Область изобретения
Настоящее изобретение относится к области биотехнологии, и, в частности, относится к высокостабильному и растворимому Т-клеточному рецептору (Т cell receptor, TCR), мутантному в своем гидрофобном коровом домене. Настоящее изобретение также относится к получению и применению рецептора.
Предшествующий уровень техники
Существует только два типа молекул, которые могут распознавать антигены специфическим образом. Один из типов представляет собой иммуноглобулин или антитело, а другой представляет собой Т-клеточный рецептор (TCR), который представляет собой α/β или γ/δ гетеродимерный гликопротеин на клеточной мембране. Физический репертуар TCR иммунной системы формируется в тимусе путем V(D)J-рекомбинации, а затем положительной и отрицательной селекции. В периферическом окружении TCR опосредуют распознавание специфических комплексов Главный комплекс гистосовместимости (Major Histocompatibility Complex, МНС)-пептид Т-клетками, и в этом качестве являются важными для функционирования иммунной системы на клеточном уровне.
TCR является единственным рецептором для антигенов конкретных пептидов в главном комплексе гистосовместимости (МНС). Экзогенные или эндогенные пептиды могут являться единственным признаком аномалии в клетке. В иммунной системе, как только антигенспецифические TCR связываются с комплексами рМНС (пептид-МНС), происходит прямой физический контакт Т-клетки и антигенпрезентирующей клетки (antigen presenting cell, АРС). Затем происходит взаимодействие других мембранных молекул в Т-клетке и АРС, и инициируются последующая передача сигнала в клетке и другие физиологические ответы таким образом, что диапазон различных антигенспецифических Т-клеток демонстрирует иммунные действия в отношении своих мишеней.
На мембране Т-клетки TCR ассоциируется с инвариантными белками CD3, вовлеченными в опосредованную передачу сигнала, с образованием комплекса. TCR существуют во множестве форм, которые являются сходными по структуре, но экспрессирующие их Т-клетки обладают весьма отличающимися анатомическими расположениями и, вероятно, функциями. Внеклеточная часть TCR состоит из двух проксимальных относительно мембраны константных доменов, и двух дистальных относительно мембраны вариабельных доменов. Вариабельные домены содержат полиморфные петли, аналогичные участкам антител, определяющим комплементарность (от англ. complementary determining region - участки, определяющие комплементарность, CDR). Именно эти петли образуют связывающий сайт молекулы TCR и определяют пептидную специфичность. Лиганды МНС класса I и класса II, соответствующие TCR, также относятся к белкам суперсемейства иммуноглобулинов, но специализированы на презентации антигенов, и обладают полиморфным сайтом связывания пептидов, позволяющим осуществлять презентацию различных наборов коротких пептидных фрагментов на поверхности клеток АРС.
Подобно иммуноглобулину (антителу), представляющему собой разновидность молекулы, распознающей антиген, TCR могут быть разработаны для применения в диагностике и терапии. Тем не менее, сложно получить белки в (водо)растворимой форме, образованные из более чем одной полипептидной субъединицы и содержащие трансмембранный домен, поскольку во многих случаях белок стабилизируются своей трансмембранной областью. Последнее относится и к TCR и отражено в научной литературе. Сообщается о том, что усеченная форма TCR, содержащая только внеклеточные домены или только внеклеточные и цитоплазматические домены, может распознаваться TCR-специфическими антителами, что свидетельствует о том, что часть рекомбинантного TCR, распознанная антителами, уложена правильным образом. Однако, ее не удается получить с хорошим выходом, она не стабильна и/или не может распознавать комплексы пептид-МНС в низкой концентрации.
Растворимый TCR полезен не только для исследования взаимодействий TCR-pMHC, но также в качестве потенциального диагностического средства для обнаружения инфекции или в качестве маркера аутоиммунных заболеваний. Аналогично, растворимые TCR могут быть использованы для доставки терапевтического агента, например, цитотоксического соединения или иммуностимулирующего соединения, в клетки, которые осуществляют презентацию определенного антигена, или для ингибирования Т-клеток, например клеток, взаимодействующих с аутоиммунным пептидным антигеном. Для этих задач важна модификация белка TCR. Она особенно важна для гетерогенной экспрессии TCR в прокариотических или эукариотических системах.
В отношении экспрессии растворимого TCR в Е. coli, когда TCR отделяют от мембраны, нестабильность и низкий выход белка представляют собой основные препятствия для разработки терапевтических или диагностических реагентов с TCR или его фрагментом. Для преодоления присущей одноцепочечным TCR нестабильности, в некоторых литературных источниках описано получение гетеродимеров TCR, содержащих нативный дисульфидный мостик, связывающий соответствующие субединицы (Garboczi, et al., (1996), Nature 384 (6605): 134-41; Garboczi, et al., (1996), J Immunol 157(12): 5403-10; Chang et al., (1994), PNAS USA 91: 11408-11412; Davodeau et al., (1993), J. Biol. Chem. 268(21): 15455-15460; Golden et al., (1997), J. Imm. Meth. 206: 163-169; патент США №6080840). Однако, хотя такие TCR могут распознаваться TCR-специфическими антителами, они могут распознавать только нативный лиганд в относительно высокой концентрации, что свидетельствует о том, что распознавание нестабильно.
Кроме того, для получения TCR с исходной антигенной специфичностью существует множество исследований на тему улучшения стабильности водорастворимых фрагментов TCR, включающих вариабельные домены одноцепочечного TCR (Novotny, et al. (1991) PNAS USA 88:8646-8650), внеклеточные домены в гетеродимерном TCR (Garcia1 et al. (1996) Science 274:209-219), или модификацию таких молекул (Shusta et al. (2000) Nature Biotechnology 18:754-759), Boulter et al. (2003) Protein Engineering 16:707-711). В этих исследованиях Novotny et al. использовали гибкий пептид для связывания вариабельных доменов для конструирования одноцепочечного TCR. Однако, стабильные молекулы могут быть получены только после замены гидрофобных остатков, представленных на поверхности, на гидрофильные остатки, содержащие водорастворимые боковые цепи. Shusta et al. модифицировали структуру вариабельного домена одноцепочечного TCR путем введения случайных мутаций в целую молекулу, отображения на поверхности дрожжей и отбора при помощи FACS (клеточный сортер с возбуждением флуоресценции). Garcia et al. сконструировали внеклеточный домен 2С в α/β TCR с сохранением в структуре нативных межцепочечных дисульфидных связей. Boulter et al. улучшили конструкцию α/β гетеродимера путем введения искусственной дисульфидной связи между двумя константными доменами.
Подход с использованием дисульфида между константными доменами использовался для векторов фагового дисплея TCR, которые использовали для получения множества высокоаффинных TCR (Li et al. (2005) Nature Biotechnology 34:349-354; Liddy et al. (2012) Nature Medicine 18:980-987). Тем не менее, авторы изобретения обнаружили, что вероятность успешного получения TCR с использованием таких конструкций все же очень низка, и сложно получать TCR, обладающие как высокой аффинностью, так и высокой стабильностью. Таким образом, необходимо разрабатывать новые стратегии для получения TCR и их фрагментов, обладающих растворимостью в воде, высокой аффинностью и высокой стабильностью.
Краткое описание изобретения
Цель настоящего изобретения состоит в разработке высокостабильного Т-клеточного рецептора (TCR).
Еще одна цель настоящего изобретения состоит в разработке способов получения и применения указанного высокостабильного Т-клеточного рецептора.
В первом аспекте изобретения предложен Т-клеточный рецептор (TCR), обладающий следующими свойствами:
(1) гидрофобный коровый домен TCR является мутантным; и
(2) TCR обладает стабильностью более высокой, чем стабильность соответствующего TCR, имеющего гидрофобное ядро дикого типа.
В одном из предпочтительных воплощений "мутантный" означает, что гидрофобный коровый домен TCR по настоящему изобретению имеет мутацию по сравнению с соответствующим гидрофобным коровым доменом TCR дикого типа.
В одном из предпочтительных воплощений "стабильностью более высокой, чем" означает, что стабильность TCR по настоящему изобретению увеличена по меньшей мере на 5%, предпочтительно по меньшей мере на 30%, и еще предпочтительней по меньшей мере на 80% по сравнению с TCR, имеющим гидрофобное ядро дикого типа.
В одном из предпочтительных воплощений "гидрофобное ядро дикого типа" относится к гидрофобному ядру, идентичному по аминокислотной последовательности гидрофобному ядру встречающегося в природе TCR, и не является мутантным.
В одном из предпочтительных воплощений "соответствующий TCR, имеющий гидрофобное ядро дикого типа" относится к TCR, идентичному по последовательности других доменов с последовательностями TCR по настоящему изобретению, за исключением того, что его гидрофобное ядро представляет собой ядро дикого типа по сравнению с TCR по настоящему изобретению, гидрофобное ядро которого является мутантным. Дополнительно или предпочтительно "соответствующий TCR, имеющий гидрофобное ядро дикого типа" относится к TCR дикого типа, который встречается в природе и не содержит какого-либо мутантного сайта, и в особенности к молекуле sTv, имеющей вариабельный домен α-цепи и вариабельный домен β-цепи дикого типа. Типичные примеры включают LC13-WT.
В одном из предпочтительных воплощений участки CDR в TCR являются участками дикого типа или содержат мутации, повышающие аффинность.
В одном из предпочтительных воплощений "аффинность" относится к аффинности связывания молекулы TCR и ее соответствующего антигена.
В одном из предпочтительных воплощений в каркасе вариабельного домена TCR и константном домене гидрофобные остатки, боковая цепь которых направлена к поверхности, являются мутантными. То есть, аминокислотные остатки, представленные на поверхности в каркасе вариабельного домена и константном домене в TCR являются мутантными. Предпочтительно, мутантные аминокислотные остатки представляют собой аминокислотные остатки, представленные на поверхности в вариабельном домене α-цепи и/или β-цепи TCR. Конкретнее, позиции аминокислот, представленных на поверхности, включают позиции 4, 12, 16, 93, 97, 100 и 105 в вариабельном домене α-цепи TCR и последнюю позицию α-цепи, кодируемой J-геном; позиции 4 и 101 в вариабельном домене β-цепи TCR, и последнюю позицию, и третью позицию с конца β-цепи, кодируемой J-геном. Нумерация позиций аминокислот представляет собой нумерацию, представленную в IMGT (международная база данных по иммуногенетике).
В одном из предпочтительных воплощений мутации гидрофобных остатков, боковая цепь которых направлена к поверхности, в каркасе вариабельного домена в TCR включают (не ограничиваясь приведенным списком) в α-цепи: I7S, A9S, A10S, V20S, А92Е, A93S; I→T во второй позиции с конца короткоцепочечного пептида, кодируемого J-геном; в β-цепи: I12S; или какая-либо их комбинация; где нумерация позиций аминокислот является нумерацией, представленной в IMGT.
В одном из предпочтительных воплощений TCR является растворимым.
В одном из предпочтительных воплощений TCR представляет собой мембранный белок.
В одном из предпочтительных воплощений TCR содержит (а) всю или часть α-цепи TCR, за исключением ее трансмембранного домена; и (б) всю или часть β-цепи TCR, за исключением ее трансмембранного домена;
и каждый из (а) и (б) независимо содержит функциональный вариабельный домен, или функциональный вариабельный домен и по меньшей мере часть константного домена цепи TCR.
В одном из предпочтительных воплощений TCR представляет собой одноцепочечный TCR, состоящий из вариабельного домена α-цепи TCR и вариабельного домена β-цепи TCR, связанных гибким линкерным пептидом.
В одном из предпочтительных воплощений мутации включают по меньшей мере одну мутацию в гидрофобном ядре.
В одном из предпочтительных воплощений TCR имеет одну или более чем одну мутацию в следующих позициях: позиции в гидрофобном ядре аминокислотной последовательности вариабельного домена α и/или β-цепи, т.е. позиции аминокислот 11, 13, 19, 21, 53, 76, 89, 91, 94 вариабельного домена и/или третья, пятая или седьмая позиции аминокислот с конца α-цепи короткоцепочечного пептида, кодируемого J-геном, и/или вторая, четвертая или шестая аминокислотные позиции с конца β-цепи короткоцепочечного пептида, кодируемого J-геном, где нумерация позиций аминокислот представляет собой нумерацию, представленную в IMGT.
В одном из предпочтительных воплощений одна или более чем одна из следующих позиций в вариабельном домене α-цепи TCR является мутантной: позиции аминокислот 11, 13, 19, 21, 53, 76, 89, 91 или 94 вариабельного домена α-цепи и/или третья, пятая или седьмая позиции аминокислот с конца α-цепи короткоцепочечного пептида, кодируемого J-геном, где нумерация позиций аминокислот представляет собой нумерацию, представленную в IMGT (международная база данных по иммуногенетике).
В одном из предпочтительных воплощений одна или более чем одна из следующих позиций в вариабельном домене α-цепи, представленном в SEQ ID NO: 9, или SEQ ID NO: 29, или SEQ ID NO: 31, или SEQ ID NO: 33 TCR, является мутантной: позиции аминокислот 11, 13, 19, 21, 53, 76, 89, 91 или 94 в вариабельном домене α-цепи, и/или третья, пятая или седьмая позиции аминокислот с конца α-цепи короткоцепочечного пептида, кодируемого J-геном, где нумерация позиций аминокислот представляет собой нумерацию, представленную в IMGT.
В одном из предпочтительных воплощений одна или более чем одна из следующих позиций в вариабельном домене β-цепи TCR является мутантной: позиции аминокислот 11, 13, 19, 21, 53, 76, 89, 91 или 94 в вариабельном домене β-цепи и/или вторая, четвертая или шестая позиции с конца β-цепи короткоцепочечного пептида, кодируемого J-геном, где нумерация позиций аминокислот представляет собой нумерацию, представленную в IMGT.
В одном из предпочтительных воплощений одна или более чем одна из следующих позиций в вариабельном домене β-цепи, представленном в SEQ ID NO: 11 или SEQ ID NO: 30, или SEQ ID NO: 32, или SEQ ID NO: 34 TCR, является мутантной: позиции аминокислот 11, 13, 19, 21, 53, 76, 89, 91 или 94 в вариабельном домене β-цепи и/или вторая, четвертая или шестая позиции с конца β-цепи короткоцепочечного пептида, кодируемого J-геном, где нумерация позиций аминокислот представляет собой нумерацию, представленную в IMGT.
В одном из предпочтительных воплощений вариабельный домен α-цепи TCR содержит один или более чем один аминокислотный остаток, выбранный из группы, состоящей из: 11L, 11М или 11Е; 13V, 13R или 13K; 19V; 21I; 91L или 91I; и 94V или 94I; и/или вариабельный домен β-цепи TCR содержит один или более чем один аминокислотный остаток, выбранный из группы, состоящей из: 11L или 11V; 13V; 19V; 89L; 91F или 91I; 94V или 94L; шестая позиция с конца β-цепи, кодируемой J-геном, представляет собой Т; и четвертая позиция с конца β-цепи, кодируемой J-геном, представляет собой М; где нумерация позиций аминокислот представляет собой нумерацию, представленную в IMGT.
В одном из предпочтительных воплощений аминокислотный остаток, находящийся в вариабельном домене α-цепи и/или β-цепи TCR и представленный на поверхности, является мутантным.
В одном из предпочтительных воплощений TCR содержит один или более чем один аминокислотный остаток вариабельного домена α-цепи, выбранный из группы, состоящей из: 4L; 12N; 16S; 93N или 93R; 97N; 100G; 105S; и последняя позиция α-цепи, кодируемой J-геном, представляет собой D; и/или TCR содержит один или более чем один аминокислотный остаток вариабельного домена β-цепи, выбранный из группы, состоящей из: 4I; 101L; последняя позиция β-цепи, кодируемой J-геном, представляет собой D; и третья позиция с конца β-цепи, кодируемой J-геном, представляет собой Е.
В одном из предпочтительных воплощений TCR содержит вариабельный домен α-цепи, аминокислотная последовательность которого выбрана из группы, состоящей из SEQ ID NO: 15, 17, 35, 37, 39, 75, 76, 77, 78, 79, 80, 81, 82, 83, 84, 85, 97, 99, 101, 103, 105 и 107.
В одном из предпочтительных воплощений TCR содержит вариабельный домен β-цепи, аминокислотная последовательность которого выбрана из группы, состоящей из SEQ ID NO: 16, 18, 36, 38, 40, 86, 87, 88, 89, 90, 91, 92, 93, 94, 95, 96, 98, 100, 102, 104, 106 и 108.
В одном из предпочтительных воплощений комбинация вариабельного домена α-цепи и вариабельного домена β-цепи TCR выбрана из группы, состоящей из:
(а) вариабельного домена α-цепи, имеющего аминокислотную последовательность SEQ ID NO: 15, и вариабельного домена β-цепи, имеющего аминокислотную последовательность SEQ ID NO: 16;
(б) вариабельного домена α-цепи, имеющего аминокислотную последовательность SEQ ID NO: 17, и вариабельного домена β-цепи, имеющего аминокислотную последовательность SEQ ID NO: 18;
(в) вариабельного домена α-цепи, имеющего аминокислотную последовательность SEQ ID NO: 15, и вариабельного домена β-цепи, имеющего аминокислотную последовательность SEQ ID NO: 18;
(г) вариабельного домена α-цепи, имеющего аминокислотную последовательность SEQ ID NO: 35, и вариабельного домена β-цепи, имеющего аминокислотную последовательность SEQ ID NO: 36;
(д) вариабельного домена α-цепи, имеющего аминокислотную последовательность SEQ ID NO: 37, и вариабельного домена β-цепи, имеющего аминокислотную последовательность SEQ ID NO: 38;
(е) вариабельного домена α-цепи, имеющего аминокислотную последовательность SEQ ID NO: 39, и вариабельного домена β-цепи, имеющего аминокислотную последовательность SEQ ID NO: 40;
(ж) вариабельного домена α-цепи, имеющего аминокислотную последовательность SEQ ID NO: 75, и вариабельного домена β-цепи, имеющего аминокислотную последовательность SEQ ID NO: 86;
(з) вариабельного домена α-цепи, имеющего аминокислотную последовательность SEQ ID NO: 76, и вариабельного домена β-цепи, имеющего аминокислотную последовательность SEQ ID NO: 87;
(и) вариабельного домена α-цепи, имеющего аминокислотную последовательность SEQ ID NO: 77, и вариабельного домена β-цепи, имеющего аминокислотную последовательность SEQ ID NO: 88;
(к) вариабельного домена α-цепи, имеющего аминокислотную последовательность SEQ ID NO: 78, и вариабельного домена β-цепи, имеющего аминокислотную последовательность SEQ ID NO: 89;
(л) вариабельного домена α-цепи, имеющего аминокислотную последовательность SEQ ID NO: 79, и вариабельного домена β-цепи, имеющего аминокислотную последовательность SEQ ID NO: 90;
(м) вариабельного домена α-цепи, имеющего аминокислотную последовательность SEQ ID NO: 80, и вариабельного домена β-цепи, имеющего аминокислотную последовательность SEQ ID NO: 91;
(н) вариабельного домена α-цепи, имеющего аминокислотную последовательность SEQ ID NO: 81, и вариабельного домена β-цепи, имеющего аминокислотную последовательность SEQ ID NO: 92;
(о) вариабельного домена α-цепи, имеющего аминокислотную последовательность SEQ ID NO: 82, и вариабельного домена β-цепи, имеющего аминокислотную последовательность SEQ ID NO: 93;
(п) вариабельного домена α-цепи, имеющего аминокислотную последовательность SEQ ID NO: 83, и вариабельного домена β-цепи, имеющего аминокислотную последовательность SEQ ID NO: 94;
(р) вариабельного домена α-цепи, имеющего аминокислотную последовательность SEQ ID NO: 84, и вариабельного домена β-цепи, имеющего аминокислотную последовательность SEQ ID NO: 95;
(с) вариабельного домена α-цепи, имеющего аминокислотную последовательность SEQ ID NO: 85, и вариабельного домена β-цепи, имеющего аминокислотную последовательность SEQ ID NO: 96;
(т) вариабельного домена α-цепи, имеющего аминокислотную последовательность SEQ ID NO: 97, и вариабельного домена β-цепи, имеющего аминокислотную последовательность SEQ ID NO: 98;
(у) вариабельного домена α-цепи, имеющего аминокислотную последовательность SEQ ID NO: 99, и вариабельного домена β-цепи, имеющего аминокислотную последовательность SEQ ID NO: 100;
(ф) вариабельного домена α-цепи, имеющего аминокислотную последовательность SEQ ID NO: 101, и вариабельного домена β-цепи, имеющего аминокислотную последовательность SEQ ID NO: 102;
(х) вариабельного домена α-цепи, имеющего аминокислотную последовательность SEQ ID NO: 103, и вариабельного домена β-цепи, имеющего аминокислотную последовательность SEQ ID NO: 104;
(ц) вариабельного домена α-цепи, имеющего аминокислотную последовательность SEQ ID NO: 105, и вариабельного домена β-цепи, имеющего аминокислотную последовательность SEQ ID NO: 106; и
(ч) вариабельного домена α-цепи, имеющего аминокислотную последовательность SEQ ID NO: 107, и вариабельного домена β-цепи, имеющего аминокислотную последовательность SEQ ID NO: 108.
