ПЕРЕКРЕСТНАЯ ССЫЛКА НА РОДСТВЕННЫЕ ЗАЯВКИ
[0001] Настоящая заявка заявляет приоритет предварительной заявки на патент США №61/605028, зарегистрированной 29 февраля 2012 г., содержание которой в полном объеме включено в данный текст посредством ссылки.
ОБЛАСТЬ ТЕХНИКИ
[0002] Настоящее изобретение относится к области техники генной экспрессии и геномного редактирования.
УРОВЕНЬ ТЕХНИКИ
[0003] Болезнь Хантингтона (БХ), также известная как хорея Хантингтона, является прогрессирующим нарушением двигательных, когнитивных функций в сочетании с психическими расстройствами. Средний возраст начала этой болезни составляет 35-44 года, хотя примерно в 10% случаев начало болезни приходится на возраст до 21 года, а средняя продолжительность жизни после диагностирования болезни составляет 15-18 лет. Частота встречаемости среди людей с западноевропейскими корнями составляет приблизительно от 3 до 7 случаев на 100000 человек.
[0004] Болезнь Хантингтона является примером нарушений, связанных с экспансией тринуклеотидных повторов, которые впервые были описаны в ранних 1990-х (см. Di Prospero and Fischbeck (2005) Nature Reviews Genetics 6:756-765). Эти нарушения включают локальную экспансию нестабильных повторов групп из трех нуклеотидов и могут привести к потере функции гена, в котором находится увеличенное количество повторов, развитию токсичной функции или к обоим этим последствиям. Тринуклеотидные повторы могут находиться в любой части гена, включая некодирующую и кодирующую области гена. Повторы, которые локализированы в пределах кодирующих областей, обычно включают повторяемый глутамин-кодирующий триплет (CAG) или аланин-кодирующий триплет (CGA). Наличие участков с увеличенным количеством повторов в пределах некодирующих последовательностей может привести к аберрантной экспрессии гена, в то время как увеличенное количество повторов в пределах кодирующих областей (также известное как нарушение повторов кодонов) может стать причиной неправильного сворачивания и агрегации белка. Зачастую настоящая причина патофизиологии, связанной с абберантными белками, остается невыясненной. Обычно в генах дикого типа, в которых может происходить экспансия тринуклеотидов, в нормальной популяции эти области содержат разное число повторяющихся последовательностей, но в пораженных популяциях число повторов может увеличиваться от удвоения до логарифмического возрастания числа повторов. При БХ эти повторы находятся в пределах N-терминальной области, кодирующей крупный цитозольный белок хантингтин (Htt). Нормальные аллели Htt содержат 15-20 повторов CAG, при этом аллели, содержащие 35 и более повторов, могут считаться аллелями, потенциально являющимися причиной БХ и несущими риск развития болезни. Аллели, содержащие 36-39 повторов, считаются частично пенетрантными, а у людей, имеющих такие аллели, болезнь может как развиться, так и не развиться (или симптомы могут появиться в более старшем возрасте), при этом аллели, содержащие 40 или более повторов, считаются полностью пенетрантными. Фактически, не сообщалось о пациентах с отсутствием симптомов среди тех, кто имеет аллели БХ с таким большим количеством повторов. У пациентов с ранним началом БХ (возрастом <21 года) часто обнаруживают 60 или более повторов CAG. Также было показано, что дополнительно к увеличению количества повторов CAG БХ может включать +1 или +2 сдвига рамки в пределах повторяющихся последовательностей, вследствие чего эта область будет кодировать полипептидный полисериновый тракт (кодируемый повторами AGC в случае +1 сдвига рамки) с большей вероятностью, чем полиглутаминовый (Davies and Rubinsztein (2006) Journal of Medical Genetics 43: 893-896).
[0005] При БХ мутантный аллель Htt обычно наследуется от одного из родителей как доминантный признак. Любой ребенок, рожденный от пациента с БХ, имеет 50% шанс развития болезни в случае, если второй из родителей не страдает этим нарушением. В некоторых случаях один из родителей может иметь промежуточный аллель БХ и не проявлять при этом симптомов заболевания, в то время как у ребенка из-за экспансии повторов болезнь проявляется. Вдобавок, аллель БХ также может характеризоваться явлением, известным как антиципация, при котором в нескольких поколениях наблюдается увеличение тяжести заболевания или снижение возраста начала заболевания, что связано с нестабильностью природы повторяемой области во время сперматогенеза.
[0006] Более того, тринуклеотидная экспансия в Htt приводит к потере нейронов среди проекционных шипиковых интернейронов стриатума, использующих в качестве нейротрансмиттеров гамма-аминомасляную кислоту (ГАМК), также потери нейронов наблюдаются в новой коре головного мозга. Шипиковые интернейроны, которые содержат энкефалин и которые проецируются на наружный сегмент бледного шара, затрагиваются чаще, чем нейроны, которые содержат субстанцию Р и проецируются на внутренний сегмент бледного шара. Другие участки мозга, которые сильно поражаются у людей с болезнью Хантингтона, включают черную субстанцию, 3, 5 и 6 слои коры головного мозга, участок СА1 гиппокампа, угловую извилину в теменной доле, клетки Пуркинье в мозжечке, боковые туберальные ядра гипоталамуса и парафасцикулярный комплекс таламуса (Walker (2007) Lancet 369:218-228).
[0007] Роль нормального белка Htt изучена плохо, но он может участвовать в нейрогенезе, апоптотической гибели клеток и миграции везикул. Вдобавок, есть свидетельства того, что Htt дикого типа стимулирует выработку нейротрофического фактора головного мозга (НФГМ) - фактора, поддерживающего развитие стриарных нейронов. Было показано, что в мышиных моделях БХ прогрессирование БХ сопровождается снижением экспрессии НФГМ (Zuccato et al (2005) Pharmacological Research 52(2): 133-139), и что доставка НФГМ или нейротрофического фактора глиальной клеточной линии (НФГК) посредством адено-ассоциированной вирусной (ААВ) вектор-опосредованной доставки генов может защитить стриарные нейроны в мышиных моделях БХ (Kells et al, (2004) Molecular Therapy 9(5): 682-688).
[0008] Возможности лечения БХ на сегодняшний день очень ограничены. Некоторые потенциальные методики, разработанные для предотвращения токсических эффектов, связанных с агрегацией белка, которая возникает при удлиненном полиглутаминовом тракте, таких как повышенная экспрессия шаперонинов или индукция ответа на тепловой шок выработкой соединения гелданамицина, продемонстрировали снижение этих токсических эффектов в in vitro моделях. Другие способы лечения нацелены на роль апоптоза в клинических проявлениях болезни. Например, замедление развития симптомов болезни в животных моделях было достигнуто путем блокирования каспазной активности у потомков пары мышей, где один из родителей имел аллель БХ, а второй из родителей имел доминантный негативный аллель для каспазы 1. Вдобавок, в патогенности болезни может играть роль расщепление мутантного БХ Htt каспазой. Было обнаружено, что у трансгенных мышей, несущих устойчивый к каспазе-6 мутантный Htt, сохраняется нормальное функционирование нейронов и не развивается стриарная нейродегенерация по сравнению с мышами, несущими неустойчивый к каспазе мутантный аллель Htt (см. Graham et al (2006) Cell 125: 1179-1191). Также было показано, что молекулы, нацеленные на участников каскада апоптотических реакций, оказывают замедляющее воздействие на симптомологию. Например, было показано, что соединения zVAD-fmk и миноциклин, которые ингибируют каспазную активность, замедляют проявление болезни у мышей. Также в клинических испытаниях на людях с нетяжелыми случаями БХ применяли лекарственный препарат ремацемид, так как считалось, что это соединение предотвращает связывание мутантного Htt с рецептором NDMA и, соответственно, предотвращает воздействие токсических эффектов на нервные клетки. Однако в этих исследованиях не наблюдали статистически значимого улучшения в функционировании нейронов. Вдобавок исследовательская группа по болезни Хантингтона (Huntington Study Group) провела рандомизированное исследование двойным слепым методом с применением кофермента Q. Хотя среди пациентов, которых лечили при помощи кофермента Q10, наблюдали тенденцию к замедлению прогрессирования болезни, никаких заметных изменений в скорости снижения общей функциональной способности не произошло. (Di Prospero and Fischbeck, там же). В патентной публикации США 2011/0082093 описываются специфические белки “цинковые пальцы”, нацеленные на Htt.
[0009] Следовательно, сохраняется потребность в составах и способах для лечения и предотвращения болезни Хантингтона.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
[0010] В данном тексте раскрыты способы и составы для лечения болезни Хантингтона. В частности, в данном тексте предложены способы и составы для модифицирования (например, модуляции экспрессии какого-либо компонента) аллеля БХ Htt таким образом, чтобы лечить болезнь Хантингтона. Также предложены способы и составы для создания животных моделей болезни Хантингтона.
[0011] Соответственно, в одном аспекте реализации изобретения предложены сконструированные ДНК-связывающие домены (например, белки цинковые пальцы или белки TAL-эффекторы (TALE)), которые модулируют экспрессию БХ аллеля (например, Htt). Сконструированные белки цинковые пальцы или TALE являются белками цинковые пальцы или TALE неприродного происхождения, чьи ДНК-связывающие домены (например, спирали узнавания или RVD (от англ. - “repeat variable diresidue”)) были изменены (например, путем селекции и/или рационального проектирования) для того, чтобы связываться с предварительно выбранным целевым участком. Любые описанные в данном тексте белки цинковые пальцы могут содержать 1, 2, 3, 4, 5, 6 или более цинковых пальцев, при этом каждый из цинковых пальцев содержит спираль узнавания, которая связывается с целевым субсайтом в выбранной последовательности(ях) (например, гене(ах)). Аналогично, любой из описанных в данном тексте белков TALE может содержать любое количество TALE RVD. В некоторых вариантах реализации изобретения по меньшей мере один RVD характеризуется неспецифическим связыванием ДНК. В некоторых вариантах реализации изобретения по меньшей мере одна спираль узнавания (или RVD) имеет неприродное происхождение. В некоторых вариантах реализации изобретения белки цинковые пальцы содержат спирали узнавания, приведенные в таблицах 1А и 1Б. В других вариантах реализации изобретения белки цинковые пальцы связываются с целевыми последовательностями, приведенными в таблицах 2А и 2Б. В некоторых вариантах реализации изобретения белки цинковые пальцы содержат спирали узнавания, приведенные в таблице 2В. В определенных вариантах реализации изобретения белки цинковые пальцы входят в фармацевтический состав, предназначенный, например, для введения пациенту.
[0012] В одном аспекте реализации изобретения предложены репрессоры (ZFP-TF или TALE-TF - транскрипционные факторы на основе цинковых пальцев и TALE (от англ. “zinc finger protein transcription factors” и “transcription activator-like effector transcription factors”)), которые связываются с последовательностями, полностью или частично находящимися снаружи повторяющегося CAG-участка Htt. В другом аспекте реализации изобретения предложены репрессоры ZFP или TALE (ZFP-TF или TALE-TF), которые связываются с последовательностями, находящимися в пределах повторяющегося CAG-участка Htt. В некоторых вариантах реализации изобретения эти ZFP-TF или TALE-TF преимущественно связываются с тринуклеотидными трактами, удлиненными относительно длины повторяемых трактов дикого типа, чем достигается преимущественная репрессия удлиненных аллелей. В некоторых вариантах реализации изобретения эти ZFP-TF или TALE-TF содержат домены белкового взаимодействия (или “димеризационные домены”), которые обеспечивают возможность мультимеризации при связывании с ДНК. В некоторых вариантах реализации изобретения эти ZFP-TF или TALE-TF достигают кооперативного связывания ДНК с повторяющейся последовательностью таким образом, что удлиненный аллель более эффективно связывается большим количеством белков ZFP или TALE, чем аллель дикого типа, что дает возможность осуществлять преимущественную репрессию мутантного аллеля. Эти ZFP-TF или TALE-TF кооперативного связывания могут дополнительно содержать или не содержать домены белкового взаимодействия, которые создают возможность мультимеризации при связывании с ДНК. В некоторых вариантах реализации изобретения ZFP-TF или TALE-TF образуют устойчивый комплекс мультимеров заданного размера и, таким образом, способны к преимущественному взаимодействию с трактом CAG, длина которого превышает определенный минимальный размер, при этом минимальный размер является большим, чем длина тракта CAG дикого типа.
[0013] В определенных вариантах реализации изобретения ZFP или белки TALE, описанные в данном тексте (например, двурукие, мультимеризующиеся и т.д.), преимущественно модифицируют экспрессию мутантного аллеля Htt. В некоторых вариантах реализации изобретения ZFP или TALE специфически связывается с мутантными аллелями Htt, при этом удлиненный тракт кодирует полиглутамин, в то время как в других вариантах реализации изобретения ZFP или TALE специфически связывается с мутантным аллелем Htt, но при этом удлиненный тракт кодирует полисерин. Таким образом, в некоторых вариантах реализации изобретения ZFP-TF или TALE-TF модулирует как дикую, так и мутантную формы аллеля Htt. В определенных вариантах реализации изобретения ZFP или TALE модулирует только аллель Htt дикого типа. В других вариантах реализации изобретения ZFP или TALE модулирует только мутантную форму Htt.
[0014] В других вариантах реализации изобретения предложены репрессирующие ZFP-TF или TALE-TF, которые преимущественно связываются с известными ОНП, связанными с удлиненными аллелями БХ Htt. В этом случае ZFP-TF или TALE-TF являются специфичными для мутантных аллелей Htt, которые содержат ОНП, что дает возможность осуществлять специфическую репрессию мутантного аллеля Htt. В другом аспекте реализации изобретения предложены ZFP-TF или TALE-TF, которые специфически активируют аллель Htt дикого типа посредством взаимодействия с ОНП, связанными с аллелями дикого типа. В этом случае активируется только аллель Htt дикого типа.
[0015] В определенных вариантах реализации изобретения белки цинковые пальцы (ZFP) или белки TALE, описанные в данном тексте, можно размещать в функциональной связи с регуляторным доменом (или функциональным доменом) как часть слитого белка. Функциональным доменом может быть, например, домен активации транскрипции, домен репрессии транскрипции и/или нуклеазный (расщепляющий) домен. Выбрав активационный домен либо репрессионный домен для сшивания с ZFP или TALE, такие слитые белки можно использовать как для активации, так и для подавления генной экспрессии. В некоторых вариантах реализации изобретения предложен слитый белок, содержащий ZFP или TALE, нацеленный на мутантный Htt, как описано в данном тексте, сшитый с доменом репрессии транскрипции, который можно применять для снижения экспрессии мутантного Htt. В некоторых вариантах реализации изобретения предложен слитый белок, содержащий ZFP или TALE, нацеленный на аллель Htt дикого типа, сшитый с доменом активации транскрипции, который может осуществлять повышающую регуляцию аллеля Htt дикого типа. В определенных вариантах реализации изобретения активность регуляторных доменов регулируется экзогенным низкомолекулярным компонентом или лигандом, таким образом, что взаимодействие с клеточным транскрипционным комплексом не будет происходить в отсутствие экзогенного лиганда. Такие внешние лиганды контролируют степень взаимодействия ZFP-TF или TALE-TF с транскрипционным комплексом. Регуляторный домен(ы) может быть функционально связан с любой частью(ями) одного или более ZFP или TALE, включая часть между одним или более ZFP или TALE, снаружи одного или более ZFP или TALE и любые комбинации этих вариантов. Любые описанные в данном тексте слитые белки могут являться частью фармацевтического состава.
[0016] В некоторых вариантах реализации изобретения описанные в данном тексте сконструированные ДНК-связывающие домены можно размещать в функциональной связи с нуклеазными (расщепляющими) доменами как часть слитого белка. В других вариантах реализации изобретения нуклеазные системы, такие как система CRISPR/Cas, можно использовать вместе со специфической одиночной “руководящей” РНК для нацеливания нуклеазы на целевой участок в ДНК. В определенных вариантах реализации изобретения такие нуклеазы и нуклеазные продукты слияния можно использовать для нацеливания на мутантные аллели Htt в стволовых клетках, таких как индуцированные полипотентные стволовые клетки (иПСК), человеческие эмбриональные стволовые клетки (чЭСК), мезенхимальные стволовые клетки (МСК) или нейрональные стволовые клетки, при этом активность нуклеазного продукта слияния приведет к появлению аллеля Htt, содержащего “дикое” количество повторов CAG. В определенных вариантах реализации изобретения предложены фармацевтические составы, содержащие модифицированные стволовые клетки.
[0017] В другом аспекте реализации изобретения предложен полинуклеотид, кодирующий любой из описанных в данном тексте ДНК-связывающих белков. В другом аспекте реализации изобретения предложены полинуклеотиды, кодирующие нуклеазу CRIPSR/Cas и одиночную “руководящую” РНК. Такие полинуклеотиды можно вводить пациенту, у которого необходимо лечить болезнь Хантингтона.
[0018] В дополнительных аспектах реализации в изобретении предложены способы и составы для создания специфических модельных систем для изучения болезни Хантингтона. В определенных вариантах реализации изобретения предложены модели, в которых создаются мутантные аллели Htt путем использования эмбриональных стволовых клеток для создания клеточных и животных линий, в которых удлиненные тринуклеотидные тракты определенной длины (50, 80, 109 и 180 повторов CAG, например) вставлены в аллель Htt дикого типа при помощи целевого внедрения, осуществляемого цинк-пальцевыми нуклеазами (ZFN), нуклеазами TALE (TALEN) или нуклеазами CRISPR/Cas. В определенных вариантах реализации изобретения модельные системы включают in vitro клеточные линии, в то время как в других вариантах реализации изобретения модельные системы включают трансгенных животных. В любой из описанных в данном тексте животных моделей животное может быть, например, грызуном (например, крысой, мышью), приматом (например, любым приматом за исключением человека) или кроликом.
[0019] В другом аспекте реализации изобретения предложен вектор доставки генов, содержащий любой из описанных в данном тексте полинуклеотидов. В определенных вариантах реализации изобретения вектор является аденовирусным вектором (например, вектором Ad5/F35), лентивирусным вектором (ЛВ), включая интеграционно-компетентные или интеграционно-дефектные лентивирусные векторы, либо аденоассоциированным вирусным вектором (ААВ). Таким образом, в данном тексте также предложены аденовирусные (Ад) векторы, ЛВ или аденоассоциированные вирусные векторы (ААВ), содержащие последовательность, кодирующую по меньшей мере одну нуклеазу (ZFN или TALEN) и/или донорную последовательность для целевой интеграции в ген-мишень. В определенных вариантах реализации изобретения аденовирусный вектор является химерным аденовирусным вектором, например, вектором Ad5/F35. В определенных вариантах реализации изобретения лентивирусный вектор является интегразо-дефектным лентивирусным вектором (ИДЛВ) или интеграционно-компетентным лентивирусным вектором. В определенных вариантах реализации изобретения вектор является псевдотипированным с VSV-G оболочкой или с другими оболочками.
[0020] В некоторых вариантах реализации изобретения предложены модельные системы болезни Хантингтона, в которых целевые аллели (например, мутантного Htt) помечены экспрессионными маркерами. В определенных вариантах реализации изобретения мутантные аллели (например, мутантного Htt) помечены. В некоторых вариантах реализации изобретения помечен аллель дикого типа (например, Htt дикого типа), а в дополнительных вариантах реализации изобретения отдельными экспрессионными маркерами помечены аллели как дикого типа, так и мутантные. В определенных вариантах реализации изобретения модельные системы включают in vitro клеточные линии, в то время как в других вариантах реализации изобретения модельные системы включают трансгенных животных.
[0021] Дополнительно предложены фармацевтические составы, содержащие нуклеиновые кислоты и/или белки (например, ZFP или TALE либо слитые белки, содержащие ZFP или TALE). Например, определенные составы содержат нуклеиновую кислоту, содержащую последовательность, которая кодирует один из описанных в данном тексте ZFP или TALE, функционально связанную с регуляторной последовательностью, соединенную с фармацевтически приемлемым носителем или разбавителем, при этом регуляторная последовательность обеспечивает возможность экспрессии нуклеиновой кислоты в клетке. В определенных вариантах реализации изобретения кодируемые ZFP или TALE являются специфическими аллелю БХ Htt. В некоторых вариантах реализации изобретения фармацевтические составы содержат ZFP или TALE, которые модулируют аллель БХ Htt, и ZFP или TALE, которые модулируют нейротрофический фактор. Составы на белковой основе содержат один или более из описанных в данном тексте ZFP или TALE и фармацевтически приемлемый носитель или разбавитель.
[0022] В другом аспекте реализации изобретения предложена также выделенная клетка, содержащая любой из белков, полинуклеотидов и/или составов, которые описаны в данном тексте.
[0023] В другом аспекте реализации изобретения предложены способы лечения и/или предотвращения болезни Хантингтона при помощи описанных в данном тексте способов и составов. В некоторых вариантах реализации изобретения способы включают применение составов, в которых доставка полинуклеотидов и/или белков может быть осуществлена при помощи вирусного вектора, невирусного вектора (например, плазмиды) и/или их комбинации. В некоторых вариантах реализации изобретения способы включают применение составов, содержащих популяции стволовых клеток, содержащих ZFP или TALE либо измененных при помощи нуклеазных систем ZFN, TALEN или CRISPR/Cas, которые являются объектами данного изобретения.
[0024] В целом эти и другие аспекты реализации станут понятны специалисту в данной области техники в свете раскрытия сущности настоящего изобретения.
КРАТКОЕ ОПИСАНИЕ ФИГУР
[0025] На фигуре 1, панелях от А до Д, приведены схематические изображения аллеля хантингтина (Htt) дикого типа и мутантного (болезни Хантингтона, БХ), а также разных типов связывание ZFP-TF с этим аллелем. На фигуре 1А показаны конструкции ZFP, которые связываются с внешней стороны CAG-повтора и, следовательно, предполагается, что они связываются как с аллелем дикого типа, так и с мутантным (БХ) аллелем. “KRAB” обозначает репрессионный домен KRAB из гена KOX1, а “ZFP” обозначает ДНК-связывающий белок «цинковые пальцы». “Стандартный ZFP TF” является слитым белком транскрипционного фактора ZFP, в котором ДНК-связывающие домены цинковых пальцев связаны с репрессионным доменом KRAB. На фигуре 1Б показаны ZFP-TF, сконструированные так, чтобы связываться в пределах участка CAG. На фигуре 1В показан “двурукий ZFP TF”, который представляет собой транскрипционный фактор ZFP, в котором два кластера из доменов цинковых пальцев разделены жесткими белковыми последовательностями. На этой фигуре функциональный (репрессионный) домен изображен снаружи одного из ZFP, но стоит понимать, что функциональный домен может находиться между двумя ZFP или снаружи ZFP, находящегося на другом конце белка. На фигуре 1Г показан “мультимеризующийся ZFP TF”, который представляет собой ZFP TF, способный к мультимеризации посредством мультимеризационного домена (изображен в виде пятнистых прямоугольников). На фигуре 1Д показана конфигурация ZFP-ZFP-KRAB, в которой два ДНК-связывающих домена цинковых пальцев связаны гибким линкером и дополнительно сшиты с доменом KRAB. Специалисту в данной области техники понятно, что во всех слитых белках функциональный домен может находиться на любом конце ДНК-связывающего домена, и что ДНК-связывающий домен может содержать большой количественный диапазон цинковых пальцев. Функциональный домен (например, активационный, репрессионный домен или расщепляющий домен) изображен на фигуре 1 в виде прямоугольника с черными ромбами. Специалисту в данной области техники понятно, что приведенные на фигурах примеры моделей применимы также к TALE TF.
[0026] Фигура 2, панели от А до Д, иллюстрирует репрессию обоих аллелей Htt, осуществляемую ZFP, как описывается фигурой 1А, при помощи ZFP TF, которые не связываются с повторяющимися последовательностями CAG. Идентификационные номера ZFP приведены в таблицах 1А и 1Б и указаны под столбиками. Фигура 2А иллюстрирует репрессию человеческих аллелей Htt в клетках HEK293 при помощи ZFP, нацеленных на пять участков в гене человека. Приведена диаграмма для человеческого гена Htt, а также показаны расположения ZFP-связывающих участков. Для каждой группы ZFP каждый из столбиков представляет независимую трансфекцию. Фигура 2Б иллюстрирует вестерн-блоттинг, показывающий уровни белка Htt в клетках HEK293, трансфицированных контрольным ЗФБ или репрессором 18856 ZFP TF (содержащим репрессионный домен KRAB KOX1), при этом для подтверждения равноценной загрузки белка использовали уровни NFκB p65 (“p65”). Результаты вестерн-блоттинга подтверждают наличие подавления экспрессии Htt, осуществляемой ZFP-TF. На фигуре 2В приведены данные, аналогичные приведенным на фигуре 2А, для ZFP, специфичного к мышиному Htt, в клетках Neuro2A. Как и на фигуре 2А, приведена диаграмма для мышиного гена Htt, а также показаны расположения ZFP-связывающих участков. Фигуры 2Г и 2Д иллюстрируют подавление экспрессии мышиного гена Htt (РНК) в иммортализованных стриарных клетках, в случаях применения разных дозировок ZFP-TF мРНК для трансфекции. Во всех случаях за исключением фигуры 2Б уровни мРНК для Htt измерены при помощи ПЦР в режиме реального времени и нормированы относительно уровней мРНК для актина.
[0027] Фигура 3, панели от А до Ж, иллюстрирует избирательную репрессию мутантного Htt, осуществляемую путем применения связывания ZFP в пределах повторяемого участка CAG, как показано на фигуре 1Б. Эта модель иллюстрирует, что более длинный повторяемый участок CAG в мутантном аллеле делает возможным повышенное связывание нацеленных на CAG репрессорных молекул ZFP. Фигура 3А иллюстрирует воздействие нацеленных на CAG ZFP, имеющих разную репрессорную активность, на эндогенный ген Htt (с нормальной длиной CAG-повторов) в клетках HEK293. Фигура 3Б иллюстрирует репрессию репортеров люциферазы, управляемую фрагментами промотора/экзона1 Htt, содержащими повторы CAG разной длины в диапазоне от 10 до 47 повторов CAG. CAG10 (крайний левый столбик для каждого из двух указанных условий) представляет результаты для 10 CAG-повторов; CAG17 (второй столбик слева для каждого из двух указанных условий) представляет результаты для 17 CAG-повторов; CAG23 (второй столбик справа для каждого из двух указанных условий) представляет результаты для 23 CAG-повторов и CAG47 (крайний правый столбик для каждого из двух указанных условий) представляет результаты для 47 CAG-повторов. На схеме, расположенной над графиком, приведена конструкция промотора, экзона1, CAG-повторов Htt и репортерного гена люциферазы, которые применяются в данной системе. Данные показывают, что увеличение количества повторов CAG приводит к сниженной экспрессии от промотора Htt под действием нацеленного на CAG ZFP. Кроме того, фигура 3В показывает, что хотя относительно слабый нацеленный на CAG ZFP не репрессирует репортер люциферазы, который содержит участок CAG-повторов нормальной длины, равно как и сильный репрессор CAG, он осуществляет такую же репрессию репортера люциферазы, который содержит удлиненный участок CAG-повторов, как и сильный нацеленный на CAG ZFP, что справедливо при всех тестируемых дозировках. “pRL-Htt-CAG23-интрон 1” (левый столбик в каждой паре) соответствует экспрессии аллеля дикого типа, в то время как “pRL-HttCAG47-интрон 1” (правый столбик в каждой паре) соответствует экспрессии мутантного удлиненного аллеля Htt (содержащего 47 повторов CAG). На фигуре 3Г приведен график, иллюстрирующий репрессию мутантного Htt (111 CAG), осуществляемую нацеленными на CAG ZFP в иммортализованных стриарных клетках, полученных от мышей с генным нокином HdH(Q111/Q7). Экспрессию дикого типа отображает левый столбик из каждой пары, а нокин-экспрессию - правый столбик из каждой пары. ZFP-TF, содержащие определенный ZFP, сшитый с репрессионным доменом KRAB, исследовали, используя для трансфекций три разных концентрации ZFP мРНК. Фигура 3Д иллюстрирует репрессию мутантного Htt, осуществляемую нацеленными на CAG ZFP, в линии фибробластов (CAG15/70), полученной от пациента с БХ. В данной линии фибробластов аллель Htt дикого типа содержит 15 повторов CAG (“099T(CAG15)”, средний столбик для каждого из указанных условий), а мутантный удлиненный аллель Htt содержит 70 повторов CAG (“099C(CAG70)”, правый столбик для каждого из указанных условий). Фигура 3F иллюстрирует избирательное подавление экспрессии мутантного Htt в фибробластных клеточных линиях, полученных от 4 разных пациентов с БХ. Числа над каждой из групп обозначают количество повторов CAG в аллеле Htt дикого типа (например, 15 или 18) и в мутантном аллеле (например, 70, 67, 45 и 44); а исследования проводили для двух разных дозировок ZFP мРНК. Левый столбик в каждой из пар отображает экспрессию Htt дикого типа, а правый столбик отображает экспрессию мутантного Htt. Фигура 3Ж иллюстрирует оцененную при помощи вестерн-блоттинга экспрессию Htt в фибробластах, полученных от пациента с БХ, в присутствии ZFP-TF 30640, 32528 и 30657. Более медленные миграционные полосы белка соответствуют тем, которые получены от удлиненных мутантных аллелей Htt. 32528 связывается с сайтом инициации транскрипции Htt (СИТ) и таким образом подавляет экспрессию обоих аллелей, в то время как 30640 и 30657 связываются с CAG-повторами (CAG).
[0028] Фигура 4, панели А и Б, иллюстрирует репрессию мутантного Htt в линии фибробластов, полученной от пациента с БХ, осуществляемую ZFP, нацеленными на CAG-повторы. Применяемый диапазон концентраций РНК составил от 0,1 до 3 мкг. На фигурах 4А и 4Б левый столбик для каждого из указанных условий отображает общую экспрессию Htt, средний столбик отображает экспрессию Htt в фибробластах, в которых аллель Htt содержит 18 повторов CAG (“099T(CAG18)”, а мутантный удлиненный аллель Htt содержит 45 повторов CAG 099T (CAG45).
