ОБЛАСТЬ ТЕХНИКИ, К КОТОРОЙ ОТНОСИТСЯ ИЗОБРЕТЕНИЕ
Изобретение относится к биотехнологии и медицине представляет собой способ культивирования гемопоэтических клеток на поддерживающих стромальных слоях, которые делаются магнитными путем инкубации с магнитными наночастицами.
УРОВЕНЬ ТЕХНИКИ
Гемопоэтические клетки, включая стволовые кроветворные клетки (СКК) и предшественники представляют большой интерес для медицины, поскольку используются в трансплантациях при лечении наследственных и онкологических заболеваний, а также перспективны для терапии ряда других патологий. Большой интерес представляют перспективы использования гемопоэтических клеток в генной терапии наследственных и приобретенных заболеваний кроветворных и иммунных систем.
При этом СКК являются чрезвычайно редкими клетками, и их низкое содержание в пуповинной крови делает препараты пуповинной крови зачастую недостаточными для трасплантации взрослым пациентам. Учитывая также, что СКК обычно являются покоящимися клетками и крайне редко делятся, что затрудняет введение в них генных конструкций, во всей остроте встает вопрос о размножении этих клеток в культуре.
До настоящего времени наиболее часто использовались подходы, связанные с попытками размножения СКК и предшественников в классической культуре с использованием различных сочетаний цитокинов. В целом, эти попытки можно считать малоудачными, поскольку они не сопровождались существенным размножением СКК в культуре.
Наиболее важным альтернативным подходом является культивирование гемопоэтических клеток на подслоях стромальных клеток. Многочисленные данные показывают, что в организме СКК находятся в костном мозге в так называемых нишах, то есть окружении, состоящем из клеток и внеклеточного матрикса. Наиболее важными типами микроокружения считается эндостальная остеобластная ниша и эндотелиальная ниша. Клетки ниши находятся в тесном контакте с СКК и предоставляют последним не только секретируемые цитокины, но также и мембрано-связанные лиганды рецепторов, задействованных в важнейших сигнальных путях, контролирующих самообновление, пролиферацию и дифференцировку СКК, а также белки клеточной адгезии, и белки внеклеточного матрикса, которые также влияют на состояние СКК. Таким образом, культивирование СКК, гемопоэтических предшественников и других гемопоэтических клеток в присутствии и в контакте со стромальными клетками, которые частично воспроизводят условия ниши in vivo, более адекватно воспроизводят условия, в которых находятся гемопоэтические клетки in vivo, и могут давать возможность достичь истинного размножения в культуре.
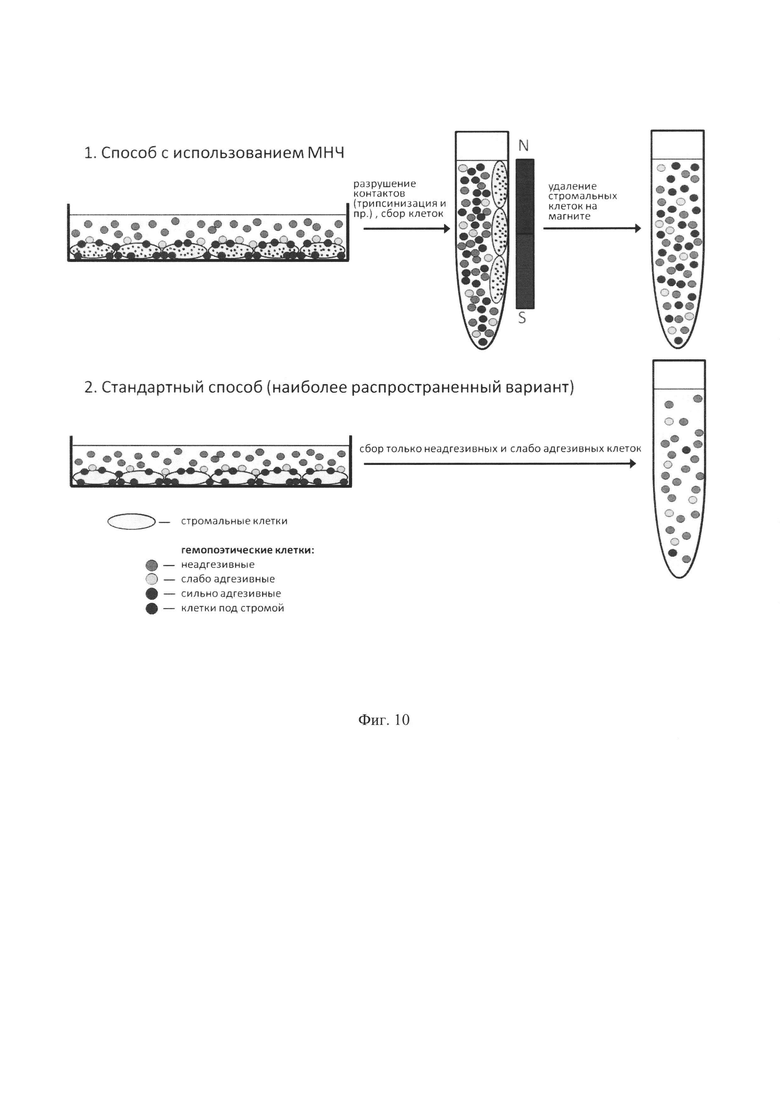
Весьма важным является исследование различных видов стромы для выяснения возможностей размножения СКК. Хотя само по себе культивирование на стромальных подслоях не представляет сложностей и является давно отработанной процедурой, сбор гемопоэтических клеток после культивирования их на строме является технической проблемой. Как правило, после культивирования гемопоэтических клеток на строме, производится сбор неадгезивных клеток, причем с минимальными воздействиями на стромальный слой. Более интенсивные воздействия, такие, как энергичное пипетирование или обработка трипсином, приводят к сильному загрязнению стромальными клетками полученных препаратов гемопоэтических клеток, что, без очистки от стромальных клеток, сильно сужает возможности анализа и дальнейшего использования полученных препаратов. Предполагается, что при таком способе сбора потери гемопоэтических клеток относительно невелики, значительных избирательных потерь каких-либо видов клеток не происходит, и собранная фракция является достаточно репрезентативной. При этом даже в случае мягких обработок происходит определенное разрушение целостности стромального слоя, и часть клеток попадает в собранную фракцию гемопоэтических клеток и загрязняет их.
Альтернативным способом сбора гемопоэтических клеток после культивирования является обработка совместных культур трипсином для разрушения взаимодействий клеток с поверхностями культивирования и между собой. Для осуществления анализа полученных препаратов и дальнейшего использования производят разделение гемопоэтических клеток и стромальных клеток. Один из вариантов решения этой проблемы состоит в использовании антител, специфических к поверхностным маркерам, чтобы разделить стромальные и гемопоэтические клетки с помощью флюоресцентно-активируемой и или магнитно-активируемой клеточных сортировок. Для того, чтобы проводить негативное удаление стромальных клеток, вначале нужно проводить исследование отличий поверхностного антигенного профиля стромальных клеток от гемопоэтических, чтобы выбрать антитела, специфически реагирующие со стромальными клетками, но не с гемопоэтическими клетками.
Описаны также подходы, основанные на позитивном выделении гемопоэтических клеток с помощью антител к маркерам Sca-1 (Butler JM, Nolan DJ, Vertes EL, Varnum-Finney B, Kobayashi H, Hooper AT, Seandel M, Shido K, White IA, Kobayashi M, Witte L, May C, Shawber C, Kimura Y, Kitajewski J, Rosenwaks Z, Bernstein ID, Rafii S. Endothelial cells are essential for the self-renewal and repopulation of Notch-dependent hematopoietic stem cells. Cell Stem Cell. 2010 6(3):251-264) или CD45 (Nakamura Y, Arai F, Iwasaki H, Hosokawa K, Kobayashi I, Gomei Y, Matsumoto Y, Yoshihara H, Suda T. Isolation and characterization of endosteal niche cell populations that regulate hematopoietic stem cells. Blood. 2010 116(9):1422-1432). При этом Sca-1 является маркером ранних гемопоэтических клеток, включая и СКК, и поэтому при использовании выделяются не все гемопоэтические клетки, а только минорная субпопуляция ранних клеток. Кроме того, Sca-1 экспрессируется некоторыми стромальными клетками, например ОР9 клетками (Ichii М, Frank MB, Iozzo RV, Kincade PW. The canonical Wnt pathway shapes niches supportive of hematopoietic stem/progenitor cells. Blood. 2012 119(7): 1683-1692), что делает невозможным отделение такой стромы от гемопоэтических клеток.
