В соответствии с настоящим изобретением предложены Т-клеточные рецепторы (TCR), связывающие рестриктированный по HLA-A*02 пептид GVYDGREHTV (SEQ ID NO: 1), полученный из ракового антигена MAGE А4 клеток зародышевой линии. Указанные TCR могут содержать неприродные мутации в вариабельных доменах альфа и/или бета-цепей относительно нативного MAGE А4 TCR. TCR согласно настоящему изобретению особенно подходят для применения в качестве новых иммунотерапевтических реагентов для лечения злокачественных заболеваний.
Уровень техники
Т-клеточные рецепторы (TCR) в природных условиях экспрессируются Т-клетками CD4+ и CD8+. TCR предназначены для распознавания коротких пептидных антигенов, которые располагаются на поверхности антигенпрезентирующих клеток в комплексе с молекулами главного комплекса гистосовместимости (МНС) (у людей молекулы МНС также называют лейкоцитарными антигенами человека или HLA) (Davis, et at., (1998), Annu Rev Immunol 16: 523-544.). Т-клетки CD8+, которые также называют цитотоксическими Т-клетками, специфически распознают пептиды, связанные с МНС класса I, и, как правило, ответственны за обнаружение и участие в уничтожении пораженных заболеванием клеток. Т-клетки CD8+ способны уничтожать раковые, а также инфицированные вирусом клетки; однако аффинность естественного репертуара TCR, экспрессируемых опухолеспецифическими Т-клетками, обычно является низкой в результате селекции в тимусе, что означает, что злокачественные клетки часто избегают обнаружения и уничтожения. Новые иммунотерапевтические подходы, направленные на содействие распознаванию рака Т-клетками, обеспечивают возможность применения многообещающей стратегии для разработки эффективных противораковых средств.
MAGE А4 принадлежит к семейству раковых антигенов MAGE, кодируемых в клетках зародышевой линии (De Plaen, et al., (1994), Immunogenetics 40 (5): 360-369) и имеет номер доступа в базе Uniprot Р43358. Было обнаружено, что такие антигены часто экспрессируются во многих раковых тканях, а в нормальных тканях их экспрессия ограничена семенниками взрослых организмов и другими иммунологически «привилегированными» областями, такими как плацента. Опухолеспецифический характер этих генов делает их идеальными мишенями для противораковой терапии. Точная функция MAGE А4 остается неизвестной, но считается, что он играет роль в эмбриональном развитии. Сообщалось о высокой степени экспрессии MAGE А4 в опухолях нескольких типов, включая меланому, карциному пищевода, головы и шеи, легких, молочной железы и мочевого пузыря (Bergeron, (2009), Int J Cancer 125(6): 1365-1371; Cabezon, et al., (2013), Mol Cell Proteomics 12(2): 381-394; Cuffel, et al., (2011), Int J Cancer 128(11): 2625-2634. Forghanifard, et al., (2011), Cancer Biol Ther 12(3): 191-197; Karimi, et al., (2012), din Lung Cancer 13(3): 214-219; Svobodova, et al., (2011), Eur J Cancer 47(3): 460-469). Указанный пептид из 10-ти аминокислот (10-мер) GVYDGREHTV (SEQ ID NO 1) соответствует аминокислотам 230-239 полноразмерного белка MAGE А4. Указанный пептид связывается с HLA-A*02, и было показано, что комплекс пептид-HLA стимулирует цитотоксические Т-клетки, приводя к лизису положительных по MAGE А4, положительных по HLA-A*02 опухолевых клеток (Duffour, et al., (1999), Eur J Immunol 29 (10): 3329-3337 и WO 2000020445). Таким образом, комплекс GVYDGREHTV HLA-A*02 представляет собой целевой антиген, который можно применять для иммунотерапевтического вмешательства.
Выявление конкретных последовательностей TCR, которые связываются с комплексом GVYDGREHTV-HLA-A*02 с высокой специфичностью, является эффективным подходом для разработки новых иммунотерапевтических средств. Терапевтические TCR могут быть использованы, например, в качестве растворимых агентов целенаправленного действия для доставки цитотоксических или иммунных эффекторных агентов в опухоль (Lissin, et al., (2013). "High-Affinity Monocloncal T-cell receptor (mTCR) Fusions. Fusion Protein Technologies for Biophamaceuticals: Applications and Challenges". S.R. Schmidt, Wiley; Boulter, et al., (2003), Protein Eng 16(9): 707-711; Liddy, et al., (2012), Nat Med 8: 980-987), или, в другом варианте, они могут быть использованы для конструирования Т-клеток для адоптивной терапии (June, et al., (2014), Cancer Immunol lmmunother 63(9): 969-975). Однако такие последовательности TCR пока не известны в данной области техники, а способы выявления TCR со специфическими характеристиками, пригодных для терапевтического применения, имеют высокий процент негативных результатов и поэтому не дают квалифицированному специалисту достаточных оснований надеяться на успех.
В первую очередь квалифицированный специалист должен выявить подходящую исходную или каркасную последовательность. Как правило, такие последовательности получают из природных источников, например, из отвечающих на антиген Т-клеток, выделенных из донорской крови. Учитывая редкость опухолеспецифических Т-клеток в естественном репертуаре, часто необходимо провести скрининг многих доноров, например, 20 или более, прежде чем найти отвечающую (реактивную) Т-клетку. Процесс скрининга может занять несколько недель или месяцев, и даже если отвечающая Т-клетка обнаружена, она может быть непригодна для иммунотерапевтического применения. Например, ответ может быть слишком слабым и/или не быть специфичным для антигена-мишени, или в качестве альтернативы, может оказаться невозможным создать клональную популяцию Т-клеток, а также размножать или поддерживать данную линию Т-клеток для получения достаточного материала для идентификации правильных последовательностей цепи TCR. Кроме того, поскольку TCR являются вырожденными и, как было спрогнозировано, способны связывать приблизительно один миллион различных HLA-пептидов (Wooldridge, et al., (2012), J Biol Chem 287(2): 1168-1177), даже опытным специалистам исключительно сложно определить, имеет ли конкретный TCR профиль специфичности, который обеспечил бы возможность его применения для конструирования молекулы для терапевтического применения.
Последовательности TCR, которые являются подходящими в качестве исходных или каркасных последовательностей, должны иметь хорошую аффинность к целевому комплексу пептид-HLA, например 200 мкМ или более сильную аффинность, демонстрировать высокий уровень специфичности к распознаванию мишени, например, относительно слабое связывание или отсутствие связывания с альтернативными комплексами пептид-HLA, поддаваться модификации для использования в библиотеках дисплеев, таких как фаговый дисплей, и допускать рефолдинг и очистку с высоким выходом.
TCR, существующие в природе, имеют слабую аффинность к антигену-мишени (низкий микромолярный диапазон) по сравнению с антителами, a TCR против раковых антигенов обычно способны к более слабому распознаванию антигена, чем вирус-специфические TCR (Aleksic, et al. (2012). Eu J Immunol., 42 (12), 3174-3179). Эта слабая аффинность в сочетании с снижением количества HLA на раковых клетках означает, что терапевтические TCR для иммунотерапии рака требуют генноинженерной модификации для увеличения их аффинности к антигену-мишени и, таким образом, обеспечения более сильного ответа. Желательной для растворимых целевых агентов на основе TCR является аффинность связывания антигена TCR в диапазоне от наномолярного до пикомолярного, с периодом полужизни в связанном состоянии, составляющим несколько часов. Такая повышенная активность, генерируемая распознаванием антигена с высокой аффинностью при низких количествах эпитопов, проиллюстрирована на Фигурах 1е и 1f в работе Liddy et al. (Liddy, et al. (2012), Nat Med, 18 (6), 980-987). При созревании аффинности квалифицированный специалист обычно должен идентифицировать конкретные мутации и/или комбинации мутаций, включая, но не ограничиваясь указанным, замены, инсерции и/или делеции, которые можно внести в исходную последовательность TCR для увеличения силы распознавания антигена. В данной области техники известны способы идентификации мутаций любого заданного TCR, увеличивающих аффинность, например, использование библиотек дисплеев. (Li et al., (2005) Nat Biotechnol. 23(3):349-354; Holler et al., (2000). Proc Natl Acad Sci USA; 97(10): 5387-5392). Однако для обеспечения значительного увеличения аффинности данного TCR к данной мишени специалист должен подобрать конкретные мутации и/или комбинации мутаций из большого числа возможных альтернатив. Конкретные мутации и/или комбинации мутаций, которые приведут к значительному увеличению аффинности, непредсказуемы, и при отборе наблюдается высокий уровень неудачных результатов. Во многих случаях может оказаться невозможно достичь значительного увеличения аффинности с заданной исходной последовательностью TCR.
Процесс созревания аффинности должен также включать необходимость сохранения специфичности антигена TCR. Увеличение аффинности TCR к его антигену-мишени приводит к значительному риску выявления перекрестной реактивности с другими непредусмотренными мишенями в результате неустранимой вырожденности распознавания антигена TCR. (Wooldridge, et al., (2012), J Biol Chem 287(2): 1168-1177; Wilson, et al., (2004), Mol Immunol 40(14-15): 1047-1055; Zhao et al., (2007) J. Immunol, 179; 9, 5845-5854). При естественном уровне аффинности распознавание перекрестно-реактивного антигена может быть слишком низким, чтобы вызвать ответ. Если перекрестно реактивный антиген презентируется на нормальных здоровых клетках, существует высокая вероятность нецелевого связывания in vivo, которое может проявляться токсичности в клинической практике. Таким образом, в дополнение к увеличению силы связывания антигена специалист должен также выбрать мутации и/или комбинации мутаций, которые позволяют TCR сохранять высокую специфичность к антигену-мишени и демонстрировать хороший профиль безопасности при доклинических испытаниях. К тому же, такие мутации и/или комбинации мутаций не являются предсказуемыми. Уровень неудач на этой стадии еще выше, и во многих случаях получение целевой молекулы из заданной исходной последовательности TCR может быть вообще невозможно.
Мутации, необходимые для обеспечения высокой аффинности и высокой специфичности, должны также обеспечивать получение TCR, который можно экспрессировать, с которым можно осуществлять рефолдинг и очистку с приемлемым выходом и который является высокостабильным в очищенной форме.
Несмотря на описанные выше трудности идентификации последовательностей TCR с подходящими характеристиками для терапевтического применения, авторы настоящего изобретения неожиданно обнаружили последовательность TCR, которая представляет собой идеальную отправную точку или каркас для получения терапевтических TCR. Кроме того, авторы настоящего изобретения неожиданно выявили подходящие мутации, которые могут быть введены в вариабельные домены альфа- и бета-цепи указанного каркаса для получения последовательностей TCR с идеальными характеристиками для направленной основанной на TCR иммунотерапии- раковых заболеваний, при которых экспрессируется MAGE А4.
Краткое описание изобретения
В первом аспекте настоящее изобретение относится к Т-клеточному рецептору (TCR), обладающему свойством связывания с GVYDGREHTV (SEQ ID NO: 1) в комплексе с HLA-А*02 и содержащему вариабельный домен альфа-цепи TCR и/или вариабельный домен бета-цепи TCR, при этом:
указанный вариабельный домен альфа-цепи содержит аминокислотную последовательность, которая по меньшей мере на 90% идентична последовательности аминокислотных остатков 1-113 SEQ ID NO: 2, и/или
указанный вариабельный домен бета-цепи содержит аминокислотную последовательность, которая по меньшей мере на 90% идентична последовательности аминокислотных остатков 1-116 SEQ ID NO: 3.
Вариабельный домен альфа-цепи может содержать аминокислотную последовательность, которая по меньшей мере на 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотам 1-113 SEQ ID NO: 2, и/или вариабельный домен бета-цепи может содержать аминокислотную последовательность, которая имеет по меньшей мере на 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотам 1-116 SEQ ID NO: 3.
Во втором аспекте изобретение относится к TCR, который связывается с комплексом HLA-А*02 GVYDGREHTV (SEQ ID NO: 1) с аффинностью более 200 мкМ, причем: CDR 1, 2 и 3 альфа-цепи содержат SEQ ID NO 6, 7 и 8 соответственно и/или CDR 1, 2 и 3 бета-цепи содержат SEQ ID NO 11, 12 и 13 соответственно; и/или по меньшей мере один из CDR содержит одну или несколько консервативных замен относительно SEQ ID NO: 6-8 и 11-13; и/или по меньшей мере один из CDR содержит до трех допустимых замен относительно SEQ ID NO: 6-8 и 11-13.
Аффинность TCR по настоящему изобретению к комплексу GVYDGREHTV HLA-A*02 может находиться в диапазоне от 200 мкМ до 1 пМ. Предпочтительно указанные замены не изменяют аффинность связывания более чем на +/- 50% или, более предпочтительно, не более чем на +/- 20% относительно TCR без замен. Предпочтительно указанные замены не увеличивают аффинность связывания с альтернативными комплексами пептид-HLA.
Каркасный TCR имеет следующие варианты использования вариабельных доменов альфа-и бета-цепей:
Альфа-цепь: TRAV10*01/TRAJ6*01
Бета-цепь: TRBV28*01/TRBD1*01/TRBJ2-7*01/ (Обратите внимание, что элемент «*01» обозначает аллельный вариант для этой последовательности, в соответствии с номенклатурой IMGT.) и следующие последовательности CDR3 альфа- и бета-цепи:
Альфа-цепь: WNHSGGSYIPTF (SEQ ID NO: 8)
Бета-цепь: ASSFLMTSGDPYEQYF (SEQ ID NO: 13)
Термин «каркасный TCR» или «исходный TCR» используется в данной заявке как синоним терминов «TCR дикого типа» или «WT TCR», или «немутантный (немутированный) TCR», или «нативный TCR», или «родительский TCR», и означает TCR, имеющий вариабельный домен альфа-цепи, содержащий остатки 1-113 последовательности SEQ ID NO: 2, и вариабельный домен бета-цепи, содержащий остатки 1-116 последовательности SEQ ID NO: 3. Константный домен TCR WT может быть полноразмерным или может быть укороченным и/или мутированным с целью получения растворимого TCR. В любом случае в области TRAC и TRBC могут быть введены остатки цистеина так, что может образоваться ненативная межцепочечная дисульфидная связь. Подходящие положения для расположения указанных участков цистеина описаны в WO 03020763. На Фигуре 2 прилагаемых чертежей показаны внеклеточные последовательности альфа- и бета-цепей TCR дикого типа соответственно в растворимом формате. SEQ ID NO: 4 идентична внеклеточной последовательности нативной альфа-цепи SEQ ID NO: 2, за исключением того, что цистеин в положении 48 константного домена был заменен треонином. Аналогично, SEQ ID NO: 5 идентична внеклеточной последовательности нативной бета-цепи SEQ ID NO: 3, за исключением того, что цистеин в положении 57 константного домена был заменен серином, цистеин в положении 75 константного домена был заменен аланином, и аспарагин в положении 89 константного домена был заменен аспарагиновой кислотой. Растворимый TCR дикого типа можно использовать в качестве эталона для сравнения с ним профиля связывания мутированных TCR по настоящему изобретению.
Определенные в настоящей заявке последовательности TCR описаны со ссылкой на номенклатуру IMGT, которая широко известна и доступна для тех, кто работает в области TCR. Например, см.: LeFranc and LeFranc, (2001). "Т cell Receptor Factsbook", Academic Press; Lefranc, (2011), Cold Spring Harb Protoc 2011(6): 595-603; Lefranc, (2001), Curr Protoc Immunol Appendix 1: Appendix 10O; и Lefranc, (2003), Leukemia 17(1): 260-266. В кратком изложении, αβ TCR состоят из двух цепей, связанных дисульфидными связями. Каждая цепь (альфа и бета) обычно рассматривается как имеющая два домена, а именно вариабельный и константный домен. Короткий J-сегмент соединяет вариабельные и константные домены и обычно считается частью вариабельной области альфа. Кроме того, бета-цепь обычно содержит короткий D-сегмент рядом с J-сегментом, который также обычно считается частью вариабельной области бета.
Вариабельный домен каждой цепи расположен на N-конце и содержит три определяющих комплементарность участки (CDR), встроенные в каркасную последовательность. CDR содержат сайт распознавания для связывания пептид-МНС. Существует несколько генов, кодирующих вариабельные области альфа-цепи (Vα), и несколько генов, кодирующих вариабельные области бета-цепи (Vβ), которые различаются своим каркасом, последовательностями CDR1 и CDR2 и частично определенной последовательностью CDR3. Гены Vα и Vβ обозначены в номенклатуре IMGT префиксом TRAV и TRBV соответственно (Folch and Lefranc, (2000), Exp Clin Immunogenet 17(1): 42-54; Scaviner and Lefranc, (2000), Exp Clin Immunogenet 17(2): 83-96; LeFranc and LeFranc, (2001), "T cell Receptor Factsbook", Academic Press). Аналогично, существует несколько соединяющих или J-генов, называемых TRAJ или TRBJ, для альфа- и бета-цепи соответственно, и для бета-цепи - генов разнообразия или D-генов, называемых TRBD. (Folch and Lefranc, (2000), Exp Clin Immunogenet 17(2): 107-114; Scaviner and Lefranc, (2000), Exp Clin Immunogenet 17(2): 97-106; LeFranc and LeFranc, (2001), "T cell Receptor Factsbook", Academic Press). Огромное разнообразие цепей рецепторов Т-клеток является результатом комбинаторных реаранжировок между различными генами V, J и D, которые включают аллельные варианты, и разнообразия J-сегментов (Arstila, et al., (1999), Science 286(5441): 958-961; Robins et al., (2009), Blood 114(19): 4099-4107.) Константные или С-области альфа- и бета-цепей TCR обозначаются как TRAC и TRBC, соответственно. (Lefranc, (2001), Curr Protoc Immunol Appendix 1: Appendix 10).
Вариабельный домен альфа-цепи первого или второго аспекта может содержать мутацию по меньшей мере в одном из следующих положений по отношению к положениям остатков 1-113 последовательности SEQ ID NO: 2: М50, Т51, F52, S53, Е54, Н94, S95, G96, S98. Мутации могут быть выбраны из следующих аминокислот по отношению к положениям остатков 1-113 последовательности SEQ ID NO: 2:


Дополнительно или в другом варианте, вариабельный домен бета-цепи по первому или второму аспекту может содержать мутацию по меньшей мере в одном из следующих положений по отношению к положениям остатков 1-116 последовательности SEQ ID NO: 3: М27; D28; Н29, Е30, N31, Y50, D51, V52, К53, М54, F95, L96, М97, Т98. Мутации могут быть выбраны из следующих аминокислот по отношению к положениям остатков 1-116 SEQ ID NO: 3:
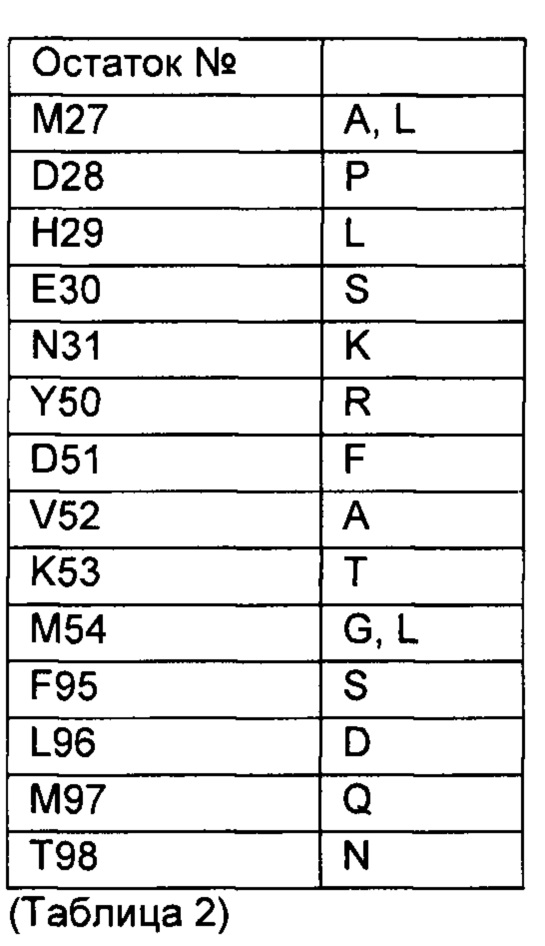
Вариабельный домен альфа-цепи может содержать 1, 2, 3, 4, 5, 6, 7, 8 или 9 мутаций, показанных в Таблице 1, и/или вариабельный домен бета-цепи может содержать 1, 2, 3, 4, 5 6, 7, 8, 9 или 10 мутаций, показанных в Таблице 2.
Вариабельный домен альфа-цепи может содержать по меньшей мере одну из следующих групп мутаций:
Группа 1: M50L, T51D, F52Y, S53A, E54I
Группа 2: H94S, S95A, G96N, S98L
Группа 3: H94R S95A, G96D, S98L
Группа 4: M50L, T51D, F52Y, S53A, E54I, H94R S95A, G96D, S98L
Группа 5: M50L, H94S, S95A, G96S, S98L
Группа 6: M50L, H94S, S95A, G96Q, S98L
и/или вариабельный домен бета-цепи может содержать по меньшей мере одну из следующих групп мутаций:
Группа 1: М27А, D28P, H29L, E30S, N31K F95S, L96D, M97Q, T98N
Группа 2: Y50R, D51F, V52A, К53Т, M54G, F95S, L96D, M97Q, T98N
Группа 3: F95S, L96D, M97Q, T98N
Группа 4: M27L, Y50R, D51F, V52A, К53Т, M54G, F95S, L96D, M97Q, T98N
Группа 5: М27А, D28P, H29L, E30S, N31К, M54L, F95S, L96D, M97Q, T98N
Например, вариабельный домен альфа-цепи может содержать мутации группы 4, а вариабельный домен бета-цепи может содержать мутации группы 1; вариабельный домен альфа-цепи может содержать мутации группы 4, а вариабельный домен бета-цепи может содержать мутации группы 5; вариабельный домен альфа-цепи может содержать мутации группы 2, а вариабельный домен бета-цепи может содержать мутации группы 2; вариабельный домен альфа-цепи может содержать мутации группы 6, а вариабельный домен бета-цепи может содержать мутации группы 4; вариабельный домен альфа-цепи может содержать мутации группы 5, а вариабельный домен бета-цепи может содержать мутации группы 4.
Мутации могут дополнительно или в другом варианте быть выполнены вне CDR; такие мутации могут улучшать связывание, и/или специфичность, и/или стабильность, и/или полезный выход очищенной растворимой формы TCR. Например, TCR по настоящему изобретению может дополнительно или в другом варианте содержать вариабельный домен альфа-цепи, имеющий следующие мутации по отношению к положениям остатков 1-113 SEQ ID NO: 2:

В вариабельном домене альфа-цепи последовательность аминокислотных остатков 27-32, 50-56 и 91-103 может быть выбрана из следующих последовательностей:


Вариабельный домен альфа-цепи TCR может содержать аминокислотную последовательность, которая по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентична любой из последовательностей SEQ ID NO: 16-24 или 46-64.
В вариабельном домене бета-цепи последовательность аминокислотных остатков 27-31, 49-54 и 92-107 может быть выбрана из следующих последовательностей:

Вариабельный домен бета-цепи TCR может содержать аминокислотную последовательность, которая по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентична любой из последовательностей SEQ ID NO: 25-29 или 65-81.
Последовательность вариабельного домена альфа-цепи аминокислотных остатков 27-32, 50-56 и 91-103 и последовательность вариабельного домена бета-цепи аминокислотных остатков 27-31, 49-54 и 92-107 могут быть выбраны из следующих:
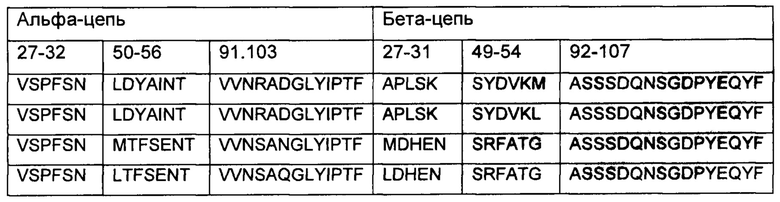

Вариабельный домен альфа-цепи может содержать аминокислотную последовательность любой из SEQ ID NO: 16-24 или 46-64, и/или вариабельный домен бета-цепи может содержать аминокислотную последовательность любой из SEQ ID NO: 25-29 или 65-81.
Например, вариабельный домен альфа-цепи может содержать аминокислотную последовательность SEQ ID NO: 22, а вариабельный домен бета-цепи может содержать аминокислотную последовательность SEQ ID NO: 26; вариабельный домен альфа-цепи может содержать аминокислотную последовательность SEQ ID NO: 22, а вариабельный домен бета-цепи может содержать аминокислотную последовательность SEQ ID NO: 27; вариабельный домен альфа-цепи может содержать аминокислотную последовательность SEQ ID NO: 20, а вариабельный домен бета-цепи может содержать аминокислотную последовательность SEQ ID NO: 28; вариабельный домен альфа-цепи может содержать аминокислотную последовательность SEQ ID NO: 24, а вариабельный домен бета-цепи может содержать аминокислотную последовательность SEQ ID NO: 29; вариабельный домен альфа-цепи может содержать аминокислотную последовательность SEQ ID NO: 23, а вариабельный домен бета-цепи может содержать аминокислотную последовательность SEQ ID NO: 29.
TCR по настоящему изобретению может представлять собой альфа-бета-гетеродимер, имеющий последовательность константного домена TRAC альфа-цепи и последовательность константного домена TRBC1 или TRBC2 бета-цепи.
TCR по настоящему изобретению может быть в одноцепочечном формате, включая, но не ограничиваясь указанным, Vα-L-Vβ, Vβ-L-Vα, Vα-Cα-L-Vβ, Vα-L-Vβ-Cβ, Vα-Cα-L-Vβ-Cβ, где Vα и Vβ представляют собой вариабельные области цепи α и β TCR соответственно, Сα and Сβ представляют собой константные области цепи α и β TCR соответственно, a L представляет собой линкерную последовательность.
TCR по настоящему изобретению может быть ассоциирован с детектируемой меткой, терапевтическим агентом или модификатором фармакокинетики (PK modifying moiety).
TCR по настоящему изобретению может содержать антитело против CD3, ковалентно связанное с С- или N-концом альфа- или бета-цепи TCR. Такой TCR может содержать вариабельный домен альфа-цепи, выбранный из любой из последовательностей SEQ ID NO: 16-24 или 46-64, и вариабельный домен бета-цепи, выбранный из любой из последовательностей SEQ ID NO: 25-29 или 65-81, гибридизованый с антителом против CD3. Бета-цепь может быть связана с последовательностью антитела против CD3 с помощью линкерной последовательности; такая линкерная последовательность может быть выбрана из группы, состоящей из GGGGS (SEQ ID NO: 30), GGGSG (SEQ ID NO: 31), GGSGG (SEQ ID NO: 32), GSGGG (SEQ ID NO: 33), GSGGGP (SEQ ID NO: 34), GGEPS (SEQ ID NO: 35), GGEGGGP (SEQ ID NO: 36) и GGEGGGSEGGGS (SEQ ID NO: 37).
Предпочтительные варианты гибридных молекул TCR-анти-СD3 содержат аминокислотную последовательность альфа-цепи, выбранную из SEQ ID NO: 38-41, или последовательность, которая по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности, представленной в SEQ ID NO: 38-41, и аминокислотную последовательность бета-цепи, выбранную из SEQ ID NO: 42-45, или последовательность, которая по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности, представленной в SEQ ID NO: 42-45.
TCR по настоящему изобретению может быть включен в библиотеку частиц. Для таких целей TCR может презентирован, например, на поверхности бактериофага, дрожжевой клетки, клетки млекопитающего или рибосомы. TCR может быть выделенным, бесклеточным и/или растворимым, то есть он может не встречаться в своем естественном состоянии в Т-клетке человеческого организма.
TCR по настоящему изобретению могут быть не встречающимися в природных условиях и/или очищенными и/или сконструированным. TCR по настоящему изобретению могут иметь более одной мутации, присутствующей в вариабельном домене альфа-цепи и/или вариабельном домене бета-цепи относительно нативного TCR MAGE А4.
TCR по настоящему изобретению может содержать каркасную область 2 (FR2) альфа-цепи и каркасную область 3 (FR3) альфа-цепи, причем области FR2 и FR3 содержат SEQ ID NO: 9 и 10 соответственно и/или содержат один или несколько например, одну, две или три, консервативные замены и/или до трех допустимых замен.