[0029] Фигура 5 иллюстрирует воздействие нацеленных на CAG ZFP репрессоров на экспрессию Htt и других CAG-содержащих генов в фибробластах, полученных от пациента с БХ. Левый столбик под каждым из указанных условий отображает результаты для 30640; средний столбик под каждым из указанных условий отображает результаты для 30675 и правый столбик отображает ложные трансфекции.
[0030] Фигура 6, панели А и Б, иллюстрирует эксперимент, в котором исследовали общегеномную специфичность трех нацеленных на CAG ZFP. На фигуре 6А приведены результаты количественного ПЦР-анализа репрессии Htt, проведенного для шести биологических образцов (шести отдельных трансфекций) БХ фибробластов (CAG18 (средние столбики)/CAG45, правые столбики) с применением 30640, 30645 или 33074. Четыре наиболее схожих согласно данным количественного ПЦР-анализа образца были отобраны для микроматричного анализа, а данные представлены на фигуре 6Б.
[0031] Фигура 7 иллюстрирует репрессию Htt в нейрональных стволовых клетках (НСК) CAG17/69. Клетки трансфицировали ZFP мРНК в указанных дозировках. Левые столбики под каждой из указанных дозировок отображают результаты для клеток CAG17, средние столбики отображают результаты для клеток дикого типа, правые столбики отображают результаты для клеток CAG69.
[0032] Фигура 8 иллюстрирует экспрессию Htt в нейронах, дифференцировавших из БХ эмбриональных стволовых клеток (ЭСК) (CAG 17/48), обработанных ZFP TF. Клетки трансфицировали ZFP мРНК в указанных дозировках.
[0033] Фигура 9 иллюстрирует подавление экспрессии мутантного трансгена Htt в R6/2 мышах с последующей обработкой ZFP TF 30640.
[0034] На Фигуре 10, панели от А до Г, показаны ZFP с мультимеризационными доменами, которые специфически нацелены на удлиненные повторы CAG, как показано на фигуре 1Г. На фигуре 10А показан одиночный ZFP, который содержит четыре компонента: (i) репрессорный домен KOX (овал, подписанный как “репрессор”); (ii) группу из 2-6 пальцев (показано два, маленькие овалы, помеченные как “Z”), которая связывается с (CAG)N или пермутацией этой последовательности, и (iii) два димеризационных домена (прямоугольники, подписанные как “д1” и “д2”), которые взаимодействуют в антипараллельной конфигурации. Эти домены обеспечивают возможность полимеризации ZFP в пределах большой бороздки CAG-тракта. На фигуре 10Б приведено схематическое изображение связывания с мультимером из 3 ZFP. Следует понимать, что можно использовать любое количество мультимеров, и что функциональный домен может размещаться в любом месте на одном или более из отдельных ZFP, и что данные диаграммы применимы также к TALE-TF. На фигуре 10В приведены белковые последовательности четырех мономерных каркасов ZFP, которые сконструированы так, чтобы мультимеризоваться через взаимодействия между димеризующимися цинковыми пальцами (DZ). Каркасы имеют названия DZ1 (SEQ ID NO:180), DZ2 (SEQ ID NO:181), DZ3 (SEQ ID NO:182) и DZ4 (SEQ ID NO:183). Димеризующиеся домены цинковых пальцев выделены подчеркиванием, а репрессионный домен и последовательность ядерной локализации выделены, соответственно, жирным подчеркнутым и курсивным текстом. На фигуре 10Г приведены белковые последовательности семи мономерных каркасов ZFP, которые сконструированы так, чтобы мультимеризоваться через взаимодействия между суперспиралями (СС). Каркасы имеют названия CC1 (SEQ ID NO:184), CC2 (SEQ ID NO:185), CC3 (SEQ ID NO:186), CC4 (SEQ ID NO:187), CC5 (SEQ ID NO:188), CC6 (SEQ ID NO:189) и CC7 (SEQ ID NO:190). Последовательности суперспиралей выделены подчеркиванием, а репрессионный домен и последовательность ядерной локализации выделены, соответственно, жирным подчеркнутым и курсивным текстом. Расположение участка ZFP каждого из каркасов, которое будет отличным в разных конструкциях, обозначено как “[ZFP]”. Расположение (ДНК-связывающего) участка ZFP каждого из каркасов, которое будет отличным в разных конструкциях, обозначено как “[ZFP]”.
[0035] Фигура 11, панели А и Б, иллюстрирует активность ZFP-TF с димеризационными доменами. На фигуре 11А ZFP-TF с “суперспиральными” (СС) доменами исследовали вместе с репортерами люциферазы. pRL-Htt CAG17 (левый столбик в каждой паре) представляет репортер люциферазы Renilla, управляемый фрагментом человеческого промотора/экзона1 Htt с 17 CAG; pGL3-Htt-CAG47 (правый столбик в каждой паре) представляет репортер люциферазы светляков, управляемый фрагментом человеческого промотора/экзона1 Htt с 47 повторами CAG. См. текст примера 10, в котором описаны разные димеризационные домены. На фигуре 11Б ZFP с димеризующимися доменами цинковых пальцев “DZ” исследовали вместе с такими же репортерами люциферазы и наблюдали повышенную репрессию некоторыми димеризационными доменами ZFP-TF. Левый столбик в каждом дублете отображает экспрессию аллеля Htt с 17 CAG-повторами, а правый столбик отображает экспрессию аллеля Htt с 47 CAG-повторами.
[0036] Фигура 12, панели А и Б, иллюстрирует репрессию Htt белками ZFP-ZFP-KOX. Фигура 12А иллюстрирует репрессию Htt одиночными 33088 и 33084 ZFP-TF и репрессию белками 33088-33088 и 33088-33084 ZFP-ZFP-KOX в БХ фибробластах дикого типа (левый столбик), CAG18 (средний столбик) и CAG45 (правый столбик) (фигура 12А); фигура 12Б иллюстрирует репрессию Htt 33088-33088 и 33088-33084 ZFP-ZFP-KOX в БХ фибробластах дикого типа (левый столбик), CAG20 (средний столбик) и CAG41 (правый столбик).
[0037] Фигура 13, панели от А до Д, иллюстрирует активацию мышиного Htt. На фигуре 13А показана ZFP-TF-стимулированная повышающая регуляция генов мышиного Htt на уровне РНК в клетках Neuro2A при применении ZFP, сшитого с активационным доменом р65. Двойные столбики обозначают дублирующие трансфекции. На фигуре 13Б приведены результаты вестерн-блоттинга, демонстрирующие повышенную выработку белка Htt, стимулированную ZFP. На фигуре 13В показан мышиный аллель Htt дикого типа и “нокин” аллель Htt, в котором мышиная последовательность (большая часть экзона 1 и часть интрона 1, линия над схематическим изображением аллеля дикого типа) была заменена соответствующей человеческой последовательностью с экспансией CAG (линия над схематическим изображением нокин-аллеля). На фигуре 13Г приведено выравнивание между мышиной последовательностью (SEQ ID NO:191), которая была заменена соответствующей человеческой последовательностью (SEQ ID NO:192), таким образом, что нокин-аллель обладает достаточным расхождением в последовательности для того, чтобы дать возможность сконструировать ZFP (показаны на А и Б) так, чтобы специфически связывать мышиную последовательность. Фигура 13Д иллюстрирует специфическую активацию мышиного аллеля Htt дикого типа в иммортализованных стриарных клетках, полученных от мышей с генным нокином HdhQ111/Q7. Левый столбик отображает результаты для клеток дикого типа, а правый столбик отображает результаты для клеток с мутантным нокин-аллелем.
[0038] На фигуре 14, панели А и Б, приведены результаты анализа ошибок спаривания Cel-I (Surveyor™, Transgenomics) с последующей обработкой клеток К562 Htt-специфическими ZFN парами. Процент активности НГСК (вставок-делеций) для активного ZFN приведен внизу соответствующей линии. “ЗФБ” обозначает клетки, которые были трансфицированы кодирующей ЗФБ плазмидой. На фигуре 14А приведены результаты по ZFN, которые расщепляют ранние экзоны Htt, а на фигуре 14Б приведены результаты по ZFN, которые расщепляют вблизи стоп-кодона. Также наблюдали неактивные ZFN пары (линии, не подписанные процентным содержанием вставок-делеций).
[0039] На фигуре 15 приведены графики результатов репрессии Htt для нескольких представляющих интерес белков TALE-TF. TALE-TF исследовали в фибробластах, полученных от пациента с БХ (CAG 20/41). Результаты показывают, что некоторые из TALE TF оказались активны в плане полного подавления экспрессии Htt, в то время как другие демонстрировали репрессию предпочтительно мутантного Htt.
ДЕТАЛЬНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
[0040] В данном тексте раскрыты составы и способы для лечения болезни Хантингтона (БХ). В частности, предложены Htt-модулирующие транскрипционные факторы, содержащие белки «цинковые пальцы» (ZFP TF) или TALE (TALE-TF), и способы, в которых используются такие белки, для применения в лечении и предотвращении болезни Хантингтона. Например, предложены ZFP-TF или TALE-TF, которые подавляют экспрессию мутантного аллеля Htt или активируют экспрессию аллеля Htt дикого типа. Дополнительно предложены цинк-пальцевые нуклеазы (ZFN), нуклеазы TALE (TALEN) или нуклеазные системы CRISPR/Cas, которые модифицируют геномную структуру гена, связанного с БХ. Например, предложены ZFN, TALEN или нуклеазные системы CRISPR/Cas, которые способны специфически изменять фрагменты мутантной формы Htt. Вышеупомянутое включает составы и способы, в которых применяют сконструированные белки «цинковые пальцы» или сконструированные белки TALE, т.е. белки неприродного происхождения, которые связываются с заданной целевой нуклеотидной последовательностью.
[0041] Таким образом, способы и составы, описанные в данном тексте, предлагают способы для лечения и предотвращения болезни Хантингтона, и эти способы и составы могут содержать транскрипционные факторы цинковых пальцев или транскрипционные факторы TALE, которые способны модулировать гены-мишени, а также сконструированные цинк-пальцевые и TALE нуклеазы и нуклеазные системы CRISPR/Cas, которые способны модифицировать или редактировать Htt.
Общая информация
[0042] В практическом применении способов, а также изготовлении и применении составов, раскрытых в данном тексте, используются, если не указано иное, общепринятые методы молекулярной биологии, биохимии, изучения структуры и анализа хроматина, вычислительной химии, клеточного культивирования, генной инженерии и родственных направлений в пределах области техники изобретения. Суть этих методов в полном объеме раскрыта в литературе. См., например, Sambrook et al. MOLECULAR CLONING: A LABORATORY MANUAL, Second edition, Cold Spring Harbor Laboratory Press, 1989 and Third edition, 2001; Ausubel et al., CURRENT PROTOCOLS IN MOLECULAR BIOLOGY, John Wiley & Sons, New York, 1987 и периодические обновления; серии METHODS IN ENZYMOLOGY, Academic Press, San Diego; Wolffe, CHROMATIN STRUCTURE AND FUNCTION, Third edition, Academic Press, San Diego, 1998; METHODS IN ENZYMOLOGY, Vol. 304, “Chromatin” (P.M. Wassarman and A. P. Wolffe, eds.), Academic Press, San Diego, 1999 и METHODS IN MOLECULAR BIOLOGY, Vol. 119, “Chromatin Protocols” (P.B. Becker, ed.) Humana Press, Totowa, 1999.
ОПРЕДЕЛЕНИЯ
[0043] Термины “нуклеиновая кислота”, “полинуклеотид” и “олигонуклеотид” употребляются взаимозаменяемо и обозначают дезоксирибонуклеотидный или рибонуклеотидный полимер, находящийся в линейной или кольцевой конформации и как в одно-, так и в двухцепочечной форме. Применительно к целям настоящего изобретения эти термины не должны толковаться как ограничивающие в отношении длины полимера. Данные термины могут включать известные аналоги нуклеотидов природного происхождения, а также те нуклеотиды, в которых были модифицированы компоненты оснований, сахаров и/или фосфатов (например, тиофосфатные остовы). В общем случае аналог конкретного нуклеотида обладает аналогичной специфичностью спаривания оснований, т.e. аналог А будет спариваться с Т.
[0044] Термины “полипептид”, “пептид” и “белок” употребляются взаимозаменяемо для обозначения полимера из аминокислотных остатков. Также данный термин применяется к аминокислотным полимерам, в которых одна или более из аминокислот представлены химическими аналогами или модифицированными производными соответствующих аминокислот природного происхождения.
[0045] “Связывание” обозначает специфическое к последовательности нековалентное взаимодействие между макромолекулами (например, между белком и нуклеиновой кислотой). Не все компоненты связывающего взаимодействия должны быть специфическими к последовательности (например, контакты с фосфатными остатками в остове ДНК), так как взаимодействие в целом является специфическим к последовательности. Такие взаимодействия в общем случае характеризуются константой диссоциации (Kd), составляющей 10-6 M-1 или меньше. “Аффинность” обозначает силу связывания: более высокая аффинность связывания соответствует более низкому значению Kd.
[0046] “Связывающий белок” представляет собой белок, который способен нековалентно связываться с другой молекулой. Связывающий белок может связываться, например, с молекулой ДНК (ДНК-связывающий белок), молекулой РНК (РНК-связывающий белок) и/или белковой молекулой (белоксвязывающий белок). В случае белоксвязывающего белка он может связываться сам с собой (для образования гомодимеров, гомотримеров и т.д.) и/или он может связываться с одной или более молекулами другого белка или белков. Связывающий белок может обладать более чем одним типом связывающей активности. Например, белки «цинковые пальцы» обладают ДНК-связывающей, РНК-связывающей и белоксвязывающей активностью.
[0047] “ДНК-связывающий белок «цинковые пальцы»” (или связывающий домен) представляет собой белок или домен в составе более крупного белка, который связывает ДНК специфическим к последовательности образом при помощи одного или более цинковых пальцев, которые являются участками аминокислотной последовательности в пределах связывающего домена и чья структура стабилизируется при помощи координации ионом цинка. Термин ДНК-связывающий белок «цинковые пальцы» часто сокращают до белка «цинковые пальцы» или аббревиатуры ZFP (от англ. “zinc finger protein”).
[0048] “ДНК-связывающий домен TALE” или “TALE” представляет собой полипептид, содержащий один или более повторяемых доменов/структурных единиц TALE. Повторяемые домены участвуют в связывании TALE с родственной ему целевой последовательностью ДНК. Одиночная “единица повтора” (называемая также “повтором”) обычно имеет длину в 33-35 аминокислот и обладает по меньшей мере некоторой гомологией последовательностей с другими повторяемыми последовательностями TALE в составе белка TALE природного происхождения.
[0049] Связывающие домены цинковых пальцев или ДНК-связывающие домены TALE можно “сконструировать” таким образом, чтобы они связывались с заданной нуклеотидной последовательностью, например, путем конструирования (изменения одной или более из аминокислот) спирального участка узнавания белка «цинковые пальцы» природного происхождения или путем конструирования RVD белка TALE. Следовательно, сконструированные белки «цинковые пальцы» или TALE являются белками неприродного происхождения. Неограничивающими примерами способов конструирования белков «цинковые пальцы» или TALE являются проектирование и селекция. Спроектированным белком «цинковые пальцы» или TALE является белок неприродного происхождения, конструкция/состав которого определяется преимущественно критерием целесообразности. В проектировании критерий целесообразности включает применение правил замещения и компьютеризированных алгоритмов для обработки информации из базы данных, в которой хранится информация по существующим конструкциям ZFP и данным связывания. См., например, патенты США 6140081; 6453242 и 6534261; также см. WO 98/53058; WO 98/53059; WO 98/53060; WO 02/016536 и WO 03/016496 и патентную публикацию США №20110301073.
[0050] “Селектированным” белком «цинковые пальцы» или TALE является белок неприродного происхождения, получение которого, главным образом, является результатом эмпирического процесса, такого как фаговое отображение, применение основанной на взаимодействии “ловушки” или гибридная селекция. См., например, патенты США 5789538; 5925523; 6007988; 6013453; 6200759; WO 95/19431; WO 96/06166; WO 98/53057; WO 98/54311; WO 00/27878; WO 01/60970 WO 01/88197, WO 02/099084 и WO 2011/146121 (патентную публикацию США №20110301073).
[0051] “Рекомбинация” обозначает процесс обмена генетической информацией между двумя полинуклеотидами. Применительно к целям настоящего изобретения “гомологичная рекомбинация (ГР)” обозначает специализированную форму такого обмена, который происходит, например, во время репарации двухцепочечных разрывов в клетках при помощи гомологически направленных механизмов репарации. В этом процессе необходима гомология нуклеотидных последовательностей, используется “донорная” молекула для матричной репарации “молекулы-мишени” (т.e. той, в которой присутствует двухцепочечный разрыв), и он известен под разными названиями, таким как “некроссоверная конверсия генов” или “конверсия генов с коротким трактом”, так как приводит к передаче генетической информации от донора к мишени. Не привязываясь к какой-либо определенной теории, можно сказать, что такая передача может включать исправление ошибок спаривания оснований гетеродуплексной ДНК, которое происходит между содержащей разрывы целью и донором, и/или “синтез-зависимый отжиг цепей”, в котором донор используется для повторного синтеза генетической информации, которая должна стать частью мишени, и/или другие подобные процессы. Подобная специализированная ГР часто приводит к изменению последовательности молекулы-мишени таким образом, что часть последовательности или вся последовательность донорного полинуклеотида оказывается включенной в полинуклеотид-мишень.
[0052] В способах настоящего изобретения одна или более из описанных в данном тексте нацеленных нуклеаз создают двухцепочечный разрыв в целевой последовательности (например, клеточного хроматина) на заданном участке, и после этого в клетку может быть внесен “донорный” полинуклеотид, обладающий гомологией с нуклеотидной последовательностью на участке разрыва. Было показано, что наличие двухцепочечного разрыва способствует интеграции донорной последовательности. Донорную последовательность можно интегрировать физически либо, в другом варианте, донорный полинуклеотид используется в качестве матрицы для репарации разрыва путем гомологичной рекомбинации, которая приводит к внесению всей или части нуклеотидной последовательности, аналогичной донорной, в клеточный хроматин. Таким образом, первую последовательность в клеточном хроматине можно изменить или, в некоторых вариантах реализации изобретения, преобразовать в последовательность, которая присутствует в донорном полинуклеотиде. Следовательно, употребление терминов “заместить” или “замещение” стоит трактовать как замещение одной нуклеотидной последовательности другой (т.e. замещение последовательности в информационном смысле), при этом замещение не обязательно требует физического или химического замещения одного полинуклеотида другим.
[0053] В любом из описанных в данном тексте способов можно применять дополнительные пары белков «цинковые пальцы» или TALE для дополнительного двухцепочечного расщепления дополнительных целевых участков в клетке.
[0054] В определенных вариантах реализации способов осуществления нацеленных рекомбинации, и/или замещения, и/или изменения последовательности на представляющем интерес участке в клеточном хроматине хромосомную последовательность изменяют путем гомологичной рекомбинации с экзогенной “донорной” нуклеотидной последовательностью. Такая гомологичная рекомбинация стимулируется в присутствии двухцепочечного разрыва в клеточном хроматине, в случае наличия последовательностей, гомологичных участку разрыва.
[0055] В любом из описанных в данном тексте способов первая нуклеотидная последовательность (“донорная последовательность”) может содержать последовательности, которые являются гомологичными, но не идентичными геномным последовательностям на представляющем интерес участке, стимулируя тем самым внесение при помощи гомологичной рекомбинации неидентичной последовательности в представляющий интерес участок. Следовательно, в некоторых вариантах реализации изобретения части донорной последовательности, которые гомологичны последовательностям на представляющем интерес участке, имеют от около 80 до 99% (или любое численное значение между этими величинами) идентичности с замещаемой геномной последовательностью. В других вариантах реализации изобретения гомологичность между донорной и геномной последовательностью превышает 99%, например, в случае, когда донорная и геномная последовательность протяженностью более 100 последовательных пар оснований различаются только на 1 нуклеотид. В отдельных случаях негомологичная часть донорной последовательности может содержать последовательности, которые не присутствуют на представляющем интерес участке, и таким образом в представляющий интерес участок вносятся новые последовательности. В этих случаях негомологичная последовательность, как правило, фланкируется последовательностями из 50-1000 пар оснований (или любое численное значение между этими величинами) или любого количества пар оснований, большего, чем 1000, которые гомологичны или идентичны последовательностям на представляющем интерес участке. В других вариантах реализации изобретения донорная последовательность является негомологичной первой последовательности и вносится в геном при помощи механизмов негомологичной рекомбинации.
[0056] Любой из описанных в данном тексте способов можно применять для частичной или полной инактивации одной или более целевых последовательностей в клетке путем нацеленной интеграции донорной последовательности, которая препятствует экспрессии представляющего интерес гена(ов). Также предложены клеточные линии с частично или полностью инактивированными генами.
[0057] Более того, описанные в данном тексте способы нацеленной интеграции можно применять для интегрирования одной или более экзогенных последовательностей. Экзогенная последовательность нуклеиновой кислоты может содержать, например, один или более генов или молекул кДНК, или любой тип кодирующих или некодирующих последовательностей, а также один или более контрольных элементов (например, промоторов). Вдобавок, экзогенная последовательность нуклеиновой кислоты может продуцировать одну или более молекул РНК (например, малую шпилечную РНК (мшРНК), ингибиторную РНК (РНКи), микроРНК (миРНК) и т.д.).
[0058] “Расщепление” обозначает создание разрыва в ковалентном остове молекулы ДНК. Расщепление можно инициировать при помощи большого количества методов, включая, но не ограничиваясь этим, ферментативный или химический гидролиз фосфодиэфирной связи. Возможны как одноцепочечное расщепление, так и двухцепочечное расщепление, а двухцепочечное расщепление может являться результатом двух отдельных актов одноцепочечного расщепления. Расщепление ДНК может привести к образованию как тупых концов, так и ступенчатых концов. В некоторых вариантах реализации изобретения для нацеленного двухцепочечного расщепления ДНК используют слитые полипептиды.
[0059] “Расщепляющий полудомен” представляет собой полипептидную последовательность, которая совместно со вторым полипептидом (как идентичным, так и отличным) образует комплекс, обладающий расщепляющей активностью (предпочтительно двухцепочечной расщепляющей активностью). Термины “первый и второй расщепляющие полудомены”, “+ и - расщепляющие полудомены” и “правый и левый расщепляющие полудомены” употребляются взаимозаменяемо для обозначения димеризующихся пар расщепляющих полудоменов.
[0060] “Сконструированный расщепляющий полудомен” представляет собой расщепляющий полудомен, который был модифицирован так, чтобы образовывать облигатные гетеродимеры с другим расщепляющим полудоменом (например, другим сконструированным расщепляющим полудоменом). См. также патентные публикации США №2005/0064474, 20070218528, 2008/0131962 и 2011/0201055, которые в полном объеме включены в данный текст посредством ссылки.
[0061] Термин “последовательность” обозначает нуклеотидную последовательность любой длины, которая может являться ДНК или РНК; может быть линейной, кольцевой или разветвленной, а также может быть как одноцепочечной, так и двухцепочечной. Термин “донорная последовательность” относится к нуклеотидной последовательности, которая вставлена в геном. Донорная последовательность может иметь любую длину, например, длину от 2 до 10000 нуклеотидов (или любое численное значение между этими величинами или превышающее их), предпочтительно - длину от около 100 до 1000 нуклеотидов (или любое численное значение между этими величинами), более предпочтительно - длину от около 200 до 500 нуклеотидов.
[0062] “Хроматин” представляет собой структуру нуклеопротеина, содержащую клеточный геном. Клеточный хроматин содержит нуклеиновую кислоту, преимущественно ДНК, и белок, включая гистоны и негистонные хромосомные белки. Большая часть эукариотического клеточного хроматина существует в форме нуклеосом, при этом ядро нуклеосомы содержит приблизительно 150 пар оснований ДНК, связанной с октамером, содержащим по два гистона из H2A, H2B, H3 и H4; а линкерная ДНК (различной длины в зависимости от организма) располагается между ядрами нуклеосом. Молекула гистона H1 в общем случае связана с линкерной ДНК. Применительно к целям настоящего изобретения подразумевается, что термин “хроматин” включает в себя все типы клеточного нуклеопротеина, как прокариотические, так и эукариотические. Клеточный хроматин включает в себя как хромосомный, так и эписомный хроматин.
[0063] “Хромосома” представляет собой хроматиновый комплекс, содержащий весь или часть клеточного генома. Клеточный геном часто характеризуется кариотипом, который представляет собой полный набор хромосом, которые содержат клеточный геном. Клеточный геном может содержать одну или более хромосом.
[0064] “Эписома” представляет собой реплицирующуюся нуклеиновую кислоту, комплекс нуклеопротеинов или другую структуру, содержащую нуклеиновую кислоту, которая не является частью хромосомного кариотипа клетки. Примеры эписом включают плазмиды и отдельные вирусные геномы.
[0065] “Целевой участок” или “целевая последовательность” (“участок-мишень” или “последовательность-мишень”) представляют собой последовательность нуклеиновой кислоты, с которой будет связываться связывающая молекула при наличии условий, достаточных для связывания.
[0066] “Экзогенная” молекула представляет собой молекулу, которая в обычном состоянии не присутствует в клетке, но которую можно внести в клетку при помощи одного или более генетических, биохимических или других методов. “Нормальное присутствие в клетке” определяется в отношении конкретного этапа развития клетки и окружающей клетку среды. Таким образом, к примеру, молекула, которая присутствует в клетке только во время эмбрионального развития мускулатуры, является экзогенной молекулой в отношении к зрелой мышечной клетке. Аналогично, молекула, индуцированная тепловым шоком, является экзогенной молекулой в отношении к клетке, не испытавшей тепловой шок. Экзогенная молекула может содержать, например, функционирующую версию неправильно функционирующей эндогенной молекулы или неправильно функционирующую версию нормально функционирующей эндогенной молекулы.
[0067] Экзогенная молекула может являться, среди всего прочего, небольшой молекулой, такой, которая образуется при помощи комбинаторного химического процесса, или макромолекулой, такой как белок, нуклеиновая кислота, углевод, липид, гликопротеин, липопротеин, полисахарид, любое модифицированное производное вышеупомянутых молекул или любой комплекс, содержащий одну или более из вышеупомянутых молекул. Нуклеиновые кислоты включают ДНК или РНК, могут быть одно- или двухцепочечными; могут быть линейными, кольцевыми или разветвленными и могут иметь любую длину. Нуклеиновые кислоты включают те, которые способны образовывать двойные спирали, а также нуклеиновые кислоты, образующие тройные спирали. См., например, патенты США №5176996 и 5422251. Белки включают, но не ограничиваются этим, ДНК-связывающие белки, транскрипционные факторы, хроматин-реконструирующие факторы, метилированные ДНК-связывающие белки, полимеразы, метилазы, деметилазы, ацетилазы, деацетилазы, киназы, фосфатазы, интегразы, рекомбиназы, лигазы, топоизомеразы, гиразы и геликазы.
[0068] Экзогенная молекула может являться молекулой того же типа, что и эндогенная молекула, например, экзогенным белком или нуклеиновой кислотой. Например, экзогенная нуклеиновая кислота может содержать инфицирующий вирусный геном, плазмиду или эписому, внесенную в клетку, или хромосому, которая в обычных условиях не присутствует в клетке. Способы внесения экзогенных молекул в клетки известны специалистам в данной области техники и включают, но не ограничиваются этим, опосредованный липидами перенос (т.e. липосомами, включая нейтральные и катионные липиды), электропорацию, прямую инъекцию, клеточное слияние, бомбардировку частицами, осаждение фосфата кальция, ДЭАЭ-декстран-опосредованный перенос и опосредованный вирусным вектором перенос. Также экзогенная молекула может являться молекулой того же типа, что и эндогенная молекула, но может быть получена от вида, отличного от того, от которого получена клетка. Например, человеческую последовательность нуклеиновой кислоты можно внести в клетку, изначально полученную от мыши или хомяка.
[0069] И наоборот, “эндогенной” молекулой является такая молекула, которая в обычном состоянии присутствует в конкретной клетке на конкретной стадии развития в конкретных окружающих условиях. Например, эндогенная нуклеиновая кислота может содержать хромосому, геном митохондрии, хлоропласта или другой органеллы, или эписомную нуклеиновую кислоту природного происхождения. Дополнительные эндогенные молекулы могут включать белки, например, транскрипционные факторы и ферменты.
[0070] “Слитой” молекулой является молекула, в которой соединены, предпочтительно - ковалентно, две или более субъединицы. Субъединичные молекулы могут быть молекулами одного и того же химического типа или могут быть молекулами разного химического типа. Примеры первого типа слитых молекул включают, но не ограничиваются этим, слитые белки (например, результат слияния между ДНК-связывающим доменом ZFP или TALE и одним или более активационными доменами) и слитые нуклеиновые кислоты (например, нуклеиновая кислота, кодирующая слитый белок, описанный выше). Примеры второго типа слитых молекул включают, но не ограничиваются этим, слияние между образующей тройную спираль нуклеиновой кислотой и полипептидом и слияние между белком, связывающим малую бороздку ДНК, и нуклеиновой кислотой.
[0071] Экспрессия слитого белка в клетке может являться следствием доставки в клетку слитого белка или доставки в клетку полинуклеотида, кодирующего слитый белок, при этом полинуклеотид транскрибируется, а транскрипт транслируется для выработки слитого белка. В экспрессию белка в клетке также могут быть вовлечены транс-сплайсинг, полипептидное расщепление и полипептидное лигирование. Способы доставки в клетки полинуклеотидов и полипептидов представлены в другом месте данного описания.
