотя совокупная последовательность-мишень из 18 нуклеотидов, вероятно, является уникальной в геномах млекопитающих и растений, любой конкретный 9-нуклеотидный сайт-мишень встречается, в среднем, приблизительно 23000 раз в геноме человека. Таким образом, может иметь место неспецифическое расщепление вследствие сайт-специфического связывания одной половины домена. Соответственно, в описываемых здесь способах предусматривается использование доминантно-негативного мутанта половины домена для расщепления, например, FokI (или кодирующей его нуклеиновой кислоты), который экспрессируется в клетке вместе с двумя составными белками. Доминантно-негативный мутант способен к димеризации, но неспособен к расщеплению, и также блокирует активность расщепления половины домена, с которой он димеризуется. При обеспечении доминантно-негативного мутанта в избыточном молярном количестве относительно составных белков только участки, в которых связываются оба составных белка, будут характеризоваться достаточно высокой локальной концентрацией функционального расщепляющего полдомена, чтобы происходила димеризация и расщепление. В сайтах, в которых связывается только один из двух составных белков, его половины домена для расщепления образуют димер с половиной домена доминантно-негативного мутанта, и нежелательное, неспецифическое расщепление не происходит.
Экспрессионные векторы
Нуклеиновую кислоту, кодирующую один или более составных белков (например, ZFN), описываемых здесь, можно клонировать в вектор для трансформации прокариотических или эукариотических клеток для репликации и/или экспрессии. Вектора могут быть прокариотическими векторами (например, пдазмидами, или «челночными» векторами, векторами насекомых) или эукариотическими векторами. Нуклеиновую кислоту, кодирующую составной белок, можно также клонировать в экспрессионный вектор для введения в клетку.
Для экспрессии составных белков (например, ZFN) последовательности, кодирующие составные белки, типично субклонируют в экспрессионный вектор, который содержит промотор, управляющий транскрипцией. Подходящие прокариотические и эукариотические промоторы хорошо известны в данной области техники и описаны, например, в Sambrook et al., Molecular Cloning, A Laboratory Manual (2nd ed. 1989; 3rd ed., 2001); Kriegler, Gene Transfer and Expression: A Laboratory Manual (1990); и Current Protocols in Molecular Biology (Ausubel et al., выше). Бактериальные экспрессионные системы для экспрессии белков применимы, например, в E. coli, Bacillus sp. и Salmonella (Palva et al., Gene 22:229-235 (1983)). Наборы для таких экспрессионных систем имеются в продаже. Эукариотические экспрессионные системы для клеток млекопитающих, дрожжей и клеток насекомых хорошо известны квалифицированным в данной области техники специалистам и также имеются в продаже.
Промотор, используемый для управления экспрессией кодирующей составной белок нуклеиновой кислоты, зависит от конкретного применения. Например, сильный конститутивный промотор, подобранный в зависимости от клетки-хозяина, типично используется для экспрессии и очистки составных белков.
Напротив, когда составной белок вводят in vivo для регуляции гена растения (см. приведенный ниже раздел «Доставка нуклеиновых кислот в клетки растений»), или конститутивный, регулируемый (например, во время развития, в соответствии с типом ткани или клеток или под действием окружающей среды), или индуцируемый промотор используется в зависимости от конкретного применения составного белка. Неограничивающие примеры промоторов растений включают промоторные последовательности, происходящие из гена убиквитина-3 (ubi-3) A. thaliana (Callis, et al., 1990, J. Biol. Chem. 265-12486-12493); маннопинсинтазы (Δmas) A. tumifaciens (Petolino et al., патент США №6730824); и/или вируса мозаики прожилок листьев маниоки (CsVMV) (Verdaguer et al., 1996, Plant Molecular Biology 31:1129-1139). См. также раздел «Примеры».
Помимо промотора, экспрессионный вектор типично содержит транскрипционную единицу или экспрессионную кассету, которая содержит все дополнительные элементы, необходимые для экспрессии нуклеиновой кислоты в клетках-хозяевах, или прокариотических, или эукариотических. Таким образом, типичная экспрессионная кассета содержит промотор (включающий участки связывания рибосомы), функционально связанный, например, с последовательностью нуклеиновой кислоты, кодирующей составной белок, и сигналы, необходимые, например, для эффективного полиаденилирования транскрипта, терминации транскрипции или терминации трансляции. Дополнительные элементы кассеты могут включать, например, энхансеры, гетерологичные сигналы сплайсинга, последовательность 2A из вируса Thosea asigna (Mattion et al. (1996) J. Virol. 70:8124-8127), и/или сигнал ядерной локализации (NLS).
Конкретный экспрессионный вектор, используемый для переноса генетической информации в клетку, выбирают, принимая во внимание предполагаемое использование составных белков, например, экспрессию в растениях, животных, бактериях, грибах, простейших и т.д. (см. экспрессионные вектора, описанные ниже). Стандартные вектора для экспрессии в бактериях и клетках животных известны в данной области техники и описаны подробно, например, в публикации заявки на патент США №20050064474 A1 и публикациях международных заявок на патенты WO 05/084190, WO 05/014791 и WO 03/080809.
Стандартные методы трансфекции могут использоваться для получения линий бактериальных клеток, клеток растений, млекопитающих, дрожжей или насекомых, которые экспрессируют большие количества белка, который можно затем очистить, используя стандартные методы (см., например, Colley et al., J. Biol. Chem. 264:17619-17622 (1989); Guide to Protein Purification, in Methods in Enzymology, vol. 182 (Deutscher, ed., 1990)). Трансформацию эукариотических и прокариотических клеток выполняют в соответствии со стандартными методами (см., например, Morrison, J Bact. 132:349-351 (1977); Clark-Curtiss & Curtiss, Methods in Enzymology 101:347-362 (Wu et al., eds., 1983)).
Любая из хорошо известных процедур для введения чужеродных нуклеотидных последовательностей в такие клетки-хозяева может использоваться. Они включают использование трансфекции с использованием кальция фосфата, полибрена, слияния протопластов, электропорации, ультразвуковых методов (например, сонопорации), липосом, микроинъекции, «голой» ДНК, плазмидных векторов, вирусных векторов, трансформации с использованием Agrobacterium, трансформации с использованием карборунда (например, WHISKERS™), как эписомной, так и интегративной, и любого из других хорошо известных способов для введения клонированной геномной ДНК, кДНК, синтетической ДНК или другого чужеродного генетического материала в клетку-хозяина (см., например, Sambrook et al., выше). Лишь необходимо, чтобы конкретная используемая процедура генетической инженерии была способна к успешному введению по крайней мере одного гена в клетку-хозяина, способную экспрессировать предпочтительный белок.
Доставка нуклеиновых кислот в клетки растений
Как отмечалось выше, ДНК-конструкции можно ввести в желаемую клетку растения (например, в ее геном) с помощью множества традиционных методов. Для обзоров таких методов см., например, Weissbach & Weissbach Methods for Plant Molecular Biology (1988, Academic Press, N.Y.) Section VIII, pp. 421-463; и Grierson & Corey, Plant Molecular Biology (1988, 2d Ed.), Blackie, London, Ch. 7-9. См. также публикации заявок на патенты США №№20090205083; 20100199389; 20110167521 и 20110189775, которые включены сюда посредством ссылки в их полном объеме.
Например, ДНК-конструкцию можно ввести непосредственно в геномную ДНК клетки растения, используя такие методы, как электропорация и микроинъекция в протопласты клеток растения, или ДНК-конструкции можно ввести непосредственно в ткань растения, используя методы баллистической трансфекции, такие как бомбардировка частицами ДНК (см., например, Klein et al. (1987) Nature 327:70-73). Альтернативно, ДНК-конструкцию можно ввести в клетку растения посредством трансформации с использованием наночастиц (см., например, публикацию заявки на патент США №20090104700, которая включена сюда посредством ссылки в ее полном объеме). Альтернативно, ДНК-конструкции можно объединить с подходящими пограничными районами Т-ДНК/фланкирующими ее районами и ввести в вектор обычного хозяина Agrobacterium tumefaciens. Методы трансформации с использованием Agrobacterium tumefaciens, в том числе нейтрализация онкогенов и разработка и применение бинарных векторов, полностью описаны в научной литературе. См., например, Horsch et al. (1984) Science 233:496-498, и Fraley et al. (1983) Proc. Natl. Acad. Sci. USA 80:4803.
Кроме того, переноса генов можно достичь, используя не относящиеся к Agrobacterium бактерии или вирусы, такие как Rhizobium sp. NGR234, Sinorhizoboium meliloti, Mesorhizobium loti, картофельный вирус X, вирус мозаики цветной капусты и вирус мозаики прожилок листьев маниоки, и/или вирус мозаики табака. См., например, Chung et al. (2006) Trends Plant Sci. 11(1):1-4.
Связанные с вирулентностью функции хозяина Agrobacterium tumefaciens будут предписывать вставку T-цепи, содержащей конструкцию и расположенный рядом маркер, в ДНК клетки-хозяина при инфицировании клетки бактериями, используя бинарный Т-ДНК-вектор (Bevan (1984) Nuc. Acid Res. 12:8711-8721) или процедуры сокультивирования (Horsch et al. (1985) Science 227:1229-1231). Как правило, система для трансформации с использованием Agrobacterium используется для создания двудольных растений (Bevan et al. (1982) Ann. Rev. Genet 16:357-384; Rogers et al. (1986) Methods Enzymol. 118:627-641). Система для трансформации с использованием Agrobacterium может также использоваться для трансформации, а также переноса, ДНК в однодольные растения и клетки растений. См. патент США №5591616; Hernalsteen et al. (1984) EMBO J 3:3039-3041; Hooykass-Van Slogteren et al. (1984) Nature 311:763-764; Grimsley et al. (1987) Nature 325:1677-179; Boulton et al. (1989) Plant Mol. Biol. 12:31-40; и Gould et al. (1991) Plant Physiol. 95:426-434.
Альтернативные методы переноса генов и трансформации включают, но без ограничения, трансформацию протопластов через поглощение «голой ДНК», опосредованное кальцием, полиэтиленгликолем (ПЭГ) или электропорацией (см. Paszkowski et al. (1984) EMBO J 3:2717-2722, Potrykus et al. (1985) Molec. Gen. Genet. 199: 169-177; Fromm et al. (1985) Proc. Nat. Acad. Sci. USA 82:5824-5828; и Shimamoto (1989) Nature 338:274-276), и электропорацию тканей растений (D'Halluin et al. (1992) Plant Cell 4:1495-1505). Дополнительные методы трансформации клеток растений включают микроинъекцию, опосредованное карборундом (например, WHISKERS™) поглощение ДНК (Kaeppler et al. (1990) Plant Cell Reporter 9:415-418), и бомбардировку микрочастицами (см. Klein et al. (1988) Proc. Nat. Acad. Sci. USA 85:4305-4309; и Gordon-Kamm et al. (1990) Plant Cell 2:603-618).
Описываемые способы и композиции могут использоваться для вставки экзогенных последовательностей в ген MDH. Они применимы, поскольку экспрессия введенного трансгена в геном растения зависит критически от сайта интеграции. Соответственно, гены, кодирующие, например, устойчивость к гербициду, устойчивость к насекомым, питательные вещества, антибиотики или терапевтические молекулы, могут быть вставлены посредством направленной рекомбинации в участках генома растений, подходящих для их экспрессии.
Трансформированные клетки растений, которые получают с помощью любого из вышеупомянутых методов трансформации, можно подвергнуть культивированию для регенерации целого растения, которое обладает измененным генотипом и, соответственно, желаемым фенотипом. Такие методы регенерации основаны на обработке определенными фитогормонами в среде для роста культуры тканей, типично опираясь на маркер устойчивости к биоциду и/или гербициду, который был введен вместе с желаемыми нуклеотидными последовательностями. Регенерация растений из культивируемых протопластов описана в Evans, et al., «Protoplasts Isolation and Culture» in Handbook of Plant Cell Culture, pp. 124-176, Macmillian Publishing Company, New York, 1983; и Binding, Regeneration of Plants, Plant Protoplasts, pp. 21-73, CRC Press, Boca Raton, 1985. Регенерации можно также достичь, исходя из каллуса, эксплантов, органов, пыльцы, завязей или частей растения. Такие методы регенерации описаны в общем в Klee et al. (1987) Ann. Rev. of Plant Phys. 38:467-486.
