Настоящее изобретение относится к ингибиторам сигнального пути CD95 для применения при лечении миелодиспластического синдрома (МДС), где МДС выбирают из подгруппы МДС с низким риском согласно IPSS и/или подгруппы МДС с промежуточным-1 (int-1) риском, а также к способу диагностики МДС.
Миелодиспластические синдромы (МДС) являются расстройствами клональных гематопоэтических стволовых клеток, характеризуемых неэффективным гематопоэзом, ведущим к цитопениям, особенно анемии, и зачастую развивающихся в острый миелоидный лейкоз (ОМЛ). В частности, МДС могут характеризоваться нарушенным ростом эритроидных предшественников. Соответственно, МДС зачастую классифицируют на основании морфологии и процента бластных клеток в крови и костном мозге (классификации French American British (FAB) и WHO) (Bennett et al., 1982; Vardiman et al., 2009).
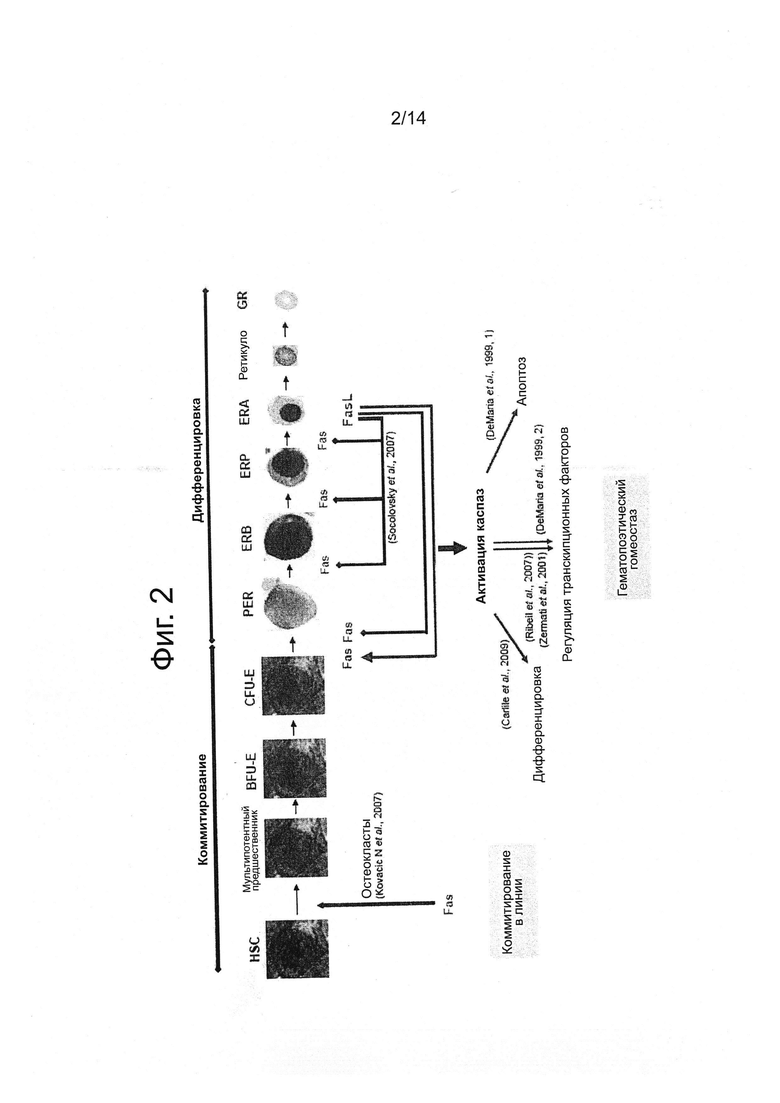
Эритропоэз контролируется балансом между положительными и отрицательными сигналами, в которые вовлечены межклеточное взаимодействие и растворимые факторы в эритробластных островках костного мозга. Обзор эритропоэза показан на фигуре 1. Коммитирование CD34+ гематопоэтической стволовой клетки (ГСК) в эритроидную линию под контролем важного транскрипционного фактора, подобного GATA-1 и цитокинов, подобных лиганду c-Kit, фактору стволовых клеток (SCF). Размер эритроидного компартмента положительно регулируется эритропоэтином, который стимулирует созревание CFU-E и проэритробластов и предотвращает их апоптоз. Эритроидные клетки приобретают экспрессию мембранного CD71 (рецептор переноса), а затем гликофорина A (GPA) на более зрелых клетках (фигура 1).
Отрицательная регуляция эритропоэза зависит от Fas/FasL, которые вносят вклад в апоптоз незрелых эритробластов, которые экспрессируют Fas, путем взаимодействия с FasL, который экспрессируется на зрелых эритроидных предшественниках (De Maria et al., 1999) и также путем авторегуляции эритробластов в той же стадии дифференцировки (Socolovski et al., 2008). Созревание нормальных эритроидных предшественников требует активации каспазы-3, несмотря на то, что активность каспазы-8 не подтверждена (De Maria et al., 1999; Zermati et al., 2001) Расщепление субстратов каспазы-3 ограничено. Например, GATA-1 является субстратом для каспазы-3 в условиях депривации эритропоэтином, хотя в присутствии Еро GATA-1 защищен от расщепления взаимодействием с Hsp70, который транслоцируется из цитозоля в ядро (Ribeil et al., 2007).
Дизэритропоэз МДС ассоциирован с эктопической активацией каспазы-8 в нисходящем направлении сигнала от Fas. Группа заявителя и другие ранее демонстрировали, что Fas сверхэкспрессирован на поверхности незрелых предшественников CD34+ и в эритроидных коммитированных предшественниках. Экспрессия Fas повышается вместе с эритроидной дифференцировкой, с началом экспрессии Fas-лиганда в GPA-положительных эритроидных предшественниках, приводящей к эктопической активации каспазы-8 в эритроидной линии. Это наблюдалось в клетках свежего костного мозга (Bouscary et al., 1997), а также в эритроидных клетках, полученных в 2-х стадийной жидкой культуре (Claessens et al, 2002). Ингибирование передачи сигналов Fas эктопической экспрессией лентивирусно экспрессируемого доминантно-негативного мутанта адаптера FADD снижает активацию каспазы-8 и ингибирование апоптоза в эритроидных предшественниках МДС (Claessens et al., 2005). Fas/FasL могут вносить вклад в предотвращение нормальной эритроидной дифференцировки и индуцировать апоптоз в эритроидных клетках МДС.
Недавние данные продемонстрировали, что созревание эритроидных клеток серьезно нарушается в МДС с низкой степенью тяжести. Предшественники эритроидных клеток выражали количественно с помощью мечения CD71/GPA, описанного Socolovski et al. (2007). Было отмечено, что в культурах клеток МДС по сравнению с нормальными культурами по прошествии 7 дней фракция клеток с высоким уровнем CD71/низким уровнем GPA повышена, а со средним уровнем CD71/высоким уровнем GPA - понижена. Транскриптомные исследования эритроидных предшественников 14 дня продемонстрировали 2-кратное снижение экспрессии GYP А, кодирующего гликофорин A (GPA). Кроме того, несколько других эритроидных генов были отрицательно регулированы.