[0072] “Мультимеризационный домен” (также называемый “димеризационным доменом” или “доменом белкового взаимодействия”) представляет собой домен, инкорпорированный в амино-, карбокси- либо амино- и карбокситерминальные участки ZFP TF или TALE TF. Эти домены обеспечивают возможность мультимеризации множественных единиц ZFP TF или TALE TF и, таким образом, более протяженные тракты доменов тринуклеотидных повторов оказываются преимущественно связанными мультимеризованными ZFP TF или TALE TF по сравнению с более короткими трактами с длиной повторов, соответствующей дикому типу. Примеры мультимеризационных доменов включают лейциновые молнии. Мультимеризационные домены также могут регулироваться небольшими молекулами, при этом мультимеризационный домен принимает надлежащую конформацию, которая обеспечивает возможность взаимодействия с другим мультимеризационным доменом только в присутствии небольшой молекулы или внешнего лиганда. Таким образом, экзогенные лиганды можно использовать для регуляции активности этих доменов.
[0073] Применительно к целям настоящего изобретения “ген” включает в себя участок ДНК, кодирующий генный продукт (см. ниже), а также все участки ДНК, которые регулируют выработку генного продукта, вне зависимости от того, прилегают ли такие регуляторные последовательности к кодирующим и/или транскрибируемым последовательностям. Соответственно, гены включают, но необязательно ограничиваются этим, промоторные последовательности, терминаторы, трансляционные регуляторные последовательности, такие как участки связывания рибосом и внутренние участки посадки рибосом, энхансеры, сайленсеры, инсуляторы, граничные элементы, участки начала репликации, участки присоединения матрицы и локусные контрольные участки.
[0074] “Генная экспрессия” означает преобразование информации, содержащейся в гене, в генный продукт. Генный продукт может являться прямым продуктом транскрипции гена (например, мРНК, тРНК, рРНК, антисмысловой РНК, рибозимом, структурной РНК или РНК любого другого типа) или белком, вырабатываемым в результате трансляции мРНК. Генные продукты также включают РНК, которые модифицированы при помощи таких процессов, как кэпирование, полиаденилирование, метилирование и редактирование, и белки, модифицированные при помощи, например, метилирования, ацетилирования, фосфорилирования, убиквитинирования, АДФ-рибозилирования, миристилирования и гликозилирования.
[0075] “Модуляция” генной экспрессии означает изменение в активности гена. Модуляция экспрессии может включать, но не ограничиваться этим, активацию гена и репрессию гена. Для модуляции экспрессии можно использовать геномное редактирование (например, расщепление, изменение, инактивацию, случайную мутацию). Инактивацией гена называется любое снижение генной экспрессии по сравнению с клеткой, которая не содержит описанный в данном тексте белок ZFP или TALE. Следовательно, инактивация гена может быть частичной либо полной.
[0076] “Представляющий интерес участок” представляет собой любой участок клеточного хроматина, такой как, к примеру, ген или некодирующая последовательность в пределах гена или прилегающая к гену, в котором необходимо связать экзогенную молекулу. Связывание может осуществляться с целью нацеленного расщепления ДНК и/или нацеленной рекомбинации. Представляющий интерес участок может находиться, например, в хромосоме, эписоме, геноме органеллы (например, митохондрии, хлоропласта) или инфицирующем вирусном геноме. Представляющий интерес участок может находиться в пределах кодирующей области гена, в пределах транскрибируемых некодирующих областей, таких как, например, лидерные последовательности, трейлерные последовательности или интроны, или в пределах нетранскрибируемых областей, расположенных как выше, так и ниже кодирующей области. Представляющий интерес участок может быть небольшим - до одной пары нуклеотидов, или длиной до 2000 пар нуклеотидов, или длиной в любое промежуточное количество пар нуклеотидов.
[0077] “Эукариотические” клетки включают, но не ограничиваются этим, клетки грибов (такие как дрожжи), клетки растений, клетки животных, клетки млекопитающих и клетки человека (например, Т-клетки).
[0078] Термины “функциональная связь” или “функционально связанный” употребляются взаимозаменяемо в отношении непосредственного соседства двух или более компонентов (таких как элементы последовательности), при котором компоненты расположены таким образом, что оба компонента функционируют нормально и существует возможность, что по меньшей мере один из компонентов может опосредовать функцию, которая влияет на по меньшей мере один из других компонентов. В качестве иллюстрации можно привести транскрипционную регуляторную последовательность, такую как промотор, функционально связанную с кодирующей последовательностью в том случае, если транскрипционная регуляторная последовательность регулирует уровень транскрипции кодирующей последовательности в ответ на присутствие или отсутствие одного или более транскрипционных регуляторных факторов. В общем случае транскрипционная регуляторная последовательность функционально связана в цис-форме с кодирующей последовательностью, но необязательно должна прилегать к ней. Например, энхансер является транскрипционной регуляторной последовательностью, которая функционально связана с кодирующей последовательностью, даже если они не являются смежными.
[0079] В случае слитых полипептидов термин “функционально связанный” может обозначать тот факт, что каждый из компонентов в связи оказывает такое же действие на другой компонент, какое оказывал бы в случае, если бы не был связан таким образом. Например, в случае слитого полипептида, в котором ДНК-связывающий домен ZFP или TALE сшит с активационным доменом, ДНК-связывающий домен ZFP или TALE и активационный домен являются функционально связанными, если в слитом белке часть ДНК-связывающего домена ZFP или TALE способна связывать свой целевой участок и/или свой связывающий участок, в то время как активационный домен способен повышающе регулировать генную экспрессию. Сшитые с доменами ZFP, способные регулировать генную экспрессию, обобщенно называют “ZFP-TF” или “транскрипционными факторами цинковых пальцев”, в то время как сшитые с доменами TALE, способные регулировать генную экспрессию, обобщенно называют “TALE-TF” или “транскрипционными факторами TALE”. В случае слитого полипептида, в котором ДНК-связывающий домен ZFP сшит с расщепляющим доменом (“ZFN” или “цинкпальцевой нуклеазой”), ДНК-связывающий домен ZFP и расщепляющий домен являются функционально связанными, если в слитом белке часть ДНК-связывающего домена ZFP способна связывать свой целевой участок и/или свой связывающий участок, в то время как расщепляющий домен способен расщеплять ДНК вблизи целевого участка. В случае слитого полипептида, в котором ДНК-связывающий домен TALE сшит с расщепляющим доменом (“TALEN” или “TALE-нуклеазой”), ДНК-связывающий домен TALE и расщепляющий домен являются функционально связанными, если в слитом белке часть ДНК-связывающего домена TALE способна связывать свой целевой участок и/или свой связывающий участок, в то время как расщепляющий домен способен расщеплять ДНК вблизи целевого участка.
[0080] “Функциональным фрагментом” белка, полипептида или нуклеиновой кислоты является белок, полипептид или нуклеиновая кислота, чья последовательность не идентична полноразмерным белку, полипептиду или нуклеиновой кислоте, но при этом он сохраняет такие же функции, что и полноразмерные белок, полипептид или нуклеиновая кислота. Функциональный фрагмент может содержать большее, меньшее или такое же самое количество остатков, что и соответствующая нативная молекула, и/или может содержать одну или более аминокислотных или нуклеотидных замен. Способы определения функции нуклеиновой кислоты (например, кодирующей функции, способности гибридизироваться с другой нуклеиновой кислотой) хорошо известны в данной области техники. Аналогично, способы определения функции белка также хорошо известны. Например, ДНК-связывающую функцию полипептида можно определить методами связывания на фильтрах, изменения электрофоретической подвижности или иммунопреципитации. Расщепление ДНК можно проанализировать при помощи гель-электрофореза. См. Ausubel et al., выше. Способность белка взаимодействовать с другим белком можно определить, например, при помощи совместной иммунопреципитации, двухгибридного анализа или комплементации, как генетической, так и биохимической. См., например, Fields et al. (1989) Nature 340:245-246; патент США №5585245 и PCT WO 98/44350.
[0081] “Вектор” способен переносить последовательности генов в целевые клетки. Как правило, “векторная конструкция”, “экспрессионный вектор” и “вектор для переноса генов” обозначают любую конструкцию нуклеиновой кислоты, способную управлять экспрессией представляющего интерес гена, и которая может переносить последовательности генов в целевые клетки. Следовательно, этот термин включает клонирующие и экспрессионные векторы, а также интегрирующие векторы.
[0082] “Репортерный ген” или “репортерная последовательность” обозначает любую последовательность, которая вырабатывает белковый продукт, который легко определяется, предпочтительно, но необязательно, при помощи общепринятых методов. Подходящие репортерные гены включают, но не ограничиваются этим, последовательности, кодирующие белки, которые опосредуют устойчивость к антибиотикам (например, устойчивость к ампициллину, устойчивость к неомицину, устойчивость к G418, устойчивость к пуромицину), последовательности, кодирующие окрашенные, или флуоресцентные, или люминесцентные белки (например, зеленый флуоресцентный белок, усиленный зеленый флуоресцентный белок, красный флуоресцентный белок, люциферазу) и белки, которые опосредуют повышенный рост клеток и/или генную амплификацию (например, дигидрофолатредуктазу). Эпитопные метки включают, например, одну или более копий FLAG, His, myc, Tap, HA или любой регистрируемой аминокислотной последовательности. “Экспрессионные метки” включают последовательности, которые кодируют репортеры, которые могут быть функционально связаны с требуемой последовательностью гена, с целью отслеживания экспрессии представляющего интерес гена.
ДНК-связывающие домены
[0083] В данном тексте описаны составы, содержащие ДНК-связывающие домены, которые специфически связываются с целевой последовательностью в любом гене, содержащем тринуклеотидные повторы, включая, но не ограничиваясь этим, Htt. Любой ДНК-связывающий домен можно применять в раскрытых в данном тексте составах и способах.
[0084] В некоторых вариантах реализации изобретения ДНК-связывающий домен содержит белок «цинковые пальцы». Предпочтительно белок «цинковые пальцы» имеет неприродное происхождение и сконструирован для того, чтобы связываться с выбранным целевым участком. См., например, Beerli et al. (2002) Nature Biotechnol. 20:135-141; Pabo et al. (2001) Ann. Rev. Biochem. 70:313-340; Isalan et al. (2001) Nature Biotechnol. 19:656-660; Segal et al. (2001) Curr. Opin. Biotechnol. 12:632-637; Choo et al. (2000) Curr. Opin. Struct. Biol. 10:411-416; патенты США №6453242; 6534261; 6599692; 6503717; 6689558; 7030215; 6794136; 7067317; 7262054; 7070934; 7361635; 7253273 и патентные публикации США №2005/0064474; 2007/0218528; 2005/0267061, которые в полном объеме включены в данный текст посредством ссылки.
[0085] Сконструированный связывающий домен цинковых пальцев может обладать новой связывающей специфичностью по сравнению с белком «цинковые пальцы» природного происхождения. Методы конструирования включают, но не ограничиваются этим, рациональное конструирование и различные типы селекции. Рациональное конструирование включает, например, использование баз данных, содержащих триплетные (или квадруплетные) нуклеотидные последовательности и индивидуальные аминокислотные последовательности цинковых пальцев, в которых каждая триплетная или квадруплетная нуклеотидная последовательность связана с одной или более аминокислотными последовательностями цинковых пальцев, которые связывают конкретную триплетную или квадруплетную последовательность. См., например, совместные патенты США 6453242 и 6534261, в полном объеме включенные в данный текст посредством ссылки.
[0086] Примеры селекционных методов, включая фаговое отображение и двухгибридные системы, раскрыты в патентах США 5789538; 5925523; 6007988; 6013453; 6410248; 6140466; 6200759 и 6242568; а также в WO 98/37186; WO 98/53057; WO 00/27878; WO 01/88197 и патенте Великобритании 2338237. Вдобавок, было описано повышение специфичности связывания для связывающих доменов цинковых пальцев, например, в совместной заявке WO 02/077227.
[0087] Вдобавок, в этих и других ссылках раскрывается, что домены цинковых пальцев и/или белки «цинковые пальцы», содержащие большое количество “пальцев”, могут быть связаны вместе посредством любых подходящих линкерных последовательностей, включая, например, линкеры длиной в 5 или более аминокислот. Также см. патенты США №6479626, 6903185 и 7153949, в которых приведены примеры линкерных последовательностей длиной в 6 или более аминокислот. Описанные в данном тексте белки могут содержать любую комбинацию подходящих линкеров между отдельными цинковыми пальцами белка. Вдобавок, было описано повышение специфичности связывания для связывающих доменов цинковых пальцев, например, в совместной заявке WO 02/077227.
[0088] Выбор целевых участков; ZFP и способы проектирования и конструирования слитых белков (и кодирующих их полинуклеотидов) известны специалистам в данной области техники и детально описаны в патентах США №61400815; 789538; 6453242; 6534261; 5925523; 6007988; 6013453; 6200759; WO 95/19431; WO 96/06166; WO 98/53057; WO 98/54311; WO 00/27878; WO 01/60970 WO 01/88197; WO 02/099084; WO 98/53058; WO 98/53059; WO 98/53060; WO 02/016536 и WO 03/016496.
[0089] Вдобавок, в этих и других ссылках раскрывается, что домены цинковых пальцев и/или белки «цинковые пальцы», содержащие большое количество “пальцев”, могут быть связаны вместе посредством любых подходящих линкерных последовательностей, включая, например, линкеры длиной в 5 или более аминокислот. Также см. патенты США №6479626, 6903185 и 7153949, в которых приведены примеры линкерных последовательностей длиной в 6 или более аминокислот. Описанные в данном тексте белки могут содержать любую комбинацию подходящих линкеров между отдельными цинковыми пальцами белка.
[0090] В определенных вариантах реализации изобретения ДНК-связывающий домен представляет собой сконструированный белок «цинковые пальцы», который связывается (специфическим к последовательности образом) с целевым участком в гене Htt и модулирует экспрессию Htt. ZFP могут выборочно связываться с мутантным аллелем Htt или последовательностью Htt дикого типа. Нацеленные на Htt участки обычно содержат по меньшей мере один цинковый палец, но могут содержать несколько цинковых пальцев (например, 2, 3, 4, 5, 6 или больше пальцев). Обычно ZFP содержат по меньшей мере три пальца. Определенные ZFP содержат четыре, пять или шесть пальцев, при этом некоторые ZFP содержат 8, 9, 10, 11 или 12 пальцев. ZFP, которые содержат три пальца, обычно распознают целевой участок, который содержит 9 или 10 нуклеотидов; ZFP, которые содержат четыре пальца, обычно распознают целевой участок, который содержит от 12 до 14 нуклеотидов; в то время как ZFP, содержащие шесть пальцев, могут распознавать целевые участки, которые содержат от 18 до 21 нуклеотида. ZFP также могут представлять собой слитые белки, которые содержат один или более регуляторных доменов, и эти домены могут являться транскрипционными активационными или репрессионными доменами. В некоторых вариантах реализации изобретения слитый белок содержит два связанных вместе ДНК-связывающих домена ZFP. Такие белки «цинковые пальцы» могут содержать 8, 9, 10, 11, 12 или более пальцев. В некоторых вариантах реализации изобретения два ДНК-связывающих домена связаны посредством удлиняемого гибкого линкера таким образом, что один ДНК-связывающий домен содержит 4, 5 или 6 цинковых пальцев, а второй ДНК-связывающий домен содержит дополнительно 4, 5 или 6 цинковых пальцев. В некоторых вариантах реализации изобретения линкер является стандартным междупальцевым линкером так, что группа пальцев содержит ДНК-связывающий домен, содержащий 8, 9, 10, 11 или 12 или более пальцев. В других вариантах реализации изобретения линкер является нетипичным линкером, таким как гибкий линкер. ДНК-связывающие домены сшиты с по меньшей мере одним регуляторным доменом, и можно считать, что они обладают архитектурой ‘ZFP-ZFP-TF’. Конкретные примеры этих вариантов реализации можно называть “ZFP-ZFP-KOX”, которая содержит два ДНК-связывающих домена, связанных с гибким линкером и сшитых с репрессором КОХ и “ZFP-KOX-ZFP-KOX”, где два слитых белка ZFP-KOX сшиты вместе посредством линкера.
[0091] В альтернативном варианте ДНК-связывающий домен можно получить из нуклеазы. Например, известны распознающие последовательности хоминг-эндонуклеаз и мегануклеаз, такие как I-SceI, I-CeuI, PI-PspI, PI-Sce, I-SceIV, I-CsmI, I-PanI, I-SceII, I-PpoI, I-SceIII, I-CreI, I-TevI, I-TevII и I-TevIII. См. также патент США №5420032, патент США №6833252; Belfort et al. (1997) Nucleic Acids Res. 25:3379-3388; Dujon et al. (1989) Gene 82:115-118; Perler et al. (1994) Nucleic Acids Res. 22, 1125-1127; Jasin (1996) Trends Genet. 12:224-228; Gimble et al. (1996) J. Mol. Biol. 263:163-180; Argast et al. (1998) J. Mol. Biol. 280:345-353 и каталог New England Biolabs. Вдобавок, ДНК-связывающую специфичность хоминг-эндонуклеаз и мегануклеаз можно подобрать так, чтобы связывать целевые последовательности неприродного происхождения. См., например, Chevalier et al. (2002) Molec. Cell 10:895-905; Epinat et al. (2003) Nucleic Acids Res. 31:2952-2962; Ashworth et al. (2006) Nature 441:656-659; Paques et al. (2007) Current Gene Therapy 7:49-66; патентную публикацию США №20070117128.
[0092] “Двурукими” белками «цинковые пальцы» являются такие белки, в которых два кластера ДНК-связывающих доменов цинковых пальцев разделены промежуточными аминокислотами таким образом, что два домена цинковых пальцев связываются с двумя несмежными целевыми участками. Примером связывающего белка «цинковые пальцы» двурукого типа является SIP1, в котором кластер из четырех цинковых пальцев расположен на амино-конце белка, а кластер из трех пальцев расположен на карбоксильном конце (См. Remacle et al, (1999) EMBO Journal 18 (18): 5073-5084). Каждый кластер из цинковых пальцев в этих белках способен связываться с уникальной целевой последовательностью, а участок между двумя целевыми последовательностями может содержать много нуклеотидов. Двурукие ZFP могут содержать функциональный домен, например, сшитый с одним или более из ZFP. Таким образом, стоит понимать, что функциональный домен можно присоединить к внешней части одного или обоих ZFP (см. фигуру 1В) или можно поместить между ZFP (присоединен к обоим ZFP) (см. фигуру 4).
[0093] Конкретные примеры Htt-нацеленных ZFP приведены в таблицах 1А и 1Б. В первой колонке этой таблицы представлено внутреннее наименование ссылки (номер) для ZFP, которое соответствует аналогичному наименованию в колонке 1 таблиц 2А и 2Б. “П” обозначает палец, а номер, следующий за “П”, обозначает какой именно это палец (например, “П1” относится к 1 пальцу).
[0094] Последовательность и расположение целевых участков этих белков приведены в таблицах 2А и 2Б. В таблицах 2А и 2Б приведены целевые последовательности указанных белков «цинковые пальцы». Нуклеотиды на целевых участках, которые контактируют со спиралями узнавания ZFP, обозначены большими буквами; неконтактирующие нуклеотиды обозначены малыми буквами.
[0095] В определенных вариантах реализации изобретения ДНК-связывающий домен содержит ДНК-связывающий домен TAL эффектора (TALE) природного происхождения или сконструированный (неприродного происхождения). См., например, патентную публикацию США №20110301073, в полном объеме включенную в данный текст посредством ссылки. Известно, что фитопатогенные бактерии рода Xanthomonas являются причиной многих заболеваний важных сельскохозяйственных культур. Патогенность Xanthomonas зависит от консервативной секреторной системы III типа (T3S), которая инжектирует в клетку растения более 25 разных эффекторных белков. Среди этих инжектированных белков присутствуют транскрипционные эффекторы активаторного типа (TALE), которые имитируют растительные транскрипционные активаторы и управляют транскриптомом растений (см. Kay et al (2007) Science 318:648-651). Эти белки содержат ДНК-связывающий домен и транскрипционный активационный домен. Одним из наиболее полно описанных TALE является AvrBs3 из Xanthomonas campestgris pv. Vesicatoria (см. Bonas et al (1989) Mol Gen Genet 218: 127-136 и WO 2010079430). TALE содержат централизованный домен тандемных повторов, где каждый повтор содержит приблизительно 34 аминокислоты, которые являются ключевыми для ДНК-связывающей специфичности этих белков. Вдобавок, они содержат последовательность ядерной локализации и кислый транскрипционный активационный домен (для ознакомления см. Schornack S, et al (2006) J Plant Physiol 163(3): 256-272). Дополнительно было обнаружено, что в фитопатогенных бактериях Ralstonia solanacearum два гена, обозначаемые как brg11 и hpx17, являются гомологичными семейству AvrBs3 Xanthomonas, а именно в R. solanacearum биовар 1 штамм GMI1000 и биовар 4 штамм RS1000 (См. Heuer et al (2007) Appland Envir Micro 73(13): 4379-4384). Эти гены на 98,9% идентичны друг другу по нуклеотидной последовательности, но отличаются делецией в 1575 п.н.о. в повторяемом домене hpx17. При этом оба генных продукта обладают менее чем 40% идентичности последовательности с белковым семейством AvrBs3 Xanthomonas.
[0096] Специфичность этих TALE зависит от последовательностей, которые находятся в тандемных повторах. Повторяемые последовательности содержат приблизительно 102 п.н.о., а повторы обычно на 91-100% гомологичны друг другу (Bonas et al, там же). Полиморфизм повторов обычно локализован в позициях 12 и 13 и, по-видимому, существует однозначное соответствие между идентичностью гипервариабельных двойных остатков в позициях 12 и 13 и идентичностью смежных нуклеотидов в целевой последовательности TALE (см. Moscou and Bogdanove, (2009) Science 326:1501 и Boch et al (2009) Science 326:1509-1512). Экспериментально был определен код для распознавания ДНК этих TALE, в котором последовательность БХ в позициях 12 и 13 приводит к связыванию с цитозином (C), NG связывается с T, NI с A, C, G или T, NN связывается с A или G, а IG связывается с T. Эти ДНК-связывающие повторы вносили в белки с новыми комбинациями и количествами повторов для создания искусственных транскрипционных факторов, которые способны взаимодействовать с новыми последовательностями и активировать экспрессию неэндогенного репортерного гена в клетках растений (Boch et al, там же). Сконструированные белки TAL связывали с полудоменом расщепления FokI, чтобы получить продукт слияния эффекторного домена TAL с нуклеазой (TALEN), обладающий активностью по данным анализа дрожжевого репортерного гена (мишень на основе плазмид). Christian et al ((2010)<Genetics epub 10.1534/genetics.110.120717). См. также патентную публикацию США №20110301073, в полном объеме включенную в данный текст посредством ссылки.
[0097] Конкретные примеры сконструированных димеризационных доменов для применения с белками ZFP или TALE приведены в таблице 3. Приведены аминокислотные последовательности двух типов доменов - суперспирального (СС) и димеризующихся цинковых пальцев (DZ).
Слитые белки
[0098] Также предложены слитые белки, содержащие описанные в данном тексте ДНК-связывающие белки (например, ZFP или TALE) и гетерологичный регуляторный (функциональный) домен (или его функциональный фрагмент). Стандартные домены включают, например, домены транскрипционных факторов (активаторы, репрессоры, ко-активаторы, ко-репрессоры), сайленсеры, онкогены (например, представителей семейств myc, jun, fos, myb, max, mad, rel, ets, bcl, myb, mos и т.д.); ДНК-репарационные ферменты и их ассоциированные факторы и модификаторы; хроматин-ассоциированные белки и их модификаторы (например, киназы, ацетилазы и деацетилазы) и ДНК-модифицирующие ферменты (например, метилтрансферазы, топоизомеразы, геликазы, лигазы, киназы, фосфатазы, полимеразы, эндонуклеазы) и их ассоциированные факторы и модификаторы. См. патентные заявки США №20050064474; 20060188987 и 2007/0218528, в которых детально описаны продукты слияния ДНК-связывающих доменов и нуклеазных доменов расщепления, все эти заявки в полном объеме включены в данный текст посредством ссылки.
[0099] Домены, подходящие для достижения активации, включают активационный домен HSV VP16 (см., например, Hagmann et al., J. Virol. 71, 5952-5962 (1997)) ядерные гормональные рецепторы (См., например, Torchia et al., Curr. Opin. Cell. Biol. 10:373-383 (1998)); субъединицу p65 ядерного фактора каппа B (Bitko & Barik, J. Virol. 72:5610-5618 (1998) и Doyle & Hunt, Neuroreport 8:2937-2942 (1997)); Liu et al., Cancer Gene Ther. 5:3-28 (1998)), или искусственные химерные функциональные домены, такие как VP64 (Beerli et al., (1998) Proc. Natl. Acad. Sci. USA 95:14623-33), и дегрон (Molinari et al., (1999) EMBO J. 18, 6439-6447). Дополнительные примеры активационных доменов включают Oct 1, Oct-2A, Sp1, AP-2 и CTF1 (Seipel et al., EMBO J. 11, 4961-4968 (1992)), а также p300, CBP, PCAF, SRC1 PvALF, AtHD2A и ERF-2. См., например, Robyr et al. (2000) Mol. Endocrinol. 14:329-347; Collingwood et al. (1999) J. Mol. Endocrinol. 23:255-275; Leo et al. (2000) Gene 245:1-11; Manteuffel-Cymborowska (1999) Acta Biochim. Pol. 46:77-89; McKenna et al. (1999) J. Steroid Biochem. Mol. Biol. 69:3-12; Malik et al. (2000) Trends Biochem. Sci. 25:277-283 и Lemon et al. (1999) Curr. Opin. Genet. Dev. 9:499-504. Дополнительные примеры активационных доменов включают, но не ограничиваются этим, OsGAI, HALF-1, C1, AP1, ARF-5, -6, -7 и -8, CPRF1, CPRF4, MYC-RP/GP и TRAB1. См., например, Ogawa et al. (2000) Gene 245:21-29; Okanami et al. (1996) Genes Cells 1:87-99; Goff et al. (1991) Genes Dev. 5:298-309; Cho et al. (1999) Plant Mol. Biol. 40:419-429; Ulmason et al. (1999) Proc. Natl. Acad. Sci. USA 96:5844-5849; Sprenger-Haussels et al. (2000) Plant J. 22:1-8; Gong et al. (1999) Plant Mol. Biol. 41:33-44 и Hobo et al. (1999) Proc. Natl. Acad. Sci. USA 96:15,348-15,353.
[0100] Для специалистов в данной области техники очевидно, что при образовании слитого белка (или кодирующей его нуклеиновой кислоты) между ДНК-связывающим доменом и функциональным доменом в качестве функционального домена подходящими являются активационный домен или молекула, которая взаимодействует с активационным доменом. Практически любую молекулу, способную привнести активационный комплекс и/или активационную активность (такую как, например, ацетилирование гистонов) в целевой ген, можно применять в качестве активационного домена слитого белка. Инсуляторные домены, локализационные домены и хроматин-реконструирующие белки, такие как ISWI-содержащие домены и/или белки, содержащие метилсвязывающие домены, подходят для их применения в качестве функциональных доменов в слитых молекулах, как описано, например, в совместных патентных заявках США 2002/0115215 и 2003/0082552 и в совместной публикации WO 02/44376.
[0101] Примеры репрессионных доменов включают, но не ограничиваются этим, KRAB A/B, KOX, ТФР-бета-индуцибельный ранний ген (TIEG), v-erbA, SID, MBD2, MBD3, представителей семейства DNMT (например, DNMT1, DNMT3A, DNMT3B), Rb и MeCP2. См., например, Bird et al. (1999) Cell 99:451-454; Tyler et al. (1999) Cell 99:443-446; Knoepfler et al. (1999) Cell 99:447-450 и Robertson et al. (2000) Nature Genet. 25:338-342. Дополнительные примеры репрессионных доменов включают, но не ограничиваются этим, ROM2 и AtHD2A. См., например, Chem et al. (1996) Plant Cell 8:305-321 и Wu et al. (2000) Plant J. 22:19-27.
[0102] Слитые молекулы конструируют при помощи методов клонирования и биохимической конъюгации, которые хорошо известны специалистам в данной области техники. Слитые молекулы содержат ДНК-связывающий домен и функциональный домен (например, транскрипционный активационный или репрессионный домен). В некоторых случаях слитые молекулы содержат также сигналы ядерной локализации (такие как, например, сигналы Т-антигена среды SV40) и эпитопные метки (такие как, например, FLAG и гемагглютинин). Слитые белки (и кодирующие их нуклеиновые кислоты) сконструированы таким образом, что трансляционная рамка считывания сохраняется в составе компонентов продукта слияния.
[0103] Продукты слияния между полипептидным компонентом функционального домена (или его функциональным фрагментом) с одной стороны и небелковым ДНК-связывающим доменом (например, антибиотиком, интеркалятором, компонентом, связывающим малую бороздку ДНК, нуклеиновой кислотой) с другой стороны конструируют при помощи методов биохимической конъюгации, известных специалистам в данной области техники. См., например, каталог Pierce Chemical Company (Рокфорд, Иллинойс). Были описаны способы и составы для получения продуктов слияния между компонентом, связывающим малую бороздку ДНК, и полипептидом. Mapp et al. (2000) Proc. Natl. Acad. Sci. USA 97:3930-3935.