Хотя CD45 считается почти универсальным маркером гемопоэтических клеток, его экспрессия не характерна для ранних стадий гемопоэтической дифференцировки, из, например, эмбриональных стволовых клеток или гемангиобластов, и может снижаться при культивировании гемопоэтических клеток на стромальных слоях (Vodyanik MA, Thomson JA, Slukvin II. Leukosialin (CD43) defines hematopoietic progenitors in human embryonic stem cell differentiation cultures. Blood. 2006 108(6): 2095-2105; Hara T, Nakano Y, Tanaka M, Tamura K, Sekiguchi T, Minehata K, Copeland NG, Jenkins NA, Okabe M, Kogo H, Mukouyama Y, Miyajima A. Identification of podocalyxin-like protein 1 as a novel cell surface marker for hemangioblasts in the murine aorta-gonad-mesonephros region. Immunity. 1999 11 (5): 567-578).
Таким образом, при использовании маркера CD45 выделение репрезентативной популяции гемопоэтических клеток также не гарантировано.
Еще одним способом, который может приводить к желаемому результату, является стабильное мечение стромальных клеток путем введения в них интегрирующих конструкций, экспрессирующих маркерный белок (например, зеленый флюоресцентный белок), который можно использовать для разделения гемопоэтических и стромальных клеток с помощью флюоресцентно-активируемой и или магнитно-активируемой клеточных сортировок. Однако это предполагает проведение значительной предварительной работы по получению генетически модифицированных линий стромальных клеток, экспрессирующий такой белок. Кроме того, данный метод малопригоден для работы с первичными клетками, такими, как мезенхимальные стромальные/стволовые клетки (МСК), которые способны проходить только ограниченное число делений в культуре. При этом высокая стоимость антител ограничивает возможность массового применения такого рода подходов. Производительность флюоресцентно-активируемой сортировки также мешает рутинному использованию подобных подходов.
Таким образом, в настоящее время не существует подхода, который позволял бы без значительных потерь получать после совместного культивирования со стромальными клетками очищенные препараты гемопоэтических клеток, адекватно представляющих набор гемопоэтических субпопуляций, образующихся в результате культивирования на строме, в том числе и ранних клеток, а также клеток, плотно ассоциированных со стромой, или находящихся под стромальным слоем, и при этом не требовал бы предварительных этапов сравнения поверхностных маркеров гемопоэтических и стромальных клеток или получения генетически модифицированных линий стромальных клеток.
Данная техническая проблема решена в заявке тем, что препараты стромальных клеток, в дополнение к стандартной обработке для необратимой остановки их пролиферации с помощью известных в технике подходов, например, митомицина С или гамма-облучения, также инкубируют с магнитными наночастицами (МНЧ), в результате чего стромальные клетки поглощают МНЧ и становятся магнитными. В качестве МНЧ используют наночастицы Fe3O4, как поверхностно модифицированные, так и без дополнительной модификации поверхности. Для выделения чистой фракции магнитных стромальных клеток проводят их выделение путем осаждения на стенки сосуда с помощью магнита, и полученную фракцию клеток используют для культивирования с гемопоэтическими клетками. Для снижения нагрузки магнитной стромы наночастицами и повышения времени ее жизни возможно также проведение предварительного титрования связывания стромальными клетками магнитных частиц, при этом для очистки магнитной стромы используют препараты магнитных стромальных клеток, в которых доля магнитных клеток не превышает 90%. После окончания совместного культивирования выделяют очищенную фракцию гемопоэтических клеток путем обработки культур трипсином или, в других вариантах исполнения, коллагеназой, диспазой или другими протеолитическими ферментами, пригодными для использования в этих целях, или же без применения протеаз, с использованием хелаторов двухвалентных ионов металлов и интенсивным пипетированием и перемешиванием для разрушения контактов между стромальными и гемопоэтическими клетками, после чего удаляют магнитные стромальные клетки путем их осаждения на стенки сосуда с помощью магнита.
Наиболее близким по техническому уровню к настоящему изобретению является работа A. Ito et al. (Ito A, Jitsunobu H, Kawabe Y, Ijima H, Kamihira M. Magnetic separation of cells in coculture systems using magnetite cationic liposomes. Tissue Eng Part С Methods. 2009 15(3): 413-423), в которой применено магнитное мечение клеток с помощью магнитных липосом, которые потом используются для сокультивирования с немечеными клетками, с целью последующего получения очищенных препаратов сокультивируемых клеток.
В отличие от работы Ito et al., в настоящем изобретении применены МНЧ, которые имеют значительное преимущество перед магнитными липосомами с точки зрения хранения. Кроме того, использованные МНЧ, как далее показано в примерах 2 и 3, обладают весьма высокой скоростью связывания с клетками, обеспечивая оптимальную эффективность мечения уже за 15-30 мин инкубации (по сравнению с несколькими часами в работе Ito et al.).
Наиболее важным отличием настоящего изобретения от работы Ito et al. является то, что возможность использования магнитно меченых клеток в качестве стромальных слоев при культивировании гемопоэтических клеток в работе Ito et al. не была показана и даже не предлагалась. Тем более, в работе Ito et al. не было показано, что обработка магнитными нанолипосомами не вызывает снижения поддерживающей активности стромальных клеток. В то же время, очевидно, что стромальные линии, используемые для совместного культивирования с гемопоэтическими клетками, должны обладать поддерживающей кроветворные клетки активностью, в противном случае их использование лишено практического смысла. Поэтому для применения магнитных стромальных слоев принципиально важно, чтобы после магнетизации стромальных клеток их поддерживающая активность оставалась бы на прежнем уровне или, если падала, то незначительно. Без информации о том, что это действительно так, возможность успешного применения магнитных стромальных слоев для совместного культивирования с гемопоэтическими клетками не является очевидной для специалиста. В примере 7 настоящей заявки приводится доказательство того, что после магнетизации стромальных клеток с помощью МНЧ их поддерживающая активность не падает, а даже, возможно, увеличивается.
В примерах 5 и 6 настоящей заявки приводятся данные о том, что применение магнитной стромы для культивирования гемопоэтических клеток с последующим сбором клеток с помощью трипсина приводит к значительному увеличению выхода тотальных гемопоэтических клеток, и еще более значительному увеличению выхода ранних гемопоэтических клеток, включая колониеобразующие клетки и клетки с фенотипом предшественников и стволовых кроветворных клеток Lin-, kit+, Scal+, по сравнению с наиболее распространенным способом с использованием немагнитной стромы и сбора неадгезивных клеток. В примере 8 настоящей заявки показано, что при культивировании гемопоэтической CD34+ популяции клеток из пуповинной крови человека на магнитной строме увеличивается как общее число собранных клеток, так и, в гораздо большей степени, число ранних клеток с фенотипом CD34+CD38-, по сравнению с контрольной немагнитной стромой.
В работе Ito et al. таких сведений не содержится. Таким образом, хотя в работе Ito et al. применен способ магнитного мечения для сокультивирования клеток в принципе, в этой работе не содержится очевидных для специалиста указаний, что данный способ может быть успешно применен для размножения гемопоэтических клеток, и что данный способ, при обеспечении существенной чистоты получаемых фракций гемопоэтических клеток, может давать значительное увеличение выхода как тотальных ядерных клеток, так и ранних гемопоэтических клеток, по сравнению с использованием немагнитной стромы и сбора неадгезивных клеток. По сравнению с другими способами отделения стромальных от гемопоэтических клеток, использование магнитной стромы позволяет получать репрезентативные препараты гемопоэтических клеток, а не только специфические субпопуляции их, и не требует проведения дополнительных работ по генетической модификации стромальных клеток или идентификацию поверхностных маркеров, дискриминирующих стромальные и гемопоэтические клетки. Магнитное мечение не вызывает снижения поддерживающей активности исследованных видов стромальных клеток, однако при необходимости уровень мечения стромальных клеток может быть понижен для дальнейшего снижения эффекта МНЧ на функционирование стромы, при этом 100% меченая строма может быть получена путем предварительной очистки частично меченых МНЧ клеток на магните. Набор для проведения культивирования гемопоэтических клеток на магнитной строме включает в себя препарат магнитных наночастиц, эффективно магнетизирующих стромальные клетки, а также магнит для удаления магнитной стромы после культивирования. Некоторые варианты набора могут также включать в себя стромальные клетки, митомицин С и среды для инкубации клеток с магнитными наночастицами.
