Настоящее изобретение относится к новым, специфично связывающимся терапевтическим и/или диагностическим белкам, направленным против гепсидина, предпочтительно представляющим собой мутеины белка липокалина. Изобретение также относится к молекулам нуклеиновых кислот, кодирующим такие белки, и к способам получения и применения таких белков и молекул нуклеиновых кислот. Соответственно, изобретение также направлено на фармацевтические и/или диагностические композиции, содержащие такие липокалиновые белки, включая применения этих белков.
ПРЕДШЕСТВУЮЩИЙ УРОВЕНЬ ТЕХНИКИ
Гепсидин, пептидный гормон, обычно существующий в двух формах, состоящих из 20 или 25 аминокислот, экспрессируемый и секретируемый рядом клеток в ответ на повышенный уровень железа и воспаление. Гепсидин образуют преимущественно гепатоциты печени, он играет центральную роль в регуляции гомеостаза железа, действует как антимикробный пептид и непосредственно или косвенно вовлечен в развитие большинства железодефицитных синдромов/синдромов перегрузки железом. Основное действие гепсидина заключается в интернализации и деградации экспортера железа ферропортина, экспрессируемого на всех железо-экспортирующих клетках. Гепсидин непосредственно связывается с ферропортином. Таким образом, высокий уровень гепсидина приводит к подавлению всасывания железа в кишечнике и высвобождения железа из макрофагов и гепатоцитов, в то время как низкая концентрация гепсидина приводит к ускорению высвобождения железа из этих клеток.
Также полагают, что гепсидин играет роль в патогенезе анемии воспалительных заболеваний и железодефицитной анемии. Анемия воспалительных заболеваний, также известная как анемия хронических заболеваний (ACD) или анемия хронических расстройств, в настоящее время является наиболее распространенной анемией у госпитализированных пациентов и частым синдромом, осложняющим многие инфекционные, неинфекционные, воспалительные и опухолевые расстройства. ACD представляет собой нормоцитарную, нормохромную анемию, характеризующуюся сниженным уровнем железа и сниженной железосвязывающей способностью (трансферрином), повышенным уровнем ферритина и присутствием железа в макрофагах костного мозга, что указывает на нарушение мобилизации железа из его депо. В то время как при анемии воспалительных заболеваний уровни гепсидина повышены, при железодефицитной анемии определяют низкие уровни гепсидина. Таким образом, гепсидин можно использовать в качестве маркера для различения этих заболеваний. Гепсидин может также быть полезным маркером для скрининга, прогнозирования и мониторинга наследственного гемохроматоза и анемий при высоком уровне железа. Уровни гепсидина могут также быть полезны при мониторинге лечения эритропоэтином (ЕРО) и прогнозировании ответа на ЕРО.
Способы выделения, анализа и определения количества гепсидина, а также агенты для лечения заболеваний и состояний, связанных с гепсидином, описаны в международных заявках на патенты WO 2008/011158, WO 2008/097461, WO 2009/094551A1, WO 2009/139822, WO 2009/058797 и WO 2010/017070. Тем не менее, белки, связывающиеся с гепсидином, обладающие свойствами белков, предложенных согласно настоящему изобретению, ранее описаны не были.
КРАТКОЕ ИЗЛОЖЕНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
Одно воплощение настоящего изобретения относится к мутеину липокалина, способному связываться с гепсидином с аффинностью, определенной по константе диссоциации (KD), приблизительно 10 нМ или менее. Более предпочтительно, липокалины могут иметь аффинность, определенную по KD, приблизительно 1 нМ или менее. В другом воплощении мутеин липокалина способен нейтрализовать биологическую активность человеческого гепсидина-25, предпочтительно со значением средней ингибирующей концентрации (IC50) приблизительно 80 нМ или менее, как определено клеточным анализом индуцированной гепсидином интернализации и деградации ферропортина.
В определенных воплощениях мутеин липокалина по настоящему изобретению содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO:1-14. В другом воплощении мутеин по меньшей мере на 75% идентичен последовательности человеческого липокалина дикого типа, включая человеческий липокалин-2.
В другом воплощении мутеин по настоящему изобретению конъюгирован с соединением, выбранным из группы, состоящей из органической молекулы, ферментной метки, радиоактивной метки, окрашенной метки, флуоресцентной метки, хромогенной метки, люминесцентной метки, гаптена, дигоксигенина, биотина, цитостатического агента, токсинов, металлического комплекса, металла и коллоидного золота. Мутеин может быть слит на его N-конце и/или его С-конце с партнером слияния, представляющим собой белок, белковый домен или пептид.
В другом воплощении мутеин конъюгирован с соединением, увеличивающим период полувыведения мутеина из сыворотки. Более предпочтительно, мутеин конъюгирован с соединением, выбранным из группы, состоящей из молекулы полиалкиленгликоля, гидроэтилкрахмала, Fc-части иммуноглобулина, CH3-домена иммуноглобулина, CH4-домена иммуноглобулина, альбуминсвязывающего пептида и альбуминсвязывающего белка.
В другом воплощении мутеин по настоящему изобретению представляет собой антагонист гепсидина. Гепсидин может представлять собой зрелый человеческий гепсидин.
В другом воплощении настоящее изобретение относится к молекуле нуклеиновой кислоты, содержащей нуклеотидную последовательность, кодирующую мутеин по настоящему изобретению.
В другом воплощении мутеин липокалина по настоящему изобретению выбран из группы, состоящей из мутеинов ретинолсвязывающего белка (RBP), билинсвязывающего белка (ВВР), аполипопротеина D (АРО D), липокалина, ассоциированного с желатиназой нейтрофилов (NGAL), липокалина слезы (TLPC), α2-микроглобулин-родственного белка (A2m), 24р3/угерокалина (24р3), белка желез Эбнера-1 (VEGP 1), белка желез Эбнера-2 (VEGP 2) и предшественника главного аллергена Can f1 (ALL-1). В родственных воплощениях мутеин липокалина выбран из группы, состоящей из человеческого липокалина, ассоциированного с желатиназой нейтрофилов (hNGAL), человеческого липокалина слезы (hTLPC), человеческого аполипопротеина D (АРО D) и билинсвязывающего белка Pien’s brassicae.
В другом воплощении изобретение относится к мутеину липокалина, предотвращающему индуцированное человеческим гепсидином-25 снижение уровней железа в сыворотке субъекта.
Изобретение также включает способ лечения заболевания или расстройства, связанного с измененным уровнем гепсидина, включающий введение фармацевтической композиции, содержащей мутеин, как описано здесь, субъекту, нуждающемуся в этом. В родственных воплощениях заболевание или расстройство включает расстройство гомеостаза железа или воспалительное состояние, связанное с повышенным уровнем гепсидина.
ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
На Фиг.1 показан способ сборки с применением полимеразной цепной реакции (PCR, PCR-сборки) для одновременного случайного мутагенеза 20 аминокислотных положений 36, 40, 41, 49, 52, 68, 70, 72, 73, 77, 79, 81, 96, 100, 103, 106, 125, 127, 132 и 134 (подчеркнутых и пронумерованных) аминокислотной последовательности зрелого Lcn 2 (липокалина-2). Эти 20 положений были разделены на четыре подгруппы в последовательности. Для рандомизации аминокислот каждой подгруппы синтезировали олигодезоксинуклеотид (SEQ ID NO:16, SEQ ID NO:17, SEQ ID NO:18, SEQ ID NO:19), в мутированных кодонах которого использовали NNK-смеси нуклеотидов. N обозначает смесь всех четырех оснований А, С, G и Т, в то время как К обозначает смесь только двух оснований G и Т; поэтому такой триплет кодирует все 20 природных аминокислот, а также амбер-стоп-кодон TAG, транслируемый как глутамин в supE-штаммах E. coli XLI-blue (Bullock et al., BioTechniques 5 (1987), 376-378) или TG1 (Sambrook et al., Molecular Cloning. A Laboratory Manual (1989), Cold Spring Harbor Press), которые использовали для получения фагмид и экспрессии генов. В реакции сборки также использовали четыре дополнительных олигодезоксинуклеотида (SEQ ID NO:20, SEQ ID NO:21, SEQ ID NO:22, SEQ ID NO:23) с постоянными нуклеотидными последовательностями, соответствующими некодирующей цепи (указанными под последовательностью двойной цепи ДНК в 3’-5’-направлении), и заполнение промежутков между указанными выше олигодезоксинуклеотидами. Два более коротких фланкирующих олигодезоксинуклеотида (SEQ ID NO:24 и SEQ ID NO:25), добавляемых в избытке и несущих биотиновые группы, служили в качестве праймеров для PCR-амплификации собранного, полностью синтетического фрагмента гена. Каждый из двух фланкирующих праймеров включал сайт рестрикции BstXI (CCANNNNNNTGG), приводя к образованию несовместимых друг с другом липких концов при ферментативном расщеплении. Такое специальное расположение сайтов рестрикции обеспечивало особенно эффективное лигирование и клонирование синтетического гена. Для введения первого сайта BstXI была необходима замена аминокислоты GIn28 на His относительно исходной последовательности Lcn2, в то время как второй сайт присутствует в кДНК (комплементарной ДНК) Lcn2 естественным образом. Кроме того, при сборке гена неспаренный остаток Cys87 заменяли на Ser. После первой PCR полученный фрагмент гена вводили в вектор, обеспечивавший недостающие части структурного гена Lcn2. На данном графическом материале также показаны два коротких праймера (SEQ ID NO:32 и SEQ ID NO:33), ближе к 3’- и 5’-концу, соответственно, относительно кассеты, фланкированной двумя сайтами рестрикции BstXI, используемыми для секвенирования двухцепочечной ДНК.
На Фиг.2 показана нуклеотидная последовательность библиотеки синтетических генов Lcn2 (показана только центральная кассета, фланкированная двумя сайтами рестрикции BstXI, как на Фиг.1). Этот фрагмент гена был получен Sloning BioTechnology GmbH. По сравнению с ДНК-библиотекой, показанной на Фиг.1, присутствуют два отличия. Во-первых, по возможности в немутированных аминокислотных положениях использовали кодоны, оптимизированные для экспрессии в Е. coli. Во-вторых, использовали смесь 19 разных триплетов (GAC, ТТС, CTG, САС, ААТ, AGC, АСС, GCA, ATG, ССТ, GTT, TGG, GAG, САА, АТС, GGA, CGT, GCA, TAC), кодирующих разные аминокислоты, за исключением Cys, в 20 рандомизированных положениях, идентичных показанным на Фиг.1. Нумерация аминокислот в данном случае соответствует внутренней схеме, использованной Sloning BioTechnology GmbH, в соответствии с которой Gly №1 является первым аминокислотным кодоном сразу после расположенного ближе к 3’-концу сайта рестрикции BsfXI.
На Фиг.3А показано выравнивание определенных аминокислотных последовательностей мутеинов липокалина на основе NGAL, специфичных в отношении человеческого гепсидина, в сравнении с полипептидной последовательностью липокалина NGAL дикого типа. Имеющие происхождение от NGAL мутеины, связывающиеся с гепсидином, содержат остатки 1-178, и это означает, что их длина равна длине зрелых белков дикого типа. Остатки 179-186 представляют собой последовательность метки, связывающейся со стрептавидином, Strep-tag™, использованной при выделении полученных мутеинов.
На Фиг.3Б показано выравнивание определенных аминокислотных последовательностей мутеинов липокалина на основе NGAL, специфичных в отношении человеческого гепсидина, в сравнении с полипептидной последовательностью липокалина NGAL дикого типа. Имеющие происхождение от NGAL мутеины, связывающиеся с гепсидином, содержат остатки 1-178, и это означает, что их длина равна длине зрелых белков дикого типа. Остатки 179-186 представляют собой последовательность метки, связывающейся со стрептавидином, Strep-tag™, использованной при выделении полученных мутеинов.
На Фиг.4А показана аминокислотная последовательность мутеина липокалина SEQ ID NO:1, слитая через линкер (показанный серым полужирным курсивом) с альбуминсвязывающим доменом (ABD) (показанным полужирным шрифтом) и меткой, связывающейся со стрептавидином, Strep-tag™ (показанной курсивом) (SEQ ID NO:15).
На Фиг.4Б показана аминокислотная последовательность липокалина hNGAL, кодируемая вектором phNGAL 98, слитая с меткой, связывающейся со стрептавидином, Strep-tag™ (показанной курсивом), и N-концевой Т7-меткой (показанной полужирным курсивом) (SEQ ID NO:34). Данный полипептид кодируется phNGAL 101.
На Фиг.5 показаны результаты прямого ELISA выбранных мутеинов Lcn2.
На Фиг.6 показаны результаты конкурентно-связывающего анализа выбранных мутеинов Lcn2.
На Фиг.7 приведены показатели аффинности выбранных мутеинов в отношении человеческого гепсидина-25 и гепсидина-25 яванского макака, определенные поверхностным плазменным резонансом (SPR).
На Фиг.8 показана нейтрализующая активность мутеинов липокалина против гепсидина-25 in vitro.
На Фиг.9 показано, что мутеин липокалина, направленный против гепсидина, нейтрализует человеческий гепсидин, вводимый мышам. SD -стандартное отклонение.
На Фиг.10 показаны фармакокинетические параметры SEQ ID NO:14-PEG и SEQ ID NO:1-ABD (равноценной SEQ ID NO:15). T1/2 - период полувыведения; в/в - внутривенно.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
В одном аспекте настоящее изобретение относится к новым, специфично связывающимся белкам, направленным против или специфичным в отношении гепсидина. Белки по изобретению могут быть использованы в терапевтических и/или диагностических целях. При использовании здесь белок по изобретению «специфично связывается» с мишенью (здесь, с гепсидином), если он способен отличать эту мишень от одной или более контрольных мишеней, поскольку специфичность связывания является не абсолютным, а относительным свойством. «Специфичное связывание» можно определить, например, вестерн-блоттингом, твердофазным иммуноферментным анализом (ELISA), радиоиммунным анализом (RIA), электрохемилюминесцентным анализом (ECL), иммунорадиометрическим анализом (IRMA), сортировкой клеток с возбуждением флуоресценции (FACS), иммуногистохимическим исследованием (IHC) и пептидным сканированием.
Белки по изобретению, направленные против или специфичные в отношении гепсидина, включают любое количество специфично связывающихся мутеинов белков на основе определенного белкового каркаса. При использовании здесь «мутеин», «мутированная» группировка (белок или нуклеиновая кислота) или «мутант» относятся к замене, делеции или вставке одного или более нуклеотидов или аминокислот, соответственно, по сравнению с встречающимся в природе нуклеотидным или белковым «эталонным» каркасом (каркасом дикого типа).
Белок по изобретению может представлять собой мутеин липокалина, предпочтительно липокалина, выбранного из группы, состоящей из ретинолсвязывающего белка (RBP), билинсвязывающего белка (ВВР), аполипопротеина D (АРО D), липокалина, ассоциированного с желатиназой нейтрофилов (NGAL), липокалина слезы (TLPC), α2-микроглобулин-родственного белка (A2m), 24р3/угерокалина (24р3), белка желез Эбнера-1 (VEGP 1), белка желез Эбнера-2 (VEGP 2) и предшественника главного аллергена Can f1 (ALL-1). При использовании здесь «липокалин» определяют как мономерный белок массой приблизительно 18-20 кДа, имеющий цилиндрическую β-складчатую область супервторичной структуры, содержащую множество β-цепей (предпочтительно восемь), соединенных попарно множеством петель (предпочтительно четырьмя) на одном конце с образованием посредством этого связывающего кармана. Именно разнообразие петель в остальном ригидном каркасе липокалинов приводит к множеству различных вариантов связывания у представителей семейства липокалинов, способных связываться с мишенями различного размера, формы и с разными химическими свойствами (обзор, например, в Flower, D.R. (1996), см. выше; Flower, D.R. et al. (2000), см. выше, или Skerra, A. (2000) Biochim. Biophys. Acta 1482, 337-350). Действительно, белковое семейство липокалинов естественным образом эволюционировало с развитием связывания с широким спектром лигандов при необычно низких уровнях общей консервативности последовательности (часто при идентичности последовательности менее 20%), сохраняя, однако, высококонсервативную общую пространственную структуру. Соответствие положений различных липокалинов хорошо известно специалисту в данной области техники. См., например, патент США №7250297.
В предпочтительном воплощении белок по изобретению представляет собой мутеин липокалина-2 (Lcn2; также известного как человеческий липокалин, ассоциированный с желатиназой нейтрофилов, hNGAL, или как сидерокалин). При использовании здесь термин «человеческий липокалин, ассоциированный с желатиназой нейтрофилов», или «hNGAL», или «липокалин-2», или «Lcn2» относится к зрелому hNGAL с регистрационным номером в базе данных SWISS-PROT/UniProt P80188 или зрелому hNGAL, показанному в SEQ ID NO:35. Зрелая форма данного белка имеет аминокислоты 21-198 полной последовательности ввиду отщепления сигнального пептида из аминокислот 1-20. Данный белок также имеет дисульфидную связь, образованную между аминокислотными остатками в положениях 76 и 175 зрелого белка.
В более предпочтительном воплощении изобретение относится к мутеину липокалина, имеющему цилиндрическую β-складчатую область супервторичной структуры, содержащую восемь β-цепей, соединенных попарно четырьмя петлями на одном конце с образованием посредством этого связывающего кармана, где по меньшей мере одна аминокислота каждой из по меньшей мере трех из указанных четырех петель мутирована и где указанный липокалин эффективно связывается с гепсидином в качестве данной искусственной мишени с поддающейся определению аффинностью. Предпочтительно, указанный мутеин липокалина имеет одну или более, как, например, 1, 2, 3, 4, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19 или 20 аминокислотных замен в положении, соответствующем положению 36, 40, 41, 49, 52, 68, 70, 72, 73, 77, 79, 81, 96, 100, 103, 106, 125, 127, 132 и/или 134 линейной полипептидной последовательности NGAL.
В данном контексте авторы изобретения идентифицировали определенную группу мутеинов липокалина-2 с мутациями в определенных положениях, демонстрирующих поддающуюся определению аффинность, а также специфичность в отношении гепсидина. Подходящие аминокислотные положения для мутаций включают положения 96, 100 и 106 линейной полипептидной последовательности человеческого липокалина-2. Настоящее изобретение также относится к нуклеиновым кислотам, кодирующим эти белки.
Другие белковые каркасы, которые могут быть сконструированы согласно настоящему изобретению для получения мутеинов белков, связывающихся с гепсидином, с поддающейся определению аффинностью, включают: домен, подобный эпидермальному фактору роста (EGF-подобный домен), домен «двойная петля», фибронектиновый домен I типа, фибронектиновый домен II типа, фибронектиновый домен III типа, домен PAN, домен G1a, домен SRCR, домен Кунитца/домен-ингибитор трипсина поджелудочной железы крупного рогатого скота, тендамистат, домен-ингибитор сериновых протеаз типа Казал (Kazal), домен-трилистник (Trefoil-домен Р-типа), домен фактора фон Виллебранда типа С, анафилатоксиноподобный домен, домен CUB, тиреоглобулиновый повтор I типа, домен рецептора липопротеинов низкой плотности (LDL) класса А, домен Sushi, домен Link, тромбоспондиновый домен I типа, домен иммуноглобулина или иммуноглобулиноподобный домен (например, однодоменные антитела или верблюжьи антитела из тяжелых цепей), пектиновый домен С-типа, домен МАМ, домен фактора фон Виллебранда типа А, домен соматомедина В, домен WAP-типа с четырьмя дисульфидными связями в сердцевине (WAP-type four disulfide core domain), домен F5/8 типа С, гемопексиновый домен, домен SH2, домен SH3, EGF-подобный домен ламининового типа, домен С2, «каппатела» ("Kappabodies") (III. et al. "Design and construction of a hybrid immunoglobulin domain with properties of both heavy and light chain variable regions" Protein Eng 10:949-57 (1997)), «минитела» ("Minibodies") (Martin et al. "The affinity-selection of a minibody polypeptide inhibitor of human interleukin-6" EMBO J 13:5303-9 (1994)), «диатела» ("Diabodies") (Holliger et al. "Diabodies": small bivalent and bispecific antibody fragments" PNAS USA 90:6444-6448 (1993)), «янусины» ("Janusins") (Traunecker et al. "Bispecific single chain molecules (Janusins) target cytotoxic lymphocytes on HIV infected cells" EMBO J 10:3655-3659 (1991) и Traunecker et al. "Janusin: new molecular design for bispecific reagents" Int J Cancer SuppI 7:51-52 (1992)), нанотело (nanobody), аднектин, тетранектин, микротело, аффилин, аффитело, анкирин, кристаллин, ноттин, убиквитин, белок типа «цинковый палец», автофлуоресцентный белок, анкирин или белок с анкириновыми повторами, или белок с повторами, богатыми лейцином, авимер (Silverman, Lu Q, Bakker A, To W, Duguay A, Alba BM, Smith R, Rivas A, Li P, Le H, Whitehorn E, Moore KW, Swimmer C, Perlroth V, Vogt M, Kolkman J, Stemmer WP 2005, Nat Biotech, Dec;23(12): 1556-61, E-Publication in Nat Biotech. 2005 Nov 20 edition); а также мультивалентные авимерные белки, эволюционировавшие перетасовкой экзонов в семействе человеческих рецепторных доменов (также описанные в Silverman J, Lu Q, Bakker А, То W, Duguay A, Alba BM, Smith R, Rivas A, Li P, Le H, Whitehorn E, Moore KW, Swimmer C, Perlroth V, Vogt M, Kolkman J, Stemmer WP, Nat Biotech, Dec;23(12):1556-61, E-Publication in Nat. Biotechnology. 2005 Nov 20 edition).
Белок по изобретению может содержать аминокислотную последовательность «исходного» белкового каркаса (такого как липокалин) дикого типа (природную последовательность) вне мутированных положений аминокислотной последовательности; альтернативно, мутеин липокалина может также содержать аминокислотные мутации вне положений последовательности, подвергнутых мутагенезу, не нарушающие связывающую активность и пространственную структуру мутеина. Такие мутации могут быть проведены на уровне ДНК с применением известных стандартных способов (Sambrook, J. et al. (1989) Molecular Cloning: A Laboratory Manual, 2nd Ed., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY). Возможные изменения аминокислотной последовательности представляют собой вставки или делеции, а также аминокислотные замены.
Такие замены могут быть консервативными, то есть аминокислотный остаток заменяют на химически сходный аминокислотный остаток. Примерами консервативных замен являются замены среди представителей следующих групп: (1) аланин, серин и треонин; (2) аспарагиновая кислота и глутаминовая кислота; (3) аспарагин и глутамин; (4) аргинин и лизин; (5) изолейцин, лейцин, метионин и валин; и (6) фенилаланин, тирозин и триптофан. С другой стороны, также возможно проведение неконсервативных изменений аминокислотной последовательности. Кроме того, вместо замены отдельных аминокислотных остатков также возможны вставка или делеция одной или более смежных аминокислот первичной структуры исходного белкового каркаса, где эти делеции или вставки приводят к образованию стабильного свернутого/функционального мутеина, который может быть легко исследован специалистом в данной области техники.