TCR по настоящему изобретению может содержать область FR2 бета-цепи и область FR3 бета-цепи, причем области FR2 и FR3 содержат SEQ ID NO: 14 и 15 соответственно и/или содержат одну или несколько, например одну, две или три, консервативные замены и/или до трех допустимых замен.
TCR по настоящему изобретению может содержать аминокислоты 1-113 последовательности SEQ ID NO: 2 и/или аминокислоты 1-116 последовательности SEQ ID NO: 3, каждая из которых может содержать одну или несколько консервативных замен и/или до трех допустимых мутаций и/или одну или несколько мутаций, указанных в Таблицах 1, 2 и 3.
«Сконструированный TCR» и «мутантный TCR» используются в настоящей заявке как синонимы для обозначения TCR, имеющего одну или несколько введенных мутаций относительно нативного TCR MAGE А4, в частности в вариабельном домене альфа-цепи и/или в вариабельном домене бета-цепи. Мутация (и) обычно улучшает аффинность связывания TCR с комплексом HLA-A*02 GVYDGREHTV (SEQ ID NO: 1), но дополнительно или в другом варианте может придавать ему другие преимущества, такие как улучшенная стабильность в выделенной форме и улучшенная специфичность. Мутации в одном или нескольких положениях могут дополнительно или в другом варианте влиять на взаимодействие смежного положения с когнатным комплексом рМНС, например, путем обеспечения более благоприятного угла для взаимодействия. Чтобы улучшить связывание TCR с комплексом HLA-A*02 GVYDGREHTV (SEQ ID NO: 1), мутации предпочтительно проводят в одном или нескольких участках CDR.
В некоторых вариантах реализации в CDR альфа-цепи имеется 1, 2, 3, 4, 5, 6, 7, 8 или 9 мутаций, например, 4, 5 или 9 мутаций, и/или 1, 2, 3, 4, 5 6, 7, 8, 9 или 10 мутаций в CDR бета-цепи, например, 4, 9 или 10 мутаций.
В некоторых вариантах реализации вариабельный домен а-цепи TCR по настоящему изобретению может содержать аминокислотную последовательность, которая по меньшей мере на 90%, по меньшей мере на 91%, по меньшей мере на 92%, по меньшей мере на 93%, по меньшей мере на 94%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98% или по меньшей мере на 99% идентична последовательности аминокислотных остатков 1-113 последовательности SEQ ID NO: 2, при условии, что вариабельный домен α-цепи имеет по меньшей мере одну из мутаций, описанных выше, например, в Таблице 1 или в Таблице 3. В некоторых вариантах реализации вариабельный домен В-цепи TCR по настоящему изобретению может содержать аминокислотную последовательность, которая по меньшей мере на 90%, по меньшей мере на 91%, при по меньшей мере на 92%, по меньшей мере на 93%, по меньшей мере на 94%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98% или, по меньшей мере на 99% идентична последовательности аминокислотных остатков 1-116 из последовательности SEQ ID NO: 3, при условии, что вариабельный домен В-цепи имеет по меньшей мере одну из мутаций, описанных выше, например, в Таблице 2.
Мутации в TCR родительского (или дикого типа) могут включать мутации, которые способны увеличивать аффинность связывания (Kd и/или период полужизни связывания) TCR с GVYDGREHTV. Мутации могут включать мутации, которые способны уменьшить количество неспецифического связывания, то есть уменьшить связывание с антигенами кроме связывания с GVYDGREHTV. Мутации могут включать мутации, которые увеличивают эффективность фолдинга и/или получения. Некоторые мутации могут способствовать каждой из этих характеристик, другие могут способствовать, например, аффинности, но не специфичности, или способствовать специфичности, но не аффинности и т.д.
В объем изобретения входят фенотипически молчащие варианты любого TCR по настоящему изобретению, раскрытого в настоящей заявке. Используемый в настоящей заявке термин «фенотипически молчащие варианты» предназначен для обозначения TCR, включающего одну или несколько дополнительных аминокислотных изменений, включая замещения, инсерции и делеции, в дополнение к тем, которые изложены выше, причем TCR имеет фенотип, сходный с соответствующим TCR без указанных изменений. Для целей настоящей заявки фенотип TCR включает антигенсвязывающую аффинность (Ко и/или период полужизни связывания связывания) и антигенную специфичность. Фенотипически молчащий вариант может иметь KD и/или период полужизни связывания для комплекса HVA-А*02 GVYDGREHTV (SEQ ID NO: 1) в пределах 50% или более предпочтительно в пределах 20% от измеренной KD и/или периода полужизни связывания соответствующего TCR без указанного изменения (изменений) при измерении в идентичных условиях (например, при 25°С и/или на одном и том же SPR-чипе). Подходящие условия дополнительно представлены в Примере 3. Антигенная специфичность дополнительно определена ниже. Как известно специалистам в данной области техники, существует возможность получать TCR, которые включают изменения в своих вариабельных доменах по сравнению с TCR, которые подробно описаны выше, без изменения аффинности взаимодействия с комплексом GVYDGREHTV (SEQ ID NO: 1) HLA-A*02. В частности, такие молчащие мутации могут быть включены в части последовательности, о которых известно, что они не принимают непосредственного участия в связывании антигена (например, CDR или части CDR, которые не контактируют с пептидным антигеном). Такие тривиальные варианты включены в объем данного изобретения.
Фенотипически молчащие варианты могут содержать одну или несколько консервативных замен и/или одну или несколько допустимых замен. Допустимые и консервативные замены могут привести к изменению KD и/или времени полужизни связывания для комплекса HVA-А*02 GVYDGREHTV (SEQ ID NO: 1) в пределах 50% или, более предпочтительно, в пределах 20%, еще более предпочтительно - в пределах 10% от измеренной KD и/или периода полужизни связывания соответствующего TCR без указанной консервативной и/или допустимой замены (замен), при измерении в идентичных условиях (например, при 25°С и/или на одном и том же SPR-чипе) при условии, что это изменение KD не приведет к тому, что аффинность окажется меньше (то есть слабее) 200 мкМ. Под допустимыми заменами подразумеваются те замены, которые не подпадают под определение консервативных, как указано ниже, но, тем не менее, являются фенотипически молчащими.
TCR по настоящему изобретению могут включать одну или несколько консервативных замен, которые имеют сходную аминокислотную последовательность и/или которые сохраняют ту же функцию (то есть являются фенотипически молчащими, как определено выше). Специалисту известно, что различные аминокислоты имеют сходные свойства и, следовательно, являются «консервативными». Одну или несколько таких аминокислот белка, полипептида или пептида часто можно заменить одной или несколькими другими такими аминокислотами без потери желаемой активности этого белка, полипептида или пептида.
Соответственно, аминокислоты глицин, аланин, валин, лейцин и изолейцин часто могут заменять друг друга (аминокислоты, имеющие алифатические боковые цепи). Из этих возможных замен предпочтительно, чтобы глицин и аланин использовались для замены друг друга (поскольку они имеют относительно короткие боковые цепи), и чтобы валин, лейцин и изолейцин использовались для замены друг друга (поскольку они имеют более крупные алифатические боковые цепи, которые являются гидрофобными). Другие аминокислоты, которые часто могут заменять друг друга, включают: фенилаланин, тирозин и триптофан (аминокислоты, имеющие ароматические боковые цепи); лизин, аргинин и гистидин (аминокислоты, имеющие основные боковые цепи); аспартат и глутамат (аминокислоты, имеющие кислотные боковые цепи); аспарагин и глутамин (аминокислоты, имеющие амидные боковые цепи); и цистеин и метионин (аминокислоты, имеющие серосодержащие боковые цепи). Следует учитывать, что аминокислотные замены в рамках настоящего изобретения могут быть выполнены с использованием встречающихся в природных условиях или не встречающихся в природных условиях аминокислот. Например, в настоящей заявке предполагается, что метильная группа на аланине может быть заменена этильной группой, и/или что незначительные изменения могут быть внесены в основную цепь пептида. Независимо от того, используются природные или синтетические аминокислоты, предпочтительно, чтобы присутствовали только L-аминокислоты.
Замены такого характера часто называют «консервативными» или «полуконсервативными» аминокислотными заменами. Следовательно, настоящее изобретение распространяется на использование TCR, содержащего аминокислотную последовательность, описанную выше, но с одной или несколькими консервативными заменами и/или одной или несколькими допустимыми заменами в последовательности, так что аминокислотная последовательность TCR идентична по меньшей мере на 90%, например идентична на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% TCR, содержащему аминокислоты 1-113 SEQ ID NO: 2, 16-24 или 46-64 и/или аминокислоты 1-116 SEQ ID NO: 3, 25-29 или 65-81.
Термин «идентичность», применяемый в данной области техники, обозначает отношение между двумя или более полипептидными последовательностями или двумя или более полинуклеотидными последовательностями, определяемую путем сравнения последовательностей. В данной области техники идентичность также означает степень родства последовательностей между полипептидными или полинуклеотидными последовательностями, соответственно, определяемую совпадением между строками таких последовательностей. Хотя существует ряд методов измерения идентичности между двумя полипептидными или двумя полинуклеотидными последовательностями, методы, обычно используемые для определения идентичности, кодифицированы в компьютерных программах. Предпочтительные компьютерные программы для определения идентичности между двумя последовательностями включают, помимо прочего, пакет программ GCG (Devereux, et al., Nucleic Acids Research, 12, 387 (1984), BLASTP, BLASTN, and FASTA (Atschul et al., J. Molec. Biol. 215, 403 (1990)).
Для сравнения аминокислотных последовательностей можно использовать такую программу, как программа CLUSTAL. Эта программа сравнивает аминокислотные последовательности и подбирает оптимальное выравнивание, вставляя пробелы в любую последовательность при необходимости. Для оптимального выравнивания можно рассчитать идентичность или сходство аминокислот (идентичность плюс консервативность типа аминокислот). Программа, подобная BLASTx, способна выровнять самый длинный отрезок похожих последовательностей и присвоить такому подбору численную оценку. Таким образом, можно получить сравнение, в котором найдено несколько областей сходства, каждая из которых имеет различную оценку. В настоящем изобретении предусмотрены оба типа анализа идентичности.
Процент идентичности двух аминокислотных последовательностей или двух последовательностей нуклеиновых кислот определяют путем выравнивания последовательностей в целях оптимального сравнения (например, в первую последовательность могут быть введены пробелы для лучшего выравнивания сданной последовательностью) и сравнения аминокислотных остатков или нуклеотидов на соответствующих позициях. «Лучшее выравнивание» представляет собой выравнивание двух последовательностей, приводящее к наибольшему проценту идентичности. Процент идентичности определяется количеством идентичных аминокислотных остатков или нуклеотидов в сравниваемых последовательностях (то есть % идентичности = количество идентичных положений/общее количество положений ×100).
Определение процента идентичности между двумя последовательностями может быть выполнено с использованием математического алгоритма, известного специалистам в данной области техники. Примером такого математического алгоритма сравнения двух последовательностей является алгоритм Карлина и Альтшуля (1990) Proc. Natl. Acad. Sci. USA 87:2264-2268, модифицированный в работе Karlin and Altschul (1993) Proc. Natl. Acad. Sci. USA 90:5873-5877. Программы NВLAST и XBLAST, описанные Altschul, et al. (1990) J. Mol. Biol. 215:403-410, включили в себя такой алгоритм. Чтобы получить нуклеотидные последовательности, гомологичные молекулам нуклеиновой кислоты, может быть выполнен поиск нуклеотидов согласно алгоритмам BLAST с помощью программы NBLAST, оценка = 100, длина слова = 12. Чтобы получить аминокислотные последовательности, гомологичные молекулам белка для использования в изобретении, поиск белков согласно алгоритмам BLAST может быть выполнен с помощью программы XBLAST, оценка = 50, длина слова = 3 слова. Для получения выравнивания с пробелами для сравнения можно использовать алгоритм Gapped BLAST, как описано в работе Altschul et al. (1997) Nucleic Acids Res. 25: 3389-3402. В другом варианте, можно использовать PSI-Blast для выполнения итеративного поиска, который обнаруживает отдаленные соответствия между молекулами (там же). При использовании программ BLAST, Gapped BLAST и PSI-Blast можно использовать параметры по умолчанию соответствующих программ (например, XBLAST и NBLAST). См. Http://www.ncbi.nlm.nih.gov. Другим примером математического алгоритма, используемого для сравнения последовательностей, является алгоритм Майерса и Миллера, Myers and Miller, CABIOS (1989). Программа ALIGN (версия 2.0), которая является частью пакета для выравнивания последовательностей CGC, содержит такой алгоритм. Другие алгоритмы анализа последовательности, известные в данной области техники, включают ADVANCE и ADAM, описанные в работе Torellis and Robotti (1994) Comput. Appl. Biosci., 10: 3-5; и FASTA, описанные в работе Pearson and Lipman (1988) Proc. Natl. Acad. Sci. 85: 2444-8. В программе FASTA контрольный параметр ktup устанавливает чувствительность и скорость поиска.
Мутации, включая консервативные и допустимые замены, инсерции и делеции, могут быть введены в последовательности по настоящему изобретению любым подходящим способом, включая, но не ограничиваясь указанным, методы, основанные на полимеразной цепной реакции (ПЦР), клонировании на основе рестрикционных ферментов или процедурах безлигазного клонирования (LIC). Эти методы подробно описаны во многих стандартных текстах по молекулярной биологии. Для получения дополнительной информации о полимеразной цепной реакции (ПЦР) и клонировании на основе рестрикционных ферментов см. Sambrook & Russell, (2001) Molecular Cloning - A Laboratory Manual (3rd Ed.) CSHL Press. Дополнительную информацию о процедурах безлигазного клонирования (LIC) можно найти в работе Rashtchian, (1995) Curr Opin Biotechnol 6(1): 30-6. Последовательности TCR по настоящему изобретению могут быть получены с помощью твердофазного синтеза или любого другого подходящего способа, известного в данной области техники.
TCR по настоящему изобретению обладают свойством связывать комплекс GVYDGREHTV (SEQ ID NO: 1) HLA-A*02. Было обнаружено, что TCR по настоящему изобретению строго распознают этот эпитоп по сравнению с другими неподходящими эпитопами и, таким образом, особенно пригодны в качестве направленных векторов для доставки терапевтических агентов или детектируемых меток в клетки и ткани, презентирующие эти эпитопы. Специфичность в отношении TCR по настоящему изобретению связана с их способностью распознавать клетки-мишени HLA-A*02, которые являются антиген-положительными, при этом обладая минимальной способностью распознавать клетки-мишени HLA-A*02, которые являются антиген-отрицательными.
Специфичность может быть измерена in vitro, например, в клеточных анализах, таких как описанные в Примере 6. Для тестирования специфичности TCR могут находиться в растворимой форме и/или могут быть гибридизованы с иммунным эффектором, и/или могут быть экспрессированы на поверхности клеток, таких как Т-клетки. Распознавание может быть определено путем измерения уровня активации Т-клеток в присутствии TCR по настоящему изобретению и клеток-мишеней. Минимальное распознавание антиген-отрицательных клеток-мишеней определяется как уровень активации Т-клеток, составляющий менее 20%, предпочтительно менее 10%, предпочтительно менее 5% и более предпочтительно менее 1% от уровня, получаемого в присутствии антиген-положительных клеток-мишеней при измерении в тех же условиях и при терапевтически значимой концентрации TCR. Для растворимых TCR по настоящему изобретению терапевтически значимая концентрация может быть определена как концентрация TCR 10-9 М или ниже и/или концентрация в 100, предпочтительно до 1000 раз больше, чем соответствующее значение ЕС 50. Антиген-положительные клетки могут быть получены с помощью пептидного примирования с использованием подходящей концентрации пептида для получения уровня презентации антигена, сравнимого с раковыми клетками (например, 10-9 М peptide, как описано в работе Bossi et al., (2013) Oncoimmunol. 1; 2 (11):е26840) или они могут по своей природе презентировать указанный пептид. Предпочтительно как антиген-положительные, так и антиген-отрицательные клетки являются клетками человека. Предпочтительно антиген-положительные клетки представляют собой раковые клетки человека. Антиген-отрицательные клетки предпочтительно включают клетки, полученные из здоровых тканей человека.
Специфичность может дополнительно или в другом варианте относиться к способности TCR связываться с комплексом HLA-A*02 GVYDGREHTV (SEQ ID NO: 1), а не с панелью альтернативных комплексов пептид-HLA. Ее можно, например, определить методом Biacore из Примера 3. Указанная панель может содержать по меньшей мере 5, а предпочтительно -по меньшей мере 10 альтернативных комплексов пептид-НLА-А*02. Альтернативные пептиды могут иметь низкий уровень идентичности последовательности с SEQ ID NO: 1 и могут быть представлены в естественных условиях. Альтернативные пептиды могут быть получены из белков, экспрессируемых в здоровых тканях человека. Связывание с комплексом GVYDGREHTV-HLA-A*02 может быть по меньшей мере в 2 раза больше, чем в других HLA-пептидных комплексах, представленных в естественных условиях, более предпочтительно - по меньшей мере в 10 раз, или по меньшей мере в 50 раз или по меньшей мере в 100 раз больше, или, даже более предпочтительно по меньшей мере в 400 раз больше.
Альтернативным или дополнительным подходом для определения специфичности TCR может быть идентификация мотива распознавания пептида TCR с использованием последовательного мутагенеза, например, аланинового сканирования. Остатки, составляющие часть связующего мотива, представляют собой остатки, которые не допустимы для замены. Недопустимые замены могут быть определены как те положения пептидов, в которых аффинность связывания TCR снижается по меньшей мере на 50% или предпочтительно по меньшей мере на 80% относительно аффинности связывания для немутантного пептида. Такой подход подробнее описан в работе Cameron et al., (2013), Sci Transl Med. 2013 Aug 7; 5 (197): 197ra103 и WO 2014096803. Специфичность TCR в этом случае можно определить путем идентификации альтернативных мотивов, содержащих пептиды, в частности альтернативных мотивов, содержащих пептиды в протеоме человека, и тестирования этих пептидов на связывание с TCR. Связывание TCR с одним или несколькими альтернативными пептидами может указывать на отсутствие специфичности. В этом случае может потребоваться дальнейшее тестирование специфичности TCR с помощью клеточных анализов.
Как известно специалистам в данной области техники, пептиды, полученные из членов семейства MAGE, могут иметь высокий уровень идентичности последовательностей с пептидами, полученными из других членов семейства MAGE. Например, существуют пептиды, полученные из MAGE-A8 и MAGE-B2, которые отличаются только двумя остатками от SEQ ID NO 1 (GVYDGREHTV). Указанные пептиды и клетки, экспрессирующие указанные члены семейства MAGE, могут быть исключены из вышеприведенного определения специфичности, особенно если известно, что указанные члены семейства MAGE являются антигенами рака, такими как MAGE-A8 и MAGE-B2. Следовательно, TCR по настоящему изобретению могут распознавать пептиды с высокой процентной идентичностью последовательности, которые получены из других членов семейства MAGE, включая MAGE-А8 и MAGE-B2, и презентированы в контексте HLA А*02. Распознавание указанных пептидов с помощью TCR по настоящему изобретению может быть на том же или более низком уровне, чем распознавание GVYDGREHTV (SEQ ID NO: 1) HLA-A*02.
Некоторые TCR по настоящему изобретению могут иметь идеальный профиль безопасности для применения в качестве терапевтических реагентов. В этом случае TCR могут быть в растворимой форме и предпочтительно могут быть гибридизованы с иммунным эффектором. Идеальный профиль безопасности означает, что помимо демонстрации хорошей специфичности TCR по настоящему изобретению, возможно, прошли дополнительные доклинические испытания безопасности. Примеры таких тестов включают анализы цельной крови для подтверждения минимального высвобождения цитокинов в присутствии цельной крови и, таким образом, низкого риска возникновения потенциального синдрома высвобождения цитокинов in vivo, и тесты аллореактивности для подтверждения низкой потенциальной вероятности для распознавания альтернативных типов HLA.
Некоторые растворимые TCR по настоящему изобретению могут поддаваться очистке с высоким выходом. "Высокий выход" означает выход более 1% или более предпочтительно более 10% или более высокий выход.
TCR по настоящему изобретению могут иметь KD для комплекса GVYDGREHTV-HLA-A*02 превышающую (то есть соответствующую более сильному связыванию) 200 мкМ, например, от 1 пМ до 200 мкМ. Некоторые TCR по настоящему изобретению могут иметь KD для этого комплекса от приблизительно 1 до приблизительно 400 нМ, от приблизительно 1 до приблизительно 200 пМ, от приблизительно 1 до приблизительно 100 пМ. Некоторые TCR по настоящему изобретению могут иметь KD для этого комплекса приблизительно 20-80 пМ. TCR по настоящему изобретению могут иметь период полужизни связывания  для комплекса в диапазоне от приблизительно 1 с до приблизительно 60 ч, от 1 мин до приблизительно 60 ч, от приблизительно 20 мин до приблизительно 50 ч или от приблизительно 2 до приблизительно 35 часов. Определенные TCR по настоящему изобретению могут иметь
для комплекса в диапазоне от приблизительно 1 с до приблизительно 60 ч, от 1 мин до приблизительно 60 ч, от приблизительно 20 мин до приблизительно 50 ч или от приблизительно 2 до приблизительно 35 часов. Определенные TCR по настоящему изобретению могут иметь  для комплекса от приблизительно 8 до приблизительно 35 часов. TCR, которые предназначены для применения в виде растворимых терапевтических средств и/или средств для диагностики, когда они связаны с детектируемой меткой или терапевтическим агентом, предпочтительно имеют KD для этого комплекса от приблизительно 1 до приблизительно 100 пМ или от приблизительно 20 до приблизительно 80 пМ, и/или период полужизни связывания для данного комплекса от приблизительно 2 ч до приблизительно 60 ч или от приблизительно 8 ч до приблизительно 35 ч. Некоторые TCR по настоящему изобретению могут быть пригодны для применения в адоптивной терапии; такие TCR могут иметь Kd для данного комплекса от приблизительно 50 нМ до приблизительно 200 мкМ или от приблизительно 100 нМ до приблизительно 1 мкМ и/или период полужизни связывания для комплекса от приблизительно 3 с до приблизительно 12 мин.
для комплекса от приблизительно 8 до приблизительно 35 часов. TCR, которые предназначены для применения в виде растворимых терапевтических средств и/или средств для диагностики, когда они связаны с детектируемой меткой или терапевтическим агентом, предпочтительно имеют KD для этого комплекса от приблизительно 1 до приблизительно 100 пМ или от приблизительно 20 до приблизительно 80 пМ, и/или период полужизни связывания для данного комплекса от приблизительно 2 ч до приблизительно 60 ч или от приблизительно 8 ч до приблизительно 35 ч. Некоторые TCR по настоящему изобретению могут быть пригодны для применения в адоптивной терапии; такие TCR могут иметь Kd для данного комплекса от приблизительно 50 нМ до приблизительно 200 мкМ или от приблизительно 100 нМ до приблизительно 1 мкМ и/или период полужизни связывания для комплекса от приблизительно 3 с до приблизительно 12 мин.
Некоторые предпочтительные TCR способны генерировать высокоэффективный Т-клеточный ответ in vitro против антиген-положительных клеток, в частности, клеток, которые имеют низкие уровни антигена, типичные для раковых клеток (т.е. приблизительно 50 антигенов на клетку (Bossi et al., (2013) Oncoimmunol) 1; 2 (11): c. 26840; Purbhoo et al. (2006). J Immunol 176 (12): 7308-7316.)). Такие TCR могут быть в растворимой форме и связаны с иммунным эффектором, таким как антитело против CD3. Измеряемым ответом Т-клеток может быть высвобождение маркеров активации Т-клеток, таких как интерферон у или гранзим В, или уничтожение клетки, или другой критерий активации Т-клеток. Предпочтительно высокоэффективный ответ представляет собой ответ со значением ЕС50 в диапазоне пМ, например, 100 пМ или ниже.
Некоторые предпочтительные TCR по настоящему изобретению имеют аффинность связывания и/или период полужизни связывания для комплекса GVYDGREHTV-HLA-A*02, существенно более высокие, чем у нативного TCR. Увеличение аффинности связывания нативного TCR часто снижает специфичность этого TCR к его пептид-МНС-лиганду, и это продемонстрировано в работе Zhao et al., (2007) J. Immunol, 179: 9, 5845-5854. Однако такие TCR по настоящему изобретению остаются специфичными к комплексу GVYDGREHTV-HLA-А*02, несмотря на то, что они имеют существенно более высокую аффинность связывания, чем нативный TCR.
Аффинность связывания (обратно пропорциональная константе равновесия KD) и период полужизни связывания (выраженный 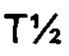 ) могут быть определены с использованием поверхностного плазмонного резонанса (BIAcore) и/или метода Octet из Примера 3 в настоящем документе. Понятно, что удвоение аффинности TCR приводит к уменьшению Kd вдвое. рассчитывается как In2, деленный на скорость диссоциации (koff). Следовательно, удвоение
) могут быть определены с использованием поверхностного плазмонного резонанса (BIAcore) и/или метода Octet из Примера 3 в настоящем документе. Понятно, что удвоение аффинности TCR приводит к уменьшению Kd вдвое. рассчитывается как In2, деленный на скорость диссоциации (koff). Следовательно, удвоение 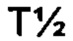 приводит к уменьшению koff вдвое. Значения Kd и koff для TCR обычно измеряют для растворимых форм TCR, то есть тех форм, которые укорочены для удаления остатков цитоплазматического и трансмембранного доменов. Предпочтительно аффинность связывания или период полужизни связывания TCR измеряют несколько раз, например, 3 или более раз, используя один и тот же протокол анализа, и получают среднее значение результатов.
приводит к уменьшению koff вдвое. Значения Kd и koff для TCR обычно измеряют для растворимых форм TCR, то есть тех форм, которые укорочены для удаления остатков цитоплазматического и трансмембранного доменов. Предпочтительно аффинность связывания или период полужизни связывания TCR измеряют несколько раз, например, 3 или более раз, используя один и тот же протокол анализа, и получают среднее значение результатов.
Для применения в качестве направленного агента для доставки терапевтических агентов к антигенпрезентирующей клетке TCR может быть в растворимой форме (то есть не иметь трансмембранного или цитоплазматического доменов). Для стабильности TCR по настоящему изобретению и предпочтительно растворимые ар гетеродимерные TCR могут содержать введенную дисульфидную связь между остатками соответствующих константных доменов, как описано, например, в WO 03/020763. Один или оба внеклеточных константных домена, присутствующих в α-гетеродимере по настоящему изобретению, могут быть укорочены на С-конце или С-концах, например, на до 15, или до 10, или до 8 или менее аминокислот. С-конец внеклеточного константного домена альфа-цепи может быть укорочен на 8 аминокислот. Один или оба внеклеточных константных домена могут содержать одну или несколько мутаций. Внеклеточная константа альфа-цепи может содержать аспарагиновый (N) или лизиновый (K) остаток в положении 4 вследствие естественного полиморфизма. Для применения в адоптивной терапии αβ гетеродимерный TCR может, например, быть трансфицирован как полноразмерные цепи, имеющие как цитоплазматический, так и трансмембранный домены. TCR для применения в адоптивной терапии могут содержать дисульфидную связь, соответствующую той, которая встречается в природе между соответствующими константными доменами альфа и бета; дополнительно или в другом варианте может присутствовать ненативная дисульфидная связь.
TCR по настоящему изобретению могут быть αβ-гетеродимерами. TCR по настоящему изобретению могут быть в одноцепочечном формате. Одноцепочечные форматы включают, но не ограничиваются указанным, αβ TCR полипептиды типов Vα-L-Vβ, Vβ-L-Vα, Vα-Cα-L-Vβ, Vα-L-Vβ-Cβ или Vα-Cα-L-Vβ-Cβ, где Vα и Vβ представляют собой вариабельные области TCRα и β соответственно, Сα и Cβ представляют собой константные области α и β TCR соответственно, a L представляет собой последовательность линкера (Weidanz et al., (1998) J. Immunol Methods. Dec 1; 221 (1-2): 59-76; Epel et al., (2002), Cancer Immunol Immunother. Nov; 51 (10): 565-73; WO 2004/033685; WO 9918129). Один или оба константных домена могут быть полноразмерными, или они могут быть укороченными, как описано выше, и/или содержать мутации. Внеклеточная константа альфа-цепи может содержать аспарагиновый (N) или лизиновый (K) остаток в положении 4 вследствие естественного полиморфизма. В некоторых вариантах реализации одноцепочечные TCR по настоящему изобретению могут иметь введенную дисульфидную связь между остатками соответствующих константных доменов, как описано в WO 2004/033685. Одноцепочечные TCR дополнительно описаны в WO 2004/033685; WO 98/39482; WO 01/62908; Weidanz et al. (1998) J Immunol Methods 221(1-2): 59-76; Hoo et al. (1992) Proc Natl Acad Sci U S A 89(10): 4759-4763; Schodin (1996) Mol Immunol 33(9): 819-829).