КРАТКОЕ ОПИСАНИЕ ФИГУР
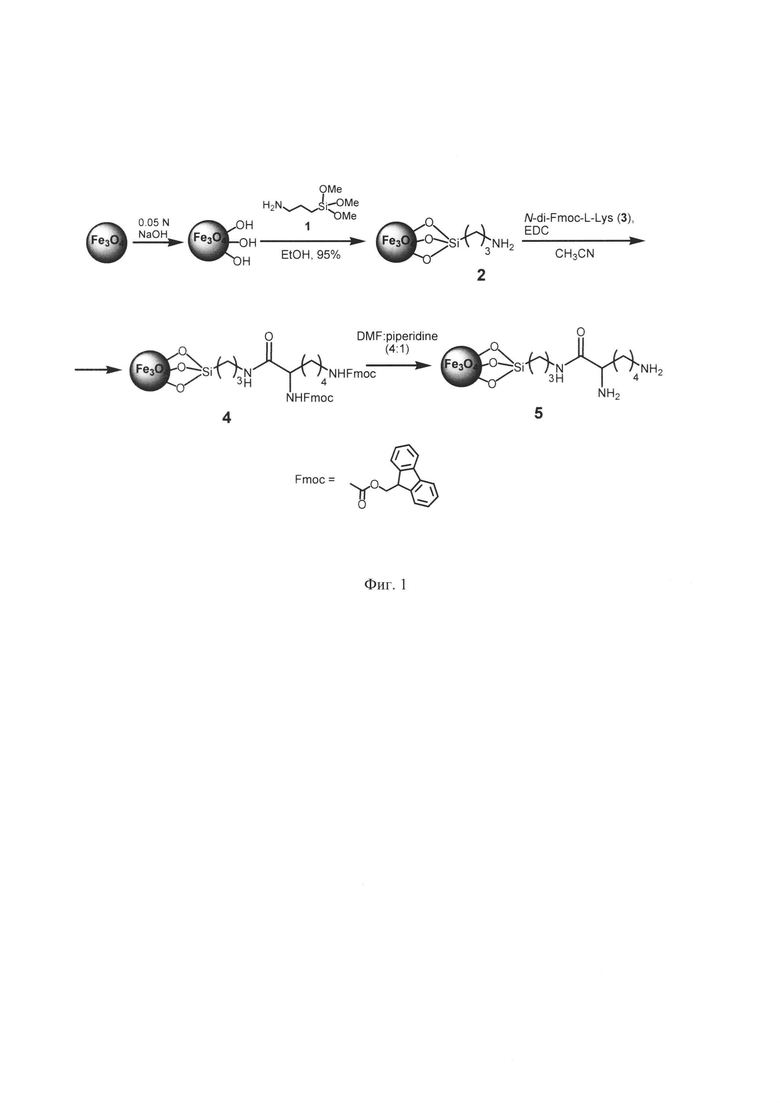
Фиг. 1. Схема, описывающая получение МНЧ.
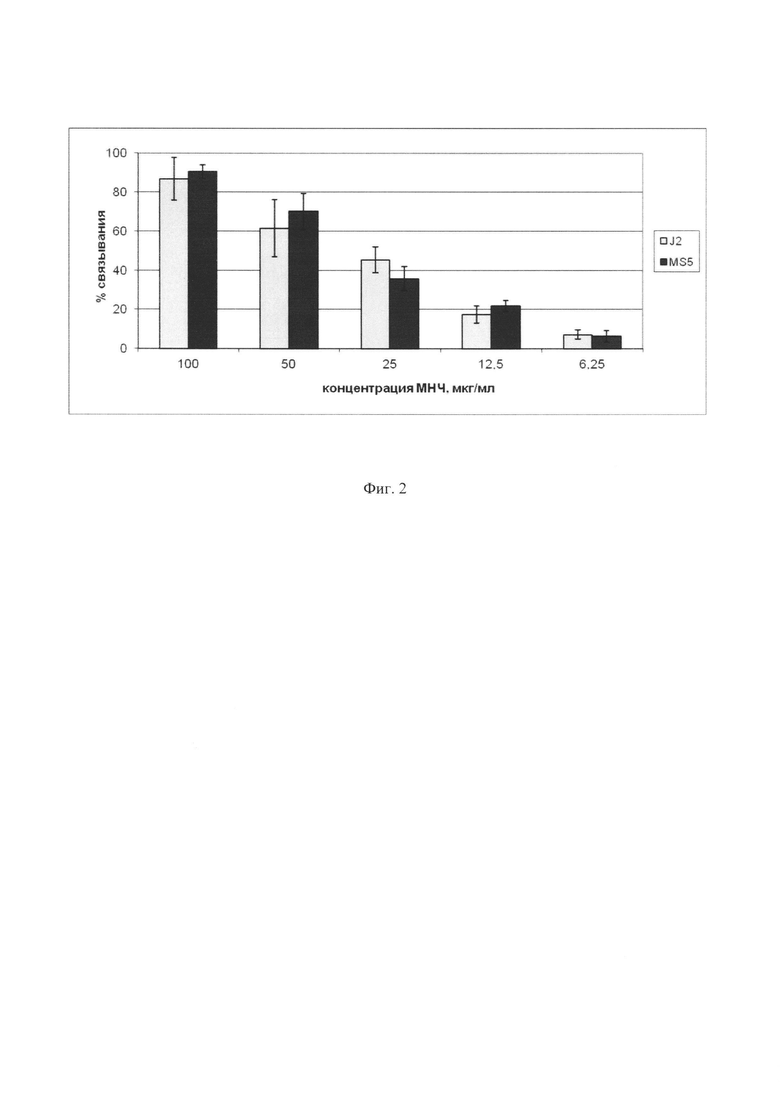
Фиг. 2. Зависимость эффективности связывания МНЧ с двумя типами стромальных клеток, MS5 и J2/3T3, от концентрации МНЧ. Инкубация клеток с МНЧ проводилась в течение 10 мин. По оси абсцисс показан процент клеток, выделяемых с помощью магнита.
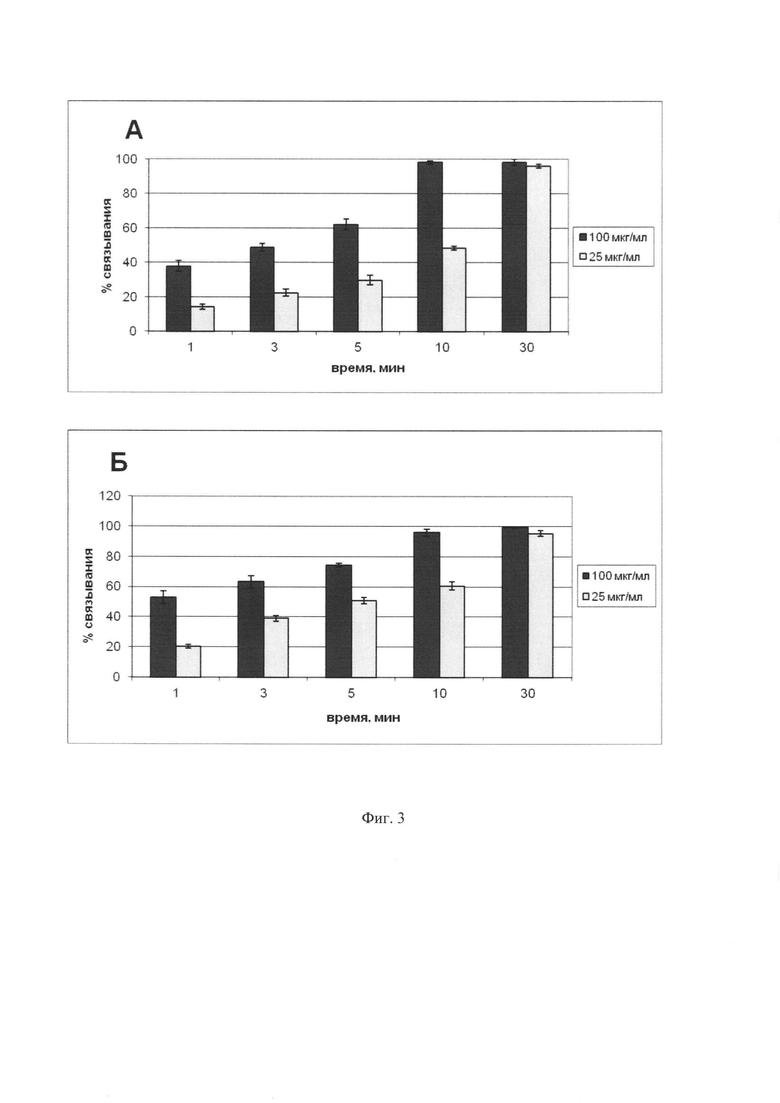
Фиг. 3. Зависимость эффективности связывания МНЧ со стромальными клетками от времени при двух концентрациях МНЧ. А - связывание с ОР9 клетками, Б - связывание с J2/3T3 клетками. МНЧ использовались в концентрациях 25 и 100 мкг/мл. По оси абсцисс показан процент клеток, выделяемых с помощью магнита.