В одном из предпочтительных воплощений гидрофобный коровый домен вариабельного домена α-цепи в TCR имеет по меньшей мере одну из следующих мутаций: аминокислота в позиции 19 мутирована до V, аминокислота в позиции 21 мутирована до I, аминокислота в позиции 91 мутирована до L и/или гидрофобный коровый домен вариабельного домена β-цепи имеет по меньшей мере одну из следующих мутаций: аминокислота в позиции 91 мутирована до F или I, и/или аминокислота в четвертой позиции относительно последней аминокислоты последовательности короткого пептида J-гена β-цепи мутирована до М. В одном из предпочтительных воплощений мутации выбраны из группы, состоящей из:
(1) в вариабельном домене α-цепи аминокислота в позиции 19 мутирована до V, аминокислота в позиции 21 мутирована до I, аминокислота в позиции 91 мутирована до L; в вариабельном домене β-цепи аминокислота в позиции 91 мутирована до F, аминокислота в четвертой позиции с конца β-цепи короткого пептида, кодируемого J-геном, мутирована до М; или
(2) в вариабельном домене α-цепи аминокислота в позиции 19 мутирована до V, аминокислота в позиции 21 мутирована до I, и в вариабельном домене β-цепи аминокислота в позиции 91 мутирована до I; или
(3) в вариабельном домене α-цепи аминокислота в позиции 19 мутирована до V, аминокислота в позиции 21 мутирована до I, аминокислота в позиции 91 мутирована до L; в вариабельном домене β-цепи аминокислота в позиции 91 мутирована до F.
В одном из предпочтительных воплощений гидрофобный коровый домен вариабельного домена α-цепи TCR имеет по меньшей мере одну из следующих мутаций: L19V, L21I, 191L; и/или гидрофобный коровый домен вариабельного домена β-цепи имеет по меньшей мере одну из следующих мутаций: V91F или V91I; и/или аминокислота в четвертой позиции с конца β-цепи короткого пептида, кодируемого J-геном, мутирована с L до М.
В одном из предпочтительных воплощений мутации выбраны из группы, состоящей из:
(1) L19V, L21I, 191L в вариабельном домене α-цепи, V91F в вариабельном домене β-цепи, и аминокислота в четвертой позиции с конца β-цепи короткого пептида, кодируемого J-геном, мутирована с L до М; или
(2) L19V, L21I в вариабельном домене α-цепи, и V91I в вариабельном домне β-цепи; или
(3) L19V, L21I, 191L в вариабельном домене α-цепи, и V91I в вариабельном домне β-цепи;
где нумерация аминокислот представлена в IMGT.
В одном из предпочтительных воплощений TCR дополнительно имеет дисульфидную связь, связывающую константный домен α-цепи и константный домен β-цепи.
В одном из предпочтительных воплощений дисульфидная связь представлена в природном TCR или введена искусственно.
В одном из предпочтительных воплощений искусственно введенная дисульфидная связь располагается между константными доменами α- и β-цепей TCR.
В одном из предпочтительных воплощений искусственно введенные цистеиновые остатки, образующие межцепочечную дисульфидную связь, замещают по меньшей мере одну пару аминокислотных остатков α- и β-цепей, включающих, но не ограниченных приведенным списком:
(а) Т в константном домене α-цепи в позиции 48 и S в константном домене β-цепи в позиции 57; или
(б) Т в константном домене α-цепи в позиции 45 и S в константном домене β-цепи в позиции 77; или
(в) Т в константном домене α-цепи в позиции 10 и S в константном домене β-цепи в позиции 17; или
(г) Т в константном домене α-цепи в позиции 45 и D в константном домене β-цепи в позиции 59; или
(д) S в константном домене α-цепи в позиции 15 и Е в константном домене β-цепи в позиции 15; или
(е) S в константном домене α-цепи в позиции 61 и S в константном домене β-цепи в позиции 57; или
(ж) L в константном домене α-цепи в позиции 50 и S в константном домене β-цепи в позиции 57; или
(з) S в константном домене α-цепи в позиции 15 и V в константном домене β-цепи в позиции 13; или
(и) L в константном домене α-цепи в позиции 12 и S в константном домене β-цепи в позиции 17; или
(к) S в константном домене α-цепи в позиции 61 и R в константном домене β-цепи в позиции 79; или
(л) L в константном домене α-цепи в позиции 12 и F в константном домене β-цепи в позиции 14; или
(м) V в константном домене α-цепи в позиции 22 и F в константном домене β-цепи в позиции 14; или
(н) Y в константном домене α-цепи в позиции 43 и L в константном домене β-цепи в позиции 63; или
(o) Y в константном домене α-цепи в позиции 10 и S в константном домене β-цепи в позиции 17.
Используемая здесь нумерация позиций аминокислот, заменяемых в константном домене α-цепи и β-цепи, представляет собой нумерацию позиций, представленную в источнике Stable, souble T-cell receptor молекул for crystallization and therapeutics (Jonathan M. Boulter et al., 2003, Protein Engineering 16 (9): 707-711).
В одном из предпочтительных воплощений TCR отбирают при помощи технологии фагового дисплея.
В одном из предпочтительных воплощений TCR связан с конъюгатом (ковалентно или другими способами).
В одном из предпочтительных воплощений конъюгат представляет собой один или более чем один конъюгат, выбранный из группы, состоящей из:
(1) обнаруживаемого маркера;
(2) терапевтического агента; и
(3) группировки, модифицирующей PK (от англ. pharmacokinetic profile - фармакокинетический профиль).
Предпочтительно, обнаруживаемый маркер содержит: флуоресцентную или люминесцентную метку, радиоактивную метку, контрастирующий агент для МРТ (магнитно-резонансная томография) или КТ (рентгеновская компьютерная томография), или фермент, способный продуцировать обнаруживаемый продукт.
Предпочтительно, терапевтический агент содержит: радионуклид, биотоксин, цитокин (например, ИЛ-2 и т.п.), антитело, Fc (кристаллизующийся фрагмент) антитела, scFv (одноцепочечный фрагмент вариабельной области) антитела, золотую наночастицу/наностержень, вирусную частицу, липосому, наномагнитную частицу, фермент, активирующий пролекарство (например, DT-диафораза (DTD) или дифенилгидролаза-подобный белок (BPHL)), химиотерапевтический агент (например, цисплатин) или наночастицу в какой-либо форме.
В одном из предпочтительных воплощений конъюгат представляет собой антитело против CD3, связанное с С- или N-концом α- и/или β-цепей TCR.
Во втором аспекте изобретения предложена молекула нуклеиновой кислоты, содержащая последовательность, кодирующую TCR в соответствии с первым аспектом изобретения, или комплементарную ей последовательность.
В третьем аспекте изобретения предложен вектор, содержащий молекулу нуклеиновой кислоты в соответствии со вторым аспектом изобретения.
В четвертом аспекте изобретения предложена клетка-хозяин, которая содержит вектор в соответствии с третьим аспектом изобретения, или в которой экзогенная молекула нуклеиновой кислоты в соответствии со вторым аспектом изобретения интегрирована в хромосоме.
В одном из предпочтительных воплощений клетка-хозяин выбрана из группы, состоящей из: прокариотической и эукариотической клетки, такой как Escherichia coli, дрожжей, клетки СНО (яичника китайского хомячка) и т.п.
В пятом аспекте изобретения предложен способ получения TCR в соответствии с первым аспектом изобретения, включающий:
(1) инкубацию клетки хозяина в соответствии с четвертым аспектом изобретения, таким образом экспрессирующей TCR в соответствии с первым аспектом изобретения; и
(2) выделение или очистку TCR.
В шестом аспекте изобретения предложен комплекс TCR, содержащий одну или более чем одну молекулу TCR в соответствии с первым аспектом изобретения.
В одном из предпочтительных воплощений комплекс содержит комплекс, образуемый TCR по изобретению, связанным с терапевтическим агентом или обнаруживаемым маркером.
В одном из предпочтительных воплощений комплекс содержит две или более чем две молекулы TCR.
В седьмом аспекте изобретения предложено применение TCR в соответствии с первым аспектом изобретения для производства лекарственного средства для лечения опухоли, вирусной инфекции или аутоиммунного заболевания.
В восьмом аспекте изобретения предложена фармацевтическая композиция, содержащая фармацевтически приемлемый носитель и безопасную и эффективную дозу какого-либо TCR в соответствии с первым аспектом изобретения.
В девятом аспекте изобретения предложен способ лечения заболевания, включающий введение субъекту, нуждающемуся в таком введении, TCR в соответствии с первым аспектом изобретения, или комплекса TCR в соответствии с шестым аспектом изобретения, или фармацевтической композиции в соответствии с восьмым аспектом изобретения.
В одном из предпочтительных воплощений заболевание включает опухоль, аутоиммунное заболевание и вирусную инфекцию.
В десятом аспекте изобретения предложен способ получения TCR в соответствии с первым аспектом изобретения, включающий:
(1) введение мутации аминокислотного остатка в гидрофобный коровый домен TCR; и
(2) отбор тех TCR, стабильность которых значительно увеличивается, таким образом получая TCR в соответствии с первым аспектом изобретения.
В одном из предпочтительных воплощений способ отбора включает метод фагового дисплея, но не ограничивается им.
В одном из предпочтительных воплощений на стадии (2) TCR, имеющий мутантный гидрофобный коровый домен, представляется при помощи метода фагового дисплея, и затем отбирается.
В одном из предпочтительных воплощений способ дополнительно включает стадию определения последовательности, активностей и/или других признаков отбираемых TCR.
Следует понимать, что в настоящем изобретении технические признаки, особо описанные выше и ниже (такие как примеры), могут комбинироваться друг с другом, таким образом составляя новое или предпочтительное техническое решение, и не обязательно должны быть описаны друг за другом.
ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
Фиг. 1 демонстрирует структурную диаграмму типичного вариабельного домена TCR. Такой TCR представляет собой TCR дикого типа, специфический к опухолевому антигену MAGE A3 HLA А1.
Фиг. 2а и 2б соответственно демонстрируют аминокислотную последовательность и последовательность нуклеиновой кислоты (SEQ ID NO: 9 и 10) вариабельного домена α-цепи TCR после сайт-направленной мутации. Аминокислотная последовательность представляет собой оптимизацию аминокислотной последовательности вариабельного домена α-цепи TCR, раскрытой в патентной литературе (WO 2012/013913). В частности, гидрофобные остатки в вариабельном домене, имеющие боковые цепи, направленные к поверхности, мутированы в гидрофильные или полярные остатки. Жирные и подчеркнутые буквы представляют аминокислотные остатки после мутации.
Фиг. 3а и 3б соответственно демонстрируют аминокислотную последовательность и последовательность нуклеиновой кислоты (SEQ ID NO: 11 и 12) вариабельного домена β-цепи TCR после сайт-направленной мутации. Аминокислотная последовательность представляет собой оптимизацию аминокислотной последовательности вариабельного домена β-цепи TCR, раскрытой в патентной литературе (WO 2012/013913). В частности, гидрофобные остатки в вариабельном домене, имеющие боковые цепи, направленные к поверхности, мутированы в гидрофильные или полярные остатки. Жирные и подчеркнутые буквы представляют аминокислотные остатки после мутации.
Фиг. 4 демонстрирует способ лигирования каждого праймера в процессе конструирования MAGE-sTv-WT.
Фиг. 5а и 5б соответственно демонстрируют аминокислотную последовательность и последовательность нуклеиновой кислоты (SEQ ID NO: 13 и 14) линкера между α- и β-цепями в процессе конструирования библиотек мутантных штаммов sTv.
Фиг. 6а и 6б соответственно демонстрируют аминокислотную последовательность вариабельного домена α- и β-цепи (SEQ ID NO: 15 и 16) мутантного штамма sTv MG29. Мутантные остатки по сравнению с референсными MAGE-sTv-WT выделены жирным и подчеркнуты.
Фиг. 7а и 7б соответственно демонстрируют аминокислотную последовательность вариабельного домена α- и β-цепей (SEQ ID NO: 17 и 18) мутантного штамма sTv P8F1. Мутантные остатки по сравнению с референсными MAGE-sTv-WT выделены жирным и подчеркнуты.
Фиг. 8а и 8б соответственно демонстрируют аминокислотную последовательность вариабельного домена α- и β-цепи (SEQ ID NO: 15 и 18) мутантного штамма sTv P8F2. Мутантные остатки по сравнению с референсным MAGE-sTv-WT выделены жирным и подчеркнуты.
Фиг. 9 демонстрирует значения OD (оптической плотности) для различных отобранных мутантных штаммов и MAGE-sTv-WT в ИФА (иммуноферментном анализе) против антигена MAGEA3, EBV (вирус Эпштейна-Барр), вируса гриппа, NY-ESO.
Фиг. 10а и 10б соответственно демонстрируют аминокислотные последовательности вариабельного домена α-цепи (SEQ ID NO: 29) и вариабельного домена β-цепи (SEQ ID NO: 30) в LC13-WT.
Фиг. 11а и 11б соответственно демонстрируют аминокислотные последовательности вариабельного домена α-цепи (SEQ ID NO: 31) и вариабельного домена β-цепи (SEQ ID NO: 32) в JM22-WT.
Фиг. 12а и 12б соответственно демонстрируют аминокислотные последовательности вариабельного домена α-цепи (SEQ ID NO: 33) и вариабельного домена β-цепи (SEQ ID NO: 34) в 1G4-WT.
Фиг. 13а и 13б соответственно демонстрируют аминокислотные последовательности вариабельного домена α-цепи (SEQ ID NO: 35) и вариабельного домена β-цепи (SEQ ID NO: 36) в LC13-sTv.
Фиг. 14а и 14б соответственно демонстрируют аминокислотные последовательности вариабельного домена α-цепи (SEQ ID NO: 37) и вариабельного домена β-цепи (SEQ ID NO: 38) в JM22-sTv.
Фиг. 15а и 15б соответственно демонстрируют аминокислотные последовательности вариабельного домена α-цепи (SEQ ID NO: 39) и вариабельного домена β-цепи (SEQ ID NO: 40) в 1G4-sTv.
Фиг. 16 демонстрирует аминокислотную последовательность линкера (SEQ ID NO: 41), используемого для конструирования одноцепочечной молекулы sTV.
Фиг. 17 демонстрирует результат SDS-PAGE (электрофореза в полиакриламидном геле с додецилсульфатом натрия) очищенных белков LC13-WT и LC13-sTv. Дорожка 1: маркеры молекулярной массы, дорожка 2: LC13-WT, дорожка 3: LC13-sTv.
Фиг. 18а и 18б соответственно демонстрируют профиль SEC (гель-фильтрационной хроматографии) очищенных белков LC13-WT и LC13-sTv.
Фиг. 19 демонстрирует результат SDS-PAGE очищенного белка JM22-WT и JM22-sTv. Дорожка 1: маркеры молекулярной массы, дорожка 2: JM22-WT, дорожка 3: JM22-sTv.
Фиг. 20а и 20б соответственно демонстрируют профиль SEC очищенного белка JM22-WT и JM22-sTv.
Фиг. 21 демонстрирует результат SDS-PAGE очищенного белка 1G4-WT и 1G4-sTv. Дорожка 1: маркеры молекулярной массы, дорожка 2: 1G4-WT, дорожка 3: 1G4-sTv.
Фиг. 22а и 22б соответственно демонстрируют профиль SEC очищенного белка 1G4-WT и 1G4-sTv.
Фиг. 23 демонстрирует аминокислотную последовательность 1G4-sTv (SEQ ID NO: 42).
Фиг. 24 демонстрирует значения OD мутантного штамма 1G4-sTv против различных антигенов.
Фиг. 25 демонстрирует аминокислотную последовательность вариабельного домена α-цепи отобранных мутантных штаммов 1G4-sTv, обладающих высокой стабильностью (SEQ ID NO: 75-85).
Фиг. 26 демонстрирует аминокислотную последовательность вариабельного домена в β-цепи отобранных мутантных штаммов 1G4-sTv, обладающих высокой стабильностью (SEQ ID NO: 86-96).
Фиг. 27 демонстрирует график ДСК (дифференциальной сканирующей калориметрии) мутантных штаммов 1G4-sTv, обладающих высокой стабильностью.
Фиг. 28 демонстрирует график ДСК для 1G4-WT.
Фиг. 29а и 29б соответственно демонстрируют аминокислотные последовательности вариабельного домена α-цепи (SEQ ID NO: 97) и вариабельного домена β-цепи (SEQ ID NO: 98) G15, обладающего высокой стабильностью.
Фиг. 30 демонстрирует результат SDS-PAGE очищенного белка 1G4-WT, 1G4-sTv, G13, G15, G9. Дорожка 1: маркеры молекулярной массы, дорожка 2: 1G4-WT, дорожка 3: 1G4-sTv, дорожка 4: G13, дорожка 5: G15, дорожка 6: маркеры молекулярной массы, дорожка 7: G9.
Фиг. 31а, 31б и 31в соответственно демонстрируют профиль SEC очищенных белков G9, G13 и G15.
Фиг. 32а и 32б соответственно демонстрируют аминокислотные последовательности вариабельного домена α-цепи (SEQ ID NO: 99) и вариабельного домена β-цепи (SEQ ID NO: 100) LC13-G9.
Фиг. 33а и 33б соответственно демонстрируют аминокислотные последовательности вариабельного домена α-цепи (SEQ ID NO: 101) и вариабельного домена β-цепи (SEQ ID NO: 102) LC13-G15.
Фиг. 34а и 34б соответственно демонстрируют аминокислотные последовательности вариабельного домена α-цепи (SEQ ID NO: 103) и вариабельного домена β-цепи (SEQ ID NO: 104) JM22-G9.
Фиг. 35а и 35б соответственно демонстрируют аминокислотные последовательности вариабельного домена α-цепи (SEQ ID NO: 105) и вариабельного домена β-цепи (SEQ ID NO: 106) JM22-G15.
Фиг. 36 демонстрирует результат SDS-PAGE очищенных белков LC13-WT, LC13-sTv, LC13-G15 и LC13-G9. Дорожка 1: маркеры молекулярной массы, дорожка 2: LC13-WT, дорожка 3: LC13-sTv, дорожка 4: LC13-sTv, дорожка 5: маркеры молекулярной массы, дорожка 6: LC13-G9.
Фиг. 37 демонстрирует профиль SEC очищенного белка LC13-G9.
Фиг. 38 демонстрирует профиль SEC очищенного белка LC13-G15.
Фиг. 39 демонстрирует результат SDS-PAGE очищенных белков JM22-WT, JM22-sTv, JM22-G15 и JM22-G9. Дорожка 1: маркеры молекулярной массы, дорожка 2: JM22-WT, дорожка 3: JM22-sTv, дорожка 4: JM22-G15, дорожка 5: JM22-G9.
Фиг. 40 демонстрирует профиль SEC очищенного белка JM22-G9.
Фиг. 41 демонстрирует профиль SEC очищенного белка JM22-G15.
Фиг. 42а и 42б соответственно демонстрируют аминокислотные последовательности вариабельного домена α-цепи (SEQ ID NO: 107) и вариабельного домена β-цепи (SEQ ID NO: 108) MAGE-G15.
Фиг. 43 демонстрирует результат SDS-PAGE очищенного белка MAGE-G15. Дорожка 1: маркеры молекулярной массы, дорожка 2: MAGE-G15.
Фиг. 44 демонстрирует профиль SEC очищенного белка MAGE-G15.
Фиг. 45 демонстрирует график ДСК очищенного белка MAGE-G15.
Фиг. 46 демонстрирует график ДСК очищенного белка G15.
Фиг. 47 демонстрирует график ДСК очищенного белка LC13-sTv.
Фиг. 48а и 48b соответственно демонстрируют графики ДСК очищенных белков JM22-WT и JM22-sTv.
Фиг. 49а и 49b соответственно демонстрируют графики ДСК очищенных белков LC13-G9 и LC13-G15.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Путем обширных и интенсивных исследований авторы изобретения неожиданно и первыми обнаружили, что мутантный TCR, обладающий высокой стабильностью и, в особенности, обладающий растворимостью, может быть получен после направленной мутации гидрофобного корового домена TCR. На основе данного открытия авторы изобретения выполнили настоящее изобретение.
Авторы настоящего изобретения использовали оптимизированную белковую структуру TCR для конструирования молекулы TCR, обладающей высокой стабильностью, путем замены гидрофобного ядра TCR. В настоящем изобретении сконструирован новый тип одноцепочечного вариабельного домена TCR при помощи способа направленной молекулярной эволюции, и выделено наилучшее гидрофобное ядро. Для фрагмента TCR, имеющего такое новое гидрофобное ядро, гидрофобные остатки, расположенные на поверхности вариабельного домена TCR, могут для дополнительного улучшения быть заменены гидрофильными или полярными остатками.
