Перекрестная ссылка на родственные заявки
Настоящее заявка испрашивает преимущество приоритета Европейских патентных заявок Nos EP18158169.5, поданной 22 февраля 2018 года; EP18189562.4, поданной 17 августа 2018 года; EP18194549.4, поданной 14 сентября 2018 года, и заявки на патент США US16/269572. Содержание вышеупомянутых патентных заявок полностью включено в настоящую заявку путем ссылки.
Область техники, к которой относится изобретение
Настоящее изобретение относится к неагонистическому лиганду, в частности к антителу, которое специфически связывается с ГМ-КСФ (CSF-2) или его рецепторным комплексом, состоящим из CSF2Rβ (CD131) и CSF2Rα (CD116), для применения при лечении гемобластоза у пациента, перенесшего алло-ТГК.
Уровень техники
Для пациентов, страдающих гемобластозом, аллогенная трансплантация гемопоэтических клеток (алло-ТГК) является потенциально излечивающим и спасающим жизнь вмешательством; однако у 40-60% всех пациентов развивается клинически значимое острая или хроническая реакция «трансплантат против хозяина» (РТПХ), что в совокупности приводит к смертности на уровне примерно 50%. Донорские Т-клетки опосредуют оба процесса: алло-реактивные Т-клетки из трансплантата атакуют злокачественные клетки хозяина, оказывая благоприятное действие «трансплантат против лейкоза» (ТПЛ); в то время как та же самая аллореактивность может быть нацелена на здоровые ткани (обычно кожу, кишечник и печень), что приводит к РТПХ. Хотя истощение Т-клеток из донорского материала до алло-ТГК может предотвратить/уменьшить РТПХ, соответственно, это происходит за счет снижения активности трансплантат против лейкоза (ТПЛ) и увеличения частоты рецидивов. Следовательно, существует острая необходимость понять, как механизмы ТПЛ и РТПХ могут быть разделены на уровне Т-клеток и модулированы для получения благоприятного клинического эффекта.
Было сделано много попыток определить на мышиных моделях ключевые субпопуляции Т-клеток и цитокины, которые лежат в основе РТПХ, но на сегодняшний день сделано мало последовательных выводов. Хотя изначально предполагалось, что РТПХ является TH1-опосредованной патологией, исследования на мышах показали, что донорские Т-клетки, дефицитные по TH1-цитокину ИФНγ, могут усугубить заболевание. Клетки TH2 могут как подавлять экспериментальную РТПХ, так и вызывать РТПХ, поражая печень и кожу. Аналогичным образом, в то время как есть доказательства того, что IL-17A-продуцирующие Т-клетки являются известными медиаторами повреждения тканей при воспалительных заболеваниях в целом, их роль в РТПХ остается спорной, так как в мышиных моделях IL-17, по-видимому, может или способствовать или облегчать РТПХ, в зависимости от условий эксперимента. Взятые вместе, поляризованные TH-клетки явно участвуют в возникновении и сохранении РТПХ, но до сих пор не удалось идентифицировать какой-либо конкретный растворимый медиатор, который имеет воспроизводимую и неизбыточную функцию в патогенезе заболевания, как у мышиных моделей, так и у людей.
Основываясь на вышеупомянутом уровне техники, целью настоящего изобретения является создание средств и способов для ослабления РТПХ при сохранении полезного эффекта ТПЛ. Эта цель достигается предметом независимых пунктов формулы настоящего описания.
Сущность изобретения
В первом аспекте настоящее изобретения относится к способу лечения пациента, страдающего от реакции «трансплантат против хозяина», или к способу предотвращения, ингибирования или уменьшения тяжести развития реакции «трансплантат против хозяина» у пациента, перенесшего аллогенную трансплантацию.
Во втором аспекте настоящее изобретения относится к способу лечения пациента, страдающего онкологическим заболеванием, в частности гемобластозом, и перенесшего аллогенную трансплантацию гемопоэтических стволовых клеток (алло-ТГК).
В соответствии с любым аспектом настоящего изобретения способы включают нарушение передачу сигналов иммунного цитокина ГМ-КСФ, запускающего РТПХ. Это может быть достигнуто путем ингибирования ГМ-КСФ лигандами, в частности антителами, более конкретно нейтрализующими антителами. Цель изобретения также может быть достигнута путем нарушения связывания ГМ-КСФ с его природным рецептором или путем ингибирования сигнального каскада при помощи введения неагонистических лигандов к этим рецепторам, в частности, при помощи введения нейтрализующих антител к рецептору ГМ-КСФ. Другой возможный механизм заключается во временном или более длительном подавлении экспрессии одного из ГМ-КСФ или его рецепторов путем интерференции нуклеиновых кислот (РНК-интерференция, антисмысловая РНК).
В конкретных вариантах осуществления настоящее изобретение включает введение пациенту неагонистического лиганда, в частности антитела, в частности нейтрализующего неагонистического антитела, реагирующего с ГМ-КСФ или с одним из CD116, CD131 и рецептором ГМ-КСФ, состоящим из CD116 и CD131.
В качестве альтернативы этот аспект может быть сформулирован как предоставление агента, выбранного из
- нейтрализующего полипептидного лиганда, в частности нейтрализующего антитела, реагирующего с одним из ГМ-КСФ, CD116, CD131 или рецептором ГМ-КСФ, состоящим из CD116 и CD131, и
- нуклеотидного агента, ингибирующего экспрессию ГМ-КСФ, CD116 и/или CD131,
для применения в лечении или предотвращении РТПХ у пациента, страдающего гемобластозом.
Краткое описание чертежей
Фигура 1A-1H Донорские Т-клетки секретируют ГМ-КСФ и ИФНγ во время аллогенных ответов
Фигура 1A: Выживаемость летально облученных мышей CD45.2+ BALB/c после алло-ТГК с использованием CD45.1+ ДТ C57BL/6 БТК-КМ отдельно или в комбинации со спленоцитами CD45.1+ ДТ C57BL/6. Данные объединены из 4 отдельных экспериментов, в каждом n=5 мышей на группу. Для сравнения кривых выживаемости использовали логарифмический ранговый критерий (критерий Кокса-Мантеля), ***p<0,001. Фигура 1B: Частота CD45.1+ клеток в живых синглетах в селезенке, печени и коже через 3 и 6 дней после алло-ТГК. Данные объединены, чтобы получить n=10 мышей на группу. Фигура 1C: Частота ИФНγ-, ГМ-КСФ- и IL-17A- продуцирующих CD4+ и CD8+ Т-клеток в популяции CD45.1+ из печени мышей через 3 дня после алло-ТГК. Репрезентативные графики показаны для 3 независимых экспериментов. Фигура 1D: Частоты ИФНγ-, ГМ-КСФ- и IL-17A- продуцирующих CD4+ и CD8+ Т-клеток в популяциях CD45.1+ из печени, селезенки и кожи через 3 и 6 дней после алло-ТГК. Данные объединены из 3 индивидуальных экспериментов, всего n=10 мышей в группе. Фигура 1E: Уровни ИФНγ и ГМ-КСФ в сыворотке крови у мышей через 6 дней после алло-ТГК. Данные объединены из 3 индивидуальных экспериментов, n=2-5 мышей на группу. Для сравнения средних значений использовали непарный двусторонний t-критерий с поправкой Уэлча, *p<0,05, **p<0,01. Фигура 1F: ИФНγ, ГМ-КСФ и IL-17A в супернатантах от совместных культур Т-клеток мышей C57BL/6 дикого типа с сингенными (C57BL/6) или аллогенными (BALB/c) селезеночными CD11c+ ДК. Данные объединены из 3 индивидуальных экспериментов, n=3-5 мышей на группу. Для сравнения средних значений использовался непарный двусторонний t-критерий с поправкой Уэлча, *p<0,05, **p<0,01, ***p<0,001. Фигура 1G: Включение меченого тритием тимидина Т-клетками мышей C57BL/6 ДТ, Ifng-/-, Csf2-/- или Il17a-/-, совместно культивированных с сингенными (C57BL/6) или аллогенными (BALB/c) селезеночными CD11c+ ДК. Данные объединены из 3 индивидуальных экспериментов, n=2-5 мышей на группу. Для сравнения средних значений использовали однофакторный дисперсионный анализ с пост-тестом Бонферрони. Данные приводятся как среднее +/- СО на Фигуре 1B и как среднее +/- СОС на (Фигурах 1D-1G). Фигура 1H: Частоты ИФНγ и ГМ-КСФ-продуцирующих CD4+ и CD8+ Т-клеток в популяциях CD45.1+ из печени и селезенки через 3 и 6 дней после алло-ТГК. Показаны репрезентативные данные, объединенные из 1 из 3 экспериментов, n=3-4 мышей на группу. Сокращения: БТК: без Т-клеток, КМ: костный мозг, ТГК: трансплантация гемопоэтических клеток, ДТ: дикий тип, имп/мин: число импульсов в минуту.
Фигура 2A-2K ГМ-КСФ имеет решающее значение для острой РТПХ после алло-ТГК с полной MHC-несовместимостью
Фигура 2A: Выживаемость летально облученных мышей BALB/c после алло-ТГК с использованием ДТ C57BL/6 БТК-КМ отдельно или в комбинации со спленоцитами от мышей C57BL/6 ДТ, Ifng-/-, Csf2-/- или Il17a-/-. Данные объединены из 4 отдельных экспериментов, каждый с n=5 мышей на группу. Для сравнения кривых выживаемости (ДТ по сравнению с другими группами) использовали логарифмический ранговый критерий (критерий Кокса-Мантеля), *p<0,05, **p<0,01, ***p<0,001. Фигура 2В: Выживаемость летально облученных мышей BALB/c после алло-ТГК с использованием ДТ C57BL/6 БТК-КМ отдельно или в комбинации с Т-клетками, очищенными из селезенки мышей C57BL/6 ДТ или Csf2-/-. Данные объединены из 3 индивидуальных экспериментов, каждый с n=5 мышей на группу. Для сравнения кривых выживаемости (ДТ по сравнению с Csf2-/-) использовали логарифмический ранговый критерий (критерий Кокса-Мантеля), ***p<0,001. Фигура 2C: Комплексная гистопатологическая оценка печени, тонкого кишечника и кожи мышей BALB/c через 6 дней после алло-ТГК. Данные объединены из 3 индивидуальных экспериментов, n=5 мышей на группу. Для сравнения средних значений (ДТ по сравнению с Csf2-/-) использовали непарный двусторонний t-критерий *p<0,05. Фигура 2D: Репрезентативные изображения окрашенных гематоксилином и эозином срезов кожи и тонкого кишечника мышей через 6 дней после алло-ТГК с использованием ДТ C57BL/6 БТК-КМ в комбинации со спленоцитами от мышей C57BL/6 ДТ или Csf2-/-, n=4-5 мышей на группу (масштабная шкала: 100 мкм). Фигуры 2E-2F: Репрезентативные изображения меченных p22phox срезов печени (Фигура 2E) и тонкого кишечника (Фигура 2F) (масштабная шкала: 100 мкм) мышей через 6 дней после алло-ТГК с использованием ДТ C57BL/6 БТК-КМ в комбинации со спленоцитами от мышей C57BL/6 ДТ или Csf2-/-, n=4-5 мышей на группу (верхние изображения). Количественная оценка среднего % общей площади, положительно меченной p22phox, на поле зрения. Для сравнения средних значений (ДТ по сравнению с Csf2-/-) использовали непарный двусторонний t-критерий, **p<0,01. Фигура 2G: Выживаемость летально облученных мышей BALB/c после алло-ТГК с использованием ДТ C57BL/6 БТК-КМ отдельно или в комбинации со спленоцитами от мышей C57BL/6 ДТ. Мышам вводили ФСБ, изотипическое контрольное антитело или антителом против ГМ-КСФ 3 раза в неделю в течение эксперимента, начиная за 2 дня до ТГК. Данные объединены из 2 индивидуальных экспериментов, в каждом n=5 мышей на группу. Для сравнения кривых выживаемости (ГМ-КСФ по сравнению с изотипическим контролем) использовали логарифмический ранговый критерий (критерий Кокса-Мантеля), ***p<0,001. Данные приводятся как среднее +/- СОС. Фигура 2H: Длина толстой кишки в см у мышей BALB/c через 6 дней после алло-ТГК, как описано в a. Данные объединены от 2 индивидуальных экспериментов. Фигура 2I: Репрезентативные изображения и количественная оценка апоптозных клеток (окрашивание TUNEL) в толстой кишке мышей BALB/c через 6 дней после алло-ТГК, как описано в а. Данные объединены из 3 индивидуальных экспериментов (верхняя масштабная шкала 50 мкм, нижняя 20 мкм). Фигура 2J: Уровни щелочной фосфатазы (AP), аланинаминотрансферазы (ALT), азота мочевины крови (BUN) и альбумина в сыворотке мышей BALB/c через 6 дней после алло-ТГК. Данные являются репрезентативными для 2 индивидуальных экспериментов, n=3-5 мышей на группу. Фигура 2K: Выживаемость летально облученных мышей BALB/c, подвергшихся алло-ТГК с использованием ДТ C57BL/6 БТК-КМ отдельно или в комбинации с CD4 и CD8 Т-клетками, очищенными из селезенок мышей C57BL/6 ДТ или Csf2-/- (CD4ДТCD8ДТ, CD4ДТCD8Csf2-/-, CD4Csf2-/-CD8ДТ и CD4Csf2-/-CD8Csf2-/-). Мышей, получавших только БТК-КМ, использовали в качестве контроля. Данные являются репрезентативными для 2 индивидуальных эксперимента с n=5 мышей на группу. Для сравнения кривых выживаемости (CD4ДТCD8ДТ по сравнению с другими группами) использовали логарифмический ранговый критерий (критерий Кокса-Мантеля), нз (не значимо), **p<0,01. Сокращения: БТК: без Т-клеток, КМ: костный мозг, ТГК: трансплантация гемопоэтических клеток, ДТ: дикий тип.
Фигура 3A-3H ГМ-КСФ опосредует патологию РТПХ после частично MHC-несовместимой алло-ТГК
Фигура 3A: Выживаемость летально облученных мышей B6D2F1 после частично MHC-несовместимой алло-ТГК с использованием ДТ C57BL/6 БТК-КМ отдельно или в комбинации со спленоцитами от мышей C57BL/6 ДТ, Csf2-/- или Ifng-/-. Данные объединены из 5 индивидуальных экспериментов, в каждом n=5 мышей на группу. Для сравнения кривых выживаемости (ДТ по сравнению с другими группами) использовали логарифмический ранговый критерий (критерий Кокса-Мантеля), *p<0,05, ***p<0,001. Фигура 3B: Выживаемость летально облученных мышей B6D2F1 после алло-ТГК с использованием ДТ C57BL/6 БТК-КМ отдельно или в комбинации с Т-клетками, очищенными из селезенок мышей C57BL/6 ДТ, Csf2-/- или Ifng-/-. Данные объединены из 2 индивидуальных экспериментов, в каждом n=5 мышей на группу. Для сравнения кривых выживаемости (ДТ по сравнению с другими группами) использовали логарифмический ранговый критерий (критерий Кокса-Мантеля), *p<0,05, ***p<0,001. Фигура 3C: Комплексная гистопатологическая оценка срезов печени, тонкой кишки и кожи мышей B6D2F1 через 11 дней после алло-ТГК. Данные объединены из 2 индивидуальных экспериментов, n=5 мышей на группу. Для сравнения средних значений (ДТ, Csf2-/- и Ifng-/-) использовался однофакторный дисперсионный анализ с пост-тестом Бонферрони. *p<0,05 ***p<0,001. Фигура 3D: Репрезентативные изображения окрашенных гематоксилином и эозином срезов кожи мышей через 1 день после алло-ТГК с использованием ДТ C57BL/6 БТК-КМ в комбинации со спленоцитами мышей C57BL/6 ДТ, Ifng-/- или Csf2-/-, n=4-5 мышей на группу (масштабная шкала: 100 мкм). Фигуры 3E-3F: репрезентативные изображения меченных p22phox (верхние изображения) и F4/80 срезов печени (Фигура 3E) и тонкой кишки (Фигура 3F) (масштабная шкала: 100 мкм) мышей через 9 дней после алло-ТГК с использованием ДТ C57BL/6 БТК-КМ в комбинации со спленоцитами мышей C57BL/6 ДТ или Csf2-/-, n=4-5 мышей на группу. Количественное определение % общей площади, положительно меченной p22phox или F4/80 на поле зрения. Для сравнения средних значений (ДТ по сравнению с Csf2-/-) использовали непарный двусторонний t-критерий ***p<0,001, **p<0,01. Фигура 3G: Уровни ИФНγ и ГМ-КСФ в сыворотке крови у мышей через 9 дней после алло-ТГК с использованием ДТ C57BL/6 БТК-КМ в комбинации со спленоцитами мышей C57BL/6 ДТ, Ifng-/- или Csf2-/-. Данные объединены из 2-3 индивидуальных экспериментов, n=4-5 мышей на группу. Для сравнения средних значений (ДТ по сравнению с Ifng-/- или ДТ по сравнению с Csf2-/-) использовали непарный двусторонний t-критерий с поправкой Уэлча, *p<0,05, **p<0,01. Данные показаны как среднее +/- СОС. Фигура 3H: Комплексная гистопатологическая оценка срезов печени мышей B6D2F1 через 9 или 28 дней после алло-ТГК, как описано в a. Данные получены от 2 независимых экспериментов, n=4-6 мышей на группу. Сокращения: БТК: без Т-клеток, КМ: костный мозг, ТГК: трансплантация гемопоэтических клеток, ДТ: дикий тип.
Фигура 4A-4H ГМ-КСФ не является обязательным для противоопухолевой активности после алло-ТГК
Фигуры 4A-4D: Летально облученным мышам BALB/c внутривенно вводили опухолевые клетки A20, экспрессирующие GFP и люциферазу, одновременно с MHC-несовместимой алло-ТГК с использованием ДТ C57BL/6 БТК-КМ отдельно или в комбинации с Т-клетками, очищенными из селезенки мышей C57BL/6 ДТ или Csf2-/-. Мышей, получавших только БТК-КМ, использовали в качестве контроля. Фигура 4A: Рост опухоли отслеживали с помощью биолюминесцентной визуализации in vivo. Показаны изображения одного репрезентативного эксперимента из двух. Фигура 4B: Интенсивность сигнала в интересующей области (ИО) отслеживалась во времени. Выживаемость с течением времени (Фигура 4C) и причина смерти (Фигура 4D) показаны как процент мышей в каждой экспериментальной группе со смертельной РТПХ (серый), опухолями (черный) или выживших (белый). Данные объединены из 2 индивидуальных экспериментов с n=5 мышей в группе. Для сравнения кривых выживаемости (ДТ по сравнению с другими группами) использовали логарифмический ранговый критерий (критерий Кокса-Мантеля), нз (не значимо), **p <0,01. Фигуры 4E-4H: Летально облученным мышам BALB/c внутривенно вводили опухолевые клетки A20, экспрессирующие GFP и люциферазу, одновременно с MHC-несовместимой алло-ТГК с использованием ДТ C57BL/6 БТК-КМ отдельно или в комбинации с Т-клетками, очищенными из селезенки мышей C57BL/6 ДТ. Мышей, получавших только БТК-КМ, использовали в качестве контроля. Мышам вводили изотипический контроль или антитело против ГМ-КСФ 3 раза в неделю в течение эксперимента, начиная за 2 дня до ТГК. Фигура 4E: Рост опухоли отслеживали с помощью биолюминесцентной визуализации in vivo. Показаны изображения из одного эксперимента с n=6-7 мышей на группу. Фигура 4F: Интенсивность сигнала в интересующей области (ИО) отслеживалась во времени. Выживаемость с течением времени (Фигура 4G) и причина смерти (Фигура 4H) показаны как процент мышей в каждой экспериментальной группе со смертельной РТПХ (серый), опухолями (черный) или выживших (белый). Для сравнения кривых выживаемости (ДТ по сравнению с Csf2-/-) использовали логарифмический ранговый критерий (критерий Кокса-Мантеля), *p<0,05. Данные показаны как среднее +/- СОС. Сокращения: БТК: без Т-клеток, КМ: костный мозг, ТГК: трансплантация гемопоэтических клеток, ДТ: дикий тип, БЛВ: биолюминесцентная визуализация, ИО: интересующая область.
Фигура 5A-5D Пациенты с тяжелой РТПХ экспрессируют высокие уровни ГМ-КСФ в биоптатах кишечника
Фигура 5A: Относительная экспрессия ГМ-КСФ на уровне мРНК в биоптатах желудочно-кишечного тракта от пациентов с различными степенями РТПХ, см. Таблицу 3. Для сравнения средних значений использовали однофакторный дисперсионный анализ с пост-тестом Бонферрони **p<0,01. Данные показаны как среднее +/- СОС. Фигура 5B: Изображения мечения ГМ-КСФ в контрольной биопсии и биопсии РТПХ IV степени из тонкой кишки пациентов с алло-ТГК, см. Таблицы 1 и 2. Коричневый: анти-человеческий ГМ-КСФ антитело, синий: гематоксилин. Масштабные шкалы - 100 мкм (20 мкм в увеличенных изображениях). Показаны репрезентативные изображения 3 отдельных контрольных образцов и образцов пациентов. Фигура 5C: Иммунофлуоресцентное окрашивание на CD3 (розовый), CD68 (зеленый) и ГМ-КСФ (красный) биоптатов желудочно-кишечного тракта от пациентов с РТПХ IV степени. Показано репрезентативное изображение 3 отдельных образцов пациентов. Ядра изображены синим цветом (DAPI) (шкала вверху 50 мкм, внизу 20 мкм). Фигура 5D: проточно-цитометрический анализ МКПК, полученных от здоровых доноров (HD) и пациентов с РТПХ (GP), стимулированных в течение 4 часов PMA/иономицином. Показан реперезентативный FACS-график Т-клеток, экспрессирующих цитокины. ГМ-КСФ, ИФН и IL-17, продуцирующие CD4+ и CD8+ Т-клетки, представлены как отдельные частоты. n=10-9 мышей на группу *P<0,05, **P<0,01. Сокращения: БТК: без Т-клеток, КМ: костный мозг, ТГК: трансплантация гемопоэтических клеток.