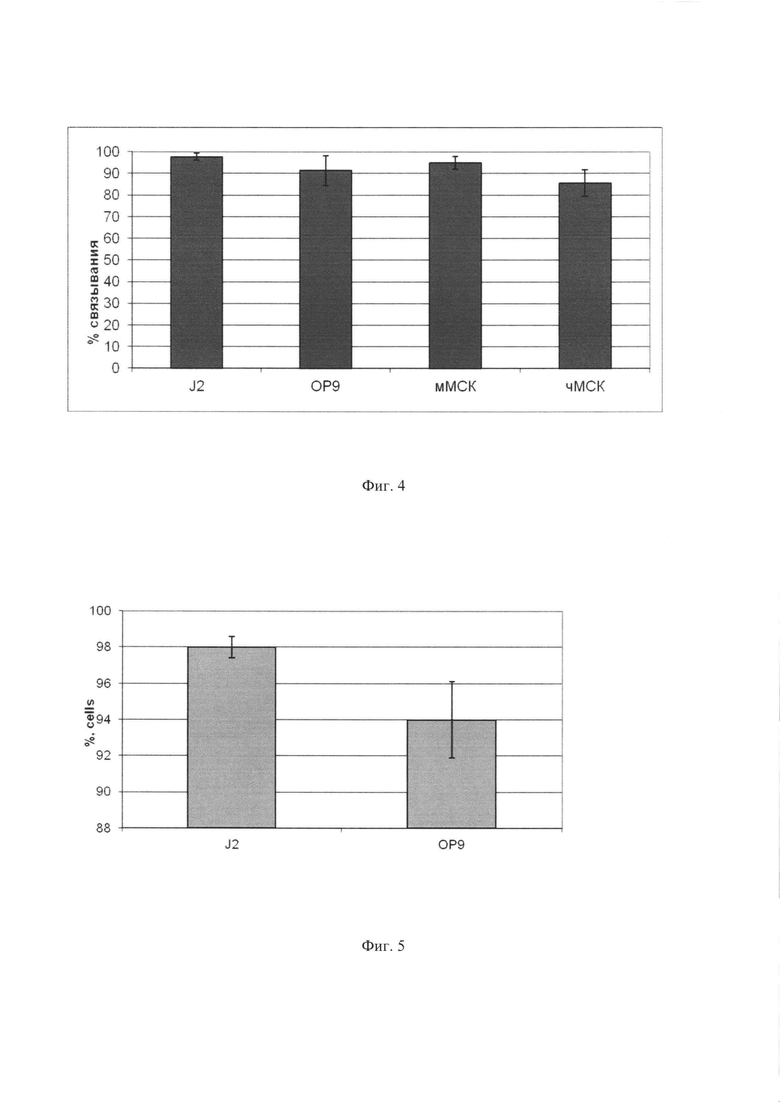
Фиг. 4. Различные типы стромальных клеток могут быть эффективно нагружены МНЧ. 106 стромальных клеток разных типов были инкубированы с МНЧ (25  g/ml) в течение 15 мин при комнатной температуре. По оси абсцисс показан процент клеток, выделяемых с помощью магнита.
g/ml) в течение 15 мин при комнатной температуре. По оси абсцисс показан процент клеток, выделяемых с помощью магнита.
Фиг. 5. Сохранение МНЧ клетками стромы после их нагрузки МНЧ и культивирования в течение периода, характерного для совместного культивирования гемопоэтических и стромальных клеток (7 дней). Стромальные клетки J2/3T3 и ОР9 были инкубированы с МНЧ (25 g/ml) в течение 15 мин, связавшие МНЧ клетки были выделены на магните, обработаны митомицином С для остановки их пролиферации и культивированы в течение 7 дней, после чего было проведено повторное выделение клеток на магните. Показан процент клеток, сохранивших магнитную метку.
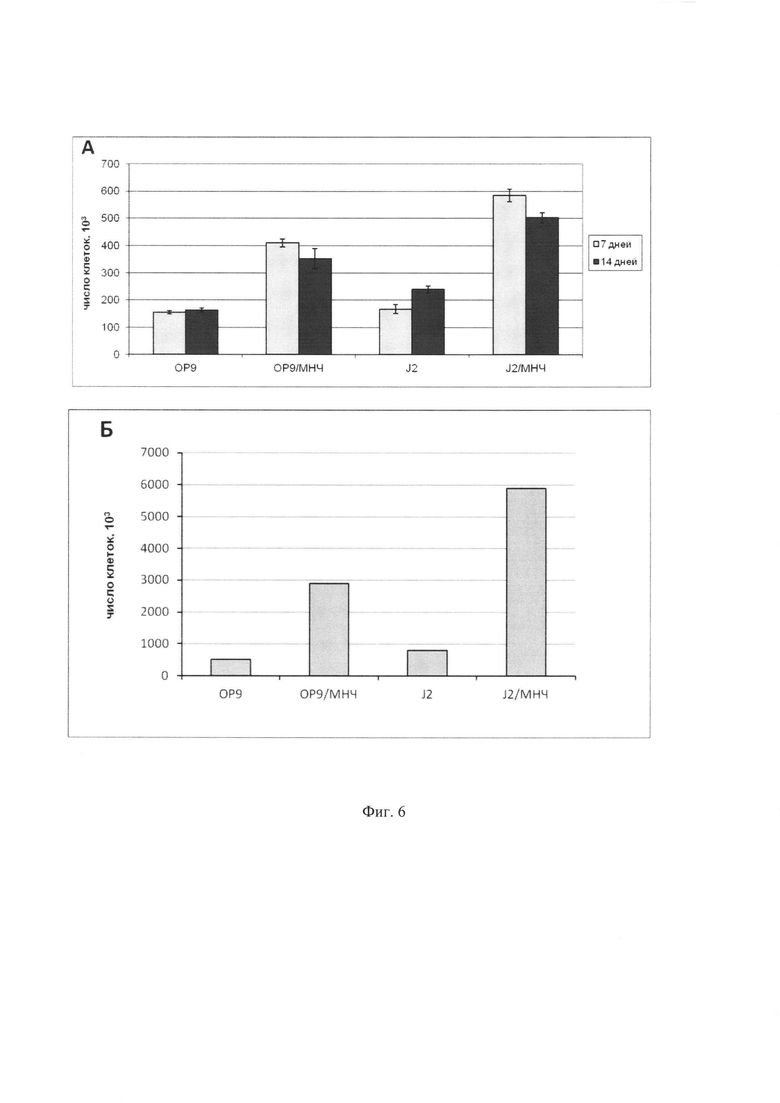
Фиг. 6. Продукция ядерных клеток после культивирования Lin- клеток на стромальных слоях. 15×103 Lin- клеток из костного мозга мыши были культивированы без добавления цитокинов в течение 7 дней на стромальных слоях ОР9 и J2/3T3. С контрольных слоев ОР9 и J2/3T3 собирали неадгезивную фракцию, в то время как с магнитных слоев ОР9+МНЧ и J2/3T3+МНЧ клетки собирали с помощью обработки трипсином и удаления стромальных клеток на магните. 50×103 клеток после первого раунда культивирования засевали на соответствующие стромальные слои, после чего культивировали еще 7 дней и собирали так же, как и в первом раунде. (А) Число собранных клеток после одного (7 дней) и двух (14 дней) раундов культивирования на строме. (Б) Кумулятивная продукция клеток после двух раундов культивирования на стромальных слоях.