Как будет понятно для специалиста в данной области техники, можно укоротить представленные последовательности на С-конце и/или N-конце, на 1, 2, 3, 4, 5 или более остатков без существенного влияния на связывающие характеристики TCR. Все такие тривиальные варианты охватываются настоящим изобретением.
Альфа-бета-гетеродимерные TCR по настоящему изобретению обычно содержат последовательность константного домена альфа-цепи TRAC и/или последовательность константного домена бета-цепи TRBC1 или TRBC2. Последовательности константного домена альфа- и бета-цепи могут быть модифицированы путем укорочения или замены для удаления нативной дисульфидной связи между Cys4 экзона 2 TRAC и Cys2 экзона 2 TRBC1 или TRBC2. Последовательность (последовательности) константного домена альфа- и/или бета-цепи может быть модифицирована путем замены остатков Thr 48 TRAC и Ser 57 TRBC1 или TRBC2 на цистеин, причем указанные цистеины образуют дисульфидную связь между константными доменами альфа- и бета-цепи TCR. TRBC1 или TRBC2 могут дополнительно включать мутацию цистеина на аланин в положении 75 константного домена и мутацию аспарагина на аспарагиновую кислоту в положении 89 константного домена. Константный домен может дополнительно или в другом варианте содержать дополнительные мутации, замены или делеции относительно нативных последовательностей TRAC и/или TRBC1/2. Термин TRAC и TRBC1/2 охватывают природные полиморфные варианты, например, от N до К в положении 4 домена TRAC (Bragado et al Int Immunol. 1994 Feb; 6(2):223-30).
Также в объем изобретения включены варианты, фрагменты и производные TCR, предусмотренных изобретением.
Изобретение также включает частицы, презентирующие TCR по настоящему изобретению, и включение указанных частиц в библиотеку частиц. Такие частицы включают, но не ограничиваются указанным, фаг, рибосомы дрожжей или клетки млекопитающих. Способ получения таких частиц и библиотек известен в данной области техники (например, см. WO 2004/044004; WO 01/48145, Chervin et al. (2008) J. Immuno. Methods 339.2: 175-184).
В дополнительном аспекте настоящее изобретение относится к нуклеиновой кислоте, кодирующей TCR по настоящему изобретению. В некоторых вариантах реализации нуклеиновая кислота представляет собой кДНК. В некоторых вариантах реализации изобретение относится к нуклеиновой кислоте, содержащей последовательность, кодирующую вариабельный домен α-цепи TCR по настоящему изобретению. В некоторых вариантах реализации изобретение относится к нуклеиновой кислоте, содержащей последовательность, кодирующую вариабельный домен β-цепи TCR по настоящему изобретению. Нуклеиновая кислота может быть не встречающейся в природных условиях и/или очищенной и/или сконструированной. Последовательность нуклеиновой кислоты может быть кодон-оптимизированной в соответствии с используемой системой экспрессии.
В другом аспекте изобретение относится к вектору, содержащему нуклеиновую кислоту по настоящему изобретению. Предпочтительно такой вектор представляет собой вектор для экспрессии TCR.
Изобретение также относится к клетке, несущей вектор по настоящему изобретению, предпочтительно вектор для экспрессии TCR. Такой вектор может содержать нуклеиновую кислоту по настоящему изобретению, кодирующую одну открытую рамку считывания или две отдельные открытые рамки считывания, кодирующие альфа-цепь и бета-цепь соответственно. В другом аспекте предлагается клетка, несущая первый вектор экспрессии, который содержит нуклеиновую кислоту, кодирующую альфа-цепь TCR по настоящему изобретению, и второй вектор экспрессии, который содержит нуклеиновую кислоту, кодирующую бета-цепь TCR по настоящему изобретению. Такие клетки особенно полезны в адоптивной терапии. Клетки по настоящему изобретению могут быть выделенными и/или рекомбинантными и/или не встречающимися в природных условиях и/или сконструированными.
Поскольку TCR по настоящему изобретению полезны в адоптивной терапии, изобретение включает не встречающиеся в природных условиях и/или очищенные и/или сконструированные клетки, в частности Т-клетки, презентирущие TCR по настоящему изобретению. Изобретение также относится к размноженной популяции Т-клеток, презентирущих TCR по настоящему изобретению. Существует ряд способов, подходящих для трансфекции Т-клеток нуклеиновой кислотой (такой как ДНК, кДНК или РНК), кодирующей TCR по настоящему изобретению (см., например, Robbins et al., (2008) J Immunol. 180: 6116-6131). Т-клетки, экспрессирующие TCR по настоящему изобретению, будут пригодны для применения при основанном на адоптивной терапии лечении рака. Как известно специалистам в данной области техники, существует ряд подходящих способов, с помощью которых можно проводить адоптивную терапию (см., например, Rosenberg et al., (2008) Nat Rev Cancer 8(4): 299-308).
Растворимые TCR по настоящему изобретению полезны для доставки детектируемых меток или терапевтических агентов в антигенпрезентирующие клетки и ткани, содержащие антигенпрезентирующие клетки. Поэтому они могут быть ассоциированы (ковалентно или иным образом) с детектируемой меткой (для диагностических целей, когда TCR используется для обнаружения присутствия клеток, презентирующих комплекс GVYDGREHTV-HLA-A*02); терапевтическим агентом; или группой, модифицирующей фармакокинетику (ФК).
Примеры модифицирующих ФК групп включают, но не ограничиваются указанным, ПЭГ
(Dozier et al., (2015) Int J Mol Sci. Oct 28; 16(10):25831-64 and Jevsevar et al., (2010) Biotechnol J. Jan; 5(1):113-28), ПАСилирование (Schlapschy et al., (2013) Protein Eng Des Sel. Aug; 26(8):489-501), альбумин (Dennis et al., (2002) J Biol Chem. Sep 20; 277(38):35035-43) и/или неструктурированные полипептиды (Schellenberger et al., (2009) Nat Biotechnol. Dec; 27(12): 1186-90).
Детектируемые метки для диагностических целей включают, например, флуоресцентные метки, радиоактивные метки, ферменты, зонды на основе нуклеиновой кислоты и контрастные реагенты.
Терапевтические агенты, которые могут быть ассоциированы с TCR по настоящему изобретению, включают иммуномодуляторы, радиоактивные соединения, ферменты (например, перфорин) или химиотерапевтические агенты (например, цис-платин). Чтобы гарантировать, что токсические эффекты проявляются в желаемом месте, токсин может находиться внутри липосомы, связанной с TCR, так чтобы соединение высвобождалось медленно. Это предотвратит повреждающие эффекты во время транспорта в организме и обеспечит максимальный эффект токсина после связывания TCR с соответствующими антигенпрезентирующими клетками.
Другие подходящие терапевтические агенты включают:
• низкомолекулярные цитотоксические агенты, то есть соединения, обладающие способностью убивать клетки млекопитающих, имеющие молекулярную массу менее 700 дальтон. Такие соединения могут также содержать токсичные металлы, способные оказывать цитотоксическое действие. Кроме того, следует понимать, что эти низкомолекулярные цитотоксические агенты также включают пролекарства, то есть соединения, которые в физиологических условиях подвергаются разложению или превращению с высвобождением цитотоксических агентов. Примеры таких агентов включают цис-платин, производные майтансина, рахельмицин, калихеамицин, доцетаксел, этопозид, гемцитабин, ифосфамид, иринотекан, мелфалан, митоксантрон, сорфимер натрийфотофрин II, темозоломид, топотекан, триметреат глюкуронат, ауристатин Е, винкристин и доксорубицин;
• пептидные цитотоксины, то есть белки или их фрагменты, способные убивать клетки млекопитающих. Например, рицин, дифтерийный токсин, псевдомонас, экзотоксин синегнойной палочки А, ДНКазу и РНКазу;
• радионуклиды, то есть нестабильные изотопы элементов, которые распадаются при одновременном испускании одной или нескольких α- или β-частиц, или γ-лучей. Например, йод 131, рений 186, индий 111, иттрий 90, висмут 210 и 213, актиний 225 и астат 213; для облегчения ассоциации этих радионуклидов с высокоаффинными TCR или их мультимерами могут быть использованы хелатирующие агенты;
• Иммуностимуляторы, то есть иммунные эффекторные молекулы, которые стимулируют иммунный ответ. Например, цитокины, такие как IL-2 и IFN -γ,
• Суперантигены и их мутанты;
• гибридные молекулы TCR-HLA, например, гибрид с комплексом пептид-HLA, где указанный пептид получен из распространенного человеческого патогена, такого как вирус Эпштейна-Барра (EBV);
• хемокины, такие как IL-8, тромбоцитарный фактор 4, белок, стимулирующий рост меланомы, и т.д.;
• антитела или их фрагменты, включая детерминантные антитела против Т-клеток или NK-клеток (например, анти-СD3, анти-СD28 или анти-СD16);
• альтернативные белковые каркасы с антителоподобными характеристиками связывания
• активаторы комплемента;
• ксеногенные белковые домены, аллогенные белковые домены, вирусные/бактериальные белковые домены, вирусные/бактериальные пептиды.
Один предпочтительный вариант реализации включает TCR по настоящему изобретению, ассоциированный (обычно путем гибридизации с N- или С-концом альфа- или бета-цепи) с анти-СD3 антителом или функциональным фрагментом или вариантом указанного анти-СD3 антитела (такие гибриды TCR-анти-СD3 можно назвать молекулами ImmTACTM). Используемый в настоящей заявке термин «антитело» охватывает такие фрагменты и варианты. Примеры анти-СD3 антител включают, но не ограничиваются указанным, ОКТ3, UCHT-1, ВМА-031 и 12F6. Фрагменты и варианты/аналоги антител, которые подходят для применения • описанных а настоящей заявке композициях и способах, включают мини-тела, Fab-фрагменты, F(аb')2 фрагменты, dsFv- и scFv-фрагменты, нанотела Nanobodies™ (эти конструкции, поставляемые на рынок фирмой Ablynx (Бельгия), содержат синтетический одиночный вариабельный домен тяжёлой цепи иммуноглобулина, полученного из антител верблюдовых (например, верблюда или ламы)), и антитела Domain Antibodies (Domantis (Бельгия), содержащие одиночный вариабельный домен тяжёлой цепи иммуноглобулина с созревшей аффинностью или вариабельный домен легкой цепи иммуноглобулина, или каркасы альтернативных белков, которые демонстрируют антителоподобные характеристики связывания, такие как Affibodies (Affibody (Швеция), содержащие каркас сконструированного белка А) или Anticalins (Pieris (Германия), содержащие сконструированные антикалины), среди многих прочих.
Связывание TCR и анти-СD3 антитела может быть через ковалентное или нековалентное присоединение. Ковалентное присоединение может быть прямым или косвенным через линкерную последовательность. Линкерные последовательности обычно являются гибкими, поскольку они состоят в основном из аминокислот, таких как глицин, аланин и серии, которые не имеют громоздких боковых цепей, которые могут ограничивать гибкость. Пригодные для применения или оптимальные длины линкерных последовательностей легко определимы. Часто линкерная последовательность будет иметь длину менее 12, например, менее 10 или 2-10 аминокислот. Подходящие линкеры, которые можно использовать в TCR по настоящему изобретению, включают, но не ограничиваются указанным: GGGGS (SEQ ID NO: 30), GGGSG (SEQ ID NO: 31), GGSGG (SEQ ID NO: 32), GSGGG (SEQ ID NO: 33), GSGGGP (SEQ ID NO: 34), GGEPS (SEQ ID NO: 35), GGEGGGP (SEQ ID NO: 36) и GGEGGGSEGGGS (SEQ ID NO: 37) (как описано в WO 2010/133828).
Характерные варианты гибридных конструкций анти-СD3-TCR по настоящему изобретению включают такие пары альфа- и бета-цепей, в которых альфа-цепь состоит из вариабельного домена, содержащего аминокислотную последовательность SEQ ID NO: 16-24 или 46-64, и/или бета-цепь состоит из вариабельного домена, содержащего аминокислотную последовательность SEQ ID NO: 25-29 или 65-81. Указанные альфа- и бета-цепи могут дополнительно содержать константную область, содержащую ненативную дисульфидную связь. N- или С-конец альфа- и/или бета-цепи может быть гибридизован с фрагментом scFv анти-СD3-антитела через линкер, выбранный из SEQ ID NO: 30-37. Некоторые предпочтительные варианты таких гибридных конструкций анти-CD3-TCR представлены ниже:

Каждый линкер с последовательностью SEQ ID NO: 30-37 можно использовать с каждым или любым из предпочтительных вариантов гибридных конструкций CD3-TCR. Например, в изобретение включен гибрид TCR-CD3, содержащий альфа-цепь SEQ ID NO: 24 и бета-цепь SEQ ID NO: 29, где бета-цепь гибридизована с анти-СD3 scFv через линкер любой из последовательностей SEQ ID NO: 31-37.
Для некоторых целей TCR по настоящему изобретению могут быть аггрегированы в комплекс, содержащий несколько TCR, с образованием поливалентного TCR-комплекса. Существует ряд человеческих белков, содержащих домен мультимеризации, который может быть использован при получении поливалентных TCR-комплексов. Например, домен тетрамеризации гена р53, который был использован для получения тетрамеров scFv-фрагментов антител, проявлявших повышенную устойчивость в сыворотке и значительно сниженную скорость диссоциации по сравнению с мономерным scFv-фрагментом (Willuda et al. (2001) J. Biol. Chem. 276 (17) 14385-14392). Гемоглобин также содержит домен тетрамеризации, который может быть использован для такого применения. Поливалентный TCR-комплекс по настоящему изобретению может обладать улучшенной способностью связывания с комплексом GVYDGREHTV-HLA-A*02 по сравнению с немультимерным гетеродимером рецептора дикого типа или Т-клеточного рецептора по настоящему изобретению. Таким образом, поливалентные TCR-комплексы по настоящему изобретению также включены в изобретение. Такие поливалентные TCR-комплексы согласно изобретению особенно полезны для отслеживания или нацеливания на клетки, презентирующих конкретные антигены in vitro или in vivo, и также полезны в качестве промежуточных продуктов для получения дальнейших поливалентных комплексов TCR, имеющих такие применения.
Как хорошо известно в данной области техники, TCR могут подвергаться посттрансляционным модификациям. Одной из таких модификаций является ликозилирование, включающее ковалентное присоединение олигосахаридных фрагментов к определенным аминокислотам в цепи TCR. Например, хорошо известными местами присоединения олигосахаридов являются остатки аспарагина или остатки серина/треонина. Статус гликозилирования конкретного белка зависит от ряда факторов, включая последовательность белка, конформацию белка и наличие определенных ферментов. Кроме того, статус гликозилирования (то есть тип олигосахарида, ковалентная связь и общее количество точек присоединения) может влиять на функцию белка. Поэтому при получении рекомбинантных белков часто желательно контролировать гликозилирование. Контролируемое гликозилирование уже использовалось для улучшения терапии на основе антител. (Jefferis et al., (2009) Nat Rev Drug Discov Mar; 8 (3): 226-34.). Для растворимых TCR по настоящему изобретению гликозилирование можно контролировать in vivo, используя, например, специальные клеточные линии, или in vitro, путем химической модификации. Такие модификации желательны, поскольку гликозилирование способно улучшать фармакокинетику, снижать иммуногенность и более точно имитировать нативный человеческий белок (Sinclair and Elliott, (2005) Pharm Sci. Aug; 94(8): 1626-35).
Для введения пациентам TCR по настоящему изобретению (предпочтительно ассоциированные с детектируемой меткой или терапевтическим средством или экспрессируемые на трансфицированных Т-клетках), гибридные молекулы TCR-анти-СD3, нуклеиновые кислоты, векторы экспрессии или клетки по настоящему изобретению могут быть предоставлены в фармацевтической композиции вместе с одним или несколькими фармацевтически приемлемыми носителями или вспомогательными веществами. Терапевтические или визуализирующие TCR или клетки в соответствии с изобретением, как правило, будут поставляться в составе стерильной фармацевтической композиции, которая обычно будет включать фармацевтически приемлемый носитель. Эта фармацевтическая композиция может быть в любой подходящей форме (в зависимости от желаемого способа введения ее пациенту). Она может предоставляться в единичной дозированной форме, как правило, будет предоставляться в герметичном контейнере и может предоставляться в составе набора. Такой набор обычно (хотя и не обязательно) будет включать инструкции по применению. Он может содержать множество указанных единичных дозированных форм.
Такая фармацевтическая композиция может быть адаптирована для введения любым подходящим путем, таким как парентеральный (включая подкожный, внутримышечный или внутривенный), энтеральный (включая пероральный или ректальный), ингаляционный или интраназальный способ введения. Такие композиции могут быть получены любым способом, известным в области фармацевтики, например, путем смешивания активного ингредиента с носителем(-ями) или вспомогательным веществом(-ами) в стерильных условиях.
Дозировки веществ по настоящему изобретению могут варьировать в широких пределах, в зависимости от заболевания или расстройства, подлежащего лечению, возраста и состояния лица, подлежащего лечению, и т.д.: подходящий диапазон доз для растворимого TCR по настоящему изобретению, ассоциированного с анти-СD3 антителом, может составлять от 25 нг/кг до 50 мкг/кг. В конечном итоге подходящие дозы для применения будут определены врачом.
TCR, фармацевтические композиции, векторы, нуклеиновые кислоты и клетки по настоящему изобретению могут быть представлены в по существу чистой форме, например, с чистотой по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 91%, по меньшей мере 92%, по меньшей мере 93%, по меньшей мере 94%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или 100%.
Изобретение также предусматривает:
• TCR, гибридную молекулу TCR-анти-CD3, нуклеиновую кислоту, фармацевтическую композицию или клетку по настоящему изобретению для применения в медицине, предпочтительно для применения в способе лечения рака или опухоли;
• применение TCR, гибридной молекулы TCR-анти-СВ3, нуклеиновой кислоты, фармацевтической композиции или клетки по настоящему изобретению при получении лекарственного средства для лечения рака или опухоли;
• способ лечения рака или опухоли у пациента, включающий введение пациенту TCR, гибридной молекулы TCR-анти-СВ3, нуклеиновой кислоты, фармацевтической композиции или клетки по настоящему изобретению;
• инъецируемую композицию для введения человеку, содержащую TCR, гибридную молекулу TCR-анти-СВ3, нуклеиновую кислоту, фармацевтическую композицию или клетку по настоящему изобретению.
Указанный рак может представлять собой рак молочной железы, пищевода, головы и шеи, легких, яичников или мочевого пузыря. Опухоль может экспрессировать MAGE А4 и/или может быть солидной опухолью. TCR, гибридную молекулу TCR-анти-СВ3, нуклеиновую кислоту, фармацевтическую композицию или клетку по настоящему изобретению можно вводить инъекцией, такой как внутривенная или непосредственная внутриопухолевая инъекция. Субъект, представляющий собой человека, может иметь подтип НLА-А*02.
Предпочтительные признаки каждого аспекта изобретения являются такими же, как для каждого из других аспектов, с соответствующими поправками. Документы предшествующего уровня техники, упомянутые в настоящей заявке, включены в максимальном объеме, разрешенном законом.
Описание чертежей
Фигура 1: аминокислотная последовательность внеклеточных областей альфа- и бета-цепи нативного MAGE А4 TCR.
Фигура 2: аминокислотная последовательность нативных внеклеточных областей альфа- и бета-цепи растворимого MAGE А4 TCR.
Фигура 3: примеры аминокислотных последовательностей мутированных вариабельных областей альфа-цепи MAGE А4 TCR.
Фигура 4: примеры аминокислотных последовательностей мутированных вариабельных областей бета-цепи MAGE А4 TCR.
Фигура 5: примеры аминокислотных последовательностей альфа-цепи гибридных молекул MAGE A4TCR-анти-CD3.
Фигура 6: примеры аминокислотных последовательностей бета-цепи гибридных молекул MAGE A4TCR-анти-CD3.
Фигура 7: клеточные данные, демонстрирующие активность и специфичность гибридных молекул MAGE А4 TCR-анти-СD3.
Фигура 8: клеточные данные, демонстрирующие активность и специфичность дополнительных гибридных молекул MAGE А4 TCR-анти-СD3.
Фигура 9: дополнительные доказательства специфичности гибридных молекул MAGE А4 TCR-анти-СD3
Фигура 10: дополнительные данные о специфичности для гибридных молекул MCR А4 TCR-анти-СD3
Фигура 11: доказательства того, что гибридные молекулы MAGE А4 TCR-анти-СD3 приводят к гибели раковых клеток
Изобретение далее описано в следующих неограничивающих примерах.
Примеры
Пример 1. Экспрессия, рефолдинг и очистка растворимых TCR
Методика
Последовательности ДНК, кодирующие внеклеточные области альфа и бета растворимых TCR по настоящему изобретению, клонировали по отдельности в плазмиды экспрессии на основе pGMT7, используя стандартные методы (как описано в Sambrook et al. Molecular cloning. Vol. 2. (1989) New York: Cold spring harbour laboratory press). Плазмиды экспрессии трансформировали отдельно в штамм E.coli Rosetta (BL21pLysS), и отдельные устойчивые к ампициллину колонии выращивали при 37°С в среде TYP (+ампициллин 100 мкг/мл) до достижения плотности OD600 ~ 0,6-0,8 перед индукцией экспрессии белка с использованием 0,5 мМ IPTG. Клетки собирали через три часа после индукции центрифугированием. Клеточный осадок лизировали реагентом для экстракции белка BugBuster (Merck Millipore) в соответствии с инструкциями производителя. Гранулы внутриклеточных телец извлекали центрифугированием. Гранулы дважды промывали в буфере Triton (50 мМ Tris-HCl, рН 8,1, 0,5% Triton-Х100, 100 мМ NaCl, 10 мМ NaEDTA) и в заключение ресуспендировали в буфере, не содержащем детергента (50 мМ Tris-HCl, рН 8,1, 100 мМ NaCl, 10 мМ NaEDTA). Выход белка внутриклеточных телец определяли количественно путем солюбилизации с 6 М гуанидин-HCl и измерения ОD280. Затем рассчитывали концентрацию белка с использованием коэффициента экстинкции. Чистоту внутриклеточных телец измеряли путем солюбилизации с 8М мочевины и загрузки ~2 мкг в 4-20% гель ДНС-ПААГ в восстанавливающих условиях. Затем оценивали или рассчитывали чистоту с помощью программного обеспечения для денситометрии (Chemidoc, Biorad). Внутриклеточные тельца хранили при +4°С для кратковременного хранения и при -20°С или -70°С для длительного хранения.
Для рефолдинга растворимого TCR, содержащие а-и В-цепи внутриклеточные тельца сперва смешивали и разбавляли в 10 мл буфера солюбилизации/денатурации (6 М гуанидин-гидрохлорида, 50 мМ Трис-HCl рН 8,1, 100 мМ NaCl, 10 мМ ЭДТА, 20 мМ ДТТ) с последующей инкубацией в течение 30 мин при 37°С. Затем инициировали рефолдинг дальнейшим разбавлением в 1 л буфера для рефолдинга (100 мМ Трис, рН 8,1, 400 мМ L-аргинина HCL, 2 мМ ЭДТА, 4 М мочевины, 10 мМ гидрохлорида цистеамина и 2,5 мМ дигидрохлорида цистамина), и раствор хорошо перемешивали. Подвергнутую рефолдингу смесь диализировали 10 л Н2О в течение 18-20 часов при 5±3°С. По истечении этого времени диализный буфер дважды заменяли 10 мМ Трис, рН 8,1 (10 л) и продолжали диализ в течение еще 15 часов. Затем подвергнутую рефолдингу смесь фильтровали через целлюлозные фильтры 0,45 мкм.
Очистку растворимых TCR инициировали, нанося диализированный рефолд на анионообменную колонку POROS® 50HQ и элюируя связанный белок с градиентом 0-500 мМ NaCl в 20 мМ Трис рН 8,1 в 50 объемах колонки с использованием пурификатора Akta® (GE Healthcare). Пиковые фракции TCR идентифицировали с помощью ДНС-ПААГ перед объединением в пул и концентрированием. Затем концентрированный образец наносили на гель-фильтрационную колонку Superdex® 75HR (GE Healthcare), предварительно уравновешенную в фосфатно-солевом буфере Дульбекко. Пиковые фракции TCR объединяли и концентрировали, затем рассчитывали конечный выход очищенного материала.
Пример 2. Экспрессия, рефолдинг и очистка молекул ImmTAC (растворимые гибридные молекулы TCR-анти-СD3)
Методика
ImmTAC получали, как описано в Примере 1, за исключением того, что бета-цепь TCR подвергали гибридизации через линкер с одноцепочечным анти-СD3-антителом. Кроме того, во время очистки после анионного обмена выполняли стадию катионного обмена. В этом случае пиковые фракции из анионного обмена разбавляли в 20 раз в 20 мМ MES (рН 6,5) и наносили на катионообменную колонку POROS® 50HS. Связанный белок элюировали с градиентом 0-500 мМ NaCl в 20 мМ MES. Пиковые фракции ImmTAC объединяли и доводили до 50 мМ Трис рН 8,1, а затем концентрировали и наносили непосредственно на матрицу для гель-фильтрации, как описано в примере 1.
Пример 3. Исследование связывания
Анализ связывания очищенных растворимых TCR и молекул ImmTAC с соответствующим комплексом пептид-HLA выполняли с помощью поверхностного плазмонного резонанса с использованием прибора BIAcore 3000 или BIAcore Т200 или с помощью биослойной интерферометрии с использованием прибора ForteBio Octet. Биотинилированные молекулы НLА-А*02 класса I подвергали рефолдингу с рассматриваемым пептидом и очищали с применением методов, известных специалистам в данной области техники (O'Callaghan et al. (1999). Anal Biochem 266(1): 9-15; Garboczi, et al. (1992). Proc Natl Acad Sci USA 89(8): 3429-3433). Все измерения проводили при 25°C в фосфатно-солевом буфере Дульбекко с добавлением 0,005% Р20.
Методика BIAcore
Биотинилированные мономеры пептид-HLA иммобилизовали на покрытых стрептавидином сенсорных чипах СМ-5. Равновесные константы связывания определяли с помощью серийных разведений растворимого TCR/ImmTAC, вводимого при постоянной скорости потока 30 мкл/мин и проходящего над проточной ячейкой, покрытой ~ 200 единицами ответа (RU) комплекса пептид-НLА-А*02. Равновесные ответы нормировали для каждой концентрации TCR путем вычитания ответа буфера на контрольной проточной ячейке, содержащей посторонний пептид-HLA. Значение KD получали подбором нелинейной кривой с использованием программного обеспечения Prism и изотермы связывания Ленгмюра, bound = С * Мах/(С+KD), где «bound» - это равновесное связывание в RU при концентрации С введенного TCR, а Мах - максимальное связывание.
Для высокоаффинных взаимодействий параметры связывания определяли с помощью анализа кинетики одного цикла. Пять различных концентраций растворимого TCR/ImmTAC вводили пропуская через проточную ячейку, покрытую ~100-200 RU комплекса пептид-HLA, при скорости потока 50-60 мкл/мин. Как правило, 60-120 мкл растворимого TCR/ImmTAC вводили при максимальной концентрации 100-200 нМ, с последующими 2-кратными разведениями для других четырех инъекций. Самую низкую концентрацию вводили первой. Затем для измерения фазы диссоциации вводили буфер до достижения диссоциации ≥ 10%, обычно через 1-3 часа. Кинетические параметры рассчитывали с использованием программного обеспечения BIAevaluation®. Фаза диссоциации аппроксимировали одним уравнением экспоненциального спада, позволяющим рассчитать время полужизни связывания. Константу равновесия KD рассчитывали из koff//kon.