Фигура 6A-6D Сравнительный фенотипический анализ популяций Т-клеток от мышей ДТ и Csf2-/-
Проточно-цитометрический анализ частот популяций Т-клеток селезенки (Фигура 6A) и частот экспрессии маркеров активации у иммунологически наивных мышей C57BL/6 ДТ и Csf2-/- (Фигура 6B). Данные были объединены из 2 индивидуальных экспериментов, в каждом n=3 мыши на группу. Фигура 6C: Проточно-цитометрический анализ инфильтратов донорских Т-клеток в лимфатических узлах (ЛУ), селезенке, печени и коже реципиента через 2 и 5 дней после алло-ТГК летально облученных мышей BALB/c, восстановленных ДТ C57BL/6 БТК-КМ в комбинации со спленоцитами мышей C57BL/6 ДТ или Csf2-/-. Клетки были гейтированы по H2-Db и CD45; CD4 и CD8 Т-клетки были предварительно гейтированы по CD3. Показан один эксперимент с n=4 мыши в группе. Данные показаны как среднее +/- СОС. Фигура 6D: Проточно-цитометрический анализ инфильтратов донорских Т-клеток в ЛУ и селезенке реципиента через 5 дней после алло-ТГК летально облученных мышей BALB/c, восстановленных ДТ C57BL/6 БТК-КМ в комбинации со спленоцитами мышей C57BL/6 ДТ или Csf2-/-. Клетки были гейтированы по H2-Db и CD45; CD4 и CD8 Т-клетки. Показан один эксперимент с n=4 мыши в группе. Данные показаны как среднее +/- СОС. Сокращения: TCD: БТК: без Т-клеток, КМ: костный мозг, ТГК: трансплантация гемопоэтических клеток, ДТ: дикий тип.
Фигуры 7A и 7B Киллинг-анализ с использованием опухолевых клеток A20 и Т-клеток, выделенных из селезенки и лимфатических узлов мышей C57BL/6 ДТ или Csf2-/-, соответственно. Фигура 7A: Проточно-цитометрический анализ мертвых клеток путем окрашивания T0-PR03, показаны репрезентативные графики. Фигура 7B: Специфичный лизис опухолевых клеток A20 указанными Т-клетками при соотношениях мишень:эффектор 1:1, 1:20 и 1:50. Данные представляют собой два эксперимента с n=3 мыши на группу в каждом. Данные показаны как среднее +/- СОС. Сокращения: ДТ: дикий тип.
Фигуры 8A и 8B. Фигура 8A: выживаемость летально облученных мышей C57BL/6 ДТ или Csf2rb-/-, после алло-ТГК с использованием без Т-клеток-КМ мышей Balb/c ДТ отдельно или в комбинации со спленоцитами мышей Balb/c ДТ. Данные объединены из 2 отдельных экспериментов, в каждом n=5-7 мышей на группу. Фигура 8В: Выживаемость летально облученных мышей Balb/c ДТ после алло-ТГК с использованием БТК-КМ мышей C57BL/6 ДТ или Csf2rb-/- отдельно или в комбинации со спленоцитами мышей C57BL/6 ДТ. Данные объединены из 2 отдельных экспериментов, в каждом n=5-7 мышей на группу.
Фигура 9A-9H. ГМ-КСФ вызывает РТПХ через миелоидные клетки донорского происхождения
Фигура 9A: Выживаемость летально облученных мышей C57BL/6 ДТ и Csf2rb-/-, после MHC-несовместимой алло-ТГК с использованием ДТ Balb/c БТК-КМ отдельно или в комбинации со спленоцитами мышей Balb/c ДТ. Данные объединены из 2 отдельных экспериментов, в каждом n=5 мышей в группе. Фигура 9В: Выживание летально облученных мышей Balb/c ДТ после MHC-несовместимой алло-ТГК с использованием БТК-КМ мышей C57BL/6 ДТ или и Csf2rb-/- отдельно или в комбинации со спленоцитами мышей C57BL/6 ДТ. Данные объединены из 2 отдельных экспериментов, в каждом n=5 мышей на группу. Для сравнения кривых выживаемости (ДТ по сравнению с Csf2rb-/- реципиенты/доноры) использовали логарифмический ранговый критерий (критерий Кокса-Мантеля) в a и b, **p<0,01. Фигура 9C: Аннотированная карта t-SNE, отображающая 200000 случайно отобранных клеток из костного мозга мышей ДТ C57BL/6 и Csf2rb-/-, демонстрирующих фосфорилирование STAT5 (градиент от черного к желтому) при стимуляции ГМ-КСФ, проанализировано с помощью проточно-цитометрического анализа. Данные представляют собой два независимых эксперимента с n=3 мыши на группу. Фигура 9D: Частоты индуцированной ГМ-КСФ активации pSTAT5 в моноцитах и нейтрофилах мышей ДТ C57BL/6 и Csf2rb-/-, как показано на Фигуре 9B. Фигура 9E: Проточно-цитометрический анализ различных популяций миелоидных клеток (ДК, нейтрофилов, моноцитов и ЗДК) после ТГК. Показан пример гейтирования для печени. Фигура 9F: Количественная оценка популяций миелоидных клеток (из Фигуры 9E) в селезенке (верхний ряд) и печени (нижний ряд). Показан один представитель из 3 индивидуальных экспериментов, n=4-5 мышей на группу. (g) Частота про-IL-1β-продуцирующих нейтрофилов и моноцитов в популяции H2Db+, CD45+ из селезенки мышей через 6 дней после алло-ТГК. Репрезентативные графики показаны для нейтрофилов, моноцитов и ЗДК (гейтирование, как в e). Данные представляют два независимых эксперимента n=5 мышей на группу Фигура 9H: Проточно-цитометрический анализ АФК (реагент CellROX) у мышей через 6 дней после алло-ТГК. Репрезентативные гистограммы СКП для нейтрофилов, моноцитов и ЗДК (гейтирование, как на Фигуре 9E). Данные представляют собой один эксперимент где n=5-7 мышей на группу. Для сравнения средних значений использовался непарный двусторонний t-критерий в f-h ***p<0,001, **p<0,01, *p<0,05. Данные показаны как среднее +/- СОС. Сокращения: БТК: без Т-клеток, КМ: костный мозг, ТГК: трансплантация гемопоэтических клеток, ДТ: дикий тип.
Краткое описание упомянутых последовательностей
Представленные здесь нуклеотидные последовательности показаны с использованием стандартных буквенных сокращений для нуклеотидных оснований, как определено в разделе 37 параграфе 1.822 Свода федеральных нормативных актов США. Показана только одна цепь каждой последовательности нуклеиновой кислоты, но комплементарная цепь предполагается как включенная путем любой ссылки на отображаемую цепь. Список последовательностей представлен в виде текстового файла ASCII с названием «Протокол последовательностей uz349wo_ST25.txt», размером около 2 КБ, который включен в настоящее описание путем ссылки.
SEQ ID NO: 1 и 2: прямой и обратный праймеры для ПЦР-амплификации для гена CSF2.
SEQ ID NO: 3 и 4: прямой и обратный праймеры для ПЦР-амплификации для гена GAPDH.
Подробное описание изобретения
Термины и определения
Термин «РТПХ» в контексте настоящего описания относится к реакции «трансплантат против хозяина», осложнению, возникающему в результате трансплантации иммунных клеток генетически другому пациенту. РТПХ обычно связана с трансплантацией стволовых клеток, особенно в контексте терапии гемобластоза, но может возникать в контексте других трансплантаций.
Термин «алло-ТГК» в контексте настоящего описания относится к трансплантации аллогенных гемопоэтических клеток.
Термин «ГМ-КСФ» в контексте настоящего описания относится к гранулоцитарно-макрофагальному колониестимулирующему фактору (Uniprot P04141; CAS No. 83869-56-1).
Термин «CD116» в контексте настоящего описания относится к кластеру дифференцировки 116, также известному как альфа-цепь рецептора ГМ-КСФ (Uniprot P15509). Рецептор ГМ-КСФ состоит из специфичной по отношению к ГМ-КСФ альфа-цепи (CD116) и бета-цепи (CD131), которая также присутствует в рецепторах интерлейкина IL-3R и IL-5R.
Лиганд ГМ-КСФ или один из CD116, CD131 и рецептор ГМ-КСФ, состоящий из CD116 и CD131 в соответствии с настоящим изобретением, способен подавлять или нейтрализовать передачу сигнала, возникающую, когда ГМ-КСФ связывается с его рецептором.
Термин «ХЛЛ» в контексте настоящего описания относится к острому хроническому лимфоцитарному или лимфобластному лейкозу.
Термин «ОЛЛ» в контексте настоящего описания относится к острому лимфобластному лейкозу.
Термин «ХМЛ» в контексте настоящего описания относится к хроническому миелогенному или миелоидному лейкозу.
Термин «ОМЛ» в контексте настоящего описания относится к острому миелогенному лейкозу.
Термин «ОМоЛ» в контексте настоящего описания относится к острому моноцитарному лейкозу.
В контексте настоящего описания термин «антитело» используется в его значении, известном в области клеточной биологии и иммунологии; он относится к полным антителам, включая, помимо прочего, иммуноглобулин типа G (IgG), типа A (IgA), типа D (IgD), типа E (IgE) или типа M (IgM), любому антигенсвязывающему фрагменту или его отдельным цепям и родственным или производным конструкциям. Полное антитело представляет собой гликопротеин, содержащий, по меньшей мере, две тяжелые (H) цепи и две легкие (L) цепи, соединенные дисульфидными связями. Каждая тяжелая цепь состоит из вариабельной области тяжелой цепи (VH) и константной области тяжелой цепи (CH). Константная область тяжелой цепи состоит из трех доменов: СН1, СН2 и СН3. Каждая легкая цепь состоит из вариабельной области легкой цепи (сокращенно VL) и константной области легкой цепи (CL). Константная область легкой цепи состоит из одного домена CL. Вариабельные области тяжелой и легкой цепей содержат связывающий домен, который взаимодействует с антигеном. Константные области антител могут опосредовать связывание иммуноглобулина с тканями или факторами хозяина, включая различные клетки иммунной системы (например, эффекторные клетки) и первый компонент классической системы комплемента. Так же данный термин включает так называемое нанотело или однодоменное антитело, фрагмент антитела, состоящий из одного мономерного вариабельного домена антитела.
Термин «антитело» включает антитело верблюдовых, в частности гуманизированное антитело верблюдовых.
Термин «антителоподобная молекула» в контексте настоящего описания относится к молекуле, способной специфично связываться с другой молекулой или мишенью с высоким сродством/a Kd≤10E-8 моль/л. Антителоподобная молекула, связывается со своей мишенью аналогично специфичному связыванию антитела. Термин «антителоподобная молекула» включает белок с повторами, такой как сконструированный белок с анкириновым повтором (Molecular Partners, Zurich), сконструированные белки-миметики антител, демонстрирующие высокоспецифичное и высокоаффинное связывание с целевым белком (см. заявки на патент США US2012142611, US2016250341, US2016075767 и US2015368302 (все включены в настоящую заявку путем ссылки). Термин «антителоподобная молекула» дополнительно включает без ограничений полипептид, полученный из белков с армадиловым повтором, полипептид, полученный из белков с богатыми лейцином повторами, и полипептид, полученный из белков с повторами тетратрикопептида.
Термин «антителоподобная молекула» дополнительно включает полипептид, полученный из доменов белка A, полипептид, полученный из домена фибронектина FN3, полипептид, полученный из консенсусных доменов фибронектина, полипептид, полученный из липокалинов, полипептид, полученный из цинковых пальцев, полипептид, полученный из Src гомологичного домена 2 (SH2), полипептид, полученный из Src гомологичного домена 3 (SH3), полипептид, полученный из доменов PDZ, полипептид, полученный из гамма-кристаллина, полипептид, полученный из убиквитина, полипептид, полученный из полипептида с цистиновым узлом и полипептид, полученный из узелтина, полипептид, полученный из цистатина, полипептид, полученный из Sac7d, суперспираль с тройной спиралью (также известный как альфа-тела), полипептид, полученный из домена Куница ингибитора протеазы типа Куница, и полипептид, полученный из модуля связывания углеводов 32-2.
Термин «полипептид, полученный из доменов белка A» относится к молекуле, которая является производной белка A и способна специфично связывать область Fc и область Fab иммуноглобулинов.
Термин «белок с армадиловым повтором» относится к полипептиду, содержащему, по меньшей мере, один армадиловый повтор, при этом армадиловый повтор характеризуется парой альфа-спиралей, которые образуют шпилечную структуру.
В контексте настоящего описания термин «гуманизированное антитело» используется в его значении, известном в области клеточной биологии и биохимии; он относится к антителу, первоначально продуцируемому иммунными клетками не относящихся к человеку видов, белковые последовательности которых были модифицированы для увеличения их сходства с вариантами антител, вырабатываемых естественным образом у людей.
Термин «гуманизированное антитело верблюдовых» в контексте настоящего описания относится к антителу, состоящему только из тяжелой цепи или вариабельного домена тяжелой цепи (домена VHH), чья аминокислотная последовательность была изменена для увеличения его сходства с антитела, вырабатываемые естественным образом у людей, и поэтому оно демонстрирует пониженную иммуногенность при введении человеку. Общая стратегия гуманизации антител верблюдовых описана в Vincke et al. «General strategy to humanize a camelid single-domain antibody and identification of a universal humanized nanobody scaffold», J Biol Chem. 2009 Jan 30; 284(5): 3273-3284, и в заявке на патент США US2011165621 A1.
В контексте настоящего описания термин «химерное антитело» используется в его значении, известном в области клеточной биологии и иммунологии; он относится к молекуле антитела, в которой константная область или ее часть изменена, заменена или заменена так, что сайт связывания антигена (вариабельная область) связан с константной областью другого или измененного класса, эффекторной функции и/или вида, или совершенно другой молекулой, которая придает новые свойства химерному антителу, например ферментом, цитокином, токсином, гормоном, фактором роста, лекарственным средством и т.д. Например, антитело можно модифицировать, заменив его константную область на цитокин. Благодаря замене на цитокин химерное антитело может сохранять свою специфичность в распознавании антигена, одновременно выполняя функцию или ее часть исходной молекулы цитокина.
В контексте настоящего описания термин «константа диссоциации (KD)» используется в его значении, известном в области химии и физики; он относится к константе равновесия, которая измеряет склонность более крупного объекта к обратимой диссоциации на более мелкие компоненты, когда комплекс распадается на составляющие его молекулы. KD выражается в молярных единицах [M] и соответствует концентрации [Ab], при которой занята половина сайтов связывания [Ag]. Другими словами, концентрация несвязанного [Ab] равна концентрации комплекса [AbAg]. Константу диссоциации можно рассчитать по следующей формуле:
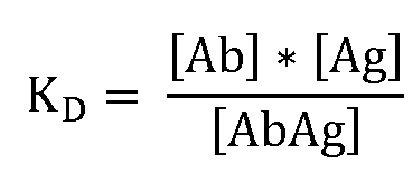
[Ab]: концентрация антитела; [Ag]: концентрация антигена; [AbAg]: концентрация комплекса антитело-антиген.
В контексте настоящего описания термины «скорость диссоциации» (Koff; [1/сек]) и «скорость ассоциации» (Kon; [1/сек*M]) используются в их значениях, известных в области химии и физики. Они относятся к константе скорости, которая измеряет диссоциацию (Koff) или ассоциацию (Kon) антитела с его антигеном-мишенью. Koff и Kon могут быть определены экспериментально с использованием методов, хорошо известных в данной области техники. В способе определения Koff и Kon антитела используется поверхностный плазмонный резонанс. Это принцип, лежащий в основе биосенсорных систем, таких как Biacore® или ProteOn®. Их также можно использовать для определения константы диссоциации KD по следующей формуле:
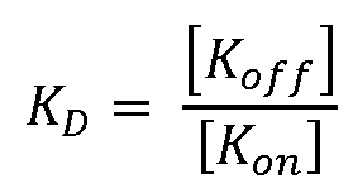
Используемый здесь термин «лечить» или «лечение» любого заболевания или расстройства (например, рака или реакции трансплантат против хозяина) относится в одном варианте осуществления изобретения к облегчению заболевания или расстройства (например, замедлению, остановке или уменьшению развития заболевания или, по меньшей мере, одного из его клинических симптомов). В другом варианте осуществления изобретения термин «лечить» или «лечение» относится к облегчению или улучшению, по меньшей мере, одного физического параметра, включая те, которые могут быть не различимы пациентом. В еще одном варианте осуществления изобретения термин «лечить» или «лечение» относится к модуляции заболевания или расстройства либо физически (например, стабилизация различимого симптома), либо физиологически (например, стабилизация физического параметра), или и тем, и другим способом. Способы оценки лечения и/или предотвращения заболевания общеизвестны в данной области техники, если специально не описано ниже.
В контексте настоящего описания термин «аптамер» относится к олигонуклеотиду или пептиду, способному специфично связываться с другой молекулой или мишенью с высоким сродством, имеющим Kd≤10E-8 моль/л. Аптамер связывается со своей мишенью аналогично специфичному связыванию антитела. Термин «аптамер» включает молекулы РНК или ДНК, аналоги нуклеиновых кислот или молекулы пептидов. Аптамер также может быть связан с саморасщепляющейся молекулой РНК, так называемым рибозимом. Известны способы получения аптамеров de novo; к ним относятся так называемый подход «SELEX» и другие методы, основанные на эволюции конкретных связующих из случайного выбранных молекул.
Используя мышиные модели ТГК, полностью или частично несовместимой с MHC, авторы настоящего изобретения показали, что ГМ-КСФ (Csf-2), цитокин, играющий важную роль в ряде воспалительных заболеваний, в большом количестве продуцируется донорскими Т-клетками сразу после трансплантации. Когда донорские Т-клетки не содержали ГМ-КСФ, РТПХ значительно уменьшалась. Важно и неожиданно, что отсутствие ГМ-КСФ не влияло на способность донорских Т-клеток контролировать рост опухоли у мышей (эффект ТПЛ), и этот контроль был достигнут без возникновения РТПХ даже в контексте полной MHC-несовместимости. Авторы настоящего изобретения также обнаружили высокие уровни ГМ-КСФ в биоптатах желудочно-кишечного тракта от пациентов с РТПХ, что согласуется с их параллельной ролью у человека. Таким образом, авторы настоящего изобретения предлагают ГМ-КСФ в качестве новой терапевтической мишени для ослабления РТПХ при сохранении ТПЛ у пациентов, получающих алло-ТГК.
Хотя использование антител против ГМ-КСФ или против рецептора ГМ-КСФ было предложено в контексте РТПХ, данные, представленные в настоящем изобретении, впервые показывают, что РТПХ можно лечить или подавлять, не нарушая терапевтический эффект аллогенного трансплантата, и что существует клиническое обоснование применения лигандов настоящего изобретения для обеспечения возможности организма формировать реакцию трансплантат против лейкоза при подавлении РТПХ (опосредованной фагоцитами). Это позволит проводить алло-ТГК у пациентов, которым в настоящее время не назначается алло-ТГК из-за риска, связанного с РТПХ, и может дополнительно ограничить использование иммунодепрессантов после алло-ТГК.
В первом аспекте настоящее изобретения относится к неагонистическому лиганду, специфично связывающемуся с ГМ-КСФ или с одним из CD116, CD131 и рецептором ГМ-КСФ, состоящим из CD116 и CD131, для применения при лечении лейкоза у пациента, перенесшего аллогенную трансплантацию гемопоэтических стволовых клеток (алло-ТГК). Другими словами, лиганд применяется для лечения лейкоза после аллогенной трансплантации гемопоэтических стволовых клеток.
Неагонистический лиганд настоящего изобретения подавляет биологический сигнал, подаваемый ГМ-КСФ на свой рецептор, что приводит к последующим эффектам взаимодействия ГМ-КСФ. Соответственно, понятно, что описанные здесь способы можно использовать для ингибирования или уменьшения тяжести РТПХ в контексте любого аллогенного трансплантата, в котором может возникнуть РТПХ, но без значительного уменьшения полезного эффекта аллогенного трансплантата. В конкретных вариантах осуществления изобретения, описанных, в настоящей заявке описываются способы лечения гемобластоза, включая лейкоз, лимфому или множественную миелому, путем введения пациенту описанного неагонистического лиганда одновременно с или после проведения лечения злокачественного новообразования при помощи алло-ТГК. В других вариантах осуществления изобретения способы ингибирования или уменьшения тяжести РТПХ позволяют получить аллогенный трансплантат, в котором существует риск индукции РТПХ за счет достаточного количества продуцирующих или индуцирующих ГМ-КСФ клеток, трансплантированных вместе с аллотрансплантатом. Трансплантация печени является одним из неограничивающих примеров такой трансплантации.
В некоторых вариантах осуществления изобретения неагонистический анти-ГМ-КСФ, анти-CD116, анти-CD131 или анти-рецептор ГМ-КСФ лиганд представляет собой антитело, фрагмент антитела, антителоподобную молекулу, аптамер или производный полипептид доменов белка A. Хотя применение антител, в частности моноклональных антител, является стандартным для терапевтических целей у пациентов-людей, и в действительности ГМ-КСФ-специфичные антитела и CD116-специфические антитела были разработаны для других терапевтических целей, квалифицированному специалисту в данной области техники понятно, что другие молекулы, такие как как дарпины, аптамеры или молекулы, производные от антител, могут применятся, по существу, для той же цели.
В некоторых вариантах осуществления изобретения неагонистический анти-ГМ-КСФ, анти-CD116, анти-CD131 или анти-рецептор ГМ-КСФ полипептидный лиганд представляет собой иммуноглобулин, состоящий из двух тяжелых цепей и двух легких цепей. В некоторых вариантах осуществления изобретения неагонистический анти-ГМ-КСФ, анти-CD116, анти-CD131 или анти-рецептор ГМ-КСФ полипептидный лиганд представляет собой однодоменное антитело, состоящее из изолированного вариабельного домена тяжелой или легкой цепи. В некоторых вариантах осуществления изобретения неагонистический анти-ГМ-КСФ, анти-CD116, анти-CD131 или анти-рецептор ГМ-КСФ полипептидный лиганд представляет собой антитело тяжелой цепи, состоящее только из тяжелых цепей, такое как антитела, обнаруженные у верблюдовых.
В некоторых вариантах осуществления неагонистический анти-ГМ-КСФ, анти-CD116, анти-CD131 или анти-рецептор ГМ-КСФ полипептидный лиганд представляет собой фрагмент антитела. В некоторых вариантах осуществления изобретения неагонистический анти-ГМ-КСФ, анти-CD116, анти-CD131 или анти-рецептор ГМ-КСФ полипептидный лиганд представляет собой Fab-фрагмент, то есть антигенсвязывающий фрагмент антитела или одноцепочечный вариабельный фрагмент, то есть слитый белок вариабельной области тяжелой и легкой цепи антитела, соединенных пептидным линкером.