Специалисту в данной области техники будут ясны способы, полезные для получения мутеинов белков, предполагаемых согласно настоящему изобретению, но белковые или нуклеотидные последовательности которых не раскрыты здесь в прямой форме. В общих чертах, такие модификации аминокислотной последовательности включают, например, направленный мутагенез отдельных аминокислотных положений для упрощения субклонирования мутированного гена липокалина или его частей включением сайтов расщепления для определенных рестриктаз. Кроме того, эти мутации могут также быть включены для дополнительного улучшения аффинности мутеина липокалина в отношении данной мишени. Более того, при необходимости могут быть введены мутации для изменения определенных свойств мутеина, как, например, для улучшения стабильности пространственной структуры, стабильности в сыворотке, устойчивости белка или растворимости в воде или для снижения склонности к агрегации. Например, природные остатки цистеина могут быть заменены на другие аминокислоты для предотвращения образования дисульфидных связей.
Соответственно, изобретение также включает функциональные варианты белков, раскрытых здесь, имеющие пороговую идентичность последовательности или гомологию последовательности относительно эталонного белка. Под «идентичностью» или «идентичностью последовательности» понимают свойство последовательностей, характеризующее их сходство или родство. При использовании в настоящем изобретении термин «идентичность последовательности» или «идентичность» обозначает процент попарно идентичных остатков, после (гомологичного) выравнивания последовательности полипептида по изобретению и рассматриваемой последовательности, относительно числа остатков в более длинной из этих двух последовательностей. Процент идентичности определяют делением числа идентичных остатков на общее число остатков и умножением полученного числа на 100. Термин «гомология» использован здесь в его обычном значении, и включает идентичные аминокислоты, а также аминокислоты, рассматриваемые как консервативные замены (например, замену остатка глутамата на остаток аспартата) в эквивалентных положениях линейной аминокислотной последовательности двух белков. Наиболее предпочтительно, аминокислотная последовательность, показанная в SEQ ID NO:35, является предпочтительной «эталонной последовательностью». В SEQ ID NO:35 показан зрелый hNGAL. Термины «эталонная последовательность» и «последовательность дикого типа» (NGAL) использованы здесь взаимозаменяемо. Альтернативно, в качестве эталонной последовательности может быть использована аминокислотная последовательность с регистрационным номером в базе данных SWISS-PROT/UniProt P80188.
Процент гомологии последовательности или идентичности последовательности может, например, быть определен здесь с использованием программы BLASTP, версии blastp 2.2.5 (16 ноября 2002 г.; cf. Altschul, S.F. et al. (1997) Nucl. Acids Res. 25, 3389-3402). В данном воплощении процент гомологии основан на выравнивании полноразмерных полипептидных последовательностей (матрица: BLOSUM 62; значения разрыва: 11,1; установленное значение отсечения 10-3), включая пропептидные последовательности, предпочтительно с использованием белкового каркаса дикого типа в качестве эталона при попарном сравнении. Его вычисляют как процент числа «положительных результатов» (гомологичных аминокислот), указанных как результат в программе BLASTP, разделенного на общее число аминокислотных остатков, выбранных программой для выравнивания.
Также возможна преднамеренная замена других положений аминокислотной последовательности на цистеин для введения новых реакционно-способных групп, например, для конъюгации с другими соединениями, такими как полиэтиленгликоль (PEG), гидроксиэтилкрахмал (HES), биотин, пептиды или белки, или для образования искусственных дисульфидных связей. Относительно мутеина человеческого липокалина-2, примерами таких возможных мутаций с введением остатка цистеина в аминокислотную последовательность липокалина, включая мутеин человеческого липокалина-2, является введение остатка цистеина (Cys) по меньшей мере в одно из положений последовательности, соответствующих положениям 14, 21, 60, 84, 88, 116, 141, 145, 143, 146 или 158 последовательности hNGAL дикого типа. В некоторых воплощениях, где мутеин человеческого липокалина-2 по изобретению имеет последовательность, в которой, по сравнению с последовательностью с регистрационным номером в базе данных SWISS-PROT/UniProt P80188, цистеин был заменен на другой аминокислотный остаток, соответствующий цистеин может быть повторно введен в последовательность. В качестве иллюстративного примера, в таком случае возможно введение остатка цистеина в аминокислотном положении 87 с возвращением цистеина, исходно присутствовавшего в последовательности с регистрационным номером в SWISS-PROT P80188. Полученная тиоловая группировка в боковой цепи любого из аминокислотных положений 14, 21, 60, 84, 88, 116, 141, 145, 143, 146 и/или 158 может быть использована для пегилирования мутеина или его связывания с HES, например, с целью увеличения периода полувыведения соответствующего мутеина человеческого липокалина-2 из сыворотки.
При использовании согласно изобретению термин «положение» обозначает положение аминокислоты в аминокислотной последовательности, показанной здесь, или положение нуклеотида в последовательности нуклеиновой кислоты, показанной здесь. При использовании здесь термин «соответствующий» также включает, что положение определено не только числом предшествующих нуклеотидов/аминокислот. Соответственно, положение данной аминокислоты согласно изобретению, которая может быть заменена, может варьировать из-за делеции или добавления аминокислот в другой части липокалина (мутантного или дикого типа). Сходным образом, положение данного нуклеотида согласно настоящему изобретению, который может быть заменен, может варьировать из-за делеций или дополнительных нуклеотидов в другой части 5’-нетранслируемой области (UTR) мутеина или липокалина дикого типа, включая промоторные и/или любые другие регуляторные последовательности гена (включая экзоны и интроны).
Таким образом, под «соответствующим положением» согласно изобретению предпочтительно понимают, что нуклеотиды/аминокислоты могут отличаться по указанному числу, но все еще могут иметь сходные соседние нуклеотиды/аминокислоты. Указанные нуклеотиды/аминокислоты, которые могут быть заменены, удалены или добавлены также включают в термин «соответствующее положение». При использовании здесь «в положении, соответствующем положению», означает, что положение в «запрашиваемой» аминокислотной (или нуклеотидной) последовательности соответствует положению в «данной» аминокислотной (или нуклеотидной) последовательности.
Конкретно, для определения того, соответствует ли нуклеотидный остаток или аминокислотный остаток аминокислотной последовательности липокалина, отличного от мутеина липокалина NGAL по изобретению, определенному положению в нуклеотидной последовательности или аминокислотной последовательности описанного мутеина липокалина NGAL, в частности любой из SEQ ID NO:1-14 или последовательности, имеющей одну или более аминокислотных замен, как, например, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19 или 20 замен, в положении 36, 40, 41, 49, 52, 68, 70, 72, 73, 77, 79, 81, 96, 100, 103, 106, 125, 127, 132 и/или 134 линейной полипептидной последовательности NGAL, специалист в данной области техники может применять средства и способы, хорошо известные в данной области техники, например, выравнивания, ручные или с использованием компьютерных программ, таких как BLAST2.0, что расшифровывается как Basic Local Alignment Search Tool, или ClustalW, или любой другой подходящей программы, подходящей для проведения выравнивания последовательностей. Соответственно, мутеин липокалина по любой из SEQ ID NO:1-14 или мутеин липокалина, имеющий одну или более аминокислотных замен, как, например, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19 или 20 замен, в положении 36, 40, 41, 49, 52, 68, 70, 72, 73, 77, 79, 81, 96, 100, 103, 106, 125, 127, 132 и/или 134 или любом другом положении, описанном здесь, линейной полипептидной последовательности NGAL, может служить в качестве «данной последовательности», в то время как аминокислотная последовательность липокалина, отличного от NGAL, служит в качестве «запрашиваемой последовательности».
С учетом описанного выше, специалист в данной области техники может, таким образом, легко определить, какое аминокислотное положение, мутированное в Lcn2, как описано здесь, соответствует аминокислоте каркаса, отличного от Lcn2, предпочтительно такого, как один из описанных здесь. Конкретно, специалист в данной области техники может провести выравнивание аминокислотной последовательности мутеина, описанного здесь, в частности мутеина NGAL (или антикалина) по изобретению, с аминокислотной последовательностью другого липокалина для определения того, какая(ие) аминокислота(ы) указанного мутеина соответствует(ют) соответствующей(им) аминокислоте(ам) аминокислотной последовательности указанного другого липокалина. Конкретнее, специалист в данной области техники может таким образом определить, какая аминокислота аминокислотной последовательности указанного другого липокалина соответствует аминокислоте в положении(ях) 36, 40, 41, 49, 52, 68, 70, 72, 73, 77, 79, 81, 96, 100, 103, 106, 125, 127, 132 и/или 134 или аминокислоте в любом другом положении линейной полипептидной последовательности NGAL, как описано здесь.
Белки по изобретению, направленные против или специфичные в отношении гепсидина, включают любое количество специфично связывающихся мутеинов белков на основе определенного белкового каркаса. Предпочтительно, каркас представляет собой hNGAL. При использовании здесь «мутеин», «мутированная» группировка (белок или нуклеиновая кислота) или «мутант» относятся к замене, делеции или вставке одного или более нуклеотидов или аминокислот, соответственно, по сравнению с встречающимся в природе нуклеотидным или белковым «эталонным» каркасом (каркасом дикого типа). Предпочтительно, количество замененных, удаленных или вставленных нуклеотидов или аминокислот, соответственно, составляет 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20 или более, как, например, 25, 30, 35, 40, 45 или 50. Тем не менее, предпочтительно, что мутеин по изобретению все еще способен связываться с гепсидином.
В некоторых воплощениях белок по изобретению связывается с гепсидином с KD 100 мкМ или менее, включая 5 мкМ или менее, приблизительно 500 нМ, приблизительно 200 нМ или менее, 100 нМ или менее, 1 нМ или менее или 0,1 нМ или менее. Белок по изобретению может специфично связываться с одним или более чем одним непрерывным, прерывистым или конформационным эпитопом зрелой, свернутой, биологически активной формы гепсидина.
Белок по изобретению способен связываться с гепсидином с поддающейся определению аффинностью, то есть с константой диссоциации по меньшей мере 200 нМ, то есть KD приблизительно 200 нМ или менее. В некоторых воплощениях белок по изобретению связывается с гепсидином с константой диссоциации по меньшей мере приблизительно 100 нМ, приблизительно 50 нМ, приблизительно 25 нМ, приблизительно 15 нМ, приблизительно 5 нМ, приблизительно 2 нМ, приблизительно 0,5 нМ, приблизительно 0,25 нМ, приблизительно 0,1 нМ, приблизительно 0,05 нМ или еще меньше. Белок по изобретению предпочтительно связывается с молекулой зрелого человеческого гепсидина с аффинностью, определенной по KD, приблизительно 10 нМ или сильнее. Авторы изобретения часто определяли аффинности связывания по KD менее приблизительно 1 нМ и, в некоторых случаях, приблизительно 0,1 нМ и менее.
Аффинность связывания белка по изобретению (например, мутеина липокалина) в отношении выбранной мишени (в данном случае, гепсидина) может быть измерена (и, таким образом, значения KD комплекса мутеин-лигнад могут быть определены) множеством способов, известных специалистам в данной области техники. Такие способы включают, без ограничения, флуоресцентное титрование, конкурентный ELISA, калориметрические способы, такие как изотермическая титрационная калориметрия (ITC), и поверхностный плазменный резонанс (BIAcore). Такие способы хорошо известны в данной области техники, и их примеры также подробно описаны ниже.
Аминокислотная последовательность белка по изобретению может иметь высокую идентичность последовательности в отношении зрелого человеческого липокалина-2 или других липокалинов. В данном контексте белок по изобретению может быть по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 82%, по меньшей мере на 85%, по меньшей мере на 87%, по меньшей мере на 90%, включая по меньшей мере 95%, идентичен белку, выбранному из группы, состоящей из последовательностей SEQ ID NO:1-14. Предпочтительно, что структурный гомолог все еще имеет аминокислотную замену в одном или более, как, например, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19 или 20 положениях, соответствующих положению 36, 40, 41, 49, 52, 68, 70, 72, 73, 77, 79, 81, 96, 100, 103, 106, 125, 127, 132 и/или 134 линейной полипептидной последовательности NGAL.
Изобретение также включает структурные гомологи белков, выбранных из группы, состоящей из последовательностей SEQ ID NO:1-14, имеющие гомологию аминокислотной последовательности или идентичность последовательности более приблизительно 60%, предпочтительно более 65%, более 70%, более 75%, более 80%, более 85%, более 90%, более 92% и наиболее предпочтительно более 95%, по сравнению с указанными последовательностями. Предпочтительно, что структурный гомолог все еще имеет аминокислотную замену в одном или более, как, например, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19 или 20 положениях, соответствующих положению 36. 40, 41, 49, 52, 68, 70, 72, 73, 77, 79, 81, 96, 100, 103, 106, 125, 127, 132 и/или 134 линейной полипептидной последовательности NGAL.
Термин «гепсидин» относится к белку, также называемому экспрессируемым в печени антимикробным пептидом-1 (liver-expressed antimicrobial peptide 1) или предполагаемым регрессором опухолей печени (putative liver tumor regressor), человеческая форма которого имеет регистрационный номер в UniProtKB/Swiss-Prot P81172. Обычно термин «гепсидин» относится к любой форме белка гепсидина, о которой известно, что она присутствует у видов позвоночных, включая млекопитающих. Длина человеческого непроцессированного белка составляет 84 аминокислоты, и он кодируется геном «НАМР», также известным как «НЕРС» или «LEAP1». Происходит его расщепление на две цепи, которые здесь также включены в термин «гепсидин». Эти две цепи представлены аминокислотами 60-84, составляющими гепсидин-25 (Нерс25), и аминокислотами 65-84, составляющими гепсидин-20 (Нерс20). Гепсидин-25 имеет форму изогнутой шпильки, стабилизированной четырьмя дисульфидными связями. Природные варианты, также включенные в термин «гепсидин», имеют, например, аминокислотную замену 59 R → G (VAR_0425129); аминокислотную замену 70 С → R (VAR_042513); аминокислотную замену 71 G → D (VAR_026648) или аминокислотную замену 78 С → Y (VAR_042514). Другим природным вариантом является гепсидин-22, еще одна (наряду с гепсидином-20) укороченная с N-конца изоформа гепсидина-25.
При использовании здесь термин «зрелый гепсидин» относится к любой зрелой биологически активной форме белка гепсидина, экспрессируемой у позвоночного, такого как млекопитающее. Термин «человеческий гепсидин» относится к любой форме белка гепсидина, присутствующей у людей. Выражение «человеческий гепсидин-25» относится к зрелой форме человеческого гепсидина с аминокислотной последовательностью, показанной в SEQ ID NO:28. Согласно настоящему изобретению предложены мутеины липокалина, способные связываться с каждой заданной формой гепсидина, включая его протеолитические фрагменты, независимо от того, проявляет ли соответствующая молекула гепсидина биологическую/физиологическую активность. Таким образом, достаточно лишь присутствия молекулы гепсидина в биологическом образце без какого-либо измеримого физиологического значения. См., например, гепсидин-22, который на настоящий момент был обнаружен только в моче и предположительно является лишь продуктом деградации гепсидина-25 в моче (обзор в Kemna et al., Haematologica. 2008 Jan;
93:(1)90-97). Конечно, мутеин по изобретению может также связываться с физиологически активными разновидностями, такими как зрелый, биологически активный гепсидин-25. Соответственно, мутеин по изобретению может быть использован в качестве диагностического и/или фармацевтического агента, в зависимости от формы гепсидина, выбранной для распознавания.
В соответствии с описанным выше, белок по изобретению предпочтительно действует как антагонист молекулы гепсидина. В некоторых воплощениях белок по изобретению (например, мутеин человеческого липокалина-2) может действовать как антагонист молекулы гепсидина, ингибируя способность молекулы гепсидина связываться или иным образом взаимодействовать с ферропортином. Гепсидин может представлять собой форму зрелого человеческого гепсидина, такую как гепсидин-25 или гепсидин-20. Связывание зрелого гепсидина с ферропортином приводит к интернализации и деградации ферропортина, стандартные процессы для белка, расположенного на поверхности/мембране клетки.
В другом аспекте настоящее изобретение включает различные мутеины липокалина, включая мутеины человеческого липокалина-2, специфично связывающиеся с гепсидином. В этом смысле, гепсидин можно рассматривать как искусственный лиганд человеческого липокалина-2 дикого типа, где «искусственный лиганд» относится к соединению, не связывающемуся с липокалинами дикого типа, включая человеческий липокалин-2, в физиологических условиях. Конструированием липокалинов дикого типа, таких как человеческий липокалин-2, с мутациями в определенных положениях, авторы настоящего изобретения продемонстрировали, что высокая аффинность и высокая специфичность возможны для искусственного лиганда. В одном аспекте по меньшей мере в 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19 и/или 20 нуклеотидных триплетах, кодирующих любые из положений 36, 40, 41, 49, 52, 68, 70, 72, 73, 77, 79, 81, 96, 100, 103, 106, 125, 127, 132 и/или 134 линейной полипептидной последовательности человеческого Lcn2 (hLcn2) или других параллельных положений липокалинов, можно проводить случайный мутагенез, осуществляя в этих положениях замены с использованием подгруппы нуклеотидных триплетов.
Аминокислотные замены в мутеинах липокалинов по изобретению, как описано здесь, предпочтительно расположены в одной, двух, трех или четырех петлевых областях липокалина, предпочтительно hNGAL Петлевые области представлены положениями 33-54 (петля 1), 66-83 (петля 2), 94-106 (петля 3) и 123-136 (петля 4) hNGAL. 24-36, 53-66, 79-84 и 103-110.
Кроме того, липокалины могут быть использованы для получения мутеинов, имеющих мутированный аминокислотный остаток в любом одном или более, включая по меньшей мере два или все три, из положений последовательности, соответствующих положениям 96, 100 и 106 линейной полипептидной последовательности зрелого человеческого липокалина-2. Замена в положении последовательности 96 может, например, представлять собой замену Asn 96 → Arg, Asp, GIn, Gly, Lys, Ser, Thr или Val. Замена в положении последовательности 100 может, например, представлять собой замену Tyr 100 → Ala, Arg, Glu, GIn, Gly, Ser и Val. Замена в положении последовательности 106 может, например, представлять собой замену Tyr 106 → IIe, Gly, Phe, Val или Arg. В некоторых воплощениях мутеин по изобретению может иметь группу аминокислотных замен, по сравнению с линейной полипептидной последовательностью зрелого человеческого липокалина-2, Asn 96 → Val и Tyr 100 → GIn. В таком воплощении тирозин в положении 106 может быть сохранен. В некоторых воплощениях мутеин по изобретению может иметь группу аминокислотных замен, по сравнению с линейной полипептидной последовательностью зрелого человеческого липокалина-2, Asn 96 → Arg, Tyr 100 → Glu и Tyr 106 → Phe. В некоторых воплощениях мутеин по изобретению может иметь группу аминокислотных замен Asn 96 → Asp, Tyr 100 → Ser и Tyr 106 → Gly. В некоторых воплощениях мутеин по изобретению может иметь группу аминокислотных замен Asn 96 → Gly, Tyr 100 → Gly и Tyr 106 → Gly. В некоторых воплощениях мутеин по изобретению может иметь группу аминокислотных замен Asn 96 → Lys, Tyr 100 → Ala и Tyr 106 → lle. В некоторых воплощениях мутеин по изобретению может иметь группу аминокислотных замен Asn 96 → Ser, Tyr 100 → Arg и Tyr 106 → Val. В некоторых воплощениях мутеин по изобретению может иметь группу аминокислотных замен Asn 96 → Ser, Tyr 100 → Val и Tyr 106 → Arg. В некоторых воплощениях мутеин по изобретению может иметь группу аминокислотных замен Asn 96 → Thr, Tyr 100 → Val и Tyr 106 → Gly. В некоторых воплощениях мутеин по настоящему изобретению дополнительно содержит мутированный аминокислотный остаток в положении 134 линейной полипептидной последовательности зрелого человеческого липокалина-2. В одном воплощении эта замена представляет собой Lys 134 → Trp.
В некоторых воплощениях мутеин по настоящему изобретению содержит, обычно в дополнение к мутации в одном или более из положений последовательности 96, 100 и 106 (описанных выше), мутированный аминокислотный остаток в любом одном или более из положений последовательности, соответствующих положениям 52, 68, 81, 127 линейной полипептидной последовательности зрелого человеческого липокалина-2. Мутеин может, например, содержать в линейной полипептидной последовательности зрелого человеческого липокалина-2 замену Tyr 52 → His, Leu, Phe или Trp. Мутеин может также содержать в линейной полипептидной последовательности зрелого человеческого липокалина-2 замену Ser 68 → Arg, Gly или IIe. Мутеин может также содержать замену Arg 81 → Glu, Gly или GIn. Мутеин может, например, содержать в линейной полипептидной последовательности зрелого человеческого липокалина-2 замену Ser 127 → Thr или Trp. В некоторых воплощениях мутеин по изобретению может иметь группу аминокислотных замен, по сравнению с линейной полипептидной последовательностью зрелого человеческого липокалина-2, Tyr 52 → His, Ser 68 → Arg, Arg 81 → Ser и Ser 127 → Trp.В некоторых воплощениях мутеин по изобретению может иметь группу аминокислотных замен Tyr 52 → Leu, Ser 68 → Arg, Arg 81 → Glu и Ser 127 → Trp. В некоторых воплощениях мутеин по изобретению может иметь группу аминокислотных замен Tyr 52 → Phe, Ser 68 → Gly, Arg 81 → Gly and Ser 127 → Trp.В некоторых воплощениях мутеин по изобретению может иметь группу аминокислотных замен Tyr 52 → Trp, Ser 68 → IIe, Arg 81 → GIn and Ser 127 → Trp.В некоторых воплощениях мутеин по изобретению может иметь группу аминокислотных замен Tyr 52 → Trp, Ser 68 → Arg, Arg 81 → Glu and Ser 127 → Trp.В некоторых воплощениях мутеин по изобретению может иметь группу аминокислотных замен, по сравнению с последовательностью зрелого человеческого липокалина-2, Tyr 52 → Trp, Ser 68 → Arg, Arg 81 → Glu и Ser 127 → Thr. В некоторых воплощениях мутеин по изобретению может иметь группу аминокислотных замен Tyr 52 → Trp, Ser 68 → Arg, Arg 81 → Glu и Ser 127 → Trp.
