ПЕРЕКРЕСТНАЯ ССЫЛКА НА РОДСТВЕННЫЕ ЗАЯВКИ
В настоящей заявке, поданной 22 февраля 2013 как Международная патентная заявка PCT, испрашивается приоритет предварительной заявки на патент США No. 61/603231, поданной 24 февраля 2012, содержание которой во всей своей полноте вводится в настоящее описание посредством ссылки.
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
Настоящая заявка включает список последовательностей, представленных в электронном формате в виде текстового файла, под именем «Список последовательностей 54428.0006WOU1_ST25», который был создан 22 февраля 2013 и имеет размер 174 килобайт (KB). Содержание текстового файла «Список последовательностей 54428.0006WOU1_ST25» вводится в настоящее описание посредством ссылки.
ПРЕДШЕСТВУЮЩИЙ УРОВЕНЬ ТЕХНИКИ
Область, к которой относится изобретение
В общих чертах, настоящее изобретение относится к лечению генетических заболеваний. Более конкретно, настоящее изобретение относится к композициям на основе эндонуклеаз и к способам, включающим композиции на основе хоминг-эндонуклеаз и на основе эндонуклеаз Cas9, а также к способам изменения уровней экспрессии генов глобина, где указанные композиции и способы могут быть применены для лечения талассемии, серповидно-клеточной болезни, и других гемоглобинопатий.
Описание прототипов
Гемоглобинопатии, такие как талассемия и серповидно-клеточная болезнь, представляют собой наиболее часто встречающиеся генетические заболевания крови, связанные с поражением эритроцитов и представляющие серьезную угрозу для здоровья населения во всем мире. Каждый год регистрируется свыше 1300000 случаев тяжелых расстройств, связанных с аномалиями структуры гемоглобина. Такими заболеваниями страдает 5% населения во всем мире, а коэффициенты рождаемости детей с клинически тяжелыми формами талассемии и серповидно-клеточной болезни (СКА) составляют 0,44 и 1,96 на тысячу человек соответственно.
У здорового млекопитающего, гемоглобины, присутствующие в эритроидных клетках, преимущественно состоят из гетеротетрамеров двух α-подобных цепей (полипептидов) и двух β-подобных цепей. Пять генов локуса β-глобина присутствуют в кластере на хромосоме 11. Эти гены экспрессируются в эритроидных клетках и на конкретной стадии развития индивидуума; при этом, гены ε, Αγ и Gγ и δ и β экспрессируются, главным образом, на стадиях развития эмбриона, плода и в постнатальной стадии, соответственно. Во время рождения, 95% β-подобных цепей представляют собой цепи γ, а остальные цепи представляют собой цепи β. На первом году жизни, такое соотношение постепенно меняется в обратную сторону, и этим можно объяснить, почему фенотипы заболеваний, ограниченных геном β-глобина, таких как серповидно-клеточная болезнь и большинство β-талассемий, не проявляются в течение нескольких месяцев. Уровни экспрессии α-подобных генов на хромосоме 16 отличаются друг от друга, эмбриональный ξ-ген экспрессируется одновременно с ε-геном, а двойные α-гены экспрессируются после развития плода. Таким образом, α-аномалии проявляется при внутриутробном развитии, что может приводить к тяжелым последствиям (например, к развитию водянки плода). Полученные α-, β-гетеротетрамеры экспрессируются на стадии развития, а именно на стадии развития эмбриона: Hb Gower1 (ξ2,ε2), Hb Gower2 (α2,ε2) и Hb Portland (ξ2,γ2); на стадии развития плода: HbF (Fetal) (α2,γ2) и у взрослых: HbA2 (α2,δ2) и HbA (взрослых) (α2,β2).
β-Талассемия обусловлена аномалиями в локусе β-глобина у взрослых, которые приводят к нарушению стехиометрического отношения β-подобных цепей глобина к α-подобным цепям и, тем самым, к преципитации неспаренных α-подобных цепей. Тяжесть талассемии непосредственно зависит от степени дисбаланса цепей глобина. В результате этого, поражения, опосредуемые несколькими путями, включая окисление клеточных и мембранных белков, приводят к неэффективному эритропоэзу, апоптозу и низкой выживаемостью эритроцитов. Было описано свыше 200 мутаций, ответственных за развитие β-талассемии.
Серповидно-клеточная болезнь вызывается одной нуклеотидной заменой в гене β-глобина, которая приводит к замене глутаминовой кислоты на валин в аминокислотном положении 6 пептида и к образованию βS. В отличие от нормального гемоглобина взрослых (HbA), гемоглобин S (α2,βS2), который имеет эту мутацию, обозначается HbS. В условиях низкой концентрации кислорода, HbS подвергается аллостерической модификации, и на этом уровне он может полимеризоваться. Дезокси-форма гемоглобина имеет гидрофобный «пэтч» на белке между спиралями E и F. Гидрофобный валин в положении 6 β-цепи гемоглобина образует гидрофобный «пэтч», который может ассоциироваться с гидрофобным «пэтчем» других молекул гемоглобина S, что может приводить к агрегации молекул гемоглобина S и к образованию нитевидного осадка, и, в свою очередь, к приобретению эритроцитами серповидной формы, а также к изменению многочисленных путей, которые вызывают поражение ткани в результате закупорки сосудов и гемолиза.
Хотя β-талассемия и серовидно-клеточная болезнь (СКА) представляют собой количественные и качественные нарушения в локусе β-глобина, соответственно, однако, экспрессия нормальных β-подобных генов глобина может приводить к ослаблению симптомов обоих заболеваний. В случае талассемии, любое снижение дисбаланса цепей глобина дает селективное преимущество для каждой клетки и позволяет достичь желаемого клинического результата. В случае серповидно-клеточной болезни, присутствие нормальных или некоторых мутантных β-подобных цепей может ослаблять клинический фенотип благодаря более эффективной конкуренции в пользу α-подобных цепей, а не мутантных цепей серповидных клеток, что приводит к снижению количества HbS в результате образования гемоглобинов (например, HbF), блокирующих полимеризацию Hbs, и, тем самым, к увеличению количества не-серповидного гемоглобина на клетку. Так, например, при серповидно-клеточной болезни, присутствие лишь 8% уровня фетального гемоглобина (HbF) приводит к ингибированию полимеризации HbS и, тем самым, к повышению выживаемости клеток, а 20% уровень HbF обеспечивает почти полную коррекцию фенотипа. Важно отметить, что потомство донорных эритроидных клеток, содержащих нормальный HbA, обладает существенным селективным преимуществом при трансплантации гемопоэтических стволовых клеток (HSCT) по сравнению с эндогенными клетками, содержащими HbS. У пациента, имеющего 11% донорных клеток в костном мозге, присутствует 35% донорных BFUe (эритроидных «взрывообразующих» единиц) и 73% донорных эритроцитов, и такому пациенту не требуется переливание крови. Таким образом, коррекция относительно небольшой фракции трансплантированных HSC дает нужный клинический эффект.
При талассемии в тяжелой форме необходимо непрерывное переливание крови, что приводит к избыточному поступлению железа в организм. Выживаемость непосредственно коррелирует с эффективностью лечения хелатообразующими соединениями, хотя эффективность такого лечения значительно ограничивается затратами на такое лечение, побочными эффектами и необходимостью строгого соблюдения режима и схемы лечения. Единственным лекарственным средством, разрешенным Управлением по контролю за качеством пищевых продуктов, медикаментов и косметических средств (FDA) для применения в терапии СКА, является гидроксимочевина, которая может приводить к ослаблению тяжести заболевания и к уменьшению случаев летальных исходов. Однако такое лечение является малоэффективным, и его назначение врачом плохо соблюдается пациентами и не дает адекватного протективного эффекта.
Трансплантация гемопоэтических клеток (HCT) представляет собой важную терапевтическую процедуру, которой каждый год подвергается несколько тысяч пациентов, страдающих злокачественными гематологическими заболеваниями и родственными расстройствами. В 2009 году, в Международном исследовательском центре по трансплантации крови и костного мозга (CIBMTR) было проведено приблизительно 60000 трансплантаций, то есть на 15000 трансплантаций в год больше, чем за последние десятилетия. Эффективность трансплантации также увеличивается, причем, полученные в последнее время результаты указывали на значительное снижение риска развития рецидивирующих и нерецидивирующих заболеваний и, в целом, на снижение смертности. Gooley et al., N. Engl. J. Med. 363:2091-101 (2011).
Трансплантация аллогенных гемопоэтических клеток (HCT), взятых от HLA-совместимых сибсов или неродственных доноров предлагается в качестве лечения пациентов с гемоглобинопатиями, но такое лечение имеет некоторые ограничения, связанные с необходимостью поиска подходящих родственных или неродственных доноров, и осложняется развитием реакции «трансплантат против хозяина» (GVHD) и инфекций. Кроме того, значительным ограничением является большое число случаев отторжения трансплантатов, которое наблюдается чаще, чем при HCT, применяемой для лечения злокачественных заболеваний. Альтернативные способы включают HCT с использованием донорных клеток крови пупочного канатика, поскольку доноры клеток крови пупочного канатика могут быть найдены почти для всех пациентов. Другими экспериментальными способами являются использование собственных гемопоэтических стволовых клеток пациента (HSC) и индуцирование экспрессии эндогенных генов глобина или добавление экзогенного β-подобного гена глобина.
Для многих пациентов, для которых невозможно найти донора, а в частности, для пациентов, составляющих этническое меньшинство или пациентов смешанной расы, самые большие надежды возлагают на лечение путем трансплантации клеток крови пупочного канатика (ПК). Источник донорных стволовых клеток (которые могут быть легко собраны при рождении ребенка без риска для матери или ребенка), то есть ПК, также имеет то преимущество, что он является легко доступным и безопасным при его использовании для HLA-несовместимых пациентов, поскольку такая трансплантация не приводит к увеличению риска развития GVHD.
К сожалению, некоторые факторы, включая низкую дозу клеток, имеющихся во многих элементах крови пупочного канатика, дают низкую приживляемость трансплантата, что повышает смертность при трансплантации у взрослых, а особенно у детей. Существенно замедленное восстановление гемопоэтических клеток, таких как нейтрофилы и тромбоциты, является известным фактором риска у реципиентов с трансплантатом клеток крови пупочного канатика (ТПК) и ассоциируется с низким общим числом ядросодержащих клеток (TNC) и CD34+-клеток, присутствующих в одинарном или двойном трансплантате ПК. Аналогичным образом, такое низкое число клеток коррелирует с более высоким риском отторжения трансплантата, и это в значительной степени относится к гемоглобинопатиям, при которых уже существует высокий риск отторжения трансплантата. Фактически, недавно проведенный анализ взрослых реципиентов с ТПК от одного донора продемонстрировал, что введенная доза CD34+-клеток является наиболее ценным прогностическим фактором приживления миелоидного трансплантата.
Случаи смертности при не-рецидивирующих заболеваниях (НРЗ) чаще всего наблюдаются у реципиентов с двойным ТПК (дТПК) по сравнению с реципиентами, которые являются неродственными реципиентами, совместимыми и не совместимыми с донорами. Brunstein et al., Blood 116:4693-9 (2010). Большинство случаев смертности при НРЗ наблюдается в первые 100 дней после трансплантации, причем, наиболее распространенной причиной летального исхода является инфекция. Важно отметить, что анализ факторов риска летального исхода при НРЗ у реципиентов с дТПК выявил повышенный риск у пациентов с замедленным восстановлением миелоидных клеток (время до достижения абсолютного числа нейтрофилов (ANC) >500/мл), то есть если такое восстановление происходит за период ≥26 дней, соответствующий среднему времени приживления трансплантата у реципиентов с дТПК. Однако если анализ факторов риска развития НРЗ был проведен только у реципиентов с дТПК, у которых приживление наблюдалось на 26-й день, каких-либо различий между донорными источниками не обнаруживалось, что еще раз указывает на важную роль замедления приживления трансплантата в повышении риска летального исхода при НРЗ.
Кроме того, недавно было показано, что абсолютное число нейтрофилов (ANC) >100 в любой день после трансплантации стволовых клеток является критическим порогом для снижения риска летального исхода через 100 дней после трансплантации (Offner et al., Blood 88:4058-62 (1996)). Таким образом, значительное замедление восстановления миелоидных клеток, наблюдающееся у реципиентов с ТПК, остается главным недостатком, препятствующим успешному осуществлению ТПК. Возможность увеличения не только абсолютного числа клеток-предшественников ПК, доступных для трансплантации, но также и клеток, которые будут способствовать более быстрому восстановлению миелоидных клеток после трансплантации, должно повысить общую выживаемость пациентов, которым была проведена ТПК. Для решения проблемы, связанной с низкими дозами клеток, присутствующих в трансплантате крови пупочного канатика, и для улучшения восстановления гемопоэтических клеток и повышения общей выживаемости при ТПК были разработаны стратегии, в которых применятся ex vivo размножение стволовых клеток/клеток-предшественников крови пупочного канатика.
Для предотвращения значительного замедления восстановления нейтрофилов, которое наблюдается после трансплантации клеток крови пупочного канатика (ПК), было проведено исследование роли путей передачи сигнала Notch в регуляции ex vivo размножения гемопоэтических стволовых клеток/клеток-предшественников в целях увеличения числа клеток-предшественников, способных быстро образовывать новые популяции in vivo. Была разработана клинически приемлемая методика с использованием сконструированного лиганда Notch (Delta 1), позволяющая достичь мультилогарифмического увеличения абсолютного числа CD34+-клеток, и клеточная терапия, способствующая быстрой репопуляции клеток in vivo.
Введение размноженных частично HLA-совместимых клеток приводит к значительному сокращению среднего времени для достижения начального абсолютного числа нейтрофилов (ANC), составляющего 500/мл всего лишь за 11 дней, тогда как у параллельной группы из 29 пациентов, прошедших аналогичное лечение, проводимое путем введения двух необработанных единиц ПК, такое среднее время составляло 25 дней (p<0,0001). Хотя число пациентов, проходивших лечение, было небольшим (то есть n=14), но при этом, уже наблюдался значительный эффект в отношении времени восстановления миелоидных клеток, что указывало не то, что данный способ является безопасным и клинически целесообразным.
Несмотря на огромные средства, которые были затрачены многими лабораториями [на исследования] в течение более 30 лет, какого-либо заметного прогресса в разработке терапевтических схем лечения гемоглобинопатий не было достигнуто, что, в основном, обусловлено отсутствием идентифицированных мишеней для лекарственных средств и необходимостью конструирования векторов для генотерапии в целях достижения стабильной экспрессии на очень высоких уровнях, которая, при этом, не приводила бы к инсерционному мутагенезу. Хотя повышенный уровень экспрессии фетального гемоглобина (HbF) способствует ослаблению симптомов обеих гемоглобинопатий, однако, обширные исследования, проведенные на основе такого наблюдения, не привели к получению новых приемлемых терапевтических агентов. Генотерапия с применением гемопоэтических стволовых клеток (HSC), включающих интегрированные в них лентивирусные векторы, была проведена несколькими исследователями. Однако генотерапия с применением HSC требует высоких уровней стабильной экспрессии и связана с высоким риском инсерционного мутагенеза и развития лейкоза.
В настоящее время особенно назрела необходимость в разработке композиций и способов, которые были бы более эффективными для лечения гемоглобинопатий, включая талассемию и серповидно-клеточную болезнь, но, при этом, были бы более безопасными по сравнению с уже существующими методами терапии.
ОПИСАНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
Настоящее изобретение направлено на решение указанных и других задач, стоящих перед специалистами, путем разработки, inter alia, композиций и способов лечения гемоглобинопатии. В описанных здесь композициях и способах используются один или более полинуклеотидов, кодирующих одну или более эндонуклеаз или эндонуклеазных гибридных белков, включая одну или более хоминг-эндонуклеаз и/или гибридных белков, содержащих хоминг-эндонуклеазу, и/или одну или более эндонуклеаз CRISPR (то есть эндонуклеазы Cas9 в комбинации с одной или более «руководящими» цепями РНК), и/или гибридных белков, содержащих эндонуклеазу CRISPR (то есть гибридных белков, содержащих эндонуклеазу Cas9 в комбинации с одной или более «руководящими» цепями РНК), где указанные композиции и способы используются: (a) для дизрупции кодирующей области Bcl11a или регуляторной области гена Bcl11a; (b) для дизрупции регуляторного элемента или пути сайленсинга ДНК HbF, такого как Bcl11a-регулируемая область сайленсинга HbF; (c) для мутации одного или более промоторов гена γ-глобина в целях повышения уровня экспрессии гена γ-глобина; (d) для мутации одного или более промоторов гена δ-глобина в целях повышения уровня экспрессии гена δ-глобина; и/или (e) для коррекции одной или более мутаций гена β-глобина.
В своем первом варианте, настоящее изобретение относится к композициям и способам, в которых используется полинуклеотид, кодирующий одну или более эндонуклеаз, таких как хоминг-эндонуклеаза (ХЭ) и/или эндонуклеазы CRISPR (то есть эндонуклеазы Cas9 в комбинации с одной или более «руководящими» цепями РНК), где указанные композиции и способы используются для достижения направленной дизрупции последовательности в кодирующей области Bcl11a или в регуляторной области гена Bcl11a и, тем самым, повышения терапевтических уровней экспрессии эндогенного гена, такого как ген γ- или ε-глобина. В родственных аспектах изобретения, композиции согласно этим вариантам осуществления изобретения включают полинуклеотид, кодирующий один или более TALEN, один или более гибридных белков TALE-ХЭ и/или один или более белков TREX2.
Во втором своем варианте, настоящее изобретение относится к композициям и способам, которые включают полинуклеотид, кодирующий одну или более эндонуклеаз, таких как хоминг-эндонуклеаза (ХЭ) или эндонуклеазы CRISPR (то есть эндонуклеазы Cas9 в комбинации с одной или более «руководящими» цепями РНК), где указанные композиции и способы используются для достижения направленной дизрупции ключевой регуляторной последовательности в локусе гена β-глобина и, тем самым, повышения терапевтических уровней экспрессии эндогенного гена, такого как ген γ- или δ-глобина. В родственных аспектах изобретения, композиции согласно этим вариантам осуществления изобретения включают полинуклеотид, кодирующий один или более TALEN, один или более гибридных белков TALE-ХЭ и/или один или более белков TREX2.
В некоторых аспектах этого варианта, настоящее изобретение относится к эндонуклеазам ХЭ и CRISPR, которые нацелены на 3,6 т.п.н.-область (SEQ ID NO: 1) в локусе гена β-глобина (chr11:5212342-5215944 в HG18), который содержит сайт связывания с регуляторным белком Bcl11a.
Описанные здесь хоминг-эндонуклеазы и эндонуклеазы CRISPR имеют уникальные преимущества по сравнению со стандартными нуклеазами, нецеленными на ген. Поскольку эти эндонуклеазы обладают широким спектром действия, независимо от генотипа, то описанные здесь хоминг-эндонуклеазы и эндонуклеазы Cas9 в комбинации с описанными здесь одной или более «руководящими» цепями РНК, не являются специфичными для пациента, то есть они дают клинический эффект в гетерозиготном состоянии, что позволяет избежать инсерции векторных последовательностей.
В своем третьем варианте, настоящее изобретение относится к композициям и способам, предназначенным для рекапитуляции, посредством редактирования генома, одной или более природных мутаций в геноме пациента, включая, например, делеционные или не-делеционные персистентные формы наследственного фетального гемоглобина (HPFH), в целях достижения нужного клинического эффекта. Более конкретно, настоящее изобретение относится к композициям и способам, предназначенным для достижения прямой коррекции мутации при талассемии и/или серповидно-клеточной болезни (СКА) посредством редактирования генома.
В некоторых аспектах этого варианта осуществления изобретения, одна или более хоминг-эндонуклеаз используются в комбинации с нормальной полинуклеотидной последовательностью или полинуклеотидной последовательностью дикого типа (корректирующей матрицы) для редактирования и/или репарации одной или более генетических последовательностей, таких как ген(ы) β-подобного глобина. Эти хоминг-эндонуклеазы позволяют осуществлять модификацию ключевых регуляторных и/или кодирующих последовательностей в генном локусе, представленном здесь как локус человеческого гена β-глобина, посредством транзиентной экспрессии полинуклеотида, включающего одну или более природных мутаций. В родственных аспектах изобретения, композиции согласно этим вариантам включают полинуклеотид, кодирующий один или более TALEN, один или более гибридных белков TALE-ХЭ и/или один или более белков TREX2.
Более конкретно, настоящее изобретение относится к композициям и способам, предназначенным для «редактирования» генома, где указанные композиции и способы включают один или более полинуклеотидов, каждый из которых кодирует ХЭ и корректирующую матрицу, и могут быть использованы для введения природных мутаций в стволовые клетки, включая, например, гемопоэтические стволовые клетки (HSC), эмбриональные стволовые (ES) клетки и индуцированные плюрипотентные стволовые клетки (iPSC). Редактированные по геному клетки HSC, ES и iPSC, включая аутологичные HSC и iPSC, могут быть трансплантированы пациенту для лечения одной или более гемоглобинопатий, таких как талассемия и/или серповидно-клеточная болезнь.
С применением описанных здесь композиций и способов можно осуществлять эффективную модификацию HSC, ES и iPSC посредством транзиентной экспрессии полинуклеотида, кодирующего ХЭ в присутствии или в отсутствии нацеливающей матрицы, эндонуклеазы Cas9 и/или «руководящей» цепи РНК, не прибегая, при этом, к стабильной экспрессии или инсерции экзогенного гена для ослабления гемоглобинопатий в зрелых эритроидных клетках и в клетках пациента in vivo. Поскольку при осуществлении таких способов терапии не требуется интеграция и/или стабильная экспрессия трансгена, то это позволяет решить проблемы безопасности, которые возникают в случае применения современных методов генотерапии.
