Перекрестная ссылка на родственные заявки
[0001] В настоящей заявке испрашивается приоритет предварительной заявки на патент США № 62/542511, поданной 8 августа 2017; предварительной заявки на патент США № 62/581290, поданной 3 ноября 2017; предварительной заявки на патент США № 62/613258, поданной 3 января 2018 и предварительной заявки на патент США № 62/652672, поданной 4 апреля 2018, содержание которых включено в настоящее описание посредством ссылки в полном объеме.
Область техники
[0002] Настоящее изобретение относится к области геномной модификации человеческих клеток, включая лимфоциты и стволовые клетки.
Уровень техники
[0003] Генная терапия представляет широкие перспективы, открывающие новую эру в медицине. Такая методика позволяет осуществлять лечение состоний, которые не поддаются лечению стандартными методами, применяемыми в медицине. Генотерапия может включать множество вариантов методов редактирования генома, таких как дизрупция или коррекция генного локуса и инсерция экспрессируемого трансгена, который может регулироваться под действием специфического экзогенного промотора, присоединенного к трансгену или под действием эндогенного промоторго, присутствующего в сайте инсерции в геноме, а также экспрессия трансгена посредством эписомной векторной системы.
[0004] Примерами проблем, которые должны быть решены для реализации этой технологии, являются доставка и инсерция трансгена. Так, например, хотя для терапии могут быть применены различные способы доставки генов, однако, важно найти компромисс между безопасностью, продолжительностью и уровнем экспрессии. Способы, в которых используется трансген в виде эписомы (например, системы на основе аденовируса (Ad), аденоассоциированного вируса (AAV) и плазмиды) являются, в основном, безопасными и могут давать высокие начальные уровни экспрессии, однако, эти способы не позволяют осуществлять надежную эписомную репликацию, что может ограничивать продолжительность экспрессии в митотически активных тканях. В противоположность этому, способы доставки, которые приводят к рандомизированной интеграции нужного трансгена (например, интеграции лентивируса (LV)), обеспечивают более длительную экспрессию, но из-за неспецифической природы рандомизированной инсерции, могут стимулировать нерегулируемый рост клеток-реципиентов, что может приводить к развитию злокачественных новообразований посредством активации онкогенов вблизи от случайно интегрированного трансгенного кластера. Кроме того, хотя интеграция трансгена исключает потери от репликации, однако, она, в конечном счете, не предотвращает сайленсинг экзогенного промотора, присоединенного к трансгену. По истечении этого времени, такой сайленсинг приводит к снижению уровня экспрессии трансгенов для большинства неспецифических событий инсерции. Кроме того, интеграция трансгена редко происходит в каждой клетке-мишени, что может затруднять обеспечение достаточно высокого уровня экспрессии представляющего интерес трансгена для достижения желаемого терапевтического эффекта.
[0005] В последние годы была разработана новая стратегия интеграции трансгенов, в которой используется расщепление под действием сайт-специфических нуклеаз (например, нуклеаз «цинковые пальцы» (ZFN), нуклеаз эффекторного домена, подобных активатору транскрипции (TALEN), система CRISSPR/Cas, в которой используется сконструированная cr-РНК/tracr-РНК («одноцепочечная руководящая РНК») для нацеленного специфического расщепления и т.п.) для обеспечения нацеленного смещения инсерции в сторону выбранного геномного локуса. См. например, патенты США №№ 9937207; 9873894; 9567573; 9394545; 9255250; 9045763; 9005973; 8956828; 8945868; 8703489; 8586526; 6534261; 6599692; 6503717; 6689558; 7067317; 7262054; 7888121; 7972854; 7914796; 7951925; 8110379; 8409861; публикации эаявок на патенты США №№ 2017/0211075; 2003/0232410; 2005/0208489; 2005/0026157; 2005/0064474; 2006/0063231; 2008/0159996; 2010/00218264; 2012/0017290; 2011/0265198; 2013/0137104; 2013/0122591; 2013/0177983, 2013/0177960 и 2015/0056705. Кроме того, были разрабатаны нацеленные нуклеазы на основе системы Argonaute (например, от T. thermophilics, известной как «TtAgo», см. Swarts et al. (2014) Nature 507 (7491): 258-261), которая может быть также использована для редактирования генома и генотерапии. Этот способ интеграции трансгенов, опосредованной нуклеазой, в будущем позволит повысить уровень экспрессии трансгена, повысить безопасность и увеличить длительность экспрессии по сравнению с классическими методами интеграции, поскольку этот метод позволяет выбрать точное положение трансгена с минимальным риском сайленсинга гена или активации находящихся поблизости онкогенов.
[0006] ACTR (связанные с антителом Т-клеточные рецепторы) представляют собой сконструированные Т-клеточные компоненты, способные связываться с экзогенно продуцируемым антителом. Связывание антитела с компонентом ACTR позволяет Т-клетке взаимодействовать с антигеном, распознаваемым антителом, а в случае присутствия антигена, ACTR, содержащийся в Т-клетке, стимулирует взаимодействие с антигеном (см. публикацию заявки на патента США 20150139943).
[0007] Т-клеточный рецептор (TCR) представляет собой основную часть селективной активации Т-клеток. При некотором сходстве с антителом, антиген-распознающая часть TCR обычно состоит из двух цепей α и β, которые подвергаются сборке с образованием гетеродимера. Сходство антител лежит в основе способа, в котором один ген, кодирующий комплекс цепей TCR-альфа и -бета, объединяет их вместе. Каждая из цепей TCR-альфа (TCR-α) и TCR-бета (TCR-β) состоит из двух областей, C-концевой константной области и N-концевой вариабельной области. Геномные локусы, которые кодируют цепи TCR-альфа и TCR-бета, напоминают антитело-кодирующие локусы, в которых ген TCR-α содержит V- и J-сегменты помимо константной области (часто называемые TRAC), а локус бета-цепи содержит, помимо V- и J-сегментов, D-сегменты. В случае локуса TCR-β, присутствуют две дополнительных различных константных области, которые были выбраны в процесса отбора. В процессе развития Т-клеток, различные сегменты подвергаются рекомбинации, в результате чего каждая Т-клетка содержит уникальную вариабельную область TCR в альфа- и бета-цепях, называемую гипервариабельной областью (CDR), и такой организм имеет большой репертуар Т-клеток, которые вследствие их уникальных CDR, способны взаимодействовать с уникальными антигенами, представленными антигенпрезентирующими клетками. После реаранжировки генов TCR-α или TCR-β, экспрессия второго соответствующего TCR-α или TCR-β подавляется, в результате чего каждая T-клетка экспрессирует только одну уникальную структуру TCR по механизму, называемому «аллельным исключением антигенных рецепторов» (см. Brady et al. (2010) J. Immunol 185: 3801-3808).
[0008] Во время активации Т-клеток, TCR взаимодействует с антигенами, представленными как пептиды на главном комплексе гистосовместимости (MHC) антигенпрезентирующей клетки. Распознавание комплекса антиген-МНС посредством TCR приводит к стимуляции Т-клеток, и, в свою очередь, к дифференцировке хелперных T-клеток (CD4+) и цитотоксических Т-лимфоцитов (CD8+), входящих в состав лимфоцитов памяти и эффекторных лимфоцитов. Затем эти клетки могут подвергаться клональному размножению с образованием активированной субпопуляции во всей популяции T-клеток, способных реагировать с одним конкретным антигеном. Человеческий МНС также называется HLA-комплексом (человеческих лейкоцитарных антигенов). Семейство генов МНС подразделяется на три подгруппы: класс I, класс II и класс III. Молекулы МНС класса I имеют β2-субъединицы, а поэтому они могут распознаваться только ко-рецепторами CD8. Молекулы МНС I встречаются в α-цепи, состоящей из трех доменов, α1, α2 и α3. α1 находится на звене β2-микроглобулина, не являющегося молекулой МНС (кодируемого на человеческой хромосоме 15, сокращенно обозначаемой B2M). α3-домен является трансмембранным и заякоривает молекулу МНС класса I на клеточной мембране. Презентированнй пептид удерживается на дне пептид-связывающей бороздки в центральной области α1/α2-гетеродимера (молекулы, состоящей из двух неидентичных субъединиц). Генетически кодируемая и экспрессируемая последовательность аминокислот, последовательность остатков на дне пептид-связывающей бороздки определяет какие именно конкретные пептидные остатки являются связанными.
[0009] Природные клетки-киллеры (NK) представляют собой специализированные клетки, которые являются частью врожденной иммунной системы. Они представляют собой один из трех типов лимфоцитов, обнаруженных в организме: Т-клеток, В-клеток и NK-клеток. Эти клетки играют главную роль в уничтожении опухолевых клеток и инфицированных бактериями и вирусами клеток в организме. См., например, публикацию заявки на патент США № 20140301990. Их цитотоксическая активность опосредована, главным образом, двумя преобладающими путями. Белок, разрушающий мембрану, перфорин, и семейство структурно родственных сериновых протеаз, гранзимов, секретируются посредством экзоцитоза, что приводит к индуцированию апоптоза клетки-мишени. Во втором пути происходит зависимый от каспазы апоптоз, включающий ассоциацию рецепторов гибели (например, Fas/CD95) на клетках-мишенях с их эквивалентными лигандами, такими как FasL, и с апоптоз-индуцирующим лигандом, связанным с фактором некроза опухоли (TRAIL), на NK-клетках, что приводит к каспазо-зависимому апоптозу. Антителозависимая клеточная цитотоксичность (ADCC) может также представлять собой механизм уничтожения опухолевых клеток NK-клетками, поскольку они экспрессируют низкоаффинный Fc-рецептор для IgG, FcγRIII (CD16) (Mandal and Viswanhan (2015) Hemoat/Onc Stem Cell Ther 8(2): 47-55).
[0010] Функционирование NK-клеток регулируется рецепторами широкого ряда, которые экспрессируются на клеточной поверхности. По своей природе, эти рецепторы являются либо ингибирующими, либо активирующими. Семейство ингибирующих рецепторов состоит из рецепторов-киллеров, подобных иммуноглобулинам (KIR), или Ig-подобных рецепторов (CD158), рецепторов лектина C-типа (CD94-NKG2A) и рецепторов, ингибирующих лейкоциты (LIR1, LAIR-1). Активирующими рецепторами являются природные цитотоксические рецепторы (NKp46, NKp44), рецепторы лектина типа С (NKG2D, CD94-NKG2C) и Ig-подобные рецепторы (2В4). Цитотоксичность NK-клеток жестко регулируется балансом между активирующими и ингибирующими сигналами, хотя ингибирующие сигналы могут быть преобладающими по сравнению с активирующими сигналами. Ингибирующие рецепторы NK-клеток распознают сами молекулы МНС класса I, что предотвращает активацию NK-клеток и объясняет аутотолерантность и предотвращение гибели клеток-хозяев. Было также показано, что NK-клетки могут активироваться, если они сталкиваются с клетками, в которых отсутствует сама молекула МНС класса I («HLA-I»). Экспрессия рецепторов, ингибирующих связывание с HLA-I (например, KIR, NKG2A/CD94, IL2 и LIR1), приводит к вырабатыванию толерантности NK-клеток к нормальным клеткам. NK-клетки обычно атакуют аномальные клетки, которые ингибируют поверхностные молекулы HLA-I, и такой механизм называется «отсутствием самораспознавания». Однако, «отсутствие самораспознавания» приводит к вырабатыванию чувствительности к хорошо известным NK-клеткам, которые лизируют клетки, не имеющие HLA-I (Crux and Elahi (2017), Front Immunol 8: 832).
[0011] HLA-E и HLA-G принадлежат к неклассическим паралогам тяжелой цепи HLA класса I, также называемым классом HLA-Ib. Эта молекула класса I представляет собой гетеродимер, состоящий из тяжелой цепи, кодируемой геном HLA-E или HLA-G, соответственно, и легкой цепи, кодируемой микроглобулином бета-2, B2M. Человеческий HLA-E представляет собой неклассическую молекулу МНС класса I, которая характеризуется ограниченным полиморфизмом и более низкой экспрессией на клеточной поверхности по сравнению с классическими паралогами. Гаплотипы HLA-E включают HLA-E*0101 (также обозначаемую как ER или HLA-E0101), которая отличается от HLA-E*0103 (также обозначаемой как EG или HLA-E0103) только в одном положении аминокислоты, а именно, где в положении 107 в HLA-E0101 присутствует аргинин, а в HLA-E0103 в этом положении, аргинин заменен глицином (Celik et al. (2016) Immunogenetics 68 (1):29-41). В 2017 году, 52 аллеля/гаплотипа HLA-G были официально внесены в Международную базу данных «International Immunogenetics Project/human majoy histocompatibility complex (IMGT/HLA)» (версия 3.23.0). Большинство полиморфизмов по одному нуклеотиду (SNP) в HLA-G-кодирующей области представляет собой кодирующие синонимичные мутации или интронные варианты. Из 52 аллелей, которые официально известны как IMGT/HLA, только 13 аллелей кодируют 4 различных полноразмерных молекулы HLA-G и часто наблюдаются в организмах во всем мире (Prakash et al. (2017) Antropol Open J. 2(1):1-9.doi:10.17140/ANT-POJ-2-106). Одним из наиболее распространенных аллелей HLA-G (часто обозначаемых как аллель дикого типа) является аллель HLA-G*01:01 (Metcalfe et Al. (2013) Int. J. Circular Health 72. Doi: 10.3402/ijch.vy2i0,21350).
[0012] HLA-E играет узкоспециализированную роль в распознавании клеток природными клетками-киллерами (NK-клетками). NK-клетки распознают пептидный комплекс HLA-E+ посредством гетеродимерного ингибирующего рецептора CD94/NKG2 A/B/C. При связывании CD94/NKG2A или CD94/NKG2B, они ингибируют цитотоксическую активность NK-клеток, что предотвращает лизис клеток, однако связывание HLA-E с CD94/NKG2C приводит к активации NK-клеток. Было показано, что такое взаимодействие стимулирует размножение субпопуляций NK-клеток при противовирусных ответах. Человеческий лейкоцитарный антиген-Е представляет собой неклассический HLA-Ib с широким распределением в тканях, но из всех молекул МНС класса I, он является наименее полиморфным. Такой антиген активируется в результате стрессов, вызываемых микроокружением, таким как гипоксия и дефицит глюкозы в опухолях, но транскрибируется на более низких уровнях, чем молекулы HLA-Ia (Crux and Elahi, ibid).
[0013] HLA-G обычно экспрессируется на мембранах клеток плаценты плода и может играть определенную роль в вырабатывании иммунологической толерантности при беременности. Ген HLA-G очень похож на гены HLA-А, B и C, но белки, кодируемые HLA-G, отличаются от продуктов других 3 генов тем, что в них отсутствует большинство внутриклеточных сегментов. С помощью Нозерн-блот-анализа и ОТ-ПЦР-анализа, проводимых с использованием зонда и праймеров, специфичных к локусу HLA-G, было продемонстрировано, что этот ген транскрибируется в различных клетках и тканях взрослых, а также в тканях плода (Onno et al. 1994) Hum. Immun. 41: 79-86, 1994). В большинстве тканей, уровень мРНК был на порядок ниже уровня классических генов класса I в тех же самых тканях. Альтернативный сплайсинг первичного транскрипта HLA-G отличается в различных тканях и может регулироваться по тканеспецифическому механизму. HLA-G представляет собой лиганд для рецептора KIR2DL4, ингибирующего NK-клетки, а поэтому, экспрессия этого HLA трофобластами теоретически будет защищать их от гибели, опосредуемой NK-клетками.
[0014] Адоптивная клеточная терапия (ACT) представляет собой находящуюся на стадии разработки форму противораковой терапии, основанную на доставке опухолеспецифических иммунных клеток пациенту для направленной атаке на клетки и лечения рака у пациента. ACT может включать использование опухоль-инфильтрирующих лимфоцитов (TIL), которые представляет собой Т-клетки, выделенные из собственной опухолевой массы пациента и размноженные vivo для повторного их введения пациенту. Такой подход является перспективным для лечения метастазирующей меланомы, где в одном исследовании наблюдалась длительность ответа >50% (см. например, Rosenberg et al. (2011) Clin Cans Res 17 (13): 4550). TIL являются перспективным источником клеток, поскольку они представляют собой смешанную популяцию собственных клеток пациента, которые имеют Т-клеточные рецепторы (TCR), специфичные к опухолеассоциированным антигенам (TAA), присутствующим на опухоли (Wu et al (2012) Cancer J. 18 (2): 160). Другие подходы включают редактирование Т-клеток, выделенных из крови пациента, для конструирования этих клеток, так, чтобы они были чувствительными к опухоли в соответствии с определенным механизмом (Kalos Et al. (2011) Sci Transl Med 3 (95): 95ra73).
[0015] Химерные антигенные рецепторы (CAR) представляют собой молекулы, сконструированные для нацеливания на иммунные клетки, специфичные к молекулярным мишеням, экспрессируемым на клеточных поверхностях. В своей основной форме, эти рецепторы, вводимые в клетку, где они специфически связываются с доменом, экспрессируются на внешней поверхности клетки и передают сигналы по определенным путям вовнутрь клетки, так, что, если домен специфически взаимодействует с мишенью, то клетка становится активированной. В большинстве случаев, CAR получают из функциональных доменов T-клеточных рецепторов (TCR), где антигенспецифический домен, такой как одноцепочечный фрагмент антитела (scFv) или рецептор какого-либо другого типа (например, рецептор цитокина), присоединяется к сигнал-передающему домену, такому как мотив активации иммунорецептора на основе тирозина (ITAM) и другие костимулирующие домены. Эти конструкции вводят в Т-клетку ex vivo, так, чтобы при повторном введении T-клеток пациенту, они активировались в присутствии клетки, экспрессирующей антиген-мишень, что будет приводить к атаке на клетку-мишень посредством активированной T-клетки по механизму, не зависящему от МНС (см., Chicaybam et al. (2011) Int Rev Immunol 30: 294-311). Могут быть проведены различные способы переноса генов в Т-клетки, и такими способами являются способы с использованием ретровирусных и лентивирусных векторов, позволяющих осуществлять перманентную модификацию генома, а также способы на основе РНК, позволяющие осуществлять временную экспрессию гена. Преимущество способов на основе ретровирусов и лентивирусов заключается в длительной экспрессии генов, а следовательно, это дает возможность осуществлять подавление заболевания на длительное время путем одного введения сконструированных T-клеток (если эти T-клетки сохраняются). Недостатками перманентной модификации являются длительная цитотоксичность по отношению к мишени и теоретически риск трансформации, если такая инсерция гена приводит к нарушению регуляции находящегося рядом онкогена. Перенос генов с использованием матричной РНК позволяет осуществлять временную экспрессию без интеграции в геном, что позволяет устранить любую проблему, связанную с трансформацией после рандомизированной интеграции. Т-клетки, сконструированные путем введения РНК посредством электропорации, обладают выраженной репликативной способностью и могут продуцировать значительные противопухолевые ответы. Однако, экспрессия обычно продолжается 7 дней или менее, что позволяет осуществлять подавление заболевания на длительное время, хотя, все же возможно, что это потребует многократных инфузий при проведении такого способа (Maude et al.. (2015) Blood 125 (26): 4017-4023).
[0016] Адоптивная клеточная терапия с использованием Т-клеток, модифицированных ex vivo с использованием сконструированного TCR или CAR, представляет собой перспективный клинический подход для лечения заболеваний нескольких типов. Так, например, раковыми опухолями и их антигенами-мишенями являются фолликулярная лимфома (CD20 или GD2), нейробластома (CD171), неходжкинская лимфома (CD19 И CD20), лимфома (CD19), глиобластома (IL13Rα2), хронический лимфоцитарный лейкоз или ХЛЛ и острый лимфоцитарный лейкоз или ОЛЛ (в обоих случаях CD19). Вирус-специфические CAR были также сконструированы для атаки на клетки, содержащие вирус, такой как ВИЧ. Так, например, клиническое испытание было начато с использованием CAR, специфичного к Gp100 для лечения ВИЧ (Chicaybam, ibid), и несколько испытаний были продолжены с использованием CD19-специфических CAR (Maude (2015) ibid) для лечения ХЛЛ (см., Kalos et al. (2011) Sci. Transl. Med. 3(95):95Ra73 и Kochenderfer JN and Rosenberg SA (2013) Nat Rev Clin Oncol. (5): 267-276), и лечения ОЛЛ (См., Maude et al. (2014) N Engl J Med. 371 (16): 1507-1517 и Lee et al. (2015) Lancet 385 (9967): 517-528).
[0017] Несмотря на недавние успехи адоптивной клеточной терапии, остаются некоторые значительные проблемы, которые необходимо решить, прежде чем широко применять эту технологию. Так, например, эндогенный αβ-Т-клеточный рецептор (TCR), присутствующий на вводимых аллогенных (донорных) сконструированных T-клетках, может распознавать основные и вспомогательные антигены гистосовместимости у реципиента, что будет приводить к развитию заболевания «трансплантат против хозяина» (GVHD). В результате этого, большинство современных клинических испытаний по инфузии аутологичных CAR+-T-клеток (T-клеток, содержащих экспрессированную CAR-кодирующую последовательность), основано на иммунологической толерантности для предотвращения TCR-опосредованного нежелательного распознавания нормальных тканей после адоптивного переноса. Этот подход обеспечил начальный клинический успех по нацеливанию на CD19+-злокачественные новообразования, но был ограничен по времени и требовал больших затрат на получение T-клеточных продуктов, специфичных для пациента (Torikai et al. (2012) Blood 119: 5697-5705). Другой проблемой, которая возникает при лечении пациентов этими сконструированными T-клетками, является один из рецидивов, когда было обнаружено, что у пациентов, у которых вначале была диагностирована опухоль, появляется только в отношении опухолевых клеток, в которых ингибировался антиген-мишень, и такой рецидив называется «ускользанием от иммунного ответа» (Sotillo et al. (2015) Cancer Dis 5(12): 1282-1295 и Gardner et al. (2016) Blood; 127(20): 2406-2410).
[0018] Таким образом, несмотря на возможности применения адоптивной клеточной терапии, необходимо разработать способы и композиции, которые могут быть использованы для повышения эффективности и специфичности «вооруженных» T-клеток. В частности, необходимо разработать способы для безопасного введения и поддержания экспрессии антигенспецифических комплексов, включающих CAR и сконструированные TCR. Кроме того, необходимо разработать способы для получения универсальных сконструированных T-клеток, которые не содержат маркеров, распознаваемых иммунной системой пациента {например, HLA-комплексами), и которые могут быть использованы для лечения большого числа пациентов без необходимости получения модифицированных клеток для лечения каждого пациента.
Сущность изобретения
[0019] В настоящей заявке раскрываются композиции и способы для введения химерного антигенного рецептора в сконструированную T-клетку. Способы согласно изобретению включают нацеленное введение гена, кодирующего один или более донорных трансгенов (например, CAR), в клетку, включая нацеленную интеграцию одного или более CAR в один или более генов TCR и/или ген B2M (также называемую CAR+-клеткой). Полученные таким образом клетки согласно изобретению содержат один или более CAR, специфичных к выбранному антигену (например, CD20, CD171, CD19) и/или к IL13Rα2, а также не содержат функционального эндогенного TCR и/или комплекса HLA. В некоторых вариантах осуществления изобретения, клетки дополнительно модифицируют в другом локусе (например, в гене контрольной точки, таком как CISH или PD1), так, чтобы полученные клетки содержали отредактированные модификации в TCR, B2M и в любых других генах (например, CISH), и чтобы полученные клетки не содержали нативного TCR, комплекса HLA и не имели нокаута в дополнительных генах, таких как ген контрольной точки CISH, а также содержали донорный трансген (например, CAR). Эти клетки могут быть использованы для удаления других клеток, несущих антиген-мишень. В предпочтительных вариантах осуществления изобретения, сконструированные клетки CAR+-Т-клетки получают для адоптивной клеточной терапии. В некоторых вариантах осуществления изобретения, сконструированные клетки CAR+-Т-клетки экспрессируют модифицированный комплекс HLA-E и/или HLA-G, что позволяет этим клеткам избежать NK-опосредуемого уничтожения. Кроме того, настоящее изобретение относится к способам и композициям для экспрессии антигена на клетках несоответствующих типов в целях анализа активности сконструированных T-клеток in vitro.
[0020] В одном из своих аспектов, настоящее изобретение относится к генетически модифицированной Т-клетке (и к популяциям Т-клеток, содержащих множество таких генетически модифицированных Т-клеток), где генетически модифицированная Т-клетка включает: первый полинуклеотид, кодирующий химерный антигеннный рецептор (CAR), и второй полинуклеотид, содержащий последовательности, кодирующие гибридный белок, содержащий белок бета-2-микроглобулин (B2M), и белок HLA-E и/или HLA-G, где первый и второй полинуклеотиды интегрированы в ген Т-клеточного рецептора-α (TCRA). В некоторых вариантах осуществления изобретения, один или более дополнительных генов являются инактивированными в клетке, например, ген B2M и/или один или более генов иммунологической контрольной точки (например, PD1, CISH и т.п.). В любых описанных здесь T-клетках, первый и второй полинуклеотиды могут быть присоединены посредством саморасщепляющегося пептида (например, пептида 2А, также называемого «Р2А» или пептидом «Р2А»). Кроме того, в некоторых вариантах осуществления изобретения, второй полинуклеотид также содержит последовательность, кодирующую линкер (например, 1, 2, 3, 4, 5, 6 или более копий линкеров G4S) между B2M и HLA-Е-или HLA-G-кодирующими последовательностями.
[0021] Настоящее изобретение также относится к способам получения описанных здесь генетически модифицированных Т-клеток, где указанные способы включают: расщепление гена TCR-α в выделенной Т-клетке под действием нуклеазы, содержащей ДНК-связывающий домен, который связывается с сайтом-мишенью в гене TCR-α; и введение одного или более доноров, содержащих первый и второй полинуклеотиды, в T-клетку так, чтобы первый и второй полинуклеотиды интегрировались в расщепленный ген TCR-α. В некоторых вариантах осуществления изобретения, донор содержит первый и второй полинуклеотиды, связанные посредством саморасщепляющегося пептида 2А. Кроме того, в некоторых вариантах осуществления изобретения, второй полинуклеотид также содержит последовательность, кодирующую линкер (например, 1, 2, 3, 4, 5, 6 или более копий линкеров G4S) между B2M и HLA-Е-или HLA-G-кодирующими последовательностями. В любом из описанных здесь способов, один или более доноров могут также содержать один или более из следующих компонентов: (1) гомологичные ветви (сцепленные с геном TCR-α), фланкирующие первый и второй полинуклеотиды; (2) один или более промоторов, которые инициируют экспрессию первого и/или второго полинуклеотида, (3) один или более последовательностей энхансеров TCR-α, (4) 3'- нетранслируемую область (3’UTR) и/или 5'-нетранслируемую область (5’UTR); (5) последовательность WPRE (например, в 3’UTR); и/или (5) последовательность гена бета-глобина Xenopus (например, в 5’UTR).
[0022] В другом аспекте изобретения, описанными здесь клетками являются выделенные клетки (например, эукариотические клетки, такие как клетки млекопитающих, включая лимфоидные клетки, стволовые клетки (например, iPSC, эмбриональные стволовые клетки, MSC или HSC) или клетки-предшественники), где один или более экзогенных CAR экспрессируются после сайт-специфической интеграции гена CAR в геном клетки, включая, например, эндогенный ген. В некоторых вариантах осуществления изобретения, CAR встраивают в ген TCR (TCRA и/или TRBC) и/или, необязательно, в ген контрольной точки (например, CISH, PD1 и т.п.) и/или в ген B2M с использованием нуклеаз, нацеленных на эти гены, как описано в настоящей заявке или в литературе. В других вариантах осуществления изобретения, CAR встраивают в ген TCRA, а в других вариантах, его встраивают в ген TRBC. В некоторых вариантах осуществления изобретения, CAR встраивают в ген B2M, а в других вариантах осуществления изобретения, его встраивают в ген контрольной точки (например, CISH, PD1 и т.п.) помимо гена TCR. В клетках, полученных этим способом, введение гена, кодирующего CAR, приводит к нокауту эндогенного TCR, гена контрольной точки (например, CISH) и/или гена B2M и к прекращению образования функционального эндогенного TCR или комплекса HLA, и/или к нокауту гена контрольной точки (например, CISH). В любой из описанных клеток, если CAR интегрируется в ген B2M, то один или более генов TCR в клетке могут быть инактивированы в этой клетке (например, посредством инактивации, опосредуемой нуклеазой), что будет приводить к нокауту множества генов (например, гена B2M и одного или более генов TCR и гена контрольной точки (например, CISH)). Аналогичным образом, при интеграции CAR в ген TCR, один или более различных генов TCR, ген контрольной точки (например, CISH) и/или ген B2M могут инактивироваться в этой клетке (например, посредством инактивации, опосредуемой нуклеазой), что будет приводить к нокауту множества генов (например, одного или более генов TCR, гена CISH и гена B2M).
[0023] В других вариантах осуществления изобретения, описанные здесь генетически модифицированные клетки (Т-клетки) также содержат модифицированный ген HLA-E или HLA-G (трансген), который встраивается в клетки, кодирующие CAR, например, в ген TCR (TCRA) вместе с CAR. В некоторых вариантах осуществления изобретения, ген HLA-Е или HLA-G (например, экзогенный трансген) совместно вводят в сконструированные клетки вместе с геном CAR. В некоторых вариантах осуществления изобретения, трансген HLA-Е или HLA-G вводят до или после введения гена CAR. В других вариантах осуществления изобретения, трансген содержит последовательность гена HLA-Е или HLA-G, которая присоединяется к последовательности, кодирующей линкер, который затем присоединяется к последовательности гена B2M (то есть, трансгена, содержащего последовательность, кодирующую HLA-E или HLA-G, линкер и B2M-кодирующую последовательность), что приводит к кодированию модифицированного комплекса HLA-E или HLA-G, где ген B2M является частью открытой рамки считывания вместе с геном HLA-E или HLA-G, в результате чего после экспрессии может образовываться стабильный комплекс HLA-E или HLA-G вместе с B2M, экспрессируемым из трансгена. В этом варианте осуществления изобретения, благодаря присутствию линкера, субъединица B2M может ассоциироваться только с белком HLA-E или HLA-G с образованием стабильного комплекса HLA-E или HLA-G на клеточной поверхности и не может ассоциироваться с любыми другими HLA-комплексами. В других вариантах Осуществления изобретения, трансген HLA-E или HLA-G-линкер-B2M присоединен к последовательности, кодирующей представляющий интерес CAR, где такая конструкция также включает последовательность саморасщепляющегося пептида 2А между геном CAR и конструкцией HLA-E или HLA-G-линкер-B2M. В других вариантах осуществления изобретения, этот комплекс CAR-HLA-G или CAR-HLA-E также содержит гомологичные ветви, что позволяет комплексу интегрироваться в геном в определенном сайте. В некоторых вариантах осуществления изобретения, сайт интеграции может индуцировать транскрипцию, инициируемую эндогенным промотором в сайте интеграции, или конструкция CAR-HLA-E или CAR-HLA-G может быть присоединена к промотору, который также является интегрированым, что позволяет экспрессионной конструкции интегрироваться в геном, но инициироваться гетерологичным промотором. В некоторых вариантах осуществления изобретения, комплекс CAR-HLA-E или CAR-HLA-G (трансген) функционально присоединен к промотору, что обеспечивает его экспрессию из эписомной нуклеиновой кислоты (например, кДНК). В донорную конструкцию могут быть также включены дополнительные последовательности, включая, но не ограничиваются ими, последовательности WPRE и/или энхансерные последовательности TCRa. В других вариантах осуществления изобретения, промотор представляет собой тканеспецифический промотор. Кроме того, любая из этих экспрессионных конструкций может дополнительно содержать AAV-ITR для упаковки в AAV-частицу. В некоторых случаях, AAV представляет собой AAV5, AAV6, AAV9 или AAV любого другого типа.
[0024] В некоторых аспектах изобретения, ген TCR модулируют путем модификации экзона c2 гена TCRA. Ген B2M модулируют путем модификации экзона 1 гена B2M. В некоторых вариантах осуществления изобретения, модификация находится внутри последовательности и/или рядом с последовательностью, содержащей 12-25 (включая сайты-мишени 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25) или более нуклеотидов, как показано в сайтах-мишенях Таблицы 1, внутри сайта-мишена и/или рядом с сайтом-мишенью, как показано в Таблице 1 (SEQ ID NO: 1 и 2; SEQ ID NO 3 и 4, SEQ ID NO: 5 и 6; SEQ ID NO: 39 и 40; SEQ ID NO: 50 и 51); в пределах 1-5, в пределах 1-10 или в пределах 1-20 пар оснований на любой стороне (фланкирующих геномные последовательности) сайтов-мишеней, показанных в Таблице 1 (SEQ ID NO: 1-6, 39, 40, 50 или 51); или в TTGAAA для пары TCRA ZFN или в GCCTTA для пары B2M ZFN. Альтернативно или дополнительно, модификации могут быть также внесены в последовательности (например, геномные последовательности) между описанными здесь спаренными сайтами-мишенями (например, между сайтами-мишенями для пар нуклеаз, представленных в Таблице 1, включая модификации между сайтами-мишенями для 55266 и 53853 (между SEQ ID NO: 1 и SEQ ID NO: 2) или для 68813 и 68812 (между SEQ ID NO: 39 и SEQ ID NO: 40) в гене TCRA; и/или между сайтами-мишенями для 57071 и 57531 (между SEQ ID NO: 3 и SEQ ID NO: 4) в гене B2M; между сайтами-мишенями для 59488 и 59489 в гене CISH (между SEQ ID NO: 50 и SEQ ID NO: 51). В дополнительных вариантах осуществления изобретения, дополнительные геномные модификации вводят между сайтами-мишенями для 37706 и 48407 в гене HPRT (SEQ ID NO: 5 и 6). Генетическая модификация может быть осуществлена путем введения в клетку экзогенной гибридной молекулы, содержащей функциональный домен (например, домен регуляции транскрипции, домен нуклеазы) и ДНК-связывающий домен, включая, но не ограничиваясь ими, домен(ы), связывающийся(еся) с ДНК ZFP, домены, связывающиеся с ДНК TALE и домены, связывающиеся с ДНК и оцрРНК системы CRISPR/Cas. Модифицированными клетками могут быть, но не ограничиваются ими, (i) клетка, содержащая экзогенный фактор транскрипции, включающий ДНК-связывающий домен, который связывается с сайтом-мишенью, как показано в Таблице 1, и домен регуляции транскрипции, где фактор транскрипции модулирует экспрессию гена B2M и/или TCRA и/или (ii) клетка, содержащая инсерцию и/или делецию в одном или более сайтах-мишенях или возле этих сайтов (внутри этих сайтов, между ними или рядом с ними), как показано в Таблице 1; в пределах 1-5, 1-10 или 1-20 пар оснований на любой стороне (фланкирующей геномной последовательности) сайтов-мишеней, показанных в Таблице 1; или в пределах TTGAAA для TCRA или GCCTTQ для B2M; и/или между описанными здесь спаренными сайтами-мишенями (например, сайтами-мишенями для пар нуклеаз, показанных в Таблице 1). Клетка может также включать дополнительные модификации, например дополнительный инактивированный ген TCR или B2M, инактивированный ген ULA, ген CISH, PD1 и/или CTLA4 и/или трансген, кодирующий связанный с антителом Т-клеточный рецептор (ACTR) и/или трансген, кодирующий CAR, и/или трансген, кодирующий антитело. Клетка может также включать экспрессионную конструкцию для модифицированного комплекса HLA-E или HLA-G. Настоящее изобретение также относится к фармацевтическим композициям, содержащим любые описанные здесь клетки, а также к способам применения клеток и фармацевтических композиций в терапии ex vivo для лечения расстройства (например, рака) у индивидуума.
[0025] Таким образом, в одном из аспектов изобретения описаны генетически модифицированные клетки, в которых, помимо интеграции CAR-кодирующего гена, экспрессия гена TCR и/или B2M является модулированной (например, активированной, ингибированной или инактивированной). В некоторых вариантах осуществления изобретения, клетки также содержат модифицированный комплекс HLA-Е или HLA-G. В предпочтительных вариантах осуществления изобретения, экзон с2 гена TCRA является модулированным, и/или экзон 1 гена B2M является модулированным. Модуляция может быть осуществлена посредством экзогенной молекулы (например, сконструированного фактора транскрипции, содержащего ДНК-связывающий домен и домен активации или ингибирования транскрипции), которая связывается с геном TCR и/или B2M и регулирует экспрессию TCR и/или B2M, и/или посредством модификации последовательности гена TCR и/или B2M (например, с использованием нуклеазы, которая расщепляет ген TCR или B2M и модифицирует генную последовательность в результате инсерций и/или делеций). В некоторых вариантах осуществления изобретения описаны клетки, которые содержат сконструированную нуклеазу (например, ZFN, TALEN, CRISPR/Cas-нуклеазу) для инициации нокаута гена TCR и/или B2M и необязательно дополнительных генов, таких как гены иммунологической контрольной точки (например, CISH (нуклеазы Таблицы 1), PD1 (патент США № 8563314, CTLA-4 и т.п.). В других вариантах осуществления изобретения описаны клетки, которые содержат один или более сконструированных факторов транскрипции (TF), так, чтобы это приводило к модуляции экспрессия гена TCR и/или B2M. В некоторых вариантах осуществления изобретения, клетками являются Т-клетки. Также описаны клетки, в которых экспрессия гена TCR и/или B2M является модулированной, и клетки, которые были дополнительно сконструированы так, чтобы они содержали по меньшей мере один экзогенный трансген и/или имели дополнительный нокаут по меньшей мере одного эндогенного гена (например, гена бета-2-микроглобулина (B2M) или гена TCR, и/или гена иммунологической контрольной точки, такого как PD1 и/или CTLA4), или их комбинации. Экзогенный(е) трансген(ы) может(могут) быть интегрирован(ы) в ген TCR или B2M (например, в случае нокаута гена TCR или B2M), а также в один или более генов, включая, но не ограничиваясь ими, ген, не являющийся ни TCR, ни B2M, такой как ген контрольной точки (например, CISH, PD1), или безопасный якорный ген. В некоторых вариантах осуществления изобретения, один или более трансгенов CAR (например, CD19-специфических CAR) интегрированы в ген TRAC, ген CISH и/или ген B2M, а также в безопасный якорный ген (например, AAVS1, Rosa, альбумина, HPRT и т.п.). Одинаковые или различные трансгены CAR могут быть интегрированы в одинаковые или различные локусы. В тех вариантах осуществления изобретения, в которых один или более CAR интегрированы в один или более безопасных якорных генов, помимо гена TRAC, CISH и/или B2M, также предпочтительно подвергают инактивации (например, путем инактивации, опосредуемой нуклеазой, инсерции и/или делеции). В некоторых случаях, экзогенный трансген кодирует ACTR. Трансгенная конструкция может быть встроена по механизмам, инициируемым HDR или NHEJ. В некоторых аспектах изобретения, CAR-содержащие клетки с модулированной экспрессией TCR и/или B2M содержат по меньшей мере экзогенный ACTR. Некоторые клетки, содержащие модулятор TCR и/или B2M, также имеют нокаут одного или более генов-ингибиторов контрольной точки. В некоторых вариантах осуществления изобретения, геном-ингибитором контрольной точки является CISH (см., Таблицу 1 ниже и предварительную заявку на патент США 62/583724), а в других вариантах осуществления изобретения, геном-ингибитором контрольной точки является PDl (см. патент США № 8563314). В других вариантах осуществления изобретения, ингибитором контрольной точки является CTLA4. См., например, патент США № 9597357. В других аспектах изобретения, CAR-содержащая и TCR- и/или B2M-модулированная клетка также имеет нокаут PDl и/или нокаут CTLA4 и/или нокаут CISH (см., ниже Таблицу 1). В некоторых вариантах осуществления изобретения, модулированным геном TCR является ген, кодирующий TCR-β (TCRB). В некоторых вариантах осуществления изобретения, это может быть достигнуто посредством нацеленного расщепления константной области этого гена (константной области TCR-β или TRBC). В некоторых вариантах осуществления изобретения, модулированным геном TCR является ген, кодирующий TCRα (TCRA). В дополнительных вариантах осуществления изобретения, инсерция достигается посредством нацеленного расщепления константной области гена TCRα, включая нацеленное расщепление константной области гена TCRα (называемого далее последовательностью «TRAC»). В некоторых вариантах осуществления изобретения, клетки с модифицированным геном TCR, также имеют модифицированный ген B2M, ген CISH (в сайтах для нацеливания, раскрытых в Таблице 1), ген PDl и/или ген CTLA4, гены HLA-A,-B,-C или ген ТАР или любые их комбинации. В других вариантах осуществления изобретения, регулятор HLA класса II, CIITA (см., патент США № 8945868), также является модулированным.
[0026] В некоторых вариантах осуществления изобретения, описанные здесь CAR-содержащие клетки также содержат модификацию (например, делецию и/или инсерцию, связывание сконструированного TF для ингибирования экспрессии TCR) в гене TCRA (например, модификацию экзона c2). В некоторых вариантах осуществления изобретения, модификация находится внутри любых сайтов-мишеней и/или рядом с этими сайтами, показанными в Таблице 1 (SEQ ID NO: 1 или 2) и/или между спаренными сайтами-мишенями (например, сайтами-мишенями пар нуклеаз, показанными в Таблице 1), включая модификацию путем связывания, расщепления, инсерции и/или делеции одного или более нуклеотидов в любой из этих последовательностей и/или в пределах 1-50 пар оснований (включая любые пары оснований между ними, такие как 1-5, 1-10 или 1-20 пар оснований) геннных (геномных) последовательностей, фланкирующих эти последовательности в гене TCRA. В некоторых вариантах осуществления изобретения, клетки содержат модификацию (связывание, расщепление, инсерции и/или делеции) в пределах одной или более из следующих последовательностей: TTGAAA в гене TCRA (например, экзона c2) или GCCTTA в гене B2M (например, экзона 1). В некоторых вариантах осуществления изобретения, модификация включает связывание описаного здесь сконструированного TF так, чтобы экспрессия гена TCRA модулировалась, например, ингибировалась или активировалась. В других вариантах осуществления изобретения, модификация представляет собой генетическую модификацию (изменение нуклеотидной последовательности) в сайте(ах) или рядом с сайтами связывания с нуклеазой (мишенью) и/или сайтами расщепления, включая, но не ограничиваясь ими, модификации в последовательностях в пределах 1-300 пар оснований (или любых пар оснований между ними), расположенных выше или ниже и/или включая 1 или более пар оснований сайта(ов) расщепления и/или связывания; модификации в пределах 1-100 пар оснований (или любых пар оснований между ними) включительно, и/или с любой стороны сайта(ов) связывания и/или расщепления; модификации в пределах 1-50 пар оснований (или любых пар оснований между ними) включительно, и/или с любой стороны (например, от 1 до 5, от 1 до 10, от 1 до 20 или более пар оснований) сайта(ов) связывания и/или расщепления; и/или модификации в одной или более парах оснований в сайте связывания с нуклеазой и/или в сайте расщепления нуклеазой. В некоторых вариантах осуществления изобретения, модификация присутствует в спаренных сайтах-мишенях или возле этих сайтов (например, 1-300 пар оснований, 1-50, 1-20, 1-10 или 1-5 или любое число пар оснований между ними) и/или между спаренными сайтами-мишенями (см., например, Таблицу 1) последовательности гена TCRA внутри, вокруг или между любыми из SEQ ID NO: 1 и SEQ ID NO: 2 и/или SEQ ID NO: 39 и 40. В некоторых вариантах осуществления изобретения, модификация включает модификации гена TCRA, находящегося рядом с одной или более последовательностями, представленными в SEQ ID NO: 1 и SEQ ID NO: 2, SEQ ID NO: 39 и 40, или в TTGAAA гена TCRA (например, экзона c2), например, модификацию одной или более пар оснований в одной или более из этих последовательностей. В некоторых вариантах осуществления изобретения, опосредованные нуклеазой генетические модификации присутствуют между спаренными сайтами-мишенями (если для расщепления мишени используется димер). Опосредованные нуклеазой генетические модификации могут включать инсерции и/или делеции любого числа пар оснований, включая инсерции некодирующих последовательностей любой длины и/или трансгенов любой длины и/или делеции от 1 пары оснований до более 1000 т.п.о. (или любое число пар оснований между ними, включая, но не ограничиваясь ими, 1-100 пар оснований, 1-50 пар оснований, 1-30 пар оснований, 1-20 пары оснований, 1-10 пар оснований или 1-5 пар оснований).
[0027] В некоторых вариантах осуществления изобретения, модификация находится внутри любых сайтов-мишеней и/или рядом с этими сайтами, показанными в Таблице 1 (SEQ ID NO: 3 и SEQ ID NO: 4) и/или между спаренными сайтами-мишенями (например, сайтами-мишенями пар нуклеаз, показанными в Таблице 1) гена В2М (например, модификация экзона 1), включая модификацию путем связывания, расщепления, инсерции и/или делеции одного или более нуклеотидов в любой из этих последовательностей и/или в пределах 1-50 пар оснований (включая любые пары оснований между ними, такие как 1-5, 1-10 или 1-20 пар оснований) генных (геномных) последовательностей, фланкирующих эти последовательности в гене В2М. В некоторых вариантах осуществления изобретения, клетки содержат модификацию (связывание, расщепление, инсерции и/или делеции) в пределах GCCTTA в экзоне 1 гена B2M. В некоторых вариантах осуществления изобретения, модификация включает связывание описаного здесь сконструированного TF так, чтобы экспрессия гена В2М модулировалась, например, ингибировалась или активировалась. В других вариантах осуществления изобретения, модификация представляет собой генетическую модификацию (изменение нуклеотидной последовательности) в сайте(ах) или рядом с сайтами связывания с нуклеазой (мишенью) и/или сайтами расщепления, включая, но не ограничиваясь ими, модификации в последовательностях в пределах 1-300 пар оснований (или любых пар оснований между ними), расположенных выше или ниже и/или включая 1 или более пар оснований сайта(ов) расщепления и/или связывания; модификации в пределах 1-100 пар оснований (или любых пар оснований между ними) включительно, и/или с любой стороны сайта(ов) связывания и/или расщепления; модификации в пределах 1-50 пар оснований (или любых пар оснований между ними) включительно, и/или с любой стороны (например, от 1 до 5, от 1 до 10, от 1 до 20 или более пар оснований) сайта(ов) связывания и/или расщепления; и/или модификации в одной или более парах оснований в сайте связывания с нуклеазой и/или в сайте расщепления нуклеазой. В некоторых вариантах осуществления изобретения, модификация присутствует в спаренных сайтах-мишенях или возле этих сайтов (например, 1-300 пар оснований, 1-50, 1-20, 1-10 или 1-5 или любое число пар оснований между ними) и/или между спаренными сайтами-мишенями (см., например, Таблицу 1) последовательности гена В2М внутри, вокруг или между любыми из SEQ ID NO: 3 и SEQ ID NO: 4. В некоторых вариантах осуществления изобретения, модификация включает модификации гена В2М в одной или более последовательностях, представленных в SEQ ID NO: 3 и SEQ ID NO: 4, или в GCCTTA в экзоне 1 гена В2М, например, модификацию одной или более пар оснований в одной или более из этих последовательностей. В некоторых вариантах осуществления изобретения, опосредованные нуклеазой генетические модификации присутствуют между спаренными сайтами-мишенями (если для расщепления мишени используется димер). Опосредованные нуклеазой генетические модификации могут включать инсерции и/или делеции любого числа пар оснований, включая инсерции некодирующих последовательностей любой длины и/или трансгенов любой длины и/или делеции от 1 пары оснований до более 1000 т.п.о. (или любое число пар оснований между ними, включая, но не ограничиваясь ими, 1-100 пар оснований, 1-50 пар оснований, 1-30 пар оснований, 1-20 пары оснований, 1-10 пар оснований или 1-5 пар оснований).