[0104] В определенных вариантах реализации изобретения целевой участок, связываемый ДНК-связывающим доменом, расположен в доступной области клеточного хроматина. Доступные области были определены и описаны, например, в совместной международной публикации WO 01/83732. В случае если целевой участок не расположен в доступной области клеточного хроматина, можно создать одну или более доступных областей, как описано в совместной публикации WO 01/83793. В дополнительных вариантах реализации изобретения ДНК-связывающий домен слитой молекулы способен связываться с клеточным хроматином вне зависимости от того, находится ли его целевой участок в доступной области или нет. Например, такие ДНК-связывающие домены способны связываться с линкерной ДНК и/или нуклеосомной ДНК. Примеры ДНК-связывающих доменов подобного “передового” типа можно обнаружить в некоторых стероидных рецепторах и в гепатоцитарном ядерном факторе 3 (ГЯФ3). Cordingley et al. (1987) Cell 48:261-270; Pina et al. (1990) Cell 60:719-731 и Cirillo et al. (1998) EMBO J. 17:244-254.
[0105] Слитые молекулы можно смешивать с фармацевтически приемлемым носителем, как известно специалистам в данной области техники. См., например, Remington's Pharmaceutical Sciences, 17th ed., 1985 и совместную публикацию WO 00/42219.
[0106] Функциональный компонент/домен слитой молекулы можно выбрать из множества разных компонентов, способных влиять на транскрипцию гена в случае, если слитая молекула связывается с целевой последовательностью посредством своего ДНК-связывающего домена. Следовательно, функциональный компонент может включать, но не ограничиваться этим, различные домены транскрипционных факторов, такие как активаторы, репрессоры, ко-активаторы, ко-репрессоры и сайленсеры.
[0107] Дополнительные примеры функциональных доменов раскрыты, например, в совместном патенте США №6534261 и патентной заявке США №2002/0160940.
[0108] Также можно выбрать функциональные домены, которые регулируются экзогенными небольшими молекулами или лигандами. Например, можно применять технологию RheoSwitch®, в которой функциональный домен принимает свою активную конформацию только в присутствии внешнего лиганда RheoChem™ (см., например, США 20090136465). Таким образом, ZFP или TALE могут быть функционально связанными с регулируемым функциональным доменом, при этом результирующая активность ZFP-TF или TALE-TF контролируется внешним лигандом.
Нуклеазы
[0109] В некоторых вариантах реализации изобретения слитый белок содержит ДНК-связывающий домен и расщепляющий (нуклеазный) домен. Таким образом, модификацию гена можно осуществить при помощи нуклеазы, например, сконструированной нуклеазы. Технология конструирования нуклеаз основана на конструировании ДНК-связывающих белков природного происхождения. Например, было описано конструирование хоминг-эндонуклеаз с заданной ДНК-связывающей специфичностью. (см., Chames et al. (2005) Nucleic Acids Res 33(20):e178; Arnould et al. (2006) J. Mol. Biol. 355:443-458). Дополнительно было описано также конструирование ZFP. См., например, патентные заявки США №6534261; 6607882; 6824978; 6979539; 6933113; 7163824 и 7013219.
[0110] Вдобавок, ZFP и TALE были сшиты с нуклеазными доменами для получения ZFN и TALEN - функциональных единиц, которые способны распознавать свою цель в виде нуклеиновой кислоты при помощи своих сконструированных (ZFP или TALE) ДНК-связывающих доменов и приводить к разрезанию ДНК вблизи ДНК-связывающего участка ZFP или TALE вследствие нуклеазной активности. См., например, Kim et al. (1996) Proc Natl Acad Sci USA 93(3):1156-1160. В последнее время ZFN стали применять для модификации генома большого количества организмов. См., например, патентные публикации США 20030232410; 20050208489; 20050026157; 20050064474; 20060188987; 20060063231 и международную публикацию WO 07/014275.
[0111] Таким образом, описанные в данном тексте способы и составы широко применяются и в них могут быть задействованы любые представляющие интерес нуклеазы. Неограничивающие примеры нуклеаз включают мегануклеазы, TALEN и цинк-пальцевые нуклеазы. Нуклеаза может содержать гетерологичный ДНК-связывающий и расщепляющий домены (например, ДНК-связывающие домены цинк-пальцевой нуклеазы; TALEN; мегануклеазы с гетерологичными расщепляющими доменами) или, в альтернативном варианте, ДНК-связывающий домен нуклеазы природного происхождения можно изменить так, чтобы он связывался с выбранным целевым участком (например, мегануклеаза, которая была сконструированы, чтобы связываться с участком, отличным от когнатного связывающего участка).
[0112] В определенных вариантах реализации изобретения нуклеаза является мегануклеазой (хоминг-эндонуклеазой). Мегануклеазы природного происхождения распознают сайты расщепления в 15-40 пар оснований и обычно их сгруппировывают в четыре семейства: семейство LAGLIDADG, семейство GIY-YIG, семейство His-Cyst box и семейство HNH. Примеры хоминг-эндонуклеаз включают I-SceI, I-CeuI, PI-PspI, PI-Sce, I-SceIV, I-CsmI, I-PanI, I-SceII, I-PpoI, I-SceIII, I-CreI, I-TevI, I-TevII и I-TevIII. Их распознающие последовательности известны. См. также патент США №5420032; патент США №6833252; Belfort et al. (1997) Nucleic Acids Res. 25:3379-3388; Dujon et al. (1989) Gene 82:115-118; Perler et al. (1994) Nucleic Acids Res. 22, 1125-1127; Jasin (1996) Trends Genet. 12:224-228; Gimble et al. (1996) J. Mol. Biol. 263:163-180; Argast et al. (1998) J. Mol. Biol. 280:345-353 и каталог New England Biolabs.
[0113] ДНК-связывающие домены мегануклеаз природного происхождения, преимущественно принадлежащих семейству LAGLIDADG, использовали для проведения сайт-специфической геномной модификации в растениях, дрожжах, дрозофилах, клетках млекопитающих и мышей, но этот эксперимент ограничивался модификацией гомологичных генов, при которой сохраняется распознающая последовательность мегануклеазы (Monet et al. (1999), Biochem. Biophysics. Res. Common. 255:88-93) или геномами до конструирования, в которые вносилась распознающая последовательность (Route et al. (1994), Mol. Cell. Biol. 14:8096-106; Chilton et al. (2003) Plant Physiology 133:956-65; Puchta et al. (1996), Proc. Natl. Acad. Sci. USA 93: 5055-60; Rong et al. (2002), Genes Dev. 16: 1568-81; Gouble et al.(2006), J. Gene Med. 8(5):616-622). Соответственно, были предприняты попытки сконструировать мегануклеазы, которые проявляли бы новую связывающую специфичность в участках, важных с точки зрения медицины или биотехнологии (Porteus et al. (2005), Nat. Biotechnol. 23: 967-73; Sussman et al. (2004), J. Mol. Biol. 342: 31-41; Epinat et al. (2003), Nucleic Acids Res. 31: 2952-62; Chevalier et al. (2002) Molec. Cell 10:895-905; Epinat et al. (2003) Nucleic Acids Res. 31:2952-2962; Ashworth et al. (2006) Nature 441:656-659; Paques et al. (2007) Current Gene Therapy 7:49-66; U.S. патентные публикации США №20070117128; 20060206949; 20060153826; 20060078552 и 20040002092). Вдобавок, ДНК-связывающие домены мегануклеаз, как природного происхождения, так и сконструированные, были также функционально связаны с расщепляющим доменом гетерологичной нуклеазы (например, FokI).
[0114] В других вариантах реализации изобретения нуклеаза является цинк-пальцевой нуклеазой (ZFN). ZFN содержат белок «цинковые пальцы», который был сконструирован так, чтобы связываться с целевым участком в выбранном гене, и расщепляющий домен или расщепляющий полудомен.
[0115] Как детально описано выше, связывающие домены цинковых пальцев можно сконструировать так, чтобы они связывались с выбранной последовательностью. См., например, Beerli et al. (2002) Nature Biotechnol. 20:135-141; Pabo et al. (2001) Ann. Rev. Biochem. 70:313-340; Isalan et al. (2001) Nature Biotechnol. 19:656-660; Segal et al. (2001) Curr. Opin. Biotechnol. 12:632-637; Choo et al. (2000) Curr. Opin. Struct. Biol. 10:411-416. Сконструированный связывающий домен цинковых пальцев может обладать новой связывающей специфичностью по сравнению с белком «цинковые пальцы» природного происхождения. Методы конструирования включают, но не ограничиваются этим, рациональное конструирование и различные типы селекции. Рациональное конструирование включает, например, использование баз данных, содержащих триплетные (или квадруплетные) нуклеотидные последовательности и индивидуальные аминокислотные последовательности цинковых пальцев, в которых каждая триплетная или квадруплетная нуклеотидная последовательность связана с одной или более аминокислотными последовательностями цинковых пальцев, которые связывают конкретную триплетную или квадруплетную последовательность. См., например, совместные патенты США 6453242 и 6534261, в полном объеме включенные в данный текст посредством ссылки.
[0116] Примеры селекционных методов, включая фаговое отображение и двухгибридные системы, раскрыты в патентах США 5789538; 5925523; 6007988; 6013453; 6410248; 6140466; 6,200,759 и 6242568; а также в WO 98/37186; WO 98/53057; WO 00/27878; WO 01/88197 и патенте Великобритании 2338237. Вдобавок, было описано повышение специфичности связывания для связывающих доменов цинковых пальцев, например, в совместной заявке WO 02/077227.
[0117] Вдобавок, в этих и других ссылках раскрывается, что домены цинковых пальцев и/или белки «цинковые пальцы», содержащие большое количество “пальцев”, могут быть связаны вместе посредством любых подходящих линкерных последовательностей, включая, например, линкеры длиной в 5 или более аминокислот (например, TGEKP (SEQ ID NO:135), TGGQRP (SEQ ID NO:136), TGQKP (SEQ ID NO:137) и/или TGSQKP (SEQ ID NO:138)). См., например, патенты США №6479626; 6903185 и 7153949, в которых приведены примеры линкерных последовательностей длиной в 6 или более аминокислот. Описанные в данном тексте белки могут содержать любую комбинацию подходящих линкеров между отдельными цинковыми пальцами белка. См. также предварительную патентную публикацию США №20110287512.
[0118] [Нуклеазная система CRISPR (от англ. “Clustered Regularly Interspaced Short Palindromic Repeats“ - кластерные короткие палиндромные повторы, разделенные регулярными промежутками)/Cas (от англ. “CRISPR Associated“ - CRISPR-ассоциированный) представляет собой недавно сконструированную на основе бактериальной системы нуклеазную систему, которую можно применять для геномного конструирования. Частично она основана на адаптивном иммунном ответе многих бактерий и архей. Когда вирус или плазмида проникают в бактерию, сегменты ДНК внедряющегося организма преобразуются в CRISPR РНК (crРНК) в ходе ‘иммунного’ ответа. Затем эта crРНК объединяется посредством области частичной комплементарности с РНК другого типа, называемой tracrРНК, для того, чтобы направлять нуклеазу Cas9 к участку, гомологичному crРНК в целевой ДНК и называемому “протоспейсером”. Cas9 расщепляет ДНК для получения тупых концов в месте ДЦР на участках, определяемых 20-нуклеотидной руководящей последовательностью, которая содержится в транскрипте crРНК. Для Cas9 необходимы как crРНК, так и tracrРНК, для сайт-специфического распознавания и расщепления ДНК. Сейчас данная система была сконструирована таким образом, чтобы crРНК и tracrРНК можно было объединить в одной молекуле (“одиночной руководящей РНК”), а эквивалентная часть crРНК одиночной руководящей РНК может быть сконструирована таким образом, чтобы направлять нуклеазу Cas9 к любой целевой последовательности (см. Jineket al (2012) Science 337, p. 816-821, Jinek et al (2013) eLife 2:e00471, и David Segal, (2013) eLife 2:e00563). Таким образом, систему CRISPR/Cas можно сконструировать таким образом, чтобы создавать ДЦР в требуемом месте генома, а на репарацию ДЦР можно воздействовать при помощи ингибиторов репарации для того, чтобы вызвать увеличение неточности репарации.
[0119] Нуклеазы, такие как ZFN, TALEN и/или мегануклеазы также содержат нуклеазу (расщепляющий домен, расщепляющий полудомен). Как указывалось выше, расщепляющий домен может быть гетерологичным ДНК-связывающему домену, например, в случае ДНК-связывающего домена цинковых пальцев и расщепляющего домена из нуклеазы или ДНК-связывающего домена мегануклеазы и расщепляющего домена из другой нуклеазы. Гетерологичные расщепляющие домены можно получить из любой эндонуклеазы или экзонуклеазы. Примеры эндонуклеаз, из которых можно получить расщепляющие домены, включают, но не ограничиваются этим, рестрикционные эндонуклеазы хоминг-эндонуклеазы. См., например, каталог 2002-2003, New England Biolabs, Beverly, MA и Belfort et al. (1997) Nucleic Acids Res. 25:3379-3388. Известны также дополнительные ферменты, которые расщепляют ДНК (например, нуклеаза S1; нуклеаза золотистой фасоли; панкреатическая ДНаза I; микрококковая нуклеаза; дрожжевая НОэндонуклеаза; см. также Linn et al. (eds.) Nucleases, Cold Spring Harbor Laboratory Press, 1993). Один или более из этих ферментов (или их функциональных фрагментов) можно использовать в качестве источника расщепляющих доменов и расщепляющих полудоменов.
[0120] Аналогично, расщепляющий полудомен можно получить из любой нуклеазы или ее части, как установлено выше, для расщепляющей активности которой требуется димеризация. В общем случае, если слитые белки содержат расщепляющие полудомены, то для расщепления необходимы два слитых белка. В альтернативном варианте можно использовать один белок, содержащий два расщепляющих полудомена. Два расщепляющих полудомена могут быть получены из одной и той же эндонуклеазы (или ее функциональных фрагментов) либо каждый расщепляющий полудомен может быть получен из разных эндонуклеаз (или их функциональных фрагментов). Вдобавок, целевые участки для двух слитых белков предпочтительно расположены по отношению друг к другу таким образом, что связывание двух слитых белков с соответствующими целевыми участками приводит к такой взаимной пространственной ориентации расщепляющих полудоменов, которая создает возможность образования расщепляющими полудоменами функционального расщепляющего домена, например, путем димеризации. Таким образом, в некоторых вариантах реализации изобретения близлежащие края целевых участков разделены 5-8 нуклеотидами или 15-18 нуклеотидами. При этом между двумя целевыми участками может находиться любое соответствующее целому числу количество нуклеотидов или пар нуклеотидов (например, от 2 до 50 нуклеотидных пар или более). В общем случае участок расщепления находится между целевыми участками.
[0121] Рестрикционные эндонуклеазы (рестрикционные ферменты) присутствуют у многих видов и способны к специфическому к последовательности связыванию с ДНК (в участке узнавания) и расщеплению ДНК в участке или вблизи участка связывания. Определенные рестрикционные ферменты (например, типа IIS) расщепляют ДНК в участках, удаленных от участка узнавания, и имеют отделяемые связывающий и расщепляющий домены. Например, принадлежащий типу IIS фермент Fok I катализирует двухцепочечное расщепление ДНК на расстоянии 9 нуклеотидов от своего сайта узнавания на одной цепи и на расстоянии 13 нуклеотидов от своего сайта узнавания на другой. См., например, патенты США 5356802; 5436150 и 5487994; а также Li et al. (1992) Proc. Natl. Acad. Sci. USA 89:4275-4279; Li et al. (1993) Proc. Natl. Acad. Sci. USA 90:2764-2768; Kim et al. (1994a) Proc. Natl. Acad. Sci. USA 91:883-887; Kim et al. (1994b) J. Biol. Chem. 269:31,978-31,982. Таким образом, в одном варианте реализации изобретения слитые белки содержат расщепляющий домен (или расщепляющий полудомен) из по меньшей мере одного рестрикционного фермента, принадлежащего типу IIS, и один или более связывающих доменов цинковых пальцев, которые могут быть или не быть сконструированными.
[0122] Примером рестрикционного фермента типа IIS, чей расщепляющий домен отделим от связывающего домена, является Fok I. Этот конкретный фермент является активным в виде димера. Bitinaite et al. (1998) Proc. Natl. Acad. Sci. USA 95: 10,570-10,575. Соответственно, применительно к целям настоящего изобретения, та часть фермента Fok I, которая используется в описанных слитых белках, считается расщепляющим полудоменом. Таким образом, для нацеленного двухцепочечного расщепления и/или нацеленного замещения клеточных последовательностей при помощи продуктов слияния «цинковые пальцы»- или TALE-Fok I, можно использовать два слитых белка, каждый из которых содержит расщепляющий полудомен FokI, для того, чтобы воссоздать каталитически активный расщепляющий домен. В альтернативном варианте можно также использовать одну молекулу полипептида, содержащую связывающий домен цинковых пальцев и два расщепляющих полудомена Fok I. Параметры для нацеленного расщепления и нацеленного изменения последовательностей при помощи продуктов слияния «цинковые пальцы»- или TALE-Fok I приведены в другом месте данного текста.
[0123] Расщепляющий домен или расщепляющий полудомен могут являться любой частью белка, которая сохраняет расщепляющую активность или которая сохраняет способность мультимеризоваться (например, димеризоваться) для образования функционального расщепляющего домена.
[0124] Примеры рестрикционных ферментов типа IIS описаны в международной публикации WO 07/014275, в полном объеме включенной в данный текст посредством ссылки. Дополнительные рестрикционные ферменты также содержат отделяемые связывающий и расщепляющий домены, которые рассматриваются в настоящем изобретении. См., например, Roberts et al. (2003) Nucleic Acids Res. 31:418-420.
[0125] В определенных вариантах реализации изобретения расщепляющий домен содержит один или более сконструированных расщепляющих полудоменов (также называемых мутантными димеризационными доменами), которые минимизируют или предотвращают гомодимеризацию, как описано, например, в патентных публикациях США №20050064474 и 20060188987 и в заявке США №11/805850 (зарегистрированной 23 мая 2007 г.), описания которых в полном объеме включены в данный текст посредством ссылки. Аминокислотные остатки в позициях 446, 447, 479, 483, 484, 486, 487, 490, 491, 496, 498, 499, 500, 531, 534, 537 и 538 Fok I являются мишенями для оказания влияния на димеризацию расщепляющих полудоменов Fok I. Примеры сконструированных расщепляющих полудоменов Fok I, которые образуют облигатные гетеродимеры, включают пару, в которой первый расщепляющий полудомен содержит мутации в аминокислотных остатках в позициях 490 и 538 и второй расщепляющий полудомен Fok I содержит мутации в аминокислотных остатках 486 и 499.
[0126] Таким образом, в одном варианте реализации изобретения, мутация в 490 замещает Glu (E) на Lys (K); мутация в 538 замещает Iso (I) на Lys (K); мутация в 486 замещает Gln (Q) на Glu (E) и мутация в позиции 499 замещает Iso (I) на Lys (K). В частности, описанные в данном тексте сконструированные расщепляющие полудомены получали при помощи изменения позиций 490 (E→K) и 538 (I→K) в одном расщепляющем полудомене для того, чтобы получить сконструированный расщепляющий полудомен, обозначаемый “E490K:I538K”, и при помощи изменения позиций 486 (Q→E) и 499 (I→L) в другом расщепляющем полудомене для того, чтобы получить сконструированный расщепляющий полудомен, обозначаемый “Q486E:I499L”. Описанные в данном тексте сконструированные расщепляющие полудомены являются облигатными мутантными гетеродимерами, в которых аберрантное расщепление минимизировано или исключено. См., например, патентные публикации США №2008/0131962 и 2011/0201055, описания которых в полном объеме включены в данный текст посредством ссылки. В определенных вариантах реализации изобретения сконструированные расщепляющие полудомены содержат мутации в позициях 486, 499 и 496 (пронумерованные относительно FokI дикого типа), например, мутации которые замещают остаток Gln (Q) дикого типа в позиции 486 остатком Glu (E), остаток Iso (I) дикого типа в позиции 499 остатком Leu (L) и остаток Asn (N) дикого типа в позиции 496 остатком Asp (D) или остатком Glu (E) (также называемые соответственно доменами “ELD” и “ELE”). В других вариантах реализации изобретения сконструированные расщепляющие полудомены содержат мутации в позициях 490, 538 и 537 (пронумерованные относительно FokI дикого типа), например, мутации которые замещают остаток Glu (E) дикого типа в позиции 490 остатком Lys (K), остаток Iso (I) дикого типа в позиции 538 остатком Lys (K) и остаток His (H) дикого типа в позиции 537 остатком Lys (K) или остатком Arg (R) (также называемые соответственно доменами “KKK” и “KKR”). В других вариантах реализации изобретения сконструированные расщепляющие полудомены содержат мутации в позициях 490 и 537 (пронумерованные относительно FokI дикого типа), например, мутации которые замещают остаток Glu (E) дикого типа в позиции 490 остатком Lys (K) и остаток His (H) дикого типа в позиции 537 остатком Lys (K) или остатком Arg (R) (также называемые соответственно доменами “KIK” и “KIR”). (См. патентную публикацию США №20110201055).
[0127] Описанные в данном тексте сконструированные расщепляющие полудомены могут быть изготовлены при помощи любого подходящего способа, например, путем сайт-направленного мутагенеза расщепляющих полудоменов (Fok I) дикого типа, как описано в патентных публикациях США №20050064474 и 20080131962.
[0128] В альтернативном варианте нуклеазы могут быть собраны in vivo на целевом участке нуклеиновой кислоты при помощи так называемой технологии “расщепленных ферментов” (см., например, патентную публикацию США №20090068164). Компоненты таких расщепленных ферментов могут экспрессироваться на отдельных экспрессионных конструкциях или могут быть связаны в одной открытой рамке считывания, где разделены индивидуальные компоненты, например, при помощи саморасщепляющегося пептида 2А или последовательности УВПР. Компоненты могут представлять собой индивидуальные связывающие домены цинковых пальцев или связывающие нуклеиновую кислоту домены мегануклеазы.
[0129] В некоторых вариантах реализации изобретения ДНК-связывающий домен представляет собой сконструированный домен из TAL-эффектора, схожий с теми, которые были получены из фитопатогенов Xanthomonas (см. Boch et al, (2009) Science 326: 1509-1512 and Moscou and Bogdanove, (2009) Science326: 1501) и Ralstonia (см. Heuer et al (2007) Applied and Environmental Microbiology 73(13): 4379-4384). Также см. публикацию согласно РСТ WO 2010/079430.
[0130] Перед применением нуклеазы (например, ZFN или TALEN) можно проверить на предмет активности, например, в хромосомной системе на основе дрожжей, как описано в WO 2009/042163 и 20090068164. Нуклеазные экспрессионные конструкции можно легко сконструировать при помощи известных в данной области техники методов. См., например, патентные публикации США 20030232410; 20050208489; 20050026157; 20050064474; 20060188987; 20060063231 и международную публикацию WO 07/014275. Экспрессия нуклеазы может находиться под управлением конститутивного промотора или индуцибельного промотора, например, промотора галактокиназы, который активируется (де-репрессируется) в присутствии раффинозы и/или галактозы и репрессируется в присутствии глюкозы.
Доставка
[0131] Описанные в данном тексте белки (например, ZFP, TALE, CRISPR/Cas), кодирующие их полинуклеотиды и составы, содержащие белки и/или полинуклеотиды, могут быть доставлены в целевую клетку при помощи любых подходящих средств, включая, например, инъекцию белков ZFP-TF, TALE-TF или использование мРНК, кодирующей ZFN или TALEN. Подходящие клетки включают, но не ограничиваются этим, эукариотические и прокариотические клетки и/или клеточные линии. Неограничивающие примеры таких клеток или клеточных линий, полученных из таких клеток, включают клетки COS, CHO (например, CHO-S, CHO-K1, CHO-DG44, CHO-DUXB11, CHO-DUKX, CHOK1SV), VERO, MDCK, WI38, V79, B14AF28-G3, BHK, HaK, NS0, SP2/0-Ag14, HeLa, HEK293 (например, HEK293-F, HEK293-H, HEK293-T) и perC6, а также клетки насекомых, таких как Spodoptera fugiperda (Sf), или клетки грибов, таких как Saccharomyces, Pichia и Schizosaccharomyces. В определенных вариантах реализации изобретения клеточная линия является клеточной линией CHO-K1, MDCK или HEK293. Подходящие клетки также включают стволовые клетки, такие как, например, эмбриональные стволовые клетки, индуцированные полипотентные стволовые клетки, гематопоэтические стволовые клетки, нейрональные стволовые клетки и мезенхимальные стволовые клетки.
[0132] Способы доставки описанных в данном тексте белков, содержащих белки «цинковые пальцы», описаны, например, в патентах США №6453242; 6503717; 6534261; 6599692; 6607882; 6689558; 6824978; 6933113; 6979539; 7013219 и 7163824, описания которых в полном объеме включены в данный текст посредством ссылки.
[0133] Описанные в данном тексте белки «цинковые пальцы», TALE или CRISPR/Cas также можно доставлять при помощи векторов, содержащих последовательности, кодирующие один или более из белков «цинковые пальцы», TALE или CRISPR/Cas. Можно использовать любые векторные системы, включая, но не ограничиваясь этим, плазмидные векторы, ретровирусные векторы, лентивирусные векторы, аденовирусные векторы, поксвирусные векторы; герпесвирусные векторы и аденоассоциированные вирусные векторы и т.д. См. также патенты США №6534261; 6607882; 6824978; 6933113; 6979539; 7013219 и 7163824, в полном объеме включенные в данный текст посредством ссылки. Кроме того, стоит понимать, что любой из этих векторов может содержать одну или более последовательностей, кодирующих белок «цинковые пальцы» или TALE. Таким образом, когда в клетку вносят один или более белков ZFP, TALE или CRISPR/Cas, тот же самый вектор или отличные векторы могут нести последовательности, кодирующие белки ZFP, TALE или CRISPR/Cas. Когда используются множественные векторы, каждый из векторов может содержать последовательность, кодирующую одну из множественных систем ZFP, TALE или CRISPR/Cas.
[0134] Для внесения нуклеиновых кислот, кодирующих сконструированные системы ZFP, TALE или CRISPR/Cas, в клетки (например, клетки млекопитающих) и целевые ткани можно использовать традиционные вирусные и невирусные методы переноса генов. Такие методы также можно применять для введения нуклеиновых кислот, кодирующих системы ZFP, TALE или CRISPR/Cas, в клетки in vitro. В определенных вариантах реализации изобретения нуклеиновые кислоты, кодирующие системы ZFP, TALE или CRISPR/Cas, вводят в рамках in vivo или ex vivo генной терапии. Невирусные векторные системы доставки включают ДНК-плазмиды, оголенные нуклеиновые кислоты и нуклеиновые кислоты, комплексированные со средством доставки, таким как липосома или полоксамер. Вирусные векторные системы доставки включают ДНК и РНК вирусы, которые после доставки в клетку имеют эписомные или интегрированные геномы. Для ознакомления с процедурами генной терапии см. Anderson, Science 256:808-813 (1992); Nabel & Felgner, TIBTECH 11:211-217 (1993); Mitani & Caskey, TIBTECH 11:162-166 (1993); Dillon, TIBTECH 11:167-175 (1993); Miller, Nature 357:455-460 (1992); Van Brunt, Biotechnology 6(10):1149-1154 (1988); Vigne, Restorative Neurology and Neuroscience 8:35-36 (1995); Kremer & Perricaudet, British Medical Bulletin 51(1):31-44 (1995); Haddada et al., в Current Topics in Microbiology and Immunology Doerfler and Böhm (eds.) (1995) и Yu et al., Gene Therapy 1:13-26 (1994).
[0135] Способы невирусной доставки нуклеиновых кислот включают электропорацию, липофекцию, микроинъекцию, баллистическую трансфекцию, виросомы, липосомы, иммунолипосомы, поликатионы или конъюгаты липид:нуклеиновая кислота, оголенные ДНК, оголенные РНК, искусственные вирионы и агент-усиленное поглощение ДНК. Также для доставки нуклеиновых кислот можно использовать сонопорацию, например, при помощи системы Sonitron 2000 (Rich-Mar). В предпочтительном варианте реализации изобретения одну или более нуклеиновых кислот доставляют в виде мРНК. Также предпочтительным является применение кэпированных мРНК для повышения трансляционной эффективности и/или стабильности мРНК. В особенности предпочтительными являются кэпы ARCA (от англ. “anti-reverse cap analog”) или их варианты. См. патенты США US7074596 и US8153773, включенные в данный текст посредством ссылки.
[0136] Дополнительные примеры систем доставки нуклеиновой кислоты включают те, которые предоставляют Amaxa Biosystems (Кельн, Германия), Maxcyte, Inc. (Роквилль, Мэрилэнд), BTX Molecular Delivery Systems (Холлистон, Массачусетс) и Copernicus Therapeutics Inc, (см., например, US6008336). Липофекция описана, к примеру, в патентах США №5049386; 4946787 и 4897355, а реагенты для липофекции имеются в продаже (например, Трансфектам™, и Липофектин™, и Липофектамин™ RNAiMAX). Катионные и нейтральные липиды, которые подходят для эффективной рецептор-распознающей липофекции полинуклеотидов, включают те, что описаны у Felgner, WO 91/17424, WO 91/16024. Доставка может осуществляться в клетки (ex vivo введение) или в целевые ткани (in vivo введение).
[0137] Технологии изготовления комплексов липид:нуклеиновая кислота, включая нацеленные липосомы, такие как иммунолипидные комплексы, хорошо известны специалисту в данной области техники (см., например, Crystal, Science 270:404-410 (1995); Blaese et al., Cancer Gene Ther. 2:291-297 (1995); Behr et al., Bioconjugate Chem. 5:382-389 (1994); Remy et al., Bioconjugate Chem. 5:647-654 (1994); Gao et al., Gene Therapy 2:710-722 (1995); Ahmad et al., Cancer Res. 52:4817-4820 (1992); патенты США №4186183, 4217344, 4235871, 4261975, 4485054, 4501728, 4774085, 4837028 и 4946787).
