Предшествующий уровень техники
Лиганд CD30 (CD30L, CD153), встречающийся в природе лиганд CD30, представляет собой мембранный гликопротеин типа II, специфично связывающий CD30, в результате чего запускается передача сигнала CD30 посредством его цитоплазматического домена. CD30 и CD30L представляют собой взаимодействующие гликопротеины клеточной поверхности, являющиеся членами надсемейств рецептора фактора некроза опухоли (TNFR; от англ. "tumor necrosis factor receptor") и фактора некроза опухоли (TNF; от англ. "tumor necrosis factor"), соответственно (Durkop et al., Cell, 68:421, 1992; Smith et al., Cell, 73:1349, 1993; патенты US №: 5480981; 5677430; 6143869 и 6652854). Экспрессия CD30 и CD30L ограничена клетками иммунной системы и строго регулируется. CD30 экспрессируется, прежде всего, на активированных В-клетках и подгруппах Т-клеток с активированным фенотипом/фенотипом памяти (Ellis et al., J. Immun., 151:2380, 1993; Falini et al., Blood, 85:1, 1995). CD30L экспрессируется на высоких уровнях на активированных Т-клетках мыши и человека (Shimozato et al., Biochem. & Biophys. Res. Comm., 256:519, 1999; Armitage, J. Biological Regulators & Homeostatic Agents, 14:142, 2000). Резкие изменения в относительной генной экспрессии CD30L происходят по мере перехода В-клеток через герминативные центры (Klein et al., Proc. Nat. Acad. Sci., USA, 100:2639, 2003). Таким образом, экспрессия CD30L на В-клетках может быть стадиеспецифичной и контекстоспецифичной. CD30L также, по-видимому, является маркером уникальной популяции дендритоподобных антигенпрезентирующих клеток мыши, присутствующих в мальпигиевых тельцах, где Т-клетки взаимодействуют с В-клетками (Kim et al., Immunity, 18:643, 2003).
Взаимодействия между клетками, экспрессирующими CD30/CD30L, по-видимому, важны для генерирования сильных вторичных ответов антител, зависимых от Т-клеток, или ответов антител с переключением изотипа. Данные, свидетельствующие о такой роли, включают данные исследований in vitro (Shanebeck et al., Eur. J. Immunol., 25:2147-53, 1995) и in vivo (Gaspal et al., J. Immunol., 174:3891-6, 2005) на мышиных системах. Кроме того, показано, что обработка мышей in vivo блокирующим, но не истощающим моноклональным антителом (mAb; от англ. "monoclonal antibody") к CD30L мыши ингибирует развитие или прогрессирование заболевания в ряде моделей аутоиммунных заболеваний, зависимых от Τ- и/или В-клеток (патент US №6667039). В модели с сильным гуморальным иммунным компонентом ингибирование заболевания коррелирует с ингибированием ответа антител, обусловленного заболеванием.
Следовательно, было бы полезно иметь композиции, содержащие антитела и/или антигенсвязывающие участки человека, связывающиеся с CD30L, для применения в терапевтических и диагностических областях применения.
Сущность изобретения
Предложены белки, связывающие антиген - лиганд CD30 (CD30L), в частности, CD30L человека. Белки, связывающие антиген CD30L человека, могут уменьшать, ингибировать, нарушать и/или модулировать по меньшей мере один из биологических ответов, относящихся к взаимодействию CD30L/CD30, и как таковые полезны для ослабления действий заболеваний или расстройств, обусловленных CD30L. Белки, связывающие антиген CD30L, можно применять, например, чтобы уменьшать, ингибировать, нарушать и/или модулировать взаимодействия CD30L/CD30.
В одном воплощении изобретения изолированные белки, связывающие антиген, связывают С-концевой участок CD30L, включающий аминокислоты (АА) 201-234. В следующем воплощении изобретения предложен антигенсвязывающий белок, связывающий С-концевой участок CD30L, включающий АА 201-234, и дополнительный участок CD30L, расположенный в N-концевой части внеклеточной области, определенный АА 75-95. В следующих воплощениях изобретения антигенсвязывающий белок обладает по меньшей мере одним свойством, выбранным из группы, состоящей из следующих свойств: а) ингибирования взаимодействия CD30/CD30L; b) ингибирования СD30L-индуцированной индукции интерлейкина (ИЛ)-8; с) перекрестной конкуренции с любым из антител A-F за связывание с CD30L человека; d) константы диссоциации к CD30L человека, составляющей максимум 70 пикомоль (пМ), и е) связывания с CD30L человека по существу с таким же или с более высоким сродством (более низкой константой диссоциации (KD)) по сравнению с любым из антител A-F. Сродство (или KD) может быть определено, как известно специалистам в данной области техники, например, с помощью поверхностного плазмонного резонанса (SPR; от англ. "Surface Plasmon Resonance") или с помощью сортировки флуоресцентно-активированных клеток (FACS; от англ. "Fluorescence Activated Cell Sorting"), как описано в Примере 4 данного изобретения.
В следующем воплощении изобретения антигенсвязывающий белок связывает CD30L и конкурирует с Fab-фрагментом одного или более из антител А, В, С, D, Ε и F за связывание с CD30L Альтернативно данный антигенсвязывающий белок, характеризующийся как антигенсвязывающий белок, который связывается с CD30L человека (hCD30L), ингибируется связыванием Fab одного или более из антител А, В, С, D, Ε и F. В следующем конкретном воплощении изобретения Fab представляет собой Fab антитела А, включающего каждое из антител А1, А2, A3, A4, А5 и А6.
В одном воплощении изобретения предложен изолированный антигенсвязывающий белок, связывающий CD30L и включающий по меньшей мере одну вариабельную область тяжелой цепи, содержащую участки определения комплементарности (CDR; от англ. "complementarity determining region") Н1, CDRH2 и CDRH3, выбранные из группы, состоящей из следующих участков: a) CDRH1, отличающегося не более чем четырьмя, тремя, двумя или одной аминокислотной заменой, инсерцией или делецией от участка CDRH1, представленного в таблице 3; b) CDRH2, отличающегося не более чем семью, шестью, пятью, четырьмя, тремя, двумя или одной аминокислотной заменой, инсерцией и/или делецией от участка CDRH2, представленного в таблице 3; с) CDRH3, отличающегося не более чем одиннадцатью, десятью, девятью, восемью, семью, шестью, пятью, четырьмя, тремя, двумя или одной аминокислотной заменой, инсерцией и/или делецией от участка CDRH3, представленного в таблице 3; и включающий по меньшей мере одну вариабельную область легкой цепи, содержащую участок CDRL1, CDRL2 и CDRL3, выбранный из группы, состоящей из следующих участков: d) CDRL1, отличающегося не более чем четырьмя, тремя, двумя или одной аминокислотной заменой, инсерцией и/или делецией от участка CDRL1, представленного в таблице 3; е) CDRL2, отличающегося не более чем двумя или одной аминокислотной заменой, инсерцией или делецией от участка CDRL2, представленного в таблице 3; f) CDRL3, отличающегося не более чем двумя или одной аминокислотной заменой, инсерцией или делецией от участка CDRL3, представленного в таблице 3. В родственном воплощении изобретения предложен участок CDRH1, выбранный из группы, состоящей из последовательностей SEQ ID NO: 14, 15, 16, 17, 18 и 33; участок CDRH2, выбранный из группы, состоящей из последовательностей SEQ ID NO: 19, 20, 21, 22, 23, 24 и 34; участок CDRH3, выбранный из группы, состоящей из последовательностей SEQ ID NO: 25, 26, 27, 28, 29 и 35; участок CDRL1, выбранный из группы, состоящей из последовательностей SEQ ID NO: 1, 2, 3, 4, 5, 6 и 30; участок CDRL2, выбранный из группы, состоящей из последовательностей SEQ ID NO: 7, 8, 9, 10 и 31; и участок CDRL3, выбранный из группы, состоящей из последовательностей SEQ ID NO: 11, 12, 13 и 32.
В другом воплощении изобретения предложен изолированный белок, связывающий антиген, который связывает CD30L человека, содержащий по меньшей мере одну вариабельную область легкой цепи, включающую CDR1, CDR2 и CDR3, где участок CDR1 содержит аминокислотные остатки 23-36, CDR2 содержит аминокислотные остатки 52-58, и CDR3 содержит аминокислотные остатки 91-100 SEQ ID NO: 36, 28, 40, 42 или 44; либо b) вариабельную область легкой цепи, включающую CDR1, CDR2 и CDR3, где участок CDR1 содержит аминокислотные остатки 25-36, CDR2 содержит аминокислотные остатки 52-58, и CDR3 содержит аминокислотные остатки 91-100 SEQ ID NO: 46; и по меньшей мере одну вариабельную область тяжелой цепи, включающую CDR1, CDR2 и CDR3, где участок CDR1 содержит аминокислотные остатки 31-35, CDR2 содержит аминокислотные остатки 50-65, и CDR3 содержит аминокислотные остатки 98-113 SEQ ID NO: 48, 50, 52, 54, 56, 58, 60, 62, 64, 66, 68, 70 или 72. В родственном воплощении изобретения предложен белок, связывающий антиген, содержащий по меньшей мере одну вариабельную область тяжелой цепи и по меньшей мере одну вариабельную область легкой цепи. В другом воплощении изобретения предложен белок, связывающий антиген, включающий по меньшей мере две вариабельные области тяжелой цепи и по меньшей мере две вариабельные области легкой цепи.
В другом воплощении изобретения предложен изолированный белок, связывающий антиген, который связывает CD30L, включающий вариабельную область тяжелой цепи и вариабельную область легкой цепи, где последовательность вариабельной области тяжелой цепи отличается не более чем 31, 30, 29, 28, 27, 26, 25, 24, 23, 22, 21, 20, 19, 18, 17, 16, 15, 14, 13, 12, 11, 10, 9, 8, 7, 6, 5, 4, 3, 2 или 1 аминокислотной заменой, добавлением и/или делецией от последовательности вариабельной области тяжелой цепи, представленной в таблице 2; и где последовательность вариабельной области легкой цепи отличается не более чем 30, 29, 28, 27, 26, 25, 24, 23, 22, 21, 20, 19, 18, 17, 16, 15, 14, 13, 12, 11, 10, 9, 8, 7, 6, 5, 4, 3, 2 или 1 аминокислотной заменой, добавлением и/или делецией от последовательности вариабельной области легкой цепи, представленной в таблице 1.
В другом воплощении изобретения предложен изолированный антигенсвязывающий белок, связывающий CD30L и включающий вариабельную область тяжелой цепи, содержащую аминокислотную последовательность, обладающую по меньшей мере 80% идентичностью последовательности SEQ ID NO: 48, 50, 52, 54, 56, 58, 60, 62, 64, 66, 68, 70 и 72; и вариабельную область легкой цепи, содержащую аминокислотную последовательность, обладающую по меньшей мере 88% идентичностью последовательности SEQ ID NO: 36, 38, 40, 42, 44 и 46.
В одном воплощении изобретения предложен изолированный антигенсвязывающий белок, связывающий CD30L, выбранный из группы, состоящей из следующих участков: а) вариабельной области тяжелой цепи, выбранной из группы, состоящей из SEQ ID NO: 48, 50, 52, 54, 56, 58, 60 и 62, и вариабельной области легкой цепи SEQ ID NO: 36; b) вариабельной области тяжелой цепи SEQ ID NO: 64 и вариабельной области легкой цепи SEQ ID NO: 38; с) вариабельной области тяжелой цепи SEQ ID NO: 66 и вариабельной области легкой цепи SEQ ID NO: 40; d) вариабельной области тяжелой цепи SEQ ID NO: 68 и вариабельной области легкой цепи SEQ ID NO: 42; е) вариабельной области тяжелой цепи SEQ ID NO: 70 и вариабельной области легкой цепи SEQ ID NO: 44; и f) вариабельной области тяжелой цепи SEQ ID NO: 72 и вариабельной области легкой цепи SEQ ID NO: 46. В родственном воплощении изобретения изолированный антигенсвязывающий белок представляет собой антитело. В другом родственном воплощении изобретения антитело представляет собой моноклональное антитело, рекомбинантное антитело, человеческое антитело, гуманизированное антитело, химерное антитело, мультиспецифическое антитело или его фрагмент антитела. В следующем воплощении изобретения фрагмент антитела представляет собой Fab фрагмент, Fab' фрагмент, F(ab')2 фрагмент, Fv фрагмент, диатело или одноцепочечную молекулу антитела. В родственном воплощении изобретения антигенсвязывающий белок представляет собой человеческое антитело. В другом родственном воплощении изобретения антигенсвязывающий белок представляет собой моноклональное антитело. В следующем родственном воплощении изобретения антигенсвязывающий белок относится к IgG1-, IgG2- IgG3- или IgG4-типу. В родственном воплощении изобретения антигенсвязывающий белок относится к IgG1- или IgG2-типу.
В другом воплощении изобретения предложены изолированные молекулы нуклеиновой кислоты, кодирующие антигенсвязывающий белок, как раскрыто выше.
В родственном воплощении изобретения по меньшей мере одна вариабельная область тяжелой цепи кодируется изолированной молекулой нуклеиновой кислоты, выбранной из группы, состоящей из SEQ ID NO: 49, 51, 53, 55, 57, 59, 61, 63, 65, 67, 69, 71 и 73, и по меньшей мере одна вариабельная область легкой цепи кодируется изолированной молекулой нуклеиновой кислоты, выбранной из группы, состоящей из SEQ ID NO: 37, 39, 41, 43, 45 и 47. В другом родственном воплощении изобретения молекула нуклеиновой кислоты функционально связана с контролирующей последовательностью. В следующих родственных воплощениях изобретения предложены векторы, содержащие нуклеиновую кислоту, раскрытую выше, и клетки-хозяева, содержащие такие векторы. Еще в одном следующем воплощении изобретения предложен изолированный полинуклеотид, достаточный для применения в качестве зонда гибридизации, праймера полимеразной цепной реакции (ПЦР) или секвенирующего праймера, представляющий собой фрагмент молекулы нуклеиновой кислоты, как раскрыто выше, или комплементарной ей молекулы.
В другом воплощении изобретения предложен способ получения антигенсвязывающего белка, как раскрыто выше, включающий стадию получения препарата антигенсвязывающего белка из клетки-хозяина, секретирующей антигенсвязывающий белок.
В другом воплощении изобретения предложен изолированный антигенсвязывающий белок, как раскрыто выше, где антигенсвязывающий белок обладает по меньшей мере одним свойством, выбранным из группы, состоящей из следующих свойств: а) ингибирования взаимодействия CD30/CD30L; b) ингибирования СD30L-индуцированной индукции ИЛ-8; с) перекрестной конкуренции с любым из антител A-F за связывание с CD30L человека; d) константой диссоциации, равной не более 70 пМ, и е) связывания с CD30L человека по существу с такой же Kd, как любого из антител A-F.
Еще в одном другом воплощении изобретения предложена фармацевтическая композиция, содержащая по меньшей мере один антигенсвязывающий белок, как раскрыто выше, и фармацевтически приемлемый эксципиент. В родственном воплощении изобретения дополнительно предложены такие фармацевтические композиции, дополнительно содержащие метящую группу или эффекторную группу. В другом родственном воплощении изобретения метящая группа выбрана из группы, состоящей из следующих групп: изотопных меток, магнитных меток, редокс-активных группировок, оптических красителей, биотинилированных групп и предопределенных полипептидных эпитопов, распознаваемых вторичным репортером. В следующем родственном воплощении изобретения эффекторная группа выбрана из группы, состоящей из следующих групп: радиоизотопа, радионуклида, токсина, терапевтической группы и химиотерапевтической группы. В другом родственном воплощении изобретения белок, связывающий антиген, связан с метящей группой.
В другом воплощении изобретения предложен способ лечения или предупреждения состояния, связанного с CD30L, у пациента, включающий введение пациенту, нуждающемуся в этом, эффективного количества по меньшей мере одного изолированного белка, связывающего антиген, как раскрыто выше. В другом родственном воплощении изобретения изолированный антигенсвязывающий белок вводят отдельно или в виде комбинированной терапии.
В другом воплощении изобретения предложен способ снижения активности CD30L у пациента, включающий введение эффективного количества по меньшей мере одного белка, связывающего антиген, как раскрыто выше.
Еще в одном другом воплощении изобретения предложены белки, связывающие антиген, конкурирующие по меньшей мере с одним белком, связывающим антиген, как раскрыто выше.
В другом воплощении изобретения предложен белок, связывающий антиген, как раскрыто выше, полностью или частично афукозилированный.
Описание графических материалов
Фиг.1А. Клетки Ramos и эффекторы, представляющие собой естественные киллерные клетки (NK; от англ. "natural killers"), Пустые квадраты: Ритуксан (IgG1), Сплошные ромбы: Антитело (Аb; от англ. Antibody) А1 IgG1f, Сплошные квадраты: Антитело А1 IgG1, Пустые круги: Антитело А1 IgG2.
Фиг.1В: Клетки JD38 и эффекторы NK, Пустые квадраты: Ритуксан (IgG1), Сплошные ромбы: Антитело А1 IgG1f, Сплошные квадраты: Антитело А1 IgG1, Пустые круги: Антитело А1 IgG2.
Фиг.1С. Клетки DS179 и эффекторы NK, Пустые квадраты: Ритуксан (IgG1), Сплошные ромбы: Антитело А1 IgG1f, Сплошные квадраты: Антитело А1 IgG1, Пустые круги: Антитело А1 IgG2.
Фиг.1D: Клетки EW36 и эффекторы NK, Пустые квадраты: Ритуксан (IgG1), Сплошные ромбы: Антитело А1 IgG1f, Сплошные квадраты: Антитело А1 IgG1, Пустые круги: Антитело А1 IgG2.
Сведения, подтверждающие возможность осуществления изобретения
В настоящем изобретении предложены композиции, наборы и способы, относящиеся к белкам, связывающим антиген CD30L, включающим антигенсвязывающие белки, которые блокируют взаимодействие между CD30L и CD30, такие как антитела против CD30L, фрагменты антител и производные антител, например, нейтрализующие антитела против CD30L, фрагменты антител или производные антител. Также предложены полинуклеотиды и их производные и фрагменты, содержащие последовательности нуклеиновых кислот, кодирующие весь полипептид или участок полипептида, связывающийся с CD30L, например, полинуклеотид, кодирующий все антитело или участок антитела против CD30L, фрагмент антитела или производное антитела, плазмиды и векторы, содержащие такие нуклеиновые кислоты, и клетки или линии клеток, содержащие такие полинуклеотиды и/или векторы или плазмиды. Предложенные способы включают, например, способы получения, идентификации или выделения белков, связывающих антиген CD30L, таких как антитела против CD30L, способы определения блокирования молекулой взаимодействия между CD30L и CD30, способы определения антагонизма молекулы CD30L, способы получения композиций, таких как фармацевтические композиции, содержащие белок, связывающий антиген CD30L, и способы введения белка, связывающего антиген CD30L, субъекту, например, способы лечения состояния, опосредованного CD30L, и блокирования взаимодействия между CD30L и CD30, in vivo или in vitro.
Если в данном изобретении не определено иное, научные и технические термины, используемые в связи с настоящим изобретением, имеют значение, обычно понимаемое обычными специалистами в данной области техники. Кроме того, если контекст не требует иного, термины в единственном числе включают термины во множественном числе, а термины множественного числа включают термины в единственном числе. Как правило, методы культивирования клеток и тканей, молекулярной биологии, иммунологии, микробиологии, генетики и химии белков и нуклеиновых кислот и гибридизации, описанные в данном изобретении, и номенклатуры, используемые в связи с ними, хорошо известны и общеприняты в данной области техники. Способы и методы по настоящему изобретению, как правило, выполняют в соответствии с традиционными способами, хорошо известными в данной области техники и описанными в различных общих и более конкретных ссылках, цитируемых и обсуждаемых на протяжении всего настоящего описания, если не указано иное. См., например, следующие ссылки: Sambrook et al., Molecular Cloning: A Laboratory Manual, 3rd ed., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y. (2001) и Ausubel et al., Current Protocols in Molecular Biology, Greene Publishing Associates (1992), а также Harlow and Lane Antibodies: A Laboratory Manual Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y. (1990). Методы ферментативных реакций и очистки выполняют в соответствии со спецификациями изготовителя, которые обычно выполняют в данной области техники, или как раскрыто в данном изобретении. Лабораторные работы и методы аналитической химии, химии органического синтеза и медицинской и фармацевтической химии, раскрытые в данном изобретении, а также терминология, используемая в связи с ними, хорошо известны и общеприняты в данной области техники. Для химических синтезов, химических анализов, получения, приготовления, и доставки фармацевтических препаратов и лечения пациентов можно использовать стандартные способы.
Все документы или части документов, цитируемые в данном изобретении, включающие, но не ограниченные ими, патенты, заявки на патенты, статьи, книги и монографии, явным образом полностью включены в данное изобретение посредством ссылки в целях описания и раскрытия, например, методологии, описанные в таких публикациях, которые могут быть использованы в сочетании с информацией, раскрытой в данном изобретении.
Термин "СD30-лиганд" (CD30L) относится к роду полипептидов, способных к связыванию с CD30, как раскрыто в статье Smith et al., Cell 73:1349-1360, 1993 и в патенте US №5480981, включая его СD30-связывающие мутеины; такие полипептиды включают мембраносвязанные белки (содержащие цитоплазматический домен, трансмембранную область и внеклеточный домен), а также укороченные белки, сохраняющие СD30-связывающее свойство. Такие укороченные белки включают, например, растворимый CD30L, содержащий только внеклеточный (рецептор-связывающий) домен. Включены также фрагменты CD30L, включающие участки полноразмерного полипептида CD30L, сохраняющие способность к связыванию с CD30, или способные вызывать антитело, специфично связывающееся с полипептидом CD30 или с участком полноразмерного CD30, способным передавать биологический сигнал, такой как активация фактора транскрипции каппа В (NF-κB).
Термин "CD30" относится к рецептору, являющемуся членом надсемейства рецепторов фактора некроза опухоли (TNF; от англ. "tumor necrosis factor")/фактора роста нервной ткани (NGF; от англ. "nerve growth factor"), клонирование которого раскрыто в статье Durkop et al. (Cell 68:421, 1992). Выражение "растворимый CD30" (sCD30) относится к растворимым молекулам, включающим весь внеклеточный домен или часть внеклеточного домена белка CD30, и сохраняющим способность к специфичному связыванию с CD30L. Растворимые полипептиды CD30 охватывают рекомбинантные SCD30 и встречающиеся в природе белки sCD30 в высокоочищенной форме.
Как используют в настоящем изобретении, выражение "фрагмент CD30" относится к участку полноразмерного полипептида CD30, сохраняющему способность к связыванию с CD30L, или способному вызывать антитело, специфично связывающееся с полипептидом CD30 или с участком полноразмерного CD30, способным передавать биологический сигнал, такой как активация NF-κB.
Выражение "взаимодействие CD30/CD30L", как используют в настоящем изобретении, относится к специфичному связыванию CD30 с CD30L, приводящему в результате к преобразованию сигнала CD30. Это связывание включает случаи, в которых по меньшей мере один партнер связывания представляет собой фрагмент либо CD30, либо CD30L, то есть этот термин может относиться к связывающему взаимодействию фрагмента CD30 с CD30L, CD30 с фрагментом CD30L или фрагмента CD30 с фрагментом CD30L. Кроме того, во взаимодействие CD30/CD30L может быть вовлечен аналог CD30 (такой как аллельный вариант или мутеин), способный к специфичному связыванию с CD30L, либо в это взаимодействие может быть вовлечен аналог CD30L (такой как аллельный вариант или мутеин), специфично связывающийся с CD30. Кроме того, во взаимодействие CD30/CD30L могут быть вовлечены либо эндогенные белки CD30 или CD30L, либо рекомбинантные белки CD30 или CD30L, экспрессируемые клеткой, трансфицированной нуклеиновой кислотой, кодирующей рекомбинантный белок.
Термин "полинуклеотид" включает как однонитевые, так и двунитевые нуклеиновые кислоты, и включает ДНК, РНК, мРНК, кДНК геномного или синтетического происхождения или некоторые их комбинации, не связанные с последовательностями, в норме обнаруживаемыми в природе. Изолированные полинуклеотиды, содержащие указанные последовательности, могут содержать, кроме указанных последовательностей, последовательности, кодирующие вплоть до десяти или даже вплоть до двенадцати других белков или их участков, либо могут включать функционально связанные регуляторные последовательности, контролирующие экспрессию кодирующей области указанных нуклеиново-кислотных последовательностей, и/или могут включать векторные последовательности. Нуклеотиды, составляющие полинуклеотид, могут представлять собой рибонуклеотиды или дезоксирибонуклеотиды, либо модифицированную форму любого из типов нуклеотидов. Модификации включают модификации оснований, такие как производные, представляющие собой бромуридин и инозин, модификации рибозы, такие как 2',3'-дидезоксирибоза, и модификации межнуклеотидной связи, такие как фосфоротиоат, фосфородитиоат, фосфороселеноат, фосфородиселеноат, фосфороанилотиоат, фосфоранилидат и фосфороамидат.
Термин "олигонуклеотид" означает полинуклеотид, содержащий 100 или менее нуклеотидов. В некоторых воплощениях изобретения олигонуклеотиды составляют от 10 до 60 оснований в длину. В других воплощениях изобретения олигонуклеотиды составляют 12, 13, 14, 15, 16, 17, 18, 19 или от 20 до 40 нуклеотидов в длину. Олигонуклеотиды могут быть однонитевыми или двунитевыми, например, для применения при конструировании мутантного гена. Олигонуклеотиды могут представлять собой смысловые или антисмысловые олигонуклеотиды. Олигонуклеотид может включать обнаружимую метку, такую как радиоактивная метка, флуоресцентная метка, гаптен или антигенная метка, для анализов определения. Олигонуклеотиды можно применять, например, в качестве праймеров ПЦР, праймеров клонирования или зондов гибридизации.
Термины "полипептид" или "белок" означает макромолекулу, имеющую аминокислотную последовательность нативного белка, то есть белка, продуцируемого встречающейся в природе и нерекомбинантной клеткой; либо продуцируемого клеткой, сконструированной генно-инженерными методами, или рекомбинантной клеткой, и включает молекулы, имеющие аминокислотную последовательность нативного белка, или молекулы, имеющие одну или более делеций, инсерций и/или замен аминокислотных остатков нативной последовательности. Данный термин также включает полимеры аминокислот, в которых одна или более аминокислот представляют собой химические аналоги соответствующих встречающихся в природе аминокислот и полимеров. Термины "полипептид" и "белок" охватывают белки, связывающие антиген CD30L (такие как антитела), и последовательности, имеющие одну или более делеций, добавлений и/или замен аминокислотных остатков последовательности белка, связывающего антиген. Термин "полипептидный фрагмент" относится к полипептиду, имеющему амино-концевую делецию, карбокси-концевую делецию и/или внутреннюю делецию по сравнению с полноразмерным нативным белком. Такие фрагменты могут также содержать модифицированные аминокислоты по сравнению с нативным белком. В определенных воплощениях изобретения фрагменты имеют длину от примерно пяти до 500 аминокислот. Например, длина фрагментов может составлять по меньшей мере 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 50, 70, 100, 110, 150, 200, 250, 300, 350, 400 или 450 аминокислот. Полезные полипептидные фрагменты включают иммунологически функциональные фрагменты антител, включая связывающие домены. В случае белка, связывающего антиген CD30L, такого как антитело, полезные фрагменты включают, но не ограничены ими, один или более участков CDR, вариабельный домен тяжелой или легкой цепи, участок цепи антитела, участок вариабельной области, включающий менее трех CDR, и тому подобное.
"Аминокислота" включает ее обычное значение в данной области техники. Двадцать встречающихся в природе аминокислот и их сокращения следуют стандартному использованию. См.: Immunology-Α Synthesis, 2nd Edition, (Ε.S. Golub and D.R. Gren, eds.), Sinauer Associates: Sunderland, Mass. (1991). Стереоизомеры (например, D-аминокислоты) двадцати стандартных аминокислот, неприродные аминокислоты, такие как [альфа]-,[альфа]-двузамещенные аминокислоты, N-алкиламинокислоты и другие нестандартные аминокислоты могут также быть подходящими компонентами для полипептидов. Примеры нестандартных аминокислот включают следующие аминокислоты: 4-гидроксипролин, [гамма]-карбоксиглутамат, [эпсилон]-N,N,N-триметиллизин, [эпсилон]-N-ацетиллизин, О-фосфосерин, N-ацетилсерин, N-формилметионин, 3-метилгистидин, 5-гидроксилизин, [сигма]-1Ч-метиларгинин и другие подобные аминокислоты и иминокислоты (например, 4-гидроксипролин). В обозначении полипептидов, используемом в данном изобретении, направление влево представляет собой амино-концевое направление, а направление вправо представляет собой карбокси-концевое направление в соответствии со стандартным использованием и правилом.
Термин "изолированный белок" относится к белку, такому как белок, связывающий антиген (примером которого может быть антитело), очищенный от белков или полипептидов, либо от других загрязняющих веществ, которые препятствовали бы его терапевтическому, диагностическому, профилактическому, исследовательскому или другому применению. Как используют в настоящем изобретении, "по существу чистый" означает, что молекула описываемого соединения является преобладающей из присутствующих соединений, то есть на молярной основе она является более многочисленной, чем любое другое индивидуальное соединение в той же смеси. В определенных воплощениях изобретения по существу чистая молекула представляет собой композицию, где молекула-объект составляет по меньшей мере 50% (на молярной основе) всех присутствующих макромолекулярных соединений. В других воплощениях изобретения по существу чистая композиция содержит по меньшей мере 80%, 85%, 90%, 95% или 99% всех макромолекулярных соединений, присутствующих в композиции. В определенных воплощениях изобретения по существу гомогенное вещество очищено до такой степени, что загрязняющие соединения не могут быть обнаружены в композиции традиционными способами обнаружения, и, следовательно, композиция состоит из единственного обнаружимого макромолекулярного соединения.
"Вариант" полипептида (например, белок, связывающий антиген, такой как антитело) содержит аминокислотную последовательность, где один или более аминокислотных остатков вставлено, делетировано и/или заменено в пределах аминокислотной последовательности относительно другой полипептидной последовательности. Варианты включают слитые белки. "Производное" полипептида представляет собой полипептид, который был химически модифицирован определенным образом, отличающимся от вариантов инсерции, делеции или замены, например, посредством конъюгации с другой химической группировкой.
Термины "встречающийся в природе" или "нативный", как используют на протяжении всего описания в сочетании с биологическими материалами, такими как полипептиды, нуклеиновые кислоты, клетки-хозяева и тому подобное, относятся к веществам, обнаруживаемым в природе. В данном контексте "рекомбинантный белок" представляет собой белок, полученный с использованием рекомбинантных методов, то есть посредством экспрессии рекомбинантной нуклеиновой кислоты, как раскрыто в данном изобретении. Способы и методы получения рекомбинантных белков хорошо известны в данной области техники.
Термин "антитело" относится к интактному иммуноглобулину любого изотипа или к его фрагменту, который может конкурировать с интактным антителом за специфичное связывание с антигеном-мишенью, и включает, например, химерные, гуманизированные, полностью человеческие и биспецифические антитела. Антитело как таковое представляет собой вид белка, связывающего антиген. Если не указано иное, термин "антитело" включает, кроме антител, содержащих две полноразмерных тяжелых цепи и две полноразмерных легких цепи, его производные, варианты, фрагменты и мутеины, примеры которых раскрыты ниже. Интактное антитело, как правило, содержит по меньшей мере две полноразмерных тяжелых цепи и две полноразмерных легких цепи, но в некоторых случаях может содержать меньшее количество цепей, как, например, антитела, в природе встречающиеся у верблюдовых, которые могут содержать только тяжелые цепи. Антитела могут иметь происхождение только из одного источника или могут быть "химерными", то есть различные участки антитела могут иметь происхождение от двух различных антител, как дополнительно описано ниже. Белки, связывающие антиген, антитела или связывающие фрагменты могут быть получены в гибридомах, методами рекомбинантных ДНК или путем ферментативного или химического расщепления интактных антител.
Термин "функциональный фрагмент" (или просто "фрагмент") антитела или цепи иммуноглобулина (тяжелой или легкой цепи), как используют в настоящем изобретении, представляет собой белок, связывающий антиген, содержащий участок (независимо от того, каким образом получен или синтезирован этот участок) антитела, в котором отсутствуют по меньшей мере некоторые из аминокислот, присутствующие в полноразмерной цепи, но способный к специфичному связыванию с антигеном. Такие фрагменты биологически активны в том, что они специфично связываются с антигеном-мишенью и могут конкурировать с другими белками, связывающими антиген, включая интактные антитела, за специфичное связывание с данным эпитопом. В одном аспекте такой фрагмент сохраняет по меньшей мере один CDR, присутствующий в полноразмерной легкой или тяжелой цепи, и в некоторых воплощениях изобретения содержит одну тяжелую цепь и/или легкую цепь, либо ее участок. Эти биологически активные фрагменты могут быть получены методами рекомбинантных ДНК, либо могут быть получены путем ферментативного или химического расщепления белков, связывающих антиген, включая интактные антитела. Фрагменты включают, но не ограничены ими, иммунологически функциональные фрагменты, такие как Fab, Fab', F(ab')2, Fv, доменные антитела и одноцепочечные антитела, и могут иметь происхождение из любого источника млекопитающего, включающего, но не ограниченного ими, человека, мышь, крысу, верблюда или кролика. Кроме того, рассматривают, что функциональный участок белков, связывающих антиген, раскрытых в данном изобретении, например, один или более CDR, может быть ковалентно связан со вторым белком или с малой молекулой для создания терапевтического агента, направленного на определенную мишень в организме, обладающего бифункциональными терапевтическими свойствами или обладающего пролонгированным периодом полувыведения в сыворотке.
Термин "конкурировать" при использовании в контексте белков, связывающих антиген (например, нейтрализующих белков, связывающих антиген, или нейтрализующих антител), означает конкуренцию между белками, связывающими антиген, которую определяют с помощью анализа, в котором белок, связывающий антиген (например, антитело или его иммунологически функциональный фрагмент), в условиях теста предотвращает или ингибирует специфичное связывание референсного белка, связывающего антиген (например, лиганда или референсного антитела) с общим антигеном (например, с белком CD30L или его фрагментом). Можно использовать многочисленные типы анализов конкурентного связывания, например, следующие анализы: твердофазный прямой или непрямой радиоиммунологический анализ (РИА), твердофазный прямой или непрямой иммуноферментный анализ (ИФА), конкурентный сэндвич-анализ (см., например, Stahli et al., 1983, Methods in Enzymology 92:242-253); твердофазный прямой ИФА с биотином-авидином (см., например, Kirkland et al., 1986, J. Immunol. 137:3614-3619) твердофазный анализ с прямым мечением, твердофазный сэндвич-анализ с прямым мечением (см., например, Harlow and Lane, 1988, Antibodies, A Laboratory Manual, Cold Spring Harbor Press); твердофазный РИА с прямым мечением с использованием метки 1-125 (см., например, Morel et al., 1988, Molec. Immunol. 25:7-15); твердофазный прямой ИФА с биотином-авидином (см., например, Cheung, et al., 1990, Virology 176:546-552); и РИА с прямым мечением (Moldenhauer et al., 1990, Scand. J. Immunol. 32:77-82). В характерном случае такой анализ включает использование очищенного антигена, связанного с твердой поверхностью, или клеток, несущих любой из них, немеченый тестируемый белок, связывающий антиген, и меченый референсный белок, связывающий антиген. Могут быть предприняты различные подходы, известные специалистам в данной области техники. Альтернативная возможность включает получение референсного белка, связывающего антиген, связанного с планшетом, необязательно посредством гибкого матрикса. Дополнительные варианты могут быть основаны на порядке добавления, то есть, смешивают ли с референсным белком, связывающим антиген, связанным с планшетом, сначала антиген или тестируемый белок, связывающий антиген. Во всех случаях необходимо насыщение антигена белком, связывающим антиген, чтобы избежать ложного неконкурентного результата.
Конкурентное ингибирование измеряют путем определения количества метки, связанной с твердой поверхностью или с клетками, в присутствии тестируемого белка, связывающего антиген. Обычно тестируемый белок, связывающий антиген, присутствует в избытке. Белки, связывающие антиген, идентифицированные с помощью конкурентного анализа (конкурирующие белки, связывающие антиген), включают антигенсвязывающие белки, связывающиеся с тем же эпитопом, что и референсные антигенсвязывающие белки, а также антигенсвязывающие белки, связывающиеся с примыкающим эпитопом, достаточно приближенным к эпитопу, связанному референсным антигенсвязывающим белком, в результате чего происходит стерическое затруднение. Обычно, когда конкурирующий белок, связывающий антиген, присутствует в избытке, он ингибирует специфичное связывание референсного белка, связывающего антиген, с общим антигеном по меньшей мере на 40%, 45%, 50%, 55%, 60%, 65%, 70% или 75%.В некоторых случаях связывание ингибировано по меньшей мере на 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более.
Эксперименты по конкуренции можно проводить с различными типами молекул, которые могут обладать различной чувствительностью. Если белок, связывающий антиген, представляет собой антитело, молекула будет бивалентной. Если антигенсвязывающая молекула представляет собой Fab, она является моновалентной и имеет существенно меньший размер, чем полноразмерное антитело, что приводит в результате к меньшему стерическому затруднению. В конкурентных экспериментах для определенных белков, связывающих антиген, связывание с hCD30L ингибируется связыванием Fab антитела, связывающего антиген, как определено в данном изобретении, такого как любое из антител А, В, С, D, Ε и F.
Термин "эпитоп" или "антигенная детерминанта" относится к сайту на антигене, с которым связывается белок, связывающий антиген. Эпитопы могут быть образованы либо из смежных аминокислот, либо из не смежных аминокислот, сближенных в результате третичного свертывания белка. Эпитопы, образованные из смежных аминокислот, в характерном случае сохраняются при воздействии денатурирующих растворителей, тогда как эпитопы, образованные в результате третичного свертывания, в характерном случае утрачиваются при обработке денатурирующими растворителями. Эпитопные детерминанты могут включать химически активные поверхностные группировки молекул, такие как аминокислоты, боковые цепи Сахаров, фосфорильные или сульфонильные группы, и могут обладать специфичными характеристиками трехмерной структуры и/или специфичными характеристиками заряда. Эпитоп в характерном случае включает 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 25, 30, 35 аминокислот в уникальной пространственной конформации. Эпитопы можно определить, используя способы, известные в данной области техники.
Связывание белка, связывающего антиген, может быть также описано областью или областями антигена, с которыми взаимодействует белок, связывающий антиген. Такая область (области) взаимодействия может быть определена различными способами, известными в данной области техники, например, путем проведения анализов связывания, используя варианты молекул антигена, или с помощью масс-спектрометрии водородного обмена (ВО-МС), пример которой приведен в данном изобретении (см. Пример 6), в результате чего определяют область (области) взаимодействия антигена с белками, связывающими антиген.
В технологии ВО-МС используют тот факт, что за обменом водорода (ВО) белка можно легко следить с помощью масс-спектрометрии (МС). Путем замены водного растворителя, содержащего водород, водным растворителем, содержащим дейтерий, включение атома дейтерия в определенный сайт в белке вызовет увеличение массы на 1 Да. Мониторинг этого увеличения массы можно проводить в зависимости от времени с помощью масс-спектрометрии в погашенных образцах реакции обмена. Информация о мечении дейтерием может быть сублокализована в областях в белке с помощью расщепления пепсином в условиях гашения, и можно следить за увеличением массы полученных в результате пептидов. ВО-МС можно использовать для зондирования сайтов, вовлеченных в молекулярные взаимодействия, путем идентификации областей сниженного водородного обмена при образовании белок-белкового комплекса, включая взаимодействия антитело-антиген. Обычно границы связывания выявляют на основании заметных снижений водородного обмена за счет стерического исключения растворителя. Образование белок-белкового комплекса может быть обнаружено с помощью ВО-МС просто путем измерения суммарного количества дейтерия, включенного в какой-либо из белков-участников в присутствии и в отсутствие соответствующего партнера связывания в зависимости от времени. В методе ВО-МС используют нативные компоненты, то есть белок и антитело или Fab фрагмент, и его выполняют в растворе. Таким образом, ВО-МС обеспечивает возможность имитации условий in vivo (обзор по технологии ВО-МС см., например, в статье Wales and Engen, Mass Spectrom. Rev. 25, 158 (2006)).
Некоторые белки, связывающие антиген, как раскрыто в данном изобретении, взаимодействуют с CD30L или связывают его посредством С-концевого участка CD30L человека, определенного АА201-234. Может быть так, что белок, связывающий антиген, связывает С-концевой участок CD30L человека, определенный АА 201-234, или участок меньшего размера, такой как АА 205-230 или АА 211-226. Как раскрыто выше, эпитоп может не быть непрерывным, как и область взаимодействия. В одном воплощении изобретения белок, связывающий антиген, согласно изобретению связывается по меньшей мере с двумя областями CD30L. Такие дополнительные белки, связывающие антиген, согласно изобретению могут взаимодействовать с С-концевым участком, как определено выше, и дополнительной областью CD30L человека, локализованной в N-концевом участке внеклеточного домена, такой как область, определенная АА70-100, полноразмерного hCD30L. Такие белки, связывающие антиген, могут связываться с АА 75-95 или с более коротким участком, таким как АА 80-90 или АА 82-88, в дополнение к С-концевому участку, определенному АА 201-234, АА 205-230 или АА 211-226.
Белки, связывающие антиген CD30L
"Белок, связывающий антиген", как используют в настоящем изобретении, означает белок, специфично связывающий указанный антиген-мишень; антиген, предложенный в данном изобретении, представляет собой CD30L, в частности, CD30L человека. Белки, связывающие антиген, включают, например, белки, блокирующие или ингибирующие взаимодействие CD30L и CD30. Такие "блокирующие" белки, связывающие антиген, можно разрабатывать в направлении CD30L, либо его фрагмента, варианта или производного и подвергать скринингу в традиционных анализах на способность препятствовать взаимодействию CD30L и CD30. Примеры подходящих анализов, представляющих собой анализы, тестирующие способность белков, связывающих антиген, к ингибированию взаимодействия CD30L и CD30, раскрыты в данном изобретении. Белки, связывающие антиген, также включают белки, ингибирующие CD30L или активирующие CD30L. Такое ингибирование или нейтрализация нарушает биологический ответ в присутствии белка, связывающего антиген, по сравнению с ответом в отсутствие белка, связывающего антиген, что можно определить, используя анализы, известные в данной области техники и раскрытые в данном изобретении. Белки, связывающие антиген, предложенные в данном изобретении, например, индуцируют продуцирование ИЛ-8 клетками CD30+. Белки, связывающие антиген, как раскрыто в данном изобретении, также не связывают другие члены надсемейства TNF, в частности, 4-1 ВВL, OX-40L, TNFα, ΤΝΡβ, RANKL, Trail, CD40L или CD27L.
Различные белки, связывающие антиген CD30L, могут связываться с различными доменами или эпитопами CD30L или действовать посредством различных механизмов действия. Белок, связывающий антиген CD30L, необязательно должен полностью ингибировать активность, индуцированную CD30L, чтобы найти применение, раскрытое в данном изобретении; вероятнее, для применения также рассмотрены белки, связывающие антиген, снижающие конкретную активность CD30L. Белки, связывающие антиген, также включают белки, перекрестно конкурирующие за связывание с CD30L человека; связываются с тем же эпитопом CD30L человека; связываются с CD30L человека по существу с одинаковой Kd; связываются с CD30L по существу с одинаковой скоростью диссоциации по сравнению с любым из референсных белков, связывающих антиген, раскрытых в данном изобретении. Различные способы измерения таких характеристик известны в данной области техники и раскрыты в данном изобретении.
Также предложены белки, связывающие антиген CD30L, истощающие клетки CD30L+. При таком истощении белками, связывающими антиген CD30L, связывание антигенсвязывающего белка с клеткой, содержащей его антиген-мишень, приводит в результате к ингибированию антигена или клеточной функции, либо приводит в результате к гибели клетки. В одном воплощении изобретения истощающее антитело по изобретению связывает CD30L и может блокировать или не блокировать связывание лиганда CD30L с CD30. Таким образом, истощающие белки, связывающие антиген, конкретно включают блокирующие и не блокирующие белки, связывающие антиген. В одном аспекте белки, связывающие антиген CD30L, которые истощают клетки CD30L+, могут индуцировать апоптоз или программируемую клеточную гибель, как определено с помощью анализов на апоптоз, известных в данной области техники. Такие белки, связывающие антиген CD30L, могут обладать иммуномодулирующим эффектом за счет следующих механизмов: 1) элиминации клеток, взаимодействующих с клетками CD30L+ для индукции иммунного ответа, и/или 2) элиминации клеток, в которых CD30L является маркером клеточной поверхности для клеточного типа, участвующего в определенном иммунном ответе, но который необязательно должен для этого взаимодействовать с клеткой CD30+. В таком случае CD30L является маркером клеточного типа, ассоциированного с конкретным заболеванием, и взаимодействие CD30/CD30L может не быть вовлечено в патогенез самого этого заболевания. Другое воплощение изобретения включает истощающие белки, связывающие антиген, включающие конъюгированный токсин или цитотоксический агент, где этот токсин или цитотоксический агент индуцирует истощение клеток, связывающих конъюгат антигенсвязывающего белка.
Белок, связывающий антиген, может включать участок, связывающийся с антигеном, и необязательно структурный или каркасный участок, дающий возможность антигенсвязывающему участку принимать конформацию, способствующую связыванию белка, связывающего антиген, с антигеном. Примеры белков, связывающих антиген, включают антитела, фрагменты антител (например, антигенсвязывающий участок антитела), производные антител и аналоги антител. Белок, связывающий антиген, может включать альтернативную белковую структуру или искусственную структуру с привитыми участками CDR или производными участков CDR. Такие структуры включают, но не ограничены ими, структуры, производные от антитела, содержащие мутации, введенные, например, для стабилизации структуры белка, связывающего антиген, а также полностью синтетические структуры, включающие, например, биосовместимый полимер. См., например, Korndorfer et al., Proteins: Structure, Function, and Bioinformatics (2003) Volume 53, Issue 1:121-129; Roque et al., Biotechnol. Prog., 2004, 20:639-654. Кроме того, можно использовать пептидомиметики антител ("ПМА"), а также структуры на основе миметиков антител, используя в качестве структуры фибропектиновые компоненты.
Некоторые белки, связывающие антиген, раскрытые в данном изобретении, представляют собой антитела или образованы от антител. Такие белки, связывающие антиген, включают, но не ограничены ими, моноклональные антитела, биспецифические антитела, минитела, доменные антитела, синтетические антитела, миметики антител, химерные антитела, гуманизированные антитела, человеческие антитела, слитые антитела, конъюгаты антител, одноцепочечные антитела, а также их фрагменты, соответственно. В некоторых случаях белок, связывающий антиген, представляет собой иммунологический фрагмент антитела (например, Fab, Fab', F(ab')2 или scFv). Различные структуры дополнительно описаны и определены в данном изобретении.
Некоторые предложенные белки, связывающие антиген, могут содержать один или более CDR, как раскрыто в данном изобретении (например, 1, 2, 3, 4, 5, 6 или более CDR). В некоторых случаях белок, связывающий антиген, содержит (а) полипептидную структуру и (b) один или более CDR, встроенных в полипептидную структуру и/или соединенных с ней. Полипептидная структура может принимать ряд различных форм. Например, она может представлять собой или включать каркас встречающегося в природе антитела или его фрагмента или варианта, либо может быть полностью синтетической по природе. Примеры различных полипептидных структур дополнительно описаны ниже.
Некоторые из белков, связывающих антиген, предложенных в данном изобретении, специфично связываются с CD30L человека. "Специфично связываться", как используют в настоящем изобретении, означает, что равновесная константа диссоциации (KD) составляет от менее 10-8 до менее 10-10 М, альтернативно от менее 10-9 до менее 10-10 М, более конкретно от менее 10-11М до менее 10-12М. В одном воплощении изобретения белки, связывающие антиген, связываются с высоким сродством, выраженным равновесной константой диссоциации (KD), равной 10-8, либо такой как 5,0×10-9, либо такой как 10-9 или даже 5,0×10-10. Равновесная константа диссоциации может быть определена, как известно в данной области техники. В таких воплощениях изобретения KD в характерном случае определяют путем иммобилизации соединения при низкой плотности (это соединение может быть мультивалентным) и нанесения серии титрования моновалентного соединения (фаза ассоциации), а затем допущения фазы диссоциации, где комплексы распадаются. Константы скорости, соответствующие ассоциации и диссоциации моновалентного комплекса, называют константой скорости ассоциации kа и константой скорости диссоциации kd, соответственно. Затем полученные в результате данные приводят в соответствие с моделью связывания 1:1, соответствующей 3 параметрам: константе скорости ассоциации kа, константе скорости диссоциации kd и Rmax, которые относятся к поверхностной плотности и стехиометрии. Отношение kd/ka равно равновесной константе диссоциации KD.
KD может быть также определена с использованием анализа сортировки флуоресцентно-активированных клеток (FACS), как описано в Примере 4 в данном изобретении, где антитела связываются с CD30L, экспрессируемым на клетках Ramos.
Одно воплощение изобретения относится к изолированному белку, связывающему антиген, как раскрыто в данном изобретении, где белок, связывающий антиген, обладает сродством (или KD) к CD30L человека по меньшей мере 75 пикомоль (пМ), например, 50 пМ, например, 40 пМ. В следующих воплощениях константа диссоциации (КD) белка, связывающего антиген, с CD30L человека составляет максимум 35 пМ, например, максимум 25 пМ, например, максимум 20 пМ, например, максимум 15 пМ.
Сродство (или КD) в таком воплощении изобретения может быть определена с помощью FACS, как описано в Примере 4 в данном изобретении.
В другом аспекте предложен белок, связывающий антиген, имеющий период полувыведения по меньшей мере одни сутки in vitro или in vivo (например, при введении субъекту-человеку). В одном воплощении изобретения белок, связывающий антиген, имеет период полувыведения по меньшей мере трое суток. В другом воплощении изобретения антитело или его участок имеет период полувыведения четверо суток или дольше. В другом воплощении изобретения антитело или его участок имеет период полувыведения восемь суток или дольше. В другом воплощении изобретения антитело или его антигенсвязывающий участок дериватизирован или модифицирован таким образом, что он имеет более длительный период полувыведения по сравнению с не дериватизированным или немодифицированным антителом. В другом воплощении изобретения белок, связывающий антиген, содержит точечные мутации для увеличения сывороточного периода полувыведения, как описано в публикации WIPO №WO 00/09560.
В воплощениях изобретения, где белок, связывающий антиген, используют для терапевтических применений, где белок, связывающий антиген, может снизить, ингибировать, нарушать или модулировать одну или более биологических активностей CD30L, таких как индукция продуцирования ИЛ-8 в клетках CD30+.
Некоторые из предложенных белков, связывающих антиген, имеют структуру, в характерном случае ассоциированную с природными антителами. Структурные звенья этих антител в характерном случае содержит один или более тетрамеров, каждый из которых состоит из двух идентичных пар полипептидных цепей, хотя некоторые виды млекопитающих также продуцируют антитела, имеющие только одну тяжелую цепь. В характерном антителе каждая пара включает одну полноразмерную "легкую" цепь (в определенных воплощениях изобретения примерно 25 кДа) и одну полноразмерную "тяжелую" цепь (в определенных воплощениях изобретения примерно 50-70 кДа). Каждая отдельная цепь иммуноглобулина состоит из нескольких "иммуноглобулиновых доменов", каждый из которых состоит приблизительно из аминокислот в количестве от 90 до 110 и экспрессирует характеристический профиль свертывания. Эти домены являются основными звеньями, из которых состоят полипептиды антител. Амино-концевой участок каждой цепи в характерном случае включает вариабельную область, ответственную за распознавание антигена. Карбокси-концевой участок эволюционно более консервативен, чем другой конец цепи, и его называют "константной областью" или "С областью". Легкие цепи человека в целом классифицируют как легкие цепи каппа и лямбда, и каждая из них содержит гипервариабельную область и один константный домен. Тяжелые цепи в характерном случае классифицируют как мю-, дельта-, гамма-, альфа- или эпсилон-цепи, и они определяют изотип антитела как IgM, IgD, IgG, IgA и IgE, соответственно. IgG имеет несколько подтипов, включающих, но не ограниченных ими, IgG1, IgG2, IgG3 и IgG4. Подтипы IgM включают IgM и IgM2. Подтипы IgA включают IgА1 и IgА2. У людей изотипы IgA и IgD содержат четыре тяжелых цепи и четыре легких цепи; изотипы IgG и IgE содержат две тяжелых цепи и две легких цепи; и изотип IgM содержит пять тяжелых цепей и пять легких цепей. Константная область тяжелой цепи (СН) в характерном случае содержит один или более доменов, которые могут быть ответственными за эффекгорную функцию. Число доменов константной области тяжелой цепи зависит от изотипа. Каждая из тяжелых цепей IgG, например, содержит три домена области СН, известных как СН1, СН2 и СН3. Предложенные белки, связывающие антиген, могут иметь любой из этих изотипов и подтипов, например, белок, связывающий антиген CD30L, имеет подтип IgG1, IgG2 или IgG4. Если желателен IgG4, может быть также желательным вводить точечную мутацию (CPSCP → СРРСР) в шарнирной области, как описано в статье Bloom et al., 1997, Protein Science 6:407), чтобы ослабить склонность к образованию внутрицепных дисульфидных связей внутри цепи Н, которое может привести к гетерогенности в антителах IgG4. Антитела, предложенные в данном изобретении, относящиеся к одному типу, могут быть изменены на другой тип, используя способы переключения подкласса. См., например, Lantto et al., 2002, Methods Mol. Biol. 178:303-316.
В полноразмерных легких и тяжелых цепях вариабельные и константные области соединены областью "J", состоящей из примерно двенадцати или более аминокислот, причем, тяжелая цепь также включает область "D", состоящую из примерно десяти или более аминокислот. См., например, Fundamental Immunology, 2nd ed., Ch. 7 (Paul, W., ed.) 1989, New York: Raven Press. Вариабельные области каждой из пар легкая/тяжелая цепь в характерном случае образуют антигенсвязывающий сайт.
Вариабельные области
Различные вариабельные области (или домены) тяжелых цепей и легких цепей, предложенные в данном изобретении, показаны в таблицах 1 и 2. Каждая из этих вариабельных областей может быть присоединена, например, к константным областям тяжелой и легкой цепи, описанным выше. Кроме того, каждая из созданных таким образом последовательностей тяжелой и легкой цепи может быть объединена с образованием полноразмерной структуры белка, связывающего антиген.


Участки определения комплементарности (CDR) выделены полужирным курсивом, каркасные области (FR; от англ. "framework regions") напечатаны светлым шрифтом. Порядок элементов является следующим: FR1, CDR1, FR2, CDR2, FR3, CDR3 и FR4.



Участки определения комплементарности (CDR) выделены полужирным курсивом, каркасные области (FR; от англ. "framework regions") напечатаны светлым шрифтом. Порядок элементов является следующим: FR1, CDR1, FR2, CDR2, FR3, CDR3 и FR4.
Предложены белки, связывающие антиген, содержащие по меньшей мере одну вариабельную область тяжелой цепи (VH), выбранную из группы, состоящей из следующих участков: VH1, VH2, VH3, VH4, VH5, VH6, VH7, VH8, VH9, VH10, VH11, VH12 и VH13 и/или по меньшей мере одну вариабельную область легкой цепи (VL), выбранную из группы, состоящей из следующих участков: VL1, VL2, VL3, VL4, VL5, VL6, представленных в таблицах 1 и 2.
Каждая из вариабельных областей тяжелой цепи, перечисленных в таблице 2, может быть объединена с любой из вариабельных областей легкой цепи, представленных в таблице 1, с образованием белка, связывающего антиген. В некоторых случаях белок, связывающий антиген, включает по меньшей мере одну вариабельную область тяжелой цепи и/или одну вариабельную область легкой цепи из перечисленных в таблицах 1 и 2. В некоторых случаях белок, связывающий антиген, включает по меньшей мере две различные вариабельные области тяжелой цепи и/или две различные вариабельные области легкой цепи из перечисленных в таблицах 1 и 2. различные комбинации вариабельных областей тяжелой цепи могут быть объединены с любой из различных комбинаций вариабельных областей легкой цепи.
В других случаях белок, связывающий антиген, содержит две идентичные вариабельные области легкой цепи и/или две идентичные вариабельные области тяжелой цепи. В качестве примера, белок, связывающий антиген, может представлять собой антитело или его иммунологически функциональный фрагмент, содержащий две вариабельные области легкой цепи и две вариабельные области тяжелой цепи в комбинациях пар вариабельных областей легких цепей и пар вариабельных областей тяжелых цепей, перечисленных в таблицах 1 и 2. Примеры таких белков, связывающих антиген, включающих две идентичные вариабельные области тяжелой цепи и две идентичные вариабельные области легкой цепи, включают следующие белки: Антитело A VH2/VL1; Антитело А1 VH1/VL1; Антитело А2 VH3A/L1; Антитело A3 VH4/VL1; Антитело A4 VH5A/L1; Антитело А5 VH6A/L1; Антитело А6 VH7/VL1; Антитело А7 VH8/VL1; Антитело В VH9/VL2; Антитело С VH10/VL3; Антитело D VH11/VL4; Антитело Ε VH12A/L5 и Антитело F VH13/VL6.
Некоторые предложенные белки, связывающие антиген, включают вариабельную область тяжелой цепи и/или вариабельную область легкой цепи, содержащую последовательность аминокислот, которая отличается от последовательности вариабельной области тяжелой цепи и/или вариабельной области легкой цепи, выбранной из таблиц 1 и 2, только 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 20, 25, 30, 31 или более аминокислотными остатками, где каждое такое различие последовательностей независимо представляет собой либо делецию, либо инсерцию, либо замену одной аминокислоты. Вариабельные области легкой и тяжелой цепи в некоторых белках, связывающих антиген, содержат последовательности аминокислот, обладающие по меньшей мере 70%, 75%, 80%, 85% 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичностью последовательности с аминокислотными последовательностями, представленными в таблицах 1 и 2. Другие белки, связывающие антиген, например, антитела или их иммунологически функциональные фрагменты, также включают формы вариантов областей тяжелой цепи и/или вариантов областей легкой цепи, как раскрыто в данном изобретении.
Термин "идентичность" относится к отношению между последовательностями двух или более молекул полипептидов или двух или более полинуклеотидов, определенному путем выравнивания и сравнения последовательностей. "Процент идентичности" означает процент идентичных остатков между аминокислотами или нуклеотидами в сравниваемых молекулах, который вычисляют на основании размера наименьшей из сравниваемых молекул. Для этих вычислений гэпы в выравниваниях (если они есть) должны относиться к конкретной математической модели или компьютерной программе (то есть "алгоритму"). Способы, которые можно использовать для вычисления идентичности выровненных нуклеиновых кислот или полипептидов, включают способы, описанные в следующих книгах и статьях: Computational Molecular Biology (Lesk, A.M., ed.), 1988, New York: Oxford University Press; Biocomputing Informatics and Genome Projects (Smith, D.W., ed.), 1993, New York: Academic Press; Computer Analysis of Sequence Data, Part I (Griffin, Α.M., and Griffin, H.G., eds.), 1994, New Jersey: Humana Press; von Heinje, G., 1987, Sequence Analysis in Molecular Biology, New York: Academic Press; Sequence Analysis Primer, (Gribskov, M. and Devereux, J., eds.), 1991, New York: M. Stockton Press; и Carillo et al., 1988, SIAM J. Applied Math. 48:1073.
При вычислении процента идентичности сравниваемые последовательности выравнивают таким путем, который дает наибольшее совпадение между последовательностями. Компьютерная программа, используемая для определения процента идентичности, представляет собой пакет программ GCG, включающий программу GAP (Devereux et al., 1984, Nucl. Acid Res. 12:387; Genetics Computer Group, University of Wisconsin, Madison, Wl). Компьютерный алгоритм GAP используют для выравнивания двух полипептидов или полинуклеотидов, для которых нужно определять процент идентичности последовательности. Последовательности выравнивают для максимального совпадения их соответствующих аминокислот или нуклеотидов ("совпадающего интервала", как определено алгоритмом). Штраф на открытие гэпа (который вычисляют как 3х среднюю диагональ, где "средняя диагональ" представляет собой среднее значение диагонали используемой матрицы сравнений; "диагональ" представляет собой балл или число, присваиваемые каждому точному совпадению аминокислот конкретной матрицей сравнений) и штраф на удлинение гэпа (который обычно равен 1/10 штрафа на открытие гэпа), а также матрица сравнений, такая как РАМ 250 или BLOSUM 62, используют в соответствии с алгоритмом. В определенных воплощениях изобретения стандартная матрица сравнений (см. Dayhoff et al., 1978, Atlas of Protein Sequence and Structure 5:345-352 для матрицы сравнений РАМ 250; Henikoff et al., 1992, Proc. Natl. Acad. Sci. U.S.A. 89:10915-10919 для матрицы сравнений BLOSUM 62) также используется алгоритмом.
Рекомендуемые параметры для определения процента идентичности для полипептидов или нуклеотидных последовательностей с использованием программы GAP являются следующими: алгоритм: Needleman et al., 1970, J. Mol. Biol. 48:443-453; матрица сравнений: BLOSUM 62 из статьи Henikoff et al., 1992, см. выше; штраф на гэп: 12 (но без штрафа на концевые гэпы), штраф на длину гэпа: 4, порог идентичности: 0. Некоторые схемы выравнивания для выравнивания двух аминокислотных последовательностей могут привести в результате к совпадению только короткого участка двух последовательностей, и этот короткий выровненный участок может иметь очень высокую идентичность последовательности, даже, несмотря на отсутствие значимого отношения между двумя полноразмерными последовательностями. Соответственно, выбранный способ выравнивания (программа GAP) может быть при желании отрегулирован, чтобы привести в результате к выравниванию, распространяющемуся по меньшей мере на 50 непрерывных аминокислот целевого полипептида.
Участки определения комплементарности
Участки определения комплементарности или "CDR" заключены внутри каркасного участка в вариабельных областях тяжелой и легкой цепи, где они составляют участки, ответственные за связывание и распознавание антигена. Вариабельные домены цепей иммуноглобулина одного и того же вида, например, как правило, проявляют сходную общую структуру, включающую относительно консервативные каркасные участки (FR), соединенные гипервариабельными участками CDR. Белок, связывающий антиген, может иметь 1, 2, 3, 4, 5, 6 или более CDR. Вариабельные области, обсуждаемые выше, например, в характерном случае содержат три CDR. CDR из вариабельных областей тяжелой цепи и вариабельных областей легкой цепи в характерном случае выровнены каркасными участками с образованием структуры, специфично связывающейся на антигене-мишени (например, CD30L). От N-конца к С-концу вариабельные области как легких, так и тяжелых цепей, встречающихся в природе, в характерном случае соответствуют следующему порядку этих элементов: FR1, CDR1, FR2, CDR2, FR3, CDR3 и FR4. Участки CDR и FR иллюстративных вариабельных доменов легкой цепи и вариабельных доменов тяжелой цепи выделены в таблицах 1 и 2. Признано, что границы областей CDR и FR могут варьировать по сравнению с выделенными границами. Разработаны системы нумерации для присвоения номеров аминокислот, занимающих положения в каждом из этих доменов. Участки определения комплементарности и каркасные участки данного белка, связывающего антиген, могут быть идентифицированы с использованием этих систем. Системы нумерации определены в книгах Kabat et al., Sequences of Proteins of Immunological Interest, 5th Ed., US Dept. of Health and Human Services, PHS, NIH, NIH Publication No. 91-3242, 1991, или Chothia & Lesk, 1987, J. Mol. Biol. 196:901-917; Chothia et al., 1989, Nature 342:878-883. Другие системы нумерации для аминокислот в иммуноглобулиновых цепях включают IMGT® (международную информационную систему ImMunoGeneTics; Lefranc et al., Dev. Сотр. Immunol. 2005, 29:185-203) и АНо (Honegger and Pluckthun, J. Mol. Biol. 2001, 309(3):657-670). CDR, предложенные в данном изобретении, можно использовать не только для определения антигенсвязывающего домена традиционной структуры антитела, но можно включать в ряд других полипептидных структур, описанных в данном изобретении.
Белки, связывающие антиген, раскрытые в данном изобретении, представляют собой полипептиды, в которых может быть привит, встроен, заключен и/или присоединен один или более CDR. Белок, связывающий антиген, может иметь, например, один CDR1 тяжелой цепи ("CDRH1"), и/или один CDR2 тяжелой цепи ("CDRH2"), и/или один CDR3 тяжелой цепи ("CDRH3"), и/или один CDR1 легкой цепи ("CDRL1"), и/или один CDR2 легкой цепи ("CDRL2"), и/или один CDR3 легкой цепи ("CDRL3"). Некоторые белки, связывающие антиген, включают оба участка, CDRH3 и CDRL3. В конкретных воплощениях, как правило, используют комбинации CDR, которые являются не повторяющимися, например, белки, связывающие антиген, как правило, не состоят из двух участков CDRH2 в одной вариабельной области тяжелой цепи, и т.д. Белки, связывающие антиген, могут содержать одну или более аминокислотных последовательностей, идентичных аминокислотным последовательностям или отличающихся от аминокислотных последовательностей одного или более участка CDR, представленного в таблице 3, только 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16 или более аминокислотных остатков, где каждое такое различие последовательностей независимо представляет собой либо делецию, либо инсерцию, либо замену одной аминокислоты. CDR в некоторых белках, связывающих антиген, содержат последовательности аминокислот, обладающие по меньшей мере 80%, 85%, 90%, 91%, 92, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичностью последовательности с последовательностями CDR, перечисленными в таблице 3. В некоторых белках, связывающих антиген, CDR заключены в "каркасной" области, которая ориентирует участок (участки) CDR таким образом, чтобы достичь правильных антигенсвязывающих свойств участка (участков) CDR.


В данном изобретении предложены участки CDR1, содержащие аминокислотные остатки 23-36 SEQ ID NO: 36, 38, 40, 42 и 44; аминокислотные остатки 25-36 SEQ ID NO: 46 и аминокислотные остатки 31-35 SEQ ID NO: 48, 50, 52, 54, 56, 58, 60, 62, 64, 66, 68, 70 и 72. Предложены участки CDR2, содержащие аминокислотные остатки 52-58 SEQ ID NO: 36, 38, 40, 42, 44 и 46 и аминокислотные остатки 50-65 SEQ ID NO: 48, 50, 52, 54, 56, 58, 60, 62, 64, 66, 68, 70 и 72. Участки CDR3 содержат аминокислотные остатки 91-100 SEQ ID NO: 36, 38, 40, 42, 44 и 46 и аминокислотные остатки 98-113 SEQ ID NO: 48, 50, 52, 54, 56, 58, 60, 62, 64, 66, 68, 70 и 72.
Участки CDR, раскрытые в данном изобретении, содержат консенсус-последовательности, выведенные из групп родственных последовательностей. Консенсус-последовательность CDRL1 состоит из TGX1SSDX2GX3YX4YVS (SEQ ID NO: 30), где Х1 представляет собой треонин или серии, Х2 представляет собой валин или изолейцин, Х3 представляет собой валин, треонин или лейцин, и Х4 представляет собой аспарагиновую кислоту или аспарагин.
Консенсус-последовательность CDRL2 состоит из EVX1X2RPS (SEQ ID NO: 31), где Х1 представляет собой серии, аспарагин или изолейцин, и Х2 представляет собой аспарагин или лизин.
Консенсус-последовательность CDRL3 включает SSYX1SX2STWV (SEQ IDN NO: 32), где Х1 представляет собой треонин или серии, и Х2 представляет собой аргинин или серии.
Консенсус-последовательность CDRH1 состоит из X1X2X3WX4 (SEQ ID NO: 33), где Х1 представляет собой серии или аспарагин, Х2 представляет собой тирозин или аспарагин, Х3 представляет собой изолейцин, тирозин или серии, и Х4 представляет собой треонин или серии. В другом воплощении изобретения консенсус-последовательность CDRH1 состоит из SYX3WX5 (SEQ ID NO: 75), где Х3 представляет собой I, S или Y, и Х5 представляет собой Τ или S.
Консенсус-последовательность CDRH2 состоит из
RX1X2X3SGX4X5NYX6PSLX7S (SEQ ID NO: 34), где Х1 представляет собой изолейцин, валин или треонин, Х2 представляет собой тирозин, фенилаланин или серии, Х3 представляет собой треонин, серии или аланин, Х4 представляет собой изолейцин, лейцин, аспарагин, серии, аргинин или глутамин, Х5 представляет собой треонин или аспарагин, Х6 представляет собой аспарагин или лизин, и Х7 представляет собой лизин или аргинин.
Консенсус-последовательность CDRH3 состоит из
Х1Х2Х3Х4Х5X6X7X8YX9YX10GX11DV (SEQ ID NO: 35), где Х1 представляет собой глутаминовую кислоту или аспарагиновую кислоту, Х2 представляет собой аргинин, тирозин или фенилаланин, Х3 представляет собой валин, аланин, аргинин или треонин, Х4 представляет собой валин, треонин, глицин или изолейцин, Х5 представляет собой валин, аланин или глицин, Х6 представляет собой аланин, треонин, глицин или глутамин, Х7 представляет собой треонин, аспарагиновую кислоту, аргинин или серии, Х8 представляет собой аргинин или тирозин, Х9 представляет собой тирозин или гистидин, Х10 представляет собой тирозин, аспарагиновую кислоту или серии, и Х11 представляет собой метионин, лейцин или валин.
Моноклональные антитела
Предложенные белки, связывающие антиген, включают моноклональные антитела, связывающиеся в CD30L. Моноклональные антитела можно продуцировать, используя любой метод, известный в данной области техники, например, путем иммортализации клеток селезенки, собранной от трансгенного животного после выполнения режима иммунизации. Клетки селезенки могут быть иммортализованы, используя любой метод, известный в данной области техники, например, путем их слияния с клетками миеломы с получением гибридом. Клетки миеломы, применяемые в методах получения гибридом путем слияния, предпочтительно не являются клетками, продуцирующими антитела, обладают высокой эффективностью слияния и дефицитами ферментов, которые делают их неспособными к росту в определенной селективной среде, поддерживающей рост только желаемых слитых клеток (гибридом). Примеры подходящих клеточных линий, применяемых при слияниях клеток мыши, включают следующие линии: Sp-20, P3-X63/Ag8, P3-X63-Ag8.653, NS1/1.Ag 4 1, Sp210-Ag14, FO, NSO/U, MPC-11, MPC11-X45-GTG 1.7 и S194/5XXO Bul; примеры клеточных линий, применяемых при слиянии клеток крысы, включают следующие линии: R210.RCY3, Y3-Ag 1.2.3, IR983F и 4 В210. Другими клеточными линиями, полезными для слияний клеток, являются следующие линии: U-266, GM1500-GRG2, LICR-LON-HMy2 и UC729-6.
В некоторых случаях линию клеток гибридомы получают путем иммунизации животного (например, трансгенного животного, имеющего последовательности иммуноглобулинов человека) иммуногеном CD30L; сбора спленоцитов от иммунизированного животного; слияния собранных спленоцитов с линией клеток миеломы, получая, таким образом, клетки гибридомы; основания линий клеток гибридомы из клеток гибридомы и идентификации линии клеток гибридомы, продуцирующей антитело, которое связывает полипептид CD30L. Такие линии клеток гибридомы и моноклональные антитела против CD30L, продуцируемые этими клетками, представляют собой аспекты настоящего изобретения.
Моноклональные антитела, секретируемые линией клеток гибридомы, могут быть очищены с использованием любого метода, известного в данной области техники. Гибридомы или mab можно дополнительно подвергать скринингу, чтобы идентифицировать mAb с определенными свойствами, такими как способность к снижению, ингибированию, нарушению или модулированию взаимодействия CD30L с CD30.
Химерные и гуманизированные антитела
Предложены также химерные и гуманизированные антитела на основе приведенных выше последовательностей. Моноклональные антитела, применяемые в качестве терапевтических средств, могут быть модифицированы различными путями перед применением. Одним из примеров является химерное антитело, которое представляет собой антитело, состоящее из сегментов белка из различных антител, ковалентно связанных с получением функциональных легких или тяжелых цепей иммуноглобулина или их иммунологически функциональных участков. Как правило, участок тяжелой цепи и/или легкой цепи идентичен или гомологичен соответствующей последовательности в антителах, имеющих происхождение от конкретного вида или принадлежащих к конкретному классу или подклассу антител, тогда как остальная часть цепи (цепей) идентична или гомологична соответствующей последовательности в антителах, имеющих происхождение от другого вида, или принадлежащих к другому классу или подклассу антител. Способы, относящиеся к химерным антителам, см., например, в патенте US №4816567; и в статье Morrison et al., 1985, Proc. Natl. Acad. Sci. USA 81:6851-6855. Прививание CDR описано, например, в патентах US №№6180370, 5693762, 5693761, 5585089 и 5530101.
Одним из полезных типов химерного антитела является "гуманизированное" антитело. Как правило, гуманизированное антитело получают из моноклонального антитела, первоначально вырабатываемого у животного, отличающегося от человека. Определенные аминокислотные остатки в данном моноклональном антителе, в характерном случае от участков антитела, не распознающих антиген, модифицированы таким образом, что являются гомологичными соответствующим остаткам в человеческом антителе соответствующего изотипа. Гуманизация может быть выполнена, например, с использованием различных способов путем замещения по меньшей мере участка вариабельной области грызуна соответствующими участками человеческого антитела (см., например, патенты US №№5585089 и 5693762; Jones et al., 1986, Nature 321:522-525; Riechmann et al., 1988, Nature 332:323-27; Verhoeyen et al., 1988, Science 239:1534-1536).
В определенных воплощениях изобретения константные области от видов, отличающихся от человека, можно использовать параллельно с вариабельной областью (областями) человека для получения гибридных антител.
Полностью человеческие антитела
Предложены также полностью человеческие антитела. Доступны способы получения полностью человеческих антител, специфичных к данному антигену, не подвергая людей воздействию антигена ("полностью человеческих антител"). Одним из конкретных средств, предложенных для выполнения продуцирования полностью человеческих антител, является "гуманизация" гуморальной иммунной системы мыши. Введение локусов иммуноглобулина (Ig) человека в мышей, у которых эндогенные гены lg инактивированы, является одним из средств продуцирования полностью человеческих моноклональных антител (mAb) у мышей, то есть животных, которых можно иммунизировать любым желаемым антигеном. Применение полностью человеческих антител может свести к минимуму иммуногенные и аллергические ответы, которые иногда могут быть вызваны введением людям мышиных или производных от мышиных mAb в качестве терапевтических агентов.
Полностью человеческие антитела можно продуцировать путем иммунизации трансгенных животных (обычно мышей), способных продуцировать набор человеческих антител в отсутствие продуцирования эндогенных иммуноглобулинов. Антигены для этой цели в характерном случае имеют шесть или более непрерывных аминокислот и необязательно конъюгированы с носителем, таким как гаптен. См., например, Jakobovits et al., 1993, Proc. Natl. Acad. Sci. USA 90:2551-2555; Jakobovits et al., 1993, Nature 362:255-258; и Bruggermann et al., 1993, Year in Immunol. 7:33. В одном примере такого способа трансгенных животных получают путем выведения из строя эндогенных локусов иммуноглобулина мыши, кодирующих тяжелые и легкие цепи иммуноглобулина мыши, и встраивания в геном мыши больших фрагментов геномной ДНК человека, содержащих локусы, кодирующие белки тяжелой и легкой цепи человека. Затем этих частично модифицированных животных, имеющих менее чем полный комплемент локусов иммуноглобулина человека, скрещивают, чтобы получить животное, имеющее все желаемые модификации иммунной системы. При введении иммуногена эти трансгенные животные продуцируют антитела, являющиеся иммуноспецифичными к иммуногену, но вероятнее имеют аминокислотные последовательности человека, чем мыши, включая вариабельные области. Дополнительные подробности таких способов см., например, в заявках на патенты WIPO WO 96/33735 и WO 94/02602. Дополнительные способы, относящиеся к трансгенным мышам, для получения человеческих антител раскрыты в патентах US №№5545807; 6713610; 6673986; 6162963; 5545807; 6300129; 6255458; 5877397; 5874299 и 5545806; в патентных публикациях WIPO WO 91/10741, WO 90/04036 и в ЕР 546073 В1 и ЕР 546073А1.
Описанные выше трансгенные мыши содержат минилокус гена иммуноглобулина человека, кодирующий неперестроенные последовательности тяжелой ([мю] и [гамма]) и легкой [каппа] цепей иммуноглобулина вместе с направленными мутациями, инактивирующими эндогенными локусами [мю] и [каппа] цепей (Lonberg et al., 1994, Nature 368:856-859). Соответственно, эти мыши проявляют сниженную экспрессию IgM или [каппа] мыши, и в ответ на иммунизацию введенные трансгены тяжелой и легкой цепи человека претерпевают переключение класса и соматическую мутацию с получением моноклональных антител IgG [каппа] человека с высоким сродством (Lonberg et al., см. выше; Lonberg and Huszar, 1995, Intern. Rev. Immunol. 13: 65-93; Harding and Lonberg, 1995, Ann. N.Y Acad. Sci. 764:536-546). Получение таких мышей подробно описано в следующих статьях: Taylor et al., 1992, Nucleic Acids Research 20:6287-6295; Chen et al., 1993, International Immunology 5:647-656; Tuaillon et al., 1994, J. Immunol. 152:2912-2920; Lonberg et al., 1994, Nature 368:856-859; Lonberg, 1994, Handbook of Exp. Pharmacology 113:49-101; Taylor et al., 1994, International Immunology 6:579-591; Lonberg and Huszar, 1995, Intern. Rev. Immunol. 13:65-93; Harding and Lonberg, 1995, Ann. N.Y Acad. Sci. 764:536-546; Fishwild et at., 1996, Nature Biotechnology 14:845-85. Дополнительно см. патенты US №№5545806; 5569825; 5625126; 5633425; 5789650; 5877397; 5661016; 5814318; 5874299 и 5770429; а также в патенте US №5545807; в публикациях WIPO №№WO 93/1227; WO 92/22646 и WO 92/03918. Технологии, используемые для продуцирования человеческих антител в этих трансгенных мышах, также раскрыты в публикации WIPO №WO 98/24893 и в статье Mendez et аl., 1997, Nature Genetics 15:146-156. Например, линии трансгенных мышей НСо7 и НСо12 можно использовать для генерирования антител против CD30L.
Используя гибридомную технологию, антигенспецифические mAb человека с желаемой специфичностью можно продуцировать и подвергать селекции у трансгенных мышей, таких как описано выше. Такие антитела можно клонировать и экспрессировать, используя подходящий вектор и клетку-хозяина, либо эти антитела можно собирать из культивируемых гибридомных клеток.
Полностью человеческие антитела могут быть также выделены из библиотек фагового дисплея (таких как раскрыто в следующих публикациях: Hoogenboom et al., 1991, J. Mol. Biol. 227:381; Marks et al., 1991, J. Mol. Biol. 222:581; публикации WIPO №WO 99/10494). Методы фагового дисплея имитируют иммунную селекцию посредством дисплея набора антител на поверхности нитевидного бактериофага и последующей селекции фага на основании его связывания с выбранным антигеном.
Биспецифические или бифункциональные белки, связывающие антиген
"Биспецифический", "двойной специфичности" или "бифункциональный" антигенсвязывающий белок или антитело представляет собой гибридный антигенсвязывающий белок или гибридное антитело, соответственно, имеющий два различных антигенсвязывающих сайта, таких как один или более CDR или одна или более вариабельных областей, как раскрыто выше. В некоторых случаях эти белки представляют собой искусственное гибридное антитело, имеющее две различные пары тяжелая/легкая цепь и два различных связывающих сайта. Мультиспецифический антигенсвязывающий белок или "мультиспецифическое антитело" представляет собой белок или антитело, направленные на более чем один антиген или эпитоп.Биспецифические антигенсвязывающие белки и антитела представляют собой виды мультиспецифических антигенсвязывающих белков и антител, и их можно продуцировать с помощью ряда способов, включающих, но не ограниченных ими, слияние гибридом или сшивание Fab' фрагментов. См., например, Songsivilai and Lachmann, 1990, Clin. Exp. Immunol. 79:315-321; Kostelny et al., 1992, J. Immunol. 148:1547-1553.
Иммунологические фрагменты
Белки, связывающие антиген, также включают иммунологические фрагменты антитела (например, Fab, Fab', F(ab')2 или scFv). "Fab фрагмент" состоит из одной легкой цепи (вариабельной области легкой цепи (VL) и ее соответствующего константного домена (CL)) и одной тяжелой цепи (вариабельной области тяжелой цепи (VH) и первого константного домена (СH1)). Тяжелая цепь Fab молекулы не может образовать дисульфидную связь с другой молекулой тяжелой цепи. "Fab' фрагмент" содержит одну легкую цепь и часть одной тяжелой цепи, которая также содержит область между доменами СH1 и Сн2, так что межцепная дисульфидная связь может быть образована между двумя тяжелыми цепями двух Fab' фрагментов с образованием молекулы F(ab')2. "F(ab')2 фрагмент", таким образом, состоит из двух Fab' фрагментов, удерживаемых вместе дисульфидной связью между двумя тяжелыми цепями. "Fv фрагмент" состоит из вариабельной области легкой цепи и вариабельной области тяжелой цепи одного плеча антитела. Одноцепочечные антитела "scFv" представляют собой молекулы Fv, в которых вариабельные области тяжелой и легкой цепей соединены гибким линкером с образованием единой полипептидной цепи, образующей антигенсвязывающий участок. Одноцепочечные антитела подробно обсуждаются в публикации WIPO №WO 88/01649, в патентах US №№4946778 и 5260203; в статьях Bird, 1988, Science 242:423; Huston et al., 1988, Proc. Natl. Acad. Sci. U.S.A. 85:5879; Ward et al., 1989, Nature 334:544, de Graaf et al., 2002, Methods Mol Biol. 178:379-387; Kortt et al., 1997, Prot. Eng. 10:423; Kortt et al., 2001, Biomol. Eng. 18:95-108 и Kriangkum et al., 2001, Biomol. Eng. 18:31-40. Участок "Fc" содержит два фрагмента тяжелой цепи, включающие домены СH2 и СH3 антитела. Два фрагмента тяжелой цепи удерживаются вместе двумя или более дисульфидными связями и гидрофобными взаимодействиями доменов СH3.
Также включены доменные антитела, иммунологически функциональные фрагменты иммуноглобулина, содержащие только вариабельную область тяжелой цепи или вариабельную область легкой цепи. В некоторых случаях две или более областей VH ковалентно соединены пептидным линкером с получением бивалентного доменного антитела. Две области VH бивалентного доменного антитела могут быть направлены на один и тот же или на различные антигены. Диатела представляют собой бивалентные антитела, содержащие две полипептидные цепи, где каждая полипептидная цепь содержит домены VH и VL, соединенные линкером, который является слишком коротким, чтобы дать возможность спаривания между двумя доменами на одной и той же цепи, таким образом, давая возможность каждому домену спариваться с комплементарным доменом на другой полипептидной цепи (см., например, Holliger et al., Proc. Natl. Acad. Sci. USA 90:6444-48, 1993 и Poljak et al., Structure 2:1121-23, 1994). Аналогично, тритела и тетратела представляют собой антитела, содержащие три и четыре полипептидных цепи, соответственно, и образующие три и четыре антигенсвязывающих сайта, соответственно, которые могут быть одинаковыми или разными. Макситела содержат бивалентные scFv, ковалентно соединенные с участком Fc IgG1 (см., например, Fredericks et al., 2004, Protein Engineering, Design & Selection, 17:95-106; Powers et al., 2001, Journal of Immunological Methods, 251:123-135; Shu et al., 1993, Proc. Natl. Acad. Sci. USA 90:7995-7999; Hayden et al., 1994, Therapeutic Immunology 1:3-15).
Различные другие формы
Предложены также вариантные формы белков, связывающих антиген, раскрытых выше, где некоторые из белков, связывающих антиген, имеют, например, одну или более консервативных аминокислотных замен в одной или более из тяжелых и легких цепей, вариабельных областей или CDR, перечисленных в таблицах 1 и 2.
Встречающиеся в природе аминокислоты могут быть разделены на классы на основании общих свойств боковых цепей: гидрофобные (норлейцин, Met, Ala, Val, Leu, Ile); нейтральные гидрофильные (Cys, Ser, Thr, Asn, Gin); кислые (Asp, Glu); основные (His, Lys, Arg); остатки, влияющие на ориентацию цепи (Gly, Pro), и ароматические (Тrр, Туr, Phe).
Консервативные аминокислотные замены могут включать замену члена одного из этих классов на другой член того же класса. Консервативные аминокислотные замены могут охватывать неприродные аминокислотные остатки, которые в характерном случае вводят вероятнее путем химического пептидного синтеза, чем путем синтеза в биологических системах. Эти остатки включают пептидомиметики и другие обращенные или инвертированные формы аминокислотных группировок. Такие существенные модификации в функциональных и/или биохимических характеристиках белков, связывающих антиген, раскрытых в данном изобретении, могут быть достигнуты путем создания замен в аминокислотных последовательностях тяжелой и легкой цепей, которые значительно отличаются по их действию на сохранение (а) структуры молекулярного остова в области замены, например, в виде складчатой или спиральной конформации, (b) заряда или гидрофобности молекулы в сайте-мишени или (с) объемности боковой цепи.
Неконсервативные замены могут включать замену члена одного из описанных выше классов членом другого из описанных выше классов. Такие замененные остатки можно вводить в области антитела, гомологичные с антителами человека или в негомологичные области молекулы.
При получении таких замен согласно определенным воплощениям изобретения можно учитывать гидропатический индекс аминокислот. Гидропатический профиль белка вычисляют путем присвоения каждой аминокислоте числового значения ("гидропатического индекса"), а затем повторяющегося усреднения этих значений по всей полипептидной цепи. Каждой аминокислоте присвоен гидропатический индекс на основании характеристик ее гидрофобности и заряда. Эти индексы составляют: изолейцин (+4,5); валин (+4,2); лейцин (+3,8); фенилаланин (+2,8); цистеин/цистин (+2,5); метионин (+1,9); аланин (+1,8); глицин (-0,4); треонин (-0,7); серии (-0,8); триптофан (-0,9); тирозин (-1,3); пролин (-1,6); гистидин (-3,2); глутамат (-3,5); глутамин (-3,5); аспартат (-3,5); аспарагин (-3,5); лизин (-3,9) и аргинин (-4,5).
Важность гидропатического профиля при придании интерактивной биологической функции белку понятна в данной области техники (см., например, Kyte et al., 1982, J. Mol. Biol. 157:105-131). Известно, что некоторые аминокислоты могут быть заменены другими аминокислотами, имеющими подобный гидропатический индекс или балл и все еще сохраняющими подобную биологическую активность. При создании замен на основании гидропатического индекса в определенных воплощениях изобретения включена замена аминокислот, гидропатические индексы которых находятся в пределах ±2. В некоторых аспектах включены те аминокислоты, гидропатические индексы которых находятся в пределах ±1, а в других аспектах включены те аминокислоты, гидропатические индексы которых находятся в пределах ±0,5.
В данной области техники также понятно, что замена подобных аминокислот может быть эффективно получена на основании гидрофильности, в частности, где биологически функциональный белок или пептид, созданный таким путем, предназначен для применения в иммунологических воплощениях изобретения, как в настоящем случае. В определенных воплощениях изобретения самая высокая локальная средняя гидрофильность белка, зависящая от гидрофильности его примыкающих аминокислот, коррелирует с его иммуногенностью и связыванием антигена или иммуногенностью, то есть с биологическим свойством белка.
Этим аминокислотным остаткам присвоены следующие значения гидрофильности: аргинин (+3,0); лизин (+3,0); аспартат (+3,0±1); глутамат (+3,0±1); серии (+0,3); аспарагин (+0,2); глутамин (+0,2); глицин (0); треонин (-0,4); пролин (-0,5±1); аланин (-0,5); гистидин (-0,5); цистеин (-1,0); метионин (-1,3); валин (-1,5); лейцин (-1,8); изолейцин (-1,8); тирозин (-2,3); фенилаланин (-2,5) и триптофан (-3,4). При создании замен на основании подобных значений гидрофильности в определенных воплощениях изобретения включена замена аминокислот, значения гидрофильности которых находятся в пределах ±2, в других воплощениях изобретения включены те аминокислоты, значения гидрофильности которых находятся в пределах ±1, и в других воплощениях изобретения включены те аминокислоты, значения гидрофильности которых находятся в пределах ±0,5. В некоторых случаях можно также идентифицировать эпитопы из первичных аминокислотных последовательностей на основании гидрофильности. Эти области также называют "эпитопными коровыми участками".
Иллюстративные консервативные аминокислотные замены представлены в таблице 4.
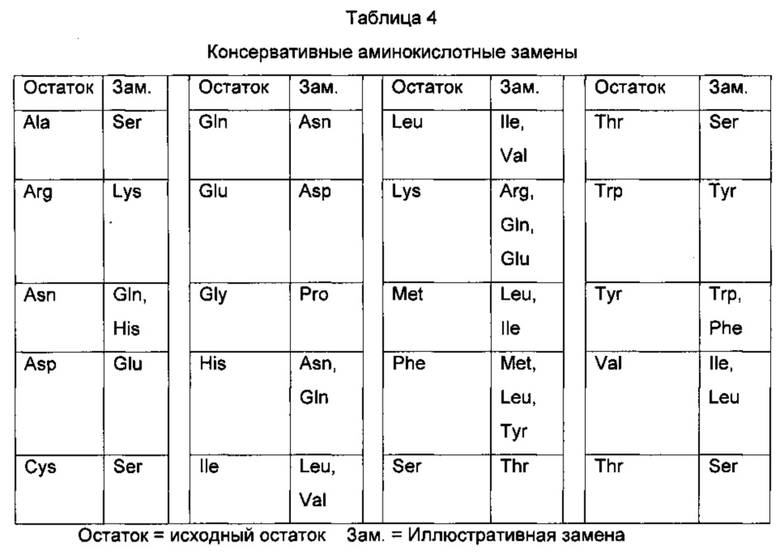
Специалист в данной области техники способен определить подходящие варианты полипептидов, как раскрыто в данном изобретении. Используя хорошо известные методы. Специалист в данной области техники может идентифицировать подходящие области молекулы, которые могут быть изменены без нарушения активности путем направленности на области, которые не считают важными для активности. Специалист в данной области техники также способен идентифицировать остатки и участки молекул, которые являются консервативными среди подобных полипептидов. В следующих воплощениях изобретения даже области, которые могут быть важны для биологической активности или для структуры, можно подвергать консервативным аминокислотным заменам без разрушения биологической активности или без отрицательного влияния на структуру полипептида.
Кроме того, специалист в данной области техники может провести обзор структурно-функциональных исследований, идентифицирующих остатки в подобных полипептидах, важные для активности или структуры. В свете такого сравнения можно прогнозировать важность аминокислотных остатков в белке, соответствующих аминокислотным остаткам, важным для активности или структуры в подобных белках. Специалист в данной области техники может выбрать замены химически подобных аминокислот для таких прогнозируемых важных аминокислотных остатков.
Специалист в данной области техники может также анализировать 3-мерную структуру и аминокислотную последовательность по отношению к этой структуре в подобных полипептидов. В свете такой информации специалист в данной области техники может прогнозировать выравнивание аминокислотных остатков антитела по отношению к его трехмерной структуре. Специалист в данной области техники может сделать выбор, чтобы не проводить радикальных изменений аминокислотных остатков, которые, как предсказано, находятся на поверхности белка, поскольку такие остатки могут быть вовлечены в важные взаимодействия с другими молекулами. Кроме того, специалист в данной области техники может создать экспериментальные варианты, содержащие единственную аминокислотную замену в каждом желаемом аминокислотном остатке. Затем эти варианты можно подвергать скринингу, используя анализы на активность CD30L (см. раздел Примеры ниже), с получением, таким образом, информации относительно того, какие аминокислоты могут быть изменены, а какие не должны быть изменены. Иными словами, на основании информации, собранной в результате таких рутинных экспериментов, специалист в данной области техники может легко определить положения аминокислот, где дополнительных замен следует избегать, как отдельно, так и в комбинации с другими мутациями.
Ряд научных публикаций посвящен прогнозированию вторичной структуры. См. Moult, 1996, Curr. Op. in Biotech. 7:422-427; Chou et al., 1974, Biochem. 13:222-245; Chou et al., 1974, Biochemistry 113:211-222; Chou et al., 1978, Adv. Enzymol. Relat. Areas Mol. Biol. 47:45-148; Chou et al., 1979, Ann. Rev. Biochem. 47:251-276; и Chou et al., 1979, Biophys. J. 26:367-384. Кроме того, в настоящее время доступны компьютерные программы, помогающие прогнозировать вторичную структуру. Один из способов прогнозирования вторичной структуры основан на моделировании гомологии. Например, два полипептида или белка, обладающие идентичностью последовательности более 30% или подобием более 40%, часто имеют сходные структурные топологии. В последнее время рост базы данных белковых структур (PDB; Protein Database) обеспечил повышенную прогнозируемость вторичной структуры, включая потенциальное число складок в пределах структуры полипептидов или белков. См. Holm et al., 1999, Nucl. Acid. Res. 27:244-247. Предположили (Brenner et al., 1997, Curr. Op. Struct. Biol. 7:369-376), что существует ограниченное число складок в данном полипептиде или белке, и что, как только разрешено критическое число структур, прогнозирование структуры станет значительно более точным.
Дополнительные способы прогнозирования вторичной структуры включают "метод протягивания" (Jones, 1997, Curr. Opin. Struct. Biol. 7:377-387; Sippl et al., 1996, Structure 4:15-19), "профильный анализ" (Bowie et al., 1991, Science 253:164-170; Gribskov et al., 1990, Meth. Enzym. 183:146-159; Gribskov et al., 1987, Proc. Nat. Acad. Sci. 84:4355-4358) и "эволюционное сцепление" (см. Holm, 1999, выше; и Brenner, 1997, выше).
В некоторых воплощениях изобретения получают аминокислотные замены, которые: (1) снижают чувствительность к протеолизу, (2) снижают чувствительность к окислению, (3) изменяют связывающее сродство к образованию белковых комплексов, (4) изменяют связывающее сродство к лиганду или антигену и/или (4) придают или модифицируют другие физико-химические или функциональные свойства таких полипептидов, например, сохранение структуры молекулярного остова в области замены, например, в виде складчатой или спиральной конформации; сохранение или изменение заряда или гидрофобности молекулы в сайте-мишени, либо сохранение или изменение объемности боковой цепи.
Например, одиночные или множественные аминокислотные замены (в определенных воплощениях изобретения консервативные аминокислотные замены) можно осуществить в природной последовательности. Замены могут быть получены в той части антитела, которая находится за пределами домена (доменов), образующего межмолекулярные контакты. В таких воплощениях изобретения можно использовать консервативные аминокислотные замены, по существу не изменяющие структурные характеристики исходной последовательности (например, одна или более заменяющих аминокислот, не нарушающих вторичную структуру, которая характеризует исходный или нативный белок, связывающий антиген). Примеры известных в данной области техники вторичных и третичных полипептидных структур описаны в кн. Proteins, Structures and Molecular Principles (Creighton, Ed.), 1984, W.H. New York: Freeman and Company; Introduction to Protein Structure (Branden and Tooze, eds.), 1991, New York: Garland Publishing; и в статье Thornton et al., 1991, Nature 354:105.
Дополнительные варианты включают цистеиновые варианты, где один или более остатков цистеина в исходной или нативной аминокислотной последовательности делетировано или заменено другой аминокислотой (например, серином). Цистеиновые варианты полезны, среди прочего, когда антитела (например) должны быть подвергнуты рефолдингу с образованием биологически активной конформации. Цистеиновые варианты могут иметь меньше остатков цистеина, чем нативный белок, и в характерном случае имеют их равное число, чтобы свести к минимуму взаимодействия в результате неспаренных цистеинов.
Вариабельную область и CDR тяжелой и легкой цепи, раскрытые в данном изобретении, можно использовать для получения антигенсвязывающих белков, содержащих антигенсвязывающий участок, которые могут специфично связываться с полипептидом CD30L. "Антигенсвязывающий участок" означает белок или участок белка, специфично связывающийся с указанным антигеном, такой как участок, содержащий аминокислотные остатки, которые взаимодействуют с антигеном и придают белку, связывающему антиген, его специфичность и сродство к антигену-мишени. Антигенсвязывающий участок может включать один или более CDR, и определенные антигенсвязывающие участки также включают одну или более "каркасную" область. Например, один или более из CDR, перечисленных в таблице 3, может быть ковалентно или нековалентно включен в молекулу (например, в полипептид) с получением иммуноадгезии. Иммуноадгезия может включать участок (участки) CDR как часть полипептидной цепи большего размера, может ковалентно связывать участок (участки) CDR с другой полипептидной цепью или может включать участок (участки) CDR нековалентно. Участок (участки) CDR дает возможность для иммуноадгезии для специфичного связывания с конкретным интересующим антигеном (например, с полипептидом CD30L).
Другие белки, связывающие антиген, включают миметики (например, "миметики пептидов" или "пептидомиметики"), основанные на вариабельных участках и CDR, раскрытых в данном изобретении. Эти аналоги могут представлять собой пептиды, непептиды или комбинации пептидных и непептидных участков. Fauchere, 1986, Adv. Drug Res. 15:29; Veber and Freidinger, 1985, TINS p.392 и Evans et al., 1987, J. Med. Chem. 30:1229. Пептидные миметики, структурно подобные терапевтически полезным пептидам, можно использовать для получения подобного терапевтического или профилактического эффекта. Такие соединения часто разрабатывают с помощью компьютерного молекулярного моделирования. Как правило, пептидомиметики представляют собой белки, структурно подобные белку, связывающему антиген, проявляющие желаемую биологическую активность, такую как способность к ингибированию или блокированию взаимодействия CD30 и CD30L, но пептидомиметики имеют одну или более пептидных связей, необязательно замещенных связью, выбранной, например, из следующих связей: -CH2NH-, -CH2S-, -CH2-CH2-, -СН-СН- (цис и транс), -СОСН2-, -СН(ОН)СН2- и -CH2SO-, способами, известными в данной области техники. Систематическую замену одной или более аминокислот консенсус-последовательности D-аминокислотой того же типа (например, D-лизин вместо L-лизина) можно использовать в определенных воплощениях изобретения для создания более стабильных белков. Кроме того, пептиды с ограниченной конформационной свободой, содержащие консенсус-последовательность или по существу идентичный вариант консенсус-последовательности, могут быть получены способами, известными в данной области техники (Rizo and Gierasch, 1992, Ann. Rev. Biochem. 61:387), например, путем добавления внутренних остатков цистеина, способных к образованию внутримолекулярных дисульфидных связей, которые замыкают пептид в цикл.
Предложены также производные белков, связывающих антиген, раскрытых в данном изобретении. Дериватизированные белки, связывающие антиген, могут включать любую молекулу или вещество, придающие желаемое свойство белку, связывающему антиген, или его фрагменту, такое как увеличенный период полувыведения при конкретном применении. Дериватизированный белок, связывающий антиген, может содержать, например, обнаружимую (или метящую) группировку (например, радиоактивную, колориметрическую, антигенную группировку или молекулу фермента, обнаружимую гранулу (такую как магнитная или электронно-плотная (например, золотая) гранула) или молекулу, связывающуюся с другой молекулой (например, биотин или стрептавидин)), терапевтическую или диагностическую группировку (например, радиоактивную, цитотоксическую или фармацевтически активную группировку) или молекулу, повышающую пригодность белка, связывающего антиген, для конкретного применения (например, введения субъекту, такому как субъект-человек, или других применений in vivo или in vitro). Примеры молекул, которые можно использовать для дериватизации белка, связывающего антиген, включают альбумин (например, человеческий сывороточный альбумин) и полиэтиленгликоль (ПЭГ). Связанные с альбумином или пегилированные производные белков, связывающих антиген, могут быть получены, используя методы, хорошо известные в данной области техники. В одном воплощении изобретения белок, связывающий антиген, конъюгируют или другим путем сшивают с транстиретином (TTR; от англ. "transthyretin") или с вариантом TTR. TTR или вариант TTR может быть химически модифицирован, например, химическим веществом, выбранным из группы, состоящей из следующих веществ: декстрана, поли(н-винилпирролидон), полиэтиленгликолей, гомополимеров пропиленгликоля, сополимеров пропиленоксида/этиленоксида, полиоксизтилированных полиолов и поливиниловых спиртов.
Другие производные включают ковалентные или агрегационные конъюгаты белков, связывающие антиген CD30L, с другими белками или полипептидами, например, путем экспрессии рекомбинантных слитых белков, содержащих гетерологичные полипептиды, слитые с N-концом или С-концом белка, связывающего антиген CD30L. Например, конъюгированный пептид может представлять собой гетерологичный сигнальный (или лидерный) полипептид, например, лидерный пептид альфа-фактора дрожжей, или пептид, такой как эпитопная метка. Белок, связывающий антиген CD30L, содержащий слитые белки, может содержать пептиды, присоединенные для облегчения очистки или идентификации белка, связывающего антиген CD30L (например, поли-His). Белок, связывающий антиген CD30L, может быть также сшит с пептидом FLAG, как описано в статье Норр et al., 1988, Bio/Technology 6:1204; и в патенте US №5011912. пептид FLAG является высоко антигенным и обеспечивает эпитоп, обратимо связываемый специфичным моноклональным антителом (mAb), что дает возможность быстрого анализа и облегчает очистку экспрессируемых рекомбинантных белков. Реагенты, полезные для получения слитых белков, в которых пептид FLAG слит с данным полипептидом, имеются в продаже (Sigma, St. Louis, МО).
Олигомеры, содержащие один или более белков, связывающих антиген CD30L, могут применяться в качестве антагонистов CD30L. Олигомеры могут находиться в форме ковалентно связанных или нековалентно связанных димеров, триммеров или высших олигомеров. Олигомеры, содержащие два или более белков, связывающих антиген CD30L, рассмотрены для применения, где одним из примеров является гомодимер. Другие олигомеры включают гетеродимеры, гомотримеры, гетеротримеры, гомотетрамеры, гетеротетрамеры и т.д. Также включены олигомеры, содержащие множественные белки, связывающие CD30L, соединенные посредством ковалентных или нековалентных взаимодействий между пептидными группировками, слитые с белками, связывающими антиген CD30L. Такие пептиды могут представлять собой пептидные линкеры (спейсеры) или пептиды, обладающие свойством стимуляции олигомеризации. Среди подходящих пептидных линкеров находятся линкеры, раскрытые в патентах US №№4751180 и 4935233. Среди пептидов, которые могут стимулировать олигомеризацию присоединенных к ним белков, связывающих антиген CD30L, находятся лейциновые молнии и некоторые полипептиды, образованные от антител. Примеры доменов типа лейциновой молнии, подходящих для продуцирования растворимых олигомерных белков, раскрыты в публикации WIPO №WO 94/10308 и в статьях Норре et al., 1994, FEBS Letters 344:191 и Fanslow et al., 1994, Semin. Immunol. 6:267-278. При одном подходе рекомбинантные слитые белки, содержащие белок, связывающий антиген CD30L, их фрагмент или производное, слитые с пептидом типа лейциновой молнии, экспрессируют в подходящих клетках-хозяевах, и подходящие растворимые олигомерные белки, связывающие антиген CD30L, их фрагменты или производные, которые при этом образуются, выделяют из супернатанта культуры.
Такие олигомеры могут содержать от двух до четырех белков, связывающих антиген CD30L. Группировки олигомера, представляющие собой белок, связывающий антиген CD30L, могут находиться в любой из раскрытых выше форм, например, вариантов или фрагментов. Предпочтительно олигомеры содержат белки, связывающие антиген CD30L, обладающие способностью к связыванию CD30L. Олигомеры могут быть получены, используя полипептиды, образованные от иммуноглобулинов. Получение слитых белков, содержащих определенные гетерологичные полипептиды, слитые с различными участками полипептидов, образованных от антитела (включая домен Fс) раскрыто, например, в следующих публикациях: Ashkenazi et al., 1991, Proc. Natl. Acad. Sci. USA 88:10535; Byrn et al., 1990, Nature 344:677 и Hollenbaugh et al., 1992 "Construction of Immunoglobulin Fusion Proteins", in Current Protocols in Immunology, Suppl. 4, pages 10.19.1-10.19.11.
Также включены димеры, содержащие два слитых белка, полученных путем слияния белка, связывающего антиген CD30L, с областью Fc антитела. Этот димер может быть получен, например, путем встраивания слитого гена, кодирующего слитый белок, в подходящий экспрессионный вектор, экспрессии этого слитого гена в клетках-хозяевах, трансформированных рекомбинантным экспрессионным вектором, и предоставления возможности для сборки экспрессированного слитого белка, в значительной степени подобного молекуле антитела, причем, межцепные дисульфидные связи образуются между группировками Fc с образованием димера. Такие Fc-полипептиды включают нативные и мутеиновые формы полипептидов, образованных от участка Fc антитела. Также включены укороченные формы таких полипептидов, содержащих шарнирную область, стимулирующую димеризацию. Слитые белки, содержащие группировки Fc (и олигомеры, образованные из них) дают преимущество, состоящее в простой очистке путем аффинной хроматографии на колонке с белком А или с белком G. Один из подходящих полипептидов Fc, раскрытых в публикации WIPO №WO 93/10151 и в патентах US №№5426048 и 5262522, представляет собой одноцепочечный полипептид, продолжающийся от N-концевого шарнирного участка до С-конца участка Fc антитела IgG1 человека. Другим полезным полипептидом Fc является мутеин Fc, раскрытый в патенте US №5457035 и в статье Baum et al., 1994, EMBO J. 13:3992-4001. Аминокислотная последовательность этого мутеина идентична нативной последовательности Fc, представленной в публикации WIPO №WO 93/10151, за исключением того, что аминокислота 19 заменена с Leu на Ala, аминокислота 20 заменена с Leu на Glu, и аминокислота 22 заменена с Gly на Ala. Этот мутеин проявляет сниженное сродство к рецепторам Fc.
Гликозилирование
Белок, связывающий антиген, может иметь профиль гликозилирования, отличающийся или измененный по сравнению с профилем, обнаруживаемым в нативных соединениях. Как известно в данной области техники, профили гликозилирования могут зависеть как от последовательности белка (например, от присутствия или отсутствия конкретных аминокислотных остатков гликозилирования, обсуждаемых ниже), так и от клетки или организма-хозяина, в которой продуцируют белок. Конкретные экспрессионные системы обсуждаются ниже.
Гликозилирование полипептидов в характерном случае является либо N-связанным, либо О-связанным. N-связанное гликозилирование относится к присоединению углеводной группировки к боковой цепи остатка аспарагина. Трипептидные последовательности аспарагин-Х-серин и аспарагин-Х-треонин, где X представляет собой любую аминокислоту за исключением пролина, являются последовательностями распознавания для ферментативного присоединения углеводной группировки к боковой цепи аспарагина. Таким образом, присутствие любой из этих трипептидных последовательностей в полипептиде создает потенциальный сайт гликозилирования. О-связанное гликозилирование относится к присоединению одного из следующих сахаров: N-ацетилгалактозамина, галактозы или ксилозы, к гидроксиаминокислоте, чаще всего к серину или треонину, хотя можно также использовать 5-гидроксипролин или 5-гидроксилизин.
Присоединение сайтов гликозилирования к белку, связывающему антиген, традиционно осуществляют путем изменения аминокислотной последовательности таким образом, чтобы она содержала один или более из вышеописанных трипептидных последовательностей (для N-связанных сайтов гликозилирования). Это изменение может быть также получено путем добавления или замены одного или более остатков серина или треонина к исходной последовательности (для О-связанных сайтов гликозилирования). Для простоты аминокислотная последовательность белка, связывающего антиген, может быть изменена посредством изменения на уровне ДНК, в частности, путем мутирования ДНК, кодирующей целевой полипептид, по предварительно выбранным основаниям, таким образом, что образуются кодоны, которые будут транслироваться в желаемые аминокислоты.
Другие способы увеличения числа углеводных группировок на белке, связывающем антиген, представляют собой химическое или ферментативное сочетание гликозидов с белком. Эти методы обладают преимуществом, состоящим в том, что они не требуют продуцирования белка в клетке-хозяине, обладающей способностями к гликозилированию путем N- и О-связанного гликозилирования. В зависимости от используемого режима сочетания сахар (сахара) может быть присоединен к следующим группам: (а) к аргинину и гистидину, (b) к свободным карбоксильным группам, (с) к свободным сульфгидрильным группам, таким как группы цистеина, (d) к свободным гидроксильным группам, таким как группы серина, треонина или гидроксипролина, (е) к ароматическим остаткам, таким как остатки фенилаланина, тирозина или триптофана, или (f) к амидной группе глутамина. Эти способы раскрыты в публикации РСТ №WO 87/05330 и в статье Aplin and Wriston, 1981, CRC Crit. Rev, Biochem., pp.259-306.
Удаление углеводных группировок, присутствующих на исходном белке, связывающем антиген, можно осуществить химическим или ферментативным путем. Для химического дегликозилирования необходимо воздействие на белок соединения трифторметансульфоновой кислоты или эквивалентного соединения. Данная обработка приводит в результате к отщеплению большинства или всех сахаров за исключением связывающего сахара (N-ацетилглюкозамина или N-ацетилгалактозамина), причем, полипептид остается интактным. Химическое дегликозилирование раскрыто в статьях Hakimuddin et al., 1987, Arch. Biochem. Biophys. 259:52 и Edge et al., 1981, Anal. Biochem. 118:131. Ферментативное отщепление углеводных группировок на полипептидах может быть достигнуто путем использования ряда эндо- и экзо-гликозидаз, как раскрыто в статье Thotakura et al., 1987, Meth. Enzymol. 138:350. Гликозилирование в потенциальных сайтах гликозилирования можно предотвратить путем использования соединения туникамицина, как раскрыто в статье Duskin et al., 1982, J. Biol. Chem. 257:3105. туникамицин блокирует образование связей белок-N-гликозид.
Следовательно, аспекты включают варианты гликозилирования белков, связывающих антиген, где число и/или тип сайта (сайтов) гликозилирования изменен по сравнению с аминокислотными последовательностями исходного полипептида. В определенных воплощениях изобретения варианты белка, связывающего антиген, содержат большее или меньшее число N-связанных сайтов гликозилирования по сравнению с исходным полипептидом. Замены, элиминирующие или изменяющие эту последовательность, предотвращают присоединение N-связанной углеводной цепи, присутствующей в исходном полипептиде. Например, гликозилирование может быть снижено путем делеции Asn или путем замены Asn другой аминокислотой. Антитела в характерном случае имеют N-связанный сайт гликозилирования в области Fс.
Метки и эффекторные группы
Белки, связывающие антиген, могут содержать одну или более меток. Термин "метка" или "метящая группа" относится к любой обнаружимой метке. Как правило, метки попадают в ряд следующих классов в зависимости от анализа, в котором они должны быть обнаружены: а) изотопные метки, которые могут представлять собой радиоактивные или тяжелые изотопы; b) магнитные метки (например, магнитные частицы); с) редокс-активные группировки; d) оптические красители; ферментные группы (например, пероксидаза хрена, β-галактозидаза, люцифераза, щелочная фосфатаза); е) биотинилированные группы; и f) предопределенные полипептидные эпитопы, распознаваемые вторичным репортером (например, парные последовательности в виде лейциновой молнии, сайты связывания вторичных антител, домены, связывающие металлы, эпитопные метки и т.д.). В некоторых воплощениях настоящего изобретения метящая группа связана с белком, связывающим антиген, посредством спейсерных плеч различной длины, чтобы снизить потенциальное стерическое затруднение. Различные способы мечения белков известны в данной области техники. Примеры подходящих метящих групп включают, но не ограничены ими, следующие группы: радиоизотопы или радионуклиды (например, 3Н, 14С, 15N, 35S, 90Y, 99Тс, 111ln, 125Ι, 131I), флуоресцентные группы (например, флуоресцеинизотиоцианат (ФИТЦ), родамин, люминофоры на основе комплексов лантанидов), ферментные группы (например, пероксидазу хрена, β-галактозидазу, люциферазу, щелочную фосфатазу), хемилюминесцентные группы, биотинильные группы или предопределенные полипептидные эпитопы, распознаваемые вторичным репортером (например, парные последовательности в виде лейциновой молнии, сайты связывания вторичных антител, домены, связывающие металлы, эпитопные метки). В некоторых воплощениях изобретения метящая группа, связана с белком, связывающим антиген, посредством спейсерных плеч различной длины, чтобы снизить потенциальное стерическое затруднение. Различные способы мечения белков известны в данной области техники и могут быть использованы, если их считают подходящими.
Термин "эффекторная группа" означает любую группу, связанную с белком, связывающим антиген, которая действует как цитотоксический агент. Примерами подходящих эффекторных групп являются радиоизотопы или радионуклиды (например, 3Н, 14С, 15N, 35S, 90Y, 99Тс, 111ln, 125Ι, 131I). Другие подходящие группы включают токсины, терапевтические группы или химиотерапевтические группы. Примеры подходящих групп включают калиехамицин, ауристатины, гелданамицин и майтансин. В некоторых воплощениях изобретения эффекторная группа связана с белком, связывающим антиген, посредством спейсерных плеч различной длины, чтобы снизить потенциальное стерическое затруднение.
Полинуклеотиды, кодирующие белки, связывающие антиген CD30L
Предложены также полинуклеотиды, кодирующие белки, связывающие антиген, раскрытые в данном изобретении, или их участки, включая полинуклеотиды, кодирующие одну или обе цепи антитела или его фрагмента, производного, мутеина или варианта, полинуклеотиды, кодирующие вариабельные области или только CDR тяжелой цепи, полинуклеотиды, достаточные для использования в качестве зондов гибридизации, праймеров ПЦР или праймеров секвенирования, для идентификации, анализа, мутирования или амплификации полинуклеотида, кодирующего полипептид, антисмысловые нуклеиновые кислоты для ингибирования экспрессии полинуклеотида и последовательности, комплементарные описанным выше. Полинуклеотиды могут иметь любую длину. Они могут иметь, например, 5, 10, 15, 20, 25, 30, 35, 40, 45, 50, 75, 85, 95, 100, 125, 150, 175, 200, 250, 300, 350, 400, 450, 500, 750, 1000, 1500, 3000, 5000 или более нуклеиновых кислот в длину, включая все значения между ними, и/или могут содержать одну или более дополнительных последовательностей, например, регуляторные последовательности, и/или составлять участок полинуклеотида большего размера, например, вектора. Полинуклеотиды могут быть однонитевыми или двунитевыми и могут включать нуклеиновые кислоты РНК и/или ДНК и их искусственные варианты (например, пептидные нуклеиновые кислоты).
Полинуклеотиды, кодирующие определенные белки, связывающие антиген, или их участки (например, полноразмерное антитело, тяжелую или легкую цепь, вариабельный домен, либо CDRH1, CDRH2, CDRH3, CDRL1, CDRL2 или CDRL3) могут быть выделены из В-клеток мышей, которые были иммунизированы CD30L или его иммуногенным фрагментом. Полинуклеотид может быть выделен общепринятыми методами, такими как полимеразная цепная реакция (ПЦР). Фаговый дисплей является другим примером известного метода, где могут быть получены производные антител и других белков, связывающих антиген. При одном подходе полипептиды, являющиеся компонентами интересующего белка, связывающего антиген, экспрессируют в любой подходящей рекомбинантной экспрессионной системе, и экспрессированным полипептидам предоставляют возможность сборки с образованием молекул белка, связывающего антиген. Фаговый дисплей также используют для образования белков, связывающих антиген, обладающих различными свойствами (то есть варьирующими значениями сродства к антигену, с которым они связываются), посредством перестановки цепей, см. Marks et al., 1992, BioTechnology 10:779.
Вследствие вырожденности генетического кода каждая из полипептидных последовательностей, раскрытых в данном изобретении, также кодируется большим числом других полинуклеотидных последовательностей, кроме предложенных. Например, вариабельные домены тяжелой цепи, предложенные в данном изобретении, могут кодироваться полинуклеотидными последовательностями SEQ ID NO: 49, 51, 53, 55, 57, 59, 61, 63, 65, 67, 69, 71 и 73. Вариабельные домены легкой цепи могут кодироваться полинуклеотидными последовательностями SEQ ID NO: 37, 39, 41, 43, 45 и 47. Обычному специалисту в данной области техники понятно, что в настоящем изобретении, таким образом, предложено адекватное письменное описание и внедрение для каждой вырожденной нуклеотидной последовательности, кодирующей каждый белок, связывающий антиген.
В аспекте изобретения дополнительно предложены полинуклеотиды, гибридизующиеся с другими полинуклеотидными молекулами в определенных условиях гибридизации. Способы гибридизации нуклеиновых кислот, основные параметры, влияющие на выбор условий гибридизации, и руководства для разработки подходящих условий хорошо известны в данной области техники. См., например, Sambrook, Fritsch, and Maniatis (2001, Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y. и Current Protocols in Molecular Biology, 1995, Ausubel et al., eds., John Wiley & Sons, Inc. Как определено в данном изобретении, при условиях гибридизации средней жесткости используют раствор для предварительной отмывки, содержащий 5х хлорид натрия/цитрат натрия (SSC), 0,5% додецилсульфат натрия (ДСН), 1,0 мМ этилендиаминтетрауксусную кислоту (ЭДТА) (рН 8,0), буфер гибридизации, состоящий из примерно 50% формамида, 6х SSC, и температуру гибридизации 55°С (или другие подобные условия гибридизации, такие как буфер, содержащий примерно 50% формамид, при температуре гибридизации 42°С), и условия отмывки при 60°С в 0,5х SSC, 0,1% ДСН. При жестких условиях гибридизации гибридизацию проводят в 6х SSC при 45°С с последующей одной или более отмывок в 0,1х SSC, 0,2% ДСН при 68°С. Кроме того, специалист в данной области техники может управлять условиями гибридизации и/или отмывки, чтобы повысить или понизить жесткость гибридизации, так что полинуклеотиды, включающие нуклеиново кислотные последовательности, являющиеся по меньшей мере на 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичными друг другу, включая все значения между ними, в характерном случае остаются гибридизованными друг с другом.
В полинуклеотид можно вводить изменения путем мутации, приводящие, таким образом, к изменению аминокислотной последовательности полипептида (например, белка, связывающего антиген, или производного белка, связывающего антиген), который он кодирует. Мутации можно вводить, используя любой метод, известный в данной области техники, такой как сайт-направленный мутагенез и случайный мутагенез. Мутантные полипептиды можно экспрессировать и отбирать на желаемое свойство. В полинуклеотид можно вводить мутации без значительного изменения биологической активности полипептида, который он кодирует, например, замены в неэссенциальных аминокислотных остатках. Альтернативно в полинуклеотид можно вводить одну или более мутаций, селективно изменяющих биологическую активность полипептида, который он кодирует. Например, мутация может количественно или качественно изменять биологическую активность, например, повышать, снижать или элиминировать активность, и изменять антигенную специфичность белка, связывающего антиген.
В другом аспекте изобретения предложены полинуклеотиды, которые подходят для использования в качестве праймеров или зондов гибридизации для определения нуклеиново-кислотных последовательностей. Полинуклеотид может содержать только участок нуклеиново-кислотной последовательности, кодирующей полноразмерный полипептид, например, фрагмент, который можно использовать в качестве зонда или праймера, или фрагмент, кодирующий активный участок (например, участок связывания CD30L) полипептида. Зонды, основанные на последовательности нуклеиновой кислоты, можно использовать для определения нуклеиновой кислоты или подобных нуклеиновых кислот, например, транскриптов, кодирующих полипептид. Зонд может содержать метящую группу, например, радиоизотоп, флуоресцентное соединение, фермент или кофактор фермента. Такие зонды можно использовать для идентификации клетки, экспрессирующей полипептид.
Способы экспрессии белков, связывающих антиген
Белки, связывающие антиген, предложенные в данном изобретении, могут быть получены с помощью любого из многочисленных традиционных способов. Например, белки, связывающие антиген CD30L, могут быть получены с помощью рекомбинантных экспрессионных систем, используя любой метод, известный в данной области техники. См., например, Monoclonal Antibodies, Hybridomas: A New Dimension in Biological Analyses, Kennet et al.(eds.) Plenum Press, New York (1980); и Antibodies: A Laboratory Manual, Harlow and Lane (eds.), Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y. (1988).
Экспрессионные системы и конструкции в форме плазмид, экспрессионных векторов, транскрипционные или экспрессионные кассеты, содержащие по меньшей мере один полинуклеотид, как раскрыто выше, также предложены в данном изобретении, а также клетки-хозяева, содержащие такие экспрессионные системы или конструкции. Как используют в настоящем изобретении, "вектор" означает любую молекулу или частицу (например, нуклеиновую кислоту, плазмиду, бактериофаг или вирус), подходящую для применения для переноса информации, кодирующей белок, в клетку-хозяина. Примеры векторов включают, но не ограничены ими, плазмиды, вирусные векторы, неэписомные векторы млекопитающих и экспрессионные векторы, например, рекомбинантные экспрессионные векторы. Экспрессионные векторы, такие как рекомбинантные экспрессионные векторы, полезны для трансформации клетки-хозяина и содержат нуклеиново-кислотные последовательности, направляющие и/или контролирующие (в сочетании с клеткой-хозяином) экспрессию одного или более гетерологичного кодирующего участка, функционально связанные с ним. Экспрессионная конструкция может включать, но не ограничена ими, последовательности, оказывающие воздействие на транскрипцию, трансляцию, или контролирующие их, и, если присутствуют интроны, оказывающие воздействие на сплайсинг РНК кодирующего участка, функционально связанные с ним. "Функционально связанный" означает, что компоненты, к которым применяют данный термин, находятся в таком взаимоотношении, что дают возможность осуществлять присущие им функции. Например, контролирующая последовательность, например, промотор, в векторе, "функционально связанный" с кодирующей последовательностью белка, расположен таким образом, что нормальная активность контролирующей последовательности приводит к транскрипции кодирующей последовательности белка, приводящей в результате к рекомбинантной экспрессии кодируемого белка.
В другом аспекте предложены клетки-хозяева, в которые введен экспрессионный вектор, такой как рекомбинантный экспрессионный вектор. Клетка хозяин может представлять собой любую прокариотическую клетку (например, Е. coli) эукариотическую клетку (например, клетки дрожжей, насекомых или млекопитающих (например, клетки яичника китайского хомячка (СНО))). Векторную ДНК можно вводить в прокариотические или эукариотические клетки общепринятыми методами трансформации или трансфекции. Для стабильной трансфекции клеток млекопитающих известно, что в зависимости от используемого экспрессионного вектора и метода трансфекции только небольшая доля клеток может интегрировать чужеродную ДНК в свой геном. В целях идентификации и селекции этих интегрантов в клетки-хозяева параллельно с интересующим геном обычно вводят ген, кодирующий селективный маркер (например, для устойчивости к антибиотикам). Предпочтительные селективные маркеры включают маркеры, придающие устойчивость к лекарственным средствам, таким как G418, гигромицин и метотрексат. Клетки, стабильно трансфицированные введенным полинуклеотидом, могут быть идентифицированы, среди прочих способов, путем селекции лекарственным средством (например, клетки, включившие селективный маркерный ген, выживут, тогда как другие клетки погибнут).
Белки, связывающие антиген, можно экспрессировать в линиях клеток гибридомы (например, в частности, антитела можно экспрессировать в гибридомах) или в других линиях клеток, отличающихся от гибридом. Экспрессионные конструкции, кодирующие белки, связывающие антиген, можно использовать для трансформации клетки-хозяина млекопитающего, насекомого или микроорганизма. Трансформацию можно проводить, используя любой известный способ введения полинуклеотидов в клетку-хозяина, включая, например, упаковку полинуклеотида в вирус или бактериофаг и трансдукцию клетки-хозяина конструкцией с помощью методов трансфекции, известных в данной области техники, примеры которых приведены в патентах US №№4399216; 4912040; 4740461; 4959455. Оптимальный используемый метод трансформации зависит от того, какой тип клетки-хозяина трансформируют. Способы введения гетерологичных полинуклеотидов в клетки млекопитающих хорошо известны в данной области техники и включают, но не ограничены ими, трансфекцию, опосредованную декстраном, кальцийфосфатную преципитацию, трансфекцию, опосредованную полибреном, слияние протопластов, электропорацию, инкапсуляцию полинуклеотида (полинуклеотидов) в липосомы, смешивание нуклеиновой кислоты с положительно заряженными липидами и прямую микроинъекцию ДНК в ядра.
Рекомбинантные экспрессионные конструкции в характерном случае содержат полинуклеотид, кодирующий полипептид. Полипептид может содержать один или более из следующих участков: один или более участков CDR, таких как предложено в данном изобретении; вариабельную область легкой цепи; вариабельную область тяжелой цепи; константную область легкой цепи; константную область тяжелой цепи (например, СH1, СH2 и/или СH3); и/или другой каркасный участок белка, связывающего антиген CD30L. Эти нуклеиново-кислотные последовательности встраивают в подходящий экспрессионный вектор, используя стандартные методы лигирования. В одном воплощении изобретения константная область тяжелой или легкой цепи присоединяют к С-концу вариабельной области тяжелой или легкой цепи, предложенной в данном изобретении, и лигируют в экспрессионный вектор. В характерном случае выбирают такой вектор, чтобы он был функциональным в конкретной используемой клетке-хозяине (то есть вектор, совместимый с механизмом клетки-хозяина, дающий возможность, чтобы могла происходить амплификация и/или экспрессия гена). В некоторых воплощениях изобретения используют векторы, для которых применяют анализы комплементации белка-фрагмента, используя белки-репортеры, такие как дигидрофолатредуктаза (см., например, патент US №6270964). Подходящие экспрессионные векторы могут быть приобретены, например, в фирме Invitrogen Life Technologies (Carlsbad, CA) или BD Biosciences (San Jose, CA). Другие полезные векторы для клонирования и экспрессии антител и их фрагментов включают векторы, описанные в статье Bianchi and McGrew, 2003, Biotech. Biotechnol. Bioeng. 84:439-44. Дополнительные подходящие экспрессионные векторы обсуждают, например, в кн. Methods Enzymol., vol. 185 (D.V. Goeddel, ed.), 1990, New York: Academic Press.
В характерном случае экспрессионные векторы, используемые в любой из клеток-хозяев, содержат последовательности для поддержания плазмид и для клонирования и экспрессии экзогенных нуклеотидных последовательностей. Такие последовательности, которые все вместе называют "фланкирующими последовательностями", в определенных воплощениях изобретения в характерном случае включают одну или более из следующих нуклеотидных последовательностей: промотора, одной или более энхансерных последовательностей, точки начала репликации, последовательности терминации транскрипции, полноразмерной интронной последовательности, содержащей донорный и акцепторный сайт сплайсинга, последовательности, кодирующей лидерную последовательность для секреции полипептида, сайта связывания рибосомы, последовательности полиаденилирования, полилинкерного участка для встраивания полинуклеотида, кодирующего полипептид, который нужно экспрессировать, и селективного маркерного элемента. Предложенные экспрессионные векторы можно конструировать из исходного вектора, такого как имеющийся в продаже вектор. Такие векторы могут содержать или не содержать все из желаемых фланкирующих последовательностей. Где одна или более из фланкирующих последовательностей, раскрытых в данном изобретении, уже отсутствуют в векторе, их можно получить отдельно и лигировать в векторе. Способы, используемые для получения каждой из фланкирующих последовательностей, хорошо известны специалистам в данной области техники.
Необязательно вектор может кодировать последовательность, кодирующую "метку", то есть олигонуклеотидную молекулу, локализованную на 5' или 3' конце кодирующей последовательности белка, связывающего антиген CD30L; эта олигонуклеотидная последовательность кодирует поли-His (такой как гекса-His) или другую "метку", такую как FLAG®, гемагглютинин вируса гриппа (НА; от англ. "hemaglutinin influenza virus") или myc, для которой существуют имеющиеся в продаже антитела. Эту метку в характерном случае сливают с полипептидом при экспрессии полипептида, и она может служить как средство аффинной очистки или обнаружения белка, связывающего антиген CD30L, из клетки-хозяина. Аффинную очистку можно осуществлять, например, путем колоночной хроматографии, используя антитела против метки в качестве аффинной матрицы. Необязательно эту метку можно впоследствии удалить из очищенного белка, связывающего антиген CD30L, различными способами, например, путем использования определенных пептидаз для отщепления.
Фланкирующие последовательности могут быть гомологичными (то есть из того же вида и/или штамма, что и клетка-хозяин), гетерологичными (то есть из вида, отличающегося от вида или штамма клетки-хозяина), гибридными (комбинация фланкирующих последовательностей более чем из одного источника), синтетическими или нативными. Как таковой, источник фланкирующей последовательности может представлять собой любой прокариотический или эукариотический организм, любой позвоночный или беспозвоночный организм или любое растение, при условии, что фланкирующая последовательность функциональна в клетке-хозяине или может быть активирована ее механизмом.
Фланкирующие последовательности, полезные в векторах, могут быть получены любым из нескольких способов, хорошо известных в данной области техники. В характерном случае фланкирующие последовательности, полезные в данном изобретении, идентифицированы ранее путем картирования и/или путем расщепления эндонуклеазами рестрикции и, следовательно, могут быть выделены из соответствующего источника ткани, используя подходящие эндонуклеазы рестрикции. В некоторых случаях может быть известна полноразмерная нуклеотидная последовательность фланкирующей последовательности. В данном изобретении фланкирующая последовательность может быть синтезирована, используя способы синтеза или клонирования нуклеиновых кислот, раскрытые в данном изобретении.
Если известна вся фланкирующая последовательность или только ее часть, она может быть получена с использованием полимеразной цепной реакции (ПЦР) и/или путем скрининга геномной библиотеки подходящим зондом, таким как олигонуклеотид и/или фрагмент фланкирующей последовательности из того же или другого вида. Если фланкирующая последовательность неизвестна, фрагмент ДНК, содержащий фланкирующую последовательность, может быть выделена из большего участка ДНК, который может содержать, например, кодирующую последовательность или даже другой ген или гены. Выделение может быть выполнено путем расщепления эндонуклеазами рестрикции с получением соответствующего фрагмента ДНК с последующим выделением с использованием очистки из агарозного геля, колоночной хроматографии Qiagen® (Qiagen, Chatsworth, СА) или других способов, известных специалистам в данной области техники. Выбор подходящих ферментов для осуществления этой цели легко очевиден обычным специалистам в данной области техники.
Точка начала репликации в характерном случае составляет часть прокариотических экспрессионных векторов, приобретенных коммерческим путем, и данная точка начала репликации способствует амплификации вектора в клетке-хозяине. Если выбранный вектор не содержит точку начала репликации, ее можно синтезировать химическим путем, основываясь на известной последовательности, и лигировать в векторе. Например, точка начала репликации из плазмиды pBR322 (New England Biolabs, Beverly, MA) подходит для большинства грамотрицательных бактерий, а точки начала репликации различных вирусов (например, SV40, полиомы, аденовируса, вируса везикулярного стоматита (VSV) или папилломавирусов, таких как HPV или BPV) полезны для клонирования векторов в клетках млекопитающих. Как правило, компонент, представляющий собой точку начала репликации, необязателен для экспрессионных векторов млекопитающих (например, точку начала репликации SV40 часто используют только в связи с тем, что она также содержит ранний промотор вируса).
Последовательность терминации транскрипции в характерном случае локализована в 3' положении к концу кодирующей области полипептида и служит для терминации транскрипции. Обычно последовательность терминации транскрипции в прокариотических клетках представляет собой G-C богатый фрагмент, за которым следует последовательность поли-Т. Хотя эту последовательность легко клонировать из библиотеки или даже приобрести коммерческим путем в виде части вектора, ее можно также легко синтезировать, используя способы синтеза нуклеиновых кислот, такие как раскрыто в данном изобретении.
Селективный маркерный ген кодирует белок, необходимый для выживания и роста клетки-хозяина, выращиваемой в селективной культуральной среде. Характерные селективные маркерные гены кодируют белки, которые: (а) придают устойчивость к антибиотикам или другим токсинам, например, к ампициллину, тетрациклину или канамицину для прокариотических клеток-хозяев; (b) комплементируют ауксотрофные дефициты для клетки; или (с) обеспечивают критические питательные вещества, недоступные из комплексной среды или из среды определенного состава. Конкретными селективными маркерами являются следующие гены: ген устойчивости к канамицину, ген устойчивости к ампициллину и ген устойчивости к тетрациклину. Предпочтительно ген устойчивости к неомицину можно также использовать для селекции и в прокариотических, и в эукариотических клетках-хозяевах.
Другие селективные гены можно использовать для амплификации экспрессируемого гена. Амплификация представляет собой процесс, где гены, требующиеся для продуцирования белка, критичного для роста или выживания клетки, повторяются в тандеме в пределах хромосом последующих поколений of рекомбинантных клеток. Примеры подходящих селективных маркеров для клеток млекопитающих включают гены дигидрофолатредуктазы (DHFR) и беспромоторной тимидинкиназы. Трансформанты клеток млекопитающих помещают в условия селекционного давления, в которых только трансформанты уникально адаптированы к выживанию за счет селективного гена, присутствующего в векторе. Селекционное давление налагают путем культивирования трансформированных клеток в условиях, в которых концентрация селективного агента в среде последовательно возрастает, что, таким образом, приводит к амплификации, как селективного гена, так и ДНК, кодирующей другой ген, такой как ген белка, связывающего антиген, который связывается с CD30L. В результате, с амплифицированной ДНК синтезируются увеличенные количества полипептида, такого как белок, связывающий антиген.
Сайт связывания рибосомы обычно необходим для инициации трансляции мРНК и характеризуется последовательностью Шайна-Дальгарно (прокариоты) или последовательностью Козака (эукариоты). Этот элемент в характерном случае локализован в 3' положении к промотору и в 5' положении к кодирующей последовательности полипептида, который нужно экспрессировать.
В некоторых случаях, например, где в экспрессионной системе эукариотической клетки-хозяина желательно гликозилирование, можно манипулировать различными пре- или πрο-последовательностями для повышения гликозилирования или выхода. Например, можно изменить сайт расщепления пептидазы конкретного сигнального пептида или присоединять про-последовательности, которые также могут влиять на гликозилирование. Конечный белковый продукт может иметь в положении -1 (относительно первой аминокислоты зрелого белка) одну или более дополнительных аминокислот, случайных для экспрессии, которые могут быть не полностью удалены. Например, в конечном белковом продукте один или два аминокислотных остатка, обнаруживаемых в сайте расщепления пептидазы, присоединены к амино-концу. Альтернативно использование нескольких сайтов ферментативного расщепления может привести в результате к несколько укороченной форме желаемого полипептида, если фермент расщепляет в такой области в пределах зрелого полипептида.
Экспрессионные и клонирующие векторы в характерном случае содержат промотор, распознаваемый организмом-хозяином и функционально связанный с молекулой, кодирующей белок, связывающий антиген CD30L. Промоторы представляют собой нетранскрибируемые последовательности, локализованные выше (то есть в 5' положении) стартового кодона структурного гена (обычно в пределах от 100 до 1000 п.о.), контролирующие транскрипцию структурного гена. Промоторы традиционно сгруппированы в один или два класса: индуцибельные промоторы и конститутивные промоторы. Индуцибельные промоторы инициируют повышенные уровни транскрипции с ДНК, находящейся под его контролем, в ответ на некоторое изменение в условиях культивирования, такое как присутствие или отсутствие питательного вещества или изменение температуры. Конститутивные промоторы, с другой стороны, однородно транскрибируют ген, с которым они функционально связаны, то есть с небольшим контролем или отсутствием контроля экспрессии гена. Хорошо известно большое число промоторов, распознаваемых разнообразными клетками-хозяевами. Подходящий промотор функционально связан с ДНК, кодирующей вариабельную область тяжелой цепи или вариабельную область легкой цепи белка, связывающего антиген CD30L, путем удаления промотора из источника ДНК путем расщепления ферментом рестрикции и встраивания желаемой промоторной последовательности в вектор.
Подходящие промоторы, используемые при дрожжевых клетках-хозяевах, также хорошо известны в данной области техники. Дрожжевые энхансеры предпочтительно используют с дрожжевыми промоторами. Промоторы, подходящие для использования с клетками-хозяевами млекопитающих, хорошо известны и включают, но не ограничены ими, промоторы, полученные из геномов вирусов, таких как вирус полиомы, вирус оспы птиц, аденовирус (такой как аденовирус 2), вирус бычьей папилломы, вирус саркомы птиц, цитомегаловирус, ретровирусы, вирус гепатита В и вирус обезьян 40 (SV40). Другие подходящие промоторы млекопитающих включают гетерогенные промоторы млекопитающих, например, промоторы теплового шока и промотор актина.
Дополнительные промоторы, которые могут представлять интерес, включают, но не ограничены ими, следующие промоторы: ранний промотор SV40 (Benoist and Chambón, 1981, Nature 290:304-310); промотор CMV (Thornsen et al., 1984, Proc. Natl. Acad. U.S.A. 81:659-663); промотор, содержащийся в 3' длинном концевом повторе вируса саркомы Рауса (Yamamoto et al., 1980, Cell 22:787-797); промотор тимидинкиназы герпеса (Wagner et al., 1981, Proc. Natl. Acad. Sci. U.S.A. 78:1444-1445); промотор и регуляторные последовательности из гена металлотионеина (Prinster et al., 1982, Nature 296:39-42); и прокариотические промоторы, такие как промотор бета-лактамазы (Villa-Kamaroff et al., 1978, Proc. Natl. Acad. Sci. U.S.A. 75:3727-3731); или промотор tac (DeBoer et al., 1983, Proc. Natl. Acad. Sci. U.S.A. 80:21-25). Также представляют интерес следующие участки транскрипционного контроля животных, проявляющие тканеспецифичность и используемые у трансгенных животных: контролирующий участок гена эластазы I, активный в ацинарных клетках поджелудочной железы (Swift et al., 1984, Cell 38:639-646; Ornitz et al., 1986, Cold Spring Harbor Symp. Quant. Biol. 50:399-409; MacDonald, 1987, Hepatology 7:425-515); контролирующий участок гена инсулина, активный в бета-клетках поджелудочной железы (Hanahan, 1985, Nature 315:115-122); контролирующий участок гена иммуноглобулина, активный в лимфоидных клетках (Grosschedl et al., 1984, Cell 38:647-658; Adames et al., 1985, Nature 318:533-538; Alexander et al., 1987, Mol. Cell. Biol. 7:1436-1444); контролирующий участок гена вируса опухоли молочной железы мыши, активный в клетках семенников, молочной железы, лимфоидных клетках и мастоцитах (Leder et al., 1986, Cell 45:485-495); контролирующий участок гена альбумина, активный в печени (Pinkert et al., 1987, Genes and Devel. 1:268-276); контролирующий участок гена альфа-фетопротеина, активный в печени (Krumlauf et al., 1985, Mol. Cell. Biol. 5:1639-1648; Hammer et al., 1987, Science 253:53-58); контролирующий участок гена альфа-1-антитрипсина, активный в печени (Kelsey et al., 1987, Genes and Devel. 1:161-171); контролирующий участок гена бета-глобина, активный в миелоидных клетках (Mogram et al., 1985, Nature 315:338-340; Kollias et al., 1986, Cell 46:89-94); контролирующий участок гена основного белка миелина, активный в олигодендроцитах головного мозга (Readhead et al., 1987, Cell 48:703-712); контролирующий участок гена легкой цепи-2 миозина, активный в скелетных мышцах (Sani, 1985, Nature 314:283-286); и контролирующий участок гена гормона, высвобождающего гонадотропин, активный в гипоталамусе (Mason et al., 1986, Science 234:1372-1378).
Энхансерная последовательность может быть встроена в вектор для повышения транскрипции высшими эукариотами. Энхансеры представляют собой цис-активные элементы ДНК, обычно примерно 10-300 п.о. в длину, действующие на промотор, усиливая его транскрипцию. Энхансеры относительно независимы от ориентации и положения и находятся как в 5', так и в 3' положении относительно транскрипционной единицы. Известно несколько энхансерных последовательностей, доступных для генов млекопитающих (например, глобина, эластазы, альбумина, альфа-фето-протеина и инсулина). Тем не менее, в характерном случае используют энхансер от вируса. Энхансер SV40, энхансер раннего промотора цитомегаловируса, энхансер полиомы и энхансеры аденовируса, известные в данной области техники, являются иллюстративными энхансерными элементами для активации эукариотических промоторов. Хотя энхансер может быть расположен в векторе либо в 5', либо в 3' положении к кодирующей последовательности, в характерном случае он локализован в 5-сайте от промотора. Последовательность, кодирующая подходящую нативную или гетерологичную сигнальную последовательность (лидерную последовательность или сигнальный пептид), может быть включена в экспрессионный вектор, для стимуляции внеклеточной секреции антитела. Выбор сигнального или лидерного пептида зависит от типа клеток-хозяев, в которых нужно получать антитело, и гетерологичная сигнальная последовательность может замещать нативную сигнальную последовательность. Примеры сигнальных пептидов, функциональных в клетках-хозяевах млекопитающих, включают следующие сигнальные пептиды: сигнальную последовательность интерлейкина-7, раскрытую в патенте US №4965195; сигнальную последовательность рецептора интерлейкина-2, раскрытую в статье Cosman et al., 1984, Nature 312:768; сигнальный пептид рецептора интерлейкина-4, раскрытый в патенте ЕР №0367566; сигнальный пептид рецептора интерлейкина-1 типа I, раскрытый в патенте US №4968607; сигнальный пептид рецептора интерлейкина-1 типа II, раскрытый в патенте ЕР №0460846.
После того, как вектор сконструирован, полноразмерный вектор может быть встроен в подходящую клетку-хозяина для амплификации и/или экспрессии полипептида. Трансформация экспрессионного вектора для белка, связывающего антиген, в выбранной клетке-хозяине может быть выполнена хорошо известными способами, включающими трансфекцию, инфекцию, кальцийфосфатную копреципитацию, электропорацию, микроинъекцию, липофекцию, трансфекцию, опосредованную ДЭАЭ-декстраном, или другими известными методами. Выбранный способ отчасти зависит от используемой клетки-хозяина. Эти способы и другие подходящие способы хорошо известны специалистам в данной области техники и представлены, например, в Sambrook et al., Molecular Cloning: A Laboratory Manual, 3rd ed., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y. (2001).
Клетка-хозяин при культивировании в подходящих условиях синтезирует белок, который затем можно собрать из культуральной среды (если клетка-хозяин секретирует его в среду) или непосредственно из клетки-хозяина, продуцирующей этот белок (если он не секретируется). Выбор подходящей клетки-хозяина зависит от различных факторов, таких как уровни экспрессии, модификации полипептида, желательные или необходимые для активности (такие как гликозилирование или фосфорилирование) и простота свертывания в биологически активную молекулу.
Линии клеток млекопитающих, доступные в качестве хозяев для экспрессии, хорошо известны в данной области техники и включают, но не ограничены ими, иммортализованные линии клеток, доступные из Американской коллекции типовых культур (АТСС; от англ. "American Type Culture Collection"), включающие, но не ограниченные ими, клетки яичника китайского хомячка (СНО; от англ. "Chinese hamster ovary"), клетки HeLa, клетки почек новорожденного хомяка (ВНК; от англ. "baby hamster kidney"), клетки почек обезьяны (COS), клетки печеночно-клеточного рака человека (например, Hep G2) и ряд других линий клеток. В определенных воплощениях изобретения линии клеток могут быть выбраны путем определения, какие из линий клеток обладают более высокими уровнями экспрессии и конститутивно продуцируют белки, связывающие антиген, обладающие свойствами связывания CD30L. В другом воплощении изобретения линия клеток может быть также выбрана из В-клеточной линии, не продуцирующей собственное антитело, но обладающей способностью к продуцированию и секреции гетерологичного антитела. В другом воплощении изобретения нокаут-линии клеток по фукозилтрансферазе продуцируют полностью или частично афукозилированные антитела (патенты US 6946292, US 7425446, US 7846725, US 8067232, клетки Potelligent®, BioWa, Princeton, NJ).
Биологические анализы CD30L In Vitro
Предложены также способы идентификации антагонистов, ингибирующих взаимодействие CD30/CD30L. При данных способах используют индукцию маркерного цитокина, продуцируемого клетками CD30+ в ответ на CD30L. Этот способ включает стадии объединения интересующего тестируемого соединения с источником CD30L; добавление комбинации тестируемое соединение/CD30L к клеткам CD30+, способным к экспрессии маркерного цитокина в ответ на взаимодействие CD30 и CD30L; сбор супернатанта клеток и определение, ингибирует ли тестируемое соединение взаимодействие CD30/CD30L, путем измерения количества маркерного цитокина, продуцируемого и/или высвобождаемого в супернатант клеток. Например, в таком CD30L-зависимом биологическом анализе in vitro можно использовать экспрессию эндогенного CD30 в линии клеток анапластической крупноклеточной лимфомы человека (ALCL; от англ. "Anaplastic Large Cell Lymphoma") Karpas-299 (K299) и высвобождение ИЛ-8 из этих клеток в ответ на взаимодействие CD30 и CD30L. Дополнительные примеры этих способов приведены в данном описании в разделе Примеры.
Тестируемые соединения включают белки, связывающие антиген CD30L, включающие, но не ограниченные ими, антитела, их антигенсвязывающие фрагменты и их производные. Тестируемые соединения могут также включать рекомбинантные растворимые соединения CD30 (такие как внеклеточный домен CD30, слитый с меткой Fc, меткой поли-His, меткой FLAG® и тому подобное), а также малые молекулы. Подходящие линии клеток CD30+ экспрессируют маркерный цитокин в ответ на взаимодействие CD30 и CD30L; такие линии клеток включают линию клеток анапластической крупноклеточной лимфомы человека CD30+ К299 (Karpas-299, DSMZ, Germany).
Источники CD30L включают растворимые конструкции CD30L и мембраносвязанный CD30L. Растворимый CD30L включает химерные композиции, содержащие фрагмент CD30L, предпочтительно фрагмент внеклеточной области CD30L. Такие растворимые конструкции включают мультимерные конструкции в виде лейциновой или изолейциновой молнии. Такие конструкции включают 33-аминокислотный мотив лейциновой молнии, присоединенный к N-концу внеклеточного домена CD30L человека или яванского макака. Растворимые рекомбинантные конструкции CD30L также имеются в продаже (R&D Systems, Minneapolis, MN). Другой растворимый CD30L также включает внеклеточный домен CD30L, слитый с меткой FLAG® (Axxora LLC, San Diego, СА). Мембраносвязанный CD30L включает нативные и трансфицированные клетки и линии клеток CD30L. Такие линии клеток включают, но не ограничены ими, такие клетки человека, как линия В-клеток человека CD30L+ Ramos (АТСС, Manassas, VA). Рассмотрены также Т-клетки крови яванского макака и линии дендритных клеток мыши.
В данном изобретении предложен способ определения индукции лигандом CD30L продуцирования ИЛ-8 клетками CD30+, включающий следующие стадии: объединение тестируемого соединения с источником CD30L; добавление комбинации тестируемое соединение-СD30L к необлученным клеткам CD30+; сбор супернатанта клеток и определение количества ИЛ-8, высвобождаемого в супернатант клеток. В данном СD30L-зависимом биологическом анализе in vitro используют экспрессию эндогенного CD30 в линии клеток анапластической крупноклеточной лимфомы человека (ALCL) Karpas-299 (К299) и высвобождение ИЛ-8 из этих клеток в ответ на взаимодействие CD30 и CD30L. Высвобождаемый ИЛ-8 можно количественно определить, например, с помощью твердофазного иммуноферментного анализа (ELISA) на ИЛ-8. Антагонисты CD30L, такие как белки, связывающие антиген, раскрытые в данном изобретении, инкубируют либо с растворимым CD30L (анализ на одной линии клеток), либо с мембраносвязанным CD30L (анализ на двух линиях клеток), а затем культивируют с клетками К299. Затем супернатанты клеток собирают и определяют ингибирование высвобождения ИЛ-8 из клеток К299 в ответ на блокирование антагонистами CD30L такими как белки, связывающие антиген, раскрытые в данном изобретении, используя сэндвич-ELISA на ИЛ-8.
Для одинарного клеточного анализа антагонисты CD30L титруют на 96-луночных микротитрационных планшетах, таких как 96-луночные планшеты фирмы Costar® (Corning; Acton, MA), до получения желаемого диапазона конечных концентраций, например, от 1 мкг/мл до 10 пг/мл. Белки, связывающие антиген CD30L, такие как раскрыто в данном изобретении, можно использовать в качестве положительных контролей. Планшеты инкубировали в течение примерно 45 минут при комнатной температуре. Затем в каждую лунку добавляли источник клеток CD30+. Планшеты инкубировали в течение 24 часов при 37°С и 5% СO2.
Для двойного клеточного анализа источник CD30L связан с мембраной и облучен. Клетки, экспрессирующие CD30L, включают, но не ограничены ими, такие клетки, как линия В-клеток человека CD30L человека+ Ramos (АТСС, Manassas, VA), активированные Т-клетки крови яванского макака или линии дендритных клеток (DC) мыши. Клетки, экспрессирующие CD30L, можно подвергать сортировке флуоресцентно-активированных клеток (FACS; от англ. "Fluorescence Activated Cell Sorting") перед использованием для селекции клеток, имеющих самый высокий уровень экспрессии CD30L клеточной поверхности. Клетки, экспрессирующие CD30L, объединяют с антагонистом CD30L и инкубируют в течение 45 минут, после чего добавляют к источнику клеток CD30+. Планшеты снова инкубируют в течение 24 часов при 37°С, 5% СO2.
Для обоих анализов высвобожденный ИЛ-8 можно обнаруживать и определять количественно любым способом определения ИЛ-8, таким как, например, сэндвич-ELISA на ИЛ-8. Такие анализы имеются в продаже, например, предложенные фирмой R&D Systems (Minneapolis, MN). Уровни ИЛ-8 можно интерполировать из стандартной кривой ELISA и определить значения IC50 с использованием имеющегося в продаже программного обеспечения, такого как DeltaSoft (DeltaSoft, Inc., Hillsborough, NJ) и GraphPad PRISM (GraphPad Software Inc., San Diego, CA).
Применение белков, связывающих антиген CD30L человека, в диагностических и терапевтических целях
Белки, связывающие антиген, предложенные в данном изобретении, полезны для определения CD30L в биологических образцах и идентификации клеток или тканей, продуцирующих CD30L. Белки, связывающие антиген, которые специфично связываются с CD30L, можно применять при диагностике и/или лечении заболеваний, обусловленных CD30L, у пациента, нуждающегося в этом, например, белки, связывающие антиген CD30L, можно применять в диагностических анализах, например, индукции ИЛ-8 в клетках CD30+. Белки, связывающие антиген, которые связываются с CD30L, могут иметь терапевтическое применение при ослаблении заболеваний, обусловленных CD30L.
Показания
Настоящее изобретение также относится к применению белков, связывающих антиген CD30L, для применения при предупреждении или терапевтическом лечении клинических расстройств, как раскрыто в данном изобретении. Белки, связывающие антиген CD30L, полезны для лечения ряда состояний, при которых CD30L связан с ними или играет роль при внесении вклада в лежащее в основе заболевание или расстройство, либо вносит иной вклад в негативный симптом.
Предложены способы лечения ряда заболеваний, таких как аутоиммунные и хронические воспалительные заболевания и рак, путем введения пациенту, нуждающемуся в этом, эффективного количества композиции, содержащей один или более из белков, связывающих антиген CD30L, раскрытых в данном изобретении. Такие композиции полезны для модулирования иммунных ответов in vivo путем блокирования взаимодействий между клетками CD30+ и CD30L+; истощения клеток CD30L+; или посредством антагонистической активности в отношении клеток CD30L+. CD30L может проявлять "обратную передачу сигнала". CD30L, экспрессируемый на нейтрофилах и Т-клетках периферической крови, может быть активирован путем сшивания, в результате чего стимулирует метаболические активности в этих клетках (Wiley et al., J Immunol 157: 3235-39, 1996; Cerutti et al., J. Immunol. 165: 786, 2000; Cerutti et al., Nat. Immunol. 2: 150, 2001). Как таковые, антигенсвязывающие белки, раскрытые в данном изобретении, которые связывают CD30L, можно применять для обратной передачи сигнала CD30L или стимуляции клеток CD30L+.
Клетки CD30+ тесно связаны с болезнью Ходжкина, и CD30 является широко применяемым клиническим маркером ряда гематологических злокачественных опухолей (обзор см. в статье Horie and Watanabe, Immuno. 10:457-470, 1998). Композиции, содержащие один или более из белков, связывающих антиген CD30L раскрытых в данном изобретении, либо отдельно, либо в комбинации с другими терапиями, могут быть полезны при лечении таких состояний.
CD30L экспрессируется на активированных Т-клетках и некоторых В-клетках и популяциях дендритных клеток. CD30 экспрессируется на части активированных Т- и В-клеток. Данный экспрессионный профиль позволяет предположить, что нацеливание на CD30L было бы полезно для модулирования взаимодействия между Т-клетками, В-клетками и дендритными клетками. Показано, что CD30L влияет на гуморальный иммунитет. Белки, связывающие антиген, раскрытые в данном изобретении, могли бы иметь применение в схемах лечения, относящихся к воспалительным заболеваниям, в частности, направляемым зависимыми от Т-клеток ответами В-клеток.
Артрит можно лечить способами и композициями, раскрытыми в данном изобретении. Как используют в данном описании, термин "артрит" относится к хроническим воспалительным состояниям, которые в первую очередь поражают суставы или соединительную ткань, окружающую суставы, хотя могут также поражаться различные органы организма. Артрит может быть аутоиммунным или травматическим по происхождению, либо он может инициироваться под действием чужеродного антигена, приводя затем к хроническому состоянию, которое больше не зависит от продолжения присутствия инициирующего антигена. Термин "артрит", как используют в настоящем изобретении, включает следующие заболевания: деформирующий артрит; остеоартрит; ревматоидный артрит (взрослый и ювенильный); артрит Лайма; реактивный артрит, включая болезнь Рейтера; псориатический артрит; узловой артрит; серонегативные спондилоартропатии, включающие, но не ограниченные ими, анкилозирующий спондилоартрит.
Белки, связывающие антиген, раскрытые в данном изобретении, полезны при лечении разнообразных ревматических расстройств, которые определены в данном изобретении как любое хроническое расстройство, включающее болезненные и часто множественные локализованные воспаления в суставах, мышцах, нервах, сухожилиях, коже, глазах, соединительных тканях или различных других системах органов. Эти расстройства включают, но не ограничены ими, следующие расстройства: артрит; склеродерму; подагру; системную красную волчанку; ревматическую полимиалгию; болезнь Стилла; хронический увеит; расстройства, приводящие в результате к воспалению скелетной мышцы, включающие дерматомиозит и полимиозит, включающий спорадичный миозит с включенными тельцами. Системная красная волчанка может вызывать воспаление суставов, кожи, почек, сердца, легких, кровеносных сосудов и головного мозга. При запущенных формах системной красной волчанки это состояние может привести к почечной недостаточности.
Также предложены способы применения белков, связывающих антиген, раскрытых в данном изобретении, при терапиях для лечения различных расстройств эндокринной системы, включающих, но не ограниченных ими, следующие расстройства: ювенильный диабет или диабет зрелого возраста (включая аутоиммунные, инсулинозависимые типы диабета; инсулиннезависимые типы и опосредованный ожирением диабет); идиопатическую атрофию надпочечников; болезнь Аддисона; гипотиреоз; диффузный токсический зоб; аутоиммунный тиреоидит, такой как тиреоидит Хасимото; и полигландулярные аутоиммунные синдромы (типов I и II).
Белки, связывающие антиген, раскрытые в данном изобретении, также полезны при терапиях для лечения патологических состояний желудочно-кишечного тракта, включающих, но не ограниченных ими, следующие состояния: аутоиммунный склерозирующий холангит; глютеновую болезнь; воспалительные кишечные заболевания, включая болезнь Крона и неспецифический язвенный колит; аутоиммунный панкреатит, включая хронический панкреатит; идиопатический парез желудка и идиопатические язвы, включая язвы желудка и двенадцатиперстной кишки.
Также включены способы применения белков, связывающих антиген, раскрытых в данном изобретении, при терапиях для лечения расстройств мочеполовой системы, таких как аутоиммунный и идиопатический гломерулонефрит; и хронический идиопатический простатит (небактериальный), включая доброкачественную гипертрофию простаты.
В данном изобретении также предложены способы применения белков, связывающих антиген, раскрытых в данном изобретении, при терапиях для лечения различных гематологических расстройств, включающих, но не ограниченных ими, следующие расстройства: анемии и гематологические расстройства, включающие пернициозную анемию и апластическую анемию, а также апластическую анемию Фанкони; аутоиммунную гемолитическую анемию; идиопатическую тромбоцитопеническую пурпуру (ИТП); миелодиспластические синдромы (включающие рефрактерную анемию, рефрактерную анемию с кольцевидными сидеробластами, рефрактерную анемию с избытком бластов, рефрактерную анемию с избытком бластов на стадии трансформации); и аутоиммунный лимфопролиферативный синдром (ALPS; от англ. "autoimmune lymphoproliferative syndrome").
Кроме того, раскрытые белки, связывающие антиген, полезны для лечения патологических состояний, поражающих печень, таких как аутоиммунный или хронический воспалительный гепатит.
Кроме того, раскрытые белки, связывающие антиген, и комбинации применяют для лечения различных аутоиммунных или хронических воспалительных расстройств, в которые вовлечена потеря слуха. Одним из них является потеря слуха, обусловленная внутренним ухом или улитковым нервом, которую считают результатом аутоиммунного процесса, то есть аутоиммунной потери слуха. В настоящее время данное состояние лечат стероидами, метотрексатом и/или циклофосфамидом, которые можно вводить одновременно с ингибитором или блокатором взаимодействия CD30/CD30L.
Ряд воспалительных легочных расстройств также можно лечить раскрытыми белками, связывающими антиген, включая следующие расстройства: идиопатический лимфангиолейомиоматоз; хроническую обструктивную болезнь легких (ХОБЛ), обусловленную хроническим неинфекционным бронхитом или эмфиземой; и фибротические легочные заболевания, такие как муковисцидоз и идиопатический легочный фиброз.
Расстройства, обусловленные трансплантацией, также поддаются лечению раскрытыми белками, связывающими антиген, включая болезнь трансплантат-против-хозяина. Для предупреждения или ослабления болезни трансплантат-против-хозяина композиции, содержащие один или более раскрытых белков, связывающих антиген, можно вводить до, во время или после трансплантации костного мозга или солидного органа, включая трансплантацию сердца, печени, легкого, кожи, почки или других органов.
Раскрытые белки, связывающие антиген, также полезны для лечения хронических воспалительных глазных заболеваний, включая аутоиммунный увеит.
Такие соединения также были бы полезны для лечения заболеваний, обусловленных воспалением дыхательных путей, таких как астма.
Белки, связывающие антиген, раскрытые в данном изобретении, также полезны для лечения воспалительных расстройств, поражающих женскую половую систему, включающих следующие расстройства: множественные неудачи имплантации/бесплодие; синдром потери плода или потери эмбриона IV недели (самопроизвольный аборт); и эндометриоз.
Другие клинические расстройства, поддающиеся лечению белками, связывающими антиген, раскрытыми в данном изобретении, включают хронические воспалительные и/или дегенеративные заболевания центральной нервной системы. Они включают, например, заболевания, обусловленные демиелинизацией, такие как рассеянный склероз, системный склероз и синдромы Гийена-Барре (включая острую воспалительную демиелинизирующую полинейропатию, острую моторную аксональную нейропатию, острую моторно-сенсорную аксональную нейропатию и синдром Фишера). Рассеянный склероз является репрезентативным хроническим дегенеративным заболеванием центральной нервной системы, которое можно лечить агентом, способным к ингибированию или блокированию взаимодействия CD30 и CD30L. См., например, патент US №6652854.
Другие хронические воспалительные состояния, поддающиеся лечению раскрытыми белками, связывающими антиген, включают синдром Холодовой агглютинации; синдром Бехчета; синдром Шегрена; и идиопатический тендовагинит, а также различные хронические воспалительные расстройства, обусловленные наследственными дефектами. Заявленные ингибиторы, композиции и комбинированные терапии, кроме того, полезны для лечения паралича Белла (идиопатического паралича лицевого нерва); синдрома хронической утомляемости (не связанного с текущей инфекцией); хронического дегенеративного заболевания позвоночного диска; синдрома войны в Персидском заливе; и тяжелую миастению, где данные заболевания можно лечить одновременно с кортикостероидами.
Расстройства, в которые вовлечена кожа или слизистые оболочки, также поддаются лечению белками, связывающими антиген, раскрытыми в данном изобретении. Они включают следующие расстройства: акантолитические заболевания, включающие дискоидную волчанку, подострую кожную красную волчанку, кожный васкулит, болезнь Дарье, фолликулярный дискератоз, вульгарную пузырчатку и паранеопластическую пузырчатку; розовые угри; гнездную алопецию; буллезный пемфигоид; экзему; эритему, включающую полиморфную эритему и буллезную полиморфную эритему (синдром Стивенса-Джонсона); воспалительное заболевание кожи; плоский лишай; линейный IgA дерматоз (хронический буллезный дерматоз у детей); потерю эластичности кожи; острый фебрильный нейтрофильный дерматоз (синдром Свита); красный волосистый питириаз; псориаз; гангренозную пиодермию; потерю эластичности кожи и токсикодермальный некролиз.
Другие заболевания, которые можно лечить раскрытыми белками, связывающими антиген, включают следующие заболевания: аутоиммунно-ассоциированный хронический кандидоз кожи и слизистых; аллергии; саркоидоз; многополюсный ретикулогистиоцитоз; гранулематоз Вегенера; артериит, включающий гигантоклеточный артериит; васкулит и хронический аутоиммунный миокардит.
Для лечения клинического расстройства с применением белков, связывающих антиген, раскрытых в данном изобретении, терапевтически эффективное количество терапевтического агента вводят млекопитающему, нуждающемуся в этом. Этот агент вводят согласно схеме дозы и частоты введения, адекватной для индукции пролонгированного улучшения по меньшей мере одного показателя, отражающего тяжесть расстройства. Улучшение считают "пролонгированным", если пациент проявляет улучшение по меньшей мере при двух случаях, разделенных по меньшей мере одними сутками, но предпочтительно разделенных одной неделей, двумя неделями, тремя неделями или четырьмя или более недель. Тяжесть расстройства определяют на основании признаков или симптомов, либо ее можно определить с помощью анкетирования, назначенного пациенту, такого как анкетирование качества жизни, часто применяемое врачами для оценки статуса хронических заболеваний.
Один или более показателей, отражающих тяжесть заболевания пациента, можно оценивать для определения, достаточна ли частота или продолжительность лечения лекарственным средством. Базовое значение выбранного показателя устанавливают путем обследования пациента перед введением первой дозы терапевтического агента. Предпочтительно базовое обследование проводят в пределах примерно 60 суток введения первой дозы.
Если состояние, подлежащее лечению, представляет собой системную красную волчанку, показатель определения достаточности лечения может состоять в наблюдаемом улучшении одного или более из следующих показателей: утомляемости; лихорадки; язв рта и носа; высыпаний на лице ("сыпь в форме бабочки"); светочувствительности (СКВ часто внезапно обостряется под действием солнечного света); плеврита; перикардита; феномена Рейно (сниженное кровообращение пальцев рук и ног под действием холода); функции почек; и числа лейкоцитов (пациенты с СКВ часто имеют сниженные количества лейкоцитов).
Предложены фармацевтические композиции, содержащие белки, связывающие антиген, раскрытые в данном изобретении, включенные в препараты с фармацевтически приемлемым носителем. В некоторых воплощениях изобретения фармацевтически приемлемый носитель подходит для введения субъектам-людям.
Диагностические способы
Раскрытые белки, связывающие антиген, можно применять в диагностических целях для обнаружения, диагностики или мониторинга заболеваний и/или патологических состояний, обусловленных взаимодействием CD30/CD30L. Примеры способов, полезных при обнаружении присутствия CD30L, включают иммунологические анализы, такие как твердофазный иммуноферментный анализ (ELISA; от англ. "enzyme linked immunosorbent assay") и радиоиммунологический анализ (РИА).
Для диагностических применений белок, связывающий антиген, в характерном случае помечен обнаружимой метящей группой. Подходящие метящие группы включают, но не ограничены ими, следующие группы: радиоизотопы или радионуклиды (например, 3Н, 14С, 15N, 35S, 90Y, 99Тс, 111ln, 125Ι, 131I), флуоресцентные группы (например, ФИТЦ, родамин, люминофоры на основе комплексов лантанидов), ферментные группы (например, пероксидазу хрена, β-галактозидазу, люциферазу, щелочную фосфатазу), хемилюминесцентные группы, биотинильные группы или предопределенные полипептидные эпитопы, распознаваемые вторичным репортером (например, парные последовательности в виде лейциновой молнии, сайты связывания вторичных антител, домены, связывающие металлы, эпитопные метки). В некоторых воплощениях изобретения метящую группу связывают с белком, связывающим антиген, посредством спейсерных плеч различной длины, чтобы уменьшить потенциальное стерическое затруднение. Различные способы мечения белков известны в данной области техники и могут быть использованы.
Другие диагностические способы предложены для диагностики клетки или клеток, экспрессирующих CD30L. В конкретном воплощении изобретения белок, связывающий антиген, метят метящей группой и определяют связывание меченого белка, связывающего антиген, с CD30L. В следующем конкретном воплощении изобретения связывание белка, связывающего антиген, с CD30L определяют in vivo. В следующем конкретном воплощении изобретения белок, связывающий антиген CD30L, выделяют и измеряют, используя методы, известные в данной области техники. См., например, Harlow and Lane, 1988, Antibodies: A Laboratory Manual, New York: Cold Spring Harbor (ed. 1991 и периодические дополнения); John Ε. Coligan, ed., 1993, Current Protocols In Immunology New York: John Wiley & Sons.
Другие способы предложены для определения присутствия тестируемой молекулы, конкурирующей за связывание с CD30L с предложенными белками, связывающими антиген. Пример одного такого анализа включает определение количества свободного белка, связывающего антиген, в растворе, содержащем количество CD30L, в присутствии или в отсутствие тестируемой молекулы. Увеличение количества свободного белка, связывающего антиген (то есть белка, связывающего антиген, не связанного с CD30L) указывает на то, что тестируемая молекула способна конкурировать за связывание CD30L с белком, связывающим антиген. В одном воплощении изобретения белок, связывающий антиген, метят метящей группой. Альтернативно метят тестируемую молекулу и проводят мониторинг свободной тестируемой молекулы в присутствии и в отсутствие белка, связывающего антиген.
Способы лечения: фармацевтические препараты, пути введения
Предложены фармацевтические композиции, содержащие терапевтически эффективное количество одного или множества белков, связывающих антиген, и фармацевтически приемлемый эксципиент, разбавитель, носитель, солюбилизатор, эмульгатор, консервант и/или адъювант.Кроме того, включены способы лечения пациента путем введения такой фармацевтической композиции. Термин "пациент" включает пациентов-людей. Термины "лечить" и "лечение" включает ослабление или предупреждение по меньшей мере одного симптома или другого аспекта расстройства, либо снижение тяжести заболевания и тому подобное. Термин "терапевтически эффективное количество" или "эффективное количество" относится к определенному количеству белка, связывающего антиген CD30L, производящему какой-либо терапевтический ответ у млекопитающего. Такие терапевтически эффективные количества легко устанавливается обычными специалистами в данной области техники. Для целей данного описания термины "болезнь", "заболевание", "клиническое состояние", "аномальное состояние", "недомогание", "клиническое расстройство", "расстройство" и тому подобное используют взаимозаменяемо.
Белок, связывающий антиген, необязательно осуществляет полное излечение или устранение каждого симптома или проявления заболевания, чтобы представлять собой пригодный терапевтический агент. Как признано в области, к которой принадлежит изобретение, лекарственные средства, применяемые в качестве терапевтических агентов, могут уменьшить тяжесть данного болезненного состояния, но не обязательно прекращает каждое проявление заболевания, чтобы они были рассмотрены как полезные терапевтические агенты. Подобным образом, профилактически вводимое лечение необязательно должно быть полностью эффективным при предупреждении возникновения состояния, чтобы представлять собой пригодный терапевтический агент. Достаточно просто уменьшение действия заболевания (например, путем снижения числа или тяжести его симптомов, либо путем увеличения эффективности другого лечения, либо путем произведения другого полезного эффекта) или снижение вероятности того, что это заболевание возникнет или ухудшится у субъекта. Определенные способы, предложенные в данном изобретении, включают введение пациенту антагониста CD30L (такого как белки, связывающие антиген, раскрытые в данном изобретении) в таком количестве и в течение такого периода времени, которые достаточны для индукции пролонгированного улучшения по сравнению с базовым уровнем показателя, отражающего тяжесть конкретного заболевания.
Как понимают в области техники, к которой принадлежит изобретение, фармацевтические композиции, содержащие молекулы по изобретению, вводят пациенту способом, соответствующим показанию. Фармацевтические композиции можно вводить с помощью любой подходящей методики, включающими, но не ограниченными ими, парентеральное введение, местное введение или введение путем ингаляции. Если фармацевтическую композицию инъецируют, ее можно вводить, например, посредством следующих путей: внутрисуставного, внутривенного, внутримышечного, внутриочагового, интраперитонеального или подкожного, путем болюсной инъекции или непрерывной инфузии. Рассмотрено локализованное введение, например, в сайт заболевания или повреждения, такое как чрескожная доставка и пролонгированное высвобождение из имплантатов. Доставка путем ингаляции включает, например, назальную или пероральную ингаляцию, применение небулайзера, ингаляцию антагониста в аэрозольной форме и тому подобное. Другие альтернативы включают глазные капли; пероральные препараты, включающие пилюли, сиропы, лепешки или жевательную резинку; и местные препараты, такие как лосьоны, гели, спреи и мази.
Также рассмотрено применение белков, связывающих антиген, в методах ех vivo. Например, кровь или другую жидкость организма пациента можно приводить в контакт с белком, связывающим антиген, который связывает CD30L ex vivo. Белок, связывающий антиген, может быть связан с подходящим нерастворимым матриксом или твердым материалом носителя.
Предпочтительно белки, связывающие антиген, вводят в форме композиции, содержащей один или более дополнительный компонент, такой как физиологически приемлемый носитель, эксципиент или разбавитель. Необязательно композиция содержит один или более чем один физиологически активный агент для комбинированной терапии. Фармацевтическая композиция содержит белок, связывающий антиген CD30L, вместе с одним или более веществ, выбранных из группы, состоящей из следующих веществ: буфера, антиоксиданта, такого как аскорбиновая кислота, низкомолекулярного полипептида (такого как полипептиды, имеющие менее 10 аминокислот), белка, аминокислоты, углевода, такого как глюкоза, сахароза или декстрины, хелатирующего агента, такого как этилендиаминтетрауксусная кислота (ЭДТА), глутатиона, стабилизатора и эксципиента. Примерами подходящих разбавителей являются следующие вещества: нейтральный забуференный физиологический раствор или физиологический раствор, смешанный с конспецифичным сывороточным альбумином. В соответствии с надлежащими промышленными стандартами можно также добавлять консерванты, такие как бензиловый спирт. Композицию можно готовить в виде лиофилизата, используя подходящие растворы эксципиентов (например, сахарозы) в качестве разбавителей. Подходящие компоненты нетоксичны для реципиентов при применяемых дозах и концентрациях. Дополнительные примеры компонентов, которые можно применять в фармацевтических препаратах, представлены в любом издании руководства Remington's Pharmaceutical Sciences, включающем 21е издание (2005), Mack Publishing Company, Easton, PA.
Наборы, применяемые практикующими врачами, включают белок, связывающий антиген CD30L, и метку или другие инструкции по применению при лечении любого из состояний, раскрытых в данном изобретении. В одном воплощении изобретения набор включает стерильный препарат одного или более белков, связывающих антиген CD30L, который может находиться в форме композиции, как раскрыто выше, или может находиться в одном или более флаконов.
Дозы и частоту введения можно варьировать в соответствии с такими факторами, как путь введения, конкретные применяемые белки, связывающие антиген, природа и тяжесть заболевания, подлежащего лечению, независимо от того, является ли это состояние острым или хроническим, и размера и общего состояния субъекта. Подходящие дозы могут быть определены методами, известными в области техники, к которой принадлежит изобретение, например, в клинических испытаниях, которые могут включать исследования с увеличением дозы.
В характерном случае доза может находиться в диапазоне от примерно 0,1 мкг/кг до вплоть до примерно 30 мг/кг или более в зависимости от факторов, указанных выше. В конкретных воплощениях изобретения доза может находиться в диапазоне от 0,1 мкг/кг вплоть до примерно 30 мг/кг, необязательно от 1 мкг/кг вплоть до примерно 30 мг/кг, необязательно от 10 мкг/кг вплоть до примерно 10 мг/кг, необязательно от примерно 0,1 мг/кг до 5 мг/кг или необязательно от примерно 0,3 мг/кг до 3 мг/кг.
Частота дозы зависит от фармакокинетических параметров конкретного белка, связывающего антиген CD30L, в применяемом препарате. В характерном случае врач вводит композицию до достижения дозы, при которой достигается желаемый эффект. Композицию можно, таким образом, вводить в однократной дозе или в виде двух или более доз (которые могут содержать одинаковое или неодинаковое количество желаемой молекулы) за период времени, либо в виде непрерывной инфузии посредством имплантационного устройства или катетера. Подходящие дозы могут быть установлены посредством использования данных доза-ответ. Белок, связывающий антиген CD30L, по изобретению можно вводить, например, один или более чем один раз, например, через регулярные интервалы в течение периода времени. В конкретных воплощениях изобретения белок, связывающий антиген CD30L, вводят в течение периода, составляющего по меньшей мере месяц или более, например, в течение одного, двух или трех месяцев или даже неопределенного времени. Для лечения хронических состояний длительное лечение, как правило, наиболее эффективно. Тем не менее, для лечения острых состояний может быть достаточно введение в течение более коротких периодов, например, от одной до шести недель. Как правило, белок, связывающий антиген, вводят до тех пор, пока пациент не проявляет клинически релевантную степень улучшения по сравнению с базовым уровнем для выбранного показателя или показателей.
Рассмотрено, что белок, связывающий антиген CD30L, вводят пациенту в достаточном количестве и в течение достаточного периода времени, чтобы индуцировать улучшение, предпочтительно пролонгированное улучшение, по меньшей мере по одному показателю, отражающему тяжесть расстройства, подлежащего лечению. Различные показатели, отражающие степень болезни, заболевания или патологического состояния пациента, можно оценивать для определения, является ли достаточным количество и время лечения. Такие показатели включают, например, клинически признанные показатели тяжести заболевания, симптомы или проявления исследуемого расстройства. В одном воплощении изобретения улучшение рассматривают как пролонгированное, если субъект проявляет улучшение по меньшей мере в двух случаях, разделенных периодом от двух до четырех недель. Степень улучшения, как правило, определяется врачом, который может основывать это определение на признаках, симптомах, биопсиях или результатах других тестов, и который может также использовать анкетирование, которое назначают субъекту, такое как анкетирование качества жизни, разработанное для данного заболевания. Конкретные воплощения способов и композиций по изобретению включают применение белка, связывающего антиген CD30L, и одного или более дополнительных антагонистов CD30L, например, двух или более белков, связывающих антиген, по изобретению или белка, связывающего антиген, по изобретению и одного или более других антагонистов CD30L. Также предложены белки, связывающие антиген CD30L, вводимые отдельно или в комбинации с другими агентами, полезными для лечения состояния, которым страдает пациент. Примеры таких агентов включают как белковые, так и небелковые лекарственные средства. Такие агенты включают, например, антагонисты фактора некроза опухоли альфа (TNFα; от англ. "Tumor Necrosis Factor"), интерлейкина (ИЛ)-1, ИЛ-2, рецептора активатора транскрипции каппа (RANK; от англ. "Receptor Activator Of Nf Kappa") или других цитокинов, которые могут вносить вклад в аутоиммунность или хроническое воспаление. Антагонисты цитокинов включают антагонисты, нацеленные на цитокин и/или его рецептор. Антагонист, нацеленный на цитокин, может включать растворимый рецептор против цитокина и обычно включает участок внеклеточного домена или весь внеклеточный домен рецептора цитокина. Растворимый рецептор цитокина можно применять в виде мономера, либо в виде димера или более высокого мультимера (например, в виде слитой молекулы, где растворимый рецептор присоединен к участку Fc иммуноглобулина человека, стимулирующему образование димеров). В других воплощениях изобретения растворимый рецептор цитокина пегилирован, чтобы увеличить его период полувыведения в сыворотке. В некоторых воплощениях изобретения антагонист растворимого цитокина включает растворимый рецептор TNF (типа I или II). Малые органические молекулы, ингибирующие воспалительные цитокины можно также применять в комбинации с заявленными терапевтическими агентами. Более чем один белок, связывающий антиген - лиганд CD30 и/или его антигенсвязывающие участки, можно вводить одновременно для лечения аутоиммунных или хронических воспалительных заболеваний, и его можно вводить отдельно или вместе с другими лекарственными средствами, эффективными против того же аутоиммунного или хронического воспалительного состояния, или средствами, которые вводят для лечения другого состояния у того же пациента.
Когда множественные терапевтические средства вводят совместно, дозы можно регулировать соответственно, как признано или известно в области техники, к которой принадлежит изобретение. Такие средства можно вводить одновременно, последовательно, поочередно или согласно любой другой схеме, позволяющей, чтобы полный курс терапии был эффективным.
Кроме пациентов-людей, белки, связывающие антиген CD30L, полезны при лечении животных, отличающихся от людей, таких как домашние питомцы (собаки, кошки, птицы, приматы и т.д.), сельскохозяйственные животные (лошади, крупный рогатый скот, овцы, свиньи, птицы и т.д.). В таких случаях соответствующая доза может быть определена в соответствии с массой тела животного. Например, можно применять дозу 0,2-1 мг/кг. Альтернативно дозу определяют в соответствии с площадью поверхности тела животного, где иллюстративная доза находится в диапазоне 0,1-20 мг/м2 или более предпочтительно 5-12 мг/м2. Для мелких животных, таких как собаки или кошки, подходящая доза составляет 0,4 мг/кг. Белок, связывающий антиген CD30L (предпочтительно сконструированный из генов, имеющих происхождение от вида-реципиента), вводят путем инъекции или другого подходящего пути один или более раз в неделю до улучшения состояния животного, либо его можно вводить в течение неопределенного времени.
Приведенные ниже воплощения изобретения и Примеры, включающие проведенные эксперименты и достигнутые результаты, предложены только в целях иллюстрации и не должны рассматриваться как ограничивающие объем прилагаемой формулы изобретения.
Воплощения изобретения
1. Изолированный антигенсвязывающий белок, который связывает CD30L, включающий по меньшей мере одну вариабельную область тяжелой цепи, содержащую участки CDRH1, CDRH2 и CDRH3, выбранные из группы, состоящей из следующих участков:
a) CDRH1, отличающегося не более чем четырьмя аминокислотными заменами, инсерциями или делециями от CDRH1, представленного в таблице 3;
b) CDRH2, отличающегося не более чем семью аминокислотными заменами, инсерциями и/или делециями от CDRH2, представленного в таблице 3;
c) CDRH3, отличающегося не более чем одиннадцатью аминокислотными заменами, инсерциями и/или делециями от CDRH3, представленного в таблице 3; и
включающий по меньшей мере одну вариабельную область легкой цепи, содержащей участки CDRL1, CDRL2 и CDRL3, выбранные из группы, состоящей из следующих участков:
d) CDRL1, отличающегося не более чем четырьмя аминокислотными заменами, инсерциями и/или делециями от CDRL1, представленного в таблице 3;
e) CDRL2, отличающегося не более чем двумя аминокислотными заменами, инсерциями или делециями от CDRL2, представленного в таблице 3;
f) CDRL3, отличающегося не более чем двумя аминокислотными заменами, инсерциями или делециями от CDRL3, представленного в таблице 3.
2. Изолированный антигенсвязывающий белок по воплощению 1 изобретения, включающий следующие участки:
CDRH1, выбранный из группы, состоящей из следующих участков: SEQ ID NO: 14, 15, 16, 17, 18 и 33;
CDRH2, выбранный из группы, состоящей из следующих участков: SEQ ID NO: 19, 20,21,22, 23, 24 и 34;
CDRH3 выбранный из группы, состоящей из следующих участков: SEQ ID NO: 25, 26, 27, 28, 29 и 35;
CDRL1, выбранный из группы, состоящей из следующих участков: SEQ ID NO: 1,2, 3, 4, 5, 6 и 30;
CDRL2, выбранный из группы, состоящей из следующих участков: SEQ ID NO: 7, 8,9, 10 и 31; и
CDRL3, выбранный из группы, состоящей из следующих участков: SEQ ID NO: 11, 12, 13 и 32.
3. Изолированный антигенсвязывающий белок, связывающий CD30L человека, включающий по меньшей мере одну вариабельную область легкой цепи, выбранную из группы, состоящей из следующих участков:
a) вариабельную область легкой цепи, содержащую CDR1, CDR2 и CDR3, где CDR1 содержит аминокислотные остатки 23-36, CDR2 содержит аминокислотные остатки 52-58, и CDR3 содержит аминокислотные остатки 91-100 SEQ ID NO: 36, 28, 40, 42 или 44; или
b) вариабельную область легкой цепи, содержащую CDR1, CDR2 и CDR3, где CDR1 содержит аминокислотные остатки 25-36, CDR2 содержит аминокислотные остатки 52-58, и CDR3 содержит аминокислотные остатки 91-100 SEQ ID NO:46; и
по меньшей мере одну вариабельную область тяжелой цепи, содержащую участки CDR1, CDR2 и CDR3, где CDR1 содержит аминокислотные остатки 31-35, CDR2 содержит аминокислотные остатки 50-65, и CDR3 содержит аминокислотные остатки 98-113 SEQ ID NO:48, 50, 52, 54, 56, 58, 60, 62, 64, 66, 68, 70 или 72.
4. Изолированный антигенсвязывающий белок по п.1, включающий по меньшей мере одну вариабельную область тяжелой цепи и по меньшей мере одну вариабельную область легкой цепи.
5. Изолированный антигенсвязывающий белок по воплощению 1 изобретения, включающий по меньшей мере две вариабельные области тяжелой цепи и по меньшей мере две вариабельные области легкой цепи.
6. Изолированный антигенсвязывающий белок, который связывает CD30L, включающий вариабельную область тяжелой цепи и вариабельную область легкой цепи, где последовательность вариабельной области тяжелой цепи отличается не более чем 31 аминокислотной заменой, добавлением и/или делецией от вариабельной области тяжелой цепи, представленной в таблице 2; и где последовательность вариабельной области легкой цепи отличается не более чем 30 аминокислотными заменами, добавлениями и/или делециями от вариабельной области легкой цепи, представленной в таблице 1.
7. Изолированный антигенсвязывающий белок, который связывает CD30L, включающий следующие участки:
вариабельную область тяжелой цепи, состоящую из аминокислотной последовательности, обладающей по меньшей мере 80% идентичностью последовательности SEQ ID NO: 48, 50, 52, 54, 56, 58, 60, 62, 64, 66, 68, 70 и 72; и
вариабельную область легкой цепи, содержащую аминокислотную последовательность, обладающую по меньшей мере 88% идентичностью последовательности SEQ ID NO: 36, 38, 40, 42, 44 и 46.
8. Изолированный антигенсвязывающий белок, который связывает CD30L, выбранный из группы, состоящей из следующих белков:
a) изолированного антигенсвязывающего белка, включающего вариабельную область тяжелой цепи, выбранную из группы, состоящей из следующих участков: SEQ ID NO: 48, 50, 52, 54, 56, 58, 60 и 62, и вариабельную область легкой цепи SEQ ID NO: 36;
b) изолированного антигенсвязывающего белка, включающего вариабельную область тяжелой цепи SEQ ID NO:64 и вариабельную область легкой цепи SEQ ID NO: 38;
c) изолированного антигенсвязывающего белка, включающего вариабельную область тяжелой цепи SEQ ID NO: 66 и вариабельную область легкой цепи SEQ ID NO: 40;
d) изолированного антигенсвязывающего белка, включающего вариабельную область тяжелой цепи SEQ ID NO: 68 и вариабельную область легкой цепи SEQ ID NO: 42;
e) изолированного антигенсвязывающего белка, включающего вариабельную область тяжелой цепи SEQ ID NO: 70 и вариабельную область легкой цепи SEQ ID NO: 44; и
f) изолированного антигенсвязывающего белка, включающего вариабельную область тяжелой цепи SEQ ID NO: 72 и вариабельную область легкой цепи SEQ ID NO: 46.
9. Изолированный белок, связывающий антиген, по любому из предшествующих воплощений 1-8 изобретения, где данный белок, связывающий антиген, представляет собой антитело.
10. Изолированный белок, связывающий антиген, по воплощению 9 изобретения, где антитело представляет собой моноклональное антитело, рекомбинантное антитело, человеческое антитело, гуманизированное антитело, химерное антитело, мультиспецифическое антитело или фрагмент антитела.
11. Изолированный белок, связывающий антиген, по воплощению 10 изобретения, где фрагмент антитела представляет собой Fab фрагмент, Fab' фрагмент, F(ab')2 фрагмент, Fv фрагмент, диатело или одноцепочечную молекулу антитела.
12. Изолированный белок, связывающий антиген, по воплощению 10 изобретения, где белок, связывающий антиген, представляет собой человеческое антитело.
13. Изолированный белок, связывающий антиген, по воплощению 10 изобретения, где белок, связывающий антиген, представляет собой моноклональное антитело.
14. Изолированный белок, связывающий антиген, по воплощению 9 изобретения, где белок, связывающий антиген, относится к IgG1-, IgG2-, IgG3- или IgG4-типу.
15. Изолированный белок, связывающий антиген, по воплощению 14 изобретения, где белок, связывающий антиген, относится к IgG1- или IgG2-типу.
16. Изолированный белок, связывающий антиген, по любому из воплощений 1-15 изобретения, где белок, связывающий антиген, обладает по меньшей мере одним свойством, выбранным из группы, состоящей из следующих свойств:
a) ингибирования взаимодействия CD30/CD30L;
b) ингибирования СD30L-индуцированной индукции ИЛ-8;
c) перекрестной конкуренции с любым из антител A-F за связывание с CD30L человека;
d) константы диссоциации, равной не более 70 пМ, и
e) связывания с CD30L человека по существу с такой же KD, как для любого из антител A-F.
17. Изолированный антигенсвязывающий белок, который связывает CD30L, где белок, связывающий антиген, связывает С-концевой участок CD30L, определенный аминокислотами (АА) 201-234, или такими, как АА 205-230 или АА 211-226.
18. Изолированный белок, связывающий антиген, по воплощению 17 изобретения, где белок, связывающий антиген, связывает дополнительный участок CD30L, определенный АА 75-95, такими как АА 80-90 или АА 82-88.
19. Изолированный белок, связывающий антиген, по воплощению 17 или 18 изобретения, где белок, связывающий антиген, обладает по меньшей мере одним свойством, выбранным из группы, состоящей из следующих свойств:
a) ингибирования взаимодействия CD30/CD30L;
b) ингибирования СD30L-индуцированной индукции ИЛ-8;
c) перекрестной конкуренции с любым из антител A-F за связывание с CD30L человека;
d) константы диссоциации с CD30L человека, составляющей максимум 70 пМ, и
e) связывания с CD30L человека по существу с такой же или более низкой Kd, как для любого из антител A-F.
20. Изолированный белок, связывающий антиген, по любому из воплощений 17-19 изобретения, где связывание белка, связывающего антиген, с hCD30L ингибируется связыванием Fab одного или более из антител А, В, С, D, Ε и F.
21. Изолированный белок, связывающий антиген, по любому из воплощений 17-19 изобретения, где белок, связывающий антиген, конкурирует с Fab фрагментом одного или более из антител А, В, С, D, Ε и F за связывание с CD30L
22. Изолированный белок, связывающий антиген, по любому из воплощений 20-21 изобретения, где Fab фрагмент представляет собой Fab фрагмент антитела А1.
23. Изолированный белок, связывающий антиген, по любому из воплощений 17-22 изобретения, где белок, связывающий антиген, имеет константу диссоциации (KD) с CD30L человека максимум 75 пМ, как, например, максимум 50 пМ, как, например, 40 пМ.
24. Изолированный белок, связывающий антиген, по любому из воплощений 17-23 изобретения, включающий участки CDRH1, CDRH2 и CDRH3 и CDRL1, CDRL2 и CDRL3,
где CDRL1 представляет собой консенсус-последовательность CDRL1 SEQ ID NO: 30,
где CDRL2 представляет собой консенсус-последовательность CDRL2 SEQ ID NO: 31,
где CDRL3 представляет собой консенсус-последовательность CDRL3 SEQ ID NO: 32,
где CDRH1 представляет собой консенсус-последовательность CDRH1 SEQ ID NO: 33,
где CDRH2 представляет собой консенсус-последовательность CDRH2 SEQ ID NO: 34 и
где CDRH3 представляет собой консенсус-последовательность CDRH3 SEQ ID NO: 35.
25. Изолированный белок, связывающий антиген, по любому из воплощений 17-23 изобретения, включающий участки CDRH1, CDRH2 и CDRH3 и CDRL1, CDRL2 и CDRL3,
где CDRL1 представляет собой консенсус-последовательность CDRL1 SEQ ID NO: 30,
где CDRL2 представляет собой консенсус-последовательность CDRL2 SEQ IDNO: 31,
где CDRL3 представляет собой консенсус-последовательность CDRL3 SEQ ID NO: 32,
где CDRH1 представляет собой последовательность SEQ ID NO: 17 или консенсус-последовательность CDRH1 SEQ ID NO: 75,
где CDRH2 представляет собой консенсус-последовательность CDRH2 SEQ ID NO: 34 и
где CDRH3 представляет собой консенсус-последовательность CDRH3 SEQ ID NO: 35.
26. Изолированный белок, связывающий антиген, по любому из воплощений 17-23 изобретения, включающий участки CDRH1, CDRH2 и CDRH3,
где CDRL1 представляет собой консенсус-последовательность CDRL1 SEQ ID NO: 30,
где CDRL2 представляет собой консенсус-последовательность CDRL2 SEQ ID NO: 31,
где CDRL3 представляет собой консенсус-последовательность CDRL3 SEQ ID NO: 32,
и участки CDRL1, CDRL2 и CDRL3,
где CDRH1 представляет собой последовательность SEQ ID NO: 17 или консенсус-последовательность CDRH1 SEQ ID NO: 75,
где CDRH2 выбран из последовательности ID NO: 19, 20, 21, 22, 23 и 24 и
где CDRH3 представляет собой консенсус-последовательность CDRH3 SEQ ID NO: 35.
27. Изолированный белок, связывающий антиген, по любому из воплощений 17-23 изобретения, включающий участки CDRH1, CDRH2 и CDRH3,
где CDRL1 представляет собой консенсус-последовательность CDRL1 SEQ ID NO: 30,
где CDRL2 представляет собой консенсус-последовательность CDRL2 SEQ ID NO: 31,
где CDRL3 представляет собой консенсус-последовательность CDRL3 SEQ ID NO: 32,
и участки CDRL1, CDRL2 и CDRL3,
где CDRH1 представляет собой последовательность SEQ ID NO: 17 или консенсус-последовательность CDRH1 SEQ ID NO: 75,
где CDRH2 выбран из группы, состоящей из SEQ ID NO: 19, 20, 21, 22, 23 и 24, и
где CDRH3 выбран из группы, состоящей из SEQ ID NO: 25, 26, 27, 28 и 29.
28. Изолированный белок, связывающий антиген, по любому из воплощений 17-23 изобретения, включающий следующие участки:
CDRH1, выбранный из группы, состоящей из SEQ ID NO: 14, 15, 16, 17 и 18; CDRH2, выбранный из группы, состоящей из SEQ ID NO: 19, 20, 21, 22, 23 и 24;
CDRH3, выбранный из группы, состоящей из SEQ ID NO: 25, 26, 27, 28 и 29; CDRL1, выбранный из группы, состоящей из SEQ ID NO: 1, 2, 3, 4, 5 и 6; CDRL2, выбранный из группы, состоящей из SEQ ID NO: 7, 8,9 и 10; и CDRL3, выбранный из группы, состоящей из SEQ ID NO: 11, 12 и 13.
29. Изолированный белок, связывающий антиген, по любому из воплощений 17-23 изобретения, включающий следующие участки:
CDRH1, CDRH2 и CDRH3, идентифицированные как SEQ ID NO: 14, 19, 25, и CDRL1, CDRL2 и CDRL3, идентифицированные как SEQ ID NO: 1, 7 и 11,
CDRH1, CDRH2 и CDRH3, идентифицированные как SEQ ID NO: 15, 21, 26, и CDRL1, CDRL2 и CDRL3, идентифицированные как SEQ ID NO: 2, 8 и 12,
CDRH1, CDRH2 и CDRH3, идентифицированные как SEQ ID NO: 15, 21, 26, и CDRL1, CDRL2 и CDRL3, идентифицированные как SEQ ID NO: 3, 8 и 12,
CDRH1, CDRH2 и CDRH3, идентифицированные как SEQ ID NO: 16, 22, 27, и CDRL1, CDRL2 и CDRL3, идентифицированные как SEQ ID NO: 4, 9 и 12,
CDRH1, CDRH2 и CDRH3, идентифицированные как SEQ ID NO: 17, 23, 28, и CDRL1, CDRL2 и CDRL3. идентифицированные как SEQ ID NO: 5, 8 и 13, или
CDRH1, CDRH2 и CDRH3, идентифицированные как SEQ ID NO: 18, 24, 29, и CDRL1, CDRL2 и CDRL3, идентифицированные как SEQ ID NO: 6, 10 и 12.
30. Белок, связывающий антиген, конкурирующий по меньшей мере с одним белком, связывающим антиген, по любому из воплощений 1-29 изобретения.
31. Белок, связывающий антиген, по любому из воплощений 1-29 изобретения, который является полностью или частично афукозилированным.
32. Изолированная молекула нуклеиновой кислоты, кодирующая белок, связывающий антиген, по любому из воплощений 1-31 изобретения.
33. Изолированная молекула нуклеиновой кислоты по воплощению 32 изобретения, где по меньшей мере одна вариабельная область тяжелой цепи кодируется изолированный молекулой нуклеиновой кислоты, выбранной из группы, состоящей из SEQ ID NO: 49, 51, 53, 55, 57, 59, 61, 63, 65, 67, 69, 71 и 73, и по меньшей мере одна вариабельная область легкой цепи кодируется изолированной молекулой нуклеиновой кислоты, выбранной из группы, состоящей из SEQ ID NO: 37, 39, 41,43, 45 и 47.
34. Молекула нуклеиновой кислоты согласно воплощению 33 изобретения, где молекула нуклеиновой кислоты функционально связана с контролирующей последовательностью.
35. Вектор, содержащий молекулу нуклеиновой кислоты согласно воплощению 32 изобретения.
36. Клетка-хозяин, содержащая молекулу нуклеиновой кислоты согласно воплощению 32 изобретения.
37. Клетка-хозяин, содержащая вектор согласно воплощению 35 изобретения.
38. Изолированный полинуклеотид, достаточный для использования в качестве зонда гибридизации, праймера ПЦР или секвенирующего праймера, представляющий собой фрагмент молекулы нуклеиновой кислоты по воплощению 33 или 34 изобретения или его комплемент.
39. Способ получения белка, связывающего антиген, по любому из воплощений 1-31 изобретения, включающий стадию получения препарата белка, связывающего антиген, из клетки-хозяина, секретирующей данный белок, связывающий антиген.
40. Фармацевтическая композиция, содержащая по меньшей мере один белок, связывающий антиген, по любому из воплощений 1-31 изобретения и фармацевтически приемлемый эксципиент.
41. Фармацевтическая композиция по воплощению 40 изобретения, дополнительно содержащая метящую группу или эффекторную группу.
42. Фармацевтическая композиция по воплощению 41 изобретения, где метящая группа выбрана из группы, состоящей из изотопных меток, магнитных меток, редокс-активных группировок, оптических красителей, биотинилированных групп и предопределенных полипептидных эпитопов, распознаваемых вторичным репортером.
43. Фармацевтическая композиция по воплощению 42 изобретения, где эффекторная группа выбранный из группы, состоящей из радиоизотопа, радионуклида, токсина, терапевтической группы и химиотерапевтической группы.
44. Способ лечения или предупреждения патологического состояния, обусловленного CD30L, у пациента, включающий введение пациенту, нуждающемуся в этом, эффективного количества по меньшей мере одного изолированного белка, связывающего антиген, по любому из воплощений 1-31 изобретения.
45. Способ по воплощению 44 изобретения, где изолированный антигенсвязывающий белок вводят отдельно или в виде комбинированной терапии.
46. Способ снижения активности CD30L у пациента, включающий введение эффективного количества по меньшей мере одного белка, связывающего антиген, по любому из воплощений 1-31 изобретения.
47. Белок, связывающий антиген, по любому из воплощений 1-31 изобретения, применяемый в способе лечения.
48. Белок, связывающий антиген, по любому из воплощений 1-31 изобретения для лечения воспалительных заболеваний.
49. Белок, связывающий антиген, по любому из воплощений 1-31 изобретения для лечения аутоиммунных заболеваний, таких как артрит (включая ревматоидный артрит (РА)) и системная красная волчанка (СКВ), воспалительные кишечные заболевания (ВКЗ), включая болезнь Крона и неспецифический язвенный колит.
Примеры
Пример 1
Получение антител к CD30L человека Для разработки человеческих моноклональных антител, распознающих рекомбинантный CD30L человека и ингибирующих его активность, использовали технологию XenoMouse™ (Amgen, Thousand Oaks, CA). Отобранные антитела обладали перекрестной реактивностью на CD30L человека и яванского макака, но не на CD30L мыши.
Супернатанты гибридом подвергали скринингу с помощью технологии флуориметрического объемного микроанализа (FMAT; от англ. "fluorometric microvolume assay technology") на связывание с рекомбинантным CD30L человека, экспрессируемым на клетках 293Т, а также подвергали скринингу с помощью анализа сортировки флуоресцентно-активированных клеток (FACS) на связывание с нативным CD30L человека на клетках Ramos. При анализе FMAT планшеты покрывают клетками 293Т, экспрессирующими CD30L, добавляют супернатант гибридомы, а затем добавляют второе антитело против lg человека для обнаружения с помощью стандартных методов ELISA. Отрицательные контроли представляли собой соответствующие нетрансфицированные клетки 293Т, не экспрессирующие CD30L.
Супернатанты очищали, и рекомбинантные антитела экспрессировали в виде конструкций IgG1 и IgG2. Антитела подвергали повторному скринингу на связывание с нативным CD30L человека и обезьяны яванского макака. Кратко, клетки лимфомы Беркитта Ramos (3×105/лунка) инкубировали в 96-луночных микротитрационных планшетах с круглым дном (Costar® 96-луночный планшет (Corning; Acton, MA)) с титрованием (конечная концентрация от 10 мкг/мл до 3,2 нг/мл) антител IgG против CD30L в блокирующем буфере (фосфатно-солевой буферный раствор (ФСБ), содержащий 0,02% азид натрия, 2% нормальную сыворотку козы и 1% нормальную сыворотку кролика). Слитый белок CD30. Fc человека включали в качестве положительного контроля на связывание. Клетки инкубировали в течение 30 минут при 4°С, затем отмывали 2 раза ФСБ и инкубировали в течение еще 30 минут при 4°С с 30 мкл/лунка антитела против IgG человека (hu-IgG; от англ. "human"), меченого биотином (1:50 в блокирующем буфере; Jackson Immunoresearch, West Grove, PA). После отмывки 2 раза ФСБ клетки инкубировали в течение 30 минут при 4°С с 30 мкл/лунка (разведение 1:50 в ФСБ) стрептавидина - фикоэритрина (РЕ; Molecular Probes, Eugene, OR). Клетки отмывали 2 раза ФСБ и готовили для анализа FACS. Результаты можно найти в таблице 5.
Активированные Т-клетки яванского макака получали путем выделения мононуклеарных клеток периферической крови (МКПК) из гепаринизированной крови обезьян яванских макак путем центрифугирования в градиенте плотности через Isolymph (2000 об/мин в течение 20 минут). МКПК ресуспендировали в 25 мл ФСБ, добавляли к 300 мкл 50% раствора эритроцитов овцы, обработанных 2-аминоэтилизотиоурония бромидом (АЕТ), и центрифугировали примерно при 700 об/мин при 4°С в течение 10 минут. Осажденные клетки инкубировали при 4°С в течение еще 20 минут, мягко ресуспендировали и центрифугировали через Isolymph. Супернатант удаляли до осадка Т-клеток/эритроцитов. Затем этот осадок ресуспендировали в 25 мл буфера для лизиса (0,85% хлорид аммония в воде) и инкубировали при 37°С в течение 10 минут. Полученные в результате Т-клетки осаждали и отмывали 2 раза в культуральной среде (среда RPMI-1640 с добавлением 10% фетальной бычьей сыворотки, инактивированной нагреванием), а затем культивировали (1×106/мл) при 37°С в течение 18 ч в присутствии 10 нг/мл форболмиристоацетата (ФМА; Sigma Chemicals, St Louis, MO) и 500 нг/мл иономицина (Calbiochem, San Francisco, CA). По окончании периода культивирования Т-клетки яванского макака считали, промывали один раз ФСБ и инкубировали с 2% сывороткой козы, 1% сывороткой кролика и 0,1% сывороткой человека в ФСБ в течение 30 минут при 4°С. Затем клетки окрашивали антителами против CD30L человека (huCD30L), как раскрыто выше для клеток Ramos человека. Результаты можно найти в таблице 5.
Антитела также подвергали скринингу на способность к блокированию связывания CD30L человека с растворимой молекулой CD30. Fc человека. Кратко, клетки Ramos человека или активированные Т-клетки яванского макака инкубировали с IgG против CD30L при 50 мкг/мл в блокирующем буфере в течение 30 минут при 4°С. Затем добавляли растворимый huCD30.Fc-His при конечной концентрации 2,5 мкг/мл (30 мкл/лунка) и инкубировали в течение 30 минут при 4°С. Клетки отмывали 2 раза ФСБ и инкубировали с 5 мкг/мл моноклонального антитела мыши против His (30 мкл/лунка в блокирующем буфере) и инкубировали в течение 30 минут при 4°С. После отмывки 2 раза ФСБ клетки инкубировали с IgG-PE против мыши (1:50 в ФСБ; BD Pharmingen, San Diego, СА) еще в течение 30 минут при 4°С. Затем клетки отмывали 2 раза ФСБ и готовили для анализа FACS. Результаты можно найти в таблице 5.
Пример 2
Функциональный скрининг антитела к CD30L
Антитела были охарактеризованы по их способности блокировать взаимодействия CD30L/CD30 с использованием двух анализов высвобождения ИЛ-8. В этих анализах используют линию клеток анапластической крупноклеточной лимфомы человека (ALCL) Karpas-299 (К299), экспресирующую эндогенный CD30 и высвобождающую ИЛ-8 в ответ на CD30L. Кратко, очищенные антитела IgG против CD30L человека инкубировали либо с рекомбинантным CD30L (анализ на одной линии клеток), либо с клетками Ramos, экспрессирующими нативный CD30L (анализ на двух линиях клеток), и культивировали с клетками К299. После определенного периода инкубации супернатанты клеток собирали и анализировали на содержание ИЛ-8, используя сэндвич-ELISA для определения ингибирования высвобождения ИЛ-8 из клеток К299 в ответ на CD30L антителами-кандидатами.
Анализ на одной линии клеток
Антитела IgG против CD30L титровали в разбавителе культуры (среда RPMI-1640 с добавлением 10% фетальной бычьей сыворотки, инактивированной нагреванием) и помещали в 96-луночные микротитрационные планшеты с круглым дном (96-луночный планшет Costar® (Corning; Acton, MA)) с получением диапазона конечных концентраций от 1 мкг/мл до 10 пг/мл. В качестве положительного контроля использовали препарат моноклонального антитела мыши против CD30L человека, как раскрыто в патенте US №5480981. Конечный объем разбавителя и IgG к CD30L составлял 100 мкл/лунка. Затем в каждую лунку добавляли 50 мкл рекомбинантного CD30L человека с мотивом изолейциновой молнии (huCD30L LZ). Конструкция huCD30L LZ состоит из внеклеточного домена CD30L человека (см. патент US №5480981), сшитого на его N-конце с мотивом изолейциновой молнии, модифицированного путем замены остатков лейцина остатками изолейцина. Анализ на одной линии клеток также использовали для определения перекрестной реактивности с CD30L яванского макака, используя CD30L яванского макака (супо CD30L LZ; от англ. "cynomolgus") вместо huCD30L LZ. Препарат cynoCD30L LZ получали, как описано для молекулы LZ человека.
Планшеты инкубировали в течение 45 минут при комнатной температуре. В каждую лунку добавляли 50 мкл клеток К299 (4×105 клеток/мл), затем планшеты инкубировали в течение 24 часов при 37°С, 5% СO2.
Количество ИЛ-8 в супернатантах клеток определяли с помощью сэндвич-ELISA, используя набор CDCL8/IL-8 ELISA DuoSet согласно инструкциям изготовителя (R&D Systems, Minneapolis, MN). Уровни ИЛ-8 интерполировали из стандартной кривой ELISA, используя программное обеспечение DeltaSoft 3 версия 2.2 (DeltaSoft, Inc. Hillsborough, NJ). Все результаты выражали как среднее ± стандартная ошибка среднего (SEM; от англ. "standard error of mean") для 3 повторов культур, вычисленного с помощью программного обеспечения GraphPad PRISM версия 4.01 (GraphPad Software Inc., San Diego, CA), и также определяли значения IC50. Значение IC50, выведенное с использованием huCD30L LZ для антитела А1 составляло 2700 пМ. Результаты с использованием cynoCD30L LZ можно найти в таблице 5.
Анализ на двух линиях клеток
Как раскрыто выше, антитела IgG против CD30L титровали в разбавителе культуры и помещали в 96-луночные микротитрационные планшеты с круглым дном для получения диапазона конечных концентраций от 1 мкг/мл до 10 пг/мл. Конечный объем разбавителя и IgG к CD30L составлял 100 мкл/лунка. В каждую лунку добавляли 50 мкл облученных (5000 рад.) клеток Ramos CD30L+ (4×106/мл, АТСС). Клетки Ramos подвергали сортировке FACS для селекции клеток, имеющих самый высокий уровень экспрессии CD30L на клеточной поверхности.
Планшеты инкубировали в течение 45 минут при комнатной температуре. В каждую лунку добавляли 50 мкл клеток К299 (4×105 клеток/мл). Планшеты инкубировали в течение 24 часов при 37°С, 5% СO2.
Количество ИЛ-8 в супернатантах культуры определяли с помощью сэндвич-ELISA, используя набор CDCL8/IL-8 ELISA DuoSet согласно инструкциям изготовителя (R&D Systems, Minneapolis, MN). Количество ИЛ-8 и значения IC50 вычисляли, как раскрыто выше, см. таблицу 5.
Производные антитела отбирали на основании их способности ингибировать CD30L-индуцированную индукцию ИЛ-8 по сравнению с контрольным моноклональным антителом мыши против CD30L человека.
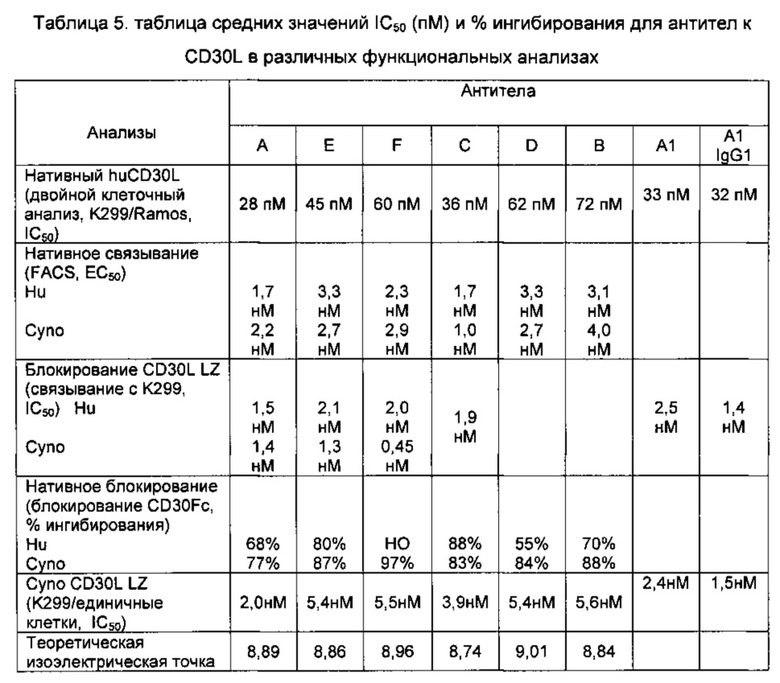
Пример 3
Биннинг антител к CD30L на основании конкурентного связывания
Количественное определение связывания проводили, используя анализ конкурентного связывания, в котором одно меченое антитело к CD30L конкурировало с избыточными количествами других немеченых антител к CD30L за связывание с rhCD30L. Антитела, конкурирующие друг с другом, определяли в одну и ту же позицию. Антитела A-F конкурировали друг с другом за связывание с rhCD30L.
Пример 4
Определение равновесной константы диссоциации на клетках (Kd) для антител к CD30L
Сродство связывания антител к CD30L с CD30L, экспрессируемым на клетках Ramos, определяли способом FACS Kd. Клетки Ramos культивировали в среде RMPI1640 с добавлением 10% фетальной бычьей сыворотки (ФБС), L-глутамина и 0,05% азида. Ставили две равновесные реакции, где антитела титровали 1:3 от 60 нМ до 338 фемтомоль (фМ), используя среду для культивирования клеток, и инкубировали с двумя различными постоянными концентрациями клеток (25000 и 250000 клеток). Планшеты герметично закрывали парафильмом и инкубировали при 37°С, 20 ч, встряхивание. Равновесные планшеты центрифугировали, и осадки клеток дважды промывали охлажденным во льду буфером FACS (1Х ФСБ Дульбекко (Д-ФСБ) с добавлением 2% ФБС). Затем клетки инкубировали со вторым антителом коза против Fc Су5 человека (Jackson Immuno Research, West Grove, PA) и с 7AAD (BD-Biosciences, Rockville, MD) в течение 1 ч на льду в темноте. После инкубации клеток дважды промывали охлажденным во льду буфером FACS, после чего проводили анализ проточной цитометрии. Значения сдвига геометрического среднего (GeoMean) FL4, открываемого популяцией живых клеток, является мерой связывания [Аb]. Затем вычисляли обратную величину GeoMean, отражающую теоретическую концентрацию свободного антигена [Аg] в равновесном растворе. Обратные величины считывания GeoMean использовали в программном обеспечении KinExA для анализа и вычисления Kd для каждого антитела. Измерение свободного [Аg] наносили на график против исходной концентрации титрующего компонента, и на основании этих графиков при двух различных концентрациях клеток значение Kd получали из соответствия кривой, используя анализ n-кривой в программном обеспечении KinExATM Pro. 95% доверительный интервал приведен в виде Kd низкой и Kd высокой.
Как видно в таблице 6, антитела могут обладать высоким сродством к CD30L, экспрессируемому на клетках Ramos. Все они имели значение Kd в низком пикомолярном диапазоне.
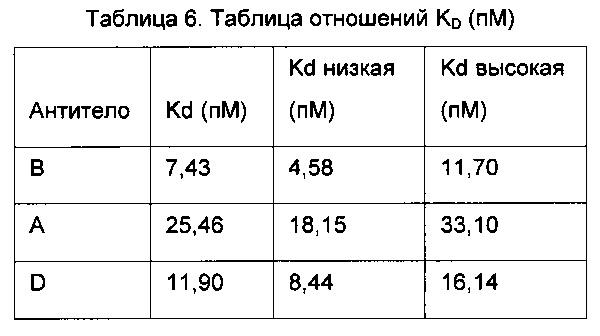

Пример 5
Анализ антителозависимой клеточной цитотоксичности (А3КЦ)
Вариабельные домены от А1 присоединяли к константному домену IgG1 человека (А1 IgG1). Данное антитело экспрессировали в линии клеток СНО с нокаутом по Fut-8, в результате чего получили афукозилированное антитело, А1 IgG1f.
Цитолиз клеток-мишеней, опосредованный антителом к CD30L, оценивали в анализе антителозависимой клеточной цитотоксичности (А3КЦ). Клетки-мишени Ramos (высокий уровень экспрессии CD30L), JD38 (средний уровень экспрессии CD30L), DS179 (низкий уровень экспрессии CD30L) и EW36 (нет экспрессии CD30L) собирали и ресуспендировали в 2×106 клеток/мл в среде cRPMI-10 (RPMI (Life Technologies, Carlsberg, CA) с добавлением 10% фетальной бычьей сыворотки). Добавляли кальцеин-АМ, 4 мМ в безводном ДМСО (Sigma-Aldrich, St. Louis, МО) до конечной концентрации 10 мкМ (разведение 1:400). Клетки инкубировали в течение 30 минут при 37°С с двумя последующими отмывками в ФСБ (1500 об/мин, 5 минут при 4°С каждая). Осадки ресуспендировали в cRPMI-10 и доводили до концентрации 0,2×106 клеток/мл и помещали на лед до использования в следующей стадии.
Антитело А1 IgG1 и антитело А1 IgG1f сначала добавляли в 96-луночные планшеты с U-образным дном (BD, Franklin Lakes, NJ) при 10-кратном серийном титровании, начиная с 10 мкг/мл. Ритуксимаб (Genentech, South San Francisco, CA) использовали в качестве положительного контроля антитела. Антитело А1 (IgG2) добавляли в качестве отрицательного контроля. Затем в каждую лунку добавляли клетки-мишени, меченые кальцеином-АМ, описанные выше (10000 клеток/лунка) и инкубировали с антителами в течение 20 минут при 37°С. После инкубации в каждую лунку добавляли 50 мкл естественных киллерных клеток (NK; от англ. "natural killers") при концентрации 4×106 клеток/мл и инкубировали в течение 4 часов при 37°С. Затем клетки NK осаждали (1500 об/мин, 5 минут при 4°С) и ресуспендировали в cRPMI-10, доводя концентрацию клеток до 4×106 клеток/мл.
После инкубации готовили контрольные лунки путем добавления 20 мкл/лунка 9% NP-40 (IGEPAL СА 630, Sigma-Aldrich) для стимуляции 100% лизиса. Во все остальные лунки добавляли 20 мкл/лунка только среды cRPMI-10. Планшеты центрифугировали при 800 об/мин в течение 4 минут при 4°С. Супернатант (150 мкл) отбирали и переносили в черный 96-луночный планшет с прозрачным дном (Corning, Lowell, MA) и считывали флуоресценцию на считывающем устройстве для планшетов (Molecular Devices, Spectra max GEMINI), возбуждение 485, испускание 535.
Процент максимального лизиса вычисляли как (значение образца - спонтанный лизис)/ (100% лизис - спонтанный лизис)*100.
Результаты цитолиза, опосредованного антителом CD30L, четырех клеток-мишеней представлены на фиг.1 (A-D). Антитело А1 IgG1 не опосредует цитолиз клеток на любой из клеток, экспрессирующих CD30L (см. фиг.1A-D). Антитело А1 IgG1f индуцировало клеточную гибель, как на клетках-мишенях с высокой экспрессией CD30L (см. фиг.1А), так и на клетках-мишенях со средней экспрессией CD30L (см. фиг.1В). Антитело А1 IgG1f опосредовало более высокий уровень истощения на клетках-мишенях с более высокой экспрессией CD30L, чем на клетках-мишенях со средними уровнями CD30L (см. фиг.1А и 1В). Антитело А1 IgG1f не лизируют клетки-мишени с низкой экспрессией (С) или без экспрессии (D) CD30L.
Пример 6
Эпитопное картирование с помощью HX-MS mAb против CD30L
Участки связывания CD30L с антителами против CD30L оценивали с помощью масс-спектрометрии водородного обмена (ВО-МС), чтобы идентифицировать участок или участки CD30L человека, вовлеченные в молекулярные взаимодействия с антителами A-F, раскрытыми в данном изобретении.
Материалы
hCD30L: Использовали два различных варианта внеклеточного домена hCD30L. His-hCD30L-EC содержит остатки 63-234 hCD30L, идентифицированные как SEQ ID NO: 74 и, как показывает название, белок продуцируется с N-концевой His-меткой (RnD Systems). FLAG-hCD30L-EC содержит остатки 66-234 SEQ ID NO: 74 и был получен из клеток млекопитающих с N-концевой меткой FLAG-His.
кДНК внеклеточного домена CD30L человека приобретали у фирмы Origene. Метку FLAG-His6 и сайт расщепления протеазы TEV сливали с N-концом кодирующей последовательности для аминокислот 66-234 с помощью двухступенчатой ПЦР с перекрывающимися праймерами и клонировали между сайтами EcoRI и BamHI плазмиды pJSV001 в одной рамке считывания с сигнальным пептидом CD33.
FLAG-hCD30L-EC, используемый ниже, включает точечную мутацию N153Q, запрещающую N-гликозилирование. Мутант N153Q был получен с помощью набора для быстрого точечного мутагенеза Qiagen quick-change point mutagenesis Kit (QIAGEN).
Клетки HEK293 6E использовали для продуцирования белка. Клетки культивировали в среде Freestyle™ (Gibco) с добавлением 25 мкг/мл генетицина 418 (Gibco) и 0,1% F-68 (Gibco). На сутки, трансфекции клетки поддерживали при плотности 1×106 клеток/мл и трансфицировали 293 Fectin (Invitrogen) согласно инструкции изготовителя. Через 24 ч после трансфекции в культуру добавляли пептон (Promega) до конечной концентрации 0,5%. Клетки продолжали выращивать в течение 5 суток, после чего собирали супернатант.
Супернатант культуры собирали центрифугированием (15000 об/мин×20 мин, 4°С), а затем осветляли фильтрованием через мембрану из нитрата целлюлозы 0,22 мкм. Осветленный супернатант наносили на колонку с Ni-хелатированной сефарозой ХК60 (20 мл) (GE Healthcare) с последующим промыванием 10 объемами колонки 20 мМ NaH2PO4 рН 7,4, 0,5 M NaCl. Затем белок элюировали 5 объемами колонки 20 мМ NaH2PO4 рН 7,4, 0,5 M NaCl, 0,5 M имидазол. Элюированные белки объединяли, концентрировали примерно до 5 мл с помощью устройств для центрифугирования Amicon ultra 15 (10000 Да MWCO, Millipore) и наносили на колонку Hiload 26/60 Superdex 200 318 мл (GE Healthcare) для удаления агрегатов и обмена буфера на ФСБ. После концентрирования конечные концентрации белка определяли путем измерения поглощения при 280 нм с помощью спектрометра NANODROP UV. Чистоту белка оценивали с помощью электрофореза с полиакриламидном геле с додецилсульфатом натрия (ДСН-ПААГ).
Дегликозилирования проводили в течение приблизительно 16 ч при 37°С, используя 1 ед. пептид-N-гликозидазы (ПНГазы) F (Roche) на 5 мкг hCD30L. Дегликозилированный белок хранили при 4°С и использовали для экспериментов в пределах одной недели. Все белки подвергали обмену буфера в ФСБ рН 7,4 перед экспериментами ВО-МС.
Эксперименты ВО-МС
Приборы и регистрация данных
Эксперименты ВО проводили на системе сверхэффективной жидкостной хроматографии (СВЭЖХ) nanoACQUITY с помощью технологии HDX (Waters Inc.), соединенной с масс-спектрометром Synapt G2 (Waters Inc.). Система Waters HDX содержала робот Leap (H/D-x PAL; Waters Inc.), управляемый программным обеспечением LeapShell (Leap Technologies Inc/Waters Inc.), выполняющий инициацию реакции дейтериевого обмена, контроль времени реакции, гашение реакции, впрыск на систему СВЭЖХ и контроль времени расщепления. Робот Leap был оборудован двумя блоками с температурным контролем, поддерживаемыми при 20°С для хранения буфера и реакций НХ и поддерживаемыми при 2°С для хранения белка и раствора для гашения, соответственно. Система Waters HDX дополнительно включает термостат, поддерживающий температуру предварительной колонки и аналитической колонки, а также трубок и переключающих клапанов для жидкостной хроматографии при 1°С. Отдельный термостат поддерживает колонку с пепсином при 25°С. Для расщепления пепсином на одной линии наносили 100 мкл погашенного образца, содержащего 300 пмоль hCD30L, и пропускали через картридж с иммобилизованным пепсином Poroszyme® Immobilized Pepsin Cartridge (2,1×30 мм (Applied Biosystems)), помещенный при 25°С, используя изократическую скорость тока 100 мкл/мин (0,1% муравьиная кислота: CH3CN 95:5). Полученные в результате пептиды улавливали и обессоливали на предварительной колонке VanGuard ВЕН С18 1,7 мкм (2,1×5 мм (Waters Inc.)). Затем клапаны переключали, чтобы подключить предварительную колонку на одной линии с аналитической колонкой, UPLC-BEH С18 1,7 мкм (1×100 мм (Waters Inc.)), и пептиды разделяли, используя 9-минутный градиент 10-50% В, подаваемый со скоростью 40 мкл/мин из системы ВЭЖХ nanoAQUITY (Waters Inc.). Подвижные фазы состояли из раствора А: 0,1% муравьиная кислота и В: 0,1% муравьиная кислота в CH3CN. Данные масс-спектрометрии с ионизацией электрораспылением (ИЭР-МС) и отдельных экспериментов высокой энергии (МСЕ) снимали в режиме положительного иона, используя масс-спектрометр Synapt G2 (Waters Inc.). Лейцин-энкефалин использовали в качестве иона фиксированной массы ([М+Н]+ при m/z 556,2771), и данные снимали в непрерывном режиме (дополнительное описание см. в Andersen and Faber, Int. J. Mass Spec., 302, 139-148 (2011)).
Анализ данных
Пепсиновые пептиды идентифицировали в отдельных экспериментах, используя стандартные способы МСЕ, где пептиды и фрагменты дополнительно выравнивали, используя свойства подвижности ионов Synapt G2 (Waters Inc.). Данные МСЕ обрабатывали, используя программу ProteinLynx Global Server версия 2.5 (Waters Inc.). Необработанные файлы данных ВО-МС обрабатывали в программе DynamX 2.0 (Waters Inc.). Программа DynamX автоматически производит коррекцию фиксированной массы и определение включения дейтерия, то есть центроидное определение дейтерированных пептидов. Кроме того, все пептиды проверяли вручную, чтобы гарантировать правильное назначение пика и дейтерирования программным обеспечением.
Эксперимент по эпитопному картированию
Обмен амидного водорода/дейтерия (ВО) инициировали 6-кратным разведением hCD30L в присутствии или в отсутствие mAb А, В, С, D, Ε или F в соответствующем дейтерированном буфере (то есть ФСБ, приготовленном в D20, 96% конечного D2O, рН 7,4 (нескорректированное значение)). Все реакции НХ проводили при 20°С, и они содержали 6 мкМ hCD30L (использовали мономерную Mw) в отсутствие или в присутствии 6,6 мкМ mAb с получением, таким образом, 2,2-кратного молярного избытка mAb. Через интервалы времени 0,25, 0,5, 1, 3 и 10 минут 50 мкл аликвоты реакции ВО гасили 50 мкл охлажденного во льду буфера гашения (1,35 M трихлорэтилфосфат (ТСЕР; от англ. "trichloroethylphosphate")) с получением в результате конечного значения рН 2,5 (нескорректированное значение).
Результаты
Белки hCD30L
Присутствие гликозилирований в hCD30L затрудняло бы анализ ВО-МС. В связи с неизвестной массой и часто гетерогенной природой гликозилирований белков полученные в результате пепсиновые пептиды невозможно идентифицировать и проанализировать в масс-спектре (МС). Тем не менее, удаление гликозилирований из hCD30L создало бы хорошо определенную аминокислотную последовательность, подходящую для анализа ВО-МС, поскольку ее можно идентифицировать и анализировать в МС.
Внеклеточный домен hCD30L содержит пять потенциальных сайтов N-гликозилирования, перечисленных в первой колонке таблицы 7 ниже. Удаление гликозилирований было предпринято и путем обработки ПНГазой F, и путем точечной мутации с получением белка, более подходящего для анализа ВО-МС.
Анализы с помощью МС и эксклюзионной хроматографии с детектированием рассеивания лазерного излучения с кратными углами (SEC-MALS; от англ. "Size-Exclusion Chromatography Combined with Multiangle Laser Light Scattering") His-hCD30L-EC показали, что четыре из пяти потенциальных сайтов N-гликозилирования действительно гликозилированы (таблица 7; данные не представлены). При обработке данного белка ПНГазой F в нативных условиях только два из гликозилирований можно удалить ферментативным путем с помощью ПНГазы F и, таким образом, два N-гликозилирования остается на белке.
При обработке FLAG-hCD30L-EC ПНГазой F в нативных условиях как МС, так и SEC-MALS продемонстрировали, что данный белок может быть полностью дегликозилирован. В заключение, мутация N153Q запрещает сайт N-гликозилирования, который не может быть ферментативно удален из His-hCD30L-ЕС, и данный белок считали оптимальным для ВО-МС. Обзор оценки сайта (сайтов) гликозилирования включен в таблицу 7.
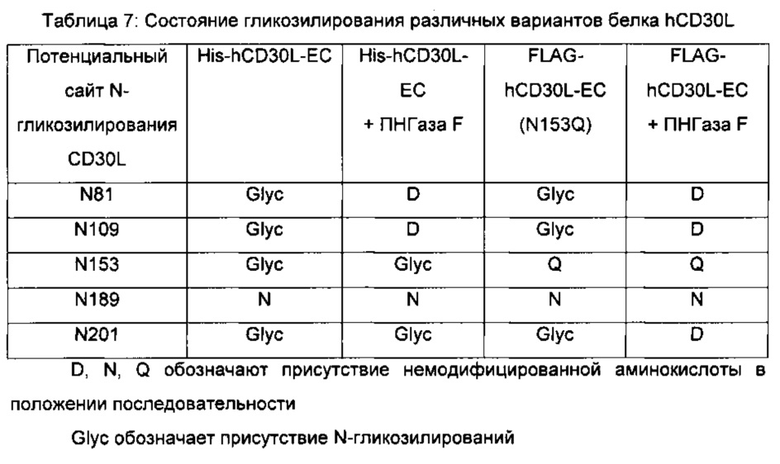
Анализ ВО-МС
После реакции дейтериевого обмена hCD30L-EC расщепляют пепсином с получением пепсиновых пептидных участков, описанных в таблице 8. нумерация остатков hCD30L следует SEQ ID NO 74, тем не менее, N-концевая метка FLAG-His DYKDDDDKHHHHHHENLYFQG не является частью последовательности hCD30L Мониторинг хода ВО 40 пепсиновых пептидов, покрывающих 83% первичной структуры FLAG-hCD30L-EC, проводили в отсутствие или в присутствии mAb А, В, С, D, Ε или F, и данные включены в таблицу 8.
Остальную последовательность, не проанализированную в настоящем исследовании, покрывали пепсиновыми пептидами; тем не менее, интенсивность сигнала была недостаточна для анализа данных. Наблюдаемый профиль обмена в ранние моменты времени (менее 10 мин) в присутствии или в отсутствие mAb А1, В, С, D, Ε или F может быть разделен на две различные группы: одна группа пепсиновых пептидов hCD30L-EC проявляет профиль обмена, на который не влияет связывание mAb. Напротив, другая группа пептидов hCD30L показывает защиту от обмена при связывании mAb (таблица 8). В случае перекрывания пепсиновых пептидов была предпринята сублокализация информации по защите от обмена в определенных остатках или участках внутри пептида, учитывая полный обратный обмен N-конца пептида и первой пептидной связи. Защита от обмена в пептиде является показателем того, что данный участок вовлечен в связывание mAb. Таким образом, эпитоп частично или полностью локализован в пределах участка, определенного специфичными пептидами. Тем не менее, поскольку разделение ВО-МС основано на расщеплении пепсином дейтерированного белка, защита от обмена в пределах данного участка не подразумевает, что каждый остаток в пределах участка, определенного пепсиновыми пептидами, необязательно вовлечен в связывание mAb.
Эпитопное картирование mAb А1
Эпитоп mAb А1 был картирован в четырех различных экспериментах, используя FLAG-hCD30L-EC три раза, и используя His-hCD30L-EC один раз. Оба белка обрабатывали ПНГазой F перед экспериментами ВО-МС. Результаты этих исследований были в высокой степени сходными независимо от используемого белка hCD30L, и из всех экспериментов была получена одна и та же информация об эпитопах. Тем не менее, покрытие последовательности было явно ниже в эксперименте His-hCD30L-EC вследствие отсутствия обнаружимых пептидов в гликозилированных участках.
Эпитопный сигнал для mAb А1 наблюдали в N-концевом участке hCD30L (таблица 8) плоть до остатка Cys88. Защита от обмена становилась сильнее и дольше для пептидов, удлиняющихся от стартовой точки (остатки 45, 62 или 66), что, таким образом, указывает на защиту от обмена в сторону С-концевого участка пептидов, а также в остатке 88. Напротив, пептиды 45-65, 45-81 и 66-81 не проявляли защиту от обмена. Кроме того, остаток D81 представляет собой сайт гликозилирования, где гликозилированный N становится D под действием ПНГазы F. Таким образом, N/D81, вероятно, не вовлечен в связывание mAb. Следовательно, эпитопный сигнал, по-видимому, возникает от остатков в пределах участка 82-88 CSEDLLC.
Эпитопный сигнал для mAb А1 также наблюдали в С-концевом участке hCD30L (таблица 8) в шести различных пептидах, покрывающих область от F211 до S226. На основании уровня наблюдаемой защиты от обмена все участки данной области, по-видимому, вовлечены в связывание или подвергаются влиянию связывания mAb А в большей или меньшей степени. Области, связываемые антителом, отмечены полужирным шрифтом в таблице 8.
В заключение, обнаружено, что участками hCD30L, вовлеченными в связывание mAb А, являются участок 82-88 CSEDLLC и участок 211-226 FQYIDTSTFPLENVLS.
Эпитопное картирование mAb В, С, D, Е и F
Эпитопы mAb В, С, D, Е и F картировали, используя FLAG-hCD30L-EC, обработанный ПНГазой F перед экспериментами ВО-МС.Результаты этих исследований эпитопов были в высокой степени сходными с mAb А (таблица 8) также в отношении величины защиты от обмена. Эпитопное картирование на mAb Е проводили два раза, тогда как А, В, С и D каждое картировали один раз.

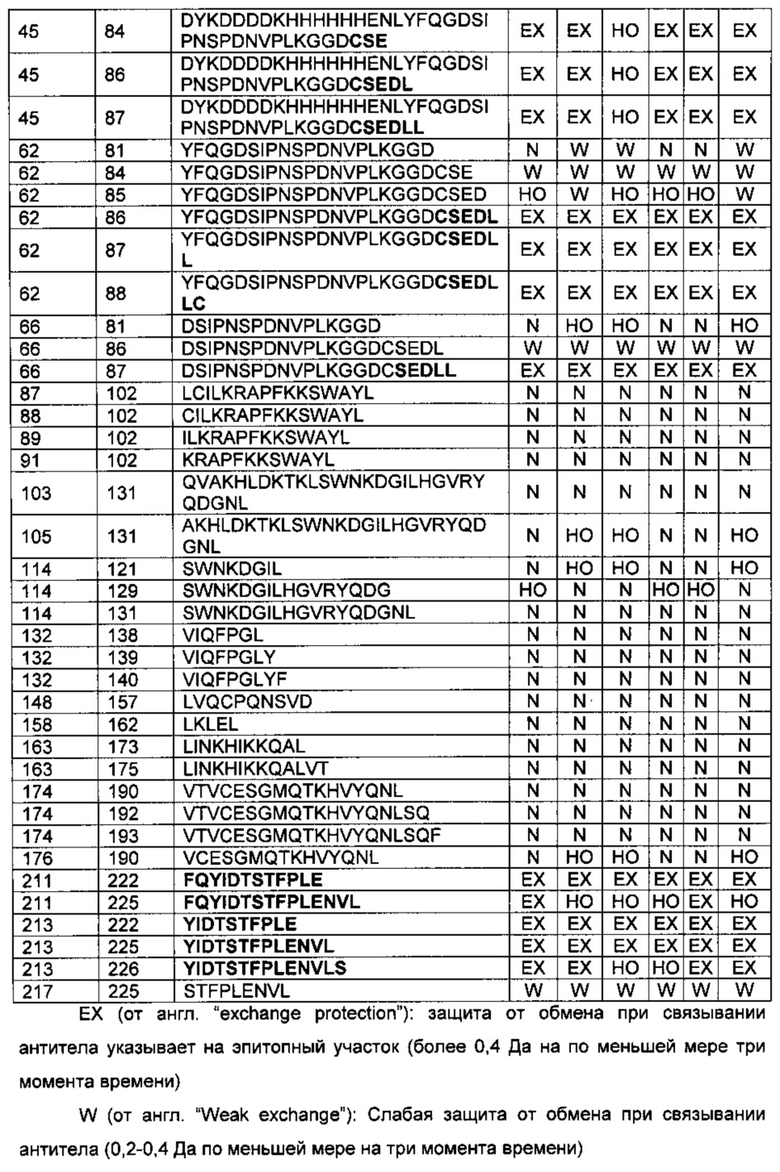

В заключение, участки hCD30L, вовлеченные в связывание mAb В, С, D, Е и F, подобны участкам, вовлеченным в связывание mAb А1, и обнаружены как участок 82-88 CSEDLLC и участок 211-226 FQYIDTSTFPLENVLS.
Пример 7
Биннинг антител к CD30L, используя иммобилизованные антитела против CD30L А-FAB
Моноклональные антитела против CD30L человека подвергали биннингу эпитопов на Biacore Т200 (GE Healthcare 28-9750-01), используя классический сэндвич-метод.
Для уменьшения стерического затруднения иммобилизованное антитело использовали в моновалентном формате. Чтобы определить, связывает ли антигенсвязывающий участок антител против hCD30L перекрывающиеся эпитопы, тестировали способность антител A-F к связыванию CD30L, уже связанного Fab антитела А1.
Fab фрагменты могут быть получены путем расщепления полноразмерных антител папаином, который расщепляет выше цистеинов шарнирного участка, или рекомбинантной экспрессии вариабельной области тяжелой цепи и вариабельной области легкой цепи. Fab фрагмент анти-hCD30L А1 (A1-FAB) был получен с использованием иммобилизованного папаина в присутствии цистеина для ферментативного расщепления IgG, находящегося непосредственно выше шарнирного участка, с получением двух Fab фрагментов и одного Fc фрагмента на молекулу антитела. Fab отделяли от Fc, используя аффинную хроматографию на белке А, а затем обмен буфера на ФСБ. Эту операцию проводили в соответствии с инструкциями изготовителя (Pierce Fab Preparation Kit #44985, US).
FAB против CD30L человека (Fab антитела A1) иммобилизовали, и на нем иммобилизовали тример CD30L человека-HiS10 (R&D Systems 1028-CL). Затем впрыскивали растворимые антитела A-F против hCD30L. Связывание второго антитела подразумевает, что два антитела находятся в различных эпитопных бинах.
A1-FAB против hCD30L разводили до 40 мкг/мл в 10 мМ, рН 5,0 и подвергали аминному сочетанию с чипом СМ5 (GE Healthcare BR-1000-12, BR-1000-50) до уровня 3566 условных единиц (УЕ), используя стандартные методы. Тример CD30L человека-His10 (R&D Systems 1028-CL) разводили до 10 нМ в HBS-P+ (GE Healthcare BR-1006-71) и впрыскивали над поверхностью A-FAB до иммобилизации примерно 175 УЕ CD30L. Второе антитело впрыскивали при 75 нМ над иммобилизованным hCD30L Контроли включали нулевую иммобилизацию (только буфер) и впрыскивания буфера вместо растворимого антитела. Все эксперименты проводили при 25°С.
Покрытие hCD30L A-FAB блокировало связывание подгруппы антител A-F. Оба контроля с использованием A-Fab и А были блокированы, как ожидали. Кроме того, связывание антител F и В с иммобилизованным hCD30L было также предотвращено, тогда как антитела Е, С и D были способны к связыванию hCD30L что демонстрирует, что их связывающий эпитоп можно отличить от связывающего эпитопа А, В и F. В заключение, антитела A, F и В конкурируют с Fab антитела А1 за связывание hCD30L.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ОТБОРА И ПОЛУЧЕНИЯ ВЫСОКОСЕЛЕКТИВНЫХ И МУЛЬТИСПЕЦИФИЧНЫХ НАЦЕЛИВАЮЩИХ ГРУПП С ЗАДАННЫМИ СВОЙСТВАМИ, ВКЛЮЧАЮЩИХ ПО МЕНЬШЕЙ МЕРЕ ДВЕ РАЗЛИЧНЫЕ СВЯЗЫВАЮЩИЕ ГРУППИРОВКИ, И ИХ ПРИМЕНЕНИЯ | 2013 |
|
RU2639287C2 |
| ВЫСОКОАФФИННЫЕ АНТИТЕЛА К CD3 И СПОСОБЫ ИХ ПОЛУЧЕНИЯ И ПРИМЕНЕНИЯ | 2020 |
|
RU2831840C2 |
| АНТИТЕЛА ПРОТИВ NGF И СВЯЗАННЫЕ С НИМИ СПОСОБЫ | 2019 |
|
RU2838175C2 |
| АНТИТЕЛО ПРОТИВ PDL-1, ЕГО ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ И ПРИМЕНЕНИЕ | 2017 |
|
RU2693661C2 |
| КАНИНИЗИРОВАННЫЕ МЫШИНЫЕ АНТИТЕЛА К ЧЕЛОВЕЧЕСКОМУ PD-1 | 2014 |
|
RU2676158C1 |
| ОДНОВАЛЕНТНЫЕ МОДУЛИ-ПЕРЕНОСЧИКИ ЧЕРЕЗ ГЕМАТОЭНЦЕФАЛИЧЕСКИЙ БАРЬЕР | 2014 |
|
RU2694659C2 |
| ОДНОВАЛЕНТНЫЕ МОДУЛИ-ПЕРЕНОСЧИКИ ЧЕРЕЗ ГЕМАТОЭНЦЕФАЛИТИЧЕСКИЙ БАРЬЕР | 2014 |
|
RU2799436C1 |
| АНТИГЕНСВЯЗЫВАЮЩИЙ БЕЛОК И ЕГО ПРИМЕНЕНИЕ В КАЧЕСТВЕ ПРОДУКТА ДЛЯ АДРЕСНОЙ ДОСТАВКИ ПРИ ЛЕЧЕНИИ РАКА | 2012 |
|
RU2659094C2 |
| ИММУНОТЕРАПИЯ С ПРИМЕНЕНИЕМ АНТИТЕЛ, СВЯЗЫВАЮЩИХ БЕЛОК 1 ПРОГРАММИРУЕМОЙ СМЕРТИ КЛЕТОК (PD-1) | 2017 |
|
RU2768404C2 |
| Иммунотерапия с применением антител, связывающих лиганд 1 белка программируемой смерти клеток (PD-L1) | 2017 |
|
RU2766582C2 |
































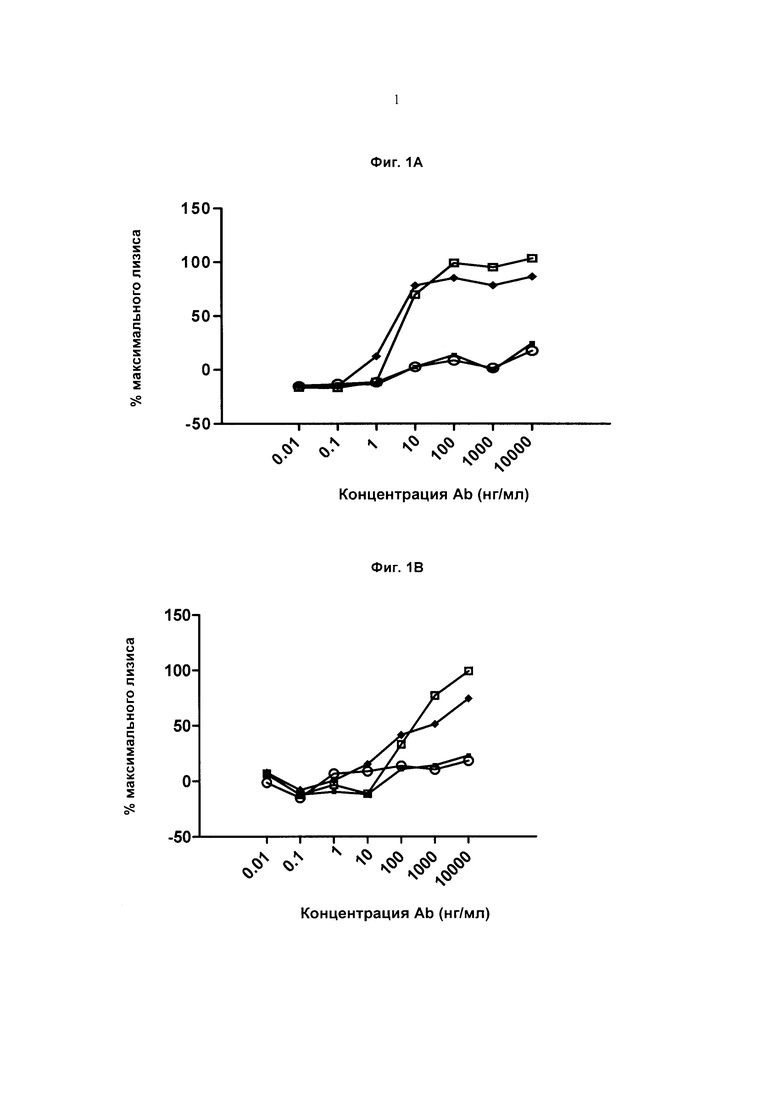
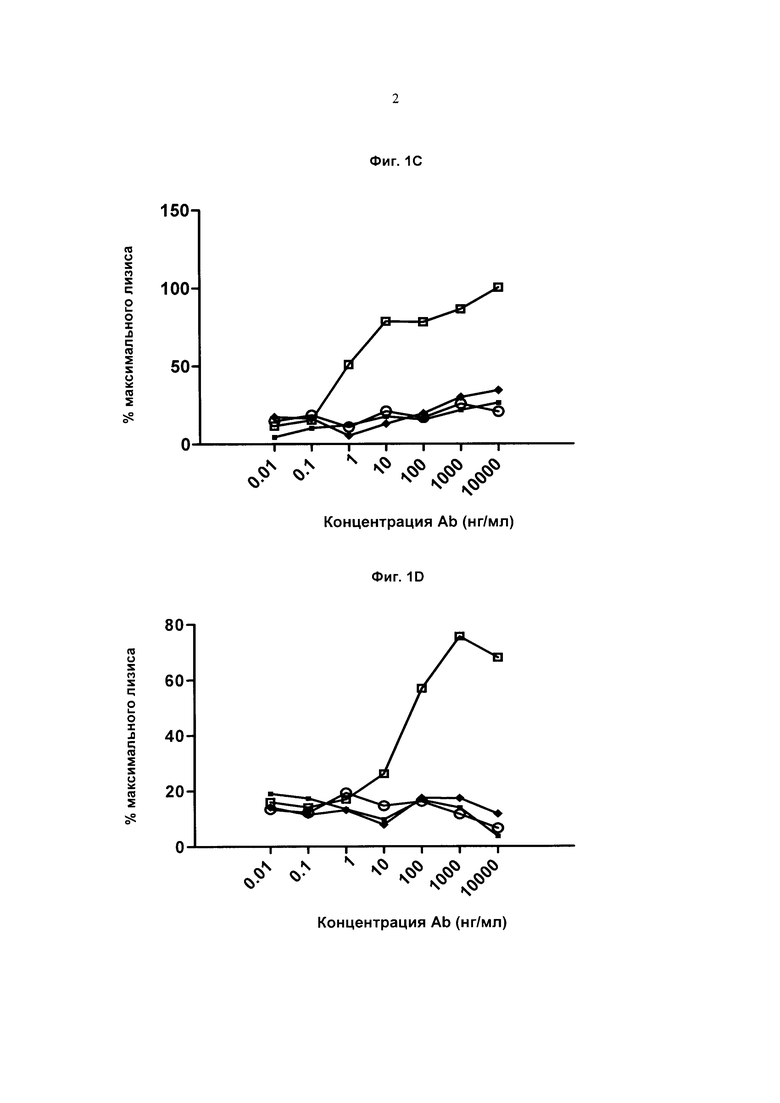
Настоящее изобретение относится к иммунологии. Предложены изолированные антитела и их антигенсвязывающие фрагменты, связывающие CD30L (CD153) и охарактеризованные аминокислотными последовательностями гипервариабельных участков (CDR) и вариабельных доменов. Рассмотрено применение антител и антигенсвязывающих фрагментов для снижения активности CD30L у пациента, а также для лечения воспалительных заболеваний, таких как аутоиммунные заболевания. Описана фармацевтическая композиция, а также способ лечения или предупреждения патологического состояния, обусловленного CD30L, и способ снижения активности CD30L у пациента. Кроме того, предложена изолированная нуклеиновая кислота, кодирующая антитело или антигенсвязывающий фрагмент, и содержащая ее клетка-хозяин. Антитела и антигенсвязывающие фрагменты по настоящему изобретению обеспечивают ингибирование взаимодействия CD30L с CD30, в связи с чем могут найти дальнейшее применение в терапии. 10 н. и 7 з.п. ф-лы, 1 ил., 8 табл., 7 пр.
1. Изолированное антитело или его антигенсвязывающий фрагмент, который связывает CD30L, где антитело или его антигенсвязывающий фрагмент включает участки CDRH1, CDRH2 и CDRH3 и CDRL1, CDRL2 и CDRL3, выбранные из группы, состоящей из:
a) CDRH1, CDRH2 и CDRH3, идентифицированные как SEQ ID NO: 14, 19, 25, и CDRL1, CDRL2 и CDRL3, идентифицированные как SEQ ID NO: 1, 7 и 11,
b) CDRH1, CDRH2 и CDRH3, идентифицированные как SEQ ID NO: 15, 21, 26, и CDRL1, CDRL2 и CDRL3, идентифицированные как SEQ ID NO: 2, 8 и 12,
c) CDRH1, CDRH2 и CDRH3, идентифицированные как SEQ ID NO: 15, 21, 26, и CDRL1, CDRL2 и CDRL3, идентифицированные как SEQ ID NO: 3, 8 и 12,
d) CDRH1, CDRH2 и CDRH3, идентифицированные как SEQ ID NO: 16, 22, 27, и CDRL1, CDRL2 и CDRL3, идентифицированные как SEQ ID NO: 4, 9 и 12,
e) CDRH1, CDRH2 и CDRH3, идентифицированные как SEQ ID NO: 17, 23, 28, и CDRL1, CDRL2 и CDRL3, идентифицированные как SEQ ID NO: 5, 8 и 13, или
f) CDRH1, CDRH2 и CDRH3, идентифицированные как SEQ ID NO: 18, 24, 29, и CDRL1, CDRL2 и CDRL3, идентифицированные как SEQ ID NO: 6, 10 и 12.
2. Изолированное антитело или его антигенсвязывающий фрагмент, который связывает CD30L, где антитело или его антигенсвязывающий фрагмент выбран из группы, состоящей из:
a) антитела, включающего вариабельную область тяжелой цепи SEQ ID NO: 48 и вариабельную область легкой цепи SEQ ID NO: 36;
b) антитела, включающего вариабельную область тяжелой цепи SEQ ID NO: 64 и вариабельную область легкой цепи SEQ ID NO: 38;
c) антитела, включающего вариабельную область тяжелой цепи SEQ ID NO: 66 и вариабельную область легкой цепи SEQ ID NO: 40;
d) антитела, включающего вариабельную область тяжелой цепи SEQ ID NO: 68 и вариабельную область легкой цепи SEQ ID NO: 42;
e) антитела, включающего вариабельную область тяжелой цепи SEQ ID NO: 70 и вариабельную область легкой цепи SEQ ID NO: 44; и
f) антитела, включающего вариабельную область тяжелой цепи SEQ ID NO: 72 и вариабельную область легкой цепи SEQ ID NO: 46.
3. Изолированное антитело или его антигенсвязывающий фрагмент по п. 1, где CDRH1, CDRH2 и CDRH3 идентифицированы как SEQ ID NO: 14, 19, 25 соответственно, a CDRL1, CDRL2 и CDRL3 идентифицированы как SEQ ID NO: 1, 7 и 11 соответственно.
4. Изолированное антитело или его антигенсвязывающий фрагмент по любому из предшествующих пунктов, включающий вариабельную область тяжелой цепи SEQ ID NO: 48 и вариабельную область легкой цепи SEQ ID NO: 36.
5. Изолированное антитело или его антигенсвязывающий фрагмент по п. 3, где антитело или его антигенсвязывающий фрагмент обладает по меньшей мере одним свойством, выбранным из группы, состоящей из следующих свойств:
a) ингибирование взаимодействия CD30/CD30L;
b) ингибирование CD30L-индуцированной индукции ИЛ-8;
c) перекрестная конкуренция с любым из антител A-F за связывание с CD30L человека;
d) константа диссоциации с CD30L человека, составляющая максимум 70 пМ.
6. Изолированное антитело или его антигенсвязывающий фрагмент по п. 3, имеющий константу диссоциации (KD) с CD30L человека, составляющую максимум 50 пМ.
7. Изолированное антитело или его антигенсвязывающий фрагмент по любому из пп. 1-6, где антигенсвязывающий белок относится к IgG1-, IgG2- IgG3- или IgG4-типу.
8. Изолированное антитело или его антигенсвязывающий фрагмент по любому из пп. 1-7 для применения в способе лечения.
9. Изолированное антитело или его антигенсвязывающий фрагмент по любому из пп. 1-7 для применения в способе лечения воспалительных заболеваний, таких как аутоиммунные заболевания.
10. Фармацевтическая композиция для лечения состояний, обусловленных CD30L, содержащая эффективное количество по меньшей мере одного антитела или его антигенсвязывающего фрагмента по любому из пп. 1-7 и фармацевтически приемлемый эксципиент.
11. Способ лечения или предупреждения патологического состояния, обусловленного CD30L, у пациента, включающий введение пациенту, нуждающемуся в этом, эффективного количества по меньшей мере одного изолированного антитела или его антигенсвязывающего фрагмента по любому из пп. 1-7.
12. Способ снижения активности CD30L у пациента, включающий введение эффективного количества по меньшей мере одного изолированного антитела или его антигенсвязывающего фрагмента по любому из пп. 1-7.
13. Применение изолированного антитела или его антигенсвязывающего фрагмента по любому из пп. 1-7 для лечения или профилактики состояния, связанного с CD30L, у пациента.
14. Применение изолированного антитела или его антигенсвязывающего фрагмента по любому из пп. 1-7 для снижения активности CD30L у пациента.
15. Применение изолированного антитела или его антигенсвязывающего фрагмента по любому из пп. 1-7 для лечения воспалительных заболеваний, таких как аутоиммунные заболевания.
16. Изолированная молекула нуклеиновой кислоты, кодирующая антитело или его антигенсвязывающий фрагмент по любому из пп. 1-7.
17. Клетка-хозяин, содержащая молекулу нуклеиновой кислоты по п. 16, для экспрессии антитела или его антигенсвязывающего фрагмента по п. 1.
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |
| US 5677430 A, 14.10.1997 | |||
| FRANKE ANJA C | |||
| et al., "Characterization of the CD30L binding domain on the human CD30 molecule using anti-CD30 antibodies." Hybridoma, 2000, 19(1): 43-48 | |||
| BLAZAR B.R | |||
| et al., "CD30/CD30 ligand (CD153) interaction regulates CD4+ T cell-mediated graft-versus-host disease." The Journal of Immunology, 2004; 173(5): 2933-2941 | |||
| GRUSS HANS-JURGEN et al., "CD30 ligand expression in nonmalignant and Hodgkin's disease-involved lymphoid tissues." The American journal of pathology, 1996; 149(2): 469-481. | |||