[0028] В некоторых вариантах осуществления изобретения, модификация находится внутри любых сайтов-мишеней и/или рядом с этими сайтами, показанными в Таблице 1 (SEQ ID NO: 5 и SEQ ID NO: 6) и/или между спаренными сайтами-мишенями (например, сайтами-мишенями пар нуклеаз, показанными в Таблице 1) гена HPRT (например, модификация интрона 1), включая модификацию путем связывания, расщепления, инсерции и/или делеции одного или более нуклеотидов в любой из этих последовательностей и/или в пределах 1-50 пар оснований (включая любое число пар оснований между ними, такие как 1-5, 1-10 или 1-20 пар оснований) генных (геномных) последовательностей, фланкирующих эти последовательности в гене В2М. В некоторых вариантах осуществления изобретения, клетки содержат инсерцию в интроне 1, так, чтобы ген HPRT не транскрибировался. Экспрессия трансгена, встроенного в ген HPRT, может быть индуцирована эндогенным промотором HPRT, либо этот ген может включать экзогенный промотор. В другом варианте осуществления изобретения, модификация представляет собой генетическую модификацию (изменение нуклеотидной последовательности) в сайте(ах) или рядом с сайтами связывания с нуклеазой (мишенью) и/или сайтами расщепления, включая, но не ограничиваясь ими, модификации в последовательностях в пределах 1-300 пар оснований (или любых пар оснований между ними), расположенных выше или ниже, и/или включая 1 или более пар оснований сайта(ов) расщепления и/или связывания; модификации в пределах 1-100 пар оснований (или любого числа пар оснований между ними) включительно, и/или с любой стороны сайта(ов) связывания и/или расщепления; модификации в пределах 1-50 пар оснований (или любого числа пар оснований между ними) включительно, и/или с любой стороны (например, от 1 до 5, от 1 до 10, от 1 до 20 или более пар оснований) сайта(ов) связывания и/или расщепления; и/или модификации в одной или более парах оснований в сайте связывания с нуклеазой и/или в сайте расщепления нуклеазой. В некоторых вариантах осуществления изобретения, модификация присутствует в спаренных сайтах-мишенях или возле этих сайтов (например, 1-300 пар оснований, 1-50, 1-20, 1-10 или 1-5 или любое число пар оснований между ними) и/или между спаренными сайтами-мишенями (см., например, Таблицу 1) последовательности гена В2М внутри, вокруг или между любыми из SEQ ID NO: 3 и SEQ ID NO: 4. В некоторых вариантах осуществления изобретения, модификация включает модификации гена В2М в одной или более последовательностях, представленных в SEQ ID NO: 3 и SEQ ID NO: 4, или в GCCTTA в экзоне 1 гена В2М, например, модификацию 1 или более пар оснований в одной или более из этих последовательностей. В некоторых вариантах осуществления изобретения, опосредованные нуклеазой генетические модификации присутствуют между спаренными сайтами-мишенями (если для расщепления мишени используется димер). Опосредованные нуклеазой генетические модификации могут включать инсерции и/или делеции любого числа пар оснований, включая инсерции некодирующих последовательностей любой длины и/или трансгенов любой длины и/или делеции от 1 пары оснований до более 1000 т.п.о. (или любое число пар оснований между ними, включая, но не ограничиваясь ими, 1-100 пар оснований, 1-50 пар оснований, 1-30 пар оснований, 1-20 пары оснований, 1-10 пар оснований или 1-5 пар оснований).
[0029] В некоторых вариантах осуществления изобретения, ген CISH был также модифицирован (например, посредством инактивации вместе с интеграцией или без нее) с использованием нуклеазы, которая связывается с любыми из этих сайтов-мишеней CISH, показанными в Таблице 1 (SEQ ID NO: 50 и SEQ ID NO: 51). В некоторых вариантах осуществления изобретения, модификация включает инсерцию и/или делецию таким образом, чтобы ген CISH не транскрибировался, или чтобы мутантный транскрипт подвергался деградации, опосредуемой несмысловой мутацией. Экспрессия одного или более трансгенов, которые были встроены в ген CISH, может быть индуцирована посредством эндогенного промотора CISH, либо этот ген может включать экзогенный промотор. В других вариантах осуществления изобретения, модификация представляет собой генетическую модификацию (изменение нуклеотидной последовательности) в сайте(ах) или рядом с сайтами связывания с нуклеазой (мишенью) и/или сайтами расщепления, включая, но не ограничиваясь ими, модификации в последовательностях в пределах 1-300 пар оснований (или любого числа пар оснований между ними), расположенных выше или ниже и/или включая 1 или более пар оснований сайта(ов) расщепления и/или связывания; модификации в пределах 1-100 пар оснований (или любого числа пар оснований между ними) включительно, и/или с любой стороны сайта(ов) связывания и/или расщепления; модификации в пределах 1-50 пар оснований (или любого числа пар оснований между ними) включительно, и/или с любой стороны (например, от 1 до 5, от 1 до 10, от 1 до 20 или более пар оснований) сайта(ов) связывания и/или расщепления; и/или модификации в одной или более парах оснований в сайте связывания с нуклеазой и/или в сайте расщепления нуклеазой. В некоторых вариантах осуществления изобретения, модификация присутствует в спаренных сайтах-мишенях или возле этих сайтов (например, 1-300 пар оснований, 1-50, 1-20, 1-10 или 1-5 или любое число пар оснований между ними) и/или между спаренными сайтами-мишенями (см., например, Таблицу 1) последовательности гена CISH внутри, вокруг или между любыми из SEQ ID NO: 50 и SEQ ID NO: 51. В некоторых вариантах осуществления изобретения, опосредованные нуклеазой генетические модификации присутствуют между спаренными сайтами-мишенями (если для расщепления мишени используется димер). Опосредованные нуклеазой генетические модификации могут включать инсерции и/или делеции любого числа пар оснований, включая инсерции некодирующих последовательностей любой длины и/или трансгенов любой длины и/или делеции от 1 пары оснований до более 1000 т.п.о. (или любое число пар оснований между ними, включая, но не ограничиваясь ими, 1-100 пар оснований, 1-50 пар оснований, 1-30 пар оснований, 1-20 пары оснований, 1-10 пар оснований или 1-5 пар оснований).
[0030] CAR+-модифицированные клетки согласно изобретению, могут представлять собой эукариотическую клетку, включая клетку млекопитающего, не являющегося человеком, и человеческую клетку, такую как лимфоидная клетка (например, Т-клетка), стволовая клетка/клетка-предшественник (например, индуцированная плюрипотентная стволовая клетка (iPSC), эмбриональная стволовая клетка (например, человеческая ES), мезенхимальная стволовая клетка (MSC) или гемопоэтическая стволовая клетка (HSC). CAR+-модифицированные клетки были также, но необязательно, модифицированы для экспрессии модифицированного комплекса HLA-Е или HLA-G. Стволовые клетки могут быть тотипотентными или плюрипотентными (например, частично дифференцированными, такими как HSC, которые представляют собой плюрипотентные миелоидные или лимфоидные стволовые клетки). В других своих вариантах, настоящее изобретение относится к способам получения CAR+-клеток, имеющих нулевой генотип по экспрессии TCR и/или B2M. Любая из описанных здесь стволовых CAR+-модифицированных клеток (модифицированных в локусе TCRA и/или B2M, необязательно экспрессирующих модифицированный HLA-E или HLA-G) может быть затем дифференцирована для получения дифференцированной (in vivo или in vitro) клетки, происходящей от описаной здесь стволовой клетки, с модифицированной экспрессией гена TCRA и/или B2M. В некоторых вариантах осуществления изобретения описана популяция CAR+-модифицированных клеток, в которых по меньшей мере 20% (или любой процент от 20% до 100%), предпочтительно, по меньшей мере 50% (или любой процент от 50% до 100%), еще более предпочтительно, по меньшей мере 60% (или любой процент от 60% до 100%), а еще более предпочтительно, по меньшей мере от 70% до 100% (или любой процент между ними) клеток являются CAR+-модифицированными.
[0031] В другом аспекте изобретения, описанные здесь композиции (CAR+-модифицированные клетки, необязательно экспрессирующие модифицированный комплекс HLA-Е или HLA-G) и способы могут быть использованы, например, для лечения или профилактики или облегчения расстройства. Эти способы обычно включают (а) интеграцию CAR-кодирующего трансгена в клетку посредством расщепления или интеграцию эндогенного гена TCR и/или B2M в выделенной клетке (например, в Т-клетке или в лимфоците) с использованием нуклеазы {например, ZFN или TALEN) или нуклеазной системы, такой как CRISPR/Cas, вместе со сконструированной cr-РНК/tracr-РНК, так, чтобы ген TCR инактивировался или подвергался негативной модуляции; и (b) введение клетки индивидууму для лечения или профилактики расстройства. В некоторых вариантах осуществления изобретения, ген, кодирующий экспрессионную конструкцию CAR или CAR-HLA-E или CAR-HLA-G, специфически интегрируют в TCRβ (TCRB), так, чтобы экспрессия TCRB была инактивирована. В некоторых вариантах осуществления изобретения, инактивацию осуществляют посредством нацеленного расщепления константной области этого гена (константной области TCRβ или TRBC) и интеграции CAR. В предпочтительных вариантах осуществления изобретения, ген, кодирующий экспрессионную конструкцию CAR или CAR-HLA-E или CAR-HLA-G, интегрируют в TCRα (TCRA), так, чтобы экспрессия TCRα была инактивирована. В других предпочтительных вариантах осуществления изобретения, инактивацию осуществляют посредством нацеленного расщепления константной области этого гена (константной области TCRα или сокращенно TRAC). В некоторых вариантах осуществления изобретения, ген, кодирующий CAR, специфически интегрируют в B2M, так, чтобы экспрессия B2M была инактивирована. В некоторых вариантах осуществления изобретения, CAR+-клетка также содержит сконструированный фактор транскрипции, который негативно модулирует ген. В других вариантах осуществления изобретения, фактор транскрипции представляет собой ZFP-TF, TALE-TF или CRISPR/dCas-TF. В некоторых вариантах осуществления изобретения, другими модулированными генами являются, но не ограничиваясь ими, ген B2M, CISH, PD1 и/или CTLA4.
[0032] В некоторых вариантах осуществления изобретения, другие гены были модулированы (посредством нокаута), например B2M, PD1, CISH и/или CTLA4, и/или в клетке присутствуют один или более терапевтических трансгенов (эписомных, случайно интегрированных или интегрированных посредством нацеленной интеграции, такой как интеграция, опосредуемая нуклеазой). По своей природе, терапевтический трансген может обладать способностью стимулировать уничтожение CAR+-клетки или он может действовать как переключатель на уничтожение самих модифицированных Т-клеток (создавая так называемую «интеллектуальную» CAR-Т-клетку, см., Hartmann et al. (2017) EMBO Mol Medicine, DOI 10. 15252/emmm.201607485). В некоторых вариантах осуществления изобретения, трансген представляет собой ген-«самоубийцу», и активация такого гена приводит к гибели CAR-T-клетки. В некоторых вариантах осуществления изобретения, ген-«самоубийца» представляет собой ген HSV-TK, который кодирует фермент тимидинкиназу, способную метаболизировать ганцикловир, то есть, противовирусное лекарственное средство, которое обычно не обладает противоопухолевым действием, с образованием токсического соединения, которое препятствует репликации ДНК и приводит к апоптозу клеток; ген цитозиндеаминазы (также сцепленный с геном урацил-фосфорибозилтрансферазы), который превращает 5-фторцитозин (обладающий противогрибковыми свойствами) в 5-фторурацил; ген нитроредуктазы, который превращает СВ1954 ([5-(азиридин-1-ил)-2,4-динитробензамид]) в токсическое соединение, 4-гидроксиламин, и ген цитохрома р450, который превращает ифосфамид в акролеин (азотный аналог горчичного газа) (Rouanet et al. (2017), Int J Mol Sci, 18(6):1231); или индуцибельный ген каспазы-9 (Jones et al. (2014) Front Pharmacol. 5:254). В дополнительных вариантах осуществления изобретения, трансген кодирует фактор, ослабляющий тяжелые побочные эффекты и/или токсические эффекты, такие как синдром высвобождения цитокинов (CRS) и/или нейротоксичность (например, scFv-конструкция, нацеленная на IL-6 или на секретируемую конструкцию IL-12, называется, например, «TRUCK» четвертого поколения (Chmielewski et al. (2014) Immunol Rev. 257 (L): 83-90).
[0033] Фактор(ы) транскрипции и/или нуклеаза(ы) может (могут) быть введен(ы) в клетку или в окружающую культуральную среду в виде мРНК, в виде белка и/или в виде последовательности нуклеиновой кислоты, кодирующей нуклеазу(ы). В некоторых вариантах осуществления изобретения, выделенная CAR+-клетка, введенная индивидууму, также включает дополнительную геномную модификацию, например, интегрированную экзогенную последовательность (в расщепленном гене TCR и/или B2M или в другом гене, например, в безопасном якорном гене или локусе) и/или инактивацию (например, опосредованную нуклеазой) дополнительных генов, например, одного или более генов HLA. В некоторых вариантах осуществления изобретения, один ген CAR вводят в ген TCR, а второй (идентичный или неидентичный) ген CAR вводят в ген B2M. В обоих этих вариантах осуществления изобретения, ген CAR может быть частью экспрессионной конструкции CAR-HLA-E или CAR-HLA-G. Экзогенная последовательность или белок могут быть введены посредством вектора (например, Ad, AAV, LV) или с применением такого метода, как электропорация. В некоторых вариантах осуществления изобретения, белки вводят в клетку путем их сжатия (см., Kollmannsperger et al. (2016) Nat. Comm. 7, 10372 doi:10.1038/ncomms10372). В некоторых аспектах изобретения, композиция может содержать выделенные клеточные фрагменты и/или дифференцированные (частично или полностью) клетки.
[0034] В некоторых аспектах изобретения, CAR+-клетки могут быть использованы для клеточной терапии, например, для адоптивного переноса клеток. В других вариантах осуществления изобретения, CAR+-клетки могут быть использованы для трансплантации T-клеток и могут содержать другую(ие) представляющую(ие) интерес модификацию(и) гена. В одном аспекте изобретения, CAR+-Т-клетки содержат CAR, специфичный к маркеру рака. В другом аспекте изобретения, встроенный CAR является специфичным к маркеру CD19, характерному для В-клеток, включая B-клеточные злокачественные новообразования. Нацеленная интеграция CAR в ген B2M позволяет создать «универсальные» клетки, подходящие для их использования в терапевтической композиции для лечения пациентов с HLA-несовместимостью, а поэтому такие клетки будут использованы в качестве коммерчески доступного терапевтического средства для лечения любого пациента, нуждающегося в этом. Нацеленная интеграция экспрессионной конструкции CAR-HLA-E или CAR-HLA-G позволит создать «коммерчески доступную универсальную» клетку, которая не будет подвергаться NK-опосредованному уничтожению у пациента, подвергаемого лечению и, таким образом, будет сохраняться в организме в течение более длительного периода времени.
[0035] В некоторых вариантах осуществления изобретения, CAR-кодирующую последовательность встраивают посредством нацеленной интеграции, где донорная последовательность CAR содержит фланкирующие гомологичные ветви, которые имеют гомологию с последовательностью, фланкирующей сайт расщепления сконструированной нуклеазы. В некоторых вариантах осуществления изобретения, донорная последовательность CAR или донорная последовательность CAR-HLA-E или CAR-HLA-G также содержат промотор и/или другие последовательности регуляции транскрипции. В других вариантах осуществления изобретения, донорная последовательность CAR или CAR-HLA-E или CAR-HLA-G не содержит промотора. В других вариантах осуществления изобретения, донорная последовательность CAR или CAR-HLA-E или CAR-HLA-G содержит переключатель, посредством которого отдельные внеклеточные антигенсвязывающие домены и внутриклеточные сигнал-передающие компоненты могут подвергаться сборке, например, посредством модуля FKBP-FRB только в присутствии гетеродимеризующихся небольших молекул (аналога рапамицина AP21967) (см., Wu et al. (2015) Science. 350 (6258): Aab4077). Любая из описанных здесь донорных конструкций может также содержать дополнительные последовательности, включая, но не ограничиваясь ими, гомологичные ветви (любой длины); последовательности WPRE и/или энхансерные последовательности TCRa.
[0036] В другом аспекте изобретения, CAR+ или CAR+-модифицированные, HLA-G+-модифицированные или HLAE+-TCR- или B2M-модулированные (модифицированные) Т-клетки также содержат встроенную донорную последовательность T-клеточного рецептора, связыванного с антителом (ACTR). В некоторых вариантах осуществления изобретения, донорную последовательность ACTR встраивают в ген TCR и/или B2M для нарушения экспрессии гена TCR и/или B2M после расщепления, индуцированного нуклеазой. В других вариантах осуществления изобретения, донорную последовательность встраивают в «безопасный якорный» локус, такой как гены AAVS1, HPRT, альбумина и CCR5.
[0037] Настоящее изобретение также относится к фармацевтическим композициям, содержащим описанные здесь модифицированные клетки (например, CAR+ или CAR+-модифицированные, HLA-E+-модифицированные или HLA-G+-модифицированные Т-клетки или стволовые клетки с инактивированным геном TCR и/или B2M), или к фармацевтическим композициям, содержащим описанные здесь одну или более молекул, связывающихся с геном TCR и/или B2M (например, сконструированные факторы транскрипции и/или нуклеазы). В некоторых вариантах осуществления изобретения, фармацевтические композиции также содержат один или более фармацевтически приемлемых наполнителей. Модифицированные клетки, молекулы, связывающиеся с геном TCR и/или B2M (или полинуклеотиды, кодирующие эти молекулы) и/или фармацевтические композиции, содержащие эти клетки или молекулы, вводят индивидууму известными методами, например, посредством внутривенного вливания, вливания в конкретный кровеносный сосуд, такой как печеночная артерия, или посредством прямой инъекции в ткань (например, в мышцу). В некоторых вариантах осуществления изобретения, индивидуумом является взрослый человек, страдающий заболеванием или состоянием, которое может быть подвергнуто лечению или ослаблению с использованием такой композиции. В других вариантах осуществления изобретения, индивидуумом является ребенок, которому вводят эту композицию для профилактики, лечения или ослабления заболевания или состояния (например, рака, заболевания «трансплантат против хозяина» и т.п.).
[0038] В некоторых аспектах изобретения, композиция (модулированные клетки, содержащие CAR, или CAR и модифицированный HLA-E или HLA-G) может содержать CAR, включающий антигенсвязывающий домен антитела. В некоторых аспектах изобретения, CAR используется в целях «оснащения» сконструированной Т-клетки для профилактики или лечения состояния. В некоторых вариантах осуществления изобретения, CAR распознает антиген, ассоциированный с опухолевой клеткой или с развитием рака, такой как BCMA, EpCAM, CEA (аденокарцинома поджелудочной железы, рак молочной железы, карцинома прямой и ободочной кишки), gpA33, муцины, TAG-72, CAIX (почечноклеточная карцинома (ПКК)), фолат-связывающие антитела, CD19 (B-клеточные злокачественные новообразования), CD20, CD22, EGFR (эпителиальная карцинома, глиома), EGFRvIII (глиобластома), EpCam, EphA2, ERBB2/HER2 (рак молочной железы, рак яичника, рак толстой кишки; остеосаркома, медуллобластома), ERBB3, FAP (злокачественная мезотелиома плевры), MET, MSLN (мезотелиома, рак яичника, аденокарцинома поджелудочной железы), Igκ, IL-1RAP, IGF1R, EPHA3, Lewis Y, лиганды NKG2D, ROR1, FR-a (рак яичника), GD2 (нейробластома, меланома), GPC3 (гепатоцеллюлярная карцинома), IL-13Rα2 (глиома), L1-САМ (нейробластома, меланома, аденокарцинома яичника), MUC1 (рак семенных канальцев), CA125 (эпителиальный рак яичника), PD-L1, PSCA, CTAG1B (меланома и рак яичника), PSMA (рак предстательной железы), VEGFR2, c-MET, CD133 (глиобластома, холангиокарцинома (CCA)), TRAILR1, TRAILR2, RANKL, FAP, VEGF, VEGFR, интегрины αVβ3 и α5β1, CD30, CD33, CD52, CD70, CD123, CD138, CD16V, CTLA4 и энасцин (Scott et al. (2012) Nat Rev Cancer 12: 278; Hartmann et al. (2017) EMBO Mol Med; DOI 10.15252/emmm. 201607485, Yu et al. (2017) J Hematol Oncol 10: 78). В других вариантах осуществления изобретения, CAR распознает антиген, ассоциированный с инфекционным заболеванием, таким как ВИЧ, HCV и т.п. (см. например, Carillo et al.. (2017) Transl Res. pii: S1931-5244(17)30233-5500. doi: 10.1016/j trsl.2017.07.002). В некоторых вариантах осуществления изобретения, сконструированная Т-клетка может содержать 2 или более неидентичных последовательностей CAR, интегрированных В геном (например, специфичных к CD19 и CD20). В некоторых вариантах осуществления изобретения, CAR содержит CAR, обозначенный как FMC63-CD8BBZ, где FMC63 представляет собой последовательность scFv против CD19 (см., патент США № 9701758), а CD8BBZ означает части CAR, за исключением scFv (шарнирный/трансмембранный домен CD8 (CD8), костимулирующий домен гена 41BB (BB), домен активации гена CD3z (Z)).
[0039] ДНК-связывающий домен фактора транскрипции или нуклеазы (например, ZFP, TALE, оцрРНК) может связываться с сайтом-мишенью в гене TCRA или B2M, включающем 9, 10, 11, 12 или более (например, 13, 14, 15, 16, 17, 18, 19, 20 или более) нуклеотидов любого из представленных здесь сайтов-мишеней, (например, как показано в Таблице 1, в SEQ ID NO: 1-4). Белки «цинковые пальцы» могут включать 1, 2, 3, 4, 5, 6 или более цинковых пальцев, где каждый цинковый палец имеет распознающую спираль, которая специфически контактирует с субсайтом-мишенью в гене-мишени. В некоторых вариантах осуществления изобретения, белки «цинковые пальцы» содержат 4, 5 или 6 пальцев (обозначенных F1, F2, F3, F4, F5 и F6 и упорядоченных как F1-F4 или F5 или F6 от N-конца до С-конца), например, как показано в Таблице 1. Описанные здесь ZFP могут также включать одну или более мутаций в контакттирующих фосфатных остатках белка ««цинковый палец»», например, мутацию nR-5Qabc, описанную в предварительных заявках на патент США № 62/378978 и 62/443981. В других вариантах осуществления изобретения, одноцепочечные руководящие РНК или домены, связывающиеся с ДНК TAL-эффектора, могут связываться с описанным здесь сайтом-мишенью (например, с сайтами-мишенями в Таблице 1, представленными в любой из SEQ ID NO: 1-4) или 12 или более парами оснований в любом из этих сайтов-мишеней или между спаренными сайтами-мишенями. Описанные здесь нуклеазы (содержащие ZFP, TALE или домен, связывающийся c ДНК оцрРНК) способны осуществлять генетические модификации в гене TCRA или в гене B2M, включающем любую из SEQ ID NO: 1-4, включая модификации (инсерции и/или делеции) в любой из этих последовательностей (SEQ ID NO: 1-4) и/или модификации в последовательностях гена TCRA и/или B2M, фланкирующих последовательности сайта-мишени, представленные в SEQ ID NO: 1-4, например, модификации в экзоне c2 гена TCRA в TTGAAA.
[0040] Любой из описанных здесь белков может также содержать домен расщепления и/или полудомен расщепления (например, полудомен расщепления дикого типа или сконструированный полудомен расщепления FokI). Таким образом, в любой из описанных здесь нуклеаз (например, ZFN, TALEN, систем CRISPR/Cas), домен нуклеазы может содержать домен нуклеазы дикого типа или полудомен нуклеазы (например, полудомен расщепления FokI). В других вариантах осуществления изобретения, нуклеазы (например, нуклеазы ZFN, TALEN, CRISR/Cas) содержат сконструированные домены или полудомены нуклеазы, например сконструированные полудомены расщепления FokI, которые образуют облигатные гетеродимеры. См. например, патенты США №№ 7914796 и 8034598. В некоторых вариантах осуществления изобретения, один или более эндонуклеазных доменов FokI описанных здесь нуклеаз могут также содержать мутации в контактирующих фосфатных остатках (например, R416S и/или K525S), как описано в публикации заявки на патент США N 20180087072.
[0041] В другом своем аспекте, настоящее изобретение относится к полинуклеотиду, кодирующему любые белки, гибридные молекулы и/или их компоненты (например, оцрРНК или другой ДНК-связывающий домен), описанные в настоящей заявке. Полинуклеотид может быть частью вирусного вектора, невирусного вектора (например, плазмиды) или может присутствовать в форме мРНК. Любые из описанных здесь полинуклеотидов могут также содержать последовательности (донорные последовательности, гомологичные ветви или последовательности-«пэтчи») для нацеленного встраивания в ген-мишень (например, в ген TCRA, TCRB, ген контрольной точки и/или ген B2M). В еще одном своем аспекте, настоящее изобретение относится к вектору для доставки генов, содержащему любые из описанных здесь полинуклеотидов. В некоторых вариантах осуществления изобретения, вектор представляет собой аденовирусный вектор (например, вектор Ad5/F35) или лентивирусный вектор (LV), включающий компетентные по интеграции или дефектные по интеграции лентивирусные векторы или аденоассоциированный вектор (AAV). Таким образом, настоящее изобретение также относится к вирусным векторам, содержащим последовательность, кодирующую нуклеазу (например, ZFN или TALEN) и/или нуклеазную систему (CRISPR/Cas или Ttago) и/или донорную последовательность для нацеленной интеграции в ген-мишень. В некоторых вариантах осуществления изобретения, донорная последовательность и последовательности, кодирующие нуклеазу, находятся на различных векторах. В других вариантах осуществления изобретения, нуклеазы представляют собой полипептиды. В предпочтительных вариантах осуществления изобретения, полинуклеотидами являются мРНК. В некоторых аспектах изобретения, мРНК может быть химически модифицирована (см., например, Kormann et al. (2011) Nature Biotechnology 29(2):154-157). В других аспектах изобретения, мРНК может содержать кэп ARCA (см., патенты США 7074596 и 8153773). В некоторых аспектах изобретения, мРНК может содержать кэп, введенный посредством ферментативной модификации. Ферментативно введенный кэп может содержать Cap0, Cap1 или Cap2 (см., например, Smietanski et al. (2014) Nature Communications 5: 3004). В других аспектах изобретения, мРНК может быть кэпирована посредством химической модификации. В других вариантах осуществления изобретения, мРНК может содержать смесь немодифицированных и модифицированных нуклеотидов (см., публикацию заявки на патент США 2012-0195936). В других вариантах осуществления изобретения, мРНК может содержать элемент WPRE (см., публикацию заявки на патент США № 20160326548). В некоторых вариантах осуществления изобретения, элемент WPRE может содержать одну или более мутаций (см., патент США №. 7419829). В некоторых вариантах осуществления изобретения, мРНК является двухцепочечной (см., например, Kariko et al. (2011) Nucl Acid Res 39:el42).
[0042] В другом своем аспекте, настоящее изобретение относится к выделенной клетке, содержащей любые из описанных здесь белков, полинуклеотидов и/или векторов. В некоторых вариантах осуществления изобретения, клетка выбрана из группы, состоящей из стволовой клетки/клетки-предшественника или Т-клетки (например, CD4+-Т-клетки). В еще одном своем аспекте, настоящее изобретение относится к CAR+ или CAR+-модифицированной HLA-Е+ и/или HLA-G+-клетке или клеточной линии, которая происходит от клетки или клеточной линии, содержащей любые из описанных здесь белков, полинуклеотидов и/или векторов, а именно, от клтеки или клеточной линии (например, в культуре), происходящей от клетки, в которой TCR и/или B2M были инактивированы посредством одной или более ZFN, и/или в которой донорный полинуклеотид (например, ACTR и/или CAR) был стабильно интегрирован в геном клетки. Таким образом, потомки описанных здесь клеток сами могут не содержать описанные здесь белки, полинуклеотиды и/или векторы, но в этих клетках, ген TCR и/или B2M является инактивированным и/или донорный полинуклеотид был интегрирован в геном и/или экспрессируется в этом геноме.
[0043] В другом аспекте изобретения, описанными здесь способами являются способы инактивации по меньшей мере одного гена TCR в клетке (например, в Т-клетке) и спсобы достижения экспрессии экзогенного CAR в клетке путем введения в клетку одного или более описанных здесь белков, полинуклеотидов и/или векторов. В любых описанных здесь способах, нуклеазы могут индуцировать нацеленный мутагенез, делеции последовательностей ДНК в клетках и/или облегчить нацеленную рекомбинацию в предварительно определенном хромосомном локусе. Таким образом, в некоторых вариантах осуществления изобретения, нуклеазы делетируют из гена-мишени и/или встраивают в ген-мишень один или более нуклеотидов. В некоторых вариантах осуществления изобретения, ген TCR и/или HLA инактивируют посредством расщепления нуклеазой с последующим присоединением негомологичных концов. В других вариантах осуществления изобретения, геномную последовательность в гене-мишени заменяют, например, с использованием описанной здесь нуклеазы (или вектора, кодирующего указанную нуклеазу), и «донорной» последовательности (например, CAR), которая встраивается в ген после нацеленного расщепления нуклеазой. Донорная последовательность может присутствовать в нуклеазном векторе, в отдельном векторе (например, в векторе AAV, Ad или LV), или альтернативно, она может быть введена в клетку по другому механизму доставки нуклеиновой кислоты. В любом из описанных здесь способов, нуклеаза(ы) и доноры могут быть совместно или последовательно введены в любом порядке. Введение нуклеазы (нуклеаз) и/или одного или более доноров может быть осуществлено повторно, например, один или более доноров (например, один или более CAR) могут быть введены до, во время и/или после введения одной нуклеазы (одной, двух, трех, четырех или более нуклеаз, нацеленных на различные гены); один или более доноров могут быть введены повторно до, во время и/или после введения одной нуклеазы; или один или более доноров могут быть введены повторно до, во время и/или после повторных введений нуклеазы. В некоторых вариантах осуществления изобретения, TRAC-, CISH- и/или B2M-специфические нуклеазы и один или более донорных CAR вводят совместно для получения клетки, экспрессирующей CAR с мультинокаутом, в способе, включающем одну стадию редактирования. В некоторых вариантах осуществления изобретения, способы также включают инактивацию одного или более дополнительных генов (например, генов иммунологической контрольной точки, таких как CISH, PD1 или CTLA4), и/или интеграцию одного или более трансгенов в геном клетки, включая, но не ограничиваясь ими, интеграцию одного или более трансгенов в инактивированный TCR (и необязательно дополнительные гены B2M и т.п.) и/или в один или более безопасных якорных генов. В некоторых вариантах осуществления изобретения, описанные здесь способы позволяют получить популяцию клеток, в которых по меньшей мере 80-100% (или любой процент в этих пределах), включая, по меньшей мере 90-100% (или любой процент в этих пределах) клеток, имеющих нокаут(ы) и/или интегрированный(е) трансген(ы).
[0044] Кроме того, любой из описанные здесь способов может быть осуществлен in vitro, in vivo и/или ex vivo. В некоторых вариантах осуществления изобретения, способы осуществляют ex vivo, например, для модификации Т-клеток в целях их использования в качестве терапевтических средств для сообщения им аутологичных или аллогенных свойств, подходящих для лечения индивидуума (например, индивидуума с раком). Неограничивающими примерами рака, который может быть подвергнут лечению и/или профилактике, являются карциномы легких, рак поджелудочной железы, рак печени, рак кости, рак молочной железы, рак прямой и ободочной кишки, лейкозы, рак яичника, лимфомы, рак головного мозга и т.п.
[0045] В другом аспекте изобретения, описанным здесь способом является способ интеграции одного или более трансгенов в геном выделенной клетки, где указанный способ включает введение в клетку: (a) одного или более донорных векторов (например, AAV, плазмид, Ad, мРНК и т.п.), содержащих один или более трансгенов, и (b) по меньшей мере одной не-природной нуклеазы в форме мРНК, где по меньшей мере одна нуклеаза расщепляет геном клетки таким образом, что один или более трансгенов интегрируются в геном клетки (например, один или более генов CAR или экспрессионных конструкций CAR=HLA-Е или CAR-HLA-G интегрируются в рецептор TCR или HLA), где донорный вектор вводят в буфер для электропорации, содержащий выделенную клетку и мРНК, непосредственно до или непосредственно после электропорации нуклеазы в клетку. В некоторых вариантах осуществления изобретения, донорный вектор вводят в буфер для электропорации после электропорации и перед переносом клеток в культуральную среду. См., например, публикации заявок на патенты США № 20150174169 и 20150110762. Эти способы могут быть применены для введения трансгена(ов) CAR или экспрессионной конструкции CAR=HLA-Е или CAR-HLA-G в любое положение генома, включая, но не ограничиваясь этим, ген TCR, ген B2M и/или безопасный якорный ген.
Краткое описание чертежей
[0046] На фигуре 1 представлено изображение гена HPRT, иллюстрирующее организацию интронов и экзонов на геномном варианте гена и конструкции трансгена, экспрессирующего CD19, для инсерции в интрон 1. Донорная конструкция трансгена имеет гомологичные ветви («гомологию»), фланкирующие трансген CD19, а также имеет акцептор сплайсинга (SA), в результате чего после созревании транскрипта, экзон 1 HPRT будет сцеплен с геном CD19. Конструкция также включает саморасщепляющийся сайт 2А между экзоном 1 HPRT и CD19. У конца CD19-кодирующей последовательности также присутствует поли-А-последовательность. При транскрибировании и созревании, эта конструкция будет, в конечном счете, транслироваться только в CD19-кодирующие последовательности. Клетки, содержащие встроенный трансгенный кластер CD19, не экспрессируют ген HPRT и, таким образом, являются резистентными к 6-тиогуанину («6-TG»), что позволяет проводить отбор только тех клеток, в которые был встроен кластер в присутствии 6-TG.
[0047] На фигурах 2А и 2В представлены графики, иллюстрирующие экспрессию CD19 в клетках К562, трансфецированнх CD19-экспрессирующим кластером, представленным на фигуре 1. Для оценки экспрессии CD19, клетки окрашивали PE-Cy7-конъюгированным антителом, нацеленным на человеческий внеклеточный домен CD19. Немодифицированные («необученные»») клетки К562 были аналогичным образом обработаны и окрашены (фигура 2А) в качестве негативного контроля. На фигуре 2В показаны трансфецированные клетки К562 после отбора на 6-TG и было продемонстрировано, что почти все эти клетки (98,3%) были позитивными по экспрессии CD19.
[0048] На фигурах 3А-3С представлены графики, иллюстрирующие результаты FACS-анализа для оценки влияния Т-клеток на клетки-мишени. CD19+-K562 (как показано на фигуре 2) смешивали в отношении 1:1 с немодифицированными нормальными клетками к562. Клетки метили с использованием красителя на пролиферацию CellTrace™, а затем культивировали вместе с T-клетками. Клетки подвергали совместному культивированию в течение трех часов, промывали, окрашивали антителом против CD19, а затем процент CD19+-K562 вычисляли с помощью FACS. В отсутствие Т-клеток, позитивная популяция CellTrace™ (то есть, CD19+-K562 и нормальные (CD19-)К562) составляли приблизительно 50%:50%, как и ожидалось (фигура 3А). При совместном культивировании с Т-клетками, которые были либо отредактированы TCRA-специфическим ZFN без донора CD19-CAR (фигура 3С, «контроль-TRAC+мишень»), либо не были обработаны (фигура 3В, «Контроль-мишень+UT»), цитотоксичность К562 не наблюдалась.
[0049] На фигурах 4А и 4В представлен ряд панелей, иллюстрирующих эффективное уничтожение клеток, содержащих антиген CD19, в отношении «эффекторные клетки : клетки-мишени» (в отношении «E:T»). Как указывалось ранее, клетки-мишени представляют собой смесь 50:50 CD19-несущих клеток К562 и «необученных» клеток К562. На фигуре 4A представлены результаты, полученные с использованием Т-клеток, где в ген B2M были интегрированы CD19-CAR, а на фигуре 4B представлены результаты, где в ген TCRA были интегрированы CD19-CAR. На самой крайней левой панели представлен набор данных, иллюстрирующий отношение E:T=2:1, где T-клетки присутствуют в количестве, в два раза превышающем количество клеток-мишеней, на самой крайней правой панели представлено отношение E:T=0,125:1, а на панелях в середине представлены отношения E:T, составляющее промежуточные значения между этими двумя значениями. Полученные данные показали, что «необученные» клетки (CD19-)К562 не уничтожались под действием T-клеток, несущих CD19-CAR, а уничтожение клеток К562 с антигеном CD19 коррелирует с количеством используемых эффекторных клеток.
[0050] На фигуре 5 представлен график, иллюстрирующий процент гибели CD19+-K562-клеток в зависимости от отношения эффектор:клетка-мишень. На графике представлены результаты для Т-клеток, содержащих CD19-CAR, интегрированный в ген TCRA или B2M. Данные показали, что интеграция в любой ген приводит к эффективному функционированию CAR и к активации T-клеток.
[0051] На фигурах 6А-6F представлены графики, иллюстрирующие FACS-анализ на экспрессию маркеров клеточной поверхности или CD19-CAR. На фигурах 6А и 6В показаны результаты сканирования клеток без обработки ZFN (Фигура 6А) или без обработки донорным CD19-CAR (фигура 6В). На фигурах 6C и 6D показаны результаты, полученные в том случае, когда клетки были обработаны нуклеазами, нацеленными на B2M и TCRA, но не были обработаны донором CAR. На фигуре 6C показаны результаты после расщепления нуклеазой и было продемонстрировано, что 80% из этих клеток имеют фенотип с двойным нокаутом. На фигурах 6Е и 6F показаны результаты, полученные при обработке клеток нуклеазами и донором CD19-CAR, содержащим гомологичные ветви TRAC. На фигуре 6F показано, что 63,4% клеток экспрессируют CD19-CAR.
[0052] На фигуре 7 представлен график, иллюстрирующий генотип и фенотип клеток после опосредованной нуклеазой инактивации (нокаута) TRAC. Как было показано, более, чем 95% клеток имеют генотип и фенотип инактивации.
[0053] На фигурах 8A и 8B показаны результаты FACS-анализа на экспрессию поверхностного рецептора CD3 в Т-клетках, обработанных нуклеазами, нацеленными на TRAC (фигура 8B) по сравнению с необработанными клетками (фигура 8A). Как было показано, по меньшей мере 99,5% обработанных нуклеазой клеток не экспрессировали поверхностного TCR по сравнению с 0,2% необработанных клеток.
[0054] На фигурах 9А-9С представлены графики, иллюстрирующие FACS-анализ на экспрессию трансгена (GFP) в Т-клетках, обработанных нуклеазами, нацеленными на TRAC (фигура 9B) или B2M (фигура 9С). Также показаны клетки, не содержащие донорного трансгена, но обработанные TRAC- и B2M-специфическими реагентами (фигура 9С). Как было показано, трансген экспрессировался по меньшей мере в 93% клеток, обработанных TRAC-специфическими нуклеазами, что указывает на эффективную опосредованную нуклеазой интеграцию в TRAC. Аналогичным образом, трансген экспрессировался по меньшей мере в 90% клеток, обработанных B2M-специфическими нуклеазами, что указывает на эффективную опосредованную нуклеазой интеграцию в B2M.
[0055] На фигурах 10А-10С проиллюстрирован FACS-анализ клеток, где показаны результаты FACS-анализа T-клеток здорового донора, обработанных TRAC-нацеленными нуклеазами (фигура 10A) и B2M-нуклеазами (фигура 10B), а также донорным CAR (фигура 10C). На фигуре 10А показано, что свыше 94% клеток с трансгеом CAR с двумя нокаутами были негативными по CD3 (имели TRAC-нокауты); на фигуре 10В показано, что свыше 88% обработанных клеток были негативными по HLA (имели B2M-нокаут), а на фигуре 10С показано, что более 76% клеток с двойным нокаутом (TRAC и B2M) экспрессировали донорный CAR.
[0056] На фигуре 11 представлен график, иллюстрирующий результаты мультиплексного эксперимента (на множество нокаутов и/или на множество нацеленных интеграций). В этом эксперименте, Т-клетки обрабатывали TRAC- и B2M-специфическими реагентами ZFN, а также ZFN и двумя донорными конструкциями AAV: донором GFP, содержащим гомологичные ветви TRAC, и донором CD19-CAR, содержащим гомологичные ветви B2M. Данные показали, что обе пары ZFN расщепляли свои мишени с эффективностью >90%, и что доноры были интегрированы на 90% в случае донора GFP и на 77% в случае донора CD19-CAR. Более 62% клеток содержали все четыре редакции (TCR k/o, B2M k/o, GFP+ и CD19-CAR+0 для клеток, обработанных парами ZFN и обеими донорными конструкциями).
[0057] На фигурах 12А и 12В представлены графики, иллюстрирующие результаты интеграции трансгенного донора в клетки, которые были подвергнуты мультиплексному редактированию генов. На фигуре 12А, T-клетки были обработаны ZFN для специфического нокаута B2M, TRAC и CISH, а также показаны результаты интеграции GFP-экспрессирующего трансгена в локус TRAC, а на фигуре 12В показаны гипотетические результаты для аналогичной обработки T-клеток трансгеном, экспрессирущим CD19- CAR.
[0058] На фигуре 13 показан сайт интеграции гибридного трансгена B2M-HLA-G в сайт инсерции «D» в экзоне с3 (SEQ ID NO: 62) в локусе TRAC (см., публикацию заявки PCT WO 2017106528). Донорная конструкция содержит правые (R-HA) и левые (L-HA) гомологичные ветви, которые гомологичны последовательностям, фланкирующим сайт расщепления TRAC-ZFN. Экспрессия гибридного трансгена инициируется промотором PGK, а донор также содержит последовательность поли-А бычьего гормона роста (BGH).
[0059] На фигуре 14А-14С представлены графики, иллюстрирующие инсерцию гибридного трансгена B2M-HLA-G в ген TRAC. На фигуре 14А показаны результаты расщепления TRAC-специфической парой ZFN SBS#68877/SBS#68876, и продемонстрировано, что ген TRAC был нокаутирован с частотой 97%, как было определено по присутствию или отсутствию субъединицы CD3 TCR (левая панель). Аналогичным образом, B2M был нокаутирован с частотой 81% парой ZFN SBS#57071/SBS#57531, как как было определено по отсутствию комплекса ULA на поверхности клеток (правая панель). На фигурах 14В и 14С проиллюстрировано внутриклеточное (фигура 14В) и поверхностное (фигура 14С) окрашивание на HLA-G и B2M (левая и правая панели, соответственно). Как было показано, HLA-G экспрессировался на высоких уровнях внутри клеток, а B2M, экспрессируемый из конструкции HLA-B2M, детектировался внутри клеток и на поверхности клеток.
[0060] На фигурах 15A и 15B показаны FACS- и MisSeq-анализ CD4- и CD8-клеток, обработанных указанными донорными конструкциями B2M (трансгеном GFP) и B2M-нацеленными нуклеазами. На фигуре 15А показан FACS-анализ; на дальней левой панели показан только донор (без нуклеаз); на второй панели слева показаны клетки, обработанные донорными конструкциями GFP, активированными промотором PGK, включая длинные гомологичные ветви (~1 т.п.о. каждая) и отсутствие молекулярной метки, для оценки эффективности нацеленной интеграции посредством секвенирования последующей генерации (то есть, метки «Miseq»), и B2M-нацеленными нуклеазами; на средней панели показаны клетки, обработанные донорными конструкциями GFP, активированными промотором PGK, включая короткие гомологичные ветви (250 п.о. для каждой ветви) и метку Miseq, и B2M-нацеленными нуклеазами; на второй панели справа показаны клетки, обработанные донорными конструкциями, включая короткие гомологичные ветви и последовательность WPRE; а на самой дальней правой панели показаны клетки, обработанные донорными конструкциями, включая короткие гомологичные ветви и энхансерную последовательность TCRa. На фигуре 15В показаны результаты молекулярного анализа Miseq и количественной оценки результатов экспрессии, полученных с помощью проточной цитометрии, как показано на 15А.
[0061] На фигуре 16 схематически представлен репрезентативный донор для доставки трансгена, содержащего гомологичные ветви (например, в TRAC или B2M), фланкирующие промоторную последовательность, и трансген (например, CAR, B2M, HLA-Е и/или HLA-G и т.п.). Дополнительными компонентами, которые могут быть включены, являются энхансерные последовательности, последовательности WPRE, 5’- и 3’-UTR, последовательности полиаденилирования и т.п.
[0062] На фигуре 17 показаны результаты молекулярного анализа Miseq на инсерции-делеции и нацеленную интеграцию в генах-мишенях TRAC и B2M после трансфекции указанных ZFN и/или доноров. Образец «01_Mock» («1») означает ложные трансфекции; образец «02_TRAC+B2M» («2») относится к клеткам, трансфецированным только ZFN TRAC и B2M; образец 3 «03_site_E_423_493_hpGK_GFP-Miseq» («3») относится к клеткам, трансфицированным донором, содержащим левую ветвь 423 п.о. и правую ветвь 393 п.о., гомологичные сайту TRAC-E, фланкирующему трансген GFP, активируемый промотором hPGK. Этот донор также содержит метку «miseq», которая позволяет количественно определить TI в локусе TRAC по метке miseq. В образце 3, этот донор был добавлен отдельно без ZFN. Образец «04 TRAC+B2M site_E_423_493_hpGK_GFP-Miseq» содержит ZFN образца 3, а также донор образца 4. Образец «05 TRAC+B2M site_E_AAV_hpGK_GFP» содержит ZFN образца 3, а также донор с левой ветвью 1000 п.о. и с правой ветвью 992 п.о., гомологичными сайту TRAC-E, фланкирующему трансген GFP, активируемый промотором hPGK. Этот донор не содержит метку «miseq». Образец «06 TRAC+B2M site_E_350-393_hpGK_GFP-WPRE-Miseq» содержит ZFN образца 3 и донор, содержащий левую ветвь 350 п.о. и правую ветвь 393 п.о., гомологичные сайту TRAC-E, фланкирующему трансген GFP, активируемый промотором hPGK, а за ними следует мутированный элемент WPRE. Этот донор также содержит метку «miseq». Образец «07 TRAC+B2M site_E_423-393_hpGK_FMC63-CD8BBZ-Miseq» содержит ZFN образца 3 и донор, содержащий левую ветвь 423 п.о. и правую ветвь 393 п.о., гомологичные сайту TRAC-E, фланкирующему трансген FM63-CD8BBZ CAR, активируемый промотором hPGK. Этот донор также содержит метку «miseq». Образец «08 TRAC+B2M site_E_423-393_hpGK_FMC63-CD8BBZ-Miseq» содержит ZFN образца 3 и донор, содержащий левую ветвь 423 п.о. и правую ветвь 393 п.о., гомологичные сайту TRAC-E, фланкирующему трансген FM63-CD8BBZ CAR, активируемый промотором hPGK, а за ними следует мутированный элемент WPRE. Этот донор также содержит метку «miseq». Образец «09 TRAC+B2M site_E_350-393_hpGK_FMC63-CD8BBZ-P2A-B2M-HLA-E-WPREm-Miseq» содержит ZFN образца 3 и донор, содержащий левую ветвь 360 п.о. и правую ветвь 393 п.о., гомологичные сайту TRAC-E, фланкирующему трансген FM63-CD8BBZ CAR, связанный посредством саморасщепляющегося пептида P2A с гибридным белком и экспрессирующий B2M и HLA-E. Этот трансген активируется промотором hPGK, и за ним следует мутированный элемент WPRE. Этот донор также содержит метку «miseq». Образец «10 TRAC+B2M site_E_350-393_hpGK_FMC63-CD8BBZ-P2A-B2M-HLA-G-WPREm-Miseq» содержит ZFN образца 3 и донор, содержащий левую ветвь 360 п.о. и правую ветвь 393 п.о., гомологичные сайту TRAC-E, фланкирующему трансген FM63-CD8BBZ CAR, связанный посредством саморасщепляющегося пептида P2A с гибридным белком и экспрессирующий B2M и HLA-G. Этот трансген активируется промотором hPGK, и за ним следует мутированный элемент WPRE. Этот донор также содержит метку «miseq».
[0063] На фигурах 18А-18D показаны результаты FACS-анализа на нацеленную интеграцию в T-клетки здорового донора, обработанные TRAC- и B2M-нацеленными нуклеазами (обозначаемыми как клетки «DKO» с двойным нокаутом), а также донором. На фигуре 18А показаны результаты экспрессии после ложных трансфекций (верхняя панель); клетки DKO с нацеленной интеграцией с использованием донора GFP, включающего длинные гомологичные ветви (образец 5 на фигуре 17) (средняя панель); и DKO-клетки с нацеленной интеграцией с использованием донорного GFP с короткими гомологичными ветвями и с последовательностью WPRE (образец 6 на фигуре 17) (нижняя панель). На фигуре 18В показаны результаты экспрессии после ложных трансфекций (верхняя панель); клетки DKO с нацеленной интеграцией с использованием донорного CAR, включающего короткие гомологичные ветви (образец 7 на фигуре 17) (средняя панель); и DKO-клетки с нацеленной интеграцией с использованием донорного CAR с короткими гомологичными ветвями и с последовательностью WPRE (образец 8 на фигуре 17) (нижняя панель). На фигуре 18С показаны результаты экспрессии в DKO-клетках с нацеленной интеграцией с использованием донорных CAR и HLA-E (образец 9 на фигуре 17) (верхняя панель); и DKO-клетках с нацеленной интеграцией с использованием донорных CAR и HLA-G (образец 10 на фигуре 17) (нижняя панель). На фигуре 18D показаны результаты после ложных трансфекций (образец 1 на фигуре 17, верхняя панель); DKO-клетки без донора (образец 2 на фигуре 17) (средняя панель); и DKO-клетки с донорными CAR и HLA-E или HLA-G (две левые панели для образца 9 или две правые панели для образца 10 на фигуре 17) (нижняя панель).