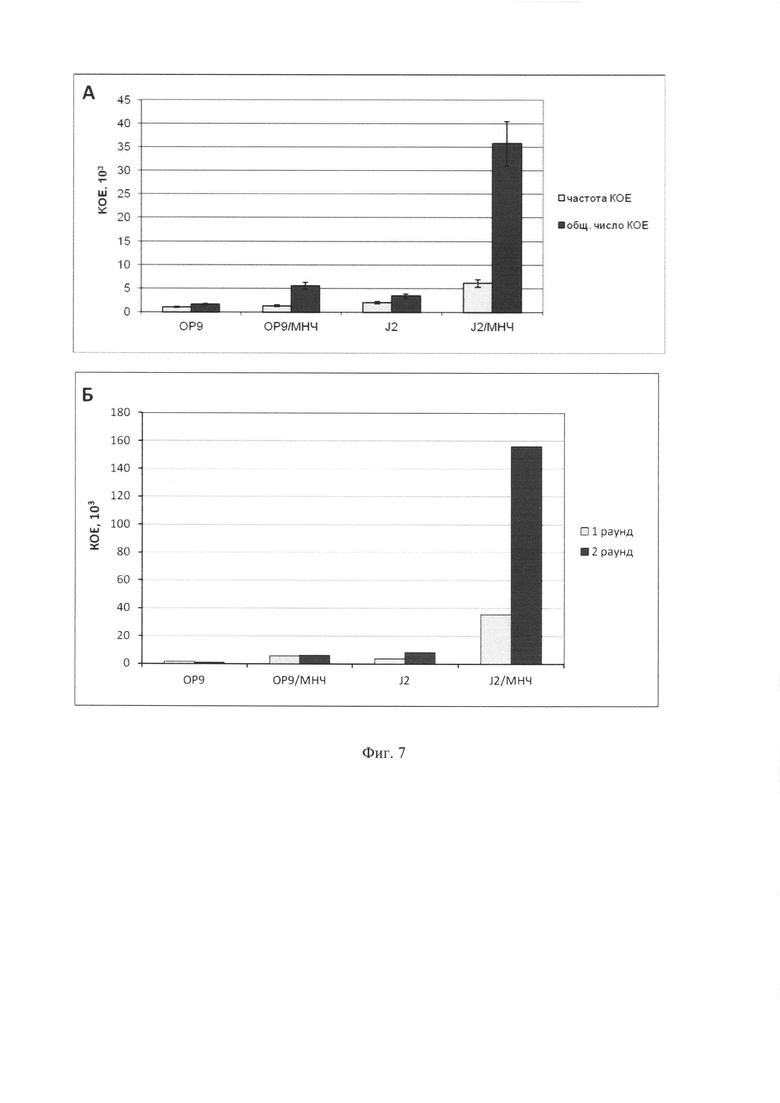
Фиг. 7. Продукция колониеобразующих клеток (КОЕ, колониеобразующих единиц) после одного (7 дней) и двух (14 дней) раундов культивирования Lin- клеток на контрольных (ОР9 и J2/3T3) и магнитных (ОР9/МНЧ и J2/3T3/ МНЧ) стромальных слоях. (А)
Анализ частоты КОЕ на 105 клеток (светло-серые столбики) и общего числа КОЕ (темно-серые столбики) после 7 дней (1 раунд) культивирования на стромальных слоях. (Б) Анализ кумулятивной продукции КОЕ после одного и двух раундов культивирования на стромальных слоях. Анализ проводился на гемопоэтических фракциях, описанных на Фиг. 6. Анализ КОЕ был проведен с помощью стандартной методики подсчета колониеобразующих клеток в метил целлюлозе. При культивировании на магнитной строме и сборе с помощью трипсинизации число КОЕ резко повышается по сравнению с немагнитной стромой, в особенности в раунде 2.
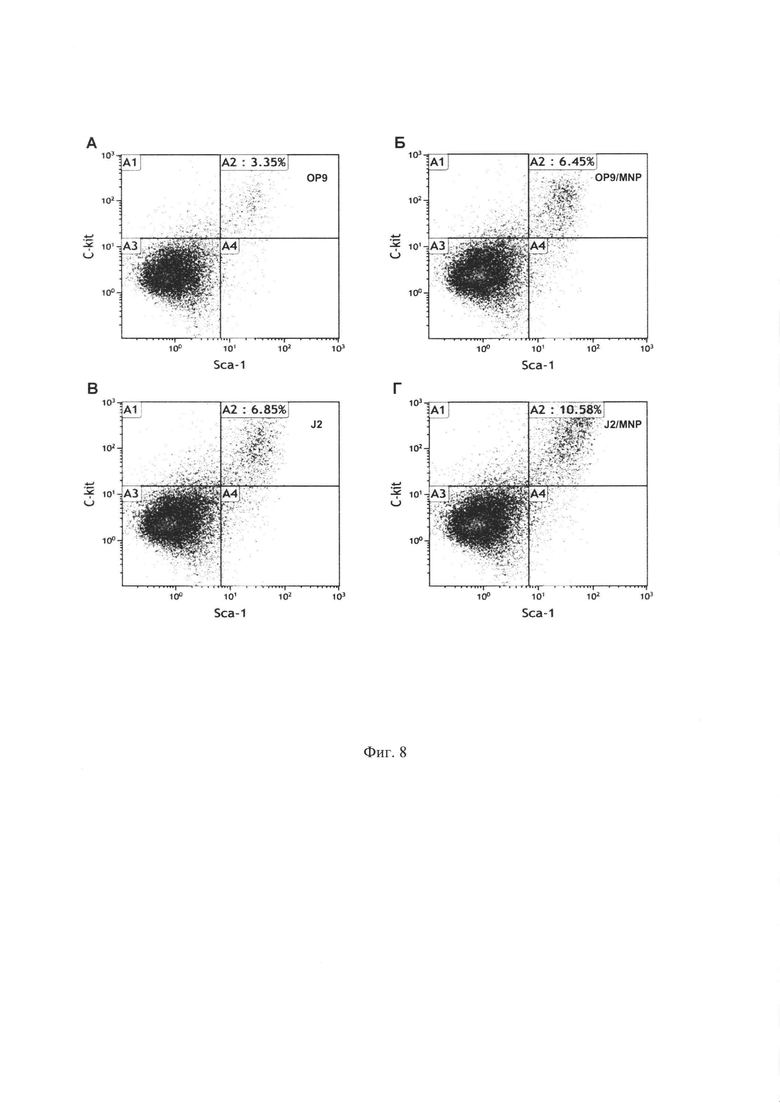
Фиг. 8. Анализ поверхностного фенотипа с помощью проточной цитофлюорометрии гемопоэтических клеток, собранных после культивирования Lin- клеток в течение 7 дней на контрольных ОР9 и J2/3T3 (А и В, соответственно) и магнитных ОР9+МНЧ и J2/3T3+ МНЧ (Б и Г, соответственно) стромальных слоях. Окрашивание проводилось с помощью PE-Lin, FITC-Sca-1 и Alexa647-c-kit антител. Клетки, изображенные на диаграммах, были гейтированы по Lin- фенотипу. Анализ проводился на гемопоэтических фракциях, описанных на Фиг. 6. При культивировании на магнитной строме и сборе с помощью трипсинизации число клеток с ранним фенотипом Lin-, kit+, Scal+ (верхние правые квадранты А2) значительно повышается по сравнению с немагнитной стромой.
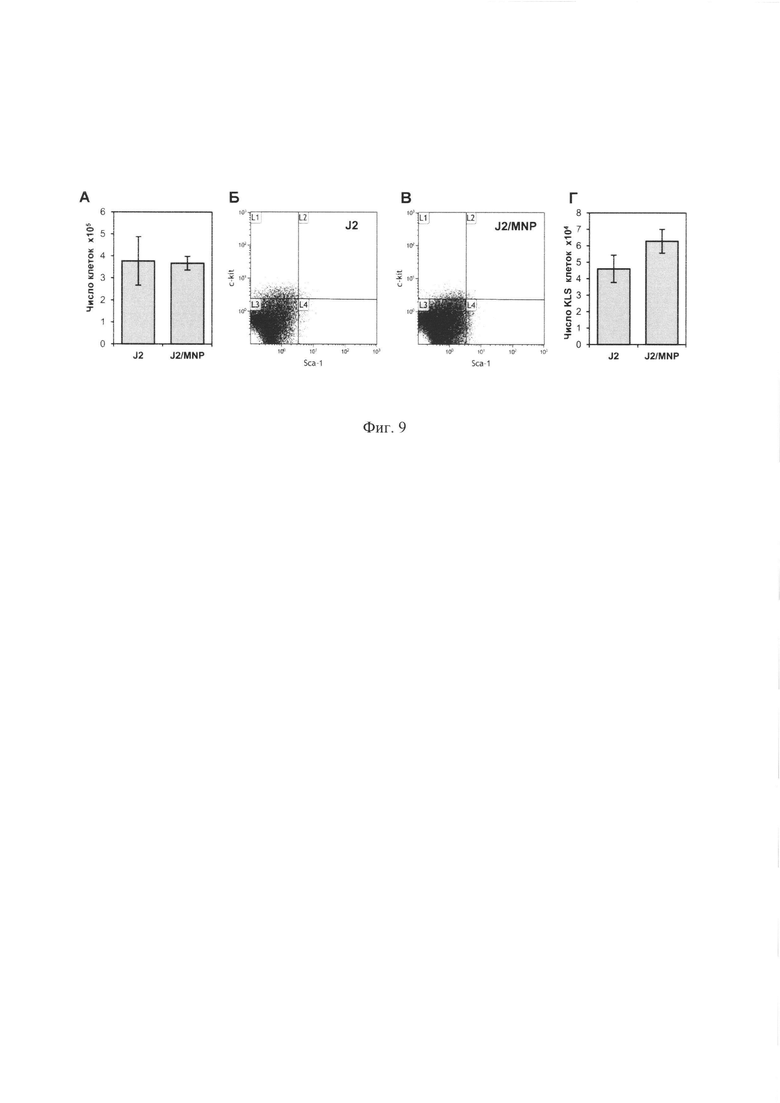
Фиг.9. Поддерживающая активность стромальных клеток не снижается после их магнетизации. (А) Общая продукция гемопоэтических клеток магнитной и обычной стромами. 30×103 Lin- клеток сокультивировали с магнитной и обычной J2/3T3 стромами, как описано в примере 5 (Фиг. 6, 7) и после 7 дней культивирования оба типа культур были трипсинизованы. Гемопоэтические клетки подсчитывали в камере Горяева, используя их морфологические отличия от стромальных клеток. (Б, В) Снятые со стромы клетки также анализировали с помощью проточной цитофлюорометрии с использованием PE-Lin, FITC-Sca-1 и Alexa647-c-kit антител. Показаны репрезентативные диаграммы для культур на контрольных (Б) и магнитных (В) J2/3T3 стромах. Клетки на диаграммах были гейтированы по Lin- фенотипу. Квадрант L3 соответствует неокрашенным клеткам, квадрант L2 - ранним клеткам фенотипа LSK. (Г) Общее число LSK клеток, собранных с обоих типов стромы.