Эффект лечения антителом к ГМ-КСФ на мышиной модели лечения лейкоза после алло-ТГК показан на Фигуре 4.
В некоторых вариантах осуществления изобретения лиганд представляет собой моноклональное антитело. В некоторых вариантах осуществления изобретения лиганд представляет собой человеческое антитело.
В некоторых вариантах осуществления изобретения лиганд для применения в способе лечения гемобластоза, такого как лейкоз, у пациента, перенесшего аллогенную трансплантацию гемопоэтических стволовых клеток (алло-ТГК), представляет собой гуманизированное антитело. В некоторых вариантах осуществления изобретения лиганд представляет собой химерное антитело.
В некоторых вариантах осуществления изобретения лиганд для применения в способе лечения гемобластоза, такого как лейкоз, у пациента, перенесшего алло-ТГК, представляет собой маврилимумаб (CAS No. 1085337-57-0).
В некоторых вариантах осуществления изобретения лиганд для применения в способе лечения гемобластоза, такого как лейкоз, у пациента, перенесшего алло-ТГК, представляет собой намилумаб (CAS No. 1206681-39-1).
В некоторых вариантах осуществления изобретения лиганд для применения в способе лечения гемобластоза, такого как лейкоз, у пациента, перенесшего алло-ТГК, представляет собой лензилумаб (CAS No. 1229575-09-0). В некоторых вариантах осуществления изобретения лиганд для применения в способе лечения гемобластоза, такого как лейкоз, у пациента, перенесшего алло-ТГК, представляет собой отилимаб (MOR103 или GSK-3196165; CAS No. 1638332-55-4). В некоторых вариантах осуществления изобретения лиганд для применения в способе лечения гемобластоза, такого как лейкоз, у пациента, перенесшего алло-ТГК, представляет собой гимсилумаб (MORAb-022; CAS No. 1648796-29-5).
Следует принимать во внимание, что в конкретных вариантах осуществления изобретения антитело, такое как маврилимумаб, намилумаб, лензилумаб, отилимаб или гимсилумаб, можно применять в способах получения аллогенного трансплантата, в которых ингибируется развитие РТПХ или уменьшается ее тяжесть.
В некоторых вариантах осуществления изобретения лиганд для применения в способе лечения лейкоза у пациента, перенесшего алло-ТГК, характеризуется KD меньшей (<) 10-7 моль/л-1, предпочтительно KD <10-8 моль/л-1, более предпочтительно KD <10-9 моль/л-1.
Во втором аспекте настоящее изобретение относится к молекуле нуклеиновой кислоты, кодирующей лиганд, описанный первом аспекте изобретения или в любом из его конкретных вариантов осуществления, для применения в лечении лейкоза у пациента, перенесшего алло-ТГК.
В некоторых вариантах осуществления изобретения молекула нуклеиновой кислоты для применения в лечении гемобластоза, такого как лейкоз, у пациента, перенесшего алло-ТГК, представляет собой одноцепочечную или двухцепочечную молекулу ДНК или одноцепочечную или двухцепочечную молекулу РНК.
В некоторых вариантах осуществления изобретения молекула нуклеиновой кислоты для применения в лечении лейкоза или другого злокачественного новообразования у пациента, перенесшего алло-ТГК в соответствии с настоящим изобретением, представляет собой экспрессионную конструкцию нуклеиновой кислоты, содержащую указанную выше последовательность нуклеиновой кислоты под контролем промотора, действующего в клетке млекопитающего.
В некоторых вариантах осуществления изобретения молекула нуклеиновой кислоты для применения в лечении гемобластоза, такого как лейкоз, у пациента, перенесшего алло-ТГК, представляет собой экспрессионную конструкцию, выбранную из плазмидной ДНК, двухцепочечной линейной ДНК, одноцепочечной РНК и вируса, в частности лентивируса, вируса герпеса, аденовируса или аденоассоциированного вируса.
Экспрессионная конструкция нуклеиновой кислоты используется для введения конкретного гена в клетку-мишень и может управлять клеточным механизмом синтеза белка для получения кодируемого геном белка. Экспрессионную конструкцию нуклеиновой кислоты конструируют таким образом, чтобы она содержала регуляторные последовательности, которые действуют как промоторные области, и при необходимости также содержала последовательности, которые действуют как энхансерные области и приводят к эффективной транскрипции гена, переносимого в экспрессионной конструкции.
Экспрессионные векторы нуклеиновых кислот для использования в описанных способах широко применяются в данной области техники. Примеры векторов для экспрессии белков млекопитающих включают векторы, доступные, среди прочего, от фирм Promega Corp (Madison, WI) и Thermo Fisher Scientific, Inc. (Waltham, MA).
В другом варианте осуществления изобретения, в качестве альтернативы введению неагонистического лиганда, описанные способы лечения включают ингибирование экспрессии ГМ-КСФ, его рецептора (CD116 и CD131) или отдельной субъединицы рецептор ГМ-КСФ. (CD1 16 и/или CD131). В таких способах нуклеиновая кислота, способная ингибировать экспрессию ГМ-КСФ или его рецепторных пептидов, вводится нуждающемуся в этом пациенту (например, после или во время алло-ТГК). В конкретных вариантах осуществления изобретения нацеливающая нуклеиновая кислота может представлять собой антисмысловую ДНК, миРНК или тому подобное. Способы определения подходящей целевой и нацеливающей последовательностей известны в данной области техники.
Альтернатива вышеуказанным аспектам настоящего изобретения относится к фармацевтической композиции для применения в лечении гемобластоза, такого как лейкоз, у пациента, перенесшего алло-ТГК. Эта композиция включает неагонистический лиганд, специфически связывающийся с ГМ-КСФ или одним из CD1 16, CD131 и рецептором ГМ-КСФ, состоящим из CD116 и CD131, или кодирующую его экспрессионную конструкцию нуклеиновой кислоты и фармацевтически приемлемый носитель, в частности изготовленная в виде формы для парентерального введения, более конкретно для внутривенного введения.
Антитела против ГМ-КСФ и против одного из CD116, CD131 и рецептора ГМ-КСФ, состоящего из CD116 и CD131, известны в данной области техники. Моноклональные антитела, специфичные к ГМ-КСФ, были разработаны и испытаны клинически на эффективность при ревматоидном артрите. Неограничивающие примеры антител для практического применения в настоящем изобретении включают антитела CD1 16/131, раскрытые в заявках на патент США US2014079708, US2012141464, US2009130093, US2014079708 и US2015376285, все авторов Cohen et al. (Маврилимумаб), включенные в настоящее изобретение путем ссылки.
Другим неограничивающим примером является анти-ГМ-КСФ антитело (отилимаб), раскрытое в заявке на патент США US2015246969, Haertle et al., и антитела, раскрытые в заявках на патент США US2011189082 и US2013071923, Kirchner et al., которые включены в настоящее изобретение путем ссылки.
Другим неограничивающим примером является анти-ГМ-КСФ антитело, раскрытое в заявках на патент США US2009053213, US2011045000, US2017218061, Steidl et al., которые включены в настоящее изобретение путем ссылки.
Другое антитело против ГМ-КСФ разрабатывается компанией Takeda для применения при ревматоидном артрите под наименованием намилумаб.
Другое антитело против ГМ-КСФ проходит клинические испытания на субъектах с ранее леченным хроническим миеломоноцитарным лейкозом (ХММЛ) под наименованием лензилумаб (CAS No. 1229575-09-0).
Аналогичным образом, в объем настоящего изобретения включен способ лечения гемобластоза, такого как лейкоз, у пациента, перенесшего алло-ТГК, включающий введение пациенту неагонистического лиганда, в частности антитела, специфично связывающегося с ГМ-КСФ или с одним из CD116, CD131 и рецептором ГМ-КСФ, состоящим из CD116 и CD131, в соответствии с приведенным выше описанием.
Аналогичным образом настоящее изобретение относится к лекарственной форме для предотвращения или лечения лейкоза у пациента, перенесшего алло-ТГК, содержащей лиганд или конструкцию нуклеиновой кислоты в соответствии с одним из вышеуказанных аспектов изобретения.
Лекарственные формы могут быть предназначены для энтерального введения, такого как назальное, буккальное, ректальное, трансдермальное или пероральное введение, или в виде ингаляционной формы или суппозитория. В качестве альтернативы можно использовать парентеральное введение, такое как формы для подкожной, внутривенной, внутрипеченочной или внутримышечной инъекции. При необходимости в рецептуре лекарственной формы может присутствовать фармацевтически приемлемый носитель и/или вспомогательное вещество.
Везде, где альтернативы для отдельных отделяемых признаков указаны в настоящей заявке как «варианты осуществления изобретения», следует понимать, что такие альтернативы могут свободно комбинироваться для образования отдельных вариантов осуществления раскрытого здесь изобретения.
Далее настоящее изобретение в качестве примера описывается следующими пунктами:
Пункт 1: Неагонистический лиганд, специфично связывающийся и способный нейтрализовать физиологическую функцию
- ГМ-КСФ или
- одного из CD116, CD131 и рецептора ГМ-КСФ, состоящего из CD116 и CD131,
для применения в лечении РТПХ.
Пункт 2: Лиганд для применения в лечении РТПХ по пункту 1, где лиганд представляет собой антитело, фрагмент антитела, аптамер или антителоподобную молекулу.
Пункт 3: Лиганд по пункту 1 или 2 для применения в лечении РТПХ, где лиганд представляет собой человеческое антитело или гуманизированное антитело.
Пункт 4: Лиганд по любому из предыдущих пунктов для применения в лечении РТПХ, выбранный из маврилимумаба, намилумаба, лензилумаба, MOR103 и MORAb-022.
Пункт 5: Лиганд по любому из предыдущих пунктов для применения в лечении РТПХ, где связывание лиганда с ГМ-КСФ или одним из CD116, CD131 и рецептором ГМ-КСФ, состоящим из CD116 и CD131, характеризуется KD меньше чем (<) 10-7, предпочтительно KD <10-8, более предпочтительно KD <10-9.
Пункт 6: Молекула нуклеиновой кислоты, кодирующая лиганд по любому из предыдущих пунктов с 1 по 4, для применения в лечении РТПХ.
Пункт 7: Молекула нуклеиновой кислоты для применения в лечении РТПХ по пункту 6, где молекула нуклеиновой кислоты представляет собой молекулу ДНК или молекулу РНК.
Пункт 8: Экспрессионная конструкция нуклеиновой кислоты, содержащая последовательность нуклеиновой кислоты по пункту 5 или 7 для лечения РТПХ.
Пункт 9: Экспрессионная конструкция нуклеиновой кислоты для применения в лечении РТПХ по пункту 8, где экспрессионная конструкция выбрана из плазмидной ДНК, двухцепочечной линейной ДНК, одноцепочечной РНК и вируса, в частности лентивируса, вируса герпеса, аденовируса или аденоассоциированного вируса.
Пункт 10: Лиганд по любому из предыдущих пунктов с 1 по 5 или молекула нуклеиновой кислоты по пунктам 6 или 7, или экспрессионная конструкция нуклеиновой кислоты по пункту 8 или 9, для применения при лечении осложнений, возникающих вследствие алло-ТГК.
Пункт 11: Лиганд по любому из предыдущих пунктов с 1 по 5 или молекула нуклеиновой кислоты по пунктами 6 или 7, или экспрессионная конструкция нуклеиновой кислоты по пункту 8 или 9 для применения при лечении осложнений, возникающих вследствие алло-ТГК, у пациента, страдающего гемобластозом или излечившегося от гемобластоза,
- предпочтительно гемобластозом, выбранным из лейкоза, лимфомы и множественной миеломы,
- более предпочтительно гемобластозом, выбранным из хронического миелоидного лейкоза (ХМЛ), острого миелогенного лейкоза (ОМЛ), хронического лимфоцитарного лейкоза (ХЛЛ), острого лимфоцитарного лейкоза (ОЛЛ) и острого моноцитарного лейкоза (ОМоЛ), лимфомы Ходжкина или неходжкинской лимфомы.
Настоящее изобретение дополнительно проиллюстрировано следующими примерами и фигурами, из которых можно понять дополнительные варианты осуществления изобретения и его преимущества. Эти примеры предназначены для иллюстрации изобретения, но не для ограничения его объема.
Примеры
Материалы и методы
Дизайн исследования
Исследование было начато для определения того, могут ли определенные цитокины, из Т-клеток, отделять РТПХ от ТПЛ и, таким образом, могут представлять собой новые многообещающие терапевтические мишени для лечения гемобластоза. Для достижения этой цели авторы настоящего изобретения использовали две разные экспериментальные модели РТПХ, экспериментальную модель ТПЛ и пациентов-людей. Для исследований на животных использовали мышей в возрасте от 8 до 12 недель. Все эксперименты на животных были одобрены местными властями (Швейцарское кантональное ветеринарное управление) и проводились в соответствии с соответствующими экспериментальными лицензиями (76/2012 и 052/2015). Животных случайным образом распределяли по экспериментальным группам, и клиническую оценку при жизни выполняли слепым методом, а также анализировали изображения срезов органов. Размер выборки и конечные сроки заболевания были выбраны на основе предыдущих исследований. Для характеристики органов-мишеней РТПХ/ТПЛ использовали проточную цитометрию, гистопатологический анализ, реакцию смешанной культуры лимфоцитов (СКЛ), киллинг-анализ и анализ цитокинов. Влияние специфичного блокирующего ГМ-КСФ антитела на клинические показатели оценивали исследователи, которые не принимали участие в лечении.
Для проведения надежного статистического анализа было проведено не менее трех независимых экспериментов для каждых данных, представленных в рукописи, если иное не указано в подписях к Фигурам. Все человеческие образцы были собраны после одобрения этическим комитетом Университета Альберта-Людвига во Фрайбурге, Германия (номер протокола: 267/1 1) после письменного информированного согласия. Авторы настоящего изобретения проводили иммуногистохимию и количественную ОТ-ПЦР на биопсиях кишечника и многопараметрическую проточную цитометрию на МКПК.
Мыши и манипуляции in vivo. Мышей содержали в лаборатории в индивидуальных вентилируемых клетках в условиях, свободных от патогенной микрофлоры. Мыши линий ДТ C57BL/6, BALB/c и B6D2F1 были приобретены в Janvier Laboratories, Франция. Конгенную линию мышей C57BL/6 CD45.1 получали в лаборатории самостоятельно. Мышей Ifng-/- получали от Jackson Laboratories; Мыши Csf2-/- были предоставлены Джеффри Уитсеттом и затем подвергнуты обратному скрещиванию с мышами C57BL/6 с использованием стратегии быстрого получения конгеников. Мыши Il17a-/- были предоставлены Y. Iwakura. Все эксперименты на животных были одобрены местными властями (Швейцарское кантональное ветеринарное управление) и проводились в соответствии с соответствующими экспериментальными лицензиями (76/2012 и 052/2015). Для экспериментов использовали самок мышей в возрасте 7-12 недель.
Индукция РТПХ . Суспензии отдельных клеток селезенки и костного мозга донорских мышей C57BL/6 или BALB/c получали, как описано ниже (в разделе «Выделение лимфоцитов»). Для истощения пула Т-клеток в костном мозге клетки инкубировали при концентрации 25×106 клеток/мл в полной среде RMPI (RPMI 1640 с 10% ФТС, 1% пенициллина/стрептомицина, 2 мМ L-глутамина и 0,5 мМ b-меркаптоэтанола) с αCD90.2-биотином (1:100, eBioscience) в течение 30 минут при температуре 4°C при перемешивании. Затем, после стадии отмывки холодным буфером MACS (ФСБ с 0,5% БСА и 2 мМ ЭДТА), клетки ресуспендировали при концентрации 100 × 106 клеток/мл в буфере MACS и инкубировали с гранулами антибиотина (1:5, Miltenyi) или гранулами стрептавидина (1:10, Miltenyi) в течение 20 минут при температуре 4°C при перемешивании. После промывки буфером MACS клетки разделяли с помощью сепаратора AutoMACS Pro, Miltenyi Biotech и программы «истощение». Отрицательную фракцию собирали, и количество клеток определяли с помощью счетной камеры. Эффективность истощения анализировали с помощью проточной цитометрии. Частота Т-лимфоцитов в костном мозге снижалась с 1-3% до 0,055 +/- 0,022%. Для выделения нетронутых Т-клеток из спленоцитов использовали набор реагентов для выделения Т-клеток Pan (II) от Miltenyi в соответствии с протоколом производителя. Клетки разделяли с помощью сепаратора AutoMACS Pro, Miltenyi Biotech и программы «истощение». Отрицательную фракцию собирали, и количество клеток определяли с помощью счетной камеры. Эффективность обогащения анализировали с помощью проточной цитометрии, и чистота Т-клеток обычно составляла 95,2 +/- 2,16%. CD4 и CD8 Т-клетки разделяли с использованием наборов реагентов для выделения CD4 Т-клеток мыши MojoSort и наборов реагентов для выделения CD8 Т-клеток мыши MojoSort в соответствии с инструкциями производителя (Biolegend).
Мышей-реципиентов BALB/c или B6D2F1 летально облучали в условиях, свободных от патогенной микрофлоры в фильтрующих клетках с разделенной дозой 850 (BALB/c), 1200 (B6D2F1) или 1100 (C57BL/6) рад, разделенных на, по меньшей мере, 5 часов. Реципиентам внутривенно вводили 5 × 106 клеток костного мозга и 0,1-10 × 106 спленоцитов или 6-7,5 * 106 Т-клеток на мышь C57BL/6 или B6D2F1. Реципиентам внутривенно вводили 7 × 106 клеток костного мозга и 20 × 106 спленоцитов на мышь BALB/c. Мышей давали 0,1% боргал (Intervet) в питьевой воде в течение трех недель для предотвращения бактериальных инфекций. Мышей также ежедневно оценивали на симптомы РТПХ, взятые из (Cooke, K. R. et al. An experimental model of idiopathic pneumonia syndrome after bone marrow transplantation: I. The roles of minor H antigens and endotoxin. Blood 88, 3230-3239 (1996)) и показаны в Таблице 4 в приложении. При оценке РТПХ исследователи не знали о распределении по группам. Контрольную массу измеряли в день 0 перед инъекцией клеток трансплантата. Для блокирования ГМ-КСФ мышам вводили ФСБ, 300 мкг изотипического контроля (2A3) или 300 мкг антитела против ГМ-КСФ (MP1) (BioXCell) 3 раза в неделю, начиная за 2 дня до ТГК.
Индукция ТПЛ : Линия клеток A20-люцифераза+GFP+ В-клеточной лимфомы (A20-luc-gfp, любезно предоставленная Эммой Свенссон, Департамент иммунологии, генетики и патологии, Университет Упсалы, Швеция) была создана на основе BALB/c путем трансдукции клеток A20 (ATCC) с лентивирусом, кодирующим CMV-GFP-(T2A)-Luc. Клетки WEHI-3-люцифераза+ были предоставлены R. Zeiser. В день ТГК мышам внутривенно инъецировали 2,5 × 105 клеток A20, 5 × 105 клеток A20 или 1 × 105 клеток WEHI-3 на мышь вместе с 5 × 106 клеток ДТ C57BL/6 КМ, или отдельно, или с 1 × 105 очищенных Т-клеток селезенки на мышь или от мыши ДТ C57BL/6 или от мыши Csf2-/- C57BL/6. В день ТГК 2,5 × 105 клеток A20 на мышь инъецировали внутривенно вместе с 5 × 106 клеток ДТ C57BL/6 BM или отдельно, или с 1 × 105 очищенных Т-клеток селезенки на мышь или от мыши ДТ C57BL/6 или от мыши Csf2-/- C57BL/6. Прогрессирование опухоли контролировали с помощью биолюминесцентной визуализации (БЛВ): мышам внутрибрюшинно вводили 150 мг/кг D-люциферина (Promega) в ФСБ за десять минут до визуализации с использованием доклинической системы визуализации in vivo Xenogen IVIS 200 (PerkinElmer, Waltham, MA.); время экспозиции 1-120 с, биннинг 2-8, поле зрения 15 см, диафрагма 1, без фильтра. Мышей анестезировали изофлураном (2% испарения в 02) до и во время визуализации. Общий поток фотонов (фотонов/сек) измеряли из фиксированной интересующей области (ИО) по всему телу с помощью программного обеспечения Living Image (PerkinElmer). Фотографии всегда делались для каждой клетки и обрезались, когда разные группы мышей смешивались в одной клетке. Смертность от опухолей и РТПХ различалась по интенсивности сигнала ВЛВ (ИО > 1 * 107 для частоты опухолей), параличу задних конечностей (указывающему на развитие опухоли) и клиническим проявлениям РТПХ (по меньшей мере, 2 степени по двум отдельным критериям РТПХ). Для блокирования ГМ-КСФ мышам вводили 300 мкг изотипического контроля (2A3) или 300 мкг антитела против ГМ-КСФ (MP1) (BioXCell) 3 раза в неделю, начиная за 2 дня до ТГК. С недели 3 доза антител снижали до 150 пг.
Выделение лимфоцитов. Мышей умерщвляли с помощью ингаляции CO2 и перфузировали 40 мл холодного ФСБ. Если ниже не указано иное, органы собирали, разрезали на мелкие кусочки и инкубировали с коллагеназой при температуре 37°C с последующим механическим разрушением путем повторного пропускания через иглу 20 калибра. Проводили лизис эритроцитов. Полученную суспензию клеток затем фильтровали через клеточное сито с размером пор 70 мкм и использовали для дальнейших процедур.