[0064] На фигуре 19 показаны результаты молекулярного анализа MiSeq на инсерции-делеции после трансфекций указанных ZFN и/или доноров. Клетки, обработанные ZFN TRAC и B2M, имели двойные нокауты (обозначаемые как DKO). Образец «01_Mock» («1») относится к ложным трансфекциям; образец «02_TRAC+B2M» («2») относится к клеткам, трансфецированным только ZFN TRAC и B2M; образец 3 «_pAAV_TRAC_E_1000_992_hPGK_HLA_G only» («3») относится к клеткам, трансфицированным донором, содержащим левую ветвь 1000 п.о. и правую ветвь 992 п.о., гомологичные сайту TRAC-E, фланкирующему трансген HLA-G, активируемый промотором hPGK. Образец 4 «_pAAV_TRAC_E_1000_992_hPGK_HLA_E0101 only» («4») относится к клеткам, трансфицированным донором, содержащим левую ветвь 1000 п.о. и правую ветвь 992 п.о., гомологичные сайту TRAC-E, фланкирующему трансген HLA-E0101, активируемый промотором hPGK. Образец 5 «_pAAV_TRAC_E_1000_992_hPGK_HLA_E0103 only» («5») относится к клеткам, трансфицированным донором, содержащим левую ветвь 1000 п.о. и правую ветвь 992 п.о., гомологичные сайту TRAC-E, фланкирующему трансген HLA-E0103, активируемый промотором hPGK. Образец 6 «_ pAAV_TRAC_E_1000_992_hPGK_HLA_G» («6») относится к клеткам, трансфицированным ZFN образца 2 и донором, содержащим левую ветвь 1000 п.о. и правую ветвь 992 п.о., гомологичные сайту TRAC-E, фланкирующему трансген HLA-G, активируемый промотором hPGK. Образец 7 «_ pAAV_TRAC_E_1000_992_hPGK_B2M_G4S_2_noSP_HLA_G» («7») относится к клеткам, трансфицированным ZFN образца 2 и донором, содержащим левую ветвь 1000 п.о. и правую ветвь 992 п.о., гомологичные сайту TRAC-E, фланкирующему трансген HLA-G В2М, активируемый промотором hPGK. Линкер между В2М и HLA-G содержит 2 копии пептида G4S. HLA-G не содержит сигнального пептида. Образец 8 «_ pAAV_TRAC_E_1000_992_hPGK_B2M_G4S_2_noSP_HLA_G» («8») относится к клеткам, трансфицированным ZFN образца 2 и донором, содержащим левую ветвь 1000 п.о. и правую ветвь 992 п.о., гомологичные сайту TRAC-E, фланкирующему связанный трансген HLA-G В2М, активируемый промотором hPGK. Линкер между В2М и HLA-G содержит 4 копии пептида G4S. HLA-G не содержит сигнального пептида. Образец 9 «_pAAV_TRAC_E_1000_992_hPGK_B2M_G4S_2_noSP_HLA_G» («9») относится к клеткам, трансфицированным ZFN образца 2 и донором, содержащим левую ветвь 1000 п.о. и правую ветвь 992 п.о., гомологичные сайту TRAC-E, фланкирующему связанный трансген HLA-G В2М, активируемый промотором hPGK. Линкер между В2М и HLA-G содержит 6 копий пептида G4S. HLA-G не содержит сигнального пептида. Образец 10 «_pAAV_TRAC_E_1000_992_hPGK_HLA_E0101» («10») относится к клеткам, трансфицированным ZFN образца 2 и донором, содержащим левую ветвь 1000 п.о. и правую ветвь 992 п.о., гомологичные сайту TRAC-E, фланкирующему трансген HLA-E0101, активируемый промотором hPGK. Образец 11 «_ pAAV_TRAC_E_1000_992_hPGK_HLA_E0101» («11») относится к клеткам, трансфицированным ZFN образца 2 и донором, содержащим левую ветвь 1000 п.о. и правую ветвь 992 п.о., гомологичные сайту TRAC-E, фланкирующему трансген HLA-E0103, активируемый промотором hPGK. Образец 12 «_ pAAV_TRAC_E_1000_992_hPGK_B2M_noSP_HLA_E0101» («12») относится к клеткам, трансфицированным ZFN образца 2 и донором, содержащим левую ветвь 1000 п.о. и правую ветвь 992 п.о., гомологичные сайту TRAC-E, фланкирующему связанный трансген HLA-E0101 В2М, активируемый промотором hPGK. Линкер между В2М и HLA-E0101 содержит 4 копии пептида G4S. HLA-E0101 не содержит сигнального пептида. Образец 13 «_pAAV_TRAC_E_1000_992_hPGK_B2MnoSP_HLA_E0103» («13») относится к клеткам, трансфицированным ZFN образца 2 и донором, содержащим левую ветвь 1000 п.о. и правую ветвь 992 п.о., гомологичные сайту TRAC-E, фланкирующему связанный трансген HLA-E0101 В2М, активируемый промотором hPGK. Линкер между В2М и HLA-E0101 содержит 4 копии пептида G4S. HLA-E0103 не содержит сигнального пептида. Образец 14 «pAAV_TRAC_E_1000_992_pgk_gfp» («14») относится к клеткам, трансфицированным ZFN образца 2 и донором, содержащим левую ветвь 1000 п.о. и правую ветвь 992 п.о., гомологичные сайту TRAC-E, фланкирующему трансген GFP, активируемый промотором hPGK. Образец 15 «pAAV_TRAC_E_350_393_hPGK_CAR_B2M_HLA_G_WPREm_Miseq» («15»), относится к клеткам, трансфицированным ZFN образца 2 и донором, содержащим левую ветвь 360 п.о. и правую ветвь 393 п.о., гомологичные сайту TRAC-E, фланкирующему трансген FM63-CD8BBZ CAR, связанный саморасщепляющимся пептидом P2A с гибридным белком и экспрессирующий B2M и HLA-G. Этот трансген активируется промотором HPGK, и за ним следует мутированный элемент WPRE. Этот донор также содержит метку «miseq».
[0065] На фигуре 20 показаны результаты FACS-анализа на нацеленную интеграцию в T-клетки здорового донора, обработанные, как было указано. Клетки, обработанные TRAC и B2M ZFN, обозначаются «DKO» (с двойным нокаутом). Ни верхних панелях показана экспрессия CD3, HLA-ABC и GFP после ложных трансфекций (образец 1 на фигуре 19); средние панели иллюстрируют экспрессию тех же самых белков в клетках DKO в отсутствии донора (образец 2 на фигуре 19); а нижняя панель иллюстрируют экспрессию в клетках DKO, которые были также обработаны донорным GFP (образец 14 на фигуре 19).
[0066] На фигуре 21 показаны результаты FACS-анализа на нацеленную интеграцию в T-клетки здорового донора, обработанные, как было указано. Клетки, обработанные TRAC и B2M ZFN, обозначаются «DKO» (с двойным нокаутом). На верхних панелях показана экспрессия указанных белков (CD3, HLA-ABC, B2M и HLA-G) после ложных трансфекций (образец 1 на фигуре 19); средние панели иллюстрируют экспрессию тех же самых белков в клетках DKO с TRAC/B2M и в отсутствии донора (образец 2 на фигуре 19), а нижняя панель иллюстрируют экспрессию в клетках, обработанных только донорным HLA-G (образец 3 на фигуре 19).
[0067] На фигуре 22 показаны результаты FACS-анализа на нацеленную интеграцию в T-клетки здорового донора, обработанные, как было указано. Клетки, обработанные TRAC и B2M ZFN, обозначаются «DKO» (с двойным нокаутом). На верхних панелях показана экспрессия указанных белков (CD3, HLA-ABC, B2M и HLA-G) после трансфекции донорного HLA-G в клетки DKO (образец 6 на фигуре 19); на второй панели сверху показана экспрессия тех же самых белков после трансфекции донора, включающего трансгены B2M и HLA-G в клетки DKO (образец 7 на фигуре 19); на второй панели снизу показана экспрессия тех же самых белков после трансфекции донора, включающего трансгены B2M и HLA-G в клетки DKO (образец 8 на фигуре 19); а на нижних панелях показана экспрессия тех же самых белков после трансфекции донора, включающего трансгены B2M и HLA-G в клетки DKO (образец 9 на фигуре 19).
[0068] На фигуре 23 показаны результаты FACS-анализа на нацеленную интеграцию в T-клетки здорового донора, обработанные, как было указано. Клетки, обработанные TRAC и B2M ZFN, обозначаются «DKO» (с двойным нокаутом). На верхних панелях показана ложная трансфекция (в отсутствии донора или ZFN, образец 1 на фигуре 19); на второй панели сверху показана экспрессия указанных белков (CD3, HLA-ABC, B2M и HLA-Е) в клетках DKO (образец 2 на фигуре 19); на второй панели снизу показана экспрессия тех же самых белков после трансфекции только донорного HLA-Е (образец 4 на фигуре 19); а на нижних панелях показана экспрессия тех же самых белков после трансфекции только донорного HLA (образец 5 на фигуре 19).
[0069] На фигуре 24 показаны результаты FACS-анализа на нацеленную интеграцию в T-клетки здорового донора, обработанные, как было указано. Клетки, обработанные TRAC и B2M ZFN, обозначаются «DKO» (с двойным нокаутом). На верхних панелях показана экспрессия указанных белков (CD3, HLA-ABC, B2M и HLA-E) в клетках DKO с донорной конструкцией HLA-E0101 (образец 10 на фигуре 19); на второй панели сверху показана экспрессия указанных белков в клетках DKO с донорной конструкцией HLA-E0103 (образец 11 на фигуре 19); на второй панели снизу показана экспрессия тех же самых белков после трансфекции донора B2M и HLA-Е в клетки DKO (образец 12 на фигуре 19); а на нижних панелях показана экспрессия тех же самых белков после трансфекции донора B2M и HLA-Е в клетки DKO (образец 13 на фигуре 19).
[0070] На фигуре 25 показаны результаты FACS-анализа на нацеленную интеграцию в T-клетки здорового донора, обработанные, как было указано. Клетки, обработанные TRAC и B2M ZFN, обозначаются «DKO» (с двойным нокаутом). На верхних панелях показана экспрессия указанных белков (CD3, HLA-ABC и белка L=CAR) после ложных трансфекций (образец 1 на фигуре 19); на средних панелях показана экспрессия тех же самых белков в клетках DKO с TRAC/B2M (образец 2 на фигуре 19) и в отсутствии донора; а на нижней панели показана экспрессия в клетках DKO, обработанных донорным CAR-B2M-HLA-G (образец 15 на фигуре 19).
[0071] На фигуре 26 показаны результаты FACS-анализа на экспрессию указанных белков (CD3, HLA-ABC, B2M и HLA-G) на клеточной поверхности (на верхних панелях, помеченных ECS) и внутри клеток (на нижних панелях, помеченных ICS) в клетках DKO с донорным CAR-B2M-HLAG (образец 15 На Фигуре 19).
Подробное описание
[0072] В настоящей заявке раскрываются композиции и способы получения Т-клеток, в которых экспрессия одного или более CAR или CAR и модифицированных HLA-E или HLA-G является эффективной для уничтожения клеток, экспрессирующих антиген, нацеленный на CAR. Кроме того, уничтожение клеток является эффективным, если CAR или экспрессионная конструкция CAR-HLA-E или CAR-HLA-G интегрированы в ген TCR и/или ген B2M. Клетки, модифицированные таким образом, могут быть использованы в качестве терапевтических средств, например, для адоптивной клеточной терапии, поскольку отсутствие комплекса TCR в результате интеграции CAR в ген TCR предотвращает нацеливание этих T-клеток на другие клетки, не несущие антигена, а нокаут HLA-комплекса посредством интеграции CAR в B2M устраняет или ослабляет иммунный ответ на основе HLA, направленный против введенных «вооруженных» T-клеток. Клетки, которые были дополнительно модифицированы для экспрессии модифицированного комплекса HLA-E или HLA-G, также могут «ускользать» от уничтожения NK-клетками-хозяевами. Кроме того, другие представляющие интерес гены могут быть встроены в клетки, несущие CAR и необязательно модифицированные HLA-E или HLA-G, и/или другие представляющие интерес гены могут быть нокаутированы.
Общее описание
[0073] Практическое применение способов, а также получение и применение раскрытых здесь композиций может быть осуществлено, если это не оговорено особо, с помощью стандартных методов молекулярной биологии, биохимии, исследования структуры хроматина и его анализа, аналитической химии, культивирования клеток, техники рекомбинантных ДНК и родственных областей, известных специалистам. Эти методы подробно описаны в литературе. См. например, Sambrook et al. MOLECULAR CLONING: A LABORATORY MANUAL, Second edition, Cold Spring Harbor Laboratory Press, 1989 and Third edition, 2001; Ausubel et al., CURRENT PROTOCOLS IN MOLECULAR BIOLOGY, John Wiley & Sons, New York, 1987 и современные периодические издания; ряд публикаций METHODS IN ENZYMOLOGY, Academic Press, San Diego; Wolffe, CHROMATIN STRUCTURE AND FUNCTION, Third edition, Academic Press, San Diego, 1998; METHODS IN ENZYMOLOGY, Vol. 304, “Chromatin” (P.M. Wassarman and A.P. Wolffe, eds.), Academic Press, San Diego, 1999; и METHODS IN MOLECULAR BIOLOGY, Vol. 119, “Chromatin Protocols” (P.B. Becker, ed.) Humana Press, Totowa, 1999.
Определения
[0074] Используемые здесь термины «нуклеиновая кислота», «полинуклеотид» и «олигонуклеотид» являются синонимами и означают дезоксирибонуклеотидный или рибонуклеотидный полимер в линейной или кольцевой конформации и в одноцепочечной или двуцепочечной форме. В целях раскрытия настоящего изобретения, эти термины не следует рассматривать как ограничение длины полимера. Эти термины могут включать известные аналоги природных нуклеотидов, а также нуклеотиды, которые были модифицированы в молекулах оснований, сахаров и/или фосфатов (например, в фосфортиоатных остовах). Вообще говоря, аналог конкретного нуклеотида имеет ту же самую специфичность спаривания оснований, то есть, аналог А будет образовывать пары оснований с Т.
[0075] Используемые здесь термины «полипептид», «пептид» и «белок» являются синонимами и означают полимер из аминокислотных остатков. Этот термин также относится к аминокислотным полимерам, в которых одна или более аминокислот представляют собой химические аналоги или модифицированные производные соответствующих природных аминокислот.
[0076] Термин «связывание» относится к последовательность-специфическому нековалентному взаимодействию между макромолекулами (например, между белком и нуклеиновой кислотой). Не все компоненты такого взаимодействия посредством связывания должны быть последовательность-специфическими (например, контакты с фосфатными остатками в остове ДНК), при условии, что такое взаимодействие в целом будет последовательность-специфическим. Такие взаимодействия обычно характеризуются константой диссоциации (Kd) 10-6 М-1 или ниже. «Аффинность» относится к силе связывания: повышенная аффинность связывания коррелирует с более низкой Kd. «Неспецифическое связывание» относится к нековалентным взаимодействиям, которые происходят между любой представляющей интерес молекулой (например, сконструированной нуклеазой) и макромолекулой (например, ДНК), и которые не зависят от последовательности-мишени.
[0077] «ДНК-связывающая молекула» представляет собой молекулу, которая может связываться с ДНК. Такая ДНК-связывающая молекула может представлять собой полипептид, домен белка, домен в более крупном белке или полинуклеотид. В некоторых вариантах осуществления изобретения, полинуклеотид представляет собой ДНК, а в других вариантах осуществления изобретения, полинуклеотид представляет собой РНК. В некоторых вариантах осуществления изобретения, ДНК-связывающая молекула является белковым доменом нуклеазы (например, доменом FokI), а в других вариантах осуществления изобретения, ДНК-связывающая молекула представляет собой компонент руководящей РНК нуклеазы, нацеленной на РНК (например, Cas9 или Cfp1).
[0078] «Связывающий белок» представляет собой белок, который способен нековалентно связываться с другой молекулой. Связывающий белок может связываться, например, с молекулой ДНК (ДНК-связывающий белок), с молекулой РНК (РНК-связывающий белок) и/или с молекулой белка (белок-связывающий белок). В случае белка, связывающегося с белком, он может связываться сам с собой (с образованием гомодимеров, гомотримеров и т.п.), и/или он может связываться с одной или более молекулами другого белка или других белков. Связывающий белок может обладать активностью связывания более, чем одного типа. Так, например, белки «цинковые пальцы» обладают ДНК-связывающей, РНК-связывающей и белок-связывающей активностью.
[0079] «ДНК-связывающий белок цинковый палец» (или его связывающий домен) представляет собой белок или домен в более крупном белке, который связывается с ДНК по последовательность-специфическому механизму посредством одного или более цинковых пальцев, которые представляют собой области аминокислотной последовательности в связывающем домене, структура которого была стабилизирована посредством координации иона цинка. Термин «ДНК-связывающий белок цинковый палец» часто сокращенно называется белком «цинковые пальцы» или ZFP. Термин «нуклеаза цинковые пальцы» включает одну ZFN, а также пару ZFN (члены пары называются «левым и правым» или «первым и вторым» или «парой»), которые димеризуются и расщепляют ген-мишень.
[0080] «ДНК-связывающий домен TALE» или «TALE» представляет собой полипептид, содержащий один или более повторяющихся доменов/звеньев TALE. Повторяющиеся домены, каждый из которых включает повторяющийся вариабельный димерный остаток (RVD), участвуют в связывании TALE с его когнатной последовательностью ДНК-мишени. Одно «повторяющееся звено» (также называемое «повтором») обычно имеет длину 33-35 аминокислот и имеет по меньшей мере некоторую гомологию последовательности с другими последовательностями повторов TALE в природном белке TALE. Белки TALE могут быть сконструированы так, чтобы они связывались с сайтом-мишенью посредством канонических или неканонически RVD в повторяющихся звеньях. См., например, патенты США № 8586526 и 9458205. Термин «TALEN» включает один TALEN, а также пару TALEN (члены пары называются «левым и правым» или «первым и вторым» или «парой»), которые димеризуются и расщепляют ген-мишень.
[0081] ДНК-связывающие домены «цинковые пальцы» и TALE могут быть «сконструированы» так, чтобы они связывались с предварительно определенной нуклеотидной последовательностью, например, путем конструирования (модификации одной или более аминокислот) области спирали распознавания природного белка «цинковый палец» или путем конструирования аминокислот, участвующих в связывании с ДНК (повторяющимся вариабельным димерным остатком или областью RVD). Следовательно, сконструированные белки «цинковые пальцы» или белки TALE представляет собой белки, которые не встречаются в природе. Неограничивающий примерами способов получения белков «цинковые пальцы» и TALE являются конструирование и отбор. Сконструированный белок представляет собой белок, который не встречается в природе, и конструкция/состав которого, в основном, соответствуют рациональным критериям. Рациональные критерии для конструирования включают применение правил замены и компьютерных алгоритмов для обработки информации в базе данных, хранящей информацию о существующих конструкциях ZFP или TALE (канонические и неканонические RVD) и данные по связыванию. См., например, патенты США №№ 9458205, 8586526, 6140081, 6453242 и 6534261; см. также WO 98/53058; WO 98/53059; WO 98/53060; WO 02/016536 и WO 03/016496.
[0082] «Выбранный» белок цинковые пальцы, белок TALE или CRISSPR/Cas-система не существуют в природе, и их получение осуществляют, главным образом, эмпирическими способами, такими как фаговое представление, взаимодействие-ловушка или гибридный отбор. См., например, патенты США 5789538; 5925523; 6007988; 6013453; 6200759; WO 95/19431; WO 96/06166; WO 98/53057; WO 98/54311; WO 00/27878; WO 01/60970; WO 01/88197 и WO 02/099084. Кроме того, белки TALE (которые обычно имеют точное соответствие между RVD и нуклеотидом, связанными в сайте-мишени) и оцрРНК систем CRISPR/Cas могут быть легко сконструированы для нацеления на сайты-мишени для ZFP, с которым связываются ZFP. См., например, патенты США № 9873894 и 8586526.
[0083] «TtAgo» представляет собой прокариотический белок Argonaute, который, как предполагается, участвует в сайленсинге гена. TtAgo получают из бактерий Thermus thermophilus. См. например, Swarts et al. ibid, G. Sheng et al. (2013) Proc Natl Acad Sci USA 111, 652). «Система TtAgo» представляет собой все необходимые компоненты, включая, например, руководящие ДНК для расщепления ферментом TtAgo.
[0084] «Рекомбинация» означает процесс обмена генетической информацией между двумя полинуклеотидами. В целях раскрытия настоящего изобретения используется термин «гомологичная рекомбинация (ГР)», который означает специализированную форму такого обмена, происходящего, например, в процессе репарации двухцепочечных разрывов в клетках по механизмам репарации в зависимости от гомологии. Для осуществления такого процесса требуется гомология нуклеотидных последовательностей, использование «донорной» молекулы для репарации матрицы молекулы-«мишени» (то есть, молекулы, имеющей двухцепочечный разрыв), и такой процесс известен как «конверсия гена по некроссинговерному механизму» или «конверсия гена по короткому пути», поскольку она приводит к переносу генетической информации от донора к мишени. Не ограничиваясь какой-либо конкретной теорией, авторы лишь отмечают, что такой перенос может включать коррекцию ошибочного спаривания гетеродуплексной ДНК, которая образуется между разрушенной мишенью и донором, и/или «зависимый от синтеза отжиг цепи», при котором донор используется для повторного синтеза генетической информации, которая будет становиться частью мишени, и/или родственные процессы. Такая специализированная ГР часто приводит к модификации последовательности молекулы-мишени таким образом, что часть или вся последовательность полинуклеотида-донора будет включена в полинуклеотид-мишень.
[0085] В способах согласно изобретению, одна или более из описанных здесь нуклеаз-мишеней создают двухцепочечный разрыв (DSB) в последовательности-мишени (например, в клеточном хроматине) в предварительно определенном сайте (например, в представляющем интерес гене или локусе), и «донорный» полинуклеотид, гомологичный нуклеотидной последовательности в области разрыва, может быть введен в клетку. Было показано, что присутствие DSB облегчает интеграцию донорной последовательности. Такая конструкция, но необязательно, гомологчна нуклеотидной последовательности в области разрыва. Донорная последовательность может быть физически интегрирована или, альтернативно, донорный полинуклеотид используется в качестве матрицы для репарации разрыва посредством гомологичной рекомбинации, что приводит к введению всей или части нуклеотидной последовательности в качестве донора в клеточный хроматин. Таким образом, первая последовательность в клеточном хроматине может быть модифицирована, а в некоторых вариантах осуществления изобретения, она может быть превращена в последовательность, присутствующую в донорном полинуклеотиде. Таким образом, использование терминов «заменить» или «замена» может быть интерпретировано как замена одной нуклеотидной последовательности другой последовательностью (то есть, замена последовательности в информационном смысле), но, при этом, необязательно, чтобы она была физической или химической заменой одного полинуклеотида другим.
[0086] В любом из описанных здесь способов, дополнительные пары белков «цинковые пальцы» могут быть использованы для дополнительного двухцепочечного расщепления дополнительных сайтов-мишеней в клетке.
[0087] В некоторых вариантах способов нацеленной рекомбинации и/или замены и/или модификации последовательности в представляющей интерес области в клеточном хроматине, хромосомную последовательность модифицируют путем гомологичной рекомбинации с экзогенной «донорой» нуклеотидной последовательностью. Такая гомологичная рекомбинация стимулируется присутствием двухцепочечного разрыва в клеточном хроматине, если присутствуют последовательности, гомологичные области разрыва.
[0088] В любом из описанных здесь способов, первая нуклеотидная последовательность («донорная последовательность») может содержать последовательности, которые являются гомологичными, но не идентичными геномным последовательностям в представляющей интерес области, что тем самым стимулирует гомологичную рекомбинацию для встраивания неидентичной последовательности в представляющую интерес область. Таким образом, в некоторых вариантах осуществления изобретения, части донорной последовательности, которые являются гомологичными последовательностям в представляющей интерес области, приблизительно на 80-99% (или любое целое число в этих пределах) идентичны геномной последовательности, которая была заменена. В других вариантах осуществления изобретения, гомология между донорной и геномной последовательностью составляет более, чем 99%, например, если только 1 нуклеотид отличается в донорных и геномных последовательностях, имеющих более чем 100 смежных пар оснований. В некоторых случаях, негомологичная часть донорной последовательности может содержать последовательности, не присутствующие в представляющей интерес области, а поэтому, в представляющую интерес область вводят новые последовательности. В этих случаях, негомологичная последовательность обычно фланкируется последовательностями, составляющими 50-1000 пар оснований (или любое целое число в этих пределах) или любое число пар оснований более 1000, которые гомологичны или идентичны последовательностям в представляющей интерес области. В других вариантах осуществления изобретения, донорная последовательность не является гомологичной первой последовательности и встраивается в геном по негомологичным механизмам рекомбинации.
[0089] Любой из описанных здесь способов может быть применен для частичной или полной инактивации одной или более последовательностей-мишеней в клетке путем нацеленной интеграции донорной последовательности, которая нарушает экспрессию представляющего(их) интерес гена(ов). Настоящее изобретение также относится к клеточным линиям с частично или полностью инактивированными генами.
[0090] Кроме того, описанные здесь способы нацеленной интеграции могут быть также применены для интеграции одной или более экзогенных последовательностей. Экзогенная последовательность нуклеиновой кислоты может содержать, например, один или более генов или молекул кДНК или кодирующих или некодирующих последовательностей любого типа, а также один или более регуляторных элементов (например, промоторов). Кроме того, экзогенная последовательность нуклеиновой кислоты может продуцировать одну или более молекул РНК (например, коротких шпилечных РНК (кшРНК), ингибирующих РНК (иРНК), микро-РНК (миPHK) и т.п.).
[0091] «Расщепление» означает разрушение ковалентного остова молекулы ДНК. Расщепление может быть инициировано различными способами, включая, но не ограничиваясь ими, ферментативный или химический гидролиз фосфодиэфирной связи. При этом, возможно как одноцепочечное расщепление, так и двухцепочечное расщепление, а двухцепочечное расщепление может происходить в результате двух отдельных событий одноцепочечного расщепления. Расщепление ДНК может приводить к образованию либо «тупых» концов, либо «ступенчатых» концов. В некоторых вариантах осуществления изобретения, гибридные полипептиды используются для нацеленного расщепления двухцепочечной ДНК.
[0092] «Полудомен расщепления» представляет собой полипептидную последовательность, которая в сочетании со вторым полипептидом (либо идентичным, либо отличающимся) образует комплекс, обладающий активностью расщепления (предпочтительно, активностью двухцепочечного расщепления). Используемые здесь термины «первый и второй полудомены расщепления»; «полудомены расщепления+и -» и «правый и левый полудомены расщепления» являются синонимами и означают пары полудоменов расщепления, которые димеризуются.
[0093] «Сконструированный полудомен расщепления» представляет собой полудомен расщепления, который был модифицирован так, чтобы он образовывал облигатные гетеродимеры с другим полудоменом расщепления (например, с другим сконструированным полудоменом расщепления). См. также патенты США № 7888121; 7914796; 3834598; 8623618 и публикацию заявки на патент США № 2011/0201055, которые включены в настоящее описание посредством ссылки в полном объеме.
[0094] Термин «последовательность» означает нуклеотидную последовательность любой длины, которая может представлять собой ДНК или РНК; может быть линейной, кольцевой или разветвленной, и может быть либо одноцепочечной, либо двуцепочечной. Термин «донорная последовательность» означает нуклеотидную последовательность, встроенную в геном. Донорная последовательность может иметь любую длину, например от 2 до 10000 нуклеотидов (или любое целое число в этих пределах или выше), предпочтительно, приблизительно от 100 до 1000 нуклеотидов (или любое целое число в этих пределах, а еще более предпочтительно, приблизительно от 200 до 500 нуклеотидов.
[0095] «Хроматин» представляет собой нуклеопротеиновую структуру, включающую клеточный геном. Клеточный хроматин содержит нуклеиновую кислоту, главным образом, ДНК, и белок, включая гистоны и негистоновые хромосомные белки. Большинство эукариотических клеточных хроматинов существует в форме нуклеосом, где нуклеосомная сердцевина содержит приблизительно 150 пар оснований ДНК, связанных с октамером, содержащим два из гистонов H2A, H2B, H3 и H4; и линкерную ДНК (длина которой может варьироваться в зависимости от организма), простирающуюся между нуклеосомными сердцевинами. Молекула гистона H1 обычно связана с линкерной ДНК. В соответствии с целями настоящего описания, термин «хроматин» охватывает клеточные нуклеопротеины всех типов как прокариотических, так и эукариотических. Клеточный хроматин включает как хромосомный, так и эписомный хроматин.
[0096] «Хромосома» представляет собой хроматиновый комплекс, содержащий всю или часть генома клетки. Геном клетки часто характеризуется его кариотипом, который представляет собой набор всех хромосом, содержащихся в геноме клетки. Геном клетки может содержать одну или более хромосом.
[0097] «Эписома» представляет собой реплицирующуюся нуклеиновую кислоту, нуклеопротеиновый комплекс или другую структуру, содержащую нуклеиновую кислоту, которая не является частью хромосомного кариотипа клетки. Примерами эписом являются плазмиды и некоторые вирусные геномы.
[0098] «Сайт-мишень» или «последовательность-мишень» представляет собой последовательность нуклеиновой кислоты, которая определяет часть нуклеиновой кислоты, с которой связывается связывающая молекула, если будут выполняться условия, достаточные для связывания. Так, например, последовательность 5’-GAATTC-3’ представляет собой сайт-мишень для рестриктирующей эндонуклеазы EcoRI. Этот термин включает смежные или несмежные пары оснований любой последовательности, а также «спаренный» сайт-мишень (например, спаренные сайты-мишени для описанных здесь ZFN и/или TALEN). После связывания одного или более ДНК-связывающих доменов (например, ZFP, TALE, одноцепочечной руководящей РНК) нуклеазы с их соответствующими сайтами-мишенями, расщепление и модификация (например, интеграция донора и/или модификации, то есть, инсерции/делеции) могут происходить в любом положении в сайте-мишени или возле этого сайта, включая, но не ограничиваясь ими, положения в последовательностях, связанных с ДНК-связывающим доменом, между спаренными и/или смежными сайтами-мишенями (например, в пределах 1-50 (включая 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25 или более) или более пар оснований со стороны 3’ или 5’ по отношению к любому из сайтов-мишеней).
[0099] «Экзогенная» молекула представляет собой молекулу, которая обычно не присутствует в клетке, но может быть введена в клетку с применением одного или более генетических, биохимических или других методов. «Нормальное присутствие в клетке» определяют в зависимости от конкретной стадии развития и условий окружающей среды клетки. Так, например, молекула, которая присутствует только во время эмбрионального развития мышц, представляет собой экзогенную молекулу по отношению к клетке мышцы взрослого. Аналогичным образом, молекула, индуцированная тепловым шоком, является экзогенной молекулой по отношению к клетке, содержащей молекулу, не подвергнутую тепловому шоку. Экзогенная молекула может содержать, например, функциональный вариант неправильно функционирующей эндогенной молекулы или неправильно функционирующий вариант природной эндогенной молекулы.
[0100] Экзогенная молекула может представлять собой, среди прочих, небольшую молекулу, такую как молекула, продуцируемая комбинаторным химическим способом, или макромолекулу, такую как белок, нуклеиновая кислота, углевод, липид, гликопротеин, липопротеин, полисахарид, любое модифицированное производное вышеуказанных молекул или любой комплекс, содержащий одну или более из вышеуказанных молекул. Нуклеиновыми кислотами являются ДНК и РНК, которые могут быть одноцепочечными или двухцепочечными; могут быть линейными, разветвленными или кольцевыми; и могут иметь любую длину. См., например, патенты США №№ 8703489 и 9255259. Нуклеиновые кислоты включают кислоты, которые способны образовывать дуплексы, а также нуклеиновые кислоты, образующие триплексы. См., например, патенты США №№ 5176996 и 5422251. Белками являются, но не ограничиваются ими, ДНК-связывающие белки, факторы транскрипции, факторы ремоделирования хроматина, метилированные ДНК-связывающие белки, полимеразы, метилазы, деметилазы, ацетилазы, деацетилазы, киназы, фосфатазы, интегразы, рекомбиназы, лигазы, топоизомеразы, гиразы и геликазы.
[0101] Экзогенная молекула может представлять собой молекулу такого же типа, как и эндогенная молекула, например, экзогенный белок или экзогенную нуклеиновую кислоту. Так, например, экзогенная нуклеиновая кислота может содержаться в инфицирующем вирусном геноме, плазмиде или эписоме, введенных в клетку, или в хромосоме, которая обычно не присутствует в клетке. Методы введения экзогенных молекул в клетки известны специалистам в данной области и включают, но не ограничиваются ими, опосредованный липидом перенос (то есть, липосомами, включая нейтральные и катионные липиды), электропорацию, прямую инжекцию, слияние клеток, бомбардировку частицами, совместное осаждение фосфатом кальция, перенос, опосредуемый DEAE-декстраном, и перенос, опосредуемый вирусным вектором. Экзогенная молекула может также представлять собой молекулу такого же типа, как и эндогенная молекула, но происходит от клеток других видов, отличающихмя от данной клетки. Так, например, человеческая последовательность нуклеиновой кислоты может быть введена в клеточную линию, первоначально полученную от мыши или хомячка.
[0102] В противоположность этому, «эндогенная» молекула представляет собой молекулу, которая обычно присутствует в конкретной клетке на определенной стадии развития в конкретных условиях окружающей среды. Так, например, эндогенная нуклеиновая кислота может содержаться в хромосоме, геноме митохондрий, хлоропласта или другой органеллы, или представлять собой природную эписомную нуклеиновую кислоту. Другие эндогенные молекулы могут включать белки, например, факторы транскрипции и ферменты.
[0103] «Гибридная» молекула представляет собой молекулу, в которой две или более молекул субъединиц являются связанными, предпочтительно, ковалентно. Молекулы субъединиц могут представлять собой химические молекулы одного и того же типа или химические молекулы различных типов. Примерами гибридных молекул первого типа являются, но не ограничиваются ими, гибридные белки (например, гибрид между ДНК-связывающим доменом ZFP или TALE и одним или более доменами активации) и гибридные нуклеиновые кислоты (например, нуклеиновая кислота, кодирующая гибридный белок, описанный выше). Примерами гибридных молекул второго типа являются, но не ограничиваются ими, гибрид между нуклеиновой кислотой, образующей триплекс, и полипептидом, и гибрид между небольшой молекулой, связывающейся с бороздкой, и нуклеиновой кислотой. Этот термин также включает системы, в которых полинуклеотидный компонент ассоциируется с полипептидным компонентом с образованием функциональной молекулы (например, системы CRISSPR/Cas, в которой одноцепочечная руководящая РНК ассоциируется с функциональным доменом и модулирует экспрессию гена).
[0104] Экспрессия гибридного белка в клетке может быть результатом доставки гибридного белка в клетку или доставки полинуклеотида, кодирующего гибридный белок, в клетку, где полинуклеотид транскрибируется, а транскрипт транслируется с образованием гибридного белка. Транс-сплайсинг, расщепление полипептида и лигирование полипептида также могут участвовать в экспрессии белка в клетке. Методы доставки полинуклеотидов и полипептидов в клетку описаны в настоящей заявке.
[0105] Термин «ген», используемый в целях раскрытия настоящего изобретения, включает область ДНК, кодирующую генный продукт (см. ниже), а также все области ДНК, которые регулируют продуцирования генного продукта, независимо от того, являются ли такие регуляторные последовательности смежными с кодирующими и/или транскрибированными последовательностями, или нет. Соответственно, ген включает, но не обязательно ограничивается ими, промоторные последовательности, терминаторы, последовательности регуляции трансляции, такие как сайты связывания с рибосомой и внутренние сайты связывания с рибосомой, энхансеры, сайленсеры, изоляторы, пограничные элементы, ориджины репликации, сайты связывания с матрицей и области регуляции локусов.
[0106] «Безопасный якорный локус» представляет собой локус в геноме, в который может быть введен ген так, чтобы он не оказывал какого-либо негативного влияния на клетку-хозяина. Наиболее предпочтительным является безопасный якорный локус, в котором экспрессия встроенной генной последовательности не нарушается в результате сквозного прочитывания экспрессии от соседних генов. Неограничивающими примерами безопасных якорных локусов, на которые нацелена(ы) нуклеаза(ы), являются CCR5, CCR5, HPRT, AAVS1, Rosa и альбумин. См., например, патенты США №№ 8771985; 8110379; 7951925; и публикации заявок на патент США №№ 20100218264; 20110265198; 20130137104; 20130122591; 20130177983; 20130177960; 20150056705 и 20150159172.
[0107] «Экспрессия генов» означает преобразование информации, содержащейся в гене, в генный продукт. Генный продукт может представлять собой продукт непосредственной транскрипции гена (например, мРНК, тРНК, рРНК, антисмысловой РНК, рибозима, структурной РНК или РНК любого другого типа РНК) или белка, продуцируемого в результате трансляции мРНК. Генные продукты также включают РНК, которые были модифицированы такими способами, как кэпирование, полиаденилирование, метилирование и редактирование, и белки, модифицированные, например, посредством метилирования, ацетилирования, фосфорилирования, убихитинизации, АДФ-рибозилирования, миристоилирования и гликозилирования. «Модуляция» или «модификация» экспрессии гена означает изменение активности гена. Модуляция экспрессии может включать, но не ограничивается ими, активацию гена и репрессию генов, включая модификацию гена посредством связывания экзогенной молекулы (например, сконструированного фактора транскрипции). Модуляция может быть также достигнута путем модификации генной последовательности посредством редактирования генома (например, расщепления, изменения, инактивации, случайной мутации). Инактивация гена означает любое снижение уровня экспрессии генов по сравнению с экспрессией в клетке, которая не была модифицирована, как описано в настоящей заявке. Таким образом, инактивация гена может быть частичной или полной.
[0108] «Представляющая интерес область» означает любую область клеточного хроматина, такую как, например, ген или некодирующая последовательность внутри или рядом с геном, в котором желательно связывание экзогенной молекулы. Связывание может быть осуществлено для нацеленного расщепления ДНК и/или нацеленной рекомбинации. Представляющая интерес область может присутствовать в хромосоме, в эписоме, в геноме органеллы (например, митохондрии, хлоропласте) или, например, в геноме инфицирующего вируса. Представляющая интерес область может находиться в кодирующей области гена, в транскрибируемых некодирующих областях, таких как, например, лидерные последовательности, последовательности-метки или интроны, или в нетранскрибируемых областях, расположенных выше или ниже кодирующей области. Представляющая интерес область иметь по меньшей мере одну пару нуклеотидов, или она может иметь длину до 2000 пар нуклеотидов или любое целое число пар нуклеотидов.
[0109] «Эукариотическими» клетками являются, но не ограничиваются ими, клетки грибов (таких как дрожжи), клетки растений, клетки животных, клетки млекопитающих и клетки человека (например, Т-клетки).
[0110] Используемые здесь термины «функциональная связь» и «функционально присоединенный» (или «функционально связанный») являются синонимами, если они относятся к юкстаположению двух или более компонентов (таких как элементы последовательности), при котором эти компоненты расположены таким образом, что оба этих компонента функционируют нормально и обеспечивают возможность того, что, по меньшей мере, один из компонентов может опосредовать функцию, которая действует, по меньшей мере, на один из других компонентов. В качестве иллюстрации, последовательность регуляции транскрипции, такая как промотор, функционально связана с кодирующей последовательностью, если последовательность регуляции транскрипции регулирует уровень транскрипции кодирующей последовательности в ответ на присутствие или отсутствие одного или более факторов регуляции транскрипции. Последовательность регуляции транскрипции обычно функционально связана в цис-положении с кодирующей последовательностью, но она необязательно должна быть непосредственно смежной с этой последовательностью. Так, например, энхансер представляет собой последовательность регуляции транскрипции, которая функционально присоединена к кодирующей последовательности, даже если они не являются смежными.
[0111] Термин «функционально присоединенный», если он относится к гибридным полипептидам, может означать, что каждый из компонентов при связывании с другим компонентом выполняет ту же самую функцию, как если бы он был не связан с ним. Так, например, что касается гибридного полипептида, в котором ДНК-связывающий домен (например, ZFP, TALE) присоединен к домену активации, то ДНК-связывающий домен и домен активации являются функционально связанными, если в гибридном полипептиде, часть ДНК-связывающего домена способна связываться с его сайтом-мишенью и/или с его сайтом связывания, а домен активации способен повышать уровень экспрессии гена. Если гибридный полипептид, в котором ДНК-связывающий домен присоединен к домену расщепления, то ДНК-связывающий домен и домен расщепления являются функционально связанными, если в гибридном полипептиде, часть ДНК-связывающего домена способна связываться с его сайтом-мишенью и/или с его сайтом связывания, а домен расщепления способен расщеплять ДНК рядом с сайтом-мишенью. Аналогичным образом, что касается гибридного полипептида, в котором ДНК-связывающий домен присоединен к домену активации или репрессии, то ДНК-связывающий домен и домен активации или репрессии являются функционально связанными, если в гибридном полипептиде, часть ДНК-связывающего домена способна связываться с его сайтом-мишенью и/или с его сайтом связывания, а домен активации способен повышать уровень экспрессии гена, либо домен репрессии способен снижать уровень экспрессии гена.
[0112] «Функциональный фрагмент» белка, полипептида или нуклеиновой кислоты представляет собой белок, полипептид или нуклеиновую кислоту, последовательность которых не идентична полноразмерным белку, полипептиду или нуклеиновой кислоте, но сохраняет такую же функцию, как и функция полноразмерного белка, полипептида или нуклеиновой кислоты. Функциональный фрагмент может иметь большее или меньшее число остатков по сравнению с соответствующей нативной молекулой или такое же число остатков, как в этой молекуле, и/или он может содержать одну или более аминокислотных или нуклеотидных замен. Методы определения функции нуклеиновой кислоты (например, кодирующей функции, способности гибридизоваться с другой нуклеиновой кислотой) хорошо известны специалистам в данной области. Аналогичным образом, методы определения функции белка хорошо известны. Так, например, ДНК-связывающая функция полипептида может быть определена, например, посредством связывания с фильтром, электрофоретического сдвига подвижности или с помощью анализов на иммунопреципитацию. Расщепление ДНК может быть проанализировано с помощью гель-электрофореза. См. Ausubel et al., см. выше. Способность белка взаимодействовать с другим белком может быть определена, например, путем совместной иммунопреципитации, анализов на наличие двух компонентов или генетическихк и биохимических анализов на комплементарность. См. например, Fields et al., (1989) Nature 340: 245-246; Патент США № 5585245 и PCT WO 98/44350.
[0113] «Вектор» способен переносить генные последовательности в клетки-мишени. Обычно, «векторная конструкция», «экспрессионный вектор» и «вектор для переноса гена» означают любую конструкцию нуклеиновой кислоты, способную направлять экспрессию представляющего интерес гена и способную переносить генные последовательности в клетки-мишени. Таким образом, этот термин включает клонирующие и экспрессирующие носители, а также интегрирующие векторы.
[0114] «Репортерный ген» или «репортерная последовательность» означают любую последовательность, продуцирующую белковый продукт, который может быть легко оценен, предпочтительно, хотя и необязательно, в обычном анализе. Подходящими репортерными генами являются, но не ограничиваются ими, последовательности, кодирующие белки, которые опосредуют резистентность к антибиотикам (например, резистентность к ампициллину, резистентность к неомицину, резистентность к G418, резистентность к пуромицину), последовательности, кодирующие окрашенные или флуоресцентные или люминесцентные белки (например, белок, флуоресцирующий в зеленом диапазоне спектра, активированный белок, флуоресцирующий в зеленом диапазоне спектра, белок, флуоресцирующий в красном диапазоне спектра, люциферазу) и белки, которые опосредуют усиленный рост клеток и/или амплификацию гена (например, дигидрофолатредуктаза). Эпитопными метками являются, например, одна или более копий FLAG, His, myc, Tap, HA или любая детектируемая аминокислотная последовательность. «Экспрессионными метками» являются последовательности, кодирующие репортеры, которые могут быть функционально присоединены к нужной генной последовательности для мониторинга экспрессии представляющего интерес гена.
[0115] Используемые здесь термины «индивидуум» и «пациент» являются синонимами и относятся к млекопитающим, таким как человек и приматы, не являющиеся человеком, а также к экспериментальным животным, таким как кролики, собаки, кошки, крысы, мыши и другие животные. Соответственно, используемые здесь термины «индивидуум» или «пациент» относятся к любому пациенту или индивидууму, такому как млекопитающее, которому могут быть введены экспрессионные кластеры согласно изобретению. Индивидуумами согласно изобретению являются индивидуумы, страдающие расстройством или имеющие риск развития такого расстройства.
[0116] Используемые здесь термины «лечение» и «терапия», относятся к снижению тяжести и/или частоты симптомов, к устранению симптомов и/или факторов, вызывающих эти симптомы, к предотвращению появления симптомов и/или факторов, вызывающих эти симптомы, а также к ослаблению или устранению повреждений. Рак и болезнь «трансплантат против хозяина» являются неограничивающими примерами состояний, которые могут быть подвергнуты лечению с использованием описанных здесь композиций и с применением описанных здесь способов. Таким образом, «лечение» и «терапия» включают:
(i) профилактику заболевания или состояния у млекопитающего, а в частности, если у такого млекопитающего имеется предрасположенность к развитию состояния, но оно еще не было диагностировано;
(ii) подавление заболевания или состояния, то есть, прекращение его развития;
(iii) ослабление заболевания или состояния, то есть, стимуляцию обратного развития этого заболевания или состояния; или
(iv) ослабление симптомов заболевания или состояния, то есть, ослабление боли без лечения основного заболевания или состояния.
[0117] Используемые здесь термины «заболевание» и «состояние» могут употребляться как синонимы, или могут различаться по смыслу, если возбудители конкретного заболевания или состояния точно неизвестны (то есть, этиология этого заболевания или состояния точно неизвестна), а поэтому это заболевание квалифицируется пока еще не как заболевание, а только как нежелательное состояние или синдром, где более или менее конкретный набор симптомов был идентифицирован клиницистами.
[0118] «Фармацевтическая композиция» означает композицию соединения согласно изобретению и среды, обычно применяемой специалистами для доставки биологически активного соединения млекопитающим, например человеку. Такая среда включает все фармацевтически приемлемые носители, разбавители или наполнители.
[0119] «Эффективное количество» или «терапевтически эффективное количество» означает количество соединения согласно изобретению, которое при его введении млекопитающему, а предпочтительно, человеку, является достаточным для эффективного лечения млекопитающего, а предпочтительно человека. Количество композиции согласно изобретению, которое входит в понятие «терапевтически эффективное количество», может варьироваться в зависимости от соединения, состояния и его тяжести, способа введения и возраста млекопитающего, подлежащего лечению, и оно может быть легко определено специалистом в данной области в соответствии с его компетенцией и исходя из данного описания.
ДНК-связывающие домены
[0120] В настоящем изобретении описаны композиции, содержащие ДНК-связывающий домен, который специфически связывается с сайтом-мишенью в любом гене, содержащем ген HLA или регулятор HLA, или ген TCR, ген иммунологической контрольной точки (например, CISH, PD1, CTLA-4 и т.п.) и/или дополнительный ген (например, безопасный якорь). В раскрытых здесь композициях и способах, может быть использован любой ДНК-связывающий домен, включая, но не ограничиваясь ими, ДНК-связывающий домен «цинковый палец», ДНК-связывающий домен TALE, ДНК-связывающую часть (оцрРНК) нуклеазы CRSPR/Cas или ДНК-связывающий домен мегануклеозы. ДНК-связывающий домен может связываться с любой последовательностью-мишенью в гене, включая, но не ограничиваясь ими, последовательность-мишень из 12 или более нуклеотидов, как показано в любом из раскрытых здесь сайтов-мишеней (например, в сайте-мишени из 12-20 или более смежных или несмежных пар оснований, как показано в Таблице 1). В некоторых вариантах осуществления изобретения, ДНК-связывающий домен связывается (по последовательность-специфическому механизму) с сайтом-мишенью в гене TCR или в регуляторном гене TCR и модулирует экспрессию гена TCR. В некоторых вариантах осуществления изобретения, ДНК-связывающий домен связывается с сайтом-мишенью в TCRA, а в других вариантах осуществления изобретения, цинковый палец связывается с сайтом-мишенью в TRBC. В других вариантах осуществления изобретения, ДНК-связывающий домен связывается по последовательность-специфическому механизму с сайтом-мишенью в гене B2M и модулирует экспрессию гена B2M. В других вариантах осуществления изобретения, ДНК-связывающий домен связывается по последовательность-специфическому механизму с геном иммунологической контрольной точки, таким как ген CISH (например, с сайтами-мишенями, представленными в Таблице 1) или ген PDl (например, с сайтами-мишенями, представленными в патенте США № 8563314). Множество ДНК-связывающих доменов (связывающихся с одинаковыми или различными генами) могут быть использованы вместе для модификации множества генов в одно и то же время (например, посредством мультиплексной реакции с использованием множества опосредуемых нуклеазой нокаутов и/или нацеленной интеграции донора).