МДС являются редкими заболеваниями (распространенность 3-5/100000 человек в год) и преобладают в пожилом возрасте (средний возраст 65-70 лет).
Обычно МДС лечат введением классических агентов, стимулирующих эритропоэз (ESA).
Однако до настоящего изобретения пациенты с МДС, которые устойчивы к ESA, с трудом подвергались лечению.
Таким образом, задача настоящего изобретения заключается в обеспечении нового варианта лечения МДС, в частности, для лечения пациентов с МДС, которые устойчивы к ESA.
Первый аспект изобретения относится к ингибитору сигнального пути CD95 для применения при лечении миелодиспластического синдрома (МДС), который выбирают из подгруппы МДС с низким риском согласно IPSS и/или подгруппы МДС с промежуточным риском (int-1) согласно IPSS).
Согласно изобретению термины CD95, CD95R и CD95-рецептор могут использоваться взаимозаменяемо. Другими синонимами являются Аро-1 или Fas, которые в данном документе могут использоваться взаимозаменяемо. Кроме того, термины CD95L, СD95-лиганд и соответствующие синонимы FasL, Apo-IL, CD178 или TNF-SF6 могут использоваться взаимозаменяемо.
«Ингибитор сигнального пути CD95» в свете настоящего изобретения могут быть любым соединением, которое мешает или блокирует, по меньшей мере, частично сигнальный путь CD95. Согласно предпочтительному воплощению «ингибитор сигнального пути CD95» блокирует сигнальный путь CD95. Способы определения и/или оценки активности сигнального пути CD95 известны специалисту в данной области и, например, описаны Lavrik et al. (Cell Death Differ. 2012 Jan; 19(1):36-41 Regulation of CD95/Fas signaling at the DISC).
Ингибитор по изобретению может действовать на уровне белка и/или уровне нуклеиновой кислоты. Ингибиторы, воздействующие на уровне белка, могут быть выбраны из антител, белков и/или малых молекул. Ингибиторы, действующие на уровне нуклеиновой кислоты, например, антисмысловые молекулы, молекулы РНКи и/или рибозимы.
Согласно особенно предпочтительным воплощениям ингибитор связывается с CD95-penenropoM (CD95) и/или СD95-лигандом (CD95L). В дополнительном воплощении взаимодействие CD95/CD95L может быть ингибировано.
В одном предпочтительном воплощении ингибитор по изобретению является антителом или его функциональным фрагментом. Ингибитор, являющийся антителом, может связаться с CD95, но, конечно, также с CD95L. Примером антитела, связывающего CD95L, является Nok-1.
Антитело может быть, например, моноклональным антителом, поликлональным антителом, рекомбинантным антителом, гуманизированным антителом, человеческим антителом, химерным антителом, мультиспецифичным антителом, или фрагментом такого антитела (например, Fab-фрагментом, Fab'-фрагментом, F(аb’)2-фрагментом, Fv-фрагментом, диателом, или молекулой одноцепочечного антитела). Антитело может быть IgG1-, IgG2-, IgG3- или IgG4-типа.
Антитело может использоваться с модификацией или без модификации, и может быть помечено, либо ковалентно или нековалентно, например, с помощью репортерной группы или эффекторной группы.
«Антительный фрагмент» согласно изобретению представляет по существу такой же эпитоп-связьшающий участок, что и на соответствующем антителе осуществляет и/или имеет фактически такую же CD95 и/или CD95L ингибирующую активность, что и у соответствующего антитела.
Способы получения антител изобретения известны специалистам в данной области.
Один тип ингибитора, охваченный настоящим изобретением, может быть ингибитором СD95-лиганда. Например, ингибиторы СD95-лиганда могут быть выбраны из (а) ингибиторного анти-СD95-лиганд антитела или его фрагмента, как указано выше;
(b) молекулы растворимого СD95-рецептора или ее части, связывающей СD95-лиганд; и
(c) ингибитор СD95-лиганда, выбранный из FLINT, DcR3 или его фрагментов.
Молекулы растворимого CD95-рецептора, например, молекула растворимого СD95-рецептора без трансмембранного домена, как описано в ЕР-А-0595659 и ЕР-А-0965637 или пептиды СD95-рецептора, как описано в WO 99/65935.
Ингибитор Fas-лиганда FLINT или DcR3 или фрагмент, например, его растворимый фрагмент, например внеклеточный домен, необязательно слитый в гетерологичный полипептид, в частности с молекулой Fc-иммуноглобулина, описан в WO 99/14330 или WO 99/50413. FLINT и DcR3 являются белками, которые способны связываться с СD95-лигандом.
В дополнительном воплощении ингибитор является химерным белком, в частности химерным белком, который связывает CD95L.
В одном воплощении ингибитор CD95L включает экстраклеточный домен молекулы CD95R, такие как аминокислоты 1-172 (MLG…SRS) зрелой последовательности CD45 согласно Пат. США N5891434. Этот внеклеточный домен молекулы CD95R может быть слит с гетерологичным полипептидным доменом, в частности молекулой Fc-иммуноглобулина, включая шарнирный участок, например, из молекулы IgG1 человека. Химерный белок, содержащий внеклеточный домен CD95 и человеческий Fc-домен описаны в WO 95/27735.
Таким образом, согласно предпочтительному воплощению, агент, который связывает CD95L, является химерным белком, содержащим внеклеточный домен CD95 и Fc-домен, в частности Fc-домен человека.
Согласно особенно предпочтительному воплощению, агент, который связывается с CD95L, является APG101 или его функциональными фрагментами, изоформами и/или производными. APG101 содержит домены CD95L (аминокислоты 26-172; ECD внеклеточный домен) и IgG1-Fc (аминокислоты 172-400) из SEQ ID NO: 1). APG101 и его производные описаны в WO 95/27735 и WO 2004/085478.
В еще одном дополнительном воплощении настоящего изобретения ингибитор является молекулой нуклеотидного эффектора. Молекула нуклеотидного эффектора может быть ДНК; РНК, ПНК или гибридом ДНК-РНК. Молекула может быть одно- или двухцепочечной. Экспрессирующие векторы, полученные из ретровирусов, аденовируса, вирусов герпеса или коровьей оспы или из различных бактериальных плазмид могут быть использованы для доставки нуклеотидных последовательностей в целевой орган, ткань или клеточную популяцию. Такие конструкции могут быть использованы для введения нетранслируемых смысловых или антисмысловых последовательностей в клетку. Даже без интеграции в ДНК такие векторы могут продолжать транскрибировать молекулы РНК до тех пор, пока они не будут повреждены эндогенными нуклеазами. Временная экспрессия может длиться в течение месяца или больше с нереплицирующимся вектором или даже дольше если соответствующие репликационные элементы являются частью векторной системы.
Молекула нуклеотидного эффектора частично может быть выбрана из антисмысловых молекул, молекул РНКи и рибозимов, которые предпочтительно способны ингибировать экспрессию гена CD95R и/или CD95L.
Как указано выше, настоящее изобретение относится, в частности, к лечению миелодиспластического синдрома (МДС), который выбран из подгруппы МДС с низким риском согласно Международной Прогностической Балльной Системе (англ. International Prognostic Scoring System, EPSS) и/или подгруппы МДС с промежуточным-1 (int-1) риском.