[0138] Дополнительные способы доставки включают применение упаковки предназначенных для доставки нуклеиновых кислот в средства доставки EnGeneIC (EDV). Эти EDV доставляются прямо в целевую ткань при помощи биспецифических антител, в которых одно плечо антитела обладает специфичностью к целевой ткани, а другое обладает специфичностью к EDV. Антитело доставляет EDV к поверхности целевой клетки, а затем EDV попадает в клетку путем эндоцитоза. Внутри клетки содержимое EDV высвобождается (см. MacDiarmid et al (2009) Nature Biotechnology 27(7):643).
[0139] Использование систем на основе вирусных РНК или ДНК для доставки нуклеиновых кислот, кодирующих сконструированные системы ZFP, TALE или CRISPR/Cas, имеет преимущество, состоящее в высокоразвитых процессах нацеливания вируса на определенные клетки тела и переноса нагрузки вируса в ядро. Вирусные векторы можно вводить пациенту прямым способом (in vivo) либо можно применять их для того, чтобы обрабатывать клетки in vitro, а модифицированные клетки вводить пациентам (ex vivo). Стандартные системы на вирусной основе для доставки систем ZFP, TALE или CRISPR/Cas включают, но не ограничиваются этим, ретровирусные, лентивирусные, аденовирусные, адено-ассоциированные векторы для переноса генов и векторы на основе вируса осповакцины и вируса простого герпеса. Интеграция в геном хозяина возможна в случае применения методов переноса генов, в которых используются ретровирусы, лентивирусы и адено-ассоциированные вирусы, которые часто приводят к долгосрочной экспрессии внесенного трансгена. Вдобавок, для многих различных типов клеток и целевых тканей наблюдали высокие эффективности трансдукции.
[0140] Тропизм ретровируса можно изменить путем внесения чужеродных оболочечных белков, расширяя потенциальную целевую популяцию целевых клеток. Лентивирусные векторы представляют собой ретровирусные векторы, которые способны преобразовывать или инфицировать неделящиеся клетки и для которых обычно наблюдают высокие вирусные титры. Выбор ретровирусной системы переноса генов зависит от целевой ткани. Ретровирусные векторы состоят из цис-действующих длинных концевых повторов с пакующей емкостью до 6-10 т.п.н. чужеродной последовательности. Минимума цис-действующих ДКП достаточно для репликации и упаковки векторов, которые затем используют для интеграции терапевтического гена в целевую клетку для обеспечения постоянной трансгенной экспрессии. Широко применяемые ретровирусные векторы включают векторы на основе вируса лейкоза мышей (MuLV), вируса лейкоза гиббонов (GaLV), вируса иммунодефицита обезьян (ВИО), вируса иммунодефицита человека (ВИЧ) и их комбинаций (см., например, Buchscher et al., J. Virol. 66:2731-2739 (1992); Johann et al., J. Virol. 66:1635-1640 (1992); Sommerfelt et al., Virol. 176:58-59 (1990); Wilson et al., J. Virol. 63:2374-2378 (1989); Miller et al., J. Virol. 65:2220-2224 (1991); PCT/US 94/05700).
[0141] В применениях, где временная экспрессия является предпочтительной, можно использовать системы на основе аденовирусов. Векторы на основе аденовирусов способны к высокоэффективной трансдукции во многих типах клеток и не требуют клеточного деления. С такими векторами были получены высокие титры и высокие уровни экспрессии. Такой вектор можно получить в большом количестве относительно простых систем. Аденоассоциированные вирусные (“ААВ”) векторы также используются для трансдукции клеток целевыми нуклеиновыми кислотами, например, при in vitro выработке нуклеиновых кислот и пептидов и для in vivo и ex vivo процедур генной (см., например, West et al., Virology 160:38-47 (1987); патент США №4797368; WO 93/24641; Kotin, Human Gene Therapy 5:793-801 (1994); Muzyczka, J. Clin. Invest. 94:1351 (1994). Конструкции рекомбинантных ААВ векторов описаны в большом количестве публикаций, включая патент США №5173414; Tratschin et al., Mol. Cell. Biol. 5:3251-3260 (1985); Tratschin, et al., Mol. Cell. Biol. 4:2072-2081 (1984); Hermonat & Muzyczka, PNAS 81:6466-6470 (1984) и Samulski et al., J. Virol. 63:03822-3828 (1989).
[0142] В настоящее время в клинических испытаниях применяют по меньшей мере шесть методов переноса генов на основе вирусных векторов, которые включают методы, которые используют комплементацию дефектных векторов генами, внесенными в клеточные линии хелперов, для получения трансдуцирующего агента.
[0143] Примерами ретровирусных векторов, которые применяли в клинических испытаниях, являются pLASN и MFG-S (Dunbar et al., Blood 85:3048-305 (1995); Kohn et al., Nat. Med. 1:1017-102 (1995); Malech et al., PNAS 94:22 12133-12138 (1997)). PA317/pLASN был первым терапевтическим вектором, который применяли в испытаниях генной терапии. (Blaese et al., Science 270:475-480 (1995)). Для упакованных векторов MFG-S наблюдали эффективность трансдукции в 50% и выше. (Ellem et al., Immunol Immunother. 44(1):10-20 (1997); Dranoff et al., Hum. Gene Ther. 1:111-2 (1997).
[0144] Рекомбинантные аденоассоциированные вирусные векторы (рААВ) на основе дефектного и непатогенного парвовируса аденоассоциированного вирусного типа 2 являются перспективными альтернативными системами генной доставки. Все векторы получены из плазмиды, в которой сохранены только ААВ инвертированные концевые повторы в 145 п.н.о., фланкирующие трансгенную экспрессионную кассету. Ключевыми характеристиками данной векторной системы являются эффективный перенос генов и стабильная трансгенная доставка благодаря интеграции в геном трансдуцированной клетки. (Wagner et al., Lancet 351:9117 1702-3 (1998), Kearns et al., Gene Ther. 9:748-55 (1996)). Также, согласно настоящему изобретению, можно использовать другие ААВ серотипы, включая AAB1, AAB3, AAB4, AAB5, AAB6, AAB8, AAB 8.2, AAB9 и AAB rh10 и псевдотипированные AAB, такие как AAB2/8, AAB2/5 и AAB2/6.
[0145] Рекомбинантные аденовирусные (Ад) векторы, дефектные по репликации, можно получить при высоких титрах и они легко инфицируют большое количество разных типов клеток. Большинство аденовирусных векторов сконструированы таким образом, чтобы трансген замещал аденовирусные гены E1a, E1b и/или E3; впоследствии вектор, дефектный по репликации, размножается в человеческих клетках 293, которые возмещают функцию удаленного гена в трансформе. Аденовирусные векторы могут трансдуцировать многие типы тканей in vivo, включая неделящиеся, дифференцировавшие клетки, такие как те, что находятся в печени, почках и мышцах. Стандартные аденовирусные векторы обладают большой переносимой емкостью. Один из примеров применения аденовирусного вектора в клиническом исследовании включал терапию полинуклеотидами путем внутримышечной инъекции для противоопухолевой иммунизации (Sterman et al., Hum. Gene Ther. 7:1083-9 (1998)). Дополнительные примеры применения аденовирусных векторов для переноса генов в клинических исследованиях включают Rosenecker et al., Infection 24:1 5-10 (1996); Sterman et al., Hum. Gene Ther. 9:7 1083-1089 (1998); Welsh et al., Hum. Gene Ther. 2:205-18 (1995); Alvarez et al., Hum. Gene Ther. 5:597-613 (1997); Topf et al., Gene Ther. 5:507-513 (1998); Sterman et al., Hum. Gene Ther. 7:1083-1089 (1998).
[0146] Пакующие клетки применяют для образования вирусных частиц, которые способны инфицировать клетку-хозяина. Такие клетки включают клетки 293, которые пакуют аденовирусы, и клетки ψ2 или клетки PA317, которые пакуют ретровирусы. Применяемые в генной терапии вирусные векторы обычно получают при помощи продуцирующей клеточной линии, которая пакует нуклеотидный вектор в вирусную частицу. Как правило, векторы содержат минимальную вирусную последовательность, необходимую для упаковки и последующей интеграции в организм хозяина (в случае применения), при этом другие вирусные последовательности заменены экспрессионной кассетой, кодирующей белок, который необходимо экспрессировать. Недостающие вирусные функции возмещаются в транс-форме пакующей клеточной линией. Например, ААВ векторы, применяемые в генной терапии, имеют, как правило, только последовательности инвертированных концевых повторов (ИКП) из ААВ генома, которые необходимы для упаковки и интеграции в геном организма-хозяина. Вирусная ДНК упакована в клеточную линию, которая содержит хелперную плазмиду, кодирующую другие ААВ гены, а именно, rep и cap, но в которой отсутствуют последовательности ИКП. Клеточная линия также инфицирована аденовирусом в качестве хелпера. Хелперный вирус стимулирует репликацию ААВ вектора и экспрессию ААВ генов из хелперной плазмиды. Хелперные плазмиды не пакуются в значительных количествах из-за отсутствия последовательностей ИКП. Заражение аденовирусом может быть снижено, например, тепловой обработкой, к которой аденовирус более чувствителен, чем ААВ.
[0147] Для многих применений генной терапии желательно, чтобы вектор генной терапии доставлялся с высокой степенью специфичности в конкретный тип ткани. Соответственно, вирусный вектор может быть модифицирован, чтобы обладать специфичностью к заданному типу клеток, путем экспрессии лиганда в виде слитого белка с белком вирусной оболочки на внешней поверхности вируса. Лиганд выбирают так, чтобы он обладал аффинностью к рецептору, который присутствует в представляющем интерес типе клеток. Например, в Han et al., Proc. Natl. Acad. Sci. USA 92:9747-9751 (1995) сообщается, что вирус мышиного лейкоза Молони можно модифицировать так, чтобы он экспрессировал человеческий херегулин, сшитый с gp70, а рекомбинантный вирус инфицировал определенные клетки рака груди человека, экспрессирующие человеческий рецептор эпидермального фактора роста. Этот принцип может быть распространен на другие пары вирус-клетка-мишень, в которых клетка-мишень экспрессирует рецептор, а вирус экспрессирует слитый белок, содержащий лиганд к рецептору клеточной поверхности. Например, нитевидный фаг можно сконструировать так, чтобы он обнаруживал фрагменты антитела (например, FAB или Fv), обладающие специфической аффинностью связывания к практически любому выбранному клеточному рецептору. Хотя вышеприведенное описание относится преимущественно к вирусным векторам, те же самые принципы применимы и к невирусным векторам. Такие векторы могут быть сконструированы так, чтобы содержать определенные последовательности поглощения, которые способствуют поглощению определенными целевыми клетками.
[0148] Векторы для генной терапии могут быть доставлены in vivo путем введения отдельному пациенту, как правило, путем систематического введения (например, внутривенной, внутрибрюшинной, внутримышечной, подкожной или внутричерепной инфузии) или местного применения, как описано ниже. В альтернативном варианте векторы могут быть доставлены в клетки ex vivo, к примеру, в клетки, эксплантированные у отдельного пациента (например, лимфоциты, пунктаты костного мозга, образцы биопсии тканей), или универсальные донорные гематопоэтические стволовые клетки, с последующей реимплантацией клеток пациенту, как правило, после отбора клеток, в которые был инкорпорирован вектор.
[0149] Ex vivo трансфекция клеток для диагностики, исследований или генной терапии (например, путем повторной инфузии трансфицированных клеток в организм-хозяин) хорошо известна специалистам в данной области техники. В предпочтительном варианте реализации изобретения клетки выделены из организма испытуемого, трансфицированы нуклеиновой кислотой системы ZFP, TALE или CRISPR/Cas (ген кДНК или мРНК) и повторно введены обратно в организм испытуемого (например, пациента). В предпочтительном варианте реализации изобретения одну или более нуклеиновых кислот доставляют в виде мРНК. Также предпочтительным является применение кэпированных мРНК для повышения трансляционной эффективности и/или стабильности мРНК. В особенности предпочтительными являются кэпы ARCA (от англ. “anti-reverse cap analog”) или их варианты. См. патенты США 7074596 и 8153773, в полном объеме включенные в данный текст посредством ссылки. Большое количество клеток, подходящих для ex vivo трансфекции, известно специалистам в данной области техники (см., например, Freshney et al., Culture of Animal Cells, A Manual of Basic Technique (3rd ed. 1994) и перечисленные в работе ссылки, в которых обсуждается, как выделять и культивировать клетки, полученные от пациентов).
[0150] В одном варианте реализации изобретения в ex vivo способах трансфекции клеток и генной терапии используют стволовые клетки. Преимуществом применения стволовых клеток является то, что они могут дифференцироваться в другие типы клеток in vitro или могут быть внесены в организм млекопитающего (такой, как донор клеток), где они внедрятся в костный мозг. Известны способы дифференцирования клеток CD34+ in vitro в клинически важные типы иммунных клеток с использованием цитокинов, таких как GM-CSF, IFN-γ и TNF-α (см. Inaba et al., J. Exp. Med. 176:1693-1702 (1992)).
[0151] Стволовые клетки выделяют для трансдукции и дифференцирования при помощи известных методов. Например, стволовые клетки выделяют из клеток костного мозга путем пэннинга клеток костного мозга антителами, которые связывают нежелательные клетки, такие как CD4+ и CD8+ (T-клетки), CD45+ (panB-клетки), GR-1 (гранулоциты) и Iad (дифференцированные антиген-презентирующие клетки) (см. Inaba et al., J. Exp. Med. 176:1693-1702 (1992)).
[0152] В некоторых вариантах реализации изобретения также могут применяться стволовые клетки, которые были модифицированы. Например, нейрональные стволовые клетки, которым была привита устойчивость к апоптозу, могут применяться в качестве терапевтических составов, в которых стволовые клетки содержат также ZFP TF, являющиеся объектами данного изобретения. Устойчивость к апоптозу может быть получена, например, путем инактивации BAX и/или BAK при помощи BAX- или BAK-специфических TALEN или ZFN (см. патентную публикацию США № 20100003756) в стволовых клетках или тех, которые были разрушены каспазой, снова при помощи специфических к каспазе-6 ZFN, к примеру. Эти клетки можно трансфицировать ZFP TF или TALE TF, которые, как известно, регулируют мутантный Htt и Htt дикого типа.
[0153] Векторы (например, ретровирусы, аденовирусы, липосомы и т.д.), содержащие терапевтические нуклеиновые кислоты ZFP, можно вводить прямо в организм для трансдукции клеток in vivo. В альтернативном варианте можно вводить оголенную ДНК. Введение проводят любым из способов, которые обычно используются для приведения молекулы в максимальный контакт с кровью или клетками ткани, включая, но не ограничиваясь этим, инъекцию, инфузию, местное применение и электропорацию. Подходящие способы введения таких нуклеиновых кислот доступны и хорошо известны специалистам в данной области техники и, хотя для введения конкретного состава можно использовать более одного способа, конкретный способ часто может обеспечить незамедлительную и более эффективную реакцию, чем другой способ.
[0154] Способы внесения ДНК в гематопоэтические стволовые клетки раскрыты, например, в патенте США №5928638. Векторы, подходящие для внесения трансгенов в гематопоэтические стволовые клетки, например, клетки CD34+, включают аденовирус типа 35.
[0155] Векторы, подходящие для внесения трансгенов в иммунные клетки (например, Т-клетки), включают неинтегрирующиеся лентивирусные векторы. См., например, Ory et al. (1996) Proc. Natl. Acad. Sci. USA 93:11382-11388; Dull et al. (1998) J. Virol. 72:8463-8471; Zuffery et al. (1998) J. Virol. 72:9873-9880; Follenzi et al. (2000) Nature Genetics 25:217-222.
[0156] Фармацевтически приемлемые носители частично определяются конкретным вводимым составом, а также конкретным способом, который применяется для введения состава. Соответственно, существует большое количество подходящих формулировок фармацевтических составов (см., например, Remington’s Pharmaceutical Sciences, 17th ed., 1989).
[0157] Как указано выше, раскрытые способы и составы можно применять в любых типах клеток, включая, но не ограничиваясь, прокариотические клетки, клетки грибов, клетки простейших, клетки растений, клетки насекомых, клетки животных, клетки позвоночных, клетки млекопитающих и клетки человека. Клеточные линии, подходящие для экспрессии белка, известны специалистам в данной области техники и включают, но не ограничиваются этим, COS, CHO (например, CHO-S, CHO-K1, CHO-DG44, CHO-DUXB11), VERO, MDCK, WI38, V79, B14AF28-G3, BHK, HaK, NS0, SP2/0-Ag14, HeLa, HEK293 (например, HEK293-F, HEK293-H, HEK293-T), perC6, клетки насекомых, такие как Spodoptera fugiperda (Sf), и клетки грибов, такие как Saccharomyces, Pischia и Schizosaccharomyces. Также можно использовать потомство, варианты и производные этих клеточных линий.
Применения
[0158] Раскрытые составы и способы можно использовать для любого применения, в котором необходимо модулировать аллель Htt, включая, но не ограничиваясь этим, терапевтические и исследовательские применения.
[0159] Болезни и болезненные состояния, при которых в качестве терапевтических средств можно применять ZFP TF или TALE TF, подавляющие Htt, включают, но не ограничиваются этим, болезнь Хантингтона. Вдобавок, способы и составы, содержащие ZFN или TALEN, специфичные к мутантным аллелям Htt, можно применять в качестве терапевтического средства для лечения болезни Хантингтона.
[0160] ZFP-TF или TALE-TF, которые подавляют БХ аллель Htt также можно использовать совместно с ZFP-TF или TALE-TF, которые активируют нейротрофические факторы, включая, но не ограничиваясь этим, НФГК и НФГМ. Данные ZFN или TALEN (или полинуклеотиды, кодирующие данные ZFN или TALEN) можно вводить одновременно (например, в одних фармацевтических составах) либо можно вводить последовательно в любом порядке.
[0161] Способы и составы для лечения болезни Хантингтона также включают составы, содержащие стволовые клетки, в которых мутантная копия аллеля Htt в стволовых клетках была модифицирована в аллель Htt дикого типа при помощи специфичных к Htt ZFN или TALEN.
[0162] Способы и составы данного изобретения также применимы для проектирования и реализации in vitro и in vivo моделей, например, животных моделей нарушений тринуклеотидных повторов, которые дают возможность для изучения данных нарушений. Неограничивающие примеры подходящих in vitro моделей включают клетки или клеточные линии, полученные из любого организма, включая фибробласты. Неограничивающие примеры подходящих животных для использования в качестве животных моделей включают беспозвоночных (C. elegans, drosophila), грызунов (например, крыс или мышей), приматов (например, любых приматов за исключением человека).
ПРИМЕРЫ
Пример 1: Проектирование и конструирование нацеленных на Htt транскрипционных факторов белков «цинковые пальцы» (ZFP-TF) и ZFN
[0163] Нацеленные на Htt белки «цинковые пальцы» конструировали главным образом так, как описано в патенте США № 6534261. В таблицах 1А и 1Б приведены примеры спиралей узнавания ДНК-связывающих доменов Htt-нацеленных ZFP, а в таблицах 2А и 2Б приведены целевые последовательности данных ZFP.
[0164] Для нацеливания на участки, полностью находящиеся в пределах области повторов CAG, проектировали ZFP с одним прилегающим комплектом цинковых пальцев (фигура 1Б). Такие ZFP могут связывать более протяженные, мутантные тракты с более высокой аффинностью и/или с большим охватом, чем достигается избирательное подавление мутантного аллеля. Также проектировали ZFP, которые нацелены на участки, которые располагаются частично или полностью снаружи CAG-области (фигура 1А), и которые, следовательно, будут одинаково связываться с аллелями дикого типа и мутантными аллелями и регулировать экспрессию из обоих аллелей с одинаковой эффективностью. При проектировании белков «цинковые пальцы» для узнавания области CAG можно использовать группу модулей, содержащих один или два цинковых пальца, в подбираемых комбинациях. Эти модули приведены ниже в таблице 2В.
[0165] Мультимеризующиеся ZFP TF конструировали тем же описанным выше способом за исключением того, что вектор также содержит последовательности, кодирующие один или более доменов белкового взаимодействия (также называемых димеризационными доменами или доменами белкового взаимодействия), которые делают возможной мультимеризацию экспрессируемого белка вдоль тракта тринуклеотидных повторов, которые функционально связаны с последовательностями, кодирующими ZFP TF. См. фигуру 1Г и фигуру 10. В таблице 3 приведены конструкции димеризационных доменов, которые применяют вместе с нацеленными на область CAG-повторов ZFP. На фигуре 10В приведены белковые последовательности четырех мономерных каркасов ZFP, которые сконструированы так, чтобы мультимеризоваться через взаимодействия между димеризующимися цинковыми пальцами (DZ), DZ1-DZ4. Основой для конструкций служит работа, описанная в Mol. Syst. Biol. (2006) 2:2006.2011. На фигуре 10Г приведены белковые последовательности семи мономерных каркасов ZFP, которые сконструированы так, чтобы мультимеризоваться через взаимодействия между суперспиралями (СС), CC1-CC7. Основой для конструкции CC#1 служит работа, описанная в (J. Am. Chem. Soc. (2001), 123:3151-3152), а CC#2, CC#3 и CC#4 основаны на (J. Am. Chem. Soc. (2000), 122:5658-5659). Домены СС и DZ обеспечивают возможность полимеризации ZFP в пределах основной бороздки CAG-тракта (проиллюстрировано на фигуре 10Б). При выборе комплекта пальцев и димеризационных доменов с соответствующими связывающими свойствами эффективное связывание будет проявляться только в случае удлиненного CAG-тракта аллеля, указывающего на заболевание.
[0166] ZFP-TF конструировали в виде слитых белков, содержащих последовательность ядерной локализации, сконструированный ДНК-связывающий домен цинковых пальцев (таблицы 1А и 1Б), нацеленный на аллель Htt, и репрессионный домен KRAB из человеческого белка KOX1. См. фиг. 1А, 1Б и 1Г. Сконструированные ДНК-связывающие домены содержат 3-6 пальцевых модулей, распознающих последовательности в 9-18 п.н.о. (таблицы 2А и 2Б). Нуклеотиды на целевых участках, которые контактируют со спиралями узнавания ZFP, обозначены большими буквами; неконтактирующие нуклеотиды обозначены малыми буквами. Также были сконструированы молекулы ZFP-ZFP-TF, в которых два ДНК-связывающих домена ZFP сшивали гибким линкером, а затем сшивали с репрессионным доменом KRAB (фигура 1Д). ДНК-связывающие домены выбирали из таблиц 2А и 2Б.
Пример 2: Репрессия двух аллелей Htt в человеческих и мышиных клетках
[0167] Для репрессии обоих аллелей Htt (не аллель-специфичных) конструировали ZFP таким образом, чтобы они связывались с промотором Htt и участком экзона 1, при этом целевой участок не полностью находился в пределах CAG-повтора. См. фигуру 1А. Для исследования активности ZFP TF, репрессирующих Htt, ZFP TF трансфицировали в клетки человека, а экспрессию Htt отслеживали при помощи ОТ-ПЦР в режиме реального времени.
[0168] Человеческие клетки HEK293 (Graham et al (1977) J Gen Virol 36:59-74) культивировали в DMEM с добавлением 10% ФБС, а 1e5 клеток трансфицировали 1 мкг плазмидной ДНК, кодирующей указанные продукты слияния ZFP-KOX от Amaxa Nucleofector®, следуя инструкциям производителя.
[0169] Трансфицированные клетки инкубировали на протяжении 2 дней, а уровни эндогенного человеческого хантингтина (Htt) и нормировочного контрольного бета-актина (ACTB) анализировали при помощи ПЦР в режиме реального времени с применением, соответственно, праймеров и зондов Hs00918176_m1 и 4352935E (Applied Biosystems) согласно стандартным протоколам. Уровни Htt выражали через соотношение Htt/ACTB, нормированное на величину, соответствующую ложнотрансфицированным образцам (обозначенную как 1).
[0170] Как показано на фигуре 2А, нацеленные на Htt ZFP подавляли экспрессию Htt. Для того чтобы подтвердить снижение уровней белка Htt, проводили вестерн-блоттинг анализ (фигура 2В) в соответствии со стандартными протоколами; в качестве загрузочного контроля использовали белок р65.
[0171] Мышиные Htt-специфические репрессоры ZFP TF временно трансфицировали в клетки Neuro2A (Klebe & Ruddle (1969) J. Cell Biol. 43: 69A) при помощи набора реагентов Lipofectamine® 2000 (Invitrogen) в соответствии с протоколами производителя. Через 48 часов после трансфекции измеряли уровни мHtt и ACTB мРНК при помощи группы праймеров/зондов ABI Taqman®, соответственно, Mm01213820 m1 и 4352933E. Соотношение мHtt/ACTB для трансфицированных ZFP образцов нормировали относительно контрольного ЗФБ (обозначенного как 1).
[0172] Как показано на фигуре 2В, ZFP подавляли экспрессию Htt. Вдобавок, мышиные Htt-специфические репрессоры ZFP-TF могут репрессировать мышиный Htt в иммортализованных стриарных клетках STHdh(Q111/Q7), полученных от мышей с Htt-нокином (Trettel et al. (2000) Hum. Mol. Genet 9: 2799-2809). См. фигуры 2Г и 2Д. мРНК для указанных ZFP создавали при помощи набора инструментов mMessage mMachine (Ambion), после чего проводили трансфекцию 0,1, 0,5 или 2 мкг этих мРНК при помощи нуклеофектора Amaxa, как описано выше. Клетки собирали через 48 часов после трансфекции для проведения анализа экспрессии мHtt и ACTB, как описано выше. Более существенную репрессию по сравнению с клетками Neuro2A наблюдали в стриарных клетках, что может являться результатом повышенной эффективности трансфекции, которая достигается путем трансфекции мРНК в стриарные клетки.
Пример 3: Избирательная репрессия мутантного Htt в человеческих и мышиных клетках
[0173] Для того чтобы достичь избирательной репрессии мутантного аллеля Htt, конструировали ZFP таким образом, чтобы связывание происходило в пределах CAG-повтора. На фигуре 1Б показан один из типов таких ZFP с прилегающим комплектом цинковых пальцев, связанным с репрессионным доменом (например, доменом KRAB из KOX1); такие ZFP можно спроектировать так, чтобы они обладали соответствующей аффинностью для того, чтобы порог занятости, необходимый для транскрипционной репрессии, мог быть установлен только в случае удлиненных CAG-повторов. На фигурах 1В, 1Г и 1Д приведены три примера других конструкций ZFP, который могут обеспечивать специфическое связывание с удлиненными CAG-повторами.
[0174] ZFP, сконструированные так, как показано на фигуре 1Б, вносили в клетки HEK293 и оценивали экспрессию Htt. Конструкции, кодирующие ZFP, трансфицировали в клетки HEK293, используя FugeneHD и придерживаясь стандартных протоколов. Через семьдесят два часа после трансфекции выделяли всю РНК и оценивали уровни человеческого хантингтина (Htt) относительно внутреннего контрольного бета-актина (ACTB) при помощи ПЦР в режиме реального времени с применением, соответственно, праймеров и зондов Hs00918176_m1 и 4352935E (Applied Biosystems). Соотношение Htt/ACTB для трансфицированных ZFP образцов нормировали относительно значений для контрольного ЗФБ (обозначенного как 1).
[0175] Как показано на фигуре 3А, репрессоры ZFP (сшитые с репрессионным доменом KRAB), сконструированные для связывания CAG-повторов (фигура 1Б) как в верхней, так и в нижней цепи, в клетках HEK293, эффективно подавляли экспрессию Htt. на фигуре 3А показаны репрессоры транскрипции, при этом экспрессию измеряли для двойных трансфекций (отдельные столбики на фигуре) и проводили множественный ПЦР-анализ в режиме реального времени (“усы” погрешностей). Разные уровни репрессии отдельными ZFP позволяют предположить, что они обладают разной аффинностью к участку CAG-повторов. Так как аллели Htt в клетках HEK293 содержат 16 и 17 CAG, данный результат также позволяет предположить, что “более слабые” ZFP, такие как 30640, эффективно не подавляют аллели Htt с длиной CAG-повторов, соответствующей дикому типу (неудлиненному).
[0176] Для того чтобы исследовать, могут ли ZFP, такие как 30640, подавлять транскрипцию аллелей Htt с удлиненными CAG-повторами, сконструировали люциферазные репортеры, управляемые фрагментом промотор/экзон 1 Htt, который содержит CAG-повторы разной длины. Сначала фрагмент промотор/экзон 1 Htt человека амплифицировали из геномной ДНК HEK293, используя прямой праймер:
5’ GAAGATCTCACTTGGGGTCCTCAGGTCGTGCCGAC (SEQ ID NO:139)
и обратный праймер:
5’ GTATCCAAGCTTCAGCTTTTCCAGGGTCGCCTAGGCGGTCT. (SEQ ID NO:140).
[0177] Прямой праймер вносит участок BglII, обратный праймер меняет первый ATG, принадлежащий Htt, на TAG и создает участок AvrII, а также содержит участок HindIII. Продукт ПЦР расщепляли BglII и HindIII и лигировали к вектору pRL-TK (Promega), который расщеплялся тем же самым ферментом, чтобы получить конструкцию pRL-Htt. Затем фрагмент экзона 1 Htt человека (кодирующая последовательность минус первый ATG) амплифицировали из геномной ДНК HEK293 или геномной ДНК пациентов с БХ с удлиненными повторами CAG, используя прямой праймер:
5’ GCCTAGGCGACCCTGGAAAAGCTGATGAAGGCC (SEQ ID NO:141)
и обратный праймер: 5’
5’ GTATCCAAGCTTGAGCTGCAGCGGGCCCAAACTCACG (SEQ ID NO:142).
[0178] Прямой праймер вносит участок AvrII, обратный праймер вносит участок HindIII. Продукт ПЦР расщепляли AvrII и HindIII и лигировали к вектору pRL-Htt, который расщеплялся теми же самыми ферментами. Клоны, содержащие 10, 17, 23 или 47 CAG-повторов (pRL-Htt-CAG(x)) идентифицировали путем секвенирования.