В другом воплощении изобретения мутеин содержит по меньшей мере 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19 или 20 мутированных аминокислотных остатков в любом из положений последовательности, соответствующих положениям 33, 36, 40, 41, 42, 43, 44, 46, 47, 48, 49, 50, 51, 52, 54, 55, 59, 65, 68, 70, 72, 73, 75, 77, 78, 79, 80, 81, 86, 87, 98, 96, 99, 100, 103, 106, 107, 110, 111, 125, 127, 132, 134, 136 и 138 линейной полипептидной последовательности hNGAL или соответствующим положениям в других липокалинах. В другом воплощении мутеин содержит по меньшей мере 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19 или 20 мутированных аминокислотных остатков в любом одном из положений 33, 36, 40, 41, 42, 43, 44, 46, 47, 48, 49, 50, 51, 52, 54, 55. 59, 65, 68, 70, 72, 73, 75, 77, 78, 79, 80, 81, 86, 87, 98, 96, 99, 100, 103, 106, 107, 110, 111, 125, 127, 132, 134, 136 и 138 линейной полипептидной последовательности hNGAL или соответствующих положений в других липокалинах. В другом воплощении мутеин содержит 18, 19 или 20 мутированных аминокислотных остатков в любом одном из положений 33, 36, 40, 41, 42, 43, 44, 46, 47, 48, 49, 50, 51, 52, 54, 55, 59, 65, 68, 70, 72, 73, 75, 77, 78, 79, 80, 81, 86, 87, 98, 96, 99, 100, 103, 106. 107, 110, 111, 125, 127, 132, 134, 136 и 138 линейной полипептидной последовательности человеческого липокалина-2 или соответствующих положений в других липокалинах.
Мутеин по изобретению может, например, по сравнению с аминокислотной последовательностью зрелого hLcn2 дикого типа, содержать одну или более аминокислотных замен, как, например, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19 или 20 замен, из следующей группы: Leu36 → Ala, Cys, Thr, Val; Ala 40 → Arg, Glu, Gly и Ser; IIe 41 → IIe, Leu, Met или Val; GIn 49 → Leu или Met; Leu 70 → Asp, Asn, GIn, Met или Phe; Arg 72 → Glu, Gly, Leu или Val; Lys 73 → Ala, Arg, Glu, Gly, Leu, Thr или Tyr; Asp 77 → Arg, Glu, Gly, Leu, Ser или Val; Trp 79 → Gly, Leu, Ser, Tyr или Val; Leu 103 → Ala, Arg, Gly или Trp; Tyr 106 → Gly, IIe, Phe или Val; Lys 125 → Arg, Leu, Met, Phe, Thr или Val; и Tyr 132 → Leu или Val. Мутеин по изобретению может, например, иметь группу аминокислотных комбинаций, по сравнению с линейной полипептидной последовательностью зрелого человеческого липокалина-2, Ala 36, Ser 40, Leu 41, Met 49, Asn 70, Gly 72, Gly 73, Ser 77, Leu 79, Leu 125 и Val 132. Мутеин по изобретению может, например, иметь группу аминокислотных комбинаций, по сравнению с последовательностью зрелого человеческого липокалина-2, Leu 36, Arg 40, Val 41, GIn 49, Asp 70, Arg 72, Thr 73, Leu 77, Ser 79, Thr 125 и Val 132. В некоторых воплощениях мутеин по изобретению может иметь группу аминокислотных комбинаций Leu 36, Glu 40, IIe 41, Leu 49, GIn 70, Gly 72, Glu 73, Gly 77, Gly 79, Phe 125 и Val 132. Мутеин по изобретению может также иметь группу аминокислотных комбинаций Leu 36, Glu 40, IIe 41, Met 49, Met 70, Leu 72, Ala 73, Glu 77, Leu 79, Val 125, Val 132 или группу аминокислотных комбинаций Leu 36, Glu 40, Val 41, Met 49, Met 70, Leu 72, Ala 73, Glu 77, Leu 79, Thr 125 и Val 132. В некоторых воплощениях мутеин по изобретению может иметь группу аминокислотных комбинаций Leu 36, Glu 40, Val 41, Met 49, Met 70, Leu 72, Ala 73, Glu 77, Leu 79, Val 125 и Val 132 или группу аминокислотных комбинаций Thr 36, Ser 40, IIe 41, GIn 49, Phe 70, Glu 72, Gly 73, Arg 77, Val 79, Val 125 и Leu 132. В качестве дополнительного примера, мутеин по изобретению может иметь группу аминокислотных комбинаций Val 36, Glu 40, Met 41, Leu 49, Met 70, Glu 72, Tyr 73, Val 77, Leu 79, Arg 125 и Val 132. Мутеин по изобретению может также иметь группу аминокислотных комбинаций Val 36, Gly 40, Leu 41, Leu 49, Leu 70, Val 72, Arg 73, Arg 77, Tyr 79, Met 125 и Val 132.
В одном воплощении настоящего изобретения мутеин содержит мутированные аминокислотные остатки по меньшей мере в любых 10, 14, 15, 20, 22, 24, 26, 28, 29, 30, 31, 32, 33, 35 или во всех 45 из перечисленных выше положений последовательности.
Мутеин по изобретению, связывающийся с гепсидином, может содержать, по сравнению с аминокислотной последовательностью зрелого человеческого липокалина-2 дикого типа (Lcn2), no меньшей мере 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16 или 17 аминокислотных замен, включающих, без ограничения, Leu36 → Val или Cys; Ala40 → Tyr, или Lys, или Val; IIe41 → Thr, или Ser, или Leu; GIn49 → Leu или Trp; Leu70 → Gly; Arg72 → Gly или Asp; Lys73 → Leu, или Thr, или Asp; Asp77 → Asn, или His, или Leu; Trp79 → Lys; Asn96 → IIe или Arg; Tyr100 → GIn, или Arg, или Glu; Leu103 → Met, или Arg, или Gly; Tyr106 → Tyr, или Ala, или Trp; Lys125 → Thr, или Val, или Glu; Ser127 → Gly, или GIn, или Ala; Tyr132 → Met, или Ser, или Thr; и Lys134 → Asn.
В одном воплощении мутеин по изобретению, связывающийся с гепсидином, содержит следующие аминокислотные замены: Leu36 → Val; Ala40 → Tyr; IIe41 → Thr; GIn49 → Leu; Leu70 → Gly; Lys73 → Leu; Asp77 → Asn; Trp79 → Lys; Asn96 → IIe; Tyr100 → GIn; Leu103 → Met; Lys125 → Thr; Ser127 → Gly; Tyr132 → Met; и Lys134 → Asn. В другом воплощении мутеин по изобретению, связывающийся с гепсидином, содержит следующие аминокислотные замены: Leu36 → Val; Ala40 → Lys; 11е41 → Ser; GIn49 → Trp; Leu70 → Gly; Arg72 → Gly; Lys73 → Thr; Asp77 → His; Trp79 → Lys; Asn96 → Arg; Tyr100 → Arg; Leu103 → Arg; Tyr106 → Ala; Lys125 → Val; Ser127 → GIn; Tyr132 → Ser; и Lys134 → Asn. В другом воплощении мутеин по изобретению, связывающийся с гепсидином, содержит следующие аминокислотные замены: Leu36 → Cys; Ala40 → Val; IIe41 → Leu; GIn49 → Leu; Leu70 → Gly; Arg72 → Asp; Lys73 → Asp; Asp77 → Leu; Trp79 → Lys; Asn96 → Arg; Tyr100 → Glu; Leu103 → Gly; Tyr106 → Trp; Lys125 → Glu; Ser127 → Ala; Tyr132 → Thr; и Lys134 → Asn.
Мутеин по настоящему изобретению может дополнительно содержать, по сравнению с аминокислотной последовательностью зрелого hLcn2 дикого типа, аминокислотную замену GIn 28 → His. Мутеин по изобретению может также содержать, по сравнению с аминокислотной последовательностью зрелого hLcn2 дикого типа, аминокислотную замену Lys 62 → Arg. Кроме того, мутеин по настоящему изобретению может содержать, по сравнению с аминокислотной последовательностью зрелого hLcn2 дикого типа, аминокислотную замену Phe 71 → Pro или Ser. Еще одна аминокислотная замена, которая может присутствовать в мутеине по настоящему изобретению, по сравнению с аминокислотной последовательностью зрелого hLcn2 дикого типа, представляет собой замену Lys 74 → Glu. Еще одна аминокислотная замена, которая может быть включена в мутеин по изобретению, представляет собой замену Lys 75 → Glu. Мутеин по изобретению может также содержать, по сравнению с аминокислотной последовательностью зрелого hLcn2 дикого типа, аминокислотную замену Cys 87 → Ser. Мутеин по изобретению может также содержать аминокислотную замену Ser 146 → Pro. Еще одна аминокислотная замена, которая может присутствовать в мутеине по настоящему изобретению, по сравнению с аминокислотной последовательностью зрелого hLcn2 дикого типа, представляет собой замену Glu 147 → Gly. Мутеин по изобретению может содержать другие аминокислотные замены. Мутеины могут дополнительно содержать такие аминокислотные замены, как Tyr52 → GIn или Val; Ser68 → Lys или Asn; или Arg81 → Trp, Asn или His.
Мутеин по изобретению обычно существует в форме мономерного белка. Тем не менее, также возможно, что мутеин липокалина по изобретению способен к спонтанной димеризации или олигомеризации. Несмотря на то, что для некоторых применений может быть предпочтительным использование мутеинов липокалина, образующих стабильные мономеры, например, ввиду более быстрой диффузии и лучшего проникновения в ткани, в других случаях может быть предпочтительным использование мутеинов липокалина, образующих стабильные гомодимеры или мультимеры, поскольку такие мультимеры могут обеспечить (дополнительное) увеличение аффинности и/или авидности в отношении данной мишени. Кроме того, олигомерные формы мутеина липокалина могут иметь меньшие скорости диссоциации или более длительный период полувыведения из сыворотки.
Также отмечено, что на образование комплекса соответствующего мутеина и его лиганда влияет множество различных факторов, таких как концентрации соответствующих партнеров связывания, присутствие конкурентов, рН и ионная сила используемой буферной системы и экспериментальный способ, применяемый для определения константы диссоциации KD (например, флуоресцентное титрование, конкурентный ELISA или поверхностный плазменный резонанс, лишь в качестве нескольких примеров), или даже математический алгоритм, используемый для оценки экспериментальных данных.
По этой причине специалисту в данной области техники также ясно, что значения KD (константы диссоциации комплекса, образованного соответствующим мутеином и его мишенью/лигандом) могут варьировать в определенном экспериментальном диапазоне, в зависимости от способа и схемы эксперимента, используемых для определения аффинности определенного мутеина липокалина в отношении данного лиганда. Это означает возможность наличия некоторого разброса измеренных значений KD или приемлемого диапазона, в зависимости, например, от того, было ли значение KD определено поверхностным плазменным резонансом (Biacore), конкурентным ELISA или «прямым ELISA».
В одном воплощении мутеины, раскрытые здесь, могут быть связаны, на N- или С-конце, с афинной меткой, такой как пентагистидиновая метка, гексагистидиновая метка или стрептавидиновая метка (например, Streptag®). Таким образом, настоящая заявка включает также все прямо и в общих чертах описанные мутеины, снабженные такими метками.
Термин «фрагмент», при использовании в настоящем изобретении в связи с признаком фрагмента мутеина липокалина, относится к белкам или пептидам, имеющим происхождение от полноразмерного зрелого Lcn 2, укороченного с N-конца и/или С-конца, то есть, не имеющим по меньшей мере одной из N-концевых и/или С-концевых аминокислот. Такие фрагменты включают предпочтительно по меньшей мере 10, более предпочтительно 20, наиболее предпочтительно 30 или более последовательных аминокислот первичной последовательности зрелого Lcn 2, и их обычно можно выявить в иммунологическом анализе на Lcn 2. При использовании здесь термин «выявлять» или «выявление» понимают как на количественном, так и на качественном уровне, а также в комбинации. Таким образом, он включает количественные, полуколичественные и качественные измерения интересующей молекулы. Соответственно, возможно определение присутствия или отсутствия молекулы, такой как гепсидин, например, в образце, а также ее концентрации или уровня.
В объем настоящего изобретения также включены описанные выше мутеины, измененные в отношении их иммуногенности для снижения любой определяемой иммуногенности с применением способов, известных специалисту в данной области техники.
Цитотоксические Т-клетки распознают пептидные антигены на клеточной поверхности антиген-презентирующей клетки в комплексе с молекулой главного комплекса гистосовместимости (МНС) I класса. Способность пептидов связываться с молекулами МНС аллель-специфична и коррелирует с их иммуногенностью. Для снижения иммуногенности данного белка очень важна возможность спрогнозировать, какие пептиды белка обладают потенциалом связывания с данной молекулой МНС. Ранее были описаны способы с применением методики компьютерного моделирования для определения возможных Т-клеточных эпитопов для прогнозирования связывания данной пептидной последовательности с молекулами МНС 1 класса (Altuvia et al. (1995) J. Mol. Biol. 249: 244-250). Такой способ может также быть применен для идентификации потенциальных Т-клеточных эпитопов в мутеинах по изобретению и для проведения, в зависимости от его предполагаемого применения, селекции специфичного мутеина на основании его прогнозируемой иммуногенности. Кроме того, может быть возможным проведение дополнительного мутагенеза в областях пептидов, которые, согласно прогнозам, содержат Т-клеточные эпитопы, для уменьшения или удаления этих Т-клеточных эпитопов и, таким образом, минимизации иммуногенности. Удаление амфипатичных эпитопов из генетически конструированных антител было описано (Mateo et al. (2000) Hybridoma 19(6):463-471), и его можно адаптировать к мутеинам по настоящему изобретению. Полученные таким образом мутеины могут обладать минимизированной иммуногенностью, что желательно для их применения в терапевтических и диагностических применениях, таких, как описанные ниже.
Для некоторых применений также полезно использовать мутеины по изобретению в конъюгированной форме. Соответственно, изобретение также направлено на мутеины липокалина, конъюгированные с соединением, которое может включать, без ограничения, органические молекулы, ферментную метку, окрашенную метку, цитостатический агент, токсин, метку, которая может быть фотоактивирована и является подходящей для применения в фотодинамической терапии, флуоресцентную метку, радиоактивную метку, хромогенную метку, люминесцентную метку, металлические комплексы, металл, такой как коллоидное золото, гаптены, дигоксигенин, биотин, химиотерапевтический металл или химиотерапевтический металл, в качестве нескольких типичных примеров. Мутеин может также быть конъюгирован с органической молекулой лекарственного средства. Конъюгация может быть проведена с применением любого обычного способа сочетания, известного в данной области техники.
Термин «органическая молекула» или «малая органическая молекула» при использовании здесь в отношении искусственной мишени обозначает органическую молекулу, содержащую по меньшей мере два атома углерода, но предпочтительно не более 7 или 12 вращающихся атомов углерода, имеющую молекулярную массу в диапазоне от 100 до 2000 дальтон, предпочтительно от 100 до 1000 дальтон и, возможно, содержащую один или два атома металла.
В целом, возможно мечение мутеина липокалина, описанного здесь, любым подходящим химическим веществом или ферментом, которое непосредственно или косвенно образует выявляемое соединение или сигнал в химической, физической, оптической или ферментативной реакции. Примером физической реакции и в тоже время оптической реакции/маркера является эмиссия флуоресценции при облучении. Щелочная фосфатаза, пероксидаза хрена и β-галактозидаза являются примерами ферментных меток (и в то же время оптических меток), катализирующих образование продуктов хромогенных реакций. В целом, все метки, обычно используемые для антител (за исключением меток, используемых исключительно с сахарной группировкой Fc-части иммуноглобулинов), могут также быть использованы для конъюгации с мутеинами по настоящему изобретению. Мутеины по изобретению могут также быть конъюгированы с любым подходящим терапевтически активным агентом, например, для направленной доставки таких агентов в данную клетку, ткань или орган или для селективного направленного воздействия на клетки, например, опухолевые клетки, без эффекта на окружающие нормальные клетки. Примеры таких терапевтически активных агентов включают радионуклиды, токсины, малые органические молекулы и терапевтические пептиды (такие как пептиды, действующие в качестве агонистов/антагонистов рецептора клеточной поверхности, или пептиды, конкурирующие за сайт связывания белка на данной клеточной мишени). Примеры подходящих токсинов включают, без ограничения, коклюшный токсин, дифтерийный токсин, рицин, сапорин, псевдомонадный экзотоксин, калихеамицин или его производное, таксоид, майтансиноид, тубулизин или аналог доластатина. Аналог доластатина может представлять собой ауристатин Е, монометилауристатин Е, ауристатин PYE и ауристатин РНЕ. Примеры цитостатического агента включают, без ограничения, цисплатин, карбоплатин, оксалиплатин, 5-фторурацил, таксотер (доцетаксел), паклитаксел, антрациклин (доксорубицин), метотрексат, винбластин, винкристин, виндезин, винорелбин, дакарбазин, циклофосфамид, этопозид, адриамицин, камптотецин, соединения, родственные комбретастатину А-4, сульфонамиды, оксадиазолины, бензо[b]тиофены, синтетические спирокетальные пираны, монотетрагидрофурановые соединения, курацин и производные курацина, производные метоксиэстрадиола и лейковорин. Мутеины липокалина по изобретению могут также быть конъюгированы с терапевтически активными нуклеиновыми кислотами, такими как антисмысловые молекулы нуклеиновых кислот, малые интерферирующие РНК, микроРНК или рибозимы. Такие конъюгаты могут быть получены способами, хорошо известными в данной области техники.
В одном воплощении мутеины по изобретению можно также сочетать с группировкой для направленной доставки, обеспечивающей направление в определенную область организма, для доставки мутеинов по изобретению в желаемую область или часть тела. Одним примером, в котором такая модификация может быть желательной, является преодоление гематоэнцефалического барьера. Для преодоления гематоэнцефалического барьера мутеины по изобретению можно сочетать с группировками, способствующими активному транспорту через данный барьер (см. Gaillard PJ, et al. (2005) International Congress Series. 1277,185-198 или Gaillard PJ, et al. (2005) Expert Opin Drug Deliv. 2(2), 299-309). Такие соединения доступны, например, под торговым наименованием 2B-Trans™ (to-BBB technologies BV, Leiden, NL). Другие примеры молекул для направленной доставки, с которыми можно сочетать мутеины по настоящему изобретению, включают антитела, фрагменты антител или мутеины липокалина с аффинностью в отношении желаемой молекулы-мишени. Молекула-мишень группировок для направленной доставки может, например, представлять собой антиген клеточной поверхности. Антигены клеточной поверхности могут быть специфичны для типа клеток или тканей, такого как, например, раковые клетки. Типичными примерами таких белков клеточной поверхности являются HER-2 или протеогликаны, такие как NEU-2.
Как указано выше, в некоторых воплощениях мутеин по изобретению может быть конъюгирован с соединением, увеличивающим период полувыведения мутеина из сыворотки (в этом отношении см. также РСТ-публикацию WO 2006/56464, где описаны такие способы конъюгации применительно к мутеинам человеческого липокалина, ассоциированного с желатиназой нейтрофилов, с аффинностью связывания в отношении CTLA-4). Соединение, увеличивающее период полувыведения из сыворотки, может представлять собой молекулу полиалкиленгликоля, такую как полиэтилен (PEG) или его активированное производное; гидроксиэтилкрахмал, молекулы жирных кислот, таких как пальмитиновая кислота (Vajo & Duckworth (2000) Pharmacol. Rev. 52, 1-9), Fc-часть иммуноглобулина, CH3-домен иммуноглобулина, CH4-домен иммуноглобулина, альбумин или его фрагмент, альбуминсвязывающий пептид, альбуминсвязывающий белок, трансферрин или метку Рго-Ala-Ser, в качестве нескольких примеров. Альбуминсвязывающий белок может представлять собой бактериальный альбуминсвязывающий белок, антитело, фрагмент антитела, включая однодоменные антитела (см., например, патент США №6696245), или мутеин липокалина со связывающей активностью в отношении альбумина. Соответственно, подходящие конъюгируемые соединения для увеличения периода полувыведения мутеина липокалина по изобретению включают альбумин (Osborn et al. (2002) J. Pharmacol. Exp.Ther. 303, 540-548) или альбуминсвязывающий белок, например, бактериальный альбуминсвязывающий домен, такой как домен стрептококкового белка G (Konig, Т. and Skerra, A. (1998) J. Immunol. Methods 218, 73-83). Другие примеры альбуминсвязывающих пептидов, которые могут быть использованы в качестве партнера по конъюгации, представляют собой, например, пептиды, имеющие консенсусную последовательность Cys-Хаа1-Хаа2-Хаа3-Хаа4-Cys, где Xaa1 представляет собой Asp, Asn, Ser, Thr или Trp; Хаа2 представляет собой Asn, GIn, His, IIe, Leu или Lys; Хаа3 представляет собой Ala, Asp, Phe, Trp или Tyr; и Xaa4 представляет собой Asp, Gly, Leu, Phe, Ser или Thr, как описано в заявке на патент США №2003/0069395 или Dennis et al. (Dennis et al. (2002) J. Biol. Chem. 277, 35035-35043).
В других воплощениях альбумин сам по себе или биологически активный фрагмент альбумина могут быть использованы в качестве соединения для мутеина липокалина по изобретению, увеличивающего период полувыведения мутеина из сыворотки. Термин «альбумин» включает альбумины всех млекопитающих, такие как человеческий сывороточный альбумин, или бычий сывороточный альбумин, или крысиный альбумин. Альбумин или его фрагмент может быть получен рекомбинантным способом, как описано в патенте США №5728553 или заявках на европейские патенты ЕР 0 330 451 и ЕР 0 361 991. Рекомбинантный человеческий альбумин (Recombumin®) для применения в качестве стабилизатора белков доступен, например, от Novozymes Delta Ltd. (Nottingham, UK).
Если альбуминсвязывающий белок представляет собой фрагмент антитела, он может представлять собой однодоменное антитело. Однодоменные антитела (dAb) сконструированы для обеспечения точного контроля биофизических свойств и периода полувыведения in vivo для получения оптимального профиля безопасности и эффективности продукта. Однодоменные антитела имеются в продаже, например, от Domantis Ltd. (Cambridge, UK и МА, USA).
При использовании трансферрина в качестве группировки для увеличения периода полувыведения мутеинов по изобретению из сыворотки мутеины могут быть генетически слиты с N- или С-концом или с обоими концами негликозилированного трансферрина. Период полувыведения негликозилированного трансферрина составляет 14-17 суток, и слитый белок трансферрина будет сходным образом иметь увеличенный период полувыведения. Трансферриновый носитель также обеспечивает высокую биодоступность, биораспределение и стабильность в циркуляции. Эта технология имеется в продаже от BioRexis (BioRexis Pharmaceutical Corporation, PA, USA). Рекомбинантный человеческий трансферрин (DeltaFemn™) для применения в качестве стабилизатора белков также имеется в продаже от Novozymes Delta Ltd. (Nottingham, UK).