IPSS хорошо известна специалистам в данной области. Основные прогностические факторы МДС для прогрессии в ОМЛ и выживаемости включают количество и существенность цитопений, процент бластных клеток костного мозга и цитогенетических аномалий костного мозга. Каждый индикатор оценивается согласно его тяжести и рейтинги объединяются в «балл», IPSS.
IPSS разделяет 4 подгруппы со значимо различным риском прогрессии в ОМЛ и выживаемостью: низкий, промежуточный-1 (int-1), промежуточный-2 (int-2) и высокий риск. Подгруппы с низким риском и риском int-1 часто группируются вместе как «благоприятные» или с низким риском МДС, а подгруппы int-2 и высокого риска являются «неблагоприятными» или МДС с высоким риском (Greenberg et al., 1997).
МДС с низким риском (с низким или int-1 IPSS) характеризуются повышенным апоптозом костномозговых предшественников, который приводит с большой вероятностью к цитопениям. У большинства пациентов эритроидная клетка демонстрирует нарушенную дифференцировку и повышенный апоптоз.
Таким образом, подгруппа МДС, подлежащая лечению согласно настоящему изобретению, т.е. с IPSS с низким риском или int-1 может характеризоваться повышенным апоптозом в ходе эритропоэза.
Согласно дополнительному воплощению подгруппа МДС подлежащая лечению может дополнительно характеризоваться тяжелым дефектом эритропоэза без избытка бластных клеток.
Другой характеризующий признак подгруппы МДС, подлежащей лечению может быть устойчивостью к классическим агентам, стимулирующим эритропоэз (ESA) и/или колониестимулирующим факторам. Агент, стимулирующий эритропоэз, согласно изобретению может быть любым соединением, которое стимулирует выработку красных кровяных клеток. Примеры ESA и/или колониестимулирующих факторов включают, без ограничения перечисленным, эритропоэтин (Еро), эпоэтин альфа (Procrit/Epogen), эпоэтин бета (NeoRecormon), дарбепоэтин альфа (Aranesp), метоксиполиэтиленгликоль-эпоэтин бета (Mircera) и/или некоторые цитокины, такие как IL-3 или IL-9.
Подгруппа МДС, подлежащая лечению, может быть охарактеризована низким значением эритроидной взрывообразующей единицы (BFU-E) и/или низким значением эритроидной колониеобразующей единицы (CFU-E).
Конечно, характеристики, обсуждаемые выше, могут быть объединены в любой возможной комбинации для описания подгруппы МДС, подлежащей лечению.
С учетом описанной выше подгруппы МДС, дополнительное воплощение изобретения относится к применению ингибиторов, которые повышают количество BFU-Е и не влияют на гранулоцитарно-макрофагальную колониеобразующую единицу (CFU-GM) и/или без повышения риска экспансии лейкозных клеток. Предпочтительно, если ингибитор также может быть соединением, которое улучшает рост эритроидных предшественников.
Ингибитор сигнального пути CD95, используемый согласно настоящему изобретению, может предлагаться в качестве фармацевтической композиции. Данная композиция может содержать фармацевтически приемлемые носители, разбавители и/или адъюванты, и т.п.
Дополнительные подробности по методам для составления и введения могут быть обнаружены в последней редакции Remington's Pharmaceutical Sciences (Maack Publishing Co., Easton, Pa.).
Фармацевтические композиции, используемые в данном изобретении, могут быть введены любыми путями, включая, без ограничения перечисленным, пероральные, внутривенные, внутримышечные, внутриартериальные, костномозговые, интратекальные, внутрижелудочковые, трансдермальные, подкожные, внутрибрюшинные, интраназальные, энтеральные, местные подъязычные и ректальные средства.
Фармацевтические композиции, подходящие для применения в изобретении включают композиции, в которых активные ингредиенты содержатся в эффективном количестве для достижения подразумеваемой цели. Определение эффективной дозы находится в пределах возможностей специалиста в данной области. Терапевтически эффективная доза относится к такому количеству активного ингредиента, например, нуклеиновой кислоты или белка изобретения или антитела, которое достаточно для лечения специфического состояния. Точная дозировка будет определена практикующим врачом в свете факторов, связанных с объектом, которому требуется лечение.
Дозировка и введение корректируются для обеспечения достаточных уровней активного составляющего или для поддержания искомого эффекта. Специалист в данной области осведомлен о дополнительных способах по обеспечению достаточных уровней активного составляющего и/или поддержания искомого эффекта. Факторы, которые могут быть учтены, включают тяжесть течения заболевания, общее состояние здоровья объекта, возраст, массу и пол объекта, диету, время и частоту введения, комбинацию(ии) лекарственного средства, чувствительности реакции и переносимость/ответ на терапию. В предпочтительном воплощении общее количество ингибитора сигнального пути CD95 согласно настоящему изобретению, вводимого пациенту, страдающему от МДС, составляет от около 50 до около 400 мг/неделю, более предпочтительно от около 100 до около 200 мг/неделю. Предпочтительная недельная доза может быть введена в виде одиночной дозы или в виде нескольких доз. Особенно предпочтительной является одиночная доза, особенно от 100 до 200 мг/неделю, которая вводится внутривенно в виде одиночной дозы.
Лечение может длиться в течение нескольких недель. В каждом индивидуальном случае длительность лечения определяется наблюдающим врачом и, например, основана на успехе лечения, встречаемости побочных эффектов и т.п.
Согласно дополнительному аспекту настоящего изобретения данная фармацевтическая композиция может включать, по меньшей мере, один дополнительный активный ингредиент, такой как агент, широко используемый для лечения МДС, такой как 5-азацитидин, децитабин или леналидомид, агент, стимулирующий эритропоэз и/или агент, ингибирующий апоптоз.
Примеры агентов, стимулирующих эритропоэз были указаны выше.
Примеры агентов, ингибирующих апоптоз включают ингибиторы каспаз, такие как xIAP, c-IAP-1, C-IAP2, сурвивин, TNF-α-ингибирующие соединения, такие как ревлимид, помалидомид, ингибиторы белков семейства Всl-2 и т.п.
Дополнительным аспектом настоящего изобретения является фармацевтическая композиция, содержащая ингибитор согласно настоящему изобретению, дополнительно содержащая агент, стимулирующий эритропоэз, в частности, агент, стимулирующий эритропоэз, определенный выше.
Дополнительный аспект изобретения относится к способу диагностики МДС, включающий стадию определения экспрессии CD95L в образце, где сверхэкспрессия CD95L является предиктивной для заболевания. Дополнительная стадия оригинального способа может быть помимо прочего сравнением определенного значения экспрессии CD95L с контрольным значением, например, из образца пациента с анемией или с происхождением отличным от МДС.
Конечно, оригинальный способ диагностики может быть объединен с известными способами, такими как система IPSS.
Подписи к чертежам
Фигура 1: Обзор эритропоэза. ГСК: гематопоэтические стволовые клетки. BFU-Е: эритроидная взрывообразующая единица, CFU-E: эритроидная колониеобразующая единица, PER: проэритробласт, ERB: базофильный эритробласт, ERP: полихроматофильный эритробласт, ERA: ацидофильный эритробласт, ретикуло: ретикулоцит, RBC: красная кровяная клетка, SCF: фактор стволовых клеток, ЕРО: эритропоэтин.