В своем четвертом варианте, настоящее изобретение относится к композициям и к способам, предназначенным для доставки одной или более хоминг-эндонуклеаз и/или одной или более эндонуклеаз Cas9 в комбинации с одной или более «руководящими» цепями РНК, каждая из которых может быть транзиентно экспрессирована в областях-мишенях, где указанные композиции и способы, как было показано, являются клинически эффективными при лечении человека. Описанные здесь последовательности, кодирующие эндонуклеазу, могут экспрессироваться в комбинации с последовательностью, кодирующей нуклеазу TAL-эффектора (TALEN), либо в виде гибрида с этой последовательностью. В настоящей заявке описаны гибридные белки TAL-эффектор-ХЭ (TALE-ХЭ) и полинуклеотиды, кодирующие указанные гибридные белки TALE-ХЭ, которые направлены на важные геномные области, влияющие на продуцирование фетального гемоглобина.
В некоторых аспектах этих вариантов осуществления изобретения, полинуклеотид, кодирующий один или более ХЭ в присутствии или в отсутствии нацеливающей матрицы, одну или более эндонуклеаз Cas9, одну или более «руководящих» цепей РНК, один или более TALEN, один или более гибридных белков TALE-ХЭ и/или один или более белков TREX2, функционально присоединяют к промоторной последовательности в вирусном векторе для доставки и транзиентной экспрессии ХЭ, Cas9, «руководящей» цепи РНК, TALEN, гибридного белка TALE-ХЭ и/или белка TREX2. Подходящие вирусные векторы, которые могут быть с успехом использованы для доставки ХЭ, TALEN, гибридного белка TALE-ХЭ и/или белка TREX2, могут быть выбраны из группы, состоящей из лентивирусного вектора, псевдотипированного в кокках; «пенистого» вирусного вектора; аденовирусного вектора и аденоассоциированного вирусного (AAV) вектора.
В своем пятом варианте, настоящее изобретение относится к композициям и к способам, включающим ex vivo размноженные модифицированные гемопоэтические стволовые клетки (HSC), что позволяет осуществлять эффективную трансплантацию «отредактированных» клеток и использовать индуцированные плюрипотентные стволовые клетки (iPSC) для скрининга и клинического применения. В некоторых аспектах этих вариантов, настоящее изобретение относится к композициям и к способам, предназначенным для эффективного размножения аутологичных HSC, аутологичных геномодифицированных HSC, HSC, происходящих от iPSC, и ES-клеток. В этих целях может быть применена методика размножения клеток крови пупочного канатика, где указанная методика включает применение Delta1 в бессывороточной среде, в которую были добавлены гемопоэтические факторы роста, с использованием мобилизованных CD34+-клеток периферической крови, взятых у здоровых доноров. Эти композиции и способы могут быть применены в комбинации с одним или более дополнительными реагентами для повышения выживаемости и пролиферации гемопоэтических стволовых клеток/клеток-предшественников. В других аспектах изобретения, с применением этих композиций и способов может быть осуществлено совместное культивирование эндотелиальных клеток для усиления размножения долгоживущих клеточных репопуляций, включая «отредактированные» HSC, происходящие от iPSC.
В своем шестом варианте, настоящее изобретение относится к композициям и способам, используемым для поддерживающей терапии, где указанные композиции и способы могут быть применены в стандартной клеточной терапии, что позволит устранить посттрансплантационную нейтропению и повысить эффективность лечения после трансплантации «отредактированных» по генам аутологичных HSC. Размноженные ex vivo криопротективные стволовые клетки/клетки-предшественники крови пупочного канатика (ПК) могут быть, например, введены как средство для поддерживающей терапии пациентам с талассемией и/или серповидно-клеточной болезнью, которые были подвергнуты миелоабляционной трансплантации гемопоэтических клеток (ТПК) с использованием аутологичных скорректированных по гену CD34+-клеток.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКОГО МАТЕРИАЛА
Для лучшего понимания некоторых аспектов настоящего изобретения ниже приводится описание графического материала.
На фиг. 1 проиллюстрировано нацеливание на мишени для повышения уровня экспрессии генов, подобных гену β-глобина, в эритроидных тканях взрослых. Представлены факторы, которые участвуют в регуляции переключения с паттерна фетальной экспрессии (двух γ-генов) на программу такой экспрессии у взрослых (δ и β). (Адаптированные технологии, описанные в публикации Wilber et al., Blood 117(15):3945-3953 (2011)).
На фиг. 2 представлены три репрезентативных метода, проводимых с использованием редко встречающейся расщепляющей нуклеазы.
На фиг. 3 представлен график, который указывает на то, что риск летального исхода при не-рецидивирующем заболевании гораздо выше у реципиентов, подвергнутых двойной трансплантации клеток крови пупочного канатика (ТПК). На этой фигуре представлены данные по смертности от не-рецидивирующего заболевания после двойной ТПК (DUCB), взятых от совместимого неродственного донора (MUD), несовместимого неродственного донора (MMUD) и совместимого родственного донора (SIB).
На фиг. 4 показано, что культивирование предшественников ПК с Delta1ext-IgG приводит к более быстрому восстановлению нейтрофилов при проведении двух миелоабляционных ТПК. Указано индивидуальное и среднее время (сплошная линия) до достижения ANC≥500/мкл у пациентов, которым трансплантировали две немодифицированных единицы клеток крови пупочного канатика (ТПК) (стандартных) и одну размноженную ex vivo единицу и одну немодифицированную («размноженную») единицу.
На фиг. 5 представлена гистограмма, на которой указано число трансплантаций клеток крови пупочного канатика, осуществляемых ежегодно в зависимости от типа заболевания.
На фиг. 6 (SEQ ID NO: 1) представлена последовательность 3,6 т.п.н.-области, в которой находится область сайленсинга HbF, охватывающая локус chr11:5212342-5215944 в HG18.
На фиг. 7 (SEQ ID NO: 2) представлена область из 350 пар оснований, начинающаяся от элемента-повтора (chr11:5213912-5214261 в HG18) и простирающаяся выше точки разрыва Френча HPFH, которая, как известно, разрушает область захвата Bcl11a в области сайленсинга HbF и включает связывающий мотив GATA-1, и из которой были сконструированы хоминг-эндонуклеазы (ХЭ) согласно изобретению.
На фиг. 8 (SEQ ID NO: 13) представлен ген человеческого бета-глобина, находящийся выше на расстоянии 1 т.п.н. от кэпа и простирающийся до сайта polyA, который начинается от chr11:5203272-5205877 в HG18 (обратная цепь).
На фиг. 9 (SEQ ID NO: 14) представлена 606 п.н.-область человеческого бета-глобина, простирающаяся от промотора и до интрона 2 (chr11:5204380-5204985 в HG18). Эта относительно небольшая область содержит большое число мутаций, приводящих к развитию тяжелой талассемии, а также мутаций, вызывающих серповидно-клеточную болезнь. Эта небольшая область легко поддается гомологичной рекомбинации с последующей коррекцией гена.
На фиг. 10 (SEQ ID NO: 24) представлена последовательность кДНК для человеческого гена Bcl11a (CCDS1862.1).
На фиг. 11 представлена рестрикрионная карта плазмиды pET-21a(+).
На фиг. 12 представлена рестрикрионная карта плазмиды pEndo (Doyon et al., J. Am. Chem. Soc. 128(7):2477-2484 (2006).
На фиг. 13 схематически представлена диаграмма прямого способа конструирования эндонуклеазы, нацеленной на ген BCL11A. Сконструированную библиотеку подвергали отбору при компартментализации in vitro (IVC) на сайт-мишень, часть которого была заменена геном-мишенью BCL11A.
На фиг. 14 (SEQ ID NO: 28) представлена нуклеотидная последовательность I-HjeMI (родительского фермента для нуклеазы, нацеленной на ген BcL11A), которая была оптимизирована по кодонам для экспрессии в E. coli.
На фиг. 15 (SEQ ID NO: 29) представлена нуклеотидная последовательность I-HjeMI (родительского фермента для нуклеазы, нацеленной на ген BCL11A), которая была оптимизирована по кодонам для экспрессии у млекопитающих.
На фиг. 16 (SEQ ID NO: 30) представлена аминокислотная последовательность хоминг-эндонуклеазы I-HjeMI.
На фиг. 17 (SEQ ID NO: 31) представлена нуклеотидная последовательность нуклеазы, нацеленной на ген BCL11A (Bcl11-Ahje), которая была получена на основе хоминг-эндонуклеазы I-HjeMI (прямым способом конструирования при IVC и в бактериях), и которая была оптимизирована по кодонам для экспрессии в E. coli.
На фиг. 18 (SEQ ID NO: 32) представлена нуклеотидная последовательность нуклеазы, нацеленной на ген BCL11A, которая была получена на основе хоминг-эндонуклеазы I-HjeMI (прямым способом конструирования при IVC и в бактериях), и которая была оптимизирована по кодонам для экспрессии у млекопитающих.
На фиг. 19 (SEQ ID NO: 33) представлена аминокислотная последовательность нуклеазы, нацеленной на ген BCL11A, которая была получена на основе хоминг-эндонуклеазы I-HjeMI (прямым способом конструирования при IVC и в бактериях).
На фиг. 20 представлена модель белка, иллюстрирующая распределение аминокислотных остатков в эндонуклеазе, нацеленной на ген BCL11A, и в родительской эндонуклеазе LHE I-HjeMI, где остатки этих ферментов отличаются друг от друга. Замененные остатки эндонуклеазы, нацеленной на ген BCL11A, были картированы на кристаллической структуре I-HjeMI, связанной с ее сайтом-мишенью (PDB ID: 3UVF). D161 был делетирован в модифицированной эндонуклеазе.
На фиг. 21 представлена гистограмма, иллюстрирующая активность эндонуклеазы, нуклеазы, нацеленной на ген BCL11A, в анализе на расщепление, проводимом с использованием двух плазмид.
На фиг. 22A (SEQ ID NO: 34) представлена нуклеотидная последовательность хоминг-эндонуклеазы I-OnuI (родительского фермента для хоминг-эндонуклеаз, нацеленных на область сайленсинга HbF), которая была оптимизирована по кодонам для экспрессии в E. coli.
На фиг. 22B (SEQ ID NO: 15) представлена аминокислотная последовательность хоминг-эндонуклеазы I-OnuI.
На фиг. 23 показан агарозный гель, иллюстрирующий активность хоминг-эндонуклеазы I-OnuI, нацеленной на область сайленсинга HbF.
На фиг. 24 (SEQ ID NO: 35) представлена нуклеотидная последовательность MegaTAL:5.5 RVD + Y2 I-AniI.
На фиг. 25 (SEQ ID NO: 36) представлена аминокислотная последовательность MegaTAL:5.5 RVD + Y2 I-AniI.
На фиг. 26 (SEQ ID NO: 37) представлена нуклеотидная последовательность эндонуклеазы Cas9 (Mali et al., Science (2013)).
На фиг. 27 (SEQ ID NO: 38) представлена нуклеотидная последовательность «руководящей» цепи РНК, используемой вместе с эндонуклеазой Cas9 (Mali et al., Science (2013)).
На фиг. 28 (SEQ ID NO: 62) представлена нуклеотидная последовательность хоминг-эндонуклеазы I-CpaMI (OPC, оптимизированной по кодонам для экспрессии у млекопитающих).
На фиг. 29 (SEQ ID NO: 63) представлена аминокислотная последовательность хоминг-эндонуклеазы I-CpaMI.
На фиг. 30 показан агарозный гель, иллюстрирующий детектирование направленного мутагенеза в эндогенном человеческом гене BCL11A, как описано в примере 4.
На фиг. 31 представлена рестрикционная карта плазмиды pCcdB wt6 (Doyon et al., J. Am. Chem. Soc. 128(7): 2477-2484 (2006).
На фиг. 32 (SEQ ID NO: 64) представлена нуклеотидная последовательность плазмиды, кодирующей нуклеазу, нацеленную на ген BcL11A (pExodusBCLIIAhje).
На фиг. 33 (SEQ ID NO: 65) представлена нуклеотидная последовательность TREX2-кодирующей плазмиды (pExodus CMV.Trex2).
На фиг. 34 представлена рестрикционная карта плазмиды pExodusBCL11Ahje.
На фиг. 35 представлена рестрикционная карта плазмиды pExodus CMV.Trex2.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
В общих чертах, настоящее изобретение относится к композициям и способам, приемлемым для лечения генетического заболевания, такого как гемоглобинопатия, посредством транзиентной или стабильной экспрессии полинуклеотида, который кодирует одну или более эндонуклеаз или эндонуклеазных гибридных белков, включая одну или более хоминг-эндонуклеаз и/или гибридных белков, содержащих хоминг-эндонуклеазу, и/или одну или более эндонуклеаз Cas9 и/или гибридных белков, содержащих эндонуклеазу Cas9, в комбинации с одной или более «руководящими» цепями РНК, где указанные композиции и способы применяются: (a) для дизрупции кодирующей области Bcl11a; (b) для дизрупции регуляторного элемента или пути ДНК сайленсинга HbF, такого как Bcl11a-регулируемая область сайленсинга HbF; (c) для мутации одного или более промоторов гена γ-глобина в целях повышения уровня экспрессии гена γ-глобина; (d) для мутации одного или более промоторов гена δ-глобина в целях повышения уровня экспрессии гена δ-глобина; и/или (e) для коррекции одной или более мутаций гена β-глобина. Описанные здесь композиции и способы могут быть применены для лечения гемоглобинопатии, включая β-талассемию и серповидно-клеточную болезнь. Описанные здесь композиции и способы могут включать, но необязательно, полинуклеотид, кодирующий один или более TALEN, один или более гибридных белков TALE-ХЭ и/или один или более белков TREX2.
Для лучшего понимания настоящего изобретения ниже приводятся определения терминов.
Определения
Используемый здесь термин «гемоглобинопатия» означает класс генетических патологий, приводящих к образованию аномальной структуры, к нарушению функции или к изменению экспрессии одной или более глобиновых цепей молекулы гемоглобина. Гемоглобинопатии представляют собой наследственные расстройства, вызываемые мутацией одного гена. Самыми распространенными гемоглобинопатиями являются талассемия и серповидно-клеточная болезнь.
Используемый здесь термин «талассемия» означает гемоглобинопатию, вызываемую изменением отношения α-подобных глобиновых цепей к β-подобным глобиновым цепям полипептида, которое приводит к недостаточному продуцированию нормальных тетрамерных белков гемоглобина и к увеличению числа свободных или неспаренных α- или β-цепей.
Используемый здесь термин «серповидно-клеточная болезнь» означает группу аутосомных рецессивных генетических болезней крови, которые вызываются мутациями в гене глобина и характеризуются приобретением эритроцитами аномальной ригидной серповидной формы. Эти клетки определяют по присутствию βS-гена, кодирующего вариант цепи β-глобина, в котором глутаминовая кислота заменена валином в положении аминокислоты 6 пептида, и второго β-гена, который имеет мутацию, способствующую кристаллизации HbS, которая приводит к образованию клинического фенотипа. Термин «серповидно-клеточная анемия» означает специфическую форму серповидно-клеточной болезни у пациентов, которые являются гомозиготными по мутации, приводящей к образованию HbS. Другими распространенными формами серповидно-клеточной болезни являются HbS/β-талассемия, HbS/HbC и HbS/HbD. В таблице 1 представлены нуклеотидные последовательности, кодирующие исходные аминокислоты цепей β-глобина дикого типа и цепей β-глобина серповидных клеток.
Используемый здесь термин «наследственное персистентное продуцирование фетального гемоглобина» или «HPFH» означает доброкачественное состояние, при котором непрерывно продуцируется значительное количество фетального гемоглобина (гемоглобина F) вплоть до достижения зрелого возраста, что приводит к игнорированию нормальной точки «отключения».
Используемый здесь термин «глобин» означает семейство гем-содержащих белков, которые участвуют в связывании и транспорте кислорода.
Используемый здесь термин «хоминг-эндонуклеаза» или «ХЭ» означает класс рестриктирующих эндонуклеаз, характеризующихся наличием последовательностей распознавания, которые являются слишком длинными, а поэтому присутствуют в геноме в единственном числе, и вероятность их встречаемости очень низка (например, одна последовательность на каждые 7×l010 п.н.).
Используемые здесь термины «нуклеаза эффектора, подобная активатору транскрипции» или «нуклеаза TAL-эффектора» или «TALEN» означают класс искусственных рестриктирующих эндонуклеаз, образующихся в результате присоединения ДНК-связывающего домена к ДНК-расщепляющему домену TAL-эффектора.
Используемые здесь термины «3ʹ-репарирующая экзонуклеаза 2» или «TREX2» означают нуклеазу, обладающую 3ʹ-экзонуклеазной активностью, которая обычно участвует в репликации, репарации и рекомбинации ДНК.
Используемый здесь термин «эндонуклеаза Cas9» означает эндонуклеазу, которая использует «руководящую» цепь РНК для нацеливания на сайт расщепления эндонуклеазой. Термин «эндонуклеаза CRISPR» означает эндонуклеазу Cas9, присутствующую в комбинации с «руководящей» цепью РНК. См. Jinek et al., Science 337:816-821 (2013); Cong et al., Science (Jan. 3, 2013) (Электронная публикация до выхода в печать); и Mali et al., Science (Jan. 3, 2013) (Электронная публикация до выхода в печать).
Если это не оговорено особо, то описанные ниже термины относятся к классу «открытых» (то есть допускающих расширение) терминов (например, термин «включающий» должен быть интерпретирован как «включающий, но не ограничивающийся»; термин «имеющий» должен быть интерпретирован как «имеющий по меньшей мере», термин «включает» должен быть интерпретирован как «включает, но не ограничивается», и т.п.). Такие выражения как «по меньшей мере один» и «один или более», а также артикли «a» или «an» употребляемые с существительными в единственном числе, могут относиться и к существительным во множественном числе.
Следует отметить, что признаки или аспекты настоящего изобретения описаны как группы Маркуша, где данное описание также относится к любому отдельному члену или к подгруппе членов группы Маркуша. Аналогично, все описанные здесь интервалы также охватывают все возможные субинтервалы и комбинации субинтервалов, и употребляемые здесь слова «между», «до», «по меньшей мере», «более, чем», «менее, чем» и т.п., указывают на ряд значений в данном интервале и на каждый отдельный член в данном интервале.
Все цитируемые выше и ниже работы, включая, но не ограничиваясь ими, патенты, патентные заявки и патентные публикации, независимо от того, являются ли они заявками США, PCT или другими иностранными заявками, а также все цитируемые технические и/или научные публикации во всей своей полноте вводятся в настоящее описание посредством ссылки.
Хотя в настоящем описании рассматриваются различные варианты осуществления изобретения, однако, для каждого специалиста очевидно, что могут быть представлены и другие варианты. Описанные здесь различные варианты осуществления изобретения приводятся лишь в иллюстративных целях и не должны рассматриваться как ограничение истинного существа и объема, описанного в формуле изобретения.
Хоминг-эндонуклеазы, используемые для достижения высокоэффективной дизрупции мультиплексного гена и обеспечения функций редактирования гена
Как описано выше и проиллюстрировано ниже, настоящее изобретение относится к композициям и к способам, в которых используется полинуклеотид, кодирующий одну или более эндонуклеаз, включая одну или более хоминг-эндонуклеаз (ХЭ), таких как одна или более хоминг-эндонуклеаз I-HjeMI, хоминг-эндонуклеаз I-CpaMI и/или хоминг-эндонуклеаз I-OnuI, и/или одну или более эндонуклеаз Cas9 в комбинации с одной или более «руководящими» цепями РНК, которые могут транзиентно или персистентно экспрессироваться в клетках-мишенях и дают, как было показано, клинический эффект при лечении человека. Репрезентативные эндонуклеазы нацелены на важные геномные области, которые влияют на продуцирование фетального гемоглобина посредством: (a) дизрупции кодирующей области Bcl11a или регуляторной области гена Bcl11a; (b) дизрупции регуляторного элемента или пути сайленсинга ДНК HbF, такого как Bcl11a-регулируемая область сайленсинга HbF; (c) мутации одного или более промоторов гена γ-глобина в целях повышения уровня экспрессии гена γ-глобина; (d) мутации одного или более промоторов гена δ-глобина в целях повышения уровня экспрессии гена δ-глобина; и/или (e) коррекции одной или более мутаций гена β-глобина. Описанные здесь композиции и способы могут быть применены для лечения гемоглобинопатии, включая β-талассемию и серповидно-клеточную болезнь.
Описанные здесь последовательности, кодирующие эндонуклеазу, могут экспрессироваться в комбинации или вместе с последовательностью, кодирующей ДНК-связывающий домен, такой как последовательность, кодирующая TAL-эффектор (TALE), или последовательностью, кодирующей нуклеазу, такой как последовательность, кодирующая 3ʹ-репарирующую экзонуклеазу 2 (TREX2). В настоящем изобретении рассматриваются гибридные белки TALE-ХЭ и полинуклеотиды, кодирующие один или более гибридных белков TALE-ХЭ.
Специалистам известны четыре каркасных белка, используемые для модификации и дизрупции гена-мишени у эукариотов, а именно нуклеазы «цинковые пальцы» (ZFN), нуклеазы TAL-эффектора (TALEN), хоминг-эндонуклеазы (ХЭ) и эндонуклеазы Cas9 в комбинации с «руководящей» цепью РНК. В настоящем изобретении используются нуклеазы TAL-эффектора, хоминг-эндонуклеазы и/или эндонуклеазы Cas9, взятые отдельно или в комбинации друг с другом. TAL-нуклеазы дает более простую модульную конструкцию и более высокую специфичность распознавания ДНК, чем нуклеаза «цинковые пальцы», тогда как хоминг-эндонуклеазы, такие как хоминг-эндонуклеазы LAGLIDADG (LHE), имеют высокоспецифичные профили расщепления, компактные структуры, и, поскольку они представляют собой компактные мономерные белки, не требующие димеризации, как это происходит в случае ZFN и TALEN, то они могут быть использованы в мультиплексных комбинациях. В соответствии с этим, эндонуклеазы ХЭ и CRISPR (то есть эндонуклеазы Cas9 в комбинации с одной или более «руководящими» цепями РНК) обладают высокой эффективностью в опосредовании дизрупции гена. Stoddard, см. выше, и Mali et al., Science (2013), см. выше.