При использовании Fc-части иммуноглобулина с целью увеличения периода полувыведения мутеинов по изобретению из сыворотки может быть применена технология SynFusion™, имеющаяся в продаже от Syntonix Pharmaceuticals, Inc (MA, USA). Применение данной технологии Fc-слияния позволяет создавать более длительно действующие биофармацевтические агенты и может, например, включать две копии мутеина, связанные с Fc-областью антитела, для улучшения фармакокинетики, растворимости и эффективности получения.
Еще одной альтернативой для увеличения периода полувыведения мутеина по изобретению является слияние N- или С-конца мутеина по изобретению с длинными, неструктурированными, гибкими, богатыми глицином последовательностями (например, полиглицином с приблизительно 20-80 последовательными остатками глицина). Этот способ, описанный, например, в WO 2007/038619, был также назван «rPEG» (рекомбинантным PEG).
При использовании полиалкиленгликоля в качестве соединения, увеличивающего период полувыведения мутеина, полиалкиленгликоль может быть замещенным или незамещенным. Он также может представлять собой активированное полиалкиленовое производное. Примерами подходящих соединений являются молекулы полиэтиленгликоля (PEG), как описано в WO 99/64016, в патенте США №6177074 или в патенте США №6403564 применительно к интерферону, или как описано применительно к другим белкам, таким как PEG-модифицированная аспарагиназа, PEG-аденозиндезаминаза (PEG-ADA) или PEG-супероксиддисмутаза (см., например, Fuertges et al. (1990) "The Clinical Efficacy of Poly(Ethylene Glycol)-Modified Proteins" J. Control. Release 11, 139-148). Молекулярная масса такого полимера, предпочтительно полиэтиленгликоля, может варьировать от приблизительно 300 до приблизительно 70000 дальтон, включая, например, полиэтиленгликоль с молекулярной массой приблизительно 10000, приблизительно 20000, приблизительно 30000 или приблизительно 40000 дальтон. Более того, например, как описано в патентах США №6500930 или 6620413, углеводные олиго- и полимеры, такие как крахмал или гидроксиэтилкрахмал (HES), могут быть конъюгированы с мутеином по изобретению с целью увеличения периода полувыведения из сыворотки.
В другом воплощении для обеспечения подходящих боковых цепей аминокислот для конъюгации одного из описанных выше соединений с мутеинами по изобретению возможно введение искусственных аминокислот посредством мутагенеза. Обычно такие искусственные аминокислоты разработаны более реакционно-способными и, таким образом, облегчают конъюгацию с желаемой группировкой. Одним примером такой искусственной аминокислоты, которая может быть введена с использованием искусственной транспортной РНК (тРНК), является пара-ацетилфенилаланин.
Для некоторых применений мутеинов, описанных здесь, может быть предпочтительным использовать их в форме слитых белков. В некоторых воплощениях мутеин по изобретению слит на его N-конце и/или его С-конце, с белком, белковым доменом или пептидом, таким как сигнальная последовательность и/или аффинная метка.
Для фармацевтических применений мутеин по изобретению может быть слит с партнером слияния, увеличивающим период полувыведения мутеина из сыворотки in vivo (снова, см. РСТ-публикацию WO 2006/56464, где описаны подходящие партнеры слияния применительно к мутеинам человеческого липокалина, ассоциированного с желатиназой нейтрофилов, с аффинностью связывания в отношении CTLA-4). Аналогично конъюгируемым соединениям, описанным выше, партнер слияния может представлять собой Fc-часть иммуноглобулина, CH3-домен иммуноглобулина, CH4-домен иммуноглобулина, альбумин, альбуминсвязывающий пептид или альбуминсвязывающий белок, лишь в качестве нескольких примеров. Снова, альбуминсвязывающий белок может представлять собой бактериальный альбуминсвязывающий белок или мутеин липокалина со связывающей активностью в отношении альбумина. Соответственно, подходящие партнеры слияния для увеличения периода полувыведения мутеина липокалина по изобретению включают альбумин (Osborn, B.L. et al. (2002) см. выше J. Pharmacol. Exp.Ther. 303, 540-548) или альбуминсвязывающий белок, например, бактериальный альбуминсвязывающий домен, такой как стрептококковый белок G (Konig, Т. and Skerra, А. (1998) см. выше J. Immunol. Methods 218, 73-83). Альбуминсвязывающие пептиды, описанные в Dennis et al, см. выше (2002) или заявке на патент США №2003/0069395, имеющие консенсусную последовательность Cys-Xaa1-Xaa2-Xaa3-Xaa4-Cys, где Xaa1 представляет собой Asp, Asn, Ser, Thr или Trp; Хаа2 представляет собой Asn, GIn, His, IIe, Leu или Lys; Хаа3 представляет собой Ala, Asp, Phe, Trp или Tyr; и Xaa4 представляет собой Asp, Gly, Leu, Phe, Ser или Thr, могут также быть использованы в качестве партнера слияния. Также возможно использование альбумина самого по себе или биологически активного фрагмента альбумина в качестве партнера слияния для мутеина липокалина по изобретению. Термин «альбумин» включает альбумины всех млекопитающих, такие как человеческий сывороточный альбумин, или бычий сывороточный альбумин, или крысиный сывороточный альбумин. Рекомбинантное получение альбумина или его фрагментов хорошо известно в данной области техники и описано, например, в патенте США №5728553, заявке на европейский патент ЕР 0 330 451 или ЕР О 361 991.
Партнер слияния может придавать новые свойства мутеину липокалина по изобретению, такие как ферментативная активность или связывающая активность в отношении других молекул. Примерами подходящих белков для слияния являются щелочная фосфатаза, пероксидаза хрена, глутатион-S-трансфераза, альбуминсвязывающий домен белка G, белок А, фрагменты антител, домены олигомеризации, мутеины липокалина с такой же или другой специфичностью связывания (что приводит к образованию «дуокалинов», Schlehuber, S., and Skerra, A. (2001), Duocalins, engineered ligand-binding proteins with dual specificity derived from the lipocalin fold (Biol. Chem. 382, 1335-1342)) или токсины.
В частности, может быть возможным слияние мутеина липокалина по изобретению с отдельным ферментативно активным сайтом, таким образом, что оба «компонента» полученного слитого белка оказывают совместное воздействие на данную терапевтическую мишень. Связывающий домен мутеина липокалина связывается с мишенью, вызывающей болезнь, позволяя ферментативному домену устранять биологическую функцию мишени.
Аффинные метки, такие как Strep-tag® или Strep-tag® II (Schmidt, T.G.M. et al. (1996) J. Mol. Biol. 255, 753-766), myc-метка, FLAG-метка, Hise-метка или НА-метка, или белки, такие как глутатион-3-трансфераза, также позволяют легко выявлять и/или очищать рекомбинантные белки, являются дополнительными примерами предпочтительных партнеров слияния. В завершение, белки с хромогенными или флуоресцентными свойствами, такие как зеленый флуоресцентный белок (GFP) или желтый флуоресцентный белок (YFP), также являются подходящими партнерами слияния для мутеина липокалина по изобретению.
При использовании здесь термин «слитый белок» также включает мутеины липокалина по изобретению, содержащие сигнальную последовательность. Сигнальные последовательности на М-конце полипептида направляют данный полипептид в определенный клеточный компартмент, например, периплазму Е. coli или эндоплазматический ретикулум эукариотических клеток. В данной области техники известно большое количество сигнальных последовательностей. Предпочтительной сигнальной последовательностью для секреции полипептида в периплазму Е. coli является сигнальная последовательность OmpA.
Настоящее изобретение также относится к молекулам нуклеиновых кислот (ДНК и РНК), содержащим нуклеотидные последовательности, кодирующие мутеины, описанные здесь. Поскольку вырожденность генетического кода позволяет заменять определенные кодоны другими кодонами, обозначающими ту же аминокислоту, изобретение не ограничено одной конкретной молекулой нуклеиновой кислоты, кодирующей мутеин по изобретению, но включает все молекулы нуклеиновых кислот, содержащие нуклеотидные последовательности, кодирующие функциональный мутеин.
Молекула нуклеиновой кислоты, раскрытая в данной заявке, может быть «функционально связана» с регуляторной последовательностью (или регуляторными последовательностями) для обеспечения экспрессии данной молекулы нуклеиновой кислоты.
Молекулу нуклеиновой кислоты, такую как ДНК, называют «способной экспрессировать молекулу нуклеиновой кислоты» или способной «обеспечивать экспрессию нуклеотидной последовательности», если она содержит элементы последовательности, содержащие информацию, относящуюся к регуляции транскрипции и/или трансляции, и такие последовательности «функционально связаны» с нуклеотидной последовательностью, кодирующей полипептид. Функциональная связь представляет собой связь, при которой регуляторные элементы последовательности и экспрессируемая последовательность соединены таким образом, который обеспечивает экспрессию генов. Конкретная природа регуляторных областей, необходимых для экспрессии генов, может варьировать у разных видов, но обычно эти области содержат промотор, который у прокариот содержит промотор сам по себе, то есть ДНК-элементы, управляющие инициацией транскрипции, а также ДНК-элементы, которые, при транскрипции с образованием РНК, будут служить сигналами инициации трансляции. Такие промоторные области обычно содержат 5’-некодирующие последовательности, вовлеченные в инициацию транскрипции и трансляции, такие как «-35/-10»-боксы и элемент Шайна-Дальгарно у прокариот или ТАТА-бокс, СААТ-последовательности и 5’-кэпирующие элементы у эукариот. Эти области могут также содержать энхансерные или репрессорные элементы, а также транслируемые сигнальные и лидерные последовательности для направления нативного полипептида в определенный компартмент клетки-хозяина.
Кроме того, 3’-некодирующие последовательности могут содержать регуляторные элементы, вовлеченные в терминацию транскрипции, полиаденилирование и тому подобное. Тем не менее, если эти последовательности терминации неудовлетворительно функционируют в определенной клетке-хозяине, они могут быть заменены сигнальными последовательностями, функционирующими в данной клетке.
Таким образом, молекула нуклеиновой кислоты по изобретению может содержать регуляторную последовательность, предпочтительно промоторную последовательность. В другом предпочтительном воплощении молекула нуклеиновой кислоты по изобретению содержит промоторную последовательность и последовательность терминации транскрипции. Подходящими прокариотическими промоторами являются, например, промотор tet, промотор /acUV5 или промотор Т7. Примерами промоторов, полезных для экспрессии в эукариотических клетках, являются промотор SV40 или промотор CMV.
Молекулы нуклеиновых кислот по изобретению могут также быть частью вектора или клонирующей конструкции любого другого типа, такой как плазмида, фагмида, фаг, бакуловирус, космида или искусственная хромосома.
Молекула ДНК, кодирующая мутеины липокалина по изобретению, и, в частности, клонирующий вектор, содержащий последовательность, кодирующую такой мутеин липокалина, может быть использована для трансформации клетки-хозяина, способной экспрессировать данный ген. Трансформация может быть проведена с применением стандартных методик (Sambrook, J. et al. (1989), см. выше).
Таким образом, изобретение также направлено на клетку-хозяина, содержащую молекулу нуклеиновой кислоты, раскрытую здесь.
Изобретение также относится к способу получения мутеина по изобретению, где мутеин, фрагмент мутеина или слитый белок мутеина и другого полипептида получают, начиная с нуклеиновой кислоты, кодирующей мутеин, способами генной инженерии. Способ может быть осуществлен in vivo, мутеин может, например, быть получен в бактериальном или эукариотическом организме-хозяине и затем концентрирован, очищен или выделен из этого организма-хозяина или его культуры. Также возможно получение белка in vitro, например, с использованием системы трансляции in vitro. Термин «концентрированно» означает, что мутеин или его функциональный фрагмент составляет значимо большую фракцию от общего количества белка, присутствующего в рассматриваемом образце или растворе, чем в образце или растворе, из которых он был получен. Концентрированно может, например, включать выделение определенной фракции из экстракта клеток. Это может быть осуществлено стандартными методиками, такими как центрифугирование. Примерами других способов концентрирования являются фильтрация или диализ, которые могут, например, быть направлены на удаление нежелательных молекул с молекулярной массой меньше определенного значения, или осаждение с использованием органических растворителей или сульфата аммония. Очистка может, например, включать хроматографическую методику, например, гель-фильтрацию, ионообменную хроматографию, аффинную очистку, хроматографию гидрофобного взаимодействия или гидрофобную хроматографию с индукцией заряда (hydrophobia charge induction chromatography). Другим примером очистки является электрофоретическая методика, такая как препаративный капиллярный электрофорез. Выделение может включать комбинацию сходных способов. При использовании здесь «по существу чистый» или «по существу очищенный» означает соединение или разновидность, являющуюся преобладающей разновидностью в образце (то есть по молярному соотношению ее в композиции больше, чем любой другой отдельной разновидности). В некоторых воплощениях по существу очищенная композиция представляет собой композицию, в которой данная разновидность включает по меньшей мере приблизительно 50 процентов (по молярному соотношению) всех присутствующих молекулярных или, если это применимо, всех макромолекулярных разновидностей. В определенных воплощениях по существу чистая композиция будет иметь более приблизительно 80%, приблизительно 85%, приблизительно 90%, приблизительно 95% или приблизительно 99% всех молекулярных или, если это применимо, всех макромолекулярных разновидностей, присутствующих в композиции.
При получении мутеина in vivo нуклеиновую кислоту, кодирующую мутеин по изобретению, вводят в подходящий бактериальный или эукариотический организм-хозяин с применением технологии рекомбинантных ДНК (как уже описано выше). С этой целью клетку-хозяина сначала трансформируют клонирующим вектором, содержащим молекулу нуклеиновой кислоты, кодирующую мутеин по изобретению, с применением общепринятых стандартных способов (Sambrook, J. et al. (1989), см. выше). Клетку-хозяина затем культивируют в условиях, обеспечивающих экспрессию гетерологичной ДНК и, таким образом, синтез соответствующего полипептида. Затем полипептид выделяют из клетки или из культуральной среды.
В одном аспекте настоящее изобретение относится к способу получения мутеина, связывающегося с гепсидином, включающему:
проведение мутагенеза молекулы нуклеиновой кислоты, кодирующей липокалин, приводящего к получению одной или более молекул нуклеиновых кислот мутеина.
Способ может дополнительно включать:
экспрессию одной или более полученных молекул нуклеиновых кислот мутеина в подходящей системе экспрессии;
приведение множества мутеинов в контакт со зрелой формой гепсидина или по меньшей мере его фрагментом; и
концентрирование одного или более мутеинов, имеющих поддающуюся определению аффинность связывания в отношении данной мишени, посредством селекции и/или выделения.
При использовании здесь термин «мутагенез» означает, что экспериментальные условия выбраны таким образом, что аминокислота, встречающаяся в природе в данном положении последовательности липокалина, включая Lcn 2 (hNGAL; регистрационный номер в базе данных Swiss-Prot P80188), может быть заменена по меньшей мере одной аминокислотой, не присутствующей в данном конкретном положении в соответствующей природной полипептидной последовательности. Термин «мутагенез» также включает (дополнительное) изменение длины сегментов последовательности делецией или вставкой одной или более аминокислот. Таким образом, объем изобретения включает, например, замену одной аминокислоты в выбранном положении последовательности на группу из трех случайный мутаций, приводящую к добавлению двух аминокислотных остатков по сравнению с длиной соответствующего сегмента белка дикого типа. Такие вставки и делеции могут быть проведены независимо друг от друга в любом из пептидных сегментов, в которых может быть проведен мутагенез согласно изобретению. В одном типичном воплощении изобретения вставка нескольких мутаций может быть проведена в петле АВ выбранного липокалинового каркаса (см. международную заявку на патент WO 2005/019256, полностью включенную сюда посредством ссылки). Термин «случайный мутагенез» означает не присутствие предопределенной одной аминокислоты (мутации) в определенном положении последовательности, но возможность включения по меньшей мере двух аминокислот с определенной вероятностью в предопределенное положение последовательности при мутагенезе.
В одном неограничивающем способе кодирующая последовательность человеческого липокалина-2 может быть использована в качестве начальной точки для мутагенеза пептидных сегментов, выбранных согласно настоящему изобретению. Для мутагенеза указанных аминокислотных остатков специалист в данной области техники может применять различные общепринятые стандартные способы сайт-направленного мутагенеза (Sambrook, J. et al. (1989), см. выше). Широко применяемой методикой является введение мутаций посредством PCR (полимеразной цепной реакции) с использованием смесей синтетических олигонуклеотидов, содержащих вырожденное сочетание оснований, в желаемых положениях последовательности. Другие сходные методики хорошо известны специалистам в данной области техники.
Молекулы нуклеиновых кислот, определенные выше, могут быть соединены лигированием с недостающими 5’- и 3’-последовательностями нуклеиновой кислоты, кодирующей полипептид липокалина, и/или вектором и могут быть клонированы в известном организме-хозяине. Для лигирования и клонирования доступно множество общепринятых способов (Sambrook, J. et al. (1989), см. выше). Например, в последовательность синтетических олигонуклеотидов могут быть включены последовательности распознавания для рестрикционных эндонуклеаз, также присутствующие в последовательности клонирующего вектора. Таким образом, после амплификации соответствующего PCR-продукта и ферментативного расщепления полученный фрагмент можно легко клонировать с использованием соответствующих последовательностей распознавания.
Также возможно проведение случайного мутагенеза более длинных сегментов последовательности в пределах гена, кодирующего белок, выбранный для мутагенеза, известными способами, например, с применением полимеразной цепной реакции в условиях повышенной частоты ошибок, химическим мутагенезом или с использованием бактериальных штаммов-мутаторов. Такие способы могут также быть применены для дополнительной оптимизации аффинности или специфичности мутеина липокалина в отношении мишени. Возможные мутации вне сегментов экспериментального мутагенеза часто приемлемы или могут даже быть полезными, например, если они способствуют повышению эффективности фолдинга или стабильности пространственной структуры мутеина липокалина.
В другом воплощении способ включает проведение мутагенеза молекулы нуклеиновой кислоты в нуклеотидных триплетах, кодирующих по меньшей мере любые 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19 или 20 из положений последовательности, соответствующих положениям 33, 36, 40, 41, 42, 43, 44, 46, 47, 48, 49, 50, 51, 52, 54, 55, 59, 65, 68, 70, 72, 73, 75, 77, 78, 79, 80, 81, 86, 87, 98, 96, 99, 100, 103, 106, 107, 110, 111, 125, 127, 132, 134, 136 и/или 138 линейной полипептидной последовательности липокалина или, например, человеческого липокалина-2. Возможно проведение мутагенеза такой нуклеиновой кислоты и ее введение в подходящий бактериальный или эукариотический организм-хозяин с применением технологии рекомбинантных ДНК. Получение библиотеки нуклеиновых кислот липокалина может быть проведено с применением любой подходящей методики, известной в данной области техники, для получения мутеинов липокалина с антителоподобными свойствами, то есть мутеинов, обладающих аффинностью в отношении данной мишени. Примеры таких комбинаторных способов подробно описаны в международных заявках на патенты, например, WO 99/16873, WO 00/75308, WO 03/029471, WO 03/029462, WO 03/029463, WO 2005/019254, WO 2005/019255, WO 2005/019256 или WO 2006/56464. Содержание каждой из этих заявок на патенты полностью включено сюда посредством ссылки. После экспрессии последовательностей нуклеиновых кислот, подвергнутых мутагенезу, в подходящем хозяине, возможна селекция клонов, несущих генетическую информацию о множестве соответствующих мутеинов липокалина, связывающихся с данной мишенью, из полученной библиотеки. Для селекции этих клонов могут быть применены хорошо известные методики, такие как фаговый дисплей (обзор в Кау, В.К. et al. (1996) см. выше; Lowman, H.B. (1997) см. выше или Rodi, D.J. and Makowski, L. (1999) см. выше), скрининг колоний (обзор в Pini, A. et al. (2002) Comb. Chem. High Throughput Screen. 5, 503-510), рибосомный дисплей (обзор BAmstutz, P. et al. (2001) Curr. Opin. Biotechnol. 12, 400-405) или мРНК-дисплей, как описано в Wilson, D.S. et al. (2001) Proc. Natl. Acad. Sci. USA 98, 3750-3755, или способы, подробно описанные в WO 99/16873, WO 00/75308, WO 03/029471, WO 03/029462, WO 03/029463, WO 2005/019254, WO 2005/019255, WO 2005/019256 или WO 2006/56464.
Согласно данному описанию другое воплощение описанных выше способов включает:
обеспечение, например, по меньшей мере фрагмента гепсидина в качестве данной мишени/лиганда;
приведение множества мутеинов в контакт с указанными мишенью/лигандом для образования комплексов указанного лиганда и мутеинов, обладающих аффинностью связывания в отношении указанных мишени/лиганда; и
удаление мутеинов, не обладающих аффинностью связывания или не обладающих существенной аффинностью связывания.
В одном воплощении способов по изобретению селекцию по аффинности связывания проводят в конкурентных условиях. При использовании здесь термин «конкурентные условия» означает, что селекция мутеинов включает по меньшей мере одну стадию, на которой мутеины и фрагмент гепсидина или зрелый гепсидин, такой как гепсидин-25 (мишень), приводят в контакт в присутствии дополнительного лиганда, конкурирующего с мишенью (гепсидином) за связывание с мутеинами. Эта дополнительная мишень может представлять собой другую форму гепсидина, например, гепсидин-20 (в случае, если необходимо проводить селекцию мутеинов, селективно связывающихся с гепсидином-25 или даже с пятью N-концевыми остатками гепсидина-25 (как указано выше, в настоящее время полагают, что железорегулирующая биологическая активность обусловлена почти исключительно формой из 25 аминокислот, гепсидином-25, показывая, что пять N-концевых аминокислот необходимы для данной активности, Kenma et al., см. выше)), избыток мишени самой по себе или любой другой нефизиологический лиганд гепсидина, связывающийся с эпитопом, по меньшей мере перекрывающимся с эпитопом, распознаваемым мутеинами по изобретению, и, таким образом, препятствующий связыванию мутеинов с мишенью (гепсидином). Альтернативно, дополнительный лиганд конкурирует с мутеинами за связывание, образуя комплекс с эпитопом, отличающимся от сайта связывания мутеинов с мишенью аллостерическими эффектами. Соответственно, любые фрагмент, предшественник или зрелая форма гепсидина могут быть использованы при получении мутеинов по изобретению.
Другое воплощение способов по изобретению включает функциональное связывание нуклеиновой кислоты, кодирующей множество мутеинов по изобретению и полученной в результате мутагенеза, на 3’-конце с геном, кодирующим белок оболочки pIII нитчатого бактериофага семейства М13 или фрагмент этого белка оболочки, с целью селекции по меньшей мере одного мутеина по связыванию сданным лигандом.