Фигура 2: Отрицательная регуляция нормального и МДС-эритропоэза. Нормальный эритропоэз контролируется отрицательной регуляцией пролиферации через Fas-зависимый апоптоз незрелых предшественников, опосредованный FasL-экспрессирующими зрелыми клетками (голубые стрелки). При МДС Fas и FasL сверхэкспрессируются, что приводит к избыточному и неприемлемому апоптозу (красные стрелки).
Фигура 3: Роль/Концентрация Fas и FasL в ходе эритропоэза.
Фигура 4: Экспрессия FasL в МДС, вторичном ОМЛ и контролях. А. Распределение FasL RFI. В. Анализ ROC, сравнивающий МДС и контроли.
Фигура 5: Влияние экспрессии Fas на CD45lo/CD34+ клетки в момент постановки диагноза на общую выживаемость.
Фигура 6: Влияние APG101 на рост BFU-E и CFU-GM в 14 день метилцеллюлозной культуры.
Фигура 7: Влияние APG101 на рост CFU-E и CFU-L в 7 день метилцеллюлозной культуры.
Фигура 8: Влияние APG101 на BFU-E и CFU-GM, полученных из жидкой культуры на 5 день.
Фигура 9: Влияние APG101, добавленного в фазе дифференцировки в жидкую культуру эритроидных предшественников.
Фигура 10: Влияние APG101 на дифференцировку эритроидных клеток. Слева: окрашенные MGG клетки, центрифугированные на предметное стекло. Справа: Анализ проточной цитометрией двойного мечения CD71/GPA.
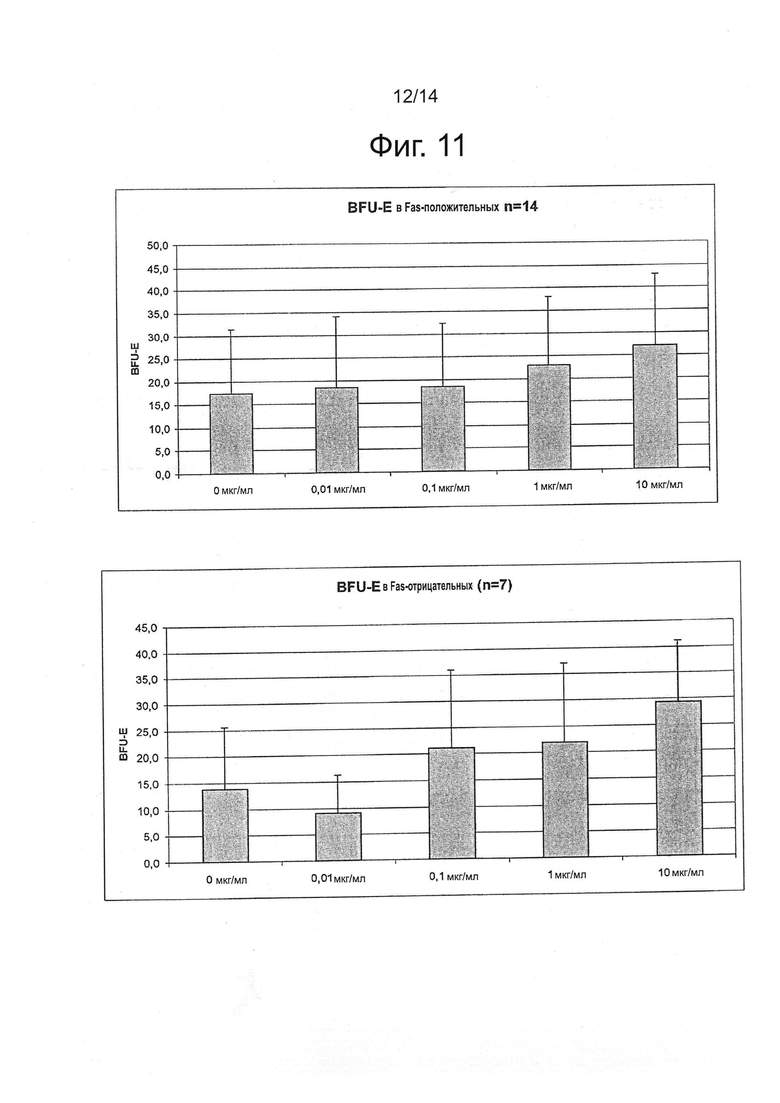
Фигура 11: Влияние APG101 на рост BFU-E. Сравнение между Fas-положительным и Fas-отрицательным МДС.
Фигура 12: Влияние APG101 согласно FasL-экспрессии на CD45low-клетки.
Фигура 13: Влияние APG101 на рост BFU-E относительно роста CFU-L и BFU-E на исходном уровне.
Материалы и методы
Лабораторные материалы

Образцы костного мозга
Нормальные и МДС-образцы костного мозга собирали грудинной аспирацией у пациентов госпиталя Cochin, Париж, и из смежных департаментов, которые ассоциированы с многоцетровыми биологическими исследованиями, координируемыми проф. М. Fontenay. Перед аспирацией костного мозга согласно рекомендациям местного этического комитета пациенты давали информированное согласие на исследования клеточной биологии и генетики.
Оборудование

Способы
Релевантные стандартные экспериментальные процедуры
Текущие стандарты проведения экспериментальных процедур в лаборатории упоминается и удостоверяется в Guide de Bonne Execution des Analyses de Bioloige medicale by M. Fontenay and C. Lacombe in 2002.

Инструменты и платформы
- APG101 Fas-Fc химера (Apogenix): ингибитор Fas-зависимого апоптоза; предоставлено Apogenix GmbH.
- CD95L-T4: тримерный лиганд для Fas поставляемый «Apogenix"
- Культуры эритроидных клеток получали, как описано: СD34+-клетки костного мозга выделяли на колонке «МIDI Macs» и культивировали с SCF, Еро, IGF-1 и дексаметазоном для индукции коммитирования СD34+-клеток в эритроидную линию и целевую пролиферацию эритроидных клеток. Через 10 дней конечное созревание эритроидных клеток получали путем переноса клеток в среду, содержащую Еро и инсулин.
- Проточную цитометрию использовали для количественной оценки созревания эритроидных клеток в ходе жидкой культуры in vitro.
Протоколы
Проточная цитометрия
Субпопуляции клеток идентифицировали двойным мечением с использованием моноклональных антител к CD71 (рецептор трансферрина) и гликопротеину A (GPA CD235a). Различными стадиями созревания были клетки CD71-/GPA-, CD71+/GPA-, CD71+/GPA+ и, наконец, CD71-/GPA+. Экспрессию мембранного Fas количественно оценивали мечением CD95 и экспрессией как соотношение средней интенсивности флуоресценции к изотопическому контролю RFI). Анализ кривой соотношений правильного и ложного обнаружения сигналов (ROC, Receiver operating curve), который является графическим представлением зависимости между истинным положительным соотношением и ложным положительным соотношением для диапазона предельных значений, использованный для сравнения 132 МДС с 25 контролями, продемонстрировал, что порог положительности составил RFI 1,8. Апоптоз измеряли мечением Аннексии v/7-AAD. Образцы анализировали на устройстве FC500 (Beckman Coulter).