Определение
TCR
Нативные α/β гетеродимерные TCR имеют α-цепь и β-цепь. Обобщенно, каждая цепь содержит вариабельные, связанные и константные области, и бета-цепь также обычно содержит короткую область разнообразия между вариабельными и связывающими областями, но эта область разнообразия часто рассматривается как часть связывающей области. Три CDR (участка, определяющих комплементарность) каждой вариабельной области встроены в каркас вариабельной области, и гидрофобное ядро также расположено в каркасе вариабельной области. Существует несколько типов вариабельных областей α-цепи (Vα) и несколько типов вариабельных областей β-цепи (Vβ). В международной базе данных по иммуногенетике (IMGT) типы Vα и типы Vβ обозначаются в номенклатуре по отдельности уникальными номерами TRAV и TRBV. TRAJ и TRBJ относятся к связывающим областям TCR. Используемые в настоящем изобретении J-ген α-цепи обозначается TRAJ и J-ген β-цепи обозначается TRBJ. Каждая из α- и β-цепей TCR в общем рассматривается как имеющая два "домена", а именно вариабельный и константный домены. Вариабельный домен состоит из объединения вариабельной области и связывающей области. Таким образом, в описании и формуле настоящего изобретения термин "вариабельный домен α TCR" относится к объединению TRAV и TRAJ, и термин "вариабельный домен β TCR" относится к объединению TRBV и TRBJ.
Аминокислотные последовательности TCR и каркаса его вариабельного домена, включающие специфический номер расположения позиции гидрофобного ядра в IMGT, устанавливаемый IMGT, широко известны и доступны для специалистов, работающих в области TCR. Например, их можно найти в публичной базе данных IMGT. Если не указано иное, в настоящем изобретении нумерация позиции аминокислоты представляет собой нумерацию, определяемую в IMGT. Если нумерация позиции, приведенная в IMGT, меняется, тогда приоритетом должна обладать нумерация позиции аминокислоты в TCR, представленная в IMGT в версии от 1 января 2013 года.
Используемый здесь термин "гидрофобное ядро" (также называемое "гидрофобный центр") относится к коровой области, в основном состоящей из гидрофобных аминокислот, которые, как правило, расположены внутри молекулярной структуры белковых доменов, когда какой-либо белок растворен в воде. В TCR гидрофобное ядро вариабельного домена α-цепи TCR представляет собой позиции аминокислот 11, 13, 19, 21, 53, 76, 89, 91, 94 в вариабельном домене, и третью, пятую или седьмую позиции аминокислот с конца α-цепи короткого пептида, кодируемого J-геном (TRAJ); гидрофобное ядро вариабельного домена β-цепи TCR представляет собой позиции аминокислот 11, 13, 19, 21, 53, 76, 89, 91, 94 в вариабельном домене, и вторую, четвертую или шестую позиции аминокислот с конца β-цепи короткого пептида, кодируемого J-геном (TRBJ). Вышеприведенная нумерация позиций аминокислот представляет собой нумерацию, представленную в IMGT.
Фиг. 1 демонстрирует структурную диаграмму вариабельного домена дикого типа TCR, специфического к раковому антигену MAGE A3 HLA А1, и выделенные жирным шрифтом аминокислотные остатки, представленные в двух эллипсах снизу слева и справа, представляют собой гидрофобное ядро, расположенное в каркасной области вариабельного домена α- и β-цепи, соответственно. Антигенсвязывающие сайты TCR расположены в областях CDR, и области CDR определяют связывающую аффинность между TCR и соответствующим его антигеном. Как представлено на фигуре, гидрофобное ядро не находится в области CDR, и мутации гидрофобного ядра не будут влиять на аффинность связывания между TCR и соответствующим антигеном. Тем не менее, мутации гидрофобного ядра влияют на стабильность TCR в соответствии с исследованием авторов изобретения.
Следует понимать, что структурная диаграмма TCR использована только для объяснения настоящего изобретения, а не для ограничения объема настоящего изобретения каким-либо образом.
Термин "sTv" относится к одноцепочечному TCR с функциональным доменом, состоящим из вариабельных доменов α- и β-цепей TCR, связанных гибким линкерным пептидом. Гибкая пептидная цепь может представлять собой любую пептидную цепь, подходящую для связывания вариабельных доменов α- и β-цепей TCR, и количество аминокислотных остатков в гибкой пептидной цепи может составлять от 1 до 50, но не ограничиваться этим количеством.
Термин "стабильность" относится к любому выражению стабильности белка. По сравнению с исходным белком дикого типа отобранный высокостабильный белок обладает одной или более чем одной из следующих характеристик: более устойчив к анфолдингу, более устойчив к неправильному или нежелательному фолдингу, повышенной способностью к ренатурации, повышенной способностью к экспрессии, более высоким выходом при ренатурации белка, увеличенной термостабильностью, и увеличенной стабильностью в различных условиях (например, при различной величине рН, концентрации соли, в присутствии детергентов или денатурирующих агентов).
Система фагового дисплея и отбор TCR, обладающих высокой стабильностью
Когда систему фагового дисплея используют для выделения рецептора, окончательный рецептор отбирают на основе двух важных свойств. Первое свойство является силой связывания или аффинности к своему лиганду, а второе является плотностью дисплея на поверхности фага. Первое свойство представляет собой основу для эволюции аффинности белка, на основании которого развиваются все методологии для получения высокоаффинных рецепторов. Может быть приведено следующее простое описание: когда библиотеку, демонстрирующую рецептор, применяют в отношении лиганда, рецепторы с более высокой силой связывания будут связываться с лигандом с большей скоростью и/или с большим временем удерживания, и они будут устойчивы к более жестким процессам отмывания, таким образом, что эти рецепторы с кодирующими их генами будут обнаружены и затем амплифицированы с использованием последующих способов. С другой стороны, когда аффинность взаимодействия рецептор-лиганд не была изменена или изменялась незначительно или даже меньше, аффинность не играет роли для отбора, и плотность дисплея управляет результатами эволюции. Последнее означает, что когда на единичной фаговой частице демонстрируются молекулы рецепторов, свернутые более правильным образом, или большее количество фаговых частиц демонстрирует один или более чем один такой рецептор, тогда рецептор и кодирующий ген обладают большими возможностями для связывания с лигандом, и при определенных условиях отмывания такой рецептор должен удерживаться дольше, и, таким образом, обнаруживаться и амплифицироваться в течение последующих стадий отбора. На основе второго свойства более стабильный белок может быть выделен путем использования фагового дисплея или других направленных способов молекулярного дисплея. Авторы изобретения сконструировали библиотеки гидрофобного ядра белка TCR для направленной эволюции с целью выделения более стабильных белков или TCR. Подтверждено, что такие гидрофобные ядра не оказывают никакого воздействия на силу связывания TCR с его лигандом рМНС или pHLA(пептид-Человеческий лейкоцитарный антиген), поскольку TCR связывается с рМНС через свои CDR.
В настоящем изобретении технологию фагового дисплея используют для выделения более стабильных белковых конструкций. В одном из предпочтительных воплощений для проверки гипотезы использовали внеклеточный домен TCR, обладающий специфичностью к раковому антигену рМНС MAGE A3 HLA А1. Внеклеточный домен синтезируют в соответствии с последовательностью, представленной в ссылочных патентных документах. Когда он экспрессируется на поверхности нитевидных бактериофагов, связывание TCR с рМНС может быть обнаружено при помощи ИФА, и может быть определена сила взаимодействия. Тем не менее, при использовании опубликованных способов, таких как мутирование гидрофобных остатков в вариабельных доменах и представленных на поверхности в гидрофильных или полярных остатках, одноцепочечная форма TCR (sTv), состоящая из вариабельных доменов α- и β-цепи TCR, демонстрируемых фагами, не может демонстрировать какую-либо функцию связывания, обнаруживаемую при помощи ИФА. Тем не менее, когда библиотеку, содержащую мутацию гидрофобного ядра вариабельного домена в одноцепочечном TCR (sTv), подвергшуюся обработке рестриктазами, клонировали в вектор фагового дисплея, то после нескольких раундов отбора неожиданно получали несколько клонов, обладающих высокой стабильностью. Затем связывание с соответствующим рМНС обнаруживали при помощи ИФА.
Активные полипептиды
В настоящем изобретении термины "полипептид(ы) по настоящему изобретению", "TCR по настоящему изобретению", "Т-клеточный(е) рецептор(ы) по настоящему изобретению" используются взаимозаменяемо, и все обозначают Т-клеточный рецептор (TCR), имеющий мутации в области своего гидрофобного ядра, и обладающий стабильностью, значительно более высокой, чем стабильность соответствующего TCR с гидрофобным ядром дикого типа.
Дополнительно, полипептид по настоящему изобретению дополнительно может содержать другие мутации за пределами области гидрофобного ядра, в особенности мутации, которые могут повышать аффинность, и мутации аминокислотных остатков, представленных на поверхности в вариабельном домене TCR.
Эти дополнительные вариации за пределами области гидрофобного ядра включают, не ограничиваясь приведенным списком, делеции, вставки и/или замены 1-6 (как правило, 1-5, предпочтительно 1-3, предпочтительней 1-2, и наиболее предпочтительно 1) аминокислот, и добавление одной или более чем одной (как правило, меньше чем 5, предпочтительно меньше чем 3, и более предпочтительно меньше чем 2) аминокислот по С-концу и/или N-концу. Например, функции белка обычно не меняются, когда аминокислотный остаток заменяется на похожий или аналогичный из известных в области техники. Кроме того, добавление одной или нескольких аминокислот по С-концу и/или N-концу как правило не меняет структуру и функцию белка. Кроме того, эти термины также включают полипептид по настоящему изобретению в мономерной и полимерной форме.
Необходимо понимать, что используемые здесь названия аминокислот представляют собой общепринятые в мире однобуквенные идентификаторы и соответствующие им сокращения названий аминокислот из трех букв английского алфавита. Они представляют собой Ala (A), Arg (R), Asn (N), Asp (D), Cys (C), Gln (Q), Glu (E), Gly (G), His (H), Ile (I), Leu (L), Lys (K), Met (M), Phe (F), Pro (P), Ser (S), Thr (T), Trp (W), Tyr (Y), и Val (V). В этом применении выражение для замены аминокислоты, такое как L19V, означает, что в позиции 19, в соответствии с нумерацией позиций в IMGT, L (лейцин) заменен на V (валин). Значения других выражений для замены аминокислот, записанных аналогичным образом, могут опираться на данный пример.
Настоящее изобретение дополнительно включает активные фрагменты, производные и аналоги полипептида по настоящему изобретению. Используемые здесь термины "фрагменты", "производные" и "аналоги" относятся к полипептидам, которые могут связываться с молекулой лиганда. Фрагменты, производные или аналоги полипептида по настоящему изобретению могут представлять собой (1) полипептид с одним или более чем одним замещенным консервативным или неконсервативным аминокислотным остатком (предпочтительно консервативные аминокислотные остатки), или (2) полипептид, имеющий замещенную(ые) группу(ы) в одном или более чем одном аминокислотном остатке, или (3) полипептид, образуемый путем слияния TCR по настоящему изобретению с другим соединением (таким как соединение, которое увеличивает период полураспада полипептида, такое как полиэтиленгликоль), или (4) полипептид с дополнительной аминокислотной последовательностью, слитой с указанной полипептидной последовательностью, такой как слитые белки, образованные путем слияния с лидерной последовательностью, последовательностью секреции или маркерной последовательностью, такой как 6His. В соответствии с применением изобретения эти фрагменты, производные и аналоги находятся в объеме, обычно известном специалистам в данной области техники.
Класс предпочтительных активных производных представляет собой полипептиды, образованные путем замены максимально 5, предпочтительно максимально 3, предпочтительней максимально 2, и наиболее предпочтительно 1 аминокислоты в аминокислотной последовательности полипептида по настоящему изобретению на аминокислоту, обладающую похожим или аналогичным свойством. Эти консервативные варианты полипептидов предпочтительно образуются путем осуществления замен аминокислот в соответствии с таблицей А.
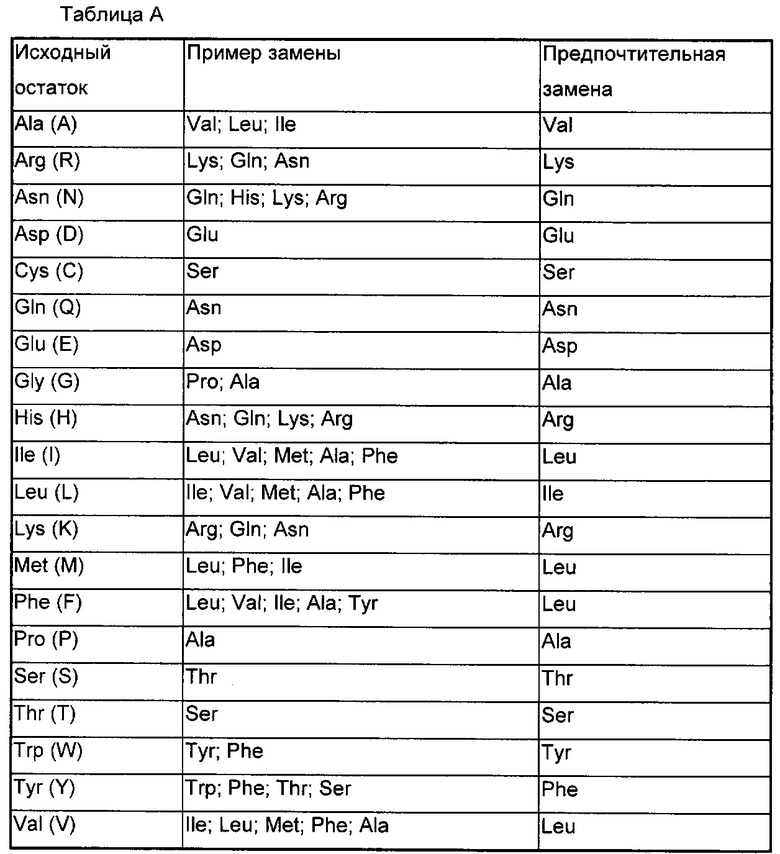
В настоящем изобретении также предложены аналоги TCR по настоящему изобретению. Эти аналоги отличаются от TCR по настоящему изобретению аминокислотной последовательностью или модификациями, не влияющим на последовательность, или и тем, и тем. Также включены аналоги, содержащие остатки, отличающиеся от встречающихся в природе L-аминокислот (например, D-аминокислоты), или не встречающиеся в природе или синтетические аминокислоты (например, бета- или гамма-аминокислоты). Понятно, что полипептиды по настоящему изобретению не ограничены типичными полипептидами, перечисленными выше.
Модификации (которые обычно не изменяют первичную последовательность) включают химическую дериватизацию полипептидов in vivo или in vitro, например ацетилирование или карбоксилирование. Гликозилирование также включено в модификацию, например полипептиды, получаемые путем модификации гликозилированием во время их синтеза и процессинга, или в течение дополнительных стадий процессинга. Эти модификации могут быть осуществлены путем воздействия на полипептид ферментов для гликозилирования (например, ферменты гликозилирования или дегликозилирования у млекопитающих). Также включены последовательности, которые имеют фосфорилированные аминокислотные остатки, например фосфотирозин, фосфосерин, фосфотреонин, а также последовательности, модифицированные для улучшения их устойчивости к протеолитической деградации или для оптимизации свойств растворимости.
Полипептиды по настоящему изобретению могут быть использованы в форме фармацевтически или физиологически приемлемой соли, полученной из кислоты или основания. Такие соли включают, не ограничиваясь приведенным списком, соли, образованные со следующими кислотами: соляной кислотой, бромистоводородной кислотой, серной кислотой, лимонной кислотой, винной кислотой, ортофосфорной кислотой, молочной кислотой, пировиноградной кислотой, уксусной кислотой, янтарной кислотой, щавелевой кислотой, фумаровой кислотой, малеиновой кислотой, щавлево-уксусной кислотой, метансульфоновой кислотой, этилсульфоновой кислотой, бензенсульфоновой кислотой или изэтионовой кислотой. Другие соли включают соли, образованные с щелочными металлами или щелочно-земельными металлами (такими как натрий, калий, кальций или магний), и сложные эфиры, карбаматы или другие традиционные формы "пролекарства".
Полипептиды по настоящему изобретению могут быть предложены в форме поливалентных комплексов. Поливалентный комплекс TCR по настоящему изобретению содержит две, три, четыре или более молекулы TCR, связанные с другой молекулой.
Кодирующая последовательность
Настоящее изобретение также относится к полинуклеотидам, кодирующим TCR по настоящему изобретению.
Полинуклеотиды по настоящему изобретению могут находиться в форме ДНК или РНК. ДНК может представлять собой кодирующую цепь или не кодирующую цепь. Например, кодирующая последовательность, кодирующая зрелый полипептид, может быть идентична кодирующей последовательности, указанной в SEQ ID NO: 10, или может представлять собой ее вырожденный вариант. Используемый здесь термин "вырожденный вариант" относится к последовательности нуклеиновой кислоты, кодирующей белок, имеющий аминокислотную последовательность SEQ ID NO: 9, но отличается от соответствующей кодирующей последовательности SEQ ID NO: 10.
Полноразмерная нуклеотидная последовательность по настоящему изобретению или ее фрагмент обычно могут быть получены при помощи амплификации путем ПЦР (полимеразной цепной реакции), рекомбинации или способов синтеза, но не ограничиваясь ими. В настоящее время, последовательности ДНК, кодирующие полипептиды по настоящему изобретению (или их фрагменты, или их производные) могут быть Полностью получены путем химического синтеза. Затем последовательности ДНК могут быть введены в различные существующие молекулы ДНК (например, векторы) и клетки, известные в области техники.
Настоящее изобретение также включает вектор, содержащий полинуклеотид по настоящему изобретению, и клетку хозяина, сконструированную при помощи вектора или кодирующей последовательности по настоящему изобретению.
Кроме того, настоящее изобретение дополнительно содержит поликлональные антитела или моноклональные антитела, специфические к полипептиду TCR по настоящему изобретению, в особенности, моноклональные антитела.
Способ получения
Один из способов получения TCR по настоящему изобретению заключается в отборе высокостабильных TCR из разнообразной библиотеки фаговых частиц, демонстрирующих такие TCR.
Мутации могут быть осуществлены с использованием любых подходящих способов, включающих, не ограничиваясь приведенным списком, способы, основанные на полимеразной цепной реакции (ПЦР), клонирование, основанное на ферментах рестриктазах, или способы безлигазного клонирования (LIC). Эти способы подробно изложены во множестве стандартных руководств по молекулярной биологии. Дополнительную подробную информацию о мутагенезе путем полимеразной цепной реакции (ПЦР) и клонировании, основанном на ферменте рестриктазе, можно найти в Sambrook & Russell, (2001) Molecular Cloning - A laboratory Manual (3rd Ed) CSHL press. Больше информации о способе LIC можно найти в Rashtchian, (1995) Curr Opin Biotechnol 6 (1): 30-6.
Полипептид по настоящему изобретению может представлять собой рекомбинантный или синтетический полипептид. Полипептид по настоящему изобретению может представлять собой химически синтезированный или рекомбинантный полипептид. Соответственно, полипептид по настоящему изобретению может быть искусственно синтезирован при помощи традиционного способа, или может быть получен при помощи рекомбинантного способа.
При помощи традиционной технологии рекомбинантной ДНК полинуклеотид по настоящему изобретению могут быть использован для экспрессии или продукции рекомбинантных полипептидов по настоящему изобретению. Как правило, способ включает следующие стадии:
(1) Трансформацию или трансфекцию подходящей клетки хозяина полинуклеотидом или его вариантом, кодирующим полипептид TCR по настоящему изобретению, или рекомбинантным экспрессирующимся вектором, содержащим указанный полинуклеотид;
(2) Выращивание клетки хозяина в подходящей культуральной среде;
(3) Выделение и очистку полипептида TCR по настоящему изобретению из культуральной среды или клетки.
Рекомбинантный полипептид может быть экспрессирован в клетках или на клеточной мембране, или секретирован из клеток. При желании физические, химические и другие свойства могут быть использованы в различных способах выделения для выделения и очистки рекомбинантного белка. Эти способы хорошо известны специалистам в данной области техники и включают, не ограничиваясь приведенным списком, традиционную обработку для ренатурации, обработку осадителем белка (такую как осаждение солью), центрифугирование, осмотический клеточный лизис, ультразвуковую обработку, ультрацентрифугирование, хроматографию на молекулярных ситах (гель-хроматографию), адсорбционную хроматографию, ионообменную хроматографию, высокоэффективную жидкостную хроматографию (ВЭЖХ) и какую-либо другую жидкостную хроматографию и их комбинацию.