Нуклеиновые кислоты, вводимые в клетку растения, могут использоваться для придания желаемых особенностей по существу любому растению. Широкий ряд растений и систем клеток растений можно создать ради желаемых физиологических и агрономических характеристик, описываемых здесь, используя НК-конструкции настоящего описания и различные вышеупомянутые методы трансформации. В предпочтительных вариантах осуществления являющиеся мишенями для инженерии растения или клетки растений включают, но без ограничения, однодольные и двудольные растения, такие как культуры, в том числе зерновые культуры (например, пшеница, кукуруза, рис, просо, ячмень), плодовые культуры {например, помидор, яблоня, грушевое дерево, клубника, апельсиновое дерево), фуражные культуры (например, люцерна), корнеплоды (например, морковь, картофель, сахарная свекла, ямс), листовые овощи (например, салат-латук, шпинат); цветковые растения (например, петуния, роза, хризантема), хвойные деревья и сосны (например, сосна, ель); растения, используемые в фиторемедиации (например, растения, накапливающие тяжелые металлы); масличные культуры (например, подсолнечник, семена рапса) и растения, используемые для экспериментальных целей (например, Arabidopsis). Таким образом, описываемые способы и композиции имеют применение в пределах широкого ряда растений, включающих, но без ограничения, виды из родов Asparagus, Avena, Brassica, Citrus, Citrullus, Capsicum, Cucurbita, Daucus, Erigeron, Glycine, Gossypium, Hordeum, Lactuca, Lolium, Lycopersicon, Malus, Manihot, Nicotiana, Orychophragmus, Oryza, Persea, Phaseolus, Pisum, Pyras, Prunus, Raphanus, Secale, Solanum, Sorghum, Triticum, Vitis, Vigna и Zea.
Введение нуклеиновых кислот, вводимых в клетку растения, может использоваться для придания желаемых особенностей по существу любому растению. В некоторых вариантах осуществления изменение экспрессии/функции MDH в клетках растений приводит к растениям, характеризующимся увеличенной величиной урожая плодов, имеющим увеличенную биомассу растения (или плода растения), большее содержание мякоти плода, концентрированное завязывание плода, являющимся растениями большего размера, имеющим увеличенный сырой вес, увеличенный сухой вес, увеличенное твердое содержимое, более высокий общий вес при сборе урожая, увеличенную интенсивность и/или однородность цвета культуры, измененные химические характеристики (например, имеющие отношение к маслам, жирным кислотам, углеводам, белкам) и т.д.
Квалифицированный в данной области техники специалист признает, что экзогенную последовательность можно временно включить в клетку растения. Для включения экзогенной полинуклеотидной последовательности может использоваться клеточный аппарат клетки растения, в которую введена последовательность. Экспрессию экзогенной полинуклеотидной последовательности, включающей ZFN, которая временно включена в клетку растения, можно исследовать посредством анализа геномной ДНК последовательности-мишени для идентификации и определения вставок или делеций, инверсий или вставок. Эти типы реаранжировок являются следствием расщепления сайта-мишени в последовательности геномной ДНК и последующей репарации ДНК. Кроме того, экспрессию экзогенной полинуклеотидной последовательности можно исследовать, используя способы, которые позволяют проверить экспрессию гена-маркера, известные специалистам со средним уровнем компетентности в данной области техники. О транзиторной экспрессии генов-маркеров сообщалось при использовании ряда растений, тканей и систем доставки ДНК. Системы анализов транзиторной экспрессии включают, но без ограничения, прямую доставку гена с помощью электропорации или бомбардировки частицами тканей в любом анализе транзиторной экспрессии в растении, используя любой вид растений, представляющий интерес. Такие системы для транзиторной экспрессии будут включать, но без ограничения, электропорацию в протопласты клеток из ряда источников тканей или бомбардировку частицами конкретных представляющих интерес тканей. Настоящее описание охватывает использование любой транзиторной экспрессионной системы для оценки сайт-специфической эндонуклеазы (например, ZFN) и для введения мутаций в ген MDH-мишень. Примеры тканей растений, предусмотренных для проверки при транзиторных экспрессиях посредством соответствующей системы доставки, будут включать, но без ограничения, основные ткани листа, каллус, семядоли, корни, эндосперм, завязи, ткань цветка, пыльцу и эпидермальную ткань.
Квалифицированный в данной области техники специалист признает, что экзогенную полинуклеотидную последовательность можно прочно включить в трансгенные растения. После подтверждения того, что экзогенная полинуклеотидная последовательность является функционирующей, ее можно ввести в другие растения с помощью полового скрещивания. Может использоваться любой из ряда стандартных методов селекции, в зависимости от скрещиваемого вида.
Трансформированную клетку, каллус, ткань растения или растение можно идентифицировать и изолировать посредством отбора или скрининга созданного растительного материала в отношении признаков, кодируемых генами-маркерами, присутствующими в трансформирующей ДНК. Например, можно выполнить отбор посредством выращивания созданного растительного материала на средах, содержащих ингибиторные количества антибиотика или гербицида, к которому трансформирующая генная конструкция придает устойчивость. Кроме того, трансформированные растения и клетки растений можно также идентифицировать посредством скрининга на предмет активностей любых генов визуальных маркеров (например, генов β-глюкуронидазы, люциферазы, B или C1), которые могут присутствовать в рекомбинантных НК-конструкциях. Методологии такого отбора и скрининга хорошо известны квалифицированным в данной области техники специалистам.
Физические и биохимические методы также могут использоваться для идентификации трансформированных растений или клеток растений, содержащих прочно вставленные генные конструкции, или клетки растения, содержащей измененную геномную ДНК гена-мишени, которая является следствием транзиторной экспрессии сайт-специфической эндонуклеазы (например, ZFN). Эти способы включают, но без ограничения: 1) анализ по Саузерну или амплификацию с использованием ПЦР для обнаружения и определения структуры вставки рекомбинантной ДНК; 2) Нозерн-блот, защиту от РНКазы S1, удлинение праймера или амплификацию с помощью ПЦР с использованием обратной транскриптазы для обнаружения и исследования РНК-транскриптов с генных конструкций; 3) ферментативные анализы для обнаружения ферментативной или рибозимной активности, когда такие продукты генов кодируются генной конструкцией; 4) гель-электрофорез белков, методы Вестерн-блоттинга, иммунопреципитацию или иммуноферментный твердофазный анализ (ELISA), когда продуктами генных конструкций являются белки. Дополнительные методы, такие как in situ гибридизация, окрашивание с использованием ферментов и иммуноокрашивание, также могут использоваться для обнаружения присутствия или экспрессии рекомбинантной конструкции в конкретных органах и тканях растений. Способы проведения всех этих анализов хорошо известны квалифицированным в данной области техники специалистам.
Эффекты манипуляции генами, используя описываемые здесь способы, можно наблюдать с использованием, например, Нозерн-блотов РНК (например, мРНК), выделенной из представляющих интерес тканей. Типично, если мРНК присутствует или количество мРНК увеличилось, можно предположить, что соответствующий трансген экспрессируется. Другие способы измерения активности гена и/или кодируемого полипептида могут использоваться. Различные типы ферментативных анализов могут использоваться в зависимости от используемого субстрата и способа обнаружения увеличения или уменьшения продукта реакции или побочного продукта. Кроме того, уровни экспрессируемого полипептида можно измерить иммунохимически, т.е. с помощью ELISA, RIA, EIA и других анализов на основе антител, хорошо известных квалифицированным в данной области техники специалистам, например, с помощью электрофоретических способов обнаружения (с использованием или окрашивания, или Вестерн-блоттинга). В качестве одного неограничивающего примера, обнаружение белков AAD-1 и PAT, используя анализ ELISA, описано в публикации заявки на патент США №20090093366, которая таким образом включена посредством ссылки в ее полном объеме. Трансген можно избирательно экспрессировать в некоторых тканях растения или на некоторых стадиях развития, или трансген можно экспрессировать по существу во всех тканях растения, по существу на протяжении всего жизненного цикла. Однако также применим любой комбинаторный способ экспрессии.
Настоящим описанием также охватываются семена трансгенных растений, описанных выше, причем семя содержит трансген или генную конструкцию. Настоящим описанием, кроме того, охватывается потомство, клоны, линии клеток или клетки трансгенных растений, описанных выше, причем указанное потомство, клоны, линия клеток или клетка содержат трансген или генную конструкцию.
Составные белки (например, ZFN) и экспрессионные вектора, кодирующие составные белки, могут вводиться непосредственно в растение для регуляции генов, направленного расщепления и/или рекомбинации. В некоторых вариантах осуществления растение содержит множество паралогичных генов MDH-мишеней. Таким образом, один или более различных составных белков или экспрессионных векторов, кодирующих составные белки, могут вводиться в растения, чтобы воздействовать на один или более из этих паралогичных генов в растении.
Введение эффективных количеств осуществляют с помощью любого из способов, обычно используемых для приведения составных белков в максимальный контакт с клеткой растения, подвергаемой обработке. ZFP вводят любым подходящим способом, предпочтительно с использованием приемлемых носителей. Подходящие способы введения таких модуляторов имеются в распоряжении и хорошо известны квалифицированным в данной области техники специалистам, и, хотя более чем один способ может использоваться для введения конкретной композиции, конкретный способ может часто обеспечить более немедленную и более эффективную реакцию, чем другой способ.
Носители могут также использоваться и определяются отчасти конкретной вводимой композицией, а также конкретным способом, используемым для введения композиции. Соответственно, существует широкий ряд подходящих препаратов носителей, которые имеются в распоряжении.
ПРИМЕРЫ
Пример 1: Характеристика гена-мишени митохондриальной малатдегидрогеназы (mMDH) из помидора (Solanum lycopersicum)
Ген малатдегидрогеназы (MDH) обычно присутствует в виде множества паралогичных копий в организме. Несмотря на присутствие схожих паралогичных последовательностей генов малатдегидрогеназы в организме, одна последовательность гена митохондриальной малатдегидрогеназы (mMDH) была идентифицирована и выделена из Solarium lycopersicum. В начале последовательность гена митохондриальной малатдегидрогеназы (mMDH) была идентифицирована, исходя из нескольких баз данных, касающихся последовательностей полинуклеотидов, посредством сравнения частей последовательностей известных генов малатдегидрогеназы, которые были описаны в литературе, с множеством различных полинуклеотидных последовательностей, содержащихся в базах данных, касающихся последовательностей полинуклеотидов. После многочисленных сравнений совмещений последовательностей были идентифицированы полные последовательности двух различных генов малатдегидрогеназы: последовательность mMDH (описанная ниже) и последовательность гликосомальной MDH (gMDH) (идентификационный номер: AY725476). Попытки скринировать базы данных, касающихся последовательностей, привели к идентификации локуса mMDH в виде идентификационного номера: Solyc07g062650 (SEQ ID NO: 1) в базе данных, касающихся геномных последовательностей помидора, доступной в Интернете в Solgenomics (см. также Bombarely et al. (2011) Nuc Acids Res. (Database issue): D1149-55).
Последовательность гена mMDH, идентифицированную посредством виртуального скрининга, использовали для подтверждения последовательности гена mMDH из двух различных генотипов помидора, M82 и Moneymaker. Праймеры для полимеразной цепной реакции (ПЦР) были разработаны на основе идентифицированной посредством виртуального скрининга последовательности гена mMDH. Фрагмент, амплифицированный с помощью ПЦР, размером приблизительно 6 т.п.о. был выделен из каждого генотипа помидора, клонирован и секвенирован. Неожиданно, поскольку незначительные различия обычно ожидаются, секвенированные гены mMDH из генотипов помидора не продемонстрировали различие с локусом mMDH, который был первоначально идентифицирован и выделен в результате виртуального скрининга.
Выделенные новые последовательности гена mMDH были использованы для дальнейшей разработки реагента «цинковые пальцы».
Пример 2: Получение белков с цинковыми пальцами, предназначенных для связывания с геном митохондриальной малатдегидрогеназы (mMDH)
Белки с цинковыми пальцами, направленные против последовательностей ДНК, кодирующих различные функциональные домены, в кодирующей области гена mMDH S. lycopersicum v. M82 mMDH, (см. фиг. 1) были разработаны, как описывалось ранее. См., например, Urnov et al. (2005) Nature 435:646-651. Приводимые в качестве примера последовательность-мишень и распознающие спиральные участки продемонстрированы в таблице 1A (конструкции распознающих спиральных участков) и таблице 1B (сайты-мишени). В таблице 1B нуклеотиды в сайте-мишени, с которыми контактируют распознающие спиральные участки ZFP, указаны с использованием заглавных букв; неконтактирующие нуклеотиды указаны с использованием строчных букв.