Выделение и культивирование клеток
Образцы костного мозга из пациентов и здоровых контролей собирали стернальной аспирацией и очищали клетки CD34+ (>85% CD34+ клеток) на MIDI-MACS иммуноаффинных колонках (Miltenyi Biotech, Бергиш-Гладбах, Германия) (Claessens et al., 2002). Очищенные СD34+-клетки культивировали в среде Игла, модифицированной по Дульбекко (DMEM), содержащей 20% BIT (бычьего сывороточного альбумина, инсулина, голотрансферрина), и соответствующих цитокинов: 50 нг/мл фактора стволовых клеток (SCF), 1 МЕ/мл эритропоэтина (Еро), 40 нг/мл инсулино-подобного фактора роста 1 (IGF-1) и 10-6 M дексаметазона вплоть до 10 дней в культуре, и 1 МЕ/мл Еро и 10-6 инсулина после этого. Обзор стадий культивирования показан на фигуре 3. Клетки активно делились вплоть до 10 дней и демонстрировали незначительное созревание эритроидов. После замены цитокинов большинство клеток стали дифференцированными эритробластами на 14 день.
Индукция апоптоза с помощью CD95L-T4 в нормальных эритроидных клетках
Нормальные эритроидные клетки обрабатывали повышающимися концентрациями CD95L-T4. В данной концентрации измеряли влияние, оказываемое CD95L-T4, в ранней фазе коммитирования эритроидных клеток на рост эритроидных клеток (BFU-E и общая амплификация эритроидных предшественников). Тестировали влияние CD95L-T4, добавленного до возникновения активации каспазы-3, предшествующей созреванию эритроидов. За каспазой-3 и мишенями следовали проточная цитометрия и Вестерн-блоттинг.
Что касается стехиометрии 1/1 комплекса CD95L-T4/APG101 с 11 нМ связывающей аффинностью в людях, то концентрации тестируемых CD95L-T4 и APG101 в культуре были в одинаковых диапазонах (0,01-10 мкг/мл).
Ингибирование апоптоза, индуцированного CD95L-T4, с помощью APG101 в нормальных эритроидных клетках.
Нормальные эритроидные клетки предварительно обрабатывали с помощью APG101 при повышенной концентрации перед лечением с помощью предварительно определенной концентрации CD95L-T4 либо в ранней фазе амплификации эритроидных клеток, либо в поздней фазе дифференцировки, либо в обеих фазах). Важно отметить, что избегали добавлять CD95L-T4 в образцы с высокими уровнями апоптоза на исходном уровне. Эффекты APG101 оценивали на общей амплификации эритроидных клеток, росте BFU-E, апоптозе, активности каспазы-3, целевом расщеплении каспазы-3, и дифференцировке (GPA, цитология).
Ингибирование апоптоза эритроидных клеток МДС с помощью APG101
Тестировали повышенные концентрации APG101 (0,001-100 мкг/мл) на количество BFU-E в клоногенных тестах
- на скорости амплификации
- на апоптоз (активность каспазы-3, экспонирование фосфатидилсерина, мишени каспазы-3)
- на дифференцировку эритроидных клеток (GPA/CD71, цитологическая проверка, экспрессия эритроидных генов в микрочипах).
Ранее было показано, что Fas сверхэкспрессируется в фракции СD34+-клеток костного мозга. Поэтому- было проведено сравнение между эритроидными клетками и клетками предшественниками гранулоцитов в образцах МДС в условиях специфической культуры для каждой линии. Протестировали возможность восстановления нормальной кинетики активности каспазы-3 и предотвращения расщепления мишеней каспазы-3 при обработке APG101.
Метилцеллюлозный Анализ
Мононуклеарные клетки, выделенные из аспиратов костного мозга после градиента фиколла, рассевали в 0,8% метилцеллюлозе, содержащей фетальную телячью сыворотку, BSA и цитокины (IL3 0,1 МЕ/мл, IL6 10 нг/мл, GM-CSF 5 нг/мл, ЕРО 1 МЕ/мл и SCF 20 нг/мл в концентрации 106 клеток/мл). CFU-E и CFU-L подсчитывали на 7 день культуры, а BFU-E и CFU-GFM подсчитывали на 14 день культуры. Добавляли возрастающие концентрации APG101.
Статистика
Биологические данные анализировали в виде медианных значений ± стандартная ошибка среднего. Чувствительность и специфичность проточной цитометрии для анализа Fas и FasL оценивали с помощью кривой соотношений правильного и ложного обнаружения сигналов (ROC) и выводили пороговое значение. Непрерывные переменные сравнивали с помощью t-критерия Стьюдента (программа Excel 2003, Microsoft). Оценку Каплана-Майера использовали для оценки влияния Fas и FasL на общую выживаемость с течение времени, в двух подгруппах популяции в исследовании, и сравнивали с помощью логарифмического рангового критерия. Все статистические анализы были двухсторонними и р-значения меньше, чем 0,05 считались значимыми. Статистические анализы осуществляли с помощью программного обеспечения «GraphPad».
Результаты
Экспрессия FasL и диагноз МДС
Экспрессию FasL измеряли как в CD45low/CD34+ так и в CD71+ популяциях клеток костного мозга в 84 МДС, 21 вторичного ОМЛ и 17 контролях. FasL был значимо более повышен в МДС, чем во вторичном ОМЛ или в контролях (фигура 4А) и согласно ROC, порог положительности был равен 6. При сравнении МДС с контролями, предиктивное значение экспрессии FasL для различения между МДС и контролями было хорошим (площадь под кривой: 0,73; Р=0,002; фигура 4 В).
Для анализа влияния экспрессии Fas или FasL на прогноз собирали клиническую информацию 166 пациентов с МДС или пациентов с вторичным ОМЛ с известным значением экспрессии Fas в момент постановки диагноза и 42 пациентов с МДС/вторичным ОМЛ с известным значением экспрессии FasL в момент постановки диагноза. В группе из 166 пациентов с МДС 41% были положительными по Fas и не демонстрировали какого-либо различия по показателям возраста, пола, параметрам гемограммы, % бластных клеток костного мозга, мультилинейной дисплазии, кариотипу, IPSS и обработкам. Экспрессия Fas не оказывала влияния на общую выживаемость (фигура 5). Кроме того, экспрессия Fas не была предиктивной относительно ответа на ЕРО
В группе из 42 пациентов с МДС/вторичным ОМЛ 18 пациентов были положительными по FasL (RFI≥5). Уровень Hb и медиана общей выживаемости были эквивалентны у FasL-положительных и FasL-отрицательных пациентов (Р=0,57 и Р=0,97, соответственно). Кроме того, экспрессия FasL в малой когорте пациентов (n=22) не была предиктивной для ответа на лечение.
В общем, эти данные демонстрируют, что Fas и FasL были сверхэкспрессированы в МДС, а сверхэкспрессия FasL является предиктивной для МДС. Ни экспрессия Fas, ни экспрессия FasL не являются прогностическими параметрами для общей выживаемости.