Фиг. 10. Сравнение схемы заявляемого и стандартного наиболее распространенного способов сбора гемопоэтических клеток после культивирования на стромальных слоях. Заявляемый способ дает возможность собирать сильно адгезивные и подстромальные клетки, в отличие от стандартного подхода. Поскольку ранние гемопоэтические клетки (колониеобразующие клетки, фенотипически ранние KSL клетки и гемопоэтические стволовые клетки) обладают высокой адгезией к стромальным слоям и активной миграцией в подстромальные области, заявляемый способ дает возможность существенно повышать выход ранних клеток, представляющих наибольший интерес для трансплантации, регенеративно медицины и других клинических применений.
Таблица 1. Общее число снятых со стромы ядерных клеток. Показано число снятых с контрольной (немагнитной) и магнитной стром. В случае с контрольной стромой проводилось снятие неадгезивных клеток, в случае с магнитной стромой - полная диссоциация культуры путем трипсинизации с последующим удалением магнитных клеток на магните.
Таблица 2. Фенотип снятых со стромы клеток. Показано число снятых с контрольной (немагнитной) и магнитной стром. CD34+ и CD34+CD38- клетки представляют собой фракции, обогащенные гемопоэтическими предшественниками.
ОСУЩЕСТВЛЕНИЕ ИЗОБРЕТЕНИЯ
Пример 1. Получение магнитных наночастиц (МНЧ) с поверхностными модификациями
В качестве основы для получения модифицированных МНЧ использовались магнитные частицы на основе Fe3O4 (20-40 нм), полученные методом газофазного синтеза. ИК-спектры регистрировались на инфракрасном Фурье-спектрометре Nicolet 6700 FT-IR (Thermo Scientific). Регистрация спектров проводилась в диапазоне 400-4000 см-1. Спектрофотометрические исследования проводились на УФ спектрофотометре UV2401PC, Shimadzu в диапазоне 190-450 нм. Морфологические исследования частиц проводились при использовании просвечивающего электронного микроскопа Philips СМ30.
Получение 3-аминопропилсилан-модифицированных МНЧ
Схема получения модифицированных МНЧ показана на Фиг. 1. 3-Аминопропилсилан-модифицированные МНЧ 2 (МНЧ-APS) получали по аналогии с ранее разработанной методикой (Фиг. 1). Для этого 0,2 г исходных МНЧ Fe3O4 предварительно диспергировали в 50 мл 0.05 N водного раствора NaOH при использовании ультразвуковой бани (УЗ-бани) в течение 10 мин, и нагревали на водяной бане до 60°C в течение 5 ч, полученную суспензию оставляли на ночь. Через сутки МНЧ отделяли с использованием внешнего магнитного поля, промывали водой до нейтрального pH, после чего упаривали досуха, полученный порошок МНЧ использовали для дальнейшей модификации.
Для проведения поверхностной модификации 0.2 г полученных наночастиц диспергировали при обработке ультразвуком (УЗ-баня, 20 мин.) в 100 мл 95% этанола. К полученной суспензии добавляли 117 мкл 3-аминопропилтриэтоксисилана (APTES)1 (2.5 ммоль в расчете на 1 г МНЧ) и перемешивали в течение 16 ч. Модифицированные МНЧ осаждали при использовании внешнего магнитного поля. Растворитель декантировали, МНЧ промывали этанолом (двумя порциями по 30 мл) и ацетоном (тремя порциями по 35 мл). После декантации МНЧ 2 сушили в вакууме, получали 0,19 г МНЧ-APS.
Получение N-di-Fmoc-L-лизин-модифицированных МНЧ
0.100 г MH4-APS 2 диспергировали в 50 мл CH3CN с использованием УЗ-бани в течение 10 мин. Затем при перемешивании добавляли 0.046 г Nα,Nε-di-Fmoc-L-лизина 3 и 0.016 г 1-этил-3-(3-диметиламинопропил)карбодиимида (EDC) в качестве конденсирующего агента. Перемешивали 20 ч, после чего модифицированные МНЧ осаждали при использовании внешнего магнитного поля. Растворитель декантировали, МНЧ промывали ацетоном (пятью порциями по 15 мл). После промывки МНЧ 4 сушили под вакуумом, получали 0,085 г коричневого порошка.
Методом УФ-спектроскопии определяли количество не вступившего в реакцию N-di-Fmoc-L-лизина 1 и, исходя из полученных данных, рассчитывали количество иммобилизированного на МНЧ производного аминокислоты. Для этого был построен калибровочный график содержания N-di-Fmoc-L-лизина 1 в растворе CH3CN. Количество N-di-Fmoc-L-лизина 1 в растворе определяли спектрофотометрически по максимуму поглощения (λ 265 нм). Растворитель и промывные растворы после проведения реакции собирали и упаривали. Остаток вновь растворяли в точном объеме CH3CN и проводили фотометрическое определение количества не вступившей в реакцию N-Fmoc-аминокислоты 1 в анализируемом растворе. Количество аминокислотных остатков на поверхности модифицированных МНЧ 4 находили по разнице загруженного в реакцию и непрореагировавшего N-di-Fmoc-L-лизина. В результате, было рассчитано, что полученные модифицированные МНЧ 4 содержали 0.29 ммоль N-di-Fmoc-L-лизина в расчете на 1 г МНЧ.
Получение L-лизин-модифицированных МНЧ
0.064 г N-di-Fmoc-L-лизин-модифицированных МНЧ 4 диспергировали в 3 мл смеси диметилформамид : пиперидин (4:1) с использованием УЗ-бани в течение 10 мин, и через 20 мин МНЧ осаждали при использовании внешнего магнитного поля. Растворитель декантировали, МНЧ промывали водой (тремя порциями по 15 мл) до нейтральной pH и ацетоном (двумя порциями по 15 мл), сушили досуха. Получали 0.064 г коричневого порошка МНЧ 5.
Количество удаленных Fmoc-групп оценивали методом УФ-спектроскопии по аналогии с методикой определения количества Fmoc-групп в пептидах и аминокислотах, иммобилизированных на полимерные носители и используемых в твердофазном синтезе. В результате, было рассчитано, что полученные МНЧ 5 содержали 0.23 ммоль L-лизина в расчете на 1 г МНЧ. Средний размер частиц составлял 20 нм.
Пример 2. Кинетика связывания МНЧ с двумя типами стромальных клеток, ОР9 и J2/3T3.
Для выяснения способности МНЧ связываться со стромальными клетками были использованы стромальные линии ОР9 и J2/3T3. Проводили связывание этих клеточных линий с МНЧ, описанными в примере 1. Вначале проверяли зависимость эффективности связывания от концентрации МНЧ. Стромальные клетки обрабатывали митомицином С для остановки их пролиферации, и после инкубации с разными концентрациями МНЧ в течение 10 мин связавшие МНЧ клетки выделяли на магните и подсчитывали в камере Горяева. Полученные данные, которые приведены на Фиг. 2, свидетельствуют о том, концентрация МНЧ 100 g/ml достаточна, чтобы около 90% клеток стали магнитными и могли быть выделены на магните.
Для выяснения времени, необходимого для эффективного связывания клеток с МНЧ стромальные клетки обрабатывали митомицином С и инкубировали с двумя концентрациями МНЧ, 25 и 100 g/ml в течение различного времени, после чего магнетизированные клетки выделяли на магните. Полученные данные, которые приведены на Фиг. 3, свидетельствуют о том, 10 и 30 мин достаточны, чтобы при концентрациях МНЧ 100 g/ml и 25 g/ml, соответственно, около 90% клеток приобрели магнитные свойства и могли быть выделены на магните.
Пример 3. Различные типы стромальных клеток эффективно накапливают МНЧ и приобретают магнитные свойства.