[0179] Репортеры pRL-Htt-CAG(x) (300 нг) и репортер с промотором pGL3 (100 нг, используемый в качестве контроля для нормировки, Progema) трансфицировали в клетки HEK293 с или без 100 нг экспрессионного вектора ZFP 30640. Активности люциферазы светляков (репортер pGL) и Renilla (репортер pRL) измеряли через 24 часа после трансфекции. Уровни люциферазы Renilla нормировали относительно уровней люциферазы светляков из одного трансфицированного образца и дополнительно нормировали относительно соотношения Renilla/светляки образца, содержащего только репортеры.
[0180] Как показано на фигуре 3Б, репрессия репортеров люциферазы ZFP-TF 30640 возрастает с увеличение длины CAG-повторов, что позволяет предположить, что ZFP с ДНК-связывающими аффинностями, схожими с 30640, могут подавлять активность промотора Htt через удлиненные CAG-повторы, а уровень репрессии зависит от длины CAG-повторов.
[0181] На фигуре 3В приведен эксперимент, аналогичный приведенному на фигуре 3Б, за исключением того, что в нем исследовали также “сильный” ZFP-TF 30657, и 30640 и 30657 исследовали, как указано, при разных дозировках. При каждом уровне дозирования 30640 демонстрировал более сильное подавление репортера pRL-Htt-CAG47 по сравнению с репортером pRL-Htt-CAG23 (зависимое от длины CAG-повторов подавление), в то время как 30657 подавлял оба репортера до одинаковых уровней. В экспериментах с репортером pRL-Htt-CAG23 30640 демонстрировал при каждом уровне дозирования менее сильное подавление, чем 30657, повторяя разницу в их воздействии на эндогенный аллель Htt с нормальной длиной CAG-повторов (клетки HEK293, Фигура 3А); но с репортером pRL-Htt-CAG47 30640 и 30657 демонстрировали одинаковое подавление при каждом уровне дозирования, что позволяет предположить, что “более слабые” ZFP, такие как 30640, могут эффективно подавлять промотор Htt с удлиненным участком CAG-повторов, скорее всего потому, что только удлиненная CAG-мишень может обеспечить порог занятости, необходимый для начала репрессии такими ZFP.
[0182] На фигуре 3Г показано, что ZFP-TF 30640 и 30657 (сшитые с репрессионным доменом KRAB из KOX1) могут репрессировать нокин-аллель Htt (CAG111) в иммортализованных стриарных клетках, полученных из Hdh(Q111/Q7) нокин-мышей, демонстрируя, что ZFP, такие как 30640, которые осуществляют зависимую от длины CAG-повторов репрессию репортеров люциферазы, также могут подавлять экспрессию из эндогенного аллеля Htt, который и привел к удлинению участке CAG-повторов. мРНК для указанных ZFP создавали при помощи набора инструментов mMessage mMachine (Ambion) и трансфицировали клетки Hdh(Q111/Q7) в указанных дозировках при помощи нуклеофектора Amaxa. Для определения экспрессии из аллеля Htt дикого типа при проведении ОТ-ПЦР в режиме реального времени использовали прямой праймер CAGGTCCGGCAGAGGAACC (SEQ ID NO:193) и обратный праймер TTCACACGGTCTTTCTTGGTGG (SEQ ID NO:194); для определения экспрессии из нокин-аллеля Htt использовали прямой праймер GCCCGGCTGTGGCTGA (SEQ ID NO:195) и обратный праймер TTCACACGGTCTTTCTTGGTGG (SEQ ID NO:196).
[0183] На фигуре 3Д приведены результаты исследования ZFP-TF 30640 и 30657 в линии фибробластов, полученной от пациента с БХ (GM21756, Coriell), которые содержат 15 и 70 CAG-повторов в, соответственно, нормальном и мутантном аллелях Htt. Сначала проводили аллель-специфический ПЦР-анализ на основе ОНП в режиме реального времени для специфического определения экспрессии из аллеля Htt дикого типа или мутантного. Фазирование ОНП (rs363099 T/C) определяли по Carroll et al. (Mol Ther. (2011) 19:2178-85); “T” обозначает нормальный аллель, а “C” обозначает мутантный аллель. Для того чтобы определить экспрессию из мутантного аллеля (099С), кДНК из фибробластов амплифицировали при помощи ПЦР в режиме реального времени (SsoFast EvaGreen Supermix, Bio-Rad), используя прямой праймер 099C.F (5’AGTTTGGAGGGTTTCTC, SEQ ID NO:143) и обратный праймер 099.R5 (5’ TCGACTAAAGCAGGATTTCAGG, SEQ ID NO:144); температура отжига/удлинения составляла 55,6°C. Для того чтобы определить экспрессию из аллеля дикого типа (099T), проводили ПЦР кДНК фибробластов в режиме реального времени, используя прямой праймер 099T.F (5’AGTTTGGAGGGTTTCTT, SEQ ID NO:145), обратный праймер 099.R5 и 3’фосфорилированный блокатор олиго 099T.BL (5’AGGGTTTCTCCGCTCAGC-3’фос, SEQ ID NO:146); температура отжига/удлинения составляла 58,3°C. Уровни общего человеческого хантингтина (чHtt, аллели дикого типа и мутантные) и уровни нормировочного контрольного бета-актина (ACTB) анализировали при помощи ПЦР в режиме реального времени с применением, соответственно, праймеров/зондов Hs00918176_m1 и 4352935E (Applied Biosystems). В экспериментах, показанных на фигуре 3Д, мРНК для указанных ZFP создавали при помощи набора инструментов mMessage mMachine (Ambion), трансфицировали 1 мкг мРНК при помощи нуклеофектора Amaxa, как указано выше. Через 48 часов после трансфекции клетки собирали; уровни мРНК из нормального (CAG15, 099T), мутантного (CAG70, 099C) аллеля Htt и общий Htt (чHtt) оценивали, как описано выше, и нормировали относительно уровней ACTB; соотношение Htt/ACTB для каждого образца дополнительно нормировали относительно ложнотрансфицированного образца. “Сильный” CAG-нацеленный ZFP 30657 репрессировал оба аллеля, как и ожидалось (на основании его активности в клетках HEK293, фиг. 3А). ZFP 30640, который проявлял зависимую от длины CAG-повторов репрессию репортеров, демонстрировал <10% подавление аллеля дикого типа и подавлял мутантный аллель >90%. Уровни общего Htt в каждом образце согласовались с уровнями Htt дикого типа и мутантного Htt в том же образце.
[0184] ZFP-30640 исследовали также в нормальной линии фибробластов, а также в других линиях БХ фибробластов, которые содержали в гене Htt CAG-повторы разной длины (см. фигуру 3Е). Экспрессию Htt из каждого аллеля определяли способом, описанным выше. Для нормальной линии фибробластов (CAG18/18) не наблюдали экспрессии Htt. И наоборот, для линий CAG 15/67 и CAG15/70 наблюдали четкую дискриминацию аллелей как при высокой, так и при низкой дозировках трансфицируемой мРНК 30640; аналогичные результаты были получены для двух линий БХ фибробластов, содержащих в мутантном аллеле CAG-повторы средней длины (CAG 18/44 и CAG 18/45) - при этом удлиненный аллель репрессировался на ~80% как при высокой, так и при низкой дозировках 30640, а аллель CAG18 оставался не подверженным какому-либо влиянию. Вместе эти данные указывают на то, что аллель-специфические репрессоры, такие как 30640, могут сохранять избирательность по длине CAG-повторов в аллеле в контексте преобладающих генотипов болезни, таких как CAG18/44 и CAG18/45.
[0185] Вестерн-блоттинг применяли для того, чтобы показать, что ZFP, такие как 30640, избирательно подавляли уровни мутантного белка Htt в двух линиях фибробластов, полученных от пациентов, подтверждая наличие аллель-специфической регуляции, которое показал кПЦР-анализ (см. фигуру 3Ж). ZFP доставляли при помощи трансфекции мРНК (Amaxa nucleofection) в дозировке 300 нг в 4 реплики по 1,5e5 клеток и смешивали перед высеванием в 12-луночные планшеты. Через 48 часов клетки собирали для получения белкового экстракта. Приблизительно 2,5 мкг экстракта загружали в 5% Трис-ацетат гель и фиксировали при помощи MAB2166 (Millipore). Те же самые образцы дополнительно загружали в 4-15% Трис-HCl гель (Bio-Rad) и переносили при помощи стандартных методов фиксации анти-В-актином (1:20000, Sigma) в качестве загрузочного контроля. На основании кПЦР-анализа, при помощи которого измеряли Htt мРНК, можно заключить, что 30640 является аллель-специфическим репрессором, нацеленным на CAG-повторы; 32528 является биаллельным репрессором, нацеленным на сайт инициации транскрипции (СИТ), а 30657 является CAG-нацеленным репрессором, который подавляет оба аллеля Htt при всех применяемых дозировках. Вестерн-блоттинг показал, что 30640 специфически снижает уровни мутантного Htt (верхняя полоса) в обоих полученных от пациентов с БХ клеточных линиях, в то время как 32528 и 30657 одинаково подавляют оба аллеля.
Пример 4: Дополнительные конструкции CAG-нацеленных ZFP, которые осуществляют аллель-специфическую репрессию Htt
[0186] На фигуре 4А приведены результаты исследования ZFP-TF 30640, 30643, 30645 и 33074 (все нацелены на CAG-повторы и задействуют репрессионный домен KRAB) в линии БХ фибробластов CAG18/45. Для трансфекции при помощи нуклеофектора Amaxa указанным выше способом использовали разные количества ZFP мРНК, экспрессию мутантного Htt (правый столбик), Htt дикого типа (средний столбик) и общего Htt (обоих аллелей, левый столбик) измеряли, как описано выше, через 24 часа после трансфекции. ZFP 30640, 30645 и 33074 осуществляли аллель-специфическую репрессию в полном диапазоне дозировок ZFP мРНК, составляющем 3 мкг - 10 нг; при этом 30643 способен существенно подавлять оба аллеля в дозировках 30 нг или выше и начинает проявлять избирательность в отношении аллелей при дозировке в 10 нг.
[0187] На фигуре 4Б приведены результаты исследования ZFP 30643, 30648, 30657 и 30658 (все нацелены на CAG-повторы и задействуют репрессионный домен KRAB) в линии БХ фибробластов CAG15/70. Для трансфекции при помощи нуклеофектора Amaxa указанным выше способом использовали разные количества ZFP мРНК, экспрессию мутантного Htt (правый столбик), Htt дикого типа (средний столбик) и общего Htt (обоих аллелей, левый столбик) измеряли, как описано выше, через 24 часа после трансфекции. По сравнению с ZFP, результаты исследований которых приведены на предыдущей фигуре (30640, 30645 и 33074), данные ZFP осуществляют специфическую к мутантному Htt репрессию при более низких дозировках. Эти результаты позволяют предположить, что в зависимости от уровней экспрессии ZFP, которые могут быть достигнуты in vivo (например, в мозгу пациентов с БХ), можно достичь аллель-специфической репрессии мутантного Htt, используя соответствующие конструкции ZFP.
Пример 5: Репрессия чередующихся CAG-содержащих генов
[0188] Используя РНК, выделенные в примере 3 (фиг. 3Д), анализировали репрессию генов, содержащих CAG-повторы, результаты приведены на фигуре 5. При помощи ПЦР в режиме реального времени исследовали и нормировали относительно актина уровни экспрессии следующих генов: атаксина 2 (“ATXN2”); динамина (“DNM1”); белка, содержащего только F-бокс 11 (“FBXO11”), нитратредуктазы (“NAP”); субъединицы пререпликационного комплекса 4 (“ORC4”); фосфокиназы (“PHK”); белка OCT3/4 (“POU3”); содержащего домен THAP апоптоз-ассоциированного белка 2 (“THAPII”); связывающего белка TATA (“TBP”) и станниокальцина 1 (“STC1”). Дополнительно определяли расположение последовательности CAG-повторов относительно сайта инициации транскрипции (СИТ) и обозначили его на фигуре 3Е как “СИТ@”, где “+” обозначает положение оснований CAG-повторов, которые расположены ниже СИТ, а “-” обозначает положение оснований CAG-повторов, которые расположены выше СИТ. Также для каждого гена указывали количество CAG-повторов (“#CAG”).
[0189] Данные демонстрируют, что репрессия мутантного удлиненного аллеля Htt, осуществляемая 30640, является высокоспецифической, и только группа генов, содержащих CAG-повторы, в которых CAG-повторы расположены относительно близко к соответствующему сайту инициации транскрипции, могут являться мишенями для ZFP, таких как 30640.
Пример 6: Полногеномная специфичность аллель-специфических ZFP-репрессоров Htt
[0190] Для изучения полногеномной специфичности CAG-нацеленных ZFP при помощи микроматричного анализа трансфицировали БХ фибробласты (CAG18/45) (фигура 6). ZFP доставляли путем трансфекции мРНК (Amaxa nucleofection) в указанных дозировках - ZFP 30640, 30645 и 33074 в шести параллельных испытаниях трансфицировали, соответственно, в дозировках 75 нг, 15 нг и 15 нг; GFP-Kox мРНК (150 нг) трансфицировали в качестве контроля и использовали как носитель для того, чтобы довести общее количество трансфицированной мРНК во всех образцах до 150 нг. Экспрессию из аллелей CAG18 (099T, средние столбики) и CAG45 (099C, правые столбики) определяли при помощи аллель-специфических реагентов кПЦР через 24 часа после трансфекции, как описано выше, при этом все образцы (1-6) являлись биологическими репликами (проводили отдельные трансфекции). Уровни Htt нормировали относительно уровней GAPDH. Для всех трех ZFP наблюдали специфическую к мутантному аллелю репрессию Htt. Затем четыре наиболее схожих реплики отбирали для микроматричного анализа (Affymetrix HGU133plus2.0), а именно: для микроматричного анализа использовали реплики ЗФБ 1, 3, 4 и 6; реплики 30640 2, 3, 5 и 6; реплики 30645 2, 3, 5 и 6 и реплики 33074 1, 3, 4 и 5. Робастное среднее по мультиматрице (РСМ) использовали для нормирования исходных сигналов от каждой группы зондов; ZFP-трансфицированные образцы сравнивали с ЗФБ-трансфицированными образцами при помощи Т-теста; сигналы об “изменении” поступали от генов (групп зондов) с >2-кратной разницей по отношению к контрольным образцам и величиной Р для Т-теста <0,05. На основании этого критерия 30640 репрессировал только два гена, станниокальцина 1 (STC1) и удлиненного синаптотагминподобного белка 1 (ESYT1); 30645 и 33074 репрессировали только по одному гену каждый, STC1 и рецептор A интерлейкина 17 (ILR17RA), соответственно. Htt не зафиксировали как репрессируемый (>2-кратная репрессия) ген, потому что группа зондов Htt на матрице фиксирует мРНК Htt как дикого типа, так и мутантного. Этот эксперимент демонстрирует, что когда ZFP, специфичные к мутантному Htt, экспрессируются на уровнях, которые обеспечивают эффективную аллель-специфическую репрессию Htt, они могут действовать в пределах полного генома с высокой специфичностью.
Пример 7: Аллель-специфическая репрессия в нейральных стволовых клетках (НСК) с БХ
[0191] БХ iPSC/ESCs перевивали с аккутазой и культивировали на покрытых матригелем планшетах в среде E8 (Life Technologies). Нейральные стволовые клетки получали, используя среду для нейральной индукции StemPro (Life Technologies). Вкратце, iPSC/ESCs высевали на покрытый geltrex 6-луночный планшет с 200000 клеток/лунка и меняли среду на среду для нейральной индукции StemPro, когда происходило слияние 10-20% клеток. Среду меняли каждые 2 дня, а на 7-й день НСК собирали и выращивали. Бессывороточную среду для НСК StemPro (Life Technologies) использовали для культивирования НСК. НСК с БХ (CAG17/69, полученные из Coriell GM23225 iPSC) трансфицировали 1,5 или 0,5 мкг ZFP мРНК при помощи нуклеофекции. Через сорок восемь часов после трансфекции клетки собирали и оценивали экспрессию при помощи ОТ-ПЦР. Аллель-специфическую детекцию экспрессии Htt проводили при помощи генотипирования #4351376 на основе ОНП (rs1143646) (Applied Biosystems). В диапазоне исследуемых дозировок ZFP 30640 проявлял аллель-специфическую репрессию мутантного Htt, 30643 проявлял ~50% репрессию Htt дикого типа и ~90% репрессию мутантного Htt, а 30648 репрессировал оба аллеля (фигура 7); поведение этих ZFP согласуется с соответствующим поведением в БХ фибробластах (фигура 4). Уровни общего Htt (средние столбики) для каждого образца согласуются с уровнями мутантного Htt и уровнями Htt дикого типа.
Пример 8: Репрессия Htt в дифференцированных БХ нейронах
[0192] НСТ с БХ перевивали с аккутазой на покрытые geltrex планшеты. Дифференцирование нейронов индуцировали путем смены среды на среду для дифференцирования нейронов, содержащую бессывороточную среду для НСК StemPRO без оФРФ и ЭФР. Среду меняли каждые 3-4 дня на протяжении 21 дня. Нейроны получали из НСТ (CAG17/48, полученных из БХ ЭСК) путем культивирования в среде для нейронального дифференцирования. На 15-й день после нейрональной индукции клетки трансфицировали 1,0 или 0,5 мкг ZFP мРНК при помощи нуклеофекции. Через сорок восемь часов после трансфекции клетки собирали и оценивали экспрессию генов при помощи ОТ-ПЦР. Так как эта, полученная от пациентов линия не содержит ОНП, которые бы делали возможной аллель-специфическую детекцию на основе кПЦР Htt дикого типа и мутантного Htt, можно измерить только уровни общего Htt. Так как было показано, что 30640 и 33074 не репрессируют аллель CAG18 или CAG17 в БХ фибробластах и НСК, уровни общего Htt, которые наблюдали в образцах, обработанных 30640 и 33074, согласуются с аллель-специфической репрессией мутантного аллеля (CAG48). Более сильная репрессия, осуществляемая 30643 и 30648 при исследуемых дозировках ZFP, также согласуется с поведением этих ZFP в БХ фибробластах (фигура 8).
Пример 9: CAG-нацеленный репрессор репрессирует мутантный трансген Htt в R6/2 мышах
[0193] R6/2 мышам (которые несли в себе трансген мутантного человеческого экзона 1 Htt с ~120 CAG-повторами, см. Mangiarini et al, (1996) Cell 15:197) проводили стереотаксические, билатеральные стриарные инъекции 3e10 векторного генома рекомбинантного AAB2/6, кодирующего ZFP 30640-KOX или ЗФБ, приводимого в действие промотором ЦМВ. Инъекцию проводили мышам в возрасте 5 недель, затем в возрасте 8 недель их умерщвляли для молекулярного анализа. Левый и правый стриатум вырезали из каждой полусферы и быстро замораживали. Для оценки репрессии мутантного трансгена Htt из каждого стриатума извлекали всю РНК при помощи TRIzol Plus (Life Technologies) с последующим синтезом кДНК при помощи High Capacity RT (Life Technologies). После этого экспрессию трансгена R6/2 измеряли при помощи кПЦР и нормировали относительно среднего геометрического по трем контрольным генам (Atp5b, Eif4a2, UbC), как описано ранее у Benn et al. ((2008) Molecular Neurodegeneration: 3, 17). Наблюдали статистически существенную репрессию (P <0,001) мутантного трансгена Htt в четырех обработанных ZFP стриатумах по сравнению с четырьмя обработанными ЗФБ стриатумами (фигура 9). Средняя репрессия R6/2 составила 64,9% от обработанных ЗФБ контрольных образцов. Так как при помощи одной стереотаксической инъекции не было достигнуто полного покрытия стриатума, а ААВ2/6 трансдуцирует преимущественно нейрональные клетки, наблюдаемое увеличение репрессии (~35%) является, вероятно, недооценкой реальной репрессии в клетках, которые были трансдуцированы ААВ вектором.
Пример 10: Избирательная репрессия мутантного Htt при применении ZFP с димеризованными/мультимеризованными доменами
[0194] Для того чтобы сконструировать транскрипционные факторы цинковых пальцев для лучшей дискриминации между короткими CAG и длинными GAG-повторами, авторы обратили внимание как на снижение аффинности связывания ДНК индивидуальных транскрипционных факторов цинковых пальцев, так и на увеличение силы взаимодействия между разными копиями слитого белка, связанными с дополнительными субсайтами в пределах GAG-повторов. Для того чтобы понизить аффинность связывания ДНК индивидуальных транскрипционных факторов цинковых пальцев, авторы создали домены цинковых пальцев, содержащие меньшее количество цинковых пальцев и/или содержащие аминокислотные последовательности, для которых предполагается связывание ДНК с аффинностью меньшей, чем оптимальная. Для того чтобы увеличить силу взаимодействия между разными копиями слитого белка, связанными с дополнительными субсайтами в пределах GAG-повторов, авторы сшили разные димеризационные домены с транскрипционными факторами цинковых пальцев. Димеризационные домены могут взаимодействовать “параллельным” способом и образовывать димеры “голова-к-голове” или “хвост-к-хвосту” содержащих их слитых белков. В одной из потенциальных димеризационных методик требуется матрица из одинаковых продуктов слияния между ZFP и транскрипционными факторами, которые связаны в ориентации “голова-к-хвосту”, и, таким образом, в этой методике требуются димеризационные домены, которые взаимодействуют “антипараллельным” способом. См., например, McClain et al. (2001) J. Am. Chem. Soc. 123:3151-3152) и информацию по димеризующимся пептидам «цинковые пальцы» (Giesecke et al. (2006), Molecular Systems Biology 2:2006.2011).
[0195] Димеризационные конструкции СС1 и СС2 основаны на парах антипараллельных суперспиралей (McClain et al, там же, Ghosh et al. (2000) J Am Chem Soc 122:5658-5659). Димеризационные конструкции СС3 и СС4 представляли собой процессированные версии СС2, в которых отсутствовали, соответственно, 4 остатка или 7 остатков. Димеризационные конструкции DZ1, DZ2, DZ3 и DZ4 основаны на парах димеризующихся доменов цинковых пальцев (Giesecke et al, там же). В каждом случае один представитель из пары был сшит с N-концом ДНК-связывающего домена цинковых пальцев, а второй представитель из пары был сшит с С-концом ДНК-связывающего домена цинковых пальцев. Для сшивания димеризационных доменов со связывающим доменом цинковых пальцев использовали короткие линкеры, богатые остатками глицина и серина. В дополнительных вариантах реализации изобретения используют линкеры с переменной длиной и/или аминокислотным составом. Линкеры с удаленным одним или более остатком или с одной или более заменой остатками глицина или серина других аминокислотных остатков будут обладать меньшей гибкостью, что может привести к улучшенной дискриминации между длинными и короткими CAG-повторами.
[0196] Для того чтобы достичь избирательной репрессии мутантного аллеля Htt, ZFP конструировали так, как проиллюстрировано на фигуре 1Г и фигуре 10А и 4Б. на фигурах 10В и 10Г показаны последовательности мультимеризационных доменов. на фигурах 11А и 11Б приведены результаты экспериментов, в которых оценивали способность ZFP-TF, содержащих домены CC и DZ, соответственно, репрессировать свои мишени. В этих экспериментах указанные конструкции ZFP (50 нг) трансфицировали совместно с pRL-Htt-CAG17 (200 нг), pGL3-Htt-CAG47 (200 нг) и pVax-SEAP (секретируемой щелочной фосфатазой, 10 нг, используемой в качестве нормировочного контроля) в клетки HEK293. Через 24 часа после трансфекции измеряли активность люциферазы и активность щелочной фосфатазы. Соотношения люцифераза Renilla (CAG17)/SEAP и люцифераза светляков (CAG47)/SEAP для каждого образца нормировали относительно соответствующих величин для образцов, содержащих только репортеры. Репортер pGL3-Htt-CAG47 конструировали таким же способом, как и репортер pRL-Htt-CAG47 (см. пример 3), за исключением того, что вместо конструкции pRL-TK использовали конструкцию pGL-промотора (Promega).
[0197] Как показано на фигуре 11А, при исследовании 3 ZFP в виде конструкций, содержащих один или более доменов СС, наблюдали усиленную репрессию одного или обоих репортеров по сравнению с конструкциями из тех же ZFP, но в отсутствие доменов СС. на фигуре 11Б показано, что домены DZ1 и DZ3 усиливают репрессию обоих репортеров, осуществляемую 32220.
[0198] Эти результаты позволяют предположить, что домены СС и DZ в общем случае могут повысить аффинность мультимеризованных ZFP и что конструирование ДНК-связывающего домена и димеризационного домена может обеспечить оптимальную дискриминацию длины CAG-повторов.
Пример 11: Избирательная репрессия мутантного Htt, осуществляемая конструкциями ZFP-ZFP-Kox
[0199] В БХ фибробластах исследовали ZFP TF, которые имели конструкцию ZFP-ZFP-KOX. В этих экспериментах два ДНК-связывающих домена ZFP соединяли вместе посредством гибкого линкера (LRQKDAARGSAAMAERPFQ, SEQ ID NO:179) и сшивали с репрессионным доменом КОХ. Линкер размещали между консервативными гистидинами и цистеинами. Белки исследовали, как описано выше, используя ZFP мРНК в указанных дозировках. Результаты для БХ линий фибробластов CAG18/45 (фигура 12A) и CAG20/41 (фигура 12Б) демонстрируют, что путем связывания таким образом менее активных ДНК-связывающих доменов ZFP можно получить ZFP, которые осуществляют аллель-специфическую репрессию.
Пример 12: Активация Htt в мышиных клетках
[0200] Описанные в данном тексте ZFP оценивали на предмет их способности активировать экспрессию Htt. ZFP, нацеленные на участок от +200 до +467 п.н.о. (относительно сайта инициации транскрипции) мышиного Htt, сшивали с транскрипционным активационным доменом субъединицы NFκB p65. Этот целевой участок был выбран потому, что данный фрагмент был заменен соответствующей последовательностью (последовательностью большей части экзона 1 и части интрона 1) из человеческого Htt в разных мышиных нокин-моделях БХ (Menalled et al. (2003) J. Comp. Neurol 4651:11-26; Wheeler et al. (2000) Hum Mol Genet 8:115-122), следовательно, нацеленные на этот участок ZFP могут избирательно активировать в этих животных аллель дикого типа, но не нокин-аллель.
[0201] ZFP трансфицировали в клетки Neuro2A (в этих клетках оба аллеля Htt дикого типа), уровни мышиного Htt и ACTB мРНК измеряли, как описано в примере 2 (двойные трансфекции и множественный анализ).
[0202] Как показано на фигуре 13, повышение уровней Htt мРНК по сравнению с ложно-трансфицированными образцами было зафиксировано для обоих ZFP-TF. См. фигуру 13А. Повышение уровней белка Htt подтверждали при помощи вестерн-блоттинга. См. фигуру 13Б.
[0203] Создание нокин-аллеля Htt проиллюстрировано на фигуре 13В; выравнивание последовательностей (фигура 13Г) показывает расхождения между мышиной последовательностью, которая была заменена, и соответствующей человеческой последовательностью. На фигуре 13Д показано, что когда такие ZFP-активаторы трансфицировали в иммортализованные стриарные клетки, полученные из HdhQ111/Q7 нокин-мышей, происходила избирательная активация только Htt дикого типа.
Пример 13: Управление экспрессией Htt in vivo
[0204] Для того чтобы исследовать эффективность Htt-специфических ZFP TF in vivo, создавали AAB2 векторы, кодирующие ZFP. Затем эти конструкции на основе ААВ2 доставляли в мозг мышей. В случае человеческих Htt-специфических ZFP TF, AAB векторы доставляли R6.2 мышам или BAC мышам с БХ (штаммы C57B1/6 или FVB/N) для того, чтобы оценить репрессию человеческого трансгена, а также изменение БХ-подобного фенотипа. В случае мышиных Htt-специфических ZFP TF (активаторов или репрессоров), AAB векторы доставляли мышам дикого типа (C57B1/6 или FVB/N) или мышам с нокином человеческого Htt (HdhQ111/Q7, HdhQ140/Q7 или HdhQ175/Q7) для того, чтобы оценить активацию или подавление экспрессии эндогенного мышиного Htt. В случае ZFP, которые нацелены преимущественно на аллель с удлиненным CAG-повтором, AAB векторы доставляли R6.2 мышам или мышам с нокином человеческого Htt (HdhQ111/Q7, HdhQ140/Q7 или HdhQ175/Q7) для того, чтобы исследовать избирательную репрессию аллеля дикого типа в сравнении с удлиненным аллелем Htt. После умерщвления ткани мозга анализировали на предмет экспрессии Htt при помощи ОТ-ПЦР Такмана в режиме реального времени и демонстрировали, что гены Htt модулированы ZFP-TF.
Пример 14: Совместная трансфекция нейротрофического фактора и БХ Htt аллель-специфического ZFP TF
[0205] Описанные выше Htt-специфические ZFP TF трансфицировали совместно с ZFP TF, специфическими к нейротрофическому фактору мозга. Применяемые ZFP TF, специфические к нейротрофическому фактору мозга, являются специфическими к НФГК или НФГМ.
Пример 15: Проектирование и конструирование Htt-нацеленных цинк-пальцевых нуклеаз (ZFN)
[0206] ZFN, нацеленные на человеческий Htt и мышиный Htt, конструировали для нацеливания на последовательности, фланкирующие CAG-повторы, последовательности, расположенные вблизи первого кодирующего ATG, стоп-кодона, а также в ранних экзонах. ZFN конструировали и вносили в плазмиды или аденовирусные векторы преимущественно так, как описано в Urnov et al. (2005) Nature 435(7042):646-651, Perez et al (2008) Nature Biotechnology 26(7): 808-816, и патентной публикации США 2008/0131962.