Влияние APG101 на рост гематопоэтических предшественников
Исследования показали, что блокирование FasL избытком растворимого Fas (Fas: Fc) а также разрушение сигнального пути Fas через эктопическую экспрессию доминантно-негативной формы адаптера FADD восстанавливает рост BFU-E. При анализе эффекта повышенных концентраций APG101 (0, 0,01, 0,1, 1, 10 μг/мл) на эритроидные и грунулоцитарно-моноцитарные предшественники в тестах с метилцеллюлозой становится очевидно, что APG101 не стимулирует рост CFU-GM хотя был отмечен умеренный положительный эффект на рост BFU-E (фигура 6).
В дополнение к эффекту APG101 на рост зрелых эритроидных предшественников (CFU-E) ответ лейкозных бластных клеток анализировали путем подсчета количества CFU-L (неэритроидных кластеров, по меньшей мере, из 50 клеток) на 7 день метилцеллюлозной культуры (фигура 7). Было установлено, что APG101 не восстанавливает рост CFU-E и не повышает количество CFU-L даже в образцах из пациентов с RAEB2 (МДС с более чем 10% бластных клеток в момент постановки диагноза), что подтверждает, что APG101 не стимулирует рост лейкозных бластных клеток.
Чтобы получить более глубокое представление о влиянии APG101 на эритропоэз при МДС, СD34+-клетки костного мозга из пациента с МДС выделяли и высевали в «эритроидных условиях» для коммитирования. Клетки собирали на 5 день жидкой культуры и затем рассевали в метилцеллюлозе для оценки количества BFU-E и CFU-GM. Как показано на фигуре 7, изначальное количество BFU-E было снижено в МДС по сравнению с контролями. APG101 индуцировало 3-х кратное увеличение количества BFU-Е без достижения нормального уровня. CFU-GM были менее тяжело повреждены, чем BFU-E и не были поражены APG101. Эти результаты подтверждают специфическое влияние APG101 на эритроидную линию.
Для тестирования влияния APG101 на дифференцировку клеток в течение эритропоэза к клеточной культуре добавляли различные концентрации APG101, а затем культуру переводили на Еро и инсулин, что совпадало с началом экспрессии FasL. На 14 день культуры общее количество клеток увеличивалось в среднем на 60%, а апоптоз оказался сниженным вплоть до 33% (фигура 9).
APG101 не оказывал влияния на дифференцировку эритроидных клеток, а что более важно - не блокировал созревание клеток (фигура 10).
Для описания группы пациентов, которые могли бы получить пользу от лекарственного средства, данные тестов на метилцеллюлозе повторно анализировали в соответствии с исходными клиническими и биологическими характеристиками. Тесты на метилцеллюлозе ранее были признаны в качестве полезного инструмента для прогноза ответа на Еро и были признаны коррелирующими с качеством эритропоэза (Frisan et al, 2010).
Во-первых, анализировали клоногенные тесты согласно исходной экспрессии Fas. Как показано на фигуре 11, улучшение роста BFU-E в присутствии 10 мкг/мл APG101 значимо не отличалось у Fas-положительных и Fas-отрицательных пациентов.
Во-вторых, анализировали влияние APG101 согласно экспрессии FasL на CD71-положительной популяции. В этих сериях порог положительности составил 2,8. FasL-положительные МДС имели среднее значение fasL RFI равное 3,7, а FasL-отрицательные пациенты имели среднее значение FasL RFI равное 2,2. Влияние APG101 не отличалось между FasL-положительными пациентами, демонстрирующими высокую экспрессию FasL, и FasL-отрицательными пациентами, демонстрирующими низкую экспрессию FasL.
В-третьих, заметили положительную корреляцию между влиянием APG101 на рост BFU-E и изначальным ростом CFU-L в момент постановки диагноза. В пациентах с избытком лейкозных кластеров, APG101 не мог восстановить рост BFU-E, при этом APG101 значительно повышал количество BFU-E у пациентов с низким изначальным ростом CFU-L (фигура 13, слева).
И наконец, APG101 более эффективно улучшает рост BFU-E в клетках костного мозга из пациентов с МДС с тяжелым изменением эритропоэза на исходной линии, чем у пациентов с сохраненным эритропоэзом (фигура 13, справа).
Резюме
Было продемонстрировано, что в культуре in vitro гематопоэтические стволовые - клетки (CD34+клетки) из пациентом с МДС с низкой или INT-1 степенью с тяжелым нарушением эритропоэза APG101 восстанавливает рост эритроидных клеток. APG101 не стимулирует рост лейкозных клеток и таким образом не увеличивает риск экспансии лейкозных клеток.
Сверхэкспрессия FasL была прогностической относительно заболевания. Однако ни экспрессия Fas, ни экспрессия FasL не продемонстрировали явное влияние на общую выживаемость.
Fas является одним из основных участников апоптотической клеточной смерти гепатопоэтических клеток предшественников приводящей к анемии. Таким образом, ингибирование взаимодействия Fas и FasL с использованием растворимого рецептора Fas (т.е. APG1010) могло восстановить эритропоэз у пациентов с МДС.
APG101 восстанавливает рост эритроидных клеток в подгруппе пациентов с МДС с тяжелым дефектом эритропоэза и без избытка лейкозных бластных клеток. APG101 не стимулирует рост лейкозных клеток согласно оценке подсчетом CFU-L, однако APG101 не способен восстановить рост BFU-E у пациентов, демонстрирующих избыток CFU-L на момент постановки диагноза. В совокупности APG101 является эффективным при восстановлении роста BFU-E в гематопоэтических стволовых клетках из пациентов с МДС с тяжелым нарушением эритропоэза на момент постановки диагноза и без избытка бластных клеток. Эти пациенты ранее были идентифицированы как устойчивые к агентам, стимулирующим эритропоэз (ESA) частично из-за сверхэкспрессии Fas и высокого уровня апоптоза (Frisan et al., 2010).
Литература
1. Bennett JM, Catovsky D, Daniel MT, Flandrin G, Galton DA, Gralnick HR, et al. Proposals for the classification of the myelodysplastic syndromes. Br. J. Haematol. 1982 Jun; 51(2):189-199.
2. Bouscary D, De Vos J, Guesnu M, Jondeau K, Viguier F, Melle J, Picard F, Dreyfus F, Fontenay-Roupie M. Fas/Apo-1 (CD95) expression and apoptosis in patients with myelodysplastic syndromes.
3. Leukemia. 1997 Jun; 11(6):839-45.
4. Carlile GW, Smith DH, Wiedmann M. Caspase-3 has a nonapoptotic function in ervthroid maturation. Blood. 2004 Jun l; 103(11):4310-6.
5. Claessens YE, Bouscary D, Dupont JM, et al. In vitro proliferation and differentiation of erythroid progenitors from patients with myelodysplastic syndromes: evidence for Fas-dependent apoptosis. Blood. 2002; 99:1594-1601.
6. Claessens YE, Park S, Dubart-Kupperschmitt A, Mariot V, Garrido С, Chrétien S, Dreyfus F, Lacombe C, Mayeux P, Fontenay M. Rescue of early-stage myelodysplastic syndrome-deriving ervthroid precursors by the ectopic expression of a dominant-negative form of FADD. Blood. 2005 May 15; 105(10):4035-42.