В процессе разработки настоящего изобретения была идентифицирована важная область в локусе β-глобина, которая ингибирует функцию HbF. Эта область имеет множество мишеней для ХЭ- и Cas9-опосредуемого расщепления. Специально сконструированные нуклеазы могут быть протестированы на активность, направленную против когнатного сайта-мишени, и на нежелательную активность, направленную против любых близкородственных геномных мишеней. Эти эндонуклеазы ХЭ и Cas9, используемые в комбинации с одной или более «руководящими» цепями РНК, могут быть сконструированы в целях предотвращения нежелательного геномного расщепления методами, описанными Stoddard, Structure 19:7-15 (2011) и Mali et al., Science (2013). Эндонуклеазы ХЭ и Cas9, используемые в комбинации с одной или более «руководящими» цепями РНК, описанными в настоящей заявке, обладают способностью нацеливаться непосредственно на γ- и δ-промоторы и осуществлять замену 606 п.н-области (SEQ ID NO: 14), которая охватывает большинство мутаций, ассоциированных с талассемией, а также мутацию HbS.
Для облегчения введения большого числа делеций, охватывающих область сайленсинга HbF или ее подпоследовательность, одна или более эндонуклеаз ХЭ и/или одна или более эндонуклеаз Cas9 в комбинации с одной или более «руководящими» цепями РНК могут быть котрансдуцированы с мостиковым олигонуклеотидом, который простирается от сайта расщепления эндонуклеазой до конца области-мишени. Chen et al., Nat. Methods 8(9):753-5 (2011). Редактирование генома с более высокой частотой может быть достигнуто с использованием одной или более ХЭ, которые связываются с последовательностью и расщепляют последовательность, фланкирующую каждую сторону данной области-мишени. Аналогичным образом, ХЭ и мутагенный олигонуклеотид могут быть использованы для введения мутаций в промоторную область, что будет приводить к повышению уровня экспрессии гена гамма или дельта.
Описанные здесь ХЭ могут быть сначала оценены в эритроидной клеточной линии и в человеческих CD34+-клетках, которые были индуцированы для их дифференцировки в эритроидные клетки, а затем была подтверждена способность этих клеток изменять уровень экспрессии гена глобина. Одна, две, три или более ХЭ, в зависимости от цели их применения, могут быть доставлены для облегчения введения более крупных делеций. Эффективность отдельных HE может быть оценена в дополнительной культуре и в анализах на животных-моделях для подтверждения способности ХЭ нацеливаться на HSCs без нарушения потенциала плюрипотентности и размножения и для оценки желаемого клинического эффекта у моделей с гемоглобинопатией. Одна или более ХЭ и одна или более экзонуклеаз, таких как экзонуклеаза TREX2 или экзонуклеаза TAL-эффектора, могут быть введены в CD34+-HSC для индуцирования целевых генетических делеций в важных областях HbF.
Отдельные нуклеазы могут быть протестированы на серию мишеней в 350 п.н.-области (SEQ ID NO: 2), определенной как область, начинающаяся от конца элемента повтора и простирающаяся выше точки разрыва HPFH Френча, которая, как известно, разрушает область захвата Bcl11a в области сайленсинга HbF и включает связывающий мотив GATA-1. В предварительных анализах было идентифицировано семь мишеней, каждая из которых равномерно распределена по всей области и включает модули последовательности ДНК, для которых были выделены и секвенированы пулы в высокой степени активных вариантов эндонуклеазы. Интересно отметить, что одна мишень перекрывается с потенциальным Bcl11a-связывающим мотивом и является смежной с мотивом GATA-1. Последовательное введение более крупных делеций в области-мишени может быть достигнуто путем трансдукции двух, трех или более ХЭ. Альтернативно, для элиминации генной функции, множество мишеней для дизрупции гена Bcl11a было идентифицировано на 5ʹ-конце гена. Аналогично, множество оптимальных мишеней для дизрупции, опосредуемой Cas9/«руководящей» цепью РНК, было идентифицировано по всему геному, и эти мишени могут быть использованы отдельно или в комбинации друг с другом, что будет приводить к образованию более крупных делеций.
Отдельные ХЭ могут быть протестированы в трансфецированных человеческих клеточных линиях с использованием интегрированных геномных репортеров и могут быть подвергнуты дополнительным стадиям отбора для последующей оптимизации расщепления и активности конверсии генов в соответствии с протоколами, описанными Stoddard, см. выше. После идентификации и доставки отдельных нужных ХЭ, которые обладают активностью, направленной против мишеней в локусе глобина, может быть осуществлена векторизация нуклеаз в экспрессионных системах. Так, например, в целях достижения высокоэффективной усиленной дизрупции гена в трандуцированных клетках может быть использована экспрессионная система, которая будет присоединять каждую ХЭ для коэкспрессии нуклеазы, такой как экзонуклеаза TALEN и/или TREX.
Настоящее изобретение также относится к гибридным белкам TALE-ХЭ и к полинуклеотидам, кодирующим гибридные белки TALE-ХЭ, которые обладают нужными свойствами, такими как ограничение активности и рекрутинга сконструированных ХЭ в нужный сайт-мишень, например, в локус глобина, посредством синергического распознавания смежных ДНК-мишеней под действием каркасных TALE и ХЭ. Такие гибриды TALE-ХЭ объединяют в себе самые ценные свойства каждого каркаса (то есть модульную сборку TALE и нуклеазную специфичность ХЭ), и при этом, снижают неспецифическую нуклеазную активность, которой обладают традиционные нуклеазы TALEN или «цинковые пальцы» (ZFN).
Кристаллические структуры высокого разрешения были определены для десяти отдельных хоминг-эндонуклеаз LAGLIDADG (LHE) в комплексе с их когнатными сайтами ДНК-мишени. Stoddard, Structure 19:7-15 (2011) и Takeuchi et al., Proc. Natl. Acad. Sci. U.S.A. 108:13077-13082 (2011). Были сконструированы химерные «гибриды» этих LHE, которые, в совокупности, представляют собой широкий ряд геноспецифических нацеливающих белков LHE. Baxter et al., Nucl. Acids Res. 40(16):7985-8000 (2012).
ХЭ, имеющие соответствующую специфичность к последовательности-мишени, могут быть идентифицированы с применением стратегии представления на поверхности дрожжей в комбинации с высокоэффективным клеточным сортингом на нужную специфичность к расщеплению ДНК. Серия «модулей белок-ДНК», которая соответствует последовательно расположенным «карманам» контактов, простирающихся по всему сайту-мишени, могут быть системно рандомизированы в отдельных библиотеках. Каждая библиотека может быть затем системно отсортирована на популяции ферментов, которые могут специфически расщеплять каждый возможный вариант ДНК в каждом модуле, после чего каждая отсортированная популяция может быть подвергнута глубокому секвенированию и архивированию для последующей сборки и конструирования ферментов. ХЭ, которые могут быть соответствующим образом использованы в композициях и способах согласно изобретению, являются коммерчески доступными (Pregenen, Seattle, WA).
В некоторых аспектах изобретения, в описанных здесь композициях и способах может быть использована коэкспрессия одной или более ХЭ, включая, например, одну или более LHE вместе с 3ʹ-экзонуклеазой TREX2. В отличие от выступающего 5ʹ-конца, находящегося слева у имеющихся в настоящее время вариантов ZFN и TALEN, ХЭ продуцируют выступающие 3ʹ-концы в сайте целевых двухцепочечных разрывов, что приводит к усилению процессинга концевых последовательностей после ХЭ-расщепления. Почти полная модификация сайта двухцепочечного разрыва в первичных клетках может быть достигнута посредством коэкспрессии ХЭ/TREX2. Поскольку путь коэкспрессии ХЭ/TREX2 влияет на процессинг с образованием разрывов, то эта комбинация позволяет вводить множество нужных делеций в одну область и повышает безопасность индуцированной нуклеазой дизрупции гена-мишени благодаря уменьшению скорости образования разрывов и снижению вероятности крупномасштабных транслокаций, опосредуемых альтернативными путями присоединения по концам.
Недавно была определена кристаллическая структура TAL-эффектора (PthXol), связанного с его сайтом ДНК-мишени. Mak et al., Science 335(6069):716-9 2012; e-pub 05Janl2 PubMed PMID: 22223736. Данные такой кристаллической структуры позволяют точно определить границы области распознавания ДНК и облегчают проведение стратегий по созданию конструкции TALEN-ХЭ с нужными свойствами или других конструкций гибридов «TALEN-нуклеаза», которые могут быть применены для осуществления ряда сложных геномных манипуляций.
Дизрупция генома для экспрессии гена Bcl11a
Отсутствие Bcl11a у мышей с моделью серповидно-клеточной болезни приводит к ослаблению симптомов данного заболевания, что подтверждает клиническую релевантность такого подхода. Xu et al., Science 334:993-6 (2011). Кроме того, мыши, содержащие трансген YAC, охватывающий локус человеческого β-глобина, были использованы для моделирования перестановок в области Bcl11a- опосредуемого сайленсинга HbF. У этих мышей, гетерозиготных и гомозиготных по отсутствию эндогенного гена Bcl11a, количество мРНК γ-глобина составляет 20 и 76% от общего количества β-подобной мРНК, соответственно, по сравнению с контролем, у которого такое количество составляет 0,24%. Sankaran et al., Nature 460:1093-7 (2009). Это позволяет предположить, что Bcl11a действует как реостат, модулирующий степень ингибирования HbF. В соответствии с этим, снижение уровня функциональных мутаций в Bcl11a приводит к повышению уровней HbF и к ослаблению клинического фенотипа талассемии и/или серповидно-клеточной болезни. Galanello et al., Blood 114:3935-7 (2009).
В некоторых своих вариантах, настоящее изобретение относится к композициям, включающим одну или более эндонуклеаз, и к способам, в которых используется указанные одна или более эндонуклеаз, включая одну или более хоминг-эндонуклеаз (ХЭ), таких как одна или более хоминг-эндонуклеаз I-HjeMI, хоминг-эндонуклеаз I-CpaMI и/или хоминг-эндонуклеаз I-OnuI, и/или одна или более эндонуклеаз Cas9, в целях достижения дизрупции последовательности, кодирующей Bcl11a или его ключевые регуляторные последовательности. Как подробно описано и проиллюстрировано в настоящей заявке, в композициях и в способах, в которых используются одна или более эндонуклеаз Cas9, также используются одна или более «руководящих» цепей РНК, специфичных к гену Bcl11a, для опосредованного нацеливания эндонуклеазы Cas9 на последовательность гена Bcl11a.
Ген Bcl11a имеет множество экзонов, охватывающих 100 т.п.н., и несколько вариантов сплайсинга, что приводит к образованию белков с различными активностями. В процессе разработки настоящего изобретения было идентифицировано несколько ДНК-мишеней, которые были транскрибированы во множество вариантов сплайсинга Bcl11a. Все указанные белки разрывают длинные (L) и очень длинные (XL) формы, которые ассоциируются с наибольшей активностью сайленсинга HbF, тогда как один из этих белков разрывает все формы Bcl11a. Эти мишени содержат модули последовательности ДНК, для которых были выделены и секвенированы пулы в высокой степени активных вариантов эндонуклеазы. Последовательность кДНК человеческого Bcl11a (CCDS1862.1) представлена здесь как SEQ ID NO: 24 (фиг. 10).
Таким образом, в некоторых своих аспектах, настоящее изобретение относится к композициям, используемым для достижения терапевтических уровней HbF, где указанные композиции содержат полинуклеотид, кодирующий одну или более хоминг-эндонуклеаз (ХЭ), способных опосредовать дизрупцию нуклеотидной последовательности в 1,3 т.п.н.-области, и тем самым, предупреждать связывание с Bcl11a и образование соответствующего репрессивного комплекса, а также стимулировать восстановление экспрессии γ-глобина.
Дизрупция генома для блокирования Bcl11a-опосредуемого сайленсинга HbF
Как было вкратце описано выше, в некоторых своих вариантах, настоящее изобретение относится к композициям и способам, которые могут быть применены для лечения и/или ослабления симптомов генетического заболевания, такого как гемоглобинопатия, включая талассемию и/или серповидно-клеточную болезнь. Некоторые аспекты этих вариантов изобретения включают осуществление транзиентной экспрессии полинуклеотида, кодирующего одну или более хоминг-эндонуклеаз, для дизрупции элемента или пути сайленсинга HbF в локусе гена β-глобина или в локусе гена δ-глобина в целях повышения терапевтических уровней экспрессии эндогенного гена, такого как ген γ- или δ-глобина.
Описанные здесь композиции включают полинуклеотид, кодирующий одну или более хоминг-эндонуклеаз (ХЭ) и/или одну или более эндонуклеаз Cas9 в комбинации с одной или более «руководящими» цепями РНК и, необязательно, с одним или более эффекторами, подобными активатору транскрипции (TAL) для достижения целевой дизрупции ключевых регуляторных последовательностей в локусе гена β-глобина. Более конкретно, описанные здесь композиции и способы могут быть применены для повышения уровней экспрессии гена γ-глобина и последующего продуцирования белка HbF посредством удаления элементов, играющих важную роль в связывании Bcl11a с областью(ями) сайленсинга HbF в локусе гена β-глобина.
При нормальном развитии организма, экспрессия эмбрионального β-подобного гена (ε-глобина) заменяется экспрессией пары генов γ-глобина в плоде и генов δ- и β-глобина у взрослых. В эритроидных тканях взрослых, белок «цинковый палец» Bcl11a связывается с областью, расположенной между генами γ-глобина и δ-глобина в локусе гена β-глобина, что приводит к сайленсингу продуцирования HbF. Важная роль Bcl11a-опосредуемого сайленсинга HbF подтверждается отсутствием мРНК Bcl11a в человеческих CD34-клетках, которое приводит к увеличению уровней HbF до 24-36% от общего уровня β-подобных белков. Sankaran et al., Science 322:1839-42 (2008). Удаление этой области в делеционной форме HPFH, а также отсутствие Bcl11a приводит к блокированию Bcl11a-опосредуемого сайленсинга HbF и к повышению уровня экспрессии гена γ-глобина и продуцирования белка HbF в эритроидных тканях взрослых (Sankaran et al., N. Engl. J. Med. 365:807-14 (2011)).
В повышении уровней белка HbF участвует множество механизмов, причем, было показано, что ключевую роль в сайленсинге HbF играет 3,6 т.п.н.-область (SEQ ID NO: 1). Sankaran et al., N. Engl. J. Med. 365:807-14 (2011). В локусе β-глобина имеется несколько пиков с большим количеством Bcl11a, при этом один пик в 3,6 т.п.н.-области сайленсинга HbF соответствует белкам, которые, как известно, образуют репрессивный комплекс со связанным Bcl11a в этой области (GATA-1 и HDAC-1) и хроматином, обогащенным триметильными группами репрессивного маркерного гистона, то есть гистона H3 на лизине 27.
Интересно отметить, что эта 3,6 т.п.н.-область содержит один пик Bcl11a, связывающейся на участке, расположенном ниже γ-гена. В промоторах γ-глобина было идентифицировано множество точковых мутаций, которые дают уровни HbF, составляющие 20-30% для гетерозиготов, что приводит к ослаблению симптомов талассемии и СКЗ. Эти точковые мутации ассоциируются с тремя областями, которые, в целом, охватывают 200 п.н. сайтов γ-кэпа: (1) -200, GC-богатую область, связанную с SP1 и стадиеспецифическим белковым комплексом; (2) -175, область, связанную с GATA-1; и (3) Octl и CCAAT-мотив в положении -117, связанный с несколькими дополнительными факторами. Forget, Ann. NY Acad. Sci. 850:38-44 (1998).
Мутации в этих трех областях блокируют связывание репрессивного комплекса в эритроидных клетках у взрослых. Следовательно, эти области представляют собой подходящие мишени для ХЭ-опосредуемой дизрупции и направленной мутации, осуществляемых с применением описанных здесь композиций и способов. Дизрупция этих областей приводит к снижению уровня продуцирования репрессивных комплексов, и тем самым, к повышению уровня экспрессии гена γ-глобина и к соответствующему повышению уровня продуцирования белка HbF до уровней, достаточных для достижения терапевтической эффективности способов лечения гемоглобинопатий, включая β-талассемию и серповидно-клеточную болезнь.
Один пик захвата Bcl11a имеется в 3,6 т.п.н.-области сайленсинга HbF (Sankaran et al., N. Engl. J. Med. 365:807-14 (2011)) (SEQ ID NO: 1). Эта область захвата Bcl11a разрывается в расположенной выше точке разрыва HPFH Френча, Sankaran et al., N. Engl. J. Med. 365:807-14 (2011). В настоящей заявке описана 350 п.н.-область, начинающаяся от края элемента повтора и простирающаяся выше точки разрыва HPFH Френча, которая, как известно, разрушает область захвата Bcl11a в области сайленсинга HbF и включает связывающий мотив GATA-1 (SEQ ID NO: 2). Основанием, находящимся перед вышерасположенной делецией HPFH Френча, является HG18 chr11:5.214.023. Мотив GATA-1 охватывает chr11:5.214.200-5.214.206. Не ограничиваясь какой-либо конкретной теорией, авторы отмечают, что GATA-1 и HDAC-1 образуют репрессивный комплекс с Bcl11a, если Bcl11a связан в этой 350 п.н.-области, что приводит к образованию репрессивного комплекса, ингибирующего экспрессию генов γ-глобина и, тем самым, к снижению уровней белка HbF в клетках.
ХЭ-опосредуемая дизрупция, которая может быть достигнута с применением описанных здесь композиций и способов, является высокоэффективной. В отличие от способов ингибирования короткой РНК, известных специалистам, высокий уровень последовательность-специфической дизрупции области сайленсинга HbF, которая опосредуется описанными здесь хоминг-эндонуклеазами, предотвращает возникновение нежелательных эффектов в других сайтах связывания Bcl11a в геноме и в клетках других типов, а в частности, в В-клетках, где для нормального развития необходимо связывание Bcl11a.
Таким образом, описанные здесь хоминг-эндонуклеазы имеют уникальные преимущества по сравнению со стандартными нуклеазами, нацеленными на ген. Описанные здесь хоминг-эндонуклеазы, благодаря широкому спектру их действия независимо от генотипа, не являются специфичными для какого-либо конкретного пациента, и дают нужный клинический эффект в гетерозиготном состоянии.
Рекапитуляция генетических модификаций для коррекции мутаций, ассоциированных с талассемией или серповидно-клеточной болезнью
В других своих вариантах, настоящее изобретение относится к композициям и способам, применяемым для рекапитуляции одной или более природных мутаций в геноме пациента, осуществляемой посредством «редактирования» генома в целях достижения нужных клинических эффектов. Более конкретно, настоящее изобретение относится к композициям и способам, которые могут быть применены для прямой коррекции мутаций при талассемии и/или серповидно-клеточной болезни (СКА) посредством «редактирования» генома.
В описанных здесь композициях и способах используется корректирующая матрица для «редактирования» и коррекции гена в целях ослабления симптомов гемоглобинопатий, включая талассемию и серповидно-клеточную болезнь, посредством повышения скорости гомологичной рекомбинации (HR) между корректирующей матрицей и соответствующей мутированной последовательностью в геноме пациента. В настоящей заявке описаны репрезентативные композиции и способы для коррекции уже имеющейся мутации β-глобина, с применением которых можно достичь клинического эффекта в гетерозиготном состоянии, не прибегая к инсерции векторных последовательностей. Эти композиции и способы могут быть применены независимо или в комбинации с описанными выше композициями и способами, применяемыми для дизрупции Bcl11a-опосредуемого сайленсинга гена.
Настоящее изобретение относится к ряду надежных технологий «редактирования» генома, в которых используются преимущества ХЭ по сравнению с альтернативными методами, известными специалистам. Эти ХЭ могут быть объединены с TAL-эффектором с применением модульного метода связывания ДНК для достижения дополнительных преимущественных терапевтических свойств.
Гомологичная рекомбинация (HR), применяемая для «редактирования» геномов, является мощным, но неэффективным средством. Введение двухцепочечного разрыва в модифицируемую область приводит к значительному повышению эффективности гомологичной рекомбинации (HR). Одновременное введение полинуклеотида, кодирующего ХЭ, и корректирующей матрицы, которая содержит по меньшей мере 100 п.н. фланкирующей гомологичной области, позволяет повысить частоту гомологичной рекомбинации и, тем самым, осуществлять «редактирование» генома после введения корректирующей матрицы.
Клеточная трансдукция с помощью короткой синтезированной корректирующей матрицы может быть также применена для эффективного введения определенных мутаций одной пары оснований. В таких подходах обычно используется одна ХЭ. Альтернативно, могут быть трансдуцированы ХЭ, фланкирующие область, предназначенную для модификации. Корректирующие матрицы могут быть трансдуцированы оптимизированными методами, описанными в настоящей заявке. Конструирование, трансдукция и оценка ХЭ могут быть осуществлены как подробно обсуждается ниже с применением методики, описанной Certo et al., Nat. Methods 8:671-6 (2011) и Jarjour et al., Nucleic Acids Res 37:6871-80 (2009).
В некоторых аспектах этих вариантов осуществления изобретения, одна или более хоминг-эндонуклеаз используются в комбинации с нормальной полинуклеотидной последовательностью или полинуклеотидной последовательностью дикого типа, что позволяет осуществлять «редактирование» и/или репарацию одного или более генов β-подобного глобина. Так, например, настоящее изобретение относится к композициям и способам, применяемым для лечения гемоглобинопатий, где указанные композиции и способы могут быть применены для модификации ключевых регуляторных и/или кодирующих последовательностей в локусе гена, представленном здесь как локус человеческого гена β-глобина, посредством транзиентной экспрессии полинуклеотида, включающего одну или более природных мутаций.