[0121] В некоторых вариантах осуществления изобретения, ДНК-связывающий домен содержит белок «цинковый палец». При этом предпочтительно, чтобы белок «цинковый палец» не встречался в природе, то есть, чтобы он был сконструирован для связывания с выбранным сайтом-мишенью. См. например, Beerli et al. (2002) Nature Biotechnol. 20: 135-141; Pabo et al., (2001) Ann. Rev. Biochem. 70: 313-340; Isalan et al.,l., (2001) Nature Biotechnol. 19: 656-660; Seal et al. (2001) Curr. Opin. Biotechnol. 12: 632-637; Choo et al. (2000) Curr. Opin. Struct. Biol. 10: 411-416; патенты США №№ 6453242; 6534261; 6599692; 6503717; 6689558; 7030215; 6794136; 7067317; 7262054; 7070934; 7361635; 7253273; и публикациях заявок на патент США №№ 2005/0064474; 2007/0218528; 2005/0267061, которые включены в настоящее описание посредством ссылки в полном объеме.
[0122] Сконструированный домен, связывающийся с цинковым пальцем, может обладать новой специфичностью связывания по сравнению с природным белком «цинковый палец». Способы конструирования включают, но не ограничиваются ими, рациональное конструирование и различные типы отбора. Рациональное конструирование включает, например, использование баз данных, содержащих триплетные (или квадруплетные) нуклеотидные последовательности и отдельные аминокислотные последовательности цинкового пальца, где каждая триплетная или квадруплетная нуклеотидная последовательность ассоциирована с одной или более аминокислотными последовательностями цинковых пальцев, которые связываются с конкретной триплетной или квадруплетной последовательностью. См., например, патенты США 6453242 и 6534361, которые включены в настоящее описание посредством ссылки в полном объеме.
[0123] Примеры методов отбора, включая системы фагового представления и двухкомпонентные системы, описаны в патентах США 5789538; 59255523; 6007988; 6013453; 6410248; 6140466; 6200759 и 6242568; а также в WO 98/37186; WO 98/53057; WO 00/27878; WO 01/88197 и в патенте Великобритании 2338237. Кроме того, повышение специфичности связывания с доменами, связывающимися с цинковым пальцем, описано, например, в патенте США № 6794136.
[0124] Кроме того, как описано в этих и других документах, домены «цинковые пальцы» и/или множество белков «цинковые пальцы» могут быть связаны друг с другом посредством любых подходящих линкерных последовательностей, включая, например, линкеры длиной в 5 или более аминокислот. См. также патенты США №№ 6479626; 6903185 и 7153949, где описаны репрезентативные линкерные последовательности длиной в 6 или более аминокислот. Описанные здесь белки могут включать любую комбинацию из подходящих линкеров, расположенных между отдельными белками «цинковые пальцы». Кроме того, повышение специфичности связывания с доменами, связывающимися с цинковыми пальцами, описано, например, в патенте США № 6794136.
[0125] Методы выбора сайтов-мишеней; ZFP и способы получения и конструирования гибридных белков (и полинуклеотидов, кодирующих эти белки) известны специалистам в данной области и подробно описаны в патентах США №№ 6140081; 5789538; 6453242; 6534261; 59255523; 6007988; 6013453; 6200759; WO 95/19431; WO 96/06166; WO 98/53057; WO 98/54311; WO 00/27878; WO 01/60970; WO 01/88197; WO 02/099084; WO 98/53058; WO 98/53059; WO 98/53060; WO 02/016536 и WO 03/016495.
[0126] В некоторых вариантах осуществления изобретения, ДНК-связывающий домен представляет собой сконструированный белок «цинковый палец», который связывается (по последовательность-специфическому механизму) с сайтом-мишенью в гене TCR или в регуляторном гене TCR и модулирует экспрессию гена TCR. В некоторых вариантах осуществления изобретения, белок «цинковый палец» связывается с сайтом-мишенью в TCRA, а в других вариантах осуществления изобретения, цинковый палец связывается с сайтом-мишенью в TRBC. В других вариантах осуществления изобретения, ДНК-связывающий домен представляет собой сконструированный белок «цинковый палец», который связывается по последовательность-специфическому механизму с сайтом-мишенью в гене B2M и модулирует экспрессию гена B2M. В других вариантах осуществления изобретения, ДНК-связывающий домен связывается по последовательность-специфическому механизму с геном иммунологической контрольной точки, таким как ген CISH или PD1.
[0127] Обычно, ZFP включают по меньшей мере три «пальца». Некоторые из ZFP включают четыре, пять или шесть пальцев. ZFP, которые включают три пальца, обычно распознают сайт-мишень, который включает 9 или 10 нуклеотидов; ZFP, которые включают четыре пальца, обычно распознают сайт-мишень, который включает от 12 до 14 нуклеотидов; а ZFP, имеющий шесть пальцев, может распознавать сайты-мишени, которые включают от 18 до 21 нуклеотидов. ZFP также могут представлять собой гибридные белки, которые включают один или более регуляторных доменов, где указанные домены могут представлять собой домены активации или ингибирования транскрипции. ZFP могут также содержать модификации в областях основа, описанные в публикации заявки на патент США № 20180087072.
[0128] В некоторых вариантах осуществления изобретения, ДНК-связывающий домен может происходить от нуклеазы. Так, например, последовательности распознавания хоминг-эндонуклеаз и мегануклеаз, таких как I-SceI, I-CeuI, PI-PspI, PI-Sce, I-SceIV, I-CsmI, I-PanI, I-SceII, I-PpoI, I-SceIII, I-CreI, I-TevI, I-TevII и I-TevIII являются известными. См., также патент США No. 5420032; патент США No. 6833252; Belfort et al. (1997) Nucleic Acids Res. 25:3379-3388; Dujon et al. (1989) Gene 82:115-118; Perler et al. (1994) Nucleic Acids Res. 22, 1125-1127; Jasin (1996) Trends Genet. 12:224-228; Gimble et al. (1996) J. Mol. Biol. 263:163-180; Argast et al. (1998) J. Mol. Biol. 280:345-353 и каталог New England Biolabs. Кроме того, специфичность связывания ДНК с хоминг-эндонуклеазами и мегануклеазами может быть создана для связывания с неприродными сайтами-мишенями. См., например, Chevalier et al. (2002) Molec. Cell 10:895-905; Epinat et al. (2003) Nucleic Acids Res. 31:2952-2962; Ashworth et al. (2006) Nature 441:656-659; Paques et al. (2007) Current Gene Therapy 7:49-66; публикацию заявки на патент США No. 20070117128.
[0129] В других вариантах осуществления изобретения, ДНК-связывающий домен содержит сконструированный домен эффектора TAL, аналогичный домену, происходящему от растительных патогенов Xanthomonas (см. Boch et al, (2009) Science 326: 1509-1512 и Moscou & Bogdanove, (2009) Science326: 1501) и Ralstonia (см. Heuer et al (2007) Applied and Environmental Microbiology 73(13): 4379-4384); публикации заявок на патенты США 20110301073 и 20110145940). Известно, что патогенные бактерии растений рода Xanthomonas вызывают множество заболеваний ценных сельскохозяйственных культур. Патогенность Xanthomonas зависит от системы секреции консервативного типа III (T3S), которая вводит более чем 25 различных эффекторных белков в клетку растения. Среди этих инъецированных белков присутствуют эффекторы, подобные активатору транскрипции (TALE), которые имитируют растительные активаторы транскрипции и модифицируют растительный транскриптом (см. Kay et al. (2007) Science 318: 648-651). Эти белки содержат ДНК-связывающий домен и домен активации транскрипции. Одним из наиболее хорошо охарактеризованных TALE является AvrBs3 от Xanthomonas campestgris pv. Vesicatoria (см. Bonas et al (1989) Mol Gen Genet 218: 127-136 и WO2010079430). TALE содержат центральный домен тандемных повторов, каждый из которых содержит приблизительно 34 аминокислоты, которые играют ключевую роль в специфичности связывания ДНК с этими белками. Кроме того, они содержат последовательность локализации в ядре и кислотный домен активации транскрипции (см. обзор Schornack S, et al (2006) J Plant Physiol 163(3): 256-272). Кроме того, было обнаружено, что в фитопатогенных бактериях Ralstonia solanacearum, два гена, обозначенные brg11 и hpx17, являются гомологичными белку AvrBs3 семейства Xanthomonas в биологической разновидности 1 R. solanacearum GMI1000 и в биологической разновидности 4 RS1000 (см. Heuer et al (2007) Appl and Envir Micro 73(13): 4379-4384). Эти гены имеют нуклеотидные последовательности, которые на 98,9% идентичны друг другу, но отличаются делецией 1575 п.о. в повторяющемся домене hpxl7. Однако, оба эти генных продукта имеют последовательности, которые менее, чем на 40% идентичны последовательностям белков AvrBs3 семейства Xanthomonas.
[0130] Специфичность этих эффекторов TAL зависит от последовательностей, обнаруженных в тандемных повторах. Повторяющаяся последовательность содержит приблизительно 102 пары оснований, и эти повторы обычно на 91-100% гомологичны друг другу (Bonas et al. ibid). Полиморфизм повторов обычно находится в положениях 12 и 13, и, по-видимому, существует точное соответствие между идентичностью гипервариабельных димерных остатков (повторяющегося вариабельного димерного остатка или область RVD) в положениях 12 и 13 и идентичностью смежных нуклеотидов в последовательности-мишени эффектора TAL (см. Moscou and Bogdanve, (2009) Science 326: 1501 и Boch et al. (2009) Science 326: 1509-1512). Был экспериментально определен природный код для распознавания ДНК этих эффекторов TAL, и было обнаружено, что последовательность HD в положениях 12 и 13 (повторяющийся вариабельный димерный остаток или RVD) способствует связыванию с цитозином (C), NG связывается с T, NI связывается с А, C, G или T, NN связывается с A или G, а ING связывается с Т. Эти ДНК-связывающиеся повторы были собраны в белки с новыми комбинациями и количествами повторов для получения искусственных факторов транскрипции, которые способны взаимодействовать с новыми последовательностями и активировать экспрессию не-эндогенного репортерного гена в растительных клетках (Boch et al. ibid). Сконструированные белки TAL были присоединены к полудомену расщепления FokI с получением гибрида нуклеазы и эффекторного домена TAL (TALEN), включая TALEN с атипичными RVD. См. например, патент США № 8586526.
[0131] В некоторых вариантах осуществления изобретения, TALEN содержит домен расщепления эндонуклеазой (например, FokI) или полудомен расщепления. В других вариантах осуществления изобретения, TALE-нуклеаза представляет собой мега-TAL. Эти нуклеазы мега-TAL представляют собой гибридные белки, содержащие ДНК-связывающий домен TALE и домен расщепления мегануклеазой. Домен расщепления мегануклеазой является активным в качестве мономера и для своей активности не требует димеризации. (См. Boissel et al. (2013) Nucl Acid Res. 1-13, doi:10. 1093/nar/gktl224).
[0132] В других вариантах осуществления изобретения, нуклеаза содержит компактную TALEN. Они представляют собой одноцепочечные гибридные белки, соединяющие ДНК-связывающий домен TALE с доменом нуклеазы TevI. Гибридный белок может действовать либо в качестве никазы, локализованной в области TALE, либо он может создать двухцепочечной разрыв, в зависимости от того, где расположен ДНК-связывающий домен TALE по отношению к домену нуклеазы TevI (см. Beurdeley et Al. (2013) Nat Comm 4: 1762 DOI:10.1038/ncomms2782). Кроме того, домен нуклеазы также может обладать функциональной активностью связывания с ДНК. Любые TALEN могут быть использованы в комбинации с дополнительными TALEN (например, с одним или более TALEN (cTALEN или FokI-TALEN) с одним или более мега-TALE.
[0133] Кроме того, как описано в этих и других документах, домены «цинковые пальцы» и/или множество белков «цинковые пальцы» или TALE могут быть связаны друг с другом посредством любых подходящих линкерных последовательностей, включая, например, линкеры длиной в 5 или более аминокислот. См. также патенты США №№ 6479626; 6903185 и 7153949, где описаны репрезентативные линкерные последовательности длиной в 6 или более аминокислот. Описанные здесь белки могут включать любую комбинацию из подходящих линкеров, расположенных между отдельными белками «цинковые пальцы». Кроме того, повышение специфичности связывания с доменами, связывающимися с цинковыми пальцами, описано, например, в патенте США № 6794136.
[0134] В некоторых вариантах осуществления изобретения, ДНК-связывающий домен представляет собой часть нуклеазной системы CRISPR/Cas, включающей одноцепочечную руководящую РНК (оцрРНК), которая связывается с ДНК. См., например, патенты США №№ 9873894 и 8697359, публикацию заявки на патент США № 20150159172. Локус CRISPR (кластеризованных регулярно перемежающихся коротких палиндромных повторов), который кодирует компоненты РНК системы, и локус cas (CRISPR-ассоциированный), который кодирует белки (Jansen et al., 2002. Mol. Microbiol. 43: 1565-1575; Makarova et al., 2002. Nucleic Acids Res. 30: 482-496; Makarova et al., 2006. Biol. Direct 1: 7; Haft et al., 2005. PLoS Comput. Biol. 1:e60), составляют генные последовательности нуклеазной системы CRISPR/Cas. Локусы CRISPR в микробных хозяевах содержат комбинацию CRISPR-ассоциированных (Cas) генов, а также некодирующие элементы РНК, способные программировать специфичность CRISPR-опосредуемого расщепления нуклеиновой кислоты.
[0135] CRISPR типа II является одной из наиболее хорошо охарактеризованных систем и осуществляет нацеленный двухцепочечный разрыв ДНК в четыре последовательных стадии. Во-первых, две некодирующие РНК, массив пре-cr-РНК и tracr-РНК транскрибируются из локуса CRISPR. Во-вторых, tracr-РНК гибридизуется с повторяющимися областями пре-crРНК и опосредует процессинг пре-cr-РНК в зрелые cr-РНК, содержащие отдельные спейсерные последовательности. В-третьих, зрелый комплекс cr-РНК:tracr-РНК направляет функциональный домен (например, нуклеазу, такую как Cas) к ДНК-мишени посредством спаривания оснований Уотсона-Крика между спейсером на cr-РНК и протоспейсером на ДНК-мишени рядом с соседним мотивом протоспейсера (PAM), что является дополнительным требованием для распознавания мишени. И наконец, Cas9 опосредует расщепление ДНК-мишени с образованием двухцепочечного разрыва в протоспейсере. Активность системы CRISPR/Cas включает три стадии: (i) инсерцию чужеродных последовательностей ДНК в массив CRISPR для предотвращения будущих атак в процессе, называемом «адаптацией», (ii) экспрессию релевантных белков, а также экспрессию и процессинг массива, а затем (iii) РНК-опосредованную интерференцию чужеродной нуклеиновой кислоты. Таким образом, в бактериальной клетке, несколько так называемых белков «Cas» участвуют в природной функции системы CRISPR/Cas и играют определенную роль в таких функциях, как инсерция чужеродной ДНК и т.п.
[0136] В некоторых вариантах осуществления изобретения, белок Cas может представлять собой «функциональное производное» природного белка Cas. «Функциональное производное» полипептида с нативной последовательностью представляет собой соединение, которое вместе с полипептидом с нативной последовательностью обладает ценным биологическим свойством. «Функциональными производными» являются, но не ограничиваются ими, фрагменты нативной последовательности и производные полипептида с нативной последовательностью и его фрагментов при условии, что они, вместе с соответствующим полипептидом с нативной последовательностью, будут обладать биологической активностью. Рассматриваемая здесь биологическая активность представляет собой способность функционального производного гидролизовать субстрат ДНК на фрагменты. Термин «производное» охватывает варианты аминокислотной последовательности полипептида, ковалентные модификации и их гибриды, такие как производные белков Cas. Подходящими производными полипептида Cas или его фрагмента являются, но не ограничиваются ими, мутанты, гибриды, ковалентные модификации белка Cas или его фрагмента. Белок Cas, который включает белок Cas или его фрагмент, а также производные белка Cas или его фрагмента, могут быть получены из клетки или синтезированы химическим способом или комбинацией этих двух процедур. Клетка может представлять собой клетку, которая в природе продуцирует белок Cas, или клетку, которая в природе продуцирует белок Cas и была генетически сконструирована для продуцирования эндогенного белка Cas с более высоким уровнем экспрессии или для продуцирования белка Cas из экзогенно введенной нуклеиновой кислоты, где эта нуклеиновая кислота кодирует Cas, который является таким же как и эндогенный Cas или отличается от него. В некоторых случаях, если клетка не продуцирует природного белка в природе, то она была генетически сконструирована для продуцирования белка Cas. В некоторых вариантах осуществления изобретения, белок Cas представляет собой небольшой ортолог Cas9 для доставки посредством AAV-вектора (Ran et al. (2015) Nature 510, p.186). Таким образом, ДНК-связывающий домен может содержать оцрРНК, которая связывается с сайтом-мишенью или по меньшей мере с 9 нуклеотидами, как показано в Таблице 1.
[0137] В некоторых вариантах осуществления изобретения, ДНК-связывающий домен является частью системы TtAago (см. Swarts et al., ibid; Sheng et al. ibid). В эукариотах, сайленсинг гена опосредуется белками семейства Argonaute (Ago). В этой парадигме, Ago связывается с небольшими РНК (из 19-31 нуклеотидов). Этот комплекс «белок-РНК» для сайленсинга распознает РНК-мишени посредством спаривания оснований Уотсона-Крика между небольшой РНК и мишенью и эндонуклеолитически расщепляет РНК-мишень (Vogel (2014) Science 344: 972-973). В противоположность этому, прокариотические белки Ago связываются с небольшими одноцепочечными фрагментами ДНК и, вероятно, их функции заключаются в выявлении и удалении чужеродной (часто вирусной) ДНК (Yuan et al., (2005) Mol. Cell 19, 405; Olovnikov, et al. (2013) Mol. Cell 51, 594; Swarts et al., ibid). Репрезентативными прокариотическими белками Ago являются белки, происходящие от Aquiflex aeolicus, Rhodobacter sphaeroides и Thermus thermophilus.
[0138] Одним из наиболее хорошо охарактеризованных прокариотических белков Ago является белок, происходящий от T. thermophilics (TtAgo; Swarts et al. ibid). TtAgo ассоциируется либо с 15 нуклеотидами, либо с 13-25 нуклеотидами одноцепочечных фрагментов ДНК с 5’-фосфатными группами. Эта «руководящая ДНК», связанная с TtAgo, служит для нацеливания комплекса белок-ДНК на связывание с комплементарной последовательностью ДНК Уотсона-Крика в молекуле ДНК третьего поколения. После получения информации о том, что последовательность этих руководящих ДНК позволяет идентифицировать ДНК-мишень, комплекс «TtAgo-руководящая ДНК» расщепляет ДНК-мишень. Такой механизм также поддерживается структурой комплекса «TtAgo-руководящая ДНК», который связывается с его ДНК-мишенью (G. Sheng et al., ibid). Ago от Rhodobacter sphaeroides (RsAgo) обладает аналогичными свойствами (Olivnikov et al., ibid).
[0139] Экзогенные руководящие ДНК с произвольной последовательностью ДНК могут быть загружены в белок TtAgo (Swarts et al., ibid). Поскольку специфичность расщепления TtAgo опосредуется руководящей ДНК, то комплекс TtAgo-ДНК, образованный экзогенной руководящей ДНК, определяемой исследователем, будет направлять расщепление ДНК-мишени посредством TtAgo на комплементарную определенную исследователем ДНК-мишень, определенную исследователем. Таким образом, можно специально создать двухцепочечный разрыв в ДНК. Применение системы TtAgo-руководящая ДНК (или ортологичных систем Ago-руководящая ДНК от других организмов) позволяет осуществлять нацеленное расщепление геномной ДНК в клетках. Такое расщепление может быть либо одноцепочечным или двухцепочечным. Для расщепления геномной ДНК млекопитающих предпочтительно использовать вариант кодона TtAgo, оптимизированный для экспрессии в клетках млекопитающих. Кроме того, может оказаться предпочтительым обрабатывать клетки комплексом TtAgo-ДНК, образованным in vitro, где белок TtAgo был присоединен к пептиду, проникающему в клетку. Кроме того, может оказаться предпочтительным использование варианта белка TtAgo, который был модифицирован посредством мутагенеза и обладал повышенной активностью при 37°C. Расщепление ДНК, опосредуемое Ago-РНК, может быть использовано для подтверждения результатов, включая нокаут гена, добавление гена-мишени, коррекцию гена, делецию гена-мишени с применение методов, известных специалистам в области, относящейся к разрывам ДНК. Таким образом, любой ДНК-связывающий домен может быть использован для связывания и модификации (например, модуляции экспрессии и/или генетической модификации посредством инсерций и/или делеций) описанных здесь клеток. В некоторых вариантах осуществления изобретения, ДНК-связывающий(е) домен(ы) молекулы (молекул), которая(ые) дает (дают) модификации гена(ов)-мишени, связывается(ются) с сайтом-мишенью, содержащим по меньшей мере 9 нуклеотидов сайта-мишени, показанного в Таблице 1.
Гибридные молекулы
[0140] Настоящее изобретение также относится к гибридным молекулам, содержащим ДНК-связывающие домены (например, ZFP или TALE, компоненты CRISPR/Cas, такие как одноцепочечные руководящие РНК), описанные в настоящей заявке и ассоциированные с гетерологичным регуляторным (функциональным) доменом (или его функциональным фрагментом). Общими доменами являются, например, домены фактора транскрипции (активаторы, репрессоры, коактиваторы, ко-репрессоры), сайленсеры, онкогены (например, члены семейства myc, jun, fos, myb, max, mad, rel, ets, bcl, myb, mos и т.п.); ДНК-репарирующие ферменты и ассоциированные с ними факторы и модификаторы; ферменты реаранжировки ДНК и ассоциированные с ними факторы и модификаторы: белки, ассоциированные с хроматином и их модификаторы (например, киназы, ацетилазы и деацетилазы); и ДНК-модифицирующие ферменты (например, метилтрансферазы, топоизомеразы, геликазы, лигазы, киназы, фосфатазы, полимеразы, эндонуклеазы) и ассоциированные с ними факторы и модификаторы. Такие гибридные молекулы включают факторы транскрипции, содержащие описанные здесь ДНК-связывающие домены, и домен регуляции транскрипции, а также нуклеазы, содержащие ДНК-связывающие домены и один или более нуклеазных доменов.
[0141] Подходящими доменами для достижения активации (доменами активации транскрипции) являются домен активации HSV VP16 (см. например, Hagmann et al., J. Virol. 71, 5952-5962 (1997)), рецепторы гормона ядра (см., например, Torchia et al., Curr. Opin. Cell. Biol. 10:373-383 (1998)); субъединица p65 ядерного фактора каппа B (Bitko & Barik, J. Virol. 72:5610-5618 (1998) и Doyle & Hunt, Neuroreport 8:2937-2942 (1997)); Liu et al., Cancer Gene Ther. 5:3-28 (1998)), или искусственные химерные функциональные домены, такие как VP64 (Beerli et al., (1998) Proc. Natl. Acad. Sci. USA 95:14623-33), и дегрон (Molinari et al., (1999) EMBO J. 18, 6439-6447). Другими репрезентативными доменами активации являются Oct 1, Oct-2A, Sp1, AP-2 и CTF1 (Seipel et al., EMBO J. 11, 4961-4968 (1992), а также p300, CBP, PCAF, SRC1 PvALF, AtHD2A и ERF-2. См, например, Robyr et al. (2000) Mol. Endocrinol. 14:329-347; Collingwood et al. (1999) J. Mol. Endocrinol. 23:255-275; Leo et al. (2000) Gene 245:1-11; Manteuffel-Cymborowska (1999) Acta Biochim. Pol. 46:77-89; McKenna et al. (1999) J. Steroid Biochem. Mol. Biol. 69:3-12; Malik et al. (2000) Trends Biochem. Sci. 25:277-283; и Lemon et al. (1999) Curr. Opin. Genet. Dev. 9:499-504. Другими репрезентативными доменами активации являются, но не ограничиваются ими, OsGAI, HALF-1, C1, AP1, ARF-5,-6,-7 и -8, CPRF1, CPRF4, MYC-RP/GP и TRAB1. См, например, Ogawa et al. (2000) Gene 245:21-29; Okanami et al. (1996) Genes Cells 1:87-99; Goff et al. (1991) Genes Dev. 5:298-309; Cho et al. (1999) Plant Mol. Biol. 40:419-429; Ulmason et al. (1999) Proc. Natl. Acad. Sci. USA 96:5844-5849; Sprenger-Haussels et al. (2000) Plant J. 22:1-8; Gong et al. (1999) Plant Mol. Biol. 41:33-44; и Hobo et al. (1999) Proc. Natl. Acad. Sci. USA 96:15348-15353.
[0142] Специалистам в данной области очевидно, что при образовании гибридного белка (или нуклеиновой кислоты, кодирующей этот белок) между ДНК-связывающим доменом и функциональным доменом, в качестве функционального домена подходящим является домен активации или молекула, которая взаимодействует с доменом активации. В качестве домена активации гибридного белка может быть использована, по существу, любая молекула, способная осуществлять рекрутинг активирующего комплекса и/или активирующей активности (такой как, например, ацетилирование гистона) в ген-мишень. Домены-изоляторы, домены локализации и белки ремоделирования хроматина, такие как ISWI-содержащие домены и/или белки с метил-связывающим доменом, которые являются подходящими для их использования в качестве функциональных доменов в гибридных молекулах, описаны, например, в патенте США № 7053264.
[0143] Репрезентативными доменами репрессии являются, но не ограничиваются ими, KRAB A/B, KOX, индуцибельный ранний ген TGF-бета (TIEG), v-erbA, SID, MBD2, MBD3, члены семейства DNMT (например, DNMT1, DNMT3A, DNMT3B), Rb и MeCP2. См., например, Bird et al. (1999) Cell 99:451-454; Tyler et al. (1999) Cell 99:443-446; Knoepfler et al. (1999) Cell 99:447-450; и Robertson et al. (2000) Nature Genet. 25:338-342. Другими репрезентативными доменами репрессии являются, но не ограничиваются ими, ROM2 и AtHD2A. См., например, Chem et al. (1996) Plant Cell 8:305-321; и Wu et al. (2000) Plant J. 22:19-27.
[0144] Гибридные молекулы конструируют методами клонирования и биохимического конъюгирования, хорошо известными специалистам в данной области. Гибридные молекулы содержат ДНК-связывающий домен (например, ZFP, TALE, оцрРНК), связанный с функциональным доменом (например, доменом активации или ингибирования транскрипции). Гибридные молекулы также содержат, но необязательно, сигналы локализации в ядре (такие как, например, сигналы, происходящие от Т-антигена среды SV40) и эпитопные метки (такие как, например, FLAG и гемагглютинин). Гибридные белки (и кодирующие их нуклеиновые кислоты) сконструированы таким образом, чтобы рамка считывания при трансляции сохранялась среди компонентов гибрида.
[0145] Гибриды между полипептидным компонентом функционального домена (или его функционального фрагмента), с одной стороны, и небелковым ДНК-связывающим доменом (например, антибиотиком, интеркалирующим агентом, агентом, связывающимся с небольшой бороздкой, нуклеиновой кислотой), с другой стороны, конструируют методами биохимического конъюгирования, известными специалистам в данной области. См. например, Pierce Chemical Company (Rockford, IL) Catalogue. Также описаны методы и композиции для получения гибридов между агентом, связывающимся с небольшой бороздкой, и полипептидом. Mapp et al. (2000) Proc. Natl. Acad. Sci. USA 97:3930-3935. Кроме того, одноцепочечные руководящие РНК CRISPR/Cas-системы ассоциируются с функциональными доменами и образуют активные регуляторы транскрипции и нуклеазы.
[0146] В некоторых вариантах осуществления изобретения, сайт-мишень присутствует в доступной области клеточного хроматина. Доступные области могут быть определены, как описано, например, в патентах США №№ 7217509 и 7923542. Если сайт-мишень отсутствует в доступной области клеточного хроматина, то одна или более доступных областей могут быть получены как описано в патентах США №№ 7785792 и 8071370. В дополнительных вариантах осуществления изобретения, ДНК-связывающий домен гибридной молекулы способен связываться с клеточным хроматином независимо от того, находится ли его сайт-мишень в доступной области или нет. Так, например, такие ДНК-связывающие домены способны связываться с линкерной ДНК и/или нуклеосомной ДНК. Примерами недавно открытого ДНК-связывающего домена такого типа являются домены, обнаруженные в некоторых стероидных рецепторах и в факторе ядра гепатоцитов 3 (HNF3) (Cordingley et al. (1987) Cell 48:261-270; Pina et al. (1990) Cell 60:719-731; и Cirillo et al. (1998) EMBO J. 17:244-254).
[0147] Гибридная молекула может быть получена вместе с фармацевтически приемлемым носителем, как известно специалистам в данной области. См. например, Remington's Pharmaceutical Sciences, 17 th ed. 1985; и патенты США №№ 6453242 и 6534261.
[0148] Функциональный компонент/домен гибридной молекулы может быть выбран из любого множества различных компонентов, способных влиять на транскрипцию гена после связывания гибридной молекулы с последовательностью-мишенью посредством ДНК-связывающего домена. Таким образом, функциональный компонент может включать, но не ограничиваться ими, различные домены факторов транскрипции, такие как активаторы, репрессоры, ко-активаторы, ко-репрессоры и сайленсеры.
[0149] Дополнительные репрезентативные функциональные домены описаны, например, в патентах США №№ 6543261 и 6933113.
[0150] Могут быть также отобраны функциональные домены, которые регулируются экзогенными небольшими молекулами или лигандами. Так, например, в том случае, когда функциональный домен принимает свою активную конформацию в присутствии внешнего лиганда RheoChem™, может быть применена технология RheoSwitch® (см. например, заявку на патент США 20090136465). Таким образом, ZFP может быть функционально присоединен к регуляторному функциональному домену, где конечная активность ZFP-TF регулируется внешним лигандом.
Нуклеазы
[0151] В некоторых вариантах осуществления изобретения, гибридная молекула содержит ДНК-связывающий домен, связанный с доменом расщепления (нуклеазой). По существу, модификация гена может быть достигнута с использованием нуклеазы, например, сконструированной нуклеазы. Технология конструирования нуклеазы основана на конструировании встречающихся в природе ДНК-связывающих белков. Так, например, было описано конструирование хоминг-эндонуклеаз с адаптированными специфичностями связывания с ДНК. Chames et al. (2005) Nucleic Acids Res 33 (20): el78; Arnuld Et Al. (2006) J Mol. Biol. 355: 443-458. Кроме того, было описано конструирование ZFP. См. например, патенты США №№ 6534261; 6607882; 6824978; 6997539; 6933113; 7163824 и 7013219.
[0152] Кроме того, ZFP и/или TALE могут быть присоединены к нуклеазным доменам для получения ZFN и TALEN, то есть, функциональной молекулы, способной распознавать нуклеиновую кислоту-мишень посредством сконструированного ДНК-связывающего домена (ZFP или TALE) и стимулировать разрезание ДНК возле ДНК-связывающего сайта посредством нуклеазной активности.
[0153] Таким образом, описанные здесь способы и композиции находят широкое применение и могут включать любую представляющую интерес нуклеазу. Неограничивающими примерами нуклеаз являются мегануклеазы, TALEN и нуклеазы «цинковые пальцы». Нуклеаза может содержать гетерологичные ДНК-связывающие домены и домены расщепления (например, нуклеазы «цинковые пальцы»; ДНК-связывающие домены мегануклеазы с гетерологичными доменами расщепления) или альтернативно, ДНК-связывающий домен природной нуклеазы может быть модифицирован так, чтобы он связывался с выбранным сайтом-мишенью {например, с мегануклеазой, которая была сконструирована так, чтобы она связывалась с сайтом, отличающимся от когнатного сайта связывания}.
[0154] Любая описанная здесь нуклеаза может содержать сконструированнуый ДНК-связывающий домен TALE и домен нуклеазы {например, домен эндонуклеазы и/или мегануклеазы), также известной как TALEN. Методы и композиции для конструирования этих белков TALEN в целях обеспечения надежного, сайт - специфического взаимодействия с последовательностью-мишенью, выбранной специалистом, были опубликованы (см. патент США № 8586526). В некоторых вариантах осуществления изобретения, TALEN содержат домен расщепления эндонуклеазой {например, FokI) или полудомен расщепления. В других вариантах осуществления изобретения, нуклеаза TALE представляет собой мега-TAL. Эти нуклеазы мега-TAL представляют собой гибридные белки, содержащие ДНК-связывающий домен TALE и домен расщепления мегануклеазой. Домен расщепления мегануклеазой является активным в качестве мономера и не требует димеризации для своей активности (см. Boissel et al., (2013) Nucl Acid Res: 1-13, doi:10.1093/nar/gktl224). Кроме того, домен нуклеазы может также обладать функциональной активностью связывания с ДНК.
[0155] В других вариантах осуществления изобретения, нуклеаза содержит компактные TALEN (cTALEN). Они представляют собой одноцепочечные гибридные белки, соединяющие ДНК-связывающий домен TALE с доменом нуклеазы TevI. Гибридный белок может действовать либо в качестве никазы, локализованной в области TALE, либо он может создавать двухцепочечной разрыв в зависимости от того, где расположен ДНК-связывающий домен TALE по отношению к домену нуклеазы TevI (см. Beurdeley et Al. (2013) Nat Comm 4: 1762 DOI:10.1038/ncomms2782). Любые TALEN могут быть использованы в комбинации с дополнительными TALEN (например, с одним или более TALEN (cTALEN или FokI-TALEN) вместе с одним или более мега-TAL) или с другими ферментами расщепления ДНК.
[0156] В некоторых вариантах осуществления изобретения, нуклеаза содержит мегануклеазу (хоминг-эндонуклеазу) или ее часть, обладающие расщепляющей активностью. Природные мегануклеазы распознают сайты расщепления в 15-40 пар оснований и обычно подразделяются на группы по четырем семействам: семейство LAGLIDADG («LAGLIDADG», представленное как SEQ ID NO: 63, семейство GIY-YIG, семейство боксов His-Cyst и семейство HNH. Репрезентативными хоминг-эндонуклеазами являются I-SceI, I-CeuI, PI-PspI, PI-Sce, I-SceIV, I-CsmI, I-PanI, I-SceII, I-PpoI, I-SceIII, I-CreI, I-TevI, I-TevII и I-TevIII. Их последовательности распознавания являются известными. См. также патент США No. 5420032; патент США No. 6833252; Belfort et al. (1997) Nucleic Acids Res. 25:3379-3388; Dujon et al. (1989) Gene 82:115-118; Perler et al. (1994) Nucleic Acids Res. 22, 1125-1127; Jasin (1996) Trends Genet. 12:224-228; Gimble et al. (1996) J. Mol. Biol. 263:163-180; Argast et al. (1998) J. Mol. Biol. 280:345-353 и каталог New England Biolabs.
[0157] ДНК-связывающие домены из природных мегануклеаз, главным образом, семейства LAGLIDADG («LAGLIDADG» представленного как SEQ ID NO: 63), были использованы для стимуляции сайт-специфической модификации генома в растениях, в дрожжах, у дрозофил, в клетках млекопитающих и мышей, но этот метод был ограничен модификацией либо гомологичных генов, которые сохраняют последовательность распознавания мегануклеазой (Monet et al. (1999), Biochem. Biophysics. Res. Common. 255: 88-93), либо предварительно сконструированными геномами, в которые была введена последовательность распознавания (Route et al. (1994), Mol. Cell. Biol. 14: 8096-106; Chilton et al. (2003), Plant Physiology. 133: 956-65; Puchta et al. (1996), Proc. Natl. Acad. Sci. USA 93: 5055-60; Rong et al. (2002), Genes Dev. 16: 1568-81; Gouble et al. (2006), J. Gene Med. 8(5):616-622). В соответствии с этим, были предприняты попытки сконструировать мегануклеазы так, чтобы они обладали новой специфичностью связывания в фармацевтически или биотехнологически релевантных сайтах (Porteus et al. (2005), Nat. Biotechnol. 23: 967-73; Sussman et al. (2004), J. Mol. Biol. 342:31-41; Epinat et al. (2003), Nucleic Acids Res. 31:2952-62; Chevalier et al. (2002) Molec. Cell 10:895-905; Epinat et al. (2003) Nucleic Acids Res. 31:2952-2962; Ashworth et al. (2006) Nature 441:656-659; Paques et al. (2007) Current Gene Therapy 7:49-66; публикации заявок на патент США №№ 20070117128; 20060206949; 20060153826; 20060078552 и 20040002092). Кроме того, природные или сконструированные ДНК-связывающие домены мегануклеаз могут быть функционально присоединены к домену расщепления гетерологичной нуклеазы (например, FokI), и/или домены расщепления мегануклеаз могут быть функционально присоединены к гетерологичному ДНК-связывающему домену (например, ZFP или TALE).
[0158] В других вариантах осуществления изобретения, нуклеаза представляет собой нуклеазу «цинковый палец» (ZFN) или гибрид ДНК-связывающего домена-нуклеазы TALE (TALEN). ZFN и TALEN содержат ДНК-связывающий домен (белок «цинковый палец» или ДНК-связывающий домен TALE), который был сконструирован так, чтобы он связывался с сайтом-мишенью в выбранном гене и в домене расщепления или в полудомене расщепления {например, в результате рестрикции и/или под действием мегануклеазы, как описано в настоящей заявке).
[0159] Как подробно описано выше, связывающие домены «цинковые пальцы» и ДНК-связывающие домены TALE могут быть сконструированы для связывания с выбранной последовательностью. См., например, Beerli et al. (2002) Nature Biotechnol. 20:135-141; Pabo et al. (2001) Ann. Rev. Biochem. 70:313-340; Isalan et al. (2001) Nature Biotechnol. 19:656-660; Segal et al. (2001) Curr. Opin. Biotechnol. 12:632-637; Choo et al. (2000) Curr. Opin. Struct. Biol. 10:411-416. Сконструированный домен, связывающийся с цинковым пальцем или с белком TALE, может обладать новой специфичностью связывания по сравнению с природным белком «цинковый палец». Способы конструирования включают, но не ограничиваются ими, рациональное конструирование и различные типы отбора. Рациональное конструирование включает, например, использование баз данных, содержащих триплетные (или квадруплетные) нуклеотидные последовательности и отдельные аминокислотные последовательности «цинкового пальца» или аминокислотные последовательности TALE, где каждая триплетная или квадруплетная нуклеотидная последовательность ассоциирована с одной или более аминокислотными последовательностями цинковых пальцев или повторяющихся звеньев TALE, которые связываются с конкретной триплетной или квадруплетной последовательностью. См., например, патенты США 6453242 и 6534261, которые включены в настоящее описание посредством ссылки в полном объеме.
[0160] Выбор сайтов-мишеней и методы получения и конструирования гибридных белков (и кодирующих их полинуклеотидов) известны специалистам в данной области и подробно описаны в патентах США №№ 7888121 и 8409861, содержание которых включено в настоящее описание посредством ссылки в полном объеме.
[0161] Кроме того, как описано в этих и других документах, домены «цинковые пальцы», TALE и/или множество белков «цинковые пальцы» могут быть связаны друг с другом посредством любых подходящих линкерных последовательностей, включая, например, линкеры длиной в 5 или более аминокислот. См., например, патенты США №№ 6479626; 6903185 и 7153949, где описаны репрезентативные линкерные последовательности длиной в 6 или более аминокислот. Описанные здесь белки могут включать любую комбинацию из подходящих линкеров, расположенных между отдельными белками «цинковые пальцы». См. также патент США № 8772453.
[0162] Таким образом, нуклеазы, такие как ZFN, TALEN и/или мегануклеазы могут содержать любой ДНК-связывающий домен и любой домен нуклеазы (домен расщепления, полудомен расщепления). Как уже отмечалось выше, домен расщепления может быть гетерологичным ДНК-связывающему домену, например, домену «цинковый палец» или ДНК-связывающему домену эффектора TAL и домену расщепления, происходящему от ДНК-связывающего домена нуклеазы или мегануклеазы, и домену расщепления, происходящему от другой нуклеазы. Гетерологические домены расщепления могут быть получены из любой эндонуклеазы или экзонуклеазы. Репрезентативными эндонуклеазами, от которых происходит домен расщепления, являются, но не ограничиваются ими, рестриктирующие эндонуклеазы и хоминг-эндонуклеазы. См., например, 2002-2003 Catalogue, New England Biolabs, Beverly, MA; и Belfort et al., (1997) Nucleic Acids Res. 25:3379-3388. Известны и другие ферменты, которые, расщепляют ДНК (например, нуклеаза S1; нуклеаза фасоли золотистой; панкреатическая ДНКаза I; микрококковая нуклеаза; дрожжевая эндонуклеаза HO; см. также Linn et al., (eds.) Nucleases, Cold Spring Harbor Laboratory Press, 1993). Один или более из этих ферментов (или их функциональных фрагментов) могут быть использованы в качестве источника доменов расщепления и полудоменов расщепления.
[0163] Аналогичным образом, полудомен расщепления может происходить от любой нуклеазы или ее части, как было указано выше, и для активности расщепления требует димеризации. Вообще говоря, для расщепления необходимы два гибридных белка, если эти гибридные белки содержат полудомены расщепления. Альтернативно, может быть использован один белок, содержащий два полудомена расщепления. Два полудомена расщепления могут происходить от одной и той же эндонуклеазы (или ее функциональных фрагментов), либо каждый полудомен расщепления может происходить от различных эндонуклеаз (или их функциональных фрагментов). Кроме того, сайты-мишени для двух гибридных белков предпочтительно расположены по отношению друг к другу так, чтобы связывание двух гибридных белков с их соответствующими сайтами-мишенями позволяло располагать полудомены расщепления в пространственной ориентации по отношению к друг к другу, что, в свою очередь, позволит полудоменам расщепления образовывать функциональный домен расщепления, например, посредством димеризации. Таким образом, в некоторых вариантах осуществления изобретения, близко расположенные края сайтов-мишеней разделены 5-8 нуклеотидами или 15-18 нуклеотидами. Однако, между двумя сайтами-мишенями может присутствовать любое целое число нуклеотидов или пар нуклеотидов (например, от 2 до 50 пар нуклеотидов или более). В целом, сайт расщепления расположен между сайтами-мишенями, но он может находиться на расстоянии 1 или более тысяч пар оснований от сайта расщепления, включая 1-50 пар оснований (или любое число пар оснований в этих пределах, включая 1-5, 1-10 и 1-20 пар оснований), 1-100 пар оснований (или любое число пар оснований в этих пределах), 100-500 пар оснований (или любое число пар оснований в этих пределах), 500-1000 пар оснований (или любое число пар оснований в этих пределах), или даже более, чем 1 т.п.о. от сайта расщепления.
[0164] Рестриктирующие эндонуклеазы (рестриктирующие ферменты) присутствуют у многих видов и способны осуществлять последовательность-специфическое связывание с ДНК (в сайте распознавания) и расщеплять ДНК в сайте или возле сайта связывания. Некоторые рестриктирующие ферменты (например, типа IIS) расщепляют ДНК в сайтах, удаленных от сайта распознавания и имеют разделяемые домены связывания и расщепления. Так, например, фермент типа IIS FokI катализирует двухцепочечное расщепление ДНК на расстоянии 9 нуклеотидов от сайта распознавания на одной цепи и 13 нуклеотидов от сайта распознавания на другой цепи. См., например, патенты США 5356802; 5436150 и 5487994; а также Li et al. (1992) Proc. Natl. Acad. Sci. USA 89:4275-4279; Li et al. (1993) Proc. Natl. Acad. Sci. USA 90:2764-2768; Kim et al. (1994a) Proc. Natl. Acad. Sci. USA 91:883-887; Kim et al. (1994b) J. Biol. Chem. 269:31978-31982. Таким образом, в одном варианте осуществления изобретения, гибридные белки содержат домен расщепления (или полудомен расщепления) по меньшей мере одного рестриктирующего фермента типа IIS, и один или более доменов, связывающихся с цинковми пальцами, которые могут быть, а могут и не быть сконструироваными.
[0165] Примером рестриктирующего фермента типа IIS, в котором домен расщепления отделен от связывающего домена, является FokI. Этот конкретный фермент является активным в качестве димера. Bitinaite et al. (1998) Proc. Natl. Acad. Sci. USA 95: 10570-10575. В соответствии с настоящим изобретением, часть фермента FokI, используемого в описанных гибридных белках, рассматривается как полудомен расщепления. Таким образом, для нацеленного двухцепочечного расщепления и/или для нацеленной замены клеточных последовательностей с использованием гибридов «цинковый палец»-FokI, два гибридных белка, каждый из которых содержит полудомен расщепления FokI, может быть использован для восстановления каталитически активного домена расщепления. Альтернативно, может быть также использована одна полипептидная молекула, содержащая домен, связывающийся с цинковым пальцем и два полудомена расщепления FokI. Параметры для нацеленного расщепления и нацеленной модификации последовательности с использованием гибридов «цинковый палец»-FokI описаны в других разделах настоящего описания.
[0166] Домен расщепления или полудомен расщепления могут представлять собой любую часть белка, которая сохраняет активность расщепления или способность к мультимеризации (например, димеризации) с образованием функционального домена расщепления.
[0167] Репрезентативные рестриктирующие ферменты типа IIS описаны в публикации Международной заявки WO 07/014275, которая включена в настоящее описание посредством ссылки в полном объеме. В настоящем описании рассматриваются и другие рестриктирующие ферменты, которые также содержат разделяемые связывающие домены и домены расщепления. См., например, Roberts et al. (2003) Nucleic Acids Res. 31:418-420.
[0168] В некоторых вариантах осуществления изобретения, домен расщепления содержит один или более сконструированных полудоменов расщепления (также называемых мутантами домена димеризации), которые минимизируют или предотвращают гомодимеризацию, как описано, например, в патентах США №№ 7914796, 8034598 и 8623618; и в публикации заявки на патент США № 20110201055, содержание которых включено в настоящее описание посредством ссылки в полном объеме. Аминокислотные остатки в положениях 446, 447, 479, 483, 484, 486, 487, 490, 491, 496, 498, 499, 500, 531, 534, 537 и 538 FokI представляют собой мишени для влияния на димеризацию полудоменов расщепления FokI.
[0169] Репрезентативные сконструированные полудомены расщепления FokI, которые образуют облигатные гетеродимеры, включают пару, в которой первый полудомен расщепления включает мутации в аминокислотных остатках в положениях 490 и 538 FokI, а второй полудомен расщепления включает мутации в аминокислотных остатках 486 и 499.
[0170] Таким образом, в одном варианте осуществления изобретения, мутацией в положении 490 является замена Glu (E) на Lys (К); мутацией в положении 538 является замена Iso (I) на Lys (К); мутацией в положении 486 является замена Gin (Q) на Glu (E); а мутацией в положении 499 является замена Iso (I) на Lys (K). В частности, описанные здесь сконструированные полудомены расщепления были получены путем замен в положениях 490 (Е→К) и 538 (I→K) в одном полудомене расщепления с получением сконструированного полудомена расщепления, обозначенного «E490K: I538K» и путем замен в положениях 486 (Q→Е) и 499 (I→L) в другом полудомене расщепления с получением сконструированного полудомена расщепления, обозначенного «Q486E:I499L». Описанные здесь сконструированные полудомены расщепления представляют собой облигатные гетеродимерные мутанты, в которых аберрантное расщепление было сведено к минимуму или отменено. См., например, патенты США № 7914796 и 8034598, содержание которых включено в настоящее описание посредством ссылки в полном объеме и во всех целях. В некоторых вариантах осуществления изобретения, сконструированные полудомены расщепления содержат мутации в положениях 486, 499 и 496 (пронумерованных относительно FokI дикого типа), например, мутации, которые представляют собой замену остатка Gln (Q) дикого типа в положении 486 на остаток Glu (E), замену остатка Iso (I) дикого типа в положении 499 на остаток Leu (L) и замену остатка Asn (N) дикого типа в положении 496 на остаток Asp (D) или Glu (E) (также обозначаемые как домены «ELD» и «ELE», соответственно). В других вариантах осуществления изобретения, сконструированные полудомены расщепления содержат мутации в положениях 490, 538 и 537 (пронумерованных относительно FokI дикого типа), например, мутации, которые представляют собой замену остатка Glu (E) дикого типа в положении 490 на остаток Lys (К), замену остатка Iso (I) дикого типа в положении 538 на остаток Lys (К), и замену остатка His (Н) дикого типа в положении 537 на остаток Lys (К) или остаток Arg (R) (эти домены также обозначаются как домены «ККК» и «KKR», соответственно). В других вариантах осуществления изобретения, сконструированный полудомен расщепления содержит мутации в положениях 490 и 537 (пронумерованных относительно FokI дикого типа), например, мутации, которые представляют собой замену остатка Glu (E) дикого типа в положении 490 на остаток Lys (К) и замену остатка His (Н) дикого типа в положении 537 на остаток Lys (К) или остаток Arg (R) (эти домены также обозначаются как домены «КIК» и «KIR», соответственно). См., например, патенты США №№ 7914796, 8034598 и 8623618, содержание которых включено в настоящее описание посредством ссылки в полном объеме и во всех целях. В других вариантах осуществления изобретения, сконструированный полудомен расщепления содержит замены «Шарки» и/или мутации «Шарки» (см., Guo et al, (2010) J. Mol. Biol. 400(1):96-107).