Метод Octet
Биотинилированные пептид-HLA-мономеры захватывали до достижения 1 нМ на биосенсорах (SA) на основе стрептавидина (Pall ForteBio), предварительно иммобилизованных стрептавидином. Сенсоры блокировали свободным биотином (2 мкМ) в течение 2 минут. Равновесные константы связывания определяли погружением загруженных биосенсоров в растворимый TCR/ImmTAC, последовательно разбавленный в 96-луночном или 384-луночном планшете для образцов. Встряхивание планшета устанавливали на 1000 об/мин. Для низкокоаффинных взаимодействий (диапазон мкМ) использовали малое время ассоциации (~2 минуты) и диссоциации (~2 минуты). Кривые связывания обрабатывали с помощью двойного вычитания эталонных биосенсоров, загруженных посторонним pHLA, с использованием программного обеспечения для анализа данных Octet (Pall ForteBio). Ответы (нм) в равновесии использовали для оценки значения KD из графиков стационарного состояния, подогнанных к уравнению Ответ = Rmax * конц/(KD + конц), где «ответ» - это равновесное связывание в нм при каждой концентрации TCR (конц), a Rmax - максимальный ответ связывания при насыщении pHLA.
Для высокоаффинных взаимодействий (в диапазоне нМ - пМ) кинетические параметры определяли по кривым связывания при ≥ 3 концентрациях TCR/ImmTAC, обычно 10 нМ, 5 нМ и 2,5 нМ. Время ассоциации составляло 30 минут, а время диссоциации - 1-2 часа. Кривые связывания обрабатывали с помощью двойного вычитания эталонных биосенсоров, загруженных посторонним pHLA и блокированных биотином. Кинетические параметры kon и koff рассчитывали путем глобального подбора непосредственно к кривым связывания с использованием программного обеспечения для анализа данных Octet (Pall ForteBio). KD рассчитывали по koff/kon, а период полудиссоциации рассчитывали по t1/2 = 0.693/koff.
Пример 4. Исследование связывания нативного TCR
Получали растворимый нативный TCR в соответствии со способами, описанными в Примере 1, и анализировали связывание с pHLA в соответствии с Примером 3. Аминокислотные последовательности альфа- и бета-цепей соответствовали показанным на Фиг. 2. Растворимый биотинилированный HLA-A*02 получали с помощью пептида MAGE А4 GVYDGREHTV и иммобилизовали на сенсорном чипе BIAcore.
Результаты
Связывание определяли при различных концентрациях, и значение KD для взаимодействия составило 142 мкМ. Перекрестную реактивность (специфичность) оценивали по панели из 15 посторонних пептидных комплексов HLA-A*02, используя метод определения равновесного связывания BIAcore из Примера 3. Пятнадцать посторонних pHLA объединяли в три группы и загружали в одну из трех проточных ячеек, получая приблизительно 1000 RU каждого pHLA на каждую проточную ячейку. Вводили 20 мкл растворимого TCR дикого типа в концентрации 73 мкМ во все проточные ячейки со скоростью 20 мкл/мин. При обеих концентрациях значительного связывания обнаружено не было, что указывает на специфичность нативного TCR к комплексу GVYDGREHTV-HLA-A*02.
Полученные данные показывают, что этот TCR связывается с мишенью с подходящей аффинностью и специфичностью и, следовательно, обеспечивает полезную исходную последовательность для получения терапевтических TCR.
Пример 5. Исследование связывания растворимых мутированных TCR и молекул ImmTAC по настоящему изобретению
Растворимые мутированные TCR и молекулы ImmTAC получали на основе последовательностей, представленных на Фиг. 2. Образцы готовили, как описано в Примерах 1 и 2, а характеристики связывания определяли в соответствии с Примером 3.
Результаты
Было обнаружено, что одиночная точечная мутация с заменой цистеина на валин в положении 19 альфа-цепи (SEQ ID NO: 6) улучшает рефолдинг и выход при очистке, не влияя на аффинность или специфичность (было зарегистрировано значение KD 145 мкМ, и перекрестного реагирования с той же панелью из 15 альтернативных пептидных комплексов HLA, которые были протестированы с WT, не наблюдалось).
Были выявлены альфа- и/или бета-цепи TCR, которые содержат мутации по меньшей мере в одном участке CDR относительно последовательностей CDR, показанных на Фиг. 2 (SEQ ID NO: 4 и 5). Эти последовательности TCR распознавали комплекс GVYDGREHTV HLA-A*02 с наиболее подходящей аффинностью и/или периодом полужизни связывания. В некоторых случаях были выявлены дополнительные мутации, которые улучшали стабильность и/или выход TCR, включая мутацию альфа-цепи K1А (по отношению к положениям SEQ ID NO: 4). Аминокислотные последовательности некоторых мутированных вариабельных областей альфа- и бета-цепи TCR по настоящему изобретению представлены на Фиг. 4 и 5 соответственно. В приведенной ниже таблице представлены характеристики связывания для растворимых TCR или молекул ImmTAC (растворимых гибридных молекул TCR анти-СD3), содержащих указанные альфа- и бета-вариабельные области.
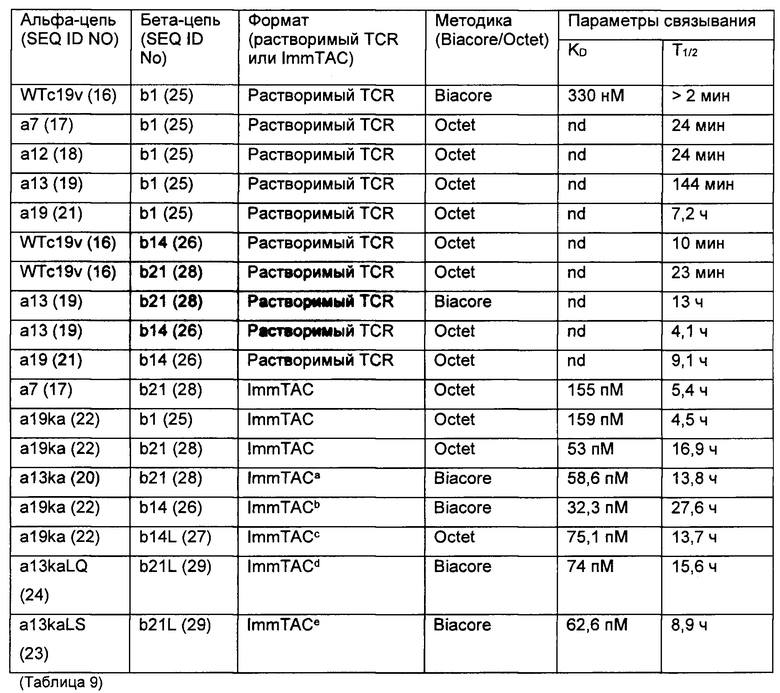
nd = не определено
а Соответствует ImmTAC3 из Примера 6, полные последовательности альфа- и бета-цепи представлены в SEQ ID NO: 39 и SEQ ID NO: 44 соответственно. Значения основаны на среднем из 7 независимых измерений
b Соответствует ImmTAC1 из Примера 6, полные последовательности альфа- и бета-цепи представлены в SEQ ID NO: 38 and SEQ ID NO: 42 соответственно. Значения основаны на среднем из 7 независимых измерений
c Соответствует ImmTAC2 из Примера 6, полные последовательности альфа- и бета-цепи представлены в SEQ ID NO: 38 and SEQ ID NO: 43 соответственно
d Соответствует ImmTAC4 из Примера 6, полные последовательности альфа- и бета-цепи представлены в SEQ ID NO: 40 and SEQ ID NO: 45 соответственно. Значения основаны на среднем из 4 независимых измерений
e Соответствует lmmTAC5 из Примера 6, полные последовательности альфа- и бета-цепи представлены в SEQ ID NO: 41 and SEQ ID NO: 45 соответственно На связывание с комплексом GVYDGREHTV HLA-A*02 также тестировали другие комбинации вариабельных областей альфа- и бета-цепи, содержащие мутации по настоящему изобретению. Данные, представленные в таблице ниже, были получены с использованием методики Biacore, как описано выше. Указанные последовательности вариабельных доменов альфа- и бета-цепи получали в виде молекул ImmTAC.
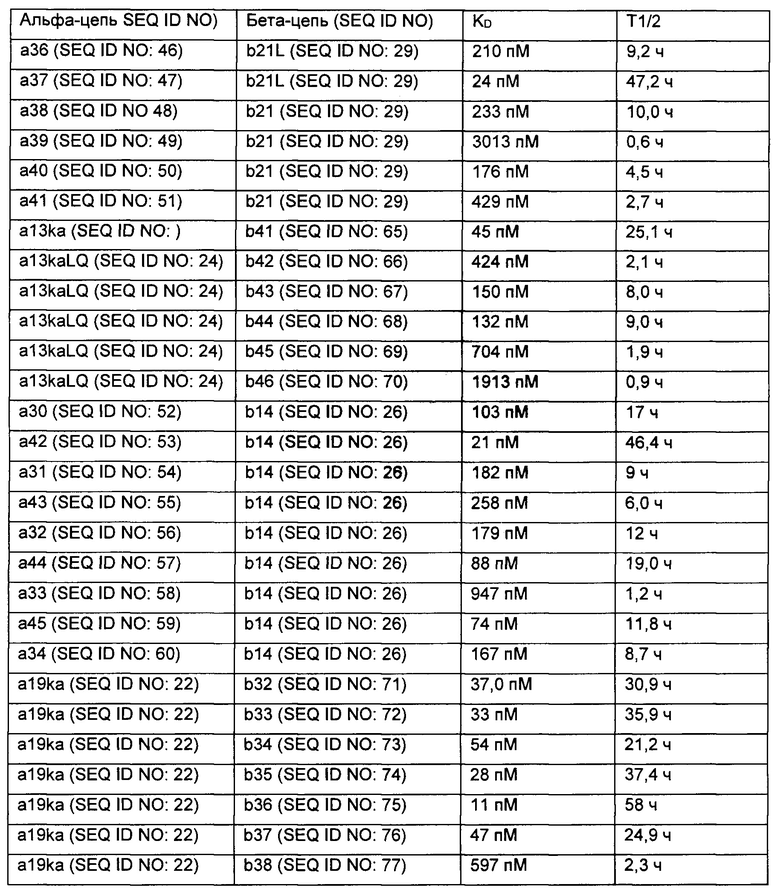
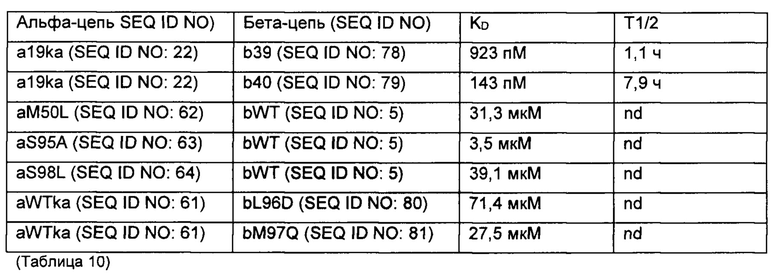
nd - не определено
Данные, представленные в Таблицах 9 и 10, показывают, что некоторые вариабельные последовательности TCR по настоящему изобретению имеют высокую аффинность связывания и большое время полужизни связывания для комплекса GVYDGREHTV HLA-А*02 и, следовательно, особенно подходят для применения в качестве растворимых терапевтических реагентов.
Помимо связывания когнатного комплекса GVYDGREHTV HLA-A*02, TCR по настоящему изобретению также оценивали на связывание со сходными пептидами, полученными из MAGE А8 и MAGE В2 и презентируемыми HLA-A*02. В таблице ниже приведены данные Biacore по связыванию для трех молекул ImmTAC, содержащих указанные последовательности вариабельного домена альфа и бета. Все три молекулы ImmTAC распознают пептид MAGE-A8 на том же уровне, что и когнатный пептид, и пептид MAGE-B2 на более слабом уровне.
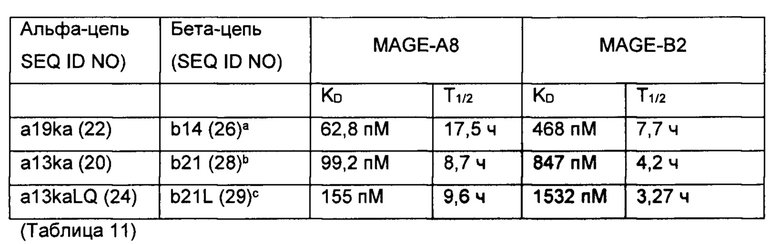
а Соответствует ImmTAC1 из Примера 6, полные последовательности альфа- и бета-цепи представлены в SEQ ID NO: 38 и SEQ ID NO: 42 соответственно.
b Соответствует ImmTAC3 from из Примера 6, полные последовательности альфа- и бета-цепи представлены в SEQ ID NO: 39 и SEQ ID NO: 44 соответственно.
с Соответствует ImmTAC4 из Примера 6, полные последовательности альфа- и бета-цепи представлены в SEQ ID NO: 40 и SEQ ID NO: 45 соответственно.
Пример 6. Эффективное и специфическое перенаправление Т-клеток молекулами ImmTAC
Молекулы ImmTAC, содержащие мутированные последовательности вариабельных областей альфа- и бета-цепи с особенно высокой аффинностью к антигену-мишени, были протестированы на их способность опосредовать эффективное и специфическое перенаправление CD3+ Т-клеток с помощью анализа ELISPOT, используя секрецию интерферона-γ (IFN-γ) в качестве регистрируемого показателя активации Т-клеток.
В этом примере последовательность вариабельной области альфа-цепи была выбрана из SEQ ID NO: 20-24, а последовательность вариабельной области бета-цепи была выбрана из SEQ ID NO: 26-29. Последовательности вариабельного домена были гибридизованы с соответствующими последовательностями внеклеточного константного домена альфа или бета и содержали ненативную дисульфидную связь. В каждом случае бета-цепь была гибридизована через линкер с анти-СD3 scFv; линкер был выбран из SEQ ID NO: 30-37. Полные последовательности протестированных молекул ImmTAC представлены в SEQ ID NO, приведенных в следующей таблице:

Методика
Анализы выполняли с использованием набора ELISPOT IFN-γ человека (BD Biosciences). Клетки-мишени готовили при плотности 1×106/мл в среде для анализа (RPM 11640, содержащей 10% инактивированной нагреванием FBS и 1% пенициллин-стрептомицин-L-глутамина) и высевали при плотности 50000 клеток на лунку в объеме 50 мкл. Мононуклеарные клетки периферической крови (РВМС), выделенные из свежей донорской крови, использовали в качестве эффекторных клеток и высевали при плотности 10000-50000 клеток на лунку в объеме 50 мкл (точное количество клеток, используемых в каждом эксперименте, зависит от донора и может быть скорректировано для получения ответа в подходящем диапазоне для анализа). Использовали различные концентрации ImmTAC, охватывающие ожидаемый клинически значимый диапазон, и добавляли их в лунку в объеме 50 мкл.
Планшеты были предварительно подготовлены в соответствии с инструкциями производителя. Клетки-мишени, эффекторные клетки и молекулы ImmTAC добавляли в соответствующие лунки и доводили до конечного объема 200 мкл средой для анализа. Все реакции выполняли в трех повторах. Также готовили контрольные лунки, исключая ImmTAC, эффекторные клетки или клетки-мишени. Затем планшеты инкубировали в течение ночи (37°С/5% СО2). На следующий день планшеты трижды промывали промывочным буфером (1 пакетик с фосфатно-солевым буфером, содержащий 0,05% Р20, разведенный в деионизированной воде). Затем в каждую лунку добавляли первичное детектирующее антитело в объеме 50 мкл. Планшеты инкубировали при комнатной температуре в течение 2 часов, а затем снова трижды промывали. Вторичное детектирование выполняли путем добавления 50 мкл разведенного конъюгата стрептавидина с пероксидазой хрена в каждую лунку и инкубирования при комнатной температуре в течение 1 часа, и повторяли стадию промывки. Не более чем за 15 минут до использования к каждому 1 мл субстрата АЕС добавляли одну каплю (20 мкл) хромогена АЕС и перемешивали, и 50 мкл добавляли в каждую лунку. Проявление пятен регулярно контролировали, и чтобы остановить реакцию проявления, чашки промывали в водопроводной воде. Затем планшетам давали высохнуть при комнатной температуре в течение по меньшей мере 2 часов до подсчета пятен с использованием анализатора CTL с программным обеспечением Immunospot (Cellular Technology Limited).
Результаты
Данные, представленные на Фигурах 7 и 8, на верхних панелях, показывают, что молекулы ImmTAC 1-2 и 3-5, соответственно, способны опосредовать эффективное (т.е. с EC50 менее 100 пМ) перенаправление Т-клеток против раковых клеток, экспрессирующих антиген-мишень (NCl-H1703 - клеточная линия рака легких человека). Активации Т-клеток против антиген-отрицательных раковых клеток (клеточная линия папиллярной аденокарциномы человека NCl-H441 для молекул 1-2 ImmTAC и клеточная линия рака молочной железы человека САМА-1 для молекул ImmTAC 3-5) в пределах клинически значимого диапазона концентраций 1 нМ) обнаружено не было, что демонстрирует специфичность ответа.
Молекулы ImmTAC были протестированы на специфичность с использованием в качестве клеток-мишеней клеток, полученных из нормальных тканей здорового человека. Нижняя панель на Фиг. 7 показывает, что молекулы 1-2 ImmTAC обладают минимальной реактивностью при клинически значимой концентрации в отношении клеток сосудистой сети кожи человека. Аналогично, нижние панели на Фиг. 8 показывают, что молекулы 3-5 ImmTAC обладают минимальной реактивностью при клинически значимой концентрации в отношении клеток сосудистой сети кожи человека и почечных клеток человека.
Молекулы ImmTAC 1 и 3 были дополнительно протестированы на специфичность в отношении панели человеческих клеток, полученных из нормальных здоровых тканей, с использованием той же методологии ELISPOT, описанной выше. Данные, приведенные на Фигуре 9, показывают ограниченную активацию Т-клеток в пределах клинически значимого диапазона концентраций 1 нМ) для здоровых тканей, включая клетки сосудистой сети кожи, сердца, скелета, печени и легких.
Молекулы ImmTAC 1 и 4 были дополнительно протестированы на реакционноспособность в отношении расширенной панели из > 10 типов нормальных клеток с использованием той же методологии ELISPOT, описанной выше, и с более мелким шагом диапазона концентраций ImmTAC (0,01 нМ, 0,1 нМ, 0,2 нМ, 0,3 нМ, 0,5 нМ и 1 нМ). На Фигуре 10 показаны репрезентативные данные, полученные от скелетных, сердечных и почечных клеток. В каждом случае в качестве контролей были включены антиген-положительные клетки (NCl-Н1703) и антиген-отрицательные клетки (NCl-H441). Полученные данные демонстрируют незначительную реакционноспособность в отношении нормальных клеток относительно антиген-положительных клеток в пределах клинически значимого диапазона концентраций (< 1 нМ).
Эти данные показывают, что эти молекулы ImmTAC демонстрируют высокий уровень активности и специфичности и поэтому особенно подходят для терапевтического применения.
Пример 7. Эффективное уничтожение опухолевых клеток перенаправленными Т-клетками ImmTAC
Способность молекул ImmTAC по настоящему изобретению опосредовать эффективное перенаправленное уничтожение Т-клетками антиген-положительных опухолевых клеток исследовали с использованием платформы IncuCyte (Essen Bioscience). Этот анализ позволяет в реальном времени микроскопически обнаружить высвобождение каспазы-3/7, маркера апоптоза.
Методика
Анализы выполняли с использованием набора для анализа апоптоза с помощью реагента Caspase-3/7 и 96-луночного планшета CellPlayer (Essen Bioscience, Cat. No. 4440) и осуществляли в соответствии с протоколом производителя. В кратком изложении клетки-мишени (NCl-H1703 - антиген-положительные HLA А*02-положительные и NCl-H441 -антиген-отрицательные HLA А*02-положительные) высевали по 5000 клеток на лунку и инкубировали в течение ночи, чтобы дать им прикрепиться. Растворы ImmTAC готовили в концентрациях от 0,5 нМ до 0,01 нМ, и 25 мкл каждой концентрации добавляли в соответствующую лунку. Эффекторные клетки использовали при соотношении эффекторов:клеток-мишеней 10:1 (50000 клеток на лунку). Также готовили контрольный образец без ImmTAC. Реагент для анализа NucView готовили при концентрации 30 мкМ, и 25 мкл добавляли в каждую лунку, а конечный объем доводили до 150 мкл (получая конечную концентрацию 5 мкМ). Планшет помещали в прибор IncuCyte, и регистрировали изображения каждые 2 часа (1 изображение на лунку) в течение 3 дней. Затем определяли количество апоптотических клеток на каждом изображении и выражали как количество апоптотических клеток на мм2. Анализы выполняли в трех повторах.
Результаты
Данные, представленные на Фигуре 11, показывают уничтожение опухолевых клеток в режиме реального времени Т-клетками, перенаправленными молекулами ImmTAC. Результаты представлены для ImmTAC1 и ImmTAC4. Обе молекулы ImmTAC демонстрируют перенаправленное уничтожение Т-клетками антиген-положительных опухолевых клеток при концентрации 0,01 нМ. Уничтожения антиген-отрицательных клеток не наблюдается даже при самой высокой концентрации (0,5 нМ).