7. De Maria R, Zeuner A, Eramo A, Domenichelli C, Bonci D, Grignani F, Srinivasula SM, Alnemri ES, Testa U, Peschle C. Negative regulation of erythropoiesis by caspase-mediated cleavage of GATA-1. Nature. 1999 Sep 30; 401(6752):489-93.
8. Frisan Ε, Pawlikowska Ρ, Pierre-Eugène С, Viallon V, Gibault L, Park S, Mayeux P, Dreyfus F, Porteu F, Fontenay M. p-ERKl/2 is a predictive factor of response to erythropoiesis-stimulating agents in low/int-1 myelodysplastic syndromes. Haematologica. 2010 Nov; 95(11): 1964-8.
9. Greenberg P, Cox C, LeBeau MM, Fenaux P, Morel P, Sanz G, Sanz M, Vallespi T, Hamblin T, Oscier D, Ohyashiki K, Toyama K, Aul C, Mufti G, Bennett J. International scoring system for evaluating prognosis in myelodysplastic syndromes. Blood. 1997 Mar 15; 89(6):2079-88.
10. Liu Y, Pop R, Sadegh C, Brugnara C, Haase VH, Socolovsky M. Suppression of Fas-FasL coexpression by erythropoietin mediates ervthroblast expansion during the erythropoietic stress response in vivo. Blood. 2006 Jul 1; 108(1): 123-33.
11. Ribeil JA, Zermati Y, Vandekerckhove J, Cathelin S, Kersual J, Dussiot M, Coulon S, Moura 1С, Zeuner A, Kirkegaard-Sørensen T, Varet B, Solary E, Garrido С, Hermine O. Hsp70 regulates erythropoiesis by preventing caspase-3-mediated cleavage of GATA-1. Nature. 2007 Jan4; 445(7123):102-5.
12. Socolovsky M., Murrell M., Liu Y., Pop R., Porpiglia E., Levchenko A. Negative autoregulation by FAS mediates robust fetal erythropoiesis. PLoS Biol. 2007 Oct; 5(10):e252.
13. Tehranchi R., Fadeel B., Forsblom A.M. et al. Granulocyte colony-stimulating factor inhibits spontaneous cytochrome с release and mitochondria-dependent apoptosis of myelodysplastic syndrome hematopoietic progenitors. Blood. 2003; 101: 1080-1086.
14. Vardiman JW, Thiele J, Arber DA, Brunning RD, Borowitz MJ, Porwit A, Harris NL, Le Beau MM, Hellström-Lindberg Ε, Tefferi A, Bloomfield CD. The 2008 revision of the World Health Organization (WHO) classification of myeloid neoplasms and acute leukemia: rationale and important changes. Blood. 2009 Jul 30; 114(5):937-51.
15. Zermati Y, Garrido С, Amsellem S et al. Caspase activation is required for terminal erythroid differentiation. J Exp Med. 2001; 1 93: 247-254.
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Apogenix GmbH
<120> Improved APG101 Drug and Substance Formulation
<130> 51510P WO
<160> 1
<170> PatentIn version 3.5
<210> 1
<211> 400
<212> PRT
<213> Artificial Sequence
<220>
<223> Recombinant fusion protein consisting of human CD95 extracellular
domain with human IgG1 FC-part to its C-terminus
<220>
<221> SIGNAL
<222> (1)..(16)
<223> Variable cleavage sites
<220>
<221> SIGNAL
<222> (1)..(20)
<223> Variable cleavage sites
<220>
<221> SIGNAL
<222> (1)..(25)
<223> Variable cleavage sites
<220>
<221> DOMAIN
<222> (26)..(172)
<223> Human CD95 extracellular domain
<220>
<221> MOD_RES
<222> (26)..(26)
<223> PYRROLIDONE CARBOXYLIC ACID
<220>
<221> DISULFID
<222> (59)..(73)
<220>
<221> DISULFID
<222> (63)..(82)
<220>
<221> DISULFID
<222> (85)..(101)
<220>
<221> DISULFID
<222> (104)..(119)
<220>
<221> DISULFID
<222> (107)..(127)
<220>
<221> CARBOHYD
<222> (118)..(118)
<223> N-linked Glycosylation at Position N118
<220>
<221> DISULFID
<222> (129)..(143)
<220>
<221> DISULFID
<222> (135)..(140)
<220>
<221> CARBOHYD
<222> (136)..(136)
<223> N-linked Glycosylation at Position N136
<220>
<221> DISULFID
<222> (146)..(157)
<220>
<221> DISULFID
<222> (149)..(165)
<220>
<221> DOMAIN
<222> (172)..(400)
<223> Human IgG1-FC domain
<220>
<221> DISULFID
<222> (173)..(173)
<223> Interchain cystine forming residue of the APG101 homodimer.
<220>
<221> DISULFID
<222> (179)..(179)
<223> Interchain cystine forming residue of the APG101 homodimer.
<220>
<221> DISULFID
<222> (182)..(182)
<223> Interchain cystine forming residue of the APG101 homodimer.
<220>
<221> DISULFID
<222> (214)..(274)
<220>
<221> CARBOHYD
<222> (250)..(250)
<223> N-linked Glycosylation at Position N250
<220>
<221> DISULFID
<222> (320)..(378)
<400> 1
Met Val Gly Ile Trp Thr Leu Leu Pro Leu Val Leu Thr Ser Val Ala
1 5 10 15
Arg Leu Ser Ser Lys Ser Val Asn Ala Gln Val Thr Asp Ile Asn Ser
20 25 30
Lys Gly Leu Glu Leu Arg Lys Thr Val Thr Thr Val Glu Thr Gln Asn
35 40 45
Leu Glu Gly Leu His His Asp Gly Gln Phe Cys His Lys Pro Cys Pro
50 55 60
Pro Gly Glu Arg Lys Ala Arg Asp Cys Thr Val Asn Gly Asp Glu Pro
65 70 75 80
Asp Cys Val Pro Cys Gln Glu Gly Lys Glu Tyr Thr Asp Lys Ala His
85 90 95
Phe Ser Ser Lys Cys Arg Arg Cys Arg Leu Cys Asp Glu Gly His Gly
100 105 110
Leu Glu Val Glu Ile Asn Cys Thr Arg Thr Gln Asn Thr Lys Cys Arg
115 120 125
Cys Lys Pro Asn Phe Phe Cys Asn Ser Thr Val Cys Glu His Cys Asp
130 135 140
Pro Cys Thr Lys Cys Glu His Gly Ile Ile Lys Glu Cys Thr Leu Thr
145 150 155 160
Ser Asn Thr Lys Cys Lys Glu Glu Gly Ser Arg Ser Cys Asp Lys Thr
165 170 175
His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser
180 185 190
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
195 200 205
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
210 215 220
Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
225 230 235 240
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val
245 250 255
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
260 265 270
Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr
275 280 285
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
290 295 300
Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys
305 310 315 320
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
325 330 335