Селезенка: для выделения миелоидных клеток кусочки органов инкубировали в 2 мл коллагеназы D (0,4 мг/мл, Roche) и растворе 0,1 мг/мл ДНКазы I (Sigma) в RPMI в течение 30 минут при температуре 37°C. Для выделения Т-клеток селезенки гомогенизировали механическим разрушением и фильтровали через клеточное сито с размером пор 70. КМ: бедренные кости, большеберцовые кости и таз промывали ФСБ для получения стволовых клеток костного мозга. Печень: кусочки органов инкубировали в растворе 1,6 мг/мл коллагеназы типа IV (из Clostridium histolyticum, Sigma) в HBSS, содержащем 10% ФТС, в течение приблизительно 45 минут при температуре 37°C. Клетки ресуспендировали в 10 мл перколла (непрерывный градиент, 27%, GE) и центрифугировали в течение 30 минут при 1700 об/мин при комнатной температуре. Жир и супернатант удаляли, а осадок подвергали лизису красных кровяных телец. Кожа: кусочки органов инкубировали в растворе 1 мг/мл коллагеназы типа IV (из Clostridium histolyticum, Sigma) и 0,1 мг мл ДНКазы I (Sigma) в RPMI в течение 1,5-2 часов при температуре 37°C. Тонкий кишечник: органы отделяли от мезентериального жира до механического удаления слизи из просвета; Затем органы инкубировали в растворе HBSS, не содержащем кальция и магния, содержащем 2% ФТС, 1 мМ DTT и 1,35 мМ ЭДТА в течение 15 минут при температуре 37°C. После дополнительной инкубации в HBSS, дополненном ЭДТА, в течение 30 минут при температуре 37°C, толстый кишечник вырезали и расщепляли с использованием 0,4 мг/мл коллагеназы IV (Sigma Aldrich) в течение 45 минут при температуре 37°C. Затем образцы гомогенизировали с помощью шприца с иглой 18 калибра и фильтровали через клеточное сито с размером пор 70 мкм.
Проточная цитометрия. Проточный цитометрический анализ выполняли с помощью стандартных методов, рассмотренных в (Perfetto, S. P., Chattopadhyay, P. K. & Roederer, M. Seventeen-colour flow cytometry: unravelling the immune system. Nat Rev Immunol 4, 648-655 (2004)). Оптимальные концентрации всех антител, конъюгированных с флуорохромом, были определены с помощью экспериментов по титрованию. Клоны антител, специфичных по отношению к мышиным CD4 (GK1.5), CD8 (53-6.7), CD3 (17A2), CD45 (30F11), CD45.1 (A20), CD45.1 (104), CD44 (IM7), CD62L (MEL-14), CD69 (H1.2F3) MHC Класс I H2-Dd (34-2-12) и H2-Db (KH95), ГМ-КСФ (MP1-22E9), ИФНγ (TC11-18H10), IL-17A (XMG1.2) и FoxP3 (FJK-16s) были приобретены или у BD, BioLegend, или у eBioscience. Для окрашивания поверхности клетки инкубировали с соответствующими антителами в течение 20-30 минут при температуре 4°C. Во всех экспериментах мертвые клетки исключали из анализа с использованием фиксируемого окрашивающего реагента Aqua или Near-IR Live/Dead (Invitrogen/BioLegend); дублеты были исключены с помощью гейтирования на гистограмме области FSC против высоты FSC. Для окрашивания внутриклеточных цитокинов Т-клетки инкубировали в течение 4-5 часов при температуре 37°C в среде RPMI, содержащей 10% ФТС с ФМА (50 нг/мл), лономицином (500 нг/мл) и GolgiPlug (содержащим брефелдин A, BD, разведение 1:1000). После окрашивания поверхности набор реагентов Cytofix/Cytoperm (BD) использовали в соответствии с инструкциями производителя, и буфер для пермеабилизации/промывки готовили непосредственно в лаборатории (ФСБ, содержащий 0,5% сапонина и 5% БСА). Для внутриклеточного окрашивания клетки инкубировали с соответствующими антителами в течение 20-30 минут при температуре 4°C. Для внутриядерного окрашивания FoxP3 после окрашивания поверхности использовали буфер фиксации/пермеабилизации (eBioscience), а затем буфер для пермеабилизации/промывки, приготовленный непосредственно в лаборатории. В целом, клетки были отобраны на основе гейтирования области FSC и области SSC, чтобы исключить дебрис, дублеты были исключены с помощью гейтирования на гистограмме области FSC против высоты FSC. Мертвые клетки исключали из анализа с использованием фиксируемого окрашивающего реагента Aqua или Near-IR Live/Dead (Invitrogen/BioLegend). Там, где это применимо, клетки CD45.1 гейтировали перед гейтированием CD4 или CD8 Т-клеток. Все маркеры, используемые для гейтирования (CD4, CD8, CD3, CD44, CD62L, CD69), показали четкое разделение отрицательной и положительной популяции. Для киллинг-анализа синглеты гейтировали, как описано выше, затем опухолевые клетки идентифицировали по фиолетовому следу клеток и строили гистограмму, показывающую поглощение TOPRO-3. Проточный цитометрический анализ проводили с использованием проточного цитометра FACSCanto II (BD) или LSR II Fortessa (продукт для исследований по специальному заказу, BD и оснащенный лазерными линиями 405 нм, 488 нм, 561 нм и 640 нм) с программным обеспечением FACS Diva. Анализ данных выполняли с помощью программного обеспечения FlowJo 10.0.x (Treestar).
Проточно-цитометрический анализ фосфорилирования STAT5
Суспензии единичных клеток костного мозга подвергали окрашиванию поверхности клеток в течение 20 минут, после чего к образцам добавляли среду, содержащую ГМ-КСФ (20 нг/мл). Клетки инкубировали при температуре 37°C в течение 30 минут для индукции фосфорилирования STAT5 перед добавлением 4% раствора ПФА (pH 7,4) до конечной концентрации 2%. Клетки фиксировали в течение 20 минут, промывали и ресуспендировали в 1 мл метанола при температуре 4°C. Через 40 минут клетки дважды промывали и ресуспендировали в 45 мл буфера FACS. Добавляли 5 мл анти-pSTAT5 АПК (BD Biosciences), чтобы конечное соотношение окрашивания составляло 1:10. Клетки инкубировали в течение 45 минут при температуре 4°C перед последним этапом промывки и сбором. Проточно-цитометрический анализ проводили с использованием проточного цитометра LSR II Fortessa (продукт для исследований по специальному заказу, BD и оснащенный лазерными линиями 405 нм, 488 нм, 561 нм и 640 нм) с программным обеспечением FACS Diva. Анализ данных выполняли с помощью программного обеспечения FlowJo 10.0.x (Treestar).
Фенотипический анализ МКПК человека методом проточной цитометрии
Криоконсервированные МКПК хранили в жидком азоте до оттаивания на водяной бане при температуре 37°C. Клетки осторожно ресуспендировали в 1 мл предварительно нагретой среды для культивирования клеток (CCM; RPMI-1640 [PAN biotech], 10% ФТС [Biochrom]), 1 × l-глутамин и 1 × пенициллин/стрептомицин [оба Life Technologies]) с добавлением бензоназы 1:10000 (Invitrogen). Затем клетки переносили в 5 мл пробирки и промывали CCM. Клетки подсчитывали и доводили до концентрации 20 × 106 клеток/мл в CCM. Для определения продукции цитокинов с помощью проточной цитометрии клетки стимулировали раствором 50 нг/мл форбол 12-миристат 13-ацетата (ФМА; Sigma-Aldrich) и 1 мкг/мл иономицина (Sigma-Aldrich) в присутствии реактива GolgiPlug (содержащего брефельдин A, BD, разведение 1:1000) и реактива GolgiStop (содержащего монензин, BD, разведение 1:1000) в течение 4 часов при температуре 37°C. Неспецифичное связывание блокировали с помощью Human TruStain FcX (Biolegend).
Реакция смешанной культуры лимфоцитов (СКЛ). Иммунореактивные Т-клетки C57BL/6 ДТ, Csf2-/-, Ifng-/- или Il17a-/- культивировали совместно с ДК мышей BALB/c (аллогенные) или C57BL/6 (сингенные). Альтернативно, иммунореактивные Т-клетки- BALB/c ДТ совместно культивировали с ДК мышей BALB/c (сингенные) или ДК мышей C57BL/6 ДТ и Csf2-/- (аллогенные). Т-клетки и ДК очищали от спленоцитов при помощи положительной селекции с использованием анти-CD4 и анти-CD8 (для Т-клеток) или анти-CD11c (для ДК) магнитных частиц (Miltenyi Biotec), соответственно, и сепаратора AutoMACS Pro. Клетки высевали в трех повторностях в 96-луночные планшеты с U-образным дном при соотношении стимулятор/респондер 1/10 (2 * 104 ДК, 2 * 105 Т-клетки) в течение 3 дней в полной среде RPMI 1640 температуре 37°C и в атмосфере 5% СО2. После этого собирали супернатанты культур и заменяли разбавленным 3H-тимидином (1/100, 50 миКи) в течение 16 часов. Клетки собирали, используя 96-луночный коллектор клеток (PerkinElmer), и сцинтилляцию измеряли с помощью бета-счетчика (1450 Microbeta Plus, Wallac).
Киллинг-анализ. Т-клетки селезенки и лимфатических узлов выделяли с использованием набора реагентов Pan T Cell Isolation Kit II, мыши (MACS Miltenyi Biotec). Клетки A20-люцифераза+ GFP+ или WEHI-3-люцифераза+ окрашивали с помощью набора реагентов CellTrace™ Violet Cell Proliferation Kit (ThermoFisher Scientific, C34557). Клетки инкубировали при соотношении эффектор/мишень 1:1, 20:1 и 50:1 в присутствии растворимых α-CD3 (1 мкг/мл) и α-CD28 (0,5 мкг/мл) в среде RPMI с добавлением 10% ФТС в течение 24 часов при температуре 37°C и в атмосфере 5% СО2. После удаления среды к клеткам добавляли Topro (0,8 мкМ) и клетки анализировали на проточном цитометре LSRII Fortessa (BD). Данные анализировали с помощью программного обеспечения FlowJo Version X (Tree Star). Процент специфичного лизиса рассчитывали следующим образом: [(экспериментальный лизис - спонтанный лизис) / (максимальный лизис - спонтанный лизис)] × 100%.
Анализ подавления Treg
Т-клетки выделяли из лимфатических узлов и селезенки мышей C57BL/6 ДТ и Csf2-/-, как описано ранее. CD4 Т-клетки сначала обогащали с использованием набора реагентов для выделения CD4 (MojoSort™ mouse CD4 Nanobeads) с последующим окрашиванием CD45, CD25, CD62L и CD4. Клетки CD4+CD25-CD62L+ (Tconv) и CD4+CD25high (Treg) сортировали методом проточной цитометрии с использованием сортировщика клеток BD FACS ARIA II с чистотой 98-99%. АПК облучали > 3400 рад (4 мин, 12,2 Гр/мин, 225 кВ, 17,7 мА) и разводили до концентрации 4 × 106 клеток/мл (или 2 × 106 клеток/мл) в 10% среде для клонов. Клетки АПК и Tconv совместно культивировали с аутологичными клетками Treg при различных концентрациях в 96-луночном круглодонном планшете (4 × 104 Tconv на лунку). Клетки стимулировали αCD3 в концентрации 1 мкг/мл при температуре 37°C в течение 2 дней, а затем инкубировали в течение 16 часов с 3H-тимидином. Клетки собирали с помощью аппарата FilterMate (PerkinElmer) и анализировали с помощью бета-счетчика с программой Microbeta. Процент подавления Treg рассчитывали среди живых клеток как [((cpm Tconv - cpm Treg:Tconv) / cpmTconv отдельно) * 100], где cpmTconv - число импульсов в минуту, рассчитанное как % подавления Tconv клеток.
Анализ цитокинов. Сыворотку получали от мышей-реципиентов алло-ТГК в указанные сроки. Культуральные супернатанты СКЛ собирали после 3 дней совместного культивирования. Измерение ГМ-КСФ, ИФНγ и IL-17A в сыворотке или супернатантах клеточных культур проводили с помощью цитокин-специфичного ИФА в соответствии с инструкциями производителя (BD Biosciences и Biolegend).
Параметры функции печени
Ферменты печени (аланинаминотрансфераза (ALT), щелочная фосфатаза (AP)), альбумин и азот мочевины крови (BUN) измеряли в сыворотках крови мышей с использованием набора реагентов Piccolo Liver Panel Plus (Abaxis).
Гистология. Для гистопатологической оценки органы фиксировали в HOPE (DCS, Гамбург, Германия) и заливали парафином. Срезы размером 3-5 мкм вырезали с помощью микротома (Micro HM 325, Thermo Scientific) и затем окрашивали гематоксилином и эозином. Для получения гистологических оценок РТПХ использовали полуколичественную тканеспецифическая система оценки: параметры тонкой кишки, включая притупление ворсинок, регенерацию крипт, апоптоз клеток крипт, прямое разрушение крипт и лимфоцитарные инфильтраты собственной пластинки, оценивались следующими баллами: 0=нормально; 0,5=очаговый и редкий; 1=очаговый и легкий; 2=диффузный и мягкий; 3=диффузный и умеренный; 4=диффузный и тяжелый. Эта система оценки была заимствована из Hill, G. R. et al. Total body irradiation and acute graft-versus-host disease: the role of gastrointestinal damage and inflammatory cytokines. Blood 90, 3204-3213 (1997). Для кожи оценивали следующие параметры: вычисленное количество волосяных фолликулов: 0=нормально (относительно близко расположенные), 0,5=количество слегка уменьшено (обычно очагово); вычисленное количество сальных желез: 0=нормально (относительно большие клетки, небольшие группы, на уровне шейки волос), 1,0=немного уменьшено (количество и размер, обычно очагово), 2,0=явно уменьшено (присутствует только несколько клеток); апоптоз и дегенерация сальных желез: 0=отсутствует, 0,5=редко, 1=затронуто небольшое количество желез, 2=затронуто умеренное количество желез; апоптоз и дегенерация волосяных фолликулов: 0=отсутствует, 0,5=редко, 1=затронуто небольшое количество фолликулов, 2=затронуто умеренное количество фолликулов, 3=затронуто большое количество фолликулов; лейкоцитарная инфильтрация (экстра-придаточная): 0=низкая, 1=слегка повышенная, 2=умеренно повышенная; лейкоцитарная инфильтрация (придаточная): 0=отсутствует, 1=слабая, 2=умеренная; эпидермис (апоптозные клетки, лейкоцитарная инфильтрация): 0=отсутствует, 0,5=редко, 1=небольшое количество, 2=умеренное количество, 3=большое количество; подкожный жир: 0=непрерывно, 1=не непрерывно, 2=отсутствует, система оценки была заимствована из Kaplan, D. H. et al. Target antigens determine graft-versus-host disease phenotype. J. Immunol. 173, 5467-5475 (2004). Для параметров печени, включая расширение и инфильтраты портального тракта, инфильтраты желчных протоков, многослойность ядер желчных протоков, пикнотические клетки желчных протоков, внутриэпителиальные клетки желчных протоков, эндотелиалит сосудов, гепатоцеллюлярный пан-лобулярный некроз, ацидофильные тельца, микроабсцессы, митотические фигуры и гепатоцеллюлярный стеатоз, использовали следующие оценки: 0=нормально; 0,5=очаговый и редкий; 1=очаговый и легкий; 2=диффузный и мягкий; 3=диффузный и умеренный; 4=диффузный и тяжелый. Эта система оценки была заимствована из Hill, G. R. et al. Interleukin-1 1 promotes T cell polarization and prevents acute graft-versus-host disease after allogeneic bone marrow transplantation. J. Clin. Invest. 102, 1 15-123 (1998). Все слайды были закодированы и прочитаны слепым методом на микроскопе Olympus BX41 TF.
Люди. Все человеческие образцы были собраны после одобрения этическим комитетом Университета Альберта-Людвига во Фрайбурге, Германия (номер протокола: 267/11) после письменного информированного согласия. Биоптаты кишечной ткани были собраны проспективным образом у людей с РТПХ или здоровых людей в контрольной группе в период с 2007 по 2014 год. Оценка кишечной РТПХ проводилась опытным патологом на основе гистопатологии в соответствии с опубликованной системой стадирования Lerner, K. G. et al. Histopathology of graft-vs.-host reaction (GvHR) in human recipients of marrow from HL-A-matched sibling donors. Transplant. Proc. 6, 367-371 (1974). У контрольных индивидуумов не было никаких отклонений от нормы при биопсии кишечника. Характеристики пациентов, включая возраст реципиента, пол, основной диагноз, тип донора и трансплантата, режим подготовки к трансплантации, режим иммуносупрессии и степень РТПХ, подробно описаны в дополнительных Таблицах 1 и 3 для пациентов с РТПХ и в дополнительной Таблице 2 для контрольных субъектов.
Иммуногистохимия. Иммуногистохимию проводили на фиксированных залитых парафином срезах тканей с использованием набора реагентов для двухэтапного ИГХ окрашивания EnVision+System HRP DAKO (Glostrup, Дания) в соответствии с инструкциями производителя. Срезы подвергали тепловому извлечению антигена с помощью раствора для извлечения мишеней Dako (1 раз) в течение 10 мин. Активность эндогенной пероксидазы блокировали с помощью набора для блокирования эндогенной пероксидазы DAKO в течение 30 мин при комнатной температуре. Первичные антитела разводили в ФСБ+5% нормальной козьей сыворотки (NGS). Одно иммуноокрашивание состояло из инкубации в течение ночи при температуре 4°C с неконъюгированным mAb против человеческого ГМ-КСФ антитела (клон 3209.1; R&D Systems) или изотипическим контролем IgG для биоптатов от людей и крысиным α-F4/80 (клон CI: A3-1, BioRad) и кроличьим α-p22phox (клон FL-195, Santa Cruz) для срезов тканей мыши. В качестве хромогена использовали DAB. Затем срезы контрастировали гематоксилином, закрепляли с помощью DPX и анализировали с помощью светового микроскопа Olympus BX41, соединенного с цветной камерой ColorViwerlllu (Olympus), с использованием пакета программ Fiji/ImageJ (Стандартная общественная лицензия GNU).
Иммунофлюоресценция
Ткани подвергали криосрезу (толщиной 10 мкм) для иммуногистохимии с использованием криостата Hyrax C60 (Zeiss) и хранили при температуре -80°C. Срезы фиксировали в 4% ПФА, промывали ФСБ и блокировали ФСБ с добавлением 0,1% Тритона X-100 и 4% нормальной козьей сыворотки. Затем срезы инкубировали со следующими первичными антителами, крысиными анти-ГМ-КСФ антителом (BD Pharmingen, клон BVD2-21C11, 1:50), кроличьим α-CD3 (Novus, клон SP7, 1:200) и мышиным анти-CD68 антителом (DAKO, клон EMB11, 1:50), разбавленными в блокирующем растворе, в течение ночи при температуре 4°C. Затем срезы промывали ФСБ и инкубировали с AF647-меченным козьим антикрысиным, AF488-меченными козьим антимышиным и AF555-меченными ослиным антикроличьим вторичными антителами (Life Technologies, 1:500) в течение ночи при температуре 4°C или при комнатной температуре в течение 1 часа. Срезы фиксировали антибактериальным реагентом SlowFade Gold с ДАПИ (Invitrogen). Флуоресцентные микрофотографии получали с помощью конфокального лазерного сканирующего микроскопа Leica SP5 (SP5; Leica, Heerbrug, Швейцария), оснащенного аргоновым и гелиевым лазерами, с использованием объектива с 40-кратным увеличением (масляная иммерсия, NA1.25). Изображения обрабатывали и объединяли с помощью программного обеспечения Imaris imaging (Bitplane, Цюрих, Швейцария).
TUNEL анализ
Срезы, зафиксированные в 4% ПФА, обрабатывали для TUNEL анализа для обнаружения фрагментированных ядер в толстой кишке. Набор реагентов для обнаружения апоптоза in situ ApopTag Red использовали в соответствии с инструкциями производителя. В кратком изложении, срезы предварительно обрабатывали H2O2 и инкубировали с реакционной смесью, содержащей TdT и dUTP, конъюгированный с дигоксигенином, в течение 1 ч при температуре 37°C. Меченую ДНК визуализировали вторичным антителом, конъюгированным с AF488. Флуоресцентные микрофотографии получали с помощью конфокального лазерного сканирующего микроскопа Leica SP5 (SP5; Leica, Heerbrug, Швейцария), оснащенного аргоновым и гелиевым лазерами, с использованием объектива с 40-кратным увеличением (масляная иммерсия, NA1.25). Изображения обрабатывали и объединяли с помощью программного обеспечения Imaris imaging (Bitplane, Цюрих, Швейцария).
Выделение РНК и количественная ОТ-ПЦР. РНК выделяли из фиксированных залитых парафином срезов тканей с использованием набора реагентов RNeasy FFPE (Qiagen, депарафинизация с использованием ксилола) в соответствии с инструкциями производителя. Концентрацию РНК оценивали с помощью спектрофотометра NanoDrop 1000 (Thermo Scientific). Для синтеза первой цепи кДНК использовали обратную транскриптазу MLV (Invitrogen) в соответствии с инструкциями производителя. Экспрессию генов измеряли количественным ПЦР-анализом в реальном времени с использованием системы обнаружения Real-Time CFX 384 (Bio-Rad, Геркулес, Калифорния, США) с SYBR Green Supermix (Bio-Rad). Последовательности использованных ПЦР для праймеров приведены в таблице ниже. Экспрессию транскрипта нормализовали по гену домашнего хозяйства GAPDH и выражали как 2-ΔΔT (ΔΔCT = ΔCT-ΔCКонтроль).
Автоматическая идентификация популяции при анализе высокоразмерных данных
Данные, полученные в анализаторе клеток BD FACSymphony™, были скомпенсированы, экспортированы в FlowJo версии v10.4 (TreeStar), нормализованы с использованием Cyt MATLAB (версия 2017b), а объективный анализ был выполнен, как описано в Nowickaet al. FIOOORes 6, 748 (2017); Hartmann et al., J. Exp. Med. 213, 2621-2633 (2016).
Статистика. Графики строили с помощью программы GraphPad Prism (пакет программ GraphPad). Кривые выживаемости строили методом Каплана-Мейера, и для сравнения кривых выживаемости использовали логарифмический ранговый критерий (критерий Кокса-Мантеля). Данные приводятся как отдельные значения данных или как среднее +/- стандартное отклонение (СО) или стандартная ошибка среднего (СОС), как показано в подписях к Фигурам. Сравнение средних значений проводилось с использованием непарного двустороннего t-критерия Стьюдента (с поправкой Уэлча, где это применимо) или однофакторного и двухфакторного дисперсионного анализа с пост-тестом Бонферрони, соответственно. Никакие статистические методы не использовались для предварительного определения размера выборки, но используемые размеры выборки были аналогичны тем, которые обычно используются в этой области техники. Метод рандомизации не использовался.
Доступность данных. Наборы данных, созданные во время и/или проанализированные в ходе текущего исследования, доступны у соответствующих авторов по разумному запросу.