Эти данные подтверждают, что ImmTAC1 и immTAC4 опосредуют эффективное перенаправленное уничтожение Т-клетками антиген-положительных опухолевых клеток.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Immunocore Limited
<120> Т-клеточные рецепторы
<130> P101450WO
<150> 1606009.7
<151> 2016-04-08
<160> 81
<170> PatentIn версия 3.5
<210> 1
<211> 10
<212> БЕЛОК
<213> Homo sapiens
<400> 1
Gly Val Tyr Asp Gly Arg Glu His Thr Val
1 5 10
<210> 2
<211> 207
<212> БЕЛОК
<213> Homo sapiens
<400> 2
Lys Asn Gln Val Glu Gln Ser Pro Gln Ser Leu Ile Ile Leu Glu Gly
1 5 10 15
Lys Asn Cys Thr Leu Gln Cys Asn Tyr Thr Val Ser Pro Phe Ser Asn
20 25 30
Leu Arg Trp Tyr Lys Gln Asp Thr Gly Arg Gly Pro Val Ser Leu Thr
35 40 45
Ile Met Thr Phe Ser Glu Asn Thr Lys Ser Asn Gly Arg Tyr Thr Ala
50 55 60
Thr Leu Asp Ala Asp Thr Lys Gln Ser Ser Leu His Ile Thr Ala Ser
65 70 75 80
Gln Leu Ser Asp Ser Ala Ser Tyr Ile Cys Val Val Asn His Ser Gly
85 90 95
Gly Ser Tyr Ile Pro Thr Phe Gly Arg Gly Thr Ser Leu Ile Val His
100 105 110
Pro Tyr Ile Gln Lys Pro Asp Pro Ala Val Tyr Gln Leu Arg Asp Ser
115 120 125
Lys Ser Ser Asp Lys Ser Val Cys Leu Phe Thr Asp Phe Asp Ser Gln
130 135 140
Thr Asn Val Ser Gln Ser Lys Asp Ser Asp Val Tyr Ile Thr Asp Lys
145 150 155 160
Thr Val Leu Asp Met Arg Ser Met Asp Phe Lys Ser Asn Ser Ala Val
165 170 175
Ala Trp Ser Asn Lys Ser Asp Phe Ala Cys Ala Asn Ala Phe Asn Asn
180 185 190
Ser Ile Ile Pro Glu Asp Thr Phe Phe Pro Ser Pro Glu Ser Ser
195 200 205
<210> 3
<211> 246
<212> БЕЛОК
<213> Homo sapiens
<400> 3
Asp Val Lys Val Thr Gln Ser Ser Arg Tyr Leu Val Lys Arg Thr Gly
1 5 10 15
Glu Lys Val Phe Leu Glu Cys Val Gln Asp Met Asp His Glu Asn Met
20 25 30
Phe Trp Tyr Arg Gln Asp Pro Gly Leu Gly Leu Arg Leu Ile Tyr Phe
35 40 45
Ser Tyr Asp Val Lys Met Lys Glu Lys Gly Asp Ile Pro Glu Gly Tyr
50 55 60
Ser Val Ser Arg Glu Lys Lys Glu Arg Phe Ser Leu Ile Leu Glu Ser
65 70 75 80
Ala Ser Thr Asn Gln Thr Ser Met Tyr Leu Cys Ala Ser Ser Phe Leu
85 90 95
Met Thr Ser Gly Asp Pro Tyr Glu Gln Tyr Phe Gly Pro Gly Thr Arg
100 105 110
Leu Thr Val Thr Glu Asp Leu Lys Asn Val Phe Pro Pro Glu Val Ala
115 120 125
Val Phe Glu Pro Ser Glu Ala Glu Ile Ser His Thr Gln Lys Ala Thr
130 135 140
Leu Val Cys Leu Ala Thr Gly Phe Tyr Pro Asp His Val Glu Leu Ser
145 150 155 160
Trp Trp Val Asn Gly Lys Glu Val His Ser Gly Val Ser Thr Asp Pro
165 170 175
Gln Pro Leu Lys Glu Gln Pro Ala Leu Asn Asp Ser Arg Tyr Cys Leu
180 185 190
Ser Ser Arg Leu Arg Val Ser Ala Thr Phe Trp Gln Asn Pro Arg Asn
195 200 205
His Phe Arg Cys Gln Val Gln Phe Tyr Gly Leu Ser Glu Asn Asp Glu
210 215 220
Trp Thr Gln Asp Arg Ala Lys Pro Val Thr Gln Ile Val Ser Ala Glu
225 230 235 240
Ala Trp Gly Arg Ala Asp
245
<210> 4
<211> 207
<212> БЕЛОК
<213> Homo sapiens
<400> 4
Lys Asn Gln Val Glu Gln Ser Pro Gln Ser Leu Ile Ile Leu Glu Gly
1 5 10 15
Lys Asn Cys Thr Leu Gln Cys Asn Tyr Thr Val Ser Pro Phe Ser Asn
20 25 30
Leu Arg Trp Tyr Lys Gln Asp Thr Gly Arg Gly Pro Val Ser Leu Thr
35 40 45
Ile Met Thr Phe Ser Glu Asn Thr Lys Ser Asn Gly Arg Tyr Thr Ala
50 55 60
Thr Leu Asp Ala Asp Thr Lys Gln Ser Ser Leu His Ile Thr Ala Ser
65 70 75 80
Gln Leu Ser Asp Ser Ala Ser Tyr Ile Cys Val Val Asn His Ser Gly
85 90 95
Gly Ser Tyr Ile Pro Thr Phe Gly Arg Gly Thr Ser Leu Ile Val His
100 105 110
Pro Tyr Ile Gln Lys Pro Asp Pro Ala Val Tyr Gln Leu Arg Asp Ser
115 120 125
Lys Ser Ser Asp Lys Ser Val Cys Leu Phe Thr Asp Phe Asp Ser Gln
130 135 140
Thr Asn Val Ser Gln Ser Lys Asp Ser Asp Val Tyr Ile Thr Asp Lys
145 150 155 160
Cys Val Leu Asp Met Arg Ser Met Asp Phe Lys Ser Asn Ser Ala Val
165 170 175
Ala Trp Ser Asn Lys Ser Asp Phe Ala Cys Ala Asn Ala Phe Asn Asn
180 185 190
Ser Ile Ile Pro Glu Asp Thr Phe Phe Pro Ser Pro Glu Ser Ser
195 200 205
<210> 5
<211> 246
<212> БЕЛОК
<213> Homo sapiens
<400> 5
Asp Val Lys Val Thr Gln Ser Ser Arg Tyr Leu Val Lys Arg Thr Gly
1 5 10 15
Glu Lys Val Phe Leu Glu Cys Val Gln Asp Met Asp His Glu Asn Met
20 25 30
Phe Trp Tyr Arg Gln Asp Pro Gly Leu Gly Leu Arg Leu Ile Tyr Phe
35 40 45
Ser Tyr Asp Val Lys Met Lys Glu Lys Gly Asp Ile Pro Glu Gly Tyr
50 55 60
Ser Val Ser Arg Glu Lys Lys Glu Arg Phe Ser Leu Ile Leu Glu Ser
65 70 75 80
Ala Ser Thr Asn Gln Thr Ser Met Tyr Leu Cys Ala Ser Ser Phe Leu
85 90 95
Met Thr Ser Gly Asp Pro Tyr Glu Gln Tyr Phe Gly Pro Gly Thr Arg
100 105 110
Leu Thr Val Thr Glu Asp Leu Lys Asn Val Phe Pro Pro Glu Val Ala
115 120 125
Val Phe Glu Pro Ser Glu Ala Glu Ile Ser His Thr Gln Lys Ala Thr
130 135 140
Leu Val Cys Leu Ala Thr Gly Phe Tyr Pro Asp His Val Glu Leu Ser
145 150 155 160
Trp Trp Val Asn Gly Lys Glu Val His Ser Gly Val Cys Thr Asp Pro
165 170 175
Gln Pro Leu Lys Glu Gln Pro Ala Leu Asn Asp Ser Arg Tyr Ala Leu
180 185 190
Ser Ser Arg Leu Arg Val Ser Ala Thr Phe Trp Gln Asp Pro Arg Asn
195 200 205
His Phe Arg Cys Gln Val Gln Phe Tyr Gly Leu Ser Glu Asn Asp Glu
210 215 220
Trp Thr Gln Asp Arg Ala Lys Pro Val Thr Gln Ile Val Ser Ala Glu
225 230 235 240
Ala Trp Gly Arg Ala Asp
245
<210> 6
<211> 6
<212> БЕЛОК
<213> Homo sapiens
<400> 6
Val Ser Pro Phe Ser Asn
1 5
<210> 7
<211> 7
<212> БЕЛОК
<213> Homo sapiens
<400> 7
Met Thr Phe Ser Glu Asn Thr
1 5
<210> 8
<211> 13
<212> БЕЛОК
<213> Homo sapiens
<400> 8
Val Val Asn His Ser Gly Gly Ser Tyr Ile Pro Thr Phe
1 5 10
<210> 9
<211> 17
<212> БЕЛОК
<213> Homo sapiens
<400> 9
Leu Arg Trp Tyr Lys Gln Asp Thr Gly Arg Gly Pro Val Ser Leu Thr
1 5 10 15
Ile
<210> 10
<211> 34
<212> БЕЛОК
<213> Homo sapiens
<400> 10
Lys Ser Asn Gly Arg Tyr Thr Ala Thr Leu Asp Ala Asp Thr Lys Gln
1 5 10 15
Ser Ser Leu His Ile Thr Ala Ser Gln Leu Ser Asp Ser Ala Ser Tyr
20 25 30
Ile Cys
<210> 11
<211> 5
<212> БЕЛОК
<213> Homo sapiens
<400> 11
Met Asp His Glu Asn
1 5
<210> 12
<211> 6
<212> БЕЛОК
<213> Homo sapiens
<400> 12
Ser Tyr Asp Val Lys Met
1 5
<210> 13
<211> 16
<212> БЕЛОК
<213> Homo sapiens
<400> 13
Ala Ser Ser Phe Leu Met Thr Ser Gly Asp Pro Tyr Glu Gln Tyr Phe
1 5 10 15
<210> 14
<211> 17
<212> БЕЛОК
<213> Homo sapiens
<400> 14
Met Phe Trp Tyr Arg Gln Asp Pro Gly Leu Gly Leu Arg Leu Ile Tyr
1 5 10 15
Phe
<210> 15
<211> 37
<212> БЕЛОК
<213> Homo sapiens
<400> 15
Lys Glu Lys Gly Asp Ile Pro Glu Gly Tyr Ser Val Ser Arg Glu Lys
1 5 10 15
Lys Glu Arg Phe Ser Leu Ile Leu Glu Ser Ala Ser Thr Asn Gln Thr
20 25 30
Ser Met Tyr Leu Cys
35
<210> 16
<211> 113
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> мутированная альфа-цепь (c19v)
<400> 16
Lys Asn Gln Val Glu Gln Ser Pro Gln Ser Leu Ile Ile Leu Glu Gly
1 5 10 15
Lys Asn Val Thr Leu Gln Cys Asn Tyr Thr Val Ser Pro Phe Ser Asn
20 25 30
Leu Arg Trp Tyr Lys Gln Asp Thr Gly Arg Gly Pro Val Ser Leu Thr
35 40 45
Ile Met Thr Phe Ser Glu Asn Thr Lys Ser Asn Gly Arg Tyr Thr Ala
50 55 60
Thr Leu Asp Ala Asp Thr Lys Gln Ser Ser Leu His Ile Thr Ala Ser
65 70 75 80
Gln Leu Ser Asp Ser Ala Ser Tyr Ile Cys Val Val Asn His Ser Gly
85 90 95
Gly Ser Tyr Ile Pro Thr Phe Gly Arg Gly Thr Ser Leu Ile Val His
100 105 110
Pro
<210> 17
<211> 113
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> мутированная альфа-цепь (a7)
<400> 17
Lys Asn Gln Val Glu Gln Ser Pro Gln Ser Leu Ile Ile Leu Glu Gly
1 5 10 15
Lys Asn Val Thr Leu Gln Cys Asn Tyr Thr Val Ser Pro Phe Ser Asn
20 25 30
Leu Arg Trp Tyr Lys Gln Asp Thr Gly Arg Gly Pro Val Ser Leu Thr
35 40 45
Ile Leu Asp Tyr Ala Ile Asn Thr Lys Ser Asn Gly Arg Tyr Thr Ala
50 55 60
Thr Leu Asp Ala Asp Thr Lys Gln Ser Ser Leu His Ile Thr Ala Ser
65 70 75 80
Gln Leu Ser Asp Ser Ala Ser Tyr Ile Cys Val Val Asn His Ser Gly
85 90 95
Gly Ser Tyr Ile Pro Thr Phe Gly Arg Gly Thr Ser Leu Ile Val His
100 105 110
Pro
<210> 18
<211> 113
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> мутированная альфа-цепь (a12)
<400> 18
Lys Asn Gln Val Glu Gln Ser Pro Gln Ser Leu Ile Ile Leu Glu Gly
1 5 10 15
Lys Asn Val Thr Leu Gln Cys Asn Tyr Thr Val Ser Pro Phe Ser Asn
20 25 30
Leu Arg Trp Tyr Lys Gln Asp Thr Gly Arg Gly Pro Val Ser Leu Thr
35 40 45
Ile Met Thr Phe Ser Glu Asn Thr Lys Ser Asn Gly Arg Tyr Thr Ala
50 55 60
Thr Leu Asp Ala Asp Thr Lys Gln Ser Ser Leu His Ile Thr Ala Ser
65 70 75 80
Gln Leu Ser Asp Ser Ala Ser Tyr Ile Cys Val Val Asn Arg Ala Asp
85 90 95
Gly Leu Tyr Ile Pro Thr Phe Gly Arg Gly Thr Ser Leu Ile Val His
100 105 110
Pro
<210> 19
<211> 113
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> мутированная альфа-цепь (a13)
<400> 19
Lys Asn Gln Val Glu Gln Ser Pro Gln Ser Leu Ile Ile Leu Glu Gly
1 5 10 15
Lys Asn Val Thr Leu Gln Cys Asn Tyr Thr Val Ser Pro Phe Ser Asn
20 25 30
Leu Arg Trp Tyr Lys Gln Asp Thr Gly Arg Gly Pro Val Ser Leu Thr
35 40 45
Ile Met Thr Phe Ser Glu Asn Thr Lys Ser Asn Gly Arg Tyr Thr Ala
50 55 60
Thr Leu Asp Ala Asp Thr Lys Gln Ser Ser Leu His Ile Thr Ala Ser
65 70 75 80
Gln Leu Ser Asp Ser Ala Ser Tyr Ile Cys Val Val Asn Ser Ala Asn
85 90 95
Gly Leu Tyr Ile Pro Thr Phe Gly Arg Gly Thr Ser Leu Ile Val His
100 105 110
Pro
<210> 20
<211> 113
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> мутированная альфа-цепь (a13ka)
<400> 20
Ala Asn Gln Val Glu Gln Ser Pro Gln Ser Leu Ile Ile Leu Glu Gly
1 5 10 15
Lys Asn Val Thr Leu Gln Cys Asn Tyr Thr Val Ser Pro Phe Ser Asn
20 25 30
Leu Arg Trp Tyr Lys Gln Asp Thr Gly Arg Gly Pro Val Ser Leu Thr
35 40 45
Ile Met Thr Phe Ser Glu Asn Thr Lys Ser Asn Gly Arg Tyr Thr Ala
50 55 60
Thr Leu Asp Ala Asp Thr Lys Gln Ser Ser Leu His Ile Thr Ala Ser
65 70 75 80
Gln Leu Ser Asp Ser Ala Ser Tyr Ile Cys Val Val Asn Ser Ala Asn
85 90 95
Gly Leu Tyr Ile Pro Thr Phe Gly Arg Gly Thr Ser Leu Ile Val His
100 105 110
Pro
<210> 21
<211> 113
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> мутированная альфа-цепь (a19)
<400> 21
Lys Asn Gln Val Glu Gln Ser Pro Gln Ser Leu Ile Ile Leu Glu Gly
1 5 10 15
Lys Asn Val Thr Leu Gln Cys Asn Tyr Thr Val Ser Pro Phe Ser Asn
20 25 30
Leu Arg Trp Tyr Lys Gln Asp Thr Gly Arg Gly Pro Val Ser Leu Thr
35 40 45
Ile Leu Asp Tyr Ala Ile Asn Thr Lys Ser Asn Gly Arg Tyr Thr Ala
50 55 60
Thr Leu Asp Ala Asp Thr Lys Gln Ser Ser Leu His Ile Thr Ala Ser
65 70 75 80
Gln Leu Ser Asp Ser Ala Ser Tyr Ile Cys Val Val Asn Arg Ala Asp
85 90 95
Gly Leu Tyr Ile Pro Thr Phe Gly Arg Gly Thr Ser Leu Ile Val His
100 105 110
Pro
<210> 22
<211> 113
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> мутированная альфа-цепь (a19ka)
<400> 22
Ala Asn Gln Val Glu Gln Ser Pro Gln Ser Leu Ile Ile Leu Glu Gly
1 5 10 15
Lys Asn Val Thr Leu Gln Cys Asn Tyr Thr Val Ser Pro Phe Ser Asn
20 25 30
Leu Arg Trp Tyr Lys Gln Asp Thr Gly Arg Gly Pro Val Ser Leu Thr
35 40 45
Ile Leu Asp Tyr Ala Ile Asn Thr Lys Ser Asn Gly Arg Tyr Thr Ala
50 55 60
Thr Leu Asp Ala Asp Thr Lys Gln Ser Ser Leu His Ile Thr Ala Ser
65 70 75 80
Gln Leu Ser Asp Ser Ala Ser Tyr Ile Cys Val Val Asn Arg Ala Asp
85 90 95
Gly Leu Tyr Ile Pro Thr Phe Gly Arg Gly Thr Ser Leu Ile Val His
100 105 110
Pro
<210> 23
<211> 113
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> мутированная альфа-цепь (a13kaLS)
<400> 23
Ala Asn Gln Val Glu Gln Ser Pro Gln Ser Leu Ile Ile Leu Glu Gly
1 5 10 15
Lys Asn Val Thr Leu Gln Cys Asn Tyr Thr Val Ser Pro Phe Ser Asn
20 25 30
Leu Arg Trp Tyr Lys Gln Asp Thr Gly Arg Gly Pro Val Ser Leu Thr
35 40 45
Ile Leu Thr Phe Ser Glu Asn Thr Lys Ser Asn Gly Arg Tyr Thr Ala
50 55 60
Thr Leu Asp Ala Asp Thr Lys Gln Ser Ser Leu His Ile Thr Ala Ser
65 70 75 80
Gln Leu Ser Asp Ser Ala Ser Tyr Ile Cys Val Val Asn Ser Ala Ser
85 90 95
Gly Leu Tyr Ile Pro Thr Phe Gly Arg Gly Thr Ser Leu Ile Val His
100 105 110
Pro
<210> 24
<211> 113
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> мутированная альфа-цепь (a13kaLQ)
<400> 24
Ala Asn Gln Val Glu Gln Ser Pro Gln Ser Leu Ile Ile Leu Glu Gly
1 5 10 15
Lys Asn Val Thr Leu Gln Cys Asn Tyr Thr Val Ser Pro Phe Ser Asn
20 25 30
Leu Arg Trp Tyr Lys Gln Asp Thr Gly Arg Gly Pro Val Ser Leu Thr
35 40 45
Ile Leu Thr Phe Ser Glu Asn Thr Lys Ser Asn Gly Arg Tyr Thr Ala
50 55 60
Thr Leu Asp Ala Asp Thr Lys Gln Ser Ser Leu His Ile Thr Ala Ser
65 70 75 80
Gln Leu Ser Asp Ser Ala Ser Tyr Ile Cys Val Val Asn Ser Ala Gln
85 90 95
Gly Leu Tyr Ile Pro Thr Phe Gly Arg Gly Thr Ser Leu Ile Val His
100 105 110
Pro
<210> 25
<211> 116
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> мутированная бета-цепь (b1)
<400> 25
Asp Val Lys Val Thr Gln Ser Ser Arg Tyr Leu Val Lys Arg Thr Gly
1 5 10 15
Glu Lys Val Phe Leu Glu Cys Val Gln Asp Met Asp His Glu Asn Met
20 25 30
Phe Trp Tyr Arg Gln Asp Pro Gly Leu Gly Leu Arg Leu Ile Tyr Phe
35 40 45
Ser Tyr Asp Val Lys Met Lys Glu Lys Gly Asp Ile Pro Glu Gly Tyr
50 55 60
Ser Val Ser Arg Glu Lys Lys Glu Arg Phe Ser Leu Ile Leu Glu Ser
65 70 75 80
Ala Ser Thr Asn Gln Thr Ser Met Tyr Leu Cys Ala Ser Ser Ser Asp
85 90 95
Gln Asn Ser Gly Asp Pro Tyr Glu Gln Tyr Phe Gly Pro Gly Thr Arg
100 105 110
Leu Thr Val Thr
115
<210> 26
<211> 116
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> мутированная бета-цепь (b14)
<400> 26
Asp Val Lys Val Thr Gln Ser Ser Arg Tyr Leu Val Lys Arg Thr Gly
1 5 10 15
Glu Lys Val Phe Leu Glu Cys Val Gln Asp Ala Pro Leu Ser Lys Met
20 25 30
Phe Trp Tyr Arg Gln Asp Pro Gly Leu Gly Leu Arg Leu Ile Tyr Phe
35 40 45
Ser Tyr Asp Val Lys Met Lys Glu Lys Gly Asp Ile Pro Glu Gly Tyr
50 55 60
Ser Val Ser Arg Glu Lys Lys Glu Arg Phe Ser Leu Ile Leu Glu Ser
65 70 75 80
Ala Ser Thr Asn Gln Thr Ser Met Tyr Leu Cys Ala Ser Ser Ser Asp
85 90 95
Gln Asn Ser Gly Asp Pro Tyr Glu Gln Tyr Phe Gly Pro Gly Thr Arg
100 105 110
Leu Thr Val Thr
115
<210> 27
<211> 116
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> мутированная бета-цепь (b14L)
<400> 27
Asp Val Lys Val Thr Gln Ser Ser Arg Tyr Leu Val Lys Arg Thr Gly
1 5 10 15
Glu Lys Val Phe Leu Glu Cys Val Gln Asp Ala Pro Leu Ser Lys Met
20 25 30
Phe Trp Tyr Arg Gln Asp Pro Gly Leu Gly Leu Arg Leu Ile Tyr Phe
35 40 45
Ser Tyr Asp Val Lys Leu Lys Glu Lys Gly Asp Ile Pro Glu Gly Tyr
50 55 60
Ser Val Ser Arg Glu Lys Lys Glu Arg Phe Ser Leu Ile Leu Glu Ser
65 70 75 80
Ala Ser Thr Asn Gln Thr Ser Met Tyr Leu Cys Ala Ser Ser Ser Asp
85 90 95
Gln Asn Ser Gly Asp Pro Tyr Glu Gln Tyr Phe Gly Pro Gly Thr Arg
100 105 110
Leu Thr Val Thr
115
<210> 28
<211> 116
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> мутированная бета-цепь (b21)
<400> 28
Asp Val Lys Val Thr Gln Ser Ser Arg Tyr Leu Val Lys Arg Thr Gly
1 5 10 15
Glu Lys Val Phe Leu Glu Cys Val Gln Asp Met Asp His Glu Asn Met
20 25 30
Phe Trp Tyr Arg Gln Asp Pro Gly Leu Gly Leu Arg Leu Ile Tyr Phe
35 40 45
Ser Arg Phe Ala Thr Gly Lys Glu Lys Gly Asp Ile Pro Glu Gly Tyr
50 55 60
Ser Val Ser Arg Glu Lys Lys Glu Arg Phe Ser Leu Ile Leu Glu Ser
65 70 75 80
Ala Ser Thr Asn Gln Thr Ser Met Tyr Leu Cys Ala Ser Ser Ser Asp
85 90 95
Gln Asn Ser Gly Asp Pro Tyr Glu Gln Tyr Phe Gly Pro Gly Thr Arg
100 105 110
Leu Thr Val Thr
115
<210> 29
<211> 116
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> мутированная бета-цепь (b21L)
<400> 29
Asp Val Lys Val Thr Gln Ser Ser Arg Tyr Leu Val Lys Arg Thr Gly
1 5 10 15
Glu Lys Val Phe Leu Glu Cys Val Gln Asp Leu Asp His Glu Asn Met
20 25 30
Phe Trp Tyr Arg Gln Asp Pro Gly Leu Gly Leu Arg Leu Ile Tyr Phe
35 40 45
Ser Arg Phe Ala Thr Gly Lys Glu Lys Gly Asp Ile Pro Glu Gly Tyr
50 55 60
Ser Val Ser Arg Glu Lys Lys Glu Arg Phe Ser Leu Ile Leu Glu Ser
65 70 75 80
Ala Ser Thr Asn Gln Thr Ser Met Tyr Leu Cys Ala Ser Ser Ser Asp
85 90 95
Gln Asn Ser Gly Asp Pro Tyr Glu Gln Tyr Phe Gly Pro Gly Thr Arg
100 105 110
Leu Thr Val Thr
115
<210> 30
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Линкерная последовательность
<400> 30
Gly Gly Gly Gly Ser
1 5
<210> 31
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Линкерная последовательность
<400> 31
Gly Gly Gly Ser Gly
1 5
<210> 32
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Линкерная последовательность
<400> 32
Gly Gly Ser Gly Gly
1 5
<210> 33
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Линкерная последовательность
<400> 33
Gly Ser Gly Gly Gly
1 5
<210> 34
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Линкерная последовательность
<400> 34
Gly Ser Gly Gly Gly Pro
1 5
<210> 35
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Линкерная последовательность
<400> 35
Gly Gly Glu Pro Ser
1 5
<210> 36
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Линкерная последовательность
<400> 36
Gly Gly Glu Gly Gly Gly Pro
1 5
<210> 37
<211> 12
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Линкерная последовательность
<400> 37
Gly Gly Glu Gly Gly Gly Ser Glu Gly Gly Gly Ser
1 5 10
<210> 38
<211> 199
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> альфа-цепь (a19ka)
<400> 38
Ala Asn Gln Val Glu Gln Ser Pro Gln Ser Leu Ile Ile Leu Glu Gly
1 5 10 15
Lys Asn Val Thr Leu Gln Cys Asn Tyr Thr Val Ser Pro Phe Ser Asn
20 25 30
Leu Arg Trp Tyr Lys Gln Asp Thr Gly Arg Gly Pro Val Ser Leu Thr
35 40 45
Ile Leu Asp Tyr Ala Ile Asn Thr Lys Ser Asn Gly Arg Tyr Thr Ala
50 55 60
Thr Leu Asp Ala Asp Thr Lys Gln Ser Ser Leu His Ile Thr Ala Ser
65 70 75 80
Gln Leu Ser Asp Ser Ala Ser Tyr Ile Cys Val Val Asn Arg Ala Asp
85 90 95
Gly Leu Tyr Ile Pro Thr Phe Gly Arg Gly Thr Ser Leu Ile Val His
100 105 110
Pro Tyr Ile Gln Lys Pro Asp Pro Ala Val Tyr Gln Leu Arg Asp Ser
115 120 125
Lys Ser Ser Asp Lys Ser Val Cys Leu Phe Thr Asp Phe Asp Ser Gln
130 135 140
Thr Asn Val Ser Gln Ser Lys Asp Ser Asp Val Tyr Ile Thr Asp Lys
145 150 155 160
Cys Val Leu Asp Met Arg Ser Met Asp Phe Lys Ser Asn Ser Ala Val
165 170 175
Ala Trp Ser Asn Lys Ser Asp Phe Ala Cys Ala Asn Ala Phe Asn Asn
180 185 190
Ser Ile Ile Pro Glu Asp Thr
195
<210> 39
<211> 199
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> альфа-цепь (a13ka)
<400> 39
Ala Asn Gln Val Glu Gln Ser Pro Gln Ser Leu Ile Ile Leu Glu Gly
1 5 10 15
Lys Asn Val Thr Leu Gln Cys Asn Tyr Thr Val Ser Pro Phe Ser Asn
20 25 30
Leu Arg Trp Tyr Lys Gln Asp Thr Gly Arg Gly Pro Val Ser Leu Thr
35 40 45
Ile Met Thr Phe Ser Glu Asn Thr Lys Ser Asn Gly Arg Tyr Thr Ala
50 55 60
Thr Leu Asp Ala Asp Thr Lys Gln Ser Ser Leu His Ile Thr Ala Ser
65 70 75 80
Gln Leu Ser Asp Ser Ala Ser Tyr Ile Cys Val Val Asn Ser Ala Asn
85 90 95
Gly Leu Tyr Ile Pro Thr Phe Gly Arg Gly Thr Ser Leu Ile Val His
100 105 110
Pro Tyr Ile Gln Lys Pro Asp Pro Ala Val Tyr Gln Leu Arg Asp Ser
115 120 125
Lys Ser Ser Asp Lys Ser Val Cys Leu Phe Thr Asp Phe Asp Ser Gln
130 135 140
Thr Asn Val Ser Gln Ser Lys Asp Ser Asp Val Tyr Ile Thr Asp Lys
145 150 155 160
Cys Val Leu Asp Met Arg Ser Met Asp Phe Lys Ser Asn Ser Ala Val
165 170 175
Ala Trp Ser Asn Lys Ser Asp Phe Ala Cys Ala Asn Ala Phe Asn Asn
180 185 190
Ser Ile Ile Pro Glu Asp Thr
195
<210> 40
<211> 199
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> альфа-цепь (a13kaLQ)
<400> 40
Ala Asn Gln Val Glu Gln Ser Pro Gln Ser Leu Ile Ile Leu Glu Gly
1 5 10 15
Lys Asn Val Thr Leu Gln Cys Asn Tyr Thr Val Ser Pro Phe Ser Asn
20 25 30
Leu Arg Trp Tyr Lys Gln Asp Thr Gly Arg Gly Pro Val Ser Leu Thr
35 40 45
Ile Leu Thr Phe Ser Glu Asn Thr Lys Ser Asn Gly Arg Tyr Thr Ala
50 55 60
Thr Leu Asp Ala Asp Thr Lys Gln Ser Ser Leu His Ile Thr Ala Ser
65 70 75 80
Gln Leu Ser Asp Ser Ala Ser Tyr Ile Cys Val Val Asn Ser Ala Gln
85 90 95
Gly Leu Tyr Ile Pro Thr Phe Gly Arg Gly Thr Ser Leu Ile Val His
100 105 110
Pro Tyr Ile Gln Lys Pro Asp Pro Ala Val Tyr Gln Leu Arg Asp Ser
115 120 125
Lys Ser Ser Asp Lys Ser Val Cys Leu Phe Thr Asp Phe Asp Ser Gln
130 135 140
Thr Asn Val Ser Gln Ser Lys Asp Ser Asp Val Tyr Ile Thr Asp Lys
145 150 155 160
Cys Val Leu Asp Met Arg Ser Met Asp Phe Lys Ser Asn Ser Ala Val
165 170 175
Ala Trp Ser Asn Lys Ser Asp Phe Ala Cys Ala Asn Ala Phe Asn Asn
180 185 190
Ser Ile Ile Pro Glu Asp Thr
195
<210> 41
<211> 199
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> альфа-цепь (a13kaLS)
<400> 41
Ala Asn Gln Val Glu Gln Ser Pro Gln Ser Leu Ile Ile Leu Glu Gly
1 5 10 15
Lys Asn Val Thr Leu Gln Cys Asn Tyr Thr Val Ser Pro Phe Ser Asn
20 25 30
Leu Arg Trp Tyr Lys Gln Asp Thr Gly Arg Gly Pro Val Ser Leu Thr
35 40 45
Ile Leu Thr Phe Ser Glu Asn Thr Lys Ser Asn Gly Arg Tyr Thr Ala
50 55 60
Thr Leu Asp Ala Asp Thr Lys Gln Ser Ser Leu His Ile Thr Ala Ser
65 70 75 80
Gln Leu Ser Asp Ser Ala Ser Tyr Ile Cys Val Val Asn Ser Ala Ser
85 90 95
Gly Leu Tyr Ile Pro Thr Phe Gly Arg Gly Thr Ser Leu Ile Val His
100 105 110
Pro Tyr Ile Gln Lys Pro Asp Pro Ala Val Tyr Gln Leu Arg Asp Ser
115 120 125
Lys Ser Ser Asp Lys Ser Val Cys Leu Phe Thr Asp Phe Asp Ser Gln
130 135 140
Thr Asn Val Ser Gln Ser Lys Asp Ser Asp Val Tyr Ile Thr Asp Lys
145 150 155 160
Cys Val Leu Asp Met Arg Ser Met Asp Phe Lys Ser Asn Ser Ala Val
165 170 175
Ala Trp Ser Asn Lys Ser Asp Phe Ala Cys Ala Asn Ala Phe Asn Asn
180 185 190
Ser Ile Ile Pro Glu Asp Thr
195
<210> 42
<211> 504
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> бета-цепь (b14)
<400> 42
Ala Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Ile Arg Asn Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Tyr Thr Ser Arg Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Tyr Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Gly Asn Thr Leu Pro Trp
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Gly Gly Gly Gly Ser
100 105 110
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
115 120 125
Gly Gly Ser Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln
130 135 140
Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Ser Phe
145 150 155 160
Thr Gly Tyr Thr Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu
165 170 175
Glu Trp Val Ala Leu Ile Asn Pro Tyr Lys Gly Val Ser Thr Tyr Asn
180 185 190
Gln Lys Phe Lys Asp Arg Phe Thr Ile Ser Val Asp Lys Ser Lys Asn
195 200 205
Thr Ala Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val
210 215 220
Tyr Tyr Cys Ala Arg Ser Gly Tyr Tyr Gly Asp Ser Asp Trp Tyr Phe
225 230 235 240
Asp Val Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Gly Gly Gly
245 250 255
Gly Ser Asp Val Lys Val Thr Gln Ser Ser Arg Tyr Leu Val Lys Arg
260 265 270
Thr Gly Glu Lys Val Phe Leu Glu Cys Val Gln Asp Ala Pro Leu Ser
275 280 285
Lys Met Phe Trp Tyr Arg Gln Asp Pro Gly Leu Gly Leu Arg Leu Ile
290 295 300
Tyr Phe Ser Tyr Asp Val Lys Met Lys Glu Lys Gly Asp Ile Pro Glu
305 310 315 320
Gly Tyr Ser Val Ser Arg Glu Lys Lys Glu Arg Phe Ser Leu Ile Leu
325 330 335
Glu Ser Ala Ser Thr Asn Gln Thr Ser Met Tyr Leu Cys Ala Ser Ser
340 345 350
Ser Asp Gln Asn Ser Gly Asp Pro Tyr Glu Gln Tyr Phe Gly Pro Gly
355 360 365
Thr Arg Leu Thr Val Thr Glu Asp Leu Lys Asn Val Phe Pro Pro Glu
370 375 380
Val Ala Val Phe Glu Pro Ser Glu Ala Glu Ile Ser His Thr Gln Lys
385 390 395 400
Ala Thr Leu Val Cys Leu Ala Thr Gly Phe Tyr Pro Asp His Val Glu
405 410 415
Leu Ser Trp Trp Val Asn Gly Lys Glu Val His Ser Gly Val Cys Thr
420 425 430
Asp Pro Gln Pro Leu Lys Glu Gln Pro Ala Leu Asn Asp Ser Arg Tyr
435 440 445
Ala Leu Ser Ser Arg Leu Arg Val Ser Ala Thr Phe Trp Gln Asp Pro
450 455 460
Arg Asn His Phe Arg Cys Gln Val Gln Phe Tyr Gly Leu Ser Glu Asn
465 470 475 480
Asp Glu Trp Thr Gln Asp Arg Ala Lys Pro Val Thr Gln Ile Val Ser
485 490 495
Ala Glu Ala Trp Gly Arg Ala Asp
500
<210> 43
<211> 504
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> бета-цепь (b14L)
<400> 43
Ala Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Ile Arg Asn Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Tyr Thr Ser Arg Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Tyr Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Gly Asn Thr Leu Pro Trp
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Gly Gly Gly Gly Ser
100 105 110
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
115 120 125
Gly Gly Ser Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln
130 135 140
Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Ser Phe
145 150 155 160
Thr Gly Tyr Thr Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu
165 170 175
Glu Trp Val Ala Leu Ile Asn Pro Tyr Lys Gly Val Ser Thr Tyr Asn
180 185 190
Gln Lys Phe Lys Asp Arg Phe Thr Ile Ser Val Asp Lys Ser Lys Asn
195 200 205
Thr Ala Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val
210 215 220
Tyr Tyr Cys Ala Arg Ser Gly Tyr Tyr Gly Asp Ser Asp Trp Tyr Phe
225 230 235 240
Asp Val Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Gly Gly Gly
245 250 255
Gly Ser Asp Val Lys Val Thr Gln Ser Ser Arg Tyr Leu Val Lys Arg
260 265 270
Thr Gly Glu Lys Val Phe Leu Glu Cys Val Gln Asp Ala Pro Leu Ser
275 280 285
Lys Met Phe Trp Tyr Arg Gln Asp Pro Gly Leu Gly Leu Arg Leu Ile
290 295 300
Tyr Phe Ser Tyr Asp Val Lys Leu Lys Glu Lys Gly Asp Ile Pro Glu
305 310 315 320
Gly Tyr Ser Val Ser Arg Glu Lys Lys Glu Arg Phe Ser Leu Ile Leu
325 330 335
Glu Ser Ala Ser Thr Asn Gln Thr Ser Met Tyr Leu Cys Ala Ser Ser
340 345 350
Ser Asp Gln Asn Ser Gly Asp Pro Tyr Glu Gln Tyr Phe Gly Pro Gly
355 360 365
Thr Arg Leu Thr Val Thr Glu Asp Leu Lys Asn Val Phe Pro Pro Glu
370 375 380
Val Ala Val Phe Glu Pro Ser Glu Ala Glu Ile Ser His Thr Gln Lys
385 390 395 400
Ala Thr Leu Val Cys Leu Ala Thr Gly Phe Tyr Pro Asp His Val Glu
405 410 415
Leu Ser Trp Trp Val Asn Gly Lys Glu Val His Ser Gly Val Cys Thr
420 425 430
Asp Pro Gln Pro Leu Lys Glu Gln Pro Ala Leu Asn Asp Ser Arg Tyr
435 440 445
Ala Leu Ser Ser Arg Leu Arg Val Ser Ala Thr Phe Trp Gln Asp Pro
450 455 460
Arg Asn His Phe Arg Cys Gln Val Gln Phe Tyr Gly Leu Ser Glu Asn
465 470 475 480
Asp Glu Trp Thr Gln Asp Arg Ala Lys Pro Val Thr Gln Ile Val Ser
485 490 495
Ala Glu Ala Trp Gly Arg Ala Asp
500
<210> 44
<211> 504
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> бета-цепь (b21)
<400> 44
Ala Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Ile Arg Asn Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Tyr Thr Ser Arg Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Tyr Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Gly Asn Thr Leu Pro Trp
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Gly Gly Gly Gly Ser
100 105 110
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
115 120 125
Gly Gly Ser Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln
130 135 140
Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Ser Phe
145 150 155 160
Thr Gly Tyr Thr Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu
165 170 175
Glu Trp Val Ala Leu Ile Asn Pro Tyr Lys Gly Val Ser Thr Tyr Asn
180 185 190
Gln Lys Phe Lys Asp Arg Phe Thr Ile Ser Val Asp Lys Ser Lys Asn
195 200 205
Thr Ala Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val
210 215 220
Tyr Tyr Cys Ala Arg Ser Gly Tyr Tyr Gly Asp Ser Asp Trp Tyr Phe
225 230 235 240
Asp Val Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Gly Gly Gly
245 250 255
Gly Ser Asp Val Lys Val Thr Gln Ser Ser Arg Tyr Leu Val Lys Arg
260 265 270
Thr Gly Glu Lys Val Phe Leu Glu Cys Val Gln Asp Met Asp His Glu
275 280 285
Asn Met Phe Trp Tyr Arg Gln Asp Pro Gly Leu Gly Leu Arg Leu Ile
290 295 300
Tyr Phe Ser Arg Phe Ala Thr Gly Lys Glu Lys Gly Asp Ile Pro Glu
305 310 315 320
Gly Tyr Ser Val Ser Arg Glu Lys Lys Glu Arg Phe Ser Leu Ile Leu
325 330 335
Glu Ser Ala Ser Thr Asn Gln Thr Ser Met Tyr Leu Cys Ala Ser Ser
340 345 350
Ser Asp Gln Asn Ser Gly Asp Pro Tyr Glu Gln Tyr Phe Gly Pro Gly
355 360 365
Thr Arg Leu Thr Val Thr Glu Asp Leu Lys Asn Val Phe Pro Pro Glu
370 375 380
Val Ala Val Phe Glu Pro Ser Glu Ala Glu Ile Ser His Thr Gln Lys
385 390 395 400
Ala Thr Leu Val Cys Leu Ala Thr Gly Phe Tyr Pro Asp His Val Glu
405 410 415
Leu Ser Trp Trp Val Asn Gly Lys Glu Val His Ser Gly Val Cys Thr
420 425 430
Asp Pro Gln Pro Leu Lys Glu Gln Pro Ala Leu Asn Asp Ser Arg Tyr
435 440 445
Ala Leu Ser Ser Arg Leu Arg Val Ser Ala Thr Phe Trp Gln Asp Pro
450 455 460
Arg Asn His Phe Arg Cys Gln Val Gln Phe Tyr Gly Leu Ser Glu Asn
465 470 475 480
Asp Glu Trp Thr Gln Asp Arg Ala Lys Pro Val Thr Gln Ile Val Ser
485 490 495
Ala Glu Ala Trp Gly Arg Ala Asp
500
<210> 45
<211> 504
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> бета-цепь (b21L)
<400> 45
Ala Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Ile Arg Asn Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Tyr Thr Ser Arg Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Tyr Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Gly Asn Thr Leu Pro Trp
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Gly Gly Gly Gly Ser
100 105 110
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
115 120 125
Gly Gly Ser Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln
130 135 140
Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Ser Phe
145 150 155 160
Thr Gly Tyr Thr Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu
165 170 175
Glu Trp Val Ala Leu Ile Asn Pro Tyr Lys Gly Val Ser Thr Tyr Asn
180 185 190
Gln Lys Phe Lys Asp Arg Phe Thr Ile Ser Val Asp Lys Ser Lys Asn
195 200 205
Thr Ala Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val
210 215 220
Tyr Tyr Cys Ala Arg Ser Gly Tyr Tyr Gly Asp Ser Asp Trp Tyr Phe
225 230 235 240
Asp Val Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Gly Gly Gly
245 250 255
Gly Ser Asp Val Lys Val Thr Gln Ser Ser Arg Tyr Leu Val Lys Arg
260 265 270
Thr Gly Glu Lys Val Phe Leu Glu Cys Val Gln Asp Leu Asp His Glu
275 280 285
Asn Met Phe Trp Tyr Arg Gln Asp Pro Gly Leu Gly Leu Arg Leu Ile
290 295 300
Tyr Phe Ser Arg Phe Ala Thr Gly Lys Glu Lys Gly Asp Ile Pro Glu
305 310 315 320
Gly Tyr Ser Val Ser Arg Glu Lys Lys Glu Arg Phe Ser Leu Ile Leu
325 330 335
Glu Ser Ala Ser Thr Asn Gln Thr Ser Met Tyr Leu Cys Ala Ser Ser
340 345 350
Ser Asp Gln Asn Ser Gly Asp Pro Tyr Glu Gln Tyr Phe Gly Pro Gly
355 360 365
Thr Arg Leu Thr Val Thr Glu Asp Leu Lys Asn Val Phe Pro Pro Glu
370 375 380
Val Ala Val Phe Glu Pro Ser Glu Ala Glu Ile Ser His Thr Gln Lys
385 390 395 400
Ala Thr Leu Val Cys Leu Ala Thr Gly Phe Tyr Pro Asp His Val Glu
405 410 415
Leu Ser Trp Trp Val Asn Gly Lys Glu Val His Ser Gly Val Cys Thr
420 425 430
Asp Pro Gln Pro Leu Lys Glu Gln Pro Ala Leu Asn Asp Ser Arg Tyr
435 440 445
Ala Leu Ser Ser Arg Leu Arg Val Ser Ala Thr Phe Trp Gln Asp Pro
450 455 460
Arg Asn His Phe Arg Cys Gln Val Gln Phe Tyr Gly Leu Ser Glu Asn
465 470 475 480
Asp Glu Trp Thr Gln Asp Arg Ala Lys Pro Val Thr Gln Ile Val Ser
485 490 495
Ala Glu Ala Trp Gly Arg Ala Asp
500
<210> 46
<211> 113
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> мутированная альфа-цепь(a36)
<400> 46
Ala Asn Gln Val Glu Gln Ser Pro Gln Ser Leu Ile Ile Leu Glu Gly
1 5 10 15
Lys Asn Val Thr Leu Gln Cys Asn Tyr Thr Val Ser Pro Phe Ser Asn
20 25 30
Leu Arg Trp Tyr Lys Gln Asp Thr Gly Arg Gly Pro Val Ser Leu Thr
35 40 45
Ile Met Thr Phe Ser Glu Asn Thr Lys Ser Asn Gly Arg Tyr Thr Ala
50 55 60
Thr Leu Asp Ala Asp Thr Lys Gln Ser Ser Leu His Ile Thr Ala Ser
65 70 75 80
Gln Leu Ser Asp Ser Ala Ser Tyr Ile Cys Val Val Asn Ser Ala Gln
85 90 95
Gly Leu Tyr Ile Pro Thr Phe Gly Arg Gly Thr Ser Leu Ile Val His
100 105 110
Pro
<210> 47
<211> 113
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> мутированная альфа-цепь(a37)
<400> 47
Ala Asn Gln Val Glu Gln Ser Pro Gln Ser Leu Ile Ile Leu Glu Gly
1 5 10 15
Lys Asn Val Thr Leu Gln Cys Asn Tyr Thr Val Ser Pro Phe Ser Asn
20 25 30
Leu Arg Trp Tyr Lys Gln Asp Thr Gly Arg Gly Pro Val Ser Leu Thr
35 40 45
Ile Leu Thr Tyr Ser Glu Asn Thr Lys Ser Asn Gly Arg Tyr Thr Ala
50 55 60
Thr Leu Asp Ala Asp Thr Lys Gln Ser Ser Leu His Ile Thr Ala Ser
65 70 75 80
Gln Leu Ser Asp Ser Ala Ser Tyr Ile Cys Val Val Asn Ser Ala Gln
85 90 95
Gly Leu Tyr Ile Pro Thr Phe Gly Arg Gly Thr Ser Leu Ile Val His
100 105 110
Pro
<210> 48
<211> 113
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> мутированная альфа-цепь(a38)
<400> 48
Ala Asn Gln Val Glu Gln Ser Pro Gln Ser Leu Ile Ile Leu Glu Gly
1 5 10 15
Lys Asn Val Thr Leu Gln Cys Asn Tyr Thr Val Ser Pro Phe Ser Asn
20 25 30
Leu Arg Trp Tyr Lys Gln Asp Thr Gly Arg Gly Pro Val Ser Leu Thr
35 40 45
Ile Leu Thr Phe Ser Glu Asn Thr Lys Ser Asn Gly Arg Tyr Thr Ala
50 55 60
Thr Leu Asp Ala Asp Thr Lys Gln Ser Ser Leu His Ile Thr Ala Ser
65 70 75 80
Gln Leu Ser Asp Ser Ala Ser Tyr Ile Cys Val Val Asn His Ala Gln
85 90 95
Gly Leu Tyr Ile Pro Thr Phe Gly Arg Gly Thr Ser Leu Ile Val His
100 105 110
Pro
<210> 49
<211> 113
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> мутированная альфа-цепь (a39)
<400> 49
Ala Asn Gln Val Glu Gln Ser Pro Gln Ser Leu Ile Ile Leu Glu Gly
1 5 10 15
Lys Asn Val Thr Leu Gln Cys Asn Tyr Thr Val Ser Pro Phe Ser Asn
20 25 30
Leu Arg Trp Tyr Lys Gln Asp Thr Gly Arg Gly Pro Val Ser Leu Thr
35 40 45
Ile Leu Thr Phe Ser Glu Asn Thr Lys Ser Asn Gly Arg Tyr Thr Ala
50 55 60
Thr Leu Asp Ala Asp Thr Lys Gln Ser Ser Leu His Ile Thr Ala Ser
65 70 75 80
Gln Leu Ser Asp Ser Ala Ser Tyr Ile Cys Val Val Asn Ser Ser Gln
85 90 95
Gly Leu Tyr Ile Pro Thr Phe Gly Arg Gly Thr Ser Leu Ile Val His
100 105 110
Pro
<210> 50
<211> 113
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> мутированная альфа-цепь(a40)
<400> 50
Ala Asn Gln Val Glu Gln Ser Pro Gln Ser Leu Ile Ile Leu Glu Gly
1 5 10 15
Lys Asn Val Thr Leu Gln Cys Asn Tyr Thr Val Ser Pro Phe Ser Asn
20 25 30
Leu Arg Trp Tyr Lys Gln Asp Thr Gly Arg Gly Pro Val Ser Leu Thr
35 40 45
Ile Leu Thr Phe Ser Glu Asn Thr Lys Ser Asn Gly Arg Tyr Thr Ala
50 55 60
Thr Leu Asp Ala Asp Thr Lys Gln Ser Ser Leu His Ile Thr Ala Ser
65 70 75 80
Gln Leu Ser Asp Ser Ala Ser Tyr Ile Cys Val Val Asn Ser Ala Gly
85 90 95
Gly Leu Tyr Ile Pro Thr Phe Gly Arg Gly Thr Ser Leu Ile Val His
100 105 110
Pro
<210> 51
<211> 113
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> мутированная альфа-цепь(a41)
<400> 51
Ala Asn Gln Val Glu Gln Ser Pro Gln Ser Leu Ile Ile Leu Glu Gly
1 5 10 15
Lys Asn Val Thr Leu Gln Cys Asn Tyr Thr Val Ser Pro Phe Ser Asn
20 25 30
Leu Arg Trp Tyr Lys Gln Asp Thr Gly Arg Gly Pro Val Ser Leu Thr
35 40 45
Ile Leu Thr Phe Ser Glu Asn Thr Lys Ser Asn Gly Arg Tyr Thr Ala
50 55 60
Thr Leu Asp Ala Asp Thr Lys Gln Ser Ser Leu His Ile Thr Ala Ser
65 70 75 80
Gln Leu Ser Asp Ser Ala Ser Tyr Ile Cys Val Val Asn Ser Ala Gln
85 90 95
Gly Ser Tyr Ile Pro Thr Phe Gly Arg Gly Thr Ser Leu Ile Val His
100 105 110
Pro
<210> 52
<211> 113
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> мутированная альфа-цепь(a30)
<400> 52
Ala Asn Gln Val Glu Gln Ser Pro Gln Ser Leu Ile Ile Leu Glu Gly
1 5 10 15
Lys Asn Val Thr Leu Gln Cys Asn Tyr Thr Val Ser Pro Phe Ser Asn
20 25 30
Leu Arg Trp Tyr Lys Gln Asp Thr Gly Arg Gly Pro Val Ser Leu Thr
35 40 45
Ile Met Asp Tyr Ala Ile Asn Thr Lys Ser Asn Gly Arg Tyr Thr Ala
50 55 60
Thr Leu Asp Ala Asp Thr Lys Gln Ser Ser Leu His Ile Thr Ala Ser
65 70 75 80
Gln Leu Ser Asp Ser Ala Ser Tyr Ile Cys Val Val Asn Arg Ala Asp
85 90 95
Gly Leu Tyr Ile Pro Thr Phe Gly Arg Gly Thr Ser Leu Ile Val His
100 105 110
Pro
<210> 53
<211> 113
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> мутированная альфа-цепь(a42)
<400> 53
Ala Asn Gln Val Glu Gln Ser Pro Gln Ser Leu Ile Ile Leu Glu Gly
1 5 10 15
Lys Asn Val Thr Leu Gln Cys Asn Tyr Thr Val Ser Pro Phe Ser Asn
20 25 30
Leu Arg Trp Tyr Lys Gln Asp Thr Gly Arg Gly Pro Val Ser Leu Thr
35 40 45
Ile Leu Thr Tyr Ala Ile Asn Thr Lys Ser Asn Gly Arg Tyr Thr Ala
50 55 60
Thr Leu Asp Ala Asp Thr Lys Gln Ser Ser Leu His Ile Thr Ala Ser
65 70 75 80
Gln Leu Ser Asp Ser Ala Ser Tyr Ile Cys Val Val Asn Arg Ala Asp
85 90 95
Gly Leu Tyr Ile Pro Thr Phe Gly Arg Gly Thr Ser Leu Ile Val His
100 105 110
Pro
<210> 54
<211> 113
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> мутированная альфа-цепь(a31)
<400> 54
Ala Asn Gln Val Glu Gln Ser Pro Gln Ser Leu Ile Ile Leu Glu Gly
1 5 10 15
Lys Asn Val Thr Leu Gln Cys Asn Tyr Thr Val Ser Pro Phe Ser Asn
20 25 30
Leu Arg Trp Tyr Lys Gln Asp Thr Gly Arg Gly Pro Val Ser Leu Thr
35 40 45
Ile Leu Asp Phe Ala Ile Asn Thr Lys Ser Asn Gly Arg Tyr Thr Ala
50 55 60
Thr Leu Asp Ala Asp Thr Lys Gln Ser Ser Leu His Ile Thr Ala Ser
65 70 75 80
Gln Leu Ser Asp Ser Ala Ser Tyr Ile Cys Val Val Asn Arg Ala Asp
85 90 95
Gly Leu Tyr Ile Pro Thr Phe Gly Arg Gly Thr Ser Leu Ile Val His
100 105 110
Pro
<210> 55
<211> 113
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> мутированная альфа-цепь(a43)
<400> 55
Ala Asn Gln Val Glu Gln Ser Pro Gln Ser Leu Ile Ile Leu Glu Gly
1 5 10 15
Lys Asn Val Thr Leu Gln Cys Asn Tyr Thr Val Ser Pro Phe Ser Asn
20 25 30
Leu Arg Trp Tyr Lys Gln Asp Thr Gly Arg Gly Pro Val Ser Leu Thr
35 40 45
Ile Leu Asp Tyr Ser Ile Asn Thr Lys Ser Asn Gly Arg Tyr Thr Ala
50 55 60
Thr Leu Asp Ala Asp Thr Lys Gln Ser Ser Leu His Ile Thr Ala Ser
65 70 75 80
Gln Leu Ser Asp Ser Ala Ser Tyr Ile Cys Val Val Asn Arg Ala Asp
85 90 95
Gly Leu Tyr Ile Pro Thr Phe Gly Arg Gly Thr Ser Leu Ile Val His
100 105 110
Pro
<210> 56
<211> 113
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> мутированная альфа-цепь(a32)
<400> 56
Ala Asn Gln Val Glu Gln Ser Pro Gln Ser Leu Ile Ile Leu Glu Gly
1 5 10 15
Lys Asn Val Thr Leu Gln Cys Asn Tyr Thr Val Ser Pro Phe Ser Asn
20 25 30
Leu Arg Trp Tyr Lys Gln Asp Thr Gly Arg Gly Pro Val Ser Leu Thr
35 40 45
Ile Leu Asp Tyr Ala Glu Asn Thr Lys Ser Asn Gly Arg Tyr Thr Ala
50 55 60
Thr Leu Asp Ala Asp Thr Lys Gln Ser Ser Leu His Ile Thr Ala Ser
65 70 75 80
Gln Leu Ser Asp Ser Ala Ser Tyr Ile Cys Val Val Asn Arg Ala Asp
85 90 95
Gly Leu Tyr Ile Pro Thr Phe Gly Arg Gly Thr Ser Leu Ile Val His
100 105 110
Pro
<210> 57
<211> 113
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> мутированная альфа-цепь(a44)
<400> 57
Ala Asn Gln Val Glu Gln Ser Pro Gln Ser Leu Ile Ile Leu Glu Gly
1 5 10 15
Lys Asn Val Thr Leu Gln Cys Asn Tyr Thr Val Ser Pro Phe Ser Asn
20 25 30
Leu Arg Trp Tyr Lys Gln Asp Thr Gly Arg Gly Pro Val Ser Leu Thr
35 40 45
Ile Leu Asp Tyr Ala Ile Asn Thr Lys Ser Asn Gly Arg Tyr Thr Ala
50 55 60
Thr Leu Asp Ala Asp Thr Lys Gln Ser Ser Leu His Ile Thr Ala Ser
65 70 75 80
Gln Leu Ser Asp Ser Ala Ser Tyr Ile Cys Val Val Asn His Ala Asp
85 90 95
Gly Leu Tyr Ile Pro Thr Phe Gly Arg Gly Thr Ser Leu Ile Val His
100 105 110
Pro
<210> 58
<211> 113
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> мутированная альфа-цепь(a33)
<400> 58
Ala Asn Gln Val Glu Gln Ser Pro Gln Ser Leu Ile Ile Leu Glu Gly
1 5 10 15
Lys Asn Val Thr Leu Gln Cys Asn Tyr Thr Val Ser Pro Phe Ser Asn
20 25 30
Leu Arg Trp Tyr Lys Gln Asp Thr Gly Arg Gly Pro Val Ser Leu Thr
35 40 45
Ile Leu Asp Tyr Ala Ile Asn Thr Lys Ser Asn Gly Arg Tyr Thr Ala
50 55 60
Thr Leu Asp Ala Asp Thr Lys Gln Ser Ser Leu His Ile Thr Ala Ser
65 70 75 80
Gln Leu Ser Asp Ser Ala Ser Tyr Ile Cys Val Val Asn Arg Ser Asp
85 90 95
Gly Leu Tyr Ile Pro Thr Phe Gly Arg Gly Thr Ser Leu Ile Val His
100 105 110
Pro
<210> 59
<211> 113
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> мутированная альфа-цепь(a45)
<400> 59
Ala Asn Gln Val Glu Gln Ser Pro Gln Ser Leu Ile Ile Leu Glu Gly
1 5 10 15
Lys Asn Val Thr Leu Gln Cys Asn Tyr Thr Val Ser Pro Phe Ser Asn
20 25 30
Leu Arg Trp Tyr Lys Gln Asp Thr Gly Arg Gly Pro Val Ser Leu Thr
35 40 45
Ile Leu Asp Tyr Ala Ile Asn Thr Lys Ser Asn Gly Arg Tyr Thr Ala
50 55 60
Thr Leu Asp Ala Asp Thr Lys Gln Ser Ser Leu His Ile Thr Ala Ser
65 70 75 80
Gln Leu Ser Asp Ser Ala Ser Tyr Ile Cys Val Val Asn Arg Ala Gly
85 90 95
Gly Leu Tyr Ile Pro Thr Phe Gly Arg Gly Thr Ser Leu Ile Val His
100 105 110
Pro
<210> 60
<211> 113
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> мутированная альфа-цепь(a34)
<400> 60
Ala Asn Gln Val Glu Gln Ser Pro Gln Ser Leu Ile Ile Leu Glu Gly
1 5 10 15
Lys Asn Val Thr Leu Gln Cys Asn Tyr Thr Val Ser Pro Phe Ser Asn
20 25 30
Leu Arg Trp Tyr Lys Gln Asp Thr Gly Arg Gly Pro Val Ser Leu Thr
35 40 45
Ile Leu Asp Tyr Ala Ile Asn Thr Lys Ser Asn Gly Arg Tyr Thr Ala
50 55 60
Thr Leu Asp Ala Asp Thr Lys Gln Ser Ser Leu His Ile Thr Ala Ser
65 70 75 80
Gln Leu Ser Asp Ser Ala Ser Tyr Ile Cys Val Val Asn Arg Ala Asp
85 90 95
Gly Ser Tyr Ile Pro Thr Phe Gly Arg Gly Thr Ser Leu Ile Val His
100 105 110
Pro
<210> 61
<211> 113
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> мутированная альфа-цепь(aWTka)
<400> 61
Ala Asn Gln Val Glu Gln Ser Pro Gln Ser Leu Ile Ile Leu Glu Gly
1 5 10 15
Lys Asn Val Thr Leu Gln Cys Asn Tyr Thr Val Ser Pro Phe Ser Asn
20 25 30
Leu Arg Trp Tyr Lys Gln Asp Thr Gly Arg Gly Pro Val Ser Leu Thr
35 40 45
Ile Met Thr Phe Ser Glu Asn Thr Lys Ser Asn Gly Arg Tyr Thr Ala
50 55 60
Thr Leu Asp Ala Asp Thr Lys Gln Ser Ser Leu His Ile Thr Ala Ser
65 70 75 80
Gln Leu Ser Asp Ser Ala Ser Tyr Ile Cys Val Val Asn His Ser Gly
85 90 95
Gly Ser Tyr Ile Pro Thr Phe Gly Arg Gly Thr Ser Leu Ile Val His
100 105 110
Pro
<210> 62
<211> 113
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> мутированная альфа-цепь(aM50L)
<400> 62
Ala Asn Gln Val Glu Gln Ser Pro Gln Ser Leu Ile Ile Leu Glu Gly
1 5 10 15
Lys Asn Val Thr Leu Gln Cys Asn Tyr Thr Val Ser Pro Phe Ser Asn
20 25 30
Leu Arg Trp Tyr Lys Gln Asp Thr Gly Arg Gly Pro Val Ser Leu Thr
35 40 45
Ile Leu Thr Phe Ser Glu Asn Thr Lys Ser Asn Gly Arg Tyr Thr Ala
50 55 60
Thr Leu Asp Ala Asp Thr Lys Gln Ser Ser Leu His Ile Thr Ala Ser
65 70 75 80
Gln Leu Ser Asp Ser Ala Ser Tyr Ile Cys Val Val Asn His Ser Gly
85 90 95
Gly Ser Tyr Ile Pro Thr Phe Gly Arg Gly Thr Ser Leu Ile Val His
100 105 110
Pro
<210> 63
<211> 113
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> мутированная альфа-цепь(aS95A)
<400> 63
Ala Asn Gln Val Glu Gln Ser Pro Gln Ser Leu Ile Ile Leu Glu Gly
1 5 10 15
Lys Asn Val Thr Leu Gln Cys Asn Tyr Thr Val Ser Pro Phe Ser Asn
20 25 30
Leu Arg Trp Tyr Lys Gln Asp Thr Gly Arg Gly Pro Val Ser Leu Thr
35 40 45
Ile Met Thr Phe Ser Glu Asn Thr Lys Ser Asn Gly Arg Tyr Thr Ala
50 55 60
Thr Leu Asp Ala Asp Thr Lys Gln Ser Ser Leu His Ile Thr Ala Ser
65 70 75 80
Gln Leu Ser Asp Ser Ala Ser Tyr Ile Cys Val Val Asn His Ala Gly
85 90 95
Gly Ser Tyr Ile Pro Thr Phe Gly Arg Gly Thr Ser Leu Ile Val His
100 105 110
Pro
<210> 64
<211> 113
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> мутированная альфа-цепь(aS98L)
<400> 64
Ala Asn Gln Val Glu Gln Ser Pro Gln Ser Leu Ile Ile Leu Glu Gly
1 5 10 15
Lys Asn Val Thr Leu Gln Cys Asn Tyr Thr Val Ser Pro Phe Ser Asn
20 25 30
Leu Arg Trp Tyr Lys Gln Asp Thr Gly Arg Gly Pro Val Ser Leu Thr
35 40 45
Ile Met Thr Phe Ser Glu Asn Thr Lys Ser Asn Gly Arg Tyr Thr Ala
50 55 60
Thr Leu Asp Ala Asp Thr Lys Gln Ser Ser Leu His Ile Thr Ala Ser
65 70 75 80
Gln Leu Ser Asp Ser Ala Ser Tyr Ile Cys Val Val Asn His Ser Gly
85 90 95
Gly Leu Tyr Ile Pro Thr Phe Gly Arg Gly Thr Ser Leu Ile Val His
100 105 110
Pro
<210> 65
<211> 116
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> мутированная бета-цепь (b41)
<400> 65
Asp Val Lys Val Thr Gln Ser Ser Arg Tyr Leu Val Lys Arg Thr Gly
1 5 10 15
Glu Lys Val Phe Leu Glu Cys Val Gln Asp Met Asp His Glu Asn Met
20 25 30
Phe Trp Tyr Arg Gln Asp Pro Gly Leu Gly Leu Arg Leu Ile Tyr Phe
35 40 45
Ser Arg Phe Ala Thr Gly Lys Glu Lys Gly Asp Ile Pro Glu Gly Tyr
50 55 60
Ser Val Ser Arg Glu Lys Lys Glu Arg Phe Ser Leu Ile Leu Glu Ser
65 70 75 80
Ala Ser Thr Asn Gln Thr Ser Met Tyr Leu Cys Ala Ser Ser Ser Asp
85 90 95
Gln Asn Ser Gly Asp Pro Tyr Glu Gln Tyr Phe Gly Pro Gly Thr Arg
100 105 110
Leu Thr Val Thr
115
<210> 66
<211> 116
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> мутированная бета-цепь (b42)
<400> 66
Asp Val Lys Val Thr Gln Ser Ser Arg Tyr Leu Val Lys Arg Thr Gly
1 5 10 15
Glu Lys Val Phe Leu Glu Cys Val Gln Asp Leu Asp His Glu Asn Met
20 25 30
Phe Trp Tyr Arg Gln Asp Pro Gly Leu Gly Leu Arg Leu Ile Tyr Phe
35 40 45
Ser Tyr Phe Ala Thr Gly Lys Glu Lys Gly Asp Ile Pro Glu Gly Tyr
50 55 60
Ser Val Ser Arg Glu Lys Lys Glu Arg Phe Ser Leu Ile Leu Glu Ser
65 70 75 80
Ala Ser Thr Asn Gln Thr Ser Met Tyr Leu Cys Ala Ser Ser Ser Asp
85 90 95
Gln Asn Ser Gly Asp Pro Tyr Glu Gln Tyr Phe Gly Pro Gly Thr Arg
100 105 110
Leu Thr Val Thr
115
<210> 67
<211> 116
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> мутированная бета-цепь (b43)
<400> 67
Asp Val Lys Val Thr Gln Ser Ser Arg Tyr Leu Val Lys Arg Thr Gly
1 5 10 15
Glu Lys Val Phe Leu Glu Cys Val Gln Asp Leu Asp His Glu Asn Met
20 25 30
Phe Trp Tyr Arg Gln Asp Pro Gly Leu Gly Leu Arg Leu Ile Tyr Phe
35 40 45
Ser Arg Asp Ala Thr Gly Lys Glu Lys Gly Asp Ile Pro Glu Gly Tyr
50 55 60
Ser Val Ser Arg Glu Lys Lys Glu Arg Phe Ser Leu Ile Leu Glu Ser
65 70 75 80
Ala Ser Thr Asn Gln Thr Ser Met Tyr Leu Cys Ala Ser Ser Ser Asp
85 90 95
Gln Asn Ser Gly Asp Pro Tyr Glu Gln Tyr Phe Gly Pro Gly Thr Arg
100 105 110
Leu Thr Val Thr
115
<210> 68
<211> 116
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> мутированная бета-цепь (b44)
<400> 68
Asp Val Lys Val Thr Gln Ser Ser Arg Tyr Leu Val Lys Arg Thr Gly
1 5 10 15
Glu Lys Val Phe Leu Glu Cys Val Gln Asp Leu Asp His Glu Asn Met
20 25 30
Phe Trp Tyr Arg Gln Asp Pro Gly Leu Gly Leu Arg Leu Ile Tyr Phe
35 40 45
Ser Arg Phe Val Thr Gly Lys Glu Lys Gly Asp Ile Pro Glu Gly Tyr
50 55 60
Ser Val Ser Arg Glu Lys Lys Glu Arg Phe Ser Leu Ile Leu Glu Ser
65 70 75 80
Ala Ser Thr Asn Gln Thr Ser Met Tyr Leu Cys Ala Ser Ser Ser Asp
85 90 95
Gln Asn Ser Gly Asp Pro Tyr Glu Gln Tyr Phe Gly Pro Gly Thr Arg
100 105 110
Leu Thr Val Thr
115
<210> 69
<211> 116
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> мутированная бета-цепь (b45)
<400> 69
Asp Val Lys Val Thr Gln Ser Ser Arg Tyr Leu Val Lys Arg Thr Gly
1 5 10 15
Glu Lys Val Phe Leu Glu Cys Val Gln Asp Leu Asp His Glu Asn Met
20 25 30
Phe Trp Tyr Arg Gln Asp Pro Gly Leu Gly Leu Arg Leu Ile Tyr Phe
35 40 45
Ser Arg Phe Ala Lys Gly Lys Glu Lys Gly Asp Ile Pro Glu Gly Tyr
50 55 60
Ser Val Ser Arg Glu Lys Lys Glu Arg Phe Ser Leu Ile Leu Glu Ser
65 70 75 80
Ala Ser Thr Asn Gln Thr Ser Met Tyr Leu Cys Ala Ser Ser Ser Asp
85 90 95
Gln Asn Ser Gly Asp Pro Tyr Glu Gln Tyr Phe Gly Pro Gly Thr Arg
100 105 110
Leu Thr Val Thr
115
<210> 70
<211> 116
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> мутированная бета-цепь (b46)
<400> 70
Asp Val Lys Val Thr Gln Ser Ser Arg Tyr Leu Val Lys Arg Thr Gly
1 5 10 15
Glu Lys Val Phe Leu Glu Cys Val Gln Asp Leu Asp His Glu Asn Met
20 25 30
Phe Trp Tyr Arg Gln Asp Pro Gly Leu Gly Leu Arg Leu Ile Tyr Phe
35 40 45
Ser Arg Phe Ala Thr Met Lys Glu Lys Gly Asp Ile Pro Glu Gly Tyr
50 55 60
Ser Val Ser Arg Glu Lys Lys Glu Arg Phe Ser Leu Ile Leu Glu Ser
65 70 75 80
Ala Ser Thr Asn Gln Thr Ser Met Tyr Leu Cys Ala Ser Ser Ser Asp
85 90 95
Gln Asn Ser Gly Asp Pro Tyr Glu Gln Tyr Phe Gly Pro Gly Thr Arg
100 105 110
Leu Thr Val Thr
115
<210> 71
<211> 116
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> мутированная бета-цепь (b32)
<400> 71
Asp Val Lys Val Thr Gln Ser Ser Arg Tyr Leu Val Lys Arg Thr Gly
1 5 10 15
Glu Lys Val Phe Leu Glu Cys Val Gln Asp Met Pro Leu Ser Lys Met
20 25 30
Phe Trp Tyr Arg Gln Asp Pro Gly Leu Gly Leu Arg Leu Ile Tyr Phe
35 40 45
Ser Tyr Asp Val Lys Met Lys Glu Lys Gly Asp Ile Pro Glu Gly Tyr
50 55 60
Ser Val Ser Arg Glu Lys Lys Glu Arg Phe Ser Leu Ile Leu Glu Ser
65 70 75 80
Ala Ser Thr Asn Gln Thr Ser Met Tyr Leu Cys Ala Ser Ser Ser Asp
85 90 95
Gln Asn Ser Gly Asp Pro Tyr Glu Gln Tyr Phe Gly Pro Gly Thr Arg
100 105 110
Leu Thr Val Thr
115
<210> 72
<211> 116
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> мутированная бета-цепь (b33)
<400> 72
Asp Val Lys Val Thr Gln Ser Ser Arg Tyr Leu Val Lys Arg Thr Gly
1 5 10 15
Glu Lys Val Phe Leu Glu Cys Val Gln Asp Ala Asp Leu Ser Lys Met
20 25 30
Phe Trp Tyr Arg Gln Asp Pro Gly Leu Gly Leu Arg Leu Ile Tyr Phe
35 40 45
Ser Tyr Asp Val Lys Met Lys Glu Lys Gly Asp Ile Pro Glu Gly Tyr
50 55 60
Ser Val Ser Arg Glu Lys Lys Glu Arg Phe Ser Leu Ile Leu Glu Ser
65 70 75 80
Ala Ser Thr Asn Gln Thr Ser Met Tyr Leu Cys Ala Ser Ser Ser Asp
85 90 95
Gln Asn Ser Gly Asp Pro Tyr Glu Gln Tyr Phe Gly Pro Gly Thr Arg
100 105 110
Leu Thr Val Thr
115
<210> 73
<211> 116
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> мутированная бета-цепь (b34)
<400> 73
Asp Val Lys Val Thr Gln Ser Ser Arg Tyr Leu Val Lys Arg Thr Gly
1 5 10 15
Glu Lys Val Phe Leu Glu Cys Val Gln Asp Ala Pro His Ser Lys Met
20 25 30
Phe Trp Tyr Arg Gln Asp Pro Gly Leu Gly Leu Arg Leu Ile Tyr Phe
35 40 45
Ser Tyr Asp Val Lys Met Lys Glu Lys Gly Asp Ile Pro Glu Gly Tyr
50 55 60
Ser Val Ser Arg Glu Lys Lys Glu Arg Phe Ser Leu Ile Leu Glu Ser
65 70 75 80
Ala Ser Thr Asn Gln Thr Ser Met Tyr Leu Cys Ala Ser Ser Ser Asp
85 90 95
Gln Asn Ser Gly Asp Pro Tyr Glu Gln Tyr Phe Gly Pro Gly Thr Arg
100 105 110
Leu Thr Val Thr
115
<210> 74
<211> 116
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> мутированная бета-цепь (b35)
<400> 74
Asp Val Lys Val Thr Gln Ser Ser Arg Tyr Leu Val Lys Arg Thr Gly
1 5 10 15
Glu Lys Val Phe Leu Glu Cys Val Gln Asp Ala Pro Leu Glu Lys Met
20 25 30
Phe Trp Tyr Arg Gln Asp Pro Gly Leu Gly Leu Arg Leu Ile Tyr Phe
35 40 45
Ser Tyr Asp Val Lys Met Lys Glu Lys Gly Asp Ile Pro Glu Gly Tyr
50 55 60
Ser Val Ser Arg Glu Lys Lys Glu Arg Phe Ser Leu Ile Leu Glu Ser
65 70 75 80
Ala Ser Thr Asn Gln Thr Ser Met Tyr Leu Cys Ala Ser Ser Ser Asp
85 90 95
Gln Asn Ser Gly Asp Pro Tyr Glu Gln Tyr Phe Gly Pro Gly Thr Arg
100 105 110
Leu Thr Val Thr
115
<210> 75
<211> 116
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> мутированная бета-цепь (b36)
<400> 75
Asp Val Lys Val Thr Gln Ser Ser Arg Tyr Leu Val Lys Arg Thr Gly
1 5 10 15
Glu Lys Val Phe Leu Glu Cys Val Gln Asp Ala Pro Leu Ser Asn Met
20 25 30
Phe Trp Tyr Arg Gln Asp Pro Gly Leu Gly Leu Arg Leu Ile Tyr Phe
35 40 45
Ser Tyr Asp Val Lys Met Lys Glu Lys Gly Asp Ile Pro Glu Gly Tyr
50 55 60
Ser Val Ser Arg Glu Lys Lys Glu Arg Phe Ser Leu Ile Leu Glu Ser
65 70 75 80
Ala Ser Thr Asn Gln Thr Ser Met Tyr Leu Cys Ala Ser Ser Ser Asp
85 90 95
Gln Asn Ser Gly Asp Pro Tyr Glu Gln Tyr Phe Gly Pro Gly Thr Arg
100 105 110
Leu Thr Val Thr
115
<210> 76
<211> 116
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> мутированная бета-цепь (b37)
<400> 76
Asp Val Lys Val Thr Gln Ser Ser Arg Tyr Leu Val Lys Arg Thr Gly
1 5 10 15
Glu Lys Val Phe Leu Glu Cys Val Gln Asp Ala Pro Leu Ser Lys Met
20 25 30
Phe Trp Tyr Arg Gln Asp Pro Gly Leu Gly Leu Arg Leu Ile Tyr Phe
35 40 45
Ser Tyr Asp Val Lys Met Lys Glu Lys Gly Asp Ile Pro Glu Gly Tyr
50 55 60
Ser Val Ser Arg Glu Lys Lys Glu Arg Phe Ser Leu Ile Leu Glu Ser
65 70 75 80
Ala Ser Thr Asn Gln Thr Ser Met Tyr Leu Cys Ala Ser Ser Phe Asp
85 90 95
Gln Asn Ser Gly Asp Pro Tyr Glu Gln Tyr Phe Gly Pro Gly Thr Arg
100 105 110
Leu Thr Val Thr
115
<210> 77
<211> 116
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> мутированная бета-цепь (b38)
<400> 77
Asp Val Lys Val Thr Gln Ser Ser Arg Tyr Leu Val Lys Arg Thr Gly
1 5 10 15
Glu Lys Val Phe Leu Glu Cys Val Gln Asp Ala Pro Leu Ser Lys Met
20 25 30
Phe Trp Tyr Arg Gln Asp Pro Gly Leu Gly Leu Arg Leu Ile Tyr Phe
35 40 45
Ser Tyr Asp Val Lys Met Lys Glu Lys Gly Asp Ile Pro Glu Gly Tyr
50 55 60
Ser Val Ser Arg Glu Lys Lys Glu Arg Phe Ser Leu Ile Leu Glu Ser
65 70 75 80
Ala Ser Thr Asn Gln Thr Ser Met Tyr Leu Cys Ala Ser Ser Ser Leu
85 90 95
Gln Asn Ser Gly Asp Pro Tyr Glu Gln Tyr Phe Gly Pro Gly Thr Arg
100 105 110
Leu Thr Val Thr
115
<210> 78
<211> 116
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> мутированная бета-цепь (b39)
<400> 78
Asp Val Lys Val Thr Gln Ser Ser Arg Tyr Leu Val Lys Arg Thr Gly
1 5 10 15
Glu Lys Val Phe Leu Glu Cys Val Gln Asp Ala Pro Leu Ser Lys Met
20 25 30
Phe Trp Tyr Arg Gln Asp Pro Gly Leu Gly Leu Arg Leu Ile Tyr Phe
35 40 45
Ser Tyr Asp Val Lys Met Lys Glu Lys Gly Asp Ile Pro Glu Gly Tyr
50 55 60
Ser Val Ser Arg Glu Lys Lys Glu Arg Phe Ser Leu Ile Leu Glu Ser
65 70 75 80
Ala Ser Thr Asn Gln Thr Ser Met Tyr Leu Cys Ala Ser Ser Ser Asp
85 90 95
Met Asn Ser Gly Asp Pro Tyr Glu Gln Tyr Phe Gly Pro Gly Thr Arg
100 105 110
Leu Thr Val Thr
115
<210> 79
<211> 116
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> мутированная бета-цепь (b40)
<400> 79
Asp Val Lys Val Thr Gln Ser Ser Arg Tyr Leu Val Lys Arg Thr Gly
1 5 10 15
Glu Lys Val Phe Leu Glu Cys Val Gln Asp Ala Pro Leu Ser Lys Met
20 25 30
Phe Trp Tyr Arg Gln Asp Pro Gly Leu Gly Leu Arg Leu Ile Tyr Phe
35 40 45
Ser Tyr Asp Val Lys Met Lys Glu Lys Gly Asp Ile Pro Glu Gly Tyr
50 55 60
Ser Val Ser Arg Glu Lys Lys Glu Arg Phe Ser Leu Ile Leu Glu Ser
65 70 75 80
Ala Ser Thr Asn Gln Thr Ser Met Tyr Leu Cys Ala Ser Ser Ser Asp
85 90 95
Gln Thr Ser Gly Asp Pro Tyr Glu Gln Tyr Phe Gly Pro Gly Thr Arg
100 105 110
Leu Thr Val Thr
115
<210> 80
<211> 116
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> мутированная бета-цепь (bL96D)
<400> 80
Asp Val Lys Val Thr Gln Ser Ser Arg Tyr Leu Val Lys Arg Thr Gly
1 5 10 15
Glu Lys Val Phe Leu Glu Cys Val Gln Asp Met Asp His Glu Asn Met
20 25 30
Phe Trp Tyr Arg Gln Asp Pro Gly Leu Gly Leu Arg Leu Ile Tyr Phe
35 40 45
Ser Tyr Asp Val Lys Met Lys Glu Lys Gly Asp Ile Pro Glu Gly Tyr
50 55 60
Ser Val Ser Arg Glu Lys Lys Glu Arg Phe Ser Leu Ile Leu Glu Ser
65 70 75 80
Ala Ser Thr Asn Gln Thr Ser Met Tyr Leu Cys Ala Ser Ser Phe Asp
85 90 95
Met Thr Ser Gly Asp Pro Tyr Glu Gln Tyr Phe Gly Pro Gly Thr Arg
100 105 110
Leu Thr Val Thr
115
<210> 81
<211> 116
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> мутированная бета-цепь (bM97Q)
<400> 81
Asp Val Lys Val Thr Gln Ser Ser Arg Tyr Leu Val Lys Arg Thr Gly
1 5 10 15
Glu Lys Val Phe Leu Glu Cys Val Gln Asp Met Asp His Glu Asn Met
20 25 30
Phe Trp Tyr Arg Gln Asp Pro Gly Leu Gly Leu Arg Leu Ile Tyr Phe
35 40 45
Ser Tyr Asp Val Lys Met Lys Glu Lys Gly Asp Ile Pro Glu Gly Tyr
50 55 60
Ser Val Ser Arg Glu Lys Lys Glu Arg Phe Ser Leu Ile Leu Glu Ser
65 70 75 80
Ala Ser Thr Asn Gln Thr Ser Met Tyr Leu Cys Ala Ser Ser Phe Leu
85 90 95
Gln Thr Ser Gly Asp Pro Tyr Glu Gln Tyr Phe Gly Pro Gly Thr Arg
100 105 110
Leu Thr Val Thr
115
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| Т-КЛЕТОЧНЫЕ РЕЦЕПТОРЫ, СПЕЦИФИЧНЫЕ В ОТНОШЕНИИ КОМПЛЕКСА ОПУХОЛЕВЫЙ АНТИГЕН NY-ESO-1/HLA-A*02 | 2016 |
|
RU2775394C2 |
| КОНСТРУКТЫ Т-КЛЕТОЧНОГО РЕЦЕПТОРА И ИХ ПРИМЕНЕНИЕ | 2019 |
|
RU2785954C2 |
| T-КЛЕТОЧНЫЕ РЕЦЕПТОРЫ, РАСПОЗНАЮЩИЕ МУТАЦИЮ R175H ИЛИ Y220C В P53 | 2020 |
|
RU2830061C2 |
| Т-КЛЕТОЧНЫЕ РЕЦЕПТОРЫ | 2018 |
|
RU2762255C2 |
| HPV-СПЕЦИФИЧЕСКИЕ СВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ | 2018 |
|
RU2804664C2 |
| СПЕЦИФИЧНЫЕ СВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ | 2020 |
|
RU2825837C2 |
| ВЫСОКОАФФИННЫЙ Т-КЛЕТОЧНЫЙ РЕЦЕПТОР ДЛЯ РАСПОЗНАВАНИЯ АНТИГЕНА AFP | 2019 |
|
RU2836365C2 |
| УЛУЧШЕННЫЙ СИНТЕТИЧЕСКИЙ Т-КЛЕТОЧНЫЙ РЕЦЕПТОР И АНТИГЕННЫЙ РЕЦЕПТОР | 2021 |
|
RU2820497C1 |
| CD3-СПЕЦИФИЧЕСКИЕ СВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ | 2020 |
|
RU2826453C2 |
| НОВЫЕ БИСПЕЦИФИЧНЫЕ ПОЛИПЕПТИДНЫЕ КОМПЛЕКСЫ ПРОТИВ CD3/CD19 | 2018 |
|
RU2788127C2 |












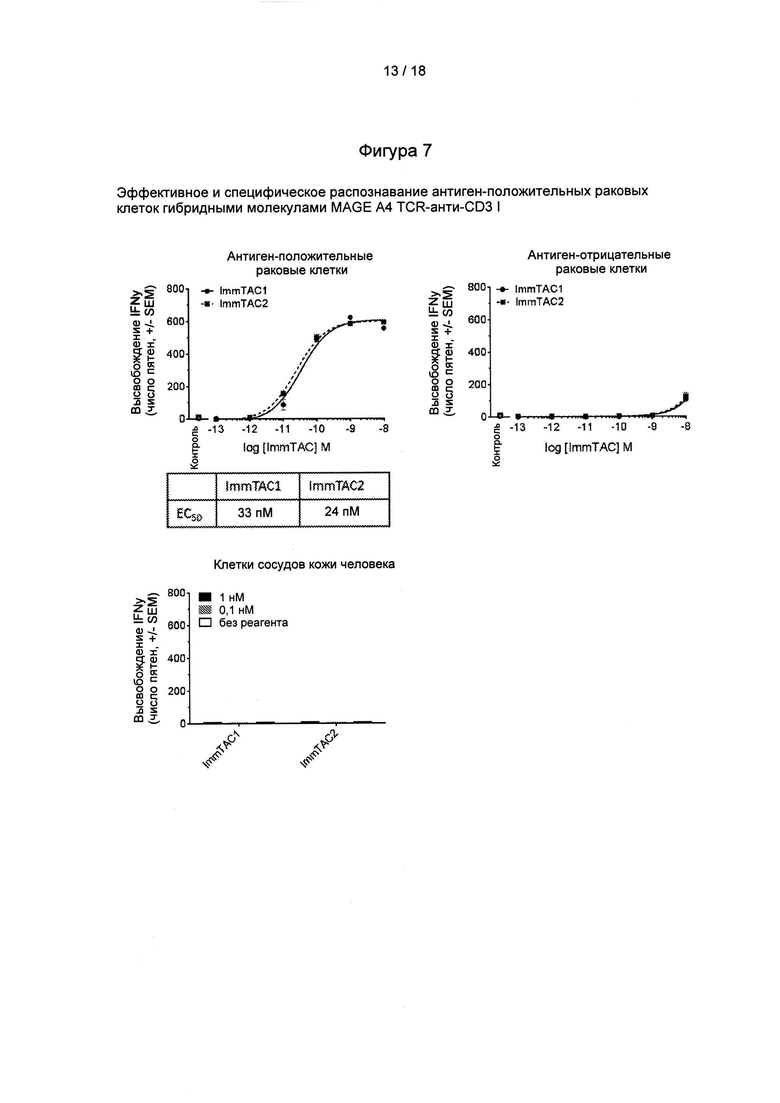
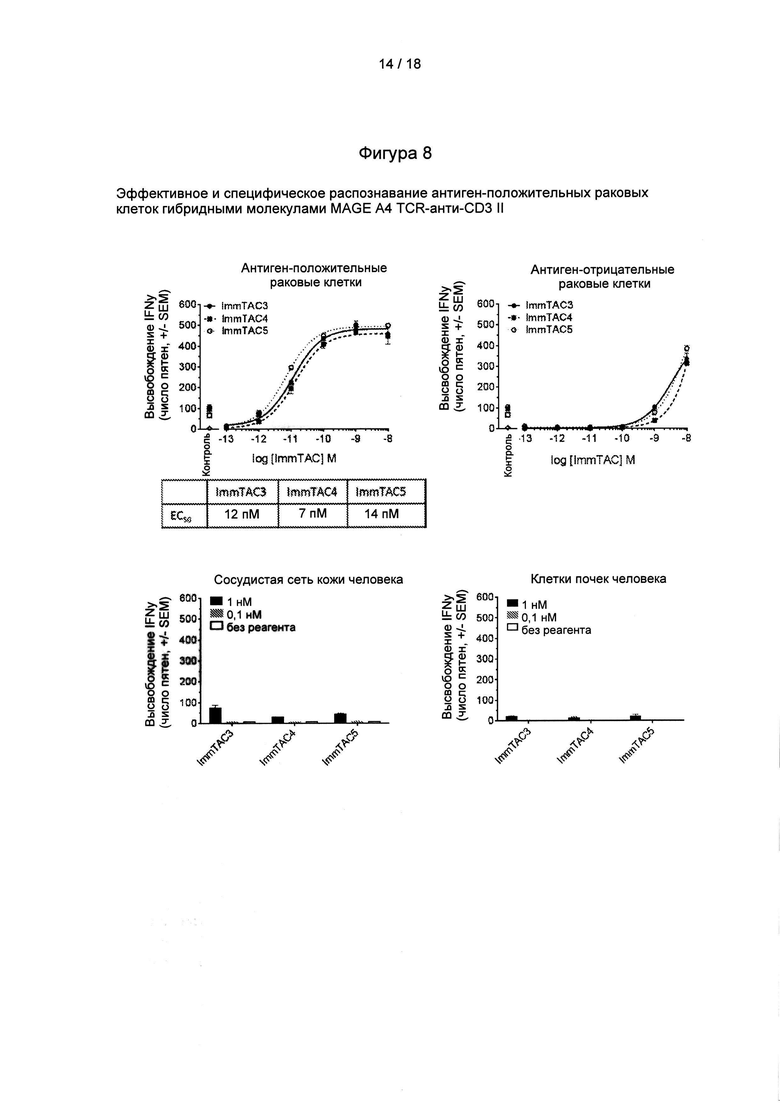
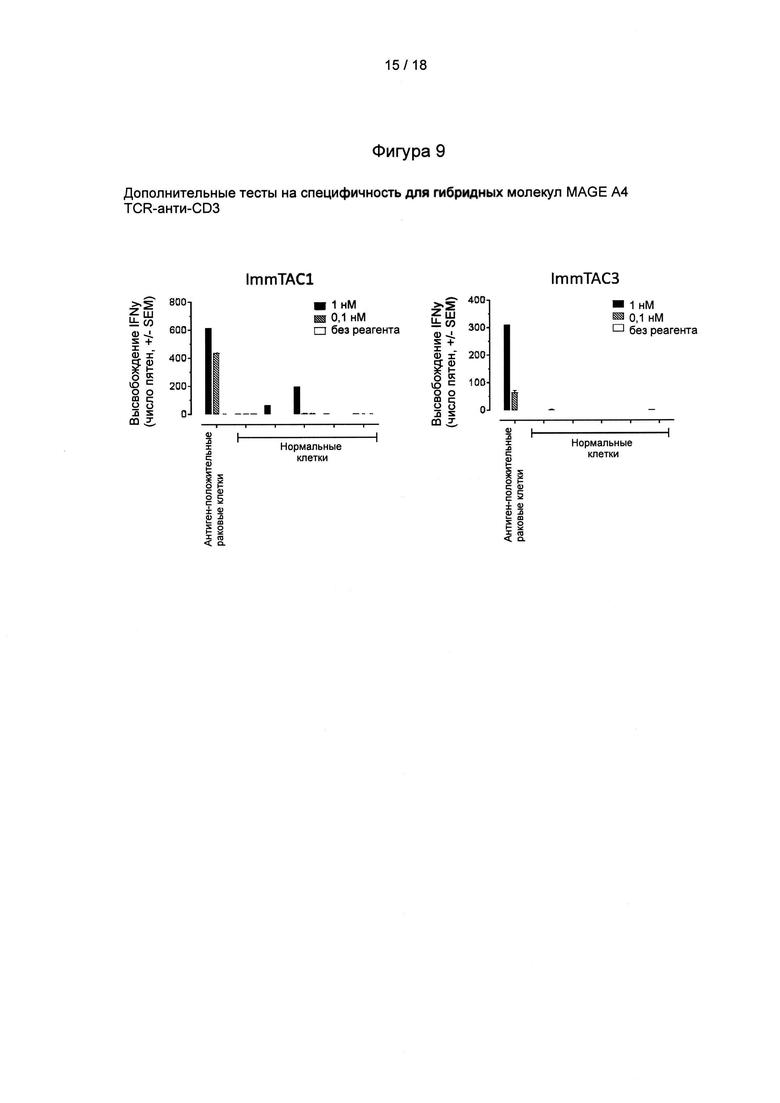
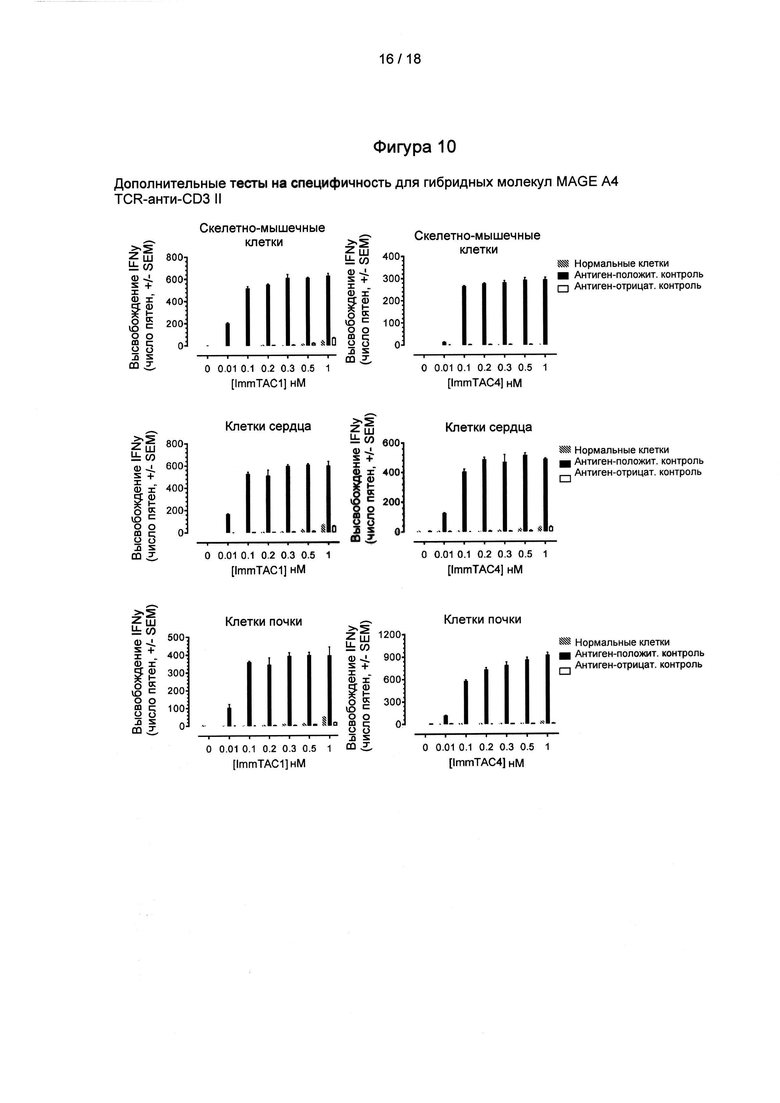
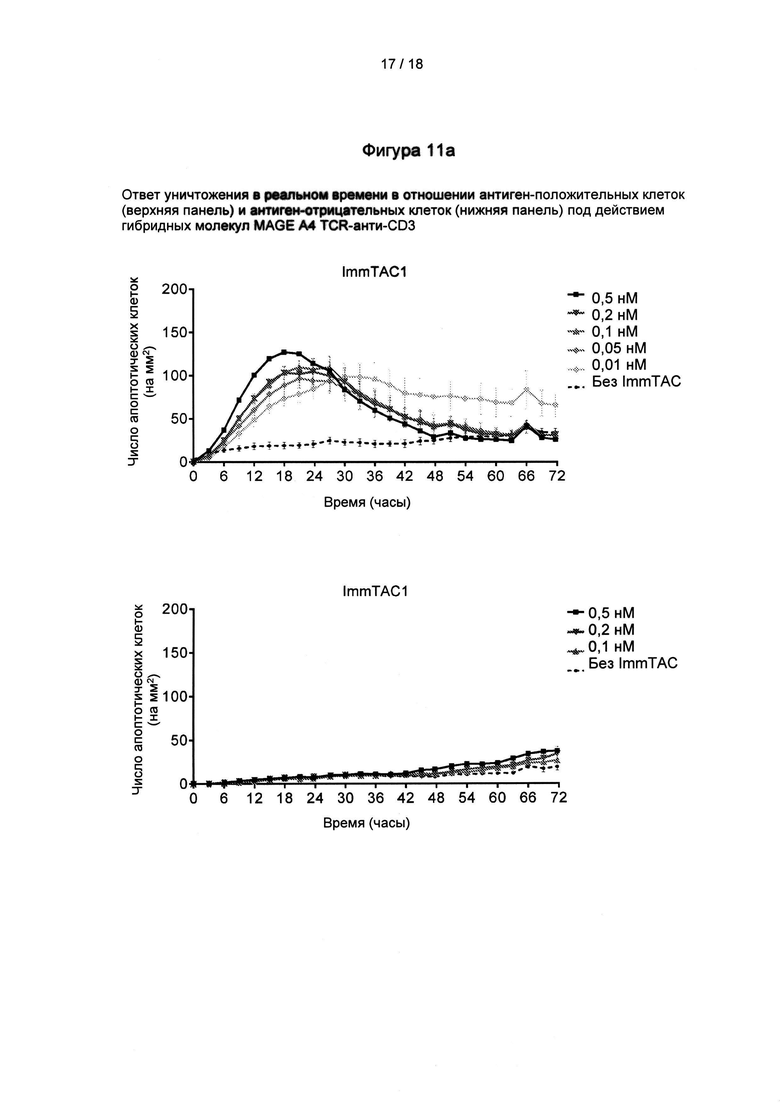

Изобретение относится к области биотехнологии, конкретно к Т-клеточным рецепторам (TCR), которые связываются с комплексом HLA-A*02 c раковым антигеном MAGE A4 GVYDGREHTV (SEQ ID NO: 1), и может быть использовано в медицине. Полученные TCR могут входить в состав гибридной молекулы TCR-анти-CD3, содержащей TCR и антитело против CD3, для перенаправления Т-клеток на клетки, презентирующие антиген MAGE-A4, и могут быть использованы для эффективной терапии MAGE A4 экспрессирующих опухолей. 19 н. и 26 з.п. ф-лы, 11 ил., 12 табл., 7 пр.
1. Т-клеточный рецептор (TCR), обладающий свойством связываться с комплексом HLA-A*02 c раковым антигеном MAGE A4 GVYDGREHTV (SEQ ID NO: 1) и содержащий гетеродимер альфа- и бета-цепей, где указанный гетеродимер включает альфа-цепь, содержащую вариабельный домен альфа-цепи TCR, и бета-цепь, содержащую вариабельный домен бета-цепи TCR, причем:
(а) вариабельный домен альфа-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 17; и вариабельный домен бета-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 28;
(b) вариабельный домен альфа-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 22; и вариабельный домен бета-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 25;
(c) вариабельный домен альфа-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 22; и вариабельный домен бета-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 28;
(d) вариабельный домен альфа-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 20; и вариабельный домен бета-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 28;
(e) вариабельный домен альфа-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 22; и вариабельный домен бета-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 26;
(f) вариабельный домен альфа-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 22; и вариабельный домен бета-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 27;
(g) вариабельный домен альфа-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 24; и вариабельный домен бета-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 29;
(h) вариабельный домен альфа-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 23; и вариабельный домен бета-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 29;
(i) вариабельный домен альфа-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 46; и вариабельный домен бета-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 29;
(j) вариабельный домен альфа-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 47; и вариабельный домен бета-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 29;
(k) вариабельный домен альфа-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 48; и вариабельный домен бета-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 29;
(l) вариабельный домен альфа-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 49; и вариабельный домен бета-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 29;
(m) вариабельный домен альфа-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 50; и вариабельный домен бета-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 29;
(n) вариабельный домен альфа-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 51; и вариабельный домен бета-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 29;
(o) вариабельный домен альфа-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 20; и вариабельный домен бета-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 65;
(p) вариабельный домен альфа-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 24; и вариабельный домен бета-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 66;
(q) вариабельный домен альфа-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 24; и вариабельный домен бета-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 67;
(r) вариабельный домен альфа-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 24; и вариабельный домен бета-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 68;
(s) вариабельный домен альфа-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 24; и вариабельный домен бета-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 69;
(t) вариабельный домен альфа-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 24; и вариабельный домен бета-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 70;
(u) вариабельный домен альфа-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 52; и вариабельный домен бета-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 26;
(v) вариабельный домен альфа-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 53; и вариабельный домен бета-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 26;
(w) вариабельный домен альфа-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 54; и вариабельный домен бета-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 26;
(x) вариабельный домен альфа-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 55; и вариабельный домен бета-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 26;
(y) вариабельный домен альфа-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 56; и вариабельный домен бета-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 26;
(z) вариабельный домен альфа-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 57; и вариабельный домен бета-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 26;
(аа) вариабельный домен альфа-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 58; и вариабельный домен бета-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 26;
(bb) вариабельный домен альфа-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 59; и вариабельный домен бета-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 26;
(cc) вариабельный домен альфа-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 60; и вариабельный домен бета-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 26;
(dd) вариабельный домен альфа-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 22; и вариабельный домен бета-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 71;
(ee) вариабельный домен альфа-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 22; и вариабельный домен бета-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 72;
(ff) вариабельный домен альфа-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 22; и вариабельный домен бета-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 73;
(gg) вариабельный домен альфа-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 22; и вариабельный домен бета-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 74;
(hh) вариабельный домен альфа-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 22; и вариабельный домен бета-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 75;
(ii) вариабельный домен альфа-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 22; и вариабельный домен бета-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 76;
(jj) вариабельный домен альфа-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 22; и вариабельный домен бета-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 77;
(kk) вариабельный домен альфа-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 22; и вариабельный домен бета-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 78; или
(ll) вариабельный домен альфа-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 22; и вариабельный домен бета-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 79.
2. TCR по п. 1, отличающийся тем, что указанный вариабельный домен альфа-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 24, и указанный вариабельный домен бета-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 29.
3. TCR по п. 1, отличающийся тем, что указанный вариабельный домен альфа-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 22, и указанный вариабельный домен бета-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 26.
4. TCR по любому из предыдущих пунктов, который представляет собой альфа-бета-гетеродимер, имеющий последовательность константного домена альфа-цепи TRAC и последовательность константного домена бета-цепи TRBC1 или TRBC2.
5. TCR по п. 4, отличающийся тем, что последовательности константного домена альфа- и бета-цепи модифицированы путем усечения или замены для удаления нативной дисульфидной связи между Cys4 экзона 2 TRAC и Cys2 экзона 2 TRBC1 или TRBC2.
6. TCR по п. 4 или 5, отличающийся тем, что последовательность(-ти) константного домена альфа- и/или бета-цепи модифицирована(-ы) путем замены остатков Thr 48 TRAC и Ser 57 TRBC1 или TRBC2 на цистеин, причем указанные цистеины образуют ненативную дисульфидную связь между константными доменами альфа- и бета-цепи TCR.
7. Гибридная молекула TCR для направленной доставки агента к клеткам, которые презентируют антиген MAGE A4, или к ткани, которая содержит клетки, которые презентируют антиген MAGE A4, включающая TCR по любому из предыдущих пунктов, который ковалентным образом связан с указанным агентом.
8. Гибридная молекула TCR по п. 7, отличающаяся тем, что указанный агент представляет собой детектируемую метку, терапевтический агент или группу, модифицирующую фармакокинетику (ФК),
при этом указанная детектируемая метка предпочтительно выбрана из флуоресцентной метки, радиоактивной метки, фермента, зонда на основе нуклеиновой кислоты или контрастного реагента;
при этом указанный терапевтический агент предпочтительно выбран из низкомолекулярного цитотоксического агента, пептидного цитотоксина, радионуклида, цитокина, суперантигена или его мутантной формы, комплекса пептид-HLA, хемокина, антитела или его антигенсвязывающего фрагмента, белкового каркаса, активатора комплемента или вирусного или бактериального белкового домена или пептида; и/или
при этом указанная группа, модифицирующая ФК, предпочтительно выбрана из ПЭГ, ПАС или альбумина.
9. Гибридная молекула TCR по п. 8, отличающаяся тем, что указанный агент представляет собой анти-CD3 антитело, ковалентно связанное с C- или N-концом альфа- или бета-цепи TCR.
10. Гибридная молекула TCR по п. 9, отличающаяся тем, что указанный агент представляет собой анти-CD3 антитело, ковалентно связанное с C- или N-концом бета-цепи TCR через линкерную последовательность.
11. Гибридная молекула TCR по п. 10, отличающаяся тем, что указанная линкерная последовательность выбрана из группы, состоящей из GGGGS (SEQ ID NO: 30), GGGSG (SEQ ID NO: 31), GGSGG (SEQ ID NO: 32), GSGGG (SEQ ID NO: 33), GSGGGP (SEQ ID NO: 34), GGEPS (SEQ ID NO: 35), GGEGGGP (SEQ ID NO: 36) и GGEGGGSEGGGS (SEQ ID NO: 37).
12. Гибридная молекула TCR по любому из пп. 9-11, отличающаяся тем, что указанное анти-CD3 антитело представляет собой антитело в формате одноцепоченого вариабельного фрагмента (scFv).
13. Гибридная молекула TCR-анти-CD3, содержащая TCR, обладающий свойством связывания с комплексом HLA-A*02 с раковым антигеном MAGE-A4 GVYDGREHTV (SEQ ID NO: 1), и антитело против CD3 для перенаправления Т-клеток на клетки, презентирующие антиген MAGE-A4, или ткань, которая содержит клетки, которые презентируют антиген MAGE A4, где указанный TCR содержит гетеродимер альфа- и бета-цепей, включающий альфа-цепь, которая содержит вариабельный домен альфа-цепи TCR, и бета-цепь, которая содержит вариабельный домен бета-цепи TCR, при этом указанное антитело против CD3 ковалентным образом связано с N-концом или C-концом бета-цепи TCR, и в которой
(а) вариабельный домен альфа-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 17; и вариабельный домен бета-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 28;
(b) вариабельный домен альфа-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 22; и вариабельный домен бета-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 25;
(c) вариабельный домен альфа-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 22; и вариабельный домен бета-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 28;
(d) вариабельный домен альфа-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 20; и вариабельный домен бета-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 28;
(e) вариабельный домен альфа-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 22; и вариабельный домен бета-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 26;
(f) вариабельный домен альфа-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 22; и вариабельный домен бета-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 27;
(g) вариабельный домен альфа-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 24; и вариабельный домен бета-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 29;
(h) вариабельный домен альфа-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 23; и вариабельный домен бета-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 29;
(i) вариабельный домен альфа-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 46; и вариабельный домен бета-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 29;
(j) вариабельный домен альфа-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 47; и вариабельный домен бета-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 29;
(k) вариабельный домен альфа-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 48; и вариабельный домен бета-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 29;
(l) вариабельный домен альфа-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 49; и вариабельный домен бета-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 29;
(m) вариабельный домен альфа-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 50; и вариабельный домен бета-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 29;
(n) вариабельный домен альфа-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 51; и вариабельный домен бета-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 29;
(o) вариабельный домен альфа-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 20; и вариабельный домен бета-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 65;
(p) вариабельный домен альфа-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 24; и вариабельный домен бета-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 66;
(q) вариабельный домен альфа-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 24; и вариабельный домен бета-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 67;
(r) вариабельный домен альфа-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 24; и вариабельный домен бета-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 68;
(s) вариабельный домен альфа-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 24; и вариабельный домен бета-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 69;
(t) вариабельный домен альфа-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 24; и вариабельный домен бета-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 70;
(u) вариабельный домен альфа-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 52; и вариабельный домен бета-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 26;
(v) вариабельный домен альфа-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 53; и вариабельный домен бета-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 26;
(w) вариабельный домен альфа-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 54; и вариабельный домен бета-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 26;
(x) вариабельный домен альфа-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 55; и вариабельный домен бета-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 26;
(y) вариабельный домен альфа-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 56; и вариабельный домен бета-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 26;
(z) вариабельный домен альфа-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 57; и вариабельный домен бета-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 26;
(аа) вариабельный домен альфа-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 58; и вариабельный домен бета-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 26;
(bb) вариабельный домен альфа-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 59; и вариабельный домен бета-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 26;
(cc) вариабельный домен альфа-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 60; и вариабельный домен бета-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 26;
(dd) вариабельный домен альфа-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 22; и вариабельный домен бета-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 71;
(ee) вариабельный домен альфа-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 22; и вариабельный домен бета-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 72;
(ff) вариабельный домен альфа-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 22; и вариабельный домен бета-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 73;
(gg) вариабельный домен альфа-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 22; и вариабельный домен бета-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 74;
(hh) вариабельный домен альфа-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 22; и вариабельный домен бета-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 75;
(ii) вариабельный домен альфа-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 22; и вариабельный домен бета-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 76;
(jj) вариабельный домен альфа-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 22; и вариабельный домен бета-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 77;
(kk) вариабельный домен альфа-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 22; и вариабельный домен бета-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 78; или
(II) вариабельный домен альфа-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 22; и вариабельный домен бета-цепи TCR содержит аминокислотную последовательность SEQ ID NO: 79;
причем указанное анти-CD3 антитело ковалентно связано с N-концом или C-концом бета-цепи TCR через линкерную последовательность, выбранную из SEQ ID NO: 30-37.
14. Гибридная молекула TCR-анти-CD3 по п. 13, содержащая
(a) аминокислотную последовательность альфа-цепи, соответствующую SEQ ID NO: 38, и аминокислотную последовательность бета-цепи, соответствующую SEQ ID NO: 42;
(b) аминокислотную последовательность альфа-цепи, соответствующую SEQ ID NO: 38, и аминокислотную последовательность бета-цепи, соответствующую SEQ ID NO: 43;
(c) аминокислотную последовательность альфа-цепи, соответствующую SEQ ID NO: 39, и аминокислотную последовательность бета-цепи, соответствующую SEQ ID NO: 44;
(d) аминокислотную последовательность альфа-цепи, соответствующую SEQ ID NO: 40, и аминокислотную последовательность бета-цепи, соответствующую SEQ ID NO: 45; или
(e) аминокислотную последовательность альфа-цепи, соответствующую SEQ ID NO: 41, и аминокислотную последовательность бета-цепи, соответствующую SEQ ID NO: 45.
15. Гибридная молекула TCR-анти-CD3 по п. 14, содержащая аминокислотную последовательность альфа-цепи, соответствующую SEQ ID NO: 40, и аминокислотную последовательность бета-цепи, соответствующую SEQ ID NO: 45.
16. Гибридная молекула TCR-анти-CD3 по п. 14, содержащая аминокислотную последовательность альфа-цепи, соответствующую SEQ ID NO: 38, и аминокислотную последовательность бета-цепи, соответствующую SEQ ID NO: 42.
17. Нуклеиновая кислота, кодирующая TCR, который обладает свойством связываться с комплексом HLA-A*02 c раковым антигеном MAGE A4 GVYDGREHTV (SEQ ID NO: 1) и содержит гетеродимер альфа- и бета-цепей, где указанная нуклеиновая кислота кодирует альфа-цепь TCR и бета-цепь TCR по любому из пп. 1-6.
18. Нуклеиновая кислота, кодирующая гибридную молекулу TCR, для доставки агента к клеткам, которые презентируют раковый антиген MAGE A4, или к ткани, которая содержит клетки, которые презентируют антиген MAGE A4, где указанная нуклеиновая кислота кодирует гибридную молекулу TCR, как заявлено в любом из пп. 7-12, при этом указанный агент представляет собой пептид или полипептид.
19. Нуклеиновая кислота, кодирующая гибридную молекулу TCR-анти-CD3, содержащую TCR, обладающий свойством связывания с комплексом HLA-A*02 с раковым антигеном MAGE-A4 GVYDGREHTV (SEQ ID NO: 1), и антитело против CD3, где указанная нуклеиновая кислота кодирует альфа-цепь и бета-цепь гибридной молекулы TCR-анти-CD3, как заявлено в любом из пп. 13-16, и при этом указанная нуклеиновая кислота дополнительно кодирует антитело против CD3, ковалентным образом соединенное с N-концом или C-концом бета-цепи TCR, через линкерную последовательность, выбранную из SEQ ID NO: 30-37.
20. Вектор экспрессии, кодирующий TCR, обладающий свойством связываться с комплексом HLA-A*02 c раковым антигеном MAGE A4 GVYDGREHTV (SEQ ID NO: 1) и содержащий гетеродимер альфа- и бета-цепей, где указанный вектор экспрессии содержит нуклеиновую кислоту по п. 17.
21. Вектор экспрессии по п. 20, отличающийся тем, что указанный вектор содержит первую открытую рамку считывания, кодирующую альфа-цепь TCR, и вторую открытую рамку считывания, кодирующую бета-цепь TCR.
22. Вектор экспрессии, кодирующий гибридную молекулу TCR для доставки агента к клеткам, которые презентируют раковый антиген MAGE A4, или к ткани, которая содержит клетки, которые презентируют раковый антиген MAGE A4, причем указанный вектор экспрессии содержит нуклеиновую кислоту по п. 18.
23. Вектор экспрессии по п. 22, где указанный вектор содержит первую открытую рамку считывания, кодирующую альфа-цепь TCR, и вторую открытую рамку считывания, кодирующую бета-цепь TCR.
24. Вектор экспрессии, кодирующий гибридную молекулу TCR-анти-CD3, содержащую TCR, обладающий свойством связываться с комплексом HLA-A*02 с раковым антигеном MAGE-A4 GVYDGREHTV (SEQ ID NO: 1), и антитело против CD3, где указанный вектор экспрессии содержит нуклеиновую кислоту по п. 19.
25. Вектор экспрессии по п. 24, где указанный вектор содержит первую открытую рамку считывания, кодирующую альфа-цепь TCR, и вторую открытую рамку считывания, кодирующую бета-цепь TCR.
26. Клетка для экспрессии TCR, обладающего свойством связываться с комплексом HLA-A*02 c раковым антигеном MAGE A4 GVYDGREHTV (SEQ ID NO: 1) и содержащего гетеродимер альфа и бета цепей, несущая
(a) вектор экспрессии по п. 20, содержащий одну открытую рамку считывания или вектор экспрессии по п. 21, содержащий две различных открытых рамки считывания; или
(b) первый вектор экспрессии, который содержит нуклеиновую кислоту, кодирующую альфа-цепь TCR по любому из пп. 1-6, и второй вектор экспрессии, который содержит нуклеиновую кислоту, кодирующую бета-цепь TCR по любому из пп. 1-6.
27. Клетка для экспрессии гибридной молекулы TCR для доставки агента к клеткам, которые презентируют антиген MAGE A4, или к ткани, которая содержит клетки, которые презентируют антиген MAGE A4, несущая
(a) вектор экспрессии по п. 22, содержащий одну открытую рамку считывания или вектор экспрессии по п. 23, содержащий две различных открытых рамки считывания; или
(b) первый вектор экспрессии, который содержит нуклеиновую кислоту, кодирующую альфа-цепь гибридной молекулы TCR, как заявлено в любом из пп. 9-12, и второй вектор экспрессии, который содержит нуклеиновую кислоту, кодирующую бета-цепь гибридной молекулы TCR, как заявлено в любом из пп. 9-12.
28. Клетка для экспрессии гибридной молекулы TCR-анти-CD3, содержащей TCR, обладающий свойством связываться с комплексом HLA-A*02 с раковым антигеном MAGE-A4 GVYDGREHTV (SEQ ID NO: 1), и антитело против CD3, несущая
(a) вектор экспрессии по п. 24, содержащий одну открытую рамку считывания или вектор экспрессии по п. 25, содержащий две различных открытых рамки считывания; или
(b) первый вектор экспрессии, который содержит нуклеиновую кислоту, кодирующую альфа-цепь гибридной молекулы TCR-анти-CD3, как заявлено в любом из пп. 13-16, и второй вектор экспрессии, который содержит нуклеиновую кислоту, кодирующую бета-цепь гибридной молекулы TCR-анти-CD3, как заявлено в любом из пп. 13-16.
29. Фармацевтическая композиция для диагностирования или лечения рака или опухоли, экспрессирующих комплекс HLA-A*02 c раковым антигеном MAGE A4 GVYDGREHTV (SEQ ID NO: 1), содержащая в эффективном количестве TCR по любому из пп. 1-6, гибридную молекулу TCR по любому из пп. 7-12 или гибридную молекулу TCR-анти-CD3 по любому из пп. 13-16 вместе с одним или более фармацевтически приемлемым носителем или вспомогательным веществом.
30. TCR по любому из пп. 1-6 или фармацевтическая композиция по п. 29, которая содержит TCR, для применения в медицине, предпочтительно у субъекта, представляющего собой человека.
31. Гибридная молекула TCR по любому из пп. 7-12 или фармацевтическая композиция по п. 29, которая содержит гибридную молекулу TCR, для применения в медицине, предпочтительно у субъекта, представляющего собой человека.
32. Гибридная молекула TCR-анти-CD3 по любому из пп. 13-16 или фармацевтическая композиция по п. 29, которая содержит гибридную молекулу TCR-анти-CD3, для применения в медицине, предпочтительно у субъекта, представляющего собой человека.
33. TCR по любому из пп. 1-6 или фармацевтическая композиция по п. 29, которая содержит TCR, для применения в способе лечения рака или опухоли, предпочтительно у субъекта, представляющего собой человека.
34. Гибридная молекула TCR по любому из пп. 7-12 или фармацевтическая композиция по п. 29, которая содержит гибридную молекулу TCR, для применения в способе лечения рака или опухоли, предпочтительно у субъекта, представляющего собой человека.
35. Гибридная молекула TCR-анти-CD3 по любому из пп. 13-16 или фармацевтическая композиция по п. 29, которая содержит гибридную молекулу TCR-анти-CD3, для применения в способе лечения рака или опухоли, предпочтительно у субъекта, представляющего собой человека.
36. Гибридная молекула TCR-анти-CD3 или фармацевтическая композиция для применения по п. 35, при этом указанный субъект, представляющий собой человека, имеет опухоль, экспрессирующую MAGE A4.
37. Гибридная молекула TCR-анти-CD3 или фармацевтическая композиция для применения по пп. 35,36, причем указанная опухоль представляет собой солидную опухоль.
38. Гибридная молекула TCR-анти-CD3 или фармацевтическая композиция для применения по любому из пп. 35-37, причем указанный субъект, представляющий собой человека, принадлежит к подтипу HLA-A*02.
39. Гибридная молекула TCR-анти-CD3 или фармацевтическая композиция для применения по любому из пп. 35-38, которые вводят путем инъекции, такой как внутривенная инъекция или инъекция напрямую в опухоль.
40. Способ лечения нуждающегося в этом субъекта, представляющего собой человека, включающий введение указанному субъекту фармацевтически эффективной дозы фармацевтической композиции по п. 29, где у указанного субъекта, представляющего собой человека, присутствует опухоль, которая экспрессирует MAGE A4.
41. Способ лечения нуждающегося в этом субъекта, представляющего собой человека, включающий введение указанному субъекту фармацевтически эффективной дозы фармацевтической композиции по п. 29, и антинеопластического средства, где у указанного субъекта, представляющего собой человека, присутствует опухоль, которая экспрессирует MAGE A4, причем указанную композицию и средство вводят по отдельности, в комбинации или последовательно.
42. Композиция для диагностирования или лечения рака или опухоли, экспрессирующих комплекс HLA-A*02 c раковым антигеном MAGE A4 GVYDGREHTV (SEQ ID NO: 1) для введения путем инъекции субъекту, представляющему собой человека, содержащая в эффективном количестве TCR по любому из пп. 1-6, гибридную молекулу TCR по любому из пп. 7-12 или гибридную молекулу TCR-анти-CD3 по любому из пп. 13-16.
43. Способ получения TCR, обладающего свойством связываться с комплексом HLA-A*02 c раковым антигеном MAGE A4 GVYDGREHTV (SEQ ID NO: 1) и содержащего гетеродимер альфа- и бета-цепей, включающий: а) поддержание клетки по п. 26 в оптимальных условиях для экспрессии цепей TCR, и b) выделение указанных цепей TCR.
44. Способ получения гибридной молекулы TCR для доставки агента к клеткам, которые презентируют антиген MAGE A4, или к ткани, которая содержит клетки, которые презентируют антиген MAGE A4, включающий: а) поддержание клетки по п. 27 в оптимальных условиях для экспрессии указанной гибридной молекулы TCR, и b) выделение указанной гибридной молекулы TCR.
45. Способ получения гибридной молекулы TCR-анти-CD3, содержащей TCR, обладающий свойством связывания с комплексом HLA-A*02 с раковым антигеном MAGE-A4 GVYDGREHTV (SEQ ID NO: 1), и антитело против CD3, включающий: а) поддержание клетки по п. 28 в оптимальных условиях для экспрессии указанной гибридной молекулы TCR-анти-CD3, и b) выделение указанной гибридной молекулы TCR-анти-CD3.
| WO 2014118263 A1, 07.08.2014 | |||
| WO 2016007570 A2, 14.01.2016 | |||
| WO 2004074322 A1, 02.09.2004 | |||
| НАЗАРОВ В | |||
| И., Обзор методов анализа, моделей генерации и моделей селекции репертуаров иммунных рецепторов, Новые информационные технологии в автоматизированных системах, 2015, N | |||
| Способ использования делительного аппарата ровничных (чесальных) машин, предназначенных для мериносовой шерсти, с целью переработки на них грубых шерстей | 1921 |
|
SU18A1 |
| DE LA HERA A | |||
| et al., Structure of the T cell antigen receptor | |||