Более конкретно, настоящее изобретение относится к композициям и способам, применяемым для «редактирования» генома и для введения мутаций, обеспечивающих рекапитуляцию природных мутаций в стволовых клетках, включая, например, гемопоэтические стволовые клетки (HSC), эмбриональные стволовые клетки (ES) и индуцированные плюрипотентные стволовые клетки (iPSC). Редактированные по геному клетки HSC, ES и iPSC, включая аутологичные HSC и iPSC, могут быть трансплантированы пациенту для лечения одной или более гемоглобинопатий, таких как талассемия и/или серповидно-клеточная болезнь. С применением описанных здесь композиций и способов можно проводить эффективную модификацию HSC, ES и iPSC без осуществления стабильной экспрессии или инсерции экзогенного гена для ослабления гемоглобинопатии в зрелых эритроидных клетках и в клетках пациента in vivo.
Поскольку при осуществлении таких способов терапии не требуется интеграция и/или стабильная экспрессия трансгена, то это позволяет решить проблемы безопасности, которые возникают в случае применения современных методов генотерапии. В некоторых аспектах этих вариантов осуществления изобретения, в описанных здесь композициях и способах используются один или более полинуклеотидов для целевой дизрупции области Bcl11a-опосредуемого сайленсинга HbF.
С применением описанных здесь репрезентативных композиций и способов можно осуществлять рекапитуляцию генетических модификаций в одной или более областях сайленсинга HbF, ответственных за наследственную персистентность HbF (HPFH). Поскольку такие генетические модификации приводят к повышению уровня экспрессии терапевтически эффективного гена, то для достижения терапевтического эффекта, рекапитулированные генетические модификации должны присутствовать только как гетерозиготы.
С помощью описанных здесь композиций и способов, применяемых для ослабления симптомов талассемии и серповидно-клеточной болезни, может быть достигнута терапевтическая эффективность путем введения одной или более мутаций, приводящих к повышению уровня продуцирования белка HbF и/или HbA2 и/или HbA. В настоящей заявке описаны репрезентативные композиции и способы, применяемые для рекапитуляции одной или более природных делеций гена и/или областей гена β-глобина, где такая рекапитуляция активирует экспрессию гена γ-глобина и способствует повышению уровней фетального гемоглобина. Поскольку умеренное повышение уровня продуцирования белка HbF и/или HbA2 является достаточным для улучшения фенотипов этих заболеваний, то для достижения заметного терапевтического эффекта будет достаточным введение гетерозиготных мутаций.
В некоторых аспектах изобретения, доставка корректирующей матрицы может быть осуществлена вместе с доставкой селективного маркерного гена, что позволит проводить отбор скорректированных клеток ex vivo и in vivo, хотя такой подход требует проведения продолжительной экспрессии посредством интеграции селективного маркерного гена. Beard et al., J. Clin. Invest. 120:2345-54 (2010) и Munoz et al., Nucleic Acids Res. 39{2):729-743 (2011).
Активация экспрессии β-глобина в тканях взрослых зависит от связывания KLF-1 в CACCC-боксе в его промоторе. Промотор δ-глобина не содержит интактного CACCC-бокса, KLF-1 не является связанным, а экспрессия ограничена 2% β-глобина. Мутации δ-промотора, обеспечивающие рекапитуляцию промотора β-глобина, например, посредством введения интактного CACCC-бокса, приводят к связыванию с KLF-1, и тем самым, к терапевтически эффективному повышению уровня экспрессии δ-глобина.
В некоторых аспектах этих вариантов осуществления изобретения может быть введена не-делеционная мутация в промотор γ-глобина HPFH. Для достижения эффективности должна быть модифицирована только одна пара оснований. Так, например, мутация -175 T→C (SEQ ID NO: 21) может быть рекапитулирована для максимизации уровней HbF. Мутация любого из четырех генов γ-глобина будет давать нужный эффект, а именно повышение уровней потенциальных мишеней.
Доставка хоминг-эндонуклеаз, эндонуклеазы Cas9, нуклеаз TAL-эффектора и экзонуклеаз TREX2
В других своих вариантах, настоящее изобретение относится к системам, а в частности, к неинтегрирующим векторным системам для доставки одной или более из описанных здесь нуклеаз ХЭ, Cas9, TALEN и/или TREX2. Для «редактирования» терапевтического гена в гемопоэтических стволовых клетках (HSC) должно выполняться три главных условия: (1) нуклеазные реагенты должны быть транзиентно доставлены в HSC; (2) эффективность «редактирования» гена в клетках, в которые была введена нуклеаза, должна быть высокой; и (3) отредактированные по гену клетки HSC должны быть трансплантированы на уровне, достаточном для достижения терапевтического эффекта. Эти требования могут быть удовлетворены с применением различных методов векторизации.
В настоящей заявке рассматриваются репрезентативные лентивирусные векторы, псевдотипированные в кокках, и «пенистые» вирусные векторы, используемые для эффективного переноса гена в HSC. Trobridge et al., Mol. Ther. 18:725-33 (2008). Альтернативно, аденовирусные векторы могут быть модифицированы, как описано ранее, для их использования в целях переноса гена в HSC. Wang et al., Exp. Hematol. 36:823-31 (2008) и Wang et al., Nat. Med. 17:96-104 (2011). В других аспектах этих вариантов осуществления изобретения, векторные системы на основе AAV могут быть также использованы для доставки нуклеаз ХЭ, Cas9 (и/или «руководящих» цепей РНК), TALE-ХЭ, TALEN и/или TREX2.
Рекомбинантные AAV-векторы серотипа AAV6 дают полезную нагрузку 4,5 т.п.н., достаточную для доставки кластера «промотор-ХЭ-экзонуклеаза» или кластера «промотор-TAL-ХЭ-экзонуклеаза», а также небольшой матрицы для рекомбинации. Альтернативно, эта матрица несет небольшой полипептид Cas9 и «руководящие» РНК. AAV6 обеспечивает эффективную трансдукцию человеческих CD34+-клеток крови пупочного канатика в капсиды всех известных AAV и обладает способностью опосредовать значительные уровни транзиентной экспрессии гена в HSC. Аутокомплементарные и одноцепочечные векторы AAV6 могут быть использованы для ингибирования и «редактирования» гена посредством рекомбинации в клеточных линиях HSC и в первичных CD34+-клетках.
Аденовирусные векторы с гибридными капсидами обладают способностью эффективно трансдуцировать гемопоэтические клетки многих типов, включая CD34+-клетки. Для улучшения трансдукции гемопоэтических клеток может быть использован химерный аденовирусный вектор, имеющий волоконный белок серотипа 35 (Ad5-F35) и серотипа 11 (Ad5-F11). Хелпер-зависимые аденовирусные векторы дают полезную нагрузку до 30 т.п.н. и обеспечивают транзиентную экспрессию гена в HSC, а также могут быть использованы для доставки множества кластеров «ХЭ/экзонуклеаза», гибридов «ХЭ-TAL» и очень крупных матриц для рекомбинации. Альтернативно, эти векторы могут содержать небольшой полипептид Cas9 и «руководящие» РНК. Поэтому, эти модифицированные химерные аденовирусные векторы могут быть использованы для ингибирования гена и «редактирования» гена посредством рекомбинации в клеточных линиях HSC.
Дефицитные по интеграции лентивирусные векторы и «пенистые» вирусные векторы (IDLV и IDFV) дают полезную нагрузку 6 т.п.н. (IDLV) - 9 т.п.н. (IDFV) и, как было хорошо задокументировано, обладают способностью к трансдукции в человеческие HSC. В некоторых аспектах изобретения, оба вектора IDLV и IDFV могут быть использованы для ингибирования гена и для «редактирования» гена посредством рекомбинации в клеточных линиях HSC. IDLV с альтернативными кластерами «промотор-GFP» обеспечивают эффективную экспрессию в CD34+-HSC на высоком уровне. Высокие исходные титры могут быть достигнуты путем проведения стадии очистки TFF. Векторы с серией кластеров «промотор/GFP» могут быть использованы для достижения эффективной экспрессии ХЭ в CD34+-HSC на высоком уровне и для экспрессии одной ХЭ, ХЭ/Trex2, комбинации «мультиплекс-ХЭ» (то есть двух, трех или четырех ХЭ, которые были коэкспрессированы) и комбинации мультиплекс-ХЭ/TREX2. Экспрессия гибрида «мультиплекс/ХЭ» позволяет осуществлять множество событий расщепления в представляющей интерес области, и такая экспрессия, в зависимости от цели ее применения, может оказаться желательной для повышения уровня дерепрессии HbF. Такие мультиплексные стратегии следует проводить с использованием LHE, поскольку они функционируют автономно и могут быть с успехом использованы в комбинации с коэкспрессией TREX2, что позволяет осуществлять высокоэффективный и синхронный процессинг с одновременным образованием двухцепочечного разрыва. Альтернативно, эти векторы могут содержать небольшой полипептид Cas9 и «руководящие» РНК.
Эффективность нацеливания на ген-мишень; уровни экспрессии гена глобина в отдельных клетках-мишенях, а также в популяциях клеток и в их потомстве; влияние такого нацеливания на эритропоэз и на функцию стволовых клеток, а также на гематологические параметры и на функцию органов могут быть подтверждены в организмах-моделях.
После трансдукции может быть проведена оценка эффективности модификации одной клетки и всей клеточной популяции и оценка экспрессии β-подобных генов на уровни РНК и белка. Могут быть оценены изменения уровня связывания с фактором и изменения в структуре хроматина, а также морфология, степень неэффективного эритропоэза и апоптоза. Кандидаты, которые получили хорошую оценку в предварительном скрининге, могут быть дополнительно оценены на их влияние на плюрипотентность HSC, а также на способность ослаблять заболевание-специфические фенотипы in vitro и in vivo.
Предварительный скрининг ХЭ-кандидатов и систем доставки может быть осуществлен на клеточных линиях мышиного эритролейкоза, содержащих одну интактную человеческую хромосому 11 (N-MEL), и на нормальных человеческих CD34+-HSC клинической категории, где конечными точками такого скрининга является оценка эффективности введения целевой мутации и экспрессии гена глобина. Клетки обоих типов могут быть индуцированы для дифференцировки по эритроидному пути, в процессе которого сильно индуцируется экспрессия β-подобных генов с высокими отношениями β-глобина к γ- и δ-глобинам, что позволяет осуществлять количественную оценку эффектов регуляции гена глобина на уровне одной клетки и клеточной популяции. Оценки второго уровня могут включать проведение анализа на плюрипотентность трансдуцированных CD34+-клеток и на эритропоэз. Подходящие аналитические системы могут включать культивирование для оценки длительного пролиферативного потенциала, оценку миелоидных и эритроидных колоний для клонального анализа и трансплантацию этих колоний NOD-мышам SCID gamma (NSG) с последующей оценкой приживления первичных и вторичных трансплантатов клеток с множественной линией дифференцировки у реципиентов. Клиническая эффективность может быть одновременно оценена in vitro и in vivo.
Дефицит мышиного Bcl11a приводит к резкому дозозависимому увеличению уровня γ-глобина у мышей, содержащих локус человеческого β-глобина, и к улучшению серповидно-клеточного фенотипа у гуманизованных мышей-молелей. Хотя обе эти системы позволяют проводить анализ на экспрессию гена глобина, однако, мыши с серповидно-клеточной болезнью могут быть также оценены на улучшение фенотипа болезни у этих мышей, причем, особое внимание следует уделить оценке гематологических параметров, патологии печени и легких, функции почек и размера селезенки. Улучшение фенотипа может быть скоррелировано по числу HbF-содержащих клеток, по отношению HbF/HbS и по профилям экспрессии в анализах одной клетки.
Кроме того, продолжительность жизни и морфология эритроцитов могут быть оценены путем трансдукции человеческих CD34+-HSC у пациентов с гемоглобинопатией. Культивированные клетки при талассемии, в отличие от нормальных клеток, обнаруживают минимальный уровень размножения, отсутствие гемоглобинизации, неэффективный эритропоэз и усиленный апоптоз. По этим признакам может быть осуществлена количественная оценка уровней экспрессии и степени эритропоэза после нацеливания. Может быть также оценена степень серповидности потомства эритроидных CD34+-клеток в условиях гипоксии. CD34+-клетки, взятые у пациентов, могут быть трансплантированы мышам NSG, после чего, некоторые признаки аномального эритропоэза могут быть рекапитулированы, что позволит проводить оценку эффекта направленного мутагенеза.
Размножение аутологичных клеток HSC, ES и HSC, происходящих от iPSC
В других своих вариантах, настоящее изобретение относится к композициям и способам, применяемым для ex vivo размножения модифицированных гемопоэтических стволовых клеток (HSC) в целях осуществления эффективной трансплантации скорректированных клеток и использования индуцированных плюрипотентных стволовых клеток (iPSC) для скрининга и клинического применения. В некоторых аспектах этих вариантов, настоящее изобретение относится к композициям и способам, применяемым для эффективного размножения аутологичных HSC, аутологичных геномодифицированных HSC, ES и HSC, происходящих от iPSC. Может быть применена методика размножения клеток крови пупочного канатика, включающая использование Delta1 в бессывороточной среде, в которую были добавлены гемопоэтические факторы роста, и мобилизованных CD34+-клеток периферической крови, взятых у нормальных доноров. Эти композиции и способы могут быть применены в комбинации с одним или более дополнительными реагентами для повышения выживаемости и пролиферации гемопоэтических стволовых клеток/клеток-предшественников. В других аспектах изобретения, в этих композициях и способах могут быть использованы эндотелиальные сокультуры клеток для усиления размножения репопуляции долгоживущих клеток, включая скорректированные HSC, происходящие от iPSC.
Для эффективной клинической трансляции описанной здесь стратегии коррекции гена могут быть применены описанные здесь способы ex vivo размножения абсолютного числа скорректированных аутологичных HSC. Процедуры коррекции генов, по существу, являются более эффективными, если их проводят в небольших масштабах, и часто они ограничены лишь числом клеток HSC, которые могут быть использованы для коррекции. Таким образом, в настоящем изобретении рассматриваются способы размножения, которые могут быть применены для клинически приемлемого размножения ex vivo скорректированных HSC, ES и/или HSC, происходящих от индуцированных плюрипотентных стволовых клеток (iPSC). В некоторых своих аспектах, настоящее изобретение относится к способам размножения гемопоэтических стволовых клеток/клеток-предшественников, используемых в терапевтических целях, с учетом роли передачи сигнала Notch в определении дальнейшего метаболического пути стволовых клеток. Dahlberg et al., Blood 117:6083-90 (2010); Delaney et al., Nat. Med. 16:232-6 (2010); и Varnum-Finney et al., Nat. Med. 6:1278-81 (2000).
Эти способы позволяют осуществлять клинически релевантное ex vivo размножение клеток крови пупочного канатика/клеток-предшественников и терапию с применением размноженных клеток для лечения миелосупрессии у пациентов, которым была проведена трансплантация клеток крови пупочного канатика сначала с использованием частично HLA-совместимого свежего продукта (собранного после культивирования и введенного непосредственно в организм) и/или с использованием уже размноженного и криопротективного продукта в качестве стандартной клеточной терапии, проводимой на основе клеток, не совместимых с HLA.
Ex vivo размножение скорректированных по гену аутологичных HSC повышает безопасность и эффективность генотерапии на основе HSC благодаря трансплантации большего числа соответствующим образом скорректированных клеточных репопуляций, что позволяет достичь быстрой репопуляции и обеспечить преобладание скорректированных по генам клеток in vivo. В соответствии с этим, настоящее изобретение относится к композициям и способам, применяемым для поддерживающей терапии с использованием клеток, взятых у третьего лица, то есть HLA-несовместимых донорных ex vivo размноженных стволовых клеток/клеток-предшественников, которые способны обеспечивать быстрое, но транзиентное восстановление миелоидных клеток, значительно снижать риск раннего ассоциированного с трансплантацией летального исхода, вызванного инфекциями, появляющимися после трансплантации миелоалбяционных аутологичных клеток, не содержащих Т-клеток. Delaney et al., Nat. Med. 16:232-6 (2010).
Агенты, которые ингибируют дифференцировку (например, лиганд Notch), могут быть объединены с композициями и введены одновременно со способами усиления пролиферации и повышения выживаемости недифференцированных стволовых клеток/клеток-предшественников и таким образом может быть достигнуто улучшение Notch-опосредуемого разложения ex vivo. Повышенный уровень пролиферации стволовых клеток/клеток-предшественников крови пупочного канатика может быть достигнут путем объединения лиганда Notch, Delta1, с ингибитором арилуглеводородного рецептора (SRI) (Boitano et al., Science 329:1345-8 (2011)) или HoxB4 (Watts et al., Blood 116:5859-66 (2010) и Zhang et al., PLoS Med. 3:e173 (2006)) в целях повышения уровня пролиферации и ускорения самообновления гемопоэтических предшественников, и с ангиопоэтин-подобным рецептором 5 в целях повышения их выживаемости. Важным фактором клинического применения генотерапии является возможность размножения долгоживущих репопуляций клеток и гарантия долговечности трансплантата скорректированных клеток.
Akt-активированные эндотелиальные клетки могут быть использованы в системах совместного культивирования для подтверждения размножения клеток, скорректированных по гену. Butler et al., Cell Stem Cell 6:251-64 (2011). Размножение скорректированных по гену клеток зависит от индуцированной эндотелиальными клетками активации передачи Notch-сигнала в гемопоэтических предшественниках. Вторым важным аспектом клинического применения генотерапии является генетическая и эпигенетическая точность генерированных клеток по сравнению с их нормальными аналогами, что обеспечивает приобретение клеткой соответствующих свойств и отсутствие онкогенного потенциала in vivo. Важно отметить, что широкомасштабный анализ генома размноженных стволовых клеток/клеток-предшественников крови пупочного канатика, по сравнению с первичными выделенными CD34+-клетками, выявил наличие транскриптомы, хроматиновой структуры и ДНК-метиломы.
Стратегии размножения нормальных CD34+-клеток могут быть проведены в комбинации с определенными методами, в которых используются CD34+-клетки, взятые у пациентов с гемоглобинопатиями. В методике размножения клеток крови пупочного канатика могут быть использованы лиганд Delta1 в бессывороточной среде с добавленными гемопоэтическими факторами роста и мобилизованные CD34+-клетки периферической крови, взятые у нормальных доноров. Условия размножения ex vivo, оптимизированные с использованием разработанных анализов in vitro (иммунофенотипирования, культивирования и т.п.), и способность к репопуляции in vivo могут быть оценены на мышах-моделях NSG. Оптимизированные условия могут быть использованы в комбинации с композициями, которые включают SR1 (ингибитор арилуглеводородного рецептора), белки Hox или ангиопоэтины, в целях повышения уровня пролиферации и повышения выживаемости недифференцированных стволовых клеток/клеток-предшественников. Представляющие интерес комбинации могут быть оценены в анализах клеток-предшественников in vitro и у мышей с моделью иммунодефицита (NSG-мышей), а затем получены после размножения CD34+-клеток, взятых у здоровых индивидуумов для оценки способов размножения CD34+-клеток, взятых у пациентов с талассемией (и с другими гемоглобинопатиями).
Транскрипционная, генетическая и эпигенетическая точность клеток, размноженных вместе с их нормальными аналогами HSC, может быть оценена с помощью широкомасштабных анализов генома для выявления онкогенного потенциала генерированных клеток. После культивирования клеток in vivo (после инфузии) можно определить, имеют ли эти клетки функционально значимые аберрации, которые усиливают рост in vivo любого(ых) измененного(ых) клона(ов) и, тем самым, позволяют осуществлять селективное размножение и детектирование редко встречающихся клеток.
Клеточная терапия, направленная на устранение посттрансляционной нейтропении и улучшение состояния после трансплантации скорректированных по гену аутологичных HSC
В другом своем варианте, настоящее изобретение относится к композициям и к способам, применяемым для поддерживающей терапии, где указанные композиции и способы включают традиционную клеточную терапию, которая устраняет посттрансляционную нейтропению и улучшает состояние после трансплантации скорректированных по гену аутологичных HSC. Ex vivo размноженные криопротективные стволовые клетки/клетки-предшественники крови пупочного канатика (ПК) могут быть, например, введены как средство для поддерживающей терапии пациентам с талассемией и/или серповидно-клеточной болезнью, которым была проведена миелоабляционная HCT с использованием аутологичных скорректированных по гену CD34+-клеток.
Благодаря исследованиям, проводимым в целях получения экономически выгодного «готового к применению» источника клеток-предшественников, способных к быстрому восстановлению нейтрофилов, был создан банк продуктов предварительно размноженных криопротективных гемопоэтических клеток/клеток-предшественников, каждый из которых был получен из отдельной клеточной единицы ПК, которая может быть положена на хранение для будущих клинических исследований.
Безопасность введения этого «готового» и не совместимого с HLA продукта взрослым была продемонстрирована сразу после первой восстановительной химиотерапии рецидивирующего/не поддающегося лечению ОМЛ, а также после миелоабляционной ТПК, проводимой детям и взрослым со злокачественными гематологическими заболеваниями.
Была высказана гипотеза, что такой продукт размноженных клеток, который не содержат Т-клеток, может быть введен путем инфузии в качестве стандартной клеточной терапии для быстрой, но временной трансплантации миелоидных клеток и потенциального ускорения восстановления аутологичных гемопоэтических клеток у пациентов, которым была проведена миелоабляционная HCT с использованием аутологичных трансплантатов стволовых клеток, скорректированных по гену, и для предотвращения инфекционных осложнений и снижения риска летального исхода.
Очень важно определить, необходима ли HLA-совместимость для безопасного введения «готового» не совместимого с HLA продукта, который не содержит Т-клеток. Если нет необходимости в HLA-совместимости, то свежие единицы ПК могут быть собраны для немедленного размножения ex vivo, и конечный продукт может быть заморожен до тех пор, пока он не потребуется. Доступность «готового» продукта с размноженными клетками ПК для пациента может значительно возрасти, если все продукты с размноженными клетками будут депонированы в банк и будут доступны для любого пациента, независимо от HLA-типирования, расовой/этнической принадлежности или местонахождения пациента.