Пример 1:
Донорские Т-клетки, инфильтрирующие ткань, продуцируют ГМ-КСФ и ИФНγ после MHC-несовместимой ТГК
Чтобы понять вклад ГМ-КСФ из Т-клеток в острую РТПХ, авторы настоящего изобретения сначала использовали модель MHC-несовместимой ТГК. Авторы настоящего изобретения летально облучили мышей BALB/c CD45.2+ дикого типа (ДТ) (гаплотип MHC H2d), а затем внутривенно вводили им клетки костного мозга (КМ) без Т-клеток (БТК) от мышей ДТ CD45.1 C57BL/6 (B6) (гаплотип MHC H2b) со спленоцитами B6 CD45.1+ или без них, которые служили источником зрелых Т-клеток. У реципиентов, получавших КМ и спленоциты, развилась фатальная РТПХ между 3 и 6 днями после алло-ТГК (Фигура 1A), что совпадало с восстановлением и размножением донорских CD45.1+ клеток в селезенке, печени и коже (Фигура 1B). В донорском компартменте CD45.1+ популяции Т-клеток содержали высокие частоты ГМ-КСФ- и ИФНγ-продуцирующих клеток, которые сначала наблюдались в печени (Фигура 1C) и селезенке (Фигура 1D) с 3-го дня после алло-ТГК, затем в коже с 6-го дня (Фигура 1D). IL-17A практически не обнаруживался в этих популяциях (Фигура 1C, 1D). Соответственно, авторы настоящего изобретения обнаружили значительно более высокие количества ГМ-КСФ и ИФНγ в сыворотке мышей, получавших КМ и спленоциты, по сравнению мышами, получавшими только КМ (Фигура 1E). Авторы настоящего изобретения подтвердили продуцирование ГМ-КСФ и ИФНγ во время аллогенного ответа in vitro, используя реакции смешанной культуры лимфоцитов (СКЛ) (Фигура 1F). Кроме того, ДТ, Csf2-/-, Il17a-/-, и Ifng-/- Т-клетки пролиферировали одинаково после совместного культивирования с аллогенными ДК, за исключением априорных различий в пролиферативном потенциале этих клеток (Фигура 1G). Взятые вместе, ГМ-КСФ и ИФНγ, но не IL-17A, продуцируются Т-клетками, инфильтрирующими органы-мишени во время РТПХ.
Пример 2:
Продуцирование ГМ-КСФ аллогенными Т-клетками существенно для патологии РТПХ
Учитывая продуцирование ГМ-КСФ и ИФНγ алло-реактивными клетками TH, авторы настоящего изобретения оценили значимость этих цитокинов для определения тяжести острой РТПХ. Как указано выше, авторы настоящего изобретения летально облучили мышей ДТ BALB/c, а затем сравнили результаты инъекции клеток БТК-КМ от мышей ДТ B6 со спленоцитами или без них от мышей ДТ B6 или от мышей B6, лишенных ГМ-КСФ (Csf2-/-), ИФНγ (Ifng-/-) или IL-17 (Il17a-/-). Мыши, получавшие КМ плюс спленоциты Csf2-/-, были защищены от летальной РТПХ до 20 дней после алло-ТГК, в то время как все мыши, получавшие КМ плюс спленоциты ДТ, погибли на 15 день после ТГК (Фигура 2A). Введение мышам КМ и спленоцитов Ifng-/- или Il17a-/- также значительно повлияло на кинетику фатальной РТПХ, но все же почти 100% этих мышей погибли к 20 дню (Фигура 2A).
Поскольку препараты донорских спленоцитов содержат несколько различных типов иммунных клеток, авторы настоящего изобретения затем решили выяснить, действительно ли Т-клетки являются биологически значимым источником ГМ-КСФ в данной модели РТПХ. Авторы настоящего изобретения вводили летально облученным мышам-реципиентам BALB/c клетки БТК-КМ от доноров B6 ДТ вместе с очищенными Т-клетками, выделенными из селезенок мышей B6 ДТ или Csf2-/-, или без них; этот подход подтвердил, что мыши, получавшие Csf2-/- T-клетки, были полностью защищены от летальной РТПХ в течение 70 дней после алло-ТГК, в отличие от мышей, получавших ДТ T-клетки (Фигура 2B). Не было доказательств того, что этот защитный эффект связан с врожденными различиями между Т-клетками селезенки от мышей дикого типа и мышей Csf2-/-, поскольку две популяции демонстрировали сопоставимые клеточный состав (Фигура 6A), статус активации (Фигура 6B), способность РТПХ проникать в органы-мишени после ТГК (Фигура 6C), частоты активированных (CD44high) Т-клеток в селезенке или периферических лимфатических узлах (ЛУ) (Фигура 6D), частоты регуляторных T-клеток (Treg) и продукцию гранзима (данные не показаны) или их способность продуцировать провоспалительные цитокины, такие как ИФНγ, ФНОα или IL-17A. Затем авторы настоящего изобретения исследовали относительный вклад ГМ-КСФ от CD4 и CD8 Т-клеток в патологию РТПХ. С этой целью авторы настоящего изобретения провели «перекрестный» эксперимент с использованием 4 комбинаций: CD4 ДТ T-клетки+CD8 ДТ T-клетки, CD4 ДТ T-клетки+CD8 Csf2-/- T-клетки, CD4 Csf2-/- T-клетки+CD8 ДТ T-клетки и CD4 Csf2-/- T-клетки+CD8 Csf2-/- T-клетки. Только комбинации, в которых в CD4 T-клетках отсутствовал ГМ-КСФ, обеспечивали защиту от летальной РТПХ (Фигура 2K), подтверждая представление о том, что аллореактивные TH-клетки являются важным источником ГМ-КСФ в патогенезе РТПХ. Таким образом, авторы настоящего изобретения пришли к выводу, что ГМ-КСФ, полученный из Т-клеток, ответственен за запуск летальной РТПХ после MHC-несовместимой ТГК.
Преимущество выживания, обеспечиваемое алло-ТГК Т-клеток, лишенных ГМ-КСФ, также было подтверждено на уровне отдельных тканей-мишеней РТПХ. Гистопатологический анализ через 6 дней после алло-ТГК показал, что перенос Csf2-/- спленоцитов вызвал значительно меньшее повреждение тканей кожи и тонкой кишки по сравнению с переносом спленоцитов дикого типа (Фигура 2C, 2D). Более подробно, кожа мышей, получавших Csf2-/- клетки, показывала снижение клеточного апоптоза и клеточной инфильтрации вокруг и внутри волосяных фолликулов и эпидермиса, тогда как в тонком кишечнике наблюдалось уменьшение прямого разрушения крипт (Фигура 2D). Авторы настоящего изобретения далее установили уменьшении патологии толстой кишки при переносе Csf2-/- клеток, о чем свидетельствует увеличение длины толстой кишки и снижение индекса апоптоза (Фигура 2H, 2I). Кроме того, скрининг субклинических признаков дисфункции печени показал снижение сывороточных уровней аланинаминотрансферазы (ALT), щелочной фосфатазы (AP) и азота мочевины крови (BUN) у мышей, которым трансплантировали Csf2-/- спленоциты, по сравнению с аналогами ДТ (Фигура 2G). Кроме того, системные уровни ФНОα в сыворотке крови снижались в случае отсутствия ГМ-КСФ, тогда как количества IL-6 не изменялись (данные не показаны).
Поскольку активные формы кислорода (АФК) вовлечены в повреждение тканей при РТПХ, авторы настоящего изобретения решили выяснить, связано ли это уменьшение повреждения ткани с более низкими уровнями продукции АФК. Авторы настоящего изобретения обнаружили, что срезы печени и тонкой кишки мышей через 6 дней после переноса Csf2-/- спленоцитов содержали значительно меньше клеток, экспрессирующих p22phox, критический компонент механизма продуцирования АФК фагоцитами, чем срезы мышей, получавших спленоциты ДТ (Фигура 2E, 2F).
Чтобы дополнительно подтвердить функциональную роль ГМ-КСФ в РТПХ, авторы настоящего изобретения вводили мышам-реципиентам антителами против ГМ-КСФ перед MHC-несовместимой ТГК мышей BALB/c с использованием или только БТК-КМ мышей B6 ДТ или КМ плюс ДТ спленоциты. Системное блокирование действия ГМ-КСФ успешно воспроизводило преимущество выживания от переноса Csf2-/- спленоцитов этим мышам (Фигура 2G).
Так как мышиные модели MHC-несовместимой ТГК являются информативными, авторы настоящего изобретения затем решили выяснить, сохраняется ли роль ГМ-КСФ в патологии РТПХ в более клинически значимых условиях модели острой РТПХ с гаплонесовместимостью. Авторы настоящего изобретения объединили ДТ B6 БТК-КМ со спленоцитами ДТ, Csf2-/- или Ifng-/- и перенесли клетки летально облученным мышам B6D2F1, которые являются гибридом мышей линий B6 и DBA и, таким образом, несут гаплотип MHC H2b/d. Также в этом эксперименте перенос Csf2-/- спленоцитов давал значительное преимущество в выживаемости по сравнению с мышами, которые получали спленоциты ДТ, защищая мышей от фатальной РТПХ до уровня почти сравнимого с уровнем при переносе только БТК-КМ (Фигура 3A), который снова воспроизводился при переносе очищенных Т-клеток селезенки Csf2-/- (Фигура 3B). Как в экспериментах по переносу спленоцитов, так и Т-клеток, авторы настоящего изобретения также отметили заметное увеличение смертности после переноса клеток Ifng-/- (Фигура 3A, 3B), что согласуется с известной защитной иммунорегуляторной функцией этого цитокина при РТПХ. На 9-й день после ТГК кожа была наиболее пораженным органом с самыми высокими баллами гистологической оценки; здесь авторы настоящего изобретения увидели меньшее повреждение ткани у мышей, получавших Csf2-/- по сравнению со спленоцитами ДТ, и большее повреждение ткани у реципиентов Ifng-/- спленоцитов (Фигура 3C, 3D). Это совпало с изменениями количества сальных желез, тканевой организации и клеточных инфильтратов вокруг и внутри волосяных фолликулов и эпидермиса (Фигура 3D). На более поздней стадии (28 дней) после алло-ТГК перенос Csf2-/- спленоцитов также привел к уменьшению патологии печени (Фигура 3H), характеризующейся изменениями инфильтратов портального тракта и дезорганизацией желчных протоков. В этой модели частичной несовместимости кожа была наиболее пораженным органом с самыми высокими показателями РТПХ; здесь авторы настоящего изобретения увидели значительно меньшее повреждение ткани у мышей, получавших Csf2-/- по сравнению со спленоцитами ДТ, и значительно большее повреждение ткани у реципиентов Ifng-/- спленоцитов (Фигура 3C, 3D). На клеточном уровне авторы настоящего изобретения также наблюдали признаки уменьшения РТПХ в печени и тонком кишечнике мышей, получавших клетки Csf2-/-, у которых было значительно меньше клеток p22phox+, что сопровождалось уменьшением инфильтрации миелоидных клеток F4/80+ (Фигура 3E, 3F). Учитывая защитную роль ИФНγ при РТПХ, наблюдаемую у этих мышей, авторы настоящего изобретения затем решили выяснить, связано ли отсутствие ГМ-КСФ с уровнями этого цитокина: через десять дней после гаплонесовместимой ТГК авторы настоящего изобретения обнаружили значительно более высокие уровни ИФНγ у мышей, которые получили Csf2-/- спленоцитов, по сравнению с мышами, получавшими спленоциты дикого типа (Фигура 3G). Интересно, что авторы настоящего изобретения также обнаружили доказательства увеличения ГМ-КСФ в сыворотке крови мышей, получавших Ifng-/- спленоциты (Фигура 3G), что позволяет предположить, что ГМ-КСФ может дополнительно опосредовать усиление РТПХ, индуцированную отсутствием ИФНγ.
В целом авторы настоящего изобретения пришли к выводу, что ГМ-КСФ, полученный из Т-клеток, играет жизненно важную роль в развитии иммунопатологии РТПХ в двух мышиных моделях алло-ТГК.
Пример 3:
ГМ-КСФ опосредует летальность РТПХ через миелоидные клетки донорского происхождения
После летального облучения радиочувствительные антигенпрезентирующие клетки хозяина (АПК) теряются в течение первых дней после ТГК и заменяются АПК донора. В то время как АПК хозяина необходимы для фазы инициации РТПХ, донорские АПК играют меньшую роль в индукции заболевания, но могут быть вовлечены в сохранение повреждения тканей. Чтобы определить, имеют ли патологически релевантные ГМ-КСФ-чувствительные клетки донорское или хозяйское происхождение, авторы настоящего изобретения использовали мышей Csf2rb-/-, лишенных бета-субъединицы рецептора ГМ-КСФ, в качестве доноров или реципиентов ТГК. Отсутствие ГМ-КСФ в реципиентном компартменте не влияло на выживаемость при РТПХ (Фигура 9A). В противоположность этому, перенос КМ от мышей Csf2rb-/- приводил к резкому уменьшению смертности (Фигура 9B), фенокопируя перенос Csf2-/- спленоцитов (Фигура 2A). Авторы настоящего изобретения использовали неконтролируемое нелинейное сокращение размерности (t-SNE) для идентификации и визуализации ГМ-КСФ-чувствительных клеток в донорском трансплантате. С помощью этого подхода авторы настоящего изобретения обнаружили, что моноциты и нейтрофилы были особенно чувствительны к ГМ-КСФ, что они оценили с помощью анализа отдельных клеток фосфорилированного STAT-5, фактора транскрипции, расположенного после рецепторного комплекса ГМ-КСФ. Мышей Csf2rb-/- использовали в качестве источника нечувствительных к ГМ-КСФ клеток (Фигура 9C, 9D). Подтвердив, что клетки, чувствительные к ГМ-КСФ, ограничены миелоидным компартментом, авторы настоящего изобретения провели тщательную характеристику субпопуляций миелоидных клеток, обнаруженных в воспаленных органах-мишенях через 5 дней после алло-ТГК. Перенос Csf2-/- спленоцитов приводил к снижению частоты нескольких миелоидных субпопуляций, включая дендритные клетки (ДК), нейтрофилы, моноциты и клетки моноцитарного происхождения (MDC) (Фигура 9E, 9F). Уменьшение количества нейтрофилов и моноцитов/MDC также было обнаружено в органах-мишенях после лечения анти-ГМ-КСФ, тогда как системные количества этих субпопуляций миелоидных клеток оставались неизменными. Авторы настоящего изобретения наблюдали, что донорские моноциты/MDC продуцируют меньше IL-1β в отсутствие ГМ-КСФ (Фигура 9G). Важно отметить, что IL-1β является сигнатурным цитокином патогенной программы, вызываемой ГМ-КСФ, и было показано, что он играет критическую роль в развитии РТПХ. Авторы настоящего изобретения также исследовали экспрессию АФК донорскими миелоидными клетками после алло-ТГК, учитывая их низкие уровни в срезах ткани мышей, получавших аллогенные Csf2-/- T-клетки, и их ранее подтверденную роль в управлении повреждением ткани. Авторы настоящего изобретения обнаружили, что перенос Csf2-/- спленоцитов приводит к снижению продуцирования АФК как субпопуляциями моноцитов/MDC, так и субпопуляциями нейтрофилов (Фигура 9H). В совокупности эти данные демонстрируют, что ГМ-КСФ-лицензированные миелоидные клетки донорского происхождения имеют решающее значение для патологии РТПХ.
Пример 4
Эффективная ТПЛ сохраняется при отсутствии ГМ-КСФ
Считается, что иммунные ответы, приводящие к повреждению тканей при РТПХ и элиминации опухоли при ТПЛ, опосредуются одним и тем же механизмом. Поскольку ГМ-КСФ, полученный из донорских Т-клеток, важен для летальной РТПХ, авторы настоящего изобретения исследовали, опосредует ли он также ТПЛ в модели MHC-несовместимой ТГК. Чтобы проверить это, авторы настоящего изобретения внутривенно вводили A20 клетки В-клеточной лимфомы (происхождения BALB/c), коэкспрессирующие GFP и люциферазу (A20-GFP-Luc), летально облученным реципиентам BALB/c вместе с клетками ДТ B6 БТК-КМ или отдельно или с очищенными ДТ или Csf2-/- Т-клетками селезенки мышей линии B6. Рост опухоли контролировали с помощью биолюминесцентной визуализации (БЛВ). Когда в клетки A20 вводили только с БТК-КМ, все реципиенты погибли от растущей опухоли в течение 35 дней (Фигура 4A-4C), что соответствует критической роли Т-клеток в эффекте ТПЛ. Соответственно, добавление к переносу КМ T-клеток ДТ или Csf2-/- приводило к эффективному контролю роста опухоли (Фигура 4A-4C), хотя только мыши, получавшие T-клетки Csf2-/-, показали значительное улучшение выживаемости (Фигура 4C). В то время как мыши, несущие лимфому, получавшие БТК-КМ плюс Т-клетки ДТ, в основном умирали от тяжелой РТПХ, мыши, получавшие БТК-КМ плюс Т-клетки Csf2-/-, проявляли неуменьшенный эффект ТПЛ, но в то же время были защищены от РТПХ (Фигура 4C, 4D). Это улучшение выживаемости мышей, получавших T-клетки Csf2-/-, также наблюдалось в альтернативной модели ТПЛ с использованием линии мономиелоцитарных клеток WEHI-3. Кроме того, когда авторы настоящего изобретения увеличили количество инокулированных клеток лимфомы A20, T-клетки Csf2-/- осуществляли ТПЛ даже более эффективно, чем T-клетки дикого типа, что приводило к значительному увеличению общей выживаемости. Соответственно, T-клетки Csf2-/- и ДТ, выделенные у интактных мышей или мышей, получавших алло-ТГК, были одинаково способны уничтожать лимфому A20 и клетки WEHI-3 ex vivo, одинаково размножались в селезенке и ЛУ и демонстрировали аналогичные частоты активацированых (CD44high), пролиферативных (Ki67 +) и гранзим B-продуцирующих клеток. Чтобы проверить возможность применения результатов в клинических условиях, авторы настоящего изобретения использовали нейтрализующие mAb против ГМ-КСФ. Поразительно, что лейкозные мыши, получившие анти-ГМ-КСФ антитела, получавшие БТК-КМ плюс Т-клетки, показали сильный эффект ТПЛ и имели значительно лучшую выживаемость, чем контрольные мыши, получившие изотипические антитела (Фигура 4E-4H). Лучшая выживаемость была связана со снижением заболеваемости РТПХ (Фигура 4H). Следует отметить, что нейтрализация ГМ-КСФ не оказала никакого влияния на рост опухоли у мышей, которые не подвергались ТГК.
Авторы настоящего изобретения сравнили причину гибели (опухоль или РТПХ) в разных группах: мыши с лимфомой, которые не подвергались ТГК, достигли критериев отмены и нуждались в эвтаназии. У мышей с лимфомой, получавших БТК-КМ плюс Т-клетки ДТ, наблюдалось замедленное прогрессирование опухоли, но вместо этого они погибали от тяжелой РТПХ, тогда как мыши, получавшие БТК-КМ плюс T-клетки Csf2-/-, проявляли сильный ТПЛ-эффект и были защищены от РТПХ (Фигура 4D). В соответствии с этими данными, T-клетки Csf2-/- и ДТ были в равной степени способны уничтожать клетки лимфомы A20 ex vivo (Фигура 7A, 7B). Чтобы проверить применимость полученных результатов к клиническим условиям, авторы настоящего изобретения использовали нейтрализующие mAb против ГМ-КСФ. Поразительно, что лейкозные мыши, получившие анти-ГМ-КСФ антитела, получавшие БТК-КМ плюс Т-клетки, показали сильный эффект ТПЛ и имели значительно лучшую выживаемость, по сравнению с контрольными мышами, получившими изотипические антитела (Фигура 4E-4H). Лучшая выживаемость была связана со снижением заболеваемости РТПХ (рис. 4H). Следует отметить, что нейтрализация ГМ-КСФ не оказала никакого влияния на рост опухоли у мышей, которые не подвергались ТГК.
Следует отметить, что мыши, получавшие алло-ТГК от Csf2-/- доноров, показали значительное улучшение выживаемости по сравнению с донорами ДТ (Фигура 8A и 8B), демонстрируя, что нейтрализация рецептора ГМ-КСФ эффективна для облегчения РТПХ.
Таким образом, при отсутствии ГМ-КСФ донорские Т-клетки являются эффективными медиаторами терапевтического ответа ТПЛ у мышей, не вызывая летальной РТПХ, даже в высокоиммуногенном контексте MHC-несовместимой алло-ТГК.
Пример 5:
Уровень ГМ-КСФ повышен в биоптатах человека с РТПХ
Затем авторы настоящего изобретения решили выяснить, имеются ли доказательства того, что ГМ-КСФ играет аналогичную роль в РТПХ у реципиентов ТГК человека. Авторы настоящего изобретения оценили экспрессию гена ГМ-КСФ и количество белка в биоптатах желудочно-кишечного тракта от пациентов с различными степенями клинически подтвержденной РТПХ после ТГК для ряда различных состояний (Таблицы 1-3). На уровне транскрипции авторы настоящего изобретения обнаружили значительно более высокую экспрессию ГМ-КСФ в биоптатах желудочно-кишечного тракта от пациентов с IV степенью РТПХ по сравнению с РТПХ I степени (Фигура 5A). Более того, в образцах от пациентов с РТПХ IV степени авторы настоящего изобретения наблюдали сильную иммунореактивность в отношении ГМ-КСФ в стромальном компартменте; в то время как авторы настоящего изобретения не обнаружили ГМ-КСФ в биоптатах пациентов без РТПХ (Фигура 5B). Вместе эти результаты указывают на корреляцию между ГМ-КСФ и наличием тяжелой РТПХ после алло-ТГК у людей.
Обсуждение:
Несмотря на то, что существует большое количество литературы, в которой обсуждается потенциальный вклад различных субпопуляций TH-клеток и их сигнатурных цитокинов в РТПХ, механизмы, лежащие в основе воспаления и последующего разрушения ткани, до конца не поняты. Кроме того, пластичность коммитирования линий TH-клеток, а также манипуляции in vitro с перенесенными поляризованными Т-клетками ограничивают интерпретацию этих результатов. Авторы настоящего изобретения показывают здесь, что ГМ-КСФ играет ранее недооцененную роль в управлении летальной РТПХ у мышей и аналогичным образом повышен у перенесших алло-ТГК пациентов людей с тяжелой РТПХ. Более того, ГМ-КСФ, по-видимому, незаменим для терапевтического эффекта ТПЛ и, таким образом, может представлять собой многообещающую новую терапевтическую мишень для предотвращения и лечения РТПХ. Наши результаты также исключают роль IL-17A, полученного из Т-клеток, в острой РТПХ, но демонстрируют защитный эффект ИФНγ.