[0171] Альтернативно, нуклеазы могут быть подвергнуты сборке in vivo в сайте-мишени нуклеиновой кислоты с применением так называемой технологии «ферментативного расщепления» (см., например, публикацию заявки на патент США 20090068164). Компоненты таких расщепляющих ферментов могут быть экспрессированы либо в отдельных экспрессионных конструкциях, либо они могут быть присоединены в одной открытой рамке считывания, где отдельные компоненты разделены, например, саморасщепляющимся пептидом 2А или последовательностью IRES. Компоненты могут представлять собой отдельные домены, связывающиеся с цинковым пальцем, или домены, связывающиеся с нуклеиновой кислотой, кодирующей мегануклеазу.
[0172] Нуклеазы (например, ZFN и/или TALEN) могут быть подвергнуты скринингу на активность перед их использованием, например, в хромосомной системе на основе дрожжей, как описано в патенте США № 8563314.
[0173] В некоторых вариантах осуществления изобретения, нуклеаза содержит систему CRISPR/Cas. Локус CRISPR (кластеризованных регулярно перемежающихся коротких палиндромных повторов), который кодирует компоненты РНК системы, и локус Cas (CRISPR-ассоциированный), который кодирует белки (Jansen et al., 2002. Mol. Microbiol. 43: 1565-1575; Makarova et al., 2002. Nucleic Acids Res. 30: 482-496; Makarova et al., 2006. Biol. Direct 1: 7; Haft et al., 2005. PLoS Comput. Biol. 1:e60), составляют генные последовательности нуклеазной системы CRISPR/Cas. Локусы CRISPR в микробных хозяевах содержат комбинацию CRISPR-ассоциированных (Cas) генов, а также некодирующие элементы РНК, способные программировать специфичность CRISPR-опосредуемого расщепления нуклеиновой кислоты.
[0174] CRISPR типа II является одной из наиболее хорошо охарактеризованных систем и осуществляет нацеленный двухцепочечный разрыв ДНК в четыре последовательных стадии. В первой стадии, две некодирующие РНК, массив пре-cr-РНК и tracr-РНК транскрибируются из локуса CRISPR. Во второй стадии, tracr-РНК гибридизуется с повторяющимися областями пре-crРНК и опосредует процессинг пре-cr-РНК в зрелые cr-РНК, содержащие отдельные спейсерные последовательности. В третьей стадии, зрелый комплекс cr-РНК:tracr-РНК направляет Cas9 к ДНК-мишени посредством спаривания оснований Уотсона-Крика между спейсером на cr-РНК и протоспейсером на ДНК-мишени рядом с соседним мотивом протоспейсера (PAM), что является дополнительным требованием для распознавания мишени. И наконец, Cas9 опосредует расщепление ДНК-мишени с образованием двухцепочечного разрыва в протоспейсере. Активность системы CRISPR/Cas включает три стадии: (i) инсерцию чужеродных последовательностей ДНК в массив CRISPR для предотвращения будущих атак в процессе, называемом «адаптацией», (ii) экспрессию релевантных белков, а также экспрессию и процессинг массива, а затем (iii) РНК-опосредованную интерференцию чужеродной нуклеиновой кислоты. Таким образом, в бактериальной клетке, несколько так называемых белков «Cas» участвуют в природной функции системы CRISPR/Cas и играют определенную роль в таких функциях, как инсерция чужеродной ДНК и т.п.
[0175] В некоторых вариантах осуществления изобретения, белок Cas может представлять собой «функциональное производное» природного белка Cas. «Функциональное производное» полипептида с нативной последовательностью представляет собой соединение, которое вместе с полипептидом с нативной последовательностью обладает ценным биологическим свойством. «Функциональными производными» являются, но не ограничиваются ими, фрагменты нативной последовательности и производные полипептида с нативной последовательностью и его фрагментов при условии, что они, вместе с соответствующим полипептидом с нативной последовательностью, будут обладать биологической активностью. Рассматриваемая здесь биологическая активность представляет собой способность функционального производного гидролизовать субстрат ДНК на фрагменты. Термин «производное» охватывает варианты аминокислотной последовательности полипептида, ковалентные модификации и их гибриды. Подходящими производными полипептида Cas или его фрагмента являются, но не ограничиваются ими, мутанты, гибриды, ковалентные модификации белка Cas или его фрагмента. Белок Cas, который включает белок Cas или его фрагмент, а также производные белка Cas или его фрагмента, могут быть получены из клетки или синтезированы химическим способом или комбинацией этих двух процедур. Клетка может представлять собой клетку, которая в природе продуцирует белок Cas, или клетку, которая в природе продуцирует белок Cas и была генетически сконструирована так, чтобы она для продуцировала эндогенный белок Cas с более высоким уровнем экспрессии или белок Cas из экзогенно введенной нуклеиновой кислоты, где эта нуклеиновая кислота кодирует Cas, который является таким же как и эндогенный Cas или отличается от него. В некоторых случаях, если клетка не продуцирует природного белка Cas, то она была генетически сконструирована для продуцирования белка Cas.
[0176] Репрезентативные нуклеазные системы CRISPR/Cas, нацеленные на гены TCR и другие гены, раскрываются, например, в публикации заявки на патент США № 20150056705. Нуклеаза(ы) может (могут) создавать один или более двухцепочечных и/или одноцепочечных разрывов в сайте-мишени. В некоторых вариантах осуществления изобретения, нуклеаза включает каталитический неактивный домен расщепления (например, FokI и/или белок Cas). См., например, патенты США №№ 9200266; 8703489 и Guillinger et al.,. (2014) Nature Biotech. 32(6):577-582. Каталитический неактивный домен расщепления может, в сочетании с каталитический активным доменом, действовать как никаза и создавать одноцепочечный разрыв. Таким образом, две никазы могут быть использованы в комбинации с образованием двухцепочечного разрыва в специфической области. Специалистам известны также и другие никазы, см., например, McCaffrey et al., (2016) Nucleic Acids Res. 44(2): e11.doi: 10.1093/nar/gkv878. Epub 2015 Oct.19
Сайты-мишени
[0177] Как было подробно описано выше, ДНК-связывающие домены могут быть сконструированы так, чтобы они связывались с любой выбранной последовательностью. Сконструированный ДНК-связывающий домен может иметь новую специфичность связывания по сравнению с природным ДНК-связывающим доменом.
[0178] Описанные здесь нуклеазы нацелены на ген TCR или B2M, например, в сайте-мишени по меньшей мере из 9 нуклеотидов (например, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21 или более смежных или несмежных нуклеотидов), как показано в Таблице 1. Таким образом, описанные здесь композиции включают клетки, в которых: ген TCR является модифицированным (например, посредством инсерций и/или делеций, включая инсерцию трансгена CAR или CAR-HLA-Е или CAR-HLA-G) в экзоне c2 гена TCRA: ген B2M является модифицированным (например, посредством инсерций и/или делеций, включая инсерцию трансгена CAR) экзона 1 гена B2M; и/или ген HPRT является модифицированным (например, посредством инсерций и/или делеций, включая инсерцию трансгена CAR или CAR-HLA-Е или CAR-HLA-G). Модификации могут присутствовать в сайте-мишени или, альтернативно, поблизости (например, в пределах 1-5, 1-10 или 1-20 нуклеотидов) от 3’- или 5’-конца сайта-мишени. В других вариантах осуществления изобретения, модификации может присутствовать между спаренными сайтами-мишенями, как показано в Таблице 1.
Доноры
[0179] Любой донор может быть интегрирован путем встраивания в геном после описанного здесь расщепления, опосредованного нуклеазой. Донор может включать одну или более кодирующих последовательностей (например, CAR); одну или более РНК (РНКи, кшРНК и т.п.); одну или более некодирующих последовательностей и/или другие последовательности. Донорная конструкция может быть доставлена в клетку или индивидууму в виде мРНК или с использованием вирусного или невирусного ДНК-вектора, как описано в настоящей заявке. В некоторых вариантах осуществления изобретения, донор доставляют в виде мРНК или в виде одного или более AAV-векторов, включая, но не ограничиваясь ими, AAV1, AAV3, AAV4, AAV5, AAV6, AAV8, AAV8.2, AAV9 и AAVrh10 и/или псевдотипированные AAV, такие как AAV2/8, AAV2/5 и AAV2/6.
[0180] В некоторых вариантах осуществления изобретения, донор содержит один или более трансгенов, включая, но не ограничиваясь ими, одну или более CAR-кодирующих последовательностей, а также одну или более последовательностей, кодирующих B2M; одну или более HLA-G- и/или HLA-Е-кодирующих последовательностей; один или более репортеров (например, GFP) и т.п.
[0181] CAR-кодирующие последовательности (используемые для получения CAR-позитивных (+) Т-клеток) могут включать внеклеточный одноцепочечный вариабельный фрагмент (scFv) со специфичностью к конкретному опухолевому антигену, связанному с внутриклеточной сигнал-передающей частью, содержащей костимулирующий домен и домен активации. Костимулирующий домен может происходить, например, от CD28, а домен активации может происходить, например, от CD3-дзета. Трансгены CAR могут включать два, три, четыре или более костимулирующих доменов. scFv CAR может быть сконструирован для нацеливания, например, на CD19, который представляет собой трансмембранный белок, экспрессируемый клетками в В-клеточной линии дифференцировки, включая все нормальные В-клетки и В-клеточные злокачественные новообразования, включая, но не ограничиваясь ими, НХЛ, ХЛЛ и не-Т-клеточную ОЛЛ. См., например, патент США № 9855298. В некоторых вариантах осуществления изобретения, CAR содержит CAR, обозначенный FMC63-CD8BBZ, где FMC63 представляет собой последовательность анти-CD19 scFv (см., патент США № 9701758), а CD8BBZ означает части CAR, за исключением scFv (шарнирный/трансмембранный домен CD8 (CD8), костимулирующий домен гена 41BB (BB), домен активации гена CD3z (Z)).
[0182] В описанных здесь композициях и способах может быть использована любая HLA-Е- и/или HLA-G-кодирующая последовательность, включая, но не ограничиваясь ими, их аллельные варианты и/или функциональные фрагменты. В некоторых вариантах осуществления изобретения, трансген содержит последовательность, кодирующую один из двух основных аллельных вариантов HLA-E: HLA-E*0101 (также называемый здесь как ER или HLA-E0101) или HLA-E*0103 (также называемый здесь как EG или HLA-E0103). HLA-E0101 и HLA-E0103 различаются только в одном положении аминокислоты, где в положении 107 в HLA-E0101 присутствует аргинин, а в HLA-E0103 в этом положении, аргинин заменен глицином (Celik et al., ibid). В другом варианте осуществления изобретения, донор содержит гибридный белок, например, гибридные белки B2M-HLA-G и/или HLA-E. Компоненты трансгена, кодирующего гибридный белок, могут присутствовать в любом порядке, включая, но не ограничиваясь ими: последовательности гена B2M, расположенные перед последовательностями гена HLA-G и/или HLA-E; или последовательности гена HLA-G и/или HLA-E, расположенные перед последовательностями гена B2M.
[0183] Для введения трансгенов могут быть использованы один или более доноров, включая, например, один донор для последовательности CAR, последовательности B2M, последовательности HLA-E и/или HLA-G (например, вместе с саморасщепляющимся пептидом 2A и/или линкером между одной или более последовательностями). В некоторых вариантах осуществления изобретения, донор содержит последовательность 2А между CAR-кодирующей(ими) последовательностью(ями) и последовательностью, кодирующей B2M/HLA-G или E (например, последовательностью, кодирующей гибридный белок B2M-HLA-G/E). Альтернативно, могут быть использованы один донор для последовательности CAR и один или более отдельных доноров для генов B2M, HLA-E и/или HLA-G, например, первый донор CAR и второй донор для гибридного белка B2M/HLA-E/G (который также может дополнительно содержать одну или более последовательностей CAR). В некоторых вариантах осуществления изобретения, донор содержит трансген, кодирующий CAR и гибридный белок B2M-HLA-Е и/или G, вместе с саморасщепляющимся пептидом 2A между CAR и гибридным белком, и дополнительно, один или более линкеров (например, 1, 2, 3, 4, 5, 6 или более линкеров GS4) между белок-кодирующими последовательностями гибридных белков.
[0184] Любые из описанных здесь доноров могут включать гомологичные ветви (для гена, на который нацелена нуклеаза) любой длины. См., например, патенты США № 8822221; 7972854. «Длинные гомологичные ветви» имеют длину ~1 т.п.о., а «короткие гомологичные ветви» имеют длину приблизительно от 250 п.о. до 750 п.о.
[0185] Кроме того, любой из доноров может включать любую комбинацию компонентов, включая, но не ограничиваясь ими, гомологичные ветви; один или более трансгенов (экспрессия которых инициируется одинаковыми или различными регуляторными элементами), таких как репортер, B2M, HLA-G, HLA-E и/или один или более CAR; а также дополнительные последовательности, такие как один или более конститутивных или индуцибельных промоторов (например, PGK), одну или более энхансерных последовательностей (например, энхансерных последовательностей TCR); последовательности 2A; сигнал(ы) полиаденилирования; последовательность IRES, области 5’-UTR и/или 3’-UTR; и/или один или более (G4S)4-линкеров, а в некоторых вариантах осуществления изобретения, последовательность бета-глобина Xenopus (например, в 5’-UTR).
[0186] Любой из доноров может содержать последовательность WPRE. Для осуществления настоящего изобретения может быть использована любая последовательность WPRE. Неограничивающие примеры подходящих последовательностей раскрыты в публикации заявки на патент США 20160326548 и в патентах США № 6136597; 6284469; 6312912 и 6287814. В некоторых вариантах осуществления изобретения, последовательность WPRE содержит мутацию по сравнению с последовательностью дикого типа. См., например, патент США № 7419829 и Zanta-Boussif et al., (2009) Gene Therapy 16:605-619, или усеченную последовательность (Choi et al.,, ibid). Могут быть также использованы одна или более одинаковых или различных последовательностей WPRE. WPRE может присутствовать в 3’-UTR и/или 5’-UTR донора. В некоторых вариантах осуществления изобретения, последовательности WPRE включают:
AATCAACCTCTGGATTACAAAATTTGTGAAAGATTGACTGGTATTCTTAACTATGTTGCTCCTTTTACGCTATGTGGATACGCTGCTTTAATGCCTTTGTATCATGCTATTGCTTCCCGTATGGCTTTCATTTTCTCCTCCTTGTATAAATCCTGGTTGCTGTCTCTTTATGAGGAGTTGTGGCCCGTTGTCAGGCAACGTGGCGTGGTGTGCACTGTGTTTGCTGACGCAACCCCCACTGGTTGGGGCATTGCCACCACCTGTCAGCTCCTTTCCGGGACTTTCGCTTTCCCCCTCCCTATTGCCACGGCGGAACTCATCGCCGCCTGCCTTGCCCGCTGCTGGACAGGGGCTCGGCTGTTGGGCACTGACAATTCCGTGGTGTTGTCGGGGAAGCTGACGTCCTTTCCATGGCTGCTCGCCTGTGTTGCCACCTGGATTCTGCGCGGGACGTCCTTCTGCTACGTCCCTTCGGCCCTCAATCCAGCGGACCTTCCTTCCCGCGGCCTGCTGCCGGCTCTGCGGCCTCTTCCGCGTCTTCGCCTTCGCCCTCAGACGAGTCGGATCTCCCTTTGGGCCGCCTCCCCGCCTG (SEQ ID NO:64).
[0187] Кроме того, доноры могут включать один или более энхансеров Т-клеточного рецептора, в том числе последовательность энхансера TCRa. См., например, Ho & Leiden (1990) Mol.Cell. Biol. 10 (9): 4720-4727; Kappes et al.,. (1991), Proc. Natl. Acad. Sci. USA 88: 2204-2208. В некоторых вариантах осуществления изобретения, энхансер Т-клеточного рецептора содержит последовательность:
GATATCATTGGTCAATTGCAGCAGACATGTCTCTTGTGGTGTAAAGCCTCATAGCATCTTCCTGCTGGAGTCCACCTGGCAGCCCAAAGCCTCAGCAAGTGCTGGGGACAGAGAGATAGAAAGGCTTTGCTGTGGCTCAACTCAGTTGTACTTTCTCAAGCTCCTTTTGGTTATCTGCTCTGTCTATGTAGTTGCTTCCTGGAAAATGGTTAAAGTCAAACCTCCTCTGTTTGGGGCAGCACAAAATGTTGCTAAGGCGTTAGAAGACAACATAGGAGCAGTTAAACCACAGCCAGTTTTTGCTCGCTTTCGAAAGACCACAGCTATTAGACAGAAAACCTCCTGTACTGGAAGTGGCCCTGAAACAGCATCTGTGCTGACAGACACTGGGCTTGCTATGGTTGCTGCTCATCATGCTCAGCTG (SEQ ID NO: 65). В других вариантах осуществления изобретения, энхансерная последовательность TCRα представлена ниже: CTCCCATTTCCATGACGTCATGGTTAAACCTCTTTTACCTGCTTTAACCCTTTGAAGCTCTCCCGCAGAAGCCACATCCTCTG (SEQ ID NO:66).
[0188] Таким образом, неограничивающими примерами доноров являются:
(1) донор с длинными гомологичными ветвями (длинными ветвями), содержащими следующие последовательности: последовательность промотора PGK, функционально присоединенную к трансгену (GFP, CAR и т.п.), фланкированному гомологичными ветвями (нацеленными на B2M) размером ~1 т.п.о. каждая;
(2) донор с короткими гомологичными ветвями (короткими ветвями), содержащими следующие последовательности: последовательность промотора PGK, функционально присоединенную к трансгену, фланкированному гомологичными ветвями (нацеленными на B2M) размером 250 п.о. каждая;
(3) донор (2), дополнительно содержащий последовательность WPRE в 3’-UTR, например, последовательность WPRE, включающую:
AATCAACCTCTGGATTACAAAATTTGTGAAAGATTGACTGGTATTCTTAACTATGTTGCTCCTTTTACGCTATGTGGATACGCTGCTTTAATGCCTTTGTATCATGCTATTGCTTCCCGTATGGCTTTCATTTTCTCCTCCTTGTATAAATCCTGGTTGCTGTCTCTTTATGAGGAGTTGTGGCCCGTTGTCAGGCAACGTGGCGTGGTGTGCACTGTGTTTGCTGACGCAACCCCCACTGGTTGGGGCATTGCCACCACCTGTCAGCTCCTTTCCGGGACTTTCGCTTTCCCCCTCCCTATTGCCACGGCGGAACTCATCGCCGCCTGCCTTGCCCGCTGCTGGACAGGGGCTCGGCTGTTGGGCACTGACAATTCCGTGGTGTTGTCGGGGAAGCTGACGTCCTTTCCATGGCTGCTCGCCTGTGTTGCCACCTGGATTCTGCGCGGGACGTCCTTCTGCTACGTCCCTTCGGCCCTCAATCCAGCGGACCTTCCTTCCCGCGGCCTGCTGCCGGCTCTGCGGCCTCTTCCGCGTCTTCGCCTTCGCCCTCAGACGAGTCGGATCTCCCTTTGGGCCGCCTCCCCGCCTG (SEQ ID NO:64);
(4) донор (2), дополнительно содержащий энхансерную T-клеточную последовательность, расположенную выше промотора PGK, например, последовательность, представленную ниже:
GATATCATTGGTCAATTGCAGCAGACATGTCTCTTGTGGTGTAAAGCCTCATAGCATCTTCCTGCTGGAGTCCACCTGGCAGCCCAAAGCCTCAGCAAGTGCTGGGGACAGAGAGATAGAAAGGCTTTGCTGTGGCTCAACTCAGTTGTACTTTCTCAAGCTCCTTTTGGTTATCTGCTCTGTCTATGTAGTTGCTTCCTGGAAAATGGTTAAAGTCAAACCTCCTCTGTTTGGGGCAGCACAAAATGTTGCTAAGGCGTTAGAAGACAACATAGGAGCAGTTAAACCACAGCCAGTTTTTGCTCGCTTTCGAAAGACCACAGCTATTAGACAGAAAACCTCCTGTACTGGAAGTGGCCCTGAAACAGCATCTGTGCTGACAGACACTGGGCTTGCTATGGTTGCTGCTCATCATGCTCAGCTG (SEQ ID NO:65); или
(5) донор (2), дополнительно содержащий энхансерную последовательность TCRα, например, последовательность, представленную ниже:
CTCCCATTTCCATGACGTCATGGTTAAACCTCTTTTACCTGCTTTAACCCTTTGAAGCTCTCCCGCAGAAGCCACATCCTCTG (SEQ ID NO:66);
(6) донор (2), дополнительно включающий последовательность 5’-нетранслируемой области гена бета-глобина Xenopus, необязательно последовательность, представленную ниже: TGCTTGTTCTTTTTGCAGAAGCTCAGAATAAACGCTCAACTTTGGCAGAT (SEQ ID NO:67);
(7) донор, содержащий короткие гомологичные ветви (левую ветвь 423 п.о. и правую ветвь 393 п.о.), нацеленные на сайт TRAC-E, фланкирующий трансген (например, GFP, CAR, B2M, HLA-E и/или HLA-G), активируемый промотором hPGK. Донор также содержит, но необязательно, «метку Miseq», позволяющую количественно определить уровень TI в локусе TRAC по miseq;
(8) донор, содержащий длинные гомологичные ветви (левую ветвь 1000 п.о. и правую ветвь 992 п.о.), нацеленные на сайт TRAC-E, фланкирующий трансген (например, GFP, CAR, B2M, HLA-E и/или HLA-G), активируемый промотором hPGK. Этот донор не содержит «метку miseq»;
(9) донор, содержащий короткие гомологичные ветви (левую ветвь 350 п.о. и правую ветвь 393 п.о.), нацеленные на сайт TRAC-E, фланкирующий трансген (например, GFP, CAR, B2M, HLA-E и/или HLA-G), активируемый промотором hPGK, и мутированный элемент WPRE (после трансгена). Этот донор также содержит, но необязательно, метку «miseq»;
(10) донор, содержащий короткие гомологичные ветви (левую ветвь 423 п.о. и правую ветвь 393 п.о.), нацеленные на сайт TRAC-E, фланкирующий трансген FMC63-CD8BBZ CAR, активируемый промотором hPGK. Этот донор также содержит, но необязательно, метку «miseq», позволяющую количественно определить уровень TI в локусе TRAC по miseq;
(11) донор (10), дополнительно содержащий последовательность WPRE после последовательности трансгена;
(12) донор, содержащий короткие гомологичные ветви (левую ветвь 360 п.о. и правую ветвь 393 п.о.), нацеленные на сайт TRAC-E, фланкирующий трансген FMC63-CD8BBZ CAR, связанный посредством саморасщепляющегося пептида P2A с гибридным белком и экспрессирующий B2M и HLA-E. Трансген активируется промотором hPGK, и за этим трансгеном следует мутированный элемент WPRE. Этот донор также, но необязательно, содержит метку «miseq»;
(13) донор (12), в котором HLA-Е заменен на HLA-G;
(14) донор (13), содержащий HLA-E и HLA-G;
(15) донор, содержащий длинные гомологичные ветви (левую ветвь 1000 п.о. и правую ветвь 992 п.о.), нацеленные на сайт TRAC-E, фланкирующий трансген HLA-G, активируемый промотором hPGK;
(16) донор (15), в котором трансген HLA-G заменен трансгеном HLA-E (например, HLA-E0101 или E0103 (Celik et al., ibid));
(17) донор (15), дополнительно содержащий трансген HLA-G;
(18) донор по любому из (15)-(17), дополнительно содержащий B2M-кодирующую последовательность, связанную с трансгеном HLA-E и/или HLA-G, необязательно посредством линкера с 1, 2, 3, 4, 5, 6 или более копиями пептида G4S, расположенного между любыми из трансгенов (например, между B2M и HLA-G, между B2M и HLA-E, между B2M, HLA-G и HLA-E, между HLA-G и HLA-E, и т.п.);
(19) донор по любому из (15)-(18), дополнительно содержащий, но необязательно, сигнальный пептид, расположенный перед одним или более трансгенами (например, B2M и HLA-E/G или только B2M, например донор, который не содержит один пептид для HLA-G);
(20) донор по любому из (15)-(19), дополнительно содержащий трансген CAR (например, FMC63-CD8BBZ), необязательно связанный посредством саморасщепляющегося пептида P2A с гибридным белком и экспрессирующий B2M и HLA-G и/или HLA-Е; и
(21) донор (20), дополнительно содержащий мутированный элемент WPRE и необязательно метку miseq.
Доставка
[0189] Белки (например, факторы транскрипции, нуклеазы, молекулы TCR и CAR), полинуклеотиды и/или композиции, содержащие описанные здесь белки и/или полинуклеотиды, могут быть доставлены в клетку-мишень любыми подходящими способами, включая, например, инжекцию белка и/или компонентов мРНК. В некоторых аспектах изобретения, нуклеазы и/или доноры доставляются в виде мРНК, а трансген доставляется посредством других молекул, таких как вирусные векторы, миникольцевая ДНК, плазмидная ДНК, одноцепочечная ДНК, линейная ДНК, липосомы, наночастицы и т.п. См., например, патент США № 20140335063. В некоторых вариантах осуществления изобретения, белки вводят в клетку путем сжатия клеток (см., Kollmannsperger et al. (2016) Nat Comm 7, 10372 doi:10.1038/ncomms10372).
[0190] Подходящими клетками являются, но не ограничиваются ими, эукариотические и прокариотические клетки и/или клеточные линии. Неограничивающими примерами таких клеток или клеточных линий, полученных из таких клеток, являются Т-клетки, COS, CHO (например, CHO-S, CHO-K1, CHO-DG44, СНО-DUXB11, СНО-DUKX, CHOK1SV), VERO, MDCK, WI38, V79, B14AF28-G3, ВНК, Hak, NS0, SP2/0-Agl4, HeLa, HEK293 (например, НЕК293-F, НЕК293-H, HEK293-Т) и клетки perC6, а также клетки насекомых, таких как Spodoptera fugiperda (Sf), или клетки грибов, таких как Saccharomyces, Pichia и Schizosaccharomyces. В некоторых вариантах осуществления изобретения, клеточная линия представляет собой клеточную линию СНО-К1, MDCK или НЕК293. Подходящими клетками также являются стволовые клетки, такие как, например, эмбриональные стволовые клетки, индуцированные плюрипотентные стволовые клетки (iPS-клетки), гемопоэтические стволовые клетки, нейронные стволовые клетки и мезенхимальные стволовые клетки.
[0191] Методы доставки описанных здесь белков, содержащих ДНК-связывающие домены, описаны, например, в патентах США №№ 6453242, 6503717; 6534261; 6599692; 6607882; 6689558; 6824978; 6933113; 6979539; 7013219 и 7163824, содержание которых включено в настоящее описание посредством ссылки в полном объеме.
[0192] ДНК-связывающие домены и гибридные белки, содержащие эти описанные здесь ДНК-связывающие домены, могут быть также доставлены с помощью векторов, содержащих последовательности, кодирующие один или более ДНК-связывающих белков. Кроме того, посредством этих векторов могут быть доставлены и другие нуклеиновые кислоты (например, доноры). При этом, могут быть использованы любые векторные системы, включая, но не ограничиваясь ими, плазмидные векторы, ретровирусные векторы, лентивирусные векторы, аденовирусные векторы, поксвирусные векторы; герпесвирусные векторы и аденоассоциированные вирусные векторы и т.п. См. также патенты США №№ 6534261, 6607882; 6824978; 6933113; 6979539; 7013219 и 7163824, содержание которых включено в настоящее описание посредством ссылки в полном объеме. Кроме того, очевидно, что любой из этих векторов может содержать одну или более последовательностей, кодирующих ДНК-связывающий белок, и/или дополнительные нуклеиновые кислоты, если это необходимо. Таким образом, при введении в клетку одного или более описанных здесь ДНК-связывающих белков и дополнительных ДНК, если это необходимо, они могут быть перенесены на один и тот же вектор или на различные векторы. При использовании множества векторов, каждый вектор может содержать последовательность, кодирующую один или более ДНК-связывающих белков и дополнительные нуклеиновые кислоты, если это необходимо.
[0193] Обычные вирусные и невирусные методы переноса генов могут быть использованы для введения нуклеиновых кислот, кодирующих сконструированные ДНК-связывающие белки, в клетки (например, в клетки млекопитающих) и ткани-мишени, и для совместного введения дополнительных нуклеотидных последовательностей, если это необходимо. Такие методы могут быть также применены для введения нуклеиновых кислот (например, кодирующих ДНК-связывающие белки и/или доноры) в клетки in vitro. В некоторых вариантах осуществления изобретения, нуклеиновые кислоты вводят с применением генотерапии in vivo или ex vivo. Системами доставки на основе невирусных векторов являются плазмидные ДНК, «оголенная» нуклеиновая кислота и нуклеиновая кислота, образующая комплекс с носителем для доставки, таким как липосомы, липидные наночастицы или полоксамер. Системы доставки на основе вирусных векторов включают ДНК- и РНК-вирусы, которые имеют либо эписомный, либо интегрированный геном после доставки в клетку. Обзор процедур генотерапии см., Anderson, Science 256:808-813 (1992); Nabel & Felgner, TIBTECH 11:211-217 (1993); Mitani & Caskey, TIBTECH 11:162-166 (1993); Dillon, TIBTECH 11:167-175 (1993); Miller, Nature 357:455-460 (1992); Van Brunt, Biotechnology 6(10):1149-1154 (1988); Vigne, Restorative Neurology and Neuroscience 8:35-36 (1995); Kremer & Perricaudet, British Medical Bulletin 51(1):31-44 (1995); Haddada et al., in Current Topics in Microbiology and Immunology Doerfler and  (eds.) (1995); и Yu et al., Gene Therapy 1:13-26 (1994).
(eds.) (1995); и Yu et al., Gene Therapy 1:13-26 (1994).
[0194] Способы не-вирусной доставки нуклеиновых кислот включают электропорацию, липофекцию, микроинжекцию, биобаллистику, использование виросом, липосом, липидных наночастиц, иммунолипосом, поликатионов или конъюгатов «липид: нуклеиновая кислота», «оголенных» ДНК, мРНК, искусственных вирионов, и поглощение ДНК, индуцируемое агентом. Для доставки нуклеиновых кислот может быть также применена сонопорация, проводимая с использованием, например, системы Sonitron 2000 (Rich-Mar). В предпочтительном варианте осуществления изобретения, одну или более нуклеиновых кислот доставляют в виде мРНК. Также предпочтительным является использование кэпированных мРНК для повышения эффективности трансляции и/или стабильности мРНК. Особенно предпочтительными являются кэпы ARCA (антитело против кэпированного аналога обратной транскриптазы) или их варианты. См. патенты США №№ 7074596 и 8153773, которые включены в настоящее описание посредством ссылки.
[0195] Дополнительные репрезентативные системы доставки нуклеиновых кислот включают системы, поставляемые Amaxa Biosystems (Cologne, Germany), Maxcyte, Inc. (Rockville, Maryland), системы молекулярной доставки BTX (Holliston, MA) и Copernicus Therapeutics Inc, (см., например, US6008336). Липофекция описана, например, в патентах США 5049386, 4946787 и 4897355, а реагенты для липофекции являются коммерчески доступными (например, Трансфектам™, липофектин™ и липофектамин™ RNAiMAX). Катионными и нейтральными липидами, подходящими для эффективной липофекции полинуклеотидов с распознаванием рецептора, являются липиды Felgner, WO 91/17424, WO 91/16024. Доставка может быть осуществлена в клетки (введение ex vivo) или в ткани-мишени (введение in vivo).
[0196] Получение комплексов «липид:нуклеиновая кислота», включая липосомы-мишени, такие как иммунолипидные комплексы, хорошо известно специалистам в данной области (см., например, Crystal, Science 270:404-410 (1995); Blaese et al., Cancer Gene Ther. 2:291-297 (1995); Behr et al., Bioconjugate Chem. 5:382-389 (1994); Remy et al., Bioconjugate Chem. 5:647-654 (1994); Gao et al., Gene Therapy 2:710-722 (1995); Ahmad et al., Cancer Res. 52:4817-4820 (1992); патенты США №№ 4186183, 4217344, 4235871, 4261975, 4485054, 4501728, 4774085, 4837028 и 4946787).
[0197] Другими способами доставки является упаковка нуклеиновых кислот, доставляемых в носительи для доставки EnGeneIC (EDV). Эти EDV, в частности, доставляются в ткани-мишени с использованием биспецифических антител, где одна ветвь антитела является специфичной к ткани-мишени, а другая к EDV. Антитело переносит EDV на поверхность клетки-мишени, в результате чего EDV проникает в клетку посредством эндоцитоза. После этого, содержимое высвобождается в клетку (см. MacDiarmid et al. (2009) Nature Biotechnology 27(7) p. 643).
[0198] При применении вирусных систем на основе РНК или ДНК для доставки нуклеиновых кислот, кодирующих сконструированные ДНК-связывающие белки и/или доноры (например, CAR или ACTR), желательно использовать в высокой степени эволюционные процессы для нацеливания вируса на конкретные клетки в организме и доставки вирусной полезной нагрузки в ядро. Вирусные векторы могут быть введены непосредственно пациентам (in vivo), либо они могут быть использованы для обработки клеток in vitro с последующим введением модифицированных клеток пациентам {ex vivo). Стандартные вирусные системы доставки нуклеиновых кислот включают, но не ограничиваются ими, ретровирусные векторы, лентивирусные векторы, аденовирусные векторы, векторы на основе аденоассоциированного вируса, векторы на основе вируса осповакцины и векторы на основе вируса простого герпеса для переноса генов. Интеграция в геном хозяина может быть осуществлена с применением методов переноса генов на основе ретровируса, лентивируса и аденоассоциированного вируса, и это часто приводит к более длительной экспрессии встроенного трансгена. Кроме того, высокая эффективность трансдукции наблюдалась во многих клетках различных типов и в тканях-мишенях.
[0199] Тропизм ретровируса может быть изменен путем введения чужеродных оболочечных белков, что позволяет расширить популяцию клеток-мишеней. Лентивирусными векторами являются ретровирусные векторы, способные к трансдукци или инфицированию неделящихся клеток и обычно продуцирующие высокие вирусные титры. Выбор системы переноса ретровирусного гена зависит от ткани-мишени. Ретровирусные векторы состоят из цис-действующих длинных концевых повторров с емкостью упаковки до 6-10 т.п.о. чужеродной последовательности. Минимальные цис-действующие LTR является достаточными для репликации и упаковки векторов, которые затем используются для интеграции терапевтического гена в клетку-мишень для достижения перманетной экспрессии трансгена. Широко используемыми ретровирусными векторами являются векторы на основе вируса мышиного лейкоза (MuLV), вируса лейкоза гиббонов (GaLV), вируса обезьяньего иммунодефицита (SIV), вируса иммунодефицита человека (ВИЧ) и их комбинации (см., например, Buchscher et al., J. Virol. 66:2731-2739 (1992); Johann et al., J. Virol. 66:1635-1640 (1992); Sommerfelt et al., Virol. 176:58-59 (1990); Wilson et al., J. Virol. 63:2374-2378 (1989); Miller et al., J. Virol. 65:2220-2224 (1991); PCT/US94/05700).
[0200] В тех случаях, когда предпочтительной является временная экспрессия, то могут быть использованы системы на основе аденовирусов. Аденовирусные векторы имеют очень высокую эффективность трансдукции в клетках многих типов и не требуют деления клеток. При использовании таких векторов достигаются высокие титры и высокие уровни экспрессии. Этот вектор может быть получен в больших количествах в относительно простой системе. Векторы на основе аденоассоциированного вируса («AAV») также используется для инфицирования клеток нуклеиновыми кислотами-мишенями, например, при продуцировании in vitro нуклеиновых кислот и пептидов, а также для генотерапии in vivo и ex vivo (см, например, West et al., Virology 160:38-47 (1987); патент США No. 4797368; WO 93/24641; Kotin, Human Gene Therapy 5:793-801 (1994); Muzyczka, J. Clin. Invest. 94:1351 (1994). Конструирование рекомбинантных AAV-векторов описано в ряде публикаций, включая патент США No. 5173414; Tratschin et al., Mol. Cell. Biol. 5:3251-3260 (1985); Tratschin, et al., Mol. Cell. Biol. 4:2072-2081 (1984); Hermonat & Muzyczka, PNAS USA 81:6466-6470 (1984); и Samulski et al., J. Virol. 63:03822-3828 (1989).
[0201] При клинических испытаниях, для переноса генов в настоящее время применяется по меньшей мере шесть методов с использованием вирусных векторов, и в этих методах используются подходы, которые включают комплементацию дефектных векторов генами, встроенными в хелперные клеточные линии для получения трансдуцирующего агента.
[0202] pLASN и MFG-S являются примерами ретровирусных векторов, которые были использованы в клинических испытаниях (Dunbar et al., Blood 85:3048-305 (1995); Kohn et al., Nat. Med. 1:1017-102 (1995); Malech et al., PNAS USA 94:22 12133-12138 (1997)). PA317/pLASN представляет собой первый терапевтический вектор, используемый в испытаниях по генотерапии (Blaese et al., Science 270:475-480 (1995)). Эффективность трандукции для MFG-S-упакованных векторов составляет 50% или более (Ellem et al., Immunol Immunother. 44(1):10-20 (1997); Dranoff et al., Hum. Gene Ther. 1:111-2 (1997).
[0203] Векторы на основе рекомбинантного аденоассоциированного вируса (rAAV) представляют собой перспективную альтернативную систему доставки генов на основе дефектного и непатогенного аденоассоциированного парвовируса типа 2. Все векторы были получены из плазмиды, которая сохраняет только 145 п.о. инвертированных концевых повторов AAV, фланкирующих кластер для экспрессии трансгена. Эффективный перенос генов и стабильная доставка трансгена благодаря интеграции в геномы трансдуцированных клеток являются ключевыми свойствами этой векторной системы. (Wagner et al., Lancet 351:9117 1702-3 (1998), Kearns et al., Gene Ther. 9:748-55 (1996)). В настоящем изобретении могут быть также использованы AAV других серотипов, включая AAV1, AAV3, AAV4, AAV5, AAV6, AAV8, AAV8.2, AAV9 и AAVrh10 и псевдотипированные AAV, такие как AAV2/8, AAV2/5 и AAV2/6.
[0204] Дефицитные по репликации рекомбинантные аденовирусные векторы (Ad) могут быть получены с высоким титром и могут легко инфицировать ряд клеток различных типов. Большинство аденовирусных векторов сконструированы так, чтобы трансген заменял гены (Ad) Ela, E1b и/или E3, в результате чего дефектный по репликации вектор будет размножаться в человеческих клетках 293, которые будут выполнять функцию делетированного гена in trans. Векторы Ad могут трансдуцировать ткани множества типов in vivo, включая неделящиеся дифференцированные клетки, такие как клетки, присутствующие в печени, почках и в мышцах. Стандартные векторы Ad имеют большую емкость переноса частиц. Примером использования вектора Ad в клинических испытаниях является полинуклеотидная терапия для противоопухолевой иммунизации путем внутримышечной инъекции (Sterman et al., Hum. Gene Ther. 7:1083-9 (1998)). Другие примеры использования аденовирусных векторов для переноса генов в клинических испытаниях описаны Rosenecker et al., Infection 24:1 5-10 (1996); Sterman et al., Hum. Gene Ther. 9:7 1083-1089 (1998); Welsh et al., Hum. Gene Ther. 2:205-18 (1995); Alvarez et al., Hum. Gene Ther. 5:597-613 (1997); Topf et al., Gene Ther. 5:507-513 (1998); Sterman et al., Hum. Gene Ther. 7:1083-1089 (1998).
[0205] Упаковывающие клетки используют для получения вирусных частиц, способных инфицировать клетку-хозяина. Такими клетками являются клетки 293, которые упаковывают аденовирус и клетки ψ2 или клетки PA317, которые упаковывают ретровирус. Вирусные векторы, используемые в генотерапии, обычно генерируются клеточной линией-продуцентом, которая упаковывает вектор нуклеиновой кислоты в вирусную частицу. Векторы обычно содержат минимальные вирусные последовательности, необходимые для упаковки и последующей интеграции в клетку хозяина (если это необходимо), а другие вирусные последовательности заменяются экспрессионным кластером, кодирующим экспрессируемый белок. Недостающие вирусные функции поставляются in trans упаковывающей клеточной линией. Так, например, AAV-векторы, используемые в генотерапии, обычно имеют только последовательности инвертированных концевых повторов (ITR) генома AAV, которые необходимы для упаковки и интеграции в геном хозяина. Вирусная ДНК упакована в клеточной линии, которая содержит хелперную плазмиду, кодирующую другие гены AAV, а именно, rep и cap, но не содержит последовательностей ITR. Клеточная линия также инфицируется аденовирусом в качестве вируса-помощника. Вирус-помощник стимулирует репликацию вектора AAV и экспрессию генов AAV из хелперной плазмиды. Хелперная плазмида не упаковывается в значительных количествах из-за отсутствия последовательностей ITR. Контаминация аденовирусом может быть снижена, например, путем тепловой обработки, к которой аденовирус, по сравнению с AAV, является более чувствительным. Кроме того, AAV может быть получен с использованием бакуловирусной системы (см., например, патенты США 6723551 и 7271002).
[0206] Очистка частиц AAV из системы 293 или бакуловирусной системы обычно включает культивирование клеток, продуцирующих вирус, с последующим сбором вирусных частиц из клеточного супернатанта или лизисом клеток и сбором вируса из неочищенного лизата. AAV затем очищают известными методами, включая ионообменную хроматографию {см., например, патенты США 7419817 и 6989264), ионообменную хроматографию и центрифугирование в градиенте плотности CsCl {см., например, публикацию заявки РСТ WO2011094198A10), иммуноаффинную хроматографию {например, WO2016128408) или очистку с использованием сефарозы AVB (например, GE Healthcare Life Sciences).
[0207] Во многих случаях применения генотерапии, желательно, чтобы вектор для генотерапии доставлялся с высокой степенью специфичности в ткань конкретного типа. В соответствии с этим, вирусный вектор может быть модифицирован для сообщения специфичности к клетке данного типа путем экспрессии лиганда в виде гибридного белка вместе с белком вирусной оболочки на внешней поверхности вируса. Лиганд выбирают так, чтобы он обладал аффинностью к известному рецептору, присутствующему на клетке представляющего интерес типа. Так, например, Han et al., (Proc. Natl. Acad. Sci., USA 92:9747-9751 (1995)) сообщали, что вирус мышиного лейкоза Молони может быть модифицирован так, чтобы он экспрессировал человеческий герегулин, присоединенный к gp70, а рекомбинантный вирус инфицировал определенные клетки рака молочной железы человека, экспрессирующие человеческий рецептор эпидермального фактора роста. Этот принцип может быть применен и к другим парам «вирус-клетка-мишень», где клетка-мишень экспрессирует рецептор, а вирус экспрессирует гибридный белок, содержащий лиганд для рецептора клеточной поверхности. Так, например, нитчатый фаг может быть сконструирован так, чтобы он представлял фрагменты антител (например, Fab или Fv), обладающие специфической аффинностью связывания фактически с любым выбранным клеточным рецептором. Хотя приведенное выше описание относится прежде всего к вирусным векторам, однако, одни и те же принципы могут быть применены и к невирусным векторам. Такие векторы могут быть сконструированы так, чтобы они содержали специфические последовательности, которые поглощаются, предпочтительно, специфическими клетками-мишенями.
[0208] Векторы для генотерапии могут быть доставлены in vivo путем их введения отдельному пациенту, обычно путем системного введения (например, внутривенного, внутрибрюшинного, внутримышечного, субдермального или внутричерепного вливания) или путем местного применения, как описано ниже. Альтернативно, векторы могут быть доставлены в клетки ex vivo, такие как клетки, эксплантированные у отдельного пациента (например, лимфоциты, аспираты костного мозга, биоптаты ткани), или универсальные донорные гемопоэтические стволовые клетки, с последующей повторной имплантацией клеток пациенту, обычно после отбора клеток, которые имеют включенный в них вектор.
[0209] Ex vivo трансфекция клеток для диагностики, исследования, трансплантации или генотерапии (например, путем повторной инфузии трансфецированных клеток в организм хозяина) хорошо известна специалистам в данной области. В предпочтительном варианте осуществления изобретения, клетки выделяют из организма индивидуума, трансфицируют нуклеиновой кислотой ДНК-связывающего белка (геном или кДНК), и повторно вводят в организм индивидуума (например, пациента). Различные типы клеток, подходящие для трансфекции ex vivo, хорошо известны специалистам в данной области (см., например, Freshney et al., Culture of Animal Cells, A Manual of Basic Technique (3rd ed. 1994)) и цитируемые там материалы, в которых обсуждается выделение клеток у пациентов и их культивирование).
[0210] В одном варианте осуществления изобретения, стволовые клетки используют в процедурах ex vivo для трансфекции клеток и генотерапии. Преимущество использования стволовых клеток заключается в том, что они могут быть дифференцированы в клетки других типов in vitro, или могут быть введены млекопитающему (такому, как донор клеток), где они будут трансплантированы в костный мозг. Методы дифференцировки CD34+-клеток in vitro в клинически ценные иммунные клетки с использованием цитокинов, таких как GM-CSF, IFN-γ и TNF-α являются известными (см. Inaba et al., J. Exp. Med. 176:1693-1702 (1992)).
[0211] Стволовые клетки выделяют для трансдукции и дифференцировки известными методами. Так, например, стволовые клетки выделяют из клеток костного мозга путем пэннинга клеток костного мозга с использованием антител, которые связываются с нежелательными клетками, такими как CD4+- и CD8+-клетки (Т-клетки), CD45+-клетки (клетки panB), GR-1 (гранулоциты) и Iad (дифференцированные антигенпрезентирующие клетки) (см., Inaba et al., J. Exp. Med. 176:1693-1702 (1992)).
[0212] Стволовые клетки, которые были модифицированы, могут быть также использованы в некоторых вариантах осуществления изобретения. Так, например, нейронные стволовые клетки, которым была сообщена резистентность к апоптозу, могут быть использованы в качестве терапевтических композиций, где стволовые клетки также содержат ZFP TF согласно изобретению. Резистентность к апоптозу может быть сообщена, например, путем нокаута BAX и/или BAK с использованием BAX- или BAK-специфических ZFN (см., публикацию заявки на патент США № 20100003756) в стволовых клетках, или в клетках, которые были подвергнуты дизрупции в каспазе, и в этом случае, с использованием, например, ZFN, специфичных к каспазе-6. Эти клетки могут быть трансфицированы ZFP TF, которые, как известно, регулируют TCR.
[0213] Векторы {например, ретровирусы, аденовирусы, липосомы и т.п.), содержащие терапевтические ДНК-связывающие белки (или нуклеиновые кислоты, кодирующие эти белки), могут быть также введены непосредственно в организм для трансдукции клеток in vivo. Альтернативно, может быть введена «оголенная» ДНК. Введение осуществляют любымм способами, обычно применяемыми для ее контактирования, в конечном счете, с клетками крови или ткани, включая, но не ограничиваясь ими, инъекцию, инфузию, местное применение и электропорацию. Подходящие способы введения таких нуклеиновых кислот являются доступными и хорошо известны специалистам в данной области, и, хотя для введения конкретной композиции может быть применен более, чем один способ, однако, какой-либо конкретный способ, часто может давать более быструю и более эффективную реакцию, чем другой способ.
[0214] Способы введения ДНК в гемопоэтические стволовые клетки, описаны, например, в патенте США № 5928638. Векторы, подходящие для введения трансгенов в гемопоэтические стволовые клетки, например, CD34+-клетки, включают аденовирус типа 35.