Кроме того, возможность разработки терапии на основе уже «готовых» размноженных универсальных донорных клеток является перспективной не только с точки зрения снижения продолжительности тяжелой нейтропении после HCT, но также, вероятно, с точки зрения расширения областей исследования по трансплантации не только стволовых клеток, но также, например, временной трансплантации миелоидных клеток для лечения тяжелой нейтропении, индуцированной химиотерапией; любой приобретенной тяжелой нейтропении; или случайного радиоактивного облучения.
Ex vivo размножение клеток позволяет снизить риск при ТПК благодаря предотвращению замедленного восстановления гемопоэтических клеток, и значительно повысить общую выживаемость. Такой пониженный риск развития рецидивов у пациентов, подвергнутых двойной ТПК, и пациентов с хронической реакцией «трансплантат против хозяина» (GVHD) будет еще ниже, несмотря на высокую степень несовместимости трансплантатов. Если риск летального исхода, связанного с трансплантацией недифференцированных клеток, может быть снижен путем инфузии ex vivo размноженных клеток-предшественников крови пупочного канатика, вводимых для усиления восстановления гемопоэтических клеток, то, вероятно, что общая выживаемость будет выше, чем выживаемость, наблюдаемая при введении клеток, взятых у обычных неродственных доноров.
В других своих вариантах, настоящее изобретение относится к клеточной терапии, осуществляемой в целях предотвращения посттрансляционной нейтропении и улучшения состояния после трансплантации скорректированных по гену аутологичных HSC. У пациентов с талассемией, которые были подвергнуты миелоабляционной HCT с использованием скорректированных по гену аутологичных клеток, наблюдался повышенный риск инфицирования и летального исхода, вызванного ограниченным числом CD34+-клеток во введенном трансплантате (до тех пор, пока не будет достигнуто ex vivo размножение этих скорректированных по гену клеток до клинически приемлемого количества). Инфузия продукта предварительно размноженных и криопротективных клеток-предшественников крови пупочного канатика, проводимая в качестве стандартной поддерживающей терапии, может быть осуществлена для снижения риска летального исхода благодаря быстрому, но временному восстановлению миелоидных клеток вплоть до введения трансплантата долгоживущих клеток, обеспечивающих восстановление гемопоэтических клеток.
У пациентов, которые были подвергнуты миелоабляционной HCT, наблюдалась тяжелая панцитопения, непосредственно вызванная соответствующим курсом лечения, и у всех пациентов во время такого лечения наблюдались повышенный риск инфицирования и кровотечения. Время восстановления гемопоэтических клеток (нейтрофилов и тромбоцитов) непосредственно зависит от дозы CD34+-клеток, и, таким образом, у пациентов, которым была проведена миелоабляционная HCT с использованием клеток крови пупочного канатика, где доза стволовых клеток составляла 1/10 от всего стандартного донорного трансплантата, или с использованием трансплантатов с низкой дозой аутологичных CD34-богатых клеток, риск летального исхода после трансплантации, вызываемый замедленным восстановлением гемопоэтических клеток, еще больше увеличивался.
Для устранения риска при применении таких методов HCT и повышения безопасности при проведении этих методов крайне необходимо разработать такие новые методы терапии, которые могли бы способствовать предотвращению пролонгированной панцитопении и более быстрому восстановлению гемопоэтических клеток. Как обсуждалось выше, была разработана стратегия, при которой абсолютное число репопуляций гемопоэтических стволовых клеток/клеток-предшественников крови пупочного канатика (ПК) в костном мозге (HSPC) может быть увеличено путем культивирования с лигандом Notch Delta1. Было продемонстрировано, что инфузия этих частично HLA-совместимых ex vivo размноженных клеток ПК детям или взрослым, которым была проведена трансплантация клеток крови пупочного канатика (ТПК), является безопасной и позволяет значительно сократить время до достижения первоначального абсолютного числа нейтрофилов, равного 500, с 26 до 11 дней в результате быстрой трансплантации миелоидных клеток, обеспечиваемой размноженными клетками.
Благодаря исследованиям, проводимым совсем недавно в целях получения экономически выгодного «готового к применению» источника клеток-предшественников, способных к быстрому восстановлению нейтрофилов, был создан банк продуктов предварительно размноженных криопротективных гемопоэтических клеток/клеток-предшественников, где каждый из этих продуктов был получен из отдельной клеточной единицы ПК, которая может быть положена на хранение для будущих клинических применений. Авторами настоящего изобретения была также продемонстрирована безопасность введения этого «готового» не совместимого с HLA продукта взрослым сразу после первой восстановительной химиотерапии рецидивирующего/не поддающегося лечению ОМЛ, а также после миелоабляционной ТПК, проводимой детям и взрослым со злокачественными гематологическими заболеваниями. Авторами настоящего изобретения была высказана гипотеза, что такой продукт размноженных клеток, который не содержит Т-клеток, может быть введен путем инфузии в качестве стандартной клеточной терапии для быстрой, но временной трансплантации миелоидных клеток и потенциального ускорения восстановления аутологичных гемопоэтических клеток у пациентов, которым была проведена миелоабляционная HCT с использованием аутологичных трансплантатов скорректированных по гену стволовых клеток и, тем самым, для предотвращения инфекционных осложнений и снижения риска летального исхода.
С применением определенных оптимальных методов получения ex vivo размноженных стволовых клеток/клеток-предшественников крови пупочного канатика был создан банк готовых продуктов размноженных клеток для оценки безопасности введения этих клеток в качестве поддерживающей терапии, проводимой путем трансплантации HCT аутологичных скорректированных по гену гемопоэтических клеток.
Для лучшего понимания настоящего изобретения ниже приводятся нижеследующие неограничивающие примеры.
ПРИМЕРЫ
Пример 1
Отбор хоминг-эндонуклеаз, нацеленных на ген Bcl11a, по I-HjeMI, I-CpaMI и I-OnuI с применением компартментализации in vitro (IVC)
Открытую рамку считывания (OPC) родительской хоминг-эндонуклеазы LAGLIDADG (LHE), I-HjeMI (фиг.14; SEQ ID NO: 28; Jacoby et al., Nucl. Acids Res. 40(11):4954-4964 (2012); Taylor et al., Nucl. Acids Res. 40 (Web Server issue): Wl 10-6 (2012)), оптимизированную по кодонам для экспрессии в E. coli, клонировали между рестрикционными NcoI- и NotI-сайтами pET21-a(+) (фиг.11; EMD Millipore (Novagen) division of Merck KGaA). Для осуществления сайт-направленного насыщающего мутагенеза в ОРС I-HjeMI, ДНК-фрагменты, содержащие неполную ОРС приблизительно с 20 парами оснований в области, перекрывающейся с фланкирующими фрагментами на обеих сторонах, подвергали ПЦР-амплификации с использованием праймеров, содержащих вырожденный кодон 5ʹ-NNK-3ʹ (кодирующий все 20 аминокислот). Аминокислотные остатки, мутированные с использованием таких ПЦР-праймеров, представлены в таблице 2. ПЦР-продукты очищали путем экстракции из агарозного геля и подвергали сборке в последующем раунде ПЦР с использованием последовательности, содержащей 2 копии сайтов-мишеней, для отбора вариантов эндонуклеаз. Собранный ДНК-фрагмент снова очищали посредством гель-экстракции и использовали в качестве библиотеки в методе компартментализации in vitro (IVC).
После каждого раунда сайт-направленного насыщающего мутагенеза было проведено три раунда IVC для получения большого количества вариантов генов нуклеазы с измененной специфичностью. Смесь «масло-поверхностно-активное вещество» (2% ABIL EM 90 (Evonik Industries AG Personal Care, Essen, North Rhine- Westphalia, Germany), 0,05% тритона X-100 в легком минеральном масле) тщательно смешивали с насыщающим буфером (100 мМ глутамата калия (pH 7,5), 10 мМ ацетата магния (pH 7,5), 1 мМ дитиотреитола и 5 мг/мл альбумина бычьей сыворотки), инкубировали при 37°C в течение 20 минут, и центрифугировали при 16000×g в течение 15 минут при 4°C. 500 микролитров верхней фазы использовали для эмульгирования 30 мкл смеси для синтеза белка in vitro (25 мкл PURExpress (New England Biolabs, Ipswich, MA), 20 единиц ингибитора РНКазы, 1 мг/мл альбумина бычьей сыворотки и 8 нг библиотеки ДНК) при непрерывном перемешивании при 1400 об/мин в течение 3,5 минут на льду. Эмульсию инкубировали при 30°C в течение 4 часов, а затем нагревали при 75°C в течение 15 минут. Эмульгированные капли собирали путем центрифугирования при 16000×g в течение 15 минут при 4°C, и разрушали путем добавления фенола/хлороформа/изоамилового спирта. Нуклеиновые кислоты регенерировали путем осаждения этанолом и обрабатывали смесью РНКаз (Life Technologies Corporation (Invitrogen), Grand Island, New York). После очистки с использованием набора для ПЦР-очистки QIAquick (Qiagen, Hilden, Germany), библиотеку ДНК лигировали с ДНК-адаптором, содержащим 3ʹ-выступающую последовательность из 4 оснований, комплементарную липкому концу сайта-мишени, генерированного эндонуклеазными вариантами, экспрессированными в эмульгированных каплях, а затем добавляли к ПЦР-смеси, содержащей пару праймеров, один из которых является специфичным к лигированному ДНК-адаптору, в целях получения множества генов эндонуклеазных вариантов, присоединенных к расщепленному сайту-мишени. ПЦР-ампликон подвергали гель-очистке, и ОРС вариантов гена снова подвергали ПЦР-амплификации с получением библиотеки ДНК, используемой в последующем раунде IVC.
Во втором раунде IVC, эмульсию приготавливали с использованием 1 нг реконструированной библиотеки и инкубировали при 42°C в течение 75 минут, а затем реакцию транскрипции/трансляции in vitro гасили путем нагревания при 75°C. Библиотеку ДНК регенерировали, и гены активной эндонуклеазы специфически обогащали путем с помощью ПЦР после лигирования с ДНК-адаптором, описанным выше.
В третьем раунде IVC, смесь для синтеза белка in vitro, содержащую 0,5 нг фрагмента библиотеки, эмульгировали в 4,5% Span 80/0,5% тритона X-100/легком минеральном масле. Реакцию проводили при 42°C в течение 45 минут, и смесь термоинактивировали при 75°C. После экстракции из эмульсии, гены эндонуклеазы, ассоциированные с расщепленным сайтом-мишенью, подвергали ПЦР-амплификации и второму раунду сайт-направленного мутагенеза.
Для реконструирования вариантов I-HjeMI, которые распознавали половину (-)- и (+)-сайтов гена-мишени BCL11A, проводили 4 и 2 раунда сайт-направленного насыщающего мутагенеза, соответственно (фиг. 13). Пул нуклеазных вариантов, нацеленных на первую половину сайта, подвергали дополнительному (пятому) раунду мутагенеза на поверхности, противоположной пограничной области белок-ДНК, а затем проводили 3 раунда IVC (таблица 2).
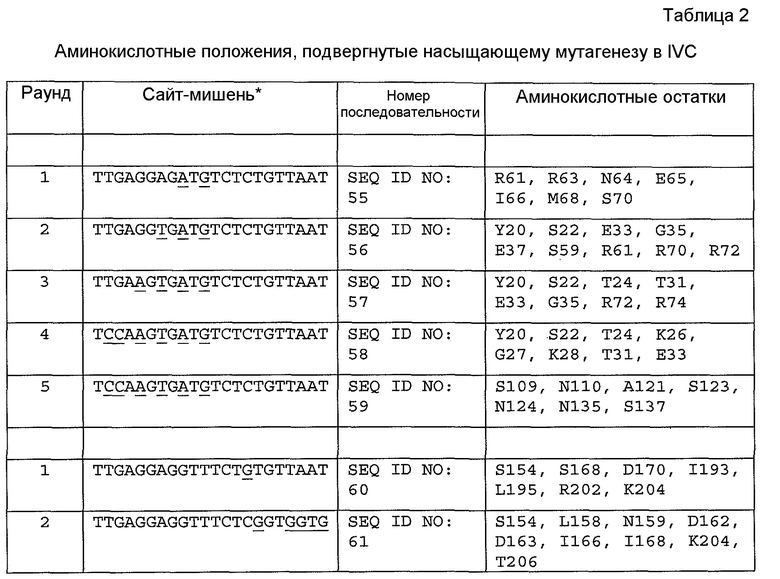
* Подчеркнутые нуклеотиды отличаются от нуклеотидов, присутствующих в сайте-мишени для родительской LHE I-HjeMI.
Была проведена сборка ДНК-фрагментов, кодирующих N-концевую и С-концевую половину доменов вариантов эндонуклеазы I-HjeMI, ответственных за распознавание половины (-)- и (+)-сайтов гена-мишени BCL11A, после чего, пул нуклеаз, которые расщепляли полноразмерный сайт-мишень гена BCL11A, отбирали за 3 раунда IVC (фиг.13).
В таблице 3 представлены репрезентативные последовательности-мишени BCL11a, содержащие модули последовательности ДНК, для которых были выделены пулы в высокой степени активных вариантов эндонуклеазы (на основе хоминг-эндонуклеаз I-CpaMI и I-OnuI) и определены их последовательности.
Chr2:60, 542, 847-60, 542, 868
Дизрупция форм Bcl11a-Х и XL
Chr2:60, 542, 630-60, 542, 651
Дизрупция форм Bcl11a-Х и XL
Chr2: 60, 543, 005-60, 543, 026
Дизрупция всех форм Bcl11a
Пример 2
Оптимизация активности вариантов I-HjeMI, нацеленных на ген Bcl11a, с помощью анализа на расщепление, проводимого в бактериях с использованием двух плазмид с элиминацией гена
Активность вариантов I-HjeMI, полученных путем отбора с применением селективных методов представления в IVC (описанных выше в примере 1), оптимизировали с использованием двухплазмидной системы отбора в бактериальных клетках методом Doyon et al., J. Am. Chem. Soc. 128(7): 2477-2484 (2006). ОРС эндонуклеазных генов встраивали между NcoI- и NotI-сайтами экспрессионной плазмиды pENDO (фиг. 12, Doyon et al., J. Am. Chem. Soc. 128(7): 2477-2484 (2006)). Компетентные клетки NovaXGF (EMD Millipore (Novagen)), содержащие репортерную плазмиду pCcdB (фиг. 31, Doyon et al., J. Am. Chem. Soc. 128(7): 2477-2484 (2006); Takeuchi et al., Nucl. Acids Res. 37(3): 877-890 (2009); и Takeuchi et al., Proc. Natl. Acad. Sci. U.S.A. 10.1073/pnas.1107719108 (2011)), включающую 4 копии гена-мишени BCL11A, трансформировали пулом плазмиды pEndo, кодирующей варианты I-HjeMI. Трансформанты культивировали в среде 2×YT (16 г/л триптона, 10 г/л дрожжевого экстракта и 5 г/л NaCl) при 37°C в течение 30 минут, а затем 10-кратно разводили средой 2×YT, в которую было добавлено 100 мкг/мл карбенициллина и 0,02% L-арабинозы (для предварительного индуцирования экспрессии вариантов I-HjeMI). После выращивания культуры при 30°C в течение 15 часов, клетки собирали, ресуспендировали в стерильной воде и распределяли по неселективным (1×M9 соль, 1% глицерин, 0,8% триптон, 1 мМ MgSO4, 1 мМ CaCl2, 2 мкг/мл тиамина и 100 мкг/мл карбенициллина) и селективным планшетам (то есть неселективным планшетам, в которые добавляли 0,02% L-арабинозы и 0,4 мМ IPTG для индуцирования экспрессии токсичного белка CcdB). После инкубирования при 30°C в течение 30-40 часов, плазмиду pEndo экстрагировали из выживших колоний на селективных планшетах.
ОРС, кодирующие активные варианты I-HjeMI, амплифицировали посредством ПЦР с вероятностью ошибки с использованием набора для неспецифического мутагенеза (Gene Morph II Random Mutagenesis Kit) (Agilent Technologies, Santa Clara, CA). После гидролиза ферментами NcoI, NotI и DpnI, полученные фрагменты снова клонировали в вектор pEndo. Плазмиду подвергали 2 раундам отбора в условиях, при которых варианты эндонуклеаз экспрессировали при 30°C в течение 4 часов, а затем переносили в планшет. N-концевую половину и C-концевую половину доменов выбранных генов подвергали перестановке с помощью перекрывающейся ПЦР, а затем снова клонировали в вектор pEndo. Трансформированные клетки, несущие плазмиду pEndo и репортерную плазмиду pCcdB, культивировали в среде 2×YT, содержащей 0,02% L-арабинозы при 37°C в течение одного часа, а затем распределяли по селективным планшетам и оставляли на 16-20 часов при 37°C. После 2 раундов отбора, проводимых в тех же самых условиях жесткости, плазмиду pEndo экстрагировали из выживших колоний на селективных планшетах, и секвенировали ОРС вариантов генов, находящихся на этой плазмиде.
Пример 3
Активность эндонуклеаз, нацеленных на ген BCL11A, протестированная в анализе на расщепление с использованием двух плазмид
Активность репрезентативной эндонуклеазы, нацеленной на ген BCL11A (BCL11Ahje; фиг. 17, SEQ ID NO: 31), ее каталитически неактивного варианта (BCL11-Ahje D18N) и родительского фермента LHE I-HjeMI (фиг. 14, SEQ ID NO: 28) измеряли в бактериальных клетках, несущих репортерную плазмиду pCcdB (Doyon et al., J. Am. Chem. Soc. 128(7): 2477-2484 (2006)), содержащую 4 копии сайта-мишени для I-HjeMI (мишени для I-HjeMI) или гена-мишени BCL11A (TCCAAGTGATGTCTCGGTGGTG (SEQ ID NO: 39; нуклеотиды, отличающиеся от нуклеотидов, присутствующих в сайте-мишени для родительского LHE I-HjeMI, подчеркнуты). Репортерная плазмида pCcdB кодирует «белок, ответственный за предотвращение гибели клеток В» («ccdB», токсичный белок в бактериях, индуцируемый добавлением IPTG). Расщепление сайтов-мишеней в репортерной плазмиде приводит к RecBCD-опосредуемой деградации репортерной плазмиды и соответствующей выживаемости клеток в селективной среде, содержащей IPTG. Степень выживаемости определяли путем деления числа колоний, присутствующих на селективных планшетах, на число колоний, присутствующих на неселективных планшетах. Величины ошибок указаны как ± ср.кв.откл. для 3 независимых экспериментов.
Пример 4
Детектирование направленного мутагенеза в эндогенном человеческом гене BCL11A
Клетки HEK 293T (1,6×105) высевали в 12-луночные планшеты за 24 часа до трансфекции и трансфецировали 0,5 мкг экспрессионной плазмиды для нуклеазы, нацеленной на ген BCL11A, и нуклеазы TREX2. Через 48 часов после трансфекции, трансфецированные клетки подвергали лизису, и геномную ДНК экстрагировали с использованием набора Quick-gDNA MiniPrep kit (Zymo Research). Приблизительно 500 п.н.-фрагмент, охватывающий ген-мишень BCl11A, подвергали ПЦР-амплификации из 50 нг экстрагированных геномов с использованием пары следующих праймеров: BCl11A_up1, 5ʹ- GCT GGA ATG GTT GCA GTA AC -3ʹ (SEQ ID NO: 66); BCL11A_down1, 5ʹ- CAA ACA GCC ATT CAC CAG TG -3ʹ (SEQ ID NO: 67). ПЦР-ампликон инкубировали в 1×NEB буфере 4 плюс 1×BSA (New England Biolabs) в отсутствии или в присутствии 0,5 мкМ нуклеазы, нацеленной на ген BCL11A, которая была выделена из E. coli, сверхэкспресирующих рекомбинантный белок, и оставляли на 2 часа при 37°C. Реакцию завершали добавлением 5 × терминирующего раствора (50 мМ трис-HCl (pH7,5), 5 мМ EDTA, 0,5% ДСН, 25% глицерина, 0,1 оранжевого G и 0,5 мг/мл протеиназы K). После инкубирования при комнатной температуре в течение 15 минут, половину каждого образца разделяли в 1,6% агарозном геле, содержащем этидийбромид в TAE (верхние панели). Другую половину каждого образца очищали с помощью набора DNA Clean & Concentrator-5 kit (Zymo Research) и использовали в качестве матрицы во втором раунде ПЦР, проводимом с применением пары следующих праймеров: BCl11A_up2, 5ʹ- CTG CCA GCT CTC TAA GTC TCC -3ʹ (SEQ ID NO: 68); BCl11A_down2, 5ʹ- TGC AAC ACG CAC AGA ACA CTC -3ʹ (SEQ ID NO: 69). ПЦР-продукт снова гидролизовали нуклеазой, нацеленной на ген BCL11A, в условиях, описанных выше, и анализировали на 1,6% агарозном геле (нижние панели) (см. фиг. 30).
Пример 5
Отбор эндонуклеаз, нацеленных на область сайленсинга фетального гемоглобина, на основе I-OnuI посредством компартментализации in vitro
Репрезентативные последовательности-мишени для хоминг-эндонуклеазы (ХЭ), которые равномерно распределены по всей 350 п.н.-области (SEQ ID NO: 2), включающей область захвата Bcl11a, присутствующую в области сайленсинга HbF в эритроидных клетках взрослых, которая имеет разрыв в точке делеции HPFH Френча, представлены в таблице 4. Эти последовательности-мишени содержат модули последовательности ДНК, для которых были выделены и секвенированы пулы в высокой степени активных вариантов эндонуклеазы.