Слитый белок может содержать дополнительные компоненты, такие как аффинная метка, позволяющие иммобилизовать, выявлять и/или очищать слитый белок или его части. Кроме того, между областями последовательности, кодирующими липокалин или его мутеины, и геном капсида фага или его фрагментами может быть расположен стоп-кодон, предпочтительно амбер-стоп-кодон, по меньшей мере частично транслируемый до аминокислоты при трансляции в подходящем штамме-супрессоре.
Например, плазмидный вектор pTLPC27, сейчас также называемый рТ1с27, описанный здесь, может быть использован для получения фагмидной библиотеки, кодирующей мутеины по изобретению. Молекулы нуклеиновых кислот по изобретению, кодирующие мутеины по изобретению, могут быть введены в вектор с использованием двух сайтов рестрикции BstXI. После лигирования подходящий штамм-хозяин, такой как Е. coli XLI-Blue, трансформируют полученной смесью нуклеиновых кислот с получением большого числа независимых клонов. Если желательно, может быть получен соответствующий вектор для получения гиперфагмидной библиотеки.
По завершении селекции мутеина с аффинностью в отношении данной мишени дополнительно возможно проведение еще одного мутагенеза такого мутеина с целью последующей селекции вариантов с еще большей аффинностью или вариантов с улучшенными свойствами, такими как большая термостабильность, улучшенная стабильность в сыворотке, термодинамическая стабильность, улучшенная растворимость, улучшенные мономерные свойства, улучшенная устойчивость к термической денатурации, химической денатурации, протеолизу или детергентам и так далее. Этот дополнительный мутагенез, который, в случае, если он направлен на повышение аффинности, можно рассматривать как «созревание аффинности» in vitro, может быть проведен сайт-направленным мутагенезом, основанным на рациональном планировании или случайной мутации. Другим возможным способом повышения аффинности или улучшения свойств является применение PCR пониженной точности, приводящей к точковым мутациям в выбранном диапазоне положений последовательности мутеина липокалина. PCR пониженной точности может быть проведена в соответствии с любым известным протоколом, таким как описанный Zaccolo et al. (1996) J. Mol. Biol. 255, 589-603. Другие способы случайного мутагенеза, подходящие для таких целей, включают мутагенез случайной вставкой/делецией (random insertion/deletion mutagenesis, RID), как описано Murakami et al. (2002) Nat. Biotechnol. 20, 76-81, или негомологичную случайную рекомбинацию (nonhomologous random recombination, NRR) как описано Bittker et al. (2002) Nat. Biotechnol. 20,1024-1029. Если желательно, созревание аффинности может также быть проведено способом, описанным в WO 00/75308 или Schlehuber et al. (2000) J. Mol. Biol. 297, 1105-1120, где были получены мутеины билинсвязывающего белка, обладающие высокой аффинностью в отношении дигоксигенина. Другим способом улучшения аффинности является проведение сайт-насыщающего мутагенеза. В этом способе возможно получение «малых» библиотек нуклеиновых кислот, где аминокислотные замены/мутации вводят только в единичных положениях в любом из четырех петлевых сегментов. Затем проводят стадию непосредственной селекции (скрининга аффинности) этих библиотек без дополнительных раундов пэннинга. Этот способ позволяет определить остатки, способствующие улучшенному связыванию с желаемой мишенью, и позволяет определить «горячие точки», важные для связывания.
В одном воплощении описанный выше способ модификации мутеина дополнительно включает введение остатка Cys в по меньшей мере одно из любых положений последовательности, соответствующих положениям 14, 21, 60, 84, 88, 116, 141, 145, 143, 146 или 158 последовательности человеческого липокалина-2 дикого типа, и сочетание с группировкой, способной изменять период полувыведения указанного мутеина из сыворотки, через тиоловую группу остатка Cys, введенного в по меньшей мере одно из любых положений последовательности, соответствующих положениям 14, 21, 60, 84, 88, 116, 141, 145, 143, 146 или 158 последовательности DNGAL дикого типа. Группировка, способная изменять период полувыведения указанного мутеина из сыворотки, может быть выбрана из группы, состоящей из молекулы полиалкиленгликоля и гидроксиэтилкрахмала.
Там, где белок по изобретению представляет собой мутеин человеческого липокалина-2 по изобретению, встречающаяся в природе дисульфидная связь между Cys 76 и Cys 175 может быть удалена. Соответственно, такие мутеины (или любой другой мутеин человеческого липокалина-2, не содержащий внутримолекулярную дисульфидную связь) могут быть получены в клеточном компартменте, имеющем восстановительную среду, например, в цитоплазме грамотрицательных бактерий.
В случае если мутеин липокалина содержит внутримолекулярные дисульфидные связи, может быть предпочтительным направить получаемый полипептид в клеточный компартмент, имеющий окислительную среду, используя подходящую сигнальную последовательность. Такая окислительная среда может быть обеспечена в периплазме грамотрицательных бактерий, таких как E. coli, во внеклеточной среде грамположительных бактерий или в просвете эндоплазматического ретикулума эукариотических клеток, и она обычно способствует образованию структурных дисульфидных связей.
Тем не менее, также возможно получение мутеина по изобретению в цитозоле клетки-хозяина, предпочтительно E. coli. В этом случае полипептид может быть получен непосредственно в растворимом и свернутом состоянии или выделен в форме телец включения с последующей ренатурацией in vitro. Еще одним вариантом является использование определенных штаммов-хозяев, имеющих окислительную внутриклеточную среду, способных, таким образом, обеспечивать образование дисульфидных связей в цитозоле (Venturi et al. (2002) J. Mol. Biol. 315, 1-8.).
Тем не менее, мутеин по изобретению не обязательно получают или производят с применением только генной инженерии. Наоборот, мутеин липокалина может также быть получен химическим синтезом, таким как твердофазный синтез полипептидов по Меррифилду, или транскрипцией и трансляцией in vitro. Например, возможно определение перспективных мутаций с применением молекулярного моделирования и затем синтез желаемого (конструированного) полипептида in vitro и исследование связывающей активности в отношении данной мишени. Способы твердофазного и/или жидкофазного синтеза белков хорошо известны в данной области техники (обзор, например, в Lloyd-Williams et al. (1997) Chemical Approaches to the Synthesis of Peptides and Proteins. CRC Press, Boca Raton, Fields, GB, и Colowick (1997) Solid-Phase Peptide Synthesis. Academic Press, San Diego, или Bruckdorfer et al. (2004) Curr. Pharm. Biotechnol. 5, 29-43).
В другом воплощении мутеины по изобретению могут быть получены транскрипцией/трансляцией in vitro с применением общепринятых способов, известных специалистам в данной области техники.
Изобретение также относится к фармацевтической композиции, содержащей по меньшей мере один мутеин по изобретению, указанный в формуле изобретения, или его слитый белок или конъюгаты, и, возможно, фармацевтически приемлемый эксципиент.
Мутеины липокалина по изобретению могут быть введены любым парентеральным или непарентеральным (например, энтеральным) способом введения, терапевтически эффективным для белковых лекарственных средств.
Соответственно, мутеины по настоящему изобретению могут быть включены в композиции с применением фармацевтически приемлемых ингредиентов, а также общепринятых способов изготовления (Gennaro and Gennaro (2000) Remington: The Science and Practice of Pharmacy, 20th Ed., Lippincott Williams & Wilkins, Philadelphia, PA). Для изготовления фармацевтических композиций могут быть использованы фармацевтически инертные неорганические или органические эксципиенты.
Белок по изобретению может также быть использован для направленной доставки соединения в предварительно выбранное место. В одном таком воплощении белок по изобретению используют для направленной доставки фармацевтически активного соединения в предварительно выбранное место в организме или ткани, включая:
а) конъюгацию белка с указанным соединением; и
б) доставку комплекса белок/соединение в предварительно выбранное место.
С этой целью мутеин приводят в контакт с интересующим соединением для образования комплекса. Затем осуществляют доставку комплекса, содержащего мутеин и интересующее соединение, в предварительно выбранное место. Это может, например, быть осуществлено сочетанием мутеина с группировкой для направленной доставки, такой как антитело, фрагмент антитела, мутеин липокалина или фрагмент мутеина липокалина с аффинностью связывания в отношении выбранной мишени.
В частности, это применение является подходящим, без ограничения, для (селективной) доставки лекарственного средства в предварительно выбранное место в организме, такое как инфицированная часть тела, ткань или орган, на который следует воздействовать лекарственным средством. Кроме образования комплекса мутеина и интересующего соединения, мутеин можно также подвергать взаимодействию с данным соединением с получением конъюгата мутеина и соединения. Аналогично описанному выше комплексу, такой конъюгат может быть подходящим для доставки соединения в предварительно выбранное целевое место. Такой конъюгат мутеина и соединения может также содержать линкер, ковалентно связывающий мутеин и соединение друг с другом. Возможно, такой линкер стабилен в кровотоке, но подвержен расщеплению в клеточной среде.
Мутеины, раскрытые здесь, и их производные могут, таким образом, быть использованы во многих областях, аналогично антителам или их фрагментам. Помимо их применения для связывания с подложкой, позволяющего иммобилизовать или выделить мишень данного мутеина или конъюгат или слитый белок данной мишени, мутеины могут быть использованы для мечения ферментом, антителом, радиоактивным веществом или любой другой группой, обладающей биохимической активностью или определенными связывающими свойствами. Посредством этого можно выявлять и приводить в контакт с ними их соответствующие мишени или конъюгаты или слитые белки этих мишеней. Например, мутеины по изобретению можно использовать для выявления химических структур общепринятыми аналитическими способами (например, ELISA или вестерн-блоттингом) или микроскопией или иммуносенсорными способами. В данном случае, выявляемый сигнал может быть получен непосредственно, применением подходящего конъюгата или слитого белка мутеина, или косвенно, иммунохимическим выявлением связанного мутеина с использованием антитела.
Также существует множество возможных применений мутеинов по изобретению в медицине. Помимо их применения в диагностике и доставке лекарственных средств, может быть получен мутантный полипептид по изобретению, связывающийся, например, с ткане- или опухольспецифичными молекулами клеточной поверхности. Такой мутеин может, например, быть использован в конъюгированной форме или в форме слитого белка для «визуализации опухолей» или непосредственно для лечения рака.
В другом аспекте настоящее изобретение также включает применение мутеина по изобретению для изготовления фармацевтической композиции. Изготовленная таким образом фармацевтическая композиция может быть подходящей для снижения уровня гепсидина. Фармацевтическая композиция может быть использована в форме монотерапии или в форме комбинированной терапии. Соответственно, изобретение также относится к мутеину, как описано выше, для лечения заболевания или расстройства, связанного с измененным, например, повышенным или сниженным, уровнем гепсидина. Заболевания, связанные с гепсидином
Анемия представляет собой заболевание, связанное с уменьшением количества железа в сыворотке, приводящим к снижению таких гематологических показателей, как количество эритроцитов (RBC), гематокрит (Ht), гемоглобин (Hb), уровень железа в сыворотке и насыщение трансферрина (Tf). Это приводит к снижению уровня кислорода в крови и связано со снижением качества жизни (QOL), описываемым слабостью, недостаточной концентрацией, одышкой и диспноэ. Тяжелая анемия может приводить к учащенному сердечному ритму, увеличению сердца и сердечной недостаточности. Анемия часто связана с хронической болезнью почек (СКО)/хронической почечной недостаточностью, раковой анемией (АС), анемией, вызванной химиотерапией (CIA) и анемией хронических заболеваний (ACD).
Эффективное лечение анемии оказывает значительный эффект на качество жизни и может влиять на выживаемость пациентов. Сниженное качество жизни может быть описано слабостью, утомлением, недостаточной концентрацией, одышкой вплоть до диспноэ. Тяжелая анемия связана с частым сердечным ритмом и может приводить к увеличению сердца и сердечной недостаточности.
Стандартным лечением являются трансфузии и введение агентов, стимулирующих эритропоэз (ESA), и железа. Тем не менее, необходимы новые способы лечения, поскольку стандартные способы лечения связаны со следующими недостатками или возможными недостатками. Трансфузия несет риск гемолиза, инфекций и аллергических реакций из-за несовместимой группы крови. Лечение железом может приводить к перегрузке железом при длительном лечении, и его не рекомендуют для лечения воспалительной анемии, поскольку железо способствует воспалительным ответам (например, воспалительному заболеванию суставов). Что касается ESA, приблизительно 40-50% пациентов с анемией не отвечают на ESA без Hb-ответа или с отсроченным НЬ-ответом только после лечения высокими дозами ESA, связанными с проблемами безопасности, такими как худшая выживаемость и меньшее время выживаемости без прогрессирования у пациентов с раком.
Железодефицитная анемия представляет собой расстройство гомеостаза железа, легко поддающееся лечению введением железа, в отличие от анемии, связанной с воспалительным заболеванием. Гепсидин является параметром, позволяющим различить эти два расстройства, поскольку в сочетании с воспалением происходит только повышающая регуляция уровня гепсидина.
Анемии, связанные с хроническим воспалительным заболеванием, таким как хронические инфекции, ревматические и системные аутоиммунные расстройства и воспалительное заболевание кишечника, называют воспалительной анемией (Al) или анемией хронических заболеваний (ACD). Экспрессию гепсидина индуцирует воспалительный цитокин интерлейкин-6 (IL-6), как часть воспалительного ответа, что приводит к железодефицитной анемии и слабому ответу на ESA.
У пациентов с хронической почечной недостаточностью (CRF) развивается уремическая анемия, как один из наиболее очевидных признаков заболевания. Этот симптом обусловлен сниженной продукцией эритропоэтина (ЕРО) в почках. ЕРО контролирует образование эритроцитов (RBC), стимулируя выживание, пролиферацию и дифференцировку эритроидных предшественников в костном мозге. Эффективное лечение анемии при хронической почечной недостаточности (CRF) оказывает значительный эффект на качество жизни и может влиять на выживаемость. Заместительная терапия рекомбинантным человеческим эритропоэтином (rhEPO) является в настоящее время стандартным лечением анемии у этих пациентов. В клинических исследованиях с пациентами с CRF наблюдаемая частота ответа на различные ESA (агенты, стимулирующие эритропоэз) составляла 70-90%. При анемии, связанной с CKD, гепсидин играет важную роль только у пациентов с сопутствующим воспалительным заболеванием.
Анемия распространена у пациентов с раком и имеет многофакторную этиологию. Она может быть связана со злокачественным новообразованием самим по себе и степенью его развития, а также с типом, продолжительностью и интенсивностью химиотерапии, направленной на подавление функции костного мозга. Более того, было показано, что у большинства пациентов с раком уровни циркулирующего ЕРО неадекватно низки для степени их анемии, отражая изменение этого гомеостатического механизма. При определенных типах опухолей частота анемии, тяжесть которой требует переливания крови, может достигать 60%. У пациентов с анемией и раком могут быть такие симптомы, как утомление, головокружение, одышка и сердечно-сосудистые симптомы, такие как учащенное сердцебиение и сердечная недостаточность. Такие клинические последствия могут снижать качество жизни этих пациентов. Кроме того, в последнее время обсуждается возможная взаимосвязь коррекции анемии и повышенной выживаемости у пациентов, получающих химиотерапию. В настоящее время возможным лечением анемии у пациентов с раком являются RBC-трансфузии или ESA. RBC-трансфузии могут быть связаны с негемолитическими и гемолитическими трансфузионными реакциями, перегрузкой железом у пациентов, получающих массивные трансфузии, или передачей инфекций. Требования по безопасности и скринингу при трансфузионной терапии усложнили логистику и повысили стоимость трансфузионной терапии, ограничив, таким образом, трансфузии случаями тяжелой и/или симптоматической анемии. ESA обеспечили альтернативу переливанию крови при лечении симптоматической анемии, тяжесть которой, по современным стандартам, еще не требует трансфузий. Тем не менее, четкого соотношения «доза-ответ» для ESA установлено не было, и от 40% до 50% пациентов демонстрируют полное отсутствие Hb-ответа или отсроченный ответ. В последние годы появились серьезные опасения о влиянии ESA на выживаемость пациентов с раком, а также их способности увеличивать риск тромбоэмболии (в марте 2007 г. Управление по контролю за продуктами и лекарственными средствами США (FDA) ввело предостережение в черной рамке (black-box warning) о возможной связи ESA со стимуляцией опухоли и тромбоэмболическими событиями). В литературе все больше данных о том, что прогностическим фактором резистентности к ESA у пациентов с раком является не только отсутствие повышения Hb-ответа за 4 недели введения ESA, но также повышенный уровень гепсидина, предположительно как часть воспалительного ответа.
Как объяснено выше, гепсидин является центральным отрицательным регулятором гомеостаза железа. Образование гепсидина усиливается при избытке железа и воспалении и снижается в условиях недостатка железа и гипоксии. Гепсидин действует, связываясь с единственным известным клеточным экспортером железа млекопитающих, ферропортином, и приводит к его интернализации и деградации. Поскольку ферропортин экспрессирован на энтероцитах двенадцатиперстной кишки, в селезенке и печени, повышение уровня гепсидина и последующее снижение уровня ферропортина приводят к ингибированию всасывания железа в двенадцатиперстной кишке, высвобождения рециркулирующего железа из макрофагов и мобилизации железа из депо в печени. Полагают, что гепсидин играет ключевую роль в развитии анемии, связанной с воспалительным заболеванием. Острые или хронические воспалительные состояния приводят к повышающей регуляции экспрессии гепсидина, вызывая дефицит железа, который может приводить к анемии, связанной с воспалительным заболеванием (ACD), раком (AC, CIA) и хронической болезнью почек (CKD) (CKD-анемии).
Мутеин липокалина по изобретению может быть использован в качестве антагониста гепсидина (см. выше). В этом отношении мутеин липокалина по изобретению, обычно выделенный мутеин липокалина, может быть использован в лечении, таком как лечение человека. Соответствующий мутеин способен образовывать комплекс с гепсидином, например, человеческим гепсидином, обычно с высокой аффинностью. Посредством этого мутеин липокалина обычно блокирует взаимодействие с рецептором гепсидина ферропортином. В результате это приводит к предотвращению интернализации и деградации ферропортина. Посредством этого мутеин липокалина поддерживает эритропоэз, обеспечивая мобилизацию железа из депо и улучшенное всасывание железа в кишечнике. В качестве иллюстративного примера, субъектом, нуждающимся в применении соответствующего антагониста гепсидина по изобретению, является субъект, слабо отвечающий на ESA-терапию (приблизительно 40-50% пациентов), что, как полагают, обусловлено сниженной доступностью железа для синтеза гемоглобина из-за повышающей регуляции гепсидина. Термин «субъект» относится к позвоночному животному, включая млекопитающее, и, в частности к человеку, в таком случае может также быть использован термин «пациент». В некоторых воплощениях у субъекта может быть расстройство, при котором будут полезны сниженный уровень гепсидина, такого как гепсидин-25, снижение биологической активности гепсидина (например, биологической активности гепсидина-25) и/или повышение уровня железа в сыворотке, увеличение количества ретикулоцитов, количества эритроцитов, повышение гемоглобина и/или гематокрита.
Мутеин липокалина по изобретению может быть использован для повышения уровней железа в биологической жидкости, такой как сыворотка. Он может также быть использован для увеличения количества ретикулоцитов, количества эритроцитов, повышения гемоглобина и/или гематокрита у субъекта, например, человека. В этом отношении может быть использована фармацевтическая композиция, содержащая мутеин липокалина по изобретению.
Другой аспект настоящего изобретения относится к способу лечения субъекта, страдающего от заболевания или расстройства, связанного с измененным уровнем гепсидина, таким как повышенный или сниженный уровень гепсидина. Соответствующее заболевание или расстройство может включать наследственное или ненаследственное заболевание/расстройство, приводящее к дефициту или избытку железа. Болезненное состояние или расстройство может включать инфекционное заболевание, включающее, например, бактерии, грибы, дрожжи или вирусы. Как объяснено выше, в некоторых воплощениях заболевание или расстройство представляет собой анемию, включая, без ограничения, анемию, являющуюся результатом инфекции, воспаления, хронического заболевания и/или рака. В некоторых воплощениях оно может включать воспалительное заболевание, такое как артрит, и определенные типы рака, заболевание печени или гематологическое заболевание. В некоторых воплощениях заболевание, связанное с повышенным уровнем гепсидина, представляет собой анемию, или хроническую болезнь почек, или анемию, связанную с хронической болезнью почек. Как уже объяснено выше, такой способ включает введение соответствующего мутеина по изобретению или фармацевтической композиции, содержащей мутеин по изобретению, субъекту, нуждающемуся в этом.
Мутеин липокалина по изобретению может, например, быть использован для лечения субъекта, имеющего повышенный уровень гепсидина, расстройство, связанное с гепсидином, расстройство гомеостаза железа, анемию или воспалительное состояние, связанное с повышенным уровнем гепсидина. Субъект может, например, представлять собой млекопитающее, такое как человек, страдающее от африканской перегрузки железом, альфа-талассемии, болезни Альцгеймера, анемии, раковой анемии, анемии хронических заболеваний, воспалительной анемии, артериосклероза или атеросклероза (включая ишемическую болезнь сердца, цереброваскулярную болезнь или окклюзионное заболевание периферических артерий), атаксий, атаксий, связанных с железом, атрансферринемии, рака, дефицита церулоплазмина, анемии, вызванной химиотерапией, хронической болезни почек (в частности анемии, связанной с хронической болезнью почек), включая терминальную почечную недостаточность или хроническую почечную недостаточность, цирроза печени, классического гемохроматоза, коллаген-индуцированного артрита (С1А), состояния, включающего избыток гепсидина (повышенный уровень гепсидина), врожденной дизэритропоэтической анемии, застойной сердечной недостаточности, болезни Крона, диабета, расстройства биораспределения железа, расстройства гомеостаза железа, расстройства метаболизма железа, ферропортинового заболевания, гемохроматоза с мутацией ферропортина, дефицита фолатов, атаксии Фридриха, фуникулярного миелоза, GRACILE-синдрома, бактериальной инфекции, такой как инфекция, вызванная Н. pyelori, болезни Галлервордена-Шпатца, гемохроматоза, гемохроматоза, являющегося результатом мутаций рецептора трансферрина 2 типа, гемоглобинопатий, гепатита, гепатита (Brock), гепатита С, печеночно-клеточного рака, наследственного гемохроматоза, вирусной инфекции, такой как инфекция, вызванная вирусом иммунодефицита человека (H1V), болезни Гентингтона, гиперферритинемии, гипохромной микроцитарной анемии, пониженного уровня железа в крови, инсулинорезистентности, железодефицитной анемии, железодефицитного расстройства, расстройства с избытком железа, железодефицитного состояния с избытком гепсидина, ювенильного гемохроматоза (HFE2), рассеянного склероза, мутации гена, вовлеченного в метаболизм железа, например, экспрессирующего белок, вовлеченный в метаболизм железа, такой как рецептор трансферрина 2 типа, HFE, гемоювелин или ферропортин, неонатального гемохроматоза, нейродегенеративного заболевания, связанного с железом, остеопении, остеопороза, панкреатита, нейродегенерации, связанной с пантотенаткиназой, болезни Паркинсона, пеллагры, геофагии, порфирии, поздней порфирии кожи, псевдоэнцефалита, гемосидероза легких, эритроцитарного расстройства, ревматоидного артрита, сепсиса, сидеробластной анемии, системной красной волчанки, талассемии, промежуточной талассемии, трансфузионной перегрузки железом, опухоли, васкулита, дефицита витамина B6, дефицита витамина B12, болезни Вильсона или воспалительного расстройства, связанного с повышенным уровнем гепсидина.