Фармацевтическая композиция и способы введения
TCR по настоящему изобретению и Т-клетки, трансфицированные TCR по настоящему изобретению, могут быть представлены в фармацевтической композиции вместе с фармацевтически приемлемым носителем. TCR, поливалентные комплексы TCR и клетки по настоящему изобретению обычно поставляются как часть стерильной фармацевтической композиции, которая обычно включает фармацевтически приемлемый носитель. Фармацевтическая композиция может находиться в любых подходящих формах (в зависимости от желаемого способа введения пациенту). Она может быть представлена в единичной лекарственной форме, как правило, представлена в герметичном контейнере, и может быть представлена как часть набора. Набор обычно (хотя и не обязательно) включает инструкции по использованию. Он может включать множество указанных единичных лекарственных форм.
Дополнительно, полипептиды по настоящему изобретению могут быть использованы сами по себе или в ассоциации, или связи с другими терапевтическими агентами (например, агентами, находящимися в составе той же самой фармацевтической композиции).
Терапевтические агенты, которые могут быть ассоциированы с или связаны с TCR по настоящему изобретению, включают, не ограничиваясь приведенным списком: 1. Радиоактивный нуклид (Koppe, et al., 2005, Cancer metastasis reviews 24, 539); 2. Биологический токсин (Chaudhary et al., 1989, Nature, 339, 394; Epel et al., 2002, Cancer immunology and immunotherapy 51, 565); 3. Цитокин (Gillies, et al., 1992, PNAS, 89, 1428; Card, et al., 2004, Cancer immunology and immunotherapy 53, 345; Halin, et al., 2003, Cancer research 63, 3202); 4. Фрагмент Fc антитела (Mosquera et al., 2005, The journal of immunology 174, 4381); 5. scFv антитела (Zhu, et al., 1995, International journal of cancer 62, 319); 6. Золотую наночастицу/наностержень (Lapotko, et al., 2005, Cancer letters 239, 36; Huang, et al., 2006, Journal of the American chemical society 128, 2115); 7. Вирусные частицы (Peng, et al., 2004, Gene therapy, 11, 1234); 8. Липосому (Mamot, et al., 2005, Cancer research 65, 11631); 9. Магнитные наночастицы; 10. Ферменты, активизирующие пролекарство (такие как DT-диафораза (DTD) или дифенилгидролаза-подобный белок (BPHL)); 11. Химиотерапевтический агент (например, цисплатин) и т.п.
Фармацевтическая композиция дополнительно может содержать фармацевтически приемлемый носитель. Термин "фармацевтически приемлемый носитель" относится к носителю для использования при введении терапевтических агентов. Термин относится к таким медицинским носителям, которые сами не индуцируют продукцию антитела, вредного для субъекта, которому вводят композицию, и не обладают чрезмерной токсичностью после введения. Эти носители хорошо известны специалистам в данной области техники. Подробное обсуждение фармацевтически приемлемого эксципиента можно найти в Remington's Pharmaceutical Sciences (Mack Pub. Co., N.J., 1991). Такие носители включают, не ограничиваясь приведенным списком, физиологический раствор, буферный раствор, глюкозу, воду, глицерин, этанол, адъювант или их комбинацию.
Фармацевтически приемлемый носитель в терапевтической композиции может содержать жидкость, такую как вода, физиологический раствор, глицерин и этанол. Кроме того, эти носители могут содержать вспомогательное(ые) вещество(а), такое(ие) как увлажнитель или эмульгатор, буферное вещество, регулирующее рН и т.п.
Как правило, терапевтическая композиция может быть приготовлена в виде инъецируемой формы, такой как жидкий раствор или суспензия; или она может находиться в твердой форме, подходящей для введения в раствор, или суспензию, или жидкий носитель перед инъекцией.
После приготовления композиция по настоящему изобретению может быть введена обычными путями, которые включают, не ограничиваясь приведенным списком, внутриглазное, внутримышечное, внутривенное, подкожное, внутрикожное или местное введение. Субъект, в отношении которого проводят лечение или профилактику, может представлять собой животное, в особенности человека.
При использовании фармацевтической композиции по настоящему изобретению используют в реальном лечении лекарственная форма фармацевтической композиции может варьироваться в соответствии с применениями. Предпочтительно, в качестве примера, лекарственная форма может включать инъекцию, композицию для перорального введения и т.п.
Фармацевтическая композиция может быть приготовлена путем смешивания, разбавления или растворения в соответствии с традиционными способами. Иногда могут быть добавлены подходящие медицинские добавки, такие как эксципиенты, разрыхлители, адгезивные средства, смазывающие вещества, разбавители, буферные агенты, агенты, придающие изотоничность, консерванты, увлажнители, эмульгаторы, диспергаторы, стабилизаторы и усилители растворимости. Приготовление может быть осуществлено обычным образом в соответствии с лекарственной формой.
Фармацевтическая композиция по настоящему изобретению дополнительно может быть введена в форме композиции с замедленным высвобождением. Например, пептид по настоящему изобретению может быть включен в пилюлю или микрокапсулу, в которой полимер с замедленным высвобождением использован в качестве носителя, и затем пилюлю или микрокапсулу имплантируют оперативным путем в ткань, которая подвергается лечению. Примеры полимера с замедленным высвобождением включают этилен-этиленацетатный сополимер, полигидроксиметилакрилат, полиакриламид, поливинилпирролидон, метилцеллюлозу, полимер молочной кислоты, сополимер молочной кислоты и гликолевой кислоты и т.п. Предпочтительные примеры включают биоразлагаемые полимеры, такие как полимер молочной кислоты и сополимер молочной кислоты и гликолевой кислоты.
При использовании фармацевтической композиции по настоящему изобретению в реальном лечении, дозировка пептида по настоящему изобретению или его фармацевтически приемлемой соли в качестве активного ингредиента может быть определена подходящим образом для каждого пациента в соответствии с массой его тела, возрастом, полом, симптомами.
Применение TCR по изобретению
TCR по настоящему изобретению может быть использован в качестве лекарственного средства или диагностического агента. Признаки, подходящие для применения в качестве лекарственного средства или диагностического агента, могут быть получены путем модификаций или других улучшений. Такие лекарственные средства или диагностические агенты могут быть использованы для лечения или диагностики различных заболеваний, включающих, не ограничиваясь приведенным списком, рак (такой как рак почки, рак яичника, рак головы и шеи, рак яичка, рак легкого, рак желудка, рак шейки матки, рак мочевого пузыря, карциномы предстательной железы или меланомы), аутоиммунное заболевание, вирусное инфекционное заболевание, отторжение трансплантата и болезнь «трансплантат против хозяина».
Локализация лекарственного средства или направленная доставка лекарственного средства могут быть реализованы на основе специфичности TCR по изобретению, таким образом, усиливая терапевтические или диагностические эффекты в отношении различных заболеваний.
Для рака локализация вокруг опухолей или метастаза может усилить действие токсинов или иммуностимуляторов. При аутоиммунных заболеваниях иммунная реакция в отношении нормальных клеток или тканей может специфическим образом подавляться, или иммуносупрессорные лекарственные средства могут медленно высвобождаться для обеспечения более локального действия в течение более длительного периода времени при минимальном влиянии на общую иммунологическую активность субъекта. Для предотвращения отторжения трансплантата действие иммуносупрессии может быть оптимизировано тем же самым образом. При вирусных заболеваниях, для которых существуют лекарственные средства, например ВИЧ (вирус иммунодефицита человека), SIV (вирус иммунодефицита африканских обезьян), EBV (вирус Эпштейна-Барр), CMV (цитомегаловирус), HCV (вирус гепатита С), HBV (вирус гепатита В), благоприятно высвобождение лекарственного средства или выполнение им активизирующей функции вокруг инфицированных клеток.
TCR по изобретению могут быть использованы для модулирования Т-клеточной активации путем связывания с определенными рМНС и, таким образом, подавления активации Т-клеток. Этот подход может применяться в отношении аутоиммунных заболеваний, связанных с опосредованным Т-клетками воспалением и/или повреждением ткани, например сахарного диабета 1 типа.
TCR по изобретению также могут быть использованы для доставки цитотоксических агентов в опухолевые клетки, или могут быть трансформированы в Т-клетки, таким образом, придавая им способность повреждать опухолевые клетки, презентирующие комплексы HLA, таким образом, что они могут быть введены пациенту в процессе лечения, называемом адоптивной иммунотерапией.
TCR по изобретению также может использоваться в качестве терапевтического агента. TCR по изобретению могут быть мечены обнаруживаемой меткой, например меткой, подходящей для диагностических целей, для обнаружения связывания МНС-пептида с TCR по изобретению, который является специфическим к МНС-пептиду. Флуоресцентно меченый мультимерный TCR подходит для применения в анализе FACS для обнаружения антигенпрезентирующих клеток, несущих пептид, к которому TCR обладает специфичностью.
TCR по настоящему изобретению, связанный с конъюгатом (конъюгат, включающий, но не ограниченный приведенным примером, антитело против CD3), может перенаправлять Т-клетки, таким образом, что Т-клетки направлены против клеток, презентирующих специфический антиген, таких как раковые клетки.
Промышленная применимость
Высокостабильный TCR по настоящему изобретению полезен не только в исследовании взаимодействия между TCR и рМНС (пептид-главный комплекс гистосовместимости), но также в диагностике и лечении заболеваний.
Основные преимущества настоящего изобретения включают:
(а) Полипептиды TCR по настоящему изобретению обладают высокой стабильностью.
(б) Высокостабильные полипептиды TCR могут быть отобраны эффективным и удобным образом.
(в) Дополнительно могут быть отобраны высокостабильные и обладающие высокой аффинностью полипептиды TCR.
Настоящее изобретение будет дополнительно проиллюстрировано ниже со ссылкой на конкретные примеры. Необходимо понимать, что целью этих примеров является только иллюстрация изобретения, а не ограничение объема изобретения. Экспериментальные способы без конкретных условий, описанные в следующих примерах, как правило проводят в обычных условиях (например, в условиях, описанных в Sambrook and Russell et al., Molecular Cloning-A Laboratory Manual (3rd Ed) CSHL Press), или в соответствии с указаниями производителя. Если не указано иное, то части и проценты рассчитаны по отношению к массе.
Пример 1. Конструирование и оптимизация последовательности исходного одноцепочечного вариабельного домена TCR (sTv)
Для сайт-направленной мутации аминокислотной последовательности вариабельного домена α- и β-цепей синтетического TCR (см. WO 2012/013913) были разработаны праймеры, представленные в таблице 1. Вследствие этих мутаций гидрофобный остатки, расположенные на поверхности каркаса вариабельного домена α- и β-цепи TCR, были мутированы до гидрофильных или полярных остатков для получения матриц для библиотек мутаций гидрофобного ядра. Мутацию поверхностного гидрофобного остатка V в позиции 20 α-цепи до гидрофильного остатка S завершали путем направленного мутагенеза в процессе конструирования библиотеки мутантного гидрофобного ядра.


YW800, YW801, YW802, YW803 и YW804 представляли собой праймеры, разработанные для сайт-направленной мутации вариабельного домена α-цепи; YW806 и YW807 представляли собой праймеры, разработанные для сайт-направленной мутации вариабельного домена β-цепи; и YW805 представлял собой линкер, используемый для введения гибкого пептидного фрагмента. sTv конструировали при помощи ПЦР, осуществляемой в соответствии со способом лигирования праймера, представленным на фиг. 4, и этот sTv назван MAGE-sTv-WT.
Специфические стадии мутагенеза при помощи ПЦР были следующими:
Первая стадия ПЦР. ПЦР осуществляли с использованием синтетической α- или β-цепи в качестве матрицы, соответственно, и следующих пар праймеров: YW801/YW803 (α-цепь), YW802/YW804 (α-цепь), YW806/YW807 (β-цепь). Процесс реакции представлял собой: однократную денатурацию при 98°С в течение 30 секунд; и при 94°С в течение 5 секунд, при 55°С в течение 10 секунд, и при 72°С в течение 20 секунд, в общем 25 циклов.
Вторая стадия ПЦР. Способ ПЦР с перекрывающимися праймерами осуществляли на второй стадии ПЦР с использованием очищенного продукта в соответствии с первой стадией ПЦР и химически синтезированной одноцепочечной ДНК, кодирующей гибкий пептидный фрагмент в качестве матрицы и YW800/YW807 в качестве праймеров. Процесс реакции представлял собой: однократную денатурацию при 98°С в течение 30 секунд; при 94°С в течение 5 секунд, при 55°С в течение 10 секунд, при 72°С в течение 30 секунд, в общем 30 циклов; и однократно при 72°С в течение 5 минут. Продукт после второй стадии PCR очищали, расщепляли и лигировали в вектор фагового дисплея.
Пример 2. Клонирование последовательности MAGE-sTv-WT в основанную на рЕТ-28а экспрессирующую плазмиду
MAGE-sTv-WT клонировали в основанную на рЕТ-28а экспрессирующую плазмиду при помощи стандартных способов, описанных в Molecular Cloning: а Laboratory Manual (Third edition by Sambrook and Russell). Плазмиду секвенировали с использованием анализатора ДНК 3730 от ABI.
Последовательность ДНК, кодирующую MAGE-sTv-WT, расщепленную при помощи Ncol и Notll, лигировали в вектор рЕТ-28а (Novagen).
Лигированную плазмиду трансформировали в компетентные клетки штамма Escherichia coli BL21(DE3) (приобретенного в Merck) и высевали в планшеты со средой LB(Луриа-Бертани)-агар, содержащие 50 мкг/мл канамицина. После инкубации на протяжении ночи при 37°С отбирали единичную колонию и выращивали в 5 мл LB, содержащей 50 мкг/мл канамицина, в на протяжении ночи при 37°С при встряхивании. Клонированную плазмиду очищали с использованием набора Zyppy Plasmid Midiprep Kit от Zymo (Zyppy Plasmid Midiprep Kit, Zymo), и вставку сексвенировали с использованием секвенатора ДНК 3730 от ABI.
Фиг. 2а и 3а соответственно демонстрируют аминокислотную последовательность вариабельных доменов α- и β-цепи MAGE-sTv-WT (SEQ ID NO: 9 и 11), где оптимизированные аминокислотные остатки выделены жирным шрифтом и подчеркнуты.
Фиг. 2b и 3б соответственно демонстрируют нуклеотидную последовательность вариабельных доменов α- и β-цепи MAGE-sTv-WT (SEQ ID NO: 10 и 12).
Пример 3. Экспрессия, ренатурация и очистка MAGE-sTv-WT
Экспрессирующую плазмиду, содержащую MAGE-sTv-WT, трансформировали в штамм E. coli Rosetta (DE3) (Merck) на планшете с культуральной средой. После выращивания при 37°С на протяжении ночи единичную колонию отбирали и выращивали при 37°С в среде, содержащей канамицин, до тех пор, пока OD600 (оптическая плотность при 600 нм) не достигала 0,6-0,8. Затем экспрессию белка индуцировали 0,5 мМ IPTG (изопропилтиогалактозид) в течение 4 ч. Клетки собирали путем центрифугирования при 5000 об/мин в течение 15 минут с использованием центрифуги FisherThermo Sovall R6+. Клеточный осадок лизировали при помощи Bugbuster MasterMix (Merck). Осадок внутриклеточного тельца выделяли после центрифугирования при 6000 g в течение 15 минут с использованием центрифуги FisherThermo Sovall X1R. Внутриклеточное тельце промывали 3 раза раствором Bugbuster (10-кратное разведение) для удаления клеточного детрита и компонентов мембран. Затем внутриклеточное тельце растворяли в следующем буфере: 20 мМ Tris, рН 9,0, 8М мочевина. Его разделяли на аликвоты по 10 мг на пробирку после количественного анализа с использованием способа на основе ВСА (бицинхониновой кислоты) и замораживали при -80°С.
10 мг солюбилизированного белка внутриклеточного тельца MAGE-sTv-WT оттаивали, добавляли в 200 мл буфера для ренатурации, содержащего 100 мМ Tris, рН 9,0, 400 мМ L-аргинина и 2 мМ ЭДТА (этилендиаминтетрауксусная кислота). Окислительно-восстановительнную пару из окисленного и восстановленного глутатиона добавляли в раствор до конечных концентраций в 1 мМ и 10 мМ, соответственно. Раствор перемешивали в течение 10 минут при 10°С и встряхивали в течение 1-2 суток при 100 об/мин. Ренатурированный MAGE-sTv-WT диализировали с использованием контейнера для диализа с целлюлозной мембраной с отсечением по молекулярной массе 4 кДа и 5 л 20 мМ Tris (рН 9,0) при 4-8°С в течение 8 часов. Буфер для диализа дважды заменяли на свежий буфер.
После центрифугирования подвергнутые диализу и ренатурированные MAGE-sTv-WT наносили на анионообменную колонку Q HP (GE, 5 мл) и связанные белки элюировали с линейным градиентом 0-1М NaCl, приготовленного с 20 мМ Tris с рН 9,0, для 10 объемов колонки с использованием установки для очистки AKTA (GE). Элюированный пик (при относительной молекулярной массе приблизительно 28 кДа) собирали и затем анализировали при помощи SDS-PAGE (Bio-Rad). Компонент, содержащий MAGE-sTv-WT, концентрировали и дополнительно очищали при помощи колонки для гель-фильтрации (Superdex 75 10/300, GE Healthcare). Если целевой компонент получали после очистки, его анализировали при помощи SDS-PAGE и затем хранили при 4°С. Целевые компоненты пика объединяли и концентрировали, и затем переносили в буфер HEPES (10 мМ, рН 7,4).
Элюированные фракции дополнительно тестировали в отношении чистоты при помощи способа гель-фильтрации. Условия: колонки Agilent Bio SEC-3 (300 А, ϕ7,8×300 мм), подвижная фаза, состоящая из 150 мМ фосфатного буфера, скорость потока 0,5 мл/мин, температура колонки 25°С, и длина волны для УФ(ультрафиолетового)-детектирования составляла 214 нм.
Пример 4. Получение высокостабильных вариантов MAGE-sTv-WT
Фаговый дисплей использовали для формирования библиотеки вариантов гидрофобных ядер MAGE-sTv-WT, которые использовали для скрининга и идентификации высокостабильных мутантов. Сайты гидрофобного ядра MAGE-sTv-WT мутировали для конструирования библиотеки мутантного гидрофобного ядра, библиотеку разделяли методом пэннинга и производили отбор. Способ конструирования и отбора из библиотеки гидрофобного ядра представлен в Li et al. ((2005) Nature Biotech 23 (3): 349-354), в которой описан фаговый дисплей и способ конструирования и скрининга фаговой библиотеки высокоаффинного TCR. Различие заключалось в том, что при конструировании библиотеки мутантного гидрофобного ядра, праймеры должны быть разработаны в соответствии с сайтом гидрофобного ядра матричной цепи, тогда как при конструировании библиотеки высокоаффинного TCR праймеры разрабатывают на основе областей CDR матричной цепи. Праймеры, разработанные для конструирования библиотеки мутантного гидрофобного ядра, представлены в таблице 2 ниже.


Вырожденные основания, используемые в настоящем изобретении, как хорошо известно специалистам в данной области техники, могут представлять типы оснований в соответствии со следующими: В=С или G или Т; D=А или G или Т; Н=А или С, или Т; K=G или Т; М=А или С; N=А или С, или G, или Т; R=А или G; S=С или G; V=А или С, или G; W=А или Т; Y=С или Т.
Для устранения мутантных штаммов с плохой стабильностью и отбора более стабильных мутантных штаммов в способе фагового дисплея были приняты три следующих способа обработки: 1. демонстрация sTv при 37°С; 2. Добавление индуктора (например, IPTG) для индуцирования sTv, демонстрируемых на поверхности фага; 3. инкубация фага, демонстрирующего sTv, при 55°С в течение 60 минут до отбора.
После идентификации путем секвенирования все гидрофобные ядра высокостабильных мутантных штаммов sTv, отобранных при помощи вышеприведенного способа, имели мутации. Отобранные высокостабильные мутантные штаммы были названы MG29, P8F1 и P8F2. В соответствии с нумерацией позиций аминокислот в IMGT вариабельный домен α-цепи имел одну или более чем одну аминокислотную мутацию в гидрофобном ядре: позиция 19, позиция 21, позиция 91; и/или вариабельный домен β цепи имел одну или более чем одну аминокислотную мутацию в гидрофобном ядре: позиция 91, 4-я позиция относительно последнего короткого пептида J-гена β-цепи. Конкретней, в соответствии с нумерацией позиций аминокислот в IMGT они имели один или более чем один из следующих аминокислотных остатков в вариабельном домене α-цепи: 19V, 21I, 91L; и/или они имели один или более чем один из следующих аминокислотных остатков в вариабельном домене β-цепи: 91F или 91I, 4-я позиция относительно последнего J-гена β-цепи представляла собой М. Конкретнее, аминокислотная последовательность вариабельного домена α-цепи представляла собой SEQ ID NO: 15 или 17; и аминокислотная последовательность вариабельного домена β-цепи представляла собой SEQ ID NO: 16 или 18. Аминокислотные последовательности вариабельного домена α- и β-цепи, образующего мутантный штамм MG29, представляли собой SEQ ID NO: 15 и 16, соответственно, как представлено на фиг. 6а и 6б; аминокислотные последовательности вариабельного домена α- и β-цепи, образующего мутантный штамм P8F1, представляли собой SEQ ID NO: 17 и 18, соответственно, как представлено на фиг. 7а и 7б; и аминокислотные последовательности вариабельного домена α- и β-цепи, образующего мутантный штамм P8F2, представляли собой SEQ ID NO: 15 и 18, соответственно, как представлено на фиг. 8а и 8б.