Пример 16: Расщепляющая активность Htt-специфических ZFN
[0207] Для того чтобы исследовать расщепляющую активность, плазмиды, кодирующие описанные выше пары человеческих Htt-специфических ZFN, трансфицировали в клетки К562. Клетки К562 были получены из американской коллекции типовых культур и выращены согласно рекомендациям в среде F-12 (Invitrogen) с добавлением 10% разбавленной фетальной телячьей сыворотки (ФТС, Cyclone). Клетки отделяли от пластиковой посуды при помощи протеазы TrypLE Select™ (Invitrogen). Для проведения трансфекции один миллион клеток К562 смешивали с 2 мкг плазмид, содержащих цинк-пальцевые нуклеазы, и 100 мкл Amaxa Solution T. Клетки трансфицировали в Amaxa Nucleofector II™, используя программу U-23, и восстанавливали в 1,4 мл теплой среды F-12 + 10% ФТС.
[0208] Геномную ДНК собирали, а часть локуса Htt, включающую необходимый сайт расщепления, амплифицировали при помощи ПЦР. ПЦР, в которой использовали Accuprime HiFi полимеразу от InVitrogen, проводили следующим образом: после первичной 3-минутной денатурации при 94°C, проводили 30 циклов ПЦР с 30-секундным этапом денатурации при 94°C, с последующим 30-секундным этапом отжига при 58°C, с последующим 30-секундным этапом удлинения при 68°C. После завершения 30 циклов реакцию инкубировали при 68°C на протяжении 7 минут, затем хранили при 10°C на протяжении произвольного времени.
[0209] Геномную ДНК из клеток К562, обработанных Htt-специфическим ZFN, изучали при помощи нуклеазы Surveyor™ (Transgenomic), как описано, например, в патентных публикациях США №20080015164; 20080131962 и 20080159996.
[0210] Плазмиды, кодирующие пары мышиных Htt-специфических FN, исследовали тем же способом в клетках Neuro-2a.
[0211] Фигуры 14А и Б демонстрируют, что ZFN способны к нацеливанию на гены Htt с эффективностью генной модификации между 8-40%, оцененной, как ранее описывалось, по количеству наблюдаемых вставок-делеций.
Пример 17: Целевая интеграция разных длин тринуклеотидных повторов
[0212] Htt-специфические ZFN, обладающие наибольшей расщепляющей активностью по отношению к последовательностям, фланкирующим CAG-повторы, как описано выше, применяли в методике целевой интеграции для внесения разных длин CAG-повторов в копию Htt дикого типа. Конструировали доноры, содержащие 50, 80, 109 и 180 единиц CAG. Затем эти доноры трансфицировали в клетки К562 вместе с плазмидами, кодирующими Htt-специфические ZFN, как описано выше. Подтверждение интеграции донора получали путем выделения геномной ДНК, ПЦР-амплификации (как описано выше) с последующим секвенированием представляющего интерес участка.
[0213] Выявленные в клетках К562 ZFN, которые привели к нацеленной интеграции донорных аллелей в аллель Htt, применяли для вставки донорных нуклеиновых кислот переменной длины в человеческие эмбриональные стволовые клетки (чЭСК). Успешную интеграцию донора подтверждали путем выделения геномной ДНК, ПЦР-амплификации и секвенирования, как описано выше.
Пример 18: Разрушение/нокаут Htt дикого типа и/или мутантного Htt
[0214] ZFN, которые расщепляют в области ранних экзонов, могут привести к появлению небольших вставок или делеций (вставок-делеций) в результате негомологичного соединения концов (НГСК), с помощью чего можно создать клеточные модели с разрушенным одним или обоими аллелями Htt.
[0215] Указанные пары ZFN изготавливали, как описано выше, и исследовали на предмет расщепляющей активности, используя ошибку спаривания Cel I, как описано для примера 8. Данные пары ZFN нацелены на ранние экзоны человеческого Htt, и, следовательно, могут быть использованы для нокаута аллеля Htt дикого типа или мутантного Htt.
[0216] Как показано на фигуре 14А, пары ZFP 29627/29628, 29631/29632 (экзон 12) и 29637/29638 (экзон 18) расщепляли ген Htt и, следовательно, могут быть использованы для создания клеточных линий с генным нокаутом.
Пример 19: Экспрессия меченых аллелей Htt дикого типа и БХ Htt аллелей
[0217] ZFN, обладающие наибольшей расщепляющей активностью в отношении первого или последнего кодирующего экзона, использовали для мечения аллеля Htt дикого типа и мутантного Htt разными репортерными белками. Донорные ДНК для каждого репортера (А и Б) конструировали на основе сайта расщепления главной пары (пар) ZFN для того, чтобы обеспечить возможность целевой интеграции репортерного гена для получения слияния с Htt внутри рамки считывания. Донорные ДНК трансфицировали совместно с главной парой(ами) ZFN в клетки К562 для того, чтобы выбрать донорную конструкцию ДНК, которая обладает наивысшей частотой интеграции.
[0218] Пары ZFN изготавливали, как описано выше, и исследовали на предмет расщепляющей активности, используя ошибку спаривания Cel I, как описано для примера 8. Применяемые пары ZFN нацелены на 3’ конец кодирующей последовательности Htt, и, следовательно, могут быть использованы для нацеливания на аллель Htt дикого типа или мутантного Htt. Как показано на фигуре 14Б, пары ZFP 25917/25916, 25920/25921 и 25923/25922 были способны расщеплять ген Htt и, следовательно, могут быть использованы для внесения репортерной метки.
[0219] Выбранную донорную конструкцию ДНК для репортера А вместе с соответствующими ZFN доставляют в клетки, полученные от испытуемых, несущих мутантный ген Htt (например, фибробласты, индуцированные полипотентные клетки). Клоны выделяли и исследовали на предмет целевой интеграции репортера А. Гетерозиготные проявления считали желательными, а целевые аллели определяли при помощи ПЦР. Клоны, содержащие одиночный меченный репортером аллель Htt и немодифицированную целевую последовательность ZFN на втором аллеле, отбирали; донорную конструкцию для репортера Б и соответствующие ZFN трансфицировали для того, чтобы пометить второй аллель репортером Б.
[0220] Полученный клон мышиной эмбриональной стволовой клетки содержит аллель Htt дикого типа и мутантный аллель, меченный двумя разными маркерами, что позволяет проследить экспрессию из каждого аллеля; эти клетки использовали для создания мышиных моделей нарушений тринуклеотидных повторов, используя стандартные протоколы.
Пример 20: Конструирование активных белков TALE-TF против Htt
[0221] ДНК-связывающие домены TALE соединяли с репрессионным доменом KRAB из белка Кох1 (TALE TF) и использовали для исследования репрессии гена Htt в фибробластах, полученных от пациентов с БХ (CAG 20/41). Конструирование белков TALE осуществляли так, как было описано ранее (см. совместную патентную публикацию США 20110301073 и совместную патентную заявку США 13/679684, которые включены в данный текст посредством ссылки), а белки конструировали с тремя различными С-конечными архитектурами: +63, +231 и +278, как описано в US 20110301073. Для того чтобы сконструировать экспрессионные плазмиды TALE TF, использовали описанные ранее (см. US 20110301073) экспрессионные плазмиды TALEN, за исключением того, что используемый для TALEN домен FokI был заменен репрессионным доменом KRAB. Соединения C-конца белка TALE и домена KRAB приведены ниже, при этом последовательность домена KRAB выделена подчеркиванием. Жирным курсивом выделена тройная метка, жирным текстом выделена последовательность ядерной локализации, “[повторы]” обозначает месторасположение группы повторов единиц TALE (полный набор повторов плюс C-концевой полуповтор), а часть, выделенная волнистым подчеркиванием, представляет последовательность домена KRAB:
TALE-C63-Kox1:
MDYKDHDGDYKDHDIDYKDDDDKMAPKKKRKVGIHGVPMVDLRTLGYSQQQQEKIKPKVRSTVAQHHEALVGHGF
THAHIVALSQHPAALGTVAVKYQDMIAALPEATHEAIVGVGKQWSGARALEALLTVAGELRGPPLQLDTGQLLKI
AKRGGVTAVEAVHAWRNALTGAPLN[повторы]GGRPALESIVAQLSRPDPALAALTNDHLVALACLGGRPALD
AVKKGLPHAPALIKRTNRRIPERTSHRVAGSGMDAKSLTAWSRTLVTFKDVFVDFTREEWKLLDTAQQIVYRNVM
LENYKNLVSLGYQLTKPDVILRLEKGEEPWLVEREIHQETHPDSETAFEIKSSV (SEQ ID NO:197)
TALE-C231-Kox1:
MDYKDHDGDYKDHDIDYKDDDDKMAPKKKRKVGIHGVPMVDLRTLGYSQQQQEKIKPKVRSTVAQHHEALVGHGF
THAHIVALSQHPAALGTVAVKYQDMIAALPEATHEAIVGVGKQWSGARALEALLTVAGELRGPPLQLDTGQLLKI
AKRGGVTAVEAVHAWRNALTGAPLN[повторы]GGRPALESIVAQLSRPDPALAALTNDHLVALACLGGRPALD
AVKKGLPHAPALIKRTNRRIPERTSHRVADHAQVVRVLGFFQCHSHPAQAFDDAMTQFGMSRHGLLQLFRRVGVT
ELEARSGTLPPASQRWDRILQASGMKRAKPSPTSTQTPDQASLHAFADSLERDLDAPSPTHEGDQRRASSRKRSR
SDRAVTGPSAQQSFEVRAPEQRDALHLPLSWRVKRPRTSIGGGLPDPGSGMDAKSLTAWSRTLVTFKDVFVDFTR
EEWKLLDTAQQIVYRNVMLENYKNLVSLGYQLTKPDVILRLEKGEEPWLVEREIHQETHPDSETAFEIKSSV (SEQ ID NO:198)
TALE-C278-Kox1
MDYKDHDGDYKDHDIDYKDDDDKMAPKKKRKVGIHGVPMVDLRTLGYSQQQQEKIKPKVRSTVAQHHEALVGHGF
THAHIVALSQHPAALGTVAVKYQDMIAALPEATHEAIVGVGKQWSGARALEALLTVAGELRGPPLQLDTGQLLKI
AKRGGVTAVEAVHAWRNALTGAPLN[повторы]GGRPALESIVAQLSRPDPALAALTNDHLVALACLGGRPALD
AVKKGLPHAPALIKRTNRRIPERTSHRVADHAQVVRVLGFFQCHSHPAQAFDDAMTQFGMSRHGLLQLFRRVGVT
ELEARSGTLPPASQRWDRILQASGMKRAKPSPTSTQTPDQASLHAFADSLERDLDAPSPTHEGDQRRASSRKRSR
SDRAVTGPSAQQSFEVRAPEQRDALHLPLSWRVKRPRTSIGGGLPDPTPTAADLAASSTVMREQDEDPFAGAADD
FPAFNEEELAWLMELLPQGSGMDAKSLTAWSRTLVTFKDVFVDFTREEWKLLDTAQQIVYRNVMLENYKNLVSLG
YQLTKPDVILRLEKGEEPWLVEREIHQETHPDSETAFEIKSSV (SEQ ID NO:199)
[0222] Распознавания оснований достигали, используя канонические соответствия RVD-основание (“код TALE”: NI соответствует A, HD соответствует C, NN соответствует G (NK в полуповторе), NG соответствует T). В некоторых TALE TF белок сконструирован так, чтобы связывать смысловую (5’-3’) нить ДНК, в то время как в других TALE TF сконструирован так, чтобы связывать антисмысловую (3’-5’) нить. Группу TALE TF сконструировали для нацеливания на CAG-повторы гена Htt. ДНК-связывающие белки TALE часто взаимодействуют преимущественно с нуклеотидным основанием ‘T’ в 5’ конце мишени, и так как мишенями являются участки CAG-повторов, можно предсказать, что белки, которые связываются с антисмысловой нитью ДНК и, следовательно, последовательности CTG-повтора с основанием ‘T’ в 5’ конце мишени, могут обладать лучшей аффинностью связывания и специфичностью, и, следовательно, репрессорной активностью.