В качестве еще одного иллюстративного примера, мутеин по настоящему изобретению может в некоторых воплощениях быть использован в комбинации с эритропоэтином. Анемия у пациентов с раком (АС) и анемия хронических заболеваний (ACD) связаны с высокими концентрациями гепсидина (приблизительно 30 нмоль/л), приводящими к дефициту железа в сыворотке и, таким образом, к ослабленному эритропоэзу. Было сообщено, что субъекты с исходными концентрациями гепсидина в сыворотке менее 13 нмоль/л демонстрируют лучший ответ на терапию эритропоэтином (ЕРО), чем субъекты с концентрациями более 13 нмоль/л. По этой причине лечение пациентов с раком и анемией антагонистом гепсидина может улучшить их ответ на эритропоэтин.
Кроме того, широко распространенным явлением у пациентов с анемией является резистентность к рекомбинантному человеческому эритропоэтину (rhEPO), и эта терапевтическая проблема может быть преодолена комбинированной терапией с использованием мутеина по настоящему изобретению. Вероятно, гепсидин играет значительную роль в такой резистентности к rhEPO. Sasu et al. (Blood (2010) 115, 17, 3616-3624) показали отчетливую корреляцию повышенного уровня гепсидина и резистентности к агентам, стимулирующим эритропоэз (ESA), у мышей. Им также удалось восстановить чувствительность к ESA введением гепсидинспецифичного антитела.
В еще одном аспекте изобретение относится к применению мутеина по изобретению в диагностике. Обычно мутеин по изобретению применяют для диагностики заболевания или расстройства, связанного с измененным уровнем гепсидина, а также в соответствующем способе диагностики. В некоторых воплощениях применение может включать оценку уровня гепсидина в биологической жидкости субъекта. С этой целью от соответствующего субъекта может быть получена биологическая жидкость. Уровень гепсидина может быть сопоставлен с контрольным образцом с известным нормальным уровнем гепсидина. Посредством этого можно определить наличие нефизиологичных уровней гепсидина у субъекта.
Соответственно, изобретение также относится к мутеину, как определено выше, для диагностики заболевания или расстройства, связанного с измененным, например, повышенным или сниженным, уровнем гепсидина. В некоторых воплощениях заболевание представляет собой анемию, включая, без ограничения, анемию, являющуюся результатом инфекции, воспаления, хронического заболевания и/или рака. Заболевание или расстройство может, например, быть связано со сниженными уровнями гепсидина, как, например, наследственный гемохроматоз, анемия при избытке железа или гепатит С. Заболевание или расстройство может также быть связано со сниженными уровнями гепсидина, например, воспалительная анемия, железорефрактерная железодефицитная анемия или хроническая болезнь почек. Например, гепатит С обычно включает избыток железа в печени, в большинстве случаев в результате подавления синтеза гепсидина. В контексте диагностики мутеин по изобретению может быть использован для оценки уровней гепсидина в биологической жидкости субъекта. С учетом того наблюдения, что пациенты с раком и анемией с низкими концентрациями гепсидина (менее 13 нмоль/л) демонстрировали лучший ответ на терапию эритропоэтином, чем пациенты с высокими концентрациями гепсидина (более 13 нмоль/л), концентрации гепсидина в сыворотке могут, например, быть использованы для прогнозирования ответа на терапию эритропоэтином (приблизительно 50% пациентов резистентны к ЕРО).
В еще одном аспекте настоящее изобретение относится к диагностическому или аналитическому набору, содержащему мутеин по настоящему изобретению.
Субъект, нуждающийся в таком лечении, может представлять собой млекопитающее, такое как человек, собака, мышь, крыса, свинья, человекообразная обезьяна, такая как яванские макаки, в качестве лишь нескольких иллюстративных примеров.
В еще одном аспекте настоящее изобретение относится к способу in vivo-визуализации у субъекта, включающему введение указанному субъекту мутеина по изобретению или фармацевтической композиции, содержащей мутеин по изобретению. Субъект может представлять собой субъекта, определенного выше.
Изобретение дополнительно проиллюстрировано следующими неограничивающими Примерами и приложенными графическими материалами.
Если не указано иное, были применены общепринятые способы рекомбинантной генной инженерии, например, как описано в Sambrook et al. (см. выше).
Пример 1: Конструирование библиотеки для фагового дисплея мутантного Lcn2
Комбинаторную библиотеку вариантов Lcn2 получали на основании клонированной кДНК (Breustedt et al. (2006) Biochim. Biophys. Acta 1764, 161-173), содержавшей аминокислотные замены Cys87Ser для удаления одной неспаренной тиоловой боковой цепи (Goetz et al. (2000) Biochemistry 39, 1935-1941), а также GIn28His для введения второго сайта рестрикции BsfXI. Мутагенез и сборку данной области полимеразной цепной реакцией (PCR) проводили по существу согласно опубликованному способу (Beste et al. (1999) Proc. Natl. Acad. Sci. USA 96, 1898-1903; Skerra (2001) J. Biotechnol. 74, 257-275), в данном случае с использованием одной реакции амплификации с олигодезоксинуклеотидами (последовательности SEQ ID NO:16-SEQ ID NO:25), как показано на Фиг.1. Олигодезоксинуклеотиды были разработаны таким образом, что праймеры с последовательностями SEQ ID NO:16-SEQ ID NO:19 соответствовали кодирующей цепи и содержали вырожденные кодоны в аминокислотных положениях 36, 40, 41, 49, 52, или 68, 70, 72, 73, 77, 79, 81, или 96, 100, 103, 106, или 125, 127, 132, 134, соответственно, в то время как праймеры с последовательностями SEQ ID NO:20-SEQ ID NO:23 соответствовали некодирующей цепи и не содержали вырожденных кодонов или антикодонов. Два фланкирующих праймера SEQ ID NO:24 и SEQ ID NO:25 использовали в избытке, и они служили для амплификации собранного рандомизированного фрагмента гена. Все PCR-стадии проводили с использованием ДНК-полимеразы с «горячим стартом» Go-Taq (Promega, Mannheim, Germany), как описано Schlehuber et al. (2000) J. Mol. Biol. 297, 1105-1120.
Олигодезоксинуклеотиды без вырожденных кодонов со степенью очистки высокоэффективной жидкостной хроматографией (HPLC) приобретали у Metabion (Munich, Germany). NNK-содержащие Олигодезоксинуклеотиды приобретали обессоленными у того же поставщика и дополнительно очищали электрофорезом в полиакриламидном геле (PAGE) с мочевиной. Полученную ДНК-библиотеку обрабатывали BstXI (Promega, Mannheim, Germany) и клонировали на фагмидный вектор phNGAL102 (SEQ ID NO:26), основанный на типовом векторе экспрессии pASK111 (Vogt and Skerra (2001) J. Mol. Recognit. 14 (1), 79-86) и кодирующий слитый белок, состоящий из сигнального пептида ОтрА, модифицированного зрелого Lcn2 с последующим амбер-кодоном и С-концевого фрагмента гена белка оболочки III нитчатого бактериофага М13, то есть аналогично тому, как описано ранее для билинсвязывающего белка (Beste et а1., см. выше’, Skerra, см. выше). После электропорации Е. coli XLI-Blue (Bullock et аl. (1987) Biotechniques 5, 376-378) лигазной смесью из 8,4 мкг расщепленного PCR-продукта и 94 мкг расщепленной плазмидной ДНК получали 1×1010 трансформантов.
Альтернативно, случайную библиотеку клонированных синтетических Lcn2, описанную на Фиг.2, получали от Sloning BioTechnoIogy GmbH (Puchheim, Germany). Центральную генную кассету, фланкированную двумя сайтами рестрикции BstXI, амплифицировали посредством PCR за 20 циклов с использованием подходящих праймеров (SEQ ID NO:24 и SEQ ID NO:25) и субклонировали на phNGAL108 (SEQ ID NO:27), основанный на типовом векторе экспрессии pASK75 (Skerra (1994) Gene 151, 131-135) и содержащий по существу те же элементы, что и phNGAL102 (SEQ ID NO:26), но опосредующий резистентность к ампициллину вместо резистентности к хлорамфениколу, таким же образом с получением библиотеки, сложность которой соответствовала 1,7×1010 независимых трансформантов.
Последующие стадии получения библиотек проводили одинаково для обеих библиотек Lcn2. 100 мл культуры, содержащей клетки, трансформированные плазмидными векторами на основе phNGAL102 или phNGAL108, соответственно, кодирующими библиотеку мутеинов липокалина в форме слитых белков фагового белка pIII, переносили в стерильную колбу Эрленмейера и инкубировали в течение одного часа при 37°С, 160 об/мин в среде 2YT без селекционного давления антибиотиками. Перед инфицированием фагом-помощником VCS-M13 культуру разводили средой 2YT до оптической плотности при 550 нм (OD550) 0,1 с добавлением соответствующего антибиотика и выращивали далее в идентичных условиях с до достижения OD550 0,6. После инфицирования фагом-помощником VCS-M13 (Agilent Technologies, La Jolla, USA) при множественности инфицирования приблизительно 10, культуру перемешивали на протяжении еще 30 минут при 37°С, 100 об/мин. Затем температуру в инкубаторе снижали до 26°С и скорость качалки снова увеличивали до 160 об/мин, через 10 минут добавляли канамицин (70 мкг/мл) с последующей индукцией экспрессии генов добавлением ангидротетрациклина (ACROS Organics, Geel, Belgium) в концентрации 25 мкг/л (125 мкл исходного раствора с концентрацией 200 мкг/мл в диметилформамиде (DMF) на литр культуры). Инкубацию продолжали еще 12-15 часов при 26°С, 160 об/мин.
Клетки из полной культуры осаждали центрифугированием (30 минут, 18000 g, 4°С). Супернатант, содержащий фагмидные частицы, стерилизовали фильтрацией (0,45 мкм), смешивали с 1/4 объема 20% масс./об. PEG 8000, 15% масс./об. NaCl и инкубировали на льду в течение по меньшей мере 2 часов. После центрифугирования (30 минут, 18000 g, 4°С) осажденные фагмидные частицы из 1 литра культуры растворяли в 30 мл холодного BBS/E (200 мМ борат натрия, 160 мМ NaCl, 1 мМ EDTA (этилендиаминтетрауксусная кислота), рН 8,0), содержащего 50 мМ бензамидина (Sigma) и 1 мкг/мл Pefabloc (Roth, Karlsruhe, Germany). Раствор инкубировали на льду в течение 1 часа. После осаждения нерастворенных компонентов центрифугированием (10 минут, 43000 g, 4°C) каждый супернатант перемещали в новую реакционную емкость.
Добавление 1/4 объема 20% масс./об. PEG 8000, 15% масс./об. NaCl и инкубацию в течение 60 минут на льду использовали для повторного осаждения фагмидных частиц перед разделением фагмид на аликвоты и их заморозкой при -80°С для хранения. Для первого цикла селекции фагмиды размораживали и центрифугировали (30 минут, 34000 g, 4°С), супернатант удаляли и осажденные фагмидные частицы растворяли и смешивали в 400 мкл PBS (фосфатно-солевой буферный раствор), содержащего 50 мМ бензамидина. После инкубации в течение 30 минут на льду раствор центрифугировали (5 минут, 18500 g, 4°С) для удаления остаточных агрегатов и супернатант использовали непосредственно для селекции фаговым дисплеем.
Пример 2: Получение растворимых пептидов гепсидина-25
Синтетический немодифицированный гепсидин-25 (человеческий, DTHFPICIFCCGCCHRSKCGMCCKT, SEQ ID NO:28, 2789,4 г/моль; мышиный, DTNFPICIFCCKCCNNSQCGICCKT, SEQ ID NO:29, 2754,2 г/моль; крысиный, DTNFPICLFCCKCCKNSSCGLCCIT, SEQ ID NO:30, 2711,9 г/моль) и биотинилированный на С-конце крысиный гепсидин-25 (DTNFPICLFCCKCCKNSSCGLCCIT (SEQ ID NO:30)-мини-РЕС-линкер-К-биотин, 3210,5 г/моль) получали от PeptaNova GmbH (Sandhausen, GE).
Человеческий и мышиный гепсидин-25, биотинилированный на С-конце, получали от Bachem AG (Bubendorf, CM). Аналогично крысиному гепсидину-25, эти мишени были биотинилированы через остаток лизина, присоединенный к С-концу через мини-РЕС-линкер.
Пример 3: Получение библиотеки с 10 миллиардами независимых мутеинов NGAL
Случайную библиотеку липокалина NGAL (Lcn2) с высокой сложностью получали по существу, как описано в Примере 1 выше. Реакция амплификации показана на Фиг.1, фагмидный вектор phNGAL102 имеет последовательность SEQ ID NO:26.
Пример 4: Представление фагмид и селекция мутеинов NGAL с аффинностью в отношении человеческого гепсидина
Фагмидный дисплей и селекцию проводили с использованием фагмид, полученных в Примере 1, по существу, как описано в международной заявке на патент WO/2005/019256. Библиотеку подвергали 3 циклам селекции фаговым дисплеем против растворимого пептида-мишени человеческого гепсидина-25, биотинилированного на С-конце.
Использовали от 2×1012 до 1×1013 фагмид библиотеки, полученной в Примере 1. Кратко, фагмиды центрифугировали (21460 × g, 4°С, 20 минут) и ресуспендировали в 1 мл PBS (4 мМ КН2Р04, 16 мМ Na2BP04, 115 мМ NaCl, pH 7,4) содержащего 50 мМ бензамидина. PBS, содержащий 1% масс./об. казеина (Sigma) и 0,1% Tween 20®, использовали в качестве блокирующего буфера. Перед инкубацией с белком-мишенью фагмиды из библиотеки инкубировали с блокированными казеином стрептавидиновыми гранулами в течение 30 минут для удаления фагмид с мультиреактивными мутеинами липокалина, мутеинами липокалина с неправильной пространственной структурой или мутеинами, специфичными в отношении стрептавидиновых гранул.
В различных способах пэннинга либо проводили захват мишени из 1 мкМ раствора на покрытых Streptavidin™ блокированных 1% казеином магнитных гранулах перед инкубацией фагмид (способ «твердое тело в растворе»), либо 500 нМ гепсидин-25 инкубировали в растворе с 3×1012 фагмид из библиотеки NGAL, блокированной 1% казеином («растворный» способ). В «растворном» способе проводили захват связанных с пептидом фагмид с использованием магнитных гранул, покрытых Streptavidin™ (Invitrogen/Dynal), за 20 минут, с последующими 8 циклами отмывки и элюированием с использованием либо 300 мкл 70 мМ триэтиламина в течение 10 минут и нейтрализации подходящим количеством 1 М трис-HCl, рН 7,4 (основное элюирование), либо с использованием 300 мкл 0,1 М глицина/HCl, рН 2,2, в течение 10 минут и нейтрализации подходящим количеством 0,5 М трис-основания (кислотное элюирование).
В способе «твердое тело в растворе» блокированные фагмиды инкубировали с мишенью, нанесенной на стрептавидиновые гранулы, с последующими 8 циклами отмывки и элюированием, как описано выше.
Начиная со второго цикла обогащения, для амплификации фагмид использовали только половину объединенных фагмидных растворов.
Амплификацию фагмид между всеми циклами пэннинга проводили, как описано в Schlehuber, S. et al. (J. Mol. Biol. (2000), 297, 1105-1120).
Таким же образом проводили еще два раунда селекции против гепсидина-25 с использованием препарата амплифицированных фагмид от соответствующего предыдущего цикла обогащения, за исключением того, что, начиная со второго цикла обогащения, использовали только 1×1011 фагмид.
Пример 5: Идентификация мутеинов. специфичных в отношении человеческого гепсидина, с применением высокоэффективного ELISA-скрининга
Скрининг мутеинов, отобранных в Примере 4, проводили по существу, как описано в Примере 3 международной заявки на патент WO 2006/56464.
Селекцию липокалинов проводили с применением высокоэффективного ELISA-скрининга. При этом варианты NGAL с выявляемой Т7-меткой (Novagen), а также аффинной меткой Strep-tag II (IBA) экспрессировали в растворенной форме в 96-луночном титрационном микропланшете с использованием штамма Е. coli TG1/F- с phNGAL 101. Этот вектор соответствует phNGAL 98 (SEQ ID NO:31) с N-концевой Т7-меткой, состоящей из 11 аминокислот (MASMTGGQQMG) (SEQ ID NO:34, см. также Фиг.4Б). Экспрессию мутеинов липокалина индуцировали в течение ночи при 22°С и 700 об/мин с ангидротетрациклином (0,2 мкг/мл) при OD550 0,6. Затем клетки лизировали (100 мМ борат натрия, рН 8,0, 80 мМ NaCl, 1 мМ EDTA, 0,025% масс./об. лизоцим) в течение 1 часа при перемешивании. Для минимизации неспецифичного связывания при последующем ELISA-скрининге в необработанные лизаты клеток добавляли 2% масс./об. BSA и 0,1% об./об. Tween 20 и исследовали их в ELISA на предмет связывания с человеческим гепсидином-25. Таким образом, растворимый биотинилированный на С-конце человеческий гепсидин-25 иммобилизовали на лунках черных ELISA-планшетов Fluotrac 600 (Greiner; 384-луночные) в концентрации 1 мкг/мл захватом на нейтравидин (5 мкг/мл, Thermo Scientific). Нейтравидин, стрептавидин, каждый в концентрации 5 мкг/мл, и 3% молоко использовали в качестве отрицательного контроля. Планшеты блокировали РВ8Т(фосфатно-буферный солевой раствор/Tween)/0,1, содержащим 2% масс./об. BSA (бычий сывороточный альбумин), и затем инкубировали с экстрактом бактериальных клеток в течение 1 часа при комнатной температуре, планшеты отмывали пять раз и связанные мутеины липокалина выявляли моноклональным антителом против Т7, конъюгированным с пероксидазой хрена (HRP) (Novagen), разведенным 1:10.000 в PBST/0,1. Таким образом, QuantaBlu™ (Pierce; разведенный 1:2 в PBS/T 0,1%) использовали в качестве флуорогенного субстрата для HRP. Через 45 минут проявления сигнала при комнатной температуре проводили возбуждение флуоресценции при длине волны 320 нм (±12,5 нм) и измеряли ее при 430 нм (±17,5 нм) в спектрофотометре для прочтения планшетов GENiosPlus (Tecan).
В способе обратного ELISA проводили захват растворимых экспрессированных мутеинов из необработанного лизата клеток в ELISA-планшетах с использованием их Т7-метки после инкубации с различными количествами биотинилированного на С-конце человеческого гепсидина для создания условий ограниченного количества мишени с целью различения мутеинов по их аффинности. Связывание с мишенью выявляли конъюгатом экстравидин-HRP (Sigma). Конкуренция за связывание с мутеином была возможна при добавлении 100 нМ небиотинилированного человеческого гепсидина-25, показывая, что мутеины связываются и с немодифицированным человеческим гепсидином-25.
Скрининг 2160 колоний, отобранных, как описано в Примере 4, привел к идентификации более 1000 первичных удачных вариантов, продемонстрировав успешное выделение мутеинов, специфичных в отношении мишени. Способ обратного ELISA в условиях ограниченного количества мишени и конкурентный ELISA позволили различать гепсидинспецифичные мутеины по их аффинности в отношении мишени. С применением этих ELISA-способов были идентифицированы клоны, имеющие последовательности SEQ ID NO:1, SEQ ID NO:2, SEQ ID NO:3, SEQ ID NO:4, SEQ ID NO:5, SEQ ID NO:6, SEQ ID NO:7 SEQ ID NO:8, SEQ ID NO:9, SEQ ID NO:10. SEQ ID NO:11 и SEQ ID NO:12. Последовательности этих мутеинов показаны на Фиг.3.
Пример 6: Получение мутеинов NGAL. связывающихся с гепсидином
Рекомбинантный Lcn2 и варианты Lcn2, специфичные в отношении человеческого гепсидина, получали секрецией в периплазму штамма Е. coli К12 JM83 (Yanisch-Perron et al. (1985) Gene 33, 103-119), штамма Е. coli supE TG1-F-(производного Е. coli К12 TG1 [Kirn et al. (2009) J. Am. Chem. Soc. 131, 3565-3576], эписома которого была удалена с использованием акридинового оранжевого), Е. coli BL21 (Studier and Moffat (1986) J. Mol. Biol. 189, 113-130) или Е. coli W3110 (Bachmann (1990) Microbiol. Rev. 54, 130-197).
Для мелкомасштабной экспрессии растворимого белка использовали плазмиду phNGAL.98 (SEQ ID NO:31), кодирующую сигнальный пептид OmpA, слитый с соответствующим мутеином и С-концевой меткой Strep-tag II, содержащую два несовместимых сайта рестрикции BstXI для однонаправленного субклонирования мутированной генной кассеты. Выращивание проводили в культуре во встряхиваемой колбе объемом 2 л в присутствии среды LB (Лурия-Бертани) с ампициллином согласно протоколу, описанному в Schlehuber, S. et al. (J. Mol. Biol. (2000), 297, 1105-1120). Для больших количеств белка получение в периплазме проводили с использованием того же вектора, экспрессированного в штамме Е. coli W3110, культивацией в настольном ферментере в емкости объемом 1 л или 10 л на основании протокола, описанного в Schiweck, W., and Skerra, A. Proteins (1995) 23, 561-565).
Для увеличения периода полувыведения in vivo выбранные мутеины липокалина модифицировали, например, следующими способами.
Для мутеина по SEQ ID NO:1 конструировали ABD-слитый белок и экспрессировали его в периплазме. Альбуминсвязывающий домен стрептококкового белка G сливали с С-концом мутеина через оригинальный линкер, имеющий происхождение от стрептококкового белка G, как описано в SEQ ID NO:15.