Вышеприведенные отобранные высокостабильные мутантные штаммы MG29, P8F1, P8F2 с высокими значениями OD и MAGE-sTv-WT с немутантным гидрофобным ядром подвергали эксперименту ИФА для сравнения значений OD, и проверяли специфичность мутантных штаммов.
Стадии экспериментов ИФА:
1. Штаммы MAGE-sTv-WT, MG29, P8F1 и P8F2 в глицерине соответственно инокулировали в 5 мл 2xTY (100 мкг/мл ампициллина, 2% глюкозы), 250 об/мин, 37°С в течение ночи.
2. После выращивания в течение ночи 50 мкл бульона соответственно инокулировали в 5 мл свежей 2xTY (100 мкг/мл ампициллина, 2% глюкозы), 250 об/мин, и выращивали при 37°С до тех пор, пока OD600 не достигла 0,4. После инфицирования 5 мкл (6,5×1010) хелперного фага KM13 (Source Bioscience) и выдерживания при 37°С в течение 30 мин его встряхивали при 200 об/мин при 37°С в течение 30 мин и центрифугировали. Осадок ресуспендировали в 30 мл 2xTY (100 мкг/мл ампициллина, 50 мкг/мл канамицина, 0,1% глюкозы), и выращивали при 30°С в течение ночи при 250 об/мин.
3. Иммуносорбентный планшет (NUNC) покрывали 10 мкг/мл стрептавидина (PBS (натрий-фосфатный буфер), рН=7,4), 100 мкл на лунку, и выдерживали на протяжении ночи при 4°С. 4. Супернатант культурального бульона на следующий день собирали после центрифугирования. Фаг в супернатанте осаждали ПЭГ (полиэтиленгликоль)/NaCl, имеющим объемное отношение 1/4, помещали на лед на 1 ч и центрифугировали для сбора осадков. Осадки ресуспендировали в 3 мл PBS.
5. Планшет трижды промывали 0,1% TBST (Tris-Buffered Saline with Tween 20), добавляли в каждую лунку по 400 мкл 3% Marvel-PBS (Cadbury Schweppes), и блокировали планшет при 37°С в течение 2 ч. Планшет трижды промывали PBST (Phosphate Buffered Saline with Tween 20), добавляли в каждую лунку по 100 мкл 10 мкг/мл рМНС, и выдерживали планшет при комнатной температуре в течение 1 ч. После трехкратного промывания планшета 100 мкл фаговых образцов (10 мкл осажденного ПЭГ образца с 3% Marvel-PBS инкубировали при комнатной температуре в течение 1 ч) добавляли в каждую лунку, и планшет выдерживали при комнатной температуре в течение 1 ч; после трехкратного промывания планшета 100 мкл конъюгата антитела анти-M13-HRP (пероксидаза хрена) (GE Healthcare) (разведение 1:5000 в 3% Marvel-PBS) добавляли в каждую лунку, и планшет выдерживали при комнатной температуре в течение 1 ч. Планшет промывали шесть раз, и в каждую лунку добавляли 100 мкл TMD (тетраметил-1,2-диоксетан) и оставляли на 5 мин. 100 мкл 1М серной кислоты добавляли в каждую лунку для остановки реакции.
6. Регистрировали поглощение при 450 нм и 650 нм
Значения OD для мутантных штаммов в эксперименте ИФА представлены на фиг. 9, и результат демонстрирует, что специфичность sTv, имеющих оптимизированное гидрофобное ядро поддерживается, и является такой же, как для MAGE-sTv-WT, имеющего гидрофобное ядро дикого типа. В этом эксперименте значение OD для MAGE-sTv-WT было очень низким вследствие плохой демонстрации, свидетельствуя о том, что даже если гидрофобные остатки, демонстрируемые на поверхности вариабельного домена α-цепи и β-цепи, заменить на гидрофильные или полярные остатки, то стабильность белка все равно остается сравнительно плохой, и, таким образом, гидрофобное ядро все еще нуждается в оптимизации. Клоны с оптимизированным гидрофобным ядром могут демонстрировать sTv в отличающейся степени и специфически связываться с антигеном исходного лиганда MAGE A3 pHLA-A1, но не могут связываться с другими нерелевантными антигенами, такими как антигены EBV, вируса гриппа и NY-ESO-1. Связывание молекул sTv, имеющих мутантное гидрофобное ядро (MG29, P8F1, P8F2 и другие, обнаруженные при помощи фагового дисплея), со специфическими антигенами, не происходило, поскольку они обладали более высокой аффинностью, чем TCR дикого типа, как показано в примере 15.
Пример 5. Конструирование высокостабильных sTv молекул с мутацией в гидрофобном ядре
Часть гидрофобного ядра высокостабильного мутанта, отобранного в примере 4, вводили в несколько других молекул TCR для построения высокостабильных молекул sTv с использованием хорошо известного в области техники способа направленного мутагенеза.
Несколько вышеприведенных молекул в одноцепочечной форме конструировали в соответствии с вариабельными доменами α- и β-цепей TCR дикого типа соответственно против коротких пептидов антигена HLA-B8/FLRGRAYGL (выделен из вирусного антигена ЕВ EBNA3A), HLA-A2/GILGFVFTL (выделен из белка матрикса вируса гриппа) и HLA-A2/SLLMWITQC (опухолеспецифический антиген NY-ESO-1), и соответственно назвали как LC13-WT, JM22-WT и 1G4-WT. Аминокислотные последовательности вариабельного домена α-цепи и β-цепи LC13-WT представляли собой SEQ ID NO: 29 и SEQ ID NO: 30, соответственно, как представлено на фиг. 10а и 10б; аминокислотные последовательности вариабельного домена α-цепи и β-цепи JM22-WT представляли собой SEQ ID NO: 31 и SEQ ID NO: 32, соответственно, как представлено на фиг. 11а и 11б; и аминокислотные последовательности вариабельного домена α-цепи и β-цепи 1G4-WT представляли собой SEQ ID NO: 33 и SEQ ID NO: 34, соответственно, как представлено на фиг. 12а и 12б.
Часть гидрофобного ядра высокостабильного варианта, отобранного в примере 4, соответственно вводили в LC13-WT, JM22-WT и 1G4-WT в соответствии с хорошо известным в области техники способам направленного мутагенеза. Молекулы, полученные после введения мутации, соответственно называли как LC13-sTv, JM22-sTv и 1G4-sTv, и введенное гидрофобное ядро выделено жирным шрифтом и подчеркнуто. Аминокислотные последовательности вариабельного домена α-цепи и β-цепи LC13-sTv представляли собой SEQ ID NO: 35 и SEQ ID NO: 36, соответственно, как представлено на фиг. 13а и 13б, и гидрофобное ядро, введенное в α-цепь, содержало 11L, 13V, 21I и 91I, и гидрофобное ядро, введенное в β-цепь, содержало 94L. Аминокислотные последовательности вариабельного домена α-цепи и β-цепи JM22-sTv представляли собой SEQ ID NO: 37 и SEQ ID NO: 38, соответственно, как представлено на фиг. 14а и 14б, и гидрофобное ядро, введенное в α-цепь, содержало 19V и 21I, и гидрофобное ядро, введенное в β-цепь, содержало 91I и 94L. Аминокислотные последовательности вариабельного домена α-цепи и β-цепи 1G4-sTv представляли собой SEQ ID NO: 39 и SEQ ID NO: 40, соответственно, как представлено на фиг. 15а и 15б, и гидрофобное ядро, введенное в α-цепь, содержало 19V и 21I, и гидрофобное ядро, введенное в β-цепь, содержало 19V, 91I, и 94L, и шестая позиция с конца β-цепи, кодируемой J-геном, представляла собой Т. Вышеприведенная нумерация позиций аминокислот является нумерацией, представленной в IMGT. Короткий пептид для связывания (линкер), полезный в конструировании вышеприведенных одноцепочечных молекул, может иметь любую подходящую последовательность, и предпочтительная аминокислотная последовательность в настоящем изобретении представляла собой SEQ ID NO: 41, как представлено на фиг. 16.
Пример 6. Тестирование стабильности белка LC13-WT и LC13-sTv
С использованием способа, описанного в примере 3, белки LC13-WT и LC13-sTv экспрессировали, подвергали рефолдингу и очищали. После очистки на колонке для гель-фильтрации проводили разделение SDS-PAGE, и получали профили SEC для двух белков с использованием гель-фильтрации. Экспрессируемое количество, количество очищенного белка и выход белка после рефолдинга рассчитывали одновременно. Среди них экспрессируемое количество представляло собой выход очищенного внутриклеточного тельца, индуктивно экспрессированного в 1 л бульона Е. coli. Количество очищенного белка представляло собой количество белка, полученное после очистки подвергнутого рефолдингу внутриклеточного тельца, которое очищали из 1 л бульона Е. coli с индуктивно экспрессированным внутриклеточным тельцем.
Расчетная формула для выхода белка после рефолдинга представляла собой: выход белка после рефолдинга (%)=100 * количество очищенного белка (мг)/количество внутриклеточного тельца, использованного в рефолдинге (мг). Если не указано иное, в настоящем изобретении экспрессируемое количество и выход белка после рефолдинга рассчитывали в соответствии с вышеприведенным способом расчета.
Величину Tm очищенных белков LC13-WT и LC13-sTv измеряли при помощи дифференциального сканирующего калориметра (Nano DSC) от US ТА company (Waters). Диапазон сканирования составлял 10-90°С, скорость нагревания составляла 1°С/мин, и объем образца составлял 900 мкл. Величину Tm получали с использованием модели подгонки TwostateScaled в программном обеспечении Nanoanalyze.
Таблица 3 демонстрирует данные об экспрессируемом количестве, количестве очищенного белка и выходе белка LC13-WT и LC13-sTv после рефолдинга.
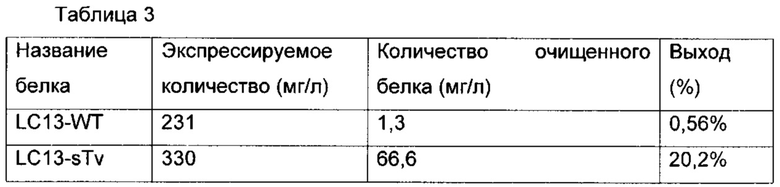
Данные в вышеприведенной таблице демонстрируют, что после очистки выход после рефолдинга белка LC13-sTv, имеющего мутации, введенные в гидрофобное ядро, увеличивается в 35 раз по сравнению с выходом после рефолдинга белка LC13-WT, не имеющего мутаций в гидрофобном ядре.
Фиг. 17 демонстрирует результаты SDS-PAGE для белков LC13-WT и LC13-sTv, очищенных при помощи колонки для гель-фильтрации (Superdex 75 10/300, GE Healthcare) в соответствии с примером 3. Фотография геля демонстрирует, что полоса, образуемая очищенным белком LC13-WT, была гетерогенной, a LC13-sTv образовал единичную полосу и обладал высокой чистотой. Последнее свидетельствует о том, что рефолдинг LC13-sTv был гораздо лучше, чем рефолдинг LC13-WT.
Фиг. 18а и 18б соответственно демонстрируют профили SEC очищенных белков LC13-WT и LC13-sTv. В соответствии с представленным очищенный белок LC13-WT не демонстрирует пика, тогда как LC13-sTv образует единичный и симметричный пик элюции. Последнее свидетельствует о том, что рефолдинг LC13-sTv был значительно лучше, чем рефолдинг LC13-WT.
Белок с правильной конформацией, полученный в результате рефолдинга LC13-WT, был очень небольшим, явный эндотермический пик анфолдинга белка отсутствовал. Величина Tm для LC13-WT не была обнаружена с использованием программного обеспечения для анализа Nanoanalyze. Тогда как величина Tm для LC13-sTv с мутациями гидрофобного ядра составляла 43,6°С, и его кривая ДСК представлена на фиг. 47. Последнее свидетельствует о том, что по сравнению с LC13-WT, LC13-sTv обладал более сильной способностью к ренатурации, был более устойчив к анфолдингу, был более устойчив к неподходящему или нежелательному фолдингу, и обладал существенно улучшенной температурной стабильностью.
После сравнительного анализа экспрессируемого количества, количества очищенного белка, выхода белка после рефолдинга, карт SDS-PAGE, профилей SEC и величин Tm белков LC13-WT и LC13-sTv, можно видеть, что по сравнению с LC13-WT, имеющим немодифицированное гидрофобное ядро, LC13-sTv, имеющий модифицированное гидрофобное ядро, обладал более сильной способностью к ренатурации, был более устойчив к анфолдингу, был более устойчив к неподходящему или нежелательному фолдингу, обладал более высоким выходом белка после рефолдинга и обладал существенно улучшенной термической стабильностью. Таким образом, по сравнению с LC13-WT, LC13-sTv обладал существенно улучшенной стабильностью. В настоящем изобретении стабильность LC13-sTv увеличилась в 35 раз по сравнению со стабильностью LC13-WT, рассчитанной на основе данных для выхода белка после рефолдинга.
Пример 7. Тестирование стабильности белков JM22-WT и JM22-sTv
С использованием способа, описанного в примере 3, белки JM22-WT и JM22-sTv экспрессировали, подвергали рефолдингу и очищали. После очистки на колонке для гель-фильтрации, проводили разделение методом SDS-PAGE, и получали профили SEC для двух белков с использованием гель-фильтрации. Экспрессируемое количество, количество очищенного белка и выход белка после рефолдинга рассчитывали одновременно. Величины Tm определяли с использованием способа в примере 6.
Таблица 4 демонстрирует данные об экспрессируемом количестве, количестве очищенного белка и выходе белков JM22-WT и JM22-sTv после рефолдинга.
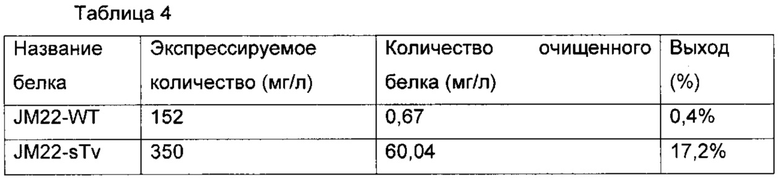
Данные в вышеприведенной таблице демонстрируют, что после очистки выход после рефолдинга белка JM22-sTv, имеющего мутации, введенные в гидрофобное ядро, увеличивался в 42 раза по сравнению с выходом белка JM22-WT, не имеющим мутаций в гидрофобном ядре.
Фиг. 19 демонстрирует результаты SDS-PAGE для белков JM22-WT и JM22-sTv, очищенных при помощи колонки для гель-фильтрации (Superdex 75 10/300, GE Healthcare) в соответствии с примером 3. Фотография геля демонстрирует, что мономерные полосы, образованные вследствие рефолдинга белка JM22-WT были гетерогенными, и имелись три полосы, а JM22-sTv образует единичную полосу мономера с высокой чистотой. Последнее свидетельствует о том, что рефолдинг JM22-sTv был гораздо лучше рефолдинга JM22-WT.
Фиг. 20а и 20б соответственно демонстрируют профили SEC для очищенных белков JM22-WT и JM22-sTv. Как представлено, пик элюции, образованный очищенным белком JM22-WT, не был единичным, и сигнал был очень низким, хотя очищенный LC13-sTv образовывал единичный и симметричный пик элюции. Последнее свидетельствует о том, что рефолдинг JM22-sTv был значительно лучше, чем рефолдинг JM22-WT.
Фиг. 48а и 48б соответственно демонстрируют графики ДСК для очищенных белков JM22-WT и JM22-sTv. Белок с правильной конформацией, полученный в результате рефолдинга JM22-WT, был очень небольшим, явный эндотермический пик анфолдинга белка отсутствовал. Величина Tm для JM22-WT не может быть обнаружена с использованием программного обеспечения для анализа Nanoanalyze. Тогда как величина Tm для JM22-sTv с мутацией гидрофобного ядра составляла 43,7°С. Вышеприведенный график ДСК демонстрирует, что по сравнению с JM22-WT, JM22-sTv обладал большей способностью к ренатурации, был более устойчив к анфолдингу, был более устойчив к неподходящему или нежелательному фолдингу и обладал существенно улучшенной термической стабильностью.
После сравнительного анализа экспрессируемого количества, количества очищенного белка, выхода белка после рефолдинга, карт SDS-PAGE, графиков ДСК и профилей SEC для белков JM22-WT и JM22-sTv, можно видеть, что по сравнению с JM22-WT, имеющим немодифицированное гидрофобное ядро, JM22-sTv, имеющий модифицированное гидрофобное ядро, обладал большей способностью к ренатурации, был более устойчив к анфолдингу, был более устойчив к неподходящему или нежелательному фолдингу, обладал более высоким выходом белка после рефолдинга и обладал существенно улучшенной температурной стабильностью. Таким образом, по сравнению с JM22-WT, JM22-sTv обладал существенно улучшенной стабильностью. В настоящем изобретении стабильность JM22-sTv увеличилась на 4200% по сравнению со стабильностью JM22-WT, рассчитанной на основе данных для выхода белка после рефолдинга.
Пример 8. Тестирование стабильности белка 1G4-WT и 1G4-sTv
С использованием способа, описанного в примере 3, белки 1G4-WT и 1G4-sTv экспрессировали, подвергали рефолдингу и очищали. После очистки на колонке для гель-фильтрации проводили разделение методом SDS-PAGE, и профили SEC для двух белков получали с использованием способа гель-фильтрации. Экспрессируемое количество, количество очищенного белка и выход белка после рефолдинга рассчитывали одновременно.
Таблица 5 демонстрирует данные об экспрессируемом количестве, количестве очищенного белка и выходе белков 1G4-WT и 1G4-sTv после рефолдинга.
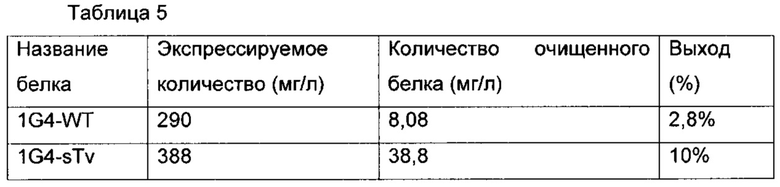
Данные в вышеприведенной таблице демонстрируют, что после очистки выход после рефолдинга белка 1G4-sTv, имеющего мутации, введенные в гидрофобное ядро, увеличился в 2,6 раза по сравнению с выходом белка 1G4-WT, не имеющим мутаций в гидрофобном ядре.
Фиг. 21 демонстрирует результаты SDS-PAGE для белков 1G4-WT и 1G4-sTv, очищенных при помощи колонки для гель-фильтрации (Superdex 75 10/300, GE Healthcare) в соответствии с примером 3. Фотография геля демонстрирует, что полосы, образованные очищенным белком 1G4-WT, были гетерогенными и имелись две полосы, a 1G4-sTv образует единичную полосу мономера с высокой чистотой. Последнее свидетельствует о том, что рефолдинг 1G4-sTv был гораздо лучше рефолдинга 1G4-WT.
Фиг. 22а и 22б соответственно демонстрируют профили SEC очищенных белков 1G4-WT и 1G4-sTv. Как показано, пик элюции, образованный очищенным белком 1G4-WT, не был единичным, и сигнал был очень низким, хотя очищенный 1G4-sTv образовывал единичный и симметричный пик элюции. Последнее свидетельствует о том, что рефолдинг 1G4-sTv был значительно лучше рефолдинга 1G4-WT.
После сравнительного анализа экспрессируемого количества, количества очищенного белка, выхода белка после рефолдинга, карт SDS-PAGE и профилей SEC для белков 1G4-WT и 1G4-sTv, можно видеть, что по сравнению с 1G4-WT, имеющим немодифицированное гидрофобное ядро, 1G4-sTv, имеющий модифицированное гидрофобное ядро, обладал большей способностью к ренатурации, более высоким экспрессируемым количеством и более высоким выходом белка после рефолдинга. Таким образом, по сравнению с 1G4-WT, 1G4-sTv обладал существенно улучшенной стабильностью. В настоящем изобретении стабильность 1G4-sTv увеличилась на 260% по сравнению со стабильностью 1G4-WT, рассчитанной на основе данных для выхода белка после рефолдинга.