[0215] Векторы, подходящие для введения трансгенов в иммунные клетки (например, Т-клетки), включают не-интегрирующиеся лентивирусные векторы. См., например, Ory et al. (1996) Proc. Natl. Acad. Sci. USA 93:11382-11388; Dull et al. (1998) J. Virol. 72:8463-8471; Zuffery et al. (1998) J. Virol. 72:9873-9880; Follenzi et al. (2000) Nature Genetics 25:217-222.
[0216] Фармацевтически приемлемые носители определяются отчасти в зависимости от конкретно вводимой композиции, а также конкретного метода, применяемого для введения композиции. Соответственно, существует широкий ряд подходящих и доступных препаратов фармацевтических композиций, описанных ниже (см., например, Remington’s Pharmaceutical Sciences, 17th ed., 1989).
[0217] Как было отмечено выше, описанные здесь способы и композиции могут быть применены к клеткам любого типа, включая, но не ограничиваясь ими, прокариотические клетки, клетки грибов, клетки архебактерий, клетки растений, клетки насекомых, клетки животных, клетки позвоночных, клетки млекопитающих и клетки человека, включая Т-клетки и стволовые клетки любого типа. Подходящие клеточные линии для экспрессии белка известны специалистам в данной области и включают, но не ограничиваются ими, COS, CHO (например, СНО-S, СНО-К1, СНО-DG44, CHO-DUXB11), VERO, MDCK, WI38, V79, B14AF28-G3, ВНК, Hak, NS0, SP2/0-Agl4, HeLa, HEK293 (например, НЕК293-F, НЕК293-Н, НЕК293-Т), perC6, клетки насекомых, такие как Spodoptera fugiperda (Sf) и клетки грибов, такие как Saccharomyces, Pichia и Schizosaccharomyces. Могут быть также использованы потомство, варианты и производные этих клеточных линий.
Применение
[0218] Описанные здесь композиции и способы могут быть применены в любых целях, включая, но не ограничиваясь ими, применение в терапевтических и исследовательских целях, если желательно получить T-клетки («вооруженные» Т-клетки), отвечающие на конкретные антигены клеточной поверхности.
[0219] Так, например, описанные здесь композиции могут быть использованы in vivo и/или ex vivo (в клеточной терапии) для нарушения экспрессии функциональных эндогенных TCR, и/или комплексов HLA в Т-клетках, модифицированных для адоптивной клеточной терапии, так, чтобы они экспрессировали один или более экзогенных CAR, экзогенных TCR или других молекул рецепторов, специфичных к раку, и тем самым, для лечения и/или профилактики рака. Кроме того, в таких случаях, отмена функциональной экспрессии TCR и/или экспрессии HLA в клетке позволяет устранять или значительно уменьшить риск нежелательной перекрестной реакции со здоровыми тканями, которые не являются мишенями (т.е., в случае ответа на реакцию «трансплантат против хозяина»). Кроме того, включение модифицированного трансгена HLA-Е или HLA-G так, чтобы клетка экспрессировала модифицированный комплекс HLA-Е или HLA-G, может способствовать предотвращению их цитолиза NK-клетками-хозяевами. Таким образом, универсальные донорные Т-клетки могут быть получены так, чтобы они содержали CAR, необязательно вместе с модифицированным комплексом HLA-Е или HLA-G, и так, чтобы эти универсальные клетки могли быть введены любому пациенту, нуждающемуся в этом.
[0220] Кроме того, сконструированные CAR-содержащие Т-клетки могут включать дополнительные трансгены, которые могут быть подходящими для адоптивной клеточной терапии. Так, например, эти клетки могут включать гены-«самоубийцы», регулируемые экзогенным сигналом, для стимуляции апоптоза сконструированных Т-клеток после активации экспрессии гена-«самоубийцы» (Wang et al. (2017) J. Hematol Oncol 10:53). Было также показано, что сверхэкспрессия CD40L в CAR-Т-клеток может оказаться предпочтительной для сообщения активности CAR-T (см., Curran et al (2015) Mol Ther. 23:769-78). В настоящее время, несколько групп специалистов советуют получить CAR-T-клетки «четвертого поколения», в которых секреция IL-12 объединена с экспрессией CAR. Koneru и его коллеги (Oncoimmunology (2015);4:e994446) использовали так называемые «Т-клетки, перенацеленные на универсальный цитолиз, опосредуемый цитокинами (TRUCK), для лечения рака яичнника у ортотропной модели трансплантата опухоли яичника.
[0221] Другие трансгены могут содержать дополнительные последовательности CAR, которые являются специфичными для других представляющих интерес антигенов, или трансген, экспрессирующий модифицированные HLA-Е или HLA-G. Было показано, что адоптивный перенос Т-клеток, экспрессирующих антитела против химерных антигенных рецепторов CD19 (CAR), имеет значительный лечебный потенциал против прогрессирующих В-клеточных злокачественных новообразований, но многочисленные исследования также показали, что у пациентов наблюдаются рецидивы из-за появления CD19-негативных клеток лейкоза. Таким образом, использование двух CAR, экспрессирующих, но необязательно, модифицированный комплекс HLA-Е или HLA-G, где один CAR является специфичным к CD19, а другой к CD20, может предотвратить рецидив. Кроме того, вместо использования двух отдельных конструкций CAR, может быть использован сам трансген CAR, который может кодировать биспецифический CAR, способный распознавать CD19 и CD20 (Zah et al (2016) Cancer Immunol Res, 4(6): 498-508). T-клетки, снабженные CD19-специфическими CAR и необязательно содержащие модифицированный комплекс HLA-Е или HLA-G, могут быть использованы в терапии для лечения В-клеточных злокачественных новообразований, таких как В-клеточные лимфомы, и других раковых заболеваний крови, таких как ОЛЛ, ОМЛ и НХЛ (Scheuermann and Racila (1995) Leuk Lymphoma. 18(5-6):385-97). Другими раковыми заболеваниями, которые могут быть подвергнуты лечению с использованием CAR-содержащих T-клеток, являются злокачественные глиомы (см., идентификатор клинических испытаний: NCT01454596), плоскоклеточный рак головы и шеи (SCCHN) (см. идентификатор клинических испытаний: NCT01818323), нейробластома (см. идентификатор клинических испытаний: NCT01822652), злокачественная мезотелиома плевры (см. идентификатор клинических испытаний: NCT01722149), прогрессирующая саркома (см. идентификатор клинических испытаний: NCT00902044), почечные карциномы (Kakarla & Gottschalk, (2014) Cancer J. 20(2):151-155). Таким образом, описанные здесь композиции и способы могут быть использованы для лечения рака любого типа, включая, но не ограничиваясь ими, рак яичника, рак головного мозга, рак предстательной железы, рак печени, рак почек, лейкоз, лимфомы, рак спинного мозга и рак ЦНС, и т.п.
[0222] Другие антигены, которые имеют терапевтическую ценность для потенциального нацеливания с использованием CAR, включает антигены, ассоциированные с опухолевыми клетками или с раковыми процессами, такие как BCMA, EpCAM, CEA, HER-2, gpA33, муцины, TAG-72, CAIX, PSMA, антитела, связывающиеся с фолатом, фолатный рецептор FR-α, CD19, CD20, CD22, EGFR, EGFRvIII, EpCam, EphA2, ERBB2/HER2, ERBB3, FAP, MET, Igκ, IL-1RAP, IGF1R, EPHA3, Lewis Y, лиганды NKG2D, ROR1, FR-a, GD2, GPC3, IL-13Ra2, L1-CAM, MUC1, PD-L1, PSCA, PSMA, VEGFR2, c-MET, CD133, TRAILR1, TRAILR2, RANKL, FAP, VEGF, VEGFR, интегрины αVβ3 и α5β1, CD30, CD33, CD52, CD70, CD123, CD138, CD171, CD16V, CTLA4 и энасцин.
[0223] Другие возможные области применения описанных здесь способов и композиций включают дополнительные способы конструирования, такие как модуляция других представляющих интерес генов. Эти другие гены включают гены ингибитора контрольной точки, такие как PD1 и/или CTLA4, где нокаут этих генов может предотвращать опухолеопосредованную супрессию Т-клеточного ответа. Кроме того, Т-клетки, содержащие инсерцию представляющго интерес CAR в TCR-родственном гене, могут быть дополнительно сконструированы для нокаута гена B2M, а Т-клетки, включающие CAR, интегрированный в ген B2M, могут быть дополнительно сконструированы для нокаута TCR-родственного гена. Эти модифицированные Т-клетки могут также содержать модифицированный комплекс HLA-E или HLA-G, что позволяло бы сконструированным Т-клеткам «ускользать» от уничтожения NK-клетками-хозяевами. Эти дополнительные мутации могут быть подходящими для создания универсальных Т-клеток, «готовых для применения».
[0224] Способы и композиция также включают композиции стволовых клеток, в которых гены TCRA и/или TCRB и/или B2M внутри стволовых клеток были модулированы (модифицированы), и эти клетки также содержат один или более CAR. Так, например, аллогенные гемопоэтические стволовые клетки, модулированные путем нокаута или нокдауна TCR, могут быть введены HLA-совместимому пацциенту после операции по удалению костного мозга. Эти модифицированные CAR-содержащие HSC позволяют осуществлять реколонизацию у пациента, но не вызывают возможной реакции «трансплантат против хозяина» (GvHD). Введенные клетки также могут иметь другие модификации для облегчения проведения последующей терапии (например, при резистентности к химиотерапии) для лечения основного заболевания. Клетки, не содержащие TCR и/или HLA и необязательно содержащие модифицированный комплекс HLA-E или HLA-G, также используются в качестве терапии, подходящей для применения в отделении неотложной помощи у пациентов с травмой.
[0225] Способы и композиции согласно изобретению могут быть также использованы для разработки и реализации моделей in vitro и in vivo, например, у животных с моделями TCR и/или ассоциированных с ними расстройств, что позволяет проводить изучение этих расстройств.
[0226] Все упомянутые здесь патенты, патентные заявки и публикации включены в настоящее описание посредством ссылки в полном объеме.
[0227] Несмотря на то, что раскрытие изобретения представлено здесь в некоторых деталях, например, для иллюстрации и для лучшего понимания изобретения, однако, специалистам в данной области очевидно, что в настоящее изобретение могут быть внесены различные изменения и модификации, не выходящие за рамки существа и объема настоящего изобретения. В соответствии с этим, вышеприведенное описание и последующие примеры не должны быть истолкованы как ограничивающие. В частности, хотя в качестве примера представлены ДНК-связывающие домены белка «цинковый палец», однако, для нацеленной модификации может быть использован любой ДНК-связывающий домен, включая, но не ограничиваясь ими, ДНК-связывающие домены эффекторного домена TAL, ДНК-связывающие домены оцрРНК (систем CRISPR/Cas) и/или Ttago, включая любой ДНК-связывающий домен, который связывается с 12-25 нуклеотидами сайтов-мишеней, представленных ниже в качестве примеров.
ПРИМЕРЫ
Пример 1: Конструирование нуклеаз
[0228] TCRA-специфические, B2M-специфические, CISH- специфические, PD1-специфические, CTLA-4- и HPRT-специфические ZFN были получены и сконструированы для сайт-специфического введения двухцепочечноых ДНК-разрывов, в основном, как описано Urnov et al. (2005) Nature 435(7042):646-651, Lombardo et al (2007) Nat Biotechnol. Nov;25(11):1298-306, в публикациях заявок на патенты США 20150164954, 20140301990, в патентах США 8563314, 8956828, 9402879 и 9597357; и в заявке на патент США No. 62/583724. См., также патент США No. 9873894, где описаны оцрРНК-системы CRISPR/Cas для связывания и модификации этих и других генов. Кроме того, спирали распознавания для репрезентативных пар ZFN, а также для последовательности-мишени представлены ниже в Таблице 1. Сайты-мишени конструкций ««цинковые пальцы» представлены в первом столбце. Нуклеотиды в сайте-мишени, на который нацелены спирали распознавания ZFP, обозначены прописными буквами; а нуклеотиды, не являющиеся мишенями, обозначены строчными буквами. Для TRAC-специфических ZFN, см. также заявку на патент США 16/009975. Для B2M- специфической ZFN, см. также публикацию заявки на патент США 20170173080. Для CISH-специфических ZFN см. предварительную заявку на патент США 62/583724.
[0229] Также описаны линкеры, используемые для связывания нуклеазного домена FokI и ДНК-связывающего домена ZFP (см., публикацию заявки на патент США 20150132269). Так, например, аминокислотная последовательность линкерного домена L0 представляет собой ДНК-связывающий домен-QLVKS-нуклеазный домен FokI (SEQ ID NO: 5). Аналогичным образом, аминокислотные последовательности для линкерного домена N7a представляет собой нуклеазный домен FokI-SGTPHEVGVYTL-ДНК-связывающий домен (SEQ ID NO:37), а N7c представляет собой нуклеазный домен FokI-SGAIRCHDEFWF-ДНК-связывающий домен (SEQ ID NO: 38). Кроме того, в соответствующих последовательностях показаны модификации остова «цинковый палец», описанного в публикации заявки на патент США № 20180087072. В обозначениях, используемых в Таблице 1, «Qm5» означает положение минус 5 (по отношению к спирали, которая пронумерована от -1 до +6) указанного пальца, где аргинин в этом положении заменен на глутамин (Q), а «Qml4» означает, что аргинин (R), обычно присутствующий в положении минус 14, был заменен на глутамин (Q). Аббревиатура «n», как в nQm5, означает, что в N-концевом пальце модуля из двух пальцев, используемого для конструирования белка с 5 или 6 пальцами, присутствует мутация. «Нет» означает отсутствие изменений за пределами области спирали распознавания.
Таблица 1: Конструкции ««цинковые пальцы»
R416E
ctCCTGAAAGTGGCCGGg tttaatctgc
(SEQ ID NO:40)
R416E
agGATTCGGAACCCAATC ACtgacaggt (SEQ ID NO:39)
K525S
(SEQ ID NO:40)
ggAAGGCCcCAGCAGGC AAGGgctgcat (SEQ ID NO:50)
gaGGAGGTgGCAGAGGG TACCccagccc (SEQ ID NO:51)
[0230] Были протестированы все ZFN на их связывание и было обнаружено, что они связываются с сайтами-мишенями и являются активными как нуклеазы.
[0231] Описанные здесь ZFP могут также включать одну или более мутаций в фосфатных контактирующих остатках белка «цинковый палец» и/или домена FokI, например, мутации NR-5Qabc (в остове ZFP) и/или мутации R416S и/или K525S (в FokI), описанные в публикации заявки на патент США № 20180087072. Так, например, ZFN SBS#68812 и SBS#68813 содержат мутации в остове ZFP и в домене FokI, снижающие неспецифические фосфатные контакты с остовом ДНК, и эти мутации указаны в Таблице 1.
[0232] Таким образом, описанные здесь нуклеазы (например, нуклеазы, содержащие ZFP) связываются с их сайтами-мишенями и расщепляют ген TCRA с образованием генетических модификаций в гене TCRA, содержащем любые из SEQ ID NO:1-2, включая модификации (инсерции и/или делеции) в любой из этих последовательностей и/или рядом с ними (например, в последовательностях-мишенях, представленных в любой из SEQ ID NO: 1-2 и/или между спаренными сайтами-мишенями) и/или модификации в TTGAAA экзона 2. Описанные здесь нуклеазы (например, нуклеазы, содержащие ZFP) связываются с их сайтами-мишенями и расщепляют ген B2M с образованием генетических модификаций в гене B2M, содержащем любые из SEQ ID NO: 3-4, включая модификации (инсерции и/или делеции) в любой из этих последовательностей и/или рядом с ними (например, в последовательностях-мишенях, представленных в любой из SEQ ID NO: 3-4 и/или между спаренными сайтами-мишенями) и/или модификации в GCCTTA экзона 1. Описанные здесь нуклеазы (например, нуклеазы, содержащие ZFP) связываются с их сайтами-мишенями и расщепляют ген HPRT с образованием генетических модификаций (например, инсерции CAR-кодирующего трансгена) в гене HPRT, содержащем любые из SEQ ID NO: 5-6, включая модификации (инсерции и/или делеции) в любой из этих последовательностей и/или рядом с ними (например, в последовательностях-мишенях, представленных в любой из SEQ ID NO: 5 или 6 и/или между спаренными сайтами-мишенями) и/или модификации в интроне 1, включая модификацию, в которой трансген экспрессируются из эндогенного промотора HPRT (но ген HPRT не экспрессируется). См. фигуру 1.
[0233] Кроме того, ДНК-связывающие домены (ZFP), которые связываются с их сайтами-мишенями, и ДНК-связывающие домены ZFP, TALE и оцРНК, которые распознают эти сайты-мишени (сайты-мишени, которые имеют длину 12 или более нуклеотидов в последовательностях-мишенях, как показано в Таблице 1), также вводят в активные сконструированные факторы транскрипции, если они связаны с одним или более доменами регуляции транскрипции.
Пример 2: Конструирование CD19-несущих клеток К562
[0234] Вкратце, нормальные клетки K562 культивировали в среде RPMI с 10% FBS, а затем подвергали электропорации в устройстве для электропорации Amaxa в присутствии 40 мкг/мл мРНК, кодирующей HPRT-специфические ZFN (37706/48407), нацеленные на интрон 1 человеческого гена HPRT, вместе с 50 мкг/мл плазмиды, содержащей кластер для экспрессии человеческого CD19 (ген NCBI ID:930, оптимизированный по кодонам), активируемый промотором EF1a, и гомологичные ветви, фланкирующие сайт расщепления ZFN в гене HPRT. Клетки оставляли на ночь для восстановления в среде после электропорации, а затем добавляли 6-тиогуанин (6-TG) в концентрации 6 мкМ, и клетки культивировали в течение 27 дней. Для оценки экспрессии CD19, клетки окрашивали РЕ-Cy7-конъюгированным антителом, нацеленным на человеческий внеклеточный домен CD19. Немодифицированные («необученные») K562 культивировали аналогичным образом и окрашивали как негативный контроль. Результаты (фигура 2) показали, что клетки, которые имели трансген CD19, окрашивались антителом.
Пример 3: Антиген-специфическое уничтожение клеток под действием CD19-CAR-Т-клеток
[0235] Затем был проведен эксперимент для получения клеток с нацеленной инсерцией экспрессионного кластера CAR GFP или CD19 CAR (Kochenderfer et al., (2009) J. Immunother 32 (7): 689-702) в локус TCRA или B2M. TCRA-специфическую пару ZFN SBS#55266/SBS# 53853 или пару B2M SBS#7071/SBS#57531 (все они показаны в Таблице 1) вводили в Т-клетки или клетки К562 в виде мРНК посредством электропорации, вместе с векторами AAV6, кодирующими кластер для экспрессии трансгена (GFP или CD19-CAR), фланкированный гомологичными ветвями в локусах расщепления TRAC или B2M для нацеленной инсерции, опосредуемой гомологией.
[0236] Вкратце, человеческие CD4:CD8-Т-клетки в отношении 1:1 оттаивали и активировали сферами CD3/28 Dynabeads® (в отношении клетки:сферы=1:3) в среде для культивирования клеток X-vivo15 (день 0). После культивирования в течение 3 дней (день 3), клетки концентрировали до 37 клеток/мл в буфере для электропорации Maxcyte в присутствии мРНК ZFN (TCRA или B2M ZFN) и подвергали электропорации на устройстве Maxcyte. Затем концентрированные клетки, подвергнутые электропорации, помещали в лунку для культивирования ткани, и к концентрированным клеткам добавляли AAV6-векторы, содержащие трансгенные доноры в совместимом локусе (например, донорные AAV6-векторы с TCRA-гомологичными ветвями добавляли к TCRA ZFN-обрабатанным клеткам). Кластеры для экспрессии трансгенов состояли из гомологичных ветвей (HA), специфичных к сайту расщепления (TCRA или B2M), промотора hPGK, активирующего трансген (либо GFP, либо CD19 CAR), и поли-А-последовательности BGH. После добавления AAV6-векторов, клетки оставляли для восстановления на 20 минут при 37°C. Затем клетки разводили в культуральной среде до плотности 36 клеток/мл, и культивировали при 30°C в течение ночи (см. публикацию заявки на патент США № 20170137845). На следующее утро, клетки разводили до 0,56 клеток/мл в дополнительной культуральной среде. Были получены клеточные популяции, включающие нижеследующие комбинации ZFN и доноров:
(а) Нетрансфецированные: клетки, подвергнутые электропорации без добавления мРНК ZFN или донора AAV6;
(b) клетки, подвергнутые электропорации с использованием только мРНК B2M ZFN, без добавления донора AAV6;
(с) клетки, подвергнутые электропорации с использованием только мРНК TCRA ZFN, без добавления донора AAV6;
(d) клетки, подвергнутые электропорации с использованием мРНК B2M ZFN и AAV6 B2M HA; c добавлением донора hPGK-eGFP-BGH-поли-А;
(е) клетки, подвергнутые электропорации с использованием мРНК TCRA ZFN и AAV6 TCRA HA, c добавлением донора hPGK-eGFP-BGH-поли-А;
(f) клетки, подвергнутые электропорации с использованием мРНК B2M ZFN и AAV6 B2M HA, c добавлением донора hPGK-FMC63-CD8BBZ (CD-19 CAR)-BGH-поли-А;
(е) клетки, подвергнутые электропорации с использованием мРНК TCRA ZFN и AAV6 TCRA HA, c добавлением донора hPGK-FMC63-CD8BBZ (CD-19 CAR)-BGH-поли-А.
[0237] Как было отмечено выше, указанные выше аббревиатуры означают: HA=гомологичная ветвь; B2M=ген B2M; PGK=промотор PGK; BGH=последовательность поли-А гена бычьего гормона роста (BGH); FMC63 представляет собой последовательность анти-CD19 ScFv, взятую из NCI (см. патент США № 9701758); CD8BBZ означает части CAR, за исключением scFv=шарнирный/трансмембранный домен CD8 (CD8), костимулирующего домена гена 41BB (BB), домена активации гена CD3z (Z).
[0238] Все эксперименты проводили при плотности 37 клеток/мл в процессе электропорации. На следующий день (день 4), клетки разводили до 0,56 клеток/мл и переносили в культуры при 37°C. Через три дня (на день 7), клетки снова разводили до 0,56 клеток/мл. Еще через три и семь дней культивирования (дни 10 и 14, соответственно), клетки собирали для проведения FACS- и MiSeq-анализа (клетки разводили до 0,56 клеток/мл). Данные представлены ниже в Таблице 2.
Таблица 2: Активность ZFN
[0239] Затем была измерена функциональная активность. Сначала, смесь 50:50 получали с использованием «необученных» клеток К562, смешанных с CD19-экспрессирующими клетками К562 Примера 2. Эту смесь обрабатывали либо нетрансфецированными Т-клетками, либо T-клетками с нокаутом TCRA. Результаты показаны на фигуре 3, где на фигурах 3А-3C показана только смесь, обработанная «необученными» T-клетками или T-клетками с нокаутом TCRA. Клетки окрашивали CD19-специфическими антителами, и было обнаружено, что добавление T-клеток не оказывало какого-либо влияния на популяцию клеток К562.
[0240] Затем, аналогичную 50:50-смесь клеток-мишеней обрабатывали Т-клетками, несущими трансген CAR, встроенный в локус B2M или TCRA. Популяция «необученных» клеток и CD19+-мишеней К562 обрабатывали смесью «эффектор:клетка-мишень» в пределах от 2:1 до 0,125, включительно. Результаты показали, что встраивание CAR в гены B2M или TCRA приводит к экспрессии функционального CAR (см. 4А-4В). Для обеих популяций CAR-T-клеток, взятых в отношении «эффектор:мишень» = 2:1, почти все CD19+-клетки К562 были уничтожены (0,4% и 0,1% CD19+-клеток К562 оставались после обработки Т-клетами, в которые был встроен CAR в гене B2M или TCRA, соответственно).
[0241] И наконец, был построен график зависимости процента оставшихся CD19+-клеток К562 от отношения «эффектор:клетка-мишень» (фигура 5), и на этом графике продемонстрировано, что CAR-Т-клетки обоих типов были активными и эффективными для уничтожения клеток, экспрессирующих антиген-мишень.
[0242] Второй эксперимент проводили, как описано выше, и условия этого эксперимента были такими же, за исключением того, что клетки обрабатывали обеими TCRA- и B2M-специфическими парами ZFN (с двойным нокаутом) и трансгенным донором CD19-CAR, содержащим TCRA-специфические гомологичные ветви. Данные представлены ниже в Таблице 3 и на фигуре 6.
Таблица 3: Двойное расщепление TCRA и B2M и ZFN-опосредованное встраивание CD19-CAR в локус TCRA
[0243] Третий эксперимент проводили как описано выше, и в этом эксперименте, генотип и фенотип Т-клеток, обработанных TRAC-специфическими нуклеазами, также оценивали после введения различных доз TRAC-нуклеаз. Как показано на фигуре 7 и в Таблице 4, опосредуемая нуклеазой модификация TRAC давала >95% клеток с генотипом TRAC-нокаута (инсерции/делеции TRAC) и CD3--фенотипом (CD3-негативным).
Таблица 4: Титрование дозы TRAC с нокаутом
[0244] Кроме того, как показано на фигуре 8, FACS-анализ показал, что опосредуемая нуклеазой инактивация (нокаут) TRAC последовательно достигалась по меньшей мере при 99,5% потере поверхностного TCR (CD3) без проведения отбора. Аналогичным образом, как показано на фигуре 9, трансгены (GFP), интегрированные в TRAC (левая панель) или в B2M (правая панель) посредством специфического расщепления этих генов нуклеазой, давали высокоэффективную экспрессию трансгена (93% для TRAC и 91% для B2M) как было определено с помощью FACS-анализа. В этом эксперименте, в отсутствие донора, 93% клеток были нокаутированы в обоих локусах TRAC и B2M.
[0245] Т-клетки здорового донора также обрабатывали TRAC- и B2M-нуклеазами (для инактивации этих генов) в комбинации с донором AAV, содержащим трансген CAR (CD19), нацеленный на ген TRAC (гомологичные ветви), за одну стадию (совместного введения TRAC и B2M и донора). Как показано на фигуре 10, высокоэффективная экспрессия CAR (>75% клеток) и TRAC/B2M-нокаута (>94% и >88%, соответственно) была достигнута за одну стадию редактирования.
[0246] Четвертая серия экспериментов была проведена для оценки эффективности «мультиплексного» нокаута множества генетических локусов в Т-клетках в комбинации с введением донора GFP и/или CD19 CAR. В первом эксперименте была протестирована эффективность интеграции двух донорных трансгенов посредством расщепления в TRAC (с использованием SBS68812/SBS68813) и B2M (с использованием SBS57071/SBS57531). В этом эксперименте, донор CD19 CAR, содержащий гомологичные ветви TRAC, был введен в клетки посредством трансдукции AAV. Донор GFP состоял из гомологичных ветвей B2M и был введен в клетки посредством трансдукции AAV.
[0247] Как показано на фигуре 11, гены-мишени расщеплялись с эффективностью более, чем 90%, и при добавлении доноров, нацеленная интеграция достигалась на уровне 90% для донора GFP и 77% для донора CD19 CAR. Результаты по интеграции CD19 CAR соответствовали результатам, представленным на фигуре 10С, где был использован только трансген CD19 CAR. Таким образом, эффективность интеграции CD19 CAR в расщепленном локусе TRAC не зависит от одновременной интеграции трансгена GFP в расщепленном локусе В2В, и наоборот.
[0248] В другом эксперименте были использованы реагенты TRAC SBS68812/SBS68813 и реагенты B2M ZFN SBS57071/SBS57531. Был также использован третий набор реагентов ZFN (SBS59488/SBS59489), нацеленных на локус CISH (ген контрольной точки, см. предварительную заявку на патент США 62/583724), как показано выше в Таблице 1.
[0249] CISH-специфические реагенты были использованы в комбинации с TRAC- и B2M-специфическими ZFN в описанных выше способах. Клетки были также обработаны, помимо различных пар ZFN, донорным ААВ-GFP, и полученные данные по нокауту и интеграции приводятся на фигуре 12А. Для клеток, обработанных донором GFP, содержащим гомологичные ветви TRAC, и множеством различных нуклеаз, данные по нокауту и интеграции донора представлены ниже в Таблице 5. С использованием данных по эффективности донора трансгена CD19 CAR, полученные в предыдущем эксперименте (как показано на фигуре 11), была оценена предполагаемая эффективность интеграции трансгена CD19 CAR в локус TRAC, и данные этой оценки представлены на фигуре 12В и в Таблице 5.
Таблица 5: Мультиплексное редактирование генов и нацеленная интеграция
k/o
k/o
k/o
(оцененный)
* Предполагаемая эффективность для CD19 CAR исходя из достигнутой ранее эффективности нацеленной интеграции CD19 CAR в TRAC, как показано на фигуре 11.
[0250] Таким образом, как показано в настоящем описании, CAR+-модифицированные клетки были получены в больших количествах с применением описанных здесь способов.
Пример 4: Конструирование клеток так, чтобы они экспрессировали CAR и модифицированные HLA-E и/или HLA-G
[0251] Клетки, экспрессирующие CD19 CAR и модифицированный комплекс HLA-E и/или HLA-G, получали методами, описанными в Примере 3. Вкратце, TCRA-специфическую пару ZFN SBS#55266/SBS# 53853 или пару B2M SBS#7071/SBS#57531 (все они показаны в Таблице 1) вводили в Т-клетки в виде мРНК посредством электропорации, вместе с векторами AAV6, кодирующими кластер для экспрессии трансгена (GFP или CD19-CAR или один или оба CD19-HLA-E и CD19-HLA-G)), фланкированного гомологичными ветвями в локусах расщепления TRAC или B2M для нацеленной инсерции, опосредуемой гомологией.
[0252] Эти клетки охарактеризовывали на CD19-специфическую экспрессию CAR и экспрессию HLA-Е или HLA-G с помощью FACS-анализа и с использованием меченных антител, специфичных к этим двум белкам клеточной поверхности. FACS-анализ показал, что эти клетки экспрессируют CD19-специфический CAR и модифицированный комплекс HLA-E и/или HLA-G. Активность CD19-специфического CAR измеряли как описано выше, и было обнаружено, что клетки подвергались CD19 CAR-опосредуемому уничтожению. Затем клетки анализировали для мониторинга уничтожения клеток NK-клетками с применением известных методов (например, с помощью анализов на высвобождение хрома и т.п.). Клетки, содержащие CD19 CAR и модифицированный комплекс HLA-E или HLA-G, были более резистентными к NК-опосредуемому уничтожению, чем клетки, не содержащие модифицированного комплекса HLA-E или HLA-G.
[0253] Были использованы пары ZFN, нацеленные на белок- кодирующие области локусов TRAC и B2M (SBS#68877/SBS#68876 для TRAC и SBS#7071/SBS#57531 для B2M). Вкратце, человеческие CD4:CD8-Т-клетки в отношении 1:1 оттаивали и активировали сферами CD3/28 Dynabeads® (в отношении клетки;сферы=1:3) в среде для культивирования клеток X-vivo15 (день 0). После культивирования в течение 3 дней (день 3), клетки концентрировали до 37 клеток/мл в буфере электропорации Maxcyte в присутствии мРНК ZFN и подвергали электропорации на устройстве Maxcyte. Затем концентрированные клетки, подвергнутые электропорации, помещали в лунку для культивирования ткани, и к концентрированным клеткам добавляли AAV6, содержащий трансгенный донор, и эти клетки оставляли для восстановления и инкубировали при 37°C в течение 20 минут. Затем клетки разводили в культуральной среде до плотности 36 клеток/мл и культивировали при 30°C в течение ночи. На следующее утро, клетки разводили до 0,56 клеток/мл в дополнительной культуральной среде.
[0254] Ниже приводится описание групп (все ZFN подвергали электропорации при 60 мкг/мл мРНК на ZFN, и добавляли донор AAV6 при 105 вирусных геномов/клетку). Гибридный трансген AAV B2M-HLA-G был активирован промотором PGK в присутствии гомологичных ветвей TRAC, экспрессирующих гибридный трансген B2M-HLA-G. Вкратце, трансген B2M-HLA-G включает полноразмерный ген B2M, содержащий лидерную последовательность сигнального пептида, но не имеющий стоп-кодона у своего конца, а за этой последовательностью находится (G4S)4-линкер, за которым следует полноразмерная последовательность HLA-G*01:01 без лидерного сигнального пептида. Весь гибридный кластер B2M-HLA-G был оптимизирован по кодонам перед синтезом (фигура 13).
[0255] Эксперимент проводили при плотности 37 клеток/мл в процессе электропорации, как описано в публикации заявки на патент США 20170137845, и подвергали холодовому шоку при 30°C в течение ночи после электропорации. На следующий день (день 4), клетки разводили до 0,56 клеток/мл и переносили в культуры при 37°C. Через три дня (день 7), клетки снова разводили до 0,56 клеток/мл. На день 10, клетки собирали для проведения FACS-анализа (клетки разводили до 0,56 клеток/мл).
[0256] Для проведения FACS-анализа, окрашивание клеточной поверхности осуществляли в 100 мкл окрашивающего буфера (PBS с 1% BSA и 0,02% NaN3), содержащего антитела, конъюгированные с флуорохромом и специфичные к каждому оцениваемому антигену клеточной поверхности. «Коктейли» антител получали как подробно показано ниже в Таблице 6 на панели 1 или 2. Клетки ресуспендировали в любом коктейле антител, инкубировали в темноте в течение 30 минут и два раза промывали PBS, а затем осуществляли сбор данных на проточном цитометре.
[0257] Для внутриклеточного окрашивания, клетки сначала ресуспендировали в 100 мкл раствора Cytofix/Cytoperm™ и инкубировали в течение 20 минут при 4°C. После двухкратной промывки клеток 1× раствором для сообщения проницаемости/промывочным раствором, фиксированные клетки ресуспендировали в любом коктейле антител в течение 30 минут в темноте. Клетки два раза промывали 1 × раствором для сообщения проницаемости/промывочным раствором™, и проводили сбор данных на цитометре.
Таблица 6: Используемые коктейли антител
[0258] Как показано на фигуре 14, гибридный белок B2M-HLA-G экспрессировался только внутри клеток, а на поверхности Т-клеток в эндогенных локусах B2M и TRAC - не экспрессировался. На фигуре 14A показано, что ZFN-опосредованный нокаут (КО) TRAC приводил к 97% потере экспрессии поверхностного CD3 (левая панель), а нокаут B2M приводил к 81% потере молекул HLA класса I на поверхности (HLA-ABC, правая панель), на что указывала проточная цитометрия. Светло-серые пики соответствовали ложнообработанным или необработанным Т-клеткам, а темно-серые пики соответствовали ZFN-обработанным клеткам. Т-клетки с двойным нокаутом были трансдуцированы донорным AAV6, экспрессирующим описанный выше трансген B2M-HLA-G. На фигурах 14В и 14С проиллюстрирована успешная экспрессия B2M и HLA-G в Т-клетках с двойным нокаутом, которые были трансдуцированы донорным AAV6, экспрессирующим гибридный трансген B2M-HLA-G, для нацеленной интеграции в локус TRAC. В частности, поскольку HLA-G обычно не экспрессируется на Т-клетках, то наличие темно-серого пика HLA-G на фиг. 14В и 14С продемонстрировало позитивную интеграцию трансгена B2M-HLA-G и его экспрессию, как внутри клеток, так и на клеточной поверхности (левые панели). Светло-серые пики соответствуют клеткам, которые имели двойной нокаут для CD3 и В2М, но не были трансдуцированы донорным AAV. Экспрессия трансгена B2M-HLA-G, также подтверждалась наличием светло-серых пиков B2M на фиг. 14В и 14С, на правых панелях, где B2M экспрессировался в результате интеграции конструкции B2M-HLA-G в клетках, не содержащих эндогенного B2M, а сигнал детектировался как внутри клеток, так и на их поверхности. Серые пики с более темной окраской соответствуют клеткам, не экспрессирующим B2M.
[0259] Таким образом, была достигнута эффективная модификация клеток, опосредуемая нуклеазой.
Пример 5: Нацеленная интеграция
А. B2M
[0260] Была использована пара ZFN, нацеленная на B2M, SBS#57071/SBS#57531 и на белок-кодирующую область локуса В2М, для нацеленной интеграции различных донорных конструкций, описанных ниже. Вкратце, человеческие CD4:CD8-Т-клетки в отношении 1:1 оттаивали и активировали сферами CD3/28 Dynabeads® (в отношении клетки:сферы=1:3) в среде для культивирования клеток X-VIVO15™, содержащей IL-2 (день 0, Lonza BioWhittaker). После культивирования в течение 3 дней (день 3), клетки концентрировали до плотности 37 клеток/мл в буфере для электропорации Maxcyte в присутствии мРНК ZFN и подвергали электропорации на устройстве Maxcyte. Затем концентрированные клетки, подвергнутые электропорации, помещали в лунку для культивирования ткани, и к концентрированным клеткам добавляли AAV6, содержащий трансгенный донор, а затем, эти клетки оставляли для восстановления и инкубировали при 37°C в течение 20 минут. Используемые донорные конструкции представлены ниже:
(1) донор с длинными гомологичными ветвями (длинными ветвями), содержащими следующие последовательности: последовательность промотора PGK, функционально присоединенную к трансгену GFP, фланкированному гомологичными ветвями (нацеленными на B2M) размером 1 т.п.о. каждая;
(2) донор с короткими гомологичными ветвями (короткими ветвями), содержащими следующие последовательности: последовательность промотора PGK, функционально присоединенную к трансгену GFP, фланкированному гомологичными ветвями (нацеленными на B2M) размером 250 п.о. каждая;
(3) донор (2), дополнительно содержащий последовательность WPRE в 3’-UTR, например, последовательность WPRE, включающую:
AATCAACCTCTGGATTACAAAATTTGTGAAAGATTGACTGGTATTCTTAACTATGTTGCTCCTTTTACGCTATGTGGATACGCTGCTTTAATGCCTTTGTATCATGCTATTGCTTCCCGTATGGCTTTCATTTTCTCCTCCTTGTATAAATCCTGGTTGCTGTCTCTTTATGAGGAGTTGTGGCCCGTTGTCAGGCAACGTGGCGTGGTGTGCACTGTGTTTGCTGACGCAACCCCCACTGGTTGGGGCATTGCCACCACCTGTCAGCTCCTTTCCGGGACTTTCGCTTTCCCCCTCCCTATTGCCACGGCGGAACTCATCGCCGCCTGCCTTGCCCGCTGCTGGACAGGGGCTCGGCTGTTGGGCACTGACAATTCCGTGGTGTTGTCGGGGAAGCTGACGTCCTTTCCATGGCTGCTCGCCTGTGTTGCCACCTGGATTCTGCGCGGGACGTCCTTCTGCTACGTCCCTTCGGCCCTCAATCCAGCGGACCTTCCTTCCCGCGGCCTGCTGCCGGCTCTGCGGCCTCTTCCGCGTCTTCGCCTTCGCCCTCAGACGAGTCGGATCTCCCTTTGGGCCGCCTCCCCGCCTG (SEQ ID NO:64);
(4) донор (2), дополнительно содержащий энхансерную T-клеточную последовательность, расположенную выше промотора PGK, то есть, последовательность, представленную ниже:
GATATCATTGGTCAATTGCAGCAGACATGTCTCTTGTGGTGTAAAGCCTCATAGCATCTTCCTGCTGGAGTCCACCTGGCAGCCCAAAGCCTCAGCAAGTGCTGGGGACAGAGAGATAGAAAGGCTTTGCTGTGGCTCAACTCAGTTGTACTTTCTCAAGCTCCTTTTGGTTATCTGCTCTGTCTATGTAGTTGCTTCCTGGAAAATGGTTAAAGTCAAACCTCCTCTGTTTGGGGCAGCACAAAATGTTGCTAAGGCGTTAGAAGACAACATAGGAGCAGTTAAACCACAGCCAGTTTTTGCTCGCTTTCGAAAGACCACAGCTATTAGACAGAAAACCTCCTGTACTGGAAGTGGCCCTGAAACAGCATCTGTGCTGACAGACACTGGGCTTGCTATGGTTGCTGCTCATCATGCTCAGCTG (SEQ ID NO:65); или
(5) донор (2), дополнительно включающий энхансерную последовательность TCRα, представленную ниже:
CTCCCATTTCCATGACGTCATGGTTAAACCTCTTTTACCTGCTTTAACCCTTTGAAGCTCTCCCGCAGAAGCCACATCCTCTG (SEQ ID NO:66);
(6) донор (2), дополнительно включающий последовательность 5’-нетранслируемой области гена бета-глобина Xenopus:
TGCTTGTTCTTTTTGCAGAAGCTCAGAATAAACGCTCAACTTTGGCAGAT (SEQ ID NO:67).
[0261] Затем клетки разводили в культуральной среде до 36 клеток/мл, и культивировали при 30°C в течение ночи.
[0262] Все ZFN подвергали электропорации при 60 мкг/мл мРНК на ZFN, и все доноры AAV6 добавляли при плотности 105 вирусных геномов/клетку. Все эксперименты проводили при плотности 37 клеток/мл в соответствии с протоколом, описанным в публикации заявки на патент США 20170137845 (в экстремальных условиях холодового шока), и культивировали в условиях холодового шока при 30°C в течение ночи после электропорации.
[0263] На следующий день (день 4), клетки разводили до 0,56 клеток/мл и переносили в культуры при 37°C. Через три дня (день 7), клетки снова разводили до 0,56 клеток/мл. Еще через три и семь дней культивирования (дни 10 и 14, соответственно), клетки собирали для проведения FACS- и MiSeq-анализа (клетки разводили до 0,56 клеток/мл).
[0264] Как указано на фигурах 15A и B, экспрессия GFP, показала, что нацеленная интеграция была проведена успешно, и были получены генетически модифицированные клетки, содержащие геномные модификации B2M (инсерции/делеции и TI) в описанном здесь сайте-мишени ZFN.
[0265] Самая высокая средняя интенсивность флуоресценции (MFI) наблюдалась в конструкциях, содержащих энхансер, причем, в этих конструкциях, такая интенсивность повышалась на 65% по сравнению со стандартной конструкцией, содержащей короткие гомологичные ветви.
B. Нацеленная интеграция донора в клетки с двойным нокаутом B2M/TRAC
[0266] Кроме того, эксперименты были также осуществлены как описано выше (за исключением того, что в качестве культуральной среды была использована среда RPMI с добавлением 10% человеческой сыворотки и IL-2) с использованием B2M и TRAC ZFN и различных доноров для создания TRAC/B2M с двумя нокаутами (DKO) и со встроенными трансгенами, включая интегрированные трансгены HLA-Е или HLA-G. Доноры подробно описаны выше на фигурах 17 и 19 и указаны ниже:
(1) донор, содержащий левую ветвь 423 п.о. и правую ветвь 393 п.о., гомологичные сайту TRAC-E, фланкирующему трансген GFP, активируемый промотором hPGK. Донор также содержит метку «miseq», позволяющую количественно определить уровень TI в локусе TRAC по miseq;
(2) донор, содержащий левую ветвь 1000 п.о. и правую ветвь 992 п.о., гомологичные сайту TRAC-E, фланкирующему трансген GFP, активируемый промотором hPGK. Донор не содержит метку «miseq»;
(3) донор, содержащий левую ветвь 350 п.о. и правую ветвь 393 п.о., гомологичные сайту TRAC-E, фланкирующему трансген GFP, активируемый промотором hPGK, и за этим трансгеном находится мутированный элемент WPRE. Донор также содержит метку «miseq»;
(4) донор, содержащий левую ветвь 423 п.о. и правую ветвь 393 п.о., гомологичные сайту TRAC-E, фланкирующему трансген FMC63-CD8BBZ CAR, активируемый промотором hPGK. Донор также содержит метку «miseq»;
(5) донор, содержащий левую ветвь 423 п.о. и правую ветвь 393 п.о., гомологичные сайту TRAC-E, фланкирующему трансген FMC63-CD8BBZ CAR, активируемый промотором hPGK, и за этим трансгеном находится мутированный элемент WPRE. Донор также содержит метку «miseq»;
(6) донор, содержащий левую ветвь 360 п.о. и правую ветвь 393 п.о., гомологичные сайту TRAC-E, фланкирующему трансген FMC63-CD8BBZ CAR, связанный посредством саморасщепляющегося пептида P2A с гибридным белком и экспрессирующий B2M и HLA-E. Трансген активируется промотором hPGK, и за этим трансгеном следует мутированный элемент WPRE. Донор также содержит метку «miseq»;
(7) донор, содержащий левую ветвь 360 п.о. и правую ветвь 393 п.о., гомологичные сайту TRAC-E, фланкирующему трансген FMC63-CD8BBZ CAR, связанный посредством саморасщепляющегося пептида P2A с гибридным белком и экспрессирующий B2M и HLA-G. Трансген активируется промотором hPGK, и за этим трансгеном следует мутированный элемент WPRE. Донор также содержит метку «miseq»;
(8) донор, содержащий левую ветвь 1000 п.о. и правую ветвь 992 п.о., гомологичные сайту TRAC-E, фланкирующему трансген HLA-G, активируемый промотором hPGK;
(9) донор, содержащий левую ветвь 1000 п.о. и правую ветвь 992 п.о., гомологичные сайту TRAC-E, фланкирующему трансген HLA-E0101, активируемый промотором hPGK;
(10) донор, содержащий левую ветвь 1000 п.о. и правую ветвь 992 п.о., гомологичные сайту TRAC-E, фланкирующему трансген HLA-E0103, активируемый промотором hPGK;
(11) донор, содержащий левую ветвь 1000 п.о. и правую ветвь 992 п.о., гомологичные сайту TRAC-E, фланкирующему трансген HLA-G, активируемый промотором hPGK;
(12) донор, содержащий левую ветвь 1000 п.о. и правую ветвь 992 п.о., гомологичные сайту TRAC-E, фланкирующему связанный трансген В2М HLA-G, активируемый промотором hPGK. Линкер между В2М и HLA-G содержит 2 копии пептида G4S. HLA-G не содержит сигнального пептида; донор, содержащий левую ветвь 1000 п.о. и правую ветвь 992 п.о., гомологичные сайту TRAC-E, фланкирующему связанный трансген В2М HLA-G, активируемый промотором hPGK. Линкер между В2М и HLA-G содержит 4 копии пептида G4S. HLA-G не содержит сигнального пептида;
(13) донор, содержащий левую ветвь 1000 п.о. и правую ветвь 992 п.о., гомологичные сайту TRAC-E, фланкирующему связанный трансген В2М HLA-G, активируемый промотором hPGK. Линкер между В2М и HLA-G содержит 6 копий пептида G4S. HLA-G не содержит сигнального пептида;
(14) донор, содержащий левую ветвь 1000 п.о. и правую ветвь 992 п.о., гомологичные сайту TRAC-E, фланкирующему трансген HLA-E0101, активируемый промотором hPGK;
(15) донор, содержащий левую ветвь 1000 п.о. и правую ветвь 992 п.о., гомологичные сайту TRAC-E, фланкирующему трансген HLA-E0103, активируемый промотором hPGK;
(16) донор, содержащий левую ветвь 1000 п.о. и правую ветвь 992 п.о., гомологичные сайту TRAC-E, фланкирующему связанный трансген В2М HLA-E0101, активируемый промотором hPGK. Линкер между В2М и HLA-E0101 содержит 4 копии пептида G4S. HLA-E0101 не содержит сигнального пептида (Celik et al., ibid);
(17) донор, содержащий левую ветвь 1000 п.о. и правую ветвь 992 п.о., гомологичные сайту TRAC-E, фланкирующему связанный трансген В2М HLA-E0101, активируемый промотором hPGK. Линкер между В2М и HLA-E0101 содержит 4 копии пептида G4S. HLA-E0103 не содержит сигнального пептида;
(18) донор, содержащий левую ветвь 1000 п.о. и правую ветвь 992 п.о., гомологичные сайту TRAC-E, фланкирующему трансген GFP, активируемый промотором hPGK;
(19) донор, содержащий левую ветвь 360 п.о. и правую ветвь 393 п.о., гомологичные сайту TRAC-E, фланкирующему трансген FMC63-CD8BBZ CAR, связанный посредством саморасщепляющегося пептида P2A с гибридным белком и экспрессирующий B2M и HLA-G. Трансген активируется промотором hPGK, и за этим трансгеном следует мутированный элемент WPRE. Донор также содержит метку «miseq».
[0267] Результаты, полученные с использованием указанных репрезентативных доноров, представлены на фигурах 17 и 19, где проиллюстрирована опосредуемая нуклеазой нацеленная интеграция, позволяющая эффективно встраивать трансген донора. На фигуре 18 показаны результаты проточного цитометрического анализа указанных образцов, представленных на фигуре 17. На фигурах 20-26 показаны результаты для указанных доноров (фигура 19).
[0268] Как уже указывалось ранее, нацеленая интеграция приводила к экспрессии белка, включая экспрессию в CAR+-клетках (нацеленных на ген TCR), также экспрессирующих гибридные белки B2M-HLA-E или HLA-G.