[0223] Исследуемые мишени и численные обозначения TALE TF приведены ниже в таблице 4. Численные обозначения помечены “SBS#”, специфичность к смысловой или антисмысловой нити обозначена как (“С/A”), также указаны мишень, количество единиц повторов или RVD и тип С-конца.
[0224] Затем приведенные в таблице TALE TF исследовали на предмет репрессии Htt в фибробластах, полученных от пациентов с БХ (CAG 20/41), результаты приведены на фигуре 15. В этом эксперименте клетки трансфицировали 1000, 100 или 10 нг мРНК, кодирующей TALE-TF. Результаты для каждого исследуемого TALE TF собраны в группы по три, соответствующие трем количественным значениям трансфицируемой мРНК. В каждой группе содержится по три образца: левый столбик представляет экспрессию общего Htt, средний столбик представляет экспрессию из аллеля Htt CAG20, а правый столбик представляет экспрессию из аллеля Htt CAG41. Эти данные показывают, что некоторые TALE TF были способны репрессировать оба аллеля (см., например, 102454), в то время как другие TALE TF были способны избирательно ингибировать мутантный Htt с удлиненным CAG-повтором (см., например, 102451 и 102472).
[0225] Все патенты, патентные заявки и публикации, упоминаемые в данном тексте, в полном объеме включены в этот текст посредством ссылки.
[0226] Хотя для четкости понимания описание данного изобретения сопровождается достаточно подробными иллюстрациями и примерами, для специалистов в данной области техники очевидно, что на практике можно вносить различные изменения и модификации без отступления от сущности и объема настоящего изобретения. Соответственно, вышеприведенные описания и примеры не должны расцениваться как ограничивающие.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ ЛЕЧЕНИЯ БОЛЕЗНИ ХАНТИНГТОНА | 2014 |
|
RU2693891C1 |
| СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ ЛЕЧЕНИЯ БОЛЕЗНИ ХАНТИНГТОНА | 2015 |
|
RU2691102C2 |
| МОДУЛЯТОРЫ ЭКСПРЕССИИ ГЕНА ОТКРЫТОЙ РАМКИ СЧИТЫВАНИЯ 72 ХРОМОСОМЫ 9 И ИХ ПРИМЕНЕНИЯ | 2020 |
|
RU2828216C2 |
| МОДУЛЯТОРЫ ТАУ И СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ ИХ ДОСТАВКИ | 2017 |
|
RU2789459C2 |
| КОНСТРУИРУЕМЫЕ БЕЛКИ С ЦИНКОВЫМИ ПАЛЬЦАМИ, НАПРАВЛЕННЫЕ НА ГЕНЫ РАСТЕНИЙ, ВОВЛЕЧЕННЫЕ В БИОСИНТЕЗ ЖИРНЫХ КИСЛОТ | 2010 |
|
RU2712833C2 |
| КОНСТРУИРУЕМЫЕ БЕЛКИ С ЦИНКОВЫМИ ПАЛЬЦАМИ, НАПРАВЛЕННЫЕ НА ГЕНЫ РАСТЕНИЙ, ВОВЛЕЧЕННЫЕ В БИОСИНТЕЗ ЖИРНЫХ КИСЛОТ | 2010 |
|
RU2568056C2 |
| НАПРАВЛЕННАЯ МОДИФИКАЦИЯ МАЛАТДЕГИДРОГЕНАЗЫ | 2013 |
|
RU2658437C2 |
| НАЦЕЛИВАНИЕ НА КЛЕТКИ, ОПОСРЕДУЕМОЕ ХИМЕРНЫМ АНТИГЕННЫМ РЕЦЕПТОРОМ | 2018 |
|
RU2783316C2 |
| СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ ЛЕЧЕНИЯ ГЕМОФИЛИИ В | 2011 |
|
RU2608643C2 |
| ФУНКЦИОНАЛЬНЫЕ ЛОКУСЫ FAD2 И СООТВЕТСТВУЮЩИЕ СПЕЦИФИЧНЫЕ ДЛЯ САЙТА-МИШЕНИ СВЯЗЫВАЮЩИЕСЯ БЕЛКИ, СПОСОБНЫЕ ИНДУЦИРОВАТЬ НАПРАВЛЕННЫЕ РАЗРЫВЫ | 2013 |
|
RU2656158C2 |


















































































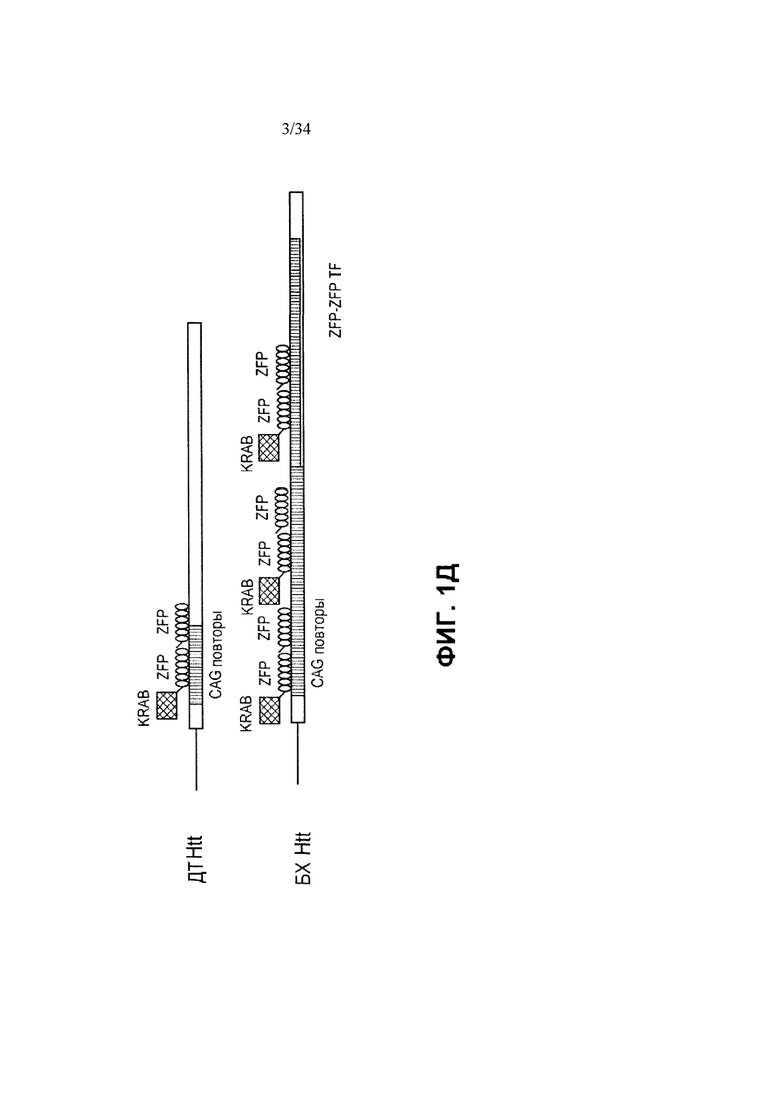


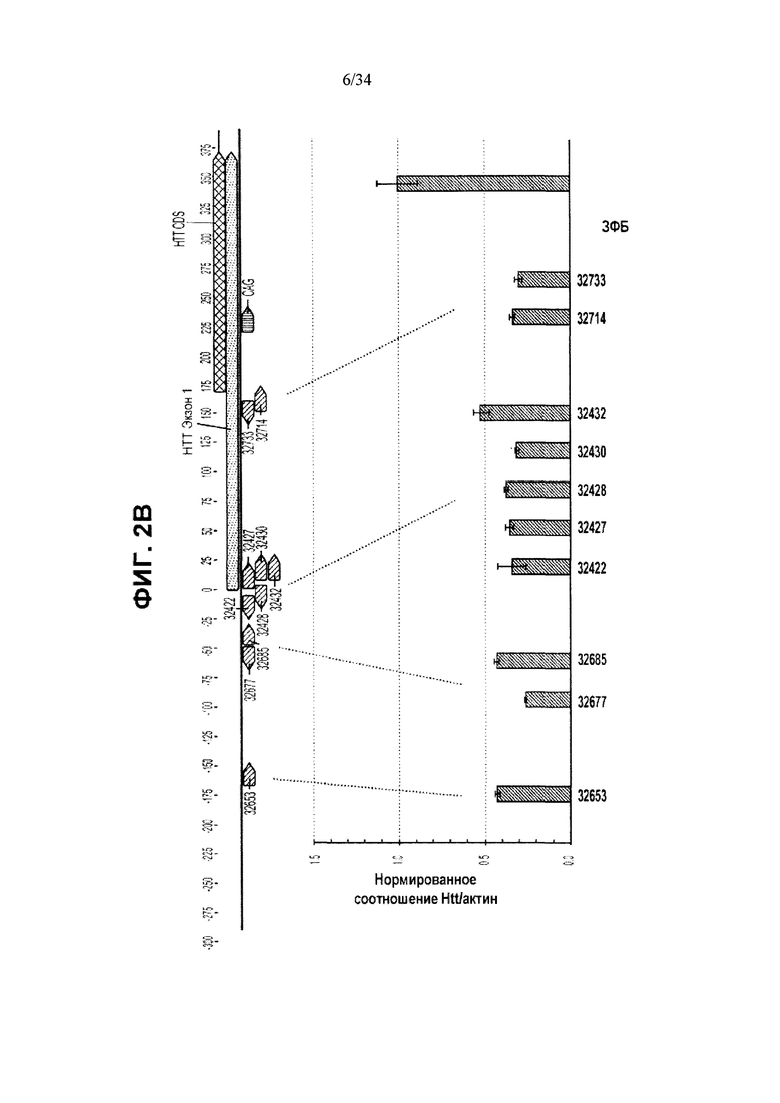

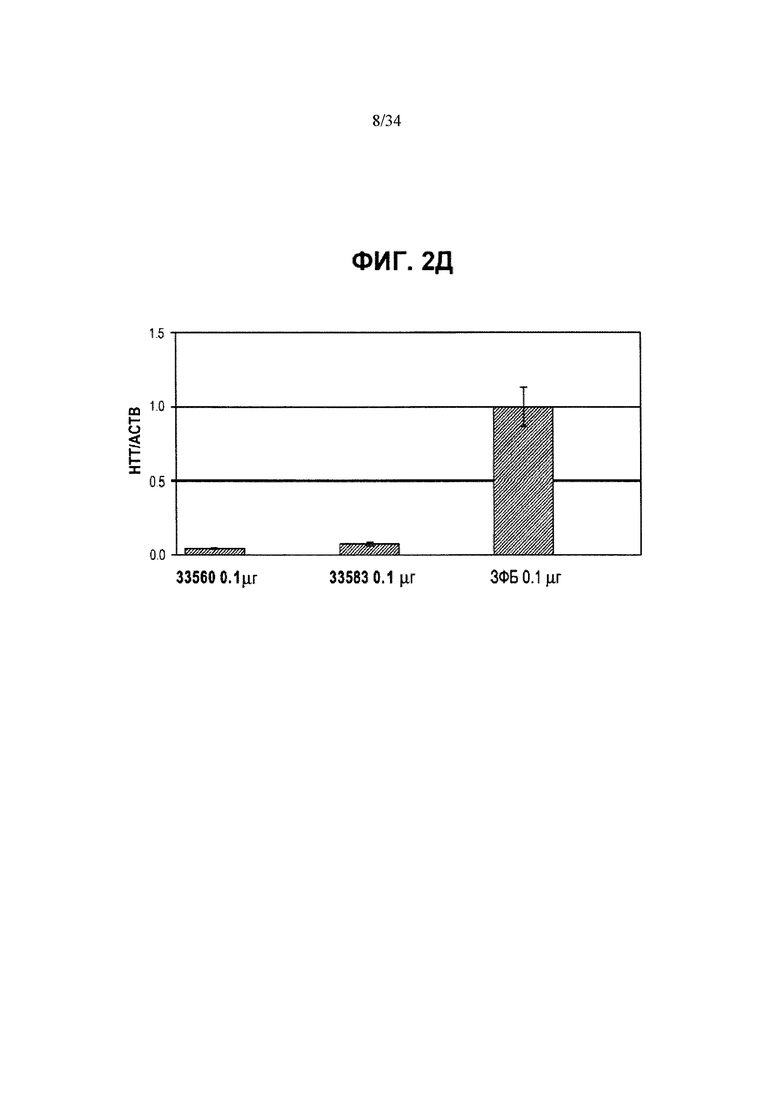
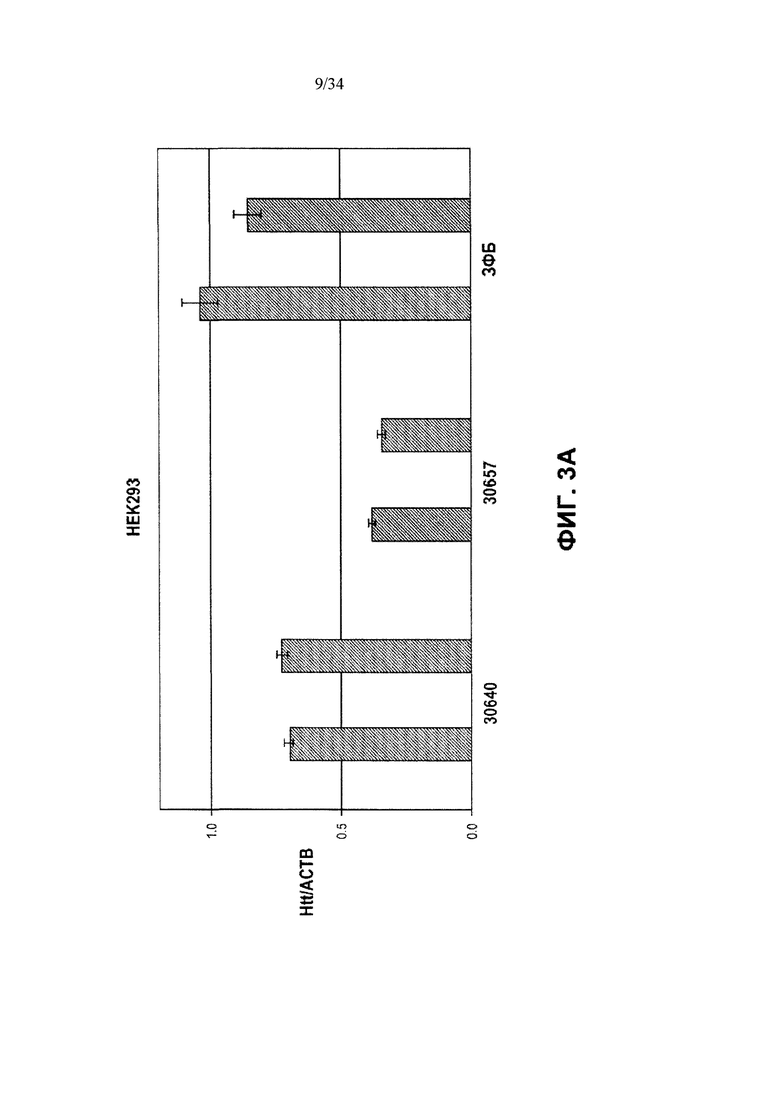

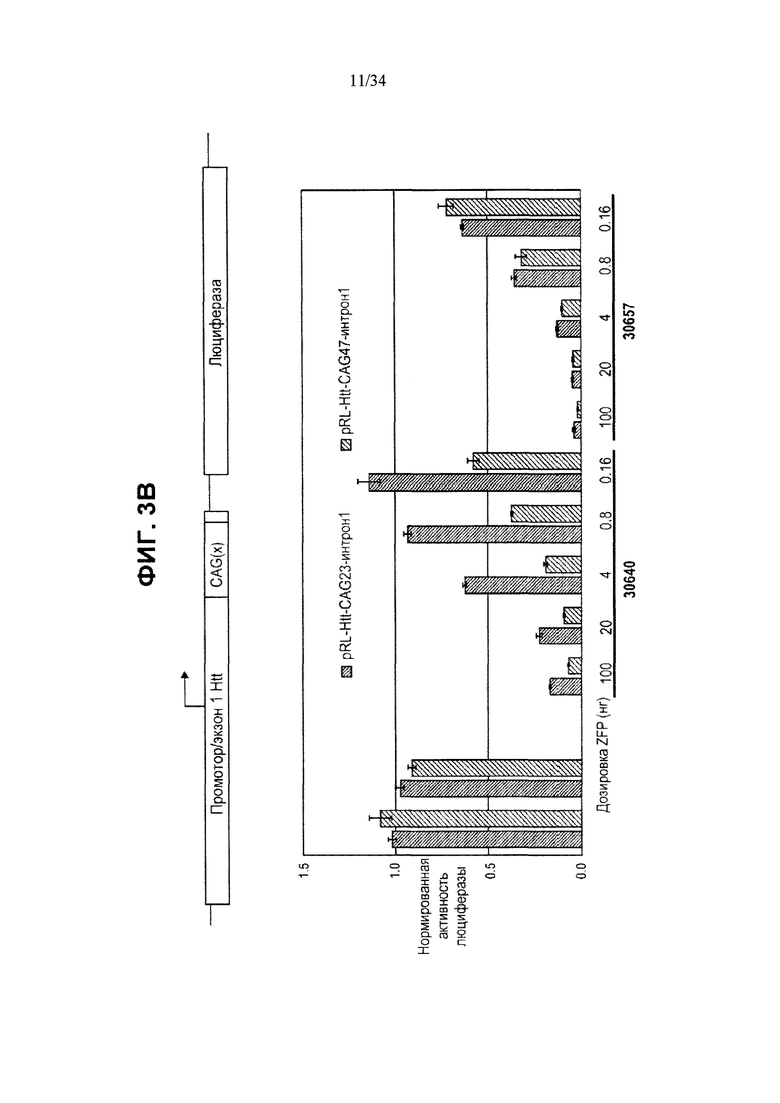



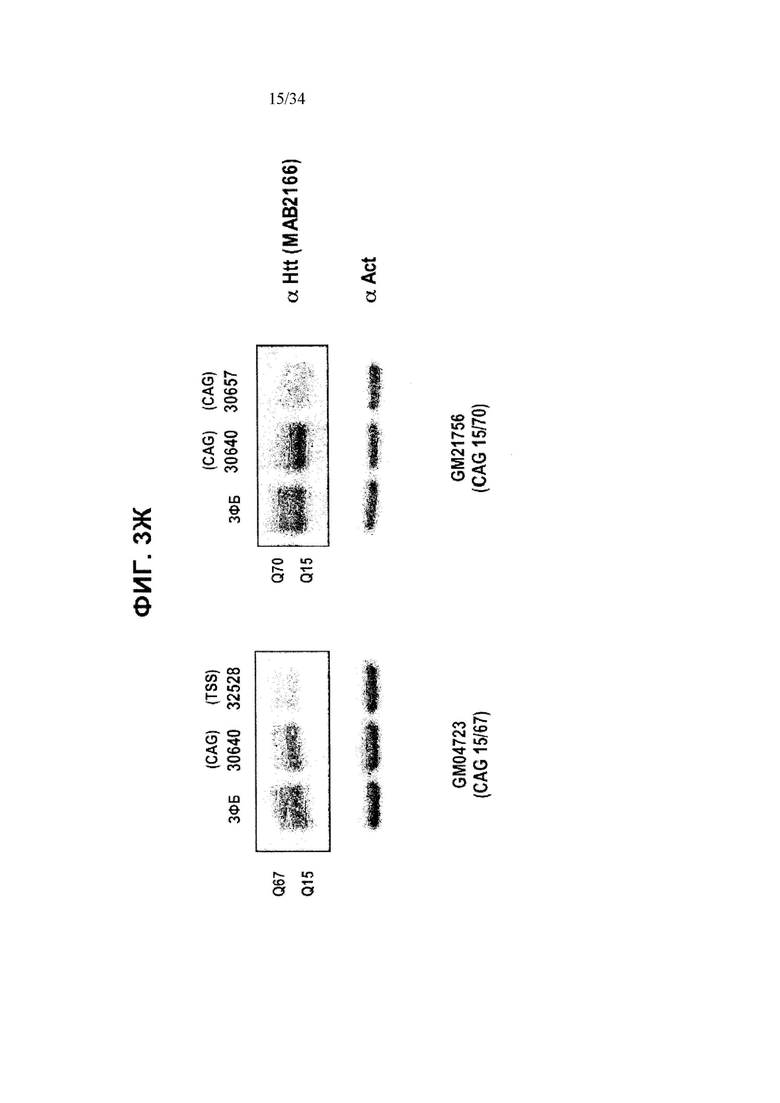

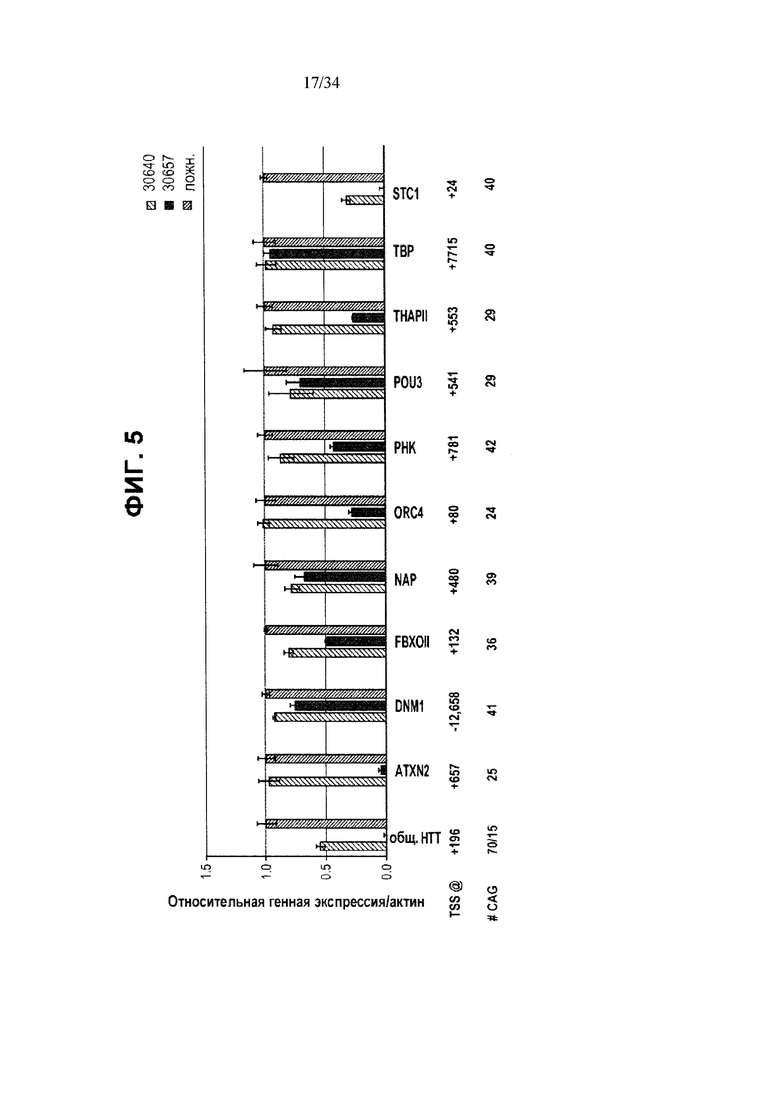



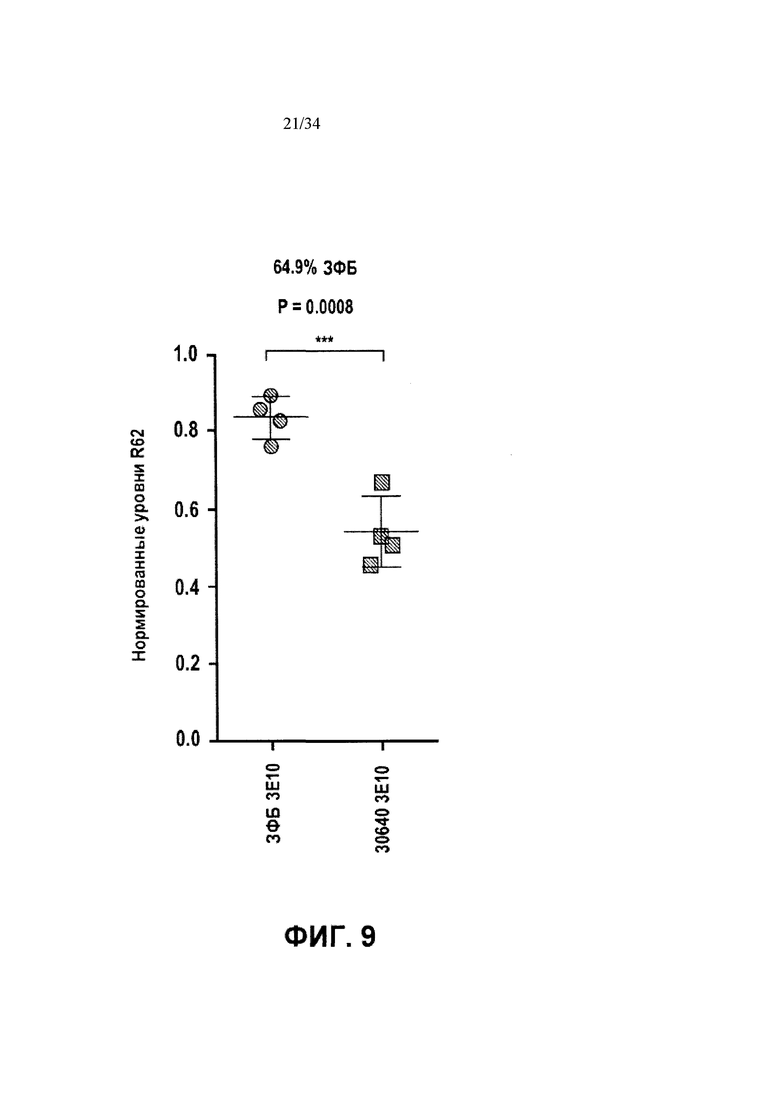





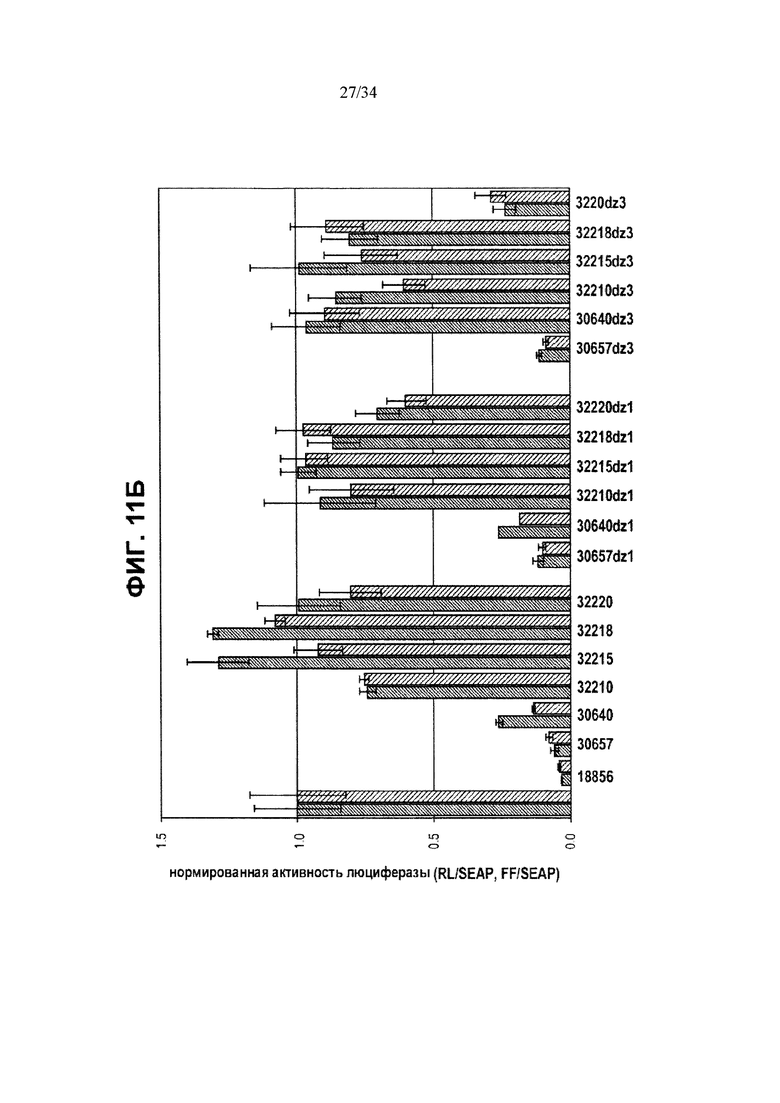
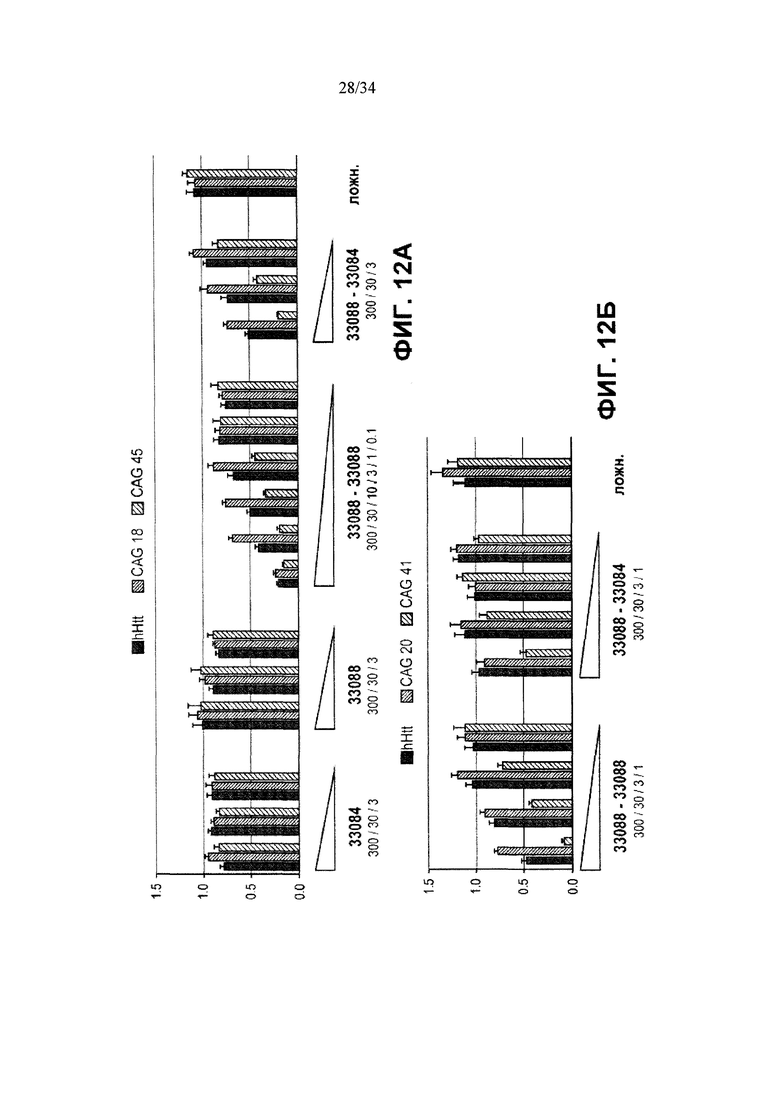

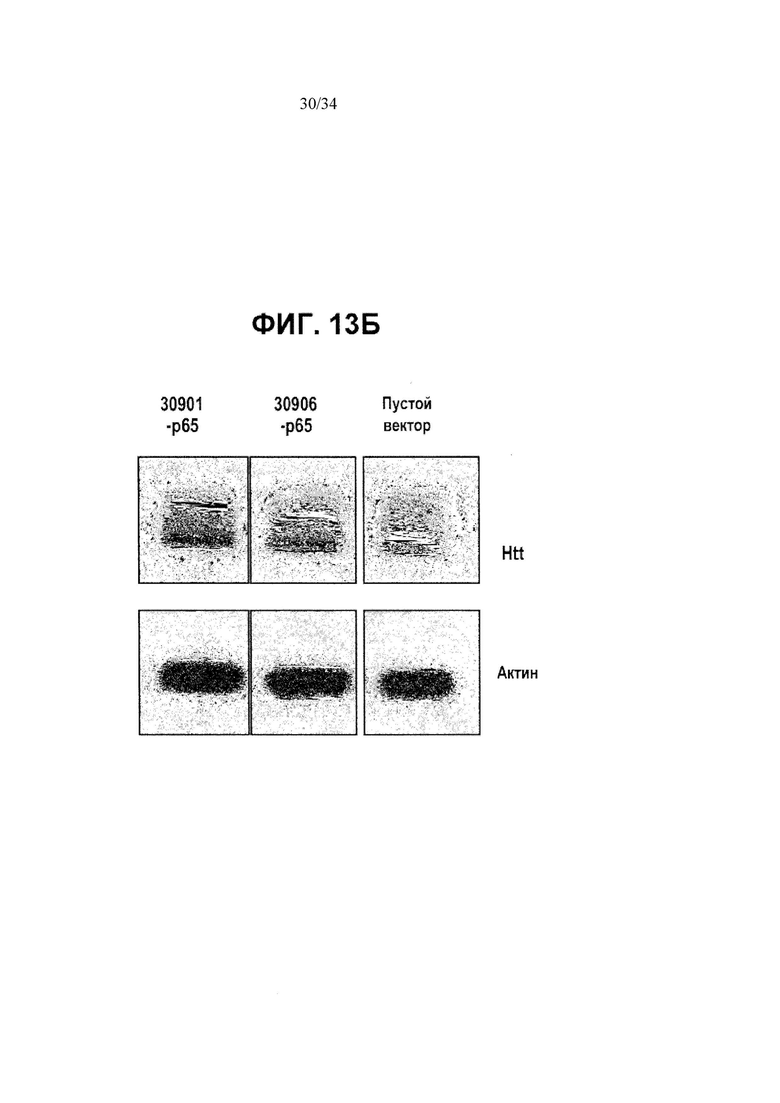
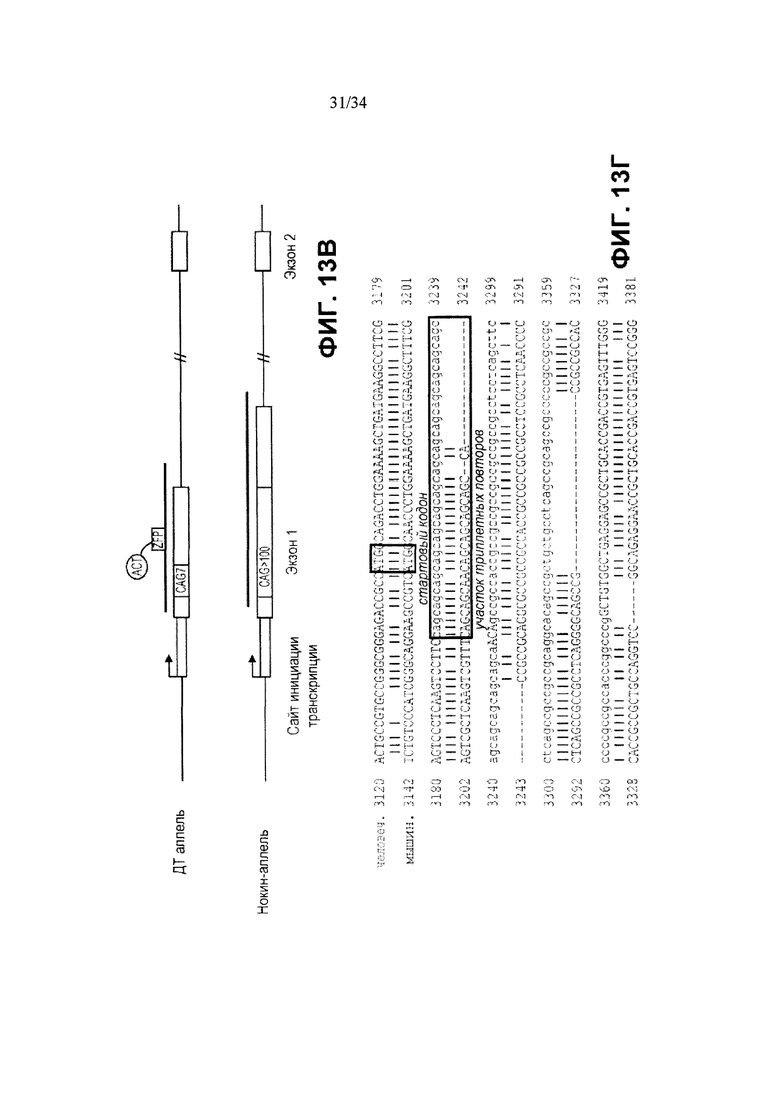


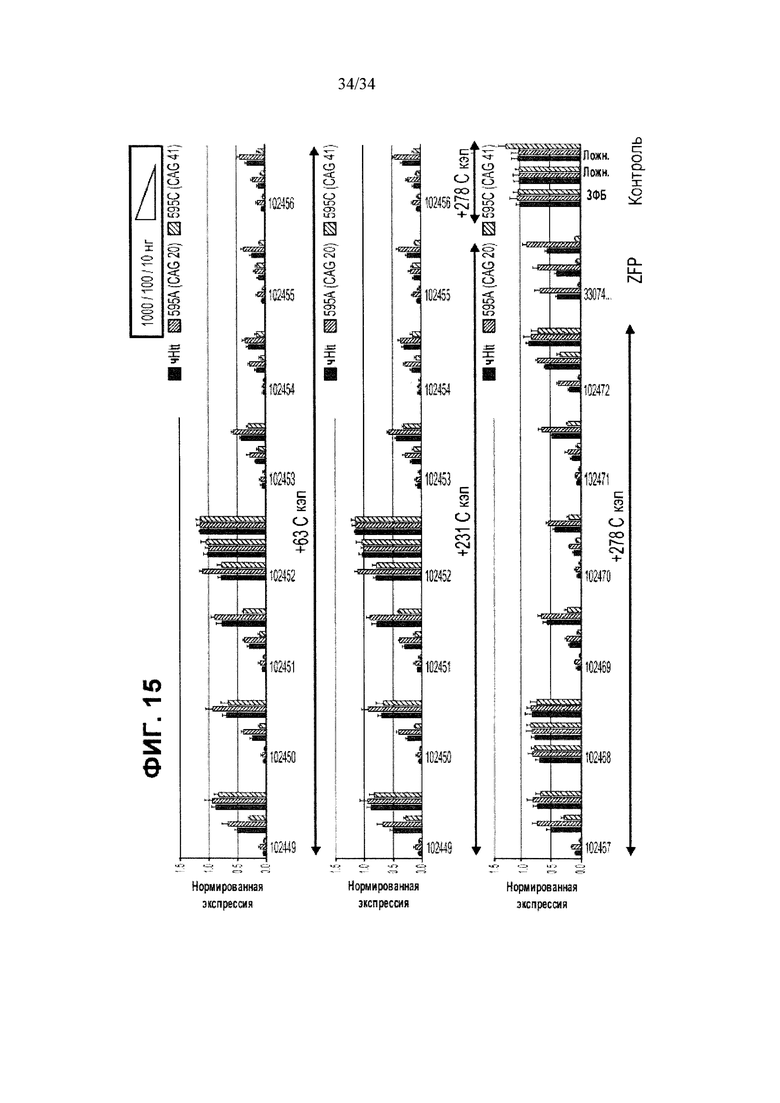
Изобретение относится к области биотехнологии, конкретно к белку «цинковые пальцы» неприродного происхождения, который связывается с геном Htt, что может быть использовано в медицине. Получают указанный белок «цинковые пальцы», а также слитый белок, включающий указанный белок, функционально связанный с нуклеазным доменом, полинуклеотид, кодирующий указанный белок и указанный слитый белок, клетку-хозяина для получения указанного белка и указанного слитого белка, фармацевтическую композицию для подавления экспрессии гена Htt в клетке. Также изобретение описывает способ подавления экспрессии гена Htt, способ лечения болезни Хантингтона. Изобретение позволяет эффективно подавлять экспрессию гена Htt, что позволяет успешно лечить болезнь Хантингтона. 15 н. и 9 з.п. ф-лы, 15 ил., 7 табл., 20 пр.
1. Белок «цинковые пальцы» неприродного происхождения, который связывается с геном Htt, содержащим последовательность, представленную SEQ ID NO: 107, при этом указанный белок «цинковые пальцы» содержит 3, 5 или 6 доменов цинковых пальцев в порядке от П1 до П4, от П1 до П5 и от П1 до П6, и где домены цинковых пальцев белка связывают целевые субсайты в SEQ ID NO: 107 и включают следующие участки спиралей узнавания:
(i) П1: QSSDLSR (SEQ ID NO: 31);
П2: QWSTRKR (SEQ ID NO: 63);
П3: QSSDLSR (SEQ ID NO: 31);
П4: QWSTRKR (SEQ ID NO: 63); и
П5: QSGDLTR (SEQ ID NO: 18); или
(ii) П1: RSDNLSE (SEQ ID NO: 60);
П2: KRCNLRC (SEQ ID NO: 61);
П3: QSSDLSR (SEQ ID NO: 31);
П4: QWSTRKR (SEQ ID NO: 63); и
П5: QSGDLTR (SEQ ID NO: 18); или
(iii) ni: QSGDLTR (SEQ ID NO: 18);
П2: QSGDLTR (SEQ ID NO: 18);
П3: RSDNLSE (SEQ ID NO: 60);
П4: KRCNLRC (SEQ ID NO: 61);
П5: QSGDLTR (SEQ ID NO: 18); и
П6: QSGDLTR (SEQ ID NO: 18); или
(iv) П1: RSDNLSE (SEQ ID NO: 60);
П2: KRCNLRC (SEQ ID NO: 61);
П3: QSSDLSR (SEQ ID NO: 31);
П4: QWSTRKR (SEQ ID NO: 63);
П5: QSSDLSR (SEQ ID NO: 31); и
П6: QWSTRKR (SEQ ID NO: 63); или
(v) П1: QSGDLTR (SEQ ID NO: 18);
П2: QSSDLSR (SEQ ID NO: 31);
П3: QWSTRKR (SEQ ID NO: 63);
П4: QSSDLSR (SEQ ID NO: 31); и
П5: QWSTRKR (SEQ ID NO: 63); или
(vi) П1: RSDNLSE (SEQ ID NO: 60);
П2: KRCNLRC (SEQ ID NO: 61);
П3: QSGDLTR (SEQ ID NO: 18);
П4: QSSDLSR (SEQ ID NO: 31); и
П5: QWSTRKR (SEQ ID NO: 63); или
(vii) П1: QSGDLTR (SEQ ID NO: 18);
П2: QSSDLSR (SEQ ID NO: 31); и
П3: QWSTRKR (SEQ ID NO: 63), необязательно содержащий димеризационный домен, который обеспечивает возможность мультимеризации белков «цинковые пальцы» при связывании с ДНК.
2. Белок «цинковые пальцы» по п. 1, отличающийся тем, что белок «цинковые пальцы» связывается полностью или частично снаружи участка CAG-повторов гена Htt.
3. Белок «цинковые пальцы» по п. 1, отличающийся тем, что белок «цинковые пальцы» связывается с последовательностями в пределах участка CAG-повторов гена Htt.
4. Слитый белок, включающий белок «цинковые пальцы» по любому из пп. 1-3, функционально связанный с нуклеазным доменом, где слитый белок расщепляет и подавляет экспрессию гена Htt.
5. Слитый белок, содержащий белок «цинковые пальцы» по любому из пп. 1-3, функционально слитый с транскрипционным регуляторным доменом, где слитый белок расщепляет и подавляет экспрессию гена Htt.
6. Слитый белок по п. 5, отличающийся тем, что транскрипционный регуляторный домен представляет собой репрессионный домен и экспрессия гена Htt подавлена.
7. Полинуклеотид, кодирующий белок «цинковые пальцы» по пп. 1-3.
8. Клетка-хозяин для получения белка «цинковые пальцы» по п. 1, где клетка трансфицирована экспрессионным вектором, кодирующим указанный белок «цинковые пальцы», и где клетка не является эмбриональной линией клеток человека.
9. Фармацевтический композиция для подавления экспрессии гена Htt, содержащая эффективное количество белка «цинковые пальцы» по пп. 1-3.
10. Способ подавления экспрессии гена Htt в клетке, включающий введение в клетку полинуклеотида, кодирующего слитый белок по п. 5.
11. Способ по п. 10, отличающийся тем, что ген Htt содержит по меньшей мере один мутантный аллель.
12. Способ по п. 10, отличающийся тем, что ген Htt является геном дикого типа.
13. Способ по п. 10, отличающийся тем, что слитый белок содержит нуклеазный домен, а экспрессия гена Htt является инактивированной.
14. Способ подавления гена Htt в клетке, включающий введение в клетку полинуклеотида, кодирующего слитый белок по п. 5, где слитый белок содержит нуклеазный домен и где последовательность гена Htt является модифицированной.
15. Способ лечения болезни Хантингтона, включающий введение нуждающемуся в этом пациенту эффективного количества полинуклеотида, кодирующего слитый белок по п. 5.
16. Способ создания модельной системы для изучения болезни Хантингтона, включающий подавление гена Htt с использованием слитого белка по п. 4.
17. Способ по п. 16, отличающийся тем, что ген Htt является модифицированным таким образом, чтобы содержать один или более мутантных аллелей.
18. Способ по п. 17, отличающийся тем, что мутантные аллели содержат удлиненные тринуклеотидные повторы.
19. Способ по любому из пп. 16-18, отличающийся тем, что клетка содержит эмбриональную стволовую клетку, не принадлежащую человеку.
20. Полинуклеотид, кодирующий слитый белок по п. 5.
21. Клетка-хозяин для получения белка «цинковые пальцы», который подавляет экспрессию гена Htt, где клетка-хозяин содержит один или несколько полинуклеотидов по п. 7, где клетка не является эмбриональной линией клеток человека.
22. Клетка-хозяин для получения слитого белка по п. 5, где клетка трансфицирована экспрессионным вектором, кодирующим указанный слитый белок, и где клетка не является эмбриональной линией клеток человека.
23. Фармацевтическая композиция для подавления экспрессии гена Htt, содержащая эффективное количество одного или нескольких полинуклеотидов по п. 7.
24. Фармацевтическая композиция для подавления экспрессии гена Htt, содержащая эффективное количество одного или нескольких слитых белков по п. 5.
| US 20110082093 A1, 07.04.2011 | |||
| MITTELMAN D | |||
| et al | |||
| Zinc-finger directed double-strand breaks within CAG repeat tracts promote repeat instability in human cells, PNAS, 2009, v.106, no.24, p.9607-9612 | |||
| US 6534261 B1, 18.03.2003 | |||
| РЕГУЛИРУЮЩЕЕ ДИФФЕРЕНЦИРОВКУ СРЕДСТВО, СОДЕРЖАЩЕЕ ГЕН, КОТОРЫЙ РЕГУЛИРУЕТ ДИФФЕРЕНЦИРОВКУ ИЗ СТВОЛОВЫХ КЛЕТОК В КЛЕТКИ-ПРИРОДНЫЕ КИЛЛЕРЫ, В КАЧЕСТВЕ ЭФФЕКТИВНОГО ИНГРЕДИЕНТА | 2005 |
|
RU2350649C2 |