Пример 9. Дополнительная оптимизация стабильности молекул с использованием 1G4-sTv в качестве матрицы
При использовании 1G4-sTv в качестве матрицы его гидрофобное ядро и поверхностные аминокислотные остатки в вариабельном домене мутировали для конструирования библиотеки для отбора высокостабильных молекул. Сайты гидрофобного ядра, подвергаемые мутации, представлены в последовательности SEQ ID NO: 42, они выделены жирным шрифтом и подчеркнуты. Поверхностные аминокислотные остатки, подвергаемые мутации, выделены жирным шрифтом, как представлено на фиг. 23.
Основной способ, используемый для конструирования библиотеки, уже описан в примере 4. В этом примере конструировали три библиотеки для подвергаемых мутации сайтов. Все сайты гидрофобного ядра, подвергаемые мутации, были в библиотеке 1. Библиотеку 2 и библиотеку 3 конструировали для поверхностных аминокислотных остатков. Конкретнее, осуществляли ПЦР с перекрывающимися праймерами с использованием плазмиды 1G4-sTv в качестве матрицы, и мутантные праймеры разрабатывали таким образом, чтобы получать мутантные фрагменты ДНК. Мутантные фрагменты ДНК расщепляли при помощи Ncol/Notl, и фрагменты клонировали в фаговый плазмидный вектор pLitmus28 (NEB), основанный на скелете pUC19. После электротрансфекции ДНК в компетентные клетки TG1 (Lucigen) в общей сложности получали три библиотеки фагового плазмидного вектора, емкость которых составляла 1×109-3×109 на основе количества колоний. «Газон», выращенный в этих трех библиотеках, соскребали, помещали в глицерин с конечной концентрацией 20% и хранили при -80°С. В следующих таблицах 8, 9 и 10 соответственно продемонстрированы праймеры, разработанные для библиотеки 1, библиотеки 2 и библиотеки 3.




Для получения высокостабильных клонов sTv фаги, выращенные в библиотеках, подвергали тепловой шоковой обработке при 65°С после осаждения и концентрирования, и инкубировали вместе с 0,02% SDS (додецилсульфатом натрия), добавляемым для дополнительного увеличения мощности отбора. Затем обработанный фаг подвергали последующему отбору. Клоны с более высоким значением OD и отобранные из трех библиотек, комбинировали, и в итоге получили 11 клонов.
Пример 10. Проверка стабильности клонов, отобранных в примере 9
Величины OD для 11 клонов, отобранных в примере 9, обнаруживали при помощи ИФА, экспериментальная процедура которого описана в примере 4, и проверяли их антигенную специфичность. Результаты представлены на фиг. 24. Эти результаты демонстрируют, что величины OD для 11 клонов были высокими, и эти клоны могли специфически связываться с антигеном исходного лиганда HLA-A2/SLLMWITQC (опухолеспецифический антиген NY-ESO-1), и по существу не связываться с другими нерелевантными антигенами. Связывание молекул sTv, обнаруженных при помощи фагового дисплея, с антигеном HLA-A2/LLMWITQC, не происходило, поскольку они более высокой аффинностью, чем TCR дикого типа, как показано в примере 11.
В соответствии с нумерацией позиций аминокислот в IMGT, вышеприведенные 11 клонов имели одну или более чем одну мутацию в позициях гидрофобного ядра, выбранных из группы, состоящей из: позиция 11, позиция 13 или позиция 94 вариабельного домена α-цепи; и/или позиция 11, позиция 13 или позиция 94 вариабельного домена β-цепи. Конкретнее, они имели один или более чем один из следующих аминокислотных остатков гидрофобного ядра в вариабельном домене α-цепи: 11М, 11Е, 13R, 13K, 94V или 94I; и/или аминокислотных остатков гидрофобного ядра в вариабельном домене β-цепи: 11L, 11V, 13V или 94V. Дополнительно к гидрофобному ядру клоны, отобранные авторами изобретения, дополнительно содержали один или более чем один из следующих аминокислотных остатков в вариабельном домене α-цепи: 4L, 12N, 16S, 93N, 93R, 97N, 100G, 105S или последняя позиция α-цепи, кодируемой J-геном, представляет собой D, и/или содержали один или более чем один из следующих аминокислотных остатков в вариабельном домене β-цепи: 4I, 101L, или последняя позиция β-цепи, кодируемой J-геном, представляет собой D, или третья позиция с конца β-цепи, кодируемой J-геном, представляет собой Е.
Аминокислотные последовательности вариабельного домена α-цепи (SEQ ID NO: 75-85) и аминокислотная последовательность вариабельного домена β-цепи (SEQ ID NO: 86-96) вышеприведенных отобранных высокостабильных клонов соответственно представлены на фиг. 25 и фиг. 26.
С использованием способов, описанных в примере 2 и примере 3, каждый из 11 отобранных клонов связывали, экспрессировали, подвергали рефолдингу и очищали. Величины Tm для вышеприведенных 11 клонов измеряли с использованием дифференциального сканирующего калориметра (Nano DSC) от US ТА company (Waters). Диапазон сканирования составлял 10-90°С, скорость нагревания составляла 1°С/мин, и объем образца составлял 900 мкл. Величину Tm получали с использованием модели подгонки TwostateScaled в программном обеспечении для анализа Nanoanalyze.
Результаты представлены на фиг. 27 и в таблице 9, которые демонстрируют, что все величины Tm были не меньше чем 37,9°С, и обладали явным эндотермическим пиком анфолдинга белка. Результаты ДСК для 1G4-WT представлены на фиг. 28, где процессы экспрессии, рефолдинга, очистки и экспериментальные условия ДСК были такими же, как для вышеприведенных клонов. На фигуре видно, что не было показано явного эндотермического пика, свидетельствующий о том, что белок с правильной конформацией был очень небольшим. Сравнение графиков ДСК для вышеприведенных 11 клонов с графиком ДСК для 1G4-WT демонстрирует, что отобранные клоны были более устойчивыми к анфолдингу, были более устойчивыми к неправильному или нежелательному фолдингу, обладали более сильной способностью к ренатурации, и обладали существенно улучшенной термической стабильностью по сравнению с 1G4-WT. Таким образом, стабильность отобранных клонов была гораздо выше стабильности 1G4-WT без мутаций в гидрофобном ядре.
Величина Tm не может быть получена при помощи программного обеспечения, поскольку вышеприведенный 1G4-WT с правильной конформацией после рефолдинга и очистки был очень небольшим и не имел явного эндотермического пика анфолдинга белка. Тогда как вышеприведенный TCR с мутантным гидрофобным ядром обладал величинами Tm, составляющими приблизительно 38°С или выше, что свидетельствует о том, что стабильность 11 мутантных TCR вышеприведенных G3-G7 и G9-G14 по настоящему изобретению увеличилась очень значительно (по меньшей мере однократное увеличение).


Специалист в данной области техники может повторно собрать вышеприведенные отобранные высокостабильные мутации для построения новых стабильных мутантов. Авторы изобретения повторно собрали вышеприведенные сайты мутации для построения нового вариабельного домена α-цепи (SEQ ID NO: 97) и вариабельного домена β-цепи (SEQ ID NO: 98), аминокислотные последовательности которых представлены на фиг. 29а и 29б, соответственно. Молекулы sTv, сконструированные с указанным вариабельным доменом α-цепи и вариабельным доменом β-цепи, названы G15.
Пример 11. Дополнительное тестирование стабильности мутантов 1G4-WT
С использованием способа, описанного в примере 3, мутанты G9, G13 и G15 примера 10 экспрессировали, подвергали рефолдингу и очищали. После очистки на колонке для гель-фильтрации проводили разделение способом SDS-PAGE и профили SEC трех белков получали с использованием способа гель-фильтрации. Экспрессируемое количество, количество очищенного белка и выход белка после рефолдинга рассчитывали одновременно, и сравнивали с 1G4-WT.
Таблица 10 демонстрирует данные об экспрессируемом количестве, количестве очищенного белка и выходе белка после рефолдинга 1G4-WT, G9, G13 и G15.


Данные в вышеприведенной таблице демонстрируют, что после очистки выход белка после рефолдинга мутантов G9, G13 и G15 значительно увеличился по сравнению с выходом 1G4-WT, и соответственно увеличился в 4,5 раза, 15,2 раза и 15,6 раз.
Фиг. 30 демонстрирует результаты SDS-PAGE белков 1G4-WT, G9, G13 и G15, очищенных при помощи колонки для гель-фильтрации (Superdex 75 10/300, GE Healthcare), как описано в примере 3. Фотография геля демонстрирует, что полоса, образованная путем рефолдинга белка 1G4-WT, была гетерогенной, и G9, G13 и G15 образуют единичную полосу мономера с высокой чистотой. Последнее свидетельствует о том, что рефолдинг G9, G13 и G15 был лучше рефолдинга 1G4-WT.
Фиг. 31а, 31б и 31в демонстрируют соответственно профили SEC белков G9, G13 и G15. Как представлено на фиг. 22а в профиле SEC для 1G4-WT, пик элюции, образованный очищенным белком 1G4-WT, не был единичным, и сигнал был очень низким, тогда как очищенные G9, G13 и G15 образовывали единичный и симметричный пик элюции. Последнее свидетельствует о том, что рефолдинг G9, G13 и G15 был значительно лучше рефолдинга 1G4-WT.
Величина Tm для G15, измеренная с использованием способа в примере 10, составляла 46,6°С, и кривая ДСК для него представлена на фиг. 46. В соответствии с результатами, измеренными как в примере 10, величины Tm мутантных штаммов G9 и G13 были относительно высокими, и составляли 49,55°С и 49,63°С, соответственно.
Связывание белков G9, G13 и G15 с их лигандами определяли при помощи системы анализа в реальном времени BIAcore Т200. Результат показал, что аффинность трех белков sTv к антигену HLA-A2/SLLMWITQC была не лучше, чем связывание 1G4 TCR дикого типа с антигеном. Константа диссоциации при связывании 1G4 TCR дикого типа с антигеном HLA-A2/SLLMWITQC составляла 32 мкМ (в соответствии с Li et al. ((2005) Nature Biotech 23 (3): 349-354)).
После сравнительного анализа экспрессируемого количества, количества очищенного белка, выхода белка после рефолдинга, карт SDS-PAGE и профилей SEC белков 1G4-WT и G9, G13 и G15, можно видеть, что способность к ренатурации, термическая стабильность и выход после рефолдинга белка мутантов, имеющих модифицированное гидрофобное ядро, были гораздо выше, чем для 1G4-WT, имеющего немодифицированное гидрофобное ядро. Таким образом, по сравнению с 1G4-WT, мутанты, имеющие модифицированное гидрофобное ядро, обладали существенно увеличенной стабильностью. В настоящем изобретении стабильность G9, G13 и G15 соответственно увеличивалась на 450%, 1520% и 1560% по сравнению со стабильностью 1G4-WT, рассчитанной на основе данных выхода белка после рефолдинга.
Пример 12. Конструирование высокостабильных sTv молекул с мутацией в гидрофобном ядре
Высокостабильные молекулы sTv конструировали в соответствии с гидрофобным ядром и аминокислотными остатками на поверхности скелета вариабельных доменов высокостабильного мутанта, отобранного в примере 9.
Часть гидрофобного ядра и аминокислотных остатков на поверхности скелета вариабельных доменов высокостабильного мутанта, отобранного в примере 9, вводили в молекулы LC13-WT, JM22-WT и MAGE-sTv-WT в соответствии с хорошо известными в области техники способами направленного мутагенеза. Молекулы, имеющие введенные мутации, соответственно назвали как LC13-G9, LC13-G15, JM22-G9, JM22-G15 и MAGE-G15, введенное гидрофобное ядро выделено жирным шрифтом и подчеркнуто.
Аминокислотные последовательности вариабельного домена α-цепи и β-цепи LC13-G9 представляли собой SEQ ID NO: 99 и SEQ ID NO: 100, соответственно, как представлено на фиг. 32а и 32б, и введенное гидрофобное ядро в α-цепи содержало 13V, 21I, 91I и 94I а введенное гидрофобное ядро в β-цепи содержало 11V, 13V и 94V. Аминокислотные последовательности вариабельного домена α-цепи и β-цепи LC13-G15 представляли собой SEQ ID NO: 101 и SEQ ID NO: 102, соответственно, как представлено на фиг. 33а и 33б, и введенное гидрофобное ядро в α-цепи содержало 11L, 13V, 21I, 91I и 94I, а введенное гидрофобное ядро в β-цепи содержало 11L, 13V и 94V. Аминокислотные последовательности вариабельного домена α-цепи и β-цепи JM22-G9 представляли собой SEQ ID NO: 103 и SEQ ID NO: 104, соответственно, как представлено на фиг. 34а и 34б, и введенное гидрофобное ядро в α-цепи содержало 11М, 13V, 19V, 21I и 94I, а введенное гидрофобное ядро в β-цепи содержало 11V, 13V, 91I и 94V. Аминокислотные последовательности вариабельного домена α-цепи и β-цепи JM22-G15 представляли собой SEQ ID NO: 105 и SEQ ID NO: 106, соответственно, как представлено на фиг. 35а и 35б, и введенное гидрофобное ядро в α-цепи содержало 13V, 19V, 21I и 94I, а введенное гидрофобное ядро в β-цепи содержало 13V, 91I и 94V. Аминокислотные последовательности вариабельного домена α-цепи и β-цепи MAGE-G15 представляли собой SEQ ID NO: 107 и SEQ ID NO: 108, соответственно, как представлено на фиг. 42а и 42б, и введенное гидрофобное ядро в α-цепи содержало 19V, 21I и 94I, а введенное гидрофобное ядро в β-цепи содержало 13V, 89L, 91I и 94V.
Вышеприведенная нумерация позиций аминокислот является нумерацией, представленной в IMGT. Короткий пептид для связывания (линкер), используемый для конструирования вышеприведенной одноцепочечной молекулы, может иметь любую подходящую последовательность, и предпочтительная аминокислотная последовательность в настоящем изобретении представляла собой SEQ ID NO: 41, как показано на фиг. 16.
Пример 13. Тестирование стабильности белков LC13-G9 и LC13-G15
С использованием способа, описанного в примере 3, белки LC13-G9 и LC13-G15 экспрессировали, подвергали рефолдингу и очищали. После очистки на колонке для гель-фильтрации проводили разделение методом SDS-PAGE, и получали профили SEC для двух белков с использованием способа гель-фильтрации. Экспрессируемое количество, количество очищенного белка и выход белка после рефолдинга рассчитывали одновременно.
Таблица 11 демонстрирует данные об экспрессируемом количестве, количестве очищенного белка и выходе белка после рефолдинга LC13-G9 и LC13-G15. Релевантные данные для LC13-WT также приведены для анализа.

Данные в вышеприведенной таблице демонстрируют, что после очистки выход белка после рефолдинга белков LC13-G9 и LC13-G15, имеющих мутации, введенные в гидрофобное ядро, соответственно увеличивался на 5,4% и в 57,9 раз по сравнению с выходом белка 1G4-WT, не имеющего мутаций в гидрофобном ядре.
Фиг. 36 демонстрирует результаты SDS-PAGE белков LC13-G9 и LC13-G15, очищенных при помощи колонки для гель-фильтрации (Superdex 75 10/300, GE Healthcare), как описано в примере 3. Фотография геля демонстрирует, что полоса, образуемая очищенным белком LC13-WT, была гетерогенной, и как модифицированный LC13-G9, так и LC13-G15, образуют единичную полосу мономера с высокой чистотой. Последнее свидетельствует о том, что рефолдинг LC13-G9 и LC13-G15 был гораздо лучше рефолдинга LC13-WT.
Фиг. 37 и 38 демонстрируют профили SEC белков LC13-G9 и LC13-G15, соответственно. Профиль SEC для LC13-WT не демонстрировал пик, тогда как белки LC13-G9 и LC13-G15 формировали единичные и симметричные пики элюции. Последнее свидетельствует о том, что рефолдинг LC13-G9 и LC13-G15 был значительно лучше рефолдинга LC13-WT.
Величины Tm для LC13-G9 и LC13-G15, измеренные с использованием способа в соответствии с примером 10, составляли 43°С и 50,5°С, соответственно, и их кривые ДСК представлены на фиг. 49а и 49б. Белок с правильной конформацией, полученный в результате рефолдинга LC13-WT, был очень небольшим и не обладал явным эндотермическим пиком анфолдинга белка. Величина Tm для LC13-WT не может быть обнаружена. Последнее свидетельствует о том, что термическая стабильность LC13-G9 и LC13-G15 по настоящему изобретению увеличилась по меньшей мере однократно по сравнению со стабильностью LC13-WT. В то же самое время, последнее свидетельствует о том, что по сравнению с LC13-WT, LC13-G9 и LC13-G15 были более устойчивыми к анфолдингу, были более устойчивыми к неправильному или нежелательному фолдингу и обладали более сильной способностью к ренатурации.
После сравнительного анализа экспрессируемого количества, количества очищенного белка, выхода белка после рефолдинга, карт SDS-PAGE, графиков ДСК и профилей SEC белков LC13-G9 и LC13-G15, и релевантных данных для LC13-WT, можно видеть, что, по сравнению с LC13-WT, имеющим немодифицированное гидрофобное ядро, LC13-G9 и LC13-G15, имеющие модифицированное гидрофобное ядро, обладали более высокой способностью к ренатурации, были более устойчивыми к анфолдингу, были более устойчивыми к неправильному или нежелательному фолдингу, обладали улучшенной термической стабильностью и обладали более высоким выходом белка после рефолдинга. Таким образом, по сравнению с LC13-WT, LC13-G9 и LC13-G15 по настоящему изобретению обладали существенно увеличенной стабильностью. В настоящем изобретении стабильность LC13-G9 и LC13-G15 увеличивалась на 5,4% и в 57,9 раз по сравнению со стабильностью LC13-WT, рассчитанной на основе данных для выхода белка после рефолдинга.
Пример 14. Тестирование стабильности белков JM22-G9 и JM22-G15
С использованием способа, описанного в примере 3, белки JM22-G9 и JM22-G15 экспрессировали, подвергали рефолдингу и очищали. После очистки на колонке для гель-фильтрации проводили разделение способом SDS-PAGE, и профили SEC для двух белков получали с использованием способа гель-фильтрации. Экспрессируемое количество, количество очищенного белка и выход белка после рефолдинга рассчитывали одновременно.
Таблица 12 демонстрирует данные об экспрессируемом количестве, количестве очищенного белка и выходе белка после рефолдинга JM22-G9 и JM22-G15. Релевантные данные для JM22-WT также приведены для анализа.


Данные в вышеприведенной таблице демонстрируют, что после очистки выход после рефолдинга белков JM22-G9 и JM22-G15 увеличился в 28,5 раз и 127,25 раз, соответственно, по сравнению с выходом для белка JM22-WT, не имеющего мутации гидрофобного ядра.
Фиг. 39 демонстрирует результаты SDS-PAGE для белков JM22-G9 и JM22-G15, очищенных при помощи колонки для гель-фильтрации (Superdex 75 10/300, GE Healthcare), как описано в примере 3. Фотография геля демонстрирует, что мономерные полосы, образованные путем рефолдинга белка JM22-WT, были гетерогенными, и имелись три полосы, и модифицированные JM22-G9 и JM22-G15 образовывали единичную полосу мономера с высокой чистотой. Последнее свидетельствует о том, что рефолдинг JM22-G9 и JM22-G15 был гораздо лучше рефолдинга JM22-WT.
Фиг. 40 и 41 демонстрируют профили SEC для очищенных белков JM22-G9 и JM22-G15, соответственно. Как представлено, пик элюции, образующийся очищенным белком JM22-WT, не был единичным, и сигнал был очень низким, хотя очищенные JM22-G9 и JM22-G15 образовывали единичный и симметричный пик элюции. Последнее дополнительно свидетельствует о том, что рефолдинг JM22-G9 и JM22-G15 был значительно лучше, чем рефолдинг JM22-WT.
После сравнительного анализа экспрессируемого количества, количества очищенного белка, выхода белка после рефолдинга, карт гелей SDS-PAGE, графиков ДСК и профилей SEC белков JM22-G9 и JM22-G15, можно видеть, что по сравнению с JM22-WT, имеющим немодифицированное гидрофобное ядро, JM22-G9 и JM22-G15, имеющие модифицированное гидрофобное ядро, обладали более сильной способностью к ренатурации, имели более высокое экспрессируемое количество и более высокий выход белка после рефолдинга. Таким образом, по сравнению с JM22-WT, JM22-G9 и JM22-G15 обладали существенно увеличенной стабильностью. В настоящем изобретении стабильность JM22-G9 и JM22-G15 увеличивалась в 28,5 раз и 127,25 раз соответственно по сравнению со стабильностью JM22-WT, рассчитанной на основе данных для выхода белка после рефолдинга.