[0269] Все упомянутые здесь патенты, патентные заявки и публикации включены в настоящее описание посредством ссылки в полном объеме.
[0270] Хотя раскрытие изобретения представлено здесь в некоторых деталях, например, для иллюстрации и для лучшего понимания изобретения, однако, специалистам в данной области очевидно, что в настоящее изобретение могут быть внесены различные изменения и модификации, не выходящие за рамки существа и объема настоящего изобретения. В соответствии с этим, вышеприведенное описание и последующие примеры не должны быть истолкованы как ограничивающие.
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> SANGAMO THERAPEUTICS, INC.
<120> НАЦЕЛИВАНИЕ НА КЛЕТКИ, ОПОСРЕДУЕМОЕ ХИМЕРНЫМ
АНТИГЕННЫМ РЕЦЕПТОРОМ
<130> 8325-0165.40
<140> PCT/US2018/045759
<141> 2018-08-08
<150> 62/652,672
<151> 2018-04-04
<150> 62/613,258
<151> 2018-01-03
<150> 62/581,290
<151> 2017-11-03
<150> 62/542,511
<151> 2017-08-08
<160> 70
<170> PatentIn version 3.5
<210> 1
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 1
tcaagctggt cgagaaaagc tttgaaac 28
<210> 2
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 2
aacaggtaag acaggggtct agcctggg 28
<210> 3
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 3
gccacggagc gagacatctc ggcccgaa 28
<210> 4
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 4
gagtagcgcg agcacagcta aggccacg 28
<210> 5
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 5
acccagatac aaacaatgga tagaaaac 28
<210> 6
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 6
tgcaaaaggt aggaaaagga ccaaccag 28
<210> 7
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 7
Gln Ser Ser Asp Leu Ser Arg
1 5
<210> 8
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 8
Gln Ser Gly Asn Arg Thr Thr
1 5
<210> 9
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 9
Arg Ser Ala Asn Leu Ala Arg
1 5
<210> 10
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 10
Asp Arg Ser Ala Leu Ala Arg
1 5
<210> 11
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 11
Arg Ser Asp Val Leu Ser Glu
1 5
<210> 12
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 12
Lys His Ser Thr Arg Arg Val
1 5
<210> 13
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 13
Thr Met His Gln Arg Val Glu
1 5
<210> 14
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 14
Thr Ser Gly His Leu Ser Arg
1 5
<210> 15
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 15
Arg Ser Asp His Leu Thr Gln
1 5
<210> 16
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 16
Asp Ser Ala Asn Leu Ser Arg
1 5
<210> 17
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 17
Gln Ser Gly Ser Leu Thr Arg
1 5
<210> 18
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 18
Ala Lys Trp Asn Leu Asp Ala
1 5
<210> 19
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 19
Arg Ser Asp Val Leu Ser Ala
1 5
<210> 20
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 20
Gln Asn Ala Thr Arg Ile Asn
1 5
<210> 21
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 21
Gln Ser Gly Asp Leu Thr Arg
1 5
<210> 22
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 22
Thr Ser Gly Asn Leu Thr Arg
1 5
<210> 23
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 23
Gln Ser Asn Asp Leu Asn Ser
1 5
<210> 24
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 24
Arg Ser Asp Asp Leu Ser Lys
1 5
<210> 25
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 25
Asp Ser Ser Ala Arg Lys Lys
1 5
<210> 26
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 26
Asp Arg Ser Asn Leu Ser Arg
1 5
<210> 27
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 27
Gln Arg Thr His Leu Arg Asp
1 5
<210> 28
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 28
Gln Ser Gly His Leu Ala Arg
1 5
<210> 29
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 29
Asp Ser Ser Asn Arg Glu Ala
1 5
<210> 30
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 30
Ala Gln Cys Cys Leu Phe His
1 5
<210> 31
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 31
Asp Gln Ser Asn Leu Arg Ala
1 5
<210> 32
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 32
Arg Ser Ala Asn Leu Thr Arg
1 5
<210> 33
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 33
Arg Ser Asp Asp Leu Thr Arg
1 5
<210> 34
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 34
Arg Ser Asp Asn Leu Ser Val
1 5
<210> 35
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 35
Arg Ser Asp Tyr Leu Ala Lys
1 5
<210> 36
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 36
Gln Asn Ala Asn Arg Ile Thr
1 5
<210> 37
<211> 12
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 37
Ser Gly Thr Pro His Glu Val Gly Val Tyr Thr Leu
1 5 10
<210> 38
<211> 12
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 38
Ser Gly Ala Ile Arg Cys His Asp Glu Phe Trp Phe
1 5 10
<210> 39
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотида
<400> 39
aggattcgga acccaatcac tgacaggt 28
<210> 40
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотида
<400> 40
ctcctgaaag tggccgggtt taatctgc 28
<210> 41
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 41
Thr Ser Ser Asn Arg Lys Thr
1 5
<210> 42
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 42
Leu Gln Gln Thr Leu Ala Asp
1 5
<210> 43
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 43
Gln Ser Gly Asn Leu Ala Arg
1 5
<210> 44
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 44
Arg Arg Glu Asp Leu Ile Thr
1 5
<210> 45
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 45
Thr Ser Ser Asn Leu Ser Arg
1 5
<210> 46
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 46
Arg Ser Asp His Leu Ser Thr
1 5
<210> 47
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 47
Asp Arg Ser His Leu Ala Arg
1 5
<210> 48
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 48
Leu Lys Gln His Leu Asn Glu
1 5
<210> 49
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 49
His Asn Ser Ser Leu Lys Asp
1 5
<210> 50
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотида
<400> 50
ggaaggcccc agcaggcaag ggctgcat 28
<210> 51
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотида
<400> 51
gaggaggtgg cagagggtac cccagccc 28
<210> 52
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 52
Arg Ser Asp His Leu Ser Gln
1 5
<210> 53
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 53
Gln Asn Ala Thr Arg Thr Lys
1 5
<210> 54
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 54
Arg Ser Asp Asn Leu Ser Glu
1 5
<210> 55
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 55
Lys Arg Cys Asn Leu Arg Cys
1 5
<210> 56
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 56
Asp Arg Ser Thr Arg Thr Lys
1 5
<210> 57
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 57
Arg Arg Asp Asn Leu His Ser
1 5
<210> 58
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 58
Gly His Thr Ser Leu Lys Arg
1 5
<210> 59
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 59
Arg Ser Asp Asn Leu Ala Arg
1 5
<210> 60
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 60
Gln Asn Val Ser Arg Pro Arg
1 5
<210> 61
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 61
Gln Ser Gly His Leu Ser Arg
1 5
<210> 62
<211> 100
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полинуклеотид
<400> 62
aagcccataa ccgctgtggc ctcttggttt tacagatacg aacctaaact ttcaaaacct 60
gtcagtgatt gggttccgaa tcctcctcct gaaagtggcc 100
<210> 63
<211> 9
<212> PRT
<213> Неизвестный
<220>
<223> Описание неизвестного элемента: последовательность
мотива пептидного семейства "LAGLIDADG"
<400> 63
Leu Ala Gly Leu Ile Asp Ala Asp Gly
1 5
<210> 64
<211> 592
<212> ДНК
<213> Вирус гепатита сурка
<400> 64
aatcaacctc tggattacaa aatttgtgaa agattgactg gtattcttaa ctatgttgct 60