Конструкции связывающихся с mMDH помидора цинковых пальцев
субъединица
(F5)
(F6)
28492
(SEQ ID NO:11)
(SEQ ID NO:12)
(SEQ ID NO:13)
(SEQ ID NO:14)
(SEQ ID NO:15)
(SEQ ID NO:16)
28491
(SEQ ID NO:17)
(SEQ ID NO:18)
(SEQ ID NO:19)
28536
(SEQ ID NO:27)
(SEQ ID NO:18)
28535
(SEQ ID NO:31)
(SEQ ID NO:33)
28550
28549
(SEQ ID NO:40)
28564
28563
(SEQ ID NO:40)
Последовательности-мишени для белков с цинковыми пальцами
Конструкции цинковых пальцев, связывающихся с mMDH, были включены в вектора, кодирующие белок, имеющий по крайней мере один палец со структурой CCHC. См. публикацию заявки на патент США № 2008/0182332. В частности, последний палец в каждом белке имел остов CCHC для распознающего спирального участка. Кодирующие неканонические цинковые пальцы последовательности были слиты с нуклеазным доменом рестрикционного фермента типа IIS-FokI (аминокислотами 384-579 последовательности Wah et al. (1998) Proc. Natl. Acad. Sci. USA 95:10564-10569) посредством ZC линкера из четырех аминокислот и сигнала ядерной локализации opaque-2, происходящего из Zea mays, с образованием нуклеаз с доменами «цинковые пальцы», специфическими в отношении mMDH (ZFN). Экспрессией составных белков в бицистронной экспрессионной конструкции, используя рибосомальный прерывистый сигнал 2A, описанной в Shukla et al. (2009) Nature 459:437-441, управлял относительно сильный, конститутивный и эктопический промотор, такой как промотор CsVMV или промотор, происходящий из промотора Solarium lycopersicum AA6 (AA6).
Расщепляющую активность оптимальных цинковых пальцев подтверждали, используя систему на основе почкующихся дрожжей, которая, как было ранее установлено, позволяет идентифицировать активные нуклеазы. См., например, публикацию заявки на патент США №20090111119; Doyon et al. (2008) Nat Biotechnol. 26:702-708; Geurts et al. (2009) Science 325:433. Цинковые пальцы для различных функциональных доменов были выбраны для in-vivo использования. Из многочисленных ZFN, которые были сконструированы, продуцированы и, как проверено, связываются с предполагаемыми полинуклеотидными сайтами-мишенями в геномном локусе mMDH, были идентифицированы четыре ZFN, обладающие in vivo активностью на высоком уровне, и они были выбраны для дальнейших экспериментальных работ. См. таблицу 1A. Эти четыре ZFN были охарактеризованы как способные эффективно связывать и расщеплять четыре уникальных полинуклеотидных сайта-мишени в геномном локусе mMDH в полевых условиях.
На фиг. 2A и 2B (SEQ ID NO: 2) продемонстрирована геномная организация локуса mMDH относительно полинуклеотидных сайтов связывания ZFN/сайтов-мишеней четырех пар ZFN. Первая пара ZFN (107830L/107830R; называемых, соответственно, «830L» и «830R» на фиг.2A) связывается в экзоне 1, вторая пара ZFN (107832L/107832R; называемых, соответственно, «832L» и «832R» на фиг.2A) связывается в экзоне 3, третья пара ZFN (107833L/107833R; называемых, соответственно, «833L» и «833R» на фиг. 2A) связывается в экзоне 4, и четвертая пара ZFN (107835L/107835R; называемых, соответственно, «835L» и «835R» на фиг. 2A) связывается в экзоне 6.
Пример 3: Конструкции для экспрессии нуклеаз с доменами «цинковые пальцы» в помидоре
Плазмидные вектора, содержащие конструкции для экспрессии четырех, приводимых в качестве примера нуклеаз с доменами «цинковые пальцы» - ZFN, которые были идентифицированы, используя анализ на основе дрожжей, и описаны в примере 2, были разработаны и укомплектованы, используя навыки и методы, общеизвестные в данной области техники. Каждая кодирующая цинковые пальцы последовательность была слита с последовательностью, кодирующей сигнал ядерной локализации opaque-2 (Maddaloni et al. (1989) Nuc. Acids Res. 17(18):7532), который был расположен на N-конце нуклеазы c доменами «цинковые пальцы».
Затем, слитая последовательность сигнал ядерной локализации opaque-2::нуклеаза с доменами «цинковые пальцы» была объединена в пару с комплементарной слитой последовательностью сигнал ядерной локализации opaque-2::нуклеаза с доменами «цинковые пальцы». Как таковая, каждая конструкция включала одну открытую рамку считывания, состоящую из двух слитых последовательностей сигнал ядерной локализации opaque-2::нуклеаза с доменами «цинковые пальцы», разделенных последовательностью 2A из вируса Thosea asigna (Mattion et al. (1996) J. Virol. 70:8124-8127). Экспрессией кодирующей ZFN последовательности управлял приводящий к экспрессии на высоком уровне конститутивный промотор AA6 (публикация заявки на патент США №2009/0328248), и эта кодирующая последовательность была фланкирована нетранслируемым участком nos 3' поли-A (Bevan et al. (1983) Nucl. Acid Res. 11:369-385).
Результирующие четыре плазмидных конструкции, pKG7479 (содержащая конструкцию ZFN 107830L/R), pKG7480 (содержащая конструкцию ZFN 107832L/R), pKG7481 (содержащая конструкцию ZFN 107833L/R) и pKG7482 (содержащая конструкцию ZFN 107835L/R), были подтверждены с помощью расщепления рестрикционными ферментами и посредством секвенирования ДНК. См. фиг. 3-6.
Пример 4: Крупномасштабное выделение плазмид
Протокол крупномасштабного выделения плазмидной ДНК был использован для получения больших количеств ДНК для трансфекции в протопласты, используя следующий протокол. Сначала в 250 мл среды LB, содержащей 100 мкг/мл карбенициллина, засевали штамм Escherichia coli TOP 10 (Invitrogen, Carlsbad, CA), содержащий одну из конструкций для экспрессии нуклеазы с доменами «цинковые пальцы», описанную выше в примере 3, с последующим культивированием при 37°C в течение ночи. Затем культуру центрифугировали при 6000 об/мин в течение 15 минут и результирующий осадок ресуспендировали в 20 мл стерильного буфера GTE (на литр: 10 г глюкозы, 25 мл 1M Tris при pH 8,0 и 20 мл 0,5 M EDTA при pH 8,0) и 20 мг лизоцима (Duchefa, St. Louis, Mo). Затем добавляли 30 мл буфера NaOH-SDS (200 мМ NaOH, 1% SDS (в отношении веса к объему)), осуществляли интенсивное перемешивание без перемешивания на вортексе и инкубацию на льду в течение 3 минут. Наконец, добавляли 22,5 мл калийацетатного буфера (294,5 г/л) и образец инкубировали в течение 10 минут на льду. Результирующую суспензию подвергали центрифугированию (6000 об/мин в течение 25 минут при 5°C) и супернатант собирали после его пропускания через стерильную фильтровальную бумагу.
Затем к прошедшему через фильтр супернатанту добавляли 60 мл изопропанола (1 объем) и осуществляли инкубацию при комнатной температуре в течение 10 минут. Затем смесь подвергали центрифугированию (6000 об/мин в течение 30 минут при комнатной температуре), супернатант отбрасывали и осадок промывали в 70% этаноле. После второго цикла центрифугирования (6000 об/мин в течение 5 минут при комнатной температуре) супернатант удаляли и осадок растворяли в 4 мл TE (10 мМ Tris, 1 мМ EDTA при pH 8,0), добавляли 4 мкл раствора РНКазы и раствор инкубировали в течение 20 минут при комнатной температуре. Затем образцы переносили в 12-мл пробирку и добавляли 400 мкл 3M NaOAc при pH 5,2 и 4 мл фенола, осуществляли перемешивание на вортексе, а затем центрифугирование (4000 об/мин в течение 10 минут при комнатной температуре). Верхнюю часть подвергнутого центрифугированию образца собирали в новую пробирку и добавляли 4 мл смеси хлороформ/изоамиловый спирт (24:1), осуществляли перемешивание на вортексе, а затем центрифугирование (4000 об/мин в течение 10 минут при комнатной температуре). Снова верхнюю часть подвергнутого центрифугированию образца собирали и добавляли 8 мл абсолютного этанола, и осуществляли инкубацию в течение 30 минут при -20°C.
После дополнительного цикла центрифугирования (4000 об/мин в течение 30 минут при 5°C) осадок промывали 70% этанолом и центрифугировали (4000 об/мин в течение 5 минут при комнатной температуре), а затем сушили на воздухе в ламинаре. Осадок растворяли в 4 мл MILLI-Q™ очищенной воды и разделяли на аликвоты с использованием 1,5-мл пробирок Eppendorf. Затем добавляли 0,5 мл раствора ПЭГ (40% полиэтиленгликоля 6000 (в отношении веса к объему), MgCl2:6H2О, фильтр (0,2 мкм)), и после инкубации в течение 30 минут при комнатной температуре пробирки подвергали центрифугированию (14000 об/мин в течение 10 минут). Осадок промывали 70% этанолом, а затем центрифугировали (14000 об/мин в течение 5 минут); этот шаг повторяли дважды. Супернатант отбрасывали и осадок растворяли в конечном объеме 0,5 мл MILLI-Q™ воды. Концентрацию плазмидной ДНК определяли, используя аппарат NANO DROP™ (Thermo Scientific, Wilmington, DE).
Пример 5: Изоляция протопластов клеток помидора и их трансфекция
Изоляция и регенерация протопластов клеток листьев помидора были описаны ранее (Shahin (1985) Theor. Appl. Genet. 69:235-240; Tan et al. (1987) Theor. Appl. Genet. 75:105-108; Tan et al. (1987) Plant Cell Rep. 6:172-175), и приводимые в качестве примера растворы и среды, которые могут использоваться для этого протокола, встречаются в этих публикациях. Вкратце, семена Solanum lycopersicum подвергали стерилизации с использованием 0,1% гипохлорида натрия и выращивали in vitro на стерильной среде MS20 с использованием фотопериода 16/8 часов при 2000 люкс при 25°C и 50-70% относительной влажности. Затем 1 г свежесорванных листьев помещали в чашку с 5 мл жидкой среды CPW9M. Используя лезвие скальпеля, сорванные листья разрезали перпендикулярно основному стеблю. Сегменты листа были шириной ~1 мм.
Сегменты листа переносили в новую чашку, содержащую 25 мл раствора фермента (CPW9M, содержащего 2% целлюлозы ONOZUKA RS™, 0,4% мацерозима ONOZUKA R10™, 2,4-D (2 мг/мл), NAA (2 мг/мл), BAP (2 мг/мл) при pH 5,8), и расщепление осуществляли в течение ночи при 25°C в темноте. Затем протопласты освобождали посредством помещения их на круговую качалку (40-50 об/мин) на 1 час. Протопласты отделяли от клеточного дебриса с помощью их пропускания через 50-мкм сито и промывки сита дважды CPW9M. Затем протопласты подвергали центрифугированию при 85×g, супернатант отбрасывали и осадок размещали в два раза меньшем объеме раствора CPW9M.
Наконец, протопласты размещали в 3 мл раствора CPW9M и затем осторожно добавляли 3 мл CPW18S, чтобы создавалась разделенная на слои поверхность раздела, и не было смешивания между двумя растворами. Протопласты подвергали центрифугированию при 85×g в течение 10 минут, и жизнеспособные протопласты, плавающие в слое границы раздела, собирали, используя длинную пастеровскую пипетку. Объем протопластов увеличивали до 10 мл посредством добавления большего количества среды CPW9M и количество изолированных протопластов определяли в счетной камере. Суспензию протопластов центрифугировали при 85×g в течение 10 минут при 5°C. Супернатант отбрасывали и осадок протопластов ресуспендировали до конечной концентрации, составляющей 106∙⋅ мл-l в среде для промывки CPW9M.
Затем выполняли трансфекцию изолированных протопластов конструкциями для экспрессии ZFN. В 10-мл пробирке 250 мкл суспензии протопластов и плазмидную ДНК (используемую в концентрации = 20 мкг или 30 мкг), и 250 мкл раствора ПЭГ (40% ПЭГ 4000, 0,1 M Ca(NO3)2, 0,4 M маннит) осторожно, но интенсивно перемешивали. После инкубации в течение 20 минут при комнатной температуре по каплям добавляли 5 мл холодного 0,275 М Ca(NO3)2. Суспензию протопластов центрифугировали в течение 10 минут при 85×g при 4°C. После центрифугирования супернатант отбрасывали и протопласты ресуспендировали в 4 мл жидкой среды K8p. Протопласты инкубировали в течение 48 часов при 28°C в темноте и затем собирали с помощью центрифугирования для выделения геномной ДНК.