Пример 15. Тестирование стабильности белков MAGE-sTv-WT и MAGE-G15
С использованием способа, описанного в примере 3, белок MAGE-G15 экспрессировали, подвергали рефолдингу и очищали. После очистки на колонке для гель-фильтрации проводили разделение способом SDS-PAGE, и профиль SEC получали с использованием способа гель-фильтрации. Экспрессируемое количество, количество очищенного белка и выход белка после рефолдинга рассчитывали одновременно.
Таблица 13 демонстрирует данные об экспрессируемом количестве, количестве очищенного белка и выходе белка после рефолдинга MAGE-sTv-WT и MAGE-G15.

Целевой компонент из белка MG-sTv-WT не может быть получен с использованием колонки для гель-фильтрации, как описано в примере 3. Таким образом, карта SDS-PAGE, профиль SEC и график ДСК (величина Tm) не могут быть получены.
Фиг. 43 демонстрирует результат SDS-PAGE для белка MAGE-G15, имеющего модифицированное гидрофобное ядро и очищенного при помощи колонки для гель-фильтрации (Superdex 75 10/300, GE Healthcare). Фотография геля демонстрирует, что MAGE-G15 образует единичную полосу с высокой чистотой. Она свидетельствует о том, что рефолдинг MAGE-G15 был гораздо лучше рефолдинга MG-sTv-WT.
Фиг. 45 демонстрирует график ДСК для MAGE-G15. Величина Tm для него, полученная с использованием модели подгонки TwostateScaled программного обеспечения для анализа Nanoanalyze, составляла 46,7°С.
Связывание MAGE-G15 со своим лигандом определяли при помощи системы анализа в реальном времени BIAcore Т200. Результат демонстрирует, что аффинность белка MAGE-G15 со своим лигандом была не лучше, чем аффинность соответствующего ему TCR дикого типа, для которого величина KD составляла 30,4 мкМ.
Величину увеличенной стабильности рассчитывали на основе данных выхода белка после рефолдинга. Из таблицы 13 можно видеть, что по сравнению с MAGE-sTv-WT стабильность MAGE-G15 по настоящему изобретению была увеличена в бесконечно большое количество раз (по меньшей мере в 10000 раз).
Вышеприведенные данные демонстрируют, что по сравнению с MAGE-sTv-WT, способность к ренатурации, выход белка после рефолдинга и термическая стабильность MAGE-G15 по настоящему изобретению демонстрируют весьма значимое улучшение. Таким образом, стабильность MAGE-G15 по настоящему изобретению демонстрирует значимое улучшение по сравнению с MAGE-sTv-WT.
Пример 16. Анализ путем масс-спектрометрии
Конструированные белки очищали путем гель-фильтрации на колонке (Superdex 75 10/300, GE Healthcare). Молекулярную массу целого белка определяли при помощи масс-спектрометрии и анализировали, согласуется ли она с теоретической молекулярной массой, для того, чтобы проверить, идентична ли последовательность очищенного белка последовательности исходной конструкции.
Молекулярную массу полноразмерного белка определяли при помощи масс-спектрометрии (Eksigent nano LC (nanoflex) - Triple TOF 5600 LC-MS system) от компании AB SCIEX из США. Образец разбавляли 10% ацетонитрилом (Fisher А955-4), 1% муравьиной кислотой (Fisher А11750) и водой (Sigma 39253-1L-R) до проведения масс-спектрометрического. Условия анализа системы были следующими:
Модуль ЖХ (жидкостная хроматография)
Eksigent nano LC (nanoflex) от AB SCIEX
Колонка Guard: C4-3 мкм 300 200 мкм × 0,5 мм; Партия 804-00019
200 мкм × 0,5 мм; Партия 804-00019
Колонка для анализа: С4; 3 мкм, 300; 75 мкм * 15 см, партия 804-00018
Подвижная фаза А: 2% ацетонитрил, 0,1% муравьиная кислота
Подвижная фаза В: 98% ацетонитрил, 0,1% муравьиная кислота
Скорость потока: 300 нл/мин
Градиент: отношение жидкости В увеличивалось с 5% до 90% в течение 10 минут, и общее время анализа составляло 30 минут.
Модуль масс-спектрометра
Элюент с колонки анализировали при помощи Triple TOF 5600 от Nanospray
Способы сбора данных: МС (масс-спектрометрия) в режиме регистрации положительных ионов
Диапазон сбора данных: 400-200 m/z.
Молекулярную массу целого белка в образце получали в результате сбора данных МС после процесса обратной свертки с использованием программного обеспечения Bioanalyst.
После анализа молекулярная масса целого белка (молекулы, сконструированные в настоящем изобретении после экспрессии, рефолдинга и очистки), определенная при помощи МС, согласовывалась с теоретической молекулярной массой, что свидетельствует о том, что последовательность белка, полученного после очистки, была идентична последовательности белка исходной конструкции.
Гидрофобное ядро, отобранное в настоящем изобретении, способно существенно улучшить стабильность молекул TCR. В то же самое время, вышеприведенные примеры продемонстрировали, что введение гидрофобного ядра, отобранного в настоящем изобретении, в другие молекулы TCR, может также играть роль усиления стабильности.
Все документы, цитированные в настоящем изобретении, включены путем ссылки таким образом, как если бы каждая ссылка была цитирована сама по себе как ссылка на применение по настоящему изобретению. Дополнительно, понятно, что после прочтения настоящего изобретения, описанного выше, специалист в данной области техники может осуществить различные изменения или модификации по изобретению, и эти эквивалентные формы также входят в объем, определенный формулой изобретения.
| название | год | авторы | номер документа |
|---|---|---|---|
| Т-КЛЕТОЧНЫЕ РЕЦЕПТОРЫ | 2017 |
|
RU2775623C2 |
| УЛУЧШЕННЫЙ СИНТЕТИЧЕСКИЙ Т-КЛЕТОЧНЫЙ РЕЦЕПТОР И АНТИГЕННЫЙ РЕЦЕПТОР | 2021 |
|
RU2820497C1 |
| СПЕЦИФИЧНЫЕ СВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ | 2020 |
|
RU2825837C2 |
| ВЫСОКОАФФИННЫЙ Т-КЛЕТОЧНЫЙ РЕЦЕПТОР ДЛЯ РАСПОЗНАВАНИЯ АНТИГЕНА AFP | 2019 |
|
RU2836365C2 |
| Т-КЛЕТОЧНЫЕ РЕЦЕПТОРЫ | 2018 |
|
RU2762255C2 |
| МЕЧЕНЫЕ ХИМЕРНЫЕ ЭФФЕКТОРНЫЕ МОЛЕКУЛЫ И ИХ РЕЦЕПТОРЫ | 2014 |
|
RU2729463C2 |
| Т-КЛЕТОЧНЫЕ РЕЦЕПТОРЫ, СПЕЦИФИЧНЫЕ В ОТНОШЕНИИ КОМПЛЕКСА ОПУХОЛЕВЫЙ АНТИГЕН NY-ESO-1/HLA-A*02 | 2016 |
|
RU2775394C2 |
| СПОСОБЫ ПОЛУЧЕНИЯ КЛЕТОК, ЭКСПРЕССИРУЮЩИХ РЕКОМБИНАНТНЫЙ РЕЦЕПТОР, И РОДСТВЕННЫЕ КОМПОЗИЦИИ | 2019 |
|
RU2835579C2 |
| ХИМЕРНЫЕ КОНСТРУКЦИИ АНТИТЕЛО/Т-КЛЕТОЧНЫЙ РЕЦЕПТОР И ИХ ПРИМЕНЕНИЯ | 2016 |
|
RU2767209C2 |
| МОДИФИЦИРОВАННЫЙ ЭКЗОТОКСИН А ПСЕВДОМОНАД | 2014 |
|
RU2687143C2 |















































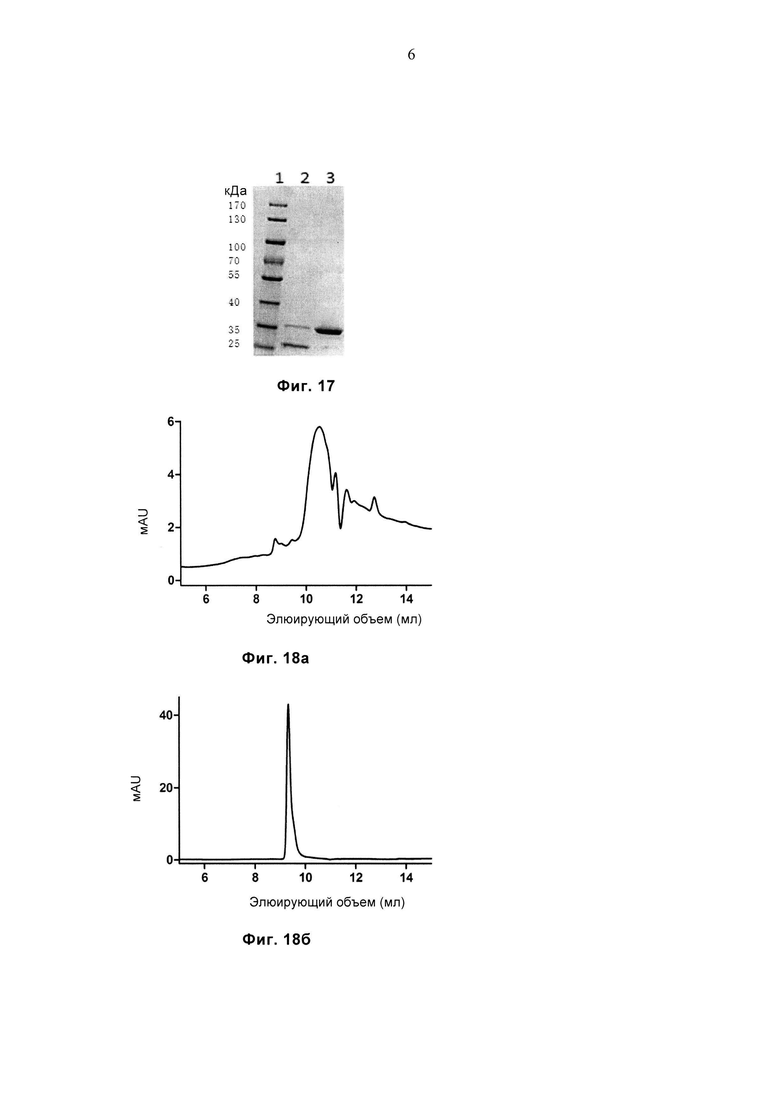
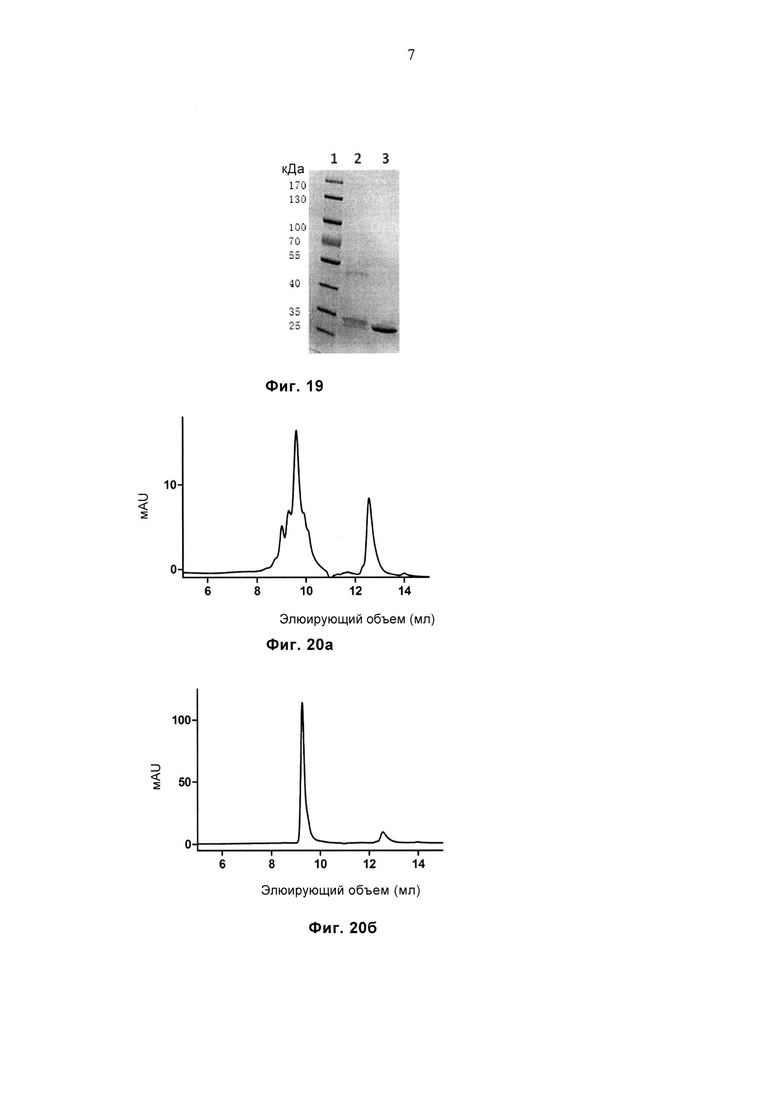





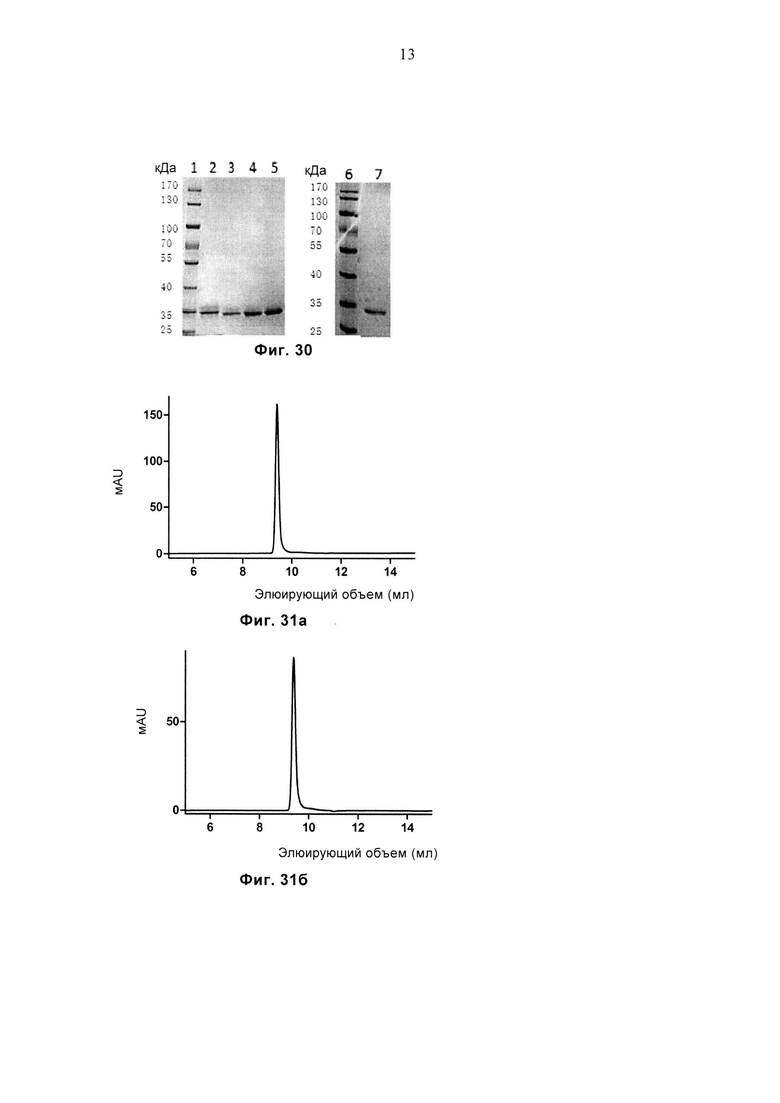

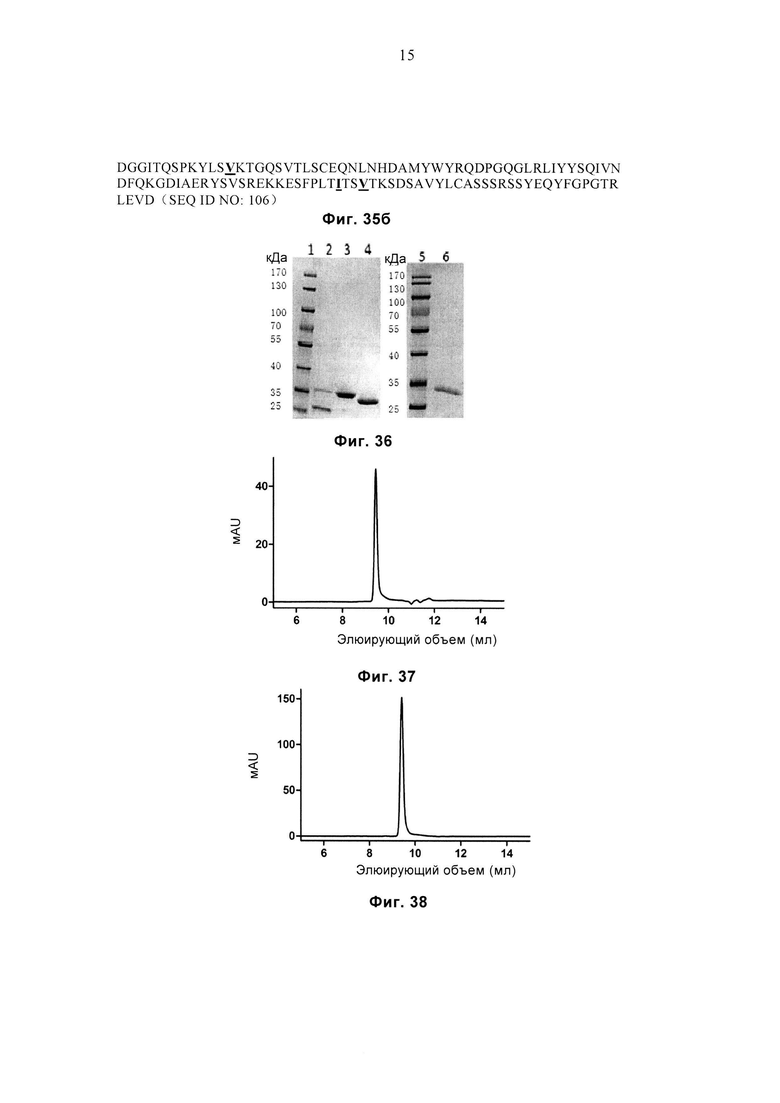
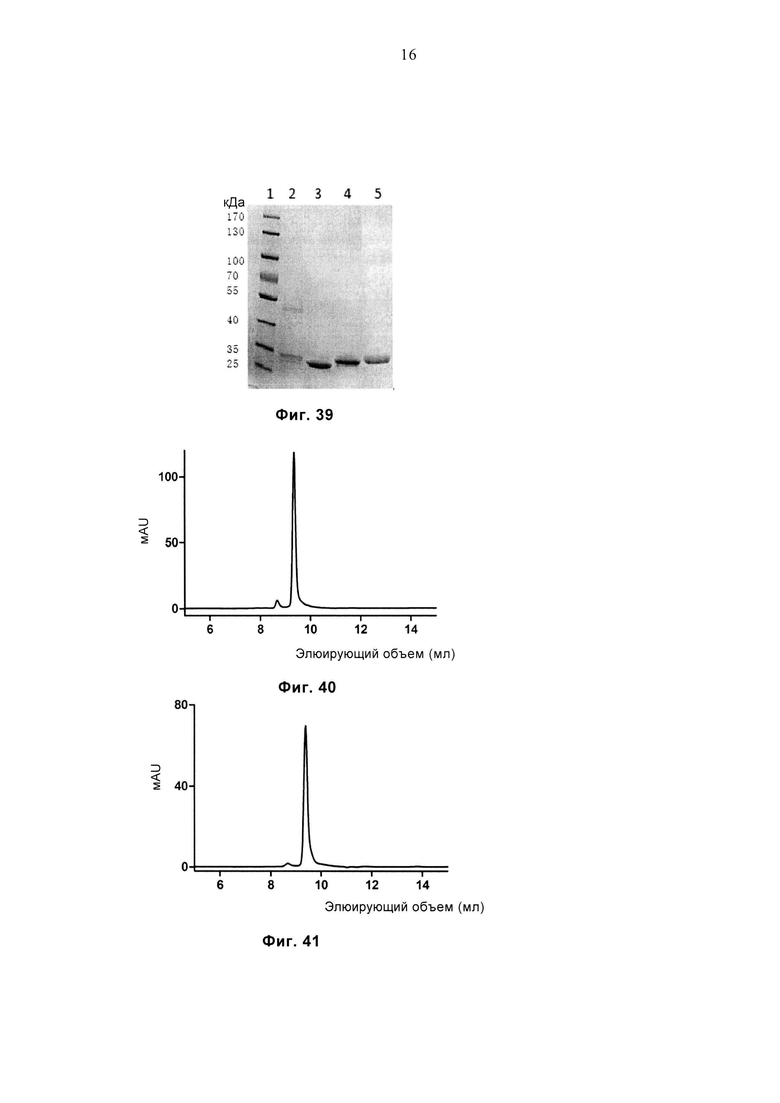
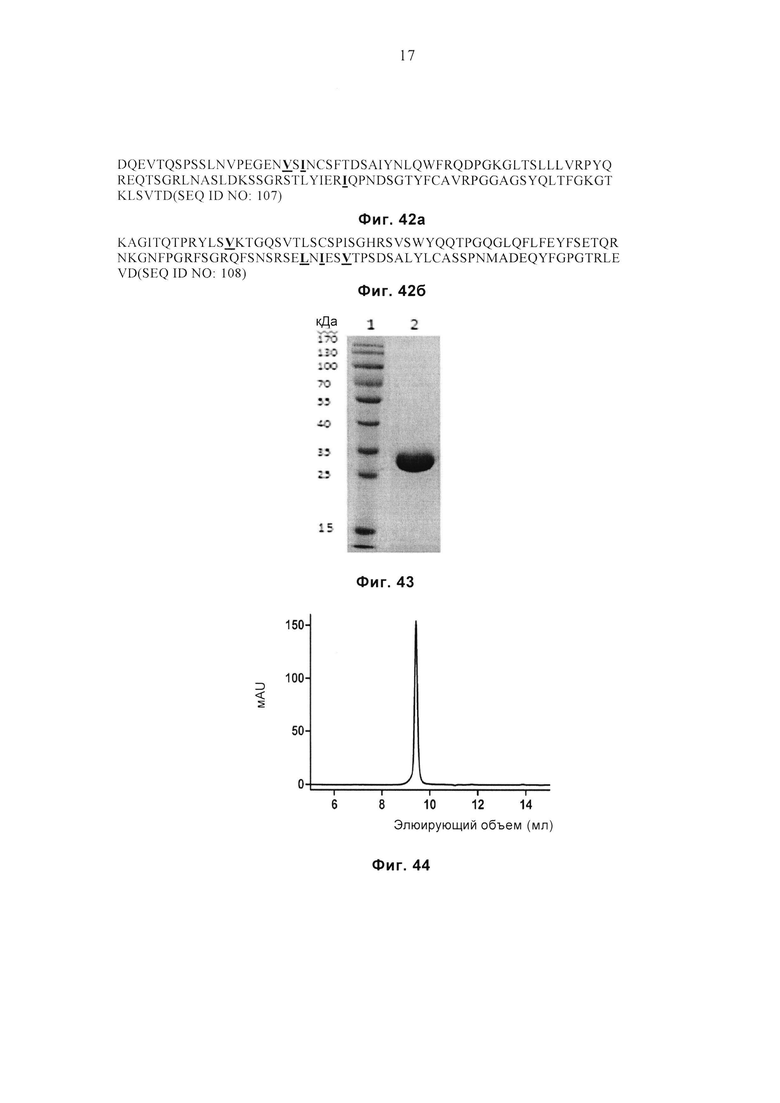
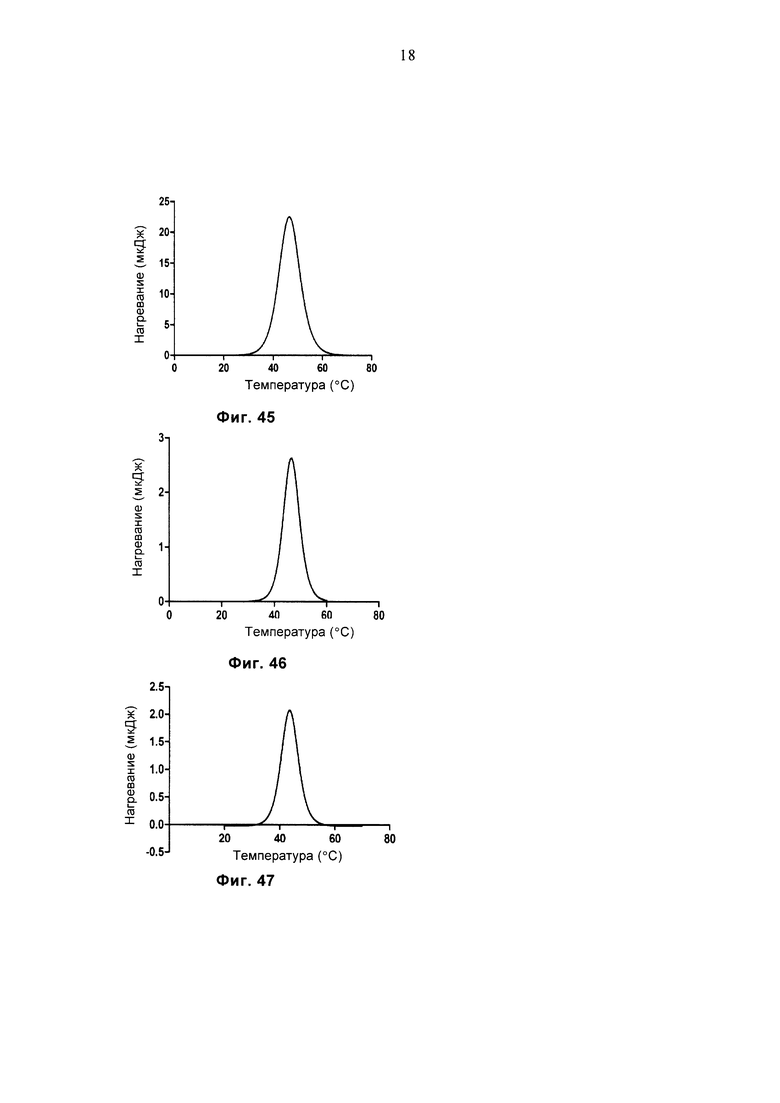


Изобретение относится к области биотехнологии, конкретно предложен высокостабильный Т-клеточный рецептор, что может быть использовано в медицине. Полученный Т-клеточный рецептор, имеющий мутации в своем гидрофобном коровом домене, используют в составе фармацевтической композиции для лечения опухоли. Настоящее изобретение позволяет осуществлять эффективную терапию заболеваний, связанных с образованием опухолей. 9 н. и 16 з.п. ф-лы, 49 ил., 13 табл., 16 пр.
1. Т-клеточный рецептор (TCR), где:
(1) гидрофобный коровый домен TCR является мутантным и
(2) TCR обладает стабильностью более высокой, чем стабильность соответствующего TCR, имеющего гидрофобное ядро дикого типа; и
где одна или более чем одна из следующих позиций в вариабельном домене α-цепи TCR является мутантной: аминокислота в позициях 11, 13, 19, 21, 91 или 94 вариабельного домена α-цепи, где нумерация позиций аминокислот является нумерацией, представленной в IMGT (международная база данных по иммуногенетике); и
где одна или более чем одна из следующих позиций в вариабельном домене β-цепи TCR является мутантной: аминокислота в позиции 11, 13, 19, 89, 91 или 94 вариабельного домена β-цепи и/или аминокислота в четвертой или шестой позиции с конца β-цепи короткого пептида, кодируемого J-геном, где нумерация позиций аминокислот является нумерацией, представленной в IMGT; и
где "стабильность" относится к более высокому выходу при ренатурации белка и/или увеличенной термостабильности и "стабильность более высокая, чем" относится к стабильности TCR, увеличенной по меньшей мере на 30% по сравнению со стабильностью TCR, имеющего гидрофобное ядро дикого типа; и
где вариабельный домен α-цепи и вариабельный домен β-цепи необязательно связаны гибким линкерным пептидом; и
где TCR необязательно связан с конъюгатом.
2. TCR по п. 1, где участки CDR (участки, определяющие комплементарность) TCR являются такими же, как участки CDR TCR дикого типа, или содержат мутацию, повышающую аффинность.
3. TCR по п. 1, где TCR является растворимым.
4. TCR по п. 1, где TCR содержит (а) всю или часть α-цепи TCR, за исключением ее трансмембранного домена; и (б) всю или часть β-цепи TCR, за исключением ее трансмембранного домена;
и каждый из (а) и (б) независимо содержит функциональный вариабельный домен или функциональный вариабельный домен и по меньшей мере часть константного домена цепи TCR.
5. TCR по п. 1, где TCR представляет собой одноцепочечный TCR, состоящий из вариабельного домена α-цепи TCR и вариабельного домена β-цепи TCR, связанных гибким линкерным пептидом.
6. TCR по любому из пп. 1-5, где одна или более чем одна из следующих позиций в вариабельном домене α-цепи, представленном в SEQ ID NO: 9, или SEQ ID NO: 29, или SEQ ID NO: 31, или SEQ ID NO: 33 TCR, является мутантной: аминокислота в позиции 11, 13, 19, 21, 91 или 94 вариабельного домена α-цепи, где нумерация позиций аминокислот является нумерацией, представленной в IMGT.
7. TCR по любому из пп. 1-5, где одна или более чем одна из следующих позиций в вариабельном домене β-цепи, представленном в SEQ ID NO: 11, или SEQ ID NO: 30, или SEQ ID NO: 32, или SEQ ID NO: 34 в TCR, является мутантной: аминокислота в позиции 11, 13, 19, 89, 91 или 94 вариабельного домена β-цепи и/или аминокислота в четвертой или шестой позиции с конца β-цепи короткого пептида, кодируемого J-геном, где нумерация позиций аминокислот является нумерацией, представленной в IMGT.