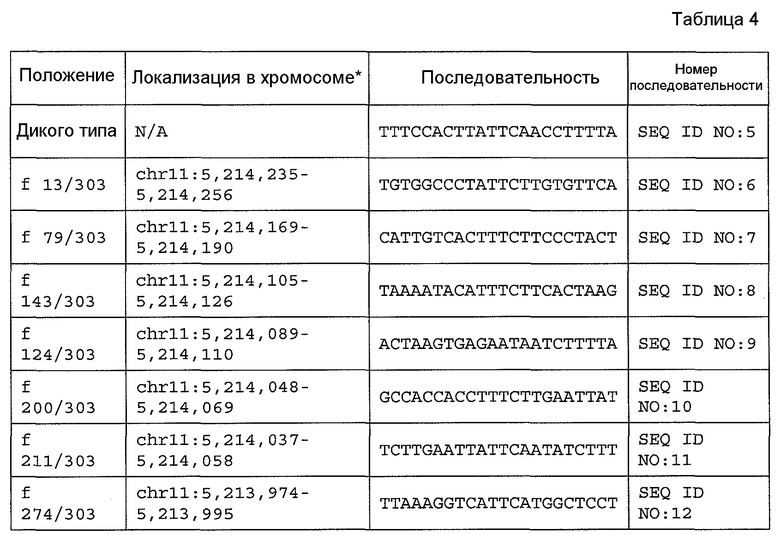
В таблице 5 представлена область, которая простирается на -100 п.н. - 210 п.н. выше генов глобина, и которая идентична обоим генам Aγ- и Gγ-глобина и содержит множество не-делеционных мутаций HPFH. Редактирование гена по этим мутациям приводит к снижению степени репрессии и, тем самым, к активации гена гамма и к повышению уровня HbF.
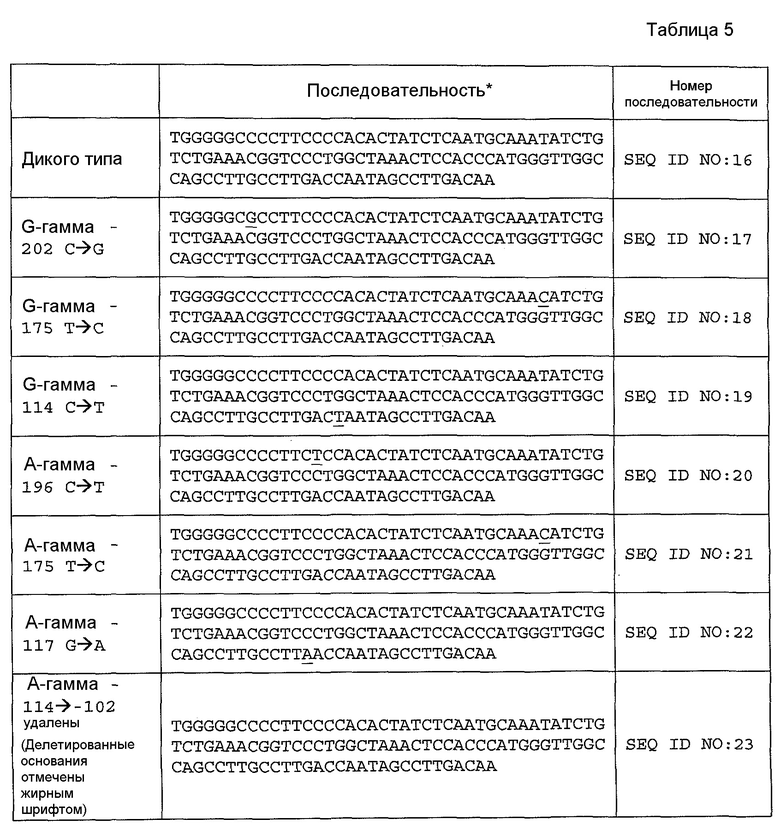
В таблице 6 представлены аминокислотные положения в родительской хоминг-эндонуклеазе LHE I-OnuI (фиг. 22A, SEQ ID NO: 34), которая была подвергнута насыщающему мутагенезу в IVC (как описано выше в примере 1) с получением хоминг-эндонуклеаз, нацеленных на область сайленсинга человеческого фетального глобина.


* Подчеркнутые нуклеотиды отличаются от нуклеотидов, присутствующих в сайте-мишени для родительской LHE I-OnuI.
На фиг.23 представлены результаты анализа на расщепление, проводимого с использованием «половины мишеней», происходящей от области сайленсинга человеческого фетального глобина. Амплифицированные полосы содержат обе расщепленные половины сайтов (захваченные посредством лигирования с комплементарными дуплексными олигонуклеотидами и соответствующими выступающими оцДНК) и последовательности вариантов фермента, ответственных за генерирование расщепленных продуктов ДНК. Конечная стадия после завершения обогащения библиотек «половинными сайтами» эндонуклеазы включает сборку фрагментов ДНК, кодирующих N-концевые и C-концевые половины доменов хоминг-эндонуклеазы I-OnuI, ответственные за распознавание левых (L) и правых (R) половин сайтов области-мишени сайленсинга гамма-глобина. Активные эндонуклеазы I-OnuI были отобраны из пула эндонуклеаз, расщепляющих полноразмерную область-мишень сайленсинга человеческого фетального глобина.
Пример 6
Хоминг-эндонуклеазы MegaTAL с N-концевыми гибридами TAL-якорей, используемые для повышения специфичности и активности эндонуклеазы, нацеленной на ген
N-концевые гибриды TAL-якорей могут быть использованы для повышения специфичности и активности эндонуклеазы, нацеленной на ген, включая одну или более хоминг-эндонуклеаз, таких как одна или более хоминг-эндонуклеаз I-HjeMI, I-CpaMI и I-OnuI. MegaTAL конструировали с применением стратегии сборки Golden Gate, описанной Cermak et al., Nucl. Acids Res. 39:e82-e82 (2011), с использованием плазмидной библиотеки RVD и специально сконструированного вектора (см., фиг.24, SEQ ID NO: 35, и фиг.25, SEQ ID NO: 36, для нуклеотидной и аминокислотной последовательностей MegaTAL:5.5 RVD + Y21-AniI).
Плазмиды модифицировали для сборки TAL-эффектора, содержащего 1,5-10,5 TAL-повторов и их соответствующих RVD («вариабельных повторяющихся остатков», которые контактируют со смежными основаниями ДНК в 5ʹ-области сайта-мишени и, тем самым, определяют когнатную последовательность ДНК в этой области). Специально сконструированный вектор pthX01 модифицировали так, чтобы он включал гемаглютининовую (HA) метку, расположенную непосредственно за NLS, и получали каркасную молекулу TALEN, начинающуюся в положении остатка 154 (по сравнению с TAL-эффектором PthXo1 дикого типа) и заканчивающуюся 63 остатками, расположенными за конечной последовательностью «половины TAL-повтора».
TAL-эффекторы конструировали с использованием нижеследующих RVD для нацеливания на каждый специфический нуклеотид: A - NI, C - HD, G - NN и T - NG. После клонирования повторов TAL-эффектора в специально сконструированный вектор, отдельный белковый линкер («Zn4»; VGGS) и сконструированные варианты хоминг-эндонуклеазы клонировали в каталитический домен нуклеазы FokI между сконструированными рестрикционными Xba-I- и Sal-I-сайтами.
Эта конструкция представляет собой «набор тест-моделей» MegaTAL (гибрид TAL-эффектора в N-концевой области одной цепи белка, полученный путем лигирования гибкого линкера с хоминг-эндонуклеазой Y2 I-AniI дикого типа, и впервые описанный Takeuchi et al., Nucl. Acids Res. 37(3): 877-890 (2009)).
Пример 7
Эндонуклеазная система на основе Cas9, сконструированая для дизрупции Bcl11a-регулируемой области сайленсинга фетального гемоглобина (HbF)
Недавно высказанная механистическия трактовка кластеризованной системы коротких регулярно чередующихся палиндромных повторов (CRISPR), которые используются бактериями для адаптивного иммунитета, позволила разработать мощный инструмент, с помощью которого можно осуществлять «редактирование» генома клеток млекопитающих, и который может быть применен в композициях и способах для лечения описанных здесь гемоглобинопатий, где указанные композиции и способы применяются: (a) для дизрупции кодирующей области Bcl11a; (b) для дизрупции регуляторного элемента или пути сайленсинга ДНК HbF, такого как Bcl11a-регулируемая область сайленсинга HbF; (c) для мутации одного или более промоторов гена γ-глобина в целях повышения уровня экспрессии гена γ-глобина; (d) для мутации одного или более промоторов гена δ-глобина в целях повышения уровня экспрессии гена δ-глобина; и/или (e) для коррекции одной или более мутаций гена β-глобина. Бактериальная система CRISPR описана в публикациях Jinek et al., Science 337:816-821 (2013); Cong et al., Science (Jan. 3, 2013) (Электронная публикация до выхода в печать); и Mali et al., Science (Jan. 3, 2013) (Электронная публикация до выхода в печать).
Белок Cas9 продуцирует двухцепочечный разрыв в специфическом сайте, локализация которого определяется последовательностью «руководящей» РНК. Все «руководящие» РНК содержат одну и ту же каркасную последовательность, связывающуюся с Cas9, а также вариабельную нецеливающую последовательность, имеющую структуру G-N20-GG, которая специфически расщепляет комплекс Cas9-РНК. Коэкспрессия белка Cas9 и «руководящей» РНК приводит к эффективному расщеплению и дизрупции в последовательность-специфическом участке человеческого генома, где такое последовательность-специфическое расщепление определяется последовательностью «руководящей» РНК. Коэкспрессия Cas9 и «руководящих» РНК, которые являются специфичными к множеству мишеней, позволяет осуществлять эффективную делецию в промежуточной области, находящейся между сайтами-мишенями.
Таким образом, в некоторых аспектах настоящего изобретения, Cas9-опосредуемое «редактирование» генома применяется: (1) для дизрупции сайта связывания Bel11a в области сайленсинга HbF; (2) для нарушения функции гена Bcl11a и (3) для делеции всей области сайленсинга HbF. Были идентифицированы области-мишени, а затем были сконструированы «руководящие» РНК исходя из информации об оптимальных последовательностях-мишенях для «руководящей» РНК. В настоящей заявке описаны репрезентативные «руководящие» РНК, нацеленные на Bcl11a-связывающую область в области сайленсинга HbF, а также на один связывающий мотив GATA-1. Эти «руководящие» РНК используются отдельно или в комбинации для направленной дизрупции области сайленсинга HbF. Cas9 с «руководящими» РНК, присоединенная к дополнительным областям в областях сайленсинга HbF, соответствующих пикам захвата Bcl11a, также используются отдельно и в комбинации друг с другом. Несколько пар «руководящих» РНК, фланкирующих Bcl11a-связывающий сайт и мотив GATA-1, а также полноразмерный участок футпринтинга, могут быть также коэкспрессированы с Cas9 для генерирования делеций в области сайленсинга HbF.
Последовательность человеческого оптимизированного по кодонам Cas9, описанная Mali et al., Science (Jan. 3, 2013), представлена на фиг.26, SEQ ID NO: 37. Родовая последовательность «руководящей» РНК (Mali et al.) представлена на фиг. 27, SEQ ID NO: 38, а ключевые элементы этой последовательности представлены в таблице 7. Репрезентативные последовательности Cas9 с «руководящими» РНК, специфически связывающиеся с мишенью и расщепляющие область сайленсинга человеческого фетального гемоглобина (HbF) (фиг. 6, SEQ ID NO: 1 и фиг. 7, SEQ ID NO: 2) представлены в таблице 8.
AAGTTAAAATAAGGCTAGTC
CGTTATCAACTTGAAAAAGT
GGCACCGAGTCGGTGCT
TCAGACTTGG
TGCAGTAGGG
AAATAGCTGG
AAGGTGGTGG
ATCACAAGGG
GGCCATGAGG
Пример 8
Векторные системы для экспрессии эндонуклеаз
Для обогащения клеток перед трансплантацией, мышам с моделью NSG, серповидно-клеточной болезни и талассемии были перенесены человеческие CD34-клетки или мышиные ядросодержащие клетки костного мозга вместе с флуоресцентным маркером с помощью проточной цитометрии. Подходящими методами трансдукции являются нижеследующие методы.
Векторы AAV6. Рекомбинантные AAV-векторы серотипа AAV6 давали полезную нагрузку в 4,5 т.п.н., достаточную для доставки кластеров «промотор-ХЭ-экзонуклеаза» или «промотор-гибрид TAL-ХЭ-экзонуклеаза» вместе с небольшой рекомбинантной матрицей. Альтернативно, эти векторы могут содержать Cas9 и «руководящую» РНК. Кроме того, авторами были получены предварительные данные, которые показали, что AAV6 позволяет осуществлять наиболее эффективную трансдукцию человеческих CD34+-клеток крови пупочного канатика в капсиды всех известных AAV и может опосредовать транзиентную экспрессию гена в HSC на значительном уровне.
Модифицированные аденовирусные векторы. Аденовирусные векторы с гибридными капсидами обладают способностью эффективно трансдуцировать гемопоэтические клетки многих типов, включая CD34+-клетки. Более эффективная трансдукция с использованием химерного вектора, содержащего волоконный белок серотипа 35 (Ad5-F35), была продемонстрирована Dr. Rawlings (SCH), а полученные совсем недавно данные позволяют предположить, что волоконный белок серотипа 11 (Ad5-F11) может быть даже более эффективным в гемопоэтических клетках. Хелпер-зависимые аденовирусные векторы дают полезную нагрузку до 30 т.п.н. и обеспечивают транзиентную экспрессию гена в HSC, а поэтому они могут быть использованы для доставки множества кластеров ХЭ/экзонуклеаза, гибридов ХЭ-TAL, а также очень крупных рекомбинантных матриц или Cas9-экспрессирующего кластера и множества «руководящих» РНК.
Дефицитные по интеграции лентивирусные векторы и векторы на основе «пенистых» вирусов (IDLV и IDFV). Эти векторы дают полезную нагрузку от 6 т.п.н. (IDLV) до 9 т.п.н. (IDFV) и обладают хорошо подтвержденной способностью трансдуцировать человеческие HSC. Оба вектора IDLV и IDFV были использованы для ингибирования гена и «редактирования» гена в HSC на основе рекомбинации. Dr. Rawlings (SCH) и Kiem (FHCRC) были получены и оценены серии IDLV с альтернативными кластерами «промотор-GFP», и было установлено, что они имеют конструкции, обеспечивающие эффективную экспрессию CD34+-HSC на высоком уровне.
Прямая нуклеофекция плазмиды и мРНК. Условия для эффективной трансдукции клеток N-MEL и CD34-клеток были определены с использованием системы нуклеофекции Amaxa. Преимуществами являются отсутствие интеграции и способность трансдуцировать множество экспрессионных плазмид или молекул РНК одновременно.
Одновременно с этим, отсортированные и неотсортированные клетки могут быть трансплантированы отдельным мышам. Хотя более поздние трансплантаты могут содержать небольшое число модифицированных клеток, однако, исследования посттрансплантационных химер, проводимые с участием человека, показали, что эти клетки имеют селективное преимущество в отношении выживания и присутствуют в большом количестве на периферии. Несмотря на это, анализ РНК одиночных ретикулоцитов и анализ F-клеток позволяет выявить дизрупцию генов в клетках, даже если они присутствуют в небольшом количестве.
Может быть также проведен анализ второго уровня на плюрипотентность трансдуцированных CD34+-клеток и эритропоэз. Эти анализы могут включать культивирование клеток для оценки способности к длительной пролиферации, анализ миелоидных и эритроидных колоний на присутствие клонов и трансплантацию этих колоний мышам NOD scid gamma (NSG), а затем оценку приживления клеток множественных линий дифференцировки первичного и вторичного трансплантата у реципиентов.
Обычно, стволовые клетки вводят путем инъекции в хвостовую вену после облучения всего организма (275 рад для NSG-мышей или 1000 рад для мышей C57). Хотя такая доза является эффективной, однако, при проведении большинства исследований, авторами была проведена прямая инъекция мышам непосредственно в бедренную вену стволовых клеток, количество которых в 50 раз меньше необходимой дозы, а в идеальном случае, для оценки эффективности такой инъекции, возможно введение еще более ограниченного числа клеток, отсортированных методом проточной цитометрии, и/или модифицированных клеток. После анестезии и местной аналгезии и после определения анатомических ориентиров, в пространство мозгового вещества бедренной кости непосредственно вводили 0,5-1 миллион клеток.
Пример 9
Характеризация хоминг-эндонуклеаз и эндонуклеазы Cas9 для эффективного нацеливания на ген
Клинический эффект нацеливания на ген был продемонстрирован путем оценки уровней экспрессии гена глобина в отдельных клетках-мишенях и в популяциях клеток, а также оценки влияния нацеливания на эритропоэз, на функцию стволовых клеток, на гематологические параметры и на функцию органов в организмах-моделях.
После трансдукции проводили оценку одиночных клеток и популяций всех клеток на эффективность нацеливания на ген и экспрессию всех β-подобных генов на уровне РНК и белка. Изменения в связывании с фактором и в структуре хроматина оценивали по морфологии клетки и по степени неэффективного эритропоэза и апоптоза. Эндонуклеазы-кандидаты, которые имели хорошую оценку в предварительном скрининге, дополнительно оценивали на их влияние на плюрипотентность HSC, а также на способность ослаблять заболевание-специфические фенотипы in vitro и in vivo.
Предварительный скрининг эндонуклеаз-кандидатов и систем доставки осуществляли в клеточной линии мышиного эритролейкоза, содержащей одну интактную человеческую хромосому 11 (N-MEL) и на нормальных человеческих CD34+-HSC клинической категории, где конечными точками такого скрининга является оценка эффективности введения целевой мутации и экспрессии гена глобина. Клетки обоих типов могут быть индуцированы для дифференцировки по эритроидному пути, в процессе которого экспрессия β-подобных генов в высокой степени индуцируется с низкими отношениями РНК □-глобина/□- + β-глобина. С использованием HbF-специфического антитела может быть количественно оценен процент «F-клеток». Такие низкие отношения являются идеальными, поскольку указанные системы являются чувствительными для детектирования и оценки даже небольшого увеличения уровня мРНК □-глобина, а также экспрессии HbF на уровне одной клетки и на уровне их популяций.
Клетки N-MEL представляют собой производные клеток мышиного эритролейкоза, содержащие одну интактную человеческую хромосому 11, которая имеет локус β-глобина. Такая эритроидная клеточная линия обычно экспрессирует низкие уровни мышиных и человеческих β-подобных генов глобина, но она может быть индуцирована, в результате чего происходит дифференцировка, во время которой уровень экспрессии глобина значительно возрастает.
Клетки N-MEL эффективно трансдуцировали с использованием системы нуклеофекции Amaxa. 25% клеток трансдуцировали с использованием 2 мкг плазмидной ДНК, набора «L» и программы A20. В результате инфицирования при множественности заражения (m.o.i.) 20 лентивирусным вектором на основе RSCS-MCS-PG-WZ, содержащим хоминг-эндонуклеазу, сконструированную в целях дизрупции гена Bcl11a, получали приблизительно 40% трансдуцированных клеток N-MEL.
Эффективность направленной дизрупции оценивали в анализе Cel-1. Область-мишень амплифицировали с помощью ПЦР, подвергали тепловой денатурации, повторному отжигу и обработке ферментом Cel-1, который эффективно расщепляет пузырьки, происходящие от несоответствующих областей размером по меньшей мере в одну пару оснований. Если область-мишень была мутирована любым способом, то гетеродуплексы цепей дикого типа и мутантных цепей расщепляли и детектировали с помощью гель-электрофореза. Этот анализ может быть проведен на популяциях всех клеток для оценки эффективности мутации или на отдельных клетках, отсортированных с помощью проточной цитометрии, и такой анализ множества клеток давал точную оценку частоты мутации.
С помощью рутинных количественных анализов ОТ-ПЦР Taqman (кол. ОТ-ПЦР) на РНК, полученные от популяции всех клеток, было установлено, что экспрессия β-глобина 11-кратно индуцируется, в результате чего происходит дифференцировка с отношением □-глобина/□- + β-глобина = 0,1%. После инфицирования Bcl11a-дефицитным вектором может наблюдаться 30-кратное увеличение такого отношения, что указывает на то, что эта система клеточных культур позволяет получить точные данные по дизрупции Bcl11a-опосредуемого пути сайленсинга HbF.
Поскольку изменение путей Bcl11a может приводить к относительному повышению уровня РНК □-глобина, но к снижению уровня экспрессии гена глобина, может быть проведена проточная цитометрия для сортинга 1000 клеточных пулов, которые были лизированы, а также могут быть проведены вышеописанные анализы с помощью кол.ОТ-ПЦР. Поскольку РНК непосредственно сравнивают для одного и того же числа клеток, то любое снижение количества РНК глобина зависит от увеличения уровня Ct, что позволяет проводить прямое измерение уровня экспрессии β- и □-глобина, а также отношения □-глобина/□- + β-глобина.
Для определения процента клеток, которые имеют измененное отношение □-глобина/□- + β-глобина после модификации, и для определения уровней изменения наблюдаемой экспрессии проводили анализы на одиночных клетках. Отдельные клетки, отсортированные с помощью проточной цитометрии, подвергали одновременно нескольким циклам рутинной ОТ-ПЦР РНК □- и β-глобина, и после получения адекватного материала, позволяющего осуществлять точное расщепление образца, □- и β-глобин анализировали с помощью кол. ОТ-ПЦР, как описано выше.
В конечном счете проводили анализ отдельных клеток и всех клеток на присутствие HbF, но не на уровне РНК, а на уровне белка, который является терапевтическим. Популяции всех клеток оценивали с помощью ВЭЖХ после клеточного лизиса и элюирования на колонке с гемоглобином, а затем определяли отношения HbF к HbA и HbA2. Число клеток, экспрессирующих HbF, сравнивали до и после трансдукции с использованием клеток, фиксированных на глутаральдегиде, после чего эти клетки делали проницаемыми с помощью детергента и добавляли HbF-специфическое антитело, а затем проводили количественную оценку с помощью проточной цитометрии.
Эффекты от введения мутаций оценивали в модифицированных клеточных клонах после выделения отдельных клеток с помощью проточной цитометрии. Для этого проводили описанные выше анализы. Кроме того, для того, чтобы показать, что целевые мутации нарушают связывание Bcl11a-репрессивного комплекса, клетки фиксировали в формальдегиде, выделяли хроматин и фрагментировали путем обработки ультразвуком. Иммунную преципитацию на хроматине (ChIP) осуществляли с использованием коммерчески доступных антител против Bcl11a, GATA-1 и HDAC-1. Связывание этих белков с областью-мишенью в клетках N-MEL, а также в эритроидных дифференцированных CD34-клетках описано ниже. После направленной дизрупции проводили анализ на отсутствие связывания.
Пример 10
Гемопоэтические стволовые CD34+-клетки клинической категории
CD34+-клетки, взятые от нормальных доноров, подвергали адгезии к культуральным чашкам под действием фибронектинового пептида CH-296, и два раза, c перерывами в 8 часов, инфицировали описанными выше лентивирусами на основе RSCS-MCS-PG-WZ при m.o.i. 20 в среде, содержащей G-CSF, SCF, IL-3, IL-6, FLT-3 и TPO. В результате этой процедуры было инфицировано ~80% клеток. Трансдуцированные клетки подвергали дифференцировке в эритроидные клетки в соответствии с протоколом Douy, Giarratana et al., Nat. Biotechnol 23(1):69-74 (2005). Описанные выше кол.ОТ-ПЦР-анализы показали, что уровень □-глобина по отношению к уровню □- + β-глобина составляет 4%. Такое низкое отношение позволяет проводить чувствительное детектирование увеличения уровня глобина после «редактирования» генома.
Для оценки изменений на стадии дифференцировки осуществляли проточную цитометрию с использованием множества маркеров клеточной поверхности, включая CD34, CD71 и гликофорин, а также проводили анализ роста клеток и морфологии на цитоцентрифуге после окрашивания по Райту-Гимзе. Эффективность дизрупции оценивали с помощью анализов Cel-1 на популяциях всех клеток и на одиночных клетках, как описано выше. Дополнительную оценку уровней экспрессии гена глобина и HbF осуществляли как описано выше с помощью кол.ОТ-ПЦР на 1000 клеточных пулах и на отдельных клетках, а также с помощью ВЭЖХ и анализов F-клеток. ChIP проводили для оценки связывания Bcl11a, GATA-1 и HDAC-1 с областью-мишенью.
Для оценки клинической эффективности в человеческих CD34+-клетках, HSC, взятые у пациентов с гемоглобинопатией, трансдуцировали и культивировали теми же самыми методами. Помимо рутинного анализа для нормальных клеток проводили дополнительные заболевание-специфические анализы. Культивированные клетки, взятые у пациента с талассемией, обнаруживали минимальное размножение, не вырабатывали гемоглобин, обнаруживали эффективный эритропоэз и подвергались повышенному апоптозу по сравнению с нормальными клетками. Это дало возможность провести количественную оценку повышения уровня экспрессии и эритропоэза после нацеливания. Аналогичным образом оценивали степень серповидности эритроидного потомства CD34+-клеток в условиях гипоксии. Эти трансдуцированные CD34+-клетки, взятые у пациентов с гемоглобинопатией, трансплантировали мышам NSG, после чего осуществляли рекапитуляцию нескольких признаков аномального эритропоэза, что позволяло проводить оценку эффекта направленного мутагенеза.
Клиническую эффективность оценивали in vivo у мышей с моделью гемоглобинопатии. Ингибирование мышиного Bcl11a приводило к резкому дозозависимому увеличению уровня □-глобина у мышей, содержащих локус человеческого β-глобина на трансгене, и к уменьшению серповидного фенотипа у гуманизованных мышей-моделей. Хотя анализ на экспрессию гена глобина может быть проведен с использованием обеих систем, однако, для мышей с серповидным фенотипом клеток может быть проведен анализ на улучшение фенотипа, причем, особое внимание следует уделить таким гематологическим параметрам, как гематокрит, патология печени и легких, функция почек и размер селезенки. Эти данные могут коррелировать с числом HbF-содержащих клеток, отношением HbF/S и паттернами экспрессии в анализах на одиночных клетках, проводимых методами, аналогичными методам, описанным выше.
Для анализа РНК использовали цельную кровь, поскольку она содержит достаточное количество РНК-содержащих ретикулоцитов, подходящих для анализа. Для анализов, проводимых на одиночных клетках, кровь окрашивали тиазоловым оранжевым и собирали РНК-содержащие клетки в соответствии с профилями прямого и бокового рассеяния эритроцитов. Время жизни эритроцитов у мышей с серповидными клетками является довольно низким, и увеличение времени жизни было оценено после внутрибрюшинной инъекции 100% эритроцитов, помеченных NHS-биотином. На этой стадии, один микролитр крови окрашивали стрептавидином-ФИТЦ и оставшийся процент меченых эритроцитов оценивали с помощью проточной цитометрии. Этот эксперимент проводили на мышах, выведенных от мышей с серповидными клетками BERK, имеющихся у авторов настоящего изобретения. Paszty et al., Science 278(5339}: 876-878 (1997). Кроме того, отношение □-глобина/□- + β-глобина оценивали у мышей, содержащих человеческие YAC с β-глобином A25 и A85. Porcu et al., Blood 90(11):4602-4609 (1997).
Были оценены две модели с талассемией, то есть мыши, гетерозиготные по Th-3 (Yang et al., Proc. Natl. Acad. Sci. U.S.A. 92(25):11608-11612 (1995)), и мыши, гетерозиготные по делеции LCR (Bender et al., Mol Cell. 5(2):387-393 (2000)). В каждом случае, в анализе мышей с талассемией определяли гематокрит, проводили оценку неэффективного эритропоэза, определяли размер селезенки, оценивали избыточный уровень железа и проводили анализ морфологии органов.
| название | год | авторы | номер документа |
|---|---|---|---|
| КОМПОЗИЦИИ И СПОСОБЫ ДЛЯ ЛЕЧЕНИЯ ГЕМОГЛОБИНОПАТИЙ | 2015 |
|
RU2707540C2 |
| КОМПОЗИЦИИ И СПОСОБЫ, ПРЕДНАЗНАЧЕННЫЕ ДЛЯ ЛЕЧЕНИЯ ГЕМОГЛОБИНОПАТИЙ | 2018 |
|
RU2834231C2 |
| КОМПОЗИЦИИ И СПОСОБЫ ЛЕЧЕНИЯ ГЕМОГЛОБИНОПАТИЙ | 2016 |
|
RU2812491C2 |
| ИММУНОЛОГИЧЕСКИ ОТЛИЧИМЫЕ ВАРИАНТЫ ПОВЕРХНОСТИ КЛЕТОК ДЛЯ ПРИМЕНЕНИЯ В КЛЕТОЧНОЙ ТЕРАПИИ | 2017 |
|
RU2793803C2 |
| ИНСЕРЦИЯ ТАРГЕТНОГО ГЕНА ДЛЯ УЛУЧШЕННОЙ КЛЕТОЧНОЙ ИММУНОТЕРАПИИ | 2017 |
|
RU2824204C2 |
| КОМПОЗИЦИИ, ПОВЫШАЮЩИЕ ЧИСЛО КОПИЙ ВЕКТОРА (ЧКВ), И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2017 |
|
RU2744603C2 |
| НАЦЕЛИВАНИЕ НА КЛЕТКИ, ОПОСРЕДУЕМОЕ ХИМЕРНЫМ АНТИГЕННЫМ РЕЦЕПТОРОМ | 2018 |
|
RU2783316C2 |
| Направляемое РНК уничтожение вируса JC человека и других полиомавирусов | 2015 |
|
RU2747722C2 |
| СПОСОБ СОЗДАНИЯ Т-КЛЕТОК, ПРИГОДНЫХ ДЛЯ АЛЛОГЕННОЙ ТРАНСПЛАНТАЦИИ | 2015 |
|
RU2752933C2 |
| МОДУЛЯТОРЫ ТАУ И СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ ИХ ДОСТАВКИ | 2017 |
|
RU2789459C2 |





































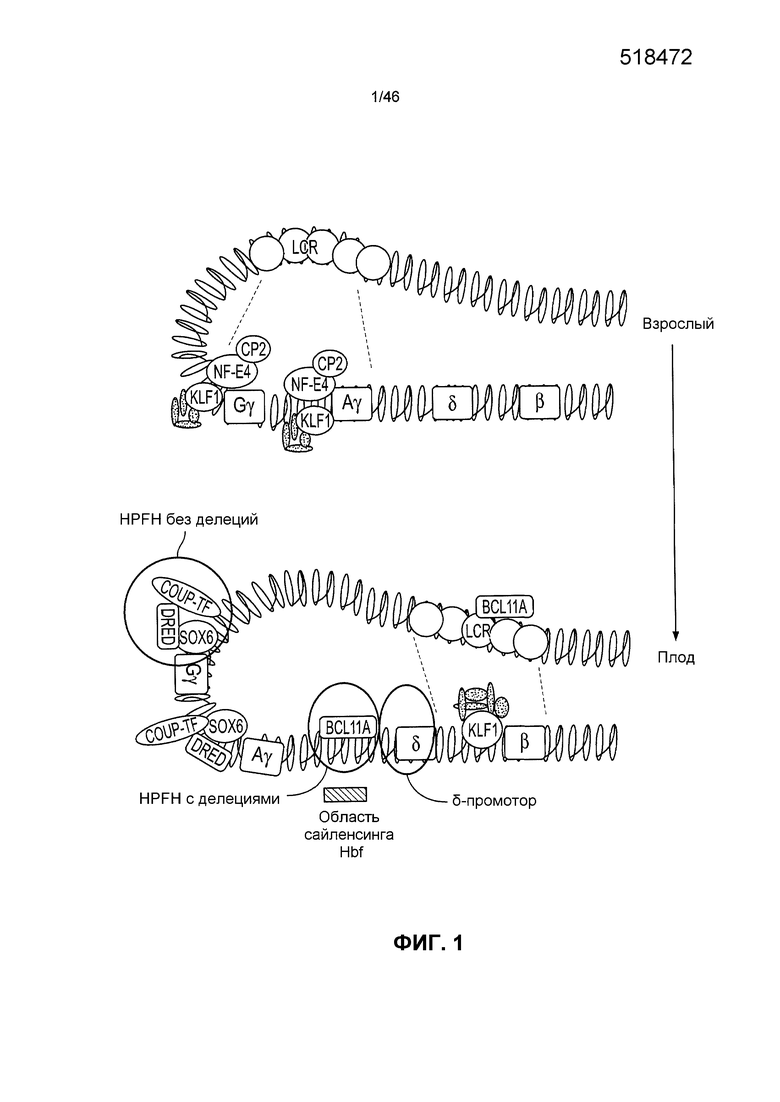

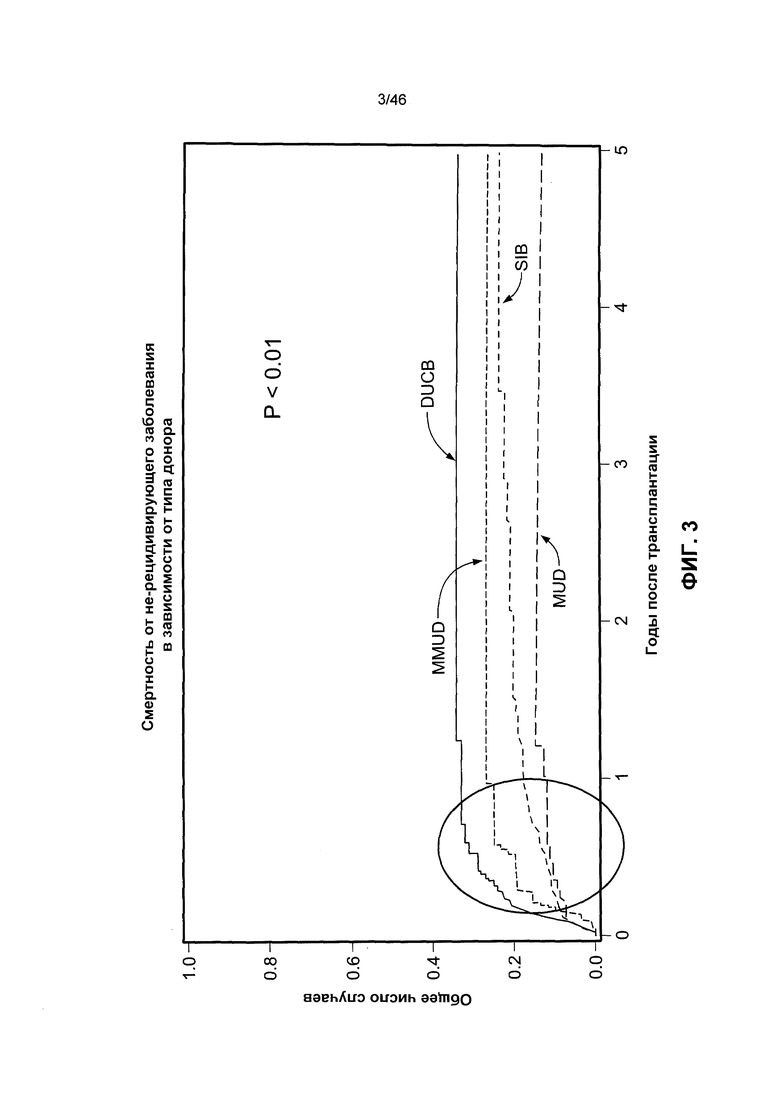