Активность ZFN в протопластах клеток помидора проверяли посредством трансфекции протопластов клеток помидора, используя описанный выше протокол. Протопласты клеток помидора изолировали в больших количествах и плазмидную ДНК вводили в клетку протопласта, используя протокол трансфекции с использованием ПЭГ. Типично, при использовании репортерных конструкций, таких как репортерная конструкция, которая содержит ген зеленого флуоресцентного белка (GFP), отмечаются степени трансфекции вплоть до 80%. Большие количества копий плазмид, введенные в клетки помидора, порождают большое количество белка, кодируемого плазмидой. В итоге, экспрессируемый в клетках помидора белок GFP является обнаруживаемым вплоть до 48 часов после трансфекции. Однако экспрессия с введенной конструкцией является транзиторной, поскольку в пределах 48 часов плазмидная ДНК в клетке ликвидируется.
Пример 6: Активность ZFN в протопластах клеток помидора
Для измерения экспрессии с конструкций для экспрессии ZFN в протопластах клеток помидора и присущей ZFN активности расщепления локуса mMDH геномной ДНК помидора футпринтинг был выполнен, как указано ниже. Протопласты клеток помидора были изолированы и трансфецированы плазмидной ДНК, содержащей одну из четырех приводимых в качестве примера конструкций для экспрессии ZFN; pKG7479 (содержащей конструкцию ZFN 107830L/R); pKG7480 (содержащей конструкцию ZFN 107832L/R); pKG7481 (содержащей конструкцию ZFN 107833L/R); или pКG7482 (содержащей конструкцию ZFN 107835L/R). После введения в клетку ZFN экспрессировалась, и фермент ZFN индуцировал двухцепочечные разрывы (DSB) в специфическом сайте-мишени в mMDH. DSB ДНК подвергался репарации белками, включенными в путь негомологичного соединения концов (NHEJ), этот процесс репарации иногда склонен к ошибкам и приводит к небольшим вставкам или делециям («вставкам или делециям») в месте индукции DSB. После обработки ZFN сайты-мишени в mMDH были амплифицированы с помощью ПЦР, и результирующие вставки или делеции были детектированы и количественно определены, используя анализ кривой высокоразрешающего плавления (HRM) и/или секвенирование.
Протопласты (~250000) клеток помидора сорта M82 были изолированы и трансфецированы с использованием варьирующих концентраций (20 или 30 мкг) плазмид pKG7479, pKG7480, pKG7481 и pKG7482, и клетки собирали через 48 часов. В качестве отрицательного контроля, выполняли трансфекцию протопластов клеток помидора, используя плазмиду, содержащую конструкцию AA6::GFP. Затем геномную ДНК выделяли из каждого образца, используя набор DNEASY MINI KIT™ (Qiagen, Valencia, CA), и определяли концентрацию ДНК. Соответствующие сайты-мишени, которые соответствуют введенной конструкции для экспрессии ZFN, амплифицировали, используя набор HERCULASE II FUSION KIT™ в соответствии с инструкциями производителя (Agilent Technologies, Santa Clara, CA), из каждого образца трасфецированных протопластов. Все из сайтов-мишеней были амплифицированы из образца отрицательного контроля, который был трансфецирован контрольной конструкцией для экспрессии GFP, используя набор HERCULASE II FUSION KIT™ в соответствии с инструкциями производителя. Праймеры, используемые для амплификаций с помощью ПЦР, описаны в таблицах 2A (для анализа с помощью ПЦР) и 2B (для анализа HRM).
Специфические для сайтов-мишеней праймеры для анализа с помощью ПЦР
Праймеры, используемые для анализа HRM
Амплифицированные с помощью ПЦР продукты очищали, используя набор PCR PURIFICATION KIT™ (Qiagen), и 50 нг очищенной ДНК использовали для клонирования в ZERO BLUNT CLONING KIT™ (Invitrogen) в соответствии с инструкциями производителя. Затем 2 мкл смеси для лигирования трансформировали в компетентные клетки Escherichia coli ONE-SHOT TOP 10 (Invitrogen) и колонии отбирали на среде LB, дополненной 100 мкг/мл канамицина. ПЦР-реакцию HERCULASE II FUSION KIT™ (конечный объем = 50 мкл) выполняли в соответствии с инструкциями от производителя, используя HRM праймеры, описанные в таблице 2. Каждый набор праймеров для ПЦР амплифицировал фрагмент размером = приблизительно 200 п.о. Результирующие продукты ПЦР использовали для анализа HRM.
Выполнение анализа HRM было осуществлено, используя принятые в данной области техники процедуры. Вкратце, продукты ПЦР смешивали в соотношении 1:1 с продуктом ПЦР дикого типа, созданным с использованием тех же праймеров на основе необработанной (отсутствие ZFN) геномной ДНК. Добавляли краситель SYBR™-зеленый и профиль кривой плавления продуктов ПЦР определяли в датчике циклов ROCHE LIGHT CYCLER™ (Roche Diagnostics, Indianapolis, IN). Продукты ПЦР с кривыми плавления, которые значимо отличались от продукта ПЦР дикого типа в отсутствие обработки, были идентифицированы и сохранены. ПЦР-реакцию, используя праймеры M13F и M13R, затем выполняли непосредственно на бактериальных клонах, которые были идентифицированы, используя процедуру HRM, и результирующий продукт ПЦР очищали и секвенировали. Результаты этого анализа представлены в таблице 3 и на фиг. 7.
Итоговая сводка анализа HRM и анализа последовательности
Как показано, образование вставок или делеций было обнаружено во всех сайтах-мишенях для ZFN, что свидетельствует о том, что все четыре ZFN являются активными в протопластах клеток помидора. Оценку эффективности конкретной конструкции ZFN можно сделать с помощью расчета количества продуктов ПЦР, образуемых при обработке, которые содержат вставки или делеции. ZFN 107830 и 107832 дали наибольшее количество продуктов ПЦР со вставками или делециями (4,1% и 5,5%, соответственно). Тогда как в случае ZFN 107833 и 107835 количество продуктов ПЦР со вставками или делециями было значительно меньше (2% и 0,7%, соответственно). Не было обнаружено каких-либо вставок или делеций в продуктах ПЦР, полученных на основе контрольных протопластов клеток помидора, обработанных кодирующей GFP плазмидой.
Пример 7: Изоляция растений, содержащих мутации в локусе mMDH
Протопласты клеток помидора были изолированы из сортов помидора M82 и Moneymaker, и трансфекции с использованием pKG7479, pKG7480, pКG7481 и pKG7482 были выполнены, как описывалось ранее. Для регенерации растений помидора, исходя из трансфекции, протопласты, в конце концов, ресуспендировали в 2 мл раствора альгината (маннит 90 г/л, CaCl2⋅2H2O 140 мг/л, альгинат Na 20 г/л (Sigma-Aldrich, St. Louis, MO)) и интенсивно перемешивали с помощью перевертывания. Из этой ресуспензии 1 мл клеток наслаивали равномерно на чашку с Ca-агаром (72,5 г/л маннита, 7,35 г/л CaCl2⋅2H2O, 8 г/л агара) и допускали полимеризацию. Затем альгинатные диски переносили в 4-см чашки Петри, содержащие 4 мл среды для культивирования K8p, и выращивали при 28°C в темноте в течение 7 дней. Спустя отведенное для инкубации время альгинатные диски разрезали на тонкие полоски и помещали в чашки, содержащие среду GM-ZG, на 3 недели при 28°C в темноте для стимуляции развития каллуса. После инкубации в течение этого периода времени отдельные каллусы затем отбирали с пололок и располагали на свежей среде GM-ZG, и выращивали при 25°C при свете. Спустя 3 недели каллусы переносили на свежую среду GM-ZG. Этот этап повторяли дважды до достижения каллусами размера = приблизительно 2 см.
Затем каллусы переносили на среду для стимуляции образования ростков (MS20-ZI; MS20 + 2 мг/л зеатина + 0,1 мг/л IAA) и перенос повторяли до образования ростков. Один лист от каждого ростка затем срывали для выделения ДНК и соответствующие сайты-мишени в mMDH амплифицировали, используя праймеры, описанные в таблице 2, и ПЦР-реакции, описанные ранее. Продукты ПЦР затем очищали и секвенировали для идентификации ростков, которые содержали мутации в локусе mMDH. Эти ростки переносили в среду для укоренения (MS20+0,5 мг/л IBA) и, в конце концов, в теплицу.
Итоговая сводка мутаций, которые были порождены в геноме растений помидора в случае этих экспериментов, представлена в таблице 4. Сайты связывания ZFN подчеркнуты и указаны делетированные нуклеотиды (-). Идентификационные номера растений, помеченные звездочками, означают растения, которые имели идентичные мутации в обеих копиях mMDH и, как было определено, являются гомозиготными по описанным мутациям.
Итоговая сводка ZFN-индуцированных мутаций в mMDH
Мутантные линии растений, которые были фертильными, затем скрещивали с родительскими растениями и собирали семена F1. Проростки F1 подвергали скринингу для обнаружения мутации в виде вставки или делеции, которая присутствовала в родительской мутантной линии, и эти растения выращивали до достижения зрелости и подвергали самоопылению с получением семени F2. Растения помидора, которые выращивают из семени F2, могут использоваться для определения эффектов мутаций на урожай помидоров.
Пример 8: Молекулярное и биохимическое подтверждение мутаций в локусе mMDH
Семя F2 от каждой мутантной линии растений проращивали и 12 проростков генотипировали посредством амплификации сайта мутации и секвенирования продуктов ПЦР для определения того, обладал ли проросток последовательностью дикого типа, или был ли он или гетерозиготным, или гомозиготным по мутации в виде вставки или делеции. Двум проросткам каждого типа (всего 6) затем предоставляли возможность расти до достижения зрелости и завязи плода. От каждого растения отбирали образцы тканей и определяли эффекты различных мутаций на уровни белка mMDH и активность mMDH. Уровни белка mMDH определяли полуколичественно, используя Вестерн-блоттинг, а активность mMDH определяли посредством определения урожая плодов растения помидора.
Для определения уровней белка mMDH поликлональное антитело против mMDH, продуцированное благодаря прививке кроликов с использованием эпитопа mMDH (RSEVAGFAGEEQLGQA, SEQ ID NO: 62) и проверенное с использованием рекомбинантного белка mMDH, сверхэкспрессируемого в E. coli, использовали для детектирования белков растительного происхождения. mMDH детектировали в образцах листьев, полученных от трансгенных и контрольных растений. Экстракты из трансгенных и контрольных растений анализировали вместе со стандартами белков mMDH, которые инкубировали с буфером для образца с LDS NUPAGE® (Invitrogen, Carlsbad, CA), содержащим DTT, при 90°C в течение 10 минут и электрофоретически разделяли в готовом полиакриламидном геле с использованием подвижного буфера MES (Invitrogen). Затем белки электропереносили на нитроцеллюлозную мембрану, используя протокол производителя Invitrogen. После блокирования с использованием смеси для блокирования Superblock® (Invitrogen) и отмывки 0,1% PBST белок mMDH детектировали с использованием антисыворотки против mMDH, а затем козьего антикроличьего антитела, конъюгированного с фосфатазой. Детектируемый белок визуализировали посредством хемилюминесценции, используя SuperSignal West Pico Luminol Enhancer™ и Stable Peroxide Solution™, смешенные в равных объемах (Pierce, Rockford, IL).
Анализ с использованием Вестерн-блота выполняли на семействах мутантных растений F2 для обнаружения присутствия и определения молекулярной массы белка mMDH. Полноразмерный белок mMDH смогли обнаружить в контрольных растениях F2, у которых отсутствовала мутация в виде вставки или делеции, и в растениях F2, гетерозиготных по мутациям в виде вставок или делеций. Однако сигнал для белка mMDH не смогли обнаружить в растениях F2, которые были гомозиготными по мутации в виде вставки или делеции, которая нарушала открытую рамку считывания для mMDH. Растения F2, которые содержали гомозиготную вставку или делецию, синтезировали усеченную последовательность белка mMDH. Последовательность белка mMDH была усеченной в результате введения преждевременных стоп-кодонов или укорочения белка mMDH. Ожидалось, что усеченные белки mMDH будут присутствовать на Вестерн-блоте в виде полосы с меньшей молекулярной массой. Однако такие полосы с меньшей М.м. не были обнаружены. Эти результаты говорят о том, что усеченные белки подверглись деградации в клетке. Мутантные растения F2, 107832_8-3 и 107832_9-6, которые содержали гомозиготные мутации в виде вставок или делеций, которые приводили к делеции или изменению одной, двух или более аминокислот (т.е. белок mMDH не был усеченным) в консервативном НАД-связывающем домене, синтезировали полноразмерный белок mMDH, на что указывал анализ с помощью Вестерн-блота.