8. TCR по любому из пп. 1-5, где вариабельный домен α-цепи TCR содержит один или более чем один аминокислотный остаток, выбранный из группы, состоящей из: 11L, 11М или 11Е; 13V, 13R или 13K; 19V; 21I; 91L или 91I; и 94V или 94I; и/или вариабельный домен β-цепи TCR содержит один или более чем один аминокислотный остаток, выбранный из группы, состоящей из: 11L или 11V; 13V; 19V; 89L; 91F или 91I; 94V или 94L; шестая позиция с конца β-цепи, кодируемой J-геном, представляет собой Т; и четвертая позиция с конца β-цепи, кодируемой J-геном, представляет собой М; где нумерация позиций аминокислот является нумерацией, представленной в IMGT.
9. TCR по любому из пп. 1-5, где аминокислотный остаток, находящийся в вариабельном домене α-цепи и/или вариабельном домене β-цепи TCR и расположенный на поверхности, является мутантным.
10. TCR по п. 9, где TCR содержит один или более чем один аминокислотный остаток вариабельного домена α-цепи, выбранный из группы, состоящей из: 4L; 12N; 16S; 93N или 93R; 97N; 100G; 105S; и последняя позиция α-цепи, кодируемой J-геном, представляет собой D; и/или TCR содержит один или более чем один аминокислотный остаток вариабельного домена β-цепи, выбранный из группы, состоящей из: 4I; 101L; последняя позиция β-цепи, кодируемой J-геном, представляет собой D; и третья позиция с конца β-цепи, кодируемой J-геном, представляет собой Е; где нумерация позиций аминокислот является нумерацией, представленной в IMGT.
11. TCR по любому из пп. 1-5, где TCR содержит вариабельный домен α-цепи, аминокислотная последовательность которого выбрана из группы, состоящей SEQ ID NO: 15, 17, 35, 37, 39, 75, 76, 77, 78, 79, 80, 81, 82, 83, 84, 85, 97, 99, 101, 103, 105 и 107; и/или
TCR содержит вариабельный домен β-цепи, аминокислотная последовательность которого выбрана из группы, состоящей из SEQ ID NO: 16, 18, 36, 38, 40, 86, 87, 88, 89, 90, 91, 92, 93, 94, 95, 96, 98, 100, 102, 104, 106 и 108.
12. TCR по любому из пп. 1-5, где комбинация вариабельного домена α-цепи и вариабельного домена β-цепи TCR выбрана из группы, состоящей из:
(а) вариабельного домена α-цепи, имеющего аминокислотную последовательность SEQ ID NO: 15, и вариабельного домена β-цепи, имеющего аминокислотную последовательность SEQ ID NO: 16;
(б) вариабельного домена α-цепи, имеющего аминокислотную последовательность SEQ ID NO: 17, и вариабельного домена β-цепи, имеющего аминокислотную последовательность SEQ ID NO: 18;
(в) вариабельного домена α-цепи, имеющего аминокислотную последовательность SEQ ID NO: 15, и вариабельного домена β-цепи, имеющего аминокислотную последовательность SEQ ID NO: 18;
(г) вариабельного домена α-цепи, имеющего аминокислотную последовательность SEQ ID NO: 35, и вариабельного домена β-цепи, имеющего аминокислотную последовательность SEQ ID NO: 36;
(д) вариабельного домена α-цепи, имеющего аминокислотную последовательность SEQ ID NO: 37, и вариабельного домена β-цепи, имеющего аминокислотную последовательность SEQ ID NO: 38;
(е) вариабельного домена α-цепи, имеющего аминокислотную последовательность SEQ ID NO: 39, и вариабельного домена β-цепи, имеющего аминокислотную последовательность SEQ ID NO: 40;
(ж) вариабельного домена α-цепи, имеющего аминокислотную последовательность SEQ ID NO: 75, и вариабельного домена β-цепи, имеющего аминокислотную последовательность SEQ ID NO: 86;
(з) вариабельного домена α-цепи, имеющего аминокислотную последовательность SEQ ID NO: 76, и вариабельного домена β-цепи, имеющего аминокислотную последовательность SEQ ID NO: 87;
(и) вариабельного домена α-цепи, имеющего аминокислотную последовательность SEQ ID NO: 77, и вариабельного домена β-цепи, имеющего аминокислотную последовательность SEQ ID NO: 88;
(к) вариабельного домена α-цепи, имеющего аминокислотную последовательность SEQ ID NO: 78, и вариабельного домена β-цепи, имеющего аминокислотную последовательность SEQ ID NO: 89;
(л) вариабельного домена α-цепи, имеющего аминокислотную последовательность SEQ ID NO: 79, и вариабельного домена β-цепи, имеющего аминокислотную последовательность SEQ ID NO: 90;
(м) вариабельного домена α-цепи, имеющего аминокислотную последовательность SEQ ID NO: 80, и вариабельного домена β-цепи, имеющего аминокислотную последовательность SEQ ID NO: 91;
(н) вариабельного домена α-цепи, имеющего аминокислотную последовательность SEQ ID NO: 81, и вариабельного домена β-цепи, имеющего аминокислотную последовательность SEQ ID NO: 92;
(о) вариабельного домена α-цепи, имеющего аминокислотную последовательность SEQ ID NO: 82, и вариабельного домена β-цепи, имеющего аминокислотную последовательность SEQ ID NO: 93;
(п) вариабельного домена α-цепи, имеющего аминокислотную последовательность SEQ ID NO: 83, и вариабельного домена β-цепи, имеющего аминокислотную последовательность SEQ ID NO: 94;
(р) вариабельного домена α-цепи, имеющего аминокислотную последовательность SEQ ID NO: 84, и вариабельного домена β-цепи, имеющего аминокислотную последовательность SEQ ID NO: 95;
(с) вариабельного домена α-цепи, имеющего аминокислотную последовательность SEQ ID NO: 85, и вариабельного домена β-цепи, имеющего аминокислотную последовательность SEQ ID NO: 96;
(т) вариабельного домена α-цепи, имеющего аминокислотную последовательность SEQ ID NO: 97, и вариабельного домена β-цепи, имеющего аминокислотную последовательность SEQ ID NO: 98;
(у) вариабельного домена α-цепи, имеющего аминокислотную последовательность SEQ ID NO: 99, и вариабельного домена β-цепи, имеющего аминокислотную последовательность SEQ ID NO: 100;
(ф) вариабельного домена α-цепи, имеющего аминокислотную последовательность SEQ ID NO: 101, и вариабельного домена β-цепи, имеющего аминокислотную последовательность SEQ ID NO: 102;
(х) вариабельного домена α-цепи, имеющего аминокислотную последовательность SEQ ID NO: 103, и вариабельного домена β-цепи, имеющего аминокислотную последовательность SEQ ID NO: 104;
(ц) вариабельного домена α-цепи, имеющего аминокислотную последовательность SEQ ID NO: 105, и вариабельного домена β-цепи, имеющего аминокислотную последовательность SEQ ID NO: 106; и
(ч) вариабельного домена α-цепи, имеющего аминокислотную последовательность SEQ ID NO: 107, и вариабельного домена β-цепи, имеющего аминокислотную последовательность SEQ ID NO: 108.
13. TCR по п. 4, где TCR представляет собой α/β гетеродимерный TCR, имеющий последовательность константного домена α-цепи и последовательность константного домена β-цепи, и цистеиновые остатки образуют дисульфидную связь между константными доменами α- и β-цепей.
14. TCR по п. 13, где цистеиновые остатки образуют искусственную дисульфидную связь между константными доменами α- и β-цепей TCR.
15. TCR по любому из пп. 1-5, где С-конец или N-конец α-цепи и/или β-цепи TCR связан с конъюгатом.
16. TCR по п. 15, где конъюгат, связанный с TCR, представляет собой обнаруживаемый маркер, терапевтический агент, группировку, модифицирующую PK (фармакокинетический профиль), или их комбинацию; и предпочтительно терапевтический агент, связанный с TCR, представляет собой антитело против CD3, связанное с С- или N-концом α-цепи или β-цепи TCR.
17. TCR по п. 1, где мутация в TCR отобрана при помощи технологии фагового дисплея.
18. Молекула нуклеиновой кислоты, кодирующая TCR по любому из пп. 1-17.
19. Вектор экспрессии, содержащий молекулу нуклеиновой кислоты по п. 18.
20. Клетка-хозяин для экспрессии Т-клеточного рецептора по любому из пп. 1-17, содержащая вектор экспрессии по п. 19, или в которой экзогенная молекула нуклеиновой кислоты по п. 18 интегрирована в хромосому, где клетка-хозяин представляет собой прокариотическую или эукариотическую клетку.
21. Способ получения TCR по любому из пп. 1-17, включающий:
(1) инкубацию клетки-хозяина по п. 20, экспрессирующей TCR по п. 1,
(2) выделение или очистку TCR;
где способ выделения для выделения и очистки TCR выбран из группы, состоящей из обработки для ренатурации, обработки осадителем белка, центрифугирования, осмотического клеточного лизиса, ультразвуковой обработки, ультрацентрифугирования, хроматографии на молекулярных ситах, адсорбционной хроматографии, ионообменной хроматографии, высокоэффективной жидкостной хроматографии (ВЭЖХ) и их комбинации.
22. Применение TCR по любому из пп. 1-17 для производства лекарственного средства для лечения опухоли.
23. Фармацевтическая композиция для лечения опухоли, содержащая фармацевтически приемлемый носитель и безопасную и эффективную дозу TCR по любому из пп. 1-17.
24. Способ лечения заболевания, включающий введение эффективного количества TCR по любому из пп. 1-17 или эффективного количества фармацевтической композиции по п. 23 субъекту, нуждающемуся в таком введении; заболевание представляет собой опухоль.
25. Способ получения TCR по любому из пп. 1-17, включающий:
(1) введение мутации аминокислотного остатка в гидрофобный коровый домен TCR и
(2) отбор TCR, стабильность которых существенно увеличена, таким образом получая TCR по любому из пп. 1-17,
где TCR отбирают при помощи технологии фагового дисплея.
| Способ приготовления лака | 1924 |
|
SU2011A1 |
| Изложница с суживающимся книзу сечением и с вертикально перемещающимся днищем | 1924 |
|
SU2012A1 |
| Колосоуборка | 1923 |
|
SU2009A1 |
| SHUSTA E.V., Directed evolution of a stable scaffold for T-cell receptor engineering, Nat | |||
| Biotechnol., 2000, v.18, is.7, p.754-759 | |||
| ОДНОЦЕПОЧЕЧНЫЕ РЕКОМБИНАНТНЫЕ Т-КЛЕТОЧНЫЕ РЕЦЕПТОРЫ | 2003 |
|
RU2355703C2 |
| PAKULA A.A | |||
| et al., Genetic analysis of protein stability and function | |||
| Anna | |||
| Rev | |||
| Genet | |||
| Механизм для сообщения поршню рабочего цилиндра возвратно-поступательного движения | 1918 |
|
SU1989A1 |
| FRANKEL A.E | |||
| et al., Characterization of diphtheria fusion proteins targeted to the human interleukin-3 receptor, Protein Eng., 2000, v.13, n.8, p.575-581. | |||

