В настоящем изобретении предложены T-клеточные рецепторы (TCR), которые связываются с рестриктированным по HLA-A*02 пептидом SLLQHLIGL (SEQ ID NO: 1), происходящим из ракового антигена зародышевой линии PRAME. Указанные TCR могут содержать не встречающиеся в природе мутации в пределах вариабельных доменов их альфа- и/или бета-цепей по сравнению с нативным TCR, специфичным по отношению к PRAME. TCR согласно настоящему изобретению, в частности, подходят для применения в качестве новых иммунотерапевтических реагентов для лечения злокачественного заболевания.
Уровень техники
T-клеточные рецепторы (TCR) в естественных условиях экспрессируются CD4+ и CD8+ T-клетками. TCR предназначены для распознавания коротких пептидных антигенов, представленных на поверхности антиген-презентирующих клеток в комплексе с молекулами главного комплекса гистосовместимости - MHC (у людей молекулы MHC также известны как человеческие лейкоцитарные антигены, или HLA, Davis, et al., Annu Rev Immunol 1998;16:523-44). CD8+ T-клетки, которые также называют цитотоксическими T-клетками, содержат TCR, специфично распознающие пептиды, связанные с MHC I класса. CD8+ T-клетки, в целом, отвечают за обнаружение и содействие в уничтожении больных клеток, включая раковые инфицированные вирусом клетки. Вследствие селекции в тимусе, аффинность природного репертуара специфичных к раку TCR к соответствующему антигену, как правило, очень низкая, что означает, что раковые клетки нередко избегают обнаружения и уничтожения. Новые иммунотерапевтические подходы, направленные на содействие распознаванию рака Т-клетками, предлагают очень многообещающую стратегию для разработки эффективных противоопухолевых способов лечения.
PRAME (Preferentially Expressed Antigen In Melanoma, антиген, преимущественно экспрессирующийся в меланоме), был впервые идентифицирован как антиген, который сверхэкспрессируется в меланоме (Ikeda et al. Immunity. 1997 Feb; 6 (2): 199-208); он также известен как CT130, MAPE, OIP-4 и имеет номер доступа в базе данных Uniprot P78395. Данный белок функционирует в качестве репрессора передачи сигналов от рецептора ретиноевой кислоты (Epping et al., Cell. 2005 Sep 23; 122 (6): 835-47). PRAME принадлежит к семейству антигенов, кодируемых генами зародышевой линии, известных как раково-тестикулярные антигены. Раково-тестикулярные антигены являются привлекательными мишенями для иммунотерапевтического вмешательства, так как они обычно имеют ограниченную или нулевую экспрессию в нормальных тканях взрослого человека. PRAME эспрессируется в ряде солидных опухолей, а также при лейкозах и в лимфомах (Doolan et al Breast Cancer Res Treat. 2008 May;109(2):359-65; Epping et al Cancer Res. 2006 Nov 15;66(22):10639-42; Ercolak et al Breast Cancer Res Treat. 2008 May;109(2):359-65; Matsushita et al Leuk Lymphoma. 2003 Mar;44(3):439-44; Mitsuhashi et al Int. J Hematol. 2014;100(1):88-95; Proto-Sequeire et al Leuk Res. 2006 Nov;30(11):1333-9; Szczepanski et al Oral Oncol. 2013 Feb;49(2):144-51; Van Baren et al Br J Haematol. 1998 Sep;102(5):1376-9). Терапия, нацеленная на PRAME, согласно настоящему изобретению, может быть особенно подходящей для лечения рака, включая, но не ограничиваясь перечисленным, рака легких (NSCLC и SCLC), рака молочной железы (включая тройной негативный), рака яичников, рака эндометрия, рака пищевода, рака мочевого пузыря и рака головы и шеи.
Пептид SLLQHLIGL (SEQ ID NO: 1) соответствует аминокислотам 425-433 полноразмерного белка PRAME и процентируется на клеточной поверхности в комплексе с HLA-A*02 (Kessler et al., J Exp Med. 2001 Jan 1; 193 (1): 73-88). Данный комплекс пептид-HLA служит удобной мишенью для иммунотерапевтического вмешательства на основе TCR.
Идентификация конкретных последовательностей TCR, которые связываются с комплексом SLLQHLIGL (SEQ ID NO: 1) HLA-A*02 с высокой аффинностью и специфичностью, является перспективной для разработки новых иммунотерапевтических средств. Терапевтические TCR могут быть использованы, например, в качестве растворимых агентов с возможностью нацеливания для доставки цитотоксических агентов к месту опухоли или активации иммунных эффекторных функций против опухолевых клеток ((Lissin, et al., “High-Affinity Monocloncal T-cell receptor (mTCR) Fusions” в Fusion Protein Technologies for Biophamaceuticals: Applications and Challenges. 2013. S. R. Schmidt, Wiley; Boulter et al., Protein Eng. 2003 Sep;16(9):707-11; Liddy, et al., Nat Med. 2012 Jun;18(6):980-7), или, в качестве альтернативы, их можно применять для конструирования Т-клеток для адоптивной терапии (Fesnak et al., Nat Rev Cancer. 2016 Aug 23; 16 (9): 566-81).
Ранее сообщали о TCR, которые связываются с SLLQHLIGL (SEQ ID NO: 1) в комплексе с HLA-A*02 (Amir et al., Clin Cancer Res. 2011 Sep 1; 17 (17): 5615-25; Griffioen et al.. Clin Cancer Res. 2006 15 мая; 12 (10): 3130-6; WO2016142783). Однако эти TCR не были сконструированы так, чтобы связываться с повышенной аффинностью с антигеном-мишенью по сравнению с нативным TCR. Как поясняется ниже, супра-физиологическая аффинность антигена является желательной характеристикой для терапевтического TCR, получение которого не является простым, особенно когда оно сочетается с другими желательными признаками, такими как специфичность.
Последовательности TCR, определенные в настоящей заявке, описаны в соответствии с номенклатурой IMGT, которая широко известна и доступна специалистам, работающим в области изучения TCR. См., например: LeFranc and LeFranc, (2001). «T-cell Receptor Factsbook», Academic Press; Lefranc, (2011), Cold Spring Harb Protoc 2011(6): 595-603; Lefranc, (2001), Curr Protoc Immunol Appendix 1: Appendix 1O; Lefranc, (2003), Leukemia 17(1): 260-266. Вкратце, αβ-TCR состоят из двух связанных дисульфидными связями цепей. В целом считается, что каждая цепь (альфа и бета) содержит два домена, а именно, вариабельный и константный домен. Короткий соединяющий участок соединяет вариабельный и константный домены и, как правило, считается частью вариабельного участка альфа. Кроме того, бета-цепь обычно содержит короткую область вариабельности рядом с соединяющим участком, которая, как правило, считается частью вариабельного участка бета.
Вариабельный домен каждый цепи расположен на N-конце и содержит три определяющих комплементарность участка (CDR), встроенных в каркасную последовательность (FR). CDR содержат сайт распознавания связывания пептид-MHC. Несколько генов кодируют вариабельные участки альфа-цепи (Vα), и несколько генов кодирует вариабельные участки бета-цепи (Vβ), указанные гены различаются по их каркасным последовательностям, CDR1 и CDR2 и по частично определенной CDR3-последовательности. Гены Vα и Vβ в соответствии с номенклатурой IMGT обозначаются с помощью приставки ‘TRAV’ и ‘TRBV’ соответственно (Folch and Lefranc, (2000), Exp Clin Immunogenet 17(1): 42-54; Scaviner and Lefranc, (2000), Exp Clin Immunogenet 17(2): 83-96; LeFranc and LeFranc, (2001), “T-cell Receptor Factsbook”, Academic Press). Подобным образом, существует несколько соединительных или J генов, называемых ‘TRAJ’ или ‘TRBJ’ для альфа и бета-цепи соответственно, и ген вариабельности или ген D, для бета-цепи, называемый ‘TRBD’ (Folch and Lefranc, (2000), Exp Clin Immunogenet 17(2): 107-114; Scaviner and Lefranc, (2000), Exp Clin Immunogenet 17(2): 97-106; LeFranc and LeFranc, (2001), «T-Cell Receptor Factsbook», Academic Press). Огромное разнообразие цепей Т клеточного рецептора является результатом комбинаторных реаранжировок различных генов V, J и D, которые включают аллельные варианты, и дополнительного разнообразия J-сегментов (Arstila, et al., (1999), Science 286(5441): 958-961; Robins et al., (2009), Blood 114(19): 4099-4107.) Константные участки, или C-участки, альфа- и бета-цепей TCR называются ‘TRAC’ и ‘TRBC’ соответственно (Lefranc, (2001), Curr Protoc Immunol Appendix 1: Appendix 10).
Авторы настоящей заявки неожиданно обнаружили новые TCR, которые способны связываться с комплексом SLLQHLIGL-HLA-A*02 с высокой аффинностью и специфичностью. Указанные TCR сконструированы на основе подходящей скелетной (“scaffold”) последовательности, в которую введен ряд мутаций. TCR согласно настоящему изобретению имеют особенно подходящий профиль для терапевтического применения. В целом, идентификация таких TCR не является простой и обычно имеет высокую вероятность получения отрицательных результатов.
В первую очередь, специалист должен идентифицировать подходящую исходную или «скелетную» последовательность. Обычно такие последовательности получают из природных источников, например, отвечающих на антиген Т-клеток, выделенных из донорской крови. Учитывая редкость онкоспецифичных Т-клеток в естественных условиях, часто необходимо провести скрининг многих доноров, например, 20 или более, прежде чем можно будет найти отвечающую на антиген Т-клетку. Процесс скрининга может занять несколько недель или месяцев, и даже если обнаружена отвечающая на антиген Т-клетка, она может быть непригодна для применения в иммунотерапии. Например, ответ может быть слишком слабым и/или может не быть специфичным для целевого антигена. Кроме того, может оказаться невозможным ни создать клональную популяцию Т-клеток, ни размножить или поддерживать данную линию Т-клеток для получения достаточного материала для идентификации корректных последовательностей цепи TCR. Последовательности TCR, которые подходят в качестве исходных или скелетных последовательностей, должны иметь одно или несколько из следующих свойств: высокую аффинность к комплексу целевой пептид-HLA, например, 200 мкМ или более; высокий уровень целевой специфичности, например, относительно слабое связывание или отсутствие связывания с альтернативными комплексами пептид-HLA; быть пригодным для использования в дисплейных библиотеках, таких как фаговый дисплей; и быть способными к рефолдингу и очистке с высоким выходом. Принимая во внимание вырожденную природу распознавания TCR, даже опытным специалистам исключительно трудно определить, имеет ли конкретная скелетная последовательность TCR профиль специфичности, который сделал бы ее пригодной для инженерии с целью их применения в терапии (Wooldridge, et al., J Biol Chem. 2012 Jan 6;287(2):1168-77).
Следующая проблема состоит в том, чтобы сконструировать TCR с более высокой аффинностью к антигену-мишени при сохранении желательных характеристик, таких как специфичность и выход. TCR, в таком виде как они существуют в природе, имеют слабую аффинность к антигену-мишени (низкий микромолярный диапазон) по сравнению с антителами, а TCR обычно более слабо распознают раковые антигены, чем вирусные антигены (Aleksic, et al. Eur J. Immunol. 2012 Dec; 42 (12): 3174-9). Указанная слабая аффинность в сочетании с отрицательной регуляцией HLA на раковых клетках означает, что терапевтические TCR для иммунотерапии рака, как правило, требуют конструирования для увеличения их аффинности к антигену-мишени с целью получения более мощного ответа. Такое увеличение аффинности необходимо для растворимых реагентов на основе TCR. В таких случаях желательны аффинности связывания антигена в диапазоне от наномолярного до пикомолярного, с периодами полувыведения в связанной форме в несколько часов. Повышенная активность, генерируемая высокой степенью аффинности к распознаваемому антигену при небольшом количестве эпитопов, показана на Фиг.1е и 1f работы Liddy et al. (Liddy et al.., Nat Med. 2012 Jun; 18 (6): 980-7). Процесс созревания аффинности требует привлечения квалифицированного специалиста, которому необходимо спроектировать специфичные мутации и/или комбинации мутаций, включая, но не ограничиваясь перечисленным, заменами, вставками и/или делециями в исходной последовательности TCR, чтобы увеличить силу распознавания антигена. В данной области известны способы конструирования мутаций, увеличивающих аффинность для выбранного TCR, например, использование дисплейных библиотек (Li et al., Nat Biotechnol. 2005 Mar;23(3):349-54; Holler et al., Proc Natl Acad Sci U S A. 2000 May 9;97(10):5387-92). Тем не менее, для получения значительного увеличения аффинности выбранного TCR к заданной мишени, специалисту в данной области может потребоваться разработка комбинаций мутаций из большого числа возможных вариантов. Конкретные мутации и/или комбинации мутаций, которые приводят к значительному увеличению аффинности, непредсказуемы, и существует высокая вероятность получения отрицательного результата. Во многих случаях может оказаться невозможным достичь значительного увеличения аффинности, используя выбранную стартовую последовательность TCR.
Процесс созревания аффинности должен также учитывать необходимость поддержания антигенной специфичности TCR. Увеличение аффинности TCR к его антигену-мишени приводит к значительному риску выявления кросс-реактивности с другими непреднамеренными мишенями в результате присущей вырожденности распознавания антигена (Wooldridge, et al., J Biol Chem. 2012 Jan 6;287(2):1168-77; Wilson, et al., Mol Immunol.2004 Feb;40(14-15):1047-55; Zhao et al., J Immunol. 2007 Nov 1;179(9):5845-54). При естественном уровне аффинности распознавание кросс-реактивного антигена может быть слишком слабым, чтобы вызвать ответ. Если перекрестно-реактивный антиген отображается на нормальных здоровых клетках, существует высокая вероятность нецелевого связывания in vivo, которое может проявляться в клинической токсичности. Таким образом, в дополнение к увеличению силы связывания антигена специалист должен также разработать мутации и/или комбинации мутаций, которые позволяют TCR сохранять высокую специфичность по отношению к антигену-мишени и демонстрировать хороший профиль безопасности при доклинических испытаниях. Опять же, подходящие мутации и/или комбинации мутаций не являются предсказуемыми. Возможность получения отрицательного результата на этой стадии еще выше, и во многих случаях это может быть недостижимо для выбранной исходной последовательности TCR.
Несмотря на трудности, описанные выше, авторы настоящего изобретения идентифицировали мутированные TCR с особенно высокой аффинностью (пикомолярный диапазон) и высокой степенью антигенной специфичности. Указанные TCR демонстрируют эффективное уничтожение PRAME-положительных раковых клеток, когда их получают в виде растворимых реагентов, гибридизованных с группой, перенаправляющей Т-клетки.
Описание изобретения
В первом аспекте настоящего изобретения предложен T-клеточный рецептор (TCR), обладающий свойством связывания с SLLQHLIGL (SEQ ID NO: 1) в комплексе с HLA-A*02 и содержащий вариабельный домен альфа-цепи TCR и/или вариабельный домен бета-цепи TCR, каждый из которых включает в себя FR1-CDR1-FR2-CDR2-FR3-CDR3-FR4, где FR представляет собой каркасный участок, а CDR представляет собой область, определяющую комплементарность, при этом
(а) CDR альфа-цепи имеют следующие последовательности:
CDR1 - TISGTDY(SEQ ID NO: 39)
CDR2 - GLTSN (SEQ ID NO: 40)
CDR3 - CILILGHSGAGSYQLTF(SEQ ID NO: 41)
необязательно с одной или более мутациями в них,
и/или
(b) CDR бета-цепи имеют следующие последовательности:
CDR1 - LNHDA (SEQ ID NO: 42)
CDR2 - SQIVNDF (SEQ ID NO: 43)
CDR3 - CASSPWTSGSREQYF (SEQ ID NO: 44)
необязательно с одной или более мутациями в них.
В TCR согласно первому аспекту каркасные участки вариабельного домена альфа-цепи могут содержать следующие каркасные последовательности:
FR1 - аминокислоты 1-25 SEQ ID NO: 2
FR2 - аминокислоты 33-49 SEQ ID NO: 2
FR3 - аминокислоты 55-87 SEQ ID NO: 2
FR4 - аминокислоты 105-114 SEQ ID NO: 2
или соответствующие последовательности, имеющие по меньшей мере 90, 91, 92, 93, 94, 95, 96, 97, 98 или 99% идентичности с указанными последовательностями, и/или
каркасные участки вариабельного домена бета-цепи могут содержать следующие последовательности:
FR1 - аминокислоты 1-26 of SEQ ID NO: 3
FR2 - аминокислоты 32-48 of SEQ ID NO: 3
FR3 - аминокислоты 56-90 of SEQ ID NO: 3
FR4 - аминокислоты 106-114 of SEQ ID NO: 3
или соответствующие последовательности, имеющие по меньшей мере 90, 91, 92, 93, 94, 95, 96, 97, 98 или 99% идентичности с указанными последовательностями.
Термин «мутации» охватывает замены, вставки и делеции. Мутации в исходном TCR (или дикого типа, или скелетном) могут включать те, которые увеличивают аффинность связывания (kD и/или период полувыведения в связанной форме) TCR с комплексом SLLQHLIGL-HLA-A*02.
Обычно остаток бета-цепи F55 считается частью каркасной области 3. Однако для целей настоящего изобретения остаток бета-цепи F55 считается частью CDR2.
Каркасные участки альфа-цепи FR1, FR2 и FR3 могут содержать аминокислотные последовательности, соответствующие цепи TRAV 26-2, и/или каркасные участки бета-цепи FR1, FR2 и FR3 могут содержать аминокислотные последовательности, соответствующие таковым в цепи TRBV19.
Область FR4 может содержать соединительный участок альфа- и бета-вариабельных цепей (TRAJ и TRBJ, соответственно).
В вариабельной области альфа-цепи TCR может присутствовать по меньшей мере одна мутация. В CDR3 альфа-цепи могут быть одна, две, три, четыре или пять мутаций. Одна или более из указанных мутаций могут быть выбраны из следующих мутаций в соответствии с нумерацией последовательности SEQ ID NO: 2:
Таким образом, могут присутствовать любые или все мутации, перечисленные в приведенной выше таблице, необязательно в сочетании с другими мутациями.
CDR3 альфа-цепи может содержать одну из следующих групп мутаций (в соответствии с нумерацией последовательности SEQ ID NO: 2):
Группа 1 является предпочтительной группой мутаций. Другой предпочтительной группой мутаций является группа 2.
CDR3 альфа-цепи может иметь последовательность, выбранную из:
CILILGHSRAGNYIATF (SEQ ID NO: 45)
CILILGHSRLGNYIATF (SEQ ID NO: 46)
CILILGHSRLGNYQATF (SEQ ID NO: 47)
Предпочтительным CDR3 альфа-цепи является CILILGHSRAGNYIATF (SEQ ID NO: 45). Предпочтительным CDR3 альфа-цепи является CILILGHSRLGNYIATF (SEQ ID NO: 46).
В вариабельной области бета-цепи TCR может присутствовать по меньшей мере одна мутация. Может присутствовать одна, две, три, четыре, пять, шесть, семь, восемь, девять, 10 или более, мутаций в CDR бета-цепи. Может присутствовать одна, две, три, четыре, пять, шесть, семь, восемь, девять или десять мутаций в CDR3 бета-цепи. Одна или более из указанных мутаций могут быть выбраны из следующих мутаций в соответствии с нумерацией последовательности SEQ ID NO: 3:
Таким образом, могут присутствовать любые или все мутации, перечисленные в приведенной выше таблице, необязательно в сочетании с другими мутациями.
CDR2 и CDR3 бета-цепи могут включать одну из следующих групп мутаций (в соответствии с нумерацией последовательности SEQ ID NO: 3):
Группа 1 является предпочтительной группой мутаций. Группа 9 является предпочтительной группой мутаций.
CDR2 бета-цепи может иметь последовательность, выбранную из:
SQIMGDE (SEQ ID NO: 48)
SQIMNDE (SEQ ID NO: 49)
SQIVGDE (SEQ ID NO: 50)
Предпочтительным CDR2 бета-цепи является SQIMGDE (SEQ ID NO: 48).
CDR3 бета-цепи может иметь последовательность, выбранную из:
CASSWWTGGASPISF (SEQ ID NO: 51)
CASSWWTSGASPISF (SEQ ID NO: 52)
CASSWWTGGSAPIRF (SEQ ID NO: 53)
CASSWWTSGSAPIRF (SEQ ID NO: 54)
CASSWWTGGSAEIRF (SEQ ID NO: 55)
CASSWWTGGSAPIYF (SEQ ID NO: 56)
CASSWWTGGAAPISF (SEQ ID NO: 57)
CASSWWTGGASPIRF (SEQ ID NO: 58)
CASSWWTGGAAPIRF (SEQ ID NO: 59)
CASSWWTGGASEISF (SEQ ID NO: 60)
CASSWWTGGSAPISF (SEQ ID NO: 61)
CASSWWTGGSSPISF (SEQ ID NO: 62)
CASSPWTGGSAPIRF (SEQ ID NO: 63)
CASSWWTGGSSPIRF (SEQ ID NO: 64)
Предпочтительным CDR3 бета-цепи является CASSWWTGGASPISF (SEQ ID NO: 51). Предпочтительным CDR3 бета-цепи является CASSWWTGGASPIRF (SEQ ID NO: 58).
Предпочтительными комбинациями CDR2 и CDR3 бета-цепи являются следующие:
Предпочтительной комбинацией является комбинация 1. Другой предпочтительной комбинацией является комбинация 9.
В предпочтительном варианте реализации последовательности CDR альфа- и бета-цепи TCR выбраны из:
(SEQ ID NO: 40)
(SEQ ID NO: 40)
(SEQ ID NO: 40)
(SEQ ID NO: 40)
(SEQ ID NO: 40)
(SEQ ID NO: 40)
(SEQ ID NO: 40)
(SEQ ID NO: 40)
(SEQ ID NO: 47)
(SEQ ID NO: 40)
(SEQ ID NO: 50)
(SEQ ID NO: 40)
(SEQ ID NO: 49)
(SEQ ID NO: 40)
(SEQ ID NO: 40)
(SEQ ID NO: 40)
(SEQ ID NO: 40)
(SEQ ID NO: 40)
(SEQ ID NO: 40)
(SEQ ID NO: 40)
(SEQ ID NO: 40)
(SEQ ID NO: 40)
(SEQ ID NO: 40)
Предпочтительной комбинацией является комбинация 1. Предпочтительной комбинацией является комбинация 17.
Мутация(и) в CDR предпочтительно улучшает аффинность связывания TCR с комплексом SLLQHLIGL-HLA-A*02, но может дополнительно или в качестве альтернативы обеспечивать другие преимущества, такие как повышенная стабильность в выделенной форме и улучшенная специфичность. Мутации в одном или более положениях могут дополнительно или в качестве альтернативы влиять на взаимодействие расположенного в соседнем положении остатка с родственным комплексом pMHC, например, обеспечивая более благоприятный угол для взаимодействия. Мутации могут включать те, которые способны уменьшить количество неспецифичного связывания, то есть уменьшить связывание с альтернативными антигенами по отношению к SLLQHLIGL-HLA-A*02. Мутации могут включать те, которые увеличивают эффективность фолдинга и/или продукции. Некоторые мутации могут влиять на каждую из этих характеристик; другие могут влиять, например, на аффинность, но не специфичность, или на специфичность, но не аффинность, и т.д.
Как правило, для получения TCR с пкM аффинностью к антигену-мишени требуется по меньшей мере 5, по меньшей мере 10, по меньшей мере 15 или более мутаций CDR. По меньшей мере 5, по меньшей мере 10 или по меньшей мере 15 мутаций CDR в целом могут быть необходимы для получения TCR с пкM аффинностью к антигену-мишени. TCR с пкM аффинностью к целевому антигену особенно подходят в качестве растворимых терапевтических средств. TCR, подходящие для применения в адоптивной терапии, могут иметь более низкую аффинность к антигену-мишени и, следовательно, меньше мутаций CDR, например, до 1, до 2, до 5 или более мутаций CDR в целом.
Мутации можно дополнительно или в качестве альтернативы производить вне CDR, внутри каркасных участков; такие мутации могут улучшать связывание, и/или специфичность, и/или стабильность, и/или выход очищенной растворимой формы TCR. Например, TCR согласно настоящему изобретению может дополнительно или в качестве альтернативы содержать вариабельный домен альфа-цепи, где вариабельная область FR1 альфа-цепи имеет остаток G в положении -1 в соответствии с нумерацией SEQ ID NO: 2, т.е. вставлена перед положением 1. Было обнаружено, что G в положении -1 улучшает эффективность отщепления N-концевого метионина во время продукции в E.coli. Неэффективное отщепление может иметь негативные последствия для терапевтического применения, так как оно может привести к гетерогенному белковому продукту, или присутствие инициирующего метионина может быть иммуногенным для человека.
Предпочтительно, согласно настоящему изобретению, вариабельный домен α-цепи TCR может содержать соответствующие каркасные аминокислотные последовательности, которые имеют, по меньшей мере, 90%, по меньшей мере, 91%, по меньшей мере, 92%, по меньшей мере, 93%, по меньшей мере, 94%, по меньшей мере, 95%, по меньшей мере, 96%, по меньшей мере, 97%, по меньшей мере, 98% или, по меньшей мере, 99% идентичности с каркасными аминокислотными остатками 1-25, 33-49, 55-87, 105-114 SEQ ID NO: 2. Согласно настоящему изобретению, вариабельный домен бета-цепи TCR может содержать соответствующие каркасные аминокислотные последовательности, которые имеют, по меньшей мере, 90%, по меньшей мере, 91%, по меньшей мере, 92%, по меньшей мере, 93%, по меньшей мере, 94%, по меньшей мере, 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98% или по меньшей мере 99% идентичности с каркасными аминокислотными остатками 1-26, 32-48, 56-90, 106-114 SEQ ID NO: 3. В качестве альтернативы, указанная процентная идентичность может быть вне каркасных последовательностей при рассмотрении в целом.
Вариабельный домен альфа-цепи может содержать любую из аминокислотных последовательностей SEQ ID NO: 6-8, и вариабельный домен бета-цепи может содержать любую одну из аминокислотных последовательностей SEQ ID NO: 9-24.
Например, TCR может содержать следующие пары альфа- и бета-цепей.
Предпочтительным сочетанием цепей TCR является SEQ ID NO: 6 и SEQ ID NO: 9. Предпочтительным сочетанием цепей TCR является SEQ ID NO: 7 и SEQ ID NO: 17.
Фенотипически не проявляющиеся («молчащие») варианты любого TCR согласно настоящему изобретению, описанные в настоящей заявке, включены в объем изобретения. При использовании в настоящей заявке термин «фенотипически не проявляющийся («молчащий») вариант» относится к вариабельному домену TCR, который содержит одну или более дополнительных аминокислотных замен, включая замены, вставки и делеции, в дополнение к тем, которые изложены выше, где фенотип указанного TCR подобен фенотипу соответствующего TCR, не содержащего указанную замену (замены). В рамках настоящей заявки фенотип TCR включает аффинность связывания антигена (KD и/или период полувыведения в связанной форме) и специфичность. Предпочтительно, фенотип растворимого TCR, связанного с иммунным эффектором, включает в себя эффективность иммунной активации и выход при очистке, в дополнение к аффинности связывания и специфичности. Фенотипически не проявляющийся вариант может иметь KD и/или период полувыведения в связанной форме с комплексом SLLQHLIGL-HLA-A*02 в пределах 50% или, более предпочтительно, в пределах 30%, 25% или 20% от измеренного KD и/или периода полувыведения в связанной форме соответствующего TCR, не содержащего указанной замены (замен), при измерении в идентичных условиях (например, при температуре 25°C и на одной и той же SPR-матрице). Подходящие условия дополнительно определены в Примере 3. Как известно специалисту в данной области техники, возможно получать TCR, содержащие замены в вариабельных доменах по отношению к TCR, подробно описанным выше, без изменения аффинности взаимодействия с комплексом SLLQHLIGL-HLA-A*02 и/или других функциональных характеристик. В частности, такие «молчащие» мутации могут быть включены в части последовательности, которые, как известно, непосредственно не вовлечены в связывание антигена (например, каркасные участки и части CDR, не контактирующие с антигеном). Такие варианты включены в объем настоящего изобретения.
Фенотипически «молчащие» варианты могут содержать одну или более консервативных замен и/или одну или более допустимых замен. Под допустимыми заменами подразумевают те замены, которые не попадают под определение консервативности, как представлено ниже, но, тем не менее, фенотипически не проявляются. Специалисту в данной области техники известно, что различные аминокислоты обладают сходными свойствами и, таким образом, являются «консервативными». Одна или более таких аминокислот белка, полипептида или пептида часто могут быть замещены на одну или более других таких аминокислот без устранения желаемой активности указанного белка, полипептида или пептида.
Таким образом, аминокислоты глицин, аланин, валин, лейцин и изолейцин часто можно заменить друг на друга (аминокислоты, содержащие алифатические боковые цепи). Из указанных возможных замен глицин и аланин предпочтительно использую для замещения друг друга (поскольку они имеют относительно короткие боковые цепи), и валин, лейцин и изолейцин используют для замещения друг друга (поскольку они имеют более объемные алифатические боковые цепи, которые являются гидрофобными). Другие аминокислоты, которые могут часто замещать друг друга, включают: фенилаланин, тирозин и триптофан (аминокислоты, имеющие ароматические боковые цепи); лизин, аргинин и гистидин (аминокислоты, имеющие основные боковые цепи); аспартат и глутамат (аминокислоты, имеющие кислотные боковые цепи); аспарагин и глутамин (аминокислоты, имеющие амидные боковые цепи); и цистеин и метионин (аминокислоты, имеющие сера-содержащие боковые цепи). Следует понимать, что в пределах настоящего изобретения возможны аминокислотные замены с использованием природных или не встречающихся в природе аминокислот. Например, в настоящей заявке предполагается, что метильная группа аланина может быть замещена на этильную группу, и/или что минорные изменения могут быть внесены в пептидный остов. Независимо от использования природных или синтетических аминокислот, предпочтительно, чтобы присутствовали только L- аминокислоты.
Замены такого типа часто называются «консервативными» или «полуконсервативными» аминокислотными заменами. Следовательно, настоящее изобретение распространяется на применение TCR, содержащего любую аминокислотную последовательность, описанную выше, но с одной или более консервативными заменами и/или одной или более допустимыми заменами в последовательности, таким образом, что аминокислотная последовательность TCR имеет, по меньшей мере, 90 % идентичности, например 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности, по отношению к TCR, содержащему аминокислоты 1-114 SEQ ID NO: 2, 6-8 и/или аминокислоты 1-114 из SEQ ID NO: 3, 9-24.
«Идентичность», как известно в данной области техники, представляет собой сходство между двумя или более полипептидными последовательностями или двумя или более полинуклеотидными последовательностями по результатам сравнения указанных последовательностей. В данной области техники идентичность также означает степень родства последовательностей между полипептидными или полинуклеотидными последовательностями (в зависимости от каждого случая) по результатам определения совпадений между цепями таких последовательностей. В то время как существует ряд способов для измерения идентичности между двумя полипептидными или двумя полинуклеотидными последовательностями, способы, которые обычно используются для определения идентичности, закодированы в компьютерных программах. Предпочтительные компьютерные программы для определения идентичности между двумя последовательностями включают пакет программ GCG, но не ограничиваются указанным (Devereux, et al., Nucleic Acids Research, 12, 387 (1984), BLASTP, BLASTN, и FASTA (Atschul et al., J. Molec. Biol. 215, 403 (1990)).
Можно использовать программу, такую как программа CLUSTAL, для сравнения аминокислотных последовательностей. Указанная программа сравнивает аминокислотные последовательности и находит оптимальное выравнивание путем введения промежутков в любую последовательность при необходимости. Возможно рассчитать идентичность или сходство по аминокислотам (идентичность плюс консервативность типа аминокислот) для оптимального выравнивания. Программа типа BLASTx выравнивает сходные последовательности с максимальной протяженностью и присваивает значение соответствию. Таким образом, возможно получать результаты сравнения, в которых обнаруживается несколько областей подобия, все из которых имеют разную оценку. Оба типа анализа идентичности входят в объем настоящего изобретения.
Процентную идентичность двух аминокислотных последовательностей или двух последовательностей нуклеиновых кислот определяют путем выравнивания последовательностей для оптимального сравнения (например, можно вводить разрывы (гэпы) в первую последовательность для наилучшего выравнивания с последовательностью) и сравнения аминокислотных остатков или нуклеотидов в соответствующих положениях. «Наилучшее выравнивание» представляет собой выравнивание двух последовательностей, которое приводит к максимальной процентной идентичности. Процентную идентичность определяют на основе числа идентичных аминокислотных остатков или нуклеотидов в сравниваемых последовательностях (т.е. % идентичности = число идентичных положений/общее число положений x 100).
Определение процентной идентичности между двумя последовательностями можно осуществлять с применением математического алгоритма, известного специалистам в данной области техники. Пример математического алгоритма для сравнения двух последовательностей представляет собой алгоритм Карлина и Алтшуля (Karlin and Altschul (1990) Proc. Natl. Acad. Sci. USA 87:2264-2268), модифицированный в соответствии с источником Karlin and Altschul (1993) Proc. Natl. Acad. Sci. USA 90:5873-5877). Программы NBLAST и XBLAST по Алтшулю и др. (Altschul, et al. (1990) J. Mol. Biol. 215:403-410) включают указанный алгоритм. Определение процентной идентичности
между двумя нуклеотидными последовательностями можно осуществлять с использованием программы BLASTn. Определение процентной идентичности между двумя нуклеотидными последовательностями можно осуществлять с использованием программы BLASTp. Для получения выровненных последовательностей с разрывами (гэпами) для их сравнения можно применять Gapped BLAST, как описано в источнике Altschul et al. (1997) Nucleic Acids Res. 25:3389-3402. В качестве альтернативы, PSI-Blast можно использовать для осуществления поиска с итерациями, который выявляет отдаленные сходства между молекулами (тот же источник). При использовании программ BLAST, Gapped BLAST и PSI-Blast можно использовать параметры, установленные по умолчанию в соответствующих программах (например, BLASTp и BLASTp). См. http://www.ncbi.nlm.nih.gov. Общие параметры по умолчанию могут включать, например, Размер слова = 3, Ожидаемый порог = 10. Данные параметры могут быть выбраны для автоматической настройки коротких вводимых последовательностей. Другим примером математического алгоритма, используемого для сравнения последовательностей, является алгоритм Майерса и Миллера, CABIOS (1989). Программа ALIGN (версия 2.0), которая является частью пакета программного обеспечения для выравнивания последовательностей CGC, содержит такой алгоритм. Общие параметры, установленные по умолчанию, могут включать, например, Размер слова = 3, Ожидаемый порог = 10. Данные параметры могут быть выбраны для автоматической настройки коротких вводимых последовательностей. Другим примером математического алгоритма, используемого для сравнения последовательностей, является алгоритм Майерса и Миллера, CABIOS (1989). Программа ALIGN (версия 2.0), которая является частью пакета программного обеспечения для выравнивания последовательностей CGC, содержит такой алгоритм. Другие алгоритмы для анализа последовательностей, известные в данной области техники, включают ADVANCE и ADAM, как описано в источнике Torellis and Robotti (1994) Comput. Appl. Biosci., 10 :3-5; и FASTA, описанный в источнике Pearson and Lipman (1988) Proc. Natl. Acad. Sci. 85:2444-8. В случае алгоритма FASTA ktup представляет собой контрольный параметр, который устанавливает чувствительность и скорость поиска. В целях оценки процентной идентичности согласно настоящему изобретению, BLASTp используется в качестве способа сравнения с параметрами по умолчанию. Кроме того, когда указанный процент идентичности дает значение не целого числа аминокислот (то есть последовательность из 25 аминокислот, имеющая идентичность последовательности 90%, дает значение «22,5», полученное значение округляется до следующего целого числа, в данном случае, «22»). Соответственно, в представленном примере последовательность, имеющая 22 совпадения из 25 аминокислот, имеет идентичность в пределах 90%.
Как будет понятно для специалиста в данной области техники, можно укоротить или удлинять последовательности, представленные на С-конце и/или N-конце, на 1, 2, 3, 4, 5 или более остатков, существенно не влияя на функциональные характеристики TCR. Последовательности, представленные на С-конце и/или его N-конце, могут быть укорочены или удлинены на 1, 2, 3, 4 или 5 остатков. Все такие варианты охватываются настоящим изобретением.
Мутации, включая консервативные и допустимые замены, вставки и делеции, могут быть введены в указанные последовательности при условии применения подходящего способа, включая, но не ограничиваясь указанными, способы на основе способов полимеразной цепной реакции (ПЦР), рестрикционного клонирования на основе фермента или независимого от лигирования клонирования (НЛК). Указанные способы подробно описаны во многих стандартных работах по молекулярной биологии. Дополнительные подробности относительно полимеразной цепной реакции (ПЦР) и рестрикционного клонирования на основе фермента можно найти в источнике Sambrook & Russell, (2001) Molecular Cloning - A Laboratory Manual (3rd Ed.) CSHL Press. Дополнительную информацию по способам независимого от лигирования клонирования (LIC) можно найти в источнике Rashtchian, (1995) Curr Opin Biotechnol 6(1): 30-6. Последовательности TCR, раскрытые в настоящем изобретении, могут быть получены путем твердофазного синтеза или любого другого подходящего способа, известного в данной области техники.
Согласно настоящему изобретению, TCR обладают свойством связывать комплекс SLLQHLIGL-HLA-A*02. Согласно настоящему изобретению, TCR демонстрируют высокую степень специфичности по отношению к комплексу SLLQHLIGL-HLA-A*02 и, таким образом, особенно подходят для терапевтического применения. Согласно настоящему изобретению, специфичность в контексте TCR относится к их способности распознавать HLA-A*02 клетки-мишени, которые являются антиген-положительными, при этом обладая минимальной способностью распознавать HLA-A*02 клетки-мишени, которые являются антиген-отрицательными.
Специфичность можно измерять in vitro, например, в клеточных тестах, таких как описаны в Примерах 6, 7 и 8. Для оценки специфичности, TCR могут быть представлены в растворимой форме и ассоциированы с иммунным эффектором, и/или могут экспрессироваться на поверхности клеток, таких как T-клетки. Специфичность может быть определена путем измерения уровня активации Т-клеток в присутствии антиген-положительных и антиген-отрицательных клеток-мишеней. Минимальное распознавание антиген-отрицательных клеток-мишеней определяют как уровень активации Т-клеток, составляющий менее 20%, предпочтительно менее 10%, предпочтительно менее 5% и более предпочтительно менее 1% от уровня активации, наблюдаемого в присутствии антиген-положительных клеток-мишеней при измерении в тех же условиях и при терапевтически значимой концентрации TCR. Для растворимых TCR, связанных с иммунным эффектором, терапевтически значимая концентрация может быть определена как концентрация TCR, составляющая 10-9 М или менее, и/или концентрация TCR, до 100 или до 1000 раз превышающая соответствующее значение EC50. Предпочтительно для растворимых TCR, связанных с иммунным эффектором, существует по меньшей мере 100-кратное различие в концентрации, необходимой для активации Т-клеток антиген-положительными клетками по сравнению с антиген-отрицательными клетками. Антиген-положительные клетки могут быть получены путем периодической активации пептидом с использованием подходящей концентрации пептида для получения уровня презентации антигена, сравнимого с раковыми клетками (например, 10-9 М пептида, как описано в источнике Bossi et al., (2013) Oncoimmunol. 1 ; 2 (11): e26840) или данные клетки могут естественно презентировать указанный пептид. Предпочтительно как антиген-положительные, так и антиген-отрицательные клетки принадлежат к клеткам человека. Предпочтительно антиген-положительными клетками являются раковые клетки человека. Антиген-отрицательные клетки предпочтительно включают в себя клетки, полученные из здоровых тканей человека.
Специфичность может дополнительно или в качестве альтернативы относиться к способности TCR связываться с комплексом SLLQHLIGL (SEQ ID NO: 1) HLA-A*02, а не с панелью альтернативных комплексов пептид-HLA, что можно определить, например, используя способ Biacore в соответствии с Примером 3. Указанная панель может содержать, по меньшей мере, 5, а предпочтительно, по меньшей мере, 10 альтернативных комплексов пептид-HLA-A*02. Альтернативные пептиды могут иметь низкий уровень идентичности последовательности с SEQ ID NO: 1 и могут быть презентированы в естественных условиях. Альтернативные пептиды предпочтительно получают из белков, экспрессируемых в здоровых тканях человека. Связывание TCR с комплексом SLLQHLIGL-HLA-A*02 может быть по меньшей мере в 2 раза, по меньшей мере в 10 раз, или по меньшей мере в 50 раз, или по меньшей мере в 100 раз, еще более предпочтительно по меньшей мере в 400 раз сильнее, чем с другими природными пептидными комплексами HLA,
Альтернативный или дополнительный подход для определения специфичности TCR может заключаться в идентификации мотива распознавания пептида TCR с использованием последовательного мутагенеза, например, аланинового сканирования. Остатки, которые составляют часть связующего мотива, не допускаются к замене. Недопустимыми заменами называются те положения пептидов, в которых аффинность связывания TCR снижена по меньшей мере на 50% или предпочтительно по меньшей мере на 80% относительно аффинности связывания немутантного пептида. Такой подход подробно описан в источнике Cameron et al., (2013), Sci Transl Med. 2013 Aug 7; 5 (197): 197ra103 и в публикации международной заявки WO2014096803. Специфичность TCR в этом случае может быть определена путем идентификации пептидов, содержащих альтернативные мотивы, в частности пептидов, содержащих альтернативные мотивы в протеоме человека, и тестирования этих пептидов на связывание с TCR. Связывание TCR с одним или более альтернативными пептидами может указывать на отсутствие специфичности. В этом случае может потребоваться дальнейшее тестирование специфичности TCR с помощью клеточных анализов.
Согласно настоящему изобретению, TCR могут иметь идеальный профиль безопасности для использования в качестве терапевтических реагентов. В этом случае TCR могут находиться в растворимой форме и предпочтительно могут быть гибридизованы с иммунным эффектором. Подходящие иммунные эффекторы включают, но не ограничиваются цитокинами, такими как IL-2 и IFN-γ; суперантигенами и их мутантными формами; хемокинами, такими как IL-8, тромбоцитарный фактор 4, белок, стимулирующий рост меланомы; антителами, включая фрагменты, производные и их варианты, которые связываются с антигенами на иммунных клетках, таких как Т- или NK-клетки (например, анти-CD3, анти-CD28 или анти-CD16); и активаторами системы комплемента. Идеальный профиль безопасности означает, что в дополнение к демонстрации хорошей специфичности, TCR согласно данному изобретению, возможно, прошли дополнительные доклинические испытания безопасности. Примеры таких испытаний включают анализы цельной крови, чтобы подтвердить минимальное высвобождение цитокинов в присутствии цельной крови и, таким образом, показать низкий риск потенциального возникновения синдрома высвобождения цитокинов in vivo, и тесты аллореактивности, чтобы подтвердить низкий потенциал распознавания альтернативных типов HLA.
Согласно настоящему изобретению, TCR, особенно TCR в растворимой форме, могут поддаваться очистке с высоким выходом. Выход может быть определен на основе количества материала, удерживаемого в процессе очистки (т.е. количества правильно сложенного материала, полученного в конце процесса очистки относительно количества солюбилизированного материала, полученного перед рефолдингом), и/или выход может быть основан на количестве правильно сложенного материала, полученного в конце процесса очистки, относительно исходного объема культуры. Высокий выход означает более 1% или более предпочтительно более 5% или более высокий выход материала в процессе очистки. Высокий выход означает более 1 мг/мл или более предпочтительно более 3 мг/мл или более 5 мг/мл или более высокий выход материала в процессе очистки.
TCR согласно настоящему изобретению предпочтительно характеризуются KD для комплекса SLLQHLIGL -HLA-A*02, больше (то есть быть сильнее, чем) KD немутантного или скелетного TCR, например, может находиться в диапазоне от 1 пМ до 100 мкМ. Согласно одному аспекту настоящего изобретения KD конкретных TCR для указанного комплекса может составлять от приблизительно (то есть +/- 10%) 1 пкМ до приблизительно 400 нМ, от приблизительно 1 пМ до приблизительно 1000 пкМ, от приблизительно 1 до приблизительно 500 пкМ. Указанные TCR могут дополнительно или в качестве альтернативы иметь период полувыведения в связанной форме (T½) с комплексом в диапазоне от приблизительно 1 мин до приблизительно 60 ч, от приблизительно 20 мин до приблизительно 50 ч или от приблизительно 2 ч до приблизительно 35 ч. Согласно особенно предпочтительному варианту реализации настоящего изобретения, KD TCR для комплекса SLLQHLIGL -HLA-A*02 составляет от приблизительно 1 пкМ до приблизительно 500 пкМ и/или период полувыведения в связанной форме от приблизительно 2 ч до приблизительно 35 ч. Такая высокая аффинность является предпочтительной для TCR, находящегося в растворимой форме, при ассоциации с терапевтическими агентами и/или с метками с возможностью обнаружения.
Согласно одному аспекту настоящего изобретения KD TCR согласно настоящему изобретению для указанного комплекса может составлять от приблизительно 50 нМ до приблизительно 200 мкМ или от приблизительно 100 нМ до приблизительно 1 мкМ и/или период полувыведения в связанной форме для комплекса может составлять от приблизительно 3 сек до приблизительно 12 мин. Такие TCR могут быть предпочтительными для применения в области адоптивной терапии.
Способы определения аффинности связывания (обратно пропорциональной константе равновесия KD) и периода полувыведения в связанной форме (выраженного как T½) известны специалистам в данной области техники. Согласно предпочтительному варианту реализации, аффинность связывания и периода полувыведения в связанной форме определяют с помощью поверхностного плазмонного резонанса (SPR) или биослойной интерферометрии (BLI), например, с помощью способов BIAcore или Octet, соответственно. Предпочтительный способ представлен в Примере 3. Следует понимать, что удвоение аффинности TCR приводит к уменьшению KD в два раза. T½ рассчитывается как ln2, деленный на скорость диссоциации (koff). Таким образом, удвоение T½ приводит к уменьшению koff в два раза. Значения KD и koff для TCR обычно измеряют для растворимых форм TCR, т.е. для форм TCR, укороченных с удалением остатков цитоплазматического и трансмембранного домена. Чтобы учесть разницу между независимыми измерениями и, в частности, для взаимодействий с временем диссоциации, превышающим 20 часов, аффинность связывания или период полувыведения данного TCR измеряют несколько раз, например, 3 или более, с использованием одного и того же аналитического протокола и берут среднее от полученных результатов. Для сравнения данных связывания между двумя образцами (то есть двумя различными TCR и/или двумя препаратами одного и того же TCR) предпочтительно, чтобы измерения проводились с использованием одних и тех же условий анализа (например, температуры), например, таких как описаны в Примере 3.
Согласно настоящему изобретению, некоторые предпочтительные TCR имеют значительно более высокую аффинность связывания и или период полувыведения в связанной форме для комплекса SLLQHLIGL-HLA-A*02, чем нативные TCR. Увеличение аффинности связывания нативного TCR часто снижает специфичность TCR для его лиганда пептид-MH-, что было продемонстрировано в источнике Zhao et al., (2007) J.Immunol, 179:9, 5845-5854. Однако такие TCR, согласно настоящему изобретению, остаются специфичными для комплекса SLLQHLIGL-HLA-A*02, несмотря на то, что они имеют существенно более высокую аффинность связывания, чем нативные TCR.
Некоторые предпочтительные TCR способны генерировать высокоэффективный Т-клеточный ответ in vitro против антиген-положительных клеток, в частности, тех клеток, которые презентируют антиген на низком уровне, что типично для раковых клеток (т.е. порядка 5-100, например 50, антигенов на клетку). (Bossi et al., (2013) Oncoimmunol. 1;2 (11) :e26840; Purbhoo et al.,(2006). J Immunol 176(12): 7308-7316.)). Такие TCR могут находиться в растворимой форме и могут быть связаны с иммунным эффектором, таким как антитело против CD3. Измеряемый ответ Т-клеток может характеризоваться высвобождением маркеров активации Т-клеток, таких как интерферон γ или гранзим В, или уничтожением клеток-мишеней, или другим показателем активации Т-клеток, таким как пролиферация Т-клеток. Предпочтительно высокоэффективный ответ представляет собой ответ со значением ЕС50 в диапазоне пкМ, наиболее предпочтительно, 100 пкМ или ниже.
Согласно настоящему изобретению, TCR могут представлять собой αβ гетеродимеры. Альфа/бета-гетеродимерные TCR согласно настоящему изобретению обычно содержат последовательность константного домена альфа-цепи TRAC и/или последовательность константного домена бета-цепи TRBC1 или TRBC2. Константные домены могут быть полноразмерными, что предполагает наличие внеклеточных, трансмембранных и цитоплазматических доменов, или они могут находиться в растворимой форме (то есть не иметь трансмембранных или цитоплазматических доменов). Один или оба константных домена могут содержать мутации, замены или делеции по сравнению с нативными последовательностями TRAC и/или TRBC1/2. Термины TRAC и TRBC1/2 также включают естественные полиморфные варианты, например, от N до K в положении 4 TRAC (Bragado et al International immunology. 1994 Feb;6(2):223-30).
Согласно настоящему изобретению, для растворимых TCR последовательности константных доменов альфа- и бета-цепи можно модифицировать путем их укорочения или введения замены с удалением нативной дисульфидной связи между Cys4 экзона 2 TRAC и Cys2 экзона 2 TRBC1 или TRBC2. Последовательность (последовательности) константного домена альфа- и/или бета-цепи может иметь введенную дисульфидную связь между остатками соответствующих константных доменов, как описано, например, в публикации международной заявки WO 03/020763. В предпочтительном варианте реализации, альфа- и бета-константные домены могут быть модифицированы путем замены остатков на цистеин в положении Thr 48 TRAC и положении Ser 57 TRBC1 или TRBC2, причем указанные цистеины образуют дисульфидную связь между альфа- и бета-константными доменами TCR. TRBC1 или TRBC2 могут дополнительно включать мутацию цистеина в аланин в положении 75 константного домена и мутацию аспарагина на аспарагиновую кислоту в положении 89 константного домена. Один или два из константных доменов, присутствующих в αβ-гетеродимере согласно настоящему изобретению, могут быть укорочены на C-конце или C-концах, например, укорочены на вплоть до 15 или до 10, или до 8 или менее аминокислот. Один или оба из внеклеточных константных доменов, присутствующих в αβ-гетеродимере согласно настоящему изобретению, могут быть укорочены на C-конце или C-концах, укорочены, например, на вплоть до 15, или до 10, или до 8 аминокислот. C-конец внеклеточного константного домена альфа-цепи может быть укорочен на 8 аминокислот. Растворимые TCR предпочтительно связаны с терапевтическими агентами и/или с метками с возможностью обнаружения.
Константные домены α/β гетеродимерного TCR могут быть полноразмерными, и иметь как трансмембранный, так и цитоплазматический домены. Такие TCR могут содержать дисульфидную связь, которая соответствует встречающейся в естественных условиях связи между соответствующими альфа- и бета-константными доменами. Кроме того, или в качестве альтернативы, между внеклеточными константными доменами может находится ненативная дисульфидная связь. Указанные ненативные дисульфидные связи дополнительно описаны в публикации международной заявки WO03020763 и WO06000830. Ненативная дисульфидная связь может находиться между положением Thr 48 TRAC и положением Ser 57 TRBC1 или TRBC2. Один или оба константных домена могут содержать одну или более мутаций, замен или делеций по сравнению с нативными последовательностями TRAC и/или TRBC1/2. TCR, содержащие полноразмерные константные домены, являются предпочтительными для использования в адоптивной терапии.
TCR согласно настоящему изобретению могут быть представлены в одноцепочечной форме. Одноцепочечные формы αβ TCR полипептидов включают, но не ограничиваются такими как Vα-L-Vβ, Vβ-L-Vα, Vα-Cα-L-Vβ, Vα-L-Vβ-Cβ или Vα-Сα-L-Vβ-Cβ типами, где Vα и Vβ представляют собой вариабельные участки α- и β-цепей TCR соответственно, Cα и Cβ представляют собой константные области α- и β-цепей TCR соответственно, и L представляет собой линкерную последовательность (Weidanz et al., (1998) J Immunol Methods. Dec 1;221(1-2):59-76; Epel et al., (2002), Cancer Immunol Immunother. Nov;51(10):565-73; WO 2004/033685; WO9918129). Там, где они присутствуют, один или оба константных домена могут быть полноразмерными или они могут быть укорочены и/или содержать мутации, как описано выше. Предпочтительно одноцепочечные TCR являются растворимыми. Согласно некоторым вариантам реализации настоящего изобретения, одноцепочечные TCR могут иметь введенную дисульфидную связь между остатками соответствующих константных доменов, как описано в публикации международной заявки WO 2004/033685. Одноцепочечные TCR дополнительно описаны в публикации международной заявки WO 2004/033685; WO98/39482; WO01/62908; Weidanz et al. (1998) J Immunol Methods 221(1-2): 59-76; Hoo et al. (1992) Proc Natl Acad Sci U S A 89(10): 4759-4763; Schodin (1996) Mol Immunol 33(9): 819-829).
Настоящее изобретение также включает частицы, экспонирующие TCR согласно настоящему изобретению, и включение указанных частиц в библиотеку частиц. Такие частицы включают, но не ограничиваются такими частицами как фаговые частицы, дрожжевые клетки, рибосомы или клетки млекопитающих. Способ получения таких частиц и библиотек известен в данной области техники (например, см. публикацию международной заявки WO 2004/044004; WO 01/48145, Chervin et al. (2008) J. Immuno. Methods 339.2: 175-184).
Согласно настоящему изобретению, растворимые TCR могут применяться для доставки поддающихся выявлению меток или терапевтических агентов в антигенпрезентирующие клетки и ткани, содержащие антигенпрезентирующие клетки. Поэтому они могут быть связаны (ковалентно или иным образом) с меткой с возможностью обнаружения (для диагностических целей, где TCR используется для обнаружения клеток, презентирующих распознанный антиген); и/или c терапевтическим агентом; и/или группой, модифицирующей фармакокинетические (ФК) свойства.
Примеры групп, модифицирующих фармакокинетические (ФК) свойства включают, но не ограничиваются, ПЭГ ((Dozier et al., (2015) Int J Mol Sci. Oct 28;16(10):25831-64 and Jevsevar et al., (2010) Biotechnol J.Jan;5(1):113-28), ПАСилирование (Schlapschy et al., (2013) Protein Eng Des Sel. Aug; 26 (8): 489-501), альбумин и альбумин-связывающие домены (Dennis et. al., (2002) J Biol Chem. Sep 20; 277 (38): 35035-43) и/или неструктурированные полипептиды (Schellenberger et al., (2009) Nat Biotechnol. Dec;27(12):1186-90). Другие группы, модифицирующие фармакокинетические (ФК) свойства, включают Fc-фрагменты антител.
Метки с возможностью обнаружения для диагностических целей включают, например, флуоресцентные метки, радиоактивные метки, ферменты, нуклеиновокислые зонды и контрастирующие реагенты.
Для некоторых целей TCR согласно настоящему изобретению можно объединять в комплекс, содержащий несколько TCR, с получением мультивалентного TCR-комплекса. Существует ряд белков человека, которые содержат домен мультимеризации, который можно использовать для получения мультивалентных комплексов TCR. Например, существует домен тетрамеризации р53, который был использован для получения тетрамеров scFv-фрагментов антитела, обладающих повышенной стабильностью в сыворотке и значительно сниженной скоростью диссоциации по сравнению с мономерным scFv-фрагментом (Willuda et al. (2001) J. Biol. Chem. 276 (17) 14385-14392). Гемоглобин также имеет домен тетрамеризации, который можно использовать для такого применения. Согласно настоящему изобретению, мультивалентный комплекс TCR может иметь улучшенную способность к связыванию комплекса по сравнению с немультимерным рецептором дикого типа или гетеродимером Т-клеточного рецептора согласно настоящему изобретению. Таким образом, мультивалентные комплексы TCR, согласно настоящему изобретению, также включены согласно настоящему изобретению. Такие мультивалентные TCR-комплексы согласно настоящему изобретению, в частности, применимы для отслеживания или нацеливания на клетки, презентирующие конкретные антигены in vitro или in vivo, и также полезны в качестве промежуточных соединений для получения дополнительных мультивалентных TCR-комплексов, которые можно применять таким образом.
Терапевтические агенты, которые могут быть связаны с TCR согласно настоящему изобретению, включают иммуномодуляторы, радиоактивные соединения, ферменты (например, перфорин) или химиотерапевтические агенты (например, цисплатин). Для гарантии того, что токсическое действие развивается в желаемой области, токсин может быть помещен внутрь липосомы, связанной с TCR, таким образом, чтобы происходило медленное высвобождение соединения. Это позволяет предотвращать повреждающее действие во время транспорта в организме и гарантировать, что указанный токсин оказывает максимальное воздействие после связывания TCR с релевантными антиген-презентирующими клетками.
Примеры подходящих терапевтических агентов включают, но не ограничиваются:
низкомолекулярными цитотоксическими агентами, т.е. соединениями, обладающими способностью уничтожать клетки млекопитающих и имеющие молекулярную массу, составляющую менее 700 Дальтон. Такие соединения также могут содержать токсичные металлы, способные оказывать цитотоксическое действие. Более того, необходимо понимать, что указанные низкомолекулярные цитотоксичные агенты также включают про-лекарственные средства, т.е. соединения, которые разрушаются или преобразуются при физиологических условиях с высвобождением цитотоксических агентов. Примеры таких агентов включают цисплатин, производные майтансина, рахелмицин, калихеамицин, доцетаксел, этопозид, гемцитабин, ифосфамид, иринотекан, мелфалан, митоксантрон, порфимер натрий (фотофрин II), темозоломид, топотекан, триметреат глюкуронат, ауристатин E, винкристин и доксорубицин;
 пептидные цитотоксины, т.е. белки или их фрагменты, способные уничтожать клетки млекопитающих. Например, рицин, дифтерийный токсин, экзотоксин А бактерий рода Pseudomonas, ДНКаза и РНКаза;
пептидные цитотоксины, т.е. белки или их фрагменты, способные уничтожать клетки млекопитающих. Например, рицин, дифтерийный токсин, экзотоксин А бактерий рода Pseudomonas, ДНКаза и РНКаза;
радионуклиды, т.е. нестабильные изотопы элементов, распад которых сопровождается испусканием одной или более α- или β- частиц или γ-лучей. Например, йод-131, рений 186, индий 111, иттрий 90, висмут 210 и 213, актиний 225 и астат 213; хелатирующие агенты можно использовать для облегчения соединения указанных радионуклидов с высокоаффинными TCR или их мультимерами;
иммуностимуляторы, т.е. эффекторные молекулы иммунной системы, стимулирующие иммунный ответ. Например, цитокины, такие как IL-2 и IFN-γ,
суперантигены и их мутантные формы;
гибриды TCR/HLA, например, слияние в комплекс пептид/HLA, где указанный пептид происходит из типичного патогена человека, такого как вирус Эпштейна-Барра (EBV);
хемокины, такие как IL-8, тромбоцитарный фактор 4, стимулирующий рост меланомы белок и т.д.;
антитела или их фрагменты, включая антитела против детерминанты T-клеток или NK-клеток (например, антитела против CD3, CD28 или CD16);
альтернативные белковые «скелеты», обладающие подобными антителу характеристиками связывания;
активаторы комплемента;
ксеногенные белковые домены, аллогенные белковые домены, вирусные/бактериальные белковые домены, вирусные/бактериальные пептиды.
Один предпочтительный вариант реализации обеспечивает растворимый TCR согласно настоящему изобретению, связанный (как правило, путем слияния с N- или C-концом альфа- или бета-цепи) с иммунным эффектором. В частности, предпочтительным иммунным эффектором является антитело против CD3 или его функциональный фрагмент или вариант указанного антитела против CD3 (такие гибриды TCR/анти-CD3 могут называться молекулами ImmTAC™). При использовании в настоящей заявке, термин «антитело» включает указанные фрагменты и варианты. Примеры антител против CD3 включают, но не ограничиваются указанными, OKT3, UCHT-1, BMA-031 и 12F6. Фрагменты антител и варианты/аналоги, подходящие для применения в композициях и способах, описанных в настоящей заявке, включают минитела, Fab-фрагменты, F(ab')2-фрагменты, dsFv- и scFv-фрагменты, нанотела (nanobodies™ - конструкции, которые поставляются компанией Ablynx (Бельгия) и содержат один синтетический вариабельный домен тяжелой цепи иммуноглобулина, полученный из антитела верблюдовых (например, верблюда или ламы), и доменные антитела (Domantis, Бельгия), содержащие один вариабельный домен тяжелой цепи иммуноглобулина или вариабельный домен легкой цепи иммуноглобулина с созревшей аффинностью), или альтернативные белковые «скелеты», обладающие подобными антителу характеристиками связывания, такие как аффитела (Affibody, Швеция, содержащие сконструированный скелет белка A), или антикалины (Pieris, Германия, содержащие сконструированные антикалины), среди прочих.
Связывание TCR и антитела против CD3 может происходить через ковалентное или нековалентное присоединение. Ковалентное присоединение может быть прямым или не прямым - через линкерную последовательность. Линкерные последовательности, как правило, являются гибкими, поскольку они состоят главным образом из таких аминокислот, как глицин, аланин и серин, которые не имеют объемных боковых цепей, вероятно, ограничивающих подвижность. В качестве альтернативы, линкеры с большей жесткостью могут быть желательны. Применимую или оптимальную длину линкерных последовательностей легко определить. Длина линкерной последовательности часто составляет менее чем приблизительно 12, например, менее 10 или от 2 до 10 аминокислот. Подходящие линкеры, которые можно использовать в TCR согласно настоящему изобретению, включают, но не ограничиваются указанными: GGGGS (SEQ ID NO: 31), GGGSG (SEQ ID NO: 32), GGSGG (SEQ ID NO: 33), GSGGG (SEQ ID NO: 34), GSGGGP (SEQ ID NO: 35), GGEPS (SEQ ID NO: 36), GGEGGGP (SEQ ID NO: 37), и GGEGGGSEGGGS (SEQ ID NO: 38) (как описано в публикации международной заявки WO2010/133828).
Конкретные варианты реализации конструктов гибридов анти-CD3/TCR согласно настоящему изобретению включают пары альфа- и бета-цепей, в которых альфа-цепь состоит из вариабельного домена TCR, содержащего аминокислотную последовательность SEQ ID NO: 6-8, и/или бета-цепь состоит из вариабельного домена TCR, содержащего аминокислотную последовательность SEQ ID NO: 9-24. Указанные альфа- и бета-цепи могут дополнительно содержать константную область, содержащую ненативную дисульфидную связь. Константный домен альфа-цепи может быть укорочен на восемь аминокислот. N или С-конец альфа- и/или бета-цепи может быть гибридизован с scFv-фрагментом антитела против CD3 через линкер, выбранный из SEQ ID NO: 31-38. Некоторые предпочтительные варианты реализации таких конструктов, представляющих собой гибридные молекулы анти-CD3/TCR, представлены ниже:
Также в объем настоящего изобретения включены функциональные варианты указанных гибридных молекул TCR/анти-CD3. Указанные функциональные варианты предпочтительно идентичны по меньшей мере на 90%, например, на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичны референсной последовательности, но тем не менее функционально эквивалентны.
Согласно дополнительному аспекту, настоящее изобретение относится к нуклеиновой кислоте, кодирующей TCR или гибридную молекулу TCR/анти-CD3 согласно настоящему изобретению. В некоторых вариантах реализации нуклеиновая кислота представляет собой кДНК. В некоторых вариантах реализации нуклеиновая кислота может представлять собой мРНК. В некоторых вариантах реализации изобретение относится к нуклеиновой кислоте, содержащей последовательность, кодирующую вариабельный домен α-цепи TCR согласно настоящему изобретению. В некоторых вариантах реализации изобретение относится к нуклеиновой кислоте, содержащей последовательность, кодирующую вариабельный домен β-цепи TCR согласно настоящему изобретению. Нуклеиновая кислота может быть не встречающейся в природе и/или очищенной и/или сконструированной. Последовательность нуклеиновой кислоты может быть кодон-оптимизирована в соответствии с используемой системой экспрессии. Как известно специалистам в данной области техники, системы экспрессии могут включать бактериальные клетки, такие как Е.coli, или дрожжевые клетки, или клетки млекопитающих, или клетки насекомых, или могут представлять собой бесклеточные системы экспрессии.
Согласно другому аспекту, изобретение обеспечивает вектор, который содержит нуклеиновую кислоту согласно настоящему изобретению. Предпочтительно, вектор представляет собой вектор экспрессии TCR. Подходящие векторы экспрессии TCR включают, например, гамма-ретровирусные векторы или, более предпочтительно, лентивирусные векторы. Более подробную информацию можно найти в источнике Zhang 2012 и в процитированных в нем источниках (Zhang et al,. Adv Drug Deliv Rev. 2012 Jun 1; 64(8): 756-762).
Изобретение также обеспечивает клетку, содержащую вектор согласно настоящему изобретению, предпочтительно, вектор экспрессии TCR. Подходящие клетки включают клетки млекопитающих, предпочтительно иммунные клетки, еще более предпочтительно, Т-клетки. Вектор может содержать нуклеиновую кислоту согласно настоящему изобретению, кодирующую в одной открытой рамке считывания или двух отдельных открытых рамках считывания альфа-цепь и бета-цепь соответственно. Согласно другому аспекту, изобретение обеспечивает клетку, содержащую первый вектор экспрессии, который содержит нуклеиновую кислоту, кодирующую альфа-цепь TCR согласно настоящему изобретению, и второй вектор экспрессии, который содержит нуклеиновую кислоту, кодирующую бета-цепь TCR согласно настоящему изобретению. Такие клетки, в частности, применимы для адоптивной терапии. Клетки, согласно настоящему изобретению, могут быть выделенными и/или рекомбинантными и/или не встречающимися в природе и/или сконструированными.
Поскольку TCR согласно настоящему изобретению применимы в адоптивной терапии, изобретение включает не встречающуюся в природе и/или очищенную и/или сконструированную клетку, в частности, T-клетку, презентирующую TCR согласно настоящему изобретению. Изобретение также обеспечивает размноженную популяцию T-клеток, презентирующих TCR согласно настоящему изобретению. Существует ряд способов, подходящих для трансфекции T-клеток нуклеиновой кислотой (такой как, ДНК, кДНК или РНК), кодирующей TCR согласно настоящему изобретению (см., например, Robbins et al., (2008) J Immunol. 180: 6116-6131). T-клетки, экспрессирующие TCR согласно настоящему изобретению, подходят для применения в лечении рака на основе адоптивной терапии. Как известно специалисту в данной области техники, существует ряд подходящих способов, с помощью которых можно проводить адоптивную терапию (см., например, Rosenberg et al., (2008) Nat Rev Cancer 8(4): 299-308).
Как хорошо известно в данной области техники, TCR можно подвергать посттрансляционным модификациям. Гликозилирование представляет собой одну из таких модификаций, которая включает ковалентное присоединение олигосахаридных групп к определенным аминокислотам в цепи TCR. Например, аспарагиновые остатки или серин/треониновые остатки представляют собой хорошо известные положения присоединения олигосахарида. Статус гликозилирования конкретного белка зависит от ряда факторов, включая белковую последовательность, конформацию белка и доступность конкретных ферментов. Более того, статус гликозилирования (т.е. тип олигосахарида, ковалентная связь и общее количество присоединений) может влиять на функцию белка. Таким образом, при получении рекомбинантных белков часто является желательным контролировать гликозилирование. Контролируемое гликозилирование использовали для улучшения терапевтических средств на основе антител (Jefferis R., Nat Rev Drug Discov. 2009 Mar;8(3):226-34.). Для растворимых TCR согласно настоящему изобретению гликозилирование можно контролировать, например, с использованием конкретных клеточных линий, например (включая, но не ограничиваясь клеточными линиями млекопитающих, такими как клетки яичников китайского хомячка (CHO) или клетки эмбриональной почки человека (HEK)), или путем химической модификации. Такие модификации могут быть желательны, поскольку гликозилирование может улучшать фармакокинетику, уменьшать иммуногенность и более точно имитировать нативный белок человека (Sinclair and Elliott, (2005) Pharm Sci.Aug; 94 (8): 1626-35).
Для введения пациентам, TCR согласно настоящему изобретению (предпочтительно связанные с меткой с возможностью обнаружения или терапевтическим агентом или экспрессируемые на трансфицированной T-клетке), гибридные молекулы TCR/анти-CD3, нуклеиновые кислоты, векторы экспрессии или клетки согласно настоящему изобретению могут быть представлены в составе стерильной фармацевтической композиции вместе с одним или более фармацевтически приемлемыми носителями или вспомогательными веществами. Указанная фармацевтическая композиция может быть представлена в любой подходящей форме (в зависимости от желаемого способа ее введения пациенту). Указанная композиция может обеспечиваться в единичной форме дозирования и ее в целом обеспечивают в запечатанном контейнере и она может быть представлена как часть набора. Такой набор обычно (но не обязательно) содержит инструкции по применению. Указанный набор может включать множество указанных единичных форм дозирования.
Фармацевтическая композиция может быть адаптирована для введения с помощью любого подходящего способа, такого как парентеральное (включая подкожное, внутримышечное, интратекальное или внутривенное), кишечное (включая пероральное или ректальное), ингаляционное или интраназальное введение. Такие композиции могут быть получены с помощью любого способа, известного в области фармацевтики, например, путем смешивания активного ингредиента с носителем (носителями) или вспомогательным веществом (веществами) в стерильных условиях.
Дозы веществ согласно настоящему изобретению могут варьировать в широких пределах в зависимости от заболевания или расстройства, подлежащего лечению, возраста и состояния субъекта, подлежащего лечению, и т.д. Подходящий диапазон доз для гибридных молекул TCR/анти-CD3, согласно настоящему изобретению, может составлять между 25 нг/кг и 50 мкг/кг или между 1 мкг и 1 г. В конечном итоге, подходящие дозы для применения определяет врач.
TCR, гибридные молекулы TCR/анти-CD3, фармацевтические композиции, векторы, нуклеиновые кислоты и клетки согласно настоящему изобретению могут быть представлены по существу в чистой форме, например, по меньшей мере на 80%, по меньшей мере на 85 %, по меньшей мере на 90%, по меньшей мере на 91%, по меньшей мере на 92%, по меньшей мере на 93%, по меньшей мере на 94%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99% или на 100% чистой форме.
Изобретение также обеспечивает:
TCR, гибридную молекулу TCR/анти-CD3, нуклеиновую кислоту, фармацевтическую композицию или клетку согласно настоящему изобретению для применения в медицине, предпочтительно для применения в способе лечения рака или опухоли;
применение TCR, гибридной молекулы TCR/анти-CD3, нуклеиновой кислоты, фармацевтической композиции или клетки согласно настоящему изобретению в изготовлении лекарственного средства для лечения рака или опухоли;
способ лечения рака или опухоли у пациента, включающий введение указанному пациенту TCR, гибридной молекулы TCR/анти-CD3, нуклеиновой кислоты, фармацевтической композиции или клетки согласно настоящему изобретению;
состав для инъекции для введения субъекту, представляющему собой человека, содержащий TCR, гибридную молекулу TCR/анти-CD3, нуклеиновую кислоту, фармацевтическую композицию или клетку согласно настоящему изобретению.
Рак, подлежащий лечению, может представлять собой солидную опухоль или гемобластоз (“liquid tumor”). Предпочтительно опухоль экспрессирует PRAME. Рак может представлять собой рак молочной железы (включая тройной отрицательный), яичника, эндометрия, пищевода, легкого (NSCLC и SCLC), мочевого пузыря или головы и шеи. В качестве альтернативы или дополнительно рак может представлять собой лейкоз или лимфому. Из этих видов рака предпочтительными являются рак молочной железы (включая тройной негативный), яичника и эндометрия. TCR, гибридную молекулу TCR-анти-CD3, нуклеиновую кислоту, фармацевтическую композицию или клетку, согласно настоящему изобретению, можно вводить путем инъекции, такой как внутривенная или прямая внутриопухолевая инъекция. Субъект, представляющий собой человека, может иметь подтип HLA-A*02.
Способ лечения может дополнительно включать введение отдельно, в комбинации или последовательно дополнительного антинеопластического агента. Примеры таких агентов известны в данной области и могут включать иммуноактивирующие агенты и/или агенты, модулирующие Т-клетки.
Предпочтительные признаки каждого аспекта настоящего изобретения соответствуют предпочтительным признакам каждого из других аспектов с соответствующими изменениями. Документы согласно предшествующему уровню техники, упомянутые в настоящей заявке, включены посредством ссылки в максимальном объеме, разрешенном законом.
Описание чертежей
На Фигуре 1 представлены аминокислотные последовательности внеклеточных участков альфа- и бета-цепей «скелетного» TCR, специфичного к PRAME.
На Фигуре 2 представлены аминокислотные последовательности внеклеточных участков растворимых вариантов нативных альфа- и бета-цепей «скелетного» TCR, специфичного к PRAME.
На Фигуре 3 представлены аминокислотные последовательности мутантных вариабельных участков альфа-цепи TCR, специфичного к PRAME.
На Фигуре 4 представлены аминокислотные последовательности мутантных вариабельных участков бета-цепи TCR, специфичного к PRAME.
На Фигуре 5 представлены аминокислотные последовательности молекул ImmTAC, (гибриды TCR/анти-CD3) содержащие определенные мутантные вариабельные домены TCR, специфичного к PRAME, как показано на фигурах 3 и 4.
На Фигуре 6 представлены данные анализа на клетках, демонстрирующие активность и специфичность молекул ImmTAC, представленных на фигуре 5, содержащих мутантные вариабельные домены TCR, специфичного к PRAME, как показано на фигурах 3 и 4.
На Фигуре 7 (панели a и b) представлены данные анализа на клетках, демонстрирующие специфичность молекул ImmTAC, представленных на фигуре 5, содержащих мутантные вариабельные домены TCR, специфичного к PRAME, как показано на фигурах 3 и 4.
На фигуре 8 представлены данные анализа на клетках, демонстрирующие уничтожение PRAME положительных раковых клеток меланомы молекулами ImmTAC, представленными на фигуре 5, включающими мутантные вариабельные домены TCR, специфичного к PRAME, как показано на фигурах 3 и 4.
Фигура 9 представлены данные анализа на клетках, демонстрирующие уничтожение PRAME положительных клеток рака легкого молекулами ImmTAC представленными на фигуре 5, включающими мутантные вариабельные домены TCR, специфичного к PRAME, как показано на фигурах 3 и 4.
Изобретение описано далее в следующих неограничивающих примерах:
Примеры
Пример 1 - Экспрессия, рефолдинг и очистка растворимых TCR
Способ
Последовательности ДНК, кодирующие внеклеточные участки альфа- и бета-цепей растворимых TCR согласно настоящему изобретению клонировали раздельно в экспрессионные плазмиды на основе pGMT7 с применением стандартных способов (как описано в источнике Sambrook, et al. Molecular cloning. Vol. 2. (1989) New York: Cold spring harbor laboratory press). Экспрессионые плазмиды трансформировали раздельно в штамм E. coli Rosetta (BL21pLysS) или Т7 Express и одиночные ампициллин-устойчивые колонии растили при температуре 37°C в среде TYP (+ ампициллин, 100 мкг/мл) до достижения OD600 ~0,6-0,8 с последующей индукцией экспрессии белка с помощью 0,5 мМ IPTG. Клетки собирали через три часа после индукции посредством центрифугирования. Клеточные осадки лизировали с помощью реагента для белковой экстракции BugBuster (Merck Millipore) в соответствии с инструкциями производителя. Осадки, содержащие тельца включения, восстанавливали путем центрифугирования. Осадки промывали два раза в Тритон-содержащем буфере (50 мМ Трис-HCI pH 8,1, 0,5% Тритон-X100, 100 мМ NaCl, 10 мМ Na-ЭДТА) и, наконец, ресуспендировали в не содержавшем детергент буфере (50 мМ Трис-HCl pH 8,1, 100 мМ NaCl, 10 мМ Na-ЭДТА). Белковый выход телец включения определяли количественно путем солюбилизирования с помощью 6 М гуанидина-HCl и измерения OD280. Затем рассчитывали концентрацию белка с использованием коэффициента экстинкции. Чистоту телец включения измеряли путем солюбилизирования с помощью 8 М мочевины и нанесения ~2 мкг на 4-20% ДСН-ПААГ при восстанавливающих условиях. Затем оценивали или рассчитывали чистоту с применением денситометрической программы (Chemidoc, Biorad). Тельца включения хранили при +4°C для кратковременного хранения и при -20°C или -70°C для более длительного хранения.
Для рефолдинга растворимых TCR содержащие α- и β-цепи тельца включения сначала перемешивали и разводили в 10 мл буфера для солюбилизирования/денатурирования (6 М гидрохлорид гуанидина, 50 мМ Трис HCl pH 8,1, 100 мМ NaCl, 10 мМ ЭДТА, 20 мМ DTT) с последующим инкубированием в течение 30 мин при 37°C. Затем инициировали рефолдинг путем дополнительного разведения в 1 л буфера для рефолдинга (100 мМ Трис pH 8,1, 800 или 1000 мМ L-аргинин-HCL, 2 мМ ЭДТА, 4 M мочевина, 10 мМ гидрохлорид цистеамина и 2,5 мМ дигидрохлорид цистамина) и раствор хорошо перемешивали. Подвергнутую рефолдингу смесь диализовали против 10 л H2O в течение 18-20 часов при 5°C ± 3°C. После этого диализный буфер дважды замещали на 10 мМ Трис, (pH 8,1, 10 л) и продолжали проводить диализ в течение дополнительных 15 часов. Подвергнутую рефолдингу смесь затем фильтровали через 0,45 мкм целлюлозные фильтры.
Очистку растворимых TCR инициировали путем нанесения диализированного продукта рефолдинга на анионообменную колонку POROS® 50HQ и элюирования связанного белка с помощью градиента 0-500 мМ NaCl в 20 мМ Трис pH 8,1 с использованием 6 колоночных объемов с применением очистителя Akta® (GE Healthcare). Пиковые фракции TCR идентифицировали с помощью электрофореза в полиакриламидном геле в присутствии додецил-сульфата натрия (ДСН-ПААГ) с их последующим объединением и концентрированием. Концентрированный образец затем наносили на колонку для гель-фильтрации Superdex® 75HR (GE Healthcare), предварительно уравновешенную в ФСБ-буфере Дульбекко. Пиковые фракции TCR объединяли и концентрировали и рассчитывали конечный выход очищенного материала.
Пример 2 - Экспрессия, рефолдинг и очистка молекул ImmTAC (растворимых гибридных молекул TCR/анти-CD3)
Способ
ImmTAC получали, как описано в Примере 1, за исключением того, что бета-цепь TCR гибридизовали через линкер с одноцепочечным антителом против CD3. Кроме того, этап катионного обмена осуществляли во время очистки после анионного обмена. В этом случае пиковые фракции, полученные в результате анионного обмена, разводили в 20 раз в 20 мМ MES (pH 6,5) и наносили на катионообменную колонку POROS® 50HS. Связанный белок элюировали с помощью градиента 0-500 мМ NaCl в 20 мМ MES. Пиковые фракции ImmTAC объединяли и доводили до pH 8,1 с помощью 50 мМ раствора Tris, затем концентрировали и наносили непосредственно на матрицу для гель-фильтрации, как описано в Примере 1.
Пример 3 - Характеристика связывания
Анализ связывания очищенных растворимых TCR и молекул ImmTAC с комплексом релевантный пептид/HLA проводили с помощью поверхностного плазмонного резонанса с применением инструмента BIAcore 3000 или BIAcore T200 или с помощью биослойной интерферометрии с применением инструмента ForteBio Octet). Биотинилированные молекулы HLA-A*02 I класса подвергали рефолдингу с интересующим пептидом и очищали с помощью способов, известных специалистам в данной области техники (O'Callaghan et al. (1999). Anal Biochem 266(1): 9-15; Garboczi, et al. (1992). Proc Natl Acad Sci USA 89(8): 3429-3433). Все измерения проводили при температуре 25°C в фосфатно-солевом буфере (ФСБ) Дульбекко с добавлением 0,005% P20.
Анализ BIAcore
Биотинилированные мономеры пептид-HLA иммобилизировали на связанных со стрептавидином сенсорных чипах CM-5. Равновесные константы связывания определяли путем пропускания растворимых TCR/ImmTAC в серийных разведениях при постоянной скорости потока, составляющей 30 мкл/мин-1, через проточную кювету, покрытую комплексом пептид/HLA-A*02 (~200 единиц ответа, RU). Равновесные ответы нормировали для каждой концентрации TCR путем вычитания общего ответа для буфера в контрольной проточной кювете, содержащей нерелевантный пептид-HLA. Значение KD получали с помощью нелинейной аппроксимации кривой с использованием программы Prism и изотермы связывания Ленгмюра: связывание = C*Max/(C + KD), где «связывание» представляет собой равновесное связывание в RU, C представляет собой концентрацию вводимого TCR и Max представляет собой максимальное связывание.
Для высокоаффинных взаимодействий параметры связывания определяли с помощью кинетического анализа с одним циклом. Растворимые TCR/ImmTAC в пяти различных концентрациях пропускали через проточную кювету, покрытую комплексом пептид-HLA (~100 - 200 RU), при скорости потока, составляющей 50-60 мкл/мин-1. Как правило, 60-120 мкл растворимых TCR/ImmTAC вводили в максимальной концентрации, составляющей 50-100 нМ, с последующими 2-кратными разведениями, которые использовали для других четырех инъекций. Минимальную концентрацию вводили первой. Затем вводили буфер для измерения фазы диссоциации до тех пор, пока не происходило ≥ 10% диссоциации, как правило, через 1 - 3 часа. Кинетические параметры рассчитывали с помощью программы BIAevaluation®. Фазу диссоциации приводили к единому уравнению экспоненциального распада, обеспечивая расчет полувыведения. Константу равновесия KD рассчитывали из koff/kon.
Анализ Octet
Биотинилированные мономеры пептид/HLA захватывали на стрептавидиновые (SA) биосенсоры размером 1 нм (Pall ForteBio), предварительно иммобилизированные со стрептавидином. Сенсоры блокировали свободным биотином (2 мкМ) в течение 2 минут. Равновесные константы связывания определяли путем помещения нагруженных биосенсоров в серийные разведения растворимого TCR/ImmTAC в 96-луночный или 384-луночный планшет для образцов. Скорость перемешивания планшета устанавливали на 1000 об./мин. Для низкоаффинных взаимодействий (мкМ диапазон) использовали короткое время ассоциации (~2 минуты) и короткое время диссоциации (~2 минуты). Кривые связывания обрабатывали путем двукратного вычитания референсного значения для референсных биосенсоров, нагруженных нерелевантным pHLA, с применением программы анализа данных Octet (Pall ForteBio). Ответы (нм) при равновесном состоянии использовали для оценки значения KD из графиков для состояния покоя, приведенных к уравнению: ответ = Rmax*конц./(KD + конц.), где «ответ» представляет собой равновесное связывание в нм для каждой концентрации TCR (конц.), и Rmax представляет собой ответ, соответствующий максимальному связыванию, при насыщении pHLA.
Для взаимодействий с высокой аффинностью (диапазон нМ - пкМ) кинетические параметры определяли из кривых связывания при концентрации ≥ 3 TCR/ImmTAC, составляющей, как правило, 10 нМ, 5 нМ и 2,5 нМ. Время ассоциации составляло 30 минут, и время диссоциации составляло 1 - 2 часа. Кривые связывания обрабатывали путем двукратного вычитания референсных значений для референсных биосенсоров, нагруженных нерелевантным pHLA и блокированных биотином. Кинетические параметры kon и koff рассчитывали путем общей аппроксимации непосредственно к кривым связывания с применением программы анализа данных Octet (Pall ForteBio). KD рассчитывали из koff/kon и период полудиссоциации рассчитывали из уравнения t1/2 = 0,693/koff.
Пример 4 - Характеристика связывания нативного TCR
Растворимый нативный TCR готовили в соответствии со способами, описанными в Примере 1, и связывание с pHLA анализировали в соответствии с Примером 3. Аминокислотные последовательности альфа- и бета-цепей соответствовали последовательностям, показанным на Фигуре 2. Растворимый биотинилированный HLA-A*02 готовили с пептидом PRAME SLLQHLIGL (SEQ ID NO: 1) и иммобилизировали на сенсорном чипе BIAcore.
Результаты
Связывание определяли при различных концентрациях, и значение KD взаимодействия составляло 141 мкМ. Перекрестную реактивность (специфичность) оценивали по панели, состоящей из 14 нерелевантных пептидных комплексов HLA-A*02 с использованием равновесного способа BIAcore как описано в Примере 3. 14 нерелевантных pHLA разделили на три группы и загрузили в одну из трех проточных кювет для получения приблизительно 1000 RU каждого pHLA на проточную кювету. 30 мкл растворимого TCR дикого типа пропускали в концентрациях 130 и 488 мкМ через все проточные кюветы при скорости потока 20 мкл/мин. При обеих концентрациях не было обнаружено значительного связывания, что указывает на то, что нативный TCR специфичен для комплекса SLLQHLIGL-HLA-A*02.
Эти данные указывают на то, что данный нативный TCR обладает характеристиками, подходящими для его использования в качестве исходной последовательности для конструирования высокоаффинных терапевтических TCR.
Пример 5 - Характеристика связывания определенных мутантных TCR согласно настоящему изобретению
Мутантные аминокислотные последовательности вариабельного домена альфа- и бета-цепи TCR, представленные на Фигурах 3 и 4 соответственно (SEQ ID NO: 6-24), использовали для получения молекул ImmTAC. Следует отметить, что включение остатка глицина в начале альфа-цепи (положение -1 относительно нумерации SEQ ID NO: 2), как было обнаружено, улучшает эффективность отщепления N-концевого метионина во время продуцирования в E.coli. Неэффективное отщепление может быть вредным для терапевтического средства, поскольку оно может привести к образованию гетерогенного белкового продукта и/или присутствие инициирующего метионина может быть иммуногенным для людей. Полные аминокислотные последовательности молекул ImmTAC, включающие следующие альфа- и бета-цепи, представлены на Фигуре 5
a28b50 - ImmTAC1
a79b74 - ImmTAC2
a79b46 - ImmTAC3
Молекулы получали, как описано в примере 2, и связывание с комплексом SLLQHLIGL-HLA-A*02 определяли согласно примеру 3.
Результаты
Данные, представленные в таблице ниже, показывают, что молекулы ImmTAC, содержащие указанные последовательности вариабельного домена TCR, распознают комплекс SLLQHLIGL-HLA-A*02 с особенно подходящей аффинностью и/или периодом полувыведения.
Пример 6 - Характеристика эффективности и специфичности некоторых мутантных TCR согласно настоящему изобретению
Молекулы ImmTAC, содержащие те же последовательности вариабельного домена TCR, которые представлены в Примере 5, оценивали по их способности опосредовать активное и специфичное перенаправление CD3+ T-клеток против PRAME-положительных раковых клеток. Секрецию интерферона-γ (IFN- γ) использовали в качестве регистрируемого показателя активации T-клеток. Полноразмерные аминокислотные последовательности альфа и бета-цепей исследованных молекул ImmTAC представлены на Фигуре 5.
a28b50 - ImmTAC1
a79b74 - ImmTAC2
a79b46 - ImmTAC3
Анализы проводили с применением набора для ELISPOT-анализа IFN-γ человека (BD Biosciences). Вкратце, клетки-мишени готовили в плотности 1×106/мл в аналитической среде (RPMI 1640, содержащей 10% термоинактивированной ЭБС и 1% смеси пенициллин-стрептомицин-L-глутамин) и высевали в плотности 50,000 клеток на лунку в объеме 50 мкл. Мононуклеарные клетки периферической крови (МКПК), выделенные из свежей крови донора, использовали в качестве эффекторных клеток и помещали в концентрации 50,000 клеток на лунку в объеме 50 мкл (точное количество клеток, используемых в каждом эксперименте, зависит от донора и может быть скорректировано для получения ответа в пределах подходящего диапазона). Молекулы ImmTAC титровали для получения конечных концентраций в 10 нМ, 1 нМ, 0,1 нМ, 0,01 нМ и 0,001 нМ, охватывая предполагаемый клинически значимый диапазон, и добавляли в лунку в объеме 50 мкл.
Планшеты готовили в соответствии с инструкциями производителя. Клетки-мишени, эффекторные клетки и молекулы ImmTAC добавляли в соответствующие лунки и доводили до конечного объема, составляющего 200 мкл аналитической среды. Все реакции проводили в трех повторениях. Также готовили контрольные лунки с отсутствием ImmTAC, эффекторных клеток или клеток-мишеней. Планшеты затем инкубировали в течение ночи (37°C/5% CO2). На следующий день планшеты промывали три раза промывочным буфером (пакетик для приготовления 1xPBS, содержащий 0,05% Tween-20, разведенный в деионизированной воде). Затем в каждую лунку добавляли первичные антитела для выявления в объеме 50 мкл. Планшеты инкубировали при комнатной температуре в течение 2 часов до повторной трехкратной промывки. Вторичное выявление осуществляли путем добавления 50 мкл разведенного конъюгата стрептавидин-HRP в каждую лунку и инкубирования при комнатной температуре в течение 1 часа и повторяли этап промывки. Не более чем за 15 мин до использования на каждый 1 мл субстрата AEC добавляли одну каплю (20 мкл) хромогена AEC и перемешивали. 50 мкл полученной смеси добавляли в каждую лунку. Постоянно отслеживали появление точек и планшеты промывали в водопроводной воде для остановки проявления. Планшеты высушивали при комнатной температуре в течение по меньшей мере 2 часов с последующим подсчетом точек при помощи анализатора CTL с программным обеспечением Immunospot (Cellular Technology Limited).
В этом примере в качестве клеток-мишеней использовались следующие линии раковых клеток:
Mel624 (меланома) PRAME+ve HLA-A*02+ve
Granta519 (гемо-лимфоцитарный рак) PRAME-ve HLA-A*02+ve
SW620 (рак толстой кишки) PRAME-ve HLA-A*02+ve
HT144 (меланома) PRAME+ve HLA-A*02-ve
Результаты
Каждая из молекул ImmTAC, включающая альфа- и бета-вариабельные домены, указанные в таблице ниже, продемонстрировала эффективную активацию перенаправленных Т-клеток в присутствии антиген-положительных клеток Mel624. В каждом случае значения EC50 были рассчитаны на основе данных и показаны в таблице ниже. Кроме того, каждая молекула ImmTAC продемонстрировала минимальное распознавание или отсутствие распознавания двух антиген-отрицательных, HLA-A*02-положительных клеток в концентрации до 1 нМ. Молекулы ImmTAC также продемонстрировали отсутствие распознавания PRAME-положительных клеток, которые являются HLA-A*02-отрицательными (данные не показаны). На рисунке 6 показаны репрезентативные данные из четырех молекул ImmTAC, перечисленных в таблице ниже.
Эти данные показывают, что молекулы ImmTAC, содержащие мутантные последовательности вариабельного домена TCR, согласно настоящему изобретению, могут опосредовать активное и специфичное перенаправление Т-клеток против PRAME-положительных, HLA-A*02-положительных раковых клеток в диапазоне концентраций, подходящих для терапевтического применения.
Пример 7. Дальнейшая характеристика специфичности некоторых мутантных TCR согласно настоящему изобретению.
Чтобы дополнительно продемонстрировать специфичность молекул ImmTAC, содержащих мутантные последовательности TCR, было проведено дополнительное тестирование с использованием того же способа ELISPOT, как описано в Примере 6, с использованием панели нормальных клеток, полученных из здоровых тканей человека, в качестве клеток-мишеней.
Нормальные ткани включали сердечно-сосудистые, почечные, скелетные мышцы, легочную, сосудистую сеть, печеночную и ткань мозга. В каждом случае антиген-положительные раковые клетки Mel624 использовали в качестве положительного контроля.
Данные, представленные в этом примере, включают молекулы ImmTAC, содержащие следующие альфа- и бета-цепи TCR
a28b50
a79b74
a79b46
a79b77
Полные аминокислотные последовательности молекул ImmTAC, включающие a28b50, a79b74 и a79b46, представлены на Фигуре 5 (ImmTAC 1-3 соответственно).
Результаты
Данные, представленные на фиг.7 (панель а), указывают, что молекулы ImmTAC, содержащие мутантные альфа- и бета-цепи a28b50 и a79b46, демонстрируют минимальную реактивность по отношению к панели из 8 нормальных клеток по сравнению с антиген-положительными раковыми клетками в концентрации до 1 нМ. Аналогично, данные на фиг.7 (панель b) демонстрируют, что молекулы ImmTAC, содержащие a28b57 и a79b46, проявляют минимальную реактивность по отношению к панели, состоящей из 4 нормальных клеток по сравнению с антиген-положительными раковыми клетками в концентрации до 1 нМ.
Пример 8 - Уничтожение раковых клеток, опосредованное некоторыми мутантными TCR согласно настоящему изобретению
Способность молекул ImmTAC, содержащих мутантные последовательности TCR, опосредовать активное перенаправленное уничтожение T-клетками антиген-положительных опухолевых клеток исследовали с использованием платформы IncuCyte (Essen BioScience). Указанный анализ позволяет выявлять высвобождения каспазы-3/7 - маркера апоптоза - в реальном времени с помощью микроскопии.
Способ
Анализы проводили с использованием набора для анализа апоптоза на основе каспазы-3/7 CellPlayer для 96-луночго планшета (Essen BioScience, каталожный номер: 4440) в соответствии с протоколом производителя. Вкратце, клетки-мишени (Mel624 (PRAME+ve HLA-A*02+ve) или NCI-H1755) высевали в концентрации 10000 клеток на лунку и инкубировали в течение ночи, чтобы позволить им прикрепиться. Растворы ImmTAC готовили в различных концентрациях и 25 мкл раствора в каждой концентрации добавляли в соответствующую лунку с получением конечной концентрации от 1 пкМ до 100 пкМ. Эффекторные клетки использовали при соотношении эффекторных клеток-мишеней 10: 1 (100000 клеток на лунку). Контрольный образец, не содержавший ImmTAC, также готовили вместе с образцами, содержащими либо только эффекторные клетки, либо только клетки-мишени. 25 мкл реагента для анализа NucView, приготовленного в концентрации 30 мкМ добавляли в каждую лунку и доводили до 150 мкл (с получением конечной концентрации 5 мкМ). Планшет помещали в прибор IncuCyte и изображения получали каждые 2 часа (1 изображение на лунку) в течение 3 дней. Количество апоптотических клеток на каждом изображении определяли и записывали как количество апоптотических клеток на мм2. Анализы проводили в трех повторениях.
Данные, представленные в этом примере, включают молекулы ImmTAC, содержащие следующие альфа- и бета-цепи TCR
a28b50
a79b74
a79b46
Полные аминокислотные последовательности молекул ImmTAC, включающие a28b50, a79b74 и a79b46, представлены на Фигуре 5 (ImmTAC 1, 2 и 3 соответственно).
Результаты
Данные, представленные на Фигурах 8 и 9, показывают уничтожение в реальном времени антиген-положительных раковых клеток (клеточные линии меланомы Mel624 на Фигуре 8 и клеточной линии рака легкого NCI-H1755 на Фигуре 9) в присутствии молекул ImmTAC, содержащих мутантные последовательности TCR, в концентрации 100 пМ или ниже. Уничтожение не наблюдали в отсутствие молекул ImmTAC.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> IMMUNOCORE LIMITED
<120> T-КЛЕТОЧНЫЕ РЕЦЕПТОРЫ
<130> P111234WO
<160> 64
<170> PatentIn версия 3.5
<210> 1
<211> 9
<212> Белок
<213> Homo sapiens
<400> 1
Ser Leu Leu Gln His Leu Ile Gly Leu
1 5
<210> 2
<211> 200
<212> Белок
<213> Homo sapiens
<400> 2
Asp Ala Lys Thr Thr Gln Pro Asn Ser Met Glu Ser Asn Glu Glu Glu
1 5 10 15
Pro Val His Leu Pro Cys Asn His Ser Thr Ile Ser Gly Thr Asp Tyr
20 25 30
Ile His Trp Tyr Arg Gln Leu Pro Ser Gln Gly Pro Glu Tyr Val Ile
35 40 45
His Gly Leu Thr Ser Asn Val Asn Asn Arg Met Ala Ser Leu Ala Ile
50 55 60
Ala Glu Asp Arg Lys Ser Ser Thr Leu Ile Leu His Arg Ala Thr Leu
65 70 75 80
Arg Asp Ala Ala Val Tyr Tyr Cys Ile Leu Ile Leu Gly His Ser Gly
85 90 95
Ala Gly Ser Tyr Gln Leu Thr Phe Gly Lys Gly Thr Lys Leu Ser Val
100 105 110
Ile Pro Asn Ile Gln Asn Pro Asp Pro Ala Val Tyr Gln Leu Arg Asp
115 120 125
Ser Lys Ser Ser Asp Lys Ser Val Cys Leu Phe Thr Asp Phe Asp Ser
130 135 140
Gln Thr Asn Val Ser Gln Ser Lys Asp Ser Asp Val Tyr Ile Thr Asp
145 150 155 160
Lys Thr Val Leu Asp Met Arg Ser Met Asp Phe Lys Ser Asn Ser Ala
165 170 175
Val Ala Trp Ser Asn Lys Ser Asp Phe Ala Cys Ala Asn Ala Phe Asn
180 185 190
Asn Ser Ile Ile Pro Glu Asp Thr
195 200
<210> 3
<211> 244
<212> Белок
<213> Homo sapiens
<400> 3
Asp Gly Gly Ile Thr Gln Ser Pro Lys Tyr Leu Phe Arg Lys Glu Gly
1 5 10 15
Gln Asn Val Thr Leu Ser Cys Glu Gln Asn Leu Asn His Asp Ala Met
20 25 30
Tyr Trp Tyr Arg Gln Asp Pro Gly Gln Gly Leu Arg Leu Ile Tyr Tyr
35 40 45
Ser Gln Ile Val Asn Asp Phe Gln Lys Gly Asp Ile Ala Glu Gly Tyr
50 55 60
Ser Val Ser Arg Glu Lys Lys Glu Ser Phe Pro Leu Thr Val Thr Ser
65 70 75 80
Ala Gln Lys Asn Pro Thr Ala Phe Tyr Leu Cys Ala Ser Ser Pro Trp
85 90 95
Thr Ser Gly Ser Arg Glu Gln Tyr Phe Gly Pro Gly Thr Arg Leu Thr
100 105 110
Val Thr Glu Asp Leu Lys Asn Val Phe Pro Pro Glu Val Ala Val Phe
115 120 125
Glu Pro Ser Glu Ala Glu Ile Ser His Thr Gln Lys Ala Thr Leu Val
130 135 140
Cys Leu Ala Thr Gly Phe Tyr Pro Asp His Val Glu Leu Ser Trp Trp
145 150 155 160
Val Asn Gly Lys Glu Val His Ser Gly Val Ser Thr Asp Pro Gln Pro
165 170 175
Leu Lys Glu Gln Pro Ala Leu Asn Asp Ser Arg Tyr Cys Leu Ser Ser
180 185 190
Arg Leu Arg Val Ser Ala Thr Phe Trp Gln Asn Pro Arg Asn His Phe
195 200 205
Arg Cys Gln Val Gln Phe Tyr Gly Leu Ser Glu Asn Asp Glu Trp Thr
210 215 220
Gln Asp Arg Ala Lys Pro Val Thr Gln Ile Val Ser Ala Glu Ala Trp
225 230 235 240
Gly Arg Ala Asp
<210> 4
<211> 200
<212> Белок
<213> Homo sapiens
<400> 4
Asp Ala Lys Thr Thr Gln Pro Asn Ser Met Glu Ser Asn Glu Glu Glu
1 5 10 15
Pro Val His Leu Pro Cys Asn His Ser Thr Ile Ser Gly Thr Asp Tyr
20 25 30
Ile His Trp Tyr Arg Gln Leu Pro Ser Gln Gly Pro Glu Tyr Val Ile
35 40 45
His Gly Leu Thr Ser Asn Val Asn Asn Arg Met Ala Ser Leu Ala Ile
50 55 60
Ala Glu Asp Arg Lys Ser Ser Thr Leu Ile Leu His Arg Ala Thr Leu
65 70 75 80
Arg Asp Ala Ala Val Tyr Tyr Cys Ile Leu Ile Leu Gly His Ser Gly
85 90 95
Ala Gly Ser Tyr Gln Leu Thr Phe Gly Lys Gly Thr Lys Leu Ser Val
100 105 110
Ile Pro Asn Ile Gln Asn Pro Asp Pro Ala Val Tyr Gln Leu Arg Asp
115 120 125
Ser Lys Ser Ser Asp Lys Ser Val Cys Leu Phe Thr Asp Phe Asp Ser
130 135 140
Gln Thr Asn Val Ser Gln Ser Lys Asp Ser Asp Val Tyr Ile Thr Asp
145 150 155 160
Lys Cys Val Leu Asp Met Arg Ser Met Asp Phe Lys Ser Asn Ser Ala
165 170 175
Val Ala Trp Ser Asn Lys Ser Asp Phe Ala Cys Ala Asn Ala Phe Asn
180 185 190
Asn Ser Ile Ile Pro Glu Asp Thr
195 200
<210> 5
<211> 244
<212> Белок
<213> Homo sapiens
<400> 5
Asp Gly Gly Ile Thr Gln Ser Pro Lys Tyr Leu Phe Arg Lys Glu Gly
1 5 10 15
Gln Asn Val Thr Leu Ser Cys Glu Gln Asn Leu Asn His Asp Ala Met
20 25 30
Tyr Trp Tyr Arg Gln Asp Pro Gly Gln Gly Leu Arg Leu Ile Tyr Tyr
35 40 45
Ser Gln Ile Val Asn Asp Phe Gln Lys Gly Asp Ile Ala Glu Gly Tyr
50 55 60
Ser Val Ser Arg Glu Lys Lys Glu Ser Phe Pro Leu Thr Val Thr Ser
65 70 75 80
Ala Gln Lys Asn Pro Thr Ala Phe Tyr Leu Cys Ala Ser Ser Pro Trp
85 90 95
Thr Ser Gly Ser Arg Glu Gln Tyr Phe Gly Pro Gly Thr Arg Leu Thr
100 105 110
Val Thr Glu Asp Leu Lys Asn Val Phe Pro Pro Glu Val Ala Val Phe
115 120 125
Glu Pro Ser Glu Ala Glu Ile Ser His Thr Gln Lys Ala Thr Leu Val
130 135 140
Cys Leu Ala Thr Gly Phe Tyr Pro Asp His Val Glu Leu Ser Trp Trp
145 150 155 160
Val Asn Gly Lys Glu Val His Ser Gly Val Cys Thr Asp Pro Gln Pro
165 170 175
Leu Lys Glu Gln Pro Ala Leu Asn Asp Ser Arg Tyr Ala Leu Ser Ser
180 185 190
Arg Leu Arg Val Ser Ala Thr Phe Trp Gln Asp Pro Arg Asn His Phe
195 200 205
Arg Cys Gln Val Gln Phe Tyr Gly Leu Ser Glu Asn Asp Glu Trp Thr
210 215 220
Gln Asp Arg Ala Lys Pro Val Thr Gln Ile Val Ser Ala Glu Ala Trp
225 230 235 240
Gly Arg Ala Asp
<210> 6
<211> 115
<212> Белок
<213> Homo sapiens
<400> 6
Gly Asp Ala Lys Thr Thr Gln Pro Asn Ser Met Glu Ser Asn Glu Glu
1 5 10 15
Glu Pro Val His Leu Pro Cys Asn His Ser Thr Ile Ser Gly Thr Asp
20 25 30
Tyr Ile His Trp Tyr Arg Gln Leu Pro Ser Gln Gly Pro Glu Tyr Val
35 40 45
Ile His Gly Leu Thr Ser Asn Val Asn Asn Arg Met Ala Ser Leu Ala
50 55 60
Ile Ala Glu Asp Arg Lys Ser Ser Thr Leu Ile Leu His Arg Ala Thr
65 70 75 80
Leu Arg Asp Ala Ala Val Tyr Tyr Cys Ile Leu Ile Leu Gly His Ser
85 90 95
Arg Ala Gly Asn Tyr Ile Ala Thr Phe Gly Lys Gly Thr Lys Leu Ser
100 105 110
Val Ile Pro
115
<210> 7
<211> 115
<212> Белок
<213> Homo sapiens
<400> 7
Gly Asp Ala Lys Thr Thr Gln Pro Asn Ser Met Glu Ser Asn Glu Glu
1 5 10 15
Glu Pro Val His Leu Pro Cys Asn His Ser Thr Ile Ser Gly Thr Asp
20 25 30
Tyr Ile His Trp Tyr Arg Gln Leu Pro Ser Gln Gly Pro Glu Tyr Val
35 40 45
Ile His Gly Leu Thr Ser Asn Val Asn Asn Arg Met Ala Ser Leu Ala
50 55 60
Ile Ala Glu Asp Arg Lys Ser Ser Thr Leu Ile Leu His Arg Ala Thr
65 70 75 80
Leu Arg Asp Ala Ala Val Tyr Tyr Cys Ile Leu Ile Leu Gly His Ser
85 90 95
Arg Leu Gly Asn Tyr Ile Ala Thr Phe Gly Lys Gly Thr Lys Leu Ser
100 105 110
Val Ile Pro
115
<210> 8
<211> 115
<212> Белок
<213> Homo sapiens
<400> 8
Gly Asp Ala Lys Thr Thr Gln Pro Asn Ser Met Glu Ser Asn Glu Glu
1 5 10 15
Glu Pro Val His Leu Pro Cys Asn His Ser Thr Ile Ser Gly Thr Asp
20 25 30
Tyr Ile His Trp Tyr Arg Gln Leu Pro Ser Gln Gly Pro Glu Tyr Val
35 40 45
Ile His Gly Leu Thr Ser Asn Val Asn Asn Arg Met Ala Ser Leu Ala
50 55 60
Ile Ala Glu Asp Arg Lys Ser Ser Thr Leu Ile Leu His Arg Ala Thr
65 70 75 80
Leu Arg Asp Ala Ala Val Tyr Tyr Cys Ile Leu Ile Leu Gly His Ser
85 90 95
Arg Leu Gly Asn Tyr Gln Ala Thr Phe Gly Lys Gly Thr Lys Leu Ser
100 105 110
Val Ile Pro
115
<210> 9
<211> 114
<212> Белок
<213> Homo sapiens
<400> 9
Asp Gly Gly Ile Thr Gln Ser Pro Lys Tyr Leu Phe Arg Lys Glu Gly
1 5 10 15
Gln Asn Val Thr Leu Ser Cys Glu Gln Asn Leu Asn His Asp Ala Met
20 25 30
Tyr Trp Tyr Arg Gln Asp Pro Gly Gln Gly Leu Arg Leu Ile Tyr Tyr
35 40 45
Ser Gln Ile Met Gly Asp Glu Gln Lys Gly Asp Ile Ala Glu Gly Tyr
50 55 60
Ser Val Ser Arg Glu Lys Lys Glu Ser Phe Pro Leu Thr Val Thr Ser
65 70 75 80
Ala Gln Lys Asn Pro Thr Ala Phe Tyr Leu Cys Ala Ser Ser Trp Trp
85 90 95
Thr Gly Gly Ala Ser Pro Ile Ser Phe Gly Pro Gly Thr Arg Leu Thr
100 105 110
Val Thr
<210> 10
<211> 114
<212> Белок
<213> Homo sapiens
<400> 10
Asp Gly Gly Ile Thr Gln Ser Pro Lys Tyr Leu Phe Arg Lys Glu Gly
1 5 10 15
Gln Asn Val Thr Leu Ser Cys Glu Gln Asn Leu Asn His Asp Ala Met
20 25 30
Tyr Trp Tyr Arg Gln Asp Pro Gly Gln Gly Leu Arg Leu Ile Tyr Tyr
35 40 45
Ser Gln Ile Met Gly Asp Glu Gln Lys Gly Asp Ile Ala Glu Gly Tyr
50 55 60
Ser Val Ser Arg Glu Lys Lys Glu Ser Phe Pro Leu Thr Val Thr Ser
65 70 75 80
Ala Gln Lys Asn Pro Thr Ala Phe Tyr Leu Cys Ala Ser Ser Trp Trp
85 90 95
Thr Ser Gly Ala Ser Pro Ile Ser Phe Gly Pro Gly Thr Arg Leu Thr
100 105 110
Val Thr
<210> 11
<211> 114
<212> Белок
<213> Homo sapiens
<400> 11
Asp Gly Gly Ile Thr Gln Ser Pro Lys Tyr Leu Phe Arg Lys Glu Gly
1 5 10 15
Gln Asn Val Thr Leu Ser Cys Glu Gln Asn Leu Asn His Asp Ala Met
20 25 30
Tyr Trp Tyr Arg Gln Asp Pro Gly Gln Gly Leu Arg Leu Ile Tyr Tyr
35 40 45
Ser Gln Ile Met Gly Asp Glu Gln Lys Gly Asp Ile Ala Glu Gly Tyr
50 55 60
Ser Val Ser Arg Glu Lys Lys Glu Ser Phe Pro Leu Thr Val Thr Ser
65 70 75 80
Ala Gln Lys Asn Pro Thr Ala Phe Tyr Leu Cys Ala Ser Ser Trp Trp
85 90 95
Thr Gly Gly Ser Ala Pro Ile Arg Phe Gly Pro Gly Thr Arg Leu Thr
100 105 110
Val Thr
<210> 12
<211> 114
<212> Белок
<213> Homo sapiens
<400> 12
Asp Gly Gly Ile Thr Gln Ser Pro Lys Tyr Leu Phe Arg Lys Glu Gly
1 5 10 15
Gln Asn Val Thr Leu Ser Cys Glu Gln Asn Leu Asn His Asp Ala Met
20 25 30
Tyr Trp Tyr Arg Gln Asp Pro Gly Gln Gly Leu Arg Leu Ile Tyr Tyr
35 40 45
Ser Gln Ile Met Asn Asp Glu Gln Lys Gly Asp Ile Ala Glu Gly Tyr
50 55 60
Ser Val Ser Arg Glu Lys Lys Glu Ser Phe Pro Leu Thr Val Thr Ser
65 70 75 80
Ala Gln Lys Asn Pro Thr Ala Phe Tyr Leu Cys Ala Ser Ser Trp Trp
85 90 95
Thr Gly Gly Ser Ala Pro Ile Arg Phe Gly Pro Gly Thr Arg Leu Thr
100 105 110
Val Thr
<210> 13
<211> 114
<212> Белок
<213> Homo sapiens
<400> 13
Asp Gly Gly Ile Thr Gln Ser Pro Lys Tyr Leu Phe Arg Lys Glu Gly
1 5 10 15
Gln Asn Val Thr Leu Ser Cys Glu Gln Asn Leu Asn His Asp Ala Met
20 25 30
Tyr Trp Tyr Arg Gln Asp Pro Gly Gln Gly Leu Arg Leu Ile Tyr Tyr
35 40 45
Ser Gln Ile Met Gly Asp Glu Gln Lys Gly Asp Ile Ala Glu Gly Tyr
50 55 60
Ser Val Ser Arg Glu Lys Lys Glu Ser Phe Pro Leu Thr Val Thr Ser
65 70 75 80
Ala Gln Lys Asn Pro Thr Ala Phe Tyr Leu Cys Ala Ser Ser Trp Trp
85 90 95
Thr Ser Gly Ser Ala Pro Ile Arg Phe Gly Pro Gly Thr Arg Leu Thr
100 105 110
Val Thr
<210> 14
<211> 114
<212> Белок
<213> Homo sapiens
<400> 14
Asp Gly Gly Ile Thr Gln Ser Pro Lys Tyr Leu Phe Arg Lys Glu Gly
1 5 10 15
Gln Asn Val Thr Leu Ser Cys Glu Gln Asn Leu Asn His Asp Ala Met
20 25 30
Tyr Trp Tyr Arg Gln Asp Pro Gly Gln Gly Leu Arg Leu Ile Tyr Tyr
35 40 45
Ser Gln Ile Met Gly Asp Glu Gln Lys Gly Asp Ile Ala Glu Gly Tyr
50 55 60
Ser Val Ser Arg Glu Lys Lys Glu Ser Phe Pro Leu Thr Val Thr Ser
65 70 75 80
Ala Gln Lys Asn Pro Thr Ala Phe Tyr Leu Cys Ala Ser Ser Trp Trp
85 90 95
Thr Gly Gly Ser Ala Glu Ile Arg Phe Gly Pro Gly Thr Arg Leu Thr
100 105 110
Val Thr
<210> 15
<211> 114
<212> Белок
<213> Homo sapiens
<400> 15
Asp Gly Gly Ile Thr Gln Ser Pro Lys Tyr Leu Phe Arg Lys Glu Gly
1 5 10 15
Gln Asn Val Thr Leu Ser Cys Glu Gln Asn Leu Asn His Asp Ala Met
20 25 30
Tyr Trp Tyr Arg Gln Asp Pro Gly Gln Gly Leu Arg Leu Ile Tyr Tyr
35 40 45
Ser Gln Ile Met Gly Asp Glu Gln Lys Gly Asp Ile Ala Glu Gly Tyr
50 55 60
Ser Val Ser Arg Glu Lys Lys Glu Ser Phe Pro Leu Thr Val Thr Ser
65 70 75 80
Ala Gln Lys Asn Pro Thr Ala Phe Tyr Leu Cys Ala Ser Ser Trp Trp
85 90 95
Thr Gly Gly Ser Ala Pro Ile Tyr Phe Gly Pro Gly Thr Arg Leu Thr
100 105 110
Val Thr
<210> 16
<211> 114
<212> Белок
<213> Homo sapiens
<400> 16
Asp Gly Gly Ile Thr Gln Ser Pro Lys Tyr Leu Phe Arg Lys Glu Gly
1 5 10 15
Gln Asn Val Thr Leu Ser Cys Glu Gln Asn Leu Asn His Asp Ala Met
20 25 30
Tyr Trp Tyr Arg Gln Asp Pro Gly Gln Gly Leu Arg Leu Ile Tyr Tyr
35 40 45
Ser Gln Ile Met Gly Asp Glu Gln Lys Gly Asp Ile Ala Glu Gly Tyr
50 55 60
Ser Val Ser Arg Glu Lys Lys Glu Ser Phe Pro Leu Thr Val Thr Ser
65 70 75 80
Ala Gln Lys Asn Pro Thr Ala Phe Tyr Leu Cys Ala Ser Ser Trp Trp
85 90 95
Thr Gly Gly Ala Ala Pro Ile Ser Phe Gly Pro Gly Thr Arg Leu Thr
100 105 110
Val Thr
<210> 17
<211> 114
<212> Белок
<213> Homo sapiens
<400> 17
Asp Gly Gly Ile Thr Gln Ser Pro Lys Tyr Leu Phe Arg Lys Glu Gly
1 5 10 15
Gln Asn Val Thr Leu Ser Cys Glu Gln Asn Leu Asn His Asp Ala Met
20 25 30
Tyr Trp Tyr Arg Gln Asp Pro Gly Gln Gly Leu Arg Leu Ile Tyr Tyr
35 40 45
Ser Gln Ile Met Gly Asp Glu Gln Lys Gly Asp Ile Ala Glu Gly Tyr
50 55 60
Ser Val Ser Arg Glu Lys Lys Glu Ser Phe Pro Leu Thr Val Thr Ser
65 70 75 80
Ala Gln Lys Asn Pro Thr Ala Phe Tyr Leu Cys Ala Ser Ser Trp Trp
85 90 95
Thr Gly Gly Ala Ser Pro Ile Arg Phe Gly Pro Gly Thr Arg Leu Thr
100 105 110
Val Thr
<210> 18
<211> 114
<212> Белок
<213> Homo sapiens
<400> 18
Asp Gly Gly Ile Thr Gln Ser Pro Lys Tyr Leu Phe Arg Lys Glu Gly
1 5 10 15
Gln Asn Val Thr Leu Ser Cys Glu Gln Asn Leu Asn His Asp Ala Met
20 25 30
Tyr Trp Tyr Arg Gln Asp Pro Gly Gln Gly Leu Arg Leu Ile Tyr Tyr
35 40 45
Ser Gln Ile Met Gly Asp Glu Gln Lys Gly Asp Ile Ala Glu Gly Tyr
50 55 60
Ser Val Ser Arg Glu Lys Lys Glu Ser Phe Pro Leu Thr Val Thr Ser
65 70 75 80
Ala Gln Lys Asn Pro Thr Ala Phe Tyr Leu Cys Ala Ser Ser Trp Trp
85 90 95
Thr Gly Gly Ala Ala Pro Ile Arg Phe Gly Pro Gly Thr Arg Leu Thr
100 105 110
Val Thr
<210> 19
<211> 114
<212> Белок
<213> Homo sapiens
<400> 19
Asp Gly Gly Ile Thr Gln Ser Pro Lys Tyr Leu Phe Arg Lys Glu Gly
1 5 10 15
Gln Asn Val Thr Leu Ser Cys Glu Gln Asn Leu Asn His Asp Ala Met
20 25 30
Tyr Trp Tyr Arg Gln Asp Pro Gly Gln Gly Leu Arg Leu Ile Tyr Tyr
35 40 45
Ser Gln Ile Met Gly Asp Glu Gln Lys Gly Asp Ile Ala Glu Gly Tyr
50 55 60
Ser Val Ser Arg Glu Lys Lys Glu Ser Phe Pro Leu Thr Val Thr Ser
65 70 75 80
Ala Gln Lys Asn Pro Thr Ala Phe Tyr Leu Cys Ala Ser Ser Trp Trp
85 90 95
Thr Gly Gly Ala Ser Glu Ile Ser Phe Gly Pro Gly Thr Arg Leu Thr
100 105 110
Val Thr
<210> 20
<211> 114
<212> Белок
<213> Homo sapiens
<400> 20
Asp Gly Gly Ile Thr Gln Ser Pro Lys Tyr Leu Phe Arg Lys Glu Gly
1 5 10 15
Gln Asn Val Thr Leu Ser Cys Glu Gln Asn Leu Asn His Asp Ala Met
20 25 30
Tyr Trp Tyr Arg Gln Asp Pro Gly Gln Gly Leu Arg Leu Ile Tyr Tyr
35 40 45
Ser Gln Ile Met Gly Asp Glu Gln Lys Gly Asp Ile Ala Glu Gly Tyr
50 55 60
Ser Val Ser Arg Glu Lys Lys Glu Ser Phe Pro Leu Thr Val Thr Ser
65 70 75 80
Ala Gln Lys Asn Pro Thr Ala Phe Tyr Leu Cys Ala Ser Ser Trp Trp
85 90 95
Thr Gly Gly Ser Ala Pro Ile Ser Phe Gly Pro Gly Thr Arg Leu Thr
100 105 110
Val Thr
<210> 21
<211> 114
<212> Белок
<213> Homo sapiens
<400> 21
Asp Gly Gly Ile Thr Gln Ser Pro Lys Tyr Leu Phe Arg Lys Glu Gly
1 5 10 15
Gln Asn Val Thr Leu Ser Cys Glu Gln Asn Leu Asn His Asp Ala Met
20 25 30
Tyr Trp Tyr Arg Gln Asp Pro Gly Gln Gly Leu Arg Leu Ile Tyr Tyr
35 40 45
Ser Gln Ile Met Gly Asp Glu Gln Lys Gly Asp Ile Ala Glu Gly Tyr
50 55 60
Ser Val Ser Arg Glu Lys Lys Glu Ser Phe Pro Leu Thr Val Thr Ser
65 70 75 80
Ala Gln Lys Asn Pro Thr Ala Phe Tyr Leu Cys Ala Ser Ser Trp Trp
85 90 95
Thr Gly Gly Ser Ser Pro Ile Ser Phe Gly Pro Gly Thr Arg Leu Thr
100 105 110
Val Thr
<210> 22
<211> 114
<212> Белок
<213> Homo sapiens
<400> 22
Asp Gly Gly Ile Thr Gln Ser Pro Lys Tyr Leu Phe Arg Lys Glu Gly
1 5 10 15
Gln Asn Val Thr Leu Ser Cys Glu Gln Asn Leu Asn His Asp Ala Met
20 25 30
Tyr Trp Tyr Arg Gln Asp Pro Gly Gln Gly Leu Arg Leu Ile Tyr Tyr
35 40 45
Ser Gln Ile Val Gly Asp Glu Gln Lys Gly Asp Ile Ala Glu Gly Tyr
50 55 60
Ser Val Ser Arg Glu Lys Lys Glu Ser Phe Pro Leu Thr Val Thr Ser
65 70 75 80
Ala Gln Lys Asn Pro Thr Ala Phe Tyr Leu Cys Ala Ser Ser Trp Trp
85 90 95
Thr Gly Gly Ser Ala Pro Ile Arg Phe Gly Pro Gly Thr Arg Leu Thr
100 105 110
Val Thr
<210> 23
<211> 114
<212> Белок
<213> Homo sapiens
<400> 23
Asp Gly Gly Ile Thr Gln Ser Pro Lys Tyr Leu Phe Arg Lys Glu Gly
1 5 10 15
Gln Asn Val Thr Leu Ser Cys Glu Gln Asn Leu Asn His Asp Ala Met
20 25 30
Tyr Trp Tyr Arg Gln Asp Pro Gly Gln Gly Leu Arg Leu Ile Tyr Tyr
35 40 45
Ser Gln Ile Met Gly Asp Glu Gln Lys Gly Asp Ile Ala Glu Gly Tyr
50 55 60
Ser Val Ser Arg Glu Lys Lys Glu Ser Phe Pro Leu Thr Val Thr Ser
65 70 75 80
Ala Gln Lys Asn Pro Thr Ala Phe Tyr Leu Cys Ala Ser Ser Pro Trp
85 90 95
Thr Gly Gly Ser Ala Pro Ile Arg Phe Gly Pro Gly Thr Arg Leu Thr
100 105 110
Val Thr
<210> 24
<211> 114
<212> Белок
<213> Homo sapiens
<400> 24
Asp Gly Gly Ile Thr Gln Ser Pro Lys Tyr Leu Phe Arg Lys Glu Gly
1 5 10 15
Gln Asn Val Thr Leu Ser Cys Glu Gln Asn Leu Asn His Asp Ala Met
20 25 30
Tyr Trp Tyr Arg Gln Asp Pro Gly Gln Gly Leu Arg Leu Ile Tyr Tyr
35 40 45
Ser Gln Ile Met Gly Asp Glu Gln Lys Gly Asp Ile Ala Glu Gly Tyr
50 55 60
Ser Val Ser Arg Glu Lys Lys Glu Ser Phe Pro Leu Thr Val Thr Ser
65 70 75 80
Ala Gln Lys Asn Pro Thr Ala Phe Tyr Leu Cys Ala Ser Ser Trp Trp
85 90 95
Thr Gly Gly Ser Ser Pro Ile Arg Phe Gly Pro Gly Thr Arg Leu Thr
100 105 110
Val Thr
<210> 25
<211> 201
<212> Белок
<213> Homo sapiens
<400> 25
Gly Asp Ala Lys Thr Thr Gln Pro Asn Ser Met Glu Ser Asn Glu Glu
1 5 10 15
Glu Pro Val His Leu Pro Cys Asn His Ser Thr Ile Ser Gly Thr Asp
20 25 30
Tyr Ile His Trp Tyr Arg Gln Leu Pro Ser Gln Gly Pro Glu Tyr Val
35 40 45
Ile His Gly Leu Thr Ser Asn Val Asn Asn Arg Met Ala Ser Leu Ala
50 55 60
Ile Ala Glu Asp Arg Lys Ser Ser Thr Leu Ile Leu His Arg Ala Thr
65 70 75 80
Leu Arg Asp Ala Ala Val Tyr Tyr Cys Ile Leu Ile Leu Gly His Ser
85 90 95
Arg Ala Gly Asn Tyr Ile Ala Thr Phe Gly Lys Gly Thr Lys Leu Ser
100 105 110
Val Ile Pro Asn Ile Gln Asn Pro Asp Pro Ala Val Tyr Gln Leu Arg
115 120 125
Asp Ser Lys Ser Ser Asp Lys Ser Val Cys Leu Phe Thr Asp Phe Asp
130 135 140
Ser Gln Thr Asn Val Ser Gln Ser Lys Asp Ser Asp Val Tyr Ile Thr
145 150 155 160
Asp Lys Cys Val Leu Asp Met Arg Ser Met Asp Phe Lys Ser Asn Ser
165 170 175
Ala Val Ala Trp Ser Asn Lys Ser Asp Phe Ala Cys Ala Asn Ala Phe
180 185 190
Asn Asn Ser Ile Ile Pro Glu Asp Thr
195 200
<210> 26
<211> 502
<212> Белок
<213> Homo sapiens
<400> 26
Ala Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Ile Arg Asn Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Tyr Thr Ser Arg Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Tyr Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Gly Asn Thr Leu Pro Trp
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Gly Gly Gly Gly Ser
100 105 110
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
115 120 125
Gly Gly Ser Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln
130 135 140
Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Ser Phe
145 150 155 160
Thr Gly Tyr Thr Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu
165 170 175
Glu Trp Val Ala Leu Ile Asn Pro Tyr Lys Gly Val Ser Thr Tyr Asn
180 185 190
Gln Lys Phe Lys Asp Arg Phe Thr Ile Ser Val Asp Lys Ser Lys Asn
195 200 205
Thr Ala Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val
210 215 220
Tyr Tyr Cys Ala Arg Ser Gly Tyr Tyr Gly Asp Ser Asp Trp Tyr Phe
225 230 235 240
Asp Val Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Gly Gly Gly
245 250 255
Gly Ser Asp Gly Gly Ile Thr Gln Ser Pro Lys Tyr Leu Phe Arg Lys
260 265 270
Glu Gly Gln Asn Val Thr Leu Ser Cys Glu Gln Asn Leu Asn His Asp
275 280 285
Ala Met Tyr Trp Tyr Arg Gln Asp Pro Gly Gln Gly Leu Arg Leu Ile
290 295 300
Tyr Tyr Ser Gln Ile Met Gly Asp Glu Gln Lys Gly Asp Ile Ala Glu
305 310 315 320
Gly Tyr Ser Val Ser Arg Glu Lys Lys Glu Ser Phe Pro Leu Thr Val
325 330 335
Thr Ser Ala Gln Lys Asn Pro Thr Ala Phe Tyr Leu Cys Ala Ser Ser
340 345 350
Trp Trp Thr Gly Gly Ala Ser Pro Ile Ser Phe Gly Pro Gly Thr Arg
355 360 365
Leu Thr Val Thr Glu Asp Leu Lys Asn Val Phe Pro Pro Glu Val Ala
370 375 380
Val Phe Glu Pro Ser Glu Ala Glu Ile Ser His Thr Gln Lys Ala Thr
385 390 395 400
Leu Val Cys Leu Ala Thr Gly Phe Tyr Pro Asp His Val Glu Leu Ser
405 410 415
Trp Trp Val Asn Gly Lys Glu Val His Ser Gly Val Cys Thr Asp Pro
420 425 430
Gln Pro Leu Lys Glu Gln Pro Ala Leu Asn Asp Ser Arg Tyr Ala Leu
435 440 445
Ser Ser Arg Leu Arg Val Ser Ala Thr Phe Trp Gln Asp Pro Arg Asn
450 455 460
His Phe Arg Cys Gln Val Gln Phe Tyr Gly Leu Ser Glu Asn Asp Glu
465 470 475 480
Trp Thr Gln Asp Arg Ala Lys Pro Val Thr Gln Ile Val Ser Ala Glu
485 490 495
Ala Trp Gly Arg Ala Asp
500
<210> 27
<211> 201
<212> Белок
<213> Homo sapiens
<400> 27
Gly Asp Ala Lys Thr Thr Gln Pro Asn Ser Met Glu Ser Asn Glu Glu
1 5 10 15
Glu Pro Val His Leu Pro Cys Asn His Ser Thr Ile Ser Gly Thr Asp
20 25 30
Tyr Ile His Trp Tyr Arg Gln Leu Pro Ser Gln Gly Pro Glu Tyr Val
35 40 45
Ile His Gly Leu Thr Ser Asn Val Asn Asn Arg Met Ala Ser Leu Ala
50 55 60
Ile Ala Glu Asp Arg Lys Ser Ser Thr Leu Ile Leu His Arg Ala Thr
65 70 75 80
Leu Arg Asp Ala Ala Val Tyr Tyr Cys Ile Leu Ile Leu Gly His Ser
85 90 95
Arg Leu Gly Asn Tyr Ile Ala Thr Phe Gly Lys Gly Thr Lys Leu Ser
100 105 110
Val Ile Pro Asn Ile Gln Asn Pro Asp Pro Ala Val Tyr Gln Leu Arg
115 120 125
Asp Ser Lys Ser Ser Asp Lys Ser Val Cys Leu Phe Thr Asp Phe Asp
130 135 140
Ser Gln Thr Asn Val Ser Gln Ser Lys Asp Ser Asp Val Tyr Ile Thr
145 150 155 160
Asp Lys Cys Val Leu Asp Met Arg Ser Met Asp Phe Lys Ser Asn Ser
165 170 175
Ala Val Ala Trp Ser Asn Lys Ser Asp Phe Ala Cys Ala Asn Ala Phe
180 185 190
Asn Asn Ser Ile Ile Pro Glu Asp Thr
195 200
<210> 28
<211> 502
<212> Белок
<213> Homo sapiens
<400> 28
Ala Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Ile Arg Asn Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Tyr Thr Ser Arg Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Tyr Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Gly Asn Thr Leu Pro Trp
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Gly Gly Gly Gly Ser
100 105 110
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
115 120 125
Gly Gly Ser Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln
130 135 140
Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Ser Phe
145 150 155 160
Thr Gly Tyr Thr Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu
165 170 175
Glu Trp Val Ala Leu Ile Asn Pro Tyr Lys Gly Val Ser Thr Tyr Asn
180 185 190
Gln Lys Phe Lys Asp Arg Phe Thr Ile Ser Val Asp Lys Ser Lys Asn
195 200 205
Thr Ala Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val
210 215 220
Tyr Tyr Cys Ala Arg Ser Gly Tyr Tyr Gly Asp Ser Asp Trp Tyr Phe
225 230 235 240
Asp Val Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Gly Gly Gly
245 250 255
Gly Ser Asp Gly Gly Ile Thr Gln Ser Pro Lys Tyr Leu Phe Arg Lys
260 265 270
Glu Gly Gln Asn Val Thr Leu Ser Cys Glu Gln Asn Leu Asn His Asp
275 280 285
Ala Met Tyr Trp Tyr Arg Gln Asp Pro Gly Gln Gly Leu Arg Leu Ile
290 295 300
Tyr Tyr Ser Gln Ile Met Gly Asp Glu Gln Lys Gly Asp Ile Ala Glu
305 310 315 320
Gly Tyr Ser Val Ser Arg Glu Lys Lys Glu Ser Phe Pro Leu Thr Val
325 330 335
Thr Ser Ala Gln Lys Asn Pro Thr Ala Phe Tyr Leu Cys Ala Ser Ser
340 345 350
Trp Trp Thr Gly Gly Ala Ser Pro Ile Arg Phe Gly Pro Gly Thr Arg
355 360 365
Leu Thr Val Thr Glu Asp Leu Lys Asn Val Phe Pro Pro Glu Val Ala
370 375 380
Val Phe Glu Pro Ser Glu Ala Glu Ile Ser His Thr Gln Lys Ala Thr
385 390 395 400
Leu Val Cys Leu Ala Thr Gly Phe Tyr Pro Asp His Val Glu Leu Ser
405 410 415
Trp Trp Val Asn Gly Lys Glu Val His Ser Gly Val Cys Thr Asp Pro
420 425 430
Gln Pro Leu Lys Glu Gln Pro Ala Leu Asn Asp Ser Arg Tyr Ala Leu
435 440 445
Ser Ser Arg Leu Arg Val Ser Ala Thr Phe Trp Gln Asp Pro Arg Asn
450 455 460
His Phe Arg Cys Gln Val Gln Phe Tyr Gly Leu Ser Glu Asn Asp Glu
465 470 475 480
Trp Thr Gln Asp Arg Ala Lys Pro Val Thr Gln Ile Val Ser Ala Glu
485 490 495
Ala Trp Gly Arg Ala Asp
500
<210> 29
<211> 201
<212> Белок
<213> Homo sapiens
<400> 29
Gly Asp Ala Lys Thr Thr Gln Pro Asn Ser Met Glu Ser Asn Glu Glu
1 5 10 15
Glu Pro Val His Leu Pro Cys Asn His Ser Thr Ile Ser Gly Thr Asp
20 25 30
Tyr Ile His Trp Tyr Arg Gln Leu Pro Ser Gln Gly Pro Glu Tyr Val
35 40 45
Ile His Gly Leu Thr Ser Asn Val Asn Asn Arg Met Ala Ser Leu Ala
50 55 60
Ile Ala Glu Asp Arg Lys Ser Ser Thr Leu Ile Leu His Arg Ala Thr
65 70 75 80
Leu Arg Asp Ala Ala Val Tyr Tyr Cys Ile Leu Ile Leu Gly His Ser
85 90 95
Arg Leu Gly Asn Tyr Ile Ala Thr Phe Gly Lys Gly Thr Lys Leu Ser
100 105 110
Val Ile Pro Asn Ile Gln Asn Pro Asp Pro Ala Val Tyr Gln Leu Arg
115 120 125
Asp Ser Lys Ser Ser Asp Lys Ser Val Cys Leu Phe Thr Asp Phe Asp
130 135 140
Ser Gln Thr Asn Val Ser Gln Ser Lys Asp Ser Asp Val Tyr Ile Thr
145 150 155 160
Asp Lys Cys Val Leu Asp Met Arg Ser Met Asp Phe Lys Ser Asn Ser
165 170 175
Ala Val Ala Trp Ser Asn Lys Ser Asp Phe Ala Cys Ala Asn Ala Phe
180 185 190
Asn Asn Ser Ile Ile Pro Glu Asp Thr
195 200
<210> 30
<211> 501
<212> Белок
<213> Homo sapiens
<400> 30
Ala Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Ile Arg Asn Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Tyr Thr Ser Arg Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Tyr Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Gly Asn Thr Leu Pro Trp
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Gly Gly Gly Gly Ser
100 105 110
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
115 120 125
Gly Gly Ser Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln
130 135 140
Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Ser Phe
145 150 155 160
Thr Gly Tyr Thr Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu
165 170 175
Glu Trp Val Ala Leu Ile Asn Pro Tyr Lys Gly Val Ser Thr Tyr Asn
180 185 190
Gln Lys Phe Lys Asp Arg Phe Thr Ile Ser Val Asp Lys Ser Lys Asn
195 200 205
Thr Ala Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val
210 215 220
Tyr Tyr Cys Ala Arg Ser Gly Tyr Tyr Gly Asp Ser Asp Trp Tyr Phe
225 230 235 240
Asp Val Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Gly Gly Gly
245 250 255
Gly Ser Asp Gly Gly Ile Thr Gln Ser Pro Lys Tyr Leu Phe Arg Lys
260 265 270
Glu Gly Gln Asn Val Thr Leu Ser Cys Glu Gln Asn Leu Asn His Asp
275 280 285
Ala Met Tyr Trp Tyr Arg Gln Asp Pro Gly Gln Gly Leu Arg Leu Ile
290 295 300
Tyr Tyr Ser Gln Ile Met Gly Asp Glu Gln Lys Gly Asp Ile Ala Glu
305 310 315 320
Gly Tyr Ser Val Ser Arg Glu Lys Lys Glu Ser Phe Pro Leu Thr Val
325 330 335
Thr Ser Ala Gln Lys Asn Pro Thr Ala Phe Tyr Leu Cys Ala Ser Ser
340 345 350
Trp Trp Thr Gly Gly Ser Ala Pro Ile Arg Phe Gly Pro Gly Thr Arg
355 360 365
Leu Thr Val Thr Glu Asp Leu Lys Asn Val Phe Pro Pro Glu Val Ala
370 375 380
Val Phe Glu Pro Ser Glu Ala Glu Ile Ser His Thr Gln Lys Ala Thr
385 390 395 400
Leu Val Cys Leu Ala Thr Gly Phe Tyr Pro Asp His Val Glu Leu Ser
405 410 415
Trp Trp Val Asn Gly Lys Glu Val His Ser Gly Val Cys Thr Asp Pro
420 425 430
Gln Pro Leu Lys Glu Gln Pro Ala Leu Asn Asp Ser Arg Tyr Ala Leu
435 440 445
Ser Ser Arg Leu Arg Val Ser Ala Thr Phe Trp Gln Asp Pro Arg Asn
450 455 460
His Phe Arg Cys Gln Val Gln Phe Tyr Gly Leu Ser Glu Asn Asp Glu
465 470 475 480
Trp Thr Gln Asp Arg Ala Lys Pro Val Thr Gln Ile Val Ser Ala Glu
485 490 495
Ala Trp Gly Arg Ala
500
<210> 31
<211> 5
<212> Белок
<213> Homo sapiens
<400> 31
Gly Gly Gly Gly Ser
1 5
<210> 32
<211> 5
<212> Белок
<213> Homo sapiens
<400> 32
Gly Gly Gly Ser Gly
1 5
<210> 33
<211> 5
<212> Белок
<213> Homo sapiens
<400> 33
Gly Gly Ser Gly Gly
1 5
<210> 34
<211> 5
<212> Белок
<213> Homo sapiens
<400> 34
Gly Ser Gly Gly Gly
1 5
<210> 35
<211> 6
<212> Белок
<213> Homo sapiens
<400> 35
Gly Ser Gly Gly Gly Pro
1 5
<210> 36
<211> 5
<212> Белок
<213> Homo sapiens
<400> 36
Gly Gly Glu Pro Ser
1 5
<210> 37
<211> 7
<212> Белок
<213> Homo sapiens
<400> 37
Gly Gly Glu Gly Gly Gly Pro
1 5
<210> 38
<211> 12
<212> Белок
<213> Homo sapiens
<400> 38
Gly Gly Glu Gly Gly Gly Ser Glu Gly Gly Gly Ser
1 5 10
<210> 39
<211> 7
<212> Белок
<213> Homo sapiens
<400> 39
Thr Ile Ser Gly Thr Asp Tyr
1 5
<210> 40
<211> 5
<212> Белок
<213> Homo sapiens
<400> 40
Gly Leu Thr Ser Asn
1 5
<210> 41
<211> 17
<212> Белок
<213> Homo sapiens
<400> 41
Cys Ile Leu Ile Leu Gly His Ser Gly Ala Gly Ser Tyr Gln Leu Thr
1 5 10 15
Phe
<210> 42
<211> 5
<212> Белок
<213> Homo sapiens
<400> 42
Leu Asn His Asp Ala
1 5
<210> 43
<211> 7
<212> Белок
<213> Homo sapiens
<400> 43
Ser Gln Ile Val Asn Asp Phe
1 5
<210> 44
<211> 15
<212> Белок
<213> Homo sapiens
<400> 44
Cys Ala Ser Ser Pro Trp Thr Ser Gly Ser Arg Glu Gln Tyr Phe
1 5 10 15
<210> 45
<211> 17
<212> Белок
<213> Homo sapiens
<400> 45
Cys Ile Leu Ile Leu Gly His Ser Arg Ala Gly Asn Tyr Ile Ala Thr
1 5 10 15
Phe
<210> 46
<211> 17
<212> Белок
<213> Homo sapiens
<400> 46
Cys Ile Leu Ile Leu Gly His Ser Arg Leu Gly Asn Tyr Ile Ala Thr
1 5 10 15
Phe
<210> 47
<211> 17
<212> Белок
<213> Homo sapiens
<400> 47
Cys Ile Leu Ile Leu Gly His Ser Arg Leu Gly Asn Tyr Gln Ala Thr
1 5 10 15
Phe
<210> 48
<211> 7
<212> Белок
<213> Homo sapiens
<400> 48
Ser Gln Ile Met Gly Asp Glu
1 5
<210> 49
<211> 7
<212> Белок
<213> Homo sapiens
<400> 49
Ser Gln Ile Met Asn Asp Glu
1 5
<210> 50
<211> 7
<212> Белок
<213> Homo sapiens
<400> 50
Ser Gln Ile Val Gly Asp Glu
1 5
<210> 51
<211> 15
<212> Белок
<213> Homo sapiens
<400> 51
Cys Ala Ser Ser Trp Trp Thr Gly Gly Ala Ser Pro Ile Ser Phe
1 5 10 15
<210> 52
<211> 15
<212> Белок
<213> Homo sapiens
<400> 52
Cys Ala Ser Ser Trp Trp Thr Ser Gly Ala Ser Pro Ile Ser Phe
1 5 10 15
<210> 53
<211> 15
<212> Белок
<213> Homo sapiens
<400> 53
Cys Ala Ser Ser Trp Trp Thr Gly Gly Ser Ala Pro Ile Arg Phe
1 5 10 15
<210> 54
<211> 15
<212> Белок
<213> Homo sapiens
<400> 54
Cys Ala Ser Ser Trp Trp Thr Ser Gly Ser Ala Pro Ile Arg Phe
1 5 10 15
<210> 55
<211> 15
<212> Белок
<213> Homo sapiens
<400> 55
Cys Ala Ser Ser Trp Trp Thr Gly Gly Ser Ala Glu Ile Arg Phe
1 5 10 15
<210> 56
<211> 15
<212> Белок
<213> Homo sapiens
<400> 56
Cys Ala Ser Ser Trp Trp Thr Gly Gly Ser Ala Pro Ile Tyr Phe
1 5 10 15
<210> 57
<211> 15
<212> Белок
<213> Homo sapiens
<400> 57
Cys Ala Ser Ser Trp Trp Thr Gly Gly Ala Ala Pro Ile Ser Phe
1 5 10 15
<210> 58
<211> 15
<212> Белок
<213> Homo sapiens
<400> 58
Cys Ala Ser Ser Trp Trp Thr Gly Gly Ala Ser Pro Ile Arg Phe
1 5 10 15
<210> 59
<211> 15
<212> Белок
<213> Homo sapiens
<400> 59
Cys Ala Ser Ser Trp Trp Thr Gly Gly Ala Ala Pro Ile Arg Phe
1 5 10 15
<210> 60
<211> 15
<212> Белок
<213> Homo sapiens
<400> 60
Cys Ala Ser Ser Trp Trp Thr Gly Gly Ala Ser Glu Ile Ser Phe
1 5 10 15
<210> 61
<211> 15
<212> Белок
<213> Homo sapiens
<400> 61
Cys Ala Ser Ser Trp Trp Thr Gly Gly Ser Ala Pro Ile Ser Phe
1 5 10 15
<210> 62
<211> 15
<212> Белок
<213> Homo sapiens
<400> 62
Cys Ala Ser Ser Trp Trp Thr Gly Gly Ser Ser Pro Ile Ser Phe
1 5 10 15
<210> 63
<211> 15
<212> Белок
<213> Homo sapiens
<400> 63
Cys Ala Ser Ser Pro Trp Thr Gly Gly Ser Ala Pro Ile Arg Phe
1 5 10 15
<210> 64
<211> 15
<212> Белок
<213> Homo sapiens
<400> 64
Cys Ala Ser Ser Trp Trp Thr Gly Gly Ser Ser Pro Ile Arg Phe
1 5 10 15
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| Т-КЛЕТОЧНЫЕ РЕЦЕПТОРЫ, СПЕЦИФИЧНЫЕ В ОТНОШЕНИИ КОМПЛЕКСА ОПУХОЛЕВЫЙ АНТИГЕН NY-ESO-1/HLA-A*02 | 2016 |
|
RU2775394C2 |
| Т-КЛЕТОЧНЫЕ РЕЦЕПТОРЫ | 2017 |
|
RU2775623C2 |
| T-КЛЕТОЧНЫЕ РЕЦЕПТОРЫ, РАСПОЗНАЮЩИЕ МУТАЦИЮ R175H ИЛИ Y220C В P53 | 2020 |
|
RU2830061C2 |
| HPV-СПЕЦИФИЧЕСКИЕ СВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ | 2018 |
|
RU2804664C2 |
| КОНСТРУКТЫ Т-КЛЕТОЧНОГО РЕЦЕПТОРА И ИХ ПРИМЕНЕНИЕ | 2019 |
|
RU2785954C2 |
| РЕКРУТИРУЮЩИЕ Т-КЛЕТКИ ПОЛИПЕПТИДЫ, СПОСОБНЫЕ СВЯЗЫВАТЬ CD123 И TCR АЛЬФА/БЕТА | 2017 |
|
RU2775063C2 |
| CD3-СПЕЦИФИЧЕСКИЕ СВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ | 2020 |
|
RU2826453C2 |
| ВЫСОКОАФФИННЫЙ Т-КЛЕТОЧНЫЙ РЕЦЕПТОР ДЛЯ РАСПОЗНАВАНИЯ АНТИГЕНА AFP | 2019 |
|
RU2836365C2 |
| УЛУЧШЕННЫЙ СИНТЕТИЧЕСКИЙ Т-КЛЕТОЧНЫЙ РЕЦЕПТОР И АНТИГЕННЫЙ РЕЦЕПТОР | 2021 |
|
RU2820497C1 |
| Композиции и библиотеки, содержащие рекомбинантные полинуклеотиды, кодирующие Т-клеточные рецепторы, и способы применения рекомбинантных Т-клеточных рецепторов | 2017 |
|
RU2752528C2 |







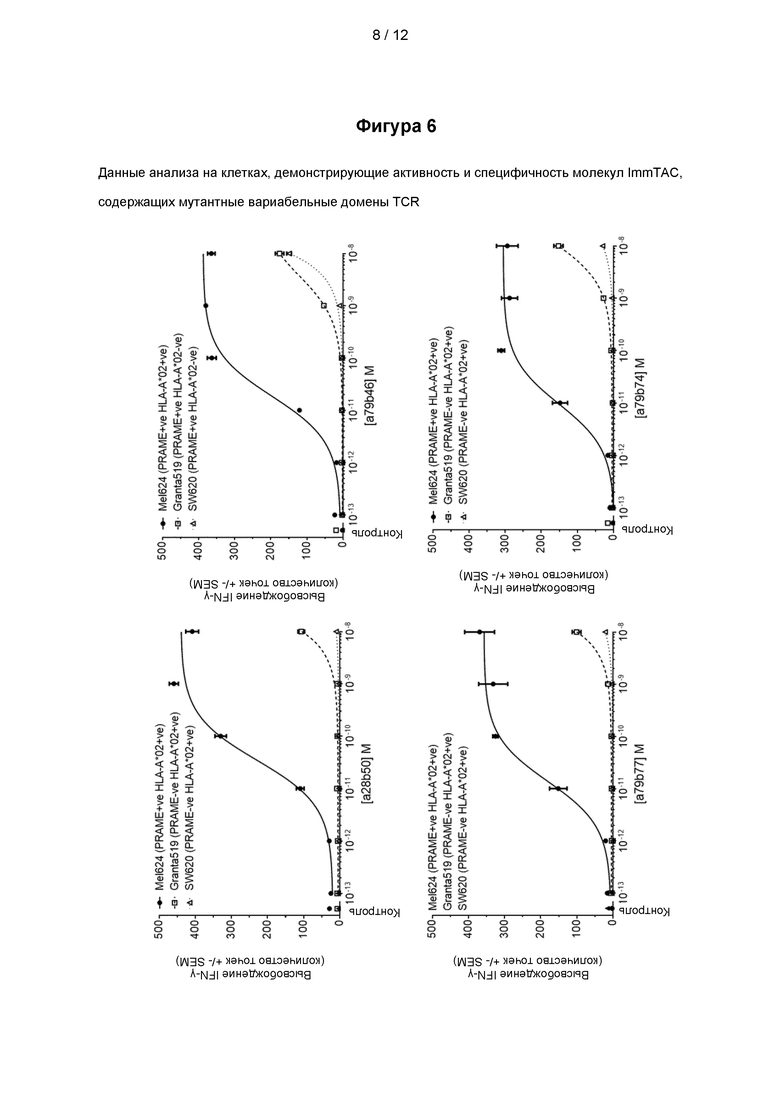
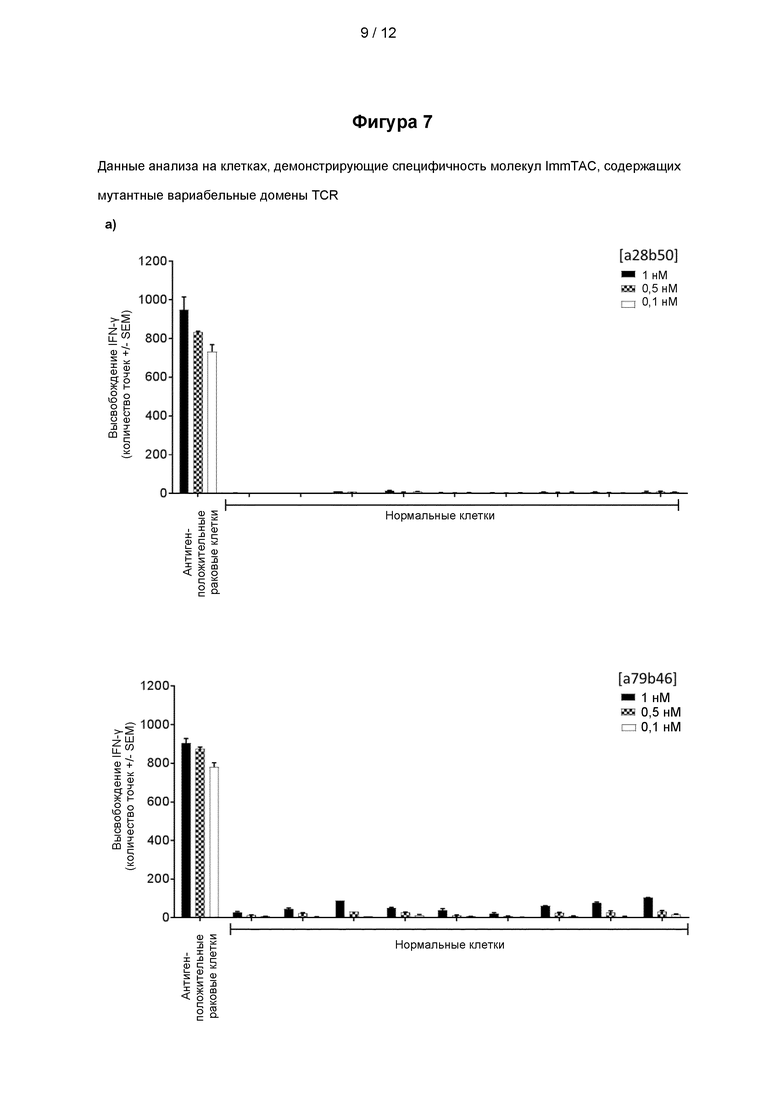
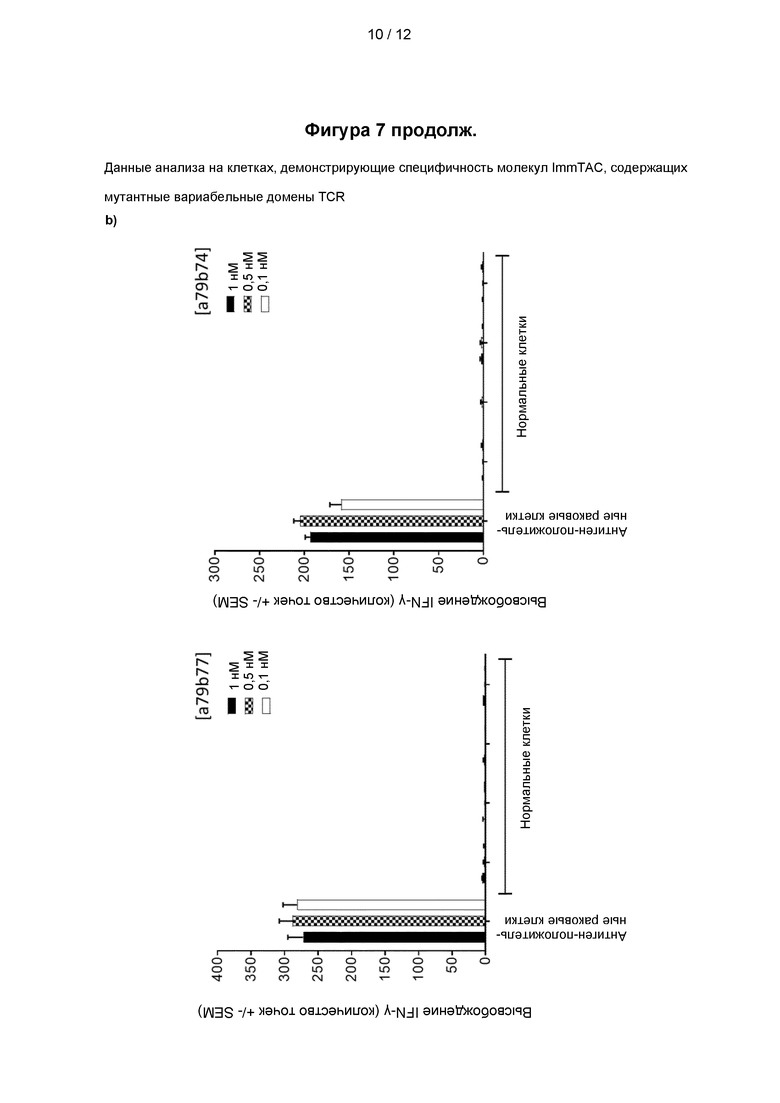
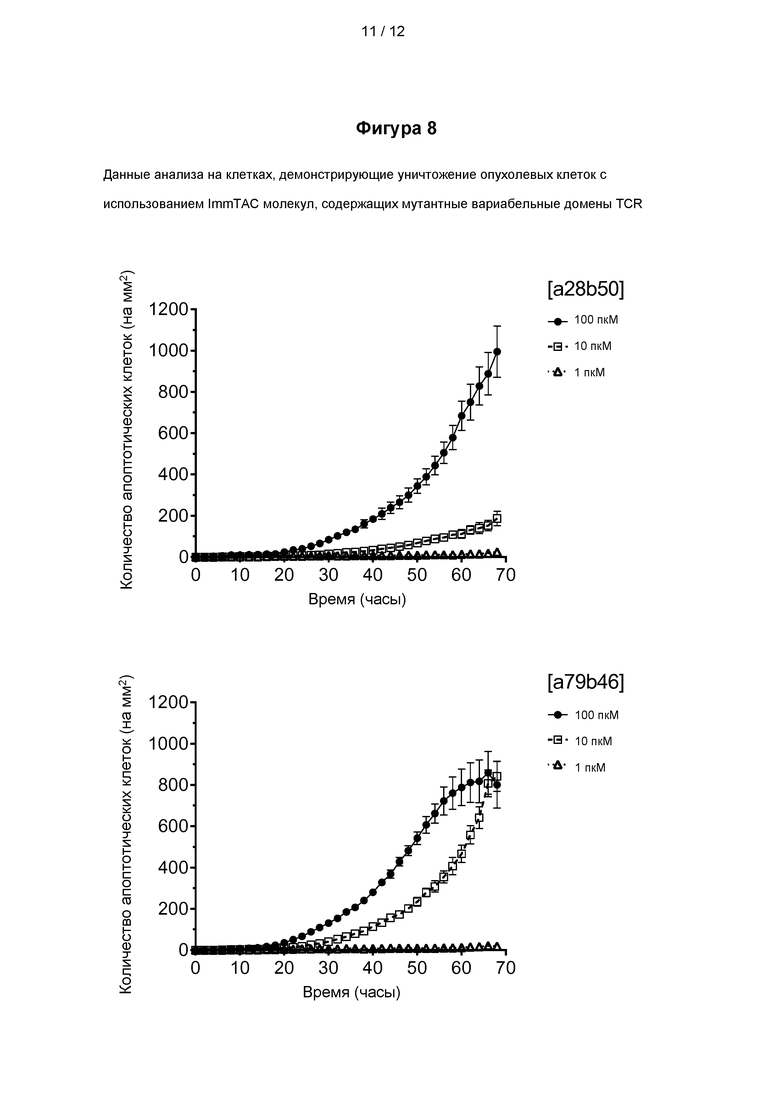
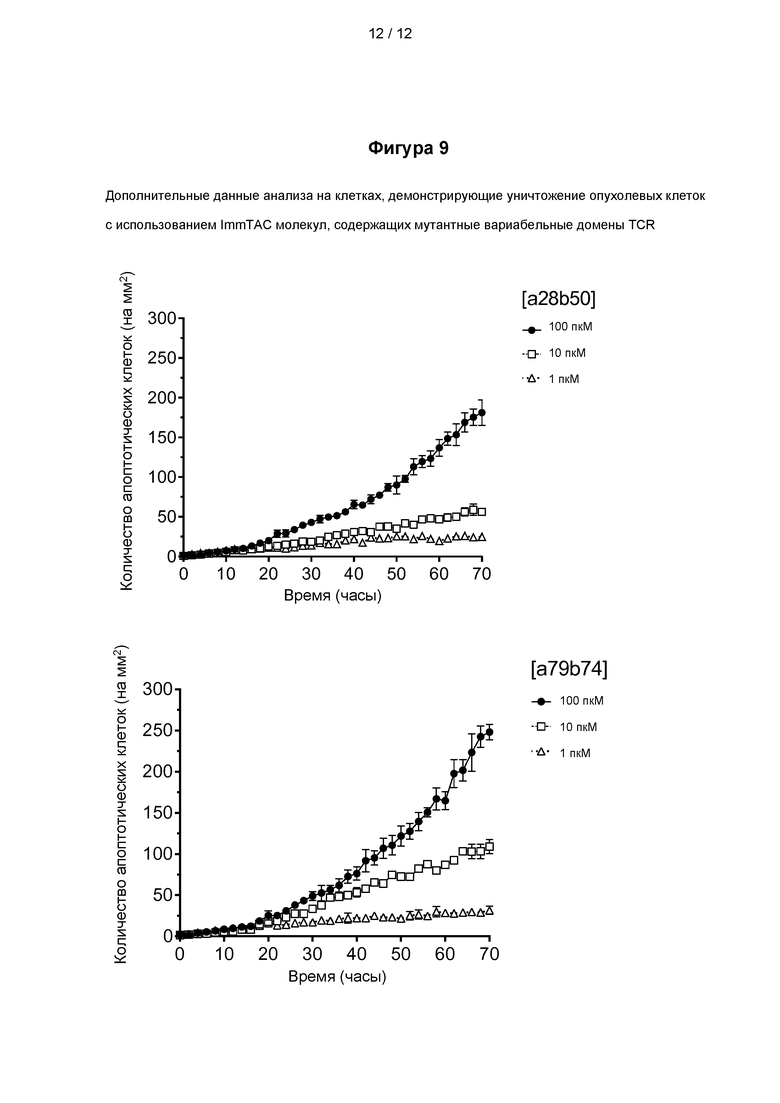
Группа изобретений относится к биотехнологии. Представлены T-клеточные рецепторы (TCR), которые связываются с рестриктированным по HLA-A*02 пептидом SLLQHLIGL (SEQ ID NO:1), гибридная молекула TCR/анти-CD3 для лечения рака, нуклеиновая кислота, вектор экспрессии, клетка для экспрессии TCR, не встречающаяся в природе и/или очищенная и/или сконструированная Т-клетка для адоптивной клеточной терапии рака, фармацевтическая композиция для лечения рака, способ лечения субъекта, состав для инъекций для введения субъекту и способ получения TCR или гибридной молекулы TCR/анти-CD3. Данная группа изобретений применяется в качестве новых иммунотерапевтических реагентов для лечения злокачественного заболевания. 10 н. и 27 з.п. ф-лы, 9 ил., 13 табл., 8 пр.
1. Т-клеточный рецептор (TCR), обладающий свойством связываться с комплексом SLLQHLIGL (SEQ ID NO: 1) HLA-A*02 и содержащий вариабельный домен альфа-цепи TCR и вариабельный домен бета-цепи TCR, каждый из которых содержит FR1-CDR1-FR2-CDR2-FR3-CDR3-FR4, где FR представляет собой каркасный участок, и CDR представляет собой определяющий комплементарность участок, при этом
(a) CDR альфа-цепи имеют следующие последовательности:
CDR1 - TISGTDY (SEQ ID NO: 39);
CDR2 - GLTSN (SEQ ID NO: 40);
CDR3 - CILILGHSGAGSYQLTF (SEQ ID NO: 41);
с одной или более мутациями (в соответствии с нумерацией последовательности SEQ ID NO: 2):
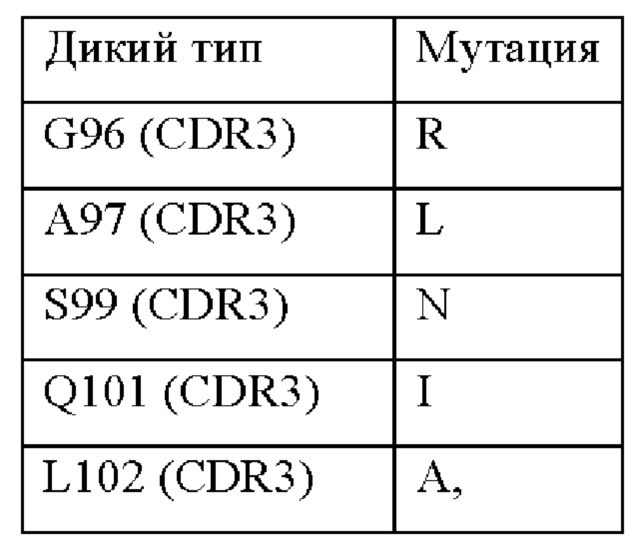
и
(b) CDR бета-цепи имеют следующие последовательности:
CDR1 - LNHDA (SEQ ID NO: 42);
CDR2 - SQIVNDF (SEQ ID NO: 43);
CDR3 - CASSPWTSGSREQYF (SEQ ID NO: 44);
с одной или более мутациями (в соответствии с нумерацией последовательности SEQ ID NO: 3):
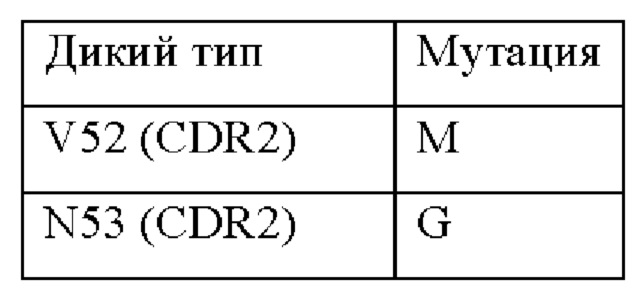

2. TCR по п. 1, отличающийся тем, что указанные каркасные участки вариабельного домена альфа-цепи содержат следующие последовательности:
FR1 - аминокислоты 1-25 SEQ ID NO: 2;
FR2 - аминокислоты 33-49 SEQ ID NO: 2;
FR3 - аминокислоты 55-87 SEQ ID NO: 2;
FR4 - аминокислоты 105-114 SEQ ID NO: 2;
или соответствующие последовательности, которые по меньшей мере на 90% идентичны указанными последовательностям, и/или
указанные каркасные участки вариабельного домена бета-цепи содержат следующие последовательности:
FR1 - аминокислоты 1-26 SEQ ID NO: 3;
FR2 - аминокислоты 32-48 SEQ ID NO: 3;
FR3 - аминокислоты 56-90 SEQ ID NO: 3;
FR4 - аминокислоты 106-114 SEQ ID NO: 3;
или соответствующие последовательности, которые по меньшей мере на 90% идентичны указанными последовательностям.
3. TCR по п. 1 или 2, отличающийся тем, что CDR альфа-цепи содержат одну, две, три, четыре или пять указанных мутаций.
4. TCR по п. 3, отличающийся тем, что CDR альфа-цепи содержат одну из следующих групп мутаций (в соответствии с нумерацией последовательности SEQ ID NO: 2):
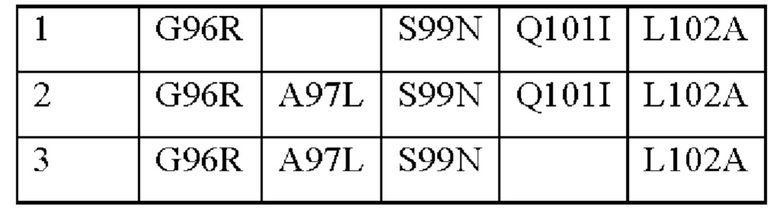
5. TCR по п. 3 или 4, отличающийся тем, что указанный CDR3 альфа-цепи имеет последовательность, выбранную из:
CILILGHSRAGNYIATF (SEQ ID NO: 45);
CILILGHSRLGNYIATF (SEQ ID NO: 46);
CILILGHSRLGNYQATF (SEQ ID NO: 47).
6. TCR по любому из пп. 1-5, отличающийся тем, что CDR бета-цепи содержат одну, две, три, четыре, пять, шесть, семь, восемь, девять или десять указанных мутаций.
7. TCR по п. 6, отличающийся тем, что CDR бета-цепи содержат одну из следующих групп мутаций (в соответствии с нумерацией последовательности SEQ ID NO: 3):
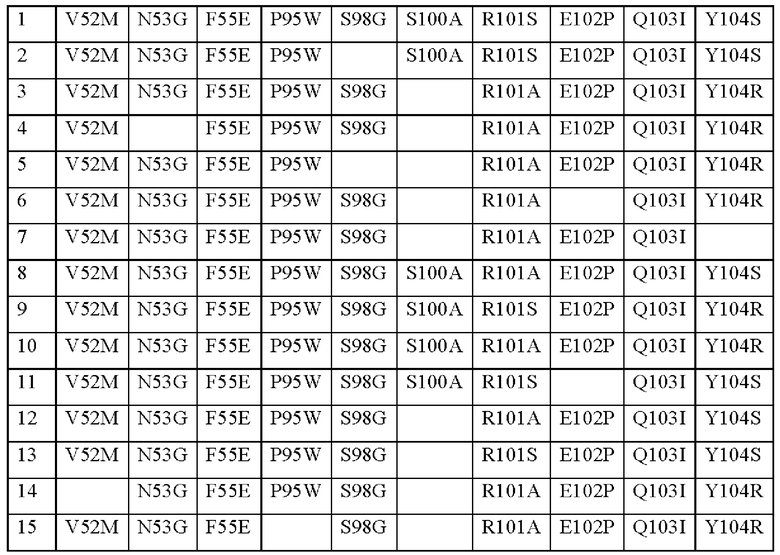

8. TCR по п. 6 или 7, отличающийся тем, что CDR2 бета-цепи имеет последовательность, выбранную из:
SQIMGDE (SEQ ID NO: 48);
SQIMNDE (SEQ ID NO: 49);
SQIVGDE (SEQ ID NO: 50).
9. TCR по пп. 6, 7 или 8, отличающийся тем, что CDR3 бета-цепи имеет последовательность, выбранную из:
CASSWWTGGASPISF (SEQ ID NO: 51);
CASSWWTSGASPISF (SEQ ID NO: 52);
CASSWWTGGSAPIRF (SEQ ID NO: 53);
CASSWWTSGSAPIRF (SEQ ID NO: 54);
CASSWWTGGSAEIRF (SEQ ID NO: 55);
CASSWWTGGSAPIYF (SEQ ID NO: 56);
CASSWWTGGAAPISF (SEQ ID NO: 57);
CASSWWTGGASPIRF (SEQ ID NO: 58);
CASSWWTGGAAPIRF (SEQ ID NO: 59);
CASSWWTGGASEISF (SEQ ID NO: 60);
CASSWWTGGSAPISF (SEQ ID NO: 61);
CASSWWTGGSSPISF (SEQ ID NO: 62);
CASSPWTGGSAPIRF (SEQ ID NO: 63);
CASSWWTGGSSPIRF (SEQ ID NO: 64).
10. TCR по п. 8 или 9, отличающийся тем, что соответствующие CDR2 и CDR3 бета-цепи являются следующими:


11. TCR по любому из предшествующих пунктов, отличающийся тем, что указанный TCR содержит одну из следующих комбинаций участков CDR альфа-цепи и бета-цепи:
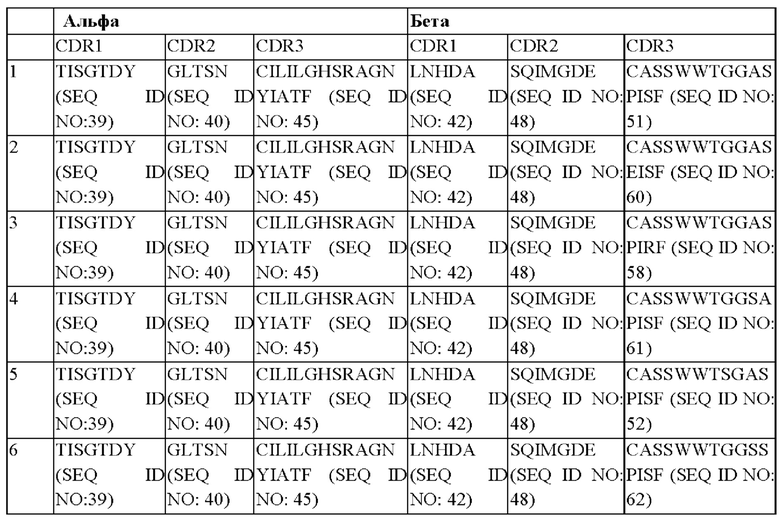
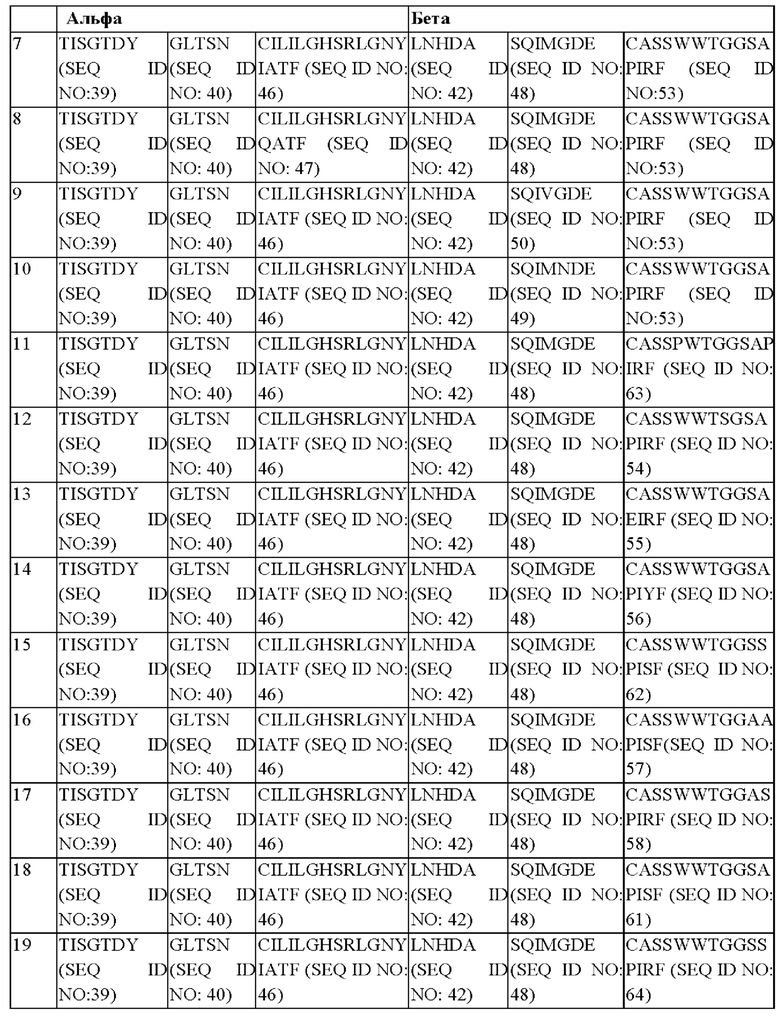

12. TCR по любому из предшествующих пунктов, отличающийся тем, что FR1 вариабельной области альфа-цепи имеет остаток G в положении -1 с использованием нумерации последовательности SEQ ID NO: 2.
13. TCR по любому из предшествующих пунктов, отличающийся тем, что вариабельный домен альфа-цепи содержит любую одну из аминокислотных последовательностей SEQ ID NO: 6-8, и вариабельный домен бета-цепи содержит любую одну из аминокислотных последовательностей SEQ ID. NO: 9-24.
14. TCR по любому из предыдущих пунктов, отличающийся тем, что вариабельный домен альфа-цепи и вариабельный домен бета-цепи выбраны из аминокислотных последовательностей:
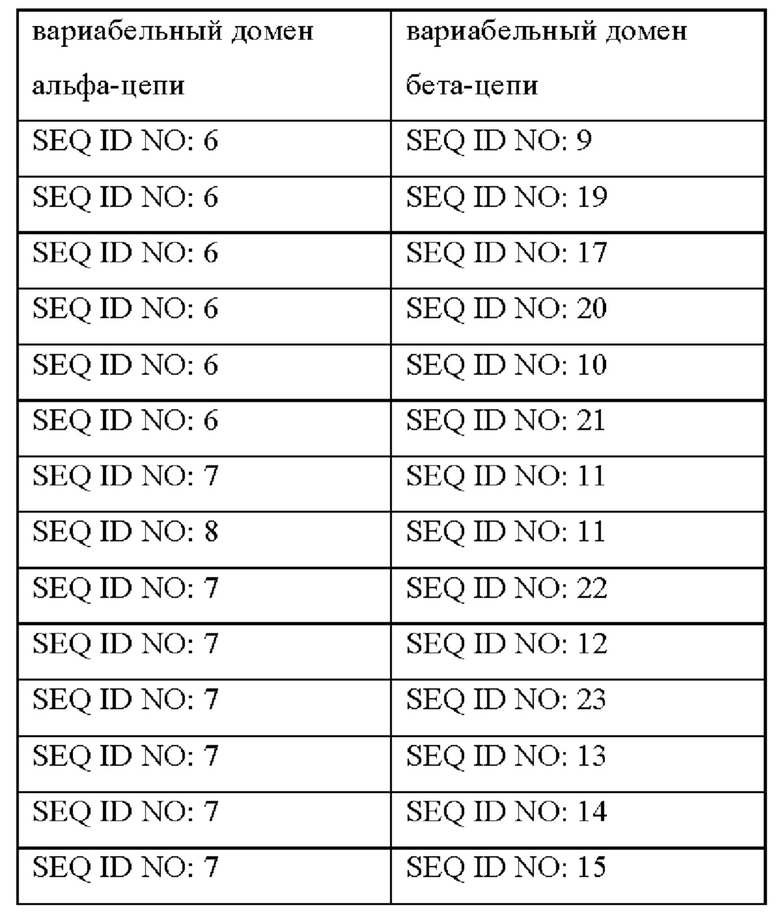

15. TCR по любому из предшествующих пунктов, представляющий собой альфа-бета-гетеродимер, имеющий последовательность константного домена TRAC альфа-цепи и последовательность константного домена TRBC1 или TRBC2 бета-цепи.
16. TCR по п. 15, отличающийся тем, что последовательности константного домена альфа- и бета-цепи модифицированы путем укорочения или замены с удалением нативной дисульфидной связи между Cys4 экзона 2 TRAC и Cys2 экзона 2 TRBC1 или TRBC2.
17. TCR по п. 15 или 16, отличающийся тем, что последовательности константного домена альфа- и/или бета-цепи модифицированы путем замены Thr 48 TRAC и Ser 57 TRBC1 или TRBC2 на остатки цистеина, где указанные остатки цистеина образуют ненативную дисульфидную связь между константными доменами альфа- и бета-цепи TCR.
18. TCR по любому из предшествующих пунктов, представленный в одноцепочечной форме типа Vα-L-Vβ, Vβ-L-Vα, Vα-Cα-L-Vβ, Vα-L-Vβ-Cβ, где Vα и Vβ представляют собой вариабельные участки α- и β-цепей TCR соответственно, Сα и Сβ представляют собой константные области α- и β-цепей TCR соответственно и L представляет собой линкерную последовательность.
19. TCR по любому из предшествующих пунктов, связанный с детектируемой меткой, терапевтическим агентом или группой, модифицирующей фармакокинетические (ФК) свойства.
20. TCR по п. 19, отличающийся тем, что с С- или N-концом альфа- или бета-цепи TCR ковалентно связано антитело против CD3, необязательно посредством линкерной последовательности.
21. TCR по п. 19, отличающийся тем, что указанная линкерная последовательность выбрана из группы, состоящей из GGGGS (SEQ ID NO: 31), GGGSG (SEQ ID NO: 32), GGSGG (SEQ ID NO: 33), GSGGG (SEQ ID NO: 34), GSGGGP (SEQ ID NO: 35), GGEPS (SEQ ID NO: 36), GGEGGGP (SEQ ID NO: 37) и GGEGGGSEGGGS (SEQ ID NO: 38).
22. Гибридная молекула TCR/анти-CD3 для лечения рака, экспрессирующего PRAME, у субъекта, представляющего собой человека с подтипом HLA-A*02, где вариабельный домен альфа-цепи содержит аминокислотную последовательность, выбранную из последовательностей SEQ ID NO: 6-8, и вариабельный домен бета-цепи содержит аминокислотную последовательность, выбранную из последовательностей SEQ ID NO: 9-24, и при этом антитело против CD3 ковалентно связано с С- или N-концом бета-цепи TCR посредством линкерной последовательности, выбранной из последовательностей SEQ ID NO: 31-38.
23. Гибридная молекула TCR/анти-CD3 по п. 22, содержащая:
аминокислотную последовательность альфа-цепи, выбранную из последовательностей SEQ ID NO: 25, 27 или 29, или аминокислотную последовательность альфа-цепи, которая по меньшей мере на 90%, например, на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотным последовательностям SEQ ID NO: 25, 27 и 29,
и аминокислотную последовательность бета-цепи, выбранную из последовательностей SEQ ID NO: 26, 28 или 30, или аминокислотную последовательность бета-цепи, которая по меньшей мере на 90%, например, на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотным последовательностям SEQ ID NO: 26, 28 и 29.
24. Гибридная молекула TCR/анти-CD3 по п. 23, содержащая:
(а) аминокислотную последовательность альфа-цепи, соответствующую SEQ ID NO: 25, и аминокислотную последовательность бета-цепи, соответствующую SEQ ID NO: 26;
(b) аминокислотную последовательность альфа-цепи, соответствующую SEQ ID NO: 27, и аминокислотную последовательность бета-цепи, соответствующую SEQ ID NO: 28; или
(c) аминокислотную последовательность альфа-цепи, соответствующую SEQ ID NO: 29, и аминокислотную последовательность бета-цепи, соответствующую SEQ ID NO: 30.
25. Нуклеиновая кислота, кодирующая альфа-цепь TCR и/или бета-цепь TCR по любому из предшествующих пунктов.
26. Вектор экспрессии, содержащий нуклеиновую кислоту по п. 25.
27. Клетка для экспрессии TCR, содержащая:
(a) вектор экспрессии по п. 26, кодирующий альфа- и бета-цепь TCR по любому из пп. 1-24, в одной открытой рамке считывания или двух отдельных открытых рамках считывания;
или
(b) первый вектор экспрессии, который содержит нуклеиновую кислоту, кодирующую альфа-цепь TCR по любому из пп. 1-24, и второй вектор экспрессии, который содержит нуклеиновую кислоту, кодирующую бета-цепь TCR по любому из пп. 1-24.
28. Не встречающаяся в природе и/или очищенная и/или сконструированная Т-клетка для адоптивной клеточной терапии рака, экспрессирующего PRAME, у субъекта, представляющего собой человека, с подтипом HLA-A*02, презентирующая TCR по любому из пп. 1-18.
29. Фармацевтическая композиция для лечения рака, который экспрессирует PRAME, у субъекта, представляющего собой человека, с подтипом HLA-A*02, содержащая в терапевтически эффективном количестве TCR по любому из пп. 1-21, или гибридную молекулу TCR/анти-CD3 по любому из пп. 22-24, или клетку по п. 27 или 28 с одним или более фармацевтически приемлемыми носителями или вспомогательными веществами.
30. TCR по любому из пп. 1-21, или гибридная молекула TCR/анти-CD3 по любому из пп. 22-24, или нуклеиновая кислота по п. 25, фармацевтическая композиция по п. 29 или клетка по п. 27 или 28 для применения в медицине, предпочтительно, у субъекта, представляющего собой человека.
31. TCR по любому из пп. 1-21, гибридная молекула TCR/анти-CD3 по любому из пп. 22-24, или нуклеиновая кислота по п. 25, фармацевтическая композиция по п. 29 или клетка по п. 27 или 28 для применения в способе лечения рака или опухоли, которые экспрессируют PRAME, предпочтительно, у субъекта, представляющего собой человека.
32. TCR, гибридная молекула TCR/анти-CD3, нуклеиновая кислота, фармацевтическая композиция или все перечисленное для применения по п. 31, отличающиеся тем, что указанная опухоль представляет собой солидную опухоль.
33. TCR, гибридная молекула TCR/анти-CD3, нуклеиновая кислота, фармацевтическая композиция или клетка для применения по любому из пп. 30-32, которые вводят путем инъекции, например внутривенной инъекции или непосредственной инъекции в опухоль.
34. Способ лечения субъекта, представляющего собой человека с подтипом HLA-A*02, имеющего рак, экспрессирующий PRAME, включающий введение указанному субъекту, нуждающемуся в указанном лечении, фармацевтически эффективной дозы фармацевтической композиции по п. 29.
35. Способ по п. 34, который дополнительно включает раздельное, комбинированное или последовательное введение антинеопластического агента.
36. Состав для инъекций для введения субъекту, представляющему собой человека с подтипом HLA-A*02, для лечения рака, экспрессирующего PRAME, содержащий терапевтически эффективное количество TCR по любому из пп. 1-21 или гибридной молекулы TCR/анти-CD3 по любому из пп. 22-24 совместно с одним или более фармацевтически приемлемым носителем или наполнителем.
37. Способ получения TCR по любому из пп. 1-21 или гибридной молекулы TCR/анти-CD3 по любому из пп. 22-24, включающий: а) содержание клетки по п. 27 при оптимальных условиях для экспрессии нуклеиновой кислоты цепей TCR и b) выделение цепей TCR.
| Токарный резец | 1924 |
|
SU2016A1 |
| AMIR, AVITAL L., et al., "PRAME-specific allo-HLA-restricted Tcells with potent antitumor reactivity useful for therapeutic T-cell receptor gene transfer" | |||
| Clinical Cancer Research, 2011, 17: 5615-5625 | |||
| KESSLER, JAN H., et al., "Efficient identification of novel HLA-A*0201-presented cytotoxic T lymphocyte epitopes | |||