Биохимический анализ выполняли для измерения активности mMDH в растениях F2. Белок mMDH катализирует превращение малата и НАД+ в оксалоацетат и НАДН и обратно. Образованный в результате превращения НАДН демонстрирует максимальные значения оптической плотности при 340 нм, тогда как при этой длине волны НАД+ обладает незначительной оптической плотностью. Следовательно, активность mMDH в образце можно измерить, следя за скоростью превращения НАДН в НАД+ в присутствии оксалоацетата. Пять листовых выколоток диметром приблизительно 2 см были сделаны из молодых листьев отдельных растений F2 и измельчены в буфере для экстракции (50 мМ имидазол-HCl буфер, содержащий 10 мМ дитиотрейтол, 20 мМ MgCl2 и 2 мМ EDTA) при 4°C. Экстракт из листьев центрифугировали при 10000×g в течение 20 минут в охлажденной центрифуге; супернатант сливали и помещали на лед до использования препарата фермента в анализе. Для анализа, 1,5 мл буфера для анализа (50 мМ Tris-HCl pH 8,0, 1 мМ EDTA) пипетировали в пробирку, добавляли 100 мкл экстракта фермента и осуществляли инкубацию при 37°C в водяной бане в течение 5 минут. Смесь наливали в кварцевую кювету и добавляли 50 мкл НАДН (6 мМ НАДН в Tris-HCl, pH 8,0), и помещали в спектрофотометр, установленный на 340 нм. Первоначальное показание прибора и последующее увеличение оптической плотности регистрировали с 15-секундными интервалами вплоть до 3 минут, и эти показания прибора служили в качестве контроля. Затем добавляли 50 мкл оксалоацетата (0,3 M оксалоацета, свежеприготовленного в дистиллированной воде) и оптическую плотность регистрировали в течение 3 минут. Активность mMDH в линиях F2, выделяющихся и по мутациям внутри рамки, и нуль-мутациям в виде вставок или делеций, продемонстрирована на фиг.8.
Растения F2, происходящие из линии 107832_9-6, выделенной по мутации в виде вставки или делеции (-3 п.о.), синтезировали полноразмерный белок mMDH, обнаруживаемый на Вестерн-блоте. В биохимическом анализе активность mMDH была схожей во всех проверенных растениях. Результаты биохимического анализа следовали одной и той же тенденции в случае всех проверенных растений. Проверенные растения включали растения с нуль-мутацией, гетерозиготной и гомозиготной мутацией, которые содержали мутацию в виде вставки или делеции и которые синтезировали белок mMDH, в котором отсутствует одна аминокислота. Следовательно, утрата аминокислоты не оказывала отрицательное влияние на активность mMDH в биохимическом анализе. Тем не менее, биохимическая эффективность может быть все же слегка сниженной, поскольку биохимический анализ может не быть достаточно чувствительным для обнаружения умеренного уменьшения активности mMDH. Растения F2, происходящие из линии 107832_10-2 и выделяющиеся по мутации в виде делеции или вставки (-2 п.о.), были также проверены, используя биохимический анализ. Растения, которые содержали нуль-мутацию в виде вставки или делеции и были гетерозиготными по этой мутации в виде вставки или делеции, приводили к схожим уровням активности mMDH. Растения, которые были гомозиготными по мутации в виде вставки или делеции, продемонстрировали значительно уменьшенную активность mMDH. На Вестерн-блоте белок mMDH не был обнаружен в этих растениях.
Пример 9: Эффект мутаций в локусе mMDH на урожай плодов помидора
Для определения эффекта ранее описанных мутаций на урожай плодов помидора все плоды первой кисти помидора были собраны и количество помидоров и сырой вес каждого помидора определяли, используя способ, описанный в Centeno et al. (2011) Plant Cell 23:162-184.
В случае растений F2, происходящих от мутанта с делецией или вставкой 107832_9-6 (-3 п.о.), определяли средний сырой вес каждого полученного помидора и общий сырой вес всех помидоров, собранных с первой кисти мутантных растений (гетерозиготных или гомозиготных). Сырой вес помидоров, полученных с гомозиготных растений помидора 107832_9-6, был выше сырого веса помидоров, полученных с контрольных растений (которые не содержат мутацию). См. фиг. 9. Мутации, которые были порождены в локусе mMDH в растениях помидора 107832_9-6, приводили к уменьшенной активности белка, которая увеличивала урожай плодов. Напротив, растения помидора, которые были гетерозиготными по какой-либо мутации в виде вставки или делеции, которая приводила к нарушению открытой рамки считывания для mMDH, продемонстрировали уменьшенный рост и меньший общий урожай плодов. Следовательно, уменьшение активности mMDH, которое было определено в растениях, гомозиготных по мутации в виде вставки или делеции, приводит к увеличению общего урожая плодов помидора.
Пример 10: Эффект мутаций в локусе mMDH на скорости ферментативной реакции и активность фермента
Мутации в ферменте mMDH в геноме помидора, которые были результатом расщепления ZFN, как ранее описывалось, были введены в выделенные полинуклеотидные последовательности, которые кодируют природные ферменты mMDH помидора (SEQ ID NO: 1), сои (SEQ ID NO: 125) и кукурузы (SEQ ID NO: 126).
Совмещение природных вариантов этих ферментов представлено на фиг.10, которая демонстрирует, что ферменты mMDH различных видов растений характеризуются высокими уровнями схожести последовательностей и имеют общие консервативные белковые мотивы. Мутации, которые были результатом расщепления ZFN в ферменте mMDH помидора, продемонстрированы на фиг.11, эти пептидные последовательности соответствуют расщепленным последовательностям ДНК, представленным в таблице 4.
В качестве варианта осуществления этого изобретения, кроме того, включены новые мутации, которые введены по всей выделенной полинуклеотидной последовательности, которая кодирует ферменты mMDH. И первый, и второй НАДН-связывающий сайт содержат большую часть введенных мутаций. Приводимые в качестве примера мутации (например, делеции и/или вставки) одной или более аминокислот, пронумерованных относительно (природных) последовательностей дикого типа, включают, но без ограничения, мутации в одном или более аминокислотных остатков 104-136, пронумерованных относительно последовательности MDH помидора, представленной в SEQ ID NO: 1, и совмещенных с ней (V, V, I, I, P, A, G, V, P, R, K, P, G, M, T, R, D, D, L, F, N, I, N, A, G, I, V, K, S, L, C, T, A), и/или аминокислотных остатков 171-220, пронумерованных относительно последовательности MDH помидора, представленной в SEQ ID NO: 1 (D, E, K, K, L, F, G, V, T, M, L, D, V, V, R, A, K, T, F, Y, A, G, K, A, K, V, N, V, A, E, V, N, L, P, V, V, G, G, H, A, G, I, T, I, L, P, L, F, S, Q); одну или более мутаций в аминокислотных остатках 104-136 (V, V, I, I, P, A, G, V, P, R, K, P, G, M, T, R, D, D, L, F, N, I, N, A, G, I, V, K, N, L, S, T, A) и/или аминокислотных остатках 171-220 (D, E, K, K, L, F, G, V, T, T, L, D, V, V, R, A, K, T, F, Y, A, G, K, A, N, L, P, V, T, D, V, N, V, P, V, V, G, G, H, A, G, I, T, I, L, P, L, F, S, Q), пронумерованных относительно последовательности MDH кукурузы, представленной в SEQ ID NO: 126, и совмещенных с SEQ ID NO: 1; и мутации в одном или более аминокислотных остатках 104-136 (V, V, I, I, P, A, G, V, P, R, K, P, G, M, T, R, D, D, L, F, N, I, N, A, G, I, V, E, T, L, C, T, A) и/или аминокислотных остатках 171-220 (D, E, K, R, L, F, G, V, T, T, L, D, V, V, R, A, K, T, F, Y, A, G, K, A, N, V, P, V, A, G, V, N, V, P, V, V, G, G, H, A, G, I, T, I, L, P, L, F, S, Q), пронумерованных относительно последовательности MDH сои, представленной в SEQ ID NO: 125, и совмещенных с SEQ ID NO: 1.
Полинуклеотидные последовательности, кодирующие такие мутации в MDH, представлены ниже (SEQ ID NO: 132-137 и SEQ ID NO: 124):




Конструирование плазмид, содержащих открытые рамки считывания для мутантных mMDH
Плазмиду pKG7495, содержащую открытую рамку считывания для mMDH помидора, слитую с 6x His-меткой, расщепляли рестрикционными ферментами HindIII и XhoI, и полосу, соответствующую вектору размером 5,8 т.п.о., выделяли из агарозного геля, используя набор QIAGEN GEL ISOLATION KIT™ (Qiagen, Carlsbad, CA). Были синтезированы (GeneArt) части последовательности mMDH помидора, фланкированные сайтами для HindIII/XhoI. В этих последовательностях отсутствовала 3 п.о. последовательность (нуклеотиды 543-545 делетированы; SEQ ID NO: 136) во втором НАДН-связывающем домене, и они соответствовали мутациям, которые вызывались в растениях помидора в результате расщепления ZFN в этих местах генома. Вторая мутация в mMDH помидора была синтезирована в HindIII/XhoI фрагменте, в котором отсутствуют 3 п.о. (нуклеотиды 353-355; SEQ ID NO: 124) в первом НАДН-связывающем домене. Все фрагменты были лигированы в вектор pKG7495, и клоны подтверждали посредством секвенирования. Эти конструкции приводят к ряду мутантных клонов mMDH помидора с делециями внутри рамки считывания в обоих НАДН-связывающих доменах, которые были сверхэкспрессированы в системе для гетерологичной продукции в E. coli.
Сверхэкспрессия и очистка митохондриальной малатдегидрогеназы (mMDH)
Конструкции для экспрессии mMDH дикого типа и мутантных mMDH были клонированы в E. coli экспрессионный вектор. Векторной ДНК трансформировали химически компетентные клетки ONE SHOT® BL21(DE3) (Invitrogen). Гены-мишени были клонированы из условия, чтобы, после сверхэкспресии, результирующие ферменты содержали N-концевые гексагистидиновые метки. Колонии выращивали в течение ночи при 37°C на чашках с LB, содержащей 100 мкг/мл карбенициллина. Одну колонию использовали для засева 50 мл культуры для засева LB той же среды. Культуру выращивали в течение ночи при 37°C и впоследствии использовали для засева в 1,2 л LB, содержащей 100 мкг/мл карбенициллина. В случае ферментов mMDH дикого типа индукцию культуры осуществляли с использованием 1 мМ IPTG при ОП600=0,6. Допускали прохождение индукции при 37°C в течение 4 часов, при достижении этого момента времени клетки собирали с помощью центрифугирования при 8000 об/мин в течение 15 минут. Все варианты помещали в ледяную баню на 10 минут и затем индуцировали с использованием 200 мкМ IPTG в течение 18 часов при низкой температуре (19°C).
Клеточные осадки ресуспендировали в 15-20 мл буфера A (50 мМ Tris-HCl pH 8,2, 300 мМ NaCl, 10 мМ имидазол и 5% глицерина). После растворения добавляли ингибитор протеазы (таблетки Roche Complete Mini) и лизоцим (1 мг/мл). Допускали перемешивание растворов в течение 20 минут при 4°C и их обрабатывали ультразвуком 4×1-минутный импульс при нахождении на льду. Клеточный дебрис удаляли с помощью центрифугирования (16500 об/мин в течение 45 минут), при достижении этого момента времени неочищенный лизат вносили в 1-мл колонку His Trap FF, уравновешенную буфером A. Белки-мишени подвергали элюированию, используя линейный градиент 0-100% буфера B (50 мМ Tris-HCl pH 8,2, 300 мМ NaCl, 10 мМ имидазол и 5% глицерина), более 20 объемов колонки. Фракции, содержащие mMDH, были идентифицированы с помощью анализа с использованием электрофореза в SDS-ПААГ и исследований активности. Позитивные фракции объединяли вместе и концентрировали с помощью устройства Amicon Ultrafiltration, оснащенного мембраной с отсечением по молекулярной массе = 10 кДа. Буфер сконцентрированных фракций меняли на буфер A, в котором отсутствует имидазол, используя гель-фильтрацию PD10. Концентрации белков определяли по теоретическим молярным коэффициентам поглощения, рассчитанным, используя сервисную программу Expasy Bioinformatics Resource Portal ProtParam. Обессоленные ферменты быстро замораживали в жидком азоте и хранили при -80°C до дальнейшего использования. Анализ с использованием электрофореза в SDS-ПААГ показал, что молекулярные массы очищенных белков соответствовали рассчитанным для них массам, равным ~37 кДа.