ГМ-КСФ участвует в нескольких патологических воспалительных заболеваниях и оказывает сильное провоспалительное действие на миелоидные клетки. Авторы настоящего изобретения использовали здесь две разные мышиные модели острой РТПХ, чтобы показать, что ГМ-КСФ, полученный из донорских Т-клеток, напрямую опосредует тяжелую РТПХ, которая связана с обильной инфильтрацией миелоидных клеток и повышенным окрашиванием тканей на компоненты механизма образования АФК, который обычно используется фагоцитами для защиты хозяина от патогенов. Те же механизмы явно оказывают пагубное действие в иммунопатологии РТПХ, где повреждение тканей и инфильтрация лейкоцитов находятся в тесной связи. Снижение окислительного стресса, вызванного миелоидными клетками, в чувствительных к РТПХ органах, наблюдаемое здесь у животных, получающих алло-реактивные Csf2-/- Т-клетки, подтверждает это мнение. Ключевая роль ГМ-КСФ в хроническом воспалении кишечника и высокое количество ГМ-КСФ, обнаруженное в биоптатах желудочно-кишечного тракта от пациентов с РТПХ, позволяет сделать вывод о том, что регулирование количества этого цитокина в этом конкретном органе может быть очень полезным для предотвращения и/или лечения РТПХ.
Авторы настоящего изобретения также обнаружили доказательства потенциальной реципрокной регуляции ГМ-КСФ и ИФНγ при РТПХ у мышей. Ранее было показано, что ИФНγ, продуцируемый активированными донорскими Т-клетками, как способствует, так и защищает от РТПХ; авторами настоящего изобретения было обнаружено, что мыши, подвергавшиеся частично MHC-несовместимой ТГК с использованием ИФН-дефицитных спленоцитов, были значительно более восприимчивы к летальной РТПХ, чем мыши, получавшие спленоциты дикого типа. Интересно, что у этих мышей наблюдались повышенные уровни сывороточного ГМ-КСФ, в то время как у мышей, получавших ГМ-КСФ-дефицитные спленоциты, в их сыворотке крови наблюдался повышенный уровень ИФНγ. Ранее сообщалось о способности ИФНγ отрицательно регулировать ГМ-КСФ в эффекторных TH-клетках; однако ГМ-КСФ не влияет напрямую на Т-клетки, которые не экспрессируют рецепторный комплекс, и поэтому механизмы, посредством которых ГМ-КСФ влияет на продуцирование ИФНγ, требуют дальнейшего изучения.
Давно существовало предположение, что РТПХ, управляемая донорскими Т-клетками, и терапевтический эффект ТПЛ опосредуются одними и теми же механизмами: данные, представленные здесь, опровергают это утверждение. Введение мышам антител, блокирующими ГМ-КСФ, перед алло-ТГК значительно защищало их от развития острой летальной РТПХ; в то время как у мышей с опухолями, получавших анти-ГМ-КСФ антитела, наблюдался благоприятный эффект от эффективного контроля роста опухоли и защиты от РТПХ. Таким образом, ГМ-КСФ не контролирует опосредованное Т-клетками уничтожение злокачественных лимфогематопоэтических клеток; вместо этого, не ограничиваясь какой-либо, теорией авторы настоящего изобретения предполагают, что при РТПХ ГМ-КСФ действует как канал связи между патогенными Т-клетками и миелоидными клетками, позволяя последним вызывать разрушение ткани. В то время как ТПЛ, вероятно, зависит от прямой цитотоксичности T-клеток, ответы миелоидных клеток, вызванные ГМ-КСФ, могут быть более актуальными для патологии РТПХ, что, таким образом, представляет собой потенциальный механизм отделения эффекта ТПЛ от РТПХ.
Патологическая роль ГМ-КСФ в аутоиммунных и воспалительных заболеваниях привела к разработке моноклональных антител, направленно воздействующих на путь ГМ-КСФ при заболеваниях человека. Учитывая наблюдаемые здесь положительные эффекты нейтрализации ГМ-КСФ при экспериментальной РТПХ, является очевидным, что блокирование ГМ-КСФ у пациентов с алло-ТГК улучшит клинический результат. Заметное повышение уровней этого цитокина, наблюдаемое в пораженных тканях пациентов с тяжелой РТПХ, еще раз подтверждает эту идею. Однако, поскольку ГМ-КСФ имеет решающее значение для гомеостаза сурфактанта и защиты легкого хозяина, важно будет тщательно контролировать параметры легочного заболевания при блокировании ГМ-КСФ в клинических условиях. Имеются полученные на мышах данные о том, что ГМ-КСФ критически влияет на иммунные ответы, опосредованные IL-23, и что терапия анти-L-23p19 антителом может облегчить сингенный колит, связанный с РТПХ. Таким образом, прямая блокада ГМ-КСФ может стать ценным дополнением к любым клиническим испытаниям, нацеленным на IL-23. Тот факт, что отсутствие ГМ-КСФ может остановить развитие РТПХ без нарушения ТПЛ-ответа, облегчает тестирование этой терапевтической стратегии для предотвращения и лечения РТПХ после алло-ТГК в клинических испытаниях. Более того, тот факт, что ГМ-КСФ был повышен в образцах пациентов с самыми тяжелыми проявлениями РТПХ, означает, что эта терапевтическая стратегия имеет особые перспективы для групп пациентов с наихудшим исходом и самым высоким риском летального исхода.
Таблица 1. Клинико-патологические характеристики I группы пациентов с РТПХ.
ОВИ: общий вариабельный иммунодефицит: вОМЛ: вторичный острый миелоидный лейкоз; Т-ОЛЛ: Т-клеточный острый лимфоидный лейкоз; МДС: миелодиспластический синдром; СРД: совместимый родственный донор; ННРД: несовместимый неродственный донор; СНРД: совместимый неродственный донор; СКПК: стволовые клетки периферической крови; КМ: костный мозг; Флу: флударабин; БХНМ: бис-хлорэтилнитрозомочевина; Мел: мелфалан; ОВТ: облучение всего тела; VP16: этопозид; ТТ: тиотепа; АТГ: антитимоцитарный глобулин; ЦиА: циклоспорин A; ММФ: микофенолата мофетил; МТР: метотрексат.
Таблица 2. Характеристики контрольных субъектов
Таблица 3. Клинико-патологические характеристики II группы пациентов с РТПХ.