Для выяснения способности МНЧ связываться с разными типами стромальных клеток, применяемых для совместного культивирования с гемопоэтическими клетками, МНЧ в концентрации (25 g/ml) были инкубированы в течение 15 мин с 106 стромальных клеток следующих типов: MS5, ОР9, J2/3T3, МСК (мезенхимальные стромальные клетки) мыши и МСК человека, после чего магнетизированные клетки выделяли на магните. Полученные данные, которые приведены на Фиг. 4, свидетельствуют о том, что все типы клеток обладали высокой, не ниже 85%, эффективностью связывания МНЧ и выделения на магните.
Пример 4. МНЧ хорошо сохраняются клетками стромы в течение времени культивирования.
Стромальные клетки J2/3T3 и ОР9 были инкубированы в течение 15 мин с МНЧ (25 g/ml). Клетки, связавшие МНЧ, были выделены на магните, обработаны митомицином С для остановки их деления и культивированы в стандартных условиях (среда DMEM с высоким содержанием глюкозы, 10% фетальной сывороткой крупного рогатого скота, 2 мМ глутамином, пенициллином и стрептомицином) при 37°C, 5% CO2 в течение 7 дней. После завершения культивирования среду отобрали, клетки обработали трипсином-ЭДТА, выделили на магните и подсчитали. Полученные результаты, представленные на Фиг. 5, показывают, что магнитная метка хорошо сохраняется стромальными клетками, причем от 94% до 98% клеток сохраняют МНЧ и могут, таким образом, быть удалены из смеси с гемопоэтическими клетками. Таким образом, МНЧ пригодны для получения очищенных препаратов гемопоэтических клеток после культивирования.
Пример 5. При культивировании на магнитной строме и сборе с помощью трипсинизации общее число ядерных клеток и число колониеобразующих клеток повышается по сравнению с культивированием на немагнитной строме и сбором неадгезивных клеток.
Для сравнения эффективности сбора гемопоэтических клеток после культивирования на магнитной и немагнитной стромах из костного мозга мышей была выделена фракция гемопоэтических клеток, обогащенная ранними клетками-предшественниками и гемопоэтическими стволовыми клетками (ГСК) (так называемая Lin- фракция). Выделение это фракции провели с помощью магнитной сортировки, основанной на удалении гемопоэтических клеток, несущих поверхностные маркеры дифференцированных клеток, согласно протоколу фирмы Miltenyi Biotec (Германия). Для этого к клеткам костного мозга мышей добавляли конъюгированные с биотином антитела к набору антигенов Lin, инкубировали с покрытыми стрептавидином магнитными шариками и разделяли на колонках MS MACS (Miltenyi Biotec), а прошедшую клеточную фракцию использовали для дальнейшей работы.
Далее было проведено культивирование клеток Lin- фракции костного мозга мышей в течение 1 недели на магнитном и немагнитном стромальных слоях клеток J2/3T3 и ОР9.
Для этого 15×103 клеток Lin- фракции культивировали на стромальных слоях клеток линий J2/3T3 и ОР9, которые были нагружены МНЧ, после чего магнитные клетки были дополнительно очищены осаждением на магните и далее обработаны митомицином С. Для контроля использовали соответствующие немагнитные слои, которые были обработаны митомицином С. Культивирование на строме проводили в CO2 инкубаторе в среде DMEM с высоким содержанием глюкозы, 10% фетальной сывороткой крупного рогатого скота, 2 мМ глутамином, пенициллином и стрептомицином. 50% смены среды проводили каждые 3 дня. После 7 дней культивирования (первый раунд культивирования) на немагнитных слоях проводили аккуратное пипетирование и собирали неадгезивные гемопоэтические клетки, в то время как на магнитных слоях проводили полную диссоциацию культур с помощью трипсинизации, после чего магнитные стромальные клетки удаляли на магните. В случае большого числа собранных клеток удаление стромальных клеток на магните проводили дважды. Собранные ядерные клетки подсчитывали, и часть собранных гемопоэтических клеток (50×103) была использована для второго раунда культивирования в течение 1 недели на соответствующей строме. Общее число собранных клеток и число колониеобразующих единиц (КОЕ) оценивали после первого и второго раунда культивирования на строме. Результаты этого эксперимента показаны на Фиг. 6 и 7. Культивирование на магнитной строме и сбор клеток с помощью трипсинизации дали на первом раунде примерно в 2,6 и в 3,5 раз больше ядерных клеток для ОР9 и J2/3T3 стромы, соответственно, по сравнению с соответствующей немагнитной стромой. Во время второго раунда культивирования эти различия в целом сохранялись, что в результате привело к кумулятивной наработке (то есть общей наработке клеток в случае, если бы все снятые после первого раунда клетки были использованы для второго раунда) ядерных клеток с использованием магнитной стромы примерно в 7 и 10 раз больше для ОР9 и J2/3T3 стромы по сравнению с соответствующей немагнитной стромой (Фиг. 6). Подсчет гемопоэтических и стромальных клеток в собранных с магнитной стромы клеточных популяциях показал, что степень контаминации собранных образцов стромальными клетками (7-10%) был значительно ниже, чем в случае с немагнитной стромы, где число стромальных клеток даже превышало число гемопоэтических. Такая степень контаминации, по-видимому, является результатом постепенного ухудшения состояния стромального слоя во время культивирования, что приводит к нарушению его целостности, отшелушиванию стромальных клеток и загрязнению ими собираемой неадгезивной фракции.
Для анализа выхода прогениторных клеток после культивирования на строме в собранных клеточных популяциях был проведен анализ содержания миелоидных КОЕ путем подсчета колоний в метил целлюлозе. Для этого 10×103 клеток после первого раунда или 50×103 клеток после второго раунда высаживали в среду IMDM с метилцеллюлозой MethoCult М3134 (StemCell Technologies, Vancouver, Canada), содержащей 15% фетальную сыворотку крупного рогатого скота, 1×BIT 9500 заменитель сыворотки, 1,4×10-4 М бета-меркаптоэтанол, фактор стволовых клеток (SCF) 100 нг/мл, интерлейкин-3 (IL-3) 10 нг/мл, интерлейкин-6 (IL-6) 10 нг/мл, эритропоэтин (ЕРО) 3 ед./мл, 2 мМ глутамин, пенициллин и стрептомицин. Клетки культивировали в течение 7 дней при 37°C в CO2 инкубаторе (5% CO2), колонии подсчитывали под инвертированным микроскопом. Полученные результаты (Фиг. 7) показывают, что число миелоидных КОЕ значительно выше во фракции, собранной с магнитной строме, чем с немагнитной. Так, после первого раунда культивирования частота КОЕ в полученной фракции гемопоэтических клеток была в 1,3 раза выше для ОР9 стромы и примерно в 3 раза выше для J2/3T3 стромы. Общее число миелоидных КОЕ было в 3,4 раза выше для магнитной ОР9 стромы и в 10,4 раза выше для магнитной J2/3T3 стромы, чем для соответствующих немагнитных стром. Второй раунд культивирования привел к дальнейшему увеличению числа КОЕ для магнитной по сравнению с немагнитной. Так, кумулятивное число КОЕ в собранных фракциях было выше в 9,7 раз в случае со стромой ОР9 и в 19,8 раз в случае со стромой J2/3T3 для магнитной стромы, чем для немагнитной.
Более высокая частота КОЕ в собранных с магнитных стром клеточных фракций, вероятно, является следствием того, что гемопоэтические предшественники локализуются в основном под стромой или тесно ассоциированы с ней (Jing D, Fonseca AV, Alakel N, Fierro FA, Muller K, Bornhauser M, Ehninger G, Corbeil D, Ordemann R. Hematopoietic stem cells in co-culture with mesenchymal stromal cells - modeling the niche compartments in vitro. Haematologica. 2010 95(4):542-550).
Пример 6. При культивировании на магнитной строме и сборе с помощью трипсинизации число клеток с ранним фенотипом Lin-, kit+, Sca1+ значительно повышается по сравнению с немагнитной стромой.
Для того, чтобы определить выход ранних гемопоэтических клеток (гемопоэтические стволовые клетки и клетки-предшественники) после культивирования на строме, мы проанализировали с помощью проточной цитофлюорометрии частоту клеток с поверхностным фенотипом Lin-Sca-+kit+(LSK клетки) в клеточных популяциях, полученных после 1 раунда культивирования на строме. Частота LSK клеток (Фиг. 8, верхние правые квадранты А2) была значительно выше в популяциях клеток, собранных с магнитной стромы, в том числе в 1,9 раз для ОР9 стромы и в 1,5 раз для стромы J2/3T3. Увеличение общего числа собранных LSK клеток для магнитной стромы, с учетом увеличения общего сбора ядерных клеток (пример 5), по сравнению с немагнитной, было еще выше.
Пример 7. Поддерживающая активность стромальных клеток не снижается после их магнетизации.
Для выяснения того, может ли магнетизация стромальных клеток негативно повлиять на их способность поддерживать гемопоэтические клетки в культуре, 15×103 клеток Lin- фракции культивировали с магнитной J2/3T3 и контрольной J2/3T3 стромами, после чего было подсчитано количество гемопоэтических клеток, а также проведена оценка фенотипа культивированных клеток с помощью проточной цитофлюорометрии. Полученные результаты представлены на Фиг. 9. Из Фиг. 9А следует, что магнетизация стромы не уменьшает количество размноженных на строме гемопоэтических клеток. Данные Фиг. 9Г показывают, что количество клеток с ранним фенотипом Lin-/Sca1+/c-kit+ даже несколько увеличивается при культивировании на магнитной строме.
Пример 8. При культивировании на магнитной строме и сборе с помощью трипсинизации число клеток из костного мозга человека с ранним фенотипом CD34+CD38- значительно повышается по сравнению с немагнитной стромой.
В качестве стромального слоя были использованы мезенхимальные стволовые клетки (МСК) из жировой ткани человека. В качестве исходного клеточного образца использовали обогащенную гемопоэтическими клетками и клетками-предшественниками фракцию CD34-позитивных клеток (CD34+ фракции) из пуповинной крови человека. Выделение CD34+ фракции проводилось с помощью магнитной селекции с использованием реагентов и оборудования фирмы Miltenyi Biotec (Германия). Было посеяно по 22 500 CD34+ клеток в каждую лунку 6-ти луночного планшета. Культивирование проводилось в течении 10 дней. Для последующего анализа три повтора были объединены. Полученные результаты показывают, что при культивировании на магнитной строме и сборе клеток с помощью трипсинизации общее число собранных ядерных клеток увеличивается в 1.66 раз, а частота клеток фенотипа CD34+CD38-, соответствующего гемопоэтическим клеткам-предшественникам и стволовым клеткам, увеличивается в несколько десятков раз (40-50 раз).


| название | год | авторы | номер документа |
|---|---|---|---|
| Способ получения ассоциатов гемопоэтических и стромальных клеток-предшественников, способных подавлять активацию и пролиферацию аллогенных лимфоцитов | 2019 |
|
RU2722669C1 |
| Способ получения МСК-ассоциированных недифференцированных гемопоэтических клеток-предшественников с фенотипов CD34+/CD133+ | 2016 |
|
RU2628092C1 |
| СПОСОБ ЭКСПАНСИИ CD34+ ГЕМАТОПОЭТИЧЕСКИХ КЛЕТОК | 2011 |
|
RU2469086C1 |
| СПОСОБ ЭКСПАНСИИ МОНОНУКЛЕАРНЫХ КЛЕТОК ПУПОВИННОЙ КРОВИ (пкМНК) ex vivo В ПРИСУТСТВИИ МУЛЬТИПОТЕНТНЫХ СТРОМАЛЬНЫХ МЕЗЕНХИМАЛЬНЫХ КЛЕТОК (ММСК) | 2013 |
|
RU2525143C1 |
| СПОСОБ ПОЛУЧЕНИЯ КУЛЬТУР МЕЗЕНХИМАЛЬНЫХ СТРОМАЛЬНЫХ КЛЕТОК ЧЕЛОВЕКА, ВЫДЕЛЕННЫХ ИЗ ЛИПОАСПИРАТА | 2007 |
|
RU2351649C1 |
| КЛЕТОЧНЫЙ ПРОДУКТ ДЛЯ ЛЕЧЕНИЯ И КОРРЕКЦИИ ПЕЧЕНОЧНОЙ НЕДОСТАТОЧНОСТИ | 2012 |
|
RU2510833C1 |
| Способ получения и ведения мезенхимальных стволовых клеток из костного материала млекопитающих | 2023 |
|
RU2821926C1 |
| Ex vivo СПОСОБ ПОВЫШЕНИЯ КАЧЕСТВА КЛЕТОК-ПРЕДШЕСТВЕННИКОВ ОТ ВОЗРАСТНЫХ ДОНОРОВ | 2012 |
|
RU2484133C1 |
| Способ прогнозирования риска опухолевой прогрессии у больных раком молочной железы | 2024 |
|
RU2838618C1 |
| Способ использования 4-((5,10-диметил-6-оксо-6,10-дигидро-5Н-пиримидо[5,4-b]тиено[3,2-e][1,4]диазепин-2-ил)амино)бензолсульфонамида (XMU-MP-1) для подавления роста клеток лимфомы Беркитта | 2023 |
|
RU2814013C1 |
Изобретение относится к биотехнологии, а именно к получению репрезентативных популяций гемопоэтических клеток и набору для их получения. Способ включает культивирование гемопоэтических клеток на магнитных стромальных слоях, полученных путем инкубации стромальных клеток с магнитными наночастицами или магнитными липосомами с последующей обработкой клеток митомицином С или облучением рентгеновским или гамма излучением для остановки их деления, при сборе гемопоэтических клеток проводят диссоциацию культуры путем обработки трипсином, коллагеназой, или другим протеолитическим ферментом, или интенсивным перемешиванием культуры, а очистку клеточных препаратов проводят путем удаления магнитной стромы на магните. Изобретение позволяет увеличить общее количество собираемых после культивирования гемопоэтических клеток. 2 н. и 3 з.п. ф-лы, 10 ил., 2 табл., 9 пр.
1. Способ получения репрезентативных популяций гемопоэтических клеток и увеличения выхода гемопоэтических клеток-предшественников и гемопоэтических стволовых клеток, включающий культивирование гемопоэтических клеток на стромальных клеточных слоях, отличающийся тем, что культивирование проводят на магнитных стромальных слоях, полученных путем инкубации стромальных клеток с магнитными наночастицами или магнитными липосомами с последующей обработкой клеток митомицином С или облучением рентгеновским или гамма излучением для остановки их деления, при сборе гемопоэтических клеток проводят диссоциацию культуры путем обработки трипсином, коллагеназой, или другим протеолитическим ферментом, или интенсивным перемешиванием культуры, а очистку клеточных препаратов проводят путем удаления магнитной стромы на магните.
2. Способ по п. 1, отличающийся тем, что стромальные слои делают магнитными с помощью инкубации с магнитными Fe3O4 наночастицами, способными захватываться клетками.
3. Способ по п. 2, отличающийся тем, что для снижения контаминации гемопоэтических клеток стромальными клетками перед культивированием проводят предварительную очистку магнитных стромальных клеток на магните.
4. Способ по п. 3, отличающийся тем, что для снижения нагрузки магнитной стромы наночастицами и повышения времени ее жизни проводят предварительное титрование связывания стромальными клетками магнитных частиц и для очистки магнитной стромы на магните используют препараты магнитных стромальных клеток, в которых доля магнитных клеток не превышает 90%.
5. Набор для получения репрезентативных популяций гемопоэтических клеток и увеличения выхода гемопоэтических клеток-предшественников и гемопоэтических стволовых клеток, включающий в себя магнитные Fe3O4 наночастицы, эффективно магнетизирующие стромальные клетки, магнит, стромальные клетки, митомицин С и среды для культивирования клеток на магнитных стромальных слоях.
| ITO A | |||
| et al | |||
| "Magnetic separation of cells in coculture systems using magnetite cationic liposomes", Tissue Eng Part C Methods | |||
| Колосоуборка | 1923 |
|
SU2009A1 |
| СПОСОБ РАЗМНОЖЕНИЯ/ПОДДЕРЖАНИЯ НЕДИФФЕРЕНЦИРОВАННЫХ ГЕМОПОЭТИЧЕСКИХ СТВОЛОВЫХ КЛЕТОК ИЛИ КЛЕТОК-ПРЕДШЕСТВЕННИКОВ (ВАРИАНТЫ), СПОСОБ ПРИГОТОВЛЕНИЯ КОНДИЦИОНИРОВАННОЙ СРЕДЫ СТРОМАЛЬНЫХ КЛЕТОК, СПОСОБ ТРАНСПЛАНТАЦИИ НЕДИФФЕРЕНЦИРОВАННЫХ ГЕМОПОЭТИЧЕСКИХ СТВОЛОВЫХ КЛЕТОК ИЛИ КЛЕТОК-ПРЕДШЕСТВЕННИКОВ (ВАРИАНТЫ) | 2000 |
|
RU2249039C2 |
| СПОСОБ КУЛЬТИВИРОВАНИЯ, ФИБРОБЛАСТ, СПОСОБ ОТДЕЛЕНИЯ КЛЕТОК | 1991 |
|
RU2164240C2 |
| US 0008940294 B2, 27.01.2015. | |||