Специфические активности mMDH дикого типа и мутантных mMDH
Анализы активности ферментов были выполнены в считывающем устройстве для микропланшета при 35°C и содержали 100 мМ Tris-HCl pH 8,2, 400 мкМ НАДН, 0,0075-0,045 мкМ mMDH или мутантного mMDH в конечном объеме = 200 мкл. Реакции проводили при 35°C в течение 1 минуты до инициирования посредством добавления оксалоацетата до конечной концентрации = 3 мМ. Начальные скорости определяли спектрофотометрически, следя за окислением НАДН (340 нм). Скорости были преобразованы из ОП×10-3/мин в мкМ мин-1, используя молярный коэффициент поглощения 6220 M-1 см-1. Результаты анализов активности ферментов представлены на фиг. 13 и в таблице 5.
Как продемонстрировано, специфические активности mMDH дикого типа помидора и двух мутантных ферментов mMDH помидора определяли, спектрофотометрически следя за окислением НАДН. Мутация в первой НАДН-связывающей последовательности, кодируемой SEQ ID NO: 124 (обозначенная как «mMDH del3 NADH BS1» в таблице 5 и на фиг. 13), является мутацией со значительным уменьшением активности и порождает приблизительно 1,5% от активности фермента дикого типа. Сравнительно, мутация во второй НАДН-связывающей последовательности, кодируемой SEQ ID NO: 136 (обозначенная как «mMDH del3» в таблице 5 и на фиг. 13), сохраняет приблизительно 23% активности фермента дикого типа. Мутации, введенные в НАДН-связывающий сайт в ферменте mMDH, приводят к уменьшению ферментативной активности, в результате растения помидора, которые содержат эти мутации в ферменте mMDH, дают плод с большей величиной сырого веса.
Кинетические исследования ферментов, которые были выполнены на мутированных ферментах mMDH, которые описаны как «mMDH del3» и «mMDH del3 NADH BS1». Мутации, приведшие к ферментам, которые обладают меньшей ферментативной активностью по сравнению с mMDH дикого типа «WT mMDH»
Введение дополнительных мутаций в конструкцию векторов для гетерологичной экспрессии и сверхэкспрессии фермента митохондриальной малатдегидрогеназы (mMDH)
Выделенные полинуклеотидные последовательности, которые кодируют природные и мутированные ферменты mMDH помидора, сои и кукурузы, встраивают в вектора и используют для трансформации микроорганизмов для гетерологичной экспрессии. Конструкцию плазмид, содержащих открытые рамки считывания для мутантных mMDH, можно слить с 6x His-меткой или другой последовательностью, используемой для выделения. Синтезируют фрагменты полинуклеотидной последовательности для mMDH. Фрагменты фланкируют сайтами для рестрикционных ферментов, и их можно без труда клонировать в плазмиду, содержащую открытую рамку считывания для mMDH. Конструируют фрагменты, содержащие мутации приблизительно 3 п.о., 4 п.о., 5 п.о., 6 п.о., 7 п.о., 8 п.о. или 9 п.о. на всем протяжении первого и второго НАДН-связывающего домена. Помимо этих различных мутаций, вводимых во второй НАДН-связывающий домен открытой рамки считывания для mMDH, мутации, которые были вызваны в растениях помидора, вводят в выделенные полинуклеотидные последовательности, которые кодируют ферменты mMDH помидора, сои и кукурузы. В результате, ряд кодирующих мутированные mMDH последовательностей, включающих делеции внутри рамки считывания в обоих НАДН-связывающих доменах, создают для ферментов, выделяемых из помидора, сои и кукурузы.
Сверхэкспрессия и очистка митохондриальной малатдегидрогеназы (mMDH)
Плазмидами, содержащими различные мутации, введенные в кодирующую последовательность помидора, сои и кукурузы, трансформируют химически компетентные клетки Escherichia coli. Выращивают трансформированные бактериальные колонии. Одну колонию используют для засева культуры для засева. Культуру выращивают и впоследствии используют для засева среды. Культуры индуцируют и за ростом культуры следят, используя спектрофотометр. Допускают протекание индукции в течение периода времени до сбора клеток.
Культуры центрифугируют и результирующие клеточные осадки ресуспендируют в буфере для лизиса. Растворы с ресуспендированными клетками подвергают обработке ультразвуком или лизируют, используя другие способы, известные квалифицированным в данной области техники специалистам. Клеточный дебрис удаляют с помощью центрифугирования. Белки подвергают элюированию. Фракции, содержащие mMDH, идентифицируют с помощью анализа с использованием электрофореза в SDS-ПААГ и исследований активности. Позитивные фракции объединяют вместе и концентрируют. Сконцентрированные фракции обессоливают. Концентрации белков определяют, используя предсказанные молярные коэффициенты поглощения, рассчитанные, используя сервисную программу Expasy Bioinformatics Resource Portal ProtParam. Обессоленные ферменты можно быстро заморозить в жидком азоте и хранить при -80°C до дальнейшего использования или сразу же исследовать.