ОМЛ: острый миелоидный лейкоз; B-ОЛЛ: В-клеточный острый лимфоидный лейкоз; ОВИ: общий вариабельный иммунодефицит, МДС: миелодиспластический синдром; РАИБ: рефрактерная анемия с избытком бластов; Т-НХЛ: Т-клеточная неходжкинская лимфома; вОМЛ: вторичный острый миелоидный лейкоз; МПН: миелопролиферативные новообразования; ММ: множественная миелома; ННРД: несовместимый неродственный донор; СРД: совместимый родственный донор; СНРД: совместимый неродственный донор; СКПК: стволовые клетки периферической крови; Флу: флударабин; БХНМ: бис-хлорэтилнитрозомочевина; Мел: мелфалан; Треос: треосульфан; ОВТ: облучение всего тела; VP16: этопозид; ТТ: тиотепа; БС: бусульфан; ЦФ: циклофосфамид; ЦиА: циклоспорин A; МТР: метотрексат; ММФ: микофенолата мофетил; АТГ: антитимоцитарный глобулин.
Таблица 4. Система оценки РТПХ для мышей с алло-ТГК
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Universitaet Zuerich
<120> Лиганды ГМ-КСФ или рецептора ГМ-КСФ для применения при лечении
гемобластоза у пациента, перенесшего алло-ТГК
<130> uz349wo
<160> 4
<170> Патентная версия 3.5
<210> 1
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ПЦР праймер
<400> 1
cactgctgct gagatgaatg aaa 23
<210> 2
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ПЦР праймер
<400> 2
gtctgtaggc aggtcggctc 20
<210> 3
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ПЦР праймер
<400> 3
gaaggtgaag gtcggagtca ac 22
<210> 4
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ПЦР праймер
<400> 4
tgattttgga gggatctcgc tc 22
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ПРОФИЛАКТИКИ И ЛЕЧЕНИЯ РЕАКЦИИ ТРАНСПЛАНТАТ ПРОТИВ ХОЗЯИНА СТИМУЛЯЦИЕЙ АНТИИДИОТИПИЧЕСКОЙ СУПРЕССИИ ДОНОРСКИХ ЛИМФОЦИТОВ НА ЭКСПЕРИМЕНТАЛЬНОЙ МОДЕЛИ | 1999 |
|
RU2175246C2 |
| АНТИТЕЛО ПРОТИВ КСФ-1R | 2012 |
|
RU2621859C2 |
| МАТЕРИАЛЫ И СПОСОБЫ ЛЕЧЕНИЯ РАКА | 2019 |
|
RU2787828C2 |
| ТЕРАПЕВТИЧЕСКОЕ ПРИМЕНЕНИЕ МЫШЬЯКА И ВОЗМОЖНЫЕ МЕХАНИЗМЫ ДЕЙСТВИЯ | 2020 |
|
RU2815051C2 |
| Т-КЛЕТКИ ЦЕНТРАЛЬНОЙ ПАМЯТИ ПРОТИВ ТРЕТЬЕЙ СТОРОНЫ, СПОСОБЫ ИХ ПОЛУЧЕНИЯ И ИХ ПРИМЕНЕНИЕ В ТРАНСПЛАНТАЦИИ И ЛЕЧЕНИИ ЗАБОЛЕВАНИЙ | 2009 |
|
RU2506311C2 |
| ПОСЛЕДОВАТЕЛЬНОЕ ЛЕЧЕНИЕ РАКА С ИСПОЛЬЗОВАНИЕМ 6-ТИО-dG, ИНГИБИТОРОВ КОНТРОЛЬНЫХ ТОЧЕК И ЛУЧЕВОЙ ТЕРАПИИ | 2021 |
|
RU2833475C1 |
| МЕДИКАМЕНТЫ, ПРИМЕНЕНИЯ И СПОСОБЫ | 2015 |
|
RU2712283C2 |
| СПОСОБ ПОЛУЧЕНИЯ ФИБРОБЛАСТОВ И Г-КСФ-ПОЛОЖИТЕЛЬНАЯ ФИБРОБЛАСТНАЯ МАССА | 2019 |
|
RU2800644C2 |
| ПРИМЕНЕНИЕ РАЗМНОЖЕННЫХ ПОПУЛЯЦИЙ ГЕМАТОПОЭТИЧЕСКИХ СТВОЛОВЫХ КЛЕТОК/КЛЕТОК-ПРЕДШЕСТВЕННИКОВ | 2016 |
|
RU2747728C2 |
| СПОСОБ ПОДАВЛЕНИЯ ИММУННЫХ РЕАКЦИЙ | 2008 |
|
RU2393481C1 |
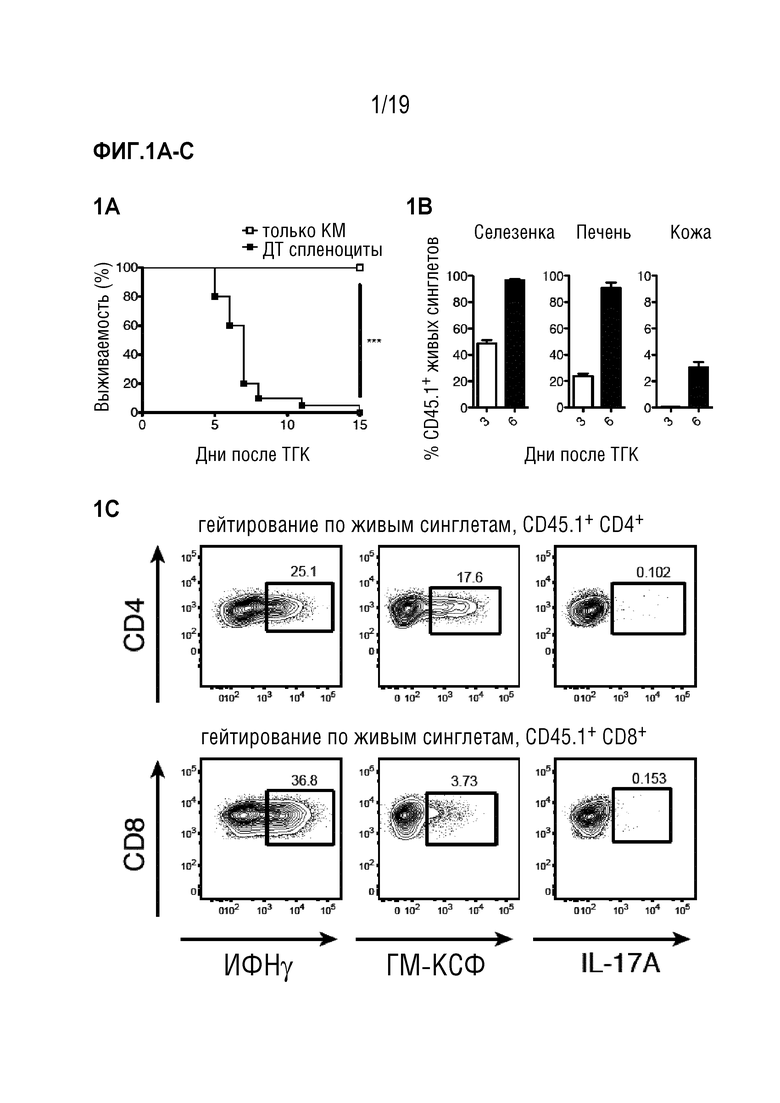
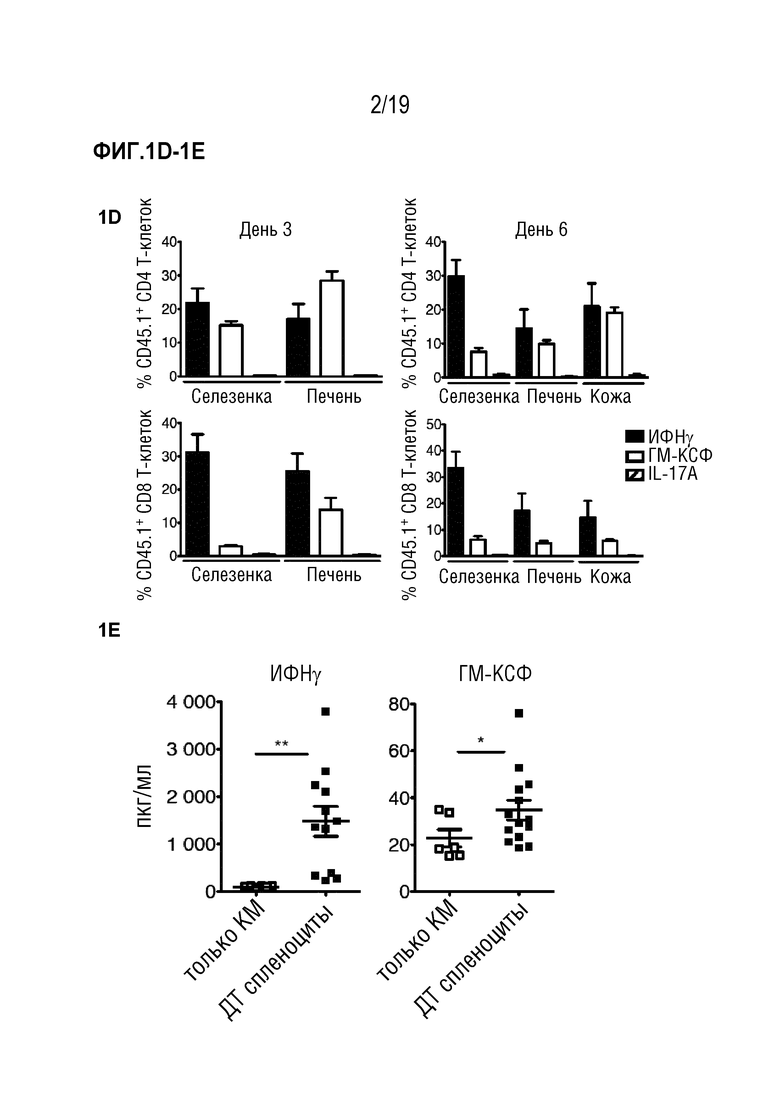
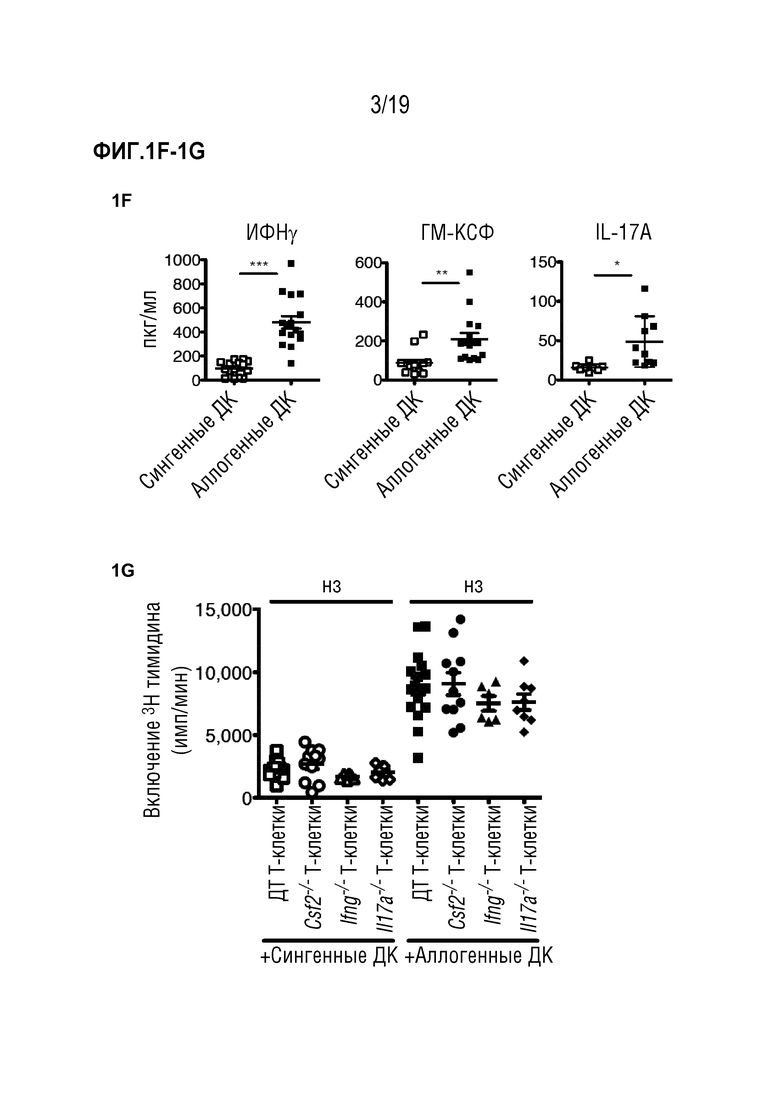
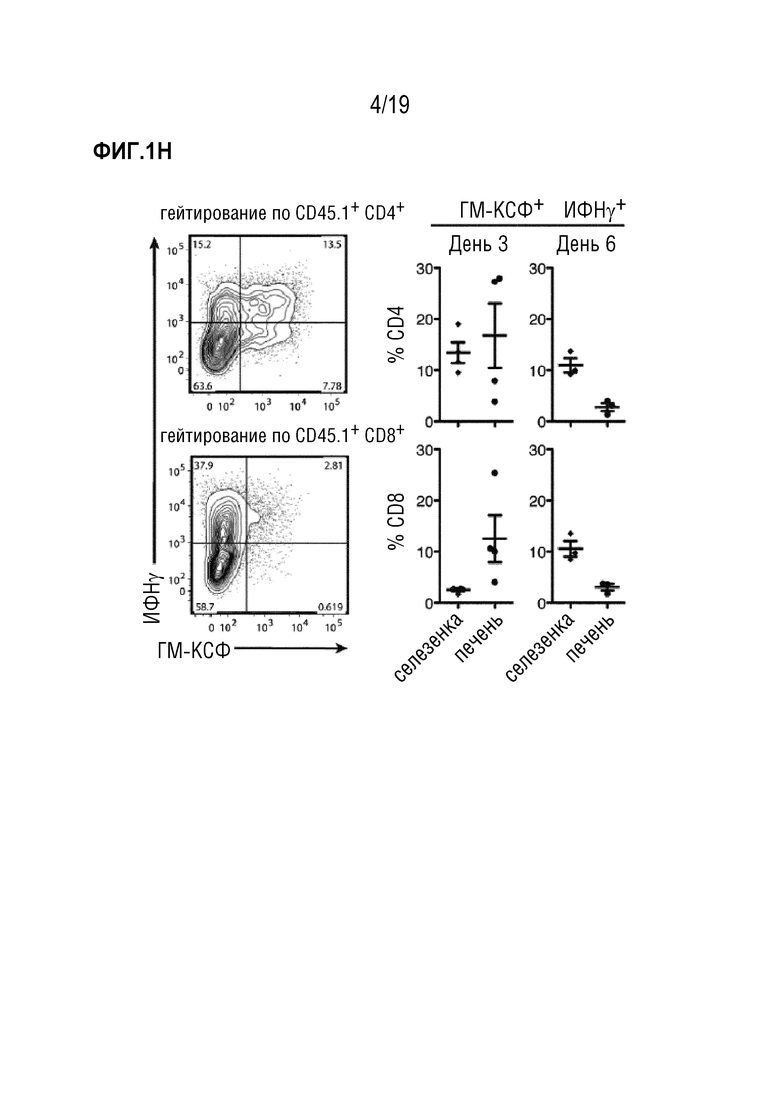
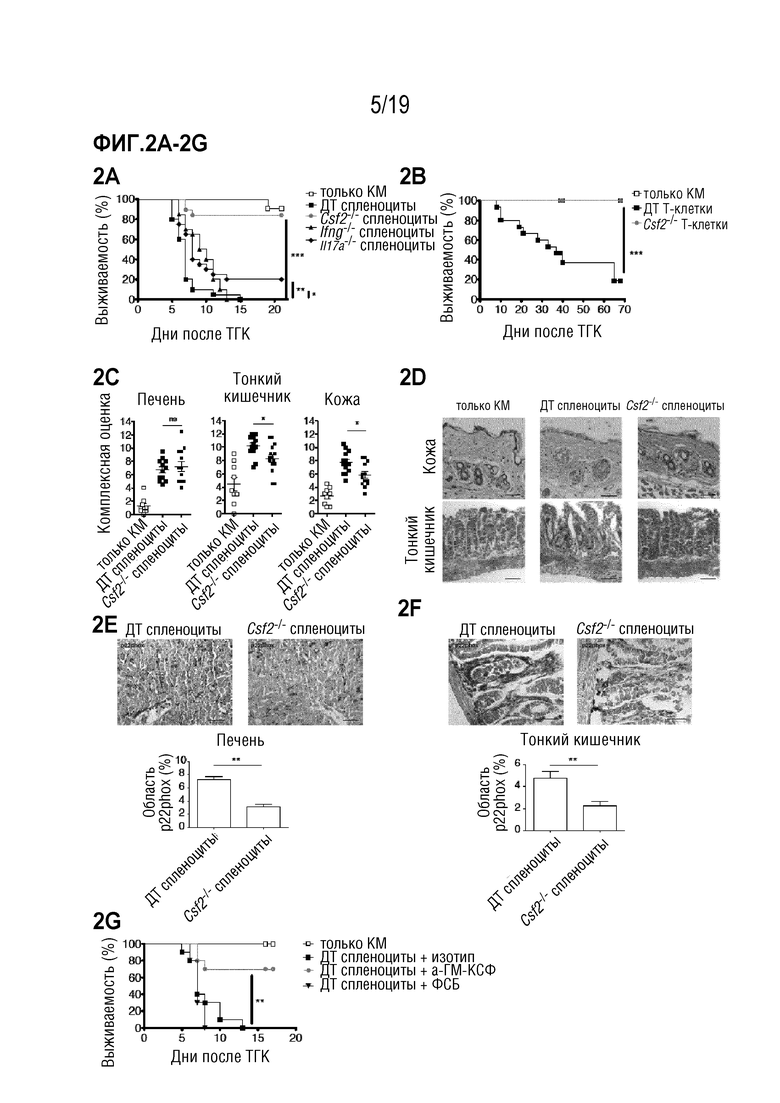
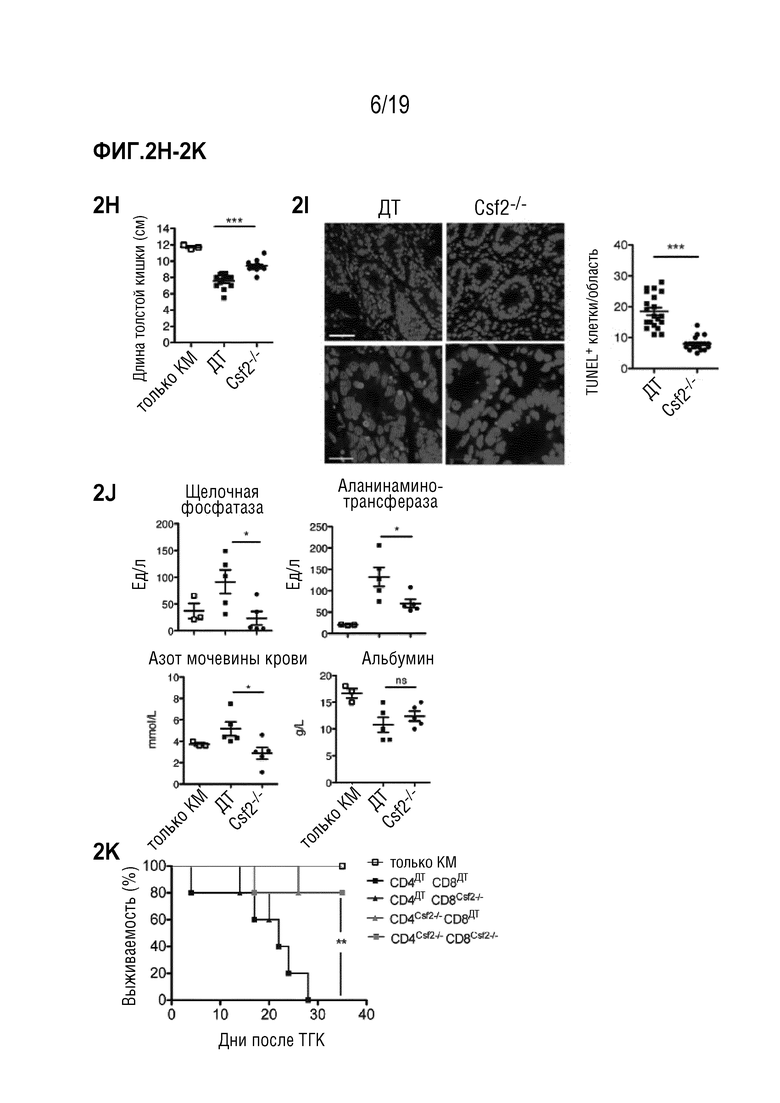
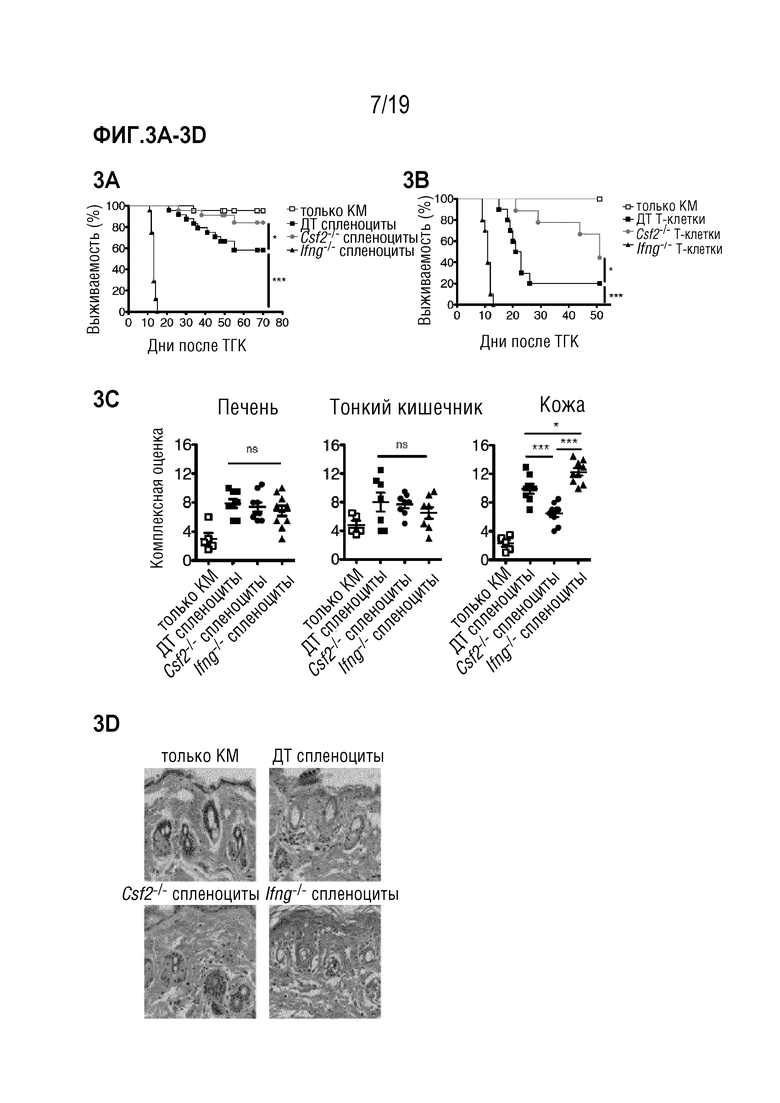
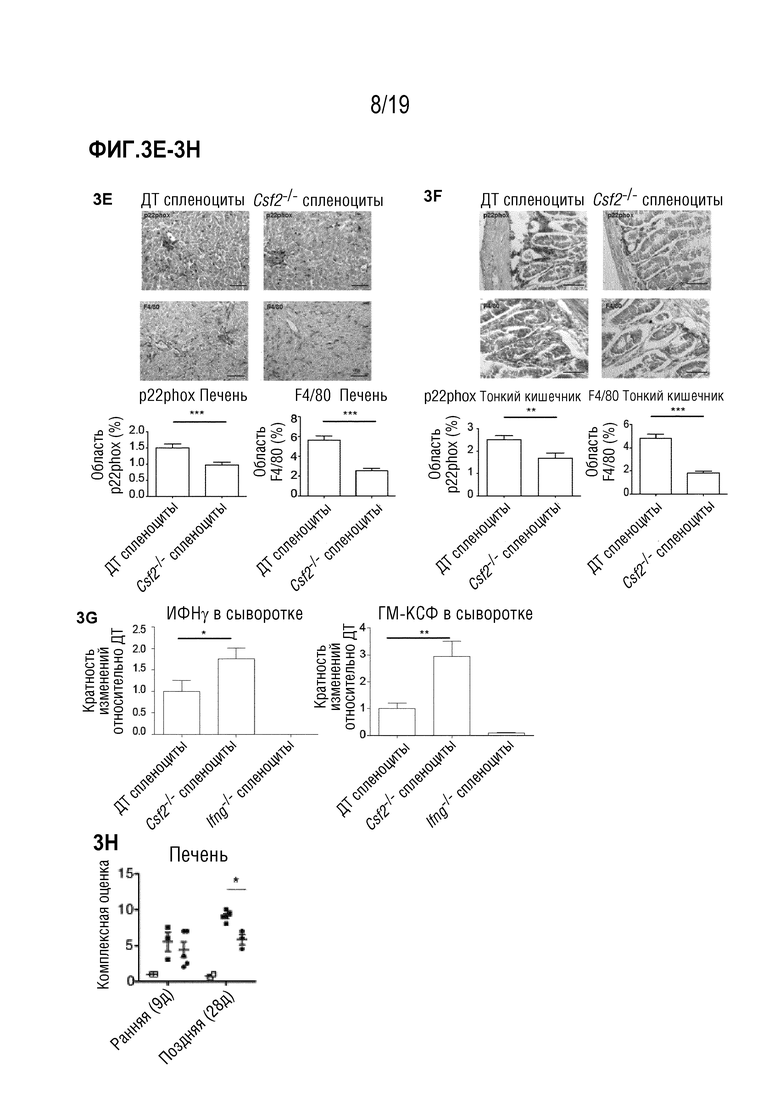
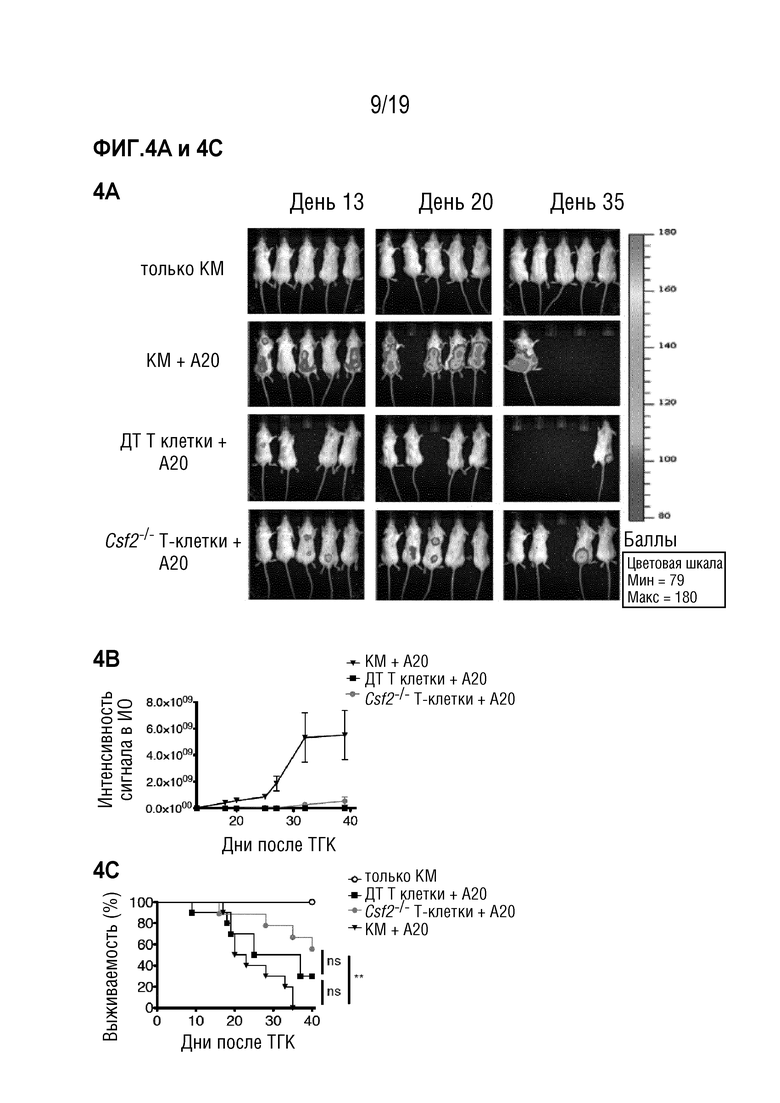
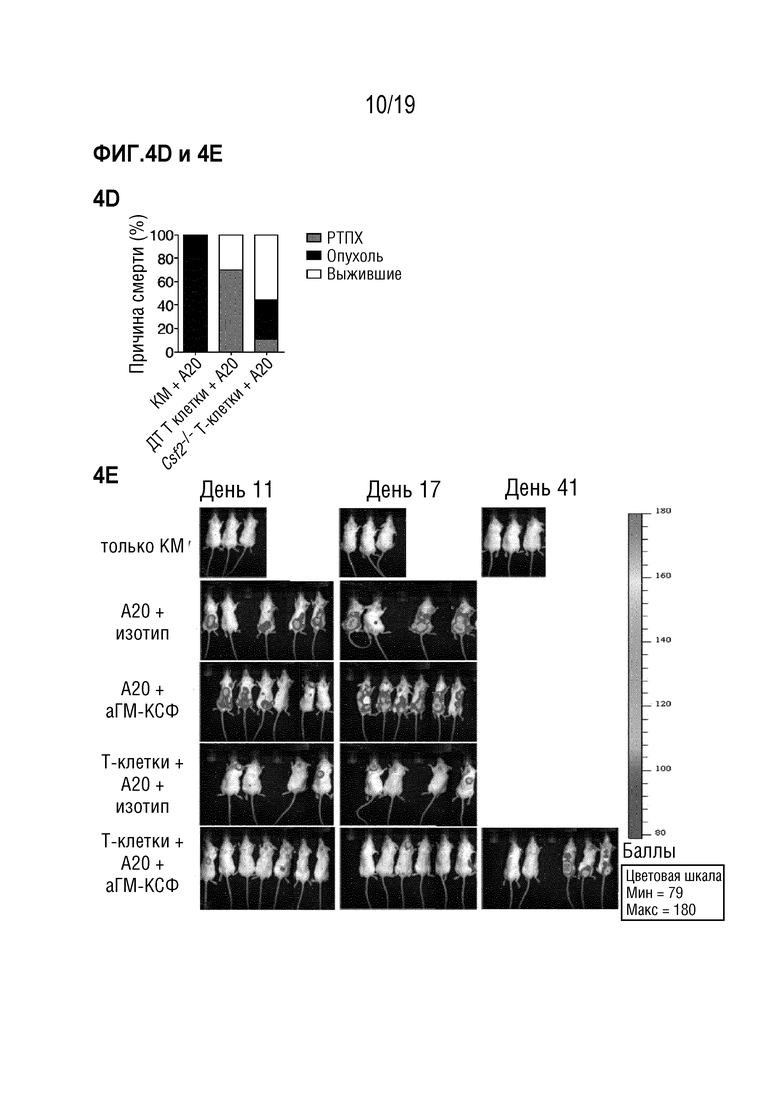


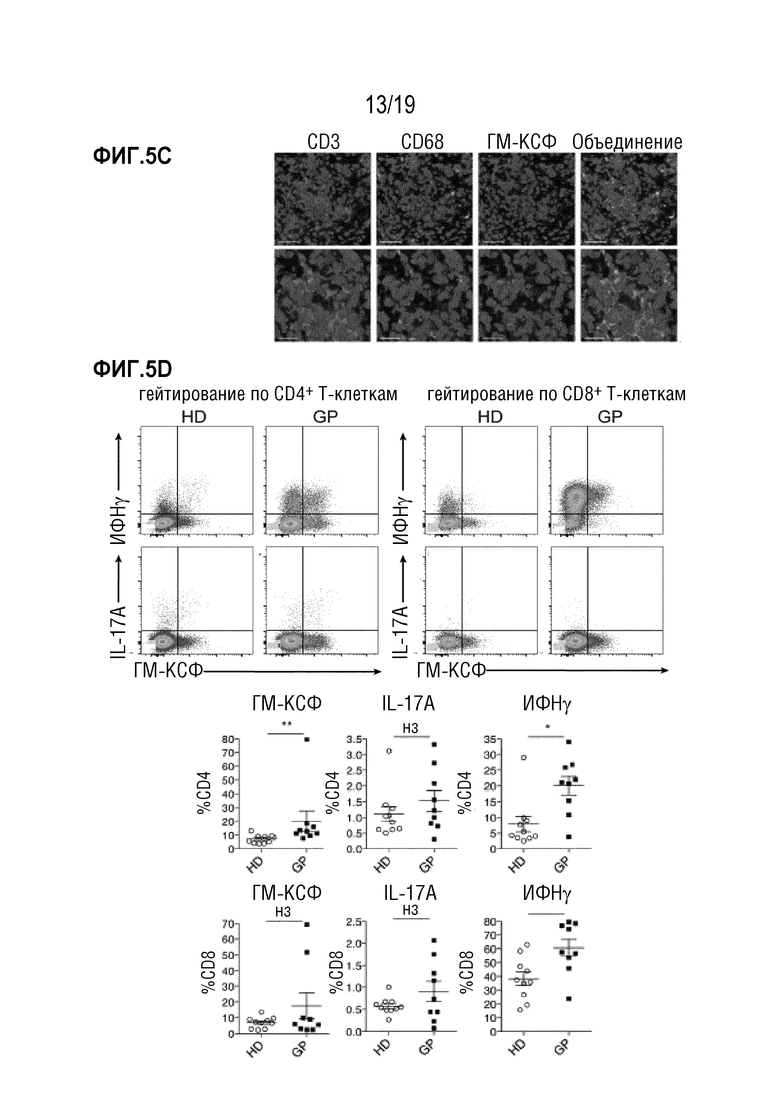
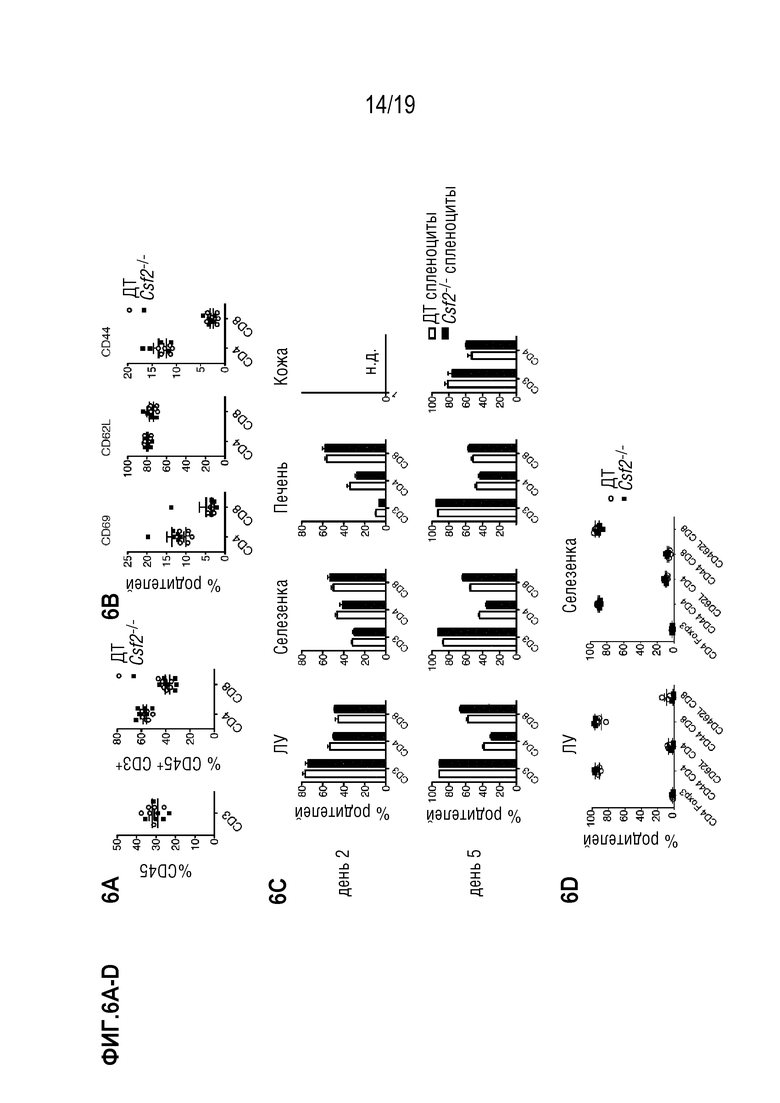
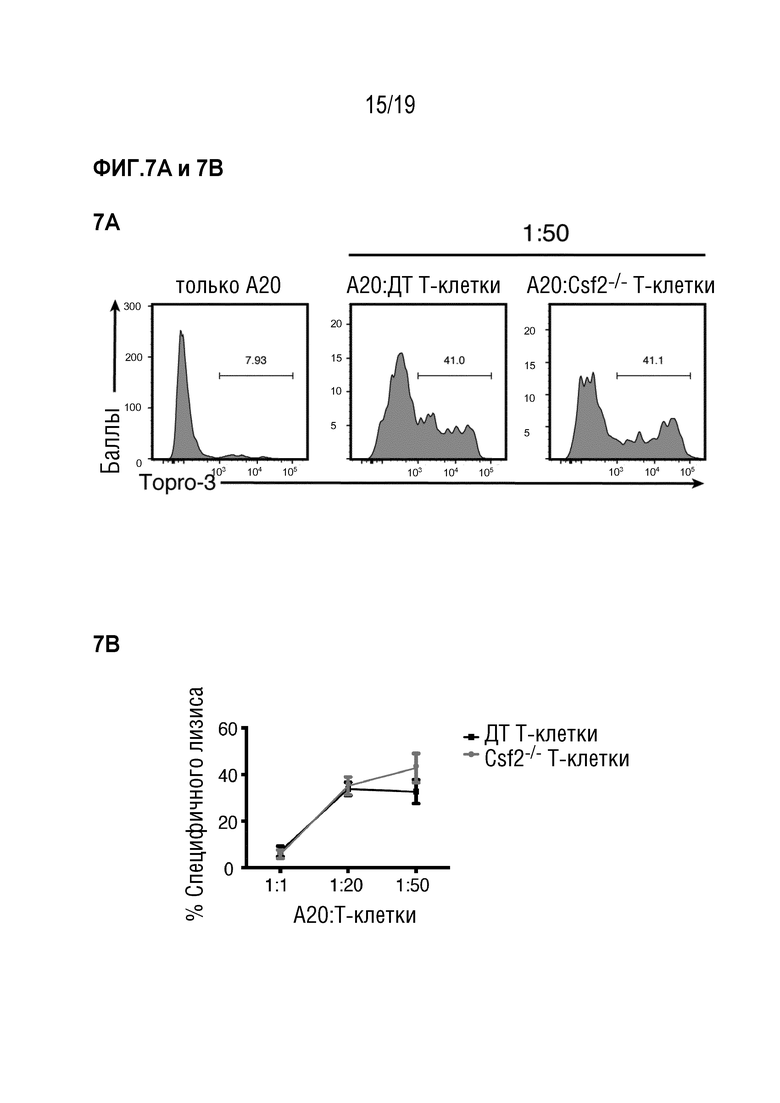
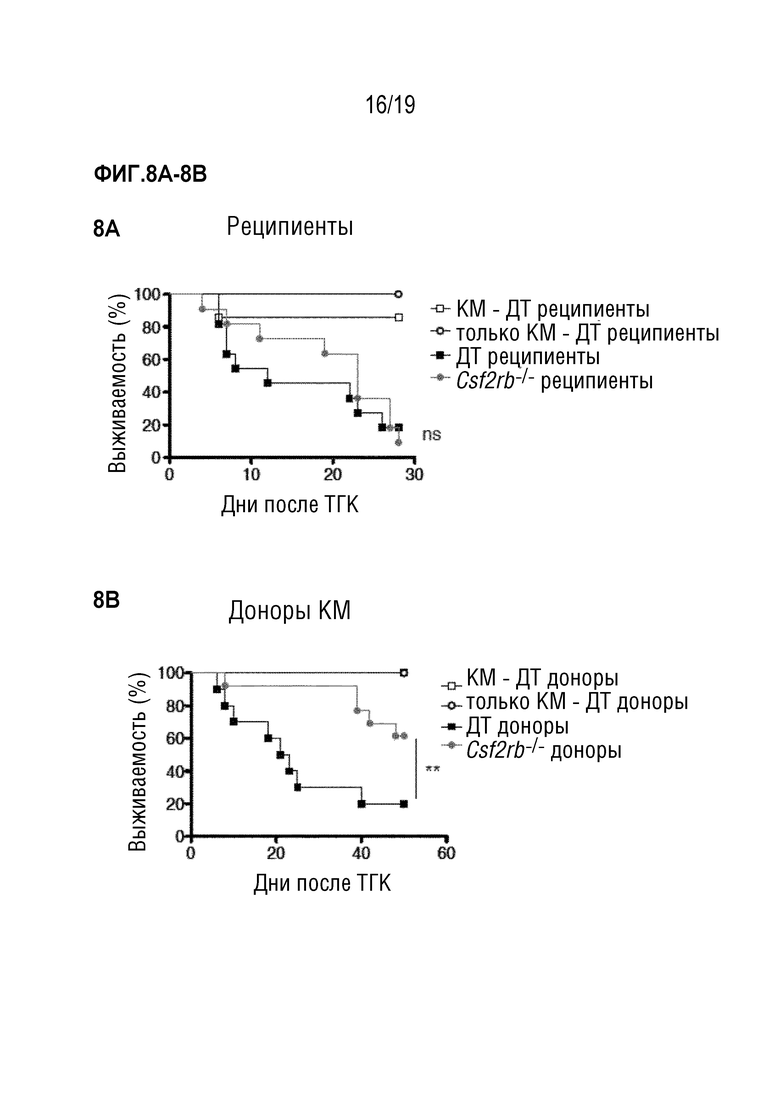
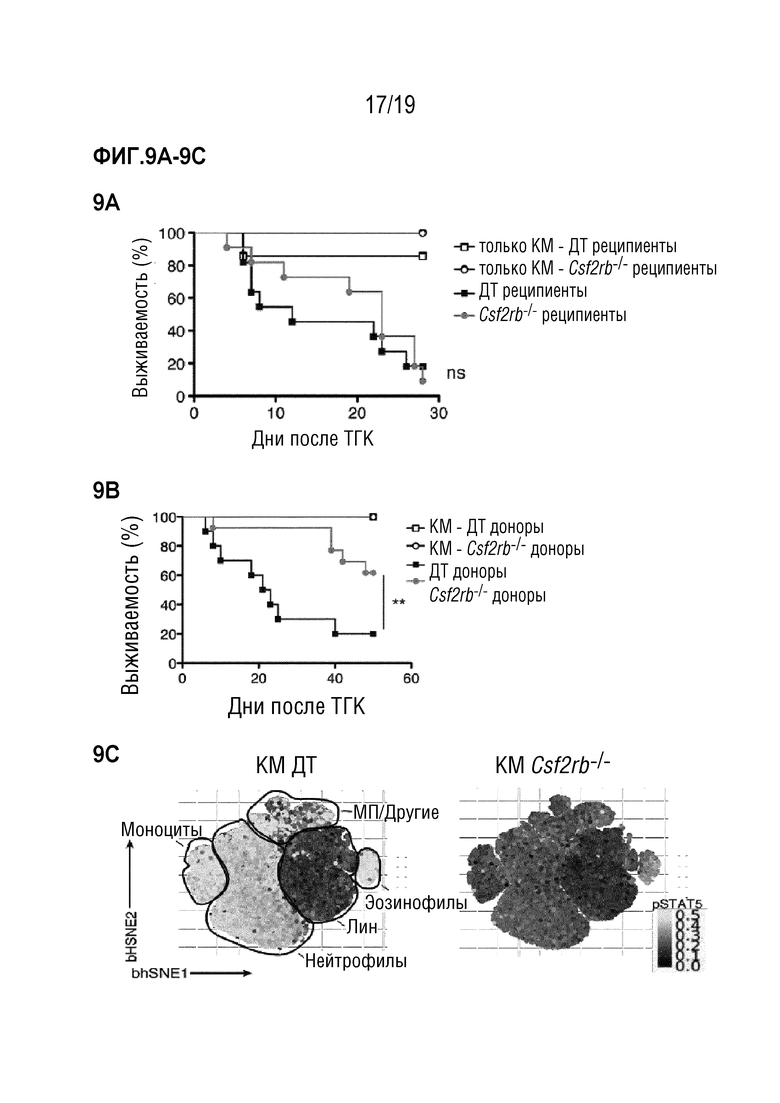
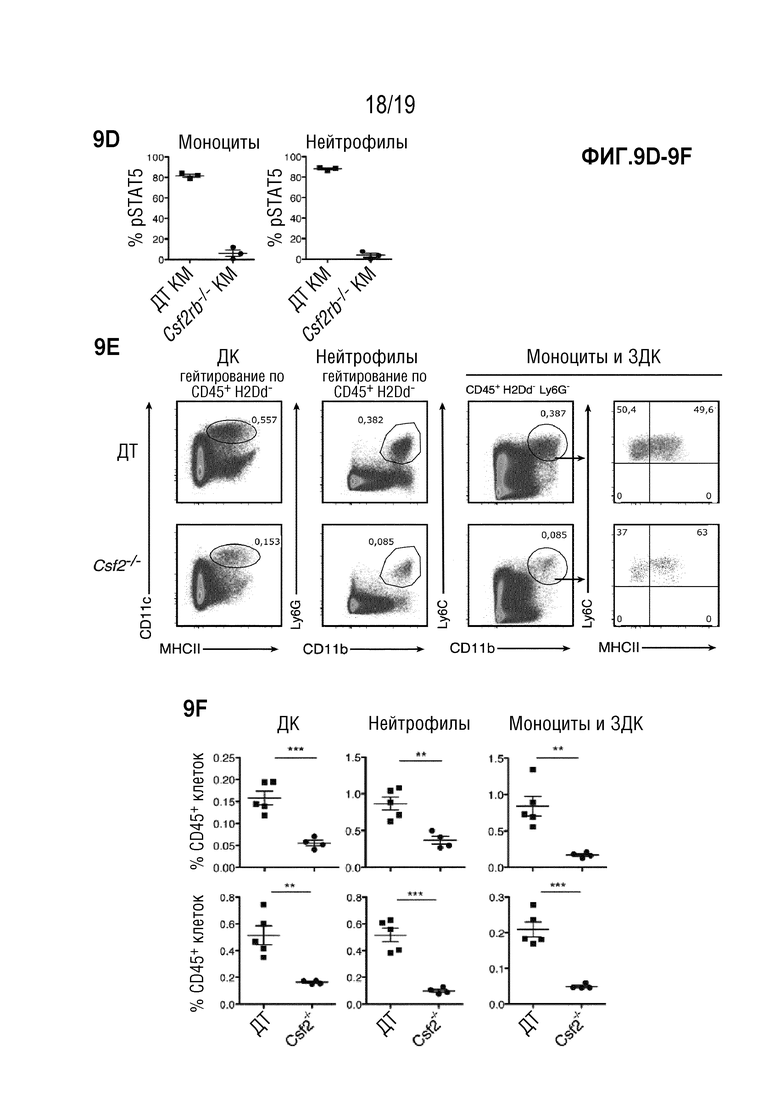
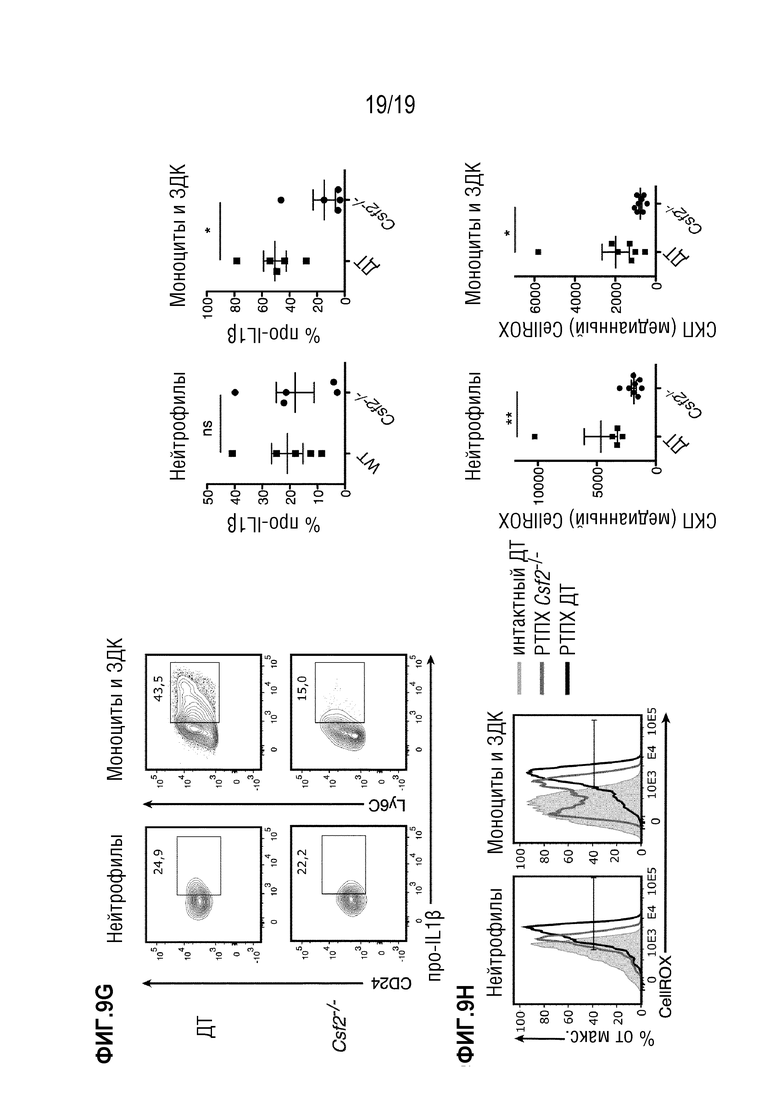
Настоящее изобретение относится к области клеточной биологии, трансплантологии и медицины, в частности к способу лечения пациента, страдающего или подверженного риску реакции «трансплантат против хозяина» (РТПХ). Способ включает введение пациенту нейтрализующего антитела к рецептору ГМ-КСФ, специфично связывающегося с гранулоцитарно-макрофагальным колониестимулирующим фактором (ГМ-КСФ) или по меньшей мере с одним из CD116, CD131 или рецептором ГМ-КСФ, состоящим из CD116 и CD131. Изобретение также раскрывает применение нейтрализующего антитела к рецептору ГМ-КСФ, применение молекулы нуклеиновой кислоты, кодирующей указанное антитело, экспрессионной конструкции нуклеиновой кислоты, содержащей указанную молекулу нуклеиновой кислоты, а также фармацевтической композиции, содержащей указанное антитело, нуклеиновую кислоту или экспрессионную конструкцию. Настоящее изобретение позволяет ослабить РТПХ при сохранении полезного эффекта лечения. 7 н. и 22 з.п. ф-лы, 9 ил., 4 табл., 5 пр.
1. Способ лечения пациента, страдающего или подверженного риску реакции «трансплантат против хозяина», включающий:
введение пациенту нейтрализующего антитела к рецептору ГМ-КСФ, специфично связывающегося с гранулоцитарно-макрофагальным колониестимулирующим фактором (ГМ-КСФ) или по меньшей мере с одним из CD116, CD131 или рецептором ГМ-КСФ, состоящим из CD116 и CD131,
благодаря чему осуществляется лечение и/или ингибирование развития реакции «трансплантат против хозяина».
2. Способ по п.1, где пациент получает аллогенный трансплантат.
3. Способ по п.2, где аллогенный трансплантат представляет собой аллогенную трансплантацию гемопоэтических стволовых клеток (алло-ТГК).
4. Способ по любому из пп.1-3, где пациент страдает лейкозом, лимфомой или множественной миеломой.
5. Способ по п.4, где лейкоз выбран из группы, состоящей из хронического миелоидного лейкоза (ХМЛ), острого миелогенного лейкоза (ОМЛ), хронического лимфоцитарного лейкоза (ХЛЛ), острого лимфоцитарного лейкоза (ОЛЛ) и острого моноцитарного лейкоза (ОМоЛ).
6. Способ по п.4, где лимфома представляет собой лимфому Ходжкина или неходжкинскую лимфому.
7. Способ по любому из пп.1-6, где нейтрализующее антитело к рецептору ГМ-КСФ представляет собой человеческое антитело или гуманизированное антитело.
8. Способ по любому из пп.1-7, где нейтрализующее антитело к рецептору ГМ-КСФ представляет собой маврилимумаб, намилумаб, лензилумаб, отилимаб или гимсилумаб.
9. Способ по любому из пп.1-8, где связывание нейтрализующего антитела к рецептору ГМ-КСФ с ГМ-КСФ или одним из CD116, CD131 и рецептором ГМ-КСФ, состоящим из CD116 и CD131, характеризуется KD меньшей чем 10-7 М, KD меньшей чем 10-8 М или KD меньшей чем 10-9 М.
10. Способ лечения пациента, перенесшего аллогенную трансплантацию, чтобы подавить развитие и/или уменьшить тяжесть связанной с ней реакции «трансплантат против хозяина», где способ включает:
введение пациенту аллогенного трансплантата и
введение пациенту нейтрализующего антитела к рецептору ГМ-КСФ, специфично связывающегося с гранулоцитарно-макрофагальным колониестимулирующим фактором (ГМ-КСФ) или по меньшей мере с одним из CD116, CD131, рецептором ГМ-КСФ, состоящим из CD116 и CD131,
благодаря чему происходит подавление развития или уменьшение тяжести реакции «трансплантат против хозяина».
11. Способ по п.10, где аллогенный трансплантат представляет собой аллогенную трансплантацию гемопоэтических стволовых клеток (алло-ТГК) и где алло-ТГК проводится пациенту при лечении лейкоза, лимфомы или множественной миеломы.
12. Способ по п.10 или 11, где нейтрализующее антитело к рецептору ГМ-КСФ вводят пациенту одновременно с аллогенной трансплантацией или после нее.
13. Способ по любому из пп.10-12, где лейкоз выбирают из группы, состоящей из хронического миелоидного лейкоза (ХМЛ), острого миелогенного лейкоза (ОМЛ), хронического лимфоцитарного лейкоза (ХЛЛ), острого лимфоцитарного лейкоза (ОЛЛ) и острого моноцитарного лейкоза (ОМоЛ).
14. Способ по любому из пп.10-13, где нейтрализующее антитело к рецептору ГМ-КСФ представляет собой маврилимумаб, намилумаб, лензилумаб, отилимаб или гимсилумаб.
15. Применение нейтрализующего антитела к рецептору ГМ-КСФ, специфично связывающегося с гранулоцитарно-макрофагальным колониестимулирующим фактором (ГМ-КСФ) или по меньшей мере с одним из CD116, CD131 или рецептором ГМ-КСФ, состоящим из CD116 и CD131, для лечения реакции «трансплантат против хозяина» у пациента, перенесшего аллогенную трансплантацию гемопоэтических стволовых клеток (алло-ТГК).
16. Применение по п.15, где нейтрализующее антитело к рецептору ГМ-КСФ нейтрализует физиологическую функцию ГМ-КСФ или одного из CD116, CD131 и рецептора ГМ-КСФ, состоящего из CD116 и CD131 соответственно.
17. Применение по п.15 или 16, где нейтрализующее антитело к рецептору ГМ-КСФ представляет собой человеческое антитело или гуманизированное антитело.
18. Применение по любому из предшествующих пп.15-17, где нейтрализующее антитело к рецептору ГМ-КСФ представляет собой маврилимумаб, намилумаб, лензилумаб, отилимаб или гимсилумаб.
19. Применение по любому из пп.15-18, где связывание нейтрализующего антитела к рецептору ГМ-КСФ с ГМ-КСФ или одним из CD116, CD131 и рецептором ГМ-КСФ, состоящим из CD116 и CD131, характеризуется KD меньшей чем 10-7 М, KD меньшей чем 10-8 М или KD меньше чем 10-9 М.
20. Применение молекулы нуклеиновой кислоты, кодирующей нейтрализующее антитело к рецептору ГМ-КСФ, специфично связывающееся с гранулоцитарно-макрофагальным колониестимулирующим фактором (ГМ-КСФ) или по меньшей мере с одним из CD116, CD131 и/или рецептором ГМ-КСФ, состоящим из CD116 и CD131, для лечения реакции «трансплантат против хозяина»у пациента, перенесшего алло-ТГК.
21. Применение по п.20, где молекула нуклеиновой кислоты представляет собой молекулу ДНК или молекулу РНК.
22. Применение экспрессионной конструкции нуклеиновой кислоты, содержащей молекулу нуклеиновой кислоты, кодирующую нейтрализующее антитело к рецептору ГМ-КСФ, специфично связывающееся с гранулоцитарно-макрофагальным колониестимулирующим фактором (ГМ-КСФ) или по меньшей мере с одним из CD116, CD131 и/или рецептором ГМ-КСФ, состоящим из CD116 и CD131, для лечения реакции «трансплантат против хозяина» у пациента, перенесшего алло-ТГК.
23. Применение по п.22, где экспрессионная конструкция выбрана из группы, состоящей из плазмидной ДНК, двухцепочечной линейной ДНК, одноцепочечной РНК и вируса, выбранного из лентивируса, вируса герпеса, аденовируса и аденоассоциированного вируса.
24. Применение нейтрализующего антитела к рецептору ГМ-КСФ для нейтрализации физиологической функции ГМ-КСФ или одного из CD116, CD131 и рецептора ГМ-КСФ, состоящего из CD116 и CD131 соответственно.
25. Применение по любому из пп.20-24, где нейтрализующее антитело к рецептору ГМ-КСФ представляет собой человеческое антитело или гуманизированное антитело.
26. Применение по любому из предшествующих пп.20-25, где нейтрализующее антитело к рецептору ГМ-КСФ представляет собой маврилимумаб, намилумаб, лензилумаб, отилимаб или гимсилумаб.
27. Применение по любому из пп.21-26, где связывание нейтрализующего антитела к рецептору ГМ-КСФ с ГМ-КСФ или одним из CD116, CD131 и рецептором ГМ-КСФ, состоящим из CD116 и CD131, характеризуется KD меньшей чем 10-7 М, KD меньшей чем 10-8 М или KD меньше чем 10-9 М.
28. Применение фармацевтической композиции для лечения реакции «трансплантат против хозяина» у пациента, перенесшего алло-ТГК, где фармацевтическая композиция содержит нейтрализующее антитело к рецептору ГМ-КСФ, специфично связывающееся с гранулоцитарно-макрофагальным колониестимулирующим фактором (ГМ-КСФ) или по меньшей мере с одним из CD116, CD131 и/или рецептором ГМ-КСФ, состоящим из CD116 и CD131, молекулу нуклеиновой кислоты, кодирующую нейтрализующее антитело к рецептору ГМ-КСФ, или экспрессионную конструкцию нуклеиновой кислоты, содержащую молекулу нуклеиновой кислоты, кодирующую нейтрализующее антитело к рецептору ГМ-КСФ, и фармацевтически приемлемый носитель и изготовленна в виде формы для парентерального введения.
29. Применение по п.28, где нейтрализующее антитело к рецептору ГМ-КСФ представляет собой маврилимумаб, намилумаб, лензилумаб, отилимаб или гимсилумаб.
| WO 2009064399 A1, 22.05.2009 | |||
| MILITANO O | |||
| et al | |||
| Sequential administration of sargramostim (GM-CSF) and filgrastim (G-CSF) in pediatric allogeneic stem cell transplant (allosct) recipients undergoing myeloablative (MA) conditioning: Cost-effective and more rapid platelet recovery in UCB recipients, POSTER SESSION I ALLOGENEIC, Vol | |||
| Способ гальванического снятия позолоты с серебряных изделий без заметного изменения их формы | 1923 |
|
SU12A1 |
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |