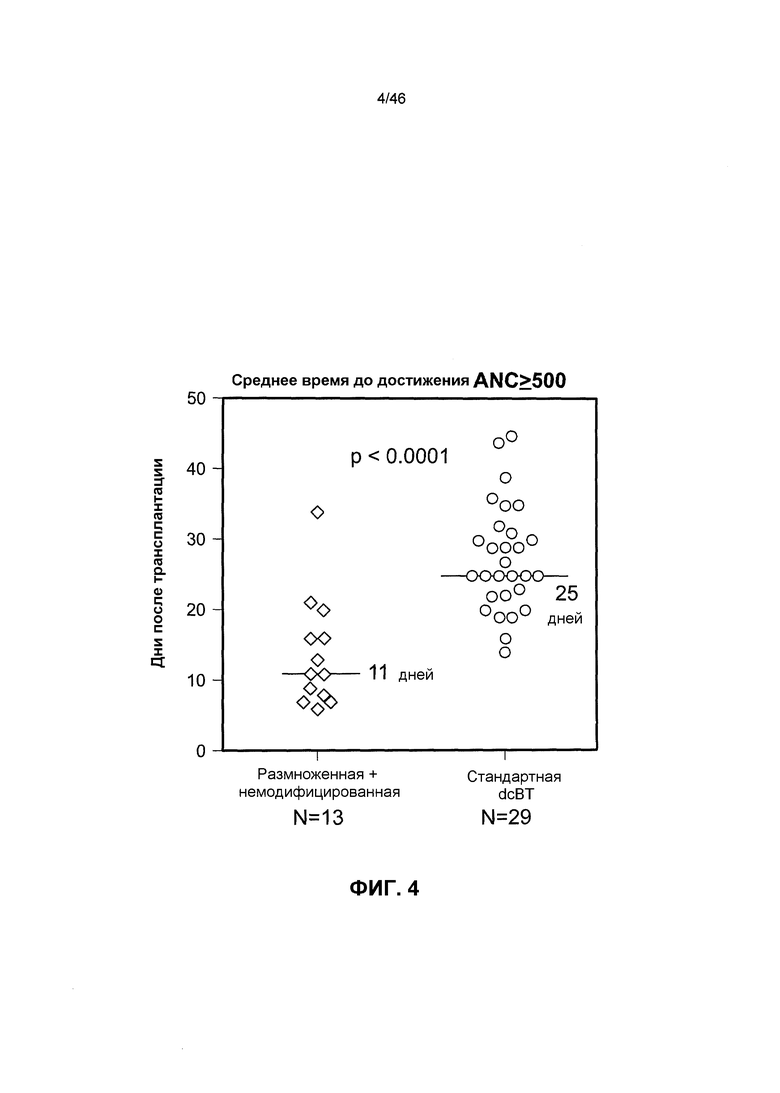
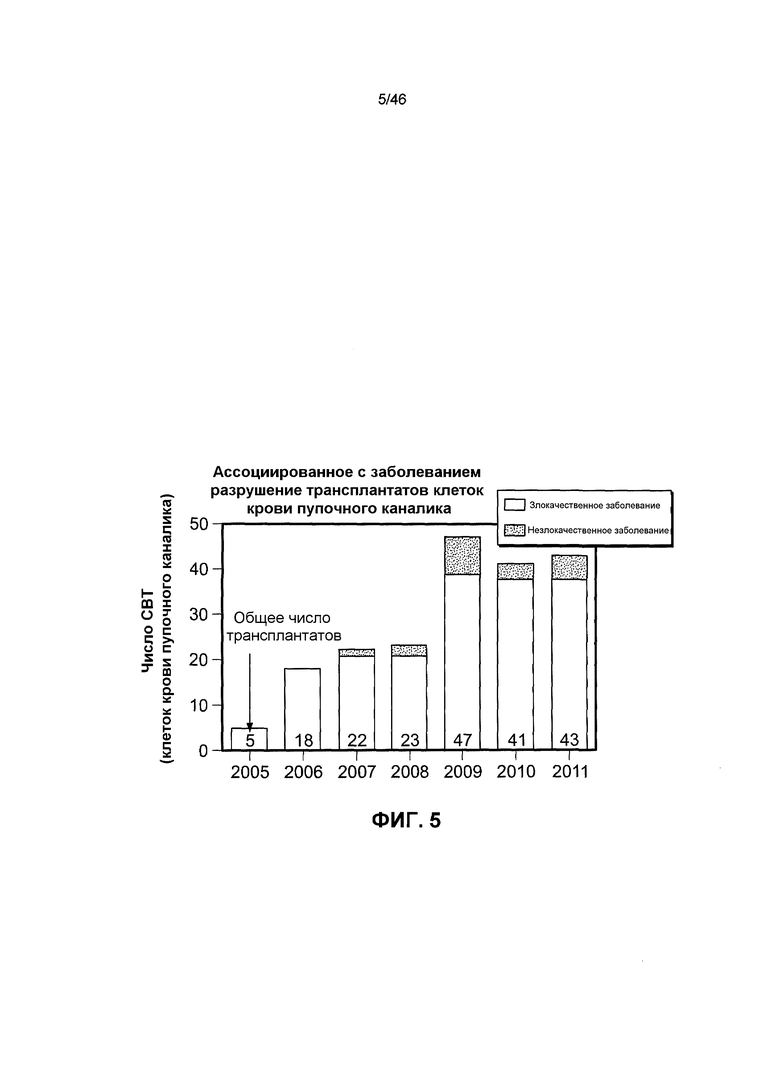









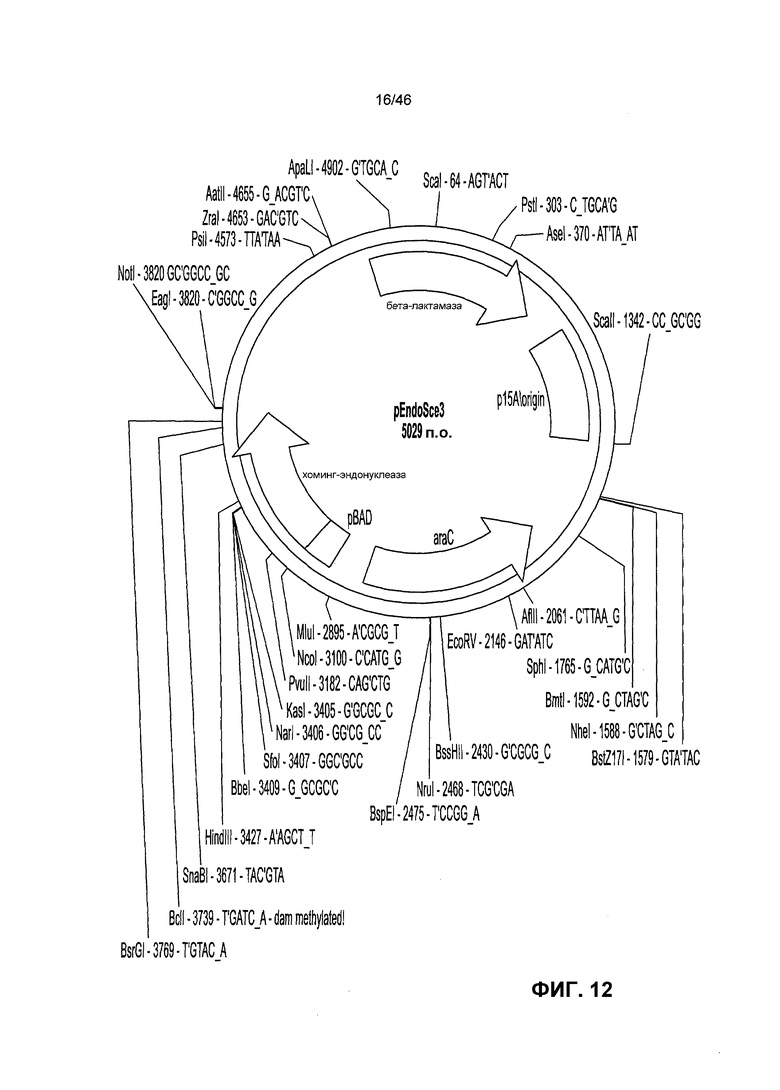
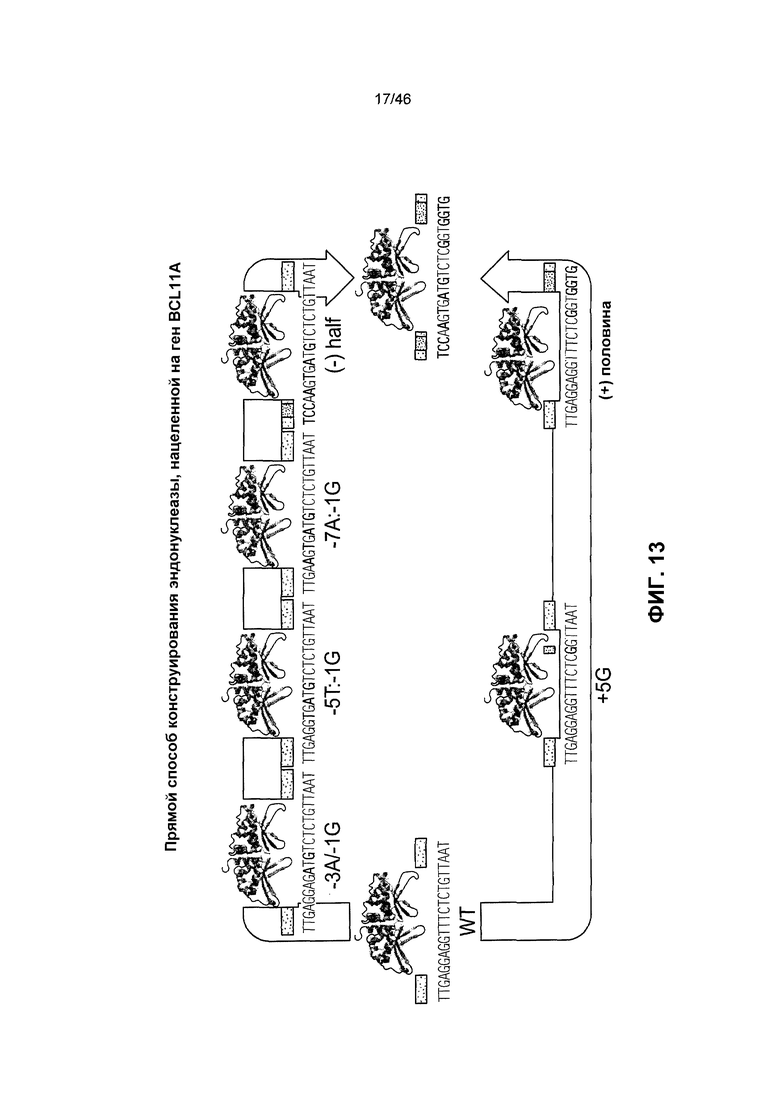






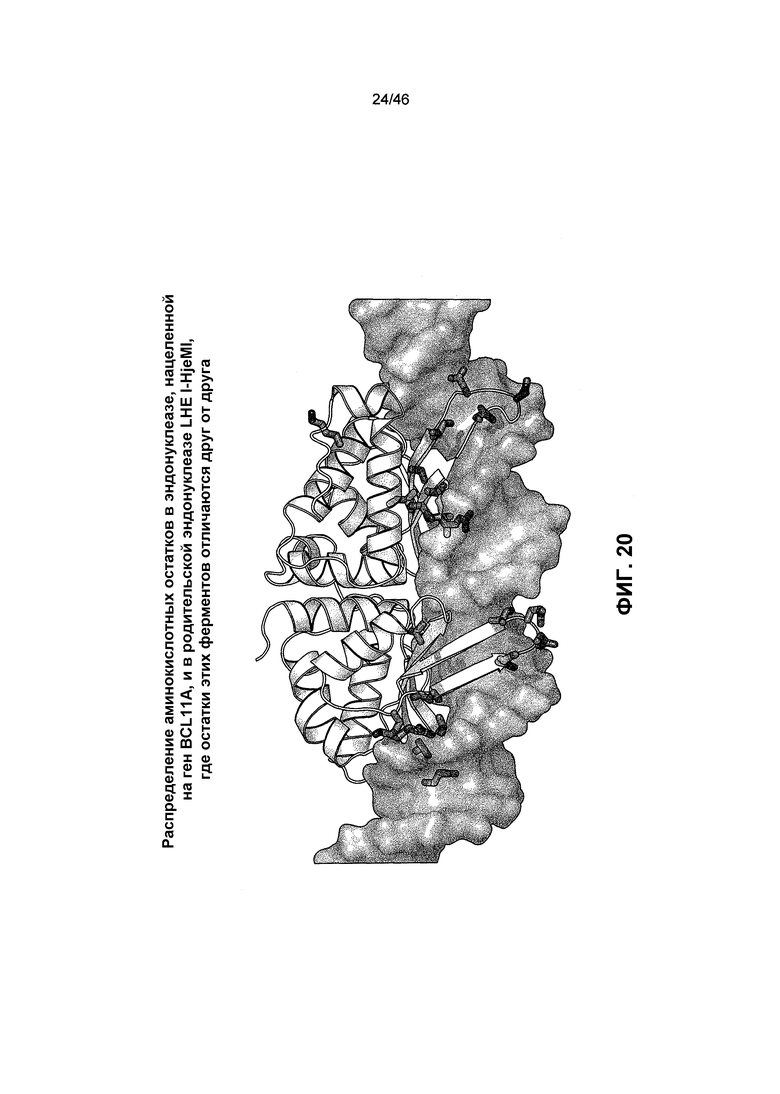
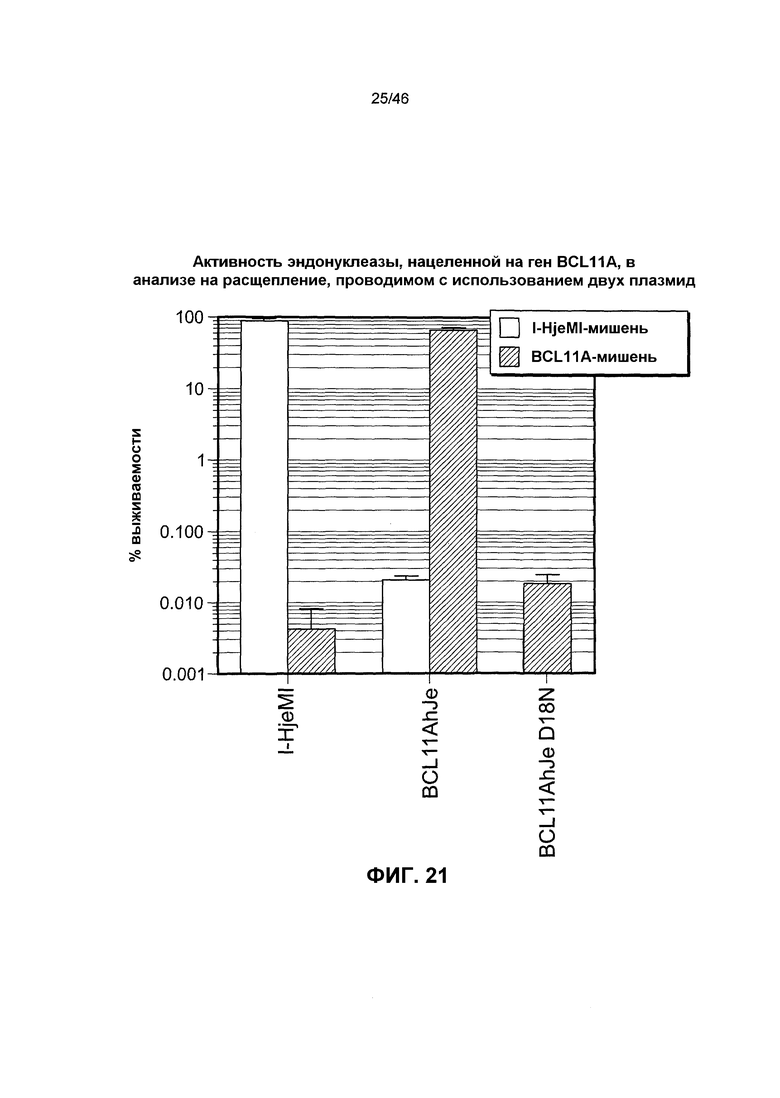


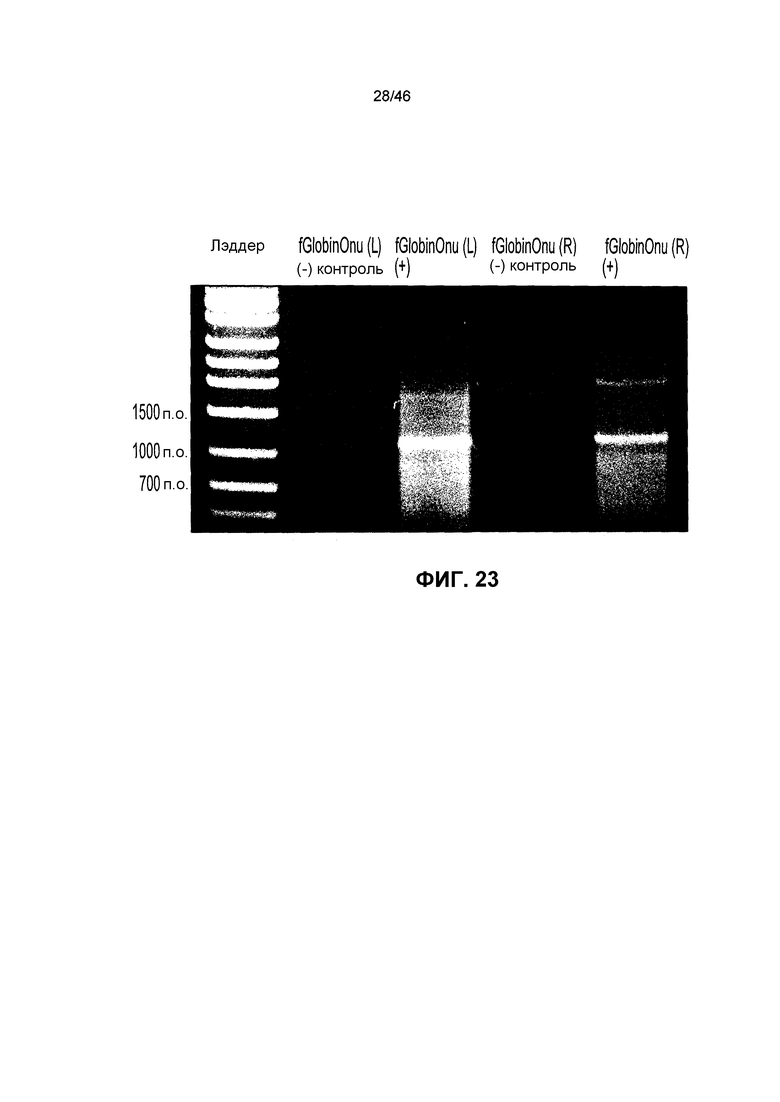









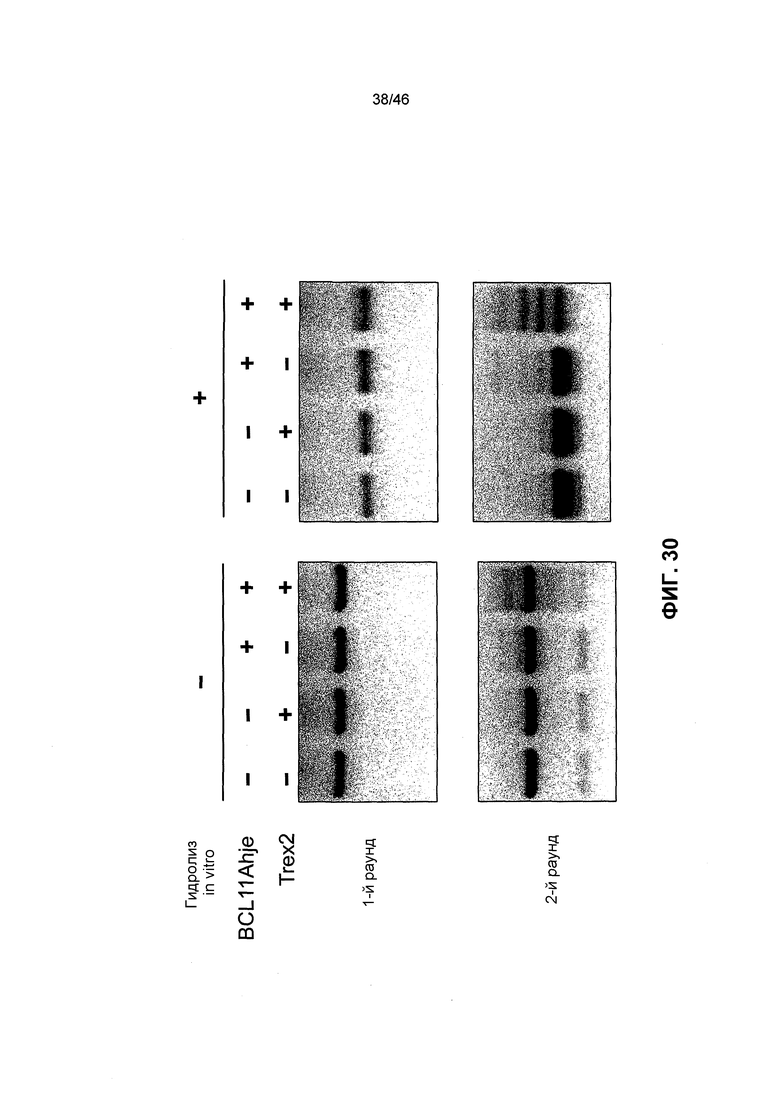





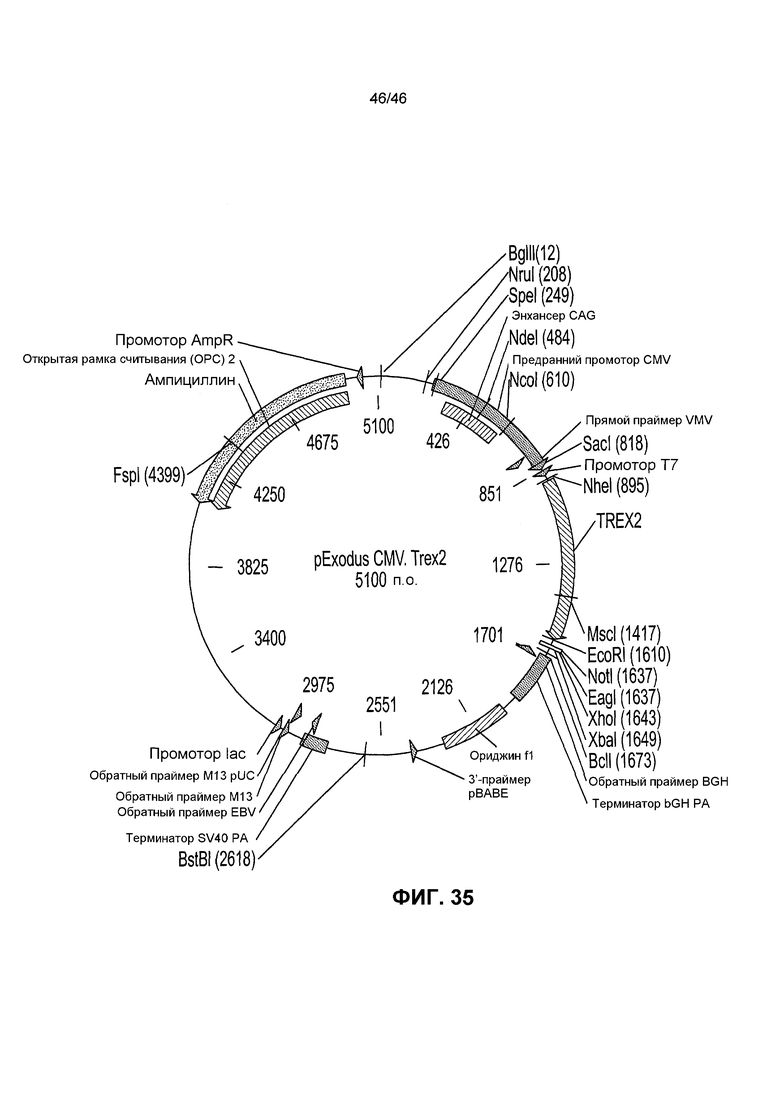
Представленная группа изобретений касается полинуклеотида, кодирующего вариант хоминг-эндонуклеазы (ХЭ) I-OnuI, экспрессионного вектора, содержащего такой полинуклеотид, полипептида эндонуклеазы, композиции для лечения гемоглобинопатии и клетки, содержащей указанные полинуклеотид, вектор или полипетид эндонуклеазы. Охарактеризованный полинуклеотид имеет последовательность SEQ ID NO: 34 с мутациями, соответствующими одной или более аминокислотным заменам, выбранными из группы, состоящей из L26, R28, R30, N32, S40, E42, G44, Q46, A70, S72, S78, K80, T82, F182, N184, I186, S190, K191, Q197, V199, S201, K225, K227, D236, V238 и T240. Получаемый вариант ХЭ I-OnuI связывается с нуклеотидной последовательностью области сайленсинга фетального гемоглобина (HbF) или Bсl11а-регулируемой области сайленсинга HbF, представленной в любой из SEQ ID NO: 5-12 и 40-43, и расщепляет указанную нуклеотидную последовательность. Изобретения могут быть использованы для лечения талассемии, серповидно-клеточной болезни и других гемоглобинопатий. 6 н. и 5 з.п. ф-лы, 35 ил., 8 табл., 10 пр.
1. Полинуклеотид, кодирующий вариант хоминг-эндонуклеазы (ХЭ) I-OnuI,
где указанный полинуклеотид имеет последовательность SEQ ID NO: 34 с мутациями, соответствующими одной или более аминокислотным заменам, выбранными из группы, состоящей из L26, R28, R30, N32, S40, E42, G44, Q46, A70, S72, S78, K80, T82, F182, N184, I186, S190, K191, Q197, V199, S201, K225, K227, D236, V238 и T240,
где указанный вариант ХЭ I-OnuI связывается с нуклеотидной последовательностью области сайленсинга фетального гемоглобина (HbF) или Bсl11а-регулируемой области сайленсинга HbF, представленной в любой из SEQ ID NO: 5-12 и 40-43, и расщепляет указанную нуклеотидную последовательность.
2. Полинуклеотид по п.1, который дополнительно содержит полинуклеотид, кодирующий ДНК-связывающий домен TAL-эффектора (TALE) и/или домен нуклеазы TREX2.
3. Экспрессионный вектор, содержащий полинуклеотид по п.1 или 2.
4. Вектор по п.3, где вектор выбран из группы, состоящей из AAV6, модифицированного аденовирусного вектора, дефицитного по интеграции лентивирусного вектора (IDLV) и дефицитного по интеграции вектора на основе «пенистого» вируса (IDFV).
5. Полипептид эндонуклеазы, кодируемый полинуклеотидом по любому из пп.1-3.
6. Композиция для лечения гемоглобинопатии, включающая эффективное количество:
а) полинуклеотида по п.1 или 2;
b) вектора по п.3 или 4 или
с) полипептида по п.5.
7. Отредактированная по геному клетка, содержащая полинуклеотид по п.1 или 2, вектор по п.3 или 4 или полипептид эндонуклеазы по п.5, где указанная клетка не является эмбриональной клеткой человека, где геном клетки отредактирован (1) для дизрупции сайта связывания Bel11a в области сайленсинга HbF; (2) для нарушения функции гена Bсl11а или (3) для делеции всей области сайленсинга HbF.
8. Отредактированная по геному клетка по п.7, где клетка представляет собой:
а) стволовую клетку;
b) стволовую клетку, выбранную из группы, состоящей из гемопоэтической стволовой клетки (HSC), индуцированной плюрипотентной стволовой клетки (iPSC), эмбриональной стволовой клетки (ES) и эритроидной клетки-предшественника, или
с) стволовую клетку, выбранную из группы, состоящей из гемопоэтической стволовой клетки (HSC), индуцированной плюрипотентной стволовой клетки (iPSC) и эритроидной клетки-предшественника.
9. Отредактированная по геному стволовая клетка, где указанную отредактированную по геному стволовую клетку получают путем введения полинуклеотида, кодирующего вариант ХЭ I-OnuI,
где указанный полинуклеотид имеет последовательность SEQ ID NO: 34 с мутациями, соответствующими одной или более аминокислотным заменам, выбранными из группы, состоящей из L26, R28, R30, N32, S40, E42, G44, Q46, A70, S72, S78, K80, T82, F182, N184, I186, S190, K191, Q197, V199, S201, K225, K227, D236, V238 и T240,
где указанный вариант ХЭ I-OnuI специфично связывается с нуклеотидной последовательностью области сайленсинга HbF или Bсl11а-регулируемой области сайленсинга HbF, представленной в любой из SEQ ID NO: 5-12 и 40-43, и расщепляет указанную нуклеотидную последовательность,
где указанная стволовая клетка не является эмбриональной клеткой человека,
где геном указанной стволовой клетки отредактирован (1) для дизрупции сайта связывания Bcl11a в области сайленсинга HbF; (2) для нарушения функции гена Bсl11а или (3) для делеции всей области сайленсинга HbF.
10. Отредактированная по геному стволовая клетка по п.9, где вариант ХЭ I-OnuI слит с ДНК-связывающим доменом TAL-эффектора (TALE) и/или c доменом нуклеазы TREX2.
11. Отредактированная по геному стволовая клетка по п.9 или 10, где:
а) корректирующая матрица содержит нуклеотидную последовательность, которая позволяет вносить модификации в ключевые регуляторные или кодирующие последовательности, присутствующие в локусе гена глобина;
b) стволовая клетка выбрана из группы, состоящей из гемопоэтической стволовой клетки (HSC), индуцированной плюрипотентной стволовой клетки (iPSC), эмбриональной стволовой клетки (ES) и эритроидной клетки-предшественника, или
с) стволовая клетка выбрана из группы, состоящей из гемопоэтической стволовой клетки (HSC), индуцированной плюрипотентной стволовой клетки (iPSC) и эритроидной клетки-предшественника.
| Способ приготовления лака | 1924 |
|
SU2011A1 |
| В.С | |||
| БРОК-ВОЛЧАНСКАЯ, Хоминг-эндонуклеаза SegB бактериофага T4: биохимические свойства, Вестник биотехнологии и физико-химической биологии имени Ю.А | |||
| Овчинникова, Научно-практический журнал, 2007, т.3, No | |||
| Переносная печь для варки пищи и отопления в окопах, походных помещениях и т.п. | 1921 |
|
SU3A1 |