Оценка активности mMDH дикого типа и мутантных mMDH
Анализы активности ферментов выполняют в конечной смеси, которая включает буфер, НАДН и очищенный фермент mMDH (дополнительные реагенты включают по месту). Реакцию инициируют посредством добавления оксалоацетата (0-50 мМ). Начальные скорости ферментативных реакций определяют, спектрофотометрически следя за уменьшением при 340 нм (окислением НАДН). Скорости в случае различных мутаций, введенных в фермент mMDH, определяют посредством преобразования в мкМ мин-1, используя молярный коэффициент поглощения 6220 M-1 см-1. Изменения ферментов в результате введения мутаций приводят к модифицированной субстратной специфичности фермента mMDH. Фермент mMDH катализирует биохимическую реакцию, представленную на фиг. 12, причем фермент mMDH обратимо катализирует окисление малата до оксалоацетата, используя восстановление НАД+ до НАДН. Исходя их различных проверенных ферментов mMDH, идентифицируют мутации, которые порождают самые подходящие общие кинетические параметры (значения kcat/Km). Эти мутации приводят к ферменту mMDH с уменьшенной активностью, в результате растения, которые содержат мутации в этих ферментах mMDH, дают семя и плод с большими значениями сырого веса.
Все патенты, заявки на патенты и публикации, упомянутые здесь, таким образом, включены посредством ссылки в их полном объеме.
Хотя описание было предоставлено довольно подробно в качестве иллюстрации и примера с целью ясности понимания, для квалифицированных в данной области техники специалистов будет очевидно, что различные изменения и модификации могут быть осуществлены на практике без отступа от сущности или объема настоящего изобретения. Соответственно, предшествующее описание и примеры не следует рассматривать как ограничение.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ ЛЕЧЕНИЯ БОЛЕЗНИ ХАНТИНГТОНА | 2014 |
|
RU2693891C1 |
| НАЦЕЛИВАНИЕ НА КЛЕТКИ, ОПОСРЕДУЕМОЕ ХИМЕРНЫМ АНТИГЕННЫМ РЕЦЕПТОРОМ | 2018 |
|
RU2783316C2 |
| СПОСОБЫ И СОСТАВЫ ЛЕЧЕНИЯ БОЛЕЗНИ ХАНТИНГТОНА | 2013 |
|
RU2639277C2 |
| ЛОКУСЫ FAD3 ДЛЯ ВЫПОЛНЕНИЯ ОПЕРАЦИЙ И СООТВЕТСТВУЮЩИЕ СВЯЗЫВАЮЩИЕСЯ СО СПЕЦИФИЧЕСКИМИ САЙТАМИ-МИШЕНЯМИ БЕЛКИ, СПОСОБНЫЕ К ВЫЗОВУ НАПРАВЛЕННЫХ РАЗРЫВОВ | 2013 |
|
RU2665811C2 |
| МОДУЛЯТОРЫ ТАУ И СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ ИХ ДОСТАВКИ | 2017 |
|
RU2789459C2 |
| КОНСТРУИРУЕМЫЕ БЕЛКИ С ЦИНКОВЫМИ ПАЛЬЦАМИ, НАПРАВЛЕННЫЕ НА ГЕНЫ РАСТЕНИЙ, ВОВЛЕЧЕННЫЕ В БИОСИНТЕЗ ЖИРНЫХ КИСЛОТ | 2010 |
|
RU2568056C2 |
| СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ ВСТРАИВАНИЯ ЭКЗОГЕННОЙ ПОСЛЕДОВАТЕЛЬНОСТИ В ГЕНОМ РАСТЕНИЙ | 2014 |
|
RU2723130C2 |
| КОНСТРУИРУЕМЫЕ БЕЛКИ С ЦИНКОВЫМИ ПАЛЬЦАМИ, НАПРАВЛЕННЫЕ НА ГЕНЫ РАСТЕНИЙ, ВОВЛЕЧЕННЫЕ В БИОСИНТЕЗ ЖИРНЫХ КИСЛОТ | 2010 |
|
RU2712833C2 |
| ФУНКЦИОНАЛЬНЫЕ ЛОКУСЫ FAD2 И СООТВЕТСТВУЮЩИЕ СПЕЦИФИЧНЫЕ ДЛЯ САЙТА-МИШЕНИ СВЯЗЫВАЮЩИЕСЯ БЕЛКИ, СПОСОБНЫЕ ИНДУЦИРОВАТЬ НАПРАВЛЕННЫЕ РАЗРЫВЫ | 2013 |
|
RU2656158C2 |
| СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ ЛЕЧЕНИЯ ГЕМОФИЛИИ В | 2011 |
|
RU2608643C2 |





































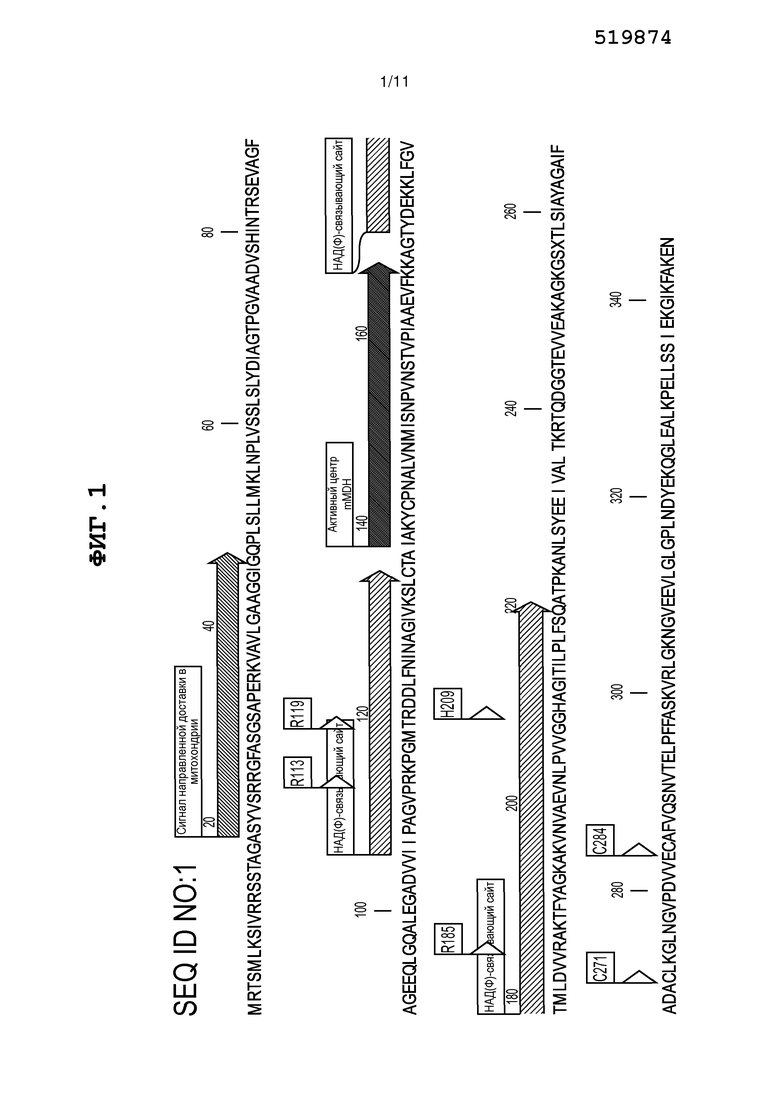
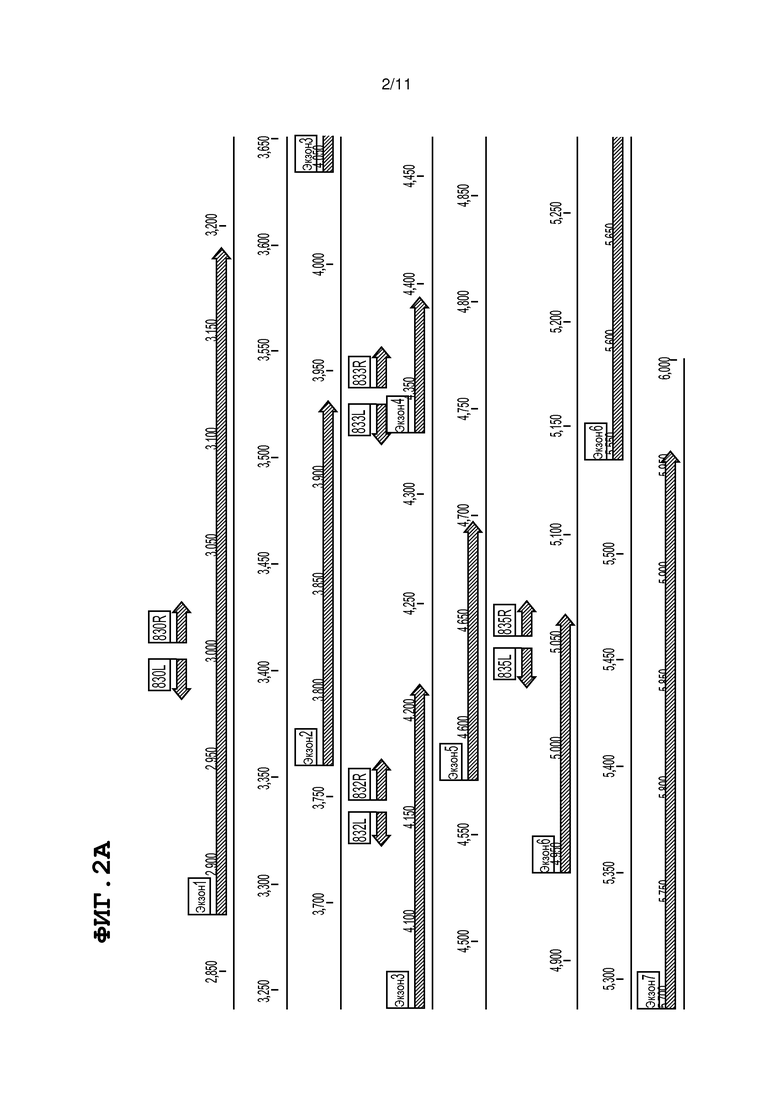

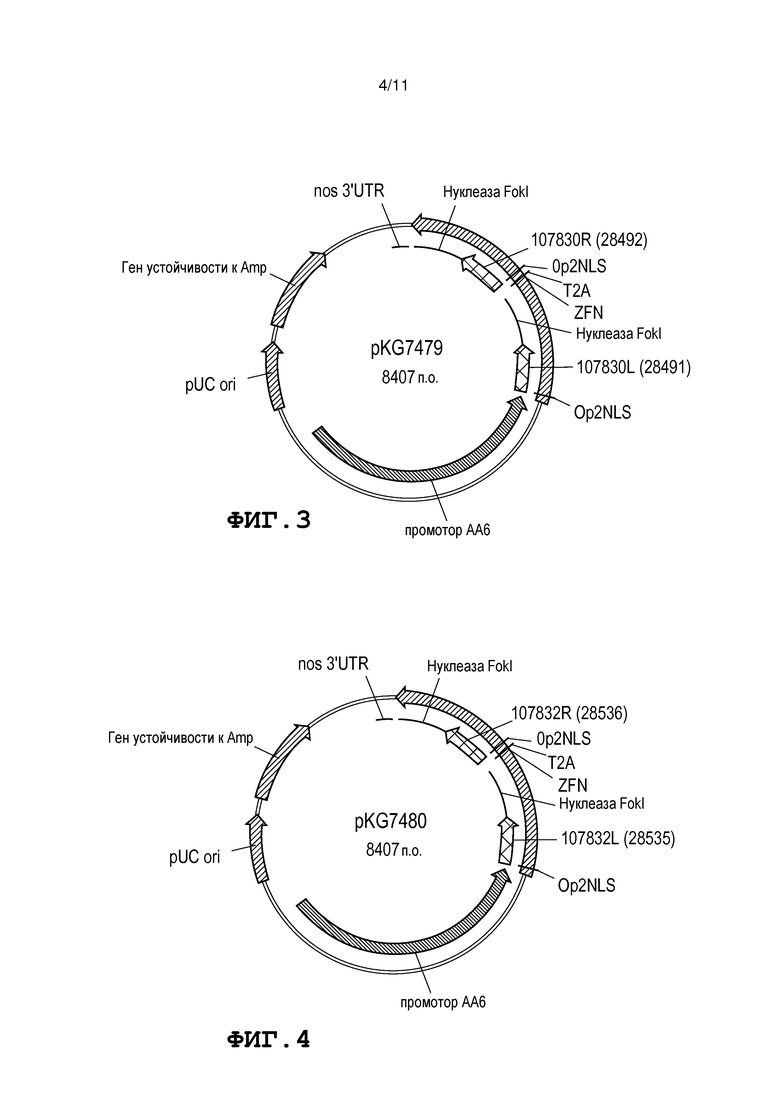
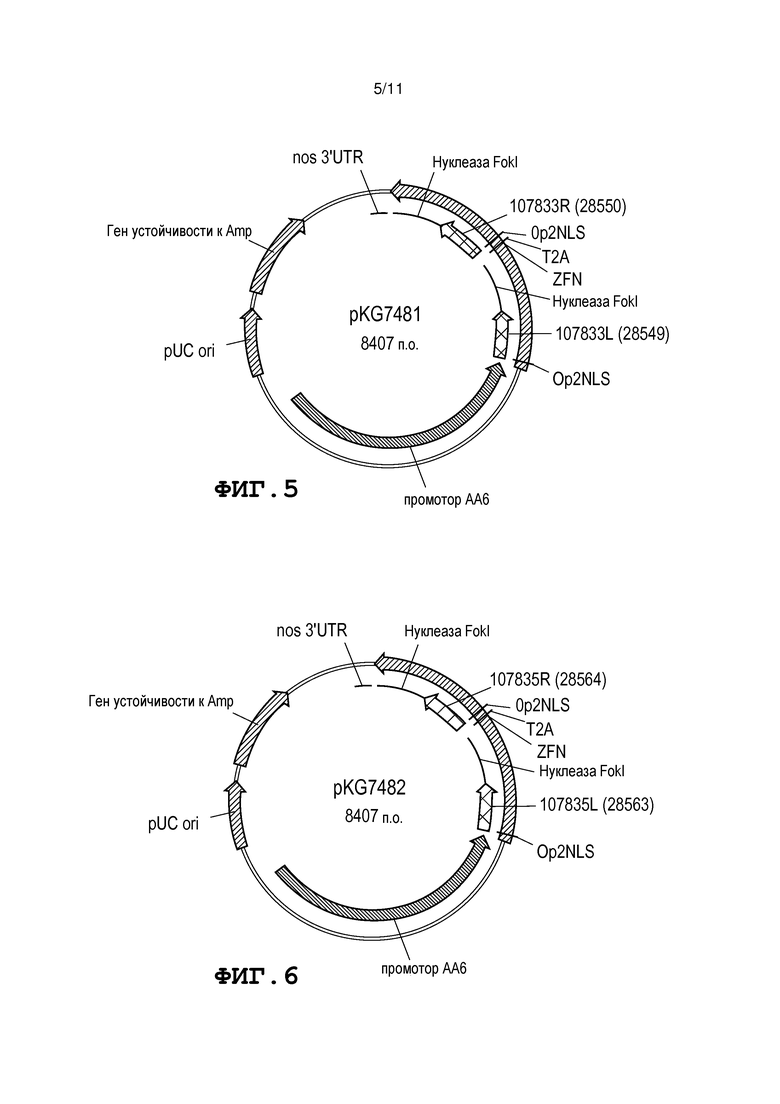

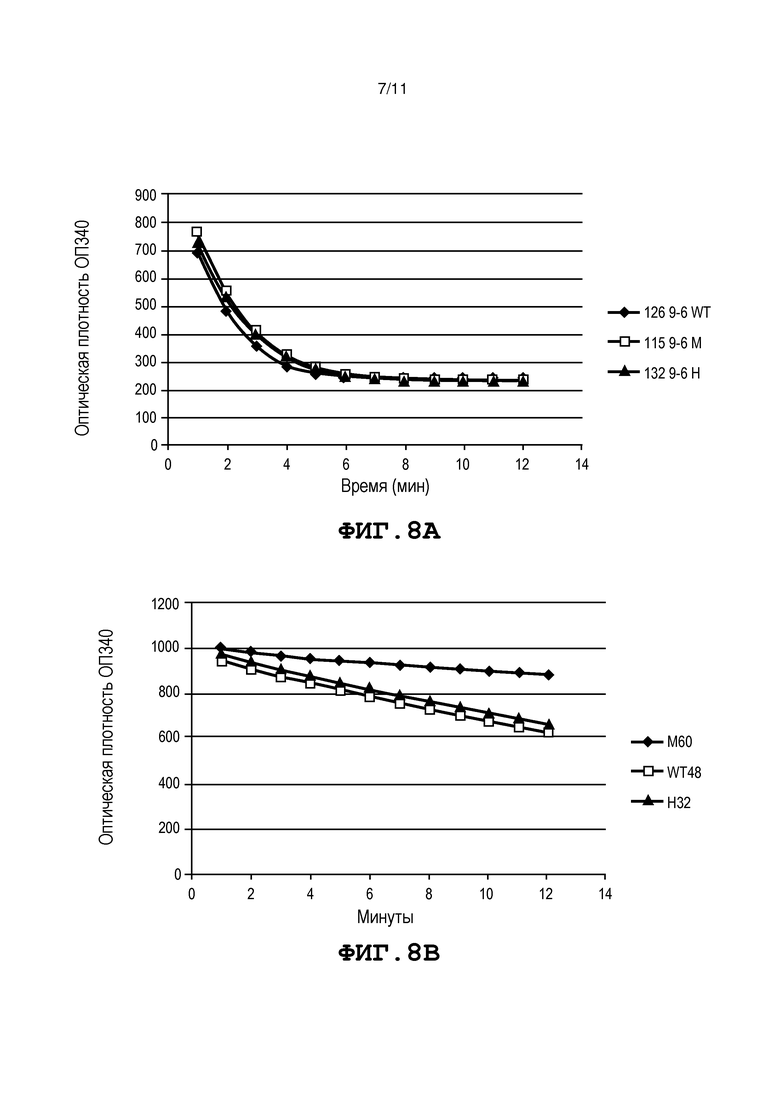
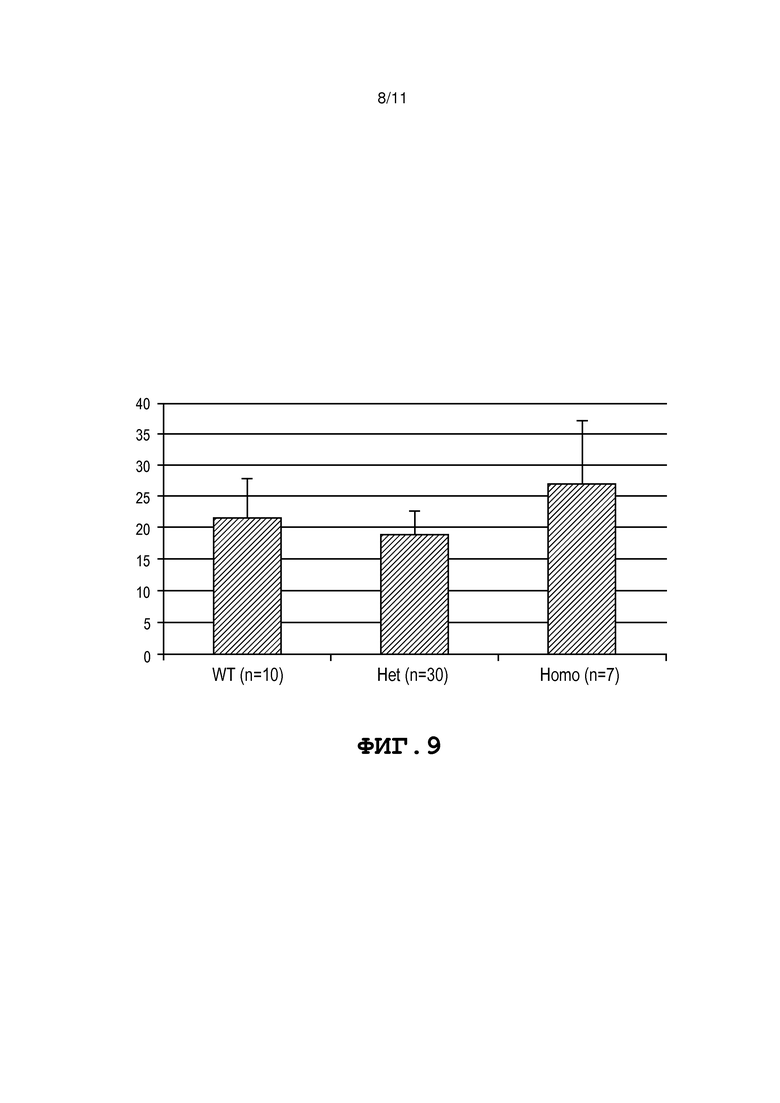

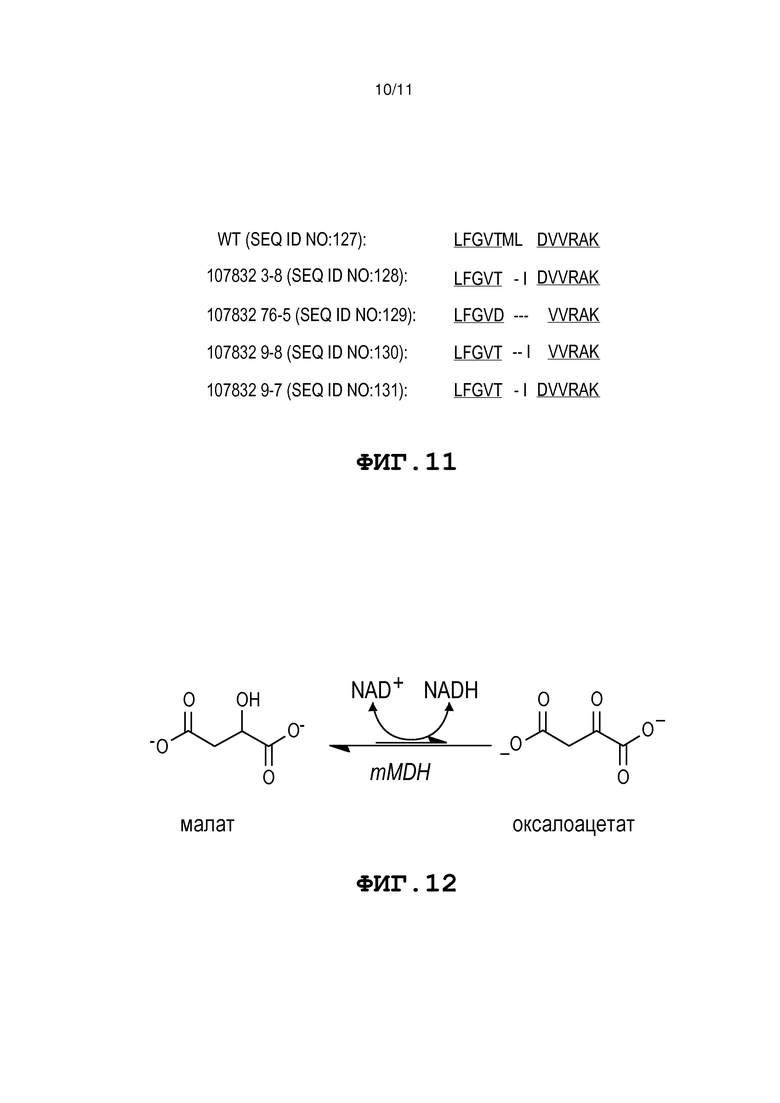
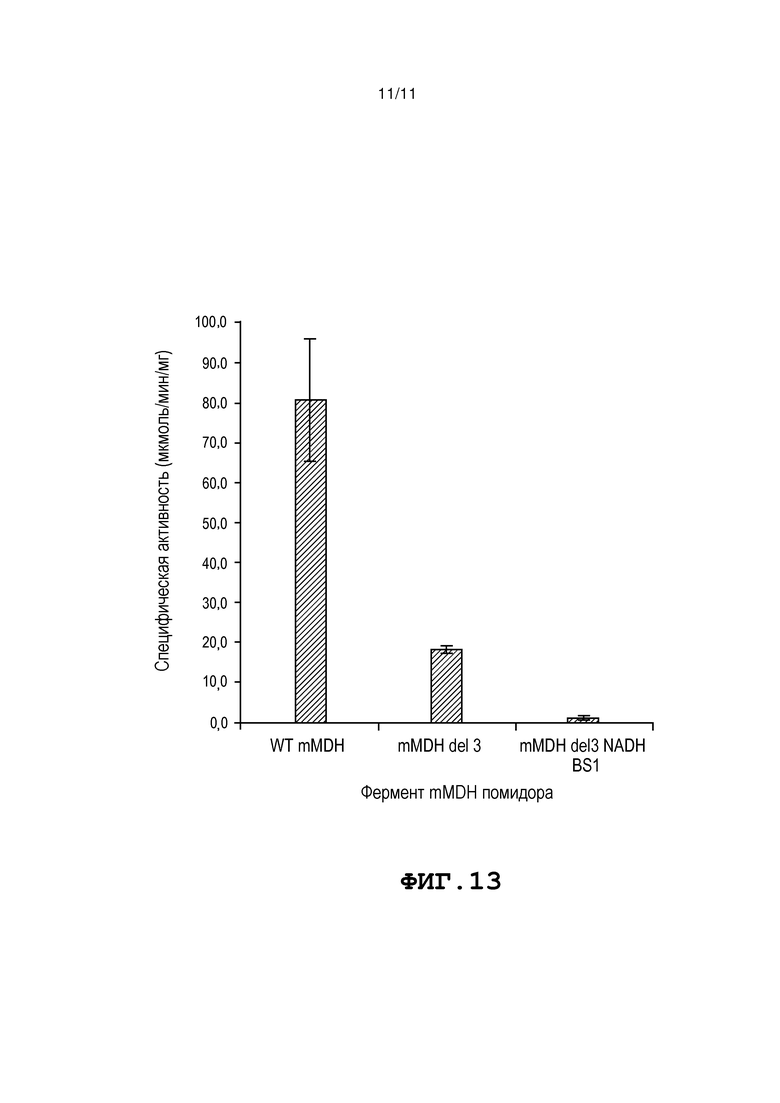
Изобретение относится к области биохимии, в частности к растению помидора для применения в увеличении производства помидоров, включающему одну или более клеток растения, в которых последовательность эндогенного гена митохондриальной малатдегидрогеназы (mMDH) модифицирована путем вставки и/или делеции таким образом, что активность любого белка mMDH, экспрессируемого из модифицированного гена, снижается, а также к его части. Также раскрыт способ получения растения помидора для применения в увеличении урожая помидоров, включающий введение пары нуклеаз с доменами «цинковые пальцы» (ZFN) в клетку растения помидора, так что ген mMDH модифицируется в клетке растения. Изобретение позволяет эффективно увеличивать производство помидоров. 3 н. и 9 з.п. ф-лы, 13 ил., 5 табл., 10 пр.