ccttttacgc tatgtggata cgctgcttta atgcctttgt atcatgctat tgcttcccgt 120
atggctttca ttttctcctc cttgtataaa tcctggttgc tgtctcttta tgaggagttg 180
tggcccgttg tcaggcaacg tggcgtggtg tgcactgtgt ttgctgacgc aacccccact 240
ggttggggca ttgccaccac ctgtcagctc ctttccggga ctttcgcttt ccccctccct 300
attgccacgg cggaactcat cgccgcctgc cttgcccgct gctggacagg ggctcggctg 360
ttgggcactg acaattccgt ggtgttgtcg gggaagctga cgtcctttcc atggctgctc 420
gcctgtgttg ccacctggat tctgcgcggg acgtccttct gctacgtccc ttcggccctc 480
aatccagcgg accttccttc ccgcggcctg ctgccggctc tgcggcctct tccgcgtctt 540
cgccttcgcc ctcagacgag tcggatctcc ctttgggccg cctccccgcc tg 592
<210> 65
<211> 424
<212> ДНК
<213> Неизвестный
<220>
<223> Описание неизвестного элемента: последовательность
энхансера T-клеточного рецептора
<400> 65
gatatcattg gtcaattgca gcagacatgt ctcttgtggt gtaaagcctc atagcatctt 60
cctgctggag tccacctggc agcccaaagc ctcagcaagt gctggggaca gagagataga 120
aaggctttgc tgtggctcaa ctcagttgta ctttctcaag ctccttttgg ttatctgctc 180
tgtctatgta gttgcttcct ggaaaatggt taaagtcaaa cctcctctgt ttggggcagc 240
acaaaatgtt gctaaggcgt tagaagacaa cataggagca gttaaaccac agccagtttt 300
tgctcgcttt cgaaagacca cagctattag acagaaaacc tcctgtactg gaagtggccc 360
tgaaacagca tctgtgctga cagacactgg gcttgctatg gttgctgctc atcatgctca 420
gctg 424
<210> 66
<211> 83
<212> ДНК
<213> Неизвестный
<220>
<223> Описание неизвестного элемента: альфа-последовательность
энхансера Т-клеточного рецептора
<400> 66
ctcccatttc catgacgtca tggttaaacc tcttttacct gctttaaccc tttgaagctc 60
tcccgcagaa gccacatcct ctg 83
<210> 67
<211> 50
<212> ДНК
<213> Xenopus sp.
<400> 67
tgcttgttct ttttgcagaa gctcagaata aacgctcaac tttggcagat 50
<210> 68
<211> 5
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 68
Gly Gly Gly Gly Ser
1 5
<210> 69
<211> 20
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 69
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
1 5 10 15
Gly Gly Gly Ser
20
<210> 70
<211> 5
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 70
Gln Leu Val Lys Ser
1 5
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ СОЗДАНИЯ Т-КЛЕТОК, ПРИГОДНЫХ ДЛЯ АЛЛОГЕННОЙ ТРАНСПЛАНТАЦИИ | 2015 |
|
RU2752933C2 |
| МОДИФИКАЦИЯ B-КЛЕТОК | 2018 |
|
RU2783116C2 |
| МНОГОЦЕПОЧЕЧНЫЙ ХИМЕРНЫЙ АНТИГЕННЫЙ РЕЦЕПТОР И ЕГО ПРИМЕНЕНИЯ | 2013 |
|
RU2663725C2 |
| СПОСОБЫ ПОЛУЧЕНИЯ КЛЕТОК, ЭКСПРЕССИРУЮЩИХ РЕКОМБИНАНТНЫЙ РЕЦЕПТОР, И РОДСТВЕННЫЕ КОМПОЗИЦИИ | 2019 |
|
RU2835579C2 |
| ДНК-СОДЕРЖАЩИЕ ПОЛИНУКЛЕОТИДЫ И РУКОВОДЯЩИЕ ПОСЛЕДОВАТЕЛЬНОСТИ ДЛЯ СИСТЕМ CRISPR ТИПА V И СПОСОБЫ ИХ ПОЛУЧЕНИЯ И ПРИМЕНЕНИЯ | 2021 |
|
RU2832314C1 |
| СПОСОБ КОНСТРУИРОВАНИЯ АЛЛОГЕННЫХ И УСТОЙЧИВЫХ К ЛЕКАРСТВЕННЫМ ПРЕПАРАТАМ Т-КЛЕТОК ДЛЯ ИММУНОТЕРАПИИ | 2014 |
|
RU2689558C1 |
| НЕ РЕСТРИКТИРОВАННЫЕ ПО HLA T-КЛЕТОЧНЫЕ РЕЦЕПТОРЫ И ИХ ПРИМЕНЕНИЕ | 2019 |
|
RU2812917C2 |
| СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ ЛЕЧЕНИЯ БОЛЕЗНИ ФАБРИ | 2017 |
|
RU2788133C2 |
| СКОНСТРУИРОВАННАЯ СПОСОБАМИ ИНЖЕНЕРИИ ПЛАТФОРМА ДЛЯ ВСТРАИВАНИЯ ТРАНСГЕНА (ETIP) ДЛЯ НАЦЕЛИВАНИЯ ГЕНОВ И СТЭКИНГА ПРИЗНАКОВ | 2013 |
|
RU2666916C2 |
| ФУНКЦИОНАЛЬНЫЕ ЛОКУСЫ FAD2 И СООТВЕТСТВУЮЩИЕ СПЕЦИФИЧНЫЕ ДЛЯ САЙТА-МИШЕНИ СВЯЗЫВАЮЩИЕСЯ БЕЛКИ, СПОСОБНЫЕ ИНДУЦИРОВАТЬ НАПРАВЛЕННЫЕ РАЗРЫВЫ | 2013 |
|
RU2656158C2 |
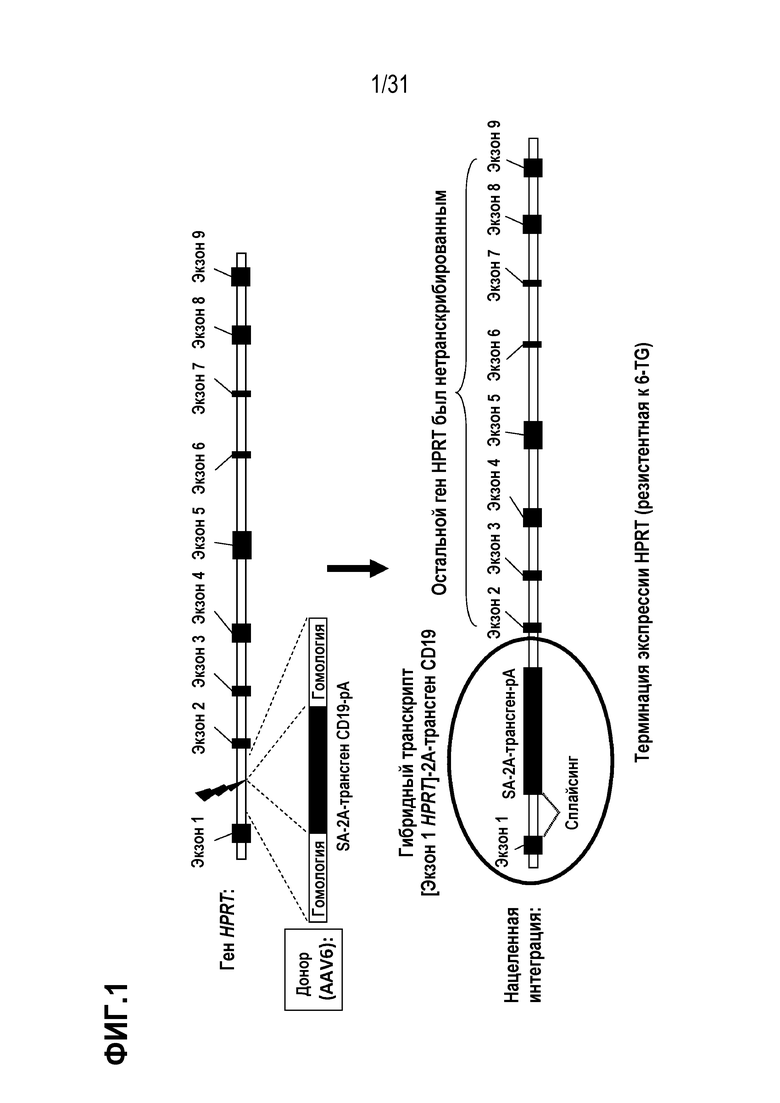
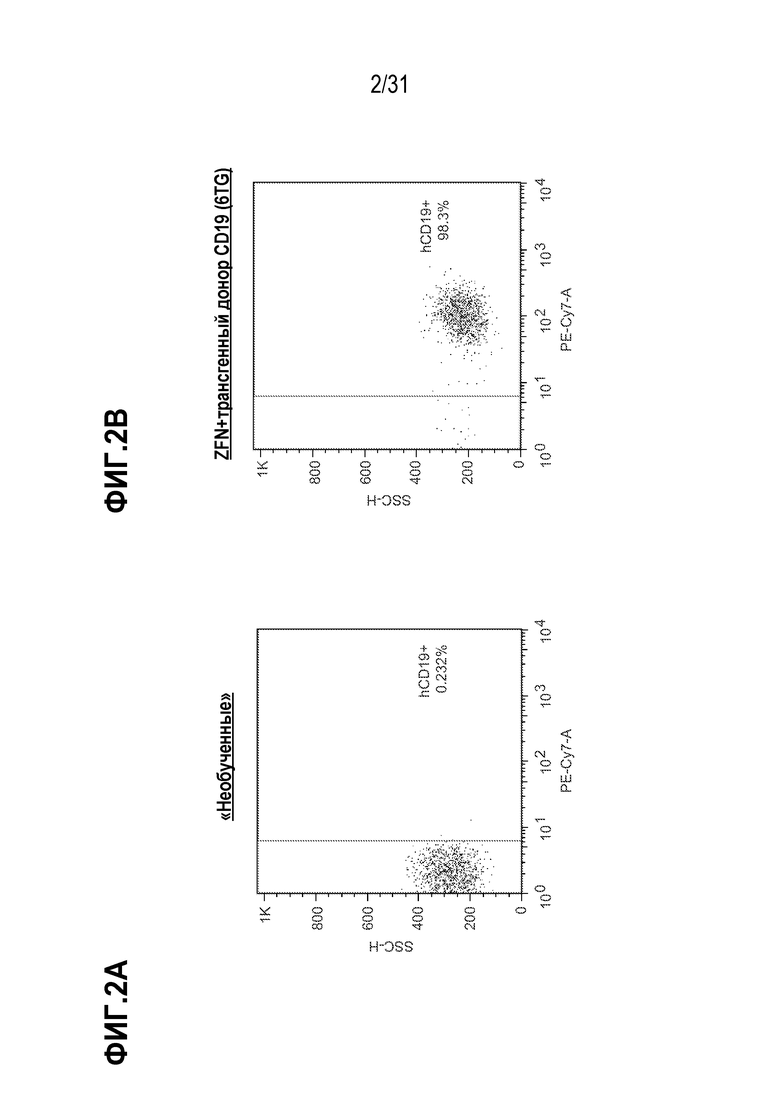
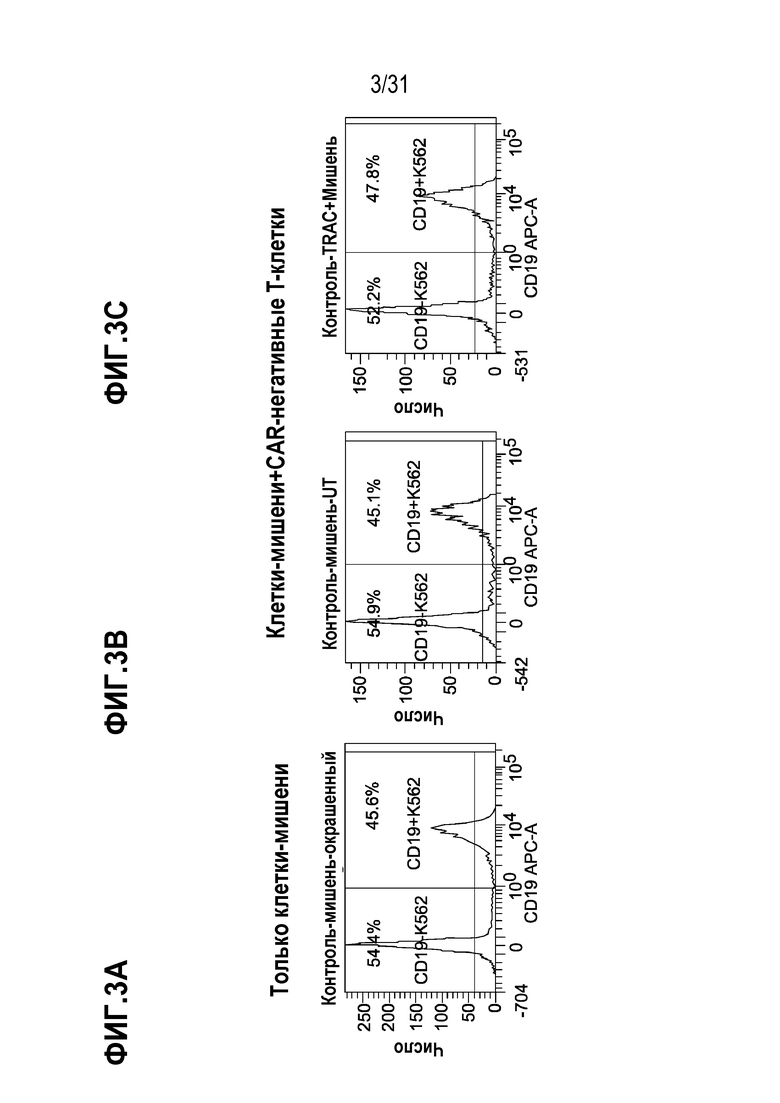
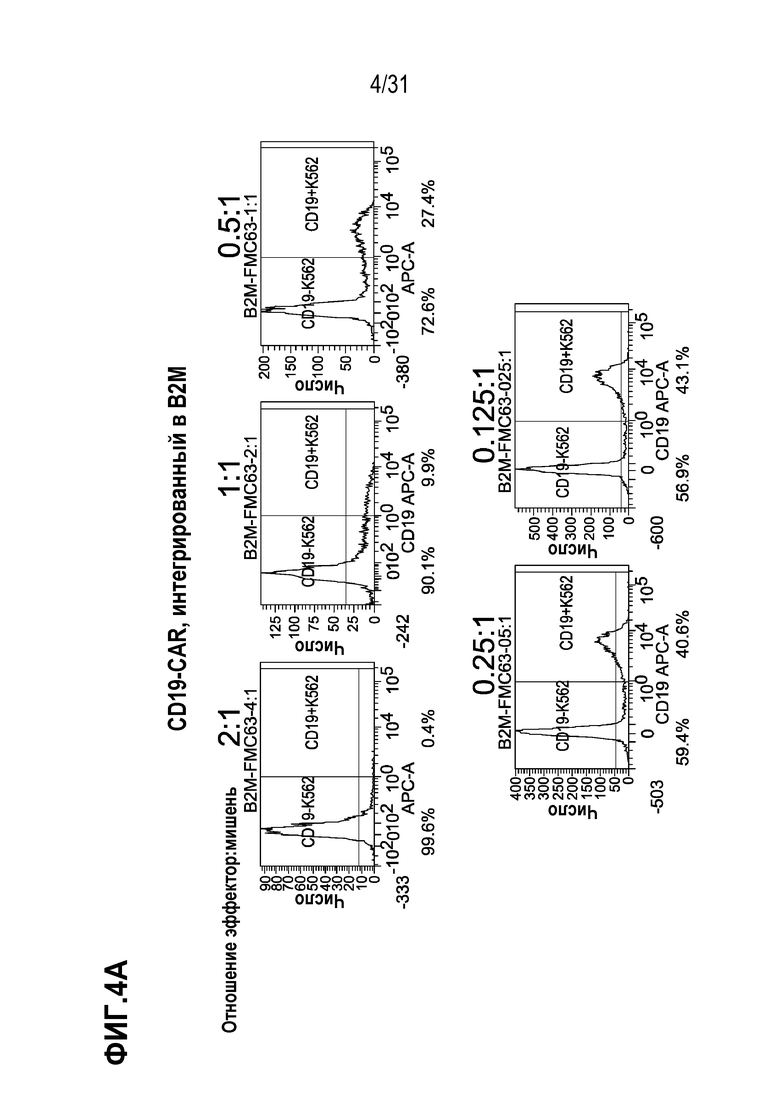
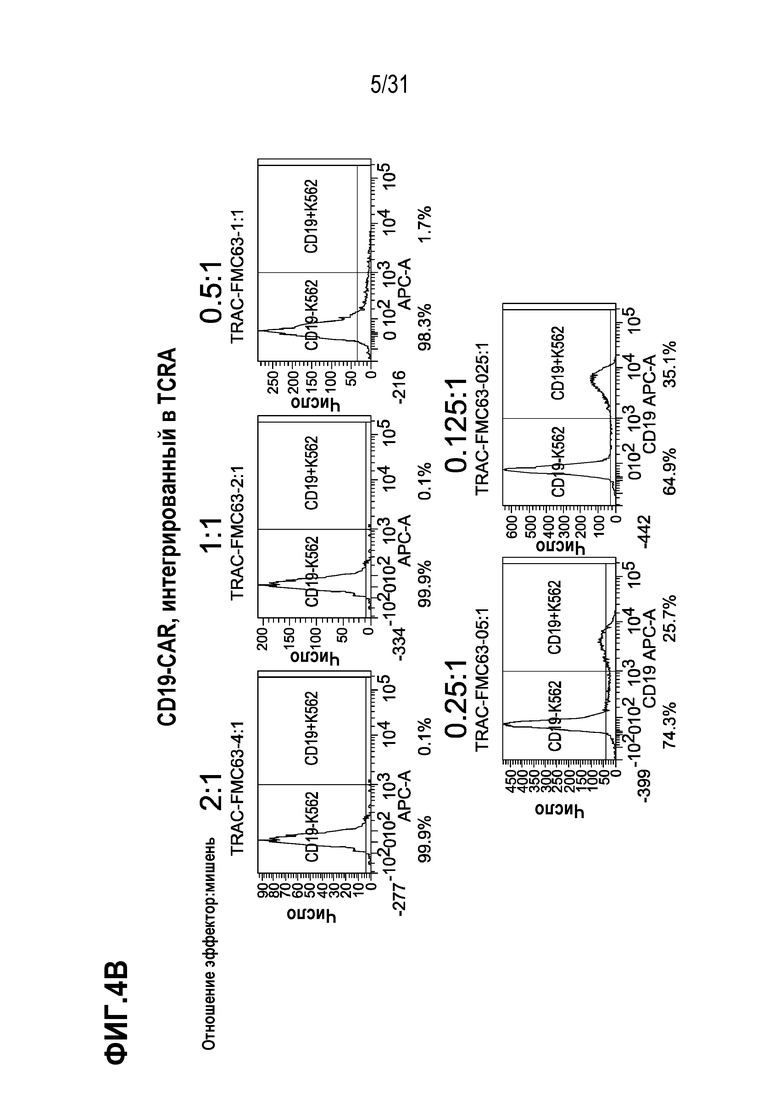
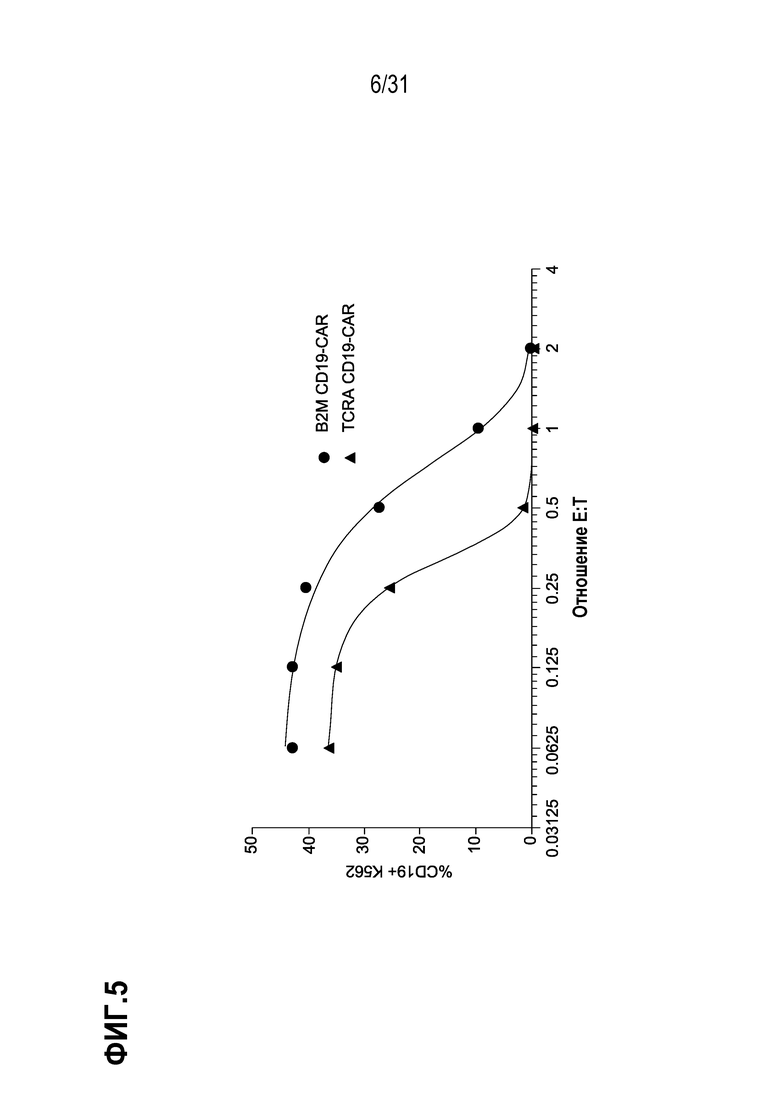
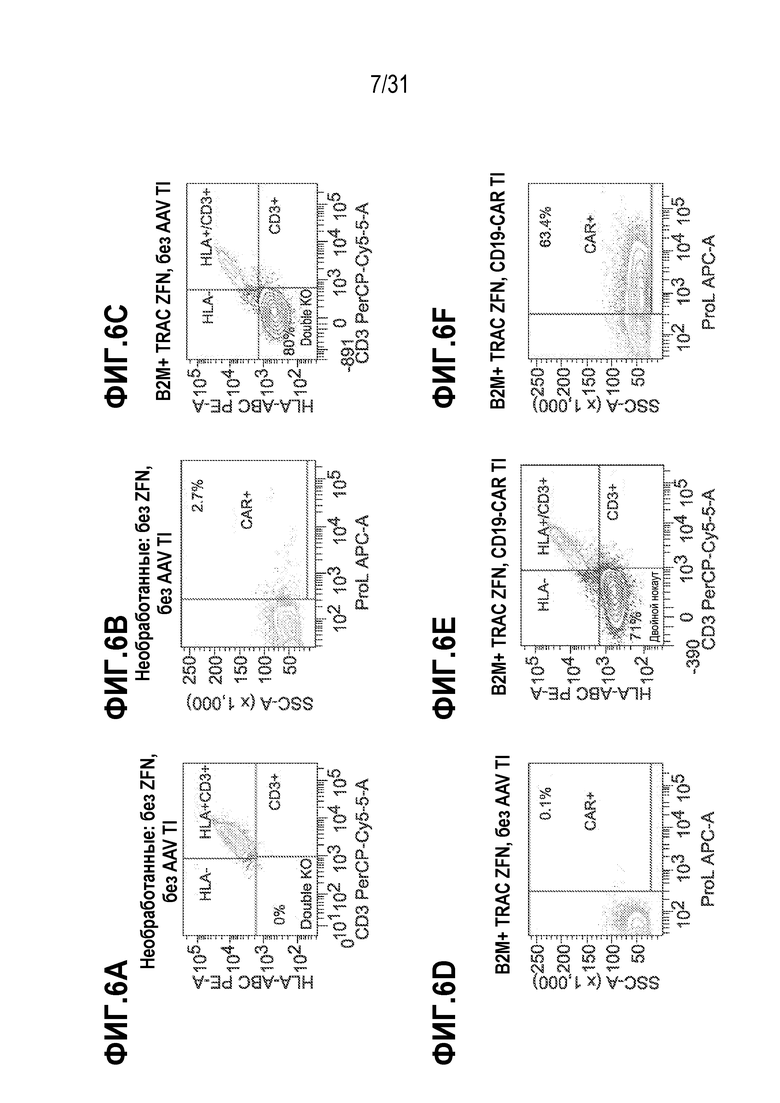
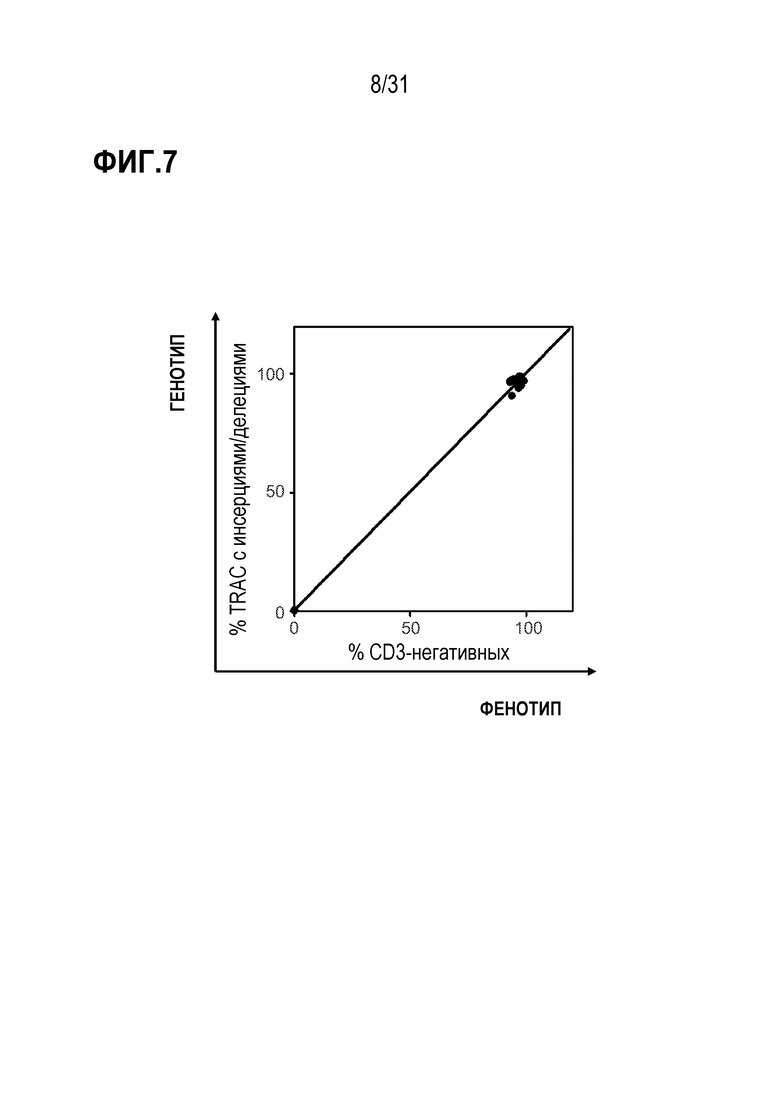
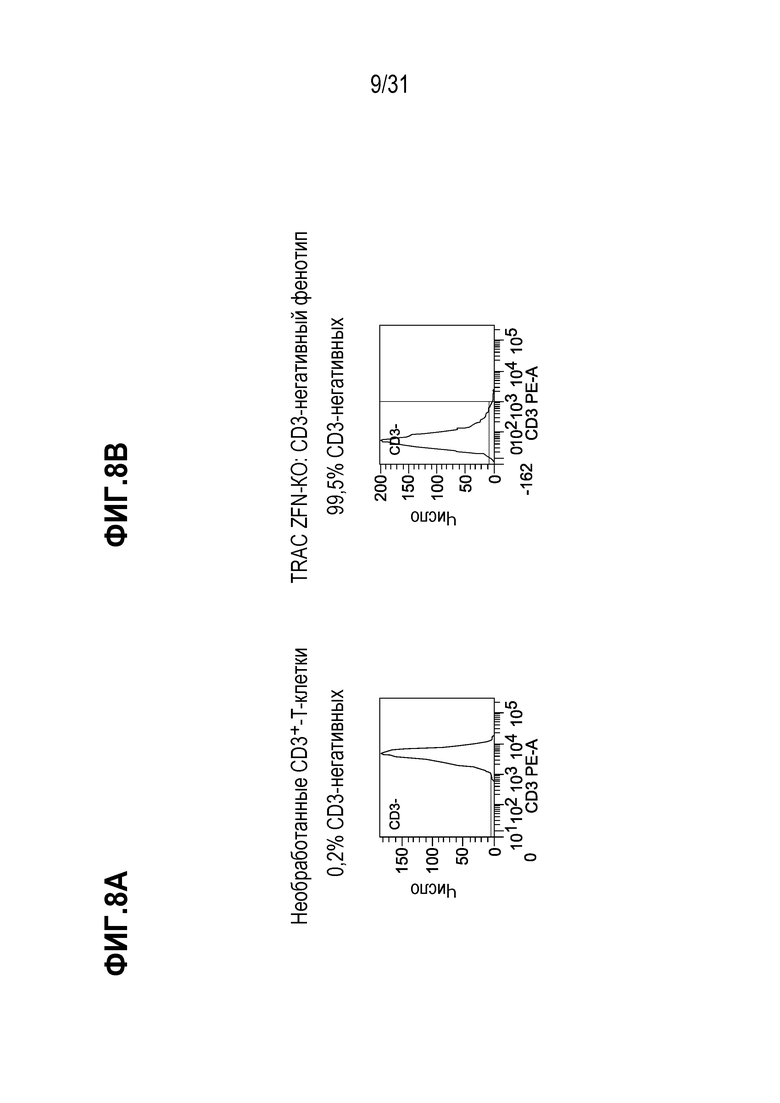
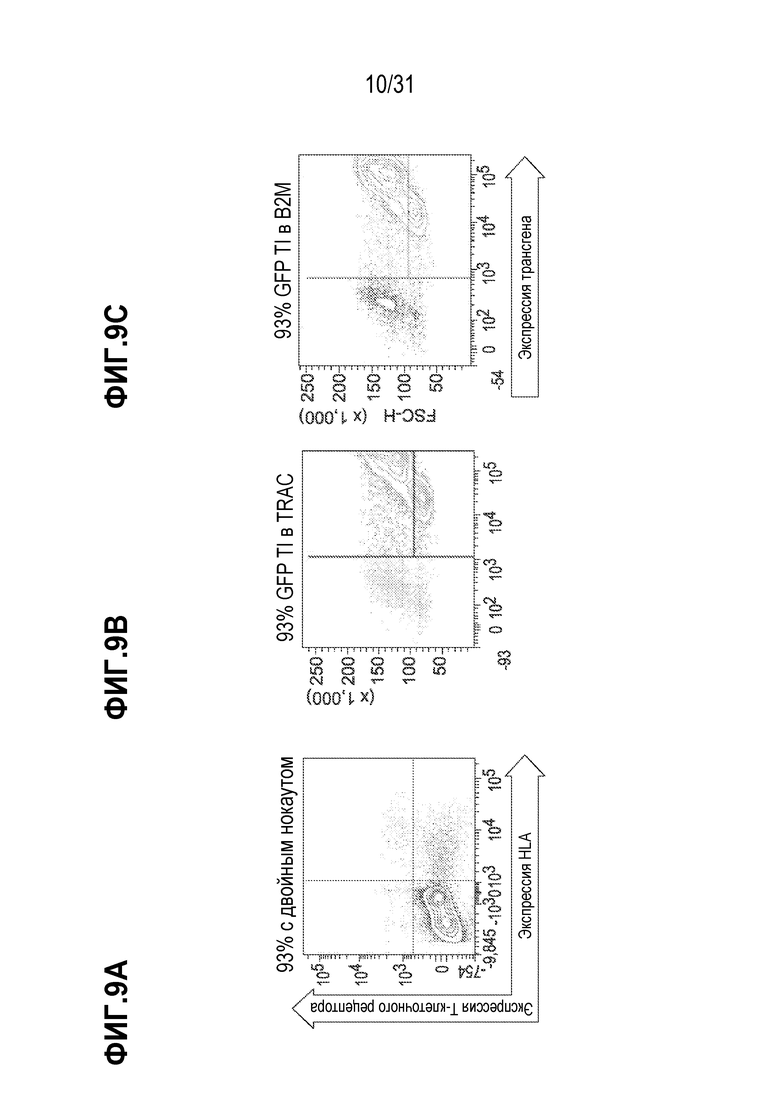
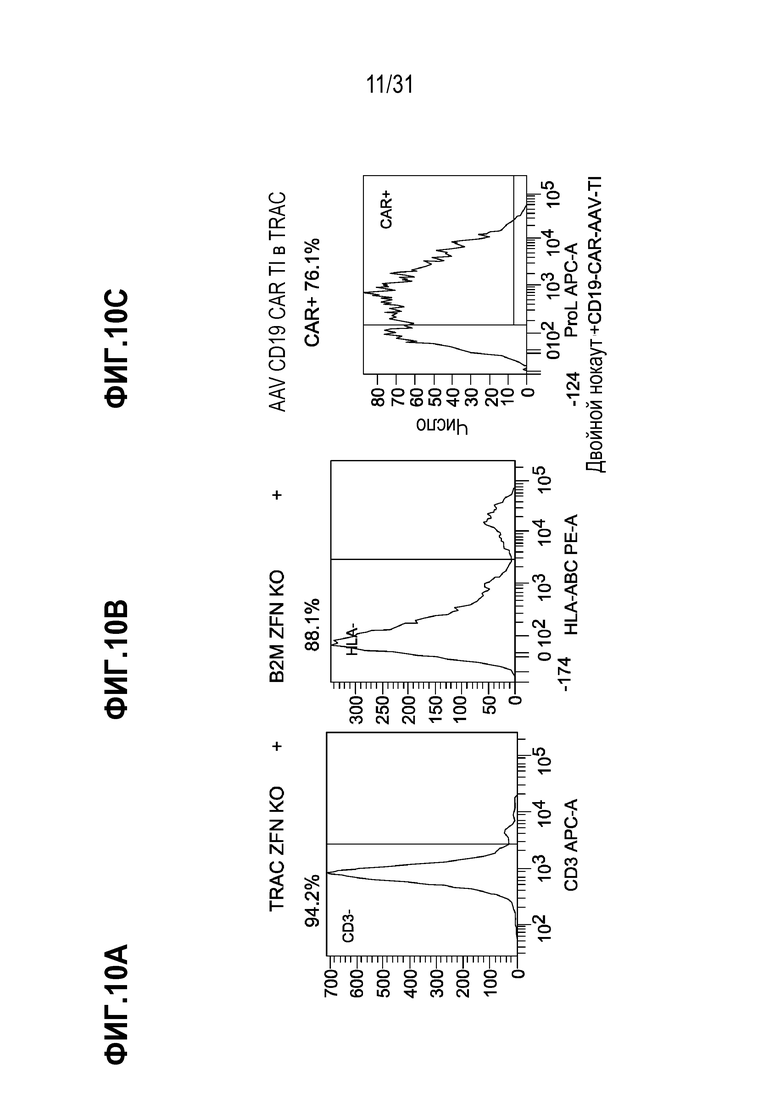
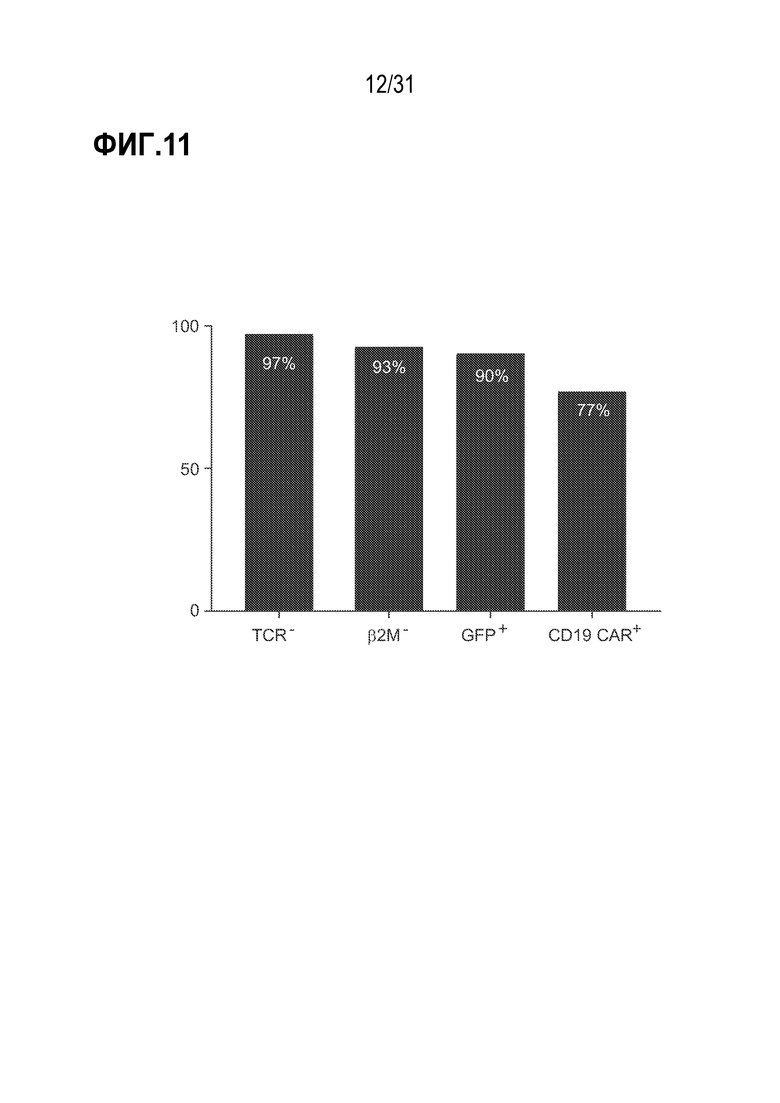
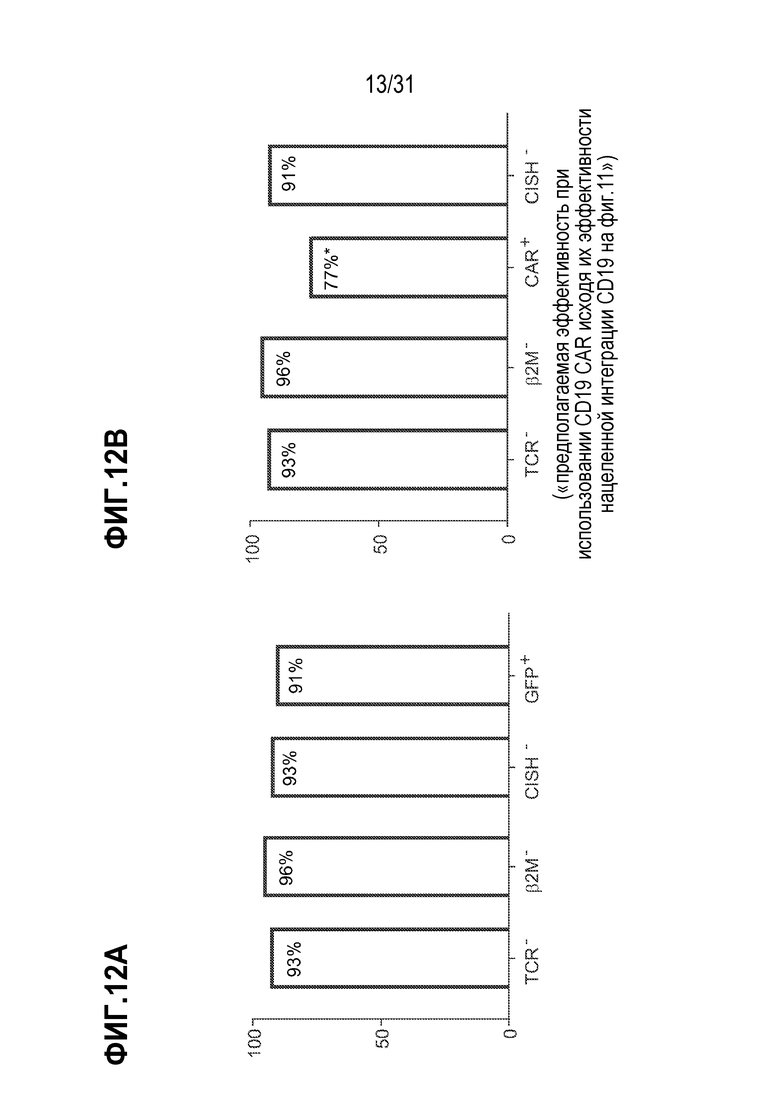
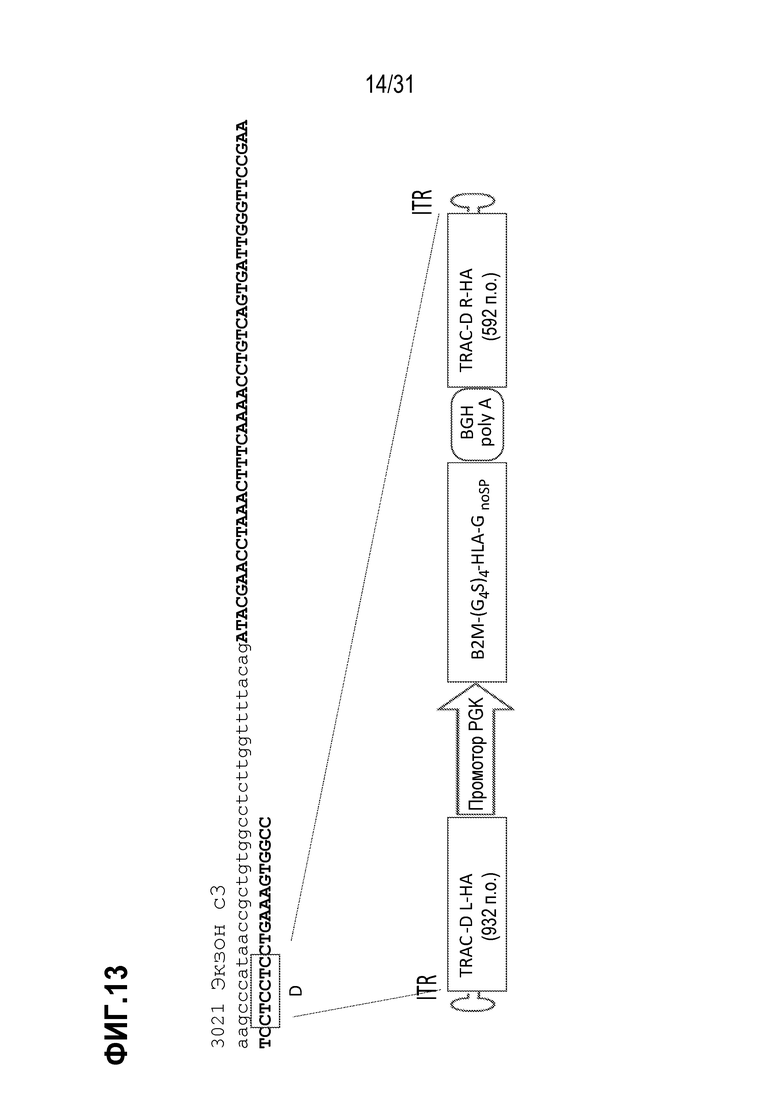
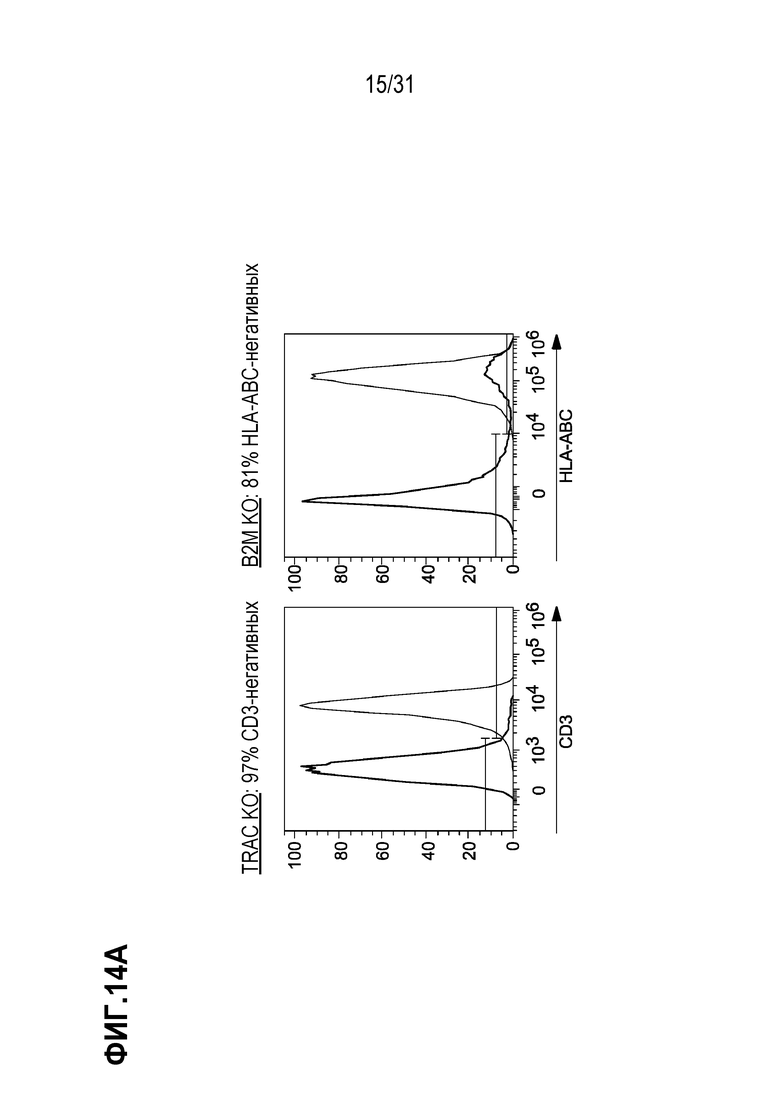
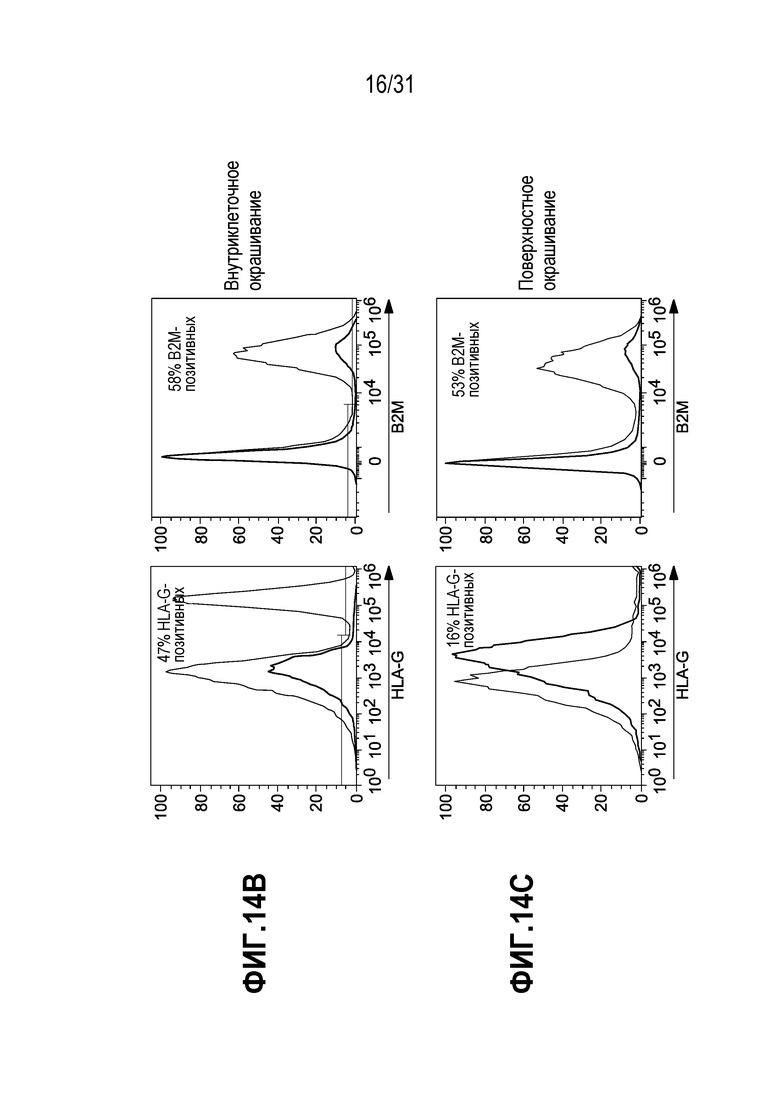
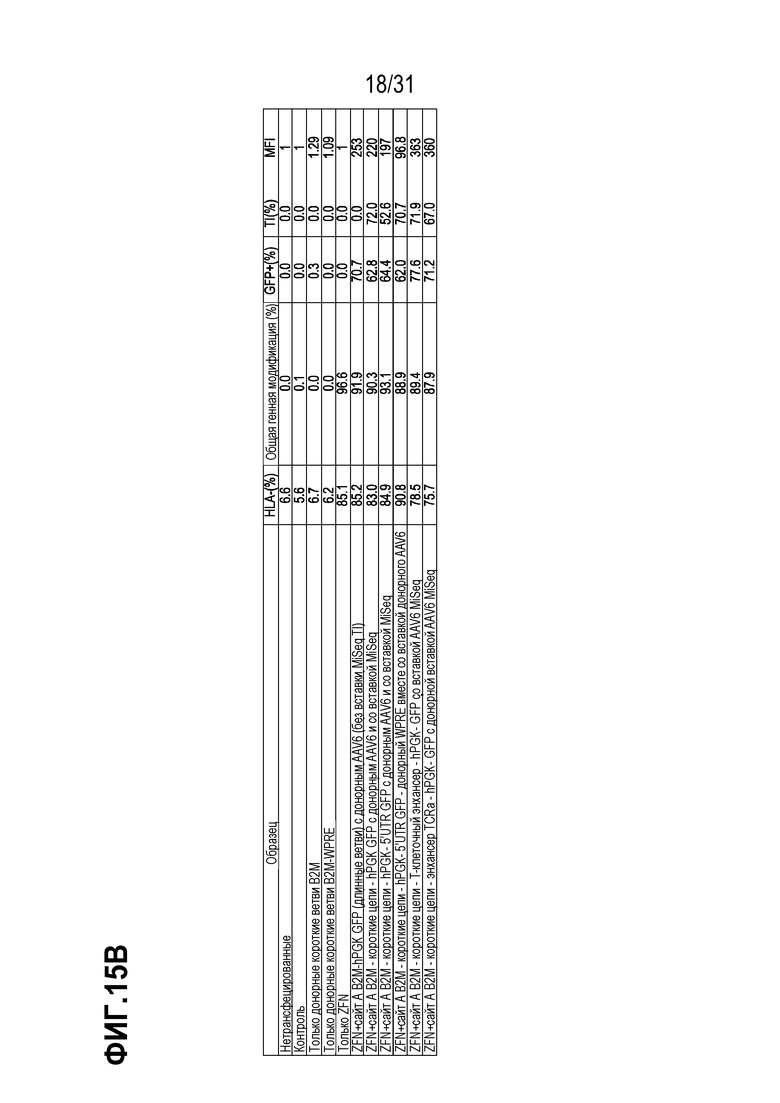
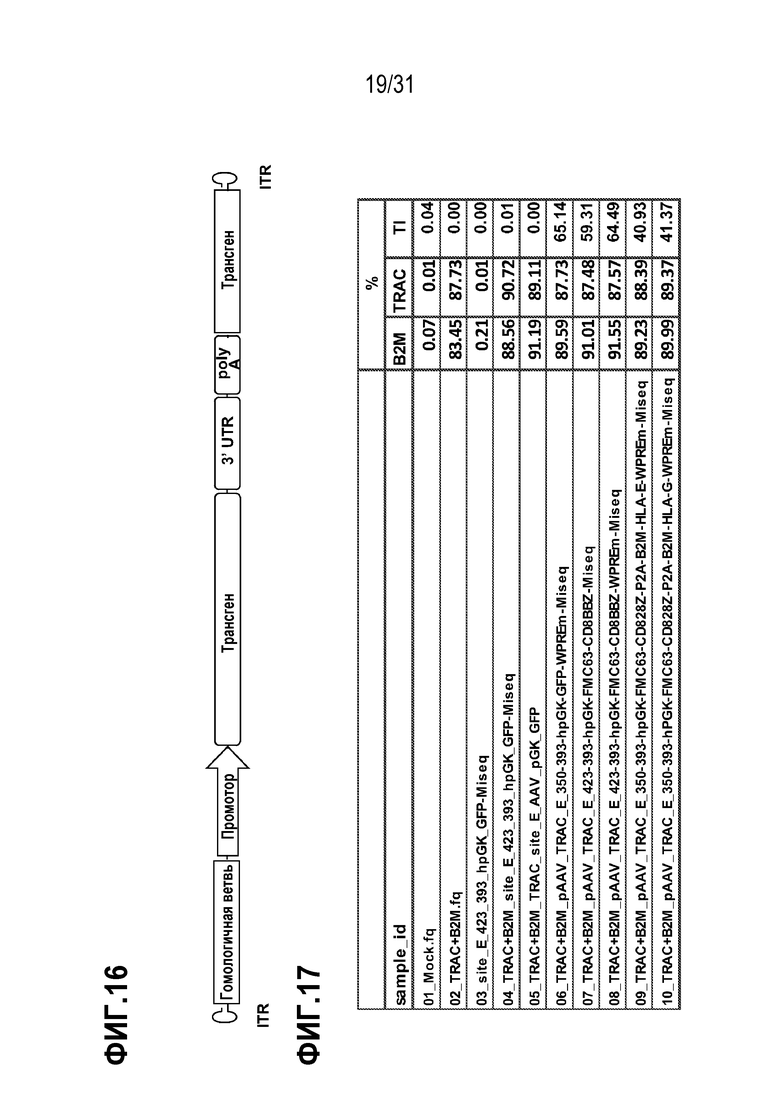
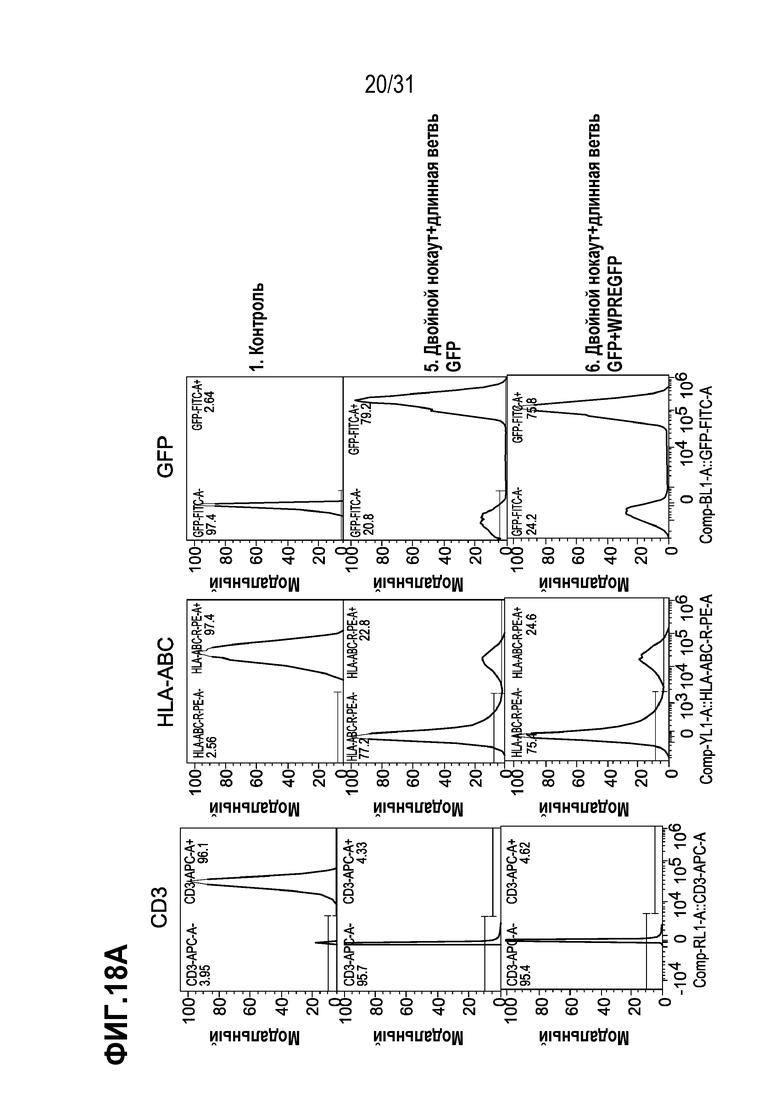
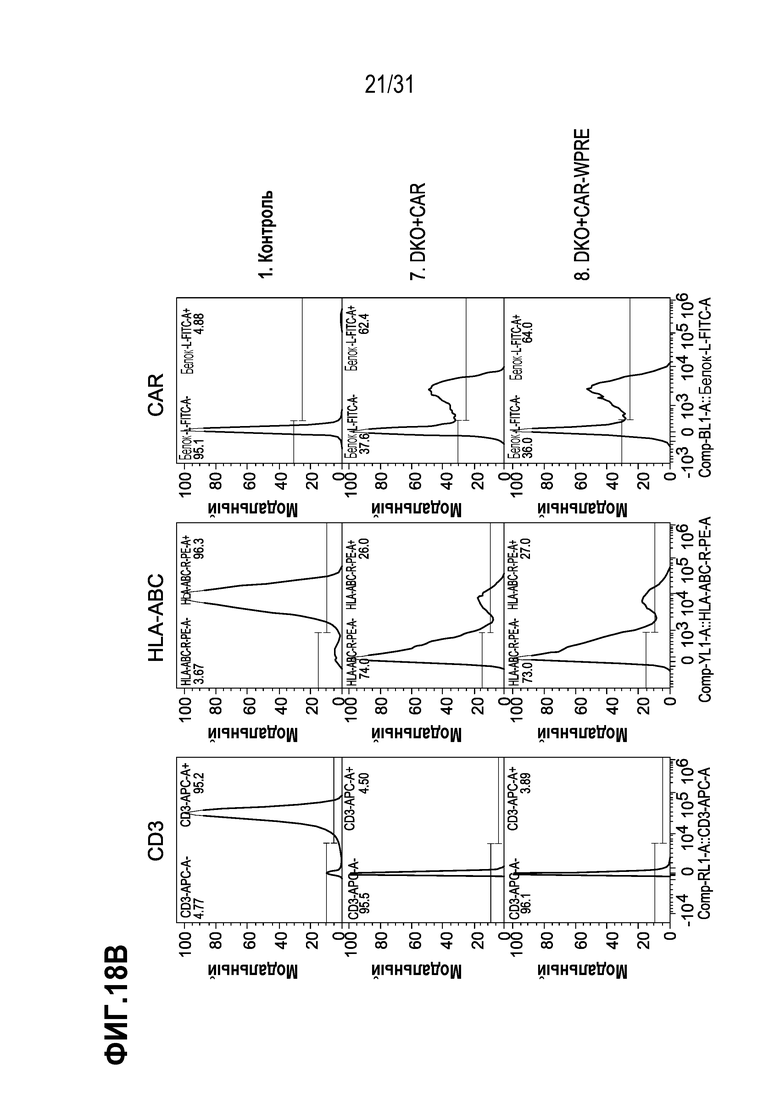
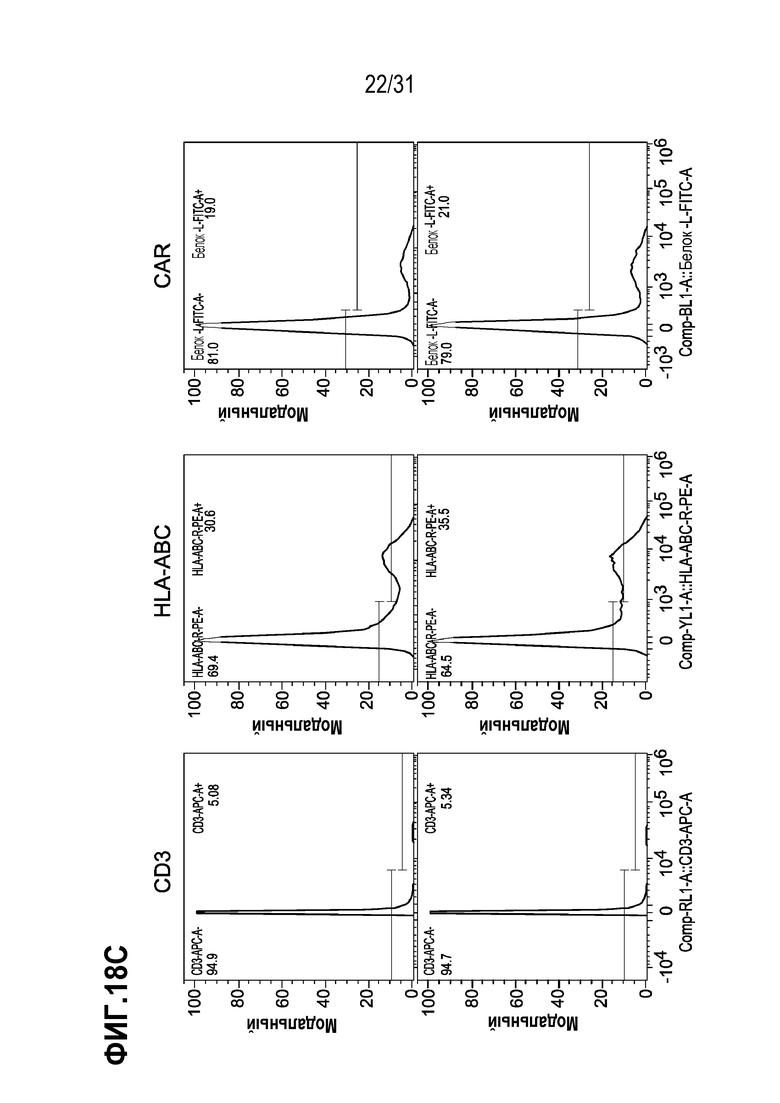
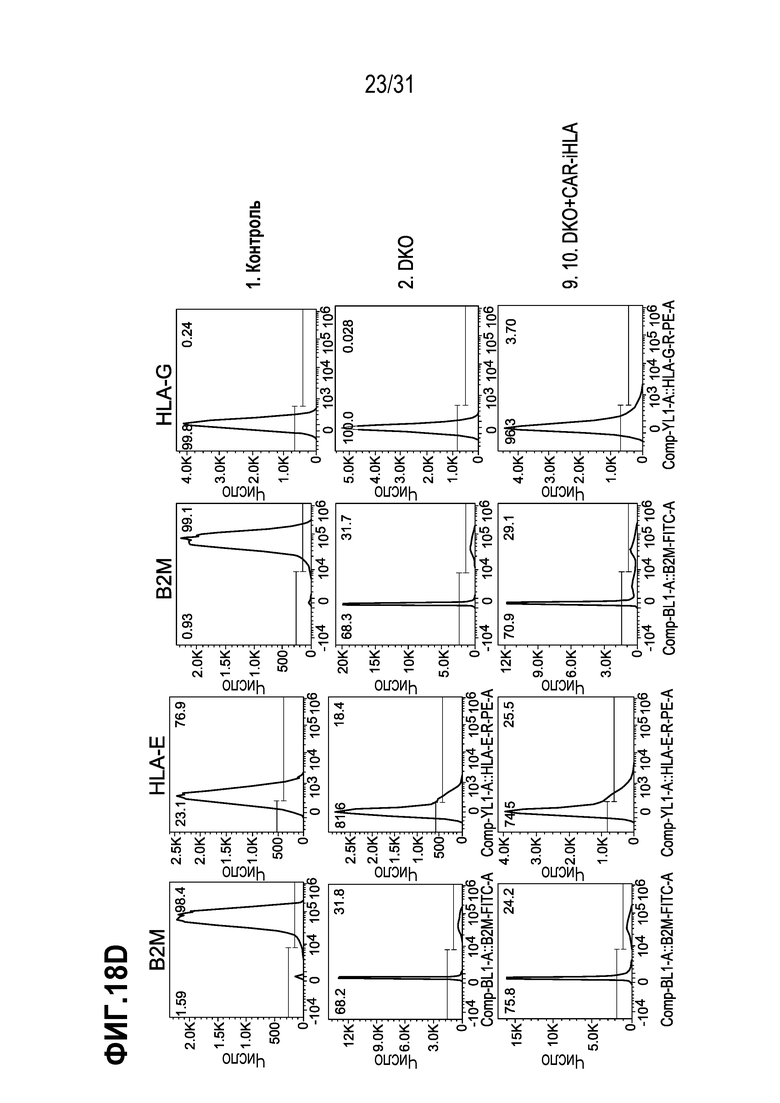
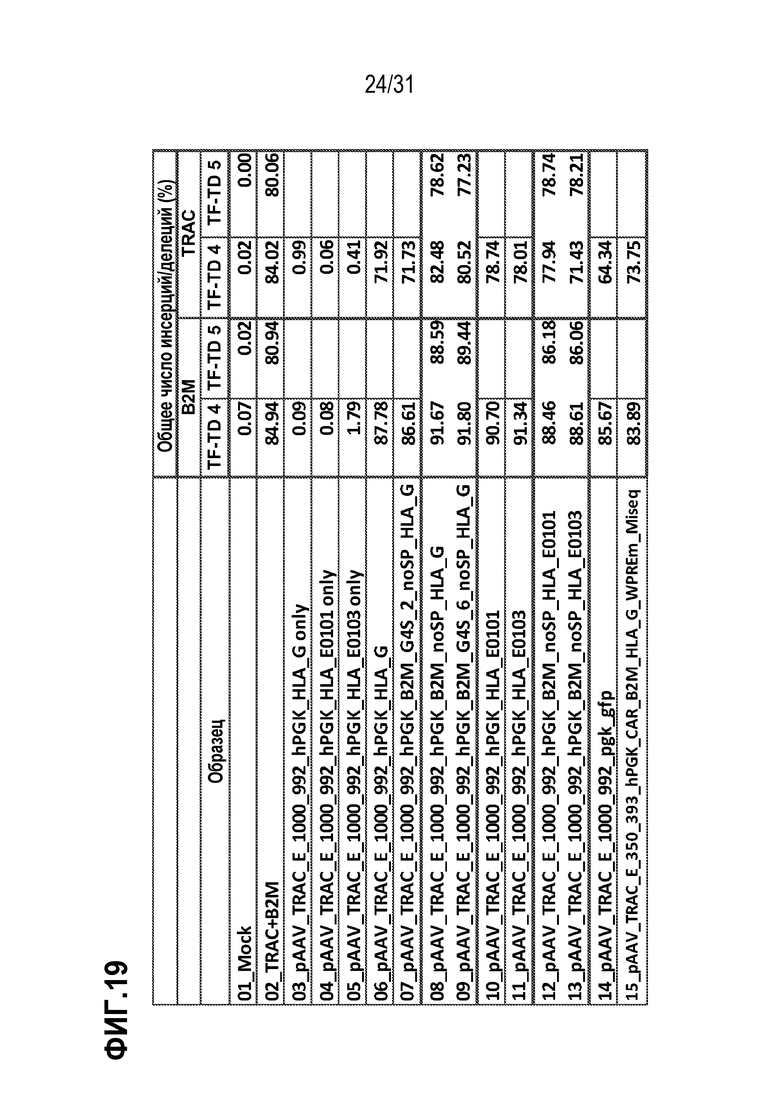
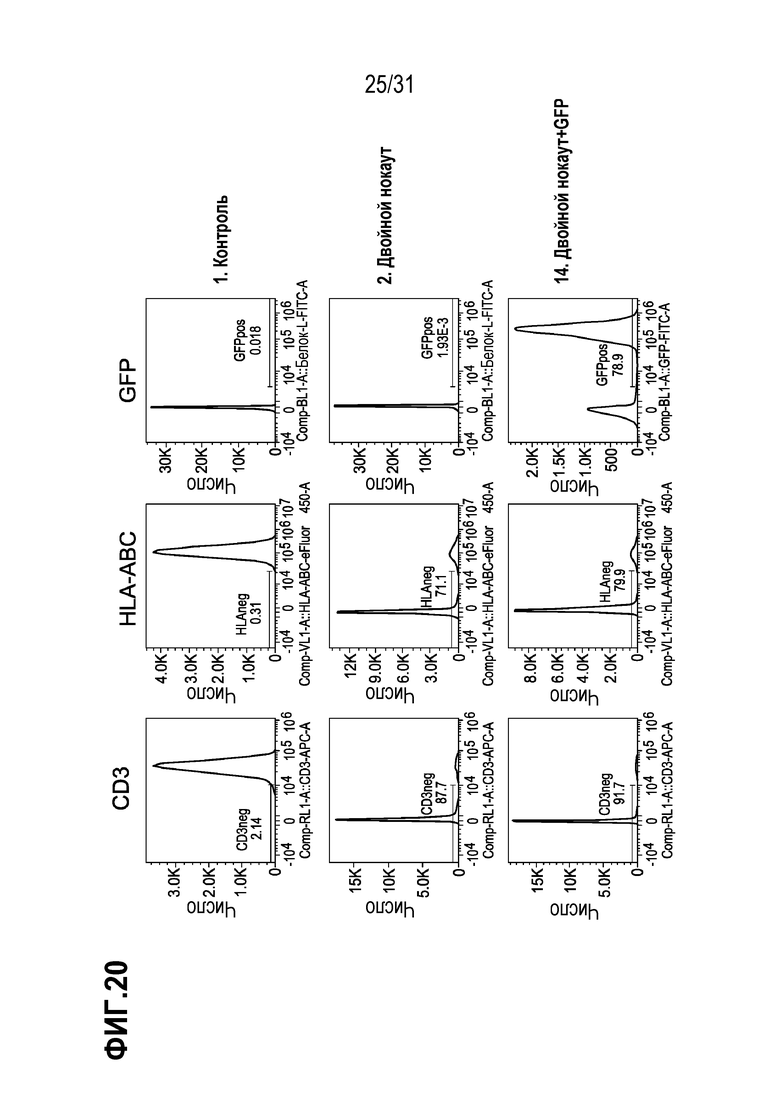
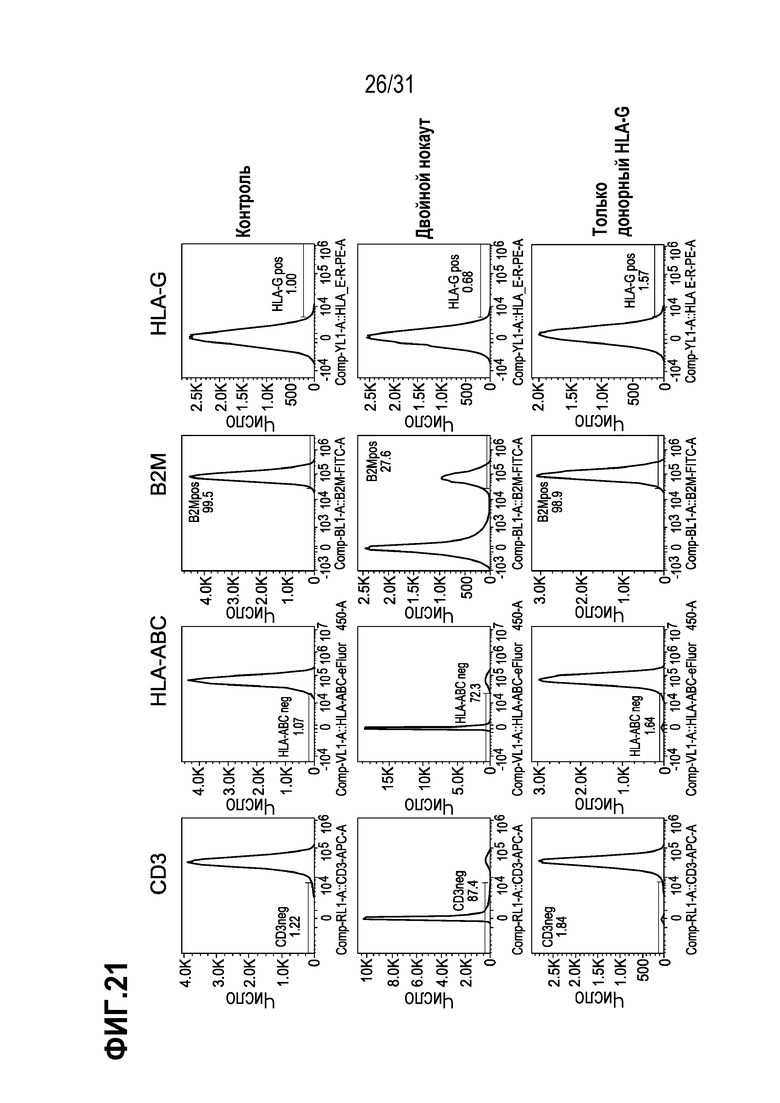
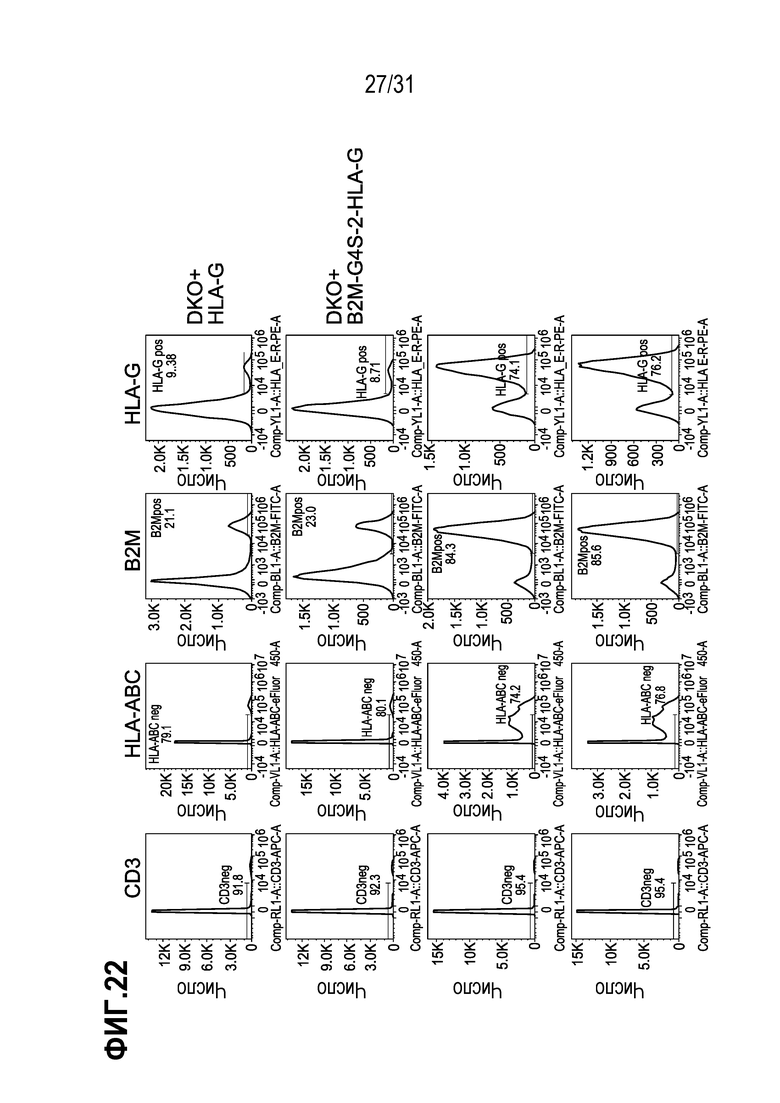
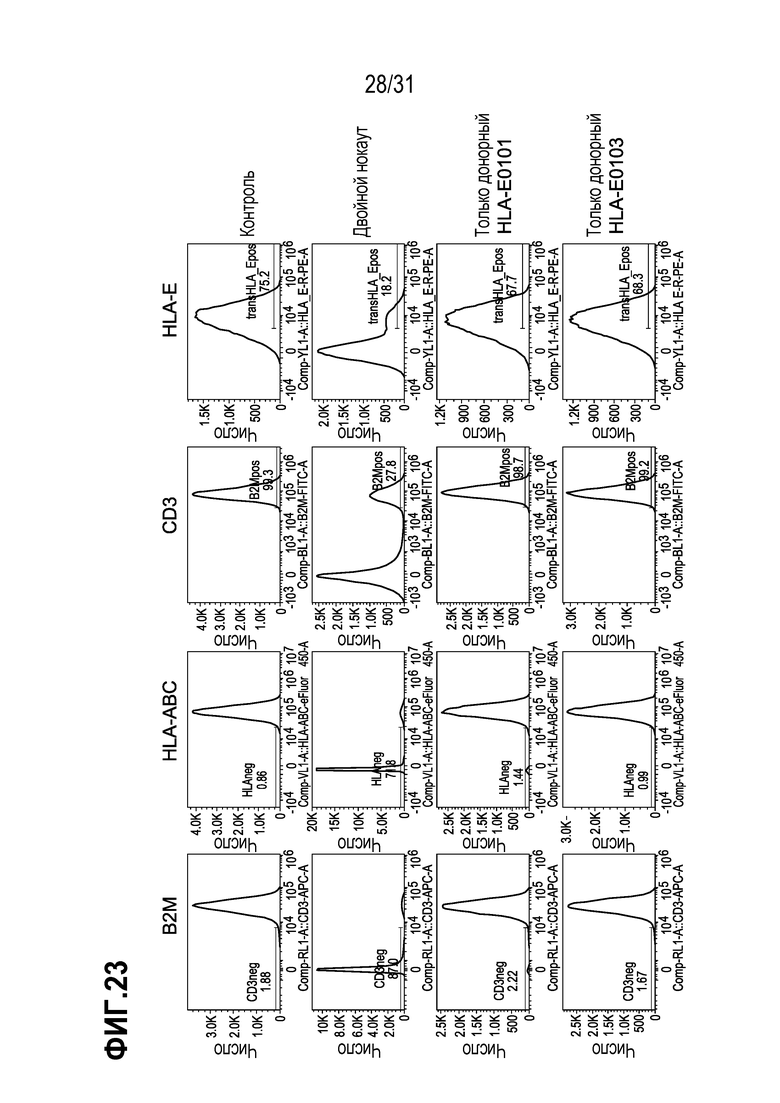
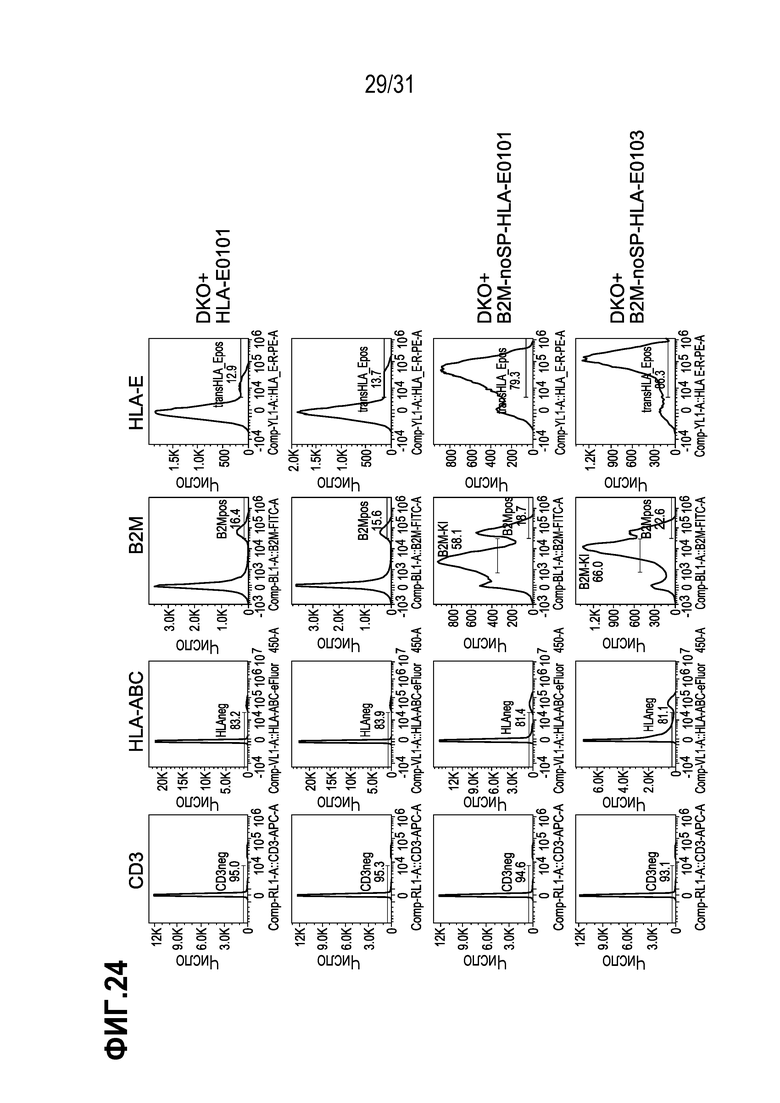
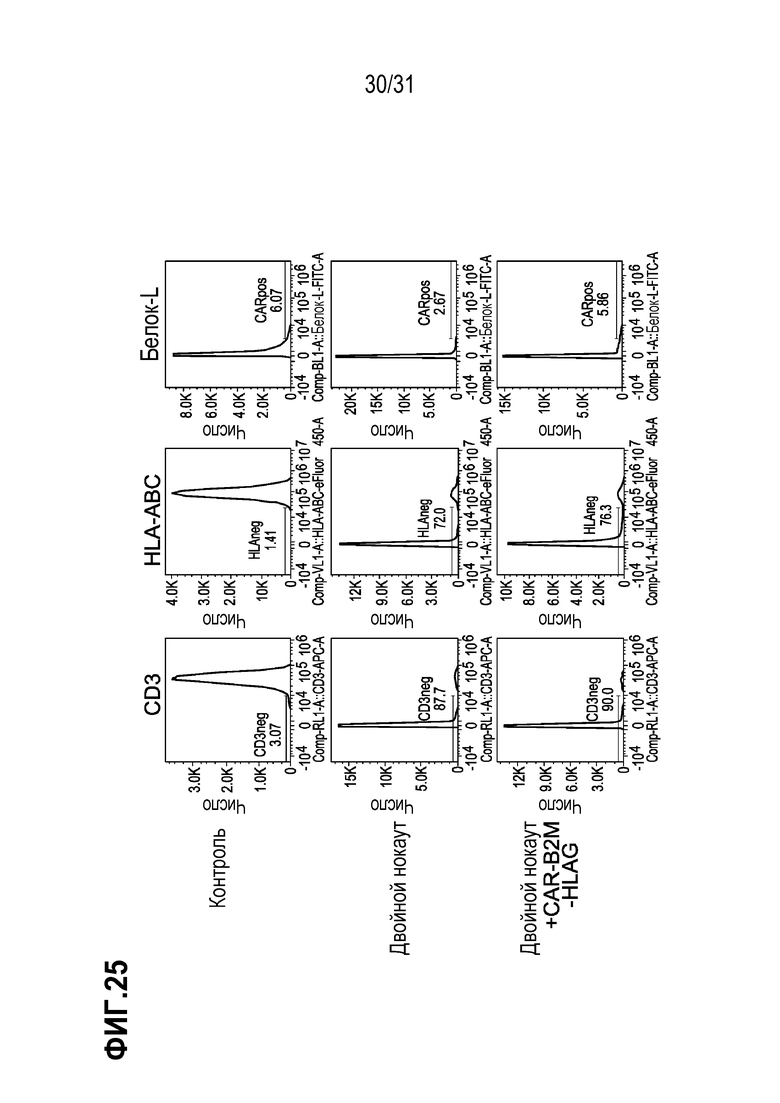
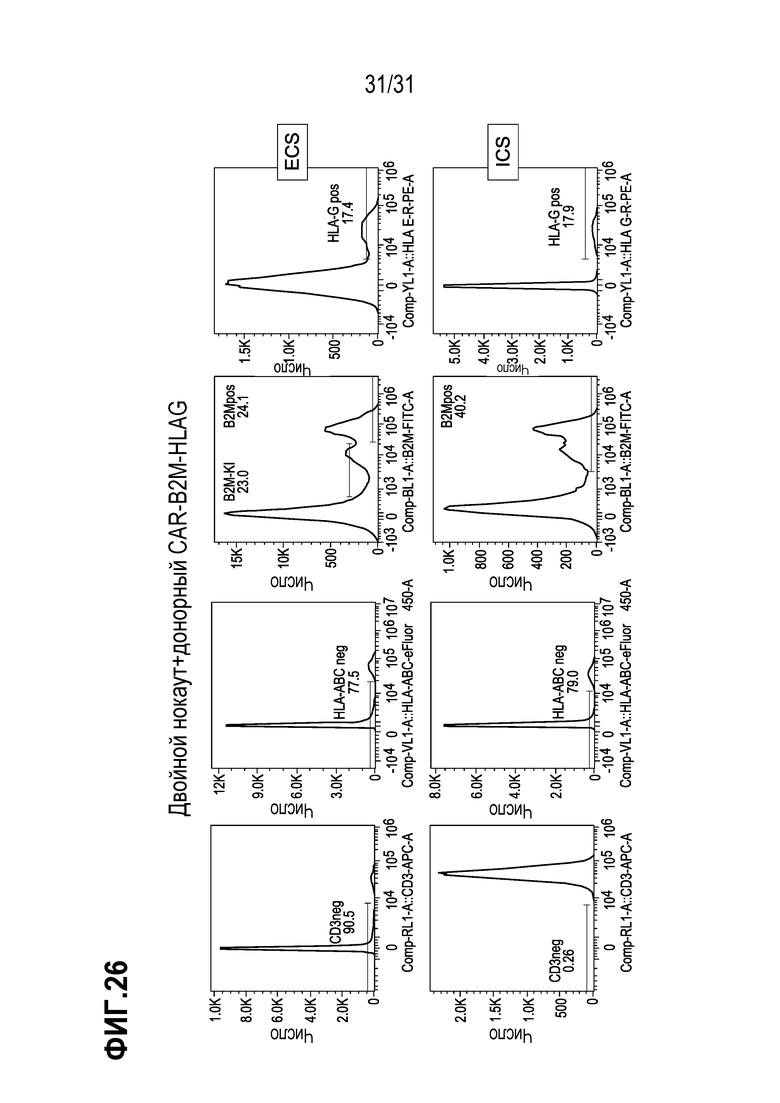
Изобретение относится к области биотехнологии, в частности к выделенной генетически модифицированной Т-клетке. Указанная клетка включает первый полинуклеотид, кодирующий химерный антигенный рецептор (CAR), и второй полинуклеотид, содержащий последовательности, кодирующие гибридный белок, содержащий белок бета-2-микро-глобулин (B2M) и белок HLA-E и/или HLA-G, где первый и второй полинуклеотиды интегрированы в ген Т-клеточного рецептора-альфа (TCRA), а также к способу ее получения. Изобретение эффективно в адоптивной клеточной терапии. 2 н. и 11 з.п. ф-лы, 26 ил., 6 табл., 5 пр.
1. Выделенная генетически модифицированная Т-клетка, обладающая устойчивостью к NK-опосредованному уничтожению, включающая: первый полинуклеотид, кодирующий химерный антигенный рецептор (CAR), и второй полинуклеотид, содержащий последовательности, кодирующие гибридный белок, содержащий белок бета-2-микро-глобулин (B2M) и белок HLA-E и/или HLA-G, где первый и второй полинуклеотиды интегрированы в ген Т-клеточного рецептора-альфа (TCRA).
2. Генетически модифицированная Т-клетка по п. 1, где эндогенный ген B2M является инактивированным в Т-клетке.
3. Генетически модифицированная Т-клетка по п. 1 или 2, где ген иммунологической контрольной точки является инактивированным в Т-клетке.
4. Генетически модифицированная Т-клетка по любому из пп. 1-3, где первый и второй полинуклеотиды связаны посредством саморасщепляющегося пептида P2A.
5. Генетически модифицированная Т-клетка по любому из пп. 1-4, где второй полинуклеотид дополнительно содержит последовательность, кодирующую линкер между B2M и последовательностями, кодирующими HLA-Е или HLA-G.
6. Генетически модифицированная Т-клетка по п. 4, где линкер содержит 1, 2, 3, 4, 5 или 6 копий G4S.
7. Способ получения генетически модифицированной Т-клетки по любому из пп. 1-6, где указанный способ включает:
расщепление гена TCR-α в выделенной Т-клетке с использованием нуклеазы, содержащей ДНК-связывающий домен, который связывается с сайтом-мишенью в гене TCR-α; и
введение одного или более доноров, содержащих первый полинуклеотид, кодирующий химерный антигенный рецептор (CAR), и второй полинуклеотид, кодирующий гибридный белок, содержащий белок бета-2-микро-глобулин (B2M) и белок HLA-E и/или HLA-G, в T-клетку так, чтобы первый и второй полинуклеотиды интегрировались в расщепленный ген TCR-α.
8. Способ по п. 7, где донор также включает гомологичные ветви, сцепленные с геном TCR-α, фланкирующим первый и второй полинуклеотиды.
9. Способ по п. 7 или 8, где донор дополнительно содержит промотор, который стимулирует экспрессию первого и/или второго полинуклеотида.
10. Способ по любому из пп. 7-9, где донор также содержит энхансерную последовательность TCR-α.
11. Способ по любому из пп. 7-10, где донор также содержит 3’-нетранслируемую область (3’UTR) и/или 5’-нетранслируемую область (5’UTR).
12. Способ по п. 11, где 3’UTR дополнительно содержит последовательность WPRE.
13. Способ по п. 11 или 12, где 5’UTR дополнительно содержит последовательность гена бета-глобина Xenopus.
| WO 2017078807 A1, 11.05.2017 | |||
| WO 2017079673 A1, 11.05.2017 | |||
| WO 2017106528 A2, 22.06.2017 | |||
| WO 2017093969 A1, 08.06.2017 | |||
| RU 2015139874 A, 24.03.2017. |