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
340 345 350
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser
355 360 365
Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
370 375 380
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
385 390 395 400
| название | год | авторы | номер документа |
|---|---|---|---|
| КОМБИНИРОВАННОЕ ПРИМЕНЕНИЕ ЛОВУШЕК GDF И АКТИВАТОРОВ РЕЦЕПТОРОВ ЭРИТРОПОЭТИНА ДЛЯ ПОВЫШЕНИЯ СОДЕРЖАНИЯ ЭРИТРОЦИТОВ | 2020 |
|
RU2814047C2 |
| ЛЕКАРСТВЕННЫЕ СРЕДСТВА ДЛЯ ЛЕЧЕНИЯ ПЕМФИГУСА, СОДЕРЖАЩИЕ АНТИТЕЛА К Fas-ЛИГАНДУ | 2008 |
|
RU2558260C2 |
| СРЕДСТВА ДЛЯ ЛЕЧЕНИЯ ПУЗЫРЧАТКИ, СОДЕРЖАЩИЕ АНТИТЕЛА ПРОТИВ ЛИГАНДА Fas | 2009 |
|
RU2556818C2 |
| ИСКУССТВЕННЫЕ АНТИГЕНПРЕЗЕНТИРУЮЩИЕ КЛЕТКИ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2018 |
|
RU2763798C1 |
| ФУНКЦИОНАЛИЗИРОВАННЫЕ ЭРИТРОИДНЫЕ КЛЕТКИ | 2018 |
|
RU2779315C2 |
| ИММУНОМОДУЛИРУЮЩИЕ СЛИТЫЕ БЕЛКИ И ПУТИ ИХ ПРИМЕНЕНИЯ | 2016 |
|
RU2755227C2 |
| СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ ПРОГНОЗИРОВАНИЯ ТЕРАПЕВТИЧЕСКОЙ ЭФФЕКТИВНОСТИ ЛЕЧЕНИЯ ОНКОЛОГИЧЕСКИХ ЗАБОЛЕВАНИЙ И ПРОГНОЗА ОНКОЛОГИЧЕСКИХ ЗАБОЛЕВАНИЙ | 2016 |
|
RU2785291C2 |
| СПОСОБЫ ГЕННОЙ МОДИФИКАЦИИ ГЕМОПОЭТИЧЕСКИХ КЛЕТОК | 2019 |
|
RU2824194C2 |
| СПОСОБЫ КУЛЬТИВИРОВАНИЯ КЛЕТОК И НАБОРЫ И УСТРОЙСТВО ДЛЯ НИХ | 2016 |
|
RU2761557C2 |
| КОМПОЗИЦИИ И СПОСОБЫ ИММУНОТЕРАПИИ | 2014 |
|
RU2798348C2 |
















Изобретение относится к медицине и предназначено для лечения миелодиспластического синдрома (МДС), где МДС выбирают из подгруппы МДС с низким риском по IPSS и/или подгруппы МДС с промежуточным-1 (int-1) риском. Используется ингибитор сигнального пути CD95, который связывает CD95-рецептор (CD95) и/или CD95-лиганд (CD95L). Способ позволяет расширить арсенал терапевтических средств. 17 з.п. ф-лы, 13 ил.
1. Способ лечения миелодиспластического синдрома (МДС), где МДС выбирают из подгруппы МДС с низким риском по IPSS и/или подгруппы МДС с промежуточным-1 риском (int-1), включающий введение ингибитора сигнального пути CD95, где ингибитор связывает CD95-рецептор (CD95) и/или CD95-лиганд (CD95L).
2. Способ по п. 1, где ингибитор является соединением, которое улучшает рост эритроидных предшественников.
3. Способ по п.1, где ингибитор является антителом, в частности антителом или его фрагментом, которое связывает CD95L.
4. Способ по п.2, где ингибитор является антителом, в частности антителом или его фрагментом, которое связывает CD95L.
5. Способ по п.1, где ингибитор является молекулой CD95-рецептора или ее частью, связывающей CD95-лиганд, и/или ингибитором CD95-лиганда.
6. Способ по п.2, где ингибитор является молекулой CD95-рецептора или ее частью, связывающей CD95-лиганд, и/или ингибитором CD95-лиганда
7. Способ по п. 1, где ингибитор является химерным белком, который связывает CD95L.
8. Способ по п. 2, где ингибитор является химерным белком, который связывает CD95L.
9. Способ по п. 7, где химерный белок содержит внеклеточный домен СD95 или его функциональный фрагмент и человеческий Fc-домен или его функциональный фрагмент.
10. Способ по п. 8, где химерный белок содержит внеклеточный домен СD95 или его функциональный фрагмент и человеческий Fc-домен или его функциональный фрагмент.
11. Способ по п. 9, где ингибитором является APG101 и/или его функциональное производное, где APG101 содержит домены CD95R (аминокислоты 26-172) из SEQ ID NO: 1 и IgG1-Fc (аминокислоты 172-400) из SEQ ID NO: 1.
12. Способ по п. 10, где ингибитором является APG101 и/или его функциональное производное, где APG101 содержит домены CD95R (аминокислоты 26-172) из SEQ ID NO: 1 и IgG1-Fc (аминокислоты 172-400) из SEQ ID NO: 1
13. Способ согласно любому из пп. 1-12, где популяция МДС характеризуется повышенным апоптозом в ходе эритропоэза.
14. Способ согласно любому из пп. 1-12, где популяция МДС характеризуется тяжелым дефектом эритропоэза без избытка бластных клеток.
15. Способ согласно любому из пп. 1-12, где популяция МДС характеризуется устойчивостью к классическим агентам, стимулирующим эритропоэз (ESA) и/или колониестимулирующим факторам.
16. Способ согласно любому из пп. 1-12, где ингибитор обеспечивается в виде фармацевтической композиции, содержащей фармацевтически приемлемые носители, разбавители и/или адъюванты.
17. Способ согласно любому из пп. 1-12, где ингибитор обеспечивается в виде фармацевтической композиции, содержащей по меньшей мере один дополнительный активный ингредиент, такой как агент, стимулирующий эритропоэз и/или агент, ингибирующий апоптоз.
18. Способ согласно любому из пп. 1-12, где ингибитор подлежит введению в общем количестве 50-400 мг/неделю, предпочтительно 100-200 мг/неделю.
| СПОСОБ ОПРЕДЕЛЕНИЯ ТАКТИКИ ИММУНОДЕПРЕССИВНОЙ ТЕРАПИИ ПРИ МИЕЛОДИСПЛАСТИЧЕСКОМ СИНДРОМЕ | 2000 |
|
RU2212037C2 |
| WO 2006055302 A2, 26.05.2006 | |||
| PARK MJ et al., Is International Prognostic Scoring System (IPSS) still standard in predicting prognosis in patients with myelodysplastic syndrome? External validation of the WHO Classification-Based Prognostic Scoring System (WPSS) and comparison with IPSS | |||
| Eur J Haematol | |||
| Станок для изготовления деревянных ниточных катушек из цилиндрических, снабженных осевым отверстием, заготовок | 1923 |
|
SU2008A1 |
| BOUSCARY D et al., Fas/Apo-1 (CD95) expression and apoptosis in patients with myelodysplastic syndromes | |||
| Leukemia | |||
| Электрическое сопротивление для нагревательных приборов и нагревательный элемент для этих приборов | 1922 |
|
SU1997A1 |
| Паровозный поршень, останавливающийся при езде без пара | 1924 |
|
SU839A1 |