1. Модифицированное растение помидора для применения в увеличении производства помидоров по сравнению с растением помидора дикого типа, включающее одну или более клеток растения, в которых последовательность эндогенного гена митохондриальной малатдегидрогеназы (mMDH) модифицирована путем вставки и/или делеции таким образом, что активность любого белка mMDH, экспрессируемого из модифицированного гена, снижается, и причем модификацию осуществляют с использованием нуклеазы, которая связывается с последовательностью-мишенью в составе любой из SEQ ID NOs: 3-10 и расщепляет ген mMDH внутри или между SEQ ID NO: 3 и 4, внутри или между SEQ ID NO: 5 и 6, внутри или между SEQ ID NO: 7 и 8 или внутри или между SEQ ID NO: 9 и 10.
2. Растение помидора по п. 1, в котором модификация включает мутацию в одном или более НАДН-связывающих доменах гена mMDH.
3. Растение помидора по п. 1 или 2, в котором модификация гена mMDH включает вставку и/или делецию в пределах последовательности mMDH, кодирующей аминокислотные остатки, соответствующие аминокислотам 104-136 и/или 171-220 SEQ ID NO: 1 в растении.
4. Растение помидора по любому из пп. 1-3, в котором один или более аллелей mMDH модифицированы опосредуемой нуклеазой вставкой и/или делецией.
5. Часть растения помидора от растения по любому из пп. 1-4 для применения в увеличении производства помидоров, включающая одну или более модифицированных клеток растения, включающих вставку и/или делецию, сделанную с использованием нуклеазы, которая связывается с последовательностью-мишенью в составе любой из SEQ ID NOs: 3-10 и расщепляет ген mMDH внутри или между SEQ ID NO: 3 и 4, внутри или между SEQ ID NO: 5 и 6, внутри или между SEQ ID NO: 7 и 8 или внутри или между SEQ ID NO: 9 и 10, причем частью растения предпочтительно является лист, корень, цветок, семя или плод.
6. Растение по любому из пп. 1-4 или часть растения по п. 5, причем модификацию осуществляют, используя слитый белок, включающий ДНК-связывающий домен и функциональный домен, причем слитым белком предпочтительно является нуклеаза с доменами «цинковые пальцы»,
причем нуклеаза с доменами «цинковые пальцы» предпочтительно включает пять или шесть доменов «цинковые пальцы», расположенных в порядке от пальца 1 к пальцу 5 или от пальца 1 к пальцу 6, при этом каждый домен «цинковые пальцы» включает распознающий спиральный участок, причем нуклеаза с доменами «цинковые пальцы» включает распознающие спиральные участки, упорядоченные и продемонстрированные в (i) SEQ ID NO: 11-16; (ii) SEQ ID NO: 17, 18, 19, 20, 18 и 22; (iii) SEQ ID NO: 23, 24, 25, 26, 27 и 18; (iv) SEQ ID NO: 29, 30, 31, 32 и 33; (v) SEQ ID NO: 32, 35, 36, 37, 38 и 29; (vi) SEQ ID NO: 40, 41, 32, 43, 29 и 45; (vii) SEQ ID NO: 46, 47, 48, 49 и 31 или (viii) SEQ ID NOs: 32, 52, 17, 40, 55 и 56.
7. Растение или часть растения по п. 6, в котором нуклеаза с доменами «цинковые пальцы» включает домен «неканонический цинковый палец».
8. Растение или часть растения по любому из пп. 6, 7, где полинуклеотид, кодирующий слитый белок, вводят в клетку растения, так что слитый белок экспрессируется в клетке.
9. Способ получения растения помидора для применения в увеличении урожая помидоров по любому из пп. 1-4, включающий введение пары нуклеаз с доменами «цинковые пальцы» (ZFN) в клетку растения помидора, так что ген mMDH модифицируется в клетке растения, причем пара ZFN включает белки с доменами «цинковые пальцы», включающие следующие распознающие спиральные участки, упорядоченные и продемонстрированные в (i) SEQ ID NO: 11-16 и SEQ ID NO: 17, 18, 19, 20, 18 и 22; (ii) SEQ ID NO: 23, 24, 25, 26, 27 и 18 и SEQ ID NO: 29, 30, 31, 32 и 33; (iii) SEQ ID NO: 32, 35, 36, 37, 38 и 29 и SEQ ID NO: 40, 41, 32, 43, 29 и 45; (iv) SEQ ID NO: 46, 47, 48, 49 и 31 и SEQ ID NO: 32, 52, 17, 40, 55, так что активность белка mMDH уменьшается по сравнению с активностью дикого типа и растение помидора демонстрирует увеличенное производство помидоров по сравнению с растением помидора дикого типа.
10. Способ по п. 9, дополнительно включающий введение экзогенной последовательности нуклеиновой кислоты в клетку, причем экзогенная последовательность интегрируется в ген mMDH с последующим расщеплением нуклеазами с доменами «цинковые пальцы».
11. Способ по п. 9 или 10, в котором модификация включает мутацию в одном или более НАДН-связывающих доменах, так что активность белка mMDH снижается.
12. Способ по любому из пп. 9-11, в котором сниженная активность белка mMDH приводит к увеличению урожая плодов.
| NUNES-NESI A | |||
| et al | |||
| Enhanced photosynthetic performance and growth as a consequence of decreasing mitochondrial malate dehydrogenase activity in transgenic tomato plants, Plant Physiol, 2005, vol.137, N.2, pp.611-622 | |||
| WO 2008056915 A1, 15.05.2008 | |||
| US 2006058190 A1, 16.03.2006 | |||
| US 2005208489 A1, 22.09.2005 | |||
| RU 2010101604 A, 27.08.2011. |