В случае сайт-направленного пегилирования мутеины hNGAL, имеющие свободный остаток цистеина в аминокислотном положении 87 (SEQ ID NO:13, SEQ ID NO:14), использовали для пегилирования разветвленным 40k PEG-малеимидом. С этой целью проводили обратную мутацию серина в положении 87 на цистеин, исходно присутствующий в hNGAL дикого типа, сайт-направленным мутагенезом (набор Quick-change mutagenesis Kit, Stratagene). Перед реакцией пегилирования свободный остаток цистеина восстанавливали при молярном соотношении антикалина и ТСЕР (трис(2-карбоксиэтил)фосфин) 1:1 в течение 3 часов при комнатной температуре. Затем проводили пегилирование, смешивая белок с молярным избытком (более 2) реагента РЕС40-малеимид в течение 1,5 часов при комнатной температуре.
Проводили очистку вариантов Lcn2 из периплазматической фракции за одну стадию посредством аффинной хроматографии со стрептавидином (Strep-Tactin™ Superflow, IBA) с использованием колонки с подходящим объемом слоя, способом, описанным Skerra, А. & Schmidt, Т. G. М. (2000) (Use of the Sfrep-tag and streptavidin for detection and purification of recombinant proteins. Methods Enzymol. 326A. 271-304). Для достижения более высокой степени очистки и полного удаления агрегированного рекомбинантного белка в завершение проводили гель-фильтрацию мутеинов на колонке Superdex 75 HR 10/30 (объем слоя 24 мл, Amersham Pharmacia Biotech, Freiburg, Germany) в присутствии PBS-буфера. Мономерные белковые фракции объединяли, анализировали их чистоту электрофорезом в полиакриламидном геле с додецилсульфатом натрия (SDS-PAGE) (Fling and Gregerson (1986) Anal. Biochem. 155, 83-88) и использовали их для дальнейшего биохимического описания.
Пегилированные варианты мутеинов hNGAL очищали хроматографией и, по необходимости, проводили дополнительную очистку от бактериальных экзотоксинов фильтрацией через мембрану MustangE (Pall Corporation, US).
Пример 7: Измерение аффинности с применением ELISA-методик
«Прямой» ELISA проводили для верификации аффинности и специфичности связывания выбранных мутеинов Lcn2. Таким образом, проводили захват биотинилированного на С-конце гепсидина (Bachem AG, СН) в постоянной концентрации 1 мкг/мл на поверхности полистиролового планшета (Greiner, GE) с использованием нейтравидина (Thermo Scientific, 5 мкг/мл). Двухступенчатую серию разведении очищенных мутеинов Lcn2 инкубировали с захваченным гепсидином в течение 1 часа при комнатной температуре и проводили выявление либо посредством метки Strep-tag II с использованием кроличьего поликлонального антитела против метки Strep-tag II (GenScript, USA), либо с использованием кроличьего поликлонального антитела, специфичного в отношении каркаса. В обоих случаях в качестве вторичного антитела для выявления использовали антитело против кроличьего IgG, конъюгированное с HRP (Abeam, UK).
Оптическую плотность ДА при 320 нм измеряли с использованием спектрофотометра для ELISA (Tecan, GE) и данные подгоняли с применением программного обеспечения Graphpad Prism (Statcom, USA).
Результаты измерений с использованием мутеинов, имеющих последовательности SEQ ID NO:1-SEQ ID NO:12, а также SEQ ID NO:14, связанную с PEG40, SEQ ID NO:13, связанную с PEG40, и SEQ ID NO:1, связанную с альбуминсвязывающим доменом (ABD) (SEQ ID NO:15), представлены на Фиг.5.
Значения Ко выбранных мутеинов Lcn2 варьируют от 220 пМ до 6,8 нМ. Все мутеины связывались с человеческим гепсидином-25 и гепсидином-25 яванского макака с сопоставимой аффинностью. Увеличение периода полувыведения мутеина липокалина по SEQ ID NO:1 из сыворотки С-концевым слиянием с альбуминсвязывающим доменом не оказывало существенного эффекта на аффинность связывания мутеина, в то время как пегилирование значительно снижало аффинности связывания в данном ELISA-формате, в 5 раз для SEQ ID NO:8 и в 8 раз для мутеина по SEQ ID NO:1.
Аффинность связывания мутеинов Lcn2 в отношении немодифицированного гепсидина-25 в растворе оценивали способом конкурентного ELISA. Таким образом, проводили захват человеческого гепсидина, биотинилированного на С-конце (Bachem AG, СН), в постоянной концентрации 1 мкг/мл на поверхности полистиролового планшета (Greiner, GE) с использованием нейтравидина (Thermo Scientific, 5 мкг/мл, GE). Одновременно двухступенчатую серию разведении небиотинилированного человеческого гепсидина, начиная с 1 мкМ, инкубировали с постоянной концентрацией гепсидинспецифичного мутеина в течение 1 часа при комнатной температуре в 96-луночном полипропиленовом планшете, не связывающем белки (Nunc, GE). Постоянная концентрация мутеинов липокалина соответствует средней эффективной концентрации (EC50) соответствующих мутеинов, определенной в прямом ELISA, как описано выше в данном примере. Затем смесь немодифицированного человеческого гепсидина и мутеина липокалина переносили в планшет с нейтравидином и захваченным гепсидином. Биотинилированному на С-конце гепсидину позволяли конкурировать с немодифицированным гепсидином за связывание с антикалином на протяжении 20 минут при комнатной температуре. В течение этих 20 минут свободный мутеин липокалина связывался с захваченным гепсидином, и его выявляли с использованием кроличьего поликлонального антитела против метки Strep-tag II (GenScript, USA). В качестве вторичного антитела для выявления использовали козье антитело против кроличьего IgG, конъюгированное с HRP (Abeam, UK). Одновременно с конкурентным анализом связывание антикалинов определяли в том же планшете в «прямом» ELISA для получения стандартной кривой, связывавшей значения в относительных единицах флуоресценции (RFU) с концентрацией антикалинов. Эту кривую затем использовали для нормализации данных конкурентного анализа относительно уровня связывания антикалинов с планшетом, и проводили анализ с использованием программного обеспечения Graphpad. Значения IC50 соответствуют половине наибольшего количества мутеина липокалина, связанного с планшетом.
Результаты измерений с использованием мутеинов, имеющих последовательности SEQ ID NO:1-SEQ ID NO:12, а также SEQ ID NO:14, связанную с PEG40, SEQ ID NO:13, связанную с PEG40, и SEQ ID NO:1, связанную с ABD (SEQ ID NO:15), представлены на Фиг.6.
Значения IC50 выбранных мутеинов Lcn2 варьировали от 100 пМ до 10,8 нМ. Увеличение периода полувыведения из сыворотки с использованием альбуминсвязывающего домена не оказывало эффекта на аффинность связывания мутеина по SEQ ID NO:1, в то время как пегилирование снижало аффинности связывания в 2 раза для SEQ ID NO:13-PEG40 и в 4 раза для SEQ ID NO:14-PEG40, соответственно.
Пример 8: Измерение аффинности с применением поверхностного плазмонного резонанса (SPR)
Поверхностный плазменный резонанс применяли для оценки кинетики и аффинности связывания мутеинов липокалина, раскрытых здесь.
Мутеины липокалина иммобилизовали на сенсорном чипе СМ5 с применением стандартной аминной химии. Поверхность чипа активировали с использованием EDC (1-этил-3-(3-диметиламинопропил)карбодиимид) и NHS (N-гидроксисукцинимид). Затем вносили растворы мутеинов липокалина в концентрации 20 мкг/мл в 10 мМ ацетате натрия, рН 4,5 (60 мкг/мл в 10 мМ ацетате натрия рН 4,0 для пегилированного мутеина липокалина) при скорости потока 5 мкл/мин до достижения поверхностной плотности 500-700 резонансных единиц (RU) для немодифицированных мутеинов липокалина и приблизительно 1600 RU для пегилированного липокалина с последовательностью SEQ ID NO:13. Оставшиеся активированные группы насыщали этаноламином. Контрольные каналы обрабатывали EDC/NHS с последующей обработкой этаноламином (отрицательный контроль иммобилизации). Все реагенты и вещества приобретали у GE Healthcare.
Серийные разведения человеческого гепсидина-25 и гепсидина-25 яванского макака в буфере для проведения анализа (HBS-EP+, GE Healthcare, BR-1006-68) наносили на подготовленную поверхность. Для анализа связывания использовали следующие параметры: время контакта 60 с, время диссоциации 180-360 с, скорость потока 30 мкл/мин. Все измерения проводили на аппарате Biacore T100 (GE Healthcare) при 25°С. Восстановление поверхностей с иммобилизованными на них липокалинами проводили последующими инъекциями 2 М/4 М гуанидина-HCl (120-600 с) и 10 мМ глицина-HCl рН 1,5/2,0 (40-240 с) с последующей дополнительной отмывкой буфером для проведения анализа и периодом стабилизации 120 с.
Данные оценивали с использованием программного обеспечения Biacore T100 Evaluation (V 2.0.1). Применяли двойной контроль. Для анализа первичных данных использовали модель связывания 1:1 (Langmuir).
Парные анализы были воспроизводимы, и связывания в контрольном канале выявлено не было. Показатели связывания мутеинов липокалина, имеющих последовательности SEQ ID NO:1, SEQ ID NO:8, а также SEQ ID NO:13, связанную с PEG40, с человеческим гепсидином-25 и гепсидином-25 яванского макака представлены на Фиг.7.
Аффинность связывания гепсидина-25 яванского макака с иммобилизованными мутеинами липокалина была приблизительно в 2 раза выше, по сравнению с аффинностью связывания с человеческой мишенью. Кинетический анализ человеческого гепсидина-25 на иммобилизованном пегилированном варианте, имеющем последовательность SEQ ID NO:13, выявил высокую аффинность, составлявшую 40 пМ.
Пример 9: Клеточный анализ индуцированной гепсидином интернализации и деградации ферропортина
Клеточный анализ in vitro использовали для измерения нейтрализующей активности мутеинов липокалина по настоящему изобретению, направленных против человеческого гепсидина. Анализ основан на индуцированной гепсидином интернализации и деградации его рецептора, ферропортина, и его проводили в основном, как описано в Nemeth et al. 2004, 2006.
Кратко, получали стабильную клеточную линию НЕК-293, обеспечивавшую индуцируемую экспрессию мышиного ферропортина (FPN), слитого на С-конце с зеленым флуоресцентным белком (GFP). Индуцируемую экспрессию слитого белка FPN-GFP контролировали доксициклином с использованием имеющейся в продаже системы экспрессии T-REx, регулируемой тетрациклином (Invitrogen, Karlsruhe, Germany). Последовательность, кодирующую FPN-GFP, клонировали в вектор pcDNA 4/ТО, содержащий индуцируемый промотор и маркер резистентности к зеоцину. Полученную конструкцию использовали для стабильной трансфекции клеток Т-REx-293, экспрессирующих регуляторный белок, необходимый для индуцируемой доксициклином экспрессии.
Анализ индуцированной гепсидином интернализации рецептора гепсидина проводили следующим образом. Клетки стабильной линии T-REx-293::FPN-GFP высевали в культуральные флаконы Т75 с конфлюэнтностью 80%. Вечером экспрессию FPN-GFP индуцировали 4 нг/мл доксициклина и стабилизировали 10 мкМ цитратом аммония-железа (III) в течение 16 часов при 37°С. На следующее утро клетки трипсинизировали и высевали на 24-луночный планшет в количестве 0,3 миллиона клеток на лунку в объеме 450 мкл. Перед добавлением гепсидина клетки оставляли на 1 ч при 37°С для их прикрепления. Клетки инкубировали при 37°С в течение 24 часов и флуоресценцию GFP в суспензии свободных клеток анализировали проточной цитометрией.
В анализах нейтрализации использовали ЕС80 (40 нМ) опосредованной гепсидином деградации слитого белка FPN-GFP. Для этого антикалины инкубировали с гепсидином при комнатной температуре в течение 30 минут перед добавлением клеток. После периода инкубации продолжительностью 24 часа флуоресценцию измеряли, как описано выше.
Мутеины липокалина против гепсидина, имеющие последовательности SEQ ID NO:1, SEQ ID NO:2, SEQ ID NO:4, SEQ ID NO:6, SEQ’ID NO:7, SEQ ID NO:8, SEQ ID NO:10, SEQ ID NO:11, а также SEQ ID NO:14, связанную с PEG40, SEQ ID NO:13, связанную с PEG40, и SEQ ID NO:1, связанную cABD (SEQ ID NO:15), нейтрализовали биологическую активность человеческого гепсидина-25 со значениями IC50, показанными на Фиг.8.
Пример 10: Мутеины липокалина против гепсидина нейтрализуют человеческий гепсидин у мышей
Активность мутеинов липокалина против человеческого гепсидина оценивали in vivo у мышей, которым вводили человеческий гепсидин в количестве, достаточном для развития ответа в виде снижения уровня железа в крови, как описано Nemeth et al. (2006) Blood, 107:328-333.
За две недели до эксперимента мышей C57BL/6 переводили на железодефицитную диету для подавления эндогенного гепсидина. Перед экспериментом 3-хкратному молярному избытку мутеина липокалина позволяли связываться с синтетическим человеческим гепсидином-25 на протяжении 30 минут. Одновременно липокалин дикого типа (NGAL 98) предварительно инкубировали с гепсидином-25 в том же молярном соотношении в качестве изотипического контроля. Мышам проводили одну внутрибрюшинную (i.p.) инъекцию PBS (наполнителя), или 2 мг/кг гепсидина, или 2 мг/кг гепсидина, предварительно инкубированного либо с мутеином липокалина, либо с липокалином дикого типа (отрицательный контроль). Через два часа под изофлурановой анестезией получали образцы крови, и уровни общего железа сыворотки определяли колориметрическим анализом на клиническом анализаторе KoneLab XTi.
Результаты показаны на Фиг.9 в виде уровней общего железа сыворотки в форме концентраций в мкМ. Введение гепсидина приводило к значимому снижению уровней железа в сыворотке у мышей, переведенных на железодефицитную диету. Гепсидин, предварительно инкубированный с липокалином дикого типа, также приводил к снижению уровня железа в крови. Предварительное образование комплекса гепсидина с мутеином липокалина защищало животных от ответа в виде снижения уровня железа в крови.
Пример 11: Определение Фармакокинетического (РК) параметра мутеинов липокалина против гепсидина-25
Фармакокинетические (РК) параметры (период полувыведения, концентрацию в плазме) мутеина Lcn 2, имеющего последовательность SEQ ID NO: 14, связанную с PEG40, и SEQ ID NO:1, связанную с ABD (SEQ ID NO:15), определяли после однократного внутривенного болюсного введения у мышей NMRI и яванских макаков (Масаса fascicularis) в дозах, показанных на Фиг.10. Плазму получали из терминальных образцов крови, полученных в предопределенных временных точках, и концентрации мутеина липокалина определяли посредством ELISA. Константу скорости элиминации вычисляли линейной регрессией методом наименьших квадратов по терминальной части логарифмически трансформированной кривой «концентрация в плазме-время». Начало терминальной фазы элиминации каждого отдельного профиля определяли визуально, и оно представляло собой первую точку, в которой не было систематического отклонения от линейно-логарифмического снижения концентраций в сыворотке. Период полувыведения (Т1/2) вычисляли по следующей формуле:
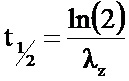
Т1/2 SEQ ID NO:14-PEG (мышь): 27,9 ч; T1/2 SEQ ID NO:1-ABD (мышь): 30 ч; T1/2 SEQ ID NO:14-PEG (яванский макак): 88 ч.
Специалисту в данной области техники будет ясно, что настоящее изобретение хорошо адаптировано для выполнения задач и получения результатов и преимуществ, упомянутых в настоящей заявке или являющихся ее неотъемлемым признаком. Кроме того, специалисту в данной области техники будет ясно, что возможно проведение различных изменений и модификаций изобретения, раскрытого здесь, без выхода за рамки объема и сущности изобретения. Композиции, способы, методики, способы лечения, молекулы и конкретные соединения, описанные здесь, являются необходимыми примерами предпочтительных воплощений и подразумевают, что они не ограничивают объем изобретения. Специалистам в данной области техники будут ясны изменения настоящего изобретения и другие применения, входящие в сущность изобретения и определенные объемом формулы изобретения. Упоминание или обсуждение опубликованного ранее документа в данном описании не следует обязательно толковать, как признание того, что документ представляет собой часть предшествующего уровня техники или является общеизвестным.
Следует отметить, что при использовании здесь формы единственного числа включают множественное число, если контекстом ясно не указано иное. Таким образом, например, ссылка на «антитело» включает одно или более таких различных антител, ссылка на «способ» включает ссылку на эквивалентные стадии и способы, известные специалистам в данной области техники, которые могут быть модифицированы или которыми могут быть заменены способы, описанные здесь.
Все публикации и патенты, упомянутые в данном описании, полностью включены в него посредством ссылки. В той степени, в которой материал, включенный посредством ссылки, противоречит или не соответствует данному описанию, описание будет заменять собой любой такой материал.
Если не указано иное, подразумевают, что термин «по меньшей мере», расположенный перед группой элементов, относится к каждому элементу группы. Специалистам в данной области техники будут ясны многие эквиваленты конкретных воплощений изобретения, описанных здесь, или специалисты в данной области техники смогут установить такие эквиваленты с применением не более чем рутинных экспериментов. Подразумевают, что настоящее изобретение включает такие эквиваленты.
Подразумевают, что при использовании здесь союзный термин «и/или» между несколькими перечисленными элементами включает как отдельные, так и комбинированные варианты. Например, там, где два элемента соединены союзным термином «и/или», первый вариант относится к возможности использования первого элемента без второго. Второй вариант относится к возможности использования второго элемента без первого. Третий вариант относится к возможности использования первого и второго элемента совместно. Подразумевают, что любой из этих вариантов включен в значение данного термина и, следовательно, соответствует требованию термина «и/или», как он использован здесь. Подразумевают, что возможность одновременного использования более чем одного варианта также включена в значение данного термина и, следовательно, соответствует требованию термина «и/или», как он использован здесь.
Изобретения, иллюстративно описанные здесь, могут быть подходящим образом применены на практике в отсутствии любого элемента или элементов, ограничения или ограничений, не раскрытых здесь конкретно. Таким образом, например, термины «включающий», «включая», «содержащий» и так далее следует толковать широко и без ограничения. В каждом случае в настоящей заявке любой из терминов «включающий», «состоящий по существу из» и «состоящий из» может быть заменен любым из двух других терминов.
Кроме того, термины и выражения, использованные здесь, были использованы как описательные и не ограничивающие термины, и использование таких терминов и выражений не направлено на исключение каких-либо эквивалентов показанных и описанных признаков или частей, при этом предполагают, что различные модификации возможны в рамках объема заявленного изобретения. Таким образом, следует понимать, что, несмотря на то, что настоящее изобретение было раскрыто конкретно посредством типичных воплощений и возможных признаков, специалист в данной области техники может прибегнуть к модификации и изменению описанных в них изобретений, раскрытых здесь, и подразумевают, что такие модификации и изменения входят в объем данного изобретения.
Изобретение было описано здесь широко и в общем. Каждая более конкретная разновидность и менее общая группа, входящая в общее описание, также является частью изобретения. Это включает общее описание изобретения, при условии или отрицательном ограничении, исключающем любой предмет из общего ряда, независимо от конкретного упоминания исключаемого предмета в настоящей заявке.
Другие воплощения входят в следующую формулу изобретения. Кроме того, там, где признаки и аспекты изобретения описаны с применением групп Маркуша, специалистам в данной области техники будет ясно, что посредством этого изобретение также описано с применением любого отдельного представителя или подгруппы представителей группы Маркуша.
| название | год | авторы | номер документа |
|---|---|---|---|
| НОВЫЕ БЕЛКИ, СПЕЦИФИЧЕСКИЕ В ОТНОШЕНИИ АНГИОГЕНЕЗА | 2016 |
|
RU2720688C2 |
| НОВЫЙ СЛИТЫЙ БЕЛОК, СПЕЦИФИЧНЫЙ ДЛЯ CD137 И PD-L1 | 2019 |
|
RU2818349C2 |
| НОВЫЕ СЛИТЫЕ БЕЛКИ, СПЕЦИФИЧЕСКИЕ В ОТНОШЕНИИ CD137 И GPC3 | 2020 |
|
RU2814653C2 |
| МУТЕИНЫ ЛИПОКАЛИНА 2 ЧЕЛОВЕКА С АФФИННОСТЬЮ В ОТНОШЕНИИ ГЛИПИКАНА-3 (GPC3) | 2016 |
|
RU2756318C2 |
| БЕЛКИ, СПЕЦИФИЧНЫЕ В ОТНОШЕНИИ CD137 | 2016 |
|
RU2736312C2 |
| НОВЫЕ ПОЛИПЕПТИДЫ СО СПЕЦИФИЧЕСКИМ СВЯЗЫВАНИЕМ И ПУТИ ИХ ПРИМЕНЕНИЯ | 2015 |
|
RU2723034C2 |
| СЛИТЫЙ ПОЛИПЕПТИД С ПРОТИВОРАКОВОЙ АКТИВНОСТЬЮ | 2016 |
|
RU2754466C2 |
| МУТЕИНЫ ЛИПОКАЛИНА 2 ЧЕЛОВЕКА (Lcn2, hNGAL) С АФФИННОСТЬЮ ДЛЯ ОПРЕДЕЛЕННОЙ МИШЕНИ | 2019 |
|
RU2804336C2 |
| МУТЕИНЫ ЛИПОКАЛИНА 2 ЧЕЛОВЕКА (Lcn2, hNGAL) С АФФИННОСТЬЮ ДЛЯ ОПРЕДЕЛЕННОЙ МИШЕНИ | 2010 |
|
RU2707126C2 |
| МУТЕИНЫ ЛИПОКАЛИНА СЛЕЗНОЙ ЖИДКОСТИ, ОБЛАДАЮЩИЕ АФФИННОСТЬЮ К С-МЕТ РЕЦЕПТОРНОЙ ТИРОЗИНКИНАЗЕ ЧЕЛОВЕКА, И СПОСОБЫ ИХ ПОЛУЧЕНИЯ | 2009 |
|
RU2515063C9 |












Изобретение относится к области биотехнологии, конкретно к мутеинам белка липокалина, а также к полученным на их основе специфично связывающимся терапевтическим или диагностическим белкам, направленным против гепсидина, и может быть использовано в медицине. Получают мутеин липокалина, по меньшей мере на 90% идентичный последовательности белка, выбранной из группы, состоящей из последовательностей SEQ ID NO: 1-14. Изобретение позволяет получить мутеин липокалина, способный связываться с гепсидином с аффинностью, определенной по константе диссоциации (KD), равной приблизительно 10 нМ или менее. 19 н. и 54 з.п. ф-лы, 12 ил., 11 пр.
1. Мутеин липокалина, способный связываться с гепсидином с аффинностью, определенной по константе диссоциации (KD), равной приблизительно 10 нМ или менее, содержащий в положениях, соответствующих линейной полипептидной последовательности зрелого hNGAL, представленного в SEQ ID NO: 35:
(i) один из перечисленных наборов аминокислотных замен:
(a) Asn 96 → Val; Tyr 100 → Gln;
(b) Asn 96 → Arg; Tyr 100 → Glu; Tyr 106 → Phe;
(c) Asn 96 → Asp; Tyr 100 → Ser; Tyr 106 → Gly;
(d) Asn 96 → Gly; Tyr 100 → Gly; Tyr 106 → Gly;
(e) Asn 96 → Lys; Tyr 100 → Ala; Tyr 106 → Ile;
(f) Asn 96 → Ser; Tyr 100 → Arg; Tyr 106 → Val;
(g) Asn 96 → Ser; Tyr 100 → Val; Tyr 106 → Arg; и
(h) Asn 96 → Thr; Tyr 100 → Val; Tyr 106 → Gly;
и
(ii) 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16 или 17 аминокислотных замен выбранных из группы, состоящей из:
Leu 36 → Ala, Cys, Thr, или Val; Ala 40 → Arg, Glu, Gly, или Ser; Ile 41 → Ile, Leu, Met, или Val; Gln 49 → Leu или Met; Tyr 52 → His, Leu, Phe, или Trp; Ser 68 → Arg, Gly, или Ile; Leu 70 → Asp, Asn, Gln, Met или Phe; Arg 72 → Glu, Gly, Leu, or Val; Lys 73 → Ala, Arg, Glu, Gly, Leu, Thr или Tyr; Asp 77 → Arg, Glu, Gly, Leu, Ser или Val; Trp 79 → Gly, Leu, Ser, Tyr, или Val; Arg 81 → Glu, Gly, Ser или Gln; Leu 103 → Ala, Arg, Gly или Trp; Ser 127 → Thr или Trp; Lys 125 → Arg, Leu, Met, Phe, Thr или Val; Tyr 132 → Leu или Val; и Lys 134 → Trp;
и
(iii) необязательно, одну или несколько замен, выбранных из:
Gln 28 → His; Lys 59 → Glu; Lys 62 → Arg; Phe 71 → Pro или Ser; Lys 74 → Glu; Lys 75 Glu; Ile 80 → Phe; Cys 87 → Ser; Ile 135 → Val; Ser 146 → Pro и Glu 147 → Gly, причем данный мутеин липокалина по меньшей мере на 90% идентичен последовательности белка, выбранного из группы, состоящей из последовательностей SEQ ID NO: 1-14.
2. Мутеин по п. 1, который имеет аффинность к гепсидину, определенную по KD, приблизительно 1 нМ или менее.
3. Мутеин по п. 2, который имеет аффинность к гепсидину, определенную по KD, приблизительно 0,1 нМ или менее.
4. Мутеин по п. 1, который способен нейтрализовать биологическую активность человеческого гепсидина-25.
5. Мутеин по п. 4, который имеет значение средней ингибирующей концентрации IC50, равное примерно 80 нМ или менее, как определено клеточным анализом индуцированной гепсидином интернализации и деградации ферропортина.
6. Мутеин по п. 5, который имеет значение IC50, равное приблизительно 50 нМ или менее.
7. Мутеин по п. 5, который имеет значение IC50, равное приблизительно 25 нМ или менее.
8. Мутеин по п. 1, включающий аминокислотную последовательность, на не менее чем 95% идентичную последовательности, выбранную из группы, состоящей из SEQ ID NO: 1-14.
9. Мутеин по п. 1, который связывает зрелый человеческий гепсидин.
10. Мутеин по п. 1, который по меньшей мере на 75% идентичен последовательности зрелого hNGAL.
11. Мутеин по п. 1, который содержит мутированный аминокислотный остаток в одном или нескольких положениях последовательности, соответствующих положениям 52, 68, 81, 127 и 134 в линейной полипептидной последовательности зрелого hNGAL.
12. Мутеин по п. 1, который содержит в положениях, соответствующих линейной полипептидной последовательности зрелого hNGAL, замену Lys 134 → Trp.
13. Мутеин по п. 1, который включает в линейной полипептидной последовательности зрелого hNGAL одну или несколько замен, Tyr 52 → His, Leu, Phe или Trp; Ser 68 → Arg, Gly, или Ile; Arg 81 → Glu, Gly, и Gln; и Ser 127 → Thr или Trp.
14. Мутеин по п. 1, в котором аминокислотная последовательность данного мутеина содержит в положениях, соответствующих линейной полипептидной последовательности зрелого hGAL, один из следующих наборов аминокислотных замен:
(a) Tyr 52 → His; Ser 68 → Arg; Arg 81 → Ser; и Ser 127 → Trp;
(b) Tyr 52 → Leu; Ser 68 → Arg; Arg 81 → Glu; и Ser 127 → Trp;
(c) Tyr 52 → Phe; Ser 68 → Gly; Arg 81 → Gly; и Ser 127 → Trp;
(d) Tyr 52 → Trp; Ser 68 → Ile; Arg 81 -→ Gln; и Ser 127 → Trp;
(e) Tyr 52 → Trp; Ser 68 → Arg; Arg 81 → Glu; и Ser 127 → Trp;
(f) Tyr 52 → Trp; Ser 68 → Arg; Arg 81 → Glu; и Ser 127 → Thr; и
(g) Tyr 52 → Trp; Ser 68 → Arg; Arg 81 → Glu; и Ser 127→ Trp.
15. Мутеин по п. 1, дополнительно содержащий по меньшей мере 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15 или 16 мутированных аминокислотных остатков в любом из положений последовательности, соответствующих положениям 36, 40, 41, 49, 52, 68, 70, 72, 73, 77, 79, 103, 125 и 132 в линейной полипептидной последовательности зрелого hNGAL.
16. Мутеин по п. 15, который содержит в положениях, соответствующих линейной полипептидной последовательности зрелого hNGAL дикого типа, одну или более аминокислотных замен, выбранных из группы, состоящей из: Leu 36 → Ala, Cys, Thr, или Val; Ala 40 → Arg, Glu, Gly или Ser; Ile 41 → Ile, Leu, Met или Val; Gln 49 → Leu или Met; Leu 70 → Asp, Asn, Gln, Met, или Phe; Arg 72 → Glu, Gly, Leu или Val; Lys 73 → Ala, Arg, Glu, Gly, Leu, Thr или Tyr; Asp 77 → Arg, Glu, Gly, Leu, Ser, или Val; Trp 79 → Gly, Leu, Ser, Tyr, или Val; Leu 103 → Ala, Arg, Gly или Trp; Lys 125 → Arg, Leu, Met, Phe, Thr, или Val; и Tyr 132 → Leu или Val.
17. Мутеин по п. 1, в котором аминокислотная последовательность данного мутеина содержит один из следующих наборов комбинаций аминокислот:
(a) Ala 36, Ser 40; Leu 41; Met 49; Asn 70; Gly 72; Gly 73; Ser 77; Leu 79; Leu 125; Val 132;
(b) Leu 36, Arg 40; Val 41; Gln 49; Asp 70; Arg 72; Thr 73; Leu 77; Ser 79; Thr 125; Val 132;
(c) Leu 36, Glu 40; Ile 41; Leu 49; Gln 70; Gly 72; Glu 73; Gly 77; Gly 79; Phe 125; Val 132;
(d) Leu 36, Glu 40; Ile 41; Met 49; Met 70; Leu 72; Ala 73; Glu 77; Leu 79; Val 125; Val 132;
(e) Leu 36, Glu 40; Val 41; Met 49; Met 70; Leu 72; Ala 73; Glu 77; Leu 79; Thr 125; Val 132;
(f) Leu 36, Glu 40; Val 41; Met 49; Met 70; Leu 72; Ala 73; Glu 77; Leu 79; Val 125; Val 132;
(g) Thr 36, Ser 40; Ile 41; Gln 49; Phe 70; Glu 72; Gly 73; Arg 77; Val 79; Val 125; Leu 132;
(h) Val 36, Glu 40; Met 41; Leu 49; Met 70; Glu 72; Tyr 73; Val 77; Leu 79; Arg 125; Val 132; и
(i) Val 36, Gly 40; Leu 41; Leu 49; Leu 70; Val 72; Arg 73; Arg 77; Tyr 79; Met 125; Val 132.
18. Мутеин по п. 17, содержащий в положениях, соответствующих линейной полипептидной последовательности зрелого hNGAL из следующих мутаций: Leu36 → Thr, Leu36 → Val, Leu36 → Ala; Ala40 → Ser, Ala40 → Gly, Ala40 → Glu, Ala40 → Arg; Ile41 → Val, Ile41 → Leu, Ile41 → Met; Gln49 → Leu, Gln49 → Met; Tyr52 → His, Tyr52 → Leu, Tyr52 → Phe, Tyr52 → Trp; Ser68 → Arg, Ser68 → Gly, Ser68 → Ile; Leu70 → Phe, Leu70 → Asn, Leu 70 → Gln, Leu70 → Met, Leu70 → Asp; Arg72 → Glu, Arg72 → Val, Arg72 → Gly, Arg72 → Leu 72; Lys73 → Gly, Lys73 → Arg, Lys73 → Ala, Lys73 → Thr, Lys73 → Tyr, Lys73 → Glu; Asp77 → Arg, Asp77 → Val, Asp77 → Ser, Asp77 → Glu, Asp77 → Gly, Asp77 → Leu; Trp79 → Val, Trp79 → Tyr, Trp79 → Ser, Trp79 → Gly, Trp79 → Leu 79; Arg81 → Glu, Arg81 → Ser, Arg81 → Gly, Arg81 → Gln; Asn96 → Lys, Asn96 → Arg, Asn96 → Thr, Asn96 → Ser, Asn96 → Asp 96, Asn 96 → Val; Tyr100 → Ala, Tyr 100 → Glu, Tyr100 → Val, Tyr100 → Arg, Tyr100 → Ser 100, Tyr100 → Gln; Leu103 → Gly, Leu103 → Arg, Leu103 → Ala, Leu103 → Gly, Leu103 → Arg, Leu103 → Trp; Tyr106 → Ile, Tyr106 → Phe, Tyr 106 → Val, Tyr106 → Gly, Tyr106 → Arg; Lys125 → Val, Lys125 → Met, Lys 125 → Arg, Lys 125 → Leu, Lys 125 → Thr; Lys 125 → Phe; Ser127 → Trp, Ser127 → Thr; Tyr132 → Leu, Tyr132 → Val; Lys134 → Trp.
19. Мутеин по п. 17, содержащий в положениях, соответствующих линейной полипептидной последовательности зрелого hNGAL из следующих мутаций: Ala40 → Glu; Ile41 → Val; Gln49 → Met; Tyr52 → Trp, Ser68 → Ile; Leu70 → Met; Arg72 → Leu 72, Lys73 → Ala; Asp77 → Glu, Trp79 → Leu 79; Arg81 → Gln; Asn96 → Asp 96, Asn 96 → Gly; Tyr100 → Ser 100, Tyr100 → Gly; Leu103 → Arg; Tyr106 → Gly; Lys125 → Thr, Lys 125 → Val; Ser 127 → Trp; Tyr 132 → Val; Lys134 → Trp.
20. Мутеин по п. 18, в положениях, соответствующих линейной полипептидной последовательности зрелого hNGAL, включающий в себя один из следующих наборов комбинаций аминокислот:
(a) Leu 36; Glu 40; Val41; Met 49; Trp 52; Ile 68; Met 70; Leu 72; Ala 73; Glu 77; Leu 79; Gln 81; Asp 96; Ser 100; Arg 103; Gly 106; Thr 125; Trp 127; Val 132; Trp 134;
(b) Leu 36, Glu 40, Val41, Met 49, Trp 52, Ile 68, Met 70; Leu 72, Ala 73, Glu 77, Leu 79, Gln 81, Gly 96, Gly 100, Arg 103, Gly 106, Val 125, Trp 127, Val 132, Trp 134;
(c) Leu 36, Glu 40, Val41, Met 49, Trp 52, Ile 68, Met 70, Leu 72, Ala 73, Glu 77, Leu 79; Gln 81, Asp 96, Ser 100, Arg 103, Gly 106, Val 125, Trp 127, Val 132, Trp 134;
(d) Leu 36, Glu 40, Ile41, Met 49, Trp 52, Ile 68, Met 70, Leu 72, Ala 73, Glu 77; Leu 79; Gln 81, Asp 96, Ser 100, Arg 103, Gly 106, Val 125, Trp 127, Val 132, Trp 134;
(e) Leu 36, Glu 40, Ile41, Met 49, Trp 52, Ile 68, Met 70, Leu 72, Ala 73, Glu 77, Leu 79, Gln 81, Asp 96, Ser 100, Arg 103, Gly 106, Val 125, Trp 127, Val 132, Trp 134 и
(f) Leu 36, Glu 40, Val41, Met 49, Trp 52, Ile 68, Met 70, Leu 72, Ala 73, Glu 77, Leu 79, Gln 81, Asp 96, Ser 100, Arg 103, Gly 106, Val 125, Trp 127, Val 132, Trp 134.
21. Мутеин по п. 19, дополнительно содержащий по меньшей мере 1, 2, 3 или 4 из следующих мутаций: Lys59 → Glu; Lys62 → Arg; Phe71 → Pro; Phe 71 → Ser; Lys74 →Glu; Lys75 → Glu; Ile80 → Phe; Ile135 → Val; Ser146 → Pro, и Glu147 → Gly в положениях, соответствующих линейной полипептидной последовательности зрелого hNGAL.
22. Мутеин по п. 1, который дополнительно содержит в положениях, соответствующих аминокислотной полипептидной последовательности зрелого hNGAL дикого типа, одну или более аминокислотных замен, выбранных из группы, состоящей из 28 → His; Lys 62 → Arg; Phe 71 → Pro or Ser; Lys 74 → Glu; Lys 75 → Glu; Cys 87 → Ser; lie 135 Val; Ser 146 → Pro и Glu 147 → Gly.
23. Мутеин по п. 1, который обладает аффинностью к зрелому человеческому гепсидину с KD 10 нМ или менее.
24. Мутеин по п. 23, который обладает аффинностью к зрелому человеческому гепсидину, определенной по KD 1 нМ или менее.
25. Мутеин по п. 1, который имеет аминокислотную последовательность, представленную в SEQ ID NO: 8.
26. Мутеин по п. 1, который имеет аминокислотную последовательность, представленную в SEQ ID NO: 9.
27. Мутеин по п. 1, который имеет аминокислотную последовательность, представленную в SEQ ID NO: 10.
28. Мутеин по п. 1, который имеет аминокислотную последовательность, представленную в SEQ ID NO: 11.
29. Мутеин по п. 1, который имеет аминокислотную последовательность, представленную в SEQ ID NO: 12.
30. Мутеин по п. 1, который имеет аминокислотную последовательность, представленную в SEQ ID NO: 13.
31. Конъюгированный мутеин липокалина, способный связываться с гепсидином с аффинностью, определенной по константе диссоциации (KD), равной приблизительно 10 нМ или менее, содержащий мутеин по любому из пп. 1-30, который конъюгирован с соединением, выбранным из группы, состоящей из ферментной метки, радиоактивной метки, цветной метки, флуоресцентной метки, хромогенной метки, люминесцентной метки, гаптена, дигоксигенина, биотина, цитостатического средства, токсинов, комплексов металлов, металла и коллоидного золота.
32. Конъюгированный мутеин липокаина, способный связываться с гепсидином с аффинностью, определенной по константе диссоциации (KD), равной приблизительно 10 нМ или менее, содержащий мутеин по любому из пп. 1-30, который конъюгирован с соединением, которое продлевает время полужизни мутеина в сыворотке.
33. Конъюгированный мутеин по п. 32, в котором соединение, которое увеличивает время полужизни в сыворотке, выбрано из группы, состоящей из молекулы полиалкиленгликоля, гидроэтилкрахмала, части Fc иммуноглобулина, домена СН3 иммуноглобулина, домена СН4 иммуноглобулина, альбумин-связывающего пептида и альбумин-связывающего белка.
34. Конъюгированный мутеин по п. 33, в котором полиалкиленгликоль представляет собой полиэтиленгликоль или его активированное производное.
35. Конъюгированный мутеин по п. 31, который имеет аминокислотную последовательность, представленную в SEQ ID NO: 8.
36. Конъюгированный мутеин по п. 31, который имеет аминокислотную последовательность, представленную в SEQ ID NO: 9.
37. Конъюгированный мутеин по п. 31, который имеет аминокислотную последовательность, представленную в SEQ ID NO: 10.
38. Конъюгированный мутеин по п. 31, который имеет аминокислотную последовательность, представленную в SEQ ID NO: 11.
39. Конъюгированный мутеин по п. 31, который имеет аминокислотную последовательность, представленную в SEQ ID NO: 12.
40. Конъюгированный мутеин по п. 31, который имеет аминокислотную последовательность, представленную в SEQ ID NO: 13.
41. Гибридный белок для повышения времени полужизни мутеина липокаина в сыворотке, содержащий мутеин по любому из пп. 1-30, который на своем N-конце и/или С-конце функционально слит с партнером слияния, который является белком, доменом белка или пептидом.
42. Гибридный белок по п. 41, в котором партнер слияния мутеина является доменом белка.
43. Гибридный белок по п. 42, в котором белковый домен представляет собой Fc-часть иммуноглобулина, домен СН3 иммуноглобулина, домен СН4 иммуноглобулина, альбумин-связывающий пептид или альбумин-связывающий белок.
44. Применение мутеина по любому из пп. 1-30 в качестве антагониста гепсидина.
45. Применение мутеина по п. 44, где антагонист представляет собой антагонист зрелого человеческого гепсидина ферропортином.
46. Применение мутеина по любому из пп. 1-30 в качестве терапевтического средства для лечения анемии: анемии вследствие хронического заболевания, анемии воспаления, хронической воспалительной анемии, анемии вследствие воспалительного заболевания, железодефицитной анемии, анемии на почве перегрузки организма железом, сепсиса, наследственного гемохроматоза, анемии, связанной с хроническим заболеванием почек, анемии рака, анемии, индуцированной химиотерапией, анемии, связанной с устойчивостью к агентам, стимулирующим эритропоэз.
47. Применение мутеина по п. 46 в лечении анемии вследствие хронического заболевания.
48. Применение мутеина по п. 47, где данное хроническое заболевание характеризуется повышенным уровнем гепцидина.
49. Применение мутеина по п. 46 в лечении анемии вследствие хронического заболевания почек.
50. Применение мутеина по п. 49, в котором данное хроническое заболевание почек является почечным расстройством в конечной стадии.
51. Применение мутеина по п. 46 в лечении анемии вследствие рака.
52. Применение мутеина по п. 46 в лечении анемии вследствие воспалительного заболевания.
53. Применение мутеина по п. 52, где воспалительным заболеванием является ревматоидный артрит.
54. Применение мутеина по п. 46 в лечении железодефицитной анемии.
55. Применение мутеина по любому из пп. 1-30 при мониторинге лечения эритропоэтином (ЭПО) или в прогнозировании ответа на ЭПО.
56. Молекула нуклеиновой кислоты, имеющая нуклеотидную последовательность, кодирующую мутеин по п. 1.
57. Молекула нуклеиновой кислоты, имеющая нуклеотидную последовательность, кодирующую гибридный белок по п. 41.
58. Вектор экспрессии, содержащий молекулу нуклеиновой кислоты по п. 56.
59. Вектор экспрессии, содержащий молекулу нуклеиновой кислоты по п. 57.
60. Клетка-хозяин для производства мутеина по п. 1, содержащая вектор экспрессии по п. 58.
61. Клетка-хозяин для производства гибридного белка по п. 41, содержащая вектор экспрессии по п. 59.
62. Способ получения мутеина по п. 1-30, включающий культивирование клетки-хозяина по п. 60 в условиях, благоприятных для экспрессии мутеина.
63. Способ по п. 62, в котором мутеин получают в бактериальном или эукариотическом организме-хозяине и выделяют из этого организма-хозяина или его культуры.
64. Способ получения гибридного белка по пп. 41-43, включающий культивирование клетки-хозяина по п. 61, в условиях, благоприятных для экспрессии гибридного белка.
65. Фармацевтическая композиция для лечения анемии: анемии вследствие хронического заболевания, анемии воспаления, хронической воспалительной анемии, анемии вследствие воспалительного заболевания, железодефицитной анемии, анемии на почве перегрузки организма железом, анемии, связанной с хроническим заболеванием почек, анемии рака, анемии, индуцированной химиотерапией, или анемии, связанной с устойчивостью к агентам, стимулирующим эритропоэз, содержащая в эффективном количестве мутеин по любому из пп. 1-30, или конъюгированный мутеин по любому из пп. 31-40, или гибридный белок по любому из пп. 41-43 и фармацевтически приемлемый наполнитель, причем данная фармацевтическая композиция необязательно содержит стимулятор эритропоэза.
66. Фармацевтическая композиция по п. 65, дополнительно содержащая стимулятор эритропоэза.
67. Фармацевтическая композиция по п. 66, в которой стимулятором эритропоэза является один из: эритропоэтин, вариант эритропоэтина, антитело, которое связывает эритропоэтин, и мутеин липокалина, связывающий эритропоэтин.
68. Применение мутеина по любому из пп. 1-30 для связывания/обнаружения гепсидина, включающее:
(a) приведение данного мутеина в контакт с исследуемым образцом, в котором предполагается наличие гепсидина, способствуя тем самым образованию комплекса между мутеином и гепсидином, и
(b) обнаружение указанного комплекса между мутеином и гепсидином с помощью соответствующего сигнала.
69. Применение мутеина по п. 68, где гепсидин является зрелым человеческим гепсидином.
70. Аналитический набор для обнаружения гепсидина, содержащий в качестве одного из компонентов набора мутеин по любому из пп. 1-30.
71. Способ обнаружения присутствия зрелого человеческого гепсидина в биологическом образце, включающий приведение образца в контакт с мутеином по любому из пп. 1-30 в условиях, обеспечивающих образование комплекса мутеина и гепсидина, где образец включает жидкость организма.
72. Способ по п. 71, дополнительно содержащий определение комплекса мутеина и гепсидина.
73. Способ по п. 71 или 72, в котором биологический образец изолируют от человека.
| WO 03029463 A2, 10.04.2003 | |||
| Приспособление для графления и штрихования | 1928 |
|
SU11816A1 |
| MALYSZKO J | |||
| ET AL., Neutrophil Gelatinase-Associated Lipocalin and Hepcidin: What Do They Have in Common and Is There a Potential Interaction?, KIDNEY AND BLOOD PRESSURE RESEARCH, 2010, v.33, n.2, p.157 - 165 | |||
| TOKURIKI N | |||
| ET AL., Stability effects of mutations and protein evolvability, Curr | |||
| Opin | |||
| Struct | |||
| Biol., 2009, v.19, n.5, p.596-604. | |||

